Embed Size (px)
Citation preview

16°
TÍTULO: PAPEL DA NEUROGÊNESE HIPOCAMPAL NA EXTINÇÃO DA DEFESA PREDATÓRIACONDICIONADA: ESTUDO PARA A PADRONIZAÇÃO DO PROTOCOLO DE ABLAÇÃO DANEUROGÊNESE HIPOCAMPAL
TÍTULO:
CATEGORIA: CONCLUÍDOCATEGORIA:
ÁREA: CIÊNCIAS BIOLÓGICAS E SAÚDEÁREA:
SUBÁREA: MEDICINASUBÁREA:
INSTITUIÇÃO: UNIVERSIDADE CIDADE DE SÃO PAULOINSTITUIÇÃO:
AUTOR(ES): MARJORIE ORQUISA CARLOSAUTOR(ES):
ORIENTADOR(ES): CLÁUDIA DE BRITO FATURIORIENTADOR(ES):

1. Resumo
Dentre os critérios para elaboração de um modelo animal de psicopatologia a
validade de constructo tem grande relevância, uma vez que para preenchê-lo o
modelo deve apresentar similaridade com a neurobiologia do transtorno em
questão. De forma complementar, a psiquiatria tem buscado endofenótipos que
permitam identificar de forma mais precisa e precoce o aparecimento de
psicopatologias. Vários trabalhos indicam que a alteração na neurogênese
hipocampal seria um possível endofenótipo para depressão e transtorno de
estresse pós traumático (TEPT). A neurogênese está envolvida, por exemplo, no
processo de padrão de separação. Neste fenômeno, os novos neurônios recém-
inseridos na camada granular do giro denteado melhorariam a resolução de novas
memórias permitindo que elas sejam mais facilmente distinguíveis de estímulos
semelhantes. Dentre os sintomas apresentados pelos pacientes diagnosticados
com o TEPT está a vivência repetida do evento traumático quando o indivíduo se
depara com pistas que o relembrem do trauma. Assim, é possível que tais
sintomas estejam relacionados com um déficit no padrão de separação. Tendo isso
em vista, a utilização de testes comportamentais que envolvam o estudo dos
mecanismos neurais subjacentes ao condicionamento e extinção da memória de
medo têm se tornado uma ferramenta importante para investigar e inferir sobre os
mecanismos neurais do TEPT. Vários estudos têm sido realizados com esse
propósito, sendo que a maioria deles utiliza o choque nas patas como estímulo
aversivo. No entanto, vários trabalhos mostram que os circuitos neurais recrutados
em resposta ao choque nas patas e um estímulo natural, como a presença do
predador, são diferentes. A exposição ao predador é potencialmente o estímulo
que mais se assemelha ao evento desencadeador de uma patologia como o TEPT.
Este é, em geral, provocado por um evento que atenta contra a vida e/ou
integridade física dos portadores do transtorno, que é coerente com as reações do
rato frente à um dos seus predadores naturais, o gato. Assim, o presente trabalho
teve como objetivo iniciar a padronização das respostas comportamentais de
extinção do comportamento de defesa. Os resultados indicam que no 6º dia de
reexposição o comportamento de defesa já está totalmente extinto. À partir desses

dados será possível compreender melhor o efeito da ablação da neurogênese
hipocampal na extinção do comportamento de defesa condicionado.
2. Introdução
A elaboração e validação de um modelo experimental de transtornos
psiquiátricos pode se dar pelos critérios de validade de face, preditiva e de
constructo. O primeiro leva em consideração o fenótipo clínico da patologia, e o
segundo diz respeito à capacidade do teste de predizer, por exemplo, a eficácia
de um agente farmacológico (Treit 1985). Estes dois critérios são considerados
superficiais, uma vez que não levam em consideração as bases neurobiológicas
do comportamento. Já a validade de constructo mede a similaridade do modelo
animal com os processos fisiológicos e etiologia da patologia em questão (Treit
1985). Esse tipo de validade parte do princípio que animais e humanos
apresentam similaridades na estrutura cerebral e função (Homberg 2013). A
psiquiatria tem se engajado também na investigação de endofenótipos, definidos
como características herdáveis que estariam conectadas com as alterações
comportamentais, neurofisiológicas, bioquímicas, endócrinas ou
neuroanatômicas apresentados numa dada psicopatologia (Andreatini et al
2006).
A investigação de endofenótipos é importante para fundamentar a conclusão
diagnóstica em várias áreas da medicina, no entanto a psiquiatria é uma área em
que o estudo da neurobiologia dos transtornos ainda requer avanços para
possibilitar a utilização dos mesmos (Gould and Gottesman 2006). A
determinação de endofenótipos permitiria ainda um diagnóstico precoce e início
do tratamento tão logo os primeiros sintomas da doença se manifestassem (Flint
and Munafo 2007).
Vários estudos têm apontado que alterações na plasticidade sináptica
hipocampal possam estar estreitamente relacionadas com a gênese de
transtornos psiquiátricos (McEwen 1999; McEwen and Magarinos 2001;
Lehmann et al. 2013; Schloesser et al. 2010; Kheirbek et al. 2012). Além de ter
papel fundamental em processos mnemônicos o hipocampo também participa da
organização de comportamentos de defesa, tendo uma contribuição importante

para a percepção da ameaça predatória haja vista que, principalmente a porção
ventral, recebe projeções de setores da amigdala envolvidos com a interpretação
de informação olfatória (Wang et al. 2013; Canteras and Swanson 1992;
Kemppainen et al. 2002) e regiões envolvidas com a regulação da resposta de
estresse, como o núcleo intersticial da estria terminal (Blanchard et al. 2005;
Herman et al. 2005; Dong and Swanson 2006). Tais funções têm motivado
alguns autores a relacionar alterações no fenômeno de padrão de separação
com a gênese do Transtorno de Estresse Pós-Traumático (TEPT). De acordo
com Kheirbek et al (2012) prejuízos nos processos de padrão de separação seria
substrato neural para a supergeneralização frequentemente vista em transtornos
de ansiedade. A fundamentação teórica do fenômeno de padrão de separação
indica que a ablação da neurogênese no giro denteado pode prejudicar
processos responsáveis pela codificação de uma memória que apresente
características parcialmente semelhantes com um traço mnemônico apresentado
anteriormente, prejudicando assim a resolução de tais memórias (Aimone et al.
2011). Tal prejuízo poderia favorecer respostas fisiológicas de estresse quando
um dado estímulo outrora relacionado com o trauma vivido é apresentado num
contexto diferente.
O comportamento é o produto final do funcionamento do SNC e a facilidade
em observá-lo em modelos animais faz dos modelos de medo
condicionado/extinção um dos protocolos mais usados para o estudo do TEPT
(Bouton 2002). Em sua maioria, os estudos que investigam a neurobiologia
subjacente ao condicionamento de medo/extinção utilizam o choque nas patas
como estímulo aversivo. O choque nas patas provoca reações comportamentais
de defesa como a imobilidade, em que o animal permanece parado,
apresentando apenas os movimentos respiratórios. No entanto, vários autores
discutem se o circuito neural responsável por tais respostas de defesa
provocadas por um estímulo doloroso como o choque nas patas seriam os
mesmos recrutados por um estímulo que represente uma real ameaça à vida do
animal, como por exemplo, a exposição ao predador. No trabalho de revisão
bibliográfica de (Gross and Canteras 2012) os autores apontam que circuitos
neurais diferentes medeiam as mesmas respostas comportamentais de defesa
nas duas situações. É importante notar que vários trabalhos indicam que a
resposta de defesa frente o odor do predador recruta as mesmas estruturas

neurais observadas durante à exposição ao predador (do Monte et al. 2008;
Kincheski et al. 2012). Assim, é possível que a utilização da defesa predatória
para elaboração de um modelo animal de TEPT tenha maior validade de
constructo do que a utilização do choque nas patas, tendo maior probabilidade
de que os resultados obtidos à partir deste modelo possibilite a validação de um
endofenótipo (Matar et al. 2013). (Quirk et al. 2010) destaca ainda que o
desenvolvimento de um estratégica terapêutica que possibilite realçar os
processos de extinção poderia reduzir a evocação espontânea de memórias de
medo observada por exemplo no TEPT, possibilitando ainda uma redução de
sessões de terapia necessárias para a alta do paciente.
Embora a extinção tenha sido vista por muito tempo como um processo em
que a memória de medo é simplesmente apagada, vários trabalhos indicam que
se trata na verdade de um novo aprendizado. (Quirk et al. 2010). Como em
qualquer outro processo de aprendizagem ele ocorre em três fases: aquisição,
consolidação e expressão (Quirk and Mueller 2007). O processo de extinção é
altamente adaptativo e permite que o indivíduo altere suas estratégicas para lidar
com dificuldades impostas pelas mudanças no ambiente. Atualmente o circuito
hipotético envolvido nos processos de extinção inclui o córtex pré-frontal, córtex
cingulado anterior rostral, córtex orbitofrontal e hipocampo (Rauch et al. 2005;
Rauch et al. 2006). No entanto, tais estudos foram realizados utilizando
principalmente o paradigma que envolve choque nas patas, e ainda não se sabe
se a extinção da memória de medo provocada pelo odor do predador recruta
essas mesmas regiões.
3. OBJETIVOS
Assim, levando em consideração a fundamentação teórica apresentada o
objetivo geral deste projeto foi padronizar o protocolo de extinção da defesa
condicionada.
de ablação da neurogênese hipocampal.

4. Metodologia
Animais. Em todos os estudos foram utilizados ratos albinos (Rattus
norvegicus, linhagem Wistar), machos, adultos, com aproximadamente 90 dias de
idade. Os animais foram alojados em gaiolas de polipropileno (30X40X18cm), em
salas com sistema de ventilação (23±2oC) e ciclo de luz de 12h (06:00-18:00). Água
e comida foram oferecidas ad libitum durante todos os procedimentos experimentais.
Os animais foram obtidos no CEDEME (UNIFESP) e os experimentos foram
realizados no Núcleo de Pesquisa em Neurociências (NUPEN) da UNICID. Os
procedimentos experimentais foram submetidos à apreciação das CEUAs de ambas
instituições e foram aprovados sob número 1376230314 na UNIFESP e 002/2015 na
UNICID.
5. Desenvolvimento
Os dados apresentados neste relatório consistem de um primeiro experimento
piloto realizado para conhecermos o padrão de resposta comportamental de defesa
ao longo do processo de extinção. Neste, apenas um grupo de animais foi testado.
Os mesmos foram expostos ao predador por 10 minutos e posteriormente
reexpostos ao contexto por cinco dias consecutivos. Após a análise desses dados
fizemos algumas alterações no protocolo inicialmente proposto. As alterações
consistiram na diminuição do tempo de reexposição ao contexto de dez para cinco
minutos. Assim, apresentamos também dados preliminares obtidos com a realização
deste experimento.
No segundo experimento foram testados 3 grupos: experimental, controle 1
(CTL1) e controle 2 (CTL2). O grupo experimental foi inicialmente habituado,
exposto ao gato por dez minutos, e posteriormente à sucessivas exposições ao
aparato até que os comportamentos de defesa fossem extintos. O grupo CTL1,
também chamado controle positivo, foi habituado, exposto ao predador e exposto ao
contexto no dia seguinte, mas durante o período de extinção ficou alocado na caixa
moradia dentro do biotério e foi reexposto ao aparato apenas no último dia de
extinção do grupo experimental. O grupo CTL2, denominado controle negativo, foi
submetido ao procedimento de habituação durante todo o período em que o grupo
experimental foi testado.

Aparato experimental. O aparato experimental consiste de 3 compartimentos:
caixa-moradia, um corredor de acesso e a caixa 2, onde é colocada ração durante o
procedimento de habituação e o gato no dia do experimento (Figura 1).
Figura 1. Esquema do aparato experimental.
Durante 10 dias, os animais passaram por um período de habituação a este
aparato, sendo que todos os dias, 3 horas antes do início do período de escuro, os
animais foram privados de ração. No início do período de escuro, quando ocorre o
período de maior atividade e o primeiro pico alimentar do rato, a porta da caixa-
moradia foi aberta permitindo que os animais explorassem o corredor e se
deslocassem até a caixa 2, para obterem a ração ali depositada.
Teste comportamental. No décimo primeiro dia, os animais do grupo
experimental e do grupo CTL1 foram expostos ao gato, que foi colocado na caixa 2,
onde a comida era oferecida durante o procedimento de habituação, e a resposta
comportamental foi observada por 10 minutos, sendo que em nenhum momento
houve contato físico entre eles. No estudo piloto, no décimo segundo dia do
procedimento experimental (1ª reexposição), os animais foram avaliados por dez
minutos durante a exposição ao aparato previamente associado à presença do
predador. Esse procedimento foi repetido nos 4 dias subsequentes.
Em função dos resultados obtidos no experimento piloto, o tempo de
exposição ao contexto no segundo experimento foi reduzido a cinco minutos e a
reexposição ao contexto foi realizada por 6 dias consecutivos.
Todos os procedimentos (exposição ao gato e exposição ao contexto) foram
filmados e as respostas comportamentais foram analisadas utilizando o programa
específico para análise comportamental “Anymaze” (Stoelting Co).

As seguintes respostas comportamentais foram categorizadas:
Congelamento o animal permanece imóvel, num estado de congelamento motor.
Exploração animal se locomove mais de 1cm sem estar com o corpo estendido.
Avaliação de risco que incluem:
“Crouch sniff” o animal permanece parado, com dorso arqueado, fazendo
movimentos com a cabeça para cheirar e esquadrinhar o ambiente.
“Stretch Attend Posture” o animal estende a cabeça e parte do corpo para frente,
mantém a cauda elevada, porém não se desloca.
“Stretch Approach” o animal mantém a mesma postura anterior, mas desloca-se
para frente.
Após o período de observação comportamental, os animais dos grupos
controle e experimental foram profundamente anestesiados e perfundidos por via
trans-aórtica. Os encéfalos foram retirados e processados para posterior realização
de imunohistoquímica para proteína deltaFosB, cujo aumento sinaliza ativação
neuronal crônica.
Analise Estatística. Temos ainda uma amostra pequena, apenas 3
animais/grupos, e por isso nenhum teste foi realizado. Mas de acordo com o
desenho experimental proposto os dados comportamentais serão realizadas
utilizando a ANOVA de duas vias (fator medidas repetidas X fator grupo) seguido do
teste a posteriori de Newman-Keuls quando necessário, considerando a diferença
significativa sempre que p < 0,05. Os resultados estão expressos em média ± erro
padrão.
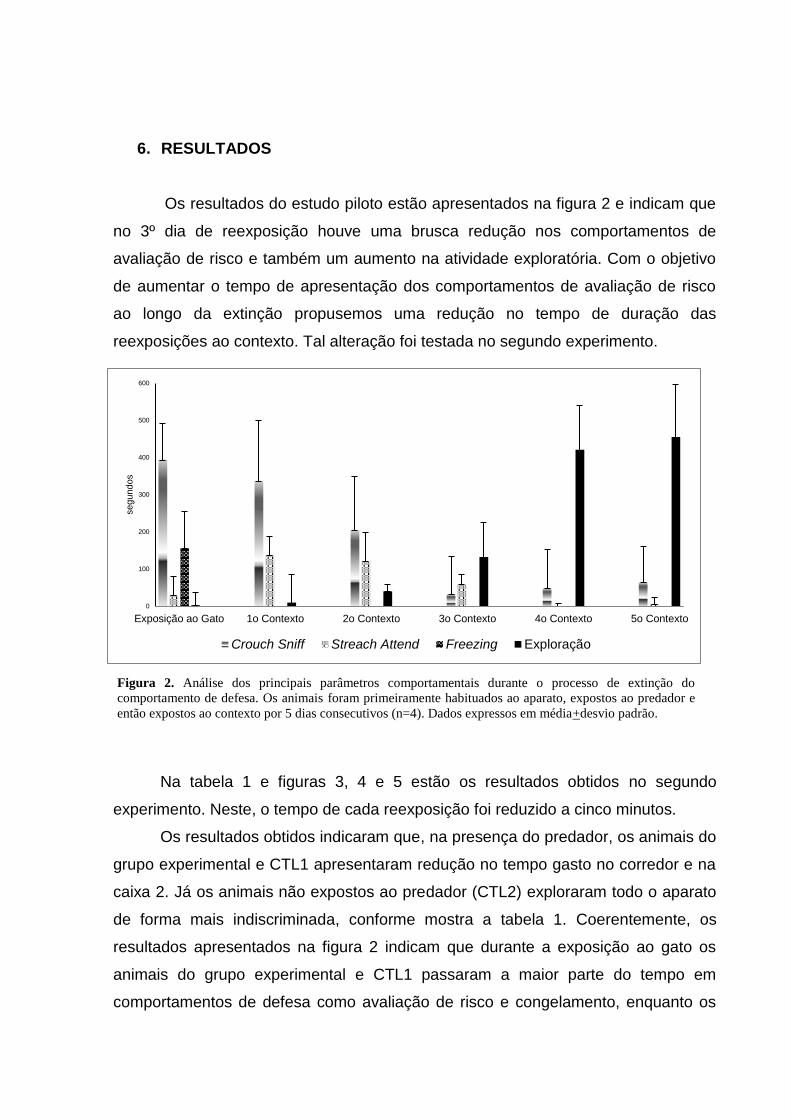
6. RESULTADOS
Os resultados do estudo piloto estão apresentados na figura 2 e indicam que
no 3º dia de reexposição houve uma brusca redução nos comportamentos de
avaliação de risco e também um aumento na atividade exploratória. Com o objetivo
de aumentar o tempo de apresentação dos comportamentos de avaliação de risco
ao longo da extinção propusemos uma redução no tempo de duração das
reexposições ao contexto. Tal alteração foi testada no segundo experimento.
Na tabela 1 e figuras 3, 4 e 5 estão os resultados obtidos no segundo
experimento. Neste, o tempo de cada reexposição foi reduzido a cinco minutos.
Os resultados obtidos indicaram que, na presença do predador, os animais do
grupo experimental e CTL1 apresentaram redução no tempo gasto no corredor e na
caixa 2. Já os animais não expostos ao predador (CTL2) exploraram todo o aparato
de forma mais indiscriminada, conforme mostra a tabela 1. Coerentemente, os
resultados apresentados na figura 2 indicam que durante a exposição ao gato os
animais do grupo experimental e CTL1 passaram a maior parte do tempo em
comportamentos de defesa como avaliação de risco e congelamento, enquanto os
0
100
200
300
400
500
600
Exposição ao Gato 1o Contexto 2o Contexto 3o Contexto 4o Contexto 5o Contexto
segundos
Crouch Sniff Streach Attend Freezing Exploração
Figura 2. Análise dos principais parâmetros comportamentais durante o processo de extinção do
comportamento de defesa. Os animais foram primeiramente habituados ao aparato, expostos ao predador e
então expostos ao contexto por 5 dias consecutivos (n=4). Dados expressos em média+desvio padrão.
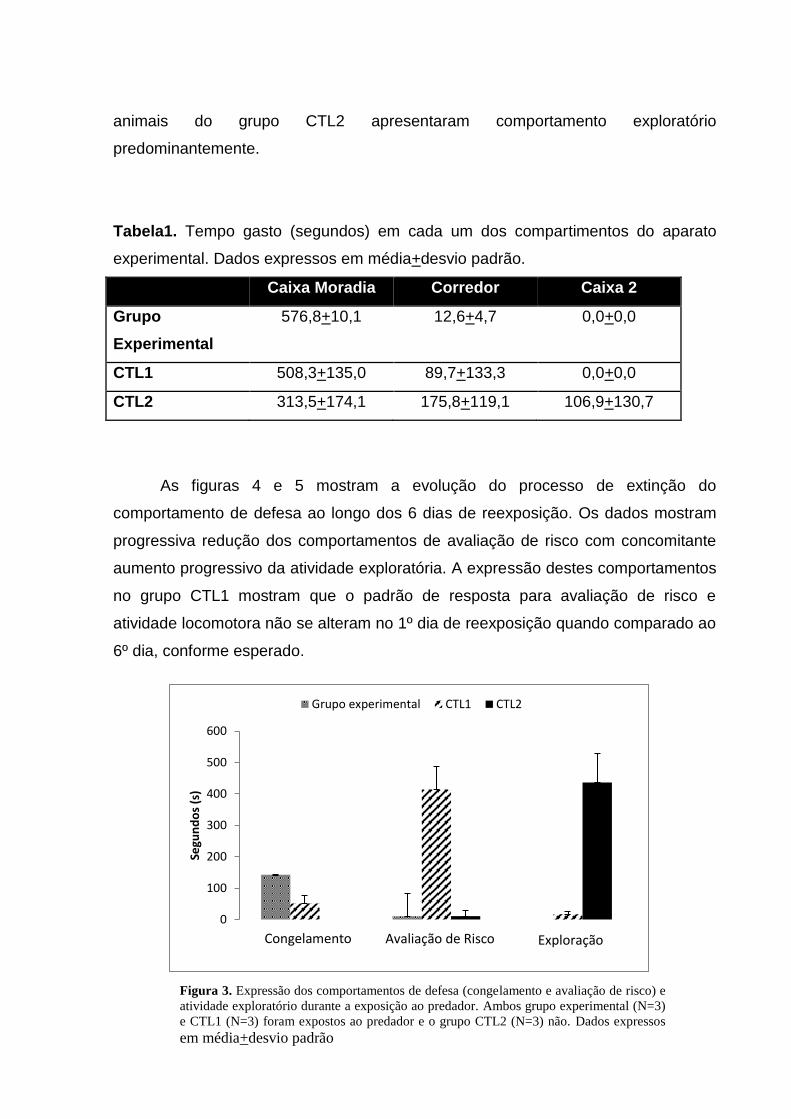
animais do grupo CTL2 apresentaram comportamento exploratório
predominantemente.
Tabela1. Tempo gasto (segundos) em cada um dos compartimentos do aparato
experimental. Dados expressos em média+desvio padrão.
Caixa Moradia Corredor Caixa 2
Grupo
Experimental
576,8+10,1 12,6+4,7 0,0+0,0
CTL1 508,3+135,0 89,7+133,3 0,0+0,0
CTL2 313,5+174,1 175,8+119,1 106,9+130,7
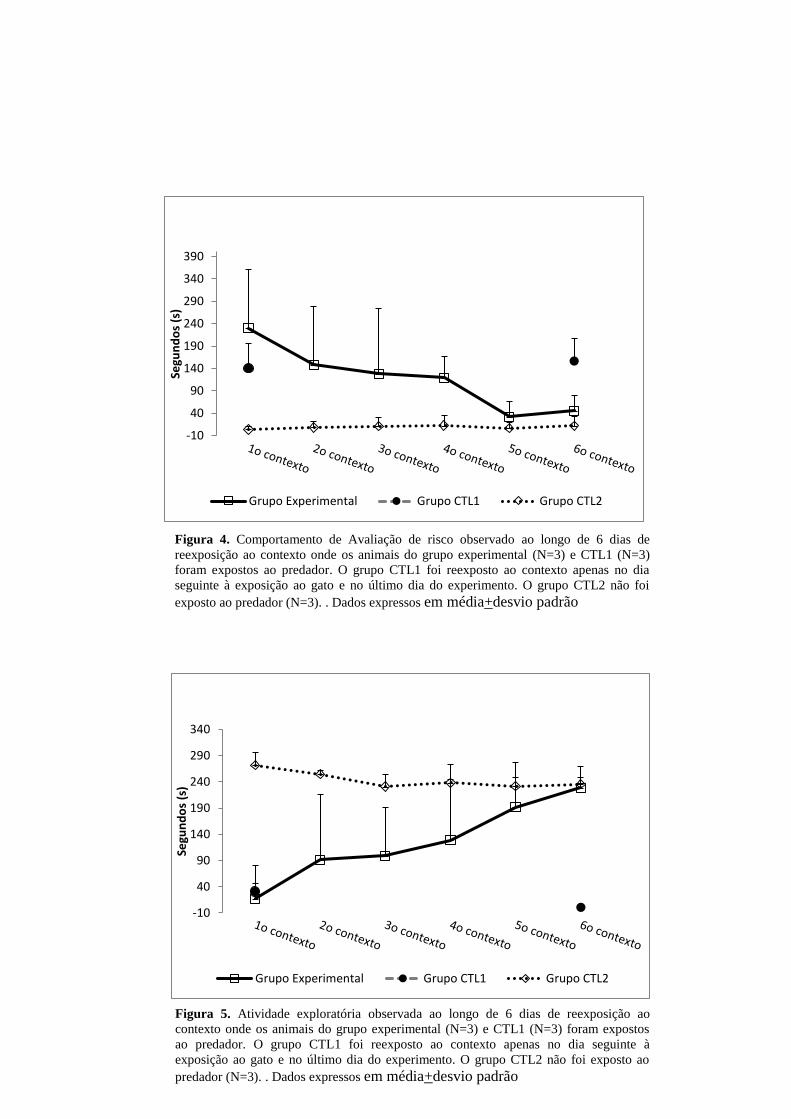
As figuras 4 e 5 mostram a evolução do processo de extinção do
comportamento de defesa ao longo dos 6 dias de reexposição. Os dados mostram
progressiva redução dos comportamentos de avaliação de risco com concomitante
aumento progressivo da atividade exploratória. A expressão destes comportamentos
no grupo CTL1 mostram que o padrão de resposta para avaliação de risco e
atividade locomotora não se alteram no 1º dia de reexposição quando comparado ao
6º dia, conforme esperado.
0
100
200
300
400
500
600
Segu
nd
os
(s)
Grupo experimental CTL1 CTL2
Congelamento Avaliação de Risco Exploração
Figura 3. Expressão dos comportamentos de defesa (congelamento e avaliação de risco) e
atividade exploratório durante a exposição ao predador. Ambos grupo experimental (N=3)
e CTL1 (N=3) foram expostos ao predador e o grupo CTL2 (N=3) não. Dados expressos
em média+desvio padrão
-10
40
90
140
190
240
290
340
390
Segu
nd
os
(s)
Grupo Experimental Grupo CTL1 Grupo CTL2
Figura 4. Comportamento de Avaliação de risco observado ao longo de 6 dias de
reexposição ao contexto onde os animais do grupo experimental (N=3) e CTL1 (N=3)
foram expostos ao predador. O grupo CTL1 foi reexposto ao contexto apenas no dia
seguinte à exposição ao gato e no último dia do experimento. O grupo CTL2 não foi
exposto ao predador (N=3). . Dados expressos em média+desvio padrão
-10
40
90
140
190
240
290
340
Segu
nd
os
(s)
Grupo Experimental Grupo CTL1 Grupo CTL2
Figura 5. Atividade exploratória observada ao longo de 6 dias de reexposição ao
contexto onde os animais do grupo experimental (N=3) e CTL1 (N=3) foram expostos
ao predador. O grupo CTL1 foi reexposto ao contexto apenas no dia seguinte à
exposição ao gato e no último dia do experimento. O grupo CTL2 não foi exposto ao
predador (N=3). . Dados expressos em média+desvio padrão

7. Considerações finais
Os resultados obtidos com o primeiro experimento piloto indicou que os
comportamentos de defesa condicionada utilizando o gato como estímulo aversivo,
predador natural da espécie estudada, poderia ser extinguido após 3 exposições ao
aparato experimental. É possível observar que os comportamentos de avaliação de
risco reduzem drasticamente já no terceiro dia de reexposição. Durante as análises
comportamentais observamos que nos primeiros minutos da reexposição o animal
apresentava um pouco de comportamento de defesa, que logo dava lugar ao
comportamento exploratório (dados não mostrados). Assim, era difícil determinar o
padrão de comportamento de defesa de cada dia de reexposição. Dado que o
objetivo do presente projeto é investigar o papel da neurogênese hipocampal no
processo de extinção da memória de medo decidimos propor uma alteração no
protocolo comportamental inicialmente apresentado. Assim, no segundo
experimento, reduzimos o tempo de reexposição para cinco minutos. Com isso, em
cada reexposição os animais apresentaram um padrão de comportamento mais
característico. Essa alteração permitirá determinar mais facilmente o momento mais
oportuno para intervirmos no processo de neurogênese ao longo da extinção da
memória de medo.
No segundo experimento os animais do grupo experimental apresentaram um
crescente aumento na atividade exploratória que culminou no 6º dia de extinção. Um
padrão coerente se observa para o comportamento de avaliação de risco, que
diminui paulatinamente, sendo totalmente extinto no 5º dia de reexposição. O
comportamento do grupo CTL1 observados no 1º e no último dia de reexposição
indicam que a redução no comportamento de defesa observado no grupo
experimental é de fato decorrente do processo de extinção da memória de medo, e
não apenas esquecimento. Os animais desse grupo apresentaram níveis muito
semelhantes de atividade locomotora e de avaliação de risco no 1º e 6º dia de
reexposição.
À partir dos encéfalos coletados dos animais do segundo experimento
faremos um estudo de ativação neuronal para conhecer as principais áreas
recrutadas para a extinção da memória de medo condicionado ao predador e melhor
compreender como este modelo animal pode ser útil para esclarecer as alterações

neuronais que ocorrem em pacientes que apresentam o transtorno de estresse pós-
traumático.
8. REFERÊNCIAS BIBLIOGRÁFICAS
Andreatini R, Boerngen-Lacerda R, Vital MABF (2006). Modelos animais em
psicofarmacologia. In: Almeida, R. N. Psicofarmacologia: fundamentos
práticos (1ª edição). Rio de Janeiro: Guanabara Koogan.
Aimone JB, Deng W, Gage FH (2011) Resolving new memories: a critical look at the
dentate gyrus, adult neurogenesis, and pattern separation. Neuron 70 (4):589-
596. doi:10.1016/j.neuron.2011.05.010
Blanchard D, Canteras N, Markham C, Pentkowski N, Blanchard R (2005) Lesions of
structures showing FOS expression to cat presentation: effects on responsivity
to a Cat, Cat odor, and nonpredator threat. Neuroscience & Biobehavioral
Reviews 29 (8):1243-1253
Bouton ME (2002) Context, ambiguity, and unlearning: sources of relapse after
behavioral extinction. Biol Psychiatry 52 (10):976-986
Canteras N, Swanson L (1992) Projections of the ventral subiculum to the amygdala,
septum, and hypothalamus: A PHAL anterograde tract tracing study in the rat.
The Journal of Comparative Neurology 324 (2):180-194
Couillard-despres S, Winner B, Schaubeck S, Aigner R, Vroemen M, Weidner N,
Bogdahn U, Winkler J, Kuhn HG, Aigner l (2005) Doublecortin expression
levels in adult brain reflect neurogenesis. European Journal of Neuroscience
21:1–14
Dong HW, Swanson LW (2006) Projections from bed nuclei of the stria terminalis,
anteromedial area: cerebral hemisphere integration of neuroendocrine,
autonomic, and behavioral aspects of energy balance. The Journal of
Comparative Neurology 494 (1):142-178
Flint J, Munafo MR (2007) The endophenotype concept in psychiatric genetics.
Psychological medicine 37 (2):163-180. doi:10.1017/S0033291706008750
Gould TD, Gottesman, II (2006) Psychiatric endophenotypes and the development of
valid animal models. Genes, brain, and behavior 5 (2):113-119.
doi:10.1111/j.1601-183X.2005.00186.x

Gross CT, Canteras NS (2012) The many paths to fear. Nature Reviews
Neuroscience 13 (9):651-658
Herman JP, Ostrander MM, Mueller NK, Figueiredo H (2005) Limbic system
mechanisms of stress regulation: hypothalamo-pituitary-adrenocortical axis.
Progress in neuro-psychopharmacology & biological psychiatry 29 (8):1201-
1213. doi:10.1016/j.pnpbp.2005.08.006
Homberg JR (2013) Measuring behaviour in rodents: towards translational
neuropsychiatric research. Behav Brain Res 236 (1):295-306.
doi:10.1016/j.bbr.2012.09.005
Kemppainen S, Jolkkonen E, Pitkanen A (2002) Projections from the posterior
cortical nucleus of the amygdala to the hippocampal formation and
parahippocampal region in rat. Hippocampus 12 (6):735-755.
doi:10.1002/hipo.10020
Kheirbek MA, Klemenhagen KC, Sahay A, Hen R (2012) Neurogenesis and
generalization: a new approach to stratify and treat anxiety disorders. Nat
Neurosci 15 (12):1613-1620. doi:10.1038/nn.3262
Lehmann ML, Brachman RA, Martinowich K, Schloesser RJ, Herkenham M (2013)
Glucocorticoids orchestrate divergent effects on mood through adult
neurogenesis. The Journal of neuroscience : the official journal of the Society
for Neuroscience 33 (7):2961-2972. doi:10.1523/JNEUROSCI.3878-12.2013
Matar MA, Zohar J, Cohen H (2013) Translationally relevant modeling of PTSD in
rodents. Cell and tissue research. doi:10.1007/s00441-013-1687-6
McEwen BS (1999) Stress and hippocampal plasticity. Annual review of
neuroscience 22:105-122. doi:10.1146/annurev.neuro.22.1.105
McEwen BS, Magarinos AM (2001) Stress and hippocampal plasticity: implications
for the pathophysiology of affective disorders. Human psychopharmacology 16
(S1):S7-S19. doi:10.1002/hup.266
Nacher J, Crespo C, Mcewen BS (2001) Doublecortin expression in the adult rat
telencephalon. European Journal of Neuroscience 14: 629–644
Quirk G, Mueller D (2007) Neural mechanisms of extinction learning and retrieval.
Neuropsychopharmacology 33 (1):56-72
Quirk GJ, Pare D, Richardson R, Herry C, Monfils MH, Schiller D, Vicentic A (2010)
Erasing fear memories with extinction training. The Journal of neuroscience :

the official journal of the Society for Neuroscience 30 (45):14993-14997.
doi:10.1523/JNEUROSCI.4268-10.2010
Rauch SL, Milad MR, Orr SP, Quinn BT, Fischl B, Pitman RK (2005) Orbitofrontal
thickness, retention of fear extinction, and extraversion. Neuroreport 16
(17):1909-1912
Rauch SL, Shin LM, Phelps EA (2006) Neurocircuitry models of posttraumatic stress
disorder and extinction: human neuroimaging research--past, present, and
future. Biol Psychiatry 60 (4):376-382. doi:10.1016/j.biopsych.2006.06.004
Schloesser RJ, Lehmann M, Martinowich K, Manji HK, Herkenham M (2010)
Environmental enrichment requires adult neurogenesis to facilitate the
recovery from psychosocial stress. Mol Psychiatry 15 (12):1152-1163.
doi:10.1038/mp.2010.34
Treit D (1985) Animal models for the study of anti-anxiety agents: a review.
Neuroscience and biobehavioral reviews 9 (2):203-222
Wang ME, Fraize NP, Yin L, Yuan RK, Petsagourakis D, Wann EG, Muzzio IA (2013)
Differential roles of the dorsal and ventral hippocampus in predator odor
contextual fear conditioning. Hippocampus 23 (6):451-466.
doi:10.1002/hipo.22105





























