Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE NUTRIÇÃO
AÇÃO INSULÍNICA NO HIPOTÁLAMO DE RATOS
SUBMETIDOS À RESTRIÇÃO PROTEICA INTRAUTERINA
E RECUPERADOS COM DIETA À BASE DE SOJA
ALINE SOUZA DE MORAES HEREDIA
Cuiabá-MT, Setembro de 2017

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE NUTRIÇÃO
AÇÃO INSULÍNICA NO HIPOTÁLAMO DE RATOS
SUBMETIDOS À RESTRIÇÃO PROTEICA INTRAUTERINA
E RECUPERADOS COM DIETA À BASE DE SOJA
ALINE SOUZA DE MORAES HEREDIA
Trabalho de Graduação apresentado ao Curso de
Nutrição da Universidade Federal de Mato
Grosso como parte dos requisitos exigidos para
obtenção do título de Bacharel em Nutrição, sob a
orientação da professora Dra. Marise Auxiliadora
de Barros Reis.
Cuiabá-MT, Setembro de 2017

Dados Internacionais de Catalogação na Fonte.
Ficha catalográfica elaborada automaticamente de acordo com os dados fornecidos pelo(a) autor(a).
Permitida a reprodução parcial ou total, desde que citada a fonte.
S729a Heredia, Aline Souza de Moraes.
Ação insulínica no hipotálamo de ratos submetidos à restrição proteica intrauterina e recuperados com dieta à base de soja / Aline Souza de Moraes Heredia. -- 2017
32 f. : il. color. ; 30 cm.
Orientadora: Marise Auxiliadora de Barros Reis. TCC (graduação em Nutrição) - Universidade Federal de Mato Grosso, Faculdade
de Nutrição, Cuiabá, 2017. Inclui bibliografia.
1. Hipotálamo. 2. Desnutrição. 3. Soja. 4. Akt. I. Título.

UNIVERSIDADE FEDERAL DE MATO GROSS

RESUMO
A desnutrição intrauterina tem sido associada ao desenvolvimento de diversas Doenças Crônicas Não
Transmissíveis na vida adulta e o consumo de alimentos à base de soja parece ser benéfico na prevenção e
tratamento destas doenças. Estudos referem que animais recuperados após desnutrição intrauterina apresentam
alterações hipotalâmicas consistentes com resistência à insulina neste órgão. Assim, analisamos os efeitos da
recuperação nutricional com dieta à base de soja na via de sinalização da insulina no hipotálamo de animais
submetidos à restrição proteica na vida intrauterina e recuperados com dieta à base de soja. Prole de mães
alimentadas com 17% ou 6% proteína durante a prenhez e lactação foram mantidas com dieta à base de caseína
(CC e RC) ou de soja (CS e RS), desde o desmame até os 90 dias de vida. O consumo relativo foi maior nos
grupos recuperados. O peso corporal final foi diminuído pelo efeito do estado nutricional (RC e RS) e da dieta
(RS e CS). Os animais recuperados apresentaram peso hipotalâmico menor. Quanto a pAkt, o grupo RS não
apresentou difereça em relação a CC, enquanto os grupos CS e RC tiveram expressão dimuída em comparação
ao grupo CC, porém similar ao RS. Os dados do presente estudo sugerem que a dieta à base de farinha de soja
gerou redução do peso corporal final e hipotalâmico e aumentou o consumo relativo nesses grupos. A farinha de
soja não exerceu efeito de recuperação no peso corporal. A recuperação e a dieta à base de soja alteraram
negativamente a expressão pAkt.
PALAVRAS-CHAVES: Hipotálamo; Soja; Desnutrição; pAkt.

ABSTRACT
The Intrauterine malnutrition has been related to the development of several Chronic non-communicable
diseases in adult life and the soy food consumption seems benefic for prevention and treatment of those
diseases. Studies point out that animals recovered after intrauterine malnutrition present
consistent hypothalamic alterations with insulin resistance in this organ. Thus, we analyzed the effects of
nutritional recovery with soy-based diet on the insulin signaling pathway in the hypothalamus of animals
submitted to protein restriction in intrauterine life and recovered with soy-based diet. Offspring of mothers fed
with 17% or 6% protein during pregnancy and lactation were maintained with diet based on caseins (CC and
RC) or soybean (CS and RS), from weaning to 90 days of life. The relative consumption was higher in the
recovered groups. The final body weight was decreased by the effect of nutritional status (CR and RS) and diet
(RS and CS). The animals recovered had lower hypothalamic weight. As for pAkt, the RS group did not differ
from the CC, whereas the CS and RC groups had decreased expression in comparison to the CC group, but
similar to the RS group. The data from the present study suggest that the soybean meal diet led to a reduction in
final and hypothalamic body weight and increased relative consumption in these groups. Soybean meal had no
recovery effect on body weight. Recovery and the soybean diet negatively altered the expression of Akt and pAkt.
KEYWORDS: Hypothalamus; Soybeans; Malnutrition; pAkt.

SUMÁRIO
1. INTRODUÇÃO ............................................................................................................................. 7
2. REVISÃO DE LITERATURA .................................................................................................... 9
2.1 DESNUTRIÇÃO ..................................................................................................................... 9
2.2 INSULINA ............................................................................................................................ 10
2.3 HIPOTÁLAMO .................................................................................................................... 11
2.4 SOJA ........................................................................................................................................... 14
3. OBJETIVOS: ............................................................................................................................... 17
3.1 OBJETIVO GERAL: ............................................................................................................ 17
3.2 OBJETIVOS ESPECÍFICOS: ............................................................................................... 17
4. PROCEDIMENTOS METODOLÓGICOS ............................................................................. 18
4.2 DIETAS ....................................................................................................................................... 19
4.3. PROCEDIMENTOS EXPERIMENTAIS .................................................................................. 21
4.4. ANÁLISES ESTATÍSTICAS .................................................................................................... 22
5. RESULTADOS E DISCUSSÃO ................................................................................................ 23
6. CONCLUSÃO ............................................................................................................................. 27
7. REFERÊNCIAS .......................................................................................................................... 28

7
1. INTRODUÇÃO
A desnutrição continua a ser uma das causas de morbidade e mortalidade mais comuns
entre crianças de todo o mundo. Embora no Brasil a prevalência da desnutrição na infância
tenha caído nas últimas décadas, o percentual de óbitos por desnutrição grave em nível
hospitalar se mantém em torno de 20%, valores muito acima dos recomendados pela
Organização Mundial da Saúde (2000), sendo considerado um problema de saúde pública.
Entretanto, 45,9% da taxa mundial de doenças atualmente são atribuídas às Doenças
Crônicas Não Transmissíveis (DCNT), estimando-se dois terços dessa taxa no ano de 2020,
com um possível deslocamento da epidemia de doenças crônicas para países menos
desenvolvidos (Campos & Neto, 2009).
Estudos mostram que além de predisposição genética, os fatores ambientais, tais
como, condições de desnutrição na vida intrauterina, podem contribuir para o aumento do
número de DCNT. Isso tem sido explicado pela hipótese do “fenótipo da economia” (thrifty
phenotype), formulada em 1992, por Hales & Baker.
De acordo com o fenótipo da economia, o feto sofre adaptações metabólicas para
economizar energia, no caso de uma má nutrição, que desaceleram o crescimento das
vísceras, o que reduz as necessidades nutricionais e preserva o crescimento do cérebro. Como
consequência desse fenótipo, após o nascimento, a prole fica metabolicamente econômica,
estocando energia na forma de gordura (Hales & Baker 1992).
Dessa forma, a restrição proteica em fases iniciais da vida, pode produzir efeitos
distintos sobre o metabolismo da glicose e consequentemente sobre a ação da insulina,
incluindo alterações na regulação da produção de glicose hepática e na supressão da lipólise
no tecido adiposo, situações essas que são consistentes com o estado de resistência à insulina
(Feres et al., 2010).
O papel crucial da sinalização da insulina, para o metabolismo humano, tem sido
conhecido há muito tempo para os órgãos sensíveis à insulina, como músculo, fígado e tecido
adiposo. No entanto, a contribuição da ação insulínica no cérebro só ganhou atenção nas
últimas décadas. Dado o aumento dos casos de obesidade e diabetes mellitus, uma melhor
compreensão de como o comportamento alimentar, peso corporal, distribuição de gordura
corporal e os fluxos metabólicos em todo o corpo são regulados, é necessária para
desenvolver novas abordagens para combater estas doenças (Heni et al., 2015).
Segundo Vogt & Bruning (2013), a recuperação nutricional após desnutrição proteica
é capaz de alterar a sinalização de insulina no hipotálamo.

8
Ademais, existem hipóteses de que prejuízos na sinalização de insulina no sistema
nervoso, devido tanto à secreção ou ação da insulina, contribuem para a patogênese de
distúrbios metabólicos comuns, incluindo diabetes e obesidade (Porte, Baskin & Schwartz,
2005).
Diante dessas alterações metabólicas, intervenções nutricionais têm se mostrado úteis
no tratamento e na prevenção de DCNT, como o diabetes mellitus; e a soja (Glycinemax) tem
sido associada a esses benefícios (Bhathena & Velasquez, 2002).
A proteína da soja é rica em glicina e arginina que estão envolvidas na secreção de
insulina (Fajans et al., 1997) além de melhorar a glicemia de jejum e a sensibilidade periférica
à insulina em ratos (Sanchez & Hubbard, 1991).
Estudos experimentais têm utilizado uma dieta à base de farinha de soja como fonte
proteica alternativa de baixo custo e alta qualidade para a recuperação de animais desnutridos
na vida intrauterina, na tentativa de prevenir e/ou tratar as consequências tardias dessa
desnutrição. Esta escolha foi motivada por evidências científicas que mostram que, pelo
menos um dos componentes da soja, a genisteína, aumenta a secreção de insulina (Sorenson,
Brelje & Roth, 1994).
Animais desnutridos quando recuperados com dieta à base de soja apresentaram
aumento da concentração sérica de insulina e moderada resistência hepática à insulina, uma
situação que poderia interferir no metabolismo lipídico e na função hepática (Oliveira, 2006).
Assim, visto que estudos já comprovam que animais recuperados com farinha de soja
apresentaram resistência hepática à insulina e alteração na via de sinalização desse hormônio,
e sabendo-se que a resistência insulínica pode ser tecido específica (Carvalheira, Zecchin &
Saad, 1995), objetivou-se investigar se a soja como uma alternativa alimentar, poderia
também alterar uma das vias de sinalização da insulina, a Akt, no hipotálamo, já que este
órgão quando recuperado também apresenta prejuízos em suas funções (Vogt & Bruning,
2013).

9
2. REVISÃO DE LITERATURA
2.1 DESNUTRIÇÃO
O crescimento intrauterino compreende a fase de multiplicação, crescimento e
diferenciação celular, e sua importância se deve ao fato de que se o crescimento fetal for
deficiente, geralmente manifesto pelo baixo peso ao nascer, gera consequências que poderão
apresentar efeito imediato como aumento da mortalidade e morbidade perinatais, ou tardio,
como as deficiências nutricionais, processos patológicos e distúrbios de crescimento
(Guimarães, 1982).
A desnutrição pode ser definida como uma condição clínica decorrente de uma
deficiência ou excesso, relativo ou absoluto, de um ou mais nutrientes essenciais (Mahan &
Arlin, 1995), sendo a desnutrição intrauterina aquela que acomete o feto durante o período
gestacional.
Não se sabe ao certo que tipo de deficiência nutricional tornaria o feto mais
susceptível a desenvolver doenças na idade adulta, porém grande atenção é dada a restrição
proteica, pois estudos demonstram que o crescimento do embrião e ou do feto é altamente
relacionado com o ambiente materno (Mc Millen & Robinson, 2005) e que a restrição de
proteínas nesse período institui-se um fator que predispõe tanto ao baixo peso ao nascer
(Lucas et al., 1999) como ao surgimento de várias doenças DCNT (Godfrey & Barber, 2000)
sendo os aminoácidos importantes para o desenvolvimento das células β pancreáticas e
secreção de insulina (Hales & Barker, 1992).
Hales & Barker (1992) relacionaram o retardo do crescimento durante a vida fetal e a
infância com o aumento das taxas de morbidades como as doenças cardiovasculares, o risco
de desenvolver uma intolerância à glicose e a síndrome metabólica durante a vida adulta.
Assim, o organismo que foi metabolicamente programado para sobreviver em
ambiente escasso de nutrientes, não terá mecanismos suficientes para lidar com as
modificações ambientais fora do útero, como por exemplo, a exposição a uma dieta
desequilibrada, marcada pelo excessivo consumo de produtos com alto valor energético e
baixo valor nutritivo, ou mesmo se utilizar uma dieta balanceada que também será uma
sobrecarga para esse animal. Sendo assim, essa sobrecarga pode, então, acarretar o
desenvolvimento de doenças como diabetes, obesidade e co-morbidades muito mais
facilmente do que indivíduos com a mesma dieta, mas que não sofreram desnutrição no
período intrauterino (Bismarck-Nasr, Frutuoso & Gamabardella, 2008).

10
Uma dieta materna hipoproteica durante as fases de prenhez e lactação é um modelo
utilizado para representar a programação nutricional. Prole de ratas alimentadas ad libitum
com uma dieta com baixo teor de proteínas contendo de 5 a 8% de proteína à base de caseína,
apresentaram menor peso corporal do que a prole de mães que consumiram uma dieta
normoproteica (Langley-Evans, 2000). Além disso, animais submetidos à restrição proteica
em fases críticas do desenvolvimento apresentaram um aumento da expressão dos receptores
de insulina e resposta alterada da insulina (Ozanne et al., 1999)
A manutenção dessa dieta hipoproteica durante o período de lactação aumenta a diferença
de peso e limita o crescimento do animal, entretanto, o desenvolvimento não esta limitado
somente ao ambiente intrauterino, uma vez que o sistema fisiológico continua a se
desenvolver e as condições nutricionais podem ter impactos consideráveis na vida adulta
(Vickers & Sloboda, 2012).
2.2 INSULINA
Muitos estudos têm sido dedicados ao entendimento do mecanismo molecular de ação
da insulina devido à alta prevalência de resistência ao hormônio e pelo fato desta estar
envolvida na síndrome metabólica (Haber et al., 2001).
A insulina é um hormônio de natureza peptídica, essencial para o desenvolvimento,
crescimento, diferenciação celular, desempenha importante papel na manutenção da
homeostase glicêmica e controle metabólico. Produzida pelas células beta do pâncreas e
secretada tonicamente e, com incrementos durante as refeições, o principal estímulo para sua
secreção é o aumento da glicemia (Woods & D’Alessio, 2008).
A insulina é transportada através da barreira hematoencefálica e age em receptores
expressos predominantemente em neurônios no núcleo arqueado do hipotálamo, mas também
em outras regiões do cérebro. Após a ligação da insulina à subunidade α extracelular do seu
receptor (IR), a subunidade β intracelular, que possui atividade tirosina quinase intrínseca, se
autofosforila. Em seguida, ela promove o recrutamento e a fosforilação em tirosina dos
substratos do receptor de insulina (IRS), e, entre os membros da família das proteínas IRS, o
IRS-2 é o que possui maior expressão no núcleo arqueado e está implicado na mediação dos
efeitos centrais da insulina (Lin et al., 2004).
O IRS-2 fosforilado se liga à subunidade regulatória (p85) da enzima
fosfatidilinositol-3-quinase (PI3K), ativando a subunidade catalítica (p110) que, por sua vez,
fosforila o fosfatidilinositolbifosfato (PIP2) para gerar o fosfatidilinositoltrifosfato (PIP3).

11
A PI3-quinase é uma serina/treonina quinase, identificada como um dímero composto
de uma subunidade catalítica (p110) e uma subunidade regulatória (p85) (Biddinger & Khan,
2006; Stein & Waterfield, 2000). A ligação da subunidade p85 da PI3-K ao IRS-2 estimula a
atividade quinase da subunidade catalítica p110. Como resultado ocorre fosforilação de uma
serina quinase denominada Akt/PKB, que dentre outras funções, participa diretamente do
transporte de glicose dependente de insulina (Kim et al., 1999). A Akt/PKB também catalisa a
fosforilação em serina da glicogênio sintetase quinase (GSK3), inativando-a e,
consequentemente, ativando a glicogênio sintetase. O resultado dessas reações é o aumento da
síntese de glicogênio (Saad, 1994).
As quinases Akt/PKB (proteína quinase B), que incluem Akt1, Akt2 e Akt3, são
intermediárias das vias de sinalização que regulam os processos celulares que controlam o
tamanho e crescimento das células, proliferação, sobrevivência, metabolismo da glicose,
estabilidade do genoma e neovascularização (Saad, 1994).
Além da função clássica na regulação do metabolismo da glicose, a insulina possui
ações centrais no controle do balanço energético, ou seja, em contraste com seus efeitos
anabólicos sobre tecidos periféricos, sua ação hipotalâmica produz efeitos catabólicos (Porte,
Baskin & Schwartz, 2005).
2.3 HIPOTÁLAMO
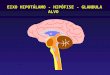
O hipotálamo é a porção do diencéfalo (Figura 1) localizada abaixo do tálamo (Figura
2) e entre a lâmina terminal e os corpos mamilares, formando as paredes e o assoalho do
terceiro ventrículo (Raff & Levitzky, 2012), estando envolvido na coordenação das respostas
fisiológicas dos diferentes órgãos que mantêm, de forma conjunta, a homeostasia. Ele integra
sinais provenientes do ambiente, de outras regiões encefálicas e de aferentes viscerais para,
então produzir as respostas neuroendócrinas apropriadas. Fazendo isso, o hipotálamo
influencia muitas funções do dia a dia, inclusive na ingestão de alimentos, no consumo
energético, no peso corporal, na ingestão e no balanço hídrico, na pressão arterial, na sede, na
temperatura corporal e no ciclo de sono (Raff & Levitzky, 2012).

12
Figura 1. Corte sagital do encéfalo.
Figura 2. Diencéfalo. O diencéfalo localiza-se entre o tronco encefálico e o cérebro. Ele
consiste em tálamo, hipotálamo, glândula pineal e glândula hipófise.
Fonte: Silverthon, 2010.
Fonte: Valdearcos, Xu & Koliwad, 2015.

13
O Hipotálamo Médio Basal (HMB) controla uma variedade de funções metabólicas
como a fome e saciedade, metabolismo energético, metabolismo lipídico periférico,
homeostase da glicose e a capacidade termogênica (Morton et al., 2006).
Um desequilíbrio na dieta poderia alterar o controle do HMB e juntamente com
excesso de citocinas pró-inflamatórias levar a um estresse celular entre os neurônios
envolvidos no controle metabólico (Morton et al., 2006).
Sinais metabólicos gerados pela insulina e leptina exercem ações recíprocas no HMB
revelando outra via de entrada que no controle metabólico dependente do Sistema Nervoso
Central (Sarper, Chou & Elmquist, 2002).
A detecção de glicose hipotalâmica também faz parte do mecanismo de regulação que
mantém a homeostase glicêmica no organismo de mamíferos. Por exemplo, as mudanças nas
concentrações hipotalâmicas de glicose modulam a ação de eferentes vagais que regulam a
produção hepática de glicose (Pocai et al., 2005).
Desordens hipotalâmicas observadas na sinalização da insulina têm sido atribuídas ao
desenvolvimento de alterações metabólicas relacionadas à reabilitação após a desnutrição
proteica (Vogt & Bruning, 2013).
Inúmeros estudos experimentais contribuíram para o avanço no conhecimento sobre os
mecanismos envolvidos na resistência hipotalâmica à ação da insulina. Os principais achados
revelaram a indução de um processo inflamatório especificamente no hipotálamo, que resulta
na ativação de vias de sinalização intracelular que atenuam os efeitos biológicos locais da
insulina (Carvalheira et al., 2001; Vogt & Bruning, 2013; Valdearcos, Xu & Koliwad, 2015).
Ao longo da última década tem sido demonstrado o papel central da sinalização
hipotalâmica da insulina no controle da produção hepática de glicose. A ativação da via da
Akt pela insulina no hipotálamo leva a supressão da produção de glicose hepática,
independentemente de mudanças na concentração sérica de insulina e outros hormônios
glicorreguladores (Obici et al., 2002).
As vias subjacentes a esta comunicação interórgãos ainda estão sendo investigadas,
mas os dados recolhidos até o momento indicam que a ativação hipotalâmica da PI3K/Akt
transmite mensagens de nervos parassimpáticos ao fígado que podem conduzir a supressão da
gliconeogênese (Pocai et al., 2005).
Como a inflamação hipotalâmica induzida pela dieta precede ou ocorre
simultaneamente com disfunção hipotalâmica, há um crescente interesse pela hipótese de que
a inflamação hipotalâmica é uma causa e não uma consequência da doença metabólica
induzida pela dieta (Kang et al., 2009).

14
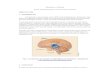
Esses esforços mostram que a ativação inflamatória no HMB exerce amplos efeitos
sobre os tecidos periféricos (Figura 3). Por exemplo, a inflamação hipotalâmica,
independentemente do peso corporal, é associada à diminuição da liberação de insulina por
células β pancreáticas com ação de insulina nos tecidos alvo e com o desenvolvimento de
disfunção renovascular que leva à hipertensão (Purkayastha, 2011).
Figura 3. A inflamação no HMB afetando tecidos periféricos. Uma inflamação no hipotálamo
resulta em alterações dos mecanismos de controles hipotalâmicos que regulam a termogênese,
metabolismo hepático e secreção de insulina pancreática.
2.4 SOJA
A soja é uma leguminosa que surgiu como importante alimento no primeiro milênio e
tem sido cultivada na China há séculos. O valor nutritivo da soja varia de acordo com a
composição de suas partes, onde 8% de seu peso correspondem à casca, sendo o restante
composto por 40,2% de proteínas, 21,0% de lipídeos, 33,9% de carboidratos, 6 a 7% de fibras
e 4,9% de cinzas. Observa-se que a proteína da soja contém maiores quantidades dos
aminoácidos arginina e glicina, porém é deficiente em aminoácidos sulfurados (metionina e
cisteína) quando comparada à proteína animal (Moraes & Silvia, 2000).
Fonte: Valdearcos, Xu & Koliwad, 2015.

15
Além do seu alto valor nutricional, a soja apresenta ação preventiva e terapêutica sobre
uma série de DCNT como diabetes mellitus, obesidade, enfermidades cardiovasculares e
câncer, sendo considerado um alimento funcional. Esse termo é aplicado para alimentos ou
ingredientes que além de ter funções nutricionais básicas, produzem efeitos fisiológicos e
metabólicos que são benéficos à saúde, quando consumidos como parte de uma dieta habitual,
sendo seguros para o consumo sem a necessidade de uma supervisão médica (Pelletier,
Kundrat & Hasler, 2002). Grande parte dos efeitos biológicos benéficos atribuídos à soja se
deve à composição de aminoácidos de sua proteína e às isoflavonas (Zhan & Ho, 2005).
As isoflavonas são fitoestrógenos da classe das flavonas, e atribui-se a elas possível
ação na regulação de atividade de proteínas e do ciclo celular, efeitos estrogênicos, anti-
estrogênicos e antioxidantes (Szkudelska et al., 2000).
Uma das principais isoflavonas bioativas encontradas na soja é a genisteína, que afeta
favoravelmente a homeostase glicêmica por seus efeitos sobre a secreção de insulina
(Szkudelska et al., 2000).
Estudos sugerem que a genisteína tem papel complacente na regulação da secreção de
insulina por competir com os sítios de ligação do ATP das proteínas tirosina-quinase,
inibindo-as (Fajans et al., 1997).
Em adipócitos, a genisteína inibiu a oxidação de glicose estimulada por insulina e
bloqueou o efeito antilipolítico da insulina na presença de isoproterenol, porém, não bloqueou
a autofosforilação do receptor ou fosforilação dos substratos do receptor de insulina e não
apresentou efeito sobre a atividade da glicogênio sintase estimulada por insulina. No fígado, a
genisteína aumenta a produção de ácidos graxos (Nogowski, Macrowiak & Kandulska, 1998).
A proteína da soja pode afetar também a homeostase glicêmica por aumentar a
secreção de insulina e alterar a sensibilidade dos tecidos periféricos a esse hormônio. Esse
aumento da secreção de insulina tem sido atribuído à composição de aminoácidos da proteína
da soja: devido às altas concentrações de arginina, um potente liberador da insulina (Fajans et
al., 1997).
Os aminoácidos compostos na proteína da soja produzem um aumento da
sensibilidade à insulina e melhoram a tolerância à glicose, por modularem a captação de
glicose pelo tecido muscular (Lavigne, Marette & Jacques, 2000). Estudos têm mostrado que
a composição de aminoácidos pode alterar o transporte de glicose em tecidos periféricos. Em
humanos, a infusão de aminoácidos de cadeia ramificada (leucina, isoleucina e valina) que são
predominantemente metabolizados no músculo esquelético, inibe a captação de glicose
mediada pela insulina (Schwenk & Haymond, 1987). A quantidade de aminoácidos de cadeia

16
ramificada é menor na proteína da soja em comparação à caseína (Bhathena & Velasquez,
2002). Por seus efeitos benéficos sobre a secreção e ação da insulina, a soja tem sido
recomendada como uma fonte alternativa na prevenção e tratamento de desordens
metabólicas.

17
3. OBJETIVOS:
3.1 OBJETIVO GERAL:
Analisar a ação insulínica no hipotálamo de ratos recuperados com dieta à base de
soja.
3.2 OBJETIVOS ESPECÍFICOS:
Avaliar os efeitos da dieta à base de farinha de soja em ratos adultos recuperados da
desnutrição sobre:
Peso corporal;
Consumo alimentar;
Peso do hipotálamo.
Avaliar a expressão da proteína AKT.
Avaliar o nível de atividade da AKT pela sua fosforilação.

18
4. PROCEDIMENTOS METODOLÓGICOS
O presente estudo foi conduzido no Laboratório de Avaliação Biológica de Alimentos
(LABA) do Departamento de Alimentos e Nutrição da Faculdade de Nutrição da
Universidade Federal de Mato Grosso.
4.1 ANIMAIS E DELINEAMENTO EXPERIMENTAL
Foram utilizadas ratas adultas virgens, da linhagem Wistar, com 90 dias de idade,
provenientes do Biotério Central da Universidade Federal de Mato Grosso. As ratas foram
mantidas com machos da mesma linhagem, por uma noite (4 fêmeas e 1 macho); em gaiolas
coletivas de polipropileno, medindo 37,0 x 31,0 x 16,0 cm, com tampa de material
galvanizado. A prenhez foi confirmada pela presença de espermatozóides no esfregaço
vaginal.
As fêmeas prenhes foram aleatoriamente divididas em 2 grupos: controle e desnutrido
e receberam dietas isocalóricas à base de caseína com 17% de proteína (normoproteica) e 6%
de proteína (hipoproteica) respectivamente, durante a prenhez e lactação.
No terceiro dia de vida, as ninhadas foram reduzidas a oito filhotes, com a finalidade
de padronizar as condições experimentais. O desmame aconteceu aos 28 dias de vida e os
machos recém-desmamados foram distribuídos aleatoriamente nos seguintes grupos, e
mantidos por 60 dias como apresenta a Tabela 1.
Durante todo o período experimental, os animais foram mantidos sob condições
controladas de umidade, temperatura (24ºC - 26ºC), e ciclo de luz claro/escuro (12h/12h),
tendo livre acesso à água e às respectivas dietas.
Todos os experimentos foram dirigidos por princípios de boas práticas de laboratório e
procedimentos científicos, seguindo o guia prático do manual de biotério do International
Committeeon Laboratory Animals (ICLA).
Este projeto foi certificado pela Comissão de Ética no Uso de Animais (CEUA) n°
23108.123676/2015-62.

19
Tabela1- Distribuições dos grupos e as dietas após o desmame.
Grupo Hipoproteico (HP) Prole de mães alimentadas com dieta
hipoproteica durante a prenhez e a
lactação.
Grupo Controle (CC) Prole de mães alimentadas com dieta
normoproteica durante a prenhez e a
lactação e subsequentemente alimentada
com a mesma dieta após o desmame.
Grupo Controle Soja (CS) Prole de mães alimentadas com dieta
normoproteica durante a prenhez e a
lactação e alimentadas com dieta à base
de farinha de soja com 17% de proteína,
após o desmame.
Grupo Recuperado com Caseína (RC) Prole de mães alimentadas com dieta
hipoproteica durante a prenhez e a
lactação, e recuperada com dieta
normoproteica à base de caseína após o
desmame.
Grupo Recuperado com Soja (RS) Prole de mães alimentadas com dieta
hipoproteica durante a prenhez e a
lactação, e recuperada com dieta
normoproteica à base de farinha de soja
após o desmame.
4.2 DIETAS
As dietas, cujas composições encontram-se descritas na Tabela 2, foram utilizadas nas
fases de prenhez, lactação e recuperação e preparadas no LABA.
Os grupos controle CC e RC receberam dieta à base de caseína, contendo 17% de
proteína, adequada para alimentação de roedores nas fases de crescimento, prenhez e lactação,
segundo as recomendações da AIN-93G (American Institute of Nutrition) enquanto que o
grupo hipoproteico foi alimentado com dieta hipoproteica (6% de proteína), que teve a
caseína substituída por carboidratos, sendo mantidas as proporções de amido, amido
dextrinizado e sacarose, bem como a de L-cistina em relação ao teor de caseína, e as
concentrações dos demais nutrientes. A dieta à base de farinha integral de soja micronizada
inativa seguiu as recomendações do American Institute of Nutrition (AIN-93G), sendo a
caseína, o óleo de soja e fibra substituídos pela farinha integral de soja, mantendo-se o teor de
proteína a 17%.

20
A farinha integral de soja micronizada inativa foi obtida por processamento industrial
(tratamento térmico, descascamento, pré-moagem e micronização), resultando na redução do
sistema enzimático e fator anti-tripsina, atingindo 80% do valor nutricional da caseína animal
(DINAL 4.3260.0003.01-2).
Todas as dietas passaram por processo de peletização, sendo os ingredientes secos
pesados em balança analítica da marca Marte, modelo A 500, com precisão de 0,01 g,
peneirados (malha 200) e homogeneizados. As dietas foram preparadas em quantidade
suficiente para todo o estudo, acondicionadas em recipientes de polipropileno,
hermeticamente fechados e armazenadas a 5ºC.
Tabela 2 – Composição das dietas controle, hipoproteica e à base de farinha de soja utilizadas
(g/Kg).
Ingredientes (g/Kg) Controle (17%
proteína)
Hipoproteica(6%
proteína)
Farinha de soja (17%
proteína)
Caseína (84%
proteína)
202,0 71,5 -
Farinha Integral de
soja
- - 415,0
Amido de milho 397,0 480,0 312,2
Amido de milho
dextrinizado
130,5 159,0 103,7
Sacarose 100,0 121,0 78,6
Óleo de soja 70,0 70,0 -
Fibras 50,0 50,0 40,0
Mistura de minerais
(AIN-93)G*
35,0 35,0 35,0
Mistura de vitaminas
(AIN-93)G*
10,0 10,0 10,0
L-cistina 3,0 1,0 3,0
Bitartarato de colina 2,5 2,5 2,5
*Reeves et al. 1993

21
4.3. PROCEDIMENTOS EXPERIMENTAIS
4.3.1. Perfusão e coleta de tecido
Após laparotomia mediana, foram injetadas 10 UI de insulina regular na via porta do
animal e foram aguardados 10 minutos. Em seguida, o crânio foi aberto, o cérebro removido e
o hipotálamo rapidamente retirado e congelado em nitrogênio líquido e posteriormente
armazenado a -80 °C.
4.3.2. Imunobloting
O hipotálamo congelado foi homogeneizado em tampão de extração [100 mmol/L
Tris-HCl (pH 7.4), 10 mmol/L EDTA, 100 mmol/L SDS 10%, 100 mmol/L pirosfosfato de
sódio, 100 mmol/L fluoreto de sódio e 10 mmol/L ortovanadato de sódio] usando “Polytron”.
O material insolúvel foi removido por centrifugação a 12000 rpm durante 20 minutos a 4°C.
A concentração de proteína foi determinada pelo método de Bradford (Bradford, 1976).
Amostras contendo 125μg de proteína foram incubadas em tampão concentrado de
Laemmli 4x e 15mg de DTT, e a corrida eletroforética realizada em gel de poliacrilamida a
120 V por 30 minutos (gel para Akt e pAkt). As proteínas foram transferidas para membranas
de nitrocelulose (Bio-Rad) por 90 minutos a 120 V, em tampão contendo metanol e SDS. Para
verificar a eficiência da transferência das proteínas, as membranas foram coradas com
Ponceau. Em seguida, as membranas foram bloqueadas com leite desnatado a 5% com
solução Tris-salina (10mmol Tris/l, 150 mmolNaCl/l, 0,5% Tween 20) por 2 horas à
temperatura ambiente, sob agitação constante.
Posteriormente, as membranas foram lavadas com solução Tris-salina em três sessões
de 10 minutos e incubadas com anticorpos específicos (para Akt e pAkt), sob agitação
constante a 4°C “overnight”.
A seguir, e as membranas foram lavadas com solução Tris-salina, como descrito
anteriormente, e incubadas em solução contendo anticorpo secundário “anti-rabbit
peroxidase” ou “anti-goat peroxidase” (diluição 1/5000) e 3% de albumina, por duas horas em
temperatura ambiente. A detecção das proteínas foi feita por quimiluminescência (Super
Signal West Pico, Pierce).
A intensidade das bandas foi determinada através da leitura das autoradiografias,
reveladas por densitometria óptica, utilizando um scanner (Epson Stylus CX 3500) e o
programa ScionImage (Scion Corporation). Os dados numéricos obtidos, correspondentes às
bandas proteicas, foram comparados estatisticamente como descritos em análises estatísticas.

22
4.4. ANÁLISES ESTATÍSTICAS
Os resultados foram apresentados como média e erro padrão da média, com o número
de animais indicado entre parênteses.
Para atestar a homogeneidade dos dados foi aplicado o Teste de Levene e para
comparar os grupos CC, CS, RC e RS, utilizou-se a análise de variância (ANOVA) a dois
fatores (estado nutricional e dieta). Quando necessário utilizou-se o teste de comparação
múltipla de médias LSD. Na comparação entre os grupos CC e HP, utilizou-se teste t de
Student. Em todos os casos foi estabelecido um nível de significância para a rejeição da
hipótese de nulidade de 5% (p<0,05).

23
5. RESULTADOS E DISCUSSÃO
Este estudo avaliou o efeito da recuperação nutricional com duas fontes proteicas
sobre a ação insulínica no hipotálamo de ratos adultos. Como esperado, a restrição proteica
durante a prenhez produziu uma redução significativa no peso corporal dos filhotes ao nascer
(p <0,001) (Tabela 3). Conforme Mc Millen & Robinson (2005), a baixa ingestão de proteínas
durante a vida intrauterina resultaria em animais com déficit corporal.
Durante a fase de recuperação, o consumo absoluto foi menor nos grupos
recuperados (RC e RS) em comparação com os grupos controles (CC e CS) (p <0,001)
(Tabela 3). A dieta à base de soja também produziu efeito no consumo absoluto, uma vez que
os grupos CS e RS apresentaram valores menores em relação os CC e RC (P <0,05). Quanto
ao estado nutricional prévio, o consumo relativo maior foram observados nos grupos RC e RS
em relaçao ao grupo CC e CS; os animais alimentados com dieta à base de soja também
tiveram um consumo relativo maior em comparação aos grupos que receberam caseína
(Tabela 3).
Ainda na Tabela 3, pode-se observar que o peso corporal no final do período
experimental foi significativamente menor nos grupos RC e RS do que nos grupos CC e CS
(p<0,001). Os ratos mantidos com a dieta de farinha de soja após o desmame (grupos RS e
CS) também apresentaram menor peso corporal final em relação aos que foram alimentados
com uma dieta à base de caseína (RC e CC) (P <0,001).
Tais dados corroboram com os de Orozco-Solis et al. (2011) e Orozco-Solis et al.
(2009) que relataram que uma dieta pobre em proteínas no período de gestação e lactação
resultaria em hiperfagia da prole devido a desnutriçao perinatal, alterando permanentemente a
estrutura e funcionalidade do hipotálamo, órgão responsável pelo controle da ingestão
alimentar e peso corporal.
A β-conglicinina, uma proteína de soja, e peptídios ricos em resíduos de arginina
apresentam alta atividade de supressão alimentar e potencialmente atrasam o esvaziamento
gástrico (Torres et al., 2006). A partir desse mecanismo, a expectativa da soja para diminuir o
peso corporal é criada, no entanto, o efeito oposto observado poderia estar relacionado, pelo
menos, à presença de fatores antinutricionais na soja, que prejudicariam a digestibilidade e a
biodisponibilidade das proteínas afetando negativamente o peso corporal (Cheim et al., 2009).

24
Tabela 3- Peso corporal ao nascer, peso corporal após 90 dias de vida, e consumo alimentar
absoluto e relativo de ratos adultos mantidos com dieta a base de caseína (grupos CC e RC)
farinha de soja (grupos CS e RS) após o desmame até 90 dias de idade.
GRUPOS
VARIÁVEIS CC CS RC RS HP
Peso
corporal ao
nascer (g)
5,62±0,34
(12)
- - - 5,11 ± 0,31+
(12)
Consumo
Absoluto(g)
1056 ± 82
(8)
994 ± 53 #
(8)
840 ± 51*
(8)
768 ± 70 * #
(8)
Consumo
Relativo(g)
2,45±0,14
(8)
2,82±0,16#
(8)
2,66±0,18*
(8)
3,09±0,11*#
(8)
Peso
corporal
Final (g)
468 ± 25
(12)
394 ± 19 #
(12)
357 ± 21*
(12)
280 ± 11* #
(12)
Valores expressos em média ± DP do número de ratos indicado entre parênteses. + valores médios foram
sifnificativamente diferentes em relaçao aos ratos controles (p <0,05, teste t de Student).*Os valores médios
foram significativamente diferentes dos ratos controle (p <0,05, ANOVA de duas vias). # Os valores médios
foram significativamente diferentes dos ratos alimentados com uma dieta de caseína (p <0,05, ANOVA de duas
vias).
Ao avaliar o peso do hipotálamo, os grupos CS e RS apresentaram peso absoluto, e
também relativo, menores em relação aos grupos CC e RC (P <0,05) (Figura 4).
Embora fosse observado que a soja reduziu o peso do hipotálamo (Figura 4), esse dado
não é suficiente para comprovar uma alteração da função hipotalâmica, porém, como citado
anteriormente (Orozco-Solis et al., 2011), alterações de ordem química e morfológica no
hipotálamo podem prejudicar a funcionalidade desse órgão, o que justificaria a hiperfagia
apresentada por esse grupo (Tabela 3).

25
Figura 4. Peso absoluto e relativo do hipotálamo (A e B, respectivamente) de ratos adultos
dos grupos controle caseína (CC), controle soja (CS), recuperado caseína (RC) e recuperado
soja (RS).
Valores expressos em média (g) ± desvio padrão (n= 7). # Os valores médios foram significativamente diferentes
dos ratos alimentados com uma dieta de caseína (p <0,05, ANOVA de duas vias).
O estado nutricional exerceu influência na expressão de Akt, sendo que os grupos
recuperados apresentaram menor expressão dessa proteína em relação aos controles (Figura
5). Observou-se ainda, influência da dieta neste parâmetro, sendo que os grupos alimentados
com dieta à base de soja tiveram menor expressão de Akt, porém, sem interação da dieta com
o estado nutricional. Contrapondo estes resultados, Feres et al. (2010) não observou nenhuma
alteração na expressão de Akt nos grupos CC, CS, RC e RS no tecido hepático.
Figura 5. Representação de “immunoblot” da expressão de Akt (A) e Akt fosforilada (B) no
hipotálamo de ratos adultos mantidos com dieta controle (grupos CC e RC) e farinha de soja
(grupos CS e RS).
(A) (B)
Valores expressos em média ± desvio padrão (n= 3). *Os valores médios foram significativamente diferentes dos
ratos controle (p <0,05, ANOVA de duas vias). #Os valores médios foram significativamente diferentes dos
ratos alimentados com uma dieta de caseína (p <0,05, ANOVA de duas vias). Letras minúsculas indicam
diferenças significativas entre os grupos (p<0,05, teste de comparação múltipla de médias LSD).
Akt
CC CS RC RS0
10000
20000
30000CC
CS
RC
RS
# # #
#
(B) (A)
# #* *
Co
nte
úd
o d
e A
kt
(Un
idad
e ar
bit
rári
a)
Co
nte
úd
o d
e p
Ak
t
(Un
idad
e ar
bit
rári
a)
pAkt
CC CS RC RS0
50000
100000
150000CC
CS
RC
RS
a
b
ab
b
Pes
o h
ipo
tála
mo
(g
)
Pes
o h
ipo
tála
mo
(g
/100
g
pes
o c
orp
ora
l)

26
A pAkt é a forma ativa da Akt, sendo a insulina responsável pela sua fosforilação. Em
relação à expressão da pAkt, os grupos CS e RC apresentaram valores significativamente
menores em relação ao grupo CC. O grupo RS não apresentou diferença estatística em relação
aos demais grupos.
Feres et al. (2010) verificou que animais alimentados com dieta à base de soja
apresentaram fosforilação da Akt diminuída no fígado, assim como foi possível observar no
hipotálamo do grupo controle soja em nosso trabalho.
Tanto a insulina quanto a leptina ativam a PI3K/Akt no hipotálamo e esta via é
importante tanto no controle do comportamento de ingestão alimentar quanto no controle da
homeostasia glicêmica (Niswender et al., 2003; Obici et al., 2002). Assim, o menor sinal
insulínico observado no grupo CS poderia justificar o consumo relativo maior desses animais,
porém não justificaria o maior consumo relativo nos grupos recuperados (Tabela 3), uma vez
que só o grupo RC apresentou menor fosforilação (Figura 5), sugerindo que outra via poderia
estar alterada.

27
6. CONCLUSÃO
Os dados do presente estudo sugerem que a dieta à base de farinha de soja gerou
redução do peso corporal final e hipotalâmico e aumentou o consumo relativo nesses grupos. A farinha de soja não exerceu efeito de recuperação no peso corporal. A recuperação e a dieta
à base de soja alteraram negativamente a expressão de Akt e pAkt.
A dieta à base de soja não alterou a sensibilidade hipotalâmica à insulina quando
utilizada na recuperação de ratos desnutridos durante a vida intrauterina e lactação e a dieta
citada foi mais prejudicial em animais que não sofreram uma interferência nutricional na fase
intrauterina e passaram a alimentar-se com a dieta à base de soja após o desmame.

28
7. REFERÊNCIAS
Biddinger S, Khan CR. From Mice to Mem: Insight sinto the Insulin Resistance Syndromes.
Annu. Rev Physiol. 2006; 68: 20.1–20.
Bismarck-Nasr EM, Frutuoso MFP, Gamabardella AMD. Efeitos tardios do baixo peso ao
nascer. Rev Bras Crescimento Desenv Hum. 2008; 18(1): 98-103
Bhathena SJ, Velasquez MT. Beneficial role of dietary phytoestrogens in obesity and
diabetes. J Clin Nutrition. 2002; 6: 1191-01.
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of
protein utilizing the principle of protein-dye binding. Anal Biochem. 1976; 72: 248-54.
Campos MO, Neto JFR. Doenças Crônicas Não Transmissíveis: Fatores de risco e
repercussão na qualidade de vida. Rev Baiana de Saúde Pública. 2009; 33(4): 561-581.
Carvalheira JBC, Siloto RM, Ignacchitti I, Brenelli SL, Carvalho CR, Leite A, et al. Insulin
modulates leptin-induced STAT3 activation in rat hypothalamus. FEBS Lett. 2001; 500(3):
119-24.
Carvalheira JBC, Zecchin HG, Saad MJA. Vias de sinalização da insulina. Arq Bras
Endocrinol Metab. 2002; 46.
Cheim LM, Oliveira EA, Arantes VC, Veloso RV, Reis MA, Gomes-da-Silva MH, Carneiro
EM, Boschero AC, Latorraca MQ. Effect of nutritional recovery with soybean flour diet on
body composition, energy balance and serum leptin concentration in adult rats. Nutr Metab.
2009; 24: 6-34.
DINAL 4.3260.0003.01-2. Divisão Nacional de Vigilância Sanitária de alimentos/DINAL da
Secretaria Nacional de Vigilância Sanitária.
Fajans SS, Floyd JC, Knof RF Jr, Conn FW. Effect of amino acids and proteins on insulin
secretion in man. Recent Prog Horm. 1997; 23: 617-62.
Feres NH, Reis SR, Veloso RV, Arantes VC, Souza LM, Carneiro EM, Boschero AC, Reis
MA, Latorraca MQ. Soybean diet alters the insulin-signaling pathway in the liver of rats
recovering from early-life malnutrition. Nutrition. 2010; 26: 441-448.
Guimarães CM. Estudo da desnutrição materna e da patologia da gestação sobre o
crescimento do concepto e da criança no ano de vida. São Paulo, 1982. [Tese de
Doutoramento - Faculdade de Saúde Pública da USP].
Godfrey KM & Barker DJP. Fetal nutrition and adult disease. Am J Clin Nutr. 2000; 71:1344-
1352.
Haber RC, Curi R, Carvalho CRO, Carpinelli AR. Secreção da Insulina: Efeito Autócrino da
Insulina e Modulação por Ácidos Graxos. Arq Bras Endocrinol Metab. 2001; 45: 219-27.
Hales CN, Barker DJ. Type 2 (non-insulin-dependent) diabetes mellitus: the thrifty phenotype
hypothesis. Diabetologia. 1992; 35(7):595-601.

29
Heni M, Kullmann S, Preissl H, Fritsche A, Haring HU. Impaired insulin action in the human
brain: causes and metabolic consequences. Nat Rev Endocrinol. 2015; 11:701–711.
Kang YM, Ma Y, Zheng JP, Elks C, Sriramula S, et al. Brain nuclear factor-κB activation
contributes o neuro humoral excitation in angiotensin II–induced hypertension. Cardiovasc
Res. 2009; 82:503–12.
Kim YB, Nikoulina SE, Ciaraldi TP, Henry RR, Kahn BB. Normal insulin dependent
activation of Akt/protein kinase B, with diminished activation of phosphoinositide 3- kinase,
in muscle in type 2 diabetes. J Clin Invest. 1999; 104: 733-41.
Langley-Evans SC. Critical differences between two low protein diet protocol in the
programming of hypertension in the rat. Food Sci Nutr. 2000; 51:11-17.
Lavigne C, Marette A, Jacques H. Cod and soy proteins compared with casein improve
glucose tolerance and insulin sensitivity in rats. Am J Physiol Endocrinol Metab. 2000; 278:
E491-E00.
Lin X, Taguchi A, Park S, Kushner JA, Li F, Li Y, et al. Dysregulation of insulin receptor
substrate 2 in beta cells and brain causes obesity and diabetes. J Clin Invest. 2004;
114(7):908-16.
Lucas A, Fewtrell MS & Cole TJ. Fetal origins of adult disease- the hypothesis revisted. BMJ.
1999; 19:245-9.
Manual para Técnicos em Laboratórios. Comissão de Ensino do Colégio Brasileiro de
Experimentação Animal (COBEA), 2a ed. 1996.
Mahan LK, Arlin MT. Krause: alimentos, nutrição e dietoterapia. 8a ed. São Paulo: Roca,
1995.
Mc Millen IC & Robinson JS. Development al origins of the metabolic syndrome: prediction,
plasticity and programming. Physiol Rev. 2005; 85: 571-633.
Morais AAC, Silva AL. Soja e suas aplicações. 3a ed. Medsi Editora Médica e Científica
Ltda: 2000.
Morton GJ, Cummings DE, Baskin DG, Barsh GS, Schwartz MW. Central nervous system
control of food intake and body weight. Nature. 2006; 443:289–95.
Niswender KD, Morrison CD, Clegg DJ, Olson R, Baskin DG, Myers MG Jr, et al. Insulin
activation of phosphatidylinositol 3-kinase in the hypothalamic arcuate nucleus: a key
mediator of insulin-induced anorexia. Diabetes. 2003; 52(2):227-31.
Nogowski L, Mackowiak P, Kandulska K. Genistein-induced changes in lipid metabolism of
ovariectomized rats. Ann Nutr Metab. 1998; 42: 360-6.
Obici S, Feng Z, Karkanias G, Baskin DG, Rossetti L. Decreasing hypothalamic insulin
receptors causes hyperfhagia and insulin resistance in rats. Nat Neuro sci. 2002; 5:566-572.
Organização Mundial de Saúde. Manejo da desnutrição grave: um manual para profissionais
de saúde de nível superior (médicos, enfermeiros, nutricionistas e outros) e seus auxiliares.
Genebra/Brasília: 2000.

30
Oliveira EA. Homeostase glicêmica de ratos expostos à restrição proteica durante a fase
intrauterina e lactação e recuperados com dieta à base de farinha integral de soja [Dissertação]
Cuiabá (MT): Universidade Federal de Mato Grosso – Faculdade de Ciências Médicas; 2006.
Orozco-Solis R, Lopes de Souza S, Barbosa Matos RJ, Grit I, Le Bloch J, Nguyen P, et al.
Perinatal undernutrition-induced obesity is independent of the developmental programming of
feeding. Physiol Behav. 2009; 96(3):481-92. doi: 10.1016/ j.physbeh.2008.11.016.
Orozco-Solis R, Matos RJ, Lopes de Souza S, Grit I, Kaeffer B, Manhaes de Castro R, et al.
Perinatal nutrient restriction induces long-lasting alterations in the circadian expression
pattern of genes regulating food intake and energy metabolism. Int J Obes. 2011; 35(7):990-
1000. doi: 10.10 38/ijo.2010.223.
Ozanne SE, Wang CL, Dorling MW, Petry CJ. Dissection of the metabolic actions of insulin
in adipocytes from early growth-retarded male rats. J Endocrinol. 1999; 162: 313-9.
Pelletier S, Kundrat S, Hasler CM. Effects of an educational program on intent to consume
functional foods. J Am Diet Assoc. 2002; 102:1297-300.
Pocai A, Lam TK, Gutierrez-Juarez R, Obici S, Schwartz GJ, Bryan J, Aguilar-Bryan L,
Rossetti L. Hypothalamic K (ATP) channels control hepatic glucose production. Nature.
2005; 434: 1026-1031.
Porte D Jr, Baskin DG, Schwartz MW. Insulin signaling in the central nervous system: a
critical role in metabolic homeostasis and disease from C. elegans to humans. Diabetes. 2005;
54(5):1264-76.
Purkayastha S, Zhang H, Zhang G, Ahmed Z, Wang Y, Cai D. Neural dysregulation of
peripheral insulin action and blood pressure by brain endoplasmic reticulum stress.PNAS.
2011; 108:2939–44.
Raff H, Levitzky M. Fisiologia Médica: uma abordagem integrada; tradução Luciano Sturmer
de Fraga…[et al.]; revisão técnica: Luciano Sturmer de Fraga, Renata Padilha Guedes.-Porto
Alegre: AMGH. 2012.
Reeves PG, Nielsen FH, Fahey CG Jr. AIN-93 purified diets for laboratory rodents: reports of
the American Institute of Nutrition ad hoc working committee on the reformulation of the
AIN-76 rodent diet. J Nutr. 1993; 123:1939-51.
Saad MJA. Molecular mechanisms of insulin resistance. Braz J Med Biol Res. 1994; 27: 941-
57.
Sanchez A & Hubbard RW. Plasma amino acids and the insulin/glucagon ratio as na
explanation for the dietary protein modulation of atherosclerosis. Med Hypotheses. 1991; 35:
324-9.
Saper CB, Chou TC, Elmquist JK. The need to feed: homeostatic and hedonic control of
eating. Neuron. 2002; 36:199–211.
Schwenk WF, Haymond MW. Effects of leucine, isoleucine, or threonine infusion on leucine
metabolism in humans. Am J Physiol Endocrinol Metab. 1987; 253: E428-E34.
Stein RC, Waterfield MD. PI3-kinase inhibition: A targer for drug development? Mol Med
Today. 2000; 6: 347-57.

31
Silverthorn DU. Fisiologia humana [recurso eletrônico]: uma abordagem integrada; tradução
Ivana Beatrice Mânica da cruz...[et al.].- 5. Ed.- Dados eletrônicos- Porto alegre: Artmed,
2010.
Sorenson R, Brelje CT, Roth C. Effect of tyrosine kinase inhibitors on islets of Langerhans:
evidence for tyrosine kinases in the regulation of insulin secretion.Endocrinology. 1994;
4:1975-8.
Szkudelska K, Nogowski L, Szkudelski T. Genistein affects lipogenesis and lipolysis in
isolated rat adipocytes. J Steroid Biochem Mol Biol. 2000; 75: 265-71.
Torres N, Torre-Villalvazo I, Tovar AR. Regulation of lipid metabolism by soy protein and its
implication in diseases mediated by lipid disorders. Nutr Biochem. 2006; 17(6):365-73.
Valdearcos M, Xu AW, Koliwad SK. Hypothalamic Inflammation in the Control of
Metabolic Function. Annu Rev Physiol. 2015; 77:131–60.
Vickers MH & Sloboda DM. Strategies for reversing the effects of metabolic disorders
induced as a consequence of developmental programming. Frontiers in Physiology. 2012;
3:242.
Vogt MC, Bruning JC. CNS insulin signaling in the control of energy homeostasis and
glucose metabolism – from embryo to old age. Trends in Endocrinology and Metabolism.
2013; 24: 2.
Zhan S, Ho SC. Meta-analysis of the effects of soy protein containing isoflavonas on the lipid
profile. Am J Clin Nutr. 2005; 81:397-08.
Woods SC, D’Alessio DA. Central control of body weight and appetite. J Clin Endocrinol
Metab. 2008; 93(11 Suppl 1):S37-50.