Embed Size (px)
Citation preview

8/3/2019 Apoptose Trabalho de Bio Cell
http://slidepdf.com/reader/full/apoptose-trabalho-de-bio-cell 1/2
Apoptosis
Apoptosis, or programmed cell death, is a normal component of the development and health of multicellular organisms. Cells die in response to a variety of stimuli and during apoptosis they do so ina controlled, regulated fashion. This makes apoptosis distinct from another form of cell death callednecrosis in which uncontrolled cell death leads to lysis of cells, inflammatory responses and,potentially, to serious health problems. Apoptosis, by contrast, is a process in which cells play anactive role in their own death (which is why apoptosis is often referred to as cell suicide).
Upon receiving specific signals instructing the cells to undergo apoptosis a number of distinctivechanges occur in the cell. A family of proteins known as caspases are typically activated in the earlystages of apoptosis. These proteins breakdown or cleave key cellular components that are required for normal cellular function including structural proteins in the cytoskeleton and nuclear proteins such asDNA repair enzymes. The caspases can also activate other degradative enzymes such as DNases,which begin to cleave the DNA in the nucleus.
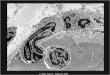
Apoptotic cells display distinctive morphology during the apoptotic process. This can be seen in theimage below which shows a trophoblast cell undergoing apoptosis.
Typically, the cell begins to shrink following thecleavage of lamins and actin filaments in thecytoskeleton (A). The breakdown of chromatin in thenucleus often leads to nuclear condensation and inmany cases the nuclei of apoptotic cells take on a"horse-shoe" like appearance (B). Cells continue to
shrink (C), packaging themselves into a form thatallows for their removal by macrophages. Thesephagocytic cells are responsible for clearing theapoptotic cells from tissues in a clean and tidyfashion that avoids many of the problemsassociated with necrotic cell death. In order topromote their phagocytosis by macrophages,apoptotic cells often ungergo plasma membranechanges that trigger the macrophage response. Onesuch change is the translocation of phosphatidylserine from the inside of the cell to theouter surface. The end stages of apoptosis are often
characterised by the appearance of membrane blebs (D) or blisters process. Small vesicles calledapoptotic bodies are also sometimes observed (D, arrow).
There are a number of mechanisms through which apoptosis can be induced in cells. The sensitivity of
cells to any of these stimuli can vary depending on a number of factors such as the expression of pro-and anti-apoptotic proteins (eg. the Bcl-2 proteins or the Inhibitor of Apoptosis Proteins), the severity of the stimulus and the stage of the cell cycle. Some of the major stimuli that can induce apoptosis are
outlined in the illustration below.
In some cases the apoptotic stimuli compriseextrinsic signals such as the binding of deathinducing ligands to cell surface receptorscalled death receptors. These ligands caneither be soluble factors or can be expressed
on the surface of cells such as cytotoxic Tlymphocytes. The latter occurs when T-cellsrecognise damaged or virus infected cells andinitiate apoptosis in order to prevent damaged
cells from becoming neoplastic (cancerous)or virus-infected cells from spreading theinfection. Apoptosis can also be induced bycytotoxic T-lymphocytes using the enzymegranzyme.
In other cases apoptosis can be initiatedfollowing intrinsic signals that are producedfollowing cellular stress. Cellular stress may
occur from exposure to radiation or chemicals or to viral infection. It might also be a consequence of growth factor deprivation or oxidative stress caused by free radicals. In general intrinsic signals initiate

8/3/2019 Apoptose Trabalho de Bio Cell
http://slidepdf.com/reader/full/apoptose-trabalho-de-bio-cell 2/2
apoptosis via the involvement of the mitochondria. The relative ratios of the various bcl-2 proteins canoften determine how much cellular stress is necessary to induce apoptosis.
THE ROLE OF CASPASE-3 IN APOPTOSIS The term apoptosis is derived from the Greek word that signifies the dropping of leaves from the trees.This term is applied to a group of characteristic structural and molecular events that separate this type of cell deletion from necrosis. In contrast to necrosis that involves a group of cells simultaneously, apoptosismay occur in a single cell surrounded by a group of viable cells. There is a distinct and precisely localizedcontrol over the fate of specific cells in a mixed cell population that undergo apoptosis. Apoptosis is aselective process for deletion of cells in various biological systems. This event similar to proliferation istightly regulated with both processes playing essential roles in the homeostasis of renewable tissues.
Diverse groups of molecules are involved in the apoptosis pathway. One set of mediators implicated inapoptosis belong to the asparate-specific cysteinyl proteases or caspases. A member of this family,caspase-3 (CPP32, apopain, YAMA) has been identified as being a key mediator of apoptosis of mammalian cells. Kothakotaet al , screened the translation products of a murine protein library to find thesubstrates that are susceptible to cleavage by caspase-3. They found that in cells exposed to Fas, gelsolinwas cleaved in vivo in a caspase dependent manner. The cleaved fragments of gelsolin led to thecleavage of the actin filaments in a Ca
2+independent manner. The expression of the gelsolin fragments
also led to the apoptosis of cells. Additional evidence for the role of gelsolin in apoptosis was provided byshowing that, as compared with wild-type neutrophis, those from mice lacking gelsolin exhibited a delayedonset of induced apoptosis. Therefore, the authors suggested that cleaved fragments of gelsolin may be
implicated in apoptosis.
Apoptosis-inducing factor Apoptosis inducing factor is a flavoprotein.
[2]
Apoptosis inducing factor is involved in initiating a caspase-independent pathway of apoptosis
(positive intrinsic regulator of apoptosis) by causing DNA fragmentation and chromatin
condensation. It also acts as an NADH oxidase. Another AIF function is to regulate the
permeability of the mitochondrial membrane upon apoptosis. Normally it is found behind the
outer membrane of the mitochondria and is therefore secluded from the nucleus. However,
when the mitochondria is damaged, it moves to the cytosol and to the nucleus. Inactivation of
AIF leads to resistance of embryonic stem cells to death following the withdrawal of growthfactors indicating that it is involved in apoptosis.