Embed Size (px)
Citation preview

CECÍLIA OLIVEIRA DE AZEVEDO
A FAMÍLIA ORCHIDACEAE NO PARQUE MUNICIPAL DE
MUCUGÊ, BAHIA, BRASIL
Feira de Santana, Bahia
Fevereiro - 2004

A FAMÍLIA
ORIE
CO-ORIENTA
UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA
DEPARTAMENTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
ORCHIDACEAE NO PARQUE MUNICIPAL DE
MUCUGÊ, BAHIA, BRASIL
CECÍLIA OLIVEIRA DE AZEVEDO
Dissertação apresentada ao Programa de Pós-
Graduação em Botânica da Universidade
Estadual de Feira de Santana como requisito
parcial para obtenção do grau de Mestre em
Botânica.
NTADOR: PROF. DR. CÁSSIO VAN DEN BERG (UEFS)
DOR: PROF. DR. ANTONIO L. V. TOSCANO DE BRITO (UEFS)
Feira de Santana, Bahia
Fevereiro - 2004

Banca Examinadora
_________________________________________________________________
Prof. Dr.
_________________________________________________________________
Prof. Dr.
_________________________________________________________________
Prof. Dr. Cássio van den Berg
Orientador e Presidente da Banca
Feira de Santana - Bahia
Fevereiro – 2004

AGRADECIMENTOS
A todos do Parque Municipal de Mucugê: Oremildes, Euvaldo, Joaab, Elaine, Ilza,
Gilvan, Baúla, Nilton e Adriano por terem me recebido com tanto carinho, sempre
cuidando para que nada me faltasse, pela ajuda e atenção.
Aos meus orientadores Cássio van den Berg e Toscano de Brito por todos os
ensinamentos. Ao Professor Eduardo Borba por ter lido e corrigido cuidadosamente as
descrições das espécies. Obrigada pelas sugestões e comentários críticos.
Aos colegas de curso: Alexa, Adilva, Carlianne, Cosme, Elisângela, Êlvia, Eric,
Hilder, Jomar, Lia, Moema, Paty, Rosineide, Silvana e Viviane pelos momentos passados
juntos, a Jorge, Cris e Tati, grandes amigos, pelos momentos compartilhados desde a época
da graduação.
Aos curadores dos Herbários ALCB, CEPEC, HRB pelo empréstimo de material. A
todos do HUEFS, especialmente a Téo, Kelly, Elaine e Zezé. A dona Vanilda e o pessoal
do laboratório de Taxonomia Vegetal. A Adriana e Dione pela preocupação e empenho. A
Ivan pela grande ajuda nas ilustrações.
A Rogério por me iniciar no mundo da pesquisa e das orquídeas, pelos
ensinamentos em campo nas muitas viagens. A Maria Clara pelo apoio e incentivo sempre,
desde a orientação na Iniciação Científica e sobretudo pela amizade.
Ao Dr. Fábio de Barros por disponibilizar seu artigo da Flora de Grão-Mogol, ainda
inédito.
Aos Professores Luciano e Flávio pela paciência no estágio em docência. Aos
demais professores, colegas e técnicos com quem convivi durante esses dois anos de UEFS.
Ao prof. Caio Graco pelas caronas a Mucugê, e a todos do LORMA pela companhia nas
viagens ao Parque.
A Universidade Estadual de Feira de Santana, a Fundação de Amparo à Pesquisa da
Bahia pela bolsa de pesquisa, e a CAPES e CADCT pelo financiamento do curso.
A minha família, especialmente minha mãe e meu pai, exemplos de força de
vontade e dedicação, a minha irmã pela amizade e constante ajuda e a Peri pelo
companheirismo e paciência.

ÌNDICE
AGRADECIMENTOS
ÌNDICE DE FIGURAS .......................................................................................................... 6
INTRODUÇÃO GERAL ....................................................................................................... 8
CAPÍTULO 1 - A FAMÍLIA ORCHIDACEAE NO PARQUE MUNICIPAL DE
MUCUGÊ, BAHIA, BRASIL.............................................................................................. 10
CAPÍTULO 2 - ANÁLISE COMPARATIVA DA FAMÍLIA ORCHIDACEAE EM
ÁREAS DE CAMPO RUPESTRE DA CADEIA DO ESPINHAÇO, BAHIA E MINAS
GERAIS, BRASIL ............................................................................................................... 97
CAPÍTULO 3 - A NEW COMBINATION IN THE GENUS ANATHALLIS
(ORCHIDACEAE), AND A NEW RECORD FOR BAHIA STATE, BRAZIL................ 112
CONCLUSÃO GERAL ..................................................................................................... 116
REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................... 117
APÊNDICE I - ÍNDICE DOS ESPÉCIMES EXAMINADOS: COLETOR, SEGUIDO
PELO NÚMERO DE COLETA E PELA NUMERAÇÃO DO TÁXON.......................... 124
APÊNDICE II - DISTRIBUIÇÃO DAS ESPÉCIES DE ORCHIDACEAE NAS ÁREAS
COMPARADAS, UTILIZADA PARA ANÁLISE DE SIMILARIDADE....................... 126
APÊNDICE III - ESPÉCIMES EXAMINADOS REFERENTES AO MATERIAL
UTILIZADO PARA A ANÁLISE DE SIMILARIDADE DEPOSITADAS NOS
HERBÁRIOS ALCB, CEPEC, HRB E HUEFS ................................................................ 130
APÊNDICE IV - ESPÉCIMES EXAMINADOS QUE SOFRERAM ALTERAÇÕES.... 134
RESUMO ........................................................................................................................... 135
ABSTRACT ....................................................................................................................... 136

ÌNDICE DE FIGURAS
CAPÍTULO 1
Figura I: Ilustração de Acianthera hamosa e Acianthera ochreata subsp. ochreata. .......... 79
Figura II: Ilustração de Anathallis microphyta e Anathallis montipelladensis. ................... 80
Figura III: Ilustração de Bulbophyllum ×cipoense, Bulbophyllum cribbianum, Bulbophyllum
involutum e Bulbophyllum weddellii. ........................................................................... 81
Figura IV: Ilustração de Brassavola tuberculata e Campylocentrum micranthum.............. 82
Figura V: Ilustração de Cattleya elongata e Cleistes exilis.................................................. 83
Figura VI: Ilustração de Cyrtopodium edmundoi e Cyrtopodium polyphyllum. .................. 84
Figura VII: Ilustração de Epidendrum cristatum, Epidendrum orchidiflorum, Epidendrum
secundum e Epidendrum warasii.................................................................................. 85
Figura VIII: Ilustração de Encyclia alboxanthina e Epistephium lucidum. ......................... 86
Figura IX: Ilustração de Habenaria fluminensis e Maxillaria cerifera................................ 87
Figura X: Ilustração de Octomeria alexandrii, Octomeria flabellifera e Octomeria stellaris.
...................................................................................................................................... 88
Figura XI: Ilustração de Oncidium blanchetii e Polystachya micrantha. ............................ 89
Figura XII: Ilustração de Prescottia leptostachya, Prescottia montana e Prescottia
stachyodes..................................................................................................................... 90
Figura XIII: Ilustração de Prosthechea moojenii. ................................................................ 91
Figura XIV: Ilustração de Scaphyglottis cf. modesta e Sobralia sessilis. ............................ 92
Figura XV: Ilustração de Sophronitis bahiensis e Thelyschista ghillanyi............................ 93
Figura XVI: Fotografia de A. Acianthera hamosa, B. Acianthera ochreata subsp. ochreata,
C-D. Anathallis microphyta, E-F. Anathallis montipelladensis, G. Brassavola
tuberculata, H-I. Bulbophyllum ×cipoense, J. Bulbophyllum involutum, K.
Bulbophyllum weddellii e L. Campylocentrum micranthum. ....................................... 94
Figura XVII: Fotografia de A. Cattleya elongata, B. Cleistes exilis, C. Cyrtopodium
edmundoi, D. Cyrtopodium polyphyllum, E. Encyclia alboxanthina, F. Epidendrum
cristatum, G. Epidendrum orchidiflorum, H. Epidendrum secundum, I. Epidendrum
warasii, J. Epistephium lucidum, K. Habenaria fluminensis e L. Maxillaria cerifera.95
Figura XVIII: Fotografia de A. Octomeria alexandrii, B. Octomeria flabellifera, C.
Octomeria stellaris, D. Oncidium blanchetii, E. Polystachya micrantha, F. Prescottia

montana, G-H. Prescottia stachyodes, I. Prosthechea moojenii, J. Sobralia sessilis, K.
Sophronitis bahiensis e L-M. Thelyschista ghillanyi. .................................................. 96
CAPÍTULO 2
Figura I. Dendrograma mostrando a similaridade entre áreas de campo rupestre da Bahia e
Minas Gerais, utilizando coeficiente de Jaccard com UPGMA. ................................ 106
Figura II. Dendrograma mostrando a similaridade entre áreas de campo rupestre da Bahia e
Minas Gerais, utilizando coeficiente de Dice com UPGMA. .................................... 106

8
INTRODUÇÃO GERAL
A família Orchidaceae é uma das mais numerosas e especializadas do Reino
Vegetal, com cerca de 20.000 espécies distribuídas em aproximadamente 850 gêneros
(Atwood, 1986; Dressler, 1993), encontradas em praticamente todos os continentes, com
predominância nas regiões tropicais e subtropicais (Dressler, 1993). As Orchidaceae estão
bem representadas no Brasil, com aproximadamente 2.350 espécies distribuídas em 200
gêneros (Pabst & Dungs, 1975, 1977). Na Bahia foram registradas 285 espécies (Pabst &
Dungs, 1975, 1977; Harley & Mayo, 1980; Harley & Simmons, 1986; Toscano de Brito,
1995; Toscano de Brito, 1998; Toscano de Brito & Queiroz, 2003).
Estudos recentes têm demonstrado que a melhor estratégia de investigação da
dinâmica e distribuição geográfica das espécies dos campos rupestres da Cadeia do
Espinhaço é a realização de inventários florísticos detalhados de áreas reduzidas,
escolhidas devido à sua alta diversidade e estado de conservação (Harley, 1995). Seguindo
essa estratégia, inventários foram executados em várias áreas da Chapada Diamantina
(Harley & Simmons, 1986; Toscano de Brito, 1995; Toscano de Brito, 1998; Toscano de
Brito & Queiroz, 2003). Estes servem não somente para propiciar um melhor
conhecimento sobre a composição florística dos campos rupestres, mas também fornecem
subsídios importantes para uma análise comparativa da flora desse ecossistema presente
tanto na porção baiana, como na porção mineira da Cadeia do Espinhaço.
Como forma de contribuir para o conhecimento desta área o presente trabalho teve
como objetivo estudar as espécies da família Orchidaceae do Parque Municipal de
Mucugê, localizado no município de Mucugê, Chapada Diamantina, integrante da Cadeia
do Espinhaço. Além de comparar levantamentos florísticos realizados em outras áreas de
campo rupestre da Bahia e de Minas Gerais.
A Cadeia do Espinhaço, constituída por dois blocos principais, a porção mineira, e
a baiana, conhecida como Chapada Diamantina, estende-se da Serra de Jacobina, ao norte,
atingindo a Serra de Ouro Branco, próximo a Ouro Preto, ao sul. No seu extremo sul, a
Chapada Diamantina divide-se em duas cadeias independentes, a Serra do Rio de Contas e
a Serra das Almas, a oeste, e a Serra do Sincorá, a leste (Harley, 1995). Em Minas Gerais a
Serra de Grão-Mogol localiza-se em posição intermediária e disjunta entre as serras de
Minas Gerais e Bahia (Pirani et al., 2003). Na porção sul da Cadeia do Espinhaço está
situada a Serra do Cipó (Giulietti et al., 1987).
Inventários florísticos desenvolvidos na Cadeia do Espinhaço revelaram a presença

9
de 139 espécies da família para a Chapada Diamantina (Harley & Simmons, 1986;
Toscano de Brito, 1995; Toscano de Brito, 1998; Toscano de Brito & Queiroz, 2003),
enquanto que estudos feitos para a Serra do Cipó (Barros, 1987) e para a Serra de Grão-
Mogol (Barros & Pinheiro, no prelo) mostraram a presença de cerca de 105 espécies de
orquídeas para esta região.
A fisionomia vegetal predominante na Cadeia do Espinhaço é o campo rupestre,
que ocorre principalmente em regiões acima de 1.000 m de altitude, podendo ser
encontrado a partir de 700 m (Giulietti et al., 1987). É um importante centro de diversidade
da flora brasileira, destacando-se por apresentar um grande número de espécies endêmicas
(Harley & Simmons, 1986; Harley, 1995). Ocorre principalmente nos estados da Bahia e
Minas Gerais. Longe de formar um tipo de vegetação homogênea, o campo rupestre é um
conjunto de comunidades predominantemente herbáceo-arbustivas, floristicamente
relacionadas, mas fisionomicamente distintas em função da topografia, natureza do
substrato, profundidade do solo e do microclima (Giulietti et al., 1996). Nas encostas e
topos de morros, onde predominam afloramentos rochosos, a diversificação florística e a
multiplicidade de microambientes atingem grau máximo, onde as famílias Velloziaceae,
Orchidaceae, Bromeliaceae e Eriocaulaceae apresentam maior destaque (Pinto et al.,
1990).
A cidade de Mucugê, sede municipal, leva o nome do rio que a cerca, que, por sua
vez, se refere a uma fruta utilizada como alimento pelos índios, outrora nativos da região.
Fundada em 1844, época em que foram descobertos diamantes nos rios Mucugê e
Cumbuca (Moraes, 1991), é uma das cidades mais antigas da Chapada Diamantina, tendo
preservado grande parte do seu conjunto arquitetônico e paisagístico, tombado pelo
Instituto do Patrimônio Histórico e Artístico Nacional - IPHAN (IPAC, 1980).
O Parque Municipal de Mucugê, localizado no município de Mucugê, teve origem a
partir de um projeto denominado Sempre Viva, executado entre 1996 e 1998, que previu a
implantação de uma Unidade de Conservação, criando a Unidade de Manejo Sustentável
de Mucugê, com área de 270 ha. Em 15 de março de 1999, a Prefeitura de Mucugê criou o
Parque Municipal de Mucugê, incorporando uma área, situada em seu entorno, de 180 ha,
elevando sua área total para 450 ha (Stradmann, 1998). Seus principais objetivos são a
conservação, reprodução e utilização racional de espécies popularmente conhecidas como
sempre-vivas, algumas ameaçadas de extinção, a exemplo de Syngonanthus mucugensis
(Eriocaulaceae), espécie endêmica dos arredores de Mucugê (Giulietti, 1996).

10
CAPÍTULO 1 - A FAMÍLIA ORCHIDACEAE NO PARQUE MUNICIPAL DE
MUCUGÊ, BAHIA, BRASIL
Este capítulo é um artigo expandido, formatado para publicação na revista Sitientibus série
Ciências Biológicas.

11
RESUMO
O levantamento da família Orchidaceae no Parque Municipal de Mucugê foi realizado
através de coletas mensais durante o período de um ano. Foram encontradas 35 espécies
em 22 gêneros. Destas, 11 são citadas pela primeira vez para o município de Mucugê; três
para a região da Chapada Diamantina e sete para o estado da Bahia. Os gêneros
Bulbophyllum, Epidendrum, Octomeria e Prescottia foram os mais representativos.
Lectótipos foram selecionados para Acianthera hamosa, Cattleya elongata, Maxillaria
cerifera, Octomeria stellaris e Prescottia montana. Todos os táxons são descritos e
ilustrados e chaves para identificação dos gêneros e espécies são apresentadas.
Palavras-chave: Chapada Diamantina, Orchidaceae, Orquídea, Parque Municipal de
Mucugê, Taxonomia.
ABSTRACT
This study presents a survey of the Orchidaceae occurring in the Parque Municipal de
Mucugê, Mucugê, Bahia, Brazil. It was carried out trough monthly random sampling
during 12 months. 35 species of orchids were recorded in 22 genera, of which 11 species
are for the first time reported for Mucugê; three for the Chapada Diamantina and seven for
the state of Bahia. The main genera in number of species are Bulbophyllum, Epidendrum,
Octomeria and Prescottia. Lectotypes for Acianthera hamosa, Cattleya elongata,
Maxillaria cerifera, Octomeria stellaris e Prescottia montana are chosen here. All taxa are
described and illustrated, as well as identification keys for species and genera are provided.
Keywords: Chapada Diamantina, Orchidaceae, Orchids, Parque Municipal de Mucugê,
Taxonomy.

12
INTRODUÇÃO
Com cerca de 2.350 espécies distribuídas em 200 gêneros (Pabst & Dungs, 1975,
1977), a família Orchidaceae encontra-se bem representada no Brasil. Estudos sobre a flora
baiana indicaram a presença de cerca de 285 espécies de orquídeas no estado (Pabst &
Dungs, 1975, 1977; Harley & Mayo, 1980; Harley & Simmons, 1986; Toscano de Brito,
1995; Toscano de Brito, 1998; Toscano de Brito & Queiroz, 2003).
A Chapada Diamantina, setor baiano da Cadeia do Espinhaço, apresenta uma
grande diversidade florística. Levantamentos realizados na área detectaram, até o
momento, 139 espécies de orquídeas nessa região. Catolés apresentou 76 espécies
(Toscano de Brito & Queiroz, 2003), o Pico das Almas, 45 espécies (Toscano de Brito,
1995); a Serra da Chapadinha possui 27 espécies (Toscano de Brito, 1998);
Andaraí/Mucugê, 26 espécies (Harley & Simmons, 1986) e o Morro do Pai Inácio
apresentou 18 espécies (Toscano de Brito, 1998). Além disso, monografias e dissertações
realizadas na Chapada Diamantina revelaram a presença de oito espécies do gênero
Oncidium (Silva, 1999), nove espécies do gênero Cleistes (Silva, 2002) e 21 espécies da
subtribo Spiranthinae (Smidt, 2003).
O Parque Municipal de Mucugê, objeto desta pesquisa, localiza-se no município de
Mucugê, nas imediações do Parque Nacional da Chapada Diamantina, na Serra do Sincorá,
a cerca de 4 km da cidade de Mucugê, com coordenadas 12o 59’ 02”-13o 00’ 18”S e 41o 19’
40”-41o 21’ 33”W, a uma altitude em torno de 1.000 m acima do nível do mar e área de 4,5
km2. A vegetação predominante na área é de campo rupestre, que é um importante centro
de diversidade da flora brasileira, destacando-se por apresentar um grande número de
espécies endêmicas (Harley & Simmons, 1986; Harley, 1995), onde Orchidaceae apresenta
uma grande importância florística, estando sempre entre as dez famílias de maior riqueza
específica (Harley & Simmons, 1986; Giulietti et al., 1987; Stannard, 1995; Guedes &
Orge, 1998; Pirani et al., 2003; Zappi et al., 2003).
O presente trabalho teve como objetivo inventariar, descrever e ilustrar as espécies
da família Orchidaceae do Parque Municipal de Mucugê; apresentar comentários sobre a
morfologia, hábito, aspectos fenológicos e distribuição geográfica das várias espécies dessa
família presentes na área de estudo; elaborar chaves para identificação dos gêneros e
espécies de Orchidaceae presentes no Parque; contribuir para o conhecimento da flora da
Bahia e para o processo de aprimoramento da gestão dos recursos naturais do Parque
Municipal de Mucugê.

13
MATERIAL E MÉTODOS
Excursões mensais à área foram realizadas durante o período de junho de 2002 a
maio de 2003, visando a coleta de amostras vegetais de forma aleatória, em toda a área do
Parque. O material coletado foi fotografado, desidratado e herborizado de acordo com
Mori et al. (1989), além da conservação de suas flores em solução alcoólica (álcool 70%:
água 25%: glicerina 5%). As exsicatas foram depositadas no acervo do herbário da
Universidade Estadual de Feira de Santana (HUEFS), e as duplicatas serão enviadas para a
coleção do Parque Municipal de Mucugê. Os dados apresentados neste trabalho foram
baseados em revisão bibliográfica e análise morfológica dos espécimes coletados e das
coleções depositadas nos seguintes herbários: ALCB, CEPEC, HRB e HUEFS (acrônimos
de acordo com Holmgren et al., 1990).
As descrições morfológicas foram realizadas através da análise das porções
vegetativas e florais dos vários espécimes. Flores conservadas em solução alcoólica ou
retiradas das exsicatas, foram examinadas utilizando-se um microscópio estereoscópico,
com o auxílio de uma câmara clara. As medidas das porções florais foram feitas com o
auxílio de régua e paquímetro eletrônico.
Foram elaboradas chaves dicotômicas para a identificação dos gêneros e espécies
presentes na área estudada, utilizando-se principalmente os caracteres mais evidentes para
a delimitação dos táxons, sem levar em consideração suas afinidades filogenéticas. A
classificação das Orchidaceae seguiu, em sua maior parte, o sistema de Dressler (1993),
porém informações contidas em estudos recentes, baseados em análises de DNA (e.g.
Cameron et al. 1999; Cameron & Chase, 2000; Pridgeon, Solano & Chase, 2001; van den
Berg et al., 2000), também foram utilizadas.
A determinação das espécies seguiu os padrões de taxonomia clássica e foi
realizada através de consulta a obras especializadas, entre elas Barbosa Rodrigues (1877,
1882), Cogniaux (1893-1896, 1898-1902, 1904-1906), Hoehne (1940, 1942, 1945, 1953),
Pabst & Dungs (1975, 1977), Sprunger et al. (1996) e Toscano de Brito (1995), assim
como através de consulta de obras príncipes dos vários táxons. Nomes dos autores das
espécies estão abreviados de acordo com Brummitt & Powell (1992), periódicos
encontram-se abreviados de acordo com Bridson e Smith (1991) e outras obras de acordo
com Stafleu & Cowan (1976-1978).
O material examinado de cada táxon encontra-se listado em ordem alfabética dos
coletores e número de coleta, além de um índice destes espécimes em ordem alfabética de

14
coletor seguido pelo número de coleta e pela numeração de cada táxon (Apêndice I).
As ilustrações foram baseadas em material conservado em líquido, além do material
herborizado. A origem desse material encontra-se indicada nas legendas dos respectivos
táxons. Para os gêneros com mais de uma espécie, apenas um hábito foi ilustrado, e
diagramas florais são apresentados para todas as espécies. Gêneros com uma única espécie
foram ilustrados aos pares, respeitando apenas ordem alfabética.
Dados sobre distribuição geográfica e fenologia foram obtidos de informações
contidas nas etiquetas dos espécimes herborizados examinados e complementados com
trabalhos de campo. Dados sobre a distribuição geral de cada táxon foram obtidos através
de consulta à bibliografia especializada, sobretudo Pabst & Dungs (1975, 1977) e Sprunger
et al. (1996). Sinônimos só foram apresentados para casos recentes, que não foram citados
anteriormente por Pabst & Dungs (1975, 1977), ou para casos duvidosos que ainda hoje
apresentam problemas.
Algumas coleções estão referidas pelas seguintes siglas: Coleção Flora dos Campos
Rupestres (Universidade de São Paulo) - CFCR, Projeto Chapada Diamantina
(Universidade Federal da Bahia) - PCD e Unidade de Manejo Sustentável de Mucugê -
UMS. É importante salientar que as denominações Projeto Sempre-Viva, Unidade de
Manejo Sustentável de Mucugê e Parque Municipal de Mucugê, se referem à mesma área,
objeto de estudo deste capítulo.
RESULTADOS E DISCUSSÃO
Foram encontradas no Parque Municipal de Mucugê 35 espécies de orquídeas
(Quadro I), distribuídas em 22 gêneros, das quais uma é um híbrido natural. Os gêneros de
maior representatividade foram Bulbophyllum, Epidendrum, Octomeria e Prescottia,
enquanto 68% dos gêneros apresentaram apenas uma espécie.
Algumas espécies de Orchidaceae encontradas no Parque Municipal de Mucugê
possuem ampla distribuição geográfica, podendo ser encontradas em outros países, e
apresentando-se bem distribuídas no Brasil. Das 35 espécies coletadas no Parque, sete são
novos registros para o estado da Bahia, anteriormente citadas para outras regiões,
sobretudo Sul e Sudeste do país. Três são novos registros para a região da Chapada
Diamantina e outras 11 são novos registros para o município de Mucugê. É importante
ressaltar que a área apresenta espécies como Encyclia alboxanthina, Sophronitis bahiensis
e Thelyschista ghillanyi, que são endêmicas da Chapada Diamantina.

15
O Parque Municipal de Mucugê está compreendido na área onde foi realizada a
Flórula de Mucugê (Harley & Simmons, 1986). Esta envolveu partes dos municípios de
Andaraí e Mucugê, abrangendo 896 km2 (Harley & Simmons, 1986). Nela foram
encontradas 26 espécies da família Orchidaceae, distribuídas em 13 gêneros. Destas,
somente 10 espécies são comuns aos dois levantamentos, enquanto 16 espécies foram
encontradas apenas na Flórula e não no Parque. O número de espécies exclusivas da
Flórula era esperado, devido ao maior tamanho da área amostrada, porém o fato de o
Parque apresentar 26 espécies, que não foram encontradas na Flórula, é bastante
significativo. Este caso representa um exemplo claro de como a diferença na metodologia e
no esforço de amostragem pode influenciar nos resultados, gerando uma falsa impressão da
situação. É importante ressaltar que as Orchidaceae do Parque Municipal de Mucugê foram
inventariadas através de coletas mensais durante o período de um ano, onde os focos
estavam voltados unicamente para esta família.
As espécies encontradas na área de estudo, descritas por Barbosa Rodrigues,
Acianthera hamosa, Cattleya elongata, Maxillaria cerifera, Octomeria stellaris e
Prescottia montana, foram lectotipificadas, uma vez que o holótipo destas espécies, assim
como a quase totalidade da coleção de Barbosa Rodrigues, foram destruídos durante uma
enchente na cidade do Rio de Janeiro (Cribb & Toscano de Brito, 1996). Desta forma, as
ilustrações originais, preparadas por este autor e ilustradas em sua Iconographie des
orchidées du Brésil, recentemente publicada por Sprunger et al. (1996), constituem o único
material original disponível, e por esse motivo foram selecionadas como lectótipos. Dos
seis volumes de ilustração de Barbosa Rodrigues, cinco estão no Rio de Janeiro e o volume
4, está depositado no Herbário Oakes Ames, na Universidade de Harvard.

16
Quadro I. Lista das espécies encontradas no Parque Municipal de Mucugê.
no ESPÉCIE 01 Acianthera hamosa (Barb.Rodr.) Pridgeon & M.W.Chase 02 Acianthera ochreata (Lindl.) Pridgeon & M.W.Chase subsp. ochreata 03 Anathallis microphyta (Barb.Rodr.) C.O.Azevedo & Van den Berg (Capítulo 3) 04 Anathallis montipelladensis (Hoehne) F.Barros 05 Brassavola tuberculata Hook. 06 Bulbophyllum ×cipoense Borba & Semir 07 Bulbophyllum cribbianum Toscano 08 Bulbophyllum involutum Borba, Semir & F.Barros 09 Bulbophyllum weddellii (Lindl.) Rchb.f. 10 Campylocentrum micranthum (Lindl.) Rolfe 11 Cattleya elongata Barb.Rodr 12 Cleistes exilis Hoehne 13 Cyrtopodium edmundoi Pabst 14 Cyrtopodium polyphyllum (Vell.) Pabst ex F.Barros 15 Encyclia alboxanthina Fowlie 16 Epidendrum cristatum Ruiz & Pav. 17 Epidendrum orchidiflorum Salzm. ex Lindl. 18 Epidendrum secundum Jacq. 19 Epidendrum warasii Pabst 20 Epistephium lucidum Cogn. 21 Habenaria fluminensis Hoehne 22 Maxillaria cerifera Barb.Rodr. 23 Octomeria alexandrii Schltr. 24 Octomeria flabellifera Pabst 25 Octomeria stellaris Barb.Rodr. 26 Oncidium blanchetii Rchb.f. 27 Polystachya micrantha Schltr. 28 Prescottia leptostachya Lindl. 29 Prescottia montana Barb.Rodr. 30 Prescottia stachyodes (Sw.) Lindl. 31 Prosthechea moojenii (Pabst) W.E.Higgins 32 Scaphyglottis cf. modesta Schltr. 33 Sobralia sessilis Lindl. 34 Sophronitis bahiensis (Schltr.) Van den Berg & M.W.Chase 35 Thelyschista ghillanyi (Pabst) Garay
Aspectos morfológicos e ecológicos das Orchidaceae ocorrentes no Parque Municipal de
Mucugê
A família Orchidaceae apresenta uma grande variação de tamanho, podendo ser
encontgradas no Parque espécies de 10-50 mm de altura, como Anathallis microphyta e
Anathallis montipelladensis, até espécies com cerca de 1,5 a 2 m de altura, como
Epidendrum cristatum, Epidendrum warasii, Epistephium lucidum e Sobralia sessilis. As

17
orquídeas podem apresentar dois padrões de crescimento, sendo eles simpodial ou
monopodial. Nas simpodiais cada broto tem crescimento limitado e, geralmente, a cada
ano um novo pseudobulbo ou caule surge da base do mesmo. Novos pseudobulbos também
podem crescer do ápice do pseudobulbo anterior, como ocorre em Scaphyglottis modesta.
Nas monopodiais, o caule cresce continuamente a partir de um único broto apical, que tem
crescimento indeterminado. Entre as espécies estudadas, a grande maioria é simpodial,
com exceção do gênero Campylocentrum.
As espécies encontradas no Parque apresentam hábito terrestre, epífito e rupícola,
sendo aí enquadrada a grande maioria. Algumas espécies são terrestres, como Cleistes
exilis, Epistephium lucidum, Habenaria fluminensis, Oncidium blanchetii, Sobralia sessilis
e Thelyschista ghillanyi, o gênero Prescottia, e algumas espécies do gênero Epidendrum,
sendo que apenas Campylocentrum micranthum é epífita. Na área de estudo, as orquídeas
são encontradas nos diferentes ecossistemas dos campos rupestres, crescendo diretamente
sobre os afloramentos rochosos, nas encostas e em matas de grotão, em áreas
caracterizadas por possuírem uma vegetação aberta de baixo porte, dominada por espécies
herbáceas, subarbustivas e arbustivas, crescendo sobre solo arenoso, e em áreas com uma
fisionomia um pouco mais fechada, dominada por arbustos e árvores, onde encontram-se
espécies terrestres crescendo na serrapilheira.
As raízes podem variar muito em sua morfologia, diferindo em sua espessura de
acordo com a espécie. Em muitas terrestres, as raízes tornam-se órgãos de armazenamento
ou tuberóides, em algumas a raiz inteira pode ser engrossada, como em Prescottia e
Thelyschista, em outras, como em Cleistes, alguns segmentos são espessos e o resto mais
fino. Raízes carnosas armazenadoras não estão limitadas a orquídeas terrestres. Epífitas e
rupícolas, como Sobralia e algumas espécies de Epidendrum, podem ter raízes carnosas,
sendo que estas plantas geralmente apresentam caules e folhas finos.
Pseudobulbos são caules espessados, essencialmente armazenadores de água, que
podem sustentar folhas em todo o seu comprimento ou só no ápice. Os pseudobulbos
podem apresentar bainhas, que envolvem sua base e podem sustentar uma folha, como em
Maxillaria cerifera e Oncidium blanchetii.
A maioria das folhas de orquídeas é paralelinérvea. Nervação reticulada é rara em
orquídeas, embora ocorra em alguns gêneros como em Epistephium. A maioria apresenta
filotaxia dística e algumas estão arranjadas em roseta, como nos gêneros Prescottia e
Thelyschista. As folhas podem ser conduplicadas ou plicadas, como em Sobralia.
Geralmente são dorsiventrais e em ambientes secos freqüentemente são coriáceas ou

18
carnosas e nos casos extremos podem ser triangulares ou cilíndricas (teretiformes), como
em algumas espécies de Acianthera e Octomeria. Plantas terrestres com brotos anuais,
como Cleistes, Habenaria, Prescottia e Thelyschista, normalmente apresentam folhas
membranáceas. A forma das folhas é relativamente uniforme. A maioria tem folha simples,
linear, lanceolada, oblonga, ovada, obovada ou elíptica.
Os tipos de inflorescência mais comuns são racemo, panícula e espiga. Algumas
sustentam apenas uma flor como em Maxillaria. A inflorescência pode formar-se de
qualquer parte do caule, aqui se usou a terminologia lateral quando se forma do lado ou na
base do caule/pseudobulbo ou do rizoma, e terminal quando esta é uma continuação do
caule ou desenvolve-se do ápice do pseudobulbo. A grande maioria dos gêneros apresenta
este tipo, embora Bulbophyllum, Campylocentrum, Cyrtopodium, Maxillaria e Oncidium
apresentem inflorescência lateral. Campylocentrum apresenta inflorescência saindo ao
longo do caule, e os outros gêneros supracitados apresentam inflorescências produzidas da
base do pseudobulbo ou rizoma.
As flores apresentam brácteas, que podem ser conspícuas e coloridas, como em
Cyrtopodium, ou inconspícuas e paleáceas. Os botões florais podem estar envolvidos por
uma ou duas grandes bainhas, chamadas de espata, que suportam e protegem as flores
enquanto estas se desenvolvem, podendo ser observadas em Cattleya elongata,
Prosthechea moojenii e Sophronitis bahiensis. Neste trabalho, usou-se a seguinte
terminologia para a inflorescência: escapo, que começa da base da inflorescência até a
primeira bráctea floral, e raque, que tem início na primeira bráctea floral até a última flor
ou botão floral.
Um aspecto interessante no desenvolvimento da maioria das flores de orquídeas é o
fenômeno da ressupinação. No botão, o labelo está normalmente voltado para cima, mas na
maioria das flores, este está voltado para baixo, pois o pedicelo sofre uma torção de 180o
durante o desenvolvimento do botão. O termo ressupinado é usado para a flor que
apresente o labelo voltado para baixo, em alguns casos a flor é ressupinada simplesmente
pela posição pendente da inflorescência. No Parque, os gêneros Polystachya, Prescottia,
Prosthechea e Epidendrum secundum, apresentam flores não ressupinadas. Alguns
gêneros, como Epistephium, apresentam uma estrutura de origem desconhecida na base do
perianto, chamada de calículo ou epicálice.
As flores de orquídeas são trímeras, o cálice é formado por uma sépala dorsal e
duas laterais, que podem estar parcialmente ou completamente unidas, como nos gêneros
Acianthera e Anathallis, que apresentam diferentes graus de concrescimento. A corola é

19
formada por três pétalas que são geralmente mais finas que as sépalas. Elas podem estar
adnadas à sépala dorsal, como em Thelyschista ghillanyi, ser bastante reduzidas, como em
Bulbophyllum cribbianum, ou partidas, como em Habenaria fluminensis. A pétala mediana
é geralmente diferente das outras duas e é chamada de labelo. O labelo é normalmente
maior e mais complexo que as outras pétalas e é um dos principais elementos diagnósticos
de uma orquídea. O labelo pode estar unido em diferentes graus à coluna, como nos
gêneros Encyclia e Epidendrum, sendo geralmente trilobado e podendo apresentar calos
variados.
Suas flores podem apresentar nectários florais a partir da extensão de um de seus
segmentos, geralmente o labelo, chamados de esporão ou cálcar, como é o caso de
Campylocentrum e Habenaria. A base da coluna forma, em muitas orquídeas, uma
extensão ventral a qual o labelo está ligado, chamado de pé de coluna. Em plantas com pés
de coluna longos, a base das sépalas laterais geralmente está unida ao mesmo, formando
um mento.
Taxonomia
Chave de identificação para os gêneros de Orchidaceae ocorrentes no Parque Municipal de
Mucugê.
1. Folhas em roseta.
2. Flores não ressupinadas, verdes, com labelo verde e cuculado. ..........................................
......................................................................................................................... Prescottia (17)
2’. Flores ressupinadas, verdes, com labelo branco e oblongo. ...............................................
..................................................................................................................... Thelyschista (22)
1’. Folhas dísticas.
3. Pseudobulbo ou caule com folhas apenas no ápice.
4. Inflorescência terminal.
5. Pseudobulbos sobrepostos, emergindo da base e do ápice do pseudobulbo anterior. ...
.............................................................................................................. Scaphyglottis (19)
5’. Pseudobulbos ou caule não sobrepostos, emergindo apenas da base do
pseudobulbo anterior.
6. Labelo livre.
7. Inflorescência fasciculada. ............................................................. Octomeria (14)

20
7’. Inflorescência não fasciculada.
8. Antera com 8 polínias. ............................................................... Sophronitis (21)
8’. Antera com 2 a 4 polínias.
9. Sépalas laterais livres.
10. Pseudobulbos 10-20 mm comp., ovóides, flores amarelo-esverdeadas,
antera com 2 polínias. ............................................................. Polystachya (16)
10’. Pseudobulbos 10-32 cm comp., cilíndricos, flores vermelho-
amarronzadas, labelo magenta, antera com 4 polínias. ................ Cattleya (06)
9’. Sépalas laterais concrescidas ao menos na base.
11. Plantas 60-170 mm alt., inflorescência com c. 5-9 flores, sépalas laterais
concrescidas da base até no mínimo 2/3 do comprimento. ................................
.................................................................................................. Acianthera (01)
11’. Plantas 10-50 mm alt., inflorescência com 1-2 flores, sépalas laterais
concrescidas na base até 1/3 do comprimento. .......................... Anathallis (02)
6’. Labelo concrescido em alguma extensão da coluna.
12. Caule cilíndrico, 1-foliado, folha terete, polínias 8. ..............................................
........................................................................................................... Brassavola (03)
12’. Pseudobulbo ovóide, 2-4-foliado, folha lanceolada a oblonga, polínias 4.
13. Inflorescência 110-130 cm comp., 8-30 flores ressupinadas. .............................
............................................................................................................ Encyclia (09)
13’. Inflorescência 47-68 mm comp., 4-6 flores não ressupinadas. .........................
...................................................................................................... Prosthechea (18)
4’. Inflorescência lateral.
14. Pseudobulbo angulado. ................................................................. Bulbophyllum (04)
14’. Pseudobulbo não angulado.
15. Flores amarelas com máculas marrons, labelo marcadamente 3-lobado, lobos
laterais 4,5-5,0 mm comp., polínias 2. ................................................... Oncidium (15)
15’. Flores esverdeadas com nuances vinácea no ápice, labelo obscuramente 3-
lobado, lobos laterais inconspícuos 1,5 mm comp., polínias 4. ........... Maxillaria (13)
3’. Pseudobulbo ou caule com folhas ao longo de sua extensão.
16. Plantas de crescimento monopodial. .......................................... Campylocentrum (05)
16’. Plantas de crescimento simpodial.
17. Planta com pseudobulbo fusiforme, inflorescência lateral. ........... Cyrtopodium (08)
17’. Plantas com caule cilíndrico, inflorescência terminal.

21
18. Flores esverdeadas, labelo tripartido, base do labelo prolongada em cálcar,
antera completamente fundida com o resto da coluna. ...................... Habenaria (12)
18’. Flores coloridas (amarelas, marrom-esverdeadas, róseas ou purpúreas),
labelo inteiro a 3-lobado, base do labelo não prolongada em cálcar, antera
claramente distinguível do resto da coluna.
19. Labelo livre.
20. Folhas 7-12 cm comp., plicadas, coriáceas, labelo 1-lobado. .........................
......................................................................................................... Sobralia (20)
20’. Folhas 1-2 cm comp., não plicadas, membranáceas, labelo distintamente
3-lobado. .......................................................................................... Cleistes (07)
19’. Labelo concrescido em alguma extensão com a coluna.
21. Folhas de nervação reticulada, presença de epicálice, labelo concrescido
com a porção lateral da coluna. ............................................. Epistephium (11)
21’. Folhas de nervação não reticulada, sem epicálice, labelo concrescido à
face adaxial da coluna. ........................................................... Epidendrum (10)

22
01. ACIANTHERA (Scheidw.) C.A.Luer
Recentemente, a subtribo Pleurothallidinae sofreu uma série de mudanças
nomenclaturais (Pridgeon & Chase, 2001), baseadas em análise filogenética molecular
(Pridgeon, Solano & Chase, 2001). Dentre elas o gênero Anathallis Barb.Rodr. foi
restabelecido; o subgênero Acianthera foi elevado para gênero; três novos gêneros foram
estabelecidos, Anthereon, Diodonopsis e Echinella; e outros três, Phloeophila,
Pleurothallopsis e Stelis, sofreram redefinições e expansões. Espécies brasileiras,
principalmente do gênero Pleurothallis R. Br. sensu lato, foram transferidas para outros
gêneros (Pridgeon & Chase, 2001). Posteriormente, Barros (2002) apresentou outras
alterações nomenclaturais para os gêneros Acianthera, Anathallis, Anthereon (= Pabstiella)
e Specklinia.
Chave de identificação para as espécies de Acianthera ocorrentes no Parque Municipal de
Mucugê.
1. Caule compresso no ápice, folha patente, ovada, comprimento c. 2 vezes a largura,
verde a arroxeada, flores vinácea, sépalas laterais concrescidas da base até 2/3 do
comprimento. ................................................................................ Acianthera hamosa (01.1)
1’. Caule não compresso no ápice, folha ereta, lanceolada, comprimento 9-10 vezes a
largura, amarelo-alaranjada, flores amarelo-alaranjadas, sépalas laterais concrescidas da
base ao ápice formando uma navícula. ............. Acianthera ochreata subsp. ochreata (01.2)
01.1. Acianthera hamosa (Barb.Rodr.) Pridgeon & M.W.Chase, Lindleyana 16(4): 243,
2001.
Pleurothallis hamosa Barb.Rodr., Gen. spec. Orchid. 2: 31, t.774, 1882. Tipo: Brasil,
Barbosa Rodrigues s.n. (perdido). Lectótipo aqui designado: ilustração original
preparada por João Barbosa Rodrigues, t.774, que consta de sua Iconographie des
orchidées du Brésil, depositada na biblioteca do Jardim Botânico do Rio de Janeiro,
reproduzida por Sprunger et al. (1996) sob o número t.177B do volume 3.
Erva rupícola. Rizoma 5,0-10 mm comp. entre pseudobulbos, raízes emergindo ao longo
de todo o rizoma. Caule 30-120 x 1,5-2,0 mm, 1-foliado, ereto, cilíndrico, compresso no
ápice, verde a arroxeado, coberto por bainhas. Folha 35-60 x 15-30 mm, séssil, patente,

23
ovada, verde a arroxeada, coriácea a carnosa, ápice agudo apiculado, base cordada.
Inflorescência racemo terminal, adpressa à folha; escapo 4,5-7,0 mm comp.; brácteas
escapais 1,5-2,0 x 1,0-1,5 mm, ovadas, membranáceas, ápice agudo apiculado, base
amplexicaule; raque 13-15 mm comp., 5-8 flores; brácteas florais 1,5-2,0 x 1,5-2,0 mm,
ovadas, membranáceas, ápice agudo apiculado, base amplexicaule. Flores ressupinadas;
pedicelo incluindo o ovário 2,5-3,0 mm comp. Sépalas vinácea, membranáceas, papilosas,
ápice agudo, base truncada; dorsal 4,0-4,5 x 1,5 mm, ereta, fundida na base com as sépalas
laterais, lanceolada; laterais 4,5-5,0 x 1,0-1,5 mm, eretas, concrescidas da base até 2/3 do
comprimento formando uma navícula. Pétalas 2,0-2,5 x 0,5-1,0 mm, eretas, lanceoladas,
vinácea, membranáceas, ápice agudo, base atenuada, margem serrada. Labelo preso ao pé
de coluna, ereto, 3-lobado, obovado, vináceo, membranáceo, papiloso, base atenuada;
lobos laterais inconspícuos, erguidos, oblongos, ápice obtuso, margem serrilhada; lobo
terminal 2,5-3,0 x 1,0-1,5 mm, ereto, obovado, ápice obtuso. Coluna 2,0 x 0,5 mm, 0,5
mm alt., esverdeada; prolongada na base em pé de 0,5 mm comp.; antera 0,6 x 0,4 mm,
prolongada em bico papiloso; polínias 2, obcônicas. Fruto cápsula 10-13 x 5,5-6,5 mm.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
18.set.2002, C.Azevedo & R.Oliveira 150 (HUEFS).
Descrita originalmente a partir de material proveniente de Minas Gerais (Barbosa
Rodrigues, 1882), também foi citada para o Rio de Janeiro e São Paulo (Pabst & Dungs,
1975; Sprunger et al., 1996). Na Bahia, é conhecida da Chapada Diamantina nos
municípios de Rio de Contas (Toscano de Brito, 1995), Lençóis (Toscano de Brito, 1998) e
Palmeiras (Toscano de Brito, 1998; Conceição & Giulietti, 2002). Esta é a primeira citação
para o município de Mucugê, onde é encontrada sobre as rochas em áreas de campo
rupestre e em matas de grotão. O comprimento do caule, a textura das folhas e sua
coloração variam de acordo com o habitat. Plantas mais expostas ao sol apresentam caules
mais curtos, folhas mais carnosas e coloração arroxeada, enquanto as que crescem em
áreas mais sombreadas, nas matas de grotão, apresentam caules mais longos, folhas mais
finas e coloração esverdeada. Floresce entre agosto e setembro.
Acianthera hamosa é reconhecida por seu caule compresso no ápice, sua folha
patente, inflorescência adpressa à folha, pela coloração verde a arroxeada de seus caules e
folhas, por suas flores vináceas e por apresentar pétalas e lobos laterais do labelo de
margem serrilhada. Acianthera hamosa e Acianthera prolifera (Herb. Ex Lindl.) Pridgeon

24
& M.W.Chase apresentam a mesma distribuição geográfica (Pabst & Dungs, 1975), apesar
da última ter sido descrita originalmente do Rio de Janeiro (Lindley, 1830) e ser citada
também para a Venezuela (Dunsterville & Garay, 1961d). Assemelham-se muito em seu
hábito, apresentando caule compresso no ápice, folha patente e inflorescência adpressa à
folha, embora A. prolifera diferencie-se por ser epífita, produzir constantemente brotos nas
axilas das folhas e por apresentar labelo inteiro. Pabst (1966) diferenciou as duas espécies
pelo tamanho maior (8-10 cm comp.) e forma aconchavada das folhas de A. prolifera,
enquanto A. hamosa apresenta folhas menores (5-6 cm comp.), só um pouco côncavas.
Barbosa Rodrigues (1882) refere-se a A. hamosa, vegetativamente, como uma miniatura de
A. pectinata (Lindl.) Pridgeon & M.W.Chase. Entretanto, as flores desta diferem bastante
em sua morfologia e coloração esverdeada.
O holótipo de A. hamosa encontra-se perdido. Desta forma, a ilustração original de
Acianthera hamosa, preparada na natureza por Barbosa Rodrigues, constitui o único
material original disponível e por esse motivo é aqui selecionada como lectótipo.
01.2. Acianthera ochreata (Lindl.) Pridgeon & M.W.Chase subsp. ochreata, Lindleyana
16(4): 245, 2001.
Pleurothallis ochreata Lindl., Bot. Reg. 21 (n.s. 8): sub t. 1797, 1836.
Erva rupícola. Rizoma 5,0 mm comp. entre pseudobulbos, 3,0 mm diâm., raízes
emergindo de todo o rizoma. Caule 60-70 x 2,0-2,5 mm, 1-foliado, ereto, cilíndrico,
amarelo-alaranjado a esverdeado, coberto por bainhas. Folha 90-100 x 10 mm, séssil,
ereta, lanceolada, amarelo-alaranjada a esverdeada, carnosa, ápice agudo, base atenuada.
Inflorescência racemo terminal, ereto; escapo 30-45 mm comp., amarelado; brácteas
escapais 3,0-4,0 x 3,5-4,0 mm, ovadas, membranáceas, ápice agudo apiculado, base
amplexicaule; raque 20-35 mm comp., 5-9 flores; brácteas florais 2,5-3,0 x 4,0-4,5 mm,
ovadas, membranáceas, ápice agudo apiculado, base amplexicaule. Flores ressupinadas;
pedicelo incluindo o ovário 2,5-3,0 mm comp. Sépalas eretas, amarelo-alaranjada,
carnosas, ápice agudo, base truncada; dorsal 5,5-6,0 x 1,0-1,5 mm, lanceolada; laterais 5,5-
6,0 x 1,0-1,5 mm, concrescidas da base até o ápice formando uma navícula. Pétalas 2,0-
2,5 x 0,5-1,0 mm, eretas, lineares, amarelo-alaranjada, carnosas, ápice agudo apiculado,
base truncada, margem serrilhada. Labelo preso ao pé de coluna, ereto, 3-lobado, oblongo,
amarelo-alaranjado, carnoso, base truncada; calosidade em forma de duas quilhas erguidas
na porção mediana do lobo terminal; lobos laterais 1,0-1,5 x 0,5-1,0 mm, erguidos,

25
oblongos, ápice obtuso; lobo terminal 1,0-1,5 x 1,0 mm, ereto, oblongo, ápice obtuso.
Coluna 2,5-3,0 x 0,5-1,0 mm, 0,5-1,0 mm alt., alada, prolongada na base em pé 0,5-1,0
mm comp.; antera 0,5 x 0,4 mm, prolongada em bico; polínias 2, obovadas. Fruto cápsula
14-14,5 x 5,0-6,0 mm.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
11.jun.2002, C.Azevedo & J.Oliveira 139 (HUEFS); Unidade de Manejo Sustentável,
04.jan.1997, UMS 124 (HRB); Unidade de Manejo Sustentável, 01.nov.1996, UMS 345
(HRB).
Material adicional examinado: BRASIL, Bahia: Mucugê, 10 km N da cidade na rod.
para Andaraí, 27.jan.2000, J.G.Jardim et al. 2550 (CEPEC); 2 km al N de Mucugê, camino
a Andaraí, 20.jan.1997, M.M.Arbo et al. 7571 (CEPEC); by rio Cumbuca, about 3 km N of
Mucugê on the road to Andaraí, 15.fev.1977, R.M.Harley et al. 18712 (CEPEC).
Esta espécie foi descrita originalmente da Serra do Monte Santo, Bahia (Lindley,
1836a). Pabst & Dungs (1975) registraram sua ocorrência para Pernambuco e Bahia.
Segundo Borba & Semir (2001), Acianthera ochreata é uma espécie com ampla
distribuição no Nordeste do Brasil, especialmente no estado da Bahia, mas também é
encontrada em Pernambuco e na Paraíba. Na Bahia é conhecida da Serra do Curral Feio,
Morro do Chapéu, Serra do Sincorá, Serra do Rio de Contas, Serra das Almas (Harley &
Mayo, 1980), Catolés (Toscano de Brito & Queiroz, 2003), e dos municípios de
Mucugê/Andaraí (Harley & Simmons, 1986), Rio de Contas (Toscano de Brito, 1995),
Lençóis (Toscano de Brito, 1998) e Palmeiras (Toscano de Brito, 1998; Conceição &
Giulietti, 2002). Acianthera ochreata subsp. ochreata é abundante nos campos rupestres da
Chapada Diamantina, e muito comum na área do Parque, onde cresce em grandes
populações, diretamente sobre as rochas a pleno sol. Sua coloração e textura podem variar
de acordo com o habitat. Plantas mais expostas ao sol apresentam folhas mais carnosas e
coloração amarelo-alaranjada, enquanto as que crescem em áreas mais sombreadas
apresentam folhas menos carnosas e coloração esverdeada. Floresce entre novembro e
fevereiro. Segundo Borba & Semir (2001), suas flores possuem néctar na base do labelo,
cheiram a queijo estragado e são polinizadas por Megaselia spp., dípteros da família
Phoridae.
O próprio Pabst (1966), após examinar o holótipo de A. ochreata, concluiu que a
espécie descrita por ele para a Chapada Diamantina, entre Lençóis e Palmeiras,

26
Pleurothallis bahiensis (Pabst, 1956), era somente uma forma mais robusta daquela.
Recentemente, uma subespécie de A. ochreata foi descrita A. ochreata subsp. cylindrifolia,
baseada em diferenças morfológicas de caracteres vegetativos, diferenças químicas e de
localização geográfica. A. ochreata subsp. cylindrifolia diferencia-se vegetativamente da
subespécie tipo por apresentar folhas cilíndricas sulcadas e ocorre apenas na Cadeia do
Espinhaço em Minas Gerais. A. ochreata subsp. ochreata apresenta folhas carnosas
nitidamente conduplicadas (Borba et al., 2002).
A. ochreata subsp. ochreata é reconhecida por suas folhas carnosas de coloração
amarelo-alaranjada, e por suas flores de sépalas laterais concrescidas da base até o ápice,
formando uma navícula. Assemelha-se a A. teres, por apresentar caracteres florais bastante
similares, mas difere em caracteres vegetativos e em suas distribuições geográficas, sendo
que esta ocorre, principalmente, em Minas Gerais, e algumas populações no Rio de Janeiro
e Espírito Santo (Borba et al., 2002). Ao contrário de A. teres, A. fabiobarrosii (Borba &
Semir) F.Barros & F.Pinheiro apresenta grande similaridade vegetativa com A. ochreata
subsp. ochreata, sendo que esta ocorre apenas na região centro-norte da Cadeia do
Espinhaço, em Minas Gerais (Borba et al., 2000).
02. ANATHALLIS Barb.Rodr.
Chave de identificação para as espécies de Anathallis ocorrentes no Parque Municipal de
Mucugê.
1. Inflorescência com uma única flor em antese, flores vináceas, sépalas e pétalas de
margem ciliada. ..........................................................................................................................
..................................................................................................... Anathallis microphyta (02.1)
1’. Inflorescência com duas flores, flores amareladas, sépalas e pétalas de margem não
ciliada. ................................................................................ Anathallis montipelladensis (02.2)
02.1. Anathallis microphyta (Barb.Rodr.) C.O.Azevedo & Van den Berg (Capítulo 3)
Pleurothallis microphyta (Barb.Rodr.) Cogn., Fl. bras. 3(4): 484, 1895.
Erva rupícola. Rizoma inconspícuo. Caule 20-40 x 0,5-1,0 mm, 1-foliado, ereto,
cilíndrico, marrom, coberto por bainhas. Folha 5,0-10 x 3,0-4,0 mm, séssil, ereta,
lanceolada a orbicular, verde, coriácea, ápice agudo a obtuso retuso, base atenuada.

27
Inflorescência racemo terminal, ereto; escapo 3,5-6,0 mm comp., marrom; brácteas
escapais inconspícuas; raque 3,0-8,0 mm comp., 1 flor; brácteas florais 1,5-2,0 x 1,0-1,5
mm, orbiculares, membranáceas, ápice agudo, base amplexicaule. Flores ressupinadas;
pedicelo incluindo o ovário 1,0-1,5 mm comp. Sépalas lanceoladas, membranáceas, ápice
agudo, base truncada, margem ciliada; dorsal 3,0-3,5 x 1,0-1,5 mm, ereta, concrescida na
base às sépalas laterais, vinácea; laterais 3,0 x 1,0 mm, reflexas, concrescidas na base, até
1/3 do comprimento, vináceas com lista branca na base. Pétalas 3,0 x 0,5-1,0 mm, eretas,
lineares, vináceas, membranáceas, ápice agudo, base truncada, margem ciliada. Labelo
articulado com o pé de coluna de 1,0 mm comp., ereto, 3-lobado, orbicular-oblongo,
vináceo, carnoso, base atenuada; calo convexo na porção basal do labelo; lobos laterais
inconspícuos, eretos, semi-orbiculares, ápice obtuso, ciliado; lobo terminal 2,0 x 1,0 mm,
orbicular-oblongo, ápice obtuso. Coluna 2,0 x 0,5 mm, 0,5 mm alt., branca com manchas
vináceas; antera 0,3 x 0,4 mm; polínias 2, orbiculares, amarelas. Fruto cápsula 4,0-5,0 x
2,0-3,0 mm.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
18.set.2002, C.Azevedo & R.Oliveira 152 (HUEFS).
Descrita originalmente de espécime originário da Serra do Mar (Barbosa
Rodrigues, 1882), esta espécie é citada para Argentina, e no Brasil, para os estados do Rio
de Janeiro e Rio Grande do Sul (Pabst & Dungs, 1975; Sprunger et al., 1996), sendo aqui
registrada pela primeira vez para o estado da Bahia (CAPÍTULO 3). Anathallis microphyta
é rara na área do Parque, encontrada somente em áreas sombreadas e úmidas das matas de
grotão, crescendo sobre rochas cobertas por musgos, o que dificulta ainda mais sua
localização. Floresce entre maio e setembro.
Anathallis microphyta é reconhecida por seu pequeno porte e suas flores vináceas.
Assemelha-se muito a A. corticicola (Schltr. Ex Hoehne) Pridgeon & M.W.Chase,
diferindo por apresentar pétalas de margem inteira, lisa, enquanto A. microphyta possui
sépalas e pétalas de margem ciliada.

28
02.2. Anathallis montipelladensis (Hoehne) F.Barros, Bradea 8(43): 295, 2002.
Pleurothallis montipelladensis Hoehne, Arch. Inst. Biol. (São Paulo) 2: 36, pl. 6, fig.
1, 1929.
Erva rupícola. Rizoma 0,5-1,0 mm comp. entre pseudobulbos, 1,0-1,5 mm diâm., coberto
por bainhas, raízes emergindo ao longo de todo o rizoma. Caule (1,5-)3,0-6,0 x 0,5-1,5
mm, 1-foliado, ereto, cilíndrico, verde, coberto por bainhas. Folha 6-16 x 2,5-4,0 mm,
séssil, ereta, lanceolada a quase orbicular, verde, carnosa, ápice agudo acuminado, base
atenuada. Inflorescência racemo terminal, ereto; escapo 9-29 mm comp., amarelado;
brácteas escapais inconspícuas; raque 3,5-10 mm comp., 2 flores; brácteas florais
inconspícuas. Flores ressupinadas; pedicelo incluindo o ovário 0,5-1,5 mm comp. Sépalas
eretas, lanceoladas, amareladas, membranáceas, ápice obtuso, base truncada; dorsal 3,0-3,5
x 1,0-1,5 mm, concrescidas na base às sépalas laterais; laterais 3,0-3,5 x 0,5-1,0 mm,
concrescidas na base. Pétalas 2,5-3,0 x 0,5-1,0 mm, eretas, lineares, amareladas,
membranáceas, ápice obtuso, base atenuada, margem serrilhada. Labelo articulado com o
pé da coluna, ereto, 3-lobado, oblongo, amarelado, membranáceo, base truncada; lobos
laterais inconspícuos, erguidos, semi-orbiculares, ápice obtuso, margem ciliada; lobo
terminal 1,0-1,5 x 0,5-1,0 mm, ereto, oblongo, ápice arredondado. Coluna 1,5 x 0,5 mm,
0,6 mm alt., amarelada, alada; antera 0,3 x 0,3 mm; polínias 2. Fruto cápsula 4,0-5,0 x 2,0-
3,0 mm.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
13.jul.2002, C.Azevedo 142 (HUEFS); Parque Municipal de Mucugê, 14.ago.2002,
C.Azevedo & J.Oliveira 146 (HUEFS); Unidade de Manejo Sustentável, 06.nov.1996,
UMS 343 (HRB).
Citada para a Bahia, São Paulo, Minas Gerais e Mato Grosso (Pabst & Dungs,
1975). Na Chapada Diamantina, foi registrada para o município de Rio de Contas (Toscano
de Brito, 1995) e Mucugê (Stradmann, 1998). Cresce em paredões rochosos no campo
rupestre. Foi observada florida nos meses de abril, julho, agosto e novembro.
Anathallis montipelladensis é facilmente reconhecida pelo seu pequeno porte, 10-20
mm alt., e sua inflorescência com duas flores amareladas.

29
03. BRASSAVOLA R.Br.
03.1. Brassavola tuberculata Hook., Bot. Mag. 56: t. 2878, 1829.
Erva rupícola. Rizoma 5,0-10 mm comp. entre pseudobulbos, 4,0-5,0 mm diâm., coberto
por bainhas, raízes emergindo ao longo de todo o rizoma. Caule 60-110 x 4,0 mm, 1-
foliado, pendente, cilíndrico, marrom, coberto por bainha. Folha 220 x 5,0 mm, séssil,
pendente, terete, verde, carnosa, ápice agudo, base truncada. Inflorescência racemo
terminal, pendente; escapo 10-20 mm comp., verde; brácteas escapais 3,0-4,0 x 4,0-5,0
mm, ovadas a orbiculares, paleáceas, ápice agudo, base amplexicaule; raque 10-20 mm
comp., 3 flores; brácteas florais 3,0-5,0 x 2,0-3,0 mm, ovadas, paleáceas, ápice agudo, base
truncada. Flores ressupinadas; pedicelo incluindo o ovário 40-55 mm comp. Sépalas
patentes, lanceoladas, creme-amareladas, membranáceas, ápice agudo, base truncada;
dorsal 30-38 x 4,0-5,0 mm; laterais 30-35 x 5,0-6,0 mm. Pétalas 30-36 x 3,0-4,0 mm,
patentes, lanceoladas, creme-amareladas, membranáceas, ápice agudo, base obtusa. Labelo
30-35 x 15-20 mm, concrescido à base da coluna, ereto, 1-lobado, ovado, branco com
centro amarelo, membranáceo, ápice agudo cuspidado, base atenuada, margem
ligeiramente revoluta. Coluna 10 x 3,0 mm, 4,0 mm alt., branca; alada; antera 2,0-2,5 x 2,0
mm, 1,0 mm alt., branca; polínias 8, cerosas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
26.nov.2002, C.Azevedo & R.Oliveira 165 (HUEFS).
Espécie conhecida da Bolívia e Brasil (Sprunger, 1991), onde foi citada para os
estados de Sergipe, Rio de Janeiro, Paraná, Santa Catarina, Rio Grande do Sul e Minas
Gerais (Pabst & Dungs, 1975), neste último, para a Serra do Cipó (Barros, 1987). Na
Bahia, foi citada para Monte Santo (Harley & Mayo, 1980), sendo aqui registrada pela
primeira vez para a Chapada Diamantina. É rara no Parque, sendo encontrada crescendo
sob paredões rochosos apenas em matas de grotão. Floresce em novembro.
Brassavola tuberculata é caracterizada por seu hábito pendente, folha terete e flores
de sépalas e pétalas creme-amareladas e labelo branco com centro amarelo. Withner (1998)
considerou B. fragrans, B. perrinii e B. tuberculata espécies diferentes, apesar de afirmar
que B. perrinii, B. fragrans e B. flagellaris são parte de um complexo de espécies
proximamente relacionadas, que diferem apenas em sua localização. Aqui, preferiu-se

30
seguir Pabst & Dungs (1975) e Sprunger (1991) que consideram B. fragrans Lem. B.
gibbiana hort. e B. perrinii Lindl. sinônimos de B. tuberculata.
04. BULBOPHYLLUM Thouars
Chave de identificação para as espécies de Bulbophyllum ocorrentes no Parque Municipal
de Mucugê.
1. Inflorescência de escapo ereto e raque genuflexa, pétalas lineares. .......................................
................................................................................................... Bulbophyllum weddellii (04.4)
1’. Inflorescência sem a característica acima, pétalas não lineares.
2. Pétalas muito reduzidas, c. 0,5 mm comp., semi-orbiculares, pé de coluna longo, c. 4,0
mm comp. .......................................................................... Bulbophyllum cribbianum (04.2)
2’. Pétalas 2-5 mm comp., ovadas a triangulares, pé de coluna 1,5-2,0 mm comp.
3. Sépalas ovadas, base não falcada, pétalas ovadas, de ápice obtuso, margem ciliada
labelo púrpuro com ápice branco ......................................................................................
.................................................................................... Bulbophyllum involutum (04.3)
3’. Sépalas triangulares, base ligeiramente falcada, pétalas triangulares, de ápice
agudo, margem não ciliada, labelo vináceo com manchas e pontuações creme-
amareladas com relances metálicos. ............................ Bulbophyllum ×cipoense (04.1.)
04.1. Bulbophyllum ×cipoense Borba & Semir, Lindleyana 13: 113-120, 1998.
Erva rupícola. Rizoma 15-20 mm comp. entre pseudobulbos, 5,0 mm diâm., coberto por
bainhas, raízes emergindo principalmente da base do pseudobulbo. Pseudobulbos 22-25 x
12-15 mm, 1-foliados, eretos, ovóides-tetrágonos, amarelados, cobertos por bainhas. Folha
80-95 x 15-18 mm, séssil, semi-ereto, elíptico-oblonga, verde, coriácea, ápice agudo
acuminado, base atenuada. Inflorescência racemo lateral; escapo 50-55 cm comp., ereto,
vináceo; brácteas escapais 10-14 x 5,0-7,0 mm, orbiculares, paleáceas, ápice agudo
acuminado, base amplexicaule; raque 30-35 cm comp., pendente, 3-4 flores em antese;
brácteas florais 5,0-9,0 x 2,0-3,0 mm, triangulares a ovadas, membranáceas, ápice agudo,
base truncada. Flores ressupinadas; pedicelo incluindo o ovário 4,0 mm comp. Sépalas
triangulares, creme-arroxeadas com manchas e pontuações purpúreas na base,
membranáceas, ápice agudo; dorsal 10-15 x 3,0-4,0 mm, ereta, base truncada; laterais 10-

31
15 x 3,0-4,0 mm, reflexas, base ligeiramente falcada. Pétalas 4,0-5,0 x 1,0-2,0 mm,
patentes, triangulares, cremes com manchas e pontuações purpúreas, membranáceas, ápice
agudo, base truncada. Labelo articulado com o pé da coluna, ereto, 3-lobado, obovado,
vináceo com manchas e pontuações creme-amareladas com relances metálicos, carnoso,
base atenuada; calo convexo, piloso; lobos laterais 2,0-2,5 x 1,0 mm, eretos, auriculares,
ápice obtuso, ciliado; lobo terminal 5,0-6,0 x 3,0-4,0 mm, oblongo, ápice obtuso, ciliado.
Coluna 2,0 x 1,0 mm, 1,0 mm alt., creme-amarelada; provido de dois longos braços
cruzados voltados para frente e dois menores para baixo; prolongada na base em pé de 2,0
mm comp.; antera 1,0 x 1,0 mm, conectada a coluna, amarela, papilosa, ápice prolongado
em bico; polínias 2. Fruto cápsula 10-12 x 6,0-9,0 mm, verde com nuances vinácea.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
18.mar.2003, C.Azevedo & E.Oliveira 182 (HUEFS).
Recentemente, Bulbophyllum ×cipoense, um híbrido natural de B. weddellii e B.
involutum, foi descrito para a Cadeia do Espinhaço, Minas Gerais (Borba & Semir, 1998).
Na área em estudo, os parentais de B. ×cipoense estão presentes, ocorrendo muitas vezes
sobre a mesma rocha. Espécimes encontrados no Parque apresentam caracteres que
sugerem que o mesmo evento possa ter ocorrido.
Borba & Semir (1998), através de estudos de biologia floral e fenologia, concluíram
que as espécies parentais são simpátricas, possuem eventos fenológicos sincronizados,
caracterizados por um período de floração longo. Compartilham os mesmos polinizadores,
fêmeas de Pholeomyia, dípteros da família Milichiidae, e são intercompatíveis, o que pode
levar à formação de híbridos naturais.
Borba & Semir (1998), quando descreveram B. ×cipoense, chamaram a atenção de
que o híbrido atrai os mesmos polinizadores dos parentais, o que pode causar introgressão.
Este evento parece estar acontecendo no Parque, onde os espécimens encontrados
apresentam caracteres intermediários entre os parentais e entre si. Como o espécime
C.Azevedo & J.Oliveira 177 (HUEFS) (Figura XVI - H), que apesar de semelhante ao
descrito por Borba & Semir (1998), apresenta algumas diferenças na coloração, tamanho e
forma de seus segmentos florais. B. ×cipoense apresenta sépalas e pétalas oval-lanceoladas,
pouco falcadas, sépalas com manchas e pontuações purpúreas na base e labelo purpúreo
com terço distal e margens laterais brancas com manchas purpúreas, e o indivíduo aqui
tratado possui sépalas e pétalas ovadas, não falcadas, sépalas com manchas e pontuações

32
purpúreas em praticamente todo seu comprimento e labelo sem terço distal e margens
laterais brancas. Apesar, de Borba & Semir (1998) destacarem que o híbrido por eles
descrito possui um alto grau de esterilidade ou auto-incompatibilidade, e características do
labelo fazem com que sua polinização seja ineficiente.
O indivíduo aqui estudado e o descrito para Minas Gerais apresentam muitas
semelhanças no formato e coloração de suas peças florais, tais como sépalas ligeiramente
falcadas, com manchas e pontuações purpúreas apenas na base. Porém, o espécime
encontrado no Parque difere-se por apresentar pétala de margem não ciliada e labelo sem
terço distal e margens laterais brancas.
Rieseberg (1995) chama a atenção de que a maioria dos híbridos verdadeiros não
apresenta caracteres intermediários aos de seus parentais, da mesma maneira, caracteres
intermediários não indicam necessariamente hibridação. Este fato foi observado por Borba
et al. (2000), que inicialmente imaginaram que Acianthera fabiobarrosii tratava-se de um
híbrido entre Acianthera ochreata, devido à similaridade vegetativa, e Acianthera
johannensis, pela similaridade nos caracteres florais. Após testarem hipótese de origem por
hibridação, detectaram tratar-se de uma nova espécie e não de um híbrido como era
esperado.
Foram realizados estudos para testar hipótese de origem por hibridação (dados não
publicados), onde foram utilizados 12 loci em 10 sistemas enzimáticos. Tal hipótese não
pode ser definitivamente comprovada, devido à ausência de loci diagnósticos entre os
supostos parentais (B. weddellii e B. involutum). Porém, a mesma pode ser fortalecida por
dois motivos: o primeiro é que entre os três táxons testados, B. ×cipoense é o único que
não apresentou alelos exclusivos, e segundo é que, entre os 14 indivíduos de possíveis
híbridos, a hipótese de hibridação não pode ser descartada em nenhum, exceto dois deles,
por estes possuírem em todos os loci alelos presentes em ambos os parentais. Os outros
dois indivíduos são homozigotos em pelo menos um locus para um alelo que não está
presente em B. weddellii (alelo exclusivo de B. involutum). Isto sugere a ocorrência de
introgressão em B. involutum, que pode ser reforçada por caracteres morfológicos, como
discutido acima.
Outro fato interessante é que a variabilidade encontrada em B. ×cipoense indica que
o evento de hibridação entre B. involutum e B. weddellii não foi único na população
estudada, mas que os indivíduos foram originados por múltiplos eventos de hibridação.

33
04.2. Bulbophyllum cribbianum Toscano, Kew Bull. 47(4): 774, 1992.
Bulbophyllum micropetalum Barb.Rodr., Gen. spec. Orchid. 2: 121, 1882. (non
Lindl. 1862).
Erva rupícola. Rizoma 5,0-10 mm comp. entre pseudobulbos, 2,0 mm diâm., coberto por
bainhas, raízes emergindo apenas na base dos pseudobulbos. Pseudobulbos 10-15 x 5,0-
8,0 mm, 1-foliados, eretos, ovóides-tetrágonos, amarelados, cobertos por bainhas. Folha
25-55 x 6,0-10 mm, séssil, ereto-patente, elíptico-oblonga a oblonga, verde, coriácea, ápice
agudo apiculado, base atenuada. Inflorescência racemo lateral; escapo 70-120 mm comp.,
ereto, verde; brácteas escapais 10-15 x 2,0-3,0 mm, lanceoladas, paleáceas, ápice agudo,
base amplexicaule; raque 25-30 mm comp., pendente, 2-3 flores em antese; brácteas florais
5,0-6,0 x 3,0-4,0 mm, ovadas, membranáceas, ápice agudo, base amplexicaule. Flores
ressupinadas; pedicelo incluindo o ovário 2,0-2,5 mm comp. Sépalas eretas, amarelo
esverdeadas com veios púrpuros, membranáceas; dorsal 5,0-6,0 x 5,0-6,0 mm, ovada a
orbicular, ápice obtuso, base truncada; laterais 6,0-7,0 x 5,0-6,0 mm, fundidas lateralmente
ao pé da coluna, triangulares a ovadas, ápice obtuso, base truncada. Pétalas 0,5 x 1,0 mm,
eretas, semi-orbiculares, róseas a amareladas com veios púrpuros, membranáceas, ápice
obtuso, base truncada. Labelo 5,0-6,0 x 3,5-4,0 mm, articulado com o pé da coluna, ereto,
1-lobado, oblongo, magenta com margem vinho, membranáceo, ápice obtuso, base
atenuada. Coluna 2,5 x 2,0 mm, 1,5 mm alt., esbranquiçada; prolongada na base em pé de
4,0 mm comp.; antera 0,5 x 1,0 mm, conectada a coluna; polínias 2, orbiculares,
amareladas. Fruto cápsula 5,0-7,0 x 4,0 mm, amarelo-esverdeado.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
26.nov.2002, C.Azevedo & R.Oliveira 167 (HUEFS).
Material adicional examinado: BRASIL, Bahia: Mucugê, Gerais do Guiné,
17.nov.2001, E.L.Borba 2127 (HUEFS).
Este Bulbophyllum foi descrito originalmente do Rio de Janeiro (Barbosa
Rodrigues, 1882), e citado também para o estado do Espírito Santo (Pabst & Dungs, 1975;
Sprunger et al., 1996). Na Bahia, ocorre no município de Rio de Contas (Toscano de Brito,
1995), sendo aqui o primeiro registro para o município de Mucugê. Raro na área do
Parque, esta espécie foi encontrada crescendo em paredões rochosos de matas de grotão.
Floresce em novembro.

34
Bulbophyllum cribbianum não foi encontrado em flor. Sua identificação foi
possível, não só pela análise da porção vegetativa característica da espécie, como pelos
restos florais encontrados no fruto. É facilmente reconhecido por suas pétalas muito
pequenas, semi-orbiculares e pelo longo pé de coluna. A descrição e ilustração das flores
desta espécie foram feitas baseadas em material conservado em líquido, coletado no
município de Mucugê, por E.L.Borba, 2127. Toscano de Brito (1995) comenta que B.
cribbianum parece ser rara em herbários brasileiros, pois o espécime citado para Rio de
Contas, foi a terceira coleção por ele examinada.
Como existe uma espécie de Bulbophyllum na África com o epíteto micropetalum,
descrita por Lindley antes de Barbosa Rodrigues publicar Bulbophyllum micropetalum
Barb.Rodr., houve a necessidade de se criar um novo nome para a espécie brasileira
(Toscano de Britto, 1992).
04.3. Bulbophyllum involutum Borba, Semir & F.Barros, Novon 8(3): 225-229, 1998.
Erva rupícola. Rizoma 10-25 mm comp. entre pseudobulbos, 3,0-5,0 mm diâm., coberto
por bainhas, raízes emergindo principalmente da base do pseudobulbo. Pseudobulbos 20-
30 x 15-20 mm, 1-foliados, eretos, ovóides-tetrágonos, amarelados, cobertos por bainhas.
Folha 50-80 x 15-25 mm, séssil, semi-ereto, elíptico-oblonga a oblonga, verde, coriácea,
ápice agudo apiculado, base atenuada. Inflorescência racemo lateral; escapo 24-50 cm
comp., ereto, vináceo; brácteas escapais 10-18 x 5,0-7,0 mm, oblongas a orbiculares,
paleáceas, ápice obtuso apiculado, base amplexicaule; raque (7,0-)23-28 cm comp.,
pendente, 1-2 flores em antese; brácteas florais 5,0-7,5 x 2,5-4,0 mm, triangulares a
ovadas, paleáceas, ápice agudo, base truncada. Flores ressupinadas; pedicelo incluindo o
ovário 6,0-6,5 mm comp. Sépalas ovadas, creme com manchas e pontuações purpúreas
cobrindo quase completamente a superfície, membranáceas, ápice agudo, base truncada;
dorsal 8,0-9,0 x 3,5-4,0 mm, recurvada; laterais 8,0-8,5 x 3,5-4,0 mm, reflexas. Pétalas
2,0-2,5 x 1,0-1,5 mm, patentes, ovadas, creme com manchas e pontuações purpúreas,
membranáceas, ápice obtuso, base truncada, margem ciliada. Labelo articulado com o pé
da coluna, ereto, 3-lobado, orbicular, púrpuro com ápice branco, carnoso, base atenuada;
calo convexo, piloso; lobos laterais 1,0-1,5 x 0,5-1,0 mm, eretos, auriculares, ápice obtuso,
ciliado; lobo terminal 2,0-2,5 x 1,5-2,0 mm, orbicular, ápice obtuso ciliado. Coluna 2,5 x
1,5 mm, 1,0-1,5 mm alt., esbranquiçada; provido de dois longos braços voltados para
frente e dois menores para baixo; prolongada na base em pé 1,5-2,0 mm comp.; antera 1,0-

35
1,5 x 0,5-1,0 mm, conectada a coluna, verde, papilosa, o ápice prolongado em bico;
polínias 2. Fruto cápsula 10-11,5 x 9,0-10 mm, verde.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
17.set.2002, C.Azevedo & J.Oliveira 149 (HUEFS)
Descrito originalmente de Minas Gerais, é encontrado também nos campos
rupestres da Bahia, ao longo da Cadeia do Espinhaço (Borba et al., 1998). Em Minas
Gerais foi citado para a Serra de Grão-Mogol (Barros & Pinheiro, no prelo). No estado da
Bahia, foi citado para Rio de Contas (Harley & Mayo, 1980; Toscano de Brito, 1995) e
Mucugê (Harley & Simmons, 1986), identificado como B. ipanemensis. O espécime deste
último levantamento, CFCR 1477, foi mais tarde designado como parátipo de B. involutum
(Borba et al., 1998). Cresce diretamente sobre as rochas em áreas abertas de campo
rupestre, dividindo o mesmo habitat com B. weddellii. As duas espécies podem ser
encontradas vivendo sobre a mesma rocha. Floresce em setembro.
Bulbophyllum involutum é reconhecido no campo pela coloração púrpura de suas
flores, brácteas, escapo e raque, sempre com uma única flor em antese, diferentemente de
B. warmingianum e B. ipanemensis que têm inflorescências verdes ou amarelo-
esverdeadas, e três a cinco flores simultaneamente em antese. Estas características
auxiliam no reconhecimento de B. involutum em material de herbário, uma vez que a
coloração do escapo, brácteas e sépalas não se perde. As pétalas ovadas com ápice obtuso
são muito peculiares em comparação com as das outras duas espécies, que são triangulares
com ápice agudo (Borba et al., 1998). Sua polinização ocorre através de correntes de vento
que movimentam o labelo e pressionam o polinizador contra a coluna da flor (Borba &
Semir, 1998).
04.4. Bulbophyllum weddellii (Lindl.) Rchb.f., Ann. bot. syst. 6: 251, 1861.
Erva rupícola. Rizoma 10 mm comp. entre pseudobulbos, 4,0-5,0 mm diâm., coberto por
bainhas, raízes emergindo da base dos pseudobulbos. Pseudobulbos 20-25 x 10-15 mm, 1-
foliados, eretos, ovóides-tetrágonos, amarelados, cobertos por bainhas. Folha 40-50 x 15-
20 mm, séssil, semi-ereto, elíptico-oblonga a oblonga, verde, coriácea, ápice agudo
apiculado, base atenuada. Inflorescência racemo lateral; escapo 15-20 cm comp., ereto,
verde; brácteas escapais 10-15 x 5,0-6,0 mm, orbiculares, ápice agudo, base amplexicaule;

36
raque 5-10 cm comp., genuflexa, 6-20 flores; brácteas florais 7,0-9,5 x 2,0-3,0 mm,
ovadas. Flores ressupinadas; pedicelo incluindo o ovário 4-5 mm comp. Sépalas
triangulares, creme com manchas e pontuações purpúreas na base, membranáceas; dorsal
10-15,5 x 2,5-3,0 mm, recurvada, triangular, creme com manchas e pontuações purpúreas
na base, membranácea, ápice agudo caudado, base truncada; laterais 10-15,5 x 2,5-3,0 mm,
reflexas, ápice agudo, base falcada. Pétalas 3,0-4,5 x 0,5-1,0 mm, patentes, lineares,
falcada na base, cremes com manchas e pontuações purpúreas, membranáceas, ápice
agudo, base truncada, margem ciliada. Labelo articulado com o pé da coluna, ereto, 3-
lobado, lanceolado, creme com manchas e pontuações purpúreas, carnoso, base obtusa;
calo convexo, piloso; lobos laterais 1,0-1,5 x 0,5-1,0 mm, eretos, auriculares, ápice obtuso,
ciliado; lobo terminal 8,0-12,5 x 2,0-4,0 mm, lanceolado, ápice agudo. Coluna 3,0-3,5 x
1,5-2,0 mm, 1,5-2,0 mm alt., branca com manchas purpúreas; provido de dois longos
braços voltados para frente e dois menores para baixo; prolongada na base em pé 2,0-2,5
mm comp.; antera 1,5 mm comp., verde, papilosa, ápice prolongado em bico; polínias 2.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
12.jun.2002, C.Azevedo & O.Oliveira 140 (HUEFS); Parque Municipal de Mucugê,
13.abr.2003, C.Azevedo 188 (HUEFS).
Material adicional examinado: BRASIL, Bahia: Mucugê, 06.jul.1998, C.Azevedo 88
(HRB).
Esta espécie foi descrita originalmente de material proveniente de Minas Gerais
(Lindley, 1852). Barros (1987) citou sua presença também para a Serra do Cipó. É
conhecida também do estado do Rio de Janeiro (Pabst & Dungs, 1975), e, na Bahia, do
município de Rio de Contas (Toscano de Brito, 1995). É aqui registrada pela primeira vez
para Mucugê, onde cresce diretamente sobre as rochas dos campos rupestres. A
polinização, como em B. involutum, ocorre através de correntes de vento que movimentam
o labelo e pressionam o polinizador contra a coluna da flor (Borba & Semir, 1998).
Floresce entre abril e julho.
Bulbophyllum weddellii é facilmente reconhecido pela inflorescência genuflexa na
raque, pela coloração creme com manchas e pontuações purpúreas, além da forma de seus
segmentos florais, que apresentam sépalas laterais de base falcada e pétalas lineares.

37
05. CAMPYLOCENTRUM Bentham
05.1. Campylocentrum micranthum (Lindl.) Rolfe, Orchid Rev. 11(128): 245, 1903.
Erva epífita. Caule 6,0-12 cm comp., 3,0 mm diâm., ereto, cilíndrico, verdes, coberto
pelas bainhas das folhas, raízes emergindo ao longo de todo o caule, 4-7 folhas. Folhas
2,0-3,5 x 0,5-1,0 cm, dísticas, sésseis, patentes, lanceoladas, verdes, coriáceas, ápice
obtuso retuso, base atenuada, articulada com a bainha. Inflorescência racemo lateral,
raque 1,0-1,5 cm comp, 6-15 flores; brácteas florais 1,0-1,5 x 1,0-1,5 mm, ovadas,
membranáceas, ápice agudo, base truncada, margem serrilhada. Flores ressupinadas;
pedicelo incluindo o ovário 2,0-2,5 mm comp. Sépalas 3,5-4,0 x 1,0-1,5 mm, eretas,
concrescidas na base, lanceoladas, amareladas, membranáceas, ápice agudo reflexo, base
truncada. Pétalas 3,5-4,0 x 0,5-1,0 mm, eretas, lanceoladas, amareladas, membranáceas,
ápice agudo reflexo, base truncada. Labelo ereto, 3-lobado, ovado, amarelado,
membranáceo, base atenuada; esporão 2,0-2,5 x 0,5-1,0 mm; lobos laterais 1,5-2,0 x 0,5-
1,0 mm, eretos envolvendo a coluna, orbiculares, ápice obtuso; lobo terminal 4,0-4,5 x 2,0-
2,5 mm, lanceolado, ápice agudo. Coluna 0,5-1,0 x 0,5-1,0 mm, 0,5 mm alt., amarelada;
antera 0,5 x 0,5 mm, amarelada; polínias 2, orbiculares.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
27.jan.2003, C.Azevedo 173 (HUEFS).
Campylocentrum micranthum ocorre desde o México até o Panamá e também da
Venezuela à Bolívia, Guianas, Brasil, Trindade e Índias Ocidentais (Dunsterville & Garay,
1961b; Sprunger, 1991). É amplamente distribuído no Brasil, sendo encontrado nos estados
do Acre, Amazonas, Pará, Amapá, Maranhão, Pernambuco, Bahia, Rio de Janeiro, São
Paulo, Minas Gerais, Goiás e Mato Grosso (Pabst & Dungs, 1977). É aqui registrado pela
primeira vez para a Chapada Diamantina. É a única espécie epífita encontrada no Parque,
sendo raro na área, crescendo em galhos finos de árvores e arbustos de matas de grotão.
Foi encontrado com flor em janeiro.
Campylocentrum micranthum é facilmente reconhecido por sua forma de
crescimento, sendo o único gênero brasileiro de crescimento monopodial (Pabst & Dungs,
1975), além de sua inflorescência lateral mais curta que as folhas e pela presença de
esporão. Dunsterville & Garay (1961b) e Sprunger (1991) apresentam uma lista de

38
sinônimos para esta espécie.
06. CATTLEYA Lindl.
06.1. Cattleya elongata Barb.Rodr., Gen. spec. Orchid. 1: 72, t.363, 1877. Tipo: Brasil,
Barbosa Rodrigues s.n. (perdido). Lectótipo aqui designado: ilustração original
preparada por João Barbosa Rodrigues, t.363, que consta de sua Iconographie des
orchidées du Brésil, depositada no Herbário Oakes Ames, na Universidade de
Harvard, reproduzida por Sprunger et al. (1996) sob o número t.48 do volume 4.
Erva rupícola. Rizoma 5,0-10 mm comp. entre pseudobulbos, 7,0-8,0 mm diâm., coberto
por bainhas, raízes emergindo ao longo de todo o rizoma. Pseudobulbos 10-32 x 0,7-1,5
cm, 2-3-foliados, eretos, cilíndricos, verde-arroxeados, cobertos por bainhas. Folhas 6-11 x
3-5 cm, sésseis, semi-eretas a patentes, elípticas a oblongas, verdes, fortemente coriáceas,
ápice obtuso retuso, base atenuada. Inflorescência racemo terminal, ereto, espata 5-9 x 1
cm, oblíqua, ápice obtuso apiculado, base amplexicaule; escapo (10-)46-50 cm, vermelho-
amarronzado; brácteas escapais 5,0-7,0 x 3,0-5,0 mm, triangulares a ovadas, paleáceas,
ápice agudo, base truncada; raque 3-11 cm comp., 1-6(-12) flores; brácteas florais 4,0-5,0 x
2,0-3,0 mm, triangulares a ovadas, membranáceas, ápice agudo, base truncada. Flores
ressupinadas; pedicelo incluindo o ovário 3,5-5,0 cm comp. Sépalas patentes, oblongo-
lanceoladas, vermelho-amarronzadas, carnosas, ápice agudo acuminado, base atenuada,
margem revoluta; dorsal 40-45 x 10-15 mm; laterais 35-40 x 10-15 mm. Pétalas 40-45 x
10-15 mm, patentes, oblongo-lanceoladas, vermelho-amarronzadas, carnosas, ápice
acuminado, base atenuada, margem ondulada. Labelo ereto, 3-lobado, magenta, carnoso,
base truncada; 2 calos conspícuos centrais e 2-3 laterais menores; lobos laterais 25-30 x
10-15 mm, eretos envolvendo completamente a coluna, triangulares, ápices agudos
sobrepostos; lobo terminal 22-25 x 20-25 mm, dividido em 2 lóbulos, transversalmente
oblongo, ápice obtuso emarginado. Coluna 20-25 x 10-15 mm, 10 mm alt., branca;
estigma bilobado, antera 5,0 x 4,0 mm; polínias 4.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
20.fev.2003, C.Azevedo & T.C.Faustino 179 (HUEFS); Unidade de Manejo Sustentável,
04.jan.1997, UMS 123 (HRB); Unidade de Manejo Sustentável, 05.abr.1997, UMS 210
(HRB).

39
Material adicional examinado: BRASIL, Bahia: Serra do Sincorá, 6,5 km SW of
Mucugê on the Cascavel road, 27.mar.1980, R.M.Harley et al. 21024 (CEPEC); South of
Andaraí, along road to Mucugê near small town of Xique-Xique, 14.fev.1977, R.M.Harley
et al 18690 (CEPEC). Mucugê, Serra do Sincorá, 7 km N of Mucugê on road to Andaraí,
09.mai.1976, C.E.Calderon et al. 2422 (CEPEC); nova rodovia Mucugê/Andaraí, coletas
entre os km 0 e 10. Área do Parque Nacional da Chapada Diamantina, 19.mai.1989, L.A.
Matos Silva et al. 2788 (CEPEC); a 7 km, estrada Mucugê/Andaraí, 23.mai.1985,
W.N.Fonseca & A.P.Araújo 451 (HRB).
Descrita originalmente de material proveniente de Carangola, Minas Gerais
(Barbosa Rodrigues,1877), esta espécie foi citada também para os estados de Pernambuco
e Bahia (Pabst & Dungs, 1975; Sprunger et al., 1996). Na Chapada Diamantina, foi citada
para Morro do Chapéu, Serra do Sincorá (Harley & Mayo, 1980), Mucugê/Andaraí (Harley
& Simmons, 1986), Rio de Contas (Toscano de Brito, 1995), Lençóis (Toscano de Brito,
1998), Palmeiras (Toscano de Brito, 1998; Conceição & Giulietti, 2002) e Catolés
(Toscano de Brito & Queiroz, 2003). É comum no Parque e cresce a pleno sol, diretamente
sobre rochas. Floresce entre janeiro e maio.
Em 1892, plantas de C. elongata foram exportadas para a Inglaterra, onde foram
descritas como C. alexandrae. Só mais tarde se descobriu que era a mesma planta descrita
anteriormente por Barbosa Rodrigues (Withner, 1988). O próprio Rolfe (1894) reconheceu
que a espécie descrita por ele era sinônimo de C. elongata.
É facilmente reconhecida por apresentar pseudobulbos cilíndricos alongados, 2-3-
foliados e por suas grandes flores vermelho-amarronzadas, de labelo magenta, cujos lobos
envolvem completamente a coluna. Uma lista de sinônimos é dada em Sprunger et al.
(1996).
O holótipo desta espécie encontra-se perdido. Conseqüentemente a ilustração
original de Cattleya elongata, preparada na natureza por Barbosa Rodrigues e ilustrada em
sua Iconographie des orchidées du Brésil, e recentemente publicada por Sprunger et al.,
(1996), constitui o único material original disponível. Por esse motivo, esta ilustração é
aqui selecionada como lectótipo.

40
07. CLEISTES L.C.Rich. ex Lindl.
07.1. Cleistes exilis Hoehne, Comm. Linh. Telegr. Matto Grosso Amoz. Annexo 5 Bot. 9:
26, t. 167, f. 1, 1916.
Erva terrestre. Raízes com segmentos espessados, 10-15 x 5,0-10 mm, formando
tuberóides e o resto mais fino. Caule 450-700 x 1-5 mm, ereto, cilíndrico, verde, 3-4
folhas. Folhas 10-20 x 3,0-4,0 mm, dísticas, sésseis, eretas, oblongas a ovadas, verdes,
membranáceas, ápice agudo, base truncada. Inflorescência racemo terminal, ereto; raque
15-20 cm comp., 1-3 flores; brácteas florais foliáceas 10-20 x 2,0-3,0 mm, ovadas,
membranáceas, ápice agudo, base truncada. Flores ressupinadas; pedicelo incluindo o
ovário 15-20 mm comp. Sépalas eretas, lanceoladas, róseas, membranáceas, ápice agudo,
base truncada; dorsal 25-30 x 5,0-8,0 mm; laterais 20-25 x 4,0-5,0 mm. Pétalas 20-25 x
5,0-7,0 mm, eretas, lanceoladas, róseas, membranáceas, ápice agudo, base truncada.
Labelo ereto, 3-lobado, rósea, calosidade amarela, membranáceo, base atenuada;
calosidade (glândulas) em todo comprimento central do labelo; lobos laterais 10-15 x 3,0-
3,5 mm, envolvendo a coluna, semi-obovados, ápice obtuso; lobo terminal 6,0-7,0 x 4,0-
5,0 mm, ereto, orbicular, ápice agudo reflexo, margem ondulada. Coluna 10-13 x 2,0-3,0
mm, 1,5-2,0 mm alt., rósea; antera 3,0-3,5 x 2,0-2,5 mm; polínias 2.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
20.fev.2003, C.Azevedo et al. 178 (HUEFS).
Material adicional examinado: BRASIL, Bahia: entre Andaraí e Mucugê, 15.fev.1974,
Ghillány s.n. (HB 63141); Serra do Sincorá, 10 km S of Andaraí on the road to Mucugê,
16.fev.1977, R.M.Harley et al. 18765 (CEPEC, HB, HRB, K, NY). Mucugê: Serra do
Sincorá, c. 15 km N.W. of Mucugê, on the road to Guiné and Palmeiras, 26.mar.1980,
R.M.Harley et al. 20984 (CEPEC, K); Serra do Sincorá, c. 15 km N.W. of Mucugê, on the
road to Guiné and Palmeiras, 26.mar.1980, R.M.Harley et al. 20986 (CEPEC, K); Serra do
Sincorá, by rio Cumbuca, S of Mucugê, 04.fev.1974, R.M.Harley et al. 15941 (CEPEC,
K).
Citada por Hoehne (1940) para Minas Gerais, esta espécie também foi citada para
Goiás (Pabst & Dungs, 1975), e na Bahia para a Chapada Diamantina, nos municípios de
Mucugê/Andaraí (Harley & Mayo, 1980; Harley & Simmons, 1986), Rio de Contas

41
(Toscano de Brito, 1995), Lençóis (Toscano de Brito, 1998) e Catolés (Toscano de Brito &
Queiroz, 2003). Segundo Silva (2002), Cleistes exilis é comum na Chapada Diamantina,
ocorrendo nos municípios de Abaíra, Andaraí, Lençóis, Mucugê, Piatã e Rio de Contas. O
espécime R.M.Harley et al. 20984, citado para Mucugê (Harley & Simmons, 1986) como
C. aphylla (Barb.Rodr.) Hoehne, foi identificado como C. exilis por Silva (2002). Cresce
em áreas abertas e ensolaradas, dominadas por gramíneas, em solo arenoso. Floresce entre
fevereiro e abril.
No Parque, C. exilis é facilmente reconhecida pelo porte esguio e delicado, pelo
tamanho muito reduzido de suas folhas, e pela coloração rósea de suas flores. A morfologia
do labelo é bastante variável, principalmente na forma do lóbulo terminal. É difícil de ser
encontrada, pois a planta fenece logo após a floração e frutificação, restando apenas o
tuberóide subterrâneo, que brota novamente na floração seguinte.
08. CYRTOPODIUM R. Br.
Chave de identificação para as espécies de Cyrtopodium ocorrentes no Parque Municipal
de Mucugê.
1. Flor branca com máculas purpúreas, brácteas, sépalas e pétalas fortemente onduladas. .......
............................................................................................. Cyrtopodium edmundoi (08.1)
1’. Flor amarela esverdeada a marrom-avermelhada, brácteas, sépalas e pétalas planos ou
fracamente ondulados. .......................................................... Cyrtopodium polyphyllum (08.2)
08.1. Cyrtopodium edmundoi Pabst, Orquideologia 6(4): 207, 1971. without type; Bradea
1(8): 54-55, fig. C, 1971.
Erva rupícola. Rizoma inconspícuo. Pseudobulbos 30-33 x 4-5 cm, eretos, fusiformes,
marrons, cobertos por bainhas. Folhas 30-35 x 1,0-1,5 cm, dísticas, sésseis, eretas,
plicadas, lineares, verdes, membranáceas, ápice agudo, base truncada. Inflorescência
panícula lateral, ereta; escapo 65-75 cm comp., verde; brácteas escapais 25-30 x 10-15
mm, ovadas, paleáceas, ápice agudo, base amplexicaule; raque 35-50 cm comp., c. 30
flores; brácteas florais 20-25 x 10-15 mm, ovadas, coloridas como as sépalas,
membranáceas, ápice agudo caudado, base obtusa, margem ondulada. Flores ressupinadas;
pedicelo incluindo o ovário 20-25 mm comp. Sépalas 15-20 x 10-15 mm, ovada, branca

42
com máculas purpúreas, membranácea, ápice agudo caudado, base truncada, margem
ondulada; dorsal patente; laterais reflexa. Pétalas 15-20 x 10-15 mm, reflexas, obovadas,
brancas com máculas purpúreas, membranáceas, ápice agudo, base atenuada, margens
onduladas. Labelo articulado com o pé da coluna, ereto, 3-lobado, branco com máculas
purpúreas, calo amarelo, membranáceo, base atenuada; calo convexo, verrugoso; lobos
laterais 9,0-10 x 6,0-7,0 mm, eretos, obovado, ápice obtuso; lobo terminal 8,0-11 x 5,0-7,0
mm, obovado a orbicular, ápice obtuso caudado, margem ondulada. Coluna 5,0-7,0 x 2,0-
3,0 mm, 2,0-2,5 mm alt., rósea, prolongada na base em pé de 3,0-4,0 mm comp.; antera
2,0-3,0 x 1,5-2,0 mm, verde; polínias 4, amarelas, oblongas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
15.ago.2002, C.Azevedo & O.A.Oliveira 147 (HUEFS); Unidade de Manejo Sustentável,
02.nov.1996, UMS 342 (HRB).
Material adicional examinado: BRASIL, Bahia: Mucugê, a 3 km da cidade em direção a
Igatu, 22.ago.1986, J.D.C.Arouck Ferreira et al. 363 (HRB); 10.out.1987, L.P.Queiroz
1850 (HUEFS); a 1 km ao N, 10.out.1987, M.L.Guedes et. al 1428 (ALCB).
Cyrtopodium edmundoi foi descrito originalmente da Bahia, entre Palmeiras e
Lençóis (Pabst, 1971). Mais tarde, Pabst & Dungs (1975) também citaram sua ocorrência
para este estado. Em Minas Gerais é conhecido de Grão-Mogol (Barros & Pinheiro, no
prelo). Na Chapada Diamantina, foi citado para Mucugê/Andaraí (Harley & Simmons,
1986), Rio de Contas (Toscano de Brito, 1995), Lençóis (Toscano de Brito, 1998),
Palmeiras (Toscano de Brito, 1998; Conceição & Giulietti, 2002) e Catolés (Toscano de
Brito & Queiroz, 2003). Cresce nos campos rupestres, em áreas abertas e ensolaradas, e foi
encontrado em flor nos meses de julho, agosto e novembro.
C. edmundoi é reconhecida pela coloração branca com máculas purpúreas e forma de
seus segmentos florais de margens onduladas.
08.2. Cyrtopodium polyphyllum (Vell.) Pabst ex F.Barros, Acta Bot. Brasil. 8(1): 12,
1994.
Epidendrum polyphyllum Vell., Fl. flumin. Ic. 9:t. 17, 1827.
Cyrtopodium paranaense Schltr., Repert. Spec. Nov. Regni Veg. 16: 333, 1920.
Erva rupícola. Rizoma inconspícuo. Pseudobulbos 25-49 x 3-4 cm, eretos, fusiformes,

43
marrons, cobertos por bainhas. Folhas 20-27 x 0,5-1 cm, dísticas, sésseis, eretas, plicadas,
lineares a lanceoladas, verdes, membranáceas, ápice agudo, base atenuada. Inflorescência
panícula lateral, ereta; escapo 65-70 cm comp., verde; brácteas escapais 40-60 x 15-30
mm, ovadas, paleáceas, ápice truncado a agudo, base amplexicaule; raque 30-40 cm comp.,
40-45 flores; brácteas florais 15-25 x 5-10 mm, lanceoladas, elípticas a ovadas, coloridas
como as sépalas, membranáceas, ápice agudo acuminado, base atenuada. Flores
ressupinadas; pedicelo incluindo o ovário 20-30 mm comp. Sépalas 15-20 x 7,0-10 mm,
patentes, ovadas, amarela esverdeada a marrom-avermelhada, mais escuras que as pétalas,
membranáceas, ápice agudo caudado, base truncada a atenuada, margens ligeiramente
onduladas. Pétalas 15-17 x 7,0-11 mm, patentes, lanceoladas a orbiculares, amarelas a
amarelas com mancha marrom-avermelhadas no ápice, membranáceas, ápice agudo
reflexo, base atenuada. Labelo articulado com o pé da coluna, ereto, 3-lobado, amarelo,
calo avermelhado, membranáceo, base atenuada; calo verrugoso, convexo; lobos laterais
8,0-10 x 5,0-7,0 mm, eretos, auriculares a orbiculares, ápice obtuso; lobo terminal 8,0-10 x
10-12 mm, orbicular, ápice retuso. Coluna 5,0-6,5 x 3,0-4,0 mm, 1,5-2,5 mm alt.,
esverdeada, prolongada na base em pé de 5,0 mm comp., amarelo; antera 2,0-3,0 x 1,5-2,0
mm, verde; polínias 4, amarelas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
19.set.2002, C.Azevedo & E.Ribeiro 155 (HUEFS); Parque Municipal de Mucugê,
20.set.2002, C.Azevedo 157 (HUEFS).
Material adicional examinado: BRASIL, Bahia: Mucugê, 10.out.1987, L.P.Queiroz
1844 (HUEFS).
Cyrtopodium paranaense foi descrito originalmente para o estado do Paraná
(Schlechter, 1920), e citado por Pabst & Dungs (1975) para Pernambuco, Bahia, Distrito
Federal, Rio de Janeiro, São Paulo, Paraná e Santa Catarina. Na Chapada Diamantina,
Cyrtopodium polyphyllum é conhecido do município de Lençóis (Toscano de Brito, 1998),
sendo aqui registrado pela primeira vez para o município de Mucugê. Ocorre também nas
restingas do litoral do Brasil, crescendo sobre solo arenoso. Floresce entre setembro e
outubro.
Epidendrum polyphyllum foi por muito tempo tratado como sinônimo de Cyrtopodium
andersonii R.Br., que é na verdade endêmica das Índias Ocidentais, não ocorrendo no
Brasil. Apenas recentemente, passaram a ser consideradas espécies distintas. Por outro

44
lado, Barros (1994) considerou que o nome correto desta espécie é Cyrtopodium
polyphyllum, com base na ilustração de Epidendrum polyphyllum na Flora Fluminensis.
09. ENCYCLIA Hook.
09.1. Encyclia alboxanthina Fowlie, Orchid Digest. 54: 26-27, 1990.
Erva rupícola. Rizoma inconspícuo. Pseudobulbos 5-8 x 2-4 cm, 2-4-foliados, eretos,
ovóides, verdes, cobertos por bainhas. Folhas 20-30 x 2-3 cm, sésseis, semi-eretas a eretas,
lanceoladas, verdes, coriáceas, ápice agudo mucronado, base atenuada. Inflorescência
panícula terminal, ereta; escapo 65-80 cm comp., verde; brácteas escapais 6,0-10 x 10-12
mm, ovadas a orbiculares, paleáceas, ápice agudo, base amplexicaule; raque 45-50 cm
comp., 8-30 flores; brácteas florais 3,0-5,0 x 2,0-3,0 mm, ovadas, paleáceas, ápice agudo,
base truncada. Flores ressupinadas; pedicelo incluindo o ovário 2,5-3,0 cm comp. Sépalas
20-25 x 8,0-10 mm, patentes, amarelo-esverdeadas com estrias vinácea, membranáceas,
ápice agudo apiculado, base truncada; dorsal levemente obovada; laterais lanceoladas.
Pétalas 20-25 x 7,0-10 mm, patentes, espatuladas, amarelo-esverdeadas com estrias
vinácea, membranáceas, ápice obtuso apiculado, base atenuada. Labelo concrescido à base
da coluna, ereto, 3-lobado, branco com linhas purpúreas no lobo terminal, disco amarelo,
membranáceo, base atenuada; disco carnoso, com duas carenas; lobos laterais 8,0-11 x 5,0-
6,0 mm, envolvendo a coluna e então reflexos, obtusos, ápice obtuso; lobo terminal 15-18
x 11-13 mm, orbicular, ápice agudo, margem ligeiramente ondulada. Coluna 12-13 x 5,0-
6,0 mm, 3,0-4,0 mm alt., amarelo-esverdeada; provida de dois braços voltados para baixo,
antera 3,0 x 4,0 mm, amarela; polínias 4, amarelas.
Material examinado: BRASIL, Bahia: Parque Municipal de Mucugê, 18.out.2002,
C.Azevedo 159 (HUEFS); Mucugê, Reserva do Projeto Sempre-Viva, 24.out.2000,
E.R.Souza et al. 88 (HUEFS); sede do "Projeto Sempre-Viva", 16.dez.1999, L.P.Queiroz et
al. 5585 (HUEFS); Unidade de Manejo Sustentável, 03.jan.1997, UMS 70 (HRB); Unidade
de Manejo Sustentável, 06.abr.1997, UMS 284 (HRB); Unidade de Manejo Sustentável,
02.nov.1996, UMS 348 (HRB).
Material adicional examinado: BRASIL, Bahia: estrada entre Andaraí e Mucugê,
20.nov.1983, L.R.Noblick & A.Pinto 2883 (HUEFS); 10 km S. of Andaraí, on the road to
Mucugê, 16.fev.1977, R.M.Harley et al. 18767 (ALCB, CEPEC, HUEFS). Mucugê, Serra

45
do Capa Bode, nov.1973, G.C.P.Pinto s.n. (ALCB 477, HRB 31060); by rio Cumbuca c. 3
km S of Mucugê, near site of small dam on road to Cascavel, 04.fev.1974, R.M.Harley et
al 15962 (CEPEC).
Descrita originalmente do município de Mucugê (Fowlie, 1990), esta espécie foi
citada para Serra do Curral Feio e Serra do Sincorá, identificada como E. advena e E.
osmantha (Harley & Mayo, 1980), para Mucugê/Andaraí como E. advena (Harley &
Simmons, 1986), e para Lençóis (Toscano de Brito, 1998) e Catolés (Toscano de Brito &
Queiroz, 2003). No Plano de Manejo do Parque, UMS 70, foi identificada como E. advena.
Esta espécie foi por muito tempo confundida com E. advena e E. osmantha, até ser
descrita como espécie nova (Fowlie, 1990). E. advena foi sempre um nome de aplicação
controversa, até ser mais recentemente aplicado à espécie comumente chamada de E.
megalantha (Castro & Campacci, 2000). E. alboxanthina difere de E. osmantha por suas
plantas e flores bem maiores, labelo branco, por vezes maculado, e de textura lisa, hastes
florais muito mais longas e por ser de hábito exclusivamente rupícola na Chapada
Diamantina, enquanto que em E. osmantha existem rugosidades sobre as venulações,
sendo epífita nas matas de galeria em vegetação de cerrado em Minas Gerais, Goiás e
Sudoeste da Bahia. E. alboxanthina é abundante nos campos rupestres do Parque,
crescendo diretamente sobre as rochas a pleno sol, muitas vezes ao lado de Cattleya
elongata. Floresce entre outubro e fevereiro.
Fowlie (1990), ao descrever E. alboxanthina, comenta que por muito tempo ele e o
Dr. Denis Duveen sabiam sobre esta espécie que crescia na Serra do Sincorá, na região de
Mucugê, e que o próprio Pabst, antes de sua morte, reconheceu que esta era uma espécie
nova, mas não teve oportunidade de nomeá-la. No Parque, Encyclia alboxanthina é
reconhecida por suas longas inflorescências e grande quantidade de flores de coloração
amarelo-esverdeadas com estrias vináceas e labelo branco com linhas purpúreas.
10. EPIDENDRUM L.
Chave de identificação para as espécies de Epidendrum ocorrentes no Parque Municipal de
Mucugê.
1. Inflorescência ereta, longa, entre 36-71 cm comp., escapo mais longo que a raque,
brácteas escapais não imbricadas.

46
2. Flores ressupinadas, marrom-esverdeadas, labelo com margem inteira. ............................
........................................................................................... Epidendrum orchidiflorum (10.2)
2’. Flores não ressupinadas, róseas, labelo com margem denticulada. ...................................
................................................................................................. Epidendrum secundum (10.3)
1’. Inflorescência pendente, curta, entre 11-23 cm comp., escapo mais curto que a raque
brácteas escapais imbricadas.
3. Flores amarelas com máculas vináceas, e labelo amarelo com máculas vináceas, de
margem laciniada. ............................................................. Epidendrum cristatum (10.1)
3’. Flores marrom-esverdeadas, e labelo branco-esverdeado, de margem lisa. ................
............................................................................................... Epidendrum warasii (10.4)
10.1. Epidendrum cristatum Ruiz & Pav., Syst. veg. fl. peruv. chil. 1: 243. 1798.
Erva terrestre. Rizoma inconspícuo. Caule 150-200 x 1-2 cm, ereto, cilíndrico, verde,
coberto por 7-12 folhas. Folhas 10-15 x 1,5-2 cm, dísticas, sésseis, semi-eretas, oblongas,
verdes, semicoriáceas, ápice obtuso, base truncada, articulada com a bainha.
Inflorescência racemo terminal, pendente; escapo 8-10 cm comp., vináceo; brácteas
escapais 50-75 x 10-15 mm, oblongas, imbricadas, paleáceas, ápice agudo, base truncada;
raque 10-15 cm comp., 25-30 flores; brácteas florais 3,0-8,0 x 1,0-2,0 mm, triangulares,
paleáceas, ápice agudo, base truncada. Flores ressupinadas; pedicelo incluindo o ovário
40-45 mm comp. Sépalas reflexas, lanceoladas, amareladas com máculas vinácea,
membranáceas, ápice agudo, base truncada; dorsal 15-20 x 4,0-6,0 mm; laterais 15-17 x
5,0-6,0 mm. Pétalas 15-17 x 4,0-5,0 mm, reflexas, obovadas, amareladas com máculas
vinácea, menos máculas que as sépalas, membranáceas, ápice agudo, base truncada.
Labelo 10-15 x 10-15 mm, concrescido à face adaxial da coluna em toda sua extensão,
patente, 3-lobado, orbicular, amarelado com máculas vinácea, membranáceo, base
atenuada, margem laciniada; lobos laterais patentes, orbiculares, ápice obtusa; lobo
terminal patente e reflexo, dividido em lóbulos, lineares, ápice obtuso. Coluna 10-15 x 3,0
mm, 2,0 mm alt., branca com ápice vináceo; antera 2,0 x ,02 mm, branca; polínias 4.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
20.set.2002, C.Azevedo 156 (HUEFS).
Material adicional examinado: BRASIL, Bahia: Mucugê, 14.out.1996, M.C.Ferreira
1199 (HRB).

47
Citada para Colômbia, Venezuela, Guiana, Suriname, Brasil, Equador, Peru,
Bolívia, (Bennett & Christenson, 1993; Sprunger et al., 1996), esta espécie ocorre no
Brasil nos estados de Minas Gerais, Espírito Santo, Rio de Janeiro, São Paulo, Paraná,
Santa Catarina e Rio Grande do Sul (Pabst & Dungs, 1975; Sprunger et al., 1996). Na
Bahia, foi citado para Catolés (Toscano de Brito & Queiroz, 2003), sendo aqui registrada
pela primeira vez para o município de Mucugê. Floresce entre setembro e outubro.
Esta espécie é altamente variável, tanto vegetativamente quanto em suas partes
florais e, como conseqüência disso, foi descrita várias vezes, apresentando um grande
número de sinônimos. Pabst & Dungs (1975) consideram E. raniferum Lindl., entre outros
(E. calliferum Lem., E. hexadactyllum Barb.Rodr., E. longovarium Barb.Rodr., E.
bathyschistum Schltr., E. validum Schltr., E. alexandrii Schltr., E. loefgrenii (Cogn.)
Cogn., E. millei Schltr.) como sinônimos de E. cristatum, enquanto Dunsterville & Garay
(1961c) e Sprunger et al. (1991, 1996), além de considerarem todos os citados por Pabst &
Dungs (1975), citaram todas as variedades de E. raniferum (E. raniferum var. luteum
Lindl., E. raniferum var. obtusilobum Cogn., E. raniferum var. hexadactyllum (Barb.Rodr.)
Cogn., E. raniferum var. lutescens Lindl. ex Broadw., E. raniferum var. loefgrenii Cogn.)
como sinônimos da espécie, além de E. rantierium Lindl. ex Sanchez, Bennett &
Christenson (1993) comentam que E. raniferum, anteriormente considerado sinônimo de E.
cristatum, é agora aplicado a espécies mexicanas e mesoamericanas. As variedades aceitas
por eles como sinônimas, foram apenas aquelas baseadas em coleções da América do Sul
(E. calliferum, E. hexadactyllum, E. longovarium, E. bathyschistum, E. validum, E.
alexandrii, E. raniferum var. luteum, E. raniferum var. obtusilobum, E. raniferum var.
hexadactylum, E. raniferum var. lutescens).
Assemelha-se vegetativamente a E. warasii pelo porte alto de suas plantas, que
chegam a 2 m de altura, embora diferencie-se deste por apresentar folhas mais longas e de
textura mais fina. As duas apresentam escapo mais curto que a raque, brácteas escapais
longas e imbricadas, mas diferenciam-se na coloração e formato de suas flores, já que E.
cristatum tem flores amareladas com máculas vináceas e labelo de margem laciniada.
10.2. Epidendrum orchidiflorum Salzm. ex Lindl., Gen. sp. orchid. pl.103, 1831.
Epidendrum huebneri Schltr., Beih. Bot. Centralbl. 42, pt 2: 99, 1925.
Erva rupícola. Rizoma inconspícuo. Caule 16-40 x 0,5-1 cm, ereto, cilíndrico, marrom,
coberto pelas bainhas das folhas, 6-22 folhas. Folhas 4-5 x 1-2 cm, dísticas, sésseis, semi-

48
eretas, lanceoladas, verdes, coriáceas, ápice agudo, base atenuada, articulada com as
bainhas. Inflorescência racemo terminal, ereta; escapo 50-65 cm comp., marrom-
esverdeado; brácteas escapais 30-45 x 5,0-10 mm, oblongas, paleáceas, ápice agudo, base
amplexicaule; raque 3-6 cm comp., 5-10 flores; brácteas florais 4,0-7,0 x 1,0-2,0 mm,
triangulares, membranáceas, ápice agudo, base truncada. Flores ressupinadas; pedicelo
incluindo o ovário 20-25 mm comp. Sépalas patentes, marrom-esverdeadas,
membranáceas, ápice agudo, base atenuada; dorsal 10-15 x 5,0-6,0 mm, obovada; laterais
10-12 x 5,0-6,0 mm, oblongo-obovadas. Pétalas 10-12 x 1,0-2,0 mm, patentes, lineares,
marrom-esverdeadas, membranáceas, ápice agudo, base atenuada. Labelo 8,0-10 x 15 mm,
concrescido à face adaxial da coluna em toda sua extensão, patente, 1-lobado, orbicular,
marrom-esverdeado, carnoso, ápice obtuso retuso, base atenuada. Coluna 6,0-7,0 x 2,5-3,0
mm, 2,5-3,0 mm alt., marrom-esverdeada; antera 1,5-2,0 x 1,0-1,5 mm, orbicular, marrom-
esverdeada; polínias 4, deltóides.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
11.jun.2002, C.Azevedo & J.Oliveira 138 (HUEFS); Unidade de Manejo Sustentável,
12.jan.1997, UMS 159 (HRB); Unidade de Manejo Sustentável, 02.nov.1996, UMS 346
(HRB).
Material adicional examinado: BRASIL, Bahia: Mucugê, próximo à área do Parque da
Chapada Diamantina, 29.jun.1997, M.L.Guedes et al. 5109 (ALCB); 31.jan.1997, PCD
4745 (HUEFS).
Descrito originalmente de material proveniente da Bahia (Lindley, 1831), E.
orchidiflorum foi citado para a Venezuela e Brasil, neste para os estados de Alagoas e
Bahia (Pabst & Dungs, 1975) Amazonas, Roraima, Para, Sergipe, Bahia e Rio de Janeiro
(Sprunger et al., 1996), e países como Venezuela, Guiana, Brasil e Peru por Bennett &
Christenson (1993) e Sprunger et al. (1996). Na Chapada Diamantina, é citado para os
municípios de Lençóis (Toscano de Brito, 1998), Palmeiras (Toscano de Brito, 1998;
Conceição & Giulietti, 2002), e no município de Mucugê foi citado para o Parque
(Stradmann, 1998). Comum na área do Parque, E. orchidiflorum ocorre em vários
ambientes da reserva. Foi observado em flor entre novembro e janeiro, e no mês de julho.
Embora Pabst & Dungs (1975) considerem E. orchidiflorum e E. huebneri como
espécies distintas, Bennett & Christenson (1993) comentaram ter seguido Carnevali
(unpubl.), que considera E. huebneri sinônimo de E. orchidiflorum, além de outros

49
sinônimos: E. caespitosum Barb.Rodr., E. rectopedunculatum C.Schweinf., E. acrirachis
Pabst, E. garayanum Hawkes. A mesma delimitação foi seguida por Sprunger et al. (1996),
sendo esta a adotada neste trabalho.
10.3. Epidendrum secundum Jacq., Enum. syst. pl. 29, 1760.
Epidendrum crassifolium Lindl., Gen. et Sp. Orch. 107, 1831.
Epidendrum elongatum Jacq., Coll. 3: 260, 1789.
Epidendrum ellipticum Grah., Edinb. N. Phil. J. 1: 171, 1826.
Erva terrestre. Rizoma inconspícuo. Caule (7-)20-27 x 0,5-1 cm, ereto, cilíndrico, verde,
coberto pelas bainhas das folhas, 6-20 folhas. Folhas 5-8 x 1-2 cm, dísticas, sésseis, semi-
eretas, lanceoladas, verdes, coriáceas, ápice agudo, base atenuada, articulada com as
bainhas. Inflorescência racemo terminal, ereta; escapo (17-24-)33-48 cm comp., verde;
brácteas escapais 35-55 x 2,0-4,0 mm, oblongo-lanceoladas, paleáceas, ápice agudo, base
amplexicaule; raque 3-5 cm comp., 4-6 flores; brácteas florais 2,0-5,0 x 1,0 mm, ovadas a
triangulares, paleáceas, ápice agudo, base truncada. Flores não ressupinadas; pedicelo
incluindo o ovário 10-20 mm comp. Sépalas róseas, membranáceas, base truncada; dorsal
10-15 x 3,0-5,0 mm, reflexa, lanceolada, ápice agudo; laterais 10-14 x 4,0-5,0 mm,
patentes, oblongas a obovadas, ápice obtuso apiculado. Pétalas 10-14 x 2,0-3,0 mm,
patentes, lanceoladas, róseas, membranáceas, ápice agudo, base atenuada. Labelo
concrescido à face adaxial da coluna em toda sua extensão, ereto, 3-lobado, orbicular,
róseo, membranáceo, base atenuada, margem denticulada, calosidade branca com centro
amarelo; lobos laterais 2,0-3,0 x 2,0-3,0 mm, eretos, ovados a orbiculares, ápice obtuso,
lobo terminal 3,0-4,5 x 3,0-4,5 mm, ereto, orbicular, ápice agudo. Coluna 4,0-5,0 x 1,0-2,0
mm, 1,0-1,5 mm alt., rósea; antera 1,0-1,5 x 1,0-1,2 mm, amarela; polínias 4, obovadas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
18.out.2002, C.Azevedo 158 (HUEFS); Unidade de Manejo Sustentável, 06.abr.1997, UMS
262 (HRB); Unidade de Manejo Sustentável, 02.nov.1996, UMS 347 (HRB).
Material adicional examinado: BRASIL, Bahia: estrada entre Andaraí e Mucugê,
20.nov.1983, L.R.Noblick & A.Pinto 2900 (HUEFS). Mucugê, Guiné, 21.out.2002,
E.C.Smidt 289 (HUEFS); caminho para Guiné, 15.feb.1997, PCD 5708 (ALCB); estrada
Mucugê - Andaraí, c. 8 km de Mucugê, torre da Telebahia, 12.nov.1998, R.P.Oliveira et al.
37 (HUEFS); fazenda Paiol, 20.out.1986, R.P. Orlandi et al. 792 (HRB).

50
É uma espécie de ampla distribuição geográfica, conhecida de toda a América
tropical (Dunsterville & Garay, 1961b; Pabst & Dungs, 1975; Sprunger et al., 1996). No
Brasil, é citada para os estados da Amazonas, Amapá, Pará, Ceará, Bahia, Rio de Janeiro,
Mato Grosso, Minas Gerais, Goiás, Distrito Federal, São Paulo, Paraná, Pernambuco,
Santa Catarina e Rio Grande do Sul (Pabst & Dungs, 1975). Na Bahia, foi citada por
Harley & Mayo (1980) para Alcobaça, Ilhéus, Serra do Curral Feio, Serra do Sincorá e
Serra das Almas, identificada como E. crassifolium e E. elongatum, foi citada também para
Mucugê/Andaraí como E. crassifolium, e E. elongatum (Harley & Simmons, 1986), Rio de
Contas (Toscano de Brito, 1995), Lençóis (Toscano de Brito, 1998), Palmeiras (Toscano
de Brito, 1998; Conceição & Giulietti, 2002) e Catolés (Toscano de Brito & Queiroz,
2003), e na Cadeia do Espinhaço para a Serra do Cipó (Barros, 1987) e Grão-Mogol
(Barros & Pinheiro, no prelo). Cresce em locais ensolarados, com substratos pedregosos e
arenosos. Floresce entre outubro e fevereiro.
Esta espécie é altamente variável, tanto vegetativamente como no tamanho, forma e
cor de suas flores, e por este motivo foi aparentemente descrita várias vezes. Pabst &
Dungs (1975) consideram E. crassifolium, E. ellipticum e E. elongatum espécies distintas,
enquanto Sprunger et al. (1991, 1996) consideram E. crassifolium e E. ellipticum sinônimo
de E. elongatum. Neste trabalho seguimos Dunsterville & Garay (1961b), que consideram
todos os quatro nomes conspecíficos, além de apresentar uma longa lista com outros
sinônimos, totalizando 37 nomes, e ainda ilustração mostrando a variação de formas do
labelo e calo. Entre as outras espécies de Epidendrum da área, E. secundum pode ser
reconhecida por suas flores não ressupinadas, pequenas, róseas e pelo formato de seu
labelo, de margem denticulada.
10.4. Epidendrum warasii Pabst, Orchid. Rev. 79(941): 306, 1971.
Erva rupícola. Rizoma inconspícuo, raízes emergindo de todo o rizoma. Caule 66-200 x
1,5-2 cm, ereto, cilíndrico, verde, recoberto pelas bainhas das folhas, 8-16 folhas. Folhas
6-7 x 1,5-2 cm, dísticas, sésseis, semi-eretas, ovadas a oblongas, verdes, carnosa, ápice
obtuso, base truncada. Inflorescência racemo terminal, pendente; escapo 8-9 cm comp.;
brácteas escapais 30-40 x 10 mm, ovadas, imbricadas, paleáceas, ápice agudo, base
truncada; raque 30-40 mm comp., 10-15 flores; brácteas florais 7,0-10 x 3,0-4,0 mm,
ovadas, membranáceas, ápice agudo, base truncada. Flores ressupinadas; pedicelo
incluindo o ovário 15-33 mm comp. Sépalas patentes, lanceoladas, marrom-esverdeadas,

51
membranáceas, base truncada; dorsal 15-20 x 5,0-6,0 mm, ápice agudo; laterais 15-20 x
5,0-7,0 mm, ápice obtuso apiculado. Pétalas 10-15 x 3,0-4,0 mm, patentes, lanceoladas,
marrom-esverdeadas, membranáceas, ápice levemente obtuso apiculado, base atenuada.
Labelo concrescido à face adaxial da coluna em toda sua extensão, ereto, 3-lobado,
branco-esverdeado, carnoso, base atenuada; disco com 2 calosidades; lobos laterais 7,0-10
x 3,0-4,0 mm, patentes e reflexos, oblongos, ápice obtuso; lobo terminal 5,0-9,0 x 6,0-7,0
mm, patente, orbicular, ápice truncado mucronado. Coluna 8,0-9,0 x 3,0-4,0 mm, 4,0-5,0
mm alt., branca-esverdeada; antera 1,5-2,0 x 2,0 mm, branca; polínias 4.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
20.fev.2003, C.Azevedo & J.Oliveira 176 (HUEFS).
Citada por Pabst & Dungs (1975) apenas para o estado de Minas Gerais, e
posteriormente para a Serra do Cipó (Barros, 1987) e Grão-Mogol (Barros & Pinheiro, no
prelo), esta espécie foi também citada na Bahia para Rio de Contas (Toscano de Brito,
1995) e Catolés (Toscano de Brito & Queiroz, 2003), sendo aqui registrada para o
município de Mucugê, onde cresce em áreas abertas, diretamente sobre as rochas em
campo rupestre. Floresce em fevereiro.
É vegetativamente parecido com E. cristatum, pela altura da planta, que chega a 2
m alt., diferindo desta por seu porte mais robusto, suas folhas mais curtas e carnosas, além
da coloração e formato de suas flores, marrom-esverdeadas e labelo branco-esverdeado, de
margem lisa.
11. EPISTEPHIUM Kunth
11.1. Epistephium lucidum Cogn., Fl. bras. 3(4): 141, pl. 30, 1893.
Erva terrestre. Rizoma inconspícuo. Caule 150-200 x 0,5-1 cm, ereto, cilíndrico, verde,
coberto por 4-11 folhas. Folhas (4-)8-11 x 2-4 cm, dísticas, sésseis, semi-eretas, ovadas,
verdes, coriáceas, ápice agudo apiculado, base amplexicaule, margem ligeiramente reflexa,
nervação reticulada. Inflorescência racemo terminal, ereto; raque 15-26 cm comp., 1-3
flores; brácteas florais 3,0-6,0 x 4,0-6,0 mm, ovadas a orbiculares, membranáceas, ápice
agudo, base truncada. Flores ressupinadas; pedicelo incluindo o ovário 35-40 mm comp.,
presença de epicálice. Sépalas 40-50 x 15-20 mm, patentes, lanceoladas, lilás,

52
membranáceas, ápice agudo apiculado, base atenuada. Pétalas 40-50 x 20-25 mm,
patentes, obovadas, lilás, membranáceas, ápice obtuso, base atenuada. Labelo 50-60 x 40-
50 mm, concrescido à face lateral da coluna até 1/3 de seu comprimento, os outros 2/3
envolvem a mesma, ereto, 1-lobado, obovado, lilás, ápice mais escuro, internamente
branco com veias lilás, membranáceo, ápice obtuso retuso, base atenuada, margem
ondulada, pêlos em todo comprimento na porção central do labelo. Coluna 30-35 x 5,0
mm, 4,0-5,0 mm alt., lilás com ápice branco; estigma fechado; antera 4,0-5,0 x 4,0-5,0
mm, branca; polínias 4. Fruto cápsula 40-50 x 5,0-7,0 mm, verde.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
27.jan.2003, C.Azevedo 172 (HUEFS); c. 4 km N de Mucugê na Chapada para Andaraí,
sede do "Projeto Sempre-Viva", 23.jan.2000, L.P.Queiroz 5619 (HUEFS); Unidade de
Manejo Sustentável, 04.jan.1997, UMS 114 (HRB).
Material adicional examinado: BRASIL, Bahia, 2 km al N de Mucugê, camino a
Andaraí, 20.jan.1997, M.M.Arbo et al. 7575 (CEPEC, CTES); 10 km S of Andaraí on the
road to Mucugê, 16.fev.1977, R.M. Harley et al. 18773 (CEPEC); by rio Cumbuca c. 3 km
S of Mucugê, near site of small dam on road to Cascavel, 04.fev.1974, R.M.Harley et al.
15905 (CEPEC). Mucugê, Serra do Capa Bode, nov.1973, G.Pinto s.n. (ALCB 471).
Hoehne (1945) citou a ocorrência de E. lucidum para o Espírito Santo, Rio de
Janeiro, São Paulo e Minas Gerais, sendo também conhecido dos estados da Bahia, Distrito
Federal, Goiás e Roraima (Pabst & Dungs, 1975). Na Bahia, foi citado para a Serra de
Jacobina, Serra do Sincorá, Serra das Almas e Belmonte (Harley & Mayo, 1980), e para
Mucugê/Andaraí (Harley & Simmons, 1986), Palmeiras (Conceição & Giulietti, 2002), Rio
de Contas (Toscano de Brito, 1995) e Catolés (Toscano de Brito & Queiroz, 2003). Na
porção mineira da Cadeia do Espinhaço, foi citado para Grão-Mogol (Barros & Pinheiro,
no prelo). Cresce em áreas abertas e ensolaradas, dominadas por gramíneas, em solo
arenoso, ocorrendo também em restinga. Floresce entre janeiro e fevereiro.
Facilmente reconhecido por suas plantas grandes, chegando até a 2 m de altura,
pela nervura reticulada de suas folhas, pela presença de epicálice e por suas flores grandes
de cor lilás, que se assemelham às flores de Cattleya.

53
12. HABENARIA Willd.
12.1. Habenaria fluminensis Hoehne, Arq. Bot. Estado São Paulo n.s. 1: 41, tab. 46, 1939.
Erva terrestre. Rizoma inconspícuo. Caule 80-100 x 0,5-1 cm, até 1a flor, ereto,
cilíndrico, verde, coberto pelas bainhas das folhas, 4-6 folhas. Folhas 20-27 x 1-1,5 cm,
dísticas, sésseis, semi-eretas a eretas, linear-lanceoladas, verdes, membranáceas, ápice
agudo, base truncada. Inflorescência racemo terminal, ereto; raque 20-25 cm comp., 15-17
flores; brácteas florais 2-3 x 1 cm, obovadas, membranáceas, ápice obtuso caudado, base
atenuada. Flores ressupinadas; pedicelo incluindo o ovário 2,5-3 cm comp., sulcado.
Sépalas esverdeadas, membranáceas, base truncada; dorsal 12-15 x 7,0-9,0 mm, patente,
ovada, ápice obtuso acuminado; laterais 12-14 x 4,0-5,0 mm, reflexas, ovadas a
lanceoladas, ápice agudo. Pétalas 12-14 x 4,0-5,0 mm (base), bipartidas até perto da base,
reflexas, lineares, esverdeadas, membranáceas, ápice agudo, base truncada. Labelo
patente, tripartido, linear, esverdeado, membranáceo, base truncada; lobos laterais 15-17 x
1,0-2,0 mm, patentes, lineares, ápice agudo; lobo mediano 15-16 x 3,0-4,0 mm, patente,
linear, ápice agudo. Cálcar 20-25 x 2,0-3,0 mm, dilatado no ápice. Coluna 4,0-5,0 x 3,0-
4,0 mm, 2,0 mm alt., verde; polínias 2.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
12.jun.2002, C.Azevedo & O.Oliveira 141 (HUEFS); Parque Municipal de Mucugê, acima
da cachoeira, 13.jul.2002, C.Azevedo 144 (HUEFS).
Descrita originalmente da Serra do Mar, São Paulo e Petrópolis, Rio de Janeiro
(Hoehne, 1939), esta espécie foi citada também para Minas Gerais (Pabst & Dungs, 1975),
Bahia e Espírito Santo (Batista & Bianchetti, 2002). No estado da Bahia, Harley & Mayo
(1980) citaram sua ocorrência para o município de Morro do Chapéu. É aqui registrada
para o município de Mucugê. Cresce em solo arenoso, em área aberta, ensolarada,
dominada por gramíneas. Floresce entre junho e julho.
Habenaria fluminensis é facilmente reconhecida no Parque por suas flores
esverdeadas, pétalas bipartidas, labelo tripartido e por apresentar um longo cálcar. Planta
encontrada apenas no período de floração e frutificação, já que, logo após esta fase, a
planta fenece, perdendo toda parte vegetativa.

54
13. MAXILLARIA Ruiz & Pav.
13.1. Maxillaria cerifera Barb.Rodr., Gen. spec. Orchid. 1: 118, 1877. Tipo: Brasil,
Barbosa Rodrigues s.n. (perdido). Lectótipo aqui designado: ilustração original
preparada por João Barbosa Rodrigues, que consta de sua Iconographie des
orchidées du Brésil, depositada na biblioteca do Jardim Botânico do Rio de Janeiro,
reproduzida por Sprunger et al. (1996) sob o número t. 296 do volume 6.
Erva rupícola. Rizoma 5,0-10 mm comp. entre pseudobulbos, 2,0-3,0 mm diâm., coberto
por bainhas, raízes emergindo ao longo de todo o rizoma. Pseudobulbos 10-15 x 5,0-7,0
mm, 2-foliados, eretos, ovóides, verdes, cobertos por 2 bainhas das folhas laterais. Folhas
laterais 15-20 x 3,0-4,0 mm, sésseis, semi-eretos, lanceoladas a oblongas, verdes,
coriáceas, ápice obtuso retuso, base atenuada. Folhas apicais 40-60 x 4,0-5,0 mm, sésseis,
semi-eretos, oblongas, verdes, coriáceas, ápice obtuso, retuso, base atenuada formando
pseudopecíolo. Inflorescência uniflora, lateral; escapo 4-4,5 cm comp., ereto, marrom;
brácteas escapais 5-8, 7,0-10 x 2,0-3,0 mm, oblongas, paleáceas, ápice agudo, apiculado,
base amplexicaule. Flores ressupinadas; pedicelo incluindo o ovário 10-12 mm comp.
Sépalas 8,0-10 x 2,0-3,0 mm, eretas, esverdeadas, ápice com nuances vinácea,
membranáceas, ápice agudo, base truncada; dorsal linear; laterais triangulares. Pétalas 6,0-
7,0 x 1,5-2,0 mm, eretas, lanceoladas, esverdeadas, ápice com nuances vinácea,
membranáceas, ápice agudo, base atenuada. Labelo articulado com o pé da coluna, ereto,
3-lobado, esverdeado, ápice com nuances vinácea, calosidade amarelada, membranácea,
base atenuada; calo convexo orbicular; lobos laterais 1,5 x 0,5 mm, eretos, oblongos, ápice
obtuso; lobo terminal 6,0-7,0 x 2,0-3,0 mm, ereto, lanceolado, coberto por substância
cerosa na face adaxial, em forma de V, ápice agudo. Coluna 4,0 x 1,0 mm, 1,0 mm alt.,
prolongada na base em pé, vinácea, levemente curvada; antera 1,0 x 1,0 mm, vinácea,
orbicular, prolongada em bico granuloso; polínias 4, orbiculares, amarelas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
19.set.2002, C.Azevedo & J.Oliveira 153 (HUEFS).
Descrita originalmente de material proveniente de Petrópolis, Rio de Janeiro
(Barbosa Rodrigues, 1877), Maxillaria cerifera é citada também para os estados do
Espírito Santo, São Paulo, Paraná, Santa Catarina, Rio Grande do Sul e Minas Gerais

55
(Pabst & Dungs, 1977; Sprunger et al., 1996), sendo aqui registrada pela primeira vez para
o estado da Bahia. Rara na área do Parque, esta espécie é encontrada em paredões rochosos
verticais e floresce entre maio e setembro.
M. cerifera assemelha-se muito a M. notylioglossa Rchb.f., apesar da última ter
sido descrita a partir de material proveniente de Caracas, Venezuela (Reichenbach, 1854).
Dunsterville & Garay (1961a) indicaram seu endemismo para este país, embora Pabst &
Dungs (1977) tenham citado sua ocorrência no Brasil para Minas Gerais e São Paulo.
Toscano de Brito (1998) citou a ocorrência desta espécie para a Serra da Chapadinha,
Bahia. Infelizmente, este material (PCD 1290) encontra-se estéril, tendo sido possível
estudar apenas suas características vegetativas, que se assemelham bastante ao material do
Parque. Por outro lado, o espécime aqui estudado concorda inteiramente com as ilustrações
de Barbosa Rodrigues (Sprunger et al., 1996). Diante disso, preferiu-se considerá-lo como
M. Cerifera, até que estudos posteriores investiguem a delimitação de M. cerifera em
relação a M. notylioglossa.
O holótipo desta espécie encontra-se perdido, desta forma a ilustração original de
M. cerifera, preparada na natureza por Barbosa Rodrigues e ilustrada em sua Iconographie
des orchidées du Brésil, recentemente publicada por Sprunger et al., (1996), constitui o
único material original disponível, e, por esse motivo, é aqui selecionado como lectótipo.
14. OCTOMERIA R.Br.
Chave de identificação para as espécies de Octomeria ocorrentes no Parque Municipal de
Mucugê.
1. Folhas teretes, verdes, flores brancas com máculas vinácea a amarelas.
2. Planta 28-55 mm alt., labelo 1-lobado, flores brancas, translúcidas com máculas
vinácea. .................................................................................... Octomeria flabellifera (14.2)
2’. Planta 70-150 mm alt., labelo 3-lobado, flores amarelas. .... Octomeria alexandrii (14.1)
1’. Folhas lanceoladas, verdes a arroxeadas, flores róseas com margem alaranjada e labelo
avermelhado. ................................................................................... Octomeria stellaris (14.3)

56
14.1. Octomeria alexandrii Schltr., Mem. Inst. Butantan, Secc. Bot. 1. 4:53, pl. 12, 1922.
Erva rupícola. Rizoma inconspícuo, raízes emergindo ao longo de todo o rizoma. Caule
40-90 x 1,0-2,0 mm, 1-foliado, ereto, cilíndrico, marrom, coberto por bainhas. Folha 30-60
x 5,0 mm, séssil, ereta, terete, verde, carnosa, ápice agudo, base atenuada. Inflorescência
fasciculada terminal; raque 3,0-4,0 mm comp., 1-2 flores em antese. Flores ressupinadas;
pedicelo incluindo o ovário 5,0 mm comp. Sépalas 5,0-6,0 x 2,5-3,0 mm, patentes, ovadas,
amarelas com ápice amarronzado, membranáceas, ápice obtuso reflexo, base truncada.
Pétalas 5,5-6,0 x 2,5 mm, patentes, elíptico-lanceoladas, amarelas com ápice amarronzado,
membranáceas, ápice obtuso reflexo, base truncada. Labelo articulado com o pé da coluna,
ereto, 3-lobado; obovado, amarelo, membranáceo, base atenuada; calosidade provida de
duas quilhas erguidas; lobos laterais 1,0-1,5 x 0,5-1,0 mm, eretos, oblongos, ápice obtuso;
lobo terminal 1,5-2,0 x 2,0-2,5 mm, ereto, obovado, ápice emarginado. Coluna 2,0-3,0 x
0,5-1,0 mm, 0,5 mm alt., prolongada na base em pé de 2,0 mm comp., amarelada,
levemente curvada; antera 0,5-1,0 x 0,6 mm, amarelada; polínias 8, obovadas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
11.jun.2002, C.Azevedo & J.Oliveira 137 (HUEFS); Parque Municipal de Mucugê,
13.jul.2002, C.Azevedo 145 (HUEFS).
Descrita originalmente de São Paulo (Schlechter, 1922), O. alexandrii é citada
também para o estado do Rio de Janeiro (Pabst & Dungs, 1975). Na Bahia, foi citada para
Lençóis e Palmeiras (Toscano de Brito, 1998), sendo aqui registrada pela primeira vez para
o município de Mucugê. Esta espécie é comum na área do Parque, onde cresce diretamente
sobre as rochas em campo rupestre, e é encontrada em flor nos meses de abril, junho e
setembro.
O. alexandrii caracteriza-se por apresentar folha terete, ereta e inflorescência
uniflora ou, mais raramente, duas flores simultaneamente em antese, e pela forma e
coloração de suas flores, amarelas com ápice amarronzado.

57
14.2. Octomeria flabellifera Pabst, Bradea 2(10): 56, t. 1, fig. H, 1975.
Erva rupícola. Rizoma inconspícuo. Caule 13-20 x 0,5-1,0 mm, 1-foliado, ereto,
cilíndrico, marrom, coberto por bainhas. Folha 15-35 x 2,0-3,0 mm, séssil, ereta, terete,
verde, coriácea, ápice agudo, base truncada. Inflorescência fasciculada terminal; raque
4,0-5,0 mm comp., 1 flor em antese; brácteas florais inconspícuas. Flor ressupinada;
pedicelo incluindo o ovário 2,5-3,0 mm comp. Sépalas 7,0-8,0 x 2,0-2,5 mm, eretas,
lanceoladas, brancas translúcidas com máculas vinácea, uma maior, bem marcante na base
das sépalas laterais, membranáceas, ápice agudo, base truncada; laterais concrescidas na
base e com o pé da coluna. Pétalas 7,0-8,0 x 1,5-2,0 mm, eretas, lanceoladas, brancas
translúcidas com máculas vinácea, membranáceas, ápice agudo, base truncada. Labelo 3,5-
4,0 x 2,5-3,0 mm, articulado com o pé da coluna, ereto, 1-lobado, transversalmente
elíptico, branco translúcido, calosidade central amarela e quilhas vinácea, membranáceo,
ápice obtuso retuso, base ungüiculada; disco do labelo com calosidade central e três
quilhas erguidas. Coluna 2,0-2,5 x 0,5 mm, 1,0 mm alt., esverdeada; antera 0,5 x 0,7 mm.,
amarela; polínias 8, amarelas. Fruto cápsula 8,0-9,0 x 2,5-3,0 mm.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
18.set.2002, C.Azevedo & R.Oliveira 151 (HUEFS).
Originalmente descrita do Morro do Cambirela, Florianópolis, Santa Catarina
(Pabst, 1975), esta espécie foi citada por Pabst & Dungs (1975) apenas para o estado de
Santa Catarina e é aqui citada pela primeira vez para a Bahia. Planta rara no Parque, foi
encontrada crescendo em paredões rochosos, apenas em áreas sombreadas de matas de
grotão. Foi observada em flor nos meses de maio, setembro e outubro.
Octomeria flabellifera é facilmente reconhecida por seu labelo de base muito
estreita e lâmina largamente flabelada, característica que a diferencia da outra espécie da
Seção Kinetoglossum, O. campos-portoi Schltr. (Pabst, 1975). Diferencia-se das outras
Octomeria do Parque por seu porte delicado e por apresentar flores grandes em relação à
sua planta, sendo maior que as das outras, além do formato de suas flores, principalmente
do labelo, que apresenta um único lóbulo e formato bastante característico. Suas flores
apresentam coloração branco-translúcida com máculas vináceas, uma das quais é maior e
mais marcante na base de cada sépala lateral, calosidade central amarela e quilhas
vináceas.

58
14.3. Octomeria stellaris Barb.Rodr., Gen. spec. Orchid. 2: 99, t.580, 1882.
Tipo: Brasil, Barbosa Rodrigues s.n. (perdido). Lectótipo aqui designado: ilustração
original preparada por João Barbosa Rodrigues, t.580, que consta de sua
Iconographie des orchidées du Brésil, depositada na biblioteca do Jardim Botânico
do Rio de Janeiro, reproduzida por Sprunger et al. (1996) sob o número t.189A do
volume 3.
Erva rupícola. Rizoma 1,0-3,0 mm comp. entre pseudobulbos, 1,0 mm diâm., raízes
emergindo ao longo de todo o rizoma. Caule (4,5-)11-22 x 1,0 mm, 1-foliado, ereto,
cilíndrico, verde a arroxeado, coberto por bainhas. Folha 15-25 x 4,0-8,0 mm, séssil, ereta,
lanceolada, verde a arroxeada, carnosa, ápice agudo, base obtusa. Inflorescência
fasciculada terminal; raque 2,0-3,0 mm comp., 1 flor em antese; brácteas florais
inconspícuas. Flor ressupinada; pedicelo incluindo o ovário 1,0-2,0 mm comp. Sépalas
4,0-5,0 x 1,5-2,0 mm, ovadas, róseas com margem alaranjada, membranáceas, ápice agudo,
base truncada; dorsal patente a ereta; laterais patentes, concrescidas na base até 1/3 do
comprimento. Pétalas 3,5-4,0 x 1,0-1,5 mm, patentes, lanceoladas, róseas com margem
alaranjada, membranáceas, ápice agudo, base obtusa. Labelo articulado com o pé da
coluna, ereto, 3-lobado, obovado, avermelhado, membranáceo, base obtusa; calosidade
formada por duas quilhas erguidas; lóbulos laterais 0,5-1,0 x 0,5 mm, eretos, arredondados,
ápice obtuso; lóbulo terminal 2,5-3,0 x 1,0-1,5 mm, ereto, obovado, ápice obtuso
emarginado. Coluna 1,0-1,5 x 0,5 mm, 0,5 mm alt., amarelada; prolongada na base em pé
de 0,5-1,0 mm comp.; antera branco-amarelada; polínias 8, obovadas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
13.jul.2002, C.Azevedo 143 (HUEFS).
Descrita originalmente do Rio de Janeiro (Barbosa Rodrigues, 1882), esta espécie
foi citada também para São Paulo (Pabst & Dungs, 1975). É aqui registrada pela primeira
vez para o estado da Bahia. Na área do Parque, é encontrada em paredões rochosos,
florindo nos meses de abril, julho e setembro.
Octomeria stellaris diferencia-se das outras Octomeria pelo seu pequeno porte, por
apresentar folhas lanceoladas, carnosas, e pela coloração e formas de suas flores, róseas
com margem alaranjada, apresentando sépalas ovadas e pétalas lanceoladas, ambas de
ápice agudo, não caudado, sépalas dorsais concrescidas na base, apenas até 1/3 do

59
comprimento e labelo trilobado, obovado, avermelhado, com calosidade formada por duas
quilhas erguidas e lóbulo terminal obovado de ápice obtuso emarginado.
O holótipo desta espécie encontra-se perdido. Desta forma, a ilustração original de
Octomeria stellaris, preparada na natureza por Barbosa Rodrigues e ilustrada em sua
Iconographie des orchidées du Brésil, recentemente publicada por Sprunger et al. (1996),
constitui o único material original disponível e por esse motivo é aqui selecionado como
lectótipo.
15. ONCIDIUM Sw.
15.1. Oncidium blanchetii Rchb.f., Linnaea. 22: 845, 1849.
Erva terrestre. Rizoma inconspícuo. Pseudobulbos 5-10 x 2-3 cm, 2-foliados, eretos,
ovóides, amarelo-esverdeados, cobertos pelas bainhas das folhas laterais. Folhas laterais
10-15 x 1-2 cm, 2-3, sésseis, semi-eretas, linear-lanceoladas, verdes, coriáceas, ápice
agudo, base articulada com as bainhas 8-10 x 1,5-2 cm, lanceoladas, verdes. Folha apical
33-35 x 1,5-2 cm, séssil, semi-ereta, linear-lanceolado, verde, coriácea, ápice agudo, base
atenuada. Inflorescência panícula lateral, ereta; escapo 25-85 cm comp., verde; brácteas
escapais 8,0-10 x 5,0-7,0 mm, ovadas, paleáceas, ápice agudo, base truncada; raque 18-22
cm comp., 20-30 flores; brácteas florais 3,0-7,0 x 1,0-2,0 mm, triangulares, membranáceas,
ápice agudo, base truncada. Flores ressupinadas; pedicelo incluindo o ovário 8,0-10 mm
comp. Sépalas amarelas com máculas marrons, membranáceas, ápice agudo, base
atenuada; dorsal 6,0-7,0 x 3,0-4,0 mm, ereta, ovada, côncava; laterais 8,0-9,0 x 3,5-4,0
mm, reflexas, concrescidas na base até a metade, lanceoladas. Pétalas 6,0-7,0 x 4,0-5,0
mm, reflexas, orbiculares, amarelas com máculas marrons, membranáceas, ápice obtuso,
base atenuada. Labelo patente, 3-lobado, orbicular, amarelo com máculas marrons no calo
e na face adaxial, membranáceo, base atenuada; calo formado por uma calosidade central
de quilhas curtas e elevadas e por verrugas laterais; lobos laterais 4,5-5,0 x 1,5-2,5 mm,
patentes, oblongos, ápice obtuso; lobo terminal 7,0-11 x 15-18 mm, patente, orbicular,
ápice obtuso emarginado. Coluna 3,5-5,0 x 2,0-2,5 mm, 1,5-2,0 mm alt., marrom; tábula
infrastigmática amarela; antera 2,0-2,5 x 1,0-1,6 mm, obovada, marrom; polínias 2,
amarelas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,

60
11.jun.2002, C.Azevedo & J.Oliveira 135 (HUEFS).
Material adicional examinado: BRASIL, Bahia: Mucugê, Guiné, 19.nov.2002,
E.C.Smidt 274 (HUEFS); Serra do Capa Bode, set.1973, G.C.P.Pinto s.n. (ALCB 9076).
Pabst & Dungs (1977) citaram a ocorrência desta espécie para a Bolívia, e no Brasil
para Pernambuco, Bahia, Rio de Janeiro, São Paulo, Paraná, Santa Catarina e Minas
Gerais. Na Bahia foi citado para a Serra do Ouro - Barra da Estiva/Ituaçú (Harley & Mayo,
1980) e para os municípios de Rio de Contas, identificado como O. ramosum (Toscano de
Brito, 1995), Lençóis (Toscano de Brito, 1998), Palmeiras (Toscano de Brito, 1998;
Conceição & Giulietti, 2002) e Catolés (Toscano de Brito & Queiroz, 2003). Silva (1999)
cita sua ocorrência para Abaíra, Andaraí, Barra da Estiva, Ibicoara, Lençóis, Mucugê,
Palmeiras, Rio de Contas, Seabra e Jacobina. Cresce geralmente em solos rasos, entre as
rochas dos campos rupestres, florescendo entre junho e julho.
Oncidium blanchetii possui morfologia floral bastante variável, principalmente no
formato do labelo, dos lóbulos laterais e verrugas laterais ao calo central, as quais podem
estar ausentes ou presentes em diferentes indivíduos e em número bastante variável (Silva,
1999). As sépalas e pétalas também variam em forma e coloração. Pabst & Dungs (1977)
apresentaram três ilustrações desta espécie, que mostram as variações que esta pode
apresentar. Assemelha-se a O. hydrophilum, embora este último apresente flores maiores e
inflorescência racemosa.
16. POLYSTACHYA Hook.
16.1. Polystachya micrantha Schltr., Repert. Beih. 35: 81, 1925.
Erva rupícola. Rizoma inconspícuo, raízes emergindo ao longo de todo o rizoma.
Pseudobulbos 10-20 x 5-10 mm, eretos, ovóides, marrons, cobertos pelas bainhas das
folhas. Folhas 2-10 x 0,5-1,5 cm, sésseis, semi-eretas, oblongas, verdes, coriáceas, ápice
obtuso, base truncada. Inflorescência panícula terminal, ereta; escapo 7-10 cm comp.,
verde; brácteas escapais 40-64 x 4,0-5,5 mm, oblongas, paleáceas, ápice agudo, base
amplexicaule; raque 44-90 mm comp., ciliado, 3-12 flores; brácteas florais 2,5-4,0 x 1,0-
1,5 mm, ovadas a triangulares, membranáceas, ápice agudo, base truncada. Flores não
ressupinadas; pedicelo incluindo o ovário 3,0-4,0 mm comp. Sépalas semipatentes,
amarelo-esverdeadas, membranáceas, ápice agudo acuminado, base truncada; dorsal 2,5-

61
3,0 x 1,5-2,0 mm, ovada; laterais 3,0-3,5 x 2,0-2,5 mm, ovado-orbiculares. Pétalas 2,0-2,5
x 0,5-1,0 mm, semipatentes, linear-lanceoladas, amarelo-esverdeadas, membranáceas,
ápice obtuso, base truncada. Labelo ereto, 3-lobado, amarelo-esverdeado, membranáceo,
base obtusa, calosidade em forma de quilha erguida no centro do labelo; lobos laterais 1,0-
1,5 x 0,5-1,0 mm, erguidos, orbiculares, ápice obtuso; lobo terminal 2,5-3,0 x 1,5-2,0 mm,
ereto, oblongo, ápice agudo reflexo, porção central, da face adaxial, coberto por tricomas.
Coluna 1,0-1,5 x 1,0-1,5 mm, 0,7 mm alt., amarelo-esverdeada; antera 1,0 x 1,0 mm,
amarela; polínias 2, cilíndricas, amarelas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
18.mar.2003, C.Azevedo & J.Oliveira 181 (HUEFS); Parque Municipal de Mucugê,
13.abr.2003, C.Azevedo 189 (HUEFS); Unidade de Manejo Sustentável, 01.nov.1996,
UMS 344 (HRB).
Esta espécie foi citada por Pabst & Dungs (1975) para São Paulo e Rio Grande do
Sul, sendo aqui registrada pela primeira vez para o estado da Bahia. Cresce diretamente
sobre as rochas do campo rupestre. Floresce entre março e abril, podendo ser encontrada
florida em outros meses, como novembro.
Polystachya micrantha apresenta flores não ressupinadas, o que a diferencia de
grande parte das espécies do Parque. Possui labelo trilobado, amarelo-esverdeado e
calosidade em forma de quilha erguida no centro do labelo.
17. PRESCOTTIA Lindl.
Chave de identificação para as espécies de Prescottia ocorrentes no Parque Municipal de
Mucugê.
1. Entre 2-3 folhas, inflorescência alongada, raque 8-24 cm comp.
2. Folhas verde-prateadas de margem inteira, labelo de ápice obtuso levemente
emarginado. ........................................................................... Prescottia leptostachya (17.1)
2’. Folhas verdes com lista branca na margem, esta ligeiramente serrilhada, labelo de
ápice agudo. .............................................................................. Prescottia stachyodes (17.3)
1’. Uma única folha, inflorescência congesta, cônica, raque c. 6 cm comp. ..............................
.......................................................................................................... Prescottia montana (17.2)

62
17.1. Prescottia leptostachya Lindl., Bot. Reg. 22:sub t. 1916, 1836.
Erva terrestre. Raízes carnosas, pilosas emergindo do rizoma. Caule inconspícuo, 2-3
folhas em roseta. Folhas 4-5,5 x 2-2,5 cm, pecioladas 1,5-2 cm comp., patentes,
lanceoladas, verde-prateadas, membranáceas, ápice agudo, base obtusa. Inflorescência
racemo terminal, ereta; escapo 22-28 cm comp., verde; brácteas escapais 5,0-15 x 2,0 mm,
ovadas a lanceoladas, membranáceas, ápice agudo, base amplexicaule; raque 8-20 cm
comp., 35-40 flores; brácteas florais inconspícuas, triangulares, membranáceas, ápice
agudo, base truncada. Flores não ressupinadas; pedicelo incluindo o ovário 3,0-4,0 mm
comp. Sépalas ovadas, verdes, membranáceas, ápice obtuso, base truncada; dorsal 2,0-2,5
x 0,5-1,0 mm, completamente enrolada para trás; laterais 3,0-3,5 x 0,5-1,0 mm, reflexas,
concrescidas na base. Pétalas 2,0-2,5 x 0,3 mm, completamente enroladas para trás,
lineares, verdes, membranáceas, ápice obtuso, base truncada. Labelo 3,0-3,5 x 1,5-2,0 mm,
ereto, cuculado, na base bi-auriculado c. 1,0 mm comp., verde, membranáceo, ápice obtuso
levemente emarginado, base truncada, margem involuta, internamente glabro. Coluna 1,0
x 0,6 mm, 0,4 mm alt., verde; antera amarela; polínias 4, obovadas, granulosas, amarelas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
26.nov.2002, C.Azevedo & R.Oliveira 164 (HUEFS).
Descrita originalmente de espécime proveniente do estado da Bahia (Lindley,
1836b), esta espécie é conhecida também do Rio de Janeiro (Hoehne, 1945; Pabst &
Dungs, 1975). Na Chapada Diamantina, foi citada para Rio de Contas (Toscano de Brito,
1995), Lençóis (Toscano de Brito, 1998) e Catolés (Toscano de Brito & Queiroz, 2003),
sendo aqui registrada pela primeira vez para o município de Mucugê. Cresce em matéria
orgânica acumulada sobre as rochas em matas de grotão. Floresce em novembro.
Prescottia leptostachya é caracterizada por apresentar folhas de coloração verde-
prateada de margem inteira, inflorescência alongada com raque 8-20 cm comp., flores
diminutas e labelo de ápice obtuso levemente emarginado.

63
17.2. Prescottia montana Barb.Rodr., Gen. Spec. Orchid. 1:178, t.485, 1877.
Tipo: Brasil, Barbosa Rodrigues s.n. (perdido). Lectótipo aqui designado: ilustração
original preparada por João Barbosa Rodrigues, t.485, que consta de sua
Iconographie des orchidées du Brésil, depositada na biblioteca do Jardim Botânico
do Rio de Janeiro, reproduzida por Sprunger et al. (1996) sob o número t.89 do
volume 2.
Erva terrestre. Raízes carnosas, pilosas emergindo do rizoma. Caule inconspícuo, 1 folha
basal (em roseta). Folha 10-11 x 1,5-2 cm, peciolada 8-9 cm comp., semi-ereta,
lanceolada, verde, membranácea, ápice agudo, base obtusa. Inflorescência racemo
terminal, congesto, cônico, ereto; escapo 45-50 cm comp., verde; brácteas escapais 15-40 x
4,0-6,0 mm, orbiculares, membranáceas, ápice agudo caudado, base amplexicaule; raque c.
6 cm comp., 30-35 flores; brácteas florais 7,0-9,0 x 2,0 mm, orbiculares a lanceoladas,
membranáceas, ápice agudo caudado, base truncada. Flores não ressupinadas; pedicelo
incluindo o ovário 3,5 mm comp. Sépalas eretas, amarelo-esverdeadas, membranáceas,
base truncada; dorsal 3,0-3,5 x 1,5 mm, ovada, ápice obtuso; laterais 4,0-4,5 x 1,5 mm,
concrescidas na base, lanceoladas, ápice agudo. Pétalas 3,0-3,5 x 1,0 mm, eretas, lineares,
amarelo-esverdeadas, membranáceas, ápice obtuso, base truncada. Labelo 4,0-4,5 x 3,0
mm, ereto, cuculado, na base levemente bi-auriculado c. 0,5 mm comp., verde,
membranáceo, ápice obtuso, base truncada, margem involuta, internamente glabro. Coluna
1,5 x 1,5 mm, 1,0 mm alt., verde; antera amarela; polínias 4, obovada, granulosa, amarelas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
16.dez.2002, C.Azevedo & J.Oliveira 192 (HUEFS).
Descrita originalmente da Pedra Branca, Caldas (Barbosa Rodrigues, 1877), esta
espécie é citada para os estados do Rio de Janeiro, São Paulo, Minas Gerais (Hoehne,
1945; Pabst & Dungs, 1975; Sprunger et al., 1996), Espírito Santo, Paraná e Santa Catarina
(Pabst & Dungs, 1975; Sprunger et al., 1996), sendo aqui registrada pela primeira vez para
o estado da Bahia. É conhecida para outras áreas de campo rupestre, em Minas Gerais, na
Serra do Cipó (Barros, 1987) e Grão-Mogol (Barros & Pinheiro, no prelo). No Parque,
cresce em área fechada de mata, sobre a serrapilheira. Foi encontrada em flor em
dezembro. Embora todas as espécies deste gênero apresentem plantas terrestres com brotos

64
anuais, dificultando sua localização quando estas não estão férteis, esta espécie é menos
comum na área do Parque que as outras duas.
Prescottia montana é facilmente reconhecida por sua inflorescência congesta, de
forma cônica, por apresentar apenas uma folha e flores amarelo-esverdeadas. Uma lista de
sinônimos é fornecida por Sprunger et al. (1996).
O holótipo desta espécie encontra-se perdido, e desta forma a ilustração original de
Prescottia montana, preparada na natureza por Barbosa Rodrigues e ilustrada em sua
Iconographie des orchidées du Brésil, recentemente publicada por Sprunger et al. (1996),
constitui o único material original disponível, e por esse motivo é aqui selecionado como
lectótipo.
17.3. Prescottia stachyodes (Sw.) Lindl., Bot. Reg. 22: sub t., 1916:1, 1836.
Erva terrestre. Raízes carnosas, pilosas emergindo do rizoma. Caule inconspícuo, 2-3
folhas em roseta. Folhas (4,5-)8-11 x 2-4 cm, pecioladas 2-4 cm comp., semi-eretas,
lanceoladas, verdes com lista branca na margem, membranáceas, ápice agudo, base obtusa,
margem ligeiramente serrilhada. Inflorescência racemo terminal, ereta; escapo 36-63 cm
comp., verde; brácteas escapais 20-50 x 5-10 mm, oblongas, membranáceas, ápice agudo,
base amplexicaule; raque 15-24 cm comp., 40-50 flores; brácteas florais 6,0-7,0 x 2,0-2,5
mm, lanceoladas, membranáceas, ápice agudo caudado, base truncada. Flores não
ressupinadas; pedicelo incluindo o ovário 6,0-6,5 mm comp. Sépalas completamente
enroladas para trás, ovadas, verdes, membranáceas, ápice agudo, base truncada; dorsal 3,0-
3,5 x 1,0 mm; laterais 6,0-6,5 x 1,5 mm, concrescidas na base. Pétalas 3,0-3,5 x 1,0 mm,
completamente enroladas para trás, lineares, verdes, membranáceas, ápice agudo, base
truncada. Labelo 5,0-5,5 x 2,5-3,0 mm, ereto, cuculado, na base bi-auriculado c. 1 mm
comp., verde, membranáceo, ápice agudo, base truncada, margem involuta, internamente
glabro. Coluna 2,0 x 1,5 mm, 0,7 mm alt., verde; antera amarela; polínias 4, obovada,
granulosa, amarelas.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
16.ago.2002, C.Azevedo & J.Oliveira 148 (HUEFS).
Esta espécie foi citada desde o México até o Panamá, Venezuela, Colômbia,
Equador, Peru, Brasil (Dunsterville & Garay, 1961a; Pabst & Dungs, 1975; Sprunger et al.,

65
1996), Guianas e Índias Ocidentais (Dunsterville & Garay, 1961a). No Brasil, é citada para
os estados do Pará, Bahia, Espírito Santo, Rio de Janeiro, São Paulo, Paraná, Santa
Catarina, Rio Grande do Sul, Minas Gerais e Distrito Federal (Pabst & Dungs, 1975;
Sprunger et al., 1996). É aqui registrada pela primeira vez para a Chapada Diamantina. Na
Cadeia do Espinhaço já era conhecida da Serra do Cipó (Barros, 1987). Ocorre no Parque
apenas em área fechada, bastante sombreada, crescendo na serrapilheira, e floresce em
agosto.
Prescottia stachyodes apresenta folhas verdes de margem branca e ligeiramente
serrilhada, inflorescência alongada com raque 15-24 cm comp. e labelo de ápice agudo. As
folhas de Prescottia são muito variáveis em seu formato e dimensões, em função do
ambiente em que as plantas crescem. Plantas que vegetam sobre rochas diferem muito
daquelas que habitam a terra farta, o que é bastante notável em P. stachyodes (Hoehne,
1945), motivo pelo qual devem ter ocorrido tantos sinônimos para algumas espécies do
gênero. Sprunger et al. (1996) apresentam uma lista de sinônimos.
18. PROSTHECHEA Knowles & Westc.
18.1. Prosthechea moojenii (Pabst) W.E.Higgins, Phytologia 82(5): 379, 1998.
Encyclia moojenii (Pabst) Pabst, Orquídea (Rio de Janeiro) 29(6): 276, 1967.
Erva rupícola. Rizoma 10-25 mm comp. entre pseudobulbos, 5,0-10 mm diâm., coberto
por bainhas, raízes emergindo ao longo do rizoma. Pseudobulbos 4,5-5,0 x 1,5-2,0 cm, 1-
2-foliados, eretos, ovóides, verdes, base arredondada, coberto por bainhas. Folhas 12-14 x
1,5-2 cm, sésseis, semi-eretas, lanceoladas a oblongas, verdes, coriáceas, ápice agudo, base
atenuada. Inflorescência racemo terminal, pendente, espata 10-15 x 10 mm, ovada, ápice
agudo, base amplexicaule; escapo 40-60 mm comp., verde; brácteas escapais inconspícuas;
raque 7-8 cm comp., 4-6 flores; brácteas florais inconspícuas. Flores não ressupinadas;
pedicelo incluindo o ovário 8,0-11 mm comp. Sépalas 15-20 x 5,0 mm, patente, amarelo-
esverdeadas, carnosa, ápice agudo, base truncada; dorsal lanceolada; laterais oblongas.
Pétalas 12-15 x 5,0 mm, patentes, lanceoladas, amarelo-esverdeadas, carnosas, ápice
agudo, base atenuada. Labelo 12-15 x 6,0 mm, parcialmente concrescido à base da coluna,
ereto, 1-lobado, ovado, branco podendo apresentar linhas purpúreas, carnoso, ápice agudo,
base arredondada. Coluna 7,0-8,0 x 4,0-5,0 mm, 4,0 mm alt., amarelo-esverdeada; antera
2,0 x 2,5 mm, orbicular, amarela; polínias 4, deltóides.

66
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
26.nov.2002, C.Azevedo & J.Oliveira 163 (HUEFS); Parque Municipal de Mucugê,
26.nov.2002, C.Azevedo & R.Oliveira 166 (HUEFS).
Esta espécie é citada para os estados da Bahia e Minas Gerais (Pabst & Dungs,
1975), tendo sido descrita originalmente a partir de material da Chapada Diamantina, entre
Lençóis e Palmeiras (Pabst, 1955). É aqui registrada pela primeira vez para o município de
Mucugê, onde cresce em paredões rochosos. Floresce em novembro.
Pabst (1955) provavelmente se enganou quanto à localização desta espécie,
referindo-se ao local de coleta como município de Castro Alves, ao mesmo tempo em que
afirma ter sido entre Lençóis e Palmeiras. Prosthechea moojenii apresenta flores não
ressupinadas, amarelo-esverdeadas, labelo branco podendo apresentar linhas purpúreas,
inteiro, parcialmente concrescido à base da coluna.
19. SCAPHYGLOTTIS Poepp. & Endl.
19.1. Scaphyglottis cf. modesta Schltr., Repert. Spec. Nov. Regni Veg. 23: 46, 1926.
Tetragamestus modestus Rchb.f., Bonplandia 2, 21, 1854.
Erva rupícola. Rizoma inconspícuo. Pseudobulbos 30-90 x 2,0-3,0 mm, 2-foliados,
eretos, cilíndricos, verdes, sobrepostos, emergindo do ápice do pseudobulbo anterior,
coberto por bainhas. Folhas 40-80 x 7,0-12 mm, sésseis, semi-eretas, lanceoladas a
oblongas, verdes, coriáceas, ápice obtuso, base atenuada. Inflorescência fasciculada
terminal. Flores não observadas, restos florais Sépalas 4,0-5,0 x 2,0-2,5 mm, ápice agudo,
base truncada; dorsal elíptica; laterais ovada. Pétalas 4,0-5,0 x 1,5 mm, lanceoladas, ápice
agudo, base truncada. Labelo 5,0 x 2,0 mm, ereto, discretamente 3-lobado, oblongo, base
atenuada; calo bilobado; lobos laterais auriculares, ápice obtuso; lobo terminal orbicular,
ápice obtuso. Coluna 4,0 x 1,5 mm, 1,0 mm alt., prolongada na em pé de 2 mm comp.;
antera 1,0 x 1,0 mm, orbicular; polínias 4. Fruto cápsula 8,0-10 x 3,0-5,0 mm.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
19.out.2002, C.Azevedo & R.Oliveira 160 (HUEFS).

67
A planta tipo de Tetragamestus modestus tem origem desconhecida (Reichenbach,
1854). Esta espécie foi citada para a Guiana, Venezuela, Caribe, Trindade e no Brasil, para
os estados do Espírito Santo, Rio de Janeiro, São Paulo, Paraná, Santa Catarina, Rio
Grande do Sul e Minas Gerais (Pabst & Dungs, 1975), sendo aqui registrada pela primeira
vez para a Bahia. Rara na área do Parque, cresce somente em áreas de matas de grotão, em
paredões rochosos.
O material examinado não foi encontrado em flor, mas sua identificação foi feita
através da análise da porção vegetativa e dos restos florais persistentes no fruto.
Scaphyglottis modesta pode ser facilmente diferenciada das outras espécies do Parque por
apresentar pseudobulbos sobrepostos, onde novos pseudobulbos crescem também do ápice
do pseudobulbo antigo.
Neste trabalho, seguimos a revisão de Adams (1993), que inclui o gênero
Tetragamestus em Scaphyglottis, posição confirmada por dados moleculares (Dressler &
Whitten, no prelo). Adams (1993) propôs duas subespécies: Scaphyglottis modesta subsp.
modesta com plantas mais delgadas, folhas até 12-16 mm largura, flores bastante abertas,
sépala dorsal 5,5-6 mm comp., labelo com calo proeminente bilobado, e Scaphyglottis
modesta subsp. striolata, que caracteriza-se por apresentar plantas robustas, folhas 12-23
mm largura, flores pouco abertas, aparentemente cleistógama, sépala dorsal 4-5,5 mm
comp., labelo com calo indistintamente bilobado ou somente carnoso entre os lobos
laterais. As subespécies apresentam distribuição geográfica diferente, S. modesta subsp.
modesta ocorrendo no Suriname e sudeste do Brasil e S. modesta subsp. striolata
ocorrendo nas Índias Ocidentais e na América do Sul, desde as Guianas até o norte do
Brasil e Equador. O material aqui estudado apresenta caracteres intermediários entre as
duas subespécies, assemelhando-se a S. modesta subsp. modesta por apresentar folhas 7,0-
12 mm largura e labelo com calo bilobado, embora apresente sépala dorsal 4,0-5,0 mm
comp. e um grande número de frutos, o que pode ser resultante de possível cleistogamia, o
que é comum em S. modesta subsp. striolata. Uma conclusão definitiva sobre as afinidades
infraspecíficas do material do Parque não pode ser obtida até que se tenha material com
flores e se estude uma amostragem mais ampla desta espécie, e seus tipos.

68
20. SOBRALIA Ruiz & Pav.
20.1. Sobralia sessilis Lindl., Bot. Reg. Misc. 3. t. 17, 1841.
Erva terrestre. Rizoma inconspícuo, raízes espessas emergindo ao longo do rizoma. Caule
100-150 x 0,5-1 cm, ereto, cilíndrico, verde, coberto pelas bainhas das folhas, 6-7 folhas.
Folhas 7-12 x 1,5-2,5 cm, dísticas, sésseis, plicadas, semi-eretas, oblongo-lanceoladas,
verdes, coriáceas, ápice agudo acuminado, base atenuada. Inflorescência racemo terminal,
bráctea escapais 28-38 x 2,5-4,5 mm, imbricadas, paleáceas, purpúreas, ápice agudo; 1-2
flores em antese. Flores ressupinadas; pedicelo incluindo o ovário 26-37 mm comp.
Sépalas patentes, lanceoladas, purpúreas, membranáceas, ápice agudo apiculado reflexo;
dorsal 45-46 x 12-14 mm, base atenuada; laterais 45-50 x 13-15 mm, base truncada.
Pétalas 40-45 x 14-15 mm, patentes, lanceoladas, purpúreas, membranáceas, ápice agudo
reflexo, base atenuada. Labelo 40-45 x 25-27 mm, ereto, 1-lobado, obovado, púrpuro,
membranáceo, ápice arredondado retuso, base atenuada, margem ondulada; apresentam 2
calos na base do labelo. Coluna 22-24 x 3,5-4,0 mm, 4,0-4,5 mm alt., branca; 2 braços
voltadas para frente; antera 3,5-4,0 x 1,5-2,0 mm, conectada a coluna; polínias 4, brancas.
Fruto cápsula 70-75 x 15-20 mm, verde.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
11.jun.2002, C.Azevedo & J.Oliveira 136 (HUEFS); Mucugê, Parque Municipal de
Mucugê, 16.dez.2002, C.Azevedo & J.Oliveira 171 (HUEFS); Mucugê, Parque Municipal
de Mucugê, 20.fev.2003, C.Azevedo & J.Oliveira 180 (HUEFS).
Esta espécie é citada para a Venezuela, Colômbia, Peru (Sprunger, 1991), Guianas
e Brasil (Hoehne, 1945; Sprunger, 1991), e, neste último, do Pará até Minas Gerais
(Hoehne, 1945). Pabst & Dungs (1975) também citaram sua ocorrência para os estados do
Amazonas, Roraima, Acre, Amapá, Pará e Minas Gerais. Na Bahia, Harley & Mayo (1980)
indicaram sua presença em Morro do Chapéu, sendo aqui registrada pela primeira vez para
o município de Mucugê. Cresce sobre matéria orgânica que se acumula sobre rochas do
campo rupestre, formando densas touceiras. Foi encontrada em flor nos meses de fevereiro,
maio, junho e dezembro.
No Parque, Sobralia sessilis é reconhecida por suas folhas plicadas, brácteas
imbricadas e grandes flores purpúreas, porém bastante efêmeras, murchando em

69
aproximadamente 24 horas. Embora não tenha sido encontrada na área do Parque, outra
espécie do gênero, S. liliastrum, é comum não só no município de Mucugê, como na
Chapada Diamantina, em afloramentos rochosos e na restinga. É facilmente distinguível de
S. sessilis por apresentar folhas mais estreitas, inflorescências sem brácteas imbricadas, em
zig-zag e flores brancas com centro do labelo amarelo.
21. SOPHRONITIS Lindl.
Recente o gênero Laelia sensu lato sofreu mudanças nomenclaturais (van den Berg
& Chase, 2000) baseadas em análises de seqüências de DNA na subtribo Laeliinae, onde as
espécies brasileiras ficaram posicionadas em um clado distinto das mexicanas.
Conseqüentemente, as espécies brasileiras foram, então, transferidas para o gênero
Sophronitis (van den Berg et al., 2000).
21.1. Sophronitis bahiensis (Schltr.) Van den Berg & M.W.Chase, Lindleyana 15(2): 116,
2000.
Laelia bahiensis Schltr., Repert. Spec. Nov. Regni Veg. 17: 272, 1921.
Erva rupícola. Rizoma inconspícuo. Pseudobulbos 4-6 x 1,5-2 cm, 1-foliados, eretos,
ovóides, verdes a arroxeados, cobertos por bainhas. Folha 10-11 x 1,5-2,5 cm, séssil, semi-
ereta a ereta, oblonga a lanceolada, verde a arroxeada, coriácea, ápice obtuso, base
truncada. Inflorescência racemo terminal, ereto, espata 40-75 x 10-20 mm, ovada a
oblonga, ápice agudo, base amplexicaule; escapo 42-80 cm comp.; brácteas escapais 5,0-
10 x 5,0-9,0 mm, ovadas a orbiculares, paleáceas, ápice agudo aculeado, base
amplexicaule; raque 10-15 cm comp., 4-8 flores; brácteas florais 3,0-6,0 x 1,0-2,0 mm,
triangulares a ovadas, membranáceas, ápice agudo, base truncada. Flores ressupinadas;
pedicelo incluindo o ovário 25-40 mm comp. Sépalas 18-20 x 4,0-5,0 mm, patentes,
oblongo-lanceoladas, amarelas a laranja, membranáceas, ápice agudo apiculado reflexo,
base truncada. Pétalas 18-20 x 4,0-6,0 mm, patentes, obtusas, amarelas a laranja,
membranáceas, ápice agudo reflexo, base ligeiramente obtusa. Labelo ereto, 3-lobado,
obovado, amarelo a laranja, membranáceo, base atenuada; disco com 4 pequenas quilhas
centrais, mais ou menos até o meio do labelo; lobos laterais 5,0-6,0 x 3,0-4,0 mm, eretos
envolvendo a coluna, obtusos, ápice obtuso; lobo terminal 7,0-9,0 x 5,0-6,0 mm, obovado,
ápice agudo, ungüiculado, reflexo e ondulado. Coluna 7,0-9,0 x 3,0-4,0 mm, 3,0 mm alt.,

70
levemente curvada; antera 1,5-2,0 x 1,5-2,0 mm, orbicular; polínias 8.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
19.set.2002, C.Azevedo & E.Ribeiro 154 (HUEFS); Parque Municipal de Mucugê,
19.out.2002, C.Azevedo 161 (HUEFS).
Material adicional examinado: BRASIL, Bahia: estrada entre Andaraí e Mucugê,
20.nov.1983, L.R.Noblick & A.Pinto 2884 (CEPEC). Mucugê, Serra do Capa Bode,
nov.1973, G.C.P.Pinto s.n (ALCB 633, HRB 31062); a 7 km de Mucugê em direção a
Andaraí, 17.nov.1983, H.P.Bautista 1418 (HRB, HUEFS).
Descrita originalmente de material procedente da Bahia (Schlechter, 1921), Pabst &
Dungs (1975) também citaram sua presença no estado. S. bahiensis foi descrita como
sendo epífita, a partir de um espécime coletado pelo Dr. Philipp von Luetzelburg, sem
localidade. Em 1926, Luetzelburg publicou em Estudo Botânico do Nordeste a sua correta
localização como Serra do São José, próximo a Feira de Santana. Em 1968, ela foi
redescoberta por Anton de Ghillany na Serra do Sincorá (Toscano de Brito, A. L. V. &
Robins, S. 1992). Na Chapada Diamantina foi citada para Mucugê/Andaraí (Harley &
Simmons, 1986), Rio de Contas (Toscano de Brito, 1995) e Catolés (Toscano de Brito &
Queiroz, 2003), identificada como Laelia bahiensis. Cresce diretamente sobre as rochas do
campo rupestre a pleno sol, e floresce entre setembro e janeiro.
Sophronitis bahiensis é a única espécie do gênero com flores amarelas na Bahia.
Apresenta uma variação grande na coloração não só de suas flores, como também
vegetativamente, apresentando pseudobulbos e folhas que variam de verde a arroxeado, e
flores de amarelo a laranja. Luetzelburg provavelmente se enganou quanto à procedência
desta espécie, pois esta planta não foi mais coletada nesta serra, sendo aparentemente
encontrada, apenas, na Chapada Diamantina.
22. THELYSCHISTA Garay
22.1. Thelyschista ghillanyi (Pabst) Garay, Bot. Mus. Leafl. 28(4): 377, 1982.
Odontorrhynchus ghillanyi Pabst, Bradea 2: 166, 1977.
Erva terrestre. Rizoma inconspícuo, raízes engrossadas emergindo ao longo do rizoma.
Caule inconspícuo, 3-4 folhas em roseta. Folhas (5-)10-16 x 2,5-4,5 cm, sésseis, semi-

71
eretas, oblongo-lanceoladas, verdes, membranáceas, ápice agudo, base atenuada.
Inflorescência racemo terminal, ereto; escapo 17-40 cm comp., verde; brácteas escapais
30-35 x 5,0-10 mm, ovadas, membranáceas, ápice agudo, base atenuada; raque 13-22 cm
comp, 10-23 flores; brácteas florais 20-23 x 7,0-8,0 mm, lanceoladas, membranáceas,
ciliadas, ápice agudo, base truncada, margem ciliada. Flores ressupinadas; pedicelo
incluindo o ovário 14-15 mm comp. Sépalas verdes, membranáceas, ciliadas, base
truncada; dorsal 18-20 x 4,0-5,0 mm, ereta, lanceolada, ápice obtuso e reflexo; laterais 20-
23 x 3,0-4,0 mm, patentes, lineares, ápice agudo e reflexo, margem involuta. Pétalas 18-20
x 5,0-6,0 mm, eretas, lanceoladas, esbranquiçadas, membranáceas, ápice agudo e reflexo,
base atenuada, margem serrilhada, adnadas a sépala dorsal. Labelo preso ao pé da coluna,
ereto, 3-lobado, oblongo, esbranquiçado, membranáceo, base atenuada; lobos laterais 8,0-
10 x 2,0-3,0 mm, envolvendo a coluna e concrescido na base a sua face lateral, semi-
orbiculares, ápice obtuso; lobo terminal 7,0-7,5 x 7,0-8,0 mm, oblongo, ápice obtuso e
reflexo, margem ondulada. Coluna 6,0-7,5 x 3,0-3,5 mm, 1,5 mm alt., branca, prolongada
na base em pé de 3,0-4,0 mm comp.; antera ovada a oblonga; polínias 2, lineares.
Material examinado: BRASIL, Bahia: Mucugê, Parque Municipal de Mucugê,
27.nov.2002, C.Azevedo et al. 169 (HUEFS); Parque Municipal de Mucugê, 28.jan.2003,
C.Azevedo & J.Oliveira 174 (HUEFS); Área da UMS, 25.jan.2000, L.P.Queiroz 5681
(HUEFS); Unidade de Manejo Sustentável, 03.jan.1997, UMS 67 (HRB); Unidade de
Manejo Sustentável, 12.jan.1997, UMS 187 (HRB).
Material adicional examinado: BRASIL, Bahia: Mucugê, 2 km al N de Mucugê, camino
a Andaraí, 20.jan.1997, M.M.Arbo et al. 7574 (CEPEC, CTES); by Rio Cumbuca, about 3
km N of Mucugê on the Andaraí road, 05.fev.1974, R.M.Harley et al. 15993 (CEPEC, K);
by Rio Cumbuca, c. 3 km S of Mucugê, near site of small dam on road to Cascavel,
04.fev.1974, R.M.Harley et al. 15965 (CEPEC); a 3 km ao S de Mucugê, estrada que vai
para Jussiape, 22.dez.1979, S.A.Mori & F.P.Benton 13172 (CEPEC).
Thelyschista ghillanyi foi originalmente descrita de espécime vindo da Chapada da
Calabocaria, entre o município de Mucugê e Andaraí (Pabst, 1977). Foi citada para a Serra
do Sincorá por Harley & Mayo (1980) e para Mucugê/Andaraí por Harley & Simmons
(1986), identificada como Odontorrhynchus ghillanyi. Thelyschista é um gênero
monotípico, endêmico do estado da Bahia. Através de estudos mais detalhados, Smidt
(2003) confirmou o seu endemismo para a região central da Chapada Diamantina: Andaraí,

72
Lençóis e Mucugê, e recentemente foi citada para Catolés (Toscano de Brito & Queiroz,
2003). No Parque, T. ghillanyi cresce tanto em área mais fechada de mata, como sobre a
matéria orgânica que se acumula sobre as rochas do campo rupestre. Floresce entre
novembro e fevereiro.
Esta espécie é facilmente reconhecida por suas grandes folhas dispostas em roseta e
por suas flores ressupinadas, verdes e de labelo branco.

73
REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, B.R. 1993. A taxonomic revision of the genus Scaphyglottis Peoppig & Endl.
(Orchidaceae-Epidendroideae). Degree of Doctor of Fhilosophy. Department of
Plant Biology, Graduate School, Southern Illinois University at Carbondale. 418 p.
BARBOSA RODRIGUES, J. 1877. Genera et species Orchidearum novarum I. C. & H.
Fleiuss. Sebastianópolis, Rio de Janeiro. 191 p.
BARBOSA RODRIGUES, J. 1882. Genera et species Orchidearum novarum II. C. & H.
Fleiuss. Sebastianópolis, Rio de Janeiro. 295 p.
BARROS, F. 1987. Orchidaceae. In: A.M. GIULIETTI; N.L. MENEZES; J.R. PIRANI;
M. MEGURO & M.G.L. WANDERLEY. Flora da Serra do Cipó, Minas Gerais:
caracterização e lista de espécies. Bol. Bot. Univ. São Paulo. 9: 1-151.
BARROS, F. 1994. Novas combinações, novas ocorrências e notas sobre espécies pouco
conhecidas, para as orquídeas do Brasil. Acta Bot. Brasil. 8(1): 12.
BARROS, F. 2002. Notas Nomenclaturais em Pleurothallidinae (Orchidaceae),
principalmente brasileiras. Bradea 8(43): 293-297
BARROS, F. & F. PINHEIRO (no prelo). Flora de Grão-Mogol, Minas Gerais:
Orchidaceae. Bol. Bot. Univ. São Paulo.
BATISTA, J.A.N. & L.B. BIANCHETTI. 2002. A review of Habenaria (Orchidaceae) in
Pabst and Dungs’ Orchidaceae Brasilienses. Lindleyana 17(2): 75-84.
BENNETT Jr., D.E. & E.A. CHRISTENSON. 1993. Icones Orchidacearum Peruviarum.
Privately published by A. Pastorelli de Bennett, Lima. 600 p.
BORBA, E.L.; J.M. FELIX; J. SEMIR & V.N. SOLFERINI. 2000. Pleurothallis
fabiobarrosii, a new Brazilian species: morphological and genetic data with notes on
the taxonomy of Brazilian rupicolous Pleurothallis. Lindleyana 15(1): 2-9.
BORBA, E. L. & J. SEMIR. 2001. Pollinator specificity and convergence in fly-pollinated
Pleurothallis (Orchidaceae) species: a multiple population approach. Ann. Bot. 88: 75-
88.
BORBA, E.L.; J. SEMIR & F. BARROS. 1998. Bulbophyllum involutum Borba, Semir &
F.Barros (Orchidaceae), a New Species from the Brasilian “Campos Rupestres”.
Novon 8(3): 225-229.
BORBA, E.L. & J. SEMIR. 1998. Bulbophyllum ×cipoense (Orchidaceae), a new natural
hybrid from the Brazilian "campos rupestres": description and biology. Lindleyana 13:
113-120.

74
BORBA, E.L; G.J. SHEPHERD; C. VAN DEN BERG & J. SEMIR. 2002. Floral and
Vegetative Morphometrics of Five Pleurothallis (Orchidaceae) Species: Correlation
with Taxonomy, Phylogeny, Genetic Variability and Pollination Systems. Ann. Bot.
90: 1-12.
BRAEM, G.J. 1984. Die Bifoliaten Cattleyen Brasiliens. Hildesheim, Brucke-Verlag Kurt
Schmersow, Germany. 94 p.
BRIDSON, G.D.R. & E.R. SMITH (eds.) 1991. Botanico-Periodicum-
Huntianum/Supplementum. Hunt Institute for Botanical Documentation. Carnegie
Mellon University, Pittsburgh. 1068 p.
BRUMMIT, R.K & C.E. POWELL (eds.) 1992. Authors of Plant Names. Royal Botanic
Gardens, London. 732 p.
CAMERON, K.M.; M.W. CHASE; W.M. WHITTEN; P.J. KORES; D.C. JARRELL; V.A.
ALBERT; T. YUKAWA; H.G. HILLS & D.H. GOLDMAN. 1999. A phylogenetic
analysis of the Orchidaceae: evidence from rbcL nucleotide sequences. Amer. J. Bot.
86(2): 208-224.
CAMERON, K.M. & M.W. CHASE. 2000. Nuclear 18S rDNA sequences of Orchidaceae
confirm the subfamilial status and circumscription of Vanilloideae. In: K.L. WILSON
& D.A. MORRISON (eds.) Monocots Systematics and Envolution. CSIRO,
Melbourne, pp. 457-464.
CASTRO, V.P. & M.A. CAMPACCI. 2000. Icones Orchidacearum Brasiliensis I. Edição
do autor, São Paulo.
COGNIAUX, A. 1893-1896. Orchidaceae. In: C.F.P. MARTIUS & A.G. EICHLER (eds.)
Flora Brasiliensis. vol. 3(4). Frid. Fleischer, Lipsiae, 1-672.
COGNIAUX, A. 1898-1902. Orchidaceae. In: C.F.P. MARTIUS & A.G. EICHLER (eds.)
Flora Brasiliensis. vol. 3(5). Frid. Fleischer, Lipsiae, 1-663.
COGNIAUX, A. 1904-1906. Orchidaceae. In: C.F.P. MARTIUS & A.G. EICHLER (eds.)
Flora Brasiliensis. vol. 3(6). Frid. Fleischer, Lipsiae, 1-604.
CONCEIÇÃO, A.A. & A.M. GIULIETTI. 2002. Composição florística e aspectos
estruturais de campo rupestre em dois platôs do Morro do Pai Inácio, Chapada
Diamantina, Bahia, Brasil. Hoehnea 29(1): 37-48, 3 tab., 3 fig.
CRIBB, P. & A.L.V. TOSCANO DE BRITO. 1996. Histórico. In: S. SPRUNGER; P.
CRIBB & A.L.V. TOSCANO DE BRITO (eds.) Iconographie des Orchidées du
Brèsil. vol. 1. Reinhardt, Basle, pp. 42-47.
DRESSLER, L.R. 1993. Phylogeny and classification of the orchid Family. Dioscorides

75
Press, Portland, 314 p.
DRESSLER, R.L. & WHITTEN, W.M. (no prelo). Phylogenetic relationships of
Scaphyglottis and related genera (Laeliinae: Orchidaceae) based on ITS sequence data.
Brittonia.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961a. Venezuelan Orchids Illustrated. vol.
1. Andre Deutsch Limited, London, pp. 232, 366.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961b. Venezuelan Orchids Illustrated. vol.
2. Andre Deutsch Limited, London, pp. 52, 148.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961c. Venezuelan Orchids Illustrated. vol.
3. Andre Deutsch Limited, London, pp. 100.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961d. Venezuelan Orchids Illustrated. vol.
4. Andre Deutsch Limited, London. pp.234.
FOWLIE, J.A. 1990. A New Encyclia Species From Serra da Sincorá National Park,
Bahia, Brazil; Encyclia alboxanthina Fowl., sp. nov. Orchid Digest 54(1): 27.
GUEDES, M.L.S. & M.D. ORGE (eds.) 1998. Checklist das Espécies Vasculares do
Morro do Pai Inácio (Palmeiras) e Serra da Chapadinha (Lençóis), Chapada
Diamantina, Bahia, Brasil. UFBA, Salvador, 68 p.
HARLEY, R.M. 1995. Introdução. In: B.L. STANNARD (ed.) Flora of the Pico das
Almas: Chapada Diamantina, Bahia, Brazil. Royal Botanic Gardens, London, pp. 43-
76.
HARLEY, R.M. & S.J. MAYO. 1980. Towards a Checklist of the Flora of Bahia. Royal
Botanic Gardens, London, 245 p.
HARLEY, R.M. & N.A. SIMMONS. 1986. Florula of Mucugê: Chapada Diamantina -
Bahia, Brazil. Royal Botanic Gardens, London, 228 p.
HOEHNE, F.C. 1939. Orchidaceae. Arq. Bot. Estado São Paulo n.s. 1: 41, tab. 46.
HOEHNE, F.C. 1940. Orchidaceae. In: F.C. HOEHNE (ed.) Flora Brasilica. vol. 12(1).
Secretaria da Agricultura, Indústria e Comércio de São Paulo, São Paulo, pp. 1-254.
HOEHNE, F.C. 1942. Orchidaceae. In: F.C. HOEHNE (ed.) Flora Brasilica. vol. 12(6).
Secretaria da Agricultura, Indústria e Comércio de São Paulo, São Paulo, pp. 1-128.
HOEHNE, F.C. 1945. Orchidaceae. In: F.C. HOEHNE (ed.) Flora Brasilica. vol. 12(2).
Secretaria da Agricultura, Indústria e Comércio de São Paulo, São Paulo, pp 1-389.
HOEHNE, F.C. 1953. Orchidaceae. In: F.C. HOEHNE (ed.) Flora Brasilica. vol. 12(7).
Secretaria da Agricultura, Indústria e Comércio de São Paulo, São Paulo, pp. 1-397.
HOLMGREN, P.K.; N.H. HOLMGREN & L.C. BARNETT (eds.) 1990. Index

76
Herbariorum Part I: The Herbaria of the World (Regnum Veg. 120). New York
Botanical Garden, New York. 693 p.
LINDLEY, J. 1830. Pleurothallis prolífera. Bot. Reg. 15: t. 1298.
LINDLEY, J. 1831. Genera and Species of Orchidaceous Plants, Part I. Ridgways,
London. pp. 65.
LINDLEY, J. 1836a. Pleurothallis Gróbyi.Bot. Reg. 21 (n.s. 8) sub t. 1797.
LINDLEY, J. 1836b. Prescóttia colórans. Bot. Reg. 22 sub t. 1916.
LINDLEY, J. 1852. Folia Orchidacea. 2. London.
MORI, S.A; L.A. SILVA; G. LISBOA & L. CORADIN. 1989. Manual de Manejo do
Herbário Fanerogâmico. 2.ed. CEPLAC, Ilhéus - Bahia. 104 p.
PABST, G.F.J. 1955. Um Epidendrum novo da Bahia. Orquídea (Rio de Janeiro) 17(6):
204.
PABST, G.F.J. 1956. Additamenta ad Orchideologiam Brasiliensem - III. Orquídea (Rio
de Janeiro) 18(5): 184-185, t. 1, fig. A.
PABST, G.F.J. 1966. Additamenta ad Orchideologiam Brasiliensem - IX. Orquídea (Rio
de Janeiro) 28(3): 165.
PABST, G.F.J. 1971. Additamenta ad Orchideologiam Brasiliensem -XI. Bradea 1(8): 54-
55, fig. C.
PABST, G.F.J. 1975. Additamenta ad Orchideologiam Brasiliensem - XX. Bradea 2(10):
56, t. 1, fig. H.
PABST, G.F.J. 1977. Additamenta ad Orchideologiam Brasiliensem - XXIV. Bradea
2(24): 166-167, t. 1, fig. C.
PABST, G.F.J. & F. DUNGS. 1975. Orchidaceae Brasilienses. vol. 1. Kurt Schmersow,
Hildesheim, 408 p.
PABST, G.F.J. & F. DUNGS. 1977. Orchidaceae Brasilienses. vol. 2. Kurt Schmersow,
Hildesheim, 418 p.
PRIDGEON A.M. & M.W. CHASE. 2001. A phylogenetic reclassification of
Pleurothallidinae (Orchidaceae). Lindleyana 16(4): 235-271.
PRIDGEON, A.M.; R. SOLANO & M.W. CHASE. 2001. Phylogenetic relationships of
Pleurothallidinae (Orchidaceae): combined evidence from nuclear and plastid DNA
sequences. Amer. J. Bot. 88: 2286-2308.
REICHENBACH, H.G. 1854. Die Wagener’schen Orchideen. Bonplandia 2, 21-22.
REICHENBACH, H.G. 1854. Die Wagener’schen Orchideen. Bonplandia 2, 16.
RIESEBERG, L.H. 1995. The role of hybridization in evolution: old wine in new skins.

77
Amer. J. Bot. 82(7): 944-953.
ROLFE, R.A. 1894. Cattleya elongata. Orchid Review 2, 206 p.
SCHLECHTER, R. 1920. Cyrtopodium paranaense. Repert. Spec. Nov. Regni Veg. 16:
333.
SCHLECHTER, R. 1921. Laelia bahiensis. Repert. Spec. Nov. Regni Veg. 17: 272.
SCHLECHTER, R. 1922. Octomeria alexandrii. Mem. Inst. Butantan, Secc. Bot. 1. 4:53,
pl. 12.
SILVA, D.G. 1999. O gênero Oncidium Sw. (Orchidaceae) na Chapada Diamantina,
Bahia, Brasil. Monografia (Especialização em Botânica) - Departamento de Biologia,
Universidade Estadual de Feira de Santana. 41 p.
SILVA, D.G. 2002. O gênero Cleistes Rich. ex Lindl. (Orchidaceae: Vanilloideae:
Pogoniinae) na Chapada Diamantina, Bahia, Brasil. Dissertação (Mestrado em
Botânica) - Programa de Pós-graduação em Botânica, Universidade Estadual de Feira
de Santana. 69 p.
SMIDT, E.C. 2003. A subtribo Spiranthinae Lindl. (Orchidaceae - Orchidoideae) na
Capada Diamantina, Bahia, Brasil. Dissertação (Mestrado em Botânica) - Programa
de Pós-graduação em Botânica, Universidade Estadual de Feira de Santana. 113 p.
SPRUNGER, S. 1991. Catalogue. In: S. SPRUNGER; P. CRIBB & W.T. STEARN (eds.)
Orchids from the Botanical Register 1815-1847. The illustrations. Birkhauser, Verlag.
SPRUNGER, S.; P. CRIBB & A.L.V. TOSCANO DE BRITO (eds.) 1996. Iconographie
des Orchidées du Brèsil. vol. 1. Reinhardt, Basle. 540 p. il.
SPRUNGER, S.; P. CRIBB & A.L.V. TOSCANO DE BRITO (eds.) 1996. Iconographie
des Orchidées du Brèsil. vol. 2. Reinhardt, Basle. 182 p.
STAFLEU, F.A & R.S. COWAN. 1976-1978. Taxonomic Literature. vol. 1-7. Bohn
Schetel & Hoekma, Utrech.
STANNARD, B.L. (ed.) 1995. Flora of the Pico das Almas: Chapada Diamantina, Bahia,
Brazil. Royal Botanic Gardens, London. 853 p.
STRADMANN, M.T.S. 1998. Plano de Manejo do Parque Municipal de Mucugê.
Prefeitura Municipal de Mucugê. Projeto Sempre Viva/Convênio MMA/PNMA/PED
96 CV 00027/96. Mucugê. 164 p. il. Mapas.
TOSCANO DE BRITO, A.L.V. 1995. Orchidaceae. In: B. L. STANNARD (ed.) Flora of
the Pico das Almas: Chapada Diamantina, Bahia, Brazil. Royal Botanic Gardens,
London, pp. 725-767.
TOSCANO DE BRITO, A.L.V. 1998. Orchidaceae. In: M.L.S. GUEDES & M.D. ORGE

78
(eds.) Checklist das Espécies Vasculares do Morro do Pai Inácio (Palmeiras) e Serra
da Chapadinha (Lençóis), Chapada Diamantina, Bahia, Brasil. UFBA, Salvador, pp.
33, 53-54.
TOSCANO DE BRITO, A.L.V. 1992. Notes on Brazilian Orchidaceae I. Kew Bull. 47 (4):
774.
TOSCANO DE BRITO, A.L.V. & L.P. QUEIROZ. 2003. Orchidaceae. In: D.C. ZAPPI;
E. LUCAS; B.L. STANNARD; E.N. LUGHADHA; J.R. PIRANI; L.P. QUEIROZ;
S. ATKINS; D.J.N. HIND; A.M. GIULIETTI; R.M. HARLEY & A.M.
CARVALHO. Lista das plantas vasculares de Catolés, Chapada Diamantina, Bahia,
Brasil. Bol. Bot. Univ. São Paulo. 21(2): 396-397.
TOSCANO DE BRITO, A.L.V. & S. ROBINS. 1992. Laelia bahiensis. Kew Mag. 9 (3):
116-120.
VAN DEN BERG, C.; W.E. HIGGINS; R.L. DRESSLER; W.M. WHITTEN; M.A. SOTO
ARENAS; A. CULHAM & M.W. CHASE. 2000. A phylogenetic analysis of
Laeliinae (Orchidaceae) based on sequence data from internal transcribed spacers
(ITS) of nuclear ribosomal DNA. Lindleyana 15(2): 96:114
VAN DEN BERG, C. & M.W. CHASE. 2000. Nomenclatural notes on Laeliinae - I.
Lindleyana 15(2): 115-119
WITHNER, C.L. 1988. The Cattleyas and their relatives. The Cattleyas. vol. 1. Timber
Press, Portland. 147 p.
WITHNER, C.L. 1998. The Cattleyas and their relatives. Brassavola, Encyclia and other
genera of México and Central America. vol. 5. Timber Press, Portland. 198 p.
ZAPPI, D.C; E. LUCAS; B.L. STANNARD; E.N. LUGHADHA; J.R. PIRANI; L.P.
QUEIROZ; S. ATKINS; D.J.N. HIND; A.M. GIULIETTI; R.M. HARLEY & A.M.
CARVALHO. 2003. Lista das plantas vasculares de Catolés, Chapada Diamantina,
Bahia, Brasil. Bol. Bot. Univ. São Paulo. 21(2): 345-398.

D A
C B
1 mm
2 mm
1 cm E
F
1 mm 1 cm
2 mm
Figura I: Acianthera hamosa: A. hábito, B. diagrama floral, C. detalhe do labelo (C.Azevedo &
R.Oliveira 150) e Acianthera ochreata subsp. ochreata: D. hábito, E. diagrama floral, F.
detalhe do labelo (C.Azevedo & J.Oliveira 139).

B A
C 2 mm
2 mm
E
D
2 mm2 mm
2 mm
Figura II: Anathallis microphyta: A. hábito, B. diagrama floral, C. fruto (C.Azevedo &
R.Oliveira 152) e Anathallis montipelladensis: D. hábito, E. diagrama floral (C.Azevedo
& J.Oliveira 142).

1 cm 5 mm
A
C
5 mm D
B
3 mm
5 mmE F
2 mm
H
3 mmG
5 mm 2 mm I
Figura III: Bulbophyllum ×cipoense: A. diagrama floral, B. coluna e labelo em vista lateral
(C.Azevedo & E.Oliveira 182), Bulbophyllum cribbianum: C. hábito (C.Azevedo &
R.Oliveira 167), D. diagrama floral, E. coluna e labelo em vista lateral (E.L.Borba 2127),
Bulbophyllum involutum: F. diagrama floral, G. coluna e labelo em vista lateral
C.Azevedo & J.Oliveira 149) e Bulbophyllum weddellii: H. diagrama floral, I. coluna e
labelo em vista lateral (C.Azevedo & O.Oliveira 140).

B H
1 mm
2 mm
1 cm F G
2 mm
A 2 cm
E
D
2 mm 2 cm
C
2 mm
Figura IV: Brassavola tuberculata: A. hábito, B. diagrama floral, C. folha em corte transversal
(C.Azevedo & R.Oliveira 165) e Campylocentrum micranthum: D. hábito, E.
inflorescência em detalhe, F. flor em vista lateral, G. diagrama floral, H. coluna em vista
lateral (C.Azevedo 173).

B
A
2 cm C
2 cm
E D
4 cm
1 cm
4 cm F
1 cm
Figura V: Cattleya elongata: A. hábito, B. diagrama floral, C. coluna em vista lateral
(C.Azevedo & T.C.Faustino 179) e Cleistes exilis: D. hábito, E. diagrama floral, F. coluna
e labelo em vista lateral (C.Azevedo et al. 178).

B 5 mm
3 cm
5 mm C
A
5 mm D
E
5 mm
Figura VI: Cyrtopodium edmundoi: A. hábito, B. diagrama floral, C. coluna e labelo em vista
lateral (C.Azevedo & O.A.Oliveira 147) e Cyrtopodium polyphyllum: D. diagrama floral,
E. coluna e labelo em vista lateral (C.Azevedo 157).

A 1 cm
B
1 cm C
2 cm
5 mm 1 cm E D
Figura VII: Epidendrum cristatum: A. hábito, B. diagrama floral (C.Azevedo 156),
Epidendrum orchidiflorum: C. diagrama floral (C.Azevedo & J.Oliveira 138),
Epidendrum secundum: D. diagrama floral (C.Azevedo 158) e Epidendrum warasii: E.
diagrama floral (C.Azevedo & J.Oliveira 176).

A B 1 cm
C 4 cm
5 mm
2 cm
E 4 cm
D
2 cm 1 cm F
Figura VIII: Encyclia alboxanthina: A. hábito, B. diagrama floral, C. coluna e labelo em vista
lateral (C.Azevedo 159) e Epistephium lucidum: D. hábito, E. diagrama floral, F. coluna
em vista lateral (C.Azevedo 172).

B
C
A
5 mm
E D
2 cm
3 mm
F 1 cm
3 mm
Figura IX: Habenaria fluminensis: A. hábito, B. flor, C. diagrama floral (C.Azevedo 141) e
Maxillaria cerifera: D. hábito, E. diagrama floral, F. coluna e labelo em vista lateral
(C.Azevedo & J.Oliveira 153).

5 mm A B C
5 mm 2 mm
E 2 mm D
2 mm
F 4 mm
1 mm G
I 1 mm
1 mm H
J 2 mm
2 mm L K 2 mm
1 mm M
Figura X: Octomeria alexandrii: A. folha em corte transversal, B. diagrama floral, C.
coluna e labelo em vista lateral, D. labelo em detalhe (C.Azevedo & J.Oliveira
137), Octomeria flabellifera: E. hábito, F. folha em corte transversal, G. diagrama
floral, H. coluna e labelo em vista lateral, I. labelo em detalhe (C.Azevedo &
R.Oliveira 151) e Octomeria stellaris: J. folha em corte transversal, K. diagrama
floral, L. coluna e labelo em vista lateral, M. labelo em detalhe (C.Azevedo 143).

A
B
5 mm
2,5 cm
2,5 cm D
C
1 mm
E
1 mm
Figura XI: Oncidium blanchetii: A. hábito, B. diagrama floral (C.Azevedo & J.Oliveira 135) e
Polystachya micrantha: C. hábito, D. diagrama floral, E. coluna e labelo em vista lateral
(C.Azevedo & J.Oliveira 181).
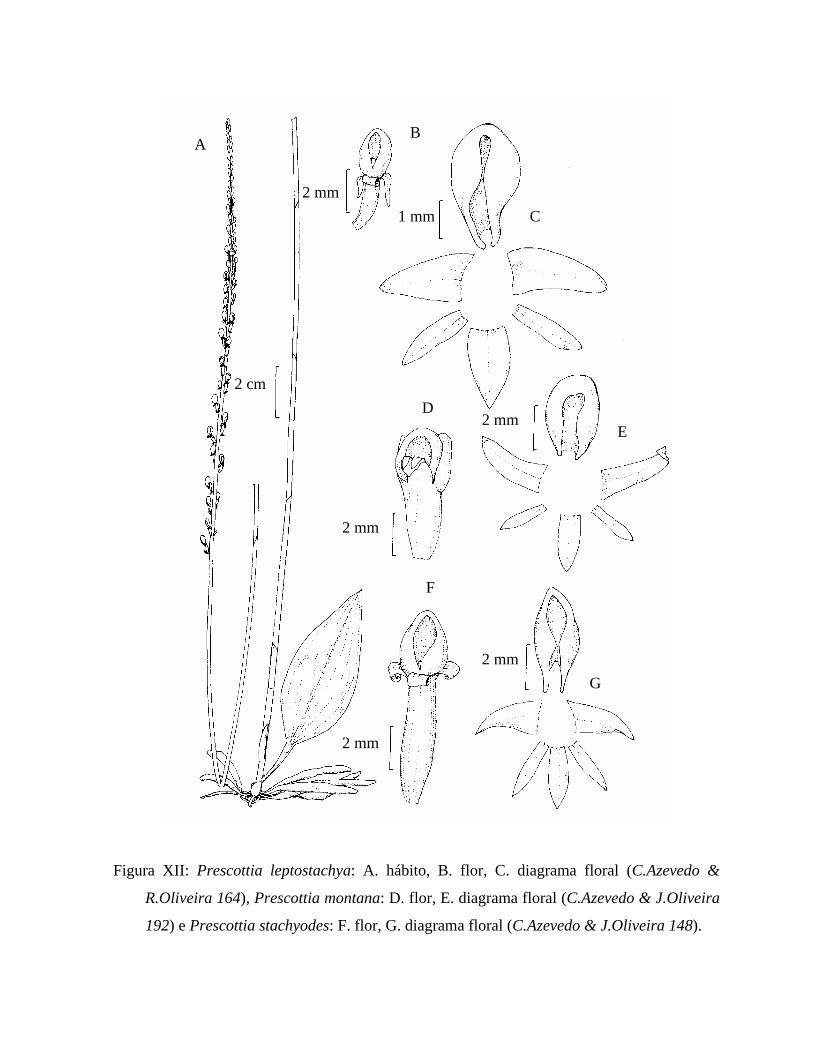
B A
2 mm 1 mm C
E
2 cm D
2 mm E
2 mm
F
2 mm G
2 mm
Figura XII: Prescottia leptostachya: A. hábito, B. flor, C. diagrama floral (C.Azevedo &
R.Oliveira 164), Prescottia montana: D. flor, E. diagrama floral (C.Azevedo & J.Oliveira
192) e Prescottia stachyodes: F. flor, G. diagrama floral (C.Azevedo & J.Oliveira 148).

D C
5 mm 5 mm
B 4 cm
A 1 cm
Figura XIII: Prosthechea moojenii: A. hábito, B. flor, C. coluna e labelo em vista frontal, D.
coluna e labelo em vista lateral (C.Azevedo & J.Oliveira 163).
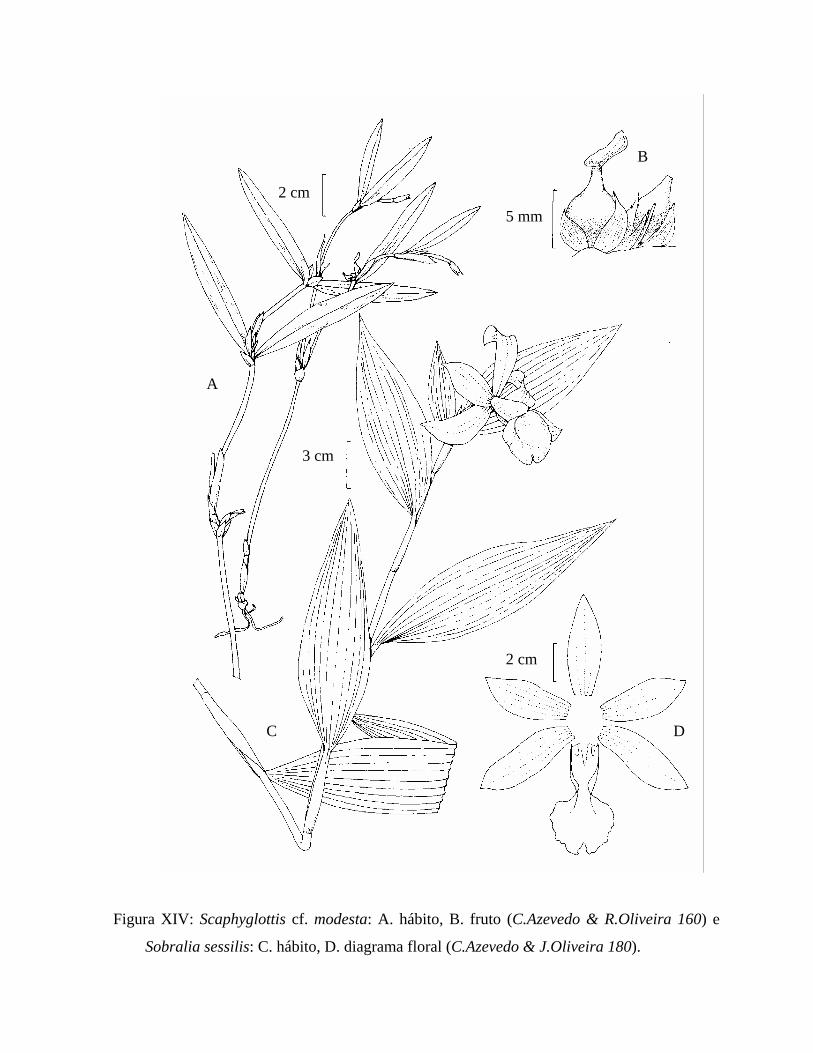
B
2 cm 5 mm
A
3 cm
2 cm
C D
Figura XIV: Scaphyglottis cf. modesta: A. hábito, B. fruto (C.Azevedo & R.Oliveira 160) e
Sobralia sessilis: C. hábito, D. diagrama floral (C.Azevedo & J.Oliveira 180).
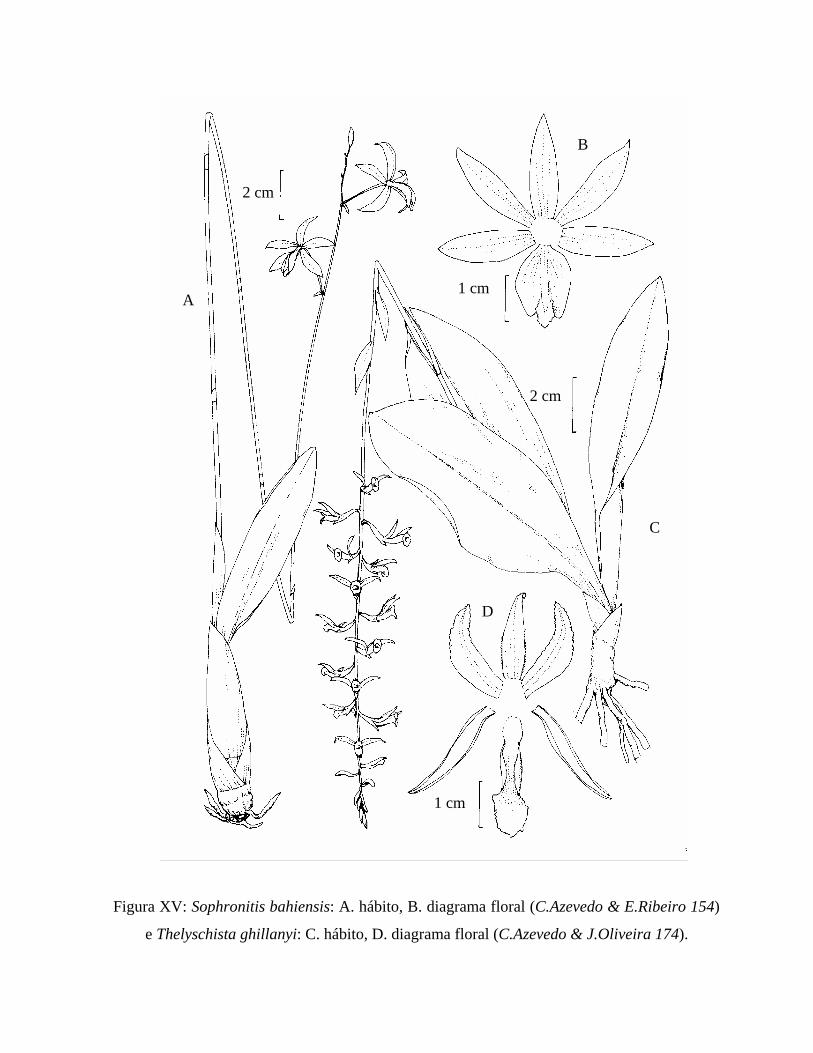
B
2 cm
1 cm A
2 cm
C
D
1 cm
Figura XV: Sophronitis bahiensis: A. hábito, B. diagrama floral (C.Azevedo & E.Ribeiro 154)
e Thelyschista ghillanyi: C. hábito, D. diagrama floral (C.Azevedo & J.Oliveira 174).

E F
G I
L
H
J K
D
CBA
Figura XVI: A. Acianthera hamosa, B. Acianthera ochreata subsp. ochreata, C-D. Anathallis microphyta, E-F. Anathallis montipelladensis, G. Brassavola tuberculata, H-I. Bulbophyllum ×cipoense, J. Bulbophyllum involutum, K. Bulbophyllum weddellii e L. Campylocentrum micranthum.

A B C
D E F
G I
L
H
J K
Figura XVII: A. Cattleya elongata, B. Cleistes exilis, C. Cyrtopodium edmundoi,D. Cyrtopodium polyphyllum, E. Encyclia alboxanthina, F. Epidendrum cristatum, G. Epidendrum orchidiflorum, H. Epidendrum secundum, I. Epidendrum warasii, J. Epistephium lucidum, K. Habenaria fluminensis e L. Maxillaria cerifera.

A B C
D F G
M
H
J
E
I
M
OLK
Figura XVIII: A. Octomeria alexandrii, B. Octomeria flabellifera, C. Octomeria stellaris, D. Oncidium blanchetii, E. Polystachya micrantha, F. Prescottia montana, G-H. Prescottia stachyodes, I. Prosthechea moojenii, J. Sobralia sessilis, K. Sophronitis bahiensis e L-M. Thelyschista ghillanyi.

97
CAPÍTULO 2 - ANÁLISE COMPARATIVA DA FAMÍLIA ORCHIDACEAE EM
ÁREAS DE CAMPO RUPESTRE DA CADEIA DO ESPINHAÇO, BAHIA E MINAS
GERAIS, BRASIL
Este capítulo é um artigo expandido, formatado para publicação na revista Sitientibus série
Ciências Biológicas.

98
RESUMO
Este trabalho apresenta uma análise comparativa da família Orchidaceae em áreas da
Cadeia do Espinhaço, nos estados de Minas Gerais e Bahia realizada por meio de análises
de similaridade e agrupamento, utilizando-se os coeficientes de Jaccard e Dice. Todas as
áreas apresentaram uma grande proporção de espécies exclusivas. Os valores de
similaridade obtidos foram relativamente baixos e três grupos foram indicados,
equivalendo respectivamente à Serra do Cipó, Grão-Mogol, e às áreas da Chapada
Diamantina. Este apresentou-se dividido em dois subgrupos, um representado pelo setor
norte da Serra do Sincorá, incluindo a Serra da Chapadinha, o Parque Municipal de
Mucugê e o Pai Inácio, e outro constituído pela porção sudoeste da Chapada Diamantina,
incluindo o Pico das Almas e Catolés.
Palavras-chave: Florística, Cadeia do Espinhaço, Chapada Diamantina, Orquídea.
ABSTRACT
In this paper we present a comparative analysis of the Orchidaceae family in seven areas at
the Espinhaço Range at the states of Minas and Bahia, Brazil. The orchid floras were
compared through similarity and clustering based on presence/absence data from species
composition. All areas included a large proportion of exclusive species, and overall
similarity among areas was low. Despite this, three groups were indicated, equivalent
respectively to Serra do Cipó, Grão-Mogol and a cluster with all the areas of the Chapada
Diamantina.
Keywords: Chapada Diamantina, Espinhaço Range, Floristics, Orchids.

99
INTRODUÇÃO
O presente trabalho pretende comparar a diversidade e composição florística da
família Orchidaceae em áreas de campo rupestre na Cadeia do Espinhaço, localizadas nos
estados de Minas Gerais e Bahia, através de análises de similaridade e agrupamento.
Os campos rupestres, formação que, no Brasil, ocorre principalmente nos estados
da Bahia e Minas Gerais, são considerados importantes centros de diversidade da flora
brasileira, destacando-se por apresentarem um grande número de espécies endêmicas
(Harley & Simmons, 1986; Giulietti & Pirani, 1988). A composição florística dos campos
rupestres da Bahia e de Minas Gerais é bastante semelhante (Zappi et al., 2003), embora,
na Bahia, este esteja predominantemente cercado por caatinga, enquanto em Minas Gerais
encontra-se inserido no cerrado (Harley & Simmons, 1986). Apesar dessa semelhança
florística, devido à abundância das mesmas famílias e gêneros, cada região possui uma
composição característica, apresentando um grande número de espécies exclusivas. Do
ponto de vista biogeográfico, os campos rupestres apresentam dois aspectos bastante
interessantes, a elevada diversidade florística e elevado número de espécies com
distribuição restrita (Harley & Simmons, 1986; Giulietti & Pirani, 1988; Harley, 1988,
1995; Giulietti et al., 1996).
A Cadeia do Espinhaço, a leste do Brasil, é constituída por dois blocos principais, a
porção mineira, composta pelas Serras da região de Ouro Preto, Serra do Cipó, Planalto de
Diamantina, Serra do Cabral e Serra de Grão-Mogol, e a porção baiana, denominada
Chapada Diamantina. Esta estende-se desde a Serra de Jacobina (norte da Bahia), até a
Serra de Ouro Branco, sul de Minas Gerais (Harley, 1995). Os estudos na Cadeia do
Espinhaço tiveram início na década de setenta, e, desde então, muitos trabalhos vêm sendo
realizados na área (e.g. Harley & Simmons, 1986; Giulietti et al., 1987; Stannard, 1995;
Guedes & Orge, 1998; Pirani et al., 2003; Zappi et al., 2003).
Orchidaceae constitui uma das maiores famílias de angiospermas, com cerca de
20.000 espécies, distribuídas em todos os continentes, exceto nos pólos, predominando nos
trópicos e subtrópicos (Dressler, 1993). Apresenta uma grande importância florística nos
campos rupestres da Bahia e Minas Gerais, estando sempre entre as dez famílias de maior
riqueza específica, juntamente com Compositae, Eriocaulaceae, Gramineae, Leguminosae,
Melastomataceae, Myrtaceae, Rubiaceae, Velloziaceae e Xyridaceae (Harley & Simmons,
1986; Giulietti et al., 1987; Stannard, 1995; Guedes & Orge, 1998; Pirani et al., 2003;
Zappi et al., 2003).

100
Alguns estudos foram realizados na Chapada Diamantina, tais como a Flórula de
Mucugê (Harley & Simmons, 1986); a Flora do Pico das Almas (Stannard, 1995); o
Checklist das Espécies Vasculares do Morro do Pai Inácio, Palmeiras e Serra da
Chapadinha, Lençóis (Guedes & Orge, 1998); a Lista das Plantas Vasculares de Catolés
(Zappi et al., 2003) e o levantamento da família Orchidaceae no Parque Municipal de
Mucugê (Capítulo 1). Em Minas Gerais foram estudadas a Flora da Serra do Cipó
(Giulietti et al., 1987) e a Flora de Grão-Mogol (Pirani et al., 2003).
MATERIAL E MÉTODOS
Dados de distribuição
Os dados de distribuição das espécies usadas neste trabalho, foram baseados em
floras publicadas para áreas de campo rupestre da Cadeia do Espinhaço, na Bahia e em
Minas Gerais, e do levantamento da família Orchidaceae para o Parque Municipal de
Mucugê (Capítulo 1). Foram incluídas a Flórula de Mucugê (Harley & Simmons, 1986); a
Flora da Serra do Cipó (Barros, 1987); a Flora do Pico das Almas (Toscano de Brito,
1995); o Checklist das Espécies Vasculares do Morro do Pai Inácio (Palmeiras) e Serra da
Chapadinha (Lençóis) (Toscano de Brito, 1998); a Lista das Plantas Vasculares de Catolés
(Toscano de Brito & Queiroz, 2003) e a Flora da Serra de Grão-Mogol (Barros & Pinheiro,
no prelo) , como pode ser observado no Apêndice II.
A Flórula de Mucugê foi retirada da análise de agrupamento, pois a área do
levantamento da família Orchidaceae no Parque Municipal de Mucugê (Capítulo 1) está
contida na área da Flórula, que não foi um trabalho de amostragem intensa. Desta forma,
apresentou menos espécies de orquídeas que o levantamento do Parque. O universo aqui
comparado se restringe a estas áreas. Não se pretende aqui abordar padrões biogeográficos,
mas comparar a distribuição das Orchidaceae apenas nas áreas citadas a cima.
Localização geográfica das áreas incluídas
Os levantamentos da porção baiana da Cadeia do Espinhaço foram realizados nas
seguintes áreas: Parque Municipal de Mucugê, localizado no setor oriental da Serra do
Sincorá; Serra da Chapadinha, no município de Lençóis e Morro do Pai Inácio, no
município de Palmeiras, localizados no setor noroeste da Serra do Sincorá; Pico da Almas,
que inclui partes dos municípios de Rio de Contas e Água Quente, e corresponde à área
mais meridional da Chapada Diamantina; Catolés, que compreende partes dos municípios

101
de Piatã e Abaíra, e corresponde a um conjunto de serras elevadas, incluindo o ponto mais
alto da Chapada Diamantina, o Pico do Barbado, com 2.050 m de altitude. Na porção
mineira as áreas analisadas foram: Grão-Mogol, situada no nordeste de Minas Gerais,
apresentando-se em posição intermediária e disjunta à Cadeia do Espinhaço de Minas e
Bahia, e Serra do Cipó, situada na porção centro-sul da Cadeia do Espinhaço.
Considerações taxonômicas
A subtribo Pleurothallidinae foi tratada segundo Pridgeon & Chase (2001), assim
como o gênero Laelia sensu lato, segundo van den Berg & Chase (2000), alterações
baseadas em análise filogenética molecular (Pridgeon, Solano & Chase, 2001; van den
Berg et al., 2000).
As coleções referentes aos levantamentos abordados neste trabalho, depositadas nos
herbários: ALCB, CEPEC, HRB e HUEFS (acrônimos de acordo com Holmgren et al.,
1990), foram examinadas (Apêndice III). O material referente ao levantamento do Parque
Municipal de Mucugê encontra-se citado no Capítulo 1. Alguns ajustes foram necessários
devido ao uso de diferentes delimitações genéricas e específicas em cada levantamento
(Apêndice IV). Sinônimos nomenclaturais foram agrupados, sendo eles: Encyclia
odoratissima a Encyclia patens; Epidendrum crassifolium e Epidendrum elongatum a
Epidendrum secundum; Habenaria vaupellii a Habenaria johannensis e Galeandra
alagoensis a Galeandra montana. Algumas coleções estão referidas pelas seguintes siglas:
Coleção Flora dos Campos Rupestres (Universidade de São Paulo) - CFCR e Projeto
Chapada Diamantina (Universidade Federal da Bahia) - PCD.
Para a análise de agrupamento, os seguintes critérios foram aplicados: espécimes
identificados apenas até gênero foram removidos quando outras espécies do gênero já
constavam na lista, como em Cleistes, Habenaria e Octomeria. Espécies que constavam
como cf. (identificação a conferir) em uma área e perfeitamente identificadas em outra área
foram consideradas conspecíficas (ex. Habenaria graciliscapa, Oncidium hydrophilum,
Pelexia parva, Sarcoglottis schwackei e Veyretia sincorensis). Espécies que estavam
identificadas como cf. ao mesmo tempo em que a espécie em questão, constando no
mesmo levantamento, foram consideradas como espécies separadas (ex. Epistephium
lucidum e Epistephium cf. lucidum; e Rodriguezia obtusifolia e Rodriguezia cf.
obtusifolia).

102
Análises de similaridade
Para comparação entre as áreas foram utilizados dois coeficientes de similaridade,
freqüentemente utilizados na literatura. O primeiro foi o de Jaccard, que é calculado pela
fórmula:
J aa b cxy = + +
onde a é o número de espécies compartilhadas entre as áreas x e y, b é o número de
espécies presentes em x e ausentes em y, e c o número de espécies presentes em y e
ausentes em x. Dessa forma, as ausências de espécies compartilhadas não são levadas em
conta como evidência de similaridade.
O outro coeficiente utilizado foi o de Dice, também conhecido como Sorensen. Este
coeficiente foi utilizado devido ao fato de que as áreas deste trabalho apresentam poucas
espécies compartilhadas em relação ao número de espécies únicas em cada área. Assim, o
coeficiente de Dice atribui um maior peso nas espécies compartilhadas utilizando a
seguinte equação:
D aa b cxy = + +
22
onde a é o número de espécies compartilhadas entre as áreas x e y, b é o número de
espécies presentes em x e ausentes em y, e c é o número de espécies presentes em y e
ausentes em x.
Análises de agrupamento
Após calcular a similaridade, foi feita uma análise de agrupamento das sete áreas da
Bahia e Minas Gerais, utilizando o algorítmo UPGMA (Unweighted Pair-Group Method
using Arithmetic Averages) para agrupamento (Sneath & Sokal, 1973), que é o de uso mais
comum na literatura. Este método procede de forma hierárquica, agrupando o par de OTUs
mais similares, e em seguida calculando uma nova matriz de similaridades com a média
das similaridades das OTUs do grupo formado a todas as outras OTUs, e assim por diante.
As análises de agrupamento foram realizadas no software PAST (Paleontological
Statistics) Versão 0.79 (Hammer & Harper, 2002).

103
RESULTADOS
Ao comparar a composição florística das oito áreas, foi possível notar que a Serra
do Cipó apresentou o maior número de espécies, seguida de Catolés, Pico das Almas,
Parque Municipal de Mucugê, Grão-Mogol, Serra da Chapadinha, a região de
Andaraí/Mucugê e Morro do Pai Inácio, como pode ser visto na Tabela I. Os gêneros de
maior riqueza foram Habenaria, Epidendrum, Oncidium, Bulbophyllum e Cleistes.
O Parque Municipal de Mucugê está compreendido na área da Flórula de Mucugê,
que envolveu partes dos municípios de Andaraí e Mucugê, abrangendo uma área de 896
km2 (Harley & Simmons, 1986). Este levantamento revelou a presença de 26 espécies da
família Orchidaceae, em 13 gêneros. As duas floras só apresentaram 10 espécies em
comum, enquanto 16 espécies foram encontradas apenas na Flórula e não no Parque. Este
número de espécies exclusivas da Flórula era esperado, pois a área em questão era
consideravelmente maior que a área do Parque, porém o fato do Parque apresentar 26
espécies exclusivas, que não foram encontradas na Flórula, é bastante significativo. Este
caso representa um exemplo de que a metodologia e o esforço amostral podem influenciar
nos resultados. As Orchidaceae do Parque Municipal de Mucugê foram acompanhadas
durante todos os meses do ano, sendo que todos os esforços estavam voltados para esta
família.
Tabela I. Comparação da composição florística da Cadeia do Espinhaço – Minas gerais e
Bahia.
Flora Espécie Gênero
Serra do Cipó (Barros, 1987) 80 32
Catolés (Toscano de Brito & Queiroz, 2003) 76 31
Pico das Almas (Toscano de Brito, 1995) 46 19
Parque Municipal de Mucugê (Capítulo 1) 35 22
Grão-Mogol (Barros & Pinheiro, no prelo) 30 17
Serra da Chapadinha (Toscano de Brito, 1998) 27 20
Andaraí/Mucugê (Harley & Simmons, 1986) 26 13
Morro do Pai Inácio (Toscano de Brito, 1998) 18 12

104
Das 191 espécies consideradas para este estudo, ocorrentes na Cadeia do
Espinhaço, 67% (128 spp.), ocorreram apenas em uma determinada área, não tendo sido
encontrada nas outras áreas comparadas. Minas Gerais e Bahia compartilham apenas 16 %
(30 spp.) das espécies. 45% (86 spp.) é compartilhada por áreas da Chapada Diamantina e
não são encontradas em Minas Gerais, enquanto que 39% (75 spp.) ocorrem apenas na
porção mineira da Cadeia do Espinhaço, e não foram encontradas na porção baiana. Esta
distinção também ocorreu em relação aos gêneros, dos 55 ocorrentes, 31% foram
encontrados apenas na porção baiana da Cadeia do Espinhaço, enquanto 24%, apenas na
porção mineira.
Na Chapada Diamantina, sete espécies são comuns a todas as suas cinco áreas:
Acianthera ochreata subsp. ochreata, Cyrtopodium edmundoi, Epidendrum secundum e
Epistephium lucidum, que também são encontradas em alguma das áreas de Minas Gerais;
e Acianthera hamosa, Cattleya elongata e Oncidium blanchetii, que não ocorrem nas áreas
de Minas aqui estudadas. Na Cadeia do Espinhaço, Grão-Mogol e Serra do Cipó
apresentam apenas nove espécies em comum: Cyrtopodium parviflorum, Epidendrum
secundum, Epidendrum warasii, Habenaria caldensis, Prescottia montana, que também
são encontradas em algumas áreas da Bahia, e Cyrtopodium eugenii, Pelexia parva,
Sophronitis mirandae e Veyretia rupicola, que não ocorrem nas áreas estudadas da Bahia.
Apenas Epidendrum secundum foi encontrado em todas as sete áreas comparadas, sendo
esta espécie de ampla distribuição geográfica, conhecida da região neotropical
(Dunsterville & Garay, 1961; Pabst & Dungs, 1975; Sprunger et al., 1996) e, no Brasil, dos
estados do Amazonas, Amapá, Pará, Ceará, Bahia, Rio de Janeiro, Mato Grosso, Minas
Gerais, Goiás, Distrito Federal, São Paulo, Paraná, Pernambuco, Santa Catarina e Rio
Grande do Sul (Pabst & Dungs, 1975). Muitas das espécies de Orchidaceae ocorrentes na
Cadeia do Espinhaço apresentam ampla distribuição, sendo encontrada também em outros
estados, principalmente nas regiões sul e sudeste do Brasil.
Todos os valores de similaridade foram relativamente baixos, tanto a partir do
coeficiente de Jaccard como de Dice (Figuras I e II). A análise de agrupamento revelou a
existência de três grupos. Os dois coeficientes apresentaram a mesma seqüência de
agrupamento. O Grupo 1 corresponde à Serra do Cipó; o Grupo 2 à Grão-Mogol e o Grupo
3 às áreas da Chapada Diamantina, apresentndo-se dividido em dois subgrupos. O
Subgrupo 1 corresponde ao setor norte da Serra do Sincorá, incluindo a Serra da
Chapadinha, o Parque Municipal de Mucugê e o Pai Inácio, e o Subgrupo 2 que
corresponde à porção sudoeste da Chapada Diamantina, incluindo o Pico das Almas e

105
Catolés.
O Grupo 1 apresentou 51 espécies exclusivas a esta área, não tendo sido
encontradas nas outras áreas comparadas, enquanto o Grupo 2 apresentou 20 espécies
restritas. O Subgrupo 1 apresenta 20 espécies restritas, das quais oito são exclusivas de
Mucugê, três da Chapadinha e duas do Pai Inácio. Epidendrum orchidiflorum, Octomeria
alexandrii são comuns às três áreas. O Parque Municipal de Mucugê se agrupou primeiro à
Serra da Chapadinha (Lençóis). Estas duas áreas apresentam três espécies exclusivas em
comum, que são Cyrtopodium polyphyllum, Maxillaria cerifera e Sobralia sessilis,
enquanto Campylocentrum linearifolium e Sarcoglottis curvisepala são exclusivas das
áreas da Chapadinha e Pai Inácio. O Subgrupo 2 apresenta 51 espécies restritas, das quais
31 são exclusivas de Catolés, e 15 do Pico das Almas. Cleistes pluriflora, Epidendrum
ochrochlorum, Habenaria johannensis, Habenaria pseudohamata e Oncidium warmingii
são comuns às duas áreas.

106
Figura I. Dendrograma mostrando a similaridade entre áreas de campo rupestre da Bahia e
Minas Gerais, utilizando coeficiente de Jaccard com UPGMA.
0,1
0,2
0,3
0,4
0,50,6
0,7
0,8
0,9
1
Sim
ilarid
ade
(Jac
card
)
Serra
do
Cip
ó
Grã
o-M
ogol
Chap
adin
ha
P.M
un. M
ucug
ê
Pai I
náci
o
Pico
das
Alm
as
Cato
lés
0,1
0,2
0,3
0,4
0,50,6
0,7
0,8
0,9
1
Sim
ilarid
ade
(Jac
card
)
Serra
do
Cip
ó
Grã
o-M
ogol
Chap
adin
ha
P.M
un. M
ucug
ê
Pai I
náci
o
Pico
das
Alm
as
Cato
lés
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
Sim
ilarid
ade
(Dic
e)
Serra
do
Cip
ó
Grã
o-M
ogol
Chap
adin
ha
P. M
un. M
ucug
ê
Pai I
náci
o
Pico
das
Alm
as
Cato
lés
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
Sim
ilarid
ade
(Dic
e)
Serra
do
Cip
ó
Grã
o-M
ogol
Chap
adin
ha
P. M
un. M
ucug
ê
Pai I
náci
o
Pico
das
Alm
as
Cato
lés
Figura II. Dendrograma mostrando a similaridade entre áreas de campo rupestre da Bahia e
Minas Gerais, utilizando coeficiente de Dice com UPGMA.

107
DISCUSSÃO
Os Grupos 1 e 2, obtidos na análise de agrupamento, equivalem à porção mineira da
Cadeia do Espinhaço, diferindo bastante do outro grupo e também entre si. A baixa
similaridade destes com a porção baiana é um padrão que foi verificado em outros grupos
de plantas. Harley (1988), ao estudar a distribuição de Eriope (Labiatae) no Brasil,
verificou que nenhuma das espécies associadas a campo rupestre é comum às porções
mineira e baiana da Cadeia do Espinhaço, sugerindo que pode ter havido algum tipo de
barreira de migração destas espécies entre as duas áreas. Ao comparar a representatividade
das famílias em áreas de campo rupestre utilizando a Flórula de Mucugê (Harley &
Simmons, 1986), a Flora do Pico das Almas (Stannard, 1995) e a Flora da Serra do Cipó
(Giulietti et al, 1987), Harley (1995), observou que as duas áreas da Chapada Diamantina
são muito semelhantes entre si, enquanto a flora da Cadeia do Espinhaço, em Minas Gerais
apresenta maiores diferenças em sua composição florística. Rapini et al. (2002), em estudo
na porção mineira da Cadeia do Espinhaço, observou que, das 47 espécies de
Asclepiadoideae que apresentam forte associação com os campos rupestres, 83% não
foram encontradas na porção baiana da Serra, embora ressalte que a quantidade de coletas
para o grupo nesta área corresponde a 10% do coletado para a porção mineira.
A Serra do Cipó apresentou maior número de Orchidaceae que Grão-Mogol,
resultado também encontrado por Rapini et al. (2002) para as Asclepiadoideae. O mesmo
ressalta que o esforço amostral foi muito maior na região sul, decrescendo
progressivamente para o norte. A relação do número de coletas por número de espécie no
sul foi de 15, enquanto na região norte este valor ficou entre 4 e 5.
O Grupo 3 reuniu todas as áreas da Chapada Diamantina. A subdivisão deste segue
um padrão geográfico previsível, pois, nesta área, a Chapada divide-se em duas cadeias
independentes, a Serra do Sincorá, a leste, que corresponde ao Subgrupo 1, e a Serra do
Rio de Contas e Serra das Almas, a oeste, que correspondem ao Subgrupo 2,
compreendendo os pontos de maior altitude da Chapada Diamantina. Este mesmo
agrupamento foi obtido por Souza (2001), apenas diferindo no agrupamento mais próximo
do Pai Inácio com Lençóis. No presente estudo, Mucugê agrupou primeiro com a
Chapadinha (Lençóis). Isto pode estar relacionado ao maior número de espécies
encontradas nestas duas áreas, aumentando a chance de apresentarem maior número de
espécies em comum, conforme verificado. O mesmo não ocorreu entre o Parque Municipal
de Mucugê e Pai Inácio, que não possuem espécies em comum restritas a estas áreas. Por

108
outro lado, a Chapadinha e o Parque são áreas de fisionomia bastante similares, próximas
de cursos d’água e com matas de grotão, enquanto o Pai Inácio é um morro tabular isolado,
de maior altitude e com predominância de afloramentos rochosos. Entretanto, os valores de
similaridade entre as áreas do Subgrupo 1 são próximos. Por este motivo, temos que levar
em consideração a hipótese de que Chapadinha-Parque Municipal de Mucugê tenham sido
agrupados primeiro devido a questões de amostragem. Baixos índices de similaridade entre
áreas de cerrado foram obtidos por Simon & Proença (2000) em análise biogeográfica para
o gênero Mimosa, bem como em áreas de campo rupestre da Chapada Diamantina obtidos
por Souza (2001), em análise baseada na distribuição de espécies do gênero Calliandra.
O fato dos valores de similaridade terem sido baixos pode expressar a grande
diversidade florística dos campos rupestres, justificado pelo número muito baixo de
espécies compartilhadas, sendo que a grande maioria é exclusiva de uma única área. Por
outro lado, esse grande número de espécies restritas pode estar relacionado a uma
subamostragem das áreas, o que pode gerar uma falsa impressão da situação, o que é
claramente verificado para a Flórula de Mucugê e o Parque Municipal de Mucugê. Outro
ponto discutido por Rapini et al. (2002) foi o período em que essas coletas vêm sendo
realizadas. Muitos grupos de plantas na Cadeia do Espinhaço apresentam um período
limitado de floração e frutificação, como em Asclepiadoideae, sendo que o mesmo foi
observado, neste trabalho, para a família Orchidaceae. Na ausência de coletas extensivas, a
fenologia pode influenciar nos resultados de levantamentos florísticos em áreas onde o
acompanhamento é realizado apenas algumas vezes durante o ano (Rapini et al., 2002).
Outro fato que pode interferir é a ausência de especialistas em certas famílias no campo,
como foi sugerido por Harley (1995) e Zappi et al. (2003), justificando o baixo número de
determinadas famílias, como Gramineae e Xyridaceae, quando comparado a outras áreas
de campo rupestre. Fatos que podem ter influenciado nos resultados obtidos para a Flórula
de Mucugê, comparada com os do Parque Municipal de Mucugê.

109
REFERÊNCIAS BIBLIOGRÁFICAS
BARROS, F. 1987. Orchidaceae. In: A.M. GIULIETTI; N.L. MENEZES; J.R. PIRANI;
M. MEGURO & M.G.L. WANDERLEY. 1987. Flora da Serra do Cipó, Minas
Gerais: caracterização e lista de espécies. Bol. Bot. Univ. São Paulo. 9: 1-151.
BARROS, F. & F. PINHEIRO (no prelo). Flora de Grão-Mogol, Minas Gerais:
Orchidaceae. Bol. Bot. Univ. São Paulo.
DRESSLER, L.R. 1993. Phylogeny and classification of the orchid Family. Dioscorides
Press, Portland, 314 p.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961. Venezuelan Orchids Illustrated. vol. 2.
Andre Deutsch Limited, London, pp. 148.
GIULIETTI, A.M & J.R. PIRANI. 1988. Patterns of geographic distribution of some plant
species from the Espinhaço Range, Minas Gerais and Bahia, Brazil. In: P.E.
VAZZOLINI & W.R. HEYER (eds.) Proceedings of a workshop on neotropical
distributions patterns. Academia Brasileira de Ciências, Rio de Janeiro, pp. 39-68.
GIULIETTI, A.M.; N.L. MENEZES; J.R. PIRANI; M. MEGURO & M.G.L.
WANDERLEY. 1987. Flora da Serra do Cipó, Minas Gerais: caracterização e lista de
espécies. Bol. Bot. Univ. São Paulo. 9: 1-151.
GIULIETTI, A.M.; L.P. QUEIROZ & R.M. HARLEY. 1996. Vegetação e Flora da
Chapada Diamantina, Bahia. In: REUNIÃO ESPECIAL DA SBPC, n. 4, 1996, Feira
de Santana. Anais... Feira de Santana, pp. 144-156.
GUEDES, M.L.S. & M.D. ORGE (eds.) 1998. Checklist das Espécies Vasculares do
Morro do Pai Inácio (Palmeiras) e Serra da Chapadinha (Lençóis), Chapada
Diamantina, Bahia, Brasil. UFBA, Salvador, 68 p.
HAMMER, O. & D.A.T. HARPER. 2002. PAST - Paleontological Statistics, Versão 0.79.
Versão eletrônica (http://folk.uio.no/ohammer/past, acesso em 12/12/2003).
HARLEY, R.M. 1988. Evolution and distribution of Eriope (Labiatae) and its relatives in
Brazil. In: P.E. VAZZOLINI & W.R. HEYER (eds.) Proceedings of a workshop on
neotropical distributions patterns. Academia Brasileira de Ciências, Rio de Janeiro,
pp. 71-120.
HARLEY, R.M. 1995. Introdução. In: B.L. STANNARD (ed.) Flora of the Pico das
Almas: Chapada Diamantina, Bahia, Brazil. Royal Botanic Gardens, London, pp. 43-
76.

110
HARLEY, R.M. & N.A. SIMMONS. 1986. Florula of Mucugê: Chapada Diamantina -
Bahia, Brazil. Royal Botanic Gardens, London, 228 p.
HOLMGREN, P.K.; N.H. HOLMGREN & L.C. BARNETT (eds.) 1990. Index
Herbariorum Part I: The Herbaria of the World (Regnum Veg. 120). New York
Botanical Garden, New York, 693 p.
PABST, G.F.J. & F. DUNGS. 1975. Orchidaceae Brasilienses. vol. 1. Kurt Schmersow,
Hildesheim, 408 p.
PIRANI, J.R.; R. MELLO-SILVA & A.M. GIULIETTI. 2003. Flora de Grão-Mogol,
Minas Gerais, Brasil. Bol. Bot. Univ. São Paulo. 21(1): 1-24.
PRIDGEON A.M. & M.W. CHASE. 2001. A phylogenetic reclassification of
Pleurothallidinae (Orchidaceae). Lindleyana 16(4): 235-271.
PRIDGEON, A.M.; R. SOLANO & M.W. CHASE. 2001. Phylogenetic relationships of
Pleurothallidinae (Orchidaceae): combined evidence from nuclear and plastid DNA
sequences. Amer. J. Bot. 88: 2286-2308.
RAPINI, A.; R. MELLO-SILVA & M.L. KAWASAKI. 2002. Richness and endemism in
Asclepiadoideae (Apocynaceae) from the Espinhaço Range of Minas Gerais, Brazil –
a conservationist view. Biodiversity and Conservation 11: 1733-1746.
SIMON, M.F. & C. PROENÇA. 2000. Phytogeographic patterns of Mimosa
(Mimosoideae, Leguminosae) in the Cerrado biome of Brazil: an indicator genus of
high-altitude centers of endemism? Biol. Conservation 96: 279-296.
SNEATH, P.H.A. & R.R. SOKAL. 1973. Numerical Taxonomy. W. H. Freeman, San
Francisco.
SOUZA, E.R. 2003. Aspectos Taxonômicos e Biogeográficos do Gênero Calliandra Benth.
(Leguminosae – Mimosoideae) na Capada Diamantina, Bahia, Brasil. Dissertação
(Mestrado em Botânica) - Programa de Pós-graduação em Botânica, Universidade
Estadual de Feira de Santana. 187 p.
SPRUNGER, S.; P. CRIBB & A.L.V. TOSCANO DE BRITO (eds.) 1996. Iconographie
des Orchidées du Brèsil. vol. 1. Reinhardt, Basle. 540 p. il.
STANNARD, B.L. (ed.) 1995. Flora of the Pico das Almas: Chapada Diamantina, Bahia,
Brazil. Royal Botanic Gardens, London, 853 p.
TOSCANO DE BRITO, A.L.V. 1995. Orchidaceae. In: B.L. STANNARD (ed.) Flora of
the Pico das Almas: Chapada Diamantina, Bahia, Brazil. Royal Botanic Gardens,
London, pp.725-767.
TOSCANO DE BRITO, A.L.V. 1998. Orchidaceae. In: M.L.S. GUEDES & M.D. ORGE

111
(eds.) Checklist das Espécies Vasculares do Morro do Pai Inácio (Palmeiras) e Serra
da Chapadinha (Lençóis), Chapada Diamantina, Bahia, Brasil. UFBA, Salvador, pp.
33, 53-54.
TOSCANO DE BRITO, A.L.V. & L.P. QUEIROZ. 2003. Orchidaceae. In: D.C. ZAPPI;
E. LUCAS; B.L. STANNARD; E.N. LUGHADHA; J.R. PIRANI; L.P. QUEIROZ;
S. ATKINS; D.J.N. HIND; A.M. GIULIETTI; R.M. HARLEY & A.M.
CARVALHO. Lista das plantas vasculares de Catolés, Chapada Diamantina, Bahia,
Brasil. Bol. Bot. Univ. São Paulo. 21(2): 396-397.
VAN DEN BERG, C.; W.E. HIGGINS; R.L. DRESSLER; W.M. WHITTEN; M.A. SOTO
ARENAS; A. CULHAM & M.W. CHASE. 2000. A phylogenetic analysis of
Laeliinae (Orchidaceae) based on sequence data from internal transcribed spacers
(ITS) of nuclear ribosomal DNA. Lindleyana 15(2): 96:114
VAN DEN BERG, C. & M.W. CHASE. 2000. Nomenclatural notes on Laeliinae - I.
Lindleyana 15(2): 115-119.
ZAPPI, D.C; E. LUCAS; B.L STANNARD; E.N. LUGHADHA; J.R. PIRANI; L.P.
QUEIROZ; S. ATKINS; D.J.N. HIND; A.M. GIULIETTI; R.M. HARLEY & A.M.
CARVALHO. 2003. Lista das plantas vasculares de Catolés, Chapada Diamantina,
Bahia, Brasil. Bol. Bot. Univ. São Paulo. 21(2): 345-398.

112
CAPÍTULO 3 - A NEW COMBINATION IN THE GENUS ANATHALLIS
(ORCHIDACEAE), AND A NEW RECORD FOR BAHIA STATE, BRAZIL.
Artigo submetido para publicação no Kew Bulletin em outubro de 2003

113
RESUMO
A subtribo Pleurothallidinae sofreu uma série de mudanças nomenclaturais, baseadas em
análise filogenética molecular, dentre elas o restabelecido do gênero Anathallis Barb.Rodr.
Durante a realização do levantamento da família Orchidaceae no Parque Municipal de
Mucugê (Capítulo 1) percebeu-se a necessidade de uma nova combinação para
Pleurothallis microphyta (Barb.Rodr.) Cogn. O exame da descrição original e ilustração de
P. microphyta, juntamente com o material coletado em Mucugê, indicaram que esta
espécie pertence ao gênero Anathallis. Esta espécie foi descrita de espécime originário da
Serra do Mar, anteriormente foi citada para Argentina e no Brasil para os estados do Rio de
Janeiro e Rio Grande do Sul, sendo aqui registrada pela primeira vez para o estado da
Bahia. No Parque Municipal de Mucugê A. microphyta é encontrada somente em áreas
sombreadas e úmidas das matas de grotão, crescendo sobre rochas cobertas por musgos. O
holótipo de A. microphyta encontra-se perdido, desta forma a ilustração original desta
espécie, preparada na natureza por Barbosa Rodrigues, constitui o único material original
disponível e por esse motivo foi aqui selecionada como lectótipo.

114
A new combination in the genus Anathallis (Orchidaceae), and a new record for Bahia
State, Brazil.
Cecília Oliveira de Azevedo1,2 & Cássio van den Berg1
Universidade Estadual de Feira de Santana, Departamento de Biologia, Km 03, BR 116. 44.031-
460, Feira de Santana, Bahia, Brasil.
2 Author for correspondence ([email protected])
Summary. The new combination Anathallis microphyta (Barb.Rodr.) C.O.Azevedo & Van
den Berg is proposed herein. This taxon is also lectotypified and recorded for the first time
in Bahia State, Brazil.
Many nomenclatural changes in the subtribe Pleurothallidinae were proposed by Pridgeon
& Chase (2001), based on molecular phylogenetic analyses (Pridgeon, Solano & Chase,
2001). Well-supported clades in these analyses warranted nomenclatural reassignments
within Pleurothallidinae, including the resurrection of Anathallis Barb.Rodr. In the course
of preparing an orchid flora of the “Parque Municipal de Mucugê”, Mucugê, Bahia, Brazil,
we realized the need of a new combination for A. microphyta. Careful examination of the
original description and illustration of Pleurothallis microphyta (Barb.Rodr.) Cogn.
together with additional herbarium material deposited at HUEFS, collected in Mucugê,
Bahia indicates this species is a member of the genus Anathallis, and it is duly transferred
here. A. microphyta was first collected in the Serra do Mar range (Barbosa Rodrigues,
1882), and it was afterwards recorded for Argentina and Brazil, in the latter only for the
states of Rio de Janeiro and Rio Grande do Sul. In the Parque Municipal de Mucugê this
species grow on rocks covered with mosses in shady positions within grooves where a thin
scrub forest has developed. In the original publication, no type specimen was designated,
and after verifying at the Herbarium of the Jardim Botânico do Rio de Janeiro (Rio de
Janeiro Botanical Gardens) that Barbosa Rodrigues’ original specimen is lost, we chose to
lectotypify the basionym upon the plate mentioned by the author in the original description
(plate 557), recently reproduced in Sprunger et al. (1996).

115
Anathallis microphyta (Barb.Rodr.) C.O.Azevedo & Van den Berg, comb. nov.
Basionym: Lepanthes microphyta Barb.Rodr., Gen. spec. Orchid. 2:67, 1882. Type: Brazil,
Serra do Mar, près de la gare du chemin de fer nomée Rodeio. Barbosa Rodrigues s.n.
(now lost). Lectotype, here chosen; Barbosa Rodrigues’ original drawing (plate 557)
in the library of the Jardim Botânico do Rio de Janeiro [reproduction printed in
“Iconographie des Orchidées du Brésil”, Sprunger, S.; Cribb, P. & Toscano de Brito,
A.L.V. (1996). 6: t.298B]
Pleurothallis microphyta (Barb.Rodr.) Cogn., Fl. bras. 3(4): 484, 1895.
REFERENCES
Barbosa Rodrigues, J. 1882. Genera et species Orchidearum novarum II. Rio de Janeiro:
C. & H. Fleiuss. Sebastianópolis. 294 p.
Pridgeon A. M. & Chase, M. W. (2001). A phylogenetic reclassification of
Pleurothallidinae (Orchidaceae). Lindleyana 16(4): 235-271.
Pridgeon, A. M., Solano, R. & Chase, M. W. (2001). Phylogenetic relationships of
Pleurothallidinae (Orchidaceae): combined evidence from nuclear and plastid DNA
sequences. Amer. J. Bot. 88: 2286-2308.
Sprunger, S., Cribb, P. J. & Toscano de Brito, A. L. V. (1996). Iconographie des Orchidées
du Brèsil. vol. 1. Reinhardt, Basle. 540 p. il.

116
CONCLUSÃO GERAL
Foram encontradas no Parque Municipal de Mucugê 35 espécies de orquídeas,
distribuídas em 22 gêneros. Os gêneros de maior riqueza de espécie foram Bulbophyllum,
Epidendrum, Octomeria e Prescottia, enquanto a maioria dos gêneros apresenta apenas
uma espécie. Das espécies coletadas no Parque, 11 são novos registros para o município de
Mucugê, três para a região da Chapada Diamantina e outras sete para o estado da Bahia. O
Parque possui espécies como Encyclia alboxanthina, Sophronitis bahiensis e Thelyschista
ghillanyi endêmicas da porção baiana da Cadeia do Espinhaço. Acianthera hamosa,
Anathallis microphyta, Cattleya elongata, Maxillaria cerifera, Octomeria stellaris e
Prescottia montana, foram lectotipificadas. Uma nova combinação para Pleurothallis
microphyta foi proposta: Anathallis microphyta.
A Serra do Cipó apresentou a maior riqueza de espécies, seguida de Catolés, Pico
das Almas, Parque Municipal de Mucugê, Grão-Mogol, Serra da Chapadinha, e Morro do
Pai Inácio. A análise de agrupamento revelou a existência de três grupos. O Grupo 1
corresponde a Serra do Cipó; o Grupo 2 a Grão-Mogol, representando à porção mineira da
Cadeia do Espinhaço, estes dois diferem bastante do outro grupo e entre si. O Grupo 3
reuniu todas as áreas da Chapada Diamantina, apresentndo-se dividido em dois subgrupos,
que segue um padrão geográfico. Um corresponde ao setor norte da Serra do Sincorá,
incluindo a Serra da Chapadinha, o Parque Municipal de Mucugê e o Pai Inácio, e o outro
corresponde à porção sudoeste da Chapada Diamantina, incluindo o Pico das Almas e
Catolés.
Os baixos valores de similaridade podem expressar a grande diversidade florística
dos campos rupestres, que apresentam muitas espécies exclusivas de uma única área. Por
outro lado, esse grande número de espécies restritas pode estar relacionado a problemas de
amostragem, sendo que muitas áreas ainda foram pouco coletadas. Além de questões
metodológicas, onde muitos inventários vêm sendo realizados a partir de excussões
concentradas apenas em alguns meses do ano, não acompanhando o ciclo reprodutivo de
muitas espécies que apresentam floração curta, outro fator que interfere é a ausência de
especialistas no campo.

117
REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, B.R. 1993. A taxonomic revision of the genus Scaphyglottis Peoppig & Endl.
(Orchidaceae-Epidendroideae). Degree of Doctor of Fhilosophy. Department of
Plant Biology, Graduate School, Southern Illinois University at Carbondale. 418 p.
ATWOOD, J. 1986. The size of the Orchidaceae and the systematic distribution of
epiphytic orchids. Selbyana 9: 171-186
BARBOSA RODRIGUES, J. 1877. Genera et species Orchidearum novarum I. C. & H.
Fleiuss. Sebastianópolis, Rio de Janeiro. 191 p.
BARBOSA RODRIGUES, J. 1882. Genera et species Orchidearum novarum II. C. & H.
Fleiuss. Sebastianópolis, Rio de Janeiro. 295 p.
BARROS, F. 1987. Orchidaceae. In: A.M. GIULIETTI; N.L. MENEZES; J.R. PIRANI;
M. MEGURO & M.G.L. WANDERLEY. Flora da Serra do Cipó, Minas Gerais:
caracterização e lista de espécies. Bol. Bot. Univ. São Paulo. 9: 1-151.
BARROS, F. 1994. Novas combinações, novas ocorrências e notas sobre espécies pouco
conhecidas, para as orquídeas do Brasil. Acta Bot. Brasil. 8(1): 12.
BARROS, F. 2002. Notas Nomenclaturais em Pleurothallidinae (Orchidaceae),
principalmente brasileiras. Bradea 8(43): 293-297
BARROS, F. & F. PINHEIRO (no prelo). Flora de Grão-Mogol, Minas Gerais:
Orchidaceae. Bol. Bot. Univ. São Paulo.
BATISTA, J.A.N. & L.B. BIANCHETTI. 2002. A review of Habenaria (Orchidaceae) in
Pabst and Dungs’ Orchidaceae Brasilienses. Lindleyana 17(2): 75-84.
BENNETT Jr., D.E. & E.A. CHRISTENSON. 1993. Icones Orchidacearum Peruviarum.
Privately published by A. Pastorelli de Bennett, Lima. 600 p.
BORBA, E.L.; J.M. FELIX; J. SEMIR & V.N. SOLFERINI. 2000. Pleurothallis
fabiobarrosii, a new Brazilian species: morphological and genetic data with notes on
the taxonomy of Brazilian rupicolous Pleurothallis. Lindleyana 15(1): 2-9.
BORBA, E. L. & J. SEMIR. 2001. Pollinator specificity and convergence in fly-pollinated
Pleurothallis (Orchidaceae) species: a multiple population approach. Ann. Bot. 88: 75-
88.
BORBA, E.L.; J. SEMIR & F. BARROS. 1998. Bulbophyllum involutum Borba, Semir &
F.Barros (Orchidaceae), a New Species from the Brasilian “Campos Rupestres”.
Novon 8(3): 225-229.
BORBA, E.L. & J. SEMIR. 1998. Bulbophyllum ×cipoense (Orchidaceae), a new natural

118
hybrid from the Brazilian "campos rupestres": description and biology. Lindleyana 13:
113-120.
BORBA, E.L; G.J. SHEPHERD; C. VAN DEN BERG & J. SEMIR. 2002. Floral and
Vegetative Morphometrics of Five Pleurothallis (Orchidaceae) Species: Correlation
with Taxonomy, Phylogeny, Genetic Variability and Pollination Systems. Ann. Bot.
90: 1-12.
BRAEM, G.J. 1984. Die Bifoliaten Cattleyen Brasiliens. Hildesheim, Brucke-Verlag Kurt
Schmersow, Germany. 94 p.
BRIDSON, G.D.R. & E.R. SMITH (eds.) 1991. Botanico-Periodicum-
Huntianum/Supplementum. Hunt Institute for Botanical Documentation. Carnegie
Mellon University, Pittsburgh. 1068 p.
BRUMMIT, R.K & C.E. POWELL (eds.) 1992. Authors of Plant Names. Royal Botanic
Gardens, London. 732 p.
CAMERON, K.M.; M.W. CHASE; W.M. WHITTEN; P.J. KORES; D.C. JARRELL; V.A.
ALBERT; T. YUKAWA; H.G. HILLS & D.H. GOLDMAN. 1999. A phylogenetic
analysis of the Orchidaceae: evidence from rbcL nucleotide sequences. Amer. J. Bot.
86(2): 208-224.
CAMERON, K.M. & M.W. CHASE. 2000. Nuclear 18S rDNA sequences of Orchidaceae
confirm the subfamilial status and circumscription of Vanilloideae. In: K.L. WILSON
& D.A. MORRISON (eds.) Monocots Systematics and Envolution. CSIRO,
Melbourne, pp. 457-464.
CASTRO, V.P. & M.A. CAMPACCI. 2000. Icones Orchidacearum Brasiliensis I. Edição
do autor, São Paulo.
COGNIAUX, A. 1893-1896. Orchidaceae. In: C.F.P. MARTIUS & A.G. EICHLER (eds.)
Flora Brasiliensis. vol. 3(4). Frid. Fleischer, Lipsiae, 1-672.
COGNIAUX, A. 1898-1902. Orchidaceae. In: C.F.P. MARTIUS & A.G. EICHLER (eds.)
Flora Brasiliensis. vol. 3(5). Frid. Fleischer, Lipsiae, 1-663.
COGNIAUX, A. 1904-1906. Orchidaceae. In: C.F.P. MARTIUS & A.G. EICHLER (eds.)
Flora Brasiliensis. vol. 3(6). Frid. Fleischer, Lipsiae, 1-604.
CONCEIÇÃO, A.A. & A.M. GIULIETTI. 2002. Composição florística e aspectos
estruturais de campo rupestre em dois platôs do Morro do Pai Inácio, Chapada
Diamantina, Bahia, Brasil. Hoehnea 29(1): 37-48, 3 tab., 3 fig.
CRIBB, P. & A.L.V. TOSCANO DE BRITO. 1996. Histórico. In: S. SPRUNGER; P.
CRIBB & A.L.V. TOSCANO DE BRITO (eds.) Iconographie des Orchidées du

119
Brèsil. vol. 1. Reinhardt, Basle, pp. 42-47.
DRESSLER, L.R. 1993. Phylogeny and classification of the orchid Family. Dioscorides
Press, Portland, 314 p.
DRESSLER, R.L. & WHITTEN, W.M. (no prelo). Phylogenetic relationships of
Scaphyglottis and related genera (Laeliinae: Orchidaceae) based on ITS sequence data.
Brittonia.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961a. Venezuelan Orchids Illustrated. vol.
1. Andre Deutsch Limited, London, pp. 232, 366.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961b. Venezuelan Orchids Illustrated. vol.
2. Andre Deutsch Limited, London, pp. 52, 148.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961c. Venezuelan Orchids Illustrated. vol.
3. Andre Deutsch Limited, London, pp. 100.
DUNSTERVILLE, G.C.K. & L.A. GARAY. 1961d. Venezuelan Orchids Illustrated. vol.
4. Andre Deutsch Limited, London. pp.234.
FOWLIE, J.A. 1990. A New Encyclia Species From Serra da Sincorá National Park,
Bahia, Brazil; Encyclia alboxanthina Fowl., sp. nov. Orchid Digest 54(1): 27.
GIULIETTI, A.M. 1996. Novas espécies no gênero Syngonanthus Ruhl. (Eriocaulaceae)
para o Brasil. Bol. Bot. Univ. S. Paulo 15: 63-71.
GIULIETTI, A.M & J.R. PIRANI. 1988. Patterns of geographic distribution of some plant
species from the Espinhaço Range, Minas Gerais and Bahia, Brazil. In: P.E.
VAZZOLINI & W.R. HEYER (eds.) Proceedings of a workshop on neotropical
distributions patterns. Academia Brasileira de Ciências, Rio de Janeiro, pp. 39-68.
GIULIETTI, A.M.; N.L. MENEZES; J.R. PIRANI; M. MEGURO & M.G.L.
WANDERLEY. 1987. Flora da Serra do Cipó, Minas Gerais: caracterização e lista de
espécies. Bol. Bot. Univ. São Paulo. 9: 1-151.
GIULIETTI, A.M.; L.P. QUEIROZ & R.M. HARLEY. 1996. Vegetação e Flora da
Chapada Diamantina, Bahia. In: REUNIÃO ESPECIAL DA SBPC, n. 4, 1996, Feira
de Santana. Anais... Feira de Santana, pp. 144-156.
GUEDES, M.L.S. & M.D. ORGE (eds.) 1998. Checklist das Espécies Vasculares do
Morro do Pai Inácio (Palmeiras) e Serra da Chapadinha (Lençóis), Chapada
Diamantina, Bahia, Brasil. UFBA, Salvador, 68 p.
HAMMER, O. & D.A.T. HARPER. 2002. PAST - Paleontological Statistics, Versão 0.79.
Versão eletrônica (http://folk.uio.no/ohammer/past, acesso em 12/12/2003).
HARLEY, R.M. 1988. Evolution and distribution of Eriope (Labiatae) and its relatives in

120
Brazil. In: P.E. VAZZOLINI & W.R. HEYER (eds.) Proceedings of a workshop on
neotropical distributions patterns. Academia Brasileira de Ciências, Rio de Janeiro,
pp. 71-120.
HARLEY, R.M. 1995. Introdução. In: B.L. STANNARD (ed.) Flora of the Pico das
Almas: Chapada Diamantina, Bahia, Brazil. Royal Botanic Gardens, London, pp. 43-
76.
HARLEY, R.M. & S.J. MAYO. 1980. Towards a Checklist of the Flora of Bahia. Royal
Botanic Gardens, London, 245 p.
HARLEY, R.M. & N.A. SIMMONS. 1986. Florula of Mucugê: Chapada Diamantina -
Bahia, Brazil. Royal Botanic Gardens, London, 228 p.
HOEHNE, F.C. 1939. Orchidaceae. Arq. Bot. Estado São Paulo n.s. 1: 41, tab. 46.
HOEHNE, F.C. 1940. Orchidaceae. In: F.C. HOEHNE (ed.) Flora Brasilica. vol. 12(1).
Secretaria da Agricultura, Indústria e Comércio de São Paulo, São Paulo, pp. 1-254.
HOEHNE, F.C. 1942. Orchidaceae. In: F.C. HOEHNE (ed.) Flora Brasilica. vol. 12(6).
Secretaria da Agricultura, Indústria e Comércio de São Paulo, São Paulo, pp. 1-128.
HOEHNE, F.C. 1945. Orchidaceae. In: F.C. HOEHNE (ed.) Flora Brasilica. vol. 12(2).
Secretaria da Agricultura, Indústria e Comércio de São Paulo, São Paulo, pp 1-389.
HOEHNE, F.C. 1953. Orchidaceae. In: F.C. HOEHNE (ed.) Flora Brasilica. vol. 12(7).
Secretaria da Agricultura, Indústria e Comércio de São Paulo, São Paulo, pp. 1-397.
HOLMGREN, P.K.; N.H. HOLMGREN & L.C. BARNETT (eds.) 1990. Index
Herbariorum Part I: The Herbaria of the World (Regnum Veg. 120). New York
Botanical Garden, New York. 693 p.
IPAC - BA. 1980. Inventário de Proteção do Acervo Cultural: Monumentos e Sítios da
Serra Geral e Chapada Diamantina. vol. 4. Secretaria da Indústria e Comércio,
Salvador.
LINDLEY, J. 1830. Pleurothallis prolífera. Bot. Reg. 15: t. 1298.
LINDLEY, J. 1831. Genera and Species of Orchidaceous Plants, Part I. Ridgways,
London. pp. 65.
LINDLEY, J. 1836a. Pleurothallis Gróbyi.Bot. Reg. 21 (n.s. 8) sub t. 1797.
LINDLEY, J. 1836b. Prescóttia colórans. Bot. Reg. 22 sub t. 1916.
LINDLEY, J. 1852. Folia Orchidacea. 2. London.
MORAES, W. 1991. Jagunços e Heróis. 4.ed. Empresa Gráfica da Bahia/IPAC, Salvador,
217 p.
MORI, S.A; L.A. SILVA; G. LISBOA & L. CORADIN. 1989. Manual de Manejo do

121
Herbário Fanerogâmico. 2.ed. CEPLAC, Ilhéus - Bahia. 104 p.
PABST, G.F.J. 1955. Um Epidendrum novo da Bahia. Orquídea (Rio de Janeiro) 17(6):
204.
PABST, G.F.J. 1956. Additamenta ad Orchideologiam Brasiliensem - III. Orquídea (Rio
de Janeiro) 18(5): 184-185, t. 1, fig. A.
PABST, G.F.J. 1966. Additamenta ad Orchideologiam Brasiliensem - IX. Orquídea (Rio
de Janeiro) 28(3): 165.
PABST, G.F.J. 1971. Additamenta ad Orchideologiam Brasiliensem -XI. Bradea 1(8): 54-
55, fig. C.
PABST, G.F.J. 1975. Additamenta ad Orchideologiam Brasiliensem - XX. Bradea 2(10):
56, t. 1, fig. H.
PABST, G.F.J. 1977. Additamenta ad Orchideologiam Brasiliensem - XXIV. Bradea
2(24): 166-167, t. 1, fig. C.
PABST, G.F.J. & F. DUNGS. 1975. Orchidaceae Brasilienses. vol. 1. Kurt Schmersow,
Hildesheim, 408 p.
PABST, G.F.J. & F. DUNGS. 1977. Orchidaceae Brasilienses. vol. 2. Kurt Schmersow,
Hildesheim, 418 p.
PINTO, G.O.; H.P. BAUTISTA & J.C. LIMA. 1990. A Chapada Diamantina, sua
Fitofisionomia e Peculiaridades Florísticas. Anais do XXXV Congresso Nacional de
Botânica, Manaus, 1984, Brasilia, 1990, pp. 256-295.
PIRANI, J.R.; R. MELLO-SILVA & A.M. GIULIETTI. 2003. Flora de Grão-Mogol,
Minas Gerais, Brasil. Bol. Bot. Univ. São Paulo. 21(1): 1-24.
PRIDGEON A.M. & M.W. CHASE. 2001. A phylogenetic reclassification of
Pleurothallidinae (Orchidaceae). Lindleyana 16(4): 235-271.
PRIDGEON, A.M.; R. SOLANO & M.W. CHASE. 2001. Phylogenetic relationships of
Pleurothallidinae (Orchidaceae): combined evidence from nuclear and plastid DNA
sequences. Amer. J. Bot. 88: 2286-2308.
RAPINI, A.; R. MELLO-SILVA & M.L. KAWASAKI. 2002. Richness and endemism in
Asclepiadoideae (Apocynaceae) from the Espinhaço Range of Minas Gerais, Brazil - a
conservationist view. Biodiversity and Conservation 11: 1733-1746.
REICHENBACH, H.G. 1854. Die Wagener’schen Orchideen. Bonplandia 2, 21-22.
REICHENBACH, H.G. 1854. Die Wagener’schen Orchideen. Bonplandia 2, 16.
RIESEBERG, L.H. 1995. The role of hybridization in evolution: old wine in new skins.
Amer. J. Bot. 82(7): 944-953.

122
ROLFE, R.A. 1894. Cattleya elongata. Orchid Review 2, 206 p.
SCHLECHTER, R. 1920. Cyrtopodium paranaense. Repert. Spec. Nov. Regni Veg. 16:
333.
SCHLECHTER, R. 1921. Laelia bahiensis. Repert. Spec. Nov. Regni Veg. 17: 272.
SCHLECHTER, R. 1922. Octomeria alexandrii. Mem. Inst. Butantan, Secc. Bot. 1. 4:53,
pl. 12.
SILVA, D.G. 1999. O gênero Oncidium Sw. (Orchidaceae) na Chapada Diamantina,
Bahia, Brasil. Monografia (Especialização em Botânica) - Departamento de Biologia,
Universidade Estadual de Feira de Santana. 41 p.
SILVA, D.G. 2002. O gênero Cleistes Rich. ex Lindl. (Orchidaceae: Vanilloideae:
Pogoniinae) na Chapada Diamantina, Bahia, Brasil. Dissertação (Mestrado em
Botânica) - Programa de Pós-graduação em Botânica, Universidade Estadual de Feira
de Santana. 69 p.
SIMON, M.F. & C. PROENÇA. 2000. Phytogeographic patterns of Mimosa
(Mimosoideae, Leguminosae) in the Cerrado biome of Brazil: an indicator genus of
high-altitude centers of endemism? Biol. Conservation 96: 279-296.
SMIDT, E.C. 2003. A subtribo Spiranthinae Lindl. (Orchidaceae - Orchidoideae) na
Capada Diamantina, Bahia, Brasil. Dissertação (Mestrado em Botânica) - Programa
de Pós-graduação em Botânica, Universidade Estadual de Feira de Santana. 113 p.
SNEATH, P.H.A. & R.R. SOKAL. 1973. Numerical Taxonomy. W. H. Freeman, San
Francisco.
SOUZA, E.R. 2003. Aspectos Taxonômicos e Biogeográficos do Gênero Calliandra Benth.
(Leguminosae – Mimosoideae) na Capada Diamantina, Bahia, Brasil. Dissertação
(Mestrado em Botânica) - Programa de Pós-graduação em Botânica, Universidade
Estadual de Feira de Santana. 187 p.
SPRUNGER, S. 1991. Catalogue. In: S. SPRUNGER; P. CRIBB & W.T. STEARN (eds.)
Orchids from the Botanical Register 1815-1847. The illustrations. Birkhauser, Verlag.
SPRUNGER, S.; P. CRIBB & A.L.V. TOSCANO DE BRITO (eds.) 1996. Iconographie
des Orchidées du Brèsil. vol. 1. Reinhardt, Basle. 540 p. il.
SPRUNGER, S.; P. CRIBB & A.L.V. TOSCANO DE BRITO (eds.) 1996. Iconographie
des Orchidées du Brèsil. vol. 2. Reinhardt, Basle. 182 p.
STAFLEU, F.A & R.S. COWAN. 1976-1978. Taxonomic Literature. vol. 1-7. Bohn
Schetel & Hoekma, Utrech.
STANNARD, B.L. (ed.) 1995. Flora of the Pico das Almas: Chapada Diamantina, Bahia,

123
Brazil. Royal Botanic Gardens, London. 853 p.
STRADMANN, M.T.S. 1998. Plano de Manejo do Parque Municipal de Mucugê.
Prefeitura Municipal de Mucugê. Projeto Sempre Viva/Convênio MMA/PNMA/PED
96 CV 00027/96. Mucugê. 164 p. il. Mapas.
TOSCANO DE BRITO, A.L.V. 1995. Orchidaceae. In: B. L. STANNARD (ed.) Flora of
the Pico das Almas: Chapada Diamantina, Bahia, Brazil. Royal Botanic Gardens,
London, pp. 725-767.
TOSCANO DE BRITO, A.L.V. 1998. Orchidaceae. In: M.L.S. GUEDES & M.D. ORGE
(eds.) Checklist das Espécies Vasculares do Morro do Pai Inácio (Palmeiras) e Serra
da Chapadinha (Lençóis), Chapada Diamantina, Bahia, Brasil. UFBA, Salvador, pp.
33, 53-54.
TOSCANO DE BRITO, A.L.V. 1992. Notes on Brazilian Orchidaceae I. Kew Bull. 47 (4):
774.
TOSCANO DE BRITO, A.L.V. & L.P. QUEIROZ. 2003. Orchidaceae. In: D.C. ZAPPI;
E. LUCAS; B.L. STANNARD; E.N. LUGHADHA; J.R. PIRANI; L.P. QUEIROZ;
S. ATKINS; D.J.N. HIND; A.M. GIULIETTI; R.M. HARLEY & A.M.
CARVALHO. Lista das plantas vasculares de Catolés, Chapada Diamantina, Bahia,
Brasil. Bol. Bot. Univ. São Paulo. 21(2): 396-397.
TOSCANO DE BRITO, A.L.V. & S. ROBINS. 1992. Laelia bahiensis. Kew Mag. 9 (3):
116-120.
VAN DEN BERG, C.; W.E. HIGGINS; R.L. DRESSLER; W.M. WHITTEN; M.A. SOTO
ARENAS; A. CULHAM & M.W. CHASE. 2000. A phylogenetic analysis of
Laeliinae (Orchidaceae) based on sequence data from internal transcribed spacers
(ITS) of nuclear ribosomal DNA. Lindleyana 15(2): 96:114
VAN DEN BERG, C. & M.W. CHASE. 2000. Nomenclatural notes on Laeliinae - I.
Lindleyana 15(2): 115-119
WITHNER, C.L. 1988. The Cattleyas and their relatives. The Cattleyas. vol. 1. Timber
Press, Portland. 147 p.
WITHNER, C.L. 1998. The Cattleyas and their relatives. Brassavola, Encyclia and other
genera of México and Central America. vol. 5. Timber Press, Portland. 198 p.
ZAPPI, D.C; E. LUCAS; B.L. STANNARD; E.N. LUGHADHA; J.R. PIRANI; L.P.
QUEIROZ; S. ATKINS; D.J.N. HIND; A.M. GIULIETTI; R.M. HARLEY & A.M.
CARVALHO. 2003. Lista das plantas vasculares de Catolés, Chapada Diamantina,
Bahia, Brasil. Bol. Bot. Univ. São Paulo. 21(2): 345-398.

124
Apêndice I - Índice dos espécimes examinados: coletor, seguido pelo número de coleta e
pela numeração do táxon.
C.E.Calderon et al. 2422 (06.1).
C.Azevedo 88 (04.4), 142 (02.2), 143 (14.3), 144 (12.1), 145 (14.1), 156 (10.1), 157 (08.2),
158 (10.3), 159 (09.1), 161 (21.1), 172 (11.1), 173 (05.1), 188 (04.4), 189 (16.1).
C.Azevedo & E.Oliveira 182 (04.1).
C.Azevedo et al. 169 (22.1), 178 (07.1).
C.Azevedo & E.Ribeiro 154 (21.1), 155 (08.2).
C.Azevedo & J.Oliveira 135 (15.1), 136 (20.1), 137 (14.1), 138 (10.2), 139 (01.2), 146
(02.2), 148 (17.3), 149 (04.3), 153 (13.1), 163 (18.1), 171 (20.1), 174 (22.1), 176 (10.4),
177 (04.1), 180 (20.1), 181 (16.1), 192 (17.2).
C.Azevedo & O.Oliveira 140 (04.4), 141 (12.1), 147 (08.1).
C.Azevedo & R.Oliveira 150 (01.1), 151 (14.2), 152 (02.1), 160 (19.1), 164 (17.1), 165
(03.1), 166 (18.1), 167 (04.2).
C.Azevedo & T.C.Faustino 179 (06.1).
E.L.Borba 2127 (04.2).
E.C.Smidt 274 (15.1), 289 (10.3).
E.R.Souza et al. 88 (09.1).
G.C.P.Pinto s.n. (ALCB 471) (11.1), (ALCB 477, HRB 31060)(09.1), (ALCB 633, HRB
31062) (21.1), (ALCB 9076) (15.1).
Ghillány s.n. (HB 63141) (07.1).
H.P.Bautista 1418 (21.1).
J.D.C.Arouck Ferreira et al. 363 (08.1).
J.G.Jardim et al. 2550 (01.2).
L.A.Matos Silva et al. 2788 (06.1).
L.P.Queiroz 1844 (08.2), 1850 (08.1), 5619 (11.1), 5681 (22.1).
L.P.Queiroz et al. 5585 (09.1).
L.R.Noblick & A.Pinto 2883 (09.1), 2884 (21.1), 2900 (10.3).
M.C.Ferreira 1199 (10.1).
M.L.Guedes et. al 1428 (08.1), 5109 (10.2).
M.M.Arbo et al. 7571 (01.2), 7574 (22.1), 7575 (11.1).
PCD 4745 (10.2), 5708 (10.3).
R.M.Harley et al. 15905 (11.1), 15941 (07.1), 15962 (09.1), 15965 (22.1), 15993 (22.1),

125
18690 (06.1), 18712 (01.2), 18765 (07.1), 18767 (09.1), 18773 (11.1), 20984 (07.1), 20986
(07.1), 21024 (06.1).
R.P.Oliveira et al. 37 (10.3).
R.P. Orlandi et al. 792 (10.3).
S.A.Mori & F.P.Benton 13172 (22.1).
W.N.Fonseca & A.P.Araújo 451 (06.1).
UMS 67 (22.1), 070 (09.1), 114 (11.1), 123 (06.1), 124 (01.2), 159 (10.2), 187 (22.1), 210
(06.1), 262 (10.3), 284 (09.1), 342 (08.1), 343 (02.2), 344 (16.1), 345 (01.2), 346 (10.2),
347 (10.3), 348 (09.1).

126
Apêndice II - Distribuição das espécies de Orchidaceae nas áreas comparadas, utilizada
para análise de similaridade.
Parque Municipal de Mucugê - PMM, Pico das Almas - PA, Chapadinha - CH, Pai Inácio -
PI, Catolés - CA, Grão-Mogol - GM e Serra do Cipó - CI.
Espécies PMM PA CH PI CA GM CI Acianthera adamantinensis x Acianthera fabiobarrosii x Acianthera hamosa x x x x x Acianthera ochreata subsp. ochreata
x x x x x x
Acianthera prolifera (aff.) x Acianthera rupestris x Acianthera saudersiana x x Amblostoma armeniacum x Anathallis imbricata (aff.) x Anathallis microphyta x Anathallis montipelladensis x x Anathallis rubens x x x x Anathallis sclerophylla x Barbosella crassifolia x Bifrenaria aureofulva x x Bifrenaria magnicalcarata x x x Bletia catenulata x Brachystele ulaei x Brassavola tuberculata x x Bulbophyllum cribbianum x x Bulbophyllum filifolium x Bulbophyllum involutum x x x Bulbophyllum ipanemensis x x Bulbophyllum mentosum x Bulbophyllum micropetaliforme x Bulbophyllum napelli x Bulbophyllum plumosum x Bulbophyllum warmingianum x Bulbophyllum weddellii x x x Campylocentrum aciculatum x Campylocentrum linearifolium x x Campylocentrum micranthum x Capanemia gehrtii x Cattleya elongata x x x x x Cattleya tenuis x Cattleya walkeriana x Centroglossa macroceras x Cleistes aphylla x Cleistes bella x Cleistes castanoides x Cleistes exilis x x x x Cleistes gracilis x Cleistes itatiaiae x Cleistes paranaensis x x Cleistes paranaensis var. bradeana x Cleistes pluriflora x x Constantia cipoensis x Constantia microscopica x Cyclopogon congestus x Cyclopogon elegans (aff.) x
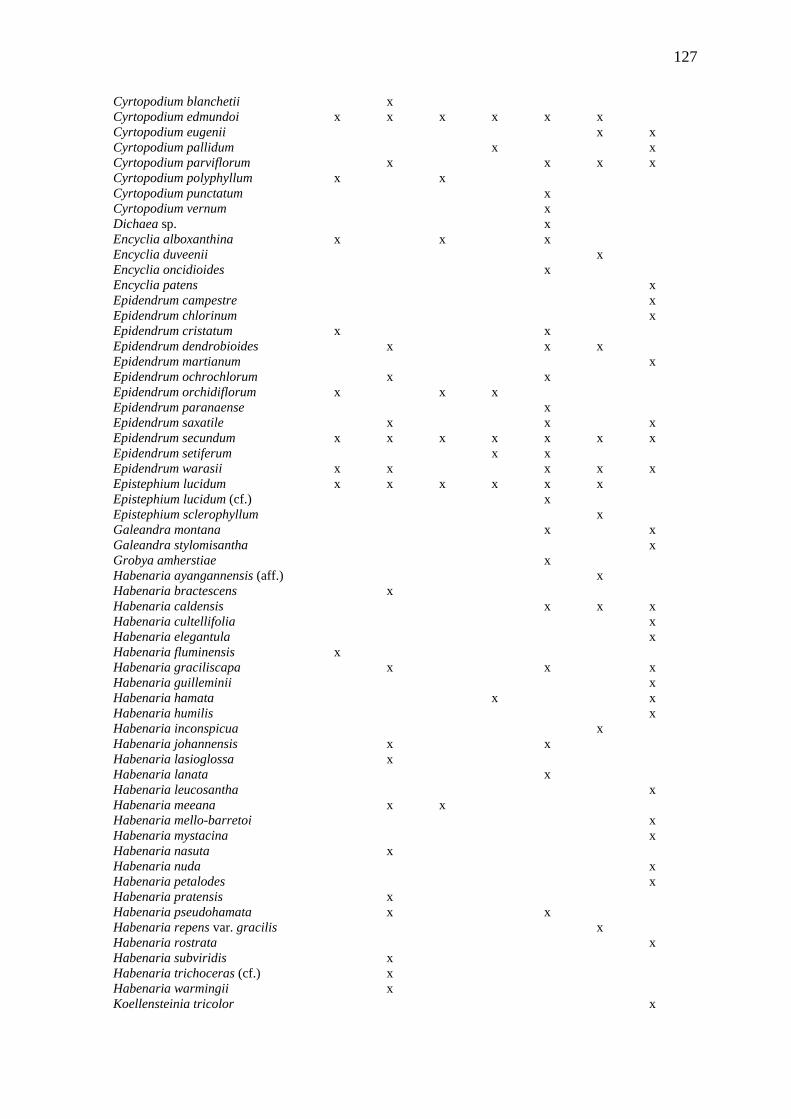
127
Cyrtopodium blanchetii x Cyrtopodium edmundoi x x x x x x Cyrtopodium eugenii x x Cyrtopodium pallidum x x Cyrtopodium parviflorum x x x x Cyrtopodium polyphyllum x x Cyrtopodium punctatum x Cyrtopodium vernum x Dichaea sp. x Encyclia alboxanthina x x x Encyclia duveenii x Encyclia oncidioides x Encyclia patens x Epidendrum campestre x Epidendrum chlorinum x Epidendrum cristatum x x Epidendrum dendrobioides x x x Epidendrum martianum x Epidendrum ochrochlorum x x Epidendrum orchidiflorum x x x Epidendrum paranaense x Epidendrum saxatile x x x Epidendrum secundum x x x x x x x Epidendrum setiferum x x Epidendrum warasii x x x x x Epistephium lucidum x x x x x x Epistephium lucidum (cf.) x Epistephium sclerophyllum x Galeandra montana x x Galeandra stylomisantha x Grobya amherstiae x Habenaria ayangannensis (aff.) x Habenaria bractescens x Habenaria caldensis x x x Habenaria cultellifolia x Habenaria elegantula x Habenaria fluminensis x Habenaria graciliscapa x x x Habenaria guilleminii x Habenaria hamata x x Habenaria humilis x Habenaria inconspicua x Habenaria johannensis x x Habenaria lasioglossa x Habenaria lanata x Habenaria leucosantha x Habenaria meeana x x Habenaria mello-barretoi x Habenaria mystacina x Habenaria nasuta x Habenaria nuda x Habenaria petalodes x Habenaria pratensis x Habenaria pseudohamata x x Habenaria repens var. gracilis x Habenaria rostrata x Habenaria subviridis x Habenaria trichoceras (cf.) x Habenaria warmingii x Koellensteinia tricolor x
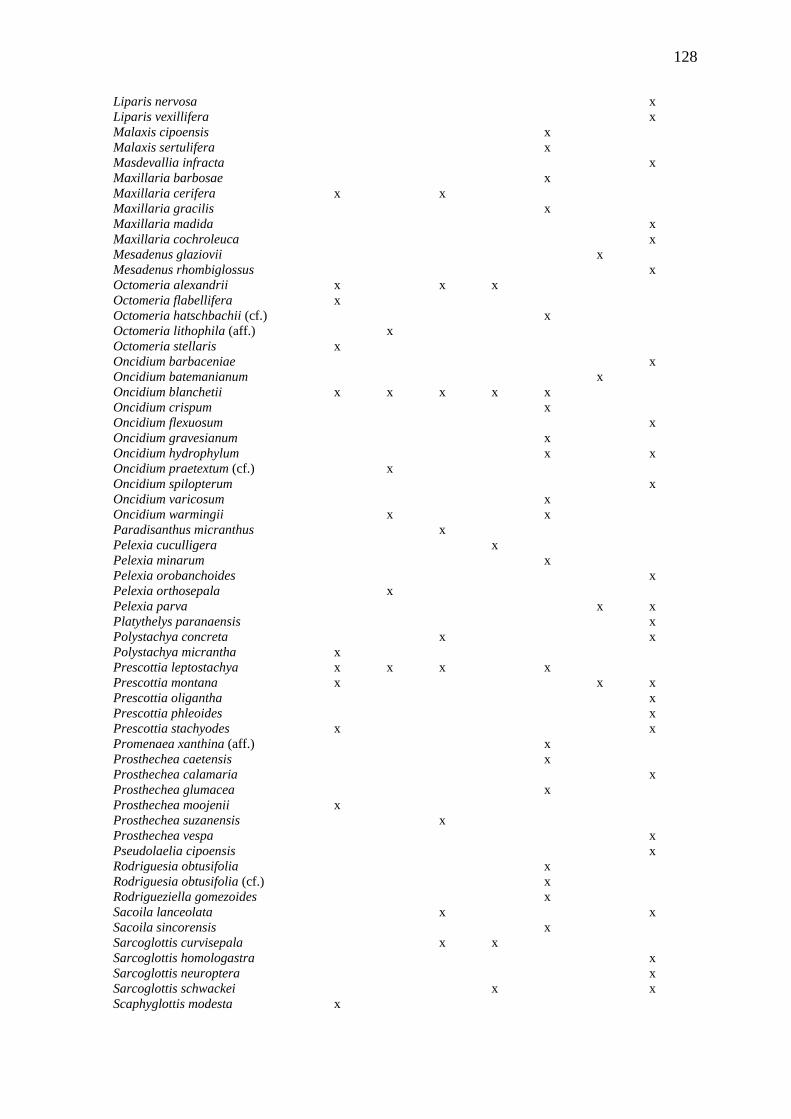
128
Liparis nervosa x Liparis vexillifera x Malaxis cipoensis x Malaxis sertulifera x Masdevallia infracta x Maxillaria barbosae x Maxillaria cerifera x x Maxillaria gracilis x Maxillaria madida x Maxillaria cochroleuca x Mesadenus glaziovii x Mesadenus rhombiglossus x Octomeria alexandrii x x x Octomeria flabellifera x Octomeria hatschbachii (cf.) x Octomeria lithophila (aff.) x Octomeria stellaris x Oncidium barbaceniae x Oncidium batemanianum x Oncidium blanchetii x x x x x Oncidium crispum x Oncidium flexuosum x Oncidium gravesianum x Oncidium hydrophylum x x Oncidium praetextum (cf.) x Oncidium spilopterum x Oncidium varicosum x Oncidium warmingii x x Paradisanthus micranthus x Pelexia cuculligera x Pelexia minarum x Pelexia orobanchoides x Pelexia orthosepala x Pelexia parva x x Platythelys paranaensis x Polystachya concreta x x Polystachya micrantha x Prescottia leptostachya x x x x Prescottia montana x x x Prescottia oligantha x Prescottia phleoides x Prescottia stachyodes x x Promenaea xanthina (aff.) x Prosthechea caetensis x Prosthechea calamaria x Prosthechea glumacea x Prosthechea moojenii x Prosthechea suzanensis x Prosthechea vespa x Pseudolaelia cipoensis x Rodriguesia obtusifolia x Rodriguesia obtusifolia (cf.) x Rodrigueziella gomezoides x Sacoila lanceolata x x Sacoila sincorensis x Sarcoglottis curvisepala x x Sarcoglottis homologastra x Sarcoglottis neuroptera x Sarcoglottis schwackei x x Scaphyglottis modesta x

129
Scuticaria hadwenii (cf.) x Skeptrostachys congestiflora x x x x Sobralia liliastrum x Sobralia sessils x x Sophronitella violacea x Sophronitis bahiensis x x x Sophronitis caulescens x Sophronitis ghillanyi x Sophronitis mirandae x x Sophronitis pumila x Stelis aprica x Stelis parvula x Sternorrynchos arechavaletanii x Thelyschista ghillanyi x x Vanilla chamissonis (cf.) x Veyretia rupicola x x Veyretia simplex x Veyretia sincorensis x x Zygopetalum mackayi x x x Zygopetalum sellowii x x Zygopetalum triste x

130
Apêndice III - Espécimes examinados referentes ao material utilizado para a análise de
similaridade depositadas nos herbários ALCB, CEPEC, HRB e HUEFS
Acianthera hamosa - Lençóis, Morro da Chapadinha, 23.nov.1994, PCD 1265 (ALCB, CEPEC,
HRB, HUEFS). Palmeiras, Pai Inácio, 30.dez.1994, PCD 1523 (ALCB); Pai Inácio, 26.out.1994,
PCD 1026 (ALCB, CEPEC, HRB)
Acianthera ochreata - Lençóis, Serra da Chapadinha, 30.jun.1995, PCD 2045 (ALCB, CEPEC);
Serra da Chapadinha, 24.nov.1994, PCD 1377 (ALCB, CEPEC, HRB); Serra da Chapadinha,
05.jul.1994, PCD 43 (ALCB, CEPEC, HUEFS). Mucugê, about 3 km N of Mucugê on the road to
Andaraí, 15.fev.1977, R.M.Harley et al. 18712 (CEPEC, NY). Palmeiras, Pai Inácio, 24.abr.1995,
PCD 1750 (ALCB, CEPEC, HRB, HUEFS). Rio de Contas, c. 25 km WNW of the town of Rio de
Contas, 22.jan.1974, R.M.Harley et al. 15414 (CEPEC, NY); near Junco, by track to Pico das
Almas, c. 15 km WNW of the town of Rio de Contas, 24.jan.1974, R.M.Harley et al. 15636
(CEPEC, NY); vale acima da Faz. Silvina, 22.dez.1988, R.M.Harley et al. 27349 (CEPEC).
Anathallis montipelladensis - Água Quente, trilho do povoado da Sta. Rosa, 23 km ao O da cidade,
01.dez.1988, R.M.Harley et al. 27050 (CEPEC, K); entre Paramirim das Crioulas e a face NNW do
Pico, 16.dez.1988, R.M.Harley et al. 27522 (CEPEC, K, NY). Rio de Contas, campo do Queiroz,
25.dez.1988, R.M.Harley et al. 27369 (CEPEC, K); 26.nov.1988, R.M.Harley et al. 26293
(CEPEC, K); campo do Queiroz, 11.nov.1988, R.M.Harley et al. 26372 (CEPEC, K)
Anathallis rubens - Palmeiras, Pai Inácio, 24.abr.1995, PCD 1775 (CEPEC). Rio de Contas, vale
ao sudeste do Campo do Queiroz, 09.dez.1988, R.M.Harley et al. 27088 (CEPEC); Pico do Campo
do Queiroz, 16.nov.1988, R.M.Harley et al. 26168 (CEPEC, NY); Pico das Almas, 24.nov.1988,
R.M.Harley et al. 26267 (CEPEC).
Anathallis sclerophylla - Palmeiras, Pai Inácio, 24.abr.1995, PCD 1775 (ALCB, HRB, HUEFS).
Barbosella crassifolia - Rio de Contas, Trilho Faz. Silvina - Campo do Queiroz, 12.dez.1988,
R.M.Harley et al. 27219 (CEPEC).
Bifrenaria magnicalcarata - Rio de Contas, Campo do Queiroz, 12.nov.1988, R.M.Harley et al.
26104 (CEPEC).
Cattleya elongata - Bahia: 10 km S. of Andaraí, on the road to Mucugê, 16.fev.1977, R.M.Harley
et al. 18767 (ALCB, CEPEC, HUEFS); Serra do Sincorá, 6,5 km SW of Mucugê on the Cascavel
road, 27.mar.1980, R.M.Harley et al. 21024 (CEPEC); South of Andaraí, along road to Mucugê
near small town of Xique-Xique, 14.fev.1977, R.M.Harley et al. 18690 (CEPEC). Lençóis, Serra da
Chapadinha, 21.fev.1995, PCD 1661 (ALCB, CEPEC, HRB, HUEFS). Mucugê, by rio Cumbuca c.
3 km S of Mucugê, near site of small dam on road to Cascavel, 04.fev.1974, R.M.Harley et al
15962 (CEPEC). Palmeiras, Pai Inácio, 24.abr.1995, PCD 1741 (CEPEC).
Cleistes sp1 - Palmeiras, Pai Inácio, 27.jun.1995, PCD 1927 (HUEFS).

131
Cleistes castanoides - Andaraí, 10 km S of Andaraí on the road to Mucugê, 16.fev.1977,
R.M.Harley et al. 18765A (CEPEC). Rio de Contas, near Junco, c. 15 km W.N.W of the town of
Rio de Contas, 22.jan.1974, R.M.Harley et al. 15613 (CEPEC, K)
Cleistes exilis - Serra do Sincorá, c. 15 km N.W. of Mucugê, on the road to Guiné and Palmeiras,
26.mar.1980, R.M.Harley et al. 20986 (CEPEC, K); Serra do Sincorá, c. 15 km N.W. of Mucugê,
on the road to Guiné and Palmeiras, 26.mar.1980, R.M.Harley et al. 20984 (CEPEC, K); Serra do
Sincorá, 10 km S of Andaraí on the road to Mucugê, 16.fev.1977, R.M.Harley et al. 18765
(CEPEC, HB, HRB, K, NY); Lençóis, Serra da Chapadinha, 05.jul.1994, PCD 37 (ALCB, CEPEC,
HRB, HUEFS).
Cleistes pluriflora - Mucugê: Serra do Sincorá, 2-3 km approximately S.W. of Mucugê on the road
to Cascavel, 17.fev.1977, R.M.Harley et al. 18813 (CEPEC, K). Rio de Contas, c. 25 km W.N.W.
of the Vila do Rio de Contas, 17.fev.1977, R.M.Harley et al. 19513 (CEPEC, K, HB).
Cyrtopodium blanchetii - Rio de Contas, entre Junco - Faz. Brumadinho, 9-14 km ao NO da cidade,
11.nov.1988, R.M.Harley et al. 27121 (CEPEC , K).
Cyrtopodium edmundoi - Lençóis, Chapadinha, 27.out.1994, PCD 1070 (ALCB); Chapadinha,
27.set.1994, PCD 933 (ALCB, HRB, HUEFS); Morro da Chapadinha, 22.nov.1994, PCD 1253
(ALCB, CEPEC). Palmeiras, Pai Inácio, 21.nov.1994, PCD 1136 (ALCB, CEPEC). Rio de Contas.
4 km ao N de Rio de Contas, 21.jul.1979, S.A.Mori et al. 12397(CEPEC).
Cyrtopodium parviflorum - Rio de Contas, trilho Faz. Silvina – Queiroz, 13.nov.1988, R.M.Harley
et al. 26139 (CEPEC, K).
Encyclia alboxanthina - Andaraí: 10 km S. of Andaraí, 16.fev.1977, R.M.Harley et al. 18767
(ALCB, CEPEC, HUEFS). Lençóis, Morro da Chapadinha, 29.dez.1994, PCD 1489 (ALCB,
CEPEC); Morro da Chapadinha, 24.nov.1994, PCD 1333 (ALCB, CEPEC, HRB, HUEFS); Serra
da Chapadinha, 21.fev.1995, PCD 1663 (ALCB, CEPEC).
Epidendrum orchidiflorum - Lençóis, Morro da Chapadinha, 29.nov.1994, PCD 1474 (ALCB,
HRB, HUEFS); Morro da Chapadinha, 27.out.1994, PCD 1133 (ALCB, HRB). Palmeiras, Pai
Inácio, 21.nov.1994, PCD 1205 (ALCB, HRB, HUEFS).
Epidendrum secundum - Lençóis, Morro da Chapadinha, 23.nov.1994, PCD 1282 (ALCB); Serra
da Chapadinha, 25.set.1994, PCD 750 (ALCB, HRB, HUEFS). Palmeiras, Pai Inácio, 29.ago.1994,
PCD 449 (ALCB); Pai Inácio, 25.out.1994, PCD 1009 (ALCB); Pai Inácio, 21.nov.1994, PCD
1164 (ALCB); Pai Inácio, 21.nov.1994, PCD 1200 (ALCB, HRB, HUEFS).
Epistephium lucidum - 10 km S of Andaraí on the road to Mucugê, 16.fev.1977, R.M.Harley et al.
18773 (CEPEC, NY); by rio Cumbuca c. 3 km S of Mucugê, 04.fev.1974, R.M.Harley et al. 15905
(CEPEC, NY). Lençóis, Morro da Chapadinha, 22.nov.1994, PCD 1209 (CEPEC, HRB); Morro da
Chapadinha, 23.nov.1994, PCD 1266 (CEPEC). Palmeiras, Pai Inácio, 27.dez.1994, PCD 1407
(ALCB, CEPEC). Rio de Contas, Trilho Faz. Silvina - Queiroz, 31.out.1988, R.M.Harley et al.

132
25805 (CEPEC); near Junco, c. 20 km WNW of the town of Rio de Contas, 22.jan.1974,
R.M.Harley et al. 15396 (CEPEC, NY).
Habenaria bractescens - Rio de Contas, between 2,5 and 5 km S of the vila of Rio de Contas on
side road to W of the road to Livramento, 28.mar.1977, R.M.Harley et al. 20099 (CEPEC, NY).
Habenaria graciliscapa - Rio de Contas, c. 25 km WNW of the vila of Rio de Contas, 17.fev.1977,
R.M.Harley et al. 19532 (CEPEC); Junco 9-11 km ao NO da cidade, 06.nov.1988, R.M.Harley et
al. 25928 (CEPEC, K).
Habenaria hamata - Palmeiras, Pai Inácio, 30.ago.1994, PCD 601 (ALCB, CEPEC, HRB).
Habenaria humilis - Serra do Sincorá, c. 15 km NW of Mucugê on road to Guiné e Palmeiras,
26.mar.1980, R.M.Harley et al. 21013 (CEPEC).
Habenaria johannensis - 2-3 km SW of Mucugê on road to Cascavel, 17.fev.1977, R.M.Harley et
al. 18787 (CEPEC); 9 km SW of Mucugê on road to Cascavel, 06.fev.1974, R.M.Harley et al.
16086 (CEPEC). Rio de Contas, c. 25 km WNW of the vila of Rio de Contas, 17.fev.1977,
R.M.Harley et al. 19597 (CEPEC).
Habenaria lasioglossa - Rio de Contas, c. 25 km WNW of the vila of Rio de Contas, 20.mar.1977,
R.M.Harley et al. 19760 (CEPEC).
Habenaria meeana - Água Quente, a 17 km a NW de Rio de Contas, 25.mar.1980, S.Mori &
F.Benton 13593 (CEPEC). Rio de Contas, ca. 25 km NW of the Vila de Rio de Contas,
17.fev.1977, R.M.Harley et al.19531 (HRB).
Habenaria mystacina - Serra do Sincorá, c. 15 km NW of Mucugê on road to Guiné e Palmeiras,
26.mar.1980, R.M.Harley et al. 20985 (CEPEC).
Habenaria nasuta - Rio de Contas, c. 25 km WNW of the vila of Rio de Contas, 17.fev.1977,
R.M.Harley et al. 19506A (CEPEC).
Habenaria pratensis - Rio de Contas, 18 km WNW along road from vila of Rio de Contas to the
Pico das Almas, 21.mar.1977, R.M.Harley et al. 19793 (CEPEC).
Habenaria repens - 9 km SW of mucugê, on road from Cascavel, 07.fev.1974, R.M.Harley et al.
16115 (CEPEC).
Habenaria subviridis - Água Quente, a 17 km ao NW de Rio de Contas, 24.mar.1980, S.Mori &
F.Benton 13580 (CEPEC). Rio de Contas, c. 25 km WNW of the vila of Rio de Contas,
18.mar.1977, R.M.Harley et al. 19626 (CEPEC).
Habenaria cf. trichoceras - Rio de Contas, c. 25 km WNW of the vila of Rio de Contas,
17.fev.1977, R.M.Harley et al. 19506B (CEPEC).
Habenaria trichoceras - Rio de Contas, c. 25 km WNW of the vila of Rio de Contas, 20.mar.1977,
R.M.Harley et al. 19760A (CEPEC).
Habenaria warmingii - Rio de Contas, c. 25 km WNW of the vila of Rio de Contas, 17.fev.1977,
R.M.Harley et al. 19506C (CEPEC).
Maxillaria notyliglossa - Lençóis, Morro da Chapadinha, 23.nov.1994, PCD 1290 (ALCB).

133
Octomeria alexandrii - Lençóis, Morro da Chapadinha, 22.nov.1994, PCD 1240 (ALCB, CEPEC).
Palmeiras, Pai Inácio, 24.abr.1995, PCD 1757 (ALCB, CEPEC, HRB, HUEFS)
Octomeria cf. decumbens - Serra do Sincorá 6,5 km SW of Mucugê on the Cascavel road,
27.mar.1980, R.M.Harley et al. 21023 (CEPEC).
Oncidium blanchetii - Lençóis, Morro da Chapadinha, Chapadinha, 22.nov.1994, PCD 1241
(ALCB).
Oncidium warmingii - c. 25 km WNW of the town of Rio de Contas, 23.jan.1974, R.M.Harley
15444 (CEPEC).
Paradisanthus micranthus - Lençóis, Chapadinha, 26.abr.1995, PCD 1824 (ALCB, CEPEC); Serra
da Chapadinha, 21.fev.1995, PCD 1652 (ALCB, CEPEC, HRB).
Pelexia orthosepala - Rio de Contas, Pico das Almas, 10.nov.1988, R.M.Harley 26332 (CEPEC,
K).
Polystachya concreta - Lençóis, Serra da Chapadinha, 21.fev.1995, PCD 1669 (ALCB).
Prescottia leptostachya - Rio de Contas, Pico das Almas, 25.dez.1988, R.M.Harley et al. 27370
(CEPEC).
Sacoila lanceolata - Lençóis, Serra da Chapadinha, 31.ago.1994, PCD 710 (ALCB).
Sarcoglottis curvisepala - Lençóis, Morro da Chapadinha, 23.nov.1994, PCD 1259 (ALCB,
CEPEC, HRB, HUEFS). Palmeiras, Pai Inácio, 21.nov.1994, PCD 1184 (ALCB).
Skeptrostachys congestiflora - Lençóis, Serra da Chapadinha, 23.fev.1995, PCD 1726 (ALCB,
CEPEC, HRB, HUEFS). Rio de Contas, Pico das Almas, 31.out.1988, R.M.Harley et al. 25811
(CEPEC, K); Pico das Almas, 09.nov.1988, R.M.Harley et al. 25997 (CEPEC, K, HUEFS); Pico
das Almas, 26.nov.1988, R.M.Harley et al. 26295 (CEPEC, K).
Sobralia liastrum - Lençóis, Chapadinha, 27.set.1994, PCD 922 (ALCB, CEPEC, HRB, HUEFS);
Morro da Chapadinha, 24.nov.1994, PCD 1328 (ALCB, CEPEC, HRB, HUEFS)
Sobralia sesilis - Lençóis, Serra da Chapadinha, 23.fev.1995, PCD 1706 (ALCB, CEPEC, HRB,
HUEFS); Morro da Chapadinha, 29.dez.1994, PCD 1478 (CEPEC).
Sophronitis bahiensis - Rio de Contas, 10-13 km ao norte da cidade na estrada para o povoado de
Mato Grosso, 27.out.1988, R.M.Harley et al. 25698 (CEPEC, K); Pico das Almas, 23.out.1988,
R.M.Harley et al. 25308 (CEPEC); Pico das Almas, 30.out.1988, R.M.Harley et al. 25770
(CEPEC).
Stelis parvula - Rio de Contas, 26.nov.1988, R.M.Harley 26294 (CEPEC, HUEFS).
Thelyschista ghillanyi - Mucugê, by Rio Cumbuca, about 3 km N of Mucugê on the Andaraí road,
05.fev.1974, R.M.Harley et al. 15993 (CEPEC, K); by Rio Cumbuca, c. 3 km S of Mucugê,
04.fev.1974, R.M.Harley et al. 15965 (CEPEC).
Veyretia simplex - Rio de Contas, Pico das Almas, 05.dez.1988, R.M.Harley et al. 26591 (CEPEC).
Veyretia sincorensis - Lençóis, Morro da Chapadinha, 22.nov.1994, PCD 1210 (ALCB, CEPEC,
HRB); Morro da Chapadinha, 21.fev.1995, PCD 1664 (ALCB).

134
Zygopetalum mackayi - Rio de Contas, c. 25 km WNW of the vila of Rio de Contas,
19.mar.1977, R.M.Harley et al. 19705 (CEPEC).
Apêndice IV - Espécimes examinados que sofreram alterações.
Nome original Alteração Fonte Anathallis rubens PCD 1775 (Toscano de Brito, 1998) Anathallis sclerophylla C. Azevedo Bulbophyllum ipanemensis R.M.Harley et al. 19498 (Toscano de Brito,1995)
Bulbophyllum involutum Toscano de Brito
Bulbophyllum ipanemensis CFCR 1477 (Harley & Simmons, 1986)
Bulbophyllum involutum Borba et al (1998)
Cleistes paranaensis var. bradeana R.M.Harley et al. 15613 (Toscano de Brito,1995)
Cleistes castanoides Silva, 2002
Cleistes aphylla R.M.Harley et al. 20984 (Harley & Simmons, 1986)
Cleistes exilis Silva, 2002
Cleistes ionoglossa R.M.Harley et al. 18813 (Harley & Simmons, 1986)
Cleistes pluriflora Silva, 2002
Encyclia advena R.M.Harley et al. 15962, 18767 (Harley & Simmons, 1986)
Encyclia alboxanthina. C. van den Berg
Epistephium mardeni PCD 1209 (Toscano de Brito, 1998) Epistephium lucidum C. van den Berg Epistephium mardeni PCD 1266 (Toscano de Brito, 1998) Epistephium lucidum C.N. Fraga Epistephium mardeni PCD 1407 (Toscano de Brito, 1998) Epistephium lucidum C. van den Berg Epistephium praestans R.M.Harley et al. 18773 (Harley & Simmons, 1986)
Epistephium lucidum C. van den Berg
Habenaria entomantha S.Mori & F.Benton 13580, R.M.Harley et al. 19626 (Toscano de Brito,1995)
Habenaria subviridis J.Batista
Habenaria pentadactyla R.M.Harley et al. 16115 (Harley & Simmons, 1986)
Habenaria repens J.Batista
Odontorrhynchus ghillanyi, R.M.Harley 15965, 15993 (Harley & Simmons, 1986)
Thelyschista ghillanyi Smidt, 2002
Oncidium ramosum R.M.Harley et al. 24441 (Toscano de Brito,1995)
Oncidium blanchetii Toscano de Brito
Sarcoglottis simplex R.M.Harley 26591 (Toscano de Brito,1995)
Veyretia simplex Smidt, 2002
Sarcoglottis sincorensis PCD 1210, PCD 1664 (Toscano de Brito, 1998)
Veyretia sincorensis Smidt, 2002
Sarcoglottis fasciculata PCD 1259 (Toscano de Brito, 1998)
Sarcoglottis curvisepala Smidt, 2002
Sarcoglottis sp. PCD 1184 (Toscano de Brito, 1998) Sarcoglottis curvisepala Smidt, 2002 Sobralia sp 1 PCD 1706 (Toscano de Brito, 1998) Sobralia sesilis C. van den Berg
Sobralia sp 2 PCD 1478 (Toscano de Brito, 1998) Sobralia sesilis C. van den Berg

135
RESUMO
Este trabalho apresenta um inventário da família Orchidaceae no Parque Municipal de
Mucugê e uma análise comparativa desta família em áreas da Cadeia do Espinhaço,
situadas nos estados de Minas Gerais e Bahia. O levantamento foi realizado através de
coletas aleatórias nessa área, a partir de excursões mensais durante o período de um ano.
Foram encontradas 35 espécies distribuídas em 22 gêneros, das quais 11 são citadas pela
primeira vez para o Município de Mucugê; três para a região da Chapada Diamantina e
sete para o estado da Bahia. Os gêneros que apresentaram maior número de espécies
foram: Bulbophyllum, Epidendrum, Octomeria e Prescottia. Lectótipos foram selecionados
para Acianthera hamosa, Anathallis microphyta, Cattleya elongata, Maxillaria cerifera,
Octomeria stellaris e Prescottia montana. Descrições e ilustrações são apresentadas para
todas as espécies, além de chaves para identificação dos gêneros e espécies. Uma nova
combinação para Pleurothallis microphyta (Barb.Rodr.) Cogn. foi proposta: Anathallis
microphyta. A comparação florística levou em consideração sete áreas da Cadeia do
Espinhaço, sendo duas em Minas Gerais e cinco na Bahia. Para comparar as floras de
Orchidaceae, foram utilizadas análises de similaridade e agrupamento, utilizando os
coeficientes de Jaccard e Dice. Todas as áreas estão representadas por um grande número
de espécies exclusivas, apresentando valores de similaridade relativamente baixos. Três
grupos foram formados, um representado pela Serra do Cipó, outro por Grão-Mogol, e o
terceiro por todas às áreas da porção baiana da Cadeia do Espinhaço, sendo que esta foi
dividida em dois subgrupos, um compreendendo o setor norte da Serra do Sincorá,
incluindo a Serra da Chapadinha, o Parque Municipal de Mucugê e o Pai Inácio, e outro a
porção sudoeste da Chapada Diamantina, abrangendo o Pico das Almas e Catolés. Os
grupos encontrados correspondem ao esperado em relação às subdivisões topogeográficas
da Cadeia do Espinhaço e dentro da Chapada Diamantina.

136
ABSTRACT
This study presents a survey of the Orchidaceae occurring in the Parque Municipal de
Mucugê, Mucugê, and a comparative analysis of this family in areas at the Espinhaço
Range at the states of Minas and Bahia. The survey consisted of random sampling within
the Parque montly during one year. We found 35 species of orchids in 22 genera, of which
11 species are for the first time reported for Mucugê; three species are new records for the
Chapada Diamantina and seven are new records for state Bahia State. The main genera in
number of species are Bulbophyllum, Epidendrum, Octomeria ande Prescottia. Lectotypes
for Acianthera hamosa, Anathallis microphyta, Cattleya elongata, Maxillaria cerifera,
Octomeria stellaris e Prescottia montana are chosen here. All taxa are described and each
genus is illustrated. Identification keys for species and genera are provided. A new
combination for Pleurothallis microphyta (Barb.Rodr.) Cogn. is proposed: Anathallis
microphyta. For the floristic comparison seven areas of the Espinhaço Range were
included, being two in Minas Gerais State and five in Bahia. In order to compare the orchid
floras, we used similarity and clustering analysis based on presence/absence data from
species composition, with the coefficients of Jaccard and Dice. All areas included
presented a large proportion of exclusive species, and overall similarity among areas was
low. Despite this, three main groups were indicated, equivalent respectively to Serra do
Cipó, Grão-Mogol and a cluster with all the areas of the Chapada Diamantina. The latter is
subdivided in two subgroups, the first is equivalent to the north of Serra do do Sincorá,
namely Serra da Chapadinha, the Parque Municipal de Mucugê and Pai Inácio, and the
second corresponds to southwest Chapada Diamantina, namely Pico das Almas and
Catolés. The groups found to the pattern that could be expected from the geography and
relief of the Espinhaço Range and within the Chapada Diamantina.

NORMAS PARA PUBLICAÇÃO
Sitientibus Série Ciências Biológicas publica trabalhos científicos originais dedicados aqualquer ramo da Biologia, além de resenhas bibliográficas, resumos de tese, comunicações,revisões, cartas ao editor, réplicas e tréplicas.
Os manuscritos devem ser enviados, no original e duas cópias (incluindo figuras), aoEditor da Série Ciências Biológicas: UEFS, Departamento de Ciências Biológicas, km 03 -BR 116, Campus, 44031-460, Feira de Santana, Bahia, Brasil.Manuscrito: Os trabalhos devem ser redigidos em Português, Inglês ou Espanhol, O aceite deoutro idioma ficará a critério da Comissão Editorial.
Os trabalhos devem ser datilografados em papel A4, espaço duplo e com margens largas(2,5 cm). Alterações de pequena monta poderão ser feitas pela Comissão Editorial. Correçõese acréscimos encaminhados pelos autores após o aceite do trabalho poderão ser recusados.
Uma cópia do manuscrito em disquete deve ser enviada apenas após o aceite do trabalho,digitado em Word para Windows, formato Rich Text (.rtf), e deve ser acompanhada de umacópia impressa.
Cada página deve ser numerada, sendo que a página de rosto deve conter: (1) título dotrabalho; (2) nome dos autores com respectiva afil iação; (3) endereço profissional,preferencialmente, incluindo: (3.1) instituição; (3.2) departamento ou unidade equivalente;(3.3) caixa postal ou logradouro; (3.4) código de endereçamento postal; (3.5) cidade; (3.6)estado ou equivalente; (3.7) país; (3.8) e-mail do autor ao qual devem ser enviadas ascorrespondências. A Segunda página deve conter Resumo (e o título do trabalho, entreparênteses), três palavras-chave, Abstract (e título em inglês, entre parênteses) e três keywords.
No texto serão escritos em itálico, os nomes dos grupos taxonômicos de gênero e deespécie, além de palavras estrangeiras. Não devem ser usados quaisquer outros sinais demarcação ou ênfase. Em trabalhos taxonômicos, a primeira citação de um táxon no texto, deveser feita com a apresentação do nome científico por extenso, paraa nomes regidos pelo ICBN,ou nome e data, para os regidos pelo ICZN.
As citações bibliográficas no texto deverão seguir as seguintes modelos: Quate (1965);(Quate, 1965); Quate (1965: 820); Erwin & Sott (1980); Guimarães et al. (1983). Os artigosdevem ser citados em ordem cronológica.
Agradecimentos e indicações de financiamentos devem ser relacionados antes do itemReferências Bibliográficas.Referências Bibliográficas: todas as mencionadas no texto devem ser arroladas no final dotrabalho. O nome dos autores deve ser digitado em Caixa Alta. A forma de citações dasreferências deve estar de acordo com os seguintes exemplos:a) Periódicos (abreviados de acordo com o World List of Scientific Periodicals ou como
indicado pelo próprio periódico):QUATE, L.W. 1965. A taxonomic study of Philipine Psychodidae. Pacif. Ins. 7(4): 815-
902.

ERWIN, T.L. & J.C. SCOTT. 1980. Seasonal and size patterns, trophic structure, andrichness of Coleoptera in the tropical arboreal ecosystem: the fauna of the treeLuchea seemannii Triana and Planch in the Canal Zone of Panama. Coleopt. Bull.34(3): 305-322.
MORI, S.A., B.M.BOOM & G.T. PRANCE. 1981. Distribution patterns and conservation ofeastern Brazilian coastal tree species. Brittonia 33 (2): 233-245.
b) Livros:HENNIG, W. 1981. Insect phylogeny. Chichester, John Wiley, 514p.POLHILL, R.M. & P.H. RAVEN (eds.) 1981. Advances em Legume Systematics. London,
Royal Botanic Gardens Kew, 1049 p.c) Capítulos de livro:
HULL, D.L. 1974. Darwinism and historiography. In: T.F. GLICK (ed.), The comparativereception of Darwinism, pp. 388-402. Austin, Univ. Texas.
d) Teses e Dissertações:SILVEIRA, L.T. 1991. Revisão taxonômica do gênero Periandra Mart. ex Benth. Univ.
Estandarte. Campinas, MSc diss.e) Publicações em eventos:FERNANDES, A. & P. BEZERRA. 1982. O gênero Moldenhawera (Leg. Caesalp.) no Brasil.Sociedade Botânica do Brasil , XXX II Congresso Nacional de Botânica, Anais. Teresina, pp.58-62.FORTUNATO, R.H. 1994. Revisión del género Collaea. Sociedad Latinoamericana deBotánica, VI Congreso Latinoamericano de Botánica, Resúmenes, Mar del Plata, p. 252.
I lustrações e tabelas: fotografias, desenhos, gráficos e mapas são denominados figuras,devem ser numerados com algarismos arábicos e chamados no texto em ordem crescente.Desenhos devem ser feitos a nanquim e fotografias devem ter nitidez e contraste. Nãomisturar fotografias e desenhos na mesma página.
As figuras devem ser montadas em cartolina branca marcada, no verso, com nome dosautores e título do trabalho, proporcionais às dimensões do espelho (19,0 cm de largura x 23,0cm de altura; coluna de 9,0 cm) e reservando um espaço para a legenda.
Tabelas devem ser numeradas em algarismos romanos e apresentadas em páginasseparadas.Legendas devem ser datilografadas em folha à parte.
Espécime-testemunha, quando apropriado, deve ser mencionado juntamente com a coleção ea instituição depositária.25 separatas serão fornecidas aos autores.
Modelo: um exemplar recente da revista deve ser usado como modelo para organização dotexto. Uma orientação para checagem está disponível na página da UEFS (www.uefs.br).

KEW BULLETIN - GUIDE FOR AUTHORS
Kew Bulletin (KB) is published by the Royal Botanic Gardens, Kew. Four parts appear each year,usually in March, June, August and November.
SCOPE
KB publishes papers in the fields of vascular plant taxonomy, nomenclature, phylogeny,systematics, plant geography, and floristics, and in morphology, palynology, cytotaxonomy,chemotaxonomy, anatomy and other relevant disciplines where the results have implications forvascular plant taxonomy. Purely biographical and bibliographic papers are not usually accepted.
In recent years about 50% of Kew Bulletin’s content has been authored or co-authored by Kewstaff, but this is not a rigid policy. Work based in a large part on Kew material, or dealing withregions or groups of major interest to Kew, is always welcome.
THE TYPESCRIPT
Two copies of all typescripts are required, on clean thick A4 paper (we will request the text ondiskette or as an e-mail attachment after the review stage). The work should be clearly typed,double spaced, with wide margins on each side. Number all pages. Do not justify text.
Initial submission may be by e-mail attachment but check first with the Editor([email protected]). At present (May 2001) Kew uses Word 97 and Pegasus Mail. Diagramsand figures can cause problems and should be discussed before submission.
Distinguish between hyphens, typed without spaces (as in brown-tomentose; 3-flowered), anden-rules, typed with spaces (as in leaves 24 – 30 � 10 – 15 cm; inflorescences 2 – 3-flowered). No full stops after common contractions (e.g., Mt, Mts), nor after points of the compass (N, S, NEetc.), not after abbreviations for units of measurement (e.g., mm, cm, km). Include, however,after other abbreviations (e.g., R., fl., fr.).
Months are listed as follows: Jan., Feb., March, April, May, June, July, Aug., Sept., Oct., Nov., Dec.Give the year in full - 1991 not /91.
Tables should be submitted separately, as should captions for illustrations.
ILLUSTRATIONS
The KB printed area is 197 � 130 mm. However, illustrations which are to be used elsewhere,such as in Kew Floras, should be drawn to the old KB print area (197 � 114 mm).
Line drawings and maps in Indian ink should be drawn on smooth white card or plastic film, onethird larger than they are to appear. No illustration or map should have its longest dimensiongreater than 270 mm long before reduction. Label figures, and indicate lettering and scale barsclearly on a good-quality photocopy. Lettering and scale bars will be added electronically afterthe original is scanned (Kew Bull. does not use "I" or "O" in lettering figures). Photographs ('Plates') for continuous tone reproduction should be of the highest quality. Theyshould show good tonal range and be absolutely sharp. They should be either at the final size or

slightly larger. Provide two copies of each. Good photocopies will suffice for the review process,but we will require transparencies, sharp prints or electronic 'pict' or 'tif' files for reproduction. Incomposite 'plates' the individual photographs will be butted together unless that would createconfusion, in which case a white space will be interposed. It is usually simpler and quicker forcomposite plates to be made up electronically at Kew, but a clear mock-up must be provided.
Line drawings and plates are numbered in one sequence as figures (Fig. 1 , etc.); maps arenumbered separately (Map 1, etc.). Scale lines may be given on the illustration (preferred for'plates') or final (reduced) magnifications given in the caption (preferred for line drawings).Full- or part-page illustrations are acceptable. Captions should be brought together on a separatesheet. All illustrations and parts of composite artwork should be referred to in the text.
Colour illustrations can only be included in very special cases; advance warning is required fortheir inclusion, and a charge may be made.
FORMAT & STRUCTURELayout
Please refer to a recent part of KB (Vol. 47 or later) for details of the typographic and layoutcharacteristics of KB. If in any doubt, do not mark anything. The recommended layout foraccounts of taxa is given below.
Authors of plant names are normally omitted from the title. If relevant, the title shouldinclude the relevant family and an indication of the geographical area.
The summary is important. Most readers will read nothing else. Some may feel they onlyneed to write for the other specialists in their field, but if you wish your paper to reach a wideraudience, make the summary interesting and comprehensive. It should indicate what theresearch set out to achieve, how it was carried out, and the degree to which the objectives havebeen reached. It should include any authors of plant names omitted from the title, and the namesof all new taxa described, and of new combinations unless the number is very large. Conclusionsshould be included, and mention made of the methods used.
Headings
A careful division of your work using headings will make it easier for the reader to follow.Plan a sensible and consistent hierarchy; the more important headings are centred (in large andsmall capitals), the less important ranged left and italicised.
For longer papers, a contents list at the beginning may be appropriate. Extensive taxonomicrevisions may be made more useful by adding an index to epithets at the end.
Keys
Keys may be bracketed (preferable) or indented, but couplets should always be numbered;please consult a recent issue of KB and follow the key layouts used therein.
Synonyms
Homotypic synonyms should be listed in chronological order after the accepted name,followed by heterotypic synonyms, also with their respective homotypic synonyms inchronological order. New synonyms should be clearly marked ‘synon. nov.’.
Types

As required by the ICBN, for newly-described taxa the herbarium in which the holotype isdeposited must be specified. Herbaria that are definitely known to hold isotypes should also belisted. The Editors consider it to be best practice for type material to be deposited in the country oforigin. Authors are reminded that in some countries, national legislation requires that theholotype be deposited in a local or national herbarium.
Citation of types, when known, is encouraged. Please indicate if lectotypes, neotypes orepitypes are being designated ('chosen here'), or have been chosen before by giving a reference:-'Papua, Boridi, Carr 12345 (lectotype K! (selected by Bloggs 1977); isolectotypes BRI, L!).' Explainwhy you need to select lectotypes, neotypes and epitypes, and also the reasons for selecting aparticular specimen.
Whenever possible, try to see and cite types when publishing new combinations.
Citation of specimens
It is not easy to make hard and fast rules for this, but the following guidelines may be useful.Remember that you are citing specimens to (inter alia) help communicate your concept of ataxon to the user, and help curation of material in other herbaria.
For new taxa, all material seen should normally be cited. In revisions, space and cost generally prohibit publication of all the label details, and of all
specimens seen. Cite specimens chosen to represent the geographical and morphologicalvariation of the taxon, preferring collections distributed to several herbaria. The complete list ofspecimens seen should preferably be deposited somewhere accessible (e.g. Kew Library), andcopies made available on request. However, revisions of whole genera or higher taxa are moreuseful to curators if accompanied by an abbreviated list of all specimens with determinations (see,e.g., KB 55(1): 38 – 40 or KB 54(4): 851 – 852).
Reproduce only those label data that add significantly to localizing the collection or to fieldknowledge. Label data are normally translated into English, but data for types can be left in theoriginal language. Remember that it may be unwise to give very precise localities for rare andhorticulturally interesting taxa.
It is useful to state the number of collections examined, and to cite at least one specimen fromeach country in the range of the taxon.
In cases of long-standing confusion, there may be a case for citing all specimens, butconsider reducing detail to a bare minimum.
Use an exclamation mark (!) to show that a specimen has been seen, or state in theintroduction "All cited specimens have been seen by the author".
Basic Sources
Herbarium codes should follow Holmgren, Holmgren & Barnett (1990) and the associatedweb site (www.nybg.org/bsci/ih).
Citations of authors of plant names should follow Brummitt & Powell (1992 – see alsowww.ipni.org). If in doubt, do not abbreviate. Joint author abbreviations are linked by '&'.
Book abbreviations should follow those in Stafleu & Cowan — Taxonomic Literature Ed. 2(1976 – 88), although KB capitalizes most words. If in doubt, do not abbreviate.
Journal abbreviations should follow Botanico-Periodicum-Huntianum (1976) and itsSupplementum (1991). The principles therein should allow the correct abbreviations to be madefor journals not included (mostly recent); if in doubt, do not abbreviate.
Spellings of place-names should follow the Times Atlas (2000 edition if possible). A usefullist of country names and subdivisions is available (Hollis & Brummitt 1992 – new edition inpreparation). Use normally accepted English usage — Ghana not Gold Coast, Zimbabwe notRhodesia — but Thailand not Muang Thai, Brazil not Brasil. Some country names can beambiguous and at present Kew Bulletin uses Congo (Brazzaville) and Congo (Kinshasa), as well asMyanmar (Burma). Several large islands have well-established English names which should beused; Madagascar (not Malagasy Republic), Sicily (not Sicilia), New Guinea (for the whole island– the eastern part is Papua New Guinea and the western part, West Papua). Old names of

localities — as used on old labels — may be given with the modern equivalent in square brackets- Stanleyville [Kisangani]; Salisbury [Harare].
Altitudes are cited in metres to the nearest 50 m. If the label gives the altitude in feet, thisshould be converted to metres (to the nearest 50 m). The original label altitude may be given insquare brackets.
The use of latitude and longitude (in this order) for obscure localities is encouraged.
Layout of accounts of taxa in revisions
In revisions, taxon accounts should be laid out as follows. Note the positions of indents, spaces, bold and ita
6. Sclerochiton boivinii (Baill.) C. B. Clarke in Fl. Trop. Afr. 5: 110 (1899); [list further works in which thehas been treated]. Type: Kenya, Mombasa, Boivin s.n. (holotype P; isotype K).
Pseudoblepharis boivinii Baill., Bull. Mens. Soc. Linn. Paris 2: 837 (1890); [further works inwhich this homotypic synonym has been used].
Pseudoblepharis heinsenii Lindau, Bot. Jahrb. Syst. 24: 320 (1897); [further works in which thisheterotypic synonym has been used]; synon. nov. Type: Tanzania, E Usambara Mts, Nderema,Heinsen 4 (holotype B†; isotypes BR, K).
Description in the order: general habit; underground parts; stem; leaves; inflorescences;flowers (calyx, corolla, androecium, gynoecium); fruits; seeds. [In long descriptions it is helpful ifmajor headings are in italics]. Fig. 4.
DISTRIBUTION. Kenya, Tanzania. Map 5.
[specimen listings either by country — if listing is complete — e.g.:
KENYA. Kilifi Distr., N of Giriama, Adu, Jan. 1937, Dale 3664 (FT, K); Kwale Distr., MweleMdogo Forest, 6 Feb. 1953, Drummond & Hemsley 1143 (BR, FT, K) & Shimba Hills, MakadaraForest, 17 Sept. 1982, Polhill & Robertson 4795 (C, K, P).
or as SELECTED COLLECTIONS. KENYA: Kilifi Distr., N of Giriama, Adu, Jan. 1937, Dale 3664 (FT, K);
Kwale Distr., Mwele Mdogo Forest, 6 Feb. 1953, Drummond & Hemsley 1143 (BR, FT, K) &Shimba Hills, Makadara Forest, 17 Sept. 1982, Polhill & Robertson 4795 (C, K, P). TANZANIA:Lushoto Distr., E Usambara Mts, Maramba, 18 Nov. 1936, Greenway 4748 (BR, FHO, K, S) & EUsambara Mts, Ndola, 17 Feb. 1954, Faulkner 1350 (BR, K, S) & W Usambara Mts, Dindira, 6Aug. 1957, Faulkner 2032 (BR, K). 62 collections studied.
HABITAT. Shrub layer in lowland and medium-altitude evergreen and semi-evergreen forest,riverine forest; 0 – 1400 ( – 1500) m. [Note that the use of vernacular terms for vegetation typesis discouraged].
VERNACULAR NAME(S). [Give name and language].
NOTE(S). Sclerochiton boivinii is an isolated species, so distinct that it deserves to be placed ina section of its own. It differs from all other species of Sclerochiton in its yellow or orange corolla,..... [discussion continues].
Literature
Literature references from the text of the paper are normally gathered together under a'References' section at the end of the paper. They are referred to in the text as Bloggs (1962),Bloggs (1962: 234), (Bloggs 1962), (Bloggs 1962: 234) or (Bloggs 1962; Another 1976)

depending on the context. Use 'et al.' for papers with more than two authors, but give all authorsin the 'References' section. Literature references which appear only in citations of names, in abbreviated form, should not berepeated in the reference list. Loc. cit. (same work, same volume, same page); tom. cit. (same work, same volume, differentpage — give page number); op.cit. ( same work; different volume; different page — give volumeand page numbers) can all be used within reason.See section 'Basic Sources' above for abbreviations of book and journal titles. Include publisherand place of publication for books.
The following examples show the layout used for various kinds of reference:
Dransfield, J. (1989). Voanioala (Arecoideae: Cocoeae: Butiinae), a new palm genus fromMadagascar. Kew Bull. 44: 191 – 198.
Uhl, N.W. & Dransfield, J. (1987). Genera Palmarum: a classification of palms based on the workof H. E. Moore Jr. The L. H. Bailey Hortorium and the International Palm Society, Lawrence,Kansas, USA.
Li, H. (1979). Arisaema. In: C. Y. Wu & H. Li (eds.), Flora Reipublicae Popularis Sinicae 13 (2):116 – 194 (in Chinese).
Gentry, A. H. (1986). Endemism in tropical versus temperate plant communities. In: M. E. Soulé(ed.), Conservation Biology - The science of scarcity and diversity, pp. 153 – 181. SinauerAssociates, Sunderland, Massachusetts, USA.
Prance, G.T. (1989). Chrysobalanaceae. Flora Neotrop. Monogr. 98.
Some notes: Separate page numbers by an en-rule (type 1 – 2 not 1-2) Do not include part numbers of volumes unless the parts are separately paginated. Plant species names should be italicised in references, whether or not they were in italics in theoriginal reference. Do not include the total numbers of pages in single works.
Kew Bulletin: Up to and including 1941 - cite as (for example):
Bull. Misc. Inform., Kew 1929: 16 – 28. (Note that this is not as in BPH).From Vol.1 (1946) - cite as (for example): Kew Bull. 44: 601 – 680.
Latin
While Latin remains the official language of description, KB prefers the spirit to the letter ofthe law. Meagre diagnoses are discouraged, especially if comparison is made with only onesupposed 'relative' which further work may show to be unrelated. Full diagnoses comparing withmore than one other taxon are preferred, with or without brief Latin descriptions. In large generathe diagnosis should mention the infra-generic group to which the new taxon belongs — if suchgroupings exist.
Full English descriptions are required. Full Latin descriptions are no longer encouraged. Note that types of new species are usually given in abbreviated form after the diagnoses, and
repeated in full amongst the cited specimens. Please use W.T. Stearn's 'Botanical Latin' as the standard.
ACKNOWLEDGEMENTS

Please keep these brief; it is not necessary to spell out full titles of any institute which has anaccepted Herbarium Code.
ADDRESS FOR SUBMISSION
Manuscripts should be submitted to the Editor, Kew Bulletin, Royal Botanic Gardens, Kew,Richmond, Surrey, TW9 3AB, U.K. e-mail: [email protected]
PROOFS
Proofs are generally sent to the first-named author. However, where one of the co-authors(but not the first author) is based at Kew then the proofs will be sent to the Kew-based authorunless an alternative arrangement is requested. Please send proofs back promptly, markedclearly, not in pencil. Ensure that your marking is comprehensible; practices vary widely indifferent countries. Particularly for overseas authors, we prefer if you can advise us of correctionsby e-mail or fax if possible. Nominate someone else to deal with proofs if you expect to be awayat the time of their arrival.
Proofs are designed for the correction of printers' errors. Please do not regard them as anopportunity to change your mind and rewrite your paper. Please advise us if you are in doubt asto the quality of illustration proofs.
OFFPRINTS
20 offprints are given to each author, joint or sole. Extra offprints (in batches of 25) can beordered when proofs are returned. A rough estimate of costs will be provided at this stage — ifnot, please ask. KB offprints are expensive, and can only be printed at the time of production ofthe journal number in which they appear.
REFERENCES
Bridson, G. D. R. (ed.) (1991). Botanico-Periodicum-Huntianum/Supplementum. Hunt Institutefor Botanical Documentation, Pittsburgh.
Brummitt, R. K. & Powell, E. (eds.) (1992). Authors of Plant Names. Royal Botanic Gardens, Kew.See also http://www.ipni.org.
Hollis, S. & Brummitt, R. K. (1992). World Geographical Scheme for Recording PlantDistributions. Hunt Institute for Botanical Documentation, Pittsburgh.
Holmgren, P. K., Holmgren, N. H & Barnett, L. C. (eds.) (1990). Index Herbariorum Part 1: TheHerbaria of the World, 8th Edn. Website: http://www.nybg.org/bsci/ih/ih.html
Holttum, R. E. & Lin, Y. X. (1990). A re-assessment of the fern genus Pseudotectaria. Kew Bull. 45:257 – 263.
Lawrence, G. H. M., Buchheim, A. F. G., Daniels, G. S. & Dolezal, H. (1968). Botanico-Periodicum-Huntianum. Hunt Botanical Library, Pittsburgh.
Stafleu, F. A. & Cowan., R. S. (1976 – 1988). Taxonomic Literature. (2nd edn.) Bohn, Scheltema &Holkema, Utrecht. Later Supplements are also available.
The Times Atlas of the World. (10th (Millennium) Edn.). (1999). Times Books, London.








![[Orchidaceae-Tese] Ventura, G. M. - Cultivo in vitro de orquídeas do grupo Cattleya, em diferentes meios de culturas e irradiâncias](https://img.document.onl/doc/110x75/557204a5497959fc0b8b5beb/orchidaceae-tese-ventura-g-m-cultivo-in-vitro-de-orquideas-do-grupo-cattleya-em-diferentes-meios-de-culturas-e-irradiancias.jpg)



















