Embed Size (px)
Citation preview

Há três tipos principais de alcalóides:
A colchicina é exemplo de um proto-alcalóide. Os Pseudo-alcalóides podem ser derivados de;
Terpenoids ou
Purines
ALCALÓIDES VERDADEIROS
A unidade básica na biogenesis dos alcalóides verdadeiros é um aminoácido. Os nitrogênios não heterocíclicos anéis ou que estão em cadeias laterais são derivados do ACETATO e/ou do TERPENO, quando o METHIONINE for responsável pela a adição de grupos metílicos aos átomos do nitrogênio.
Os alcalóides são substâncias altamente reativas com atividade biológica em doses baixos.
DEFINIÇÃO
1. Contem o nitrogênio - derivado geralmente de um aminoácido.
2. Gosto amargo, sólidos geralmente brancos (exceção - a nicotina é um líquido marrom).
3. Dão precipitados com iodetos de metal pesado.
A maioria de alcalóides são precipitados em pH neutro ou de solução ligeiramente acídica pelo reagente de Mayer (solução iodo mercuriato de potássio). Precipitate de cor creme.
O reagente de Dragendorff (solução do iodo bismutato de potassio) dá precipitado colorido de laranja com alcalóides.

A cafeína, um derivado da purine, não precipita como a maioria de alcalóides.
4. Os alcalóides são básicos - formam sais solúveis em água. A maioria de alcalóides são substâncias cristalinas bem definidas que se unem com os ácidos formando sais. Nas plantas, podem existir
no estado livre, como sais ou como N-óxidos.
5. Ocorre em um número limitado das plantas. O ácido nucléico existe em todas as plantas, já a morfina existe em somente uma espécie da planta.
Os alcalóides podem ser classificados;
nos termos de sua atividade BIOLÓGICA,
Estrutura QUÍMICA (núcleo que contem o nitrogênio),
caminho BIOSYNTHETIC (a maneira como são produzidos na planta).

Perguntas Freqüentemente Feitas Sobre A Cafeína
1. A química da cafeína e de produtos relacionados
1. Quanto cafeína há dentro [ drink/food/pill ]? 2. Quanto cafeína lá está no café de X? 3. Quimicamente falando, que é cafeína? 4. É verdadeiro que o chá não tem nenhum caffeine/What é
theine, theobromine, etc.? 5. Onde posso eu encontrar um GIF da molécula da cafeína? 6. É verdadeiro que o espresso tem menos cafeína do que o café
regular? 7. Como a cafeína prova? 8. Quanto theobromine/theophylline há...? 9. O café escuro dos roast tem menos cafeína do que roast claros? 10.Como eu meço o índice da cafeína no repouso? 11.Há um limite legal para o índice da cafeína?
2. Cafeína e sua saúde 1. Retirada Da Cafeína 2. Que acontecer quando você overdose? 3. Efeitos da cafeína em mulheres grávidas. 4. Cafeína e Osteoporosis (perda do cálcio) 5. Estudos nos side-effects da cafeína... 6. Cafeína e seu metabolism.
3. Variado 1. Como você pronuncia o mate?
4. Receitas 1. O chocolate cobriu feijões do espresso 2. Como fazer seu próprio chocolate 3. NOTA: para ver se há o café as receitas verificam o FAQ do
café 5. Recursos Eletrônicos 6. Administrivia
1. Como eu começo a cópia a mais nova deste FAQ? 2. Lista dos contribuinte 3. Copyright

1. A química da cafeína e de produtos relacionados
1. Quanto cafeína há dentro [ drink/food/pill ]?
De acordo com a associação nacional de bebidas ;eves, o seguinte é o índice da cafeína em mg por a lata de 12 onças de soda:
Sr. sugar-Free Pibb 58,8
Pepsi um da sacudida 71,2
RedBull 80 da Afri-Cola 100,0 (?) (por 250 ml) 55,5 bebida da energia da bateria da aba 46,8 do surge 51,0 do amarelo 52,8 de Mello do citrino 54 do pontapé do orvalho 55,0 da montanha da dieta do orvalho 55,0 da montanha (nenhuma cafeína em Canadá) (36mg por a lata 8oz, a cafeína do guarana) -- 140mg/l = 46.7mg/can de Barq seco grande da cola 30,0 da dieta RC 36,0 Canadá da cola 36,0 de Pepsi 35,4 RC da dieta de Aspen 36,0 da cola 37,2 de Pepsi do vermelho 38 da tempestade 38 do Dr. Pimenta 39,6 da laranja 40 de Sunkist da soda 40,5 da APROVAÇÃO do Sr. Pibb 40,8 da cola 44,4 da dieta de Shasta da cola 44,4 da cereja de Shasta da cola 44,4 de Shasta da Coca-Cola 45,6 cola seca 1,2 da dieta de Canadá da cerveja 23 da raiz 7 Acima De 0 Barq Da Dieta Da Cerveja 0 Da Raiz Da Caneca De Rite Da Dieta Colas Laranja Minuciosa 0 Da Empregada doméstica Da Laranja 0 De Sundrop Da Cerveja 0 Da Raiz De 0 Sprite 0 & Uma Cerveja 0 Da Raiz De W
Laboratório 101.6mg/500ml da amostra 97.7mg/500ml 2 da amostra 1 de Krankò: Laboratórios De Ameritech, Faculdade Pinta, NY; testado setembro 03, 96

Laboratório 96.4mg/500ml médio de Krankò: Laboratórios de Ameritech, testados agosto 29, 96
Por meio da comparação, um copo de 7 onças do café tem as seguintes quantidades da cafeína (magnésio), de acordo com o bunker e o McWilliams na dieta do J. Am.. 74:28-32, 1979:
Goteje 115-175 Espresso 100mg da cafeína 1 serving (1.5-òz)
80-135 instante brewed 65-100 Decaf, brewed 3-4 Decaf, chá do instante 2-3, congelado (12 onças.) 70 chá, chá 60 brewed, importado, brewed, E. U.. 40 chá, mate 25-150mg do instante 30
A variabilidade na quantidade de cafeína em um copo do café ou de chá é relativamente grande mesmo se preparado pela mesma pessoa usando o mesmo dia do equipamento e dos ingredientes após o dia.
Reference Variability in caffeine consumption from coffee and tea: Possible significance for epidemiological studies by B. Stavric, R. Klassen, B. Watkinson, K. Karpinski, R. Stapley, and P. Fried in "Foundations of Chemical Toxicology ", Volume 26, number 2, pp. 111-118, 1988 e um fácil de ler a vista geral, Looking for the Perfect Brew by S. Eisenberg, "Science News ", Volume 133, April 16, 1988, pp. 252-253.
Cite do manual de laboratório: A cafeína está atual no chá sae e no café à extensão de aproximadamente 4%. O chá contem também outros dois alcalóides, theobromine e theophylline. Estes últimos dois relaxam os músculos lisos onde a cafeína estimula o coração e os sistemas respiratory.
Os efeitos do theobromine são, comparado à cafeína e ao theophylline, relativamente moderados. Entretanto, a cacau contem oito vezes mais theophylline do que a cafeína. Também, a cafeína foi mostrada à

liga com outras substâncias para o potency adicionado. Assim os efeitos do theobromine puderam ser realçados pela cafeína no chocolate.
O theobromine é altamente tóxico aos cães e mata muitos canids/year através do envenenamento do chocolate . Faz exame completamente de um dose aos níveis fatais do alcance (um bodyweight de mais de 200 mg/kg) mas alguns cães têm um hábito mau de comer fora das latas do lixo e alguns proprietários têm um hábito mau de cães de alimentação doce. Alguns oreos não ferirão um cão, mas uma libra do chocolate pode fazer os danos consideráveis.
Os sinais clínicos do toxicity do theobromine nos canids geralmente manifestam 8 horas após o ingestion e podem incluir: thirst, vomiting, diarrhea, incontinence urinary, nervousness, spasms clonic do músculo, apreensões e coma. Todo o pensamento do cão para ingested uma quantidade grande do chocolate deve ser trazido a uma clínica da emergência o mais cedo possível, onde o tratamento inclui geralmente o uso do emetics e do carvão de lenha ativado. O cão necessitará assim ser monitorado para manter o contrapeso apropriado do líquido e do eletrólito.
Pathogenesis do toxicity do theobromine: evidente as quantidades grandes do theobromine têm um efeito diuretic, relaxam os músculos lisos, e estimulam o coração e o cns.
Referência: Fraser, M. de Clarence, et al, eds. O manual veterinário de Merck, 7o ed. Rahway, NJ: Merck & Co., Inc. 1991. pp. 1643-44.
Em atos da cafeína dos seres humanos particularmente no cérebro e nos músculos esqueletais quando o theophylline alvejar o coração, o bronchia, e os kidneys.
Outros dados na cafeína:
O copo da barra 30mg do chocolate da cola 30-45mg do mate 25-150mg do chá 30-70mg do café imediato 60-80mg do café 90-150mg Permaneç-acorda a tabuleta fria 30mg do relevo do pill 100mg Vivarin 200mg

A seguinte informação é de Bowes and Church's Food values of portions commonly used, by Anna De Planter Bowes. Lippincott, Phila. 1989. Pages 261-2: Caffeine. Doce:
Choc do baking da cafeína do magnésio do chocolate, unsweetened, padeiros -- 1 doce alemão de oz(28 g) 25, padeiros -- 1 onça (28 g) 8 semi-sweet, padeiros -- 1 onça (28 g) 13
Choc lasca padeiros -- 1/4 de copo (43 g) 13 doce alemão, padeiros -- 1/4 de copo (43 g) 15
Barra do chocolate, Cadbury -- 1 onça (28 g) 15 barra de 1,4 onças do choc do leite. 3-10 barra de 1,4 onças do choc branco 2-4 barra de 1,4 onças do leite de chocolate escuro do leite de chocolate 8oz do choc 28 8 vidro 2-7 de 8 onças
Desserts: PNF do pudim de Jello, Choc (47 g) 2 mousse de Choc da mistura de Jell-O (95 g) 6 mousse do fudge do choc de Jello (86 g) 12 chocolate cobriram o feijão do espresso 3-5
Bebidas3 teaspoons heaping da mistura 8 do pó do choc 2 colheres do syrup 5 do choc 1 mistura quente 5 da cacau do envelope
As fórmulas dietéticas asseguram-se, sinal de adição, choc, laboratórios de Ross -- 8 onças (259 g) 10 barra do chocolate de leite de Cadbury

Mais material:
O poder " mágico de Guarana " (completamente comum em Germany), 15 ml de álcool com 5g Guarana semeia 250,0 cápsulas do magnésio Guarana com 500 magnésio das sementes 25,0 do magnésio G./cápsula
(cafeína supondo de 5% nas sementes como indicado na literatura)O PNF da soda de Guarana está ubiquitous em Brasil e frequentemente disponível no mantimento tropical aqui. É realmente tasty e embala um wallop. Guarana acorda-o acima como louco, mas não causa tremores do café. É possível que além à cafeína, há alguma outra substância no guarana que produz também um efeito, desde que ' sentiu diferente do que o café. Mesmo vai para o mate. Retorne ao índice
2. Quanto cafeína lá está no café de X? Índice da cafeína nos feijões e nas misturas (fonte: Boletim de notícias -- Mountanos Bros. Café Co., San Francisco) Bourbons 1,20% Celebes Kalossi de VARIETALS/STRAIGHTS Brasil Colômbia 1,22 Excelso Colômbia 1,37 Supremo 1,37 Harrar-Moka Ethiopian G 1,13
*** TRANSLATION ENDS HERE ***uatemala Antigua 1.32Indian Mysore 1.37Jamaican Blue Mtn/Wallensford Estate 1.24Java Estate Kuyumas 1.20Kenya AA 1.36Kona Extra Prime 1.32Mexico Pluma Altura 1.17Mocha Mattari (Yemen) 1.01New Guinea 1.30Panama Organic 1.34

Sumatra Mandheling-Lintong 1.30Tanzania Peaberry 1.42Zimbabwe 1.10
BLENDS & DARK ROASTSColombia Supremo Dark 1.37%Espresso Roast 1.32French Roast 1.22Vienna Roast 1.27Mocha-Java 1.17
DECAFS--all @ .02% with Swiss Water Process
Return to Index
3. Chemically speaking, what is caffeine? Caffeine is an alkaloid. There are numerous compounds called alkaloids, among them we have the methylxanthines, with three distinguished compounds: caffeine, theophylline, and theobromine, found in cola nuts, coffee, tea, cacao beans, mate and other plants. These compounds have different biochemical effects, and are present in different ratios in the different plant sources. These compounds are very similar and differ only by the presence of methyl groups in two positions of the chemical structure. They are easily oxidized to uric acid and other methyluric acids which are also similar in chemical structure. Caffeine: Sources: Coffee, tea, cola nuts, mate, guarana. Effects: Stimulant of central nervous system, cardiac muscle, and respiratory system, diuretic Delays fatigue. Theophylline: Sources: Tea Effects: Cariac stimulant, smooth muscle relaxant, diuretic, vasodilator Theobromine: Sources: Principle alkaloid of the cocoa bean (1.5-3%) Cola nuts and tea

Effects: Diuretic, smooth muscle relaxant, cardiac stimulant, vasodilator. (Info from Merck Index) The presence of the other alkaloids in colas and tea may explain why these sometimes have a stronger kick than coffee. Colas, which have lower caffeine contents than coffee are, reportedly, sometimes more active. Tea seems the strongest for some. Coffee seems more lasting for mental alertness and offers fewer jitters than the others. A search in CAS and produced these names and synonyms: RN 58-08-2 REGISTRYCN 1H-Purine-2,6-dione, 3,7-dihydro-1,3,7-trimethyl- (9CI) (CA INDEX NAME)OTHER CA INDEX NAMES:CN Caffeine (8CI)OTHER NAMES:CN 1,3,7-Trimethyl-2,6-dioxopurineCN 1,3,7-TrimethylxanthineCN 7-MethyltheophyllineCN Alert-PepCN CafeinaCN CaffeinCN CafipelCN GuaranineCN KoffeinCN MateinaCN MethyltheobromineCN No-DozCN Refresh'nCN StimCN TheinCN TheineCN Tri-Aqua
MF C8 H10 N4 O2
The correct name is the first one, 1H-Purine-2,6-dione, 3,7-dihydro-1,3,7-trimethyl- (This is the "inverted name ") The "uninverted name " is 3,7-Dihydro-1,3,7-trimethyl-1H-purine-2,6-dione

Merck Index excerpt... Caffeine: 3,7-dihydro- 1,3,7-trimethyl- 1H-purine- 2,6-dione; 1,3,7-trimethylxanthine; 1,3,7-trimethyl- 2,6-dioxopurine; coffeine; thein; guaranine; methyltheobromine; No-Doz.
C8H10N4O2; mol wt 194.19. C 49.48%, H 5.19%, N 28.85%, O 16.48%.
Occurs in tea, coffee, mate leaves; also in guarana paste and cola nuts: Shuman, U.S. pat. 2,508,545 (1950 to General Foods). Obtained as a by-product from the manuf of caffeine-free coffee: Barch, U.S. pat. 2,817,588 (1957 to Standard Brands); Nutting, U.S. pat. 2,802,739 (1957 to Hill Bros. Coffee); Adler, Earle, U.S. pat. 2,933,395 (1960 to General Foods).
Crystal structure: Sutor, Acta Cryst. 11, 453, (1958). Synthesis: Fischer, Ach, Ber. 28, 2473, 3135 (1895); Gepner, Kreps, J. Gen. Chem. USSR 16, 179 (1946); Bredereck et al., Ber. 83, 201 (1950); Crippa, Crippa, Farmaco Ed. Sci. 10, 616 (1955); Swidinsky, Baizer, U.S. pats. 2,785,162 and 2,785,163 (1957 to Quinine Chem. Works); Bredereck, Gotsmann, Ber. 95, 1902 (1962).
Hexagonal prisms by sublimation, mp 238 C. Sublimes 178 C. Fast sublimation is obtained at 160-165 C under 1mm press. at 5 mm distance. d 1.23. Kb at 19 C: 0.7 x 10^(-14). Ka at 25 C: <1.0 x 10^(-14). pH of 1% soln 6.9. Aq solns of caffeine salts dissociate quickly. Absorption spectrum: Hartley, J. Chem. Soc. 87, 1802 (1905). One gram dissolves in 46 ml water, 5.5 ml water at 80 C, 1.5 ml boiling water, 66 ml alcohol,

22 ml alcohol at 60 C, 50 ml acetone, 5.5 ml chloroform, 530 ml ether, 100 ml benzene, 22 ml boiling benzene. Freely sol in pyrrole; in tetrahydrofuran contg about 4% water; also sol in ethyl acetate; slightly in petr ether. Soly in water is increased by alkali benzoates, cinnamates, citrates, or salicylates.
Monohydrate, felted needles, contg 8.5% H2O. Efflorescent in air; complete dehydration takes place at 80 C. LD50 orally in rats: 200 mg/kg.
Acetate, C8H10N4O2.(CH3COOH)2, granules or powder; acetic acid odor; acid reaction. Loses acetic acid on exposure to air. Soluble in water or alcohol with hydrolysis into caffeine and acetic acid. Keep well stoppered.
Hydrochloride dihydrate, C8H10N4O2.HCl.2H2O, crystals, dec 80-100 C with loss of water and HCl. Sol in water and in alcohol with dec.
Therap Cat: Central stimulant.
Therap Cat (Vet): Has been used as a cardiac and respiratory stimulant and as a diuretic.
Return to Index
4. Is it true that tea has no caffeine/What is theine, theobromine, etc? From "Principles of biochemistry ", Horton and al, 1993.
Caffeine is sometimes called "theine " when it's in tea. This is probably due to an ancient misconception that the active constituent is different. Theophylline is present only in

trace amounts. It is more diuretic, more toxic and less speedy.
Caffeine
1,3,7-trimethylxanthine
Theophylline
1,3-dimethylxanthine
Theobromine
3,7-dimethylxanthine
Coffee and tea contain caffeine and theophylline, respectively, which are methylated purine derivatives that inhibit cAMP phosphodiesterase. In the presence of these inhibitors, the effects of cAMP, and thus the stimulatory effects of the hormones that lead to its production, are prolonged and intensified.
Theobromine and theophylline are two dimethylxanthines that have two rather than three methyl groups. Theobromine is considerably weaker than caffeine and theophylline, having about one tenth the stimulating effect of either. Theobromine is found in cocoa products, tea (only in very small amounts) and kola nuts, but is not found in coffee. In cocoa, its concentration is generally about 7 times as great as caffeine. Although, caffeine is relatively scarce in cocoa, its mainly because of theobromine that cocoa is "stimulating ". Theophylline is found in very small amounts in tea, but has a stronger effect on the heart and breathing than caffeine. For this reason it is often the drug of choice in home remedies for treating asthma bronchitis and emphysema. The theophylline found in medicine is made from extracts from coffee or tea. Return to Index
5. Where can I find a gif of the caffeine molecule? Caffeine = 1,3,7-Trimethylxanthine A different view of the caffeine molecule.

The Department of Chemistry at Jamaica of the University of Western Indies has made available an avi and an mpeg of a rotation of the caffeine molecule, among other molecules and chemical processes. The index page contains more information and the links to the clips.
CH3 | N / \ N----C C==O || || | || || | CH C N--CH3 \ / \ / N C | || CH3 O
There is a gif picture at the wuarchive.wustl.edu ftp site or any of its mirror sites under multimedia/images/gif/c
caffeine
Theobromine is also a common component of coffee, tea, chocolate, and mate (particularly in these last two).

Theobromine
CH3 | N / \ N----C C==O || || | || || | CH C N--H \ / \ / N C | || CH3 O
Theophylline was once thought to be a major component of tea. This is not correct. Tea contains significantly more amounts of caffeine than of theophylline. Theophylline
CH3 | N / \ N----C C==O || || | || || | CH C N--CH3 \ / \ / N C | || H O
Return to Index

6. Is it true that espresso has less caffeine than regular coffee? Yes and no. An espresso cup has about as much caffeine as a cup of dark brew. But servings for espresso are much smaller. Which means that the content of caffeine per milliliter are much higher than with a regular brew. Moreover, caffeine is more quickly assimilated when taken in concentrated dosages, such as an espresso cup. The myth of lower caffeine espresso comes comes from the fact that the darker roast beans used for espresso do have less caffeine than regularly roasted beans as roasting is supposed to break up or sublimate the caffeine in the beans (I have read this quote on research articles, but found no scientific studies supporting it. Anybody out there?). But espresso is prepared using pressurized water through significantly more ground (twice as much?) than regular drip coffee, resulting in a higher percentage of caffeine per milliliter. Please refer also to Does dark roast coffee have less caffeine than light roast? Here's the caffeine content of Drip/Espresso/Brewed Coffee: Drip 115-175Espresso 100 1 serving (1-2oz)Brewed 80-135
Return to Index 7. How does caffeine taste?
Caffeine is very bitter. Barq's Root Beer contains caffeine and the company says that it has "12.78mg per 6oz " and that they "add it as a flavoring agent for the sharp bitterness "Return to Index
8. How much theobromine/theophylline there is in ...? Sources: Physicians Desk Reference and Institute of Food Technologies from Pafai and Jankiewicz (1991) DRUGS AND HUMAN BEHAVIOUR cocoa 250mg theobrominebittersweet choc. bar 130mg theobromine5 oz cup brewed coffee no theobromine

tea 5oz cup brewed 3min with teabag 3-4 mg theophyllineDiet Coke no theobromine or theophylline
Return to Index 9. Does dark roast coffee have less caffeine than light roast?
It really depends on how you measure the caffeine. If you measure by weight you actually have more caffeine in dark roast because the water loss if faster than the caffeine loss. If you measure by volume you have less caffeine because the beans expand as they roast. Return to Index
10.How do I measure caffeine content at home? To the best of my knowledge this can not be accomplished without sophisticated equipment. The Department of Energy's web page briefly explains what is involved. Return to Index
11.Is there a legal limit for caffeine content? The answer to that is it depends on the country. A few examples of laws related to caffeine content for food and drinks include the following: In the United States there is a limit of 6mg of caffeine per liquid ounce in beverages. There is also a limit of 200mg in pills such as Vivrin.
Australia has a limit of Australia 145mg of caffeine per liter.
In parts of Northern Thailand it is completely illegal. It was outlawed as a precursor to meth. Return to Index
Caffeine and your Health Important: This information was excerpted from several sources, no claims are made to its accuracy. The FAQ mantainer is not a medical doctor and cannot vouch for the accuracy of this information.

1. Caffeine Withdrawal: Procedures and Symptoms.
How to cut caffeine intake? Most people report a very good success ratio by cutting down caffeine intake at the rate of 1/2 cup of coffee a day. This is known as Caffeine Fading . Alternatively you might try reducing coffee intake in discrete steps of two-five cups of coffee less per week (depending on how high is your initial intake). If you are drinking more than 10 cups of coffee a day, you should seriously consider cutting down. The best way to proceed is to consume caffeine regularly for a week, while keeping a precise log of the times and amounts of caffeine intake (remember that chocolate, tea, soda beverages and many headache pills contain caffeine as well as coffee). At the end of the week proceed to reduce your coffee intake at the rate recommended above. Remember to have substitutes available for drinking: if you are not going to have a hot cup of coffee at your 10 minute break, you might consider having hot chocolate or herbal tea, but NOT decaff, since decaff has also been shown to be addictive. This should take you through the works without much problem. Some other people quit cold turkey. Withdrawal symptoms are quite nasty this way (see section below) but they can usually be countered with lots of sleep and exercise. Many people report being able to stop drinking caffeine almost cold-turkey while on holidays on the beach. If quitting cold turkey is proving too hard even in the beach, drinking a coke might help. What are the symptoms of caffeine withdrawal? Regular caffeine consumption reduces sensitivity to caffeine. When caffeine intake is reduced, the body becomes oversensitive to adenosine. In response to this oversensitiveness, blood pressure drops dramatically, causing an excess of blood in the head (though not necessarily on the brain), leading to a headache. This headache, well known among coffee drinkers, usually lasts from one to five days, and can be alleviated with analgesics such as aspirin. It is also alleviated with caffeine intake (in fact several analgesics contain caffeine dosages).

Often, people who are reducing caffeine intake report being irritable, unable to work, nervous, restless, and feeling sleepy, as well as having a headache. In extreme cases, nausea and vomiting has also been reported. References. Caffeine and Health. J. E. James, Academic Press, 1991. Progress in Clinical and Biological Research Volume 158. G. A. Spiller, Ed. Alan R. Liss Inc, 1984. Return to Index
2. What happens when you overdose? From Desk Reference to the Diagnostic Criteria from DSM-3-R (American Psychiatric Association, 1987):
Caffeine-Induced Organic Mental Disorder 305.90 Caffeine Intoxication
1. Recent consumption of caffeine, usually in excess of 250 mg.
2. At least five of the following signs:
1. restlessness
2. nervousness
3. excitement
4. insomnia
5. flushed face
6. diuresis
7. gastrointestinal disturbance
8. muscle twitching
9. rambling flow of thought and speech
10.tachycardia or cardiac arrhythmia
11.periods of inexhaustibility

12.psychomotor agitation
3. Not due to any physical or other mental disorder, such as an Anxiety Disorder.
Basically, overdosing on caffeine will probably be very very unpleasant but not kill or deliver permanent damage. However, People do die from it. Toxic dose
The LD_50 of caffeine (that is the lethal dosage reported to kill 50% of the population) is estimated at 10 grams for oral administration. As it is usually the case, lethal dosage varies from individual to individual according to weight. Ingestion of 150mg/kg of caffeine seems to be the LD_50 for all people. That is, people weighting 50 kilos have an LD_50 of approx. 7.5 grams, people weighting 80 kilos have an LD_50 of about 12 grams.
In cups of coffee the LD_50 varies from 50 to 200 cups of coffee or about 50 vivarins (200mg each).
One exceptional case documents survival after ingesting 24 grams. The minimum lethal dose ever reported was 3.2 grams intravenously , this does not represent the oral MLD (minimum lethal dose).
In small children ingestion of 35 mg/kg can lead to moderate toxicity. The amount of caffeine in an average cup of coffee is 50 - 200 mg. Infants metabolize caffeine very slowly.
Symptoms
Acute caffeine poisoning gives early symptoms of anorexia, tremor, and restlessness. Followed by nausea, vomiting, tachycardia, and confusion. Serious intoxication may cause delirium, seizures, supraventricular and ventricular tachyarrhythmias, hypokalemia, and hyperglycemia.

Chronic high-dose caffeine intake can lead to nervousness, irritability, anxiety, tremulousness, muscle twitching, insomnia, palpitations and hyperreflexia. For blood testing, cross-reaction with theophylline assays will detect toxic amounts. (Method IA) Blood concentration of 1-10 mg/L is normal in coffee drinkers, while 80 mg/L has been associated with death.
Treatment
Emergency Measures
Maintain the airway and assist ventilation. (See Appendix A)
Treat seizures & hypotension if they occur.
Hypokalemia usually goes away by itself.
Monitor Vital Signs.
Specific drugs & antidotes. Beta blockers effectively reverse cardiotoxic effects mediated by excessive beta-adrenergic stimulation. Treat hypotension or tachyarrhythmias with intravenous propanolol, .01 - .02 mg/kg. , or esmolol, .05 mg/kg , carefully titrated with low doses. Esmolol is preferred because of its short half life and low cardioselectivity.
Decontamination
Induce vomiting or perform gastric lavage.
Administer activated charcoal and cathartic.
Gut emptying is probably not needed if 1 2 are performed promptly.
Appendix A
Performing airway assistance.

4. If no neck injury is suspected, place in the "Sniffing " position by tilting the head back and extending the front of the neck.
5. Apply the "Jaw Thrust " to move the tongue out of the way without flexing the neck: Place thumb fingers from both hands under the back of the jaw and thrust the jaw forward so that the chin sticks out. This should also hurt the patient, allowing you to judge depth of coma. :)
6. Tilt the head to the side to allow vomit and snot to drain out.
From conversations on alt.drugs.caffeine: The toxic dose is going to vary from person to person, depending primarily on built-up tolerance. A couple people report swallowing 10 to 13 vivarin and ending up in the hospital with their stomaches pumped, while a few say they've taken that many and barely stayed awake. A symptom lacking in the clinical manual but reported by at least two people on the net is a loss of motor ability: inability to move, speak, or even blink. The experience is consistently described as very unpleasant and not fun at all, even by those very familiar with caffeine nausea and headaches. Return to Index
3. Effects of caffeine on pregnant women. Caffeine has long been suspect of causing mal-formations in fetus, and that it may reduce fertility rates. These reports have proved controversial. What is known is that caffeine does causes malformations in rats, when ingested at rates comparable to 70 cups a day for humans. Many other species respond equally to such large amounts of caffeine. Data is scant, as experimentation on humans is not feasible. In any case moderation in caffeine ingestion seems to be a prudent course for pregnant women. Recent references are Pastore and Savitz, Case-control study of caffeinated beverages and preterm delivery. American Journal of Epidemiology, Jan 1995. A recent study found a weak link between Sudden-Infant-Death-Syndrome (SIDS) and caffeine consumption by the

mother, which reinforces the recommendation for moderation -possibly even abstinence- above. On men, it has been shown that caffeine reduces rates of sperm motility which may account for some findings of reduced fertility. Return to Index
4. Caffeine and Osteoporosis (Calcium loss) From the Journal of AMA: (JAMA, 26 Jan. 1994, p. 280-3.) "There was a significant association between (drinking more) caffeinated coffee and decreasing bone mineral density at both the hip and the spine, independent of age, obesity, years since menopause, and the use of tobacco, estrogen, alcohol, thiazides, and calcium supplements [in women]. " Except when: "Bone density did not vary [...] in women who reported drinking at least one glass of milk per day during most of their adult lives. " That is, if you drink a glass of milk a day, there is no need to worry about the caffeine related loss of calcium. Return to Index
5. Studies on the side-effects of caffeine. OAKLAND, California (UPI) -- Coffee may be good for life. A major study has found fewer suicides among coffee drinkers than those who abstained from the hot black brew. The study of nearly 130,000 Northern California residents and the records of 4,500 who have died looked at the effects of coffee and tea on mortality. Cardiologist Arthur Klatsky said of the surprising results, ``This is not a fluke finding because our study was very large, involved a multiracial population, men, women, and examined closely numerous factors related to mortality such as alcohol consumption and smoking.'' The unique survey also found no link between coffee consumption and death risk.

And it confirmed a ``weak'' connection of coffee or tea to heart attack risk -- but not to other cardiovascular conditions such as stroke. The study was conducted by the health maintenance organization Kaiser Permanente and was reported Wednesday in the Annals of Epidemiology. Return to Index
6. Caffeine and your metabolism. Caffeine increases the level of circulating fatty acids. This has been shown to increase the oxidation of these fuels, hence enhancing fat oxidation. Caffeine has been used for years by runners and endurance people to enhance fatty acid metabolism. It's particularly effective in those who are not habitual users. Caffeine is not an appetite suppressant. It does affect metabolism, though it is a good question whether its use truly makes any difference during a diet. The questionable rationale for its original inclusion in diet pills was to make a poor man's amphetamine-like preparation from the non-stimulant sympathomimetic phenylpropanolamine and the stimulant caffeine. (That you end up with something very non-amphetamine like is neither here nor there.) The combination drugs were called "Dexatrim " or Dexa-whosis (as in Dexedrine) for a reason, namely, to assert its similarity in the minds of prospective buyers. However, caffeine has not been in OTC diet pills for many years per order of the FDA, which stated that there was no evidence of efficacy for such a combination. From Goodman and Gilman's The Pharmacological Basis of Therapeutics:
Caffeine in combination with an analgesic, such as aspirin, is widely used in the treatment of ordinary types of headache. There are few data to substantiate its efficacy for this purpose. Caffeine is also used in combination with an ergot alkaloid in the treatment of migrane (Chapter 39).

Ergotamine is usually administered orally (in combination with caffeine) or sublingually [...] If a patient cannot tolerate ergotamine orally, rectal administration of a mixture of caffeine and ergotamine tartarate may be attempted.
The bioavailability [of ergotamine] after sublingual administration is also poor and is often inadequate for therapeutic purposes [...] the concurrent administration of caffeine (50-100 mg per mg of ergotamine) improves both the rate and extent of absorption [...] However, there is little correspondence between the concentration of ergotamine in plasma and the intensity or duration of therapeutic or toxic effects.
Caffeine enhances the action of the ergot alkaloids in the treatment of migrane, a discovery that must be credited to the sufferers from the disease who observed that strong coffee gave symptomatic relief, especially when combined with the ergot alkaloids. As mentioned, caffeine increases the oral and rectal absorption of ergotamine, and it is widely believed that this accounts for its enhancement of therapeutic effects.
Nowadays most of researchers believe that the stimulatory actions are attributable to the antagonism of the adenosine. Agonists at the adenosine receptors produce sedation while antagonists at these sites, like caffeine and theophylline induce stimulation, and what is even more important, the latter substance also reverse agonists-induced symptoms of sedation, thus indicating that this effects go through these receptors. Another possibility, however, is that methylxanthines enhance release of excitatory aminoacids, like glutamate and aspartate, which are the main stimulatory neurotransmitters in the brain.

As to the side effects: methylxanthines inhibit protective activity of common antiepileptic drugs in exptl. animals in doses comparable to those used in humans when correction to the surface area is made. It should be underlined, that although tolerance develop to the stimulatory effects of theo or caffeine when administered on a chronic base, we found no tolerance to the above effects . This hazardous influence was even enhanced over time. Therefore, it should be emphasized that individuals suffering from epilepsy should avoid, or at least reduce consumption of coffee and other caffeine-containing beverages. Return to Index
Miscellaneous
1. How do you pronounce mate? MAH-teh. MAH like in malt, and -teh like in Gral. Patten. Return to Index
Recipes.
1. Chocolate covered espresso beans You won't get single, glossy beans, but the taste is there!
1. Put dark roast coffee beans on a waxpaper-covered baking sheet.
2. Melt some chocolate by puting a container with the chocolate in a pan of boiling water, stir the chocolate when it is getting hot. Some experimentation regarding what chocolate to use is in place. I used chocolate chips of from Girardelli. One should probably aim for dark and not too sweet chocolate.
3. Pour the chocolate over the beans and smear it so that each bean is covered - you should have a single layer of covered beans not too far apart.
4. When the beans have cooled off a little bit, put the sheet in the fridge/freezer.
5. When solid, break off a piece and enjoy. 2. How to make your own chocolate
Here's the recipe for making a real chocolate beverage. Important steps are in boldface .

Ingredients 1-2kg (2-4pounds) of cocoa beans. A manually operated grinder.
Instructions Sift through the beans removing any impurities (pieces of
grass, leaves, etc). Place the beans in a pan (no teflon) and roast them. Stir
frequently. As the beans roast they start making "pop " sounds like popcorn. Beans are ready when you estimate that approx 50-75% of the beans have popped. Do not let the beans burn, though a bit of black on each bean is ok.
Peel the beans. Peeling roasted cocoa beans is like peeling baked potatoes: The hotter they are the easier it is to peel the darn things, at the expense of third degree burns on your fingers. (Tip: Use kitchen mittens and brush the beans in your hands). If the beans are too hard to peel roast them a bit longer.
Grind the beans into a pan. They produce a dark oily paste called "cocoa paste ".
The oil in the cocoa has a bitter taste that you have to get used to. I like it this way, but not all people do. Here are the alternatives: With oil , which gives you a richer flavour: Spread aluminum foil on a table and make small pies of chocolate, about 1/4 of an inch high, and 6 inches in diameter. Let them rest overnight. The morning after they are hard tablets. Remove them from the aluminum foil and rap them in it. Store in the freezer. Without oil , some flavour is gone, less bitter, weaker (whimper) chocolate: Put the paste inside a thin cloth (like linen), close the cloth and squeeze until the oil comes out. If you manage to get most of the oil out, what is left is high quality cocoa powder, like Droste's. What is left now is either bitter tablets or bitter cocoa powder.
You can now make a nice beverage as follows:

Boil a liter of milk (or water, like in ancient Mexican style. Like water for chocolate, "Como agua para chocolate ": you know).
When the milk is warm (not hot) add a chocolate pie in pieces. Stir with a blender (but be careful! the blender's electric cord should NOT touch the pot or any other hot thing around it).
When the chocolate has dissolved add 1/2-3/4 cups of sugar (depending how sweet you like your chocolate) and blend in fast. Make sure the sugar is completely dissolved in the chocolate otherwise it would be bitter no matter how much sugar you may add afterwards .
Add a teaspoon of cinnamon or natural vanilla flavour (artificial vanilla flavour with chocolate results in an awful medicine like flavour) if you like, and blend again.
Let the mixture boil, when it starts to get bubbly quickly remove the pan from the stove top, and rest the bottom against a soaked cloth. Put again on stove top, it should get bubbly almost immediately, remove once again and repeat one last time. This aerates the chocolate which enhances flavour.
In a mug, put about 1/2-3/4 of the chocolate mixture, and add cold milk, until the temperature and/or the concentration of the flavour is right for your tastes. Accompany with French Pastries. Yum Yum!!
Enjoy! Return to Index
Electronic Resources Return to Index
Administrivia
How do I get the newest copy of this FAQ? How do I get the newest copy of this FAQ? My page at http://coffeefaq.com/ or via e-mail send a message to [email protected] or for the coffee faq: My page at http://coffeefaq.com/

or via e-mail send a message to [email protected]
List of Contributors This FAQ is a collective effort. Here's a list of most (all?) of the contributors.
Oktay Ahiska Marc Aurel Scott Austin Tom Benjamin Jennifer Beyer Steve Bliss David Alan Bozak Rajiv Trevor P. Bugera Jack Carter Richard Drapeau Jym Dyer Steve Dyer Stefan Engstrom Lemieux Francois Scott Fisher Dave Huddle Matt Humphrey Tom F Karlsson Bob Kummerfeld Dr. Robert Lancashire John Levine Alex Lopez-Ortiz Alec Muffett Dana Myers Tim Nemec Mike Oliver Jim Pailin Dave Palmer Stuart Phillips Siobhan Purcell Cary A. Sandvig Jesse T Sheidlower Stepahine da Silva

Michael A Smith Mari J. Stoddard Thom Deanna K. Tobin T.E. Nick Tsoukas Adam Turoff Ganesh Uttam David R. B. Walker Orion Wilson Piotr Wlaz Ted Young Steven Zikopoulos Susan Smith Kevin Mackie
Copyright This FAQ is Copyright (C) 1994,1995 by Alex Lopez-Ortiz. This FAQ is Copyright © 1998,2001 by Daniel Owen. This text, in whole or in part, may not be sold in any medium, including, but not limited to, electronic, CD-ROM, or published in print, without the explicit, written permission of Daniel Owen. Return to Index
Copyright (C) 1994,1995 Alex López-Ortiz. Copyright © 1998,2001 Daniel Owen.
Home Page Please send comments to Daniel Owen.
FAQ Dos Portadores Do Tryptamine
Por Petrus Pennanen

com ajuda de Michael de Melbourne . Hypertexture e esforço ©ontinuing da ilustração através de http://deoxy.org
Traduza de
Agradecimentos a muitos indivíduos para a ajuda em unir isto. Se você souber as fontes dos tryptamines que não são mencionados aqui satisfazem enviam-nos.
Última busca de julho 1999 do update do deoxy este FAQ para
Inibidores e tryptamines de MAO Síntese de derivatives de DMT Sapos De Psychedelic Os Fungos As Plantas Referências
Oral e parenterally derivatives psychotropic ativos do tryptamine Baseado em McKenna & em torres 1984
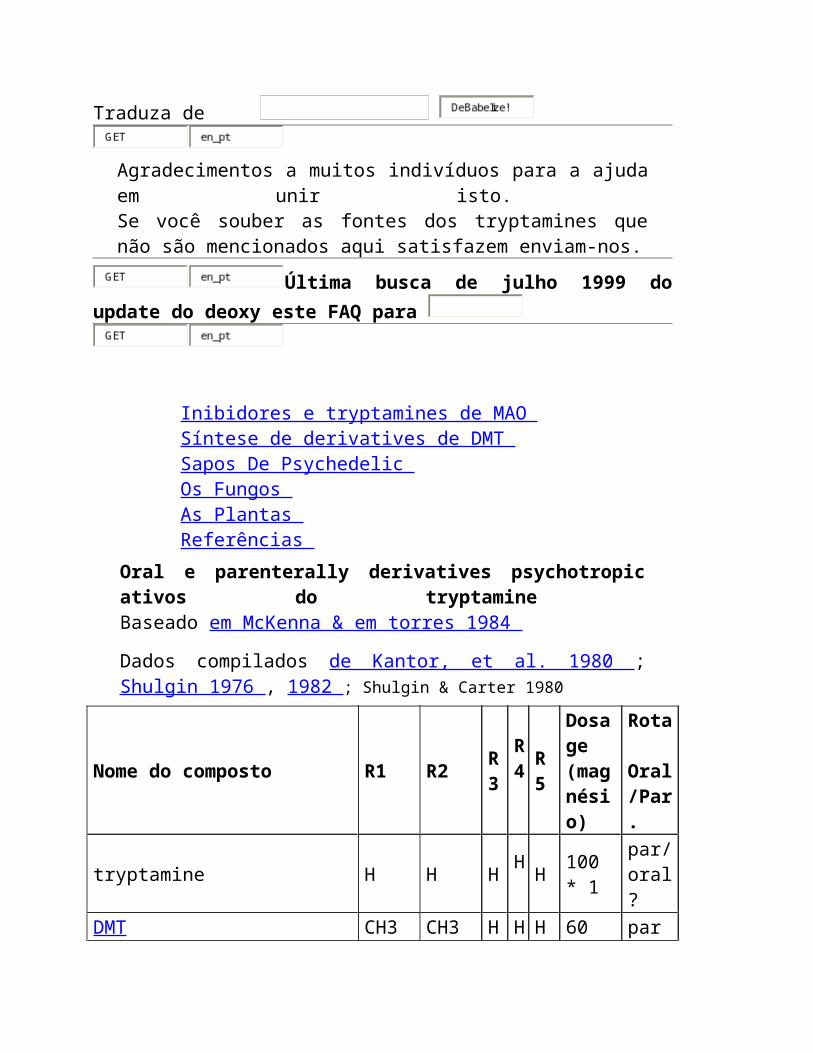
Dados compilados de Kantor, et al. 1980 ; Shulgin 1976 , 1982 ; Shulgin & Carter 1980
Nome do composto R1 R2 R3
R4
R5
Dosage (magnésio)
Rota Oral/Par.
tryptamine H H H H H 100 *
1 par/oral?
DMT (dimethyltryptamine) CH3 CH3 H H H 60 par
DET C2H5 C2H5 H H H 60 par/
oral
DPT n-suporte
n-suporte
H H H 60 par/
oral

DAT C3H5 C3H5 H H H 30 par/
oral
DIPT eu-suporte
eu-suporte
H H H 30 oral
5-MeO-DIPT eu-suporte
eu-suporte
H H
OCH3
12 oral
5-MeO-DMT CH3 CH3 H H
OCH3
6 par
psilocin CH3 CH3 H Oh
H 12 * 2 oral
Cz-74 C2H5 C2H5 H Oh
H 15 * 2 oral
serotonin H H H H
Oh
100 * 3 oral
bufotenine CH3 CH3 H H
Oh 16 * 4 par
It-290 H H CH3
H H 30 oral
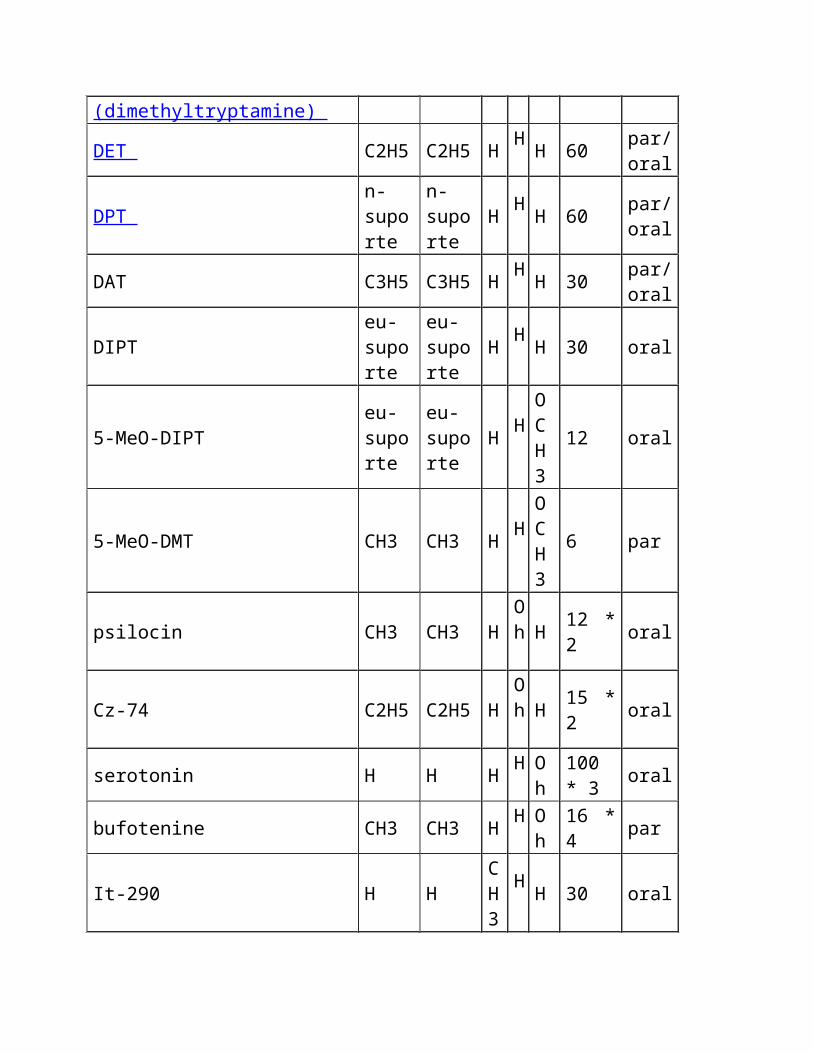
4-hydroxy-alfa-methyl-tryptamine H H CH3
Oh
H 20 * 3 oral
Mp-809 H H CH3
H
CH3
60 * 5 oral
5-fluoro-alfa-methyl-tryptamine H H CH3
H F 25 * 6 oral
5-methoxy-alfa-methyl-tryptamine
H H CH3
H
OCH
3 oral

3
4-hydroxy-diisopropyl-tryptamine eu-suporte
eu-suporte
H Oh
H 12 * 6 oral
4-hydroxy-N-isopropyl, N-methyl-n-methyl-tryptamine
eu-suporte
CH3 H Oh
H 6 * 6 oral
N-t-butyl-n-t-butyl-tryptamine H t-butylH
H H ? * 7 par?
3-(2-(2,5-dimethylpyrrolyl) ethyl)-ethyl)-indole H H H ? ? ? ?
5-alfa-DMT CH3 CH3 CH3
H H ? ?
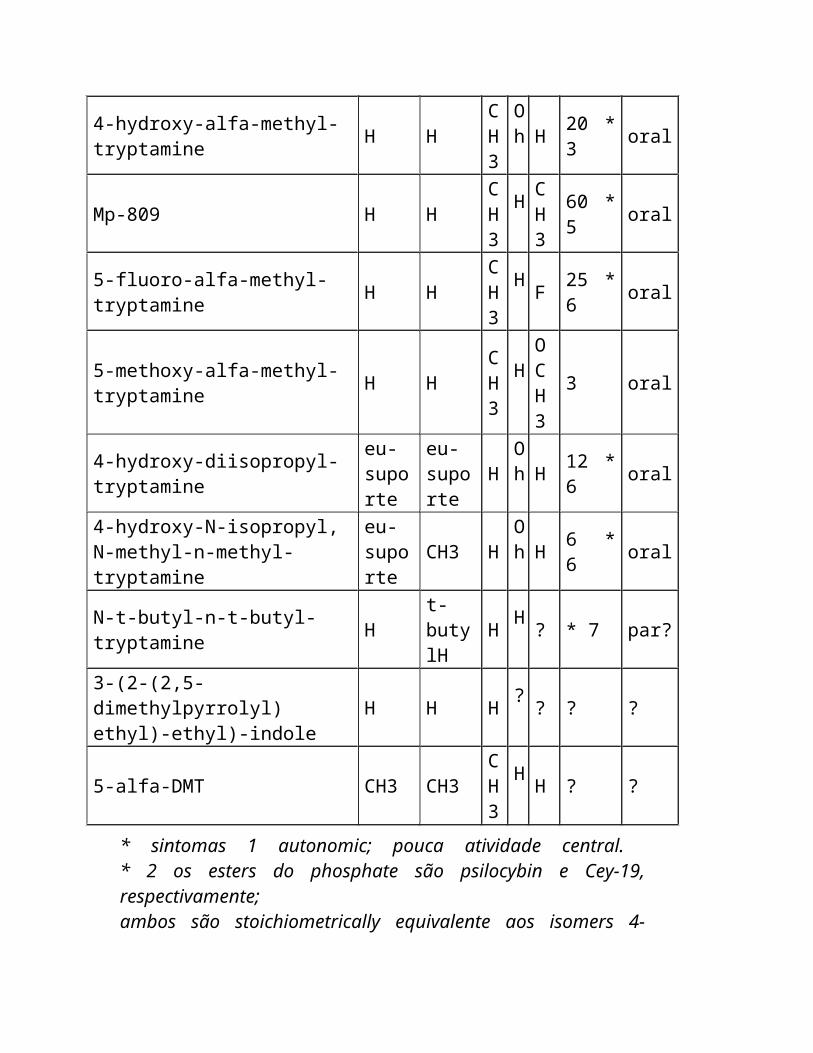
* sintomas 1 autonomic; pouca atividade central. * 2 os esters do phosphate são psilocybin e Cey-19, respectivamente; ambos são stoichiometrically equivalente aos isomers 4-hydroxy. * 3 cardiovascular e sintomas autonomic; pouca atividade central. * amine pressor de 4 A melhor que um hallucinogen no homem. * 5 um antidepressivo melhor que um hallucinogen no homem. * 6 basearam em relatórios anonymous na imprensa da configuração.
Nenhum estudo clínico foi publicado. * 7 nenhuma atividade oral com doses até magnésio 20, podem ser parenterally ativos.
Outros tryptamines potencial psychedelic incluem:
6-fluoro-alfa-methyltryptamine, 7-methyltryptamine, 5-methyltryptamine 5-fluorotryptamine, 6-fluorotryptamine e 5- e 6-fluorotryptophans.
Procurare este FAQ por índice

Inibidores e tryptamines de MAO
O oxidase de Monoamine (MAO) é o pathway preliminar do inactivation de a maioria de tryptamines. Por causa disto, os inibidores do enzyme de MAO (MAOIs) podem ser usados potentiate os efeitos dos tryptamines e fazer oral DMT e 5-MeO-DMT ativos.
Os inibidores de MAO caem em duas classes: MAOIs irreversible e reversível. Além podem inibir qualquer um ou ambos os dois tipos do enzyme, de Mao-a e de Mao-b de MAO que são associados com os neurônios serotonergic e dopaminergic respectivamente. Ligamento irreversible de MAOIs (por exemplo o iproniazid e o phenelzine dos hydrazides) permanentemente ao enzyme e à inibição da causa MAO que duram 1-2 semanas após o ingestion. São usados clìnica tratar o depression. MAOIs reversível, tal como o moclobemide, que é usado como um antidepressivo, e o harmine e o harmaline beta-carbolines-carbolines, é eficaz por um tempo muito mais curto, talvez até 24 horas. Os usuários recreacionais da droga em torno do mundo estão usando principalmente o harmine e o harmaline apesar da falta de estudos científicos em seus efeitos em seres humanos.
Os nativos de Amazon combinaram tradicional a videira do caapi de Banisteriopsis, que contem o harmine, o harmaline e beta-carbolines-carbolines relacionado, com as plantas DMT-contendo para fazer um oral ativo brew o ayahuasca chamado . Outras plantas que contêm o harmine e/ou o harmaline podem ser substituídas para o caapi do B.. ' o ayahuasca norte-americano usual ' consiste em raizes das sementes do harmala de Peganum e do illinoensis de Desmanthus, e ' no acaciahuasca australian ' sae do complanata do acacia são combinados com o material dos acacias DMT-contendo (o effectivity desta mistura não foi confirmado). MAOIs foi usado também potentiate os efeitos dos cogumelos que contêm o psilocybin. Terence McKenna mencionou o chocolate que é um MAOI fraco, que poderia ser uma razão para o hábito popular de ingesting cogumelos com cacau.

As sementes do harmala de Peganum (rue syrian) são a fonte natural a mais concentrada do harmine e harmaline - aproximadamente 3% de seu peso consiste nestes alcalóides. O caapi de Banisteriopsis foi encontrado para conter 0,18% a 1,36% beta-carbolines-carbolines, com a concentração do harmine que é 0,057% a 0,635% ( McKenna et al. 1984 ). De acordo com relatórios anecdotal um grama das sementes do harmala do P. ingested inibe MAO bastante para fazer oral DMT ativo.
Harmine e o harmaline são hallucinogenic no seus próprios com os doses que partem de magnésio ao redor 300 ( Naranjo 1967 ), mas causam frequentemente side-effects físicos tais como o nausea e os tremors neste dose variam. Têm efeitos emocionais ou ' psychedelic ' pouco, mas produzem hallucinations visuais fortes. Por causa desta os nativos de Amazon adicionam frequentemente as quantidades maiores (75-100 cm da haste por o dose) de caapi do B. ao ayahuasca brew do que é needed para a inibição de MAO ( Luna 1984 ).
Há uns perigos significativos em usar inibidores de MAO. A maioria de MAOIs potentiate os efeitos cardiovascular do tyramine e dos outros monoamines encontrados nos alimentos. O ingestion do queijo envelhecido, cerveja, vinho, pickled herring, fígado de galinha, fermento, quantidades grandes de café, frutas de citrino, figs enlatados, feijões largos, chocolate ou o creme quando MAO for inibido pode causar uma crise hypertensive including uma ascensão perigosa na pressão de sangue. Os efeitos dos amphetamines, de anestésicos gerais, de sedatives, de anti-histamines, de álcool, de analgesics potent e de agentes anticholinergic e do antidepressivo são prolongados e intensified. O overdosage de MAOIs por se é também possível com efeitos including a hiper-reflexia e os convulsions.
Procurare este FAQ por índice
Síntese de derivatives de DMT
Os derivatives do tryptamine e beta-Carbolines-Carbolines foram detectados como metabolites endogenous nos mamíferos, including seres humanos. Os transferases methyl que catalyze a síntese dos

tryptamines, including DMT, 5-MeO-DMT e bufotenine, são encontrados no pulmão humano, no cérebro, no líquido cerebrospinal, no fígado e no coração ( McKenna & torres 1984 ). Na glândula pineal MAO é o pathway preliminar do serotonin, um neurotransmitter do inactivation synthesized do tryptophan do amino-ácido. Se MAO for obstruído pelo harmine, o harmaline ou o outro serotonin dos inibidores de MAO podem ser convertidos pelos enzymes HIOMT e INMT do methyltransferase em tryptamines psychedelic (serotonin -- --> 5-MeO-trypt do (hiomt). -- (2*inmt) --> 5-MeO-DMT).
Assim, ingesting o l-l-tryptophan para aumentar níveis do serotonin, uma barra do doce para aumentar a quantidade de tryptophan que começa a seus cérebro e material de planta natural que contêm 25-50 magnésio harmine/harmaline ao bloco MAO, tudo ao mesmo tempo, pôde causar sua glândula pineal synthesize quantidades substanciais de 5-MeO-DMT ( a maioria 1986 ). Isto é extremamente perigoso para pessoas com desequilíbrio ou esquizofrenia existente do amine. Para conseqüências possíveis dos povos normais, saudáveis seja mau. NÃO TENTE ISTO.
Um inibidor potent de INMT, que é um enzyme necessário para a síntese de DMT e de 5-MeO-DMT, é encontrado em concentrações particularmente elevadas na glândula pineal. Contornear ou uma inibição da síntese deste inibidor puderam ser responsável para trances e outros estados psychedelic conseguiram "sem drogas" ( Strassman 1990 ). Veja o artigo de Strassman para mais info e speculation sobre a glândula pineal.
Procurare este FAQ por índice
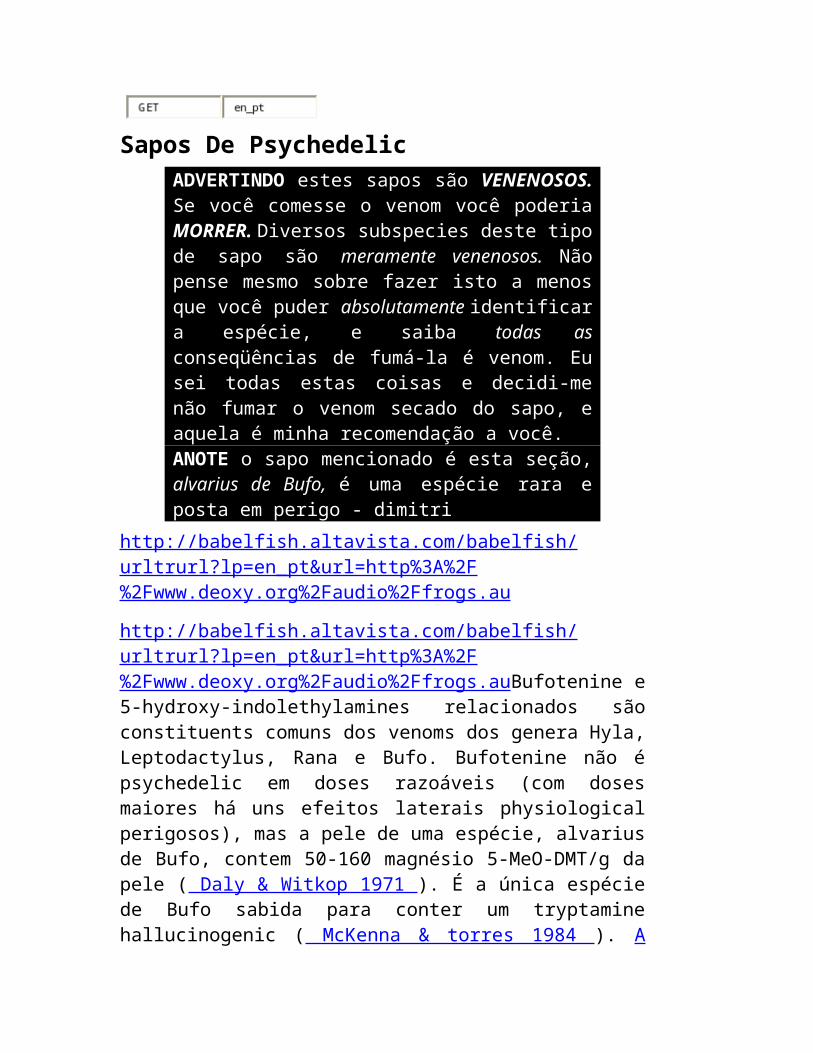
Sapos De Psychedelic ADVERTINDO estes sapos são VENENOSOS. Se você comesse o venom você poderia MORRER. Diversos subspecies deste tipo de sapo são meramente venenosos. Não pense mesmo sobre fazer isto a menos que você puder absolutamente identificar a espécie, e

saiba todas as conseqüências de fumá-la é venom. Eu sei todas estas coisas e decidi-me não fumar o venom secado do sapo, e aquela é minha recomendação a você. ANOTE o sapo mencionado é esta seção, alvarius de Bufo, é uma espécie rara e posta em perigo - dimitri
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.deoxy.org%2Faudio%2Ffrogs.au
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.deoxy.org%2Faudio%2Ffrogs.auBufotenine e 5-hydroxy-indolethylamines relacionados são constituents comuns dos venoms dos genera Hyla, Leptodactylus, Rana e Bufo. Bufotenine não é psychedelic em doses razoáveis (com doses maiores há uns efeitos laterais physiological perigosos), mas a pele de uma espécie, alvarius de Bufo, contem 50-160 magnésio 5-MeO-DMT/g da pele ( Daly & Witkop 1971 ). É a única espécie de Bufo sabida para conter um tryptamine hallucinogenic ( McKenna & torres 1984 ). A maioria (1984) dá instruções para coletar e secar o venom:
O venom fresco pode fàcilmente ser coletado sem dano ao sapo. Use uma placa de vidro lisa ou todo o outro quadrado 12-inches liso, nonporous da superfície ao menos. Prenda o sapo na frente da placa, que é reparada em uma posição vertical. Nesta maneira, o venom pode ser coletado na placa de vidro, livre da sujeira e do líquido liberados quando o sapo é segurado.
Quando você está pronto para começar, prenda o sapo firmemente com uma mão e, com o polegar e o forefinger de sua outra mão, esprema-o perto da base da glândula até que o venom esguinche fora dos pores e na placa de vidro. Use este método coletar sistematicamente o venom de cada uma das glândulas granular do sapo: aqueles no forearm, aqueles no tibia e no femur do pé hind, e, naturalmente, os parotids na garganta . Cada glândula pode ser espremida uma segunda vez para um rendimento adicional do venom se você permitir ao sapo um preiod one-hour do descanso. Depois que isto as glândulas está vazio e requer quatro a seis semanas para a regeneração.
O venom é viscous e leitoso-branco na cor quando espremido primeiramente das glândulas. Começa a secar dentro dos minutos e adquire a cor e a textura do cimento de borracha. Raspe o venom da

placa de vidro, seque-o completamente, e armazene-o em um recipiente hermético até que você esteja pronto para o fumar.
Davis e Weil (1992) fumaram o venom e descreveram o que aconteceu:
Na comparação com os compostos puros o venom do sapo parece mais por muito tempo durando e, porque um não perde completamente o contato com realidade, um sensual distante mais agradável, mais uniforme. Logo após o inhalation I experimentaram sensations nivelando mornos, um sentido da maravilha e o bem estar, os hallucinations auditory fortes, que incluíram um som da inseto-cigarra que funcionasse através de minha mente e parecesse ligar meu corpo à terra. Embora eu estava dentro, havia um sentido da sensação da terra, o pa seco do solo do deserto *** TRANSLATION ENDS HERE ***ssing through my fingers, the stars at midday, the scent of cactus and sage, the feel of dry leaves through hands. Strong visual hallucinations in orblike brilliance, diamond patterns that undulated across my visual field. The experience was in every sense pleasant, with no disturbing physical symptoms, no nausea, perhaps a slight sense of increased heart rate. Warm waves coursed up and down my body. The effects lasted only a few minutes but a pleasant afterglow continued for almost an hour. (Wade Davis, personal observation, January 12, 1991)
Profound alteration of consciousness within a few seconds of exhaling. I relax into a deep, peaceful interior awareness. There is nothing scary about the effects and no sense of toxicity. I try to describe my feelings but am unable to talk for the first five minutes and then only with some difficulty. This is a powerful psychoactive drug, one that I think would appeal to most people who like the effects of hallucinogens. For the next hour I feel slow and velvety, with a slight pressure in my head. No long-lasting effects to report. (Andrew T. Weil, personal observation, January 12, 1991).
Other animals contain DMT such as the gorgonian Paramuricea chamaeleon ( Cimino & De Stefano, 1978 ).

Search this FAQ for table of contents
F U N G I B E M U S H R Ö Ö M E D
Family: Bolbitiaceae Genus: Agrocybe Species: farinacea
Contains psilocybin ( Koike et al. 1981 ). Genus: Conocybe Species: cyanopus, kuehneriana, siligineoides, smithii
C. cyanopus ( Benedict et al. 1962 ) and C. smithii ( Benedict et al. 1967 ) contain psilocybin and psilocin while C. kuehneriana contains psilocin only ( Ohenoja et al. 1987 ). C. siligineoides may also contain these alkaloids ( Schultes & Hofmann 1979 p. 40).
Family: Coprinaceae Genus: Copelandia Species: anomala, bispora, cambodginiensis, chlorocystis,
cyanescens, tropicalis All species contain psilocin and psilocybin, for C. cyanescens ( Schultes & Hofmann 1979 p. 40) and for C. cambodginiensis as well as C. tropicalis ( Arora, 1986 ), and for C. anomala as well as C. bispora ( Merlin & Allen, 1993 ).
Genus: Panaeolina, Species: castaneifolius, foenisecii
P. foenisecii contains psilocybin ( Robbers et al. 1969 ). Genus: Panaeolus Species: africanus, antillarum, ater, campanulatus, firmicola,
microsporus, olivacens, retirugis, separatus, sphinctrinus, subbalteatus
Several Panaeolus species contain psilocybin. For P. antillarum refer to Allen et al. (1991), for P. ater refer to Bresinsky et al. (1990) , for P. papilionaceus ( Gurevich et al. 1992 ), for P. retirugis ( Fiussello et al. 1971/72 ), for P. separatus (( Miller Jr. 1972 ), for P. sphinctrinus ( Hein & Wasson, 1958 p. 322) and for P. olivacens ( Ohenoja et al. 1987 ). P. subbalteatus contains both psilocin and psilocybin ( Ohenoja et al. 1987 ) but was known to be hallucinogenic since 1959 ( Stein,

1959 ). P. firmicola is also described as hallucinogenic and probably contains the same alkaloids ( Schultes, 1979 ).
Genus: Psathyrella Species: candollenana
Contains psilocybin ( Koike et al. 1981 ) and psilocin ( Ohenoja et al. 1987 ).
Family: Cortinariaceae Genus: Galerina Species: steglichii
Contains psilocybin and psilocin ( Besl, 1993 ). Genus: Gymnopilus Species: aeruginosus, liquiritiae, luteus, purpuratus, spectabilis,
validipes, viridans Many Gymnopilus contain psilocybin, for G. aeruginosus, G. luteus, G. spectabilis, G. validipes and G.viridans refer to Hatfield et al. (1978) . For G. liquiritiae ( Koike, 1981 ) and for G. purpuratus ( Gartz 1991 ).
Genus: Inocybe Species: aeruginascens, calamistrata, coelestium, corydalna,
haemacta, tricolor These contain psilocin and psilocybin, for P. aeruginascens refer to Haeselbarth et al. (1985) and for the others Stijve et al. (1985) .
Family: Lepiotaceae Genus: Lepiota Species: humei Murrill also known as peele "Peele's Lepiota"
This mushrooms was claimed by Peele of Florida Mycology Research Center (PO Box 8104 Pensacola Florida 32505) to contain a legal tryptamine, which produces a trip with less physical symptoms and better ability of logical thinking than psilocin or psilocybin. Bioassay results didn't find psychoactivity ( Akers 1992 ).
Family: Pluteaceae Genus: Pluteus Species: atricapillus, cyanopus, nigroviridis, salicinus
P. atricapillus contains psilocybin ( Ohenoja et al. 1987 ) while both P. salicinus ( Saupe 1981 ) and P. nigroviridis ( Christiansen et al. 1984 ) contain psilocin and psilocybin.
Family: Polyporaceae Genus: Gerronema Species: fibula, swartzii
These contain psilocybin.

Genus: Hygrocybe Species: psittacina
This contains psilocybin.
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.deoxy.org%2Fmushword.htm http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.deoxy.org%2Fmushword.htm
Family: Strophariaceae Genus: Psilocybe Species: 75 Known hallucinogenic species + antioquensis,
aucklandii, beliconiae, coprophila, crobulus, cubensis (pictured) , guatapensis, samuiensis, subacutipilea,
There are at least 75 mushroom species in this genera that contain psilocin and psilocybin in Guzman 1983 , and there are several more recently discovered species such as P. aucklandii ( Guzman et al. 1993 ), P. samuiensis ( Guzman et al. 1991 ) and P. antioquensis, P. beliconiae, P. guatapensis and P. subacutipilea ( Guzman et al. 1994 ). Also P. coprophila, while lacking psilocin (making it a non-blueing psilocybe) is known to contain psilocybin ( Arora, 1986 ). P. crobulus is also known to be hallucinogenic ( Phillips, 1981 ). Images of Psilocybe cyanescens, cyanofibrillosa, azurescens, semilanceata, pelliculosa, silvatica can be seen at A Gallery of Psilocybe Mushrooms , Hyperreal Teonanacatl
Search this FAQ for table of contents

P L A N T S PLAN, PLANT, PLANET
Family: Acanthaceae Genus: Justicia Species: pectoralis (var. stenophylla)
Waikas of Orinoco headwaters in Venezuela add dried and pulverized leaves of this herb to their Virola-snuff. Intensely aromatic smelling leaves probably contain tryptamines ( Schultes 1977 ). Plants are available from from Hoti Products or ..Of the jungle .
Family: Aizoaceae Genus: Delosperma
Contains DMT and N-methyltryptamine ( Rivier & Pilet, 1971 ). Family: Alariaceae Genus: Ecklonia Species: maxima
Crouch et al. (1992) reported finding DMT in brown seaweed extract sold as Kelpak and intended to be used as a plant fertilizer. However, subsequent analysis by J. Gartz suggests none is present.
Family: Apocynaceae Genus: Prestonia Species: amazonica?
May contain DMT but the specimen used may have been misidentified ( Smith 1977 ).
Family: Cactaceae Genus: Echinocereus Species: salm-dyckianus, triglochidiatus
These cacti growing in Mexico are known to Tarahumare Indians as

peyote or hikuli and used in their festivals. E. triglochidiatus contains a tryptamine derivative, possibly 5-MeO-DMT ( Bye 1979 ). E. salm-dyckianus is also supposed to contain tryptamines according to Horus Botanicals catalog 1992.
Genus: Trichocereus Species: terscheckii "Cardon grande"
DMT has been isolated from this species growing in North-Western Argentina ( Schultes & Hofmann 1979 p. 58).
Family: Caesalpininaceae Genus: Petalostylis Species: cassiodies
Leaves and stem contain 0.4-0.5% tryptamine, DMT and other alkaloids ( Johns et al. 1966 ).
Family: Fabaceae Genus: Desmodium Species: gangetium, gyrans, tiliaefolium, triflorum,
Leaves, root, stem and seeds contain DMT and 0.06% 5-MeO-DMT of wet weight ( Banerjee & Ghosal 1969 ).
Genus: Lespedeza Species: bicolor
Leaves and root contain DMT and 5-MeO-DMT ( Smith 1977 ). Seeds of this hardy perennial shrub are available from ..Of the jungle for $5.
Genus: Mucuna Species: pruriens
Leaves, stem and fruit of this jungle vine contains DMT and 5-MeO-DMT ( Ghosal et al., 1971 ). Seeds are available from ..Of the jungle for $5.
Genus: Phyllodium Species: pulchellum
Dried plant material produced 0.2% 5-MeO-DMT and small amounts
of DMT ( Ghosal & Mukherjee 1966 ).
Family: Mimosaceae Genus: Anadenanthera (Piptandenia) Species: colubrina, contorta, excelsa, macrocarpa, peregrina
Black beans from these trees are toasted, pulverized and mixed with ashes or calcined shells to make psychedelic snuff called yopo by Indians in Orinoco basin in Colombia, Venezuela and possibly in southern part of Brasilian Amazon. Yopo is blown into the nostrils through bamboo tubes or snuffed by birdbone tubes. The trees grow in open plain areas, and leaves, bark and seeds contain DMT, 5-MeO-DMT and related compounds (Schultes 1976 , 1977 ; Pachter et al. 1959 ; Phytochem 11, 737).
Genus: Acacia Species: confusa [DMT,st], jurema, maidenni [DMT,b] niopo,
nubica [DMT, l], phlebophylla [DMT,l], polyacantha [DMT,l] subsp. campylacantha, senegal [DMT,l], simplicifolia [DMT,b,l,st], simplex, tortilis.
The letters following the tryptamine (DMT in all of these cases) refer to where the tryptamine is found in the plant: "st" is stems, "b" is bark, and "l" is leaves.
Dried A. confusa stems contain 0.04% N-methyltryptamine and 0.02% DMT ( Arthur et al. 1967 ). The dried leaves of A. phlebophylla contain 0.3% DMT ( Rovelli & Vaughan 1967 ). The bark of A. maidenii contains 0.6% of N-methyltryptamine and DMT in the proportions approx. 2:3 ( Fitzgerald & Sioumis 1965 ). A. simplex also
contains DMT ( Poupat et al. 1976 ). Several Sudanese species have DMT ( Wahba & Elkheir, 1975 ). Seeds of several acacia species are available from ..Of the jungle.
Genus: Desmanthus Species: illinoensis "Illinois Bundleflower"
Thompson et al. report that the root bark of this North American perennial shrub contains 0.34% DMT and 0.11% N-methyltryptamine. The bark accounts for about a half of the total weight of the roots. The plant should be resistant to cold and draught and easy to grow. ..Of the Jungle sells D. illinoensis seeds and dried roots (seed packet $3, 7 grams $10, oz $25; roots 4 oz $15, pound $50). Seeds are also available from more main-stream mail-order houses.
Genus: Mimosa Species: scabrella, tenuiflora (== hostilis) "tepescohuite",
verrucosa The roots of M. hostilis, which is not the common houseplant M. pudica ("sensitive plant"), contain 0.57% DMT and are used by Indians of Pernambuso State in Brazil as part of their Yurema cult ( Pachter et al. 1959 , Schultes 1977 , ( Meckes-Lozoya et al. 1990 ). M. scabrella contains DMT and N-methyltryptamine ( De Moraes et al., 1990 ). Bark of M. verrucosa also contains DMT ( Smith 1977 ).
Genus: Testulea Species: gabonensis
Contains DMT ( Leboeuf et al., 1977 ). Family: Malpighiaceae Genus: Banisteriopsis Species: muricata (=argentea), rusbyana (see Diplopterys
cabrerana) Both contain DMT ( Smith 1977 ).
Genus: Diplopterys Species: cabrerana
Natives of western Amazon add DMT- and N-methyltryptamine containing leaves of the vine D. cabrerana to a drink made from Banisteriopsis caapi, which contains beta-carbolines harmine and harmaline, to heighten and lengthen the visions ( Schultes 1977 , Smith 1977 ). D. cabrerana is also known as Banisteriopsis rusbyana.
Family: Myristicaceae Genus: Horsfieldia Species: superba
Contains 5-MeO-DMT as well as a beta-carboline ( Jossang et al. 1991 ).
Genus: Iryanthera Species: macrophylla
The bark contains 5-MeO-DMT ( Holmstedt et al. 1980 ).
Genus: Virola Species: calophylla, calophylloidea, rufula, sebifera, theiodora
The bark resin of these trees is used to prepare hallucinogenic snuffs in northwestern Brazil by boiling, drying and pulverizing it. Sometimes leaves of a Justicia are added. The snuff acts rapidly and violently, "effects include excitement, numbness of the limbs, twitching of facial muscles, nausea, hallucinations, and finally a deep sleep; macroscopia is frequent and enters into Waika beliefs about the spirits resident in the drug." Snuffs made from V. theiodora bark contain up to 11% 5-MeO-DMT and DMT. Also leaves, roots and flowers contain DMT. Amazonian Colombia natives roll small pellets of boiled resin in a evaporated filtrate of bark ashes of Gustavia Poeppigiana and ingest them to bring on a rapid intoxication ( Smith 1977 , Schultes 1977 ).
Family: Pandanaceae Genus: Pandanus "Screw pine"
DMT has been isolated from Pandanus nuts growing in New Guinea (

Barrau 1958, 1962 ). Family: Poaceae Genus: Arundo Species: donax
Leaves, flowers and rhizomes contain DMT, bufotenine and related compounds ( Ghosal et al. 1972 ).
Genus: Phalaris Species: aquatica (tuberosa), arundinacea
Leaves of P. arundinacea and leaves and seedlings of P. aquatica contain DMT, 5-MeO-DMT and related compounds ( Smith 1977 ). P. arundinacea plants are available from ..Of the jungle for $15.
Genus: Phragmites Species: australis
Rhizomes contain DMT ( Wassel et al. 1985 ).
Family: Rubiaceae Genus: Psychotria

Species: carthaginensis, viridis (psychotriaefolia) Psychotria leaves are added to a hallucinogenic drink prepared from Banisteriopsis caapi and B. rusbyana (which contain beta-carbolines) to strengthen and lengthen the effects in western Amazon. P. carthaginensis and P. viridis both contain DMT ( Rivier, 1972 ). 5 seeds of P. viridis cost $10 from ..Of the jungle.
Family: Rutaceae Genus: Dictyoloma Species: incanescens
Bark contains 0.04% 5-MeO-DMT ( Pachter et al. 1959 ). Genus: Limonia Species: acidissima
Contains DMT ( Abu Zarga, 1986 ). Genus: Melicope Species: leptococca
Contains 5-MeO-DMT as well as 5-MeO-DMT-Oxide and a beta-carboline ( Skaltsounis et al. 1983 ).
Genus: Vepris Species: ampody
The leaves and branches contains up to 0.2% DMT ( Kan-Fan 1970 ). Genus: Zanthoxylum Species: aborescens
Contains DMT ( Grina et al., 1982 ).
table of contents
References
Abu Zarga, M.H. 1986. Three new simple indole alkaloids from Limonia acidissima. Lloydia 49(5), 901-904.
Akers, B.P. 1992. Peele's Lepiota: An identification and a clarification. Mycotaxon 43(0), 461-469.
Arora, D. 1986. Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi. Ten Speed Press, Berkley.
Arthur, H.R., Loo, S.N. & Lamberton, J.A. 1967. Nb-methylated tryptamines and other constituents of Acacia confusa Merr. of Hong Kong. Aust. J Chem. 20, 811.

Banerjee, P.K. & Ghosal, S. 1969. Simple indole bases of Desmodium gangeticum. Aust. J Chem. 22, 275-277.
Barrau, J. 1958. Nouvelles observations au sujet des plantes hallucinogenes d'usage autochtone en Nouvelle-Guinee. J Agric. Trop. Bot. Appl. 5, 377-378.
Barrau, J. 1962. Observations et travaux recents sur les vegetaux hallucinogenes de la Nouvelle-Guinee. J Agric. Trop. Bot. Appl. 9, 245-249.
Benedict, R.G., Brady, L.R., Smith, A.H. & Tyler, V.E. 1962. Occurrence of psilocybin and psilocin in certain Conocybe and Psilocybe species. Lloydia 25, 156-159.
Benedict, R.G., Tyler, V.E. & Watling, R. 1967. Blueing in Conocybe, Psilocybe and a Stropharia Species and the Dectection of Psilocybin. Lloydia 30(2), 150-157.
Besl, H. 1993. Galerina steglichii spec. nov., a hallucinogenic Galerina. Zeitschrift fuer Mykologie 59(2), 215-218.
Bresinsky, A. & Besl, H. 1990. A Colour Atlas of Poisonous Fungi. Wolfe Publishing Ltd, London.
Bye, R.A. 1979. Hallucinogenic plants of the Tarahumara. J. Ethnopharmacology 1, 23-48.
Christiansen, A.L., Rasmussen, K.E. & Hoeiland, K. 1984. Detection of psilocybin and psilocin in Norwegian species of Pluteus and Conocybe. Planta Med. 50, 341-343.
Cimino, G. & De Stefano, S. 1978. Chemistry of Mediterranean Gorgonians. Simple indole derivatives from Paramuricea chamaeleon. Comptes Rendus Biochem. Physiol. Ser. C. 61, 361-362.
Crouch, I.J., Smith M.T., Van Staden J., Lewis, M.J. & Hoad, G.V. 1992. Identification of auxins in a commercial seaweed concentrate. J Plant Physiology 139(5), 590-594.

Daly, J.W. & Witkop, B. 1971. Chemistry and pharmacology of frog venoms. In: Venomous animals and their venoms. Vol II. New York: Academic Press.
Davis, W. & Weil, A.T. 1992. Identity of a New World Psychoactive Toad. Ancient Mesoamerica 3 (1992) 5, 51-59.
De Moraes, E.H.F., Alvarenga, Z.M.A., Ferreira, Z.M.G.S. & Alisue, G. 1990. Quim. Nova 13, 308.
Fitzgerald, J.S. & Sioumis, A.A. 1965. Alkaloids of Australian Leguminosae V. Aust. J Chem. 18, 433.
Fiussello, N. & Ceruti-Scarti, J. 1971/72. Presenza di psilocibina edi 5-idrossi-indolderivati in Panaeolus retirugis. Atti Acc. Sci. Torino 106, 725-735.
Gartz, J. 1991. Influence of phosphate on fruiting and secondary metabolism of mycelia of Psilocybe cubensis, Psilocybe semilanceata and Gymnopilus purpuratus. Zeitschrift fuer Mykologie 57(1), 149-154.
Ghosal, S., Chaudhuri, R.K., Dutta, S.K. & Bhattacharya, S.K. 1972. Occurrence of curaromimetic indoles in the flowers of Arundo donax. Planta Med. 21, 22.
Ghosal, S. & Mukherjee, B. 1966. Indole-3-alkylamine Bases of Desmodium pulchellum. J, Org. Chem. 31, 2284.
Ghosal, S., Singh, S. & Bhattacharya, S.K. 1971. Alkaloids of Mucuna pruriens, Chemistry and Pharmacology. Planta Med. 19, 279
Grina, J.A. et al. 1982. Constituents of Zanthoxylum aborescens. Part 7. Old & new alkaloids from Zanthoxylum aborescens. J. Organic Chemistry 47, 2648-2651.
Gurevich, L.S. 1993. Indole derivatives in certain Panaeolus species from East Europe & Siberia. Mycological Research 97(2), 251-254.

Gurevich, L.S. & Astapenko, V.V. 1992. Chromatographic study of some indole metabolites in Panaeolus basidiomycetes. Mikologiya I Fitopathologiga 26(3), 189-194.
Guzman, G. 1983. The Genus Psilocybe. Beihefte Zur Nova Hedwingia 74, 1-439.
Guzman, G., Bandala, V.M. & Allen, J.W. 1993. A New Bluing Psilocybe from Thailand. Mycotaxon 26, 155-160.
Guzman, G., Bandala, V.M. & King, C. 1991. A New Species of Psilocybe of Section Zapotecorum from New Zealand. Mycological Research 95, 507-508.
Guzman, G., Saldarriaga, Y., Pineda, F., Garcia, G. & Velazquez, L.F. 1994. New Species of Psilocybe from Colombia and Discussion on the known species. Mycotaxon , 225.
Haeselbarth, G., Michaelis, H. & Salnikow, J. 1985. Nachweis von Psilocybin in Inocybe aeruginescens. Mykol. Mitt. bl. 28(1), 59-62.
Hatfield, G.M., Valdes, L.J. & Smith, A.H. 1978. The occurrence of psilocybin in Gymnopilus species. Lloydia 41, 140-144.
Hein, R. & Wasson, R.G. 1958. Les champignons hallucinogenes du Mexique. Museum National d'Histoire Naturelle, Paris.
Holmstedt, B., Lindgren, J.E., et al. 1980. Indole alkaloids in Amazonian Myristicaceae: Field and laboratory research. Bot. Mus. Leafl., Harvard Univ. 28, 215-234.
Johns, S.R., Lamberton, J.A. & Sioumis, A.A. 1966. Alkaloids of the Australian Leguminosae VI. Aust. J Chem. 19, 893.
Jossang, A., Jossang, C., Hadi, H.A., Sevenet, T. & Bodo, B. 1991. Horsfiline, an oxindole alkaloid from Horsfieldia superba. J. Organic Chem. 56(23), 6527-6530.
Kan Fan, C. et al. 1970. Alcaloides de Vepris ampody (Rutacees). Phytochem. 9, 1283-1291.
Kantor, R.E., Dudlettes, S.D. & Shulgin, A.T. 1980. 5-Methoxy-alfa-methyl- tryptamine (alfa,O-dimethylserotonin), a hallucinogenic homolog of serotonin. Biological Psychiatry Vol 15, 349-352.
Koike, Y., Wada, K., Kusano, G., Nozoe, S., & Yokoyama, K. 1981. Isolation of Psilocybin from Psilocybe argentipes and its Determination in Specimens of some Mushrooms. Lloydia 44(3), 362-365.
Leboeuf, M. et al., 1977. Alkaloids and triterpenes of Testulea gabonensis. Plant Medicine Phytotherapy 11, 230.
Luna, L.E. 1984. The Healing Practices of a Peruvian Shaman. J. Ethnopharmacology 11, 123-133.
McKenna, D.J., Towers, G.H.N., & Abbott, F. (1984). Monoamine oxidase inhibitors in South American hallucinogenic plants: Tryptamines and Beta-carboline constituents of ayahuasca. J Ethnopharmacology 10, 195-223.
Mckenna, D.J. & Towers, G.H.N. 1984. Biochemistry and Pharmacology of Tryptamines and beta-Carbolines: A Minireview. J Psychoactive Drugs 16(4).
Meckes-Lozoya, M., Lozoya, X., Marles, R.J., Soucy-Breau, C., Sen, A. & Arnason, J.T. 1990. N,N-dimethyltryptamine alkaloid in Mimosa tenuiflora bark (tepescohuite). Arch. Invest. Med. Mex. 21(2), 175-7.
Merlin, M.D. & Allen, J.W. 1993. Species identification and chemical analysis of psychoactive fungi in the Hawaiian islands. Journal of Ethnopharmacology 40(1), 21-40.
Miller Jr., O.K. 1972. Mushrooms of North America. Dutton & Co., Springfield.
Most, Albert. 1984. Bufo Alvarius: the Psychedelic Toad of the Sonoran Desert . Venom Press Box 2863 Denton TX 76202.

Most, Albert. 1986. Eros and the Pineal: the layman's guide to cerebral solitaire . Venom Press, Denton, TX.
Naranjo, C. 1969. Psychotropic Properties of the Harmala Alkaloids. In: Efron (Ed.) The Ethnopharmacologic Search for Psychoactive Drugs.
Ohenoja, E., Jokiranta, J., Makinen, T., Kaikkonen, A. & Airaksinen, M.M. 1987. The Occurrence of Psilocybin and Psilocin in Finnish Fungi. Lloydia 50(4), 741-744.
Pachter, I.J, Zacharias, D.E & Ribeir, O. 1959. Indole Alkaloids of Acer saccharinum (the Silever Maple), Dictyoloma incanescens, Piptadenia colubrina, and Mimosa hostilis. J Org Chem 24, 1285-7.
Phillips, R. 1981. Mushrooms and other fungi of Great Britain and Europe. Pan Books, London.
Poupat, C., Ahond, A. & Sevenet, T. 1976. Alcaloides De Acaia simplicifolia. Phytochemistry 15, 2019-2120.
Rivier, L. & Lindgren, J-E. 1972. "Ayahuasca," the South American Hallucinogenic Drink: an Ethnobotanical and Chemical Investigation. Economic Botany 26, 101-129.
Rivier, L. & Pilet, P.E. 1971. Annee Biologique 10, 129.
Robbers, J.E., Tyler, V.E. & Ola'h, G.M. 1969. Additional evidence supporting the occurrence of psilocybin in Panaeolus foenisecii. Lloydia 32, 399-400.
Rovelli, B. & Vaughan, G.N. 1967. Alkaloids of Acacia I. Dimethyltryptamine in Acacia phlebophylla. Aust. J Chem. 20, 1299-1300.
Saupe, S.G. 1981. Occurence of Psilocybin/Psilocin in Pluteus salicinus (Pluteaceae). Mycologia 73, 781-784.

Schultes, R.E. 1976. Indole Alkaloids in Plant Hallucinogens. J of Psychedelic Drugs Vol 8(1), 7-25.
Schultes, R.E. 1977. The Botanical and Chemical Distribution of Hallucinogens. J of Psychedelic Drugs Vol 9(3), 247-263.
Schultes, R.E. 1979. Hallucinogenic Plants: Their Earliest Botanical Descriptions. J of Psychedelic Drugs Vol 11(1-2), 13-24.
Schultes, R.E. & Hofmann, A. 1979. Plants of the Gods. McGraw-Hill. Reprint available from Healing Arts Press, Rochester, VT.
Schultes, R.E. & Hofmann, A. 1980. The Botany and Chemistry of Hallucinogens. Springfield, Ill: Thomas.
Shulgin, A.T. 1976. Psychotomimetic agents. In: Gordon, M. (Ed.) Psychopharmacological Agents, Vol IV. New York: Academic Press.
Shulgin, A.T. 1982. Chemistry of Psychotomimetics. In: Hoffmeister, F. & Stille, G. (Eds.) Handbook of Experimental Pharmacology, Vol 55: Alcohol and Psychotomimetics, Psychotropic Effects of Central-Acting Drugs. New York: Springer-Verlag.
Skaltsounis, A.L., Tillequin, F. & Koch, M. 1983. Plantes des Nouvelle- Caledonie LXXXIII: Alcaloides des tiges feuillees de Melicope leptococca. Lloydia 46(5), 732.
Smith, T.A. 1977. Review: Tryptamine and Related Compounds in Plants. Phytochemistry 16 171-175.
Stein, S.I. 1959. Clinical Observations on the Effects of Panaeolus venenosus Versus Psilocybe caerulescens Mushrooms. Mycologica 51, 49.

Stijve, T., Klan, J. & Kugper, W. 1985. Occurrence of psilocybin and baeocystin in the genus Inocybe. Persoonia 12, 469-472.
Strassman, R.J. 1990. The Pineal Gland: Current Evidence For Its Role In Consciousness. In: Lyttle, T. (Ed.) Psychedelic Monographs and Essays Vol 5.
Thompson, A.C., Nicollier, G.F. & Pope, D.F 1987. Indolealkylamines of Desmanthus illinoensis and Their Growth Inhibition Activity. J Agric. Food Chem. 35, 361-365.
Wahba, S.K. & Elkheir, Y.M. 1975. Dimethyltyrptamine from the leaves of certain Acacia species of northern Sudan. Lloydia 38, 176-177.
Wassel, G.M. et al. 1985. Alkaloids from the rhizomes of Phragmites australis Cav. Scientia Pharmaceutica 53, 169-170.
[email protected] * Everything is perfect forever Michael from Melbourne * Ditto
RELATED SPACES DMT
N,N-DIMETHYLTRYPTAMINE H Y P E R S P A C E Terence McKenna Land Shamanism Gracie & Zarkov
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fdeoxy.orgD E O X Y . O R G
END OF FAQ Erro! O nome de arquivo não foi especificado.

FAQ Dos Portadores Do Tryptamine
por Petrus Pennanen com ajuda de Michael de Melbourne . Hypertexture e esforço ©ontinuing da ilustração através de http://deoxy.org
Traduza de
Agradecimentos a muitos indivíduos para a ajuda em unir isto. Se você souber as fontes dos tryptamines que não são mencionados aqui satisfazem enviam-nos.
Última busca de julho 1999 do update do deoxy este FAQ para
Inibidores e tryptamines de MAO Síntese de derivatives de DMT Sapos De Psychedelic Os Fungos As Plantas Referências
Oral e parenterally derivatives psychotropic ativos do tryptamine Baseado em McKenna & em torres 1984
Dados compilados de Kantor, et al. 1980 ; Shulgin 1976 , 1982 ; Shulgin & Carter 1980
Nome do composto R1 R2 R3
R4
R5
Dosage (magnésio)
Rota Oral/Par.
tryptamine H H H H H 100 *
1 par/oral?
DMT (dimethyltryptamine) CH3 CH3 H H H 60 par
DET C2H5 C2H5 H H H 60 par/

oral
DPT n-suporte
n-suporte
H H H 60 par/
oral
DAT C3H5 C3H5 H H H 30 par/
oral
DIPT eu-suporte
eu-suporte
H H H 30 oral
5-MeO-DIPT eu-suporte
eu-suporte
H H
OCH3
12 oral
5-MeO-DMT CH3 CH3 H H
OCH3
6 par
psilocin CH3 CH3 H Oh
H 12 * 2 oral
Cz-74 C2H5 C2H5 H Oh
H 15 * 2 oral
serotonin H H H H
Oh
100 * 3 oral
bufotenine CH3 CH3 H H
Oh 16 * 4 par
It-290 H H CH3
H H 30 oral
4-hydroxy-alfa-methyl-tryptamine H H CH3
Oh
H 20 * 3 oral
Mp-809 H H CH3
H
CH3
60 * 5 oral
5-fluoro-alfa-methyl-tryptamine H H CH
H
F 25 * 6 oral

3
5-methoxy-alfa-methyl-tryptamine H H
CH3
H
OCH3
3 oral
4-hydroxy-diisopropyl-tryptamine eu-suporte
eu-suporte
H Oh
H 12 * 6 oral
4-hydroxy-N-isopropyl, N-methyl-n-methyl-tryptamine
eu-suporte
CH3 H Oh
H 6 * 6 oral
N-t-butyl-n-t-butyl-tryptamine H t-butylH
H H ? * 7 par?
3-(2-(2,5-dimethylpyrrolyl) ethyl)-ethyl)-indole H H H ? ? ? ?
5-alfa-DMT CH3 CH3 CH3
H H ? ?
* sintomas 1 autonomic; pouca atividade central. * 2 os esters do phosphate são psilocybin e Cey-19, respectivamente; ambos são stoichiometrically equivalente aos isomers 4-hydroxy. * 3 cardiovascular e sintomas autonomic; pouca atividade central. * amine pressor de 4 A melhor que um hallucinogen no homem. * 5 um antidepressivo melhor que um hallucinogen no homem. * 6 basearam em relatórios anonymous na imprensa da configuração.
Nenhum estudo clínico foi publicado. * 7 nenhuma atividade oral com doses até magnésio 20, podem ser parenterally ativos.
Outros tryptamines potencial psychedelic incluem:
6-fluoro-alfa-methyltryptamine, 7-methyltryptamine, 5-methyltryptamine 5-fluorotryptamine, 6-fluorotryptamine e 5- e 6-fluorotryptophans.
Procurare este FAQ por índice

Inibidores e tryptamines de MAO
O oxidase de Monoamine (MAO) é o pathway preliminar do inactivation de a maioria de tryptamines. Por causa disto, os inibidores do enzyme de MAO (MAOIs) podem ser usados potentiate os efeitos dos tryptamines e fazer oral DMT e 5-MeO-DMT ativos.
Os inibidores de MAO caem em duas classes: MAOIs irreversible e reversível. Além podem inibir qualquer um ou ambos os dois tipos do enzyme, de Mao-a e de Mao-b de MAO que são associados com os neurônios serotonergic e dopaminergic respectivamente. Ligamento irreversible de MAOIs (por exemplo o iproniazid e o phenelzine dos hydrazides) permanentemente ao enzyme e à inibição da causa MAO que duram 1-2 semanas após o ingestion. São usados clìnica tratar o depression. MAOIs reversível, tal como o moclobemide, que é usado como um antidepressivo, e o harmine e o harmaline beta-carbolines-carbolines, é eficaz por um tempo muito mais curto, talvez até 24 horas. Os usuários recreacionais da droga em torno do mundo estão usando principalmente o harmine e o harmaline apesar da falta de estudos científicos em seus efeitos em seres humanos.
Os nativos de Amazon combinaram tradicional a videira do caapi de Banisteriopsis, que contem o harmine, o harmaline e beta-carbolines-carbolines relacionado, com as plantas DMT-contendo para fazer um oral ativo brew o ayahuasca chamado . Outras plantas que contêm o harmine e/ou o harmaline podem ser substituídas para o caapi do B.. ' o ayahuasca norte-americano usual ' consiste em raizes das sementes do harmala de Peganum e do illinoensis de Desmanthus, e ' no acaciahuasca australian ' sae do complanata do acacia são combinados com o material dos acacias DMT-contendo (o effectivity desta mistura não foi confirmado). MAOIs foi usado também potentiate os efeitos dos cogumelos que contêm o psilocybin. Terence McKenna mencionou o chocolate que é um MAOI fraco, que poderia ser uma razão para o hábito popular de ingesting cogumelos com cacau.

As sementes do harmala de Peganum (rue syrian) são a fonte natural a mais concentrada do harmine e harmaline - aproximadamente 3% de seu peso consiste nestes alcalóides. O caapi de Banisteriopsis foi encontrado para conter 0,18% a 1,36% beta-carbolines-carbolines, com a concentração do harmine que é 0,057% a 0,635% ( McKenna et al. 1984 ). De acordo com relatórios anecdotal um grama das sementes do harmala do P. ingested inibe MAO bastante para fazer oral DMT ativo.
Harmine e o harmaline são hallucinogenic no seus próprios com os doses que partem de magnésio ao redor 300 ( Naranjo 1967 ), mas causam frequentemente side-effects físicos tais como o nausea e os tremors neste dose variam. Têm efeitos emocionais ou ' psychedelic ' pouco, mas produzem hallucinations visuais fortes. Por causa desta os nativos de Amazon adicionam frequentemente as quantidades maiores (75-100 cm da haste por o dose) de caapi do B. ao ayahuasca brew do que é needed para a inibição de MAO ( Luna 1984 ).
Há uns perigos significativos em usar inibidores de MAO. A maioria de MAOIs potentiate os efeitos cardiovascular do tyramine e dos outros monoamines encontrados nos alimentos. O ingestion do queijo envelhecido, cerveja, vinho, pickled herring, fígado de galinha, fermento, quantidades grandes de café, frutas de citrino, figs enlatados, feijões largos, chocolate ou o creme quando MAO for inibido pode causar uma crise hypertensive including uma ascensão perigosa na pressão de sangue. Os efeitos dos amphetamines, de anestésicos gerais, de sedatives, de anti-histamines, de álcool, de analgesics potent e de agentes anticholinergic e do antidepressivo são prolongados e intensified. O overdosage de MAOIs por se é também possível com efeitos including a hiper-reflexia e os convulsions.
Procurare este FAQ por índice
Síntese de derivatives de DMT
Os derivatives do tryptamine e beta-Carbolines-Carbolines foram detectados como metabolites endogenous nos mamíferos, including seres humanos. Os transferases methyl que catalyze a síntese dos

tryptamines, including DMT, 5-MeO-DMT e bufotenine, são encontrados no pulmão humano, no cérebro, no líquido cerebrospinal, no fígado e no coração ( McKenna & torres 1984 ). Na glândula pineal MAO é o pathway preliminar do serotonin, um neurotransmitter do inactivation synthesized do tryptophan do amino-ácido. Se MAO for obstruído pelo harmine, o harmaline ou o outro serotonin dos inibidores de MAO podem ser convertidos pelos enzymes HIOMT e INMT do methyltransferase em tryptamines psychedelic (serotonin -- --> 5-MeO-trypt do (hiomt). -- (2*inmt) --> 5-MeO-DMT).
Assim, ingesting o l-l-tryptophan para aumentar níveis do serotonin, uma barra do doce para aumentar a quantidade de tryptophan que começa a seus cérebro e material de planta natural que contêm 25-50 magnésio harmine/harmaline ao bloco MAO, tudo ao mesmo tempo, pôde causar sua glândula pineal synthesize quantidades substanciais de 5-MeO-DMT ( a maioria 1986 ). Isto é extremamente perigoso para pessoas com desequilíbrio ou esquizofrenia existente do amine. Para conseqüências possíveis dos povos normais, saudáveis seja mau. NÃO TENTE ISTO.
Um inibidor potent de INMT, que é um enzyme necessário para a síntese de DMT e de 5-MeO-DMT, é encontrado em concentrações particularmente elevadas na glândula pineal. Contornear ou uma inibição da síntese deste inibidor puderam ser responsável para trances e outros estados psychedelic conseguiram "sem drogas" ( Strassman 1990 ). Veja o artigo de Strassman para mais info e speculation sobre a glândula pineal.
Procurare este FAQ por índice
Sapos De Psychedelic ADVERTINDO estes sapos são VENENOSOS. Se você comesse o venom você poderia MORRER. Diversos subspecies deste tipo de sapo são meramente venenosos. Não pense mesmo sobre fazer isto a menos que você puder absolutamente identificar a espécie, e

saiba todas as conseqüências de fumá-la é venom. Eu sei todas estas coisas e decidi-me não fumar o venom secado do sapo, e aquela é minha recomendação a você. ANOTE o sapo mencionado é esta seção, alvarius de Bufo, é uma espécie rara e posta em perigo - dimitri
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.deoxy.org%2Faudio%2Ffrogs.au
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.deoxy.org%2Faudio%2Ffrogs.auBufotenine e 5-hydroxy-indolethylamines relacionados são constituents comuns dos venoms dos genera Hyla, Leptodactylus, Rana e Bufo. Bufotenine não é psychedelic em doses razoáveis (com doses maiores há uns efeitos laterais physiological perigosos), mas a pele de uma espécie, alvarius de Bufo, contem 50-160 magnésio 5-MeO-DMT/g da pele ( Daly & Witkop 1971 ). É a única espécie de Bufo sabida para conter um tryptamine hallucinogenic ( McKenna & torres 1984 ). A maioria (1984) dá instruções para coletar e secar o venom:
O venom fresco pode fàcilmente ser coletado sem dano ao sapo. Use uma placa de vidro lisa ou todo o outro quadrado 12-inches liso, nonporous da superfície ao menos. Prenda o sapo na frente da placa, que é reparada em uma posição vertical. Nesta maneira, o venom pode ser coletado na placa de vidro, livre da sujeira e do líquido liberados quando o sapo é segurado.
Quando você está pronto para começar, prenda o sapo firmemente com uma mão e, com o polegar e o forefinger de sua outra mão, esprema-o perto da base da glândula até que o venom esguinche fora dos pores e na placa de vidro. Use este método coletar sistematicamente o venom de cada uma das glândulas granular do sapo: aqueles no forearm, aqueles no tibia e no femur do pé hind, e, naturalmente, os parotids na garganta . Cada glândula pode ser espremida uma segunda vez para um rendimento adicional do venom se você permitir ao sapo um preiod one-hour do descanso. Depois que isto as glândulas está vazio e requer quatro a seis semanas para a regeneração.
O venom é viscous e leitoso-branco na cor quando espremido primeiramente das glândulas. Começa a secar dentro dos minutos e adquire a cor e a textura do cimento de borracha. Raspe o venom da

placa de vidro, seque-o completamente, e armazene-o em um recipiente hermético até que você esteja pronto para o fumar.
Davis e Weil (1992) fumaram o venom e descreveram o que aconteceu:
Na comparação com os compostos puros o venom do sapo parece mais por muito tempo durando e, porque um não perde completamente o contato com realidade, um sensual distante mais agradável, mais uniforme. Logo após o inhalation I experimentaram sensations nivelando mornos, um sentido da maravilha e o bem estar, os hallucinations auditory fortes, que incluíram um som da inseto-cigarra que funcionasse através de minha mente e parecesse ligar meu corpo à terra. Embora eu estava dentro, havia um sentido da sensação da terra, o pa seco do solo do deserto *** TRANSLATION ENDS HERE ***ssing through my fingers, the stars at midday, the scent of cactus and sage, the feel of dry leaves through hands. Strong visual hallucinations in orblike brilliance, diamond patterns that undulated across my visual field. The experience was in every sense pleasant, with no disturbing physical symptoms, no nausea, perhaps a slight sense of increased heart rate. Warm waves coursed up and down my body. The effects lasted only a few minutes but a pleasant afterglow continued for almost an hour. (Wade Davis, personal observation, January 12, 1991)
Profound alteration of consciousness within a few seconds of exhaling. I relax into a deep, peaceful interior awareness. There is nothing scary about the effects and no sense of toxicity. I try to describe my feelings but am unable to talk for the first five minutes and then only with some difficulty. This is a powerful psychoactive drug, one that I think would appeal to most people who like the effects of hallucinogens. For the next hour I feel slow and velvety, with a slight pressure in my head. No long-lasting effects to report. (Andrew T. Weil, personal observation, January 12, 1991).
Other animals contain DMT such as the gorgonian Paramuricea chamaeleon ( Cimino & De Stefano, 1978 ).

Search this FAQ for table of contents
F U N G I B E M U S H R Ö Ö M E D
Family: Bolbitiaceae Genus: Agrocybe Species: farinacea
Contains psilocybin ( Koike et al. 1981 ). Genus: Conocybe Species: cyanopus, kuehneriana, siligineoides, smithii
C. cyanopus ( Benedict et al. 1962 ) and C. smithii ( Benedict et al. 1967 ) contain psilocybin and psilocin while C. kuehneriana contains psilocin only ( Ohenoja et al. 1987 ). C. siligineoides may also contain these alkaloids ( Schultes & Hofmann 1979 p. 40).
Family: Coprinaceae Genus: Copelandia Species: anomala, bispora, cambodginiensis, chlorocystis,
cyanescens, tropicalis All species contain psilocin and psilocybin, for C. cyanescens ( Schultes & Hofmann 1979 p. 40) and for C. cambodginiensis as well as C. tropicalis ( Arora, 1986 ), and for C. anomala as well as C. bispora ( Merlin & Allen, 1993 ).
Genus: Panaeolina, Species: castaneifolius, foenisecii
P. foenisecii contains psilocybin ( Robbers et al. 1969 ). Genus: Panaeolus Species: africanus, antillarum, ater, campanulatus, firmicola,
microsporus, olivacens, retirugis, separatus, sphinctrinus, subbalteatus
Several Panaeolus species contain psilocybin. For P. antillarum refer to Allen et al. (1991), for P. ater refer to Bresinsky et al. (1990) , for P. papilionaceus ( Gurevich et al. 1992 ), for P. retirugis ( Fiussello et al. 1971/72 ), for P. separatus (( Miller Jr. 1972 ), for P. sphinctrinus ( Hein & Wasson, 1958 p. 322) and for P. olivacens ( Ohenoja et al. 1987 ). P. subbalteatus contains both psilocin and psilocybin ( Ohenoja et al. 1987 ) but was known to be hallucinogenic since 1959 ( Stein,

1959 ). P. firmicola is also described as hallucinogenic and probably contains the same alkaloids ( Schultes, 1979 ).
Genus: Psathyrella Species: candollenana
Contains psilocybin ( Koike et al. 1981 ) and psilocin ( Ohenoja et al. 1987 ).
Family: Cortinariaceae Genus: Galerina Species: steglichii
Contains psilocybin and psilocin ( Besl, 1993 ). Genus: Gymnopilus Species: aeruginosus, liquiritiae, luteus, purpuratus, spectabilis,
validipes, viridans Many Gymnopilus contain psilocybin, for G. aeruginosus, G. luteus, G. spectabilis, G. validipes and G.viridans refer to Hatfield et al. (1978) . For G. liquiritiae ( Koike, 1981 ) and for G. purpuratus ( Gartz 1991 ).
Genus: Inocybe Species: aeruginascens, calamistrata, coelestium, corydalna,
haemacta, tricolor These contain psilocin and psilocybin, for P. aeruginascens refer to Haeselbarth et al. (1985) and for the others Stijve et al. (1985) .
Family: Lepiotaceae Genus: Lepiota Species: humei Murrill also known as peele "Peele's Lepiota"
This mushrooms was claimed by Peele of Florida Mycology Research Center (PO Box 8104 Pensacola Florida 32505) to contain a legal tryptamine, which produces a trip with less physical symptoms and better ability of logical thinking than psilocin or psilocybin. Bioassay results didn't find psychoactivity ( Akers 1992 ).
Family: Pluteaceae Genus: Pluteus Species: atricapillus, cyanopus, nigroviridis, salicinus
P. atricapillus contains psilocybin ( Ohenoja et al. 1987 ) while both P. salicinus ( Saupe 1981 ) and P. nigroviridis ( Christiansen et al. 1984 ) contain psilocin and psilocybin.
Family: Polyporaceae Genus: Gerronema Species: fibula, swartzii
These contain psilocybin.

Genus: Hygrocybe Species: psittacina
This contains psilocybin.
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.deoxy.org%2Fmushword.htm http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.deoxy.org%2Fmushword.htm
Family: Strophariaceae Genus: Psilocybe Species: 75 Known hallucinogenic species + antioquensis,
aucklandii, beliconiae, coprophila, crobulus, cubensis (pictured) , guatapensis, samuiensis, subacutipilea,
There are at least 75 mushroom species in this genera that contain psilocin and psilocybin in Guzman 1983 , and there are several more recently discovered species such as P. aucklandii ( Guzman et al. 1993 ), P. samuiensis ( Guzman et al. 1991 ) and P. antioquensis, P. beliconiae, P. guatapensis and P. subacutipilea ( Guzman et al. 1994 ). Also P. coprophila, while lacking psilocin (making it a non-blueing psilocybe) is known to contain psilocybin ( Arora, 1986 ). P. crobulus is also known to be hallucinogenic ( Phillips, 1981 ). Images of Psilocybe cyanescens, cyanofibrillosa, azurescens, semilanceata, pelliculosa, silvatica can be seen at A Gallery of Psilocybe Mushrooms , Hyperreal Teonanacatl
Search this FAQ for table of contents

P L A N T S PLAN, PLANT, PLANET
Family: Acanthaceae Genus: Justicia Species: pectoralis (var. stenophylla)
Waikas of Orinoco headwaters in Venezuela add dried and pulverized leaves of this herb to their Virola-snuff. Intensely aromatic smelling leaves probably contain tryptamines ( Schultes 1977 ). Plants are available from from Hoti Products or ..Of the jungle .
Family: Aizoaceae Genus: Delosperma
Contains DMT and N-methyltryptamine ( Rivier & Pilet, 1971 ). Family: Alariaceae Genus: Ecklonia Species: maxima
Crouch et al. (1992) reported finding DMT in brown seaweed extract sold as Kelpak and intended to be used as a plant fertilizer. However, subsequent analysis by J. Gartz suggests none is present.
Family: Apocynaceae Genus: Prestonia Species: amazonica?
May contain DMT but the specimen used may have been misidentified ( Smith 1977 ).
Family: Cactaceae Genus: Echinocereus Species: salm-dyckianus, triglochidiatus
These cacti growing in Mexico are known to Tarahumare Indians as

peyote or hikuli and used in their festivals. E. triglochidiatus contains a tryptamine derivative, possibly 5-MeO-DMT ( Bye 1979 ). E. salm-dyckianus is also supposed to contain tryptamines according to Horus Botanicals catalog 1992.
Genus: Trichocereus Species: terscheckii "Cardon grande"
DMT has been isolated from this species growing in North-Western Argentina ( Schultes & Hofmann 1979 p. 58).
Family: Caesalpininaceae Genus: Petalostylis Species: cassiodies
Leaves and stem contain 0.4-0.5% tryptamine, DMT and other alkaloids ( Johns et al. 1966 ).
Family: Fabaceae Genus: Desmodium Species: gangetium, gyrans, tiliaefolium, triflorum,
Leaves, root, stem and seeds contain DMT and 0.06% 5-MeO-DMT of wet weight ( Banerjee & Ghosal 1969 ).
Genus: Lespedeza Species: bicolor
Leaves and root contain DMT and 5-MeO-DMT ( Smith 1977 ). Seeds of this hardy perennial shrub are available from ..Of the jungle for $5.
Genus: Mucuna Species: pruriens
Leaves, stem and fruit of this jungle vine contains DMT and 5-MeO-DMT ( Ghosal et al., 1971 ). Seeds are available from ..Of the jungle for $5.
Genus: Phyllodium Species: pulchellum
Dried plant material produced 0.2% 5-MeO-DMT and small amounts

of DMT ( Ghosal & Mukherjee 1966 ).
Family: Mimosaceae Genus: Anadenanthera (Piptandenia) Species: colubrina, contorta, excelsa, macrocarpa, peregrina
Black beans from these trees are toasted, pulverized and mixed with ashes or calcined shells to make psychedelic snuff called yopo by Indians in Orinoco basin in Colombia, Venezuela and possibly in southern part of Brasilian Amazon. Yopo is blown into the nostrils through bamboo tubes or snuffed by birdbone tubes. The trees grow in open plain areas, and leaves, bark and seeds contain DMT, 5-MeO-DMT and related compounds (Schultes 1976 , 1977 ; Pachter et al. 1959 ; Phytochem 11, 737).
Genus: Acacia Species: confusa [DMT,st], jurema, maidenni [DMT,b] niopo,
nubica [DMT, l], phlebophylla [DMT,l], polyacantha [DMT,l] subsp. campylacantha, senegal [DMT,l], simplicifolia [DMT,b,l,st], simplex, tortilis.
The letters following the tryptamine (DMT in all of these cases) refer to where the tryptamine is found in the plant: "st" is stems, "b" is bark, and "l" is leaves.
Dried A. confusa stems contain 0.04% N-methyltryptamine and 0.02% DMT ( Arthur et al. 1967 ). The dried leaves of A. phlebophylla contain 0.3% DMT ( Rovelli & Vaughan 1967 ). The bark of A. maidenii contains 0.6% of N-methyltryptamine and DMT in the proportions approx. 2:3 ( Fitzgerald & Sioumis 1965 ). A. simplex also

contains DMT ( Poupat et al. 1976 ). Several Sudanese species have DMT ( Wahba & Elkheir, 1975 ). Seeds of several acacia species are available from ..Of the jungle.
Genus: Desmanthus Species: illinoensis "Illinois Bundleflower"
Thompson et al. report that the root bark of this North American perennial shrub contains 0.34% DMT and 0.11% N-methyltryptamine. The bark accounts for about a half of the total weight of the roots. The plant should be resistant to cold and draught and easy to grow. ..Of the Jungle sells D. illinoensis seeds and dried roots (seed packet $3, 7 grams $10, oz $25; roots 4 oz $15, pound $50). Seeds are also available from more main-stream mail-order houses.
Genus: Mimosa Species: scabrella, tenuiflora (== hostilis) "tepescohuite",
verrucosa The roots of M. hostilis, which is not the common houseplant M. pudica ("sensitive plant"), contain 0.57% DMT and are used by Indians of Pernambuso State in Brazil as part of their Yurema cult ( Pachter et al. 1959 , Schultes 1977 , ( Meckes-Lozoya et al. 1990 ). M. scabrella contains DMT and N-methyltryptamine ( De Moraes et al., 1990 ). Bark of M. verrucosa also contains DMT ( Smith 1977 ).
Genus: Testulea Species: gabonensis
Contains DMT ( Leboeuf et al., 1977 ). Family: Malpighiaceae Genus: Banisteriopsis Species: muricata (=argentea), rusbyana (see Diplopterys
cabrerana) Both contain DMT ( Smith 1977 ).
Genus: Diplopterys Species: cabrerana
Natives of western Amazon add DMT- and N-methyltryptamine containing leaves of the vine D. cabrerana to a drink made from Banisteriopsis caapi, which contains beta-carbolines harmine and harmaline, to heighten and lengthen the visions ( Schultes 1977 , Smith 1977 ). D. cabrerana is also known as Banisteriopsis rusbyana.
Family: Myristicaceae Genus: Horsfieldia Species: superba

Contains 5-MeO-DMT as well as a beta-carboline ( Jossang et al. 1991 ).
Genus: Iryanthera Species: macrophylla
The bark contains 5-MeO-DMT ( Holmstedt et al. 1980 ).
Genus: Virola Species: calophylla, calophylloidea, rufula, sebifera, theiodora
The bark resin of these trees is used to prepare hallucinogenic snuffs in northwestern Brazil by boiling, drying and pulverizing it. Sometimes leaves of a Justicia are added. The snuff acts rapidly and violently, "effects include excitement, numbness of the limbs, twitching of facial muscles, nausea, hallucinations, and finally a deep sleep; macroscopia is frequent and enters into Waika beliefs about the spirits resident in the drug." Snuffs made from V. theiodora bark contain up to 11% 5-MeO-DMT and DMT. Also leaves, roots and flowers contain DMT. Amazonian Colombia natives roll small pellets of boiled resin in a evaporated filtrate of bark ashes of Gustavia Poeppigiana and ingest them to bring on a rapid intoxication ( Smith 1977 , Schultes 1977 ).
Family: Pandanaceae Genus: Pandanus "Screw pine"
DMT has been isolated from Pandanus nuts growing in New Guinea (

Barrau 1958, 1962 ). Family: Poaceae Genus: Arundo Species: donax
Leaves, flowers and rhizomes contain DMT, bufotenine and related compounds ( Ghosal et al. 1972 ).
Genus: Phalaris Species: aquatica (tuberosa), arundinacea
Leaves of P. arundinacea and leaves and seedlings of P. aquatica contain DMT, 5-MeO-DMT and related compounds ( Smith 1977 ). P. arundinacea plants are available from ..Of the jungle for $15.
Genus: Phragmites Species: australis
Rhizomes contain DMT ( Wassel et al. 1985 ).
Family: Rubiaceae Genus: Psychotria

Species: carthaginensis, viridis (psychotriaefolia) Psychotria leaves are added to a hallucinogenic drink prepared from Banisteriopsis caapi and B. rusbyana (which contain beta-carbolines) to strengthen and lengthen the effects in western Amazon. P. carthaginensis and P. viridis both contain DMT ( Rivier, 1972 ). 5 seeds of P. viridis cost $10 from ..Of the jungle.
Family: Rutaceae Genus: Dictyoloma Species: incanescens
Bark contains 0.04% 5-MeO-DMT ( Pachter et al. 1959 ). Genus: Limonia Species: acidissima
Contains DMT ( Abu Zarga, 1986 ). Genus: Melicope Species: leptococca
Contains 5-MeO-DMT as well as 5-MeO-DMT-Oxide and a beta-carboline ( Skaltsounis et al. 1983 ).
Genus: Vepris Species: ampody
The leaves and branches contains up to 0.2% DMT ( Kan-Fan 1970 ). Genus: Zanthoxylum Species: aborescens
Contains DMT ( Grina et al., 1982 ).
table of contents
References
Abu Zarga, M.H. 1986. Three new simple indole alkaloids from Limonia acidissima. Lloydia 49(5), 901-904.
Akers, B.P. 1992. Peele's Lepiota: An identification and a clarification. Mycotaxon 43(0), 461-469.
Arora, D. 1986. Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi. Ten Speed Press, Berkley.
Arthur, H.R., Loo, S.N. & Lamberton, J.A. 1967. Nb-methylated tryptamines and other constituents of Acacia confusa Merr. of Hong Kong. Aust. J Chem. 20, 811.

Banerjee, P.K. & Ghosal, S. 1969. Simple indole bases of Desmodium gangeticum. Aust. J Chem. 22, 275-277.
Barrau, J. 1958. Nouvelles observations au sujet des plantes hallucinogenes d'usage autochtone en Nouvelle-Guinee. J Agric. Trop. Bot. Appl. 5, 377-378.
Barrau, J. 1962. Observations et travaux recents sur les vegetaux hallucinogenes de la Nouvelle-Guinee. J Agric. Trop. Bot. Appl. 9, 245-249.
Benedict, R.G., Brady, L.R., Smith, A.H. & Tyler, V.E. 1962. Occurrence of psilocybin and psilocin in certain Conocybe and Psilocybe species. Lloydia 25, 156-159.
Benedict, R.G., Tyler, V.E. & Watling, R. 1967. Blueing in Conocybe, Psilocybe and a Stropharia Species and the Dectection of Psilocybin. Lloydia 30(2), 150-157.
Besl, H. 1993. Galerina steglichii spec. nov., a hallucinogenic Galerina. Zeitschrift fuer Mykologie 59(2), 215-218.
Bresinsky, A. & Besl, H. 1990. A Colour Atlas of Poisonous Fungi. Wolfe Publishing Ltd, London.
Bye, R.A. 1979. Hallucinogenic plants of the Tarahumara. J. Ethnopharmacology 1, 23-48.
Christiansen, A.L., Rasmussen, K.E. & Hoeiland, K. 1984. Detection of psilocybin and psilocin in Norwegian species of Pluteus and Conocybe. Planta Med. 50, 341-343.
Cimino, G. & De Stefano, S. 1978. Chemistry of Mediterranean Gorgonians. Simple indole derivatives from Paramuricea chamaeleon. Comptes Rendus Biochem. Physiol. Ser. C. 61, 361-362.
Crouch, I.J., Smith M.T., Van Staden J., Lewis, M.J. & Hoad, G.V. 1992. Identification of auxins in a commercial seaweed concentrate. J Plant Physiology 139(5), 590-594.

Daly, J.W. & Witkop, B. 1971. Chemistry and pharmacology of frog venoms. In: Venomous animals and their venoms. Vol II. New York: Academic Press.
Davis, W. & Weil, A.T. 1992. Identity of a New World Psychoactive Toad. Ancient Mesoamerica 3 (1992) 5, 51-59.
De Moraes, E.H.F., Alvarenga, Z.M.A., Ferreira, Z.M.G.S. & Alisue, G. 1990. Quim. Nova 13, 308.
Fitzgerald, J.S. & Sioumis, A.A. 1965. Alkaloids of Australian Leguminosae V. Aust. J Chem. 18, 433.
Fiussello, N. & Ceruti-Scarti, J. 1971/72. Presenza di psilocibina edi 5-idrossi-indolderivati in Panaeolus retirugis. Atti Acc. Sci. Torino 106, 725-735.
Gartz, J. 1991. Influence of phosphate on fruiting and secondary metabolism of mycelia of Psilocybe cubensis, Psilocybe semilanceata and Gymnopilus purpuratus. Zeitschrift fuer Mykologie 57(1), 149-154.
Ghosal, S., Chaudhuri, R.K., Dutta, S.K. & Bhattacharya, S.K. 1972. Occurrence of curaromimetic indoles in the flowers of Arundo donax. Planta Med. 21, 22.
Ghosal, S. & Mukherjee, B. 1966. Indole-3-alkylamine Bases of Desmodium pulchellum. J, Org. Chem. 31, 2284.
Ghosal, S., Singh, S. & Bhattacharya, S.K. 1971. Alkaloids of Mucuna pruriens, Chemistry and Pharmacology. Planta Med. 19, 279
Grina, J.A. et al. 1982. Constituents of Zanthoxylum aborescens. Part 7. Old & new alkaloids from Zanthoxylum aborescens. J. Organic Chemistry 47, 2648-2651.
Gurevich, L.S. 1993. Indole derivatives in certain Panaeolus species from East Europe & Siberia. Mycological Research 97(2), 251-254.

Gurevich, L.S. & Astapenko, V.V. 1992. Chromatographic study of some indole metabolites in Panaeolus basidiomycetes. Mikologiya I Fitopathologiga 26(3), 189-194.
Guzman, G. 1983. The Genus Psilocybe. Beihefte Zur Nova Hedwingia 74, 1-439.
Guzman, G., Bandala, V.M. & Allen, J.W. 1993. A New Bluing Psilocybe from Thailand. Mycotaxon 26, 155-160.
Guzman, G., Bandala, V.M. & King, C. 1991. A New Species of Psilocybe of Section Zapotecorum from New Zealand. Mycological Research 95, 507-508.
Guzman, G., Saldarriaga, Y., Pineda, F., Garcia, G. & Velazquez, L.F. 1994. New Species of Psilocybe from Colombia and Discussion on the known species. Mycotaxon , 225.
Haeselbarth, G., Michaelis, H. & Salnikow, J. 1985. Nachweis von Psilocybin in Inocybe aeruginescens. Mykol. Mitt. bl. 28(1), 59-62.
Hatfield, G.M., Valdes, L.J. & Smith, A.H. 1978. The occurrence of psilocybin in Gymnopilus species. Lloydia 41, 140-144.
Hein, R. & Wasson, R.G. 1958. Les champignons hallucinogenes du Mexique. Museum National d'Histoire Naturelle, Paris.
Holmstedt, B., Lindgren, J.E., et al. 1980. Indole alkaloids in Amazonian Myristicaceae: Field and laboratory research. Bot. Mus. Leafl., Harvard Univ. 28, 215-234.
Johns, S.R., Lamberton, J.A. & Sioumis, A.A. 1966. Alkaloids of the Australian Leguminosae VI. Aust. J Chem. 19, 893.
Jossang, A., Jossang, C., Hadi, H.A., Sevenet, T. & Bodo, B. 1991. Horsfiline, an oxindole alkaloid from Horsfieldia superba. J. Organic Chem. 56(23), 6527-6530.
Kan Fan, C. et al. 1970. Alcaloides de Vepris ampody (Rutacees). Phytochem. 9, 1283-1291.

Kantor, R.E., Dudlettes, S.D. & Shulgin, A.T. 1980. 5-Methoxy-alfa-methyl- tryptamine (alfa,O-dimethylserotonin), a hallucinogenic homolog of serotonin. Biological Psychiatry Vol 15, 349-352.
Koike, Y., Wada, K., Kusano, G., Nozoe, S., & Yokoyama, K. 1981. Isolation of Psilocybin from Psilocybe argentipes and its Determination in Specimens of some Mushrooms. Lloydia 44(3), 362-365.
Leboeuf, M. et al., 1977. Alkaloids and triterpenes of Testulea gabonensis. Plant Medicine Phytotherapy 11, 230.
Luna, L.E. 1984. The Healing Practices of a Peruvian Shaman. J. Ethnopharmacology 11, 123-133.
McKenna, D.J., Towers, G.H.N., & Abbott, F. (1984). Monoamine oxidase inhibitors in South American hallucinogenic plants: Tryptamines and Beta-carboline constituents of ayahuasca. J Ethnopharmacology 10, 195-223.
Mckenna, D.J. & Towers, G.H.N. 1984. Biochemistry and Pharmacology of Tryptamines and beta-Carbolines: A Minireview. J Psychoactive Drugs 16(4).
Meckes-Lozoya, M., Lozoya, X., Marles, R.J., Soucy-Breau, C., Sen, A. & Arnason, J.T. 1990. N,N-dimethyltryptamine alkaloid in Mimosa tenuiflora bark (tepescohuite). Arch. Invest. Med. Mex. 21(2), 175-7.
Merlin, M.D. & Allen, J.W. 1993. Species identification and chemical analysis of psychoactive fungi in the Hawaiian islands. Journal of Ethnopharmacology 40(1), 21-40.
Miller Jr., O.K. 1972. Mushrooms of North America. Dutton & Co., Springfield.
Most, Albert. 1984. Bufo Alvarius: the Psychedelic Toad of the Sonoran Desert . Venom Press Box 2863 Denton TX 76202.

Most, Albert. 1986. Eros and the Pineal: the layman's guide to cerebral solitaire . Venom Press, Denton, TX.
Naranjo, C. 1969. Psychotropic Properties of the Harmala Alkaloids. In: Efron (Ed.) The Ethnopharmacologic Search for Psychoactive Drugs.
Ohenoja, E., Jokiranta, J., Makinen, T., Kaikkonen, A. & Airaksinen, M.M. 1987. The Occurrence of Psilocybin and Psilocin in Finnish Fungi. Lloydia 50(4), 741-744.
Pachter, I.J, Zacharias, D.E & Ribeir, O. 1959. Indole Alkaloids of Acer saccharinum (the Silever Maple), Dictyoloma incanescens, Piptadenia colubrina, and Mimosa hostilis. J Org Chem 24, 1285-7.
Phillips, R. 1981. Mushrooms and other fungi of Great Britain and Europe. Pan Books, London.
Poupat, C., Ahond, A. & Sevenet, T. 1976. Alcaloides De Acaia simplicifolia. Phytochemistry 15, 2019-2120.
Rivier, L. & Lindgren, J-E. 1972. "Ayahuasca," the South American Hallucinogenic Drink: an Ethnobotanical and Chemical Investigation. Economic Botany 26, 101-129.
Rivier, L. & Pilet, P.E. 1971. Annee Biologique 10, 129.
Robbers, J.E., Tyler, V.E. & Ola'h, G.M. 1969. Additional evidence supporting the occurrence of psilocybin in Panaeolus foenisecii. Lloydia 32, 399-400.
Rovelli, B. & Vaughan, G.N. 1967. Alkaloids of Acacia I. Dimethyltryptamine in Acacia phlebophylla. Aust. J Chem. 20, 1299-1300.
Saupe, S.G. 1981. Occurence of Psilocybin/Psilocin in Pluteus salicinus (Pluteaceae). Mycologia 73, 781-784.
Schultes, R.E. 1976. Indole Alkaloids in Plant Hallucinogens. J of Psychedelic Drugs Vol 8(1), 7-25.
Schultes, R.E. 1977. The Botanical and Chemical Distribution of Hallucinogens. J of Psychedelic Drugs Vol 9(3), 247-263.
Schultes, R.E. 1979. Hallucinogenic Plants: Their Earliest Botanical Descriptions. J of Psychedelic Drugs Vol 11(1-2), 13-24.
Schultes, R.E. & Hofmann, A. 1979. Plants of the Gods. McGraw-Hill. Reprint available from Healing Arts Press, Rochester, VT.
Schultes, R.E. & Hofmann, A. 1980. The Botany and Chemistry of Hallucinogens. Springfield, Ill: Thomas.
Shulgin, A.T. 1976. Psychotomimetic agents. In: Gordon, M. (Ed.) Psychopharmacological Agents, Vol IV. New York: Academic Press.
Shulgin, A.T. 1982. Chemistry of Psychotomimetics. In: Hoffmeister, F. & Stille, G. (Eds.) Handbook of Experimental Pharmacology, Vol 55: Alcohol and Psychotomimetics, Psychotropic Effects of Central-Acting Drugs. New York: Springer-Verlag.
Skaltsounis, A.L., Tillequin, F. & Koch, M. 1983. Plantes des Nouvelle- Caledonie LXXXIII: Alcaloides des tiges feuillees de Melicope leptococca. Lloydia 46(5), 732.
Smith, T.A. 1977. Review: Tryptamine and Related Compounds in Plants. Phytochemistry 16 171-175.
Stein, S.I. 1959. Clinical Observations on the Effects of Panaeolus venenosus Versus Psilocybe caerulescens Mushrooms. Mycologica 51, 49.

Stijve, T., Klan, J. & Kugper, W. 1985. Occurrence of psilocybin and baeocystin in the genus Inocybe. Persoonia 12, 469-472.
Strassman, R.J. 1990. The Pineal Gland: Current Evidence For Its Role In Consciousness. In: Lyttle, T. (Ed.) Psychedelic Monographs and Essays Vol 5.
Thompson, A.C., Nicollier, G.F. & Pope, D.F 1987. Indolealkylamines of Desmanthus illinoensis and Their Growth Inhibition Activity. J Agric. Food Chem. 35, 361-365.
Wahba, S.K. & Elkheir, Y.M. 1975. Dimethyltyrptamine from the leaves of certain Acacia species of northern Sudan. Lloydia 38, 176-177.
Wassel, G.M. et al. 1985. Alkaloids from the rhizomes of Phragmites australis Cav. Scientia Pharmaceutica 53, 169-170.
[email protected] * Everything is perfect forever Michael from Melbourne * Ditto
RELATED SPACES DMT
N,N-DIMETHYLTRYPTAMINE H Y P E R S P A C E Terence McKenna Land Shamanism Gracie & Zarkov
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fdeoxy.orgD E O X Y . O R G
END OF FAQ Erro! O nome de arquivo não foi especificado.
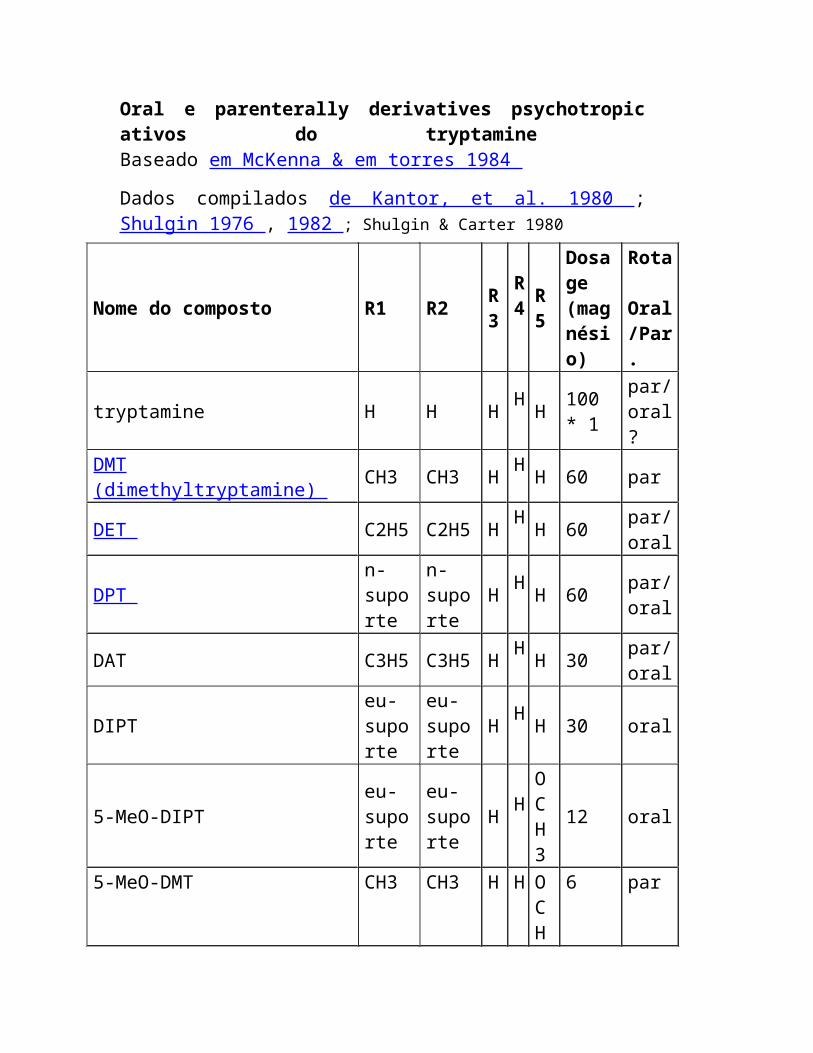
2. Alcalóides Muitos dos compostos puros o mais cedo isolados com atividade biológica eram alcalóides. Isto era devido à facilidade da isolação. O nitrogênio faz geralmente o básico composto e o composto existe na planta como um sal. Assim, os alcalóides são extraídos frequentemente com água ou ácido suave e recuperados então como o material cristalino pelo tratamento com a base.
Antes de aproximadamente 300 anos há, a malária era o scourge de Europa, provavelmente sendo introduzido através de o Oriente Médio. A malária é causada pelos protozoa do genus Plasmodium , contidos como spores no gut dos mosquitos dos anófeles, que espalha então os spores aos seres humanos quando morde. Enquanto os exploradores do espanhol e do Portugese começaram a colonize América sul, descobriram uma cura para a malária sabida aos indians nativos. Este era o bark das árvores da chinchona. O uso do bark da chinchona tratar a malária foi relatado primeiramente em Europa em 1633, e o primeiro bark alcançou Roma aproximadamente 12 anos mais tarde. Os chás feitos do bark curaram os povos que sofrem da malária, um dos scourges principais em Europa naquele tempo, e o bark tornou-se sabido como o bark de Jesuit. Por causa das diferenças filosóficas entre Protestants e catholics, muitos Protestants recusou ser tratado com o bark. Um do Protestants o mais proeminente do tempo, Oliver Cromwell, morrido reportedly da malária por causa deste stubbornness.
Isolado originalmente do succirubra da chinchona , o quinine é um de 31 alcalóides com estruturas relacionadas, e o composto antimalarial principal, na planta. (os alcalóides foram definidos em várias maneiras, mas uma definição vem razoavelmente perto da realidade. Um alcalóide é um composto planta-derivado que seja tóxico ou physiologically ativo, contenha um nitrogênio em um anel heterocyclic, seja básico, tenha uma estrutura complexa, e seja de distribuição limitada no reino de planta.) A malária é ainda um problema principal durante todo o mundo, e, embora as drogas antimalarial sintéticas suplantem pela maior parte o quinine como o tratamento para a malária durante a guerra de mundo II, o quinine é frequentemente uma vez outra vez a droga da escolha porque as
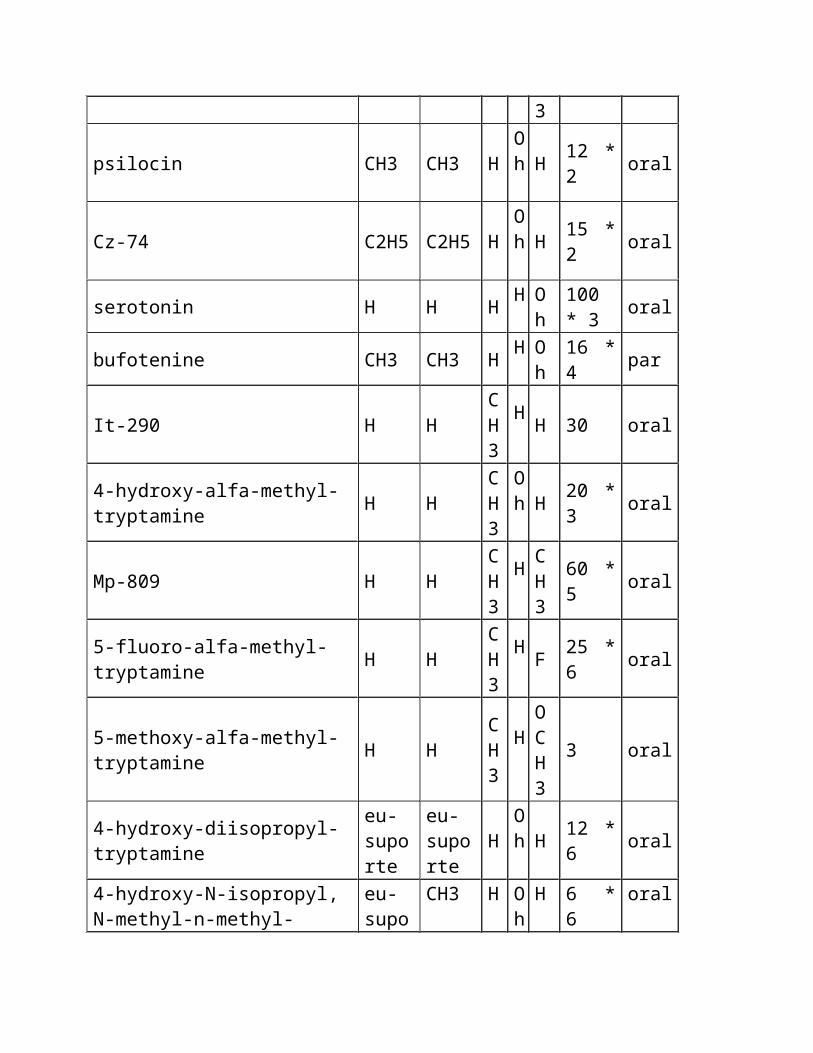
tensões da malária se tornaram resistentes às drogas sintéticas. Entretanto, a busca para outras drogas antimalarial das fontes naturais continuou também. Uma das drogas novas as mais prometedoras é qinghaosu, isolado do annua do artemisia , um sesquiterpene (veja abaixo) que contenha uma estrutura original do trioxane.
Entre o mais famoso dos alcalóides são os alcalóides de Solanaceae ou de tropane . As plantas que contêm estes alcalóides foram usadas durante todo o history gravado como venenos, mas muitos dos alcalóides têm propriedades pharmaceutical valiosas. A atropina, o formulário racemic do hyoscyamine, vem do belladonna do atropa (nightshade mortal) e é usada dilate as pupilas do olho. A atropina é também um stimulant do CNS e é usada como um tratamento para o envenenamento do gás de nervo. Scopolamine, um outro membro desta classe é usado como um tratamento para a doença de movimento. A cocaína, da coca de Erythroxylum , é relacionada pròxima na estrutura, é também um stimulant do CNS, e foi usada como um anestésico tópico no opthamology. É também uma droga do abuso. A cocaína foi encontrada em quantidades muito pequenas na fórmula original da Coca-Cola, mas não era o interesse principal do USDA naquele tempo. A cafeína foi considerada ser o problema principal com a bebida. O stramonium do datura (Jimsonweed), uma planta encontrada em Virgínia contem compostos similares.
Os alcalóides do ergot vêm de um fungo, o purpurea de Claviceps , que é um parasite no centeio e no trigo. Os alcalóides do ergot são responsáveis para o ergotismo (fogo do St. Anthony), que se manifesta como o ergotismo gangrenous, tendo por resultado a perda dos membros, ou o ergotismo convulsive, tendo por resultado hallucinations. Em ambos os casos, a morte segue geralmente e os outbreaks do ergotismo causaram 11.000 mortes em Rússia tão tarde quanto 1926. Hoje o problema é reconhecido e controlado. Alguns dos alcalóides do ergot foram usados tratar headaches do migraine e disorders sexual em aplicações clínicas. O mais famoso destes alcalóides é o diethylamide ácido lysergic, LSD, um hallucinogen poderoso que seja um derivative sintético dos produtos naturais. Os alcalóides similares, particularmente ergine, são

encontrados também em glories mexicanos da manhã, tais como Ipomeoa tricolor .
Os alcalóides do morphine , derivados do poppy de opium, somniferum do papaver , são apaziguadores da dor e narcotics poderosos. A atividade narcotic do somniferum do P. foi anotada em tabuletas sumerian em 3500 B.C., fazendo lhe um das drogas as mais velhas conhecido. O opium é o latex secado das cabeças da semente do somniferum do P. , e foi usado como um analgesic (elimina ou alivia a dor) e narcotic (induz o sono ou o drowsiness) nas preparações tais como o laudanum e paregoric. O morphine é o alcalóide principal e foi isolado primeiramente entre 1803 e 1806. Foi usado extensamente para o relevo da dor que começa nos 1830's, mas reconhecido também como sendo addictive. Em uma tentativa de fazer o morphine mais menos addictive, os químicos de Bayer acetylated os grupos do hydroxyl para produzir o diacetylmorphine. Isto foi introduzído no mercado como um apaziguador non-non-addictive da dor sob o heroin do nome de comércio por aproximadamente dois anos no 1900?s adiantado, até que se estêve reconhecido para estar mais addictive do que o morphine. Outros derivatives do morphine foram desenvolvidos e encontraram o uso como antagonistas do opiate ou como os tranquilizers animais.
Vincristine , uma das drogas antileukemic as mais potent no uso hoje, foi isolado em uma busca para tratamentos do diabetes do rosea de Vinca (agora roseus de Catharanthus ) nos 1950 junto com o vinblastine, um homologue em que o grupo n-methyl é oxidado a um moiety do aldeído. Esta é uma estrutura tão complexa que está isolado ainda da planta (o periwinkle de Madagascan) hoje melhor que preparado pela síntese. A mudança pequena na estrutura, entretanto, causa uma mudança significativa no efficacy pharmoacological. Vincristine (leurocristine, VCR) é o mais eficaz em leukemias da infância e nos lymphomas de tratamento dos non-Hodgkin?s, onde o vinblastine (vincaleukoblastine, VLB) é usado tratar a doença de Hodgkin?s. Para trás à introdução Aos Terpenes Ao ácido e ao Polyketide de Shikimic produtos naturais derivados Home

The Quinine AlkaloidsThe Cinchona alkaloids are another very important subgroup of the polycyclic -carboline alkaloids.Prior to about 300 years ago, malaria was the scourge of the Western world. Malaria was probably the most common disease in Rome and London at the time. Even today, millions of people die each year from malaria, and hundreds of millions of people suffer from the disease. The disease is caused by a protozoa of the genus Plasmodium spread as spores contained in the Anopheles mosquito which spreads them into the bloodstream of human victims. The major cure for this dread disease is quinine, theprincipal alkaloid of the Cinchona tree which is indigenous to the tropical Andes mountains. There are many tales of how the antimalarial properties of Cinchona bark was first discovered, but the first written report of its efficacy was by Father Calancha in the Chronicle of St. Augustine in 1633. It was at least twelve more years before a Father Tafur brought some of the bark back to Rome. The cures which were accomplished by drinking a "beverage" made from the powdered bark were nothing short of amazing.However, the fact that it was coming from the Catholic church made the cure suspect to Protestants, and Oliver Cromwell died in 1658 from malaria, even though the cure was available. The actual Cinchona or fever tree was not discovered until 1737. Before that time, the Jesuits who discovered the cure had to make due with trading with the Indians for the bark. Supposedly, the Indians hid the tree from the Europeans as a means of driving them from South America. Supplies of the tree were rapidly depleted in South America, and Java, under the Dutch, became the main supplier of Cinchona alkaloids until World War II when synthetic antimalarial agents became widely available. In recent years, strains of malaria which are resistant to the synthetic antimalarials have begun to make their appearance, and the value of quinine as an antimalarial agent is once again on the rise. R1 R2 (+)-isomer (-)-isomer -CH=CH2 -H cinchonine cinchonidine-CH=CH2 -OCH3 quinidine quinine -CH2CH3 -H dihydrocinchonine dihydrocinchonidine -CH2CH3 -OCH3 dihydroquinidine dihydroquinine Quinine was first isolated in 1820 by Joseph Pelletier and Joseph Caventou, but the structure determination took over 100 years. By 1884, 25 additional alkaloids related to quinine had been isolated, and an additional 6 were added between 1884 and 1941. Approximately 13 out of the 31 still have unknown structures. There

are eight major alkaloids which occur as four pairs of enantiomers, as shown above. The (+)-isomers have the 2R,3S configuration, and the (-)-isomers have the 2S,3R configuration.Quinine has typically been the most used of the alkaloids for pharmacological purposes, simply because it was the first isolated. Quinine, in addition to its antimalarial properties, is a bacteriocide, local anesthetic, cardiosvascular stimulant, and analgesic. Quinidine, in addition to its antimalarial properties, has been used to prevent certain types of cardiac arrythmias.
The Tropane AlkaloidsThe tropane alkaloids, which have the 8-azabicyclo[3.2.1]octane nucleus, are commonly found in plants of three families, the Solanaceae, Erythroxylaceae, and Convolvulaceae families. The alkaloids isolated from plants of these families, while having several legitimate medicinal uses, are probably best known for their toxic properties. This can be a major problem since the plants produce very attractive berries which are tempting to small children. As few as three berries of henbane (Hyoscyamus niger) or deadly nightshade (Atropa belladonna) can cause death in infants. Many of the plants in the Solanaceae family contain tropane alkaloids, which are responsible for the toxic effects of the plants. Cleopatra is reputed tohave tested the effects of henbane and deadly nightshade on her slaves to investigate the possibility of using these extracts to commit suicide (she found the toxic effects too painful). The wives of the Roman emperors, Augustus and Claudius, used deadly nightshade to murder large numbers of Romans. The mandrake (Mandragora officinarum) was reputed to possess aphrodisiac properties and was prized for these properties. However, the roots also contain large quantities of the tropane alkaloid hyoscine (scopolamine), making the plant highly toxic. Datura stramonium, a plant common to Virginia, and other Datura species are widely distributed throughout the world. These plants have a long history of being used as both sedatives and as poisons.Extracts of Datura metel, sometimes called Hindu Datura, were used as knockout drops to lure virgins into prostitution and then by the prostitutes to sedate their clients. The Roman army led by Mark Antony in Asia Minor was severely crippled by deaths caused by eating a Datura species. Indians in Colombia used another Datura species for infanticide. Infants died after nursing when an extract of

Datura was smeared on the nipples of the mother. Datura stramonium is also known as jimsonweed and, after nearly taking thelives of some of the Jamestown settlers, was widely used for medicinal purposes, primarily as a sedative.In the 1960’s and 1970’s, their were a number of cases of poisoning due to jimsonweed when people ingested the plant to try to induce a high similar to cocaine. The Solanaceae alkaloids derived from these plants, while very toxic, are often important medicinal agents. Atropine, the racemic form of (-)-hyoscyamine, was first isolated from Atropa belladonna in 1833. (-)-Hyoscyamine was also isolated in 1833 from Hyoscyamus niger. This tropane alkaloid is a muscarinic receptor antagonist. Muscarinic receptors are involved in constriction of the pupil, vasodilation, moderating the heartbeat, and stimulating secretions. The most familiar use of atropine, as the sulfate salt, is as a mydriatic to dilate the pupil of the eye during an opthamological examination. In fact, the juice of the berries of A. belladonna was used during the Renaissance by ladies of the Italian courts to exaggerate the size of their eyes by dilating the pupils. (The rough translation of belladonna from Italian is beautiful lady.) Atropine has also found use as a CNS stimulant and is useful in treating poisoning cases, particularly anticholinesterase poisoning induced by organophosphorous insecticides and nerve gas and poisoning induced by the toxic principles of the mushroom Amanita muscaria. Scopolamine, (-)-hyoscine, which was first isolated in 1881 from Hyoscyamus muticus and Scopolia atropides is a CNS depressant athigh doses, and has been used in combination with morphine to induce "twilight sleep" before and during labor, and in smaller doses, to prevent motion sickness.In recent years, a tropane alkaloid isolated from several Erythroxylaceae species has become more notorious. This, of course, is cocaine, which was first isolated from coca, the dried leaves of Erythroxylum coca (Bolivian coca) or Erythroxylum truxillense (Peruvian coca), in 1862 by Wohler. The anesthetic properties of cocaine have been known since the 1880's.
The hydrochloride of cocaine is used as a topical anesthetic in opthamology as 1% solution (maximum). Cocaine is toxic in large enough doses when taken internally, particularly when taken by injection, but is also a potent CNS stimulant which has led to its abuse. It is addictive but withdrawal effects are reputed to be mild.

(These effects have also been known since the late 19th (-)-hyoscyamine atropine NH3COO HC, 6H5OH OH, C6H5H OONH3C.(-)-scopolamine NH3COO HC6H5OH,Oecgonine cocaineNH3CCO2CH3 CO2HNH3CO2CC6H5 OH century. Remember that Sherlock Holmes, Sir Arthur Conan Doyle's legendary detective, was a habitual user of cocaine.)Coca has been of economic importance in Western cultures since the 1500’s. But coca was alsoimportant to the Indian cultures of South America, particularly the Incas in Peru, for at least 1000 yearsbefore the Spanish arrived in the New World. Chewing coca leaves provided a stimulatory effect forherdsmen and other workers in the Inca culture, but it was not in common usage by every Indian. Oncethe Spanish arrived, coca leaves became an important commodity, and for a while was used in place ofmoney in South America. Coca chewing increased among both natives and Europeans, but manychurchmen and missionaries were opposed to its use, particularly among the natives (who needed to beconverted to Catholicism). Since the alkaloid content of the coca leaves degraded upon long storage, itwasn’t until the 1800’s that coca made larger inroads into Europe. In the late 1800’s, a number ofbeverages based on coca began to appear, including several coca wines, for use as mild stimulants. In1886, John Styth Pemberton reformulated his coca wine into a nonalcoholic beverage, which he namedCoca-Cola, to get around a developing temperance movement. The original formulation of Coca-Colaused coca leaf from South America which contains very low amounts of cocaine. One estimate is that theoriginal formulation contained, at most, 1.5 mg of cocaine per ounce of Coca-Cola, not enough to provideany stimulation or cause addiction. (The stimulatory effect of Coca-Cola more likely comes from thecaffeine content.) By 1906, all cocaine had been eliminated from the formula of Coca-Cola, and in 1911,in a strange turn of events, the makers were sued by the government for false advertising because the
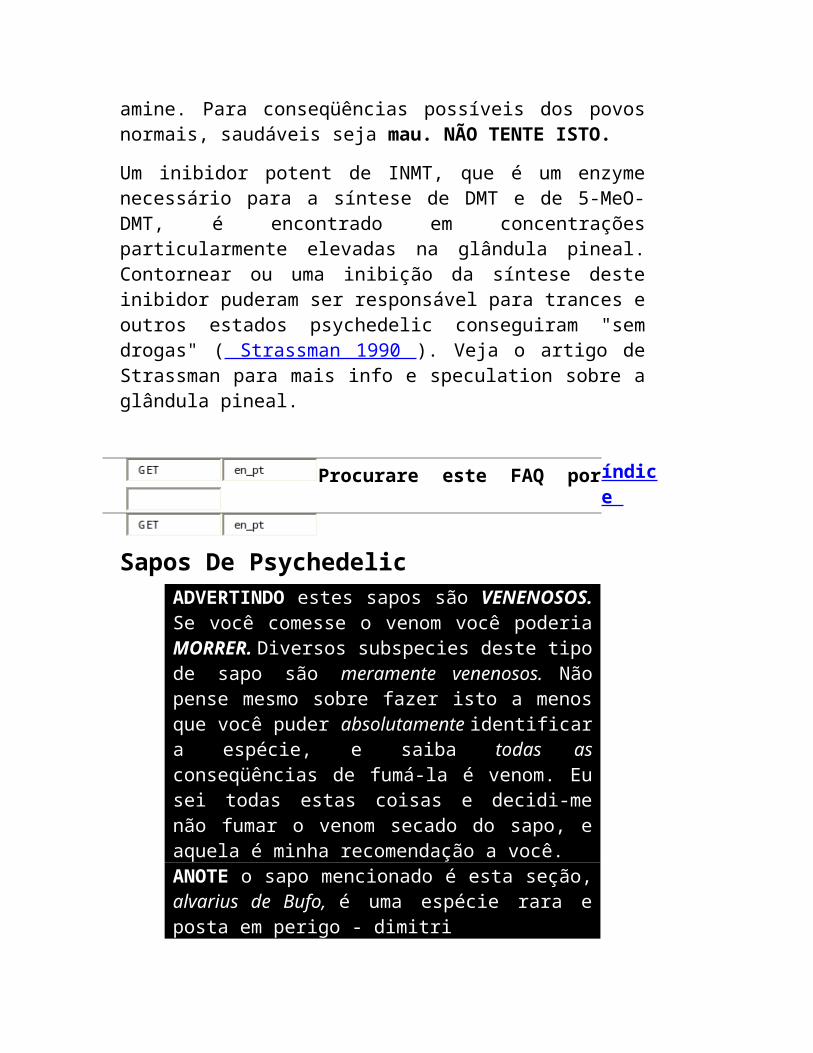
beverage did not contain cocaine!Ecgonine is the parent alkaloid of cocaine, and can be easily produced from cocaine and other estersisolated from the plant by acid hydrolysis. Cocaine can then be prepared from ecgonine by treatment withmethanol followed by benzoic acid. This is the basic procedure used in the clandestine drug labs.
The Ergot AlkaloidsOne of the pharmacologically most important groups of indole alkaloids is the ergoline, or ergot,alkaloids. These alkaloids are isolated from the dried sclerotium of the fungus Claviceps purpurea(Hypocreaceae) (ergot). This fungus is a parasite on rye and wheat and other grains. Ingestion ofcontaminated grain, most often after the grain has been made into bread, causes ergotism, also knownas the "Devil's curse" or "St. Anthony's fire," and has been a problem for centuries. It has been noted inwritings from China as early as 1100 B.C. and in Assyria in 600 B.C., and Julius Caesar's legions sufferedan epidemic of ergotism during one of the campaigns in Gaul. In 994 A.D., an epidemic in France killedbetween 20,000 and 50,000 people, and in 1926, at least 11,000 cases of ergotism occurred in Russia.Ergotism can cause convulsions, nausea, and diarrhea in mild forms, and there is some thought that anoutbreak of ergotism may have been the cause of the "bewitchings" which led to the Salem witch trials inthe United States in 1691. Ergotism may also have caused some of the extreme destruction associatedwith the French Revolution. In the Middle Ages, ergotism was described as causing victims to dies"miserably, their limbs eaten up by the holy fire that blacked like charcoal." People turned to the churchfor help, assuming that the disease was retribution for their sins. In particular, they prayed to St. Anthonyfor deliverance, giving rise to the name for the disease. Ergotism takes two forms, gangrenous ergotism,

in which tingling effects were felt in fingers and toes followed in many cases by dry gangrene of the limbsand finally loss of the limbs, and convulsive ergotism, in which the tingling was followed by hallucinationsand delerium and epileptic-type seizures. In both cases, death was slow and painful. Ergotism has nowbeen recognized as a result of infection by a mycotoxin, and the ergotism plagues have been eliminated.However, the alkaloids derived from ergot have assumed new importance for their pharmacologicalproperties, and ergot is produced commercially for the preparation of these alkaloids.There are three main groups of ergot alkaloids, the clavine type, the water-soluble lysergic acid type,and the water-insoluble lysergic acid type or peptide ergot alkaloids. The clavine type of alkaloids, suchas agroclavine and elymoclavine, are generallyregarded as precursors to the other groups of ergotalkaloids in the biogenetic pathway. These alkaloids areamong several of the ergot alkaloids also isolated fromhigher plants, particularly the seeds of Ipomoea violaceaand Rivea corymbosa ('ololiuqui," the Mexican morningglory), both members of the Convolvulaceae family.These alkaloids are not used pharmacologically, butagroclavine is a powerful uterine stimulant, and many ofthe ergot alkaloids are prolactin release inhibitors.The water-soluble lysergic acid derivatives are most oftenamide derivatives. Among the most important of these areergonovine and methysergide. Ergonovine has potent uterinecontraction activity and is used in treating postpartum hemorrhages.It has low vasoconstrictor action. Methysergide is used as a cranialvasodilator in the treatment of migraine headaches.Perhaps the most infamous of the semi-synthetic derivatives ofthe ergot alkaloids is lysergic acid diethyl amide (LSD). It was firstsynthesized by Albert Hofmann of Sandoz AG in 1938, but itshallucinogenic properties were not known until 1943. While workingwith a sample of LSD in his laboratory, Hoffmann accidentallyingested some. He described how "On a Friday afternoon, April 16,1943, while working in the laboratory, I was seized by a peculiar sensation of

vertigo and restlessness. Objects, as well as the shape of my associates inthe laboratories, appeared to undergo optical changes. I was unable toconcentrate on my work. In a dreamlike state, I left for home, where anirresistable urge to lie down and sleep overcame me. Light was so intense asto be unpleasant. I drew the curtains and immediately fell into a peculiar stateof 'drunkenness', characterized by an exaggerated imagination. With myeyes closed, fantastic pictures of extraordinary plasticity and intensive colorseemed to surge towards me. After two hours, this state gradually subsidedmethysergide ergonovineHNOHHNOHO ONHNCH3H HNCH3NCH3lysergic acid diethylamideHN CH3NHONlysergic acid agroclavine elymoclavineN

HNCH3HO2CHNHNCH3HH HHNCH3NHOHand I was able to eat dinner with a good appetite." The following Monday, to confirm that he had indeedingested some of the LSD, Hoffmann prepared a solution containing 250 µg of LSD and deliberatelyingested it. After 40 minutes, he found he had " difficulty in concentration, visual disturbances, markeddesire to laugh" and left for home. On the ride home "I had great difficulty in speaking coherently, my fieldof vision swayed before me....I had the impression of being unable to move from the spot." The symptomscontinued for six hours after Hoffmann reached his home and he described how "all objects appeared inunpleasant, constantly changing colors, the predominant shades being sickly green and blue...Aremarkable feature was the manner in which all acoustic perceptions were transformed into opticaleffects." Hoffmann had taken approximately five times the "normal" dose of LSD and had experiencedthe first "bad trip."For a while following the discovery of the pharmacological effects of LSD, it was used in psychiatry,particularly in the treatment of alcoholic schizophrenia. In recent years, evidence has come out that it was

used in the 1950s on military "volunteers" to study its effect, presumably as a chemical warfare weapon.In the early 1960s, proponents of its use as a way to achieve a state of nirvana, such as Dr. TimothyLeary, began using it heavily and distributing it throughout the high school and college age population,often adsorbed into a sugar cube. This attitude that LSD was a "good drug" was fostered by popularsongs, like the Jefferson Airplane's "White Rabbit" and the Beatles' "Lucy In The Sky With Diamonds",which supposedly described the effects of "good" LSD trips. Unfortunately, many LSD trips turned out tobe bad trips, and many heavy users of LSD experienced bad flashback trips at a later time. (One sidelightto the hallucinogenic effects of LSD is the result of finding ergot alkaloids in Mexican morning glory seeds.Some seekers of nirvana through hallucinogens began ingesting large quantities of morning glory seeds.Rather than hallucinogenic activity, these foolish people experienced primarily toxic reactions thusobtaining nirvana in an unintended way.) After a relatively brief time, the popularity of LSD as ahallucinogen diminished, and it became somewhat of a historical relic. However, it has started to make acomeback in the drug underground in recent years. This time, the target seems to be grade school agechildren, and the pushing of LSD to these children may be viewed by some as a means of hooking themon hard drugs like cocaine and heroin as they grow older.The water-insoluble lysergic acid derivatives areprimarily peptide ergot alkaloids like ergotamine.This compound was first isolated in 1918, but itsstructure was not determined until 1951. Ergotamine,as its tartrate salt, is a analgesic specifically used fortreatment of severe migraine headaches. It is oftenused in conjunction with caffeine, which constrictscerebral blood vessels, and a dose of 2 mg takenorally often results in quick relief. 2-Bromo--ergocryptine, a semisynthetic derivative, has reducedtoxicity and is now commercially available to be used

in the reduction of lactation in women. It has alsobeen used in treatment of sexual disorders, and hasbeen shown to enhance sexual libidos in both menand women.2-bromo--ergokryptine ergotamineHN CH3NHONNHNOOHOOC6H5HN OOOHNNOONHNCH3H
Morphine and Related AlkaloidsThe narcotic activity of Papaver somniferum was recorded on Sumerian tablets dating back to 3500B.C. The Ebers papyrus also cites its use in treating "colic." The first description of the collection of crudeopium appear in the writings of Dioscorides. The Greeks prescribed opium for a variety of illnesses, and it

was particularly useful as a sedative. The opium poppy was first cultivated in the Mediterranean area,then introduced into India in the 15th century, and then somewhat later into China. It is now grownthroughout the region from the Middle East to the Far East asa "cash crop." Opium is the air-dried latex of the head (orseed case) of P. somniferum, and contains primarily thealkaloids morphine (4-21%) and codeine (0.8-2.5%), alongwith other alkaloids. Codeine appears to be the precursor tomorphine. The seeds do not contain any of the morphinealkaloids. Raw opium is usually contaminated withadulterants such as fragments of the heads, pulp from figsand other fruits, tragacanth, beeswax, cumin seed, starch,and inorganic substances such as clay, sand, stone, and lead from pipe or bullets. Opium acts on theCNS, first stimulating then depressing nerve response. It has analgesic, narcotic, and antiperistalticeffects. Tinctures of opium have been used for many years. Laudanum, which is a deodorized tincture ofopium, was used extensively as an antiperistaltic treatment or sedative, particularly in Europe. Paregoric,which is a tincture of opium with added camphor, was used widely in Europe and the U.S. to treatdiarrhea. It was often used in combination with a mixture of belladona, kaolin, and pectin for this purpose.Morphine, the principal alkaloid of P. somniferum, was first isolated in either 1803 or 1806 bySerturner. The structure was first proposed in 1925 by Sir Robert Robinson, and later confirmed by thetotal synthesis of Gates and Marshall. Pure morphine was used, beginning in the 1830s, for pain relief,particularly after the advent of the hypodermic syringe in 1853, and was quickly recognized to beaddictive. However, it was widely used, particularly in military medicine, dueto its very potent analgesic properties. Other side effects include nausea,vomiting, drowsiness, mood changes, a decrease in mental accuity,respiratory arrest, and constipation. The constipation is caused by decreased

HCl secretion in the stomach which in turn increases the tone of the first partof the duodenum and delays passage of the gastric contents. Since morphinewas the best major analgesic available, pharmaceutical companies began tosearch for a nonaddictive substitute. In 1874, diacetylmorphine wassynthesized by Alder Wright of St. Mary's Hospital Medical School, and Bayermarketed this derivative from 1898 through 1910 as "Heroin," a nonaddictivemorphine substitute and cough medicine. The name heroin came from itsreputation as a "heroic" drug. Other pharmaceutical companies, such as Eli Lilly, also sold heroin over thecounter in the U.S. In 1900, heroin was toutedas "...the perfect drug, more potent yet lessharmful....Habituation has been noted in a smallpercentage of cases,....none of the patientssuffer from this habituation." In 1905, it wasnoted in a classic case of understatement that"although heroin is stated not to give rise tohabituation, a more extended knowledge of thedrug would seem to indicate that this is notentirely correct." By 1910, it was universallyrecognized that heroin was more addictive thanmorphine and it was removed from the market.It has become apparent over the years asresearchers have searched for a morphinesubstitute that small structural changes to themorphine nucleus can have major effects on theactivity. For instance, codeine is one tenth aspowerful a narcotic as morphine, and its effectdiacetylmorphine"heroin"N CH3OCH3CO2CH3CO2etorphine

naloxone RnalorphineN CH3OCH3OHON ROHOHO HOOONOHHOcodeine morphineN CH3OHOCH3O HOHOON CH3on the respiratory center is about one fourth that of morphine. In addition to being less toxic, it is lessaddictive (although it is still very addictive). The N-allyl, N-propyl, and N-cyclopropylmethyl homologues ofmorphine are all morphine (opiate) antagonists. They compete for the morphine receptor site andeliminate the respiratory depression effect of morphine. Naloxone is a pure antagonist. It is active at alower dose than the agonist and has been used to treat morphine or heroin overdoses. Etorphine, whichis prepared via a Diels-Alder reaction of methyl vinvyl ketone with thebaine followed by furthermodification, is much more powerful (5000-10000 times as powerful) than morphine as an analgesic. It isalso a hallucinogen with potency equivalent to that of LSD. The principal use of etorphine has been as atranquilizer for the capture of large wild animals.

Pharmacology de Vinblastine, de Vincristine, de Vindesine e de Vinorelbine [ Tabela Química Da Comparação | Página Do Periwinkle ]
O inblastine V e o vincristine são alcalóides encontrados no periwinkle de Madagascar , roseus de Catharanthus (classificado anteriormente como o rosea de Vinca , que conduziu a estes alcalóides chamados se tornando de Vinca dos compostos ).
E o vindesine e o vinorelbine , derivatives semisynthetic do vinblastine, todos trabalham inibindo o mitosis (divisão da pilha) no metaphase . Estes alcalóides ligam ao tubulin , assim impedindo que a pilha faça os eixos que necessita poder mover ao redor seus chromosomes enquanto se divide (esta é similar à ação do colchicine , mas é om diferente do franco a ação do paclitaxel , que interfere com a divisão da pilha mantendo os eixos de ser quebrado para baixo). Estes alcalóides parecem também interferir com a abilidade das pilhas synthesize o DNA e o RNA . Todos são administrados intravenously em seu formulário do sulfate uma vez uma semana; estas soluções são fatais se forem administradas qualquer outra maneira, e podem causar muitos da irritação do tecido se escaparem fora da veia. Embora estes três compostos sejam muito similares na estrutura e tenham a mesma ação básica, têm distintamente efeitos diferentes no corpo.
Vinblastine é administrado tipicamente em um dose de 6 milligrams por o medidor quadrado da superfície do corpo. É introduzído no mercado como Velban por Eli Lilly e tem um half-life no bloodstream de 24 horas. Vinblastine é principalmente útil para tratar a doença de Hodgkin, o lymphoma lymphocytic, o lymphoma histiocytic, cancer testicular avançado, cancer de peito avançado, sarcoma de Kaposi, e doença de Letterer-Siwe. Parece também lutar o cancer interferindo com o metabolism do ácido glutamic (especificamente, os pathways que conduzem do ácido glutamic ao ciclo de Krebs e à formação do urea). Os povos com infecções bacterianas não devem ser dados esta droga, nem devem mulheres grávidas, desde que causou defeitos severos do nascimento nos estudos animais. Os efeitos laterais incluem a perda do cabelo, o nausea, contagens abaixadas da pilha do sangue, headache, dor

do estômago, numbness, constipation e sores da boca. Os danos do marrow de osso são o fator dose-limitando típico.
Vincristine, que é introduzído no mercado como Oncovin por Eli Lilly, tem um half-life do serum de aproximadamente 85 horas. É usado principalmente tratar o leukemia agudo, o rhabdomyosarcoma, o neuroblastoma, a doença de Hodgkin e os outros lymphomas. O dose típico em 1,4 milligrams por o medidor quadrado da superfície do corpo uma vez que uma semana, e o neurotoxicity são o dose que limita o fator (ele pode causar os danos ao sistema nervoso periférico). Por causa deste, os povos com disorders neuromuscular devem dirigir claramente desta droga se possível. Do mesmo modo, os povos com alguns formulários do syndrome do Charcot-Marie-Dente devem evitar o vincristine. As mulheres grávidas não devem definitivamente fazer exame d, porque causa defeitos severos do nascimento nos testes animais. Os efeitos laterais incluem aqueles encontrados com vinblastine, mais problemas nervosos do sistema tais como o impairment sensory; alguns povos podem também desenvolver problemas ou spasms respirando do pulmão logo depois que a droga é administrada. Os povos desenvolvem ocasionalmente cancers secundários se receberem a droga junto com outras drogas anticancer que estão sabidas para ser carcinogens.
Vindesine tem um half-life do serum de aproximadamente 24 horas e é administrado em um dose de 3 milligrams por o medidor quadrado da superfície do corpo. Seus toxicity e efeitos laterais são similares àqueles do vinblastine. Vindesine, que é introduzído no mercado sob os nomes Eldisine e Fildesin, é usado principalmente tratar cancers do melanoma e de pulmão (carcinomas) e, com outras drogas, tratar cancers uterine.
Vinorelbine está atualmente em experimentações clínicas da fase II como um tratamento para o cancer ovarian. Estará introduzído no mercado como Navelbine por Glaxo Wellcome, Inc., se as experimentações forem bem sucedidas e o FDA aprovar a droga. Assim distante, o vinorelbine parece ter uma escala mais larga da atividade antitumor do que os outros alcalóides do vinca. Em experimentações preclinical, mostrou a promessa em tratar pacientes com os cancers ovarian epithelial e, em combinação com o cisplatin da droga do chemotherapy, em tratar pacientes com os cancers de pulmão da non-pequeno-pilha. Os efeitos laterais desta droga

incluem o diarrhea, o nausea, e a perda do cabelo; parece ser menos de um veneno do nervo do que o vindesine.
Para mais informação, visita:
Caderno Ovarian Da Pesquisa De Cancer: Vinorelbine (Navelbine)
Folha do fato de Vincristine do instituto nacional do cancer
O Projeto Do Acesso: Vinblastine
O Projeto Do Acesso: Vincristine
Vindesine Compreensivo: Usos e efeitos laterais
Referências:
Budavari, Susan, ed. 1989. O Índice De Merck: Uma enciclopédia dos produtos químicos, das drogas e do Biologicals . Rahway, NJ, Merck & Co.
Goodman, Louis Sanford, Alfred Gilman, e Alfred Goodman Gilman, eds. 1990. A base pharmacological do therapeutics , å edição. Elmsford, NY, Imprensa De Pergamon.
Mutschler, Ernst, e Hartmut Derendorf. 1995. Ações Da Droga: Princípios básicos e aspectos therapeutic. Estugarda, Germany, publishers científicos do medpharm.
referência 1995 & cópia da mesa do médico . Montvale, NJ, Médico Economia Dados Produção Companhia.

http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fbiotech.icmb.utexas.edu%2F http://babelfish.altavista.com/ babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fbiotech.icmb.utexas.edu%2F http://babelfish.altavista.com/ babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fbiotech.icmb.utexas.edu%2Fbotany%2F http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fbiotech.icmb.utexas.edu%2Fbotany%2F http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fbiotech.icmb.utexas.edu%2Fbotany%2Fcite.html http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fbiotech.icmb.utexas.edu%2Fbotany%2Fcite.html http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fbiotech.icmb.utexas.edu%2Fbotany%2Ftest.html http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fbiotech.icmb.utexas.edu%2Fbotany%2Ftest.html
URL: http://biotech.icmb.utexas.edu/botany/vvv.html Cyberbotanica é mantido por Lucy A. Snyder . Emita por favor seu E-mail ou encha para fora um formulário do gabarito se você manchar quaisquer erros nestas páginas, ou se você tiver quaisquer sugestões ou comentários. Todo o copyright material 1996, recursos de BioTech.
Esta página foi atualizada por último 11/6/97.
CARTA DO PHYTOCHEMISTRY

Envenenamento Da Planta, Alcalóides - Tropane Último Updated: Novembro 29, 2001 Avalie este artigo Email a um colega Synonyms e keywords relacionados: belladonna, erva daninha do jimson, erva daninha do loco, datura sacred, trombeta do angel, mandrake, henbane

& nbsp INFORMAÇÃO DO AUTOR Seção 1 de 11 Da Continuação Clínica Do Medication Do Tratamento De Workup Dos Diferenciais Da Introdução Da Informação Do Autor Bibliografia Variada Dos Retratos
Autor: Richard um Wagner, MD, PhD , equipe de funcionários consultando, departamento da medicina da emergência, hospital do EL Dorado de Tucson; Consultante, droga do arizona e centro de informação do veneno Coauthor(s): Samuel M Keim, MD , diretor do programa, professor do associado, departamento da cirurgia, divisão da medicina da emergência, universidade do centro das ciências da saúde do arizona
Richard um Wagner, MD, PhD, é um membro das seguintes sociedades médicas: Faculdade americana de médicos da emergência , e associação médica americana Editor(s): Michael S Beeson, MD, MBA , diretor do programa, departamento da medicina da emergência, sistema da saúde de Summa; Professor do associado da medicina clínica da emergência, faculdade do nordeste das universidades de Ohio; John T VanDeVoort, PharmD, DABAT , Gerente, Professor Assistente Clínico, Departamento Do Pharmacy, Hospital Das Regiões; Michael Hodgman, MD , professor clínico assistente da medicina, departamento da medicina da emergência, Bassett Healthcare; John Halamka, MD , oficial de informação principal, sistema de CareGroup Healthcare, professor assistente da medicina, departamento da medicina da emergência, centro médico de Beth Israel Deaconess; Professor assistente da medicina, escola médica de Harvard; e Raymond J Roberge, MD, MPH, FAAEM, FACMT , professor clínico do associado da medicina da emergência, universidade da escola de Pittsburgh da medicina; Atendendo à equipe de funcionários, departamento da medicina da emergência, hospital das Magee-Mulheres da universidade do centro médico de Pittsburgh & nbsp INTRODUÇÃO Seção 2 de 11 Da Continuação Clínica Do Medication Do Tratamento De Workup Dos Diferenciais Da Introdução Da Informação Do Autor Bibliografia Variada Dos Retratos

Fundo: Os alcalóides são os metabolites da planta que têm uma estrutura química nitrogen-containing do anel, alcalóide-como o reactivity químico, e a atividade pharmacologic. Os amines do alcalóide do stimulant, os amines hallucinogenic do alcalóide, e os alcalóides altamente anticholinergic do tropane (ou os alcalóides do belladonna) compreendem os 3 grupos pharmacologic principais de amines do alcalóide.
As plantas que contêm alcalóides do tropane (ou o bicyclic), atropina, scopolamine, e hyoscyamine incluem o seguinte:
Espécie do datura (erva daninha do jimson, trombeta do angel, maçã do espinho) Hyoscyamus niger (henbane) Belladonna do atropa (nightshade mortal) Officinarum de Mandragora (mandrake) Todas estas plantas têm histories longos do uso hallucinogenic e foram conectadas com o sorcery, o witchcraft, a medicina nativa, e os rites magico-religiosos que datam para trás a 1500 BC e a odyssey de Homer . (uso de Homer da planta moly como um antidote às drogas anticholinergic venenosas de Circe pode ter sido o uso primeiramente gravado de uma anticolinesterase inverter o intoxication anticholinergic central.)
As medicinas herbal chinesas que contêm alcalóides do tropane foram usadas tratar o asthma, o bronchitis crônico, a dor, e os sintomas do flu. Em México, o datura é feito exame por mulheres de Yaqui para diminuir a dor do parto. Em África, um uso comum deve fumar sae do datura para aliviar o asthma e problemas pulmonary. Muitas culturas worldwide adicionam plantas com alcalóides do tropane (particularmente espécie do datura) às bebidas alcoólicas ao intoxication do aumento.
Recentemente, o datura foi usado como um hallucinogen recreacional nos E. U., tendo por resultado casos esporádicos do envenenamento e da morte anticholinergic. Os casos numerosos do envenenamento anticholinergic também resultaram dos contaminadores do alcalóide do belladonna nos alimentos, including o chá de Paraguai comercialmente compra (um chá herbal derivado do paraguariensis de Ilex ), no hamburger, no mel, no

porridge duro feito do millet contaminado, e da "no vinho homemade da flor lua". Outros ingestions acidentais incluem o emprego errado como um vegetal e um inclusion selvagens edible no toothpaste homemade.
Pathophysiology: O toxicity das plantas que contêm alcalóides do tropane manifesta-se como o envenenamento anticholinergic clássico. Os sintomas ocorrem geralmente 30-60 minutos depois que ingestion e podem continuar por 24-48 horas porque os alcalóides do tropane atrasam esvaziar e absorption gastric.
Scopolamine, agindo como um antagonista em receptors muscarinic periféricos e centrais, é pensado para ser o composto preliminar responsável para os efeitos tóxicos destas plantas. Os alcalóides de Tropane são encontrados em todas as partes das plantas, com concentrações as mais elevadas nas raizes e nas sementes.
A atropina é um artifact do purification, produzido pelo racemization do l-hyoscyamine. A proporção de cada presente do alcalóide varia entre a espécie, a época do ano, a posição, e a parte de planta. Tão pouco quanto um meio de teaspoon da semente do datura, equivalente a 0,1 magnésio da atropina por a semente, causou a morte da apreensão cardiopulmonary. A rota usual do ingestion é como um chá, embora ingesting sementes ou a outra planta parte e fumar secado sae é também comum.
Freqüência:
Nos E. U.: A incidência é esporádica, com os conjuntos de casos do envenenamento, na maior parte entre adolescents usando plantas para seus efeitos hallucinogenic. Em 1994, um aumento 10-fold no número dos casos foi relatado em Florida. O acesso difundido à informação em plantas hallucinogenic através do Internet pode conduzir a um aumento mais adicional na incidência. Internacional: A incidência worldwide é desconhecida. Entretanto, os casos foram relatados em Austrália, Brasil, Hong Kong, México, Tanzânia, Formosa, e Venezuela, atestando à distribuição geográfica larga da espécie do datura.

Mortality/Morbidity: Os casos de Nonfatal são underreported provavelmente.
Em 1993, 318 casos do envenenamento do datura foram relatados à associação americana de centros de controle do veneno; os centros para o controle da doença (CDC) relataram 2 mortes. Em 1994, o CDC relatou 7 casos do envenenamento anticholinergic em 3 famílias que consumiram o chá de Paraguai comercial contaminado. Os relatórios de caixas ou de conjuntos esporádicos dos casos que envolvem o uso intencional como um hallucinogen são freqüentes; a maioria de pacientes recuperam uneventfully.
As mortes relacionadas de afogar-se, de exposição, e de falta do cuidado de suporte foram relatadas. Sexo: Nenhum predilection sexual existe.
Idade: Nenhum predilection da idade existe, embora o uso do datura como uma droga recreacional seja mais comum entre adolescents. & nbsp CLÍNICO Seção 3 de 11 Da Continuação Clínica Do Medication Do Tratamento De Workup Dos Diferenciais Da Introdução Da Informação Do Autor Bibliografia Variada Dos Retratos
History:
Como em algum paciente que apresenta-se com uma mudança aguda no status mental ou no envenenamento suspeitado, tentativa de obter a seguinte informação: Termine após o history médico History do medication Descrição precisa ou amostra do toxicant(s) suspeitado Rota de administração, de quantidade ingested, de tempo desde o ingestion, e de razão para o ingestion Co-ingestants-ingestants e uso do álcool ou das outras drogas da rua Os sinais e os sintomas iniciais podem incluir o seguinte: Membranas mucous e pele secas Disfagia e dysarthria

Photophobia Visão borrada Tachycardia Retenção urinary Os sinais e os sintomas iniciais podem ser seguidos pelo hyperthermia, pela confusão, pelo agitation, pelo combativeness, pelas apreensões, pelo coma, e pela morte. O amnesia a respeito dos eventos que seguem o ingestion de alcalóides do tropane é comum. A identificação de plantas ingested pode ser diagnostic do envenenamento do alcalóide do tropane. As espécies do datura, as plantas o mais geralmente encontradas que contêm alcalóides do tropane, são anuários de 3-5 pés com grosseiro-coarse-toothed saem. as flores Trombeta-dadas forma são 3-5 polegadas no comprimento, com as pétalas coloridas branco-pálido-violetas (veja o retrato 1 , retrato 2). Os mais importantes para a identificação taxonomic são os pods spiny, redondos, chambered da semente (veja o retrato 3). Exame: O "vermelho mnemônico como uma beterraba, seca como um osso, cortina porque um bastão, louco como um hatter, e por mais quente que um hare" esteja útil recordar o toxidrome anticholinergic.
Sinais vitais Tachycardia e tachypnea Hiperpirexia (em aproximadamente 20% dos casos) Hypertension e hypotension inconsistent, depression respiratory (raro) Pele seca morna (pode ser nivelado) Cabeça, orelhas, olhos, nariz, e garganta Mydriasis e cicloplegia (quase sempre ocorra e possa persistir por dias) Seque as membranas mucous Abdômen Sons diminuídos do bowel Distention da bexiga urinary Neurologic Agitation, confusão, e hallucinations Apreensões, coordenação diminuída do músculo, paralysis, depression respiratory, e coma (raro) & nbsp DIFERENCIAIS Seção 4 de 11

Da Continuação Clínica Do Medication Do Tratamento De Workup Dos Diferenciais Da Introdução Da Informação Do Autor Bibliografia Variada Dos Retratos
Delirium, dementia, e amnesia Infecção de HIV e AIDS [ hyperthyroidism, tempestade do thyroid e doença das sepulturas ]
Hypoglycemia Meningitis Envenenamento Da Planta, Glycosides - Cardiac Envenenamento Da Planta, Herbs Envenenamento Da Planta, Hypoglycemics Esquizofrenia Status Epilepticus Toxicity, Amphetamine Toxicity, Anticholinergic Toxicity, Antidepressivo Toxicity, Antidysrhythmic Toxicity, Antihistamine Toxicity, Cocaína Toxicity, Hallucinogen Toxicity, Cogumelo - Hallucinogens Syndromes Da Retirada *** TRANSLATION ENDS HERE ***
Quick Find Author Information Introduction Clinical Differentials Workup Treatment Medication Follow-up Miscellaneous

Pictures Bibliography
Click for related images.
Related Articles Delirium, Dementia, and Amnesia
HIV Infection and AIDS
[Hyperthyroidism, Thyroid Storm and Graves Disease]
Hypoglycemia
Meningitis
Plant Poisoning, Glycosides - Cardiac
Plant Poisoning, Herbs
Plant Poisoning, Hypoglycemics
Schizophrenia
Status Epilepticus
Toxicity, Amphetamine
Toxicity, Anticholinergic
Toxicity, Antidepressant
Toxicity, Antidysrhythmic
Toxicity, Antihistamine
Toxicity, Cocaine
Toxicity, Hallucinogen

Toxicity, Mushroom - Hallucinogens
Withdrawal Syndromes
Continuing Education CME available for this topic. Click here to take this CME.
Patient Education Click here for patient education.
& nbsp WORKUP Section 5 of 11 Author Information Introduction Clinical Differentials Workup Treatment Medication Follow-up Miscellaneous Pictures Bibliography
Lab Studies:
No specific diagnostic studies for tropane alkaloid poisonings exist. Tropane alkaloids are not included in standard drugs of abuse or comprehensive toxicology panels; serum drug concentrations are neither helpful nor readily available to aid in initial management. Since acetaminophen and salicylate are common co-ingestants in many poisonings, consider assays for acetaminophen and salicylate levels. In febrile patients, consider blood and urine cultures. Serum electrolyte levels may provide insight into other intoxicating agents and co-ingestants. Consider a pregnancy test for all patients capable of bearing children.

Imaging Studies:
Consider CT scan of the head for patients with altered mental status that is inadequately explained by toxicity from ingested agent or is unresponsive to appropriate intervention. Other Tests:
Obtain an electrocardiogram (ECG) at presentation for all patients with suspected toxic ingestions. Procedures:
Consider lumbar puncture (LP) for patients with fever and altered mental status. Consider invasive monitoring for hemodynamically unstable patients. & nbsp TREATMENT Section 6 of 11 Author Information Introduction Clinical Differentials Workup Treatment Medication Follow-up Miscellaneous Pictures Bibliography
Prehospital Care: Transport patient to nearest emergency facility with capabilities for advanced life support (ALS), at minimum. Primary assessment should focus on airway and respiratory, circulatory, and neurologic systems.
Unless patient is extremely agitated, obtain IV access and monitor vital signs frequently. Consider administration of naloxone and thiamine. Do not use ipecac and defer administration of activated charcoal, unless a prolonged transport time is anticipated. Assess for hypoglycemia and other causes of altered mental status. Manage seizures with benzodiazepines. Physostigmine is not recommended in prehospital setting. Emergency Department Care: As in all cases of suspected poisoning, follow the ABCDEs of emergency medicine (airway, breathing, circulation, disability, exposure), then the ABCDEs of toxicology (antidotes, basics, change absorption, change distribution, change elimination).

Provide oxygen and intubate if significant CNS or respiratory depression exists and no gag reflex is present. Assess circulation and initiate cardiac and pulse oximetry monitoring. Obtain a 12-lead ECG and evaluate for QRS prolongation, ischemia, and evidence of arrhythmia. Sinus tachycardia is common and does not require treatment for a stable patient. Obtain blood for laboratory analysis and bedside glucose measurement while obtaining IV access. Inspection after full-body exposure should be performed to assess signs of trauma or seizure. Agitated or hallucinating patients often respond to reassurance and a darkened room. If chemical restraint is required, benzodiazepines are the drugs of choice. Early consultation with a poison control center is frequently helpful. Consider GI decontamination foremost. Ipecac is contraindicated because of the potential for seizures. Gastric lavage is controversial; while it is commonly performed, no reliable data on outcomes exist to support its use, and the risk of aspiration and other complications is increased. Administer activated charcoal (1-2 g/kg) orally or per nasogastric or orogastric tube. One or 2 additional doses may be given at 1- or 2-hour intervals to ensure adequate gut decontamination. An ileus without distension is not a contraindication to a single dose of charcoal, and charcoal given alone may be as effective or more effective than emesis and lavage procedures. Use of cathartics to hasten elimination from GI tract remains controversial. Sorbitol may be used with first dose of charcoal; further use may cause serious fluid shifts to the intestine, diarrhea, dehydration, and hypernatremia. Tropane alkaloids are lipophilic and cross the blood-brain barrier; hemodialysis and hemoperfusion are generally ineffective. No effective methods of changing distribution or elimination of tropane alkaloids exist. Specific antidote for tropane alkaloid toxicity is physostigmine salicylate, a reversible acetylcholinesterase inhibitor capable of directly antagonizing CNS manifestations of anticholinergic toxicity. Physostigmine (at doses lower than those producing peripheral side effects of salivation, lacrimation, urination, defecation, emesis) can reverse central effects of the following conditions:
Coma Seizures Hallucinations Agitation

Severe dyskinesias Respiratory depression However, unless absolute certainty can be established that the toxicity present is due to tropane alkaloid poisoning and not to co-ingestants or other substances, confirmatory peripheral manifestations of anticholinergic toxicity should coexist prior to administration of physostigmine.
Physostigmine can induce a life-threatening cholinergic crisis (eg, seizures, respiratory depression, asystole). Since most patients can be safely treated without this antidote, physostigmine preferably should be used in consultation with a poison control center and generally should be used only for patients in the following states:
Unresponsive to supportive measures Tachydysrhythmias and subsequent hemodynamic compromise Intractable seizures unresponsive to benzodiazepines Extremely severe agitation or psychosis Physostigmine is contraindicated in patients receiving tricyclic antidepressants, disopyramide, quinidine, procainamide, cocaine, or other agents producing cardiac conduction abnormalities. Relative contraindications include reactive airway disease, intestinal obstruction, and administration of depolarizing paralytic agents.
Following GI decontamination, most patients rarely require more than physiologic monitoring and psychologic support.
Patients experiencing agitation and hallucinations usually respond to reassurance and benzodiazepines.
Most phenothiazines are contraindicated because of their anticholinergic properties.
If signs or symptoms of urinary retention exist, Foley catheterization should be performed for bladder decompression. Consultations:
Early consultation with a toxicologist or poison control center is frequently useful for toxic exposures or ingestions to help in decision-making with regard to decontamination and therapeutic interventions; this is particularly true with the use of physostigmine in cases of tropane alkaloid poisoning.

Psychiatric consultation is important for all intentional ingestions. Contact with primary care provider is optimal for all hospital admissions or cases of serious illness. & nbsp MEDICATION Section 7 of 11 Author Information Introduction Clinical Differentials Workup Treatment Medication Follow-up Miscellaneous Pictures Bibliography
Activated charcoal is indicated for all tropane alkaloid poisonings, with the possible exception of poisoning from smoking leaves. Benzodiazepines are first-line agents for agitation and seizures. Physostigmine should be used only for life-threatening complications.
Drug Category: GI decontaminant -- Activated charcoal is used after a drug or plant ingestion to limit adsorption of toxins. Traditionally given after the stomach has been emptied by emesis or lavage, recent evidence indicates that it may be used alone, without lavage. Drug Name Activated charcoal (Liqui-Char) -- Emergency treatment in poisoning caused by drugs and chemicals. Network of pores present in activated charcoal adsorbs 100-1000 mg of drug per gram of charcoal. An extremely important component of tropane alkaloid poisoning. May decrease severity and duration of poisoning. Does not dissolve in water. For maximum effect, administer within 30 min after ingesting poison. Adult Dose 1 g/kg PO or by gastric tube; repeat dose of 0.25-0.5 g/kg may be given in 2 h; first dose may be given with cathartic (eg, sorbitol) Pediatric Dose <2 years: Not recommended>2 years: 0.5-1 g/kg PO or by gastric tube Contraindications Documented hypersensitivity; poisoning or overdosage of mineral acids and alkalies; GI obstruction Interactions May inactivate ipecac syrup if used concomitantly; effectiveness of other medications decrease with coadministration; do not mix charcoal with sherbet, milk, or ice cream (decreases adsorptive properties of activated charcoal) Pregnancy C - Safety for use during pregnancy has not been established. Precautions Protect airway in patients with absent gag reflex; not very effective in poisonings of ethanol, methanol, and iron salts; induce emesis before giving activated charcoal; after emesis with ipecac, patient may not

tolerate activated charcoal for 1-2 h; can administer in early stages of gastric lavage; without sorbitol, gastric lavage returns are black Drug Category: Benzodiazepines -- First-line agents for treatment of tropane-alkaloid-induced seizures. Lorazepam is thought to be most effective and has a longer seizure half-life than diazepam. Drug Name Lorazepam (Ativan) -- Sedative hypnotic with short onset of effects and relatively long half-life. DOC if IV access is available. Increasing the action of gamma-aminobutyric acid (GABA), which is a major inhibitory neurotransmitter in the brain, may depress all levels of CNS, including limbic and reticular formation. Monitoring patient's blood pressure after administering dose is important. Adjust prn. Adult Dose 0.1 mg/kg IV; rate of 2 mg/min; commonly administered 1-4 mg initially, with doses up to 10 mg prn Pediatric Dose 0.1 mg/kg IV initial dose (0.15-0.2 mg/kg PR) Contraindications Documented hypersensitivity; preexisting CNS depression; hypotension; narrow-angle glaucoma Interactions Toxicity of benzodiazepines in CNS increases when used concurrently with alcohol, phenothiazines, barbiturates, and MAOIs Pregnancy D - Unsafe in pregnancy Precautions Caution with renal or hepatic impairment, myasthenia gravis, organic brain syndrome, or Parkinson disease Drug Name Midazolam (Versed) -- Used as alternative in termination of refractory status epilepticus. Because midazolam is water soluble, it takes approximately 3 times longer than diazepam to peak EEG effects. Thus clinician must wait 2-3 min to fully evaluate sedative effects before initiating procedure or repeating dose. Published reports of IM and anecdotal reports of nasal use exist (when IV access is not possible). Adult Dose 10 mg IM (when IV/PR is impossible) Refractory status: 0.2 mg/kg IV then infusion 0.1-0.4 mg/kg/h; intubation and pressor support are necessary Pediatric Dose 0.2 mg/kg IM, then obtain IV access Contraindications Documented hypersensitivity; preexisting hypotension; narrow-angle glaucoma; sensitivity to propylene glycol (the diluent) Interactions Sedative effects of midazolam may be antagonized by theophyllines; narcotics and erythromycin may accentuate sedative effects of midazolam because of decreased clearance Pregnancy D - Unsafe in pregnancy
Precautions Caution with congestive heart failure, pulmonary disease, renal impairment, and hepatic failure Drug Name Diazepam (Valium) -- Depresses all levels of CNS (eg, limbic and reticular formation), possibly by increasing activity of GABA. Frequently used in prehospital systems since refrigeration is not required. Adult Dose 0.15 mg/kg IV; not to exceed 20 mg Pediatric Dose Administer as in adults IV; 0.3-0.5 mg/kg PR Contraindications Documented hypersensitivity; narrow-angle glaucoma Interactions Increases toxicity of benzodiazepines in CNS with coadministration of phenothiazines, barbiturates, alcohols, and MAOIs Pregnancy D - Unsafe in pregnancy Precautions Caution with other CNS depressants, low albumin levels, or hepatic disease (may increase toxicity) Drug Category: Cholinergic agents -- Physostigmine is indicated only for reversal of life-threatening complications of tropane alkaloid poisoning (eg, tachydysrhythmias with hemodynamic compromise, seizures refractory to other therapeutic interventions, and severe agitation or hallucinations unresponsive to other therapy). The decision to use physostigmine ideally should be made in consultation with a toxicologist or poison control center. Drug Name Physostigmine (Antilirium) -- Reversible anticholinsterase inhibitor that increases the concentration of acetylcholine at cholinergic synapses. The only reversible anticholinesterase inhibitor that readily crosses the blood-brain barrier to produce the desired CNS effects. Some recommend repeated slow IV pushes of 0.1-0.3 mg q3min to a maximum of 2 mg to decrease potential for life-threatening cardiovascular adverse effects. Adult Dose 2 mg slow IVP; may be repeated as indicated; clinical effects last 20-60 min Pediatric Dose 0.02-0.06 mg IVP; not to exceed 0.5 mg/min or 2 mg as a single dose; clinical effects last 20-60 min; may repeat prn Contraindications Documented hypersensitivity; asthma; cardiovascular disease; diabetes; gangrene; intestinal obstruction; urogenital obstruction; patients receiving choline esters or depolarizing neuromuscular blockers Interactions May inhibit or reverse effects of nondepolarizing neuromuscular blockers (eg, vecuronium or pancuronium) Pregnancy C - Safety for use during pregnancy has not been established. Precautions Patients should be on a cardiac monitor; may precipitate a cholinergic crisis (eg, seizures, respiratory depression, asystole); atropine

should be at bedside; may precipitate bronchorrhea and bronchospasm; may induce fasciculations and muscle weakness; administer 0.5 mg of atropine IV/mg of physostigmine to reverse cholinergic crisis induced by physostigmine & nbsp FOLLOW-UP Section 8 of 11 Author Information Introduction Clinical Differentials Workup Treatment Medication Follow-up Miscellaneous Pictures Bibliography
Further Inpatient Care:
Base admission decisions on patient's signs and symptoms. Asymptomatic patients without signs of anticholinergic toxicity or altered mental status may be discharged after 6 hours of observation. Admit symptomatic patients to an ICU setting for monitoring and treatment. They may be discharged after a symptom-free period of 6 hours without use of supportive therapy or antidotes and after appropriate consultations and follow-up have been arranged. & nbsp MISCELLANEOUS Section 9 of 11 Author Information Introduction Clinical Differentials Workup Treatment Medication Follow-up Miscellaneous Pictures Bibliography
Medical/Legal Pitfalls:
Failure to make the diagnosis Failure to obtain a toxicology consult in a critically ill patient with polysubstance toxicity. & nbsp PICTURES Section 10 of 11 Author Information Introduction Clinical Differentials Workup Treatment Medication Follow-up Miscellaneous Pictures Bibliography

Caption: Picture 1. Datura stramonium (jimson weed). Note 4-5 inch long white flowers. Š 2000 Richard Wagner View Full Size Image eMedicine Zoom View (Interactive!) Picture Type: Photo Caption: Picture 2. Datura stramonium flower. Note the trumpetlike shape. Š 2000 Richard Wagner View Full Size Image eMedicine Zoom View (Interactive!) Picture Type: Photo Caption: Picture 3. Datura stramonium (close-up of unripe seed pods). Note spiny appearance of pods. Š 2000 Richard Wagner View Full Size Image eMedicine Zoom View (Interactive!) Picture Type: Photo & nbsp BIBLIOGRAPHY Section 11 of 11 Author Information Introduction Clinical Differentials Workup Treatment Medication Follow-up Miscellaneous Pictures Bibliography
Anonymous: Anticholinergic poisoning associated with an herbal tea--New York City, 1994. MMWR Morb Mortal Wkly Rep 1995 Mar 24; 44(11): 193-5 [Medline] . Anonymous: Jimson weed poisoning--Texas, New York, and California, 1994. MMWR Morb Mortal Wkly Rep 1995 Jan 27; 44(3): 41-4 [Medline] . Chan TY: Anticholinergic poisoning due to Chinese herbal medicines. Vet Hum Toxicol 1995 Apr; 37(2): 156-7 [Medline] . Chang SS, Wu ML, Deng JF: Poisoning by Datura leaves used as edible wild vegetables. Vet Hum Toxicol 1999 Aug; 41(4): 242-5 [Medline] . Dewitt MS, Swain R, Gibson LB Jr: The dangers of jimson weed and its abuse by teenagers in the Kanawha Valley of West Virginia. W V Med J 1997 Jul-Aug; 93(4): 182-5 [Medline] . Francis PD, Clarke CF, ALIA: Angel trumpet lily poisoning in five adolescents: clinical findings and management. J Paediatr Child Health 1999 Feb; 35(1): 93-5 [Medline] .

Greene GS, Patterson SG, Warner E: Ingestion of angel's trumpet: an increasingly common source of toxicity. South Med J 1996 Apr; 89(4): 365-9 [Medline] . Hayman J, ALIA: Datura poisoning--the Angel's Trumpet. Pathology 1985 Jul; 17(3): 465-6 [Medline] . Hung OL, Lewin NA, Howland MA: Herbal preparations. In: Goldfrank's Toxicologic Emergencies. 6th ed. 1998. Huxtable RJ: The toxicity of alkaloids in foods and herbs. In: Food Poisoning: Handbook of Natural Toxins. Vol 7. Marcel Dekker; 1992: 237-62. Micke MM: The case of hallucinogenic plants and the Internet. J Sch Health 1996 Oct; 66(8): 277-80 [Medline] . Olson KR: Poisoning and Drug Overdose. 3rd ed. Simon & Schuster Trade; 1998. Pereira CA, Nishioka S de D: Poisoning by the use of Datura leaves in a homemade toothpaste. J Toxicol Clin Toxicol 1994; 32(3): 329-31 [Medline] . Plaitakis A, Duvoisin RC: Homer's moly identified as Galanthus nivalis L.: physiologic antidote to stramonium poisoning. Clin Neuropharmacol 1983 Mar; 6(1): 1-5 [Medline] . Ramirez M, Rivera E, Ereu C: Fifteen cases of atropine poisoning after honey ingestion. Vet Hum Toxicol 1999 Feb; 41(1): 19-20 [Medline] . Rodgers GC Jr, Von Kanel RL: Conservative treatment of jimsonweed ingestion. Vet Hum Toxicol 1993 Feb; 35(1): 32-3 [Medline] . Rwiza HT: Jimson weed food poisoning. An epidemic at Usangi rural government hospital. Trop Geogr Med 1991 Jan-Apr; 43(1-2): 85-90 [Medline] . Schultes RE, Hoffmann A: Holy flower of the North Star: Datura. In: Plants of the Gods: Their Sacred, Healing, and Hallucinogenic Powers. 1992: 106-11. Smith EA, Meloan CE, Pickell JA: Scopolamine poisoning from homemade 'moon flower' wine. J Anal Toxicol 1991 Jul-Aug; 15(4): 216-9 [Medline] . Tiongson J, Salen P: Mass ingestion of Jimson Weed by eleven teenagers. Del Med J 1998 Nov; 70(11): 471-6 [Medline] . Urich RW, Bowerman DL, Levisky JA: Datura stramonium: a fatal poisoning. J Forensic Sci 1982 Oct; 27(4): 948-54 [Medline] . Vanderhoff BT, Mosser KH: Jimson weed toxicity: management of anticholinergic plant ingestion. Am Fam Physician 1992 Aug; 46(2): 526-30 [Medline] .

NOTE: Medicine is a constantly changing science and not all therapies are clearly established. New research changes drug and treatment therapies daily. The authors, editors, and publisher of this journal have used their best efforts to provide information that is up-to-date and accurate and is generally accepted within medical standards at the time of publication. However, as medical science is constantly changing and human error is always possible , the authors, editors, and publisher or any other party involved with the publication of this article do not warrant the information in this article is accurate or complete, nor are they responsible for omissions or errors in the article or for the results of using this information. The reader should confirm the information in this article from other sources prior to use. In particular, all drug doses, indications, and contraindications should be confirmed in the package insert. FULL DISCLAIMER
Plant Poisoning, Alkaloids - Tropane excerpt
& copy Copyright 2003, eMedicine.com, Inc. About Us | Privacy | Terms of Use | Contact Us | Advertise | Institutional Subscribers
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.inchem.org http://babelfish.altavista.com/babelfish/urltrurl? lp=en_pt&url=http%3A%2F%2Fwww.inchem.org
PROGRAMA INTERNACIONAL NA SEGURANÇA QUÍMICA

OS CRITÉRIOS de SAÚDE AMBIENTAIS 80
ALCALÓIDES do PYRROLIZIDINE
este relatório contêm as opiniões coletivas um grupo internacional dos peritos e não representam necessariamente as decisões ou a política indicada da organização da saúde do mundo ou do programa unido do ambiente das nações
Publicado sob o sponsorship comum do programa unido do ambiente das nações e da organização da saúde do mundo
Organização Genebra 1988 Da Saúde Do Mundo
ISBN 92 4 154280 2
(c) Organização 1988 Da Saúde Do Mundo
As publicações da organização da saúde do mundo aprecíam a proteção do copyright de acordo com as provisões do protocolo 2 da convenção universal do copyright. Para direitas da reprodução ou da tradução de publicações do WHO, na parte ou no toto, a aplicação deve ser feita ao escritório de publicações, organização da saúde do mundo, Genebra, Switzerland. A organização da saúde do mundo dá boas-vindas a tais aplicações.
As designações empregadas e a apresentação do material nesta publicação não implicam a expressão de nenhuma opinião qualquer na parte do secretariat da organização da saúde do mundo a respeito do status legal de

alguma país, território, cidade ou área ou de suas autoridades, ou de concernir o delimitation de seus fronteiras ou limites.
O mention de companhias específicas ou de produtos de determinados fabricantes não implica que estão endossados ou recomendados pela organização da saúde do mundo na preferência a outra de uma natureza similar que não é mencionada. Os erros e as omissões excetuadas, os nomes de produtos proprietários são distinguidos por letras importantas iniciais.
CRITÉRIOS
DE SAÚDE AMBIENTAIS DOS ÍNDICES PARA ALCALÓIDES DO PYRROLIZIDINE
PREFÁCIO
INTRODUÇÃO - ALCALÓIDES DO PYRROLIZIDINE E SAÚDE HUMANA
1. SUMÁRIO E RECOMENDAÇÕES
1,1. Sumário 1,2. Fontes e estrutura química 1,3. Mecanismos e características do toxicity 1,4. Efeitos no homem 1,4,1. Natureza e extensão dos riscos de saúde 1,5. Métodos para a prevenção 1,6. Recomendações 1,6,1. Recomendações gerais 1,6,2. Recomendações para a pesquisa
2. PROPRIEDADES E MÉTODOS ANALÍTICOS
2,1. Estrutura química e propriedades 2,2. Métodos analíticos 2,2,1. Líquidos do tecido de planta 2,2,1,2 da extração 2,2,1,1 e tecidos biológicos 2,2,2. Análise para o chromatography thin-layer

dos alcalóides 2,2,2,1 do pyrrolizidine (TLC) 2,2,2,2 chromatography de gás high-performance do chromatography líquido (HPLC) 2,2,2,3 (GC) e do resonance 2,2,2,4 magnético nuclear de spectrometry maciço (MS) spectrometry (NMR) 2,2,2,5 as tinturas do indicador da reação 2,2,2,6 de Ehrlich 2,2,2,7 2,3 pesando diretos. Determinação dos metabolites nos tecidos animais
3. FONTES E PATHWAYS DA EXPOSIÇÃO
3,1. Alcalóides hepatotóxicos do pyrrolizidine e suas fontes 3,2. Pneumotoxic e outros alcalóides tóxicos 3,3 do pyrrolizidine. Pathways da exposição 3,3,1. Contaminação das colheitas de alimento 3,3,2 do grampo. Infusions herbal 3,3,3. Uso de PA-conter plantas como o alimento 3,3,4. Mel contaminado 3,3,5. Leite 3,3,6. Carne 3,3,7. Uso de PAs como agentes chemotherapeutic para o cancer
4. METABOLISM
4,1. Absorption, excretion, e distribuição 4,1,1 do tecido. Absorption 4,1,2. Excretion e distribuição 4,2. Rotas metabolic 4,2,1. Hydrolysis 4,2,2. N - oxidação 4,2,3. Conversão aos metabolites pyrrolic 4,3. Efeitos dos tratamentos que afetam o metabolism 4,4. Outros fatores que afetam o metabolism 4,5. Outras rotas metabolic 4,6. Metabolism do pyrrolizidine N - óxidos 4,7. Metabolism no homem
5. MECANISMOS DO TOXICITY E DE OUTRAS AÇÕES BIOLÓGICAS
5,1. Metabolites responsáveis para o toxicity 5,1,1. Base metabolic do toxicity

5,1,2. Isolação dos metabolites pyrrolic 5,1,3. Os aspectos químicos do chemistry pyrrolic da preparação 5,1,3,2 dos metabolites 5,1,3,1 associaram com as ações tóxicas 5,1,4. Metabolites mais adicionais possíveis 5,2. Ações tóxicas dos metabolites pyrrolic 5,2,1. Animais 5,2,1,1 esters de Pyrrolic (dehydro-alcalóides) 5,2,1,2 álcoois de Pyrrolic (dehydro-necines) 5,2,2. Culturas de pilha 5,2,3. Participação possível do peroxidation 5,3 do lipid da membrana. Fatores químicos e metabolic que afetam o toxicity 5,3,1. Características estruturais de um alcalóide tóxico 5,3,2. Ativação e detoxication 5,3,3. Fatores que afetam o toxicity do reactivity ativo dos metabolites 5,3,3,1 do metabolite 5,3,3,2 o número dos grupos reactive 5,4. Os metabolites associaram com as ações biológicas dos alcalóides 5,4,1 do pyrrolizidine. Hepatotoxicity agudo 5,4,2. Hepatotoxicity crônico 5,4,3. Pneumotoxicity 5,4,4. Toxicity em outros tecidos 5,4,5. Carcinogenicity 5,4,6. Atividade antitumour 5,5. Prevenção e tratamento do pyrrolizidine que envenena 5,5,1. Dietas modificadas 5,5,2. Pre-treatment para realçar o detoxication dos metabolites ativos 5,5,3. Outros tratamentos
6. EFEITOS NOS ANIMAIS
6,1. Testes padrões da doença causados por genera diferentes da planta e da participação do órgão na espécie diferente 6,2. Observações do campo - outbreaks nos animais de fazenda 6,3. Estudos nos animais de fazenda 6,4. Estudos animais experimentais 6,4,1. Efeitos no hepatotoxicity do parente do fígado 6,4,1,1 de PAs diferente e de seu N - óxidos 6,4,1,2 fatores que afetam o mecanismo agudo dos efeitos 6,4,1,4 do

hepatotoxicity 6,4,1,3 dos efeitos crônicos 6,4,2 da ação 6,4,1,5 tóxicos. Efeitos nos pulmões 6,4,2,1 efeitos agudos 6,4,2,2 efeitos crônicos 6,4,2,3 mecanismos da ação tóxica 6,4,3. Efeitos no sistema nervoso central 6,4,4. Efeitos em outros órgãos 6,4,5. Teratogenicity 6,4,6. Fetotoxicity 6,4,7. Os danos 6,4,8 do chromosome do mutagenicity 6,4,7,1. O carcinogenesis 6,4,8,1 purified alcalóides 6,4,8,2 materiais de planta 6,4,8,3 metabolites do alcalóide do pyrrolizidine e o synthetic analogous combina a estrutura 6,4,8,4 molecular e a atividade carcinogenic 6,4,9. Atividade antimitotic 6,4,10. Immunosuppression 6,4,11. Efeitos no metabolism mineral 6,4,12. Métodos para a avaliação do hepatotoxicity e do pneumotoxicity crônicos 6,5. Efeitos nos animais selvagens 6,5,1. Cervos 6,5,2. Peixes 6,5,3. Insetos
7. EFEITOS NO HOMEM
7,1. Características clínicas da doença veno-veno-occlusive (VOD) 7,2. Características pathological salient da doença veno-veno-occlusive 7,3. Relatórios humanos do caso da doença veno-veno-occlusive 7,4. VOD e cirrhosis do fígado 7,5. Diferenças entre VOD e o cirrhosis indian da infância (ICC) 7,6. Doença de pulmão crônica 7,7. Trichodesma que envenena 7,8. Relacionamento entre o nível de dose e os efeitos tóxicos 7,9. Alcalóides do pyrrolizidine como um agente chemotherapeutic para o cancer 7,10. Prevenção de envenenar no homem
8. CONTROLE BIOLÓGICO

9. AVALIAÇÃO DE RISCOS E DE EFEITOS DE SAÚDE HUMANA NO AMBIENTE
9,1. Condições humanas 9,1,1 da exposição. Fontes relatadas da exposição humana 9,1,2. A espécie da planta envolveu 9,1,3. Modalidades e pathways da contaminação da exposição 9,1,3,1 de colheitas da grão 9,1,3,2 medicinas herbal 9,1,3,3 plantas PA-contendo usadas como o alimento e as bebidas 9,1,3,4 o outro alimento contaminado por PAs 9,1,4. Níveis da entrada 9,2. Efeitos agudos da exposição 9,2,1. Doença de fígado aguda 9,3. Efeitos crônicos da exposição 9,3,1. Cirrhosis do fígado 9,3,2. Mutagenicity e teratogenicity 9,3,3. Cancer do fígado 9,3,4. Efeitos em outros órgãos 9,4. Efeitos no ambiente 9,4,1. Agricultura 9,4,2. Animais selvagens 9,4,3. Insetos 9,4,4. REFERÊNCIAS do solo e
da água
APÊNDICE I. ALCALÓIDES DO PYRROLIZIDINE E SUAS FONTES DA PLANTA
APÊNDICE II.
NOTA AOS LEITORES DOS ORIGINAIS DOS CRITÉRIOS
Cada esforço foi feito apresentar tão exatamente a informação nos originais dos critérios como possível sem impropriamente retardado sua publicação. No interesse de todos os usuários dos originais ambientais dos critérios de saúde, os leitores são pedidos amavelmente comunicar todos os erros que puderem ter ocorrido ao gerente do programa internacional na segurança química, organização da saúde do mundo, Genebra, Switzerland, a fim de que possam ser incluídos nos corrigenda, que aparecerão em volumes subseqüentes.

* * *
CRITÉRIOS DE SAÚDE AMBIENTAIS PARA ALCALÓIDES DO PYRROLIZIDINE
Um grupo de tarefa do WHO em critérios de saúde ambientais para alcalóides do pyrrolizidine encontrou-se com em Tashkent, URSS, em 1 - 5 dezembro 1986. O Dr. M. Gounar abriu a reunião em nome das três organizações co-patrocinando do IPCS (UNEP/ILO/WHO). O grupo de tarefa reviu e revisou o original dos critérios do esboço e fêz uma avaliação dos riscos de saúde da exposição aos alcalóides do pyrrolizidine.
O acesso aos papéis originais no assunto publicado na URSS foi feito possível pelo PROFESSOR M. ABDULLAHODJAEVA. DR. A.R. Os MATTOCKS escreveram os primeiros esboços das seções em propriedades e em métodos analíticos, em metabolism, e em mecanismos do toxicity e de outras ações biológicas. Dr. C.C.J. CULVENOR, PROFESSOR ajudado H.D. TANDON no finalization do original após a reunião de grupo da tarefa. O Dr. J. Parizek, que era originalmente o membro da equipe de funcionários de IPCS responsável para a preparação do original, e devia ser secretária do grupo de tarefa, não poderia assistir à reunião por causa da doença repentina, e o grupo de tarefa foi ajudado em seu lugar pelo Dr. M. Gounar, membro anterior da equipe de funcionários de IPCS. O Dr. A. Prost era responsável para a versão final do original.
O secretariat reconhece a ajuda do bot
*** TRANSLATION ENDS HERE ***h Professor H.D. Tandon and Dr C.C.J. Culvenor. The Task Group meeting in Tashkent was organized by the Centre of International Projects, USSR State Committee for Science and Technology.
The efforts of all who helped in the preparation and finalization of the document are gratefully acknowledged.
* * *
Partial financial support for the publication of this criteria

document was kindly provided by the United States Department of Health and Human Services, through a contract from the National Institute of Environmental Health Sciences, Research Triangle Park, North Carolina, USA - a WHO Collaborating Centre for Environmental Health Effects.
* * *
A comprehensive data base on pyrrolizidine alkaloids has been made available by CSIRO Division of Animal Health, Private Bag No. 1, Parkville, Vic. 3052, Australia. The data base consists of alkaloid occurrence tables and keyworded bibliography readable by SCI-MATE software system (Bibliographic Manager, Institute for Scientific Information), but adaptable to other systems. It is available from CSIRO on IBM - PC diskettes; price on application to L.W. Smith.
PREFACE
A disease caused by the consumption of plants containing pyrrolizidine alkaloids (PAs) has been recognized independently as an endemic disease in certain parts of the West Indies and in Uzbekistan in the USSR. Outbreaks of the disease have affected significant segments of populations or large numbers of people in geographically confined areas in Afghanistan, India, and Uzbekistan. The outbreaks have been caused through contamination of the staple food crops with the seeds of plants containing PAs, growing among the crops; such plants are likely to thrive following periods of drought.
It is notable that the same family of plants that caused endemic disease and large-scale outbreaks in Uzbekistan also caused another outbreak of the disease in adjacent Afghanistan, long after the chemical etiology of the disease (through consumption of toxic seeds in the food) had been identified in the USSR. This happened because there was a lack of general awareness of the causal relationship between the chemical present in the plant and the disease. Sporadic cases continue to occur in different parts of the world through the consumption of seeds or plant parts

containing toxic PAs, as home remedies, beverages, or food.
The IPCS recognized that this was a health problem that might be lethal, and that it was entirely preventable, provided that it was recognized in time. It was also recognized that the dissemination of knowledge, about both the disease and the sources of the chemicals involved, would be a critical step in its prevention.
Accordingly, the IPCS invited Professor H.D. Tandon, who was responsible for establishing such a causal relationship in the outbreaks in Afghanistan and India, to prepare a draft criteria document and to assist in its further development and finalization after the Task Group meeting, which was held in Tashkent, USSR, on 1 - 5 December, 1986.
In most episodes of toxic human disease caused by PAs, the liver has been the principal target organ, except for an outbreak in the USSR caused by Trichodesma alkaloids, in which the symptoms were mostly extra-hepatic. The Environmental Health Criteria document provides comprehensive coverage of the hepatotoxic PAS, but lack of relevant documentation prevented the Task Group from analysing the role of Trichodesma alkaloids in detail.
INTRODUCTION - PYRROLIZIDINE ALKALOIDS AND HUMAN HEALTH
Pyrrolizidine alkaloids (PAs) are found in plants growing in most environments and all parts of the world. The main sources are the families Boraginaceae (all genera), Compositae (tribes Senecionae and Eupatoriae

), and Leguminosae (genus Crotalaria), and the potential number of alkaloid-containing species is as high as 6000, or 3% of the world's flowering plants (Culvenor, 1980). They have long been known to be a health hazard for livestock, at least since 1902 (Schoental, 1963), and loss of livestock in various parts of the world has been traced to their grazing on certain plants growing in pastures, especially following periods of drought or in arid climates. They have been found to be toxic for all species of animals tested (Schoental, 1963), though some species, notably the guinea-pig, are resistant (Chesney & Allen, 1973a; White et al., 1973). Human disease caused by PA toxicity has been known to be endemic in the central Asian republics of the USSR, at least since the early thirties (Ismailov, 1948a,b; Mnushkin, 1949) when several outbreaks occurred, and the cause was discovered to be the seeds of plants of Heliotropium species (Dubrovinskii, 1947, 1952; Khanin, 1948), which contaminated the staple food crops. A spate of reports followed, mostly from the West Indies, of acute and chronic liver disease (Bras et al., 1954, 1961; Bras & Hill, 1956; Stirling et al., 1962), associated with the ingestion by people of herbal infusions for the treatment of certain ailments. Schoental (1961) and Davidson (1963) suggested that, in view of the evidence of the hepatotoxicity of PAs, consumption of plants containing them could be of etiological significance in human liver disease, especially in developing countries where they are consumed as food or herbal medicines. In spite of this, and the fact that such an ubiquitous source of toxic material is capable of producing animal and human disease and that there have been more recent reports, the PAs have not attracted much attention in the world as a health hazard. In fact, a recent handbook on naturally occurring toxic agents in food (Rechicigl, Jr, 1983) refers to them only in passing and makes no mention of human disease caused by them. Veno-occlusive disease (VOD) (Bras

& Hill, 1956), which is characterized by the dominant occlusive lesion of the centrilobular veins of the liver lobule and is caused by these alkaloids, has since been reported from all parts of the world, in both man and animals (Hill, 1960; Bras, 1973). It has been attributed to the accidental contamination of food by toxic plant products or the ingestion of herbal infusions. There have been reports of stray cases and of small outbreaks from both developing and developed countries. However, in the most recent studies from Afghanistan (Tandon & Tandon, 1975; Mohabbat et al., 1976; Tandon, B.N. et al., 1978; Tandon, H.D. et al., 1978) and India (Tandon, B.N. et al., 1976; Tandon, R.K. et al., 1976; Krishnamachari et al., 1977; Tandon, H.D. et al., 1977; Tandon, B.N. et al., 1978), the disease has been reported to affect large masses of the population, resulting in high mortality, and has been attributed to the accidental contamination of their staple food crops by PA- containing seeds of plants, following periods of drought.
There is conclusive evidence from studies on experimental animals that the effects of a single exposure to PAs may progress relentlessly to advanced chronic liver disease and cirrhosis (Schoental & Magee, 1957, 1959; Nolan et al., 1966), following a long interval of apparent well-being, and without any other latent or provocative factor (Schoental & Magee, 1959). The lowest levels of such alkaloids administered thus far to experimental animals, e.g., 1 - 4 mg/kg diet, have produced chronic liver disease and tumours (Hooper & Scanlan, 1977; Culvenor & Jago, 1979). Pyrrolizidine alkaloids have also been shown to act synergistically with aflatoxin, another environmental toxin present in agricultural

products, in causing cirrhosis and hepatoma in primates (Lin et al., 1974). Though there is no conclusive evidence yet of a carcinogenic role of PAs in man, such a possibility has been suspected on the basis of experimental data (Hill, 1960; Williams et al., 1967; IARC, 1976, 1983; Huxtable, 1980; Culvenor, 1983), and experimental studies have demonstrated carcinogenicity in rats given dosages equivalent to those reported to have been ingested in human cases (Cook et al., 1950; Culvenor, 1983).
Alkaloids/toxic metabolites have been shown to be secreted in the milk of lactating dairy cattle (Dickinson et al., 1976) and rats, and the young of both sexes have been shown to suffer toxic damage, even when suckled by mothers treated with retrosine, who apparently are not affected themselves (Schoental, 1959). Such suckling animals may also be in apparent good health while the livers show toxic effects. Protein-deficient and young suckling animals are particularly vulnerable (Schoental, 1959).
Chromosomal aberrations have been demonstrated in rats and humans with veno-occlusive disease (Martin et al., 1972).
Alkaloids have been found in the honey secreted by bees feeding on the toxic plants (Deinzer et al., 1977). According to Culvenor and his co-workers, populations in some countries are exposed to low levels of alkaloids in commonly used foodstuffs, e.g., honey in Australia (Culvenor et al., 1981; Culvenor, 1983, 1985) and comfrey in many countries (Culvenor et al., 1980a; Culvenor, 1985).
Human cases of acute disease following the brief ingestion of the alkaloids have been known to progress to cirrhosis (Stuart & Bras, 1957; Braginskii & Bobokhadzaev, 1965; Stillman et al., 1977; Tandon, B.N. et al., 1977; Tandon, H.D. et al., 1977) in as short a period as 3 months from the acute phase (Stuart & Bras, 1957). The initial disease may be cryptic (Braginskii

& Bobokhadzaev, 1965) and may not be ascribed to herbal consumption, and yet may progress to cirrhosis (Huxtable, 1980). Veno-occlusive disease was stated to be the most common cause of cirrhosis in infants in Jamaica (Bras et al., 1961) and has been believed to be a significant etiological factor for adult cirrhosis, especially in developing countries (Gupta et al., 1963).
Plants known or suspected to contain toxic alkaloids are widely used for medicinal purposes as home remedies all over the world, without systematic testing for safety (Schoental, 1963; Smith &
Culvenor, 1981) and some are even used as food (Schoental & Coady, 1968; Culvenor, 1980). There are several reports of the continued use of such herbs for medicinal purposes in technically advanced countries (Culvenor, 1980). Senecio jacobaea continues to be sold at herbalists shops in the United Kingdom (Schoental, 1963; Burns, 1972), and Symphytum spp. (comfrey) are still used as a vegetables, beverages, or remedies (Mattocks, 1980). Both these herbs are known to be carcinogenic (IARC, 1976; Hirono et al., 1978). Young flower stalks of Petasites japonicus Maxim, the pre-bloom flower of coltsfoot, Tussilago farfara, the leaf and root of comfrey, Symphytum officinale, and the young leaves and stalks of Farfugium

japonicum and Senecio cannabifolius, which are all used in Japan as human food or herbal remedies, are known to be carcinogenic for rats (Hirono et al., 1983). Symphytum x uplandicum Nyman (Russian comfrey), which contains several toxic PAs (Culvenor et al., 1980b) echimidine and 7 acetylycopsamine being the main constituents, is used as a salad plant, green drink, and medicinal herb. It has been estimated that the rate of ingestion of alkaloids from this herb may, over a period of time, exceed the levels reported to have been taken during the Afghan outbreak. There is a report of at least one patient who developed toxic effects as a result of consuming a comfrey preparation (Culvenor et al., 1980a; Ridker et al., 1985). Arseculeratne et al. (1981) found that 3 of the 50 medicinal herbs commonly used in Sri Lanka contained PAs that had been proved to be hepatotoxic for animals. They suggested that consumption of such herbs might contribute to the high incidence of chronic liver disease, including primary liver cancer, in Asian and African countries, especially as they may act synergistically with aflatoxin and hepatitis B virus. The risk of toxic effects due to these alkaloids may be particularly high in children (Schoental, 1959; Jago, 1970) and protein malnutrition, which exists in some countries, may potentiate them (Schoental & Magee, 1957). Recent studies from Hong Kong (Kumana et al., 1985; Culvenor et al., 1986), the United Kingdom (McGee et. al, 1976; Ridker et al., 1985), and the USA (Stillman et al., 1977; Fox et al., 1978; Ridker et al., 1985) report instances of human disease that have been caused by the use of such herbs, resulting in fatality or the development of cirrhosis, even in countries with well-developed health services and among the higher economic and educated strata of society. Indeed, Stillman et al. (1977), from the USA, called PA toxicosis the "iceberg disease", implying that cases of this disease might be more frequent than reported in the USA, especially among populations of Mexican-American origin. In general, the use of herbal remedies is not elicited in the clinical history and

patients do not volunteer this information themselves. Furthermore, the alkaloids are eliminated within 24 h (Huxtable, 1980) and, even though methods are available for their detection in biological tissues and fluids, the suspicion cannot be confirmed, as the symptoms may take several days or months to appear.
Contamination of food crops is particularly likely to occur in parts of the world with arid climates, poor or uncertain rainfall, poor irrigation facilities, and following periods of drought, all of which promote the growth of the PA-containing plants that grow as weeds among cultivated crops, as has been found in studies on
the outbreaks in Afghanistan, India, and the USSR (Terekhov, 1939; Dubrovinskii, 1947; Ismailov, 1948a,b; Tandon & Tandon, 1975; Mohabbat et al., 1976; Tandon, B.N. et al., 1976; Tandon, R.K. et al., 1976; Tandon, H.D. et al., 1978) and in grazing pastures. The use of traditional medicines is common in these countries and there is insufficient awareness of this hazard, the disease condition, and its diagnostic pathological picture. Furthermore, health services are poorly developed. Thus, many of the cases or even outbreaks may go unnoticed or unrecorded and may even be ascribed to malnutrition (Lancet, 1984). Also, many of the reported cases of so-called "Budd-Chiari syndrome", a condition associated with obstruction of major hepatic veins and/or inferior vena cava, may actually be cases of veno-occlusive disease (Sherlock, 1968), in which only the central veins of the liver lobule or sublobular veins are occluded.
Another type of PAs, Trichodesma alkaloids, has been known to cause a human outbreak of disease in the USSR, through contamination of the staple cereal with the seeds containing these PAs; in this outbreak, the symptoms were principally extra-hepatic (Ismailov et al., 1970).
This document is aimed at focusing on a health menace that is insufficiently recognized, in order to evaluate the health risks on

the basis of published data, and to draft a set of recommendations that would help in its recognition, prevention, and control.
1. SUMMARY AND RECOMMENDATIONS
1.1. Summary
The ingestion of pyrrolizidine alkaloids (PAs) in foods and medicinal herbs results in acute and chronic effects in man, affecting mainly the liver. Data from experimental animal studies indicate that PAs represent a potential cause of cancer in man.
The alkaloids are produced by numerous plant species and occur throughout the world. In the present document, the alkaloids and their properties are described together with the sources of human exposure and the diseases that they produce in man and animals. The risks for human health are evaluated and recommendations are made for reducing such risks.
1.2. Sources and Chemical Structure
The known pyrrolizidine alkaloids, most of which are hepatotoxic, are produced by plant species within the following families: Boraginaceae ( Heliotropium, Trichodesma, Symphytum, and most other genera), Compositae ( Senecio, Eupatorium, and other genera of the tribes Senecioneae and Eupatoriae), Leguminosae
(genus

Crotalaria), and Scrophul-ariaceae (genus Castilleja). These genera are mainly herbaceous and very widely distributed, some species being found in most regions of the world. The majority of the species within these genera have not yet been investigated, but are expected to contain pyrrolizidine alkaloids.
The hepatotoxic alkaloids have a 1,2-double bond in the pyrrolizidine ring and branched chain acids, esterifying a 9-hydroxyl and preferably also the 7-hydroxyl substituent. Modified seco-pyrrolizidine alkaloids, in which the central bond between the N and C8 atoms is broken, are also hepatotoxic. Some Senecio species contain non-basic derivatives that are 5-oxopyrroles. The toxicity of these derivatives may be similar to that of the alkaloids, but this aspect has not been investigated. The alkaloids occur as free bases and N-oxides. The latter are reduced to the free bases in the gastrointestinal tract of animals and have a similar toxicity when ingested orally.
Suitable analytical procedures are available for screening plant species, including a simple field test for toxic alkaloids. Thin-layer chromatography (TLC), high-performance liquid (HPLC), gas chromatography (GC), and gas chromato-graphy-mass spectrometry (GC-MS) have been applied for separating, characterizing, and quantifying the alkaloids present. Effective use of these procedures requires authentic alkaloids for standards, few of which are available. Improved analytical methods are required for the determination of very low levels of alkaloids in some foodstuffs.
1.3. Mechanisms and Features of Toxicity
The toxic effects of pyrrolizidine alkaloids are due to

activation in the liver. Metabolism of the alkaloids by mixed- function oxidases leads to pyrrolic dehydro-alkaloids, which are reactive alkylating agents. Reaction of initial metabolites with constituents of the liver cell in which they are formed are probably the main cause of liver cell necrosis. Metabolites are released into the circulation and are believed to pass beyond the liver to the lung causing vascular lesions characteristic of primary pulmonary hypertension, especially when alkaloids, such as monocrotaline, are administered to animals.
In experimental animals, PAs are quickly metabolized and are almost completely excreted in 24 h, so that no residual products are detectable in the biological fluids or body tissues after this period.
The rate of formation of pyrrolic metabolites is influenced by the induction or inhibition of the mixed-function oxidases in the liver, but the relationship between the rate of metabolism and expression of toxicity is uncertain.
Several pyrrolizidine alkaloid-derivatives and related compounds are known to cause chromosome aberrations in plants, leukocyte cell cultures of the marsupial (Potorus tridactylus), and in hamster cell lines. Some pyrrolizidine alkaloids induce micronuclei formation in erythrocytes in the bone marrow and fetal liver in mice, sister chromatid exchanges in a Chinese hamster cell line and human lymphocytes in vitro, and repair DNA synthesis in rodent hepatocyte cell cultures. Chromosome aberrations have been reported in the blood cells of children suffering from veno- occlusive disease VOD, presumably caused by fulvine.
A number of pyrrolizidine alkaloids have been shown to be mutagenic in the Salmonella typhimurium assay, after metabolic activation. The carcinogenic activity of pyrrolizidine alkaloids

appears to parallel their mutagenic behaviour, but not their hepatotoxicity.
Heliotrine at doses of 50 mg/kg body weight or more, administered to rats during the second week of gestation, has been shown to induce several abnormalities in the fetus. Doses of 200 mg/kg body weight resulted in intrauterine deaths or resorption of fetuses. Dehydroheliotridine, the metabolic pyrrole derivative of heliotrine, was 2.5 times more effective on a molar basis than its parent PA in inducing teratogenic effects.
The ability of PAs to cross the placental barrier in the rat and to induce premature delivery or death of litters has been demonstrated. The embryo in utero appears to be more resistant to the toxic effects of pyrrolizidine alkaloids than the neonate. PAs are known to have passed through the mother's milk to the sucklings.
Megalocytosis, the presence of enlarged hepatocytes containing large, hyper-chromatic nuclei, is a characteristic feature of pyrrolizidine alkaloid-induced chronic hepatotoxicity in experimental animals. The enlarged hepatocytes arise through the powerful antimitotic action of the pyrrole metabolites of pyrrolizidine alkaloids. This change has not been observed in the human liver, though human fetal liver cells in vitro culture become enlarged when exposed to PAs, indicating susceptibility to the antimitotic effect of the alkaloids.
In experimental animals, protein-rich and sucrose-only diets have given some measure of protection against the effects of the alkaloids, as has pre-treatment of animals with thiols, anti- oxidants, or zinc chloride.
PAs are noted mainly for the poisoning of livestock due to the animals grazing on PA-containing toxic weeds, and large-scale outbreaks have been recorded. Such episodes have been reported

from most parts of the world, including those with temperate or cold climates. Studies carried out on a wide variety of farm and laboratory animals have revealed generally common features of toxicity with some species variations. The liver is the principal target organ. In small laboratory animals, doses approaching a lethal dose produce a confluent, strictly zonal haemorrhagic necrosis in the liver lobule, within 12 - 48 h of administration of PAs. Simultaneously in non-human primates, or after a short time in the rat, chicken, and swine, changes begin to occur, and later become organized, in the subintima of the central or sublobular veins in the liver resulting in their occlusion. The reticulin framework in the central zone of the lobule collapses following necrosis leading to scarring. Repeated administration of suitable doses leads to chronic liver lesion characterized by megalocytosis, and increasing fibrosis, which may result in cirrhosis. Chronic liver disease including cirrhosis has been shown to develop in the rat following administration of a single dose of a PA. In a number of animal species, the lungs develop vascular lesions characteristic of primary pulmonary hypertension with secondary hypertrophy of the right ventricle of the heart. In rats, appropriately low repeated doses of several alkaloids have been shown to induce tumours, mainly in the liver. In some studies, a single dose has been carcinogenic.
The central nervous system is the target organ of the toxic PAs contained in Trichodesma, which produce spongy degeneration of the brain.
1.4. Effects on Man
In man, PA poisoning is usually manifested as acute veno- occlusive disease characterized by a dull dragging ache in the right upper abdomen, rapidly filling ascites resulting in marked distension of the abdomen, and sometimes associated with oliguria, and massive pleural effusion. It can also manifest as subacute disease with vague symptoms and persistent hepatomegaly. Children are particularly vulnerable. Many cases progress to cirrhosis and,

in some cases, a single episode of acute disease has been demonstrated to progress to cirrhosis, in spite of the fact that the patient has been removed from the source of toxic exposure and has been given symptomatic treatment. Mortality can be high with death due to hepatic failure in the acute phase or due to hematemesis resulting from ruptured oesophageal varices caused by cirrhosis. Less severely affected cases may show clinical, or even apparently complete, recovery. The Task Group was not aware of any substantiated report of primary pulmonary hypertension resulting from PA toxicity. However, in view of the evidence in experimental animals and circumstantial evidence in one case report, the possibility of the development of toxic pulmonary disease in man cannot be ruled out. There is a report of an outbreak of Trichodesma poisoning in the USSR in which the symptoms were mainly neurological.
1.4.1. Nature and extent of health risks
The two main sources of pyrrolizidine alkaloid poisoning reported in human beings are the consumption of cereal grain contaminated by weeds containing the alkaloids and the use of alkaloid-containing herbs for medicinal and dietary purposes. A third form of exposure, with the potential to affect large populations is the possible low-level contamination of some foodstuffs, such as honey and milk, but the Task Group was not aware of any cases of human toxicity having been caused through the contamination of these foods.
Liver disease caused by the contamination of cereal grains has been reported in rural populations in Afghanistan, India, South Africa, and the USSR. A contributing factor appears to be abnormally dry weather, resulting in the growth of an exceptionally high proportion of the alkaloid-containing weeds in the crops, the seeds of which contaminate the cereal grain on harvesting. The weeds responsible for known outbreaks have been Heliotropium,

Trichodesma, Senecio, and Crotalaria species. Mortality in such outbreaks has been reported to be high. In the largest reported outbreak in northwestern Afghanistan, an estimated 8000 people were affected in a total population of 35 000 with 1600 - 2000 deaths.
Human poisoning through the medicinal use of herbs containing pyrrolizidine alkaloids has been reported from all parts of the world. PAs were responsible for a common liver disease in children in Jamaica, and individual cases in Ecuador, Hong Kong, India, the United Kingdom, and the USA. The plants involved were species of Crotalaria, Heliotropium, Senecio, Symphytum, and Gynura.
Symphytum-containing preparations present a particular hazard because of their widespread use and the generally high levels of individual exposures. The use of herbs is almost universal in traditional folk medicine and is increasing in developed countries. Some of the herbs used contain pyrrolizidine alkaloids and have a long-term toxicity that is unsuspected by the people taking them. Knowledge of the species used in herbal medicine and the frequency of such use is very limited in the scientific literature. About 40 such species are listed in this report, about one-third of which
are in use in developed countries. They are often prescribed by herbalists, naturopaths, and other non-orthodox practitioners. The extent of the contribution to acute and chronic liver disease cannot be accurately assessed. It may also constitute an etiological factor in cirrhosis of the liver and, once this stage is reached, it may not be possible to identify the cause as a PA.
PAs are known to be transmitted from the feed of dairy animals into milk and to cause toxic damage in the suckling young. One instance of large-scale contamination of honey is known to have been caused by a common weed rich in PAs, which was the source of

nectar and pollen for the honey-secreting bees. No reports of cases of acute toxicity caused by consumption of contaminated dairy products or honey were available to the Task Group. Furthermore, no information is available on the possible presence of PAs or their metabolites in the meat of animals fed toxic weeds before slaughter; however, the possibility of toxic disease being caused through this medium is considered to be low.
There are no substantial, long-term follow-up data to assess whether exposure to PAs results in increased incidence of chronic liver disease or cancer in man. Available clinical and experimental data suggest that a single episode of PA toxicity and possibly also a long-term low level exposure may lead to cirrhosis of the liver. PAs could also be possible carcinogens in man, since a number of them have been demonstrated to induce cancer in experimental animals, the main target organ being the liver. These include some which have caused episodes of human toxicity, and some others which are found in herbs traditionally used as items of food. Also, in several instances of human toxicity, the reported daily rates of intake of such PAs were in close range of those known to induce tumours in rats. However, these risks cannot be adequately assessed on a quantitative basis. There are indications that PA intoxications leading to liver disease are more prevalent than the reported frequency of cases would seem to indicate.
Because of their known involvement in human poisoning and their possible carcinogenicity, exposure to pyrrolizidine alkaloids should be kept as low as practically achievable. The setting of regulatory tolerance levels for certain food products may be required in some situations.
1.5. Methods for Prevention
The only known method of prevention is to avoid consumption of the alkaloids. In the USSR, a set of agricultural (or agrotechnical) legislative, phyto-sanitary and educational measures has prevented new outbreaks of poisoning due to Heliotropium

and Trichodesma, since 1947.
1.6. Recommendations
1.6.1. General recommendations
1. Cereal crops should be assessed throughout the world for possible contamination by weeds likely to contain pyrrolizidine alkaloids. Appropriate grain inspection systems are desirable in order to achieve near-zero levels of contamination by such weeds.
2. There is a need to create awareness, among the general population and those responsible for the delivery of health services, with regard to the hazards of consuming such plants as contaminants in food or as food, or for medicinal purposes. Advice on hazards should include mention of possible increased risks, if the alkaloid intake is associated with drug treatment, (e.g. phenobarbitone) or foods which increase the level of liver metabolizing enzymes.
3. Ethnobotanical and taxonomic studies are required in many countries to provide specific information on the use of plant species containing pyrrolizidine alkaloids for medicinal and dietary purposes. There may be a need to control the sale of some species, and their prescription by herbalists and other practitioners of traditional systems of medicine.
4. Honey and dairy products, both local and bulk supplies, should be assayed for pyrrolizidine alkaloids in all regions where a risk of contamination of these foodstuffs has been identified.
1.6.2 Recommendations for research

1. Long-term follow-up studies of the survivors of both alkaloid poisoning in human beings and animal outbreaks are required, in order to determine the possible development of chronic liver disease or cancer. Similar studies are also desirable on individuals who regularly consume comfrey or other PA- containing herbs over a substantial period of time.
2. Epidemiological studies should be carried out in countries with a high incidence of primary liver cancer, in order to determine whether there is an association with the intake of herbs containing pyrrolizidine alkaloids.
3. A network of reference laboratories is needed to assist member states in identifying plants and their seeds suspected of producing toxic effects and for the assay and identification of PAs. Provisions may be made for the easy availability of pure alkaloids for use as reference standards for assays.
4. It is necessary to develop improved assay procedures, suitable for the purposes of recommendation (4) in section 1.6.1, particularly using fluorescence and immunochemical methods.
5. There is a need for further toxicological studies, such as studies on the carcinogenicity of echimidine and the toxicity of the 5-oxopyrrole constituents of Senecio species, and for studies that would provide more quantitative information on the various adverse biological effects of PAs. A study of the carcinogenicity of the alkaloids in the pig is also indicated, since the pig exhibits a high sensitivity to acute and subacute toxicity similar to that seen in man.
6. Study is required of the possible alkaloid content of the meat, organs, and fat of animals that have recently consumed plants containing pyrrolizidine alkaloids.
7. Experimental studies are needed on the influence of nutritional status on the metabolism, and acute and chronic effects of PAs.

8. Further metabolic studies are required to define more specifically the enzymes involved in the microsomal activation and detoxification of PAs, to determine whether organelles other than microsomes are involved, and to explore further, quantitative relationships between different routes of metabolism.
9. The maximum no-observed-adverse-effect dose levels for repeated long-term administration in the rat and the pig need to be determined.
10. Experimental studies should be conducted to determine:
(a) whether pyrrolizidine alkaloid N-oxides may be metabolized directly into the pyrrolic dehydroalkaloid in mitochondria, especially in tumour cells; and
(b) which P450 enzymes are involved in the activation and N-oxidation of PAs and thence in the selective induction of N-oxidation enzymes.
11. A study might be conducted of human variability and its genetic aspects in relation to factors that influence susceptibility to PAs; for example, the study of mixed-function oxidase levels in the liver by metabolism of appropriate test substances recognized as harmless.
2. PROPERTIES AND ANALYTICAL METHODS
2.1 Chemical Structure and Properties

The chemical structure of PAs in relation to their toxic effects has been reviewed recently by Mattocks (1986). The pyrrolizidine alkaloids with which this document is concerned are those that have previously been called "hepatotoxic" or "nucleotoxic". Here it is proposed to refer to them as "toxic" PAs, because of the weight of evidence now available that they produce damage in other organs as well as the liver, and the need to avoid a restrictive term. There are other types of pyrrolizidine alkaloids, such as those that occur in the plant family Orchidaceae, which are not toxic and are not discussed here.
The toxic PAs are esters of the amino-alcohols derived from the heterocyclic nucleus. The pyrrolizidine molecule is made up of two 5-membered rings inclined to each other as shown in Fig. 1 so that geometric isomerism is possible, and which share a common nitrogen at position 4.
Most hepatotoxic alkaloids are esters of molecules similar to that shown in Fig. 1(b) (1-hydroxymethyl-1:2-dehydro- pyrrolizidine). However, a few hepatotoxic alkaloids are esters of the amino-alcohol otonecine, e.g., petasitenine (Fig. 2, No.7). The unsaturated pyrrolizidine nucleus itself is not toxic, but esters of branched-chain acids are. Ester linkages may be at positions 9, 7, or (rarely) 6. Some esters have an "open" molecule, e.g., heliotrine, whereas others are macrocyclic diesters, e.g., monocrotaline and retrosine. Examples of some pyrrolizidine alkaloid structures are shown in Fig. 2.
The ring nucleus contains a double bond at the 1:2 position, which is essential for the toxic effects of the alkaloid, but not for unrelated effects.
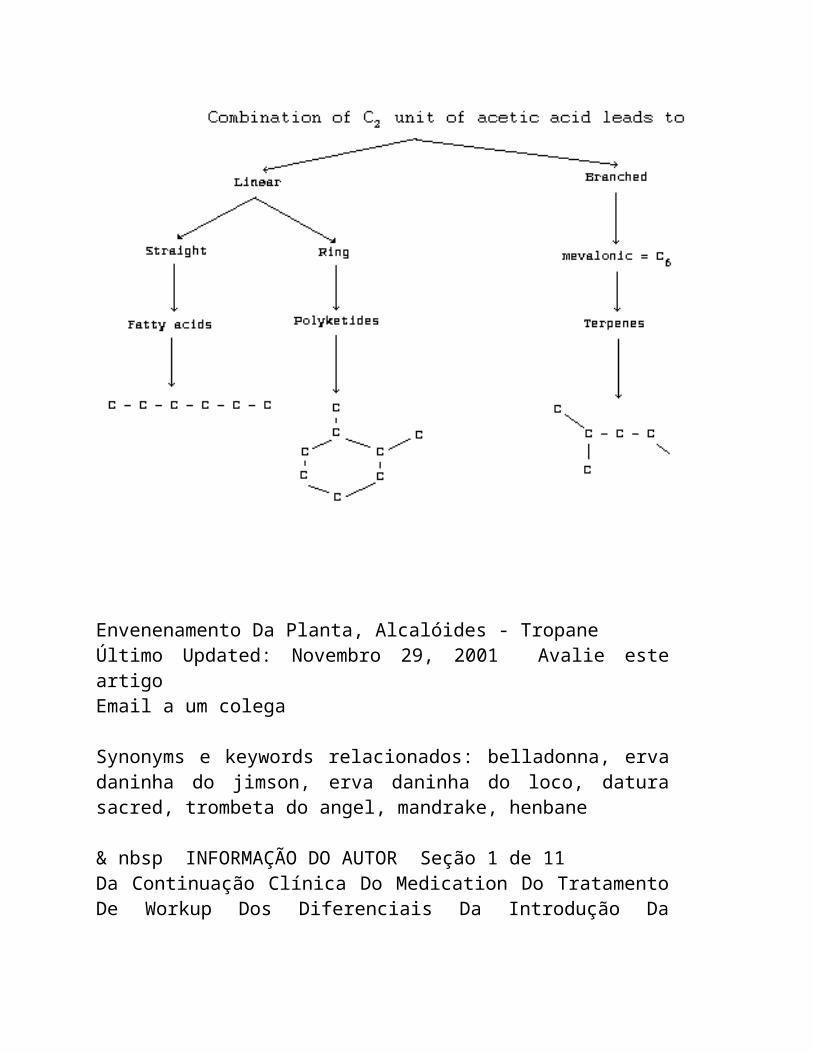
1. Echimidine
Chemical structure:
Chemical formula: C20H
31
NO7
Relative molecular mass: 397
CAS registry number: 520-68-3
2. Heliotrine
Chemical structure:
Chemical formula: C16
H

27
NO5
Relative molecular mass: 313
CAS registry number: 303-33-3
3. Indicine- N -oxide
Chemical structure:
Chemical formula: C15
H25
NO6
Relative molecular mass: 315
CAS registry number: 41708-76-3
4. Jacobine
Chemical structure:

Chemical formula: C18
H25
NO6
Relative molecular mass: 351
CAS registry number: 6870-67-3
5. Lasiocarpine
Chemical structure:
Chemical formula: C21
H33
NO7
Relative molecular mass: 411

CAS registry number: 303-34-4
6. Monocrotaline
Chemical structure:
Chemical formula: C16
H23
NO6
Relative molecular mass: 325
CAS registry number: 315-22-0
7. Petasitenine
Chemical structure:
Chemical formula: C19
H

27
NO7
Relative molecular mass: 381
CAS registry number: 60132-19-6
8. Retrorsine (retrosine N -oxide = isatidine)
Chemical structure:
Chemical formula: C18
H25
NO6
Relative molecular mass: 351
CAS registry number: 480-54-6
9. Senecionine

Chemical structure:
Chemical formula: C18
H25
NO5
Relative molecular mass: 335
CAS registry number: 130-01-8
10. Symphytine
Chemical structure:
hemical formula: C20
H31
NO6
Relative molecular mass: 381

CAS registry number: 22571-95-5
11. Trichodesmine
Chemical structure:
Chemical formula: C18
H27
NO6
Relative molecular mass: 353
CAS registry number: 548-90-3
12. Incanine
Chemical structure:
Chemical formula: C18

H27
NO5
Relative molecular mass: 337
CAS registry number: 480-77-3
As the Task Group met in Tashkent, it is of historical interest to recall that the structures of heliotrine and lasiocarpine, the main alkaloids of Heliotropium lasiocarpum, were worked out by Dr G.P. Men'shikov and associates in Moscow in the 1930s. This work included determining the structure of heliotridine, the parent compound of the amino-alcohol, heliotridane. Dr Men'shikov's studies were carried out at essentially the same time, but independently of studies by English and American authors on retronecine-based alkaloids.
The alkaloids in plants are often found together with their N-oxides, which are also toxic, when ingested orally. The pyrrolizidine alkaloids acquire their toxic properties only through the toxic pyrrolic intermediates (the general structure of which is shown in Fig. 3) formed by the mixed-function oxidases of the hepatocytes. To form these pyrrolic derivatives, the alkaloid molecule should have:
(a) a double bond at the 1:2 position of the ring nucleus;
(b) esterified hydroxyl groups in the nucleus at the C 9 and/or C 7 positions; and
(c) a branched carbon chain in at least one of the ester side- chains (McLean, 1974).

Substitution at the a position of the acid and esterification of the C-7 hydroxy group both enhance the toxicity of the alkaloid (Robins, 1982).
A group of related alkaloids, isolated from Senecio species by Bohlmann et al. (1979), have non-basic pyrrolic structures similar to those of toxic pyrrolizidine alkaloid metabolites, but they are chemically deactivated by the presence of a carbonyl group at position 3 of the pyrrolizidine nucleus, e.g., senaetnine (Fig. 4). Senaetnine does not possess the acute hepatotoxic characteristics of basic pyrrolizidine alkaloids. However, it had a direct irritant action on tissues near the site of intraperitoneal administration and caused damage to pulmonary vascular tissue when given intraveinous to rats (Mattocks & Driver, 1987).
The alkaloids are fairly stable chemically, but the ester groups may undergo hydrolysis under alkaline conditions. Some alkaloids in plant material may decompose during drying (Bull et al., 1968), but others appear to be stable under similar conditions (Pedersen, 1975; Birecka et al., 1980). The N-oxides of

unsaturated pyrrolizidines are more readily decomposed by heat than the basic alkaloids, especially when dry. However, the stability of the alkaloids and N-oxides in hot water as, for example, in cooking, is not known.
Some pyrrolizidine alkaloids have a limited water solubility, unless neutralized with acid; but others (e.g., indicine), and all the N-oxides, are readily soluble.
2.2 Analytical Methods
When analysing for PAs, it is important to recognize that this group consists of many different compounds (section 2.1) and that these often occur as mixtures in plants or in materials of plant origin. They may vary in structure, relative molecular mass, response to analytical procedures, and toxicity. Both basic alkaloids and corresponding N-oxides may be present at the same time. Thus, where such mixtures are present, analyses will inevitably be approximate, unless the individual components are separated and identified.
Nevertheless, such estimates can be useful. In particular, all hepatotoxic PAs are unsaturated in the sense that they possess a 1:2-double bond in the pyrrolizidine nucleus, and analytical methods that are specific for this structure can be of value in screening for potential toxicity. A simple qualitative field test for screening plant materials for the presence of such alkaloids and their N-oxides, without the need of high technology equipment, is described in section 2.2.2.5.

2.2.1 Extraction
2.2.1.1 Plant tissue
Pyrrolizidine alkaloids are usually extracted from dried, milled plant material with hot or cold alcohol. The alcohol is evaporated, the bases taken up in dilute acid, and fats extracted with ether or petroleum. It is usual, at this stage, to reduce any N-oxides present to the corresponding basic alkaloids with zinc, before making the solution alkaline and extracting the alkaloids with chloroform (Koekemoer & Warren, 1951). Alternatively, alcohol can be continuously circulated through the plant material and then cation exchange resin, and the alkaloids subsequently eluted from the resin (Mattocks, 1961; Deagen & Deinzer, 1977). PAs can also be extracted by soaking plant material in dilute aqueous acid (Briggs et al., 1965; Craig et al., 1984).
2.2.1.2 Biological fluids and tissues
Pyrrolizidine alkaloids have been extracted for analytical purposes from honey (Deinzer et al., 1977), milk (Dickinson et al., 1976), blood-plasma (Ames & Powis, 1978; McComish et al., 1980), urine (Mattocks, 1967a; Jago et al., 1969; Evans et al., 1979), and bile (Jago et al., 1969; Lafranconi et al., 1985).
When attempting to isolate PAs from animal tissues, it must be appreciated that the toxic alkaloids are often metabolized very rapidly in animals, so that the amounts that are recoverable (except from urine), only a few hours after alkaloid ingestion, may be extremely small. Various methods have been used to separate PAs, but some mixtures are extremely difficult to separate. On the

analytical scale, the most useful methods are thin-layer chromatography (TLC), high-performance liquid chromatography (HPLC), and gas chromatography (GC) (section 2.2.2).
2.2.2 Analysis for pyrrolizidine alkaloids
2.2.2.1 Thin-layer chromatography (TLC)
For TLC, silica plates are usually used, eluted with chloroform: methanol:aqueous ammonia mixtures (Sharma et al., 1965; Chalmers et al., 1965); solvents suitable for the N-oxides, which are more water-soluble, have been described by Mattocks (1967b) and Wagner et al. (1981). The most sensitive methods for detecting PAs on TLC are those using Ehrlich reagent (4-dimethylaminobenzaldehyde) (Mattocks, 1967b). The unsaturated alkaloids are best visualized by spraying the plates first with a solution of orthochloranil, then with Ehrlich reagent, heating after each spray (Molyneux & Roitman, 1980). The N-oxides of unsaturated pyrrolizidines are detected by spraying a solution of acetic anhydride, heating the plate, and then spraying Ehrlich reagent (Mattocks, 1967b).
Pyrrolizidine alkaloids with a saturated base moiety must be detected in other ways (which are not specific for pyrrolizidines), e.g., by exposing the dried plates to iodine vapour, or by spraying with an iodobismuth (Dragendorff) reagent (Munier, 1953).
2.2.2.2 High-performance liquid chromatography (HPLC)
Analytical or preparative scale HPLC separation of pyrrolizidine alkaloids has been described by Segall (1979a,b) and Dimenna et al. (1980), and an improved method has been reported by Ramsdell

& Buhler (1981). Alkaloids from Symphytum officinale (comfrey) have been separated on an analytical scale by Tittel et al. (1979), and partially separated on a preparative scale by Huizing et al. (1981). UV detectors are usually used for the HPLC of pyrrolizidine compounds (Mattocks, 1986).
2.2.2.3 Gas chromatography (GC) and mass spectrometry (MS)
The GC characterization of PAs using packed columns has been described by Chalmers et al. (1965) and Wiedenfeld et al. (1981). Mixtures of alkaloids from comfrey ( Symphytum sp.), normally hard to separate, were resolved by Culvenor et al. (1980a) and Frahn et al. (1980) by GC of the methylboronate derivatives.
Gas chromatography combined with mass spectrometry (GC-MS) has become a valuable and highly sensitive means for both the identification and the quantitative determination of pyrrolizidine alkaloids. Thus, alkaloids extracted from honey were separated and identified by Deinzer et al. (1977) and (as butylboronate derivatives) by Culvenor et al. (1981). Deinzer et al. (1978) described a method for the recognition (but not the individual identification) of retronecine-based pyrrolizidine alkaloids, by hydrolysing them to retronecine (the amino alcohol moiety) followed by GC-MS of its bis-trifluoroacetate. The use of capillary GC has greatly improved the sensitivity of pyrrolizidine alkaloid analysis, especially when used with MS (Luthy et al., 1981). The MS of pyrrolizidine compounds has been reviewed (Bull et al., 1968; Mattocks, 1986).
Pyrrolizidine N-oxides generally undergo thermal decomposition, when subjected to GC, but they can first be reduced to the corresponding basic alkaloids (Koekemoer &

Warren, 1951). Alternatively they may be derivatised. Thus, trimethylsilylation of indicine N-oxide or heliotrine N-oxide can lead either to the trimethylsilyl (TMS) derivative of the parent alkaloid or to the TMS derivative of the dehydro-alkaloid (pyrrolic derivative), depending on the reagents used, and these products will run successfully on GC-MS (Evans et al., 1979, 1980).
2.2.2.4 Nuclear magnetic resonance (NMR) spectrometry
A convenient, but relatively insensitive, method, specifically for the determination of unsaturated PAs, has been described by Molyneux et al. (1979). The basic alkaloids are extracted, then subjected to NMR spectrometry along with an internal standard
( p-dinitrobenzene). This enables quantitative measurements to be made of the signal(s) representing the H2 proton(s) in unsaturated pyrrolizidines, and thus the alkaloid(s) can be determined. Quantitative NMR analysis of pyrrolizidine alkaloid mixtures from Senecio vulgaris has been described by Pieters & Vlietinck (1985) and compared with an HPLC method by the same authors (1986). Qualitative aspects of the NMR spectrometry of pyrrolizidine alkaloids have been reviewed by Bull et al. (1968) and Mattocks (1986).
2.2.2.5 The Ehrlich reaction
This method (Mattocks, 1967a, 1968b) is specific for unsaturated pyrrolizidine alkaloids and is not suitable for other alkaloids. Thus, it is the most useful colorimetric method for

potentially hepatotoxic pyrrolizidine compounds. The procedure converts the alkaloid into its N-oxide, using hydrogen peroxide. The product reacts with acetic anhydride to form a pyrrolic derivative (dehydro-alkaloid) that gives a magenta colour with a specially modified Ehrlich reagent. The latter contains boron trifluoride to give maximum sensitivity. As little as 5 µg of most unsaturated pyrrolizidines can be measured by this method. If the oxidation stage is omitted, only the unsaturated pyrrolizidine N-oxides can be determined. The determination of pyrrolizidine N-oxides has also been discussed by Mattocks (1971b).
A simplification of the above colorimetric procedure was described by Mattocks (1971d) to provide a qualitative test that could be used to screen large numbers of plant samples for the presence of unsaturated pyrrolizidine alkaloid N-oxides. An improved version of this field test is now available (Mattocks & Jukes, 1987). It is suitable for any plant parts, such as leaves, stems, flowers, seeds, or roots, or materials of plant origin, such as cereals or herbal teas, but has not yet been applied to cooked food.
The plant material (0.2 - 1 g) is extracted by grinding it with aqueous ascorbic acid (5%) and a small amount of sand. The solution is filtered and divided into two equal portions ("test" and "blank"). An aqueous solution (0.2 ml) of sodium nitroprusside (5%) containing sodium hydroxide (10-3
mol) is added to the "test" sample. Both portions are heated for approximately 1 min at 70 - 80 °C; then Ehrlich reagent is added and heating is continued for

1 min. The Ehrlich reagent contains 4-dimethylaminobenzaldehyde (5 g) dissolved in a mixture of acetic acid (60 ml), water (30 ml), and 60% perchloric acid (10 ml). A magenta colour in the "test" compared with the "blank" indicates the presence of an unsaturated PA N-oxide. The "blank" may show a colour if the plant contains compounds, such as indoles or pyrroles, which can themselves give a colour with Ehrlich reagent. The intensity of colour in the "sample" compared with the "blank" can give a rough idea of the amount of alkaloids present, and indicate whether further chemical or toxicological testing of the plant material is adviseable.
In practice, the majority of PA-containing plants contain enough alkaloid in the N-oxide form (often a large proportion) to react positively in this test. The main exceptions are some seeds (Crotalaria), which may contain much alkaloid base, but little or no N-oxide. These (and any other sample not containing chlorophyll) can be tested for basic PAs by grinding them with chloroform, heating the filtered extract with a solution (0.1 ml) of orthochloranil (0.5%) in acetonitrile, and then heating it with Ehrlich reagent. A magenta colour indicates the presence of an unsaturated PA. Non-toxic pyrrolizidine alkaloids having a saturated pyrrolizidine nucleus, and pyrrolizidine alkaloids that are otonecine esters, such as petasitenine, will not respond to this test.
2.2.2.6 Indicator dyes
A method generally applicable to tertiary bases has been adapted for pyrrolizidine alkaloids by Birecka et al. (1981). It is sensitive, but is not specific for this group of alkaloids, and it does not distinguish between the saturated and unsaturated alkaloids. A chloroform solution of the alkaloid is shaken with

acidified aqueous methyl orange. The yellow alkaloid:dye complex is subsequently released from the chloroform phase, using ethanolic sulfuric acid, and measured spectrophotometrically.
2.2.2.7 Direct weighing
An insensitive way to determine the alkaloids in, for example, a plant sample, providing enough is available, is to extract the alkaloids (section 2.2.1) and weigh them. This will provide a rough measure of the total bases present in the sample; however, these may not necessarily be PAs. Nevertheless, the sample can then be subjected to further tests, e.g., GC-MC, nuclear magnetic resonance (NMR), or colorimetric analysis. Furthermore, pyrrolizidine N-oxides are generally too water soluble to be appreciably extractable from aqueous solution by chloroform. Thus, if two portions of the sample are extracted, and one of them is reduced to convert N-oxides to bases, the weight difference between the two products will represent the alkaloid existing in the form of N-oxide in the original sample.
2.3 Determination of Metabolites in Animal Tissues
Important metabolites of toxic pyrrolizidine alkaloids in animals include "pyrrolic" derivatives (dehydro-alkaloids) and N-oxides. A procedure for measuring pyrrolic metabolites in tissue samples (such as liver or lung) has been described by Mattocks & White (1970). The sample (usually 0.5 g) is homogenized in an ethanolic solution of mercuric chloride; the solids are separated

by centrifugation and heated with Ehrlich reagent to give a soluble colour that can be measured spectrophotometrically.
The measurement of pyrrolic and N-oxide metabolites, formed by the action of hepatic microsomal preparations on PAs in vitro, is an improvement described by Mattocks & Bird (1983).
3. SOURCES AND PATHWAYS OF EXPOSURE
3.1 Hepatotoxic Pyrrolizidine Alkaloids and Their Sources
Plants constitute the only natural source of pyrrolizidine alkaloids (PAs) that cause toxic reactions in man and animals. PAs occur in a number of species in the families Boraginaceae, Compositae, Leguminosae (genus Crotalaria), Ranunculaceae (genus Caltha), and Scrophulariaceae (genus Castilleja) (Table 1). The most important genera of PA-containing toxic plants are Crotalaria (Leguminosae), Senecio (Compositae), Heliotropium, Trichodesma,
Amsinckia, Echium

, and Symphytum (Boraginaceae) (Hooper, 1978). The recorded cases of human toxicity have mainly been caused by at least 12 different pyrrolizidine alkaloids, mostly derived from Heliotropium, Senecio, and Crotalaria genera. The Senecio spp. grow throughout the world; the Crotalaria spp. are mainly found in the tropics and subtropics (Culvenor, 1980). Table 1. List of plant genera containing toxic pyrrolizidine alkaloids (with number of species investigated) ------------------------------------------------------------------------------------- Family Genera ------------------------------------------------------------------------------------- Apocynaceae Fernaldia (1), Parsonsia (4),
Boraginaceae Alkanna (1), Amsinckia (4), Anchusa (2), Asperugo (1), Borago (1), Caccinia

(1), Cynoglossum (9), Echium (3), Hackelia (1), Heliotropium (25), Lappula (2), Lindelofia (7), Lithosperum (1), Macrotomia (1), Messerschmidtia (1), Myosotis (2), Paracaryum (1), Paracynoglossum (1), Rindera (5), Solenanthus (4), Symphytum (7), Tournefortia (2), Trachelanthus (2), Trichodesma

(2), Ulugbekia (1)
Compositae Adenostyles (3), Brachyglottis (1), Cacalia (4), Conoclinium (1), Crassocephalum (1), Doronicum (2), Echinacea (2), Emilia (2), Erechtites (1), Eupatorium (8), Farfugium (1), Gynura (2), Ligularia (5), Petasites (4), Senecio (142), Syneilesis (1),

Tussilago (1)
Leguminosae Crotalaria (60)
Ranunculaceae Caltha (2)
Scrophulariaceae Castilleja (1) ------------------------------------------------------------------------------------- An alphabetical list of pyrrolizidine alkaloids with their plant sources has been published by Smith & Culvenor (1981) and Mattocks (1986). An updated version is attached as Appendix I. The plant genera containing toxic PAs are listed in Table 1 indicating the number of species investigated for PAs. A comprehensive list of species of plants belonging to each of these genera, the alkaloids isolated from each, and the part of the plant containing the alkaloid are presented in Appendix II. Table 1 in Appendix II includes species known to contain alkaloids of proved hepatotoxicity, or of a molecular structure that would make them very probably hepatotoxic. Table 2 in Appendix II includes species
containing pyrrolizidine amino-alcohols or esters, which, while not having all the features of hepatotoxicity, would need only minor structural modifications to render them hepatotoxic. Plants of the same taxonomic groups as the plants of proven hepatotoxicity are listed in part (a) of the table. There is a possibility that, on further examination, hepatotoxic alkaloids may be found, as minor constituents, in strains or parts of these plants not yet investigated or under specific conditions of growth. It should be noted that the species that have been investigated and are listed are only few compared with the total number of species in each genera. It has been recommended by Smith &

Culvenor (1981) that it would be prudent to regard all species in the family Boraginaceae and the genera Crotalaria, Senecio, and Eupatorium as potentially
hepatotoxic.
It is pertinent to note that the alkaloid content in different parts of the plant (e.g., roots, leaves, stalks, flowers, and buds) varies and is subject to fluctuations according to the climate, soil conditions, and time of harvesting (Danninger et al., 1983; Hartmann & Zimmer, 1986). Mattocks (1980) demonstrated that the alkaloid content of the leaves of Symphytum spp. (Russian comfrey), which are used as an item of food, varies with their maturity. The toxic PA content is highest at the beginning of the vegetative period and declines as the leaves mature. The PA content of the roots is much higher than that of the leaves, and dried leaves contain a higher concentration than fresh leaves (Mattocks, 1986). According to Danninger et al. (1983), in some species (Symphytum asperum), relatively long storage may lead to a reduction in the alkaloid content, presumably because enzymes are released during drying. Candrian et al. (1984b) studied the stability of PAs in hay and silage containing various amounts of Senecio alpinus. The PA content of hay remained constant for several months, but the PAs in silage were mainly degraded. However, the degradation of PAs was much less complete in the lower concentration range. A quantitatively significant PA-degradation product in silage was identified as retronecine. Silage with an S. alpinus

percentage of 3.5 - 23 still contained macrocyclic PAs at a concentration of about 20 mg/kg wet weight. Such silage was not considered safe for cattle bearing in mind that a 600-kg calf eats about 30 kg silage/day, amounting approximately to a daily intake of about 1 mg PAs/kg body weight. In feeding trials with Senecio
jacobaea, Johnson (1979) found that the minimum lethal dose for cattle was between 1 and 2 mg PAs/kg body weight per day.
PAs known to have been associated with instances of human toxic liver disease in different parts of the world are listed in Table 2. Two groups of alkaloids that, according to Culvenor (1983), are consumed in significant amounts by people in different parts of the world include:
(a) Echimidine, acetyllycopsamine, and related alkaloids
(many countries)
Leaves of plants of the Symphytum sp. ( Symphytum officinale (comfrey) and Symphytum x uplandicum) are used traditionally as a salad and as a medicinal herb in Australia, many countries of
Europe, and the USA. S. officinale has been shown to be carcinogenic for rats (Hirono et al., 1978). Leaves of Russian comfrey contain a concentration of alkaloids (mainly echimidine) of 0.1 - 1.5 g/kg. The highest level of daily consumption of the alkaloids has been estimated to be 5 - 6 mg (Culvenor, 1983).

(b) Echimidine and related alkaloids (Australia)
PAs derived from Echium plantagineum, with echimidine as the major component, have been found in honey secreted by bees feeding on the plant (Culvenor et al., 1981). The plant is a major source of honey (section 3.3.4).
3.2 Pneumotoxic and Other Toxic Pyrrolizidine Alkaloids
Not all hepatotoxic alkaloids are pneumotoxic. The commonest ones used to produce experimental lung injury are fulvine (Barnes et al., 1964; Kay et al., 1971a; Wagenvoort et al., 1974a,b) and monocrotaline (Lalich & Ehrhart, 1962; Chesney & Allen, 1973b; Huxtable et al., 1977). These are also the most active (Mattocks, 1986). The seeds of Crotalaria spectabilis, which contain monocrotaline, have also been used to study pneumotoxic effects on experimental animals (Turner & Lalich, 1965; Kay & Heath, 1966; Kay et al., 1967a) and C. spectabilis has been called the pulmonary hypertension plant (Kay & Heath, 1969), because of the pulmonary hypertensionogenic properties of the PAs it contains. Culvenor et

al. (1976a) screened 62 PAs for hepatotoxicity and pneumotoxicity. Chronic lung lesions were produced by most compounds that induced chronic liver lesions, though high doses were required in some instances. It is possible that chronic lung lesions may not occur in experimental animals because of early death due to acute toxicity. However, the authors identified a number of PAs that were particularly prone to produce chronic lung damage in rats including crispatine, senecionine, seneciphylline, and usaramine (12-membered macrocyclic, retronecine diesters), anacrotine and madurensine (crotonecine esters), and the heliotridine esters, heliosupine, lasiocarpine, and rinderine.
The molecular structure-activity requirements for pneumotoxicity are the same as those for hepatotoxicity. This is consistent with their both being caused by the same toxic metabolites and by the metabolic activation of the alkaloids in the liver cells to form a reactive pyrrolic dehydro-alkaloid (Culvenor et al., 1976a).
Trichodesmine and incanine, found in the seeds of Trichodesma
incanum (Yunusov & Plekhanova, 1959), are believed to have been the causative factors of the "Ozhalangar encephalitis" that was endemic in Uzbekistan, USSR (1942 - 51), in which the symptoms and signs were related primarily to the central nervous system (Shtenberg & Orlova, 1955) (section 7.7).
Table 2. Instances of human toxicity caused by pyrrolizidine alkaloidsa
Principal Plant Country/ Cause of intake Reference alkaloid Region

Heliotrine and Heliotropium Afghanistan contamination Tandon & Tandon other alkaloids popovii (1975); Tandon, similar to B.N. et al. lasiocarpine (1978); Tandon, H.D. et al. (1978); Mohabbat et al. (1976)
Senecionine Senecio South contamination Wilmot &
illiciformis; Africa Robertson Senecio-burchelli (1920)
Senecio spp. South contamination Selzer &
Africa Parker (1951)
Alkaloids of Crotalaria Ecuador medicine Lyford et al. trichodesmine juncea (1976) and senecionine type
Heliotrine and Heliotropium Hong Kong medicine Kumana et al. lasiocarpine lasiocarpum (1985); Culvenor et al. (1986)

Table 2. (cont'd)
Principal Plant Country/ Cause of intake Reference alkaloid Region
Crotananine and Crotalaria India contamination Tandon, R.K. cronaburmine nana et al. (1976); Krishnamachari et al. (1977); Siddiqui et al. (1978a,b)
Heliotrine Heliotropium India medicine Datta et al. N-oxide eichwaldii (1978a,b)
Monocrotaline Crotalaria West Indies medicine Bras et al. fulvine retusa; (1954, 1957) Crotalaria fulva Stuart & Bras (1957)
Ilex sp. United medicine McGee et al. Kingdom (1976)
Riddelline Senecio USA medicine Stillman et al. retrorsine longilobus (1977); Fox et N-oxide al. (1978); (with others) Huxtable (1980)

Indicine N-oxide purified USA medicine Letendre et al. chemical (1984)
Symphytine, Symphytum sp. USA medicine Ridker et al. symglandine, and (1985); other symphytum Huxtable alkaloids et al (1986) Table 2. (cont'd)
Principal Plant Country/ Cause of intake Reference alkaloid Region
Lasiocarpine and Heliotropium USSR contamination Dubrovinskii heliotrine lasiocarpum (1952); Mnushkin (1952)
Trichodesmine and Trichodesma USSR contamination Shtenberg &
incanine incanum Orlova (1955); Yunosov &
Plekhanova (1959)

a
Adapted from: Culvenor (1983) and Mattocks (1986). Refer also to Table 15 for details and section 7. 3.3 Pathways of Exposure
Naturally-occurring animal disease is caused by the alkaloid- containing plants growing in fields and pastures or being fed accidentally as fodder. They are mostly herbaceous or small shrubs and many thrive in dry and arid climates. One such plant containing toxic PA alkaloids has been reported to grow in the western desert of Egypt (Hammouda et al., 1984). The growth of this group of plants is particularly prolific during, and following, periods of drought, as has been reported in association with the outbreaks of human disease in Afghanistan (Tandon & Tandon, 1975; Mohabbat et al., 1976) and India (Tandon, B.N. et al., 1976). Alkaloid-containing plants are widespread in the tropics, especially Crotalaria, of which there are over 300 species in Africa. Ordinarily, the alkaloid-containing plants have a bitter taste and grazing animals will reject them, unless their normal fodder is scarce. However, PAs often occur largely as N-oxides, which are said not to be bitter, and plants containing PAs are readily eaten by some animal species.
Human intoxication may result from the ingestion of the toxic substance in either contamined food or herbal infusion.
3.3.1 Contamination of staple food crops

The products of pyrrolizidine alkaloid-containing plants, generally seeds, may contaminate the staple food and may be eaten over long periods of time. The fact that these plants may cause disease is generally not recognized by the people and such contamination is known to have resulted in large-scale outbreaks of poisoning (Dubrovinskii, 1952; Mnushkin, 1952; Shtenberg & Orlova, 1955; Tandon & Tandon, 1975; Mohabbat et al., 1976; Tandon, B.N. et al., 1976, 1977; Tandon, R.K. et al., 1976; Krishnamachari et al., 1977; Tandon, H.D. et al., 1977) (Table 2, section 3.1).
3.3.2 Herbal infusions
Plants have been used traditionally for medicinal purposes all over the world. Herbs have been the mainstay of the indigenous systems of medicine, especially in China, Greece, and India, since ancient times. Table 3 includes a list of some plants that are suspected, or known, to contain PAs and have been used as herbal medicines in different countries (Mattocks, 1986).
Several PA-containing plants are included among the list of plants used in indigenous systems of medicine in India (Chopra, 1933). As a part of a research study on the etiological factors of chronic liver disease in Sri Lanka, Arseculeratne et al. (1981) chemically screened the first 50 plants used in Sri Lanka's traditional medicine pharmacopoaea, and found that 3 of them
contained PAs. All 3 were hepatotoxic in rats. Of the 3, the presence of alkaloids in Cassia auriculata and that of PAs in Hollarhena antidysenterica had not previously been recorded. It should be noted that the amount of experimental plant material used in this study was approximately 6.5 g/kg body weight per day, in

contrast to the approximate intake by a human being estimated to be in the range of 0.3 - 0.6 g/kg body weight per day. Some, but not all, of the plants reported to be etiological agents in human cases of veno-occlusive disease can be found in an inventory of medicinal plants used in different countries (WHO, 1980), which also indicates the countries that they are used in. The above lists may not be complete as many such plants may be used in folk medicine but have not been mentioned in the scientific literature. However, the lists do indicate the wide and varied use of such toxic herbs in all parts of the world.
Lately, there has been a growing interest in the developed countries in organically grown products for food, as well as home remedies (Table 3), and some of the PA-containing herbs have been freely available in herbal shops (Schoental, 1968; Burns, 1972). Danninger et al. (1983) listed plants containing PAs that are commonly used in the Federal Republic of Germany as medicaments (Table 4). He also listed 9 plants in which alkaloids have only been identified qualitatively, the toxicity of which has not been, or has been insufficiently, investigated (Table 5). Similarly, Roitman (1983) listed 10 plants, in which the presence of PAs is suspected or has been proved and which are used as herbal teas in the USA. The lists include 10 plants containing PAs, most of which have been proved hepatotoxic experimentally, some having highly carcinogenic promoter activity. Some of these alkaloids have been associated with human case reports of PA toxicity. The more recent reports (Table 2) of instances of PA poisoning through the use of herbal medicines are from developed countries (Lyford et al., 1976; Stillman et al., 1977; Fox et al., 1978; Kumana et al., 1985; Ridker et al., 1985). Such use of the herbs is the reason that veno-occlusive disease is endemic in Jamaica (Bras et al., 1954; Jellife et al., 1954a,b; Bras & Watler, 1955; Stuart & Bras, 1955, 1957). There are obvious difficulties in exercising any kind of control to restrict this use only to plants that have been tested and certified as safe for human use. It is impossible to identify many such herbs, as they are sold as plants or their amorphous

products in the herbal shops.
Table 3. Some plants containing (or suspected of containing) PAs, which have been used by people either as herbal medicines (M) or foods (F)
Plant Mode Country Referencea
of use or region
BORAGINACEAE
Anchusa officinalis M Europe Broch-Due & Aasen (1980) B
Borago officinalis M USA Delorme et al. (1977) A
Cynoglossum M East Africa Schoental & Coady (1968) A geometricum
Cynoglossum M Iran Coady (1973) B officinale
Heliotropium M India Gandhi et al. (1966a); B eichwaldii Datta et al. (1978a,b) A
H. europaeum M India, Greece IARC (1976) A

H. lasiocarpum M Hong Kong Kumana et al. (1985); A Culvenor et al. (1986) A
H. indicum M India, Africa, Schoental (1968a); B South America, Hoque et al. (1976) B and elsewhere
H. ramossissimum M Arabia Macksad et al. (1970); B (ramram) Coady (1973) B
H. supinum M Tanzania Schoental & Coady (1968) A
Table 3 (contd.)
Plant Mode Country Referencea
of use or region
Pulmonaria spp. M USA Delorme et al. (1977) A
Symphytum officinale F, M Japan and Hirono et al. (1978, 1979b) A M USA Furuya & Hikichi (1971); A Delorme et al. (1977) A

S. x uplandicum F, M General Hills (1976) B
USA Culvenor et al. (1980a,b) A
S. asperum M USA Pedersen (1975) A
COMPOSITAE
Cacalia decomposita M USA Sullivan (1981) B (matarique)
C. yatabei F Japan Hikichi & Furuya (1978) B
Farfugium japonicum M Japan Furuya et al. (1971) B
Ligularia dentata F Japan Asada & Furuya (1984) B
Petasites japonicus F, M Japan Hirono et al. (1973, 1979b) A
Senecio abyssinicus M Nigeria Williams & Schoental (1970) B
S. aureus M USA Wade (1977) B Table 3 (contd.)

Plant Mode Country Referencea
of use or region
S. bupleuroides M Africa Watt & Breyer-Brandwijk (1962) A
S. burchelli F, M South Africa Rose (1972) A
S. coronatus M South Africa Rose (1972) A
S. discolor M Jamaica Asprey & Thornton (1955) B
S. doronicum M Germany Roeder et al. (1980a) B
S. inaequidens F South Africa Rose (1972) B
S. jacobaea M Europe Schoental & Pullinger (1972); B (ragwort) Wade (1977) B
S. longilobus M USA Stillman et al. (1977); A (S. douglassi) Huxtable (1979a) B
S. monoensis M USA Huxtable (1980) A

S. nemorensis M Germany Habs et al. (1982) A spp. fuchsii
S. pierotti F Japan Asada & Furuya (1982) B
S. retrorsus M South Africa Rose (1972) A (S. latifolius) Table 3 (contd.)
Plant Mode Country Referencea
of use or region
S. vulgaris M Europe Watt & Breyer-Brandwijk (1962) A (common groundsel)
Netherlands Wade (1977) B
M Iran Coady (1973) B
Syneilesis palmata F Japan Hikichi & Furuya (1976) B
Trichodesma africana M Asia Omar et al. (1983) B
Tussilago farfara M Japan Culvenor et al. (1976a) A

(coltsfoot)
M China Hirono et al. (1976b) A
M Norway Borka & Onshuus (1979) B
M USA Borka & Onshuus (1979); B Culvenor et al. (1976b); B
LEGUMINOSAE
Crotalaria brevidens F East Africa Coady (1973) B
C. fulva M Jamaica Barnes et al. (1964); A McLean (1970, 1974) A Table 3 (contd.)
Plant Mode Country Referencea
of use or region
C. incana M East Africa Schoental & Coady (1968) A Watt & Breyer-Brandwijk A

(1962)
C. laburnifolia M Tanzania Schoental & Coady (1968) A
F Asia Coady (1973) B
C. mucronata M Tanzania Coady (1973) B
C. recta M, F Tanzania Schoental & Coady (1968); A Coady (1973) B
C. retusa M, F Africa IARC (1976) A
India Watt & Breyer-Brandwijk (1962) A
C. verrucosa M Sri Lanka Arseculeratne et al. (1981) A
a
A = Reference in the reference list of this document. B = Reference in Mattocks (1986). Manufactured preparations may also contain PA-containing herbs, e.g., comfrey-pepsin capsules sold as a digestive aid (Huxtable et al., 1986).
3.3.3 Use of PA-containing plants as food
Several PA-containing plants are used as food as can be seen in Table 3 (Mattocks, 1986).

Petasites japonicus Maxim, Tussilago
farfara L. (coltsfoot), and Symphytum officinale L. (comfrey or Russian comfrey) are known as edible plants in Japan, and have been proved to contain carcinogenic pyrrolizidine alkaloids (Hirono et al., 1973, 1979a,b). The young flower-stalks of P. japonicus and the buds of coltsfoot have been used in Japan as human food or herbal remedies. The leaf and root of comfrey are also used as an edible vegetable or tonic (Hirono et al., 1978) in Japan and other countries (Culvenor, 1985). The carcinogenic PAs in these plants are petasitenine (P. japonicus), senkirkine (coltsfoot), and the group including symphytine (comfrey). They were also mutagenic in the Ames system of Salmonella typhimurium and V79 hamster cell line and induced transformation in cryo-preserved hamster embryonic cells (Hirono et al., 1979b). Other such PA-containing plants, used as food in Japan, include young leaves of Syneilesis palmata, various Cacalia species, and young Senecio pierotti (Mattocks, 1986). According to Culvenor (1985), consumers of comfrey could be ingesting up to 5 mg PAs per day. Rose (1972) listed a number of plants of the genus Senecio that are used as spinach in South Africa. These include

S. burchelli, which is known to have caused an episode of PA poisoning through the ingestion of contaminated bread (Wilmot & Robertson, 1920).
3.3.4 Contaminated honey
In the USA, Deinzer et al. (1977) reported the presence of all PAs contained in Senecio jacobaea (ragwort) and proved to be hepatotoxic, in the honey secreted by bees feeding on the plant. The total alkaloid content ranged from 0.3 to 3.9 mg/kg. It has been estimated that an average annual human intake of honey (600 g) at the highest alkaloid level quoted would contain less than 3 mg of PAs (Mattocks, 1986). Culvenor et al. (1981) and Culvenor (1983, 1985) drew attention to the same potential hazard in honey from Echium plantagineum, a weed that grows widely in Southern Australia and is a major source of honey, yielding an estimated 2000 - 3000 tonnes per annum for human consumption. Echimidine is the major component of the alkaloids of Echium, which are present in concentrations of up to 1 mg/kg. Culvenor (1983) estimated that individuals may consume up to 80 g honey/day with a corresponding alkaloid intake of 80 µg/day, if only the Echium honey were used. No reports of acute human toxicity through this source are available.
Table 4. Medicinal plants containing PAs of known hepatotoxicity, reported as commonly

used in the Federal Republic of Germany, and the PAs contained in thema
Family Genus Species Pyrrolizidine alkaloids
Compositae Eupatorium E. cannabinum amabiline± (hemp agrimony) supinineb
Petasites P. hybirdus senecionineb,c
integerrimineb
senkirkineb
Senecio S. nemorensis fuchsisenecionine (groundsel) sp. fuchsii senecionineb,c
(Fuch's groundsel)
S. vulgaris senecionineb,c
(groundsel) seneciophyllineb
retrorsineb
riddellineb,c
S. Jacobaea jacobineb
(ragwort) senecionineb,c
seneciphylline

b
jacoline, jaconine chlorinated PAsd
S. aureus senecionineb,c
(American golden ragwort)
Tussilago T. farfara senkirkineb
(coltsfoot) (coltsfoot) senecionineb,c
tussilagine
Table 4 (contd.)
Family Genus Species Pyrrolizidine alkaloids
Alkanna A. tinctoria 7-angelylretronecine triangularine dihydroxytriangularine
Anchusa A. officinalis lycopsamine
Boraginaceae Borago B. officinalis lycopsamine/intermedine± (borage) acetyllycopsamine/ acetylintermedine amabiline supinine

Symphytum S. officinale symphytineb
(comfrey) (comfrey) echimidine(?) lycopsamine acetyllycopsamineb
lasiocarpineb,c
heliosupine N-oxide
S. peregrinum lycopsamineb
S. x uplandicum intermedineb
symphytineb
echimidineb
7-acetyllycopsamine 7-acetylintermedine symlandine uplandicine
S. asperum asperumine (prickly comfrey) heliosupine N-oxide echimidineb
echinatine
Table 4 (contd.)
Family Genus Species Pyrrolizidine alkaloids

Cynoglossum C. officinale heliosupine N-oxide (hound's (hound's tongue) echinatine tongue) acetyl heliosupineb
O-7-angelylhelio- tridineb
Heliotropium H. europaeum heliotrineb,c,e
(Heliotrope) (common heliotrope) lasiocarpineb,c,e
supinine heleurine europine acetyllasiocarpineb
a
Modified from: Danninger et al. (1983). b
Toxic alkaloids. c
Alkaloids known to have caused human toxicity. d
Alkaloids with highly carcinogenic promoter activity. e
Used only in homeopathy. Table 5. Medicinal plants containing PAs, reported as commonly used in the Federal Republic of Germany, the toxicity of which has not been, or has been insufficiently, investigateda

----------------------------------------------------------- Family Genus Species ----------------------------------------------------------- Compositae Eupatorium E. perforatum
Brachyglottis B. repens
Arnica A. montana (mountain arnica)
Boraginaceae Lappula L. intermedia (stickseed)
Pulmonaria P. officinalis (lungwort) ----------------------------------------------------------- a
Modified from: Danninger et al. (1983).
3.3.5 Milk
PAs have been shown to produce toxic effects via transference into the milk of dams (Schoental, 1959). Retrorsine was administered orally to 17, and intraperitoneally to 6, lactating rats weighing 185 - 350 g in 5 - 10 mg doses daily, the first dose being given during the first 24 h after parturition. The rats received from 1 to 14 doses, the total intake amounting to 21 - 335 mg/kg body weight. The litters were separated from the mothers for ´ h following the administration of PA to avoid direct contamination of the former by licking. Apparently the milk production was not affected as the stomachs of many of the young, examined postmortem, were distended with milk. All animals whose mothers had received a total dose of 138 mg PA or more died within

30 days. Many of the young whose mothers had received smaller doses survived until they were killed at 6 months. Biopsy of the liver of the young at various intervals or at autopsy showed marked changes, even in cases where the mothers did not appear to be affected. Animals dying at 18 - 30 days showed hydropic or fatty vacuolation of liver cells. In the liver of animals dying or killed later, various degrees of haemorrhagic necrosis and increase in the centrilobular reticulin of the liver, and some thickening of centrilobular veins were seen. In animals that survived 6 months, the appearance was less abnormal, but some hyperplastic nodules and bile-duct proliferation were seen. The lactating rats dosed with the PAs generally survived longer than the suckling animals and usually did not show any ill effects, suggesting that the susceptibility of the suckling rats was greater than that of the mothers.
Dickinson et al. (1976) demonstrated the presence of PAs in the milk of dairy cattle fed or dosed with ragwort (Senecio jacobaea).
When 4 cows were administered the dried plant material at levels of up to 10 g/kg body weight per day through rumen cannula, PA levels of up to 0.84 mg/kg were observed in the milk. However, only one
(jacoline) of the several PAs contained in the plant was secreted. Calves, bucket fed on the milk did not show any signs of PA toxicity.
Dickinson (1980) repeated the study on goats. Four milk goats were freshly prepared with rumen cannulae. The kids were separated from their dams and were fed milk twice a day. Dried tansy ragwort plant material with a PA content of 0.16% (dry weight) was administered through the cannulae to each goat at a dosage rate of 10 g/kg body weight per day over 125 days. During this period, each of the 4 kids received milk from their dams at approximately 125 ml/kg per day in addition to ad lib feeding on alfalfa grass hay. Six PAs were isolated from the plant material: jacobine, jaconine, jaconline, jacozine, senecionine, and seneciphylline.

Milk samples collected twice daily showed PA contents of 225 - 530 µg/litre with a mean of 381 µg/litre. No apparent health effects were noted in the kids, and only mild hepatic damage was suspected in the dams, on the basis of liver function tests. Fifty percent of the kids were killed after 10 weeks. No lesions of PA toxicity were seen. The dams were rebred and appeared normal throughout the gestation period. However, three dams aborted at almost full term, and the fetuses were born dead. One of the dams died shortly after parturition and showed evidence of severe liver damage characteristic of PA toxicity. Another, which delivered normally, also showed a lesser degree of liver damage at biopsy.
Data relating to PA secretion were compared with similar earlier data on cows. Mean secretion of PAs in cows appeared much higher, e.g., 684 µg/litre. The authors concluded that the amount of PAs secreted in the goat's milk did not cause any serious deleterious effects in the kids.
Johnson (1976) fed long-term lethal doses of Senecio jacobaea, by stomach tube, to 6 cows. Feeding started at term or within 30 days post-partum, and continued until what was considered to be a lethal dose had been fed. The daily dose of the plant ranged from 1 to 4.4 g/kg body weight, the total amount fed representing 5 - 15% of body weight over a period of 54 - 126 days. Five cows died within 98 days; one, in a moribund state, was killed on day 126. The calves suckled for 30 - 126 days. Suckling started immediately after birth in the case of 4 calves and 10 and 30 days later, respectively, in the 2 remaining calves. Three calves were killed with their dams or soon after, and 3 were retained for 1 year for observation. Milk samples from 2 cows were collected and pooled in 14- to 16-day lots during 64 days of feeding of the Senecio plant. Each pooled sample was administered intragastrically to a group of rats in daily doses of 12 ml for 15 - 30 days. A control group of rats were fed raw milk from cows not fed Senecio. Blood samples of

the dams and the calves were analysed for glutamic oxaloacetic transaminase (GOT), lactic dehydrogenase (LDH), and gamma-glutamyl transpeptidase (GGTP). Serum-enzyme levels in all cows indicated statistically significant deviations suggesting liver dysfunction, and the livers at autopsy had characteristic features of PA toxicosis. The LDH and GOT levels in calves were generally abnormal after 20 - 45 days of suckling. The abnormalities ranged
from mild to a 15- to 170-fold increase. One calf was autopsied at the peak increase of serum-enzymes and was found to have mild focal hepatitis. No significant pathological features were seen in the livers of other animals, nor of the rats, some of which were retained for up to 150 days.
Goeger et al. (1982) fed dried Senecio jacobaea (tansy ragwort) to lactating goats in a proportion of 25% of the feed. The milk contained 7.5 µg PA/kg dry weight. The milk produced by the goats was pooled and then bottle fed to appetite to 2 Jersey bull calves (1 day old) that also had access to tansy ragwort-free hay for 109 and 124 days, respectively. They were then weaned and given normal feed and observed for 6 months, after which they were killed and autopsied. In another study, rats were fed a diet containing the freeze-dried milk at 80% level for 180 days with a calculated total PA intake of 0.96 mg/rat. Other groups of rats were fed tansy ragwort at dietary levels of 0.01 - 10 g/kg (corresponding to PA intakes of 39.77, 5.04, 0.52, and 0.05 mg/rat). The calf livers only showed very mild non-specific changes, but the livers of rats fed tansy ragwort or the milk from tansy ragwort-fed goats showed definite, but mild, changes including swollen hepatocytes, megalocytosis, biliary hyperplasia, and fibrosis. Histopathological changes in milk-fed rats were similar to those in the group fed tansy ragwort in the diet at 0.01 g/kg. The authors concluded that there was evidence of PA transfer into milk, which proved hepatotoxic for rats. It was also noted that the goats had been fed high levels of tansy ragwort at the upper limit of their acceptance, and that the hepatic changes observed in rats fed high levels of milk, for extensive periods, were slight.

Luthy et al. (1983) produced direct evidence of excretion of macrocyclic esters of retronecine of the senecionine and seneciphylline-type into rat milk. 3
H-retronecine, an 3
H-necic acid-labelled senecionine, and seneciphylline were prepared biosynthetically with seedlings of Senecio vulgaris L. Two lactating rats (Ivanovas, Sprague Dawley), weighing 300 - 400 g, were fed the first of the second compound by stomach tube, in doses of 2.7 mg/kg and 5.5 mg/kg body weight, respectively. Samples of blood were examined 1, 3, and 6 h after treatment, and those of milk 1 and 3 h after. Animals were killed after 6 h. They were found to have excreted approximately 0.08% of the applied radioactivity in the milk within 3 h, mainly as unidentified retronecine-derived metabolites, and approximately 0.02% as unchanged PAs. The highest levels of PAs and metabolites in tissues were found in the liver and lungs, 6 h after administration.
Candrian et al. (1984a) also demonstrated that Drosophila
melanogaster flies fed on milk from lactating rats that had been administered an oral dose of seneciphylline showed 1.2% sex-linked recessive lethals, compared with 0.3% in controls, indicating the transfer of the mutagenic properties of the PA via milk (section 6.4.7).
The implications of the above studies on the possibility of carry-over of PAs into foodstuffs of animal origin are obvious. However, no reports of human cases of acute PA toxicity, ascribed to the consumption of contaminated milk, are available.
3.3.6 Meat

There have not been any reports of the detection of PAs in meat products from livestock exposed to them.
3.3.7 Use of PAs as chemotherapeutic agents for cancer
An alkaloid of Heliotropium indicum L. (indicine N-oxide) has been found to have antitumour activity and has been used in experimental clinical chemotherapy for cancer (section 7.9).
4. METABOLISM
4.1 Absorption, Excretion, and Tissue Distribution
4.1.1 Absorption
There have been few studies on the absorption of PAs in man, but absorption has been inferred from studies on tissue distribution and the amounts of alkaloids and their metabolites excreted in the urine, faeces, and bile of animals (section 4.1.2).
Swick et al. (1982c) measured the transfer of a mixture of pyrrolizidine alkaloids extracted from Senecio jacobaea, across isolated intestine and stomach segments from rabbits. The alkaloid mixture contained seneciphylline, jacobine, jacozine, jacoline, and senecionine. The alkaloids were transferred across the ileum and jejunum, but not the stomach. Brauchli et al. (1982) compared the oral and percutaneous absorption in rats of a crude alkaloid mixture obtained from comfrey ( Symphytum officinale

L.). The mixture consisted of N-oxides of 7 alkaloids, principally 7-acetyl- intermedine and 7-acetyl-lycopsamine. A dose of 194 mg/kg was either given by gavage, or was applied to the shaved skin and left for 44 h. After the dermal application, the excreted N-oxides in urine (up to 48 h) amounted to 0.1 - 0.4% of the dose. After oral dosage the excreted level of N-oxides and alkaloid bases was quoted as being 20 - 50 times greater.
4.1.2 Excretion and distribution
The excretion and distribution of heliotrine in rats has been reported in Bull et al., 1968. Young rats (150 g), given the LD50
of heliotrine by ip injection, were killed at intervals, bled quickly, and the organs and tissues analysed. Heliotrine was present in the liver after 2 min (3.7% of total dose), the level peaking at 5 min (6.3%), and dropping to 2.2% at 1 h and 0.5% at 2.5 h. In adult rats, the level in the liver at 5 h was 0.07% of the total dose. Five min after dosing, 30 - 40% of the initial dose remained in the peritoneal cavity, and the blood level of heliotrine was 60 mg/litre, dropping to 3 mg/litre at 1 h. The urinary excretion of base and metabolites other than pyrrolic metabolites, collected and measured 16 h after administration of several alkaloids by ip injection, is shown in Table 6. The proportion of base excreted unchanged increased with the hydrophilicity of the alkaloid, being 62% for heliotrine N-oxide, 30% for heliotrine, and only 1 - 1.5% for lasiocarpine. Heliotridine, the hydrolysis product from heliotrine and lasiocarpine, was excreted in the form of the

N-oxide in larger quantities after the administration of each of these alkaloids.
The distribution and excretion of monocrotaline was studied in rats by Hayashi (1966) who found that 50 - 70% was excreted in the urine within the first day. However, the analysis was by a non- specific chemical method that did not distinguish between the unchanged alkaloid and its metabolites. Mattocks (1968a) gave toxic pyrrolizidine alkaloids intraperitoneally to male rats and
measured the urinary excretion of the unchanged alkaloid, and of N-oxide and pyrrolic metabolites. The excretion of N-oxide and unchanged alkaloid was rapid and almost complete in the first 24 h. Excretion of pyrroles was also rapid but continued for a little longer. For example, in rats given retrosine (60 mg/kg body weight), the urine in the first 24 h contained 10.6% unchanged alkaloid, 13.3% N-oxide, and 13.4% pyrrolic metabolites. During the second day, only 0.1% alkaloid, 0.2% N-oxide, and 1.8% pyrroles were excreted. Biliary excretion also occurred. About one-quarter of an iv dose of retrosine in rats was excreted in the bile as pyrrolic metabolites, and 4% as unchanged alkaloid; most of this excretion occurred during the first hour after the injection (White, 1977).
Jago et al. (1969) gave heliotrine iv to sheep; urinary excretion of the unchanged alkaloid together with metabolites ( N-oxide, and demethylation and hydrolysis products) occurred rapidly and continued for up to 8 h. Excretion in the bile was only 2% of that in the urine.

The tissue distribution of radioactivity from a tritiated toxic pyrrolizidine alkaloid analogue was studied by Mattocks & White (1976) using synthanecine A bis- N-ethylcarbamate (40 mg/kg body weight). The highest concentrations of radioactivity were seen in the liver (where metabolism occurs), lungs, kidneys, and spleen (respectively, 3.9%, 0.19%, 0.18%, and 0.27% of the dose given), and about 69% of the dose was eliminated in the urine during the first day. Radioactivity in the expired air was negligible. The binding of radioactivity in the liver, and especially the lungs, was more persistent than in other organs. Similar results were given by the semisynthetic pyrrolizidine alkaloid analogue, retronecine bis- N-ethylcarbamate (Mattocks, 1977).
Table 6. Urinary metabolites of pyrrolizidine bases in the rat (16-h urine)a
Urine constituent (amount in percentage of dose injected)
Base Unchanged Base Heliotridine Heliotridine Heliotridine Heliotridine administered base N-oxide trachelanthate trachelanthate N-oxide (ip injection) N-oxide
Heliotrine 30 Trace 10 5 3 15

Heliotrine 62 (62) 2.7 ca. 6 ca. 1 ca. 10 N-oxide Lasiocarpine 1-1.5 1.5-3 6 Heliotridine 35 ca. 1 (35) (ca. 1) 5 20 trachelanthate Heliotridine 40 20 (40) (20)
a
From: Bull et al. (1968). The distribution of the uniformly 14
C-labelled natural pyrrolizidine alkaloid senecionine in lactating mice was studied by Eastman et al. (1982). After 16 h, 75% of the radioactivity had been recovered in the urine, 14% in the faeces, but only 0.04% was in the milk; the liver contained 1.92%. The mice were milked using teat cups. Candrian et al. (1985) studied the distribution of radioactivity in rats given small doses of senecionine or seneciphylline (0.3 - 3.3 mg/kg), tritiated in the pyrrolizidine (retronecine) moiety. Most radioactivity was eliminated in the urine and faeces within 4 days. Using mass spectrometry, Dickinson et al. (1976) found a concentration of up to 0.84 mg PAs/litre in the milk of cows fed Senecio jacobaea. Blood levels of senecionine in rats given 0.1 LD50
ip were determined by Culvenor (1978). The levels were 0.38, 0.32, and 0.14 mg/litre at 0.5, 1, and 2 h after injection, respectively.
To summarize, the available evidence suggests that ingested toxic pyrrolizidine alkaloids are rapidly metabolized and that the excretion of unchanged alkaloid and of most metabolites is also rapid. Thus, within a few hours, only a relatively small proportion of the dose remains in the body, much of this in the form of metabolites bound to tissue constituents. It appears

improbable that a significant amount of unchanged alkaloid will remain in the body after the first day.
Pyrrolizidine N-oxides are much more water soluble than their parent alkaloids. Indicine N-oxide (which is exceptionally water soluble) is very rapidly excreted, either unchanged or conjugated. Thus, indicine N-oxide given iv to mice, monkeys, or rabbits disappeared from the serum with initial half-lives ranging from 3 to 20 min (Powis et al., 1979; El Dareer et al., 1982). Over 80% of tritium-labelled indicine N-oxide given iv was excreted in the urine of mice or monkeys within 24 h (El Dareer et al., 1982); at 2 h, the highest concentrations of radioactivity were in the kidneys, liver, and intestines. Urinary excretion of indicine N-oxide was also rapid in rabbits, but somewhat slower in human beings (Powis et al., 1979).
4.2 Metabolic Routes
The major metabolic routes of unsaturated pyrrolizidine alkaloids in animals are: (a) hydrolysis (of the ester groups); (b) N-oxidation; and (c) dehydrogenation (of the pyrrolizidine nucleus) to dehydro-alkaloids (pyrrolic derivatives). Other minor routes of metabolism are known, but the three pathways account for the major known toxic effects of these alkaloids (Fig. 5). Routes (a) and (b) are believed to be detoxification mechanisms. Route (c) leads to toxic metabolites and appears to be the major

activation mechanism. Route (a) may occur in various tissues, including the liver and blood. Routes (b) and (c) are brought about in the liver by the microsomal mixed-function oxidase system.
4.2.1 Hydrolysis
The hydrolysis of a PA leads to the formation of the amino- alcohol moiety (necine base) and the acid moiety. Neither of these is hepatotoxic (Schoental & Mattocks, 1960; Culvenor et al., 1976a). The highly water-soluble necine base is readily excreted, is not accessible to the microsomal system, and is not activated to a toxic metabolite. Thus, pyrrolizidine alkaloids that are very susceptible to (enzymic) hydrolysis have low toxicity (Mattocks, 1982). A major factor contributing to resistance to esterase is the steric hindrance in the acid moiety. Thus, the chain branching near the carbonyl groups slows hydrolysis allowing the formation of relatively high levels of pyrrolic metabolites; a conformation of the basic moiety, which brings the two ester groups close together, thus leading to mutual steric hindrance, can also prevent hydrolysis (Mattocks, 1981a).
The influence of hydrolysis in vivo on alternative metabolic pathways is demonstrated by the fact that treatment of rats with an esterase inhibitor, before giving pyrrolizidine alkaloids (or synthetic analogues), can lead to greatly increased production of pyrrolic metabolites from alkaloids that are normally susceptible to hydrolysis, but little increase in those from alkaloids normally resistant to hydrolysis (Mattocks, 1981a).

4.2.2 N-oxidation
The N-oxidation of pyrrolizidine alkaloids is induced by the hepatic microsomal enzymes. The N-oxide metabolites are highly water soluble and are rapidly excreted in the urine (Mattocks, 1968a). Pyrrolizidine N-oxides are not converted to any significant extent to pyrrolic metabolites by microsomal enzymes (Jago et al., 1970; Mattocks & White, 1971a), and there is no evidence that they are toxic, unless first reduced to the corresponding basic alkaloids, which can then be activated by the microsomal system (Mattocks, 1971c). Thus, it appears that the formation of N-oxides represents a detoxification pathway.
4.2.3 Conversion to pyrrolic metabolites
In laboratory animals, toxic pyrrolizidine alkaloids are metabolized to pyrrolic derivatives, so-called because the unsaturated ring of the pyrrolizidine system loses 2 hydrogen atoms to form what is in effect a pyrrole ring (though the structure is more correctly a dihydropyrrolizidine). Pyrrolic metabolites are easily detectable in the tissues shortly after giving a toxic pyrrolizidine alkaloid to an animal, by treating the tissue with an Ehrlich reagent containing boron trifluoride, when a red colour is produced; this reaction also occurs with the urine (Mattocks, 1968a; Mattocks & White, 1970). In rats given retrosine, pyrrolic

metabolites were found principally in the liver, with highest levels associated with the microsomal and solid debris fractions and less in the mitochondrial fraction; low levels were found in the lungs, heart, spleen, and kidneys, within 4 h of giving retrosine. Rats given 60 mg retrosine/kg body weight excreted 14% of the dose in the urine, within 48 h.
Pyrrolic metabolites are formed by the hepatic mixed-function oxidase system, with a requirement for cytochrome P450, oxygen, and NADPH, as has been demonstrated in vitro (Jago et al., 1970; Mattocks & White, 1971a). Conversion of pyrrolizidine alkaloids to pyrrolic metabolites by the lung tissue of the human embryo (Armstrong & Zuckerman, 1970), rat (Mattocks & White, 1971a; Hilliker et al., 1983), or rabbit (Guengerich, 1977) was negligible. The formation of pyrrolic metabolites does not proceed via N-oxide intermediates, but appears to result from an initial hydroxylation of the unsaturated pyrrolizidine ring adjacent to the nitrogen atom (Mattocks & White, 1971a; Mattocks & Bird, 1983). This would lead to a chemically unstable intermediate that would be expected to decompose spontaneously to the pyrrolic product. The primary pyrrolic metabolites (or dehydro-alkaloid) formed by dehydrogenation of pyrrolizidine alkaloids are chemically dehydropyrrolizidine esters (Fig. 5). These are highly reactive compounds that can rapidly react with tissue constituents or hydrolyse to the corresponding pyrrolic alcohols, or dehydro- necines, which can thus be regarded as secondary metabolites. The

latter can also react with tissue constituents, but more slowly. Because of their high chemical reactivity, the primary metabolites would be expected to have a short life in the liver cell (minutes or seconds) before they are hydrolysed or react with nucleophilic tissue constituents. Some might escape into the blood stream and reach other organs, especially the lungs. Dehydro-necines are more stable and also more water soluble, and can become more widely distributed throughout the body. However, they are also capable of reacting with tissue constituents. Thus, measurements of pyrroles formed from pyrrolizidine alkaloids in tissue samples, using a colour reaction (Mattocks & White, 1970), will not represent a single metabolite, but mixtures of the metabolites together with various reaction products of these with tissue constituents. It will be seen (section 5) that pyrrolic metabolites are believed to be responsible for major toxic actions of pyrrolizidine alkaloids (Mattocks, 1972a).
A pyrrolic metabolite with reactivity midway between that of dehydromonocrotaline and dehydroretronecine has been reported to be formed from monocrotaline in isolated, perfused rat liver (Lafranconi et al., 1985). Studies on this metabolite (isolated from bile) indicated that it is a monoester, and that it is toxic in perfused rat lung. This suggests that monoester pyrrolic metabolites may play a part in the toxic actions of PAs in extra- hepatic tissues.
When rats or other laboratory animals are given a toxic pyrrolizidine alkaloid, pyrrolic metabolites accumulate rapidly in the liver (Mattocks, 1973; White et al., 1973), reaching a peak within 1 - 2 h, then falling slowly during the next 24 h; the metabolites may still be detectable after 2 days. Accumulation is especially rapid after intraperitoneal injection, a very high level
of pyrrolic metabolites being attained within 20 min; this indicates how rapid the metabolism of pyrrolizidine alkaloids can be.
The level of pyrrolic metabolites in rat liver is generally

directly related to the amount of alkaloid given 2 h previously, at least up to an acute LD50
dose. The pyrrole level depends on the alkaloid used, and is related to the acute hepatotoxicity of the alkaloid (Mattocks, 1972a).
To be converted to the type of chemically reactive, toxic pyrrolic metabolites described above, an alkaloid must possess a 1-hydroxymethyl pyrrolizidine system, unsaturated in the 1,2-position (this makes the ring susceptible to dehydrogenation), and at least one hydroxyl group must be esterified, usually by a branched-chain acid. Otonecine esters are converted to similar pyrrol metabolites by a different media involving N-demethylation. Pyrrolizidine amino-alcohols (e.g., retronecine) are not metabolized to more than small amounts of pyrroles (Jago et al., 1970; Mattocks, 1981a), possibly because they are too water soluble to reach the microsomal enzymes.
The metabolic formation of pyrroles is catalysed by cytochrome P450 and specificity exists in the various isozymes (Guengerich, 1977; Juneja et al., 1984).
A few non-toxic pyrrolizidine alkaloids (e.g., rosmarinine and hygrophylline) are converted to pyrrolic metabolites in vivo
(Mattocks, 1973). Such metabolites are chemically different from the pyrroles of toxic alkaloids, and they are neither reactive nor toxic (Mattocks & White, 1971b). The balance of structural features necessary for a pyrrolizidine alkaloid to be converted to give high concentrations of toxic pyrrolic metabolite has been discussed by Mattocks (1981a); the optimum conditions appear to be met in some alkaloids that are macrocyclic diesters, such as retrosine.

4.3 Effects of Treatments Affecting Metabolism
The formation of pyrrolic metabolites (and of N-oxides) is altered by treatments that affect the hepatic microsomal enzymes. Such effects have been studied by measuring rates of metabolism of pyrrolizidine alkaloids in vitro using microsomal preparations from animals pre-treated in various ways (Table 7). For example, microsomes from rats given the microsomal enzyme inducers phenobarbitone or DDT (but not those from rats given 3-methylcholanthrene) induce greatly increased pyrrole formation and smaller increases in N-oxide formation, from the alkaloid retrorsine (Mattocks & White, 1971a). Enzyme preparations from rats treated with inhibitors of microsomal enzymes, including SKF 525A and chloramphenicol, are much less active in converting monocrotaline to pyrroles (Chesney et al., 1974). The ability to metabolize retrorsine is diminished in microsomes from rats fed a protein-free diet, or from rats acutely poisoned with retrorsine (Mattocks & White, 1971a).
Table 7. Effect of pre-treatment of male rats on the conversion of PAs to pyrrolic derivatives and to N-oxides by liver microsomes in vitro

---------------------------------------------------------------------------- Alkaloid Pre-treatment, and Enzyme activity as Reference time before enzyme % of control values measurements for formation of:
Pyrroles N-oxides ---------------------------------------------------------------------------- retrorsine phenobarbitone, ip, 311 232 Mattocks &
3 x 100 mg/kg, White (1971a) 1 - 3 days
retrorsine DDT, ip, 75 mg/kg, 407 203 Mattocks &
3 days White (1971a)
retrorsine 3-methylcholanthrene, 95 (ns) 116 (ns) Mattocks &
ip, 3 x 20 mg/kg, White (1971a) 1 - 3 days
retrorsine retrorsine, ip, 63 - Mattocks &
35 mg/kg, 20 h White (1971a)
retrorsine protein-free diet, 39 - Mattocks &
3 days White (1971a)
monocrotaline phenobarbitone, sc, 448 165 Chesney et al. 4 x 75 mg/kg, (1974) 1 - 4 days

monocrotaline chloramphenicol, sc, 10 109 (ns) Chesney et al. 200 mg/kg, 1 h (1974)
monocrotaline SKF 525A, ip, 75 mg/kg, 10 87 (ns) Chesney et al. 1 h (1974) ---------------------------------------------------------------------------- ns = not significantly different from controls. The effects of in vivo treatment with several types of enzyme inducers on the toxicity of lasiocarpine and senecionine for primary rat hepatocyte cultures was investigated by Hayes et al. (1985). Pre-treatment with phenobarbitone potentiated the cytotoxicity of senecionine towards the cultured cells, whereas pre-treatment with 3-methylcholanthrene diminished the toxic action of senecionine, but had little effect on lasiocarpine cytotoxicity. The cytocidal effects of both alkaloids were substantially inhibited in the presence of SKF 525A.
4.4 Other Factors Affecting Metabolism
Variations between animal species have been investigated by White et al. (1973) and Shull et al. (1976). For instance, metabolism to form pyrroles is high in rats and very low in guinea- pigs, which, however, have higher rates of N-oxidation. For example, 2 h after an ip dose of retrosine (100 mg/kg body weight),
the liver-pyrrole level in male rats was 13 times higher than that in male guinea-pigs (White et al., 1973). Liver microsome preparations from male rats were 28 times more active than microsomes from male guinea-pigs in the dehydrogenation of monocrotaline (Chesney & Allen, 1973a).

The development with age of the ability of Wistar rats to metabolize retrosine was studied by Mattocks & White (1973). The ability to form pyrroles is very low in new-born rats, but, by 5 days of age, it is nearly as high as in adult males. This activity continues at a similar level in male rats, but, in females, it falls after the age of about 20 days until, by 60 days, it is about one-eighth that in males. Such a sex difference was not observed in mice (White et al., 1973).
4.5 Other Metabolic Routes
The actions of hepatic microsomal enzymes on pyrrolizidine alkaloids can produce other metabolites as well as pyrroles and N-oxides, but there are few reports of these. Eastman & Segall (1982) demonstrated hydroxylation of the acid moiety of senecionine by liver microsomes from female mice. Such metabolism should not prevent the subsequent conversion of the product to pyrrolic or N-oxide metabolites. The formation of other microsomal metabolites of senecionine has been reported by Segall et al. (1984).
The O-demethylation of the acid moiety of heliotrine has been demonstrated by Jago et al. (1969) and represents a partial detoxification mechanism, since the product is about half as toxic as heliotrine. Other detoxification mechanisms exist in the rumen of sheep (Dick et al., 1963; Lanigan & Smith, 1970a,b), which are, thus, particularly resistant to the effects of pyrrolizidine

alkaloids.
4.6 Metabolism of Pyrrolizidine N-Oxides
As mentioned in section 4.2, the N-oxides of pyrrolizidine alkaloids are not converted to pyrrolic metabolites by liver microsomes. It appears that their main route of metabolism in animals is reduction to the corresponding basic alkaloids, which may then be further metabolized as already described. This reduction has been shown to occur in the rat or rabbit gut (Mattocks, 1971c; Powis et al., 1979), and may be brought about by intestinal bacteria or possibly by gut enzymes. Such reduction can also be brought about by hepatic microsomal fractions (Powis et al., 1979) in the presence of NADH or of NADPH, and by sheep rumen fluid (Lanigan et al., 1970a, b).
The reduction of pyrrolizidine N-oxides in vivo is of great importance as a step in the bioactivation of these compounds (Mattocks, 1971c), as shown in section 4.2.2.
4.7 Metabolism in Man
Powis et al. (1979) found that indicine N-oxide given iv to 3 human patients as an antitumour drug was partially reduced to indicine base, detectable in the urine and plasma. Armstrong &

Zuckerman (1970) showed that human embryo liver slices, but not lung slices, converted the pyrrolizidine alkaloids lasiocarpine, retrorsine, and fulvine to pyrrolic metabolites in vitro.
5. MECHANISMS OF TOXICITY AND OTHER BIOLOGICAL ACTIONS
5.1 Metabolites Responsible for Toxicity
5.1.1 Metabolic basis of toxicity
The toxic effects of pyrrolizidine alkaloids are mediated through their toxic metabolites and not by the alkaloids themselves. The following observations are evidence for the above statement (Mattocks, 1972a):
(a) The alkaloids are chemically rather unreactive and it is hard to envisage reactions with cell constituents that they could undergo readily under physiological conditions. On the other hand, chemically prepared derivatives, similar or identical to known metabolites of these alkaloids, are highly reactive and are capable of causing toxic effects similar to those of PAs, often at dose levels much lower than those required by the alkaloids themselves.
(b) The liver is usually the main organ affected, whatever the route of administration of the alkaloid. The alkaloids are known to be metabolized in the liver.
(c) Direct application of these alkaloids to the skin does not cause local toxic effects (Schoental et al., 1954), nor do cytotoxic effects occur at sites of injection.
(d) The susceptibility of animals to the toxic actions of PAs is related to the ability of the animal to metabolize the

alkaloids. For example, the hepatic microsomal enzymes of rats less than 1 h old have very low activity towards retrorsine and these rats are relatively resistant to it, whereas rats aged several days have a high enzyme activity and are highly susceptible to the alkaloid (Mattocks & White, 1973). Guinea- pigs are very resistant to retrorsine, unless they have been given phenobarbitone, which potentiates the enzymes that metabolize it (White et al., 1973). Rats pre-treated with microsomal enzyme inhibitors, such as SKF 525A or chloramphenicol, have increased resistance to retrorsine or monocrotaline (Allen et al., 1972; Mattocks, 1973). In general, there is a good relationship between the rate of hepatic metabolism of PAs to pyrrole in vitro (Shull et al., 1976) and chronic toxicity. Highly resistant species, e.g., guinea-pigs, Japanese quail, and sheep, have a low rate of pyrrole formation, while susceptible species, such as the horse, cattle, and rat, have a high rate. Notable exceptions are the rabbit and hamster, which have high rates of pyrrole formation, but are resistant. It is possible that this may be due to changes in the balance between activation and the involvement of other factors, such as activity of detoxification. For example, sheep have a high epoxide hydrolase activity in the liver (Swick et al., 1983), which may affect PA detoxification (Cheeke & Pierson-Goeger, 1983).
5.1.2 Isolation of pyrrolic metabolites
There is plenty of evidence that many unsaturated PAs are converted into pyrrolic esters (dehydro-alkaloids) in the mammalian liver (section 4.2.3). These primary pyrrolic metabolites cannot be isolated, because of their high reactivity and rapid rate of hydrolysis. However, their more stable hydrolysis products (pyrrolic alcohols; dehydronecines) have been isolated and

identified. Thus dehydroheliotridine has been obtained from the in vitro incubation of both the heliotridine-based alkaloids, lasiocarpine and heliotrine, with rat liver microsomes (Jago et al., 1970) and dehydro-retronecine was found to be the main detectable pyrrolic metabolite in the liver, blood, and urine of rats injected with the retronecine-based alkaloid, monocrotaline (Hsu et al., 1973). There is evidence that these materials are identical, i.e., the (±)-form resulting from racemization during hydrolysis of the parent pyrrolic esters (Kedzierski & Buhler, 1985).
The results of these studies confirm that rat liver enzymes convert PAs into metabolites with known cytotoxic activity (section 5.2), and imply that these metabolites are formed via the yet more toxic and short-lived dehydro-alkaloids (Jago et al., 1970).
5.1.3 Chemical aspects of pyrrolic metabolites
5.1.3.1 Preparation
Chemical methods are available for converting unsaturated PAs into pyrrolic esters (dehydro-alkaloids), the putative primary toxic metabolites, enabling the physical, chemical, and toxicological properties of the latter to be studied.
Small amounts of dehydro-pyrrolizidine alkaloids are usually prepared by the reaction of the corresponding alkaloid N-oxides with either acetic anhydride (Mattocks, 1969; Culvenor et al., 1970a) or methanolic ferrous sulfate (Mattocks, 1969). The products must be protected from moisture and from acids, which can cause their immediate decomposition.
A variety of reagents can dehydrogenate the alkaloid bases to

pyrrolic derivatives, these include manganese dioxide (Culvenor et al., 1970a,b; Mattocks, 1969), potassium permanganate (Culvenor et al., 1970a), chloranil (Culvenor et al., 1970a), 2,3-dichloro-5,6- dicyanobenzoquinone (Mattocks, 1969), iodine (Culvenor et al., 1970b), and aryl thiols (Juneja et al., 1984). Some PAs are slowly oxidized to pyrroles by molecular oxygen (Bick et al., 1975).
The more stable pyrrolic alcohol, dehydroretronecine (Fig. 6), is prepared from retronecine using chloranil (Culvenor et al., 1970a) or aqueous potassium nitro-sodisulfonate (Mattocks, 1981c) or from retronecine N-oxide (isatinecine) using ferrous sulfate (Mattocks, 1969). The enantiomeric dehydroheliotridine can be prepared from heliotridine in similar ways. Racemic dehydro- heliotridine has been synthesized (Viscontini & Gilhof- Schaufelberger, 1971; Bohlmann et al., 1979).
5.1.3.2 Chemistry associated with toxic actions
Dehydro-pyrrolizidine alkaloids and dehydronecines (pyrrolic esters and alcohols) act chemically as alkylating (electrophilic) agents, i.e., they can react with compounds possessing electron- rich (nucleophilic) groups, such as amines, thiols, and some hydroxyl compounds. The products of alkylation consist of the "pyrrole" moiety covalently bonded to the substrate molecule. The mechanism of alkylation is illustrated in Fig. 7 (Mattocks, 1972a). An ester (R = COR) or hydroxyl group (R = H) attached to the pyrrole ring via one carbon atom (i.e., at C7 or C9) is highly reactive, being easily cleaved, leaving a positively charged

pyrrole moiety with a high affinity for electron-rich substrates. Pyrrolic esters are the most reactive, RCOO being a better "leaving group" than HO. When 2 oxygen functions are present (as illustrated), either (in turn) can act as an alkylating centre. Such bifunctional alkylation could lead to cross linking of macromolecules (Mattocks, 1969; White & Mattocks, 1972; Petry et al., 1984, 1986). When the groups (R) are the same or similar, C7 is the more reactive site. Examples of such alkylations using pure chemicals (amines or alcohol) have been given by Mattocks (1969) and Culvenor et al. (1970a). Mattocks & Bird (1983) showed that a variety of nucleophiles of biological interest could be alkylated by dehydroretronecine. Black & Jago (1970) demonstrated the in vitro alkylation of DNA by dehydroheliotridine, and Robertson (1982) and Wickramanayake et al. (1985) the alkylation of deoxyguanosine by dehydroretronecine. The alkylation of mouse or rat liver DNA by pyrrolizidine alkaloids has been shown in vivo by Eastman et al. (1982) and Candrian et al. (1985).
5.1.4 Possible further metabolites
The possibility that pyrrolic metabolites of PAs might themselves be metabolized by microsomal enzymes to further

cytotoxic derivatives was suggested by Guengerich & Mitchell (1980). These authors showed that the tritium-labelled model compounds 1,2,3-trimethylpyrrole and 1-methyl-3-4 bishydroxy- methylpyrrole could be metabolized in rats or by rat liver microsomes to unidentified derivatives able to bind covalently to proteins and nucleic acids. It is possible that liver damage, seen in some rats given iv injections of pneumotoxic pyrrolic esters, might have been due to metabolites of the latter formed in the liver (Mattocks & Driver, 1983). Segall et al. (1985) have identified trans-4-hydroxy-2-hexenal in an in vitro mouse liver microsomal system metabolizing the PA senecionine and suggested that it might have been formed from the alkaloid via a pyrrolic intermediate. The compound is capable of causing liver damage and might contribute to the acute hepatotoxicity of senecionine and other alkaloids. However, this has not been proved, and the highly reactive and toxic primary pyrrolic metabolites from PAs are themselves capable of causing the known hepatotoxic effects of these alkaloids.
5.2 Toxic Actions of Pyrrolic Metabolites
Pyrrolic derivatives prepared chemically from PAs, as well as some analogous compounds, have been tested in experimental animals and in vitro systems, and shown to have a variety of toxic actions.
5.2.1 Animals
5.2.1.1 Pyrrolic esters (dehydro-alkaloids)

Dehydro-pyrrolizidine alkaloids are very reactive and their effects in vivo are largely confined to the first tissues they encounter. When given orally to rats, they are destroyed almost immediately in the aqueous acid of the stomach and show no toxic action. When given ip, they cause severe local irritation and peritonitis (Mattocks, 1968a; Butler et al., 1970); subcutaneous injection leads to skin lesions (Hooson & Grasso, 1976). After iv injection of pyrroles, such as dehydromonocrotaline (monocrotaline pyrrole), into the tail veins of rats, the toxic injuries appear principally in the lungs. Depending on the dose, these include vascular lesions and pulmonary oedema (Plestina & Stoner, 1972); a progressive alveolar proliferation similar to that produced by very much larger doses of the parent alkaloid (Butler et al., 1970) and pulmonary hypertension (Hilliker et al., 1983). Dehydromonocrotaline does not require further metabolism to express its pneumotoxicity, and it is rapidly rendered inactive after exposure to aqueous media (Bruner et al., 1986). Similar pneumotoxicity is produced by totally synthetic pyrrolic esters having a simpler structure but the same type of chemical reactivity as the alkaloid derivatives (Mattocks & Driver, 1983), thus confirming the chemical mechanism of this action.
Injections of dehydro-pyrrolizidine alkaloids or synthetic analogues into mesenteric veins of rats lead to liver damage after smaller doses than the alkaloids themselves (Butler et al., 1970; Shumaker et al., 1976). The liver damage differs somewhat from the alkaloid damage, consistent with the toxin being introduced via the hepatic vascular system rather than being produced within the hepatocytes, as is the case with the alkaloids. Nevertheless, the progressive liver lesions are very similar to those produced by PAs (Butler et al., 1970). The lung damage after tail vein injections bears a closer resemblance to pyrrolizidine damage, since the

latter is also believed to be caused by metabolites entering the lungs via the bloodstream (Barnes et al., 1964).
5.2.1.2 Pyrrolic alcohols (dehydro-necines)
Dehydroheliotridine (Fig. 6), a secondary pyrrolic metabolite from heliotridine-based PAs, such as heliotrine and lasiocarpine, is less acutely toxic than its parent alkaloids; it has an LD50
(7 days) of about 250 mg/kg body weight in mice (Percy & Pierce, 1971). Its effects on 14-day-old rats were studied by Peterson et al. (1972). All rats given ip doses of 0.4 mmol/kg body weight survived, but a dose of 0.6 mmol/kg killed most animals within 10 days. Toxic effects were mainly found in rapidly developing tissues. In young rats, it caused fur loss, tooth defects, and atrophy of hair follicles, gut mucosa, spleen, thymus, testis, and bone marrow. The lungs were not affected. Pathological effects in the liver were confined to necrosis of isolated cells and antimitotic action, which was manifested as a mild megalocytosis (development of giant hepatocytes) in rats surviving 4 weeks or more. The persistent antimitotic action of dehydroheliotridine and of its parent alkaloid lasiocarpine in the liver of rats was investigated by Samuel & Jago (1975), who located the mitotic block as being either late in the DNA synthetic (S) phase or early in the post synthetic (G2) phase of the cell cycle.
Dehydroheliotridine is also carcinogenic. Peterson et al. (1983) showed that rats given 9 ip injections of this compound (60 - 76.5 mg/kg body weight) over 23 weeks had a shorter life span and suffered a significantly higher incidence of tumours than control rats. The authors concluded that dehydroheliotridine is responsible for some, or possibly all, of the carcinogenicity of its parent alkaloids.
Dehydroheliotridine was found to be teratogenic when given ip

to female hooded rats on the 14th day of pregnancy. A dose of 40 mg/kg body weight produced effects similar to those produced by the alkaloid heliotrine at a dose of 200 mg/kg (Peterson & Jago, 1980). For the immunosuppressant activity of this compound, see section 6.4.10.
The toxic actions of dehydroretronecine (DHR) (Fig. 7) when given sc to rats are similar to those of dehydroheliotridine (Hsu et al., 1973; Shumaker et al., 1976). Repeated large doses also caused ulceration of the glandular stomach. Daily sc doses (4 mg/kg body weight), administered to rats for 1 week, caused lung
damage leading to right ventricular hypertrophy (Huxtable et al., 1978). DHR was carcinogenic when applied repeatedly to mouse skin (Johnson et al., 1978; Mattocks & Cabral, 1982).
5.2.2 Cell cultures
Dehydroheliotridine and dehydrosupinidine both have an inhibitory action in cultures of KB cells (human epidermoid carcinoma of the nasopharynx) with ED50
concentrations of 10-4
mol and 10-5
mol, respectively (Culvernor et al., 1969).
Bick & Culvenor (1971) found dehydroheliotridine (DHR) to be considerably more effective than the alkaloid heliotrine in suppressing cell division and causing chromosome breaks, in cultures of leukocytes from the marsupial

Potorus tridactylus; at a concentration of 6 x 10-5
mol, the mitotic index was zero, and more than half the cells had disintegrated. In a study by Mattocks & Legg (1980), dehydroretronecine and several synthetic analogues completely inhibited cell division in a cultured rat liver cell line at a concentration of 10-4
mol. Ord et al., (1985) found that DHR induced sister chromatid exchange in human lymphocytes without the need for metabolic activation. Analogous pyrroles with only one functional (reactive) group were much less effective. DHR was also weakly active in inducing mutations in the Salmonella
typhimurium base substitution strain, TA92, and gave positive results in an in vitro cell transformation test using a culture derived from hamster kidney cells (Styles et al., 1980).
The toxicity of the pyrrolic ester, dehydromonocrotaline, for cultures of mouse fibroblasts was studied in vitro by Johnson (1981). The level of exposure was approximately 1 ng per cell. Cell death was preceded, first by the swelling and disruption of organelles, including mitochondria, and then by the rupture of plasma membranes with the release of cell components.
Bick et al., (1975) investigated whether the effects of PAs on leukocyte cultures of Potorus tridactylus were due to pyrrolic metabolites. Levels of dihydropyrrolizines, which could be

demonstrated in the culture media, were insufficient to account for the observed effects of heliotrine, lasiocarpine, and monocrotaline on the cells, but the actual amounts formed within the cells may have been higher than those observed.
5.2.3 Possible participation of membrane lipid peroxidation
Distinct increases in NADPH- and ascorbate-dependent peroxidation of microsomal membrane lipids were found in rats given heliotrine subcutaneously (300 mg/kg body weight) (Savin 1983). The primary biochemical interactions and cellular macromolecular targets for the pathogenesis of PA-induced toxicity remain unidentified.
5.3 Chemical and Metabolic Factors Affecting Toxicity
The toxicity of an alkaloid depends on the extent to which it is converted into active metabolites and on the disposition and reactivity of these metabolites, once formed.
5.3.1 Structural features of a toxic alkaloid
The essential structural features of a hepatotoxic PA (Fig. 8) are:
(a) a 1-hydroxymethylpyrrolizidine ring system unsaturated in the 1:2-position, with preferably a second hydroxyl group in the 7-position;
(b) esterification of at least one of the hydroxyls, though toxicity is much greater when both hydroxyls are esterified; and
(c) ester groups that are resistant to enzymic hydrolysis, which usually means that there is a high degree of chain branching in the acid moiety.

The above requirements apply to natural PAs but, strictly speaking, only the right hand (pyrroline) ring is essential, being the ring that is metabolized to a pyrrole derivative. Thus, esters of 2,3-bis-hydroxymethyl-1-methyl-3-pyrroline (synthanecine A) (Fig. 9) have pyrrolizidine-like hepatotoxicity (Mattocks, 1971a; Driver & Mattocks, 1984).
Structural requirements for N -oxides are the same as those for the hepatotoxic alkaloids. However, it is important to note that a PA N -oxide is not hepatotoxic itself; toxicity depends on it being reduced to the corresponding basic alkaloid, chiefly in the gut (Mattocks 1971c), but possibly in other organs, such as the liver (Powis et al., 1979).
5.3.2 Activation and detoxication
Factors affecting the proportion of an ingested alkaloid that is converted into toxic metabolites in an animal include the

following:
(a) Lipid solubility
Highly water-soluble alkaloids (such as indicine) are easily excreted and have low toxicity. Alkaloids that are more lipophilic are more open to activation by liver microsomes (Mattocks, 1981a).
(b) Subceptibility to hydrolysis
This is determined by the molecular structure and conformation of the alkaloid (Mattocks, 1981a,b). If the alkaloid is open to esterase attack, it may be largely detoxified by hydrolysis.
(c) Susceptibility to N-oxidation
The relative amounts of an alkaloid converted by hepatic microsomal enzymes to N-oxide and to pyrrolic metabolites depends on its molecular structure and conformation (Mattocks & Bird, 1983). N-oxidation represents a detoxication pathway (Mattocks, 1972b).
5.3.3 Factors affecting the toxicity of active metabolites
5.3.3.1 Reactivity of the metabolite
Toxic metabolites are formed in liver cells. Primary pyrrolic metabolites (dehydro-alkaloids) are very reactive and, thus, are

quickly hydrolysed or deactivated by reaction with cell constituents. To damage tissues other than the cells in which they are formed, active metabolites must cross the cell membrane and survive while being transported in the bloodstream. The more stable pyrrolic metabolites, such as dehydromonocrotaline from the alkaloids monocrotaline, are able to reach, and become bound to, lung tissue (Mattocks, 1973). Thus, monocrotaline frequently damages the lungs, whereas retrorsine, which yields a more reactive pyrrolic metabolite, normally does not.
Secondary metabolites (pyrrolic alcohols, e.g., dehydroretronecine), formed by the hydrolysis of primary pyrrolic metabolites, are water soluble, relatively stable compounds that can become more widely distributed throughout the body or excreted; these are not acutely toxic.
5.3.3.2 The number of reactive groups
The toxicity of a pyrrolic alkylating agent is affected by the number of reactive ester or hydroxyl groups (1 or 2) present as the following examples show:
(a) Many pyrrolic esters can cause acute lung damage when given iv to rats, but only bifunctional ones also cause delayed effects on the lungs (Mattocks & Driver, 1983).
(b) Bifunctional pyrrolic alcohols are more effective inhibitors of mitosis in cultured cells than monofunctional pyrroles (Mattocks & Legg, 1980).
(c) Bifunctional pyrrolic alcohols are much better inducers of sister chromatid exchange (SCE) in lymphocytes than monoalcohols (Ord et al., 1985).
Reasons for these differences might be that the bifunctional pyrroles are able to crosslink macromolecules or simply that they

can bind more strongly to target molecules.
5.4 Metabolites Associated with the Biological Actions of Pyrrolizidine Alkaloids
5.4.1 Acute hepatotoxicity
The following is good evidence that acute liver necrosis is caused by primary pyrrolic ester metabolites (dehydro-alkaloids):
(a) The liver, in which these metabolites are formed, is the only organ exposed to them in relatively high concentrations.
(b) There are good correlations between amounts of pyrroles bound to liver tissue and acute hepatotoxicity (Mattocks, 1973).
(c) Pyrrolic alcohols are not acutely hepatotoxic, even when given to animals in very large amounts.
(d) Pyrrolic esters injected iv into the liver are much more acutely hepatotoxic than the parent alkaloids (Butler et al., 1970).
It is possible that other metabolites, such as 4-hydroxy 2,3-unsaturated aldehydes, might also contribute to the acute hepatotoxicity of some PAs (Segall et al., 1985). However, this has still to be confirmed.
5.4.2 Chronic hepatotoxicity
The persistent antimitotic action on the liver that leads to the formation of giant hepatocytes can be produced both by pyrrolic ester metabolites, such as dehydromonocrotaline (Hsu et al., 1973), and by pyrrolic alcohols, such as dehydroheliotridine (Peterson et al., 1972). Both kinds of metabolites can lead to similar alkylation products and both are likely to be present in the liver

when the alkaloids are metabolized. Thus, either could be responsible for chronic hepatotoxic effects. However, the antimitotic action alone is not sufficient. It must be accompanied or followed by a stimulus of cell division. This may be provided by the acute necrotic effect of primary pyrrolic metabolites or by any other cause of acute liver injury that leads to tissue regeneration. In very young animals, the stimulus can be the enhanced rate of replication that already exists in them.
5.4.3 Pneumotoxicity
Characteristic pyrrolizidine lung damage is produced by iv injections of pyrrolic ester metabolites, which are effective at much lower doses than the parent alkaloids. The latter are not metabolized in lung tissue; thus, lung damage from PAs is believed to be due to pyrrolic esters reaching the lungs from the liver (Butler et al., 1970). Chronic lung damage appears to be caused by bifunctional rather than by monofunctional pyrrolic alkylating agents (Mattocks & Driver, 1983) (section 5.3.3.2).
There is some evidence that pyrrolic alcohol metabolites might also be able to contribute to chronic (but not acute) pneumotoxicity (Huxtable et al., 1978).
5.4.4 Toxicity in other tissues
Chronic heart damage including right ventricular hypertrophy is a consequence of pyrrolizidine lung damage (pulmonary hypertension) (Hayashi et al., 1967). Brain damage is attributed to ammonia intoxication secondary to severe pyrrolizidine liver injury (Hooper, 1972). This view has been contested and some PAs are known to have direct effects on the central nervous system (section 6.4.3). There is no evidence that PAs are appreciably metabolized in tissues other than the liver. Thus, damage to other organs is probably due to metabolites transported from the liver. For example, in the relatively uncommon cases of chronic kidney damage

after pyrrolizidine intoxication (Hooper, 1974; Hooper & Scanlan, 1977) megalocytosis in this organ suggests that pyrrolic metabolites (either ester or alcohol) are involved. Overall, patterns of disease, as observed in extrahepatic sites, are
consistent with a "spillover" effect of the pyrroles produced in the liver (Hooper, 1978). Toxicity of an alkaloid reflects its rate of metabolism to a pyrrole (Tuchweber et al., 1974) and so the spillover effect is likely to be more evident at higher doses. Studies of Culvenor et al. (1976a) suggest that the PAs that are hepatotoxic for rats should also be pneumotoxic when administered at higher doses. In acute poisoning, the hepatotoxic effects could outweigh the pneumotoxic effects or those on other organs, to such a degree that the latter are not manifested. Variation in expression of disease (primarily hepatic or extrahepatic) also depends on the reactions of host tissues in different species of animals, in addition to the quantities of the pyrroles (Hooper, 1978). The sensitivity of the blood vessels might explain severe interstitial pneumonias in some animals, or severe nephroses in pigs (McGrath et al., 1975).
5.4.5 Carcinogenicity
The pyrrolic alcohols dehydroretronecine and dehydroheliotridine are known carcinogens (Johnson et al., 1978; Peterson et al., 1983), whereas the pyrrolic esters dehydromonocrotaline and dehydroretrorsine are only carcinogenic in conjunction with a tumour promotor (Mattocks & Cabral, 1979, 1982). This suggests that the more persistent secondary metabolites (pyrrolic alcohols) might account for the rather weak carcinogenicity of some PAs.
5.4.6 Antitumour activity
Some PAs and their

N-oxides are active as tumour inhibitors in test systems (Culvenor, 1968; Suffness & Cordell, 1985). Indicine N-oxide, in particular, showed high activity against B16 melanoma, mammary xenograft, M5076 sarcoma, P388 leukaemia, and Walker 256 carcinoma. In clinical studies, indicine N-oxide has shown significant activity against some forms of leukaemia, with dosage limited mainly by myelosuppression and sometimes by hepatotoxicity. It is tempting to suppose that this action is related to the powerful antimitotic action of their pyrrolic metabolites, even though some of these alkaloids and derived pyrroles are themselves carcinogenic. On the other hand, there is evidence suggesting that indicine N-oxide owes it activity to a property of the compound itself rather than to the pyrrolic metabolites, which could be formed through reduction to indicine (Powis et al., 1979). The evidence, that indicine is less effective than indicine N-oxide, is not conclusive and other structure-activity data (Milkowsky, 1985) point to a need for a structural capability to form a pyrrolic metabolite. It is also possible that indicine N-oxide is converted directly to dehydroindicine by mitochondrial enzymes in liver or tumour cells, since the type of reaction required has been observed in the mitochondrial metabolism of the N-oxides of tryptamine alkaloids and certain methylated amino acids (Fish et al., 1956; Smith et al., 1962).

5.5 Prevention and Treatment of Pyrrolizidine Poisoning
There is no known way to prevent pyrrolizidine liver damage, once a hepatotoxic dose of the alkaloid has been ingested. A number of dietary regimes have been found to partially protect animals (chiefly rodents) from the acute effects of subsequent alkaloids ingestion. None of these are of any practical use for preventing pyrrolizidine intoxication in livestock. Furthermore, chronic toxic effects in the liver or in other organs are sometimes more severe in animals receiving higher doses of alkaloids after being protected against acute hepatotoxicty.
5.5.1 Modified diets
The mechanism of action of modified diets is not clear, but they may be associated with the decreased metabolic activation of the alkaloids. Some examples follow:
(a) A protein-rich diet can give some protection to rats against Senecio jacobaea alkaloids (Cheeke & Gorman, 1974). Rats fed a high casein diet survived longer than rats given a normal diet, when poisoned with retrorsine or riddelline, but the survivors were more liable to develop liver tumours (Schoental & Head, 1957). However, whether this was simply due to a prolongation of life of the animals by the diet is open to question.
(b) Male rats previously fed a sucrose-only diet for 4 days were considerably protected against the acute hepatotoxicity of retrorsine (LD50
120 mg/kg body weight compared with 34 mg/kg

in normal rats). However, lung damage, rare in control rats, was frequently seen in "protected" rats given high doses of retrorsine (Mattocks, 1973).
(c) Restriction of feed intake to 40% of normal attenuated the increase in lung weight and lavage protein concentration in cell-free bronchopulmonary lavage fluid and abolished the right ventricular hypertrophy in monocrotaline-treated rats. Furthermore, the percentage of diet-restricted animals that survived was significantly higher than that in animals that had eaten ad libitum up to day 28, but, from this time onwards, there was no difference. Alterations of dietary sodium intake alone did not affect the results of monocrotaline-induced toxicity (Ganey et al., 1985).
5.5.2 Pretreatment to enhance the detoxication of active metabolites
Treatments that have afforded some protection against pyrrolizidine hepatotoxicity (probably by increasing the liver level of sulfydryl compounds, which are known to react with pyrrolic metabolites) (White, 1976) include the following:
(a) Pre-treatment of rats with mercaptoethylamine (150 mg/kg body weight ip) partially protected rats against the acute hepatotoxicity of monocrotaline given 15 min later (Hayashi & Lalich, 1968); it gave no protection when administered 2 h after the alkaloid. Mercaptoethylamine, when given orally (300 mg/kg body weight) at the same time as the lasiocarpine, also increased the resistance of rats to the alkaloid (Rogers & Newberne, 1971).
(b) Cysteine (1% in the diet) partially protected rats against Senecio jacobaea alkaloids (Buckmaster et al., 1977) and mice

against monocrotaline (Miranda et al., 1981c).
(c) The antioxidant ethoxyquin fed at a level of 2.5 g/kg diet to female mice for 38 days, increased the liver thiol concentration and raised the acute LD50
of monocrotaline, given ip on the 10th day, to 364 mg/kg compared with 243 mg/kg in control mice (Miranda et al., 1981a).
(d) Rats or mice also had increased resistance to acute pyrrolizidine hepatotoxicity when fed the antioxidant butylated hydroxyanisole (BHA) (up to 7.5 g/kg diet) (Miranda et al., 1981c, 1982a,b; Kim & Jones, 1982).
(e) Heliotrine-induced toxicity can be modified by the co-administration of cupir (a copper-containing complex) at a level of 1 mg/kg per day for 20 days. It prevented the exit of hepatic cytosolic enzymes into the blood and improved all the energy reactions studied in the mitochondria of heliotrine- intoxicated rats (Yuldasheva & Sultanova, 1983). Inhibition of lipid peroxidation by cytoplasmic copper was shown later (Wittig & Stephen, 1964). Savin (1983) found that lethality to rats of heliotrine (300 mg/kg sc) was completely prevented by co-administration of alpha-tocopherol (6 ml/kg ip).
(f) Rats pre-treated with ip doses of zinc chloride (72 µmol/kg body weight) had increased resistance to the hepatotoxicity of Senecio jacobaea alkaloids, as assessed by histology and enzyme measurements (Miranda et al., 1982c). The zinc treatment increased the liver level of metallothionein, a sulfhydryl-rich protein that might react with pyrrolic metabolites.

Metabolic inhibitors of the microsomal P450 mixed-function oxidase system, SKF 525A, metyrapone, and allylisopropyl acetamide, which inhibit the formation of toxic pyrroles in the liver, have been tried successfully in the prevention of the toxic effects of monocrotaline in rats (Eisenstein & Huxtable, 1979). The use of P450 inhibitors was stated to show "potential therapeutic promise". However, this would seem impracticable considering that, at least in the rat, PAs undergo a high rate of metabolism commencing a few minutes after ingestion (Mattocks, 1972b). In some instances, they have been known to lead to an increase in toxicity, e.g., with lasiocarpine as reported by Tuchweber et al. (1974).
5.5.3 Other treatments
Lanigan & Whittem (1970) attempted, unsuccessfully, to protect sheep against Heliotropium europaeum poisoning by treating them with cobalt, in the hope that this would enhance the vitamin B12
-mediated detoxication of the alkaloids in the rumen (Dick et al., 1963).
Lanigan et al. (1978) found that the resistance of sheep to dietary Heliotropium europaeum was increased by giving them large daily doses of the antimethanogenic drug, iodoform. However, Swick et al., (1983) found that Senecio jacobaea alkaloids were not detoxified by incubation for 48 h with sheep rumen fluid in vitro.

6. EFFECTS ON ANIMALS
6.1 Patterns of Disease Caused by Different Plant Genera and of Organ Involvement in Different Species
The most important genera of PA-containing plants listed in section 3.1 are all hepatotoxic. Among these, Crotalaria spp. cause damage in the broadest range of tissues in most domestic species. In pigs, they are known to be severely nephrotoxic (Peckham et al., 1974; McGrath et al., 1975; Hooper & Scanlan, 1977). Some species are known to be pneumotoxic for horses (Watt & Breyer-Brandwijk, 1962; Gardiner et al., 1965), cattle (Sanders et al., 1936; Berry & Bras, 1957), sheep (Laws, 1968), and pigs (Peckham et al., 1974; Hooper & Scanlan, 1977), as well as hepatotoxic.
Although several Crotalaria spp. are known to be pneumotoxic for horses (Gardiner et al., 1965), C. retusa is an exception. It is an important cause of disease in horses in northern Australia (Hooper, 1978) and has been shown to be pneumotoxic for pigs in the same area (Hooper &

Scanlan, 1977); yet it produces only hepatic disease in horses (Rose et al., 1957a,b).
Similarly, Senecio spp. are primarily hepatotoxic, but S. jacobaea has been demonstrated to be pneumotoxic for pigs (Harding et al., 1964), though it could probably be an inconsistent change (Bull et al., 1968). This plant is also known to cause pulmonary disease in rats and mice (Hooper, 1974). However, there are no reports of its affecting the lungs in cattle, sheep, horses, or chicken. Renal megalocytosis and mild nephrosis are reported in most species poisoned with S. jacobaea (Harding et al., 1964; Bull et al., 1968). Heliotropium spp., Amsinckia spp., and Echium spp. are all mainly hepatotoxic.
Roitman (1983) summarized the pattern of organ involvement observed in man and different species of farm and experimental animals affected by pyrrolizidine alkaloids (Table 8). Even within a single species, the nature of a toxic effect, as well as the organ affected, can be altered by changing the dose rate and duration.
6.2 Field Observations - Outbreaks in Farm Animals
The veterinary problem of PA toxicity has been reviewed by Bull et al. (1968) and McLean (1970). Mattocks (1986) listed the cases of livestock poisoning and feeding trials since 1968, and cited relevant literature. Peterson &

Culvenor (1983) produced a useful and comprehensive table of the plant species known or suspected of causing natural outbreaks of poisoning in each animal species. The influence of factors such as species, age, sex, and diet, on toxicity is also reviewed in the same paper.
Table 8. Animal species and organs affected by pyrrolizidine alkaloidsa
--------------------------------------------------------------- Species Liver Lung Kidney Heart Pancreas Gastric Muscle mucosa
--------------------------------------------------------------- Man + Monkey + + + + Horse + + + Pig + + +
Home | produtos e serviços | informação médica | ordens | nutrition modular Problemas no alimento
Toxins Naturais Pesquisa De Environmed
Toxins De Alimento Nativos
Aditivos De Alimento
Contaminadores Químicos

Infecção Carregada Alimento
Sensibilidade Química
Soluções Alfa ENF
Feriado Do Alimento
Programas Do Nutrition Do Alfa
Muitas substâncias naturais do non-nutriente " em alimentos " naturais comportam-se mal. Nós podemos pensar de sua atividade como quimicamente stressful. Os stressors químicos ou molecular aumentam o caos e diminuem a função em ordem, sensible do corpo. Um stressor químico é um pouco de absurdo, uma substância com nenhum papel significativo ao jogo ou um papel tóxico que seja destrutivo. Nesta revisão, um perfil breve dos stressors e os toxins são desenvolvidos mais para ilustrar o papel de stressors químicos naturais do que para esgotar este assunto vasto e complicado.
Stephen Gislason MD
O alimento está cheio das substâncias extra que devem ser processadas no corpo e então excreted, mas que não tenha nenhum valor nutriente e possa ter a droga e efeitos tóxicos. Os stressors químicos podem ser encontrados nos alimentos como ingredientes nativos e não são necessariamente aditivos ou contaminadores. Estas substâncias têm o chemistry complexo; incluem sais inorgánicos e orgânicos, minerais tóxicos, álcoois, aldeídos, alcalóides,

compostos polyphenolic, salicylates, aminos-ácido do non-nutriente e peptides.
Um stressor do produto químico de alimento torna-se frankly tóxico quando ingested demasiado frequentemente, em um dose demasiado grande, ou quando ingested por uma pessoa que falte a maquinaria metabolic para detoxify a. Os stressors molecular podem simplesmente reque o corpo segurá-lo e excrete sem dano. O trabalho metabolic desta atividade é " o custo " de ingesting estas substâncias. Quando nós consumimos stressors no excesso, o efeito torna-se tóxico. Nossa capacidade segurar stressors químicos é limitada e varia de individual ao indivíduo.
A sensibilidade " química " é relatada quando a exposição aos produtos químicos transportados por via aérea, tais como o fumo do cigarro, a exaustão do motor, os perfumes, os detergentes da casa e os solventes causa sintomas. Os pacientes que estão recuperando das doenças allergic do alimento relatarão frequentemente a consciência aumentada de produtos químicos transportados por via aérea; isto parece representar um hypersensitivity generalizado. A parte da estratégia da recuperação deve evitar a exposição química tanto quanto possível.
Muitos stressors químicos competem para os mesmos pathways metabolic para o excretion. O fígado é responsável para remover muitos toxins. Um de seus métodos deve unir grupos do acetyl às moléculas tóxicas que as fazem mais soluble para o excretion do kidney. Alguns povos são " acetylators pobres " e relatam a intolerance ao escalas largas das drogas, dos alimentos, e de produtos químicos transportados por via aérea. Se muitas moléculas competirem para os mesmos pathways do excretion, é fácil imaginar que sobrecarregar ocorre regularmente nos cidadãos modernos que são expostos a uma escala larga dos produtos químicos no alimento, na água e no ar. Uma vez que a sobrecarga ocorre, mesmo as quantidades pequenas de stressors químicos extra transformam-se sintomas e dysfunction tóxicos e do produto.
Os stressors químicos inhaled e ingested simultaneamente. A poluição de ar reduzirá a tolerância para a poluição do alimento e o versa do visto. Um jantar típico em um restaurante agradável pode ser biochemically e metabolically stressful, como fumo inhaled e álcool, café, chá, spices, e liga ingested do açúcar com o complexo já no alimento. Você pode ser em mais problema se você estiver lidando também com os efeitos das drogas da prescrição feitas exame para aliviar seus sintomas da sobrecarga química!

As plantas são nossas fontes mais desejáveis do alimento. Nós tendemos a pensar os vegetais do e a fruta como inteiramente amigável e benigna; entretanto isto não é necessariamente verdadeiro. Os vegetais, as frutas, os herbs, e os spices são quimicamente complicados, e contêm uma variedade das substâncias químicas que seja droga-como, tóxica, e allergenic. As plantas são inerente tóxicas desde que evoluíram as defesas químicas que desanimam predadores de os comer.
Há muitas substâncias molecular nos alimentos que não oferecem nenhum benefício nutritivo, e deve ser processado e excreted. O ácido oxalic, para o exemplo, excreted no urine, e seus cristais são encontrados geralmente no urinalysis microscópico. Demasiado ácido oxalic no urine resultará em pedras do kidney ou da bexiga. O cálcio combina com o ácido oxalic para dar forma ao sal mais menos soluble, o oxalate do cálcio, que é encontrado também em pedras do kidney. A planta sae, especial rhubarb, repolho, spinach, e os altos de beterraba, contêm o ácido oxalic. O ácido oxalic é encontrado também as batatas e as ervilhas. A vitamina C metabolized ao ácido oxalic; contribui ao sobre-over-saturation do urine com cristais e para apedrejar possivelmente a formação.
Diversos produtos químicos usados como aditivos de alimento são encontrados também naturalmente em muitos alimentos. Os nitrates e os nitrites são ubiquitous nas plantas. Dão forma à parte do chemistry essencial os solos e as plantas. Como cada gardener sabe, o nitrogênio é essencial para o crescimento de planta; os fertilizantes do nitrogênio, contendo nitrates, são os produtos químicos agriculturais os mais abundantes. As beterrabas, os radishes, o spinach, e a alface contêm os níveis os mais elevados dos nitrates. O consumo diário é estimado para estar na escala de 100 mg/day.
Substâncias Aromatic
Todas as plantas contêm as moléculas que têm e impacto em nossos sentidos químicos. Provam e cheiram. Os compostos aromáticos atraem-nos a um alimento. A indústria de alimento usa quantidades grandes dos compostos aromáticos, das fontes naturais e sintéticas. O chemistry destas substâncias pode ser imaginado pensando da forma de sua estrutura molecular. O anel básico é uma molécula six-sided, benzene. As correntes laterais diferentes unidas ao anel mudam seus cor, gosto, e cheiro. Se os anéis do benzene forem ligados junto, uma variedade de estruturas do anel causa classes

diferentes das substâncias. As moléculas baseadas na estrutura do anel do benzene são comuns na natureza. Um nome ocasional para muitas das substâncias é " phenolics ".
Com estruturas aromatic do anel inclua moléculas das drogas do alimento a cafeína e ácido salicylic; a cânfora dos sabores, o ácido cinnamic, o eugenol (nutmeg e cravos-da-índia), o safrole, o anethole (anis), o tannin (chá), o ácido gallic, e o vanillin (vanilla); e as vitaminas ácido ascorbic e niacin. Os compostos phenolic estão droga-como; alguns são tóxicos e outros parecem ser benéficos.
Um outro grupo de produtos químicos interessantes da planta é os óleos essenciais das plantas, que são também aromatic. Nós tendemos como ao cheiro dos turpenes e usamo-los em nossos perfumes, garnishes, e chás. Os turpenes aromatic comuns em plantas do alimento incluem turpenes complexos, tais como o lanosterol (no lanolin), um segundo primo do cholesterol, e o squalene, encontrou no fermento, no germe do trigo, e no óleo verde-oliva. Alguns turpenes são frankly tóxicos mas remanescem em nossa fonte de alimento como sabores. Os extratos do álcool das plantas contêm frequentemente turpenes tóxicos. Outros turpenes são moléculas ou ao menos benignos positivo. A vitamina A não é uma substância; é uma família de turpenes relacionados com atividade biológica compartilhada. O beta-carotene é o pigment amarelo nas cenouras que podem ser convertidas na vitamina ativa A (Retinols) depois que nós ingest a. O lycopene é um turpene vermelho similar encontrado nos tomates. O consumo excessivo os tomates e as cenouras de pode induzir um complexion colorido que alarme amigos mas do parece fazer pouco dano e do pode ter benefícios tais como a prevenção do cancer.
Garlic e cebolas
O garlic (alium sativum) e as cebolas (allium Cepa) são ambos os membros da família do lírio. Ambos contêm as substâncias aromatic fortes que nós usamos como flavoring para o alimento. As propriedades medicinal destes alimentos foram usadas por séculos. Allicin é o princípio aromatic do garlic. Allicin é um turpene sulphur-containing. Um bulbo intato do garlic tem pouco odor. O odor forte do allicin aparece somente depois que um bulbo do garlic é cortado ou esmagado. O trauma da exposição do ar excita um enzyme qual mude uma molécula inodora do precursor, Alliin, a Allicin.

As propriedades medicinal dos produtos químicos combinados no garlic incluem a atividade antiseptic e (uma atividade anticlotting da platelet-viscosidade reduzida). O fator anticlotting é o ajoene (4,5,9-trithiadodeca-1,6,11-triene 9-oxide!). Os entusiastas do garlic devem anotar que nenhum ajoene, ou a atividade anticlotting, estiveram encontrados em preparações proprietárias do garlic, including o óleo do garlic ou as tabuletas do garlic. O ingestion do garlic recentemente esmagado parece necessário para este efeito de droga. O efeito anticlotting do ajoene é similar ao efeito da aspirina (ASA). Os respiração-odores desagradáveis que seguem o ingestion do garlic são os metabolites sulphur-containing temporários do allicin.
As cebolas são famosas para seu efeito rasgando. A substância que nos faz weep é s-oxide propanethial. O efeito rasgando pode ser reduzido chilling a cebola antes do corte, ou processando a cebola sob a água funcionando da torneira.
Spices
Todas as plantas contêm uma variedade das substâncias químicas que agem em a droga-como a maneira. Os herbs e os spices tendem a ser quimicamente complexos e droga-como em sua atividade. A maioria de culturas nativas são relutantes comer plantas non-food para a razão boa. Nós começamos provavelmente afastado com ingestion do herb e do spice somente em doses pequenos e infrequëntemente. O nutmeg, um spice comum favorecido nos desserts e bebidas, rende muitos produtos químicos tais como o eugenol, isoeugenol, safrole, myristicin, elemicin, limolene. O uso therapeutic do eugenol como um antidiarreico, anticlotting, e agente anti-inflammatory podia ser sugerido; entretanto, se nós devêssemos usar o óleo do nutmeg como a terapia, nós enfrentaríamos a droga, e efeitos tóxicos dos outros produtos químicos no nutmeg. Nós seríamos concernidos sobre os efeitos hallucinogenic do psychotropic, myristicin. Nós seríamos concernidos também sobre a abilidade do safrole de induzir o cancer do fígado nos ratos. A aproximação pharmaceutical apropriada deveria isolar a substância medicinal, eugenol, e decidir-se, após testar cuidadoso de seu efficacy contra seu toxicity, se o uso therapeutic fosse sábio.
As ervilhas e os feijões são legumes edible comuns. Estes alimentos são grampos *** TRANSLATION ENDS HERE ***world-wide and have desirable nutritional properties, but several interesting biochemical problems may arise with their use. Soybeans, for example, may be poorly tolerated

unless well prepared. The process of making tofu illustrates the effort required to process a raw food to obtain a reliable nutrient source. Soybeans contain indigestible carbohydrates and inhibitors of digestive enzymes (soybean trypsin inhibitor). Both problems contribute to difficulty digesting beans, excessive gas, and, occasionally, abdominal pain and diarrhea.
Lima and kidney beans are toxic uncooked. Both beans contain cyanide producing compounds ( cyanogenic glycosides ), which can be destroyed by adequate cooking. Small amounts of cyanogenic glycosides will be detoxified by the liver. Cyanogenic glycosides are also found in fruit pits millet, sprouts, yams, maize, chick peas, and cassava root.
Cassava (manioc) is an important vegetable of South East Asia, Africa and South America and is inherently toxic. These tuber contain linamarin which can be converted to hydrocyanic acid . They must be processed by soaking, boiling, drying, and fermentation to reduce toxic cyanide effects. Neurological disorders and thyroid enlargement occur in African peoples who eat large amounts of inadequately processed cassava.
An unusual genetic condition, "favism ", makes some people sensitive to vicine, a nucleotide in fave beans; these people develop red blood cell damage (hemolytic anemia) after eating the beans. Cooking the beans thoroughly can reduce this effect. This is a specific example of the cytotoxic mechanism of food molecules, and illustrates the advantages of cooking foods.
Nightshades
Plants of the nightshade family contain toxic substances. "Deadly " refers to the toxicity of the leaves of this plant group which includes tomato, potato, peppers, eggplant, and tobacco. All the nightshades contain nicotine; tobacco has the highest concentration, eggplant is next. Country lore tells us not to eat green potatoes and to store potatoes in the dark to avoid the greening effect of light. The green potato contains toxic compounds (glycoalkaloids) similar to those found in the leaves. Solanine poisoning from green potatoes will produce throat burning, weakness, diarrhea, and even convulsions with collapse and coma. Adverse and allergic reactions to tomatoes and peppers are common, and these vegetables are not on our most favored food list. Nightshades have often been implicated in arthritis.

The occurrence of toxic non-nutrient amino acids is not unusual in plants who produce toxic compounds to deter predators from eating them. Over a 150 non-protein amino acids derived from plant materials have been chemically characterized. One of the problems with these amino acids is their ability to imitate and replace normal amino acids in protein synthesis. Canavanine in alfalfa seeds and sprouts may a cause hypersensitivity illness. The toxic agent in inky cap mushrooms (coprinus atramentarius) which produces alcohol intolerance is the amino acid, Coprine . Toxic amino acids BMAA in cycad seeds are thought to cause a severe neurological disease in Guam; this amino acid resembles BOAA in the grass pea, lathyrus sativa, which can cause a paralytic illness. Carnosine and its methylated form, anersine , in skeletal muscle, and brain are associated with seizures and carnosinemia may lead to mental retardation.
Some vegetables also become undesirable when they are damaged or diseased. Fungal growth is a major cause of toxic alteration of plant tissue. Sweet potato , for example, supports a fungal growth (Fusarium solani), especially when the tuber's surface is damaged. The fungus alters the potatoes' metabolism, and toxic stressors are produced. Ipomeanol is one such chemical that is liver and lung toxic. Lung disease in cattle is caused by infected sweet potatoes. No similar human syndrome has been described.
Aflatoxins
A variety of fungi (Fusaria, Trichothecium, Cephalosporium, etc.) may contaminate grains, and cause illness with symptoms such as vomiting, diarrhea, headaches, chills, dizziness, and blurred vision. Aflatoxins are produced by molds which favor nuts, corn, millet, and figs. Higher concentrations of these toxins may produce acute symptoms such as loss of appetite and jaundice (hepatitis) and they are also carcinogenic if eaten in low concentrations over a longer period of time. Some of these fungal metabolites are also neurotoxins which produce tremors as a conspicuous symptom. The same fungi which produce aflatoxin produce a tremorgen, known to cause "staggers " in sheep and cattle. The common fungi which grow on our food, even in the refrigerator and cupboard, are Penicillium, Aspergillus, and Claviceps.
Over 15 tremorgenic mycotoxins have been isolated from these fungi. The role of these mycotoxins in human brain dysfunction has not been

determined and is usually not considered in medical evaluations. Aspergillus is a ubiquitous fungus that can fatally infect patients with reduced immunity.
The rules for safe storage of food to reduce fungal growth include immediate refrigeration, properly covered containers, and limits on the duration of storage. In general, proper reheating of refrigerated foods can reduce toxicity. Botulinis toxin is an example of a food poison destroyed by adequate cooking of preserved foods.
Brassicas
The common and popular cabbage or Brassica family is not without problems. The gas-producing properties of Brassica vegetables are well known. Some Brassicas (broccoli) have high vitamin K content. The therapeutic effect of anticoagulant drugs which interfere with the conversion of vitamin K to prothrombin may be reduced by brassica ingestion. Brassicas also contain high levels of chemicals that may interfere with thyroid function, promoting thyroid enlargement (goitre) . Cabbage, brussel sprouts, and kohlrabi contain progoitrin (in the range of 65-140 mg per 100 gram of fresh vegetable). Cooking reduces the goitre effect of these vegetables. Goitrogens are also found in turnips, soybeans, radishes, rapeseed, and mustard. On the plus side of the Brassica profile, there is some evidence that regular ingestion of Brassicas may offer protection against bowel cancer.
Herbs and Teas
Many patients inquire about the use of herbal teas and herbal treatments. From a medical point of view, all plant materials are potentially allergenic. From a biochemist's point of view, plant materials contain many active substances in complex combinations whose body-effects are generally not known. Beneficial effects of plant materials can be associated with negative metabolic and toxic effects which need to be considered when any plant is used with increasing intensity, especially on a daily basis.
Medicinal herbs are drug-containing plants which should not be used everyday. Like other drugs, medicinal herbs have side effects, toxic effects, and allergenic effects, and they may not be helpful. The problem with whole-plant medicines is that the active ingredients are mixed with everything else in the plant. This means that the control over the drug effect

that is achieved with purified substances is not possible with plant preparations. The appearance on the market of many "natural " and "herbal " teas is unprecedented. The safety of these products is in question. A quick review of toxicity reported to occur with popular tea-plants follows.
Cathartic teas, including those with senna leaves, flowers and bark, buckthorn bark, dock roots, or aloe leaves, do cause diarrhea. If used continuously, these herbs may induce laxative-dependence, often with abdominal discomfort, bowel dysfunction, and malabsorption of nutrients.
Laxatives, herbal allergenic teas, such as those from camomile, goldenrod, marigold, and yarrow, can cause allergic reactions in persons that are sensitive to ragweed, asters, chrysanthemums, and other related plants. Delayed allergic reactions and sun sensitivity can follow consumption of tea from the leaves of many plant products. St. John's Wort is known to be photosensitizing. Tannins in tea, including ordinary tea and peppermint tea, are surface irritants to the gastrointestinal tract and have been linked to cancer of the esophagus and stomach. Adding milk to the tea binds the plant tannins and may protect the digestive tract from irritant effects.
Diuretics are present in teas made from buchu, quack grass, and dandelion. Diuretics increase urine production with water and mineral losses. Coffee and tea are potent diuretics; the other plant teas are similar in their stressful diuretic effects. A variety of brain-active chemicals are also found in catnip, juniper, hydrangea, jimson weed, lobelia, and wormwood. Teas made from the petals of flowering plants (rose, hibiscus, hydrangea) are also neurotoxic and cause headaches, thinking disturbances, irritability, and depression. Alfalfa tea contains saponins which can disrupt digestion and respiration. While the saponins of alfalfa have been found experimentally to clear the arteries of fatty plaques in monkeys, the ingestion of alfalfa teas may have adverse effects.
Liver toxicity has been linked with a number of herbal teas.
Comfrey is a popular herb that is potentially hepatotoxic because of pyrrolizidine alkaloids, known to cause hepatocellular adenomas and increased incidence of bladder tumors in rats.
Sassafras contains safrole (as in nutmeg), another potentially hepatotoxic substance.

Ginsenghas caused breast enlargement in men (gynecomastia), due to the presence of an estrogen-like substance.
Licorice has been found to have substances that aid healing of stomach ulcers; however, it also causes sodium and water retention and loss of potassium. High blood pressure may result from excessive consumption of licorice.
Mistletoe contains alkaloids, small proteins (viscotoxins), and lectins (which collectively have hypotensive, diuretic, and antispasmodic properties). Mistletoe has been used by some herbal therapists as an anti-cancer drug. The complex of alkaloids may be cytotoxic. As well, hepatitis has been reported with mistletoe ingestion.
Pennyroyal extract has long been recommended to produce abortions, a doubtful effect, but death due to liver damage has been blamed on regular pennyroyal ingestion.
There are many possible interactions of herbal medicines and prescription drugs. Lily of the Valley, for example, contains cardiac glycosides and may lead to digitalis toxicity in a person taking adequate doses of the prescription drug. Horse chestnut contains natural anticoagulants and may increase the effect of coumarin anticoagulants. Ink Cap is a natural source of disulfiram, with the risk of an "Antabuse " reaction with alcohol. Disulfiram interferes with the metabolism of alcohol and increases the accumulation of a toxic metabolite, acetaldehyde.
The Solution
The Alpha Nutrition Program is a standardized method of diet revision that is helpful in resolving a surprising number of health problems. Food selection and preparation in the program has taken in account many of the problems associated with native food chemistry and contamination with chemicals. A food holiday on Alpha ENF is a quick way to eliminate all food contaminants and native food toxins.
Environmed Research Inc.

Contact Us | Alpha Nutrition Programs | Scope of Services | eMail Support | Answering Questions
Home | produtos e serviços | informação médica | ordens | nutrition modular Problemas no alimento
Toxins Naturais Pesquisa De Environmed

Toxins De Alimento Nativos
Aditivos De Alimento
Contaminadores Químicos
Infecção Carregada Alimento
Sensibilidade Química
Soluções Alfa ENF
Feriado Do Alimento
Programas Do Nutrition Do Alfa
Muitas substâncias naturais do non-nutriente " em alimentos " naturais comportam-se mal. Nós podemos pensar de sua atividade como quimicamente stressful. Os stressors químicos ou molecular aumentam o caos e diminuem a função em ordem, sensible do corpo. Um stressor químico é um pouco de absurdo, uma substância com nenhum papel significativo ao jogo ou um papel tóxico que seja destrutivo. Nesta revisão, um perfil breve dos stressors e os toxins são desenvolvidos mais para ilustrar o papel de stressors químicos naturais do que para esgotar este assunto vasto e complicado.
Stephen Gislason MD

O alimento está cheio das substâncias extra que devem ser processadas no corpo e então excreted, mas que não tenha nenhum valor nutriente e possa ter a droga e efeitos tóxicos. Os stressors químicos podem ser encontrados nos alimentos como ingredientes nativos e não são necessariamente aditivos ou contaminadores. Estas substâncias têm o chemistry complexo; incluem sais inorgánicos e orgânicos, minerais tóxicos, álcoois, aldeídos, alcalóides, compostos polyphenolic, salicylates, aminos-ácido do non-nutriente e peptides.
Um stressor do produto químico de alimento torna-se frankly tóxico quando ingested demasiado frequentemente, em um dose demasiado grande, ou quando ingested por uma pessoa que falte a maquinaria metabolic para detoxify a. Os stressors molecular podem simplesmente reque o corpo segurá-lo e excrete sem dano. O trabalho metabolic desta atividade é " o custo " de ingesting estas substâncias. Quando nós consumimos stressors no excesso, o efeito torna-se tóxico. Nossa capacidade segurar stressors químicos é limitada e varia de individual ao indivíduo.
A sensibilidade " química " é relatada quando a exposição aos produtos químicos transportados por via aérea, tais como o fumo do cigarro, a exaustão do motor, os perfumes, os detergentes da casa e os solventes causa sintomas. Os pacientes que estão recuperando das doenças allergic do alimento relatarão frequentemente a consciência aumentada de produtos químicos transportados por via aérea; isto parece representar um hypersensitivity generalizado. A parte da estratégia da recuperação deve evitar a exposição química tanto quanto possível.
Muitos stressors químicos competem para os mesmos pathways metabolic para o excretion. O fígado é responsável para remover muitos toxins. Um de seus métodos deve unir grupos do acetyl às moléculas tóxicas que as fazem mais soluble para o excretion do kidney. Alguns povos são " acetylators pobres " e relatam a intolerance ao escalas largas das drogas, dos alimentos, e de produtos químicos transportados por via aérea. Se muitas moléculas competirem para os mesmos pathways do excretion, é fácil imaginar que sobrecarregar ocorre regularmente nos cidadãos modernos que são expostos a uma escala larga dos produtos químicos no alimento, na água e no ar. Uma vez que a sobrecarga ocorre, mesmo as quantidades pequenas de stressors químicos extra transformam-se sintomas e dysfunction tóxicos e do produto.

Os stressors químicos inhaled e ingested simultaneamente. A poluição de ar reduzirá a tolerância para a poluição do alimento e o versa do visto. Um jantar típico em um restaurante agradável pode ser biochemically e metabolically stressful, como fumo inhaled e álcool, café, chá, spices, e liga ingested do açúcar com o complexo já no alimento. Você pode ser em mais problema se você estiver lidando também com os efeitos das drogas da prescrição feitas exame para aliviar seus sintomas da sobrecarga química!
As plantas são nossas fontes mais desejáveis do alimento. Nós tendemos a pensar os vegetais do e a fruta como inteiramente amigável e benigna; entretanto isto não é necessariamente verdadeiro. Os vegetais, as frutas, os herbs, e os spices são quimicamente complicados, e contêm uma variedade das substâncias químicas que seja droga-como, tóxica, e allergenic. As plantas são inerente tóxicas desde que evoluíram as defesas químicas que desanimam predadores de os comer.
Há muitas substâncias molecular nos alimentos que não oferecem nenhum benefício nutritivo, e deve ser processado e excreted. O ácido oxalic, para o exemplo, excreted no urine, e seus cristais são encontrados geralmente no urinalysis microscópico. Demasiado ácido oxalic no urine resultará em pedras do kidney ou da bexiga. O cálcio combina com o ácido oxalic para dar forma ao sal mais menos soluble, o oxalate do cálcio, que é encontrado também em pedras do kidney. A planta sae, especial rhubarb, repolho, spinach, e os altos de beterraba, contêm o ácido oxalic. O ácido oxalic é encontrado também as batatas e as ervilhas. A vitamina C metabolized ao ácido oxalic; contribui ao sobre-over-saturation do urine com cristais e para apedrejar possivelmente a formação.
Diversos produtos químicos usados como aditivos de alimento são encontrados também naturalmente em muitos alimentos. Os nitrates e os nitrites são ubiquitous nas plantas. Dão forma à parte do chemistry essencial os solos e as plantas. Como cada gardener sabe, o nitrogênio é essencial para o crescimento de planta; os fertilizantes do nitrogênio, contendo nitrates, são os produtos químicos agriculturais os mais abundantes. As beterrabas, os radishes, o spinach, e a alface contêm os níveis os mais elevados dos nitrates. O consumo diário é estimado para estar na escala de 100 mg/day.
Substâncias Aromatic

Todas as plantas contêm as moléculas que têm e impacto em nossos sentidos químicos. Provam e cheiram. Os compostos aromáticos atraem-nos a um alimento. A indústria de alimento usa quantidades grandes dos compostos aromáticos, das fontes naturais e sintéticas. O chemistry destas substâncias pode ser imaginado pensando da forma de sua estrutura molecular. O anel básico é uma molécula six-sided, benzene. As correntes laterais diferentes unidas ao anel mudam seus cor, gosto, e cheiro. Se os anéis do benzene forem ligados junto, uma variedade de estruturas do anel causa classes diferentes das substâncias. As moléculas baseadas na estrutura do anel do benzene são comuns na natureza. Um nome ocasional para muitas das substâncias é " phenolics ".
Com estruturas aromatic do anel inclua moléculas das drogas do alimento a cafeína e ácido salicylic; a cânfora dos sabores, o ácido cinnamic, o eugenol (nutmeg e cravos-da-índia), o safrole, o anethole (anis), o tannin (chá), o ácido gallic, e o vanillin (vanilla); e as vitaminas ácido ascorbic e niacin. Os compostos phenolic estão droga-como; alguns são tóxicos e outros parecem ser benéficos.
Um outro grupo de produtos químicos interessantes da planta é os óleos essenciais das plantas, que são também aromatic. Nós tendemos como ao cheiro dos turpenes e usamo-los em nossos perfumes, garnishes, e chás. Os turpenes aromatic comuns em plantas do alimento incluem turpenes complexos, tais como o lanosterol (no lanolin), um segundo primo do cholesterol, e o squalene, encontrou no fermento, no germe do trigo, e no óleo verde-oliva. Alguns turpenes são frankly tóxicos mas remanescem em nossa fonte de alimento como sabores. Os extratos do álcool das plantas contêm frequentemente turpenes tóxicos. Outros turpenes são moléculas ou ao menos benignos positivo. A vitamina A não é uma substância; é uma família de turpenes relacionados com atividade biológica compartilhada. O beta-carotene é o pigment amarelo nas cenouras que podem ser convertidas na vitamina ativa A (Retinols) depois que nós ingest a. O lycopene é um turpene vermelho similar encontrado nos tomates. O consumo excessivo os tomates e as cenouras de pode induzir um complexion colorido que alarme amigos mas do parece fazer pouco dano e do pode ter benefícios tais como a prevenção do cancer.
Garlic e cebolas

O garlic (alium sativum) e as cebolas (allium Cepa) são ambos os membros da família do lírio. Ambos contêm as substâncias aromatic fortes que nós usamos como flavoring para o alimento. As propriedades medicinal destes alimentos foram usadas por séculos. Allicin é o princípio aromatic do garlic. Allicin é um turpene sulphur-containing. Um bulbo intato do garlic tem pouco odor. O odor forte do allicin aparece somente depois que um bulbo do garlic é cortado ou esmagado. O trauma da exposição do ar excita um enzyme qual mude uma molécula inodora do precursor, Alliin, a Allicin.
As propriedades medicinal dos produtos químicos combinados no garlic incluem a atividade antiseptic e (uma atividade anticlotting da platelet-viscosidade reduzida). O fator anticlotting é o ajoene (4,5,9-trithiadodeca-1,6,11-triene 9-oxide!). Os entusiastas do garlic devem anotar que nenhum ajoene, ou a atividade anticlotting, estiveram encontrados em preparações proprietárias do garlic, including o óleo do garlic ou as tabuletas do garlic. O ingestion do garlic recentemente esmagado parece necessário para este efeito de droga. O efeito anticlotting do ajoene é similar ao efeito da aspirina (ASA). Os respiração-odores desagradáveis que seguem o ingestion do garlic são os metabolites sulphur-containing temporários do allicin.
As cebolas são famosas para seu efeito rasgando. A substância que nos faz weep é s-oxide propanethial. O efeito rasgando pode ser reduzido chilling a cebola antes do corte, ou processando a cebola sob a água funcionando da torneira.
Spices
Todas as plantas contêm uma variedade das substâncias químicas que agem em a droga-como a maneira. Os herbs e os spices tendem a ser quimicamente complexos e droga-como em sua atividade. A maioria de culturas nativas são relutantes comer plantas non-food para a razão boa. Nós começamos provavelmente afastado com ingestion do herb e do spice somente em doses pequenos e infrequëntemente. O nutmeg, um spice comum favorecido nos desserts e bebidas, rende muitos produtos químicos tais como o eugenol, isoeugenol, safrole, myristicin, elemicin, limolene. O uso therapeutic do eugenol como um antidiarreico, anticlotting, e agente anti-inflammatory podia ser sugerido; entretanto, se nós devêssemos usar o óleo do nutmeg como a terapia, nós enfrentaríamos a droga, e efeitos tóxicos dos outros produtos químicos no nutmeg. Nós seríamos concernidos sobre os efeitos hallucinogenic do psychotropic, myristicin. Nós seríamos

concernidos também sobre a abilidade do safrole de induzir o cancer do fígado nos ratos. A aproximação pharmaceutical apropriada deveria isolar a substância medicinal, eugenol, e decidir-se, após testar cuidadoso de seu efficacy contra seu toxicity, se o uso therapeutic fosse sábio.
As ervilhas e os feijões são legumes edible comuns. Estes alimentos são grampos *** TRANSLATION ENDS HERE ***world-wide and have desirable nutritional properties, but several interesting biochemical problems may arise with their use. Soybeans, for example, may be poorly tolerated unless well prepared. The process of making tofu illustrates the effort required to process a raw food to obtain a reliable nutrient source. Soybeans contain indigestible carbohydrates and inhibitors of digestive enzymes (soybean trypsin inhibitor). Both problems contribute to difficulty digesting beans, excessive gas, and, occasionally, abdominal pain and diarrhea.
Lima and kidney beans are toxic uncooked. Both beans contain cyanide producing compounds ( cyanogenic glycosides ), which can be destroyed by adequate cooking. Small amounts of cyanogenic glycosides will be detoxified by the liver. Cyanogenic glycosides are also found in fruit pits millet, sprouts, yams, maize, chick peas, and cassava root.
Cassava (manioc) is an important vegetable of South East Asia, Africa and South America and is inherently toxic. These tuber contain linamarin which can be converted to hydrocyanic acid . They must be processed by soaking, boiling, drying, and fermentation to reduce toxic cyanide effects. Neurological disorders and thyroid enlargement occur in African peoples who eat large amounts of inadequately processed cassava.
An unusual genetic condition, "favism ", makes some people sensitive to vicine, a nucleotide in fave beans; these people develop red blood cell damage (hemolytic anemia) after eating the beans. Cooking the beans thoroughly can reduce this effect. This is a specific example of the cytotoxic mechanism of food molecules, and illustrates the advantages of cooking foods.
Nightshades
Plants of the nightshade family contain toxic substances. "Deadly " refers to the toxicity of the leaves of this plant group which includes tomato, potato, peppers, eggplant, and tobacco. All the nightshades contain nicotine;

tobacco has the highest concentration, eggplant is next. Country lore tells us not to eat green potatoes and to store potatoes in the dark to avoid the greening effect of light. The green potato contains toxic compounds (glycoalkaloids) similar to those found in the leaves. Solanine poisoning from green potatoes will produce throat burning, weakness, diarrhea, and even convulsions with collapse and coma. Adverse and allergic reactions to tomatoes and peppers are common, and these vegetables are not on our most favored food list. Nightshades have often been implicated in arthritis.
The occurrence of toxic non-nutrient amino acids is not unusual in plants who produce toxic compounds to deter predators from eating them. Over a 150 non-protein amino acids derived from plant materials have been chemically characterized. One of the problems with these amino acids is their ability to imitate and replace normal amino acids in protein synthesis. Canavanine in alfalfa seeds and sprouts may a cause hypersensitivity illness. The toxic agent in inky cap mushrooms (coprinus atramentarius) which produces alcohol intolerance is the amino acid, Coprine . Toxic amino acids BMAA in cycad seeds are thought to cause a severe neurological disease in Guam; this amino acid resembles BOAA in the grass pea, lathyrus sativa, which can cause a paralytic illness. Carnosine and its methylated form, anersine , in skeletal muscle, and brain are associated with seizures and carnosinemia may lead to mental retardation.
Some vegetables also become undesirable when they are damaged or diseased. Fungal growth is a major cause of toxic alteration of plant tissue. Sweet potato , for example, supports a fungal growth (Fusarium solani), especially when the tuber's surface is damaged. The fungus alters the potatoes' metabolism, and toxic stressors are produced. Ipomeanol is one such chemical that is liver and lung toxic. Lung disease in cattle is caused by infected sweet potatoes. No similar human syndrome has been described.
Aflatoxins
A variety of fungi (Fusaria, Trichothecium, Cephalosporium, etc.) may contaminate grains, and cause illness with symptoms such as vomiting, diarrhea, headaches, chills, dizziness, and blurred vision. Aflatoxins are produced by molds which favor nuts, corn, millet, and figs. Higher concentrations of these toxins may produce acute symptoms such as loss of appetite and jaundice (hepatitis) and they are also carcinogenic if eaten in low concentrations over a longer period of time. Some of these fungal

metabolites are also neurotoxins which produce tremors as a conspicuous symptom. The same fungi which produce aflatoxin produce a tremorgen, known to cause "staggers " in sheep and cattle. The common fungi which grow on our food, even in the refrigerator and cupboard, are Penicillium, Aspergillus, and Claviceps.
Over 15 tremorgenic mycotoxins have been isolated from these fungi. The role of these mycotoxins in human brain dysfunction has not been determined and is usually not considered in medical evaluations. Aspergillus is a ubiquitous fungus that can fatally infect patients with reduced immunity.
The rules for safe storage of food to reduce fungal growth include immediate refrigeration, properly covered containers, and limits on the duration of storage. In general, proper reheating of refrigerated foods can reduce toxicity. Botulinis toxin is an example of a food poison destroyed by adequate cooking of preserved foods.
Brassicas
The common and popular cabbage or Brassica family is not without problems. The gas-producing properties of Brassica vegetables are well known. Some Brassicas (broccoli) have high vitamin K content. The therapeutic effect of anticoagulant drugs which interfere with the conversion of vitamin K to prothrombin may be reduced by brassica ingestion. Brassicas also contain high levels of chemicals that may interfere with thyroid function, promoting thyroid enlargement (goitre) . Cabbage, brussel sprouts, and kohlrabi contain progoitrin (in the range of 65-140 mg per 100 gram of fresh vegetable). Cooking reduces the goitre effect of these vegetables. Goitrogens are also found in turnips, soybeans, radishes, rapeseed, and mustard. On the plus side of the Brassica profile, there is some evidence that regular ingestion of Brassicas may offer protection against bowel cancer.
Herbs and Teas
Many patients inquire about the use of herbal teas and herbal treatments. From a medical point of view, all plant materials are potentially allergenic. From a biochemist's point of view, plant materials contain many active substances in complex combinations whose body-effects are generally not known. Beneficial effects of plant materials can be associated with negative

metabolic and toxic effects which need to be considered when any plant is used with increasing intensity, especially on a daily basis.
Medicinal herbs are drug-containing plants which should not be used everyday. Like other drugs, medicinal herbs have side effects, toxic effects, and allergenic effects, and they may not be helpful. The problem with whole-plant medicines is that the active ingredients are mixed with everything else in the plant. This means that the control over the drug effect that is achieved with purified substances is not possible with plant preparations. The appearance on the market of many "natural " and "herbal " teas is unprecedented. The safety of these products is in question. A quick review of toxicity reported to occur with popular tea-plants follows.
Cathartic teas, including those with senna leaves, flowers and bark, buckthorn bark, dock roots, or aloe leaves, do cause diarrhea. If used continuously, these herbs may induce laxative-dependence, often with abdominal discomfort, bowel dysfunction, and malabsorption of nutrients.
Laxatives, herbal allergenic teas, such as those from camomile, goldenrod, marigold, and yarrow, can cause allergic reactions in persons that are sensitive to ragweed, asters, chrysanthemums, and other related plants. Delayed allergic reactions and sun sensitivity can follow consumption of tea from the leaves of many plant products. St. John's Wort is known to be photosensitizing. Tannins in tea, including ordinary tea and peppermint tea, are surface irritants to the gastrointestinal tract and have been linked to cancer of the esophagus and stomach. Adding milk to the tea binds the plant tannins and may protect the digestive tract from irritant effects.
Diuretics are present in teas made from buchu, quack grass, and dandelion. Diuretics increase urine production with water and mineral losses. Coffee and tea are potent diuretics; the other plant teas are similar in their stressful diuretic effects. A variety of brain-active chemicals are also found in catnip, juniper, hydrangea, jimson weed, lobelia, and wormwood. Teas made from the petals of flowering plants (rose, hibiscus, hydrangea) are also neurotoxic and cause headaches, thinking disturbances, irritability, and depression. Alfalfa tea contains saponins which can disrupt digestion and respiration. While the saponins of alfalfa have been found experimentally to clear the arteries of fatty plaques in monkeys, the ingestion of alfalfa teas may have adverse effects.

Liver toxicity has been linked with a number of herbal teas.
Comfrey is a popular herb that is potentially hepatotoxic because of pyrrolizidine alkaloids, known to cause hepatocellular adenomas and increased incidence of bladder tumors in rats.
Sassafras contains safrole (as in nutmeg), another potentially hepatotoxic substance.
Ginsenghas caused breast enlargement in men (gynecomastia), due to the presence of an estrogen-like substance.
Licorice has been found to have substances that aid healing of stomach ulcers; however, it also causes sodium and water retention and loss of potassium. High blood pressure may result from excessive consumption of licorice.
Mistletoe contains alkaloids, small proteins (viscotoxins), and lectins (which collectively have hypotensive, diuretic, and antispasmodic properties). Mistletoe has been used by some herbal therapists as an anti-cancer drug. The complex of alkaloids may be cytotoxic. As well, hepatitis has been reported with mistletoe ingestion.
Pennyroyal extract has long been recommended to produce abortions, a doubtful effect, but death due to liver damage has been blamed on regular pennyroyal ingestion.
There are many possible interactions of herbal medicines and prescription drugs. Lily of the Valley, for example, contains cardiac glycosides and may lead to digitalis toxicity in a person taking adequate doses of the prescription drug. Horse chestnut contains natural anticoagulants and may increase the effect of coumarin anticoagulants. Ink Cap is a natural source of disulfiram, with the risk of an "Antabuse " reaction with alcohol. Disulfiram interferes with the metabolism of alcohol and increases the accumulation of a toxic metabolite, acetaldehyde.
The Solution
The Alpha Nutrition Program is a standardized method of diet revision that is helpful in resolving a surprising number of health problems. Food

selection and preparation in the program has taken in account many of the problems associated with native food chemistry and contamination with chemicals. A food holiday on Alpha ENF is a quick way to eliminate all food contaminants and native food toxins.
Environmed Research Inc. Contact Us | Alpha Nutrition Programs | Scope of Services | eMail Support | Answering Questions
" SINTA O PODER DO INCA! "
Que é Maca & o que ele faz? A ciência atrás da conversa Propriedades & Dos Nutrientes Maca Antigo History Sobre nossos growers O Vale Mágico Nossa Linha De Produto Facilidades da embalagem Sobre nós Torna-se a Distribuidor 3 níveis do poder de compra Conversa De Maca! Medicinas De Amazon Contate-nos " Estas são as RAIZES da MÁGICA de MACA... "
Todo o preto de Jerome do copyright das fotos/herbs América, 1994-2002.
Os herbs América cultivam a raiz de Maca da qualidade superior; o alimento super novo das indústrias " dos produtos naturais "...
COMPRE MACA no mart de Maca agora!
O varejo, os clientes preferidos, e todos os níveis da compra por atacado de Maca compra aqui.
Cartões de crédito aceitados em um usuário seguro

--------------------------------------------------------------------------------
A PLANTA SURPREENDENTE DE MACA Maca tem um history mais complexo do que a maioria dos herbs que nós ouvimos assim geralmente sobre hoje... sua raiz que é a fonte do poder, Maca, ( Lepidium Peruvianum Ch. ) é crescido apenas abaixo do glaciated inclinações dos Andes peruvian. Maca foi usado primeiramente pelo Incas mais de 3000 anos há para a energia e a resistência e é negociado ainda geralmente como uma medicina e uma energia que dão o herb durante todo as cidades e as vilas de Peru hoje. Os alcalóides recentemente descritos de Maca, os sterols e os vários poders surpreendentes fizeram completamente legendário durante todo o mundo, embora, até agora, remanesceram razoavelmente elusive a respeito de suas fontes. No vale mágico de Perus, os cultivars de Maca foram desenvolvidos pelo Inca para a energia e a resistência.
Embora este membro raro da família do radish fosse usado por povos indígenos para milhares dos anos , muitos consideraram Maca ser "um herb perdido" ao mundo tornando-se até recentemente. Agora, tudo está mudando enquanto Maca está sendo procurado ansiosamente em seguida para seus poders e atributos medicinal extraordinários. (PARA Um OLHAR FASCINANTE E DETALHADO No HISTORY De MACA, VEJA NOSSA PÁGINA Do HISTORY.)
No mundo moderno de hoje, Macaroot é comido até 3 vezes um o dia por Peruvians e pelos povos toda ao redor do mundo, (dos atletas profissionais às pessoas idosas), dar-lhes a energia e ajudá-la recuperar do depression, dos addictions, dos traumas, ou da doença.
POR QUE VOCÊ SENTE ASSIM BOM? É MÁGICA de MACA naturalmente! A nós, esta indicação é muito mais do que um slogan! É uma maneira de vida worth compartilhar com o outro. Você sentirá bom o dia inteiro! Cada raiz mágica do hypocotylo de Maca contem sobre 55 naturais, phyto-produtos químicos benéficos. Estes nutrientes naturais têm a abilidade

de ajudar no nourishment diário de nossos corpos e de fazer povos físicos das diferenças sentir!
QUE FAZ PARA VOCÊ? MACA MAGIC™ é considerado por investigadores superiores ser um adaptogen verdadeiro, trabalhando com os ritmos naturais dos corpos para ajudar reconstruir sistemas imunes da semana, re-re-mineralize corpos mal nourished, e aumentar a energia e a resistência. Outros exemplos de como a raiz de Maca afeta diretamente o corpo humano incluem reduzir o stress físico ou mental, irregularidades hormonal balançando, promovendo o crescimento do cabelo saudável, os ossos, e os dentes, e promover o tom do músculo. A maioria de povos sentem seu nível do modo e de energia levantar em um instante!
" O Viagra Natural " Maca é também bom conhecido para ele é fertilidade que aumenta e desempenho sexual que realça propriedades. No mundo conscious moderno de hoje da saúde e da juventude, esta é a propriedade touted de Maca mais provável e agressively introduzída no mercado a melhor. Está ganhando ràpidamente o nickname " do Viagra natural ".
A CIÊNCIA ATRÁS DA CONVERSA Maca é uma raiz com mais de 3000 anos do uso histórico; seu mérito touted pela ciência e por estudos atuais de hoje. Com seu muito a saúde que realça as propriedades que tornam-se finalmente publicized extensamente, raiz de Maca é um produto que possa fazer povos sentir, olhar, e executar mais melhor... Os doutores, os naturalopaths, e os herbalists o excesso do mundo estão estudando ativamente e aggressively esta raiz surpreendente e publicam seus muitos relatórios.
Estale sobre as ligações do interesse abaixo ou VEJA NOSSA PÁGINA da CIÊNCIA AGORA!
* Maca elevates o modo e melhora a abilidade mental (concentração) e a resistência física. * Maca aumenta a circulação do sangue na pele que dá à pele uma aparência jovem. * Os sterols naturais na ajuda de Macaroot constroem o músculo especial quando usados conjuntamente com o exercício.

* Maca dá um sentido geral do bem estar (bem de nourished.) * Maca tem a abilidade de levantar povos do depression. * Maca energiza o stimulation without excedente. * Função sexual dos enlivens de Maca em anos avançados (Viagra provado gosta do efeito, sem efeito lateral) * É non addictive. * Como anti-force a medicina homeopathic, um aura calmo acompanha o uso de MACA MAGIC™ * Maca pode ajudar regular irregularidades hormonal das mulheres.
NUTRIENTES E PROPRIEDADES Uma única raiz MÁGICA de MACA contem quase 60 phyto-nutrientes! As análises químicas de Maca Magic™ revelam um perfil cérebro-brain-powering dos aminos-ácido, dos minerais, dos sterols, e de ácidos fatty. Uma avaria destes produtos químicos é como segue:
AMINOS-ácido: alanina, argine, aspartime, glutamine, glycine, histidine, OH-oH-proline, isoleucine, leucine, lysine, methionine, phenylalanine, proline, sarcosine, serine, threonine, tyrosine, e valine. MINERAIS: cálcio, cobre, ferro, magnésio, phosphorus, potassium e zinco. STEROLS: brassicasterol, erogosterol, ergostadienol, campesterol, sitosterol, e stigmasterol. 20 ÁCIDOS FATTY: laric, myristic, palmitic, palmitoleic, linoleic, arachidic, steric, behanic, nervonic, lignoceric, tridecanoic, 7-tridecanoic, perntadecanoic, 7-pentadecanoic, heptadecanoic, 9-heptadecanoic, nonadecanoic, 11-nonadecanoic, e 15-eicosenoic.
Todos estes phytochemicals combinaram com a disposição cheia da mágica de MACA das vitaminas, including A, B1, B2, B3, B12, C, D e E; junto com saponins, os tannins, hidratos de carbono, protien, e isothiocyanate benzyl, isothiocyanate do p-methoxybenzyl, e makes de I-ecdysone a planta uma de Maca das plantas medicinal as mais poderosas na terra!
Maca é conhecido bem nas ruas de Cusco, Peru HISTORY DE NOSSO MACA

Quando o espanhol chegou a Peru em 1533, estas plantas pequenas foram escondidas afastado e remanescidas na borda do brink da extinção por séculos. Embora este membro raro da família do radish fosse usado por povos indígenos para milhares dos anos , muitos consideraram Maca ser "um herb perdido" ao mundo tornando-se até recentemente. (PARA Um OLHAR FASCINANTE E DETALHADO No HISTORY De MACA, VEJA NOSSA PÁGINA Do HISTORY AGORA.)
Os herbs-America™-America™ exploraram os highlands peruvian assim que 1992 e foram os primeiros para introduzir 20 cultivars de raizes vivas de Maca aos EUA em 1994! Estes cultivars nurtured no tipo especial de AMÉRICA dos HERBS de Maca Maca chamado raiz Mágica. É, completamente simplesmente, a qualidade a mais fina Maca disponível, e é para esta razão que nós somos orgulhosos a estar em uma posição para compartilhar finalmente desta raiz surpreendente ao mundo!
COMECE SEUS PRODUTOS DE MACA HOJE! (e comece a experimentar os benefícios de saúde bons e a energia positiva da raiz pura de Maca hoje!) Necessite mais energia ou sinta-a lento com para fora de cafeína ou outras misturas químicas deleterious? Tente nossos pós crus ou brindados da raiz de MACA MAGIC™, extratos bio-ativos puros e termine a linha de sucos inteiramente enzymatic e as barras nutrientes dos rich fizeram toda da raiz inteira de 100% MACA MAGIC™.
O *** VAI AO MART DE MACA AGORA COMPRAR MACA! * * * Para encontrar para fora mais sobre nossa linha de produto, os detalhes em nossos níveis de compra diferentes, discontos, specials, e mais, VERIFICAM PARA FORA de NOSSOS PRODUTOS WEBPAGE de MACA AGORA!
--------------------------------------------------------------------------------
CONTATO E. U. HOJE! E-mail
Telefone: 541-846-maca (6222), FAX: 541-846-9488

--------------------------------------------------------------------------------
Contato postal em PERU: HERBS América S.A.C. Peru, Avoirdupois. 28 de Julio 549, Miraflores, Lima 18 PERU Contato postal nos EUA: Caixa 446 De América Po. dos HERBS, Murphy, Oregon EUA 97533
REPOUSO MÁGICO DE MACA | GROWER DE MACA HOME | PRODUTOS DE MACA HOME
Copyright 2000- 2003, REDE de AMÉRICA dos HERBS. Todo o texto e propriedade das fotos da rede de América dos herbs e para não reprinted em algum formulário sem permissão escrita da companhia. " Os herbs América " e " a mágica de Maca " são marcas registradas internacionais registadas.
--------------------------------------------------------------------------------
Website projetado e hospedado por BASECAMP INTERNACIONAL (visto melhor nos pixels 800X600 monitore o ajuste com explorador 4,5 de Microsoft ou mais novo)

NOTAS NO MUITOS MEDICINAL E CURATIVE PROPRIEDADES DA RAIZ DE MACA Artigos, estudos, fatos, e mais...
Artigo: (nota) do médico (fonte: Dr. Garry P. Gordan, faculdade americana para o avanço na medicina.) Maca como um herb do Anti-Envelhecimento para homens e mulheres Garry P. Gordon, MD, presidente anterior da faculdade americana para o avanço na medicina, founder agora e presidente da faculdade internacional de medicina avançada do longevity, situada em Chicago, Illinois, bases sua apreciação do maca em sua própria experiência com ele. Falando com mim de Payson, arizona, o Dr. Gordon disse, " nós todos ouvimos boatos sobre vários produtos como o maca. Mas usando esta raiz peruvian eu mesmo, eu experimentei pessoalmente uma melhoria significativa na resposta erectile do tecido. Eu chamo-a ' resposta da natureza a Viagra™ ". " O que eu v no maca é meios de normalizar nossos hormones steroid como o testosterone, o progesterone, e o estrogen. Conseqüentemente tem a facilidade para prevenir as mudanças hormonal do envelhecimento, " Dr. Gordon acredita. " Age em homens para restaurá-los a um status funcional saudável em que experimentam um líbido mais ativo. Os lotes dos homens e das mulheres que acreditaram previamente seus problemas sexual eram psicológicos agora estão indo claramente procurar algo physiological para melhorar a qualidade de vida na área de sexuality, " dizem o Dr. Gordon. " Naturalmente, como alguém interessado no longevity, eu estou ciente que o mortality se aproxima muito mais logo para aqueles indivíduos cuja a atividade sexual é diminuída ou inexistente. Ou seja eu acredito que os povos que acoplam no sexo duas vezes uma semana ou mais mais longo

vivo. Eu encontrei a atividade sexual para ser um marcador de confiança para o envelhecimento total."
Artigo: (Nota) Do Médico (Fonte: Dr. Gabriel Cousens, MD) Maca dá uma resposta aos efeitos do envelhecimento no sistema do endocrine O médico americano Gabriel Cousens, MD, acredita este herb tem o potencial de uma resposta equilibrada aos efeitos do envelhecimento no sistema do endocrine. Muitos que tentaram phytoestrogens e/ou hormones do precursor tais como DHEA ou pregnenolone, ou mesmo a terapia natural da recolocação do hormone e foram descontentadas, estão começando resultados excelentes de seu uso da raiz do maca. Gabriel Cousens, MD, medicina interna praticando em Patagonia, arizona, diz, "sempre que possível, eu prefiro usar a terapia do maca melhor que a terapia da recolocação do hormone porque HRT envelhece realmente o corpo que diminui o hormone produzindo a potencialidade das glândulas. Maca provou ser muito eficaz com os pacientes menopausal em eliminar flashes e o depression quentes e em níveis de energia crescentes. _ encontr direito dosage nível, às vezes eu t começ paciente maca tratamento com um metade um teaspoon pó ou três cápsula um dia. Em alguns casos eu levantei o dosage para um teaspoon ou seis cápsulas um o dia para a eficácia cheia."
Artigo: (nota do médico e Testimonial) (fonte: Dr. Henry Campanile, MD)
O doutor oferece Maca como uma raiz balançando adrenal O henry Campanile, M.D., oferece a raiz balançando adrenal de Maca a seus pacientes. De acordo com sua modalidade de agir através do hypothalamus e pituitary, Maca tem um efeito balançando e nourishing nas glândulas adrenal. O henry Campanile, MD, um specialist 50-year velho na medicina interna e de family/complementary que pratica em St. Petersburg, Florida, relaciona-se: " Eu aconteço ter sido nascido com a uma glândula adrenal apenas como meu pai. Eu comecei fazer exame da cortisona em meus twenties atrasados para aliviar a fatiga quais eu estava sentindo já. Sabendo os perigos do uso a longo prazo da cortisona, eu olhei ao redor para uma alternativa, e esta circunstância é que começado me interessado na medicina complementar. Eu comecei usar o pregnenelone aproximadamente 10 anos há e foi razoavelmente satisfatório. Mas um de meus pacientes disse-me que sobre Maca, e mim começou fazer exame d aproximadamente um mês há. É

phenomenal! Eu não senti este bom desde que eu tinha 20 anos velho. Eu tenho assim muita energia e olho assim bem, meus pacientes observaram nela e disseram-me como descansado eu pareço. Eu have.got assim muita energia tenho começado agora um programa do exercício. " Após ter tentado o para fora nhimself, Dr.Campanile começou a usar o maca com seus pacientes. " Meu primeiro paciente para fazer exame das cápsulas do maca experimentava flashes quentes e outros sintomas menopausal. Começou sentir muito mais melhor após ter usado este herb por somente quatro dias. Eu estou empregando-o também com pacientes que têm a função adrenal baixa. "
Artigo: (Nota) Do Médico (Fonte: Dr. Jorge Um Calderon, MD) O pioneiro peruvian prescreve Maca Um outro pioneiro peruvian na aplicação therapeutic do maca integrou em uma prática médica moderna é Jorge Aguila Calderon, MD. Um intemist, Dr.Aguila Calderon é chefe anterior do departamento de ciências biológicas e de decano da faculdade da medicina humana na universidade nacional de Federico Villarreal em Lima. Como o Dr. Malaspina e outro, (veja notas em outras páginas da ciência), prescreve o maca para uma variedade larga das condições including o osteoporosis e healing de fraturas do osso no muito idoso. " Maca tem muitos do cálcio fàcilmente absorbable nele, mais o magnésio, e uma quantidade justa de aliados que nós estamos encontrando muito úteis em tratar o decalcification dos ossos nas crianças e nos adultos. " Junto com prescrever uma dieta excelente e um determinado lifestyle muda, Dr. Aguila Calderon ajudou a pacientes com impotence masculino, sterility masculino, e sterility fêmea empregando a terapia do maca. Os problemas que adicionais trata com o maca são os rickets, vários formulários do anemia, sintomas menopausal tais como flashes quentes e a noite sua, dificuldades climacteric e erectile nos homens, envelhecimento prematuro, e estados gerais da fraqueza tais como a fatiga crônica.
Artigo: (Nota) Do Médico (Fonte: Dr. Harold Clark, MD) Harold Clark, MD, faz a Maca um remédio chave Um outro doutor americano que tivesse recentemente começou a usar o maca therapeutically para alguns pacientes é de Rochelle novo, york novo. O Dr. Clark, que utiliza a terapia do chelation e a terapia do ozônio além aos herbs, vitaminas e minerais em sua prática indicou, " mim é espantado em

como rapidamente o maca trabalhou em dois pacientes que eu estive concernido aproximadamente por alguma hora. " Descreveu um paciente como 55 year-old Mary T, uma postmenopausal, mulher. Mary T foi possuída de problemas de saúde numerosos, including o açúcar um tanto elevated do sangue, o hypertension, o fibrillation atrial, e o hypomagnesemia. Tinha sido aguda doente por dois meses com osteomyelitis e o sepsis generalizado. Incapaz de trabalhar, estava sofrendo da fatiga e do depression grandes e estava sentindo ' mais má e mais má ' sobre os últimos cinco anos. " Dentro de apenas quatro dias de fazer exame das cápsulas do maca, Mary T atravessou uma rotação enorme, " disse o Dr. Clark. " Saiu comprar nas lojas; é limpeza sua casa; sente forte e vigorosa; e seu depression é ido. "
Nota e disclaimer: Esta informação é apresentada somente tão material do interesse geral e não quanto uma prescrição para toda a pessoa específica ou qualquer circunstância em um caso específico. Nós não reivindicamos que algumas destas experiências estarão duplicadas por outras, e nós incentivamos todos procurar o dae (dispositivo automático de entrada) de um practitioner qualificado da saúde para o conselho que pertence ela/suas condição e necessidades particulares.
MACAROOT HOME | MACAMAGIC HOME MACAGROWER HOME | MACAPRODUCTS HOME
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.herbs-america.com http://babelfish.altavista.com/babelfish/ urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.herbs-america.com
Copyright 2000-2002, REDE de AMÉRICA dos HERBS. Todo o texto e propriedade das fotos da rede de América dos herbs e para não reprinted em algum formulário sem permissão escrita da companhia. " Os herbs América " e " a mágica de Maca " são marcas registradas internacionais registadas.

Website projetado e hospedado por BASECAMP INTERNACIONAL (visto melhor nos pixels 800X600 monitore o ajuste com explorador 4,5 de Microsoft ou mais novo)
NOTAS EM REALÇAR SEXUAL DO DESEMPENHO PROPRIEDADES DA RAIZ DE MACA
Artigo: Viagra Fêmea! (publicado com permissão dos meios de notícia herbal scottish)
Em Scotland, um aphrodisiac natural que touted como o Viagra fêmea está sendo fornecido. O extrato líquido é dito ser do benefício em impulsionar o líbido de ambos os sexos mas do valor particular às fêmeas. Colhido 13.000 pés acima nos Andes centrais, o herb foi considerado assim potent que o Incas confinou seu uso a sua família real. Após quase tornar-se extinct, as propriedades da planta para melhorar vidas do sexo e fertilidade estão sendo apreendidas outra vez sobre na vigília da mania world-wide de Viagra. Somente de alguns acres da colheita em 1995, há agora 1500 acres com as plantas para plantar uns 5000 acres mais adicionais. O líquido Maca, Extract™ expresso, "destrava inteiramente o potencial de o que o Incas se chamou seu superfood", propionates herbal da palavra em Europa, "é material muito quente e está indo ser muito grande porque não tem nenhuns side-effects ". Aparte de aumentar o stamina sexual e a fertilidade, seus outros usos incluem do hormone a terapia da recolocação e facilitar da tensão pre-pre-menstrual. Quando algumas companhias procurarem patentes nos produtos derivados de Maca, outras consideram-no um presente ao mundo e oferecem-no sem uma patente.
Estudo e relatório chineses do laboratório de Maca: (© 2000, Elsevier Ciência Inc., publicado com permissão do jornal o urology, o volume # de 55, de 2000) Efeitos de um extrato lipidic de Lepidium Meyenii no comportamento sexual os ratos e os ratos. Este estudo revela para a primeira vez uma atividade aphrodisiac de L.

Meyenii, um herb andean da montanha.
OBJETIVOS ABSTRATOS: Para determinar o efeito de da administração oral de um extrato lipidic purified do lepidium meyenii/peruvianum (M-01 M-02) no número of intromissions completos e acoplamento em ratos normais, e no período latente da ereção (LPE) nos ratos com dysfunction erectile. MÉTODOS: Os ratos e os ratos foram divididos aleatòria em diversos grupos experimentais e de controle. De 10% do ethanol uma suspensão de M-01 e M-02 foi administrado oral por 22 dias aos grupos experimentais de acordo com o dosage especificado pelo projeto experimental. No dia 22, 30 minutos depois que o dose foi administrado aos ratos masculinos, 2 ratos fêmeas virgens foram colocados com o 1 rato masculino. O número de intromissions completos de cada rato masculino em 3 horas foi gravado. Em uma avaliação de 1 dia do acoplamento, cada rato masculino cohabited com os 5 ratos fêmeas estrous durante a noite. O número de fêmeas sperm-positivas foi gravado. O LPE foi medido aos burros a função sexual nos ratos com dysfunction erectile. Usando um instrumento multifunction de Ysd-4g, um pulso elétrico em 20 V foi aplicado para estimular o penis dos rat`s, e a duração do começo ao stimulus à ereção cheia foi medida nos segundos como o LPE. RESULTADOS: Nos ratos masculinos normais, o número de intromissions completos durante o período 3-hour era 16,33 o ± 1,78, 46,67 ± 2,39, e 67,01 o ± 2,55 para o grupo de controle, o grupo M-01, e o grupo M-02, respectivamente. Na avaliação do acoplamento, o número do increasede sperm-positivo das fêmeas de 0,6 ± 0,7 no grupo de controle 1,5 ao ± 0,5 no grupo M-01 experimental. LPE dos ratos masculinos com dysfunction erectile era o ± 112 13 segundos com uma dieta regular (grupo de controle). A administração oral de M-01 em um dose do peso de corpo de 180 ou 1800 mg/kg e do M-02 em um dose de 45, 180, peso de corpo de 1800 mg/kg reduziu o LPE ao ± 54 12 segundos, 54 ± 13 segundos, o ± 71 12 segundos, os 73 segundos do ±, e o ± 41 13 segundos, respectivamente. O LPE dos ratos cirúrgicos tratou com o M-01 no dose o mais baixo (45 mg/kg) era o ± 121 12 segundos; assim, a mudança não era significativa. CONCLUSÕES: A administração oral M-01 e M-02 realçou a função sexual os ratos e ratos, do como evidenciado por um aumento o número de intromissions completos e o número de fêmeas sperm-positivas em ratos normais, e diminuição no LPE nos ratos masculinos com dysfunction erectile.

Este estudo foi conduzido perto: Bo Lin Zheng, Kan ele, yen Huang de Calvin Hyungchan Kim, de Lingling Rogers, de Yu Shao, de Zhen, cheng Qien de Yang lu, de Sui junho Yan, de Lu, e yi Zheng de Qun na faculdade médica de Liaoning & da faculdade de Shenyang da medicina chinesa tradicional em Shenyang China, e também no academy chinês da medicina preventiva, Beijing, República Popular da China.
Referências: 1. Leon T: ' o Maca ' (meyenii de Lepidium): pouca planta conhecida do alimento de Peru. Econ Bot 18: 122-127, 1964. 2. Chacón RC: Meyenii Walp de de Lepidium do fitoquímico de Estudio. Thesis, universidade Nac. mayor de San Marcos, Lima, Peru, 1961. 3. Zheng BL, Kim CH, ele K, et al: Um processo para a isolação e purification do meyenii de Lepidium. Patente pendente, 1999. 4. Procedimento de teste clínico nacional, FDA de China, 1998. 5. Hermann M, e Heller J: Raizes e tubers andean: Ahipa, arracacha, Maca e Yacom Roma, IPGRI, 1997, pp 175-195. 6. Cobo B: Historia del Nuevo Mundo. Biblioteca de Autores Españoles 81: 430, 1956. 7. Ruiz H: Relación histórica del viaje um del Peru y o Chile dos reinos dos los, 1777-1778, Madrid Acad. De Ciene Exaetas: Fis y 1 nat: 526, 1952. 8. Pulgar VJ: Fecundante de Poderoso dos sp. de Las Maca Lepidium vegetal. Voz de Huancayo do la 24:10, 1964. 9. John T: O anu e o maca. J Ethnobiol 1: 208-212.1981.
Estudo e relatório peruvian do laboratório de Maca: (Dr. Gloria Chacon, Peru Lima- Do ©) Efeitos de Maca nas glândulas do endocrine. O Dr. Gloria Chacon isolou quatro alcalóides da raiz do maca e realizou os estudos animais com os ratos masculinos e fêmeas dados a raiz pulverizada maca ou os alcalóides isolados das raizes. Em comparação com os grupos de controle animais, aqueles que recebem ou o pó da raiz os alcalóides mostraram o maturation múltiplo do follicle do ovo nas fêmeas e, nos machos, umas taxas significativamente mais elevadas da produção e do motility do sperm do que grupos de controle. O Dr. Chacon estabeleceu em que era os alcalóides na raiz do maca, não seus hormones da planta, que produziram efeitos da fertilidade os ovaries e testes dos ratos. Estes efeitos são measurable dentro de 72 horas de dosing os animais, ' ela ofereceram em uma entrevista recente do telefone de Lima, Peru. Com as experiências, deduziu que os alcalóides estavam agindo na glândula hypothalamus-hypothalamus-pituitary, que explica porque os ratos masculinos e fêmeas afflicted em uma maneira gender-apropriada. Isto a

explica também porque os efeitos nos seres humanos não são limitados os ovaries e os testes, mas ao age também nos adrenals, dando um sentimento de uma energia mais grande e vitality, e em o pancreas e thyroid também. As implicações da descoberta do Dr. Chacon dos efeitos estimulando pituitary do maca são enormes. O que parece significar é essa terapia da recolocação do hormone, mesmo as variedades naturais, quer seja não mais por muito tempo o padrão de ouro para optimising um ponto holístico da vista.
Artigo: (Nota Dos Médicos) " Viagra natural " (notas dos doutores Malaspina, muller, e Chacon. (veja vários estudos em nossas páginas da ciência por estes doutores) Os doutores Malaspina, muller e Chacon, as.well.as doutores os os EUA e Canadá, têm a notícia boa para os homens que estão sofrendo dysfunction sexual age-related. Podem esquecer-se de Viagra caro, possivelmente perigoso. Maca trabalha extremamente bom e com segurança. O Dr. Jorge Aguila Calderon, decano da faculdade da medicina humana na universidade nacional de FedericoVillareal em Lima, prescreve o maca para uma variedade larga das condições, including o osteoporosis e healing de fraturas do osso no muito idoso. Diz, no "Maca tem muitos do cálcio fàcilmente absorbable neles, mais o magnésio, e uma quantidade justa de silicone, que nós estejamos encontrando muito útil no decalcification de tratamento dos ossos as crianças e os adultos." O Dr. Calderon ajudou também a pacientes superar o impotence masculino, o sterility masculino, e o sterility fêmea empregando a terapia do maca. Os problemas que adicionais trata com o maca são os rickets, vários formulários do anaemia, sintomas menopausal tais como flashes quentes e a noite sua, dificuldades climacteric e erectile nos homens, envelhecimento prematuro, e estados gerais da fraqueza, tais como a fatiga crônica.
Artigo: (médicos testimonial e nota) Burton Goldberg, presidente da medicina alternativa que publica em Tiburon, Califórnia, cujo o livro o mais atrasado é " uma guia definitive da medicina alternativa ao cancer " é um outro entusiasta do maca. Diz que quando tentou o maca era muito pleased com os resultados e começava a fazer exame d regularmente. " Eu sou um homem velho de 72 anos e este maca fêz exame de 25 anos fora de minha vida de sexo do envelhecimento, " declara Burton Goldberg. ' que é consideravelmente importante para mim! "

O Dr. Garry Gordon é concernido sobre problemas reproductive no mundo de hoje. " A sociedade enfrenta um problema enorme de deixar cair contagens do sperm e dificuldades do hormone do sexo. Mas o maca promete uma solução nontoxic com nenhuns efeitos downside. É uma terapia que pareça oferecer homens e mulheres a possibilidade para o rejuvenation hormonal, " concli o Dr. Gordon. " Nós vivemos atualmente em uma era em que quase todos estará fazendo algo tratar das conseqüências hormonal do envelhecimento. E Maca está agora prontamente disponível."
Nota e disclaimer: T esta informação é apresentado somente tão material do interesse geral e não quanto uma prescrição para toda a pessoa específica ou qualquer circunstância em um caso específico. Nós não reivindicamos que algumas destas experiências estarão duplicadas por outras, e nós incentivamos todos procurar o dae (dispositivo automático de entrada) de um practitioner qualificado da saúde para o conselho que pertence ela/suas condição e necessidades particulares.
MACAROOT HOME | MACAMAGIC HOME MACAGROWER HOME | MACAPRODUCTS HOME
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.herbs-america.com http://babelfish.altavista.com/babelfish/ urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.herbs-america.com
Copyright 2000-2002, REDE de AMÉRICA dos HERBS. Todo o texto e propriedade das fotos da rede de América dos herbs e para não reprinted em algum formulário sem permissão escrita da companhia. " Os herbs América " e " a mágica de Maca " são marcas registradas internacionais registadas.
Website projetado e hospedado por BASECAMP INTERNACIONAL (visto melhor nos pixels 800X600 monitore o ajuste com explorador 4,5 de Microsoft ou mais novo)

NOTAS EM REALÇAR SEXUAL DO DESEMPENHO PROPRIEDADES DA RAIZ DE MACA
Artigo: Viagra Fêmea! (publicado com permissão dos meios de notícia herbal scottish)
Em Scotland, um aphrodisiac natural que touted como o Viagra fêmea está sendo fornecido. O extrato líquido é dito ser do benefício em impulsionar o líbido de ambos os sexos mas do valor particular às fêmeas. Colhido 13.000 pés acima nos Andes centrais, o herb foi considerado assim potent que o Incas confinou seu uso a sua família real. Após quase tornar-se extinct, as propriedades da planta para melhorar vidas do sexo e fertilidade estão sendo apreendidas outra vez sobre na vigília da mania world-wide de Viagra. Somente de alguns acres da colheita em 1995, há agora 1500 acres com as plantas para plantar uns 5000 acres mais adicionais. O líquido Maca, Extract™ expresso, "destrava inteiramente o potencial de o que o Incas se chamou seu superfood", propionates herbal da palavra em Europa, "é material muito quente e está indo ser muito grande porque não tem nenhuns side-effects ". Aparte de aumentar o stamina sexual e a fertilidade, seus outros usos incluem do hormone a terapia da recolocação e facilitar da tensão pre-pre-menstrual. Quando algumas companhias procurarem patentes nos produtos derivados de Maca, outras consideram-no um presente ao mundo e oferecem-no sem uma patente.
Estudo e relatório chineses do laboratório de Maca: (© 2000, Elsevier Ciência Inc., publicado com permissão do jornal o urology, o volume # de 55, de 2000) Efeitos de um extrato lipidic de Lepidium Meyenii no comportamento sexual os ratos e os ratos. Este estudo revela para a primeira vez uma atividade aphrodisiac de L. Meyenii, um herb andean da montanha.
OBJETIVOS ABSTRATOS: Para determinar o efeito de da administração oral de um extrato lipidic purified do lepidium meyenii/peruvianum (M-01 M-02) no número of intromissions completos e acoplamento em ratos normais, e no período latente da ereção (LPE) nos ratos com dysfunction

erectile. MÉTODOS: Os ratos e os ratos foram divididos aleatòria em diversos grupos experimentais e de controle. De 10% do ethanol uma suspensão de M-01 e M-02 foi administrado oral por 22 dias aos grupos experimentais de acordo com o dosage especificado pelo projeto experimental. No dia 22, 30 minutos depois que o dose foi administrado aos ratos masculinos, 2 ratos fêmeas virgens foram colocados com o 1 rato masculino. O número de intromissions completos de cada rato masculino em 3 horas foi gravado. Em uma avaliação de 1 dia do acoplamento, cada rato masculino cohabited com os 5 ratos fêmeas estrous durante a noite. O número de fêmeas sperm-positivas foi gravado. O LPE foi medido aos burros a função sexual nos ratos com dysfunction erectile. Usando um instrumento multifunction de Ysd-4g, um pulso elétrico em 20 V foi aplicado para estimular o penis dos rat`s, e a duração do começo ao stimulus à ereção cheia foi medida nos segundos como o LPE. RESULTADOS: Nos ratos masculinos normais, o número de intromissions completos durante o período 3-hour era 16,33 o ± 1,78, 46,67 ± 2,39, e 67,01 o ± 2,55 para o grupo de controle, o grupo M-01, e o grupo M-02, respectivamente. Na avaliação do acoplamento, o número do increasede sperm-positivo das fêmeas de 0,6 ± 0,7 no grupo de controle 1,5 ao ± 0,5 no grupo M-01 experimental. LPE dos ratos masculinos com dysfunction erectile era o ± 112 13 segundos com uma dieta regular (grupo de controle). A administração oral de M-01 em um dose do peso de corpo de 180 ou 1800 mg/kg e do M-02 em um dose de 45, 180, peso de corpo de 1800 mg/kg reduziu o LPE ao ± 54 12 segundos, 54 ± 13 segundos, o ± 71 12 segundos, os 73 segundos do ±, e o ± 41 13 segundos, respectivamente. O LPE dos ratos cirúrgicos tratou com o M-01 no dose o mais baixo (45 mg/kg) era o ± 121 12 segundos; assim, a mudança não era significativa. CONCLUSÕES: A administração oral M-01 e M-02 realçou a função sexual os ratos e ratos, do como evidenciado por um aumento o número de intromissions completos e o número de fêmeas sperm-positivas em ratos normais, e diminuição no LPE nos ratos masculinos com dysfunction erectile.
Este estudo foi conduzido perto: Bo Lin Zheng, Kan ele, yen Huang de Calvin Hyungchan Kim, de Lingling Rogers, de Yu Shao, de Zhen, cheng Qien de Yang lu, de Sui junho Yan, de Lu, e yi Zheng de Qun na faculdade médica de Liaoning & da faculdade de Shenyang da medicina chinesa tradicional em Shenyang China, e também no academy chinês da medicina preventiva, Beijing, República Popular da China.

Referências: 1. Leon T: ' o Maca ' (meyenii de Lepidium): pouca planta conhecida do alimento de Peru. Econ Bot 18: 122-127, 1964. 2. Chacón RC: Meyenii Walp de de Lepidium do fitoquímico de Estudio. Thesis, universidade Nac. mayor de San Marcos, Lima, Peru, 1961. 3. Zheng BL, Kim CH, ele K, et al: Um processo para a isolação e purification do meyenii de Lepidium. Patente pendente, 1999. 4. Procedimento de teste clínico nacional, FDA de China, 1998. 5. Hermann M, e Heller J: Raizes e tubers andean: Ahipa, arracacha, Maca e Yacom Roma, IPGRI, 1997, pp 175-195. 6. Cobo B: Historia del Nuevo Mundo. Biblioteca de Autores Españoles 81: 430, 1956. 7. Ruiz H: Relación histórica del viaje um del Peru y o Chile dos reinos dos los, 1777-1778, Madrid Acad. De Ciene Exaetas: Fis y 1 nat: 526, 1952. 8. Pulgar VJ: Fecundante de Poderoso dos sp. de Las Maca Lepidium vegetal. Voz de Huancayo do la 24:10, 1964. 9. John T: O anu e o maca. J Ethnobiol 1: 208-212.1981.
Estudo e relatório peruvian do laboratório de Maca: (Dr. Gloria Chacon, Peru Lima- Do ©) Efeitos de Maca nas glândulas do endocrine. O Dr. Gloria Chacon isolou quatro alcalóides da raiz do maca e realizou os estudos animais com os ratos masculinos e fêmeas dados a raiz pulverizada maca ou os alcalóides isolados das raizes. Em comparação com os grupos de controle animais, aqueles que recebem ou o pó da raiz os alcalóides mostraram o maturation múltiplo do follicle do ovo nas fêmeas e, nos machos, umas taxas significativamente mais elevadas da produção e do motility do sperm do que grupos de controle. O Dr. Chacon estabeleceu em que era os alcalóides na raiz do maca, não seus hormones da planta, que produziram efeitos da fertilidade os ovaries e testes dos ratos. Estes efeitos são measurable dentro de 72 horas de dosing os animais, ' ela ofereceram em uma entrevista recente do telefone de Lima, Peru. Com as experiências, deduziu que os alcalóides estavam agindo na glândula hypothalamus-hypothalamus-pituitary, que explica porque os ratos masculinos e fêmeas afflicted em uma maneira gender-apropriada. Isto a explica também porque os efeitos nos seres humanos não são limitados os ovaries e os testes, mas ao age também nos adrenals, dando um sentimento de uma energia mais grande e vitality, e em o pancreas e thyroid também. As implicações da descoberta do Dr. Chacon dos efeitos estimulando pituitary do maca são enormes. O que parece significar é essa terapia da recolocação do hormone, mesmo as variedades naturais, quer seja não mais

por muito tempo o padrão de ouro para optimising um ponto holístico da vista.
Artigo: (Nota Dos Médicos) " Viagra natural " (notas dos doutores Malaspina, muller, e Chacon. (veja vários estudos em nossas páginas da ciência por estes doutores) Os doutores Malaspina, muller e Chacon, as.well.as doutores os os EUA e Canadá, têm a notícia boa para os homens que estão sofrendo dysfunction sexual age-related. Podem esquecer-se de Viagra caro, possivelmente perigoso. Maca trabalha extremamente bom e com segurança. O Dr. Jorge Aguila Calderon, decano da faculdade da medicina humana na universidade nacional de FedericoVillareal em Lima, prescreve o maca para uma variedade larga das condições, including o osteoporosis e healing de fraturas do osso no muito idoso. Diz, no "Maca tem muitos do cálcio fàcilmente absorbable neles, mais o magnésio, e uma quantidade justa de silicone, que nós estejamos encontrando muito útil no decalcification de tratamento dos ossos as crianças e os adultos." O Dr. Calderon ajudou também a pacientes superar o impotence masculino, o sterility masculino, e o sterility fêmea empregando a terapia do maca. Os problemas que adicionais trata com o maca são os rickets, vários formulários do anaemia, sintomas menopausal tais como flashes quentes e a noite sua, dificuldades climacteric e erectile nos homens, envelhecimento prematuro, e estados gerais da fraqueza, tais como a fatiga crônica.
Artigo: (médicos testimonial e nota) Burton Goldberg, presidente da medicina alternativa que publica em Tiburon, Califórnia, cujo o livro o mais atrasado é " uma guia definitive da medicina alternativa ao cancer " é um outro entusiasta do maca. Diz que quando tentou o maca era muito pleased com os resultados e começava a fazer exame d regularmente. " Eu sou um homem velho de 72 anos e este maca fêz exame de 25 anos fora de minha vida de sexo do envelhecimento, " declara Burton Goldberg. ' que é consideravelmente importante para mim! "O Dr. Garry Gordon é concernido sobre problemas reproductive no mundo de hoje. " A sociedade enfrenta um problema enorme de deixar cair contagens do sperm e dificuldades do hormone do sexo. Mas o maca promete uma solução nontoxic com nenhuns efeitos downside. É uma terapia que pareça oferecer homens e mulheres a possibilidade para o rejuvenation hormonal, " concli o Dr. Gordon. " Nós vivemos atualmente

em uma era em que quase todos estará fazendo algo tratar das conseqüências hormonal do envelhecimento. E Maca está agora prontamente disponível."
Nota e disclaimer: T esta informação é apresentado somente tão material do interesse geral e não quanto uma prescrição para toda a pessoa específica ou qualquer circunstância em um caso específico. Nós não reivindicamos que algumas destas experiências estarão duplicadas por outras, e nós incentivamos todos procurar o dae (dispositivo automático de entrada) de um practitioner qualificado da saúde para o conselho que pertence ela/suas condição e necessidades particulares.
MACAROOT HOME | MACAMAGIC HOME MACAGROWER HOME | MACAPRODUCTS HOME
http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.herbs-america.com http://babelfish.altavista.com/babelfish/ urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.herbs-america.com
Copyright 2000-2002, REDE de AMÉRICA dos HERBS. Todo o texto e propriedade das fotos da rede de América dos herbs e para não reprinted em algum formulário sem permissão escrita da companhia. " Os herbs América " e " a mágica de Maca " são marcas registradas internacionais registadas.
Website projetado e hospedado por BASECAMP INTERNACIONAL (visto melhor nos pixels 800X600 monitore o ajuste com explorador 4,5 de Microsoft ou mais novo)
HISTORY DE MACA EM PERU Artigos, histórias, fatos, e mais...

Artigo: ((c) Por Scott Wilson, Pantera Gulch Permaculture- Williams, Oregon) Segredos da raiz perdida - um history breve da mágica de Maca nas planícies elevadas. Embora se acredita que Maca estêve cultivado assim que 4000 B.C., era mais provável domesticated inteiramente entre 1200 e 100 B.C. pelo Pumpush, tribes ferozes do guerreiro que migraram acima das selvas. Continuou a ser cultivado durante todo os highlands andean e foi trazido a uma perfeição mais grande pelo Yaro, que chegou entre 1100 e 1470 A.D. Cultivaram os campos immense de Maca, altamente desejáveis para sua "fertilidade fabulous e propriedades aphrodisiacal." Após o conquest de Inca destes tribes, emitiram quantidades grandes a Cusco como o tributo a suas réguas novas. Muita dele foi alimentada às tropas para aumentar seu vitality e fortitude. Em sua volta, o Incas conquistado e seus minions payed o tributo ao espanhol Maca e outros bens. Em 1549, é alistado nos registros como o único bom emitido como o tributo ao governo colonial, um 15000-18000 whopping martela. Mesmo que o espanhol despised oficialmente alimentos nativos, podem ter mergulhado no saco de Maca para dois reasons:1. As alturas elevadas dos Andes fizeram os spaniards quase infertile. 2. Maca teve uma tradição indígena longa a fertilidade crescente e o fortitude. Dentro do conceito andean tradicional da medicina de frio e de quente, Maca é uma planta quente. As propriedades atribuídas a esta planta singular incluem o aumento na fertilidade em todos os mamíferos, aphrodisiac, revitalizor e regulador, anti-arthritic, útil em maladies respiratory. Nenhum consumidor tradicional da maravilha de Maca tem um provérbio: Maca é vida, Maca é saúde. O conhecimento do este propriedades pequenas das raizes, passado palavra da boca através das gerações desde o tempo em immemorial, diz de seu uso aumentar igualmente a fertilidade os seres humanos e os animais domésticos; sua abilidade de aliviar o frigidity as mulheres e o impotence nos homens; seus virtues adaptogenic do revitalizer órgãos e regulador do menstruation, e do apaziguador interno dos sintomas do menopause. Recomenda-se também para o malnutrition, convalescense, perda da memória, debility mental, e como um tonic geral; Suas propriedades anti-arthritic como uma planta quente; Seu uso em ailments respiratory de tratamento. Alguns herbalists recomendam não usar Maca para povos com hypertension. Entretanto, este counterindication não foi testado scientifically. Em épocas antigas, Maca era inteiro cozinhado nos poços, mergulhados com

carvões da terra charred e as raizes. Isto chamaram o "huatia". Ou fizeram o "atunca" fervendo, mashing, e rolando o em esferas e cozinhando o em uns potenciômetros de argila alinhados com palha. Hoje os usos de Maca são completamente variados. Seu uso mais popular no mercado internacional é como cápsulas e tabuletas. Mas a diversidade deve ser seu nome médio porque é também um ingrediente excelente nos concoctions doces e savory. O caráter piquant do caramelo de manteiga de Maca é a fundação dos produtos originais que variam dos liqueurs aos bens cozidos. Para toda a que history surpreendente de Maca e seu serviço indubitable aos seres humanos, é virtualmente incomprehensible encontrar tão tarde quanto 1992, ele estêve alistado como no perigo da extinção. Em 1979, o ano o mais escuro de Maca, o dept. peruvian da agricultura encontrou o hectares(about somente 25 70 acres) de Maca sob o cultivation no país inteiro! Desde os 1980 though, o cultivation de Maca tem-se levantado lentamente, e agora há um renascimento verdadeiro afoot. A vida nova está sendo respirada outra vez nos solos dos Andes elevados enquanto os segredos da raiz perdida estão sendo revelados em torno do mundo.
Artigo: (desconhecido da fonte) A importância de Maca no history de Peru O cultivation de Maca vai para trás talvez cinco millennia. Era uma parte integral da região elevada da dieta e o comércio de Andes. Quando controlaram essa determinada região americana sul, o Incas encontrou o maca assim potent que restringiram seu uso a sua corte do royalty. Em cima de overrunning os povos de Inca, conquistar spaniards tornou-se ciente do valor desta planta e coletou-se o tributo em raizes do maca para a exportação a Spain. Maca foi usado como um enhancer da energia e para o nutrition pelo royalty espanhol também. Mas eventualmente o conhecimento de qualidades especiais dos maca morreu para fora, sendo preservado somente em algumas comunidades peruvian remotas. Nos 1960s e mais tarde nos cientistas dos 1980s, os alemães e os norte-americanos que pesquisam botanicals em Peru, interesse reaceso no maca com as análises nutritivas de o que foi designado como ' as colheitas perdidas dos Andes.' A publicação de um livro por esse maca introduzido conhecido ao mundo.
MACAROOT HOME | MACAMAGIC HOME MACAGROWER HOME | MACAPRODUCTS HOME

http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.herbs-america.com http://babelfish.altavista.com/babelfish/ urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.herbs-america.com
Copyright 2000-2002, REDE de AMÉRICA dos HERBS. Toda a cópia e propriedade das fotos da rede de América dos herbs e para não reprinted em algum formulário sem permissão escrita da companhia. " Os herbs América " e " a mágica de Maca " são marcas registradas internacionais registadas.
Website projetado e hospedado por BASECAMP INTERNACIONAL (visto melhor nos pixels 800X600 monitore o ajuste com explorador 4,5 de Microsoft ou mais novo)
O Dossier De Ibogaine
Conferência de NYU sobre Ibogaine novembro 5-6, 1999
Raizes do iboga do T.
Ibogaine é extraído de o bark da raiz Cano principal | Busca | Ciência | Opinião | Literatura | Ligações | Tratamento | Livraria | Gabarito

Pharmacology Ibogaine do e alcalóides Ibogaine-Relacionados Piotr Popik , instituto do pharmacology, academy polonês das ciências, 31-343 Kraków, Poland e Phil Skolnick , descoberta do neuroscience, laboratórios de pesquisa de Lilly, centro incorporado de Lilly, Indianapolis, em 46285, EUA
Aparecido como o capítulo # 3, "NOS ALCALÓIDES", Vol.52, pp. 197-231. 1998, San Diego, ISBN 0-12-469552-3 dos EUA. Editado por G.A. Cordell. © 1998 pela imprensa academic
Requisite Os Alcalóides, Vol. 52
ÍNDICE
I. INTRODUÇÃO II. VISTA GERAL HISTÓRICA. III. ESTRUTURA QUÍMICA E PROPRIEDADES. IV. PHARMACOKINETICS . V. PHARMACOLOGICAL GERAL AÇÕES. A. Animal Estudos 1. Locomotor atividade. a. Efeitos na atividade locomotor induzida por outras drogas 2. Tremor. 3. Ansiedade e medo. 4. Efeitos no self-administration de outras drogas. 5. Efeitos na dependência de droga. 6. Dor e analgesia. 7. Aggression. 8. Interoceptive propriedades. 9. Reforçar efeitos. 10. Efeitos na aprendizagem e a memória. 11. Cardiovascular ações. B. Humano Estudos. VI. LETHALITY E EFEITOS DE NEUROTOXIC. VII. EFEITOS NO ESPECÍFICO SISTEMAS DO NEUROTRANSMITTER

A. Ibogaine Efeitos em sistemas dopaminergic. 1. Dopaminergic efeitos: Specificity Pharmacological. 2. Ibogaine altera os efeitos de drogas abusadas em sistemas dopaminergic. B. Opioid Sistemas C. Serotonergic Sistemas. D. Cálcio Regulamento. E. Cholinergic Sistemas. F. Gamma-Gamma-Aminobutyric Sistemas De Acidergic [ GABAergic ]. G. Tensão-Dependente Canaletas Do Sodium. H. Glutamatergic Sistemas. I. Sigma Receptors. J. Variado Ações de Ibogaine. VIII. CONCLUSÕES. IX. Reconhecimentos X. REFERÊNCIAS XI Tabela 1. Interações do ibogaine com sistemas do neurotransmitter: estudos radioligand do emperramento.
I. INTRODUÇÃO Ibogaine (12-methoxyibogamine, NIH 10567, Endabuse) é um dos alcalóides psychoactive do indole encontrados no shrub africano ocidental, Tabernanthe iboga . Por sobre um século, ambos os extratos de T. Iboga e seu os alcalóides constituent, including o ibogaine, foram usados como medicinals (1). O que faz este alcalóide do interesse particular ao comtemporâneo o pharmacology é observações anecdotal que indicam que o ibogaine possui propriedades "anti-anti-addictive". Assim, o ibogaine (6-25 mg/kg, nos seres humanos) tem reivindicado atenuar sintomas da dependência e da retirada a a variedade de drogas abusadas including opiates, álcool, nicotine e psychostimulants ( 2-9 ). Estudos de Preclinical que demonstram que o ibogaine reduz o self-administration a cocaína do e o morphine, e atenua os sintomas da morphine-retirada, seja consistente com estas reivindicações [ revistas dentro (Popik e Glick ( 10 ) ]. Este capítulo revê as propriedades pharmacological do ibogaine e relacionadas alcalóides. Desde nossa última revisão detalhada ( 11 ), mais de uma cem relatórios novos nas ações pharmacological do ibogaine e ibogaine-como os alcalóides apareceram. O chemistry do ibogaine foi revisto perto Alfaiate nesta série ( 12.13 ). ALTO
II. VISTA GERAL HISTÓRICA.

Ibogaine é derivado do iboga de Tabernanthe , um shrub indígeno ao Central-Oeste África. O shrub do iboga, um membro de a família Apocynaceae (ordem Contortae), é encontrada tipicamente no undergrowth das florestas tropicais ( 14 ). As raizes do iboga de Tabernanthe foram usadas nos rites tribais da iniciação ( 15.16 ). Embora os detalhes de tais ceremonies varie, ele foi acreditado que a raiz do iboga permitiu novatos para fazer o contato com os antepassados no mundo do espírito. Ibogaine foi também encontrado no crassa de Tabernanthe ( 17 ). Décimos nonos relatórios do século dos exploradores franceses e belgas descreveu primeiramente o stimulant e o aphrodisiac efeitos de comer a raiz do iboga ( 1.16 ). A primeira descrição botanical da planta, foi feito por Baillon em 1889 ( 18 ). Dybovsky e Landrin ( 19 ), as.well.as Haller e Heckel ( 20 ), era o primeiro para isolar um alcalóide cristalino de raiz do iboga, que chamaram o "ibogaine" ou o "ibogine". Em 1901 Franceses os pharmacologists encontraram o ibogaine para ter um tipo incomun de efeito excitatory nos animais ( 21-23 ). Phisalix ( 23 ) sugeriu esse ibogaine podia produzir os hallucinations baseados em observações do comportamento incomun nos cães. O alcalóide foi testado subseqüentemente em ajustes clínicos ocidentais, e foi recomendado como um stimulant para o tratamento do convalescence e neurasthenia ( 24 ). Apesar de tais recomendações, ibogaine nunca o uso clínico largo apreciado e foi negligenciado por investigadores para quase 30 anos. Nos 1940 Raymond-Hamet e os colegas de trabalho publicaram uma série de papéis que descrevem as propriedades pharmacological do ibogaine no isolado tecidos e o sistema cardiovascular ( 25-32 ). Lambarene, um extrato de as raizes do manii relativo de Tabernanthe do iboga , foram vendidas em France durante os 1930. Conteve magnésio aproximadamente 8 do ibogaine, e foi descrito como um stimulant. Iperton, um outro extrato do ibogaine, foi usado também como um tonic ou stimulant ( 33 ). Ibogaine foi usado por atletas como um desempenho realçando a droga ( 34 ). Em muitos países, including os estados unidos, o uso do ibogaine é proibido, talvez por causa do seu hallucinogenic purported efeitos (publicized extensamente nos 1960 atrasados) e sua aparência no mercado illicit da droga. Em 1970, o alimento dos estados e a administração unidos da droga ibogaine classificado como uma substância da programação I (todo o uso da non-pesquisa proibido). Começando em 1985, uma série das patentes foi emitida para o uso do ibogaine como meios rápidos de interromper o addiction aos narcotics (morphine e heroin) (3), cocaína e amphetamine (4), álcool (5), nicotine (6)

e syndrome da dependência da poly-droga ( 35 ). Reivindicação destas patentes que um dose oral ou rectal do ibogaine (4-25 mg/kg) interrompe a dependência syndrome, permitindo que os pacientes mantenham um lifestyle droga-livre para ao menos 6 meses. Baseado em estudos clínicos abertos, reivindicou-se ( 36 ) essa terapia do ibogaine resultou em 25% de pacientes restantes droga-livre sem craving por 6 meses. Este grupo incluiu aqueles que eram ambos motivated altamente para parar e tiveram ambientes home relativamente estáveis. Uns outro 40-50% dos pacientes tiveram seus addictions interrompidos com sucesso, e psychotherapy requerido. Vinte a 30% dos pacientes tinham retornado à droga uso dentro de um mês que segue o tratamento. Abaixe um tanto as taxas do sucesso (10-15%) cited por Touchette ( 37 ). Na ausência de clínico apropriadamente controlado os estudos, o efficacy do ibogaine como um agente anti-anti-addictive não podem ser avaliado rigorously no tempo atual. Nonetheless, interesse no ibogaine como um tratamento para o addiction aumentou. Em 1985 NDA Internacionais, O Inc. (console de Staten, NY, EUA) começou uma campanha a persuadir o governo de ESTADOS UNIDOS para iniciar experimentações clínicas controladas com ibogaine ( 38 ). No o mesmo tempo, o uso do ibogaine para tratar a dependência do opioid aumentou em Europa ( 39 ). Nas experimentações atuais, clínicas para avaliar a segurança do ibogaine esteja underway na universidade de Miami e são planeados dentro York Novo. Experimentações clínicas para testar o efficacy anti-anti-addictive do ibogaine está underway in os Países Baixos e Panamá ( 38.40-44 ). Concordar a Ali et a al ., ( 45 ), ao alimento de ESTADOS UNIDOS e à administração da droga e o instituto nacional para o abuso da droga aprovou o uso do ibogaine em uma base limitada para tratar o addiction da cocaína. ALTO
III. ESTRUTURA QUÍMICA E PROPRIEDADES.
Figura 1. Composto R1 R2 R3 R4 Ibogaine CH 2 CH 3 H OCH 3 H O - Desmethylibogaine CH 2 CH 3 H Oh H ()-Ibogamine do ± CH 2 CH 3 H H H ()-Coronaridine do ± CH 2 CH 3 CO 2 CH 3 H H Tabernanthine CH 2 CH 3 H H OCH 3

O-t- Butyl-O-Desmethylibogaine-O-Desmethylibogaine CH 2 CH 3 H OC(CH 3) 3 H
Embora o ibogaine primeiramente fosse isolado e identificado em 1901, ( 19-21.46 ), na estrutura do este e em alcalóides relacionados (o Fig. 1) foi estabelecido primeiramente por Alfaiate em 1957 ( 47 ) [ veja também Alfaiate ( 12.13 ) ]. A síntese total do nicotinamide foi relatada usando 13- ( 48 ) ou (um 49 ) seqüência 14-step. Os 13 C Os spectra NMR de diversos alcalóides do iboga foram publicados em 1976 ( 50 ). A síntese de ibogaine tritiated foi relatada recentemente ( 51.52 ). Ibogaine (mol. do peso. 310.44) têm um ponto de derretimento de 153° em 0,01 milímetros hectograma e um p K a de 8,1 no methylcellosolve de 80%. Os máximos do absorption no metanol são 226 (registro e 4,39) e 296 (registro e 3,93) nm. Ibogaine cristaliza-se das soluções alcoólicas em agulhas prismatic pequenas, avermelhadas; é levorotatory [ a ] D -53° (no ethanol de 95%) e é soluble no ethanol, no metanol, no clorofórmio e na acetona, mas em insoluble dentro água. Hydrochloride de Ibogaine (o ponto se congelando 299°C, [ a ] D -63° (ethanol), [ a ] D -49° (H 2 O)) é soluble na água, ethanol e metanol, é ligeiramente o soluble a acetona e o clorofórmio, e é praticamente insoluble no ether ( 53 ). Ibogaine é calor e light-sensitive ( 54 ) e lata oxide espontâneamente na solução, dando o iboluteine e o ibochine ( 16.34 ). Os alcalóides relacionados estrutural ao ibogaine incluem o tabernanthine, ibogamine, iboxigaine, gabonine, iboquine, kisantine e ibolutenine. Similaridades estruturais entre o ibogaine e o outro alcalóide do indole hallucinogens estiveram também relatado ( 55 ). A síntese de diversos derivatives do ibogaine tem recentemente publicado por Repke e por colegas de trabalho ( 56 ). ALTO
IV. PHARMACOKINETICS. Após a administração parenteral, o ibogaine foi identificado em vários materiais biológicos, including o sangue e o urine (seres humanos) e no fígado, em kidney e no cérebro dos animais de laboratório ( 54.57-59 ). Uma hora após a administração intraperitoneal, concentrações elevadas do ibogaine estavam atuais no (o fígado do rato e os kidneys 60 ). Após a injeção intravenous de 10 mg/kg aos ratos, concentrações máximas do cérebro (48 µg/g do peso molhado [ µM ~133 ]) foram conseguidos em 10 segundo ( 61 ).

Recentemente, Gallagher et o al ., ( 62 ) têm desenvolveu um método altamente sensível e específico para quantify dentro o ibogaine plasma e tecidos. Este método usa a extração orgânica, derivatization com tr *** TRANSLATION ENDS HERE ***ifluroacetic anhydride, and detection by gas chromatography-mass spectrometry (GC/MS). Similar methods were developed by Hearn et al ., ( 63 ), Alburges et al ., ( 64 ) and Ley et al ., ( 65 ). Using a GC/MS method, Pearl and colleagues ( 66 ) reported that 1, 5 and 19 hours after intraperitoneal administration of 40 mg/kg of ibogaine, the whole brain levels of ibogaine were 10, 1 and 0.7 µM in female rats and 6, 0.9 and 0.2 µM in male rats, respectively. Hough et al ., ( 67 ) studied the tissue distribution of ibogaine after i.p. and s.c. administration in rats. One hour after i.p. dosing (40 mg/kg), drug levels ranged from 106 ng/ml (~ 0.3 µM) in plasma to 11,308 ng/g (~ 36 µM) in fat, with significantly higher values after s.c. administration of the same dose. Drug levels were 10-20 fold lower 12 hours later. These data indicate that ibogaine is subject to a significant "first pass" effect after i.p. dosing, and that there is a marked propensity for ibogaine to be deposited in adipose tissue, reflecting its lipophilicity. Consistent with its lipophilicity, ibogaine levels in adipose tissue were very high for at least 12 hours after administration. Based on these data, it was suggested that a single dose of ibogaine may provide a long-acting, depot-like time course of action ( 67 ). The reported long-term effects of ibogaine (e.g. ( 68-70 )), have led to the hypothesis that this alkaloid may be metabolized to an active principle with a long half life ( 71 ). At present, there is no direct evidence to support this hypothesis. Ibogaine was reported to disappear from the rat at a rate of ~4% of the administered dose per hour with ~ 5% of the injected dose eliminated unchanged in urine. Elimination kinetics from brain yielded a half-life of 60 min in rodents ( 60,61 ) and suggest a one-compartment model. After administration of ibogaine (10 mg/kg, p.o.) to rabbits, urine concentrations reached a maximum 4-5 hours later, then decreased rapidly and disappeared after 6 hours ( 54,60 ). Taken together, these data suggest that ibogaine is extensively metabolized. Inspection of ibogaine's structure (Fig. 1) led us to hypothesize that a likely degradation pathway is O -demethylation at C12. Based on this hypothesis, O -desmethylibogaine (also known as noribogaine or 12-hydroxyibogamine), was synthesized by Dr. C. Bertha at the National Institutes of Health in 1994. At the same time, O - tert -butyl- O -desmethylibogaine was synthesized in an attempt to make an ibogaine derivative resistant to O -demethylation (Fig. 1). Thus, the first compound was synthesized to investigate the potential pharmacological actions of a likely ibogaine metabolite. The second compound permitted examination of

the pharmacological effects of an ibogaine derivative that would not be degraded by O -demethylation. The synthesis of these compounds was described by Layer et al ., ( 72 ). Recent studies have indeed demonstrated that ibogaine is metabolized, and that O -desmethylibogaine can be detected in human plasma ( 73 ) as well as in the plasma and brains of ibogaine-treated rats ( 66 ). Behavioral and neurochemical studies in rodents have established that O -desmethylibogaine is pharmacologically active (discussed later). Following an i.p. dose of ibogaine (40 mg/kg), Pearl et al ., ( 66 ) reported brain O -desmethylibogaine concentrations of 20, 10 and 0.8 µM in female rats and 13, 7 and 0.1 µM in male rats, respectively, at 1, 5, and 19 hours after administration. These data suggest that gender differences in pharmacological responses to ibogaine may be attributed to pharmacokinetic, rather than pharmacodynamic, factors. While a report of one human subject ( 73 ) indicated that O -desmethylibogaine persisted in plasma at high levels for at least 24 hours after oral ibogaine administration, it is not clear if this pattern will be representative. There is evidence indicating that the various pharmacological effects of ibogaine may be attributable, at least in part, to its metabolite(s). For example, the tremorigenic effects of ibogaine dissipate much more rapidly than its ability to attenuate the morphine withdrawal syndrome in rats ( 74 ). This finding suggests that an active principle(s) responsible for one action may be more rapidly metabolized than compound(s) involved in other actions. Alternatively, the various pharmacological effects of ibogaine may involve different neurotransmitter pathways (discussed later). TOP
V. GENERAL PHARMACOLOGICAL ACTIONS. A. Animal Studies 1. Locomotor activity. Ibogaine produces complex effects on locomotor activity in rodents. A dose of 20 mg/kg (i.p.) slightly increased locomotor activity in mice ( 75 ) while Sershen et al., ( 76 ) reported that 40 mg/kg (i.p.) decreased locomotor activity in male mice at 1, but not 24, hours after injection. The same dose inhibited locomotion in female rats during the first hour after injection, whereas one week later locomotor activity was increased ( 69 ). Recently, Pearl and colleagues ( 66 ) noted gender differences in the effects of ibogaine on locomotor activity (40 mg/kg, i.p., 5 or 19 hours before test). In control males and females the locomotor activity decreased during the second hour of observation. Ibogaine treatment in females

prevented this decrease in locomotor activity. In females, but not males, ibogaine decreased locomotor activity when given 19 hours before the test ( 66 ). Another study revealed that in male rats, a single dose of 40 mg/kg inhibited locomotor activity 4 hours after injection; a dose of 80 mg/kg decreased motor activity 24 hours after injection ( 77 ). Rats injected with doses of 20-60 mg/kg of ibogaine displayed slower response times on sensory and sensory-motor tests and were also impaired in performing specific motor reflexes at doses of 40-60 mg/kg. Furthermore, these rats exhibited a marked reduction in locomotor activity as well as in emotionality at doses ranging from 10- 40 mg/kg. At higher doses (40 mg/kg), rats appeared virtually inactive ( 78 ). In other studies, at doses above 25 mg/kg, ibogaine produced ataxia, splayed hind limbs, outstretched forelimbs, Straub tail and hyperexcitability ( 79 ). One hour after O -desmethylibogaine or 18-methoxy-coronaridine injection (40 mg/kg), locomotor activity was increased during the second hour of observation ( 66,80 ). In our studies, high doses (120 mg/kg) of O -desmethylibogaine and O - t -butyl- O -desmethylibogaine produced profound ataxia and convulsions ( 72 ). Ibogaine, O -desmethylibogaine, and O - t -butyl- O -desmethylibogaine, (80 mg/kg) did not significantly influence rotorod performance in mice ( 72 ). TOP
a. Effects on locomotor activity induced by other drugs Ibogaine has been found to affect the motor stimulant properties of amphetamine, cocaine, and morphine in rodents (hyperlocomotion induced by these drugs is believed to reflect their "psychotomimetic" qualities in man). Although the results of these studies are not uniform, in general, it has been found that in female rats this alkaloid potentiates the locomotor response to amphetamine and cocaine, whereas opposite effects were reported in male rats and mice. Sershen et al., ( 81 ) found that ibogaine (40 mg/ kg i.p., 2 or 18 hours before amphetamine) enhanced amphetamine (1 mg/kg) - induced hypermotility in female rats. In other studies, an amphetamine-induced increase in locomotor activity was potentiated in female rats pretreated with ibogaine (40 mg/kg, i.p.) 19 hours earlier ( 82 ). Cocaine-induced hypermotility in female rats was also potentiated by ibogaine ( 83,84 ). Broderick et al. , ( 85,86 ) reported that ibogaine (20-40 mg/kg, i.p.) administration to male rats for four days reduced cocaine (20 mg/kg) - induced hypermotility. Ibogaine (40 mg/kg, i.p.) administration also reduced cocaine- (25 mg/kg, s.c.) induced hypermotility in male mice ( 76 ), a

finding in agreement with the amphetamine (1 mg/kg) - ibogaine interaction ( 81 ) in this gender and species. Recent data demonstrate that the effects of ibogaine on cocaine (20 mg/kg) -induced hyperactivity in female rats are time dependent. Thus, given 1 h before cocaine, ibogaine and O -desmethylibogaine (40 mg/kg) inhibited cocaine-induced hyperactivity, but when given 19 h before cocaine they produced the opposite effect ( 80 ). Ibogaine pretreatment (40 mg/kg, i.p. 19 hours before measurement) decreased or blocked the locomotor stimulation induced by morphine (0.5-20 mg/kg) in rats ( 69,71 ). Ibogaine administered one week (but not one month) before morphine (5 mg/kg) reduced the motor stimulant effects of this opiate ( 69 ). Pearl et al ., ( 87 ) found that ibogaine (5-60 mg/kg) is more potent in inhibiting morphine-induced hyperlocomotion in rats pretreated with morphine for several (1-4) days compared to non-pretreated rats. Doses of ibogaine (5-10 mg/kg) that alone were inactive in drug-naive animals attenuated morphine-induced hyperactivity in the morphine pretreated rats. The inhibitory effects of ibogaine on morphine-induced hyperlocomotion appear gender related, because ibogaine is more potent in female rats ( 66 ). Ibogaine-induced inhibition of morphine - induced hyperlocomotion can be reversed by coadministration of a kappa antagonist (norbinaltorphine, 10 mg/kg) and an NMDA agonist (NMDA, 20 mg/kg). However, neither norbinaltorphine nor NMDA alone blocked this action of ibogaine ( 88 ). O -Desmethylibogaine (10-40 mg/kg) also inhibited morphine-induced hyperlocomotion in female rats. However in male rats, the dose of 10 mg/kg potentiated and 40 mg/kg inhibited morphine-induced hyperlocomotion ( 66,89 ). TOP
2. Tremor. Like the somewhat structurally related alkaloid harmaline, ibogaine produces tremors. In mice, ibogaine is tremorigenic both when given intracerebrally (ED 50 127 nmol/g brain, ~ 46 µ g/g with a latency to tremor of about 1 minute) ( 90 ), and systemically (ED 50 12 mg/kg, s.c.) ( 61 ). In rats, ibogaine produced fine tremors, flattening of body posture, and flaccid hind limbs up to 2 hours after administration of 40 mg/kg (i.p.) ( 91 ). Low-amplitude whole body tremors appearing within 10 min after administration of as little as 10 mg/kg of ibogaine have also been reported ( 92 ). O'Hearn et al ., ( 93 ) reported that a high dose of ibogaine (100 mg/kg) produced ataxia and high-frequency tremor of the head and trunk in rats. Ibogaine-induced tremor preferentially involves the head and upper extremity in rats

and mice ( 94 ). Ibogaine (20 mg/kg) - induced tremors in mice were blocked more potently by CCK-8 and ceruletide compared to other reference compounds, including prolyl-leucylglycine amide (MIF), atropine, haloperidol, biperiden, ethopropazine, trihexyphenidyl, methixene and clonazepam ( 95 ). Zetler et al., ( 61 ) established the tremorigenic structure-activity relationship of several ibogaine-like compounds in descending order of potency: tabernanthine > ibogaline > ibogaine > iboxygaine > O -desmethylibogaine. Glick et al., ( 96 ) found that at behaviorally effective doses (2-80 mg/kg) ibogaine, desethylcoronaridine, harmaline and tabernanthine produced tremors for at least 2-3 hours. Both the R and S enantioners of ibogamine and coronaridine were devoid of this action. The ibogaine-like alkaloids, 18-methoxycoronaridine and O -desmethylibogaine were also found to lack tremorigenic effects ( 89,97 ). The tremorigenic properties of ibogaine and related compounds have been attributed to an action on GABAergic pathways ( 98-100 ) and to the blockade of voltage-dependent sodium channels. TOP
3. Anxiety and fear. Schneider and Sigg ( 101 ) described the behavioral effects of ibogaine in cats. The authors concluded that after intravenous administration of 2-10 mg/kg, ibogaine produced fear-like reactions that persisted for 10-20 minutes with a normal appearance observed 1-2 hours after injection. The electroencephalographic pattern obtained after ibogaine administration (2-5 mg/kg) showed a typical arousal syndrome, resembling that observed after direct stimulation of the reticular formation. This arousal syndrome was inhibited by atropine (2 mg/kg) ( 101 ). Gershon and Lang ( 102 ) described the effects of ibogaine in dogs, which become more tense and alert, interpreted as the appearance of anxiety. Moreover, they observed that the dogs exhibited a lack of recognition of both their regular handlers and environment. Recently, Benwell et al ., ( 103 ) reported reductions in open arm entries in the elevated plus-maze test when rats were tested 22 hours after pretreatment with ibogaine (40 mg/kg, i.p.). In mice, ibogaine (2.5 mg/kg) exhibited anxiogenic actions, whereas a dose of 1 mg/kg had anxiolytic effects ( 104 ). These are perhaps the most compelling preclinical data that ibogaine may influence anxiety levels because anxiolytic agents (e.g. benzodiazepines) increase open arm entries in this test. TOP

4. Effects on self-administration of other drugs. Ibogaine (40 mg/kg, i.p.) inhibits the self-administration of cocaine in rodents. Cappendijk and Dzoljic ( 105 ) trained male Wistar rats to intravenously self-administer cocaine; a single dose of ibogaine (40 mg/kg) decreased cocaine intake by 40-60% for several days, and repeated treatment with ibogaine at one-week intervals decreased cocaine self-administration by 60-80%. This decrease was maintained for several weeks. Similar effects were found in mice that developed a preference for cocaine in the drinking water. Thus, ibogaine administration (two weeks after the beginning of a choice period, 2 doses of 40 mg/kg, 6 hours apart) diminished cocaine preference for five days (70). According to Vocci and London ( 106 ), some investigators have failed to replicate ibogaine's effect on cocaine self-administration in the rat ( 107 ) and rhesus monkey ( 108 ). Also Dworkin et al ., ( 109 ) reported that neither 40 mg/kg of ibogaine given 60 min prior to the session, nor 80 mg/kg given 24 hour before the session, suppressed responding maintained by intravenous cocaine infusions. In this study, cocaine self-administration was inhibited by pretreatment with ibogaine (80 mg/kg) either 60 or 90 min prior to the session ( 109 ). However, because this dose of ibogaine reduced scheduled food intake, these latter effects of ibogaine on cocaine self-administration appear to be unspecific. Glick et al., ( 96 ) demonstrated that ibogaine and several iboga alkaloids (tabernanthine, R - and S -coronaridine, R - and S - ibogamine, desethylcoronaridine, and harmaline) reduced cocaine self-administration in rats in a dose-related fashion (2.5-80 mg/kg). For some alkaloids, these effects were seen the day after injection. O -Desmethylibogaine (40 mg/kg) ( 89 ) and 18-methoxycoronaridine ( 97 ) were also reported to inhibit cocaine self-administration. Ibogaine dose dependently (2.5-40 mg/kg) reduced intravenous morphine self-administration in female Sprague-Dawley rats immediately after injection as well as on the next day ( 68 ). In some animals, a reduced morphine intake was observed for several days; other rats required several doses of ibogaine to achieve a prolonged reduction. Similar effects were demonstrated for other ibogaine-like alkaloids including O -desmethylibogaine ( 89 ), tabernanthine, R - and S -coronaridine, R - and S - ibogamine, desethylcoronaridine, harmaline ( 96 ) and 18-methoxycoronaridine ( 97 ). However, data from another study revealed somewhat different results. Thus, Dworkin et al ., ( 109 ) found that ibogaine (40 or 80 mg/kg) diminished heroin self-administration in male Fisher rats only on the day it was administered. Moreover, the same study revealed that

ibogaine treatment resulted in a 97% decrease in responding for a food reinforcement schedule, suggesting that its effects on heroin self-administration were unspecific. Ibogaine-induced inhibition of morphine self-administration has been found to be reversed by sequential administration of a kappa antagonist (norbinaltorphine, 10 mg/kg) and an NMDA agonist (NMDA, 20 mg/kg). Neither norbinaltorphine nor NMDA alone were effective in this respect (88). Ibogaine (10-60 mg/kg) reduced alcohol intake in alcohol-preferring Fawn Hooded rats, without affecting either blood alcohol concentrations or food intake ( 110,111 ). The authors concluded that a metabolite could be involved, because ibogaine was effective in this measure when administered intraperitoneally and intragastrically, but not subcutaneously ( 112 ). A recent study demonstrated an attenuation of alcohol consumption by the ibogaine congener, 18-methoxycoronaridine in rats (113). TOP
5. Effects on drug dependence. Repeated administration of ibogaine (10 or 40 mg/kg) did not produce dependence in rats as measured using the Primary Physical Dependence test ( 114 ). In morphine-dependent rats, the opioid antagonist naloxone induces a withdrawal syndrome, characterized (in rats) by increased rearing, digging, jumping, salivation and "wet-dog" head shaking. Ibogaine dose-dependently reduced the frequency of some of these withdrawal symptoms (jumping, rearing, digging, head hiding, chewing, teeth chattering, writhing, penile licking) after both intracerebroventricular (4-16 µg) ( 115 ) and i.p. administration (40 and 80 mg/kg) ( 74,116 ). However, these effects could not be replicated in other studies in either rats ( 39,117 ) or mice ( 118 ). At least the second failure to replicate can be attributed to the fact that in the Frances et al ., ( 118 ) study, ibogaine was administered to animals that developed a full withdrawal syndrome. In morphine-dependent monkeys, ibogaine (2 and 8 mg/kg, s.c.) partially suppressed the total number of withdrawal signs ( 114 ). Our studies ( 72,119 ) demonstrate that ibogaine inhibits the morphine withdrawal syndrome in mice in a dose-related fashion. This effect was reversed by combining ibogaine treatment with glycine. Structure-activity studies revealed that among various ibogaine-like compounds (including O -desmethylibogaine and O - t -butyl- O -desmethylibogaine), only ibogaine inhibited the intensity of morphine withdrawal ( 72 ). Both the ability of glycine to inhibit this effect of ibogaine

and the failure of other ibogaine derivatives to potently inhibit the binding of noncompetitive NMDA antagonists (e.g., [ 3 H]–N-[1-(2-thienyl)cyclo-hexyl]-3,4-pipenoline (TCP) and [ 3 H]–MK-801) suggests that the NMDA antagonist actions of ibogaine are responsible for its anti-withdrawal effects. This hypothesis is supported by the observation that while O -desmethylibogaine and O - t -butyl- O -desmethylibogaine had much higher affinities for kappa opioid receptors than ibogaine did, only ibogaine exhibited a significant affinity for NMDA receptors. TOP
6. Pain and analgesia. Ibogaine did not mimic the analgesic action of morphine in either the tail flick (1-40 mg/kg, i.p.) or hot plate (up to 20 mg/kg, i.p.) tests, although it exhibited analgesic activity in the phenylquinone writhing test (ED 50 9.7 mg/kg) ( 114,120,121 ). Ibogaine did not exhibit antinociceptive activity when given twice a day for 4 days ( 122 ). Ibogaine either increased ( 120,123 ) or did not affect ( 114,121 ) morphine analgesia in the tail flick test. Similarly, it did not influence analgesia produced by either a kappa opioid agonist (U-50,488H) or a delta opioid agonist [D-Pen 2 ,D-Pen 5 ]enkephalin (DPDPE) ( 121 ). Ibogaine has been reported to decrease analgesia in rats when given 19 hours prior to morphine ( 123 ), but another report indicates ibogaine is not effective when given 4-24 hours prior to morphine administration in mice ( 121 ). In addition, Cao and Bhargava ( 122 ) demonstrated that ibogaine (40-80 mg/kg) inhibited the development of analgesia to mu, but not kappa or delta, agonists in mice. O -Desmethylibogaine (40 mg/kg) potentiated morphine-induced analgesia in rats ( 123 ) and mice ( 121 ). This effect was no longer apparent 19 hours after its administration ( 123 ). The potentiation of morphine-induced analgesia may be attributed to the relatively high affinity of O -desmethylibogaine at opioid mu (K i 2.66 ± 0.62 µM) and kappa (K i 0.96 ± 0.08 µ M) receptors ( 124 ). However, this interpretation appears unlikely because O -desmethylibogaine pretreatment did not influence either kappa - or delta - opioid agonist - induced antinociception ( 121 ). Ibogaine (10-40 mg/kg) completely blocked the antinociceptive effect of (–)-epibatidine in rodents, but was ineffective when given at a dose of 40 mg/kg 24 h before epibatidine. These data suggest that this was an effect of ibogaine and not that of its putative, long-lasting metabolite ( 125 ). This blockade of the antinociceptive effect of epibatidine is not surprising, because epibatidine-induced analgesia is mediated by a mechanism fundamentally different from that of the opioids.

TOP
7. Aggression. Compared to other psychoactive compounds (e.g. psilocybin, JB-336, and bufotenine), ibogaine (10 mg/kg) had a negligible effect on the aggressiveness of isolated mice and muricidal behavior in rats ( 126 ). TOP
8. Interoceptive properties. Animals can be trained to "recognize" similarities among drugs. Such discriminative (interoceptive) properties may suggest a similar mechanism of action not necessarily related to the structure of a compound. No generalization between ibogaine and serotonergic ligands (e.g. fenfluramine, N -(3-trifluoromethylphenyl)piperazine [TFMPP], 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane [DOI], methyl-enedioxymethamphetamine [MDMA], quipazine or LSD) was found in drug-discrimination paradigms ( 127,128 ). However, Palumbo and Winter ( 129 ) did observe a generalization between ibogaine (15-20 mg/kg) and dimethoxymethylamphetamine [DOM] (0.6 mg/kg), as well as between ibogaine and LSD (0.1 mg/kg) in a two-lever discrimination task. Because pizotyline (BC-105) blocked DOM-appropriate and LSD-appropriate responses, an involvement of 5-HT 2 or 5-HT 1 receptors in the stimulus properties of ibogaine was suggested. Similarly, no generalization between ibogaine and CGS 10476B (a dopamine release-inhibiting agent) was found in a drug-discrimination paradigm ( 127 ). In contrast, ibogaine substituted as an interoceptive cue in mice trained to recognize MK-801 (dizocilpine) ( 119 ), but not to [(+)-HA-966] (a low efficacy partial agonist of the glycine site at the NMDA receptor) ( 130 ) in a T-maze drug discrimination paradigm. Helsley and colleagues ( 131 ) studied the interoceptive cue produced by ibogaine in male Fisher rats. The time course of the ibogaine (10 mg/kg) cue revealed that a maximum of ibogaine-appropriate responses were observed at a 60 min pretreatment time, and, that at the pretreatment time of 8 hours, no ibogaine-like responses were observed. These findings, together with observation that O -desmethylibogaine substituted only partially to the ibogaine cue, suggest that the subjective effects of ibogaine are not due to this putative metabolite. The same study however, revealed that harmaline completely substituted as an ibogaine cue ( 131 ). This later finding indicates that animals may recognize the tremorigenic effects of ibogaine. TOP

9. Reinforcing effects. Ibogaine does not appear to possess rewarding or aversive effects as measured in the conditioned place preference/aversion test ( 132 ), a preclinical procedure that can predict abuse potential in humans. Nonetheless, the same authors reported that ibogaine (40 mg/kg) may attenuate the acquisition, but not expression of morphine and amphetamine place-preference in male rats ( 77,132,133 ). This dose of ibogaine did not interfere with the acquisition of conditioned place aversion induced by either naloxone or lithium chloride ( 132 ). Ibogaine (40 mg/kg, 22 hours before the test) attenuated the establishment of lithium- and morphine-induced conditioned taste aversion ( 134 ). These results suggest a specific action of ibogaine on the neurochemical and behavioral (both reinforcing and aversive) actions of morphine rather than on opioid system(s), because the reinforcing effects of naloxone were unaffected. In support to these findings, it has been reported that ibogaine (20 or 40 mg/kg, 24 h before the test) neither decreased the preference for a sweet solution nor attenuated conditioned preference for a flavor previously associated with sweet taste ( 135 ). TOP
10. Effects on learning and memory. At a dose used in the majority of contemporary behavioral studies in rodents (40 mg/kg), ibogaine has been found to attenuate the acquisition of spatial memory, perhaps due to reductions in locomotor activity and in detection of sensory information ( 78 ). However, at much lower doses (0.25 - 2.5 mg/kg), ibogaine as well as O -desmethylibogaine (but not O - t -butyl- O -desmethylibogaine) facilitated spatial memory retrieval ( 136 ). Using a spatial memory task, Helsley et al ., ( 92 ) found that: 1) two doses of ibogaine (50 mg/kg, spaced by 8 hours) decreased the response rate, but did not affect acquisition rate; 2) ibogaine, even at the highest doses of 30 and 46 mg/kg given 20 min before the learning trial did not affect task acquisition; 3) 30 mg/kg of ibogaine administered just after the learning trial facilitated the consolidation of memory trace. TOP
11. Cardiovascular actions. Gershon and Lang ( 102 ) found that ibogaine produced a rise in blood pressure and increased heart rate in conscious dogs. These effects were blocked by atropine ( 137 ). However, in anesthetized dogs, ibogaine

produced a fall in blood pressure and reduced heart rate reduction, leading the authors to propose an interaction between anaesthesia and the cardiovascular effects of ibogaine ( 102 ). Schneider and Rinehart ( 137 ) postulated a centrally mediated stimulatory effect of ibogaine. Ibogaine also potentiated the pressor response to both adrenaline and noradrenaline. More recently, Hajo-Tello et al., ( 138 ) found that tabernanthine (an alkaloid closely related to ibogaine) induced a negative inotropic effect in electrically stimulated myocardial tissue and a negative chronotropic effect in the perfused rat heart. Tabernanthine also produced bradycardia and hypotension in anesthetized rats and dogs ( 139 ). Binienda et al . ( 140 ) reported that ibogaine (50 mg/kg) reduced heart rate in rats immediately after injection; this reduction persisted up to 90 minutes after injection. TOP
B. Human Studies. Numerous psychotropic actions of ibogaine have been reported. These actions seem to depend on both dose and setting. In addition, the psychoactive effects of iboga extracts (which are likely to contain additional alkaloids and are usually taken in a ritualistic setting) may be different from those of ibogaine. Thus, users of the crude extract of Tabernanthe iboga taken in sufficiently high doses have reported fantastic visions, feelings of excitement, drunkenness, mental confusion and hallucinations when ( 101 ). The total extract of iboga shrub is certainly a central stimulant, and in higher doses may lead to convulsions, paralysis and finally respiratory arrest. The psychotropic actions of the plant extract include visual sensations; objects are seen to be surrounded by specters or rainbows. In high doses it may produce auditory, olfactory and taste synesthesias. The state of mind has been reported to vary from profound fear to frank euphoria ( 141 ). When given orally, both ibogaine and the total iboga extract elicits subjective reactions that last for approximately 6 hours. Fifty percent of subjects are reported to experience dizziness, incoordination, nausea, and vomiting ( 7,33,142 ). Typically, the drug produced a state of drowsiness in which subjects did not want to move, open their eyes, or attend to the environment. Many subjects were light-sensitive, and covered their eyes or asked that the lights be turned off. Sounds or noises were disturbing. Ibogalin (0.1-1.2 mg/kg, p.o.), an alkaloid closely related to ibogaine and a constituent of the total iboga extract, did not produce psychotomimetic effects in humans ( 143 ). Ibogalin also differs from ibogaine in pharmacokinetics and tremorigenic activity ( 90 ).

The psychoactive properties of ibogaine and related compounds were studied by Naranjo ( 33,142 ) who reported that patients described the psychic state produced by ibogaine (~ 300 mg) as similar to a dream state without loss of consciousness. Ibogaine-induced fantasies [often described as a " movie run at high speed " or " slide show " ( 7 )] were reported as rich in archetypal contents, involving animals and/or the subject with or without other individuals. These fantasies were easy to manipulate by both the subjects and the psychotherapist ( 33,142 ). At higher doses, ibogaine appears to produce visual and other hallucinations associated with severe anxiety and apprehension ( 101,144,145 ). TOP
VI. LETHALITY AND NEUROTOXIC EFFECTS. The LD 50 of ibogaine has been determined in guinea pig (82 mg/kg, i.p.) and rat (327 mg/kg, intragastrically and 145 mg/kg, i.p.) ( 60,146 ). No significant pathological changes in rat liver, kidney, heart and brain following chronic ibogaine treatment (10 mg/kg, for 30 days or 40 mg/kg, for 12 days, i.p.) were reported ( 60 ). Sanchez-Ramos and Mash ( 42 ) found no evidence of gross pathology in African green monkeys given ibogaine in doses of 5-25 mg/kg, p.o. for 4 consecutive days. However, O'Hearn et al ., ( 147,148 ) and O'Hearn and Molliver, ( 93 ) reported that repeated administration of ibogaine (100 mg/kg, i.p.) to rats caused the degeneration of a subset of Purkinje cells in the cerebellar vermis. This degeneration was accompanied by a loss of microtubule-associated protein 2 (MAP-2) and calbindin. Argyrophilic degeneration, astrocytosis and microgliosis were also observed. The damage seemed to be dependent on the presence of an intact inferior olivary nucleus ( 149 ). Ibogaine-induced cerebellar toxicity seem to be independent on its action at NMDA receptors, because neither MK-801 nor phencyclidine produce the same pattern of degeneration ( 150 ). The neurotoxic effects of high doses of ibogaine were confirmed in rats, but not mice, by Scallet et al ., ( 151,152 ) and Molinari et al ., ( 153 ), who, in addition found that the "typical" dose of 40 mg/kg did not produce significant damage to female rat cerebellum. The lack of neurotoxicity after lower, behaviorally active doses of ibogaine was also demonstrated by showing that chronic administration (60 days) of 10 mg/kg of ibogaine produced no change in the number of Purkinje cerebellar cells ( 154 ). In spite of these findings, examination of cellular markers that are more sensitive toneurotoxic agents than gross histology indicates that ibogaine administration may produce significant change in many other brain

structures. Thus, O'Callaghan et al ., ( 155,156 ) examined the effects of acute and chronic administration of ibogaine on glial fibrillary acidic protein (GFAP) levels. Acutely, ibogaine increased GFAP in both sexes; whereas chronic administration (14 days) produced increases only in females. Ibogaine - induced changes in GFAP were dose-related, and, contrary to other studies, observed in other brain structures including hippocampus, olfactory bulb, brain stem and striatum. In addition, these authors reported that in females treated chronically with ibogaine, severe hippocampal damage was present as measured by increases in the cytoskeletal proteins neurofilament 68 (NF-68) and beta-tubulin. These latter markers indicate a damage-induced sprouting response ( 156 ). Ibogaine administration also produced an increase in c-fos immunostaining in several brain regions of mice and rats; the effects in rats were observed in all cortical layers while in mice the response was limited to cortical layer 2 ( 152 ). Human SK-N-SH neuroblastoma cells cultured in the presence of 3-30 µ M ibogaine (but not O -desmethylibogaine or 18-methoxycoronaridine) demonstrated concentration- and time-dependent morphological changes characterized by the loss of processes, cell rounding, detachment and ultimately cell death ( 157 ). Similar results were observed with primary cultures of rat cerebellar granulae cells. Because in this study only alkaloids that had marked affinity at sigma 2 sites were neurotoxic, Vilner et al ., ( 157 ) proposed that sigma 2 sites may be implicated in the neurotoxicity of ibogaine. The neurotoxic effects of ibogaine have been recently reviewed by Vocci and London ( 106 ). Acute treatment with the ibogaine-like alkaloid, 18-methoxycoronaridine (100 mg/kg) did not produce gross pathological changes in the cerebellum ( 97 ). In contrast, another indole alkaloid, harmaline, produced ibogaine-like degeneration of Purkinje cells in the cerebellar vermis ( 93 ). It has been reported that multiple doses of a non-NMDA antagonist (GYKI 52466) resulted in a substantially greater loss of Purkinje cells and microglial activation compared to ibogaine (50-100 mg/kg) alone ( 158 ). On the other hand, the noncompetitive NMDA antagonist MK-801 (1 mg/kg) markedly attenuated the degree of Purkinje cell loss caused by ibogaine ( 158 ). This later finding strongly supports the notion that the loss of cerebellar Purkinje cells produced by ibogaine is unrelated to its NMDA antagonist properties ( 159 ). In fact, ibogaine can also exhibit neuroprotective properties, reducing glutamate-induced neurotoxicity in primary cultures of cerebellar granule cell neurons with an EC 50 of 4-5 µM ( 119 ). These neuroprotective effects of ibogaine have recently been patented by Olney ( 160 ). Consistent with its properties as an NMDA

antagonist, ibogaine inhibited NMDA - induced lethality in mice in a dose-dependent manner ( 161 ), and also protected mice from maximal electroshock seizures (ED 50 ~ 31 mg/kg) ( 162 ). Phase I toxicity studies in drug-addicted individuals are in progress at the University of Miami ( 42,163 ). TOP
VII. EFFECTS ON SPECIFIC NEUROTRANSMITTER SYSTEMS A. Ibogaine Effects on Dopaminergic Systems. Ibogaine (at concentrations £ 100 µM) does not affect radioligand binding to dopamine receptors (D 1 , D 2 , D 3 , D 4 ) ( 164-166 ). The affinity of ibogaine for dopamine transporters as measured by inhibition of [ 3 H]WIN 35,248, [ 125 I]RTI-121 or [ 125 I]RTI-55 binding was ~ 1.5 - 4 µ M ( 73,76,166,167 ). However, in another study, ibogaine did not affect binding of [ 3 H]GBR-12935, a ligand that also appears to label dopamine transporters ( 85 ). Ibogaine inhibited [ 3 H]dopamine uptake in porcine kidney cells transfected with dopamine transporter with a K i ~86 µM ( 168 ). The in vivo and ex vivo effects of ibogaine on dopamine metabolism in mesolimbic areas of the rodent brain (striatum, nucleus accumbens) are controversial and highly inconsistent. In an attempt to reconcile several contradictory findings, one may note the following. Dopamine concentrations are reduced and dopamine metabolites dihydroxyphenyl-acetic acid (DOPAC) and homovanilic acid (HVA) are increased by ibogaine under certain experimental conditions. For example, when either measurements are taken shortly (within 2 h) after ibogaine administration or when relatively high concentrations ( £ 100 µM) are used ( 69,71,76,81,169-173 ). Reductions in extracellular dopamine concentrations were also observed after administration of a number of ibogaine derivatives, including O -desmethylibogaine ( 89 ) and 18-methoxycoronaridine ( 97 ). When dopamine is measured at longer periods after ibogaine administration (e.g., up to a week) or low concentrations (e.g., 10 µM) are applied, brain concentrations appear unchanged and metabolite concentrations are decreased ( 69,71,76,81,82,169,170,172 ). The increased levels of extracellular dopamine metabolites together with decreased or unchanged levels of dopamine suggests that ibogaine increases dopamine turnover shortly after administration. This may be followed by a decrease in turnover that may persist for some time after ibogaine administration. French et al ., ( 91 ) demonstrated that doses of ibogaine (~

1.5 mg/kg, i.v.), much lower than a "typical" dose of 40-80 mg/kg, markedly excited dopaminergic neurons in the ventral tegmental area of the rat. TOP
1. Dopaminergic effects: Pharmacological Specificity. Administration of a kappa antagonist (norbinaltorphimine, 10 mg/kg) and NMDA (10 mg/kg) (either jointly or individually) reversed ibogaine (40 mg/kg) induced decreases in striatal dopamine and increases in dopamine metabolites ( 88 ). Similarly, Reid et al ., ( 172 ) observed that the decrease in dopamine levels produced by ibogaine (100 µ M ) was reversed by either naloxone (1 µM) or norbinaltorphimine (1-10 µM). However, functionally opposite effects were observed by Sershen et al ., ( 174,175 ) who reported that the ability of the kappa opioid agonist (U-62066) to inhibit electrical- or cocaine-induced [ 3 H]dopamine release from mouse striatum was attenuated by pretreatment of mice with ibogaine (40 mg/kg, i.p., 2 hours prior; or 2 x 40 mg/kg, 6 hours apart, killed 18 hours later) ( 174,175 ). Ibogaine-induced dopamine release from the isolated mouse striatum has been studied by Harsing et al., ( 176 ). Ibogaine increased basal tritium outflow ([ 3 H]dopamine (DA) and [ 3 H]DOPAC), but was without effect on electrically stimulated tritium overflow. This dopamine releasing effect was: a) reduced by the dopamine uptake inhibitors cocaine and nomifensine, b) unaltered by omission of Ca ++ from the perfusion buffer, c) tetrodotoxin insensitive, d) unaffected by an agonist (quinpirole) or an antagonist (sulpiride) of the D 2 dopamine receptor, and e) unaffected by pretreatment with reserpine. In this study, ibogaine did not affect dopamine uptake, whereas Reid et al ., ( 172 ) found that both ibogaine and harmaline (10 µM-1 mM) inhibited it. As mentioned above, ibogaine has been reported to inhibit radioligand binding to the dopamine transporter with relatively high affinity. Sershen et al ., ( 177 ) reported an involvement of serotonin receptors in the regulation of dopamine release by ibogaine. Thus, administration of ibogaine blocked the ability of a 5HT 1B agonist (CGS-12066A [10 µM]) to increase [ 3 H]dopamine increase in striatal slices. In other studies, a concentration of ibogaine (1 µM) that was without effect on dopamine efflux inhibited both NMDA (25 µ M) and ( ± )pentazocine (100 nM) - induced dopamine release in striatal slices ( 178 ). There are few reports of the effects of ibogaine-like alkaloids on dopamine metabolism. Like ibogaine, O -desmethylibogaine acutely decreases dopamine release in the rat nucleus accumbens and striatum ( 89 ). Administration of the R - entantiomers of coronaridine and ibogamine

decreased dopamine levels in both nucleus accumbens and striatum, whereas the S -enantiomers produced no significant changes in dopamine levels in either region ( 96 ). In an attempt to reconcile several conflicting findings, Staley et al ., ( 167 ) proposed that ibogaine might promote redistribution of intraneuronal dopamine from vesicular to cytoplasmic pools. Ibogaine displays micromolar affinity for vesicular monoamine transporters labeled with [ 125 I]-tetrabenazine ( 167 ); these sites are crucial for the translocation of dopamine into synaptic vesicles. The inhibitory effect of ibogaine on vesicular monoamine transporters could result in redistribution of dopamine in the cytoplasm. Under such conditions, rapid metabolism of dopamine by monoamine oxidase would account for the decrease in tissue dopamine content and the parallel increase in its metabolites. Multiple transmitter systems have been shown to modulate dopaminergic function in the central nervous system. Because ibogaine can interact with many of these systems, including kappa opioid receptors, NMDA receptors, serotonin receptors, and dopamine transporters, it is not surprising that this alkaloid can produce complex (and sometimes apparently opposite) effects on dopaminergic function. Thus, the effects of ibogaine on dopaminergic function described in this section likely reflect the dose (or concentration) of alkaloid, preparation employed (e.g., slice versus intact animal), and brain region studied. TOP
2. Ibogaine alters the effects of abused drugs on dopaminergic systems. In general, ibogaine attenuates the increases in mesolimbic dopamine produced by drugs (e.g, nicotine, morphine) that appear to act preferentially at dopaminergic cell bodies. In the case of drugs that act at terminal regions (e.g., cocaine and amphetamine), a gender difference has been observed. In female rats, ibogaine enhances stimulant-induced increases in dopamine concentrations, whereas it decreases the effects of these stimulants in male rats and mice. Neurochemical studies were performed in male mice given two doses of ibogaine (40 mg/kg, i.p., 18 hours apart) followed by amphetamine (5 mg/kg) administered 2 hours after the second dose of ibogaine ( 81 ). Striatal levels of dopamine and dopamine metabolites [DOPAC, HVA and 3-methoxytyramine (3-MT)] measured 1 hour after amphetamine were decreased in mice that received ibogaine relative to saline-pretreated, amphetamine-treated controls. Compared to controls, levels of DOPAC and HVA were decreased in the amphetamine and ibogaine groups, and further

decreased in the group that received ibogaine and amphetamine. However, in female rats, amphetamine-induced increases in extracellular dopamine concentrations in both the striatum and the nucleus accumbens were further potentiated by ibogaine (40 mg/kg, i.p., 19 hours preceding amphetamine) ( 82 ). Similarly, Glick et al., ( 169 ) found that ibogaine potentiated amphetamine-induced increases in extracellular dopamine concentrations in female rat nucleus accumbens and striatum. In this study, however, no effect of ibogaine was seen on amphetamine-induced decreases in extracellular concentrations of dopamine metabolites. Similarly, ibogaine potentiated cocaine-induced increases in extracellular dopamine levels in striatum and nucleus accumbens of female rats ( 84 ). However, quite opposite data were obtained by Broderick et al., ( 85,86 ) who examined dopamine release in male rats using semiderivative in vivo voltametry. In these experiments, ibogaine (40 mg/kg i.p. given for four days) reduced the increase in dopamine release from nucleus accumbens induced by cocaine (20-40 mg/kg, s.c.). A presynaptic mechanism for these actions was suggested. An inhibitory effect of ibogaine on amphetamine metabolism has been proposed ( 179 ), because amphetamine levels were higher after ibogaine administration in female rats. However, ibogaine administration had no effect on brain cocaine levels ( 169 ). Ibogaine (40 mg/kg, i.p. in rats) given 19 hours before morphine (5 mg/kg) prevented the increase in extracellular dopamine concentration in the striatum, prefrontal cortex and nucleus accumbens typically observed in rats ( 71,83 ). However, in the ibogaine plus morphine group, the levels of dopamine metabolites were increased (as was observed in the morphine group), suggesting that ibogaine did not prevent morphine from activating dopamine neurons. The authors suggest that ibogaine treatment may change the properties of dopaminergic neurons in such a way that dopamine release is unaffected under normal conditions, but altered when stimulated (in this case, by morphine). Nineteen hours after placebo or ibogaine (10 mg/kg, i.p.), female rats responded similarly with increased dopamine release in nucleus accumbens following a morphine challenge ( 180 ). However, in rats that received two doses of morphine during two days preceding the experiment, ibogaine pretreatment had inhibitory effects on dopamine response to a morphine challenge. A pharmacokinetic explanation for the effects of ibogaine on morphine-induced actions is unlikely, because ibogaine (40 mg/kg, i.p. 19 hours before measurement) did not modify brain levels of morphine (10 mg/kg) in rats ( 71 ). Benwell et al ., ( 103 ) reported that ibogaine (given 22 hours before nicotine) attenuated the increase in dopamine overflow in the nucleus

accumbens evoked by nicotine administration. Similar effects were demonstrated, when ibogaine was administered 19 hours prior to nicotine infusion ( 181 ). TOP
B. Opioid Systems At concentrations of up to 100 µM, ibogaine was reported not to affect [ 3 H]carfentanil or [ 3 H]enkephalin binding indicating that this alkaloid does not affect mu or delta opioid receptors ( 124,165 ). In contrast, Pearl et al ., ( 124 ) and Sweetnam et al ., ( 166 ) demonstrated that ibogaine inhibited radioligand binding to mu opioid receptors with K i values ~ 11-20 µM. Ex vivo studies demonstrated that ibogaine and O -desmethylibogaine enhanced the inhibition of adenylyl cyclase activity by a maximally effective concentration of morphine in the rat frontal cortex, midbrain and striartum ( 182 ). This later effect is not likely mediated via a direct action at opioid receptors because it was observed at maximally effective concentration of morphine. Ibogaine inhibits (K i ~2-4 µM) [ 3 H]U-69593 binding to kappa opioid receptors ( 56,72,124,165 ). This binding is reversible, suggesting that the long-term effects of ibogaine cannot be attributed to an irreversible effect at this site. Recently, Codd ( 183 ) demonstrated that ibogaine inhibits binding to sites labeled by [ 3 H]naloxone characterized by a two-site model, with K i values of 130 nM and 4 µM. O -Desmethylibogaine had a higher affinity than ibogaine for all of the opioid receptors studied: kappa K i ~ 1 µM, mu K i ~ 2.7 µM and delta K i ~ 24.7 µM (124) (a recent study showed much higher affinity of O -desmethylibogaine at the mu receptor; K i ~ 160 nM ( 184 )). Our work ( 72 ) demonstrated that O -desmethylibogaine had a 10- to 100-fold higher affinity for kappa receptors compared to ibogaine. The magnitude of this potency difference was species-specific (e.g., in rats: IC 50 ~ 0.3 µM for O -desmethylibogaine and IC 50 ~30 µM for ibogaine). The same study demonstrated a moderate affinity of O - t -butyl- O -desmethylibogaine for kappa receptors (IC 50 ~17 µM in rat forebrain) suggesting that if any of ibogaine's in vivo actions are produced at kappa receptors, then O - t -butyl- O -desmethylibogaine would be active. In this respect, O - t -butyl- O -desmethylibogaine did not influence the morphine withdrawal syndrome ( 72 ) at doses comparable to ibogaine. TOP
C. Serotonergic Systems.

Ibogaine (at concentrations up to 1 µM) had no effect on [ 3 H]serotonin binding ( 185 ) and concentrations of up to 3.5 µM had no effect on [ 3 H]LSD binding ( 186 ). More recent studies using serotonin subtype selective ligands are discrepant. Deecher et al ., ( 165 ) reported that ibogaine did not displace ligands acting at 5-HT 1a , 5-HT 1b , 5-HT 1c , 5-HT 1d , 5-HT 2 , or 5-HT 3 receptors. However, Repke et al ., ( 56 ) reported that ibogaine inhibited binding of 5-HT 1a , 5-HT 2a , or 5-HT 3 ligands with low affinity (K i values: >100, 12.5 and >100 µM, respectively) and Sweetnam et al ., ( 166 ) reported IC 50 values of ~ 4 µM to inhibit radioligand binding to both 5-HT 2 , and 5-HT 3 receptors. Despite these discrepancies, both ex vivo and in vivo studies suggest that ibogaine can affect serotonergic transmission. Ex vivo studies indicate that ibogaine and O -desmethylibogaine enhance the inhibitory effects of serotonin on adenylyl cyclase activity in rat hippocampus ( 182 ). Broderick et al., ( 86 ) reported that ibogaine (40 mg/kg, i.p. for 4 days) increased 5-HT concentrations in rat nucleus accumbens. Consistent with this finding, Ali et al ., ( 171 ) demonstrated that ibogaine increased 5-HT levels in striatum. Sershen et al ., ( 76 ) reported that ibogaine (40-50 mg/kg) decreased levels of the serotonin metabolite 5-hydroxy-indoleacetic acid [5-HIAA] in mouse frontal cortex, hippocampus and olfactory tubercle 2 and 24 hours after injection. Ibogaine also decreased 5-HIAA levels in rat nucleus accumbens and striatum ( 103,171 ), but increased 5-HIAA and decreased 5-HT (lasting at least 7 days) in medial prefrontal cortex ( 103 ). Long and Lerrin ( 187 ) demonstrated that ibogaine is a reversible inhibitor of the active transport of serotonin into blood platelets, a finding supported by a recent observation that ibogaine inhibited serotonin transporters (in a porcine kidney cell line) with a K i ~ 10 µM (168). Sershen et al., ( 177 ) demonstrated that ibogaine inhibited the ability of a 5-HT 1b agonist (CGS-12066A) to increase stimulation-evoked [ 3 H]dopamine release from both rat and mouse striatal slices. Additionally, ibogaine increased the ability of a 5-HT 3 agonist (phenylbiguanide) to enhance stimulation-evoked [ 3 H]dopamine release from the mouse striatal slice ( 174 ). In these studies, ibogaine (40 mg/kg, i.p.) was administered 2 hours prior to slice preparation. In other studies, ibogaine (20 mg/kg) enhanced cocaine-induced reductions in serotonin concentration in the nucleus accumbens (rat), an action attributed to a presynaptic release mechanism ( 85,86 ). However, Sershen et al ., ( 175 ) reported that cocaine increased [ 3 H]serotonin efflux in striatal slices and this efflux was absent in mice pretreated with either ibogaine or a 5-HT 1b agonist. These later findings led Sershen to suggest an action of ibogaine at the HT 1b receptor

that is likely unrelated to the ability of cocaine to inhibit serotonin reuptake blockade ( 188 ). The inhibitory effect of the kappa-opioid agonist U-62066 (1µM) on [ 3 H]serotonin release in striatal slices could be blocked by in vivo ibogaine administration ( 175 ). TOP
D. Calcium Regulation. Ibogaine (80 µM) non-competitively antagonized calcium-induced contraction of rat aorta and mesenteric artery ( 138 ), which was interpreted as an action on intracellular calcium metabolism. Tabernanthine, an alkaloid related to ibogaine, inhibited depolarization-stimulated 45 Ca influx and contractions in the rat aorta ( 189 ). Ibogaine inhibited the binding of [ 3 H]isradipine (an L-type calcium channel blocker) in the mouse cerebral cortex with an IC 50 of ~28 µM ( 11 ). TOP
E. Cholinergic Systems. Ibogaine (at concentrations of up to 100 µM) was reported not to inhibit the binding of ligands acting at nicotinic or muscarinic receptors ( 165 ). However, subsequent studies demonstrated that ibogaine inhibited the binding of muscarinic M 1 , M 2 and M 3 ligands at concentrations of ~ 31, 50 and 12.5 µM, respectively ( 56 ). Sweetnam et al ., ( 166 ) showed that ibogaine inhibited radioligand binding to M 1 , and M 2 receptors with IC 50 values of 5-7 µM. These authors also reported that ibogaine did not inhibit the binding of [ 3 H]NMCI, a nonselective ligand at nicotinic receptors. Ex vivo studies have shown that neither ibogaine nor O -desmethylibogaine affect the inhibitory action of the muscarinic acetylcholine agonist, carbachol on adenylyl cyclase activity in the rat ( 182 ). In a recent study, Badio et al ., ( 125 ) demonstrated that ibogaine potently (IC 50 ~ 20 nM) blocked 22 NaCl influx through nicotinic receptor channels in rat pheochromocytoma cells. This effect was seen in the cells expressing ganglionic, but not neuromuscular, nicotinic receptor subtypes. This inhibition was noncompetitive because it was not overcome by increasing concentrations of agonist. Moreover, the blockade was not completely reversible, suggesting that ibogaine may have a long-lasting effect. O -Desmethylibogaine and O - t -butyl- O -desmethylibogaine were 75- and 20-fold less potent, respectively, than ibogaine in blocking nicotinic receptor-mediated responses. The same study demonstrated that ibogaine, as expected for a noncompetitive blocker, had a relatively low affinity (K i ~ 4 µM) as

an inhibitor of the binding of an agonist [ 3 H]nicotine. In support to these findings, Schneider et al ., ( 190 ) reported recently that ibogaine ( £ 10 µ M) had an inhibitory action on nicotinic receptor-mediated catecholamine release in bovine adrenal chromaffin cells. Consistent with the Badio et al ., ( 125 ) study, these inhibitory effects appeared to be long-lasting. TOP
F. Gamma-Aminobutyric Acidergic [GABAergic] Systems. Two independent studies ( 165,166 ) did not find any effect of ibogaine (at concentrations of up to 100 µM) on radioreceptor binding to GABA A receptors. In addition, ibogaine did not influence 36 Cl - uptake through GABA-gated channels ( 165 ) or GABA-evoked currents in rat cultured hippocampal neurons ( 162 ). TOP
G. Voltage-Dependent Sodium Channels. Ibogaine inhibited (K i ~ 8.1 µM) [ 3 H]batrachotoxin A 20-a-benzoate binding to voltage-dependent sodium channels in depolarized mouse neuronal preparations ( 165 ). Ibogaine analogs, including ibogamine, tabernanthine and coronaridine, exhibited potencies similar to ibogaine in this assay. TOP
H. Glutamatergic Systems. Our studies ( 159 ) indicate that ibogaine is a competitive inhibitor of [ 3 H]MK-801 binding (K i ~1 µM) to NMDA receptor-coupled ion channels. In contrast, ibogaine did not affect [ 3 H]( ± )- a -amino-3-hydroxy-5-methylisoxazole-4-propionic acid ([ 3 H]AMPA), [ 3 H]kainate or [ 3 H]glutamate to either the NMDA or metabotropic receptor sites, binding. These findings are consistent with a specificity of ibogaine for NMDA receptor-coupled cation channels ( 159,162,166 ). The potency of ibogaine to inhibit [ 3 H]MK-801 binding was also examined in 8 distinct brain regions of Sprague-Dawley male rats and compared with the dissociation constants for [ 3 H]MK-801 estimated using saturation analyses. A high correlation (r=0.976, p=0.0004) was obtained between the K i of ibogaine and K d of [ 3 H]MK-801 in these brain regions ( 119 ), consistent with the notion that these compounds share a common binding site. The ability of ibogaine to act as a non-competitive NMDA antagonist can also be demonstrated using [ 3 H]1-[1-(2-thienyl)cyclohexyl]piperidine ([ 3

H]TCP), a thienyl derivative of phencyclidine, resulting in a K i ~1.5 µM in rat forebrain ( 119 ). Structure-activity studies were performed using a series of ibogaine analogs, including the putative ibogaine metabolite O -desmethylibogaine, its metabolism resistant analog O - t -butyl- O -desmethylibogaine, the iboga alkaloids [( ± )-ibogamine, ( ± )-coronaridine, tabernanthine], harmaline, and indolotropanes. Ibogaine was the most potent inhibitor of [ 3 H]MK-801 binding (K i ~ 1.2 µM); the compounds with the greatest structural similarity to ibogaine, O -desmethylibogaine and O - t -butyl- O -desmethylibogaine were much less potent (K i ~ 5.5 and 179.0 µ M respectively) ( 72 ). A ~ 5 fold lower affinity of O -desmethylibogaine compared to ibogaine at [ 3 H]MK-801 binding sites was also reported by Mash et al ., ( 191 ). Consistent with these neurochemical studies, ibogaine produced a voltage-dependent block of NMDA-evoked currents in hippocampal cultures ( 119,162 ). In addition, ibogaine (100 µ M) and O -desmethylibogaine (1 mM) blocked the ability of NMDA (100 µ M, 5 sec) to depolarize frog motoneurons in a non-competitive and use-dependent manner ( 192 ). TOP
I. Sigma Receptors. In our studies (11), ibogaine inhibited [ 3 H]pentazocine (a sigma 1 receptor ligand) binding, to high (IC 50 ~86 nM) and low (IC 50 ~5.6 µM) affinity sites in mouse cerebellum. Bowen et al ., ( 193 ) demonstrated that ibogaine had high affinity for sigma 2 sites (K i ~ 200 nM) and low affinity for sigma 1 sites (K i ~ 8.5 µM), a ~ 43- fold selectivity for sigma 2 sites. The affinities of tabernanthine (13-methoxyibogamine) and ( ± )-ibogamine for sigma 2 sites were similar to that of ibogaine. O -Desmethylibogaine, had a markedly reduced affinity for sigma 2 sites (K i ~ 5 µM) and also lacked affinity for sigma 1 sites. The related alkaloids, ( ± )-coronaridine [( ± )-18-carbomethoxyibogamine] and harmaline lacked affinity for both sigma receptor subtypes. O -t-Butyl- O -desmethylibogaine inhibited radioligand binding to sigma 1 sites with a K i ~ 3.5 µM and sigma 2 sites with a K i ~ 346 nM [c.f. Bowen et al , ( 72 )]. The much higher affinity of ibogaine for sigma 2 sites compared to sigma 1 sites was also reported by Mach et al ., ( 194 ). Bowen et al ., ( 195 ) examined the ability of ibogaine and related compounds to modulate calcium release from intracellular stores in indo-1 loaded human SK-N-SH neuroblastoma cells. Consistent with its affinity at sigma 2 sites, ibogaine produced a concentration-dependent increase (13-45%) in intracellular calcium levels. O -Desmethylibogaine,

was ineffective in this measure at concentrations up to 100 µ M. These data suggest that the shared in vivo effects of ibogaine and O -desmethylibogaine are probably not mediated by sigma sites. TOP
J. Miscellaneous Actions of Ibogaine. Deecher et al., ( 165 ) reported that ibogaine (up to 100 µM) did not inhibit radioligand binding to cannabinoid receptors. Ibogaine and O -desmethylibogaine had no influence on basal or forskolin-stimulated adenylyl cyclase in the rat frontal cortex, midbrain or striatum ( 182 ). O -Desmethylibogaine, but not ibogaine, produced concentration - dependent increases in the generation of [ 3 H]inositol phosphates that were not altered by inclusion of tetrodotoxin, cadmium or omega-conotoxin ( 196 ). These results suggest that the effect of O -desmethylibogaine on phosphoinositide hydrolysis was not secondary to the release of one or more neurotransmitters. Ali et al ., ( 45 ) reported that ibogaine (0.5-250 µ M) reduced nitric oxide synthase activity in mouse brain; similar effects were noted in the striatum, hippocampus and cerebellum of mice treated parenterally with ibogaine (50 mg/kg). In radioligand binding studies, no effect of ibogaine has been found on alpha 1 , alpha 2 or beta 1 adrenergic receptors ( 165 ). Moreover, ibogaine (20 mg/kg) did not modify cerebral noradrenaline levels in rats ( 197 ). Binienda et al ., ( 140,198 ) reported that although ibogaine (50 mg/kg) challenge in rats was associated with a decrease in delta, theta, alpha and beta power spectra of cortical EEG during the first 30 min, and subsequent recovery of all except delta bands in the next 15 min, MK-801 (1 mg/kg) treatment was followed by a decrease in power of all four frequency bands for the entire time of recording. The selective power decrease in delta EEG frequency band of the cortical EEG may suggest the activation of dopamine receptors. In the anesthetized rat, ibogaine produced a slight hypoglycemia ( 60 ). After administration of 50 mg/kg of ibogaine, elevations of corticosterone levels were noted 15 - 120 min, but not 24 hours later ( 170,171,173 ). The same dose of ibogaine rapidly and transiently increased plasma prolactin levels (171,173). Bunag and Walaszek ( 199 ) reported that ibogaine antagonized the contractile responses produced in guinea pig ileum by substance P and angiotensin. Alburges and Hanson ( 200 ) reported that ibogaine administration produced increases of neurotensin-like immunoreactivity in striatum, nucleus accumbens and substantia nigra and substance P -like immunoreactivity in striatum and substantia nigra. Ibogaine or harmaline suppressed several (T-cell regulatory and effector, B-

cell, and natural killer cell) immune functions in vitro ( 201 ). Van Beek et al ., ( 17 ) reported that ibogaine showed activity against the gram-positive Bacillus subtilis . Ibogaine did not alter colonic temperature in mice, nor did it affect morphine- or kappa [U-50,488H] – opioid induced hypothermia ( 121 ). TOP
VIII. CONCLUSIONS. The renewed interest in ibogaine during the past decade stems from anecdotal clinical observations that ibogaine offers a novel means of treating drug addictions. Preclinical studies are, in general, consistent with these claims. Thus, ibogaine reduces self-administration of cocaine and morphine, attenuates morphine withdrawal, and blocks conditioned place preference produced by morphine and amphetamine. Preclinical studies also suggest there is no abuse liability associated with ibogaine. At doses that interfere with tolerance and dependence phenomena, brain concentrations of ibogaine are at levels that can affect a variety of neurotransmitter systems. Many of these effects (e.g., use dependent block of NMDA receptor-coupled cation channels, interactions with dopamine transporters and kappa opioid receptors) have previously been implicated in drug seeking phenomena. However, at the present time, the only mechanism that can be invoked to explain ibogaine's effects on drug seeking phenomena with some certainty is its ability to inhibit naloxone-precipitated jumping through blockade of NMDA receptors. Nonetheless, it is still uncertain whether the anti-addictive properties of ibogaine result from a single mechanism or are produced at multiple loci. The involvement of dopaminergic pathways in drug seeking phenomena can be considered dogma, and ibogaine undoubtedly affects these pathways. Nonetheless, based on available data no clear picture has emerged about how this interaction contributes to the anti-addictive properties of ibogaine, or any other anti-addictive medications. Additional systematic studies are obviously needed. Anecdotal reports claim long term effects of ibogaine on drug seeking following a single administration or short course of therapy. This claim has been borne out, at least in part, by preclinical studies. Based on these observations, it is unlikely that ibogaine serves simply as substitution therapy. It has been hypothesized that a long-lived metabolite is responsible for ibogaine's putative anti-addictive properties, but additional studies are required in this area. One of the central issues regarding the molecular mechanisms responsible for the anti-addictive actions of ibogaine is whether its NMDA antagonist

action is sufficient to explain these effects. Thus, there is an established body of preclinical data (and an emerging body of clinical data) demonstrating that NMDA antagonists interrupt drug seeking phenomena to a variety of addictive substances. Although it is now well established that ibogaine is a noncompetitive NMDA antagonist (albeit 1000-fold less potent than the prototype compound, dizocilpine), with the exception of its ability to block naloxone precipitated jumping in morphine-dependent mice, it is uncertain if these effects can be attributed to other mechanisms. Recent structure activity studies demonstrate that O -desmethylibogaine, which is less potent than ibogaine at NMDA receptors, appears as active as ibogaine in acutely blocking morphine and cocaine self-administration. This observation strongly suggests that other mechanisms may be operative. A similar argument can be made for harmaline, which is somewhat structurally related to ibogaine and shares some of its pharmacological actions (e.g., tremor and neurotoxic effects, reductions in cocaine and morphine self-administration), but is not an NMDA antagonist. Although inhibition of drug self-administration by harmaline may be due to unspecific effects (e.g., general malaise), these findings nonetheless raise the possibility that ibogaine's anti-addictive properties may be produced through multiple mechanisms. The involvement of sigma sites in these phenomena appears to be even more obscure because in contrast to ibogaine, harmaline has no appreciable affinity at sigma sites whereas O -desmethylibogaine lacks affinity at a sigma 2 site, yet all three block cocaine and morphine self-administration. Ibogaine can affect several aspects of serotonergic transmission at concentrations that are readily achieved in the brain following pharmacologically relevant doses [reviewed by Sershen et al ., ( 188 )]. Because multiple serotonin receptor subtypes, as well as serotonin reuptake, are modulated by ibogaine, it is not surprising that the effects of this alkaloid on steady state levels of serotonin and its metabolites (whether measured in situ or ex vivo ) are complex. Clearly, additional clinical studies are necessary to examine the efficacy of ibogaine as an anti-addictive agent. Similarly, additional preclinical studies will be required to elucidate the molecular mechanism(s) responsible for these pharmacological actions. TOP
IX. Acknowledgments The authors thank Dr. H. Sershen for helpful discussions on the effects of ibogaine on dopaminergic and serotonergic transmission. TOP

X. REFERENCES
1. A. Lecomte, Arch. Med. Navale 2, 264 (1864). 2. R. Goutarel, O. Gollnhofer, and R. Sillans, Psychedc Mono. Essays 6, 71 (1993). 3. H.S. Lotsof, U S Patent 4,499,096, (1985). 4. H.S. Lotsof, U S Patent 4,587,243, (1986). 5. H.S. Lotsof, U S Patent 4,857,523, (1989). 6. H.S. Lotsof, U S Patent 5,026,697, (1991). 7. H.S. Lotsof, Bull. MAPS 5, 16 (1995). 8. C.D. Kaplan, E. Ketzer, J. de Jong, and M. de Vries, Soc. Neurosc. Bull.n 6, 6 (1993). 9. B. Sisko, Bull. MAPS IV, 15 (1993). 10. P. Popik and S.D. Glick, Drugs Future 21, 1109 (1996). 11. P. Popik, R.T. Layer, and P. Skolnick, Pharmaco. Rev. 47, 235 (1995). 12. W.I. Taylor, in " The Alkaloids. Volume VIII. Chemistry and Physiology" (R.H.F. Manske, ed.), p. 203, Academic Press, New York, London. 1965. 13. W.I. Taylor, in " The Alkaloids. Volume XI. Chemistry and Physiology" (R.H.F. Manske, ed.), p. 79, Academic Press, New York, London. 1968. 14. R.E. Schultes and A. Hofmann, in " The Botany and Chemistry of Hallucinogens." (R.E. Schultes and A. Hofmann, eds.), p. 235 Charles C. Thomas Publisher, Sprinfield. 1980. 15. J.W. Fernandez, "Bwiti - an ethnography of religious imagination in Africa.". Princeton Press., Princeton, New Jersey, 1982. 16. H.G. Pope, Jr., Econ. Bot. 23, 174 (1969). 17. T.A. Van Beek, C. de Smidt, and R. Verpoorte, J. Ethnopharmacol. 14, 315 (1985). 18. H. Baillon, Bull. Mens. Soc. Lin. de Paris 1, 782 (1889). 19. J. Dybovsky and E. Landrin, Acad. Sci. (Paris) 133, 748 (1901). 20. A. Haller and E. Heckel Compt. Rend. Soc Biol 133, 850 (1901). 21. M. Lambert and E. Heckel, Compt. Rend. Acad. Sci. 133, 1236 (1901). 22. M. Lambert, Arch. Int. Pharmacodyn. 10, 101 (1902). 23. M.C. Phisalix, Compt. Rend. Soc. Biol. 53, 1077 (1901). 24. G. Pouchet and J. Chevalier, Bull. Gen. Ther. 149, 211 (1905). 25. E. Rothlin and M. Raymond-Hamet, Compt. Rend. Soc. Biol. 127, 592 (1938). 26. M. Raymond-Hamet and E. Rothlin, Arch. Int. Pharmacodyn. Ther. 63, 27 (1939).

27. M. Raymond-Hamet, Compt. Rend. Soc. Biol. 134, 541 (1940). 28. M. Raymond-Hamet, Compt. Rend. Soc. Biol. 133, 426 (1940). 29. M. Raymond-Hamet, Compt. Rend. Soc. Biol. 211, 285 (1940). 30. M. Raymond-Hamet, Compt. Rend. Soc. Biol. 135, 176 (1941). 31. M. Raymond-Hamet, Compt. Rend. Soc. Biol. 212, 768 (1941). 32. M. Raymond-Hamet and D. Vincent, Compt. Rend. Soc. Biol. 154, 2223 (1960). 33. C. Naranjo, Clin. Toxicol. 2, 209 (1969). 34. F. De Sio, Medicina Dello Sport 23, 362 (1970). 35. H.S. Lotsof, U S Patent 5,152,994, (1992). 36. L.R. Regan, Justicia September, 1 (1992). 37. N. Touchette, Nature Med. 1, 288 (1995). 38. FDA Advisory CMTE in " The NDA Pipeline." (E. Clarke, C. Frederick, and K. Balog, eds.), p. V-76 F-D-C Reports, Inc. Chevy Chase, MD, USA. 1993. 39. L.G. Sharpe and J.H. Jaffe, NeuroReport 1, 17 (1990). 40. N. Touchette, J. NIH Res. 5 (November), 50 (1993). 41. J. Buie, Psych. Times XI (7), 44 (1994). 42. J. Sanchez-Ramos and D. Mash, Bull. MAPS 4, 11 (1994). 43. S.G. Sheppard, J. Subst. Abuse Treat. 11, 379 (1994). 44. B.E. Judd, Ibogaine, psychotherapy, and the treatment of substance-related disorders. The Eighth International Conference on Drug-related Harm. (1994). 45. S.F. Ali, S.C. Chetty, X.M. Meng, G.D. Newport, and W. Slikker, J. Neurochem. 65, S172 (1995). 46. A. Landrin, Bull. Sci. Pharmacol. 11, 319 (1905). 47. W.I. Taylor, J. Am. Chem. Soc. 79, 3298 (1957). 48. G. Büchi, D.L. Coffen, K. Kocsis, P.E. Sonnet, and F.E. Ziegler, J. Am. Chem. Soc. 88, 3099 (1966). 49. P. Rosenmund, W.H. Haase, J. Bauer, and R. Frische, Chem. Ber. 180, 1871 (1975). 50. E. Wenkert, D.W. Cochran, H.E. Gottlieb, and E.W. Hagaman, Helv. Chim. Acta 59, 2437 (1976). 51. H.H. Seltzman, D.F. Odear, F. Ivy Carroll, and D. Wyrick, J. Chem. Soc. Chem. Commun. 1757 (1992). 52. H.H. Seltzman, D.F. Odear, and C.P. Laudeman, J. Labled. Compds. Radiopharm. 34(4), 367 (1994). 53. S. Budavari, M. O'Neil, A. Smith, P.E. Heckelman, and J.F. Kinneary, "The Merck Index. An Encyclopedia of Chemicals, Drugs and Biologicals.". 12th Ed. Merck & Co., Inc., Whitehouse Station, N.J., USA, 1996.

54. G.P. Cartoni and A. Giarusso, J. Chromatogr. 71, 154 (1972). 55. J.M. Kelley and R.H. Adamson, Pharmacol. 10, 28 (1973). 56. D.B. Repke, D.R. Artis, J.T. Nelson, and E.H. Wong, J. Org. Chem. 59, 2164 (1994). 57. H.I. Dhahir, N.C. Jain, and R.B. Forney, J. Foren. Sci. 16, 103 (1971). 58. H.I. Dhahir, N.C. Jain, and J.I. Thornton, J. Foren. Sci. 12, 309 (1972). 59. E. Bertol, F. Mari, and R. Froldi, J. Chromatogr. 117, 239 (1976). 60. H.I. Dhahir, Diss. Abstr. Int. 32/04-B, 2311 (1971). 61. G. Zetler, G. Singbarth, and L. Schlosser, Pharmacol. 7, 237 (1972). 62. C.A. Gallagher, L.B. Hough, S.M. Keefner, A. Seyed Mozaffari, S. Archer, and S.D. Glick, Biochem. Pharmacol. 49, 73 (1995). 63. W.L. Hearn, J. Pablo, G.W. Hime, and D.C. Mash, J. Anal. Toxicol. 19, 427 (1995). 64. M.E. Alburges, R.L. Foltz, and D.E. Moody, J. Anal. Toxicol. 19, 381 (1995). 65. F.R. Ley, A.R. Jeffcoat, and B. Thomas, J. Chromatogr. A 723, 101 (1996). 66. S.M. Pearl, L.B. Hough, D.L. Boyd, and S.D. Glick, Pharmacol. Biochem. Behav. 57, 809 (1997). 67. L.B. Hough, S.M. Pearl, and S.D. Glick, Life Sci. 58, PL119 (1996). 68. S.D. Glick, K. Rossman, S. Steindorf, I.M. Maisonneuve, and J.N. Carlson, Eur. J. Pharmacol. 195, 341 (1991). 69. I.M. Maisonneuve, K.L. Rossman, R.W. Keller, Jr., and S.D. Glick, Brain Res. 575, 69 (1992). 70. H. Sershen, A. Hashim, and A. Lajtha, Pharmacol. Biochem. Behav. 47, 13 (1994). 71. I.M. Maisonneuve, R.W. Keller, and S.D. Glick, Eur. J. Pharmacol. 199, 35 (1991). 72. R.T. Layer, P. Skolnick, C.M. Bertha, M.E. Kuehne, and P. Popik, Eur. J. Pharmacol. 309, 159 (1996). 73. D. Mash, J.K. Staley, M.H. Baumann, R.B. Rothman, and W.L. Hearn, Life Sci. 57, PL45 (1995). 74. S.D. Glick, K. Rossman, N.C. Rao, I.M. Maisonneuve, and J.N. Carlson, Neuropharmacology 31, 497 (1992). 75. G. Chen and B. Bohner, J. Pharmacol. Exp. Ther. 123, 212 (1958). 76. H. Sershen, A. Hashim, L. Harsing, and A. Lajtha, Life Sci. 50, 1079 (1992). 77. T. Luxton, L.A. Parker, and S. Siegel, Prog. Neuro-Psych. Biol. Psych. 20, 857 (1996).

78. R.P. Kesner, P. Jackson-Smith, C. Henry, and K. Amann, Pharmacol. Biochem. Behav. 51, 103 (1995). 79. J.G. Page, L.E. Rodan, D.R. Franell, and J.F. Martin, NIDA Contract Report SRI-CBE-94-002-7486, (1994). 80. I.M. Maisonneuve, K.E. Visker, G.L. Mann, U.K. Bandarage, M.E. Kuehne, and S.D. Glick, Eur. J. Pharmacol. 336, 123 (1997). 81. H. Sershen, L.G. Harsing, Jr., A. Hashim, and A. Lajtha, Life Sci. 51, 1003 (1992). 82. I.M. Maisonneuve, R.W. Keller, Jr., and S.D. Glick, Brain Res. 579, 87 (1992). 83. I.M. Maisonneuve, Diss. Abst. Int. 1 (1992). 84. I.M. Maisonneuve and S.D. Glick, Eur. .J Pharmacol. 212, 263 (1992). 85. P.A. Broderick, F.T. Phelan, and S.P. Berger, in " NIDA Research Monograph No. 119. Problems of Drug Dependence" (L. Harris, ed.), p. 285, U.S. Government Printing Office, Washington, D.C., 1992. 86. P.A. Broderick, F.T. Phelan, F. Eng, and R.T. Wechsler, Pharmacol. Biochem. Behav. 49, 711 (1994). 87. S.M. Pearl, D.W. Johnson, and S.D. Glick, Psychopharmacology 121, 470 (1995). 88. S.D. Glick, I.M. Maisonneuve, and S.M. Pearl, Brain Res. 749, 340 (1997). 89. S.D. Glick, S.M. Pearl, J. Cai, and I.M. Maisonneuve, Brain Res . 713, 294 (1996). 90. G. Singbarth, G. Zetler, and L. Schlosser, Neuropharmacology 12, 239 (1973). 91. E.D. French, K. Dillon, and S.F. Ali, Life Sci. 59, PL199 (1996). 92. S. Helsley, D. Fiorella, R.A. Rabin, and J. C. Winter, Pharmacol. Biochem. Behav. 58, 37 (1997). 93. E. O'Hearn and M.E. Molliver, Neuroscience 55, 303 (1993). 94. S.L. Costache "Pharmacological attenuation of the tremorigenic effects of ibogaine." Unpublished M.S. thesis. Albany Medical College, Albany, NY, USA, 1995. 95. G. Zetler, Neuropharmacology 22, 757 (1983). 96. S.D. Glick, M.E. Kuehne, J. Raucci, T.E. Wilson, D. Larson, R.W. Keller, and J.N. Carlson, Brain Res. 657, 14 (1994). 97. S.D. Glick, M.E. Kuehne, I.M. Maisonneuve, U.K. Bandarage, and H.H. Molinari, Brain Res. 719, 29 (1996). 98. S.M. King and G. Tunnicliff, Biochem. Int. 20, 821 (1990). 99. E. Roberts, E. Wong, G. Svenneby, and P. Degener, Brain Res. 152, 614 (1978).

100. J.H. Trouvin, P. Jacqmin, C. Rouch, M. Lesne, and C. Jacquot, Eur. J. Pharmacol . 140, 303 (1987). 101. J.A. Schneider and E.B. Sigg, Ann. NY Acad. Sci. 66, 765 (1957). 102. S. Gershon and W.J. Lang, Arch. Int. Pharmacodyn. Ther. 135, 31 (1962). 103. M.E.M. Benwell, P.E. Holtom, R.J. Moran, and D.J.K. Balfour, Br. J. Pharmacol. 117, 743 (1996). 104. E.S. Onaivi, S.F. Ali, and A. Chakrabarti, Ann. N. Y. Acad. Sci. in press, (1998). 105. S.L.T. Cappendijk and M.R. Dzoljic, Eur. J. Pharmacol. 241, 261 (1993). 106. F.J. Vocci and E.D. London, Ann. N. Y. Acad. Sci. 820, 29 (1996). 107. Anonymous, Presented to the Food and Drug Administration's Drug Advisory Committee #26, (1993). 108. R.S. Mansbach, R.L. Balster, M. Gregory, and E. Soenghen, NIDA Contract Report SA92.16, (1992). 109. S.I. Dworkin, S. Gleeson, D. Meloni, T.R. Koves, and T.J. Martin, Psychopharmacology 117, 257 (1995). 110. Y.W. Lee, A.H. Rezvani, and D.H. Overstreet, Soc. Neurosci. Abstr. 20, 1608 (1994). 111. A.H. Rezvani, D.H. Overstreet, and Y.W. Leef, Pharmacol. Biochem. Behav. 52, 615 (1995). 112. A.H. Rezvani, D. Mash, D.H. Overstreet, W.L. Hearn, and Y.W. Lee, Alcoholism - Clin. Exp. Res, 19, 15A (1995). 113. A.H. Rezvani, D.H. Overstreet, Y. Yang, I.M. Maisonneuve, U.K. Bandarage, M.E. Kuehne, and S.D. Glick, Pharmacol. Biochem. Behav. 58, 615 (1997). 114. M.D. Aceto, E.R. Bowman, and L.S. Harris, NIDA. Res. Monogr. 95, 578 (1990). 115. E.D. Dzoljic, C.D. Kaplan, and M.R. Dzoljic Arch Int Pharmacodyn 294, 64 (1988). 116. S.L.T. Cappendijk, D. Fekkes, and M.R. Dzoljic, Behav. Brain Res. 65, 117 (1994). 117. L.G. Sharpe and J.H. Jaffe, NIDA Res. Monogr. 105, 477 (1991). 118. B. Frances, R. Gout, J. Cros, and J.M. Zajac, Fundam. Clin. Pharmacol. 6, 327 (1992). 119. P. Popik, R.T. Layer, L. Fossom, M. Benveniste, B. Getter-Douglas, J.M. Witkin, and P. Skolnick, J. Pharmacol. Exp. Ther. 275, 753 (1995). 120. J.A. Schneider and M. McArthur, Experientia 12, 323 (1956). 121. H.N. Bhargava, Y.J. Cao, and G.M. Zhao, Brain. Res. 752, 234 (1997).

122. Y.J. Cao and H.N. Bhargava, Brain Res. 752, 250 (1997). 123. A.A. Bagal, L.B. Hough, J.W. Nalwalk, and S.D. Glick, Brain Res. 741, 258 (1996). 124. S.M. Pearl, K. Herrickdavis, M. Teitler, and S.D. Glick, Brain Res. 675, 342 (1995). 125. B. Badio, W.L. Padgett, and J.W. Daly, Mol. Pharmacol. 51, 1 (1997). 126. W. Kostowski, W. Rewerski, and T. Piechocki, Pharmacology 7, 259 (1972). 127. M.D. Schechter and T.L. Gordon, Eur. J. Pharmacol. 249, 79 (1993). 128. M.D. Schechter, Life Sci. 60, PL83 (1997). 129. P.A. Palumbo and J.C. Winter, Pharmacol. Biochem. Behav. 43, 1221 (1992). 130. J.M. Witkin, S. Brave, D. French, and B. Geterdouglass, . J Pharmacol. Exp. Ther. 275, 1267 (1995). 131. S. Helsley, R.A. Rabin, and J.C. Winter, Life Sci. 60, PL147 (1997). 132. L.A. Parker, S. Siegel, and T. Luxton, Exp. Clin, Psychopharm. 3, 344 (1995). 133. I. Moroz, L.A. Parker, and S. Siegel, Exp. Clin. Psychopharm. 5, 119 (1997). 134. L.A. Parker and S. Siegel, Learn. Motiv. 27, 170 (1996). 135. J.R. Blackburn and K.K. Szumlinski, Soc. Neurosci. Abstr. 21, 1467 (1995). 136. P. Popik, Life Sci. 59, PL379 (1996). 137. J.A. Schneider and R.K. Rinehart, Arch. Int. Pharmacodyn. Ther. CX, 92 (1957). 138. N. Hajo-Tello, C. Dupont, J. Wepierre, Y. Cohen, R. Miller, and T. Godfraind, Arch. Int. Pharmacodyn. Ther. 276, 35 (1985). 139. N. Hajo, C. Dupont, and J. Wepierre, J. Pharmacol. 12, 441 (1981). 140. Z. Binienda, M.A. Beaudoin, B.T. Thorn, D.R. Prapurna, J.R. Johnson, C.M. Fogle, W. Slikker, and S.F. Ali, Ann. NY Acad. Sci. in press, (1998). 141. E.L. Mandrile and G.M. Bongiorno de Pfirter, Acta. Farm. Bonaerense 4, 49 (1985). 142. C. Naranjo, "The healing journey. New approaches to consciousness". N.Y. Pantheon.New York, 1973. 143. P.B. Von Schmid, Arzneim.-Forsch. 17, 485 (1967). 144. R.S. Sloviter, E.G. Drust, B.P. Damiano, and J.D. Connor, J. Pharmacol. Exp. Ther. 214, 231 (1980). 145. N.R. Farnsworth, Science 162, 1086 (1968). 146. J. Delourme-Houde, Ann. Pharm. Fr. 4, 30 (1946). 147. E. O'Hearn, D.B. Long, and M.E. Molliver, NeuroReport 4, 299 (1993).

148. E. O'Hearn, P. Zhang, and M.E. Molliver, NeuroReport 6, 1611 (1995). 149. E. O'Hearn and M.E. Molliver, J. Neurosci. 17, 8828 (1997). 150. E. O'Hearn and M.E. Molliver, Soc. Neurosci. Abstr. 23, 2308 (1997). 151. A.C. Scallet, X. Ye, R. Rountree, P. Nony, and S.F. Ali, Ann. NY Acad. Sci. 801, 217 (1996). 152. A.C. Scallet, X. Ye, and S.F. Ali, Ann. NY Acad. Sci. 801, 227 (1996). 153. H.H. Molinari, I.M. Maisonneuve, and S.D. Glick, Brain Res. 737, 255 (1996). 154. S. Helsley, C.A. Dlugos, R.J. Pentney, R.A. Rabin, and J.C. Winter, Brain Res. 759, 306 (1997). 155. J.P. O'Callaghan, L.E. Rodman, T.S. Roggers, J.B. Terril, and J.G. Page, Soc. Neurosci. Abstr. 20, 1650 (1994). 156. J.P. O'Callaghan, T.S. Rogers, L.E. Rodman, and J.G. Page, Ann. NY Acad. Sci. 801, 205 (1996). 157. B.J. Vilner, U.K. Bandarage, M.E. Kuehne, C.M. Bertha, and W.D. Bowen, in " Problems of Drug Dependence. Proceedings of the 59th Annual Scientific Meeting, NIDA Res. Monograph." (L.S. Harris, ed.), U.S. Government Printing Office, Washington, D.C. 1997. 158. E. O'Hearn and M.E. Molliver, Soc. Neurosci. Abstr. 21, 1340 (1995). 159. P. Popik, R.T. Layer, and P. Skolnick, Psychopharmacology 114, 672 (1994). 160. J.W. Olney, U S Patent 5,629,307, (1997). 161. K. Chen, T.G. Kokate, S. Yamagishi, F.I. Carroll, and M.A. Rogawski, Soc. Neurosci. Abstr. 21, 1105 (1995). 162. K. Chen, T.G. Kokate, S.D. Donevan, F.I. Carroll, and M.A. Rogawski, Neuropharmacology 35, 423 (1996). 163. D.C. Mash, R. Douyon, W.L. Hearn, N.C. Sambol, and J. Sanchezramos, Biol. Psychiat. 37, 652 (1995). 164. P.M. Whitaker and P. Seeman, J. Pharm. Pharmacol. 29, 506 (1977). 165. D.C. Deecher, M. Teitler, D.M. Soderlund, W.G. Bornmann, M.E. Kuehne, and S.D. Glick, Brain Res. 571, 242 (1992). 166. P.M. Sweetnam, J. Lancaster, A. Snowman, J. Collins, S. Perschke, C. Bauer, and J. Ferkany, Psychopharmacology 118, 369 (1995). 167. J.K. Staley, Q. Ouyang, J. Pablo, W.L. Hearn, D.D. Flynn, R.B. Rothman, K.C. Rice, and D.C. Mash, Psychopharmacology 127, 10 (1996). 168. C. Messer and G. Rudnick, Soc. Neurosci. Abstr. 21, 1383 (1995). 169. S.D. Glick, K. Rossman, S. Wang, N. Dong, and R.W. Keller, Jr., Brain Res. 628, 201 (1993). 170. S.F. Ali, G.D. Newport, W. Slikker, R.B. Rothman, and M.H. Baumann, Soc. Neurosci. Abstr. 21, 2107 (1995).

171. S.F. Ali, G.D. Newport, W. Slikker, Jr., R.B. Rothman, and M.H. Baumann, Brain Res. 737, 215 (1996). 172. M.S. Reid, K. Hsu, K.H. Souza, P.A. Broderick, and S.P. Berger, J. Neural. Transm. 103, 967 (1996). 173. M.H. Baumann, R.B. Rothman, and S.F. Ali, Ann. NY Acad. Sci. in press, (1998). 174. H. Sershen, A. Hashim, and A. Lajtha, Brain Res. Bull. 36, 587 (1995). 175. H. Sershen, A. Hashim, and A. Lajtha, Pharmacol. Biochem. Behav. 53, 863 (1996). 176. L.G. Harsing, H. Sershen, and A. Lajtha, J. Neur. Trans. 96(3), 215 (1994). 177. H. Sershen, A. Hashim, and A. Lajtha, Neurochem. Res. 19, 1463 (1994). 178. H. Sershen, A. Hashim, and A. Lajtha, Brain Res. Bull. 40, 63 (1996). 179. S.D. Glick, C.A. Gallagher, L.B. Hough, K.L. Rossman, and I.M. Maisonneuve, Brain Res. 588, 173 (1992). 180. S.M. Pearl, I.M. Maisonneuve, and S.D. Glick, Neuropharmacology 35, 1779 (1996). 181. I.M. Maisonneuve, G.L. Mann, C.R. Deibel, and S.D. Glick, Psychopharmacology (Berlin) 129, 249 (1997). 182. R.A. Rabin and J. C. Winter, Eur. J. Pharmacol. 316, 343 (1996). 183. E.E. Codd, Life Sci. 57, PL315 (1995). 184. J.P. Pablo and D.C. Mash, NeuroReport, in press, (1998). 185. P.M. Whitaker and P. Seeman, Psychopharmacology 59, 1 (1978). 186. P.M. Whitaker and P. Seeman, Proc. Nat. Acad. Sci. USA 75, 5783 (1978). 187. R.F. Long and A.W. Lessin, Biochem. J. 82, 4P (1962). 188. H. Sershen, A. Hashim, and A. Lajtha, Brain Res. Bull. 42, 161 (1997). 189. R.C. Miller and T. Godfraind, Eur. J. Pharmacol. 96, 251 (1983). 190. A.S. Schneider, J.E. Nagel, and S.J. Mah, Eur. J. Pharmacol. 317, R1 (1996). 191. D. Mash, J.K. Staley, J.P. Pablo, A.M. Holohean, J.C. Hackman, and R.A. Davidoff, Neurosc. Lett. 192, 53 (1995). 192. J. Pablo, J.K. Staley, A.M. Holohean, J.C. Hackman, R.A. Davidoff, and D.C. Mash, Soc. Neurosci. Abstr. 21, 1264 (1995). 193. W.D. Bowen, B.J. Vilner, W. Williams, C.M. Bertha, M.E. Kuehne, and A.E. Jacobson, Eur. J. Pharmacol. 279, R1 (1995). 194. R.H. Mach, C.R. Smith, and S.R. Childers, Life Sci. 57, PL57 (1995). 195. W.D. Bowen, B.J. Vilner, U.K. Bandarage, and M.E. Kuehne, Soc. Neurosci. Abstr. 22, 2006 (1996).

196. R.A. Rabin and J.C. Winter, Brain Res. 731, 226 (1996). 197. E. Cretet, M. Prioux-Guyonneau, C. Jacquot, H. Sentenac, and J. Wepierre, Arch. Pharmacol. 313, 119 (1980). 198. Z. Binienda, J. Johnson, B. Thorn, and S.F. Ali, Soc. Neurosci. Abstr. 23, 1227 (1997). 199. R.D. Bunag and E.J. Walaszek, Ann. NY Acad. Sci. 104, 437 (1968). 200. M.E. Alburges and G.R. Hanson, Soc. Neurosci. Abstr. 23, 2408 (1997). 201. R.V. House, P.T. Thomas, and H.N. Bhargava, Pharmacology 51, 56 (1995). 202. Y. Itzhak and S.F. Ali, Soc. Neurosci. Abstr. 21, 2108 (1995). TOP
XI. Table 1. Interactions of ibogaine with neurotransmitter systems: radioligand binding studies.
Receptor system Ligand K i or IC 50 † [ µ M] Reference Alpha-adrenergic 1 prazosin 7.2 ± 3.0† 166 Dopamine transporter WIN 35,248 1.5† 76 Dopamine transporter WIN 35,248 3.5 ± 0.6† 166 Dopamine transporter RTI-121 2.0 73 Dopamine transporter RTI-55 4.11 ± 0.45† 167 Monoamine transporter (vesicular) tetrabenazine 2.23 ± 0.22† 167 Muscarinic M 1 pirenzepine 7.6 ± 0.7† 166 Muscarinic M 2 AF-DX384 5.9 ± 1.4† 166 Nicotinic nicotine 4.0 ± 0.6 125 Nicotinic noncompetitive carbamylcholine-induced 22 NaCl influx 0.02 ± 0.007† 125 NMDA ion channel MK-801 1.0 ± 0.1 159 NMDA ion channel MK-801 1.1 ± 0.03 72 NMDA ion channel MK-801 5.6 ± 0.8† 166 NMDA ion channel MK-801 4-10 191 NMDA ion channel MK-801 or TCP 0.01-0.05 and 2-4 202 NMDA ion channel TCP 1.5 ± 0.3 119 Opioid naloxone 0.13 ± 0.03 183 Opioid (kappa) U69,593 2.1 ± 0.2 165 Opioid (kappa) U69,593 29.8 ± 8.3† (rat) 13.8 ± 0.6† (mouse)

21.0 ± 1.1† (giunea-pig) 72 Opioid (kappa) U69,593 5.5 56 Opioid (kappa) U69,593 3.77 ± 0.81 124 Serotonin 2 ketanserin 4.8 ± 1.4† 166 Serotonin 3 GR-75558 3.9 ± 1.1† 166 Serotonin transporter RTI-55 0.55 ± 0.03 73 Serotonin transporter RTI-55 10 168 Serotonin transporter RTI-55 0.59 ± 0.09† 167 Serotonin transporter paroxetine 9.30 ± 1.70† 167 Sigma haloperidol 0.003† 164 Sigma pentazocine 0.086† 11 Sigma 1 pentazocine 9.3 ± 0.63 194 Sigma 1 pentazocine 8.6 ± 1.1 193 Sigma 1 pentazocine 1.5-3 202 Sigma 2 DTG 0.0904 ± 0.0101 194 Sigma 2 DTG 0.201 ± 0.023 193 Sigma 2 DTG 1.5-3 202 Voltage-dependent sodium channels batrachotoxin A 20-a- benzoate 8.1 ± 1.3 165
LEGEND TO TABLE 1. Presented are K i or IC 50 (†) values for various neurotransmitter systems affected by ibogaine with affinities higher than 10 µ M. The affinities of O -desmethylibogaine for the corresponding receptors are presented in footnotes.
--------------------------------------------------------------------------------
Top of page | Main | Search | Science | Opinion | Literature | Links | Treatment | Bookshop | Feedback
T. iboga flower

© 1999 The Ibogaine Dossier
O Rainforests Desaparecendo
Nós estamos perdendo os tesouros biológicos os mais grandes da terra apenas enquanto nós estamos começando a apreciar seu valor verdadeiro. Rainforests cobriu uma vez 14% da superfície da terra da terra; agora cobrem um mero 6% e os peritos estimam que os últimos rainforests restantes poderiam ser consumidos em menos de 40 anos Um e um meio de acre de mais rainforest é perdido cada
segundo com conseqüências trágicas para ambos que tornam-se e países industriais.
Rainforests está sendo destruído porque o valor da terra a mais rainforest é percieved como somente o valor de sua madeira por governos short-sighted, por companhias registrando multinacionais, e por proprietários de terra.
Quase a metade da espécie do mundo das plantas, dos animais e dos microoganisms será destruída ou ameaçada severamente sobre o século de um quarto seguinte devido ao deforestation de Rainforest.
Os peritos estimam que nós estamos perdendo a espécie de 137 planta, de animal e de inseto cada único dia devido ao deforestation o mais rainforest. Isso iguala a 50.000 espécies um o ano. Como a espécie a mais rainforest dissapear, assim que faça muitas curas possíveis para doenças life-threatening. Atualmente, 121 drogas da prescrição venderam vindo worldwide das fontes planta-derivadas. Quando 25% de pharmaceuticals ocidentais forem derivados dos ingredientes os mais rainforest, menos que 1% destas árvores e plantas tropicais foram testadas por cientistas.
A maioria de rainforests são cancelaram por chainsaws, por escavadoras e por fogos para seu valor da madeira e são seguidos então cultivando e ranching as operações, uniformes por gigantes do mundo como Mitsubishi Corporation, Gerogia o Pacífico, Texaco e Unocal.

Havia os dez milhão indians estimados que vivem no Amazonian Rainforest cinco séculos há. Hoje há menos de 200.000.
Em Brasil sozinho, os colonos europeus destruíram mais de 90 tribes indígenos desde os 1900's. Com eles tenha séculos idos do conhecimento acumulado do valor medicinal da espécie a mais rainforest. Como seus homelands continuam a ser destoyed pelo deforestation, os povos os mais rainforest dissappearing também.
A maioria homens de medicina e de shamans restantes no Rainforests têm hoje 70 anos velhos ou mais. Cada vez que um homem de medicina de Rainforest morre, é como se uma biblioteca se queimou para baixo.
Quando um homem de medicina morre sem passar suas artes sobre à geração seguinte, o tribe e o mundo perdem milhares dos anos do conhecimento insubstituível sobre plantas medicinal.
A riqueza do Rainforests As tampas de Amazonian Rainforest sobre bilhão acres, áreas
abrangendo em Brasil, Venezuela, Colômbia e a região andean oriental de Equador e de Peru. Se Amazonia fosse um país, seria o nono maior no mundo.
O Amazon Rainforest foi descrito como os "pulmões de nosso planeta" porque fornece o serviço ambiental essencial do mundo do dióxido de carbono continuamente recyling no oxigênio . Mais de 20 por cento do oxigênio do mundo são produzidos no Amazon Rainforest.
Mais do que a metade do mundo estimada 10 milhão espécies das plantas, dos animais e dos insetos vivem nos rainforests tropicais. Um quinto da água fresca do mundo estão na bacia de Amazon.
Um hectare (2,47 acres) pode conter sobre 750 tipos de árvores e de 1500 espécies de umas plantas mais elevadas.
Ao menos 80% da dieta do mundo desenvolvido originou no mais rainforest tropical. Seus presentes bountiful ao mundo incluem frutas como abacates, cocos, figs, laranjas, limões, grapefuit, bananas, guavas, pinapples, mangos e tomates; vegetais including o milho, as batatas, o arroz, o squash de inverno e os yams; os spices gostam da pimenta, de pimenta de Caiena, do chocolate, da canela, de cravos-da-índia, do ginger,

do bastão de açúcar, do tumeric, do café e do vanilla e de porcas pretos including porcas de Brasil e cajus.
Ao menos 3000 frutas são encontradas nos rainforests; destes somente 200 é agora dentro o uso no mundo ocidental. Os indians do uso o mais rainforest sobre 2.000.
As plantas de Rainforest são ricas em metabolites secundários, particularmente alcalóides. Os bioquímicos acreditam alcalóides protegem plantas dos ataques da doença e do inseto. Muitos alcalóides de umas plantas mais elevadas provaram ser do valor medicinal e beneficiam-se.
Atualmente, 121 drogas da prescrição venderam atualmente vindo worldwide das fontes planta-derivadas. E quando 25% de pharmaceuticals ocidentais forem derivados dos ingredientes os mais rainforest, menos de 1% destas árvores tropicais e as plantas foram testadas por cientistas.
O instituto nacional do cancer de ESTADOS UNIDOS identificou 3000 plantas que são ativas de encontro às pilhas do cancer. 70% destas plantas são encontradas no mais rainforest. Twenty-five por cento dos ingredientes ativos em drogas cancer-de combate de hoje vêm dos organismos encontrados somente no mais rainforest
Vincristine, extraído da planta a mais rainforest, periwinkle , é uma das drogas anticancer as mais poderosas do mundo. Aumentou dramàtica a taxa da sobrevivência para o leukemia agudo da infância desde sua descoberta.
Em 1983, não havia nenhum fabricante pharmaceutical de ESTADOS UNIDOS envolvido em programas de pesquisa para descobrir drogas novas ou curas das plantas. Hoje, sobre 100 companhias pharmaceutical e diversas filiais do governo dos E. U., including gigantes goste de Merck e o instituto nacional do cancer, é acoplado em projetos de pesquisa da planta para drogas possíveis e curas para vírus, infecções, cancer e AIDS uniformes.
Ação De Rainforest Os peritos concordam que deixando os rainforests intatos e
colhendo os são muitas porcas, frutas, plantas oil-producing, e plantas medicinal, o mais rainforest têm um valor mais econômico do que se forem reduzidos para fazer pastar a terra para o gado ou para a madeira. .

Os statistics os mais atrasados mostram que a terra a mais rainforest convertida às operações do gado rende o proprietário de terra $60 por o acre e se a madeira for colhida, a terra valem a pena $400 por o acre. Entretanto, se estes recursos renewable e sustainable forem colhidos, a terra renderá o proprietário de terra $2.400 por o acre.
Se controlada corretamente, a lata a mais rainforest fornece a necessidade do mundo para estes recursos naturais em uma base perpetual.
Promover o uso destas fontes sustainable e renewable podia parar a destruição do Rainforests. Criando uma fonte de renda nova que colhe as plantas medicinal, porcas das frutas, óleo e outros recursos sustainable, os rainforests são estejam uns vivos mais valioso do que cortados e queimados.
A demanda suficiente de sustainable e havested ecologically produtos de Rainforest é necessária para esforços da preservação suceder. Compra produtos os mais rainforest sustainable pode efetuar a mudança positiva criando um mercado para estes produtos ao suportar a economia de pessoa nativo e fornece a solução e a alternativa econômicas a cortar a floresta apenas para o valor de sua madeira.
O seguinte excerpted do livro, segredos herbal do Rainforest (Prima que publica, Rocklin, CA) por Leslie Alfaiate A destruição do Rainforest majestoso A beleza, o majesty e o timelessness de um o mais rainforest preliminar são indescribable. É impossível capturar na película, descrever nas palavras ou explicar àquelas que nunca tiveram a experiência awe-inspirando de estar no coração de um o mais rainforest preliminar. Rainforests evoluiu sobre milhões dos anos para girar nos ambientes que incredibly complexos são hoje. Um o mais rainforest representa uma loja de viver e de respirar os recursos naturais renewable, que têm para eons, pelo virtue de seu richness na espécie do animal e da planta, contribuída uma riqueza dos recursos para a sobrevivência e o bem estar do homem. Estes incluíram fontes

de alimento básicas, roupa, abrigo, combustível, spices, materiais crus industriais e medicina para o todo o aqueles que viveram sustainably no majesty da floresta. Entretanto, a dinâmica interna de um o mais rainforest tropical é um sistema intricado e frágil onde tudo seja assim interdependente que virar uma porção pode conduzir aos danos ou mesmo a destruição desconhecida do todo. Sadly, fêz exame somente de um século da intervenção do homem para destruir o que a natureza tem projetado assim intricada ao último para sempre.
Em 1950, 15% da superfície da terra da terra foi coberto por mais rainforest. Hoje, mais do que meio tem ido já acima no fumo. Um século há, a metade de India e um third de Etiópia foram cobertos pela floresta, agora somente quatorze por cento no remains de India e somente um terço são deixados em Etiópia. Oito fora de dez árvores em Ghana foram reduzidos. Três quartos das árvores da costa de marfim são idos. Mais de vinte por cento do Amazon Rainforest estão idos já e muito mais ameaçados severamente enquanto a destruição continua a escalar. Os statistics relataram em 1996 relataram o Amazon mostraram um aumento de 34 por cento no deforestation desde 1992. Um relatório novo por um comitê congressional diz o Amazon está desaparecendo em uma taxa de 20.000 milhas quadradas um o ano. Aquele é mais de três vezes a taxa de 1994, o ano passado para que as figuras oficiais estão disponíveis. "se nada é feito, o Amazon inteiro estará ido dentro de 50 anos," disse o autor do relatório 110-page, Representante Gilney Vianna do partido do trabalhador leftist no estado de Amazon de Mato Grosso. Contudo umas outras figuras novas ditas do relatório recente mostraram que no Amazon brazilian, fogos da floresta aumentados por mais de 50 por cento sobre 1996. Em menos de 50 anos, mais do que a metade dos rainforests tropicais do mundo têm a vítima caída ao fogo e a corrente viu e a taxa da destruição está acelerando ainda. Inacreditàvel, sobre 200.000 acres de mais rainforest são queimados cada dia. Isso está sobre 150 acres perdidos cada minuto de cada dia, e 78 milhão acres são perdidos cada ano! Uma floresta mais tropical queimou-se em torno do mundo em 1997 do que em toda a outra hora no history gravado, um relatório pelo fundo largo do mundo para a natureza. O fundo dito "1997 será recordado como o ano o fogo travado mundo," disse Jean-Paul Jeanrenaud, cabeça de seu programa da floresta.

O deforestation maciço traz com ele muitas conseqüências feias - poluição do ar e da água, erosão do solo, epidemias da malária, a liberação do dióxido de carbono na atmosfera, o eviction e o decimation de tribes indian indígenos, e a perda do biodiversity com a extinção das plantas e dos animais. Menos Rainforests significa menos chuva, menos oxigênio para que nós respirem, e uma ameaça mesmo mais grande de aquecer-se global. Mas quem deve realmente responsabilizar? Considere o que nós industrialized americanos fizemos a nosso próprio homeland... nós convertemos n *** TRANSLATION ENDS HERE ***inety percent of North America's virgin forests into firewood, shingles, furniture, railroad ties and paper. Other industrialized countries have done no better. Malaysia, Indonesia and Brazil, among other tropical countries with rainforests, are often branded as "environmental villains" of the world, mainly due to their reported levels of destruction of their rainforests. But despite the levels of deforestation, they are still covered by up to 60% of their territory by natural tropical forests. In fact, much of the pressure today on their remaining rainforests come from servicing the needs and markets for wood products in industrialized countries who have already depleted their own natural resources. Industrial countries would not be buying rainforest hardwoods and timber had we not cut down our own trees long ago nor would poachers in the Amazon jungle be slaughtering jaguar, ocelot, caiman and otter if we did not provide lucrative markets for their skins in Berlin, Paris and Tokyo.
The Biodiversity of the Rainforest So why should the loss of tropical forests be of any more concern to us in light of our own poor management of natural resources? The loss of tropical rainforests has a profound and devastating world impact because rainforests are so much more biologically diverse.
Consider these facts: A single pond in Brazil can sustain a greater variety of fish than
are found in all of Europe's rivers; A twenty-five acre plot of rainforest in Borneo may contain
over seven hundred species of trees - a number equal to the total tree diversity of North America;

A single rainforest reserve in Peru is home to more species of birds than the entire United States;
One single tree in Peru was found to harbor forty-three different species of ants - a total that approximates the entire ant species in the British Isles.
The biodiversity of the tropical rainforest is so immense that less than one percent of its millions of species have been studied by scientists for active constituents and their possible uses for man. When an acre of topical rainforest is lost, the impact to the number of plant, animal and insect species lost and their possible uses is staggering. Scientific experts estimate that we are losing over 137 species of plants, animals and insects every single day because of rainforest deforestation. Surprisingly, scientists have a better understanding of how many stars there are in the galaxy than how many species there are on Earth. Estimates of global species diversity have varied from 2 million to 100 million species, with a best estimate of somewhere near 10 million, and only 1.4 million have actually been named. Today, Rainforests occupy only 2% of the entire earth's surface and 6% of the world's land surface, yet these remaining lush rainforests support over half of our plants wild plants and trees and one-half of the world's wildlife. Hundreds and thousands of these rainforest species are being extinguished before they have even been identified, much less cataloged and studied. The magnitude of this loss to the world was most poignantly described by Harvard's Pulitzer Prize-winning biologist, Edward O. Wilson, over 10 years ago... "The worst thing that can happen during the 1980's is not energy depletion, economic collapses, limited nuclear war, or conquest by a totalitarian government. As terrible as these catastrophes would be for us, they can be repaired within a few generations. The one process ongoing in the 1980's that will take millions of years to correct is the loss of genetic and species diversity by the destruction of natural habitats. This is the folly that our descendants are least likely to forgive us for."
Yet still the destruction continues. If deforestation continues at current rates, scientists estimate nearly 80 to 90 percent of tropical rainforest ecosystems will be destroyed by the year 2020. This destruction is the main force driving a species extinction rate unmatched in 65 million years.

The Amazon Rainforest... The Last Frontier on Earth If Amazonia were a country, it would be the ninth largest in the world. The Amazon Rainforest, the world's greatest remaining natural resource, is the most powerful and bio-actively diverse natural phenomenon on the planet. It has as been described as the "Lungs of our Planet" because it provides the essential environmental world service of continuously recycling carbon dioxide into oxygen. It is estimated that over twenty percent of earth's oxygen is produced in this area.
The Amazon rainforest covers over 1.2 billion acres representing two-fifths of the enormous South American continent and is found in nine South American countries: Brazil, Columbia, Peru, Venezuela, Ecuador, Bolivia and the three Guyanas. With 2.5 million square miles of rainforest, the Amazon Rainforest represents 54 percent of the total rainforests left on the planet. The life force of the Amazon Rainforest is the mighty Amazon River. It starts as a trickle high in the snow-capped Andes mountains and flows over 4,000 miles across the South American continent until it enters the Atlantic ocean at Belem, Brazil where it is 200 to 300 miles across, depending on the season. Even 1,000 miles inland, it is still 7 miles in width. The river is so deep that ocean liners can travel 2,300 miles inland, up its length. The Amazon River flows through the center of the rainforest and is fed by 1,100 tributaries, seventeen of which are over 1,000 miles long. The Amazon is by far the largest river system in the world and over two-thirds of all the fresh water found on earth is in the Amazon basin's rivers, streams and tributaries. With so much water its not unusual that that the main mode of transportation throughout the area is by boat. The smallest and most common boats used today are still made out of hollowed tree trunks, whether they are powered by outboard motors or more often by man-powered paddles. Almost 14,000 miles of Amazon waterway are navigable and several million miles through swamps and forests are penetrable by canoe. The enormous Amazon River carries massive amounts of silt from run-off from the rainforest floor. Massive

amounts of silt deposited at the mouth of the Amazon river has created the largest river island in the world, Marajo Island, which is roughly the size of Switzerland. With this massive fresh water system, it not unusual that the life beneath the water is as abundant and diverse as the surrounding rainforest's plant and animal species. Over 2,000 species of fish have been identified in the Amazon Basin - more species than the entire Atlantic Ocean. The Amazon Basin was formed in the Paleozoic period, somewhere between 500 and 200 million years ago. The extreme age of the region in geologic terms has much to do with the relative infertility of the rainforest soil and the richness and unique diversity of the plant and animal life. There are more fertile areas in the Amazon River's flood plain, where the river deposits richer soil brought from the Andes, which only formed 20 million years ago. The rich diversity of plant species in the Amazon Rainforest is the highest on earth. Experts show that one hectare (2.47 acres) may contain over 750 types of trees and 1500 species of higher plants and it is estimated that one hectare of Amazon rainforest contains about 900 tons of living plants. Altogether it contains the largest collection of living plants and animal species in the world. The Andean mountain range and the Amazon jungle are home to more than half of the world's species of flora and fauna and one in five of all the birds in the world live in the rainforests of the Amazon.. To date, some 438,000 species of plants of economic and social interest have been registered in the region and many more have yet been cataloged or even discovered.
Once a vast sea of tropical forest, the Amazon rainforest today is scarred by roads, farms, ranches and dams. (See figure 1) Brazil is gifted with a full third of the world's remaining rainforests and unfortunately, it is also one of the world's great rainforest destroyers, burning or felling over 2.7 million acres each year. Today, more than 20 percent of rainforest in the Amazon has been razed and is gone forever. This ocean of green nearly as large as Australia, is the last great rainforest in the known universe and it is being decimated like the others before it. Why? Like other rainforests already lost forever, the land is being cleared for logging timber, large scale cattle ranching, mining operations, government road building and hydroelectric schemes, military operations, and the subsistence agriculture of peasants and landless settlers. Sadder still, in many places the rainforests are burnt simply to provide charcoal to power

industrial plants in the area.
The Driving Forces of Destruction Commercial logging is the single largest cause of rainforest destruction both directly and indirectly. (See figure 2) The simple fact is that they are destroying the Amazon Rainforest and the rest of the rainforests of the world because "they can't see the forest for the trees." Logging tropical hardwoods like teak, mahogany, rosewood and other timber for furniture, building materials, charcoal and other wood products is big business and big profits. Several species of tropical hardwoods are imported by developed counties, including America, just to build coffins which are then buried or burned. The demand, extraction and consumption of tropical hardwoods has been so massive that some countries which have been traditional exporters of tropical hardwoods are now importing them because they have already exhausted their supply by destroying their native rainforests in slash and burn operations. It is anticipated that The Phillippines, Malaysia, The Ivory Coast, Nigeria and Thailand will soon follow as all these countries will run out of rainforest hardwood timber for export in less than five years. Japan is the largest importer of tropical woods. Despite recent reductions, Japan's 1995 tropical timber import total of 11,695,000 cubic meters is still gluttonous; damaging to the ecological, biological and social fabric of tropical lands, and clearly unsustainable for any length of time. Behind the hardwood logger come others down the same roads built to transport the timber. The cardboard packing and the wood chipboard industries use 15 ton machines that gobble up the rainforest with 8 foot cutting discs that have eight blades revolving 320 times a minute which cut entire trees into chips half the size of a matchbox. More than 200 species of trees can be gobbled up by these machines which are currently clearing 320 square miles of rainforest in Papua New Guinea to provide a fraction of the demand of these two industries. These same land devouring machines are meeting the remaining world demand in the Amazon and Australian rainforests. Logging rainforest timber is a large economic source, and in many cases, the main source of revenues for servicing the national debt of developing countries. Logging profits are real to these countries who must service their debts, but are they are fleeting. Governments are selling their assets too cheaply, and once the rainforest is gone, their

source of income is gone. Sadly, most of the real profits of the timber trade are made not by the developing countries, but by multi-national companies and industrialists of the northern hemisphere. These huge profit driven companies pay governments a fraction of the timber's worth for large logging concessions on immense tracts of rainforest land and reap huge profits by harvesting the timber in the most economical manner feasible with little regard to the destruction left in their wake. Logging concessions in the Amazon are sold for as little as $2 per acre with logging companies felling timber worth thousands of dollars per acre. Governments are selling their natural resources, hawking for pennies, resources that soon will be worth billions of dollars. Some of these government concessions and land deals made with industrialists make the sale of Manhattan for twenty-four dollars worth of trinkets look shrewd. In 1986, a huge industrial timber corporation bought thousands of acres in the Borneo rainforest by giving 2,000 Malaysian dollars to 12 longhouses of local tribes. This sum amounted to the price of two bottles of beer for each member of the community. Since then this company and others have managed to extract and destroy about a third of the Borneo rainforest - about 6.9 million acres and the local tribes have been evicted from the area or forced to work for the logging companies at slave wages. In addition to logging for exportation, rainforest wood stays in developing countries for fuel wood and charcoal. One single steel plant in Brazil making steel for Japanese cars needs millions of tons of wood each year to produce charcoal that can be used in the manufacture of steel. Then there is the paper industry. A pulpwood project in the Brazilian Amazon consists of a Japanese power plant and pulp mill. To set up this single plant operation, 5,600 square miles of Amazon Rainforest was burned to the ground and replanted with pulpwood trees. This single manufacturing plant consumes 2,000 tons of surrounding rainforest wood every day to produce 55 megawatts of electricity to run the plant. The plant, which has been in operation since 1978, produces over 750 tons of pulp for paper every 24 hours, worth approximately $500,000 and has built 2,800 miles of roads through the Amazon rainforest to be used by its 700 vehicles. In addition to this pulp mill, the world's biggest pulp mill is the Aracruz mill in Brazil; its two units produce one million tons of pulp a year and displaced thousands of indigenous tribes harvesting the rainforest to keep the plant in business. Where does all this pulp go? Aracruz's biggest customers are the United States, Belgium, Great Britain, and

Japan. More and more rainforest is destroyed to meet the demand of developed world's paper industry which requires a staggering 200 million tons of wood each year simply to make paper. If the world continues at the present rate, 4 billion tons of wood is estimated to be consumed annually by the year 2020 in the paper industry alone. Even more rainforest is destroyed by mining operations. Brazil sits on one of the worlds largest reserves of iron ore and has ample gold, semiprecious and precious stones, natural gas and oil reserves as well. Strip mining is common in the Amazon and huge chunks of rainforest land is lost every year to mining operations. Even more lands are lost to the polution caused by these mining operations as the constant water runoff in the rainforest carries the waste oil, mercury, and other pollutinates and contaminants used. Mecury poisoning by animal and human inhabitants alike is becomming a common problem as the mecury used in strip mining and gold mining operations runs off into the rivers and streams and is carried hundreds of miles. (See figure 3.) Once an area of rainforest has been logged, even if given the rare change to re-grow, it can never became what it once was. The intricate ecosystem nature devised is lost forever. Only 1-2 percent of light at the top of a rainforest canopy manages to reach the forest floor below. Most times when timber is harvested, the plants and animals of the original forest becomes extinct, and trees and other plants that have evolved over centuries to grow in the dark, humid environment below the canopy simply cannot live out in the open. Even if only sections of land throughout an area are destroyed, these remnants change drastically. Birds and other animals cannot cross from one to another in the canopy, so plants are not pollinated, seeds are not dispersed by the animals and the plants around the edges are not surrounded by the high jungle humidity which they need to grow properly. As a result, the remnants slowly become degraded and die. Rains come and wash away the thin topsoil that was previously protected by the canopy and this barren unfertile land results in erosion. Sometimes the land is replanted in African grasses for cattle operations and other times, more virgin rainforest is destroyed for cattle operations because planting grass on recently burned land has a better change to grow. As the demand in the Western world for cheap meat increases, more and more rainforest is destroyed to provide grazing land for animals. In South America alone, there are an estimated 220 million head of cattle, 20 million goats, 60 million pigs and 700 million chickens.

Most of Central and Latin America's tropical and temperate rainforests have been lost to cattle operations to meet the world demand, and still the cattle operations continue to move southward into the heart of the South American Rainforests. To graze one steer in Amazonia, it takes two full acres. Most of the ranchers in the Amazon operate at a loss, yielding only paper profits purely as tax shelters. Rancher's fortunes are made only when ranching is supported by government giveaways. A banker or rich land owner in Brazil can slash and burn a huge tract of land in the Amazon rainforest, seed it with grass for cattle and realize millions of dollars worth of government-subsidized loans, tax-credits and write offs in return for developing the land. These government development schemes rarely make a profit actually selling cheap beef to industrialized nations. One single cattle operation in Brazil that was co-owned by British Barclays Bank and one of Brazil's wealthiest families was responsible for the destruction of almost 500,000 acres of virgin rainforest. The cattle operation never made a profit but government write-offs sheltered huge logging profits earned off of logging other land in the Brazilian rainforest owned by the same investors. These generous tax and credit incentives have created over 29 million acres of large cattle ranches in the Brazilian Amazon, even though the typical ranch could cover less that half its costs without these subsidies. This type of government-driven destruction of rainforest land is promoted by a common attitude among governments in rainforest regions that the forest is an economic resource to be harnessed to aid in the development of their countries. The same attitudes that accompanied the colonization of our own frontier are found today in Brazil and other countries with wild unharnessed rainforest wilderness. These beliefs are exemplified in a Brazilian official's public statement that "not until all Amazonas is colonized by real Brazilians, not Indians, can we truly say we own it." Were we Americans any different with our own colonization decimating the North American Indian tribes? Like Brazil, we sent out a call to all the world that America had land for the landless in an effort to increase colonization of our land at the expense of our Indigenous Indian tribes. And like the first America colonists, colonization in the rainforest really means subsistence farming. Subsistence farming has for centuries been a driving force in the loss of rainforest land and as populations explode in third world countries in South American and the Far East, the impact has been profound.

By tradition, wildlands and unsettled lands in the rainforest are free to those who clear the forest and till the soil. "Squatter's Rights" still prevail and poor, hungry people show little enthusiasm for arguments about the value of biodiversity or the plight of endangered species. The present approach to rainforest cultivation produces wealth for a few, for a short time because farming burned-off tracts of Amazon rainforest seldom works for long. Less than ten percent of Amazonian soils are suitable for sustained conventional agriculture. However lush they look, rainforests often flourish on such nutrient-poor soils that they are essentially "wet deserts," easier to damage and harder to cultivate than any other soil. Most are exhausted by the time they have produced three or four crops. Many of the thousands of homesteaders who migrated from Brazil's cities to the wilds of the rainforest, responding to the government's call of "land without men for men without land," have already had to abandon their depleted farms and move on, leaving behind fields of baked clay dotted with stagnant pools of polluted water. Experts agree that the path to conservation begins with helping these local residents meet their own daily needs. Because of the infertility of the soil, and the lack of knowledge of sustainable cultivation practices, this type of agriculture strips the soil of nutrients within a few harvests and the far-----mers continue to move farther into the rainforest in search of new land. They must be helped and educated to break free of the need to continually clear rainforest in search of fresh, fertile land if the rainforest is to be saved. Directly and indirectly, the leading threats to rainforest ecosystems are governments and their unbridled, unplanned and uncoordinated development of natural resources. Rainforest timber exports and large scale development projects go a long way in servicing national debt in many developing countries which is why governments and, international aid-lending institutions like the World Bank supports them. In the tropics, governments own or control nearly 80 percent of tropical forests, so these forests stand or fall according to government policy and in many countries, government policies lie behind the wastage of forest resources. Besides the tax incentives and credit subsidies which guarantee large profits to private investors who convert forests to pastures and farms, governments allow private concessionaires to log the national forests on terms that induce uneconomic or wasteful uses of the public domain. Massive public expenditures on highways, dams, plantations, and agricultural

settlements, too often supported by multilateral development lending, convert or destroy large areas of forest for projects of questionable economic worth. Tropical counties are among the poorest countries on Earth. Brazil alone spends 40 percent of its annual income simply servicing its loans and the per capita income of Brazil's people is less than $2,000 annually. Sadly, these numbers don't even represent an accurate picture in the Amazon because Brazil is one of the richer countries in South America. These struggling Amazonian countries must also manage the most complex, delicate and valuable forests remaining in the planet and the economic and technological resources available to them are limited. They must also endure a dramatic social and economic situation, plus deeply adverse terms of trade and financial relationships with industrial countries. Under such conditions, the possibility of them reaching sustainable models of development alone are nearly impossible. There is a clear need for industrial countries to sincerely and effectively assist the tropics in a quest for sustainable forest management and development if the remaining rainforests are to be saved. The governments of these developing countries need help in learning how to manage and protect their natural resources for long term profits while still managing to service their debts and they must be given the incentives and tools to do so. Programs to redefine the timber concessions so concessionaires have greater incentives to guard the long-term health of the forest and programs to revive and expand community-based forestry schemes, which ensure more rational use of forests and a better life for the people who live near them must be developed First-world capital must seek out opportunities to partner with organizations that have the technical expertise to guide these programs of sustainable economic development. In addition, programs teaching techniques for sustainable harvesting practices and identifying profitable, yet sustainable forest products can enable developing countries to improve the standard of living for its people, service national debt, and contribute meaningfully to the country's land use planning and conservation of natural resources.

Rainforests, Pharmacy to the World It is estimated that nearly half of the world's estimated 10 million species of plants, animals and micro-organisms will be destroyed or severely threatened over the next quarter century due to Rainforest deforestation. Harvard's Pulitzer Prize-winning biologist, Edward O. Wilson, estimates that we are losing 137 plant, animal and insect species every single day. That's 50,000 species a year! Again, why should we in the United States be concerned about the destruction of distant tropical rain forests? Because rain forest plants are complex chemical storehouses that contain many undiscovered biodynamic compounds with unrealized potential for use in modern medicine. We can gain access to these materials only it we study and conserve the species that contain them. Rainforests currently provide sources providing one-fourth of today's medicines, and 70% of the plants found to have anti-cancer properties are found only in the rainforest. The Rainforest and it's immense undiscovered biodiversity holds the key to unlocking tomorrow's cures for devastating diseases. How many cures to devastating disease have we already lost? Two drugs obtained from a rainforest plant known as the Madagascar periwinkle, now extinct in the wild due to deforestation of the Madagascar rainforest, has increased the chances of survival for children with leukemia from 20 percent to 80 percent. Think about it - 8 out of 10 children are now saved rather than 8 of 10 children dying from leukemia. How many children have been spared and how many more will continue to be spared because of this single rainforest plant? What if we failed to discover this one important plant among millions before it was extinct due to man's destruction? When our remaining rainforests are gone, the rare plants, animals will be lost forever and so will their possible cures to diseases like cancer.
No one can challenge the fact that man is still largely dependant on plants for treating his aliments. Almost 90 percent of people in developing countries still rely on traditional medicine--based largely on species of plants and animals--for their primary health care. In the United States, some 25 percent of prescriptions are filled with drugs whose active ingredients are extracted or derived from plants. Sales of these plant-based drugs in the U.S. amounted to some $4.5 billion in 1980. Worldwide sales of these plant-based drugs were estimated at

$40 billion in 1990. Still even more drugs are derived from animals and microorganisms. Currently 121 prescription drugs sold worldwide come from plant derived sources from only 90 species of plants. The U.S. National Cancer Institute has identified over 3,000 plants that are active against cancer cells, and 70% of these plants are found only in the rainforest. Today, over 25% of the active ingredients in today's cancer-fighting drugs come from organisms found only in the Rainforest. Among the thousands of species of rainforest plants that have not been analyzed, are many more thousands of unknown plant chemicals, many of which have evolved to protect the plants from pathogens. These plant chemicals may well help us in our own constant struggle with constantly evolving pathogens such as evolving bacteria-resistant pathogens in tuberculosis, measles, and HIV. Experts now believe that if there is a cure for cancer and even AIDS, it will probably be found in the rainforest. In 1983, there were no U.S. pharmaceutical manufacturers involved in research programs to discover new drugs or cures from plants. Today, over 100 pharmaceutical companies and several branches of the US government, including giants like Merck, Abbott, Bristol-Myers Squibb, Eli Lilly, Monsanto, SmithKline Beecham and the National Cancer Institute are engaged in plant-based research projects for possible drugs and cures for viruses, infections, cancer and AIDS. Most of this research is currently taking place in the rainforest in an industry that is now called "bio-prospecting." This new pharmacological industry has sprung up, drawing together an unlikely confederacy: plant-collectors and anthropologists; ecologists and conservationists; natural product companies and nutritional supplement manufacturers, AIDS and cancer researchers; executives in the world's largest drug companies, and native indigenous shamans. They are part of a radical experiment - to preserve the world's rainforests by showing how much more valuable they are standing than cut down. And it is a race against a clock whose every tick means another acre of charred forest. Yet it is also a race that pits one explorer against another, for those who score the first big hit in chemical bio-prospecting will secure wealth and a piece of scientific immortality. In November 1991, Merck Pharmaceutical Company announced a landmark agreement to obtain samples of wild plants and animals for drug-screening purposes from Costa Rica's National Biodiversity

Institute (INBio). Spurred by this and other biodiversity prospecting ventures, interest in the commercial value of plant genetic and biochemical resources is burgeoning today. While the Merck-INBio agreement provides a fascinating example of a private partnership that contributes to rural economic development, rainforest conservation, and technology transfer, virtually no precedent exists for national policies and legislation to govern and regulate what amounts to a brand new industry. Since wealth and technology are as concentrated in the North as biodiversity and poverty are in the South, the question of equity is particularly hard to answer in ways that satisfy everyone with a stake in the outcome. The interests of bioprospecting corporations are not the same as those of people who live in a biodiversity "hot spot," many of them barely eking out a living. As the search for wild species whose genes can yield new medicines and better crops gathers momentum, these rich habitats also sport more and more bio-prospectors. Like the nineteenth-century California gold rush or its present-day counterpart in Brazil, this "gene rush" could wreak havoc on ecosystems and the people living in or near them. Done properly, however, bioprospecting can bolster both economic and conservation goals while underpinning the medical and agricultural advances needed to combat disease and sustain growing populations. The majority of our current plant-derived drugs were discovered through these traditional uses of plants by the indigenous people where they grew and flourished. History has shown that the rainforest is no different, and these bioprospectors now are working side by side with rainforest tribal shamans and herbal healers to learn the wealth of their plant knowledge and many uses of indigenous plants where drugs and pharmacies are virtually unknown. Unlocking the Secrets of the Rainforest After the Ameri-Indians discovered America, about 20 millennia before Columbus, all their clothing, food, medicine and shelter were derived from the forests. Those millennia gave the Indians time to discover and learn empirically the virtues and vices of the thousands of edible and medicinal species in the rainforest. More than 80% of the developed world's diet originated from the rainforest and this empirical indigenous knowledge of the wealth of edible fruits, vegetables and nuts. Of the estimated 3,000 edible fruits found in the rainforest, only 200 are cultivated for use today, despite the fact that

the Indians use more than 1,500. Many secrets and untold treasures await discovery with the medicinal plants used by shamans, healers and the indigenous people of the rainforest Tribes. Long regarded as hocus- pocus by science, Indigenous People's empirical plant knowledge is now thought by many to be the Amazon's new gold. This indigenous use of the plants provides the bioprospector with the necessary clues to target specific species to research in the race for time before the species are lost to deforestation. More often the race is defined as to be the first company to patent a new drug utilizing a newly discovered rainforest phytochemical, and of course, profits for the pharmaceutical companies. Laboratory syntheses of new medicines is increasingly costly and not as fruitful as companies would like. In the words of one major drug company: "Scientists may be able to make any molecule they can imagine on a computer, but Mother Nature...is an infinitely more ingenuous and exciting chemist." Scientists have developed new technologies to assess the chemical makeup of plants and they realize using medicinal plants identified by Indians makes research more efficient and less expensive. With these new trends, drug development has actually returned to its roots - traditional medicine. It is now understood by bioprospectors that tribal people of the rainforest represent the key to finding new and useful tropical forest plants. The degree to which they understand and are able sustainably to use this diversity is astounding. The Barasan Indians of Amazonian Columbia can identify all of the tree species in their territory without having to refer to the fruit or flowers, a feat that no university-trained botanist is able to accomplish! A single Amazonian tribe of Indians may use over 200 species of plants for medicinal purposes alone. Of the 121 pharmaceutical drugs that are plant-derived today, 74% were discovered through follow up research to verify the authenticity of information concerning the ethnic medical uses of the plant. Nevertheless, to this day, very few rainforest tribes have been subjected to a complete ethnobotanical analysis. Robert Goodland of the World Bank wrote,
"Indigenous knowledge is essential for the use, identification and cataloguing of the [tropical] biota. As tribal groups disappear, their knowledge vanishes with them. The preservation of these groups is a significant

economic opportunity for the [developing] nation, not a luxury."
Since Amazonian Indians are often the only ones who know both the properties of these plants and how they can best be used, their knowledge is now being considered an essential component of all efforts to conserve and develop the rainforest. Since failure to document this lore would represent a tremendous economic and scientific loss to the industrialized world, the bioprospectors are now are working side by side with the rainforest tribal shamans and herbal healers to learn the wealth of their plant knowledge. But bioprospecting has a dark side. Indian knowledge that has resisted the pressure of "modernization" is being used by bioprospectors who, like oil companies and loggers destroying the forests, threaten to leave no benefits behind them. Its a noble idea, the ethnobotanist who works with the Indians seeking a cure for cancer or even AIDS, like Sean Connery in the movie, Medicine Man. Yet, behind this lurks a system that, at its worst, steals the Indian knowledge to benefit CEOs, stockholders and academic careers and reputations. The real goal of these powerful bio-prospectors is to target novel and active phytochemicals with medical applications, synthesize them in a laboratory and have them patented for subsequent drug manufacture and resulting profits. In this process, many active and beneficial plants have been found in the Shaman's medicine chest, but have been discarded when it was found that the active ingredients of the plant numbered too many to be synthesized into a patentable drug cost effectively. It doesn't matter how active or beneficial the plant was or how long the FDA process might take to patent and approve the new drug - if the bioprospector can't capitalize on it - the public will rarely hear about a newly discovered plant's benefits. The fact is, there is a lot of money at stake. In an article published in Economic Botany, Dr. Robert Mendelsohn, an economist at Yale University, and Dr. Michael J. Balick, director of the Institute of Economic Botany at the New York Botanical Gardens, estimate the minimum number of pharmaceutical drugs potentially remaining to be extracted from the rainforests. It is staggering! They estimate that there are at least 328 new drugs that still await discovery in the rainforest with a potential value of $3-4 billion to a private pharmaceutical company and as much as $147 Billion to society as a whole.

While the Indigenous Indian shamans go about their daily lives caring for the well being of their tribe, thousands of miles away in US laboratories, the Shaman's rainforest medicines are being tested, synthesized, patented and submitted for FDA approval. Soon children with viral infections, adults with herpes, cancer patients and many others may benefit from new medicines from the Amazon Rainforest. But what will the Indigenous Tribes see of these wonderful new medicines? As corporations rush to patent indigenous medicinal knowledge, the originating Indigenous communities have received few, if any benefits. Losing the Knowledge The destruction of the rainforest has followed the pattern of seeing natural land and natural world peoples as resources to be used, and seeing wilderness as idle, empty and unproductive. Destruction of our rainforests is not only causing the extinction of plant and animal species, it is also is wiping out Indigenous Peoples which live in the rainforest. Obviously, rainforests are not idle land, nor are they uninhabited. Indigenous Peoples have developed technologies and resource use systems that have allowed them to live on the land, farming, hunting and gathering in a complex sustainable relationship with the forest. But when rainforests die, so do the Indigenous Peoples. In 1500, there were an estimated six to nine million Indigenous People inhabiting the rainforests in Brazil. When Western and European cultures were drawn to Brazil's Amazon in the hopes of finding riches beyond comprehension and artifacts from civilizations that have long since expired with the passage of time, they left behind decimated cultures in their ravenous wake. By 1900 there were only one million Indigenous People left in Brazil's Amazon. Although the fabled Fountain of Youth was never discovered, many treasures in gold and gems were spirited away by the more successful invaders of the day and the Indigenous inhabitants of the rainforest bore the brunt of these marauding explorers and conquistadors. Today there are less than 250,000 Indigenous People of Brazil surviving this catastrophe and still it continues. These surviving Indigenous People still demonstrate the remarkable diversity of the rainforest because they comprise 215 ethnic groups with 170 different languages. They live in 526 territories nationwide, which together comprise an area of 190 million acres... twice the size of California.

About 188 million acres of this land is inside the Brazilian Amazon, in the states of Acre, Amapa, Amazonas, Para, Mato Grosso, Maranhao, Rondonia, Roraima, and Tocantins. There may also be 50 or more indigenous groups still living in the depths of the rainforest that have never had contact with the outside world. Throughout the rainforest, forest-dwelling peoples whose age-old traditions allow them to live in and off the forest without destroying it are losing out to cattle ranching, logging, hydroelectric projects, large-scale farms, mining, and colonization schemes. About half of the original Amazonian Tribes have already been completely destroyed. The greatest threat to Brazil's remaining tribal people, most of whom live in the Amazon Rainforest, is the invasion of their territory by these ranchers, miners, land speculators and the conflicts which follow. In Amazonia, thousands of peasants, rubber tappers, and Indigenous Tribes have been killed in the past decade in violent conflicts over forest resources and land. As their homelands continue to be invaded and destroyed, rainforest people and their cultures are disappearing. When these Indigenous Peoples are lost forever, gone too is their empirical knowledge representing centuries of accumulated knowledge of the medicinal value of rainforest plant and animal species. Very few tribes have been subjected to a complete ethnobotanical analysis of their plant knowledge and most medicine men and shamans remaining in the rainforests today are 70 years old or more. When a medicine man dies without passing his arts on to the next generation, the tribe and the world loses thousands of years of irreplaceable knowledge about medicinal plants. Each time a Rainforest medicine man dies, it is as if a library has burned down. THE SOLUTION Profits without Plunder The problem and the solution of the destruction of the rainforest are both economic. Governments need money to service their debts, squatters and settlers need money to feed their families, and companies need to make profits. The simple fact is that the rainforest is being destroyed for the income and profits it yields - however fleeting. Money still makes the world go around... even in South America and even in the rainforest. But this also means that if land owners, governments and those living in the rainforest today were given a viable economic reason NOT to

destroy the rainforest, it could and would be saved. And this viable economic alternative DOES exist and it is working today. Many organizations have demonstrated that if the medicinal plants, fruits, nuts, oils and other resources like rubber, chocolate and chicle (used to make chewing gums), were harvested sustainably, rainforest land has much more economic value today and more long term income and profits than if just timber were harvested or if it were burned down for cattle or farming operations. In fact, the latest statistics prove that rainforest land converted to cattle operations yields the land owner $60 per acre and if timber is harvested, the land is worth $400 per acre. However, if these renewable and sustainable resources are harvested, the land will yield the land owner $2,400 per acre.This value provides an income not only today, but year after year - for generations. These sustainable resources are the true wealth of the Rainforest - not the trees. This is no longer a theory. It is a fact and it is being implemented today. Just as importantly, to wild-harvest the wealth of sustainable rainforest resources effectively, local people and indigenous tribes must be employed. Today, entire communities and tribes earn 5 to 10 times more money in wild harvesting medicinal plants, fruits, nuts and oils than they can earn by chopping down the forest for subsistence crops. This much needed income source creates the awareness and economic incentive for this population in the rainforest to protect and preserve the forests for long term profits for themselves and their children and is an important solution in saving the rainforest from destruction. When the timber is harvested for short term gain and profits, the medicinal plants, nuts, oils and other important sustainable resources which thrive in this delicate ecosystem are destroyed. The real solution to saving the rainforest is to make them see the forest AND the trees by creating a consumer demand and consumer markets for these sustainable rainforest products... markets that are larger and louder than today's tropical timber market.... markets which will put as much money in their pockets and government coffers as the timber companies do.... markets which will give them the economic incentive to protect their sustainable resources for long term profits rather than short term gain. This is the only solution that makes a real impact and it can make a real difference. Each and every person here in America can take a part in this solution by helping to create this consumer market and demand

for sustainable rainforest products. By purchasing renewable and sustainable rainforest products and resources and demanding sustainable harvesting of these resources utilizing local communities and indigenous tribes of the rainforests, we all can be part of the solution and the rainforests of the world and it's people can be saved.
Help with school reports on rainforest subjects
Raintree Nutrition Products Company Mission
Online Shopping Rainforest Philosophy

http://babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.rain-tree.com%2Findex.html http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.rain-tree.com%2Findex.html http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.rain-tree.com%2Fplants.htm http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.rain-tree.com%2Fplants.htm http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.rain-tree.com%2Flinks.htm http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.rain-tree.com%2Flinks.htm http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.rain-tree.com%2Fproduct.htm http:// babelfish.altavista.com/babelfish/urltrurl?lp=en_pt&url=http%3A%2F%2Fwww.rain-tree.com%2Fproduct.htm mailto:info@rain- tree.com mailto:[email protected]
© Copyrighted 1996 - 2002 Raintree Nutrition, Inc., Austin, Texas 78758. All rights reserved. Please read the Conditions of Use , Copyright Statement and our Privacy Policy for this web page and web site.



