Embed Size (px)
Citation preview

Apostila de Citologia

1
Apresentação
Esta apostila se destina tão somente a auxiliar o aluno na compreensão
da disciplina e ao melhor acompanhamento do que foi exposto em sala de aula.
O conteúdo desta apostila, abrangendo textos e imagens, é mera
síntese da bibliografia indicada na ementa da disciplina e de artigos disponíveis
em sítios especializados na internet, sendo, portanto, um material de apoio que
não substitui a leitura e a pesquisa das fontes bibliográficas indicadas.

2
CITOLOGIA
É possível que você já tenha ouvido falar que a célula é a unidade fundamental da vida. Isso
significa dizer que para um "ser vivo" possuir vida, no mínimo, ele deverá ser formado por uma
célula.
Assim, nada mais natural, que começarmos nosso aprendizado sobre biologia (bio = vida; logia
= estudo), analisando as estruturas presentes em uma célula e suas funções, ou seja, através
da Citologia (cito = célula; logia = estudo).
Histórico
As células são geralmente muito pequenas e, dificilmente, visíveis a olho nu. Por isso, apenas
depois da invenção do microscópio é que foi possível observar uma célula pela primeira vez.
Sabe quando isso aconteceu? No ano de 1665, pelo inglês Robert Hooke, utilizando um
microscópio muito simples.
Ele fez cortes bem finos na casca de uma árvore e descobriu que estas estruturas eram
formadas por pequenas unidades semelhantes a favo de mel.
O nome "célula" foi empregado pela primeira vez por ele como diminutivo da palavra "cella",
que em latim significa espaço cercado por paredes.
seres vivos x seres brutos
Depois da descoberta da célula, tudo que existe passou a ser classificados em dois grupos:
Seres brutos ou Acelulares – que não são formados por células, portanto, sem vida.
Seres vivos ou Celulares – que são formados por células e que, portanto, possuem vida.
Classificação quanto ao tipo de célula
Todas as células possuem dois elementos essenciais: a membrana plasmática, conhecida também por plasmalema ou membrana celular, que separa os contúdos celulares do ambiente externo. E o outro é o material genético, que constitui a informação hereditária, que regula todas as atividades celulares e características que são passadas para outros descendentes.
A organização desse material genético é uma das principais características que separam as
células procarióticas das eucarióticas.
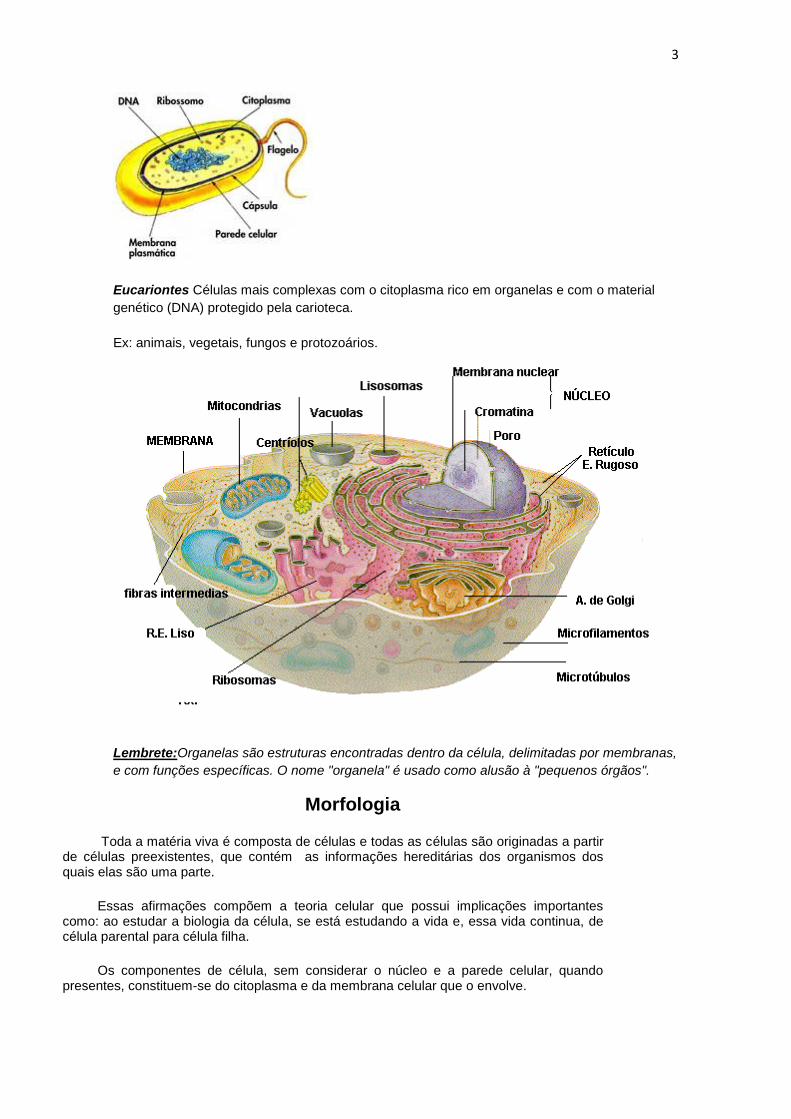
Procariontes
Células muito simples, sem organelas e com material genético (DNA) localizado em uma região
específica do citoplasma, mas não protegido por um envoltório, ou seja, sem uma membrana
nuclear (carioteca).
Ex: bactérias e algas cianofíceas, também conhecidas como algas azuis.
3
Eucariontes Células mais complexas com o citoplasma rico em organelas e com o material
genético (DNA) protegido pela carioteca.
Ex: animais, vegetais, fungos e protozoários.
Lembrete:Organelas são estruturas encontradas dentro da célula, delimitadas por membranas,
e com funções específicas. O nome "organela" é usado como alusão à "pequenos órgãos".
Morfologia
Toda a matéria viva é composta de células e todas as células são originadas a partir de células preexistentes, que contém as informações hereditárias dos organismos dos quais elas são uma parte.
Essas afirmações compõem a teoria celular que possui implicações importantes como: ao estudar a biologia da célula, se está estudando a vida e, essa vida continua, de célula parental para célula filha.
Os componentes de célula, sem considerar o núcleo e a parede celular, quando presentes, constituem-se do citoplasma e da membrana celular que o envolve.

4
No citoplasma ou citossol encontram-se todas as moléculas e organelas da célula, é onde ocorrem as reações bioquímicas. As organelas são estruturas especializadas que desempenham funções específicas dentro da célula, como por exemplo as mitocôndrias, o complexo de Golgi, vacúolos, etc. Todas as organelas serão estudadas separadamente em breve.
Composição Química das Células
MICROMOLÉCULAS
São moléculas de peso molecular entre 100 e 1000 e contém até trinta ou mais
átomos de carbono. Normalmente são encontradas livres em solução, onde algumas delas formam um conjunto de intermediárias a partir das quais as macromoléculas são formadas. As quatro micromoléculas principais são:os açúcares simples, os ácidos graxos, os aminoácidos e os nucleotídeos.
AÇUCARES
São as mais abundantes moléculas orgânicas na natureza e são primariamente moléculas que reservam energias na maioria dos organismos vivos. Os açúcares mais simples são chamados de monossacarídeos tais como ribose, glicose, frutose, que são formados com uma molécula de açúcar e possuem fórmula geral (CH2O)n, onde o n pode variar de 3 a 7. Por ex. a glicose possui fórmula C6H12O6.
Os dissacarídeos são formados por dois açúcares ligados covalentemente, por ex. a
maltose (açúcar da cana) e a lactose (açúcar do leite). Os polissacarídeos são grandes moléculas formadas por unidades de monossacarídeos, como por ex. o glicogênio, em células animais, e o amido, nos vegetais, são compostos somente de glicose.
E os oligossacarídeos que não estão livres e sim unidos a lipídeos e proteínas, de modo que são partes de glicolipídeos e de glicoproteínas, que estão na membrana plasmática.
ÁCIDOS GRAXOS
São a gorduras, óleos, fosfolipídios, cutina, suberina, ceras e esteróides. É um composto que possui um ácido carboxílico ligado a uma longa cadeia de hidrocarbonetos. São fontes valiosas de alimento, uma vez que eles podem ser quebrados para produzir o dobro de energia utilizável. Eles são estocados no citoplasma de muitas células na forma de gotas de moléculas de triglicérides (gorduras animais conhecidas da experiência diária).
A função mais importante dos ácidos graxos está na construção das membranas celulares, que envolvem as células suas organelas, que são compostas de fosfolipídios.

5
Membrana Celular Bicamada lipídica das membranas celulares
A cutina, suberina e as ceras formam barreiras contra perda de água. A superfície das células dos caules e folhas é coberta com uma cutícula, composta de cera e cutina, que protege e previne a perda de água. Os esteróides são moléculas que apresentam quatro anéis hidrocarbônicos interligados, o mais comum é o colesterol. São funcionalmente deferentes de acordo com os grupos químicos que estão unidos a sua estrutura básicos, por ex. vários hormônios importantes como estrogênio, testosterona e a vitamina A.
AMINOÁCIDOS
São compostos que apresentam um grupo de ácido carboxílico, um grupo amino e um grupo R variável, ligados a um simples átomo de carbono. São muito variados e responsáveis pela constituição de proteínas, onde os aminoácidos são ligados um ao outro por pontes peptídicas. Mas somente vinte aminoácidos são comuns em proteínas, que diferem no tamanho, carga elétrica e polaridade do grupo R. Esses aminoácidos estão distribuídos em quatro categorias: os ácidos (ácido aspártico e ácido glutâmico); básicos (histidina, lisina, arginina); neutros e polares ou hidrofílicos (serina, treonina, tirosina, asparginina, glutamina); neutros não-polares ou hidrofóbicos (glicina, alanina, triptofano, valina, cisteína, leucina, isoleucina, fenilalanina, prolina, metionina).
Alanina, Valina, Leucina, Isoleucina
Metionina, Prolina, Fenilanina, Tripotofano

6
Glicina, Serina, Treonina, Glutamina
Asparginina, Cisteína, Tirosina, Ácido glutâmico
Ácido aspártico, Arginina, Lisina, Histidina
NUCLEOTÍDEOS
São moléculas complexas, consistindo em um grupo fosfato, uma base nitrogenada e um açúcar de cinco átomos de carbono – pentose (Figura1). Eles são blocos constitutivos dos ácidos nucléicos, o ácido desoxirribonucléico (DNA) e o ácido ribonucléico (RNA), que transmitem e traduzem a informação genética. Algumas moléculas de RNA funcionam como catalisadores. Os nucleotídeos podem atuar como moléculas carregadoras de energia química, como por ex. o éster trifosfato e adenina (ATP), que participa das transferências de energia em centenas de reações celulares individuais. Eles combinam com outros para formar enzimas como a coenzima A. E são utilizadas como moléculas de sinalização específica na célula.

7
MACROMOLÉCULAS
Apresentam peso molecular entre 10.000 e 1 milhão, são construídas a partir de subunidades de baixo peso molecular (micromoléculas), que são repetidamente adicionadas para formar um longo polímero em cadeia. Como por ex. os aminoácidos ligados a outros aminoácidos para formar as proteínas.
A sua formação é mantida por ligações covalentes, as quais são fortes o suficiente para preservar a seqüência de subunidades por longos períodos de tempo. Para realizar a sua função , as macromoléculas dependem de ligação não-covalente, muito mais fraca, que se formam entre as partes distintas da mesma e entre diferentes macromoléculas. Exercendo um importante papel na determinação da estrutura tridimensional de cadeias macromoleculares e a interação com outras. As macromoléculas são: os ácidos nucléicos e as proteínas.
PROTEÍNAS
É um polímero linear de aminoácidos unidos por ligações peptídicas. Sua estrutura é formada por uma variedade de 20 aminoácidos diferentes, chamados de essenciais, que permite à proteína uma variedade de formas e funções na célula.
A estrutura da proteína é descrita em quatro níveis hierárquicos de organização. A estrutura primária é a seqüência de aminoácidos dispostos linearmente, constituindo a cadeia polipeptídica. A estrutura secundária refere-se a conformação espacial que a proteína toma, que depende da posição de certos aminoácidos, a qual é estabilizada por pontes de hidrogênio. No entanto, quando as pontes de hidrogênio estabelecem-se entre
certos aminoácidos, o esqueleto se dobra dispondo-as em formas geométrica: a -hélice, a
lâmina pregueadas, os folhetos e o giro.
A estrutura terciária resulta de interações - hidrofóbicas, ligações dissulfeto, forças de van der Waal e interações iônicas - que estabilizam a estrutura secundária, dando uma compactação ou conformação a proteína (fibrosas ou globulares). A estrutura quartenária resulta da combinação de dois ou mais polipeptídicos, chamados de subunidades, que originam moléculas com grande complexidade que se mantêm unidas e interagem, como por exemplo à hemoglobina.
A função da proteína é determinada por sua estrutura tridimensional e da capacidade de ligarem-se covalentemente à outras moléculas (ligantes). O local de fixação dos ligantes nas proteínas e os ligantes correspondentes possuem alto grau de especificidade, ou seja, são complementares. As proteínas controlam o fluxo de íons através da membrana, regulam a concentração dos metabólicos, confere rigidez a célula, catalisam uma infinidade de reações químicas, atuam como sensores e chaves, produzem movimento e controlam a função genética.

8
ENZIMAS
São proteínas catalisadoras que permitem a aceleração das reações celulares aumentando a velocidade desta. São altamente específicas para seus substratos, onde estes se ligam ao sítio ativo, ocorre a catálise formando um complexo enzima-substrato.
ÁGUA
A água é um dos compostos mais importantes, bem como o mais abundante, sendo
vital para os organismos vivos. Fora da célula os nutrientes estão dissolvidos em água, que facilita a passagem através da membrana celular e , dentro da célula, é o meio onde ocorre a maioria das reações químicas.
Ela tem propriedades estruturais e químicas que a tornam adequada para o seu papel nas células vivas como:
A água é um a molécula polar, pois tem distribuição desigual das cargas, capaz de formar quatro pontes de hidrogênio com as moléculas de água vizinhas e por isso necessita de uma grande quantidade de calor para a separação das moléculas (100
o C).
É um excelente meio de dissolução ou solvente .
A polaridade facilita a separação e a recombinação dos íons de hidrogênio (H+) e íons hidróxido (OH-), é o reagente essencial nos processos digestivos , onde as moléculas maiores são degradadas em menores e faz parte de várias reações de síntese nos organismos vivos
As pontes de hidrogênio relativamente fortes a tornam um excelente tampão de temperatura.
Estrutura de uma molécula de água
ÁCIDOS NUCLEICOS
São polímeros lineares de nucleotídeos especializados no armazenamento, na
transmissão e no uso da informação, por isso são macromoléculas de grande
importância biológica. Existem dois tipos de ácidos nucléicos: DNA e RNA, que serão
dispostos comparativamente no quadro abaixo:

9
DNA RNA
Açúcar (Pentose) Desoxirribose Ribose
Bases pirimidínicas citosina (C), timina (T) citosina (C), uracil (U)
Bases purínicas adenina (A), guanina (G) Adenina (A), guanina (G)
Localização Principalmente no núcleo
das células, também nas
mitocôndrias e
cloroplastos.
Principalmente no
citoplasma, mas também
no núcleo
Função Transmissão de
informação genética
Síntese de proteínas
Forma Formada por duas fitas
antiparalelas que são
complementares na
seqüência de
nucleotídeos. Pareadas
em dupla-hélice (figura)
Fita simples , que contém
pequenas regiões de
pareamento de bases
complementares. (Figura)

10
Membrana Celular
Estrutura da Membrana Celular
A Membrana plasmática (ou celular) engloba a célula, definindo seus limites, separa o meio intracelular do extracelular e é o principal responsável pelo controle da saída e entrada de substâncias da célula. Ela é constituída por duas camadas lipídicas fluidas e contínuas onde estão inseridas moléculas protéicas, receptores específicos, que confere o modelo mosaico fluido.
Estrutura e Composição da Membrana Celular
As moléculas lipídicas constituem 50% da massa da maioria das membranas de células
animais, sendo o restante, constituído de proteínas. As moléculas lipídicas são anfipáticas,

11
pois possuem uma extremidade hidrofílica ou polar (solúvel em meio aquoso) e uma
extremidade hidrofóbica ou não-polar (insolúvel em água).
Os três principais grupos de lipídios da membrana são os fosfolipídeos, o colesterol e
os glicolipídeos.
Os fosfolipídeos possuem uma cabeça polar e duas caudas de hidrocarboneto
hidrofóbicas (característica que confere a dupla camada lipídica). As caudas são
normalmente ácidos graxos com diferenças no comprimento, o que influi na fluidez da
membrana.
As moléculas de colesterol aumentam as propriedades de permeabilidade das duplas
camadas lipídicas. Ela torna a bicamada lipídica menos sujeita a deformações, e assim,
diminui a permeabilidade da membrana.
Os glicolipídeos auxiliam na proteção da membrana plasmática em condições
adversas, como pH baixo. Sua presença altera o campo elétrico através da membrana e das
concentrações dos íons na superfície da membrana. Acredita-se que essas moléculas

12
participem dos processos de reconhecimento celular, e alguns glicolipídeos propiciam pontos
de entrada para algumas toxinas bacterianas.
A membrana plasmática não é uma estrutura estática, os lipídios movem-se
proporcionando uma fluidez à membrana. Os lipídios podem girar em torno de seu próprio
eixo, podem difundir-se lateralmente na monocamada, migrar de uma monocamada para
outra (flip-flop) que acontece raramente, e movimentos de flexão por causa das cadeias de
hidrocarbonetos.
A fluidez da membrana plasmática depende também da temperatura e da quantidade
de colesterol, pois quanto maior a temperatura e maior quantidade de colesterol, a membrana
é menos fluida.
Proteínas da Membrana
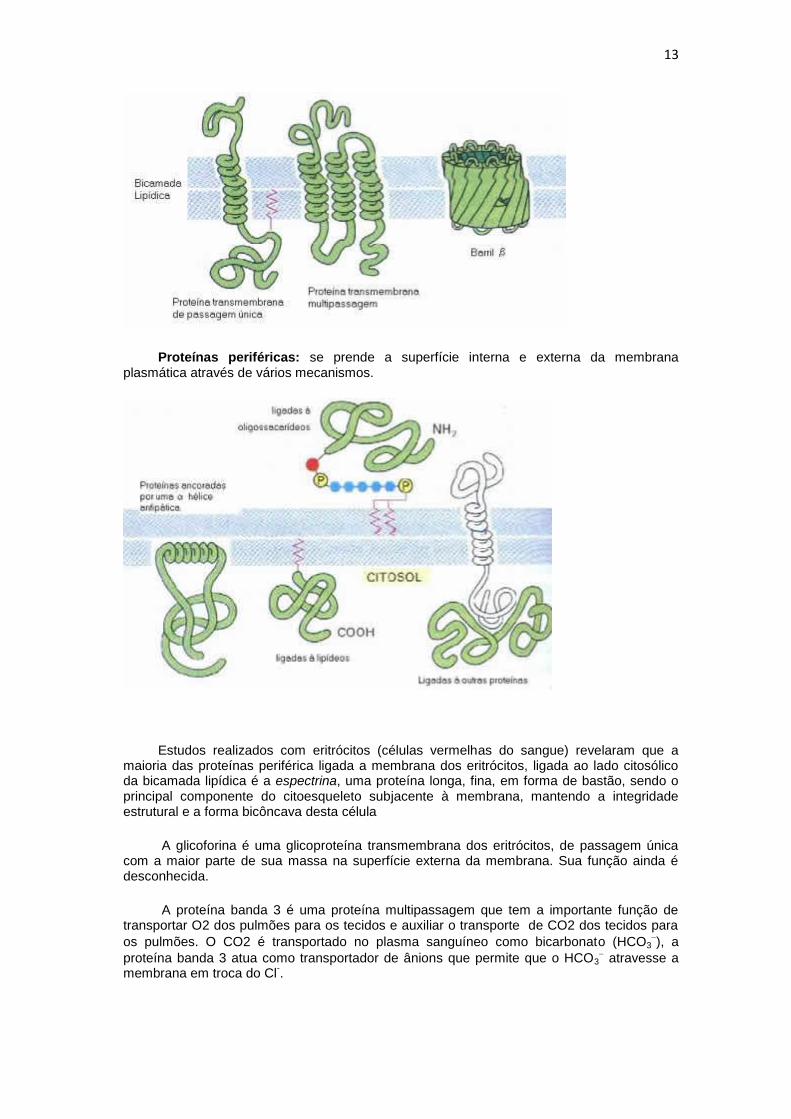
As proteínas de desempenham a maioria das funções específicas das membranas. São elas que conferem as propriedades funcionais características de cada tipo de membrana. As proteínas de membrana podem ser:
Proteínas transmembrana: atravessam a bicamada lipídica e são anfipáticas. Elas podem atravessar a membrana uma única vez (proteína transmembrana de passagem única) ou então atravessando várias vezes a membrana (proteína transmembrana multipassagem.
As proteínas transmembrana podem ser em hélice ou arranjados como barris (figura abaixo). Podem ter a função de transportar íons, funcionar como receptores ou como enzimas. A grande maioria das proteínas de membrana multipassagem das células
eucarióticas e da membrana bacteriana é formada por hélices transmembrana, enquanto
as barris se restringem principalmente às membranas externas das bactérias, das mitocôndrias e dos cloroplastos.
13
Proteínas periféricas: se prende a superfície interna e externa da membrana plasmática através de vários mecanismos.
Estudos realizados com eritrócitos (células vermelhas do sangue) revelaram que a maioria das proteínas periférica ligada a membrana dos eritrócitos, ligada ao lado citosólico da bicamada lipídica é a espectrina, uma proteína longa, fina, em forma de bastão, sendo o principal componente do citoesqueleto subjacente à membrana, mantendo a integridade estrutural e a forma bicôncava desta célula
A glicoforina é uma glicoproteína transmembrana dos eritrócitos, de passagem única com a maior parte de sua massa na superfície externa da membrana. Sua função ainda é desconhecida.
A proteína banda 3 é uma proteína multipassagem que tem a importante função de transportar O2 dos pulmões para os tecidos e auxiliar o transporte de CO2 dos tecidos para
os pulmões. O CO2 é transportado no plasma sanguíneo como bicarbonato (HCO3), a
proteína banda 3 atua como transportador de ânions que permite que o HCO3 atravesse a
membrana em troca do Cl-.

14
As proteínas de membrana estão geralmente associadas a carboidratos, que são encontrados como cadeias de oligossacarídeos ligadas covalentemente às proteínas (glicoproteínas) e a lipídeos (glicolipídeos), ou como cadeias de polissacarídeos de moléculas de proteoglicanas. O glicocálice é uma zona onde se encontra vários desses carboidratos na superfície da membrana.
Transporte Ativo e Passivo
Dizemos que a membrana plasmática seleciona a passagem destas substâncias e que
ela possui, desta forma, uma permeabilidade seletiva. A camada fosfolipídica da membrana
plasmática funciona como uma barreira fluida (maleável) e permite a passagem de substâncias
diretamente através dela.
Atravessará a barreira apenas as substâncias pequenas que consigam se entremear
através dos fosfoslipídeos. Essas substâncias precisam ter afinidades por lipídeos, senão não
conseguiriam se "misturar" com eles para atravessar a membrana.
Por outro lado não são apenas substâncias com afinidades por lipídeos que atravessam
a membrana plasmática. As substâncias que não conseguem atravessar diretamente a camada
fosfolipídica podem entrar ou sair da célula através de suas portas e janelas, que são as
proteínas.
A passagem das substâncias de pequeno porte através da membrana pode ocorrer
passivamente ou ativamente.
Antes de entender as duas formas principais de transporte na membrana, o passivo e o
ativo, é necessário que compreenda muito bem o processo de difusão.
Você já deve ter observado o que acontece quando uma pessoa passa um perfume forte
e permanece em um ambiente fechado. Em poucos instantes toda a sala fica com o cheiro do
perfume. Já parou para se perguntar por que isso ocorre?
Lembra que dois corpos não ocupam um mesmo lugar no espaço? Isso significa dizer que
quando uma "partícula" se move, ela acaba "tomando o lugar" de uma outra partícula que se
encontra ao seu lado.
O ar é composto de diversas "partículas" flutuantes diferentes, ou seja, de gases, como o
oxigênio, gás carbônico e nitrogênio, que se movem, e que estão sempre trocando de lugar uns

15
com os outros.
O perfume é feito por um líquido muito volátil, que se torna um gás facilmente. Quando
os gases perfumados são adicionados ao ar, eles também irão se mover e trocar de lugar com
os outros tipos de gases. Isso faz com que depois de um tempo tenhamos gases de perfume
espalhados por todo o ambiente fechado.
As moléculas presentes dentro de líquidos também possuem capacidade de movimento.
Dentro da célula e em seu exterior existe grande quantidade de líquidos. Então, quando uma
molécula qualquer se move dentro desses líquidos elas deverão também trocar de lugar com
as moléculas que estão ao seu redor.
Agora, responda a seguinte pergunta: Uma pessoa está parada numa estação final de
trem esperando para embarcar. O trem chega lotado de passageiros e abre as portas. A
pessoa conseguirá entrar com facilidade?
Você já deve ter percebido que esta pessoa terá grande dificuldade de entrar no trem
porque haverá um grande fluxo de passageiros saindo dele.
O mesmo acontece com as substâncias que entram e saem de uma célula. Se existe
maior quantidade de substâncias de um lado ou do outro, haverá maior fluxo de passagem
para o lado que a substância estiver em menor quantidade.
Assim, a difusão pode ser entendida como um maior fluxo de movimento de moléculas
em direção a uma região onde as mesmas se encontram em menor quantidade.
A bicamada lipídica serve como barreira, permitindo que a célula mantenha as
concentrações de solutos no citosol, que são diferentes do meio extracelular. Para isso, a
membrana desenvolveu mecanismos de transporte (proteínas carregadoras e de canal), ou
tornando-se permeável em favor do gradiente de concentração.
O transporte passivo ou difusão facilitada ocorre quando as proteínas de transporte
(canal) operam para importar moléculas específicas para dentro da célula, orientadas
somente por um gradiente de concentração.
A difusão passiva ocorre a favor de um gradiente, mas é um processo físico, que não
utiliza proteínas transportadoras e não há gasto de energia.
O transporte ativo ocorre contra um gradiente de concentração e é mediado por
carreadoras, chamadas de bombas. A atividade bombeadora consome energia (ATP).

16
As proteínas que transportam somente um tipo de soluto é denominada uniporte, outras
depende do transporte de um outro soluto, atuando como carreadores acoplados, que podem
ser simportes (transferência do segundo soluto na mesma direção) ou antiporte (transferência
do segundo soluto na direção oposta).
a) Transporte Ativo – Movimento de entrada ou saída de substâncias em uma célula com
gasto de energia. Ex: bomba de sódio e potássio. Para entender o transporte ativo, pense em
nosso exemplo acima e imagine o caso da pessoa que está do lado de fora do trem. Para
entrar no trem esta pessoa terá que "vencer" a direção natural de movimento de passageiros.
E, ao fazer isso, terá que se movimentar contra um "gradiente" de passageiros e se esforçar
bastante. Em outras palavras, ela terá que gastar energia. Para ocorrer a passagem de uma
molécula contra um gradiente de concentração também será necessário o gasto de energia.
b) Transporte Passivo – Movimento feito sem gasto de energia, ou seja, respeitando o
gradiente de condentração. Ex: osmose.
Osmose – É a difusão da água, ou seja, a passagem de água de um meio hipotônico (onde ela
se encontra em maior quantidade) para um meio hipertônico (onde ela se encontra em menor
quantidade). Em um meio hipotônico existe maior quantidade de água e menor quantidade de
sal dissolvido. O contrário ocorre em um meio hipertônico.
Difusão facilitada – É a passagem de macromoléculas através de proteínas especiais
denominadas permeases, que formam poros na membrana.
A membrana plasmática possui a capacidade de englobar substâncias de maior porte
através da endocitose
Endocitose – Transporte de moléculas em grande quantidade. Existem dois tipos de
mecanismos para esse transporte:
a) Fagocitose – Englobamento de partículas sólidas por meio da emissão de pseudópodes
pela membrana plasmática.
b) Pinocitose – Englobamento de gotículas líquidas por meio de invaginações da membrana
plasmática.

17
Modelo da Membrana Plasmática
A membrana plasmática desempenha papel fundamental na citologia. Seu estudo nao é possível sem uma visualização detalhada de sua ultra-estrutura.
Esquematizamos a ultra-estrutura da membrana plasmática em um modelo que pode ser montado por seus alunos.
Siga o passo-a-passo e depois peça que cada grupo (esse é um trabalho que deve ser realizado em grupo) faça um relatório que aponte cada uma das estruturas.
Esquema retirado do Cd interativo que acompanho o livro Junqueira - ed. 10ª
Materiais:
- Massinha para modelar (várias cores);
- Cola;
- Isopor;
- Grampo ou arame fino;
- Estilete;
- Pincel Atômico.
Passo-a-passo

18
- Com o estilete corte um retângulo de isopor, esta peça vai servir de base para a montagem da membrana;
- Lipídios: com a massinha (cor a escolha) faça pequenas bolinhas que representarão a cabeça hidrofílica dos lipídios como mostrado na foto abaixo;
- Carboidratos: usando uma outra cor de massinha, modele bolinhas menores que representarão os carboidratos da superfície da membrana e organize-os no arame ou grampo (ver foto);
- Proteínas: modele as proteínas, usando a massinha de cor diferenciada. Faça as proteínas periféricas e as integrais;
- Montagem da membrana: cole as bolinhas que represente os lipídios na base de isopor cobrindo-o completamente, exceto nas regiões que será colocada a proteína periférica. Depois de colar os "lipídios" (bolinhas maiores), coloque os arames com as bolinhas menores (carboidratos) nas bolinhas maiores (lipídio). Para finalizar, com o pincel atômico, desenhe a cabeça hidrofóbica do lipídio (as "perninhas"), em todas as laterais do isopor.
19
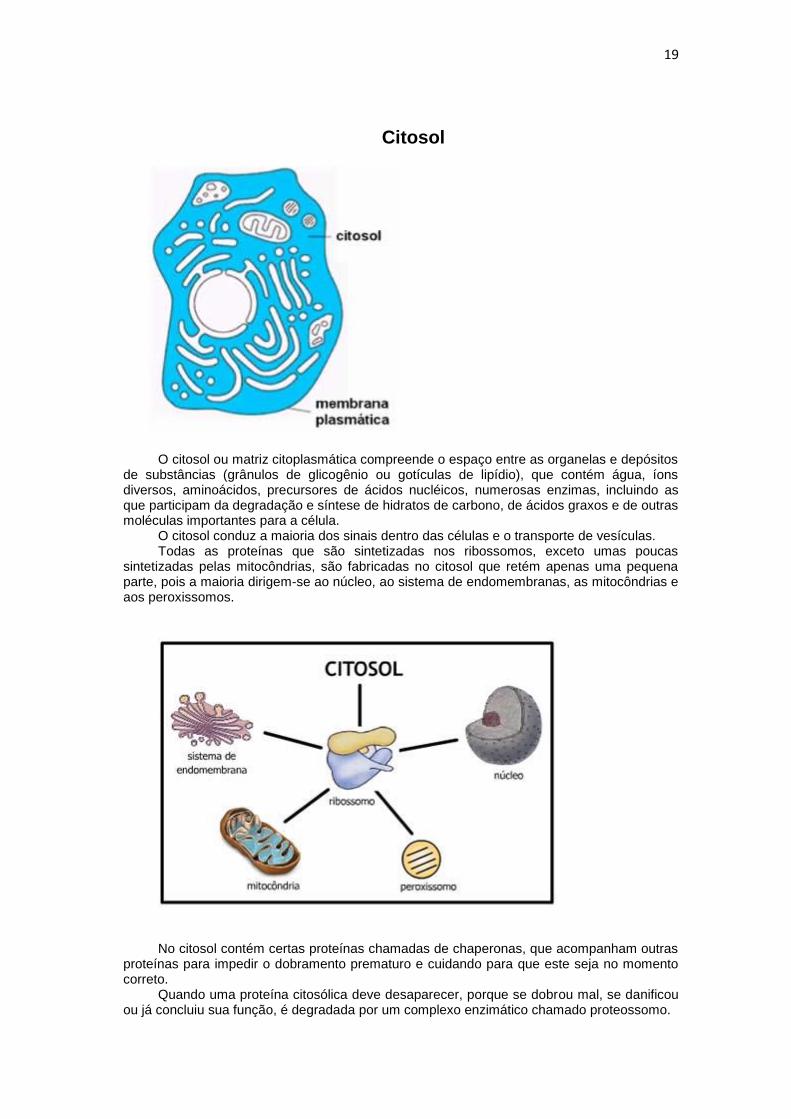
Citosol
O citosol ou matriz citoplasmática compreende o espaço entre as organelas e depósitos de substâncias (grânulos de glicogênio ou gotículas de lipídio), que contém água, íons diversos, aminoácidos, precursores de ácidos nucléicos, numerosas enzimas, incluindo as que participam da degradação e síntese de hidratos de carbono, de ácidos graxos e de outras moléculas importantes para a célula.
O citosol conduz a maioria dos sinais dentro das células e o transporte de vesículas. Todas as proteínas que são sintetizadas nos ribossomos, exceto umas poucas
sintetizadas pelas mitocôndrias, são fabricadas no citosol que retém apenas uma pequena parte, pois a maioria dirigem-se ao núcleo, ao sistema de endomembranas, as mitocôndrias e aos peroxissomos.
No citosol contém certas proteínas chamadas de chaperonas, que acompanham outras
proteínas para impedir o dobramento prematuro e cuidando para que este seja no momento correto.
Quando uma proteína citosólica deve desaparecer, porque se dobrou mal, se danificou ou já concluiu sua função, é degradada por um complexo enzimático chamado proteossomo.

20
Citoesqueleto
O esqueleto é o órgão de sustentação do corpo composto por ossos de diferentes tamanhos, que fornece proteção aos órgãos internos e ponto de apoio para a sustentação dos músculos.
As células também possuem um “esqueleto”, chamado de citoesqueleto, que mantém a forma da célula, as organizações do seu espaço interior, como também a capacidade de movimentação, são algumas das funções realizadas por esse sistema de filamentos protéicos.
citoesqueleto é composto por três tipos principais de filamentos e, cada um possui características peculiares que os diferenciam um dos outros.
Filamentos que compõem o Citoesqueleto
Filamentos Características e Função principal Esquema
Filamentos de Actina ou microfilamentos
De 7 a 9 nm de diâmetro; é uma dupla
fita helicoidal da proteína actina;
formam feixes lineares. Sustentam a
membrana plasmática e junto com
proteínas motoras, faz a locomoção
celular.
21
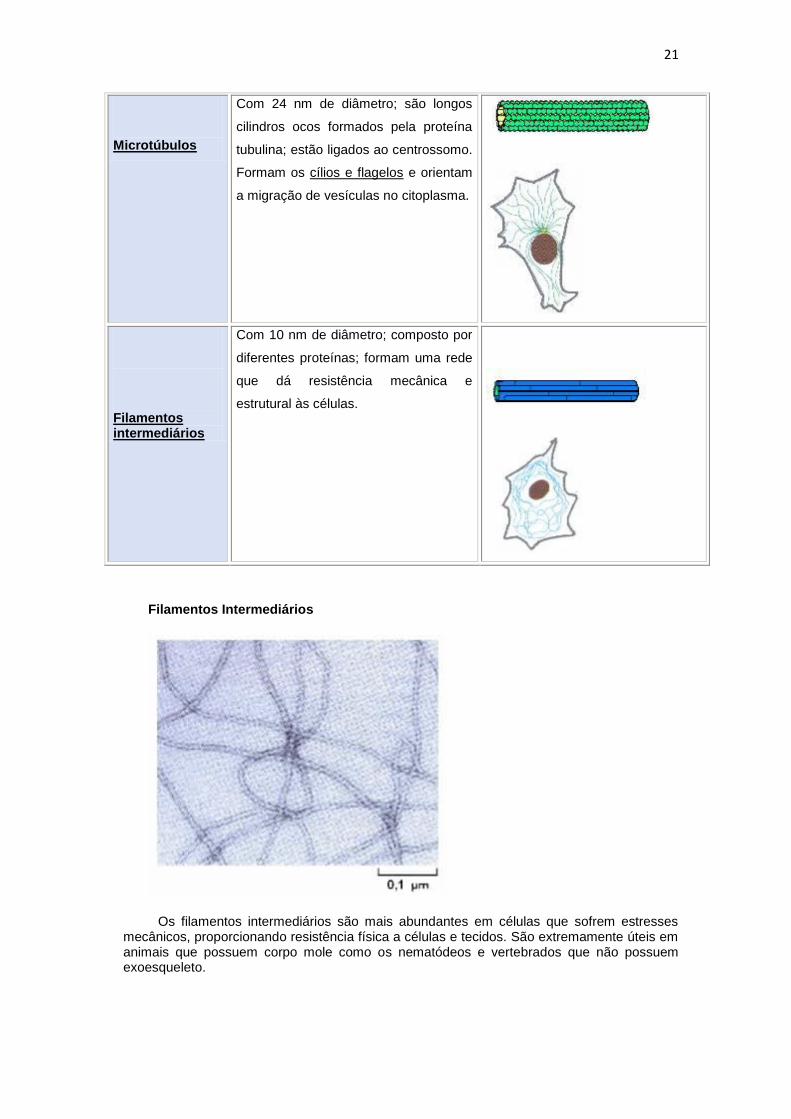
Microtúbulos
Com 24 nm de diâmetro; são longos
cilindros ocos formados pela proteína
tubulina; estão ligados ao centrossomo.
Formam os cílios e flagelos e orientam
a migração de vesículas no citoplasma.
Filamentos intermediários
Com 10 nm de diâmetro; composto por
diferentes proteínas; formam uma rede
que dá resistência mecânica e
estrutural às células.
Filamentos Intermediários
Os filamentos intermediários são mais abundantes em células que sofrem estresses mecânicos, proporcionando resistência física a células e tecidos. São extremamente úteis em animais que possuem corpo mole como os nematódeos e vertebrados que não possuem exoesqueleto.

22
Eles são formados a partir de várias proteínas que compõem as células dos tecidos, que são classificadas em seis tipos. Nos epitélios são formados a partir de queratinas ácidas (FI tipo I) e queratinas básicas (FI tipo II), proteínas tipo III são a vimentina (mesênquima), desmina (músculo), periferina (neurônios periféricos e centrais) e proteína ácida fibrilar glial (células gliais e astrócitos). As do tipo IV são neurofilamentos de baixo, médio e alto peso molecular, presentes nos neurônios maduros e a proteína internexina, distribuída no sistema nervos central em desenvolvimento. As FI tipo IV não padrão são a filensina e facinina (células das fibras do cristalino) e as do tipo V, lamina A, B e C, estão nos núcleos de todas as células.
Imagens de ME de criofratura: (A) Neurofilamentos no axônio; (B) filamentos de células da glia; (C) ME convencional transversal de axônio
O filamento intermediário possui uma estrutura em -hélice central e domínios
globulares em cada extremidade.
A organização desses filamentos, as ligações a outros filamentos e a sua função de
sustentação dependem de proteínas associadas aos filamentos intermediários (IFAP).
As redes de filamentos intermediários formam a lâmina nuclear, ao longo da superfície
interna da membrana nuclear, e estão firmemente ligados as junções celulares,
desmossomos e hemidesmossomos (serão estudadas na próxima atualização).

23
Transporte Intracelular
Para que haja transporte de vesículas ligadas à membrana, proteínas e organelas ao
longo dos microtúbulos, é necessária a participação de proteínas motoras como a cinesina e
dineína.
A cinesina movimenta-se em direção a extremidade (+) dos microtúbulos (transporte
anterógrado). Esta proteína possui dois domínios, um liga-se ao microtúbulo a ao ATP e o
domínio da cauda liga-se às vesículas. O transporte é específico porque cada vesícula possui
sua cinesina.
As dineínas movimentam-se em direção da extremidade (-) (transporte anterógrado) e são divididas em duas classes funcionais: a dineína citosólica, envolvida no movimento de vesículas e de cromossomos, e a dineína do axonema, responsável pelo batimento de cílios e flagelos.
A direção do transporte das vesículas depende da orientação dos microtúbulos, que é
fixada pelo MTOC.
Nas células pigmentosas, chamadas melanóforos, encontrada na pele de anfíbios e nas
escamas dos peixes, os microtúbulos transportam os grânulos de pigmento ao longo de toda
a célula, ajustando a cor do animal.

24
Cílios e Flagelos
Os cílios e flagelos são flexíveis prolongamentos da membrana celular, que variam de
comprimento, sendo responsáveis pelo movimento de células como o espermatozóide e
organismos unicelulares como o Paramecium.
Essas estruturas são construídas a partir de microtúbulos e proteínas motoras (dineínas).

25
O microtúbulo forma um feixe central chamado axonema, constituído de nove duplas de
microtúbulos dispostos circularmente e dois microtúbulos centrais. Moléculas de dineína ciliar
formam pontes entre os pares de microtúbulos.
O axonema é fixado por corpos basais à superfície celular, que apresenta a mesma forma do centríolo e funciona como um núcleo de montagem de microtúbulos flagelares.
As dineínas do axonema provocam o deslizamento entre os microtúbulos através da
hidrólise de ATP, proporcionando os batimentos ciliares e flagelares.
Filamentos de actina ou microfilamentos
A actina é a proteína intracelular mais abundante de uma célula eucariótica, sendo formada por subunidades globulares chamadas d0e actina G, que se polimerizam de forma helicoidal formando um filamento chamado de actina F.
O citoesqueleto de actina é dinâmico, sendo capaz de crescer e de encolher rapidamente. Primeiro ocorre a nucleação, que é a formação de um trímero estável, em
26
seguida as subunidades são adicionadas em ambas extremidades do filamento, crescendo mais rapidamente na extremidade (+) e despolarizando na extremidade (-). A estabilização é controlada por proteínas especializadas de ligação da actina que estão no citosol, como a tropomodulina e a gelsolina.
Os filamentos de actina são divididos em dois grupos:
Transcelulares: cruzam o citoplasma em todas as direções, formando feixes e redes,
interligados por proteínas de feixe (fimbrina e -actinina), que proporcionam sustentação e determinando a forma da célula.
Corticais: rede de filamentos situados abaixo da membrana plasmática (córtex), conectada a ela por proteínas de ligação (fodrina).
Algumas funções realizadas pelo filamento de actina serão apresentadas separadamente, para que não haja confusão:
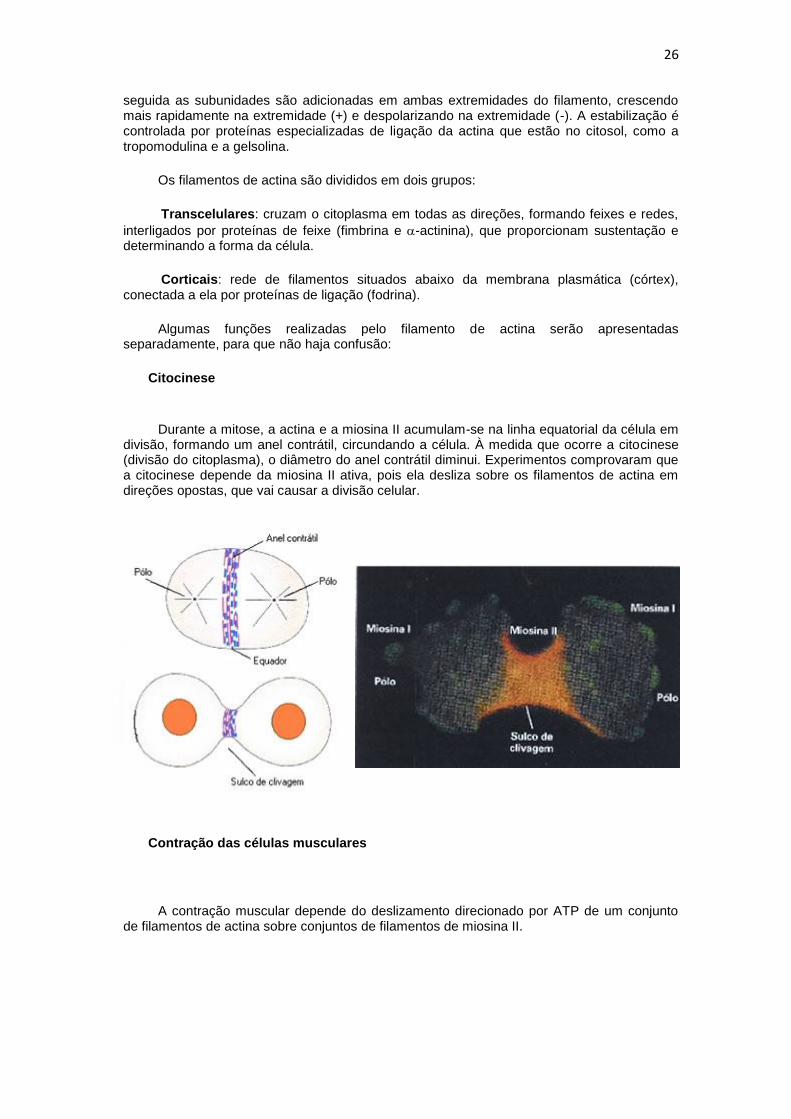
Citocinese
Durante a mitose, a actina e a miosina II acumulam-se na linha equatorial da célula em divisão, formando um anel contrátil, circundando a célula. À medida que ocorre a citocinese (divisão do citoplasma), o diâmetro do anel contrátil diminui. Experimentos comprovaram que a citocinese depende da miosina II ativa, pois ela desliza sobre os filamentos de actina em direções opostas, que vai causar a divisão celular.
Contração das células musculares
A contração muscular depende do deslizamento direcionado por ATP de um conjunto de filamentos de actina sobre conjuntos de filamentos de miosina II.

27
As células musculares esqueléticas são multinucleadas, formada por filamentos denominados de miofibrilas. As miofibrilas são formadas por unidades que se repetem, denominadas sarcômeros, que confere ao músculo esquelética, uma aparência estriada.
Cada sarcômero é formado por filamentos delgados, espessos e proteínas. Os filamentos delgados são filamentos de actina e mais duas proteínas adicionais, tropomiosina e troponina, tendo suas extremidades ligadas a uma linha elétron-densa (linha Z). Os filamentos espessos são compostos por miosina II.
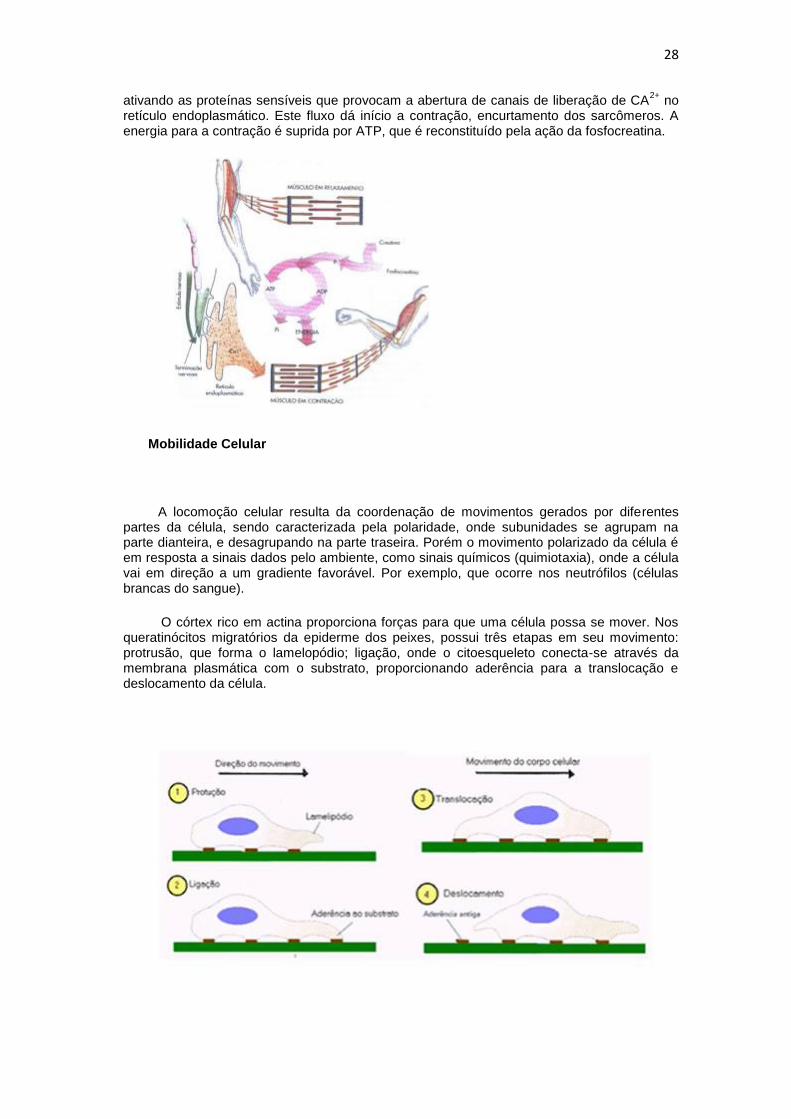
O mecanismo de contração muscular ocorre com o aumento de Ca2+
no citosol. O sinal que vem dos nervos provoca uma excitação elétrica que se espalha através dos túbulos T,
28
ativando as proteínas sensíveis que provocam a abertura de canais de liberação de CA2+
no retículo endoplasmático. Este fluxo dá início a contração, encurtamento dos sarcômeros. A energia para a contração é suprida por ATP, que é reconstituído pela ação da fosfocreatina.
Mobilidade Celular
A locomoção celular resulta da coordenação de movimentos gerados por diferentes partes da célula, sendo caracterizada pela polaridade, onde subunidades se agrupam na parte dianteira, e desagrupando na parte traseira. Porém o movimento polarizado da célula é em resposta a sinais dados pelo ambiente, como sinais químicos (quimiotaxia), onde a célula vai em direção a um gradiente favorável. Por exemplo, que ocorre nos neutrófilos (células brancas do sangue).
O córtex rico em actina proporciona forças para que uma célula possa se mover. Nos queratinócitos migratórios da epiderme dos peixes, possui três etapas em seu movimento: protrusão, que forma o lamelopódio; ligação, onde o citoesqueleto conecta-se através da membrana plasmática com o substrato, proporcionando aderência para a translocação e deslocamento da célula.
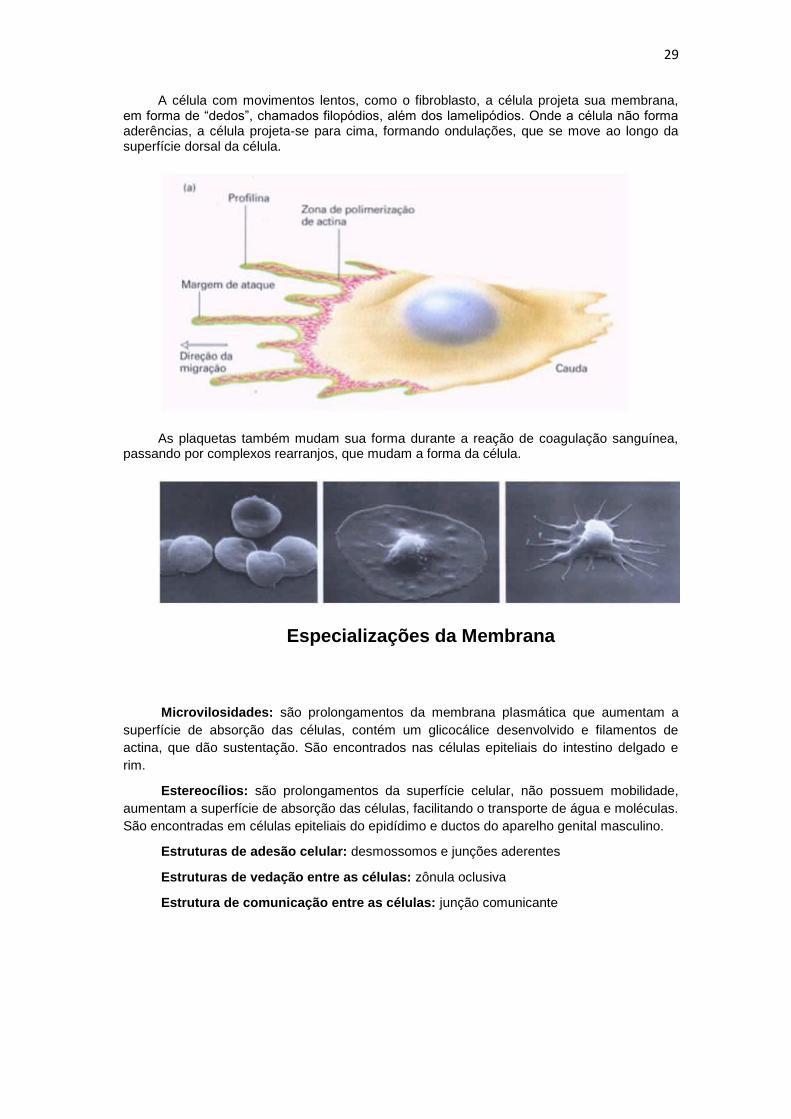
29
A célula com movimentos lentos, como o fibroblasto, a célula projeta sua membrana, em forma de “dedos”, chamados filopódios, além dos lamelipódios. Onde a célula não forma aderências, a célula projeta-se para cima, formando ondulações, que se move ao longo da superfície dorsal da célula.
As plaquetas também mudam sua forma durante a reação de coagulação sanguínea, passando por complexos rearranjos, que mudam a forma da célula.
Especializações da Membrana
Microvilosidades: são prolongamentos da membrana plasmática que aumentam a
superfície de absorção das células, contém um glicocálice desenvolvido e filamentos de
actina, que dão sustentação. São encontrados nas células epiteliais do intestino delgado e
rim.
Estereocílios: são prolongamentos da superfície celular, não possuem mobilidade,
aumentam a superfície de absorção das células, facilitando o transporte de água e moléculas.
São encontradas em células epiteliais do epidídimo e ductos do aparelho genital masculino.
Estruturas de adesão celular: desmossomos e junções aderentes
Estruturas de vedação entre as células: zônula oclusiva
Estrutura de comunicação entre as células: junção comunicante

30
Microvilosidades
As microvilosidades são projeções citoplasmáticas na superfície celular, envolta por membrana plasmática. A ponta da microvilosidade é constituída por substância amorfa, onde está imerso a extremidade (+) da actina, e a extremidade (-) está conectada ao córtex. Os feixes de filamentos de actina são dispostos paralelamente, interligados pela proteína vilina, que possui dois sítios de ligação. Os feixes laterais estão ligados a membrana plasmática através da miosina I.
31
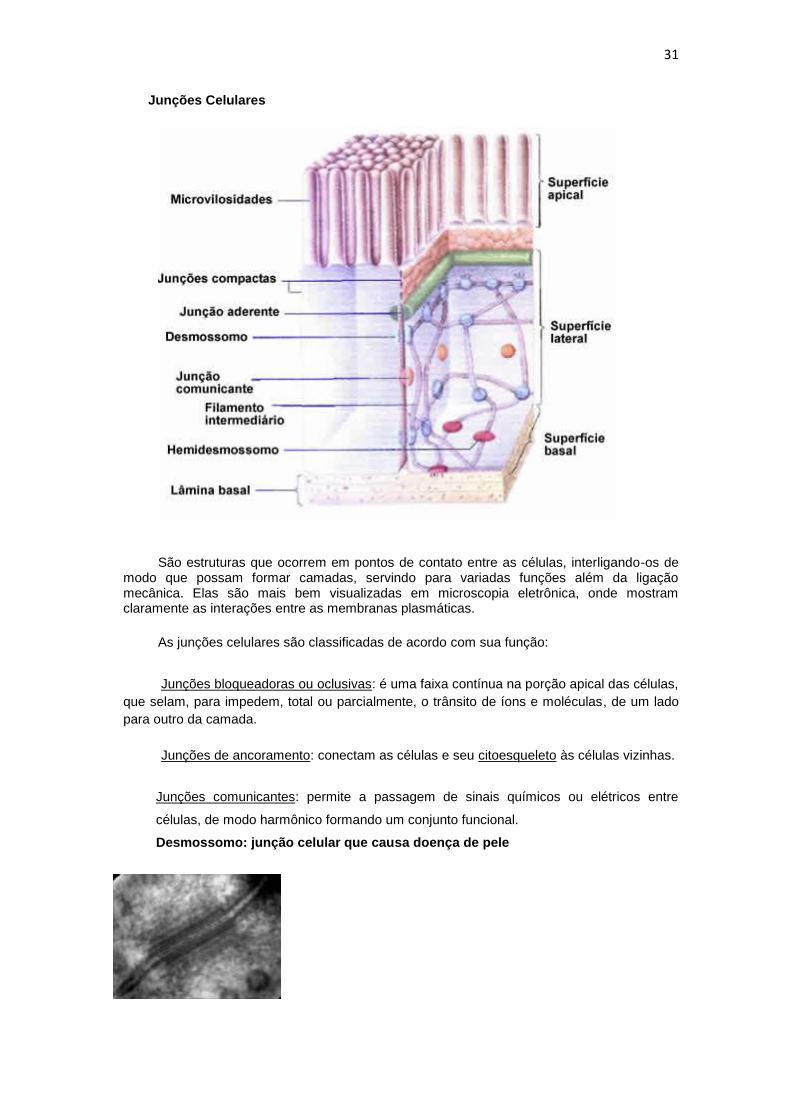
Junções Celulares
São estruturas que ocorrem em pontos de contato entre as células, interligando-os de modo que possam formar camadas, servindo para variadas funções além da ligação mecânica. Elas são mais bem visualizadas em microscopia eletrônica, onde mostram claramente as interações entre as membranas plasmáticas.
As junções celulares são classificadas de acordo com sua função:
Junções bloqueadoras ou oclusivas: é uma faixa contínua na porção apical das células,
que selam, para impedem, total ou parcialmente, o trânsito de íons e moléculas, de um lado
para outro da camada.
Junções de ancoramento: conectam as células e seu citoesqueleto às células vizinhas.
Junções comunicantes: permite a passagem de sinais químicos ou elétricos entre
células, de modo harmônico formando um conjunto funcional.
Desmossomo: junção celular que causa doença de pele
32
O Desmossomo é um tipo de junção celular muito importante encontrado na pele, cuja
função maior é unir fortemente as células umas às outras e garantir a aderência do
tecido. Neles estão inseridos placas de filamentos intermediários que são formados por
queratina garantindo a adesão e o elo de ligação do citoesqueleto com as células
vizinhas.
Muitas doenças de pele estudadas são causadas por deficiências em estruturas do desmossomos. Zdravko Peris pesquisador de dermatologia na Croácia estuda uma doença chamada Morbus Darier.
Morus Darier ou desqueratinização folicular é uma especial forma de distúrbio de
queratinização folicular a qual parece atingir clinicamente a pele, membranas e unhas
(Figuras 1, 2 e 3).
O nome da doença (Morbus Darier) é do mesmo autor que descreveu primeiramente a
histopatologia. Atinge aproximadamente 2 pessoas a cada 100 000, se expressa de
forma variável e sofre mutação aparecendo por volta dos 12 anos de idade. Esta
anomalia é devido a deficiência que ocorre no complexo de tonofilamento-desmossomo,
ou seja, nos filamentos intermediários de queratina que se ligam aos desmossomos.
Experimentos vêm mostrando que profundas raspagens de pele podem resultar na regeneração do epitélio com característica clinicas e patológicas normais. Especialistas dizem que uma provável explicação para que esse método de raspagem esteja dando certo é que durante a embriogênese os anexos epiteliais tenham sido formados antes da patologia característica de Morbus Darier começar.
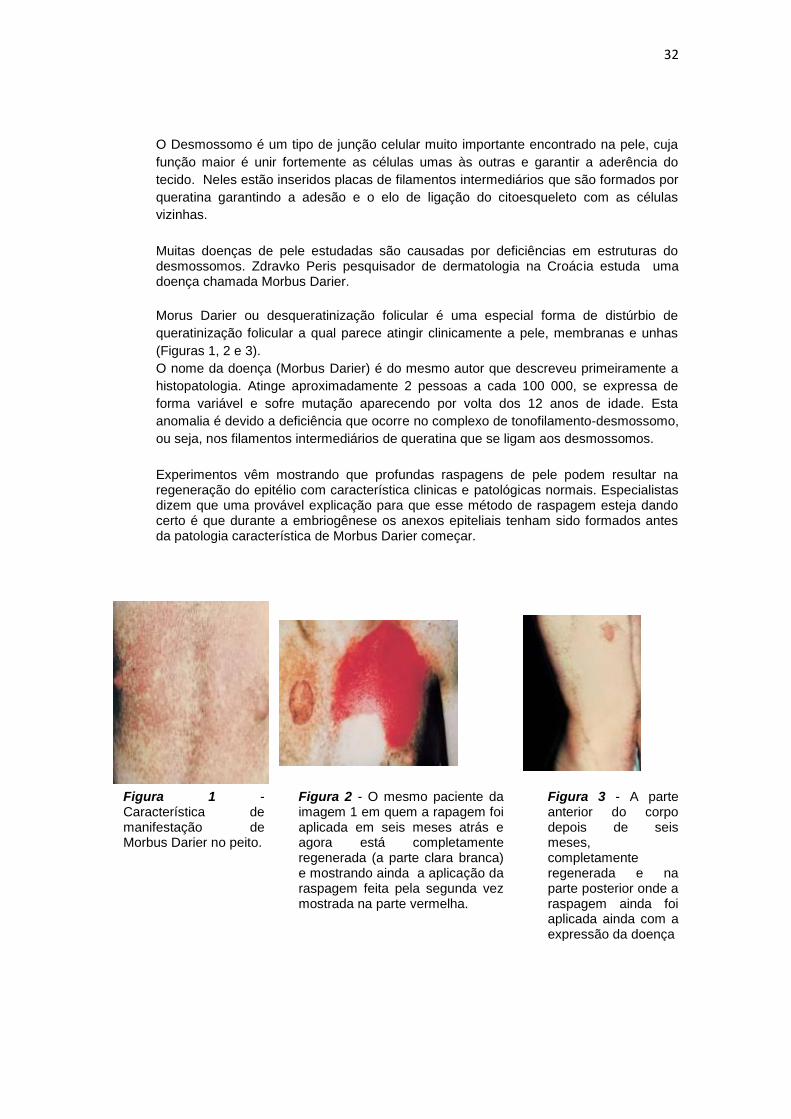
Figura 1 - Característica de manifestação de Morbus Darier no peito.
Figura 2 - O mesmo paciente da imagem 1 em quem a rapagem foi aplicada em seis meses atrás e agora está completamente regenerada (a parte clara branca) e mostrando ainda a aplicação da raspagem feita pela segunda vez mostrada na parte vermelha.
Figura 3 - A parte anterior do corpo depois de seis meses, completamente regenerada e na parte posterior onde a raspagem ainda foi aplicada ainda com a expressão da doença

33
Núcleo Celular
O núcleo é um compartimento essencial da célula eucarionte, pois é onde se localiza o
material genético, responsável pelas características que o organismo possui. Ele é delimitado pela carioteca ou envoltório nuclear, que é composto de uma
membrana nuclear externa, que é contínua com a membrana do retículo endoplasmático, e uma membrana interna, que é contínua com o lúmen do RE.
O envoltório nuclear é cheio de poros que comunicam o interior do núcleo com o citossol, e são estruturas complexas conhecidas como complexo de poro nuclear. O complexo de poro nuclear possui uma parede cilíndrica constituída por proteínas que formam um canal central com arranjo octogonal, que regula a troca de metabólitos, macromoléculas e subunidades ribossômicas entre o núcleo e o citosol.
Associada a superfície interna da carioteca encontra-se a lâmina nuclear, que constitui uma rede fibrosa de subunidades protéicas interconectadas, sendo responsável por dar forma e estabilidade ao envoltório nuclear, e liga este envoltório as fibras cromatínicas. A lâmina nuclear se despolimeriza durante a mitose, mas associam-se novamente ao seu final.
O nucleoplasma é constituído por uma solução aquosa de proteínas, RNAs, nucleosídeos, nucleotídeos e íons, onde se encontram os nucléolos e a cromatina. A maioria das proteínas da matriz nuclear são enzimas envolvidas com a transcrição e com a duplicação do DNA. O nucléolo é geralmente esférico, pode ser único ou múltiplo, é onde há transcrição de RNA ribossômico e a montagem das subunidades ribossomais.
Cromossomos
Nos organismos eucariontes, o DNA nuclear é dividido em uma série de diferentes
cromossomos. O cromossomo é constituído por uma longa molécula de DNA associado a diversas proteínas. As proteínas associadas classificam-se em dois grupos: as histonas e um grupo heterogêneo de proteínas não histônicas. O complexo formado pelo DNA, as histonas e as proteínas não histônicas chama-se cromatina. Esta organização é dinâmica, alterando-se de acordo com a fase do ciclo celular e com o grau de atividade.
No núcleo interfásico, a cromatina se apresenta compacta e/ou descompacta. No núcleo em divisão (mitose e meiose), a cromatina está altamente compactada, constituindo os cromossomos. Desta forma, a cromatina e os cromossomos representam dois aspectos morfológicos e fisiológicos da mesma estrutura.
Nos filamentos de cromatina ocorrem regiões mais claras, onde ela é menos condensada e os fios estão desenrolados, denominada de eucromatina, e outras mais escuras formadas por uma cromatina mis condensada e com fios enrolados, chamada heterocromatina.
O cromossomo possui estruturas indispensáveis para a replicação do DNA e suas proteínas associadas, que são: o centrômero (que participa da repartição para as células filhas das duas cópias cromossômicas que são geradas como conseqüência da replicação do DNA); os telômeros (que são os extremos do comossomo); pontos de origem da replicação.
Os cromossomos podem ser classificados de acordo com a posição do centrômero. Os metacêntricos são aqueles que possuem o centrômero no meio, dividindo-os em duas partes iguais. O submetacêntrico têm o centrômero um pouco afastado do centro, dividindo em duas partes com tamanhos diferentes. O acrocêntrico possui o centrômero próximo à extremidade. E o telocêntrico possui o centrômero na extremidade do cromossomo, este é ausente na espécie humana.
Cada espécie possui um conjunto de cromossomos padrão que se repete entre todos os indivíduos. Essa coleção de cromossomos própria de cada espécie é chamada cariótipo (carion = núcleo). Este conjunto pode ser identificado pelo número, pelas formas e pelos tamanhos característicos. E observa-se que os cromossomos aparecem aos pares, exceto os cromossomos sexuais masculinos, onde há um cromossomo X e um Y. Cada par de cromossomos de um determinado tipo ou forma chamamos de um par homólogo.

34
Genes
O gene é uma seqüência de nucleotídeos em uma molécula de DNA, que contêm a
informação necessária para sintetizar uma molécula de RNA messageiro para, a partir dele construir uma proteína. Eiste cerca de 100.000 genes distribuídos nos 46 cromossomos da espécie humana. Cada gene se localiza em um ponto específico do cromossomo chamado lócus.
Transcrição do DNA
A transcrição de DNA é a reprodução de uma fita de DNA em uma seqüência de RNA
complementar. Este processo é catalisado pela enzima RNA polimerase. A síntese de RNA, como a do DNA, sempre ocorre na direção 5’ para 3’. As seqüências de bases são mostradas ao longo das fitas de DNA e RNA mensageiro para ilustrar a sua natureza complementar. Devido a essa complementariedade das duas fitas do DNA, as suas seqüências de base não podem desenvolver-se independentemente.
Tradução do RNA
É o processo no qual as seqüências de nucleotídeos em uma molécula de RNA
mensageiro direciona a incorporação de aminoácidos em uma proteína, ocorrendo em um ribossomo.
Divisão Celular
A divisão celular é a separação de uma célula em suas células-filhas. Em células
eucarióticas, a divisão celular consiste da divisão do núcleo (mitose) seguida imediatamente pela divisão do citoplasma.
Mitose É a divisão do núcleo de uma célula eucariótica, envolvendo a condensação do DNA
em cromossomos visíveis e a separação dos cromossomos duplicados para formar dois conjuntos idênticos de cromossomos. Na fase M do ciclo celular, em que ocorre a mitose, é dividida em seis etapas: prófase, prometáfase, metáfase, anáfase, telófase e citocinese.
Prófase: os cromossomos replicados se condensam. No citoplasma, o fuso mitótico é formado entre os dois centrossomos, que foram replicados e separados.
Prometáfase: começa abruptamente com o rompimento do envelope nuclear. Os cromossomos podem ligar-se aos microtúbulos do fuso por meio de seus cinetócoros e sofrem movimentos ativos.
Metáfase: os cromossomos estão alinhados na região equatorial do fuso mitótico, a meio caminho entre os pólos do fuso.
Anáfase: as cromátides irmãs se separam sincronizadamente para formar dois cromossomos-filhos, e cada um é lentamente puxado em direção ao fuso polar para o qual está voltado.
Telófase: os dois conjuntos de cromátides-irmã chegam aos pólos do fuso mitótico e se descondensam. Um novo envoltório nuclear é constituído ao redor de cada grupo de
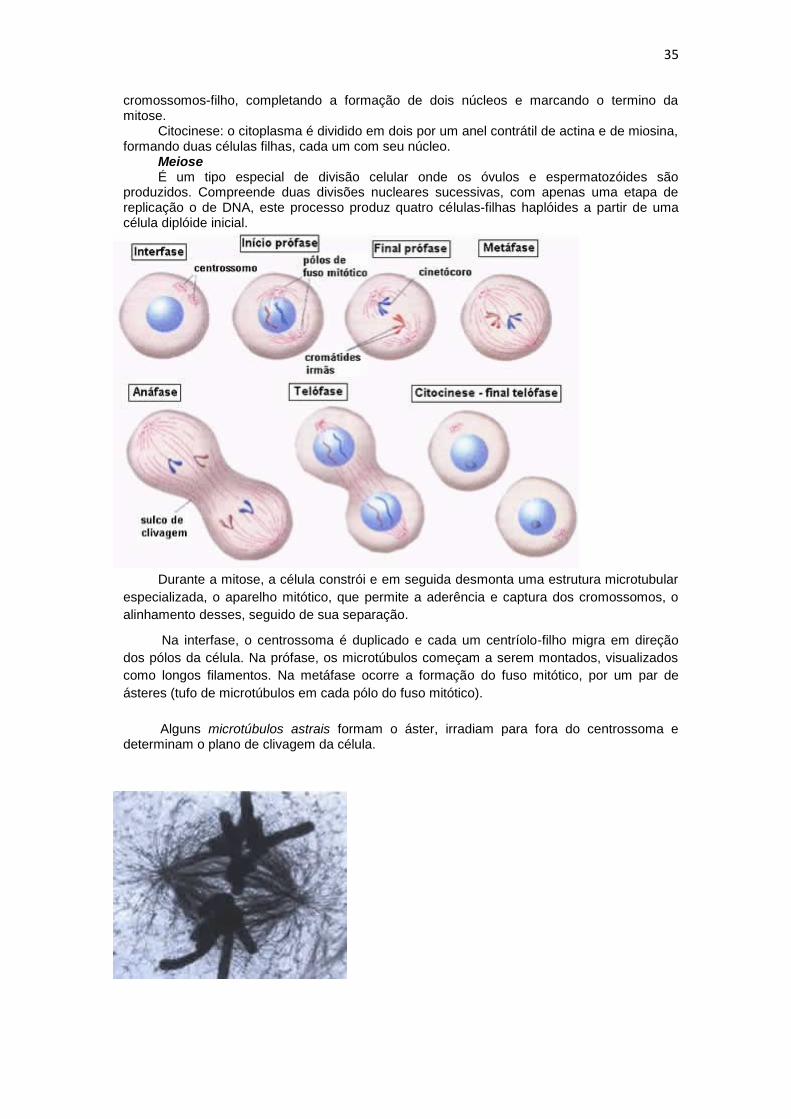
35
cromossomos-filho, completando a formação de dois núcleos e marcando o termino da mitose.
Citocinese: o citoplasma é dividido em dois por um anel contrátil de actina e de miosina, formando duas células filhas, cada um com seu núcleo.
Meiose É um tipo especial de divisão celular onde os óvulos e espermatozóides são
produzidos. Compreende duas divisões nucleares sucessivas, com apenas uma etapa de replicação o de DNA, este processo produz quatro células-filhas haplóides a partir de uma célula diplóide inicial.
Durante a mitose, a célula constrói e em seguida desmonta uma estrutura microtubular
especializada, o aparelho mitótico, que permite a aderência e captura dos cromossomos, o
alinhamento desses, seguido de sua separação.
Na interfase, o centrossoma é duplicado e cada um centríolo-filho migra em direção
dos pólos da célula. Na prófase, os microtúbulos começam a serem montados, visualizados
como longos filamentos. Na metáfase ocorre a formação do fuso mitótico, por um par de
ásteres (tufo de microtúbulos em cada pólo do fuso mitótico).
Alguns microtúbulos astrais formam o áster, irradiam para fora do centrossoma e determinam o plano de clivagem da célula.
36
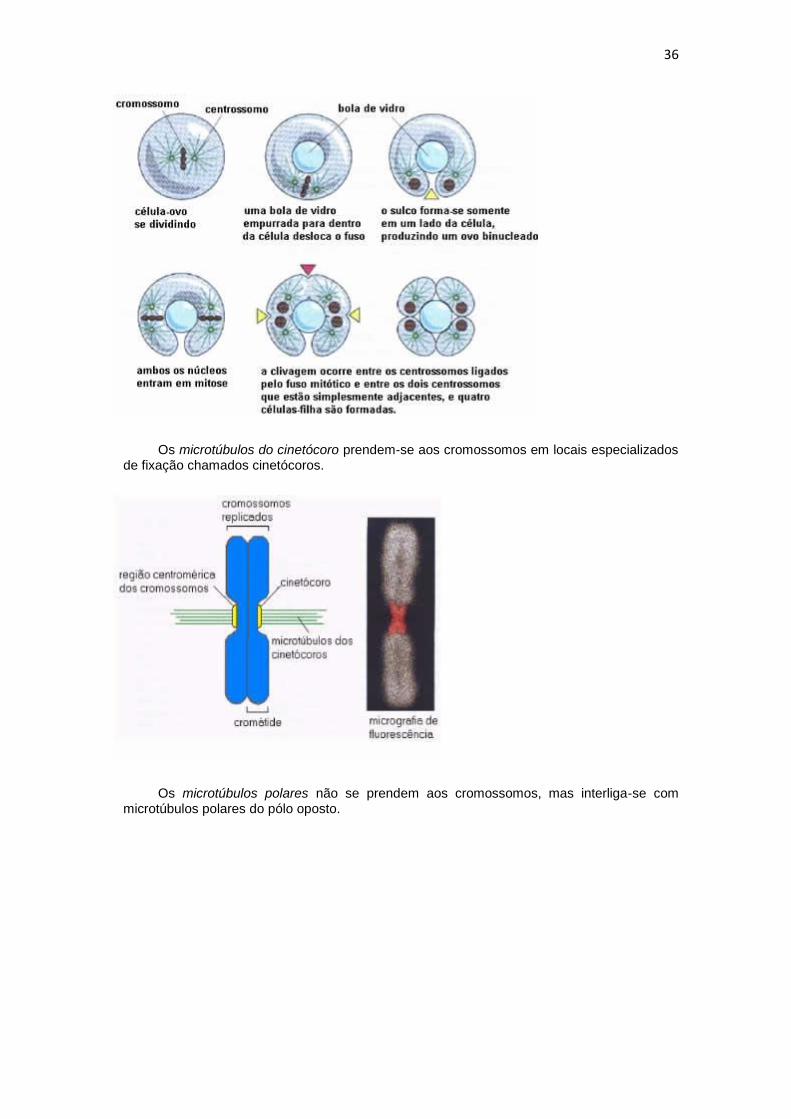
Os microtúbulos do cinetócoro prendem-se aos cromossomos em locais especializados de fixação chamados cinetócoros.
Os microtúbulos polares não se prendem aos cromossomos, mas interliga-se com microtúbulos polares do pólo oposto.

37
Com a dinâmica dos microtúbulos de polimerização e despolimerização, e agentes
motores de microtúbulos, fazem com que mantenham os cromossomos na linha equatorial da
célula, durante a metáfase e a separação e transporte dos cromossomos para os pólos
opostos, na anáfase.
Na telófase a citocinese se completa, e o fuso mitótico desaparece com a
despolimerização dos microtúbulos e de outras fibras.
Ao longo da mitose, o centríolo-filho cresce em cada pólo, até que na telófase estejam de compromisso integral.

38
CICLO CELULAR
As células passam por diversas etapas durante seu desenvolvimento, tais como, crescimento e divisão. Estas etapas são cíclicas. Assim, o ciclo celular representa o ciclo vital da célula e é dividido em duas fases: Interfase e divisão celular (mitose e meiose). 1. INTERFASE A intérfase, tal como o próprio nome diz "inter" (entre) e "fase" (período), consiste em um período entre as divisões celulares. Embora seja um momento em que a célula não está em divisão, muitos eventos importantes estão acontecendo em seu interior. Estes eventos foram divididos didaticamente em outras sub-etapas, conhecidas pelos seguintes nomes: fase G1, fase S e fase G2. Passemos agora para análise de cada uma dessas fases: G1 - É conhecida como fase de crescimento, onde ocorre a síntese intensa de várias moléculas importantes para a sobrevivência da célula, como proteínas estruturais, enzimas e RNA. S - Nesta fase o principal evento é a duplicação do material genético. G2 - É chamada de fase de preparação, pois nela ocorre a síntese de moléculas e organelas relacionadas ao processo de divisão celular. O núcleo interfásico O núcleo recebe esta denominação pois, só pode ser observado durante a interfase. componentes Membrana nuclear (cariomembrana ou carioteca): membrana dupla, porosa e apresenta ribossomos aderidos. Carioplasma (nucleoplasma ou cariolinfa): gel protéico encontrado dentro do núcleo; nele estão imersos os componentes nucleares. Nucléolo: enovelado de RNAr (ribossômico), principal componente químico dos ribossomos, de onde os mesmos surgem. Cromatina: conjunto de moléculas de DNA que se encontram na forma desespiralizada.
cromossomos, o que são? Os cromossomos nada mais são que moléculas de DNA na forma espiralizada, enroladas em proteínas, chamadas de histonas, como linha em carretel. O número de cromossomos é constante em indivíduos da mesma espécie e variável em

39
indivíduos de espécies diferentes. Ex.: homem = 46 cromossomos; macaco = 48 cromossomos; cachorro = 38 cromossomos. Quando um ser vivo possui duas cópias de cada um de seus cromossomos dentro da célula dizemos que ele é diplóide e podemos representar seu número de cromossomos como "2n". Este é o caso do ser humano e de muitos outros animais. Por outro lado, algumas plantas e algas possuem apenas uma cópia de cada cromossomo e são chamadas de haplóides, ou seja, possuem "n" cromossomos. Processo de formação de novas células.
A mitose
divisão celular onde uma célula (mãe) dá origem a duas célula.-filha com as mesmas
características e mesmo número de cromossomos, o que são? - O cromossomo nada mais é
do que moléculas de DNA na forma espiralizada, enroladas em histonas.
O número de cromossomos é constante em indivíduos da mesma espécie e variável em
indivíduos de espécies diferentes.
Observação: a mitose é chamada de divisão equacional (E!) pelo fato de manter o número de
cromossomos constante.
Ocorrência: Células Somáticas. ,
Finalidades:
-Aumentar o número de células.
-Formação do organismo.
-Reposição celular, etc.
Fases: prófase – metáfase – anáfase – telófase
Prófase
Duplicação de centríolos e migração para os pólos opostos da célula.
Durante a migração, forma-se entre os centríolos uma rede de fibras protéicas denominadas
fuso mitótico ou acromático.
Desaparecimento da carioteca e do nucléolo.

40
Metáfase
Os centríolos já estão nos pólos opostos.
O fuso mitótico está formado.
Os cromossomos estão no grau máximo de espiralização.
Os cromossomos estão alinhados na placa equatorial (região central da célula).
Os cromossomos estão ligados ao fuso mitótico pela região de maior espiralização
denominada centrômero.
Observação – É durante a metáfase que estudamos os cromossomos devido ao seu grau de
espiralização, pois ficam mais visíveis.
Anáfase
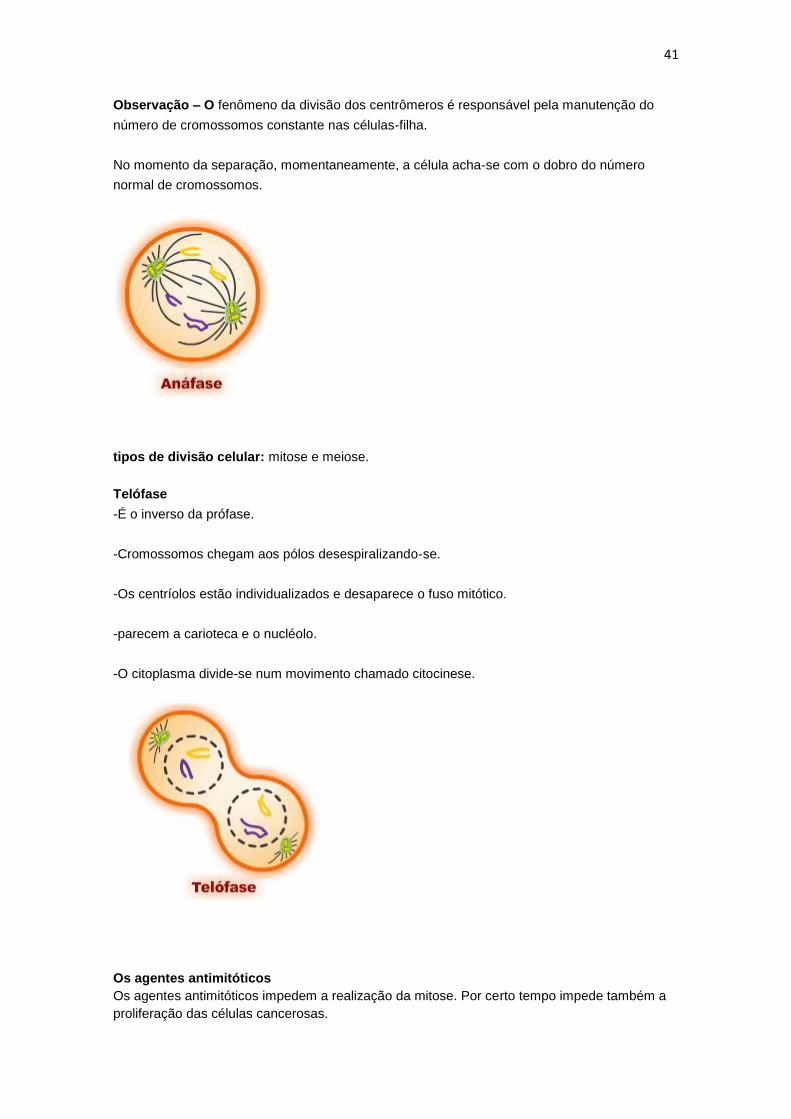
Os cromossomos são tracionados para os pólos.
O centrômero divide-se e cada cromossomo que se encontrava duplicado transforma-se em
dois cromossomos, cada um formado por uma única molécula de DNA.
41
Observação – O fenômeno da divisão dos centrômeros é responsável pela manutenção do
número de cromossomos constante nas células-filha.
No momento da separação, momentaneamente, a célula acha-se com o dobro do número
normal de cromossomos.
tipos de divisão celular: mitose e meiose.
Telófase
-É o inverso da prófase.
-Cromossomos chegam aos pólos desespiralizando-se.
-Os centríolos estão individualizados e desaparece o fuso mitótico.
-parecem a carioteca e o nucléolo.
-O citoplasma divide-se num movimento chamado citocinese.
Os agentes antimitóticos
Os agentes antimitóticos impedem a realização da mitose. Por certo tempo impede também a
proliferação das células cancerosas.

42
Certos agentes impedem somente a replicação de DNA, e alguns somente a formação do
fuso.
Os agentes que impedem a replicação do DNA são:
* aminopterina
* mercaptopurina
* fluoruracilo
* bromouracilo
Os agentes que impedem o fuso são:
* colchicina
* vincaleucoblastina
* mercaptoetanol
Os períodos da intérfase
• A interfase é o período em que a célula não esta em divisão.
• É Fase de maior atividade metabólica.
• Fase em que podemos observar o núcleo da célula.
A interfase é dividida em três fases:
G1: esta fase tem início no fim da divisão celular e termina antes da síntese do DNA. Nesta
fase ocorre a síntese de proteínas.
S: Nesta fase ocorre à replicação do DNA, está comprometida em enviar as informações
genéticas para as célula.-filhas.
G2: Nesta fase ocorre a Duplicação dos centríolos.
Mitose Divisão celular onde uma célula (mãe) dá origem a duas células-filha com as mesmas características e mesmo número de cromossomos, o que são? - Os cromossomos nada mais são que moléculas de DNA na forma espiralizada, enroladas em histonas. O número de cromossomos é constante em indivíduos da mesma espécie e variável em indivíduos de espécies diferentes.
Observação: a mitose é chamada de divisão equacional (E!) pelo fato de manter o número de cromossomos constante. Ocorrência: Células Somáticas.

43
Finalidades -Aumentar o número de células. -Formação do organismo. -Reposição celular, etc.. Fases prófase – metáfase – anáfase – telófase Prófase Duplicação de centríolos e migração para os pólos opostos da célula. Durante a migração, forma-se entre os centríolos uma rede de fibras protéicas denominadas fuso mitótico ou acromático. Desaparecimento da carioteca e do nucléolo. Cromossomos iniciando o processo de espiralização.
Ribossomos
Ribossomos são organelas citoplasmáticas encontradas em procariotos e eucariotos. Eles são amplos complexos de proteínas e moléculas de rRNA (RNA ribossômico), sendo três moléculas de rRNA nos procariotos e quatro nos eucariotos. Estes complexos de proteína e rRNA são chamados subunidades e são produzidos no nucléolo. A principal função dos ribossomos é servir de sítio para a tradução, ou seja síntese de proteínas (reunião de aminoácidos em proteínas); uma vez que duas subunidades (uma grande e uma pequena) são unidas pelo mRNA vindo do núcleo, o ribossomo traduz a seqüência do mRNA em uma seqüência específica de aminoácidos ou uma cadeia polipeptídica. Veja abaixo uma eletromicrografia mostrando uma síntese de proteínas; os ribossomos aparecem como grânulos escuros.

44
LOCALIZAÇÃO E TIPOS DE RIBOSSOMOS
Ribossomos são encontrados nas células sob duas formas: livres e associados ao retículo endoplasmático. Eles existem em várias localidades dentro da célula; entretanto esta localização depende da função da célula.
Ribossomos livres
Encontrados no citoplasma
Podem ocorrer como um único ribossomo ou em grupos conhecidos como polirribossomos ou polissomos
Ocorrem em maior número que os ribossomos associados ao retículo, em células que retém a maioria das proteínas fabricadas.
Responsáveis pelas proteínas que estão em solução no citoplasma ou formam elementos móveis ou estruturas citoplasmáticas importante
Ribossomos associados ao retículo
São encontrados associados à membrana exterior do retículo endoplasmático (RE) constituindo o RE rugoso
Ocorre em maior número que os ribossomos livre, em células que secretam suas proteínas fabricadas (ex., células pancreáticas produtoras de enzimas digestivas).
Responsáveis pelas proteínas que formam membranas ou ou são empacotadas e estocadas em vesículas no citoplasma ou são exportadas para o exterior da célula.
Ribossomos também estão localizados na mitocondria e cloroplastos de células eucariotas; eles são sempre menores que os ribossomos citoplasmáticos e são comparáveis aos ribossoomos procariotos em tamanho e sensibilidade a antibióticos; entretanto, os valores de
45
sedimentação s (s = unidade Svedberg: uma medida da taxa de sedimentação de um componentes em uma centrífuga, relacionando peso molecular e a forma 3-D do componente)varia nos diferentes filos. Ribossomos procariotos e eucariotos executam as mesmas funções pois fazem o mesmo conjunto de reações químicas; entretanto, ribossomos eucariotos são muito maiores que os procariotos e a maioria das suas proteínas são diferentes. Ribossomos mitocondriais e dos cloroplastos assemelham-se aos ribossomos bacterianos.
As células aplicam considerável esforço para a produção destas organelas essenciais; por exemplo, uma E. coli contém mais ou menos 15000 ribossomos, cada um com um peso molecular de aproximadamente 3 x 10 6 daltons constituindo 25% da massa total dessas células bacterianas.
ESTRUTURA DOS RIBOSSOMOS
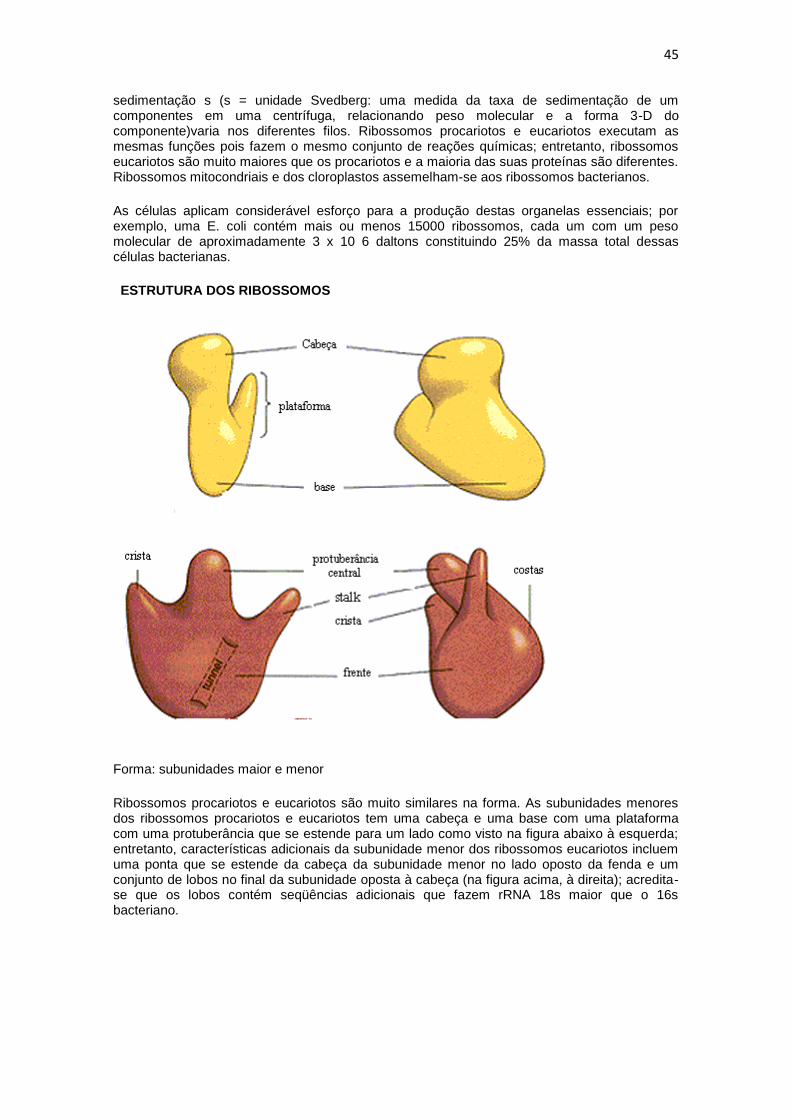
Forma: subunidades maior e menor
Ribossomos procariotos e eucariotos são muito similares na forma. As subunidades menores dos ribossomos procariotos e eucariotos tem uma cabeça e uma base com uma plataforma com uma protuberância que se estende para um lado como visto na figura abaixo à esquerda; entretanto, características adicionais da subunidade menor dos ribossomos eucariotos incluem uma ponta que se estende da cabeça da subunidade menor no lado oposto da fenda e um conjunto de lobos no final da subunidade oposta à cabeça (na figura acima, à direita); acredita-se que os lobos contém seqüências adicionais que fazem rRNA 18s maior que o 16s bacteriano.

46
A subunidade maior tem uma protuberância central proeminente, stalk, e uma crista que se estende para um lado como visto à direita na figura acima. A subunidade maior tem uma abertura de aproximadamente 10nm de comprimento e 2.5 nm de diâmetro; a abertura se estende da região que contém os sítios A (aminoacil) e P (peptidil) até a parte da subunidade maior da qual a associação da cadeia polipeptídica nascente sai do ribossomo. Pensa-se que esta abertura é um canal no qual a cadeia polipeptídica nascente atravessa no caminho de saída do ribossomo.
Componentes
Ribossomos são estruturas pequenas, mas complexas, com cerca de 20 a 30 nm de diâmetro, consistindo de duas subunidades de tamanhos desiguais, referentes à subunidades maior e menor as quais estão adaptadas intimamente como visto na figura acima. Uma subunidade é composta por um complexo formado por moléculas de RNA e proteínas; cada molécula contém pelo menos uma subunidade de RNA ribossômico (rRNA) e uma grande quantidade de proteínas ribossomais. As subunidades juntas contém mais de 82 proteínas específicas reunidas em uma seqüência precisa.
47
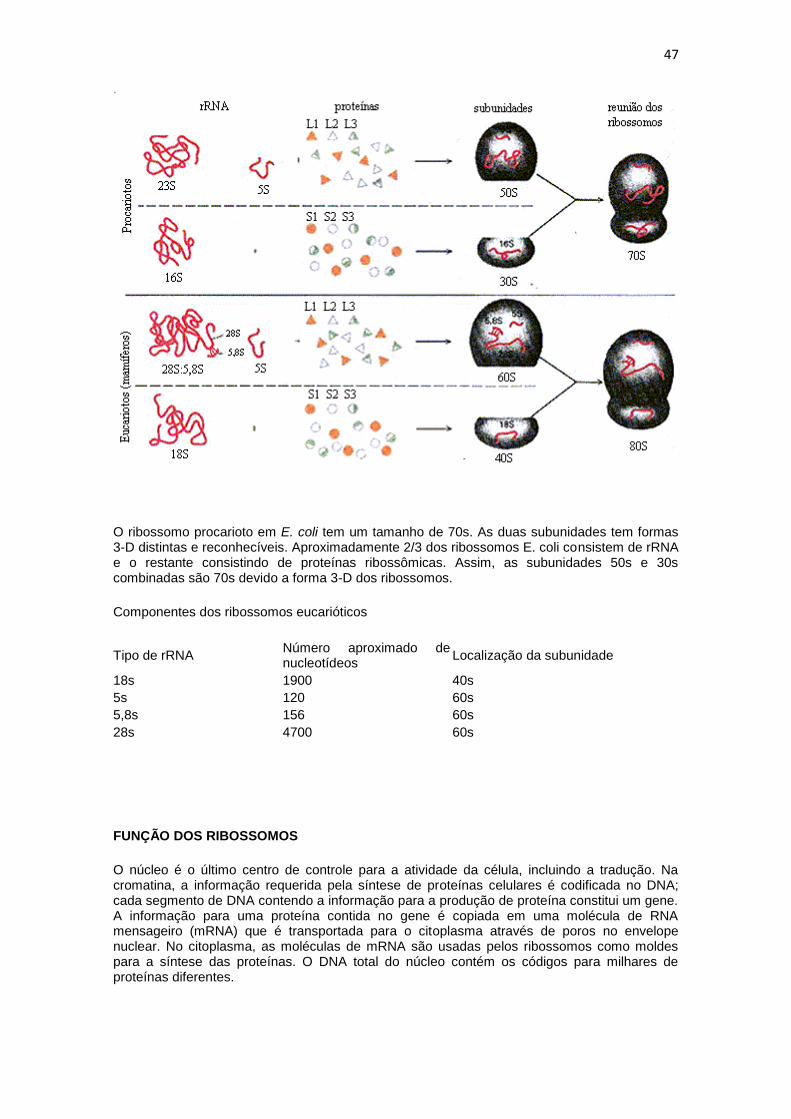
O ribossomo procarioto em E. coli tem um tamanho de 70s. As duas subunidades tem formas 3-D distintas e reconhecíveis. Aproximadamente 2/3 dos ribossomos E. coli consistem de rRNA e o restante consistindo de proteínas ribossômicas. Assim, as subunidades 50s e 30s combinadas são 70s devido a forma 3-D dos ribossomos.
Componentes dos ribossomos eucarióticos
Tipo de rRNA Número aproximado de nucleotídeos
Localização da subunidade
18s 1900 40s
5s 120 60s
5,8s 156 60s
28s 4700 60s
FUNÇÃO DOS RIBOSSOMOS
O núcleo é o último centro de controle para a atividade da célula, incluindo a tradução. Na cromatina, a informação requerida pela síntese de proteínas celulares é codificada no DNA; cada segmento de DNA contendo a informação para a produção de proteína constitui um gene. A informação para uma proteína contida no gene é copiada em uma molécula de RNA mensageiro (mRNA) que é transportada para o citoplasma através de poros no envelope nuclear. No citoplasma, as moléculas de mRNA são usadas pelos ribossomos como moldes para a síntese das proteínas. O DNA total do núcleo contém os códigos para milhares de proteínas diferentes.
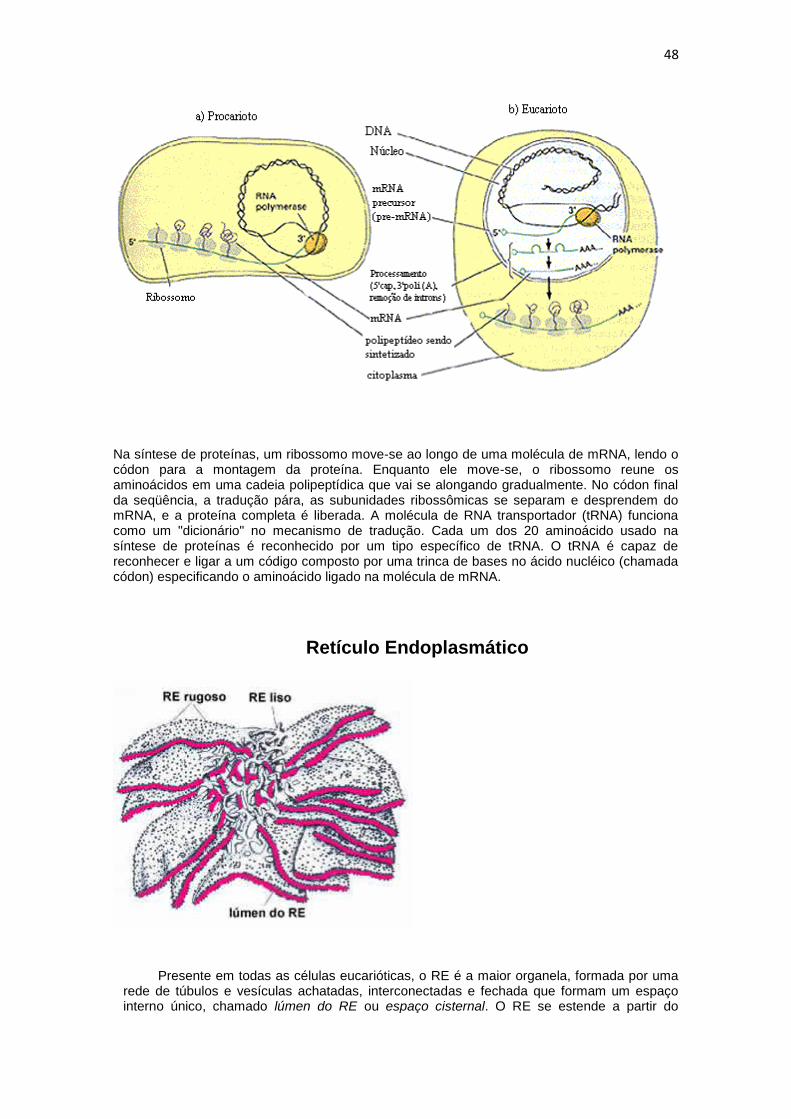
48
Na síntese de proteínas, um ribossomo move-se ao longo de uma molécula de mRNA, lendo o códon para a montagem da proteína. Enquanto ele move-se, o ribossomo reune os aminoácidos em uma cadeia polipeptídica que vai se alongando gradualmente. No códon final da seqüência, a tradução pára, as subunidades ribossômicas se separam e desprendem do mRNA, e a proteína completa é liberada. A molécula de RNA transportador (tRNA) funciona como um "dicionário" no mecanismo de tradução. Cada um dos 20 aminoácido usado na síntese de proteínas é reconhecido por um tipo específico de tRNA. O tRNA é capaz de reconhecer e ligar a um código composto por uma trinca de bases no ácido nucléico (chamada códon) especificando o aminoácido ligado na molécula de mRNA.
Retículo Endoplasmático
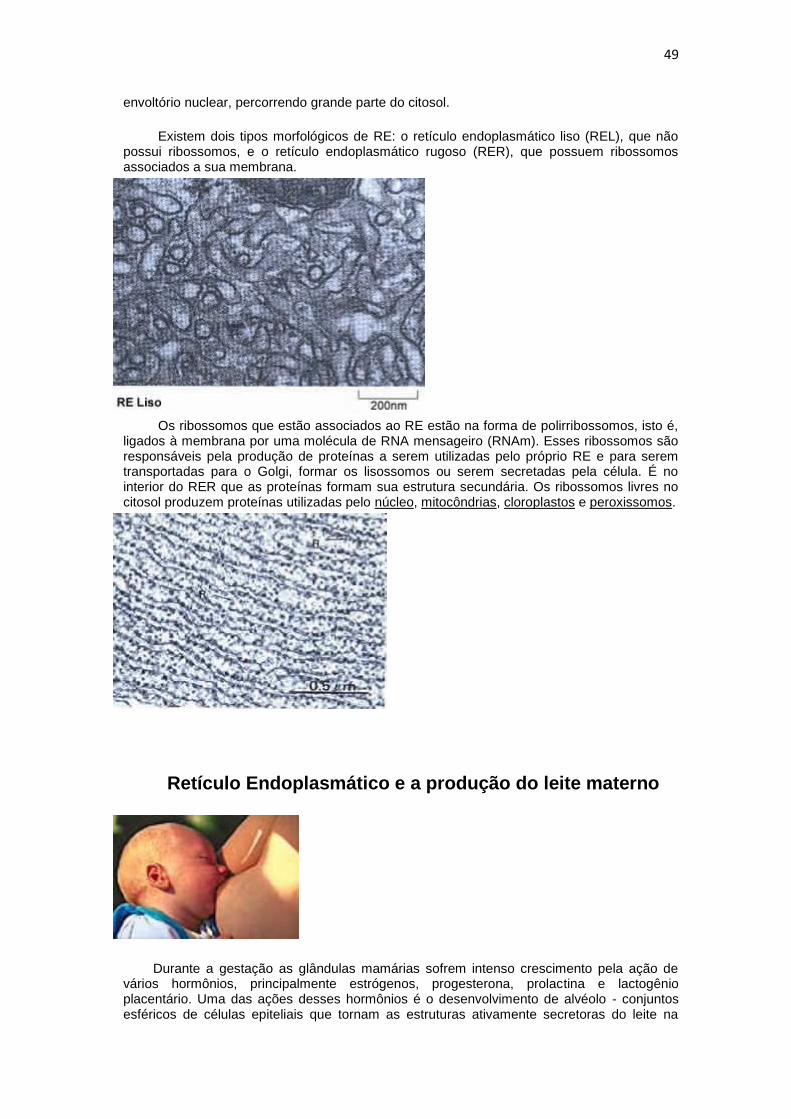
Presente em todas as células eucarióticas, o RE é a maior organela, formada por uma rede de túbulos e vesículas achatadas, interconectadas e fechada que formam um espaço interno único, chamado lúmen do RE ou espaço cisternal. O RE se estende a partir do
49
envoltório nuclear, percorrendo grande parte do citosol.
Existem dois tipos morfológicos de RE: o retículo endoplasmático liso (REL), que não possui ribossomos, e o retículo endoplasmático rugoso (RER), que possuem ribossomos associados a sua membrana.
Os ribossomos que estão associados ao RE estão na forma de polirribossomos, isto é, ligados à membrana por uma molécula de RNA mensageiro (RNAm). Esses ribossomos são responsáveis pela produção de proteínas a serem utilizadas pelo próprio RE e para serem transportadas para o Golgi, formar os lisossomos ou serem secretadas pela célula. É no interior do RER que as proteínas formam sua estrutura secundária. Os ribossomos livres no citosol produzem proteínas utilizadas pelo núcleo, mitocôndrias, cloroplastos e peroxissomos.
Retículo Endoplasmático e a produção do leite materno
Durante a gestação as glândulas mamárias sofrem intenso crescimento pela ação de vários hormônios, principalmente estrógenos, progesterona, prolactina e lactogênio placentário. Uma das ações desses hormônios é o desenvolvimento de alvéolo - conjuntos esféricos de células epiteliais que tornam as estruturas ativamente secretoras do leite na
50
lactação. Quatro a seis células mioepiteliais de forma estrelada envolvem cada alvéolo, que tem funções extremamente importantes.
Durante a lactação, o leite é produzido pelas células dos alvéolos e se acumula no lúmen destes e dentro dos ductos galactóforos (Imagens 1 e 2).
O leite é composto principalmente de lipídios, proteínas do leite e lactose (açúcar do leite). Estes componentes são sintetizados pelos retículos endoplasmáticos lisos (que sintetizam lipídios) e rugosos (que sintetizam principalmente proteínas), como detalhado na Imagem 3.
Imagem 1 - Desenho Esquemático de uma mama feminina mostrando as glândulas mamárias.
Imagem 2 - Aumento da glândula mamária em lactação. Imagem 3 - Desenho Esquemático das células secretoras da glândula mamária. Note o
acúmulo e a expulsão de lipídios e proteínas, constituintes principais do leite materno. Obs.: Imagens adaptadas do livro Junqueira e Carneiro

51
A primeira secreção das glândulas mamárias após o parto é chamada de colostro. Ele contém menos gordura e mais proteína que o leite regular e é rico em anticorpos, que fornecem algum grau de imunidade passiva ao recém-nascido, especialmente na luz intestinal.
É importante (e interessante) saber que quando uma mulher está amamentando, a ação mecânica da criança no mamilo estimula receptores táteis, resultando na liberação de oxitocina, hormônio que causa a contração das células mioepiteliais nos alvéolos e ductos, resultando a expulsão do leite (reflexo ejetor do leite). Quando a mulher sofre estímulos negativos como frustração, ansiedade ou raiva, a liberação de oxitocina pode ser inibida, e conseqüentemente a expulsão do leite pode ser prejudicada.
Retículo Endoplasmático e a contração muscular
O reticulo endoplasmatico (RE) é o maior sistema de membranas da célula, constituindo aproximadamente metade do volume total das membranas. Os processos metabólicos que ocorrem dentro do RE são síntese e modificação de proteinas, sintese de lipideos e esteróides e fabricação de todas as membranas da célula. O RE tem dois componentes: Reticulo endoplasmático liso (REL) e reticulo endoplasmático rugoso (REG).
O reticulo endoplasmático liso (REL) tornou-se especializado principalmente em células musculares, nas quais ele é denominados reticulo sarcoplasmático. Nas células musculares esqueléticas ele funciona sequestrando íons de calcio do citossol, auxiliando o controle da contração muscular. O estímulo para a contração é geralmente um impulso nervoso que se propaga pela membrana das fibras musculares, atingindo o retículo sarcoplasmático (onde há cálcio armazenado), que libera íons de cálcio no citoplasma. Ao entrar em contato com as miofibrilas, o cálcio desbloqueia os sítios de ligação de actina, permitindo que se ligue a miosina, iniciando a contração muscular. Assim que cessa o estímulo, o cálcio é rebombeado para o interior do retículo sarcoplasmático e cessa a contração muscular.
52
Retículo Endoplasmático e a Diabete
Os primeiros passos para a formação da insulina, hormônio que auxilia na absorção da
glicose e controla a quantidade de açúcar no organismo, ocorre no Retículo Endoplasmático
(RE) das células β. Estas se localizam nas Ilhotas de Langerhans, no pâncreas e são
responsáveis por apresentar um ou mais cristais de insulina. Calcula-se que as Ilhotas de
Langerhans produzam cerca de 10 mg de insulina ou aproximadamente 5 vezes a necessidade
diária.
Mutações no Retículo Endoplasmático causam profundo impacto nas células das Ilhotas de Langerhans e principalmente nas células β
o maior componentes destas ilhotas afetando o funcionamento e sobrevivência destas. Com recentes estudos Pesquisadores do Instituto Skirbal e Escola de Medicina da
Universidade de Nova York, descobriram na rara doença chamada Wolcott-Rallison, síndrome da diabete infantil, é uma desordem caracterizada por uma destruição antecipada das células β, causadas por mutações no gene que codifica a informação para a produção de insulina pelo Retículo Endoplasmático.
Assim como nesta doença, a destruição das células β, pode aumentar a concentração de glicose no sangue, causando então a diabete nas suas formas mais normais.
Vesículas Transportadoras
As vesículas de transporte brotam a partir de regiões revestidas e especializadas da membrana, podendo ser esféricas ou tubulares.
Esses revestimentos são de proteínas específicas com diferentes funções, que podem formar:
- Vesículas revestidas por clatrina: são produzidas pela membrana plasmática por endocitose ou brotam da rede trans do Golgi. Esta vesícula possui também outra proteína, a
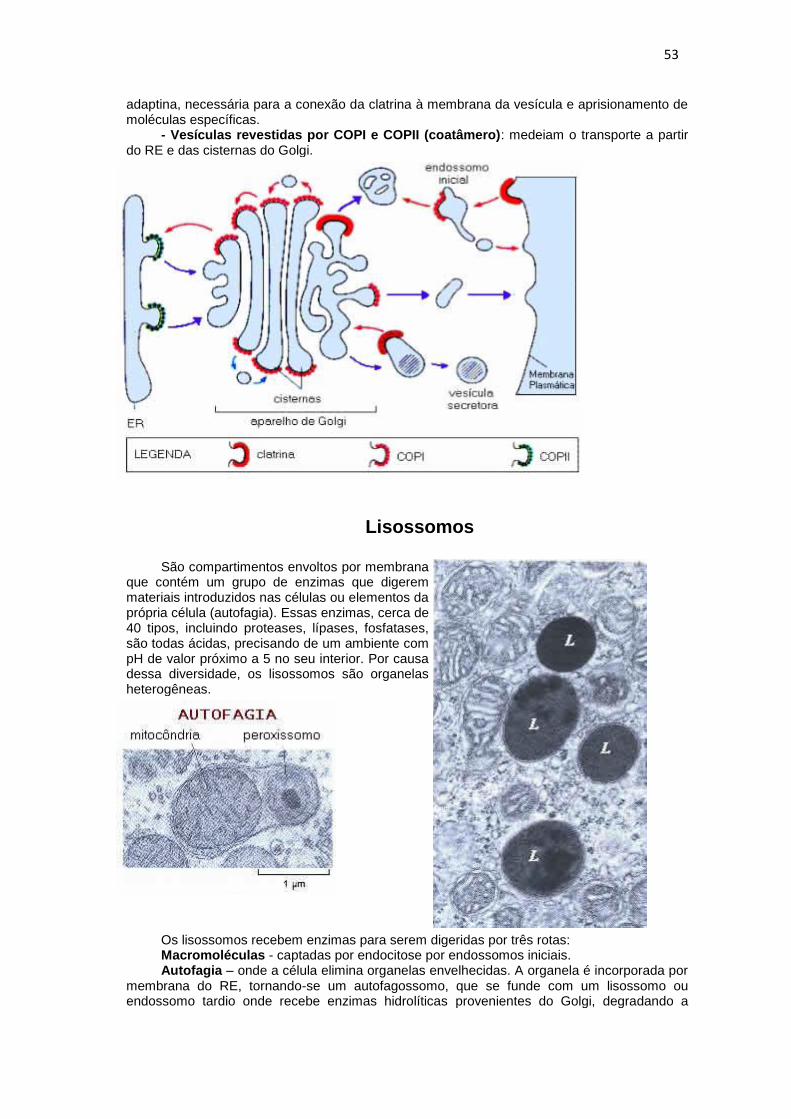
53
adaptina, necessária para a conexão da clatrina à membrana da vesícula e aprisionamento de moléculas específicas.
- Vesículas revestidas por COPI e COPII (coatâmero): medeiam o transporte a partir do RE e das cisternas do Golgi.
Lisossomos
São compartimentos envoltos por membrana que contém um grupo de enzimas que digerem materiais introduzidos nas células ou elementos da própria célula (autofagia). Essas enzimas, cerca de 40 tipos, incluindo proteases, lípases, fosfatases, são todas ácidas, precisando de um ambiente com pH de valor próximo a 5 no seu interior. Por causa dessa diversidade, os lisossomos são organelas heterogêneas.
Os lisossomos recebem enzimas para serem digeridas por três rotas: Macromoléculas - captadas por endocitose por endossomos iniciais. Autofagia – onde a célula elimina organelas envelhecidas. A organela é incorporada por
membrana do RE, tornando-se um autofagossomo, que se funde com um lisossomo ou endossomo tardio onde recebe enzimas hidrolíticas provenientes do Golgi, degradando a
54
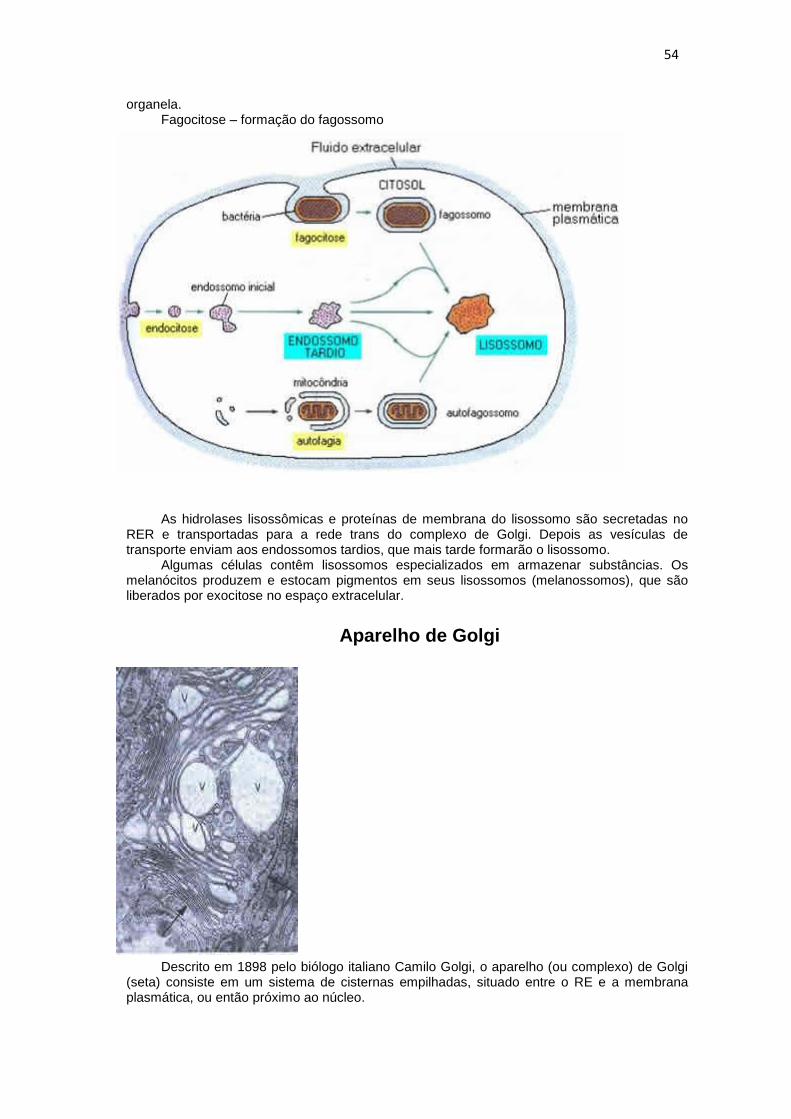
organela. Fagocitose – formação do fagossomo
As hidrolases lisossômicas e proteínas de membrana do lisossomo são secretadas no RER e transportadas para a rede trans do complexo de Golgi. Depois as vesículas de transporte enviam aos endossomos tardios, que mais tarde formarão o lisossomo.
Algumas células contêm lisossomos especializados em armazenar substâncias. Os melanócitos produzem e estocam pigmentos em seus lisossomos (melanossomos), que são liberados por exocitose no espaço extracelular.
Aparelho de Golgi
Descrito em 1898 pelo biólogo italiano Camilo Golgi, o aparelho (ou complexo) de Golgi (seta) consiste em um sistema de cisternas empilhadas, situado entre o RE e a membrana plasmática, ou então próximo ao núcleo.

55
Cada unidade do Golgi é chamada de dictiossomo, e cada pilha apresenta de 4 a 6 sáculos, embora alguns flagelados unicelulares possam ter até 60. Nas células vegetais, centenas de pilas do Golgi estão dispersas no citoplasma.
O aparelho de Golgi possui duas faces distintas: uma face cis (ou face de entrada) e uma face trans (ou face de saída), que estão intimamente associadas por compartimentos especiais de uma rede interconectada de túbulos e cisternas: rede cis de Golgi (RCG) e a rede trans de Golgi (RTG).
Esta organela é responsável pela distribuição das proteínas e lipídeos que recebe do RE, modificando-as através cisternas, para cada função específica, produzindo, por exemplo, oligossacarídeos e proteoglicanas.
A exocitose é a fusão de vesículas, que transportam substâncias (proteínas, hormônios, neurotransmissores, enzimas digestivas) secretadas para o espaço extracelular, a membrana plasmática. As vesículas secretoras formam-se a partir da rede trans do Golgi e sua formação e liberação é regulada por sinais extracelulares.
56
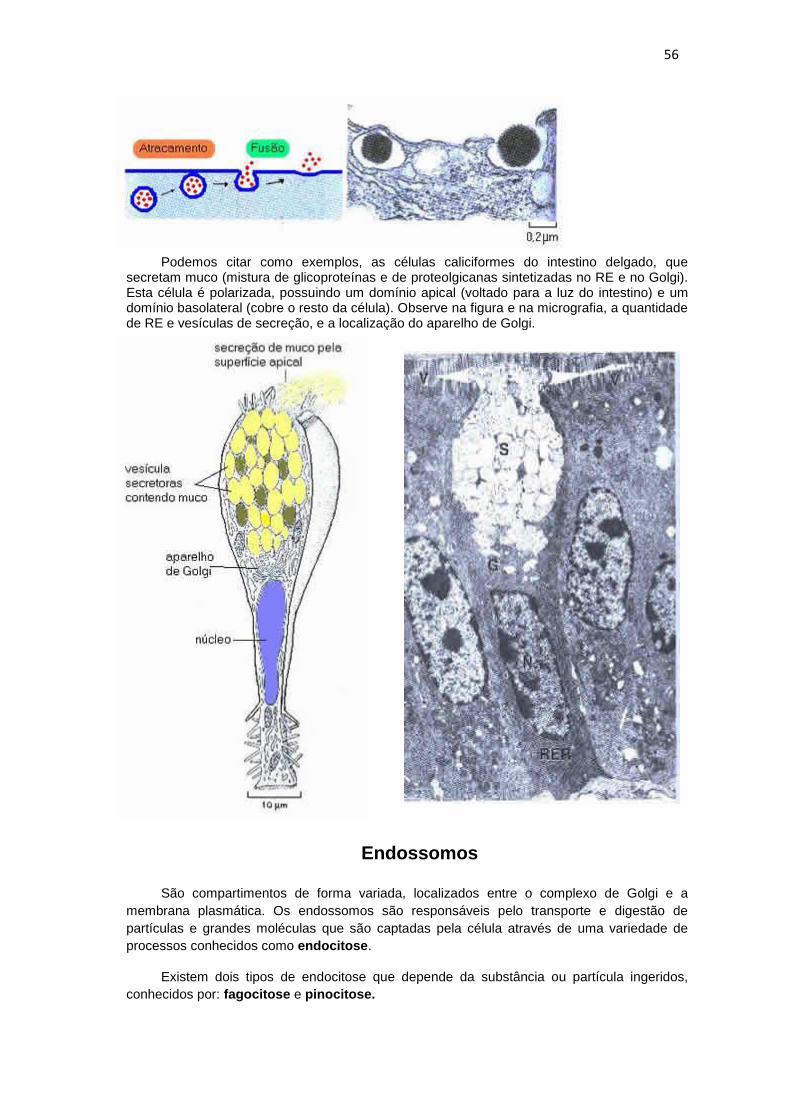
Podemos citar como exemplos, as células caliciformes do intestino delgado, que secretam muco (mistura de glicoproteínas e de proteolgicanas sintetizadas no RE e no Golgi). Esta célula é polarizada, possuindo um domínio apical (voltado para a luz do intestino) e um domínio basolateral (cobre o resto da célula). Observe na figura e na micrografia, a quantidade de RE e vesículas de secreção, e a localização do aparelho de Golgi.
Endossomos
São compartimentos de forma variada, localizados entre o complexo de Golgi e a
membrana plasmática. Os endossomos são responsáveis pelo transporte e digestão de
partículas e grandes moléculas que são captadas pela célula através de uma variedade de
processos conhecidos como endocitose.
Existem dois tipos de endocitose que depende da substância ou partícula ingeridos,
conhecidos por: fagocitose e pinocitose.
57
A fagocitose envolve a ingestão de grandes partículas, tais como parasitas, bactérias,
células prejudiciais, danificadas ou mortas, restos celulares, por meio de grandes vesículas
endocíticas chamadas fagossomos. Dependendo do tipo celular, a fagocitose é uma forma de
alimentação (nos protozoários) ou uma forma de limpeza e proteção como nos macrófagos
(foto abaixo), neutrófilos e células dendríticas.
O tamanho do fagossomo é determinado pelo tamanho da partícula a ser ingerida. Estes
se fundem com os lisossomos dentro das células, então o material ingerido é degradado.
A pinocitose envolve a entrada de líquidos e fluidos extracelulares juntamente com as
macromoléculas e os solutos dissolvidos. Neste processo, a membrana plasmática é
internalizada numa taxa que varia entre os tipos celulares. A pinocitose pode ser inespecífica,
onde as substâncias penetram na célula automaticamente, e a regulada, onde ocorre a
formação das vesículas pinocíticas quando a substância interage com um receptor específico
da membrana.
A pinocitose pode ser iniciada por cavéolos (pequenas cavidades) em regiões da membrana plasmática ricas em colesterol.
Existem dois tipos de endossomos: os endossomos primários ou iniciais, localizados nas
proximidades da membrana plasmática, e os endossomos secundários ou tardios, próximos às
redes cis e trans do Golgi.
Os endossomos iniciais formam os compartimentos de proteínas específicas
determinando seu destino: reciclagem e devolução para o mesmo domínio da membrana
plasmática; transcitose (o material interiorizado por um lado da célula atravessam o citoplasma
e saem por exocitose do lado oposto); ou destinados aos lisossomos, onde serão degradados.
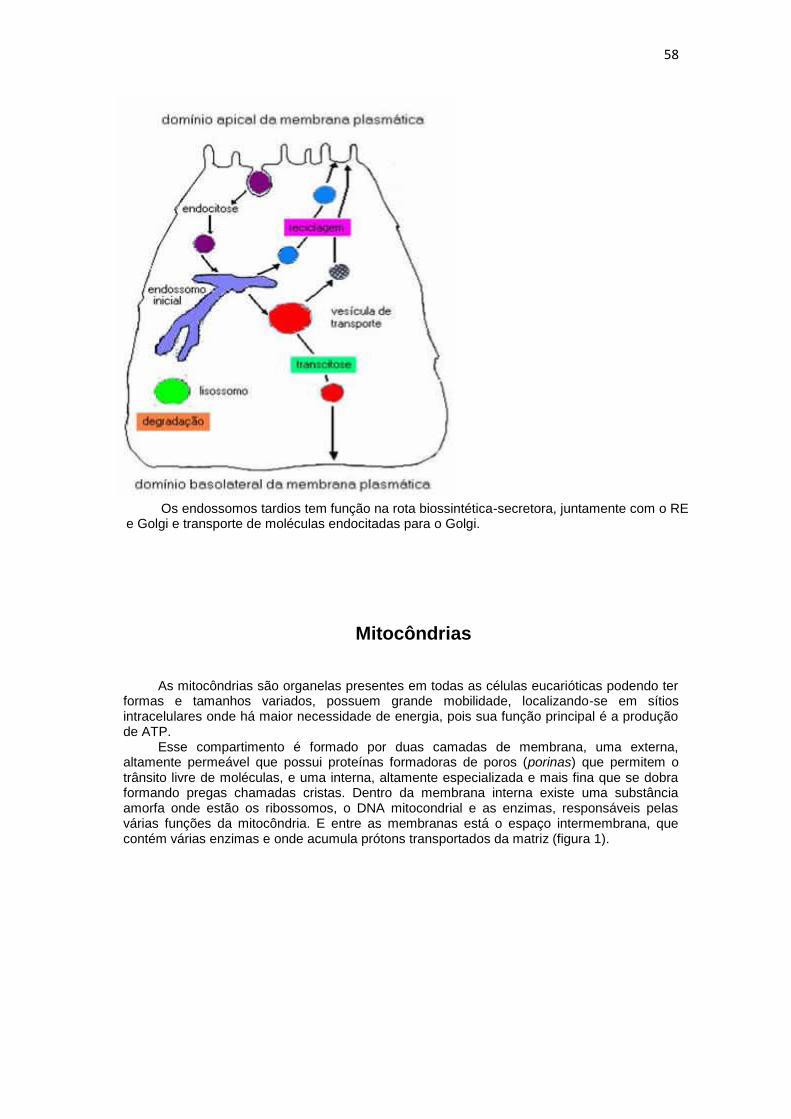
58
Os endossomos tardios tem função na rota biossintética-secretora, juntamente com o RE e Golgi e transporte de moléculas endocitadas para o Golgi.
Mitocôndrias
As mitocôndrias são organelas presentes em todas as células eucarióticas podendo ter
formas e tamanhos variados, possuem grande mobilidade, localizando-se em sítios intracelulares onde há maior necessidade de energia, pois sua função principal é a produção de ATP.
Esse compartimento é formado por duas camadas de membrana, uma externa, altamente permeável que possui proteínas formadoras de poros (porinas) que permitem o trânsito livre de moléculas, e uma interna, altamente especializada e mais fina que se dobra formando pregas chamadas cristas. Dentro da membrana interna existe uma substância amorfa onde estão os ribossomos, o DNA mitocondrial e as enzimas, responsáveis pelas várias funções da mitocôndria. E entre as membranas está o espaço intermembrana, que contém várias enzimas e onde acumula prótons transportados da matriz (figura 1).
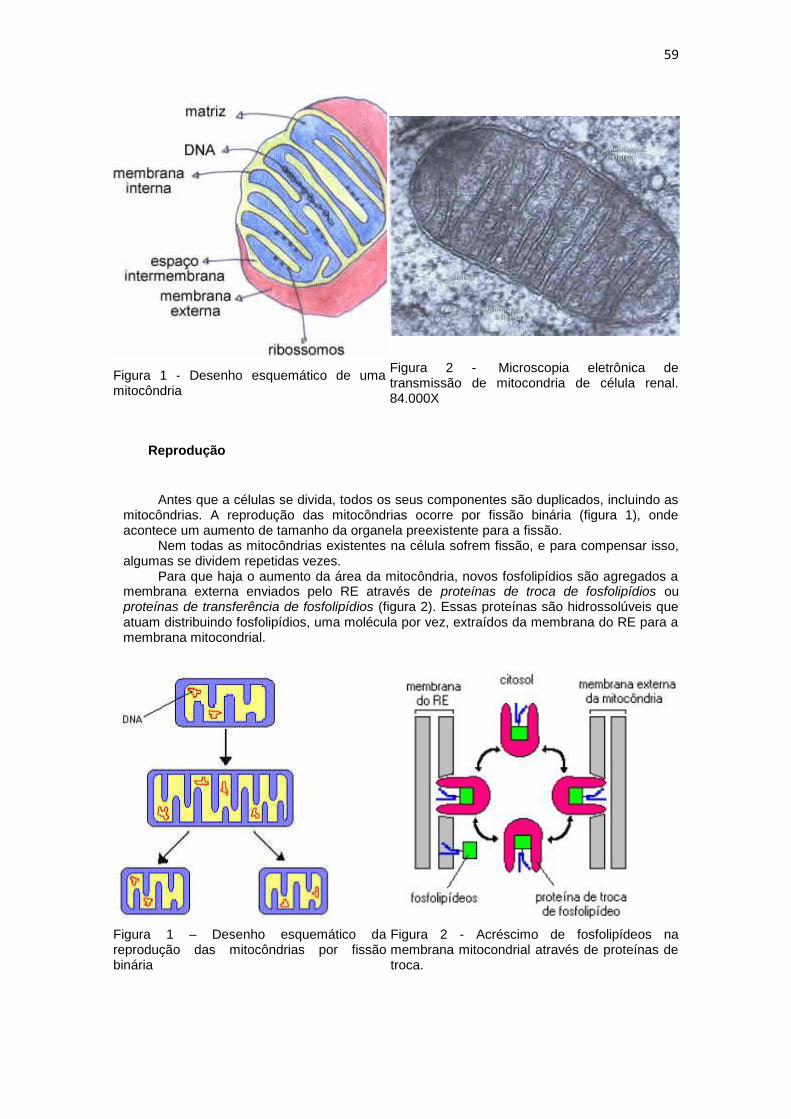
59
Figura 1 - Desenho esquemático de uma mitocôndria
Figura 2 - Microscopia eletrônica de transmissão de mitocondria de célula renal. 84.000X
Reprodução
Antes que a células se divida, todos os seus componentes são duplicados, incluindo as
mitocôndrias. A reprodução das mitocôndrias ocorre por fissão binária (figura 1), onde acontece um aumento de tamanho da organela preexistente para a fissão.
Nem todas as mitocôndrias existentes na célula sofrem fissão, e para compensar isso, algumas se dividem repetidas vezes.
Para que haja o aumento da área da mitocôndria, novos fosfolipídios são agregados a membrana externa enviados pelo RE através de proteínas de troca de fosfolipídios ou proteínas de transferência de fosfolipídios (figura 2). Essas proteínas são hidrossolúveis que atuam distribuindo fosfolipídios, uma molécula por vez, extraídos da membrana do RE para a membrana mitocondrial.
Figura 1 – Desenho esquemático da reprodução das mitocôndrias por fissão binária
Figura 2 - Acréscimo de fosfolipídeos na membrana mitocondrial através de proteínas de troca.

60
No processo de transcrição e replicação do DNA mitocondrial (figura 3) e a síntese protéica ocorrem na matriz das mitocôndrias, porém as maiorias das proteínas que medeiam esses processos são codificadas no genoma nucleares e importadas para dentro da organela. As cópias de DNA mitocondrial estão distribuídas na matriz em grupamentos chamados nucleóides.
Apesar das mitocôndrias produzirem suas proteínas, a maioria provém do citosol, onde são sintetizadas por ribossomos citosólicos.
Figura 3 - ME de moléculas circulares de DNA
Origem das Mitocôndrias
O mecanismo de reprodução por fissão semelhante ao da bactéria, a presença de DNA,
os ribossomos semelhantes ao da bactéria, sugerem que as mitocôndrias e evoluíram de bactérias endocitadas há mais de um bilhão de anos. Onde células eucariontes anaeróbicas estabeleceram relação simbiótica com bactérias aeróbicas, utilizando seu sistema de fosforilação oxidativa.
Essas bactérias teriam penetrado por fagocitose (Figura 1), escapando dos mecanismos intracelulares de destruição de organismos estranhos. Assim, a membrana do fagossomo teria tornado a membrana externa da mitocôndria e a membrana da bactéria tornou-se a membrana interna.
Com o aumento da concentração de oxigênio na atmosfera ocasionada pelas células fotossintéticas, a célula hospedeira sua atividade energética mais eficiente,m adquirindo também mecanismos de liberação deste oxigênio.
Ocorreu também a transferência de parte do DNA da organela para o DNA nuclear durante a evolução eucariótica, tornando-se dependentes das proteínas codificadas pelo núcleo celular.
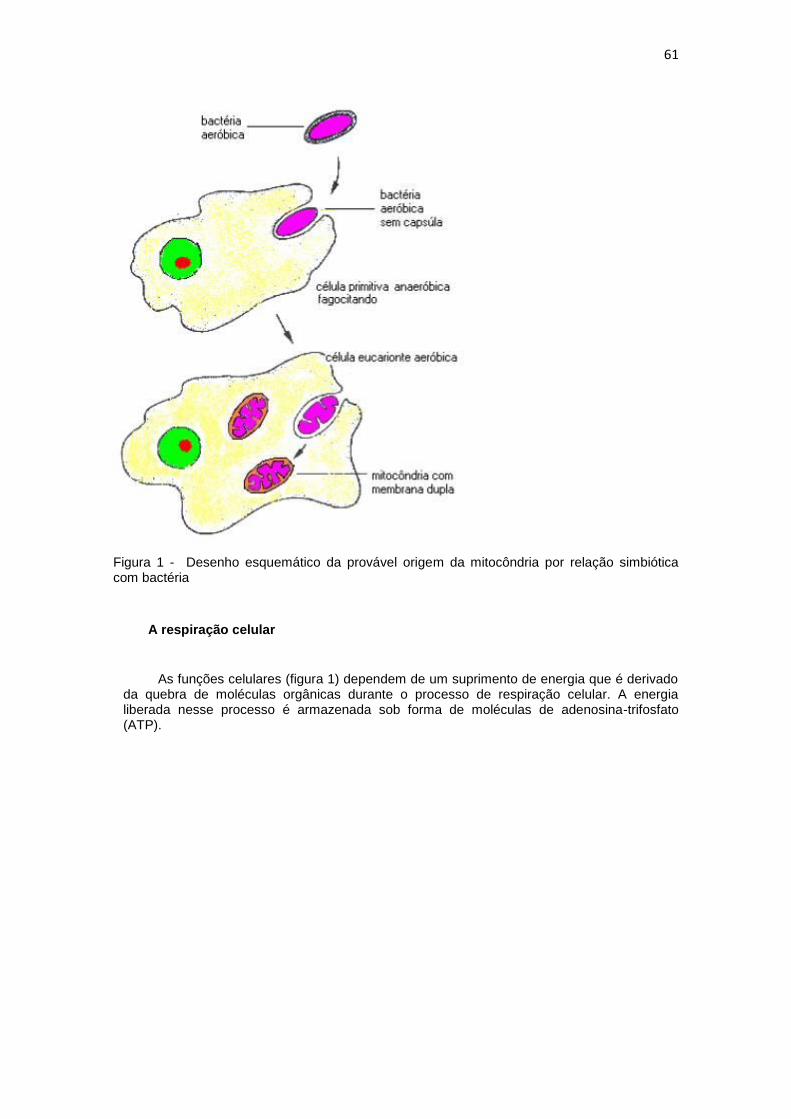
61
Figura 1 - Desenho esquemático da provável origem da mitocôndria por relação simbiótica com bactéria
A respiração celular
As funções celulares (figura 1) dependem de um suprimento de energia que é derivado
da quebra de moléculas orgânicas durante o processo de respiração celular. A energia liberada nesse processo é armazenada sob forma de moléculas de adenosina-trifosfato (ATP).

62
Figura 1 - Atividades do corpo precisam de ATP
1
a. etapa: os carboidratos e lipídeos, principalmente a glicose e os ácidos graxos, são
as principais substâncias quebradas para a respiração celular. A glicose é quebrada no citosol em um processo chamado glicólise, onde se forma duas moléculas de ácido pirúvico, liberando uma certa quantidade de energia (quatro moléculas de ATP), produz duas moléculas de NADH2 e consumindo oxigênio.
C6H12O6 2 C3H4O3 (Glicose) (Ácido pirúvico) 2
a. etapa: o ácido pirúvico entra na mitocôndria, e é convertido em acetil-coenzima A
por um sistema multienzimático da matriz mitocondrial chamado de piruvato desidrogenase, que então é metabolizada pelo ciclo do ácido cítrico (ciclo de krebs). Nesta etapa, uma quantidade de energia é liberada, tendo uma pequena parte utilizada para converter três NAD+ em três NADH.
No ciclo de Krebs (Figura 2), a acetil CoA sofre uma série de modificações que acaba produzindo ácido oxaloacético, que então recomeça o ciclo. Essas reações liberam duas moléculas de CO
2 e produzem três moléculas de NADH e uma molécula de FADH
2.
3
a. etapa: Depois os elétrons de alta energia percorrem a cadeia transportadora de
elétrons ou cadeia respiratória, que é composto por complexos enzimáticos, onde os elétrons sedem energia e produz 36 mols de ATP por mol de glicose consumida. Este processo é chamado fosforilação oxidativa, e ocorre na membrana interna da mitocôdria.
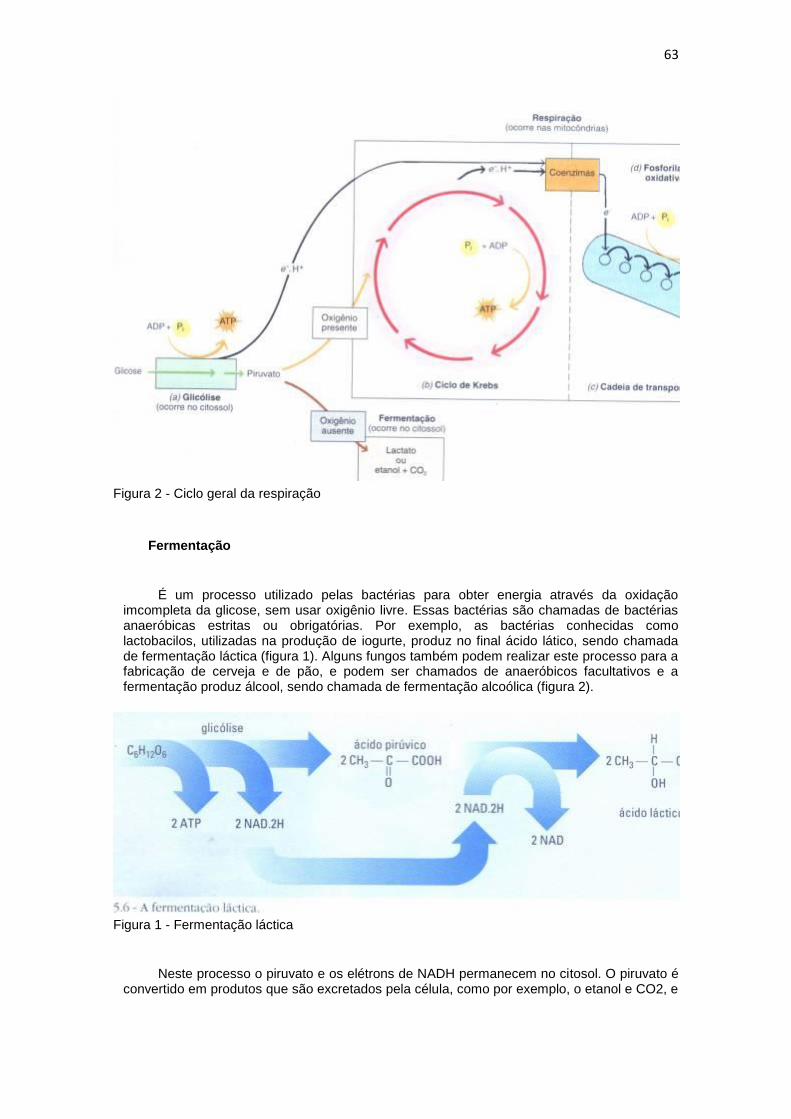
63
Figura 2 - Ciclo geral da respiração
Fermentação
É um processo utilizado pelas bactérias para obter energia através da oxidação
imcompleta da glicose, sem usar oxigênio livre. Essas bactérias são chamadas de bactérias anaeróbicas estritas ou obrigatórias. Por exemplo, as bactérias conhecidas como lactobacilos, utilizadas na produção de iogurte, produz no final ácido lático, sendo chamada de fermentação láctica (figura 1). Alguns fungos também podem realizar este processo para a fabricação de cerveja e de pão, e podem ser chamados de anaeróbicos facultativos e a fermentação produz álcool, sendo chamada de fermentação alcoólica (figura 2).
Figura 1 - Fermentação láctica
Neste processo o piruvato e os elétrons de NADH permanecem no citosol. O piruvato é
convertido em produtos que são excretados pela célula, como por exemplo, o etanol e CO2, e

64
o NADH doa os seus elétrons e é convertido novamente em NAD+, que é importante para as reações de glicólise.
Figura 2 - Fermentação alcoólica
Fermentação lática no músculo: Quando fazemos um esforço muscular intenso, a
quantidade de oxigênio que chega nos músculo, não é o suficiente para fornecer toda a energia necessária para a atividade desenvolvida. Então as células musculares passam a realizar fermentação láctica, onde o ácido láctico acumula-se no interior da fibra produzindo dores, cansaço e cãibras. Depois, uma parte desse ácido é conduzida pela corrente sanguínea ao fígado onde é convertido em ácido pirúvico.
Em repouso a célula muscular produz um excesso de ATP, que transmite sua energia para um outro composto, a creatina fosfato, que é mais estável permanecendo por mais tempo armazenada na célula. Em uma contração, este composto cede energia para produção de ATP.