Embed Size (px)
Citation preview

1
1 - INTRODUÇÃO
A região Amazônica abriga a maior bacia hidrográfica do mundo, com
aproximadamente 7 milhões de Km2, correspondendo a cerca de 20% da
vazão de todos os rios do planeta. Esta bacia é delimitada ao norte pelo maciço
das Guianas, ao sul pelo maciço do Brasil Central e a oeste pela cordilheira
dos Andes (Irion et al., 1997). Dentro desse complexo ecossistema se
destacam as várzeas, áreas situadas às margens de rios de água branca (Sioli,
1984) que ocupam cerca de 5% da Amazônia (Benchimol, 1996).
A várzea é um sistema complexo de lagos e planícies inundáveis,
remansos, canais, diques, ilhas que se estendem sobre o vale do Rio
Solimões/Amazonas e tributários de água branca, formados durante a época
glacial nos últimos 15 mil anos. A elevada carga de sedimentos do
Solimões/Amazonas, principalmente de origem andina e pré-andina, a forte
correnteza e a variação anual do nível da água, são os principais fenômenos
responsáveis pelo desenvolvimento da várzea (Irion et al., 1997). A extensão
dessas áreas alagadas depende diretamente do pulso de inundação, chegando
a ocupar um total de 300.000 km2, resultando em períodos bem definidos de
cheias e secas. Somente o complexo Solimões/Amazonas inunda
periodicamente cerca de 200.000 km2 (Junk 1993), representando o maior
sistema hidrológico do planeta. O alagamento sazonal do rio Solimões causa
uma elevação no nível da água de 10 a 12 metros todos os anos, atingindo na
Amazônia Central sua máxima inundação aproximadamente nos meses de
junho-julho e o período de vazão mínima de outubro-novembro (Ayres, 1995;
Piedade et al., 2000). O pulso de inundação é a principal força natural

2
responsável pela produtividade e pelas interações da biota nesse sistema
(Junk et al., 1989), além de produzir variações cíclicas nas condições físico-
químicas dos habitats.
Na época de enchente e cheia, o aumento da área alagada favorece a
exploração de vários habitats, aumentando a oferta de alimento para os peixes
(Santos, 1981; Almeida, 1984; Soares et al., 1986). Na vazante e seca, a retração
das águas limita o espaço, obrigando os peixes a migrarem da várzea para outros
locais como o canal principal dos rios ou para os lagos permanentes (Lowe-
McConnel, 1964; Goulding, 1980).
No período de alagação, habitats da várzea, tais como a água livre, os
bancos de macrófitas aquáticas e as florestas inundáveis, são extremamente
importantes para os peixes, pois fornecem abrigo, proteção, alimento e locais
de reprodução ao longo do ciclo hidrológico. Os bancos de macrófitas
aquáticas têm função na proteção de peixes de pequeno porte e alevinos, além
de fornecerem substratos para outros organismos (Sánchez & Araújo-Lima,
2001; Petry et al., 2003). As florestas inundáveis, por sua vez, produzem a
liteira, fonte de nutrientes, e os frutos e sementes que servem de alimento para
cerca de 200 espécies de peixes (Waldhoff & Maia, 2000), além de fornecer
para invertebrados que são ingeridos pelos peixes. Estes ambientes sustentam
populações de peixes de importância comercial, levando a pesca a ser
considerada uma das atividades econômicas de maior tradição na região.
A ictiofauna que coloniza os habitats dos lagos de várzea é abundante e
diversificada, sendo que a sua composição e distribuição estão relacionadas
com fatores abióticos como o nível da água, temperatura, oxigênio dissolvido,

3
pH e transparência. As variações sazonais do nível da água provocam
mudanças na estrutura das comunidades de peixes dos lagos e rios (Lowe-
McConnell 1964, Goulding 1980, Goulding et al. 1988; Merona & Bittencourt,
1993; Rodríguez & Lewis, 1997, Saint-Paul et al. 2000). As condições de
oxigênio e a temperatura levam os peixes a efetuarem deslocamentos entre a
vegetação flutuante, a floresta alagada e as áreas abertas (Junk et al., 1983;
Saint-Paul & Soares, 1987; Barthem, 1987). Rodríguez & Lewis (1997) e
Tejerina-Garro et al. (1998) relataram que a transparência e a profundidade da
água afetam a estrutura de comunidades de peixes em áreas alagadas no
Orinoco e no Pantanal, respectivamente. Finalmente, a cobertura de macrófitas
aquáticas e a abundância de piscívoros (Suárez et al., 2001) também
constituem importantes fatores que determinam a estrutura das comunidades
de peixes. Entretanto, muitas lacunas ainda persistem no conhecimento
integrado entre os fatores abióticos e a composição da ictiofauna ao longo do
ciclo hidrológico na Amazônia.
Estudos limnológicos realizados em corpos de água pertencentes a
sistemas rios-planícies de inundação têm demonstrado uma série de
modificações decorrentes da variação sazonal do nível da água, no potencial
hidrogeniônico, na condutividade elétrica, no material em suspensão, na
concentração de nutrientes e na comunidade biótica (da Silva, 1990; Hamilton
& Lewis Jr., 1990; Da Silva & Esteves, 1995; Hamilton et al., 1995). Além das
modificações temporais causadas pela variação do nível d’água, ambientes
aquáticos em planícies de inundação apresentam variações limnológicas
espaciais, devido à cobertura de macrófitas, vento, padrão de drenagem,

4
diversidade de ambientes terrestres no entorno, entre outros fatores, como já
constatado por Nogueira (1989), Panosso (1993) e Hamilton et al (1995). As
propriedades químicas e físicas da água são geralmente influenciadas pela
morfologia dos leitos dos rios e a estrutura das planícies inundáveis na bacia
amazônica, (Furch, 1984; Sioli, 1984), que, por sua vez, são conseqüências da
geologia dos substratos sobre os quais os rios passam (Putzer, 1984). Também
no Pantanal, Suárez et al., (2001) relatam que as características físico-
químicas, a cobertura de macrófitas aquáticas e a morfologia dos lagos
determinam a estrutura das comunidades de peixes.
Considerando que a estrutura de comunidades de peixes é regulada por
processos que ocorrem em escala temporal e espacial (Peres et al., 1995), o
presente trabalho propôs investigar a existência de relações entre a composição
da ictiofauna e alguns fatores abióticos da água ao longo do ciclo hidrológico e
entre habitats de lagos no complexo Lago Grande Manacapuru, Amazonas. Os
resultados apresentados neste estudo poderão subsidiar informações importantes
para a manutenção dos ambientes de várzea e, conseqüentemente, das
comunidades de peixes associadas a esse biótopo.

5
2- OBJETIVOS
2.1 – Geral:
Avaliar o efeito de fatores abióticos (características físico-químicos da
água) na distribuição das principais espécies de peixes em habitats de lago de
várzea, ao longo de um ciclo hidrológico.
2.2 – Específicos
√ Analisar as variações espaciais e temporais da ictiofauna em
diferentes habitats do lago na enchente, cheia, vazante e seca.
√ Avaliar as características físico-químicas da água de um lago de
várzea na enchente, cheia, vazante e seca;
√ Verificar a distribuição dos peixes em um lago de várzea conforme os
fatores abióticos.
√ Analisar as relações entre as características abióticas do lago de
várzea e a distribuição das espécies de peixes nesse ambiente.

6
3 – MATERIAL E MÉTODOS
3.1 - Área de estudo
O estudo foi realizado nos lagos Jaitêua (S 03˚13’901’’ e W 060˚44’326’’)
e São Lourenço (S 03˚17’555’’ e W 060˚43’759’’), ambos localizados no
complexo do lago grande de Manacapuru, Amazonas. O lago São Lourenço foi
utilizado como réplica do lago Jaitêua. (Fig. 1). A área total do município é de
aproximadamente 7.367,9 Km2 com uma população de 83.703 habitantes e
densidade demográfica de 11,3 hab./Km2 (IBGE, 2005).
O clima na região é do tipo equatorial úmido, com temperatura média
anual de 26,7 ˚C, com variações entre 23,3 e 31,4 ˚C e altitude de 60 metros
em relação o nível do mar. A umidade relativa do ar é de aproximadamente
80%, com média de precipitação anual de 2. 286 mm (CPRM, 2006).
Figura 1. Localização dos lagos Jaitêua e São Lourenço no complexo lago Grande de Manacapuru, Amazonas, Brasil.

7
3.1.1 – Caracterização do lago
O lago Jaitêua fica distante aproximadamente 14,8 km da sede do
município de Manacapuru (Global Mapper, 2006). Este ambiente apresenta
vegetação flutuante nos períodos de enchente e cheia com baixas ocorrências,
principalmente em suas margens. Suas florestas alagadas disponibilizam
muitos frutos e sementes para os peixes nos períodos de subida das águas.
O lago está submetido a uma flutuação média anual do nível do rio em
torno de 10 m. A enchente no rio começa em novembro e atinge a cota máxima
em julho, secando em outubro. A água começa a entrar nos lagos no final de
dezembro. Durante o período de seca, o lago permanece conectado com os
rios Manacapuru e Solimões.
3.2 – Coleta da ictiofauna
Os peixes foram coletados mensalmente na enchente (final de
dezembro, janeiro, fevereiro, março, abril e maio de 2006/2007), cheia (junho e
julho 2007), vazante (agosto, setembro e outubro de 2007) e seca (novembro e
início de dezembro de 2007). Foram amostrados três habitats: água aberta,
floresta alagada e bancos de macrófitas aquáticas. Na água aberta e floresta
alagada foram utilizadas baterias de malhadeiras, com tamanho de 25 x 2 m,
com malhas variando de 30 a 130 mm entre nós opostos, expostas durante
períodos de 24 horas e com despescas a cada 6 horas. Nos bancos de
macrófitas aquáticas os peixes foram capturados com redinha de cerco
medindo 20x5m com malhas de 10 mm entre nós opostos. Todos os pontos

8
amostrais foram georeferenciados com uso de um GPS Etrex (GARMIM
VISTA).
3.3 – Métodos de análises dos parâmetros físicos e físico-químicos do
ambiente aquático
Os valores foram obtidos concomitantes às coletas dos peixes em cada
ponto de coleta. Os fatores físicos e físico-químicos da água obtidos foram:
profundidade, transparência da água, condutividade elétrica, oxigênio
dissolvido, % de saturação do oxigênio, temperatura e potencial hidrogeniônico
(pH). Para os períodos de nível mais alto da água, foi utilizada uma garrafa
coletora de água (Van Dorn), para a obtenção de amostras de águas dos
pontos mais profundos do lago. A profundidade foi obtida com a utilização de
ecosonda ou ecobatímetro da marca Humminbird (Piranha Max 10 Fishfinder).
Para a transparência da água utilizou-se um uso de disco de Secchi de 30 cm
de diâmetro. A condutividade elétrica, o oxigênio dissolvido, a temperatura e o
pH foram medidos “in situ”, no momento das coletas de peixes, com aparelhos
do modelo YSI 30 e 55 e ORION 3 STAR/Thermo.
Os dados fluviométricos foram obtidos a partir de informações existentes
no banco de dados da CPRM Manaus.
3.5 – Triagem e identificação dos peixes
Durante as coletas os peixes foram acondicionados em sacos plásticos,
devidamente etiquetados com o número de malha, local e horário de captura.
Os exemplares foram identificados em campo com o auxílio de chaves de

9
identificação específicas (Gery, 1977; Santos, 1984; Ferreira et al., 1998), até o
menor nível taxonômico possível. Espécimes que não foram identificados em
campo foram fixados em solução de formalina a 10%, etiquetados e levados
para posterior identificação no laboratório de Bioecologia de peixes do INPA.
4 - Análises dos dados
4.1 – Atributos ecológicos da estrutura das assembléias de peixes
Nas Estimativas de riqueza da ictiofauna do lago foram calculadas com o
uso três métodos:
a) Índice de Jackknife (Krebs, 1989), que estima a riqueza absoluta
somando a riqueza observada através de um parâmetro calculado a partir do
número de espécies raras e do número de amostras. Além disso, corrige os
vícios nas estimativas, permitindo a comparação entre locais por meio do erro
padrão (Magurran, 1988), utilizando a seguinte fórmula:
Ŝ = S + (n – 1/ n) k
Onde:
Ŝ = estimativa da riqueza;
S = número total de espécies observadas na amostra “n”;
n = número total de amostras;
k = número total de espécies únicas, ou seja, coletadas uma única vez.
b) Curvas de rarefação (Krebs, 1989), a vantagem deste método é sua
independência do tamanho da amostra, além de permitir a comparação entre

10
diferentes amostras, no caso, período e ambiente, pois leva em consideração
dados sobre abundância específica e não o número absoluto de espécies.
Onde: E (Sn) = número esperado de
espécies.
S = número total de espécies na amostra.
N = número total de exemplares.
Ni = número total de exemplares da espécie i.
n = número de exemplares na amostra (número escolhido para
padronização).
c) Índice de Margalef
Para estimar a riqueza específica de cada lago foi utilizado o índice de
Margalef (Ludwig & Reynolds, 1988), que analisa a relação entre o número
total de espécies e o número total de exemplares observados, na fórmula a
seguir:
Onde: S = Número total de espécies na amostra.
n = Número total de exemplares na amostra.
Além dos valores de riqueza e abundância, foram calculados os valores de
diversidade, dominância, equitabilidade e frequência de ocorrência das
( )
−
−=Ε ∑=
n
N
n
NN
S
i
S
i
n1
1
( )n
SR
ln
11
−=

11
espécies para todos os habitats amostrados. A diversidade foi calculada com
uso do índice de Shannon-Wienner (Krebs, 1989):
H’ = - ∑ (pi) (ln pi)
Onde:
pi representa o número de exemplares da i-ésima espécie (Magurran,
1988). Dado que em uma amostra o valor de pi é desconhecido, estima-se o
mesmo pela seguinte fórmula:
pi = (ni / N)
Sendo:
ni = número de exemplares pertencentes à espécie “i”;
N = número total de exemplares coletados na amostra.
Para o cálculo de dominância foi utilizado o índice proposto por Berger-
Parker, sendo estimado pela seguinte fórmula:
d= n max / N
Onde:
n max = número de exemplares da espécie mais abundante;
N = número de exemplares da amostra.
Como o índice de Berger-Parker é um índice de dominância, foi usado
seu inverso 1/d, para indicar a diversidade.

12
Para o cálculo da abundância relativa das espécies (equitabilidade) foi
utilizado o método proposto por Pielou (1969 apud Magurran, 1988), que
propõe a relação entre a diversidade observada e a diversidade máxima. Esta
relação é representada pela seguinte fórmula:
E = H’ / ln S
Onde:
E = equitabilidade
H’ = índice de diversidade de Shannon-Wiener;
S = número de espécies presentes na amostra
4.2 - Abundância relativa das assembléias ictíicas
As medidas de abundância ou densidade relativa são consideradas
importantes ferramentas de avaliação da estrutura de assembléias. Neste
estudo o cálculo de abundância relativa foi utilizado para verificar a captura
obtida em relação a uma dada unidade de esforço (área do apetrecho), através
da CPUEn, que é definida como o somatório do número de peixes/m2/h, e
CPUEb, definido como o somatório do peso em gramas/m²/h. Ambos foram
aplicados aos dados obtidos com malhadeiras na água aberta e floresta
alagada, na enchente, cheia, vazante e seca. Este procedimento possibilita
comparações quantitativas entre espécies e locais amostrados, sendo
expresso pela seguinte equação:
Tempo de pesca Área
(“N” ou peso) CapturaTotal CPUE
X =

13
Onde:
CPUE = Captura por unidade de esforço.
N = nº da biomassa de peixes capturados para um determinado tamanho
de malha;
n = tamanhos de malha empregados (20, 30, 40, 50, 60, 70, 80, 90, 100,
110, 120 e 130);
Área = tamanho da(s) malhadeiras (m2)
Tempo de pesca = nº de horas de pesca para um dado local e período
do ciclo hidrológico
Os resultados obtidos foram analisados através de análises de variância
(ANOVA) two-way (p ≥ 0,05), aplicadas somente em dois períodos do ciclo
hidrológico (enchente e cheia) nos habitats de água aberta e floresta alagada
somente. Os períodos de vazante e seca não fizeram parte desta análise por
não apresentarem os dois habitats em seus ciclos completos.
4.3 - Variações na composição das assembléias de peixes em
relação às características ambientais do lago e características físico-
químicas da água.
Para verificar a existência de relações entre as variáveis que estruturam
as assembléias locais de peixes (riqueza de espécies, abundância relativa,
diversidade de Shannon-Wiener, dominância, equitabilidade e composição de
espécies) e variáveis ambientais (condutividade elétrica, pH, temperatura,

14
oxigênio, profundidade e transparência), foram empregadas técnicas de análise
estatística multivariada (Legendre & Legendre, 1997).
Para avaliar as relações de similaridade entre as assembléias de peixes
nos diferentes habitats estudados, foi empregado o método de Escalonamento
Multidimensional (MDS). A matriz de similaridade foi calculada a partir do índice
de Jaccard, que se baseia na presença-ausência dos habitats. Os valores do
índice variam entre O (equivalente a dissimilar) e 1 (similaridade completa)
(Krebs, 1989). O índice é expresso pela seguinte equação:
J = a / a + b + c
Onde:
J = Similaridade de Jaccard;
a = espécies comuns entre os dois ambientes;
b = espécies presentes em “a” e ausentes em “b”;
c = espécies presentes em “b” e ausentes em “a”.
Outra análise de escalonamento multidimensional (MDS) levou em
consideração os valores encontrados através do índice de Bray-Curtis que se
baseia na dissimilaridade da abundância de exemplares entre os habitats
(Krebs, 1989):
B= Σ │XiJ- XIK│/ Σ (XiJ- XIK)
Onde:
B= medida de dissimilaridade de Bray-Curtis
XiJ, XIK= Número de exemplares da espécie “i” em cada amostra
n= número de espécies nas amostras.

15
Uma análise de correlação de Pearson foi realizada entre os parâmetros
físicos e físico-químicos da água e atributos das assembléias de peixes, a fim
de verificar se eram correlacionadas positivamente ou negativamente. Como foi
encontrado um alto nível de correlações e para evitar problemas de
colinearidade nas análises, as variáveis foram reordenadas por meio de uma
Análise de Componentes Principais (PCA). A PCA foi empregada para ordenar
os habitats em função das características físico-químicas da água
(temperatura, pH, oxigênio dissolvido, porcentagem de saturação de oxigênio e
condutividade) e uma característica estrutural do lago (profundidade e
transparência). Posteriormente, para verificar as relações entre a composição
de espécies de peixes (representada pelos eixos do MDS) com os parâmetros
limnológicos e estrutural (representadas pelos eixos da PCA), foram feitas
regressões lineares utilizando os habitats do lago (MDS x PCA).
4.4 – Relação entre a estrutura das assembléias ictíicas e
parâmetros limnológicos.
As relações entre a composição de espécies das assembléias de peixes
e os parâmetros limnológicos dos habitats estudados, foram analisados por
meio de regressões lineares simples (Zar, 1999).
Para a identificação de quais parâmetros são responsáveis pelo
agrupamento das espécies foram realizadas análises de correlação, com o
intuito de se estimar quais variáveis respondem pelos agrupamentos
observados.

16
Foram aplicados testes de regressão múltipla, tendo como variável
dependente a riqueza de espécies e independentes os fatores abióticos, para
se definir quais variáveis têm influência na riqueza de espécies.
As análises utilizando-se ANOVA two-way foram realizadas
considerando menores as informações de enchente e de cheia, uma vez que
os habitats de macrófitas aquáticas e floresta alagada não ocorrem na seca,
somente no inicio da vazante.
Todas as análises estatísticas foram realizadas com a utilização dos
pacotes Statistica 6.0, SigmaStat 3.5, Past e .

17
4 - Resultados
4.1 – Fatores físico-químicos e estruturais medidos na água aberta,
floresta alagada e nos bancos de macrófitas aquáticas.
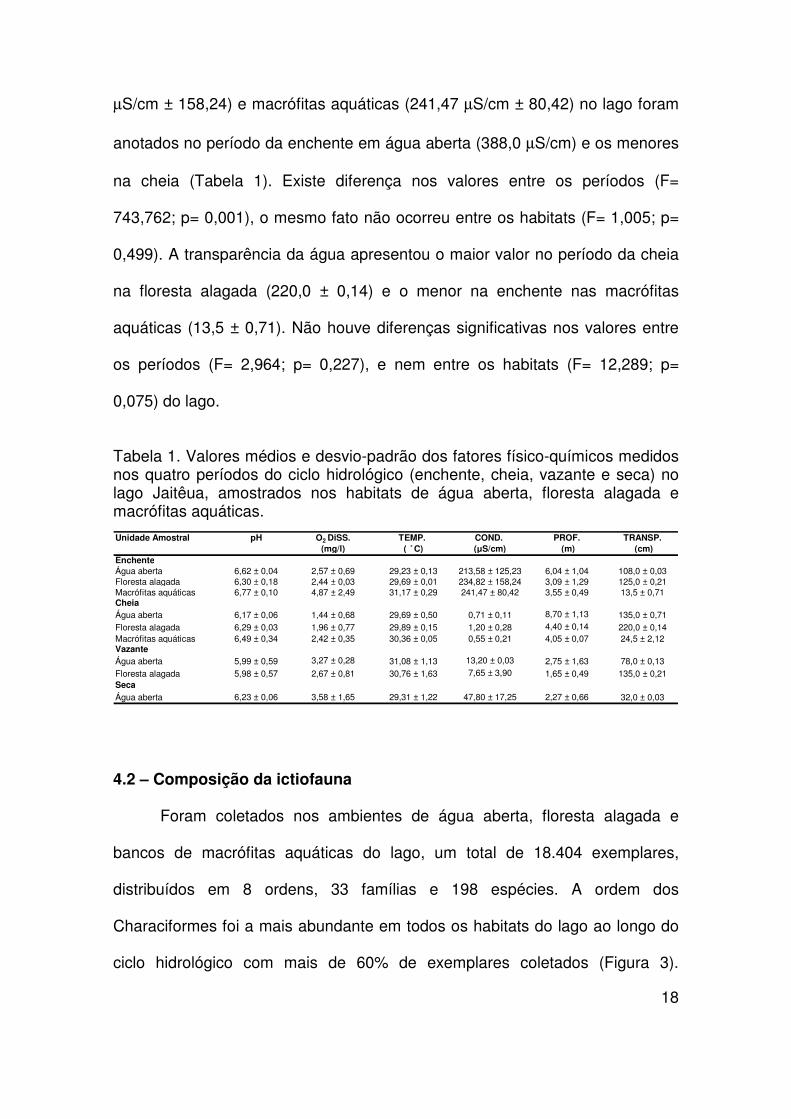
Os parâmetros limnológicos observados apresentaram valores
diferenciados entre os habitats amostrados no lago. Os valores de pH anotados
nos bancos de macrófitas aquáticas na enchente e cheia (6,8 e 6,5), foram
mais elevados em relação ao restante dos habitats durante todo o ciclo
hidrológico (Tabela 1). Não existe diferenças significativas nos valores de pH
entre os períodos (F= 3,708; p= 0,194) e habitats do lago (F= 2,403; p= 0,294).
As concentrações de oxigênio apresentaram valores muito baixos na
floresta alagada (1,96 mg/l) na cheia e valores mais elevados nas macrófitas
(4,87 mg/l) no mesmo período (Tabela 1). Não houve diferenças significativas
nos valores de oxigênio entre os períodos (F= 5,453; p= 0,145), assim como
entre habitats (F= 3,186 ; p= 0,239).
A profundidade da água do lago apresentou seu maior valor na cheia
(8,70 m) e os menores na vazante e seca (1,65 e 2,3 m), na floresta alagada e
água aberta. Não houve diferenças significativas nos valores profundidade
entre os períodos (F= 5,594; p= 0,142), e nem entre os habitats (F= 14,495; p=
0,065). A temperatura da água apresentou maior valor nas macrófitas
aquáticas na enchente (31,2 oC) e menor na água aberta (31,1 o C) na vazante.
Não existem diferenças significativas nos valores entre os períodos (F= 0,0167;
p= 0,909), assim como entre os habitats (F= 4,092; p= 0,196) do lago.
Os maiores valores de condutividade elétrica anotados no lago foram na
enchente na água aberta (213,58 µS/cm ± 125,23), floresta alagada (234,82
18
µS/cm ± 158,24) e macrófitas aquáticas (241,47 µS/cm ± 80,42) no lago foram
anotados no período da enchente em água aberta (388,0 µS/cm) e os menores
na cheia (Tabela 1). Existe diferença nos valores entre os períodos (F=
743,762; p= 0,001), o mesmo fato não ocorreu entre os habitats (F= 1,005; p=
0,499). A transparência da água apresentou o maior valor no período da cheia
na floresta alagada (220,0 ± 0,14) e o menor na enchente nas macrófitas
aquáticas (13,5 ± 0,71). Não houve diferenças significativas nos valores entre
os períodos (F= 2,964; p= 0,227), e nem entre os habitats (F= 12,289; p=
0,075) do lago.
Tabela 1. Valores médios e desvio-padrão dos fatores físico-químicos medidos nos quatro períodos do ciclo hidrológico (enchente, cheia, vazante e seca) no lago Jaitêua, amostrados nos habitats de água aberta, floresta alagada e macrófitas aquáticas.
Unidade Amostral pH O2 DiSS. TEMP. COND. PROF. TRANSP.(mg/l) ( ̊ C) (µS/cm) (m) (cm)
EnchenteÁgua aberta 6,62 ± 0,04 2,57 ± 0,69 29,23 ± 0,13 213,58 ± 125,23 6,04 ± 1,04 108,0 ± 0,03Floresta alagada 6,30 ± 0,18 2,44 ± 0,03 29,69 ± 0,01 234,82 ± 158,24 3,09 ± 1,29 125,0 ± 0,21Macrófitas aquáticas 6,77 ± 0,10 4,87 ± 2,49 31,17 ± 0,29 241,47 ± 80,42 3,55 ± 0,49 13,5 ± 0,71CheiaÁgua aberta 6,17 ± 0,06 1,44 ± 0,68 29,69 ± 0,50 0,71 ± 0,11 8,70 ± 1,13 135,0 ± 0,71Floresta alagada 6,29 ± 0,03 1,96 ± 0,77 29,89 ± 0,15 1,20 ± 0,28 4,40 ± 0,14 220,0 ± 0,14Macrófitas aquáticas 6,49 ± 0,34 2,42 ± 0,35 30,36 ± 0,05 0,55 ± 0,21 4,05 ± 0,07 24,5 ± 2,12VazanteÁgua aberta 5,99 ± 0,59 3,27 ± 0,28 31,08 ± 1,13 13,20 ± 0,03 2,75 ± 1,63 78,0 ± 0,13Floresta alagada 5,98 ± 0,57 2,67 ± 0,81 30,76 ± 1,63 7,65 ± 3,90 1,65 ± 0,49 135,0 ± 0,21SecaÁgua aberta 6,23 ± 0,06 3,58 ± 1,65 29,31 ± 1,22 47,80 ± 17,25 2,27 ± 0,66 32,0 ± 0,03
4.2 – Composição da ictiofauna
Foram coletados nos ambientes de água aberta, floresta alagada e
bancos de macrófitas aquáticas do lago, um total de 18.404 exemplares,
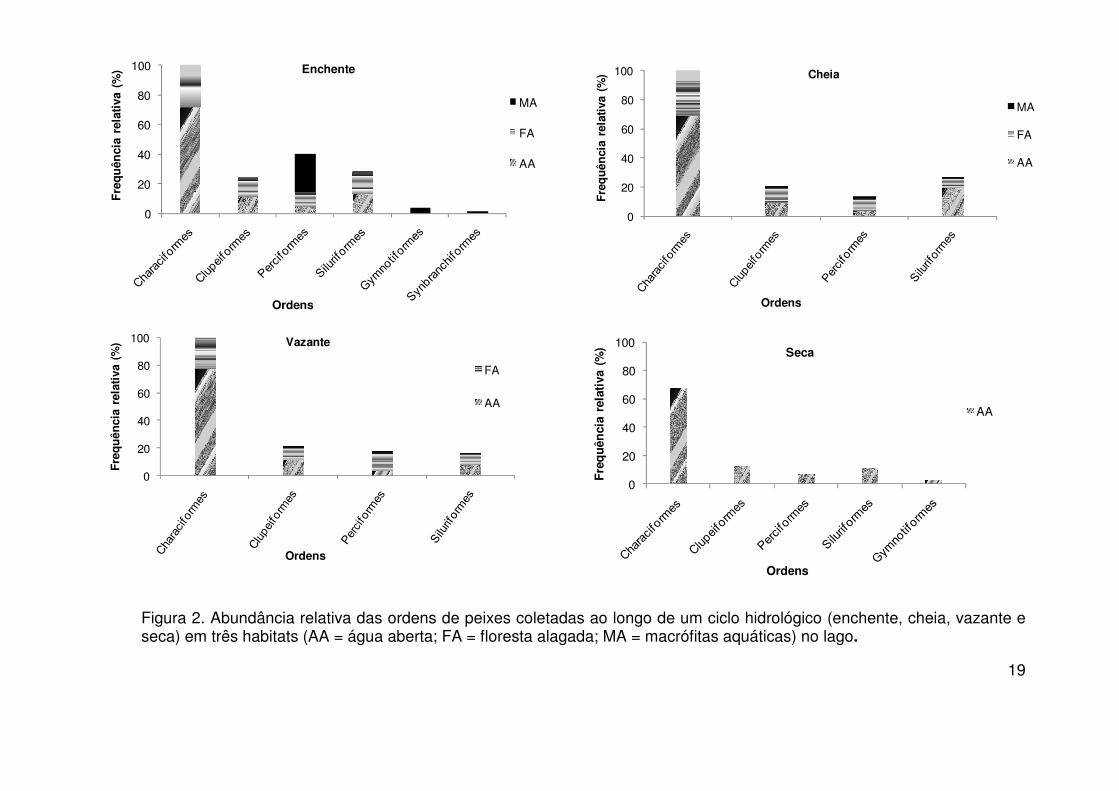
distribuídos em 8 ordens, 33 famílias e 198 espécies. A ordem dos
Characiformes foi a mais abundante em todos os habitats do lago ao longo do
ciclo hidrológico com mais de 60% de exemplares coletados (Figura 3).
19
Figura 2. Abundância relativa das ordens de peixes coletadas ao longo de um ciclo hidrológico (enchente, cheia, vazante e seca) em três habitats (AA = água aberta; FA = floresta alagada; MA = macrófitas aquáticas) no lago.
0
20
40
60
80
100
Fre
qu
ênci
a re
lati
va (
%)
Ordens
Cheia
MA
FA
AA
0
20
40
60
80
100
Fre
qu
ênci
a re
lati
va (
%)
Ordens
Enchente
MA
FA
AA
0
20
40
60
80
100
Fre
qu
ênci
a re
lati
va (
%)
Ordens
Vazante
FA
AA
0
20
40
60
80
100
Fre
qu
ênci
a re
lati
va (
%)
Ordens
Seca
AA
20
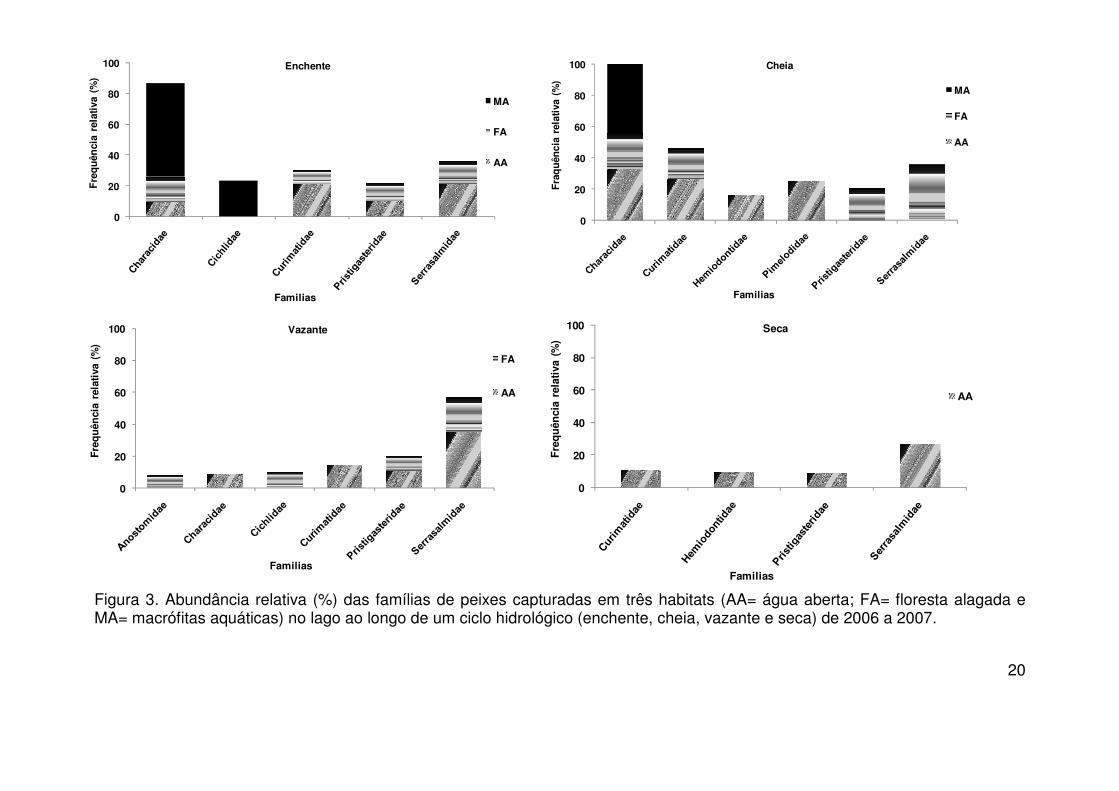
Figura 3. Abundância relativa (%) das famílias de peixes capturadas em três habitats (AA= água aberta; FA= floresta alagada e MA= macrófitas aquáticas) no lago ao longo de um ciclo hidrológico (enchente, cheia, vazante e seca) de 2006 a 2007.
0
20
40
60
80
100
Fre
qu
ênci
a re
lati
va (
%)
Familias
Enchente
MA
FA
AA
0
20
40
60
80
100
Fra
qu
ênci
a re
lati
va (
%)
Familias
Cheia
MA
FA
AA
0
20
40
60
80
100
Fre
qu
ênci
a re
lati
va (
%)
Familias
Vazante
FA
AA
0
20
40
60
80
100
Fre
qu
ênci
a re
lati
va (
%)
Familias
Seca
AA

21
4.3 - Estrutura da comunidade de peixes no lago Jaitêua: riqueza,
abundância e distribuição da ictiofauna.
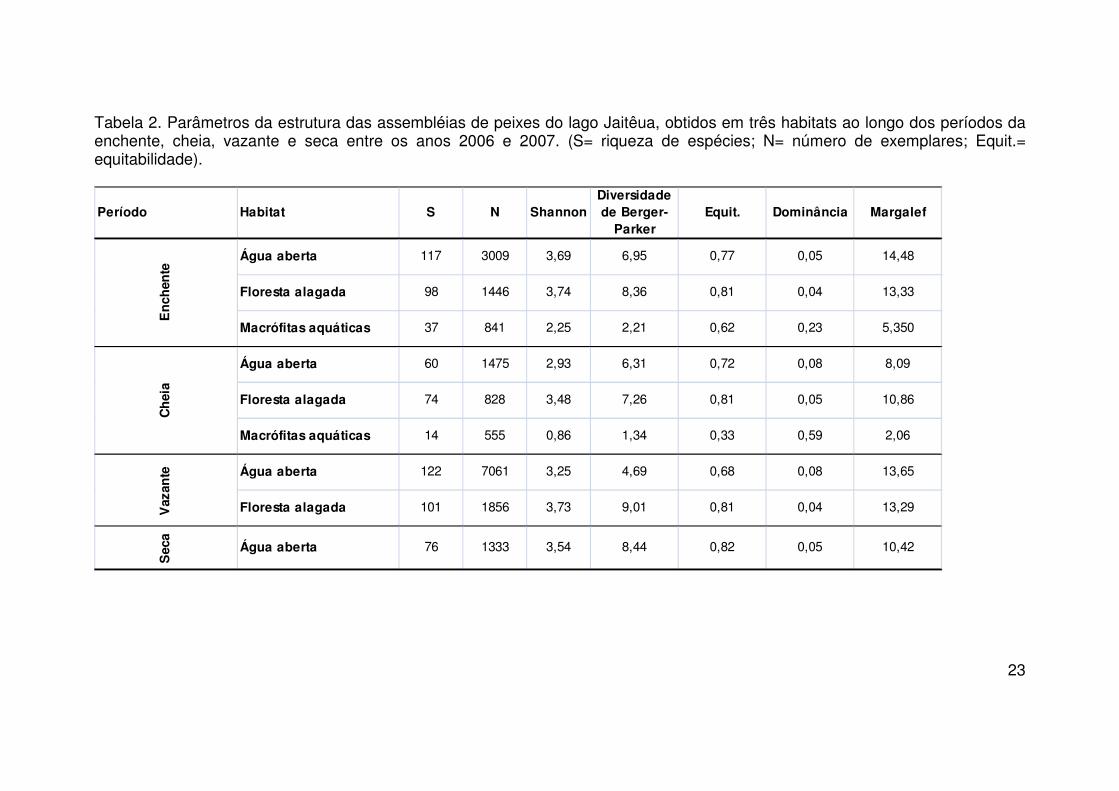
Os valores de diversidade de Shannon-Wiener obtidos no lago não
apresentaram diferenças significativas entre os períodos do ciclo hidrológico
(F= 1,735; p= 0,259). No entanto os valores entre habitats apresentaram
diferenças significativas entre (F= 5,907; p= 0,038 ). O teste Tukey a 5%,
mostrou diferenças significativas nos valores entre a água aberta e macrófitas
aquáticas (p= 0,038) do lago.
A estimativa de diversidade obtida a partir do índice de Berger-Parker
apresentou seu maior valor na floresta alagada (9,01) no período da vazante e
o menor na cheia (1,34) nas macrófitas aquáticas.
Analisando os valores de equitabilidade não foram observadas
diferenças significativas entre os períodos do ciclo hidrológico (F= 1,697; p=
0,266) e nem entre os habitats (F= 3,979; p= 0,079). Esses valores foram
maiores na água aberta (E= 0,82) na seca e floresta alagada (E= 0,81) na
enchente (Tabela 4).
A dominância apresentou seus maiores valores no habitat de macrófitas
aquáticas na cheia e enchente (0,50 e 0,23). Não foram observadas diferenças
significativas dos valores entre períodos (F= 1,283; p= 0,363) e habitats (F=
1,415; p= 0,314), respectivamente.
Os maiores valores de riqueza de espécies foram obtidos na vazante
(S= 122) e enchente (S= 117). E os menores nas macrófitas aquáticas na
enchente (S=37) e cheia (S= 14). Não foram observadas diferenças
significativas nos valores entre os períodos do ciclo hidrológico (F= 3,049; p=

22
0,114), somente entre os habitats (F= 10,036; p= 0,012). O teste post-hoc
(Tukey 5%) mostrou que a diferença foi observada entre a água aberta e
macrófitas aquáticas (p= 0,011).
117
60
122
76
98
74
101
37
14
012345678910
0
20
40
60
80
100
120
140
160
E C V S
Pro
fun
did
ade d
o lag
o (m
)
Riq
uez
a d
e es
péc
ies
Meses
AA FA MA Lago
Figura 4. Riqueza de espécies de peixes nos habitats estudados no lago ao longo do ciclo hidrológico (2006 a 2007).
23
Tabela 2. Parâmetros da estrutura das assembléias de peixes do lago Jaitêua, obtidos em três habitats ao longo dos períodos da enchente, cheia, vazante e seca entre os anos 2006 e 2007. (S= riqueza de espécies; N= número de exemplares; Equit.= equitabilidade).
Período Habitat S N ShannonDiversidade de Berger-
ParkerEquit. Dominância Margalef
Água aberta 117 3009 3,69 6,95 0,77 0,05 14,48
Floresta alagada 98 1446 3,74 8,36 0,81 0,04 13,33
Macrófitas aquáticas 37 841 2,25 2,21 0,62 0,23 5,350
Água aberta 60 1475 2,93 6,31 0,72 0,08 8,09
Floresta alagada 74 828 3,48 7,26 0,81 0,05 10,86
Macrófitas aquáticas 14 555 0,86 1,34 0,33 0,59 2,06
Água aberta 122 7061 3,25 4,69 0,68 0,08 13,65
Floresta alagada 101 1856 3,73 9,01 0,81 0,04 13,29
Sec
a
Água aberta 76 1333 3,54 8,44 0,82 0,05 10,42
Vaz
ante
En
chen
teC
hei
a

24
4.3.1 – Abundância relativa das assembléias icticas
As análises de variância mostraram que a captura por unidade de
esforço CPUEn, expressa em número de exemplares, não apresentou
diferenças significativas (F = 1,657; p= 0,3769) entre os períodos e nem entre
os habitats (F= 2,274; p= 0,271) do lago. Em relação as análises da CPUE em
biomassa, também não houve diferenças significativas nos valores amostrados
para os períodos (F= 2,786; p= 0,264) e habitats (F= 4,862; p= 0,158).
As análises de captura por unidade de esforço realizadas ao longo do
ciclo hidrológico mostram haver um aumento em mais de duas vezes no
número de exemplares e biomassa coletados na água aberta no período da
vazante em relação ao restante dos períodos do ciclo (Tabela 5).
Dentre os habitats amostrados no lago, observou-se que 16 espécies
apresentaram ocorrência única na enchente. O menor valor de ocorrências
únicas se deu no período da cheia com um total de 8 espécies amostradas.
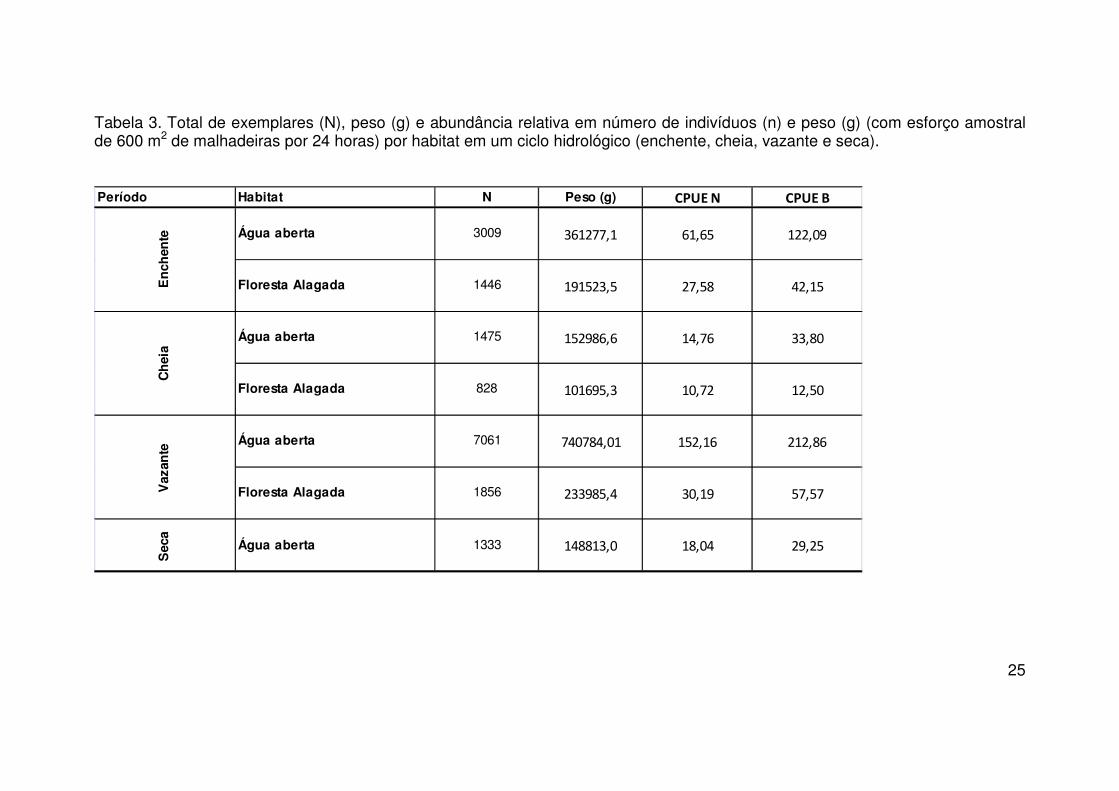
25
Tabela 3. Total de exemplares (N), peso (g) e abundância relativa em número de indivíduos (n) e peso (g) (com esforço amostral de 600 m2 de malhadeiras por 24 horas) por habitat em um ciclo hidrológico (enchente, cheia, vazante e seca).
Período Habitat N Peso (g) CPUE N CPUE B
Água aberta 3009 361277,1 61,65 122,09
Floresta Alagada 1446 191523,5 27,58 42,15
Água aberta 1475 152986,6 14,76 33,80
Floresta Alagada 828 101695,3 10,72 12,50
Água aberta 7061 740784,01 152,16 212,86
Floresta Alagada 1856 233985,4 30,19 57,57
Sec
a
Água aberta 1333 148813,0 18,04 29,25
En
chen
teC
hei
aV
azan
te

26
Figura 5. Frequência relativa das 10 espécies de peixes mais abundantes no período de enchente na água aberta (A), floresta alagada (B) e macrófitas aquáticas (C) do lago.
0 10 20 30 40 50
P. latior
P. flavipinnis
P. nattereri
T. albus
H. marginatus
S. spilopleura
M. duriventre
A. falcirostris
A. nuchalis
A. elongatus
Frequência relativa (%)
0 10 20 30 40 50
P. flavipinnis
T. angulatus
C. macropomum
T. auritus
T. galeatus
Hemiodus sp.
H. marginatus
H. microlepis
T. albus
A. falcirostris
Frequência relativa (%)
0 10 20 30 40 50
Hemigramus sp.
C. monoculus
M. duriventre
M. festivum
H. malabaricus
C. amazonarum
A. ocellatus
B. brevirostris
Serrasalmus sp.
R. myersii
Frequência relativa (%)
A B
C
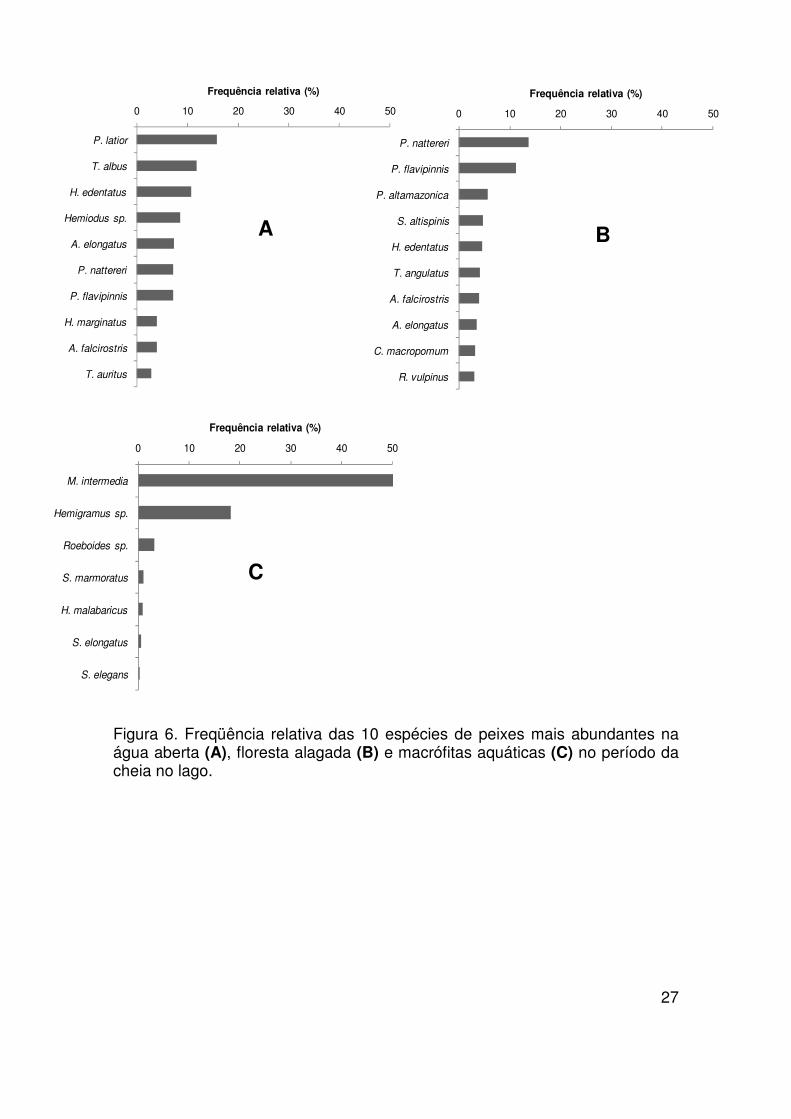
27
Figura 6. Freqüência relativa das 10 espécies de peixes mais abundantes na água aberta (A), floresta alagada (B) e macrófitas aquáticas (C) no período da cheia no lago.
0 10 20 30 40 50
P. latior
T. albus
H. edentatus
Hemiodus sp.
A. elongatus
P. nattereri
P. flavipinnis
H. marginatus
A. falcirostris
T. auritus
Frequência relativa (%)
0 10 20 30 40 50
P. nattereri
P. flavipinnis
P. altamazonica
S. altispinis
H. edentatus
T. angulatus
A. falcirostris
A. elongatus
C. macropomum
R. vulpinus
Frequência relativa (%)
0 10 20 30 40 50
M. intermedia
Hemigramus sp.
Roeboides sp.
S. marmoratus
H. malabaricus
S. elongatus
S. elegans
Frequência relativa (%)
A B
C

28
Figura 7. Freqüência relativa das 10 espécies de peixes mais abundantes na água aberta (A), floresta alagada (B) na vazante e água aberta (C) na seca no lago.
0 10 20 30 40 50
P. nattereri
P. flavipinnis
P. latior
S. spilopleura
A. falcirostris
S. insignis
A. elongatus
P. rutiloides
T. albus
H. edentatus
Frequência relativa (%)
0 10 20 30 40 50
P. nattereri
P. flavipinnis
A. falcirostris
S. insignis
Hemiodus sp.
P. squamosissimus
T. angulatus
A. elongatus
T. auritus
H. efasciatus
Frequência relativa (%)
0 10 20 30 40 50
P. nattereri
S. spilopleura
A. falcirostris
P. flavipinnis
P. latior
H. microlepis
P. squamosissimus
Hemiodus sp.
L. batesii
S. fasciatum
Frequência relativa (%)
A B
C

29
4.4 – Variação na composição da ictiofauna nos habitats do lago.
Os resultados obtidos através da análise de ordenação (MDS) realizada
com base nos dados de abundância e presença-ausência das espécies,
explicam grande parte da variação dos dados originais da composição de
espécies das assembléias, como evidenciado pelos valores de configuração do
Stress de Kruskal (0,092 e 0,166), respectivamente. As análises dos dois
primeiros eixos da MDS, com os dados quantitativos de abundância (índice de
Bray Curtis), obtidas a partir da matriz de Bray-Curtis, mostram pouca
separação (Fig. 8A) entre os períodos do ciclo hidrológico amostrados para o
lago. O habitat de macrófitas aquáticas na enchente e cheia apresenta
diferenças mais pronunciadas em relação aos demais habitats do lago em
termos de abundância por espécie. No entanto, as proximidades entre os
pontos de floresta alagada e água aberta na enchente e cheia mostram maior
homogeneidade na composição e abundância de espécies.
As análises de MDS utilizando os dados de presença-ausência e a
correspondente matriz de similaridade do índice de Jaccard (Tab. 7) mostram
uma clara separação (Fig. 8B) entre os períodos do ciclo e diferenças menores
entre os habitats do lago na enchente. Os habitats de floresta alagada e água
aberta na enchente (J = 56,9%) e cheia (J = 54,0%) foram mais semelhantes
entre si. O habitat de macrófitas aquáticas na enchente e cheia apresentou
menor similaridade com a água aberta (J = 4,8%) e floresta alagada (J = 6,7%).

30
Figura 8. Ordenação por meio de Escalonamento multidimensional (MDS) dos dados de número de indivíduos (índice de Bray Curtis) (A) e presença-ausência de espécies (índice de Jaccard) (B) nos habitats de água aberta, floresta alagada e macrófitas aquáticas do lago Jaitêua na enchente e cheia.
Água abertaEnchente
Floresta alagada
Enchente
Água abertaCheia
Floresta alagadaCheia
Macróf itasEnchente Macróf itas
Cheia
-0,8
-0,6
-0,4
-0,2
0
0,2
0,4
0,6
0,8
-1 -0,8 -0,6 -0,4 -0,2 0 0,2 0,4 0,6 0,8 1
Dim
ensã
o 2
Dimensão 1
(A) Configuração (Stress de Kruskal (1) = 0,092)
Água abertaEnchente
Floresta alagada
EnchenteÁgua aberta
Cheia
Floresta alagadaCheia
Macróf itasEnchente
Macróf itasCheia
-0,8
-0,6
-0,4
-0,2
0
0,2
0,4
0,6
0,8
-0,8 -0,6 -0,4 -0,2 0 0,2 0,4 0,6 0,8 1
Dim
ensã
o 2
Dimensão 1
(A) Configuração (Stress de Kruskal (1) = 0,166)
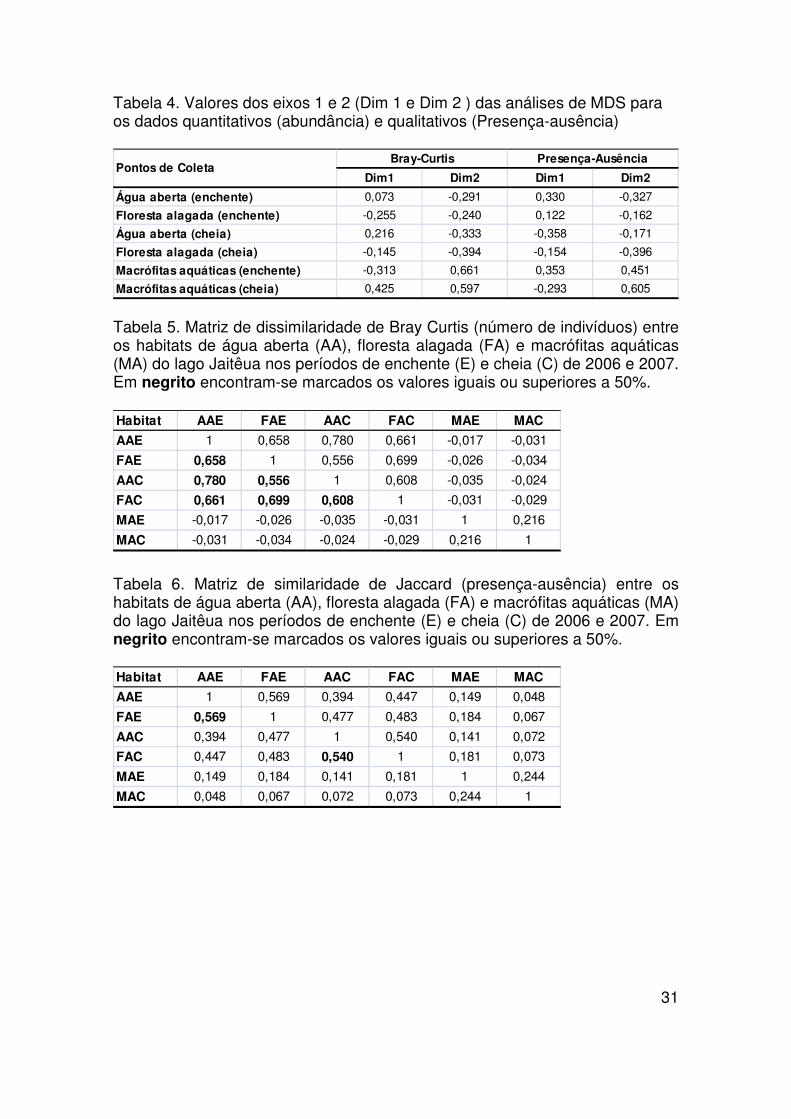
31
Tabela 4. Valores dos eixos 1 e 2 (Dim 1 e Dim 2 ) das análises de MDS para os dados quantitativos (abundância) e qualitativos (Presença-ausência)
Tabela 5. Matriz de dissimilaridade de Bray Curtis (número de indivíduos) entre os habitats de água aberta (AA), floresta alagada (FA) e macrófitas aquáticas (MA) do lago Jaitêua nos períodos de enchente (E) e cheia (C) de 2006 e 2007. Em negrito encontram-se marcados os valores iguais ou superiores a 50%.
Tabela 6. Matriz de similaridade de Jaccard (presença-ausência) entre os habitats de água aberta (AA), floresta alagada (FA) e macrófitas aquáticas (MA) do lago Jaitêua nos períodos de enchente (E) e cheia (C) de 2006 e 2007. Em negrito encontram-se marcados os valores iguais ou superiores a 50%.
Habitat AAE FAE AAC FAC MAE MAC
AAE 1 0,658 0,780 0,661 -0,017 -0,031
FAE 0,658 1 0,556 0,699 -0,026 -0,034
AAC 0,780 0,556 1 0,608 -0,035 -0,024
FAC 0,661 0,699 0,608 1 -0,031 -0,029
MAE -0,017 -0,026 -0,035 -0,031 1 0,216
MAC -0,031 -0,034 -0,024 -0,029 0,216 1
Habitat AAE FAE AAC FAC MAE MAC
AAE 1 0,569 0,394 0,447 0,149 0,048
FAE 0,569 1 0,477 0,483 0,184 0,067
AAC 0,394 0,477 1 0,540 0,141 0,072
FAC 0,447 0,483 0,540 1 0,181 0,073
MAE 0,149 0,184 0,141 0,181 1 0,244
MAC 0,048 0,067 0,072 0,073 0,244 1
Dim1 Dim2 Dim1 Dim2
Água aberta (enchente) 0,073 -0,291 0,330 -0,327
Floresta alagada (enchente) -0,255 -0,240 0,122 -0,162
Água aberta (cheia) 0,216 -0,333 -0,358 -0,171
Floresta alagada (cheia) -0,145 -0,394 -0,154 -0,396
Macrófitas aquáticas (enchente) -0,313 0,661 0,353 0,451
Macrófitas aquáticas (cheia) 0,425 0,597 -0,293 0,605
Presença-AusênciaBray-CurtisPontos de Coleta

32
4.5 – Relação entre os parâmetros limnológicos e a composição da
ictiofauna.
A análise de correlação de Pearson realizada entre os parâmetros
limnológicos (características físicas e físico-químicas da água) conseguiu
identificar 5 correlações significativas (Tabela 9). Baseado nesse número de
correlações existentes, todas as variáveis foram reordenadas em uma Análise
de Componentes Principais (PCA), gerando com isso novas variáveis (eixos
ortogonais). Após definidas as componentes principais, estas foram
relacionadas diretamente com as variáveis representativas da estrutura das
assembléias de peixes, visando-se identificar relações de dependência.
A análise de componente principal 1 (PC1) explica 42,17% da variância
total, representando as variações inversas da transparência e as variações
diretas do oxigênio dissolvido (Tabela 9 e Figura 9). Essas duas variáveis
foram às principais responsáveis pela segregação dos habitats de água aberta
e floresta alagada na cheia, e isolou os bancos de macrófitas aquáticas na
enchente, provavelmente por valores mais elevados de transparência da água
(Figura 9).
O componente principal 2 (PC2) explica 28,76% da variância total, ou
seja, representa as variações inversas da profundidade, condutividade elétrica
e pH e as variações diretas de temperatura (Figura 9 e Tabela 9). Essas quatro
variáveis foram as principais responsáveis pelo agrupamento dos habitats de
água aberta e floresta alagada na enchente e vazante, e água aberta e
macrófitas aquáticas na seca e cheia (Figura 9).
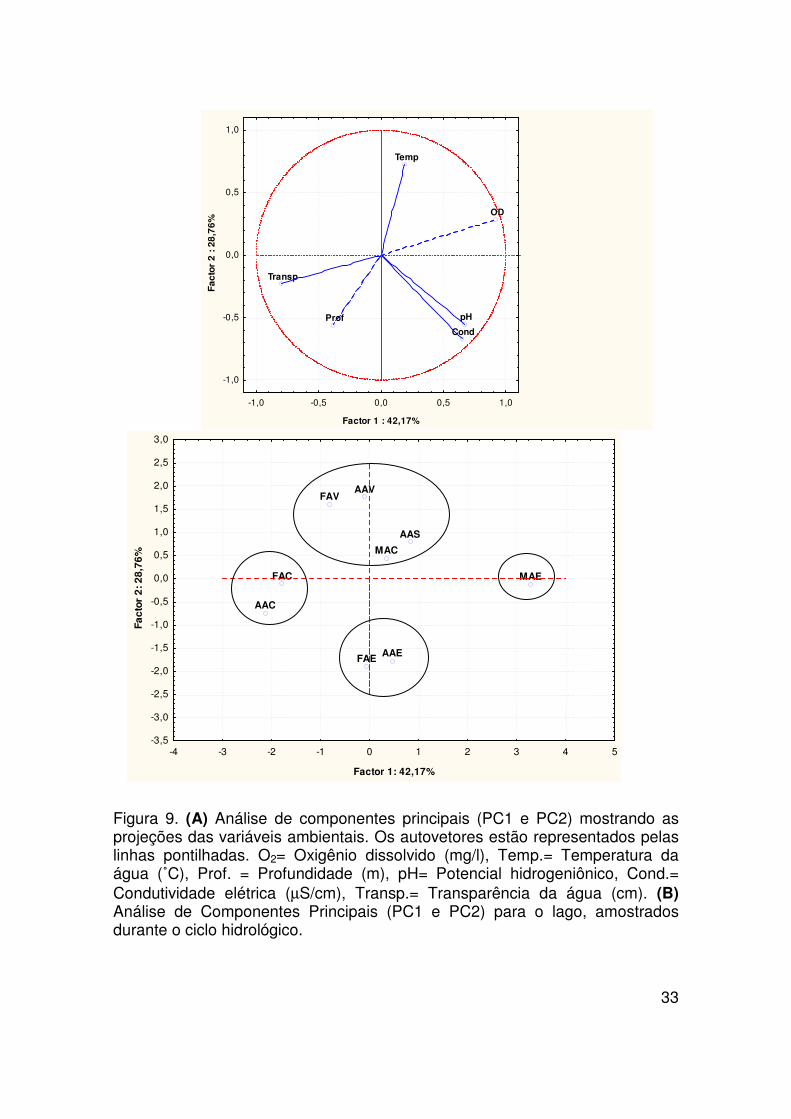
33
Figura 9. (A) Análise de componentes principais (PC1 e PC2) mostrando as projeções das variáveis ambientais. Os autovetores estão representados pelas linhas pontilhadas. O2= Oxigênio dissolvido (mg/l), Temp.= Temperatura da água (˚C), Prof. = Profundidade (m), pH= Potencial hidrogeniônico, Cond.= Condutividade elétrica (µS/cm), Transp.= Transparência da água (cm). (B) Análise de Componentes Principais (PC1 e PC2) para o lago, amostrados durante o ciclo hidrológico.
pH
OD
Temp
Cond
Prof
Transp
-1,0 -0,5 0,0 0,5 1,0
Factor 1 : 42,17%
-1,0
-0,5
0,0
0,5
1,0
Fa
cto
r 2
: 2
8,7
6%
AAEFAE
MAE
AAC
FAC
MAC
AAVFAV
AAS
-4 -3 -2 -1 0 1 2 3 4 5
Factor 1: 42,17%
-3,5
-3,0
-2,5
-2,0
-1,5
-1,0
-0,5
0,0
0,5
1,0
1,5
2,0
2,5
3,0
Fa
cto
r 2
: 2
8,7
6%

34
Para se verificar a possível relação entre as variáveis ambientais
(representadas pelos eixos 1 e 2 da PCA) com a composição de espécies
(representada pelos eixos 1 e 2 do MDS) e abundância de espécies, foram
calculadas regressões lineares entre o primeiro e o segundo eixo de ambas as
análises (Figuras 17A e 17B). Foram observadas relações positivas e
significativas entre as características ambientais dos ambientes estudados e a
composição de espécies de peixes (presença-ausência; r2= 0, 5032; r= - 0
7094 e p= 0, 0098), bem como com a abundância de espécies (r2= 0, 5215; r= -
0, 7221 e p= 0, 0080).
Tabela 7. Correlações de Pearson entre as variáveis ambientais medidas nos habitats do lago ao longo dos períodos de enchente e cheia. Os valores em negrito indicam correlações estatisticamente significativas.
Variáveis pH OD Temp Cond Prof Transp
pH 1
OD 0,869 1
Temp 0,531 0,774 1
Cond 0,593 0,661 0,098 1
Prof -0,413 -0,548 -0,471 -0,423 1
Transp -0,709 -0,674 -0,641 -0,335 0,270 1

35
Tabela 8. Autovetores dos sete parâmetros ambientais analisados para os dois primeiros componentes principais. Parâmetros PC1 PC2
pH 0,668638 -0,550810
Oxigênio dissolvido (mg/l) 0,918444 0,286241
Temperatura (˚ C) 0,193332 0,727168
Condutividade (µµµµS/cm) 0,645613 -0,669886
Profundidade (m) -0,385002 -0,557250
Transparência (cm) -0,798408 -0,228901 Tabela 9. Valores dos escores para o lago nos habitats de água aberta, floresta alagada e macrófitas, analisados na enchente e cheia.
4.6 – Relações da composição, riqueza e abundância relativa das
espécies de peixes com os parâmetros limnológicos.
A composição de espécies representada pelo eixo 1 da MDS apresentou
relações significativas com todos os parâmetros físico-químicos da água do
lago. No entanto, as relações positivas e significativas foram observadas
somente entre a transparência, temperatura, pH e oxigênio dissolvido (Tabela
11). A riqueza de espécies apresentou relações positivas e significativas com a
transparência e temperatura da água. As relações negativas foram observadas
com a transparência, condutividade, pH, profundidade e oxigênio dissolvido,
sendo que ambas apresentaram valores significativos. A captura por unidade
de esforço dada em número de exemplares (CPUEn) e biomassa (CPUEb) não
apresentou relações positivas com nenhum dos parâmetros analisados, exceto
Pontos de coletas PC1 PC2
Água aberta (Enchente) 0,46073 -1,76993
Floresta alagada (Enchente) -0,05989 -1,88996
Macrófitas aquáticas (Enchente) 3,28506 -0,11397
Água aberta (Cheia) -2,13800 -0,73488
Floresta alagada (Cheia) -1,78835 -0,10942
Macrófitas aquáticas (Cheia) 0,33643 0,44985

36
a temperatura na CPUEb. Todas as análises com relações negativas
apresentaram valores significativos.
Tabela 10. Valores de coeficiente de determinação (R2), coeficiente de correlação (R), e nível de significância (p> 0,05) das relações entre a composição, riqueza e abundância relativa (CPUEn e CPUEp) com as variáveis ambientais dos habitats do lago.
Relações R² R p
Composição de espécies Transparência 0,7201 0,8486 0,1514
Condutividade 0,1353 0,3678 0,6322
Temperatura 0,3428 0,5855 0,4145
pH 0,8416 0,9177 0,0826
Profundidade 0,0230 0,1518 0,8481
Oxigênio 0,6139 0,7835 0,2165
Riqueza Transparência 0,3225 0,5679 0,4322
Condutividade 0,0685 0,2618 0,7382
Temperatura 0,7099 0,8426 0,1574
pH 0,0055 0,0743 0,9257
Profundidade 0,00002 0,0046 0,9954
Oxigênio 0,1990 0,4461 0,5539
CPUE n Transparência 0,0441 0,2101 0,7810
Condutividade 0,1820 0,4261 0,5734
Temperatura 0,3722 0,6101 0,3899
pH 0,1084 0,3293 0,6707
Profundidade 0,0030 0,0550 0,9450
Oxigênio 0,0132 0,1150 0,8849
CPUE b Transparência 0,0042 0,0647 0,9353
Condutividade 0,2211 0,4702 0,5298
Temperatura 0,2422 0,4922 0,5078
pH 0,2165 0,4653 0,5347
Profundidade 0,0061 0,0778 0,9222
Oxigênio 0,0003 0,0162 0,9838

37
5 - Discussão
5.1 - Características ambientais e físico-químicas da água nos
habitats do lago Jaitêua.
Os ecossistemas de várzea são caracterizados por uma diversificada
quantidade de ambientes que sofrem alterações anuais de acordo com as
variações sazonais geradas pelo pulso de inundação (Junk, 1983; Barthem &
Fabré, 2004). Essas áreas possuem sistemas aquáticos que são afetados
diretamente pelo regime de alagação (Saint-Paul, et al., 2000). Segundo
Barrella (2001), um dos principais efeitos causados pela flutuação anual do
nível da água são as modificações na estrutura dos lagos, alterando
conseqüentemente as características físico-químicas da água, sendo que
estas, por sua vez, podem causar grandes modificações nas comunidades
biológicas.
Dentre os parâmetros físico-químicos anotados no lago, destacam-se as
baixas concentrações de oxigênio dissolvido no período da cheia.
Possivelmente os peixes devem apresentar adaptações específicas que
possibilitem a sobrevivência em ambientes com baixas concentrações deste
gás. Das 10 espécies dominantes no lago, apenas Potamorhina latior,
Triportheus angulatus e Colossoma macropomum possuem adaptações
morfológicas, anatômicas e fisiológicas já descritas em vários trabalhos (Saint-
Paul & Soares, 1987; Soares 1993; Fernandes et al. 1994; Parma De Croux
1994; Muusze et al. 1998; Soares & Junk, 2000). Estas espécies têm como
características o uso da respiração na superfície aquática (RSA), considerada
uma adaptação para utilização do oxigênio da camada superficial, atribuído como

38
um componente estratégico de respiração integrada, permitindo a manutenção da
vida e atividades durante os períodos de baixas concentrações de oxigênio,
aumentando o risco de morte por predação.
Fatores como o oxigênio da água e a temperatura também são
caracterizados como bons indicadores abióticos. Os habitats de floresta
alagada e água aberta do lago apresentaram déficit de oxigênio na cheia,
possivelmente por ser um período com baixa produção de fitoplâncton no
ambiente, corroborando os estudos de Junk (1983), onde cita que habitats de
água aberta têm a oxigenação influenciada pelo crescimento do fitoplâncton e
difusão com o ar. Além do que, observamos valores de transparência da água
bem mais elevados na cheia na floresta alagada, podendo ser também um
excelente indicador da baixa produção de fitoplâncton, tendo contribuição com
os processos de decomposição da matéria orgânica neste habitat. É errado
concluir que valores extremamente baixos de oxigênio são excepcionais e
dificilmente serão os mais representativos para os lagos da Amazônia (Junk et
al., 1983). O aumento dos níveis de oxigênio na vazante no lago está
relacionado diretamente com o aumento da correnteza da água, promovido
pela saída de água deste ambiente, além da redução da cobertura de
macrófitas aquáticas. Esta por sinal se ocuparem grandes áreas podem
interferir diretamente nas concentrações de oxigênio por afetarem junto com a
floresta alagada a produção de fitoplâncton no sistema Junk et al., 1980.
Os maiores valores de transparência da água observados na cheia na
floresta alagada do lago, provavelmente estão relacionados com a falta de
entrada de água com grandes quantidades de sedimentos do Solimões, além

39
de ser um habitat que não sofre grande influência dos ventos, podendo
apresentar maior eficiência na decantação das partículas em suspensão. A
flutuação do nível da água ao longo do ciclo hidrológico exerce grande
influência nos valores de profundidade da água observados no lago, mas
também promove a presença e ausência dos habitats de floresta alagada e
bancos de macrófitas aquáticas ao longo do ano.
Os maiores valores de pH e condutividade encontrados no período de
enchente indicam a intensa entrada de água proveniente do rio Solimões,
ambiente rico em sedimentos como já mencionado acima, capaz de alterar
significativamente os valores observados para estas variáveis. Os valores de
pH e condutividade anotados nos ambientes deste estudo estão de acordo com
os observados por Anjos (2007), estudando um sistema lacustre do Cururu no
município de Manacapuru, à margem direita do Solimões. Esta autora realizou
coletas em três ambientes de várzea dentro deste sistema, evidenciando
valores mais altos de pH e condutividade no período da enchente.
Os resultados obtidos neste estudo mostram a existência de variações
sazonais nos parâmetros físicos e físico-químicos da água ao longo do ciclo
hidrológico, causados principalmente pela flutuação do nível da água ao longo
do ano. Isso nos leva crer que essas mudanças alteram a estrutura da
ictiofauna de acordo com as características da água dos ambientes
observados, além de corroborar com os estudos de Hamilton & Lewis (1990) e
Rodriguez & Lewis, (1997), que também relataram tais fatos em suas
pesquisas no rio Orinoco.

40
5.2 - Composição e estrutura das assembléias de peixes no lago e
suas relações com as características ambientais.
A maioria dos estudos desenvolvidos em lagos amazônicos mostra que a
ictiofauna é caracterizada pela dominância de Otophysi (=Ostariophysi sensu
stricto) com as ordens dos Characiformes (43%), Siluriformes (36%) e
Gymnotiformes (3%) representando aproximadamente 82% dos peixes,
seguidos de Perciformes com 12% e outras ordens com 6,4% (Roberts 1972,
Lowe-McConnell, 1999). Tais valores apresentam-se semelhantes aos
encontrados nos rios (Ferreira, 1993; Ferreira et. al., 1988; Santos, 1991) e
lagos de várzea da Amazônia Central (Junk et al, 1983; Souza-Pereira, 2000;
Saint-Paul et al. 2000).
Analisando a estrutura total da ictiofauna dos lagos Jaitêua e São
Lourenço observou-se que a sequência foi bem semelhante com
Characiformes (55,70%), Siluriformes (19,35%), Perciformes (16,9%),
Clupeiformes (4,72%) e outras ordens (Gymnotiformes, Osteoglossiformes,
Pleuronectiformes e Synbranchiformes) com (3,24%). Em ambos os lagos as
famílias Serrasalmidae, Characidae, Cichlidae, Curimatidae, Pristigasteridae,
Hemiodontidae e Pimelodidae foram dominantes ao longo do ciclo hidrológico
nos diferentes habitats.
Os caracídeos, curimatídeos e pristigasterídeos são em geral espécies
formadoras de cardumes que realizam migrações de longa ou curta distância e
desenvolvem parte do seu ciclo de vida nos lagos na época de cheia, e parte
nos rios, na época de seca (Cox-Fernandes, 1988; Soares & Yamamoto, 2004).
Cox Fernandes (1988), estudando nos canais de conexão do sistema do lago

41
do Rei, e Freitas e Garcez (2004) nos canais de conexão dos lagos Cururu e
Jacaré, observaram que esses peixes realizam movimentos migratórios dos
lagos em direção ao rio principal na cheia.
Muitas dessas espécies são consideradas residentes, desenvolvendo
seu ciclo de vida nos lagos de várzea como o caso de Moenkhausia lepidura,
intermedia e Acestrorhynchus falcirostris (piabas e peixe-cachorro). No período
da seca também foi observado no lago a ocorrência de alguns exemplares de
espécies migradoras tais como Triportheus auritus, Hemiodus immaculatus,
Anodus elongatus e Hemiodus sp., pois mesmo com a conexão ainda existente
no período de seca entre os lagos e os rios Manacapuru e Solimões, esses
peixes permaneceram no lago. Segundo Lowe-McConnell (1999), a chave da
estrutura das comunidades de peixes nos trópicos é baseada em sua
mobilidade, pois muitas espécies realizam movimentos laterais dos rios em
direção aos lagos e vice-versa, além da floresta inundada e dos tributários,
tendo como fator regulador a mudança no nível da água e variáveis
limnológicas como o oxigênio. Muitos autores também observaram mudanças
sazonais na composição da ictiofauna em planícies inundáveis dos rios
Machado e Madeira (Goulding, 1980), do Rio Rupununi nas Guianas (Lowe-
McConnell, 1964), lago do Inácio (Saint-Paul et al., 2000), lago do Rei (Merona
& Bittencourt, 1993), lago Catalão (Vale, 2003), lagos Samaúma, Sacambu e
Maracá na Amazônia Central (Siqueira-Souza, 2002), lagos Comandá, Praia,
Tracajá e Acari (Yamamoto, 2004) e no complexo lacustre do Cururu (Anjos,
2007).

42
Os habitats de floresta alagada na enchente e cheia e água aberta na
vazante e seca tiveram valores mais elevados de diversidade. O elevado valor
de Shannon observado na seca pode ser explicado pelo elevado número de
espécies residentes que permaneceram no lago neste período. Com relação ao
índice de dominância de Berger- Parker, as diferenças sazonais observadas
em seus valores podem ser atribuídas ao incremento no número de espécies
dominantes. As espécies dominantes concentraram-se mais na floresta
alagada do lago na enchente e cheia, como observados nos valores de
dominância.
Os resultados de diversidade podem estar relacionados com presença
de espécies formadoras de cardumes encontradas nos lagos, ou ainda por
tratar-se de áreas impactadas pela ação antrópica (Lowe-McConnell, 1999).
Se compararmos a diversidade de Shannon-Wiener obtida nos habitats do lago
com outros valores encontrados em lagos de diferentes bacias do Brasil, tais
como: braços dos lagos do sistema lacustre do Cururu, Amazonas (H’ = 2,49 a
3,11; Anjos, 2007), lago de Itaipu, Paraná ( H’= 1,0 a 2,0; Oliveira et al., 2004),
baía do Caxiuanã, Pará (H’=2,9; Montag, 2001) e Parque Estadual Rio Doce,
Minas Gerais (H’=0,54 a 1,31; Latini & Petrere Jr., 2004), podemos considerá-
los ambientes de alta diversidade. Os valores mais elevados de riqueza de
espécies encontrados neste estudo podem ser atribuídos ao elevado esforço
de pesca aplicado, com repetições de coletas realizadas mensalmente em um
período de 12 meses, com tempo de permanência das malhadeiras na água de
24 horas de

43
Os índices de riqueza e diversidade deste estudo quando comparados
com outros trabalhos realizados também em lagos de várzea da Amazônia
Central, permitem dizer que o lago Jaitêua pode ser considerado um ambiente
que apresenta boas condições ambientais para as comunidades de peixes,
mesmo sendo áreas muito utilizadas por pescadores profissionais.
A biomassa coletada nos lagos oscilou bastante ao longo do ciclo
hidrológico, sendo a cheia o período com os menores valores. Provavelmente,
isso se deve ao aumento da área alagada dos lagos, propiciando a ampliação e
o surgimento de habitats como a floresta alagada, os bancos de macrófitas
aquáticas e capins flutuantes, capazes de disponibilizar refúgios para os
peixes, tornando-os menos vulneráveis às capturas pelas malhadeiras e
redinhas. Devemos considerar que a malhadeira é um aparelho de pesca
passivo, deixando possivelmente de capturar muitas espécies de peixes
(Merona & Bittencourt, 1988), afetando diretamente as amostragens e
conseqüentemente a caracterização da comunidade de peixe.
A retração das águas na seca ocasiona perda e redução de habitats, tais
como a floresta alagada e macrófitas aquáticas, reduzindo conseqüentemente
os valores de CPUE observados. Quando comparados os valores de CPUE em
peso e abundância nos períodos do ciclo hidrológico, notamos que a vazante
foi o período que apresentou os maiores valores. Este fato está relacionado
com o movimento dos peixes em direção aos canais mais profundos dos lagos
e até mesmo para os rios, tornando-os mais vulneráveis à pesca com
malhadeiras. Tais condições possibilitaram o aumento em mais de três vezes

44
nesses valores para este período em relação ao restante do ciclo hidrológico
nos habitats amostrados.
Os valores obtidos pela CPUE em números de exemplares encontram-se
acima dos valores já observados por outros autores em estudos realizados por
Zuanon et al., (2004) na ilha do Cantão em Tocantins (6,63), Do Valle (2003)
no lago do Catalão no Amazonas (4,42), Ferreira (1991) no rio Trombetas
(2,42) no Pará, Torrente-Vilara et al., (2003) no rio Cautário em Rondônia
(2,08), Ferreira et al., (1988) no rio Mucajaí em Roraima (1,79) e Doria et al.,
(2005) no alto Madeira em Rondônia (2,83), ambos coletando com
malhadeiras.
De acordo com alguns autores (Rodríguez & Lewis, 1997; Tejerina-
Guarro et al.,1998), as características morfológicas dos lagos (área e
profundidade) e a cobertura vegetal aquática são considerados fatores
reguladores das comunidades de peixes. Além disso, as diferenças na
composição e na riqueza de espécies estão relacionadas ao tamanho do corpo
d’água. Segundo Welcomme (1985), o número de espécies pode aumentar
proporcionalmente com o tamanho do lago, sendo este um indicador direto da
capacidade de retenção da água.
O número de espécies coletadas nos bancos de macrófitas aquáticas
neste estudo foi relativamente baixo quando comparado com outros estudos já
realizados neste mesmo tipo de ambiente por Goulding et al. (1988) em
Anavilhanas (56 espécies), Sanchez-Botero & Araújo-Lima (2001) em três
lagos de várzea do rio Solimões (91 espécies), Petry et al. (2003) na ilha da
Marchantaria (139 espécies) e Prado (2005) em quatro lagos de várzea da

45
Amazônia Central (50 espécies), ambos com esforço de pesca maior em
relação ao atual estudo.
Os maiores valores de abundância observados no lago, tiveram como
espécies mais abundantes nos períodos de enchente e cheia o tucunaré
(Cichla monoculus) e a piaba (Moenkausia intermédia) com mais de 60% dos
exemplares coletados.
A composição da ictiofauna capturada nos bancos de macrófitas
composta pelas ordens de Characiformes, Perciformes, Gymnotiformes e
Synbranchiformes, segue padrões semelhantes aos estudos realizados por
Henderson & Hamilton (1995) no lago Mamirauá em macrófitas aquáticas,
mostram também o mesmo padrão de predominância da ordem Characiformes.
Sanchez-Botero (2000), estudando três lagos da Amazônia Central, também
evidenciaram a predominância de Characiformes. Petry et al. (2003)
verificaram em seus estudos nas macrófitas, que 45% dos exemplares
capturados eram pertencentes à ordem dos Characiformes. Proporções bem
semelhantes foram observadas por Meschiatti et al, (2000), analisando a
ictiofauna associada as macrófitas aquáticas em lagos do rio Mogi-Guaçu. É
interessante ressaltar que os bancos de macrófitas do lago Jaitêua não
apresenta grande complexidade em suas raízes (obs. pessoal), por
apresentarem predominância de mureru (Eichornia crassipes) e membeca
(Paspalum repens). Este fato é muito importante se levarmos em consideração
que os peixes menores buscam os bancos mais complexos para fugirem de
seus predadores, uma vez que a baixa complexidade deste habitat, como

46
citado acima, provavelmente pode ser uma das causas para os baixos valores
de riqueza observados.
Estas afirmações corroboram as conclusões de Petry et al. (2003), que
sugerem que a diversidade de espécies esta associada diretamente à
complexidade dos bancos de macrófitas. Os caules, raízes e folhas submersas
proporcionam aos juvenis e peixes menores uma barreira visual e física contra
seus predadores (Crowder & Cooper, 1979; Savino & Stein, 1989). Segundo
Araújo-Lima et al. (1986) as espécies mais susceptíveis a predação procuram
ambientes como os de macrófitas aquáticas, compostos por raízes, caules e
folhas, considerados mais complexos, para que os peixes possam fugir de seus
predadores.
Os valores de equitabilidade mais baixos encontrados na vazante na água
aberta e floresta alagada indicam a grande dominância de espécies formadoras
de cardumes, tanto migradoras (Pellona flavipinnis, Semaprochilodus insignis,
Potamorhina latior, Triportheus albus e Triportheus angulatus), como residentes
(Pygocentrus nattereri, Serrasalmus altispinis, Serrasalmus elongatus e
Serrasalmus spilopleura).
No período da enchente a baixa dominância de P. flavipinnis e H.
marginatus resultou em uma menor diferença na distribuição do número de
exemplares por espécie, conseqüentemente elevando os valores de
equitabilidade na água aberta. O mesmo fato ocorreu para a floresta alagada,
porém com baixa de dominância de outras espécies como: A. falcirostris, C.
macropomum e P. flavipinnis. Nos bancos de macrófitas aquáticas os valores
de equitabilidade foram mais reduzidos em relação aos outros habitats, graças

47
à dominância de Hemigrammus sp. e Cichla monoculus em ambos os lagos,
porém na cheia com a baixa dominância das espécies no lago e a menor
diferença de distribuição do numero de exemplares por espécies, elevaram o
valor de equitabilidade. Ao longo da cheia nos habitats de água aberta e
floresta alagada do lago, os valores apresentaram-se bem mais elevados,
indicando a falta de dominância de espécies.
O oxigênio e a temperatura são conhecidos por afetarem diretamente a
ictiofauna. As situações de hipoxia são comumente observadas em lagos de
várzea, igapós e poças temporárias, onde as altas taxas de decomposição,
principalmente de macrófitas e liteira, acarretam em redução dos níveis de
oxigênio dissolvido. Tais condições podem levar várias espécies de peixes a
utilizarem estratégias de respiração para sobreviver nesses ambientes com
baixos níveis de oxigênio (Junk, 1983; Soares 1993; Val et al., 1995)
A variação no nível da água (profundidade) mostrou relação direta com o
aumento e redução nos valores de riqueza de espécies observadas no lago. De
acordo com Súarez et al., (2001) o aumento da profundidade em lagoas
temporárias do Pantanal, foi a responsável pelo aumento da riqueza. Anjos
(2007) também evidenciou este fato no complexo lacustre do lago cururu.
Fatores abióticos têm sido amplamente relatados como responsáveis
pelas mudanças na estrutura das comunidades de peixes em lagos. Tajerino-
Garro et al. (1998) em estudos realizados em lagos das planícies inundáveis da
bacia do rio Araguaia-Tocantins, constataram que a transparência e a
profundidade possuem relações significativas com a estrutura das
comunidades. Rodrigues & Lewis (1997), estudando áreas alagadas do rio

48
Orinoco, verificaram que as comunidades de peixes são estruturadas pela
piscivoria, sob a influência da transparência da água que é modulada pela
morfologia do lago. Junk et al. (1983) e Saint-Paul & Soares (1987) relatam que
a variação na distribuição de espécies em lagos da Amazônia Central tem
relação com ás concentrações de oxigênio dissolvido.
Ao analisarmos os parâmetros físico-químicos da água do lago,
verificamos a existência de diferenças dos habitats da cheia em relação à
enchente. As ordenações dos ambientes a partir da Análise de Componente
Principais (PCA) revelam que na cheia existe uma diminuição na variabilidade
entre os habitats do lago, diferente do que observamos na enchente. Estas
variações também podem ser evidenciadas nas análises de MDS com a
análise da composição de espécies.
Os resultados observados sugerem que os habitats estudados
apresentam composição de espécies distintas ao longo do ciclo hidrológico. As
espécies ocupam os três principais habitats do lago, a água aberta, a floresta
alagada e os bancos de macrófitas aquáticas, de acordo com a variação do
nível da água. No período de alagação, a quantidade de peixes tanto na água
aberta como na floresta alagada do lago é grande. Já no período de vazante,
muitos peixes que ocupavam os habitats de água aberta e floresta alagada do
lago passam a iniciar um processo de movimentação em direção aos canais
mais profundos do lago, poços d’água, parte central do lago Grande (ou lago
Cabaleana), canal de conexão dos lagos com os rios Solimões e Manacapuru.
Esse período caracteriza-se por grandes abundâncias de peixes nas capturas.
Mesmo com a conexão entre os rios Solimões e Manacapuru, os canais

49
menores, principalmente do lago Jaitêua, não apresentam profundidade
suficiente para que ocorra a migração dos peixes, ocorrendo isolamento de
alguns ambientes na seca, aprisionando-os os peixes em locais mais profundos
do complexo lago Grande. Dessa forma, os peixes tornam-se presas fáceis
para as malhadeiras. Na seca a abundância de recursos alimentares para os
peixes torna-se menor, com poucas opções de alimento e abrigo. No entanto,
existe o lado favorável principalmente para os predadores, como no caso de
Serrasalmus spilopleura, amplamente dominante no lago na seca.
De acordo com a distribuição das espécies de peixes ao longo do ciclo
hidrológico, observamos que os habitats de água aberta e floresta alagada
apresentaram melhores condições para as espécies de peixes, principalmente
por apresentar maiores valores de riqueza e abundância de espécies.
Esses resultados nos levam a crer que a transparência da água, a
temperatura e o oxigênio dissolvido podem constituir fatores que influenciam
mudanças nas assembléias ictíicas, como verificado através da significância de
alguns modelos lineares entre os parâmetros ambientais e estruturais da
ictiofauna. Dessa forma, a sazonalidade do habitat é considerada um fator
chave que modifica as estratégias do ciclo de vida das comunidades de peixes
tropicais, amplamente mencionada em vários estudos (Goulding, 1980; Lowe-
McConnell, 1987; Cox-Fernandes, 1988; Merona & Bittencourt, 1993 e Saint-
Paul et al., 2000).
O complexo lacustre lago Grande de Manacapuru apresenta ambientes
bastante heterogêneos, tais como lagos, paranás, igarapés, furos com grandes
áreas de floresta circundando todo o complexo, favorecendo a alta riqueza de

50
peixes, indicando que a manutenção dessas áreas é de extrema importância,
pois são consideradas locais de alimentação, abrigo e reprodução para os
peixes. Mesmo com a proximidade do lago em relação à cidade de
Manacapuru, seus ambientes ainda apresentam boa integridade, permitindo
com isso que ocorra a manutenção da ictiofauna, além da permanência dos
vários nichos ecológicos a serem explorados pelas espécies.
Os resultados apresentados e discutidos neste estudo indicam a
importância de se implementar ações de manejo e ordenamento pesqueiro
voltados principalmente para espécies de peixes como as sardinhas, o
tambaqui, apapás, branquinhas e tucunarés, amplamente comercializadas nos
mercados e feiras de Manacapuru e Manaus, e tidas neste estudo como as
mais abundantes na água aberta e floresta alagada do lago.

51
6.0 - Conclusões
As variações ocorridas nos parâmetros físicos (profundidade e
transparência da água) e físico-químicos da água (oxigênio, temperatura,
condutividade e pH) principalmente na subida e descida das águas,
apresentaram efeitos sobre a composição das comunidades de peixes nos
habitats do lago. A profundidade do lago foi um parâmetro importante na
estruturação das comunidades, exercendo grande influência sobre a
composição das comunidades, em conjunto com outros fatores, como o
oxigênio dissolvido e a temperatura da água.

52
7.0 – REFERÊNCIAS BIBLIOGRÁFICAS
Agostinho, A.A. & Zalewski, M. 1995. The dependence of fish community
structure and dynamics on floodplain and riparian ecotone zone in Paraná
River, Brazil. Hydrobiologia. Dordrecht, v 303, no. 1-3:141-148p.
Almeida, R. G. 1984. Biologia alimentar de três espécies de Triportheus
(pisces: Characidae) do lago Castanho, Amazonas. Acta Amazônica, 14
(11): 48-76.
Almeida-Val, V.M.F., Val, A.L., Duncan, W.P., Souza, F.C.A. Paula-Silva,
M.N., Land, S. 2000. Scaling effects on hypoxia tolerance in the Amazon
fish Astronotus ocellatus (Perciformes: Cichlidae): Contribution of tissue
enzyme levels. Comparative Biochemistry & Physiology - B: Comparative
Biochemistry. 125B (2): 219-226.
Anjos, C. R. Estrutura de assembléias ictíicas em sistema lacustre manejado
da Amazônia Central - 2007. 82 p. Dissertação de Mestrado – INPA/UFAM,
2007.
Ayres, J.M.1995. As matas de várzea do Mamirauá: Médio Rio Solimões. 2ed.
Brasília, DF: CNPq; Tefé, AM: Sociedade Civil Mamirauá 124p.
Barrella, W. 2001. Os peixes como indicadores da qualidade das águas dos
rios. 249-262 p. In: Maia, N. B.; Martos, H. L & Barrella, W. (orgs). 2001.
Indicadores ambientais: conceitos e aplicações. Ed. EDUC – PUC-SP: 285
pp.
Barthem, R. B. 1987. Uso de redes de espera no estudo de ritmos circadianos
de algumas espécies de peixes nos lagos de várzea do Rio Solimões.
Revista Brasileira de Zoologia, 3: 409-422.

53
Barthem, R. B.; Fabré, N. N. 2004. Biologia e diversidade dos recursos
pesqueiros na Amazônia. In: Ruffino, M. L. (Coord.) A pesca e os recursos
pesqueiros na Amazônia brasileira. Ed. Ibama/Provárzea. Manaus. 17-62p.
Braum, E. & Junk, W. J. 1982. Morphological adaptation of two Amazonian
characoids (Pisces) for surviving in oxygen deficient waters. – Int. Rev.
Ges. Hydobiol. 67: 869-886.
Cox - Fernandes, C.. 1988. Estudos de migrações laterais de peixes no sistema
lago do Rei (Ilha do Careiro) – AM, Brazil. Dissertação de Mestrado.
Manaus: PPG Instituto Nacional de pesquisas da Amazônia -INPA. 158pp.
Da Silva, C. J.; Esteves, F. A. 1995. Dinâmica das características limnológicas
das baías Porto de Fora e Acurizal (Pantanal de Mato Grosso) em função
da variação do nível da água. Oecol. Brasil. Estrutura, Funcionamento e
Manejo de Ecossistemas Brasileiros. v. 1, p.47-60.
Do Valle, J. D. 2003. Composição, diversidade e abundância da ictiofauna na
área do Catalão, Amazônia Central. Dissertação de Mestrado, INPA/UFAM.
99 pp.
Doria, C. R. C.; Torrente-Villara, G.; Zuanon, J. A. S.; Favaro, L. F.; Ruffino, M.
L. & Leite, R. G. 2005. Estudo de viabilidade das AHE’s Jirau e Santo
Antônio, localizadas no rio madeira em Rondônia, no trecho entre Porto
Velho e Abunã. Relatório Técnico Final, Porto Velho. 345 pp.
Fabré, N.N.; Saint-Paul, U. 2004. Observações sobre a ictiofauna de um lago
de Várzea da Amazônia Central. In: Cintra R (Org.). História de Vida de
Organismos Amazônicos. EDUA. Manaus-Amazonas: 179-183p.

54
Fernandes, M.N., Rantin, F.T., Kalinin, A.L. & Van Moron, S. 1994.
Comparative study of gill dimensions of three erythrinid species in relation to
their respiratory function. Can. J. Zool. 72(1):160-165.
Ferreira, E. F. G.; Santos, G. M & Jegu, M. 1988. Aspectos ecológicos da
ictiofauna do rio Mucajaí, na área da ilha Paredão, Roraima, Brasil,
Amazoniana, 10(3): 339-352.
Ferreira, F. C.; Petrere Jr., M. 2007. Anthropic effects on the fish community of
Ribeirão Claro, Rio Claro, SP, Brazil. Brazilian Journal of Biology, v. 67: 23-
32 p.
Freitas, C.E.C. & Garcez, R.C.S. 2004. Fish Communities of natural channels
between floodplain lakes and Solimões – Amazonas River (Amazon -
Brazil). Acta Limnol. Bras., 16(3):273-280 p.
Géry, J. 1977. Characoids of the World. USA, t.f.h. publications, 672 p.
Goulding, M., 1980. The Fishes and Forest. University of California.
Goulding, M.; Barthem, R.; Ferreira, E. 2003. The Smithsonian Atlas of the
Amazon. Smithsonian Institution. Hong Kong, Oakwood Arts. 254p.
Hamilton, S. K.; Lewis Jr., W. M. 1990. Basin morphology in relation to chemical
and ecological characteristics of lakes on the Orinoco River Floodplain,
Venezuela. Arch. Hydrobiol. 119 (4): 393-425.
Goulding, M.; Carvalho, M.L; Ferreira, E.G. 1988. Rio Negro: rich life in poor
water: Amazonian diversity and food chain ecology as seen through fish
communities. SPB Academic Publishing, The Hague. 200 pp.

55
Hamilton, S. K.; Sippel, S. J.; Melack, J. M. 1995. Oxygen depletion and carbon
dioxide and methane production in waters of the Pantanal wetland of Brazil.
Biogeochemistry 30: 115-141.
IBGE 2005. Censo demográfico 2005. Amazonas. IBGE. 5p. (www.ibge.gov.br)
Irion, G.; Junk, W. J.; Mello, J. A. S. N. De., 1997. The large Central Amazonian
river floodplains near Manauz: Geological, climatological, hydrological and
geomorfological aspects. In: The Central Amazon Floodplain: Ecology of a
Pulsing System. Ecological Studies. Max - Planck. Institut Für limnologie.
Junk, W. J. 1997. General Aspects of Floodplain ecology with Special reference
to Amazonian Floodplains. p. 3-17. The Central Amazon Floodplain.
Ecology of a pulsing System. Ecological Studies. Nº 126:528p.
Junk, W.J. 1999, The flood pulse concept of large rivers: learning from the
tropics.- Arch. Hydrobiol. Suppl. 115 (3): 261-280.
Junk, W.J. 1980. Áreas inundáveis. Um desafio para a limnologia. Acta
Amazônica., 20(4):775-795.
Junk, W. J. 1983. As águas da Região Amazônica. pp. 45-100. In: Salati, E.;
Junk, W. J.; Schubart, H. O. R. & Oliveira, A. E. (Eds.). Amazônia:
desenvolvimento, integração e ecologia. CNPq/Brasiliense, São Paulo.
Junk, W. J.; Soares, M.G. & Carvalho, F. M. 1983. Distribution of fish species in
a lake of the Amazon river floodplain near Manaus (Lago Camaleão), with
special reference to extreme oxygen conditions. Amazoniana, 7:397-431.
Junk, W. J.; Bayley, P. B.; Sparks, R. E. 1989. The Flood pulse concept in river-
floodplain systems. In: D. P. Dodge (ed.). Proceedings of international
Large River Symposium. Canadian Special Publication Fishery Aquatic
Sciense, Nº 106: 110-127p.

56
Krebs, C.J. 1989. Ecological methodology. New York: Krebs, C.J (ed.), 654 p.
Latini, A.O & Petrere Jr, M. 2004. Reduction of a native fish fauna by alien
species: na example from Brazilian freshwater tropical lakes. Fisheries
Management and Ecology, v. 11: 71–79p.
Lowe-McConnell, R. H. 1964. The fishes of the Rupunini savanna district of
British Guiana, Pt 1. Groupings of fish species and effects of the seasonal
cycles on the fishes. Journal of the Linnean Society (Zoology), 45, 103-
144.
____ 1999. Estudos ecológicos de comunidades de peixes tropicais.
Editora da Universidade de São Paulo, São Paulo. 5339.
Ludwig, J. A. & Reynolds, J.F. 1988. Statistical Ecology. New York. John Wiley
& Sons. Inc. Magurran, A. E. 1988. Ecological Diversity and Its
Measurement. Chapman and Hall, New York, 215p.
Magurran, A. E. 1988. Ecological Diversity and Its Measurement. Chapman and
Hall, New York, 215p.
Mérona, B. & Bittencourt, M.M. 1988. A pesca na Amazônia através dos
desembarques do mercado de Manaus: resultados preliminares.- Mem.
Soc. Cienc. Nat. La Salle (48) (Supl.): 433-453p.
Mérona, B. & Bittencourt, M. M., 1993. Fish communities of the "Lago do Rei", a
floodplain lake in the Central Amazon: a general description. Amazoniana,
v. 12: 415-441 p.
Meschiatti, A. J.; Arcifa, M. S & Fenerich-Verani, N. 2000. Fish communities
associated with macrophytes in Brazilian floodplain lakes. Environmental
Biology of Fishes, v. 58: 133–143 p.

57
Montag, L. F. A. 2001. Diversidade distribuição e estrutura da comunidade de
peixes na Estação Científica Ferreira Penna: Amazônia Oriental.
Dissertação (Mestrado) - Curso de Mestrado em Zoologia, Universidade
Federal do Pará, Museu Paraense Emílio Goeldi, Belém. 135 pp.
Muusze B., Marcon, J., Vandenthillart, G. & Almeida- Val, V.M.F. 1998. Hypoxia
tolerance of Amazon fish respirometry and energy metabolism of the cichlid
Astronotus ocellatus. Comparative Biochemistry & Physiology. Part A,
Molecular & Integrative Physiology. 120(1):151-156.
Nogueira, F. M. B. 1989. Importância das macrófitas aquáticas Eichhornia
azurea Kunt e Scirpus cubencis Poepp & Kunth na ciclagem de nutrientes
e nas principais características limnológicas da Lagoa do Infernão. 147f.
Dissertação de Mestrado. UFSCar, São Carlos.
Oliveira, P. R. S.; Ruffino, M. L. & Mitlewski, B. 1999. Levantamento Rápido
Rural (LRR) – Análise socioeconômica das comunidades de Tiningu e
Murumuru: Município de Santarém, PA – Brasil. In: Recursos Pesqueiros
do médio Amazonas: abordagem socioeconômica. Brasília : Edições
IBAMA Coleção Meio Ambiente, Série estudos pesca; 21. 187-212p.
Panosso, R. F. 1993. Influência do regime hidrológico e das características
morfométricas sobre algumas variáveis limnológicas de lago amazônico
(Lago Batata, PA) impactado por rejeito de bauxita. Dissertação de
Mestrado. UFRJ, Rio de Janeiro.
Parma De Croux, M J. 1994. Metabolic rate and oxygen consumption
requirements of some fish species from the middle Parana river. Acta Biol.
Venez. 15(2):1-10.

58
Petry, P., Bayley, P. B., Markle, D. F., Relationships between fish assemblages,
macrophytes and environmental gradients in the Amazon River floodplain.
Journal of Fish Biology (2003) 63: 547-579.
Piedade, M.T.F.; Junk, W.J.; Parolin, P. 2000. The flood pulse and
photosynthetic response of trees in a white water floodplain (várzea) of the
Central Amazon, Brazil. Verh. Internat. Verein. Limnol., (27):1-6.
Roberts, T. R. 1972. Ecology of fishes in the Amazon and Congo Basins. Bll.
Mus. Comp. Zool., 143(2): 117-147.
Rodriguez, M.A. & Lewis W.M. (1997) Structure of fish assemblages along
environmental gradients in floodplain lakes of the Orinoco River. Ecological
Monographs 67: 109- 128 p.
Ruiz, L.A & Berlanga, C. A. 1999. Modifications in coverage patterns land use in
the landscape around the Huizache-Caimanero lagoon system, Sinaloa,
México: a multitemporal analysis using Landsat images. Est. Coast. Shelf
Sci., 49: 37-44 p.
Santos, G. M. 1981. Estudos de alimentação e hábitos alimentares de
Schizodon fasciatus Agassiz, 1829, Rhytiodus microlepis Kner, 1859 e
Rhytiodus argenteofuscus Kner, 1859, do lago Janauacá-Am, Acta
Amazônica, 11: 267-283.
Santos, G. M. 1991. Pesca e ecologia dos peixes de Rondônia. Tese de
Doutorado. INPA/FUA, Manaus. 213p.
Saint-Paul, U. & M.G.M. Soares. 1987. Diurnal distribution and behavioral
responses of fishes to extreme hypoxia in an Amazon floodplain lake. Env.
Biol. Fish. 20:91-104.

59
Saint-Paul, U.; Zuanon, J.; Villacorta-Correa, M.A.; Garcia, M. Fabré, N.N.;
Berger, U.; Junk, W. J. 2000. Fish communities in Central Amazonia white-
and the blackwaters floodplains. Environmental Biology of Fishes 57:235-
250 p.
Salati, E.; Santos, A. A.; Lovejoy, T. E. & Klabin, I., 1998. Porque salvar a
Floresta Amazônica. Manaus: INPA. 114 p.
Sánchez-Botero, J. I.; Araújo-Lima, A.R.M. 2001. As macrófitas aquáticas como
berçário para a ictiofauna da várzea do rio Amazonas. Acta Amazonica,
Manaus, v.31, n.3:437-447 p.
Sioli, H., 1984. The Amazon and its main effluents: Hydrography, morphology of
the river courses, and river types. 127-166 p. In: The Amazon limnology and
landscape ecology of a mighty tropical river and its basin. The Hague, W.
Junk Publications, Netherlands.
Siqueira-Souza, F. K. 2002. Caracterização das comunidades de peixes de
lagos de várzea no Médio Rio Solimões. Monografia, UFAM, Manaus. 36p.
Soares, M. G. M.; Almeida, R. G.; Junk, W. J. 1986. The tropic status of the fish
fauna in lago Camaleão e Macrophyte dominated floodplain lake in the
middle Amazon. Amazoniana, 9 (4): 511-526.
Soares. M.G.M. 1993. Estratégias respiratórias em peixes do Lago Camaleão
(Ilha da Marchantaria) – AM, Brasil. PhD Thesis, INPA/FUA, Manaus.
Soares, M.G.M.; Junk, W.J. 2000. Commercial fishery and fish culture of the
state of Amazonas: Status and perspectives. In: The Central Amazon
Floodplain: Actual Use and Options for a Sustainable Management.
Backhuys Publishers, Leiden, The Netherlands, 433-461.

60
Soares, M.G.M & Yamamoto, K.C. 2005. Diversidade e composição da
ictiofauna do Lago Tupé. 181-197 p. In: Santos-Silva, E. N; Aprile, F. M.;
Scudeller, V. V. & Melo, S. 81 (orgs). 2005. Biotupé: Meio Físico,
Diversidade Sociocultural do Baixo Rio Negro, Amazônia Central. Ed.
INPA: 246pp.
Souza-Pereira, A. 2000. Variáveis limnológicas e sua relação com as espécies
de peixes onívoras e herbívoras do lago Camaleão, Amazonas, Brasil.
Monografia FUA, Manaus, 31 p.
Súarez, Y.R., Petrere Jr., M & Catella, A.C. 2001. Factors determining the
structure of fish communities in Pantanal lagoons (MS, Brazil). Fisheries
Management and Ecology. 8,173-186.
Tejerina-Garro F.L., Fortin, R. & Rodriguez, M.A. 1998. Fish community
structure in relation to environmental variation in floodplain lakes of the
Araguaia River, Amazon Basin. Environmental Biology of Fishes, 51: 399 –
410 p.
Torrente-Villara, G.; Araújo, T. R. & Doria, C. R. C. 2003. Base cientifica para o
manejo da pesca da região do médio Guaporé: componente Ictiofauna.
Relatório Técnico, SEDAM/TERMONORTE/ECOPORÉ, Porto Velho. 120
pp.
Vale, J. D. 2003. Composição, diversidade e abundância da ictiofauna na área
do Catalão, Amazônia Central. Dissertação de Mestrado. Instituto Nacional
de Pesquisas da Amazônia/ Fundação Universidade do Amazonas,
Manaus, Amazonas. 66 pp.

61
Waldhoff, D.; Maia, L. 2000. Production and chemical composition of fruit from
trees in floodplain forests of Central Amazonia and their importance for fish
production. In: The Central Amazon Floodplain: Actual use ands options for
a sustainable management. Ed. W. J. Junk; Ohnly; M. T. F. Piedade; M. G.
M. Soares. Backhuys Publishers. Leiden. p. 393-410.
Welcomme, R. L. 1985. River fisheries. FAO Technical Paper 262, Rome.
Yamamoto, K.C. 2004. Comparação das estruturas de comunidades de peixes
em lagos manejados da Amazônia Central. Dissertação de mestrado.
Universidade Federal do Amazonas – Instituto Nacional de Pesquisas da
Amazônia – INPA. Manaus, Amazonas. 50pp.
Zar, J.H. 1999. Biostatistical analysis. 4ª edição. Prentice-Hall, Inc., Englewood
cliffs, N.J.663p.
Zuanon, J. A. S.; Ferreira, E. J. G. Santos, G. M.; Amadio, S.; Benedito, R. M.
L. B.; Darwich, A. J.; Alves, L. F.; Almeida-Prado, R. 2004. Título:Relatório
Técnico. SEPLAN/Naturatins, Palmas. 92 pp.





























