Embed Size (px)
Citation preview

Universidade Federal de Ouro Preto – UFOP
Núcleo de Pesquisa em Ciências Biológicas – NUPEB
Laboratório de Enzimologia e Proteômica - LEP
Programa de Pós-Graduação em Biotecnologia
Alterações hepatoesplênicas associadas à infecção por Schistosoma
mansoni no modelo murino: clínica, histopatologia e proteômica
Jonatan Marques Campos
Ouro Preto
2012

Universidade Federal de Ouro Preto – UFOP
Núcleo de Pesquisa em Ciências Biológicas – NUPEB
Laboratório de Enzimologia e Proteômica - LEP
Programa de Pós-Graduação em Biotecnologia
Alterações hepatoesplênicas associadas à infecção por Schistosoma
mansoni no modelo murino: clínica, histopatologia e proteômica
Ouro Preto
2012
Dissertação apresentada ao Programa de Pós-Graduação em
Biotecnologia do Núcleo de Pesquisa em Ciências Biológicas
(NUPEB) da Universidade Federal de Ouro Preto (UFOP), como parte
integrante dos requisitos para obtenção do título de Mestre em
Biotecnologia, Área de concentração - Genômica e Proteômica.
Orientador: Prof. Dr. William de Castro Borges
Co-Orientador: Prof. Dr. Milton Hércules Guerra de Andrade

C198a Campos, Jonatan Marques.
Alterações hepatoesplênicas associadas à infecção por Schistosoma
mansoni no modelo murino [manuscrito] : clínica, histopatologia e proteômica
/ Jonatan Marques Campos - 2012.
xv, 95f.: il., color; grafs.; tabs.; mapas.
Orientador: Prof. Dr. William de Castro Borges.
Dissertação (Mestrado) - Universidade Federal de Ouro Preto.
Instituto de Ciências Exatas e Biológicas. Núcleo de Pesquisas em
Ciências Biológicas. Programa de Pós-Graduação em Biotecnologia.
Área de concentração: Genômica e Proteômica.
1. Proteômica - Teses. 2. Biotecnologia - Teses. 3. Schistosoma mansoni - Teses. 4. Biomarcadores - Teses. 5. Esquistossomose - Teses. I. Universidade Federal de Ouro Preto. II. Título.
CDU: 577.212:616.995.122
Catalogação: [email protected]


i
Colaboradores
Profa. Dra. Cláudia Martins CarneiroI
Prof. Dr. Vanderlei RodriguesII
Prof. Dr. Milton Hércules guerra de AndradeIII
Profa. Dra. Lizandra Guidi MagalhãesII
M.s. Nívia Carolina Nogueira PaivaI
Dra. Adriana Paes LemeVI
Mestranda Renata Alves de Oliveira e Castro IV
Mestranda Aline Costa LitalinoI
I – Laboratório de Imunpatologia NUPEB/UFOP
II – Laboratório de Biologia Molecular de Parasitos FMRP/USP
III – Laboratório de Enzimologia e Proteômica NUPEB/UFOP
IV – Laboratório de Imunoparasitologia NUPEB/UFOP
VI - Setor de Espectrometria de Massas do Centro Nacional de Pesquisa em Energia e
Materiais – ABTLuS)

ii
Este trabalho é dedicado a todos meus familiares;
Meus pais, Raul Antônio e Ingrid Marques;
Minha irmã, Jenifer Marques
Meus avós, Clotilde Marques e Nilo Marques (in memorian),
Tarcília Campos e Raimundo Campos.

iii
Agradecimentos
A Deus por me proporcionar paz, esperança e força durante toda minha caminhada.
Aos meus pais, irmã e familiares por serem o alicerce do meu crescimento.
Ao professor William de Castro Borges pela excelente orientação, aprendizado, confiança,
incentivo e seriedade.
Ao professor Milton Hércules Guerra de Andrade pela co-orientação, incentivo e
desenvolvimento de recursos técnicos no LEP.
Ao professor Vanderlei Rodrigues pela colaboração e oportunidade de desenvolvimento de
parte deste trabalho em seu laboratório FMRP/USP.
A professora Cláudia Martins Carneiro pela colaboração e conselhos nos experimentos.
Aos professores Ieso de Miranda e Renata Guerra por terem disponibilizados seus
laboratórios auxiliando no desenvolvimento deste trabalho.
A doutoranda Nívia Carolina pela excelente colaboração na realização da análise histológica
presente neste trabalho.
A Profa. Dra Lizandra Guidi Magalhães pela ajuda nos experimentos realizados na
FMRP/USP.
A mestranda Renata Alves de Oliveira e Castro pela ajuda nos experimentos.
Aos amigos do LEP pela amizade, apoio, aprendizado e momentos de diversão. Agradeço ao
mestrando Leandro Xavier Neves pela ajuda nos experimentos.
Ao técnico do LEP José Henrique Braga Fortes pela experiência de laboratório, por
proporcionar um ambiente de trabalho organizado e pela ajuda nos recursos técnicos.
Aos amigos da República 171 que me acolheram e foram meus parceiros de festas e futebol
durante todo este tempo. Em especial ao Daniel Discaciate (Daniboy) e ao Paulo Renato
(Marú).
Ao Programa de Pós-Graduação em Biotecnologia/NUPEB/UFOP pela oportunidade do
desenvolvimento desta dissertação.
A CAPES/REUNI pela bolsa de estudos fornecida durante a realização deste projeto.
A FAPEMIG pelo apoio financeiro para execução deste projeto.
A todos que direto ou indiretamente contribuíram para a realização deste trabalho.

iv
“Faço o melhor que sou capaz, só para viver em paz”.
(Marcelo Camelo – Los Hermanos)

v
Sumário
Lista de Figuras........................................................................................................................vii
Lista de Tabelas.........................................................................................................................ix
Lista de Abreviaturas e Símbolos...............................................................................................x
Resumo.....................................................................................................................................xii
Abstract...................................................................................................................................xiv
1 – Introdução.............................................................................................................................2
1.1 – Aspectos gerais.....................................................................................................2
1.2 - O ciclo biológico do Schistosoma mansoni..........................................................6
1.3 – Diagnóstico e Tratamento.....................................................................................9
1.4 – Proteômica..........................................................................................................10
1.5 – Modelo murino de infecção por S. mansoni.......................................................13
2 - Objetivos..............................................................................................................................15
2.1 - Objetivo geral........................................................................................................15
2.2 - Objetivos específicos............................................................................................15
3 - Metodologia.........................................................................................................................17
3.1 - Manutenção do ciclo do S. mansoni.....................................................................17
3.2 - Delineamento Experimental..................................................................................17
3.3 - Exame parasitológico de fezes..............................................................................19
3.4 - Obtenção do material biológico............................................................................19
3.5 - Razão peso do órgão e peso corporal....................................................................19
3.6 - Leucograma...........................................................................................................20
3.7 - Dosagens enzimáticas para ALT, AST e Albumina Sérica..................................20
3.8 - Análise histológica................................................................................................20
3.9 - Extração de proteínas............................................................................................21
3.10 - Eletroforese unidimensional (1D SDS-PAGE)...................................................22
3.11 - Eletroforese bidimensional (2-D SDS-PAGE)...................................................23
3.12 - Digestão in gel e preparo de amostras para espectrometria de massas...............25

vi
3.13 - Identificação das proteínas por espectrometria de massas..................................26
4 - Resultados............................................................................................................................30
4.1 - Exame parasitológico de fezes - acompanhamento da ovoposição nos animais
infectados..................................................................................................................................30
4.2 - Avaliação de esplenomegalia e hepatomegalia nos animais dos grupos I e II.....31
4.3 - Análises hematológicas e dosagens bioquímicas..................................................32
4.4 - Análises histológicas de fígado e baço.................................................................36
4.5 - Análise proteômica comparativa de fígado e baço dos animais dos grupos I e
II................................................................................................................................................40
4.6 - Perfil eletroforético unidimensional e análise densitométrica..............................40
4.7 - Perfil proteico em eletroforese bidimensional (SDS-PAGE) da fração solúvel de
fígado...................................................................................................................................41
4.8 - Perfil proteico em eletroforese bidimensional (SDS-PAGE) da fração solúvel do
baço......................................................................................................................................48
5 - Discussão.............................................................................................................................53
6 - Conclusões...........................................................................................................................64
7 - Referências..........................................................................................................................65
8 – Anexos................................................................................................................................75

vii
Lista de Figuras
Figura 1: Distribuição da Esquistossomose no mundo...............................................................3
Figura 2: As 10 principais causas de DALYs.............................................................................3
Figura 3: Distribuição da população que requer quimioterapia preventiva................................5
Figura 4: Ciclo de vida do S. mansoni........................................................................................8
Figura 5: Géis de eletroforese bidimensional (2-DE)...............................................................11
Figura 6: Métodos de ionização “soft” para análise de peptídeos e proteínas..........................12
Figura 7: Representação do método de infecção pelo anel.......................................................18
Figura 8: Diagrama esquemático para obtenção de proteínas solúveis....................................22
Figura 9: Ilustração das etapas de produção do gel bidimensional (2-DE)..............................24
Figura 10: Etapas do processo de digestão in gel.....................................................................26
Figura 11: Delineamento experimental ilustrando a identificação dos peptídeos obtidos por
digestão in gel...........................................................................................................................28
Figura 12: Gráfico demonstrando a evolução da ovoposição nos animais infectados entre o
35º e o 50º dia de infecção........................................................................................................30
Figura 13: Relação peso do órgão e peso corporal dos grupos I e II........................................31
Figura14: Análise global e diferencial de leucócitos do grupo I com 5 semanas de
infecção.....................................................................................................................................33
Figura15: Análise global e diferencial de leucócitos do grupo II com 7 semanas de
infecção.....................................................................................................................................34
Figura 16: Fotomicrografias de fígado de camundongos infectados com Schistosoma mansoni
e necropsiados com 5 e 7 semanas após a infecção..................................................................37
Figura 17: Gráfico do número de células inflamatórias na região perivascular de fígado.......38
Figura 18: Fotomicrografias de baço de camundongos infectados com Schistosoma mansoni e
necropsiados com 5 e 7 semanas após a infecção.....................................................................39
Figura 19: Gráfico comparando a área do folículo nodular do baço........................................39
Figura 20: Perfil eletroforético representativo de proteínas solúveis de fígado e baço............41
Figura 21: Géis bidimensionais representativos de proteínas solúveis de fígado de animais do
grupo I (5 semanas de infecção)...............................................................................................43

viii
Figura 22: Géis bidimensionais representativos de proteínas solúveis de fígado de animais do
grupo II (7 semanas de infecção)..............................................................................................44
Figura 23: Géis bidimensionais representativos de proteínas solúveis de fígado de animais do
grupo II (7 semanas de infecção) pH 4-7..................................................................................45
Figura 24: Géis bidimensionais representativos de proteínas solúveis de baço de animais do
grupo I (5 semanas de infecção)...............................................................................................49
Figura 25: Géis bidimensionais representativos de proteínas solúveis de baço de animais do
grupo II (7 semanas de infecção)..............................................................................................50
Figura Suplementar 1................................................................................................................91
Figura Suplementar 2................................................................................................................92
Figura Suplementar 3................................................................................................................93

ix
Lista de Tabelas
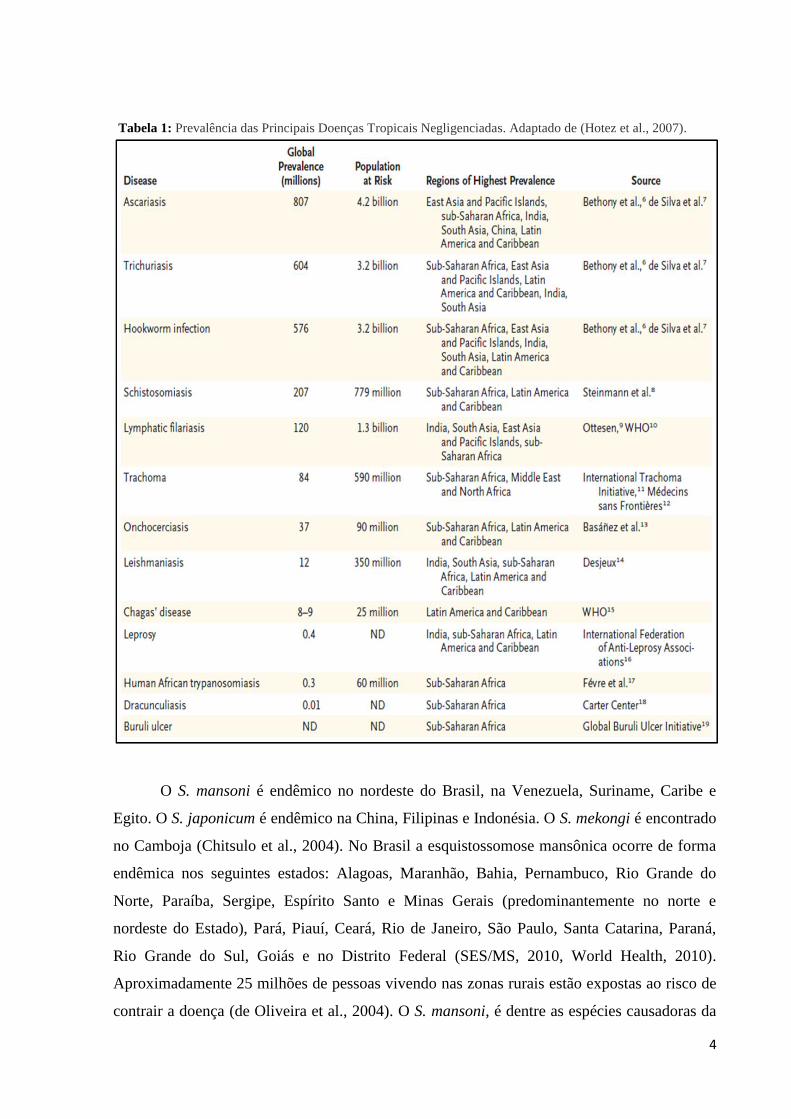
Tabela 1: Prevalência das Principais Doenças Tropicais Negligenciadas..................................4
Tabela 2: Principais metabólitos alterados em urina de camundongos infectados com S.
mansoni e em hamsters infectados com S. japonicum..............................................................14
Tabela 3: Amostragem de animais............................................................................................18
Tabela 4: Tabela de dosagens bioquímicas...............................................................................35
Tabela 5: Dosagens de proteínas solúveis de fígado e de baço................................................40
Tabela 6: Proteínas de fígado identificadas por espectrometria de massas..............................46
Tabela 7: Proteínas de baço identificadas por espectrometria de massas.................................51
Tabela Suplementar 1: Proteínas de Fígado..............................................................................76
Tabela Suplementar 2: Proteínas de Baço.................................................................................86

x
Lista de Abreviaturas e Símbolos
% - Percentual
°C – Graus Celsius
µA - Microampere
µg – Micrograma
µL - Microlitro
ACN – Acetonitrila
ADP – Adenosina difosfato
ALT – Alanino aminotrasferase
AMBIC - Bicarbonato de Amônio
AST – Aspartato aminotrasferase
ATP – Adenosina trifosfato
BCA - bicinchoninic acid
BIP - Binding immunoglobulin protein
CCA – Centro de ciência animal
CHAPS – (3-[(3-Cholamidopropil) dimetilamônio]-1- propanosulfonato)
cm – Centímetro
Da - Dalton
DNA – Ácido desoxirribonucleico
DTT - Ditiotreitol
EDTA - Ethylenediamine tetraacetic acid
EF-Tu - Elongation factor thermo unstable
ELISA – Enzyme Linked Immunosorbent Assay
emPAI – Exponentially modified protein abundance index
FDR – False discovery rate
FMRP – Faculdade de medicina de Ribeirão Preto
g - Gramas
HCL – Ácido Clorídrico
HE – Hematoxilina e Eosina
HSP – Proteína de Choque Térmico
IAA - Iodoacetamida
IEF - Isoeletrofocalização
kDa – Kilodálton
Kg – Kilograma
LEP – Laboratório de Enzimologia e Proteômica
M – Molar
m/z – Massa sobre carga
mA - Miliampere
Mb – Megabase
mg – Miligrama
min – Minutos
mm – Milímetro
mM – Milimolar
Mr – Massa relativa
N - Número
NaCl – Cloreto de Sódio
NCBI – National Center for Biotechnology Information
nL – Nanolitro
OMS – Organização Mundial de Saúde

xi
p:v – Peso por volume
pb – Pares de bases
PBS – Phosphate buffered saline
PCR – Reação em cadeia da polimerase
pH – Potencial hidrogeniônico
ROS - Reactive oxygen species
SDS – Dodecil Sulfato de Sódio
SDS-PAGE – Eletroforese em gel de poliacrilamida com SDS
TCA – Ácido tricloroacético
TFA – Ácido trifluoracético
Tris - tris(hidroximetil)aminometano
UFOP – Universidade Federal de Ouro Preto
USP – Universidade de São Paulo
V - Volts
v:v – Volume/volume
WHO – World Health Organization

xii
Resumo
A Esquistossomose é uma doença endêmica em vários países, afligindo mais de 207 milhões
de pessoas no mundo. É considerada a segunda infecção parasitária mais importante em
termos de saúde pública devido ao impacto socio-econômico, podendo ocasionar cerca de 200
mil mortes anualmente. A caracterização do genoma e transcriptoma do Schistosoma mansoni
propiciou o emprego de técnicas na área da proteômica as quais vem permitindo maior
compreensão da biologia deste parasito em todos os estágios de seu ciclo de vida. Entretanto,
até o momento, poucos estudos proteômicos abordaram a relação parasito-hospedeiro. O
entendimento desta relação é de fundamental importância para a proposição de novos
métodos de diagnóstico, avaliação de prognóstico e tratamento da doença. O presente trabalho
teve como foco a análise das alterações no proteoma solúvel de fígado e baço, utilizando o
modelo murino de infecção por S. mansoni. A primeira abordagem fundamentou-se na
discriminação dos estágios de fase aguda e fase crônica nos animais infectados. Após
realização do exame parasitológico de fezes, avaliação das alterações hepatoesplênicas através
da relação (% órgão/corpo) e histologia de fígado e baço, foi possível estabelecer que o grupo
de animais com 5 semanas de infecção desenvolveu a fase aguda da esquistossomose e
animais infectados por 7 semanas apresentaram a fase crônica da doença. A técnica de
eletroforese bidimensional comparativa, para proteínas solúveis de fígado e baço de animais
controles e infectados, demonstrou alterações nos níveis de expressão de proteínas associadas
com processos e vias bioquímicas diversas, como resposta ao estresse celular, tradução, ciclo
da uréia e via glicolítica. No fígado, a categoria de proteínas chaperoninas apresentou maior
número de representantes com alteração de expressão dentre as quais se destacam GRP-78
(proteína reguladora de glicose 78 kDa), HSP-71 (proteína de choque térmico 71 kDa), Erp-
60 (calreticulina) e PDI (proteína dissulfeto isomerase). Por outro lado, proteínas como a
CPSase I (carbamoil fosfato sintetase I), albumina, indoletilamina N-metiltransferase,
peroxirredoxina-6 e anidrase carbônica III apresentaram expressão diminuída no fígado em
decorrência da infecção. O perfil proteômico do fígado do animal infectado correlacionou-se
com a análise histológica do órgão na qual se observa uma intensa resposta imune celular. No
baço, as proteínas identificadas com alteração de expressão foram calreticulina, fosfoglicerato
quinase I, frutose-bifosfato aldolase A, gliceraldeído 3 fosfato desidrogenase, EF-Tu, (fator de
elongamento) e aspartato aminotransferase. Em geral, as proteínas diferencialmente presentes
no baço de animais infectados apresentaram aumento de expressão, sendo o perfil molecular

xiii
compatível com a demanda de produção de energia e síntese proteica. Estes dois processos
são altamente relevantes para o custeio da intensa proliferação celular observada no órgão.
Coletivamente, os resultados obtidos demonstraram que a abordagem proteômica empregada
permitiu a identificação de marcadores teciduais induzidos pela infecção por S. mansoni.
Abordagens futuras permitirão estabelecer se as alterações teciduais decorrentes da infecção
pelo parasito alteram significativamente o proteoma plasmático a ponto de indicar novos
biomarcadores da esquistossomose.

xiv
Abstract
Schistosomiasis is an endemic disease affecting 207 million people worldwide. It is
considered the second most important parasitic disease in terms of public health due to its
social and economic impact, leading to an estimated 200 thousand deaths annually. The
characterization of the S. mansoni genome and transcriptome has allowed the use of
proteomic techniques providing a deeper understanding of the parasite´s biology throughout
its life cycle stages. Nevertheless, a few proteomic studies have addressed the host-parasite
relationship. Understanding the S. mansoni parasitism is of paramount importance for the
proposal of novel diagnostic methods, evaluation of prognosis and treatment of the disease.
The present work focused on the spleen and liver soluble proteome-associated alterations in
the murine model of experimental schistosomiasis. The first approach involved discrimination
between the acute and chronic stages of the disease. After feces examination, measurement of
hepatic/spleen mass increase and tissue histology it was established that the 5 week-infected
mice developed the acute phase of schistosomiasis whereas animals infected for 7 weeks
exhibited the chronic disease. Comparative two dimensional gel electrophoresis, for cytosolic
proteins from liver and spleen, demonstrated changes in the expression profile for molecules
involved in several cellular processes such as stress response, translation, urea cycle and
glycolytic pathway. In the liver, various chaperonins such as GRP-78, HSP-71, Erp-60 and
PDI exhibited increased expression profile. In contrast, carbamoyl phosphate synthetase I,
albumin, indoethyllamine N-methyltransferase, peroxiredoxin 6 and carbonic anhydrase III
were detected at lower levels upon S. mansoni infection. The liver proteomic profile for the
infected animal correlated with the histological analysis of the organ in which an intense
cellular immune response was observed. In the spleen, the identified proteins displaying
altered expression profile were calreticulin, phosphoglycerate kinase I, fructose-biphosphate
aldolase A, glyceraldehyde-3-phosphate dehydrogenase, EF-Tu (elongation fator – Tu) and
aspartate aminotransferase. Overall, the proteins differentially expressed in the spleen of
infected animals displayed up regulation evidencing a molecular profile suitable to provide
the demand in energy production and protein synthesis. These two cellular processes are of
great relevance to maintain the intense cellular proliferation observed in the organ.
Collectively, our results demonstrated that the employed proteomic approach allowed the
identification of tissue markers induced by the S. mansoni infection. Future experiments will

xv
establish whether the observed tissue alterations promoted by the S. mansoni parasitism
significantly alter the plasma proteome to highlight novel biomarkers of schistosomiasis.

1- Introdução

2
1 - Introdução
1.1 – Aspectos gerais
O gênero Schistosoma é constituído por organismos da família Schistosomatidae,
classe Trematoda e filo Platelminto. Os representantes deste gênero possuem sexos separados
e um ciclo de vida heteroxeno. As principais espécies de Schistosoma que parasitam os seres
humanos são: Schistosoma mansoni, Schistosoma haematobium, Schistosoma japonicum,
Schistosoma intercalatum e Schistosoma mekongi (Mahmoud, 2004). Algumas destas
espécies causam a forma intestinal da esquistossomose (S. mansoni, S. japonicum, S.
intercalatum e S. mekongi); as quais, no estágio de vida adulta, localizam-se no sistema porta
hepático do hospedeiro vertebrado. O S. haematobium causa a esquistossomose urogenital por
localizar-se nas veias do trato urinário, durante a fase de vida adulta e por liberar ovos através
da urina (Utzinger et al., 2011) A Esquistossomose também conhecida como Bilharzíase é
uma doença parasitária endêmica em vários países, afligindo mais de 207 milhões de pessoas
no mundo. Estima-se ainda, que 779 milhões de pessoas possuem o risco de contrair a doença,
Figura 1 (World Health 2010); além de ser considerada a 4ª principal doença tropical
negligenciada no mundo, Tabela 1 (Hotez et al., 2008). Depois da malária é a segunda
infecção parasitária mais importante em termos de saúde pública e impacto socio-econômico,
ocasionando cerca de 200 mil mortes por ano, (Wichmann et al., 2009). As doenças tropicais
negligenciadas constituem a 6ª maior causa de DALYs (Disability-Adjusted Life Years), um
índice métrico adotado pela organização mundial de saúde, o qual avalia o impacto da doença
no indivíduo, baseando-se nos anos de vida perdidos devido à incapacidade produtiva e morte
prematura, Figura 2.
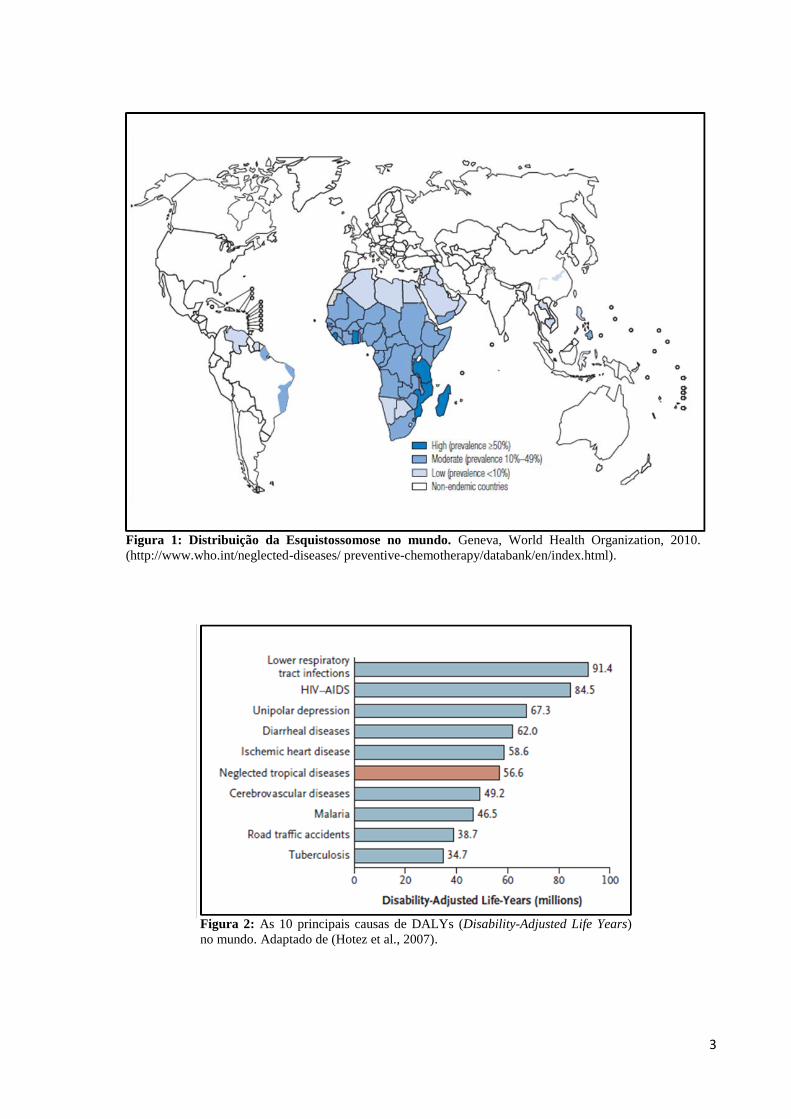
3
Figura 2: As 10 principais causas de DALYs (Disability-Adjusted Life Years)
no mundo. Adaptado de (Hotez et al., 2007).
Figura 1: Distribuição da Esquistossomose no mundo. Geneva, World Health Organization, 2010.
(http://www.who.int/neglected-diseases/ preventive-chemotherapy/databank/en/index.html).
4
O S. mansoni é endêmico no nordeste do Brasil, na Venezuela, Suriname, Caribe e
Egito. O S. japonicum é endêmico na China, Filipinas e Indonésia. O S. mekongi é encontrado
no Camboja (Chitsulo et al., 2004). No Brasil a esquistossomose mansônica ocorre de forma
endêmica nos seguintes estados: Alagoas, Maranhão, Bahia, Pernambuco, Rio Grande do
Norte, Paraíba, Sergipe, Espírito Santo e Minas Gerais (predominantemente no norte e
nordeste do Estado), Pará, Piauí, Ceará, Rio de Janeiro, São Paulo, Santa Catarina, Paraná,
Rio Grande do Sul, Goiás e no Distrito Federal (SES/MS, 2010, World Health, 2010).
Aproximadamente 25 milhões de pessoas vivendo nas zonas rurais estão expostas ao risco de
contrair a doença (de Oliveira et al., 2004). O S. mansoni, é dentre as espécies causadoras da
Tabela 1: Prevalência das Principais Doenças Tropicais Negligenciadas. Adaptado de (Hotez et al., 2007).

5
esquistossomose, a mais estudada no mundo. Acredita-se que a doença chegou às Américas
através do comércio de escravos africanos no período colonial (Campbell et al., 2000). No
Brasil, a esquistossomose foi detectada pela primeira vez em 1908, pelo médico brasileiro
Pirajá da Silva, através de exames de fezes e de vermes coletados a partir de necropsias (Katz,
2008). A transmissão da doença depende da existência de hospedeiros intermediários, e as três
principais espécies envolvidas na transmissão para o homem são: Biomphalaria glabrata,
Biomphalaria straminea e Biomphalaria tenagophila.
Clinicamente, a esquistossomose pode ser classificada em doença de fase aguda e fase
crônica. A fase aguda corresponde à etapa de penetração das cercárias através da pele,
variando desde um quadro assintomático até apresentação de um quadro clínico sintomático.
Nos seres humanos, a fase crônica inicia-se a partir do sexto mês após a infecção, podendo
durar vários anos. Está associada à postura de ovos, lesões no fígado e baço, hipertensão
portal, varizes esofagianas e hemorragias, podendo levar à morte (SES/MS, 2010, Ferrari and
Moreira, 2011, World Health, 2010).
A Assembléia Mundial de Saúde, através da resolução (WHA54. 19) realizada em
maio de 2001, recomendou aos países membros a desenvolverem atividades de controle
sustentável, garantindo o acesso à medicação e promovendo medidas preventivas para o
controle da esquistossomose. A meta mínima era a administração de quimioterapia para pelo
menos 75% de todas as crianças em risco de morbidade até 2010 (Favre et al., 2009). Porém
estimativas indicam que menos de 10% da população em risco de morbidade recebe
quimioterapia preventiva, demonstrando que a cobertura estabelecida em 2001 está longe de
ser alcançada, Figura 3.
Figura 3: Distribuição da população que requer quimioterapia preventiva. AFR – África / AMR –
Américas / EMR – leste do Mediterrâneo / SEAR – Sul e leste da Ásia / WPR – Oeste do pacífico.
Geneva, World Health Organization, 2010.

6
1.2 - O ciclo biológico do Schistosoma mansoni
O S. mansoni é o agente etiológico da esquistossomose mansônica, uma doença
inicialmente assintomática, podendo evoluir para formas clínicas extremamente graves e levar
o paciente à morte. A fêmea do S. mansoni mede aproximadamente de 1,2 a 1,6 cm, apresenta
corpo cilíndrico e encontra-se alojada em uma fenda do corpo do macho denominada de canal
ginecóforo. O macho mede cerca de 1 cm, apresenta cor mais clara que a fêmea e corpo
achatado (Loverde and Chen, 1991). O sexo do S. mansoni é determinado no zigoto, este
possui 8 pares de cromossomos (2n=16), sendo 7 pares autossômicos e 1 par sexual. A fêmea
é heterozigótica ZW e o macho é homozigoto ZZ. (Berriman et al., 2009). O genoma do
Schistosoma contém mais de 380 milhões de pares de bases divididos entre os setes pares
autossomais e o par sexual. A versão mais recente proposta para o genoma do S. mansoni
contabiliza cerca de 10.852 genes (Protasio et al., 2012).
O ser humano é o principal hospedeiro definitivo do S. mansoni. Nele, o parasito
apresenta a forma adulta, reproduz-se sexuadamente e, por meio da eliminação dos ovos,
ocasiona a contaminação das coleções hídricas. A infecção no hospedeiro vertebrado se dá
através do contato com água doce contendo cercárias, as quais são capazes de penetrar
ativamente na pele do hospedeiro, orientadas por estímulos químicos e utilizando-se do
conteúdo proteico das glândulas pré e pós acetabulares. (Collins et al., 2011, Curwen et al.,
2006). Proteínas presentes nas secreções das glândulas cercarianas possuem funções
proteolíticas e imunomoduladoras. Uma variedade de isoformas para elastase, além de outras
enzimas proteolíticas (dipeptidil peptidase e invadolisinas), constitui as secreções utilizadas
no processo de penetração. Outra proteína encontrada em secreções de cercárias é a SmKK7,
por apresentar homologia com toxinas de escorpião, acredita-se que possua a capacidade de
bloquear canais de K+, auxiliando na modulação do sistema imune (Curwen et al., 2006). O
processo de penetração das cercárias pode levar em média de 4 a 6 minutos, porém algumas
cercárias levam apenas 1.5 minutos para passar pela barreira física da pele (Haas and
Haeberlein, 2009).
Ao penetrarem na pele do hospedeiro as cercárias perdem a cauda, e após uma rápida
transformação, atingem o estágio de esquistossômulo. A transformação em esquistossômulo
dá origem ao tegumento heptalaminar, o qual permite a adaptação às alterações de
osmolaridade, nutrientes, temperatura e ação do sistema imune (Skelly and Shoemaker, 2000).
A camada mais externa do tegumento, o membranocalyx, possui a capacidade de captar

7
proteínas e moléculas do hospedeiro como CD44 e glicolipídeos provenientes de eritrócitos,
favorecendo-o na evasão do sistema imune (Castro-Borges et al., 2011a). O tegumento
também funciona como uma barreira física protegendo os componentes enzimáticos,
transportadores, canais iônicos da membrana plasmática e ainda apresenta capacidade de
renovação a cada 5 dias (Saunders et al., 1987, Braschi and Wilson, 2006).
Os esquistossômulos permanecem neste estágio por 2 a 3 dias, antes de migrarem,
através do sistema linfático e vênulas presente na derme, para o sistema circulatório. Ao
atingir a corrente sanguínea, passam pelo coração e eventualmente pelos pulmões, até
alcançarem o sistema porta-hepático. Após o 28° dia de infecção, os vermes presentes no
sistema porta-hepático se acasalam e dão início à migração até as vênulas mesentéricas para o
início da ovoposição. Através do pareamento, o macho libera os espermatozóides dentro do
canal ginecóforo, estes fecundam os ovócitos dando origem ao zigoto (Dewalick et al., 2011).
O desenvolvimento sexual da fêmea e, consequentemente, o início da ovoposição, são
dependentes do acasalamento com o macho. Acredita-se que através de vias de sinalização
envolvendo o fator de crescimento TGF-β, o macho ativa e regula a expressão gênica na
fêmea (Freitas et al., 2007, Beckmann et al., 2010).
A produção de aproximadamente 300 ovos por dia, e a consequente presença destes
ovos no tecido hepático, promove a liberação de antígenos solúveis (SEA), induzindo o
aparecimento de reações inflamatórias periovulares, originando os granulomas e, em seguida,
o processo de fibrose (Manzella et al., 2008). As glicoproteínas Omega-1 e IPSE/alpha-1
juntas constituem a fração mais abundante dos antígenos solúveis de ovos (SEA). Estas
proteínas são hepatotóxicas e imunogênicas, possuem a capacidade de estimular a liberação
de IL-4, induzindo uma resposta inflamatória do tipo Th2 no hospedeiro (Abdulla et al., 2011,
Mathieson and Wilson, 2010). Os ovos depositados no endotélio da parede das vênulas
induzem uma reação inflamatória periovular, a qual provavelmente facilita a passagem dos
mesmos pela musculatura até atingirem a luz intestinal (Lenzi et al., 1991). Ao serem
liberados juntamente com as fezes, os ovos alcançam as coleções hídricas e, em contato com a
água doce, eclodem liberando os miracídios. Os miracídios são formas larvais ciliadas,
capazes de nadar em busca do hospedeiro invertebrado, moluscos do gênero Biomphalaria.
No hepatopâncreas do molusco, as larvas transformam-se em esporocistos primários, os quais
se desenvolvem em esporocistos secundários. Estes últimos, por meio de reprodução
assexuada, dão origem às cercárias. Análises recentes de transcrição gênica por microarray
sugerem que a síntese de proteínas a serem utilizadas pelas cercárias durante a infecção do
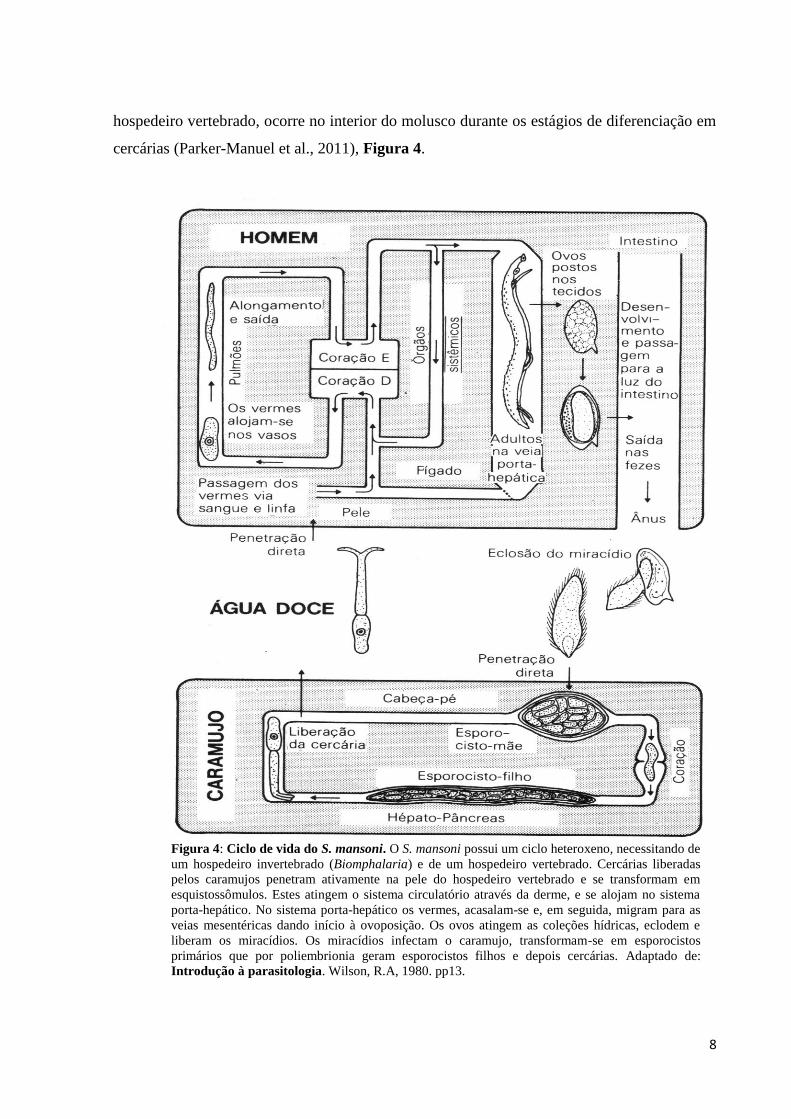
8
hospedeiro vertebrado, ocorre no interior do molusco durante os estágios de diferenciação em
cercárias (Parker-Manuel et al., 2011), Figura 4.
Figura 4: Ciclo de vida do S. mansoni. O S. mansoni possui um ciclo heteroxeno, necessitando de
um hospedeiro invertebrado (Biomphalaria) e de um hospedeiro vertebrado. Cercárias liberadas
pelos caramujos penetram ativamente na pele do hospedeiro vertebrado e se transformam em
esquistossômulos. Estes atingem o sistema circulatório através da derme, e se alojam no sistema
porta-hepático. No sistema porta-hepático os vermes, acasalam-se e, em seguida, migram para as
veias mesentéricas dando início à ovoposição. Os ovos atingem as coleções hídricas, eclodem e
liberam os miracídios. Os miracídios infectam o caramujo, transformam-se em esporocistos
primários que por poliembrionia geram esporocistos filhos e depois cercárias. Adaptado de:
Introdução à parasitologia. Wilson, R.A, 1980. pp13.

9
1.3 - Diagnóstico e Tratamento
O diagnóstico da esquistossomose pode ser realizado por métodos diretos e indiretos.
Os métodos indiretos ou sorológicos dependem de marcadores bioquímicos e imunológicos
associados à infecção, dentre os quais se destacam os testes imunológicos como a
imunofluorescência, ensaios imunoenzimáticos (ELISA), reação intradérmica e reação
periovular. Técnicas de reação em cadeia de polimerase estão sendo descritas para o
diagnóstico da doença (Pontes et al., 2002). Atualmente a técnica de Kato-Katz é o método
parasitológico recomendado para o diagnóstico da esquistossomose, por ser quantitativo e
prático (Katz et al., 1972). No entanto, a sensibilidade desta técnica diminui quando a
prevalência e intensidade da infecção são baixas (Zhou et al., 2008, Lin et al., 2008).
Os testes sorológicos para detecção de IgG e IgM no soro de pacientes, possuem uma
grande sensibilidade, podendo indicar infestação atual ou passada, principalmente em áreas de
baixa prevalência da doença, ou em pacientes com baixa parasitemia. O método de ELISA,
por exemplo, além de proporcionar ensaios quantitativos, mostra-se mais adequado para
aplicação em estudos populacionais onde se encontra baixa parasitemia, apresentando uma
sensibilidade e especificidade próximas de 98% (Oliveira et al., 2005). Teste de
imunofluorêscencia para a detecção de anticorpos contra S. mansoni demonstram alto grau de
sensibilidade 95,6% para as fases aguda e crônica (Kanamura et al., 2002). Entretanto, este
método possui especificidade em torno de 50%. Em geral os ensaios de detecção de
anticorpos demonstram ser mais sensíveis que os exames parasitológicos, mas carecem de
especificidade. Já a detecção de antígenos circulantes é um método altamente específico,
porém não tem demonstrado ser mais sensível que os exames parasitológicos em áreas de
baixa endemicidade (Attallah et al., 1999, Van Lieshout et al., 1995). Os testes de reação em
cadeia da polimerase (PCR), realizados a partir de amostras de fezes e soros de pacientes, não
demonstraram reações cruzadas com outros helmintos, atingindo sensibilidade de 96.7% e
especificidade de 88% (Pontes et al., 2002). Contudo, testes como imunofluorêscencia e PCR
não estão disponíveis na rotina do sistema público de saúde (Sorgho et al., 2005). Desta
maneira, torna-se necessário o desenvolvimento de novos métodos de diagnóstico,
especialmente para auxiliar na detecção de pacientes com baixa carga parasitária (Alan
Wilson et al., 2006).
Para o tratamento da esquistossomose, o praziquantel e a oxaminiquine são as drogas
de escolha. Os dois medicamentos se equivalem quanto à eficácia e segurança, todavia o

10
praziquantel é a droga disponível atualmente, em função do menor custo. O modo de ação do
praziquantel não é totalmente compreendido, a maioria dos estudos aponta para perturbações
na homeostase do cálcio e comprometimento das funções normais do tegumento (Kasinathan
et al., 2010). Neste sentido, o desenvolvimento de novos medicamentos para combater a
infecção também se torna necessário, uma vez que existem relatos de resistência do parasito à
ação deste fármaco (Kasinathan et al., 2010).
1.4 - Proteômica
A palavra proteoma foi primeiramente introduzida por Wilkins (1996), onde foi
utilizada para expressar a idéia do conjunto das proteínas de uma célula ou organismo. Assim,
a proteômica foi estabelecida como ferramenta para o estudo da expressão proteica e suas
alterações (Anderson and Anderson, 1998). Atualmente existe um grande interesse na
aplicação da proteômica, como ferramenta para a identificação de marcadores proteicos.
Alguns tecidos, órgãos e fluidos corporais, tais como plasma e saliva já estão sendo utilizados
na busca de possíveis biomarcadores de infecções (Bodzon-Kulakowska et al., 2007). A
proteômica também reune ferramentas que fornecem informações sobre modificações pós-
traducionais, abundância de proteínas e interações proteicas, possibilitando o estudo de
alterações metabólicas ocorridas em um organismo (Harvie et al., 2007, Manivannan et al.,
2011).
Com o melhoramento de técnicas de pré-fracionamento destes proteomas e com a
introdução da técnica de eletroforese bidimensional (2-DE), em 1975 por O'Farrell e Klose;
tornou-se possível a análise e detecção de proteínas em amostras biológicas mais complexas
(O'Farrell, 1975). Através da técnica de eletroforese bidimensional, as proteínas são
inicialmente separadas de acordo com o ponto isoelétrico e, em seguida, submetidas à
separação por massa molecular, com auxílio de uma matriz polimérica
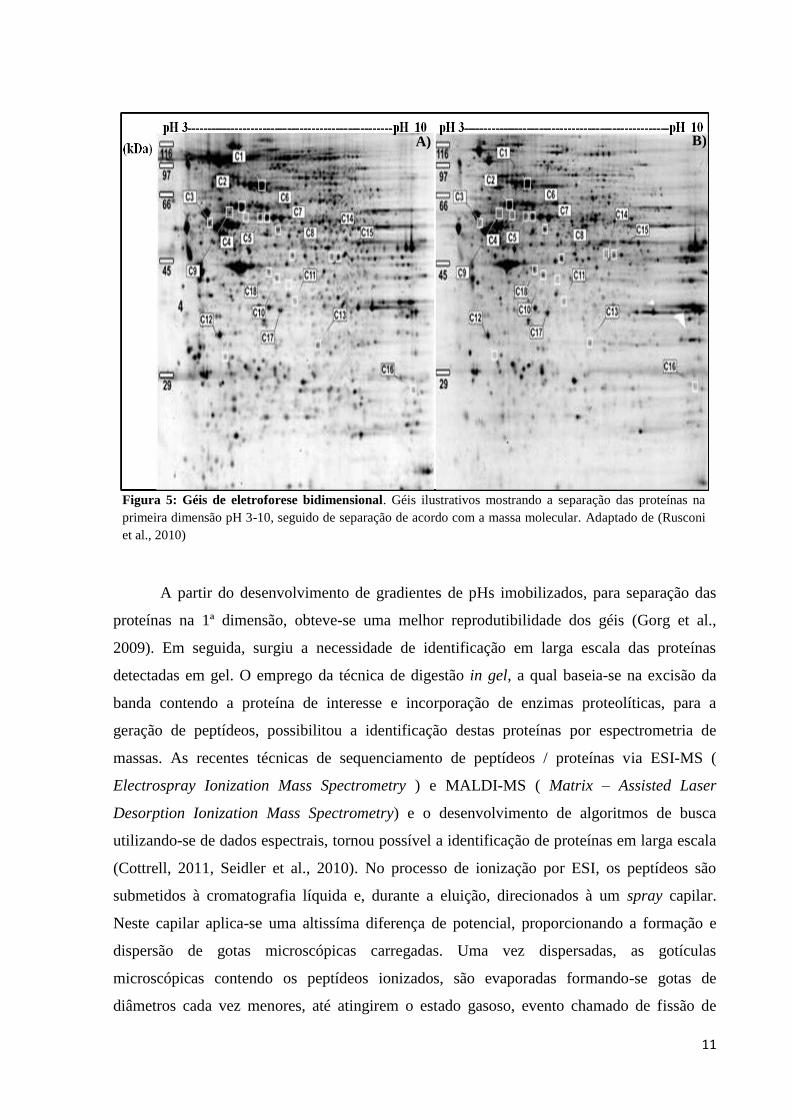
(acrilamida/bisacrilamida), Figura 5. Assim, tornou-se possível a detecção de alterações
como: aumento ou diminuição da expressão de proteínas em um proteoma específico e
modificações pós-traducionais como glicosilação e fosforilação.
11
A partir do desenvolvimento de gradientes de pHs imobilizados, para separação das
proteínas na 1ª dimensão, obteve-se uma melhor reprodutibilidade dos géis (Gorg et al.,
2009). Em seguida, surgiu a necessidade de identificação em larga escala das proteínas
detectadas em gel. O emprego da técnica de digestão in gel, a qual baseia-se na excisão da
banda contendo a proteína de interesse e incorporação de enzimas proteolíticas, para a
geração de peptídeos, possibilitou a identificação destas proteínas por espectrometria de
massas. As recentes técnicas de sequenciamento de peptídeos / proteínas via ESI-MS (
Electrospray Ionization Mass Spectrometry ) e MALDI-MS ( Matrix – Assisted Laser
Desorption Ionization Mass Spectrometry) e o desenvolvimento de algoritmos de busca
utilizando-se de dados espectrais, tornou possível a identificação de proteínas em larga escala
(Cottrell, 2011, Seidler et al., 2010). No processo de ionização por ESI, os peptídeos são
submetidos à cromatografia líquida e, durante a eluição, direcionados à um spray capilar.
Neste capilar aplica-se uma altissíma diferença de potencial, proporcionando a formação e
dispersão de gotas microscópicas carregadas. Uma vez dispersadas, as gotículas
microscópicas contendo os peptídeos ionizados, são evaporadas formando-se gotas de
diâmetros cada vez menores, até atingirem o estado gasoso, evento chamado de fissão de
Figura 5: Géis de eletroforese bidimensional. Géis ilustrativos mostrando a separação das proteínas na
primeira dimensão pH 3-10, seguido de separação de acordo com a massa molecular. Adaptado de (Rusconi
et al., 2010)
A) B)
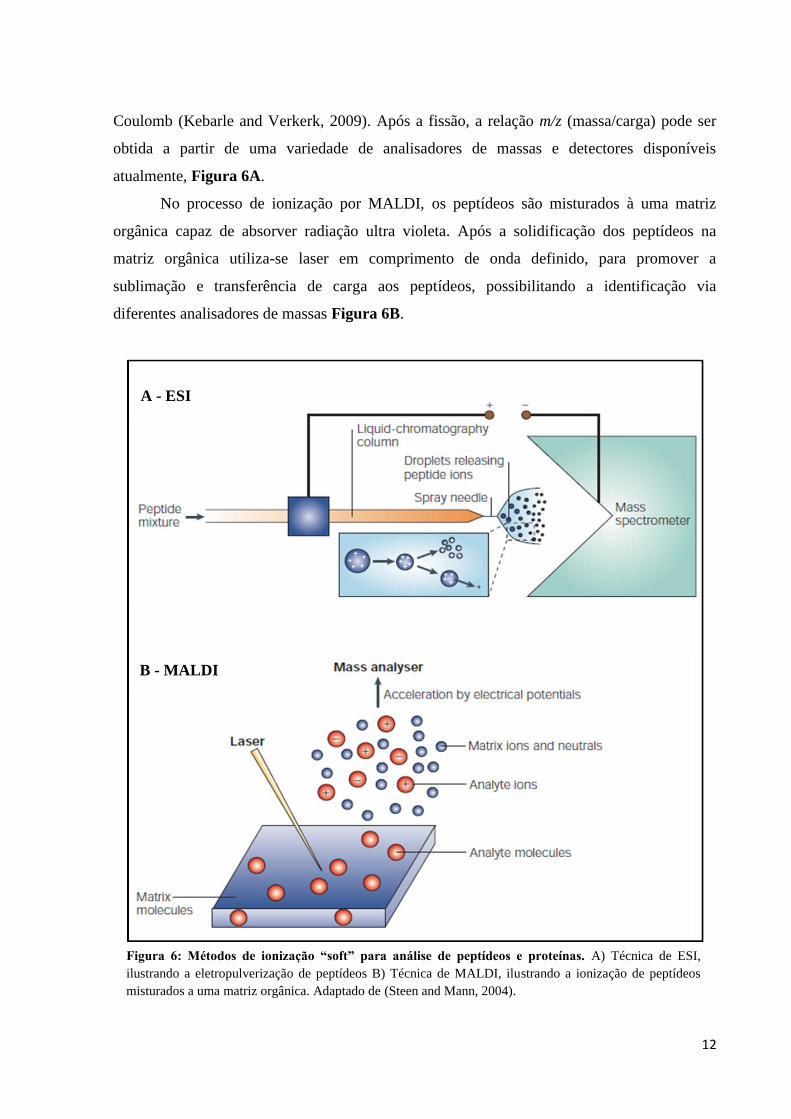
12
Coulomb (Kebarle and Verkerk, 2009). Após a fissão, a relação m/z (massa/carga) pode ser
obtida a partir de uma variedade de analisadores de massas e detectores disponíveis
atualmente, Figura 6A.
No processo de ionização por MALDI, os peptídeos são misturados à uma matriz
orgânica capaz de absorver radiação ultra violeta. Após a solidificação dos peptídeos na
matriz orgânica utiliza-se laser em comprimento de onda definido, para promover a
sublimação e transferência de carga aos peptídeos, possibilitando a identificação via
diferentes analisadores de massas Figura 6B.
A - ESI
B - MALDI
Figura 6: Métodos de ionização “soft” para análise de peptídeos e proteínas. A) Técnica de ESI,
ilustrando a eletropulverização de peptídeos B) Técnica de MALDI, ilustrando a ionização de peptídeos
misturados a uma matriz orgânica. Adaptado de (Steen and Mann, 2004).

13
1.5 - Modelo murino de infecção por S. mansoni
Considerado como o principal modelo utilizado para infecção experimental por S.
mansoni, o modelo murino vem contribuindo para o entendimento de inúmeros aspectos
ligados à esquistossomose e à biologia do parasito (Abdul-Ghani and Hassan, 2010). Dentre
os aspectos que justificam sua utilização incluem o baixo custo, a facilidade de manipulação
do animal e o tempo reduzido necessário para a evolução clínica da doença. No camundongo,
a esquistossomose de fase aguda se estabelece durante os primeiros 30 a 35 dias de infecção,
tempo necessário para a instalação dos vermes adultos no sistema porta-hepático, pareamento
e início da ovoposição (Loverde and Chen, 1991, Harvie et al., 2007). A fase crônica, por sua
vez, inicia-se a partir da postura dos ovos sendo as complicações teciduais ocasionadas pela
deposição crescente dos mesmos nos tecidos, especialmente no hepático (Manivannan et al.,
2010). Dependendo da carga parasitária, o animal dificilmente sobrevive à 8ª semana de
infecção, embora tenha sido demonstrado que a infecção com 45 cercárias garante sobrevida
por até 20 semanas em camundongos machos da linhagem CBA/J (Henderson et al., 1993).
Mais recentemente o modelo murino tem sido empregado para a identificação de
possíveis marcadores teciduais da infecção por S. mansoni, utilizando-se de abordagens na
área da proteômica (Manivannan et al., 2010). Análises do proteoma total de fígado de
camundongos infectados demonstraram alterações nos níveis de várias enzimas relacionadas
ao metabolismo energético, resposta ao estresse, dentre outras (Manivannan et al., 2012).
Espera-se que a curto prazo as inovadoras tecnologias para identificação e quantificação de
proteínas forneçam um inventário molecular do impacto da doença no hospedeiro vertebrado.
Tais investigações são de particular importância, especialmente quando aplicadas aos estágios
da doença para os quais é possível a correlação com a doença humana (Manivannan et al.,
2011, Harvie et al., 2007). Estudos de metabolômica também têm sido realizados utilizando-
se do modelo murino de infecção por S. mansoni. A identificação rápida e inequívoca de
centenas de metabólitos diferencialmente presentes no soro e urina de animais infectados vem
proporcionando a validação de métodos alternativos para a detecção dos distúrbios
metabólicos associados à infecção - Tabela 2 (Wu et al., 2010a, Balog et al., 2011). Neste
sentido, a detecção de níveis elevados do aminoácido hidroxiprolina em soros de animais
infectados tem sido avaliada como potencial indicador de fibrose em pacientes com
esquistossomose hepatoesplênica (Wang et al., 2004, Utzinger et al., 2011).

14
O presente trabalho pretende, a partir de uma coleção de dados clínicos, ensaios de
atividade enzimática, análise histológica de fígado e baço e análise comparativa de proteínas
por gel bidimensional, contribuir com o entendimento da relação parasito/hospedeiro durante
o curso da infecção por S. mansoni, utilizando camundongos isogênicos da linhagem Balb/c.
Espera-se que os resultados encontrados possam, no futuro, apontar candidatos potenciais
para melhoria das técnicas de diagnóstico, prognóstico e tratamento da esquistossomose.
Tabela 2: Principais metabólitos alterados em urina de camundongos infectados
com S. mansoni e em hamsters infectados com S. japonicum (Utzinger et al.,
2011).
a Camundongos fêmeas infectados com 80 cercárias de S. mansoni, amostras de
urina coletadas após 46 e 56 dias de infecção e analisadas por ressonância
magnética nuclear (NMR). b Camundongos fêmeas infectados com 100 cercárias de S. mansoni, amostras de
urina coletadas após 56 dias de infecção e analisadas por eletroforese em capilar
(CE). c Hamsters infectados com 100 cercárias de S. japonicum, amostras de urina
coletada após 36 dias de infecção e analisadas por ressonância magnética nuclear
(NMR).

2 - Objetivos

15
2 - Objetivos
2.1 - Objetivo geral
Identificação de proteínas solúveis diferencialmente presentes no fígado e baço de
camundongos Balb/c infectados por S. mansoni, nos estágios de fase aguda e crônica da
doença.
2.2 - Objetivos específicos
Discriminar os estágios de fase aguda e fase crônica da esquistossomose em
camundongos Balb/c infectados com cercárias de S. mansoni através de
ensaios enzimáticos, leucometria, Razão percentual (órgão/Corpo) e análise
histológica;
Obter o proteoma solúvel de fígado e baço dos animais controles e infectados
nos dois estágios distintos da infecção;
Realizar eletroforese bidimensional comparativa para a detecção de proteínas
diferencialmente presentes no fígado e baço de animais infectados;
Identificar as proteínas selecionadas por espectrometria de massas.

3- Metodologia

17
3 - Metodologia
3.1 - Manutenção do ciclo do S. mansoni
A manutenção do ciclo do S. mansoni é rotineiramente realizada no Laboratório de
Biologia Molecular de Parasitas – Faculdade de Medicina de Ribeirão Preto / USP. Cercárias
de S. mansoni da linhagem Porto Rico são recuperadas a partir de moluscos da espécie
Biomphalaria glabrata previamente infectados. Os moluscos são mantidos no escuro por 48
horas antes de serem expostos à luz artificial, pelo período mínimo de 2 horas para a liberação
das cercárias. As cercárias liberadas são contadas em microscópio óptico antes de serem
utilizadas para a infecção dos animais. O processo de infecção se dá através do contado
percutâneo de cercárias com o abdômen dos animais, simulando uma infecção natural.
3.2 – Delineamento Experimental
Fêmeas de camundongos (Mus musculus, n = 32) germ free, da linhagem Balb/c com
30 dias de idade e aproximadamente 20g foram adquiridos via Biotério Central da
Universidade de São Paulo, campus Ribeirão Preto. Os animais foram anestesiados, via
intraperitoneal, através da administração associada de dois anestésicos cloridrato de cetamina
10% (Syntec) a 8.0 mg/Kg e cloridrato de xilazina 2,3% (Vetbrands) a 4.0 mg/Kg,
proporcionando uma analgesia de aproximadamente 60 minutos. Com o auxílio de um
aparador automático de pelos, fez-se a tricotomia da região abdominal dos animais, para
facilitar a penetração das cercárias. Os animais foram colocados em decúbito dorsal,
imobilizados em um suporte de acrílico, e em seguida, os anéis de aço inoxidável de
dimensões, 1 cm de altura por 0,5 cm de raio, foram posicionados sobre o abdômen e fixados
em acrílico com o auxílio de fita adesiva. Em seguida, cerca de 1 mL de suspensão contendo
cercárias foi dispensado nos anéis para a infecção de 20 animais. Os camundongos foram
mantidos em contato com a suspensão de cercárias durante 30 minutos à temperatura
ambiente. Para os outros 12 animais, utilizados como controles não infectados, o processo de
infecção foi simulado com a utilização de água. Decorridos 30 minutos de exposição, a
suspensão remanescente de cercárias foi descartada em etanol 70%. A Figura 7 ilustra o
esquema representativo do processo de infecção pelo método de anel.

18
Após a infecção, os 32 animais foram separados em dois grupos conforme indicado na
Tabela 3: Grupo I (animais com 5 semanas de infecção) e grupo II (animais com 7 semanas
de infecção). Em seguida, os animais foram mantidos em gaiolas à temperatura ambiente com
ração e água ad libitum e ciclo claro/escuro com intervalos de 12 horas. Todos os
procedimentos realizados com os animais foram aprovados pela Comissão de Ética em Uso
Animal.
Grupo I – 5 semanas Grupo II – 7 semanas
Controle (N=6) Controle (N=6)
Infectado (N=10) Infectado (N=10)
6-Infecção por anel
5-Suspensão de
cercárias
4-Anestesia dos
animais
3-Contagem das
cercárias
2-Liberação das
cercárias
1-Biomphalaria glabrata
infectados
Figura 7: Representação do método de infecção por anel. 1 e 2- Caramujos infectados são expostos à luz
para liberação das cercárias. 3- A contagem das cercárias é realizada em microscópio óptico. 4-
Camundongos são anestesiados por via intraperitoneal. 5- A suspensão de cercárias (200 cercárias/ml) é
adicionada sobre o abdômen do animal, através do anel de infecção. 6- Decorrido 30 minutos de infecção, a
suspensão de cercárias é retirada do anel.
Tabela 3: Amostragem de animais
N – Número de animais.

19
3.3 - Exame parasitológico de fezes
Cerca de 300 mg de fezes recolhidas de cada animal infectado, foram coletadas entre o
35º e o 50º dia de infecção em intervalos de 5 dias. As amostras foram conservadas em
formol tamponado 10% até a realização dos exames. Análise das fezes foi realizada segundo o
método de sedimentação espontânea ou método de Hoffman, (Hoffman WA, 1934). O
sedimento obtido foi analisado diretamente ao microscópio óptico para a determinação do
número de ovos por lâmina. As lâminas foram confeccionadas em triplicata para cada animal,
utilizando-se 200 μl de sedimento.
3.4 - Obtenção do material biológico
Todos os animais dos grupos I e II foram eutanasiados respectivamente após 5 e 7
semanas de infecção, utilizando-se a administração intraperitoneal dos anestésicos cloridrato
de cetamina 10% (Syntec) a 24.0 mg/Kg e cloridrato de xilazina 2,3% (Vetbrands) a 12.0
mg/Kg. Após verificação de analgesia profunda, os animais foram pesados e, em seguida,
cerca de 700 L de sangue coletados por punção cardíaca. O sistema porta-hepático dos
animais foi posteriormente perfundido com solução salina citratada (NaCl 0.85%, Citrato de
sódio 0.75%).
Após a perfusão foram retirados o fígado e o baço dos animais dos grupos controle e
infectado. Os órgãos foram pesados, lavados em tampão fosfato de sódio (PBS 0,01 M pH
7,2) e segmentos de aproximadamente 0.5 cm de largura obtidos e armazenados em formol
tamponado 10% para análises histológicas. O restante dos órgãos foi congelado em freezer -
80°C para posteriores análises proteômicas.
3.5 – Razão peso do órgão e peso corporal
Para todos os animais dos grupos I e II, obteve-se a razão percentual entre o peso dos
órgãos, fígado e baço, em relação ao peso corporal dos animais (% órgão/corpo). Este
parâmetro foi obtido para avaliação de hepatoesplenomegalia dos animais infectados em
comparação com os animais controles em cada fase de infecção. As análises estatísticas foram
realizadas utilizando Student´s t-test não pareado, através do software Prism versão 5.0.

20
3.6 – Leucograma
O sangue obtido foi dividido em duas partes, sendo cerca de 400 μL mantido na
presença de EDTA a 2% para análises hematológicas e 300 μL para obtenção de soro. Para a
realização das análises hematológicas foram determinados: contagem global de leucócitos e
análise diferencial (número de linfócitos, neutrófilos segmentados, monócitos e eosinófilos).
Os leucogramas dos animais do grupo controle serviram como valores normais de referência.
Os esfregaços obtidos para a contagem diferencial foram corados utilizando-se o Kit Instant
Prov (Labor & Labor Bioclin). Para a contagem global de leucócitos, diluiu-se o sangue em
líquido de Turk (ácido acético 2%, violeta genciana 0,1%) na proporção de 1:10 e este foi
analisado em câmara de Neubauer espelhada. As análises estatísticas foram realizadas
utilizando Student´s t-test não pareado, através do software Prism versão 5.0.
3.7 - Dosagens enzimáticas para ALT, AST e Albumina Sérica
O soro obtido foi utilizado para dosagens bioquímicas de ALT (alanino
aminotransferase); AST (aspartato aminotrasferase) e Albumina sérica. As amostras de soro
foram analisadas pelo Laboratório de Análises Clínicas - Claudino / Ouro Preto, onde as
enzimas transaminases foram dosadas através de métodos cinéticos e submetidas à leitura no
ultravioleta por espectrofotometria (Bioclin). A albumina foi dosada através do método
colorimétrico verde de bromocresol (Bioclin). As dosagens enzimáticas e de albumina do
grupo controle serviram como valores normais de referência. As análises estatísticas foram
realizadas utilizando Student´s t-test não pareado, através do software Prism versão 5.0.
3.8 - Análise histológica
Durante a necropsia, amostras de fígado e baço foram fixados em formol 3,7%
tamponado (pH 7.2), processados rotineiramente e incluídos em parafina. Secções de
aproximadamente 4 μm de espessura foram obtidas em micrótomo e submetidas à coloração
com Hematoxilina & Eosina (HE) e Tricrômico de Masson.
Nos cortes de baço foi avaliado o grau de organização tecidual (distribuição da polpa
branca e polpa vermelha). Nos cortes de fígado, foram avaliados o processo inflamatório

21
perivascular e a área média dos granulomas. A coloração de Tricrômico de Masson permitiu a
avaliação qualitativa do processo de fibrose no interior dos granulomas.
Para avaliação do processo inflamatório foram quantificados os núcleos celulares
presentes no infiltrado perivascular em toda a extensão do corte histológico. As imagens
foram visualizadas pela objetiva de 40x e digitalizadas através da microcâmera Leica
DFC340FX associada ao microscópio Leica DM5000B. Todas as imagens foram analisadas
com o auxílio do software de análise e processamento de imagem Leica QwinV3 no
Laboratório Multiusuários do Núcleo de Pesquisas em Ciências Biológicas da Universidade
Federal de Ouro Preto. O processo inflamatório foi determinado pela diferença significativa
(p<0,05) entre o número de núcleos celulares presentes nos animais infectados e aquele
observado nos animais controles ± desvio padrão.
3.9 - Extração de proteínas
Para a extração de proteínas solúveis, 100 mg de tecidos foram macerados em
nitrogênio líquido com o auxílio de um gral e pistilo. O conteúdo macerado foi colocado em
eppendorf e, em seguida, adicionou-se 500 μL de tampão de extração (Tris-HCl 25 mM pH
7,5) contendo 20 μL de PIC ( coquetel inibidor de proteases, Sigma-Aldrich). A mistura foi
homogeneizada em vortex por 5 minutos, seguido de sonicação (Sonifer 250 Branson) por 5
ciclos de 20 pulsos em banho de gelo. A suspensão resultante foi centrifugada a 4°C e
100.000 x g por 1 hora em centrifuga Sorvall 5C. O sobrenadante, referente à fração solúvel,
foi recuperado e o sedimento contendo membranas, armazenado a -80°C. Uma alíquota de 50
μL do sobrenadante foi armazenada para dosagem de proteínas totais.
Após a recuperação do sobrenadante, o mesmo foi colocado em um tubo tipo Falcon
de 15 mL para a precipitação das proteínas com ácido tricloroacético (TCA) e acetona. Para
cada 1 ml de sobrenadante foram adicionados 8 mL de acetona P.A gelada e 1 mL de solução
de ácido tricloroacético (TCA) 100%. A mistura foi homogeneizada e deixada durante a noite
em freezer a -20°C para precipitação das proteínas. Após a precipitação, a mistura foi
centrifugada a 46.000 x g por 30 minutos a 4°C em centrifuga Sorvall 5C. O precipitado foi
ressuspendido e lavado por 2 vezes com 500 μL de acetona gelada e novamente centrifugado
a 4°C e 46.000 x g por 30 minutos. O precipitado de proteínas foi deixado à temperatura
ambiente para evaporação total da acetona. Finalmente, o precipitado foi ressuspendido em

22
200 μL de tampão de reidratação (uréia 7 M; tiouréia 2 M; CHAPS (3-[(3-Cholamidopropil)
dimetilamônio]-1- propanosulfonato) 4%; azul de bromofenol 0.002%), Figura 8.
Figura 8: Diagrama esquemático para obtenção de proteínas solúveis.
3.10 – Eletroforese unidimensional (1D SDS-PAGE)
Anteriormente à confecção dos géis unidimensionais, fez-se a dosagem de proteínas
solúveis, obtidas conforme o item 3.9. pra esta dosagem foi utilizando o kit BCA protein assay
(Thermo scientific). O extrato proteico da fração citosólica de cada órgão, foi analisado por
meio de eletroforese em gel de poliacrilamida em condições desnaturantes (10% SDS-PAGE)
conforme descrito por Laemmli, UK 1970. Para o preparo da amostra a ser analisada, cerca de
30 μg de proteínas foram desnaturados em 10 μL de tampão da amostra (Tris-HCl 0.125 M

23
pH 6.8; SDS 4%; glicerol 20%; azul de bromofenol 0,002%). As amostras foram fervidas em
banho-maria por 5 minutos, e, em seguida, aplicadas no gel. A eletroforese ocorreu a 200 V e
20 mA durante aproximadamente 1,5 horas. Utilizou-se solução de Coomassie Brilliant Blue
G 250 (Coomassie Brilliant Blue G 250 2%; etanol 40%; ácido acético 7%) para coloração do
gel durante 2 horas. O gel foi descorado em solução A (etanol 40%, ácido acético 7%) por 30
minutos e, em seguida, em solução B (etanol 10%, ácido acético 7%) durante a noite. As
imagens dos géis foram obtidas através do scanner ImageScanner III (GE healthcare). A
análise densitométrica, utilizando-se de canaletas referências contendo 30 g de proteínas
citossólicas, foi realizada com o auxílio do software Quantity One, versão 4.6.9 – Bio Rad.
Esta análise foi utilizada para se igualar a quantidade total de proteínas obtidas dos animais
controles e infectados e posterior confecção das réplicas técnicas nos géis bidimensionais.
3.11 - Eletroforese bidimensional (2-D SDS-PAGE)
Para a confecção de géis bidimensionais 300 μg de proteínas citosólicas foram
solubilizadas em tampão de reidratação contendo DTT a 1% e anfólitos pH 3-10 ou 4-7 0,8%.
As amostras foram acondicionadas em sarcófagos (Strip Holder 7 cm, GE Healthcare) para a
incorporação das proteínas no gel de primeira dimensão (Immobiline DryStrip Gels 7cm; pH
3-10 ou pH 4-7, Linear,GE Healthcare). A etapa de isoeletrofocalização (IEF) ocorreu à
temperatura de 20°C a 50 μA/gel no equipamento Ettan IPGphor III (GE Healthcare) de
acordo com o seguinte protocolo: reidratação por 12 horas; 300V durante 30 minutos; 1000V
por 30 minutos e 8000V por 3 horas. Após a focalização isoelétrica, as proteínas presentes nos
géis de primeira dimensão foram submetidas à redução com 1% de DTT em solução de
equilíbrio (uréia 6M; Tris-HCl pH 8.8 1.5M; glicerol 30%; SDS 2%; azul de bromofenol 1%)
e, em seguida, alquiladas com iodoacetamida (IAA) a 4% em solução de equilíbrio.
Para a separação das proteínas por massa molecular (segunda dimensão) utilizou-se
gel de acrilamida/bisacrilamida a 10%. O gel de primeira dimensão foi posicionado e
posteriormente fixado no gel de segunda dimensão com solução de agarose 1% a quente em
tampão de eletroforese. A eletroforese ocorreu nas seguintes condições: 200 V e 20 mA/gel
durante aproximadamente 1,5 horas. Para corar o gel foi utilizado solução de Coomassie Blue
G 250. Após 2 horas de coloração o gel foi descorado em solução A (etanol 40%, ácido
acético 7%) por 30 minutos e, em seguida, em solução B (etanol 10%, ácido acético 7%)
durante a noite. As imagens dos géis foram obtidas através do scanner ImageScanner III (GE

24
healthcare) e sobrepostas através do software 2-D Evolution v.2005, (GE healthcare). A
Figura 9 mostra uma representação das etapas de confecção dos géis bidimensionais.
Figura 9: Ilustração das etapas de produção do gel bidimensional (2-DE). Extratos citosólicos, obtidos de
fígado e baço de animais controle e infectados, foram adsorvidos em géis de primeira dimensão 7cm (pH3-10/4-
7) por 12 horas. Os géis reidratados foram isoeletrofocalizados no aparelho Ettan IPGphor III e as proteínas
posteriormente reduzidas e alquiladas com DTT 1% e IAA 4%, respectivamente. A segunda dimensão ocorreu
em géis de poliacrilamida 10%. Após coloração dos géis a análise de expressão diferencial foi realizada com
auxílio do software 2-D Evolution v.2005 – Non Linear Dynamics.

25
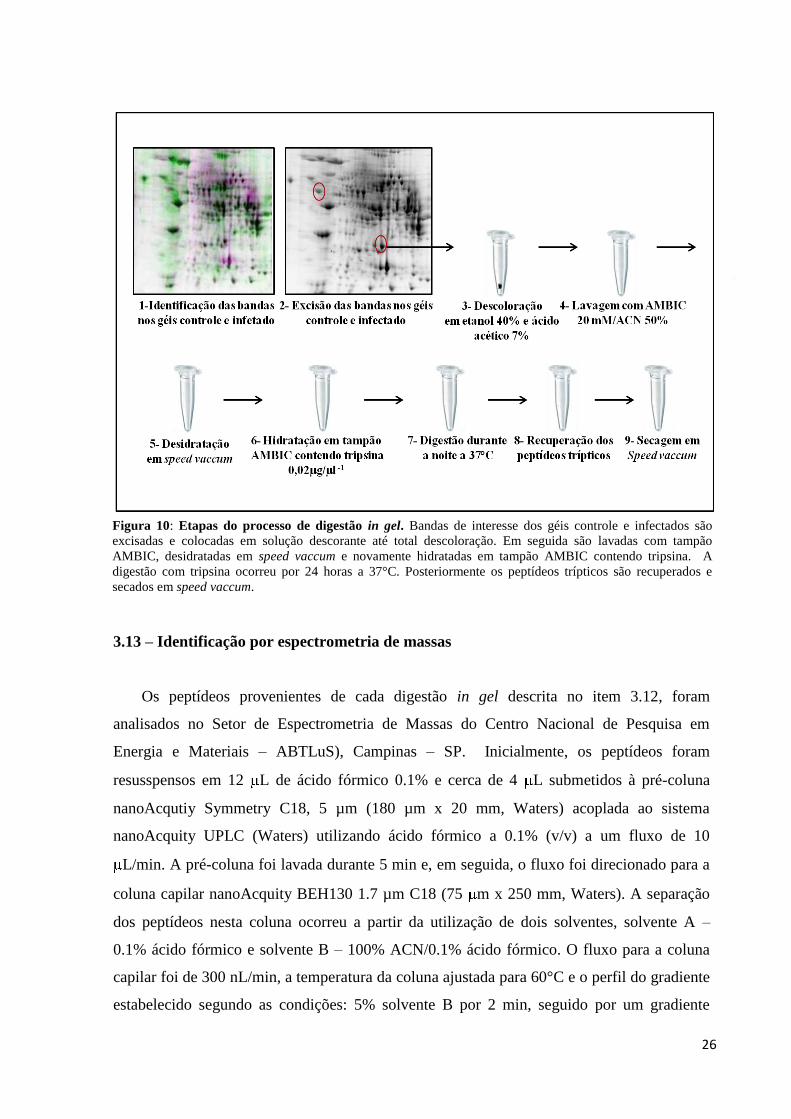
3.12 – Digestão in gel e preparo de amostras para espectrometria de massas
Após a obtenção dos géis bidimensionais, os spots de interesse (proteínas
diferencialmente presentes nos grupos controle e infectados) foram excisados e transferidos
para eppendorfs de 1,5 ml contendo 300 μL de solução descorante (etanol 40%; ácido acético
7%) e mantidas a 37°C até a completa descoloração. A solução descorante foi retirada e os
spots lavados com 500 μL de água milli-Q para remover o excesso de solução descorante.
Em seguida, os spots foram novamente lavados e desidratados por 3 vezes durante 5 min em
300 μL de solução 20 mM NH4HCO3 / Acetonitrila (ACN) 50%.
A desidratação completa dos spots ocorreu em speed vaccum durante 30 minutos. Os
spots foram reidratados com cerca de 15 μL de solução de tripsina a 0,1 μg/ μL em solução 20
mM NH4HCO3. Após 20 minutos, o excesso de solução de tripsina foi retirado e os spots
recobertos com cerca de 40 μl de solução NH4HCO3 a 20mM. A tripsinólise ocorreu a 37°C
por 24 horas. O sobrenadante de reação contendo os peptídeos trípticos foi recuperado e
transferido para um novo tubo. Para a extração dos peptídeos presentes no interior dos spots
adicionou-se ao tubo cerca de 30 μL de ácido trifluoracético (TFA) 2% / ACN 50% e o
sobrenadante recuperado após 30 minutos à temperatura ambiente. Os sobrenadantes
contendo os peptídeos de cada spot foram combinados e secos em speed vaccum. As amostras
foram mantidas a 4°C para posterior análise por espectrometria de massas. A Figura 10
ilustra as principais etapas associadas ao processo de digestão in gel.
26
3.13 – Identificação das proteínas por espectrometria de massas
3.13 – Identificação por espectrometria de massas
Os peptídeos provenientes de cada digestão in gel descrita no item 3.12, foram
analisados no Setor de Espectrometria de Massas do Centro Nacional de Pesquisa em
Energia e Materiais – ABTLuS), Campinas – SP. Inicialmente, os peptídeos foram
resusspensos em 12 L de ácido fórmico 0.1% e cerca de 4 L submetidos à pré-coluna
nanoAcqutiy Symmetry C18, 5 µm (180 µm x 20 mm, Waters) acoplada ao sistema
nanoAcquity UPLC (Waters) utilizando ácido fórmico a 0.1% (v/v) a um fluxo de 10
L/min. A pré-coluna foi lavada durante 5 min e, em seguida, o fluxo foi direcionado para a
coluna capilar nanoAcquity BEH130 1.7 µm C18 (75 m x 250 mm, Waters). A separação
dos peptídeos nesta coluna ocorreu a partir da utilização de dois solventes, solvente A –
0.1% ácido fórmico e solvente B – 100% ACN/0.1% ácido fórmico. O fluxo para a coluna
capilar foi de 300 nL/min, a temperatura da coluna ajustada para 60°C e o perfil do gradiente
estabelecido segundo as condições: 5% solvente B por 2 min, seguido por um gradiente
Figura 10: Etapas do processo de digestão in gel. Bandas de interesse dos géis controle e infectados são
excisadas e colocadas em solução descorante até total descoloração. Em seguida são lavadas com tampão
AMBIC, desidratadas em speed vaccum e novamente hidratadas em tampão AMBIC contendo tripsina. A
digestão com tripsina ocorreu por 24 horas a 37°C. Posteriormente os peptídeos trípticos são recuperados e
secados em speed vaccum.

27
linear até 35% solvente B durante 20 min. Finalmente, lavou-se a coluna utilizando 95%
solvente B por 2.5 min.
O sistema de nanocromatografia citado acima interfaceado com o espectrômetro Q-Tof
Ultima (Micromass – Waters) contendo uma fonte de nano-electrospray, constituíram o
equipamento utilizado para a detecção e fragmentação dos peptídeos. Espectros MS e
MS/MS foram adquiridos utilizando o modo AutoMSMS. Os parâmetros de espectrometria
de massas para o modo MS incluíram: voltagem do íon “spray” 1500 V, gás de secagem a
6L/min, temperatura do gás de secagem 160°C, faixa de aquisição m/z 50-2.200. No modo
AutoMSMS as condições foram as seguintes: dissociação induzida por colisão com gás N2 ,
faixa de aquisição m/z 350-1.400, carga dos íons selecionados para fragmentação +2, +3 e
+4 e exclusão de íons com carga +1.
Dados obtidos a partir dos espectros MS/MS foram submetidos à busca de identidade
utilizando uma cópia local do programa Mascot (Matrix Science Ltd., version 2.1). Os
bancos de dados utilizados foram o NCBInr (15270974 sequencias; 5234858139 resíduos) e
o NCBI_Mus musculus (139814 sequencias). As buscas levaram em consideração os
seguintes critérios: enzima – Tripsina; modificação fixa – carbamidometilação (C);
modificação variável – oxidação (M); tolerância para íon precursor - 10 ppm; tolerância para
íons provenientes de fragmentação - 0.1 Da. Considerou-se uma taxa de falsos positivos
(FDR) de 2% e os peptídeos que apresentaram expect value ≤ 0.05 foram considerados
identificações significativas. Figura 11.

28
Figura 11: Delineamento experimental ilustrando a identificação dos peptídeos obtidos por digestão in
gel. Após a obtenção dos peptídeos por digestão in gel, estes foram ressuspendidos em ácido fórmico 0.1% e
posteriormente separados por nano- UPLC, interfaceado ao espectrômetro de massas Q-Tof Ultima
(Micromass). Os dados de MS/MS foram submetidos à busca de identidade nos bancos NCBInr e Mus
Musculus.

4-Resultados
30
4 – Resultados
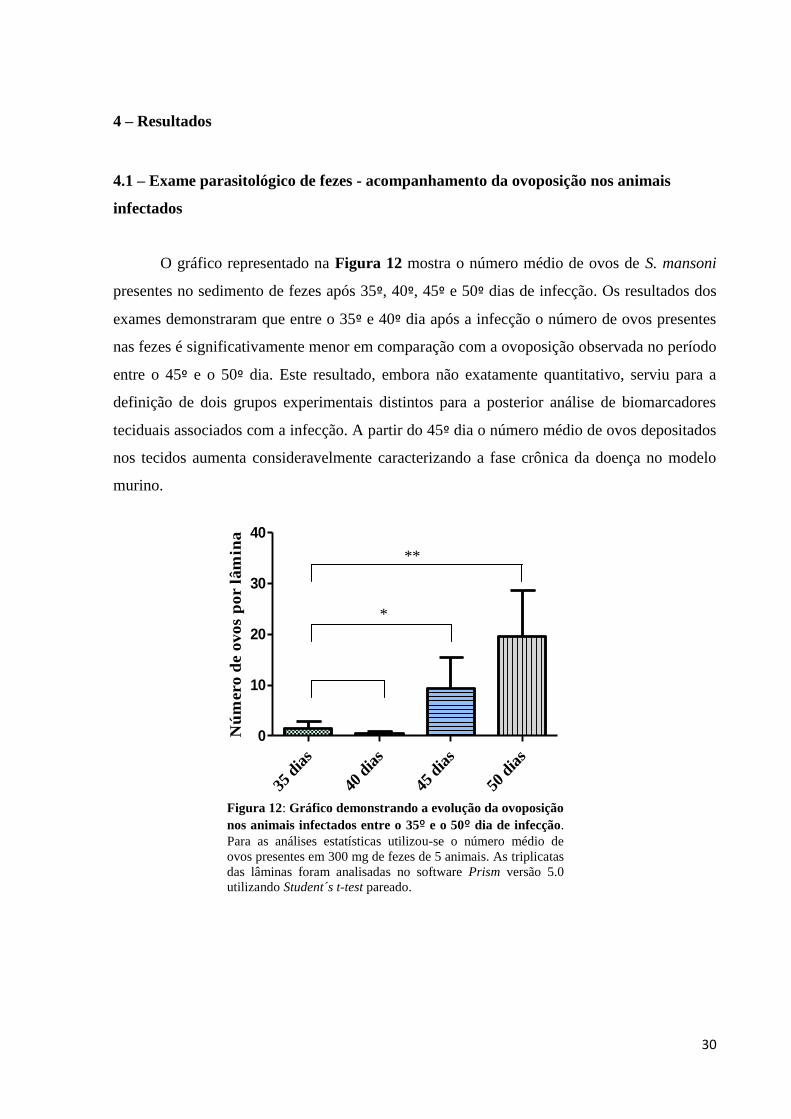
4.1 – Exame parasitológico de fezes - acompanhamento da ovoposição nos animais
infectados
O gráfico representado na Figura 12 mostra o número médio de ovos de S. mansoni
presentes no sedimento de fezes após 35º, 40º, 45º e 50º dias de infecção. Os resultados dos
exames demonstraram que entre o 35º e 40º dia após a infecção o número de ovos presentes
nas fezes é significativamente menor em comparação com a ovoposição observada no período
entre o 45º e o 50º dia. Este resultado, embora não exatamente quantitativo, serviu para a
definição de dois grupos experimentais distintos para a posterior análise de biomarcadores
teciduais associados com a infecção. A partir do 45º dia o número médio de ovos depositados
nos tecidos aumenta consideravelmente caracterizando a fase crônica da doença no modelo
murino.
Figura 12: Gráfico demonstrando a evolução da ovoposição
nos animais infectados entre o 35º e o 50º dia de infecção.
Para as análises estatísticas utilizou-se o número médio de
ovos presentes em 300 mg de fezes de 5 animais. As triplicatas
das lâminas foram analisadas no software Prism versão 5.0
utilizando Student´s t-test pareado.
35 d
ias
40 d
ias
45 d
ias
50 d
ias
0
10
20
30
40
Nú
mero
de o
vo
s p
or l
âm
ina
**
*
31
4.2 – Avaliação de esplenomegalia e hepatomegalia nos animais dos grupos I e II
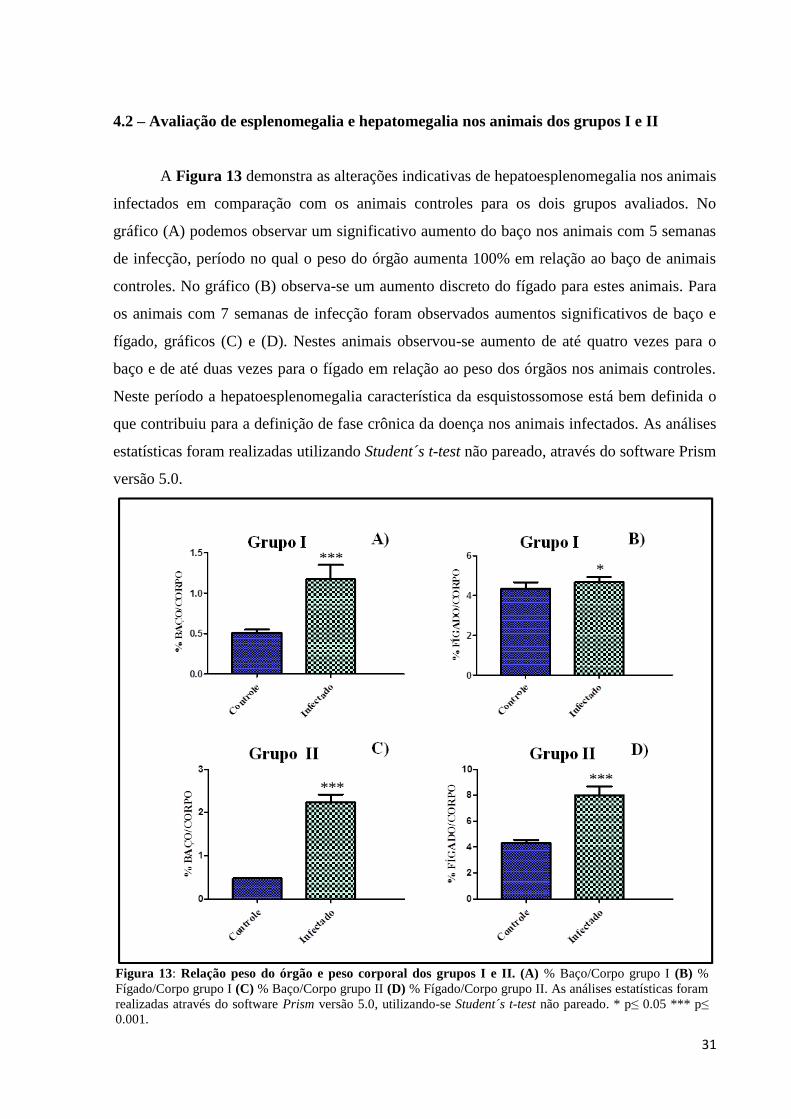
A Figura 13 demonstra as alterações indicativas de hepatoesplenomegalia nos animais
infectados em comparação com os animais controles para os dois grupos avaliados. No
gráfico (A) podemos observar um significativo aumento do baço nos animais com 5 semanas
de infecção, período no qual o peso do órgão aumenta 100% em relação ao baço de animais
controles. No gráfico (B) observa-se um aumento discreto do fígado para estes animais. Para
os animais com 7 semanas de infecção foram observados aumentos significativos de baço e
fígado, gráficos (C) e (D). Nestes animais observou-se aumento de até quatro vezes para o
baço e de até duas vezes para o fígado em relação ao peso dos órgãos nos animais controles.
Neste período a hepatoesplenomegalia característica da esquistossomose está bem definida o
que contribuiu para a definição de fase crônica da doença nos animais infectados. As análises
estatísticas foram realizadas utilizando Student´s t-test não pareado, através do software Prism
versão 5.0.
Figura 13: Relação peso do órgão e peso corporal dos grupos I e II. (A) % Baço/Corpo grupo I (B) %
Fígado/Corpo grupo I (C) % Baço/Corpo grupo II (D) % Fígado/Corpo grupo II. As análises estatísticas foram
realizadas através do software Prism versão 5.0, utilizando-se Student´s t-test não pareado. * p≤ 0.05 *** p≤
0.001.
*
***
***
***

32
4.3 – Análises hematológicas e dosagens bioquímicas
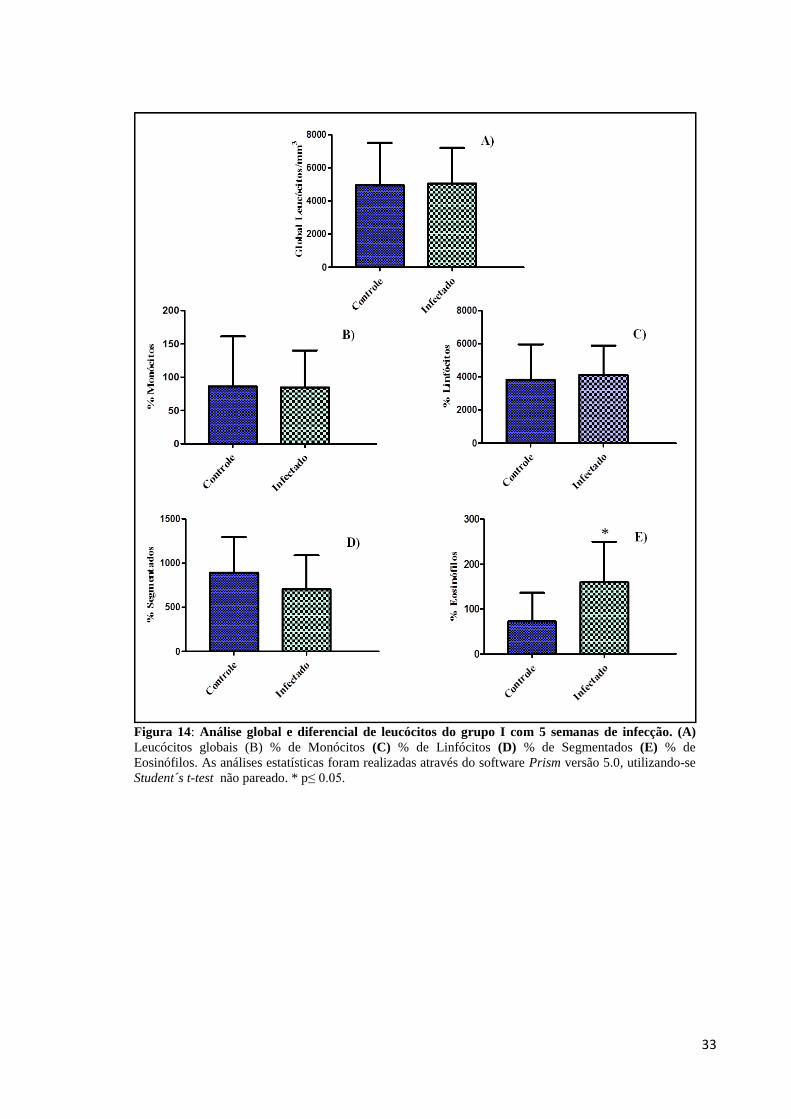
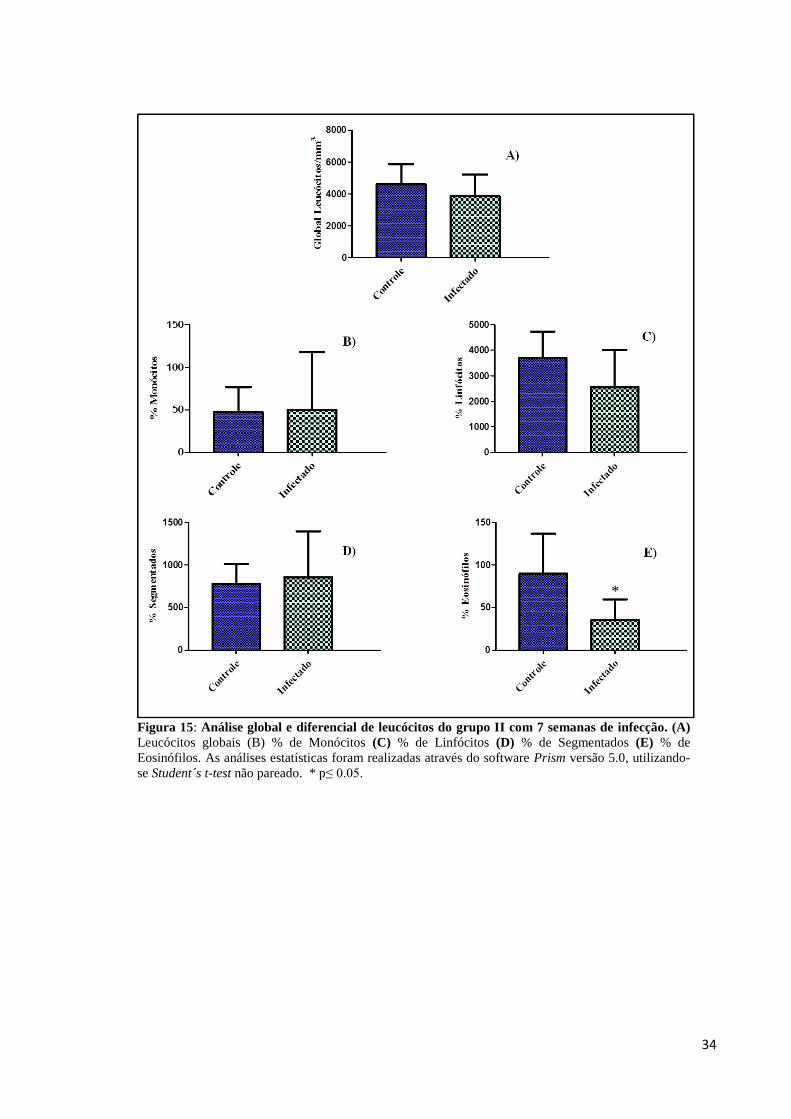
Amostras de sangue, dos animais do grupo I e do grupo II, foram coletadas através de
punção cardíaca e avaliadas quanto às alterações do número de células sanguíneas, através de
contagem global e diferencial de leucócitos. Os dados representados na Figura 14 mostram
que não houve alterações significativas na contagem global de leucócitos dos animais com 5
semanas de infecção, no entanto, a avaliação da leucometria diferencial deste grupo
demonstrou eosinofilia nos animais infectados, Figura 14E. No grupo de animais com 7
semanas de infecção, não foram observadas diferenças estatísticas nas análises de leucócitos
globais, porém a contagem diferencial revelou uma diminuição significativa de eosinófilos,
caracterizando eosinopenia associada à infecção crônica, Figura 15E.
33
*
Figura 14: Análise global e diferencial de leucócitos do grupo I com 5 semanas de infecção. (A)
Leucócitos globais (B) % de Monócitos (C) % de Linfócitos (D) % de Segmentados (E) % de
Eosinófilos. As análises estatísticas foram realizadas através do software Prism versão 5.0, utilizando-se
Student´s t-test não pareado. * p≤ 0.05.
34
*
Figura 15: Análise global e diferencial de leucócitos do grupo II com 7 semanas de infecção. (A)
Leucócitos globais (B) % de Monócitos (C) % de Linfócitos (D) % de Segmentados (E) % de
Eosinófilos. As análises estatísticas foram realizadas através do software Prism versão 5.0, utilizando-
se Student´s t-test não pareado. * p≤ 0.05.
35
As amostras de sangue recolhidas dos animais dos grupos I e II, também foram
utilizadas para avaliação de marcadores clássicos de dano tecidual, especificamente dosagem
de atividade de ALT e AST e concentração de albumina sérica. Na Tabela 4 podemos
observar as alterações das enzimas transaminases e alterações no nível de albumina sérica ao
longo da infecção. Os resultados das avaliações bioquímicas demonstraram diminuição nos
níveis das enzimas transaminases ALT e AST, entre os animais infectados e controles do
grupo I e do grupo II. A razão ALT/AST teve um aumento em torno de 50% comparando os
animais infectados dos dois grupos. A princípio esta razão poderia indicar aumento dos níveis
de ALT a partir da 5 semana de infecção, demonstrando a severidade do dano hepático. No
entanto, os valores absolutos de atividade enzimática demonstraram que os níveis de ALT
permaneceram sem grandes alterações enquanto os níveis de AST diminuíram o que
contribuiu para o aumento da razão ALT/AST. Por outro lado, os níveis séricos de albumina
diminuíram em relação ao controle nos animais com 5 semanas de infecção e,
significativamente, nos animais com 7 semanas de infecção, demonstrando o
comprometimento da função hepática associado à fase crônica da esquistossomose.
Grupo I a
(5 semanas de infecção)
Grupo II b
(7 semanas de infecção)
Controle Infectado Controle Infectado
ALT (U/L) 392,9 203,5 202,5±72,0 150,1±2,4
AST (U/L) 757,9 653,1 511,2±139,6 260,7±17,4
ALT/AST 0,52 0,31 0,40 0,58
ALB (g/dL) 3,76 3,56 3,86±0,11 2,93±0,11**
Tabela 4: Tabela de dosagens bioquímicas
a- Para os animais do grupo I, os dados mostrados correspondem a dosagens de apenas um pool
proveniente de 3 animais. b- para o grupo II foram feitas analises estatísticas utilizando triplicatas
de pools provenientes de 3 animais, Student´s t-test. **
p≤0.01.

36
4.4 – Análises histológicas de fígado e baço
As secções de fígado e baço provenientes dos animais dos grupos I e II foram
mantidas em formol, posteriormente incluídas em parafina e processadas para histologia
conforme descrito no item 3.8. A tríade portal (ramo da veia porta; ramo da artéria hepática e
ducto biliar) e a veia centro-lobular, de animais controles dos grupos I e II, estão mostradas
nas Figuras 16A/16B, respectivamente. Observou-se aspecto histológico hepático normal
nestes animais. As Figuras 16C/16D demonstraram respectivamente, inflamação moderada
perivascular presente nos animais infectados do grupo I e acentuado processo inflamatório
perivascular nos animais infectados do grupo II. A seção demonstrada na Figura 16E ilustra a
presença de vermes pareados em corte transversal nos animais com 5 semanas de infecção.
Observa-se nítido aumento de diâmetro no ramo da veia porta nestes animais e inflamação
moderada perivascular. A Figura 16F indica a presença de ovos imaturos no parênquima
hepático dos animais com 5 semanas de infecção, no entanto, observa-se discreta inflamação
granulomatosa ao redor dos ovos. Por outro lado, após 7 semanas de infecção os ovos
presentes no parênquima hepático, aparentam desenvolvimento completo, e estão circundados
por intensa inflamação granulomatosa, Figura 16G. Utilizando-se do método de coloração
Tricômico de Masson foi possível visualizar deposição de colágeno associada ao processo de
fibrose instalado para a resolução dos granulomas, Figura 16H. Avaliações quantitativas do
número de células inflamatórias da região perivascular dos animais infectados mostraram que
entre 5 e 7 semanas de infecção houve um aumento de 100% no número de células
inflamatórias, Figura 17.
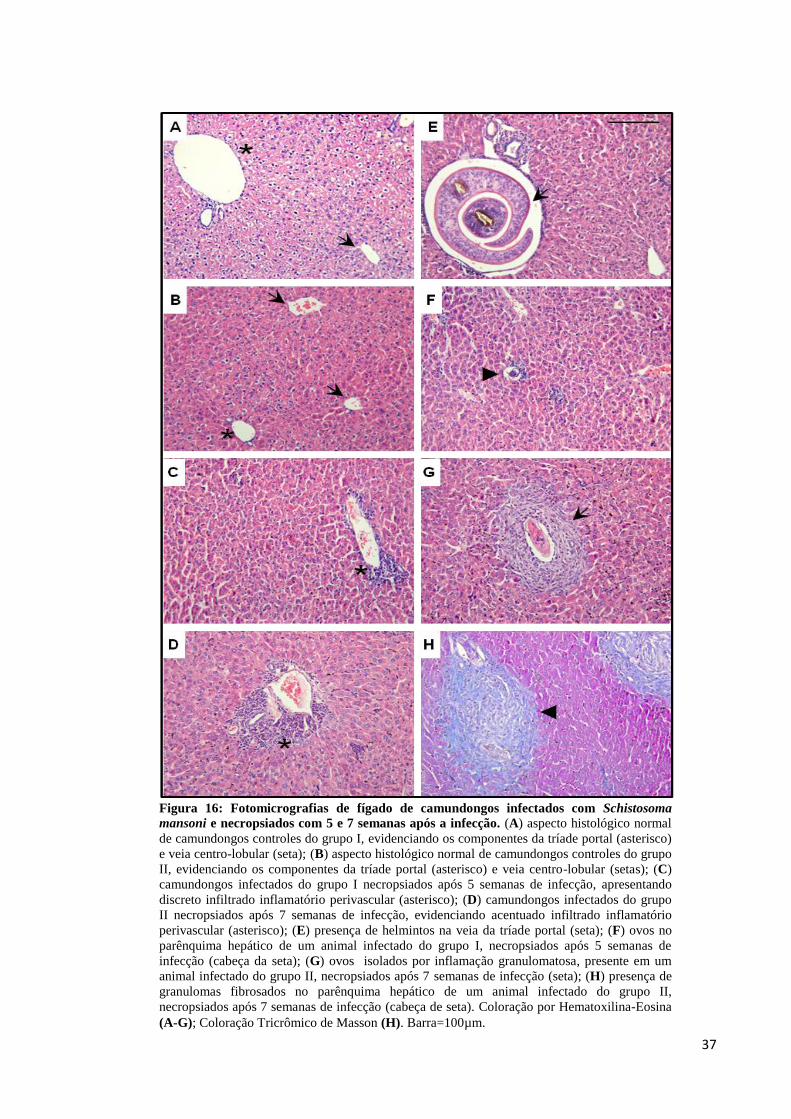
37
Figura 16: Fotomicrografias de fígado de camundongos infectados com Schistosoma
mansoni e necropsiados com 5 e 7 semanas após a infecção. (A) aspecto histológico normal
de camundongos controles do grupo I, evidenciando os componentes da tríade portal (asterisco)
e veia centro-lobular (seta); (B) aspecto histológico normal de camundongos controles do grupo
II, evidenciando os componentes da tríade portal (asterisco) e veia centro-lobular (setas); (C)
camundongos infectados do grupo I necropsiados após 5 semanas de infecção, apresentando
discreto infiltrado inflamatório perivascular (asterisco); (D) camundongos infectados do grupo
II necropsiados após 7 semanas de infecção, evidenciando acentuado infiltrado inflamatório
perivascular (asterisco); (E) presença de helmintos na veia da tríade portal (seta); (F) ovos no
parênquima hepático de um animal infectado do grupo I, necropsiados após 5 semanas de
infecção (cabeça da seta); (G) ovos isolados por inflamação granulomatosa, presente em um
animal infectado do grupo II, necropsiados após 7 semanas de infecção (seta); (H) presença de
granulomas fibrosados no parênquima hepático de um animal infectado do grupo II,
necropsiados após 7 semanas de infecção (cabeça de seta). Coloração por Hematoxilina-Eosina
(A-G); Coloração Tricrômico de Masson (H). Barra=100µm.
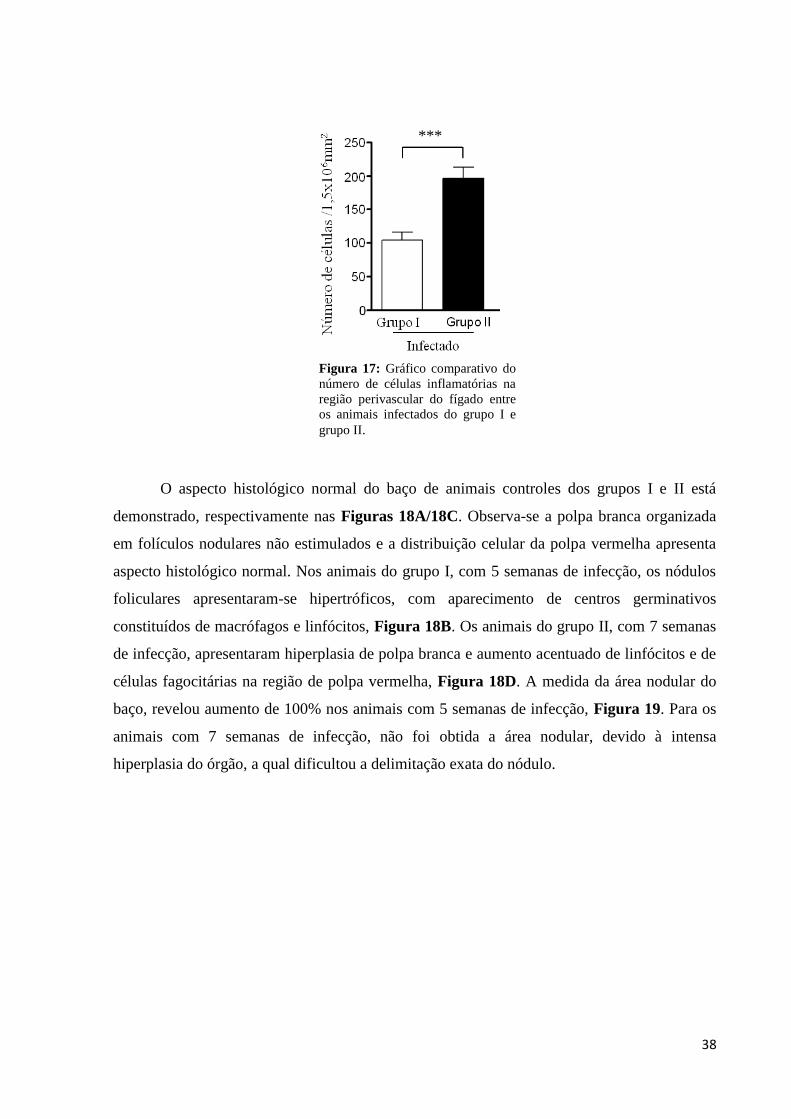
38
O aspecto histológico normal do baço de animais controles dos grupos I e II está
demonstrado, respectivamente nas Figuras 18A/18C. Observa-se a polpa branca organizada
em folículos nodulares não estimulados e a distribuição celular da polpa vermelha apresenta
aspecto histológico normal. Nos animais do grupo I, com 5 semanas de infecção, os nódulos
foliculares apresentaram-se hipertróficos, com aparecimento de centros germinativos
constituídos de macrófagos e linfócitos, Figura 18B. Os animais do grupo II, com 7 semanas
de infecção, apresentaram hiperplasia de polpa branca e aumento acentuado de linfócitos e de
células fagocitárias na região de polpa vermelha, Figura 18D. A medida da área nodular do
baço, revelou aumento de 100% nos animais com 5 semanas de infecção, Figura 19. Para os
animais com 7 semanas de infecção, não foi obtida a área nodular, devido à intensa
hiperplasia do órgão, a qual dificultou a delimitação exata do nódulo.
***
Figura 17: Gráfico comparativo do
número de células inflamatórias na
região perivascular do fígado entre
os animais infectados do grupo I e
grupo II.
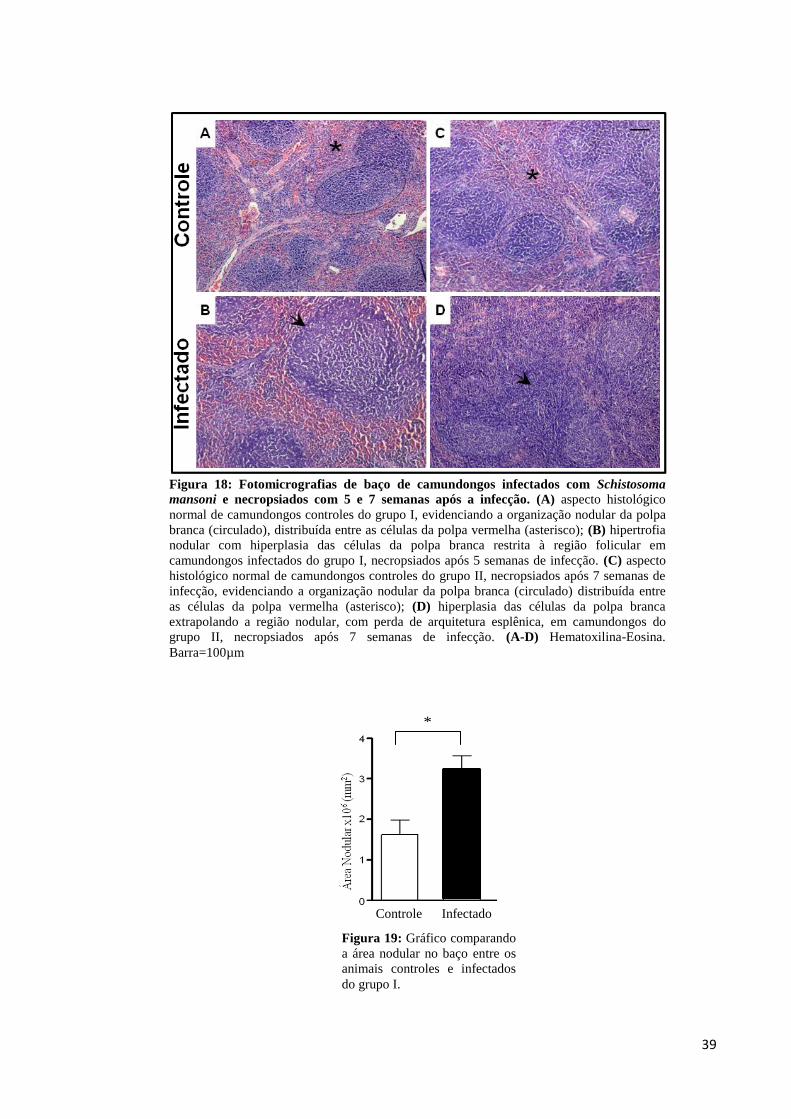
39
Figura 18: Fotomicrografias de baço de camundongos infectados com Schistosoma
mansoni e necropsiados com 5 e 7 semanas após a infecção. (A) aspecto histológico
normal de camundongos controles do grupo I, evidenciando a organização nodular da polpa
branca (circulado), distribuída entre as células da polpa vermelha (asterisco); (B) hipertrofia
nodular com hiperplasia das células da polpa branca restrita à região folicular em
camundongos infectados do grupo I, necropsiados após 5 semanas de infecção. (C) aspecto
histológico normal de camundongos controles do grupo II, necropsiados após 7 semanas de
infecção, evidenciando a organização nodular da polpa branca (circulado) distribuída entre
as células da polpa vermelha (asterisco); (D) hiperplasia das células da polpa branca
extrapolando a região nodular, com perda de arquitetura esplênica, em camundongos do
grupo II, necropsiados após 7 semanas de infecção. (A-D) Hematoxilina-Eosina.
Barra=100µm
Figura 19: Gráfico comparando
a área nodular no baço entre os
animais controles e infectados
do grupo I.
Infectado
Controle
*
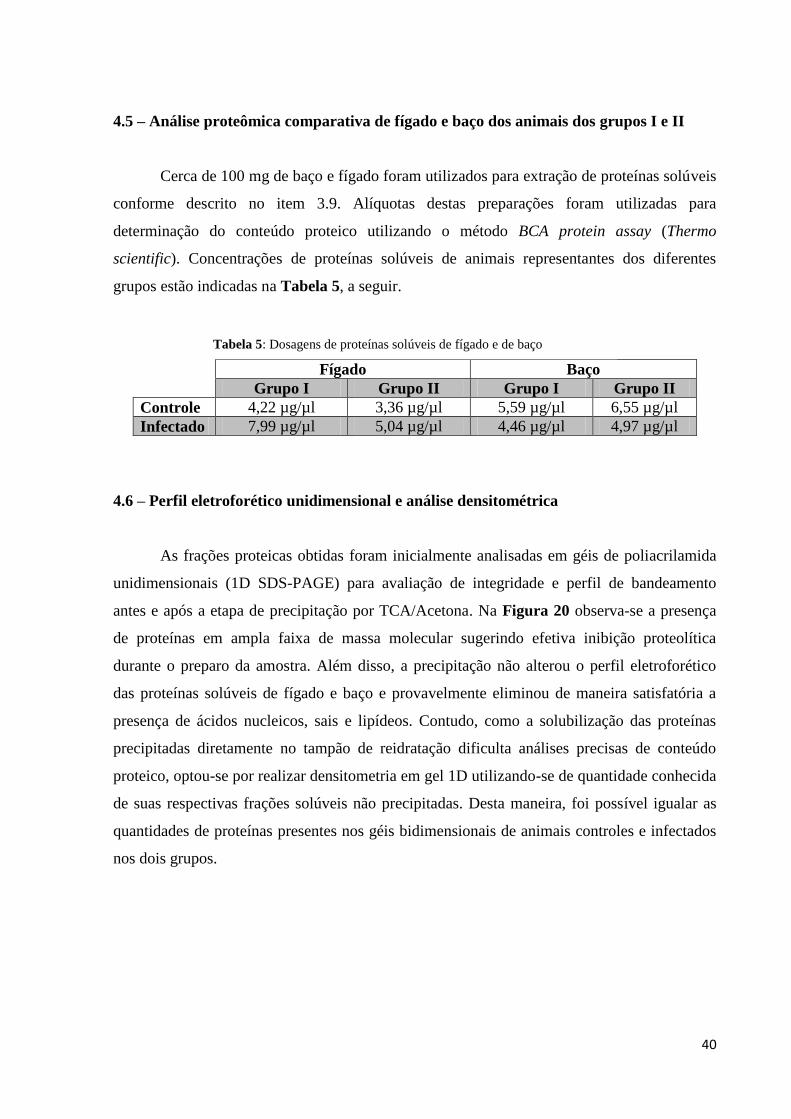
40
4.5 – Análise proteômica comparativa de fígado e baço dos animais dos grupos I e II
Cerca de 100 mg de baço e fígado foram utilizados para extração de proteínas solúveis
conforme descrito no item 3.9. Alíquotas destas preparações foram utilizadas para
determinação do conteúdo proteico utilizando o método BCA protein assay (Thermo
scientific). Concentrações de proteínas solúveis de animais representantes dos diferentes
grupos estão indicadas na Tabela 5, a seguir.
Fígado Baço
Grupo I Grupo II Grupo I Grupo II
Controle 4,22 µg/µl 3,36 µg/µl 5,59 µg/µl 6,55 µg/µl
Infectado 7,99 µg/µl 5,04 µg/µl 4,46 µg/µl 4,97 µg/µl
4.6 – Perfil eletroforético unidimensional e análise densitométrica
As frações proteicas obtidas foram inicialmente analisadas em géis de poliacrilamida
unidimensionais (1D SDS-PAGE) para avaliação de integridade e perfil de bandeamento
antes e após a etapa de precipitação por TCA/Acetona. Na Figura 20 observa-se a presença
de proteínas em ampla faixa de massa molecular sugerindo efetiva inibição proteolítica
durante o preparo da amostra. Além disso, a precipitação não alterou o perfil eletroforético
das proteínas solúveis de fígado e baço e provavelmente eliminou de maneira satisfatória a
presença de ácidos nucleicos, sais e lipídeos. Contudo, como a solubilização das proteínas
precipitadas diretamente no tampão de reidratação dificulta análises precisas de conteúdo
proteico, optou-se por realizar densitometria em gel 1D utilizando-se de quantidade conhecida
de suas respectivas frações solúveis não precipitadas. Desta maneira, foi possível igualar as
quantidades de proteínas presentes nos géis bidimensionais de animais controles e infectados
nos dois grupos.
Tabela 5: Dosagens de proteínas solúveis de fígado e de baço

41
4.7 – Perfil proteico em eletroforese bidimensional (SDS-PAGE) da fração solúvel de
fígado
Géis bidimensionais confeccionados com proteínas solúveis de fígado dos animais
com 5 e 7 semanas de infecção, foram corados por Coomassie brilliant blue G250 e
analisados com auxílio do software 2-D Evolution v.2005, para posterior comparação com
seus respectivos géis controles. Uma análise global revelou alterações pouco evidentes entre o
proteoma solúvel dos animais controles (A) e infectados (B) por 5 semanas, Figura 21.
Entretanto a análise de bandas específicas destes animais, após sobreposição dos géis,
demonstrou diferenças significativas de expressão entre animais controles e infectados
Tabela 6. Por outro lado, o perfil eletroforético bidimensional de fígado dos animais com 7
semanas de infecção, revelou nítidas alterações na abundância de proteínas compartilhadas
por animais controles (A) e infectados (B), Figura 22 e Tabela 6.
A Figura 23 mostra o perfil eletroforético bidimensional de fígado, em pH 4-7, dos
animais com 7 semanas de infecção. Esta faixa estreita de pH proporcionou uma melhor
resolução das proteínas realçando as alterações no nível de expressão de proteínas neste grupo
de animais. Os spots, diferencialmente presentes nos géis confeccionados, foram excisados e
submetidos ao processo de digestão in gel utilizando tripsina. Posteriormente os peptídeos
provenientes da digestão foram identificados por espectrometria de massas. Dos 74 spots de
interesse procedentes de cinco pares de géis réplicas de fígado (pH 3-10 e pH 4-7), foram
Figura 20: Perfil eletroforético representativo
de proteínas citosólicas de fígado e baço. FS –
Fração solúvel. FP – Fração precipitada. Alíquotas
contendo cerca de 30 μg de proteínas foram
separadas em gel SDS-PAGE 10%. Coloração por
Comassie brilliant blue G250.

42
selecionadas pelo menos 14 proteínas que apresentaram alterações de expressão comuns aos
animais de 5 e 7 semanas de infecção, Tabela 6. O aumento do poder de resolução dos géis
com o emprego do pH 4-7 não impediu com que houvesse confluência de bandas para
proteínas com massa molecular e ponto isoelétrico semelhantes. Isto resultou, na maioria dos
casos, na identificação de mais de uma proteína por spot. Contudo, a utilização da abordagem
LC-MS/MS permitiu através do parâmetro de abundância relativa, (emPAI- Exponentially
modified protein abundance index), identificar o componente de maior abundância, o qual
constitui o foco de investigação neste trabalho.
Dentre as 14 proteínas selecionadas, cinco proteínas são chaperoninas envolvidas com
estresse no retículo endoplasmático, spots 2, 3, 4, 8 e 9; duas proteínas de transporte, spots 5 e
6; duas proteínas anti-oxidantes, spots 11 e 12; uma proteína do ciclo da uréia, spot 1; uma
proteína metil-transferase, spot 10 e uma proteína de via glicolítica, spot 14, Tabela 6.
Algumas isoformas de proteínas também foram encontradas com alteração em sua expressão
nos animais infectados, como é o caso da transferrina spot 7 e anidrase carbônica spot 13.
Entre as proteínas confluentes identificadas, uma classe de proteínas estruturais chamou a
atenção, as citoqueratinas. Alterações no citoesqueleto são descritas em algumas hepatopatias,
e a presença de citoqueratinas nas mais variadas faixas de pH e massa molecular sugerem
intensa modificação pós-traducional. As chaperoninas juntamente com as proteínas
transferrina e triose-fosfato isomerase apresentaram aumento de abundância nos animais
infectados. Este aumento chegou a mais de 100% no caso das chaperoninas e mais de 200%
para a transferrina. Outras proteínas como a carbamoil fosfato sintetase I, albumina,
indoletilamina n-metiltransferase, peroxiredoxina 6 e anidrase carbônica, apresentaram
diminuição de abundância nos animais infectados, com redução de 90%; no caso da
indoletilamina n-metiltransferase e anidrase carbônica III a redução chegou a 91% e 88%
respectivamente,Tabela 6 - (vide tabela suplementar 1, em anexos).
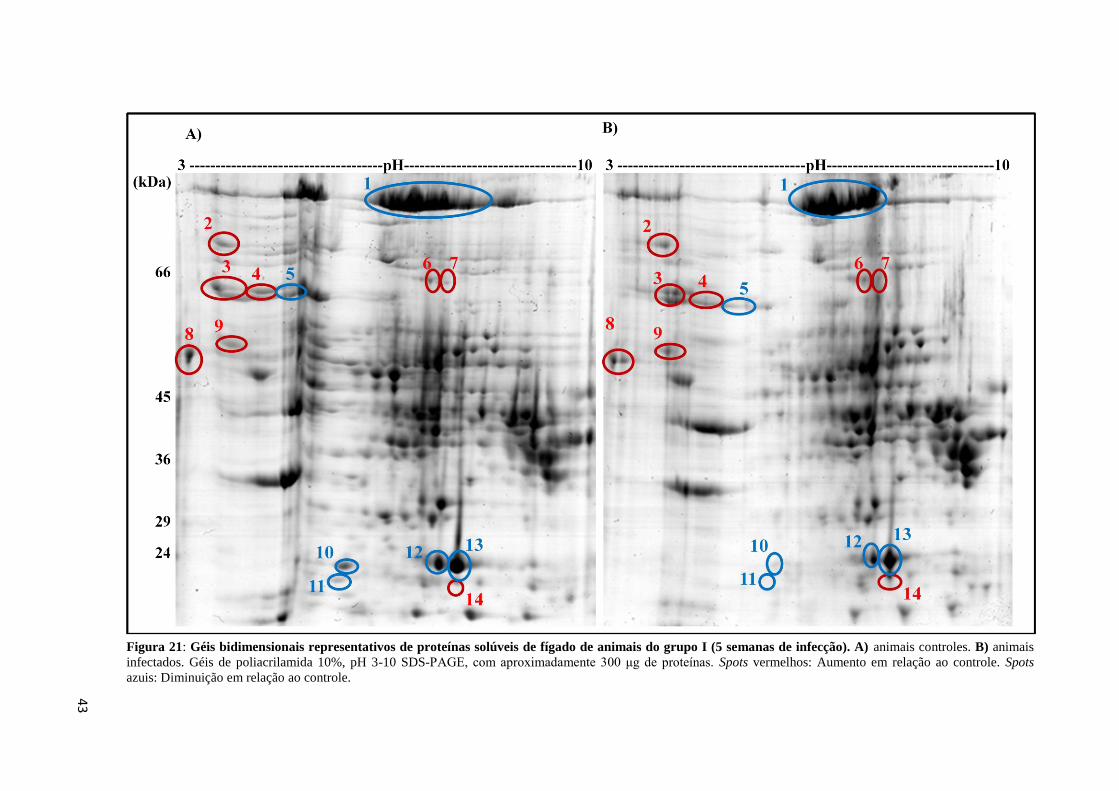
Figura 21: Géis bidimensionais representativos de proteínas solúveis de fígado de animais do grupo I (5 semanas de infecção). A) animais controles. B) animais
infectados. Géis de poliacrilamida 10%, pH 3-10 SDS-PAGE, com aproximadamente 300 μg de proteínas. Spots vermelhos: Aumento em relação ao controle. Spots
azuis: Diminuição em relação ao controle.
43
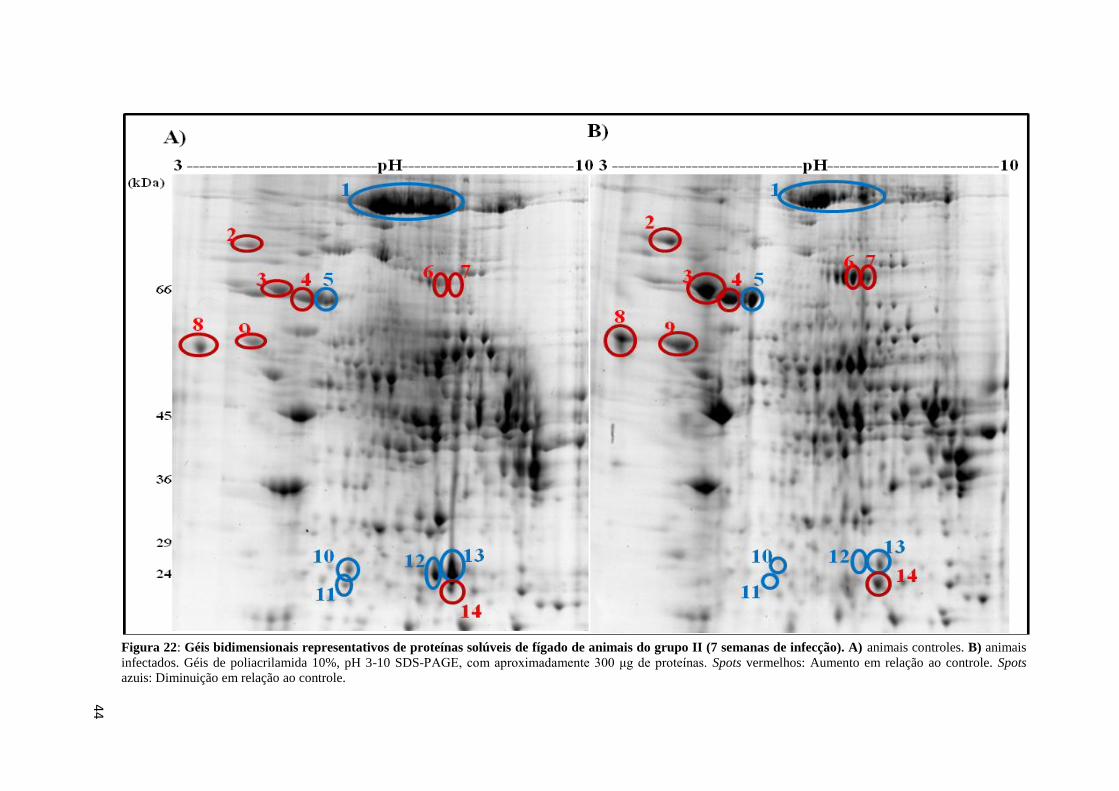
Figura 22: Géis bidimensionais representativos de proteínas solúveis de fígado de animais do grupo II (7 semanas de infecção). A) animais controles. B) animais
infectados. Géis de poliacrilamida 10%, pH 3-10 SDS-PAGE, com aproximadamente 300 μg de proteínas. Spots vermelhos: Aumento em relação ao controle. Spots
azuis: Diminuição em relação ao controle.
44
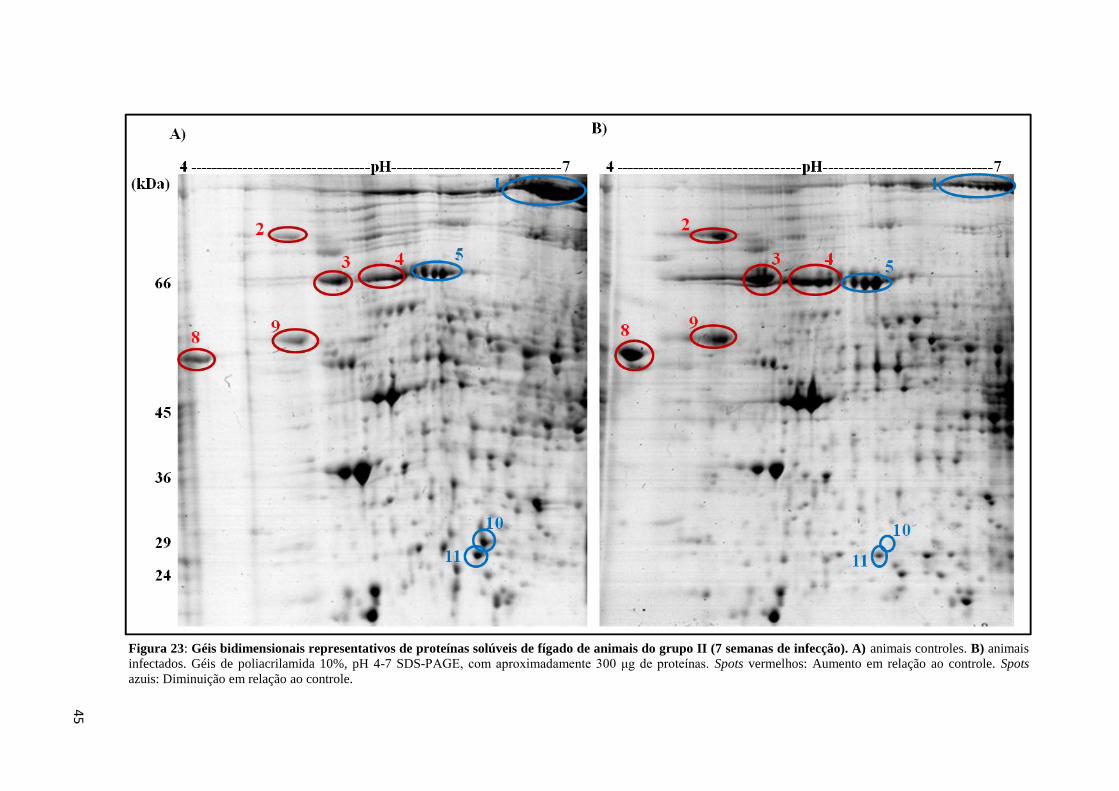
Figura 23: Géis bidimensionais representativos de proteínas solúveis de fígado de animais do grupo II (7 semanas de infecção). A) animais controles. B) animais
infectados. Géis de poliacrilamida 10%, pH 4-7 SDS-PAGE, com aproximadamente 300 μg de proteínas. Spots vermelhos: Aumento em relação ao controle. Spots
azuis: Diminuição em relação ao controle.
45
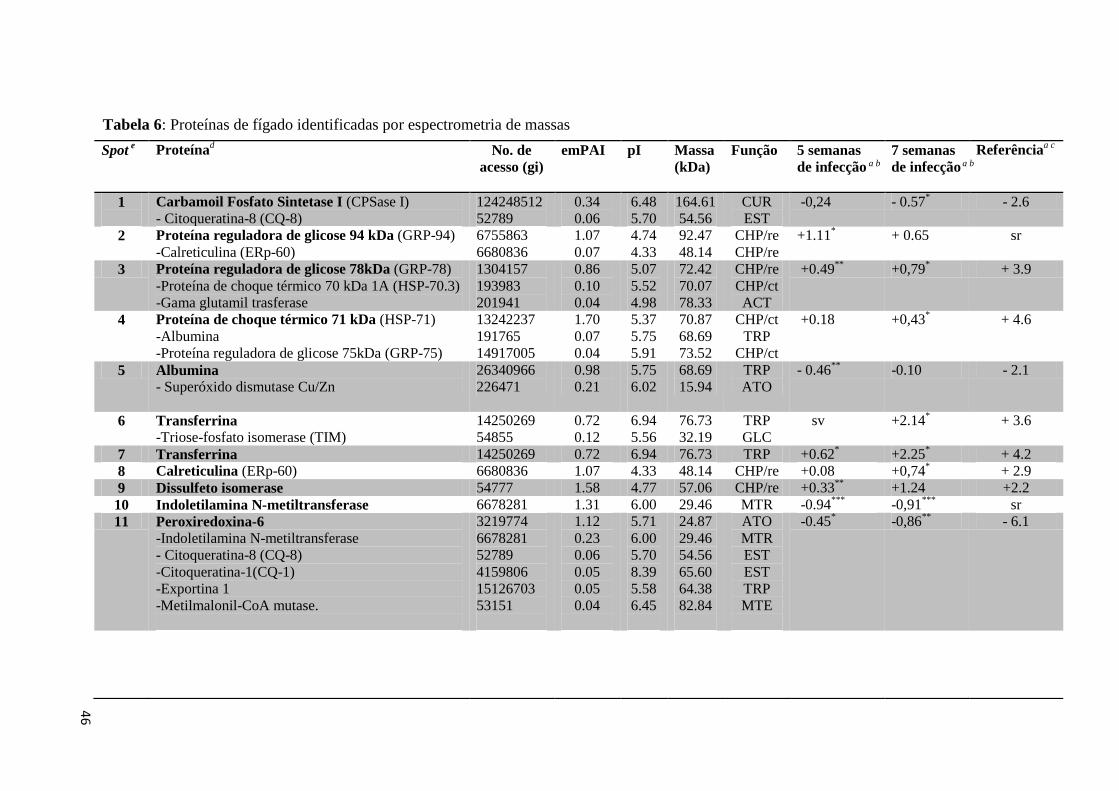
Spot e Proteína
d No. de
acesso (gi)
emPAI pI Massa
(kDa)
Função 5 semanas
de infecção
7 semanas
de infecção
Referênciaa c
1
Carbamoil Fosfato Sintetase I (CPSase I)
- Citoqueratina-8 (CQ-8)
124248512
52789
0.34
0.06
6.48
5.70
164.61
54.56
CUR
EST
-0,24 - 0.57* - 2.6
2 Proteína reguladora de glicose 94 kDa (GRP-94)
-Calreticulina (ERp-60)
6755863
6680836
1.07
0.07
4.74
4.33
92.47
48.14
CHP/re
CHP/re
+1.11* + 0.65
sr
3 Proteína reguladora de glicose 78kDa (GRP-78)
-Proteína de choque térmico 70 kDa 1A (HSP-70.3)
-Gama glutamil trasferase
1304157
193983
201941
0.86
0.10
0.04
5.07
5.52
4.98
72.42
70.07
78.33
CHP/re
CHP/ct
ACT
+0.49**
+0,79* + 3.9
4 Proteína de choque térmico 71 kDa (HSP-71)
-Albumina
-Proteína reguladora de glicose 75kDa (GRP-75)
13242237
191765
14917005
1.70
0.07
0.04
5.37
5.75
5.91
70.87
68.69
73.52
CHP/ct
TRP
CHP/ct
+0.18 +0,43* + 4.6
5 Albumina - Superóxido dismutase Cu/Zn
26340966
226471
0.98
0.21
5.75
6.02
68.69
15.94
TRP
ATO
- 0.46**
-0.10 - 2.1
6
Transferrina -Triose-fosfato isomerase (TIM)
14250269
54855
0.72
0.12
6.94
5.56
76.73
32.19
TRP
GLC
sv +2.14* + 3.6
7 Transferrina 14250269 0.72 6.94 76.73 TRP +0.62* +2.25
* + 4.2
8 Calreticulina (ERp-60) 6680836 1.07 4.33 48.14 CHP/re +0.08 +0,74* + 2.9
9 Dissulfeto isomerase 54777 1.58 4.77 57.06 CHP/re +0.33**
+1.24 +2.2
10 Indoletilamina N-metiltransferase 6678281 1.31 6.00 29.46 MTR -0.94***
-0,91***
sr
11 Peroxiredoxina-6 -Indoletilamina N-metiltransferase
- Citoqueratina-8 (CQ-8)
-Citoqueratina-1(CQ-1)
-Exportina 1
-Metilmalonil-CoA mutase.
3219774
6678281
52789
4159806
15126703
53151
1.12
0.23
0.06
0.05
0.05
0.04
5.71
6.00
5.70
8.39
5.58
6.45
24.87
29.46
54.56
65.60
64.38
82.84
ATO
MTR
EST
EST
TRP
MTE
-0.45* -0,86
** - 6.1
Tabela 6: Proteínas de fígado identificadas por espectrometria de massas
a b a b
46
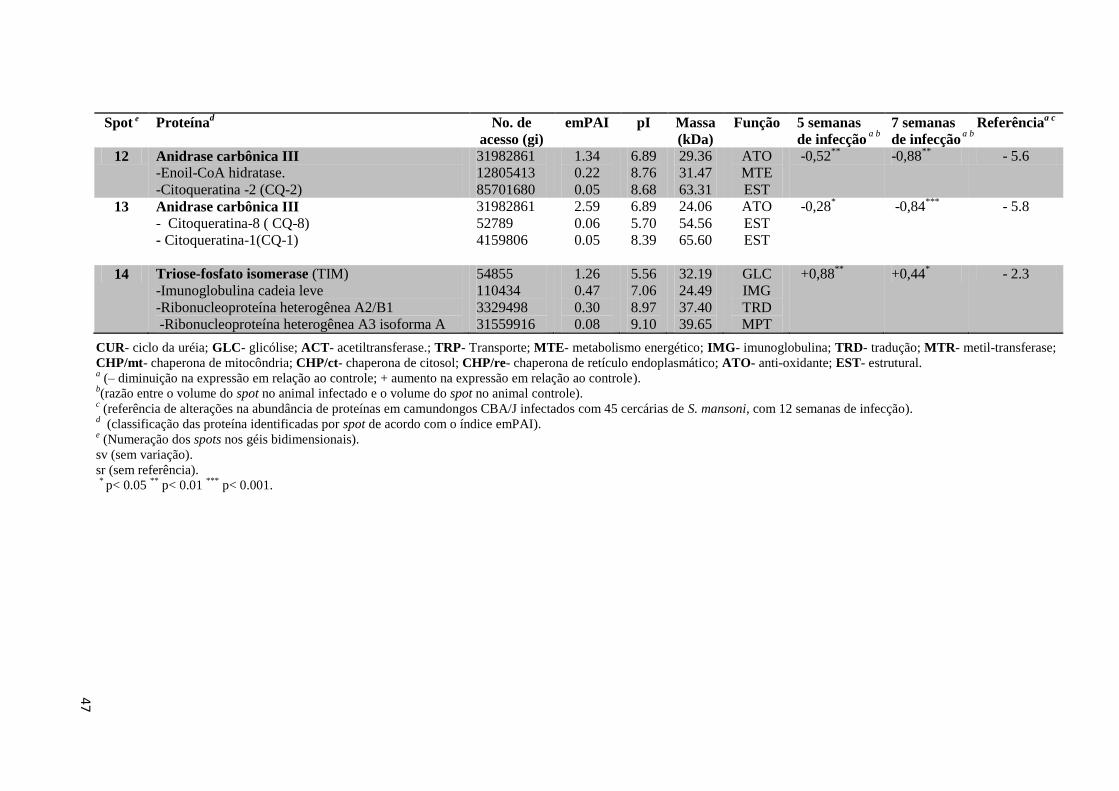
Spot e Proteínad No. de
acesso (gi)
emPAI pI Massa
(kDa)
Função 5 semanas
de infecção
7 semanas
de infecção
Referênciaa c
12 Anidrase carbônica III -Enoil-CoA hidratase.
-Citoqueratina -2 (CQ-2)
31982861
12805413
85701680
1.34
0.22
0.05
6.89
8.76
8.68
29.36
31.47
63.31
ATO
MTE
EST
-0,52**
-0,88**
- 5.6
13 Anidrase carbônica III
- Citoqueratina-8 ( CQ-8)
- Citoqueratina-1(CQ-1)
31982861
52789
4159806
2.59
0.06
0.05
6.89
5.70
8.39
24.06
54.56
65.60
ATO
EST
EST
-0,28* -0,84
*** - 5.8
14 Triose-fosfato isomerase (TIM)
-Imunoglobulina cadeia leve
-Ribonucleoproteína heterogênea A2/B1
-Ribonucleoproteína heterogênea A3 isoforma A
54855
110434
3329498
31559916
1.26
0.47
0.30
0.08
5.56
7.06
8.97
9.10
32.19
24.49
37.40
39.65
GLC
IMG
TRD
MPT
+0,88**
+0,44* - 2.3
CUR- ciclo da uréia; GLC- glicólise; ACT- acetiltransferase.; TRP- Transporte; MTE- metabolismo energético; IMG- imunoglobulina; TRD- tradução; MTR- metil-transferase;
CHP/mt- chaperona de mitocôndria; CHP/ct- chaperona de citosol; CHP/re- chaperona de retículo endoplasmático; ATO- anti-oxidante; EST- estrutural. a (– diminuição na expressão em relação ao controle; + aumento na expressão em relação ao controle).
b(razão entre o volume do spot no animal infectado e o volume do spot no animal controle).
c (referência de alterações na abundância de proteínas em camundongos CBA/J infectados com 45 cercárias de S. mansoni, com 12 semanas de infecção).
d (classificação das proteína identificadas por spot de acordo com o índice emPAI).
e (Numeração dos spots nos géis bidimensionais).
sv (sem variação).
sr (sem referência).
* p< 0.05
** p< 0.01
*** p< 0.001.
a b a b
47

48
4.8 – Perfil proteico em eletroforese bidimensional (SDS-PAGE) da fração solúvel do
baço
Géis bidimensionais de proteínas de baço dos animais com 5 e 7 semanas de infecção,
também foram confeccionados no intuito de avaliar as diferenças no proteoma solúvel dos
animais controles e infectados. Os géis foram corados por Coomassie brilliant blue G250 e
analisados com auxílio do software 2-D Evolution v.2005. A Figura 24 mostra o perfil
eletroforético bidimensional do baço dos animais com 5 semanas de infecção. Uma análise
global destes géis demonstrou significativas alterações na expressão de algumas proteínas
entre os animais controles (A) e infectados (B). A quantificação destas alterações de
abundância pode ser vista na Tabela 7. O perfil eletroforético da fração solúvel de baço dos
animais com 7 semanas de infecção é representado pela Figura 25. Estes géis nos mostra que
as alterações presentes nos animais com 5 semanas de infecção, se mantiveram nos animais
com 7 semanas de infecção, porém com um aumento na abundância de proteínas muito mais
significativo, Tabela 7. Em geral as proteínas de baço identificadas com alterações de
expressão, tiveram aumento nos animais com 5 e 7 semanas de infecção.
Dos 60 spots de interesse procedentes de três pares de géis réplicas de baço (pH 3-10),
foram selecionados pelo menos 7 proteínas que apresentaram alterações de expressão comuns
aos animais de 5 e 7 semanas de infecção, Tabela 7 – (vide tabela suplementar 2, em anexos).
Das 7 proteínas selecionadas, três proteínas são de via glicolítica, spots 3, 4 e 5; uma proteína
chaperona de retículo endoplasmático, spot 1; uma proteína de síntese proteica, spot 6 e uma
proteína de metabolismo energético, spot 7. Confluências de proteínas também foram
observadas nos géis bidimensionais de fração solúvel de baço, sendo as citoqueratinas o grupo
de proteínas confluentes com maior predominância. Algumas proteínas solúveis de baço
tiveram um aumento de mais de 600%, como é o caso da gliceraldeído 3 fosfato
desidrogenase e do fator de alongamento (EF-Tu), o qual aumentou cerca de 800% nos
animais com 7 semanas de infecção.
.
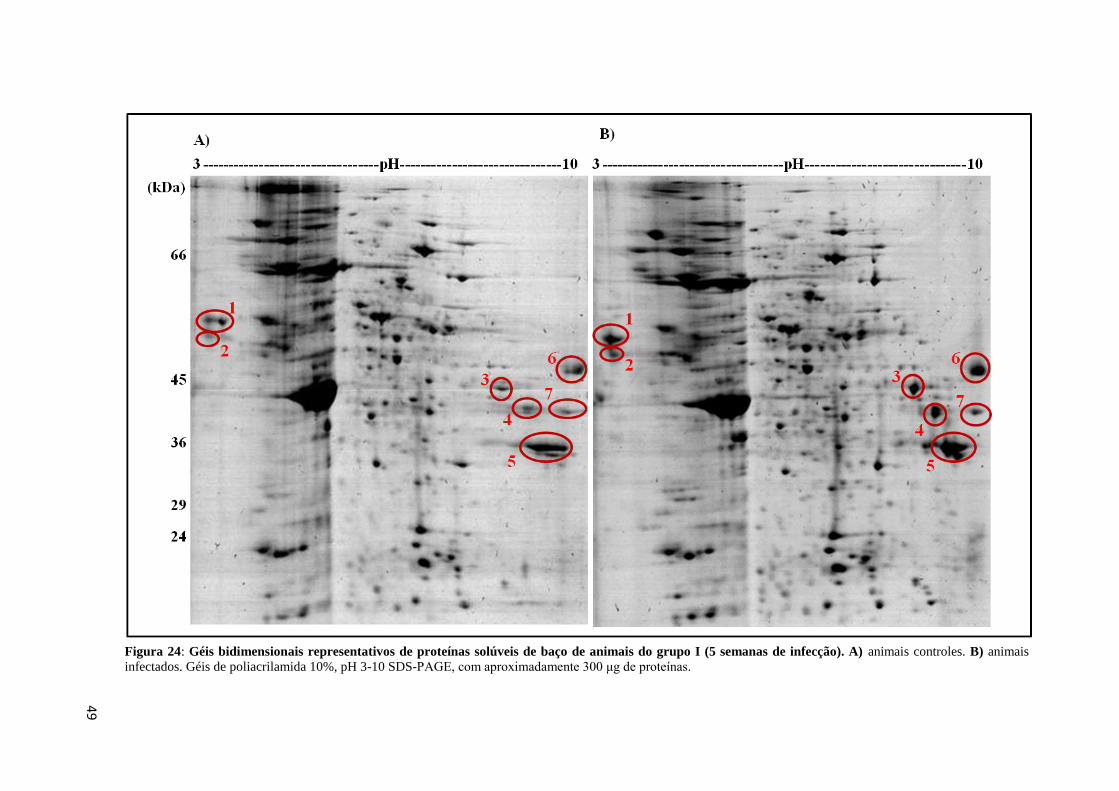
Figura 24: Géis bidimensionais representativos de proteínas solúveis de baço de animais do grupo I (5 semanas de infecção). A) animais controles. B) animais
infectados. Géis de poliacrilamida 10%, pH 3-10 SDS-PAGE, com aproximadamente 300 μg de proteínas.
49
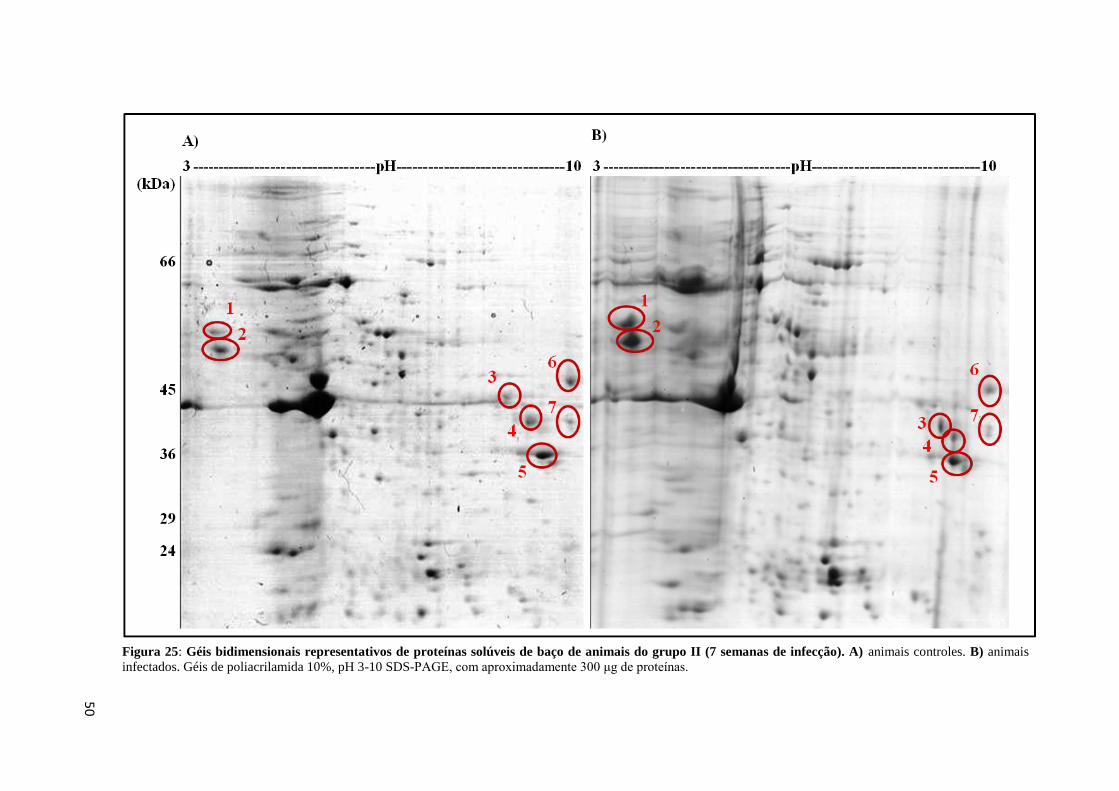
Figura 25: Géis bidimensionais representativos de proteínas solúveis de baço de animais do grupo II (7 semanas de infecção). A) animais controles. B) animais
infectados. Géis de poliacrilamida 10%, pH 3-10 SDS-PAGE, com aproximadamente 300 μg de proteínas.
50
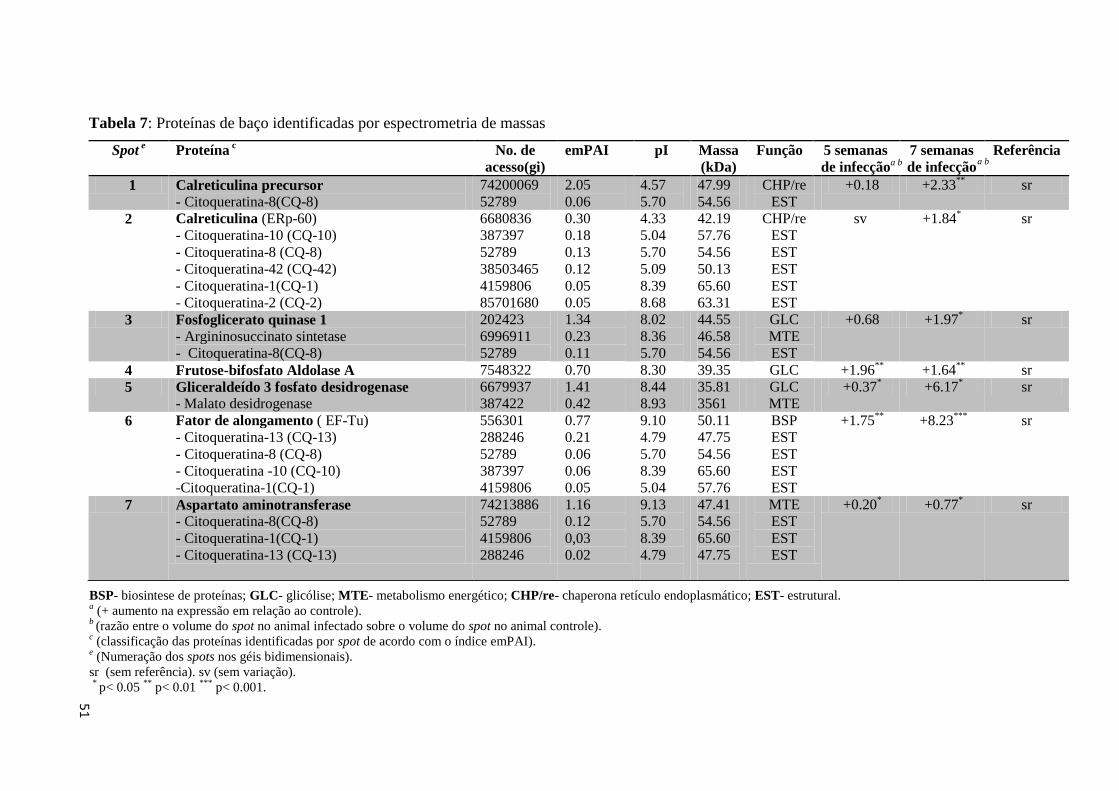
Spot e Proteína
c No. de
acesso(gi)
emPAI pI Massa
(kDa)
Função 5 semanas
de infecção
7 semanas
de infecção
Referência
1 Calreticulina precursor
- Citoqueratina-8(CQ-8)
74200069
52789
2.05
0.06
4.57
5.70
47.99
54.56
CHP/re
EST
+0.18 +2.33
** sr
2 Calreticulina (ERp-60)
- Citoqueratina-10 (CQ-10)
- Citoqueratina-8 (CQ-8)
- Citoqueratina-42 (CQ-42)
- Citoqueratina-1(CQ-1)
- Citoqueratina-2 (CQ-2)
6680836
387397
52789
38503465
4159806
85701680
0.30
0.18
0.13
0.12
0.05
0.05
4.33
5.04
5.70
5.09
8.39
8.68
42.19
57.76
54.56
50.13
65.60
63.31
CHP/re
EST
EST
EST
EST
EST
sv +1.84* sr
3 Fosfoglicerato quinase 1
- Argininosuccinato sintetase
- Citoqueratina-8(CQ-8)
202423
6996911
52789
1.34
0.23
0.11
8.02
8.36
5.70
44.55
46.58
54.56
GLC
MTE
EST
+0.68 +1.97* sr
4 Frutose-bifosfato Aldolase A 7548322 0.70 8.30 39.35 GLC +1.96**
+1.64**
sr
5 Gliceraldeído 3 fosfato desidrogenase
- Malato desidrogenase
6679937
387422
1.41
0.42
8.44
8.93
35.81
3561
GLC
MTE
+0.37* +6.17
* sr
6 Fator de alongamento ( EF-Tu)
- Citoqueratina-13 (CQ-13)
- Citoqueratina-8 (CQ-8)
- Citoqueratina -10 (CQ-10)
-Citoqueratina-1(CQ-1)
556301
288246
52789
387397
4159806
0.77
0.21
0.06
0.06
0.05
9.10
4.79
5.70
8.39
5.04
50.11
47.75
54.56
65.60
57.76
BSP
EST
EST
EST
EST
+1.75**
+8.23***
sr
7 Aspartato aminotransferase
- Citoqueratina-8(CQ-8)
- Citoqueratina-1(CQ-1)
- Citoqueratina-13 (CQ-13)
74213886
52789
4159806
288246
1.16
0.12
0,03
0.02
9.13
5.70
8.39
4.79
47.41
54.56
65.60
47.75
MTE
EST
EST
EST
+0.20* +0.77
* sr
Tabela 7: Proteínas de baço identificadas por espectrometria de massas
BSP- biosintese de proteínas; GLC- glicólise; MTE- metabolismo energético; CHP/re- chaperona retículo endoplasmático; EST- estrutural. a (+ aumento na expressão em relação ao controle).
b (razão entre o volume do spot no animal infectado sobre o volume do spot no animal controle).
c (classificação das proteínas identificadas por spot de acordo com o índice emPAI).
e (Numeração dos spots nos géis bidimensionais).
sr (sem referência). sv (sem variação).
* p< 0.05
** p< 0.01
*** p< 0.001.
a b a b
51

52
5 - Discussão

53
5 – Discussão
Abordagens proteômicas vêm sendo utilizadas nos últimos anos como ferramentas de
grande utilidade para a compreensão da biologia do S. mansoni, proporcionando um amplo
conhecimento a respeito do parasito em seus mais variados estágios de vida (Mathieson et al.,
2011, Curwen et al., 2004, Curwen et al., 2006). O emprego destas ferramentas tornou
possível o estudo detalhado de subproteomas do parasito, como aquele presente no tegumento
do verme, revelando a importância desta estrutura na obtenção de nutrientes e evasão do
sistema imune (Castro-Borges et al., 2011a, Castro-Borges et al., 2011b, Braschi et al., 2006).
Entretanto, até o momento, poucos estudos na área da protêomica abordaram a relação
parasito/hospedeiro, com o intuito de avaliar o impacto da doença nos órgãos afetados e
identificar marcadores para diagnóstico e/ou prognóstico da esquistossomose (Wilson et al.,
2007). Neste sentido, o presente trabalho refere-se à busca de marcadores proteicos,
provenientes de fígado e baço, durante as fases aguda e hepatoesplênica da esquistossomose
no modelo murino.
Nossa primeira abordagem fundamentou-se na discriminação dos estágios de fase
aguda e fase crônica nos animais infectados por S. mansoni. Para isso, inicialmente foram
realizados exames de fezes a partir do 35º dia de infecção para a comprovação do início da
ovoposição. Apesar de não terem sido realizados exames prévios, a literatura relata que a
ovoposição ocorre entre o 32º e o 35º dia de infecção no modelo murino (Brener and Chiari,
1956). Além do baixo número de ovos detectados nesta fase, observou-se que os mesmos se
encontravam em estágios iniciais de desenvolvimento embrionário. O acompanhamento da
ovoposição demonstrou aumento significativo do número de ovos nas fezes a partir do 45º dia
e a presença de até 20 vezes mais ovos por lâmina no 50º dia de infecção, quando comparados
aos números de ovos observados em 35 dias. A determinação e o acompanhamento da
ovoposição constituem ainda análises rotineiramente empregadas para o diagnóstico e
prognóstico da forma aguda da esquistossomose (Neves, 1992).
Outra abordagem utilizada para definir os estágios de fase aguda e fase crônica da
doença consistiu na avaliação da hepatoesplenomegalia desenvolvida pelos animais ao longo
da infecção. Para isso, utilizou-se a razão percentual peso do órgão (baço e fígado) / peso
corporal dos animais. Este parâmetro mostrou que os animais com 5 semanas de infecção
apresentaram aumento de 100% no tamanho do baço e um discreto aumento do fígado. Para
os animais com 7 semanas de infecção a hepatoesplenomegalia mostrou-se bem característica

54
de fase crônica da doença. Resultados similares foram encontrados em experimentos
realizados com camundongos CBA/J infectados com 45 cercárias, nos quais este aumento
chegou a aproximadamente 3 vezes para o baço e de até 2 vezes para o fígado, em animais
com 20 semanas de infecção (Manivannan et al., 2012, Henderson et al., 1993).
As análises de leucometria dos animais com 5 semanas de infecção não demonstraram
alterações no número total de leucócitos. Entretanto, a contagem diferencial mostrou aumento
significativo do número de eosinófilos, compatível com a fase aguda da esquistossomose,
caracterizando um processo de eosinofilia nos animais deste grupo. A eosinofilia
acompanhada ou não por leucocitose é característica da fase aguda da esquistossomose (Lenzi
et al., 1987, Atta, 1981). Os animais com 7 semanas de infecção também não apresentaram
alterações nos níveis de leucócitos globais. Por outro lado, observa-se nesta fase uma
diminuição significativa do número de eosinófilos. Diminuição dos níveis de leucócitos,
particularmente eosinófilos, está comumente associada à fase crônica da esquistossomose
(Vitorino RR, , 2012).
Ensaios para marcadores clássicos de danos hepáticos como a determinação das
atividades das transaminases ALT e AST, não foram suficientes para a discriminação de fase
aguda e crônica da doença. Os valores absolutos de atividade enzimática demonstraram que
os níveis de ALT permaneceram sem grandes alterações nos dois grupos de animais
infectados, enquanto os níveis de AST diminuíram; o que contribuiu para o aumento da razão
ALT/AST. Por outro lado, a dosagem de albumina sérica mostrou-se útil na identificação de
animais portadores de fase crônica da doença. Este efeito pode ser explicado pelo
comprometimento da função hepática nesta fase, devido ao intenso processo inflamatório
associado (Oliveira et al., 2008, Wu et al., 2010b). No intuito de avaliar as alterações teciduais
no fígado e baço dos animais infectados, análises histológicas foram realizadas para a
confirmação das duas fases distintas da infecção. Os animais com 5 semanas de infecção
apresentaram significativas alterações hepáticas, revelando um processo inflamatório
moderado ao redor da tríade portal e da veia-centrolobular. Sabe-se que a inflamação
periportal pode ser observada a partir da 3ª semana de infecção (Atta, 1981). Para o grupo
com 7 semanas de infecção observou-se um acentuado processo inflamatório periportal. Neste
grupo foi verificado aumento de 2 vezes no número de células inflamatórias na região
perivascular, em comparação aos animais com 5 semanas de infecção. A presença de vermes
adultos pareados foi observada nos ramos da veia porta de alguns animais, tanto no grupo
com 5 semanas de infecção quanto no grupo com 7 semanas de infecção. Ovos imaturos de S.

55
mansoni foram encontrados no parênquima dos animais com 5 semanas de infecção. Ao
serem liberados da fêmea, os ovos de S. mansoni são imaturos e continuam a se desenvolver
ainda no interior do hospedeiro vertebrado até a completa formação do miracídio (Jurberg et
al., 2009, Dewalick et al., 2011). Ovos imaturos não induzem grandes alterações teciduais,
mas após totalmente desenvolvidos estes passam a liberar secreções antigênicas solúveis
(SEA) as quais são responsáveis pelo aumento do processo inflamatório e consequente
formação de granulomas periovulares (Andrade, 2009). O fígado de animais com 7 semanas
de infecção apresentaram algumas regiões fibrosadas, nas quais ovos não foram detectados,
em consequência da resolução dos granulomas. A reação granulomatosa inicial caracteriza-se
pela presença de macrófagos, linfócitos, eosinófilos e acúmulo de colágenos dos tipos I e III
(Andrade, 2004, Silva et al., 2006). Com o passar do tempo esta reação inflamatória torna-se
fibrótica, acumulando-se no parênquima do fígado e em regiões periportais, resultando, entre
outras complicações, em hipertensão portal característica da fase crônica da esquistossomose
mansônica (Abdulla et al., 2011, Warren, 1968).
Para os animais com 5 semanas de infecção, as análises histológicas de baço revelaram
hipertrofia nodular e hiperplasia da polpa branca, restrita à região folicular. A medida da área
nodular no baço destes animais demonstrou aumento de 100% em relação ao controle, sendo
este aumento característico de nódulos sensibilizados em resposta a antígenos. Para os
animais com 7 semanas de infecção, não foi possível determinar com precisão a área nodular,
devido à intensa migração de linfócitos provenientes dos nódulos foliculares para a região de
polpa vermelha, ocasionando perda estrutural da organização folicular. Fatores diversos como
a hipertensão portal, a presença de granulomas, o alargamento de sinusóides esplênicos e a
congestão da polpa vermelha contribuem para a instalação do quadro de esplenomegalia
(Freitas et al., 1999). Coletivamente, nossos resultados permitiram concluir que os animais
com 5 semanas de infecção apresentaram quadro patológico característico de fase aguda
enquanto os animais infectados por 7 semanas apresentaram alterações celulares e teciduais
compatíveis com a fase crônica da esquistossomose.
Decorrido as etapas de diferenciação entre os grupos com infecção aguda e crônica,
foram realizadas análises proteômicas de fígado e de baço, com o intuito de determinar as
alterações no proteoma solúvel presente na fase aguda e na fase crônica da doença. A técnica
de eletroforese bidimensional demonstrou várias alterações nos níveis de expressão de
proteínas associadas com processos e vias bioquímicas diversas como resposta ao estresse
celular, tradução, ciclo da uréia, via glicolítica, entre outros.

56
Particularmente, o proteoma do fígado de animais infectados apresentou tanto
aumento quanto diminuição na abundância de algumas proteínas. Estas diferenças foram mais
evidentes no perfil bidimensional dos animais com 7 semanas de infecção, onde o dano
hepático é significativamente maior. De um total de 74 spots pré-selecionados para a
identificação por espectrometria de massas, 14 apresentaram consideráveis alterações de
abundância e foram detectadas nos animais de 5 e 7 semanas de infecção. O emprego de géis
de 7 cm pH 3-10 e pH 4-7 não impediu com que houvesse confluência de bandas para
proteínas com massa molecular e ponto isoelétrico próximos. Isto resultou, na maioria dos
casos, na identificação de mais de uma proteína por spot.
Dentre as proteínas que tiveram sua expressão aumentada no fígado dos animais
infectados, as mais abundantes são chaperoninas envolvidas com estresse no retículo
endoplasmático, proteínas carreadoras, metabolismo energético e via glicolítica. As
chaperonas identificadas apresentaram aumento de expressão, em ambos os grupos de animais
infectados. Estas moléculas constituem um grupo de proteínas que auxiliam no enovelamento
proteico, apresentação de antígeno, e estão diretamente envolvidas com o estresse celular,
termotolerância e renovação de proteínas via degradação pelo proteassoma (Hartl and Hayer-
Hartl, 2002, Moser et al., 1990). O retículo endoplasmático é uma organela intimamente
ligada com o processo de enovelamento de proteínas recém-sintetizadas, e dentre as
chaperonas que medeiam o processo de enovelamento de proteínas nesta organela estão:
GRP-94 (proteína reguladora de glicose 94 kDa); GRP-78 ou BIP (proteína reguladora de
glicose 78 kDa); PDI (proteína dissulfeto isomerase) e calreticulina (Gupta and Tuteja, 2011).
A proteína GRP-94, conhecida como endoplasmina, é uma proteína da família HSP-90,
constituída por proteínas que se ligam ao cálcio e estão intimamente envolvidas com a
manutenção da atividade do retículo endoplasmático durante o estresse celular. A proteína
GRP-94 apresentou aumento de 111% nos animais com 5 semanas de infecção e 60% nos
animais com 7 semanas de infecção. A princípio, o aumento significativo desta proteína nos
animais com 5 semanas de infecção pode estar relacionado ao início de uma resposta imune
do hospedeiro, contra a infecção pelo parasito. Com a evolução da doença e o aumento do
dano hepático, os níveis desta proteína tendem a diminuir. Estudos demonstraram que a GRP-
94 tem um papel importante na supressão de tumores (Baker-LePain et al., 2004), e,
juntamente com outras proteínas como a BIP, participam do enovelamento de
imunoglobulinas recém-sintetizadas, secreção de proteínas e apresentação de antígenos via
MHC classe I (Melnick et al., 1992). Proteínas GRP-94 e BIP promovem autofosforilação

57
dependente de ATP, regulando os processos de interação destas chaperonas com seus alvos
(Csermely et al., 1995).
As proteínas da família HSP-70 também apresentaram aumento de expressão em
decorrência da infecção por S. mansoni. As proteínas BIP e HSP-71 (proteína de choque
térmico 71 kDa) tiveram aumento significativo de expressão, chegando a 79% para a proteína
BIP e 43% para HSP-71 nos animais com 7 semanas de infecção. A proteína BIP se mostrou
significativamente aumentada, já nos animais com 5 semanas de infecção. Estudos revelaram
que a proteína BIP está intimamente ligada ao transporte de proteínas destinadas à degradação
via proteassoma. Sua síntese em altos níveis está relacionada à depleção de Ca2+
e a condições
de estresse, em paralelo ao acúmulo de proteínas não enoveladas no retículo endoplasmático
(Gething, 1999). As proteínas da família HSP-70 são capazes de interagir com reguladores de
transdução de sinais, juntamente com proteínas da família HSP-90, controlando a homeostase
e proliferação celular (Mayer and Bukau, 2005, Takashima et al., 2003). A super expressão
destas proteínas induz resistência à apoptose celular, enquanto que a baixa expressão aumenta
a sensibilidade a vários danos celulares (Jaattela et al., 1998).
Outras chaperonas como a calreticulina e a proteína dissulfeto isomerase (PDI)
também apresentaram significativo aumento nos animais infectados. Estas proteínas
desempenham papel importante no enovelamento de outras proteínas e em vias de sinalização
no retículo endoplasmático. A calreticulina é uma proteína cuja expressão aumenta no lúmen
do retículo endoplasmático em condições de estresse. Além disso, está envolvida com o
transporte de peptídeos nas células apresentadoras de antígenos auxiliando na resposta imune
contra células tumorais (Bak et al., 2008). A PDI é uma enzima que catalisa a formação de
ligações dissulfeto e, portanto, apresenta papel crucial no enovelamento de proteínas
destinadas à exportação, participa de processos redox, sinalização e homeostase celular
(Laurindo et al., 2012).
Outras proteínas que tiveram sua abundância aumentada no fígado dos animais
infectados por S. mansoni, foram a transferrina e a triose-fosfato isomerase. Duas isoformas
de transferrinas foram identificadas com alteração de expressão no fígado dos animais
infectados. De todas as proteínas que tiveram aumento de expressão no fígado, a transferrina
foi a que apresentou a maior alteração de abundância, passando do aumento de 60% nos
animais infectados por 5 semanas e culminando com aumento de 200% nos animais com 7
semanas de infecção. Estudos clássicos de padronização de cultivo in vitro de
esquistossômulos jovens demonstraram que a suplementação do meio com transferrina

58
estimula o desenvolvimento do verme especialmente durante as primeiras duas semanas e,
desta forma, justifica a necessidade de utilização de ferro pelo parasito (Clemens and Basch,
1989). A triose-fosfato isomerase participa da via glicolítica na conversão de diidroxiacetona
fosfato em gliceraldeído 3 fosfato apresentou expressão aumentada em até 80% nos animais
com 5 semanas de infecção e manteve aumento significativo de expressão (40%) com 7
semanas de infecção. Estudos realizados em fêmeas de camundongos da linhagem C57BL/6,
infectados com 45 cercárias de S. mansoni, demonstraram que proteínas de choque térmico da
família HSP-70, HSP-90, calreticulina, PDI e transferrina, tiveram expressão aumentada após
20 semanas de infecção. Estas alterações podem estar relacionadas com o aumento na síntese
de proteínas, acúmulo de proteínas não enoveladas e estresse oxidativo (Manivannan et al.,
2010).
As proteínas que apresentaram expressão diminuída nos animais infectados foram
CPSase I (carbamoil fosfato sintetase I), albumina, indoletilamina N-metiltransferase,
peroxirredoxina-6 e duas isoformas de anidrase carbônica III. A proteína CPSase I participa
do ciclo da uréia atuando na conversão de ornitina a citrulina. Esta proteína constituiu o spot
de maior volume da fração citosólica presente nos géis 2D. Observou-se uma diminuição de
expressão de 24% nos animais com 5 semanas, e de 57% nos animais com 7 semanas de
infecção. Diminuições da atividade enzimática de CPSase I e de várias outras enzimas
relacionadas ao ciclo da uréia foram previamente descritas para a infecção por S. mansoni no
modelo murino e, mais recentemente, confirmadas por análise proteômica (Tanabe et al.,
1989). Estudos de metabolômica realizados em amostras de urina de camundongos infectados
por S. mansoni revelaram intensa diminuição nos níveis de intermediários do ciclo do ácido
cítrico que participam como substratos no ciclo da uréia como o citrato e succinato (Wang et
al., 2004, Wu et al., 2010a). O mecanismo pelo qual a infecção por S. mansoni induz
diminuição de expressão de CPSase I ainda permanece por ser elucidado, no entanto, foi
demonstrado que o extrato solúvel de ovos administrado via intraperitoneal é capaz de induzir
efeito semelhante e, portanto, na ausência de alterações histopatológicas provocadas pela
formação de granulomas (Tanabe et al., 1989).
A enzima indoletilamina n-metiltransferase é uma proteína que catalisa a N-metilação
de triptaminas e outros compostos resultando com freqüência na inativação de suas atividades
biológicas (Thompson et al., 1999). A avaliação proteômica do fígado demonstrou que os
níveis desta proteína apresentam-se significativamente diminuídos após 5 semanas de
infecção e permanecem praticamente inalterados até a 7ª semana de infecção. A

59
indoletilamina n-metil transferase tem sido envolvida na participação de sinalização através
de metilação de triptaminas e em processo de desintoxicação no metabolismo de xenobióticos
(Ansher and Jakoby, 1986).
Proteínas com atividades antioxidantes também demonstraram expressão reduzida em
animais infectados. Diminuição na abundância de peroxiredoxina-6 foi verificada nos animais
com 5 e 7 semanas de infecção, no entanto, a diminuição nos níveis de peroxiredoxinas foi
pronunciada nos animais com 7 semanas de infecção, chegando a uma diminuição de 86%.
Peroxiredoxinas possuem um papel importante na detoxificação celular e em processos de
estresse oxidativo, através da eliminação de peróxidos de hidrogênio, desta forma, atuando na
prevenção de danos causados por radicais livres (Ishii et al., 2012). Estudos proteômicos de
fígado de camundongos infectados com S. mansoni, demonstraram que os níveis de
peroxiredoxinas 1 e 6 decrescem significativamente em animais infectados durante 8 e 12
semanas (Manivannan et al., 2011). Duas isoformas de anidrase carbônica III também
apresentaram diminuição de abundância nos animais infectados. A diminuição da expressão
dessas proteínas é notada já nos animais com 5 semanas de infecção, tornando-se mais
expressiva nos animais com 7 semanas de infecção (88% de redução). Anidrases carbônicas
pertencem à classe das metaloenzimas e catalisam a reação reversível de hidratação de
dióxido de carbono a ácido carbônico (Yamamoto et al., 2006, Breton, 2001). Diferentemente
das outras anidrases, a anidrase carbônica III, se distingue pelo fato de apresentar baixa
atividade de hidratase, é encontrada em altos níveis no fígado e músculos e apresenta
atividade de tirosina fosfatase através da qual desempenha papel importante na sinalização
ligada à resposta ao estresse oxidativo (Raisanen et al., 1999). A anidrase carbônica III possui
dois grupos sulfidrilas reativos em sua estrutura, capazes de formar ligações dissulfeto com a
molécula de glutationa o que contribui para a regulação de sua atividade (Thomas et al.,
1995). O mecanismo pelo qual a infecção por S. mansoni diminui a expressão de enzimas
ligadas à resposta ao estresse permanece por ser esclarecido. Por outro lado, levando-se em
consideração que a resposta celular do sistema imune contra a infecção está primariamente
baseada no recrutamento de células produtoras de espécies reativas de oxigênio, a baixa
expressão de enzimas antioxidantes poderia contribuir para a eficácia de uma resposta anti-
parasitária.
A segunda abordagem deste trabalho consistiu na avaliação de alterações no proteoma
citosólico do baço, nos animais com 5 e 7 semanas de infecção. De um total de 60 spots
selecionados nos géis bidimensionais para identificação por espectrometria de massas, 7 spots

60
apresentaram expressivas alterações de abundância no baço de animais infectados. As
proteínas que apresentaram alteração no baço foram calreticulina, fosfoglicerato quinase I,
frutose-bifosfato aldolase A, gliceraldeído 3 fosfato desidrogenase, fator de elongamento EF-
Tu e aspartato aminotransferase.
A calreticulina, pertencente ao grupo das chaperoninas, apresentou aumento discreto
nos animais com 5 semanas de infecção. Por outro lado, observou-se aumento superior a
200% para esta proteína nos animais com 7 semanas de infecção. A calreticulina é uma
proteína multifuncional pois está envolvida na captação de Ca2+
, enovelamento de proteínas,
apoptose e vias de sinalização celular (Harr and Distelhorst, 2010). Um dos principais fatores
ligados ao aumento de sua expressão está relacionado com alterações na homeostase do cálcio
e situações de estresse no retículo endoplasmático (Song et al., 2009).
Particularmente, a proteína de maior abundância encontrada no baço dos animais
infectados, foi o fator de alongamento EF-Tu. Este teve sua expressão aumentada em mais de
100% no baço dos animais com 5 semanas de infecção e em torno de 800% nos animais com
7 semanas de infecção. O EF-Tu tem o papel de se ligar à molécula de GTP formando um
complexo com o RNA transportador ligado ao aminoácido, o que propicia a ligação do RNA
transportador ao sítio A do ribossomo (Noble and Song, 2008). O aumento da abundância
desta proteína pode estar relacionado com a demanda aumentada de síntese proteica frente à
intensa resposta celular proliferativa do baço contra a infecção.
Proteínas relacionadas com o metabolismo energético como a fofosglicerato quinase I,
frutose bifosfato aldolase A, gliceraldeído 3 fosfato desidrogenase e aspartato
aminotransferase também apresentaram níveis aumentados de expressão no baço dos animais
infectados. É válido ressaltar o aumento superior a 600% para a enzima gliceraldeído 3-
fosfato desidrogenase nos animais com 7 semanas de infecção. O aumento de expressão de
proteínas ligadas ao metabolismo da glicose pode estar relacionado à demanda de produção de
energia para a multiplicação celular no baço em resposta aos estímulos antigênicos.
Avaliações metabólicas em urina de camundongos apresentaram aumento nos níveis de
piruvato, provavelmente refletindo intensa atividade da via glicolítica no fígado e baço destes
animais (Wang et al., 2004).
Outro ponto que merece atenção refere-se à identificação de citoqueratinas no fígado
e, especialmente, no baço de animais controles e infectados. As citoqueratinas compõem o
citoesqueleto celular sendo constituintes dos filamentos intermediários. A distribuição das
citoqueratinas é tecido-específica, o que tem permitido utilizá-las como importantes

61
marcadores de diferenciação tecidual (Almeida.Jr, 2004). Vários tipos de citoqueratinas foram
encontrados tanto no fígado quanto no baço dos animais infectados. Dentre elas, a CQ-8 foi
identificada com frequência em vários spots selecionados para espectrometria de massas.
(vide figura suplementar 2, em anexos). As diferentes isoformas de citoqueratinas
encontradas nos géis do fígado e do baço apresentaram pI distintos dos valores teóricos
sugerindo a existência de modificações pós traducionais. Neste trabalho não foi possível
correlacionarmos a presença de uma determinada variante de citoqueratina com a evolução da
infecção por S. mansoni, em virtude da confluência de spots para a todas as isoformas
identificadas. Entretanto, tem sido demonstrado que a citoqueratina 18 é diferencialmente
expressa no fígado de animais infectados por S. mansoni e sua presença no soro tem sido
avaliada como um potencial marcador de infecção por este parasito . Espera-se que a
confecção de géis com maior poder de resolução permita a identificação de variantes de
citoqueratina produzidas em resposta aos danos hepáticos provocados pela esquistossomose.
O estudo proteômico da fração solúvel de baço e de fígado dos animais com fase
aguda e fase crônica da doença revelou várias alterações na abundância de proteínas. No
fígado as alterações moleculares são compatíveis com o perfil histológico observado e
provavelmente explica o aumento de expressão para proteínas chaperoninas, proteínas
carreadoras e enzimas ligadas ao metabolismo energético, constituindo um perfil molecular
direcionado à montagem de uma resposta celular no combate à infecção. Em paralelo,
proteínas ligadas a funções hepáticas normais como detoxificação e resposta ao estresse
oxidativo apresentaram níveis diminuídos. Estes resultados vão de encontro ao extenso dano
hepático e perda de função tecidual em decorrência da cronicidade da infecção. No baço as
modificações caracterizaram-se por aumento de proteínas envolvidas com a síntese proteica,
enovelamento e metabolismo energético. Novamente, a análise histológica do baço é
compatível com o perfil molecular identificado e está provavelmente relacionado à intensa
proliferação celular resultando em esplenomegalia exacerbada na fase crônica da infecção.
Grande parte das alterações proteômicas hepáticas identificadas neste trabalho foram
descritas para camundongos da linhagem CBA/J infectados com 45 e 200 cercárias de S.
mansoni (Harvie et al., 2007, Manivannan et al., 2010, Manivannan et al., 2011) A estratégia
experimental utilizada mais recentemente envolveu marcação das proteínas com Cydyes
seguido da técnica de DIGE (Differential in gel electrophoresis) (Manivannan et al., 2012).
Contudo, nossos resultados demonstraram que a técnica clássica de separação bidimensional é
capaz de fornecer resultados equivalentes, embora esteja associada à maior tempo para sua

62
execução. Independentemente da técnica utilizada, o maior desafio ainda consiste no
fracionamento das amostras a serem comparadas de modo a suprimir o efeito das proteínas de
alta abundância, permitindo a identificação de biomarcadores mais específicos para a
infecção. Além disso, em face das alterações teciduais observadas é de fundamental
importância a análise detalhada do soroproteoma de animais infectados por S. mansoni. A
identificação de biomarcadores séricos poderá contribuir de maneira significativa para novos
métodos de diagnóstico, avaliação de tratamento e prognóstico da doença.

63
6 - Conclusões

64
6 – Conclusões
As análises bioquímicas, o exame parasitológico e as avaliações histopatológicas
confirmaram o desenvolvimento da fase aguda da esquistossomose nos animais
infectados por 5 semanas e o estabelecimento da fase crônica da doença com 7
semanas de infecção;
A análise proteômica da fração solúvel de fígado e baço, permitiu a identificação de
um conjunto de proteínas que sofreram alterações em sua expressão durante a infecção
por S. mansoni. A quantificação destas alterações possibilitou estabelecer quais
proteínas diferencialmente expressas durante as fases aguda e crônica da doença;
O fígado dos animais infectados apresentou aumento de expressão para proteínas
chaperoninas, proteínas carreadoras e enzimas ligadas ao metabolismo energético,
além de apresentar diminuição de proteínas envolvidas com processos de
detoxificação e resposta ao estresse oxidativo;
O baço dos animais infectados apresentou aumento de proteínas envolvidas com a
síntese proteica, enovelamento e metabolismo energético. No geral, todas as proteínas
do baço identificadas com alteração, apresentaram aumento de expressão em
comparação àquelas identificadas nos animais controles;
A identificação das alterações proteômicas de fígado e baço apontam para a
caracterização de novos marcadores para o diagnóstico e prognóstico da
esquistossomose mansônica.

65
7 - Referências

66
7-Referências
ABDUL-GHANI, R. A. & HASSAN, A. A. 2010. Murine schistosomiasis as a model for
human schistosomiasis mansoni: similarities and discrepancies. Parasitol Res, 107, 1-
8.
ABDULLA, M. H., LIM, K. C., MCKERROW, J. H. & CAFFREY, C. R. 2011. Proteomic
identification of IPSE/alpha-1 as a major hepatotoxin secreted by Schistosoma
mansoni eggs. PLoS Negl Trop Dis, 5, e1368.
ALAN WILSON, R., VAN DAM, G. J., KARIUKI, T. M., FARAH, I. O., DEELDER, A. M.
& COULSON, P. S. 2006. The detection limits for estimates of infection intensity in
schistosomiasis mansoni established by a study in non-human primates. Int J
Parasitol, 36, 1241-4.
ALMEIDA.JR 2004. Citoqueratinas. An bras Dermatol, Rio de Janeiro, 135-145.
ANDERSON, N. L. & ANDERSON, N. G. 1998. Proteome and proteomics: new
technologies, new concepts, and new words. Electrophoresis, 19, 1853-61.
ANDRADE, Z. A. 2004. Schistosomal hepatopathy. Mem Inst Oswaldo Cruz, 99, 51-7.
ANDRADE, Z. A. 2009. Schistosomiasis and liver fibrosis. Parasite Immunol, 31, 656-63.
ANSHER, S. S. & JAKOBY, W. B. 1986. Amine N-methyltransferases from rabbit liver. J
Biol Chem, 261, 3996-4001.
ATTA, A. M. E. A. 1981. Esquistossomose mansônica. I - Evolução do quadro patológico:
análise parasitológica, hematológica e histopatológica. Rev. Saúde públ., São Paulo,
72-79.
ATTALLAH, A. M., ISMAIL, H., EL MASRY, S. A., RIZK, H., HANDOUSA, A., EL
BENDARY, M., TABLL, A. & EZZAT, F. 1999. Rapid detection of a Schistosoma
mansoni circulating antigen excreted in urine of infected individuals by using a
monoclonal antibody. J Clin Microbiol, 37, 354-7.
BAK, S. P., AMIEL, E., WALTERS, J. J. & BERWIN, B. 2008. Calreticulin requires an
ancillary adjuvant for the induction of efficient cytotoxic T cell responses. Mol
Immunol, 45, 1414-23.
BAKER-LEPAIN, J. C., SARZOTTI, M. & NICCHITTA, C. V. 2004. Glucose-regulated
protein 94/glycoprotein 96 elicits bystander activation of CD4+ T cell Th1 cytokine
production in vivo. J Immunol, 172, 4195-203.
BALOG, C. I., MEISSNER, A., GORALER, S., BLADERGROEN, M. R., VENNERVALD,
B. J., MAYBORODA, O. A. & DEELDER, A. M. 2011. Metabonomic investigation
of human Schistosoma mansoni infection. Mol Biosyst, 7, 1473-80.

67
BECKMANN, S., QUACK, T., BURMEISTER, C., BURO, C., LONG, T., DISSOUS, C. &
GREVELDING, C. G. 2010. Schistosoma mansoni: signal transduction processes
during the development of the reproductive organs. Parasitology, 137, 497-520.
BERRIMAN, M., HAAS, B. J., LOVERDE, P. T., WILSON, R. A., DILLON, G. P.,
CERQUEIRA, G. C., MASHIYAMA, S. T., AL-LAZIKANI, B., ANDRADE, L. F.,
ASHTON, P. D., ASLETT, M. A., BARTHOLOMEU, D. C., BLANDIN, G.,
CAFFREY, C. R., COGHLAN, A., COULSON, R., DAY, T. A., DELCHER, A.,
DEMARCO, R., DJIKENG, A., EYRE, T., GAMBLE, J. A., GHEDIN, E., GU, Y.,
HERTZ-FOWLER, C., HIRAI, H., HIRAI, Y., HOUSTON, R., IVENS, A.,
JOHNSTON, D. A., LACERDA, D., MACEDO, C. D., MCVEIGH, P., NING, Z.,
OLIVEIRA, G., OVERINGTON, J. P., PARKHILL, J., PERTEA, M., PIERCE, R. J.,
PROTASIO, A. V., QUAIL, M. A., RAJANDREAM, M. A., ROGERS, J., SAJID,
M., SALZBERG, S. L., STANKE, M., TIVEY, A. R., WHITE, O., WILLIAMS, D.
L., WORTMAN, J., WU, W., ZAMANIAN, M., ZERLOTINI, A., FRASER-
LIGGETT, C. M., BARRELL, B. G. & EL-SAYED, N. M. 2009. The genome of the
blood fluke Schistosoma mansoni. Nature, 460, 352-8.
BODZON-KULAKOWSKA, A., BIERCZYNSKA-KRZYSIK, A., DYLAG, T., DRABIK,
A., SUDER, P., NOGA, M., JARZEBINSKA, J. & SILBERRING, J. 2007. Methods
for samples preparation in proteomic research. J Chromatogr B Analyt Technol
Biomed Life Sci, 849, 1-31.
BRASCHI, S., CURWEN, R. S., ASHTON, P. D., VERJOVSKI-ALMEIDA, S. & WILSON,
A. 2006. The tegument surface membranes of the human blood parasite Schistosoma
mansoni: a proteomic analysis after differential extraction. Proteomics, 6, 1471-82.
BRASCHI, S. & WILSON, R. A. 2006. Proteins exposed at the adult schistosome surface
revealed by biotinylation. Mol Cell Proteomics, 5, 347-56.
BRENER, Z. & CHIARI, E. 1956. [Diagnosis of Manson's schistosomiasis by the Hoffman,
Pons and Janer method of feces examination]. Rev Bras Malariol Doencas Trop, 8,
599-603.
BRETON, S. 2001. The cellular physiology of carbonic anhydrases. JOP, 2, 159-64.
CAMPBELL, G., JONES, C. S., LOCKYER, A. E., HUGHES, S., BROWN, D., NOBLE, L.
R. & ROLLINSON, D. 2000. Molecular evidence supports an african affinity of the
neotropical freshwater gastropod, Biomphalaria glabrata, say 1818, an intermediate
host for Schistosoma mansoni. Proc Biol Sci, 267, 2351-8.
CASTRO-BORGES, W., DOWLE, A., CURWEN, R. S., THOMAS-OATES, J. & WILSON,
R. A. 2011a. Enzymatic shaving of the tegument surface of live schistosomes for
proteomic analysis: a rational approach to select vaccine candidates. PLoS Negl Trop
Dis, 5, e993.
CASTRO-BORGES, W., SIMPSON, D. M., DOWLE, A., CURWEN, R. S., THOMAS-
OATES, J., BEYNON, R. J. & WILSON, R. A. 2011b. Abundance of tegument
surface proteins in the human blood fluke Schistosoma mansoni determined by
QconCAT proteomics. J Proteomics, 74, 1519-33.

68
CHITSULO, L., LOVERDE, P. & ENGELS, D. 2004. Schistosomiasis. Nat Rev Microbiol, 2,
12-3.
CLEMENS, L. E. & BASCH, P. F. 1989. Schistosoma mansoni: effect of transferrin and
growth factors on development of schistosomula in vitro. Journal of Parasitology, 75,
417-21.
COLLINS, J. J., 3RD, KING, R. S., COGSWELL, A., WILLIAMS, D. L. & NEWMARK, P.
A. 2011. An atlas for Schistosoma mansoni organs and life-cycle stages using cell
type-specific markers and confocal microscopy. PLoS Negl Trop Dis, 5, e1009.
COTTRELL, J. S. 2011. Protein identification using MS/MS data. J Proteomics, 74, 1842-51.
CSERMELY, P., MIYATA, Y., SCHNAIDER, T. & YAHARA, I. 1995.
Autophosphorylation of grp94 (endoplasmin). J Biol Chem, 270, 6381-8.
CURWEN, R. S., ASHTON, P. D., JOHNSTON, D. A. & WILSON, R. A. 2004. The
Schistosoma mansoni soluble proteome: a comparison across four life-cycle stages.
Mol Biochem Parasitol, 138, 57-66.
CURWEN, R. S., ASHTON, P. D., SUNDARALINGAM, S. & WILSON, R. A. 2006.
Identification of novel proteases and immunomodulators in the secretions of
schistosome cercariae that facilitate host entry. Mol Cell Proteomics, 5, 835-44.
DE OLIVEIRA, F. M., DA SILVA, I. C., RUMJANEK, F. D., VALADAO, A. F., FRANCO,
G. R., MESQUITA, R. D., DA SILVA-NETO, M. A. & FANTAPPIE, M. R. 2004.
Functional properties of Schistosoma mansoni single-stranded DNA-binding protein
SmPUR-alpha. Description of the interaction between SmPUR-alpha and SMYB1.
Mol Biochem Parasitol, 135, 21-30.
DEWALICK, S., TIELENS, A. G. & VAN HELLEMOND, J. J. 2011. Schistosoma mansoni:
The egg, biosynthesis of the shell and interaction with the host. Exp Parasitol.
FAVRE, T. C., PEREIRA, A. P., GALVAO, A. F., ZANI, L. C., BARBOSA, C. S. & PIERI,
O. S. 2009. A rationale for schistosomiasis control in elementary schools of the
rainforest zone of pernambuco, Brazil. PLoS Negl Trop Dis, 3, e395.
FERRARI, T. C. & MOREIRA, P. R. 2011. Neuroschistosomiasis: clinical symptoms and
pathogenesis. Lancet Neurol, 10, 853-64.
FREITAS, C. R., BARBOSA, A. A., JR., FERNANDES, A. L. & ANDRADE, Z. A. 1999.
Pathology of the spleen in hepatosplenic schistosomiasis. Morphometric evaluation
and extracellular matrix changes. Mem Inst Oswaldo Cruz, 94, 815-22.
FREITAS, T. C., JUNG, E. & PEARCE, E. J. 2007. TGF-beta signaling controls embryo
development in the parasitic flatworm Schistosoma mansoni. PLoS Pathog, 3, e52.
GETHING, M. J. 1999. Role and regulation of the ER chaperone BiP. Semin Cell Dev Biol,
10, 465-72.

69
GORG, A., DREWS, O., LUCK, C., WEILAND, F. & WEISS, W. 2009. 2-DE with IPGs.
Electrophoresis, 30 Suppl 1, S122-32.
GUPTA, D. & TUTEJA, N. 2011. Chaperones and foldases in endoplasmic reticulum stress
signaling in plants. Plant Signal Behav, 6, 232-6.
HAAS, W. & HAEBERLEIN, S. 2009. Penetration of cercariae into the living human skin:
Schistosoma mansoni vs. Trichobilharzia szidati. Parasitol Res, 105, 1061-6.
HARR, M. W. & DISTELHORST, C. W. 2010. Apoptosis and autophagy: decoding calcium
signals that mediate life or death. Cold Spring Harb Perspect Biol, 2, a005579.
HARTL, F. U. & HAYER-HARTL, M. 2002. Molecular chaperones in the cytosol: from
nascent chain to folded protein. Science, 295, 1852-8.
HARVIE, M., JORDAN, T. W. & LA FLAMME, A. C. 2007. Differential liver protein
expression during schistosomiasis. Infect Immun, 75, 736-44.
HENDERSON, G. S., NIX, N. A., MONTESANO, M. A., GOLD, D., FREEMAN, G. L.,
JR., MCCURLEY, T. L. & COLLEY, D. G. 1993. Two distinct pathological
syndromes in male CBA/J inbred mice with chronic Schistosoma mansoni infections.
Am J Pathol, 142, 703-14.
HOFFMAN WA, P. J., JANER JL 1934. Sedimentation concentration method in
Schistosomiasis mansoni. J Publ Hlth, 283-298.
HOTEZ, P. J., BRINDLEY, P. J., BETHONY, J. M., KING, C. H., PEARCE, E. J. &
JACOBSON, J. 2008. Helminth infections: the great neglected tropical diseases. J
Clin Invest, 118, 1311-21.
HOTEZ, P. J., MOLYNEUX, D. H., FENWICK, A., KUMARESAN, J., SACHS, S. E.,
SACHS, J. D. & SAVIOLI, L. 2007. Control of neglected tropical diseases. N Engl J
Med, 357, 1018-27.
ISHII, T., WARABI, E. & YANAGAWA, T. 2012. Novel roles of peroxiredoxins in
inflammation, cancer and innate immunity. J Clin Biochem Nutr, 50, 91-105.
JAATTELA, M., WISSING, D., KOKHOLM, K., KALLUNKI, T. & EGEBLAD, M. 1998.
Hsp70 exerts its anti-apoptotic function downstream of caspase-3-like proteases.
EMBO J, 17, 6124-34.
JURBERG, A. D., GONCALVES, T., COSTA, T. A., DE MATTOS, A. C., PASCARELLI,
B. M., DE MANSO, P. P., RIBEIRO-ALVES, M., PELAJO-MACHADO, M.,
PERALTA, J. M., COELHO, P. M. & LENZI, H. L. 2009. The embryonic
development of Schistosoma mansoni eggs: proposal for a new staging system. Dev
Genes Evol, 219, 219-34.
KANAMURA, H. Y., SILVA, R. M., CHIODELLI, S. G., GLASSER, C. M. & DIAS, L. C.
2002. IgM-immunofluorescence test as a diagnostic tool for epidemiologic studies of
Schistosomiasis in low endemic areas. Mem Inst Oswaldo Cruz, 97, 485-9.

70
KASINATHAN, R. S., GORONGA, T., MESSERLI, S. M., WEBB, T. R. & GREENBERG,
R. M. 2010. Modulation of a Schistosoma mansoni multidrug transporter by the
antischistosomal drug praziquantel. FASEB J, 24, 128-35.
KATZ, N. 2008. The discovery of Schistosomiasis mansoni in Brazil. Acta Trop, 108, 69-71.
KATZ, N., CHAVES, A. & PELLEGRINO, J. 1972. A simple device for quantitative stool
thick-smear technique in Schistosomiasis mansoni. Rev Inst Med Trop Sao Paulo, 14,
397-400.
KEBARLE, P. & VERKERK, U. H. 2009. Electrospray: from ions in solution to ions in the
gas phase, what we know now. Mass Spectrom Rev, 28, 898-917.
LAURINDO, F. R., PESCATORE, L. A. & FERNANDES, D. D. 2012. Protein disulfide
isomerase in redox cell signaling and homeostasis. Free Radic Biol Med.
LENZI, H. L., LENZI, J. A., KERR, I. B., ANTUNES, S. L., MOTA, E. M. & OLIVEIRA,
D. N. 1991. Extracellular matrix in parasitic and infectious diseases. Mem Inst
Oswaldo Cruz, 86 Suppl 3, 77-90.
LENZI, H. L., SOBRAL, A. C. & LENZI, J. A. 1987. "In vivo" kinetics of eosinophils and
mast cells in experimental murine schistosomiasis. Mem Inst Oswaldo Cruz, 82 Suppl
4, 67-76.
LIN, D. D., LIU, J. X., LIU, Y. M., HU, F., ZHANG, Y. Y., XU, J. M., LI, J. Y., JI, M. J.,
BERGQUIST, R., WU, G. L. & WU, H. W. 2008. Routine Kato-Katz technique
underestimates the prevalence of Schistosoma japonicum: a case study in an endemic
area of the People's Republic of China. Parasitol Int, 57, 281-6.
LOVERDE, P. T. & CHEN, L. 1991. Schistosome female reproductive development.
Parasitol Today, 7, 303-8.
MAHMOUD, A. A. 2004. Schistosomiasis (bilharziasis): from antiquity to the present. Infect
Dis Clin North Am, 18, 207-18.
MANIVANNAN, B., JORDAN, T. W., SECOR, W. E. & FLAMME, A. C. 2012. Proteomic
changes at 8 weeks after infection are associated with chronic liver pathology in
experimental schistosomiasis. J Proteomics, 75, 1838-48.
MANIVANNAN, B., RAWSON, P., JORDAN, T. W., KARANJA, D. M., MWINZI, P. N.,
SECOR, W. E. & LA FLAMME, A. C. 2011. Identification of cytokeratin 18 as a
biomarker of mouse and human hepatosplenic schistosomiasis. Infect Immun, 79,
2051-8.
MANIVANNAN, B., RAWSON, P., JORDAN, T. W., SECOR, W. E. & LA FLAMME, A.
C. 2010. Differential patterns of liver proteins in experimental murine hepatosplenic
schistosomiasis. Infect Immun, 78, 618-28.

71
MANZELLA, A., OHTOMO, K., MONZAWA, S. & LIM, J. H. 2008. Schistosomiasis of the
liver. Abdom Imaging.
MATHIESON, W., CASTRO-BORGES, W. & WILSON, R. A. 2011. The proteasome-
ubiquitin pathway in the Schistosoma mansoni egg has development- and
morphology-specific characteristics. Mol Biochem Parasitol, 175, 118-25.
MATHIESON, W. & WILSON, R. A. 2010. A comparative proteomic study of the
undeveloped and developed Schistosoma mansoni egg and its contents: the
miracidium, hatch fluid and secretions. Int J Parasitol, 40, 617-28.
MAYER, M. P. & BUKAU, B. 2005. Hsp70 chaperones: cellular functions and molecular
mechanism. Cell Mol Life Sci, 62, 670-84.
MELNICK, J., AVIEL, S. & ARGON, Y. 1992. The endoplasmic reticulum stress protein
GRP94, in addition to BiP, associates with unassembled immunoglobulin chains. J
Biol Chem, 267, 21303-6.
MOSER, D., DOUMBO, O. & KLINKERT, M. Q. 1990. The humoral response to heat shock
protein 70 in human and murine Schistosomiasis mansoni. Parasite Immunol, 12, 341-
52.
NEVES, J. 1992. [Acute or toxemic form of schistosomiasis mansoni]. Mem Inst Oswaldo
Cruz, 87 Suppl 4, 321-4.
NOBLE, C. G. & SONG, H. 2008. Structural studies of elongation and release factors. Cell
Mol Life Sci, 65, 1335-46.
O'FARRELL, P. H. 1975. High resolution two-dimensional electrophoresis of proteins. J Biol
Chem, 250, 4007-21.
OLIVEIRA, E. J., KANAMURA, H. Y. & LIMA, D. M. 2005. Efficacy of an enzyme-linked
immunosorbent assay as a diagnostic tool for schistosomiasis mansoni in individuals
with low worm burden. Mem Inst Oswaldo Cruz, 100, 421-5.
OLIVEIRA, S. A., SOUZA, B. S., GUIMARAES-FERREIRA, C. A., BARRETO, E. S.,
SOUZA, S. C., FREITAS, L. A., RIBEIRO-DOS-SANTOS, R. & SOARES, M. B.
2008. Therapy with bone marrow cells reduces liver alterations in mice chronically
infected by Schistosoma mansoni. World J Gastroenterol, 14, 5842-50.
PARKER-MANUEL, S. J., IVENS, A. C., DILLON, G. P. & WILSON, R. A. 2011. Gene
expression patterns in larval Schistosoma mansoni associated with infection of the
mammalian host. PLoS Negl Trop Dis, 5, e1274.
PONTES, L. A., DIAS-NETO, E. & RABELLO, A. 2002. Detection by polymerase chain
reaction of Schistosoma mansoni DNA in human serum and feces. Am J Trop Med
Hyg, 66, 157-62.

72
PROTASIO, A. V., TSAI, I. J., BABBAGE, A., NICHOL, S., HUNT, M., ASLETT, M. A.,
DE SILVA, N., VELARDE, G. S., ANDERSON, T. J., CLARK, R. C., DAVIDSON,
C., DILLON, G. P., HOLROYD, N. E., LOVERDE, P. T., LLOYD, C.,
MCQUILLAN, J., OLIVEIRA, G., OTTO, T. D., PARKER-MANUEL, S. J., QUAIL,
M. A., WILSON, R. A., ZERLOTINI, A., DUNNE, D. W. & BERRIMAN, M. 2012.
A systematically improved high quality genome and transcriptome of the human blood
fluke Schistosoma mansoni. PLoS Negl Trop Dis, 6, e1455.
RAISANEN, S. R., LEHENKARI, P., TASANEN, M., RAHKILA, P., HARKONEN, P. L. &
VAANANEN, H. K. 1999. Carbonic anhydrase III protects cells from hydrogen
peroxide-induced apoptosis. FASEB J, 13, 513-22.
RUSCONI, F., MANCINELLI, E., COLOMBO, G., CARDANI, R., DA RIVA, L.,
BONGARZONE, I., MEOLA, G. & ZIPPEL, R. 2010. Proteome profile in Myotonic
Dystrophy type 2 myotubes reveals dysfunction in protein processing and
mitochondrial pathways. Neurobiol Dis, 38, 273-80.
SAUNDERS, N., WILSON, R. A. & COULSON, P. S. 1987. The outer bilayer of the adult
schistosome tegument surface has a low turnover rate in vitro and in vivo. Mol
Biochem Parasitol, 25, 123-31.
SEIDLER, J., ZINN, N., BOEHM, M. E. & LEHMANN, W. D. 2010. De novo sequencing of
peptides by MS/MS. Proteomics, 10, 634-49.
SES/MS. 2010. ESQUISTOSSOMOSE MANSÔNICA [Online]. Available:
http://portalsaude.saude.gov.br [Accessed].
SILVA, L. M., RIBEIRO-DOS-SANTOS, R., SOARES, M. B. & ANDRADE, Z. A. 2006.
Characterization of the vascular changes in schistosomal portal (pipestem) fibrosis of
mice. Acta Trop, 98, 34-42.
SKELLY, P. J. & SHOEMAKER, C. B. 2000. Induction cues for tegument formation during
the transformation of Schistosoma mansoni cercariae. Int J Parasitol, 30, 625-31.
SONG, J., FINNERTY, C. C., HERNDON, D. N., BOEHNING, D. & JESCHKE, M. G.
2009. Severe burn-induced endoplasmic reticulum stress and hepatic damage in mice.
Mol Med, 15, 316-20.
SORGHO, H., BAHGAT, M., PODA, J. N., SONG, W., KIRSTEN, C., DOENHOFF, M. J.,
ZONGO, I., OUEDRAOGO, J. B. & RUPPEL, A. 2005. Serodiagnosis of
Schistosoma mansoni infections in an endemic area of Burkina Faso: performance of
several immunological tests with different parasite antigens. Acta Trop, 93, 169-80.
STEEN, H. & MANN, M. 2004. The ABC's (and XYZ's) of peptide sequencing. Nat Rev Mol
Cell Biol, 5, 699-711.
TAKASHIMA, M., KURAMITSU, Y., YOKOYAMA, Y., IIZUKA, N., TODA, T.,
SAKAIDA, I., OKITA, K., OKA, M. & NAKAMURA, K. 2003. Proteomic profiling
of heat shock protein 70 family members as biomarkers for hepatitis C virus-related
hepatocellular carcinoma. Proteomics, 3, 2487-93.

73
TANABE, M., KANEKO, N. & TAKEUCHI, T. 1989. Schistosoma mansoni: suppression of
carbamoyl phosphate synthetase (ammonia) and ornithine carbamoyltransferase
activities in the liver of infected mice. Exp Parasitol, 68, 432-42.
THOMAS, J. A., POLAND, B. & HONZATKO, R. 1995. Protein sulfhydryls and their role
in the antioxidant function of protein S-thiolation. Arch Biochem Biophys, 319, 1-9.
THOMPSON, M. A., MOON, E., KIM, U. J., XU, J., SICILIANO, M. J. &
WEINSHILBOUM, R. M. 1999. Human indolethylamine N-methyltransferase: cDNA
cloning and expression, gene cloning, and chromosomal localization. Genomics, 61,
285-97.
UTZINGER, J., N'GORAN E, K., CAFFREY, C. R. & KEISER, J. 2011. From innovation to
application: social-ecological context, diagnostics, drugs and integrated control of
schistosomiasis. Acta Trop, 120 Suppl 1, S121-37.
VAN LIESHOUT, L., PANDAY, U. G., DE JONGE, N., KRIJGER, F. W., OOSTBURG, B.
F., POLDERMAN, A. M. & DEELDER, A. M. 1995. Immunodiagnosis of
schistosomiasis mansoni in a low endemic area in Surinam by determination of the
circulating antigens CAA and CCA. Acta Trop, 59, 19-29.
VITORINO RR, S. F., COSTA AP E COL., 2012. Esquistossomose mansônica: diagnóstico,
tratamento, epidemiologia, profilaxia e controle. Rev Bras Clin Med, 10(1), 39-45.
WANG, Y., HOLMES, E., NICHOLSON, J. K., CLOAREC, O., CHOLLET, J., TANNER,
M., SINGER, B. H. & UTZINGER, J. 2004. Metabonomic investigations in mice
infected with Schistosoma mansoni: an approach for biomarker identification. Proc
Natl Acad Sci U S A, 101, 12676-81.
WARREN, K. S. 1968. Pathophysiology and pathogenesis of hepatosplenic schistosomiasis
mansoni. Bull N Y Acad Med, 44, 280-94.
WICHMANN, D., PANNING, M., QUACK, T., KRAMME, S., BURCHARD, G. D.,
GREVELDING, C. & DROSTEN, C. 2009. Diagnosing schistosomiasis by detection
of cell-free parasite DNA in human plasma. PLoS Negl Trop Dis, 3, e422.
WILSON, R. A., ASHTON, P. D., BRASCHI, S., DILLON, G. P., BERRIMAN, M. &
IVENS, A. 2007. 'Oming in on schistosomes: prospects and limitations for post-
genomics. Trends Parasitol, 23, 14-20.
WORLD HEALTH, O. 2010. Working to overcome the global impact of neglected tropical
diseases : first WHO report on neglected tropical diseases, Geneva, Department of
Reproductive health and Research, World Health Organization.
WU, J., XU, W., MING, Z., DONG, H., TANG, H. & WANG, Y. 2010a. Metabolic changes
reveal the development of schistosomiasis in mice. PLoS Negl Trop Dis, 4.
WU, J. F., HOLMES, E., XUE, J., XIAO, S. H., SINGER, B. H., TANG, H. R., UTZINGER,
J. & WANG, Y. L. 2010b. Metabolic alterations in the hamster co-infected with
Schistosoma japonicum and Necator americanus. Int J Parasitol, 40, 695-703.

74
YAMAMOTO, T., KIKKAWA, R., YAMADA, H. & HORII, I. 2006. Investigation of
proteomic biomarkers in in vivo hepatotoxicity study of rat liver: toxicity
differentiation in hepatotoxicants. J Toxicol Sci, 31, 49-60.
ZHOU, Y. B., YANG, M. X., TAO, P., JIANG, Q. L., ZHAO, G. M., WEI, J. G. & JIANG,
Q. W. 2008. A longitudinal study of comparison of the Kato-Katz technique and
indirect hemagglutination assay (IHA) for the detection of schistosomiasis japonica in
China, 2001-2006. Acta Trop, 107, 251-4.
75
8-Anexos
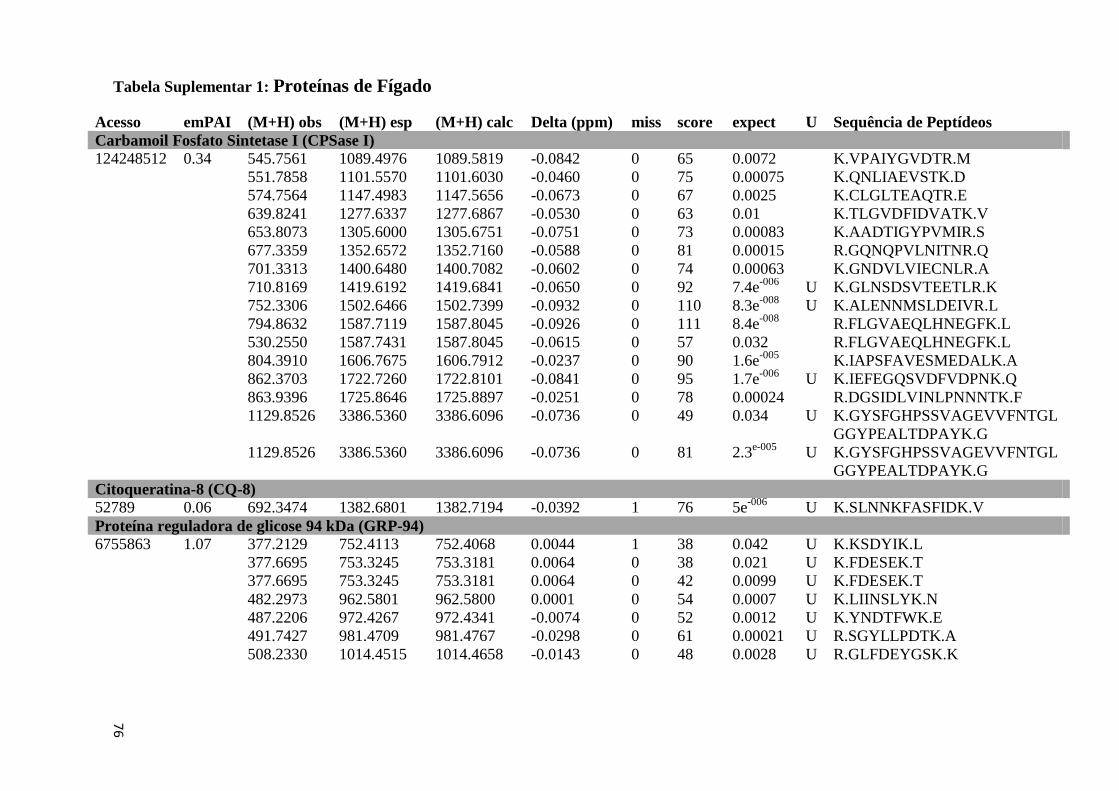
Tabela Suplementar 1: Proteínas de Fígado
Acesso emPAI (M+H) obs (M+H) esp (M+H) calc Delta (ppm) miss score expect U Sequência de Peptídeos
Carbamoil Fosfato Sintetase I (CPSase I)
124248512 0.34 545.7561 1089.4976 1089.5819 -0.0842 0 65 0.0072 K.VPAIYGVDTR.M
551.7858 1101.5570 1101.6030 -0.0460 0 75 0.00075 K.QNLIAEVSTK.D
574.7564 1147.4983 1147.5656 -0.0673 0 67 0.0025 K.CLGLTEAQTR.E
639.8241 1277.6337 1277.6867 -0.0530 0 63 0.01 K.TLGVDFIDVATK.V
653.8073 1305.6000 1305.6751 -0.0751 0 73 0.00083 K.AADTIGYPVMIR.S
677.3359 1352.6572 1352.7160 -0.0588 0 81 0.00015 R.GQNQPVLNITNR.Q
701.3313 1400.6480 1400.7082 -0.0602 0 74 0.00063 K.GNDVLVIECNLR.A
710.8169 1419.6192 1419.6841 -0.0650 0 92 7.4e-006
U K.GLNSDSVTEETLR.K
752.3306 1502.6466 1502.7399 -0.0932 0 110 8.3e-008
U K.ALENNMSLDEIVR.L
794.8632 1587.7119 1587.8045 -0.0926 0 111 8.4e-008
R.FLGVAEQLHNEGFK.L
530.2550 1587.7431 1587.8045 -0.0615 0 57 0.032 R.FLGVAEQLHNEGFK.L
804.3910 1606.7675 1606.7912 -0.0237 0 90 1.6e-005
K.IAPSFAVESMEDALK.A
862.3703 1722.7260 1722.8101 -0.0841 0 95 1.7e-006
U K.IEFEGQSVDFVDPNK.Q
863.9396 1725.8646 1725.8897 -0.0251 0 78 0.00024 R.DGSIDLVINLPNNNTK.F
1129.8526 3386.5360 3386.6096 -0.0736 0 49 0.034 U K.GYSFGHPSSVAGEVVFNTGL
GGYPEALTDPAYK.G
1129.8526 3386.5360 3386.6096 -0.0736 0 81 2.3e-005
U K.GYSFGHPSSVAGEVVFNTGL
GGYPEALTDPAYK.G
Citoqueratina-8 (CQ-8)
52789 0.06 692.3474 1382.6801 1382.7194 -0.0392 1 76 5e-006
U K.SLNNKFASFIDK.V
Proteína reguladora de glicose 94 kDa (GRP-94)
6755863 1.07 377.2129 752.4113 752.4068 0.0044 1 38 0.042 U K.KSDYIK.L
377.6695 753.3245 753.3181 0.0064 0 38 0.021 U K.FDESEK.T
377.6695 753.3245 753.3181 0.0064 0 42 0.0099 U K.FDESEK.T
482.2973 962.5801 962.5800 0.0001 0 54 0.0007 U K.LIINSLYK.N
487.2206 972.4267 972.4341 -0.0074 0 52 0.0012 U K.YNDTFWK.E
491.7427 981.4709 981.4767 -0.0298 0 61 0.00021 U R.SGYLLPDTK.A
508.2330 1014.4515 1014.4658 -0.0143 0 48 0.0028 U R.GLFDEYGSK.K
76
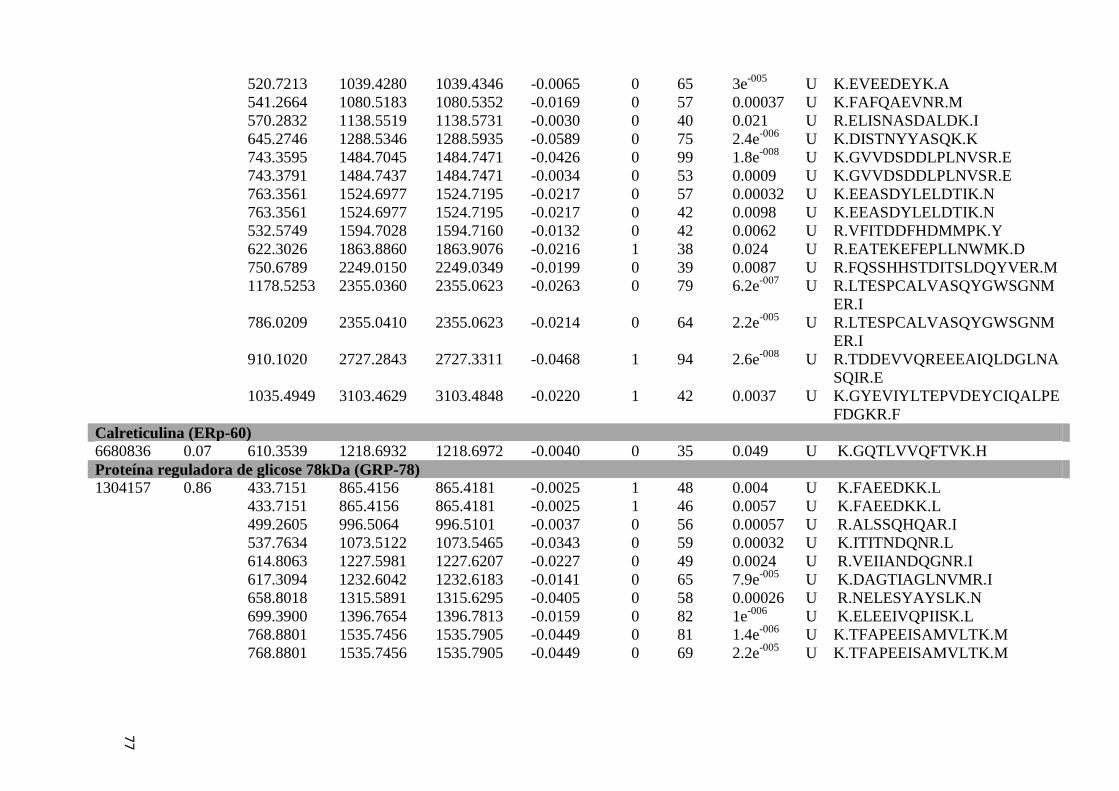
520.7213 1039.4280 1039.4346 -0.0065 0 65 3e-005
U K.EVEEDEYK.A
541.2664 1080.5183 1080.5352 -0.0169 0 57 0.00037 U K.FAFQAEVNR.M
570.2832 1138.5519 1138.5731 -0.0030 0 40 0.021 U R.ELISNASDALDK.I
645.2746 1288.5346 1288.5935 -0.0589 0 75 2.4e-006
U K.DISTNYYASQK.K
743.3595 1484.7045 1484.7471 -0.0426 0 99 1.8e-008
U K.GVVDSDDLPLNVSR.E
743.3791 1484.7437 1484.7471 -0.0034 0 53 0.0009 U K.GVVDSDDLPLNVSR.E
763.3561 1524.6977 1524.7195 -0.0217 0 57 0.00032 U K.EEASDYLELDTIK.N
763.3561 1524.6977 1524.7195 -0.0217 0 42 0.0098 U K.EEASDYLELDTIK.N
532.5749 1594.7028 1594.7160 -0.0132 0 42 0.0062 U R.VFITDDFHDMMPK.Y
622.3026 1863.8860 1863.9076 -0.0216 1 38 0.024 U R.EATEKEFEPLLNWMK.D
750.6789 2249.0150 2249.0349 -0.0199 0 39 0.0087 U R.FQSSHHSTDITSLDQYVER.M
1178.5253 2355.0360 2355.0623 -0.0263 0 79 6.2e-007
U R.LTESPCALVASQYGWSGNM
ER.I
786.0209 2355.0410 2355.0623 -0.0214 0 64 2.2e-005
U R.LTESPCALVASQYGWSGNM
ER.I
910.1020 2727.2843 2727.3311 -0.0468 1 94 2.6e-008
U R.TDDEVVQREEEAIQLDGLNA
SQIR.E
1035.4949 3103.4629 3103.4848 -0.0220 1 42 0.0037 U K.GYEVIYLTEPVDEYCIQALPE
FDGKR.F
Calreticulina (ERp-60)
6680836 0.07 610.3539 1218.6932 1218.6972 -0.0040 0 35 0.049 U K.GQTLVVQFTVK.H
Proteína reguladora de glicose 78kDa (GRP-78)
1304157 0.86 433.7151 865.4156 865.4181 -0.0025 1 48 0.004 U K.FAEEDKK.L
433.7151 865.4156 865.4181 -0.0025 1 46 0.0057 U K.FAEEDKK.L
499.2605 996.5064 996.5101 -0.0037 0 56 0.00057 U R.ALSSQHQAR.I
537.7634 1073.5122 1073.5465 -0.0343 0 59 0.00032 U K.ITITNDQNR.L
614.8063 1227.5981 1227.6207 -0.0227 0 49 0.0024 U R.VEIIANDQGNR.I
617.3094 1232.6042 1232.6183 -0.0141 0 65 7.9e-005
U K.DAGTIAGLNVMR.I
658.8018 1315.5891 1315.6295 -0.0405 0 58 0.00026 U R.NELESYAYSLK.N
699.3900 1396.7654 1396.7813 -0.0159 0 82 1e-006
U K.ELEEIVQPIISK.L
768.8801 1535.7456 1535.7905 -0.0449 0 81 1.4e-006
U K.TFAPEEISAMVLTK.M
768.8801 1535.7456 1535.7905 -0.0449 0 69 2.2e-005
U K.TFAPEEISAMVLTK.M
77
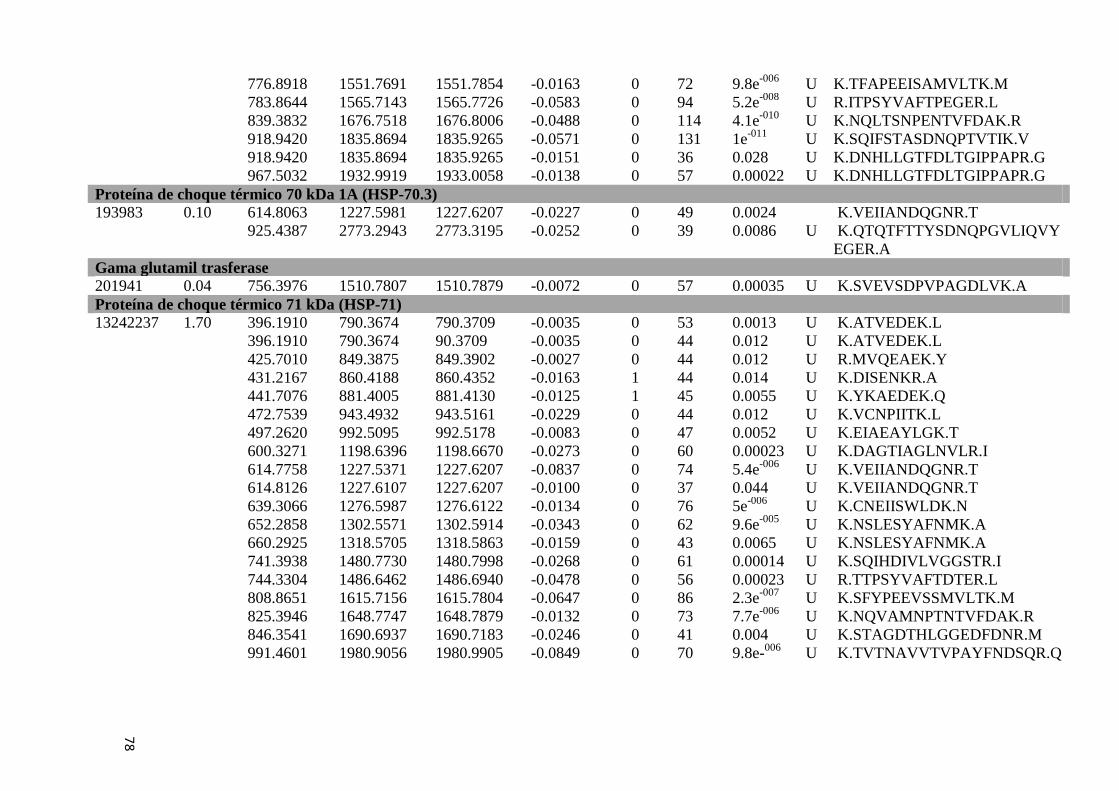
776.8918 1551.7691 1551.7854 -0.0163 0 72 9.8e-006
U K.TFAPEEISAMVLTK.M
783.8644 1565.7143 1565.7726 -0.0583 0 94 5.2e-008
U R.ITPSYVAFTPEGER.L
839.3832 1676.7518 1676.8006 -0.0488 0 114 4.1e-010
U K.NQLTSNPENTVFDAK.R
918.9420 1835.8694 1835.9265 -0.0571 0 131 1e-011
U K.SQIFSTASDNQPTVTIK.V
918.9420 1835.8694 1835.9265 -0.0151 0 36 0.028 U K.DNHLLGTFDLTGIPPAPR.G
967.5032 1932.9919 1933.0058 -0.0138 0 57 0.00022 U K.DNHLLGTFDLTGIPPAPR.G
Proteína de choque térmico 70 kDa 1A (HSP-70.3)
193983 0.10 614.8063 1227.5981 1227.6207 -0.0227 0 49 0.0024 K.VEIIANDQGNR.T
925.4387 2773.2943 2773.3195 -0.0252 0 39 0.0086 U K.QTQTFTTYSDNQPGVLIQVY
EGER.A
Gama glutamil trasferase
201941 0.04 756.3976 1510.7807 1510.7879 -0.0072 0 57 0.00035 U K.SVEVSDPVPAGDLVK.A
Proteína de choque térmico 71 kDa (HSP-71)
13242237 1.70 396.1910 790.3674 790.3709 -0.0035 0 53 0.0013 U K.ATVEDEK.L
396.1910 790.3674 90.3709 -0.0035 0 44 0.012 U K.ATVEDEK.L
425.7010 849.3875 849.3902 -0.0027 0 44 0.012 U R.MVQEAEK.Y
431.2167 860.4188 860.4352 -0.0163 1 44 0.014 U K.DISENKR.A
441.7076 881.4005 881.4130 -0.0125 1 45 0.0055 U K.YKAEDEK.Q
472.7539 943.4932 943.5161 -0.0229 0 44 0.012 U K.VCNPIITK.L
497.2620 992.5095 992.5178 -0.0083 0 47 0.0052 U K.EIAEAYLGK.T
600.3271 1198.6396 1198.6670 -0.0273 0 60 0.00023 U K.DAGTIAGLNVLR.I
614.7758 1227.5371 1227.6207 -0.0837 0 74 5.4e-006
U K.VEIIANDQGNR.T
614.8126 1227.6107 1227.6207 -0.0100 0 37 0.044 U K.VEIIANDQGNR.T
639.3066 1276.5987 1276.6122 -0.0134 0 76 5e-006
U K.CNEIISWLDK.N
652.2858 1302.5571 1302.5914 -0.0343 0 62 9.6e-005
U K.NSLESYAFNMK.A
660.2925 1318.5705 1318.5863 -0.0159 0 43 0.0065 U K.NSLESYAFNMK.A
741.3938 1480.7730 1480.7998 -0.0268 0 61 0.00014 U K.SQIHDIVLVGGSTR.I
744.3304 1486.6462 1486.6940 -0.0478 0 56 0.00023 U R.TTPSYVAFTDTER.L
808.8651 1615.7156 1615.7804 -0.0647 0 86 2.3e-007
U K.SFYPEEVSSMVLTK.M
825.3946 1648.7747 1648.7879 -0.0132 0 73 7.7e-006
U K.NQVAMNPTNTVFDAK.R
846.3541 1690.6937 1690.7183 -0.0246 0 41 0.004 U K.STAGDTHLGGEDFDNR.M
991.4601 1980.9056 1980.9905 -0.0849 0 70 9.8e-006
U K.TVTNAVVTVPAYFNDSQR.Q
78
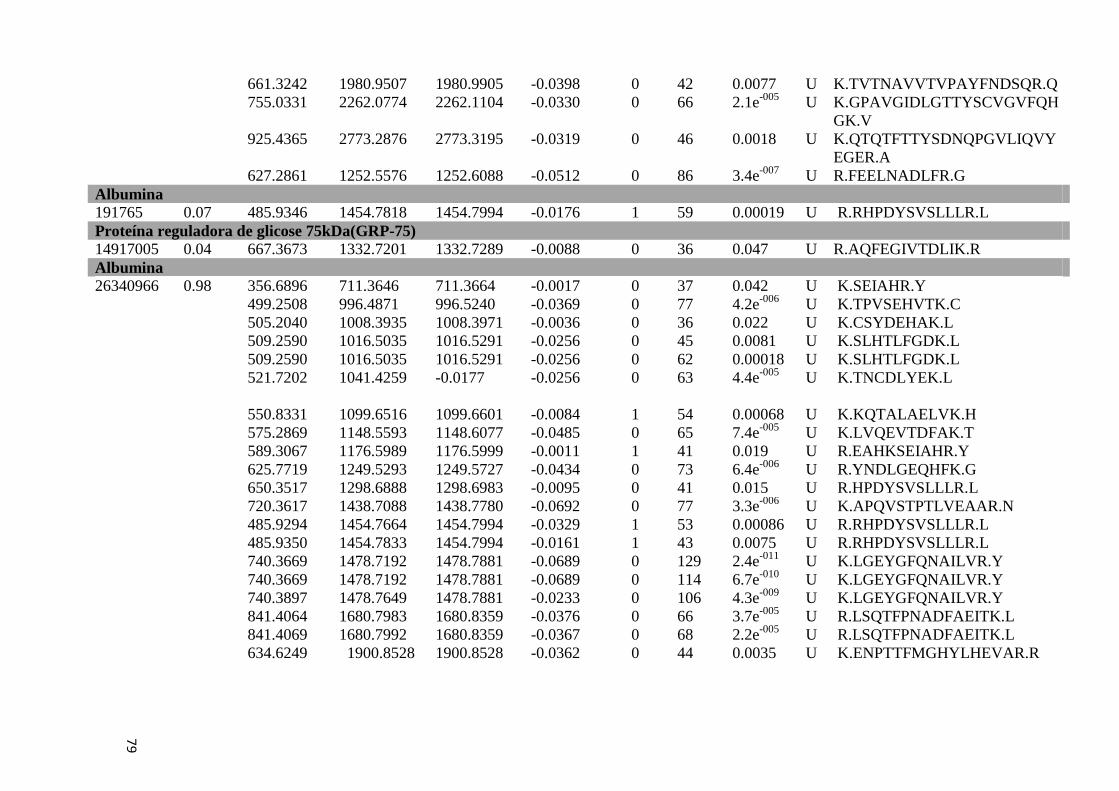
661.3242 1980.9507 1980.9905 -0.0398 0 42 0.0077 U K.TVTNAVVTVPAYFNDSQR.Q
755.0331 2262.0774 2262.1104 -0.0330 0 66 2.1e-005
U K.GPAVGIDLGTTYSCVGVFQH
GK.V
925.4365 2773.2876 2773.3195 -0.0319 0 46 0.0018 U K.QTQTFTTYSDNQPGVLIQVY
EGER.A 627.2861 1252.5576 1252.6088 -0.0512 0 86 3.4e
-007 U R.FEELNADLFR.G
Albumina
191765 0.07 485.9346 1454.7818 1454.7994 -0.0176 1 59 0.00019 U R.RHPDYSVSLLLR.L
Proteína reguladora de glicose 75kDa(GRP-75)
14917005 0.04 667.3673 1332.7201 1332.7289 -0.0088 0 36 0.047 U R.AQFEGIVTDLIK.R
Albumina 26340966 0.98 356.6896 711.3646 711.3664 -0.0017 0 37 0.042 U K.SEIAHR.Y
499.2508 996.4871 996.5240 -0.0369 0 77 4.2e-006
U K.TPVSEHVTK.C
505.2040 1008.3935 1008.3971 -0.0036 0 36 0.022 U K.CSYDEHAK.L
509.2590 1016.5035 1016.5291 -0.0256 0 45 0.0081 U K.SLHTLFGDK.L
509.2590 1016.5035 1016.5291 -0.0256 0 62 0.00018 U K.SLHTLFGDK.L
521.7202 1041.4259 -0.0177 -0.0256 0 63 4.4e-005
U K.TNCDLYEK.L
550.8331 1099.6516 1099.6601 -0.0084 1 54 0.00068 U K.KQTALAELVK.H
575.2869 1148.5593 1148.6077 -0.0485 0 65 7.4e-005
U K.LVQEVTDFAK.T
589.3067 1176.5989 1176.5999 -0.0011 1 41 0.019 U R.EAHKSEIAHR.Y
625.7719 1249.5293 1249.5727 -0.0434 0 73 6.4e-006
U R.YNDLGEQHFK.G
650.3517 1298.6888 1298.6983 -0.0095 0 41 0.015 U R.HPDYSVSLLLR.L
720.3617 1438.7088 1438.7780 -0.0692 0 77 3.3e-006
U K.APQVSTPTLVEAAR.N
485.9294 1454.7664 1454.7994 -0.0329 1 53 0.00086 U R.RHPDYSVSLLLR.L
485.9350 1454.7833 1454.7994 -0.0161 1 43 0.0075 U R.RHPDYSVSLLLR.L
740.3669 1478.7192 1478.7881 -0.0689 0 129 2.4e-011
U K.LGEYGFQNAILVR.Y
740.3669 1478.7192 1478.7881 -0.0689 0 114 6.7e-010
U K.LGEYGFQNAILVR.Y
740.3897 1478.7649 1478.7881 -0.0233 0 106 4.3e-009
U K.LGEYGFQNAILVR.Y
841.4064 1680.7983 1680.8359 -0.0376 0 66 3.7e-005
U R.LSQTFPNADFAEITK.L
841.4069 1680.7992 1680.8359 -0.0367 0 68 2.2e-005
U R.LSQTFPNADFAEITK.L
634.6249 1900.8528 1900.8528 -0.0362 0 44 0.0035 U K.ENPTTFMGHYLHEVAR.R
79
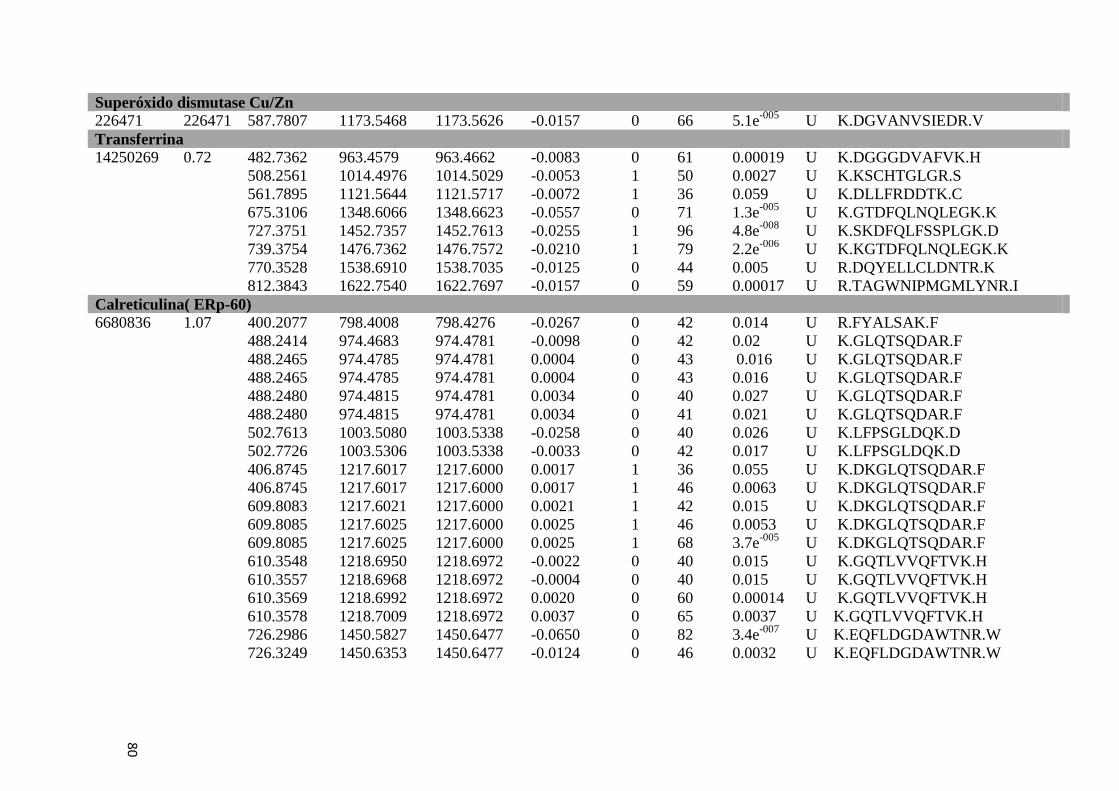
Superóxido dismutase Cu/Zn
226471 226471 587.7807 1173.5468 1173.5626 -0.0157 0 66 5.1e-005
U K.DGVANVSIEDR.V
Transferrina 14250269 0.72 482.7362 963.4579 963.4662 -0.0083 0 61 0.00019 U K.DGGGDVAFVK.H
508.2561 1014.4976 1014.5029 -0.0053 1 50 0.0027 U K.KSCHTGLGR.S
561.7895 1121.5644 1121.5717 -0.0072 1 36 0.059 U K.DLLFRDDTK.C
675.3106 1348.6066 1348.6623 -0.0557 0 71 1.3e-005
U K.GTDFQLNQLEGK.K
727.3751 1452.7357 1452.7613 -0.0255 1 96 4.8e-008
U K.SKDFQLFSSPLGK.D
739.3754 1476.7362 1476.7572 -0.0210 1 79 2.2e-006
U K.KGTDFQLNQLEGK.K
770.3528 1538.6910 1538.7035 -0.0125 0 44 0.005 U R.DQYELLCLDNTR.K
812.3843 1622.7540 1622.7697 -0.0157 0 59 0.00017 U R.TAGWNIPMGMLYNR.I
Calreticulina( ERp-60)
6680836 1.07 400.2077 798.4008 798.4276 -0.0267 0 42 0.014 U R.FYALSAK.F
488.2414 974.4683 974.4781 -0.0098 0 42 0.02 U K.GLQTSQDAR.F
488.2465 974.4785 974.4781 0.0004 0 43 0.016 U K.GLQTSQDAR.F
488.2465 974.4785 974.4781 0.0004 0 43 0.016 U K.GLQTSQDAR.F 488.2480 974.4815 974.4781 0.0034 0 40 0.027 U K.GLQTSQDAR.F 488.2480 974.4815 974.4781 0.0034 0 41 0.021 U K.GLQTSQDAR.F 502.7613 1003.5080 1003.5338 -0.0258 0 40 0.026 U K.LFPSGLDQK.D 502.7726 1003.5306 1003.5338 -0.0033 0 42 0.017 U K.LFPSGLDQK.D 406.8745 1217.6017 1217.6000 0.0017 1 36 0.055 U K.DKGLQTSQDAR.F 406.8745 1217.6017 1217.6000 0.0017 1 46 0.0063 U K.DKGLQTSQDAR.F 609.8083 1217.6021 1217.6000 0.0021 1 42 0.015 U K.DKGLQTSQDAR.F 609.8085 1217.6025 1217.6000 0.0025 1 46 0.0053 U K.DKGLQTSQDAR.F 609.8085 1217.6025 1217.6000 0.0025 1 68 3.7e
-005 U K.DKGLQTSQDAR.F
610.3548 1218.6950 1218.6972 -0.0022 0 40 0.015 U K.GQTLVVQFTVK.H 610.3557 1218.6968 1218.6972 -0.0004 0 40 0.015 U K.GQTLVVQFTVK.H 610.3569 1218.6992 1218.6972 0.0020 0 60 0.00014 U K.GQTLVVQFTVK.H 610.3578 1218.7009 1218.6972 0.0037 0 65 0.0037 U K.GQTLVVQFTVK.H 726.2986 1450.5827 1450.6477 -0.0650 0 82 3.4e
-007 U K.EQFLDGDAWTNR.W
726.3249 1450.6353 1450.6477 -0.0124 0 46 0.0032 U K.EQFLDGDAWTNR.W
80
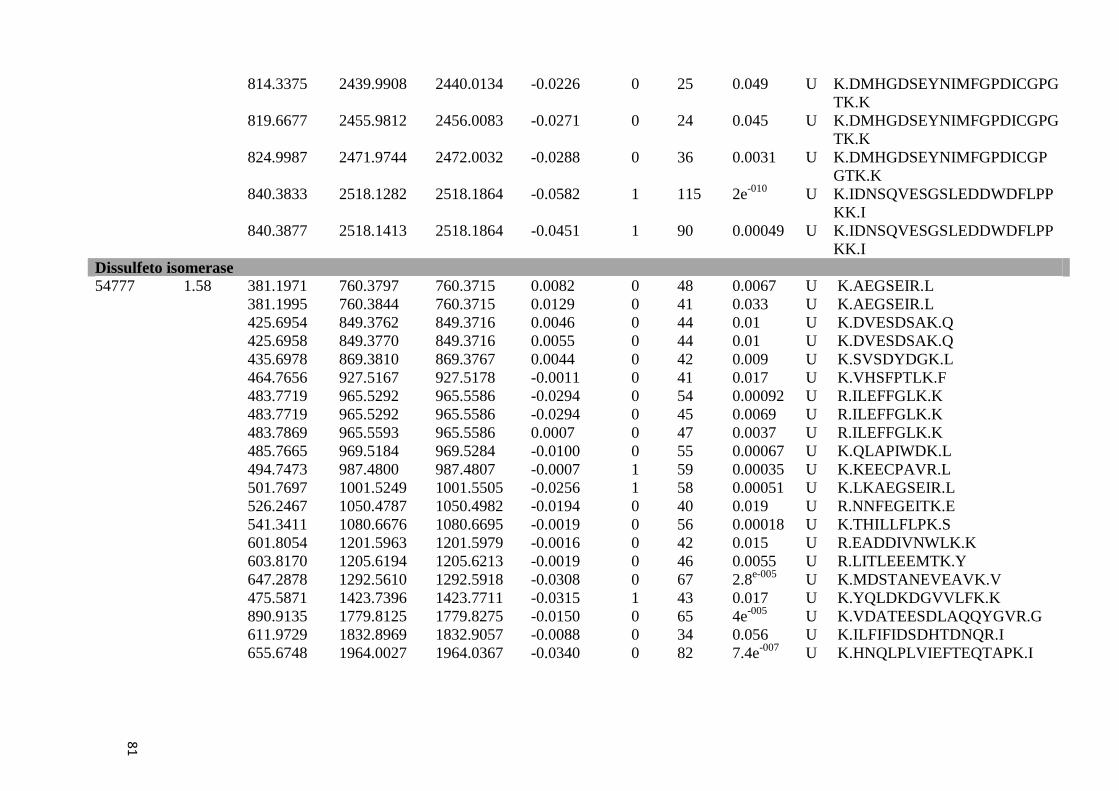
814.3375 2439.9908 2440.0134 -0.0226 0 25 0.049 U K.DMHGDSEYNIMFGPDICGPG
TK.K 819.6677 2455.9812 2456.0083 -0.0271 0 24 0.045 U K.DMHGDSEYNIMFGPDICGPG
TK.K 824.9987 2471.9744 2472.0032 -0.0288 0 36 0.0031 U K.DMHGDSEYNIMFGPDICGP
GTK.K 840.3833 2518.1282 2518.1864 -0.0582 1 115 2e
-010 U K.IDNSQVESGSLEDDWDFLPP
KK.I 840.3877 2518.1413 2518.1864 -0.0451 1 90 0.00049 U K.IDNSQVESGSLEDDWDFLPP
KK.I
Dissulfeto isomerase 54777 1.58 381.1971 760.3797 760.3715 0.0082 0 48 0.0067 U K.AEGSEIR.L
381.1995 760.3844 760.3715 0.0129 0 41 0.033 U K.AEGSEIR.L
425.6954 849.3762 849.3716 0.0046 0 44 0.01 U K.DVESDSAK.Q
425.6958 849.3770 849.3716 0.0055 0 44 0.01 U K.DVESDSAK.Q
435.6978 869.3810 869.3767 0.0044 0 42 0.009 U K.SVSDYDGK.L
464.7656 927.5167 927.5178 -0.0011 0 41 0.017 U K.VHSFPTLK.F
483.7719 965.5292 965.5586 -0.0294 0 54 0.00092 U R.ILEFFGLK.K
483.7719 965.5292 965.5586 -0.0294 0 45 0.0069 U R.ILEFFGLK.K 483.7869 965.5593 965.5586 0.0007 0 47 0.0037 U R.ILEFFGLK.K 485.7665 969.5184 969.5284 -0.0100 0 55 0.00067 U K.QLAPIWDK.L 494.7473 987.4800 987.4807 -0.0007 1 59 0.00035 U K.KEECPAVR.L 501.7697 1001.5249 1001.5505 -0.0256 1 58 0.00051 U K.LKAEGSEIR.L 526.2467 1050.4787 1050.4982 -0.0194 0 40 0.019 U R.NNFEGEITK.E 541.3411 1080.6676 1080.6695 -0.0019 0 56 0.00018 U K.THILLFLPK.S 601.8054 1201.5963 1201.5979 -0.0016 0 42 0.015 U R.EADDIVNWLK.K 603.8170 1205.6194 1205.6213 -0.0019 0 46 0.0055 U R.LITLEEEMTK.Y 647.2878 1292.5610 1292.5918 -0.0308 0 67 2.8
e-005 U K.MDSTANEVEAVK.V
475.5871 1423.7396 1423.7711 -0.0315 1 43 0.017 U K.YQLDKDGVVLFK.K 890.9135 1779.8125 1779.8275 -0.0150 0 65 4e
-005 U K.VDATEESDLAQQYGVR.G
611.9729 1832.8969 1832.9057 -0.0088 0 34 0.056 U K.ILFIFIDSDHTDNQR.I 655.6748 1964.0027 1964.0367 -0.0340 0 82 7.4e
-007 U K.HNQLPLVIEFTEQTAPK.I
81
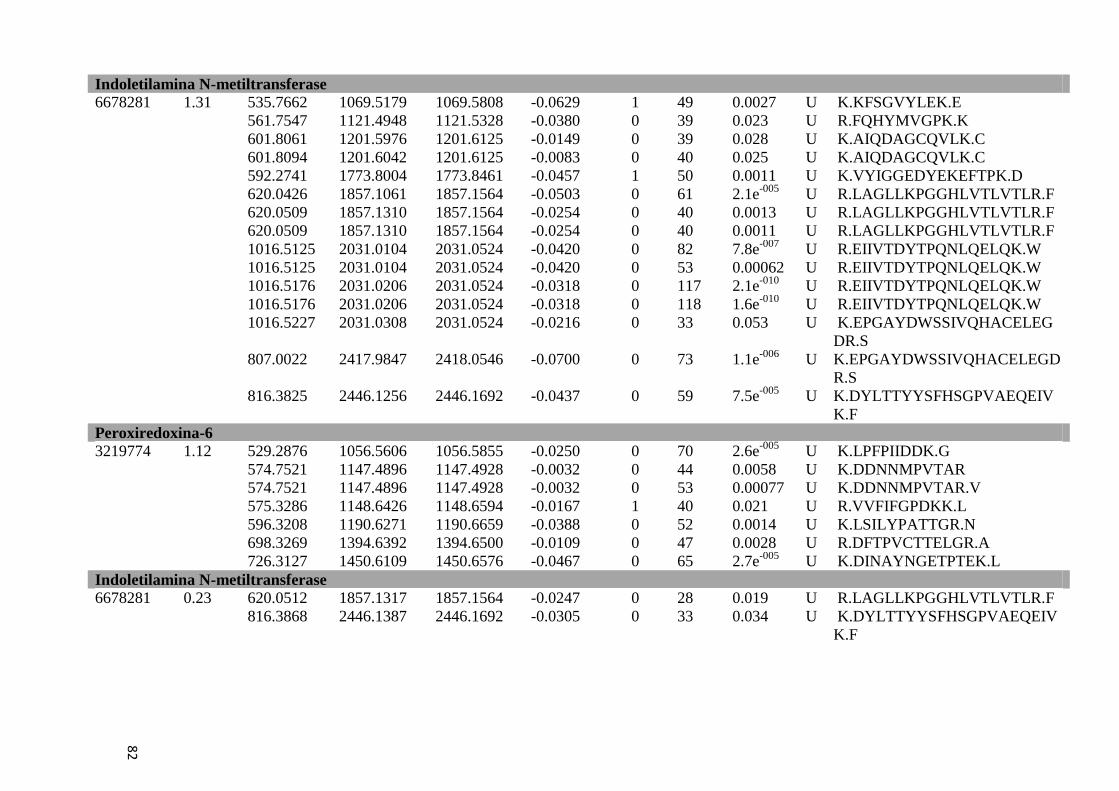
Indoletilamina N-metiltransferase 6678281 1.31 535.7662 1069.5179 1069.5808 -0.0629 1 49 0.0027 U K.KFSGVYLEK.E
561.7547 1121.4948 1121.5328 -0.0380 0 39 0.023 U R.FQHYMVGPK.K
601.8061 1201.5976 1201.6125 -0.0149 0 39 0.028 U K.AIQDAGCQVLK.C
601.8094 1201.6042 1201.6125 -0.0083 0 40 0.025 U K.AIQDAGCQVLK.C
592.2741 1773.8004 1773.8461 -0.0457 1 50 0.0011 U K.VYIGGEDYEKEFTPK.D
620.0426 1857.1061 1857.1564 -0.0503 0 61 2.1e-005
U R.LAGLLKPGGHLVTLVTLR.F
620.0509 1857.1310 1857.1564 -0.0254 0 40 0.0013 U R.LAGLLKPGGHLVTLVTLR.F
620.0509 1857.1310 1857.1564 -0.0254 0 40 0.0011 U R.LAGLLKPGGHLVTLVTLR.F
1016.5125 2031.0104 2031.0524 -0.0420 0 82 7.8e-007
U R.EIIVTDYTPQNLQELQK.W
1016.5125 2031.0104 2031.0524 -0.0420 0 53 0.00062 U R.EIIVTDYTPQNLQELQK.W
1016.5176 2031.0206 2031.0524 -0.0318 0 117 2.1e-010
U R.EIIVTDYTPQNLQELQK.W
1016.5176 2031.0206 2031.0524 -0.0318 0 118 1.6e-010
U R.EIIVTDYTPQNLQELQK.W
1016.5227 2031.0308 2031.0524 -0.0216 0 33 0.053 U K.EPGAYDWSSIVQHACELEG
DR.S 807.0022 2417.9847 2418.0546 -0.0700 0 73 1.1e
-006 U K.EPGAYDWSSIVQHACELEGD
R.S 816.3825 2446.1256 2446.1692 -0.0437 0 59 7.5e
-005 U K.DYLTTYYSFHSGPVAEQEIV
K.F
Peroxiredoxina-6 3219774 1.12 529.2876 1056.5606 1056.5855 -0.0250 0 70 2.6e
-005 U K.LPFPIIDDK.G
574.7521 1147.4896 1147.4928 -0.0032 0 44 0.0058 U K.DDNNMPVTAR
574.7521 1147.4896 1147.4928 -0.0032 0 53 0.00077 U K.DDNNMPVTAR.V
575.3286 1148.6426 1148.6594 -0.0167 1 40 0.021 U R.VVFIFGPDKK.L
596.3208 1190.6271 1190.6659 -0.0388 0 52 0.0014 U K.LSILYPATTGR.N
698.3269 1394.6392 1394.6500 -0.0109 0 47 0.0028 U R.DFTPVCTTELGR.A
726.3127 1450.6109 1450.6576 -0.0467 0 65 2.7e-005
U K.DINAYNGETPTEK.L
Indoletilamina N-metiltransferase
6678281 0.23 620.0512 1857.1317 1857.1564 -0.0247 0 28 0.019 U R.LAGLLKPGGHLVTLVTLR.F
816.3868 2446.1387 2446.1692 -0.0305 0 33 0.034 U K.DYLTTYYSFHSGPVAEQEIV
K.F
82
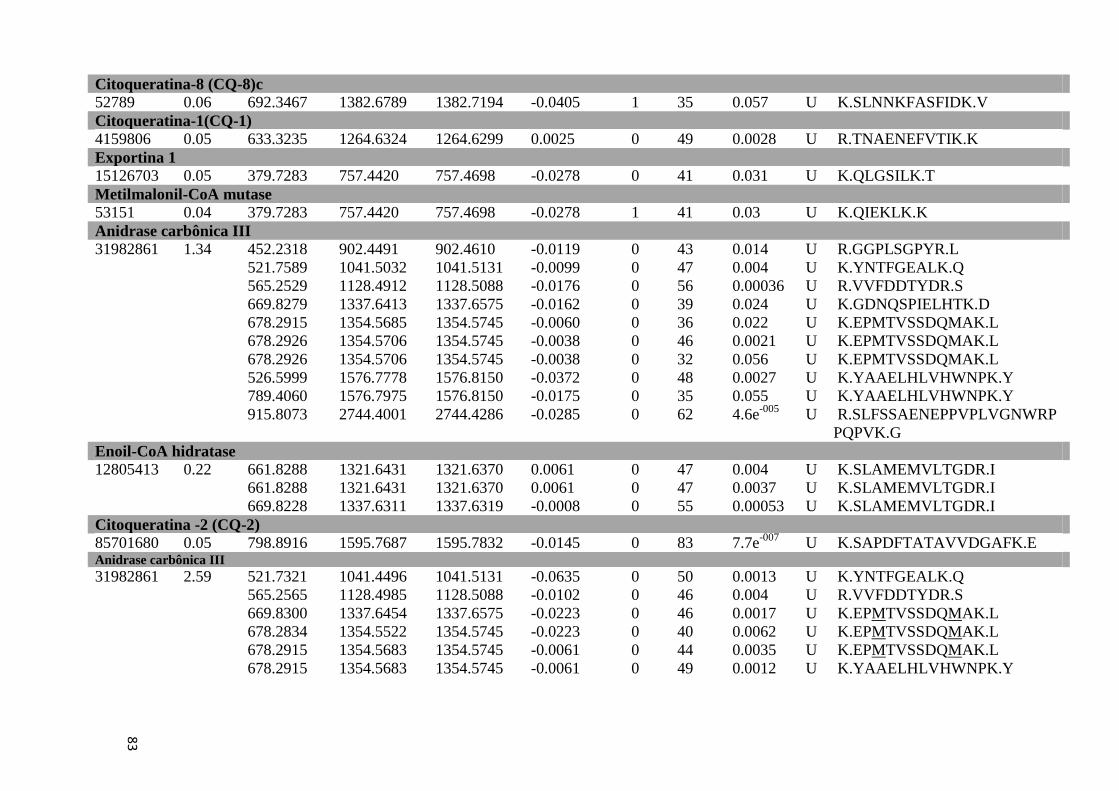
Citoqueratina-8 (CQ-8)c
52789 0.06 692.3467 1382.6789 1382.7194 -0.0405 1 35 0.057 U K.SLNNKFASFIDK.V
Citoqueratina-1(CQ-1)
4159806 0.05 633.3235 1264.6324 1264.6299 0.0025 0 49 0.0028 U R.TNAENEFVTIK.K
Exportina 1
15126703 0.05 379.7283 757.4420 757.4698 -0.0278 0 41 0.031 U K.QLGSILK.T
Metilmalonil-CoA mutase
53151 0.04 379.7283 757.4420 757.4698 -0.0278 1 41 0.03 U K.QIEKLK.K
Anidrase carbônica III 31982861 1.34 452.2318 902.4491 902.4610 -0.0119 0 43 0.014 U R.GGPLSGPYR.L
521.7589 1041.5032 1041.5131 -0.0099 0 47 0.004 U K.YNTFGEALK.Q
565.2529 1128.4912 1128.5088 -0.0176 0 56 0.00036 U R.VVFDDTYDR.S
669.8279 1337.6413 1337.6575 -0.0162 0 39 0.024 U K.GDNQSPIELHTK.D
678.2915 1354.5685 1354.5745 -0.0060 0 36 0.022 U K.EPMTVSSDQMAK.L
678.2926 1354.5706 1354.5745 -0.0038 0 46 0.0021 U K.EPMTVSSDQMAK.L
678.2926 1354.5706 1354.5745 -0.0038 0 32 0.056 U K.EPMTVSSDQMAK.L
526.5999 1576.7778 1576.8150 -0.0372 0 48 0.0027 U K.YAAELHLVHWNPK.Y
789.4060 1576.7975 1576.8150 -0.0175 0 35 0.055 U K.YAAELHLVHWNPK.Y
915.8073 2744.4001 2744.4286 -0.0285 0 62 4.6e-005
U R.SLFSSAENEPPVPLVGNWRP
PQPVK.G
Enoil-CoA hidratase
12805413 0.22 661.8288 1321.6431 1321.6370 0.0061 0 47 0.004 U K.SLAMEMVLTGDR.I
661.8288 1321.6431 1321.6370 0.0061 0 47 0.0037 U K.SLAMEMVLTGDR.I
669.8228 1337.6311 1337.6319 -0.0008 0 55 0.00053 U K.SLAMEMVLTGDR.I
Citoqueratina -2 (CQ-2)
85701680 0.05 798.8916 1595.7687 1595.7832 -0.0145 0 83 7.7e-007
U K.SAPDFTATAVVDGAFK.E Anidrase carbônica III
31982861 2.59 521.7321 1041.4496 1041.5131 -0.0635 0 50 0.0013 U K.YNTFGEALK.Q
565.2565 1128.4985 1128.5088 -0.0102 0 46 0.004 U R.VVFDDTYDR.S
669.8300 1337.6454 1337.6575 -0.0223 0 46 0.0017 U K.EPMTVSSDQMAK.L
678.2834 1354.5522 1354.5745 -0.0223 0 40 0.0062 U K.EPMTVSSDQMAK.L
678.2915 1354.5683 1354.5745 -0.0061 0 44 0.0035 U K.EPMTVSSDQMAK.L
678.2915 1354.5683 1354.5745 -0.0061 0 49 0.0012 U K.YAAELHLVHWNPK.Y
83
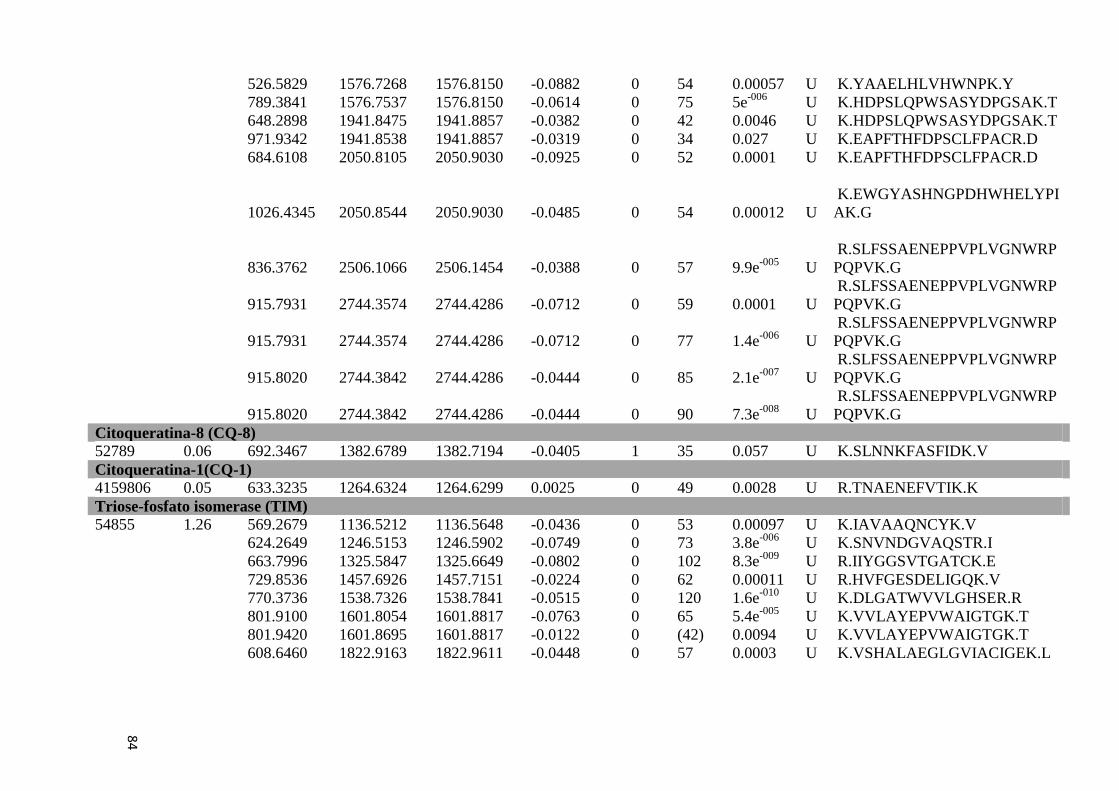
526.5829 1576.7268 1576.8150 -0.0882 0 54 0.00057 U K.YAAELHLVHWNPK.Y
789.3841 1576.7537 1576.8150 -0.0614 0 75 5e-006
U K.HDPSLQPWSASYDPGSAK.T
648.2898 1941.8475 1941.8857 -0.0382 0 42 0.0046 U K.HDPSLQPWSASYDPGSAK.T
971.9342 1941.8538 1941.8857 -0.0319 0 34 0.027 U K.EAPFTHFDPSCLFPACR.D
684.6108 2050.8105 2050.9030 -0.0925 0 52 0.0001 U K.EAPFTHFDPSCLFPACR.D
1026.4345 2050.8544 2050.9030 -0.0485 0 54 0.00012 U
K.EWGYASHNGPDHWHELYPI
AK.G
836.3762
2506.1066
2506.1454
-0.0388
0
57
9.9e-005
U
R.SLFSSAENEPPVPLVGNWRP
PQPVK.G
915.7931 2744.3574 2744.4286 -0.0712 0 59 0.0001 U
R.SLFSSAENEPPVPLVGNWRP
PQPVK.G
915.7931 2744.3574 2744.4286 -0.0712 0 77 1.4e-006
U
R.SLFSSAENEPPVPLVGNWRP
PQPVK.G
915.8020 2744.3842 2744.4286 -0.0444 0 85 2.1e-007
U
R.SLFSSAENEPPVPLVGNWRP
PQPVK.G
915.8020 2744.3842 2744.4286 -0.0444 0 90 7.3e-008
U
R.SLFSSAENEPPVPLVGNWRP
PQPVK.G
Citoqueratina-8 (CQ-8)
52789 0.06 692.3467 1382.6789 1382.7194 -0.0405 1 35 0.057 U K.SLNNKFASFIDK.V
Citoqueratina-1(CQ-1)
4159806 0.05 633.3235 1264.6324 1264.6299 0.0025 0 49 0.0028 U R.TNAENEFVTIK.K
Triose-fosfato isomerase (TIM)
54855 1.26 569.2679 1136.5212 1136.5648 -0.0436 0 53 0.00097 U K.IAVAAQNCYK.V
624.2649 1246.5153 1246.5902 -0.0749 0 73 3.8e-006
U K.SNVNDGVAQSTR.I
663.7996 1325.5847 1325.6649 -0.0802 0 102 8.3e-009
U R.IIYGGSVTGATCK.E
729.8536 1457.6926 1457.7151 -0.0224 0 62 0.00011 U R.HVFGESDELIGQK.V
770.3736 1538.7326 1538.7841 -0.0515 0 120 1.6e-010
U K.DLGATWVVLGHSER.R
801.9100 1601.8054 1601.8817 -0.0763 0 65 5.4e-005
U K.VVLAYEPVWAIGTGK.T
801.9420 1601.8695 1601.8817 -0.0122 0 (42) 0.0094 U K.VVLAYEPVWAIGTGK.T
608.6460 1822.9163 1822.9611 -0.0448 0 57 0.0003 U K.VSHALAEGLGVIACIGEK.L
84
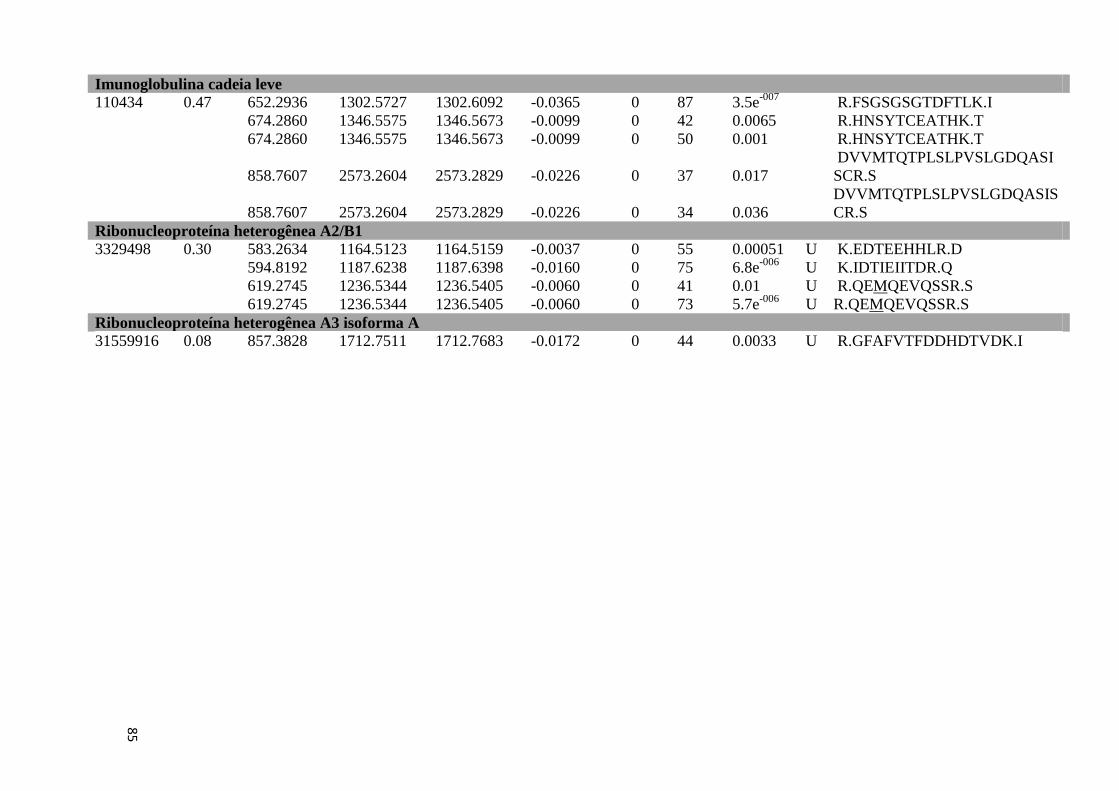
Imunoglobulina cadeia leve
110434 0.47 652.2936 1302.5727 1302.6092 -0.0365 0 87 3.5e-007
R.FSGSGSGTDFTLK.I
674.2860 1346.5575 1346.5673 -0.0099 0 42 0.0065 R.HNSYTCEATHK.T
674.2860 1346.5575 1346.5673 -0.0099 0 50 0.001 R.HNSYTCEATHK.T
858.7607 2573.2604 2573.2829 -0.0226 0 37 0.017
DVVMTQTPLSLPVSLGDQASI
SCR.S
858.7607 2573.2604 2573.2829 -0.0226 0 34 0.036
DVVMTQTPLSLPVSLGDQASIS
CR.S
Ribonucleoproteína heterogênea A2/B1
3329498 0.30 583.2634 1164.5123 1164.5159 -0.0037 0 55 0.00051 U K.EDTEEHHLR.D
594.8192 1187.6238 1187.6398 -0.0160 0 75 6.8e-006
U K.IDTIEIITDR.Q
619.2745 1236.5344 1236.5405 -0.0060 0 41 0.01 U R.QEMQEVQSSR.S
619.2745 1236.5344 1236.5405 -0.0060 0 73 5.7e-006
U R.QEMQEVQSSR.S
Ribonucleoproteína heterogênea A3 isoforma A
31559916 0.08 857.3828 1712.7511 1712.7683 -0.0172 0 44 0.0033 U R.GFAFVTFDDHDTVDK.I
85
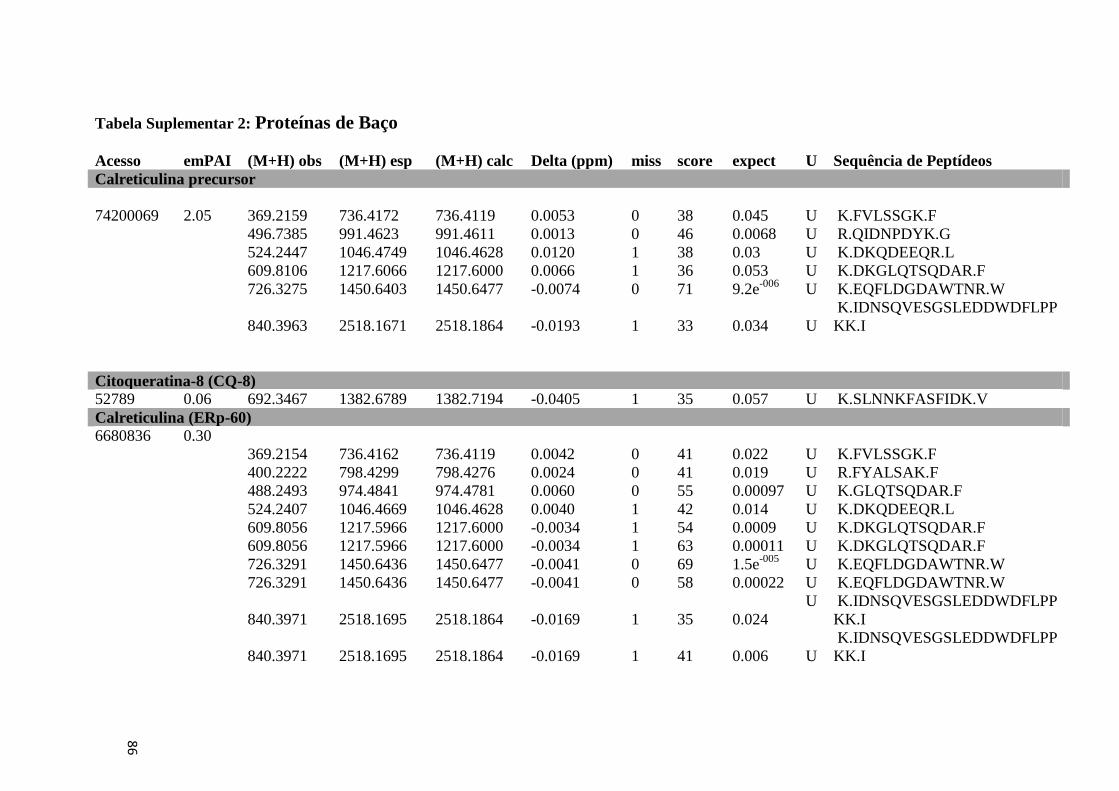
Tabela Suplementar 2: Proteínas de Baço
Acesso emPAI (M+H) obs (M+H) esp (M+H) calc Delta (ppm) miss score expect U Sequência de Peptídeos
Calreticulina precursor
74200069
2.05 369.2159 736.4172 736.4119 0.0053 0 38 0.045 U K.FVLSSGK.F
496.7385 991.4623 991.4611 0.0013 0 46 0.0068 U R.QIDNPDYK.G
524.2447 1046.4749 1046.4628 0.0120 1 38 0.03 U K.DKQDEEQR.L
609.8106 1217.6066 1217.6000 0.0066 1 36 0.053 U K.DKGLQTSQDAR.F
726.3275 1450.6403 1450.6477 -0.0074 0 71 9.2e-006
U K.EQFLDGDAWTNR.W
840.3963 2518.1671 2518.1864 -0.0193 1 33 0.034 U
K.IDNSQVESGSLEDDWDFLPP
KK.I
Citoqueratina-8 (CQ-8)
52789 0.06 692.3467 1382.6789 1382.7194 -0.0405 1 35 0.057 U K.SLNNKFASFIDK.V
Calreticulina (ERp-60)
6680836
0.30
369.2154 736.4162 736.4119 0.0042 0 41 0.022 U K.FVLSSGK.F
400.2222 798.4299 798.4276 0.0024 0 41 0.019 U R.FYALSAK.F
488.2493 974.4841 974.4781 0.0060 0 55 0.00097 U K.GLQTSQDAR.F
524.2407 1046.4669 1046.4628 0.0040 1 42 0.014 U K.DKQDEEQR.L
609.8056 1217.5966 1217.6000 -0.0034 1 54 0.0009 U K.DKGLQTSQDAR.F
609.8056 1217.5966 1217.6000 -0.0034 1 63 0.00011 U K.DKGLQTSQDAR.F
726.3291 1450.6436 1450.6477 -0.0041 0 69 1.5e-005
U K.EQFLDGDAWTNR.W
726.3291 1450.6436 1450.6477 -0.0041 0 58 0.00022 U K.EQFLDGDAWTNR.W
840.3971 2518.1695 2518.1864 -0.0169 1 35 0.024
U
K.IDNSQVESGSLEDDWDFLPP
KK.I
840.3971 2518.1695 2518.1864 -0.0169 1 41 0.006 U
K.IDNSQVESGSLEDDWDFLPP
KK.I
86
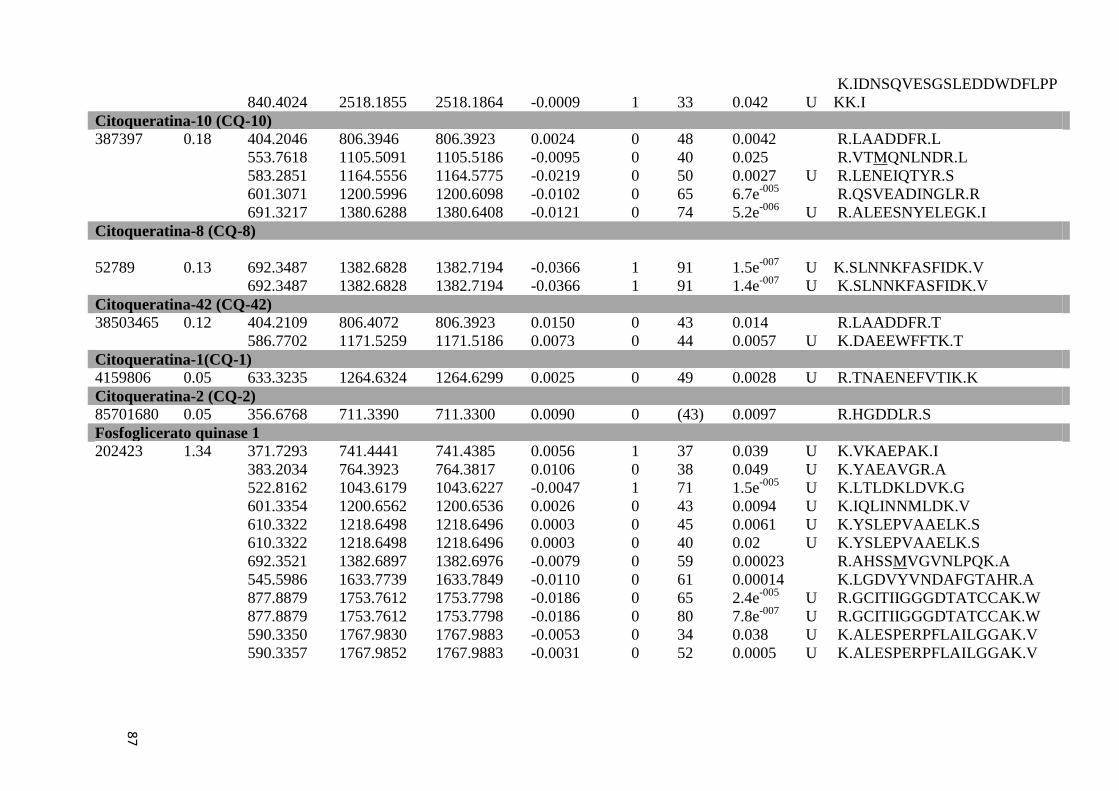
840.4024 2518.1855 2518.1864 -0.0009 1 33 0.042 U
K.IDNSQVESGSLEDDWDFLPP
KK.I
Citoqueratina-10 (CQ-10)
387397 0.18 404.2046 806.3946 806.3923 0.0024 0 48 0.0042 R.LAADDFR.L
553.7618 1105.5091 1105.5186 -0.0095 0 40 0.025 R.VTMQNLNDR.L
583.2851 1164.5556 1164.5775 -0.0219 0 50 0.0027 U R.LENEIQTYR.S
601.3071 1200.5996 1200.6098 -0.0102 0 65 6.7e-005
R.QSVEADINGLR.R
691.3217 1380.6288 1380.6408 -0.0121 0 74 5.2e-006
U R.ALEESNYELEGK.I
Citoqueratina-8 (CQ-8)
52789
0.13 692.3487 1382.6828 1382.7194 -0.0366 1 91 1.5e-007
U K.SLNNKFASFIDK.V
692.3487 1382.6828 1382.7194 -0.0366 1 91 1.4e-007
U K.SLNNKFASFIDK.V
Citoqueratina-42 (CQ-42)
38503465 0.12 404.2109 806.4072 806.3923 0.0150 0 43 0.014 R.LAADDFR.T
586.7702 1171.5259 1171.5186 0.0073 0 44 0.0057 U K.DAEEWFFTK.T
Citoqueratina-1(CQ-1)
4159806 0.05 633.3235 1264.6324 1264.6299 0.0025 0 49 0.0028 U R.TNAENEFVTIK.K
Citoqueratina-2 (CQ-2)
85701680 0.05 356.6768 711.3390 711.3300 0.0090 0 (43) 0.0097 R.HGDDLR.S
Fosfoglicerato quinase 1
202423 1.34 371.7293 741.4441 741.4385 0.0056 1 37 0.039 U K.VKAEPAK.I
383.2034 764.3923 764.3817 0.0106 0 38 0.049 U K.YAEAVGR.A
522.8162 1043.6179 1043.6227 -0.0047 1 71 1.5e-005
U K.LTLDKLDVK.G
601.3354 1200.6562 1200.6536 0.0026 0 43 0.0094 U K.IQLINNMLDK.V
610.3322 1218.6498 1218.6496 0.0003 0 45 0.0061 U K.YSLEPVAAELK.S
610.3322 1218.6498 1218.6496 0.0003 0 40 0.02 U K.YSLEPVAAELK.S
692.3521 1382.6897 1382.6976 -0.0079 0 59 0.00023 R.AHSSMVGVNLPQK.A
545.5986 1633.7739 1633.7849 -0.0110 0 61 0.00014 K.LGDVYVNDAFGTAHR.A
877.8879 1753.7612 1753.7798 -0.0186 0 65 2.4e-005
U R.GCITIIGGGDTATCCAK.W
877.8879 1753.7612 1753.7798 -0.0186 0 80 7.8e-007
U R.GCITIIGGGDTATCCAK.W
590.3350 1767.9830 1767.9883 -0.0053 0 34 0.038 U K.ALESPERPFLAILGGAK.V
590.3357 1767.9852 1767.9883 -0.0031 0 52 0.0005 U K.ALESPERPFLAILGGAK.V
87
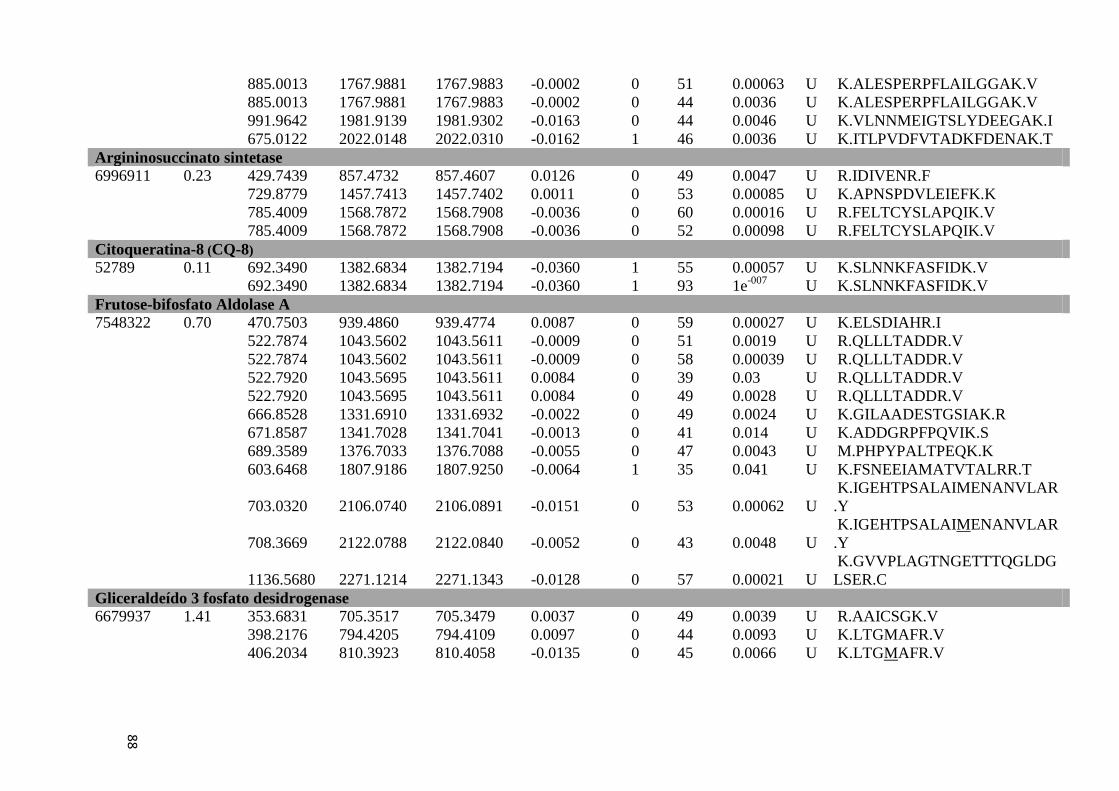
885.0013 1767.9881 1767.9883 -0.0002 0 51 0.00063 U K.ALESPERPFLAILGGAK.V
885.0013 1767.9881 1767.9883 -0.0002 0 44 0.0036 U K.ALESPERPFLAILGGAK.V
991.9642 1981.9139 1981.9302 -0.0163 0 44 0.0046 U K.VLNNMEIGTSLYDEEGAK.I
675.0122 2022.0148 2022.0310 -0.0162 1 46 0.0036 U K.ITLPVDFVTADKFDENAK.T
Argininosuccinato sintetase
6996911 0.23 429.7439 857.4732 857.4607 0.0126 0 49 0.0047 U R.IDIVENR.F
729.8779 1457.7413 1457.7402 0.0011 0 53 0.00085 U K.APNSPDVLEIEFK.K
785.4009 1568.7872 1568.7908 -0.0036 0 60 0.00016 U R.FELTCYSLAPQIK.V
785.4009 1568.7872 1568.7908 -0.0036 0 52 0.00098 U R.FELTCYSLAPQIK.V
Citoqueratina-8 (CQ-8)
52789 0.11 692.3490 1382.6834 1382.7194 -0.0360 1 55 0.00057 U K.SLNNKFASFIDK.V
692.3490 1382.6834 1382.7194 -0.0360 1 93 1e-007
U K.SLNNKFASFIDK.V
Frutose-bifosfato Aldolase A 7548322 0.70 470.7503 939.4860 939.4774 0.0087 0 59 0.00027 U K.ELSDIAHR.I
522.7874 1043.5602 1043.5611 -0.0009 0 51 0.0019 U R.QLLLTADDR.V
522.7874 1043.5602 1043.5611 -0.0009 0 58 0.00039 U R.QLLLTADDR.V
522.7920 1043.5695 1043.5611 0.0084 0 39 0.03 U R.QLLLTADDR.V
522.7920 1043.5695 1043.5611 0.0084 0 49 0.0028 U R.QLLLTADDR.V
666.8528 1331.6910 1331.6932 -0.0022 0 49 0.0024 U K.GILAADESTGSIAK.R
671.8587 1341.7028 1341.7041 -0.0013 0 41 0.014 U K.ADDGRPFPQVIK.S
689.3589 1376.7033 1376.7088 -0.0055 0 47 0.0043 U M.PHPYPALTPEQK.K
603.6468 1807.9186 1807.9250 -0.0064 1 35 0.041 U K.FSNEEIAMATVTALRR.T
703.0320 2106.0740 2106.0891 -0.0151 0 53 0.00062 U
K.IGEHTPSALAIMENANVLAR
.Y
708.3669 2122.0788 2122.0840 -0.0052 0 43 0.0048 U
K.IGEHTPSALAIMENANVLAR
.Y
1136.5680 2271.1214 2271.1343 -0.0128 0 57 0.00021 U
K.GVVPLAGTNGETTTQGLDG
LSER.C
Gliceraldeído 3 fosfato desidrogenase
6679937 1.41 353.6831 705.3517 705.3479 0.0037 0 49 0.0039 U R.AAICSGK.V
398.2176 794.4205 794.4109 0.0097 0 44 0.0093 U K.LTGMAFR.V
406.2034 810.3923 810.4058 -0.0135 0 45 0.0066 U K.LTGMAFR.V
88
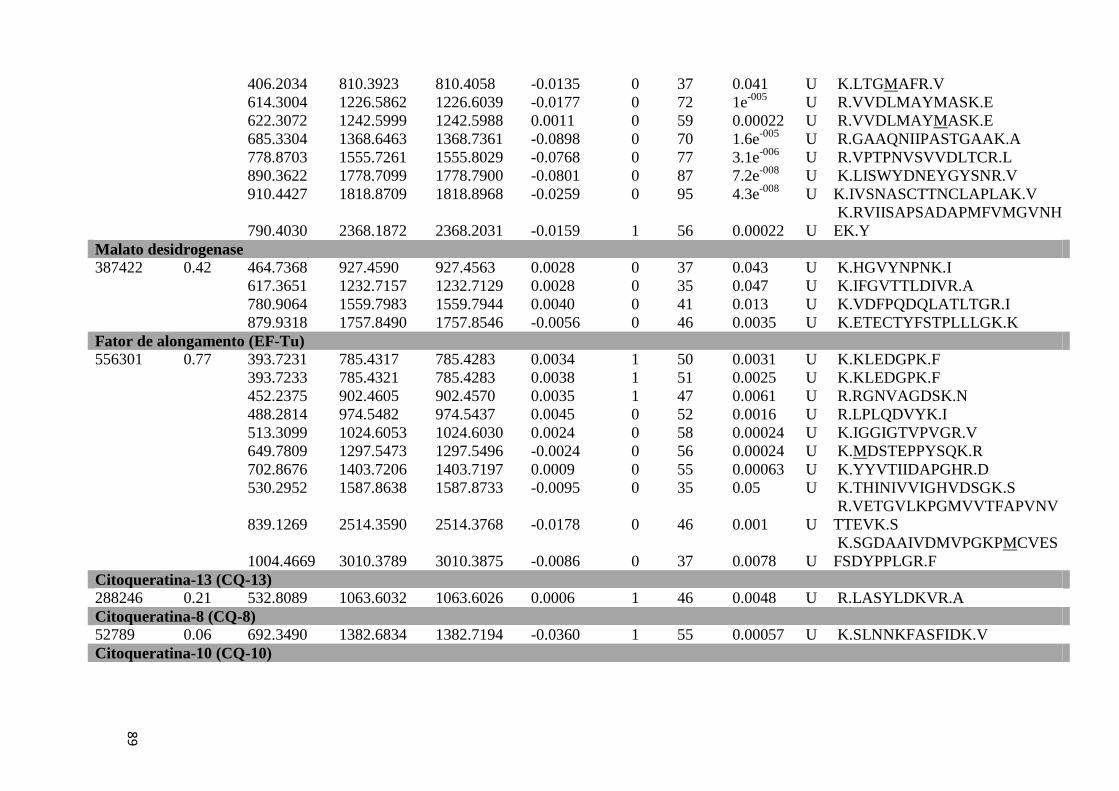
406.2034 810.3923 810.4058 -0.0135 0 37 0.041 U K.LTGMAFR.V
614.3004 1226.5862 1226.6039 -0.0177 0 72 1e-005
U R.VVDLMAYMASK.E
622.3072 1242.5999 1242.5988 0.0011 0 59 0.00022 U R.VVDLMAYMASK.E
685.3304 1368.6463 1368.7361 -0.0898 0 70 1.6e-005
U R.GAAQNIIPASTGAAK.A
778.8703 1555.7261 1555.8029 -0.0768 0 77 3.1e-006
U R.VPTPNVSVVDLTCR.L
890.3622 1778.7099 1778.7900 -0.0801 0 87 7.2e-008
U K.LISWYDNEYGYSNR.V
910.4427 1818.8709 1818.8968 -0.0259 0 95 4.3e-008
U K.IVSNASCTTNCLAPLAK.V
790.4030 2368.1872 2368.2031 -0.0159 1 56 0.00022 U
K.RVIISAPSADAPMFVMGVNH
EK.Y
Malato desidrogenase
387422 0.42 464.7368 927.4590 927.4563 0.0028 0 37 0.043 U K.HGVYNPNK.I
617.3651 1232.7157 1232.7129 0.0028 0 35 0.047 U K.IFGVTTLDIVR.A
780.9064 1559.7983 1559.7944 0.0040 0 41 0.013 U K.VDFPQDQLATLTGR.I
879.9318 1757.8490 1757.8546 -0.0056 0 46 0.0035 U K.ETECTYFSTPLLLGK.K
Fator de alongamento (EF-Tu)
556301 0.77 393.7231 785.4317 785.4283 0.0034 1 50 0.0031 U K.KLEDGPK.F
393.7233 785.4321 785.4283 0.0038 1 51 0.0025 U K.KLEDGPK.F
452.2375 902.4605 902.4570 0.0035 1 47 0.0061 U R.RGNVAGDSK.N
488.2814 974.5482 974.5437 0.0045 0 52 0.0016 U R.LPLQDVYK.I
513.3099 1024.6053 1024.6030 0.0024 0 58 0.00024 U K.IGGIGTVPVGR.V
649.7809 1297.5473 1297.5496 -0.0024 0 56 0.00024 U K.MDSTEPPYSQK.R
702.8676 1403.7206 1403.7197 0.0009 0 55 0.00063 U K.YYVTIIDAPGHR.D
530.2952 1587.8638 1587.8733 -0.0095 0 35 0.05 U K.THINIVVIGHVDSGK.S
839.1269 2514.3590 2514.3768 -0.0178 0 46 0.001 U
R.VETGVLKPGMVVTFAPVNV
TTEVK.S
1004.4669 3010.3789 3010.3875 -0.0086 0 37 0.0078 U
K.SGDAAIVDMVPGKPMCVES
FSDYPPLGR.F
Citoqueratina-13 (CQ-13)
288246 0.21 532.8089 1063.6032 1063.6026 0.0006 1 46 0.0048 U R.LASYLDKVR.A
Citoqueratina-8 (CQ-8) 52789 0.06 692.3490 1382.6834 1382.7194 -0.0360 1 55 0.00057 U K.SLNNKFASFIDK.V
Citoqueratina-10 (CQ-10)
89
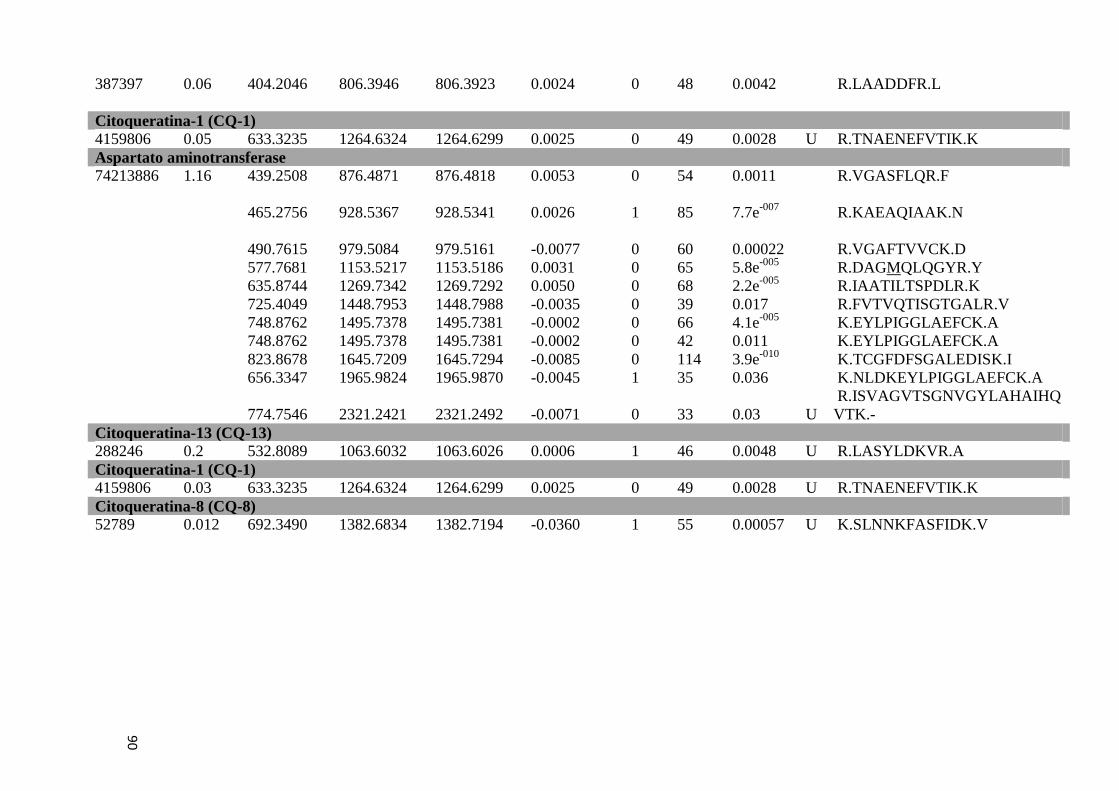
387397 0.06 404.2046 806.3946 806.3923 0.0024 0 48 0.0042 R.LAADDFR.L
Citoqueratina-1 (CQ-1)
4159806 0.05 633.3235 1264.6324 1264.6299 0.0025 0 49 0.0028 U R.TNAENEFVTIK.K
Aspartato aminotransferase 74213886 1.16 439.2508 876.4871 876.4818 0.0053 0 54 0.0011 R.VGASFLQR.F
465.2756 928.5367 928.5341 0.0026 1 85 7.7e-007
R.KAEAQIAAK.N
490.7615 979.5084 979.5161 -0.0077 0 60 0.00022
R.VGAFTVVCK.D
577.7681 1153.5217 1153.5186 0.0031 0 65 5.8e-005
R.DAGMQLQGYR.Y
635.8744 1269.7342 1269.7292 0.0050 0 68 2.2e-005
R.IAATILTSPDLR.K
725.4049 1448.7953 1448.7988 -0.0035 0 39 0.017 R.FVTVQTISGTGALR.V
748.8762 1495.7378 1495.7381 -0.0002 0 66 4.1e-005
K.EYLPIGGLAEFCK.A
748.8762 1495.7378 1495.7381 -0.0002 0 42 0.011 K.EYLPIGGLAEFCK.A
823.8678 1645.7209 1645.7294 -0.0085 0 114 3.9e-010
K.TCGFDFSGALEDISK.I
656.3347 1965.9824 1965.9870 -0.0045 1 35 0.036 K.NLDKEYLPIGGLAEFCK.A
774.7546 2321.2421 2321.2492 -0.0071 0 33 0.03 U
R.ISVAGVTSGNVGYLAHAIHQ
VTK.-
Citoqueratina-13 (CQ-13)
288246 0.2 532.8089 1063.6032 1063.6026 0.0006 1 46 0.0048 U R.LASYLDKVR.A
Citoqueratina-1 (CQ-1) 4159806 0.03 633.3235 1264.6324 1264.6299 0.0025 0 49 0.0028 U R.TNAENEFVTIK.K
Citoqueratina-8 (CQ-8) 52789 0.012 692.3490 1382.6834 1382.7194 -0.0360 1 55 0.00057 U K.SLNNKFASFIDK.V
90
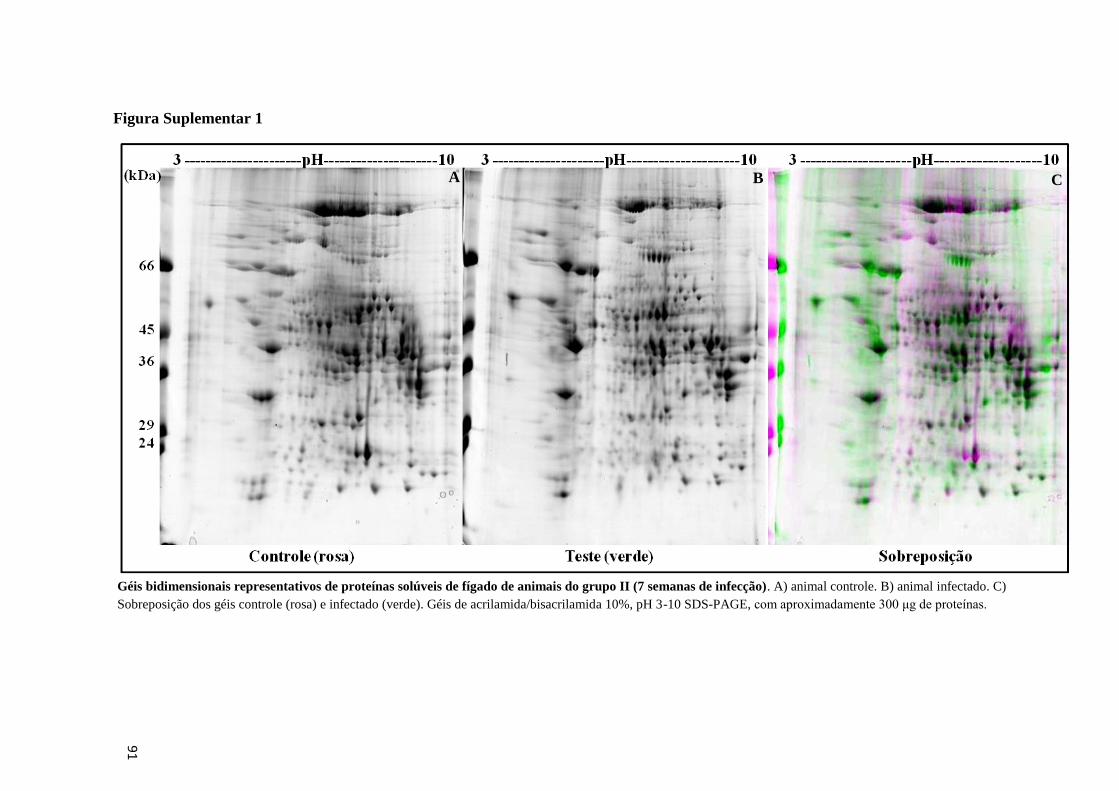
Figura Suplementar 1
Géis bidimensionais representativos de proteínas solúveis de fígado de animais do grupo II (7 semanas de infecção). A) animal controle. B) animal infectado. C)
Sobreposição dos géis controle (rosa) e infectado (verde). Géis de acrilamida/bisacrilamida 10%, pH 3-10 SDS-PAGE, com aproximadamente 300 μg de proteínas.
A B C
91
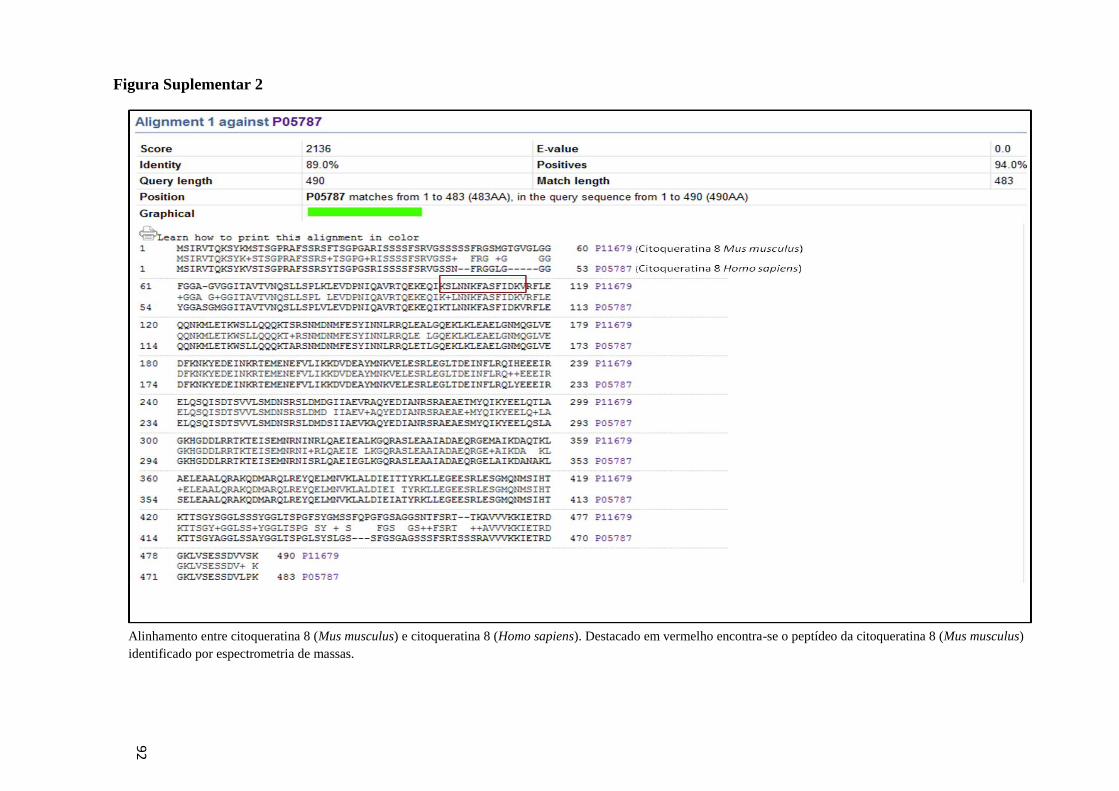
Figura Suplementar 2
92
Alinhamento entre citoqueratina 8 (Mus musculus) e citoqueratina 8 (Homo sapiens). Destacado em vermelho encontra-se o peptídeo da citoqueratina 8 (Mus musculus)
identificado por espectrometria de massas.
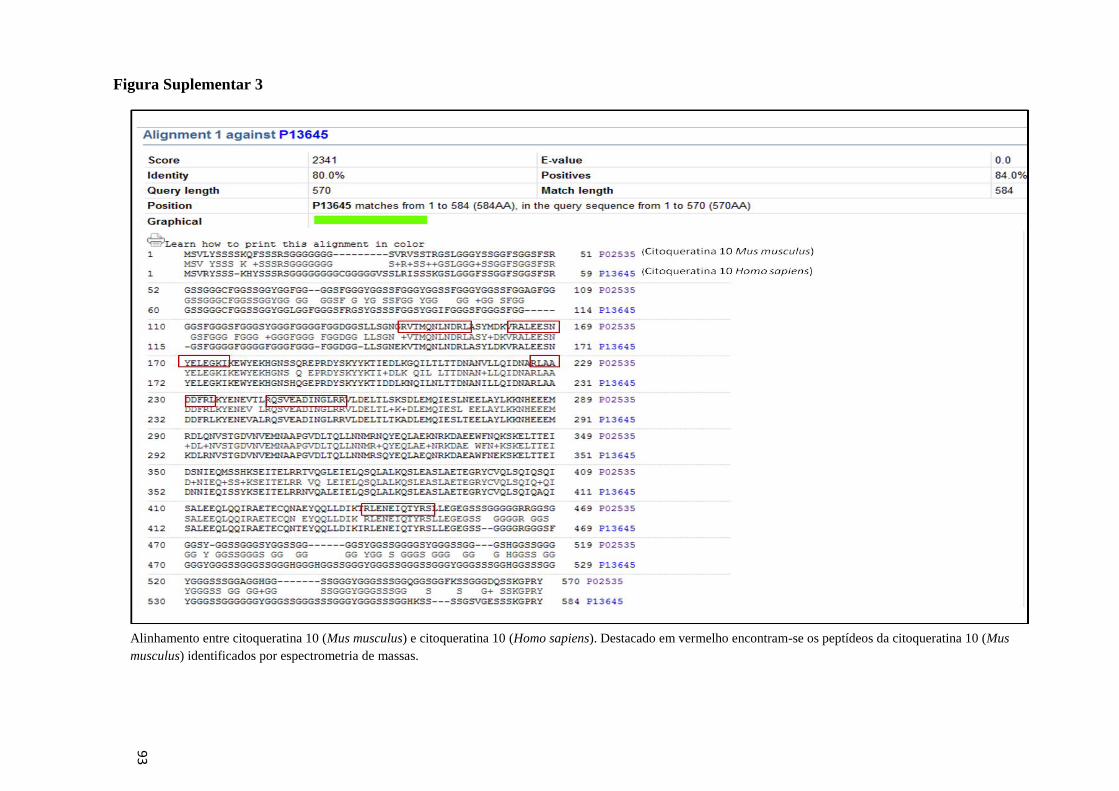
Figura Suplementar 3
93
Alinhamento entre citoqueratina 10 (Mus musculus) e citoqueratina 10 (Homo sapiens). Destacado em vermelho encontram-se os peptídeos da citoqueratina 10 (Mus
musculus) identificados por espectrometria de massas.