Embed Size (px)
Citation preview

Universidade de Aveiro Departamento de Biologia 2015
Dissertação apresentada à Universidade de Aveiro para
cumprimento dos requisitos necessários à obtenção do grau
de Mestre em Ecologia Aplicada, realizada sob a orientação
científica do Prof. Doutor António José Arsénia Nogueira do
Departamento de Biologia da Universidade de Aveiro e do
Doutor Mário Rui Canelas Boieiro do Centro de Ecologia,
Evolução e Alterações Ambientais da Universidade dos
Açores.
Ana Soraia
Pinto Lopes
Biodiversidade e ecologia de borboletas
(Lepidoptera) e libélulas (Odonata) do Planalto
Superior da Serra da Estrela
Biodiversity and ecology of butterflies
(Lepidoptera), and damselflies and dragonflies
(Odonata) from the higher plateau of Serra da
Estrela

DECLARAÇÃO
Declaro que este relatório é integralmente da minha autoria,
estando devidamente referenciadas as fontes e obras consultadas,
bem como identificadas de modo claro as citações dessas obras.
Não contém, por isso, qualquer tipo de plágio quer de textos
publicados, qualquer que seja o meio dessa publicação, incluindo
meios eletrónicos, quer de trabalhos académicos.

O Júri
presidente
Prof. Doutora Ana Maria Rodrigues
Professora Auxiliar do Departamento de Biologia da Universidade de
Aveiro
Prof. Doutora Sónia Alexandra Paiva dos Santos
Investigadora Auxiliar do Centro de Investigação de Montanha –
CIMO da Escola Superior Agrária de Bragança
Prof. Doutor António Nogueira
Professor Associado do Departamento de Biologia da Universidade
de Aveiro

Agradecimentos Ao Doutor Mário Boieiro, por proporcionar a oportunidade de
integração no projeto “Biodiversidade, endemismos e
espécies protegidas associadas às lagoas e cursos de água
da Serra da Estrela: valorização de um século de
aproveitamento hidroelétrico”, financiado pelo fundo EDP
para a biodiversidade 2011. Pelo excelente trabalho realizado
como orientador, pela disponibilidade e por todos os
conhecimentos partilhados.
Ao Professor Doutor António Nogueira pela orientação na
realização deste projeto, pela disponibilidade e pelo
conhecimento transmitido.
Aos meus colegas de trabalho de campo e amigos Sandra
Antunes e Hugo Figueiredo, pelo apoio, transmissão de
conhecimentos, risos e companheirismo.
Ao José Conde pela disponibilidade de apoio ao trabalho, pela
transmissão de conhecimentos, pelas brincadeiras e sorrisos
partilhados e claro pela amizade.
À Paula Mendes pela receção calorosa na sua casa, pelos
jantares fantásticos, pelas conversas, sorrisos e principalmente
pela amizade partilhada.
Ao Albano Soares, pelo apoio, pelos conhecimentos
transmitidos e pelo companheirismo.
À SPEN-Sociedade Portuguesa de Entomologia,
nomeadamente à Carla Rego, pelo encaminhamento para o
Doutor Mário Boieiro, que me permitiu dar o primeiro passo
para a realização desta tese.
À Fundação EDP pelo apoio financeiro cedido através do
Fundo EDP para a biodiversidade 2011.
Ao CISE-Centro de Investigação da Serra da Estrela, pelo
apoio logístico.
Aos meus pais pelo esforço realizado para me proporcionarem
a oportunidade da obtenção de um curso superior.

Palavras-chave Serra da Estrela, ecologia, Odonata, Lepidoptera,
Conservação
Resumo A Serra da Estrela apresenta um elevado valor natural
devido à presença de habitats raros e a uma assinalável
biodiversidade. O objetivo deste trabalho foi avaliar a
importância das diversas áreas do Planalto Superior da
Serra da Estrela para a conservação da natureza, através do
estudo de dois grupos bioindicadores, Odonata e
Lepidoptera. A metodologia de amostragem adotada
baseou-se na contagem de indivíduos adultos através de
transectos fixos com uma extensão de 150 m ao longo das
quatro tipologias de habitat em estudo: lagoas naturais,
barragens pequenas, barragens grandes e áreas terrestres
(pontos secos). Foram identificadas 55 espécies de
Lepidoptera nos 2301 indivíduos observados e 23 espécies
de Odonata entre os 2580 indivíduos registados. Destaca-se
a presença das espécies de Lepidoptera: Vanessa
virginiensis, Cyaniris semiargus e Coenonympha iphioides
e das espécies de Odonata: Aeshna juncea e Sympetrum
flaveolum, consideradas raras ou vulneráveis em Portugal.
Encontraram-se diferenças significativas na biodiversidade
de ambas as ordens entre as tipologias de habitat estudadas.
A riqueza e diversidade específica de borboletas foi maior
nas barragens de grandes dimensões devido,
principalmente, à existência de uma maior riqueza e
abundância de espécies de plantas nectaríferas. A
abundância e riqueza específica de libélulas foi maior nas
lagoas naturais e em algumas barragens pequenas
naturalizadas, onde a perturbação ambiental é reduzida.
Nestes habitats encontraram-se algumas espécies
estenotópicas e muito sensíveis à perturbação. Pela rica
diversidade de espécies presentes na Serra da Estrela com
estatuto de espécies vulneráveis, raras, protegidas por lei ou
endémicas, devem promover-se medidas de gestão
adequadas, sendo que a proteção dos habitats poderá ser a
de maior relevância.

Keywords Serra da Estrela, ecology, Odonata, Lepidoptera,
Conservation
Abstract Serra da Estrela has a high natural value due to the presence
of rare habitats and a remarkable biodiversity. The aim of
this study was to evaluate the importance of different areas
of the upper plateau of Serra da Estrela for the conservation
of nature through the study of two bio-indicators groups,
Odonata and Lepidoptera. The sampling methodology was
based on the count of adult individuals, using fixed
transects with a length of 150 meters along four different
habitats: natural ponds, small dams, large dams and inland
areas (dry spots). In 2301 observed individuals, 55
Lepidoptera species were identified, and in 2580
individuals, 23 species of Odonata were identified. The
presence of Lepidoptera species Vanessa virginiensis,
Cyaniris semiargus and Coenonympha iphioides, and
Odonata species Aeshna juncea and Sympetrum flaveolum,
is noteworthy, since they are considered rare or vulnerable
in Portugal. Significant differences on the biodiversity of
both orders were found between the different types of
studied habitats. The biodiversity of butterflies was higher
in large dams, mainly due to the existence of a greater
richness and abundance of nectariferous plant species,
while the diversity of Odonata species was higher in natural
lakes and some small naturalized dams, where disturbance
levels are low. In these habitats exists some stenotopic
species, and also species very sensitive to disturbance. Due
to the high diversity of species present in Serra da Estrela
with the status of vulnerable, rare, endemic or protected by
the law, appropriate management measures should be
promoted, and the protection of habitats may be the one of
greatest relevance.

i
Índice
Lista de figuras ............................................................................................................................................. ii
Lista de tabelas ............................................................................................................................................. v
Introdução ..................................................................................................................................................... 1
Material e Métodos ..................................................................................................................................... 12
Área de estudo .................................................................................................................................... 12
Trabalhos de campo ............................................................................................................................ 15
Análise estatística ............................................................................................................................... 16
Resultados e Discussão............................................................................................................................... 18
Análise faunística da ordem Lepidoptera nos diferentes habitats ....................................................... 18
Análise faunística da ordem Odonata nos diferentes habitats ............................................................ 23
Semelhanças na composição das comunidades de Lepidoptera nos diferentes habitats ..................... 28
Semelhanças na composição das comunidades de Odonata nos diferentes habitats........................... 30
Identificação de espécies de Lepidoptera indicadoras para os diferentes habitats .............................. 33
Identificação de espécies de Odonata indicadoras para os diferentes habitats ................................... 34
Identificação de áreas importantes para a conservação de borboletas e libélulas no Planalto Superior
da Serra da Estrela .............................................................................................................................. 36
Conclusões ................................................................................................................................................. 40
Referências bibliográficas ...................................................................................................................... 41
Anexo I ....................................................................................................................................................... 54
Anexo II...................................................................................................................................................... 55
Anexo III .................................................................................................................................................... 57
Anexo IV .................................................................................................................................................... 59
Anexo VI .................................................................................................................................................... 61

ii
Lista de figuras
Figura 1 Andares bioclimáticos presentes na Serra da Estrela
(adaptado de Jansen, 2002)……………………………………………………………...2
Figura 2 Exemplos de habitats naturais raros presentes na Serra da Estrela: a) Turfeiras
- Covão da Clareza; b) Lagoa natural - Lagoa Redonda. Fotos de José Conde…………3
Figura 3 Sympetrum flaveolum - espécie de Odonata presente na Serra da Estrela com
elevado valor de conservação. Foto de José Conde…………………………………….10
Figura 4 Espécies de Lepidoptera com elevado valor de conservação em Portugal e na
Serra da Estrela: a) Cyaniris semiargus (rara em Portugal); b) Euphydrias aurinia (listada
na Diretiva de Habitats) e c) Satyrus actaea (presente na Serra da Estrela, acima dos
1200m). Fotos de Albano Soares (a,c) e Hugo Figueiredo
(b)……………………………………………………………………………….………11
Figura 5 Tipologias dos habitats em estudo: a) ponto seco; b) barragem de pequenas
dimensões; c) lagoa natural e d) barragem de grandes dimensões. Fotos de José
Conde……………………………………………………………………………….…..14
Figura 6 Localização do Parque Natural da Serra da Estrela em Portugal e distribuição
espacial dos locais de amostragem. Mapa adaptado do Google Earth®, usando o software
Quantum GIS®, versão
2.10.1………………………………………………………………………...…………14
Figura 7 Espaço de contagem e identificação de indivíduos (adaptado de Pollard, &
Yates, 1993)……………………………………………………………………………15
Figura 8 Descritores de diversidade da ordem Lepidoptera para os anos de 2013 e 2014,
referentes ao índice de Shannon-Wiener (H’), ao índice de Berger-Parker (d) e à
equitabilidade (Equi) pelo índice de Pielou. Tipologia de habitat 1- pontos secos; 2-
lagoas naturais; 3- barragens pequenas; 4- barragens de
grandesdimensões.……………………………...………………………………...…….20
Figura 9 Resultados dos descritores da diversidade da ordem Odonata para os anos de
2013 e 2014, referentes ao índice de Shannon-Wiener (H’), ao índice de Berger-Parker
(d) e à equitabilidade (Equi) pelo índice de Pielou. Tipologia de habitat: 1- pontos secos;
2- lagoas naturais; 3- barragens pequenas; 4- barragens de grandes
dimensões…………………………………………………………………………...….25

iii
Figura 10 Análise de componentes principais (PCA) para determinar os principais
padrões de variação nos dados da ordem Lepidoptera. Análise da variação entre anos (a),
locais (b) e espécies que caracterizam a variação
(c)……………………………………………………………..………………………..28
Figura 11 Dendrograma da análise de ordenação dos locais de amostragem da ordem
Lepidoptera para as diferentes tipologias de habitat em estudo. Os códigos dos nomes dos
locais de amostragem (apresentados no Anexo V) incluem a indicação da tipologia de
habitat (PS - pontos secos; Nat - lagoas naturais; Bar - barragens pequenas; BarG -
barragens de grandes dimensões)……………………………………………………....29
Figura 12 Análise de componentes principais (PCA) para determinar os principais
padrões de variação nos dados da ordem Odonata. Análise da variação entre anos (a),
locais (b) e espécies que caracterizam a variação
(c)…………………………………………………………………………..…………...31
Figura 13 Dendrograma da análise de ordenação dos locais de amostragem da ordem
Odonata para as diferentes tipologias de habitat em estudo. Os códigos dos nomes dos
locais de amostragem (apresentados no Anexo V) incluem a indicação da tipologia de
habitat (PS - pontos secos; Nat - lagoas naturais; Bar - barragens pequenas; BarG -
barragens de grandes dimensões)………………………………………………………32
Figura 14 Análise IndVal para a ordem Lepidoptera. Vermelho: quartil 0-25%; Amarelo
avermelhado: quartil 25-50%; amarelo esverdeado: quartil 50-75%; Verde: quartil 75-
100%. Tipologias de habitat - 1: pontos secos; 2: lagoas naturais; 3: barragens pequenas;
4: barragens
grandes…………………………………………………….……………………………34
Figura 15 Análise IndVal para a ordem Odonata. Vermelho: quartil 0-25%; Amarelo
avermelhado: quartil 25-50%; amarelo esverdeado: quartil 50-75%; Verde: quartil 75-
100%. Tipologias de habitat - 1: pontos secos; 2: lagoas naturais; 3: barragens pequenas;
4: barragens
grandes………………………………………………………………………….………36
Figura 16 Resultados da análise de complementaridade da riqueza específica de
borboletas, libélulas e de ambos os grupos de insetos nos locais de
estudo………………………………………………………………………………..….37

iv
Figura 17 Área mínima de conservação da riqueza específica de Odonata (a vermelho) e
Lepidoptera (a azul) na área de estudo, usando o método dos polígonos. Imagem adaptada
do Google
Earth®……………………………………………………………………………...…...38

v
Lista de tabelas
Tabela 1 Média e amplitude dos valores da abundância relativa (N), da riqueza específica
(S), da equitabilidade (E) e dos índices de diversidade e de dominância (H’ e 1/d
respetivamente) de cada tipologia de habitat para a ordem
Lepidoptera…………………………………………………………………..…………19
Tabela 2 Resultados das análises de variância dos descritores da diversidade (lnN, lnS,
H’, Hmáx, E e d) entre as tipologias de habitat e os anos de amostragem para a ordem
Lepidoptera……………………………………………………………………………..21
Tabela 3 Média e amplitude dos valores da abundância relativa (N), da riqueza específica
(S), da equitabilidade (E) e dos índices de diversidade e de dominância (H’ e 1/d
respetivamente) de cada tipologia de habitat para a ordem
Odonata……………………………………………………………………….………...24
Tabela 4 Resultados das análises de variância dos descritores da diversidade (lnN, lnS,
H’, Hmáx, E e d) entre as tipologias de habitat e os anos de amostragem para a ordem
Odonata………………………………………………………………………………...27

1
Introdução
As montanhas ocupam uma área de cerca 10.000 Km2, em Portugal, o que
corresponde a 11% da superfície emersa (Aguiar et al., 2009), e são consideradas um dos
ambientes mais frágeis do mundo (Diaz et al., 2003). Elas apresentam um papel
fundamental no que concerne aos serviços do ecossistema, sendo responsáveis pela
qualidade, quantidade e regularidade da água consumida pelas populações humanas, pelo
funcionamento do ciclo dos nutrientes e pelas regulações climáticas regionais (Aguiar et
al., 2009), sabendo-se que a sua importância ultrapassa os seus limites geográficos e se
estende muito além das áreas envolventes (Spehn et al., 2011). Além disso, as montanhas
constituem centros de biodiversidade endémica, abrigando espécies e ecossistemas
ameaçados (Aguiar et al., 2009) e proporcionam também serviços com valor económico,
enquanto espaços privilegiados para o turismo e para a produção agrícola e pecuária
(Spehn et al., 2011).
Ao longo dos tempos, foram diversos os promotores responsáveis pela alteração
deste ecossistema, conduzindo a uma mudança na sua estrutura, funções e serviços
prestados. Desses fatores destacamos como principais, a agro-pastorícia, as alterações do
uso do solo, as alterações climáticas, o fogo, as políticas e a legislação ambiental e por
fim, a coesão social e a identidade cultural (Aguiar et al., 2009).
A cordilheira Central Ibérica é um maciço montanhoso da Península Ibérica,
caracterizada por ser uma morfoestrutura do tipo “montanha de blocos” (Ribeiro, 1954),
com orientação de WSW-ENE. Possui cerca de 500 Km de extensão e 40 Km de largura
(Daveau, 1969; Vegas et al., 1990). Na parte ocidental desta cordilheira encontra-se a
Serra da Estrela, a montanha mais alta de Portugal Continental, constituída por planaltos
alongados na direção SW-NE, que se estendem por 115 Km de extensão e com uma
largura média de 25 Km, desde a Guarda até à Serra da Lousã (Daveau, 1969; Lautensach,
1932), alcançando na Torre a altitude máxima de 1993 m (Ribeiro, 1954).
Genericamente, pode-se dividir a Serra da Estrela em três andares bioclimáticos
(Figura 1): o basal, o intermédio e o superior (Pinto da Silva, & Teles, 1980). Segundo
Jansen (2002), o andar basal estende-se desde o sopé da montanha até cerca de 900 m de
altitude, contendo dois macrobioclimas: o mesomediterrâneo nas áreas mais húmidas,
onde se pode encontrar bosques perenifólios ou mistos de quercíneas perenifólias e
caducifólias, como o Quercus suber e o Q. rotundifolia, e o macrobioclima
mesotemperado onde dominam os bosques de carvalhos caducifólios, constituídos

2
principalmente por Q. robur. Devido à elevada ocupação humana neste andar, a presença
destes bosques é cada vez menor, existindo uma significativa fragmentação e perturbação
dos mesmos, chegando a ser substituídos por outras formações vegetais como matos
rasteiros, prados abertos, etc. No andar intermédio, dos 900 m a 1600 m de altitude,
existem os macrobioclimas supratemperado e o supramediterrâneo. No macrobioclima
supratemperado, sob condições ideais, predominariam os bosques de Quercus pyrenaica
e de Betula celtiberica ou Taxus baccata, enquanto no macrobioclima supramediterrâneo
a vegetação natural potencial consistiria em bosques caducifólios ou mistos de quercíneas
(com Q. pyrenaica e Q. rotundifolia). Contudo, actualmente, devido à degradação dos
habitats, estas formações são escassas, encontrando-se substituídas por urzais, giestais e
caldoneirais. Por fim, acima dos 1600 m, situa-se o andar superior, onde existem os
macrobioclimas ortotemperado e criotemperado. A vegetação natural predominante
nestas áreas onde a ação do frio é mais intensa é constituída por zimbrais rasteiros,
caldoneirais pulviniformes de Echinospartum ibericum e cervunais. Verificam-se
também áreas de comunidades de rochedos, de cascalheira, turfeira, formações de
lagoachos, ribeiros e lagos. Em áreas mais degradadas, pelo fogo ou pastoreio por
exemplo, ocorrem sobretudo urzais e giestais (Jansen, 2002).
O gradiente altitudinal da Serra da Estrela possibilita a existência de um conjunto
significativo de habitats raros e protegidos por lei, alguns dos quais, no nosso país,
apresentam aqui a maior expressão (http://www.icnf.pt/portal/naturaclas/ap/p-nat/pnse).
Alguns desses habitats naturais mais raros em Portugal, como as lagoas de altitude
e as turfeiras (Figura 2), ocorrem numa área denominada Planalto Superior da Serra da
Estrela. Este planalto, segundo Rocha et al., (2004), divide-se ainda em 2 grupos
Figura 1 Andares bioclimáticos presentes na Serra da Estrela (adaptado de Jansen, 2002).

3
principais, o ocidental (Torre – Penhas Douradas) e o oriental (Alto da Pedrice – Malhada
Alta), que se estendem entre os 1450 e 1993 m.
No que concerne às lagoas de altitude, apesar de ocorrerem na Europa vários
sistemas de montanha onde estas se encontram bem representadas (Schmitt, 2009), em
Portugal (ou até mesmo na Península Ibérica) a sua representatividade é baixa, pois
apenas está representado por um conjunto de lagoas na Serra da Estrela e um pequeno
número na Serra do Gerês, facto que enaltece a singularidade das lagoas de altitude da
Serra da Estrela (www.icnf.pt).
A maioria destas lagoas originou-se durante a última glaciação, devido à erosão
provocada pelo gelo sobre a rocha matriz, sendo a sua presença favorecida pela baixa
permeabilidade dessa rocha. A localização das lagoas de altitude determina uma série de
condições ambientais que são tradicionalmente consideradas extremas para a vida. A
oligotrofia, que resulta da baixa erosão e do desenvolvimento limitado do solo, origina
águas bastante diluídas no que concerne aos nutrientes e componentes salinos, pelo que
os organismos residentes ficam expostos a condições extremas de osmorregulação. A
altitude é dos fatores que mais influencia a ocorrência de condições extremas (Catalan et
al., 2006). Esta influência na diminuição da espessura da atmosfera permite o aumento da
radiação e da incidência dos raios UV-B, expondo as lagoas, durante o período sem gelo,
a elevadas radiações prejudiciais (Catalan et al., 2006), e ainda interfere na dessecação,
na pressão atmosférica, no pH e na temperatura (Seufferheld et al., 2008).
A sensibilidade destes ecossistemas em conjunto com o seu isolamento e
afastamento das áreas de atividade humana elevada, proporcionam a estes locais
características essenciais para estudos sobre o passado e o presente relativamente a
a) b)
Figura 2 Exemplos de habitats naturais raros presentes na Serra da Estrela: a) Turfeiras - Covão da Clareza;
b) Lagoa natural - Lagoa Redonda. Fotos de José Conde.

4
mudanças ambientais em grande ou pequena escala, assim como sobre a vida em
ambientes extremos (Catalan et al., 2006).
Como referenciado anteriormente, as montanhas são cruciais para o fornecimento
de água doce para as populações, em muitas partes do mundo. Portanto, é crucial
assegurar a gestão sustentável dos ecossistemas de montanha (Viviroli, & Weingartner,
2008). O seu estudo não é fácil, pois apresentam uma elevada complexidade ao nível da
geologia, da geomorfologia e do clima. Embora se encontrem em áreas isoladas, estas
podem mostrar influências antrópicas e climáticas (Schreier, 2003; Zektser, & Everett,
2004). A sua sustentabilidade está a ser afetada por diversos fatores como a crescente
demanda de água para usos urbanos, industriais e agrícolas, as mudanças no uso da terra
para a agricultura, floresta cultivada ou urbanização, a construção de barragens para o
abastecimento de água ou produção de energia hidroelétrica, as atividades recreativas e
alterações climáticas (Schreier, 2003). Por consequência, o Programa Hidrológico
Internacional da UNESCO prevê que as montanhas de elevada altitude funcionem como
reservatórios de água para os países e considera a hidrologia das zonas de montanha como
uma das questões mais essenciais para a investigação (Aureli, 2002), o que nos encaminha
para uma crescente consciencialização sobre a importância dos sistemas hidrológicos de
montanha, como a fonte de uma fração significativa dos recursos mundiais de água
(Marques et al., 2013) e para a realização de estudos sobre a gestão e conservação dos
recursos hídricos destas regiões, a fim de colmatar a escassez de dados e a má
compreensão dos mesmos.
A geomorfologia associada ao clima, aos habitats raros, ao tipo do relevo e a
exposição das encostas, proporcionam uma paisagem singular à Serra da Estrela (Moura,
2006). Cumulativamente, também oferecem um património natural e cultural de elevado
interesse, que permitiu a classificação de parte do maciço como Parque Natural, em 1976.
Mais tarde, em 1992, adquiriu o estatuto de Reserva Biogenética, e por fim, foi feita a
inserção da maioria das áreas da Serra da Estrela na Rede Natura 2000 (Martins, &
Silveira, 2008), com o intuito de conservação dos habitats e ambientes raros (Marques,
2006).
A nível nacional, a Serra da Estrela é um local emblemático por se encontrar
estreitamente associada à história da produção de eletricidade. O sistema eletroprodutor
da Serra da Estrela é um dos mais antigos de Portugal, contabilizando mais de um século
de existência (Serrano et al., 2014). Este sistema, à exceção da subestação instalada no
vale de Unhais, encontra-se exclusivamente instalado na parte superior da bacia do rio

5
Alva – a denominada cascata da Estrela, beneficiando de diversos cursos de água que
percorrem um apreciável gradiente altitudinal para a produção de energia elétrica. Em
determinados locais, as lagoas naturais ou cursos de água existentes foram modificados
para a criação de reservatórios que originaram lagoas artificiais, permitindo o
armazenamento de água e a sua gestão para a produção de energia elétrica ao longo do
ciclo anual. Do ponto de vista ecológico, estas lagoas artificiais constituíram uma
oportunidade para a ocorrência de um número considerável de espécies, que usufrui
destes locais para a sua alimentação, repouso, reprodução ou para o desenvolvimento de
parte ou todo o seu ciclo de vida (Serrano et al., 2014).
Contudo, no que se refere às centrais hidroelétricas, sabe-se que estas acarretam
impactos não só ao nível estético, mas também na biodiversidade de montanha, pois
modificam o regime de caudais, promovem a redução da velocidade da corrente e
contribuem para o aumento da sedimentação (Bombino et al., 2014; Tortajada et al.,
2012). São ainda responsáveis pela alteração e destruição de habitats devido às
interrupções espaciais e temporais da continuidade das linhas de água, afetando tanto os
ecossistemas aquáticos como os terrestres envolventes. Por fim, causam efeito de barreira
à dispersão de muitos organismos, e aumentam a presença humana e os níveis de ruído
nas áreas em que são implantadas (Leitão et al., 2010). No entanto, atualmente não
existem estudos que estimem a importância destas lagoas para a conservação da natureza,
nem que avaliem o seu impacto nas comunidades envolventes (Serrano et al., 2014).
Apesar do potencial impacto negativo que a criação de albufeiras poderá ter tido, verifica-
se ainda a ocorrência no Planalto Superior da Serra da Estrela de um número significativo
de lagoas naturais de origem glaciária, que apresentam várias tipologias e que se
encontram em diferentes fases do processo de sucessão ecológica (Costa et al., 2004;
Serrano et al., 2014). À exceção da lagoa seca (na margem direita do rio Zêzere) que é de
natureza endorreica, as lagoas naturais integram as duas principais redes hidrográficas
existentes na serra, comunicando algumas delas entre si e com os afluentes dos rios Alva
e Zêzere (Serrano et. al., 2014). A biodiversidade associada a estes ecossistemas realçam
o seu valor para a conservação da natureza. A lagartixa-da-
montanha (Iberolacerta monticola (Boulenger, 1905)), o lagarto-d’água (Lacerta
schreiberi (Bedriaga, 1878)), o tritão-de-ventre-laranja (Lissotriton boscai (Lataste,
1879)) e a rã-ibérica (Rana iberica (Boulenger, 1879)), são alguns dos répteis e anfíbios
de grande importância que dependem destes ecossistemas. Podemos ainda observar a
petinha-dos-campos (Anthus campestris (Linnaeus,1758)), a gralha-de-bico-vermelho

6
(Pyrrhocorax pyrrhocorax (Linnaeus,1758)) e a sombria (Emberiza hortulana
(Linnaeus,1758)), espécies que se encontram listadas no Anexo I da Directiva Aves. Nos
mamíferos destaca-se a presença da lontra (Lutra lutra (Linnaeus,1758)) e do rato-de-
água (Arvicola sapidus (Miller, 1908)) (Costa et al., 2004). As ordens de insetos com
maior relevância neste ambiente são Ephemeroptera, Odonata, Plecoptera, Hemiptera,
Coleoptera, Lepidoptera e Diptera (Hynes, 1970), evidenciando-se espécies como Aeshna
juncea (Linnaeus, 1758) (Odonata), Sympetrum flaveolum (Linnaeus, 1758) (Odonata),
Leuctra estrela (Aubert, 1962) (Plecoptera), Annitella amelia (Sipahiler, 1998)
(Trichoptera), Hydraena zezerensis (Díaz Pazos and Bilton, 1995) (Coleoptera), Zabrus
estrellanus (Heyden, 1880) (Coleoptera) e Monotropus lusitanicus (Baraud, 1976)
(Coleoptera), que possuem neste local as suas principais populações ou apresentam
estatuto de espécies ameaçadas (Farminhão et al., 2014).
A destruição e fragmentação dos ecossistemas, resultante das ações exercidas pelo
Homem, atua diretamente sobre a diversidade biológica, afetando a riqueza de espécies,
alterando os padrões de abundância, a sobrevivência de endemismos e também as
propriedades estruturais do ecossistema. Deste modo, a manutenção da biodiversidade é
um desafio importante para a conservação da natureza. Para uma melhor compreensão
dos impactos decorrentes de atividades humanas, a monitorização de determinados taxa
torna-se uma ferramenta de elevado valor, permitindo a obtenção de informações sobre a
gravidade dos efeitos da destruição e fragmentação, bem como a identificação de
prioridades de gestão e a avaliação do sucesso de medidas de conservação (Didham, 1997;
Kitching, 1994). Para McGeoch (1998) um bioindicador pode ser definido como uma espécie ou
um grupo de espécies que refletem o estado biótico e abiótico do meio ambiente que
representa, o impacto das mudanças ambientais sobre o habitat, comunidade ou
ecossistema, e ainda que indiquem a diversidade de outras espécies. Assim sendo, a
mesma autora classifica-os em 3 classes: indicadores ambientais (que avaliam mudanças
no ambiente físico-químico), indicadores ecológicos (que fornecem informação sobre o
impacto de determinados fatores sobre os ecossistemas) e indicadores de biodiversidade
(que dão indicação sobre a relevância do habitat para a conservação). A maioria dos
bioindicadores ecológicos e ambientais é identificada através do estabelecimento de uma
forte relação destes com características muito particulares do ambiente onde se inserem
(Davis et al., 2001; Kitching et al., 2000). Para serem consideradas bons bioindicadores,
as espécies ou grupos de espécies devem: ter uma ecologia e a taxonomia bem conhecida,

7
incluir organismos especializados em certos requisitos do habitat, fornecer avisos
antecipados da ocorrência de mudança, ser de fácil observação e identificação, e o seu
estudo ser de baixo custo. Por outro lado, a sua resposta às alterações deverá refletir a
resposta de outras espécies da comunidade e apresentarem a capacidade de diferenciar
entre os ciclos ou tendências naturais e as provocadas por pressões antropogénicas
(McGeoch, 1998).
De entre os diversos grupos de bioindicadores, os invertebrados são bastante
solicitados, uma vez que diversas ordens e espécies possuem as características necessárias
ao estudo da qualidade ambiental. Estes podem indicar mudanças no ambiente através
das suas respostas a diferentes níveis da organização, desde o indivíduo até toda a
comunidade. Ao nível individual estes ainda podem servir como bioindicadores a curto
prazo das condições ambientais particulares, onde uma resposta a um estresse ambiental
particular pode ser demonstrada na sua fisiologia ou comportamento (Hodkinson, &
Jackson, 2005).
Independentemente dos ecossistemas em estudo, sejam montanhas ou planícies,
os princípios e as gamas no uso de invertebrados como bioindicadores para a gestão
ambiental são semelhantes (Hodkinson, & Jackson, 2005).
Nos últimos anos, verificou-se um esforço conjunto de muitos países europeus na
procura, organização e armazenamento do conhecimento sobre a biodiversidade,
incluindo a dos organismos invertebrados, e também na definição de prioridades de
conservação que orientem as estratégias para a conservação da natureza (p. ex. Borges et
al., 2010; De Jong et al., 2014).
Em Portugal, o conhecimento da biodiversidade de invertebrados dos
ecossistemas de montanha é ainda escasso. Contudo, apesar deste desconhecimento, a
Serra da Estrela, devido à sua extensão, localização, diversidade de habitats, entre outros
fatores, abrangerá seguramente o maior número de espécies conhecidas das várias áreas
protegidas existentes em Portugal (Serrano et al., 2014). Atualmente são conhecidas mais
de 1.000 espécies de plantas, 250 de vertebrados e cerca de 2.800 de invertebrados nesta
área protegida (http://www.cise.pt/). Vários grupos de organismos invertebrados apresentam
algumas espécies com estatuto de conservação desfavorável, por ostentarem raridade
local e/ou declínio populacional, justificando-se o desenvolvimento de ações de
conservação visando a sua recuperação.
Da fauna de invertebrados conhecida em Portugal, encontram-se na Serra da
Estrela diversos endemismos e algumas espécies muito raras, que apenas são conhecidas

8
nesta área ou têm aqui as suas principais populações a nível nacional. Assim, destacam-
se várias espécies de artrópodes terrestres, na sua maioria insetos, que se encontram
associados aos habitats de alta montanha, mas também a ecossistemas naturais de menor
altitude (Bivar-de-Sousa et al., 2008; Ferreira et al., 2009; Grosso-Silva, 2000; Salgueiro,
2002; Serrano et al., 2014). Muitos desses insetos são particularmente raros e apenas
alguns se encontram protegidos por lei a nível internacional (Diretiva Habitats
92/43/EEC).
Deste modo, através do projeto “Biodiversidade, endemismos e espécies
protegidas associadas às lagoas e cursos de água da Serra da Estrela: valorização de um
século de aproveitamento hidroelétrico”, financiado pelo fundo EDP para a
biodiversidade 2011, pretendeu-se caracterizar as comunidades de macroinvertebrados a
fim de obter um aumento da informação ecológica e posteriormente proceder à definição
de propostas de gestão desses habitats. Em paralelo, desenvolveu-se a recolha de
informação sobre a biologia, ecologia e estatuto de conservação não só de espécies de
macroinvertebrados aquáticos (vários grupos taxonómicos), mas também de outros
grupos de insetos como as borboletas (Lepidoptera), as libélulas (fase adulta, Odonata) e
os escaravelhos terrestres (Coleoptera), por forma a contribuir para o esforço
internacional de salvaguarda das suas populações, principalmente das espécies
endémicas, raras ou protegidas por lei.
Os objetivos gerais deste projeto foram os seguintes:
- Conhecer e valorizar a biodiversidade de macroinvertebrados associada às lagoas
naturais e artificializadas do Planalto Superior da Serra da Estrela;
- Avaliar em que medida o sistema de aproveitamento hidroelétrico da Serra da Estrela
contribui para a biodiversidade da região;
- Avaliar a importância de cada uma das lagoas selecionadas para a conservação de
espécies endémicas, raras ou protegidas por lei;
- Avaliar o estado ecológico das lagoas existentes no Planalto Superior da Serra da Estrela
e recomendar medidas de gestão para esses espaços.
Para a prossecução destes objetivos foi adotada uma metodologia centrada no
estudo de diversos grupos taxonómicos, que utilizam as lagoas de modo distinto, pois
apenas com uma perspetiva abrangente se poderá fazer uma avaliação mais precisa da
relevância dos diferentes habitats para a conservação da natureza. Deste modo, foram
aplicadas técnicas de amostragem complementares para estudar os animais que vivem no
interior das lagoas e barragens (macroinvertebrados aquáticos), os que ocorrem nas suas

9
margens ao nível do solo (artrópodes epígeos) e os que utilizam a vegetação envolvente
das massas de água (borboletas e libélulas).
Nesta tese, são apresentados e discutidos os resultados relativos a borboletas
(Lepidoptera: Rhopalocera) e libélulas (Odonata) grupos que se destacam por
apresentarem uma apreciável riqueza específica, incluindo algumas espécies raras, que
no nosso país são exclusivas da Serra da Estrela (Ferreira et al., 2009; Serrano et al.,
2014).
Sabe-se que os insetos constituem uma proporção substancial da riqueza de
espécies terrestres e da biomassa existente no planeta, desempenhando um papel
fundamental nos ecossistemas. Consequentemente, os estudos sobre estes organismos
como bioindicadores têm vindo a aumentar e a ganhar importância na comunidade
científica ( Kremen et al., 1993; Rosenberg et al., 1986). Dos diversos grupos
emblemáticos da Serra da Estrela, destacam-se as libélulas (ordem Odonata) e as
borboletas diurnas (ordem Lepidoptera; subordem Rhopalocera) devido aos valores de
riqueza específica com que ocorrem, à presença de espécies muito raras no nosso país e
aos relevantes papeis ecológicos que desempenham (Acquah-Lamptey et al., 2013a;
Čelik, 2007; Simaika, & Samways, 2011).
A ordem Odonata (Figura 3) encontra-se distribuída praticamente por todo o
Mundo, à exceção das regiões geladas permanentes, contando com aproximadamente
6.500 espécies. Em Portugal são conhecidas 65 espécies (Maravalhas, & Soares, 2013)
que se encontram, na sua maioria, associadas a zonas húmidas e de água doce, ocupando
um nicho trófico crítico nesses sistemas.

10
Algumas espécies conseguem tolerar uma maior gama de condições, enquanto
outras são extremamente sensíveis a variações no ambiente onde estão inseridas. Este
grupo de insetos é historicamente bem estudado (Acquah-Lamptey et al., 2013b),
existindo uma grande quantidade de informação ecológica sobre diversas espécies, e são
relativamente fáceis de observar e identificar, o que facilita a amostragem das suas
populações e estudos da sua diversidade (Acquah-Lamptey et al., 2013a; Simaika, &
Samways, 2011). Importa ainda mencionar que as larvas desta ordem são relativamente
sedentárias e a sua longevidade pode ser superior a um ano. Como possuem tolerâncias
específicas para algumas variáveis associadas a perturbações ambientais são também
úteis em bioindicação. Pelas razões atrás citadas, diversos autores concluíram que os
Odonata são indicadores fiáveis na avaliação dos impactos decorrentes da perturbação
humana, razão que tem motivado a sua aplicação generalizada em ecossistemas aquáticos
em diversos pontos do Mundo (Acquah-Lamptey et al., 2013a; Clark, & Samways, 1996).
As borboletas diurnas (Lepidoptera; Rhopalocera) (Figura 4) apresentam uma
elevada riqueza específica, contando cerca de 18.000 espécies conhecidas no Mundo, e
ocorrendo em quase todas as regiões e numa grande variedade de habitats (Larsen, 1988).
Figura 3 Sympetrum flaveolum (Linnaeus,
1758)- espécie de Odonata presente na Serra
da Estrela com elevado valor de conservação.
Foto de José Conde.

11
A ordem Lepidoptera apresenta um elevado valor económico, por possuir espécies
polinizadoras e indicadoras de biodiversidade e de qualidade ambiental (Chakravarthy et
al., 1997), fornecendo informação importante para a implementação de medidas de gestão
e conservação de ecossistemas. Os requisitos ecológicos para bons bioindicadores são, de
um modo geral, preenchidos por esta ordem (Čelik, 2007), uma vez que para a maioria
das espécies, a sua taxonomia e ecologia são bem conhecidas, são frequentemente
especializadas no que diz respeito às suas plantas hospedeiras e desempenham papéis
importantes nos ecossistemas. Além disso, são ativas durante o dia e, na maioria dos
casos, fáceis de identificar. Como indicadores, respondem rapidamente às mudanças
ambientais, tais como alterações climáticas ou a perturbação por atividades humanas
(Stefanescu et al., 2004). Por outro lado, o estudo da diversidade desta ordem pode servir
como um indicador da biodiversidade dessa região, uma vez que a riqueza de borboletas
está frequentemente relacionada com a diversidade de outros organismos, particularmente
com as plantas com as quais estão estreitamente associadas (Ehrlich, & Raven, 1964;
Holl, 1995; Kitahara et al., 2008). Diversos trabalhos de investigação têm sido realizados
utilizando este grupo como bioindicador, tendo sido particularmente útil na avaliação das
alterações ambientais devido a causas naturais ou ação antropogénica (Blair, & Launer,
1997; Kocher, & Williams, 2000). Estes estudos podem realizar-se ao nível de espécies
individuais, de taxa supraespecíficos ou comunidades, avaliando um ou mais grupos de
organismos numa ou em diversas tipologias de habitat, a fim de determinar prioridades
de conservação, monitorizar as alterações nas comunidades naturais ou identificar
necessidades de gestão ou recuperação dos habitats. A fiabilidade da informação
recolhida com estes bioindicadores depende da sua adequação à questão a ser investigada.
Figura 4 Espécies de Lepidoptera com elevado valor de conservação em Portugal e na Serra da Estrela: a)
Cyaniris semiargus (Rottemburg, 1775) (rara em Portugal); b) Euphydrias aurinia (Rottemburg, 1775)
(listada na Diretiva de Habitats) e c) Satyrus actaea (Esper, 1781) (presente na Serra da Estrela, acima dos
1200m). Fotos de Albano Soares (a,c) e Hugo Figueiredo (b).
a) b) c)

12
Deste modo, para determinadas tipologias de habitat existem bioindicadores ambientais
mais adequados que outros (Gerlach et al., 2013).
No nosso país, o conhecimento existente sobre a biologia de vários grupos de
insetos é ainda muito escasso, não existindo estudos elementares sobre os seus hábitos
ecológicos, distribuições e a dimensão dos seus efetivos populacionais. Este
desconhecimento biológico constitui um sério obstáculo ao desenvolvimento de medidas
de gestão ambiental que permitam a conservação das suas populações. Assim sendo, foi
desenvolvido um conjunto de objetivos para melhorar o conhecimento sobre a biologia
das espécies de alguns grupos de insetos existentes na Serra da Estrela, nomeadamente as
libélulas (Odonata) e as borboletas diurnas (Lepidoptera - Rhopalocera), bem como na
avaliação da sua utilização como potenciais bioindicadores.
Neste capítulo, pretende-se avaliar a importância de diversas áreas do planalto
superior da Serra da Estrela para a conservação da natureza, através do estudo das
comunidades de dois grupos de insetos considerados bons bioindicadores (Odonata e
Lepidoptera). Numa fase inicial procurar-se-ão avaliar os padrões de biodiversidade
associados às lagoas naturais, barragens pequenas, barragens de grande dimensão e
pontos secos. Por outro lado, pretender-se-á avaliar se existem e quais são as espécies
características de alguma das tipologias de habitat estudado. Finalmente, com base nos
resultados obtidos com os dois grupos bioindicadores, identificar-se-á um conjunto de
áreas mais relevantes para a conservação da diversidade destes grupos no Planalto
Superior da Serra da Estrela, contribuindo para uma efetiva gestão da biodiversidade
destes espaços pelas entidades competentes.
Material e Métodos
Área de estudo
O presente estudo foi desenvolvido no Parque Natural da Serra da Estrela, situado
no centro-norte de Portugal, a uma latitude de 40º 2’N e longitude de 7º 35’W (Moura,
2006), mais especificamente na área correspondente ao Planalto Superior da Serra da
Estrela, onde se encontram diversas lagoas naturais e artificiais e os cursos de água que
delas derivam e integram a cabeceira da bacia do rio Alva (Serrano et al., 2014). Este
planalto, estende-se entre os 1450 e 1993 m e constitui uma área de elevado valor
biológico, geológico e paisagístico, razões que contribuíram para a sua classificação

13
como Sítio de Interesse Comunitário, integrando a Rede Natura 2000, e apresenta também
o estatuto de Reserva Biogenética do Conselho da Europa (www.icnf.pt).
A Serra da Estrela apresenta um clima maioritariamente mediterrânico, com
influência atlântica, o que resulta em verões secos e quentes e invernos chuvosos, que
começam em outubro e prolongam-se até maio. A precipitação média anual atinge os
2500 milímetros nas áreas mais altas e é influenciada principalmente pela orientação da
vertente e a altitude, sendo menor no lado ocidental da montanha do que na parte oriental.
A média anual da temperatura do ar encontra-se abaixo dos 7 ºC na maior parte do
planalto e atinge os 4 ºC nas imediações do cume (Vieira, 2004).
As amostragens realizaram-se num conjunto diverso de lagoas naturais, barragens
(de pequena e grande dimensão) e ainda em “pontos secos” (Figura 5), isto é, áreas
afastadas de cursos de água. As lagoas naturais têm origem glaciar, resultaram da erosão
provocada pelo gelo sobre a rocha matriz e encontram-se distribuídas pelo Planalto
Superior. As lagoas artificiais resultaram da artificialização de lagoas naturais existentes
(p.ex. barragem da Lagoa Comprida) ou do emparedamento de áreas em vales (p.ex.
barragem do Vale do Rossim), diferindo entre si no grau de artificialização induzida ao
ambiente local. Dada a heterogeneidade das áreas ocupadas pelas lagoas artificiais,
optámos por dividi-las por duas categorias: “barragens pequenas” que apresentam uma
área inferior a 20 ha e “barragens grandes” quando a sua área é superior ou igual a 20 ha.
Os pontos secos correspondem a áreas terrestres afastadas de linhas de água (geralmente
> 500 m) que procuram traduzir a paisagem dominante no Planalto Superior da Serra da
Estrela, possibilitando avaliar comparativamente a relevância da biodiversidade das
lagoas no contexto da relativa homogeneidade de habitats terrestres deste planalto. A lista
dos locais amostrados é apresentada em anexo (Anexo I).
Os pontos de amostragem foram escolhidos tendo em consideração o
conhecimento existente sobre o Planalto Superior da Serra da Estrela, a diversidade
biológica que lhe está associada e a acessibilidade desses locais, encontrando-se
representados na Figura 6.

14
Figura 5 Tipologias dos habitats em estudo: a) ponto seco; b) barragem de grandes dimensões; c) lagoa natural
e d) barragem de pequenas dimensões. Fotos de José Conde.
a) b)
c) d)
N
Figura 6 Localização do Parque Natural da Serra da Estrela em Portugal e distribuição espacial dos locais de
amostragem. Mapa adaptado do Google Earth®, usando o software Quantum GIS®, versão 2.10.1

15
Trabalhos de campo
A amostragem das formas adultas das ordens Odonata e Lepidoptera foi realizada
entre os meses de junho e setembro, por ser este o período de atividade da maioria das
espécies desses grupos em áreas de elevada altitude. Nos dois anos de estudo (2013 e
2014) foi realizado um total de cinco sessões de amostragem nas diferentes áreas
selecionadas. A metodologia de amostragem adotada baseou-se na realização de
transectos fixos com uma extensão de 150 m para contagem dos insetos adultos ao longo
das margens das lagoas naturais e artificiais (pequenas e grandes) ou nas áreas terrestres
envolventes (pontos secos) dominadas por cervunal, zimbral ou matos baixos. O percurso
foi realizado a passo constante e lento, registando-se as espécies observadas num espaço
correspondente a um cubo de 5 m de lado (Figura 7), e, apenas quando ocorriam dúvidas
na identificação, os indivíduos foram capturados com o auxílio de uma rede entomológica
para identificação, sendo de imediato libertados. Esta metodologia tem sido aplicada
frequentemente em estudos de biodiversidade e ecologia de insetos voadores, pois fornece
uma boa estimativa dos valores de riqueza específica e abundância de um dado local
(Kitahara et al., 2008; Pollard, 1977; Pollard, & Yates 1993; Spitzer et al., 1997,
Stefanescu et al., 2009).
As amostragens realizaram-se entre as 10-18h, com condições climáticas
favoráveis, isto é, com uma temperatura exterior a variar entre os 15-35ºC, uma cobertura
de nuvens inferior a 75% e ausência de vento ou a existência de uma pequena brisa (até
ao máximo de 2 na escala de Beaufort (Metoffice, 2010).
Figura 7 Espaço de contagem e identificação de indivíduos (adaptado de Pollard, & Yates, 1993).

16
Análise estatística
Os dados recolhidos durante os trabalhos de campo de 2013 e 2014 sobre a
abundância e distribuição das espécies de Lepidoptera e Odonata foram organizados em
ficheiros no software Excel do Microsoft® Office 2013, antes da realização das análises
estatísticas. Numa fase preliminar foram calculados os valores de riqueza específica (S)
e de abundância relativa (N) de cada local. Com base nesta informação calcularam-se três
índices que sintetizam informação relevante sobre a diversidade local:
- o índice de diversidade de Shannon-Wiener (H’) é um dos índices de diversidade
mais frequentemente aplicados, embora não seja universalmente aceite, apresentando
forte ligação aos conceitos de entropia (Shannon, 1948). Este índice relaciona o número
de espécies de um local (riqueza específica) com a distribuição dos seus valores de
abundância através da seguinte fórmula:
Onde pi é a proporção de indivíduos da espécie i em relação ao total de indivíduos
amostrados.
- a equitabilidade (E) refere-se à regularidade na distribuição dos indivíduos pelas
espécies identificadas na comunidade em estudo. O índice utilizado para determinar a
equitabilidade foi o índice de Pielou e consiste na seguinte fórmula:
J = H' /H’max
Onde H’ é o valor máximo obtido pelo índice de Shannon-Wiener e H’max é o valor
máximo de H’ (Heip et al., 1998).
- o índice de Berger-Parker (d) é um índice de dominância e permite avaliar a
heterogeneidade das abundâncias relativas das várias espécies de um dado local. É um
índice simples, intuitivo e com significado biológico, razões que têm motivado a sua
ampla utilização (Berger, & Parker, 1970). Este índice relaciona a abundância de
indivíduos da espécie mais abundante (Nmáx) com o número de indivíduos de todas as
espécies (N) encontradas nessa área de estudo. A fórmula aplicada foi a seguinte:

17
Frequentemente os resultados da dominância são apresentados na forma recíproca do
índice de Berger-Parker, ou seja, 1/d pois a sua interpretação é mais fácil.
Os cálculos dos vários descritores de diversidade (H’, E, d) foram realizados no
programa Species Diversity and Richness®, versão 4.0.
Tomando como base as tabelas de contingência com as observações de odonatas
e lepidópteros nos diferentes locais amostrados efetuou-se a análise aglomerativa por
locais e espécies com o objetivo de identificar padrões de associação. A análise
aglomerativa média (UPGMA) das espécies foi efetuada a partir do cálculo da matriz de
distâncias euclideanas entre espécies, enquanto que a análise aglomerativa dos locais teve
como base a matriz de correlação entre locais. As análises aglomerativas foram efetuadas
através da aplicação Minitab®, versão 16. Para complementar a informação indicada com
esta análise e na tentativa de identificar e interpretar melhor as associações presentes entre
tipologias de habitat e espécies procedeu-se à aplicação de técnicas de ordenação das
matrizes de distribuição de odonatas e lepidópteros por local. Utilizou-se a análise de
componentes principais (PCA) para a construção de combinações lineares com as
variáveis originais, e determinar os principais padrões de variação nos dados iniciais
(McPeek, 1990) relativamente ao ano de amostragem e às tipologias de habitat
amostradas. A análise de componentes principais foi efetuada com a aplicação Canoco®
para Windows, versão 4.5.
Utilizando como base os valores dos diversos descritores de diversidade (Anexos II e III)
efetuou-se a análise de variância (ANOVA) para um desenho experimental fatorial,
utilizando como fatores o ano e a tipologia de habitat.
Os cálculos foram efetuados através do módulo GLM (General Linear Model) da
aplicação Minitab®, versão 16, tendo-se efetuado comparações múltiplas (utilizando o
teste de Tukey) entre todos os níveis dos fatores que apresentaram diferenças
estatisticamente significativas. Todas as análises foram efetuadas para um nível de
significância de 0.05.
Procurou-se ainda identificar quais as espécies indicadoras de cada tipologia de
habitat através da aplicação do método IndVal (Dufrêne, & Legendre, 1997). Este
consiste inicialmente em métodos de classificação das unidades amostrais, para
identificar as espécies indicadoras de um ou vários locais ou tipologias de habitat, sendo
depois feita a sua validação estatística (Borcard et al., 2008; Dufrêne & Legendre, 1997).
O IndVal expressa o grau de especificidade e fidelidade (em %) de uma espécie

18
relativamente a um local ou a um conjunto de locais de estudo e, por apresentar um claro
significado biológico, tem sido amplamente utilizado (Dai et al., 2006).
Finalmente pretendeu-se perceber qual o conjunto de áreas mais relevantes para a
conservação da diversidade das ordens em estudo no Planalto Superior da Serra da
Estrela, através de uma análise de complementaridade da riqueza específica de cada
ordem. Esta análise consiste na medida do grau de representação que uma área ou
conjunto de áreas possui para incluir a totalidade dos atributos (riqueza específica)
encontrados na área geográfica de referência (Pressey et al., 1993; Vane-Wright et al.,
1991). As análises de complementaridade foram realizadas separada e conjuntamente
para as ordens Odonata e Lepidoptera.
Resultados e Discussão
Análise faunística da ordem Lepidoptera nos diferentes habitats
Neste estudo foram assinaladas 55 espécies de borboletas dos 2301 indivíduos
registados (Anexo IV). Informação detalhada sobre a abundância e a riqueza específica
dos locais amostrados encontra-se sintetizada no relatório do projeto “Biodiversidade,
endemismos e espécies protegidas associadas às lagoas e cursos de água da Serra da
Estrela: valorização de um século de aproveitamento hidroelétrico”, financiado pelo
fundo EDP para a biodiversidade 2011 (Serrano et al., 2014). Importa destacar a presença
de um conjunto de espécies que são raras em Portugal, como as borboletas Vanessa
virginiensis (Drury, 1773), Cyaniris semiargus (Rottemburg, 1775) e Coenonympha
iphioides (Staudinger, 1870) (Maravalhas, 2003). A espécie V. virginiensis (Drury, 1773)
apenas ocorreu nas lagoas naturais, num único local de amostragem, enquanto que a
espécie C. semiargus (Rottemburg, 1775) encontrou-se em todas as tipologias, mas com
maior abundância nos pontos secos. Por fim, a espécie C. iphioides (Staudinger, 1870)
também está presente em todas as tipologias mas em baixos valores de abundância,
variando normalmente entre 1 e 2 indivíduos.
A família de Lepidoptera mais bem representada é a Nymphalidae, com 32
espécies. A sua representatividade pode dever-se ao facto de ser a família com maior
riqueza específica em Portugal e por possuir espécies com elevada capacidade de
adaptação ecológica (Jiggins et al., 1996) e de dispersão (Parandhaman et al., 2012).
Importantes, ainda que com menor representatividade, ocorreram as famílias Lycaenidae
(S=10), Pieridae (S=8), Hesperiidae (S=4) e por fim Papilionidae (S=1). Não se verificou

19
a ocorrência de dominância de nenhuma espécie nos diferentes tipos de habitat, pois o
índice de Berger-Parker apresenta valores baixos, o que é corroborado pelo cálculo da
equitabilidade, que apresenta valores elevados, isto é, as comunidades apresentam
espécies com valores de abundância homogéneos (Tabela 1).
Tabela 1 Média e amplitude dos valores da abundância relativa (N), da riqueza específica
(S), da equitabilidade (E) e dos índices de diversidade e dominância (H’ e 1/d
respetivamente) de cada tipologia de habitat para a ordem Lepidoptera.
Para entender melhor as diferenças na diversidade de borboletas entre as
tipologias de habitat em estudo realizou-se a análise dos descritores de diversidade,
equitabilidade e dominância (Tabela 1, Figura 8, Anexo II).
A diversidade de espécies nas várias tipologias de habitat não se alterou de modo
significativo de um ano para o outro, apenas existindo diferenças de uma tipologia para
outra (Figura 8). Estas diferenças são explicadas na secção sobre as semelhanças das
comunidades entre as tipologias de habitat. Os pontos secos apresentam uma maior
dispersão dos dados de biodiversidade (medidas pelo índice H’), enquanto as barragens
pequenas são as mais constantes em relação a esses valores (Figura 8). Ao nível da
dominância de espécies, é nas barragens pequenas que esta é mais evidente, embora os
valores das médias sejam semelhantes entre as quatro tipologias de habitat (Tabela 1).
Todas as tipologias apresentam um valor alto de equitabilidade em ambos os anos.
Tipologia
N S H’ E 1/d
Média Amplitude Média Amplitude Média Amplitude Média Amplitude Média Amplitude
Pontos secos 39 9-102 12 5-22 2,07 1,78-2,78 0,86 0,61-0,94 0,29 0,14-0,45
Lagoas naturais 37 20-61 13 8-17 2,17 1,46-2,64 0,86 0,70-0,94 0,28 0,14-0,50
Barragens pequenas 45 16-99 15 10-21 2,35 2,06-2,63 0,87 0,78-0,95 0,24 0,14-0,45
Barragens grandes 52 25-81 18 9-25 2,53 1,99-2,89 0,87 0,78-0,94 0,21 0,11-0,32

20
Figura 8 Descritores de diversidade da ordem Lepidoptera para os anos de 2013 e 2014, referentes
ao índice de Shannon-Wiener (H’), ao índice de Berger-Parker (d) e à equitabilidade (Equi) pelo índice de
Pielou. Tipologia de habitat 1- pontos secos; 2- lagoas naturais; 3- barragens pequenas; 4- barragens de
grandes dimensões.
A fim de complementar a informação do significado da variação entre anos e
tipologias de habitat, realizou-se uma ANOVA para os diferentes descritores de
diversidade (Tabela 2). Os resultados obtidos não permitiram detetar diferenças
significativas entre os anos de estudo mas permitiram identificar diferenças significativas
entre os pontos secos e as barragens grandes no que diz respeito à riqueza específica e à
diversidade máxima. As barragens de grandes dimensões apresentam maior riqueza e
diversidade em comparação com os pontos secos. Em nenhuma das análises se observou
existirem interações significativas entre tipologias e anos.
Tipologia
Ano
4321
20142013201420132014201320142013
3.0
2.5
2.0
1.5
H
Tipologia
Ano
4321
20142013201420132014201320142013
9
8
7
6
5
4
3
2
d
Tipologia
Ano
4321
20142013201420132014201320142013
1.0
0.9
0.8
0.7
0.6
Equi

21
Tabela 2 Resultados das análises de variância dos descritores da diversidade de
abundância relativa, riqueza específica, índice de Shannon-Weaner, valor máximo do
índice de Shannon-Weaner, equitabilidade e índice Berger-Parker (lnN, lnS, H’, Hmáx,
E e d) entre as tipologias de habitat e os anos de amostragem para a ordem Lepidoptera.
Fator DF Variância F P
ln N
Tipologia Ano Tipologia*Ano Erro
3 1 3
46
0,5932 0,0895 0,1176 0,3370
1,76 0,27 0,35
0,168 0,609
0,79
(NS) (NS) (NS)
ln S
Tipologia Ano Tipologia*Ano Erro
3 1 3
46
0,5440 0,0016 0,0614 0,1252
4,34 0,01 0,49
0,009 0,909 0,691
(S)† (NS)
(NS)
H’
Tipologia Ano Tipologia*Ano Erro
3 1 3
46
0,5827 0,0298 0,1009 0,1235
4,72 0,24 0,82
0,006 0,626 0,491
(S)† (NS) (NS)
H’max
Tipologia Ano Tipologia*Ano Erro
3 1 3
46
0,5440 0,0016 0,0614 0,1252
4,34 0,01 0,49
0,009 0,909 0,691
(S)† (NS) (NS)
E
Tipologia Ano Tipologia*Ano Erro
3 1 3
46
0,0025 0,0036 0,0029 0,0044
0,56 0,83 0,67
0,642 0,366 0,577
(NS) (NS) (NS)
D
Tipologia Ano Tipologia*Ano Erro
3 1 3
46
4,2610 0,2000 1,7440 2,5110
1,7 0,08 0,69
0,181 0,779
0,56
(NS) (NS) (NS)
Resultado das comparações efetuadas através do teste de Tukey: † 1≠4- Pontos secos ≠ barragens grandes (inconclusivo para 2 e 3- lagoas naturais e barragens pequenas).

22
O facto de as barragens de grandes dimensões possuírem maior riqueza em
Lepidoptera é um resultado inesperado, uma vez que estas áreas são mais perturbadas
pelo homem. Apesar desse aspeto negativo relacionado com a perturbação, a construção
de barragens levou à alteração da paisagem homogénea do Planalto Superior, oferecendo
uma maior heterogeneidade de recursos, proporcionando assim um impacto positivo para
a biodiversidade de borboletas. A heterogeneidade de microhabitats é considerada um
fator importante na relação da riqueza em espécies com a área (Janzen, & Schoener, 1968;
Tews et al., 2004). Spitzer et al. (1997) mostraram que a existência de perturbação
moderada pode ser positiva para as comunidades de borboletas, uma vez que os valores
de riqueza e diversidade específica foram consideravelmente maiores em locais
perturbados do que em áreas pristinas. Noutro estudo, Uehara-Prado et al. (2009)
concluíram que a fragmentação florestal também não interfere negativamente na
diversidade de espécies de borboletas, contrariamente ao que sucedia com outros grupos
taxonómicos. Ainda de acrescentar que diversos estudos mostram que a presença de
alterações na paisagem pode ser um importante mecanismo de manutenção de espécies
autóctones (Huston, 1994; Jacobs, 1988).
A maior riqueza de espécies de borboletas nas barragens grandes deve-se à
presença de uma maior diversidade e abundância de plantas nestas áreas. As plantas
nectaríferas, que se encontram na periferia das lagoas (e.g. ericáceas, leguminosas,
compostas), apresentam um papel fundamental proporcionando uma maior, mais
diversificada e estável disponibilidade de alimento para as borboletas (Holl, 1995). Por
outro lado, as plantas determinam a estrutura física do habitat, logo influenciam as
interações com as espécies animais que lhes estão associadas (sobretudo polinizadores e
herbívoros) e a distribuição destas (Lawton et al., 1987; McCoy, & Bell, 1991).
As lagoas naturais e as barragens de pequenas dimensões apresentam o mesmo
tipo de plantas nas suas margens, contudo estas ocorrem com uma abundância mais
reduzida devido ao perímetro destas também ser muito menor, quando comparadas com
as grandes barragens. Por outro lado, as lagoas naturais e as barragens pequenas
encontram-se mais isoladas, o que torna difícil a sua colonização e visitação por parte das
espécies de Lepidoptera. No que concerne aos pontos secos, estes apresentam uma maior
homogeneidade vegetal e em alguns casos, como nos cervunais, são pobres em plantas
nectaríferas. Estas razões justificam a baixa riqueza (S) e diversidade (H’) de borboletas
destes locais (Tabela 1).

23
Diversos estudos têm concluído que as diferenças observadas na diversidade de
espécies de borboletas entre locais estão geralmente relacionadas com a diversidade e
abundância de plantas, uma vez que estas determinam a estrutura dos habitats e a
disponibilidade de alimento (Camero, & Calderón, 2007; Janzen, & Schoener, 1968).
Neste contexto, algumas atividades humanas em ambientes naturais homogéneos e com
baixa riqueza específica, como as áreas de alta montanha, podem contribuir para a
diversificação de nichos ecológicos, favorecendo a presença de espécies não
características e contribuindo assim para que apresentem maior riqueza específica do que
as áreas vizinhas não perturbadas (Uehara-Prado et al., 2009).
Análise faunística da ordem Odonata nos diferentes habitats
Neste estudo foram assinaladas 23 espécies de Odonata entre os 2580 indivíduos
observados (Anexo II). Informação detalhada sobre a abundância e a riqueza específica
dos locais amostrados encontra-se sintetizada no relatório do projeto “Biodiversidade,
endemismos e espécies protegidas associadas às lagoas e cursos de água da Serra da
Estrela: valorização de um século de aproveitamento hidroelétrico”, financiado pelo
fundo EDP para a biodiversidade 2011 (Serrano et al., 2014). De mencionar que as
espécies de Odonata A. juncea (Linnaeus 1758) e S. flaveolum (Linnaeus 1758), presentes
na Serra da Estrela, encontram-se no nosso país referenciadas como espécies muito raras,
uma vez que estão associadas a zonas de altitude (Maravalhas, & Soares, 2013). Em
Portugal, ambas as espécies foram apenas citadas em áreas acima dos 1000 m da Serra da
Estrela (Ferreira et al., 2009; Maravalhas, & Soares, 2013), embora A. juncea (Linnaeus
1758) tenha sido recentemente observada no Gerês (Albano Soares, pers. comm), facto
que necessita de confirmação. As duas espécies foram observadas em todas as tipologias
mas apenas no ano de 2013.
O grupo de Odonata melhor representado foi a sub-ordem Anisoptera com 13
espécies, das quais 7 pertencem à família Libellulidae. A sub-ordem Zygoptera está
representada por 10 espécies. O número mais elevado de espécies de Anisoptera pode ser
explicado pelo facto de a maioria dos locais de amostragem ser constituída por espaços
abertos com vegetação baixa e pouco complexa, apresentando grande luminosidade. Estas
espécies são dependentes destes fatores, pois necessitam deles para o desenrolar do seu
ciclo de vida (Corbet, & May, 2008; May, 1976; Resende, 2010). A dominância da família
Libellulidae é também mencionada em diversos trabalhos (Benazzoz et al., 2009;

24
Dijkstra, 2013; Lawler, 2001; Suhling et al., 2005; Suhling et al., 2004). No que diz
respeito à sub-ordem Zygoptera, a família Coenagrionidae é a dominante (S=4). Esta
possui ampla distribuição e espécies com ciclos de vida curtos (Norma-Rashid et al.,
2001), conseguindo tolerar uma grande gama de habitats (Acquah-Lamptey et al., 2013b).
O baixo número de espécies de Zygoptera deve-se, normalmente, à sua baixa capacidade
de dispersão, à dependência de ambientes com uma estrutura de vegetação mais complexa
e ao facto de poucas espécies ocorrerem a altitudes elevadas (Acquah-Lamptey et al.,
2013b; Maravalhas, & Soares, 2013).
Os resultados faunísticos encontram-se sumarizados na tabela 3 para os dois anos
de amostragem.
Tabela 3 Média e amplitude dos valores da abundância relativa (N), da riqueza específica
(S), da equitabilidade (E) e dos índices de diversidade e dominância (H’ e 1/d
respetivamente) de cada tipologia de habitat para a ordem Odonata.
O estudo das comunidades de libélulas presentes nas diferentes tipologias de
habitat realizou-se através da análise dos descritores de diversidade, equitabilidade e
dominância (Anexo III). Os resultados são apresentados para cada tipologia (Tabela 3;
Figura 9) e ano de estudo (Figura 9).
Não se observam diferenças significativas nos descritores de diversidade dos
quatro tipos de habitat entre os anos de estudo. Os pontos secos são os que apresentam
menor diversidade e dispersão dos valores por habitat. As lagoas naturais destacam-se
pois apresentam maior diversidade, assim como as barragens pequenas, contudo estas
possuem locais com valores muito dispersos no que concerne ao ano de 2014. Por fim, as
barragens de grandes dimensões, apesar de possuírem locais com alguma diversidade, no
geral esta é baixa, comparativamente às restantes, e bastante diferente entre os locais.
Mas, no ano de 2014, existe um aumento notório da diversidade nesta tipologia ficando
mais próxima das lagoas naturais e das barragens pequenas.
Tipologia
N S H E 1/d
Média Amplitude Média Amplitude Média Amplitude Média Amplitude Média Amplitude
Pontos secos 14 1-69 3 1-5 0,55 0-1,24 0,77 0,34-1,00 0,4 0,43-1,00
Lagoas naturais 118 33-233 6 4-9 1,34 0,92-1,91 0,73 0,57-0,87 0,47 0,32-0,63
Barragens pequenas 70 6-284 6 2-9 1,23 0,33-2,07 0,72 0,44-0,99 0,53 0,21-0,90
Barragens randes 15 1-15 4 1-7 0,93 0-1,61 0,82 0,52-1,00 0,59 0,29-1,00

25
A nível de dominância de espécies, verifica-se que não existe uma dominância
clara nas diferentes tipologias para o ano de 2013, já no ano de 2014 para as lagoas
naturais e para as barragens pequenas verifica-se a dominância de espécies para alguns
locais. Por exemplo, a espécie Anax imperator (Leach, 1815) na lagoa natural da
Francelha e na barragem pequena do Covão dos Conchos e a espécie Onychogomphus
uncatus (Charpentier, 1840) na lagoa natural Seca. Por fim, a maioria dos locais das
diferentes tipologias de habitat apresentam comunidades de libélulas com valores de
abundância semelhantes (Tabela 3).
Figura 9 Descritores de diversidade da ordem Odonata para os anos de 2013 e 2014, referentes ao índice
de Shannon-Wiener (H’), ao índice de Berger-Parker (d) e à equitabilidade (Equi) pelo índice de Pielou.
Tipologia de habitat 1- pontos secos; 2- lagoas naturais; 3- barragens pequenas; 4- barragens de grandes
dimensões.
As ANOVA dos descritores de diversidade (abundância de espécies, riqueza
específica e diversidade máxima) evidenciam a existência de diferenças significativas
entre os pontos secos (que se assemelham às barragens grandes) e as lagoas naturais (mais
semelhantes às barragens pequenas). As ANOVA não evidenciaram a existência de
interações entre tipologias e anos (Tabela 4).
Os Odonata são representados por espécies predadoras que habitam os
ecossistemas de água doce e a sua diversidade está frequentemente associada à riqueza e
Tipologia
Ano
4321
20142013201420132014201320142013
2.0
1.5
1.0
0.5
0.0
H
Tipologia
Ano
4321
20142013201420132014201320142013
5
4
3
2
1
d
Tipologia
Ano
4321
20142013201420132014201320142013
1.0
0.9
0.8
0.7
0.6
0.5
0.4
0.3
Equi

26
estrutura da vegetação aquática e à qualidade ambiental dos corpos de água (Remsburg,
& Turner, 2009). Assim sendo, compreende-se a pobreza em espécies dos pontos secos,
ou seja, áreas afastadas de cursos de água. Nestes locais registaram-se sobretudo
indivíduos em movimento (dispersão, busca de alimento) de espécies com maior
capacidade de voo (por exemplo, as espécies Cordulegaster boltonii (Donovan, 1807) e
Libellula quadrimaculata (Linnaeus, 1758)). Os principais fatores para a maior riqueza
de libélulas nas lagoas naturais e nas barragens pequenas são a boa qualidade das suas
águas e o bom estado de conservação da vegetação aquática e das margens, que resultam
do facto destes locais estarem relativamente isolados e sujeitos a muito reduzida
perturbação humana. Algumas das pequenas barragens encontram-se também já
naturalizadas pois não sofrem manutenção há várias décadas e apresentam deficiências
no eficaz represamento de água. Por outro lado, alguns estudos sobre as comunidades de
odonatas mostram também uma relação positiva entre a riqueza de espécies e a duração
dos corpos de água (McCauley, 2006). A estabilidade de presença dos corpos de água nos
locais tem um efeito positivo sobre a diversidade específica de libélulas, pois são locais
de reprodução para as espécies mais abundantes e/ou frequentes dessas comunidades
(Dijkstra, & Lempert, 2003). As diferenças significativas na riqueza e abundância de
libélulas entre as lagoas naturais e as barragens grandes resultam sobretudo da maior
perturbação associada às últimas e da menor qualidade da água destas. Muitas espécies
estenotópicas ocorrem em ambientes naturais mais fechados e com vegetação mais
complexa, como se verifica nas pequenas massas de água com a espécie S. flaveolum
(Linnaeus, 1758). Por outro lado, espécies euritópicas como a C. boltonii
(Donovan, 1807) e a Libellula depressa (Linnaeus,1758), são menos sensíveis à
perturbação, conseguindo sobreviver em ambientes como as barragens de grandes
dimensões (Ilg, & Oertli, 2014). Por outro lado, a introdução de espécies exóticas de
peixes predadores (sobretudo Salmo trutta (Linnaeus,1758) e Oncorhynchus mykiss
(Walbaum, 1792)) nas grandes barragens da Serra da Estrela com o intuito de promover
o turismo da pesca desportiva e o recorrente repovoamento piscícola dessas barragens
pelos organismos estatais contribui também para o empobrecimento das comunidades de
libélulas. Diversos estudos têm demonstrado que a introdução de peixes exóticos em
sistemas lênticos contribuem para a perda de biodiversidade de macroinvertebrados
aquáticos nativos, com graves repercussões nas populações de adultos reprodutores
(Hassall et al., 2011; Knapp et al., 2009; Labaj et al., 2013; Tiberti et al.,2014).

27
Tabela 4 Resultados das análises de variância dos descritores da diversidade de
abundância relativa, riqueza específica, índice de Shannon-Weaner, valor máximo do
índice de Shannon-Weaner, equitabilidade e índice Berger-Parker (lnN, lnS, H’, Hmáx,
E e d)
entre
as
tipologias de habitat e os anos de amostragem para a ordem Odonata.
Fator DF Variância F P
ln N Tipologia Ano Tipologia*Ano Erro
3 1 3
32
107,2490 0,8673 20,556 0,8476
12,65 1,02 2,43
<0,001 0,319 0,084
(S) † (NS) (NS)
ln S Tipologia
Ano
Tipologia*Ano
Erro
3 1 3
32
0,6785 0,0124 0,2298 0,1654
4,1 0,07 1,39
0,014 0,786 0,264
(S) ‡ (NS) (NS)
H’ Tipologia Ano Tipologia*Ano Erro
3 1 3
32
0,3445 0,2264 0,0261 0,1505
2,29 1,5
0,17
0,097 0,229 0,914
(NS) (NS) (NS)
Hmáx Tipologia Ano Tipologia*Ano Erro
3 1 3
32
0,6785 0,0124 0,2298 0,1654
4,1 0,07 1,39
0,014 0,786 0,264
(S) ‡ (NS) (NS)
E Tipologia Ano Tipologia*Ano Erro
3 1 3
32
0,0161 0,0231 0,0339 0,0238
0,68 0,97 1,42
0,573 0,332 0,254
(NS) (NS) (NS)
D Tipologia Ano Tipologia*Ano Erro
3 1 3
32
0,5520 11,9530
0,4395 0,6636
0,83 1,8
0,66
0,486 0,189 0,581
(NS) (NS) (NS)
Resultado das comparações efetuadas através do teste de Tukey: † 1=4≠3≠2(pontos secos=barragens grandes≠barragens pequenas≠lagoas naturais); ‡ 1≠2 (pontos secos≠ lagoas naturais ) (inconclusivo para 3 e 4- barragens pequenas e barragens grandes).

28
Semelhanças na composição das comunidades de Lepidoptera nos diferentes
habitats
Através da Figura 10 (projeção dos dois primeiros eixos da análise de PCA)
verifica-se que o padrão de distribuição das amostragens associadas aos pontos secos e
lagoas naturais e das amostragens associadas às barragens pequenas e barragens de
grandes dimensões seguem o mesmo padrão que a distribuição do anos de 2014 e 2013,
respetivamente. As espécies que determinam a distribuição de amostragens ao longo do
eixo 1 são Brintesia circe (Fabricius, 1775), Colias crocea (Geoffroy, 1785), Plebejus
argus (Linnaeus, 1758), Satyrus actaea (Esper, 1781) e Hesperia comma (Linnaeus,
1758), e P. argus (Linnaeus, 1758), S. actaea (Esper, 1781) e H. comma (Linnaeus, 1758)
para o eixo 2. A espécie B. circe (Fabricius, 1775) encontra-se em todas as tipologias de
habitat e nos dois anos, ocorrendo com abundâncias semelhantes, mas baixas. As espécies
C. crocea (Geoffroy, 1785) e P. argus (Linnaeus, 1758) aparecem em todas as tipologias
e em quase todos os locais de amostragem em abundâncias elevadas comparativamente
com as outras espécies. Por fim, as espécies S. actaea (Esper, 1781) e a H. comma
(Linnaeus, 1758) estão em todas as tipologias, mas a sua presença é mais notória no ano
de 2014.
Figura 10 Análise de componentes principais (PCA) para determinar os principais padrões de variação nos
dados da ordem Lepidoptera. Análise da variação entre anos (a), locais (b) e espécies que caracterizam a
variação (c).
a)
b)
c)
0.0 2.0
-1.0
2.0
Ano = 2013
Ano = 2014
0.0 2.0
-1.0
2.0
Tipologi = 1
Tipologi = 2
Tipologi = 3
Tipologi = 4
-0.2 1.0
-0.6
0.8
A.io
A.urtica
A.cardam
A.adippe
A.oglae
A.pandor
A.paphiaA.cramer
B.circe
C.rubi
C.argiol
C.iphioi
C.pamphi
C.crocea
C.semiar
G.rhamni
H.comma
H.fidia
H.hermio
H.semele
H.statil
H.lycaon
I.feisth
I.lathon
L.boetic
L.maera
L.megera
L.sinapi
L.pirith
L.alciph
L.phlaea
M.jurtin
M.laches
M.russia
M.deioneM.nevadeM.parthe
N.antiop
N.polych
P.aegeri
P.brassi
P.napiP.rapae
P.argus
P.c-albu
P.icarusP.daplid
P.malvoi
P.tithon
S.actea
T.lineol
T.sylves
V.atalan
V.cardui
V.virgin

29
Analisando o dendrograma referente às tipologias em estudo verificamos que
independentemente da similaridade do grupo, estes apresentam resultados heterogéneos
(Figura 11). A aparente falta de associações entre as comunidades de lepidópteros de uma
mesma tipologia de habitat deve-se em grande medida à estrutura espacial dos locais, isto
é, locais mais próximos (mas com habitats diferentes) são mais idênticos entre si do que
de locais mais afastados da mesma tipologia de habitat. Por exemplo, a comunidade de
borboletas da Lagoa do Ângelo (lagoa natural, Ang_Nat) assemelha-se mais às das
barragens (Conchos: Con_Bar) e ponto secos (Redonda: Red_PS) que lhe estão mais
próximos, do que com as das restantes lagoas naturais. Estes resultados evidenciam a
existência de comunidades associadas a uma região (que congrega vários tipos de habitat),
em virtude da boa capacidade de dispersão de borboletas, e onde os habitats com menor
riqueza específica incluem subgrupos das espécies que ocorrem nas tipologias de habitat
com maior número de espécies. Dentro de uma mesma área existe porém maior afinidade
entre locais do mesmo tipo de habitat. Como por exemplo, a grande afinidade das
comunidades de libélulas dos pontos secos da Fonte dos Perús (Fon_PS) e do Rodeio
Grande (Rod_PS), ou das barragens das Quelhas (Que_Bar) e do Serrano (Ser_Bar), ou
ainda das lagoas naturais Redonda (Red_Nat) e Seca (Sec_Nat). Mei_Bar
Ros1_BarG
Cur_Bar
Lag2_BarG
Fom_PS
Com_PS
Red_Nat
Sec_Nat
Lag1_BarG
Ros2_BarG
Comp1_BarG
Tor_PS
Rod_PS
Fon_PS
Ser_Bar
Que_Bar
Pen_PS
Esc_Nat
Comp2_BarG
Fav_Nat
Fra_Nat
Con_Bar
Ang_Nat
Red_PS
For_Bar
Per_PS
Cor_PS
17.79
45.19
72.60
100.00
Similarity
Figura 11 Análise aglomerativa dos locais de amostragem da ordem Lepidoptera para as diferentes
tipologias de habitat em estudo. Os códigos dos nomes dos locais de amostragem (apresentados no Anexo
V) incluem a indicação da tipologia de habitat (PS - pontos secos; Nat - lagoas naturais; Bar - barragens
pequenas; BarG - barragens de grandes dimensões).

30
Semelhanças na composição das comunidades de Odonata nos diferentes habitats
Na análise de componentes principais (PCA) dos Odonata, os três primeiros eixos
explicam 89.1% da variância total das amostras Contudo, ao contrário do que aconteceu
com os Lepidoptera, não se observa nenhum padrão associando anos com tipologias
(Figura 12), observando-se uma maior dispersão dos dados em 2013 do que em 2014. As
espécies que determinam a distribuição de amostragens ao longo do eixo 1 são as espécies
A. juncea (Linnaeus, 1758), Enallagma cyathigerum (Charpentier, 1840) e Libellula
quadrimaculata (Linnaeus, 1758), apresentando um alto valor de correlação com os
mesmos. A espécie A. juncea (Linnaeus, 1758), ocorre apenas no ano de 2013 e nas
tipologias lagoas naturais, barragens pequenas e barragens grandes. Porém, enquanto que
para as duas primeiras tipologias esta espécie é abundante na maioria dos locais, para as
barragens grandes apenas ocorreu em dois locais e com um registo para cada um. No que
se refere à espécie E. cyathigerum (Charpentier, 1840), esta em 2013 apresentou uma
abundância elevada para as lagoas naturais, seguidas das barragens pequenas e por fim
pelos pontos secos. Em 2014 este padrão manteve-se, mas a abundância de espécies foi
menor. Por fim, a espécie L. quadrimaculata (Linnaeus, 1758) foi muito abundante para
as lagoas naturais e barragens pequenas. Nos pontos secos ocorre em menor número, mas
na maioria dos locais, enquanto que para as barragens grandes apenas ocorre em 2 locais,
com o registo de um e três indivíduos. No ano de 2014 não aparece nos pontos secos nem
nas lagoas naturais. Nas restantes tipologias aparece apenas num único local e com a
presença de um único individuo. A distribuição do eixo 2 é determinada pelas espécies
Lestes virens (Charpentier, 1825) e L. dryas (Kirby, 1890). A primeira, em 2013, ocorreu
em todas as tipologias de habitat, mas destacam-se os locais das lagoas naturais, uma vez
que apresentam maior abundância que as restantes. Em 2014 não aparecem nas barragens
pequenas e nas restantes tipologias a sua ocorrência consiste apenas num único local e
com um número de indivíduos baixo, variando de 1 a 4. A segunda espécie (L. dryas

31
(Kirby, 1890)) ocorre apenas no ano de 2014, num único local dos pontos secos e apenas
com um indivíduo.
Em relação ao dendrograma das tipologias, existem quatro grupos de interesse
para uma similaridade de aproximadamente 55%, sendo o primeiro o mais relevante, uma
vez que estão presentes em todas as tipologias e quase na totalidade dos pontos (Figura
13).
A ocorrência da diferenciação das últimas tipologias de habitat apresentadas no
dendrograma pode ter ocorrido devido a espécies como a L. viridis (Vander Linden, 1825)
e a L. depressa (Linnaeus, 1758), que não se encontraram nestas tipologias ou então
devido a espécies como a C. tenellum (Villers, 1789 ) que apenas ocorreu uma única vez
nas barragens pequenas. A presença ou ausência de determinadas espécies está
-0.2 1.2
-0.2
0.8
A.cyanea
A.juncea
A.impera
B.irene
C.virgo
C.xantho
C.tenell
C.bolton
E.cyathi
I.graell
L.dryas
L.virens
L.viridiL.depres
L.quadri
O.uncatu
P.latipeP.nymphu
S.flaveo
S.fonsco
S.sangui
S.sp.
S.striol
-1.0 4.0
-1.0
1.5
Tipologi = 1 Tipologi = 2
Tipologi = 3 Tipologi = 4
-1.0 4.0
-1.0
1.5
Ano = 2013 Ano = 2014
Figura 12 Análise de componentes principais (PCA) para determinar os principais padrões de variação nos
dados da ordem Odonata. Análise da variação entre anos (a), locais (b) e espécies que caracterizam a
variação (c).
a) b)
c)
c)
c)

32
geralmente relacionada com as características locais do habitat como a cobertura de
sombra (Dijkstra, & Lempert, 2003; Djikstra, & Lewington, 2006), o microclima
(Kalkman et al., 2008; Oertli, 2010) e a composição e estrutura da vegetação aquática e
das margens, pois estas fornecem não só alimento como refúgio para as comunidades de
invertebrados (Adamus et al., 2001; Zimmer et al., 2000) o que torna os pontos mais ou
menos interessantes para cada espécie. É notória a segregação de algumas das
comunidades de libélulas em função do seu habitat (Figura 13). As comunidades de
libélulas das barragens grandes e dos pontos secos apresentaram grande afinidade entre
si, caracterizando-se por apresentarem uma menor riqueza específica, na sua maioria
composta por espécies euritópicas e com boa capacidade de vôo (p. ex. O.
Uncatus (Charpentier, 1840)). Por outro lado, apesar de existir alguma afinidade entre
algumas das comunidades de lagoas naturais e de barragens pequenas (Figura 13), é
também notória a existência de comunidades muito características das lagoas naturais
(p.ex. Lagoacho das Favas: Fav_Nat). Algumas das pequenas barragens (p.ex. barragem
do Covão dos Conchos: Con_Bar) encontram-se de certo modo naturalizadas dada a
ineficiência que apresentam no represamento de água e a falta de manutenção das
estruturas edificadas. As comunidades de libélulas destas barragens apresentam uma
riqueza e composição específica comparável à das lagoas naturais (Francelha: Fra_Nat e
Escura: Esc_Nat), e acomodam diversas espécies mais exigentes do ponto de vista
ecológico (p. ex. L. quadrimaculata, E. cyathigerum, A. juncea e L.virens).
Cur_Bar
Con_Bar
Fra_Nat
Esc_Nat
Fav_Nat
Red_Nat
Sec_Nat
Ang_Nat
For_Bar
Ser_Bar
Que_Bar
Red_PS
Con_PS
Comp1_BarG
Comp2_BarG
Mei_Bar
Fom_PS
Lag1_BarG
Ros2_BarG
Lag2_BarG
Rod_PS
Fon_PS
Ros1_BarG
Tor_PS
Per_PS
33.52
55.68
77.84
100.00
Similarity
Figura 13 Análise aglomerativa dos locais de amostragem da ordem Odonata para as diferentes tipologias
de habitat em estudo. Os códigos dos nomes dos locais de amostragem (apresentados no Anexo V) incluem
a indicação da tipologia de habitat (PS - pontos secos; Nat - lagoas naturais; Bar - barragens pequenas;
BarG - barragens de grandes dimensões).

33
Identificação de espécies de Lepidoptera indicadoras para os diferentes habitats
Observando os grupos associados aos resultados do IndVal, para as espécies de
lepidópteros, existem quatro grandes grupos, com uma semelhança superior a 50%. Por
sua vez para as tipologias de habitat existem três grupos, sendo que o primeiro engloba
os pontos secos e as lagoas naturais, pela presença de espécies em baixo número. As
barragens de grandes dimensões são as que menos espécies têm em comum com as
restantes (Figura 14). As tipologias de habitat um e dois (pontos secos e lagoas naturais)
caracterizam-se por apresentarem valores de IndVal, genericamente mais baixos. A
tipologia de habitat três (barragens pequenas) exibe valores mais elevados para um
pequeno grupo de espécies (10% a 42%). Por fim a tipologia de habitat quatro é a que
contém um maior número de espécies com valores mais altos de IndVal. Em relação à
ocorrência de espécies indicadoras, não existe nenhuma que responda de modo evidente
para a avaliação das quatro tipologias de habitat. Contudo, estudando apenas um ou dois
grupos de habitat, algumas espécies aumentam o seu valor indicador, nomeadamente
espécies estenotópicas, isto é, espécies que não conseguem sobreviver numa ampla gama
de habitats, tendo uma distribuição geográfica mais restringida (Dufrêne & Legendre,
1997). São exemplos espécies como a S. actaea (Esper, 1781) e a H. comma (Linnaeus,
1758). Estas são termófilas e mais características de ambientes terrestres de montanha ou
com pequenas massas de água (Maravalhas, 2003). As espécies Gonopteryx rhamni
(Linnaeus, 1758), Hipparchia statilinus (Hufnagel, 1766) e Lampides boeticus (Linnaeus,
1767), estão mais associadas às massas de água de maior dimensão, pois nestes locais
existe uma maior abundância de plantas hospedeiras, onde se desenvolvem as lagartas e
se alimentam os adultos. Vários estudos mostram a dependência dos Lepidoptera dos
recursos fornecidos por plantas específicas, tanto para as larvas como para os adultos e
como existe uma correlação positiva entre diversidade de espécies de borboletas e a
riqueza e/ou abundância de plantas presentes, nomeadamente nectaríferas (Croxton et al.,
2005; Kitahara, & Watanabe, 2003; Kitahara et al., 2008; Simonson et al., 2001).

34
Espécie IndVal1 IndVal2 IndVal3 IndVal4
1 Iphiclides feisthamelii 5% 0% 3% 1%
2 Celastrina argiolus 6% 26% 13% 15%
3 Vanessa cardui 7% 22% 7% 6%
4 Argynnis aglaja 3% 0% 0% 4%
5 Argynnis paphia 6% 0% 0% 8%
6 Issoria lathonia 11% 33% 23% 7%
7 Thymelicus lineola 4% 0% 0% 2%
8 Melitaea nevadensis 11% 0% 0% 8%
9 Colias crocea 18% 32% 27% 24%
10 Anthocharis cardamines0% 0% 8% 0%
11 Pieris brassicae 0% 0% 11% 0%
12 Callophrys rubi 0% 0% 4% 0%
13 Melitaea deione 0% 0% 4% 0%
14 Gonepteryx rhamni 1% 2% 42% 8%
15 Leptidea sinapis 1% 2% 23% 6%
16 Argynnis adippe 3% 1% 22% 6%
17 Pieris rapae 14% 16% 39% 7%
18 Argynnis pandora 19% 14% 28% 14%
19 Hipparchia hermione 9% 26% 12% 29%
20 Lycaena phlaeas 10% 18% 11% 20%
21 Lasiommata megera 4% 4% 8% 14%
22 Pieris napi 0% 0% 6% 19%
23 Hipparchia semele 6% 5% 9% 26%
24 Pontia daplidice 4% 5% 8% 21%
25 Hyponephele lycaon 6% 8% 10% 21%
26 Coenonympha iphioides0% 3% 1% 10%
27 Coenonympha pamphilus1% 2% 0% 9%
28 Cyaniris semiargus 2% 1% 2% 6%
29 Pararge aegeria 0% 1% 2% 30%
30 Hipparchia statilinus 3% 3% 0% 45%
31 Aricia cramera 4% 4% 3% 37%
32 Lampides boeticus 0% 2% 0% 41%
33 Melitaea parthenoides0% 0% 0% 17%
34 Lasiommata maera 0% 0% 0% 25%
35 Polygonia c-album 0% 0% 0% 8%
36 Nymphalis polychloros0% 0% 0% 8%
37 Polyommatus icarus 5% 5% 5% 25%
38 Brintesia circe 14% 12% 9% 31%
39 Plebejus argus 8% 22% 26% 23%
40 Lycaena alciphron 19% 4% 1% 6%
41 Aglais urticae 24% 6% 0% 0%
42 Hesperia comma 29% 14% 11% 6%
43 Pyronia tithonus 1% 0% 9% 14%
44 Pyrgus malvoides 1% 3% 5% 7%
45 Melanargia lachesis 6% 3% 22% 21%
46 Aglais io 0% 0% 3% 3%
47 Melanargia russiae 0% 0% 4% 4%
48 Nymphalis antiopa 0% 1% 10% 10%
49 Leptotes pirithous 7% 7% 13% 13%
50 Thymelicus sylvestris 5% 0% 11% 14%
51 Hipparchia fidia 2% 5% 0% 0%
52 Maniola jurtina 1% 0% 3% 3%
53 Satyrus actea 32% 45% 7% 10%
Tipologia de habitat
Figura 14 Análise IndVal para a ordem Lepidoptera. Vermelho: quartil 0-25%; Amarelo avermelhado:
quartil 25-50%; amarelo esverdeado: quartil 50-75%; Verde: quartil 75-100%. Tipologias de habitat -
1: pontos secos; 2: lagoas naturais; 3: barragens pequenas; 4: barragens grandes

35
Identificação de espécies de Odonata indicadoras para os diferentes habitats
Na análise dos dados do IndVal, percebe-se que para as espécies de Odonata
existem quatro grupos para uma semelhança superior a 50%. Nas tipologias de habitat
observamos três grupos diferentes sendo que o primeiro engloba os pontos secos e as
barragens de grandes dimensões, com uma elevada semelhança entre eles. As tipologias
de habitat um e quatro (pontos secos e barragens grandes) são as que mais se assemelham,
uma vez que, ambas apresentam um elevado número de espécies com valores de IndVal
baixos, variando entre os 0% e os 3%. A tipologia de habitat três (barragens pequenas)
aproxima-se mais da tipologia de habitata dois (lagoas naturais), pois os valores de IndVal
aumentam para algumas espécies, atingindo os 34%. Por fim, as Lagoas naturais são as
que mais se destacam e diferenciam das restantes, devido a exibirem valores de IndVal
superiores a 15% para a maioria das espécies e alcançando um máximo de 58%. As lagoas
naturais são as que possuem menos semelhança com os restantes, mas mais próxima das
barragens pequenas. Nestas duas tipologias de habitat ocorrem as espécies mais sensíveis,
com baixa capacidade de dispersão e altamente dependentes das massas de água presentes
entre a vegetação. Para os Odonata, não se verifica a ocorrência de espécies indicadoras
para a avaliação das quatro tipologias de habitat. Contudo, apenas para um dos grupos de
habitat (lagoas naturais) algumas espécies aumentam o seu valor indicador,
nomeadamente as espécies E. cyathigerum (Charpentier, 1840), L. virens (Charpentier,
1825) e L. dryas (Kirby, 1890). Espécies como a A. juncea (Linnaeus, 1758) e S.
flaveolum (Linnaeus, 1758) aumentam o seu valor indicador devido a possuírem uma
distribuição reduzida em Portugal. Sendo na Serra da Estrela que se encontram as suas
principais populações, enaltecendo a necessidade de conservação deste local. Para as
barragens pequenas, destacam-se as espécies L. quadrimaculata (Linnaeus, 1758) e Anax
imperator (Leach, 1815), com uma percentagem elevada em relação às restantes.

36
Figura 15 Análise IndVal para a ordem Odonata. Vermelho: quartil 0-25%; Amarelo avermelhado: quartil 25-50%;
amarelo esverdeado: quartil 50-75%; Verde: quartil 75-100%. Tipologias de habitat - 1: pontos secos; 2: lagoas naturais;
3: barragens pequenas; 4: barragens grandes
Identificação de áreas importantes para a conservação de borboletas e libélulas no
Planalto Superior da Serra da Estrela
Através da análise de complementaridade (Figura 16) obteve-se o número mínimo
de sítios necessários para conservar a riqueza específica registada de borboletas, libélulas
e dos dois grupos em conjunto. Os resultados mostram que apenas são necessários seis
locais para conservar a riqueza de libélulas observada, mas já serão necessários nove se
quisermos apenas conservar a riqueza de borboletas.
IndVal1 IndVal4 IndVal3 IndVal2
Aeshnacyanea 1% 3% 12% 5%
Ceriagriontenellum 0% 0% 8% 0%
Sympetrumfonscolombii 2% 1% 28% 4%
Libellulaquadrimaculata 1% 0% 33% 18%
Lestesviridis 8% 0% 0% 0%
Sympetrumsanguineum 0% 8% 0% 9%
Calopteryxvirgo 1% 14% 4% 17%
Platycnemislatipes 1% 9% 3% 0%
Cordulegasterboltonii 3% 9% 3% 0%
Ischnuragraellsii 2% 3% 2% 2%
Libelluladepressa 0% 10% 0% 0%
Boyeriairene 1% 0% 16% 22%
Aeshnajuncea 0% 0% 21% 23%
Anaximperator 0% 0% 34% 39%
Onychogomphusuncatus 6% 1% 0% 20%
Sympetrumsp. 6% 1% 0% 23%
Pyrrosomanymphula 0% 0% 1% 7%
Calopteryxxanthostoma 0% 3% 6% 27%
Sympetrumflaveolum 2% 0% 2% 15%
Lestesdryas 0% 3% 2% 27%
Sympetrumstriolatum 0% 0% 0% 8%
Lestesvirens 0% 0% 1% 38%
Enallagmacyathigerum 1% 2% 21% 58%
Tipologia de habitat

37
Figura 16 Resultados da análise de complementaridade da riqueza específica de borboletas,
libélulas e de ambos os grupos de insetos nos locais de estudo.
A conservação da riqueza específica de ambos os grupos de insetos necessita de
um mínimo de 12 dos locais estudados. Como era de esperar esta seleção de locais deverá
incluir uma mistura de barragens (grandes e pequenas) e lagoas naturais, uma vez que os
padrões de riqueza específica das duas ordens estão associados a tipologias de habitat
diferentes. Também se verificou que três locais de cada uma das tipologias - Lagoacho
das Favas (lagoa natural), Covão do Meio (barragem pequena) e Lagoa Comprida2
(barragem grande) - são muito importantes para a conservação de borboletas e libélulas
dado apresentarem valores de riqueza específica considerável e/ou exclusividade de
algumas espécies (Anexo VI e VII).
O Lagoacho das Favas é uma área particularmente rica em libélulas (S=15),
albergando um conjunto de espécies raras como A. juncea, S. flaveolum e S. sanguineum.
Já a barragem do Covão do Meio apresentou uma considerável riqueza específica (S=24),
um pouco inferior à assinalada na barragem do Vale do Rossim, mas contando com várias
espécies pouco frequentes na Serra da Estrela como as borboletas Argynnis adippe e S.
actaea.
A zona mais importante para a conservação das libélulas encontra-se mais
localizada na envolvente da Lagoa Comprida, incluindo as lagoas naturais adjacentes do
Ângelo, Escura e Favas, e a barragem do Covão dos Conchos, estendendo-se até à
0
10
20
30
40
50
60
70
80
90
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
S_ac
um
Nº de locais de amostragem
Riqueza específica(S)_acumulada_Lepidoptera Riqueza específica(S)_acumulada_Odonata
Riqueza específica(S)_acumulada_Total

38
barragem do Covão do Meio para incluir duas outras áreas (Rodeio Grande e Covão do
Meio) relevantes para a conservação deste grupo de insetos (Figura 17). Para as
borboletas é necessário conservar um maior número de áreas e áreas mais distantes entre
si, uma vez que estas apresentam comunidades de borboletas com algumas espécies
exclusivas (p.ex. barragem do Vale do Rossim, barragem do Covão do Meio, Lagoa
Redonda (Figura 17).
Estes resultados evidenciam a necessidade de incluir áreas um pouco afastadas
entre si para conservar a riqueza específica de borboletas e libélulas, uma vez que as
comunidades destes grupos tendem a ser mais dissemelhantes quanto maior a distância
entre si. O aumento da dissemelhança na composição de comunidades com a distância é
um fenómeno bem estudado (Nekola & White, 1999; Steinitz et al., 2006), tendo sido
identificados vários fatores que contribuem para o grau de diferenciação das comunidades
com a distância, como a capacidade de dispersão e as condições ambientais.
Apesar de não existir uma grande distância entre os locais de amostragem mais
afastados (aproximadamente 10 Km) e sabendo que os dois grupos estudados apresentam,
de um modo geral, uma boa capacidade de dispersão, foi ainda assim possível identificar
Figura 17 Área de conservação correspondente a cada ordem. Legenda: Odonata
Lepidoptera
Imagem adapatada do Google Earth ®

39
alguma heterogeneidade na riqueza e composição das comunidades de borboletas e
libélulas. Os locais de amostragem identificados como mais importantes para a
conservação de borboletas e libélulas (Anexo VI) e a área que definem (Figura 16 e Figura
17) encontram-se classificados como Reserva Biogenética pelo Conselho da Europa,
Zona Húmida de Importância Internacional no âmbito da Convenção RAMSAR, e Sítio
de Interesse Comunitário integrando a Rede Natura 2000. Apesar da existência destas
figuras legais de proteção é necessário desenvolver um conjunto de medidas
especificamente direcionadas para a conservação das borboletas e libélulas do Planalto
Superior da Serra da Estrela, a par de um programa de monitorização das suas populações.
As principais causas apontadas para a perda da biodiversidade estão diretamente
relacionadas com as perturbações de origem antrópica, nomeadamente: a destruição, a
fragmentação e a degradação dos habitats; a sobre-exploração de certas espécies ou
comunidades biológicas; a introdução de espécies exóticas e a disseminação de doenças
(Pité & Avelar, 1996; Meffe et al., 1997).Assim sendo deverão ser empregues medidas
de gestão pelas entidades competentes que culminem na conservação das populações de
espécies existentes. O controlo de espécies exóticas, o ordenamento das atividades
recreativas beneficiariam direta e indiretamente as populações em estudo. Contudo, a
medida de maior importância para a conservação de invertebrados é, a proteção dos seus
habitats (Brown, 1992).
Para finalizar, é necessário promover a sensibilização das populações para a
necessidade urgente de conservar a natureza, bem como dar continuidade aos importantes
trabalhos de ecologia e de inventariação da biodiversidade, pois só com um bom
conhecimento científico, alicerçado no reconhecimento da sociedade, poderemos
promover um desenvolvilmento sustentávele compatível com a preservação da
biodiversidade (Marques, 2006).
O Parque Natural da Serra da Estrela, apesar de ser uma área protegida tem sofrido
um conjunto de diversas ameaças. A alteração das atividades agro-silvo-pastoris
tradicionais, os fogos de grande intensidade, o turismo descontrolado, a introdução de
espécies exóticas piscícolas para a realização da pesca desportiva, o corte e o arranque de
vegetação ripícola, as alterações do regime de escoamento da água e a falta de
conhecimento e educação sobre o património natural existente, são os principais
problemas que interferem para a conservação destas áreas (Marques, 2006).
O fato de determinadas espécies ocorrerem em áreas protegidas, por si só, não
garante a proteção das mesmas (Pinheiro et al., 2010). Devido à ausência de informações

40
detalhadas sobre as necessidades biológicas e ecológicas da grande maioria dos taxa de
invertebrados, a proteção do maior número possível de diferentes ambientes, torna-se a
opção mais prática para a conservação da diversidade que neles se encontra (New, 1997).
Conclusões
O presente estudo inclui-se num conjunto de trabalhos desenvolvidos no âmbito
do projeto “Biodiversidade, endemismos e espécies protegidas associadas às lagoas e
cursos de água da Serra da Estrela: valorização de um século de aproveitamento
hidroelétrico”, financiado pelo fundo “EDP para a biodiversidade 2011”, que procurou
valorizar, divulgar e aumentar o conhecimento científico sobre a biodiversidade de
macroinvertebrados da Serra da Estrela e responder a um conjunto de objetivos gerais e
específicos. Entre esses objetivos contava-se a avaliação da biodiversidade associada às
lagoas e barragens do Planalto Superior da Serra da Estrela através do estudo de vários
grupos taxonómicos, que utilizam esses espaços e as áreas envolvente de modo distinto.
Os resultados deste estudo reforçaram e complementaram as conclusões já obtidas
durante o referido projeto e tornaram evidente a necessidade de incluir grupos animais
com biologias distintas aquando da avaliação da relevância de diferentes habitats para a
conservação da natureza.
Tendo em consideração o principal objetivo desta tese – a avaliação dos padrões
de biodiversidade de borboletas e libélulas associados às lagoas naturais, barragens
pequenas, barragens de grandes e pontos secos – pudemos concluir que os padrões de
biodiversidade foram distintos para os dois grupos taxonómicos considerados. Por
exemplo, os padrões de diversidade de libélulas (que utilizam a vegetação das margens)
foram semelhantes aos obtidos com o estudo de macroinvertebrados aquáticos (que vivem
nas massas de água), onde se concluiu que as barragens de grandes dimensões são pobres
em diversidade e abundância relativa comparativamente às lagoas naturais e barragens
pequenas. Por outro lado, os padrões de diversidade de borboletas (que utilizam a
vegetação das margens para repouso, alimentação e oviposição) são idênticos aos obtidos
com os coleópteros carabídeos (que ocorrem no solo das margens, próximo da interface
terra-água) que ocorreram em maior abundância e riqueza em barragens de grandes
dimensões, sendo as barragens do Vale do Rossim e do Lagoacho, os locais de ocorrência
de muitas espécies exclusivas. O papel bioindicador destas espécies, refere-se à estrutura,
composição e grau de conservação da vegetação das margens, evidenciando aspetos
positivos das grandes albufeiras, quando a sua construção e manutenção não é

41
acompanhada da destruição do coberto natural envolvente, nem fomente uma forte
pressão humana que perturbe ou destrua (por pisoteio, etc.) a vegetação das margens.
O bom estado de conservação destes habitats e o seu isolamento são determinantes
para a manutenção de uma riqueza específica elevada e a manutenção de populações de
espécies mais raras. Importa referir que embora a biodiversidade de borboletas e libélulas
dos pontos secos seja menor do que nos restantes habitats, nestas áreas ocorreram algumas
espécies raras no nosso país e características dos ambientes de montanha, com elevado
valor para a conservação da natureza. De entre as espécies encontradas destacam-se as
borboletas Apatura ilia (Denis & Schiffermüller, 1775), C. iphioides (Staudinger, 1870),
C. semiargus (Rottemburg, 1775) e S. actaea (Esper, 1781), referenciadas como
ameaçadas no nosso país e as libélulas A. juncea (Linnaeus,1758) e S. flaveolum
(Linnaeus,1758), extremamente raras em Portugal. Deste modo pudemos constatar que a
utilização de dois grupos-alvo distintos forneceram informação complementar sobre a
relevância de cada um dos tipos de habitat para a conservação da natureza no Planalto
Superior. Hoje em dia é fundamental, que a realização deste tipo de estudos inclua uma
variedade de grupos taxonómicos uma vez que os resultados obtidos com cada um deles
podem ser diferentes, fornecendo informação científica relevante e complementar que
deverá ser tida em consideração na gestão responsável destes espaços de elevado valor
natural.
Foram ainda identificadas áreas importantes para a conservação de borboletas e libélulas
no Planalto Superior da Serra da Estrela, as quais deverão merecer especial atenção das
entidades competentes, por forma a garantir a proteção efetiva destes locais. Dado o seu
elevado valor natural, estas áreas deverão ser alvo de programas de monitorização de
biodiversidade focados no estudo de borboletas e libélulas, entre outros grupos potenciais,
por forma a identificar atempadamente alterações nas comunidades naturais que ameacem
a biodiversidade da Serra da Estrela.
Desta forma procurou-se contribuir para o esforço nacional e internacional para o
melhor conhecimento da biodiversidade de invertebrados disponibilizando informação
relevante para a melhor gestão das populações de borboletas e libélulas da Serra da
Estrela.
Referências bibliográficas
Acquah-Lamptey, D., Kyerematen, R., & Owusu, E. O. (2013a). Dragonflies (Odonata:

42
Anisoptera) as tools for habitat quality assessment and monitoring. Journal of
Agriculture and Biodiversity Research, 2(8), 178–182.
Acquah-Lamptey, D., Kyerematen, R., & Owusu, E. O. (2013b). Using Odonates as
markers of the environmental health of water and its land related ecotone.
International Journal of Biodiversity and Conservation, 5(11), 761–
769.http://doi.org/10.5897/IJBC2013.0600
Adamus, P., Danielson, T. J., & Gonyaw, A. (2001). Indicators for monitoring biological
integrity of inland, freshwater wetlands. Wetlands, 254(5035), 1178–1181.
Retrieved from http://www.sciencemag.org/content/254/5035/1178.abstract
Aguiar, C., Rodrigues, O., Azevedo, J., & Domingos, T. (2009). Montanha. Escolar
Editora. Retrieved from http://bibliotecadigital.ipb.pt/handle/10198/2925
Aureli, A. (2002). What’s ahead in UNESCO's International Hydrological Programme
(IHP VI 2002-2007)? Hydrogeology Journal, 10(3), 349–350.
http://doi.org/10.1007/s10040-002-0211-y
Benazzoz, B., Mouna, M., Amezian, M., Bensusan, K., Perez, C., & Cortes, J. (2009).
Assessment and conservation of the dragonflies and damselflies (Insecta: Odonata)
at the marshes of Smir. Bulletin Institut Scientifique, Rabat, Section Sciences de La
Vie, 31(2), 79–84. Retrieved from
http://www.israbat.ac.ma/html/PublicationsIS/BIS_SV_31_1/12-Benazzouz.pdf
Berger, W. H., & Parker, F. L. (1970). Diversity of planktonic foraminifera in deep-sea
sediments. Science, 168(3937), 1345–1347.
http://doi.org/10.1126/science.168.3937.1345Blair, R. B., & Launer, A. E. (1997).
Butterfly diversity and human land use: Species assemblages along an urban
gradient. Biological Conservation, 80(1), 113–125. http://doi.org/10.1016/S0006-
3207(96)00056-0
Bivar-De-Sousa, A., Conde, J., & Mendes, L. F. (2008). Reconfirmação da presença de
Prionotropis flexuosa (Serville,1838) em Portugal, nota sobre a espécie na Península
Ibérica e sinonimização das subespécies descritas (Orthoptera, Pamphagidae,
Prionotropisinae). Boletin Sociedad Entomológica Aragonesa, 42, 341–343.
Blair, R. B., & Launer, A. E. (1997). Butterfly diversity and human land use: Species
assemblages along an urban gradient. Biological Conservation, 80(1), 113–125.

43
http://doi.org/10.1016/S0006-3207(96)00056-0
Bombino, G., Boix-Fayos, C., Gurnell, A. M., Tamburino, V., Zema, D. A., & Zimbone,
S. M. (2014). Check dam influence on vegetation species diversity in mountain
torrents of the Mediterranean environment. Ecohydrology, 7(2), 678–691.
http://doi.org/Doi 10.1002/Eco.1389
Borcard, D., Gillet, F., & Legendre, P. (2008). Numerical Ecology with R. Applied
Spatial Data Analysis with R. http://doi.org/10.1007/978-0-387-78171-6
Borges, P. A. V., Gabriel, R., Arroz, A. M., Costa, A., Cunha, R. T., Silva, L., Mendonça,
E. P., Martins, A. M. F., Reis, F., & Cardoso, P. (2010). The Azorean Biodiversity
Portal: An internet database for regional biodiversity outreach. Systematics and
Biodiversity, 8(4), 423–434. http://doi.org/10.1080/14772000.2010.514306
Brown Jr, K. S., & Morellato, L. P. C. (1992). Borboletas da Serra do Japi: diversidade,
habitats, recursos alimentares e variação temporal. História natural da Serra do Japi.
Ecologia e preservação de uma área florestal no Sudeste do Brasil. Campinas,
Editora da Unicamp/Fapesp, 142-187.
Camero, É., & Calderón C., A. M. (2007). Comunidad de mariposas diurnas
(Lepidoptera:Rhopalocera) en un gradiente altitudinal del cañon del rio Combeima-
Tolima, Colombia. Acta Biologiaca Colombiana, 12(2), 95–110.
Catalan, J., Camarero, L., Felip, M., Pla, S., Ventura, M., Buchaca, T., Bartumeus, F., De
Mendoza, G., Miró, A., Casamayor, E. O., Medina-Sánchez, J. M., Bacardit, M.,
Altuna, M., Bartrons, M., & De Quijano, D. D. (2006). High mountain lakes:
Extreme habitats and witnesses of environmental changes. Limnetica, 25(1-2), 551–
584.
Čelik, T. (2007). Dnevni metulji ( lep .: papilionoidea in hesperioidea ) kot bioindikatorji
za ekološko in naravovarstveno vrednotenje planinskega polja. Butterflies ( lep .:
papilionoidea in hesperioidea ) as bioindicators for nature conservation evaluation
of the planins, Varstvo Narave, 20, 83–105.
Chakravarthy AK, Rajagopal D, Jagannatha R. (1997) Insect as a bioindicator of
conservation in tropics. Zoos print. ( 12): 21-25.
Clark, T. E., & Samways, M. J. (1996). Dragonflies (Odonata) as Indicators of Biotope
Quality in the Kruger National Park, South Africa. Journal of Applied Ecology, Vol.

44
33, No. 5, 33(5), 1001–1012. Retrieved from
http://www.jstor.org/stable/2404681?seq=12&uid=3738032&uid=2&uid=4&sid=2
1104560633773
Corbet, P. S., & May, M. L. (2008). Fliers and perchers among Odonata: dichotomy or
multidimensional continuum? A provisional reappraisal. International Journal of
Odonatology, 11(2), 155–171. http://doi.org/10.1080/13887890.2008.9748320
Costa, L. T., Fidalgo, J., Neves, R., & Rufino, R. (2004). Lagoas do Planalto Superior da
Serra da Estrela. Portugal.
Croxton, P. J., Hann, J. P., Greatorex-Davies, J. N., & Sparks, T. H. (2005). Linear
hotspots? The floral and butterfly diversity of green lanes. Biological Conservation,
121(4), 579–584. http://doi.org/10.1016/j.biocon.2004.06.008
Dai, X., Page, B., & Duffy, K. J. (2006). Indicator value analysis as a group prediction
technique in community classification. South African Journal of Botany, 72(4), 589–
596. http://doi.org/10.1016/j.sajb.2006.04.008
Daveau, S. (1969). Structure et relief de la Serra da Estrela. Finisterra.4(7), 31-63.
Retrieved from http://revistas.rcaap.pt/finisterra/article/view/2490
Davis, A. J., Holloway, J. D., Huijbregts, H., Krikken, J., Kirk-Spriggs, A. H., & Sutton,
S. L. (2001). Dung beetles as indicators of change in the forests of northern Borneo.
Journal of Applied Ecology, 38(3), 593–616. http://doi.org/10.1046/j.1365-
2664.2001.00619.x
De Jong, Y., Verbeek, M., Michelsen, V., Bjørn, P. D. P., Los, W., Steeman, F., & Penev,
L. (2014). Fauna Europaea - all European animal species on the web. Biodiversity
Data Journal, (2), e4034. http://doi.org/10.3897/BDJ.2.e4034
Diaz, H. F., Grosjean, M., & Graumlich, L. (2003). Climate variability and change in high
elevation regions: Past, present and future. Climatic Change, 59(1/2), 1–4.
http://doi.org/10.1023/A:1024416227887
Didham, R.K. (1997) The influence of edge efects and forest fragmentation on leaf litter
invertebrates in central Amazonia. Tropical Forest Remnants: Ecology,
Management, and Conservation of Fragmented Communities (eds W.E. Laurance &
R.O. Bierregaard), 55-70. University of Chicago Press, Chicago, IL.

45
Dijkstra, K. B. (2013). Dragonflies and Damselflies ( Odonata ) of the Atewa Range ,
Ghana. Conservation International, 50–54.
Dijkstra, K. B., & Lempert, J. (2003). Odonate assemblages of running waters in the
Upper Guinean forest. Archiv Für Hydrobiologie, 157(3), 397–412.
http://doi.org/10.1127/0003-9136/2003/0157-0397
Djikstra, K.-D. B., & Lewington, R. (2006). Field Guide to the Dragonflies of Britain and
Europe. British Wildlife Publishing.
Dufrene, M., & Legendre, P. (1997). Species assemblages and indicator species : the need
for a flexible asymmetrical approach. Ecological Monographs, 67(3), 345-366.
Ehrlich, P. R., & Raven, P. H. (1964). Butterflies and plants: a study in coevolution.
Evolution, 18(4), 586–608.Farminhão, J., Gameiro, J., Carrilho, M., & Hasan-Beigi,
Y. (2014). Ficha Técnica. Revista Ecologia, No7, 33–39.
Farminhão, J., Gameiro, J., Carrilho, M., & Hasan-Beigi, Y. (2014). Ficha Técnica.
Revista Ecologia, No7, 33–39.
Ferreira, S., Grosso-silva, J. M., & Sousa, P. (2009). The Dragonflies of Serra da Estrela
Natural Park, Portugal (Insecta, Odonata). Boletín Sociedad Entomológica
Aragonesa, 44, 417–424.
Gerlach, J., Samways, M., & Pryke, J. (2013). Terrestrial invertebrates as bioindicators:
an overview of available taxonomic groups. Journal of Insect Conservation, 17(4),
831–850. http://doi.org/10.1007/s10841-013-9565-9
Grosso-Silva, J. M. (2000) Catalogação da fauna de colóeteros do Parque Natural da Serra
da Estrela e breve abordagem sobre a distribuição altitudinal e sazonalidade de
Caraboidea. Relatório de estágio profissionalizante. Faculdade de Ciências da
Universidade do Porto, 130 pp.
Hassall, C., Hollinshead, J., & Hull, A. (2011). Environmental correlates of plant and
invertebrate species richness in ponds. Biodiversity and Conservation, 20(13), 3189–
3222. http://doi.org/10.1007/s10531-011-0142-9
Heip, C. H. R., Herman, P. M. J., & Soetaert, K. (1998). Indices of diversity and evenness.
Oceanis, 24(4), 61–87.
Hodkinson, I. D., & Jackson, J. K. (2005). Terrestrial and Aquatic Invertebrates as

46
Bioindicators for Environmental Monitoring, with Particular Reference to Mountain
Ecosystems. Environmental Management, 35(5), 649–666.
http://doi.org/10.1007/s00267-004-0211-x
Holl, K. D. (1995). Nectar Resources and Their Influence on Butterfly Communities on
Reclaimed Coal Surface Mines. Restoration Ecology. http://doi.org/10.1111/j.1526-
100X.1995.tb00080.x
Huston, M. A. (1994). Biological diversity. The coexistence of species on changing
landscapes. Journal of the Marine Biological Association of the United Kingdom,
75(01), 261. http://doi.org/10.1017/S0025315400015393
Hynes, H. B. N. (1970). The Ecology of Stream Insects. Annual Review of Entomology,
15(1), 25–42. http://doi.org/10.1146/annurev.en.15.010170.000325
Ilg, C., & Oertli, B. (2014). How can we conserve cold stenotherm communities in
warming Alpine ponds? Hydrobiologia, 723(1), 53–62.
http://doi.org/10.1007/s10750-013-1538-1
Jacobs, M. (1988). The Tropical Rain Forest. (R. Kruk, Ed.). Berlin, Heidelberg: Springer
Berlin Heidelberg. http://doi.org/10.1007/978-3-642-72793-1
Jansen, J. (2002) Guia Geobotânico da Serra da Estrela. ICN - Parque Natural da Serra
da Estrela. Manteigas.
Janzen, D. H., & Schoener, T. W. (1968). Differences in Insect Abundance and Diversity
Between Wetter and Drier Sites During a Tropical Dry Season. Ecology, 49(1), 96.
http://doi.org/10.2307/1933565
Jiggins, C. D., Mcmillan, O., Neukirchen, W., & Mallet, J. (1996). What can hybrid zones
tell us about speciation ? The case of Heliconius erato and H . himera ( Lepidoptera :
Nymphalidae ). Biological Journal of the Linnean Society, 221–242.
http://doi.org/10.1111/j.1095-8312.1996.tb01464.x
Kalkman, V. J., Clausnitzer, V., Dijkstra, K.-D. B., Orr, A. G., Paulson, D. R., & van Tol,
J. (2008). Global diversity of dragonflies (Odonata) in freshwater. Hydrobiologia,
595(1), 351–363. http://doi.org/10.1007/s10750-007-9029-x
Kitahara, M., & Watanabe, M. (2003). Diversity and rarity hotspots and conservation of
butterfly communities in and around the Aokigahara woodland of Mount Fuji,

47
central Japan. Ecological Research, 18(5), 503–522. http://doi.org/10.1046/j.1440-
1703.2003.00574.x
Kitahara, M., Yumoto, M., & Kobayashi, T. (2008). Relationship of butterfly diversity
with nectar plant species richness in and around the Aokigahara primary woodland
of Mount Fuji, central Japan. Biodiversity and Conservation, 17(11), 2713–2734.
http://doi.org/10.1007/s10531-007-9265-4
Kitching, R.L. (1994) Biodiversity political responsibilities and agendas for research and
conservation. Pacific Conservation Biology, 1, 279-283.
Kitching, R. L., Orr, a. G., Thalib, L., Mitchell, H., Hopkins, M. S., & Graham, a. W.
(2000). Moth assemblages as indicators of environmental quality in remnants of
upland Australian rain forest. Journal of Applied Ecology, 37(2), 284–297.
http://doi.org/10.1046/j.1365-2664.2000.00490.x
Knapp, R. a, Matthews, K. R., & Sarnelle, O. (2009). Resistance and Resilience of Alpine
Lake Fauna to Fish Introductions. Ecological Monographs 71:401–
421. http://dx.doi.org/10.1890/0012-9615(2001)071[0401:RAROAL]2.0.CO;2
Kocher, S. D., & Williams, E. H. (2000). The diversity and abundance of North American
butterflies vary with habitat disturbance and geography.Journal of Biogeography,
27(4), 785 – 794.
Kremen, C., Colwell, R. K., Erwin, T. L., Murphy, D. D., Noss, R. F., & Sanjayan, M. a.
(1993). Terrestrial arthropod assemblages: Their use in conservation planning.
Conservation Biology, 7(4), 796–808.
Labaj, A. L., Kurek, J., Weeber, R. C., & Smol, J. P. (2013). Long-term changes in
invertebrate size structure and composition in a boreal headwater lake with a known
minnow introduction. Journal of Limnology, 72(2), 17.
http://doi.org/10.4081/jlimnol.2013.e17
Larsen T.B. (1998). The butterflies of the Nilgiri mountains of south India Lepidoptera
Rhopalocera. Journal of the Bombay Natural History Society (86): 39-46
Lautensach, H. (1932) Estudo dos glaciares da Serra da Estrela. Memórias e Notícias,
Coimbra, 6, 1-60.

48
Lawler, S. P. (2001). Rice Fields As Temporary Wetlands: a Review. Israel Journal of
Zoology, 47(4), 513–528. http://doi.org/10.1560/X7K3-9JG8-MH2J-XGX1
Lawton, J. H., Macgarvin, M., & Heads, P. A. (1987). Effects of Altitude on the
Abundance and Species Richness of Insect Herbivores on Bracken. Journal of
Animal Ecology, 56(56), 147–160. Retrieved from http://www.jstor.org/stable/4805
Leitão, A. E., Portela, M. M., & Godinho, F. N. (2010). Produção de energia em pequenos
aproveitamentos hidroeléctricos em Portugal. Potencialidades e constrangimentos
ao seu desenvolvimento. Revista Recursos Hídricos, 31, 39–56.
Maravalhas, E (ed) (2003) As Borboletas de Portugal. Vento Norte, Porto.
Maravalhas, E & A Soares (2013) As Libélulas de Portugal. Booky Publisher, Porto.
Marques, J. E., Marques, J. M., Chaminé, H. I., Carreira, P. M., Fonseca, P. E., Monteiro
Santos, F. A., Moura, R., Samper, J., Pisani, B., Teixeira, J., Carvalho, J. M., Rocha,
F., & Borges, F. S. (2013). Conceptualizing a mountain hydrogeologic system by
using an integrated groundwater assessment (Serra da Estrela, Central Portugal): a
review. Geosciences Journal, 17(3), 371–386. http://doi.org/10.1007/s12303-013-
0019-x
Marques, P. P. (2006). Serra da Estrela: Gestão e Conservação de Habitats Prioritários
(Edição Ass). LIFE02/NAT/P/8478.
Martins, E., & Silveira, P. (2008). As glaciações e a flora da Serra da Estrela, EU, Fundo
de Desenvolvimento Regional, Departamento de Biologia da Universidade de
Aveiro, 22p.
May, M. L. (1976). Thermoregulation and Adaptation to Temperature in Dragonflies (
Odonata : Anisoptera ). Ecological Society of America- Ecological Monographs,
46(1), 1–32. Retrieved from http://www.jstor.org/stable/1942392.
McCauley, S. J. (2006). The effects of dispersal and recruitment limitation on community
structure of odonates in artificial ponds. Ecography, 29, 585–595.
http://doi.org/10.1111/j.0906-7590.2006.04787.x
McCoy, E. D., & Bell, S. S. (1991). Habitat structure: The evolution and diversification
of a complex topic. In Habitat Structure, 8: 3–27. Publicado: Springer Netherlands.
http://doi.org/10.1007/978-94-011-3076-9_1

49
McGeoch, M. a. (1998). The selection, testing and application of terrestrial insects as
bioindicators. Biological Reviews, 73, 181–201.
http://doi.org/10.1017/S000632319700515X
McPeek, M. a. (1990). Behavioral differences between Enallagma species (Odonata)
influencing differential vulnerability to predators. Ecology, 71(5), 1714–1726.
http://doi.org/10.2307/1937580
Meffe, G. K. & Carroll, C. R. (1997). Principles of Conservation Biology, 2nd edition.
Sinauer Associates, Sunderland
Metoffice. (2010). Fact sheet 6 — The Beaufort Scale. National Meteorological Library
and Archive, (version 01).
Moura, M. D. P. B. C. (2006) Diacronia e memória de uma paisagem - Análise Visual da
Paisagem do Planalto Superior da Serra da Estrela. Dissertação de Mestrado,
Faculdade e Letras, Universidade de Coimbra, Portugal. 142p.
Nekola, J. C., & White, P. S. (1999). The distance decay of similarity in biogeography
and ecology. Journal of Biogeography, 26(4), 867–878.
http://doi.org/10.1046/j.1365-2699.1999.00305.x
New, T. R. (1997). Are Lepidoptera an effective ‘umbrella group‘ for biodiversity
conservation? Journal of Insect Conservation, 1(1), 5–12.
http://doi.org/10.1023/A:1018433406701
Norma-Rashid, Y., Mohd-Sofian, A., & Zakaria-Ismail, M. (2001). Diversity and
distribution of Odonata(dragonflies and damselflies) in the freshwater swamp lake
Tasek Bera, Malaysia. Hydrobiologia, 456, 135–146. http://doi.org/10.1023/A
Oertli, B. (2010). The local species richness of Dragonflies in mountain waterbodies: an
indicator of climate warming? BIORISK – Biodiversity and Ecosystem Risk
Assessment, 5, 243–251. http://doi.org/10.3897/biorisk.5.853
Parandhaman, D., Sivasakaran, K., Meerasa, M. N., & Ignacimuthu, S. (2012). Diversity
of butterflies in different habitats from Tamilnadu part of Western Ghats
(Lepidoptera : Rhopalocera). Elixir Application Biology, 51, 10861–10865.
Pinheiro, C. E. G., Malinov, I. K., Emery, E. O., & Schmidt, K. (2010). Endemismos e
conservação de borboletas(Lepidoptera: Papilionoidea e Hesperioidea) no bioma

50
Cerrado., 223–236. In Diniz, I., Filho, J. M., & Machado, R. B. C. (2010). Cerrado
- conhecimento científico quantitativo como subsídio para ações de conservação. p.
496. Retrieved from
http://scholar.google.com.br/scholar?q=cerrado+conhecimento+cient%C3%ADfic
o+quantitativo&btnG=&hl=pt-BR&as_sdt=0%2C5#7
Pité, M.T.R., Avelar, T. (1996). Ecologia das Populações e das Comunidades: Uma
Abordagem Evolutiva do Estudo da Biodiversidade. Lisboa, Fundação Calouste
Gulbenkian.
Pinto da Silva, A.R. & Teles, A.N (1980). A flora e a vegetação da Serra da Estrela.
Parque Natural da Serra da Estrela. Lisboa, 54 p.
Pollard, E., & Yates, T. J. (1993) Monitoring butterflies for ecology and conservation.
Chapman & Hall, London. Biological Conservation, 1.
Pollard, E. (1977). A method for assessing changes in the abundance of butterflies.
Biological Conservation, 12(2), 115–134. http://doi.org/10.1016/0006-
3207(77)90065-9
Pressey, R. L., Humphries, C. J., Margules, C. R., Vane-Wright, R. I., & Williams, P. H.
(1993). Beyond opportunism: Key principles for systematic reserve selection.
Ecology & Evolution, 8(4), 124–128. http://doi.org/10.1016/0169-5347(93)90023-I
Remsburg, A. J., & Turner, M. G. (2009). Aquatic and terrestrial drivers of dragonfly
(Odonata) assemblages within and among north-temperate lakes. Journal of the
North American Benthological Society, 28(1), 44–56. http://doi.org/10.1899/08-
004.1
Resende, D. C. (2010). Residence advantage in heterospecific territorial disputes of
Erythrodiplax Brauer species (Odonata, Libellulidae). Revista Brasileira de
Entomologia, 54(1), 110–114. http://doi.org/10.1590/S0085-56262010000100014
Ribeiro, O. (1954). Estrutura e relevo da Serra da Estrela. Boletin de la Real Sociedad
Española de Historia Natural (Tomo homenaje a E. Hernández-Pacheco),[s.n.], 549-
566.
Rocha, F., Inácio, M., Marques, J. E., Gonçalves, P., Aguiar, C., & Chaminé, H. I. (2004).

51
Clay mineralogy and geochemistry of soils from Serra da Estrela Natural Park (
Central Portugal ): preliminary results, in XIV Semana de Geoquímica, VIII
Congresso de Geoquímica dos Países de Língua Portuguesa. Universidade de
Aveiro. Actas, vol. 2; p. 611-614.
Rosenberg, D. M., Danks, H. V., & Lehmkuhl, D. M. (1986). Importance of insects in
environmental impact assessment. Environmental Management, 10(6), 773–783.
http://doi.org/10.1007/BF01867730
Salgueiro, J. (2002). Variação Anual em três comunidades de formicídeos da Serra da
Estrela. Adição de um género novo e de duas espécies novas para Portugal. Boletin
de la Asociación Española de Entomología, 26(1969), 121–131.
Schreier, H., 2003, Mountain Watershed Management: Scaling-up and Scaling-out. In:
(Greminger, P. (ed.), Mountain Watershed Management, Lessons from the Past –
Lessons for the Future. Proceedings of Swiss Agency for the Environment, Forests
and Landscape (SAEFL), Environmental Documentation 165, 29–42
Schmitt, T. (2009). Biogeographical and evolutionary importance of the European high
mountain systems. Frontiers in Zoology, 6, 9. http://doi.org/10.1186/1742-9994-6-9
Serrano, A.R.M., Soares, A., Silva, A., Fonseca, A., Bívar de Sousa, A., Figueiredo, A.,
Rego, C., Aguiar, C.A.S., Monteiro, E., Figueiredo, H., Conde, J., Boieiro, M.,
Garcia-Pereira, P., Borges, P.A.V., Cardoso, P., Antunes, S., Lopes, S., Correia, T.
& Simões, V. (2014) Biodiversidade, endemismos e espécies protegidas associadas
às lagoas e cursos de água da Serra da Estrela: valorização de um século de
aproveitamento hidroeléctrico - Relatório final. Fundo EDP para a Biodioversidade
2011, Lisboa.
Seufferheld, M. J., Alvarez, H. M., & Farias, M. E. (2008). Role of polyphosphates in
microbial adaptation to extreme environments. Applied and Environmental
Microbiology, 74(19), 5867–5874. http://doi.org/10.1128/AEM.00501-08
Shannon, C. E. (1948). A Mathematical Theory of Communication. The Bell System’s
Technical Journal, 27(I), 379–423.
Simaika, J. P., & Samways, M. J. (2011). Comparative assessment of indices of
freshwater habitat conditions using different invertebrate taxon sets. Ecological
Indicators, 11(2), 370–378. http://doi.org/10.1016/j.ecolind.2010.06.005

52
Simonson, S. E., Opler, P. A., Stohlgren, T. J., & Chong, G. W. (2001). Rapid assessment
of butterfly diversity in a montane landscape. Biodiversity and Conservation, 10(8),
1369–1386. http://doi.org/10.1023/a:1016663931882
Spehn, E., Rudmann-Maurer, K., & Körner, C. (2011). Mountain biodiversity. Plant
Ecology & Diversity, 4(4), 301–302. http://doi.org/10.1080/17550874.2012.698660
Spitzer, K., Jaroš, J., Havelka, J., & Lepš, J. (1997). Effect of small-scale disturbance on
butterfly communities of an Indochinese montane rainforest. Biological
Conservation, 80(1), 9–15. http://doi.org/10.1016/S0006-3207(96)00079-1
Stefanescu, C., Herrando, S., & Paramo, F. (2004). Butterfly species richness in the north-
west Mediterranean Basin: the role of natural and human induced factors. Journal of
Biogeography, 31(6), 905–915. http://doi.org/10.1111/j.1365-2699.2004.01088.x
Stefanescu, C., Peñuelas, J., & Filella, I. (2009). Rapid changes in butterfly communities
following the abandonment of grasslands: a case study. Insect Conservation and
Diversity, 2(4), 261–269. http://doi.org/10.1111/j.1752-4598.2009.00063.x
Steinitz, O., Heller, J., Tsoar, A., Rotem, D., & Kadmon, R. (2006). Environment,
dispersal and patterns of species similarity. Journal of Biogeography, 33(6), 1044–
1054. http://doi.org/10.1111/j.1365-2699.2006.01473.x
Suhling, F., Sahlen, G., Kasperski, J., & Gaedecke, D. (2005). Behavioural and life
history traits in temporary and perennial waters: comparisons among three pairs of
sibling dragonfly species. Oikos, 108(3), 609–617. http://doi.org/10.1111/j.0030-
1299.2005.13230.x
Suhling, F., Schenk, K., Padeffke, T., & Martens, A. (2004). A field study of larval
development in a dragonfly assemblage in African desert ponds (Odonata).
Hydrobiologia, 528(1-3), 75–85. http://doi.org/10.1007/s10750-004-3047-8
Tews, J., Brose, U., Grimm, V., Tielbörger, K., Wichmann, M. C., Schwager, M., &
Jeltsch, F. (2004). Animal species diversity driven by habitat
heterogeneity/diversity: the importance of keystone structures. Journal of
Biogeography, 31(1), 79–92. http://doi.org/10.1046/j.0305-0270.2003.00994.x
Tiberti, R., von Hardenberg, A., & Bogliani, G. (2014). Ecological impact of introduced
fish in high altitude lakes: A case of study from the European Alps. Hydrobiologia,
724, 1–19. http://doi.org/10.1007/s10750-013-1696-1

53
Tortajada, C., Altinbilek, D., & Biswas, A. K. (2012). Impacts of Large Dams: A Global
Assessment. (C. Tortajada, D. Altinbilek, & A. K. Biswas, Eds.). Berlin, Heidelberg:
Springer Berlin Heidelberg. http://doi.org/10.1007/978-3-642-23571-9
Uehara-Prado, M., Fernades, J. O., Bello, A. de M., Machado, G., Santos, A. J., Vaz-de-
Mello, F. Z., & Freitas, A. V. L. (2009). Selecting terrestrial arthropods as indicators
of small-scale disturbance: a first approach in the Brazilian atlantic Forest.
Biological Conservation, 142(6), 1220-1228.
http://doi.org/10.1016/j.biocon.2009.01.008
Vane-Wright, R. I., Humphries, C. J., & Williams, P. H. (1991). What to protect?—
Systematics and the agony of choice. Biological Conservation, 55(3), 235–254.
http://doi.org/10.1016/0006-3207(91)90030-D
Vegas, R., Vázquez, J. T., Suriñach, E., & Marcos, A. (1990). Model of distributed
deformation, block rotations and crustal thickening for the formation of the Spanish
Central System. Tectonophysics, 184(3-4), 367–378. Retrieved from
http://www.scopus.com/inward/record.url?eid=2-s2.0-
0025640527&partnerID=tZOtx3y1
Vieira, G. T. (2004). Geomorfologia dos Planaltos e Altos Vales da Serra da Estrela.
Ambientes frios do Plistocénico Superior e dinâmica actual. Tese de Doutoramento
(inédita). Departamento Geografia da Faculdade de Letras, Universidade de Lisboa,
Lisboa: 724 p
Viviroli, D., & Weingartner, R. (2008). “Water towers”— a global view of the
hydrological importance of mountains. Mountains: Sources of Water, Sources of
Knowledge, 15–20.
Zektser, I. S., & Everett, L. G. (2004). Groundwater resources of the world and their use.
IhP Series on groundwater. No. 6 (Vol. 6).
Zimmer, K. D., Hanson, M. A., & Butler, M. G. (2000). Factors influencing invertebrate
communities in prairie wetlands: a multivariate approach. Canadian Journal of
Fisheries and Aquatic Sciences, 57(1), 76–85. http://doi.org/10.1139/f99-180

54
Anexo I
Anexo I.1 Locais de amostragem com indicação da tipologia e a respetiva cota de altitude
e localização (latitude e longitude no Datum Lisboa Hayford Gauss IGeoE).
Local Tipologia Altitude Latitude Longitude
Covão do Forno Artificial 1576m 377914.17 242218.00
Lag
oas
Covão do Curral Artificial 1475m 378140.37 241903.66
Covão dos Conchos Artificial 1636m 377345.75 244483.27
Lagoa do Covão das Quelhas Natural artificializada 1817m 373361.55 243032.25
Lagoa Serrano Natural artificializada 1798m 373405.87 242636.09
Lagoa do Covão do Meios Artificial 1657m 373957.64 242731.88
Vale do Rossim 1 (NE) Artificial de grandes dimensões 1435m 381341.40 246736.60
Vale do Rossim 2 (SO) Artificial de grandes dimensões 1450m 380985.00 245966.67
Lagoacho 1 (Oeste) Artificial de grandes dimensões 1433m 379571.20 243244.47
Lagoacho 2 (Sudeste) Artificial de grandes dimensões 1441m 379168.26 243443.05
Lagoa Comprida 1 Natural artificializada de grandes
dimensões
1603m 377382.13 241675.46
Lagoa Comprida 2 Natural artificializada de grandes
dimensões
1601m 376833.17 242956.74
Lagoa Seca Natural temporária 1625m 378187.31 242627.93
Lagoa Redonda Natural permanente 1616m 378070.46 243180.19
Lagoacho das Favas Natural temporária 1645m 377331.37 242198.38
Lagoa Escura Natural permanente 1670m 376381.48 242064.00
Lagoa do Ângelo Natural permanente 1738m 376158.51 242511.50
Lagoa da Francelha Natural permanente 1765m 373566.60 242474.42
Ponto
s se
cos
Corgo das Mós
1492m
381547.46
247574.14
Vale das Éguas 1477m 381373.19 247963.89
Vale de Perdiz 1425m 382289.39 245745.30
Erva da Fome 1457m 380480.91 244988.35
Lagoacho seco 1410m 380626.16 244042.90
Entre a Redonda e a Seca 1640m 378572.57 242842.79
Depois da Lagoa Seca a Oeste 1619m 378165.20 242536.28
Penha do Gato 1696m 375864.06 239825.61
Ponto seco dos Conchos 1706m 377198.01 244012.70
Torre 1927m 372219.45 243753.24
Fonte dos Perus 1867m 375156.46 243456.52
Rodeio Grande 1838m 374838.30 241684.81

55
Anexo II
Anexo II.1 Valores da abundância relativa (N), riqueza específica (S), equitabilidade (E)
e índices de diversidade e dominância ( H’ e 1/d respetivamente) por local de tipologia
para a ordem Lepidoptera.
Tipologia Ano Local N S H' E (H'/H'max) d
1 2013 Cor_PS 67 21 2.69909 0.88654 6.7
1 2013 Per_PS 42 12 1.97994 0.79679 2.47059
1 2013 Fom_PS 99 22 2.73125 0.8836 7.07143
1 2013 Red_PS 18 11 2.21226 0.92258 3.6
1 2013 Pen_PS 39 19 2.7764 0.94293 5.57143
1 2013 Con_PS 10 6 1.64342 0.91721 3.33333
1 2013 Tor_PS 25 10 2.1025 0.91311 4.16667
1 2013 Fon_PS 9 5 1.42706 0.88668 2.25
1 2013 Rod_PS 25 10 1.96333 0.85266 2.77778
2 2013 Sec_Nat 22 8 1.66633 0.80134 2.2
2 2013 Red_Nat 20 11 2.15132 0.89717 4
2 2013 Fav_Nat 61 14 2.38013 0.90189 4.35714
2 2013 Esc_Nat 60 17 2.36354 0.83423 4.61538
2 2013 Ang_Nat 33 13 2.21883 0.86506 3.3
2 2013 Fra_Nat 54 15 2.3788 0.87842 4.5
3 2013 For_Bar 34 14 2.36392 0.89574 5.66667
3 2013 Cur_Bar 97 21 2.63051 0.86401 5.38889
3 2013 Con_Bar 40 13 2.07697 0.80975 2.5
3 2013 Que_Bar 21 12 2.37565 0.95603 7
3 2013 Ser_Bar 28 17 2.63251 0.92916 4.66667
3 2013 Mei_Bar 99 17 2.44255 0.86211 4.95
4 2013 Ros1_BarG 81 25 2.78664 0.86572 5.78571
4 2013 Ros2_BarG 44 22 2.89366 0.93614 7.33333
4 2013 Lag1_BarG 36 12 2.30815 0.92887 6
4 2013 Lag2_BarG 67 14 2.23403 0.84653 3.52632
4 2013 Comp1_BarG 25 9 1.9916 0.90642 3.125
4 2013 Comp2_BarG 67 18 2.41038 0.83393 3.52632
1 2014 PerPS 55 18 2.67035 0.92388 6.11111
1 2014 PenPS 12 7 1.82008 0.93533 4
1 2014 Com PS 51 8 1.27836 0.61476 2.21739
1 2014 Tor PS 35 13 2.09848 0.81814 3.18182
1 2014 Fon PS 14 6 1.53672 0.85766 2.33333
1 2014 Rod PS 18 7 1.69041 0.8687 2.57143
1 2014 EguPS 102 18 2.16987 0.75072 3.64286
1 2014 LagPS 34 14 2.39523 0.90761 5.66667
1 2014 SecPS 38 12 2.04364 0.82242 3.16667
2 2014 Sec_Nat 34 8 1.45738 0.70085 2

56
Tipologia Ano Local N S H' E (H'/H'max) d
2 2014 RedNat 55 16 2.11925 0.76436 2.3913
2 2014 Fav_Nat 24 10 2.06883 0.89848 3.42857
2 2014 EscNat 29 16 2.6322 0.94937 7.25
2 2014 Ang_Nat 30 15 2.47089 0.91242 5
2 2014 Fra_Nat 22 10 2.16129 0.93864 5.5
3 2014 For_Bar 40 14 2.06321 0.7818 2.22222
3 2014 CurBar 54 17 2.45734 0.86733 4.5
3 2014 ConBar 40 14 2.1962 0.83219 3.33333
3 2014 QueBar 26 12 2.2913 0.92209 4.33333
3 2014 Ser_Bar 16 10 2.18732 0.94994 5.33333
3 2014 MeiBar 48 17 2.498 0.88168 4.8
4 2014 Ros1_BarG 66 22 2.80105 0.90618 5.5
4 2014 Ros2_BarG 45 20 2.77166 0.9252 9
4 2014 Lag1_BarG 45 18 2.57609 0.89127 4.5
4 2014 Lag2_BarG 70 24 2.77979 0.87468 5
4 2014 Comp1BarG 30 17 2.44212 0.86196 3.75
4 2014 Comp2BarG 45 15 2.40789 0.88916 5

57
Anexo III
Anexo III.1 Valores da abundância relativa (N), riqueza específica (S), equitabilidade
(E) e índices de diversidade e dominância ( H’ e 1/d respetivamente) por local de
tipologia para a ordem Odonata.
Tipologia Ano Local N S H' E (H'/H'max) d
1 2013 Per_PS 2 2 0.693147 1 2
1 2013 Fom_PS 17 4 0.916068 0.660803 1.416667
1 2013 Red_PS 58 5 0.549265 0.341277 1.16
1 2013 Con_PS 13 4 1.031767 0.744263 1.625
1 2013 Fon_PS 1 1 0 - 1
1 2013 Rod_PS 2 1 0 - 1
2 2013 Sec_Nat 52 4 1.006924 0.726342 1.575758
2 2013 Red_Nat 141 7 1.478879 0.759994 2.711538
2 2013 Fav_Nat 233 8 1.562554 0.751429 2.588889
2 2013 Esc_Nat 193 5 0.921851 0.572778 1.595041
2 2013 Ang_Nat 95 6 1.166258 0.650901 1.9
2 2013 Fra_Nat 205 6 1.150377 0.642038 1.708333
3 2013 For_Bar 42 5 1.198078 0.744408 1.909091
3 2013 Cur_Bar 284 8 0.921263 0.443034 1.42
3 2013 Con_Bar 186 9 1.126477 0.512682 1.55
3 2013 Que_Bar 63 4 1.002698 0.723293 1.615385
3 2013 Ser_Bar 59 6 1.605473 0.896032 2.458333
3 2013 Mei_Bar 26 6 1.444208 0.806028 2.888889
4 2013 Ros2_BarG 2 2 0.693147 1 2
4 2013 Lag1_BarG 9 3 0.848686 0.772507 1.5
4 2013 Lag2_BarG 1 1 0 - 1
4 2013 Comp1_BarG 14 4 1.19734 0.863698 2.333333
4 2013 Comp2_BarG 14 6 1.611472 0.89938 3.5
1 2014 Per_PS 1 1 0 - 1
1 2014 Con_PS 69 4 1.121454 0.808958 2.3
1 2014 Tor_PS 1 1 0 - 1
1 2014 Fon_PS 5 2 0.500402 0.721928 1.25
1 2014 Rod_PS 6 4 1.242453 0.896241 2
1 2014 Lag_PS 1 1 0 - 1
1 2014 Sec_PS 5 3 1.05492 0.96023 2.5
2 2014 Sec_Nat 118 6 1.279064 0.713859 2.36
2 2014 Red_Nat 73 8 1.659696 0.798145 3.173913
2 2014 Fav_Nat 132 9 1.537343 0.699675 2.4
2 2014 Esc_Nat 33 4 1.102593 0.795352 1.736842
2 2014 Ang_Nat 45 5 1.341999 0.833831 2.647059
2 2014 Fra_Nat 91 9 1.907632 0.868201 3.033333

58
Tipologia Ano Local N S H' E (H'/H'max) d
3 2014 For_Bar 30 2 0.325083 0.468996 1.111111
3 2014 Cur_Bar 6 2 0.450561 0.650022 1.2
3 2014 Con_Bar 88 9 1.716037 0.781002 3.142857
3 2014 Que_Bar 16 5 1.160186 0.720864 1.6
3 2014 Ser_Bar 26 6 1.768515 0.987027 4.333333
3 2014 Mei_Bar 14 9 2.069202 0.941735 4.666667
4 2014 Ros1_BarG 1 1 0 -! 1
4 2014 Ros2_BarG 11 5 1.46814 0.912207 2.75
4 2014 Lag2_BarG 8 4 1.213008 0.875 2
4 2014 Comp1_BarG 59 7 1.020223 0.524291 1.685714
4 2014 Comp2_BarG 29 6 1.222088 0.68206 1.705882

59
Anexo IV
Anexo IV.1 Lista de espécies identificadas de cada uma das ordens estudadas
Espécies de Lepidoptera Espécies de Odonata
Aglais io Aeshna cyanea
Aglais urticae Aeshna juncea
Anthocharis cardamines Anax imperator
Argynnis adippe Boyeria irene
Argynnis aglaja Calopteryx virgo
Argynnis pandora Calopteryx xanthostoma
Argynnis paphia Ceriagrion tenellum
Aricia cramera Cordulegaster boltonii
Brintesia circe Enallagma cyathigerum
Callophrys rubi Ischnura graellsii
Celastrina argiolus Lestes dryas
Coenonympha iphioides Lestes virens
Coenonympha pamphilus Lestes viridis
Colias crocea Libellula depressa
Cyaniris semiargus Libellula quadrimaculata
Gonepteryx rhamni Onychogomphus uncatus
Hesperia comma Platycnemis latipes
Hipparchia fidia Pyrrosoma nymphula
Hipparchia hermione Sympetrum flaveolum
Hipparchia semele Sympetrum fonscolombii
Hipparchia statilinus Sympetrum sanguineum
Hyponephele lycaon Sympetrum sp.
Iphiclides feisthamelii Sympetrum striolatum
Issoria lathonia
Lampides boeticus
Lasiommata maera
Lasiommata megera
Leptidea sinapis
Leptotes pirithous
Lycaena alciphron
Lycaena phlaeas
Maniola jurtina
Melanargia lachesis
Melanargia russiae
Melitaea deione
Melitaea nevadensis
Melitaea parthenoides
Nymphalis antiopa
Nymphalis polychloros
Pararge aegeria
Pieris brassicae
Pieris napi
Pieris rapae
Plebejus argus
Polygonia c-album
Polyommatus icarus
Pontia daplidice
Pyrgus malvoides
Pyronia tithonus
Satyrus actea
Thymelicus lineola
Thymelicus sylvestris
Vanessa atalanta
Vanessa cardui
Vanessa virginiensis

60
Anexo V
Anexo V.1 Lista de abreviaturas dos nomes dos locais de amostragem e da tipologia de
habitats.
Código dos
locais
Locais
Código da
Tipologia Tipologia
For Covão do Forno Bar Barragem pequena
Cur Covão do Curral Bar Barragem pequena
Con Covão dos Conchos Bar Barragem pequena
Que Covão do Quelhas Bar Barragem pequena
Ser Lagoa Serrano Bar Barragem pequena
Mei Covão do Meio Bar Barragem pequena
Ros1
Barragem do
Rossim 1 BarG Barragem de grandes dimensões
Ros2
Barragem do
Rossim 2 BarG Barragem de grandes dimensões
Lag1
Barragem do
Lagoacho 1 BarG Barragem de grandes dimensões
Lag2
Barragem do
Lagoacho 2 BarG Barragem de grandes dimensões
Comp1 Lagoa Comprida 1 BarG Barragem de grandes dimensões
Comp2 Lagoa Comprida 2 BarG Barragem de grandes dimensões
Sec Lagoa Seca Nat Lagoa natural
Red Lagoa Redonda Nat Lagoa natural
Fav Lagoacho das Favas Nat Lagoa natural
Esc Lagoa Escura Nat Lagoa natural
Ang Lagoa do Ângelo Nat Lagoa natural
Fra Lagoa da Francelha Nat Lagoa natural
Cor Corgo das Mós PS Ponto Seco
Per Vale de Perdiz PS Ponto Seco
Fom Erva da Fome PS Ponto Seco
Red Redonda Seco PS Ponto seco
Pen Penha do Gato PS Ponto seco
Con Conchos Seco PS Ponto seco
Tor Torre PS Ponto seco
Fon Fonte dos Perús PS Ponto seco
Rod Rodeio Grande PS Ponto seco
Egu Vale das Éguas PS Ponto seco
Lag Lagoacho Seco PS Ponto seco
Sec Seca seco PS Ponto seco

61
Anexo VI
Anexo VI.1 Resultados da análise de complementaridade com indicação dos locais de
amostragem mais relevantes para a conservação de cada ordem, separadamente (Odonata
e Lepidoptera) e em conjunto, mais relevantes e a sua contribuição para a riqueza
específica acumulada (S_acum).
Número
de locais
S_acum
Lepidoptera
Locais
Lepidoptera
S_acum
Odonata
Locais Odonata S_acum
Total
Locais Total
1 34 Barragem do Rossim1
15 Lagoacho das Favas
36 Barragem do covão do Curral
2 41 Barragem do Rossim2
19 Barragem do
Covão dos
Conchos
52 Barragem da lagoa Comprida1
3 45 Barragem do
Covão do Meio
20 Lagoa do Ângelo 59 Barragem do
Rossim2
4 48 Lagoa Redonda 21 Rodeio Grande 63 Barragem do Rossim1
5 50 Erva da Fome 22 Barragem do Covão do Meio
66 Lagoa Redonda
6 52 Barragem do covão do Curral
23 Barragem da lagoa Comprida2
69 Barragem do Covão do Meio
7 53 Barragem do Lagoacho1
23 71 Barragem da lagoa Comprida2
8
54 Lagoacho das Favas
23 73 Lagoa do Ângelo
9 55 Barragem da lagoa Comprida2
23 75 Barragem do Lagoacho1
10 55 23 76 Rodeio Grande
11 55 23 77 Lagoacho das Favas
12 55 23 78 Barragem do
Covão dos
Conchos
13 55 23 78
14 55 23 78
15 55 23 78
16 55 23 78
17 55 23 78
18 55 23 78
19 55 23 78
20 55 23 78
21 55 23 78
22 55 23 78
23 55 23 78
24 55 23 78
25 55 23 78
26 55 23 78
27 55 23 78

62




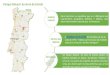


![Serra da estrela jhdf[1]](https://img.document.onl/doc/110x75/5571cc6bd8b42a3f5e8b4a78/serra-da-estrela-jhdf1.jpg)





















