Embed Size (px)
Citation preview

Programa 14
14. PROGRAMA DE MONITORAMENTO DA COMUNIDADE FITOPLANCTÔNICA E ZOOPLANCTÔNICA .......... 1
14.1. INTRODUÇÃO E OBJETIVOS ........................................................................................................................ 1
14.2. METODOLOGIA ........................................................................................................................................ 3
14.3. RESULTADOS E DISCUSSÃO ........................................................................................................................ 8
14.4. CONSIDERAÇÕES FINAIS........................................................................................................................... 69
14.5. CRONOGRAMA ...................................................................................................................................... 71
14.6. REFERÊNCIAS BIBLIOGRÁFICAS .................................................................................................................. 71
14.7. EQUIPE TÉCNICA .................................................................................................................................... 74

Programa 14 - 1
14. Programa de Monitoramento da Comunidade Fitoplanctônica e
Zooplanctônica
14.1. Introdução e Objetivos
14.1.1. Comunidade Fitoplanctônica
O fitoplâncton é importante não só por constituir uma das fontes primárias de
energia para o meio aquático, como também por desempenhar um papel
fundamental na captura do CO2 atmosférico. Por meio do processo de
fotossíntese, o fitoplâncton é capaz de fixar mais de 50 Gt de carbono por ano, o
equivalente a aproximadamente metade da produção primária global (Falkowski
et al. 1998).
Devido ao seu tamanho pequeno e ciclo de vida de alguns dias, constitui
uma ferramenta fundamental como indicador de qualidade ambiental.
A composição do fitoplâncton em um corpo de água muda continuamente
em dominância e diversidade de acordo com as mudanças físicas (luz,
temperatura, turbulência), químicas (nutrientes inorgânicos, quelantes, vitaminas)
e biológicas (competição, predação) (Hallegraeff & Reid, 1986).
Fatores físico-químicos afetam a sucessão e a dinâmica do fitoplâncton.
Descargas de água doce afetam o tempo de residência disponível para o
crescimento dos diferentes taxa do fitoplâncton, e também influencia na sucessão
do fitoplâncton marinho, estuarino e de água doce à medida que impede a
entrada da água do mar para dentro do estuário.
Os três principais grupos, Bacillariophyta, Dinophyta e Chlorophyta, estão
separados temporalmente pelas estações do ano e espacialmente, ao longo do
estuário, de acordo com o fluxo e salinidade. Bacillariophyta exibe a maior taxa de
crescimento em ambientes turbulentos. Dinophyta, é dominada por poucas
espécies de água salobra, possuem a menor taxa de crescimento e ocorrem em
locais com fluxo pequeno de água. Chlorophyta possuem taxas de crescimento
intermediarias e é restrita a ambientes de água doce. No estuário, os nutrientes

Programa 14 - 2
parecem ser menos importantes do que o fluxo e salinidade na regulação da
sucessão do fitoplâncton (Chan & Hamilton, 2001).
O fitoplâncton no estuário, também está sob a influência dos gradientes de
salinidade e mudanças entre o sedimento e a coluna de água adjacente. Muitas
das diatomáceas amostradas no plâncton são de origem bentônica como:
Entomoneis paludosa, Gyrosigma balticum, G. fasciola, G. acuminatum,
Pleurosigma strigosum, P. normanii, Nitzschia pusilla, Cylindrotheca closterium, N.
sigma ou formas meroplanctônicas Thalassiosira decipiens, Cyclotella striata, C.
meneghiniana, Paralia sulcata, Actinoptychus senarius, comuns nas camadas
superficiais do sedimento. Na presença de água doce são observadas algas do
perifiton como: Melosira varians, M. moniliformis, Bacillaria paxillifer, Achnanthes
spp. Portanto, a riqueza de espécies no plâncton é aumentada por suspensão de
microalgas epipelica-epífitas como se fosse um processo de migração do
ambiente bentônico para a coluna d’água (Cloern & Dufford, 2005).
14.1.2. Comunidade Zooplanctônica
Os organismos zooplanctônicos possuem um papel central na trama trófica
de ecossistemas costeiros e estuarinos, como consumidores, presas e
competidores, e sua atividade metabólica tem importantes implicações para a
ciclagem de compostos orgânicos e inorgânicos na coluna de água. Várias
espécies de crustáceos e de peixes utilizam os estuários como zonas de berçário
e crescimento e em muitos casos ocorrem sob a forma de ovos e larvas
compondo a comunidade zooplanctonica. Além disto, o recrutamento de muitas
espécies de importância econômica depende da disponibilidade adequada de
alimento planctônico. Várias forçantes físicas e químicas atuam sobre o
zooplâncton, com reflexos diretos (retenção, advecção, mortalidade) e indiretos
(disponibilidade de alimento, predação, competição por espaço ou recursos)
sobre a produtividade e a variabilidade espacial e temporal do ecossistema.
Alterações na qualidade ambiental dos sistemas estuarinos podem afetar os
organismos zooplanctônicos. Entre os impactos mais importantes estão as
mudanças na circulação da água nas áreas de influência de empreendimentos

Programa 14 - 3
portuários. Alterações na magnitude e nos padrões espaciais e temporais das
correntes estuarinas podem afetar o deslocamento passivo e ativo (por migração
vertical) do zooplâncton. Estas mudanças na circulação podem gerar impactos
sobre parâmetros físicos e químicos da água, como a salinidade e a turbidez.
Caso estes impactos sejam significativos, ocasionam a desestruturação de
associações de espécies nas áreas de influência direta e indireta do complexo do
porto e, em alguns casos, sua substituição por outras. A atividade portuária pode
ainda contribuir para a introdução de contaminantes orgânicos e inorgânicos no
ecossistema aquático, com consequências para os processos biogeoquímicos
envolvendo o zooplâncton. Não menos relevante é a possibilidade de introdução
de espécies exóticas pelo vetor do transporte marítimo, incluindo a água de lastro
e a bioincrustação, neste caso de organismos bentônicos que produzem larvas
meroplanctônicas.
14.2. Metodologia
Para a avaliação da Comunidade de fitoplâncton e zooplâncton, foram
realizadas coletas em janeiro (Campanha Prévia - verão) e julho de 2010
(primeira Campanha - inverno). A rede amostral do programa em questão é
composta por um total de 50 pontos de coleta, sendo 40 pontos coincidentes ao
estudo do meio biótico do EIA/RIMA (FRF, 2008) e 10 pontos adicionais na área
dragada (meio do canal), conforme a Figura 14.2-1. Assim, o monitoramento irá
ocorrer em toda a área diretamente afetada (ADA) e área de influência direta
(AID) da obra de dragagem de aprofundamento do canal de navegação do Porto
de Santos, em pontos amostrados coincidentes aos do diagnóstico do meio
biótico apresentado no EIA/RIMA (FRF, 2008).
Neste relatório são descritos e integrados os resultados das duas
campanhas executadas até o momento.

Programa 14 - 4
Figura 14.2-1. Indicação dos 50 pontos amostrais das comunidades fitoplanctônicas e zooplanctônicas. Em amarelo os pontos indicados pelo EIA/RIMA (FRF, 2008) e em azul os pontos adicionais na área dragada.

Programa 14 - 5
14.2.1. Comunidade Fitoplanctônica
As amostras foram coletadas e imediatamente fixadas com solução de
formalina neutralizada com hexametileno tetramina (0,4% concentração final).
Para a contagem e identificação dos organismos do fitoplâncton, foram utilizadas
câmaras de sedimentação com volume de 10ml, com 24 horas de sedimentação.
Após esse tempo, a cubeta foi levada ao microscópio invertido (Zeiss - axiovert),
segundo o método de Uthermöl (1931). Para as frações do nanoplâncton (< que
20μm), a contagem foi feita em transectos com aumento final de 400X e do
microfitoplâncton (> que 20μm) em toda a câmara com aumento final de 200X. Os
resultados apresentaram a densidade celular total expressa em cel.l–1. A
identificação de dinoflagelados atecados (sem placas de celulose), como exemplo
dos Gymnodiniales, foi feita após contagem em lâmina com imersão (1.600X de
magnitude) para observar detalhes da estrutura. Para os dinoflagelados com
placas de celulose, os indivíduos foram corados e as placas observadas também
em 1.600X de magnitude. A identificação foi em nível de espécie quando possível
seguindo as literaturas: Bicudo & Menezes (2006); Thomas (1997); Balech, et al.
(1984); Caljon (1983); Elster & Ohle (1983); Dodge (1982). A ordem hierárquica
seguiu a classificação no algaebase (www.algaebase.org/).
A estrutura da comunidade e a comparação entre os pontos de coleta foram
dadas por meio dos cálculos de riqueza, índice de diversidade especifica segundo
Shannon-Wiener (H´) (1948) e equitabilidade (J).
Riqueza (S): A riqueza foi estimada a partir do número de táxons
encontrados em cada amostra
Índice de Diversidade de Shanon (H’):
H’= - Σ pi . ln pi
Sendo pi = Ni/N total
Onde:
pi- Abundância relativa de gêneros/espécies;
Ni- densidade de indivíduos de um dado gênero ou espécie (fitoplâncton);

Programa 14 - 6
N total- densidade total de organismos fitoplanctônicos.
Equitabilidade (J): calculada a partir do H’ através da seguinte equação:
J= H’/ log2S
J- Equitatibilidade
H’- índice de diversidade de Shannon-Weaver;
S- Riqueza específica
Para avaliar a similaridade entre os pontos e os dois períodos de coleta,
foram aplicados os testes ANOSIM e SIMPER com a utilização do PRIMER na
matriz de dados (espécies X ponto).
14.2.2. Comunidade Zooplanctônica
Foi utilizada uma rede cônico-cilíndrica, com 40 cm de diâmetro interno, 1,76
m de comprimento e malha de 200 µm. Na boca da rede, foi acoplado um
fluxômetro (General Oceanics), devidamente calibrado, para a estimativa do
volume filtrado pela rede durante os arrastos horizontais com duração de 3
minutos cada.
As amostras foram transferidas para frascos apropriados e preservadas em
solução salina de formaldeído a 4%, tamponada.
O cálculo do volume de água filtrado nos arrastos foi feito de acordo com a
seguinte expressão:
V = a * n * c
Onde:
V = volume de água filtrada pela rede (m3);
a = área da boca da rede (m2);
n = número de rotações do fluxômetro durante o arrasto;
c = taxa de calibração do fluxômetro (rotações por metro).

Programa 14 - 7
No laboratório, as amostras de zooplâncton foram fracionadas utilizando um
quarteador do tipo Motoda (Motoda, 1959). As alíquotas, com no mínimo 300
organismos (Omori & Ikeda, 1984), foram analisadas sob estereomicroscópio
binocular até o nível de gênero ou espécie para os organismos do holoplâncton e
em grandes grupos para o meroplâncton. A identificação dos organismos do
zooplâncton foi baseada nas seguintes referências: Todd et al. (1996), Boltovskoy
(1981) e Boltovskoy (1999).
O cálculo do número de indivíduos por metro cúbico de água foi feito
dividindo-se o número total de organismos encontrados para cada táxon
identificado, pelo volume de água filtrado pela rede em cada ponto de coleta.
Os índices de diversidade de Shannon e de dominância de Simpson foram
calculados para cada ponto de coleta e separadamente para os organismos do
holoplâncton e do meroplâncton/ticoplâncton.
O índice de diversidade de Shannon (H´) mede o grau de incerteza em se
prever a qual espécie pertencerá um indivíduo escolhido ao acaso. Portanto,
quanto maior o valor, maior o grau de incerteza e em uma comunidade com
somente uma espécie não há dúvidas quanto à qual espécie pertence um
indivíduo retirado ao acaso, sendo então H´= 0. Em outras palavras, quanto maior
o valor deste índice, maior a diversidade. O valor do índice de Shannon aumenta
com o aumento do número de espécies e teoricamente pode atingir qualquer valor
máximo, mas, na prática, valores maiores do que 5 são raros. A base logarítmica
proposta inicialmente é log2 por sua relação com a Teoria da Informação (Krebs,
1999).
A obtenção deste índice para os pontos de coleta do presente estudo se deu
a partir da seguinte fórmula:
s
H´ = (pi) * (log2 pi)
i=1
Onde:
H´ = índice de diversidade (bits/indivíduo)
s = número de espécies
pi = proporção da espécie i na amostra

Programa 14 - 8
O índice de Simpson indica o grau de dominância existente em uma
comunidade. Reflete a probabilidade de dois indivíduos escolhidos ao acaso na
comunidade pertencerem à mesma espécie. Varia de 0 a 1 e quanto mais alto for,
maior a probabilidade dos dois indivíduos sorteados serem da mesma espécie, ou
seja, maior a dominância e menor a diversidade. É calculado da seguinte
maneira:
s
SI = pi2
i=1
Onde:
s = número de espécies
pi = proporção da espécie i na amostra
Para a análise de ordenação não-métrica multidimensional (N-MDS) as
matrizes de dados de densidade do zooplâncton para cada campanha foi
reduzida para 60 taxa, retirando da análise aqueles que ocorreram em 8% ou
menos dos 50 pontos de coleta. Os dados de densidade foram transformados
pela raiz quadrada e o índice de similaridade utilizado foi o de Bray-Curtis.
14.3. Resultados e Discussão
14.3.1. Comunidade Fitoplanctônica
Campanha de janeiro de 2010 (Campanha Prévia)
A composição do fitoplâncton, nos pontos de coleta amostrados no verão
(campanha prévia), foi típica de regimes costeiros com influência de água doce e
foi determinada principalmente pela estratificação salina. Nas 50 amostras
coletadas, foram observados 281 taxa pertencentes aos filos Cyanobacteria,
Chlorophyta, Bacillariophyta, Heterokontophyta, Myzozoa, Sarcomastigophora,

Programa 14 - 9
Euglenozoa, Charophyta, Chlorophyta, Ciliophora; inseridos em 16 classes e 52
ordens.
O fitoplâncton total variou entre 1,55 x 106 cels/l no Canal de São Vicente
(P37) a 22,2 x 106 cels/l Canal de Santos (P9), (Figura 14.3.1-1). Estes são
números intermediários entre os registrados por Frazão (2001) de 3,76 a
48,63 x 106 cels/l na baía de Santos e próximo ao emissário e os encontrados por
Moser (2002), num período chuvoso, de 5 x 106 cels/l para o Canal de São
Vicente e 1x 106 cels/l no Canal de Santos.
Figura 14.3.1-1. Abundância do fitoplâncton total nas 50 amostras coletadas em janeiro de 2010 (Campanha Prévia) em números de células por litro.
A fração entre 2 μm e 20μm, foi a mais abundante em todo o estuário,
variando entre 1,44 x 106 cels/l (P37) e 21 x 106 cels/l (P9), contribuindo com 89%
dentro do nanoplâncton (P38) e 74% da população total (P33).
No nanoplâncton (células entre 2μm e 20μm), o grupo dos flagelados,
juntamente com os filos Bacillariophyta e Cyanobacteria foram os mais
abundantes na região (Figura 14.3.1-2). Bacillariophyta foi o mais representativo e
abundante depois dos flagelados nesta fração de tamanho. Thalassiosira spp
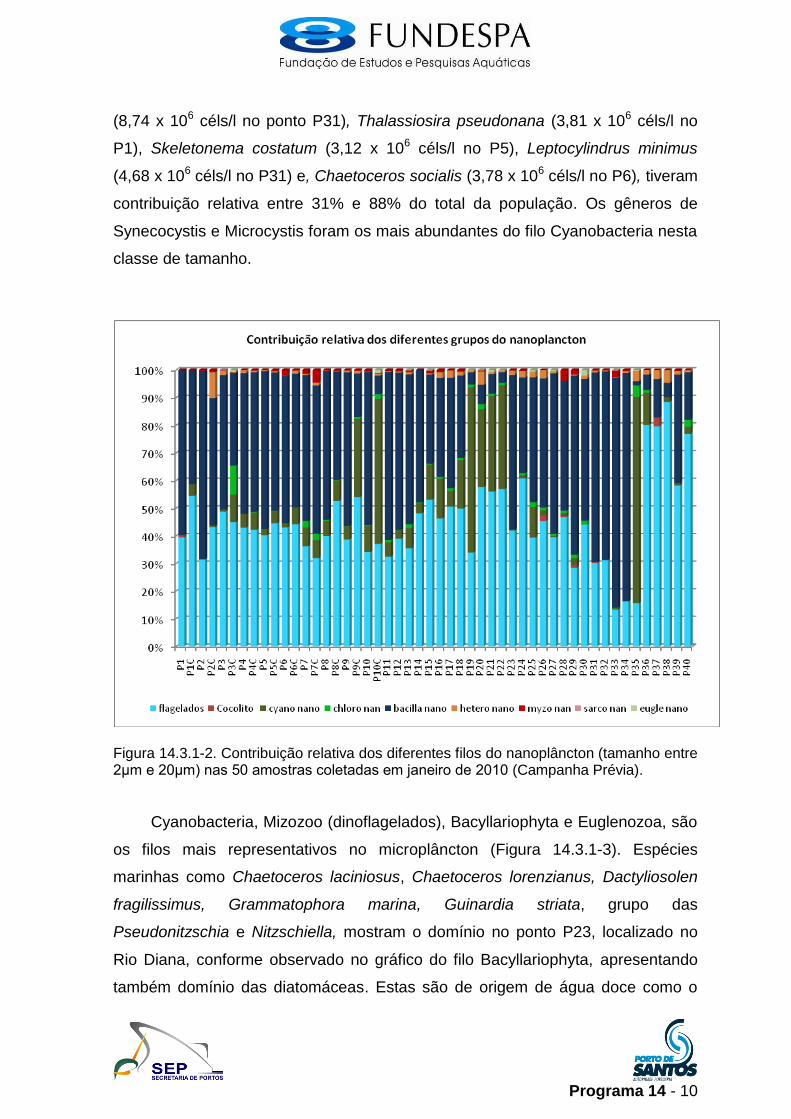
Programa 14 - 10
(8,74 x 106 céls/l no ponto P31), Thalassiosira pseudonana (3,81 x 106 céls/l no
P1), Skeletonema costatum (3,12 x 106 céls/l no P5), Leptocylindrus minimus
(4,68 x 106 céls/l no P31) e, Chaetoceros socialis (3,78 x 106 céls/l no P6), tiveram
contribuição relativa entre 31% e 88% do total da população. Os gêneros de
Synecocystis e Microcystis foram os mais abundantes do filo Cyanobacteria nesta
classe de tamanho.
Figura 14.3.1-2. Contribuição relativa dos diferentes filos do nanoplâncton (tamanho entre 2μm e 20μm) nas 50 amostras coletadas em janeiro de 2010 (Campanha Prévia).
Cyanobacteria, Mizozoo (dinoflagelados), Bacyllariophyta e Euglenozoa, são
os filos mais representativos no microplâncton (Figura 14.3.1-3). Espécies
marinhas como Chaetoceros laciniosus, Chaetoceros lorenzianus, Dactyliosolen
fragilissimus, Grammatophora marina, Guinardia striata, grupo das
Pseudonitzschia e Nitzschiella, mostram o domínio no ponto P23, localizado no
Rio Diana, conforme observado no gráfico do filo Bacyllariophyta, apresentando
também domínio das diatomáceas. Estas são de origem de água doce como o

Programa 14 - 11
genero Aulacoseira. Ao adentrarmos o Canal de Santos, em direção às águas
interiores, este domínio passa a ser das cianofíceas filamentosas, com a
presença das espécies de Cylindropermopsis raciborskii (1,3 x 106 céls/l no ponto
P9C), Planktolyngbea sp (3,5 x 105 céls/l no P9C) e Planktothrix aguardii
(5,78 x 105 céls/l no P2). No canal de Bertioga a fração do microplâncton é
dominada pelos dinoflagelados. No ponto P30, localizado na região do Canal de
Bertioga, 96% da população foram constituídas por Karenia mikimotoi, com um
número expressivo de 1,39 x 106 céls/l. Peridinium quinquecorne contribuiu com
36% da população no ponto P27. No Canal de São Vicente, o microplâncton foi
dominado pelas Euglenales, com 4,54 x 105 céls/l no P35. Ao longo de todo o
estuário, exceto para os pontos localizados na entrada do Canal de Santos, foi
registrada a presença de Mesodinium rubrum, sua maior contribuição foi no ponto
P8 com 1,02 x 105 ind./l, e maior representatividade dentro do Canal de Bertioga
no P32 (21% do microplâncton).
Figura 14.3.1-3. Contribuição relativa dos diferentes filos do microplâncton (tamanho entre 20μm e 200μm) nas 50 amostras coletadas em janeiro de 2010 (Campanha Prévia).

Programa 14 - 12
A riqueza especifica variou entre 16 espécies (ponto P37) e 62 espécies
(P2). Os menores valores foram registrados nos pontos situados no interior do
estuário, próximas e no Largo de Canéu e canal de São Vicente (Figura 14.3.1-4).
Figura 14.3.1-4. Riqueza específica do fitoplâncton registrada para as 50 amostras coletadas em janeiro de 2010 (Campanha Prévia).
O índice de diversidade específica ficou acima de 2 bits na maioria dos
pontos de coleta. Os menores valores foram obtidos nas amostras coletadas no
Canal de Bertioga nos pontos P33 e P34, nos quais foi observado valor de 0,6 bits
(Figura 14.3.1-5).

Programa 14 - 13
Figura 14.3.1-5. Diversidade específica do fitoplâncton nas 50 amostras coletadas em janeiro de 2010 (Campanha Prévia)
Valores de equitabilidade variaram entre 0,14 (P34) e 0,7 (P40). Valores
acima de 0,5 são encontrados nas amostras dentro do Canal de Santos da
entrada até a parte interna, próximo à saída do Canal de Bertioga e no Canal de
São Vicente (Figura 14.3.1-6).

Programa 14 - 14
Figura 14.3.1-6. Equitabilidade nas 50 amostras coletadas em janeiro de 2010 (Campanha Prévia).
Para as coletas de janeiro de 2010, o fitoplâncton no sistema estuarino de
Santos foi composto por espécies neríticas marinhas capazes de suportar
grandes variações de salinidade, espécies estuarinas e as encontradas em águas
interiores, lagoas e represas. Foi constituído principalmente por organismos de
pequeno tamanho denominado de nanoplâncton (entre 2μm e 20μm),
representado pelos flagelados e os filos Bacyllariophyta e Cyanobacteria.
Os índices de riqueza, diversidade e equitabilidade apontam para um
ambiente que embora, possua uma diversidade relativamente alta, não possui um
equilíbrio na sua composição (índices menores que 0,5 para equitabilidade). Isto
se deve ao fato da alternância de domínios entre as diatomáceas, dinoflagelados
e cianobacterias.
O florescimento é comum em regiões estuarinas. O fluxo de água doce
promove a ressuspensão de fundo juntamente com nutrientes e formas de
resistências (cistos) que, ao entrarem para a zona eufótica (camada de luz),
iniciam o processo de divisão. A espécie mais adaptada às condições no
momento foi a dominante.

Programa 14 - 15
Embora o florescimento seja um evento comum, o que deve ser ressaltado
neste estudo é que, as espécies dominantes, com exceção da Thalassiosira spp,
são consideradas tóxicas e ou potencialmente tóxicas a saber: Cyanobacteria:
Synechocystis sp, Microcystis sp, Anabaena sp, Cylindropermopsis raciborskii,
Planktothix agardii; Mizozoa (Dinophyta): Karenia mikimotoi, Karlodinium micrum,
Prorocentrum micans, Prorocentrum lima e Prorocentrum minutum, Scripsiella
trochoida; e dentro das Bacillariophyta as consideradas potencialmente tóxicas:
grupo Pseudonitzchia e Skeletonema costatum.
Karenia mikimotoi, causou florescimento dentro do Canal de Bertioga, com
1,39 x 106 céls/l (96% da população do microplâncton). Esta espécie causa morte
dos organismos tanto pela deplexão do oxigênio como produção de toxina
(Brevetoxina). A toxina não tem efeito sobre os seres humanos, mas em
concentração acima de 2,3 x 105 céls/l foi letal para os peixes e organismos da
macrofauna, na costa da Irlanda (McMahon & Silke, 1996).
As cianobacterias são bastante conhecidas pois os efeitos tóxicos afetam
diretamente os seres humanos. Porém dados de florescimento em ambientes
estuarinos não foram encontrados para se estabelecer uma referência.
Campanha de julho de 2010 (1ª Campanha)
A composição do fitoplâncton na campanha de inverno, realizada em julho
de 2010 (1ª campanha), foi característica de regimes costeiro-estuarinos com
pouca representatividade de espécies lacustres. Nas 50 amostras coletadas,
foram registrados 244 taxa pertencentes aos filos Cyanobacteria, Chlorophyta,
Bacillariophyta, Heterokontophyta, Myzozoa (Dinoflagellata), Sarcomastigophora,
Euglenozoa, Charophyta, Chlorophyta, Ciliophora; inseridos em 39 ordens.
Bacillariophyta foi o maior filo com 23 ordens e 153 espécies, seguido por
Dinoflagellata com 4 ordens e 27 espécies.
Numericamente, o fitoplâncton total variou entre 6,29 x 105 céls/L na Baia de
Santos próximos à entrada do canal (P3) e 7,94 x 106 céls/L, no interior do Canal
de Bertioga (P29) (Figura 14.3.1-7).

Programa 14 - 16
Figura 14.3.1-7. Abundância do fitoplâncton total nas 50 amostras coletadas em julho de 2010 (1ª Campanha) em números de células por litro.
Os flagelados juntamente com o nanoplâncton foram os mais abundantes
em todo o estuário, chegando a compor quase a totalidade da população (acima
de 99%) no Canal de Bertioga (P29 a P34) e Canal de São Vicente (P38 a P40).
(Figura 14.3.1-8).

Programa 14 - 17
Figura 14.31.-8. Contribuição relativa, das diferentes frações do fitoplâncton total, coletadas em julho de 2010 (1ª Campanha) nos 50 pontos.
Dinophyta, Mizozoo (Dinoflagelatta) e Cocolitoforídeos foram mais
representativos dentro do grupo do nanoplâncton. As diatomáceas foram
frequentes em todo o estuário, compondo 100% da fração pequena (entre 2µm e
20µm) no ponto P3. Os Cocolitoforídeos e os dinoflagelados também frequentes,
mas menos abundantes, com o máximo de representatividade nos pontos P26
(56%) e P8C (52%) respectivamente. As cianofíceas foram mais abundantes no
interior do estuário (Figura 14.3.1-9).

Programa 14 - 18
Figura 14.3.1-9. Contribuição relativa dos diferentes filos do nanoplâncton (tamanho entre 2μm e 20μm) nas 50 amostras coletadas em julho de 2010 (1ª Campanha).
Thalassiosira spp, Skeletonema costatum e Thalassiosira pseudonana foram
frequentes e abundantes principalmente nas amostras dentro do Canal de
Bertioga, enquanto que Chaetoceros socialis, Chaetoceros curvissetus e
Chaetoceros compressus, foram mais abundantes nos pontos da Baia de Santos
e entrada do Canal como pode ser observado na Figura 14.3.1-10 (A e B). Os
Gymnodiniales, e o Gymnodinium cf pygmaeum, foram os dinoflagelados mais
representativos, nesta fração de tamanho, com 65 x103 céls/L (P20) e
37 x103 céls/L (P2). Os cocolitos tiveram maior abundância no final do Canal de
Santos (P7C), com 93 x 103 indivíduos por litro.

Programa 14 - 19
Figura 14.3.1-10. Principais espécies de diatomáceas do nanoplâncton nas amostras de inverno (1ª Campanha).
Bacillariophyta foi o filo mais representativo dentro do microfitoplâncton em
todo o sistema estuarino, compondo mais de 90% nas amostras da Baia até o
final do Canal de Santos, chegando à totalidade da população no ponto P8
(100%). O ciliado autótrofo Mesodinium rubrum é o segundo mais representativo,
principalmente no Largo do Canéu e no Canal de São Vicente, com máximo de
contribuição de 34% no P36. Myzozoa (Dinoflagellata) foi mais representativo em
alguns pontos no interior do estuário (P18 e P23), Canal de Bertioga (P30) e no
Canal de São Vicente, no P39, e com o máximo de contribuição no ponto P40
(16%). (Figura 14.3.1-11).
A
B
B
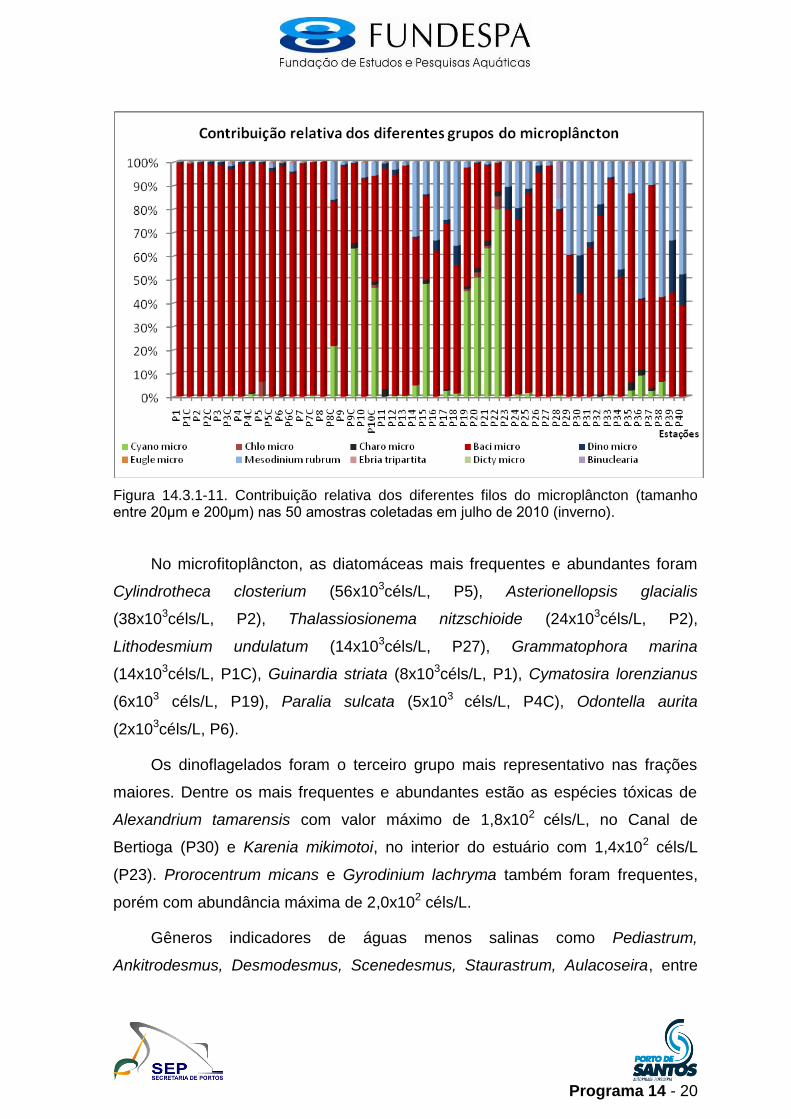
Programa 14 - 20
Figura 14.3.1-11. Contribuição relativa dos diferentes filos do microplâncton (tamanho entre 20μm e 200μm) nas 50 amostras coletadas em julho de 2010 (inverno).
No microfitoplâncton, as diatomáceas mais frequentes e abundantes foram
Cylindrotheca closterium (56x103céls/L, P5), Asterionellopsis glacialis
(38x103céls/L, P2), Thalassiosionema nitzschioide (24x103céls/L, P2),
Lithodesmium undulatum (14x103céls/L, P27), Grammatophora marina
(14x103céls/L, P1C), Guinardia striata (8x103céls/L, P1), Cymatosira lorenzianus
(6x103 céls/L, P19), Paralia sulcata (5x103 céls/L, P4C), Odontella aurita
(2x103céls/L, P6).
Os dinoflagelados foram o terceiro grupo mais representativo nas frações
maiores. Dentre os mais frequentes e abundantes estão as espécies tóxicas de
Alexandrium tamarensis com valor máximo de 1,8x102 céls/L, no Canal de
Bertioga (P30) e Karenia mikimotoi, no interior do estuário com 1,4x102 céls/L
(P23). Prorocentrum micans e Gyrodinium lachryma também foram frequentes,
porém com abundância máxima de 2,0x102 céls/L.
Gêneros indicadores de águas menos salinas como Pediastrum,
Ankitrodesmus, Desmodesmus, Scenedesmus, Staurastrum, Aulacoseira, entre
Programa 14 - 21
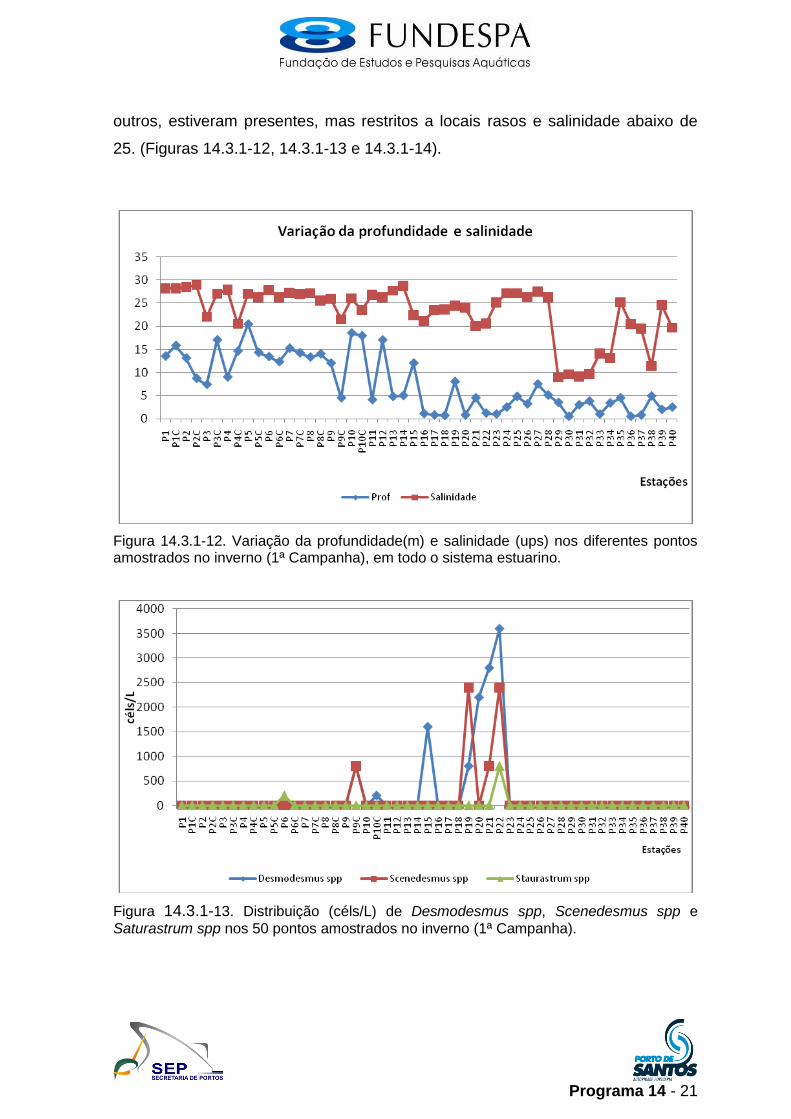
outros, estiveram presentes, mas restritos a locais rasos e salinidade abaixo de
25. (Figuras 14.3.1-12, 14.3.1-13 e 14.3.1-14).
Figura 14.3.1-12. Variação da profundidade(m) e salinidade (ups) nos diferentes pontos amostrados no inverno (1ª Campanha), em todo o sistema estuarino.
Figura 14.3.1-13. Distribuição (céls/L) de Desmodesmus spp, Scenedesmus spp e
Saturastrum spp nos 50 pontos amostrados no inverno (1ª Campanha).
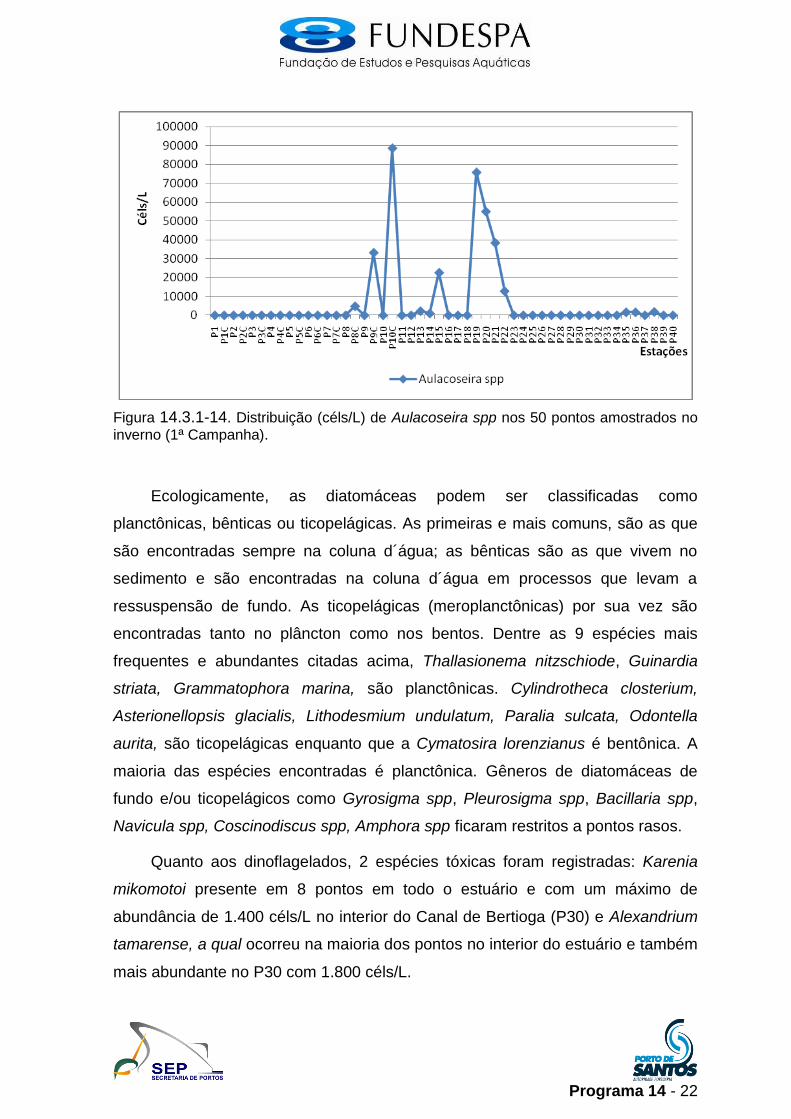
Programa 14 - 22
Figura 14.3.1-14. Distribuição (céls/L) de Aulacoseira spp nos 50 pontos amostrados no
inverno (1ª Campanha).
Ecologicamente, as diatomáceas podem ser classificadas como
planctônicas, bênticas ou ticopelágicas. As primeiras e mais comuns, são as que
são encontradas sempre na coluna d´água; as bênticas são as que vivem no
sedimento e são encontradas na coluna d´água em processos que levam a
ressuspensão de fundo. As ticopelágicas (meroplanctônicas) por sua vez são
encontradas tanto no plâncton como nos bentos. Dentre as 9 espécies mais
frequentes e abundantes citadas acima, Thallasionema nitzschiode, Guinardia
striata, Grammatophora marina, são planctônicas. Cylindrotheca closterium,
Asterionellopsis glacialis, Lithodesmium undulatum, Paralia sulcata, Odontella
aurita, são ticopelágicas enquanto que a Cymatosira lorenzianus é bentônica. A
maioria das espécies encontradas é planctônica. Gêneros de diatomáceas de
fundo e/ou ticopelágicos como Gyrosigma spp, Pleurosigma spp, Bacillaria spp,
Navicula spp, Coscinodiscus spp, Amphora spp ficaram restritos a pontos rasos.
Quanto aos dinoflagelados, 2 espécies tóxicas foram registradas: Karenia
mikomotoi presente em 8 pontos em todo o estuário e com um máximo de
abundância de 1.400 céls/L no interior do Canal de Bertioga (P30) e Alexandrium
tamarense, a qual ocorreu na maioria dos pontos no interior do estuário e também
mais abundante no P30 com 1.800 céls/L.

Programa 14 - 23
A riqueza específica mostrou gradiente, com valores mais elevados na Baia
de Santos e dentro do Canal (42 espécies na P2c) diminuindo em direção ao
interior, próximo ao Largo de Canéu, Canal de Bertioga e mostrando seu menor
índice (9 espécies) no Canal de São Vicente (P39) (Figura 14.3-15).
Figura 14.3.1-15. Riqueza específica do fitoplâncton registrada para as 50 amostras coletadas em julho de 2010 (1ª Campanha).
A diversidade (H’) é expressa em bit por indivíduo. Valores maiores do que
3bits/ind são considerados altos, e valores menores do que 1bit/ind., baixos. A
maior diversidade encontrada no inverno foi de 2,7 bits/ind. nos pontos dentro do
Largo do Canéu próximos a saída do Canal de Santos (P8C e P13). Nas
amostras da Baia e Canal de Santos, os valores ficaram acima de 2 bits/ind.
decrescendo para índices menores que 1 bit/ind. em direção ao Canal de Bertioga
onde o menor valor (0,7 bit/ind.) foi registrado no ponto P29 (Figura 14.3.1-16).

Programa 14 - 24
Figura 14.3.1-16. Diversidade específica do fitoplâncton nas 50 amostras coletadas em julho de 2010 (1ª Campanha).
A equitabilidade representa a razão entre o índice de diversidade e a
diversidade máxima que a amostra pode alcançar considerando o total de
indivíduos que possui. Seus valores variam entre 0 e 1. Valores iguais ou
superiores a 0,5 foram encontrados na Baia e Canal de Santos, valores menores
foram registrados para um ponto interno ao Largo do Canéu (P16), e no Canal de
Bertioga com o menor índice registrado também no ponto P29 de 0,2 (Figura
14.3.1-17).

Programa 14 - 25
Figura 14.3.1-17. Equitabilidade nas 50 amostras coletadas em julho de 2010 (1ª Campanha).
As diatomáceas são dependentes do processo de turbulência para
ressuspensão das células para dentro da zona eufótica, sendo mais competitivas
que outros organismos do fitoplâncton nestas condições (Reynolds, 1997). Os
dinoflagelados por sua vez se desenvolvem melhor em ambientes com baixo fluxo
e maior tempo de residência (Chan & Hamilton, 2001).
A dominância nas duas frações de tamanho (nano e micro) das
diatomáceas, indica processos de turbulências dentro da coluna d´água, que
podem ter ocorrido por condições típicas de inverno como entrada de frentes frias,
temperaturas mais amenas e ventos.
A distribuição espacial das espécies dentro do sistema estuarino foi
determinada pela interação entre a salinidade, circulação e profundidade local.
Chaetoceros spp foram mais abundantes nas amostras da baia e dentro do
canal, locais de salinidades e profundidades maiores. As Thalassiosira spp e
Coscinodiscus spp se desenvolveram melhor no Canal de Bertioga, em
profundidades rasas e favorecidas pelos pulsos de nutrientes indicados pela baixa
salinidade, típicos de habitat turbulento, raso e rico em nutriente.

Programa 14 - 26
Espécies de origem menos salina, como Desmodesmus spp, Monoraphidium
spp, Scenedesmus spp, Staurastrum spp, Aulacoseira, ficaram restritas ao interior
do estuário no Largo do Canéu. Segundo Coquemala (2005), estas espécies se
desenvolvem preferencialmente em ambientes rasos, meso-eutróficos, águas
homogêneas e são sensíveis a estratificação, o que parece ser o caso dos pontos
em questão.
Espécies bentônicas e meroplanctônicas (ticopelágicas), presentes no
plâncton também confirmam a instabilidade da coluna em alguns pontos e a
permanência na coluna vai depender do fluxo de água e profundidade local.
Karenia mikomotoi e Alexandrium tamarense merecem destaque, dentre as
espécies de dinoflagelados registrados no estuário, por produzirem toxina e
apresentarem potencial para formação de bloom. A produção de saxitoxina pode
ser cumulativa em organismos filtradores e letal para peixes e organismos
maiores da macrofauna.
A espécie de A. tamarense foi observada pela primeira vez na costa do Rio
Grande do Sul em 1996. Ocorrências de bloom na Argentina são registradas
desde o ano de 1980, 1995 (14,6 x 103céls/L) e 1998 (4,5 x 103 céls/L) (Gayoso,
2001). São diversos os mecanismos que levam à produção de toxina nas
diferentes espécies, e atualmente é objeto de estudo em todo o mundo. Embora a
abundância registrada para o inverno (1ª Campanha) de ambas as espécies
foram baixas, cabe lembrar que ocorreu um registro de boom de Karenia
mikomotoi nas coletas de verão (Campanha Prévia) (Fundespa, 2010).
A riqueza específica no sistema estuarino durante inverno (1ª Campanha) foi
menor que a registrada para o verão (Campanha Prévia). Os baixos valores de
diversidade e equitabilidade, principalmente no Canal de Bertioga (P32, P33 e
P34), são explicados pela dominância das diatomáceas, tanto no nano quanto no
microfitoplâncton, das espécies Skeletonema costatum e Thalassiosira spp, e do
ciliado autotrófico Mesodinium rubrum.

Programa 14 - 27
Análise integrada dos dados - campanhas de janeiro e julho de 2010
O teste ANOSIM, possui valores entre (-1) (0) (+1). Quando o resultado é
zero significa que as amostras são iguais; (1) que as amostras são totalmente
diferentes e (-1) que a diferença encontrada entre os pontos do mesmo período
são maiores que a diferença entre os 2 períodos. O teste aplicado para o estudo
resultou: (ANOSIM; R= 0,813, p=0,001), mostrando claramente a diferença entre
as amostras coletadas nos 2 períodos, indicando que a diferença sazonal é muito
maior que a diferença entre as amostras coletadas no mesmo período
(Figura 14.3.1-18).
Figura 14.3.1-18. Resultado da análise de ordenação (ANOSIM). Similaridade Bray Curtis. Em verde: amostras de verão; em azul amostras de inverno.
O teste Simper indicou que as espécies que mais contribuíram para a
dissimilaridade entre as amostras de verão (Campanha Prévia) e inverno
(1ª Campanha) foram as frequentes e abundantes no verão e ausente ou restrita
a poucos pontos no inverno como Chaetoceros socialis, Thalassiosira
pseudonana, Skeletonema costatum, Cylindropermopsis raciborskii,
Leptocylindrus minimus, Chaetoceros debilis, Planktothix agardhii, Karenia

Programa 14 - 28
mikimotoi, Monoraphidium contortum, Lithodesmium undullatum, Cylindrotheca
closterium, entre outras.
14.3.2. Comunidade Zooplanctônica
Campanha de janeiro de 2010 (Campanha Prévia)
Os grupos zooplanctônicos detectados nas amostras do sistema estuarino
de Santos-Bertioga, durante a campanha de janeiro de 2010, (campanha prévia)
estão listados na Tabela 14.3.2-1. As identificações, realizadas até o nível de
espécie para a maioria dos grupos holoplanctônicos, como Copepoda, Cladocera,
Appendicularia e Chaetognatha, indicam que a composição taxonômica do
zooplâncton nas áreas estudadas é similar a outros estuários do sudeste-sul do
Brasil, como a baía de Paranaguá e o complexo estuarino-lagunar de Cananéia-
Iguape (Neumann-Leitão, 1994/95; Brandini et al., 1997).
Além das espécies tipicamente estuarinas e marinho-eurihalinas, nas
amostras da campanha em análise foram encontradas espécies indicadoras de
intrusão marinha, como os copépodes Clausocalanus furcatus, Calocalanus
pavoninus e Subeucalanus pileatus, além de várias espécies indicadoras de
influência de águas continentais. Entre estas se destacaram espécies de
cladóceros não registradas anteriormente em sistemas estuarinos do litoral
paulista, como Bosmina longirostris e Diaphanosoma birgei. Foram encontrados
também muitos gêneros de copépodes de água doce ou de baixa salinidade,
incluindo Acanthocyclops, Apocyclops, Macrocyclops, Thermocyclops, entre
outros (Tabela 14.3.2-1). Estes organismos característicos de áreas oligohalinas
foram amostrados pela primeira vez em coletas de plâncton na região de Santos
porque a campanha contemplou setores internos do sistema estuarino, onde a
salinidade era provavelmente baixa. Além disto, nos estudos sobre zooplâncton
estuarino estes organismos não são geralmente identificados até espécie e
permanecem sob o nível de família ou ordem, impossibilitando sua utilização
como indicadores de águas menos salinas.

Programa 14 - 29
Tabela 14.3.2-1. Lista de táxons zooplanctônicos no sistema estuarino de Santos-Bertioga, amostragem de janeiro de 2010 (Campanha Prévia).
Foraminifera Copepoda
Radiolaria Parvocalanus crassirostris
Sticholonche zanclea Paracalanus indicus
Tintinnida Paracalanus parvus
Hydrozoa Paracalanus quasimodo
Campanulariidae Clausocalanus furcatus
Clytia sp. Calocalanus pavoninus
Obelia sp. Subeucalanus pileatus
Polychaeta (larvas e adultos) Centropages furcatus
Gastropoda (larvas) Pseudodiaptomus acutus
Bivalvia (larvas) Temora turbinata
Bryozoa (larva cyphonauta) Acartia lilljeborgi
Insecta (larvas) Acartia tonsa
Acarida Labidocera fluviatilis
Ostracoda Paracyclopina longifurca
Amphipoda Microsetella spp. copepodito
Corophiidea (adultos) Microsetella norvegica
Caprellidae (adultos) Clytemnestra rostrata
Gammaridea Euterpina acutifrons
Isopoda Macrosetella gracilis
Sphaeromatidae (adultos) Clytemnestra scutelata
Decapoda (larvas e megalopas) Metis sp.
Luciferidae Leptocaris sp.
Lucifer faxoni Tegastes sp.
Brachyura (larvas) Oithona spp. copepodito
Porcellanidae (larvas) Oithona plumifera
Stomatopoda (larvas) Oithona plumifera copepodito
Cladocera Oithona hebes
Bosmina sp. Oithona oswaldocruzi
Bosmina longirostris Oithona oculata
Ceriodaphnia sp. Oithona nana
Simocephalus sp. Oithona simplex
Moina sp. Corycaeus sp. copepodito
Moina minuta Corycaeus giesbrechti
Penilia avirostris
Diaphanosoma birgei

Programa 14 - 30
Tabela 14.3.2-1 (continuação). Lista de táxons zooplanctônicos no sistema estuarino de Santos-Bertioga, amostragem de janeiro de 2010 (Campanha Prévia).
Copepoda (continuação) Parasagitta tenuis
Clytemnestra scutelata Parasagitta friderici
Metis sp. Echinodermata (larva)
Leptocaris sp. Chordata
Tegastes sp. Ascidiacea (larva)
Oithona spp. copepodito Larvacea
Oithona plumifera Juvenis
Oithona plumifera copepodito Oikopleura (Vexillaria) dioica
Oithona hebes Oikopleura (Coecaria) longicauda
Oithona oswaldocruzi Oikopleura (Vexillaria) rufenses
Oithona oculata Vertebrata
Oithona nana Pisces (ovos e larvas)
Oithona simplex Carangidae (larvas)
Acanthocyclops sp. 1 Trachurus sp. (larva)
Apocyclops procerus Gobiidae
Halicyclops crassirostris Bathygobius soporator (larva)
Halicyclops oraeeburnensis Achiridae
Macrocyclops albidus principalis Achirus lineatus (larva)
Mesocyclops sp. 1 Sygnathidae (larvas)
Metacyclops mendocinus
Microcyclops sp. 1
Paracyclops sp. 1
Thermocyclops decipiens
Tropocyclops prasinus meridionalis
Tropocyclops priscinalis
Farranula gracilis
Oncaea spp. copepodito
Oncaea curta
Oncaea mediterranea
Oncaea venusta
Hemicyclops thalassius
Sapphirina sp. copepodito
Ergasilus sp.
Monstrilloida (adultos)
Cyclopinidae (copepoditos e adultos)
Cirripedia (larvas cypris e náuplios)
Chaetognata
Juvenis
Flaccisagitta enflata

Programa 14 - 31
Os índices de diversidade de Shannon e de dominância de Simpson para o
holoplâncton variaram de acordo com o gradiente espacial estuarino. Os valores
mais altos do índice de Shannon ocorreram nos setores externos do canal de
navegação, diminuindo gradativamente em direção aos pontos mais internos, a
tendência oposta sendo observada para o índice de Simpson (Figuras 14.3.2-1 a
14.3.2-4). Este resultado retrata a maior influência da intrusão marinha ao longo
do canal dragado, em oposição aos setores mais rasos e internos do estuário.
Figura 14.3.2-1. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o holoplâncton encontrado nos pontos de coleta no Canal de navegação em janeiro de 2010 (campanha prévia).
Figura 14.3.2-2. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o holoplâncton encontrado nos pontos de coleta na Baía de Santos e no Canal de Piaçaguera em janeiro de 2010 (campanha prévia).
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Pontos de coleta
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Pontos de coleta
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4

Programa 14 - 32
Figura 14.3.2-3. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o holoplâncton encontrado nos pontos de coleta no Canal de Piaçaguera em janeiro de 2010 (campanha prévia).
Figura 14.3.2-4. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o holoplâncton encontrado nos pontos de coleta no Canal de Bertioga em janeiro de 2010 (campanha prévia).
Ao contrário do holoplâncton, as diversidades do mero- e ticoplâncton foram
mais elevadas nos setores intermediários do sistema estuarino, particularmente
em pontos do canal de Piaçaguera (Figuras 14.3.2-5 a 14.3.2-8). Esta tendência
foi acompanhada pelo índice de Simpson, com valores inversos ao índice de
Shannon. Este resultado reflete a maior contribuição de formas larvais de
organismos bentônicos estuarinos nas áreas rasas, assim como a ocorrência de
pequenos invertebrados bentônicos nas amostras de plâncton (ticoplâncton),
Canal de Piaçaguera
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Pontos de coleta
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4

Programa 14 - 33
decorrente da influência da ressuspensão de sedimentos nestas mesmas áreas.
Considerando que o mero- e ticoplâncton puderam ser identificados somente em
níveis supragenéricos, a Figura 14.3.2-9 ilustra o efeito do maior número de
grupos nestas categorias sobre o número total de táxons, corroborando os
resultados obtidos para os índices ecológicos no sistema estuarino.
Figura 14.3.2-5. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o mero e ticoplâncton encontrados nos pontos de coleta no Canal de navegação em janeiro de 2010 (campanha prévia).
Figura 14.3.2-6. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o mero e ticoplâncton encontrados nos pontos de coleta na Baía de Santos e no Canal de Piaçaguera em janeiro de 2010 (campanha prévia).
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Pontos de coleta
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Pontos de coleta
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6

Programa 14 - 34
Figura 14.3.2-7. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o mero e ticoplâncton encontrados nos pontos de coleta no Canal de Piaçaguera em janeiro de 2010 (campanha prévia).
Figura 14.3.2-8. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o mero e ticoplâncton encontrados nos pontos de coleta no Canal de Bertioga em janeiro de 2010 (campanha prévia).
Canal de Piaçaguera
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Pontos de coleta
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6

Programa 14 - 35
Figura 14.3.2-9. Número de táxons classificados em espécies, gêneros e níveis supragenéricos na região da Baía de Santos e canais de Piaçaguera e Bertioga em janeiro de 2010 (campanha prévia).
Nas amostras coletadas em janeiro de 2010 (Campanha Prévia), a
dominância dos Copepoda foi nítida, conforme mostra a Figura 14.3.2-10. Este
padrão é comum em áreas estuarinas do Brasil (Neumann-Leitão, 1994/95), mas,
na campanha em análise, a abundância de outros grupos holoplanctônicos foi
extremamente baixa, com ocorrência significativa apenas de Chaetognatha, em
pontos da Baía de Santos. Estes resultados diferem das observações reportadas
no EIA-RIMA do empreendimento, onde outros grupos foram relatados como
subdominantes, incluindo Urochordata e Cnidaria (FRF, 2008), que contribuíram
com quase 40% da abundância em alguns pontos. Esta variabilidade na
composição e na diversidade dos grupos holoplanctônicos decorre da própria
dinâmica estuarina, em particular das mudanças na circulação durante as
diferentes fases da maré, que promovem uma alternância nos grupos dominantes
conforme o grau de intrusão marinha ou de aporte continental. No caso do
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
5
10
15
20
25Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
5
10
15
20
25
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Nú
me
ro d
e t
áxo
ns
0
5
10
15
20
25
Espécies
Gêneros
Grupos
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
5
10
15
20
25

Programa 14 - 36
meroplâncton, números relativamente elevados foram obtidos nos pontos
intermediários e internos do sistema estuarino, mas estes organismos foram
normalmente muito menos abundantes do que os Copepoda (Figura 14.3.2-10).
Figura 14.3.2-10. Abundância do zooplâncton nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga em janeiro de 2010 (campanha prévia).
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
55007500
8000
8500
9000
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
5500
7500
8000
8500
9000
9500
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Indiv
íduos.m
-3
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
55007500
8000
8500
9000
Copepoda
Holoplâncton (exceto Copepoda)
Mero/Ticoplâncton
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
5500
7500
8000
8500
9000

Programa 14 - 37
Figura 14.3.2-11. Abundância das ordens de Copepoda nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga em janeiro de 2010 (campanha prévia).
Entre os Copepoda, a ordem Calanoida foi dominante em todos os pontos e,
na maioria deles, pelo menos uma ordem de grandeza mais abundante que as
demais ordens (Figura 14.3.2-11). Os calanóides foram caracterizados por altas
abundâncias de duas famílias, Acartiidae e Paracalanidae (Figura 14.3.2-12),
representadas principalmente pelas espécies Acartia lilljeborgi, A. tonsa e
Parvocalanus crassirostris. No caso da ordem subdominante, Cyclopoida, duas
espécies foram numericamente importantes, Oithona hebes e O. oswaldocruzi,
com o predomínio da primeira (Figura 14.3.2-13). Todas as espécies citadas são
comuns em ambientes costeiros e estuarinos da costa sul-sudeste do Brasil
(Lopes, 2007), sendo Acartia lilljeborgi e Parvocalanus crassirostris indicadoras de
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
500
1000
1500
2000
2500
3000
3500
4000
4500
50006500
7000
7500
8000
8500
9000
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
6500
7000
7500
8000
8500
9000
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Indiv
íduos.m
-3
0
500
1000
1500
2000
2500
3000
3500
4000
4500
50006500
7000
7500
8000
8500
9000
Calanoida
Cyclopoida
Harpacticoida
Poecilostomatoida
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
6500
7000
7500
8000
8500
9000

Programa 14 - 38
intrusão marinha e Acartia tonsa e Oithona oswaldocruzi indicadoras de
ambientes meso- a oligohalinos. Considerando estas cinco espécies, A. lilljeborgi,
P. crassirostris e O. hebes foram as mais abundantes, indicando o predomínio da
influência marinha nos diferentes setores estudados, com exceção de parte do
canal de Piaçaguera e do Rio Casqueiro, onde ocorreram espécies tipicamente de
água doce.
Figura 14.3.2-12. Abundância das famílias da ordem Calanoida (Copepoda) nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga em janeiro de 2010 (campanha prévia).
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
500
1000
1500
2000
2500
3000
3500
4000
4500
50008500
9000
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
8500
9000
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Indiv
íduos.m
-3
0
500
1000
1500
2000
2500
3000
3500
4000
4500
50008500
9000
Acartiidae
Paracalanidae
Pseudodiaptomidae
Temoridae
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
8500
9000

Programa 14 - 39
Figura 14.3.2-13. Abundância de Oithona hebes e Oithona oswaldocruzi (Oithonidae: Cyclopoida) na região da Baía de Santos e Canais de Piaçaguera e Bertioga em janeiro de 2010 (campanha prévia).
Com relação ao mero e ticoplâncton (Figuras 14.3.2-14 e 14.3.2-15), o grupo
dominante foi o dos Cirripedia em todos os pontos, desde a Baía de Santos até as
zonas oligohalinas do sistema estuarino, com máximos nos setores intermediário
e interno do canal de Piaçaguera e no rio Casqueiro. O segundo grupo
meroplanctônico em termos de abundância foi Decapoda, formado basicamente
por larvas de Brachyura, presentes em maior número nas áreas intermediárias a
internas do estuário. Presentes em quantidades pouco expressivas, Polychaeta,
Mollusca, larvas de peixes e Amphipoda, também ocorreram principalmente
nestes setores (Figuras 14.3.2-14 e 14.3.2-15).
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
100
200
300
400
500
6001000
1100
Oithona hebes
Oithona oswaldocruzi
Baía de Santos (P1 a P4)
Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
100
200
300
400
500
6001000
1100
Canal de Piaçaguera
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Indiv
íduos.m
-3
0
100
200
300
400
500
6001000
1100 Canal de Bertioga
Pontos de coleta
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
100
200
300
400
500
6001000
1100

Programa 14 - 40
Figura 14.3.2-14. Abundância dos grupos do meroplâncton e ticoplâncton nos pontos de coleta no canal de navegação, Baía de Santos e setor externo do canal de Piaçaguera em janeiro de 2010 (campanha prévia).
A análise de ordenação N-MDS, realizada entre os 50 pontos de coleta com
base na raiz quadrada dos valores de densidade dos 111 táxons identificados,
permitiu a separação dos pontos em 5 grupos (Figura 14.3.2-16). Os resultados
do teste ANOSIM mostraram que todos os grupos formados são estatisticamente
diferentes (Tabela 14.3.2-2).
O grupo A foi formado por 17 das 24 amostras coletadas na Baía de
Santos/Canal de Navegação (área 1), que apresentaram os maiores valores de
densidade e o maior numero de táxons (ocorrência de 72 dos 111 táxons
Canal de navegação
Pontos de coleta
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
50
100
150
200
250
300350
400
Cirripedia
Decapoda
Amphipoda
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
Indiv
íduos.m
-3
0
50
100
150
200
250
300350
400
Polychaeta
Mollusca
Pisces
Baía de Santos (P1 a P4)Canal de Piaçaguera
Pontos de coleta
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
50
100150
200
Cirripedia
Decapoda
Amphipoda
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
Indiv
íduos.m
-3
0
50
100150
200
Polychaeta
Mollusca
Pisces

Programa 14 - 41
identificados). Acartia tonsa, A. lilljeborgi, copepoditos de Acartia spp. e de
Pseudodiaptomus spp., Parvocalanus crassirostris, Oithona hebes, machos de
Oithona spp., Euterpina acutifrons, Temora turbinata, larvas de Cirripedia e de
Brachyura e juvenis da família Sagittidae foram os principais representantes deste
grupo.
Figura 14.3.2-15. Abundância dos grupos do meroplâncton e ticoplâncton nos pontos de coleta no setor interno do canal de Piaçaguera e no canal de Bertioga em janeiro de 2010 (campanha prévia).
O grupo B foi composto por amostras coletadas nas áreas 1 (citada acima) e
3 (Canal de Bertioga) (Figura 14.3.2-16) e, embora a composição de táxons seja
semelhante à encontrada no grupo A, apresentou baixa densidade zooplanctônica
e uma maior contribuição de larvas de Polychaeta e dos copépodes Oithona
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16P17 P18 P19P20 P21 P22P35 P36 P37P38 P39 P40
0
50
100
150
200
250450
500
Cirripedia
Decapoda
Amphipoda
P14 P15 P16P17 P18 P19P20 P21 P22P35 P36 P37P38 P39 P40
Indiv
íduos.m
-3
0
50
100
150
200
250450
500
Polychaeta
Mollusca
Pisces
Canal de Bertioga
Pontos de coleta
P23 P24 P25 P26 P27 P29 P30 P31 P32 P33 P34
0
50
100
150
200
250300
350
400
Cirripedia
Decapoda
Amphipoda
P23 P24 P25 P26 P27 P29 P30 P31 P32 P33 P34
Indiv
íduos.m
-3
0
50
100
150
200
250300
350
400
Polychaeta
Mollusca
Pisces

Programa 14 - 42
oswaldocruzi e Ergasilus sp. Por sua vez, a composição e densidade
zooplanctônica do grupo C se mostrou bem diferente do A e mais semelhante ao
do grupo D (Tabela 14.3.2-2). Larvas e ovos de peixes mostraram uma maior
contribuição para a formação deste grupo, assim como larvas de gastrópodes e
hidromedusas.
Apesar dos pontos estarem de bem separados espacialmente, o P35 da
área 2 (Canal de Piaçaguera e Rio Casqueiro) se agrupou com outros três pontos
da área 3 devido à presença de hidromedusas, isópodes, náuplios de copépodes
e larvas de gastrópodes, que não foram identificadas nos quatro pontos que
formaram o grupo D. Este, em comparação com os demais grupos, foi o mais
diferente, com baixos valores de densidade zooplanctônica e menor número de
táxons (ocorrência de apenas 38 dos 111 táxons identificados). Larvas de peixes,
de decápodes e de cirrípedes, além dos copépodes Parvocalanus crassirostris,
Oithona oswaldocruzi, Acartia tonsa e Paracyclopina longifurca, foram os táxons
mais abundantes. No entanto, a principal característica deste grupo foi a presença
de espécies típicas de água doce, como os cladóceros Bosmina longirostris,
Bosmina sp., Diaphanosoma birgei, Simocephalus sp., Ceriodaphnia sp. e os
copépodes Thermocyclops decipiens, Acanthocyclops sp. 1, Microcyclops sp.1,
Mesocyclops sp.1, Paracyclops sp.1, Tropocyclops prasinus meridionalis, T.
priscinalis e copepoditos da família Cyclopidae.
Como pode ser visto na Tabela 14.3.2-2, apesar de estatisticamente
diferentes, os grupos A e E foram os de menor valor de R, portanto, os mais
semelhantes entre si. Agruparam as amostras mais abundantes e mais ricas, com
alta abundância relativa de Acartia tonsa, A. lilljeborgi, copepoditos de Acartia
spp., Parvocalanus crassirostris e Oithona hebes. Por outro lado, a composição
dos táxons foi um tanto diferente, uma vez que dos 88 táxons identificados
(somando ambos os grupos), 32 ocorreram apenas em pontos que formaram o
grupo A, enquanto que 16 apenas nas do grupo E. Dentre estes, destacaram-se
três cladóceros (Simocephalus sp., Moina minuta, Diaphanosoma birgei) e três
copépodes (Apocyclops procerus, Halicyclops crassirostris, Paracyclops sp. 1),
todos típicos de regiões menos salinas ou de água doce, confirmando a influência
marinha sobre a Baía de Santos e o Canal de Navegação (grupo A) em

Programa 14 - 43
contrapartida com a natureza mais estuarina do Canal de Piaçaguera e Rio
Casqueiro (grupo E) assim como do Canal de Bertioga (grupo D).
Reduzida
Resemblance: S17 Bray Curtis similarity
Áreas1
2
3
P1
P1C
P2
P2C
P3P3CP4
P4CP5
P5C
P6
P6C
P7
P7C
P8
P8C
P9
P9C
P10
P10C
P11
P12
P13
P14P15
P16P17
P18 P19
P20
P21
P22
P23
P24P25
P26
P27
P28
P29
P30P31
P32
P33
P34
P35P36
P37
P38
P39
P40
2D Stress: 0.13
A
ED
B
C
Figura 14.3.2-16. Análise de Ordenação N-MDS dos 50 pontos de coleta de janeiro de 2010 (campanha prévia) baseada na raiz quadrada dos valores de densidade dos táxons identificados e na similaridade de Bray-Curtis. Áreas: 1 – Baía de Santos e Canal de navegação; 2 – Canal de Piaçaguera e Rio Casqueiro; 3 – Canal de Bertioga.
Tabela 14.3.2-2. Resultados do teste pareado ANOSIM realizado entre os grupos obtidos na Análise de Similaridade N-MDS para a campanha de janeiro de 2010 (campanha prévia). Valores significativos: < 5%.
Nível de
significância (%)
A - B 0.880 0.1
A - C 0.987 0.1
A - D 1 0.1
A - E 0.809 0.1
B - C 0.881 0.8
B - D 1 0.8
B - E 0.904 0.1
C - D 0.865 2.9
C - E 0.818 0.2
D - E 0.983 0.2
Grupos R estatístico
R global = 0.882

Programa 14 - 44
Campanha de julho de 2010 (1ª Campanha)
Os grupos zooplanctônicos detectados nas amostras do sistema estuarino
de Santos-Bertioga, durante a campanha de julho de 2010 (primeira campanha de
monitoramento), estão listados na Tabela 14.3.2-3. As identificações, realizadas
até o nível de espécie para a maioria dos grupos holoplanctônicos, como
Copepoda, Cladocera, Appendicularia e Chaetognatha, indicam novamente que a
composição taxonômica do zooplâncton nas áreas estudadas é similar a outros
estuários do sudeste-sul do Brasil, conforme citado anteriormente.
Na campanha de inverno, não houve uma contribuição significativa de
espécies indicadoras de águas continentais. A área de estudo foi caracterizada
por espécies tipicamente estuarinas e marinho-eurihalinas, seguidas por espécies
indicadoras de intrusão marinha, como os copépodes Clausocalanus furcatus, e
Subeucalanus pileatus e algumas espécies de Oikopleura (Larvacea). Nas
imagens abaixo (Figura 14.3.2-17) estão ilustrados alguns táxons zooplanctônicos
encontrados na campanha de inverno.

Programa 14 - 45
Figura 14.3.2-17. Ilustração de alguns táxons zooplanctônicos encontrados na campanha de inverno (1ª campanha).

Programa 14 - 46
Tabela 14.3.2-3. Lista de táxons zooplanctônicos no sistema estuarino de Santos-Bertioga, amostragem de julho de 2010 (1ª campanha).
Reino Protista Classe Ostracoda
Filo Sarcomastigophora Classe Malacostraca
Subfilo Radiolaria Ordem Mysida
Sticholonche zanclea Família Mysidae
Reino Animalia Ordem Amphipoda
Subordem Corophiidea
Filo Cnidaria Subordem Gammaridea
Classe Scyphozoa
Ordem Semaeostomeae Ordem Isopoda
Família Ulmaridae
Aurelia sp. (éfira) Ordem Decapoda (larvas)
Infraordem Caridea (larvas)
Classe Hydrozoa Infraordem Brachyura (larvas)
Hidromedusas Infraordem Anomura
Ordem Leptothecata Família Porcellanidae (larvas)
Família Campanulariidae
Clytia sp. Ordem Stomatopoda (larvas)
Obelia sp.
Família Loveneliidae Classe Branchiopoda
Eucheilota sp. Ordem Diplostraca
Subordem Cladocera
Filo Annelida Família Podonidae
Classe Polychaeta (larvas) Pseudevadne tergestina
Família Sididae
Filo Mollusca Diaphanosoma birgei
Classe Gastropoda (larvas)
Ordem Thecosomata Classe Maxillopoda
Família Cavoliniidae Subclasse Copepoda
Creseis acicula Náuplios
Ordem Calanoida
Classe Bivalvia (larvas) Família Paracalanidae
Paracalanidae (copepoditos)
Filo Bryozoa (larvas cifonauta) Parvocalanus crassirostris
Paracalanus indicus
Filo Arthropoda Paracalanus parvus
Classe Arachnida Paracalanus quasimodo
Ordem Acarida (ácaros marinhos) Paracalanus spp.

Programa 14 - 47
Tabela 14.3.2-3 (continuação): Lista de táxons zooplanctônicos no sistema estuarino de Santos-Bertioga, amostragem de julho de 2010 (1ª campanha).
Copepoda (continuação) Ordem Poecilostomatoida
Família Clausocalanidae Família Corycaeidae
Clausocalanus furcatus Corycaeus spp. (copepoditos)
Família Eucalanidae Corycaeus giesbrechti
Subeucalanus pileatus Família Oncaeidae
Família Centropagidae Oncaea spp. (copepoditos)
Centropages furcatus Oncaea curta
Família Pseudodiaptomidae Oncaea mediterranea
Pseudodiaptomus spp. (copepoditos) Família Clausidiidae
Pseudodiaptomus acutus Hemicyclops thalassius
Família Temoridae
Temora stylifera Infraclasse Cirripedia
Temora turbinata Náuplios
Família Pontellidae Larvas cypris
Labidocera spp. (copepoditos)
Labidocera fluviatilis Filo Chaetognata
Família Acartiidae Classe Sagitoiidea
Acartia spp. (copepoditos) Família Sagittidae
Acartia tonsa Juvenis
Acartia lilljeborgi Parasagitta tenuis
Ordem Harpacticoida Parasagitta friderici
não identificados
Família Miraciidae Filo Echinodermata
Microsetella spp. (copepoditos) Classe Ophiuroidea (larvas)
Microsetella norvegica Classe Echinoidea (larvas)
Família Euterpinidae
Euterpina acutifrons Filo Chordata
Família Clytemnestridae Subfilo Tunicata (larvas)
Clytemnestra sp. (copepoditos) Classe Larvacea
Ordem Cyclopoida Família Oikopleuridae
não identificados Oikopleura spp.
Família Oithonidae Oikopleura (Vexillaria) dioica
Oithona spp. (copepoditos) Oikopleura (Coecaria) longicauda
Oithona plumifera Oikopleura (Vexillaria) rufenses
Oithona hebes Oikopleura (Coecaria) fusiformis
Oithona oswaldocruzi
Oithona oculata Subfilo Vertebrata
Superclasse Pisces
Larvas
Ovos
Programa 14 - 48
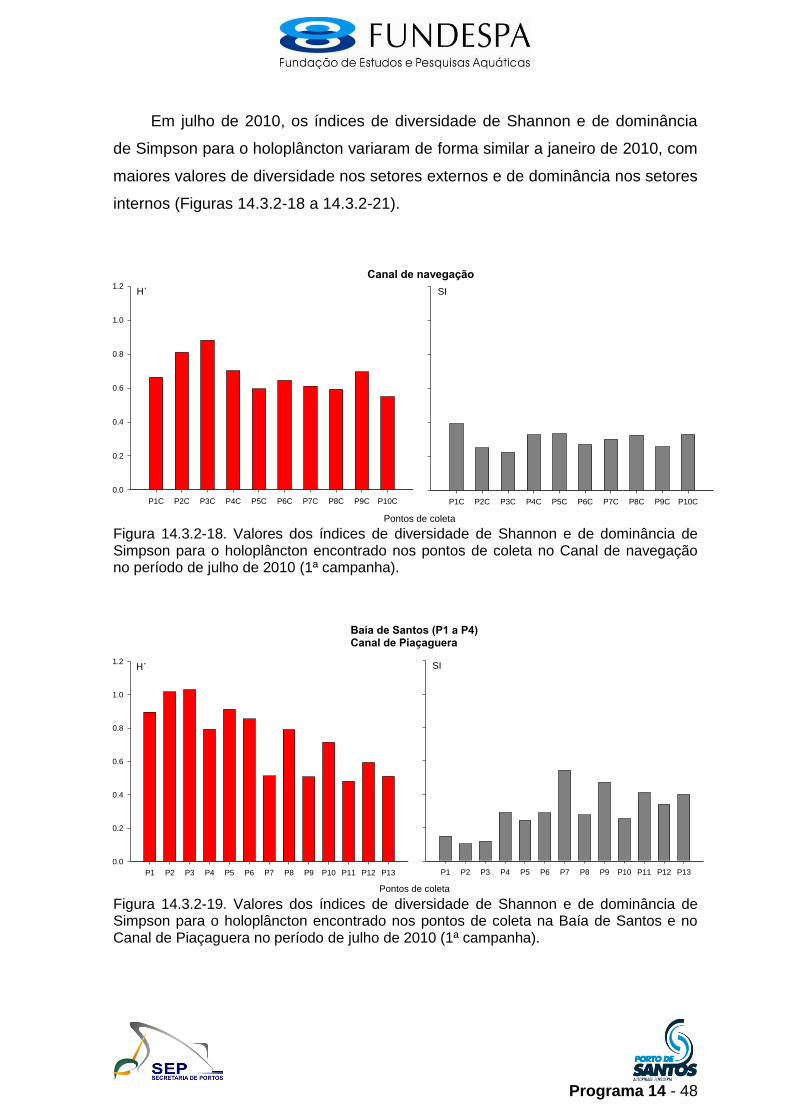
Em julho de 2010, os índices de diversidade de Shannon e de dominância
de Simpson para o holoplâncton variaram de forma similar a janeiro de 2010, com
maiores valores de diversidade nos setores externos e de dominância nos setores
internos (Figuras 14.3.2-18 a 14.3.2-21).
Figura 14.3.2-18. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o holoplâncton encontrado nos pontos de coleta no Canal de navegação no período de julho de 2010 (1ª campanha).
Figura 14.3.2-19. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o holoplâncton encontrado nos pontos de coleta na Baía de Santos e no Canal de Piaçaguera no período de julho de 2010 (1ª campanha).
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Pontos de coleta
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Pontos de coleta
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Programa 14 - 49
Canal de Piaçaguera
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
H´
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
SI
0,0
0,2
0,4
0,6
0,8
1,0
1,2
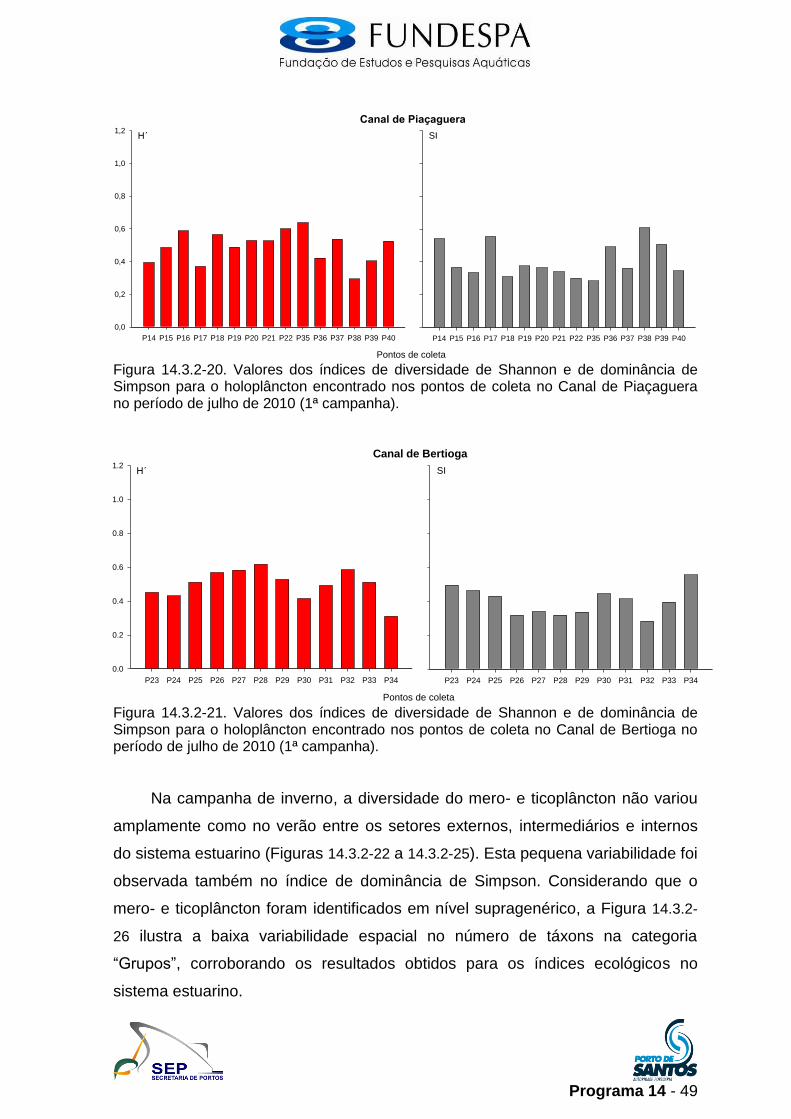
Figura 14.3.2-20. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o holoplâncton encontrado nos pontos de coleta no Canal de Piaçaguera no período de julho de 2010 (1ª campanha).
Figura 14.3.2-21. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o holoplâncton encontrado nos pontos de coleta no Canal de Bertioga no período de julho de 2010 (1ª campanha).
Na campanha de inverno, a diversidade do mero- e ticoplâncton não variou
amplamente como no verão entre os setores externos, intermediários e internos
do sistema estuarino (Figuras 14.3.2-22 a 14.3.2-25). Esta pequena variabilidade foi
observada também no índice de dominância de Simpson. Considerando que o
mero- e ticoplâncton foram identificados em nível supragenérico, a Figura 14.3.2-
26 ilustra a baixa variabilidade espacial no número de táxons na categoria
“Grupos”, corroborando os resultados obtidos para os índices ecológicos no
sistema estuarino.
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
H´
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Pontos de coleta
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
SI
0.0
0.2
0.4
0.6
0.8
1.0
1.2

Programa 14 - 50
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
H´
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Pontos de coleta
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
SI
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Figura 14.3.2-22. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o mero e ticoplâncton encontrado nos pontos de coleta no Canal de navegação no período de julho de 2010 (1ª campanha).
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
H´
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Pontos de coleta
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
SI
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Figura 14.3.2-23. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o mero e ticoplâncton encontrado nos pontos de coleta na Baía de Santos e no Canal de Piaçaguera no período de julho de 2010 (1ª campanha).

Programa 14 - 51
Canal de Piaçaguera
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
H´
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
SI
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Figura 14.3.2-24. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o mero e ticoplâncton encontrado nos pontos de coleta no Canal de Piaçaguera no período de julho de 2010 (1ª campanha).
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
H´
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Pontos de coleta
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
SI
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Figura 14.3.2-25. Valores dos índices de diversidade de Shannon e de dominância de Simpson para o mero e ticoplâncton encontrado nos pontos de coleta no Canal de Bertioga no período de julho de 2010 (1ª campanha).
Na campanha de julho de 2010 a dominância de Copepoda foi mantida,
conforme mostra a Figura 14.3.2-26, mas de forma menos evidente do que no
verão. Na campanha em análise foram observadas altas abundâncias de outros
grupos holoplanctônicos em vários pontos da Baía de Santos e do setor externo
do canal de navegação, enquanto que o meroplâncton foi abundante em pontos
mais internos dos canais de Piaçaguera e de Bertioga (Figura 14.3.2-26).

Programa 14 - 52
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
5
10
15
20
25Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
5
10
15
20
25
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Núm
ero
de t
áxons
0
5
10
15
20
25
Espécies
Gêneros
Grupos
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
5
10
15
20
25
Figura 14.3.2-26. Número de indivíduos por metro cúbico das espécies, dos gêneros e dos outros níveis taxonômicos encontrados nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga no período de julho de 2010 (1ª campanha).

Programa 14 - 53
Entre os Copepoda, a dominância foi alternada entre a ordem Calanoida e a
ordem Cyclopoida, diferindo do observado na campanha de verão (Figura 14.3.2-
28). Os calanóides foram caracterizados por altas abundâncias de duas famílias,
Acartiidae e Paracalanidae (Figura 14.3.2-29), representadas principalmente
pelas espécies Acartia lilljeborgi, A. tonsa e Parvocalanus crassirostris. A família
Pseudodiaptomidae foi também abundante em alguns pontos. No caso da ordem
Cyclopoida, duas espécies foram numericamente importantes, Oithona hebes e
O. oswaldocruzi, com o predomínio da primeira (Figura 14.3.2-30). Todas as
espécies citadas são comuns em ambientes costeiros e estuarinos da costa sul-
sudeste do Brasil, sendo Acartia lilljeborgi, Parvocalanus crassirostris e Oithona
plumifera indicadoras de intrusão marinha e Acartia tonsa e Oithona oswaldocruzi
indicadoras de ambientes meso- a oligohalinos. Considerando estas espécies, A.
lilljeborgi, P. crassirostris e O. hebes foram as mais abundantes, assim como
observado na campanha de verão, confirmando o predomínio da influência
marinha nos diferentes setores estudados. No inverno, como citado
anteriormente, não foram observadas espécies tipicamente de água doce em
parte do canal de Piaçaguera e do Rio Casqueiro, como no verão.

Programa 14 - 54
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
24000
25000
26000
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
23000
24000
25000
26000
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Ind
ivíd
uo
s.m
-3
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
23000
24000
25000
26000
Copepoda
Holoplâncton (exceto Copepoda)
Mero/Ticoplâncton
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
23000
24000
25000
26000
Figura 14.3.2-27. Número de indivíduos por metro cúbico do zooplâncton encontrados nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga no período de julho de 2010 (1ª campanha).

Programa 14 - 55
Figura 14.3.2-28. Número de indivíduos por metro cúbico da ordem de Copepoda encontrados nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga no período de julho de 2010 (1ª campanha).
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
1200014000
15000
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
14000
15000
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Indiv
íduos.m
-3
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
1200014000
15000
Calanoida
Cyclopoida
Poecilostomatoida
Harpacticoida
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
14000
15000

Programa 14 - 56
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000700080009000
Baía de Santos (P1 a P4)Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
700080009000
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Indiv
íduos.m
-3
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000700080009000
Acartiidae
Paracalanidae
Pseudodiaptomidae
Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
700080009000
Figura 14.3.2-29. Número de indivíduos por metro cúbico das famílias dominantes da ordem Calanoida (Copepoda) encontrados nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga no período de julho de 2010 (1ª campanha).

Programa 14 - 57
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
1000
2000
3000
4000
5000
6000
7000
9000
10000
Oithona oswaldocruzi
Oithona hebes
Oithona plumifera
Oithona spp. macho
Oithona spp. copepodito
Baía de Santos (P1 a P4)
Canal de Piaçaguera
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
1000
2000
3000
4000
5000
6000
7000
9000
10000
Canal de Piaçaguera
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Indiv
íduos.m
-3
0
1000
2000
3000
4000
5000
6000
7000
9000
10000 Canal de Bertioga
Pontos de coleta
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
1000
2000
3000
4000
5000
6000
7000
9000
10000
Figura 14.3.2-30. Número de indivíduos por metro cúbico das espécies da família Oithonidae (Copepoda: Cyclopoida), copepoditos e indivíduos machos do gênero encontrados nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga no período de julho de 2010 (1ª campanha).
O grupo subdominante do holoplâncton (após Copepoda) foi o dos Larvacea,
gênero Oikopleura; os demais grupos tiveram baixas abundâncias (Figura 14.3.2-
31). Espécies de Oikopleura são frequentes em ambientes estuarinos tropicais e
subtropicais (Neumann-Leitão, 1994/95), inclusive na costa sul-sudeste do Brasil
(Brandini et al., 1997). Como as espécies de Larvacea estão normalmente
associadas com a influência da intrusão marinha, a importância do grupo durante
as amostragens de inverno fornece um elemento indicativo adicional da forte
variabilidade sazonal observada na região estudada.

Programa 14 - 58
Canal de navegação
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
1000
2000
3000
4000
500014000
16000
18000
20000
22000
Oikopleura (Vexillaria) dioica
Holoplâncton (exceto Copepoda)
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
1000
2000
3000
4000
500014000
16000
18000
20000
22000
Baía de Santos (P1 a P4)
Canal de Piaçaguera
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16 P17 P18 P19 P20 P21 P22 P35 P36 P37 P38 P39 P40
Indiv
íduo
s.m
-3
0
2000
4000
14000
16000
18000
20000
22000 Canal de Bertioga
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
1000
2000
3000
4000
500014000
16000
18000
20000
22000
Figura 14.3.2-31. Número de indivíduos por metro cúbico da espécie Oikopleura (Vexillaria) dióica (Oikopleuridae: Copelata) e outros organismos do holoplâncton (exceto Copepoda) encontrados nos pontos de coleta na região da Baía de Santos e Canais de Piaçaguera e Bertioga no período de julho de 2010 (1ª campanha).
Com relação ao mero- e ticoplâncton (Figuras 14.3.2-32 a 14.3.2-35), o
grupo dominante foi o dos Cirripedia, seguidos por Brachyura em todos os pontos,
exceto em alguns localizados na Baía de Santos e pontos externos dos canais de
navegação e Piaçaguera, onde a dominância se inverteu. Os máximos
meroplanctônicos ocorreram nos setores intermediários e internos do Canal de
Piaçaguera e de Bertioga, por influência dos Cirripedia. Este resultado contrasta
com as observações do verão, quando as larvas de Brachyura estiveram
presentes em maior número nas áreas intermediárias a internas do estuário.
Presentes em quantidades pouco expressivas, Polychaeta, Mollusca, larvas de
peixes e Amphipoda, também ocorreram principalmente nestes setores (Figuras
14.3.2-32 a 14.3.2-35).

Programa 14 - 59
Figura 14.3.2-32. Número de indivíduos por metro cúbico dos organismos do meroplâncton e ticoplâncton encontrados nos pontos de coleta no canal de navegação no período de janeiro de 2010 (campanha prévia).
Figura 14.3.2-33. Número de indivíduos por metro cúbico dos organismos do meroplâncton e ticoplâncton encontrados nos pontos de coleta na Baía de Santos e no Canal de Piaçaguera no período de julho de 2010 (1ª campanha).
Canal de navegação
Pontos de coleta
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
0
1000
2000
3000
40006000
7000
8000
Cirripedia
Brachyura
Gastropoda
P1C P2C P3C P4C P5C P6C P7C P8C P9C P10C
Indiv
íduos.m
-3
0
1000
2000
3000
40006000
7000
8000
Polychaeta
Outros
Baía de Santos (P1 a P4)Canal de Piaçaguera
Pontos de coleta
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
0
1000
2000
3000
40006000
7000
8000
Cirripedia
Brachyura
Gastropoda
P1 P2 P3 P4 P5 P6 P7 P8 P9 P10 P11 P12 P13
Indiv
íduos.m
-3
0
1000
2000
3000
40006000
7000
8000
Polychaeta
Outros

Programa 14 - 60
Canal de Piaçaguera
Pontos de coleta
P14 P15 P16P17 P18 P19P20 P21 P22P35 P36 P37P38 P39 P40
0
1000
2000
3000
40006000
7000
8000
Cirripedia
Brachyura
Gastropoda
P14 P15 P16P17 P18 P19P20 P21 P22P35 P36 P37P38 P39 P40
Indiv
íduo
s.m
-3
0
1000
2000
3000
40006000
7000
8000
Polychaeta
Outros
Figura 14.3.2-34. Número de indivíduos por metro cúbico dos organismos do meroplâncton e ticoplâncton encontrados nos pontos de coleta no Canal de Piaçaguera no período de julho de 2010 (1ª campanha).
Figura 14.3.2-35. Número de indivíduos por metro cúbico dos organismos do meroplâncton e ticoplâncton encontrados nos pontos de coleta no Canal de Bertioga no período de julho de 2010 (1ª campanha).
Canal de Bertioga
Pontos de coleta
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
0
1000
2000
3000
40006000
7000
8000
Cirripedia
Brachyura
Gastropoda
P23 P24 P25 P26 P27 P28 P29 P30 P31 P32 P33 P34
Indiv
íduos.m
-3
0
1000
2000
3000
40006000
7000
8000
Polychaeta
Outros

Programa 14 - 61
A análise de ordenação N-MDS, realizada entre os 50 pontos de coleta com
base na raiz quadrada dos valores de densidade de 60 táxons identificados no
inverno (matriz reduzida), permitiu, primeiramente, a separação dos pontos em 2
grupos (Figura 14.3.2-36). O resultado do teste ANOSIM mostrou que os cinco
pontos da Baía de Santos (grupo 1) são diferentes de todas as demais (grupo 2)
(ANOSIM: R = 0,913, p<0,05). Esta separação se deu, principalmente, pelos
baixos valores de densidade zooplanctônica dos pontos do grupo 1 em
comparação com as do grupo 2. Quanto à composição de espécies, os pontos da
Baía de Santos se destacaram por apresentarem as únicas ocorrências de
Centropages furcatus e de copepoditos de Corycaceus giesbrechti e, em
comparação com os pontos mais próximos (situados no Canal de Navegação),
pela alta densidade de Euterpina acutifrons, radiolários (Sticholonche zanclea) e
larvas de Echinoidea e baixas densidades de Oikopleura (Veillaria) dioica, Acartia
spp., Oithona hebes, O. oswaldocruzi, copepoditos de Pseudodiatomus e de
larvas de caranguejos braquiúros.
Figura 14.3.2-36. Análise de Ordenação N-MDS dos 50 pontos de coleta baseada na raiz quadrada dos valores de densidade de 60 táxons identificados (no inverno, 1ª campanha) e na similaridade de Bray-Curtis. Áreas: 1 – Baía de Santos e Canal de navegação; 2 – Canal de Piaçaguera e Rio Casqueiro; 3 – Canal de Bertioga.

Programa 14 - 62
Para visualizar melhor a distribuição dos pontos do grupo 2 optou-se por
fazer uma análise de ordenação N-MDS apenas com os pontos desse grupo
(Figura 14.3.2-37). Seu resultado mostrou a formação de seis grupos diferentes
entre si (Tabela 14.3.2-4).
Apesar de semelhantes na composição de táxons e no local de amostragem
dos pontos que os formaram (pontos do canal de navegação), os grupos A e B
ficaram separados devido aos valores de densidade, uma vez que o grupo A
apresentou uma média de aproximadamente 4.000 indivíduos m-3, enquanto que
o grupo B de 30.000 indivíduos m-3. Em ambos os grupos, os larváceos
Oikopleura (Vexillaria) dioica, os copépodes Oithona hebes, Oithona spp.
(macho), Parvocalanus crassirostris, Acartia tonsa, Acartia lilljeborgi,
Pseudodiaptomus acutus e copepoditos dos gêneros Acartia e Pseudodiaptomus
e da família Paracalanidae, bem como larvas de Brachyura e de Cirripedia foram
os organismos mais abundantes. Além disso, o grupo A apresentou maior riqueza
que o grupo B (respectivamente, 52 e 44 dos 60 táxons analisados).
O grupo C, composto por pontos do final do canal de navegação/início do
canal de Piaçaguera, mostrou maior dominância e menor riqueza de táxons (40
dos 60 táxons), com poucas larvas de caranguejos braquiúros e de copépodes
dos gêneros Acartia e Pseudodiaptomus. Nele houve uma inversão de
dominância, com Oithona oswaldocruzi mais abundante que Oikopleura
(Vexillaria) dioica.

Programa 14 - 63
Figura 14.3.2-37. Análise de Ordenação N-MDS dos pontos de coleta que formaram o grupo 2 da Figura 14.3.2-36, baseada na raiz quadrada dos valores de densidade de 58 táxons identificados (no inverno, 1ª campanha) e na similaridade de Bray-Curtis. Áreas: 1 – Baía de Santos e Canal de navegação; 2 – Canal de Piaçaguera e Rio Casqueiro; 3 – Canal de Bertioga.
Apesar de distantes geograficamente, os grupos D (formado por cinco dos
seis pontos mais internos do canal de Bertioga) e E (formado por quatro dos seis
pontos do Rio Casqueiro) foram os mais similares entre si (Tabela 14.3.2-4).
Ambos foram formados por um baixo número de táxons (25 e 29 dos 60 táxons
analisados, respectivamente) e apresentaram composição zooplanctônica similar,
cujos principais representantes foram os copépodes Parvocalanus crassirostris,
Oithona oswaldocruzi e copepoditos de Paracalanidae e Acartia spp. e larvas de
Cirripedia. Por outro lado, os valores de densidade dos pontos do grupo D foram
inferiores aos do grupo E (médias de 2.850 e 6.130 indivíduos m-3,
respectivamente), além de algumas diferenças pontuais na composição e
densidade de táxons.

Programa 14 - 64
Tabela 14.3.2-4. Resultados do teste pareado ANOSIM realizado entre os grupos obtidos na Análise de Ordenação N-MDS da Figura 14.3.2-36, referentes a organismos da 1ª campanha. Valores significativos: < 5%.
Nível de
significância (%)
A-B 1 2.90
A-C 0.918 0.30
A-D 1 0.80
A-E 1 2.90
A-F 0.756 0.10
B-C 1 0.80
B-D 1 1.80
B-E 1 2.90
B-F 0.694 0.30
C-D 0.751 0.10
C-E 0.976 0.30
C-F 0.635 0.10
D-E 0.625 0.80
D-F 0.893 0.10
E-F 0.965 0.10
Grupos R estatístico
R global = 0.82
Por sua vez, o grupo F foi o único formado por pontos das três áreas de
amostragem (Baía de Santos/Canal de navegação; Canal de Piaçaguera/Rio
Casqueiro e Canal de Bertioga), apresentando maiores semelhanças com os
grupos A, B e C e menores com os grupos D e E (Tabela 14.3.2-4). Vale ressaltar
que foram realizadas algumas tentativas tanto de subdivisão deste grupo bem
como de inclusão de alguns pontos que formaram o grupo F em outros grupos,
mas nenhuma delas mostrou diferença significativa pela análise ANOSIM. Os
pontos P21, P22, P33, P35 e P37 não se enquadraram em nenhum dos grupos
formados.
Análise integrada dos dados - campanhas de janeiro e julho de 2010
Para comparar os dados de densidade e composição zooplanctônica obtidos
nas campanhas de verão e de inverno de 2010 foram realizadas quatro análises
de ordenação N-MDS; uma incluindo todas as 100 amostras coletadas em ambas

Programa 14 - 65
as campanhas (Figura 14.3.2-38) e outras três separando os pontos de coleta de
acordo com a área de amostragem (Figuras 14.3.2-39 a 14.3.2-41).
Figura 14.3.2-38. Análise de Ordenação N-MDS entre as 100 amostras coletadas nas campanhas de verão e de inverno de 2010, considerando as três áreas de amostragem, baseada na raiz quadrada dos valores de densidade de 126 táxons identificados e na similaridade de Bray-Curtis.
Nos resultados da primeira análise (Figura 14.3.2-38) nota-se uma nítida
separação entre as campanhas de verão e de inverno (Anosim: R = 0,589,
p<0,01). Esta separação ocorreu tanto em função da densidade quanto da
composição e diversidade de táxons. No geral, as amostras zooplanctônicas
coletadas na campanha de inverno foram mais densas que nas de verão (média e
desvio padrão de 7.075 ± 8.362 ind. m-3 e 1.500 ± 2.000 ind. m-3,
respectivamente). Quanto à composição dos táxons, Parvocalanus crassirostris,
Acartia tonsa, copepoditos de Acartia spp., Oithona hebes, O. oswaldocruzi,
machos de Oithona spp. e larvas de caranguejos Brachyura e de cracas
(Cirripedia) foram observadas tanto no verão quanto no inverno. Por outro lado,
Acartia lillgeborgi e ovos e larvas de peixes foram mais abundantes no verão,
enquanto que Oikopleura (Vexillaria) dioica, Paracalanus parvus, Oncaea curta,
copepoditos de Paracalanidae, Pseudodiaptomus spp. e Oithona spp., náuplios

Programa 14 - 66
de Copepoda e representantes meroplanctônicos foram mais abundantes no
inverno. Também de forma geral, os valores de diversidade dos organismos
holoplanctônicos foram maiores na campanha de inverno (máximo de 1,03 e
0,67 bits, no inverno e verão, respectivamente), enquanto que para os organismos
meroplanctônicos foi verificado o contrário (máximo de 0,68 e 1,37 bits, no inverno
e verão, respectivamente).
Outro dado interessante ilustrado na Figura 14.3.2-38 foi que os pontos da
campanha de inverno ficaram mais unidos, ou seja, foram mais similares entre si
(com exceção daquelas mais similares ao verão), enquanto que os de verão
mostraram uma separação entre as amostras coletadas na Baía de Santos/Canal
de Navegação (área 1) e as coletadas nas demais áreas (Canal de Piaçaguera e
Rio Casqueiro: área 2; Canal de Bertioga: área 3). Isto se deu tanto em função da
densidade, que foi superior nos pontos da área 1 (Mann-Whitney, p=0,0026),
quanto devido à presença de espécies indicadoras de influência de águas
continentais, como os cladóceros Bosmina longirostris e Diaphanosoma birgei e
os copépodes Acanthocyclops, Apocyclops, Macrocyclops, Thermocyclops, nos
pontos das áreas 2 e 3. Estas ocorrências não foram verificadas na campanha de
inverno.
Ao analisar apenas os pontos coletados na Baía de Santos/Canal de
Navegação de ambas as campanhas (Figura 14.3.2-39), também foi possível
verificar a separação das mesmas em dois grupos. Além das diferenças gerais já
descritas entre as duas campanhas quanto à densidade e composição de
espécies, na campanha de verão, não houve separação entre os pontos da Baía
de Santos e do Canal de navegação, enquanto que na campanha de inverno
essas áreas foram nitidamente separadas.
Comparando a composição de táxons identificados nas campanhas de verão
e inverno no Canal de Piaçaguera e Rio Casqueiro (Figura 14.3.2-40), verificou-se
que 54% deles ocorreram apenas na campanha de verão, incluindo espécies
indicadoras de águas continentais. Por outro lado, 14% dos táxons foram
identificados em maior densidade no inverno, destacando-se Oikopleura
(Vexillaria) dioica e copepoditos de Paracalanidae.

Programa 14 - 67
Figura 14.3.2-39. Análise de Ordenação N-MDS entre as 46 amostras coletadas nas campanhas de verão e de inverno, considerando apenas a Área 1 (Baía de Santos e canal de navegação), baseada na raiz quadrada dos valores de densidade de 71 táxons identificados e na similaridade de Bray-Curtis.
Figura 14.3.2-40. Análise de Ordenação N-MDS entre as 30 amostras coletadas nas campanhas de verão e de inverno, considerando apenas a Área 2 (Canal de Piaçaguera e Rio Casqueiro), baseada na raiz quadrada dos valores de densidade de 58 táxons identificados e na similaridade de Bray-Curtis.

Programa 14 - 68
Ao contrário da área 2 (Canal de Piaçaguera e Rio Casqueiro), o Canal de
Bertioga (Fig. 14.3.2-41) teve uma composição taxonômica similar no verão e no
inverno, mas os pontos do inverno tiveram maiores densidades zooplanctônicas
que os de verão (teste de Mann-Whitney: p<0,0001). Em ambas as campanhas,
Parvocalanus crassirostris, Acartia tonsa, Oithona hebes, O. oswalsocruzi,
machos de Oithona spp., copepoditos de Acartia spp.e náuplios de Copepoda
estiveram entre os táxons mais representativos. Por outro lado, Oikopleura
(Vexillaria) dioica só foi identificada na campanha de inverno nesta área e com
altos valores de densidade. Outra característica dessa área de amostragem está
no fato de que em ambas as campanhas os pontos situados na transição entre o
Canal de Piaçaguera e Canal de Bertioga (P24-P28) separaram-se dos pontos
mais internos do Canal de Bertioga (P29-P34). Tanto no verão quanto no inverno,
os pontos da área de transição foram mais ricos em táxons e com maiores
densidades de ovos e larvas de peixes, larvas de caranguejos braquiúros e
copépodes do gênero Oithona.
Figura 14.3.2-41. Análise de Ordenação N-MDS entre as 24 amostras coletadas nas campanhas de verão e de inverno, considerando apenas a Área 3 (Canal de Bertioga), baseada na raiz quadrada dos valores de densidade de 60 táxons identificados e na similaridade de Bray-Curtis.

Programa 14 - 69
14.4. Considerações Finais
14.4.1. Comunidade Fitoplanctônica
O fitoplâncton no Sistema Estuarino de Santos foi composto por espécies
marinhas, estuarinas e de água doce, dominadas pelas frações do nanoplâncton
(celulas entre 2μm e 20μm).
No verão, correspondente a Campanha Prévia realizada em janeiro de 2010,
a abundância e distribuição obedeceram os padrões halinos conferidos pelo
regime hidrográfico. Gêneros característicos de ambientes lacustres como
Desmosdemium, Scenesdesmus, Ankitrodesmus e Pediastrum, foram registrados
até nos pontos dentro do canal.
No inverno, 1ª Campanha realizada em julho de 2010, a distribuição foi
resultante de padrões halinos e fluxo de corrente. Em regiões de baixa salinidade
e águas mais calmas, como no Largo do Canéu, foram registradas espécies
características de ambientes lacustres como os citados anteriormente.
Salinidades baixas com processos de turbulências foram domínios das
diatomáceas estuarinas como Thalassiosira spp e Skeletonema costatum.
Espécies bentônicas e meroplanctonicas (ticopelágicas) foram encontradas
na coluna d´água, indicando processos de ressuspensão de fundo ocasionados
pelo aumento do fluxo de corrente em quase todo o canal. A causa pode ser as
condições de inverno, como a entrada de frentes e aumento na intensidade dos
ventos. Estas hipóteses poderão ser confirmadas após término da dragagem, a
partir dos resultados dos estudos sobre a hidrodinâmica (Programa 6 -
Monitoramento e Modelagem dos Parâmetros Hidrodinâmicos e Oceânicos).
Com a diminuição da descarga de água doce de origem continental no
inverno, houve a diminuição dos gêneros de Cyanobacteria: Synechocystis,
Anabaena, Cylindropermopsis raciborskii, Planktothix agardii; e dentro das
Bacillariophyta as consideradas potencialmente tóxicas: grupo Pseudonitzchia.
O dinoflagelado Karenia mikomotoi que formou bloom no verão, campanha
pré-dragagem, (1,8 x 106 céls/L) no interior do Canal de Bertioga (P30), esteve

Programa 14 - 70
presente, no inverno, em menor frequência e em menor número (1,4 x 103 céls/L),
no mesmo local.
A espécie de Alexandrium tamarense não foi registrada nas amostras de
verão, e no inverno ocorreram na maioria em pontos no interior do estuário sendo
mais abundante também no ponto P30 (1,8 céls/L).
14.4.2. Comunidade Zooplânctônica
Os resultados das campanhas de janeiro (Campanha Prévia - verão) e julho
(1ª Campanha - inverno) de 2010, no sistema estuarino de Santos-Bertioga,
indicam uma comunidade zooplanctônica típica de ambientes estuarinos,
dominada por espécies de copépodes marinho-eurihalinas e, secundariamente,
por espécies oligohalinas. Estas últimas tiveram importância para a composição e
a densidade do zooplâncton durante o verão, período de maior pluviosidade,
quando o aporte de água continental no estuário é mais intenso. Várias espécies
de copépodes e de cladóceros oligohalinas e de água doce foram detectadas pela
primeira vez neste complexo estuarino. As larvas meroplanctônicas,
representadas principalmente por náuplios de cirripédios e zoeas de braquiúros,
foram mais abundantes no inverno. Estas diferenças sazonais foram
representadas quantitativamente nas análises estatísticas executadas com os
dados de densidade dos táxons mais frequentes.
Os dados obtidos foram comparáveis com os resultados do EIA-RIMA (FRF,
2008) em relação à abundância total do zooplâncton e à dominância de
Copepoda, mas houve diferenças em alguns aspectos mais específicos. Entre
estes, se destacam as baixas proporções de grupos holo- e meroplanctônicos
(exceto Copepoda) em janeiro de 2010 e a presença das espécies de água doce
no mesmo período.
Os dados descritos nesta seção representam uma base importante para a
detecção de eventuais impactos da atividade de dragagem sobre esta
comunidade biológica, pois retratam tanto a variabilidade espacial (costa-estuário)
quanto sazonal (verão-inverno).

Programa 14 - 71
14.5. Cronograma
O programa de monitoramento das comunidades fitoplanctônicas e
zooplanctônicas ocorrerá no período de 18 meses, contemplando os períodos de
verão e inverno.
O cronograma de trabalho específico para o monitoramento da comunidade
planctônica está resumido conforme Tabela 14.5-1.
Tabela 14.5-1. Cronograma de atividades.
jan fev mar abr mai jun jul ago set out nov dez jan fev mar abr mai jun
Programa 14
Mobilização
Plano de Trabalho
Coletas e Atividades de Campo
Triagem das Amostras
Identificação dos organismos
Elaboração de Relatórios Semestral
ATIVIDADES
MÊS
2010 2011
14.6. Referências Bibliográficas
Balech,E.; Akselman,R.; Bernarvides,H.R. & Negri,R.M. (1984). Suplemento a los
dinoflagelados Del Atlantico Sudoccidental. Revista de Investigation y
desarrollo pesquero, 4. 309p.
Bicudo, C.E.M & Menezes, M. (2006). Generos de algas de águas continentais do
Brasil. Rima Editora, 498 p.
Boltovskoy, D. (1981). Atlas del zooplancton del Atlántico Sudoccidental y
métodos de trabajo con el zooplancton marino. Mar del Plata, INIDEP, 936 p.
Boltovskoy, D. (1999). South Atlantic zooplankton. Backhuys Publishers, Vol. 2,
1705 p.
Brandini, F.P.; Lopes, R.M.; Gutseit, K.S.; Spach, H.L. & Sassi, R. (1997).
Planctonologia na plataforma continental do Brasil: diagnose e revisão
bibliográfica. Rio de Janeiro, MMA, FEMAR, 196 p.

Programa 14 - 72
Caljon, A. (1983). Brackish-water phytoplankton of the Flemish Iowland. Dr. W.
Junk Publishers. Developments in hydrobiology, 18. 272 p.
Chan, U.T. & Hamilton, D.P. (2001). Effect of freshwater on the succession and
biomass of phytoplankton in a seasonal estuary. Mar & fresh Res. 52 (6)
869-884.
Cloern, J.E & Dufford, R. (2005). Phytoplankton community ecology: principles
applied in San Francisco Bay. Mar Ecol Prog Ser vol 285, 11-28 .
Coquemala, V. (2005). Variação anual do fitoplâncton do reservatório de
Passaúna, Paraná. Dissertação apresentada para obtenção do grau de
Mestre na Universidade do Paraná, 92 p
Dodge, J.D. (1982). Marine Dinoflagellates of the British Isles.
Elster,H.-J. & Ohle, W. (1983). Die Binnengewässe.Einzeldarstellungen aus der
limnologie und ihren Nachbargebieten. Stuttgart. Band XVI, 7.Teil, 1.Hälfte.
1044 p.
Falkowski, P.G.; Barber, R.T.; Smetacek, V. (1998). Biogeochemical controls and
feedbacks on ocean primary production. Science 281:200-206.
Frazão, L. R. (2001), Eutrofização por esgotos domésticos e sua relação com a
ecologia e fisiologia do fitoplâncton marinho no litoral do Estado de São
Paulo: Ubatuba, Praia Grande e Santos. Master of Science Dissertation.
Universidade de São Paulo, Instituto Oceanográfico. São Paulo, Brazil. 158
pp.
Fundação Ricardo Franco, (2008). Estudo de Impacto Ambiental (EIA) e Relatório
de Impacto Ambiental (RIMA): projeto de aprofundamento do canal de
navegação do Porto de Santos, Santos, SP. São Paulo.
Fundespa, (2010). Relatório Técnico Semestral do Plano Básico Ambiental da
Dragagem de Aprofundamento do Porto de Santos - RTS – 0618-
140910.1447p.
Gayoso, A.M. (2001). Observations on Alexandrium tamarense (Lebour) Balech
and other dinoflagellate populations in Golfo Nuevo, Patagonia Argentina. J.
Plank. Res. 23(5) 463-468.

Programa 14 - 73
Hallegraeff, G.M. & Reid, D.D. (1986). Phytoplankton species successions and
their hydrological environment at coastal station off Sydney. Aust. J. Mar.
Freshw. Res. 37, 361-377.
Krebs, C.J. (1999). Ecological Methodology. Addison-Welsey Educational
Publishers Inc.: California, 620 p.
Lopes, R.M. (2007) Marine zooplankton studies in Brazil: an overview. Anais da
Academia Brasileira de Ciências, 411: 161-174.
McMahon, T., Silke, J., 1996. Winter toxicity of unknown aetiology in mussels.
Harmful Algal News 14, 2.
Moser, G.A.O. Aspectos da eutrofização no sistema estuarino de Santos:
distribuição espaço-temporal da biomassa e produtividade primária
fitoplanctônica e transporte instantâneo de sal, clorofila-a, material em
suspensão e nutrientes. 2002. 410p. 2 vol. Tese (Doutorado em
Oceanografia biológica), Universidade de São Paulo, Instituto
Oceanográfico, São Paulo, 2002.
Motoda, S. (1959) Devices of simple plankton apparatus. Men. Fac. Fish.
Hokkaido Univ., 7: 73-94.
Neumann-Leitão, S. (1994/1995). Resenha literária sobre zooplâncton estuarino
no Brasil. Trab. Oceanogr. Univ. Fed. PE, 23: 25-53.
Omori, M. and Ikeda, T. (1984), Methods in marine zooplankton ecology. New
York : Wiley-Interscience Publication. 331 pp.
Reynolds, C.S. (1997). Vegetation process in the pelagic: a model for ecosystems
theory. In: Kinne, O. Excellence in ecology. Germany: Ecology Institute, 371p
Shannon, C. E. 1948. A mathematical theory of communication. Bell System
Technical Journal 27: 379-423.
Thomas, C.R. (1997). Identifying Marine Phytopplankton, Eds. Acadmic Press,
New York, 858 pp.
Tood, C.D.; Laverack, M.S.; Boxshall, G.A. (1996). Coastal marine zooplankton. A
practical manual for students. Cambridge University Press, 106 p.

Programa 14 - 74
Utermöhl, N. 1931. Neue Wege in der quantitativen Erfassung des Planktons.
Verh. Int. Verein. Theor. Angew. Linmol., 5 : 567-596.
<www.algaebase.org> Acesso em: 15 dez 2010.
14.7. Equipe Técnica
Dr. Salvador Airton Gaeta
Dr. Rubens Mendes Lopes
MSc. Daniela Rimold Cunha - Oceanógrafa
Paula Duran Nagata Perugino - Oceanógrafa
Rafael Rugna Ciglione – Oceanógrafo
Newton Dalessandro - Biólogo
Everton Kotarski – Oceanógrafo (cursando)





























