Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DO CENTRO-OESTE, UNICENTRO-PR
PROGRAMA DE PÓS-GRADUAÇÃO EM AGRONOMIA- PPGA
MESTRADO
ÁCIDO SALICÍLICO COMO INDUTOR DE
RESISTÊNCIA AO Tetranychus urticae (Koch) E
Helicoverpa armigera EM CULTIVARES DE
MORANGUEIRO
DISSERTAÇÃO DE MESTRADO
RENATA FAVARO
Guarapuava-PR
2017

RENATA FAVARO
Dissertação apresentada à Universidade
Estadual do Centro-Oeste, como parte das
exigências do Programa de Pós-Graduação em
Agronomia, área de concentração em
Produção Vegetal, para a obtenção do título de
Mestre.
Prof. Dr. Juliano Tadeu Vilela de Resende
Orientador
Dr. Rafael Gustavo Ferreira Morales
Coorientador
GUARAPUAVA-PR
2017



AGRADECIMENTOS
Primeiramente, gostaria de agradecer a Deus por me guiar, iluminar e me dar
tranquilidade para seguir em frente com os meus objetivos e não desanimar com as
dificuldades.
Agradeço em especial aos meus pais Dilio e Dilma, por serem modelos de coragem,
pelo seu apoio incondicional, incentivo, amizade e paciência demonstrada e total ajuda na
superação dos obstáculos que ao longo desta caminhada foram surgindo. A eles dedico este
trabalho.
Aos meus irmãos Carlos e Simone e a minha sobrinha Isabella pelo apoio sempre
prestado.
Ao meu orientador Juliano Tadeu Vilela de Resende pela orientação prestada, pelo seu
incentivo, disponibilidade e apoio que sempre demonstrou. Aqui a minha gratidão.
Ao meu colega e amigo Rafael Morales pelo apoio e incentivo para seguir o caminho
acadêmico, sem sua ajuda não teria seguido nessa trajetória.
A minhas companheiras de casa Ely e Ana, por me apoiarem e me aguentarem em
meus dias de desespero e angustia. Obrigada pelas palavras de incentivo sempre.
A todos os funcionários e professores, especialmente a Lucília, pela disponibilidade,
simpatia e gentileza de sempre.
Aos meus colegas do Núcleo de Pesquisa em Hortaliças por todo o auxílio e ajuda
prestada, em especial aos meus amigos Jorge Luis Favaro Jr e Ely Cristina Cordeiro por toda
ajuda a mim concedida em toda essa caminhada.
Agradeço a CAPES pela concessão da bolsa durante todo o período de realização
deste mestrado.

LISTA DE FIGURAS
Figura 1. Frequência de escolhas realizadas pelo ácaro rajado, em relação às cultivares de
morangueiro. ................................................................................................................... 22
Figura 2. Sobrevivência (A) e oviposição (B) do ácaro rajado na superfície de discos foliares
das cultivares de morangueiro Aromas e Sweet Charlie em função de diferentes doses de
ácido salicílico em teste sem chance de escolha. Guarapuava-PR, UNICENTRO, 2016.25
Figura 3. Ciclo de vida da Helicoverpa armigera em cultivares de morangueiro (Aromas e
Sweet Charlie) conduzidas sob doses de AS (0, 25, 50, 75, 100 mg L-1). Guarapuava-PR,
UNICENTRO, 2016. ...................................................................................................... 27
Figura 4. Cinco avaliações dos pesos e tamanhos das Helicoverpa armigera nas doses (0, 25,
50, 75, 100 mgL-1) de AS nas plantas de morangueiro das cultivares (Aromas e Sweet
Charlie). .......................................................................................................................... 29
Figura 5. Representação dos tipos de tricomas A) Glandulares e B) Não Glandulares, em
cultivares de morangueiro, Guarapuava-PR, UNICENTRO, 2016. ............................... 32
Figura 6. Densidade de tricomas glandulares (mm2) nas superfícies abaxial e adaxial de
folíolos das cultivares Aromas e Sweet Charlie em função de diferentes doses de ácido
salicílico. A) Tricomas glandulares na parte abaxial B) Tricomas glandulares na parte adaxial
dos folíolos. Guarapuava-PR, UNICENTRO, 2016. ..................................................... 33
Figura 7. Densidade de tricomas não glandulares (mm2) nas superfícies abaxial e adaxial de
folíolos das cultivares Aromas e Sweet Charlie em função de diferentes doses de ácido
salicílico. A) Tricomas não glandulares na parte abaxial. B) Tricomas não glandulares na
parte adaxial. Guarapuava-PR, UNICENTRO, 2016. .................................................... 34

Figura 8. Densidade de tricomas tectores, na cultivar Aromas. A - Testemunha; B - 25mgL-1
;
C – 50mgL-1
; D - 75mgL-1
; E - 100mgL-1
; de ácido salicílico e na cultivar Sweet Charlie F -
Testemunha; G - 25mgL-1
; H – 50mgL-1
; I - 75mgL-1
; J - 100mgL-1
; de ácido salicílico.
Guarapuava-PR, UNICENTRO 2016............................................................................. 34
Figura 9. Análise de parâmetros de fluorescência Fo (fluorescência inicial) em relação às
doses aplicadas de AS em três avaliações, nas cultivares Aromas e Sweet Charlie. A) Cultivar
Aromas B) Cultivar Sweet Charlie. Guarapuava-PR, UNICENTRO, 2016. ................. 37
Figura 10. Análise de parâmetros de fluorescência Fm (fluorescência máxima) em relação as
doses aplicadas em três avaliações, nas cultivares Aromas e Sweet Charlie. A) Cultivar
Aromas, B) Cultivar Sweet Charlie. Guarapuava-PR, UNICENTRO, 2016. ................ 38
Figura 11. Análise de parâmetros de fluorescência Fv/Fo (atividade potencial do PSII) em
relação as doses aplicadas em três avaliações, nas cultivares Aromas e Sweet Charlie. A)
Cultivar Aromas, B) Cultivar Sweet Charlie. Guarapuava-PR, UNICENTRO, 2016. .. 40
Figura 12. Análise de parâmetros de fluorescência Y(II) (rendimento quântico fotoquímico
efetivo do PSII, conversão fotoquímica) em relação às doses aplicadas, nas cultivares Aromas
e Sweet Charlie. A) Cultivar Aromas B) Cultivar Sweet Charlie. . Guarapuava-PR,
UNICENTRO, 2016. ...................................................................................................... 41
Figura 13. Análise dos dados não-paramétricos de fluorescência qP (coeficiente de dissipação
fotoquímica) em relação as doses aplicadas de AS, nas cultivares Aromas e Sweet Charlie. A)
Cultivar Aromas, B) Cultivar Sweet Charlie. Guarapuava-PR, UNICENTRO, 2016. .. 42
Figura 14. Análise dos dados não-paramétricos de fluorescência ETR Análise dos dados não-
paramétricos de fluorescência ETR (taxa de transferência relativa de elétrons) em relação as
doses aplicadas de AS, nas cultivares Aromas e Sweet Charlie. A) Cultivar Aromas, B)
Cultivar Sweet Charlie. Guarapuava-PR, UNICENTRO, 2016. .................................... 42

SUMÁRIO
RESUMO .................................................................................................................................................... i
ABSTRACT ................................................................................................................................................ ii
1. INTRODUÇÃO ...................................................................................................................................... 1
2. OBJETIVO ............................................................................................................................................ 3
2.1. OBJETIVO GERAL .................................................................................................................... 3
2.2. OBJETIVOS ESPECÍFICOS ....................................................................................................... 3
3. REFERENCIAL TEÓRICO ....................................................................................................................... 4
3.1. Histórico do morangueiro............................................................................................................. 4
3.2. Botânica e taxonomia ................................................................................................................... 5
3.3. Pragas do morangueiro ................................................................................................................. 5
3.4. Ácaro rajado Tetranychus urticae Koch ...................................................................................... 6
3.5. Lagarta Helicoverpa armigera ..................................................................................................... 8
3.6. Resistência induzida ..................................................................................................................... 9
3.7. Ácido salicílico como indutor de resistência .............................................................................. 10
3.8. Tricomas como fator de defesa .................................................................................................. 12
3.9. Mecanismos de defesa das plantas ............................................................................................. 14
3.10. Fluorescência ............................................................................................................................ 15
4. MATERIAL E MÉTODOS ..................................................................................................................... 17
4.1. Local do experimento ................................................................................................................. 17
4.2. Cultivares de morangueiro e cultivo das plantas ........................................................................ 17
4.3. Obtenção dos ácaros e criação ................................................................................................... 18
4.4. Bioensaio com chance de escolha .............................................................................................. 19
4.5. Bioensaio sem chance de escolha ............................................................................................... 19
4.6. Bioensaios com larvas de H. armigera ....................................................................................... 20
4.6.1. Obtenção e criação das larvas de H. Armigera.................................................................... 20
4.6.2. Bioensaios para avaliação da biologia ................................................................................. 20
4.7. Identificação e quantificação de tricomas .................................................................................. 21
4.8. Fluorescência .............................................................................................................................. 21
4.9. Análises Estatísticas ................................................................................................................... 22
5. RESULTADOS E DISCUSSÃO .............................................................................................................. 22
5.1. Bioensaio com chance de escolha – Ácaro rajado ..................................................................... 22
5.2. Bioensaio sem chance de escolha – Ácaro rajado ...................................................................... 24
5.3. Bioensaios com larvas de H. armigera ...................................................................................... 27

5.4. Identificação e quantificação de tricomas foliares ..................................................................... 31
5.5. Fluorescência .............................................................................................................................. 37
6. CONCLUSÕES ..................................................................................................................................... 44
7. REFERÊNCIAS .................................................................................................................................... 45

i
RESUMO
FAVARO, Renata. ÁCIDO SALICÍLICO COMO INDUTOR DE RESISTÊNCIA
AO Tetranychus urticae (Koch) EM MORANGUEIRO. Guarapuava: UNICENTRO,
2017. (Dissertação – Mestrado em Agronomia).
No presente trabalho, objetivou-se avaliar o efeito do ácido salicílico na indução
de resistência ao ácaro rajado e a Helicovera armigera. Para além disso, foram
avaliadas a densidade de tricomas e a fluorescência, em cultivares de morangueiro. As
cultivares avaliadas foram Aromas e Sweet Charlie. Foram utilizadas cinco doses de
ácido salicílico (AS) (0, 25, 50, 75, 100 mg L-1
). Para verificar efeitos de antixenose e
antibiose, ácaros foram submetidos aos testes com e sem chance de escolha, onde foram
avaliadas a sobrevivência e a oviposição. Foram analisados a densidade e o tipo de
tricomas foliares nas faces abaxial e adaxial dos folíolos das cultivares de morangueiro.
Foi avaliado o efeito do AS na biologia da H. armigera. Para além disso, foram
avaliadas a fluorescência das plantas sob os tratamentos e sua relação com o nível de
estresse. Nos testes sem chance de escolha ao ácaro rajado, houve redução da
sobrevivência nas doses intermediárias de ácido salicílico 25, 50 e 75mg L-1
, em ambas
as cultivares. Para o número de ovos depositados sobre folíolo, verificou-se efeito
significativo de dose, com ajuste quadrático da equação. A cultivar Sweet Charlie, foi
mais efetiva que a Aromas na redução da oviposição e na sobrevivência dos ácaros
sobre os folíolos tratados com o indutor de resistência. Para biologia da Helicoverpa
armigera, notou-se uma redução no ciclo de vida, no peso e no comprimento das larvas,
nas doses 25 e 50 mgL-1
. No teste com a fluorescência monitorando a capacidade
fotossintética detectou-se efeitos de estresse e injúrias, onde a dose 100 mgL-1
,
proporcionou maior estresse nas plantas em relação as demais. O teor de AS interferiu
na morfologia da planta, mediante alteração na densidade de tricomas glandulares e
tectores nos folíolos, demonstrando ter efeito positivo no aumento dessas estruturas,
quando aplicadas doses intermediárias. Concentrações de 25, 50 e 75 mg L-1
AS foram
as mais eficientes para as características avaliadas permitido inferir o seu potencial
como indutor de resistência em artrópodos-praga em morangueiro.
Palavras-chave: Fragaria x ananassa, Tetranychus urticae, fluorescência, tricomas
foliares.

ii
ABSTRACT
Favaro, Renata. Salicylic Acid as inductor of resistance to Tetranychus urticae
(koch) and Helicoverpa armigera in strawberry.
This work aimed to evaluate the effect of salicylic acid on induction of
resistance to spider mite (Tetranychus urticae (Koch) and Helicoverpa armigera. The
density of trichomes and fluorescence were evaluated in strawberry cultivars. The
evaluated cultivars were Aromas and Sweet Charlie. Five doses of salicylic acid (AS)
(0, 25, 50, 75, 100 mg L-1
) were used. To verify the effects of antixenosis and antibiosis,
mites were submitted to the with and without choice test, where survival and
oviposition were evaluated. The density and type of leaf trichomes were analyzed in the
abaxial and adaxial faces of the leaflets of the strawberry cultivars. The effect of AS on
the biology of H. armigera was evaluated. In addition, the fluorescence of the plants
under the treatments and their relation with the level of stress was measured. In the
without choice test, the survival mites rates were reduced of the intermediate doses of
salicylic acid 25, 50 and 75mg L-1
in both cultivars. For the number of eggs deposited
on leaflet, there was a significant dose effect, with a quadratic adjustment of the
equation, where the Sweet Charlie cultivar was more effective than Aromas in reducing
oviposition and mite survival on leaflets treated with resistance inducer. For the biology
of H. armigera, a reduction in the life cycle in weight and length was observed at doses
25 and 50 mgL-1
. In the fluorescence test monitoring the photosynthetic capacity, stress
and injury effects were detected, where the 100 mgL-1
dose provided higher stress in the
plants compared to the others. The AS doses interfered in plant morphology by altering
the density of glandular trichomes and tectors in the leaflets, showing a positive effect
on the increase of these structures when intermediate doses were applied.
Concentrations of 25, 50 and 75 mg L-1 AS were the most efficient for the evaluated
characteristics allowed to infer its potential as inducer of resistance in arthropod-pest in
strawberry.
Keywords: Fragaria x ananassa, Tetranychus urticae, fluorescence, leaf
trichomes.

1
1. INTRODUÇÃO
O morangueiro (Fragaria x ananassa Duch) é uma cultura de grande
importância econômica para produtores brasileiros, tendo maior destaque nos estados de
Minas Gerais, Rio Grande do Sul, São Paulo, Espírito Santo, Paraná, Santa Catarina e
Distrito Federal. A produção de morango no Brasil é de cerca de 105 mil toneladas
anuais, cultivadas em área de aproximadamente 4.000 hectares (ANTUNES e REISSER
JÚNIOR, 2015).
Entre as pequenas frutas, o morango é uma das mais importantes, pois
proporciona paladar apreciado em diversas regiões do mundo, versatilidade de uso, alto
retorno financeiro ao produtor e, ainda, nos últimos anos, tem despertado atenção por
ser fonte de compostos bioativos como as vitaminas C e E, β-caroteno e compostos
fenólicos, principalmente antocianinas (OSZMIANSKI e WOJDYLO, 2009).
Assim como a maioria das olerácea, o morangueiro é muito suscetível a doenças
e pragas, muitas delas de difícil analise para identificação da procedência dos sintomas,
podendo estes serem oriundos tanto de pragas como de doenças e de difícil controle, e
que podem ocasionar grandes perdas, mesmo considerando-se o ciclo de produção
relativamente curto. Por este motivo, é compreensível a grande preocupação
demonstrada pelos agricultores com a ocorrência ou infestação de pragas e doenças. De
modo geral, os produtores usam preventivamente várias aplicações de agrotóxicos,
principalmente inseticidas-acaricidas e fungicidas. No entanto, estes causam problemas
de acúmulo de resíduos nos frutos comercializados, sendo prejudiciais ao ambiente aos
aplicadores e os consumidores (LORENZETTI, 2012).
Uma das principais pragas da cultura do morangueiro é o ácaro-rajado
(Tetranychus urticae), principalmente em ambiente protegido, podendo reduzir a
produção de frutos em até 80%, no ponto máximo de desenvolvimento da população.
Elevadas infestações podem causar descoloração de folhas, perda da capacidade
fotossintética e, eventualmente, a morte das folhas (CHIAVEGATO e MISCHAN,
1981; FLECHTMANN, 1985; DEVINE et al., 2001). Um grande problema que está
associado ao controle químico do ácaro rajado é seu elevado potencial reprodutivo e seu
curto ciclo de vida, que favorecem o rápido desenvolvimento de resistência a acaricidas
(STUMPF e NAUEN, 2001).
Outra praga de grande importância que vem atacando diversas culturas é a
lagarta Helicoverpa armigera. Esta pode se alimentar tanto dos órgãos vegetativos

2
como reprodutivos de várias espécies de plantas de importância econômica. Estima-se
que a perda mundial causada por essas lagartas, nas diferentes culturas em que atacam,
chega anualmente a 5 bilhões de dólares (LAMMERS e MACLEOD, 2007).
Em plantas de morangueiro compreende-se que existem mecanismos
constitutivos e indutivos que atuam na proteção das plantas contra o ataque de pragas
(FADINI et al., 2007). Em ambos os mecanismos, em especial os constitutivos, a
produção e liberação de metabólitos secundários de defesa são de extrema importância
para a expressão das barreiras físicas e químicas das plantas. Vale salientar que grande
parte destes compostos estão contidos no interior de tricomas glandulares evidenciando
a importância dos mesmos na defesa das plantas (STEINITE e IEVINSH, 2003).
Algumas técnicas de controle aos artrópodos-praga foram estudadas por Freddi
et al. (2007). Nesse sentido, estudos com o uso do ácido salicílico (AS) concebe uma
tecnologia ambientalmente correta, sustentável, com amplo potencial para diminuir a
frequência e o uso de agrotóxicos (SILVA et al., 2010). O AS atua na regulação de
processos biológicos nas plantas, incluindo defesa (KUMAR et al., 2015). É
responsável pelo acúmulo de superóxido e peróxido de hidrogênio no apoplasto, que
ocasionam a morte de células no local da infecção, promove também a síntese de
lignina na parede celular, que dificulta a penetração de estiletes e a mastigação pelas
pragas, em decorrência do enrijecimento da estrutura de parede das células vegetais
(DATNOFF et al., 1991; EPSTEIN, 1994; MARSCHNER, 1995), atuando no
estabelecimento da resistência sistêmica adquirida (GAO et al., 2015).
A atuação do AS como indutor de resistência está relacionado á capacidade de
detectar os efeitos de estresse e injúrias nos processos fotossintéticos causados por
fatores bióticos ou abióticos (BAKER e ROSENQVIST, 2004; YUSUF et al., 2010).
Esses mecanismos de defesa podem ser explorados pelo homem para o controle de
artrópodos-praga, mediante aplicação do AS que é um indutor capaz de produzir uma
resposta de defesa nas plantas contra o ataque.

3
2. OBJETIVO
2.1. OBJETIVO GERAL
Avaliar o efeito do ácido salicílico como indutor de resistência a artrópodos-
praga em morangueiro.
2.2. OBJETIVOS ESPECÍFICOS
Avaliar os efeitos do ácido salicílico na biologia e no comportamento do ácaro
rajado em morangueiro;
Identificar e quantificar os tipos de tricomas foliares em morangueiro em função
da aplicação de AS.
Avaliar o efeito do ácido salicílico na biologia da Helicoverpa armigera.
Identificar o(s) mecanismo(s) de resistência(s) envolvido(s) na relação de
indução de resistência pelo AS entre o hospedeiro e o herbívoro.
Avaliar o efeito do AS na fluorescência e sua relação com estresses nas plantas
de morangueiro.

4
3. REFERENCIAL TEÓRICO
3.1. Histórico do morangueiro
As primeiras referências sobre o cultivo do morangueiro datam em torno do ano
1300. As plantas cultivadas nesse período pertenciam à espécie Fragaria vesca e essas
por sua vez apresentavam frutos pequenos, sendo destinadas para fins ornamentais. A
partir de 1500 as citações sobre a cultura ficaram mais constantes e seu cultivo passou a
ser frequente (COSTA et al., 2014).
O morangueiro atualmente cultivado é um híbrido proveniente do cruzamento de
duas espécies, uma do Chile e outra da América do Norte, Fragaria Chiloensis x
Fragaria virginiana, respectivamente. Esse híbrido natural (Fragaria x ananassa Duch)
surgiu há aproximadamente 250 anos (DARROW, 1966). A hibridação combinou
características dessas duas espécies, incluindo-se maior tamanho e
firmeza de frutos, vindos da F. chiloensis L. e a coloração vermelho
escuro e frutos mais aromáticos da F. virginiana M. (STEGMEIR et al.,
2010).
Apesar dessas espécies silvestres serem oriundas do continente americano, foi na
Europa, em meados de 1760, que ocorreram os cruzamentos casuais que originaram o
híbrido. Tal fato aconteceu depois que franceses cultivaram as plantas no mesmo
jardim, lado a lado (COSTA et al., 2014). De acordo com Duchesne (1766), os frutos
que se originaram tinham o gosto ácido do abacaxi, e um perfume delicioso, e devido a
essas características o nome do híbrido foi designado “morango-ananás”. A partir deste
híbrido, os ingleses iniciaram o programa de melhoramento, seguidos por franceses e
americanos. Conforme relatos este fato ocorreu somente após a Segunda Guerra
Mundial, nos Estados Unidos, onde foram desenvolvidas as principais cultivares
utilizadas, com características morfofisiológicas específicas.
A cultura do morangueiro apresentou nos últimos anos, evolução considerável
em todo o Mundo. No Brasil, a cultura começou a se expandir a partir da década de
1960, por meio do melhoramento genético, surgiram as principais cultivares, com
melhores adaptações e novas técnicas de cultivo (SILVA, 2013).

5
A cultura do morangueiro destaca-se no Brasil por sua grande importância
econômica, principalmente nas pequenas propriedades rurais, que utilizam mão-de-obra
familiar, em virtude dos valores de produção, comercialização e alta demanda de
insumos e do elevado rendimento por área (CAMARGO et al., 2000).
3.2. Botânica e taxonomia
O morangueiro é uma hortaliça que pertence à família Rosaceae, subfamília
Rosoideae, gênero Fragaria L., que abrange mais de vinte espécies, as quais diferem
tanto funcional como estruturalmente (SILVA et al., 2007). Tal gênero apresenta sete
cromossomos básicos, comuns a todas as espécies, e quatro níveis de ploidia: diploides,
tetraploides, hexaploides e octoploides. Espécies diploides, tetraploides e hexaploides
são comuns na Europa e Ásia, já as octoploides são encontradas nas Américas e Ásia
(ANTUNES e HOFFMANN, 2012).
As plantas pertencentes ao gênero Fragaria são herbáceas, rasteiras, e podem
formar touceiras que atingem de 15 a 30 cm de altura. O caule é um rizoma estolhoso,
cilíndrico e retorcido, com entrenós curtos, de onde surgem as folhas, estolhos e
inflorescências. Os frutos são secos, do tipo aquênio, e apresentam uma semente por
fruto. Após a fecundação dos óvulos, estes transformam-se em aquênios, que por sua
vez estimulam o engrossamento do receptáculo floral, tornando-o carnoso e originando
o pseudofruto ou infrutescência, chamado de morango. O pseudofruto é doce, carnoso e
suculento, possui polpa firme e coloração vermelha (SILVA et al., 2007). As sementes
são encontradas dentro dos aquênios, no exterior do pseudofruto. As sementes
estimulam o crescimento uniforme do receptáculo carnoso. Se a polinização ocorrer
bem, haverá formação de um fruto uniforme, caso contrário, poderá acontecer
deformação pela falta de estímulo hormonal. A semente serve para multiplicação
sexuada, que é útil em programas de melhoramento genético (GALVÃO et al., 2014).
3.3. Pragas do morangueiro
Existem várias pragas diferentes na cultura do morangueiro, podendo mencionar
os pulgões [(Capitophorus fragaefolii (Cockerell); Cerosipha forbesi (Weed)], lagartas-
de-solo (Agrotis sp.), Spodoptera spp.; ácaro rajado (Tetranychus urticae), ácaro-

6
rajado-do-enfezamento [Phytonemus pallidus (Banks)], broca-dos-frutos [Lobiopa
insularis (Castelnau)], tripes [Frankliniella occidentalis (Pergande)]. Estas pragas, se
não controladas, podem afetar severamente a produção de morangos no Brasil
(ANTONIOLLI et al., 2007). Outra praga de grande importância que vem atacando
diversas culturas é a lagarta Helicoverpa armigera, esta praga pode se alimentar tanto
dos órgãos vegetativos como reprodutivos de várias espécies de plantas de importância
econômica (LAMMERS e MACLEOD, 2007).
Na cultura do morangueiro é grande a incidência de pragas que atingem as várias
fases do ciclo da cultura, desde a muda recém-plantada até os frutos na fase final de
produção (OSHITA, 2012). A cultura do morangueiro tem sido uma das culturas com
maior utilização de pesticidas na agricultura nacional em consequência de práticas
agrícolas incorretas e também de sua elevada susceptibilidade a pragas. A
predominância do controle químico, além de comprometer a qualidade do produto final,
pode ser bastante impactante ao ambiente, e pode interferir na ação dos inimigos
naturais que são reguladores da densidade populacional de ácaros e insetos-praga (VAN
DE VRIE et al., 1972). Entre as pragas que prejudicam a cultura do morangueiro,
destaca-se o ácaro rajado (Tetranychus urticae Koch.), considerado como a principal
praga desta cultura (MORAES e FLECHMANN, 2008; FIGUEIREDO et al., 2010).
3.4. Ácaro rajado Tetranychus urticae Koch
Pertencente à família Tetranychidae, o ácaro rajado (Tetranychus urticae Koch)
é considerado uma das espécies mais importantes de ácaros-praga, cosmopolita e
polífaga. Ataca mais de 150 culturas de importância econômica, em meio a 1200
espécies de plantas em 70 gêneros (ZHANG, 2003). O ácaro rajado é praga importante
em frutíferas (morangueiro, mamoeiro, pessegueiro, etc.), plantas ornamentais
(crisântemo, rosa, etc.), além de várias outras culturas (CHAPMAN e MARTIN, 2005;
FADINI et al., 2006; MORAES e FLECHTMANN, 2008).
Consiste também em uma das principais pragas em cultivos protegidos. A
manutenção de condições ambientais relativamente estáveis para o desenvolvimento de
plantas geralmente favorece o rápido desenvolvimento da praga, aumentando a
população antes mesmo que o produtor perceba.

7
Frequentemente por se tratar de um ambiente protegido, dificulta o
aparecimento de inimigos naturais e favorece a rápida evolução da resistência (ZHANG,
2003). Seus ovos apresentam coloração amarelada, esféricos e de difícil visualização a
“olho nu” (FADINI et al., 2006).
A larva é incolor e translúcida, subesférica, com quatro pares de pernas e de
tamanho aproximadamente igual ao do ovo e apresentam duas manchas ocelares
vermelhas. Sua coloração muda gradativamente à medida que se alimentam (MORAES
e FLECHTMANN, 2008). As fases de protoninfa e deutoninfa difere-se entre si
somente pelo tamanho (o segundo estágio é pouco maior do que o primeiro). É na fase
de deutoninfa que se iniciam as distinções entre fêmeas (cerca de 0,46 mm de
comprimento e com manchas ocelares mais pronunciadas) e machos (aproximadamente
0,26 mm de comprimento e com o opistossoma afilado posteriormente). O dorso das
formas adultas, de coloração amarela esverdeada escura, apresenta-se coberto por
longas setas e duas manchas escuras em cada lado (FADINI et al., 2006; MORAES e
FLECHTMANN, 2008). A coloração torna-se alaranjada ou avermelhada quando
entram em diapausa. A indução ocorre quando o comprimento do dia é curto, o alimento
é escasso e a temperatura é baixa (ZHANG, 2003).
O ciclo de vida dura de 11 a 20 dias, quanto maior a temperatura, mais curto é o
tempo de desenvolvimento de ovo até adulto. Em condições ótimas de temperatura (30-
32°C), o ciclo de vida pode ser completo em menos de uma semana. Machos se
desenvolvem mais rápido que fêmeas (ZHANG, 2003). São encontrados geralmente na
face inferior das folhas, onde formam suas teias. A postura dos ovos é feita nestas teias
ou, quando diretamente nas folhas, ocorrem geralmente próximo à nervura. Uma fêmea
é capaz de ovipositar aproximadamente 10 ovos por dia, e até 100 ovos em duas
semanas, numa temperatura de 25°C (FLECHTMANN, 1985; ZHANG, 2003; FADINI
et al., 2006). A disseminação desses ácaros pode ser ativa (caminhamento) ou passiva
(vento, plantas, ferramentas ou pessoas) (ZHANG, 2003).
Quanto à alimentação, adultos e formas imaturas alimentam-se da clorofila e da
seiva, geralmente das folhas mais velhas. Eles introduzem seus estiletes nas células dos
vegetais e ingerem o conteúdo intracelular que extravasa. Em grandes infestações,
podem também atacar folhas jovens (MORAES e FLECHTMANN, 2008; VAN DER
GEEST, 1986).
Os ácaros são considerados pragas primárias para a cultura de morangueiro.
Algumas cultivares são mais preferidas pelo ácaro-rajado, sendo mais danificados

8
(LOURENÇÃO et al., 2000, FIGUEIREDO et al 2010). Esta preferência está
relacionada a fatores intrínsecos das cultivares ou mesmo da planta. Figueiredo et al.
(2010), associaram maior resistência da cultivar Camino Real ao ácaro rajado, em
decorrência da elevada densidade de tricomas glandulares, presentes na superfície foliar.
No morango, os ácaros desenvolvem-se na face inferior das folhas abertas, formando
manchas branco-prateadas, e na face superior aparecem áreas inicialmente cloróticas,
que passam a bronzeadas. As folhas podem secar e cair. Em casos de forte estresse,
pode haver redução na fotossíntese e na produtividade, afetando a qualidade e a
quantidade de frutos e folhas novas. Quando não controlado, o ataque do ácaro no
morangueiro pode reduzir a produção de frutos em até 80% (CHIAVEGATO e
MISCHAN, 1981).
A ocorrência é maior em condições de temperatura elevada e baixa umidade,
favorecida também pela presença de poeira (TANAKA et al., 2000). De uma maneira
geral, a praga determina a formação de manchas cloróticas ou descoloridas nas folhas
que depois se tornam bronzeadas e secas. Por fim, ocorre a desfolha, algumas vezes,
total (CRUZ, 2000). Outros danos causados à cultura são: redução de crescimento das
plantas e formação de teias, o que deprecia o produto final (BARBOSA, 2003).
3.5. Lagarta Helicoverpa armigera
A lagarta H. armigera é considerada uma espécie altamente polífaga, ou seja,
que apresenta a capacidade de se desenvolver em ampla gama de plantas hospedeiras.
Suas larvas têm sido registradas se alimentando causando sérios danos em mais de 100
espécies de plantas, sejam elas cultivadas ou não, incluindo cerca de 45 famílias, sendo
Asteraceae, Fabaceae, Malvaceae, Poaceae e Solanaceae (ALI e CHOUDHURY, 2009;
FITT,1989; PAWAR et al., 1986; POGUE, 2004;). No Brasil, as lagartas H. armigera já
foram constatadas se alimentando de diversas culturas de extrema importância
econômica, tais como algodão, soja, milho, tomate, feijão, sorgo, milheto, guandu, trigo
e crotalária, e também em algumas espécies de plantas daninhas. (FITT, 1989).
As lagartas se alimentam de folhas e hastes dessas plantas, mas tem preferência
pelas estruturas reprodutivas como frutos, maças, botões florais, espigas e
inflorescências, ocasionando podridões ou deformações nestas estruturas ou até mesmo
a sua queda. Essa inerente capacidade da H. armigera causar danos nas partes

9
reprodutivas das culturas, em associação à sua habilidade de atacar grande número de
hospedeiros, são fatores que abrangem o status de importância econômica desta praga
(CUNNINGHAM et al., 1999). Na Espanha, é também considerada espécie devastadora
nos cultivos de tomate para a indústria (ARNÓ et al., 1999).
3.6. Resistência induzida
Diversos estudos vêm sendo realizados a partir do sistema imune das plantas, a
fim de se entender quais são os processos celulares envolvidos na percepção dos
patógenos, assim como, as vias de sinalização que resultam na resistência do vegetal.
As plantas não apresentam um sistema imune adaptativo com células móveis, como nos
animais, pois são dotadas de um sistema inato de defesa constituído por um aparato
estrutural e bioquímico (CAVALCANTI et al., 2014). Desta maneira, elas dependem
que cada célula, individualmente, apresente a capacidade de perceber o patógeno,
sinalizar ás células vizinhas e produzir uma resposta de defesa (SPOEL e DONG, 2012;
FU e DONG, 2013).
O sistema de defesa permanece inativo ou latente, só sendo ativado após ser
exposto aos agentes de indução. Dessa maneira, a resistência é chamada de induzida, ou
seja, as plantas percebem as agressões, e sua alta capacidade de adaptação permite que
sobrevivam, mesmo tendo muitas vezes seu desenvolvimento prejudicado (BONALDO
et al., 2005). Deste modo, indutores de resistência são substâncias químicas que incitam
respostas morfofisiológicas do organismo vegetal, responsáveis por essa resistência
(POIATTI et al., 2009; JANUS et al., 2013; PO-WEN et al., 2013).
Depois de a planta apresentar a resistência induzida, esta não possui caráter
específico e se expressa por toda a planta, sendo chamada de resistência sistêmica
adquirida (SAR) (HEIDRICH et al., 2012; DAVIDSSON et al., 2013). Uma
característica evidente no SAR é o tempo no qual as defesas da planta permanecem
ativadas, podendo perdurar de semanas a meses (FU e DONG, 2013).
A resistência sistêmica nas plantas ocorre pela ação de agentes externos bióticos
ou abióticos, sem qualquer alteração do genoma da planta, sendo estes chamados, de
elicitores (HAMMERSCHMIDT et al., 2000). Segundo Campos (2009), o elicitor AS, é
um mensageiro que ativa a resistência, por meio da síntese de proteínas relacionadas à
patogênese (proteínas- RP) entre as quais as quitinases e glucanases. O aumento da
atividade das enzimas proporciona a síntese de compostos fenólicos e o acúmulo de

10
lignina em tecidos adjacentes, enrijecendo a parede das células vegetais, no local de
penetração da estrutura de ataque (DURRANT e DONG, 2004) e esse acúmulo de
lignina dificulta a mastigação e penetração de estiletes pelas pragas sugadoras (FAWE
et al., 2001; GOMES et al., 2005). Segundo Crusciol et al. (2013), além do aumento da
espessura da cutícula pela deposição de lignina nos tecidos, o acúmulo deste elicitor
promove a polimerização nos espaços extracelulares das paredes das células
epidérmicas, das folhas e dos vasos condutores (xilema e floema), fortificando essas
estruturas além de formar fitoalexinas (substâncias de defesa) e também aumentar a
capacidade fotossintética (BÉLANGER e MENZIES, 2003).
3.7. Ácido salicílico como indutor de resistência
Há compostos orgânicos que funcionam como indutores de resistência e são
chamados de reguladores vegetais. As plantas produzem uma ampla variedade de
hormônios entre os quais se incluem as auxinas, giberelinas, ácido abscísico, citocininas
e etileno. Atualmente, outros compostos que podem afetar o crescimento e o
desenvolvimento vegetal têm sido descritos, dentre eles se destaca AS (GONÇALVES
et al., 2014; PULGA, 2016).
O AS é um metabólito secundário produzido por uma vasta gama de organismo
procariotas e eucariotas incluindo as plantas (ABERG, 1981). Pertence a um grupo
bastante diverso dos compostos fenólicos, usualmente definidos como substâncias com
um anel aromático ligado a um grupo hidroxil ou ao seu derivado funcional (COLLI,
2008).
De acordo com Kerbauy (2004), Souza (2007) e Vlot et al. (2009), a biossíntese
do AS pode ocorrer por meio de duas vias enzimáticas diferentes: pela via dos
fenilpropanóides, a partir da L-fenilalanina que por ação da enzima fenilalanina-
amonialiase (FAL), é convertida em ácido trans-cinâmico que irá formar o ácido
benzóico e será convertido em AS por ação da enzima ácido benzóico-2-hidroxilase; e
pela via do isicorismato, sendo que o corismato é convertido em isocorismato por ação
da enzima isocorismato sintase (ICS) e transforma-se em AS por ação da enzima
isocorismato piruvato liase (IPL).

11
O AS produzido nas plantas pode ser convertido em SA O-β-glucosídeo (SAG),
saliciloil éster glucosa SEG, metil salicilato (MeSA) e metil salicilato O-β-glucosídeo
(MeSAG) (KERBAUY, 2004; SOUZA, 2007; VLOT et al., 2009).
Os compostos de carbono das plantas decorrem da via de síntese do ácido
chiquímico, causando grande variedade de aminoácidos aromáticos. A via do ácido
chiquímico em seu princípio forma o ácido corísmico que é um intermediário comum
para a síntese do triptofano, fenilalanina, tirosina e vários outros compostos aromáticos,
como o ácido salicílico (VELINI et al., 2009; MARTÍN, 2009), que atua como um
regulador vegetal, devido a sua implicação na regulação das plantas e induz diversos
processos fisiológicos, incluindo a fotossíntese (KUHN, 2007).
Seu transporte é realizado via floema para as partes não infectadas da planta,
especialmente na forma de metil salicilato, composto sinalizador, que regula
mecanismos de resistência no local de infecção e por meio da resistência sistêmica
adquirida, que opera na expressão (VELINI et al., 2009) e regulação de genes de defesa
nas plantas (SPOEL e DONG, 2012; FU e DONG, 2013).
O estudo do AS teve início a partir de observações de um análogo (aspirina) que
prolongava a vida pós-colheita de flores, possivelmente por interferir na biossíntese de
etileno (SOBRINHO et al., 2005). Embora o conhecimento dos diversos efeitos
fisiológicos e bioquímicos da aplicação de AS em plantas ser antigo, o seu desempenho
como regulador endógeno só foi estabelecido em um estudo de termogênese em plantas
em 1987 (RASKIN, 1992). O uso de AS no controle de perdas pós-colheita de produtos
hortícolas vêm sendo considerado uma alternativa com grande potencial. Recentemente,
o estudo do AS tem sido amplamente explorado, já que é uma molécula chave para a
expressão de resistência a estresses nas plantas, principalmente a estresses abióticos
(luz, seca, salinidade, frio, UV e choque térmico) e ao ataque de patógenos. A ação
antifúngica do ácido também vem sendo descrita (ASGHARI e AGHDAM, 2010). Os
trabalhos envolvendo compostos fenólicos já comprovaram que os mesmos
desempenham papel fundamental na regulação de diversos processos fisiológicos,
dentre eles o crescimento e desenvolvimento das plantas, a fotossíntese e a captação de
íons (POPOVA et al., 1997).
De acordo com Lee et al., (1995) e Asghari e Aghdam, (2010), o AS é essencial
na regulação das respostas ao estresse e nos processos de desenvolvimento da planta,
provocando efeitos fisiológicos e bioquímicos, incluindo a indução da floração, a
produção de calor (termogênese), a fotossíntese, a condutância estomática, a

12
transpiração, a absorção e o transporte de íons (inibição de fosfato e captação de
potássio), a germinação de sementes, a inibição da biossíntese / ação do etileno e a
resistência a doenças. O AS pode agir de modo isolado, associado ou controlar os
efeitos de outros hormônios. Os hormônios metil jasmonato e etileno induzem a
ativação genética de diversas proteínas, que por sua vez são inibidas pelo AS (SOARES
e MACHADO, 2007).
O AS é responsável pelo acúmulo de superóxido e peróxido de hidrogênio no
apoplasto, que acarretam a morte de células no local da infecção, também promovem
no local do ataque a síntese de lignina na parede celular, dificultando a penetração de
estiletes e a mastigação pelos insetos, em decorrência do enrijecimento da parede das
células vegetais (DATNOFF et al., 1991; EPSTEIN, 1994; MARSCHNER, 1995), e,
em locais distantes da infecção promovem a síntese de proteínas RP e compostos
fenólicos (GRÜNER et al., 2003).
Desta forma, a aplicação exógena de AS tem sido testada em diferentes cultivos
agrícolas a fim de investigar a capacidade deste em atenuar os efeitos adversos causados
pela deficiência hídrica (AZOOZ e YOUSSEF, 2010), como exemplo, estão as
pesquisas desenvolvidas por Singh e Usha (2003) e Horváth et al. (2007) em trigo,
Hussain et al. (2008a) em girassol, Carvalho et al. (2007) em calêndula, Mardani et al.
(2012) em pepino e Durães (2006) em feijão. Para além disso, é usada a aplicação
exógena na germinação das sementes (RAJOU et al., 2006), no estabelecimento de
plântulas (ALONSO- RAM’IREZ et al., 2009), crescimento celular (VANACKER et
al., 2001), respiração (NORMAN et al., 2004), senescência (RAO et al. 2001 e 2002) e
nodulação (STACEY et al., 2006), hormônios vegetais (ROBERT–SEILANIANTZ et
al., 2007; PIETERSE et al., 2009), como sinalizador de moléculas na resposta imune
das plantas (VLOT et al., 2009) e a resistência induzida reduzem a preferência dos
herbívoros pelas plantas (CORNELISSEN e FERNANDEZ, 2003; PULGA 2016).
3.8. Tricomas como fator de defesa
Em plantas de morangueiro sabe-se que existem mecanismos de defesas
constitutivos e indutivos que atuam na proteção das plantas contra o ataque de pragas
(FADINI et al. 2007). Nesses mecanismos há produção e liberação de metabólitos
secundários de defesa que são de importância para a expressão das barreiras físicas e

13
químicas das plantas e vale destacar que grande parte destes compostos estão contidos
no interior de tricomas glandulares evidenciando a importância dos mesmos na defesa
das plantas (STEINITE e LEVINSH, 2003).
Os tricomas são anexos epidérmicos que estão presentes sobre a superfície dos
vegetais, variando no formato, densidade, tipo e função. Em morangueiro foram
descritos dois tipos de tricomas recobrindo os seus órgãos: os não-glandulares ou
tectores e os tricomas glandulares (STEINITE e IEVINSH 2003; BRAGA et al. 2009;
FIGUEIREDO et al. 2010). Os não glandulares têm tamanho maior, unicelulares,
longos e finos podendo ser observados a “olho nu”. Já os tricomas glandulares são
pequenos, unisseriados e multicelulares com uma única célula arredondada presente no
ápice do pêlo, tendo a função de glândula, ou seja, capaz de liberar compostos químicos
presentes em seu interior quando tocados (STEINITE e LEVINSH, 2003).
Na cultura do morangueiro, grande parte da resistência das cultivares ao ácaro
rajado é atribuída à densidade de tricomas glandulares, que armazenam enzimas
oxidativas que dificultam a locomoção e alimentação do ácaro quando em contato com
o tricoma (STEINITE e LEVINSH, 2003; FIGUEIREDO et al, 2010), que funcionam
como mecanismos de defesa direta de plantas contra o ataque de artrópodo-praga
(STEINITE e LEVINSH, 2002).
Entre as funções mais estudadas dos tricomas pode-se citar o aumento da
resistência a pragas devido à formação de uma barreira física que dificulta o
caminhamento e alimentação dos artrópodos-praga (LARA, 1991). Fernandez et al.
(2006), evidenciaram o efeito inibitório da herbivoria em duas espécies de Tanchagem
(Plantago spp.) aonde os resultados comprovaram que, P. lanceolata por possuir uma
maior quantidade de tricomas apresentou taxa de herbivoria reduzida perante a P. major
que não possuía tricomas em nenhuma das faces das folhas. Logo se conclui que os
tricomas desempenham um papel importante na defesa de plantas contra o ataque de
pragas.
Quando se diz respeito aos artrópodos-pragas, são raros os trabalhos existentes,
sendo a maioria com outros reguladores vegetais que não o AS. Isso explica a
importância do presente trabalho, visando diminuir o uso dos agroquímicos utilizados
no combate a pragas.

14
3.9. Mecanismos de defesa das plantas
A resistência contra artropodo-praga é resultado de várias estratégias que as
plantas usam para tornarem-se menos atrativas a fim de dificultar a sua localização ou
se defender, tornando-se um alimento inadequado para herbívoros ou ainda sobreviver
por meio de mecanismos que podem ser denominados de tolerância ou
sobrecompensação (CORNELISSEN e FERNANDES, 2003; ALMEIDA- CORTEZ,
2005; CINGOLANI et al., 2005; MACDONALD e BACH, 2005).
A distribuição e o índice de ataque de artrópodos-pragas sofrem ampla
influência das características das plantas. Conhecer essas características permite uma
melhor compreensão das relações interespecíficas, além de determinar quais delas
afetam e, quais seus níveis de ataque, dessa forma, explicando os motivos da preferência
por determinadas espécies vegetais (PEETERS, 2002). Segundo Pizzamilgio-Gutierrez
(2009), todo o conhecimento sobre as relações entre as plantas e os herbívoros, auxilia
no desenvolvimento de cultivares resistentes e consiste em uma estratégia fundamental
para o sucesso do manejo integrado de pragas (MIP).
Algumas estruturas anatômicas são conhecidas por influenciarem o
comportamento das pragas (PEETERS, 2002). Características estruturais que
contribuem para o aumento da espessura ou a dureza dos tecidos foliares podem resultar
em obstáculo contra o ataque desses organismos (WEI et al., 2000; EDWARD; W
RATTEN, 1981; PEETERS, 2002). Os tricomas têm sido considerados uma forma de
proteção contra insetos sugadores (HOFFMAN e MCEVOY ,1985). O tamanho e a
densidade do tecido vascular também foram correlacionados com a distribuição de
fitófagos (GIBSON, 1972; COHEN et al., 1996).
A antibiose é uma modalidade de resistência, onde altera alguns dos seus
padrões biológicos, como a duração das fases imaturas, fecundidade, oviposição e
mortalidade, por exemplo. Desta forma, a ingestão pela praga destes metabólitos
deletérios produzidos pelas plantas são a causa deste tipo de resistência.
A resistência do tipo não preferência ou antixenose ocorre quando a planta
apresenta algum fator que cause uma resposta negativa na praga, deixando a planta com
menor atratividade para algumas funções básicas da praga como alimentação,
oviposição e abrigo.
Provavelmente os fatores envolvidos nessa resistência estejam relacionados aos
aspectos morfológicos, físicos e químicos apresentados pelos vegetais. Por fim, a

15
resistência do tipo tolerância acontece quando a planta não altera o comportamento ou a
biologia dos insetos, mas mantém as principais características produtivas perante outras
plantas, sobre o mesmo ataque de artrópodos-praga, compensando as perdas causadas
pelas pragas com o aumento da espessura da cutícula, aumento da área foliar, ou
mesmo, de suas brotações (GALLO et al., 2002).
3.10. Fluorescência
O sinal básico da fluorescência possui níveis característicos, que nos mostra
como está a planta naquele momento, em relação ao seu próprio metabolismo
(RIBEIRO et al., 2004; BAKER, 2008) permitindo definir o grau de tolerância a
diversos tipos de estresses bióticos e abióticos (STIRBET e GOVINDJEE, 2011).
Segundo Santos et al. (2010), a avaliação da fluorescência da clorofila a revela o nível
de excitação da energia que dirige a fotossíntese e fornece subsídios para estimar o
processo de transferência de elétrons do FSII. A energia radiante que incide sobre as
plantas pode seguir por diferentes rotas, como: a via fotoquímica, que representa a
porção energética efetivamente absorvida, e que é responsável pela assimilação do
carbono em moléculas orgânicas; e as vias de dissipação da energia que não é
consumida pela planta, que pode ocorrer na forma de calor e/ou re-emissão de fótons
pela molécula de clorofila.
A via fotoquímica é caracterizada pela fixação do carbono atmosférico em uma
molécula orgânica pela enzima Ribulose-1,5-bifosfato carboxilase/oxigenase (Rubisco)
que age no ciclo de Calvin. Para que tal ciclo ocorra, são necessários precursores
metabólicos formados na cadeia transportadora de elétrons. É nesta fase que os
pigmentos fotossintéticos exercem sua função, pois eles constituem os complexos
antena, e evitam que a energia solar incida diretamente nos centros de reação (YEOM et
al., 2014). Desta maneira, tais pigmentos, como a clorofila, atuam transferindo a energia
luminosa para os centros de reação e dissipando parte desta energia que seria danosa aos
centros de reação. Por deste motivo, tais pigmentos também são chamados de moléculas
fotoprotetoras (JOHNSON e RUBAN, 2009). É neste processo de excitação da
molécula da clorofila que ocorrem as outras duas vias de dissipação de energia:
dissipação na forma de calor e dissipação por fluorescência.

16
Quando a clorofila recebe um fóton e assume seu estado de excitação, ela torna-
se-se altamente instável, e tende a retornar rapidamente ao seu estado basal de energia.
A primeira forma dissipação é a perda de calor para o ambiente. Neste caso da
fluorescência, a clorofila excitada reemite um fóton com comprimento de onda mais
longo e menor nível energético que o absorvido (MURCHIE e LAWSON, 2013). Nesse
modo a fluorescência da clorofila a pode ser utilizada, indiretamente, como um
indicativo da eficiência no uso da radiação pela via fotoquímica e, consequentemente da
assimilação líquida de carbono (TESTER e BACIC, 2005) e sanidade do aparato
fotossintético, mais especificamente do fotossistema II.
As avaliações da fluorescência proporcionam dados que podem estimar o estado
de organização e eficiência do fotossistema II (PSII), e a taxa de transporte de elétrons
(ETR) entre os centros de reação, além da capacidade resiliente das plantas quando
expostas à pulsos de luz saturante (GENTY et al., 1989).
Nas medidas realizadas em plantas adaptadas ao escuro, é possível de avaliar a
plasticidade do PSII com relação ao estado basal da própria planta e à adaptação ao
ambiente em que se encontra. Quando plantas nestas condições são expostas a um pulso
de luz actínica fraca, é fornecido o nível mínimo de fluorescência, chamado de F0, que é
obtido quando os centros de reação do PSII estão totalmente abertos e os aceptores de
elétrons da quinona primária (QA) do PSII ficam completamente oxidados (BAKER e
ROSENQVIST, 2004; MAXWELL e JOHNSON, 2000).
Em seguida, quando se aplica um pulso saturante, é fornecido o nível de
fluorescência máxima (Fm), que informa a dissipação por re-emissão de fótons no
momento em que os centros de reação do PSII estão completamente reduzidos
(saturados). A diferença entre estes dois parâmetros determina a fluorescência variável
11 (Fv) (BAKER e ROSENQVIST, 2004) que demonstra a habilidade do PSII em
desempenhar as reações fotoquímicas primárias (fotorredução da QA).
Segundo Henschel (2016), a estimativa da fluorescência da clorofila pode servir
como uma forma rápida e não-destrutiva de mensurar a eficiência e a sanidade do
aparato fotossintético, indicando se as plantas se encontram, ou não, sob condições de
estresse. Tais avaliações tornam-se ferramentas bastante úteis para investigar a
influência da qualidade e intensidade luminosas sobre as plantas, já que estão
diretamente relacionadas ao processo fotossintético. Desse modo, a emissão da
fluorescência, permite avaliar o efeito do AS na fotossíntese, sendo que essa prática
quando combinada com outras medidas não destrutivas, permite realizar análise

17
qualitativa e quantitativa da absorção e aproveitamento da energia luminosa por meio do
fotossistema, sendo uma poderosa ferramenta para estudar o desempenho fotossintético
(BAKER, 2008).
4. MATERIAL E MÉTODOS
4.1. Local do experimento
O experimento foi realizado no Setor de Olericultura e no Laboratório de
Fisiologia Vegetal/Horticultura, localizados no Departamento de Agronomia, no
Campus Cedeteg da Universidade Estadual do Centro-Oeste, Unicentro, em
Guarapuava-PR, que está localizada nas coordenadas 25°23’00’’ de latitude Sul e
51°29’38.50’’ de longitude Oeste, a uma altitude de 1.024 metros.
4.2. Cultivares de morangueiro e cultivo das plantas
Para os bioensaios foram utilizadas mudas das cultivares Aromas e Sweet
Charlie.
A cultivar Aromas é de dias neutros, para mesa, tem bom tamanho de fruto, com
ciclo precoce, cor vermelho-brilhante, bom sabor e vigor médio. Sweet Charlie é uma
cultivar de fotoperíodo curto, de ciclo precoce, com pouca exigência ao frio e de boa
produtividade. Produz frutos grandes, com boas características qualitativas, firmeza
moderada e coloração vermelho-alaranjada externamente e alaranjada internamente.
As mudas de morangueiro foram transplantadas em vasos com capacidade para 3
dm3. O solo foi adubado com o formulado 4:14:8 em conjunto com substrato comercial
tropstrato®.
Os vasos permaneceram em casa de vegetação com irrigação diária por
microaspersão. Durante o desenvolvimento das plantas foram realizados todos os tratos
culturais rotineiros, como adubação, irrigação, toalhetes das plantas exceto o manejo
fitossanitário.
Os tratamentos foram espaçados em cerca de 3 m de distância para que não
ocorresse interferência de um tratamento sobre outro, tendo em vista que o AS, quando
convertido em metil salicilato, composto volátil, pode induzir resistência nas plantas
adjacentes.

18
Cerca de 30 dias após o transplantio iniciou-se a aplicação do ácido salicílico
(AS) com borrifador manual. As aplicações foram realizadas semanalmente totalizando
60 aplicações. O AS foi preparado e diluído em água destilada nas concentrações (0, 25,
50, 75, 100 mg L-1
).
O delineamento experimental foi em parcelas subdivididas no espaço, sendo a
parcela principal a concentração de ácido salicílico e a subparcela as cultivares de
morangueiro, em 6 repetições, com um vaso por parcela.
Com a finalidade de induzir os mecanismos de defesa das plantas, foram
adicionados em cada tratamento um vaso contendo plantas com elevada infestação de
pulgões Myzus persicae, estimulando a ocorrência da indução de resistência.
4.3. Obtenção dos ácaros e criação
Para início da criação controlada foram recebidos indivíduos de T. urticae da
Escola Superior de Agricultura Luiz de Queiroz ESALQ/USP. Com os adultos foi
estabelecida uma criação estoque, na qual os ácaros foram mantidos e multiplicados em
plantas de feijão de porco (Canavalia ensiformis DC.), cultivadas em vasos com
capacidade para 5 dm3 e acondicionadas em sala climatizada (25±2ºC, fotofase de 12
horas), sob irrigação diária. Os vasos contendo as plantas de feijão de porco e os ácaros
foram substituídos por plantas novas à medida que as plantas colonizadas pelos ácaros
apresentaram sintomas intensos de ataque ou início de senescência. Essa substituição foi
realizada colocando-se folhas com alta infestação do ácaro das plantas antigas, sobre as
plantas novas. A partir da criação estoque, deu-se início à criação programada, a qual
consistia em 10 ácaros adultos (3 machos e 7 fêmeas) mantidos em folíolos individuais
de feijão de porco, obtidos na parte mediana das plantas. Cada folíolo foi mantido em
bandejas plásticas (200 x 300 mm), com a face abaxial voltada para cima, e sobre uma
esponja umedecida em água destilada.
Os ácaros foram mantidos nos folíolos durante 24 horas, e em seguida os adultos
foram retirados, permanecendo apenas os ovos. Para a renovação e preparo de novas
bandejas, os folíolos antigos em estádio de senescência foram colocados sobre outros
folíolos, permitindo a passagem dos ácaros. A criação programada foi realizada em
câmara climatizada tipo BOD (temperatura 25±1ºC, umidade de 70±10% e fotofase de
12 horas).

19
4.4. Bioensaio com chance de escolha
Com o objetivo de avaliar a preferência das fêmeas do ácaro rajado em relação
as cultivares de morangueiro e as diferentes doses da aplicação de AS, foram realizados
experimentos com chance de escolha em arenas. Estas arenas consistiram em placas de
petri (60 mm de diâmetro) contendo uma camada de algodão, sobreposta a uma esponja
saturada em água. Nas extremidades opostas das placas foram inseridos discos foliares
(30 mm de diâmetro) de dois tratamentos distintos, sendo a testemunha com os demais
tratamentos em suas respectivas doses, os quais foram conectados por uma lamínula de
plástico (18 x 18 mm).
Todos os discos foliares foram obtidos de folhas das plantas de morangueiro
com 450 dias de desenvolvimento e dispostos em placa de petri com a face abaxial
voltada para cima. Após montadas as arenas, seis fêmeas adultas dos ácaros foram
liberadas na porção mediana da lamínula, a qual permitia a livre passagem e o acesso
dos ácaros aos folíolos de ambas as extremidades. A liberação dos indivíduos foi
realizada com auxílio de pincel de cerdas finas com auxílio de estereomicroscópio
(NIKON 1510). As placas foram mantidas durante 24 horas em câmara climatizada tipo
BOD (temperatura 25±1ºC, umidade de 70±10% e fotofase de 12 horas). Após esse
período, realizou-se a contagem do número de ácaros presentes em cada um dos discos.
O experimento foi realizado em delineamento inteiramente casualizado com fatorial 2 x
5 (duas cultivares de morangueiro e cinco doses de AS) com 10 repetições.
4.5. Bioensaio sem chance de escolha
Estes bioensaios foram realizados em placas de petri (60 mm de diâmetro),
preenchidas com uma espuma umedecida em água destilada. Sobre esta espuma foi
inserido, individualmente, um disco foliar de cada um dos tratamentos (30 mm de
diâmetro), mantendo-se a face abaxial voltada para cima. Sobre cada disco foram
transferidas seis fêmeas adultas oriundas da criação, com auxílio de um pincel de cerdas
finas e um estereomicroscópio (OLYMPUS SZ51). As fêmeas foram mantidas sobre os
discos durante 24 horas, após o qual se contabilizou o número de ácaros vivos e o
número de ovos depositados sobre cada disco foliar. O experimento foi realizado em
delineamento inteiramente casualizado em fatorial 2 x 5 com 10 repetições, sendo que o

20
conjunto de placas foi mantido em câmara climatizada do tipo BOD (temperatura
25±1ºC, umidade de 70±10% e fotofase de 12 horas.
4.6. Bioensaios com larvas de H. armigera
4.6.1. Obtenção e criação das larvas de H. Armigera
Para formação da criação estoque, foram adquiridos ovos de H. armigera de
uma criação massal (Bug Agentes Biológicos, Piracicaba, SP, Brasil). Os ovos foram
acondicionados em recipientes plásticos (250 mL) até eclosão das larvas.
As larvas recém-eclodidas foram repicadas em quantidade de 10 indivíduos em
copos plástico de 50 mL, contendo dieta artificial, preparada de acordo com a
metodologia de Greene et al. (1976), à base de feijão, germe de trigo, farelo de soja,
caseína, levedura de cerveja, antibióticos e proteínas. Os copos foram fechados com
tampas de acrílico e furados com alfinete entomológico, as larvas foram mantidas no
recipiente até fase de pupa.
As pupas foram colocadas em recipientes plásticos revestida interiormente com
papel toalha e fechados com tecido voil até a fase adulta, quando foram transferidas para
gaiolas de PVC 100 mm, forradas com papel absorvente e cobertas com tecido voil,
onde ocorria a oviposição. No interior das gaiolas foi colocada solução de mel a 10%,
embebida em algodão, para a alimentação.
Todas as fases foram mantidas em câmaras climatizadas, tipo BOD, (25±3ºC,
umidade relativa de 70 ± 5% e fotofase de 14 horas).
4.6.2. Bioensaios para avaliação da biologia
Estes bioensaios foram realizados em placas de petri (60 mm de diâmetro),
preenchidas com uma espuma umedecida em água destilada. Sobre esta espuma, foi
inserido, individualmente, uma folha de cada um dos tratamentos. Sobre cada folha foi
inserido uma lagarta, com auxílio de um pincel de cerdas finas e um estereomicroscópio
(OLYMPUS SZ51). As lagartas foram mantidas sobre os discos e a cada 72 horas,
trocava-se as folhas e contabilizava o número de lagartas vivas, pesava-se com auxilio
de uma balança de precisão (SHIMADZU AUW220D) e também media-se o diâmetro
de cada com o auxilio de um paquímetro digital (STARRETT 799). O experimento foi

21
realizado em delineamento inteiramente casualizado em fatorial 2 x 5 com 10
repetições, sendo que o conjunto de placas foi mantido em câmara climatizada do tipo
BOD (temperatura 25±1ºC, umidade de 70±10% e fotofase de 12 horas.
4.7. Identificação e quantificação de tricomas
Para identificação e quantificação dos tricomas foram coletados folíolos
expandidos das plantas de cada tratamento, com idade de 420-450 dias. Esses folíolos
foram previamente lavados com água destilada e posteriormente secos a sombra.
Para cada folíolo recém coletado foram montadas 4 amostras da face abaxial e 4
da face adaxial e fixados em stubs metálicos com auxílio de esmalte cosmético incolor
acrescido de grafite moído que são inseridos em um disco contendo sete espaços. Esse
composto atua de forma equivalente a fibra de carbono, facilitando o bombeamento de
elétrons, permitindo formação de imagens mais nítidas do tricomas e com uma boa
resolução. As amostras foram levadas para obtenção de imagens em microscopia
eletrônica de varredura (MEV) (Vega3 Tescan XM) HV a 30kv, de 2 mm2.
Foram observados o comprimento, a presença ou não de glândula na
extremidade apical e o formato de cada glândula.
4.8. Fluorescência
As leituras de fluorescência da clorofila a nas plantas mantidas em casa de
vegetação foram realizadas utilizando um fluorômetro portátil (Portable Chlorophyll
Fluorometer- PAM-2500, Walz, 2008). As avaliações foram iniciadas (16 de maio a 05
junho de 2016) totalizando três leituras em dias de pleno sol, no período das 08h ás 11h
da manhã, com adaptação ao escuro de 20 minutos, em folhas completamente
expandidas, localizadas no terço médio da planta. O experimento foi realizado em
delineamento inteiramente casualizado em fatorial 2 x 5 com 3 repetições.
A partir da resposta dos primeiros pulsos de luz da indução de curva, foram
obtidos os parâmetros: F0 (fluorescência inicial da folha); Fm (Fluorescência máxima da
folha); Y (II) (rendimento quântico fotoquímico do FSII); qP (coeficiente de dissipação
de fluorescência fotoquímica); ETR (taxa de transferência relativa de elétrons). E foram
calculado o seguinte parâmetro: Fv/Fo (atividade potencial do FSII).

22
4.9. Análises Estatísticas
O teste de Shapiro-Wilk foi utilizado para avaliar a normalidade dos dados para
comprovar a distribuição de probabilidade associada ao conjunto de dados obtidos. Foi
verificada também a homogeneidade de variâncias pelo teste Bartlett. Os dados foram
submetidos ao programa estatístico Sisvar e realizou-se análise de variância, e as médias
foram agrupadas pelo teste de Tukey, a 5% de probabilidade, (FERREIRA, 2008). Para
os dados de antixenose com chance de escolha foi analisado a tendência utilizando as
percentagens de escolha, calculadas pelos gráficos gerados pelo software Microsoft
Excel (2016). As equações foram derivadas para se determinar o ponto de máxima dose
aplicada, com melhor eficiência de indução de resistência. Os dados de densidade de
tricomas e teste de antixenose sem chance de escolha e com chance de escolha foram
transformados pela equação √(X+0,5). Os dados da fluorescência foram submetidos ao
programa estatístico Sisvar e as médias foram agrupadas pelo teste de Tukey, a 5 %.
Correlações de Pearson foram estimadas comparando as doses de AS com as fontes de
variação. A significância dos valores estimados de correlação e do R2
foram obtidas por
meio do teste de t a 5%.
5. RESULTADOS E DISCUSSÃO
5.1. Bioensaio com chance de escolha – Ácaro rajado
No bioensaio com chance de escolha houve diferença significativa na frequência
de escolha do ácaro rajado em relação às diferentes doses do AS aplicadas. Não houve
diferença significativa entre as cultivares e doses de AS aplicadas nas plantas de
morangueiro (Figura 1).
A) Cultivar Aromas sem tratamentos com AS (T0) e demais tratamentos referentes as
doses do eleicitor aplicadas (T25, T50, T75, T100), B) Cultivar Aromas sem
Figura 1. Frequência de escolhas realizadas pelo ácaro rajado, em relação às cultivares de
morangueiro.

23
tratamentos com AS (T0) e demais tratamentos referentes as doses do elicitor aplicadas
(T25, T50, T75, T100). Guarapuava-PR, UNICENTRO, 2016.
Para ambas cultivares testadas, observou-se que houve preferência dos ácaros
em se deslocar na direção da testemunha (sem aplicação de AS), quando comparada aos
demais tratamentos, ou seja, para este bioensaio o elicitor foi eficaz na repelência ao
ácaro rajado.
Entre as doses avaliadas, observou-se que a aplicação nas concentrações de 25,
50 e 75 mg L-1
foram as mais efetivas no sentido de repelir o deslocamento do ácaro
rajado. Esse efeito pode ser explicado em decorrência do mecanismo de resistência que
manifesta-se para sobrevivência da planta ao ataque, devidamente identificada ou
reconhecida. Por essa razão, tal mecanismo de defesa recebe a denominação de
Resistência Sistêmica Adquirida (SAR). A SAR é resultante da identificação do
invasor, acompanhado da indução da síntese de substâncias específicas, como quitinases
e outras enzimas hidrolíticas, devido à ação do elicitor, que funciona como um sinal
endógeno para o desencadeamento da ação de defesa. As estratégias básicas envolvidas
são barreira bioquímica e equilíbrio nutricional (FANCELLI, 2008).
Em estudos realizados por Habibi (2012) recomenda-se a quantidade de 500 μM
de ácido salicílico para aumentar o conteúdo relativo de água, a taxa fotossintética,
transpiração e condutância estomática em plantas de cevada (Hordeum vulgare L.). Isto
significa dizer que o uso de AS em concentrações adequadas aumenta a capacidade
fotossintética da cultura e que a resposta da planta a este hormônio depende das
condições ambientais, cultivar, época de aplicação dose e forma de uso
(NIVEDITHADEVI et al., 2012).
Observou-se que a dose mais elevada de AS (100 mg L-1
) não demostrou efeito
satisfatório na repelência ao Tetranychus urticae, quando comparada as doses de 25, 50
e 75 mg L-1
(Figura 1), comportando-se de forma similar a testemunha.
Uma das funções do AS na defesa é a produção de espécies reativas de oxigênio
(EROs), tais como o radical superóxido (O2•-), radical hidroxila (OH-), peróxido de
hidrogênio (H2O2) e oxigênio singleto (1O2), (JALEEL et al., 2009). Estes atuam nas
células ou tecido danificados, promovendo a sua morte e, consequentemente,
eliminando o agente danificador. As EROs formam-se naturalmente no interior das
células e a sua produção está relacionada com organelas que possuem alta atividade
oxidativa ou com uma intensa taxa de fluxo de elétrons. Mitocôndrias, cloroplastos e

24
microcorpos (peroxissomos) são considerados as principais fontes de espécies reativas,
como produtos secundários oriundos da fotossíntese e da respiração (MATAMOROS et
al., 2006; DEL RIO et al., 2006).
No entanto, o excesso de AS pode promover elevada produção de EROs,
originando o fenômeno conhecido como “explosão oxidativa” (MATAMOROS et al.,
2006; DEL RIO et al., 2006), resultando em efeito antagônico na proteção da planta.
Este fato poderia explicar os efeitos negativos da elevada dose de AS no controle do
ácaro encontrado na presente pesquisa, pois ao invés de proteger as regiões atacadas, as
EROs aceleraram a degradação do tecido, deixando a planta ainda mais vulnerável. Se o
estresse for mais severo, aumenta consideravelmente a produção de radicais livres que
podem levar a uma cascata de eventos que inicia com a peroxidação de lipídeos,
avançando para a degradação de membranas e morte celular (GREGGAINS et al.,
2000).
Outro efeito de elevadas concentrações de AS na planta, é a termogênese,
fenômeno que promove a perda de energia por entalpia, mediante reações de
oxirredução nos fotossistemas ou no aparato respiratório, como exemplo o
desacoplamento de ATP, que libera o equivalente a 15 kcal por molécula para o meio.
Essa relação do AS como regulador endógeno só foi estabelecida em um estudo de
termogênese em plantas em 1987 (RASKIN, 1992). A energia recebida do sistema por
meio dos processos endergônicos é distribuída a três processos metabólicos da planta,
fluorescência e termogênese (catabolismo) e fotossíntese (anabolismo). Quando em
altas concentrações o AS prove aumento da termogênese e fluorescência, reduzindo a
disponibilidade para a fotossíntese, diminuindo assim a quantidade de trioses P para a
própria manutenção (respiração) e para ativação de rotas metabólicas secundárias, em
especial a defesa, por meio da redução da fitoalexinas (STANGARLIN et al., 1999;
2008).
5.2. Bioensaio sem chance de escolha – Ácaro rajado
No bioensaio sem chance de escolha foram analisadas a sobrevivência de adultos
e o número de ovos em diferentes doses do AS, para as cultivares de morangueiro
Aromas e Sweet Charlie. Nas cultivares avaliadas (Aromas e Sweet Charlie), houve
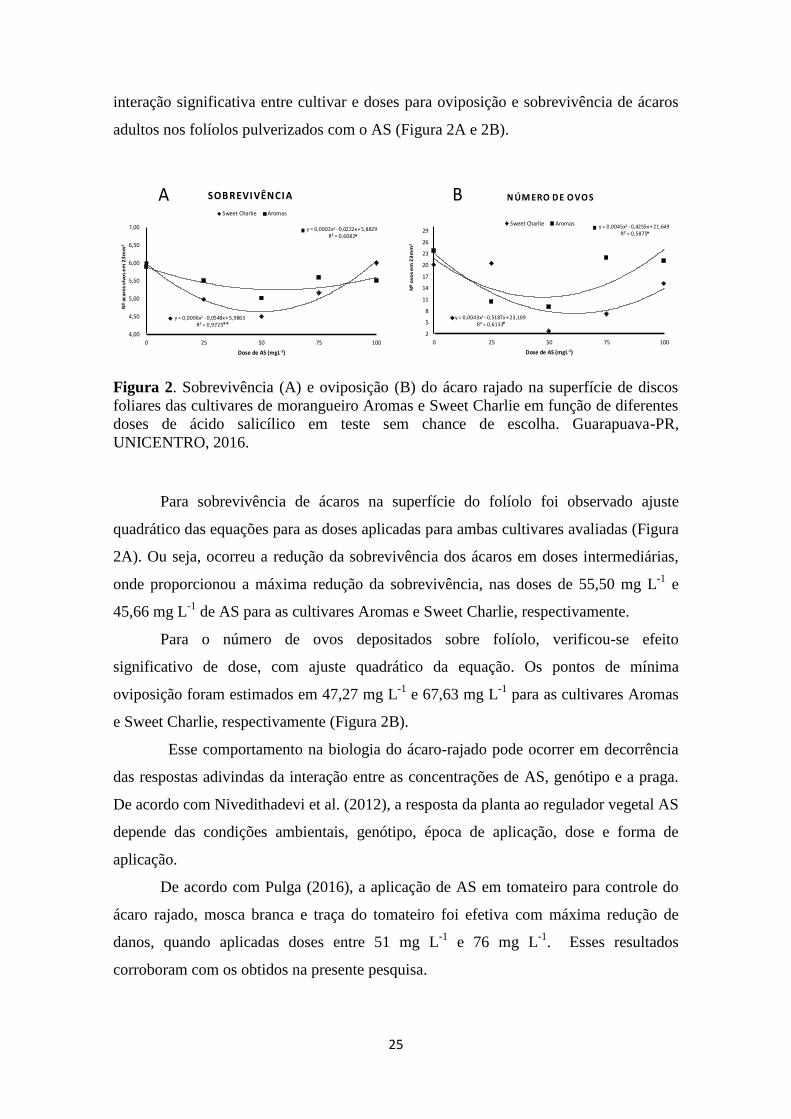
25
interação significativa entre cultivar e doses para oviposição e sobrevivência de ácaros
adultos nos folíolos pulverizados com o AS (Figura 2A e 2B).
Figura 2. Sobrevivência (A) e oviposição (B) do ácaro rajado na superfície de discos
foliares das cultivares de morangueiro Aromas e Sweet Charlie em função de diferentes
doses de ácido salicílico em teste sem chance de escolha. Guarapuava-PR,
UNICENTRO, 2016.
Para sobrevivência de ácaros na superfície do folíolo foi observado ajuste
quadrático das equações para as doses aplicadas para ambas cultivares avaliadas (Figura
2A). Ou seja, ocorreu a redução da sobrevivência dos ácaros em doses intermediárias,
onde proporcionou a máxima redução da sobrevivência, nas doses de 55,50 mg L-1
e
45,66 mg L-1
de AS para as cultivares Aromas e Sweet Charlie, respectivamente.
Para o número de ovos depositados sobre folíolo, verificou-se efeito
significativo de dose, com ajuste quadrático da equação. Os pontos de mínima
oviposição foram estimados em 47,27 mg L-1
e 67,63 mg L-1
para as cultivares Aromas
e Sweet Charlie, respectivamente (Figura 2B).
Esse comportamento na biologia do ácaro-rajado pode ocorrer em decorrência
das respostas adivindas da interação entre as concentrações de AS, genótipo e a praga.
De acordo com Nivedithadevi et al. (2012), a resposta da planta ao regulador vegetal AS
depende das condições ambientais, genótipo, época de aplicação, dose e forma de
aplicação.
De acordo com Pulga (2016), a aplicação de AS em tomateiro para controle do
ácaro rajado, mosca branca e traça do tomateiro foi efetiva com máxima redução de
danos, quando aplicadas doses entre 51 mg L-1
e 76 mg L-1
. Esses resultados
corroboram com os obtidos na presente pesquisa.
y = 0,0006x2 - 0,0548x + 5,9863R² = 0,9725
y = 0,0002x2 - 0,0222x + 5,8829R² = 0,6082
4,00
4,50
5,00
5,50
6,00
6,50
7,00
0 25 50 75 100
Nº
acar
os
vivo
s e
m 2
3m
m2
Dose de AS (mgL-1)
SOBREVIVÊNCIA
Sweet Charlie Aromas
**y = 0,0043x2 - 0,5187x + 23,109
R² = 0,6133
y = 0,0045x2 - 0,4255x + 21,649R² = 0,5875
2
5
8
11
14
17
20
23
26
29
0 25 50 75 100
Nº
ovo
s e
m 2
3m
m2
Dose de AS (mgL-1)
N ÚM ERO DE OVOS
Sweet Charlie Aromas
*
*
*
A B

26
A eficiência do AS aplicado via foliar no controle da praga em plantas de
morangueiro, provavelmente, ocorreu devido aos seus efeitos de não preferência ou
antibiose, pois o comportamento do ácaro rajado variou de acordo com a dose aplicada.
O AS, é um regulador vegetal que funciona como sinalizador, ativando as
respostas das plantas após o ataque de pragas. O acúmulo desse composto nos tecidos
das plantas induz a resistência sistêmica adquirida (SAR), uma forma de resistência
induzida (WALLING, 2000). Herbívoros que possuem uma interação próxima e de
longa duração com a planta, ativam respostas de defesa que dependem do ácido
salicílico.
O AS atua na resposta das plantas após ataque de pragas, alterando a atividade e
aumentando a expressão de genes de defesa do vegetal, potencializando sua resposta
(PONSTEIN et. al., 1994). Estudos realizados por SILVA et al. (2005), demonstram
que as pragas podem ter dificuldades de ovopositar sob células silicificadas, dessa
forma, diminuindo a herbivoria das fases imaturas desses artrópodos.
Estes resultados estão de acordo com os obtidos na presente pesquisa, tendo em
vista que o AS atuou como regulador vegetal, na indução de resistência, ativando
respostas das plantas após o ataque do ácaro rajado. Além disso, ocorreu a conversão do
AS em salicilato de metila, composto volátil, que induz resistência, nas partes não
atacadas da mesma planta, bem como em plantas adjacentes (RASKIN et al., 2000).
O salicilato de metila promove a comunicação entre uma planta estressada com
as plantas vizinhas, alertando-as da ameaça. Pesquisadores demostraram em laboratório
que uma planta pode aumentar suas defesas se estiver ligada, de alguma maneira, a
outra planta que esteja liberando essa substância. (FERNANDES et; al; 2009). Neste
trabalho, como citado na metodologia, foram adicionados em cada parcela um vaso com
elevada infestação de pulgões para induzir os mecanismos de defesa, onde observou-se
a promoção da resistência através do estresse das plantas, ocorrendo a liberação desse
composto que ativou o mecanismo de defesa dessas plantas.
Essa ativação do mecanismo de defesa da planta ocorre por meio de sucessivos
eventos e sinais que se iniciam no reconhecimento pela planta do agente agressor e
culmina com a ativação das barreiras físicas e químicas envolvidas na sua constituição
(FERNANDES et; al.; 2009).
O AS atua na alteração da atividade ou síntese de enzimas, o que aumenta a
expressão de genes de defesa, potencializando a resposta das plantas após a alimentação
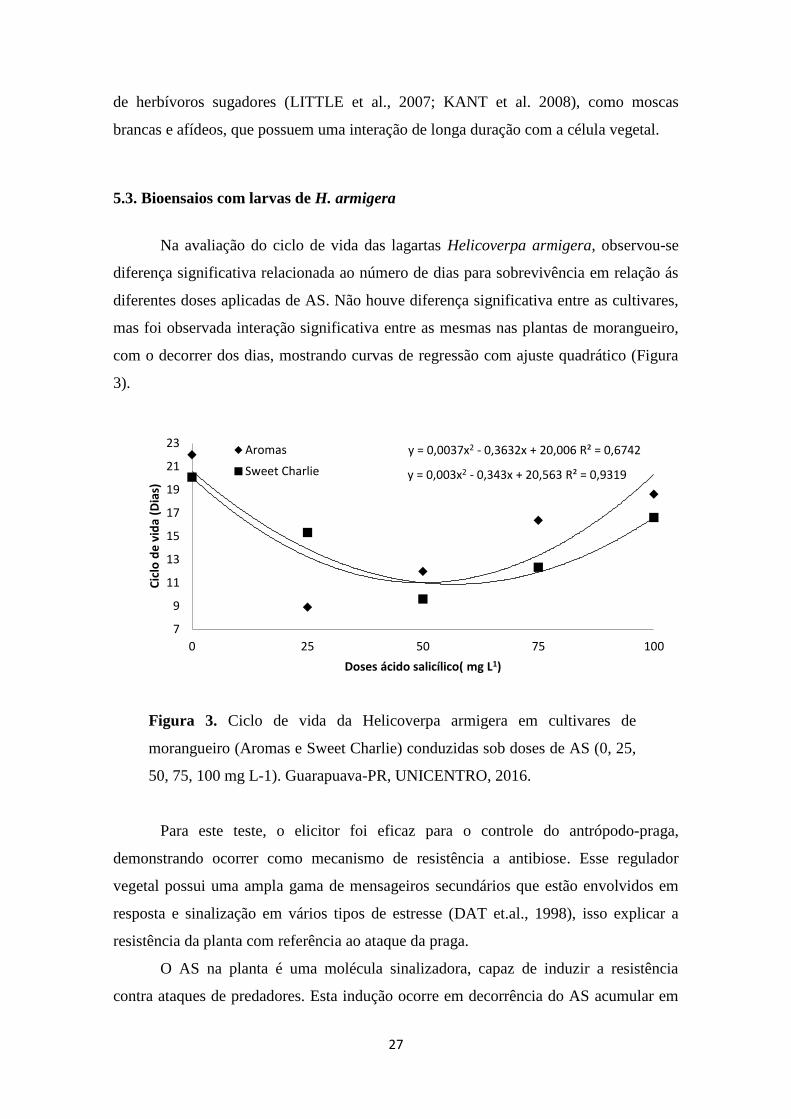
27
de herbívoros sugadores (LITTLE et al., 2007; KANT et al. 2008), como moscas
brancas e afídeos, que possuem uma interação de longa duração com a célula vegetal.
5.3. Bioensaios com larvas de H. armigera
Na avaliação do ciclo de vida das lagartas Helicoverpa armigera, observou-se
diferença significativa relacionada ao número de dias para sobrevivência em relação ás
diferentes doses aplicadas de AS. Não houve diferença significativa entre as cultivares,
mas foi observada interação significativa entre as mesmas nas plantas de morangueiro,
com o decorrer dos dias, mostrando curvas de regressão com ajuste quadrático (Figura
3).
Para este teste, o elicitor foi eficaz para o controle do antrópodo-praga,
demonstrando ocorrer como mecanismo de resistência a antibiose. Esse regulador
vegetal possui uma ampla gama de mensageiros secundários que estão envolvidos em
resposta e sinalização em vários tipos de estresse (DAT et.al., 1998), isso explicar a
resistência da planta com referência ao ataque da praga.
O AS na planta é uma molécula sinalizadora, capaz de induzir a resistência
contra ataques de predadores. Esta indução ocorre em decorrência do AS acumular em
y = 0,0037x2 - 0,3632x + 20,006 R² = 0,6742
y = 0,003x2 - 0,343x + 20,563 R² = 0,9319
7
9
11
13
15
17
19
21
23
0 25 50 75 100
Cic
lo d
e v
ida
(Dia
s)
Doses ácido salicílico( mg L1)
Aromas
Sweet Charlie
Figura 3. Ciclo de vida da Helicoverpa armigera em cultivares de
morangueiro (Aromas e Sweet Charlie) conduzidas sob doses de AS (0, 25,
50, 75, 100 mg L-1). Guarapuava-PR, UNICENTRO, 2016.

28
plantas submetidas a condições adversas, quer seja por ataque patogênicos, ou pelos
tratamentos com elicitores químicos, e por sua propriedade de induzir a expressão de
genes ligados a várias proteínas de resistência, (Pathogen ReladtiProtein-PR)
(MARTINEZ et.al., 2000).
O AS está diretamente envolvido com a defesa do vegetal, mais especificamente
contra a ação de patógenos hemi-biotrófico, biótroficos, além de ser um hormônio
indutor da resistência sistêmica adquirida (SAR) (GRANT e LAMB, 2006). Desta
forma, o aumento da concentração de AS em plantas cultivadas ocorre em resposta a
estresse causado por patógenos, pragas e estresse abiótico (GLAZEBROOK, 2005;
LORENZO e SOLANO, 2005; BROEKAERT et al., 2006; LOAKE e GRANT, 2007;
BALBI e DEVOTO, 2008). Em plantas mutantes de Arabidopsis thaliana que sofreram
ataques de patógenos notou-se o aumento na concentração de AS, entretanto plantas
deficientes na produção de AS se tornaram mais susceptíveis ao ataque de patógenos
(PARK et al., 2007).
Para os resultados obtidos, verificou-se efeito significativo de doses. Os pontos
de mínima a sobrevivência foram estimados em 49,08 mg L-1
e 57,16mg L-1
para as
cultivares Aromas e Sweet Charlie, respectivamente. Tendo em vista que o ciclo de vida
na testemunha foi de aproximadamente 23 dias, enquanto que nas doses 25 mg L-1
foi
de 9 dias, na dose 50mg L-1
de 10 dias, 75 mg L-1
13 dias e por fim, de 21 dias na dose
de 100 mg L-1
. Matthews (1999) afirma que o período larval é constituído por 5 a 6
instares e pode durar de 2 a 3 semanas, dependendo das condições climáticas e etc. Esse
resultado demostra a relevância das aplicações do AS nas doses intermediárias que fez
com que ocorresse menor ciclo do artropodo-praga Helicoverpa armigera. As medidas
dos pesos e comprimentos das larvas foram feitas a cada 72 horas totalizando cinco
avaliações para determinar os parâmetros. Nas cinco avaliações observou-se que nessas
variáveis, houve diferença entre as doses aplicadas e que a testemunha e as doses
maiores (75 e 100 mg L-1
) apresentaram maior peso das larvas comparadas com as
demais, praticamente em todas as avaliações realizadas (Figura 4).
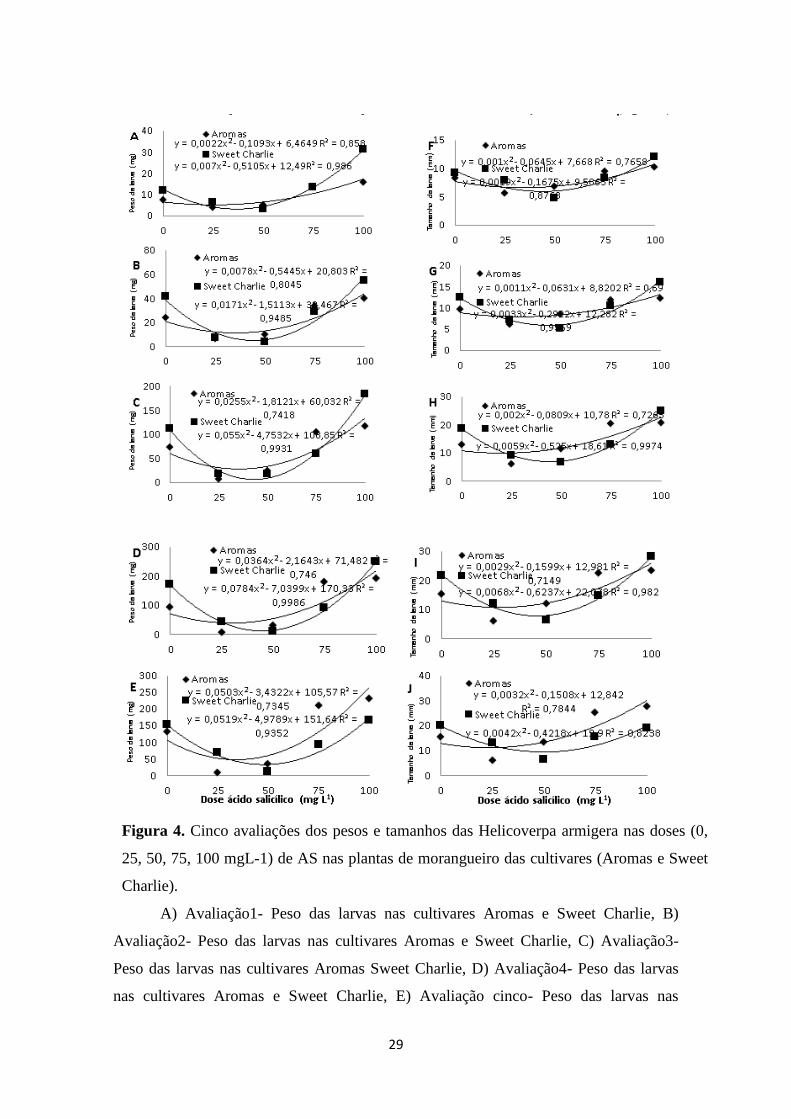
29
A) Avaliação1- Peso das larvas nas cultivares Aromas e Sweet Charlie, B)
Avaliação2- Peso das larvas nas cultivares Aromas e Sweet Charlie, C) Avaliação3-
Peso das larvas nas cultivares Aromas Sweet Charlie, D) Avaliação4- Peso das larvas
nas cultivares Aromas e Sweet Charlie, E) Avaliação cinco- Peso das larvas nas
Figura 4. Cinco avaliações dos pesos e tamanhos das Helicoverpa armigera nas doses (0,
25, 50, 75, 100 mgL-1) de AS nas plantas de morangueiro das cultivares (Aromas e Sweet
Charlie).

30
cultivares Aromas e Sweet Charlie, F) Avaliação1- Tamanho das larvas nas cultivares
Aromas e Sweet Charlie, G) Avaliação2- Tamanho das larvas nas cultivares Aromas e
Sweet Charlie, H) Avaliação3- Tamanho das larvas nas cultivares Aromas e Sweet
Charlie, I) Avaliação4- Tamanho das larvas nas cultivares Aromas e Sweet Charlie, J)
Avaliação cinco- Tamanho das larvas nas cultivares Aromas e Sweet Charlie.
Guarapuava-PR, UNICENTRO, 2016.
Foi positiva a eficiência do AS aplicado via foliar no controle de H. armigera,
os pontos de mínima no peso das larvas foi de 31,82 mg L-1
e 43,21mg L-1
. Como
Pieterse e Van Loom (1999) descrevem, o AS está envolvido em múltiplas vias de
defesa da planta, ativando independentemente sua resposta. Na presente pesquisa, o AS
deve ter atuado como regulador vegetal, na indução de resistência, ativando respostas
das plantas após o ataque da H. armigera e interferindo no peso da praga, portanto,
caracterizando o efeito de antibiose. Pickett e Poppy (2001) e Park et al. (2007) também
citam que o AS age como um composto sinalizador e amplificador de respostas
induzidas a agentes patogênicos e herbívoros. No caso do ensaio da avaliação dessa
praga os compostos produzidos pela aplicação do AS, podem ter interferido na
alimentação e com isso proporcionando as diferenças no peso do artrópodo-praga. De
acordo com Ruan e Wu (2001) e Barton e Raubenheimer (2003), a Helicoverpa
armigera precisa de condições nutricionais adequadas para seu crescimento e
proliferação, precisando de quantidade e da qualidade do alimento consumido, bem
como a eficiência de como este é utilizado.
Na quinta e última avaliação observou-se o efeito de antibiose e antixenose do
ácido salicílico em reduzir o peso e comprimento da H. armigera. Foi possível constatar
que a menor média de peso no ponto de mínima, como já mencionado foi de 31,82 mg
L-1
e 43,21 mg L-1
e no tamanho os pontos de mínima foi de 26,45 mg L-1
e 45,78 mg L-
1, respectivamente para Aromas e Sweet Charlie, enquanto a maior média foi constatada
na testemunha e nos pontos de máxima 75 e 100 mg L-1
.
Como observado, a aplicação do AS interferiu na biologia da larva e promoveu a
resistência induzida no morangueiro em quase todas as avaliações. Fu e Dong (2013),
observaram que a resistência induzida nas plantas permanece ativada por um longo
período de tempo podendo perdurar por semanas a meses. Isso evidencia a efetividade
da aplicação de ácido salicílico, como pode ser observado no presente estudo, conforme
o decorrer desse ensaio.

31
Além disso, esse regulador vegetal pode gerar, em diversas plantas, a produção
de um composto volátil, o metil-salicilato (MeSA) que, por sua vez, pode induzir
plantas próximas a sintetizar o AS, podendo torna-lás sadias, resistentes, induzindo a
produção de proteínas-PR e a ativação de genes de resistência (SHULAEV et al., 1997).
Desta forma, nesse ensaio pode ter ocorrido a conversão para o salicilato de metila que
normalmente repele os herbívoros, interferindo na evolução do artropodo-praga.
Segundo Ulland et al. (2008), o salicilato de metila inibe a oviposição da traça do
repolho (Plutella xylostella). Na literatura consta que mais de 40 variedades de insetos,
apresentam receptores olfativos para salicilato de metila (CHAMBERLAIN et al., 2000;
PICKETT et al., 2003). Se esse composto volátil foi produzido pelos morangueiros e
com os tratamentos dispostos e juntos, na mesma casa de vegetação pode ter
influenciado no comportamento biológico e de resistência da H.armigera.
5.4. Identificação e quantificação de tricomas foliares
Em relação aos tricomas foliares, os resultados indicam a existência de dois tipos
em ambas as cultivares de morangueiro analisadas. O primeiro tipo de tricoma
identificado foi o tipo I, com tamanho maior e unicelular, não glandular (Figura 5B). O
outro tipo de tricoma refere-se ao glandular, sendo pequeno, segmentado, com tamanho
variado e sempre com a glândula localizada no final da estrutura (Figura 5A).
Figueiredo et. al (2010), ao analisar dez cultivares de morangueiro, identificaram os
dois tipos de tricomas presentes na superfície foliar de todas as cultivares, conforme
obtido na presente pesquisa..

32
Quanto à densidade de tricomas foliares, verificou-se que para os glandulares,
não houve diferença estatística entre as cultivares (Tabela 2). No entanto, para os não
glandulares ou tectores, observou-se maior densidade na cultivar Aromas. As diferenças
observadas entre as cultivares, pode ocorrer em decorrência das características genéticas
inerentes a cada cultivar, ou mesmo em resposta a interação com as doses aplicadas do
AS ou ainda por ação de fatores abióticos.
Tabela 2. Densidade de tricomas glandulares e não glandulares nas cultivares de
morangueiro, Aromas e Sweet Charlie, UNICENTRO, Guarapuava, PR, 2016.
Medias seguidas da mesma letra na coluna não diferem estatisticamente pelo teste de
tukey a 95 (p<0,05) de probabilidade.
Figura 5. Representação dos tipos de tricomas A) Glandulares e B) Não Glandulares,
em cultivares de morangueiro, Guarapuava-PR, UNICENTRO, 2016.
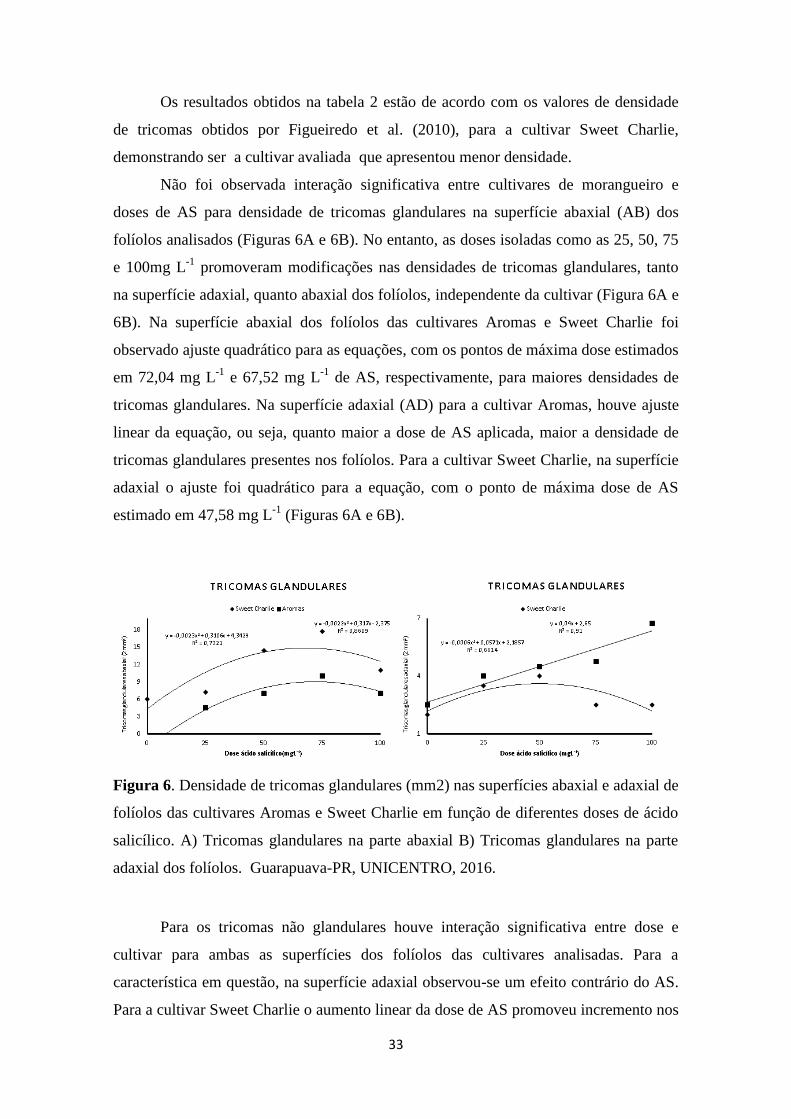
33
Os resultados obtidos na tabela 2 estão de acordo com os valores de densidade
de tricomas obtidos por Figueiredo et al. (2010), para a cultivar Sweet Charlie,
demonstrando ser a cultivar avaliada que apresentou menor densidade.
Não foi observada interação significativa entre cultivares de morangueiro e
doses de AS para densidade de tricomas glandulares na superfície abaxial (AB) dos
folíolos analisados (Figuras 6A e 6B). No entanto, as doses isoladas como as 25, 50, 75
e 100mg L-1
promoveram modificações nas densidades de tricomas glandulares, tanto
na superfície adaxial, quanto abaxial dos folíolos, independente da cultivar (Figura 6A e
6B). Na superfície abaxial dos folíolos das cultivares Aromas e Sweet Charlie foi
observado ajuste quadrático para as equações, com os pontos de máxima dose estimados
em 72,04 mg L-1
e 67,52 mg L-1
de AS, respectivamente, para maiores densidades de
tricomas glandulares. Na superfície adaxial (AD) para a cultivar Aromas, houve ajuste
linear da equação, ou seja, quanto maior a dose de AS aplicada, maior a densidade de
tricomas glandulares presentes nos folíolos. Para a cultivar Sweet Charlie, na superfície
adaxial o ajuste foi quadrático para a equação, com o ponto de máxima dose de AS
estimado em 47,58 mg L-1
(Figuras 6A e 6B).
Para os tricomas não glandulares houve interação significativa entre dose e
cultivar para ambas as superfícies dos folíolos das cultivares analisadas. Para a
característica em questão, na superfície adaxial observou-se um efeito contrário do AS.
Para a cultivar Sweet Charlie o aumento linear da dose de AS promoveu incremento nos
Figura 6. Densidade de tricomas glandulares (mm2) nas superfícies abaxial e adaxial de
folíolos das cultivares Aromas e Sweet Charlie em função de diferentes doses de ácido
salicílico. A) Tricomas glandulares na parte abaxial B) Tricomas glandulares na parte
adaxial dos folíolos. Guarapuava-PR, UNICENTRO, 2016.
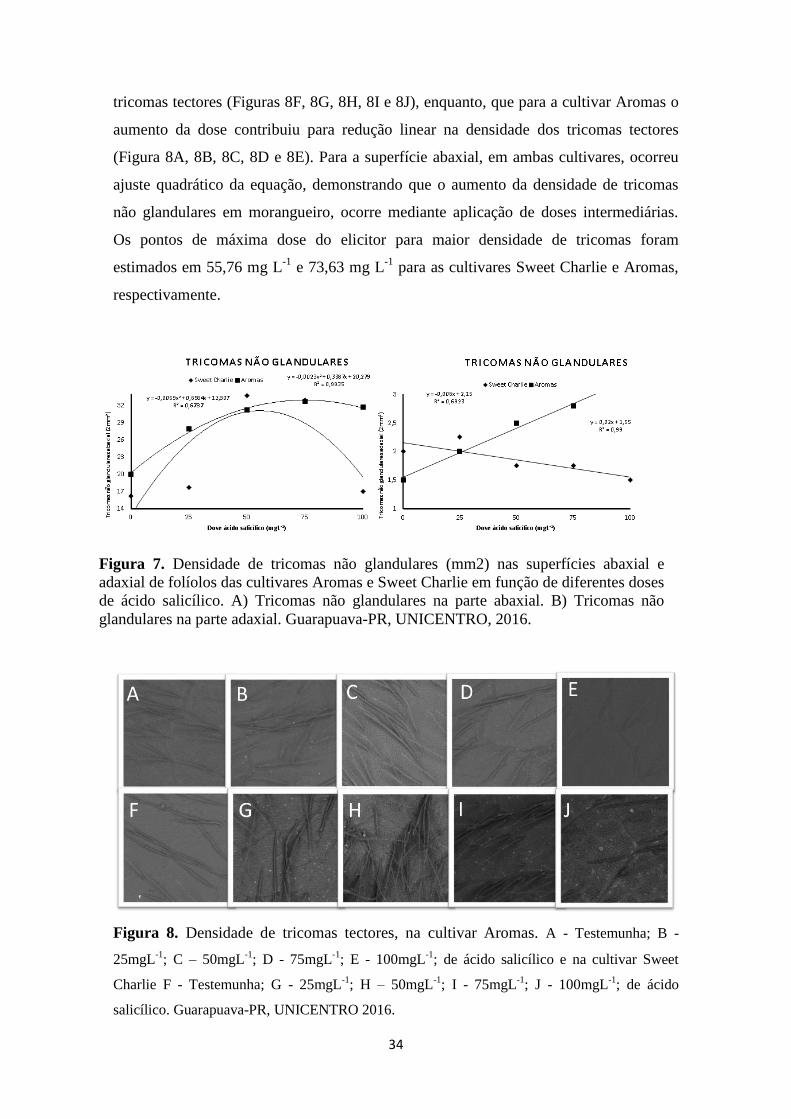
34
tricomas tectores (Figuras 8F, 8G, 8H, 8I e 8J), enquanto, que para a cultivar Aromas o
aumento da dose contribuiu para redução linear na densidade dos tricomas tectores
(Figura 8A, 8B, 8C, 8D e 8E). Para a superfície abaxial, em ambas cultivares, ocorreu
ajuste quadrático da equação, demonstrando que o aumento da densidade de tricomas
não glandulares em morangueiro, ocorre mediante aplicação de doses intermediárias.
Os pontos de máxima dose do elicitor para maior densidade de tricomas foram
estimados em 55,76 mg L-1
e 73,63 mg L-1
para as cultivares Sweet Charlie e Aromas,
respectivamente.
Figura 8. Densidade de tricomas tectores, na cultivar Aromas. A - Testemunha; B -
25mgL-1
; C – 50mgL-1
; D - 75mgL-1
; E - 100mgL-1
; de ácido salicílico e na cultivar Sweet
Charlie F - Testemunha; G - 25mgL-1
; H – 50mgL-1
; I - 75mgL-1
; J - 100mgL-1
; de ácido
salicílico. Guarapuava-PR, UNICENTRO 2016.
Figura 7. Densidade de tricomas não glandulares (mm2) nas superfícies abaxial e
adaxial de folíolos das cultivares Aromas e Sweet Charlie em função de diferentes doses
de ácido salicílico. A) Tricomas não glandulares na parte abaxial. B) Tricomas não
glandulares na parte adaxial. Guarapuava-PR, UNICENTRO, 2016.

35
Esses resultados evidenciam haver respostas diferentes das cultivares para
desenvolvimento de estruturas morfológicas de caráter físico e químico para defesa. A
maior densidade de tricomas tectores na cultivar Aromas está provavelmente,
relacionada a fatores genéticos inerentes a mesma. É salutar mencionar que houve
efeito do elicitor AS promovendo o aumento de tricomas glandulares e não glandulares,
independente da superfície e da cultivar analisada, exceto para a cultivar Sweet Charlie,
na superfície adaxial, onde o AS promoveu a redução dos tricomas tectores.
A maior relevância está no efeito do AS em proporcionar alterações
morfológicas e químicas na estrutura foliar, aumentando a densidade de tricomas
glandulares e não glandulares, responsáveis pela defesa da planta ao ataque de
artrópodos-praga.
Correlações foram estimadas para densidade de tricomas glandulares, não
glandulares e totais, de ambas as superfícies do folíolo, com o número de ovos
depositados nas duas superfícies dos folíolos de morangueiro e na sobrevivência de
ácaros adultos, nas cultivares Aromas e Sweet Charlie. Valores altos, negativos e
significativos foram obtidos ao correlacionar número de ovos depositados em todo o
folíolo com a densidade de tricomas glandulares (-0.88 e -0.70), tectores (-0.94 e -0.87)
e tricomas totais (-0.97 e -0.92), para as cultivares Sweet Charlie e Aromas,
respectivamente.
Para a sobrevivência dos ácaros na superfície dos discos foliares, foram
estimadas correlações com valores mais baixos, porém significativas e negativas. Para
tricomas glandulares foram observados valores de -0.66 e -0.70, não glandulares de -
0.72 e -0.82 e tricomas totais de -0.74 e -0.86, respectivamente para as cultivares Sweet
Charlie e Aromas. Esses valores de correlação estimados indicam que a oviposição e
sobrevivência dos ácaros foram afetadas pela presença dos tricomas, glandulares e não
glandulares na superfície dos folíolos das cultivares de morangueiro.
Na interação inseto-planta, o crescimento das populações de pragas é inibido por
características bioquímicas e morfológicas da planta hospedeira, afetando o desempenho
ou metabolismo do artrópodo-praga, reduzindo o nível de dano causado (LARA, 1991).
No presente trabalho observou-se que com a aplicação do elecitor AS houve aumento
no número de tricomas em relação às doses. Acredita-se que o efeito hormonal do AS
ativa genes responsáveis por rotas metabólicas alternativas que induzem a síntese de
substâncias que atuam como defesa química sendo repelente ao inseto e outros

36
organismos, formando assim uma combinação de defesa da planta, tanto estrutural
quanto química.
Sengundo War et al. (2012), os metabólitos secundários são os compostos que
não afetam o crescimento normal e o desenvolvimento da planta, mas reduzem a
palatabilidade dos tecidos (metabólitos secundários) e podem ser armazenados como
formas inativas ou induzida, em constitutiva resposta ao ataque de invasores. São
conhecidos como fitoalexinas. As fitoalexinas incluem isoflavonas, terpenóides,
alcalóides, etc, que influenciam o desempenho e sobrevivência dos herbívoros.
Os metabólitos secundários não apenas defendem as plantas de diferentes
tensões, mas também aumentam a aptidão das plantas. Assim constatou-se que o AS
provavelmente formam metabólitos secundários como as fitoalexinas e, com isso
repelem as pragas. Formam barreiras específicas que defendem as plantas do ataque, a
exemplo a lignificação da parede celular, aumento da espessura da epiderme,
desenvolvimento de estruturas físicas como os tricomas tectores e glandulares.
Alguns trabalhos têm demonstrado que a aplicação exógena de indutores
responsáveis por produção de proteinases (PIs) e enzimas de síntese de fitoalexinas,
aumentam a defesa estrutural, por meio da presença de pelos, espinhos, tricomas e ceras
na superfície de caules e frutos, bem como a biossíntese de flavanóides, terpenos e
compostos fenólicos (ROHWER e ERWIN, 2008; DI PIERO et al., 2005; JUNIOR,
2009; COUTO, 2006), que podem ser exsudatos pelos tricomas glandulares.
Os mecanismos estruturais atuam como barreiras físicas, promovendo o atraso
na penetração e/ou colonização pelo agressor; cutícula, estômatos, tricomas e vasos
condutores são exemplos de defesa estrutural pré-formadas e, papilas, halos, tiloses e
lignificação são pós-formados. Os mecanismos bioquímicos, por sua vez, atuam
mediante produção de substâncias tóxicas (fitoantecipinas, catecol, saponinas, espécies
reativas de oxigênio e fitoalexinas), inibindo a ação do colonizador ou criando
condições adversas ao estabelecimento do mesmo na planta (SCHWAN-ESTRADA et
al., 2008; PASCHOLATI e LEITE, 1995).
Nesse contexto, observa-se que a aplicação exógena do indutor de resistência,
propiciou o maior desenvolvimento de barreiras físicas (tricomas tectores) e físico
químicas (tricomas glandulares) na superfície dos folíolos das cultivares de
morangueiro, Aromas e Sweet Charlie.
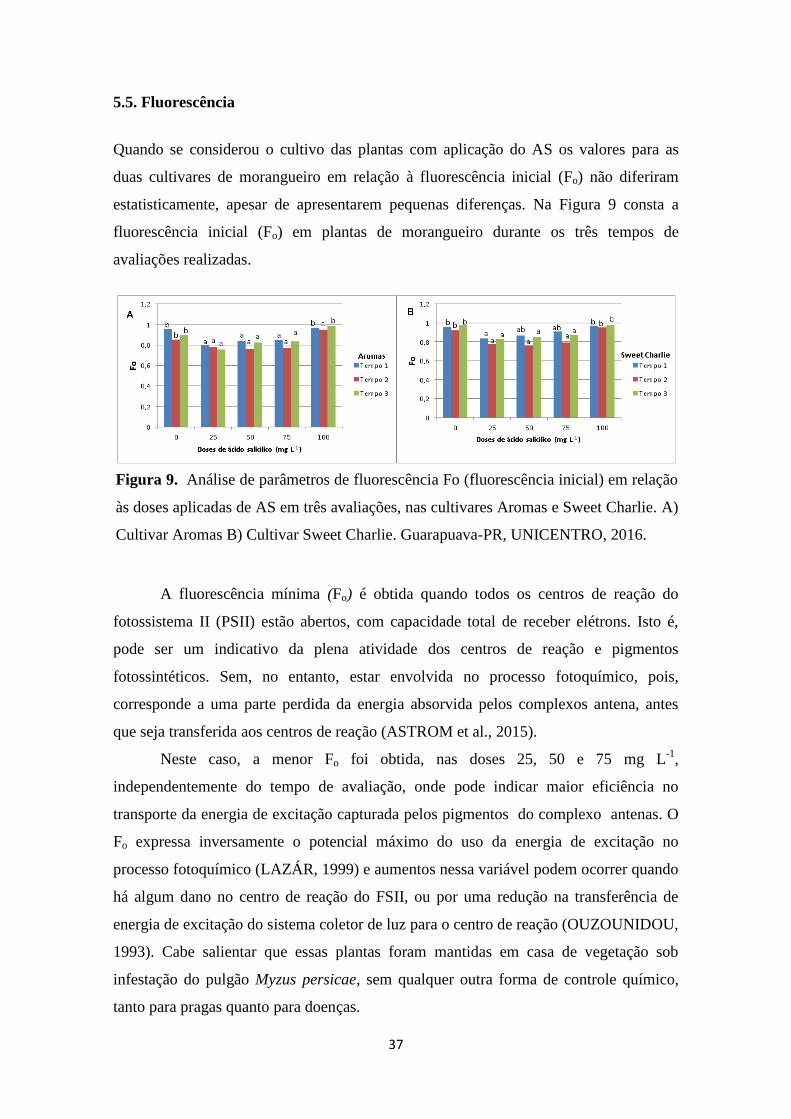
37
5.5. Fluorescência
Quando se considerou o cultivo das plantas com aplicação do AS os valores para as
duas cultivares de morangueiro em relação à fluorescência inicial (Fo) não diferiram
estatisticamente, apesar de apresentarem pequenas diferenças. Na Figura 9 consta a
fluorescência inicial (Fo) em plantas de morangueiro durante os três tempos de
avaliações realizadas.
A fluorescência mínima (Fo) é obtida quando todos os centros de reação do
fotossistema II (PSII) estão abertos, com capacidade total de receber elétrons. Isto é,
pode ser um indicativo da plena atividade dos centros de reação e pigmentos
fotossintéticos. Sem, no entanto, estar envolvida no processo fotoquímico, pois,
corresponde a uma parte perdida da energia absorvida pelos complexos antena, antes
que seja transferida aos centros de reação (ASTROM et al., 2015).
Neste caso, a menor Fo foi obtida, nas doses 25, 50 e 75 mg L-1
,
independentemente do tempo de avaliação, onde pode indicar maior eficiência no
transporte da energia de excitação capturada pelos pigmentos do complexo antenas. O
Fo expressa inversamente o potencial máximo do uso da energia de excitação no
processo fotoquímico (LAZÁR, 1999) e aumentos nessa variável podem ocorrer quando
há algum dano no centro de reação do FSII, ou por uma redução na transferência de
energia de excitação do sistema coletor de luz para o centro de reação (OUZOUNIDOU,
1993). Cabe salientar que essas plantas foram mantidas em casa de vegetação sob
infestação do pulgão Myzus persicae, sem qualquer outra forma de controle químico,
tanto para pragas quanto para doenças.
Figura 9. Análise de parâmetros de fluorescência Fo (fluorescência inicial) em relação
às doses aplicadas de AS em três avaliações, nas cultivares Aromas e Sweet Charlie. A)
Cultivar Aromas B) Cultivar Sweet Charlie. Guarapuava-PR, UNICENTRO, 2016.
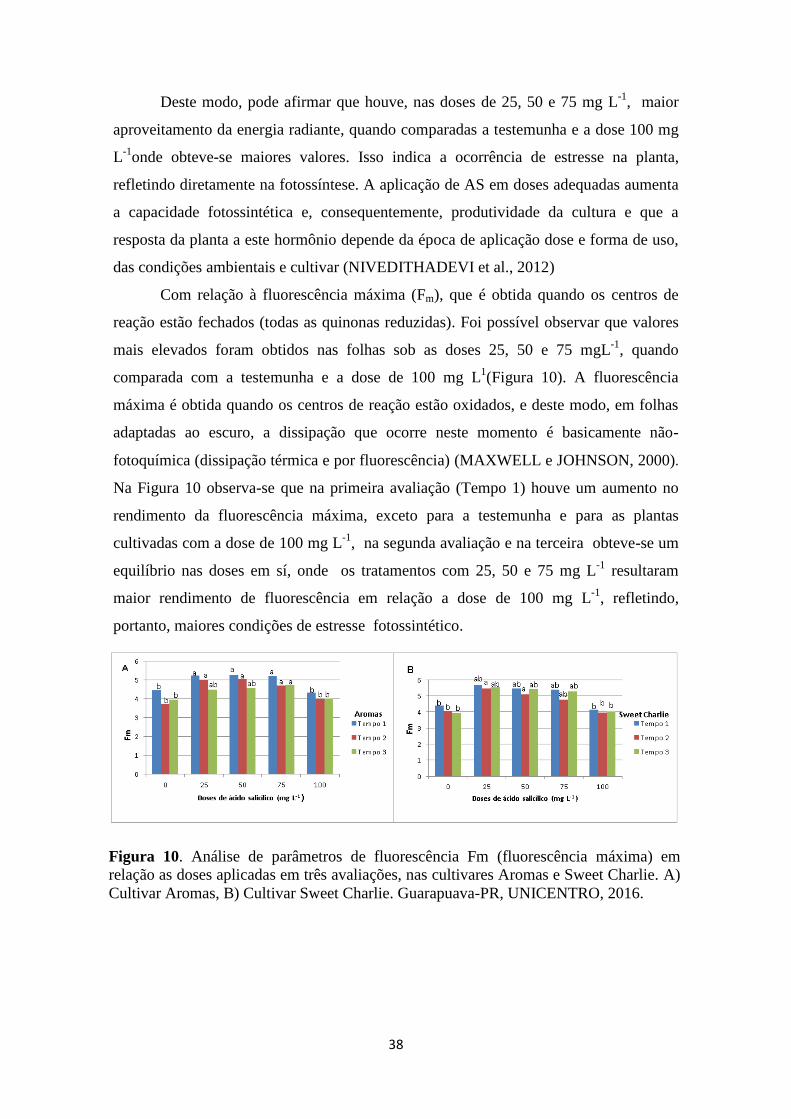
38
Deste modo, pode afirmar que houve, nas doses de 25, 50 e 75 mg L-1
, maior
aproveitamento da energia radiante, quando comparadas a testemunha e a dose 100 mg
L-1
onde obteve-se maiores valores. Isso indica a ocorrência de estresse na planta,
refletindo diretamente na fotossíntese. A aplicação de AS em doses adequadas aumenta
a capacidade fotossintética e, consequentemente, produtividade da cultura e que a
resposta da planta a este hormônio depende da época de aplicação dose e forma de uso,
das condições ambientais e cultivar (NIVEDITHADEVI et al., 2012)
Com relação à fluorescência máxima (Fm), que é obtida quando os centros de
reação estão fechados (todas as quinonas reduzidas). Foi possível observar que valores
mais elevados foram obtidos nas folhas sob as doses 25, 50 e 75 mgL-1
, quando
comparada com a testemunha e a dose de 100 mg L1(Figura 10). A fluorescência
máxima é obtida quando os centros de reação estão oxidados, e deste modo, em folhas
adaptadas ao escuro, a dissipação que ocorre neste momento é basicamente não-
fotoquímica (dissipação térmica e por fluorescência) (MAXWELL e JOHNSON, 2000).
Na Figura 10 observa-se que na primeira avaliação (Tempo 1) houve um aumento no
rendimento da fluorescência máxima, exceto para a testemunha e para as plantas
cultivadas com a dose de 100 mg L-1
, na segunda avaliação e na terceira obteve-se um
equilíbrio nas doses em sí, onde os tratamentos com 25, 50 e 75 mg L-1
resultaram
maior rendimento de fluorescência em relação a dose de 100 mg L-1
, refletindo,
portanto, maiores condições de estresse fotossintético.
Figura 10. Análise de parâmetros de fluorescência Fm (fluorescência máxima) em
relação as doses aplicadas em três avaliações, nas cultivares Aromas e Sweet Charlie. A)
Cultivar Aromas, B) Cultivar Sweet Charlie. Guarapuava-PR, UNICENTRO, 2016.

39
Quanto maior é a Fm maior a capacidade da planta em transferir a energia dos
elétrons ejetados das moléculas dos pigmentos para a formação do poder redutor
NADPH, ATP e Fd e, consequentemente, maior a capacidade de assimilação do CO2 na
fase bioquímica da fotossíntese (ROHÁČEK, 2002; BAKER, 2008). Estudos realizados
com a cultura do girassol (Helianthus annuus L) (EL-TAYEB et al., 2006) revelaram
que o uso do AS aumenta substancialmente os teores de clorofila a, b, e os teores de
carotenoides, além das proteínas solúveis nas raízes e folhas ao aplicarem a dose de 0,5
mM de AS. Por outro lado, a aplicação da dose de 100 mg L-1
de AS (DAWOOD et al.,
2012) promoveu drástica redução nos teores de carboidratos solúveis, sugerindo a
necessidade de estudos mais aprofundados no tocante a definição de doses, fontes,
formas de aplicação, sazonalidade e idade fisiológica da planta para se obter um
resultado mais consistente. O que corrobora com os resultados deste presente trabalho,
em relação à dose de 100 mg L-1
, que está demostrando baixa eficiência na fotossíntese,
em comparação como a testemunha.
Segundo Iqpal et al. (2006) e Shehata et al. (2001), o AS ocorre naturalmente na
plantas em quantidades muito baixas e participa na regulação de processos fisiológicos
dos vegetais, tais como abertura e fechamento estomático, absorção de nutrientes,
síntese de clorofila, síntese proteica. Sendo assim, a importância da aplicação exógena
desse regulador vegetal torna-se clara, contribuindo ainda mais para esses processos
fisiológicos que afetam positivamente o desenvolvimento da planta.
A Figura 11 representa os parâmetros de atividade potencial do PSII (Fv/Fo) que
tem sido recomendada para detectar mudanças induzidas pelos estresses
(LICHTENTHALER et al., 2005). O que pode-se observar na Figura 11 é que houve
uma maior atividade do PSII refletindo condições de estresses fotossintéticos e os
valores mais altos foram encontrados na maior dose (100 mg L-1
) podendo explicar que
o PSII teve que gerar mais energia, sendo assim, ocorrendo efeito de estresse mais
proeminente.

40
O AS é um regulador vegetal que participa de vários processos nas plantas
(PULGA, 2016). Neste sentido, Nivedithadevi et al. (2012), estudou os efeitos de
diferentes substâncias sobre os aspectos fisiológicos de plantas de Thuthuvalai
(Solanum trilobatum L.) (500 µg L-1
de AS, 10 mg L-1
de Paclobutrazol e 10 µg L-1 de
ácido abcísico), e constatou que o AS aumenta os teores de clorofila a, b, total e
carotenoides em plantas de Thuthuvalai, aumentando e fortalecendo a proteção das
plantas contra alguns estresses, como os provocados pelo excesso de radiação solar e
temperatura.
A análise da fluorescência da clorofila a é capaz de detectar com segurança e
confiabilidade efeitos de estresse e injúrias no processo fotossintético. As mudanças na
cinética de emissão de fluorescência a partir de organismos autotróficos são resultados
de frequentes modificações na atividade fotossintética, principalmente com relação à
eficiência quântica do transporte de elétrons através do FSII em folhas (BAKER e
ROSENQVIST, 2004; YUSUF et al., 2010).
Analisando o rendimento quântico fotoquímico efetivo do PSII Y(II) nas
cultivares (Aromas e Sweet Charlie) observa-se diferenças em relação à dose e cultivar.
(Figura 12).
Figura 11. Análise de parâmetros de fluorescência Fv/Fo (atividade potencial do PSII)
em relação as doses aplicadas em três avaliações, nas cultivares Aromas e Sweet
Charlie. A) Cultivar Aromas, B) Cultivar Sweet Charlie. Guarapuava-PR,
UNICENTRO, 2016.

41
O maior rendimento quântico fotoquímico FSII (YII) foi obtido nas doses
intermediárias para as ambas as cultivares. Fica evidente que altas doses de AS podem
causar estresse na planta, pois o rendimento quântico fotoquímico do PSII (Y(II) ou
PSII) mede a proporção de luz absorvida pelas moléculas de clorofila associadas ao
PSII que é usada fotoquimicamente, além de estimar a taxa de transporte linear de
elétrons, e, em condições de laboratório, estar fortemente relacionado à fixação de
carbono (MAXWELL e JOHNSON, 2000).
O AS exerce função central nos processos fisiológicos dos vegetais e nas
respostas de defesa contra fatores de estresse bióticos e abióticos (SHI e ZHU, 2008).
Essa resposta está diretamente relacionada a mecanismos que regulam a expressão de
genes, que controlam a síntese de proteínas e enzimas que são protagonistas na
produção de metabólitos secundários, responsáveis pelas defesas das plantas (DHONDT
et al., 2002).
Na presente pesquisa o AS potencializou as respostas de defesa do morangueiro,
para os ensaios testados com artrópodos-pragas e influenciou na fotossíntese em relação
às doses aplicadas, aumentando a interação entre as doses e os materiais testados. Além
disso, supõem-se que houve a conversão do ácido salicílico em salicilato de metila,
composto volátil, que induz resistência (RASKIN et al., 2000), o que pode ter alterado
os parâmetros avaliados de fluorescência para as doses.
Com relação ao coeficiente de dissipação de fluorescência fotoquímica (qP), este
é utilizado para estimar a fração de centros de reações (CR) do FSII abertos, cuja
estimativa não requer adaptação da folha ao escuro (KRAMER et al., 2004). Houve
Figura 12. Análise de parâmetros de fluorescência Y(II) (rendimento quântico
fotoquímico efetivo do PSII, conversão fotoquímica) em relação às doses aplicadas, nas
cultivares Aromas e Sweet Charlie. A) Cultivar Aromas B) Cultivar Sweet Charlie. .
Guarapuava-PR, UNICENTRO, 2016.
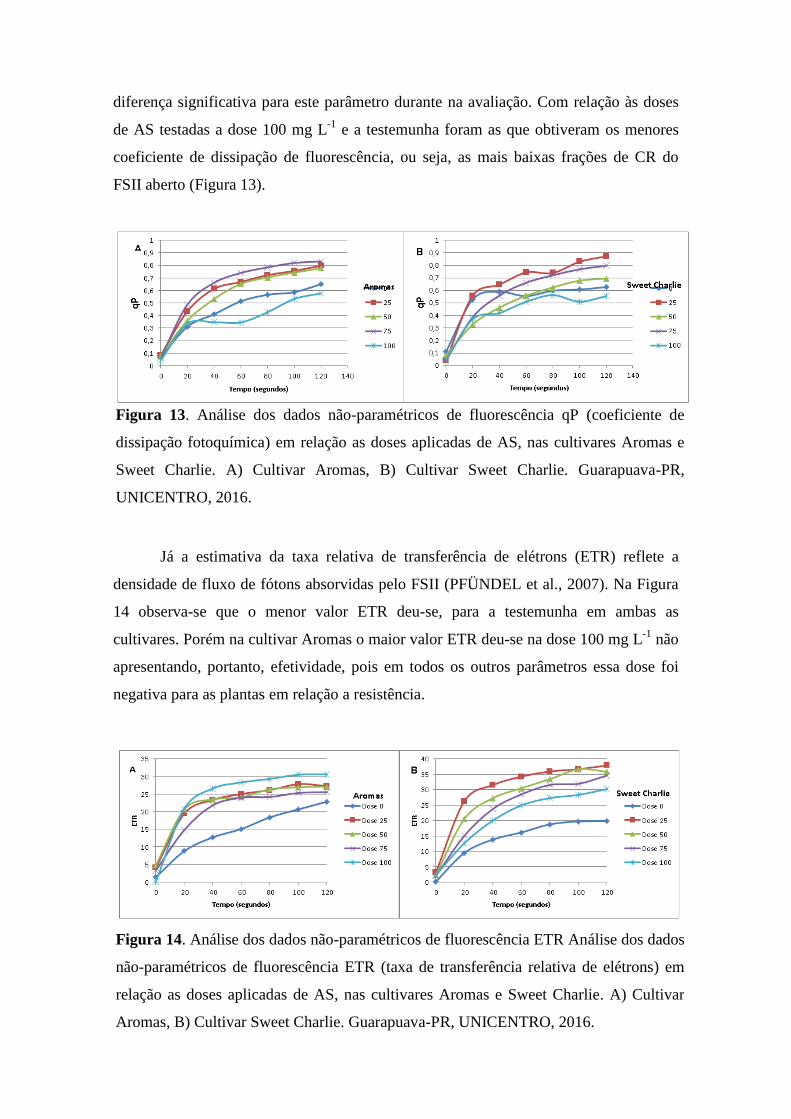
42
diferença significativa para este parâmetro durante na avaliação. Com relação às doses
de AS testadas a dose 100 mg L-1
e a testemunha foram as que obtiveram os menores
coeficiente de dissipação de fluorescência, ou seja, as mais baixas frações de CR do
FSII aberto (Figura 13).
Já a estimativa da taxa relativa de transferência de elétrons (ETR) reflete a
densidade de fluxo de fótons absorvidas pelo FSII (PFÜNDEL et al., 2007). Na Figura
14 observa-se que o menor valor ETR deu-se, para a testemunha em ambas as
cultivares. Porém na cultivar Aromas o maior valor ETR deu-se na dose 100 mg L-1
não
apresentando, portanto, efetividade, pois em todos os outros parâmetros essa dose foi
negativa para as plantas em relação a resistência.
Figura 13. Análise dos dados não-paramétricos de fluorescência qP (coeficiente de
dissipação fotoquímica) em relação as doses aplicadas de AS, nas cultivares Aromas e
Sweet Charlie. A) Cultivar Aromas, B) Cultivar Sweet Charlie. Guarapuava-PR,
UNICENTRO, 2016.
Figura 14. Análise dos dados não-paramétricos de fluorescência ETR Análise dos dados
não-paramétricos de fluorescência ETR (taxa de transferência relativa de elétrons) em
relação as doses aplicadas de AS, nas cultivares Aromas e Sweet Charlie. A) Cultivar
Aromas, B) Cultivar Sweet Charlie. Guarapuava-PR, UNICENTRO, 2016.

43
As doses de AS 25 e 50 mg L-1
proporcionaram valores médios na avaliação
para a taxa de transferência relativa de elétrons (ETR) causando possível indução de
resistência e maior eficiência na fotossíntese.
A aplicação exógena de ácidos orgânicos, como o salicílico, cítrico e ascórbico,
é utilizada como forma de combate ao estresse, sendo que destes o primeiro pode agir
como indutor de proteínas de tolerância ao estresse (McCUE et al., 2000 e BOWLER e
FLUHR, 2000), e neste contexto o trabalho nos mostra maior eficiência para a
fotossíntese em relação as doses aplicadas, pois o AS ativa os metabolitos secundários e
com isso traz defesa nas plantas e diminui o estresse causado.
Segundo Pulga (2016), a aplicação de AS em tomateiro em relação a ETR, na
dose 100 mg L-1
apresentou o maior valor, também não apresentando efetividade.
Ocorrendo que nas doses 25 e 50 mg L-1
proporcionaram valores médios para a taxa de
transferência de elétrons, estando de acordo com o presente trabalho.

44
6. CONCLUSÕES
O AS foi eficiente como indutor de resistência ao ácaro rajado e a Helicoverpa
armigera, reduzindo a sobrevivência de adultos e a oviposição nos folíolos das
cultivares de morangueiro, Aromas e Sweet Charlie, caracterizando assim os
mecanismos de resistência do tipo antibiose e antixenose.
O AS aplicado de forma exógena promoveu maior densidade de tricomas
glandulares e não glandulares nas folhas de morangueiro.
AS Doses de AS entre 41,75 e 56,75 mg L-1
foram as mais efetivas em induzir
resistência do morangueiro ao ácaro rajado.
O AS aplicado nas doses 25, 50, 75 mg L-1
resultam em eficiência e melhores
condições do aparato fotossintético, mais especificamente do fotossistema II.

45
7. REFERÊNCIAS
ABERG, B. Plant growth regulators. XLI. Monosubstituted benzoic acids. Swedish
Journal of Agricultural Research, n. 11, p. 93-105, 1981.
ALI, A.; CHOUDHURY, R. A. Some biological characteristics of Helicoverpa
armigera on chickpea. Tunisian Journal of Plant Protection, v. 4, n. 1, p. 99-106,
2009.
ALMEIDA-CORTEZ,J.S Herbivoria e mecanismos de defesa vegetal. In: NOGUEIRA,
R.J. M.C.; ARAÚJO, E.L.; WILLADINO, L.G.; CALVALCANTE, U.M.T (Eds.).
Estresses ambientais: danos e benefícios em plantas. Recife: Imprensa Universitária,
2005. P. 389-396.
ALONSO-RAMIREZ, A.; RODRIGUEZ, D.; REYES, D.; JIMENEZ, J.A.; NICOLAS,
G.; LOPEZ-CLIMENT, M.; GOMEZ-CADENAS, A.; NICOLAS, C. Crosstalk
between gibberellins and salicylic acid in early stress responses in Arabidopsis thaliana
seeds. Plant Signaling e Behavior, 750-751. 2009.
ANTONIOLLI, L.R.; MELO, G.W. de; ANTUNES, L.E.C.; BOTTON, M.; SATO,
M.E.; FERLA, N.J.; SOUZA, R.T. de; SANHUEZA, R.M.V. Boas Práticas na
Cultura do Morangueiro (Série Agronegócios). Porto Alegre, SEBRAE, 32p., 2007.
ANTUNES, L.E.C.; HOFFMANN, A. Pequenas frutas: o produtor pergunta, a
Embrapa responde. 500 perguntas, 500 respostas. Brasília: Embrapa. 2012.
ANTUNES, L.E.C.; REISSER JUNIOR, R.C. Panorama do cultivo de morango no
Brasil. Brasília: Embrapa 2015.
ANTUNES, L.E.C.; REISSER JUNIOR, R.C. Produção de morangos. Jornal da Fruta,
v.191, p.22-24, 2007.

46
ARNÓ, J.; GABARRA, R.; ROIG, J.; FOSCH, T. Integrated pest management for
processing tomatoes in the Ebro Delta (Spain). Acta Horticulturae, The Hague, v. 487,
n. 1, p. 207-212, 1999.
ÅSTRÖM, H.; METSOVUORI, E.; SAARINEN, T.; LUNDELL, R.; HÄNNINEN, H.
Morphological characteristics and photosynthetic capacity of Fragaria vesca L. winter
and summer leaves. Flora, v. 215, p. 33–39, 2015.
ASGHARI, M.; AGHDAM, M.S. Impact of salicylic acid on post-harvest physiology of
horticultural crops. Trends in Food Science e Technology. v. 21, p. 502 – 509, 2010.
AZOOZ, M.M.; YOUSSEF, M.M. Evaluation of heat shock and salycilic acid
treatments as inducers of drought stress tolerance in hassawi wheat. American Journal
of Plant Physiology, v. 5, n. 2, p. 56-70, 2010.
BALBI, V.; DEVOTO, A. Jasmonate signalling network in Arabidopsis thaliana:
crucial regulatory nodes and new physiological scenarios. New Phytologist, London, v.
177, p. 301–318, 2008.
BARBOSA, J.G. Crisântemos: produção de mudas, cultivo para corte de flor, cultivo
em vaso, cultivo hidropônico. Viçosa: Aprenda Fácil Editora. 2003. 232p.
BAKER, N.R.; ROSENQVIST, E. Applications of chlorophyll fluorescence can
improve crop production strategies: an examination of future possibilities, Journal of
Experimental Botany, 55, p. 1607-1621, 2004.
BAKER, B. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annual
Review of Plant Biology, Boca Raton, v.59, p.89-113, 2008.
BRAGA, F.T.; NUNES, C.F.; FAVERO, A.C.; PASQUAL, M.; CARVALHO, J.G.;
CASTRO, E.M. Características anatômicas de mudas de morangueiro micropropagadas
com Diferentes fontes de silício. Pesq. agropec. bras., Brasília, v.44, n.2, p.128-132,
2009.

47
BARTON, L. B.; RAUBENHEIMER, D. Ontogenetic changes in the rate of ingestion
and estimates of food consumption in fourth and fifth instar Helicoverpa armigera
caterpillars. Journal of Insect Physiology, Oxford, v. 49, n. 1, p. 63-71, 2003.
BÉLANGER, R.R.; MENZIES, J.G. Use of silicon to control diseases in vegetable
crops. In: XXXVI CONGRESSO BRASILEIRO DE FITOPATOLOGIA. Anais da
Sociedade Brasileira de Fitopatologia, v. 36, p. S42-S45, 2003.
BROEKAERT, W.F.; DELAURE, S.L.; DE BOLLE, M.F.C.; CAMMUEL, B.P.A.
2006. The role of ethylene in host-pathogen interactions. Annual Review
Phytopathology, Palo Alto, v. 44, p. 393-416, 2006.
BONALDO, S.M.; PASCHOLATI, S.F.; ROMEIRO, R.S. Indução de resistência:
noções básicas e perpectivas. In: Indução de resistência em plantas a patógenos e
insetos, p. 1128, 2005.
BOWLER, C.; FLUHR, R. The role of calcium and activated oxygens as signals for
controlling cross-tolerance. Trends Plant Science, v.5, p.241-6, 2000.
CARVALHO, S. P. Boletim do morango: cultivo convencional, segurança alimentar,
cultivo orgânico. Belo Horizonte: FAEMG, 2006. 160p.
CAMARGO, L. E. A.; BERGAMIN FILHO, A. Controle genético. In: BERGAMIN
FILHO, A.; KIMATI, H.; AMORIM, L. (Eds). Manual de fitopatologia: princípios e
conceitos, 3 ed. São Paulo, Agronômica Ceres, p. 729-758.1995.
CAMARGO, Y. R., Lima, L., Scalon, S., e SIQUEIRA, A. (2000). Efeito do cálcio
sobre o amadurecimento de morangos (Fragaria ananassa Duch.) CV.
Campineiro. Ciência e Agrotecnologia, 24(4), 968-972.

48
CARVALHO, S.P.; ZAWADNEAK, M.A.C.; ANDRADE, P.F.S.; ZANDONÁ, J.C. O
cultivo do morangueiro no Brasil. In: ZAWADNEAK, M.A.C.; SCHUBER, J.M.;
MÓGOR, A.F. Como produzir morangos. Curitiba: UFPR, 2014, p.15-68.
CAMPOS, A.D. Considerações sobre indução de resistências a patógenos em
plantas, p. 2, 2009.
CAMPOSTRINE, E. Fluorescência da clorofila a: considerações téoricas e 11
aplicações práticas. Universidade Estadual do Norte Fluminense, 2001.
CARVALHO, P.R.; MACHADO NETO, N.B.; CUSTÓDIO, C.C. Ácido salicílico em
sementes de calêndula (Calendula officinalis L.) sob diferentes estresses. Revista
Brasileira de Sementes, v. 29, n. 1, p. 114-124, 2007.
CAVALCANTI, F.R.; PEREIRA, V.F.; RIBEIRO JÚNIOR, P.M. Aspectos da
indução de resistência em plantas de videira, Embrapa Uva e Vinho, p. 20, 2014.
CHAMBERLAIN, K,; PICKETT, A.J.; WOODCOCK, M.C. Plant signalling and
induced defense in insect attack. Molecular Plant Pathology, n 1, v. 1, p. 67-72, 2000.
CHAPMAN, R.B.; MARTIN, N.A. Spider mite resistance management strategy. In:
MARTIN, N.A.; BERESFORD, R.M.; HARRINGTON, K.C. (ed.) Pesticide
Resistance: prevention & management strategies 2005. Hasting: New Zealand Plant
Protection Society. 2005. Disponível em: <
http://www.nzpps.org/resistance/pdfs/spidermite.pdf>. Acesso em 13 mar. 2008.
COHEN A.C.; HENNEBERRY T.J.; CHU C.C. Geometric relationships between
whitefly feeding behavior and vascular bundle arrangements. Emtomologia
Experimentalis et Applicata, Oxford, v.78, n. 2, p. 135-142, 1996.
CHIAVEGATO, L.G.; MISCHAN, M.M. Efeito do Tetranychus (T.) urticae (Koch,
1836) Boudreaux e Dosse, 1963 (Acari, Tetranychidae) na produção do morangueiro
(Fragaria sp) cv. ´Campinas´. Científica, v.9, n.2, p.257-266, 1981.

49
CINGOLANI, A.M.; POSSE, G.; COLLANTES, M.B. Plant functional traits, herbivore
selectivity and response to sheep grazing in Patagonian steppe grasslands. Journal of
applied Ecology, London, v. 42, n. 1, p. 50-59, 2005.
COLLI, S. Outros reguladores: Brassionosteróides, poliaminas, ácidos jasmônico e
salicílico. In: KERBAUY, G. B. Fisiologia vegetal. 2.ed. Guanabara Koogan, p. 296-
302, 2008.
COSTA, A.F.; ROSSI, D.A.; LEAL, N.R. Origem, Evolução e o Melhoramento do
Morangueriro. In: ZAWADNEAK, M. A.C.; SCHUBER, J. M.; MÓGOR, A.F. Como
produzir morangos. Curitiba: UFPR, 2014. p.15-68.
CORNELISSEN, T.G.; FERNANDES, G W. Insetos herbívoros. Ciência Hoje, v. 32,
n. 192, p. 24-30, 2013.
COUTO, E.B. Avaliação de metil jasmonato na indução de resistência de plantas de
Eucalyptus spp. ao psilídeo-deconcha Glycaspis brimblecombei moore (hemiptera:
psyllidae). 2006. 52p. Dissertação (Mestrado em Agronomia) – Faculdade de Ciências
Agronômicas – UNESP, Botucatu.
CUNNINGHAM, J. P.; ZALUCKI, M. P.; WEST, S. A. Learning in Helicoverpa
armigera (Lepidoptera: Noctuidae): a new look at the behaviour and control of a
polyphagous pest. Bulletin of Entomological Research, London, v. 89, n. 3, p. 201-
207, 1999.
CRUSCIOL, C.A.C.; SORATTO, R.P.; CASTRO, G.S.A,; DA COSTA, C.H.M.;
NETO, J.F. Aplicação foliar de ácido silícico estabilizado na soja, feijão e amendoim.
Revista Ciência Agronômica, v. 44, n. 2, p. 404-410, 2013.
CRUZ, F.Z. Pragas em plantas ornamentais. In: KÄMPF, A.N. (coord.) Produção
comercial de plantas ornamentais. Guaíba: Agropecuária, 2000. 254p.
DARROW, G. M. Holt, rinehart and winston: the strawberry history breeding and
physiology. New York: The New England Institute for Medical Research, 1966. 447 p.

50
DAT, J.F.; LOPEZ-DELGADO, H.; FOYER, C.H.; SCOTT, I.M. Parallel changes in
H2O2 and catalase during thermotolerance induced by salicylic acid or heat
acclimation in mustard seedlings. Plant Physiology, Minneapolis, v. 116, p. 1351–
1357, 1998.
DATNOFF, L.E.; RAID, R.N.; SNYDER, G.H.; JONES, D.B. Effect of calcium silicate
on blast and brown spot intensities and yields of rice. Plant Disease, v. 75, n. 7 , p. 729-
732, 1991.
DAVIDSSON P.R., KARIOLA T., NIEMI O., PALVA E.T. Pathogenicity of and plant
immunity to soft rot pectobacteria. Frontiers in Plant Science, n. 4, p. 191, 2013.
DAWOOD, M. G.; SADAK, M. S.; HOZAYEN, M. Physiological role of salicylic acid
in improving performance, yield and some biochemical aspects of sunflower plant
grown under newly reclaimed sandy soil. Australian Journal of Basic and Applied
Sciences, v. 6, n. 4, p. 82-89, 2012.
DEVINE, G.J.; BARBER, M.; DENHOLM, I. Incidence and inheritance of resistance
to METI-acaricides in European strains of the two-spotted spider mite (Tetranychus
urticae) (Acari: Tetranychidae). Pest Management Science, v.57, p.443-448, 2001.
DHONDT, S.; RUTHERFORD, S.; SPIVACK, A.J. Metabolic activity of subsurface
life in deep-sea sediments. Science, n. 295, p. 2067-2069, 2002.
DI PIERO, R.M.; KUHN, O.J.; PASCHOLATI, S.F. Indução de resistência e a
produtividade das culturas. Cap. 12. In: Indução de Resistência em plantas a
patógenos e insetos. Editores: CAVALCANTI, L. S.; DI PIERO, R. M.; CIA, P.;
PASCHOLATI, S. F.; RESENDE, M. L. V.; ROMEIRO, R. S. Piracicaba, SP, FEALQ,
2005. p. 239 – 258.
DEL RÍO, L.A.; SANDALIO, L.M.; CORPAS, F.J.; PALMA, J.M. e BARROSO, J.B.
Reactive oxygen species and reactive nitrogen species in peroxisomes: Production,
scavenging, and role in cell signaling. Physiol. Plant., 141:330-335, 2006.

51
DURÃES, M.A.B. Respostas de duas populações de feijões (Phaseolus vulgaris L.)
tratadas com ácido salicílico e submetidas a estresse hídrico. Dissertação (Mestrado
em Agronomia- Produção Vegetal) - Universidade do Oeste Paulista, p. 32, 2006.
DURRANT, W.E., DONG, X. Systemic acquired resistance. Annual Review of
Phytopathology, v. 42, p. 185-209, 2004.
DUCHESNE, N. A. Historie Naturelle des Fraisiers. A Paris, 442 p., 1766.
EL-TAYEB, M. A.; ENANY, E. A. E.; AHMED, E. N. L. Salicylic acid-induced
adaptive response to copper stress in sunflower (Helianthus annuus L.). Plant Growth
Regulation, v. 50, v. 2-3, p. 191-199, 2006.
EPSTEIN, E. A anomalia de silício em biologia vegetal. Proceedings of National
Academy of United States of America, v. 91, n. 1, p. 11-17, 1994.
EWARDS, P.J.; WRATTEN, S.D. Ecologia das interações entre insetos e plantas. São
Paulo: EPU, EDUSP, 1981.
FADINI, M. A. M.; OLIVEIRA, H. G.; VENZON, M. PALLINI, A.; VILELA, E. F.
Distribuição Espacial de Ácaros Fitófagos (Acari: Tetranychidae) em Morangueiro.
Neotropical Entomology. Londrina, v. 36, n. 5, p. 783-789, 2007.
FANCELLI, A. L. Influência da nutrição na ocorrência de doenças de plantas. In:
FANCELLI, A. L. Milho: nutrição e adubação. Piracicaba: ESALQ/USP/LPV, 2008.
p.1-35.
FACHINELLO, J.C. Produção integrada de frutas: um breve histórico. Informe
Agropecuário, v.22, n.213, p.15-18, 2001

52
FAEDI, W; MOURGUES, E; ROSATI, C. Strawberry breeding and varieties: situation
and perspectives. Acta Horticultnrae, v.567, p.51-60, 1999. IV International
Strawberry Symposium.
FAEDI, W.; MOURGUES, F.; ROSATI, C. Strawberry breeding and varieties: situation
and perspectives. Acta Horticola, Alexandria, v. 567, p. 51–60, 1999.
FADINI, M. A. M.; OLIVEIRA, H. G.; VENZON, M. PALLINI, A.; VILELA, E. F.
Distribuição Espacial de Ácaros Fitófagos (Acari: Tetranychidae) em Morangueiro.
Neotropical Entomology. Londrina, v. 36, n. 5, p. 783-789, 2007.
FADINI, M. A. M.; VENZON, M.; OLIVEIRA, H. G. de; PALLINI, A. Manejo
integrado das principais pragas do morangueiro. Boletim do morango: cultivo
convencional, segurança alimentar, cultivo orgânico. Belo Horizonte: FAEMG, 2006. p.
81-95.
FAWE, A., MENZIES, J.G., CHÉRIF, M. e BÉLANGER, R.R. Silicon and disease
resistance in dicotyledos. In: DATNOFF, L. E.; SNYDER, G.H.; KORNDÖRFER,
G.H. (Ed.). Silicon in agriculture, Elsevier Science, p. 159-169, 2001.
FERNANDES, J. M.; SERIGATTO, E. M.; LUCA, A. S; EGEWARTH, R. E. Efeito de
soluções de origem vegetal na herbívoria de duas espécies de tanchagem (plantago
major l. e plantago lanceolata l.). 2006, Revista de Biologia e Ciências da Terra, 6, 2.
FERNANDES, F.C.; VIEIRA JÚNIOR, R.J.; SILVA, S.G.D.; REIS, D.N.; ANTUNES
JÚNIOR, H. Mecanismos de defesa de plantas contra o ataque de agentes
fitopatogênicos. Embrapa, 2009
FITT, G. P. The ecology of Heliothis species in relation to agroecosystems. Annual
Review of Entomology, Palo Alto, v. 34, p. 17-52, 1989.
FLECHTMANN, C. H. W. Seringueira (Hevea sp.) um novo hospedeiro para
Oligonychus Gossypii (Zacher, 1921) (Acari: Tetranychidae). Boletim do Museu
Paraense Emílio Goeldi, Série Zoologia , Belém, v. 5, n. 1, p. 127-128, 1989.

53
FIGUEIREDO, AST; RESENDE, JTV de; SANTOS, AP dos G; CAMARGO JÚNIOR,
O; MORALES, RGF; FARIA, MV; PRECZENHAK, AP. Repelência de cultivares de
morangueiro ao ácaro rajado, mediada por tricomas presentes nos folíolos. 2010.
Horticultura Brasileira 28: S603-S609
FILGUEIRA, F.A.R. Novo Manual de Olericultura: agrotecnologia moderna na
produção e comercialização de hortaliças. 3 ed. São Paulo: UFV, 421 p. 2008.
FREDDI, O.S.; CAMPOS, A.R.; LEONAL, C.L.; FREDDI, T.S.; BARBOSA, G.F.
Período de suscetibilidade do amendoinzeiro cv. Tégua ao tripés do prateamento e seu
reflexo na produtividade. Acta Scientiarum Agronomy, v.29, n.2, p.277-282, 2007
FU Z.Q.; DONG X. Systemic Acquired Resistance: Turning Local Infection into Global
Defense. Annual Review of Plant Biology, n. 64, n. 839-863, 2013.
GALLO, D.; NAKANO, O.; SILVEIRA NETO, S.; CARVALHO, R. P. L.;
BAPTISTA, G. C. de; BERTI FILHO, E.; PARRA, J. R. P.; ZUCCHI, R. A.; ALVES,
S. B.; VENDRAMIM, J. D.; MARCHINI, L. C.; LOPES, J. R. S.; OMOTO, C.
Entomologia agrícola. Piracicaba: FEALQ, 2002, p. 920 (FEALQ. Biblioteca de
Ciências Agrárias Luiz de Queiroz, 10).
GAO, Q.-M., ZHU, S., KACHROO, P. e KACHROO, A. Signal regulators of systemic
acquired resistance. Frontiers in Plant Science, p. 228 2015.
GALVÃO, A.G.; RESENDE, L.V.; GUIMARAES, R.M.; FERRAZ, A.K.L.;
MORALES, R.G.F.; MARODIN, J.C.; CATÃO, H.C.R.M. Overcoming strawberry
achene dormancy for improved seedling production in breeding programs. IDESIA, v.
32, n. 4, p. 57-72, 2014.
GENTY, B.; BRIANTAIS, J. M.; BAKER, N. R. The relationship between the quantum
yield of photosynthetic electron transport and quenching of chlorophyll fluorescence.
Biochimica et Biophysica Acta, v. 990, p. 87-92, 1989.

54
GIBSON R.W The distribution of aphids on potato leaves in relation to vein size.
Entomologia Experimentalis et Applicata, Oxford, v. 15, n. 2, p. 213-223, 1972.
GLAZEBROOK, J. Contrasting mechanisms of defense against biotrophic and
necrotrophic pathogens. Annual Review of Phytopathology Palo Alto, v. 43, p.205–
227, 2005.
GRANT, M.; LAMB, C. Systemic immunity. Current Opinion Plant Biology,London,
v. 9, p. 414–420, 2006.
GREGGAINS, V.; FINCH-SAVAGE, W.E.; QUICK, W.P.; ATHERTON, N.M.
Metabolism-induced free radical activity does not contribute significantly to loss of
viability in moist-stored recalcitrant seeds of contrasting species. New Phytologist,
v.148, p.267-276, 2000.
GREENE, G.L.; LEPLA, N.C.; DICKERSON, W.A. Velvetbean caterpillar: a rearing
procedure and artificial medium. Journal of Economic Entomology, v.69, n.4, p.488-
497, 1976.
GOMES, F.B.; MORAES, J.C.; SANTOS, C.D.; GOUSSAIN, M.M. Resistance
induction in wheat plants by silicone and aphids. Scientia Agricola, v. 62, n. 6, p. 547-
551, 2005.
GONÇALVES, K.S.; SOUZA, A.P.; VELINI, E. D.; TRINDADE, M.L. B.; PAZ, V. P.
S. Application of potassium phosphite to eucalyptus submitted to water stress. in:
Inovagri international meeting. Anais Fortaleza Inovagri international meeting, 2014.
GRÜNER, R.; STROMPEN, G.; PFITZNER, A.P.; PFITZNER, U.M. Salicylic acid
and the hypersensitive response initiate distinct signal transduction pathways in tobacco
that converge on the as-1-like element of the PR-1 a promoter. European Journal of
Biochemistry, v. 270, n. 24, p. 4876-4886, 2003.
HABIBI, G. Exogenous salicylic acid alleviates oxidative damage of barley plants
under drought stress. Acta Biologica Szegediensis, v. 56, n. 1, p. 57-63, 2012.

55
HAMMERSCHIMIDT, R., METRAUX, J.P. e VAN LOO, L.C. Inducing resistance: a
summary of papers presented at the First International Symposium on Induced
Resistance to Plant Diseases. Europen Journal Plant Pathology, n. 107, p. 1-6, 2000.
HEIDRICH K., BLANVILLAIN-BAUFUMÉ S., PARKER J.E. Molecular and spatial
constraints on NB-LRR receptor signaling. Current Opinion in Plant Biology, v. 15 n.
4, p. 385-391, 2012.
HENSCHEL M.J.; Morangueiro cultivados sob diferentes cores de coberturas em
túnel baixo. Dissertação (Mestrado em Agronomia-Produção Vegetal) UNICENTRO,
2016.
HOFFMAN G.D.; MCEVOY P.B. The mechanism of trichomeresistance in
Anaphalismargaritaceato the meadow spittlebug Philaenusspumarius. Entomologia
Experimentalis et Applicata, Oxford v. 39, n. 2, p.123-129, 1985.
HORVÁTH, E.; PÁL, M.; SZALAI, G.; PÁLDI, E.; JANDA, T. Exogenous 4-
hydroxybenzoic acid and salicylic acid modulate the effect of short-term drought and
freezing stress on wheat plants. Biologia Plantarum, v. 51, n. 3, p. 480-487, 2007.
HUANG, J.; ZHANG, P-J.; ZHANG, J.; LU, Y-B.; HUANG, F.; LI, M-J. Chlorophyll
content and chlorophyll fluorescence in tomato leaves infested with an invasive
Mealybug, Phenacoccus solenopsis (Hemiptera: Pseudococcidae). Environmental
Entomology, 42(5):973-979, 2013.
HUSSAIN, M., M. A. MALIK, M. FARROQ, M. Y. ASHRAF, and M. A. Hussain, M.,
M. A. Malik, M. Farooq, M. Y. Ashraf, and M. A. Improving drought tolerance by
exogenous application of glycinebetaine and salicylic acid in sunflower. Journal of
Agronomy and Crop Science, Cheema v. 194, n. 3, p. 193-199, 2008a.
Iqpal M., Jamil A., ShafiqU.R. 2006. Does seed priming in-duce changes in the levels
of someendogenous plant hormones in hex-aploid wheat plant under salt stress.

56
KANT M.R.; SABELIS M. W.; HARING M.A.; SCHUURINK R.C. Intraspecific
variation in a generalista in a generalista herbivore accounts for differential induction
and impacto f host plant defenses. Proceeding of the Royal Society Biological
Sciences, n. 275, p. 443-452, 2008.
KERBAUY, G.B. Fisiologia Vegetal. 1ª ed. Editora: Guanabara Koogan S.A.. 2004.
452p.
KRAMER, D.M.; JOHNSON, G.; KIRATS, O.; EDWARDS, G. E. New fluorescence
parameters for the determination of QA redox state and excitation energy fluxes.
Photosynth Research, n. 79, . 209-218, 2004.
KUMAR, R.R., SHARMA, S.K., GOSWAMI, S., VERMA, P., SINGH, K., Dixit, N.
RAI, R.D. Salicylic acid alleviates the heat stress-induced oxidative damage of starch
biosynthesis pathway by modulating the expression of heat-stable genes and proteins in
wheat (Triticum aestivum). Acta Physiologiae Plantarum, p. 143, 2015.
KUHN, O. J. Indução de resistência em feijoeiro ( Phaseolus vulgaris) por
acibenzolar-Smetil e Bacillus cereus: aspectos fisiológicos, bioquímicos e
parâmetro de crescimento e produção. Tese (Doutorado Fitopatologia)- Escola
Superior de Agricultura “Luis de Queiroz”, Universidade de São Paulo, p. 139, 2007.
JALEEL, C. A.; RIADH, K.; GOPI, R.; MANIVANNAN, P.; INES, J.; AL-JUBURI,
H. J.; CHANG-XING, Z.; HONG-BO, S.; PANNEERSELVAM, R. Antioxidant
defense responses: physiological plasticity in higher plants under abiotic constraints.
Acta Physiology Plant, v. 31, p. 427-436, 2009.
JANUS L., MILCZAREK G., ARASIMOWICZ-JELONEK M., ABRAMOWSKI D.,
BILLERT H., FLORYSZAK-WIECZOREK J. Normoergic NO-dependent changes,
triggered by a SAR inducer in potato, create more potente defense responses to
Phytophthora infestans. Plant Science, n. 211, p. 23-34, 2013.

57
JOHNSON, M.P.; RUBAN, A.V. Photoprotetive energy dissipation in higher plants
involves alteration of the excited state energy of the emitting chlorophyll(s) in the light
harvesting antenna II (LHCII). Journal of Biological Chemestry, v. 284, n. 35, p.
23592-23601, 2009.
JUNIOR, C.G.L. Identificação e expressão de genes de biossíntese do jasmonato na
interação entre Theobroma cacao e Moniliophthora perniciosa. 2009. 89p.
Dissertação (Mestrado em Ciências) - Centro de Energia Nuclear na Agricultura – USP,
Piracicaba.
LAMMERS, J. W.; MACLEOD, A. Report of a pest risk analysis: Helicoverpa
armigera (Hübner, 1808). [S.l.: s.n.], 2007. Disponível em:
http://www.fera.defra.gov.uk/plants/plantHealth/pestsDiseases/documents/helicoverpa.p
df. Acesso em: 20 maio 2013.
LARA, F. M. Princípios de resistência de plantas a insetos. São Paulo: Ícone, 1991,
2, 336p.
LAZAR, D. Chlorophyll a fuorescence induction. Biochimica et Biophysica Acta,
Netherlands, v. 1412, p. 1 - 28, 1999.
LEE, H.; LEON, J.; RASKIN, I. Biosynthesis and metabolism of salicylic acid. Proc.
Natl. Acad. Sci. USA. v. 92, p. 4076 – 4079, May 1995.
LICHTENTHALER, H. K. et al. How to correctly determine the different chlorophyll
fluorescence parameters and the chlorophyll fluorescence decrease ratio RFd of leaves
with the PAM fluorometer. Photosynthetica, v. 43, n. 03, p. 379- 393, 2005.
LITTLE D.; GOUHIRE-DARIMONT C.; BRUESSOW F.;REYMOND P. Oviposition
by pierid butterflies triggers defense responses in Arabidopsis. Plant Physiology, n.
143, p. 784-800, 2008.
LOAKE, G.; GRANT, M. Salicylic acid in plant defence-the players and protagonists.
Currents Opinion. Plant Biology, London, v. 10, p. 466-472, 2007.

58
LORENZETTI, E.L. Controle de Doenças do morangueiro com óleos essenciais e
Trichoderma spp. Tese (Doutorado em Agronomia- Fitopatologia) – Universidade
Federal de Lavras UFLA, 2012.
LORENZO, O.; SOLANO, R. Molecular players regulating the jasmonate signalling
network. Cur rent Opinion Plant Biology London, v. 8, p. 532–540, 2005.
LOURENÇÃO, A.L. et al. Resistência de morangueiro a Tetranychus urticae Kock
(Acari: Tetranychidae). Anais da Sociedade Entomológica do Brasil, v.29, n.2, p.339-
346, 2000.
MACDONALD, K.P.; BACH, C.E. Resistance and tolerance to herbivory in Salix
cordataare affected by diferente environmental factors. Ecological Entomology, Oxford,
v.30, n.5, p. 581-589, 2005.
MARSCHINER, H. Mineral nutrition of higher plants. 2. ed. Academic, p. 889,
1995.
MARTÍN, F. J. P. Respuestas inducidas por ácido abscísico y ácido salicílico em las
simbioisis de judia y alfalfa en estrés salino. Tese (Doutorado em Ciencias
Biológicas)- Faculdade de Ciências, Universidade de Granada, p. 395, 2009.
MARDANI, H.; BAYAT, H.; SAEIDNEJAD, A.H.; REZAIE, E.E. Assessment of
salicylic acid impacts on seedling characteristic of cucumber (Cucumis sativus L.) under
water stress. Notulae Scientia Biologicae, v. 4, n. 1, p. 112-115, 2012.
McCUE, P. et al. A model for enhanced pea seedling vigour following low pH and
salicylic acid treatments. Process Biochemistry, v.35, p. 603-13, 2000.
MARSCHNER, H. Mineral nutrition of higher plants. 2. ed. New York: Academic
Press, 1995. 889 p.

59
MATAMOROS, M.A.; DALTON, D.A.; RAMOS, J.; CLEMENTE, M.R.; RUBIO,
M.C. e BACANA, M. Biochemistry and molecular biology of antioxidants in the
rhizobia-legume symbiosis. Plant Physiol., 133:499-509, 2006.
MARTÍNEZ, A.M.; DELGADO, J.V.; RODERO, A.; VEGAS-PLA, J.L. Genetic
structure of the Iberian pig breed using microsatellites. Animal Genetics, Oxford, v.31,
p. 295-301, 2000.
MATTHEWS, M. Heliothinae moths of Australia: a guide to pest bollworms and
related noctuid groups. Melbourne: CSIRO, 1999. 320 p.
MAXWELL, K.; JOHNSON, G.N. Chlorophyll fluorescence – a pratical guide.
Journal os Experimental Botany, v. 51, n. 345, p. 659-668, 2000.
MORAES, G.J. de. FLECHTMANN, C.H.W. Manual de Acarologia: acarologia
básica e ácaros de plantas cultivadas no Brasil. Ribeirão Preto: Editora Holos, 2008.
308p.
MURCHIE, E.H.; LAWSON, T. Chlorophyll fluorescence analysis: a guide to good
practice and understanding some new applications. Journal of Experimental Botany,
v.1, 2013.
NIVEDITHADEVI, D.; SOMASUNDARAM, R.; PANNERSELVAM, R. Effect of
abscisic acid, paclobutrazol and salicylic acid on the growth and pigment variation in
Solanum Trilobatum. International Journal of Drug Development e Research, n. 3,
v. 4, p. 236-246, 2012.
NORMAN C.; HOWELL K.A.; MILLAR A.H.; WHELAN J.M.; DAY D.A. Salicylic
acid is an uncoupler and inhibitor of mitochondrial electron transport. Plant
Physiology, n. 134, p. 492-501, 2004.
OSHITA, D.; JARDIM, I. C. S. F. Morango: uma preocupação alimentar, ambiental e
sanitária, monitorado por cromatografia líquida moderna. Scientia Chromatographica,
Campinas, v. 4, n. 1, p. 52-76, jan/mar 2012.

60
OSZMIANSKI, J.; WOJDYLO, A. Comparative study of phenolic content and
antioxidant activity of strawberry puree, clear, and cloudy juices. European Food
Research and Technology, v.228, p.623–631, 2009.
OUZOUNIDOU, G. Changes of photosynthetic activities in leaves as a result of Cu
treatment: dose-response relations in Silene and Thlaspi. Photosynthetica, v. 29, p. 455
– 462, 1993.
PARK S. W.; KAIOMOYO E.; KUMAR D.; MOSHER S.; KLESSIG D.F. Methyl
salicylate is a critical mobile signal for plant systemic acquired resistance. Science, n.
318, . 113-116, 2007.
PASCHOLATI, S. F.; LEITE, B. Hospedeiro: mecanismos de resistência. In:
BERGAMIN FILHO, A.; KIMATI, H.; AMORIM, L. (Ed.) Manual de fitopatologia:
princípios e conceitos. São Paulo: Ed. Agronômica Ceres, v.1, p.193-217, 1995.
PAWAR, C. S.; BHATNAGAR, V. S.; JADHAV, D. R. Heliothis species and their
natural enemies, with their potential for biological control. Proceedings Indian
Academy of Sciences, v. 95, p. 695-703, 1986.
PFÜNDEL EE, BEM GHOZLEN, N, MEYER S, CEROVIC ZG. Investigating UV
screening in leaves by two diferente types of portable UV fluorimeters reveals in vivo
screening by anthocyanins and caratenoids. Photosynthesis Research, v. 93, . 205-221,
2007.
PICKETT, J.A.; POPPY, G.M. Switching on plant genes by external chemical signals.
Trends in plant Science, v. 6, n. 4, p. 137-139, 2001.
PICKETT, J. A.; RASMUSSEN, H.B.; WOODCOCK, C.M.; MATTHES, M.;
NAPIER, J.A. Plant stress signalling: understanding and exploiting plant-plant
interactions. Biochemical Society Transactions, v. 31, n. 1, p. 123-127, 2003.

61
PIETERSE C.M.; LEON-REYES A.; VAN DER ENT S.; VAN WEES S.C.M.
Networking by small-molecule hormones in plant immunity. Nature Chemical
Biology, n. 5, p. 308-316, 2009.
PIETERSE C.M.J.; VAN LOON L.C NPR1: The spider in the web of induced
resistance signaling pathwys. Current Opinion in Plant Biology n. 7, p. 456-464,
1999.
PIZZAMIGLIO-GUTIERREZ, M.A. Interações inseto-planta.In: PANIZZI, R.A.;
PARRA J.R.P. (Eds). Bioecologia e nutrição de insetos: base para o manejo integrado
de pragas. Brasilia: EMBRAPA, 2009. P. 211-249.
PEETERS, P.J. Correlations between leaf structural traits and the densities of
herbivorous insects guilds. Biological journal of the Linnean Society, London, v. 77, n.1
p. 43-65. 2002.
POGUE, M. G. A new synonym of Helicoverpa zea (Boddie) and differentiation of
adult males of H. zea and H. armigera (Hübner) (Lepidoptera: Noctuidae: Heliothinae).
Annals of the Entomological Society of America, College Park, v. 97, n. 6, p. 1222-
1226, 2004.
POIATTI V.A., DALMAS F.R., ASTARITA L.V. Defense mechanisms of Solanum
tuberosum L. in response to attack by plant-pathogenic bacteria. Biological Research,
n. 42, p. 205-215, 2009.
PONSTEIN, A.S.; BRES-VLOEMANS, S.A.; SELA-BUURIAGE, M.B.; VAN DEN
ELZEN, P.J.M.; MELCHERS, L.S.; COMELISSEN, B.J.C. A novel pathogen- and
wound-inducible tobacco (Nicotiana tabacum) protein with antifungal activity. Plant
Physiology, n. 104, p. 109-118, 1994.
PO-WEN C., SINGH P., ZIMMERLI L. Priming of the Arabidopsis pattern-triggered
immunity response upon infection by necrotrophic Pectobacterium carotovorum
bacteria. Molecular Plant Pathology, v. 14, n.1, p. 58-70, 2013.

62
POPOVA, L.; PANCHEVA, T.; UZUNOVA, A. Salicylic Acid: Properties,
Biosynthesis and Physiological Role. Bulgarian Journal of Plant Physiology. v. 23, p.
85 – 93, 1997
PULGA P.S.; Ácido Salicílico como indutor de resistência ao Tetranychus urticae
(Kock) e Tuta absoluta (Meyrick) em genótios de tomateiro. Dissertação (Mestrado
em Agronomia- Produção Vegetal)- UNICENTRO, 2016.
ROBERT-SEILANIANTZ A.; NAVARRO L.; BARI R.; JONES J. D. G. Pathological
hormone imbalances. Current Opinion Plant Biology, n. 10, p. 372-379, 2007.
RAJOU L. BELCHAZI M.; HUGUET R. ROBIN C.; MOREAU A.; JOB C.; JOB D.
Proteomic investigation of the effect of salicylic acid on Arabidopsis seed germination
and establishment of early defense mechanisms. Plant Physiology, n. 141, p. 910-923.
2006.
RAO M.V.; DAVIS K.R. The physiology of ozone-induced cell death. Planta, n. 213,
p. 682-690, 2001.
RAO M.V.; LEE H.I.; DAVIS K.R. Ozone-induced ethylene production is dependent
on salicylic acid, and both salicylic acid ethylene act in concert to regulate ozone-
induced cell death. Plant Journal. N. 32, p. 447-456, 2002.
RASKIN, I. Role of salicylic acid in plants. Annual review of plant physiology and
plant molecular biology, Palo Alto,v. 43, n. 11, p. 439 - 463, 1992.
RASKIN, I.; SKUBATZ, H.; TANG, W.; MEEUSE, B.J.D. Salicylic acid levels in
thermogenic and non-thermogenic plants. Annals of Botany, n. 66, p. 369-373, 2000.
RIBEIRO, R.V.; SANTOS, M.G. dos; SOUZA, G.M.; MACHADO, E.C.; OLIVEIRA,
R.F. de; ANGELOCCI, L.R.; PIMENTEL, C. Environmental effects on photosynthetic
capacity of bean genotypes. Pesquisa Agropecuária Brasileira, Brasília, v.39, n.7,
p.615-623, 2004

63
ROHÁČEK, K. Chlorophyll fluorescence parameters: the definition, photosynthetic
meaning, and mutual relationship. Photosynthetica, Prague, v.40, n.1, p.13-29, 2002.
ROHWER, C.L.; ERWIN, J.E. Horticultural applications of jasmonates: A review.
Journal of Horticultural Science e Biotechnology. v. 83, n. 3, p. 283 – 304, 2008.
RUAN, Y. M.; WU, K. J. Performances of the cotton bollworm, Helicoverpa armigera,
on different food plants. Acta Entomology, Bohemoslov, v. 44, n. 2, p. 205-212, 2001.
SANTOS, C. M.; GONÇALVES, E. R.; ENDRES, L.; GOMES, T. C. A.; JADOSKI,
C. J.; NASCIMENTO, L. A.; SANTOS, E. D. Atividade fotossintética em alface
(Lactuca sativa L.) submetidas a diferentes compostagens de resíduos agroindustriais.
Pesquisa Aplicada & Agrotecnologia, v.3, p.95-102, 2010.
SHARMA, H. C.; DHILLON, M. K.; ARORA, R. Effects of Bacillus thuringiensis δδ -
endotoxin-fed Helicoverpa armigera on the survival and development of the parasitoid
Campoletis chlorideae . Entomologia Experimentalis et Applicata, Dordrecht, v. 126,
n. 1, p. 1-8, Jan. 2008.
SCHWANESTRADA,K.R.F; CARVALHO,J.B.; BONALDO, S.M.; CRUZ, M.E.S.;
CARLOS, M.M.; STANGARLIN, J.R. Fungitoxicidade de Cymbopogon
citratus e Cymbopogon martinii a Colletotrichum gloeosporioides em frutos de
pimentão. Revista Brasileira de Plantas Medicinais, v.10, p.88-93, 2008.
SHEHATA S.A.M., IBRAHIM S.I., ZAGHLOOLS. A.M. 2001. Physiological re-
sponse of flag leaf and ears of maizeplant induced by foliar applicationof Kinetin (kin)
and acetyl salicylicacid (ASA). Ann. Agric. Sci. AinShams Univ. Cairo, 46(2): 435-
449.
SHI, Q.; ZHU, Z. Effects of exogenous salicylic acid on manganese toxicity, element
contents and antioxidative system in cucumber. Environmental and Experimental
Botany, v. 63, p. 317-326, 2008.

64
SILVA, V.F.da; MORAES, J.C.; MELO, B.A. Influence of silicone on the
development, productivity and infestation by insect pests in potato crops. Ciência e
Agrotecnologia, v.34, n.6, p. 1465-1469, 2010.
SILVA, L. M.; ALQUINI, Y.; CAVALLET, V. J. Inter-relações entre a anatomia
vegetal e a produção vegetal. Acta Botanica Brasilica, v. 19, p. 183-194, 2005.
SILVA, A.F.; DIAS, M.S.C.; MARO, L.A.C. Botânica e fisiologia do morangueiro.
Informe Agropecuário, v. 28, n. 236, p.7-13, 2007.
SILVA, P. A. Qualidade de morangos cultivados na região de Lavras, MG,
armazenados em temperatura ambiente. 2013. 71p. Dissertação (Mestrado em
Agronomia). Universidade Federal de Lavras. Lavras, Minas Gerais, 2013.
SINGH, B.; USHA, K. Salicylic acid induced physiological and biochemical changes in
wheat seedlings under water stress. Plant Growth Regulation, v. 39, n. 2, p. 137-141,
2003.
SOBRINHO, C.A.; FERREIRA, P.T.O.; CAVALCANTI, L.S. Indutores Abióticos. In:
Indução de Resistência em plantas a patógenos e insetos. Editores: CAVALCANTI,
L.S.; DI PIERO, R.M.; CIA, P.; PASCHOLATI, S.F.; RESENDE, M.L.V.; ROMEIRO,
R.S. Piracicaba, SP, FEALQ, 2005. p. 51 – 80.
SOARES, A.M.S.; MACHADO, O.L.T. Defesa de plantas: Sinalização química e
espécies reativas de oxigênio. Revista Trópica – Ciências Agrárias e Biológicas. v. 1,
n. 1, p.9, 2007.
SOUZA, A.P.M. Estudo do efeito de respostas de hipersensibilidade sobre a parede
celular em cultura de células de amora-preta (Rubus fruticosus). 2007. 145p.
Dissertação (Mestrado em Ciências Farmacêuticas) – Faculdade de Ciências
Farmacêuticas – USP, Ribeirão Preto.

65
STANGARLIN, J. R.; SCHULZ, D. G.; FRANZENER, G.; ASSI, L.
SCHWANESTRADA, K. R. F.; KUHN, O. J. Indução de fitoalexinas em soja e sorgo
por preparações de Saccharomyces boulardii. Arquivos do Instituto Biológico, v.77,
n.1, p.91-98, 2010.
STEINITE, I. e IEVINSH, G. Possible role of trichomes in resistance of strawberry
cultivars against spider mite. Acta Universitatis Latviensis, v. 662, p. 59 – 65, 2003.
STEINITE, I.; IEVINSH, G. Wound-induced responses in leaves of strawberry cultivars
differing in susceptibility to spider mite. J. Plant Physiol., v.159, p. 491–497, 2002.
STADNIK, M. Indução de resistência a oídios.In: CONGRESSO PAULISTA DE
FITOPATOLOGIA, Campinas.Anais do Congresso Paulista de Fitopatologia, v. 23 , p.
176-181. 2000.
SPOEL S.H., JOHNSON J.S., DONG X. Regulation of tradeoffs between plant
defenses against pathogens with different lifestyles. PNAS, v. 104, n. 47, p. 18842-
18847, 2012.
STIRBET, A.; GOVINDJEE. On the relation between the kautsky effect (chlorophyll a
fluorescence induction) and Photosystem II: Basics and applications of the OJIP
fluorescence transient, Journal Photochem Photobiology, 2011.
STACEY, G.; LIBAULT, M.; BRECHENMACHER, L.; WAN, J.; MAY, G.D.
Genetics and functional genomics of legume nodulation. Current Opinion Plant
Biology, n. 9, p. 110-121, 2006.
STEGMEIR, T.L. et al. Performance of an elite strawberry population derived from
wild germplasm of Fragaria chiloensis and F. virginiana. Hortscience, Alexandria, v.
45, n.8, p. 1140-1145, Aug. 2010.

66
STIRBET, A.; GOVINDJEE. On the relation between the Kautsky effect (chlorophyll a
fluorescence induction) and Photosystem II. Basics and applications of the OJIP
fluorescence transient, Journal Photochem Photobiology, 2011.
STUMPF, N.; NAUEN, R. Cross-resistance, inheritance, and biochemistry of
mitochondrial electron transport inhibitor-acaricide resistance in Tetranychus urticae
(Acari: Tetranychidae). Journal of Economic Entomology, v.94, p.1.577-1.583, 2001.
TANAKA, M.A.S.; BETTI, J.A.; PASSOS, F.A.; TRANI, P.E. (coord.); MACEDO,
A.C. (coord.) Manejo Integrado de Pragas e Doenças do Morangueiro. Campinas:
Secretaria de Agricultura e Abastecimento. 2000. 61p. (Manual Técnico, Série especial
v.5).
TESTER, M.; BACIC, A. Abiotic stress tolerance in grasses. From model plants to crop
plants. Plant Physiology, v. 137, p. 791-793, 2005.
ULLAND, S.; IAN, E.; MOZURAITIS, R.; BORG-KARLSON, A.K.; MEADOW, R.;
MUSTAPARTA, H. Methyl salicylate, identified as primary odorant of a specific
receptor neuron type, inhibits ovoposition by the moth Mamestra brassicae L.
(Lepidoptera, Noctuidae). Chemical Senses, n.33, p. 35-46, 2008.
VANACKER, H.; LU, H.; RATE, D.N.; GREENBERG J.T. A role for salicylic acid
and NPR1 in regulating cell growth in Arabidopsis. Plant Journal, Oxford, v. 28, p.
209-216, 2001.
VAN de VRIE, M.; MCMURTRY, J. A.; HUFFAKER, C. B. Ecology of tetranychid
mites and their natural enemies: A review. Hilgardia, Berkeley, v. 41, p. 387-403, abr.
1972.
VELINI, E.D.; TRINDADE, M.L.B.; MESCHEDE, D.K.; CARBONARI, C.A.
Glyphosate. 1 ed. FEPAF, p. 496, 2009.

67
VLOT A.C.; DEMPSEY D.A.; KLESSIG D.F. Salicylic acid, amultifaceted hormone to
combat disease. Annual Review Phytopathology, n. 47, p. 177-206, 2009.
ZHANG, Z.Q. Mites in green house : identification, biology and control. Cambridge:
CABI Publishing, 2003. 244 p.
WALLING, L.L. The myriad plant responses to herbivores Journal Plant Growth
Regulation, n. 19, p. 195-216, 2000.
WAR, A.R.; PAULRAJ, M.G.; AHMAD, T.; BUHROO, A.A.; HUSSAIN, B.;
IGNACIMUTHU, S.; SHARMA, H.C. Mechanisms of plant defense against insect
herbivores. Plant Signaling e Behavior, 7(10): 1306-1320, 2012.
WEI, J.; ZOU, L.; RONGING, K.; LIPING, H. Influence of leaf tissue structure on host
feeding selection by Pea leafminer Liriomyzahuidobrensis (Diptera: Agromyzidae).
Zoological Studies. Taipe, v. 39, n.4, p. 295-300.
YUSUF, M.A.; KUMAR, D.; RAJWANSHI, R.; STRASSER, R.J.;
TSIMILLIMICHAEL, M.; GOVINDJEE; SARIN, N.B. Overexpression of y-totopherol
methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: 33
Physiological and chlorophyll a fluorescence measurements. Biochimica et Biophysica
Acta, v. 1797, p. 1428-1438, 2010.
YEOM, M.; KIM, H.; LIM, J.; SHIN, A. Y.; HONG, S.; KIM, J.; NAM, H. G. How do
phytochromes transmit the light quality information to the circadian clock in
Arabidopsis? Molecular Plant, v. 7, n. 11, p. 1701-1704, 2014.





























