Embed Size (px)
Citation preview

1
1. Introdução
Sendo estudante de Mestrado Integrado em ciências de Engenharia Biomédica desenvolvi o
gosto e o interesse, ao longo da minha vida académica, pela área das neurociências. Contacto com
unidades curriculares como Imagiologia e Métodos de Imagem Médica ajudaram-me a decidir, a
escolher e a procurar este campo de estudo para a realização da minha tese de mestrado.
O tema escolhido centra-se na análise de espessura cortical (EC). Esta tem bastante
relevância para o estudo do cérebro quer a nível anatómico quer do ponto de vista funcional ou
patológico, como é o caso pontual deste trabalho.
O objectivo do trabalho aqui presente é o de fornecer um estudo preliminar em como a LHON
pode afectar o córtex visual. Para atingir este fim, 16 pacientes com a patologia em questão foram
seleccionados e propostos a realizar duas sessões de ressonância magnética (Magnetization
Prepared Rapid Acquisition Gradient Echo - MPRAGE) de 9 minutos cada, uma seguida da outra.
Durante as sessões os pacientes foram sujeitos a um estímulo visual de modo a obter informação
sobre a actividade cerebral no lobo occipital (região onde está o cortex visual). Os dados
provenientes do scanner foram processados no software Brainvoyager QX v2.0. Para a obtenção dos
valores da EC do cortex visual dos pacientes, os dados foram analisados em equipa. Foi realizada
análise anatómica (feita por mim) e análise funcional (feita por um colega da equipa em que estive
inserido) no software referido. Irei explanar o processo da análise anatómica, com este software, do
ponto de vista metodológico e algorítmico.
Após a obtenção dos valores de EC, estes foram sujeitos a determinados processos
estatísticos a fim de os interpretar, atribuir significado e identificar possíveis padrões de
comportamento. Foram aqui realizados testes estatísticos de normalização, comparação de valores
médios e análise de correlação; todos com o software Statistical Package for the Social Sciences
v18.0 (SPSS).
Finalizado o processamento de dados e análise estatística, os resultados e conclusões deste
grupo com Leber estão aptos para, no decorrer da investigação sobre a doença de Leber, serem
comparados com os resultados de um trabalho equivalente realizado em um grupo de controlo.
Nos restantes sub-capítulos da Introdução falarei sobre os conceitos de: espessura cortical; o
cortex visual e seu mapeamento (análise funcional); a doença de Leber; e o software Brainvoyager
QX.

2
1.1 A importância de medir a Espessura Cortical
A Espessura Cortical (EC) é uma medida anatómica utilizada para descrever a espessura das
camadas do córtex cerebral nos cérebros dos mamíferos. A análise de espessura cortical mede a
espessura da matéria cinzenta presente no córtex. Pode referir-se à espessura global em todo o
córtex cerebral, ou à espessura local estudando o número de neurónios disponíveis por região.
Apesar das capacidades cognitivas de um indivíduo dependerem de variadíssimas condicionantes a
EC representa um papel importante no grau de capacidades cognitivas que o indivíduo pode ter.
A EC varia com alguns factores. O tamanho do cérebro, tanto dentro como entre espécies, é
preponderante nestas variações. No que diz respeito ao ser humano, a EC, pode revelar o
funcionamento e anatomia das redes neuronais [1]. As diferenças sexuais em termos de EC são
pequenas, apesar das mulheres terem valores ligeiramente superiores na região temporoparietal [2].
No cérebro humano adulto a zona mais fina é a fissura calcarina, que tem cerca de 2 mm e a mais
espessa é o giro pré-central, que tem cerca de 4 mm [3]. Globalmente, a EC varia entre 1,5 e 3 mm.
Verifica-se um processo de decaimento da EC resultante da maturação e envelhecimento dos
tecidos, o qual é conhecido como “cortical thinning” ou afinamento cortical. Esta perda ou atrofia da
EC é cerca de 10µm por ano [4]. Foi também demonstrado que, com o envelhecimento, existe um
aumento dos sulcos do córtex [5].
A EC representa uma ajuda útil. Não só no estudo do desenvolvimento e maturação do
cérebro saudável, como ainda na sua rotulagem [6 - 10]. Processos automáticos para medir a EC
humana podem fornecer ferramentas poderosas no diagnóstico de desordens neurológicas. Para fins
de diagnóstico, o conhecimento destes padrões e valores de EC, quer local quer globalmente, é
bastante vantajoso, uma vez que desvios de valores de EC podem representar a existência de uma
ou mais patologias neurológicas. São conhecidas variações da EC em doenças tais como epilepsia
[6], esquizofrenia [3], anorexia nervosa [3] e Alzheimer [3 e 10]. Quanto mais precisos estes padrões
forem, mais facilmente e precocemente se identificam patologias como estas, podendo assim
acompanhar de perto a sua evolução e aumentar a qualidade de vida dos doentes. Pretende-se
precisamente com este trabalho apresentar padrões de EC do córtex visual que possam ser
exclusivos da doença de Leber.
A EC cerebral é calculada através da segmentação cortical que separa os vários tipos de
tecidos e órgãos presentes na cabeça (crânio, dura, matéria branca, matéria cinzenta, etc…). A EC é
normalmente determinada a partir das medições de matéria cinzenta sendo esta segmentada por
métodos de neuroimagem. A EC é, então, a distância entre duas fronteiras: a fronteira entre a matéria
branca e matéria cinzenta e a fronteira entre a matéria cinzenta e o líquido cefalorraquidiano (também
conhecida como superfície pial). Os métodos de medição para este fim são trabalhosos, exigindo
vários dias de trabalho manual e de processamento. Além disso, visto o cérebro ter uma geometria
bastante irregular, existe um erro associado às medições. No entanto, hoje em dia há poderosos
algoritmos de reconstrução automatizada [11 - 14] e técnicas de alta resolução de superfícies médias

3
[15] que permitem a investigação de processos neurodegenerativos ou processos de recuperação
neurológica no que diz respeito ao córtex cerebral, globalmente ou em delimitadas áreas funcionais
como é disso exemplo o presente trabalho.
1.2 O Córtex Visual
A região do córtex responsável pela visão é o lobo occipital. O córtex visual divide-se em
córtex visual primário, também conhecido como córtex estriado ou área visual V1, e córtex extra-
estriado. O córtex extra-estriado compreende as áreas visuais V2, V3, V4 e V5. Neste trabalho serão
estudadas as áreas visuais V1, V2 e V3 excluindo as áreas visuais V4 e V5, pois são áreas pouco
definidas no cérebro humano, apesar de estudadas e conhecidas no cérebro do macaco.
A localização do córtex visual primário corresponde à região de Brodmann 17 e as áreas
extra-estriadas às regiões de Brodmann 18 e 19 (ver Figura 1.1). O córtex visual do hemisfério
esquerdo recebe sinais do campo visual direito e o córtex visual do hemisfério direito recebe sinais do
campo visual esquerdo.
Figura 1.1: Áreas de Brodmann do cerebro humano. Imagem retirada de www.mrc-
cbu.cam.ac.uk/.../brainbrodmannareas.gif
O cortex visual primário, V1, está localizado em torno da fissura calcarina situada no lobo
occipital. Em cada hemisfério, esta área visual recebe informações do seu núcleo geniculado lateral
(Lateral Geniculate Nucleos - LGN). Este último é o principal responsável pelo processamento de
informações visuais recebidas da retina no olho. Recebe assim, informações directamente das RGCs

4
através do nervo óptico, sendo que, em Leber o LGN recebe menos informação com a progressão da
doença devido à degeneração das RGCs. O LGN faz parte do sistema nervoso central pois encontra-
se dentro do tálamo, também localizado no cérebro. Os neurónios do LGN ligam-se directamente ao
córtex visual primário V1 através dos seus axónios. Permite deste modo comunicação directa com V1
recebendo também fortes ligações de feedback deste.
Acredita-se que cada área V1 transmite a informação visual para as outras áreas visuais em
duas vias: uma via chamada fluxo dorsal e outra chamada fluxo ventral. Esta hipótese foi apresentada
por Ungerleider e Mishkin em 1982 sendo amplamente aceite apesar de controversa [16 e 17]. O
fluxo ventral viaja pelo lobo temporal e é responsável por processar a informação visual do ponto de
vista da pergunta “O quê?”. Dá um significado e interpretação ao que se está a ver. O fluxo dorsal
viaja pelo lobo parietal e é responsável por processar a informação visual do ponto de vista da
pergunta “Onde?”. Está associado ao movimento, coordenação motora e a uma representação
espacial do que se está a ver. Na imagem seguinte (Figura 1.2) podemos ver uma representação
desta teoria de dois fluxos:
Figura 1.2: Fluxo dorsal (verde) e fluxo ventral (roxo). Originam-se ambos de V1. Imagem retirada de http://en.wikipedia.org/wiki/File:Ventral-dorsal_streams.svg
Os neurónios, presentes no córtex visual, disparam potenciais de acção quando recebem
estímulos visuais provenientes da zona do campo visual que interpretam. Por definição, esta zona do
campo visual é chamada de campo receptivo. No entanto os neurónios podem não responder da
mesma maneira a estímulos provenientes do seu campo receptivo. Um dado neurónio, pode
responder melhor a um subconjunto de estímulos que a outro. Esta capacidade de resposta dos
neurónios a diferentes estímulos tem o nome de “neuronal tunning” ou resposta neuronal. É
conhecido que um neurónio, da área visual V1, pode ser accionado por qualquer estímulo vertical
presente no seu campo receptivo. Já nas áreas visuais extra-estriadas os neurónios respondem de
maneira mais complexa. Por exemplo, uma resposta neuronal pode dar-se apenas quando uma certa
face ou objecto aparece no campo receptivo de um neurónio que pertença a uma área visual extra-
estriada por onde passa o fluxo ventral.

5
1.2.1 Córtex estriado - Área visual V1
O córtex visual primário ou córtex visual V1 estriado é, das áreas visuais, a maior e a melhor
definida e estudada do cérebro (quer humano quer animal). V1 é altamente especializado no
processamento de informação visual relativa a objectos estáticos e em movimento como também, no
reconhecimento de padrões. As propriedades de resposta neuronal dos neurónios presentes em V1
diferem, também, consoante o tipo de estímulo. Individualmente, neurónios em V1 têm respostas
neuronais diferentes. Podem estes descriminar pequenas mudanças nas orientações visuais,
espaciais e reconhecimento das cores. Num cérebro adulto normal a área visual V1 possui para esta
tarefa cerca de 140 milhões de neurónios em cada hemisfério [18]. Anatomicamente e
funcionalmente, V1 tem um mapa muito bem definido no que concerne à informação espacial no
campo visual. Por exemplo, no ser humano, a margem superior do sulco calcarino tem uma resposta
neuronal forte à informação visual proveniente do campo visual inferior e a margem inferior à
informação visual proveniente do campo visual superior [17]. Este tipo de mapeamento retinotópico é
uma transformação da imagem vista pela retina para V1. Nesta área visual, este mapeamento está
muito bem definido. A correspondência entre um campo receptivo e os seus neurónios em V1 é bem
conhecida e até pontos cegos estão mapeados. Noutros animais que possuem um córtex visual e
uma área visual como V1, este tipo de correspondência também é fácil de definir e básica. Verifica-se
também, que tanto em seres humanos como em animais que possuem uma fóvea na retina, existe
uma grande porção dos neurónios de V1 que processam essa pequena parte do campo visual. Este
fenómeno é conhecido como “cortical magnification” ou magnificação cortical. Entende-se que para
obter maior informação sobre o que estamos a ver, há um grande número de neurónios a processar a
pequena área correspondente ao nosso foco de visão (fóvea) e menor número de neurónios a
processar a visão periférica.
1.2.2 Córtex extra-estriado - Área visual V2
A área visual V2 é a segunda maior área do córtex visual e a primeira região da área de
associação visual. V2 tem fortes conexões com a área visual V1, trocando informação em
feedforward e feedback com este. Têm também fortes ligações com a área visual V3.
Anatomicamente V2 está dividida em quatro quadrantes. Possui duas regiões ventrais, uma em cada
hemisfério e duas regiões dorsais, também uma em cada hemisfério.
Funcionalmente, V2 tem propriedades em comum com V1. Sendo que os neurónios, também
respondem a estímulos simples como a orientação espacial, movimento e cor. Têm ainda uma maior
capacidade de modulação da atenção que V1. Aqui a resposta neuronal é ajustada segundo padrões
mais complexos e pode dividir ainda o campo receptivo em sub-regiões. Afirma-se que o fluxo ventral
tem um papel preponderante no processamento, recolha e evocação da memória visual [20 e 21].
Defende-se que V2 tem um papel importante na conversão da memória de curto prazo para memória

6
de longo prazo e também que falhas de memória visual e défices no reconhecimento de objectos
possam resultar da manipulação da área visual V2 [22].
1.2.3 Córtex extra-estriado - Área visual V3
A área visual V3 ou complexo visual V3 encontra-se imediatamente depois da área visual V2.
Denomina-se “complexo” visual visto existir ainda alguma controvérsia em relação à extensão exacta
da área visual V3. Foi proposto que V3 possa ter duas ou três subdivisões funcionais, como também
a existência de uma região V3 dorsal na parte superior de cada hemisfério e uma região V3 ventral
(ou VP de região posterior ventral) localizado na parte inferior do cérebro [23]. Supõem-se que V3
dorsal e V3 ventral têm conexões distintas com as outras áreas visuais respondendo os neurónios a
diferentes estímulos apesar de pertencerem à mesma região (neurónios de cor selectiva são os mais
comuns em V3).
V3 dorsal (ou V3 neste trabalho) é considerado como região pertencente ao fluxo dorsal,
recebendo informação de V2 dorsal e de V1, projectando-se na área de Brodman19 (ver Figura 1 e
2). Algumas conclusões por meio de ressonância magnética funcional sugerem que V3 dorsal
desempenha, ou está presente, na transformação do movimento Global [24]. Já V3 ventral (ou VP
neste trabalho) tem ligações mais fracas com V1 e mais fortes com V2. Primeiramente pensava-se
que VP processava informação visual apenas do campo visual superior mas recentemente é
evidenciado que VP é mais extenso do que se pensava e que, tal como as outras áreas visuais,
possui uma representação de todo o campo visual [25].
1.2.4 Mapeamento do cortex visual – A Retinotopia
A retinotopia é o método de mapear o córtex cerebral com dados funcionais de ressonância
magnética. Esta permite descrever e fazer corresponder a resposta neuronal dos neurónios de uma
determinada região do córtex ao estímulo visual que recebem de uma determinada região do campo
visual. Como já foi referido, os neurónios do córtex visual são estimulados por alterações no seu
campo receptivo. A retinotopia permite então saber quais os neurónios estimulados por alterações
numa determinada região do campo visual. Desta maneira é possível fazer a ligação entre neurónios
e seus respectivos campos receptivos. Os mapas retinotópicos são um caso particular de
organização topográfica e foram descobertos com o decorrer do estudo de danos neurológicos devido
a doenças ou lesões [26].
As fronteiras das áreas visuais são delimitadas, através de dados funcionais de ressonância
magnética, pois verifica-se o comportamento de simetria entre córtex e campo visual. Com o estímulo
adequado é possível verificar as transições entre as áreas visuais. Desta forma as áreas visuais são
definidas nestes mapas retinotópicos, tendo as suas fronteiras circunscritas. Sendo a área visual V1 a
mais bem conhecida, enquanto as outras áreas são alvo de intensa investigação pois não são tão

7
bem caracterizadas como V1. É usado um critério que estipula que cada área visual tem uma
representação completa do campo visual [27 - 33].
O mapeamento por dados funcionais de ressonância magnética é uma importante
ferramenta, visto ter a grande vantagem de ser um método não invasivo. Outra grande vantagem
reside na manipulação do estímulo visual que permite, para além de definir ligações entre neurónios e
seus campos receptivos, fazer a correspondência entre neurónios e conjunto de estímulos aos quais
respondem. Esta correspondência é bem conhecida para a área visual V1 [33].
Ainda relativamente a V1, a retinotopia desempenhou um papel importante na descoberta da
magnificação cortical, já referida no ponto 1.2.1. Mapeando o campo visual no córtex temos acesso
às regiões neuronais responsáveis pelo processamento do campo visual central e do campo visual
periférico. Verifica-se, no mapa retinotópico, que existe maior porção de neurónios a processar o que
a fóvea vê (visão central) e menos neurónios a processar estímulos periféricos (visão periférica). Esta
perda de número de neurónios, no processamento do estímulo visual, com o aumento da sua
distância ao centro da visão é gradual. Neste trabalho, a análise funcional, considerou 3 zonas em
cada área visual mapeada: zona da fóvea, zona intermédia e zona periférica. Na imagem seguinte
(Figura 1.3) podemos ver uma representação do olho onde está assinalada a fóvea:
Figura 1.3: O olho Imagem retirada de http://www.nei.nih.gov/health/eyediagram/
O estímulo visual usado em cada paciente consistiu numa cunha giratória no sentido anti-
horário. Durante 9 minutos os pacientes tinham de fixar o ponto vermelho no centro enquanto a cunha
girava (eu também me submeti a este exame retinotópico com o fim de saber aquilo ao qual os

8
pacientes foram sujeitos). Na imagem seguinte (Figura 1.4) está representado um snapshot do
estímulo visual:
Figura 1.4: Estímulo retinotópico
1.3 Doença hereditária de Leber (LHON)
A doença hereditária de Leber foi seguida e descrita, pela primeira vez, por Theodore Leber
no século XIX, mas só em 1980 foi entendida a sua causa e consequência. Esta doença (Leber’s
Hereditary Optic Neuropaty - LHON) é caracterizada pela degeneração das células ganglionares da
retina (Retinal Ganglion Cells - RGCs) e seus axónios que leva a uma perda aguda ou semi-aguda da
visão central. Esta degeneração celular das RGCs resulta de uma alteração no DNA mitocondrial
(DNAmt). A LHON é descrita pela perda bilateral, indolor, da visão central durante o inicio da vida
adulta. Os homens são aproximadamente quatro vezes mais susceptíveis de serem afectados do que
as mulheres. A LHON é, geralmente, assintomática até os indivíduos afectados desenvolverem a
perda da visão central. Esta surge, normalmente, entre os vinte e os trinta anos de idade, podendo
desenvolver-se em qualquer idade, tanto em homens como em mulheres [34]. Quando a perda de
visão ocorre, normalmente, é num olho e cerca de oito semanas depois no outro olho, sendo esta a
fase aguda da doença [34]. Não há registos de melhorias significativas após a fase aguda.
Com a progressão da LHON, durante a fase aguda, a acuidade visual é severamente
reduzida ao ponto de contar dedos, perde-se a capacidade de distinguir algumas cores e, em alguns
casos, formam-se regiões cegas na visão central. Contudo, após a fase aguda, a visão deixa de
piorar. Indivíduos nestas condições são considerados cegos em termos sociais e legislativos. Existem
algumas desordens neurológicas comuns em paciente com LHON. São delas exemplo tremor
postural ou distúrbios do movimento. Mulheres com LHON podem estar em risco de esclerose
múltipla [34].
Sendo uma doença hereditária transmite-se directamente por via vertical. A LHON só é
transmitida através da mãe pois é ela que fornece o genoma mitocondrial, presente no óvulo, durante
a fecundação. Os homens não podem transmiti-la aos seus filhos [34]. A LHON surge, geralmente,

9
devido a uma de três mutações patogénicas no DNAmt. Como é sabido, as estruturas mitocondriais
funcionam como as baterias das células, fornecendo energia para as mesmas. Em particular, pensa-
se que as alterações existentes no DNAmt dos doentes com LHON se reflectem numa diminuição ou
cessação da quantidade de energia disponível para as RGCs no nervo óptico e no olho. Estas células
são danificadas com a progressão da doença podendo até morrer por falta de energia. Este dano nas
RGCs provoca os sintomas, já referidos, da LHON.
O diagnóstico da LHON surge em exames oftalmológicos. Exames como o de fluorescência
em angiografia permitem identificar as características vasculares presentes no olho durante a fase
aguda. Realizam-se exames de estudo dos sinais electro-fisiológicos (potenciais evocados) no olho,
que confirmam as disfunções eléctricas presentes no nervo óptico. Posteriormente, também se
submete os pacientes a um electro-retinograma para excluir a existência de doenças na retina [36].
Na imagem seguinte (Figura 1.5) está representada a transmissão vertical da doença de Leber:
Figura 1.5: Esquema em como o DNAmt pode ser transmitido à prole. Imagem retirada de
http://en.wikipedia.org/wiki/File:Mitochondrial.jpg
Tendo em mente as características da doença de Leber, surge uma pergunta interessante do
ponto de vista das suas consequências para o cérebro, como é aliás o âmbito deste trabalho. Pode
especular-se se os danos e a morte de neurónios do nervo óptico irão causar alguma alteração na
região do cérebro responsável pela visão. É possível que a redução de estímulo visual que chega ao
córtex provoque uma atrofia neste. Espero que os resultados aqui apresentados ajudem, de algum
modo, a responder a esta pergunta.

10
1.4 O software Brainvoyager QX
O Brainvoyager QX é um software usado para a análise, processamento e visualização de
dados de ressonância magnética. É um programa de fácil aprendizagem e utilização
representando uma lufada de ar fresco neste tipo de análise. Existem softwares, como o FreeSurfer
(com o qual estou familiarizado e que aprendi antes do Brainvoyager QX) para o mesmo fim, mas que
em comparação deixa muito a desejar (elevado tempo de calculo e bastante confuso em aprender e
usar). O software Brainvoyager QX foi escolhido para integrar a metodologia desta tese devido a
vários factores. O principal foi o facto de, quando fui inserido na equipa de trabalho, todos os
investigadores trabalhavam com o Brainvoyager QX. Deste modo, para não analisar os mesmos
dados por métodos diferentes, aprendi também a funcionar com este programa. Outro, tal como já
referido, deve-se ao facto de o Brainvoyager QX ser um programa simples e agradável na óptica do
utilizador.
A análise de dados de ressonância magnética pelo Brainvoyager permitiu o desenvolvimento
de biomarcadores indirectos de efeitos do envelhecimento, efeitos de doenças em estruturas do
cérebro e compreensão da plasticidade do tecido neurológico como por exemplo em habilidades
cognitivas [37]. O Brianvoyager QX foi desenvolvido por Rainer Goebel e as suas especificidades
estão presentes em http://www.brainvoyager.com/products/brainvoyagerqx.html.
Com este programa é possível fazer toda a análise, de dados de ressonância magnética,
necessária para o trabalho aqui presente. Trata-se de trabalho de equipa em que a análise
anatómica/volumétrica foi feita por mim e a análise funcional feita por um colega. A análise
anatómica/volumétrica divide-se em duas partes: a criação de moldes para a representação 3D do
córtex e têm por nome mesh, usados para fins de visualização; a criação dos mapas corticais com os
valores de EC que são projectados nas meshes criando assim o input de dados para a análise
funcional. Quanto a esta, apesar de não ter passado por mim, trata-se do mapeamento das áreas
visuais (retinotopia). Estas, depois de definidas e delimitadas, são divididas em 3 zonas (fóvea,
intermédia e periférica) retirando de cada uma 10 pontos com valores de EC.
As principais funcionalidades usadas por mim no Brainvoyager QX são as ferramentas e
algoritmos para a segmentação do cérebro, reconstrução, visualização e modelação da superfície
cortical [38]. Estas são usadas para a criação das meshes e dos mapas de EC. Com a funcionalidade
da projecção dos mapas sobre as meshes é possível retirar informação individual anatómica de
certas regiões do córtex (análise funcional). A principal funcionalidade, para este efeito, usada na
análise funcional é a de o mapeamento cortical poder ser feito no volume e no espaço (i.e. nas
representações 3D criadas por mim).
Para o cálculo dos valores de EC o Brainvoyager QX usa o método matemático de Laplace
[39]. Este método aplica a equação diferencial de Laplace (∇2ψ = 0) onde o volume do córtex é
representado como domínio para a solução da equação diferencial, com condições de contorno nas

11
fronteiras da matéria cinzenta (com a matéria branca e com o liquido cefalorraquidiano) calculadas
pelo processo de análise anatómica/volumétrica. A normalização do gradiente ψ forma um campo
vectorial, representando vectores tangentes às linhas do campo que ligam as fronteiras da matéria
cinzenta. Desta forma e com este método a espessura é medida de uma forma tridimensional
podendo ser obtida através de qualquer ponto no córtex.

12
2. Metodologia
2.1 BrainVoyager QX
Como já referido na introdução, fui incumbido de realizar a análise anatómica dos dados
referentes aos 16 pacientes com LHON. Irei de seguida expor o meu trabalho que consistiu na
criação das meshes e dos mapas corticais para cada um dos 16 pacientes. Os mapas e as meshes
são criados por processos diferentes e começarei por relatar a criação das meshes que usa uma
ferramenta do Brainvoyager QX de nome “Segmentação Automática”. Esta última pode fornecer
dados visuais de EC, mas estes não são fiáveis para fins de medição, sendo a sua finalidade apenas
a criação de representações tridimensionais com o objectivo da sua visualização. A criação dos
mapas corticais é feita por uma outra ferramenta de nome “Segmentação Avançada”. As imagens
aqui presentes referem-se ao processo de análise completo para um dos pacientes ao qual, por
motivos de confidencialidade, chamarei de subject.
Para uma análise anatómica e tridimensional do cérebro de cada paciente, o Brainvoyager
QX cria e opera em ficheiros com a extensão VMR (Volume Magnetic Resonance). Estes ficheiros
armazenam os dados pertencentes a uma aquisição (MPRAGE) com ponderação em T1 do cérebro.
No trabalho presente foram criados estes ficheiros para cada um dos 16 pacientes. Toda a análise
levou em média 6 horas sendo constituida de análise manual e computacional, por paciente, até obter
as meshes e os mapas corticais desejados.
2.2 Análise Anatómica e Volumétrica de dados – Criação das Meshes
A ferramenta que permite a construção destes moldes 3D para fins de visualização é a
Segmentação Automática. De seguida irei relatar e explicar os vários passos presentes neste
processo.

13
2.2.1 Etapas preparatórias antes da Segmentação Automática
Antes desta fase do processo de análise usando a ferramenta “Segmentação Automática”
algumas etapas tiveram de ser levadas em consideração:
1. O conjunto de dados anatómicos 3D têm de ter uma resolução espacial de 1x1x1mm,
caso contrário terão de ser convertidos. Isto é necessário, pois o Brainvoyager QX
trabalha nesta resolução espacial para a criação dos ficheiros VMR;
2. Foram realizadas duas MPRAGE de 9 minutos cada, pelo que, foi feita uma média
entre as duas (de nome Co-registo) que permite uma melhor qualidade de imagem
melhorando a relação sinal-ruído;
3. Uma Correcção de Homogeneidade foi realizada, funcionando como uma “pré”
segmentação, definindo um bom contraste entre matéria branca e cinzenta;
4. O conjunto de dados foi transformado para o espaço Talairach, devido ao facto de o
Brainvoyager QX explorar o conhecimento anatómico deste sistema, a fim de realizar
a segmentação do cérebro.
De todos estes pontos apenas o ponto 1 não foi realizado por mim pois os dados provinham
do scanner já na resolução espacial desejada de 1x1x1mm.
2.2.1.1 O Co-registo
Cada paciente foi sujeito a duas sessões de ressonância magnética de 9 minutos cada. Para
trabalhar e analisar estes dados provenientes do scanner, o Brainvoyager QX cria um ficheiro VMR
para cada aquisição. Visto terem sido efectuadas duas sessões, é possível fazer uma média dos
dados melhorando a relação sinal-ruído. Efectuando um co-registo de dados anatómicos alinham-se
dois VMR (correspondentes a duas aquisições da mesma pessoa) resultando num VMR final, que
possui uma imagem de melhor qualidade. O co-registo de conjuntos de dados é um passo importante
na análise e visualização de dados, quer anatómicos quer funcionais, no espaço 3D.
Ao criar um ficheiro anatómico 3D (VMR), o Brainvoyager QX irá criar automaticamente um
conjunto de dados V16 correspondente (16 bits de informação referente às intensidades de cada
voxel). Este conjunto de dados é um formato com uma representação exacta da informação de
intensidades por voxel em todo o volume, em escala de cinzento de 0 a 225. Esta representação
específica é necessária para procedimentos importantes como, por exemplo, as correcções de
homogeneidade. Cada aquisição, mesmo sendo do mesmo paciente, irá ter uma distribuição de
intensidades diferente. O processo de co-registo irá criar um ficheiro VMR, com um conjunto de
dados V16 associado, que representa a média das distribuições de intensidade de ambas as
aquisições.
Seguidamente apresento o processo de criação deste VMR médio. O primeiro passo é criar
um VMR para cada aquisição. O VMR é construído seleccionando o primeiro ficheiro Dicom da
14
imagem (Dicom é o formato dos ficheiros provenientes do scanner). O Brainvoyager QX irá depois
compilar todas as fatias, criando uma imagem 2D do volume 3D. Esta imagem corresponde na
realidade a 3 imagens de três vistas: coronal, transversal e sagital. Cada uma das vistas possui 255
fatias por onde se pode “passear” e ver a anatomia do cérebro.
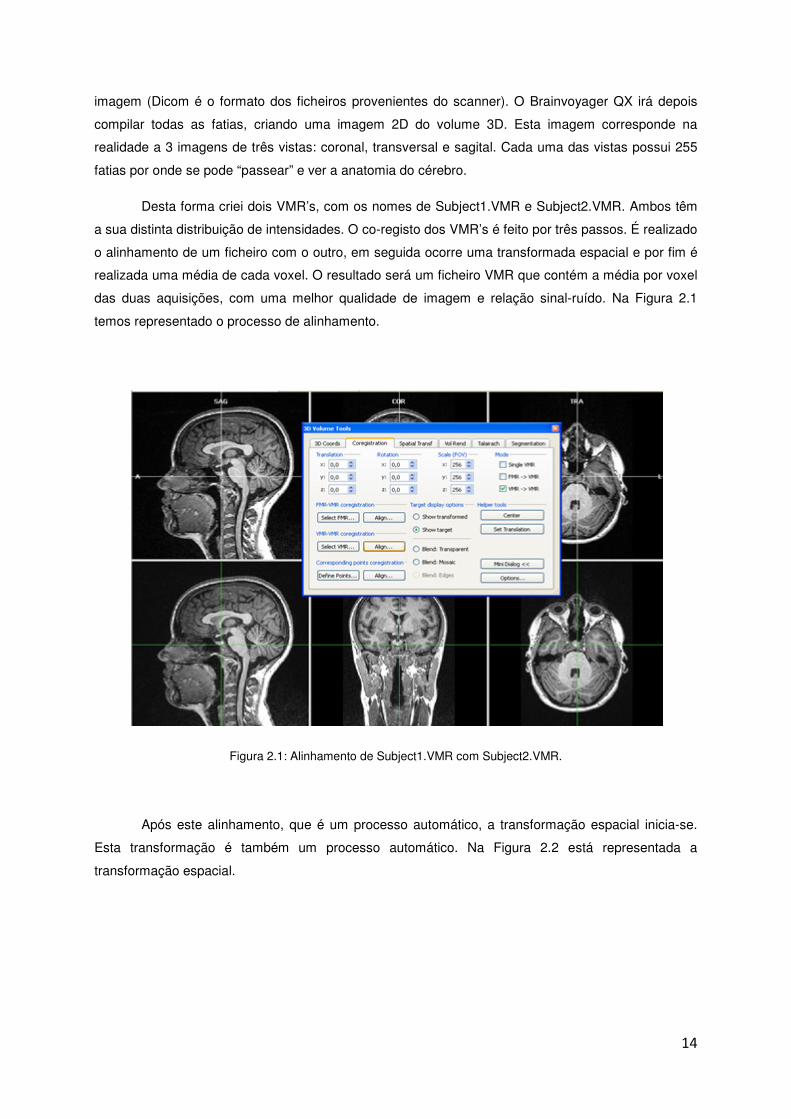
Desta forma criei dois VMR’s, com os nomes de Subject1.VMR e Subject2.VMR. Ambos têm
a sua distinta distribuição de intensidades. O co-registo dos VMR’s é feito por três passos. É realizado
o alinhamento de um ficheiro com o outro, em seguida ocorre uma transformada espacial e por fim é
realizada uma média de cada voxel. O resultado será um ficheiro VMR que contém a média por voxel
das duas aquisições, com uma melhor qualidade de imagem e relação sinal-ruído. Na Figura 2.1
temos representado o processo de alinhamento.
Figura 2.1: Alinhamento de Subject1.VMR com Subject2.VMR.
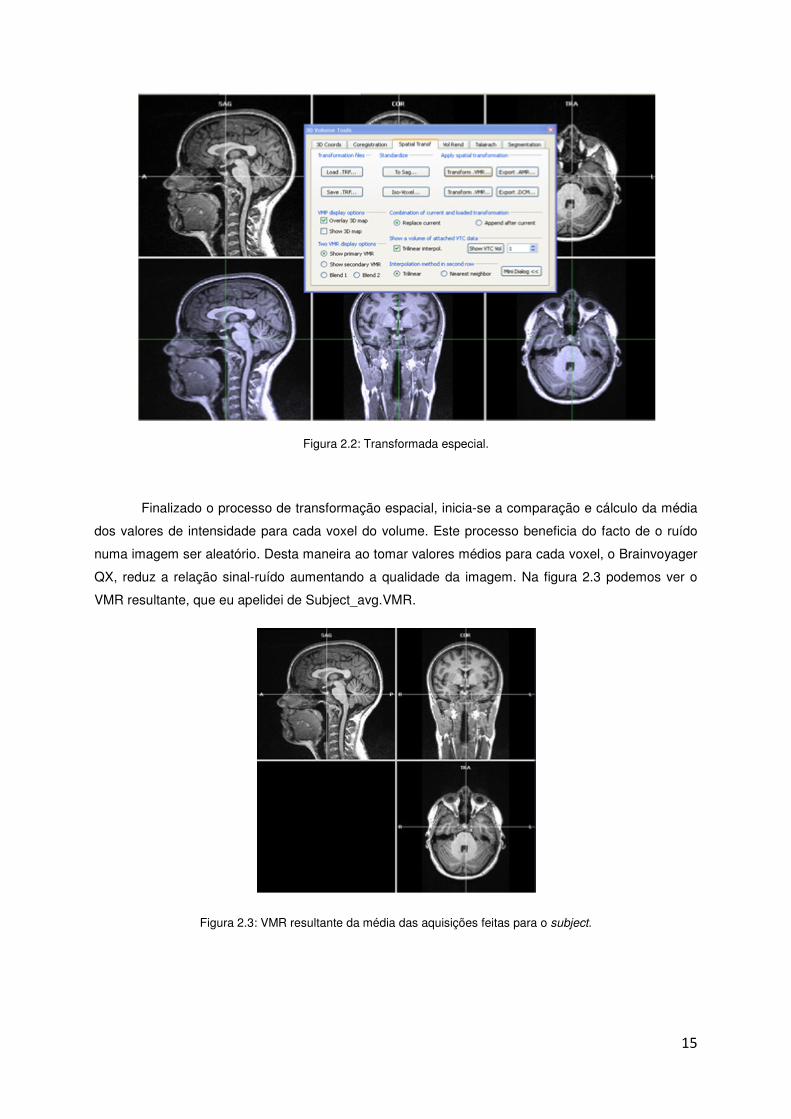
Após este alinhamento, que é um processo automático, a transformação espacial inicia-se.
Esta transformação é também um processo automático. Na Figura 2.2 está representada a
transformação espacial.
15
Figura 2.2: Transformada especial.
Finalizado o processo de transformação espacial, inicia-se a comparação e cálculo da média
dos valores de intensidade para cada voxel do volume. Este processo beneficia do facto de o ruído
numa imagem ser aleatório. Desta maneira ao tomar valores médios para cada voxel, o Brainvoyager
QX, reduz a relação sinal-ruído aumentando a qualidade da imagem. Na figura 2.3 podemos ver o
VMR resultante, que eu apelidei de Subject_avg.VMR.
Figura 2.3: VMR resultante da média das aquisições feitas para o subject.

16
2.2.1.2 As Correcções de Homogeneidade
O passo seguinte, antes de iniciar a Segmentação Automática, é o das Correcções de
Homogeneidade. Consiste, tal como o nome indica, em corrigir algumas falhas de homogeneidade
presentes na imagem. As imagens de alta resolução, de ponderação em T1, são tipicamente
corrompidas por artefactos de susceptibilidade magnética e heterogeneidades. Estes surgem como
variações na intensidade e contraste na imagem, significando que voxeis que representam o mesmo
tecido (i.e. matéria cinzenta ou matéria branca) vão ter diferentes valores de intensidade. Para um
processo de segmentação fiável estas heterogeneidades são obviamente indesejáveis, visto a
segmentação assentar em valores de intensidade para qualificar os diferentes tipos de tecidos. Para
garantir que voxeis correspondentes a um determinado tipo de tecido possuem uma gama de valores
de intensidade bastante próxima, deve esta correcção ter lugar.
Antes de iniciar a referida correcção, apliquei um pequeno passo de cosmética no meu
ficheiro Subject_avg.VMR. Como preciso apenas de informação correspondente ao cérebro (lobo
occipital), resolvi excluir uma série de voxeis referentes a estruturas como pescoço, face e parte do
cerebelo. Isto é feito seleccionando os voxels que desejo apagar. Visto estar a trabalhar em
representações 2D de um volume 3D (255x255x255), escolhi todos os voxels que estão na região a
partir de 152 (face) até 255 (pescoço) no eixo Z para serem eliminados.
Este processo de garantir que os valores dos voxeis correspondentes a um tipo de tecido são
os mesmos, pode ser encarado como uma segmentação manual. Aqui irei garantir que os voxeis,
quer de matéria branca quer de matéria cinzenta, estão dentro de intervalos conhecidos e que esses
mesmos intervalos não se sobrepõem. Seleccionando os voxeis de um intervalo, aplico um processo
de pré-segmentação que me permite alterar os valores de intensidade desse intervalo.
Para realizar esta correcção apelidei o meu VMR de Subject_avg_IC.VMR. A correcção foi
efectuada no conjunto de dados V16 desse ficheiro (pois é aqui que a informação referente às
intensidades está guardada). Usando o cursor para visualizar o estado actual do ficheiro
Subject_avg_IC.VMR quanto à sua distribuição de intensidades, posso verificar os valores de matéria
branca e matéria cinzenta. A Figura 2.4 representa esse passo na vista sagital.

17
Figura 2.4: Vista sagital de Subject_avg_IC.VMR.
Podemos ver na imagem o valor de intensidade de 163. Para o subject, a gama de
intensidades para a matéria branca era cerca de 150 valor mínimo e de 180 valor máximo. Isto
representa um intervalo bastante grande para os voxeis de matéria branca. Aplicando uma mascara à
imagem e operando nela, pude diminuir esse intervalo, seleccionando na caixa de texto o valor
mínimo e o valor máximo desejados. Após seleccionar os extremos do meu intervalo de matéria
branca basta carregar no botão de nome “Grow Region” que irá, a todos os voxeis da máscara que
estiverem nesse intervalo, atribuir uma intensidade de 255 (azul). É necessário, contudo, uma
inspecção visual por todas as fatias com o fim de verificar se não houve nenhuma zona em que
matéria branca foi mal classificada. Se houver matéria branca mal classificada é porque os intervalos
máximo ou mínimo não estão correctos. Obtendo estruturas ou tecidos classificados como matéria
branca não o sendo, terá que se realizar um reset à mascara e repetir o processo. Se houver matéria
branca por classificar, basta alterar os intervalos máximo/mínimo e carregar no botão “Expand”. O
“Expand” irá comparar as vizinhanças dos voxeis já classificados como matéria branca e “colori-los”
de azul se estes estiverem no novo intervalo definido. A Figura 2.5 tem representado o processo de
identificação de voxeis de matéria branca.

18
Figura 2.5: Subject_avg_IC.vmr
A Figura 10 representa igualmente o passo de identificação do intervalo. Agora com a matéria
branca toda definida pode-se passar à pré-segmentação. Esta irá atribuir a todos os voxeis de
intensidade 255 (azul) da máscara um valor médio (entre máximo e mínimo do intervalo definido) e
aplicá-lo aos mesmos voxeis mas no conjunto de dados V16 referente ao Subject_avg_IC.VMR.
Deste modo irá alterar os voxeis do VMR Subject_avg_IC.VMR. A Figura 2.6 representa o VMR
resultante desta etapa.
Figura 2.6: Subject_avg_IC.VMR após 1º processo de correcções de homogeneidade.

19
Estes processos de identificação do intervalo e pré-segmentação podem e devem ser
efectuados quantas vezes for necessário. Assim, é possível a obtenção de uma imagem com valores
distintos para a matéria branca, para a matéria cinzenta e um bom contraste entre ambas. Em cada
paciente realizei as Correcções de Homogeneidade duas vezes, obtendo imagens bastantes
satisfatórias para um bom processo de Segmentação Automática. No caso concreto do subject, a
matéria branca ficou toda definida num intervalo de intensidade entre 165-170 e a matéria cinzenta
entre 90-100.
2.2.1.3 A Transformada Talairach
A Transformada Talairach é a última etapa preparatória antes da Segmentação Automática.
Esta consiste em converter e gravar os dados anatómicos 3D no espaço Talairach. Este é um espaço
padrão com que contém informação sobre a localização e actividade de zonas sub-corticais.
Esta etapa preparatória é realizada em dois passos. Primeiro é definido o plano AC-PC que
corresponde ao plano definido pela comissura anterior e a comissura posterior do cérebro. Em
seguida, as fronteiras do cérebro são identificadas no AP, PP, SP, PI, PR e LP (o ponto mais anterior,
o ponto mais posterior, o ponto mais superior, o ponto mais inferior, o ponto mais à direita e o ponto
mais à esquerda, respectivamente). Este primeiro passo serve basicamente para definir as fronteiras
do cérebro. O segundo passo consiste em transformar estas fronteiras e adaptá-las ao cérebro do
espaço Talairach. A Figura 2.7 representa a definição do plano AC-PC em que podemos rodar a
imagem alterando valores nas diferentes coordenadas x, y e z.
Figura 2.7: Definição do plano AC-PC em Subject_avj_IC.VMR
Na Figura 12 podemos ver em cima o conjunto original de dados e na linha de baixo a
transformação espacial para a definição do plano AC-PC. A linha de baixo terá que ser rodada de
modo a que a comissura posterior e anterior estejam na vertical na vista transversal. A cruz verde

20
também serve para ajustar os outros ângulos suplementares se necessário. Depois do plano o AC-
PC estar definido a transformação pode ser feita. Os pontos de fronteira (AP, PP, SP, PI, PR e PL)
são usados para que o cérebro “caiba” no domínio Talairach. Estes pontos de referência são usados
para alterar o tamanho do cérebro de tal forma que ele se encaixe no tamanho do cérebro Talairach
padrão. No final, eu tenho o cérebro do meu subject no espaço Talairach, podendo tirar partido do
conhecimento de determinadas áreas e zonas de actividade do córtex para fins de segmentação. Na
Figura 2.8 podemos observar o cérebro do subject já em domínio Talairach, o qual eu apelidei de
Subject_avg_IC_TAL.VMR.
Figura 2.8: Subject_avg_IC_TAL.vmr
Com uma inspecção visual rápida posso procurar encontrar possíveis falhas no processo,
como por exemplo pontos de fronteira mal definidos. Esta busca de erros é feita com a aplicação de
uma grelha na imagem que contém, por quadrantes, a informação referente ao cérebro Talairach
padrão. Como estou interessado apenas na segmentação cortical, basta-me verificar apenas se o
cérebro cabe nas fronteiras do cérebro Talairach. A Figura 2.9 representa este processo de
troubleshooting.

21
Figure 2.9: Subject_avg_IC_TAL.vmr com a grelha Talairach sobreposta.
Após esta verificação, o VMR Subject_avg_IC_TAL.VMR está pronto para se dar início à sua segmentação e criação das meshes pela ferramenta de “Segmentação Automática.
2.2.2 A Segmentação Automática
Na Segmentação Automática, grande parte do cortex será segmentado de forma automática
pela aplicação de uma série de processos. As superfícies 3D com as representações das fronteiras
de matéria branca/matéria cinzenta (White Matter – Grey Gatter - WM-GM) são criadas neste
processo. Estas superfícies após a sua criação não estão ainda prontas e é necessário um pequeno
trabalho de correcção topológica de forma a eliminar erros comuns do processo. As meshes, após
serem criadas e corrigidas, passam por um último processo de nome “inflate” onde, por um processo
iterativo, elas são como que insufladas como um balão. Com este último passo é possível observar,
nos sulcos e giros, a actividade cerebral detalhadamente. Para os fins do trabalho aqui presente
apenas há o interesse em criar meshes representando a fronteira da WM – GM, pois é nela que
assentam os mapas de espessura cortical posteriormente construídos pelo processo da
Segmentação Avançada.
A ferramenta de Segmentação Automática tem uma série de funcionalidades que podem ser
escolhidas para terem lugar na segmentação. Em cada um dos pacientes efectuei a Segmentação
Automática duas vezes, sendo que na primeira vez não usaria todas as funcionalidades que
dispunha. Algumas falhas (i.e. uma correcção de homogeneidade mal feita ou por fazer, uma má
transformação Talairach, etc..) podem ainda existir e os resultados mais rápidos da Segmentação
Automática irão apontá-los. Após a primeira segmentação e verificação de que não existem erros
maiores a ser corrigidos, realizei simplesmente o processo de novo, desta vez com todas as opções
que me eram necessárias. Isto poupou bastante tempo de cálculo, pois certas etapas são longas e
pesadas, no que concerne aos recursos do computador em que estava a trabalhar. As
funcionalidades usadas pela Segmentação Automática que necessitei para o meu trabalho foram:
22
encontrar e preencher ventrículos com WM; aplicar uma máscara Talairach do cérebro; encontrar
picos de intensidade da WM e da GM; segmentação (region growing); expandir WM; separar os
hemisférios; remover defeitos topológicos em ambos os hemisférios; gerar superfície 3D da WM-GM.
De todos estes processos, para a primeira segmentação de verificação, não se removem os defeitos
topológicos em ambos os hemisférios por acrescentarem considerável tempo à Segmentação
Automática e não serem necessários para uma primeira correcção de eventuais erros precedentes da
Segmentação Automática. Após essa primeira verificação apenas volto a correr toda a rotina com
essa opção seleccionada.
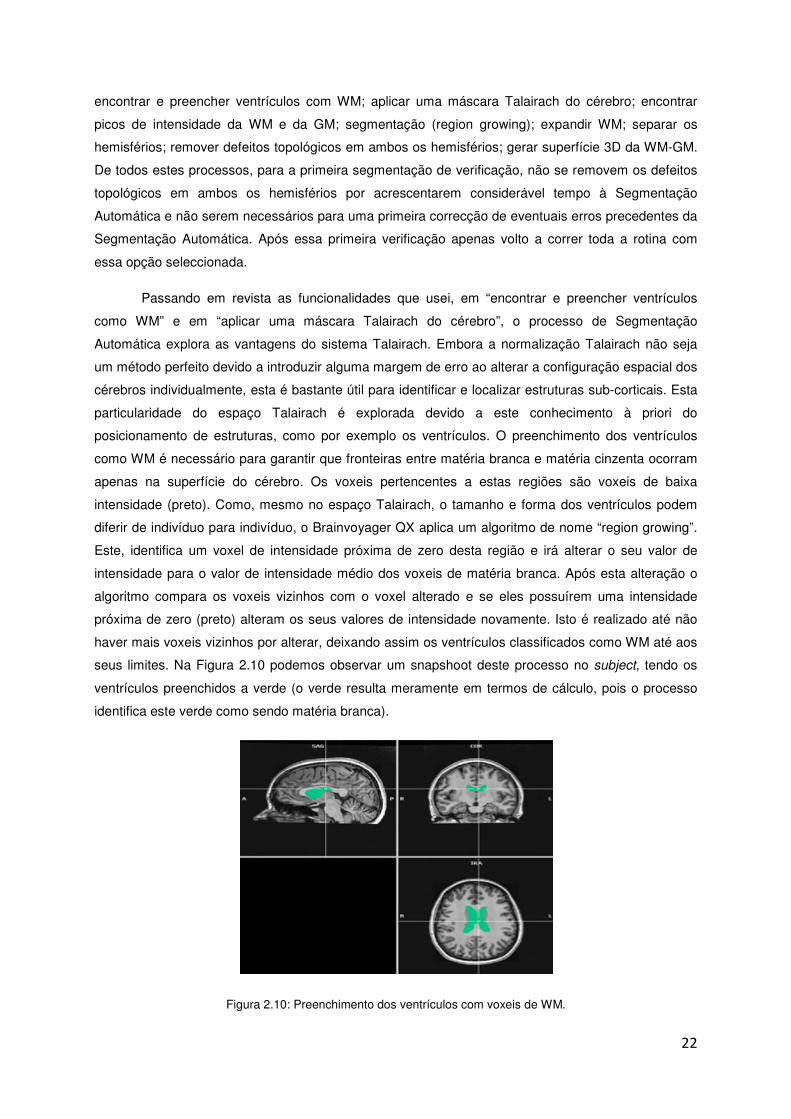
Passando em revista as funcionalidades que usei, em “encontrar e preencher ventrículos
como WM” e em “aplicar uma máscara Talairach do cérebro”, o processo de Segmentação
Automática explora as vantagens do sistema Talairach. Embora a normalização Talairach não seja
um método perfeito devido a introduzir alguma margem de erro ao alterar a configuração espacial dos
cérebros individualmente, esta é bastante útil para identificar e localizar estruturas sub-corticais. Esta
particularidade do espaço Talairach é explorada devido a este conhecimento à priori do
posicionamento de estruturas, como por exemplo os ventrículos. O preenchimento dos ventrículos
como WM é necessário para garantir que fronteiras entre matéria branca e matéria cinzenta ocorram
apenas na superfície do cérebro. Os voxeis pertencentes a estas regiões são voxeis de baixa
intensidade (preto). Como, mesmo no espaço Talairach, o tamanho e forma dos ventrículos podem
diferir de indivíduo para indivíduo, o Brainvoyager QX aplica um algoritmo de nome “region growing”.
Este, identifica um voxel de intensidade próxima de zero desta região e irá alterar o seu valor de
intensidade para o valor de intensidade médio dos voxeis de matéria branca. Após esta alteração o
algoritmo compara os voxeis vizinhos com o voxel alterado e se eles possuírem uma intensidade
próxima de zero (preto) alteram os seus valores de intensidade novamente. Isto é realizado até não
haver mais voxeis vizinhos por alterar, deixando assim os ventrículos classificados como WM até aos
seus limites. Na Figura 2.10 podemos observar um snapshoot deste processo no subject, tendo os
ventrículos preenchidos a verde (o verde resulta meramente em termos de cálculo, pois o processo
identifica este verde como sendo matéria branca).
Figura 2.10: Preenchimento dos ventrículos com voxeis de WM.

23
De seguida, no que respeita em “aplicar uma máscara Talairach do cérebro” é aplicada uma
máscara para segmentar tecidos circundantes da cabeça (i.e. crânio, cerebelo). A máscara também
rotula estruturas sub-corticais como matéria branca. A Figura 2.11 mostra a máscara usada pelo
Brainvoyager QX nas três fatias ortogonais. As regiões a verde são usadas para marcar
automaticamente estruturas sub-corticais como matéria branca. Já as regiões a vermelho e laranja
são regiões a serem segmentadas do cérebro, incluindo a remoção do cerebelo.
Figura 2.11: BrainTalMask.vmr
“The Talairach brain mask file has been created based on the segmentation of an individual
brain (dark orange color) and it was then expanded to fit many additional brains (light orange color).
This resulted in a "conservative" mask file which can be applied to any new brain without removing
relevant brain tissue.”
(retirado de: “Mask-Based Removal of Structures” user’s guide)
O Brainvoyager QX ao aplicar a máscara remove, também, todos os voxeis que não têm
representação na mesma. Os voxeis, de Subject_avg_IC_TAL.VMR, cujo valor de intensidade não é
igual a zero na máscara mantém-se. Os voxeis, de Subject_avg_IC_TAL.VMR, cujo valor de
intensidade é igual a zero na máscara são removidos. Deste modo temos apenas matéria branca e
matéria cinzenta definidas para o processo de “encontrar picos de intensidade da WM e da GM”. Na
Figura 2.12 podemos observar o resultado deste passo no subject.

24
Figura 2.12: Resultado da aplicação da mascara Talairach.
No passo “encontrar picos de intensidade da WM e da GM” da Segmentação Automática, é
determinado o limiar de intensidade entre matéria branca e matéria cinzenta. São calculados
histogramas de intensidades através de cortes axiais do córtex. Dez histogramas diferentes são
obtidos em diversas cores a partir de azul (corte mais superior) até ao vermelho (corte mais inferior).
A intensidade é representada no eixo X, tendo da esquerda (de 0 ou preto) para a direita (até 225 ou
branco) o valor da intensidade. No eixo Y é representado o número de voxeis por intensidade. É
esperado a existência de dois picos de intensidade, um para a matéria branca e outro para a matéria
cinzenta. Deste modo uma primeira observação do histograma pode revelar se há ainda, ou não,
heterogeneidades por corrigir, pela inspecção do gráfico. Se estiverem representados dois picos de
intensidade, significa que os intervalos de intensidade entre os diferentes tecidos WM e GM estão
devidamente optimizados. Após o cálculo do histograma de intensidades surge uma caixa de texto,
que nos permite definir um limiar de intensidades. Este limiar representa em valor de intensidade, a
partir do qual uma intensidade será considerada matéria branca ou matéria cinzenta. O Brainvoyager
QX calcula este valor apresentando-o como sugestão, podendo este ser alterado para fins de um
melhor resultado da Segmentação Automática (isto representa um processo de tentativa erro). Na
Figura 2.13 podemos ver o histograma de intensidades e o valor do limiar de intensidades para o
subject.

25
Figura 2.13: Limiar de intensidades e histograma de intensidades para o subject.
Podemos observar a caixa de texto com o valor 122 apresentado pelo Brainvoyager QX. Este
é também representado no histograma pela linha vertical branca. Aqui, alterei o valor do limiar de 122
para 140. O valor 140 parece um valor mais verdadeiro para a fronteira entre voxeis de matéria
branca e cinzenta pois, a 122, ainda podem estar inseridos alguns voxeis pertencentes ao pico da
matéria cinzenta.
Após a identificação dos picos de intensidade pode ter início a etapa “segmentação (region
growing)”. Aqui, serão criados os ficheiros contendo apenas a matéria branca, usados para a criação
das meshes. O processo usa o valor do limiar de intensidades determinado no passo anterior. Trata-
se de uma etapa semelhante à do preenchimento dos ventrículos. O processo aqui irá usar o valor do
limiar para classificar e alterar o valor dos voxeis. Estes serão verificados se estão entre o valor
máximo e o valor mínimo, definidos pelo histograma de intensidades. No caso do subject, a
intensidade referente à matéria branca está entre 140 e 225. Os voxeis centrais do cérebro serão
verificados se estão dentro deste intervalo. Caso isso ocorra são incluídos na segmentação da WM e
os seus voxeis vizinhos são comparados, também com o intervalo, e classificados também como
matéria branca, se possuírem uma intensidade superior à do limiar (140). Isto ocorre sucessivamente,
até não existirem mais voxeis pertencentes ao intervalo de intensidades. Os voxeis classificados
como matéria branca são guardados num ficheiro que o Brainvoyager QX cria automaticamente com
a sua representação a azul (têm uma intensidade de 240). Na Figura 2.14 está representado um
snapshoot do processo “Segmentação Automática” na etapa de segmentação da matéria branca do
subject.

26
Figura 2.14: Resultado o processo de “region growing” para a segmentação da matéria branca no subject.
Quando a matéria branca está totalmente segmentada o passo de “expandir WM” irá
melhorar a qualidade da fronteira de segmentação. Este é necessário para que haja uma boa
reconstrução da superfície 3D, visto o processo de “region growing” poder, por vezes, não classificar
finas camadas de matéria branca ao longo do córtex. Isto pode reflectir-se em pequenos buracos nas
meshes. Este passos consiste em expandir a fronteira da matéria branca em 1 voxel, eliminando um
grande número de erros topológicos, criando um suave contorno na superfície.
Após o último passo, os dois hemisférios podem ser separados pela etapa “separar os
hemisférios”. Esta, representa simplesmente um corte no meio do plano coronal (fatia 128). Este corte
ocorre apenas no corpo caloso pois, os hemisférios são assimétricos. Sobretudo, na região occipital,
um hemisfério pode cruzar a linha média (fatia 128), entrando na região de outro hemisfério. Esta
separação de hemisférios é necessário para no final deste processo o Brainvoyager QX poder
guardar a informação referente a cada hemisfério separadamente e assim poder construir as
representações 3D (meshes) para cada hemisfério individualmente. Estes ficheiros têm como
extensão no nome de WH.vmr. Na Figura 2.15 está representado o ficheiro que contém a informação
referente à segmentação da matéria branca para o subject.

27
Figura 2.15: Subject_avg_IC_TAL_LH_WM.vmr representado a material branca segmentada do hemisfério
esquerdo.
Estes ficheiros contendo a segmentação da matéria branca são automaticamente usados
para criar as meshes de cada hemisfério. Os voxeis limite (em amarelo na Figura 20) são usados
para criar uma malha 3D que representa a superfície cortical. As vistas coronal, sagital e transversal,
apesar de serem imagens em 2D, representam o volume 3D da matéria branca. Deste modo, a
reconstrução da superfície é basicamente uma conversão da fronteira 2D nas três vistas para uma
superfície 3D.
Nesta reconstrução três arquivos são criados e salvos automaticamente para cada
hemisfério. O primeiro terá uma extensão no nome RECO.srf e representa a simples transição do 2D
para 3D. A Figura 2.16 representa este ficheiro onde é possível ver a malha correspondente à
fronteira da WM – GM no hemisfério esquerdo do subject.
Figura 2.16: Subject_avg_IC_TAL_LH_RECO.srf mesh

28
Na Figura 21 é possível notar a natureza cúbica da “rendarização” proveniente dos voxeis
limite do ficheiro WM.vmr. Isto leva-nos ao segundo ficheiro criado automaticamente. Trata-se de um
ficheiro resultante de um processo de alisamento aplicado à malha RECO.srf. Este alisamento trata-
se de uma média entre voxeis para suavizar a superfície. O ficheiro é salvo com a extensão no nome
de RECOSM.srf (SM de suavização). É deste ficheiro que é criada a meshe insuflada, onde se
projectam os mapas de espessura cortical. Na Figura 2.17 está representada esta superfície.
Figura 2.17: Subject_avg_IC_TAL_LH_RECOSM.srf mesh
Finalmente, o terceiro arquivo criado para cada hemisfério contém uma superfície também
suavizada, resultante da transformação da malha RECOSM para os valores de intensidade, que
separam a matéria branca/cinzenta. Esta meshe também é utilizada para visualização dos dados
funcionais, uma vez que permite também olhar em profundidade, em sulcos. Este ficheiro é do uso
exclusivo da análise funcional e não é mais alterado por mim. A Figura 2.18 contém a sua
representação.
Figura 2.18: Subject_avg_IC_TAL_LH_WM.srf

29
Após a criação destas três malhas o processo de Segmentação Automática está concluído.
Nesta fase do trabalho é necessário procurar falhas que possam ter ocorrido. Com a verificação dos
histogramas de intensidades feita durante a sua criação, a solução de eventuais problemas reside
agora em verificar se os limites para a matéria branca foram bem delimitados para cada hemisfério.
Se tal não aconteceu as superfícies geradas podem conter erros grosseiros. Contudo, da mesma
maneira que o Brainvoyager QX transforma os ficheiros contendo a fronteira da matéria branca de 2D
para 3D, também é possível fazer o contrário. Desta maneira é pode sobrepor-se uma meshe sobre o
VMR usado na sua criação. É possível assim, procurar em todas as vistas e fatias por erros ou má
classificações existentes nas meshes. Na Figura 2.19 temos a representação da meshe RECOSM no
ficheiro Subject_avg_IC_TAL.vmr.
Figura 2.19: Superfície Subject_IC_TAL_LH_RECOSM.srf transformada espacialmente e representada em
Subject_avg_IC_TAL.vmr
Como se pode observar, a verificação é feita analisando todas as fatias em todas as vistas.
Esta procura de erros é feita também com as meshes WM.srf. Efectuei esta resolução de problemas
em todos os sujeitos e não encontrei nenhum erro nesta fase. Apesar do meu trabalho assentar num
número pequeno de pacientes -16- este resultado leva-me a concluir a robustez deste processo de
Segmentação Automática, quando as imagens contêm uma boa qualidade e homogeneidade.
Este processo de verificação dos primeiros resultados da Segmentação Automática está
concluído, podendo-se assim passar para a fase em que se corrigem os pequenos erros topológicos.
Para isto efectua-se novamente a rotina da Segmentação Automática, mas desta vez com a opção de
“remover defeitos topológicos em ambos os hemisférios” e com a confiança de que não ocorreram
erros grosseiros na definição dos histogramas de intensidade e na criação das meshes. Esta opção
irá remover pequenas pontes ou pregas na matéria branca actuando sobre o ficheiro WM.vmr,
criando e guardando um ficheiro WM_BL2.vmr onde se encontra a matéria branca com estes
pequenos erros topológicos corrigidos. O ficheiro WM_BL2.vmr será usado para reconstruir as três

30
meshes (RECO.srf, RECOSM.srf e WM.srf) ao invés do ficheiro WM.vmr como na primeira rotina da
Segmentação Automática. No entanto, e como não poderia deixar de ser, também o ficheiro
WM_BL2.vmr tem que ser submetido a uma verificação e procura de eventuais erros.
Para o caso do subject, a segunda vez que se realizou a Segmentação Automática, correu
novamente com um valor limite entre matéria branca/cinzenta de 140. A rotina continuou e após a
criação do ficheiro WM.vmr as correcções topológicas foram feitas e arquivadas no ficheiro
WM_BL2.vmr sendo este usado para a criação automática das meshes. Contudo, as correcções
topológicas não são perfeitas e para os 16 pacientes têm dificuldade em classificar a matéria branca
na zona da amígdala e recorrentemente é normal surgirem “buracos” na matéria branca. Isto é
corrigido editando o ficheiro WM_BL2.vmr e posteriormente efectuando uma reconstrução das
meshes. Para este fim usei uma funcionalidade do Brainvoyager QX que me permite observar um
VMR sobre outro VMR. O que fiz foi, por exemplo, no caso do subject, projectar o ficheiro
Subject_avg_IC_TAL.vmr (imagem secundária) sobre o ficheiro Subject_avg_IC_TAL_WM_BL2.vmr
(imagem primária). Deste modo e alterando entre imagem principal e secundária, posso editar
alterações na imagem principal. A Figura 2.20 tem uma representação, em todas as vistas, da
projecção referida e a Figura 2.21 contém a correcção de um erro comum em todos os pacientes em
que temos o nervo óptico classificado como matéria branca.
Figura 2.20: Subject_avg_IC_TAL_LH_BL2.vmr como vmr primário com Subject_avg_Ic_TAL.vmr como
secundário.
31
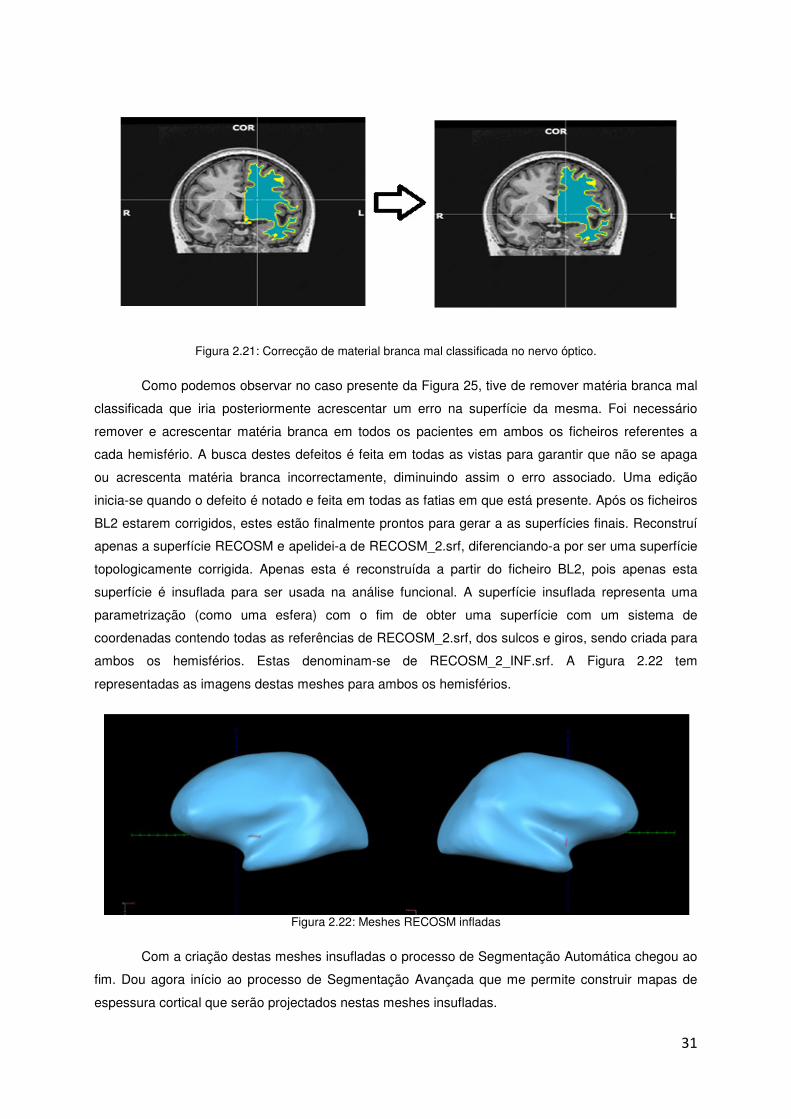
Figura 2.21: Correcção de material branca mal classificada no nervo óptico.
Como podemos observar no caso presente da Figura 25, tive de remover matéria branca mal
classificada que iria posteriormente acrescentar um erro na superfície da mesma. Foi necessário
remover e acrescentar matéria branca em todos os pacientes em ambos os ficheiros referentes a
cada hemisfério. A busca destes defeitos é feita em todas as vistas para garantir que não se apaga
ou acrescenta matéria branca incorrectamente, diminuindo assim o erro associado. Uma edição
inicia-se quando o defeito é notado e feita em todas as fatias em que está presente. Após os ficheiros
BL2 estarem corrigidos, estes estão finalmente prontos para gerar a as superfícies finais. Reconstruí
apenas a superfície RECOSM e apelidei-a de RECOSM_2.srf, diferenciando-a por ser uma superfície
topologicamente corrigida. Apenas esta é reconstruída a partir do ficheiro BL2, pois apenas esta
superfície é insuflada para ser usada na análise funcional. A superfície insuflada representa uma
parametrização (como uma esfera) com o fim de obter uma superfície com um sistema de
coordenadas contendo todas as referências de RECOSM_2.srf, dos sulcos e giros, sendo criada para
ambos os hemisférios. Estas denominam-se de RECOSM_2_INF.srf. A Figura 2.22 tem
representadas as imagens destas meshes para ambos os hemisférios.
Figura 2.22: Meshes RECOSM infladas
Com a criação destas meshes insufladas o processo de Segmentação Automática chegou ao
fim. Dou agora início ao processo de Segmentação Avançada que me permite construir mapas de
espessura cortical que serão projectados nestas meshes insufladas.

32
2.3 Análise Anatómica e Volumétrica de dados – Criação dos mapas de EC
O ponto 2.2 descreveu como é feita a segmentação de grande parte do córtex para o fim de
criação das superfícies corticais tridimensionais. Embora os métodos usados sejam adequados para
a criação de representações 3D, não o são para medidas exactas de EC, devido aos erros
associados (por exemplo a expansão da matéria branca na fronteira com a matéria cinzenta). Para
tal, o Brainvoyager QX usa outra ferramenta de nome Segmentação Avançada. Esta fornece
representações altamente precisas dos limites, WM-GM e GM-CSF, devido a calculá-las no volume e
não no espaço. São as representações destas fronteiras/limites que permitem medir, com maior
exactidão, a EC em cérebros individuais. No entanto, há uma semelhança com a Segmentação
Automática, pelo que também são necessárias etapas preparatórias antes da Segmentação
Avançada.
2.3.1 Etapas preparatórias antes da Segmentação Avançada
Antes desta fase do processo de análise usando a ferramenta “Segmentação Avançada”
algumas etapas tiveram de ser levadas em consideração:
1. Foi aplicado um processo de nome “brain peel” que remove tecido circundante do
cérebro, tal como crânio e dura;
2. Os VMRs foram convertidos para uma resolução de 0.5x0.5x0.5mm. Isto reduziu o
tamanho dos voxeis em 8 vezes;
3. Os ventrículos foram manualmente editados, sendo preenchidos com voxeis de
intensidade igual à da matéria branca;
4. Foi removido manualmente o cerebelo, dura e crânio restantes a fim de obter uma
imagem no lobo occipital apenas com cérebro presente.
Todos estes passos necessitavam que os passos presentes no ponto 2.2.1 tivessem ocorrido.
Como procedi á Segmentação Avançada depois da Segmentação Automática, todas essas condições
estavam presentes. Apesar de ter realizado estas etapas pela ordem exposta neste ponto, o processo
“brain peel” pode ser feito depois da conversão para 0.5x0.5x0.5mm, com o fim de remover tecido já
no espaço 0.5x0.5x0.5mm. Estas etapas foram feitas nos ficheiros _avg_IC_TAL.vmr e as imagens
seguintes referem-se ao ficheiro Subject_avg_IC_TAL.vmr

33
2.3.1.1 A transformação especial 0.5x0.5x0.5mm e o brain peel
A primeira etapa preparatória foi aplicar, ainda na resolução espacial 1x1x1mm, a ferramenta
brain peel. Este passo permite-me remover tecido como crânio e dura que podem introduzir erros no
cálculo posterior da espessura EC. Esta remoção resulta da aplicação do ficheiro cuja imagem está
na Figura 16. Tal como na Segmentação Automática trata-se da aplicação da máscara Talairach
usando o conhecimento “à priori” de determinadas regiões corticais, para a remoção de tecido
exterior ao cérebro. A Figura 2.23 apresenta o resultado deste passo. O ficheiro resultante apelidei de
Subject_avg_IC-TAL_PEEL.vmr
Figura 2.23: Brain peel aplicado subject
Em seguida, o conjunto de dados é convertido para uma resolução espacial de
0.5x0.5x0.5mm. Esta transformação para o espaço de 0,5 milímetros irá reduzir o tamanho dos
voxeis em 8 vezes. Aqui reside uma das grandes diferenças e vantagem da Segmentação Avançada
em relação à Segmentação Automática. Com o tamanho dos voxeis reduzido em 8 vezes, os
processos de “region growing” e “expandir WM” terão muito menos erros associados. Isto deve-se à
informação que dantes estava em apenas 1 voxel, estar agora em 8 voxeis. Desta maneira as
fronteiras WM-GM e GM-CSF serão mais precisas e terão menos erros associados. Após esta
transformação espacial o Brainvoyager QX guarda o ficheiro com uma extensão de nome
0.5mmISO.vmr. A Figura 2.24 representa o resultado da transformação espacial.

34
Figura 2.24: Transformação espacial para a resolução 0.5x0.5x0.5mm do subject
Podemos observar, como seria de esperar, o aumento do tamanho do cérebro nesta
resolução. Outro “brain peel” pode ser aplicado, desta vez com uma versão 0.5x0.5x0.5mm da
máscara Talairach, para tentar remover mais tecido excedente. Contudo, foram poucas as vezes que
recorri a este segundo “brain peel”, pois o primeiro apresentava resultados satisfatórios.
2.3.1.2 O preenchimento dos ventrículos e a remoção manual de tecido no lobo occipital
O próximo passo é identificar a forma dos ventrículos e preenche-los com voxeis de
intensidade próxima ou igual à intensidade da matéria branca circundante. Desta forma a
Segmentação Avançada usa, tal como no “brain peel”, a máscara Talairach para identificar os
ventrículos e altera todos os voxeis desta região com voxeis de uma intensidade de 150. Contudo,
pode acontecer que os valores de intensidade da matéria branca sejam diferentes de 150 devido, por
exemplo, a valores de brilho e contraste específicos. Se tal acontecer é necessário seleccionar os
voxeis presentes nos ventrículos e alterá-los para os valores da matéria branca para optimizar o
processo de segmentação. Isto é feito como no ponto 2.2.1.2 para as correcções de homogeneidade.
Pode acontecer também que os ventrículos não sejam totalmente preenchidos e aqui é necessário
editar a imagem, em todas as fatias, pintando manualmente as zonas por preencher. Isto representa
o processo de preenchimento dos ventrículos presente na Segmentação Automática mas feito e
aperfeiçoado manualmente e, também, com o objectivo de obter fronteiras apenas na superfície do
cérebro. A Figura 2.25 representa o resultado desta etapa.

35
Figure 2.25: Subject_Avg_IC_TAL_PEEL_0.5mmISO_BRAINPEEL.vmr com os ventrículos preenchidos com
voxeis de intensidade igual à da matéria branca.
Olhando atentamente para a Figura 29, verificamos que ainda persiste alguma dura e por
vezes em alguns pacientes, até partes de crânio ainda permanecem nesta fase. Estes são
indesejáveis para uma segmentação optimizada, pelo que introduzem erros durante a definição das
fronteiras devido à sua intensidade ser próxima da intensidade dos voxeis de matéria cinzenta.
Também devido a isso a fronteira entre o cerebelo e cérebro deve ser acentuada. Para se conseguir
isto é necessária uma edição manual, fatia a fatia, removendo dura e introduzindo essa
descontinuidade. Estas edições são as mais dispendiosas, em termos de tempo e de esforço, de toda
a minha análise de EC presente neste trabalho. Como o objectivo é apenas estudar o lobo occipital
(onde está a visão), estas correcções tiveram lugar apenas na metade posterior do cérebro. Isto
acarreta o aparecimento de alguns erros no mapa de EC na zona anterior do cérebro, mas poupa-me
cerca de 2-3 horas de edição. Além de, esses erros não contribuem para o trabalho em questão.
Estas edições manuais, em apenas metade do cérebro, demoraram cerca de 2-3 horas a serem
levadas a cabo em cada paciente. Na Figura 2.26 podemos ver o cérebro do subject já editado.
Salvei este ficheiro como Subject_avg_Ic_TAL_PEEL_0.5mmISO_corrigido.vmr

36
Figura 2.26: Subject_Avg_IC_TAL_PEEL_0.5mmISO_corrigido.vmr
Após estas edições é aplicado um filtro de contraste alterando o contraste e brilho entre
matéria branca e cinzenta. Deste modo as etapas preparatórias para a Segmentação Avançada
estavam concluídas.
2.3.2 A Segmentação Avançada
O primeiro passo neste processo é o cálculo da fronteira entre WM-GM. Este é feito com um
algoritmo de nome “adaptative region growing”. Este algoritmo representa outra grande diferença e
vantagem em relação à Segmentação Automática. Na Segmentação Automática é usado um valor
para o limiar de intensidade global e este por vezes pode introduzir pequenos erros pela falta de
classificação de matéria branca. Na segmentação Avançada é usado também um valor limiar de
intensidade local que funciona em várias regiões, identificando assim voxeis que são de matéria
branca mas escaparam à classificação pelo limiar global. A “adaptive region growing” analisa a
distribuição de intensidades para encontrar um limite global para começar a segmentar a matéria
branca. Imaginando o histograma de intensidades terei dois limiares representados, sendo que o
limiar local terá que ser ligeiramente menor que o limiar global, precisamente para captar matéria
branca ignorada pelo limiar global. Tal como na Segmentação Automática, posso alterar o valor
destes limiares. Antes de correr este processo é definido um valor entre 0 e 1 (para o limiar global e
outro para o limiar local) que irá “mover” estes para junto ou longe dos picos de intensidade da
matéria branca e cinzenta. Por outras palavras, o Brainvoyager QX define não um valor limiar de
intensidade mas sim um intervalo limiar de intensidades. Sendo o limite superior o limiar global e o
inferior o limiar local. Aplicando um valor normalizado ao valor dos limiares resta definir o tamanho do
intervalo e a sua magnitude. Quanto maior estes valores forem menos matéria branca será
segmentada resultando numa imagem com maior quantidade de matéria cinzenta (pois o intervalo
desloca-se para a direita no histograma de intensidades). Se estes valores forem pequenos e

37
próximos de 0 terei uma imagem com excesso de matéria branca e pouca quantidade de matéria
cinzenta (pois o intervalo desloca-se para a esquerda no histograma de intensidades). Para o subject
usei um valor para o limiar local de 0.5 e global de 0.6 bastante satisfatórios após algumas tentativas.
De paciente para paciente estes valores alteram-se, mas o tamanho do intervalo é sempre de 0.1, por
conseguir bons resultados desta maneira. Um ficheiro com a extensão no nome de WM.vmr é criado
para ser usado no cálculo da fronteira GM-CSF. A Figura 2.27 representa o resultado desta “adaptive
region growing” para o subject.
Figura 2.27: Subject_Avg_IC_TAL_PEEL_0.5mmISO_corrigido_WM.vmr representando o resultado da
segmentação pelo adaptive region growing.
A fronteira GM-CSF (superfície pial) é calculada a partir do ficheiro representado na Figura
31. A superfície pial corresponde ao limite exterior do córtex. A fronteira GM-CSF é determinada pela
expansão dos voxeis limite de matéria cinzenta de forma semelhante ao passo de “expandir WM”
presente no ponto 2.2.2. Mais uma vez temos presente uma vantagem da Segmentação Avançada
em relação à Segmentação Automática. Como este passo é feito numa resolução de 0.5x0.5x0.5mm,
ao contrário da Segmentação Automática, há uma maior exactidão na fronteira definida. Um arquivo
com o nome da extensão WM_GM.vmr é criado e salvo no disco. Na Figura 2.28 podemos ver a sua
representação.

38
Figura 2.28: Subject_Avg_IC_TAL_PEEL_0.5mmISO_corrigido_WM_GM.vmr representando o resultado do
cálculo da fronteira GM-CSF.
Ao observar a Figura 32 verificamos que a matéria branca deixou de ser representada a uma
intensidade de 255 (azul). Isto deve-se ao facto de com o cálculo da fronteira GM-CSF toda a matéria
branca é classificada com uma intensidade de 150 e toda a matéria cinzenta é classificada com uma
intensidade de 100. A análise anatómica deste paciente está quase completa faltando apenas o
cálculo do mapa de espessura cortical que é feito através deste ficheiro WM_GM.vmr. No entanto, e
como não poderia deixar de ser é necessário uma busca final de possíveis erros na segmentação.
Usando este ficheiro como imagem primária e o ficheiro base _corrigido.vmr como secundária, irei
alterar entre uma e outra e procurar erros na classificação da matéria branca e cinzenta, sobretudo no
lobo occipital. Para efectuar tais verificações descobri que a vista transversal é bastante útil para
observar o lobo occipital. Quando encontrava uma zona que teria matéria branca a mais ou a menos
simplesmente editava o vmr WM_GM.vmr manualmente. Caso se encontrem muitos erros é porque o
limiar local e limiar global foram mal escolhidos e é necessário correr o processo da Segmentação
Avançada de novo, com novos valores para os limiares. Trata-se de um processo de tentativa-erro.
Nunca consegui um resultado satisfatório à primeira vez que recorri esta rotina, devido ao facto de os
cérebros não serem iguais e chegarem ao processo de Segmentação Avançada com diferentes
valores de brilho e contraste, alterados pelos variadíssimos processos e etapas precedentes. Após
garantido que não há erros na classificação da matéria branca e cinzenta, posso finalmente calcular
os mapas de espessura cortical.
2.3.3 Calculo e construção dos mapas de EC
Para o cálculo dos valores de EC e criação dos mapas corticais no subject o Brainvoyager QX
usa o método matemático de Laplace [39]. Este método aplica a equação diferencial de Laplace (∇2ψ
= 0) onde o volume do córtex é representado como domínio para a solução da equação diferencial,
com condições de contorno nas fronteiras da matéria cinzenta (com a matéria branca e com o liquido
cefalorraquidiano) calculadas pelo processo descrito no ponto 2.3.2 e presentes no ficheiro

39
subject_avg_IC_TAL_PEEL_05mmISO_corrigido_WM_GM.vmr. A normalização do gradiente ψ
forma um campo vectorial, representando vectores tangentes às linhas do campo que ligam as
fronteiras da matéria cinzenta. Desta forma e com este método a espessura é medida de uma forma
tridimensional podendo ser obtida através de qualquer ponto no córtex. Os ficheiros estruturais de
cada paciente (WM_GM.vmr) preparados pelo processo de “Segmentação Avançada” são desta
maneira usados pelo Brainvoyager QX para, individualmente, definir mapas de EC. O ficheiro
resultante tem a extensão VMP e pode ser projectado numa mesh (para visualização 3D ou análise
funcional) ou num VMR (para visualização 2D do volume). A Figura 2.29 tem representado o mapa de
EC do subject sobreposto no seu WM_GM.vmr e na sua mesh inflada do hemisfério esquerdo.
Figure 2.29: Mapa cortical do subject sobreposto no seu WM_GM.vmr e na sua meshe inflada do hemisfério
esquerdo.
Como podemos observar na Figura 2.29, a EC varia de 0.5 mm (azul escuro) até 5 mm
(verde). O lobo frontal contém maiores zonas a verde (maior espessura cortical média), devido aos
erros deixados por corrigir durante as etapas preparatórias da Segmentação Avançada. Na Figura 34
podemos observar a meshe presente na Figura 2.29 mas de outro ângulo, deixando visível o lobo
occipital.
Figure 2.30: Hemisfério esquerdo de subject representado pela sua meshe insuflada e com o seu mapa de EC
projectado. O círculo vermelho representa a localização do lobo occipital.

40
Com a EC presente nos mapas e estes sobre as superfícies 3D insufladas, é possível olhar
profundamente para sulcos do lobo occipital e estudar a sua actividade cortical, com o fim de mapear
as áreas visuais. Dentro do círculo vermelho presente na Figura 34 está localizado o córtex visual.
Todo o processo aqui descrito, desde as etapas preparatórias da Segmentação Automática, até á
construção dos mapas de EC, foi realizado em todos os 16 pacientes. Por cada um o ciclo completo
levou cerca de 6 horas. Os vários processos de tentativa-erro poderão em alguns ser mais rápidos
que noutros, dependendo isto da qualidade dos conjuntos de dados provenientes do scanner.
Contudo, todos os 16 pacientes eram colaborantes e com isto forneceram boas imagens com bons
dados anatómicos e funcionais.
Findo o ciclo para os 16 pacientes, estavam criadas as superfícies insufladas e os mapas de
EC, para cada um. Estes foram passados por mim ao meu colega, que realizou uma análise funcional
(retinotopia) dos mesmos. Nesta análise, um mapa retinotópico foi desenhado, definindo cada área
visual (V1, V2v, V2d, V3 e VP) no córtex visual de cada hemisfério. Cada uma destas áreas foi
dividida em três sub-regiões (fóvea, intermédia e periférica) referentes às zonas do campo visual. De
cada uma foram retirados 10 pontos aleatórios, resultando em 30 pontos por área visual. Estes
valores foram usados para calcular as espessuras médias por área e realizar a análise estatística dos
resultados.
2.4 Análise de Resultados
2.4.1 Comparação de espessuras corticais em áreas homólogas
O objectivo, com esta análise estatística, foi comparar os valores de espessura cortical de
áreas homólogas no hemisfério esquerdo e direito nos pacientes com LHON.
As médias das áreas visuais entre hemisférios foram comparadas usando o programa
Statistical Package for the Social Sciences v18.0 (SPSS). O nível de significância adoptado foi de
α=0.05.
2.4.2 Análise de correlações
Este método foi também realizado com o programa SPSS. A análise por correlações é um
método estatístico que me permite determinar o tipo de relação entre duas variáveis, em termos de
direcção e grau, da relação entre elas. Usando o coeficiente de correlação (r) baseado em duas
variáveis e tendo em mente que a abordagem feita por esta análise assenta nos testes de

41
significância, tenho de escolher o nível de significância que me permite aceitar ou rejeitar as minhas
hipótese e hipótese nula:
- Hipótese: Existe uma relação entre as espessuras corticais das áreas visuais em estudo,
do córtex visual;
- Hipótese nula: Não existe uma relação entre as espessuras corticais das áreas visuais
em estudo, do córtex visual;
Os níveis de significância desta abordagem estatística foram de α=0.05 e α=0.01, que
resultam numa prova moderada e forte de que a minha hipótese é verdadeira (95% e 99% de
confiança). As correlações foram feitas entre as áreas visuais (os seus valores médios de EC) no
cortex visual. Baseado na escala Cohen para as correlações [40], decidi chamar a força de correlação
quando o coeficiente (r) se encontra nos seguintes valores (baseado no nível de significância
estatística):
- Correlação média (0.500 < r < 0.849) ou (-0.849 < r < -0.500);
- Correlação forte (r > 0.850) ou (r < - 0.850);

42
3. Resultados
3.1 Valores de Espessura Cortical
O culminar da análise anatómica e análise funcional está representado na Tabela 1. Estão
apresentados os valores das espessuras corticais por área visual por hemisfério, em milímetros. Na
análise funcional retinotópica cada área visual de cada hemisfério foi dividida em 3 zonas (fóvea,
intermédia e periférica), referentes às zonas do campo visual. De cada uma foram retirados 10
pontos. Os valores de EC para cada área visual obtiveram-se em, primeiro, calcular uma média
simples dos 10 pontos de cada zona resultando em 3 valores médios e posteriormente calcular a
média desses 3 valores médios. Os resultados obtidos estão presentes na Tabela 3.1:
Tabela 3.1 – Valores de EC em milímetros.

43
3.2 Diferenças nas espessuras corticais entre hemisférios
Com os dados presentes na tabela 3.1 foi possível determinar se existem diferenças entre as
áreas visuais homólogas relativamente às suas espessuras corticais, em ambos os hemisférios. Da
manipulação desta tabela retirei 10 variáveis, uma para cada área visual de cada hemisfério (V1-LH;
V1-RH; V2v-LH; V2v-RH; V2d-LH; V2d-RH; V3-LH; V3-RH; VP-LH e VP-RH). Cada uma delas tinha
16 medidas. Realizei testes de normalidade às variáveis, com o fim de aferir o comportamento das
suas distribuições e com isso aplicar testes de independência às suas médias. Desta forma é
possível determinar se uma dada área visual em um hemisfério é, ou não, mais espessa do que a sua
homóloga no outro hemisfério. (neste ponto de apresentação de resultados saliento que LH refere
para o hemisfério esquerdo e RH refere para o hemisfério direito)
3.2.1 Resultados estatísticos para a área visual V1
A Tabela 3.2 apresenta o resultado do teste de normalidade feito às variáveis V1 no
hemisfério esquerdo e no hemisfério direito:
Tabela 3.2 – Resultados do teste de normalidade para os resultados de V1 em ambos os hemisférios.
Hemisphere Kolmogorov-Smirnova Shapiro-Wilk
Statistic df Sig. Statistic df Sig. V1 thicknesses LH ,124 16 ,200 ,955 16 ,575
RH ,145 16 ,200 ,962 16 ,704
Da análise da Tabela 3.2 verificamos que os valores de V1-LH e V1-RH seguem uma distribuição normal para toda a amostra de 16 pacientes.
Questiono agora se a espessura da área visual V1 é diferente entre hemisférios, ou, caso contrário, se tem a mesma espessura em ambos:
0
1
: A espessura cortical é igual entre hemisférios nesta área visual
: A espessura cortical não é igual entre hemisférios nesta área visual
LH RH
LH RH
H
H
µ µ
µ µ
=
≠
A Tabela 3.3 apresenta os valores da média e desvio padrão de V1 em ambos os hemisférios. A Tabela 3.4 apresenta o resultado do teste de independência de variáveis.
Tabela 3.3 – Valores médios e desvio padrão para V1 em ambos os hemisférios.
Hemisphere N Mean Std. Deviation Std. Error Mean
V1 thicknesses LH 16 2,21 ,20 ,050
RH 16 2,40 ,19 ,048
.

44
Tabela 3.4 – Testes paramétricos para a variável V1.
Levene's Test for
Equality of Variances t-test for Equality of Means
F Sig. t df
Sig. (2-
tailed) Mean
Difference Std. Error Difference
95% Confidence Interval of the Difference
Lower Upper V1
thicknesses Equal
variances assumed
,000 ,987 -2,820 30 ,008 -,20 ,07 -,34 -,05
Equal variances
not assumed
-2,820 29,966 ,008 -,20 ,07 -,34 -,05
Da análise da Tabela 3.4, o teste paramétrico de igualdade de variâncias mostra um valor
Sig. de 0.987. Deste modo, assumo a igualdade entre as variâncias de V1-LH e V1-RH (0.987>0.05).
O valor Sig. (2-tailed) representa o nível de confiança e neste caso este valor é de 0.008. Como o
nível de confiança adoptado por mim é de α=0.05 e sabendo que 0.008 ≤ 0.05, eu rejeito a minha
hipótese nula e aceito que a espessura em V1 é diferente entre hemisférios. A espessura cortical de
V1 no hemisfério direito é superior à espessura cortical no hemisfério esquerdo (com um nível de
confiança de 95%).
3.2.2 Resultados estatísticos para a área visual V2v
A Tabela 3.5 apresenta o resultado do teste de normalidade feito às variáveis V2v no
hemisfério esquerdo e no hemisfério direito:
Tabela 3.5 – Resultados do teste de normalidade para os resultados de V2v em ambos os hemisférios.
Hemisphere Kolmogorov-Smirnova Shapiro-Wilk
Statistic df Sig. Statistic df Sig. V2v thicknesses
LH ,223 16 ,032 ,855 16 ,016
RH ,141 16 ,200 ,947 16 ,441
Da análise da Tabela 3.5 verificamos que os valores de V2v-RH seguem uma distribuição
normal para toda a amostra de 16 pacientes. Os valores de V2v-LH seguem uma distribuição não-
normal.
Questiono agora se a espessura da área visual V2v é diferente entre hemisférios, ou, caso
contrário, se tem a mesma espessura em ambos:
0
1
: A espessura cortical é igual entre hemisférios nesta área visual
: A espessura cortical não é igual entre hemisférios nesta área visual
LH RH
LH RH
H
H
µ µ
µ µ
=
≠
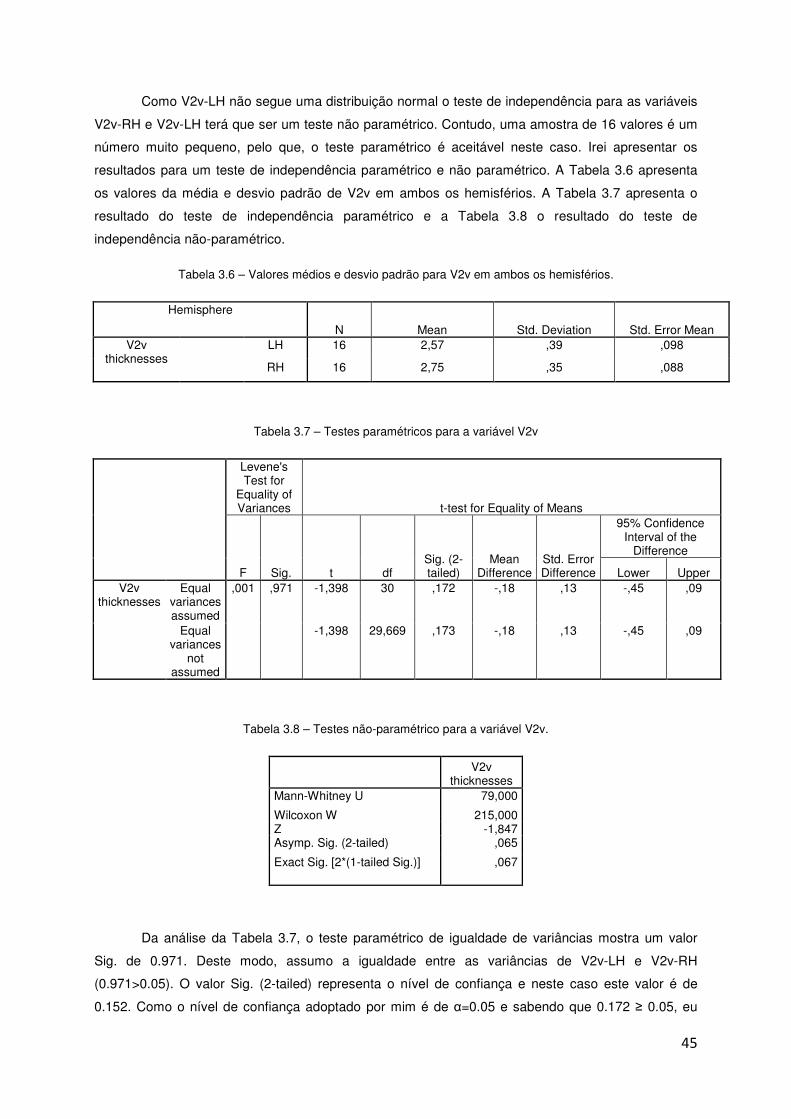
45
Como V2v-LH não segue uma distribuição normal o teste de independência para as variáveis
V2v-RH e V2v-LH terá que ser um teste não paramétrico. Contudo, uma amostra de 16 valores é um
número muito pequeno, pelo que, o teste paramétrico é aceitável neste caso. Irei apresentar os
resultados para um teste de independência paramétrico e não paramétrico. A Tabela 3.6 apresenta
os valores da média e desvio padrão de V2v em ambos os hemisférios. A Tabela 3.7 apresenta o
resultado do teste de independência paramétrico e a Tabela 3.8 o resultado do teste de
independência não-paramétrico.
Tabela 3.6 – Valores médios e desvio padrão para V2v em ambos os hemisférios.
Hemisphere
N Mean Std. Deviation Std. Error Mean V2v
thicknesses LH 16 2,57 ,39 ,098
RH 16 2,75 ,35 ,088
Tabela 3.7 – Testes paramétricos para a variável V2v
Levene's Test for
Equality of Variances t-test for Equality of Means
F Sig. t df Sig. (2-tailed)
Mean Difference
Std. Error Difference
95% Confidence Interval of the
Difference
Lower Upper V2v
thicknesses Equal
variances assumed
,001 ,971 -1,398 30 ,172 -,18 ,13 -,45 ,09
Equal variances
not assumed
-1,398 29,669 ,173 -,18 ,13 -,45 ,09
Tabela 3.8 – Testes não-paramétrico para a variável V2v.
V2v thicknesses
Mann-Whitney U 79,000
Wilcoxon W 215,000 Z -1,847 Asymp. Sig. (2-tailed) ,065
Exact Sig. [2*(1-tailed Sig.)] ,067
Da análise da Tabela 3.7, o teste paramétrico de igualdade de variâncias mostra um valor
Sig. de 0.971. Deste modo, assumo a igualdade entre as variâncias de V2v-LH e V2v-RH
(0.971>0.05). O valor Sig. (2-tailed) representa o nível de confiança e neste caso este valor é de
0.152. Como o nível de confiança adoptado por mim é de α=0.05 e sabendo que 0.172 ≥ 0.05, eu

46
aceito a minha hipótese nula de que a espessura em V2v é equivalente entre hemisférios (com um
nível de confiança de 95%). Da análise da Tabela 3.8, o teste não paramétrico apresenta um valor
Sig. (2-tailed) de 0.065. Como o nível de confiança adoptado por mim é de α=0.05 e sabendo que
0.065 ≥ 0.05, também aqui aceito a minha hipótese nula de que a espessura em V2v é equivalente
entre hemisférios (com um nível de confiança de 95%).
3.2.3 Resultados estatísticos para a área visual V2d
A Tabela 3.9 apresenta o resultado do teste de normalidade feito às variáveis V2d no
hemisfério esquerdo e no hemisfério direito:
Tabela 3.9 – Resultados do teste de normalidade para os resultados de V2d em ambos os hemisférios.
Hemisphere Kolmogorov-Smirnova Shapiro-Wilk
Statistic df Sig. Statistic df Sig. V2d thicknesses
LH ,159 16 ,200 ,968 16 ,800
RH ,131 16 ,200 ,963 16 ,718
Da análise da Tabela 3.9 verificamos que os valores de V2d-LH e V2d-RH seguem uma distribuição normal para toda a amostra de 16 pacientes.
Questiono agora se a espessura da área visual V2d é diferente entre hemisférios, ou, caso contrário, se tem a mesma espessura em ambos:
0
1
: A espessura cortical é igual entre hemisférios nesta área visual
: A espessura cortical não é igual entre hemisférios nesta área visual
LH RH
LH RH
H
H
µ µ
µ µ
=
≠
A Tabela 3.10 apresenta os valores da média e desvio padrão de V2d em ambos os hemisférios. A Tabela 3.11 apresenta o resultado do teste de independência de variáveis.
Tabela 3.10 – Valores médios e desvio padrão para V2d em ambos os hemisférios.
Hemisphere
N Mean Std. Deviation Std. Error Mean V2d
thicknesses LH 16 2,33 ,29 ,069
RH 16 2,34 ,25 ,062

47
Tabela 3.11 – Testes paramétricos para a variável V2d.
Levene's Test for
Equality of Variances t-test for Equality of Means
F Sig. t df
Sig. (2-
tailed) Mean
Difference Std. Error Difference
95% Confidence Interval of the Difference
Lower Upper V2d
thicknesses Equal
variances assumed
,218 ,644 -,040 30 ,968 -,004 ,093 -,193 ,186
Equal variances
not assumed
-,040 29,631 ,968 -,004 ,093 -,193 ,186
Da análise da Tabela 3.11, o teste paramétrico de igualdade de variâncias mostra um valor
Sig. de 0.644. Deste modo, assumo a igualdade entre as variâncias de V2d-LH e V2d-RH
(0.644>0.05). O valor Sig. (2-tailed) representa o nível de confiança e neste caso este valor é de
0.0.968. Como o nível de confiança adoptado por mim é de α=0.05 e sabendo que 0.968 ≥ 0.05, eu
aceito a minha hipótese nula de que a espessura em V2d é equivalente entre hemisférios (com um
nível de confiança de 95%).
3.2.4 Resultados estatísticos para a área visual VP
A Tabela 3.12 apresenta o resultado do teste de normalidade feito às variáveis VP no
hemisfério esquerdo e no hemisfério direito:
Table 3.12 – Resultados do teste de normalidade para os resultados de VP em ambos os hemisférios.
Hemisphere Kolmogorov-Smirnova Shapiro-Wilk
Statistic df Sig. Statistic df Sig. VP thicknesses
LH ,143 16 ,200 ,952 16 ,524
RH ,133 16 ,200 ,957 16 ,604
Da análise da Tabela 3.12 verificamos que os valores de VP-LH e VP-RH seguem uma distribuição normal para toda a amostra de 16 pacientes.
Questiono agora se a espessura da área visual VP é diferente entre hemisférios, ou, caso contrário, se tem a mesma espessura em ambos:
0
1
: A espessura cortical é igual entre hemisférios nesta área visual
: A espessura cortical não é igual entre hemisférios nesta área visual
LH RH
LH RH
H
H
µ µ
µ µ
=
≠

48
A Tabela 3.13 apresenta os valores da média e desvio padrão de VP em ambos os hemisférios. A Tabela 3.14 apresenta o resultado do teste de independência de variáveis.
Tabela 3.13 – Valores médios e desvio padrão para VP em ambos os hemisférios.
Hemisphere
N Mean Std.
Deviation Std. Error
Mean VP
thicknesses LH 16 2,53 ,22 ,055
RH 16 2,57 ,30 ,076
.
Tabela 3.14 – Testes paramétricos para a variável VP.
Levene's Test for
Equality of Variances t-test for Equality of Means
F Sig. t df Sig. (2-tailed)
Mean Differenc
e Std. Error Difference
95% Confidence Interval of the
Difference
Lower Upper VP
thicknesses
Equal variance
s assumed
1,311 ,261 -,471 30 ,641 -,04 ,09 -,24 ,15
Equal variance
s not assumed
-,471 27,167
,641 -,04 ,09 -,24 ,15
Da análise da Tabela 3.14, o teste paramétrico de igualdade de variâncias mostra um valor
Sig. de 0.261. Deste modo, assumo a igualdade entre as variâncias de VP-LH e VP-RH (0.261>0.05).
O valor Sig. (2-tailed) representa o nível de confiança e neste caso este valor é de 0.641. Como o
nível de confiança adoptado por mim é de α=0.05 e sabendo que 0.641 ≥ 0.05, eu aceito a minha
hipótese nula de que a espessura em VP é equivalente entre hemisférios (com um nível de confiança
de 95%).
3.2.5 Resultados estatísticos para a área visual V3
A Tabela 3.15 apresenta o resultado do teste de normalidade feito às variáveis V3 no
hemisfério esquerdo e no hemisfério direito:

49
Tabela 3.15 – Resultados do teste de normalidade para os resultados de V3 em ambos os hemisférios.
Hemisphere Kolmogorov-Smirnova Shapiro-Wilk
Statistic df Sig. Statistic df Sig. V3 thicknesses
LH ,171 16 ,200 ,955 16 ,571
RH ,165 16 ,200 ,936 16 ,306
Da análise da Tabela 3.15 verificamos que os valores de V3-LH e V3-RH seguem uma distribuição normal para toda a amostra de 3.16 pacientes.
Questiono agora se a espessura da área visual V3 é diferente entre hemisférios, ou, caso contrário, se tem a mesma espessura em ambos:
0
1
: A espessura cortical é igual entre hemisférios nesta área visual
: A espessura cortical não é igual entre hemisférios nesta área visual
LH RH
LH RH
H
H
µ µ
µ µ
=
≠
A Tabela 3.16 apresenta os valores da média e desvio padrão de V3 em ambos os hemisférios. A Tabela 3.17 apresenta o resultado do teste de independência de variáveis.
Tabela 3.16 – Valores médios e desvio padrão para V3 em ambos os hemisférios.
Hemisphere
N Mean Std. Deviation Std. Error Mean V3
thicknesses LH 16 2,37 ,28 ,070
RH 16 2,36 ,25 ,063
Tabela 3.17 – Testes paramétricos para a variável V3
Levene's Test for
Equality of Variances t-test for Equality of Means
F Sig. t df
Sig. (2-
tailed) Mean
Difference Std. Error Difference
95% Confidence Interval of the Difference
Lower Upper V3
thicknesses Equal
variances assumed
,059 ,809 ,171 30 ,866 ,016 ,09 -,18 ,21
Equal variances
not assumed
,171 29,709 ,866 ,016 ,09 -,18 ,21
.
Da análise da Tabela 3.17, o teste paramétrico de igualdade de variâncias mostra um valor
Sig. de 0.809. Deste modo, assumo a igualdade entre as variâncias de V3-LH e V3-RH (0.809>0.05).

50
O valor Sig. (2-tailed) representa o nível de confiança e neste caso este valor é de 0.866. Como o
nível de confiança adoptado por mim é de α=0.05 e sabendo que 0.866 ≥ 0.05, eu aceito a minha
hipótese nula de que a espessura em V3 é equivalente entre hemisférios (com um nível de confiança
de 95%).
3.2.6 Resumo
O Gráfico 3.1 apresenta uma visão geral sobre os valores de EC médios de cada área visual com seus valores máximos e mínimos.
Gráfico 3.1 – Valores de EC médios, máximos e mínimos para as áreas visuais de cada hemisfério.
3.3 Análise de Correlação entre as áreas visuais
Com base nos padrões de progressão da LHON e tendo em mente o processo de maturação
cortical, decidi criar dois grupos de estudo para este ponto: um grupo com os pacientes com mais de
21 anos e outro com os pacientes com menos de 21 anos. Com essa divisão eu pude realizar um
estudo sobre espessuras corticais tendo em mente o estado de desenvolvimento da doença LHON e
também o envelhecimento. Com este dois sub-grupos, foi possível verificar se havia ou não
correlação entre cada área visual, globalmente e em específico, nos grupos etários. Também é
possível verificar quando estas correlações são mais fortes (se na população jovem ou adulta/se no
hemisfério esquerdo ou direito).
2,41
2,60
2,80
3,11
2,61 2,592,75
2,88
2,65 2,61
2,01
2,21 2,18
2,40
2,06 2,09
2,31 2,272,09 2,10
2,21
2,402,57
2,75
2,33 2,34
2,53 2,57
2,37 2,36
-
0,50
1,00
1,50
2,00
2,50
3,00
3,50
V1-LH V1 -RH V2v-LH V2v-RH V2d-LH V2d-RH VP-LH VP-RH V3-RH V3-LH
Esp
ess
ura
(mm
)
Áreas Visuais

51
3.3.1 Correlações em todas as idades – 16 pacientes
Todas as variáveis, excepto V2V - LH seguem uma distribuição normal para esta amostra de
16 sujeitos. Os resultados da correlação são apresentados na Tabela 3.18 e 3.19.
Tabela 3.18 – Correlações paramétricas nos 16 pacientes para variáveis que seguem uma distribuição normal.
* *Correlação significativa com Sig 0,05 (2-tailed); ** Correlação significativa com Sig 0,01 (2-tailed).
Tabela 3.19 – Correlações não paramétricas nos 16 pacientes para variáveis que não seguem uma distribuição normal.
*Correlação significativa com Sig 0,05 (2-tailed); ** Correlação significativa com Sig 0,01 (2-tailed).
Legenda:
Laranja – correlação média;
Branco – sem correlação;

52
3.3.2 Correlações nos pacientes com menos de 21 anos – 7 pacientes
Todas as variáveis, excepto V2V - LH seguem uma distribuição normal para esta amostra de
7 sujeitos. Os resultados das correlações são apresentados na Tabela 3.20 e 3.21.
Tabela 3.20 – Correlações paramétricas nos pacientes com menos de 21 anos para variáveis que seguem uma distribuição normal.
* *Correlação significativa com Sig 0,05 (2-tailed); ** Correlação significativa com Sig 0,01 (2-tailed).
Tabela 3.21 – Correlações não paramétricas nos pacientes com menos de 21 anos para variáveis que não seguem uma distribuição normal.
*Correlação significativa com Sig 0,05 (2-tailed); ** Correlação significativa com Sig 0,01 (2-tailed).
Legenda:
Laranja – correlação média;
Vermelho – correlação forte;
Branco – sem correlação;

53
3.3.3 Correlações nos pacientes com mais de 21 anos – 9 pacientes
Todas as variáveis seguem uma distribuição normal para esta amostra de 9 sujeitos. Os resultados das correlações estão apresentados na Tabela 3.22.
Tabela 3.22 – Correlações paramétricas nos pacientes com mais de 21 anos para variáveis que seguem uma distribuição normal.
*Correlação significativa com Sig 0,05 (2-tailed); ** Correlação significativa com Sig 0,01 (2-tailed).
Legenda:
Laranja – correlação média;
Vermelho – correlação forte;
Branco – sem correlação;

54
4. Discussão
Os resultados técnicos aqui apresentados fornecem uma ideia preliminar e algumas pistas
sobre a organização do córtex visual em LHON. Os resultados não devem ser extrapolados para toda
a população. Como exemplo disso, os resultados da abordagem estatística entre grupos etários não
podem afirmar ser resultado da acção da LHON ou do processo normal de envelhecimento. No
entanto, estes resultados podem ser utilizados e comparados em trabalhos posteriores, quando os
dados de um grupo de controle esteja disponível. Só desta forma será possível afirmar com maior
certeza algumas hipóteses aqui por mim apresentadas.
4.1 Discussão relativa aos resultados de EC entre áreas visuais
Os resultados apresentados no ponto 3.2 “Diferenças nas espessuras corticais entre
hemisférios” são previsíveis e aceitáveis, pois parecem concordar com o conceito amplamente aceite
da existência de diferenças entre hemisférios, mesmo em regiões corticais homologas, tais como as
áreas visuais homologas.
Os resultados sugerem que, em LHON, V1 no hemisfério direito apresenta ser mais espesso
que o esquerdo. Este resultado requer uma comparação com um grupo de controlo. As outras áreas
visuais em estudo (V2v, V2d, V3 e VP) revelam, em LHON, ter uma espessura equivalente em ambos
os hemisférios.
Os resultados para V1 sugerem que as projecções retinocorticais são assimetricamente
emparelhadas em LHON, mas posterior confirmação é necessária para garantir esta hipótese. O
hemisfério direito é dominante para os aspectos visuais e espaciais das nossas funções perceptivas.
Reconhecimento de faces, auto-imagem mental, expressões faciais e memórias, são algumas das
características funcionais do hemisfério direito. Já o hemisfério esquerdo está intimamente ligado a
funções referentes ao pensamento, à moral, à linguística, à gramática e à resposta verbal. Sendo o
hemisfério direito mais talhado para processar informação visual no sentido espacial, é de algum
modo previsível que algumas áreas visuais neste apresentassem um maior valor de EC do que as
suas áreas homólogas, no hemisfério esquerdo. Se o input visual tiver mais informação a ser
processada no hemisfério direito, poderá levar a um maior número de sinapses e daí maior valor de
EC. No entanto, esta hipótese seria mais bem sustentada nas áreas visuais do cortex extra-estriado e
não no estriado, como aqui os resultados sugerem, apesar de assimetrias nesta região já terem sido
relatadas em animais vertebrados.

55
Resultados de um grupo de controle, em múltiplas comparações, irão responder de forma
definitiva a estas hipóteses preliminares tornando-as restritas da população com LHON ou, pelo
contrário, comuns à população em geral.
4.2 Discussão às correlações entre áreas visuais
Um padrão de correlações positivas foi observado entre áreas visuais. As correlações são
todas positivas e relativamente fortes. Este resultado era esperado uma vez que as áreas visuais
comunicam em regime de feedforward e feedback entre elas. É normal e de certa forma previsível
que elas se correlacionem positivamente nos seus valores de EC. O grau da correlação entre áreas
visuais refere para o nível de comunicação e estímulo entre ambas. Espera-se que uma área visual
que comunique com outra a possa influenciar. A comunicação entre áreas visuais é feita com a troca
de informação, ou seja, com sinapses. De certa forma, o valor de EC de cada área pode reflectir o
número de estímulos que ocorre. Se houver uma redução neste estímulo, por exemplo devido a uma
doença ou patologia, é previsível que possa ocorrer uma atrofia cortical nas áreas afectadas. Com
base nisto, levanto a seguinte questão: a perda de RGCs, pode causar essa atrofia devido à
diminuição do estímulo? E, se essa atrofia realmente existir, se poderá influenciar as áreas visuais
presentes no fluxo ventral e no fluxo dorsal? Mais uma vez os dados provenientes de um grupo de
controle podem reforçar esta hipótese esclarecendo se as correlações aqui presentes são exclusivas
da LHON, ou se são comuns na população geral.
Tendo em conta os sub-grupos etários, ao analisar as Tabelas 3.20 e 3.21 (pacientes com
menos de 21 anos) e as Tabela 3.22 (pacientes com mais de 21 anos), verificam-se quatro tipos de
comportamento no que concerne às correlações estabelecidas entre as áreas visuais:
1. Há áreas visuais que parecem se correlacionar sempre, independentemente da faixa
etária;
2. Há áreas visuais que parecem não se correlacionar, independentemente da faixa
etária;
3. Há áreas visuais que parecem se correlacionar apenas em grupos de adultos;
4. Há áreas visuais que parecem se correlacionar apenas em grupos de jovens;
1. Nestas condições temos: V1 do hemisfério direito, V2V do hemisfério esquerdo, V2v do hemisfério
direito, V2d do hemisfério esquerdo, VP do hemisfério direito e V3 do hemisfério direito. Este
resultado pode ser interpretado em conjunto, como uma indicação da progressão da LHON e do
processo de envelhecimento normal, que afectam os valores da espessura cortical dessas áreas.

56
2. Nestas condições temos VP do hemisfério esquerdo. As ligações fracas entre VP e o córtex
primário são reveladores, neste ponto. Este resultado sugere que esta área visual tem um
desenvolvimento separado das outras áreas visuais em estudo.
3. Nestas condições temos V1 do hemisfério esquerdo. Esta área visual não se correlacionou com
qualquer outra área visual em pacientes jovens. Ao analisarmos as Tabelas 3.20 e 3.21 verificamos
que V1-LH não se correlaciona, enquanto V1-RH o faz. Ao analisarmos a Tabela 3.22, verificamos
que agora V1-LH já se correlaciona com outras áreas e V1-RH continua a correlacionar-se, desta vez
com maior número de áreas visuais. Além das diferenças entre hemisférios, este resultado aponta
para o facto de áreas visuais do córtex extra-estriado não “dependerem” de LH-V1 para os jovens.
Isto pode ser interpretado como a progressão da LHON no córtex primário.
4. Nestas condições temos V3 do hemisfério esquerdo e V2d do hemisfério direito. Este resultado
pode ser interpretado como uma suposta semelhança apenas em fases iniciais de desenvolvimento,
independentemente de ser patológico ou não, seguindo posteriormente caminhos de
desenvolvimento distintos.
O objectivo desta tese foi o de fornecer um estudo dos padrões de correlação das medidas de
EC do córtex visual em LHON, e uma interpretação preliminar do seu significado funcional. Esta tese
abre o caminho para um trabalho futuro cheio de objectivos, que irá testar as hipóteses aqui
apresentadas e, quiçá, levantar outras relativamente a esta doença.

57
5. Referências Bibliográficas
1 - He, Yong; Zhang J. Chen & Alan C. Evans (2007), "Small-World Anatomical Networks in the Human Brain Revealed by Cortical Thickness from MRI", Cerebral Cortex 17 (10): 2407-2419, DOI:10.1093/cercor/bhl149 [e] 2 - Sowell ER, Peterson BS, Kan E, Woods RP, Yoshii J, Bansal R et al. (2007). Sex differences in cortical thickness mapped in 176 healthy individuals between 7 and 87 years of age. Cereb Cortex 17 (7): 1550-60 3 - Noor Kabani, Georges Le Goualher, David MacDonald, and Alan C. Evans. Measurement of cortical thickness using an automated 3-d algorithm: A validation study. NeuroImage,13:375–380, 2001. 4 - Salat DH, Buckner RL, Snyder AZ, Greve DN, Desikan RS, Busa E et al. (2004). Thinning of the cerebral cortex in aging. Cereb Cortex 14 (7): 721-30. 5 - Vincent A. Magnotta, Nancy C. Andreasen, Susan K. Schultz, Greg Harris, Ted Cizadlo, Dan Heckel, Peg Nopoulos, and Micheal Flaum. Quantitative in vivo measurement of gyrification in the human brain: Changes associated with aging. Cerebral Cortex, 9:151–160, mar 1999. 6 - Stephen E. Jones, Bradley R. Buchbinder, and Itzhak Aharon. Three-dimensional mapping of cortical thickness using laplace’s equation. Human Brain Mapping, 11:12–32, 2000. 7 - Georges Le Goualher, Anne Marie Argenti, Michel Duyme, William F.C. Baare, H.E. HulshoffPol, Christian Barillot, and Alan C. Evans. Statistical sulcal shape comparisons: Applicationto the detection of genetic encoding of the central sulcus shape. 8 - P.M. Thompson, J. Moussai, S. Zohoori, A. Goldkorn, A.A. Khan, M.S. Mega, G.W. Small, J.L. Cummings, and A.W. Toga. Cortical variability and asymmetry in normal aging and alzheimer’s disease. Cerebral Cortex, 8:492–509, sep 1998. 9 - Joel R. Meyer, Sudipta Roychowdhury, Eric J. Russel, Cathy Callahan, Darren Gitelman, and M. Marsel Mesulam. Location of the central sulcus via cortical thickness of the precentral and postcentral gyri on mr. American Journal of Neuroradiology, 17:1699–1706, oct 1996. 10 – K. Zilles, A. Schleicher, C. Langeman, K. Amunts, P. Morosan, N. Palomero-Gallagher, T. Schormann, H. Mohlberg, U. Burgel, H. Steinmetz, G. Schlaug, and Per E. Roland. Quantitative analysis of sulci in the human cerebral cortex: Human Brain Mapping, 5:218–221, 1997. 11 - Improved localization of cortical activity by combining EEG and MEG with MRI cortical surface reconstruction: A linear approach, Dale, A.M., and Sereno, M.I. (1993). Journal of Cognitive Neuroscience 5:162-176. 12 - Cortical Surface-Based Analysis I: Segmentation and Surface Reconstruction, Dale, A.M., Fischl, B., Sereno, M.I., (1999). NeuroImage 9(2):179-194 13 - Cortical Surface-Based Analysis II: Inflation, Flattening, and a Surface-Based Coordinate System, Fischl, B., Sereno, M.I., Dale, A.M., (1999). NeuroImage, 9(2):195-207. 14 - Fischl B, Liu A, Dale AM. Automated manifold surgery: constructing geometrically accurate and topologically correct models of the human cerebral cortex. IEEE Trans Med Imaging 2001; 20: 70-80. 15 - High-resolution inter-subject averaging and a coordinate system for the cortical surface, Fischl, B., Sereno, M.I., Tootell, R.B.H., and Dale, A.M., (1999). Human Brain Mapping, 8:272-284(1999). 16 - Goodale & Milner (1992). "Separate pathways for perception and action." Trends in Neuroscience 15: 20–25. 17 - Ungerleider and Mishkin (1982). Ingle DJ, Goodale MA and Mansfield RJW. Analysis of Visual Behavior. MIT Press. 18 - Leuba G, Kraftsik R. Changes in volume, surface estimate, three-dimensional shape and total number of neurons of the human primary visual cortex from midgestation until old age. Anat Embryol (Berl). 1994 Oct;190(4):351-66. 19 - Gazzaniga, M. S., Ivry, R. B. & Mangun, G. R. (2002). Cognitive Neuroscience: The biology of the mind (2nd ed.). New York: W.W.Norton.

58
20 - Bussey TJ, Saksida LM (2007), “Memory, perception, and the ventral visual-perirhinal-hippocampal stream: thinking outside of the boxes.” Hippocampus 17(9):898-908 21 - Winters BD, Saksida LM, Bussey TJ (2010), “Implications of animal object memory research for human amnesia.” Neuropsychologia 22 - Manuel F. López-Aranda, Juan F. López-Téllez, Irene Navarro-Lobato, Mariam Masmudi-Martín, Antonia Gutiérrez, Zafar U. Khan (2009). Role of Layer 6 of V2 Visual Cortex in Object-Recognition Memory. DOI: 10.1126/science.1170869
23 - Orban GA, Van Essen D, Vanduffel W.Trends Cogn Sci. 2004 Jul. Comparative mapping of higher visual areas in monkeys and humans., 8(7):315-24. Review.PMID: 15242691
24 - Mannion DJ, McDonald JS, Clifford CW. (2010). The influence of global form on local orientation anisotropies in human visual cortex. Neuroimage. 2010 Aug 15;52(2):600-5. 25 - Rosa MG, Tweedale R (2000) Visual areas in lateral and ventral extrastriate cortices of the marmoset monkey. J Comp Neurol 422:621-51. 26 - Ronald S. Fishman (1997). Gordon Holmes, the cortical retina, and the wounds of war. The seventh Charles B. Snyder Lecture Documental Ophthalmological 93: 9-28, 1997. 27 - Rosa MGP, 2002, Visual maps in the adult primate cerebral cortex: some implications for brain development and evolution. Braz J Med Biol Res 35(12):1485-1498. 28 - Goebel, R., Khorram-Sefat, D., Muckli, L., Hacker, H. and Singer, W. (1998) The constructive nature of vision: Direct evidence from fMRI studies of apparent motion and motion imagery. The European Journal of Neuroscience, 10, 1563-1573. 29 - Goebel, R., Muckli, L., Zanella, F.E., Singer, W. & Stoerig, P. (2001). Sustained extrastriate cortical activation without visual awareness revealed by fMRI studies of hemianopic patients. Vision Research, 41, 1459-1474. 30 - Goebel, R., Muckli, L. & Kim, D.-S. (2003). The Visual System. In: G. Paxinos & J.K. Mai (Eds.), The Human Nervous System, 2nd Edition. New York: Academic Press. 31 - Roger B.H. Tootell, Anders M. Dale, Martin I. Sereno and Rafael Malach. (1996). New images from human visual cortex. 32 - Sereno MI, Dale AM, Reppas JB, Kwong KK, Belliveau JW, Brady TJ, Rosen BR, Tootell RB. (1995). Borders of multiple visual areas in humans revealed by functional magnetic resonance imaging. Science. 1995 May 12;268(5212):889-93. 33 - DeYoe EA, Carman GJ, Bandettini P, Glickman S, Wieser J, Cox R, Miller D, Neitz J. (1996). Mapping striate and extrastriate visual areas in human cerebral cortex. 1996 Mar 19;93(6):2382-6. 34 - Leber Hereditary Optic Neuropathy .Yu-Wai-ManP,ChinneryPF. In: Pagon RA, Bird TC, Dolan CR, Stephens K, editors. GeneReviews [Internet]. Seattle (WA): University of Washington,Seattle;1993-.2000 35 - Bandelt HJ, Kong QP, Parson W, Salas A (December 2005). "More evidence for non-maternal inheritance of mitochondrial DNA?". J. Med. Genet. 42 (12): 957–60 36 - Nikoskelainen EK, Marttila RJ, Huoponen K, et al. (August 1995). "Leber's "plus": neurological abnormalities in patients with Leber's hereditary optic neuropathy". J. Neurol. Neurosurg. Psychiatr. 59 (2): 160–4. 37 - Schneider P, Sluming V, Roberts N, Bleeck S, Rupp A. Structural, functional, and perceptual differences in Heschl's gyrus and musical instrument preference. (2005). Ann N Y Acad Sci. 2005 Dec;1060:387-94 38- Goebel, R., Esposito, F. Formisano, E. (2006). Analysis of functional image analysis contest (FIAC) data with Brainvoyager QX: From single-subject to cortically aligned group general linear model analysis and self-organizing group independent component analysis. Human Brain Mapping, 27, 392-401. 39 - Jones, S.E., Buchbinder, B.R., Aharon, I. (2000). Three-Dimensional Mapping of Cortical Thickness Using Laplace’s Equation. Human Brain Mapping, 11, 12-32. 40 - Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd ed.). New Jersey: Lawrence
Erlbaum.





























