Embed Size (px)
Citation preview

0
Universidade Federal do Pará Núcleo de Ciências Agrárias e Desenvolvimento Rural
Empresa Brasileira de Pesquisa Agropecuária - Amazônia Oriental Universidade Federal Rural da Amazônia
Programa de Pós-Graduação em Ciência Animal
Karol Guimarães Oliveira
ATIVAÇÃO ESPERMÁTICA E CRIOPRESERVAÇÃO DO SÊMEN DE MACACO-PREGO (Cebus apella) EM
DILUIDORES À BASE DE ÁGUA DE COCO IN NATURA E TES-TRIS
Belém 2010

1
Karol Guimarães Oliveira
ATIVAÇÃO ESPERMÁTICA E CRIOPRESERVAÇÃO DO
SÊMEN DE MACACO-PREGO (Cebus apella) EM DILUIDORES À BASE DE ÁGUA DE COCO IN NATURA E
TES-TRIS
Dissertação apresentada para obtenção do grau de Mestre em Ciência Animal. Programa de Pós-Graduação em Ciência Animal. Núcleo de Ciências Agrárias e Desenvolvimento Rural. Universidade Federal do Pará. Empresa Brasileira de Pesquisa Agropecuária – Amazônia Oriental. Universidade Federal Rural da Amazônia. Área de concentração: Produção Animal. Orientador Profa. Dra. Sheyla Farhayldes S. Domingues
Belém 2010

2
Karol Guimarães Oliveira
ATIVAÇÃO ESPERMÁTICA E CRIOPRESERVAÇÃO DO SÊMEN DE MACACO-PREGO (Cebus apella) EM
DILUIDORES À BASE DE ÁGUA DE COCO IN NATURA E TES-TRIS
Dissertação apresentada para obtenção do grau de Mestre em Ciência Animal. Programa de Pós-Graduação em Ciência Animal. Núcleo de Ciências Agrárias e Desenvolvimento Rural. Universidade Federal do Pará. Empresa Brasileira de Pesquisa Agropecuária – Amazônia Oriental. Universidade Federal Rural da Amazônia. Área de concentração: Produção Animal.
Data da aprovação. Belém - PA: ______/_______/_______
Banca Examinadora
______________________________________ Profa. Dra. Sheyla Farhayldes S. Domingues Universidade Federal do Pará - UFPA _______________________________________ Dr. John Patrick Kastelic Agriculture and Agri-Food Canada, Lethbridge, Alberta, Canada
_______________________________________ Profa. Dra. Lúcia Daniel Machado da Silva Universidade Estadual do Ceará - UECE

3
A Deus

4
AGRADECIMENTOS
A DEUS, por permitir que eu chegasse até aqui e nunca me abandonar, dando-me força nos
momentos difíceis.
À minha mãe Eleonora, por sempre pensar em nós em primeiro lugar e na nossa felicidade
acima de tudo. Por me apoiar incondicionalmente, me ouvir e sempre ter uma palavra de
consolo nos meus momentos de aflição. Por orar por mim e pelo seu amor.
À minha família, que mesmo longe, se faz presente com seu ilimitado apoio e incentivo. E por
me fazer sentir importante a cada demonstração de carinho.
À Profa. Dra. Sheyla Farhayldes Souza Domingues pelo seu comprometimento e seriedade
que muito contribuíram para a realização deste trabalho. E ainda, por estar sempre disposta a
ajudar e superar qualquer obstáculo.
À Dra. Regiane Rodrigues dos Santos, pela valiosa contribuição durante o desenvolvimento
do artigo dessa dissertação.
Ao Dr. José Augusto Pereira Carneiro Muniz, pelo auxílio na obtenção do material necessário
para a realização desse trabalho.
Ao Prof. Dr. Otávio Mitio Ohashi e Marcela Cordeiro do Laboratório de FIV-UFPA pelos
conhecimentos repassados e pela ajuda no preparo dos diluidores.
Aos amigos do Laboratório de Biologia e Medicina de Animais Silvestres da Amazônia
(BIOMEDAM-UFPA). Em especial àqueles que incansavelmente me acompanharam durante
a realização dos experimentos: Gustavo Sales, Danuza Leão, Stefânia Miranda, Adriel Brito,
Débora Almeida.
Ao Dr. Paulo Henrique Gomes de Castro, pelo apoio e por estar sempre disposto a ajudar.

5
Aos funcionários do Centro Nacional de Primatas, especialmente: Sarah Scalercio Dra. Aline
Imbelone, Dra. Gilmara Abreu, D. Laura Ferro, D. Rosa, Obadias, Léo, Melânia, D. Socorro e
aos técnicos Sr. Oswaldo, Sr. Viana, Sr. Alfredo pelo apoio e por estarem sempre dispostos a
ajudar.
Ao Rodrigo Rodrigues, amigo e funcionário do PPGCA, pela paciência e profissionalismo.
Aos meus amigos, pelos diversos momentos de alegria e com os quais eu posso sempre
contar: Nilton Jr. Rascon, Taty, Rose, Sherlem, Danyzinha, Ramon, Suzanne, Cristina, Dani,
Shirley, Rayanne, Danigata, Natália e Cleber.
Aos animais experimentais, que mesmo sem saber, contribuíram de forma valiosa.
À CAPES, pela concessão da bolsa e ao CNPq pelo apoio financeiro.
À UFPA e ao Programa de Pós-Graduação em Ciência Animal.
Ao Centro Nacional de Primatas (IEC-SVS) pelo apoio fundamental para a realização desta
pesquisa.

6
“O Senhor é o meu pastor e nada
me faltará.”
Salmo 23

7
RESUMO
A espécie Cebus apella (macaco-prego) é amplamente utilizada como modelo experimental
na pesquisa biomédica. Entretanto, são escassos os estudos dedicados a avaliar o sêmen
desses animais, que é composto por uma fração líquida e uma coagulada de difícil
manipulação e alta concentração de espermatozóides imóveis. Portanto, objetivou-se I) avaliar
o efeito de duas concentrações de cafeína (6 e 10 mM/mL) diluídas em TES-TRIS e água de
coco in natura (ACIN) na ativação de espermatozóides de C. apella e II) testar um protocolo
de criopreservação do sêmen comparando dois diluidores (TES-TRIS e ACIN) acrescidos de
gema de ovo e glicerol. O sêmen de seis animais mantidos no Centro Nacional de Primatas foi
coletado por eletroejaculação, diluído na fração-A de TES-TRIS ou ACIN, e incubado a 35°C
até dissolução do coágulo seminal. O tempo de liquefação foi comparado. Posteriormente foi
mensurado volume, concentração, morfologia espermática e percentual de espermatozóides
vivos. No experimento I as amostras foram diluídas em TES-TRIS ou ACIN acrescidos de 6 e
10 mM de cafeína após o término da motilidade durante a liquefação e mantidas a 35°C.
Motilidade e vigor foram avaliados por 5 h. Para o experimento II, após liquefação, o sêmen
foi diluído na fração-B (fração-A + gema de ovo e glicerol) dos diluidores, envasado,
resfriado a 4°C (2 h), em seguida a -60°C (20 min), antes de ser mergulhado em nitrogênio
líquido. O sêmen foi descongelado a 35°C (5 min). Os resultados foram expressos como
média ± EP. O efeito dos diluidores foi comparado pelo teste t Student e ANOVA (p ≤ 0,05).
O coágulo liquefez em 4,5 ± 1,7 e 2,8 ± 1,1 horas (p < 0,05) em TES-TRIS e ACIN
respectivamente. O volume médio, concentração, percentual de espermatozóides normais e
vivos antes de congelação foi respectivamente 0,6 ± 0,2 mL; 1.806 ± 367 x 106
espermatozóides/mL; 81,3 ± 2, e 40,1 ± 3,3 (TES-TRIS) e 30,9 ± 4 (ACIN). A duração da
motilidade em TES-TRIS e ACIN foi, respectivamente, 5 ± 1,4 e 1 ± 0,5 h. A motilidade
média nesse período foi 38 ± 10% (TES-TRIS) e 18 ± 9% (ACIN). Foi verificado aumento da
motilidade após adição de cafeína apenas nas amostras diluídas em ACIN 6 mM (21 ± 9%) e
ACIN 10 mM (22 ± 11) (p > 0,05). O percentual de espermatozóides vivos após
descongelação foi 26,2% em TES-TRIS e 13,2% em ACIN (p < 0,05). Para a criopreservação
de sêmen de C. apella TES-TRIS é mais indicado e pode, assim como ACIN + cafeína, ser
empregado na inseminação artificial com sêmen a fresco diluído.
Palavras-chave: Cebus apella. Sêmen. Água de coco. Tes. Criopreservação. Ativação.

8
ABSTRACT
The species Cebus apella (capuchin monkey) is an important experimental model used in
biomedical research. However, there are few reports regarding their semen, which is
composed by both liquid and coagulated fractions, difficult to handle and containing a high
proportion of immobile spermatozoa. Thereafter, the aims of this study were to: I) evaluate
the effect of two caffeine concentrations (6 and 10 mM/mL) diluted in TES-TRIS and coconut
water solution (CWS) on the activation of C. apella spermatozoa and; II) test and compare
two extenders for semen cryopreservation (TES-TRIS and CWS plus egg yolk and glycerol).
Semen from six males maintained at the National Primate Center was collected by
electroejaculation, diluted in A-fraction of TES-TRIS or CWS, and incubated at 35 °C until
coagulum liquefaction. In Experiment I, the samples were diluted in TES-TRIS or CWS plus
6 and 10 mM caffeine after stopped motility during liquefaction and maintained at 35 °C.
Motility and vigor were evaluated for 5 h. In Experiment II, after liquefaction, semen was
diluted in B-fraction (A-fraction + egg yolk and glycerol) of the extenders, packed, cooled to
4 °C (2 h), then to -60 °C (20 min), before being plunged into liquid nitrogen. Semen was
thawed at 35 °C (5 min). The results were expressed as mean ± SEM. The seminal coagulum
liquefied in 4.5 ± 1.7 and 2.8 ± 1.1 hours in TES-TRIS and CWS, respectively. Volume,
concentration, percentage of normal and live sperm before freezing were, respectively, 0.6 ±
0.2 mL; 1806 ± 367 x 106 spermatozoa/mL; 81.3 ± 2, and 40.1 ± 3.3 (TES-TRIS) and 30.9 ± 4
(CWS). For TES-TRIS and CWS, the duration of sperm motility was 5 ± 1.4 and 1 ± 0.5
hours and mean motility was 38 ± 10% and 18 ± 9%, respectively. Motility increased after
caffeine addition only in samples diluted in CWS 6 mM (21 ± 9%) and CWS 10 mM (22 ±
11) (p > 0.05). Post-thaw live sperm percentage was 26.2 in TES-TRIS and 13.2 in CWS (p <
0.05). For cryopreservation of semen from C. apella TES-TRIS was more appropriate than
CWS. However, TES-TRIS and CWS + caffeine potentially may be useful in artificial
insemination of fresh diluted semen.
Key-words: Cebus apella. Semen. Coconut water. Tes. Cryopreservation. Activation.

9
SUMÁRIO
1 INTRODUÇÃO ............................................................................................................. 10
2 OBJETIVOS .................................................................................................................. 12
2.1 OBJETIVOS GERAIS ................................................................................................. .........................................................................................................
12
2.2 OBJETIVOS ESPECÍFICOS ....................................................................................... 12
3 REVISÃO DE LITERATURA .................................................................................... 13
3.1 OS PRIMATAS ........................................................................................................... 13
3.2 A ESPÉCIE Cebus apella ............................................................................................ 14
3.2.1 O sêmen de C. apella ............................................................................................... 16
3.3 O COÁGULO SEMINAL ........................................................................................... 17
3.4 MÉTODOS DE COLETA ........................................................................................... 18
3.5 CRIOPRESERVAÇÃO DE SÊMEN DE PRIMATAS ............................................... 20
3.5.1 Meio diluidor e seus componentes ......................................................................... 21
3.5.1.1 Crioprotetores ......................................................................................................... 22
3.5.1.2 Antibióticos ............................................................................................................ 23
3.5.1.3 TES-TRIS ............................................................................................................... 24
3.5.1.4 Água de coco .......................................................................................................... 24
3.6 ATIVAÇÃO ESPERMÁTICA ..................................................................................... 25
4 LIQUEFAÇÃO DO COÁGULO SEMINAL, ATIVAÇÃO ESPERMÁTICA E CRIOPRESERVAÇÃO DO SÊMEN DE MACACO-PREGO (Cebus apella) EM ÁGUA DE COCO IN NATURA (ACIN) E TES-TRIS...................................................
27 5 CONCLUSÃO GERAL................................................................................................. 44
REFERÊNCIAS..................................................................................................................
45

10
1. INTRODUÇÃO
A biodiversidade mundial, particularmente em regiões tropicais como o Brasil, está
agora mais ameaçada do que em qualquer outro período histórico (IUCN, 2008) e sua
conservação é um dos principais desafios que precisam ser atingidos para preservar a vida de
animais em risco de extinção (CHEN et al., 2004).
A conservação da variabilidade genética de animais silvestres pode ser garantida com
sua proteção em seus habitats naturais (in situ), pela manutenção das condições adequadas em
cativeiro, em criatórios conservacionistas ou de pesquisa (ex situ), bem como pela formação
de bancos de germoplasma, na qual gametas e células somáticas podem ser utilizados em
programas de biotecnologia de reprodução. A conservação in situ consiste em uma das mais
eficazes estratégias de conservação. Entretanto, é necessário considerar que a perda da
variabilidade genética vem ocorrendo de forma acelerada, provocada principalmente pela
fragmentação de habitats em decorrência de ações antrópicas como o desmatamento. Por isso,
é de suma importância lançar mão de técnicas para recuperar e preservar o potencial genético
de animais mesmo após a sua morte, através da conservação ex situ, que pode complementar
programas de conservação in situ (DOMINGUES; CALDAS-BUSSIERE, 2006).
No Brasil, existem 26 espécies de primatas neotropicais ameaçadas de extinção
(IBAMA, 1989), distribuídos nas famílias Atelidae, Pitheciidae e Cebidae. A essa última
família pertence à espécie Cebus apella, que não se encontra ameaçada e é um excelente
modelo experimental para o estudo da fisiologia da reprodução, como a biologia do oócito, a
espermatogênese (LEÃO et al., 2008) e para o desenvolvimento de biotecnologias a serviço
das espécies em extinção (DOMINGUES; CALDAS-BUSSIERE, 2006).
Entretanto, técnicas como criopreservação de gametas, inseminação artificial,
produção e transferência de embriões, ainda não são aplicadas em C. apella. Elas possuem o
seu impacto limitado devido à dificuldade de obtenção de espermatozóides viáveis
(DOMINGUES; CALDAS-BUSSIERE, 2006), pois o sêmen dessa espécie apresenta uma
porção coagulada de difícil acesso que possui concentração espermática consideravelmente
maior que o restante do ejaculado (NAGLE; DENARI, 1983; ARAÚJO et al., 2007). Por isso,
estudos para o aperfeiçoamento de técnicas para obtenção e expansão do sêmen de C. apella
são de importância inquestionável. Protocolos de coleta seminal através de eletroejaculação
por sonda retal (EEJ) já foram testados com sucesso nessa espécie (BARNABE et al., 2002;
PAZ et al., 2006; ARAÚJO et al., 2007). A EEJ é uma importante técnica de coleta em

11
homens com injúria no cordão espinhal, possuindo as vantagens de ter boa aceitação clínica
por não requerer cirurgia, além de permitir que até 90% do sêmen obtido permaneça com seu
DNA intacto, sugerindo não ser uma técnica destrutiva para o espermatozóide (SØNKSEN;
OHL, 2002).
Algumas técnicas de dissolução do coágulo seminal de C. apella foram descritas, e
substâncias naturais como a água de coco in natura constituem alternativas para sua
liquefação e diluição (ARAÚJO et al., 2009), além de apresentar potencial para ser
empregada também na criopreservação do sêmen dessa espécie, já que proporcionou
excelentes resultados para outros grupos animais (SALLES, 1989; TONIOLLI, 1989a;
CRUZ, 1994; MONTEZUMA JÚNIOR; VIANA NETO; NUNES, 1994; CARDOSO et al.,
2003; CARDOSO; SILVA; SILVA, 2006). Contudo, é imprescindível comparar sua atuação
com a de outros diluidores cuja composição e mecanismos de ação sejam bem definidos,
como o TRIS e o TES, amplamente utilizados para diversas espécies.
Para uma melhor compreensão do presente projeto de dissertação, a revisão de
literatura abordará os aspectos mais relevantes da biologia reprodutiva de C. apella, além do
estado atual da tecnologia do sêmen para essa espécie.

12
2. OBJETIVOS
2.1. OBJETIVOS GERAIS
• Avaliar a espécie Cebus apella como modelo experimental para outros
primatas neotropicais importantes para a pesquisa biomédica e/ou ameaçados de extinção;
• Desenvolver e aprimorar a tecnologia do sêmen para a espécie Cebus apella.
2.2. OBJETIVOS ESPECÍFICOS
• Analisar as características físico-químicas dos ejaculados;
• Comparar a eficiência da liquefação do coágulo seminal em diluentes à base de
TES-TRIS e ACIN;
• Investigar a influência do coágulo seminal sobre os espermatozóides inclusos
nessa porção;
• Avaliar o efeito de duas concentrações de cafeína (6 e 10 mM/mL) diluídas em
TES-TRIS e ACIN na ativação espermática após liquefação do coágulo seminal;
• Testar um protocolo de criopreservação da fração líquida e coágulo seminal
liquefeito, comparando dois diluidores (TES-TRIS e ACIN) acrescidos de gema de ovo e
glicerol.

13
3. REVISÃO DE LITERATURA
3.1. OS PRIMATAS
Devido a sua similaridade filogenética com a espécie Homo sapiens (ABEE, 2003;
WOLF et al., 2004), os primatas não-humanos se tornaram importantes modelos
experimentais utilizados na pesquisa biomédica (WRIGHT; BUSH, 1977; AURICCHIO,
1995). Estes animais são geralmente os melhores ou, às vezes, os únicos modelos para estudar
determinadas doenças em humanos, bem como para elaboração de estratégias de prevenção e
terapias (ABEE, 2003), além de serem importantes na realização de diversas pesquisas em
reprodução relacionadas à saúde humana (ABBOTT et al., 2004).
Os primatas atuais seguem a seguinte classificação taxonômica segundo Wilson e
Reeder (2005):
Ordem Primates
• Subordem Strepsirrhini
o Infra-ordem Lemuriformes (lêmures)
o Infra-ordem Chiromyiformes (ai-ai)
o Infra-ordem Lorisiformes (lóris, gálagos)
• Subordem Haplorrhini
o Infra-ordem Tarsiiformes (társios)
o Infra-ordem Simiiformes
� Parvordem Platyrrhini (macacos do Novo Mundo)
� Parvordem Catarrhini (macacos do Velho Mundo)
Dentre os Platyrrhini (= narina achatada) encontram-se os primatas Neotropicais, que
têm ampla distribuição geográfica (Américas) e são representados por um grande número de
espécies distribuídas nas famílias Cebidae, Pitheciidae e Atelidae. Dentre os primatas
neotropicais brasileiros, a espécie Cebus apella, Linnaeus, 1758, destaca-se por ser uma das
mais utilizadas como modelo biológico na pesquisa biomédica (WRIGHT; BUSH, 1977;
NAGLE et al., 1979; FRAGASZY; ADAMS-CURTIS, 1998).

14
3.2. A ESPÉCIE Cebus apella
Cebus apella é um platirrino pertencente à Superfamília Ceboidea, Família Cebidae,
Subfamília Cebinae, Tribo Cebini e Gênero Cebus (GOODMAN, 1996), tendo ampla
distribuição geográfica: Colômbia, Equador, Peru, Bolívia, Brasil, Guianas, Suriname e
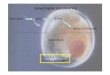
Venezuela (FRAGASZY et al., 2004) (Figura 1).
Eles são animais de porte médio, sendo que os machos têm o corpo 34% maior que o
das fêmeas e pesam em média 2,5 kg quando alcançam a maturidade sexual, por volta dos 7
anos de idade (NAGLE; DENARI, 1983). Têm um sistema poligâmico de acasalamento e as
fêmeas reproduzem-se referencialmente com o macho dominante. (EMMONS, 1990; ROWE,
1996). A gestação ocorre 1 vez ao ano e dura entre 150 e 160 dias, com uma única cria
(gêmeos são raros) que nasce com cerca de 260 g. Possuem cauda semi-preênsil e alimentam-
Fig. 1: Distribuição de espécie C. apella.
Elaborado por RASCON, N. L. (2009) baseado em FRAGASZY et al. (2004).

15
se de frutos, nozes, sementes, flores, insetos, ovos e pequenos vertebrados. Vivem em bandos
de 18 a 27 indivíduos (ROWE, 1996). Uma de suas características mais marcantes é a
presença de tufos pretos no alto da cabeça e a coloração mais escura da pelagem dos membros
e cauda (NOWAK, 1991) (Figura 2).
O sistema reprodutor do macho de C. apella é constituído por dois testículos, dois
epidídimos, dois ductos deferentes, duas vesículas seminais, próstata, duas glândulas bulbo-
uretrais (TEIXEIRA, 2005), um pênis dotado de um vestígio de osso peniano (báculo) e uma
glande muito desenvolvida em formato de prego, sendo por isso conhecido vulgarmente como
“macaco-prego” (NAPIER; NAPIER, 1986) (Figura 3).
Fig. 2: Exemplar macho da espécie Cebus apella. Fonte: Animal Park Zoo.
Fig. 3: Trato reprodutor masculino de Cebus apella. Fonte: Teixeira

16
3.2.1. O sêmen de C. apella
As glândulas acessórias de C. apella têm forma e função semelhantes às encontradas
em humanos. A vesícula seminal possui várias funções, dentre elas a mais importante é a
produção do líquido seminal, que é rico em frutose e proteínas, e serve como transporte aos
espermatozóides durante a ejaculação (TEIXEIRA, 2005).
O ejaculado dessa espécie é constituído por duas porções: a primeira é uma fração
líquida, transparente e viscosa, enquanto que a segunda é mais volumosa, opaca e consistente,
formando um coágulo seminal (Figura 4). Os valores máximos dos parâmetros espermáticos
citados na literatura para a fração líquida são: 71% de motilidade espermática; 3,5 de vigor;
pH 7,5 (PAZ, 2006); volume 1,9 ml e concentração 2,1 x 108 SPTZ/ml (ARAÚJO et al.,
2009). Poucas pesquisas se propõem a estudar a fração coagulada, sendo os melhores
resultados relatados por Araújo et al. (2009): volume 0,98 ml e concentração 1,6 x 109
SPTZ/ml. Entretanto, esses valores podem ser influenciados por variáveis como o período de
abstinência sexual, a fração seminal analisada, o método de coleta de sêmen, o anestésico
utilizado para contenção química do animal, o stress a que ele está submetido nas condições
de cativeiro, dentre outras (THOMSON et al., 1992).
Fig. 4: Coágulo e fração líquida do sêmen de Cebus apella.

17
3.3. O COÁGULO SEMINAL
A formação do coágulo seminal deve-se à reação entre as secreções provenientes do
lobo cranial da próstata e da vesícula seminal (AMBOKA; MWETHERA, 2003). O coágulo
formado tende a absorver grande número de espermatozóides do ejaculado, justificando a sua
alta concentração espermática em relação à fração líquida (HERNÁNDEZ-LÓPEZ; CERDA-
MOLINA; MONDRAGÓN-CEBALLOS, 2008). Dixson e Anderson (2002) denominam este
sêmen com aspecto gelatinoso firme de “plug copulatório”. Segundo esses autores, ele se
molda no contorno da vagina da fêmea após a cópula atuando como uma barreira que impede
a inseminação por outros machos, agindo como um mecanismo de seleção sexual. O coágulo
pode assumir diferentes consistências conforme o tipo de acasalamento da espécie, sendo
assim, em regimes monogâmicos ou poligínios, ambos em que a fêmea tem somente um
parceiro sexual, o plug tende a ser menos consistente ou até mesmo ausente, em contraste com
regimes poligâmicos (quando não há o estabelecimento de casais) em que atinge consistência
quase sólida. O plug ou coágulo seminal também pode servir como um veículo dos
espermatozóides para o interior da cérvix da fêmea, constituindo uma barreira física e
química, pois apresenta pH alcalino, aumentando o índice de sobrevivência dos
espermatozóides no meio ácido vaginal.
Várias espécies de primatas do Velho e Novo Mundo formam plug copulatório.
Hernández-López et al. (2008), analisando o ejaculado de Ateles geoffroyi durante duas
épocas do ano, verificaram aumento do volume de coágulo seminal durante as estações secas
em relação às chuvosas. Há evidências de que a produção desse coágulo seja um processo que
exija algum gasto energético pelo macho, pois ela é consideravelmente maior quando as
fêmeas encontram-se mais receptivas ao coito (HERNÁNDEZ-LÓPEZ; CERDA-MOLINA;
MONDRAGÓN-CEBALLOS, 2008). Entretanto, ainda inexistem estudos suficientes para
elucidar o processo de produção do coágulo seminal entre as espécies de primatas. (DIXSON;
ANDERSON, 2002).
Devido ao coágulo ser a porção de maior concentração espermática do sêmen (BUSH
et al., 1975; ARAÚJO et al., 2009), algumas pesquisas são realizadas com o intuito de
maximizar seu aproveitamento e impedir que ele dificulte o processo de criopreservação.
Alguns protocolos sugerem a liquefação desta fração através da imersão do tubo coletor em
banho-maria a 37 °C por 30 minutos para o sêmen de Macaca fascicularis (macaco-cauda-
longa) (LI et al., 2003; LI et al., 2005b). Outros recomendam a liquefação enzimática em

18
pronase e quimotripsina para Callithrix jacchus (sagüi) (MORRELL; HODGES, 1998) ou
com soluções contendo tripsina ou tripsina e hialuronidase para A. geoffroyi (macaco-aranha)
embora estas enzimas afetem a motilidade e o vigor espermático, além de não desfazerem
totalmente o coágulo (HERNÁNDEZ-LÓPEZ et al., 2002).
Algumas técnicas para dissolução do coágulo seminal de C. apella foram descritas.
Paz et al. (2006) utilizaram meio TCM 199 acrescido das enzimas tripsina e hialuronidase e
observaram que em ambos os casos houve uma redução significativa da motilidade e do vigor
espermático em comparação à fração líquida de acordo com o tempo de exposição, além de
não promoverem a completa dissolução do coágulo. Alguns protocolos sugerem sua
dissolução sem o uso de enzimas proteolíticas, utilizando apenas solução salina a 0,9%
(NAGLE; DENARI, 1983), diluidor a base de água de coco in natura (ARAÚJO et al., 2009),
ou a base de água de coco em pó (OLIVEIRA et al., 2010), todos associados à fragmentação
mecânica do coágulo com o uso de pipeta e incubação em banho-maria a 37 °C por 2 horas,
quando se atingiu a completa dissolução do coágulo seminal.
3.4. MÉTODOS DE COLETA
O método de coleta de sêmen mais difundido há algumas décadas em primatas tem
sido a eletroejaculação. Nos anos 60, a estimulação peniana direta foi um dos primeiros
métodos utilizados e consiste na estimulação elétrica direta do pênis do animal contido em
uma cadeira adaptada sem que haja necessidade do uso de sedativo, aplicável em espécies de
pequeno porte como C. jacchus (KUEDERLING et al., 2000; SCHNEIDERS; SONKSEN;
HODGES, 2004) e S. sciureus (YEOMAN et al., 1998). Os bons resultados alcançados com
essa metodologia podem estar relacionados à melhor estimulação do trato reprodutivo por
inteiro e diminuição das taxas de contaminação por urina, porém requer condicionamento
adequado dos animais à cadeira de contenção, se limitando a espécies de pequeno tamanho
corpóreo e fácil manipulação (contenção física) (VANDEVOORT, 2004).
Há outros métodos de coleta de sêmen utilizados em primatas não-humanos como:
vagina artificial, lavagem vaginal, punção de epidídimo, masturbação e estimulação vibratória
peniana. A vagina artificial pode ser utilizada para coleta de sêmen de algumas espécies como
Gorilla gorilla (gorila) (GOULD; MARTIN; WARNER, 1985) e Pan troglodytes
(chimpanzé) (YOUNG; SMITHWICK; GOULD, 1995). Tanto a utilização de vagina

19
artificial, quanto à masturbação, são métodos que requerem condicionamento do animal, uma
vez que é necessário realizar o estímulo pelo animal (vagina artificial) ou permitir que ele o
faça (masturbação). Ambos eliminam a necessidade de contenção química, diminuindo as
interferências na qualidade do ejaculado (VALLE, 2002). A técnica de lavagem vaginal já foi
testada em C. jacchus, fornecendo sêmen de boa qualidade (KUEDERLING; MORREL;
NAYUDU, 1996; VALLE, 2002). A micropunção de cabeça e cauda de epidídimo é um
método usualmente empregado em animais recém-mortos e já foi aplicada em M. fascicularis
(MAHONY et al., 1996), P. troglodytes (YOUNG et al., 1994), Macaca fuscata (SANKAI et
al, 1997) e C. jacchus (MORRELL et al., 1997).
Atualmente existem protocolos de coleta seminal por estimulação retal por sonda ou
eletrodo para poucas espécies de primatas, dentre elas: Gorilla gorilla (SEAGER et al., 1982),
Macaca nigra (THOMSON et al., 1992), Pan troglodytes (YOUNG; SMITHWICK;
GOULD, 1995), Alouatta caraya (MORELAND et al., 2001), C. apella (BARNABE et al.,
2002; ARAÚJO et al., 2007; 2009), A. geoffroyi (HERNÁNDEZ-LÓPEZ et al., 2002), Papio
anubis (AMBOKA; MWETHERA, 2003) e Macaca fascicularis (LI et al., 2005b).
Uma vez que a maioria das espécies neotropicais não possui um protocolo definido de
estímulos elétricos, a seqüência de estímulos utilizada para uma pode ser repetida para as
outras correlatas. Na coleta seminal de S. sciureus foram alcançados bons resultados ao
utilizar um intervalo de voltagem de 0,95-1,15 volts, obtendo tempo médio para ejaculação
entre 2 e 3 minutos (LANG, 1967). Em A. geoffroyi foram aplicados estímulos de 1 volt e 10
mA até 7 volts e 100mA, aumentado 1 volt a cada 2 ou 3 tentativas sucessivas, além de
estimulação manual do pênis (HERNÁNDEZ-LÓPEZ et al., 2002). Valle et al. (2004)
utilizaram estímulos de 0 a 8 volts para A. caraya com aumentos subseqüentes de 0,5 volt a
cada série de 30 estímulos que duravam cerca de 2 a 3 segundos com intervalo de 1 a 2
segundos.
A EEJ tornou-se uma técnica importante como método de indução da ejaculação
também na espécie C. apella devido ao seu volume corpóreo e comportamento agressivo,
fatores que predispõem contenção química para o manuseio seguro dos animais e que
dificultam a utilização de outras técnicas de coleta seminal nesta espécie. Barnabe et al.
(2002) descreveram um protocolo em que são realizadas 5 séries de 20 estímulos cada, em um
nível progressivo de intensidade, de 50 a 300 mA. Contudo, Araújo et al. (2009)
demonstraram que é possível induzir a ejaculação com estímulos que variaram de 12,5 mA a
100 mA, como o observado por Halstead, Vervoort e Seager (1987) e Ohl, Mccabe e Sønksen
(1996), com intervalos de 30 segundos entre cada série de estímulos. Esses intervalos

20
otimizaram o processo de ejaculação, sendo possível obter frações de sêmen líquidas e
coaguladas nos intervalos entre cada série. Em estudos que visam a coleta de sêmen por EEJ
em homens com disfunções ejaculatórias, tem sido descrito que os intervalos entre as séries
possibilitam o relaxamento do esfíncter uretral externo e favorecem a ejaculação em direção
anterógrada com otimização do procedimento de eletroejaculação (OHL; SØNKSEN;
BOLLING, 2000; SØNKSEN; OHL; WEDEMEYER, 2001; SØNKSEN; OHL, 2002). Esse
protocolo é vantajoso em relação aos atualmente utilizados para a espécie por oferecer menor
risco de injúrias aos indivíduos em conseqüência das baixas amperagens empregadas. Seager
et al. (1982) recomendam também a lubrificação da sonda retal com gel ou pomada de
vaselina com o objetivo de reduzir o risco de lesões no reto.
3.5. CRIOPRESERVAÇÃO DE SÊMEN DE PRIMATAS
O princípio dessa biotécnica é congelar o espermatozóide diminuindo reversivelmente
a sua atividade metabólica, permitindo o seu armazenamento por tempo indeterminado, o que
possibilita o emprego desse sêmen congelado em programas de inseminação artificial
(ENGLAND, 1993). A criopreservação inclui inicialmente uma etapa de resfriamento, que
consiste na redução progressiva da temperatura do sêmen diluído, baseado no princípio de que
o frio é o agente mais eficaz na diminuição das atividades metabólicas dos espermatozóides,
podendo ser conservado em estado líquido por várias horas com taxas de fertilidade
aceitáveis, conforme observado por Mies Filho (1987).
A importância da redução da temperatura se justifica pela ação de certos produtos do
metabolismo celular como o ácido láctico e o gás carbônico, que podem aumentar a acidez do
sêmen, desencadeando danos celulares irreversíveis (MCKINNON, 1996). A peroxidação dos
lipídios da membrana leva à perda da sua integridade, o que prejudica a função celular e,
conseqüentemente, diminui a motilidade do espermatozóide (AURICH; KUHNE; HOPPE,
1996). Quando os espermatozóides são armazenados a 5 ºC, as necessidades metabólicas
decrescem a aproximadamente 10% daquela que teriam se estivessem à temperatura de 37 ºC.
Em conseqüência, a produção de catabólitos será menor e a peroxidação da membrana
plasmática mais lenta, de modo que o desgaste da célula não ocorre de forma tão rápida
(MCKINNON, 1996).

21
A criopreservação de sêmen de alguns primatas não-humanos como: Cercopithecus
aethiops, Erythrocebus patas, Macaca speciosa, Macaca mulatta, Pan troglodytes
(ROUSSEL; AUSTIN, 1967), Papio anubis (KRAEMER; VERA CRUZ, 1969), Saimiri
sciurius (DENIS et al., 1976), Macaca fascicularis (MAHONE; DUKELOW, 1978), lêmures
(CLAVERT et al., 1986), Gorilla gorilla (LAMBERT et al., 1991), Macaca thibetana
(CHEN et al., 1994), Callithrix jacchus (MORRELL et al., 1997) e Macaca fuscata
(SANKAI et al., 1997) tem sido descrita. Contudo, somente para cinco dessas espécies foi
relatado sucesso após inseminação artificial com sêmen descongelado (LI et al., 2005a).
3.5.1. Meio diluidor e seus componentes
Para o sucesso da preservação dos espermatozóides através da criopreservação, é
necessário seguir uma série de passos que visam à redução dos danos causados às células e
que assegurem longevidade in vitro e in vivo. O meio diluente é utilizado com o intuito de
proteger os espermatozóides dos choques térmicos e osmóticos que ocorrem durante o
processo (CONCANNON; BATTISTA, 1989). A existência de diferenças na composição
lipídica da membrana plasmática do espermatozóide entre as espécies, raças e ainda entre
indivíduos da mesma espécie, pode explicar o maior ou menor efeito protetor de um diluente
aos espermatozóides de um determinado indivíduo (HOLT, 2000). A composição do diluente
é de extrema importância e deve ser específica para cada espécie. Ele deve ser constituído por
determinadas substâncias para ser completo e eficiente, tais como (ROTA, 1998):
- agentes crioprotetores;
- substâncias tampão para assegurar o controle do pH: TRIS, TES, ringer lactato, água
de coco, citrato e fosfato de sódio;
- fonte de energia: frutose ou inusitol;
- antibióticos.

22
3.5.1.1. Crioprotetores
As substâncias crioprotetoras são divididas em penetrantes e não penetrantes de
acordo com o peso molecular e a conseqüente propriedade de atravessar ou não a membrana
plasmática (RODRIGUES, 1992). Os crioprotetores permeáveis mais utilizados são o
glicerol, etilenoglicol e dimetilsulfóxido (DMSO), essenciais para minimizar ou prevenir a
formação de cristais de gelo intracelular. Os crioprotetores impermeáveis, como as
lipoproteínas da gema do ovo e as proteínas do leite, por outro lado, auxiliam na estabilização
da membrana plasmática durante o processo de congelação/descongelação (RODRIGUES,
1992). Os diluentes utilizados na criopreservação de sêmen de primatas não-humanos
geralmente contêm gema de ovo, açúcares, glicerol ou DMSO em proporções variadas
(MORREL; HODGES, 1998).
A gema de ovo é um dos protetores mais utilizados como base dos diluentes de sêmen,
sendo incorporada de forma habitual na maioria dos protocolos de conservação. Ela protege
contra o choque pelo frio devido à presença das frações lipoprotéicas de baixa densidade na
sua composição, restaurando os fosfolipídios perdidos durante o choque térmico provocado
pela mudança de temperatura que ocorre durante o resfriamento inicial do sêmen. A gema de
ovo previne também a liberação da enzima hialuronidase pela célula espermática
(HAMMERSTEDT; GRAHAM; NOLAN, 1990).
O glicerol (CH3H8O3) é o crioprotetor mais empregado na congelação do sêmen de
diferentes espécies e possui a capacidade de penetrar através das membranas celulares
(SILVA et al., 2003), permanecendo tanto na membrana quanto no citoplasma, impedindo,
dessa forma, a formação de grandes cristais de gelo intracelulares (WATSON, 2000).
Contudo, sabe-se que o glicerol exerce efeitos tóxicos sobre os espermatozóides, como
alterações físico-químicas que podem levar à ruptura da membrana plasmática, à remoção de
importantes proteínas de membrana ou provocar danos acrossomais (LOVELOCK; POLGE,
1954). A concentração final ideal de glicerol no diluente seria aquela em que há uma
predominância de seus efeitos protetores sobre os efeitos tóxicos. Essa concentração pode ser
influenciada por outros componentes do diluente, pelo padrão de resfriamento e pelos
métodos de congelação e descongelação. No entanto, os fatores determinantes seriam as
características seminais de cada espécie (WATSON, 1979). Alguns trabalhos demonstram que
concentrações excessivas desse crioprotetor podem causar diminuição da capacitação
espermática (LI et al, 2005a), demonstrando-se tóxico para espermatozóides de C. jacchus

23
quando fora dos limites da concentração ideal para essa espécie (2,5-7%) (MORREL et al.,
1997; MORREL; HODGES, 1998), intervalo que compreende a concentração adequada para
Macaca mulatta, 5% (SI et al., 2004 apud SILVA, 2005b).
A concentração ideal de glicerol varia amplamente entre as diferentes espécies de
primatas: 14% para Macaca speciosa, Erythrocebus patas e Cercopithecus aethiops
(ROUSSEL; AUSTIN, 1967), 10% para Pan troglodytes (SADLEIR, 1966), 2.5-5% para
Macaca fascicularis (CHO; HONJO, 1973), 5% para Macaca assamensis (LI; GAO; JI,
2004), 3-5% para Macaca mulatta (SI et al., 2000), 7% para Ateles paniscus e Ateles
marginatus (SILVA, 2005b). Li et al. (2005a) demonstraram que os efeitos dos crioprotetores
permeáveis são independentes do tipo do diluidor. Além disso, espermatozóides de diferentes
espécies podem reagir de forma distinta ao glicerol.
A etapa de glicerolização tem sido realizada antes do resfriamento do sêmen de
diversas espécies de primatas. Conradie, Oettle e Seier (1994) diluíram sêmen de
Cercopithecus aethiops em solução com glicerol a 32 °C antes de resfriar a 5 °C, similar a
Lanzendorf et al. (1992) com sêmen de Gorilla gorilla. Ejaculados de Macaca fascicularis
foram diluídos 20 minutos antes de resfriar a 5 °C por 2 horas. Seier et al. (1993) compararam
os efeitos do resfriamento de ejaculados de Cercopithecus aethiops a 32 °C ou 5 °C antes da
adição do glicerol a mesma temperatura. Eles observaram que motilidade era maior quando a
glicerolização ocorria a 32 °C, sugerindo que a permeabilidade do glicerol seja termo-
dependente.
3.5.1.2. Antibióticos
Os antibióticos são adicionados ao meio diluidor para prevenir contaminações das
amostras de sêmen, que podem afetar negativamente a fertilidade, devido à própria presença
das bactérias, à produção de toxinas ou à degradação dos componentes do meio. Os
antibióticos mais comumente utilizados em primatas são a penicilina e a estreptomicina
(SILVA, 2005b).

24
3.5.1.3. TES-TRIS
O TRIS (Tris-hidroximetil-aminometano - H2NC(CH2OH)3) é uma substância
facilmente solúvel em água e disponível comercialmente em um alto grau de pureza na forma
de cristais. Ele permanece estável em temperatura ambiente por diversos meses e é conhecido
por não inibir diversos sistemas enzimáticos (BATES, 1962 apud SILVA, 2005a), atuando
como tampão iônico bipolar com pH entre 7,0 e 9,0 (MCPHAIL; GOODMAN, 1984).
Rodrigues (1997) reportou que o TRIS não apenas apresenta atividade tamponante, mas que
também atua na redução do metabolismo da frutose pela célula espermática, contribuindo para
a preservação de sua energia.
Diluentes contendo TRIS (hidroximetil aminometano) e TES (ácido N-
Trishidroximetil-metil-2- aminometanosulfônico) já foram utilizados na criopreservação do
sêmen de espécies de primatas do Velho Mundo, como Clorocebus aetiops (SEIER et al.,
1993) e M. fascicularis (MAHONE; DUKELOW, 1978; TOLLNER et al., 1990; SANKAI et
al., 1994; LI et al., 2005b) e do Novo Mundo, como C. jacchus (MORRELL et al.,1998), A.
paniscus e A. marginatus (SILVA, 2005b).
3.5.1.4. Água de coco
Estudos realizados a partir da década de 80 constataram que a água de coco (Cocos
nucifera, família Palmae) pode ser uma alternativa como diluidor de sêmen de várias espécies,
já que é uma solução natural, ligeiramente ácida, estéril, composta de sais, proteínas,
açúcares, vitaminas, gorduras neutras, indutores da divisão celular, eletrólitos diversos
(NUNES, 1986; BLUME; MARQUES JR, 1994), fornecendo os nutrientes necessários para a
conservação de células espermáticas. Sua fração ativa foi identificada como um fitormônio
promotor de crescimento celular denominado ácido 3-indol-acético. Contudo, é necessária a
correção de osmolaridade e pH de acordo com as características seminais da espécie em
estudo, uma vez que a água de coco proveniente de frutos com idade de seis meses (variedade
verde-da-praia) apresenta osmolaridade em torno de 500 mOsmol/L e pH de 4,5 a 5,0
(NUNES; COBARNOUS, 1995). Para tal correção, após filtrar a água de coco, adiciona-se
água destilada e uma solução de citrato de sódio a 5% (NUNES, 1995).

25
A água de coco vem sendo amplamente utilizada para variados fins em diferentes tipos
celulares: maturação e cultivo de oócitos e embriões bovinos (BLUME et al., 1997a, BLUME
et al., 1997b); cultivo de folículos pré-antrais caprino (SILVA et al., 2000) e ovino
(ANDRADE et al., 2002 apud CORDEIRO et al., 2006); diluição do sêmen de caprinos
(SALLES, 1989; TONIOLLI, 1989a), suínos (TONIOLLI, 1989b), cães (MONTEZUMA
JÚNIOR; VIANA NETO; NUNES, 1994; CARDOSO et al., 2003; CARDOSO; SILVA;
SILVA, 2006), ovinos (CRUZ, 1994), bubalinos (VALE et al., 1999 apud CORDEIRO et al.,
2006), humano (NUNES, 1998) e do primata C. apella (ARAÚJO et al., 2009). Diluidores a
base de água de coco in natura (ACIN) forneceram excelentes melhorias relacionadas à
motilidade e vigor espermático em caprinos, mantendo a viabilidade das amostras (NUNES,
1986). Cardoso, Silva e Silva (2004) obtiveram aproximadamente 60% de espermatozóides
caninos móveis após descongelação.
Com relação à espécie C. apella, Araújo et al. (2009) demonstraram que a solução à
base ACIN é eficiente na conservação da viabilidade espermática, mantendo
aproximadamente 87% de espermatozóides vivos em banho-maria a 37 °C por até 7 horas
após a coleta do sêmen. Ela apresentou também vantagens em relação a meios diluidores
contendo enzimas proteolíticas para dissolução do coágulo, como a tripsina e a hialuronidase,
que além da queda da motilidade e do vigor espermático, podem causar danos a membrana
espermática impedindo o sucesso da criopreservação (HERNÁNDEZ-LÓPEZ et al., 2002;
PAZ et al., 2006).
3.6. ATIVAÇÃO ESPERMÁTICA
Diversos estudos relatam a baixa motilidade espermática do sêmen de primatas
coletado por eletroejaculação (MORREL et al., 1998; YEOMAN et al., 1998; SCHNEIDERS;
SONKSEN; HODGES, 2004), além do fato de que o plug mantém os espermatozóides
imóveis até que estes sejam ativados no trato reprodutor da fêmea (DIXSON; ANDERSON,
2002). Logo, surge a necessidade de administrar substâncias ao meio diluente do sêmen em
estudo, que possuem a propriedade de estimular a motilidade progressiva espermática. Dentre
essas substâncias que possuem a propriedade de estimular a motilidade, estão os derivados da
metilxantina, como a cafeína, teofilina e a pentoxifilina, pois inibem a adenosina cíclica 3''-5''
monofosfato (AMP) fosfodiesterase levando a um aumento intracelular das concentrações de

26
AMPc (BUNGE, 1973). A cafeína, especificamente, é descrita como um inibidor de
fosfodiesterase capaz de aumentar e manter a respiração e a motilidade espermática
(MAHONY et al., 1996).

27
4 LIQUEFAÇÃO DO COÁGULO SEMINAL, ATIVAÇÃO ESPERMÁTICA E
CRIOPRESERVAÇÃO DO SÊMEN DE MACACO-PREGO (Cebus apella) EM ÁGUA
DE COCO IN NATURA (ACIN) E TES-TRIS
Semen coagulum liquefaction, sperm activation and cryopreservation of capuchin
monkey (Cebus apella) semen in coconut water solution (CWS) and TES-TRIS
Karol G. Oliveira a,b, Stefânia A. Miranda a,b, Danuza L. Leão a,c, Adriel B. Brito a,b, Regiane R. Santos a,b,d, Sheyla F. S. Domingues a,b,c,*
aLaboratório de Biologia e Medicina de Animais Silvestres, Universidade Federal do Pará,
Belém, Pará, Brasil bPrograma de Pós-graduação em Ciência Animal, Universidade Federal do Pará, Belém,
Pará, Brasil cFaculdade de Medicina Veterinária, Universidade Federal do Pará, Castanhal, Pará, Brasil
dDepartment of Equine Sciences, Veterinary, Pharmacology, Pharmacy and Toxicology
Division, Utrecht University, Utrecht, The Netherlands
*Autor para correspondência: Sheyla F. S. Domingues, DMV, PhD E-mail: [email protected] Endereço: Rua Augusto Corrêa, Campus Básico, CEP 66075-110, Belém, Pará, Brasil Tel/Fax: + 55 32018011

28
Resumo
Os objetivos do presente estudo foram testar o efeito de diluidores a base de água de coco em
pó (ACIN) e TES-TRIS na liquefação do coágulo seminal, na ativação espermática do sêmen
fresco e na criopreservação do sêmen de macaco-prego (Cebus apella). O sêmen de seis
machos adultos foi coletado através de eletroejaculação, diluído em TES-TRIS ou ACIN e
incubado a 35°C até completa liquefação do coágulo seminal. No experimento I, após a
liquefação, as amostras foram diluídas em TES-TRIS ou ACIN acrescidos de 6 e 10 mM/mL
de cafeína. Motilidade espermática e vigor foram avaliados durante 5 horas. Para o
experimento II, após liquefação, as amostras de sêmen foram diluídas em TES-TRIS (3,5% de
glicerol na solução final) ou ACIN (2,5% de glicerol na solução final), criopreservado e
estocado em nitrogênio líquido por 1 semana. O coágulo seminal liquefez em 4,5 ± 1,7 e 2,8 ±
1,1 horas (média ± EPM) em TES-TRIS e ACIN, respectivamente. Os espermatozóides
mantiveram-se móveis em TES-TRIS e ACIN por 5,0 ± 1,4 e 1,0 ± 0,5 horas,
respectivamente. A motilidade média nesse período foi 38 ± 10% (TES-TRIS) e 18 ± 9%
(ACIN). Após a adição de cafeína a motilidade aumentou somente nas amostras contendo
ACIN, tanto a 6 mM (21 ± 9%) quanto a 10 mM (22 ± 11%) de cafeína. O percentual de
espermatozóides vivos após a congelação foi 26,2% em TES-TRIS e 13,2% em ACIN.
Portanto, TES-TRIS (3,5% glicerol) foi mais apropriado que ACIN (2,5% glicerol) para a
criopreservação de sêmen de C. apella. ACIN + cafeína aumentou consideravelmente a
motilidade espermática, podendo ser utilizada para melhoria da técnica de inseminação
artificial com sêmen fresco.
Palavras-chave: Cebus apella; espermatozóide; coágulo seminal; criopreservação; ativação
espermática

29
Abstract
The objectives of the present study were to test the effect of coconut water solution and TES-
TRIS on the seminal coagulum liquefaction, sperm activation in fresh diluted semen, and on
the cryopreservation of semen from capuchin monkeys (Cebus apella). Semen was collected
from six males by electro-ejaculation, diluted in TES-TRIS or coconut water solution (CWS),
and incubated at 35° C until the coagulated fraction of the semen was completely liquefied. In
the Experiment I, after liquefaction, samples were diluted in TES-TRIS or CWS, plus 6 and
10 mM/mL of caffeine. Sperm motility and vigor were evaluated during 5 hours. For
Experiment II, after liquefaction, semen samples were extended in TES-TRIS (3.5% glycerol
in the final solution) or CWS (2.5% glycerol in the final solution), cryopreserved and stored
in liquid nitrogen for 1 week. The seminal coagulum was liquefied in (mean ± SDM) 4.5 ±
1.7 and 2.8 ± 1.1 hours in TES-TRIS and CWS, respectively. Sperm were motile in TES-
TRIS and CWS for 5.0 ± 1.4 and 1.0 ± 0.5 hours, respectively. The mean motility in this
period was 38 ± 22% (TES-TRIS) and 22.0 ± 16.0 (CWS). Motility increased after caffeine
addition only in samples diluted in CWS containing 6 mM (22.5 ± 16.0) or 10 mM (28.0 ±
19.0) caffeine. Post-thaw live sperm percentage was 26.2% in TES-TRIS and 13.2% in CWS.
For cryopreservation of semen from C. apella TES-TRIS (3.5 % glycerol) was more
appropriate than CWS (2.5 % glycerol). CWS + caffeine potentially increase sperm motility
and may be useful in artificial insemination of fresh diluted semen.
Keywords: Cebus apella; spermatozoa; seminal coagulum; cryopreservation; activation

30
1. Introdução
Aquecimento global, destruição e fragmentação de habitats, introdução de espécies
exóticas, assim como caça e outras pressões de exploração, têm provocado a extinção de
muitas espécies animais (Fickel et al., 2007). Atualmente, todas as espécies de primatas do
Novo Mundo estão listadas na Convention on International Trade in Endangered Species of
Wild Fauna and Flora (CITES), mesmo que apresentem diferentes graus de vulnerabilidade.
Isso estimula o desenvolvimento e aplicação de biotécnicas reprodutivas para prevenir a
extinção de muitas espécies de primatas. Portanto, o simples estabelecimento de um banco de
sêmen poderia ajudar a manter a diversidade genética e facilitar o desenvolvimento de outras
tecnologias reprodutivas assistidas (Vidal et al., 2007).
A criopreservação do sêmen de primatas não-humanos tem sido descrita utilizando
diferentes diluidores, como glutamato de sódio ou TES-TRIS (Tabela 1).

31
Tabela 1: Composição dos diluidores usados na criopreservação do sêmen de diferentes espécies de primatas não-humanos.
NI: Não informado. * Valores médios calculados do epidídimo direito e esquerdo ** Nascimentos após inseminação artificial com sêmen descongelado
Espécie Tampão Crioprotetor Viabilidade pós-congelação (%)
Motilidade pós-congelação (%)
Referências
Callithrix jacchus TES-TRIS 3% glicerol 34 44 O’Brien et al., 2003 Cercopithecus aethiops Glutamato de sódio 14% glicerol 53 28 Roussel e Austin, 1967 Cynomolgus monkey TRIS 6% glicerol 45 29 Feradis et al., 2001 Erythrocebus patas Glutamato de sódio 14% glicerol 51 23 Roussel e Austin, 1967 Macaca assamensis TES-TRIS 5% glicerol 39 69 Li et al., 2005 Macaca fascicularis TES-TRIS 3% glicerol NI 65 Tollner et al., 1990** TRIS 5% glicerol 62 61 Li et al., 2005 Macaca fuscata TES-TRIS 5% glicerol 60 65 Sankai et al., 1997 Macaca mulatta Glutamato de sódio 14% glicerol 50 27 Roussel e Austin,1967 TES-TRIS 12% glicerol NI 27 Dong et al., 2009 TES-TRIS 3% glicerol NI 54 Dong et al., 2009 TRIS 5% glicerol 74 85 Sanchez-Partida et al., 2000** Macaca speciosa Glutamato de sódio 14% glicerol 50 25 Roussel e Austin, 1967 Papio anubis Glutamato de sódio Gema de ovo 65 44 Kraemer e Vera Cruz, 1969 TES-TRIS 3% glicerol 47 83 O’Brien et al., 2003 Pan troglodytes Glutamato de sódio 14% glicerol 54 27 Roussel e Austin, 1967 TES-TRIS 5% glicerol 21* 30* Kusunoki et al., 2001 TES-TRIS 2,5% glicerol 46* 28* Kusunoki et al., 2001** TES-TRIS 3% glicerol 28 43 O’Brien et al., 2003

32
Entretanto, somente em duas (Macaca fascicularis e Macaca mulatta) dessas espécies
de primatas há relatos de nascimentos após programas de inseminação artificial utilizando
sêmen congelado (Tollner et al., 1990; Sanchez-Partida et al., 2000).
Os primatas do Novo Mundo têm recebido comparativamente menos atenção com
relação à criopreservação do sêmen. O macaco-prego (Cebus apella) tem sido utilizado com
sucesso como modelo experimental na pesquisa biomédica (Domingues et al., 2010). Porém,
apesar da disponibilidade de métodos seguros e eficientes para coleta de sêmen dessa espécie
neotropical (Araújo et al., 2009), nenhum protocolo de criopreservação de sêmen foi relatado.
Uma das prováveis barreiras para o sucesso da preservação do sêmen de C. apella é a
formação de uma porção coagulada e consistente do ejaculado, que contém alta concentração
de espermatozóides imóveis (Nagle e Denari, 1983). Por isso se tem afirmado que a diluição
do coágulo seminal só é possível através da utilização de tripsina (Hernández-López et al.,
2002). No entanto, a tripsina danifica severamente as células espermáticas de macaco-prego
(Paz et al., 2006) que, uma vez fora do coágulo seminal, se tornam extremamente instáveis
(Hernández-López et al., 2008). Além disso, diferentemente do ejaculado humano, cujo
coágulo seminal liquefaz facilmente entre 5 e 20 minutos (Tauber et al., 1980), o plug seminal
de macaco-prego apresenta o grau mais alto de coagulação (Barnabe et al., 2002). Por isso,
é crucial otimizar o protocolo de dissolução do coágulo associado ao posterior incremento da
viabilidade e motilidade espermática.
A eficiência da água de coco in natura (ACIN) na conservação do sêmen de C. apella
foi relatada (Araújo et al., 2009), assim como na criopreservação do sêmen canino (Cardoso et
al., 2006). Além disso, a presença do fator de crescimento ácido-3-indol-acético na ACIN é
considerada um fator que promove a preservação dos espermatozóides em baixas
temperaturas (Toniolli et al., 1996). A ACIN é uma substância complexa que contém enzimas
e nutrientes, dentre outras substâncias, que também podem facilitar a liquefação do coágulo e
a sobrevivência dos espermatozóides após a diluição e a criopreservação do sêmen. Embora
seja importante salientar que a cafeína, um conhecido inibidor da fosfodiesterase, tenha sido
utilizada com sucesso a várias concentrações para aumentar a motilidade espermática em
outras espécies de primatas (Boatman e Bavister, 1984; Mahony et al., 1996), seus potenciais
efeitos e respectivas concentrações a serem administradas em espermatozóides de C. apella
após a liquefação ainda são desconhecidas.
Portanto, esse estudo foi conduzido com os objetivos de (I) avaliar a ACIN como um
diluente do coágulo seminal, (II) analisar o efeito de duas concentrações de cafeína (6 e 10
mM/mL) na ativação de espermatozóides imóveis e (III) testar um protocolo de

33
criopreservação de sêmen para C. apella após liquefação do coágulo seminal, usando ACIN e
TES-TRIS como diluidores e glicerol como crioprotetor.
2. Material e Métodos
2.1. Animais
Todos os procedimentos experimentais foram aprovados pelo Comitê de Ética em
Pesquisa Animal (nº 013/2009/CEPAN/IEC/SVS/MS). Foram utilizados seis machos
saudáveis e sexualmente adultos de C. apella, com idade média entre 10 e 20 anos, mantidos
no Centro Nacional de Primatas (CENP). Comprimento, largura, altura, circunferência e
volume dos testículos foram mensurados. O peso corporal dos animais foi aferido durante
todo o período experimental. Os animais foram alocados em gaiolas individuais de 90 x 80 x
80 cm (comprimento, largura e altura, respectivamente), sob fotoperíodo natural, no CENP,
Ananindeua, PA, Brasil (1º 22' 57"S, 48° 22' 52"N). O clima é tropical úmido, com
temperatura anual média de 28 °C. A dieta consistia de frutas e vegetais frescos, leite e ração
comercial peletizada (Foxy Junior Supreme, Curitiba, PR, Brasil). Vitaminas, minerais e ovos
eram ofertados uma vez por semana, e água ad libitum.
2.2. Diluidores
Dois diluidores foram testados: TES-TRIS (Morrell, 1997) e água de coco in natura
(ACIN) (Toniolli et al., 1996). Cada diluidor consistia de duas frações (A e B). Composição,
osmolaridade e pH das frações A (diluição) e B (criopreservação) de ambos diluidores é
apresentado na Tabela 2.

34
Tabela 2: Composição, osmolaridade e pH das frações A e B de TES-TRIS e ACIN. Composição Osmolaridade pH
Fração A ACIN Água de coco in natura Água ultrapura Solução de citrato 5% Estreptomicina Penicilina
50,00 mL 25,00 mL 25,00 mL
0,20 g 0,10 g
294 mOsm
6,5
TES-TRIS TES TRIS D-Frutose Estreptomicina Penicilina
4,90 g 1,06 g
0,20 g 0,20 g 0,10 g
327 mOsm
7,2
Fração B ACIN Fração A de ACIN Gema de ovo Glicerol
75,00 mL 20,00 mL 5,00 mL
589,2 mOsm
6,5
TES-TRIS Fração A de TES-TRIS Leite desnatado em pó a 11% Gema de ovo Glicerol
36,5 mL 36,5 mL 20,0 mL
7,0 mL
525,5 mOsm
7,0
2.3. Condicionamento animal e anestesia
Um experimento piloto foi executado com seis animais para avaliar o efeito do
condicionamento no tempo de ação da anestesia. O condicionamento foi conduzido por
cooperação, de forma que os animais eram recompensados com uvas frescas quando
permitiam a injeção do anestésico. Cada animal foi anestesiado com Cloridrato de ketamina
(10 mg/Kg; IM; Vetanarcol, Konig) e Cloridrato de xilazina (1 mg/Kg; IM; Köning S.A.,
Avellaneda, Argentina).
2.4. Coleta de sêmen e análises preliminares
O sêmen foi coletado sob condições higiênicas e no mesmo período do dia: pela
manhã, antes da 1ª refeição. Os animais foram estimulados por meio de eletroejaculação

35
(EEJ), segundo Oliveira et al (2010). A EEJ foi realizada utilizando um eletroejaculador
(Autojac-Neovet, Uberaba, Brasil) à bateria, equipado com uma probe retal bipolar (9 mm de
diâmetro e 15 cm de comprimento). A probe era lubrificada e posicionada ventralmente no
reto, estendendo-se a aproximadamente 5 cm do esfíncter anal para estimular a próstata. Cada
sessão de estímulos consistia de seis séries, compostas por 10, 15, 20, 25, 30 e 35 estímulos
elétricos (12,5 a 100 mA) com intervalos de 30 segundos entre as séries. Quando o animal não
ejaculava após a sessão, não eram realizadas tentativas adicionais no mesmo dia. As coletas
eram realizadas em intervalos mínimos de 15 dias. Todos os animais alcançaram ereção
peniana e ejacularam pelo menos uma vez. O ejaculado era coletado em tubo de
microcentrífuga (1,5 mL). Logo após a coleta, os espermatozóides eram imóveis, por isso,
somente volume e aspecto seminal eram imediatamente avaliados.
2.5. Diluição I e liquefação
O ejaculado de cada animal foi dividido em duas partes iguais; cada uma foi diluída
em 500 µL da fração A dos diluidores testados (TES-TRIS ou ACIN) e mantida a 35 °C.
Posteriormente, o coágulo seminal em cada diluidor era mecanicamente fragmentado por
meio de repetidas pipetagens a cada dez minutos, até completa liquefação, com a obtenção de
um líquido viscoso. O tempo necessário para a liquefação em ambos os meios diluidores foi
registrado. Concentração, Osmolaridade, morfologia, integridade de membrana, motilidade e
vigor espermáticos foram avaliados após a liquefação.
2.6. Avaliação do sêmen
A concentração espermática foi mensurada em câmara de Neubauer e a osmolaridade
em osmômetro (Vapro, Wescor, Utah, EUA). Morfologia espermática e percentual de
espermatozóides vivos foram avaliados através de um esfregaço preparado adicionando 5 µL
de eosina 1% (Vetec) e 5 µL nigrosina 1% (Vetec) a 5 µL de sêmen em uma lâmina pré-
aquecida (35 °C). Defeitos morfológicos identificados nos espermatozóides foram
classificados em primários e secundários (Tabela 3). A motilidade espermática foi avaliada
através de contagem de 200 células em lâmina pré-aquecida (35 °C), de acordo com Li et al.
(2005). O vigor também foi avaliado subjetivamente e classificado em um score de 0 a 5. As
análises subjetivas de todas as amostras foram realizadas pelo mesmo operador e feitas em
microscópio óptico (Nikon E400, Japão) em magnificação de 100x.

36
2.7. Delineamento experimental
Dois experimentos foram desenvolvidos neste estudo: no experimento 1, um total de
cinco ejaculados (um ejaculado por macho) foi estudado para comparar a eficiência da adição
de cafeína nos espermatozóides em ambos os diluidores. No experimento 2, um total de dez
ejaculados de seis animais (pelo menos um ejaculado por animal) foi usado para investigar a
criopreservação do sêmen usando dois diferentes diluidores (TES-TRIS e ACIN) e o glicerol
como crioprotetor.
2.8. Experimento 1: Ativação espermática
Após a liquefação do coágulo na fração A de cada um dos diluidores, vigor e
motilidade espermática foram avaliados. A duração da motilidade foi registrada. Quando
vigor e motilidade chegavam a 0 e 0%, respectivamente, duas alíquotas de cada amostra eram
retiradas e diluídas na fração A do diluidor correspondente acrescido de cafeína à
concentração final de 6 ou 10 mM/mL, de acordo com Boatman e Bavister (1984) com a
espécie Macaca mulatta. Motilidade e vigor espermáticos foram avaliados a cada 30 minutos
durante 5 horas em microscópio invertido (Motic, British Columbia, Canada).
2.9. Experimento 2: Criopreservação do sêmen
Após liquefação do coágulo seminal as amostras foram divididas em alíquotas, de
acordo com o delineamento experimental, e submetidas a uma segunda diluição (diluição II),
usando a fração B de cada diluidor separadamente na proporção 1:1. O glicerol foi usado na
concentração final de 3,5% em TES-TRIS (modificado de Morrell, 1997) e 2,5% em ACIN
(modificado de Toniolli et al., 1996). Em seguida as amostras foram envasadas em palhetas
plásticas de 0,25 mL (IMV, L’Aigle, France), seladas com esferas metálicas e estocadas
horizontalmente a 4 °C por 2 horas. Posteriormente, as esferas eram mantidas por 20 minutos
a -60 °C antes de serem mergulhadas em nitrogênio líquido (-196 °C). Após uma semana as
amostras eram descongeladas em banho maria a 35 °C por 5 minutos, e então o percentual de
espermatozóides vivos era determinado. Portanto, o percentual de espermatozóides vivos era
avaliado em quatro momentos: após diluição na fração A (diluição I), após liquefação do
coágulo seminal, após diluição na fração B (diluição II) e após congelação-descongelação.

37
2.10. Análise estatística
Os resultados foram expressos como média ± EPM. As diferenças entre parâmetros
biométricos dos testículos direito e esquerdo e as comparações entre TES-TRIS e ACIN em
todos os estágios de ambos os experimentos foram analisados pelo teste t de Student. O efeito
dos diluidores no percentual de espermatozóides vivos, motilidade e vigor espermáticos foram
avaliados por ANOVA, e as diferenças foram estabelecidas pelo teste de Fisher. P< 0,05 foi
considerado como estatisticamente significante.
3. Resultados
Largura, altura, comprimento, circunferência e volume dos testículos foram 1,6 ± 0,1
cm, 1,5 ± 0,1 cm, 2,5 ± 0,1 cm, 6,0 ± 0,3 cm, e 2,0 ± 0,2 cm3 respectivamente, sem diferença
significativa entre os testículos direitos e esquerdos. O peso corporal médio dos animais antes
do período experimental foi 3,7 ± 0,5 kg, similar ao peso final (3,6 ± 0,4 kg).
O condicionamento dos animais antes da anestesia diminuiu o stress e aumentou a
duração do efeito anestésico de 37,4 ± 3,6 para 52,3 ± 6,3 minutos (P<0,05). A coleta de
sêmen foi bem sucedida em 19 de 21 tentativas. A ereção peniana iniciou em média na
primeira série de estímulos elétricos, sendo que a ejaculação ocorria freqüentemente a partir
da terceira série, após 18 minutos. As sessões de coleta duravam em média 40 minutos.
Não houve variação individual nem variação entre os animais com relação à qualidade
do sêmen. A fração líquida do sêmen era transparente e incolor, e a fração coagulada era
levemente esbranquiçada, gelatinosa e de difícil manipulação. O volume seminal médio foi
0,6 ± 0,2 mL. O coágulo seminal liquefez em 2,8 ± 1,1 e 4,5 ± 1,7 horas (P<0,05) em ACIN e
TES-TRIS, respectivamente. A concentração espermática após total liquefação do coágulo
foi em média 1806 ± 367 x 106 espermatozóides/mL e a osmolaridade 372 mOsm. Não houve
diferença significativa (P>0,05) entre os diluidores quanto ao percentual de espermatozóides
normais e anormais (anomalias primárias e secundárias) (Tabela 3).

38
Tabela 3: Morfologia espermática após diluição nas frações A de ACIN e TES-TRIS Morfologia espermática ACIN (%)
(Média ± EPM) TES-TRIS (%)
(Média ± EPM) Espermatozóides normais 78,2 ± 2,0 83,7 ± 1,0 Anomalias primárias
Cabeça 07,8 ± 0,5 03,2 ± 0,5 Peça intermediária 01,2 ± 0,8 02,9 ± 0,7 Cauda 03,0 ± 0,4 03,0 ± 0,2 Total 12,0 ± 0,6 09,1 ± 0,7 Anomalias secundárias
Cabeça 01,4 ± 0,7 01,5 ± 0,1 Peça intermediária 01,0 ± 0,4 04,0 ± 0,5 Cauda 08,2 ± 0,2 12,7 ± 0,3 Total 10,6 ± 0,5 6,06 ± 0,4
A motilidade foi significativamente maior (entre 10 e 70%) nas amostras diluídas na
fração A de TES-TRIS que naquelas diluídas em ACIN, porém, ela não foi recuperada após a
adição de cafeína em TES-TRIS (Tabela 4). Somente nas amostras diluídas em ACIN ocorreu
ativação após a adição de cafeína (6 e 10 mM). A motilidade espermática observada após
adição de 10 mM de cafeína foi significativamente maior que a motilidade inicial nas
amostras de ACIN sem cafeína. Além disso, foi verificado aumento do percentual de amostras
exibindo espermatozóides móveis durante 5 horas de avaliação. (Tabela 4).
A comparação dos percentuais de espermatozóides vivos nos quatro diferentes
momentos de análise é apresentada na Figura 1. Não foram observadas diferenças
significativas entre os diluidores após diluição I e liquefação. Somente após a diluição II e
descongelamento, observou-se uma taxa de viabilidade espermática significativamente menor
quando ACIN foi utilizado em comparação com TES-TRIS. Sendo verificada uma redução
significativa no percentual de espermatozóides vivos após essas etapas (diluição II e
descongelamento) em ambos diluidores. Após descongelamento, o percentual de
espermatozóides nas amostras em TES-TRIS e ACIN foi 26,2 e 13,2%, respectivamente.

39
Tabela 4: Motilidade e vigor imediatamente após liquefação do coágulo, e após exposição dos espermatozóides imóveis a ACIN e ao TES-TRIS, acrescidos de 6 ou 10 mM de cafeína. Motilidade (%) Vigor Duração da motilidade (h) Liquefação do coágulo ACIN TES-TRIS
22,0 ± 16,0a 38,0 ± 22,0 b
3,0 ± 01,0 a 3,0 ± 05,0 a
1,0 ± 05,0 a 5,0 ± 01,4 b
Adição de cafeína* ACIN 6 mM 10 mM TES-TRIS 6 mM 10 mM
22,5 ± 16,0 a 28,0 ± 19,0 a
0,0 ± 0,0 c 0,0 ± 0,0 c
2,0 ± 0,8 a 2,8 ± 1,1 a
0,0 ± 0,0 c 0,0 ± 0,0 c
3,4 ± 01,1 b 4,0 ± 01,2 b
NA NA
* Cafeína foi adicionada quando a motilidade chegava a zero. NA= não se aplica
Figura 1: Percentual de espermatozóides vivos após diluição I, liquefação, diluição II e descongelamento. Letras maiúsculas diferentes (A, B) indicam diferença estatística (P < 0,05) entre diluidores em cada estágio. Letras minúsculas diferentes (a, b) indicam diferença estatística (P < 0,05) entre os estágios.
4. Discussão
No presente trabalho foi possível liquefazer o coágulo seminal usando tanto TES-
TRIS quanto ACIN, sem prejudicar a viabilidade, motilidade e vigor espermáticos. Sendo
mais lenta a liquefação em TES-TRIS (4,5 horas) que em ACIN (2,8 horas). Geralmente a
liquefação seminal é efetuada na presença de enzimas não-específicas, como a tripsina (Nagle
e Denari, 1983), para dissolver o coágulo seminal ou plug copulatório, que é composto pela
Liquefação Diluição I Diluição II Descongelamento
ACIN
TES-TRIS
P
erc
en
tua
l d
e S
PT
Z v
ivo
s

40
semenogelina I e II (Valtonen-André et al., 2005). A água de coco contém uma variedade
grande de nutrientes, incluindo vitaminas, minerais, antioxidantes, aminoácidos, fatores de
crescimento e enzimas (Gopikrishna et al., 2008), que podem auxiliar a liquefazer o coágulo
rapidamente a 35 °C.
Quando a liquefação foi seguida pela criopreservação, TES-TRIS, e não ACIN, foi o
melhor diluidor para preservar a viabilidade espermática. Embora sua eficiência tenha sido
relatada em outras espécies animais, como canina (Cardoso et al., 2006) e suína (Toniolli et
al., 1996), informações relacionadas aos efeitos crioprotetores da ACIN no sêmen de primatas
ainda são inexistentes. Em contraste, TES-TRIS tem sido usado na criopreservação de sêmen
de outros primatas não-humanos resultando em alta taxa de sobrevivência pós-congelação
(Yeoman et al., 2005) e nascimentos após IA (Morrell, 1997).
No presente estudo os melhores resultados obtidos com TES-TRIS, após
criopreservação quando comparado com ACIN, pode ser explicado por três hipóteses.
Primeiro, pela concentração mais alta de glicerol (3,5% em TES-TRIS e 2,5% em ACIN);
segundo, a presença do leite no diluidor a base de TES-TRIS aumentou a osmolaridade da
solução, o que reduziu o risco de formação de cristais de gelo intracelular; e finalmente,
componentes desconhecidos na água de coco, como enzimas, podem ter sido tóxicos ou
prejudiciais à célula após criopreservação.
A adição de cafeína para ativar espermatozóides tem sido eficiente em primatas não-
humanos (Boatman e Bavister, 1984). Todavia, mesmo quando adicionado em ambas as
concentrações ao diluidor TES-TRIS, não foi observada a recuperação da motilidade.
Diferentemente, a adição de cafeína à ACIN, em ambas as concentrações resultou ma
recuperação da motilidade espermática. A cafeína é conhecida por aumentar os níveis de
cAMP intracelular (Cascieri et al., 1976), que pode induzir a motilidade espermática
(Harayama e Miyaki, 2006). A ACIN, por sua vez, é rica em ácido 3-indol-acético (AIA),
uma auxina capaz de substituir cAMP em bactérias entéricas, restaurando vias específicas
(Bianco et al., 2006). Por isso, sugerimos que exista um efeito sinérgico entre a cafeína e os
componentes da ACIN, envolvidos na recuperação da motilidade espermática.
Deste modo, a ACIN pode conter enzimas capazes de promover a rápida liquefação do
coágulo seminal do sêmen de C. apella, o que poderia ser útil para a inseminação artificial
com sêmen fresco. Além disso, a cafeína e os componentes da ACIN, provavelmente AIA,
podem agir sinergicamente no processo de ativação espermática. Porém, para a
criopreservação do sêmen de C. apella, o diluidor a base de TES-TRIS acrescido de 3,5% de

41
glicerol assegurou melhor qualidade seminal pós-congelação que ACIN mais 2,5% de
glicerol.
Referências Bibliográficas
Araújo, L.L., Oliveira, K.G., Lima, J.S., Araújo, J.B., Domingues, S.F.S., 2009. Avaliação do uso de solução à base de água de coco a 37 °C para diluição de sêmen de Cebus apella (macaco-prego) mantido em cativeiro. Ciência Animal Brasileira 10, 588-594. Barnabe, R.C., Guimarães, M.A.B.V., Oliveira, C., Barnabe, A.H., 2002. Analysis of some normal parameters of the spermiogram of captive capuchin monkeys (Cebus apella, Linnaeus, 1758). Braz. J. Vet. Res. Anim. Sci. 39, 331-333. Bianco, C., Imperlini, E., Calogero, R., Senatore, B., Pucci, P., Defez, R., 2006. Indole-3-acetic acid regulates the central metabolic pathways in Escherichia coli. Microbiology 152, 2421–2431. Boatman, D.E., Bavister, B.D., 1984. Stimulation of rhesus monkey sperm capacitation by cyclic nucleotide mediators. J. Reprod. Fertil. 71, 357–366. Cardoso, R.C.S., Silva, A.R., Silva, L.D., 2006. Comparison of two dilution rates on canine semen quality after cryopreservation in a coconut water extender. Anim. Reprod. Sci. 92, 384–391. Cascieri, M., Amann, R.P., Hammerstedt, R.H., 1976. Adenine nucleotide changes at initiation of bull sperm motility. J. Biol. Chem. 10, 787–793 Domingues, S.F., Caldas-Bussiere, M.C., Petretski, M.D., Ohashi, O.M., Lima, J.S., Santos, R.R., Cordeiro,M.S., Gomes de Castro, P.H., 2010. Effects of follicular phase and oocyte-cumulus complexes quality on the protein profile and in vitro oocyte meiosis competence in Cebus apella. Fertil. Steril. 15, 1662–1667. Dong, Q., Correa, L.M., VandeVoort, C.A., 2009. Rhesus monkey sperm cryopreservation with TEST-yolk extender in the absence of permeable cryoprotectant. Cryobiology 58, 20–27. Feradis, A.H., Pawitri, D., Suatha, I.K., Amin, M.R., Yusuf, T.L., Sajuthi, D., Budiarsa, I.N., Hayes, E.S., 2001. Cryopreservation of epididymal spermatozoa collected by needle biopsy from cynomolgus monkeys (Macaca fascicularis). J. Med. Primatol. 30, 100–106. Fickel, J., Wagener, A., Ludwig, A., 2007. Semen cryopreservation and the conservation of endangered species. Eur. J. Wildlife Res. 53, 81-89. Gopikrishna, V., Baweja, P.S., Venkateshbabu, N., Thomas, T., Kandaswamy, D., 2008. Comparison of coconut water, propolis, HBSS, and milk on PDL cell survival. J. Endod. 34, 587–589. Harayama, H., Miyaki, M., 2006. A cyclic adenosine 3’,5’-monophosphate dependent protein kinase C activation is involved in the hyperactivation of boar spermatozoa. Mol. Reprod. Dev. 73, 1169–1178.

42
Hernández-López, L., Cerezo-Parra, G., Cerda-Molina, A.L., Pérez-Bolanõs, S.C., Mondragón-Ceballos, R., 2002. Digestion by trypsin enhances assessment of sperm parameters in the black-handed spider monkey (Ateles geoffroyi). Lab. Primate Newsl. 41, 4–6. Hernández-López, L., Cerda-Molina, A.L., Páez-Ponce, D., Mondragón-Ceballos, R., 2008. The seminal coagulum favours passage of fast-moving sperm into the uterus in the black-handed spider monkey. Reproduction 136, 411–421. Kraemer, D.C., Vera Cruz, N.C., 1969. Collection, gross characteristics and freezing of baboon semen. J. Reprod. Fertil. 20, 345–348. Kusunoki, H., Daimaru, H., Minami, S., Nishimoto, S., Yamane, K., Fukumoto, Y., 2001. Birth of a Chimpanzee (Pan troglodytes) after artificial insemination with cryopreserved epididymal spermatozoa collected postmortem. Zoo. Biol. 20, 135–143. Li, Y.H., Cai, K.J., Kovacs, A., Ji, W.Z., 2005. Effects of various extenders and permeating cryoprotectants on cryopreservation of cynomolgus monkey (Macaca fascicularis) spermatozoa. J. Androl. 26, 387–395. Mahony, M.C., Lanzendorf, S., Gordon, K., Hodgen, G.D., 1996. Effect of caffeine and dbcAMP on zona pellucida interactions by epididymal spermatozoa of cynomolgus monkeys (Macaca fascicularis). Mol. Reprod. Dev. 43, 530-535. Morrell, J.M., 1997. Cryopreservation of marmoset sperm (Callithrix jacchus). Cryo-letters
18, 45-54. Nagle, C.A., Denari, J.H., 1983. The Cebus monkey (Cebus apella). In: Hearn, J.P. (Ed.), Reproduction of New World Primates. Lancaster, pp. 281–304. O’Brien, J.K., Hollinshead, F.K., Evans, K.M., Evans, G., Maxwell, W.M., 2003. Flow cytometric sorting of frozen-thawed spermatozoa in sheep and non-human primates. Reprod. Fertil. Dev. 15, 367–375. Oliveira, K.G., Castro, P.H., Muniz, J.A., Domingues, S.F., 2010. Semen conservation and seminal clot liquefaction of capuchin monkey (Cebus apella) in powdered coconut water extender (PCW) at different temperatures. Ciência Rural 40, 1–15. Paz, R.C., Zacariotti, R.L., Teixeira, R.H., Guimarães, M.A., 2006. The effect of hyaluronidase and trypsin on the liquefaction of capuchin monkeys semen. Braz. J. Vet. Res. Anim. Sci. 43, 196–201. Roussel, J.D., Austin, C.R., 1967. Preservation of primate spermatozoa by freezing. J. Reprod. Fertil. 13, 333–335. Sanchez-Partida, L.G., Maginnis, G., Dominko, T., Martinovich, C., McVay, B., Fanton, J., Schatten, G., 2000. Live rhesus offspring by artificial insemination using fresh sperm and cryopreserved sperm. Biol. Reprod. 63, 1092–1097.

43
Sankai, T., Shimizu, K., Cho, F., Yoshikawa, Y., 1997. In vitro fertilization of follicular oocytes by frozen-thawed spermatozoa in Japanese monkeys (Macaca fuscata). Lab. Anim. Sci. 47, 58–62. Tauber, P.F., Zaneveld, L.J., Propping, D., Schumacher, G.F., 1980. A new technique to measure the liquefaction rate of human semen: the bag method. Fertil. Steril. 33, 567–570. Tollner, T.L., VandeVoort, C.A., Overstreet, J.W., Drobnis, E.Z., 1990. Cryopreservation of spermatozoa from cynomolgus monkeys (Macaca fascicularis). J. Reprod. Fertil. 90, 347–352. Toniolli, R., Bussière, J., Courot, M., Magistrini, M., Combarnous, Y., 1996. Effect of indole-3-acetic acid (plant auxin) on the preservation at 15 °C of boar semen for artificial insemination. Reprod. Nutr. Dev. 36, 503–511. Valtonen-André, C., Olsson, A.Y., Nayudu, P.L., Lundwall, A., 2005. Ejaculates from the common marmoset (Callithrix jacchus) contain semenogelin and beta-microseminoprotein but not prostate-specific antigen. Mol. Reprod. Dev. 71, 247–255. Vidal, F.D., Luz, M.S., Pinho, T.G., Pissinatti, A., 2007. Coleta de sêmen em mico-leão-de-cara-dourada (Leontopithecus chrysomelas) (Kuhl, 1820) através da eletroejaculação Callitrichidae - primates. Rev. Bras. Ci. Vet. 14, 67-71. Yeoman, R.R., Mitalipov, S., Gerami-Naini, B., Nusser, K.D., Wolf, D.P., 2005. Low temperature storage of rhesus monkey spermatozoa and fertility evaluation by intracytoplasmic injection. Theriogenology 63, 2356–2371.

44
5. CONCLUSÃO GERAL
O diluidor a base de ACIN pode ser usado para desfazer rapidamente o coágulo
seminal de C. apella. Entretanto, a motilidade espermática é mantida por mais tempo em
ACIN acrescida de cafeína e TES-TRIS. Para a criopreservação do sêmen, TES-TRIS a 3,5%
de glicerol é mais indicado que ACIN a 2,5% de glicerol.
Mais estudos in vitro e, principalmente, in vivo, são necessários para confirmar a
viabilidade do sêmen descongelado de C. apella. Contudo, as técnicas desenvolvidas nesse
estudo podem contribuir para a formação de bancos de genoma de primatas não-humanos
ameaçados de extinção.

45
REFERÊNCIAS
ABBOTT, D.H. et al. Nonhuman Primates Contribute Unique Understanding to Anovulatory Infertility in Women. ILAR Journal, v. 45, n. 2, p. 116-131, 2004. ABEE, C.R. Alternative New World Primate Models for Non-AIDS Research. ILAR
Journal, v. 44, p. 231-235, 2003. AMBOKA, J.N.O.; MWETHERA, P.G. Characterization of semen from olive baboons. Journal of Medical Primatology, v. 32, n. 6, p. 325-329, 2003. ANDRADE, E.R. et al. Evaluation of saline and coconut water solutions in the preservation of ovine preantral follicles in situ. Small Ruminant Research, v. 43, p. 235-243, 2002. ARAÚJO, L.L. et al. Avaliação do uso de solução à base de água de coco a 37 °C para diluição de sêmen de Cebus apella (macaco-prego) mantido em cativeiro. Ciência Animal Brasileira, v. 10, p. 588-594, 2009. ARAÚJO, L. L. et al. Biometria testicular e efeito de diluidor à base de água de coco na conservação a 37 ºC de sêmen de Cebus apella (macaco-prego). In: REUNIÃO ANUAL DA SBPC, 59, 2007. Belém-PA. Resumos... Belém, 2007. AURICCHIO, P. Primatas do Brasil. São Paulo: Terra Brasilis, 1995. AURICH, J.E.; KUHNE, A.; HOPPE, H. Seminal plasma effect of milk fractions on survival of equine spermatozoa. Theriogenology, v. 46, p. 791-797, 1996. BARNABE, R.C. et al. Analysis of some normal parameters of the spermiogram of captive capuchin monkeys (Cebus apella, Linnaeus, 1758). Brazilian Journal of Veterinary Research and Animal Science, v. 39, n. 6, p. 331-333, 2002. BATES, R. G. Determination of pH: Theory and Practice. 2. ed. New York: John Wiley and Sons Inc., 1962, 161p. BIANCO, C. et al. Indole-3-acetic acid regulates the central metabolic pathways in
Escherichia coli. Microbiology, v.152, p. 2421–2431, 2006. BLUME, H.; MARQUES JR., A.P. Avaliação da água de coco no cultivo e criopreservação

46
de embriões murídeos. Revista Brasileira de Reprodução Animal, v. 18, p. 97-104, 1994. BLUME, H. et al. Avaliação da água de coco na maturação de oócitos bovinos. Revista Brasileira de Reprodução Animal, v. 21, p. 72-75, 1997a. BLUME, H. et al. Uso da água de coco no cultivo de embriões bovinos. Revista Brasileira de Reprodução Animal, v. 21, p. 78-81, 1997b. BOATMAN, D.E.; BAVISTER, B.D. Stimulation of rhesus monkey sperm capacitation by cyclic nucleotide mediators. Journal of Reproduction and Fertility, v.71, p. 357–366, 1984. BUNGE, R.G. Caffeine stimulation for borderline male infertility. Fertility and Sterility, v. 51, p. 863-868, 1973. BUSH, D.F. et al. Semen evaluation in capuchin monkeys (Cebus apella). Laboratory Animal Science, v. 25, n. 5, p. 588-593, 1975. CARDOSO, R.C.S.; SILVA, A.R.; SILVA, L.D.M. Comparison of two dilution rates on canine semen quality after cryopreservation in a coconut water extender. Animal Reproduction Science, v. 92, p. 384-391, 2006. CARDOSO, R. C. S.; SILVA, A.R.; SILVA, L.D.M. Use of the alternative extender powder coconut water (PCW 106®) for canine semen freezing. In: INTERNATIONAL SYMPOSIUM ON CANINE AND FELINE REPRODUCTION, 5., 2004, Embu das Artes-SP. Anais... Embu das Artes, 2004. p 96-97. CARDOSO, R.C.S. et al. Cryopreservation of canine semen using a coconut water extender with egg yolk and three different glycerol concentrations. Theriogenology, v. 59, p. 743-751, 2003. CASCIERI, M.; AMANN, R.P.; HAMMERSTEDT, R.H. Adenine nucleotide changes at initiation of bull sperm motility. The Journal of Biological Chemistry, v.10, p. 787–793, 1976. CHEN, J.C. et al. Semen cryopreservation in the Tibetan macaque (Macaca thibetana)-comparison of different cooling programs and freezing media. Acta Zoologica Sinica., v. 40, p. 174-181, 1994.

47
CHEN, Z.J. et al. Effects of sucrose concentration on the developmental potential of human frozen-thawed oocytes at different stages of maturity. Human Reproduction, v. 19, p. 2345-2349, 2004. CHO, F.; HONJO, S. A simplified method for collecting and preserving cynomolgus macaque semen. Japanese Journal of Medical Science and Biology. v. 26, p. 261-268, 1973. CLAVERT, A. et al. A method for sperm cryopreservation of the lemur fulvus. Primate Reproduction, v. 14, p. 101, 1986. CONCANNON, P. W.; BATTISTA, M. Canine semen freezing and artificial insemination. In: KIRK, R.W. (Ed.). Current Veterinary Therapy. Philadelphia: W.B. Saunders, 1989, p. 1247-1259. CONRADIE, E.; OETTLE, E.E.; SEIER, J.V. Assessment of acrosomal integrity of vervet monkey spermatozoa after cryopreservation. Journal of Medical Primatology, v. 23, p. 315-316, 1994. CORDEIRO, M.S. et al. The use of coconut water solution (Cocos nucifera) as a holding medium for immature bovine oocytes for in vitro embryo production. Animal Reproduction, v. 3, n. 3, p. 376-379, 2006. CRUZ, J.F. Conservação e fertilidade do sêmen ovino mantido á temperatura de +4 ºC por um período de 48 horas diluído em frações ativas da água de coco. 1994. Dissertação (Mestrado em Produção e Reprodução de Pequenos Ruminantes) - Universidade Estadual do Ceará, 1994. DENIS, L.T. et al. Freeze preservation of squirrel monkey sperm for use in timed fertilization studies. Fertility and Sterility, v.27, p.723-729, 1976. DIXSON, A.F.; ANDERSON, M.J. Sexual selection, seminal coagulation and copulatory plug formation in primates. Folia Primatologica, v.73, p.63-69, 2002. DOMINGUES, S.F.S.; CALDAS-BUSSIERE, M.C. Fisiologia e biotécnicas da reprodução desenvolvidas em fêmeas de Primatas Neotropicais importantes para a pesquisa biomédica. Revista Brasileira de Reprodução Animal, v.30, n.1/2, p. 57-71, 2006. DOMINGUES, S.F.S. et al. Effects of follicular phase and oocyte-cumulus complexes quality on the protein profile and in vitro oocyte meiosis competence in Cebus apella. Fertility and Sterility, v.15, p. 1662-1667, 2010.

48
DONG, Q.; CORREA, L.M.; VANDEVOORT, C.A. Rhesus monkey sperm cryopreservation with TEST-yolk extender in the absence of permeable cryoprotectant. Cryobiology, v. 58, p. 20-27, 2009. EMMONS, L. H. Neotropical Rainforest Mammals. Chicago and London: The University of Chicago Press, 1990. ENGLAND, G.C.W. Cryopreservation of dog semen: a review. Journal of Reproduction and Fertility, v.47, p. 243-255, 1993. FERADIS, A.H. et al. Cryopreservation of epididymal spermatozoa collected by needle biopsy from cynomolgus monkeys (Macaca fascicularis). Journal of Medical Primatology, v.30, p. 100-106, 2001. FICKEL, J.; WAGENER, A.; LUDWIG, A. Semen cryopreservation and the conservation of endangered species. European Journal of Wildlife Research, v.53, p. 81-89, 2007. FRAGASZY, D.M.; ADAMS-CURTIS, L. Growth and reproduction in captive tufted capucins (Cebus apella). American Journal of Primatology, v.44, p.197-203, 1998. FRAGASZY, D. M. et al. The complete capuchin: the biology of the genus Cebus. Massachusetts: Cambridge University Press, 2004. GOODMAN, M. Epilogue: a personal account of the origins of a new paradigm. Molecular and Phylogenetic Evolution, v.5, p. 269-285, 1996. GOPIKRISHNA, V. et al. Comparison of coconut water, propolis, HBSS, and milk on PDL cell survival. Journal of Endodontics, v.34, p. 587-589, 2008. GOULD, K.G.; MARTIN, D.E.; WARNER, H. Improved Methods for Artificial Insemination in the Great Apes. American Journal of Primatology, v.8, p. 61-67, 1985. HALSTEAD, L.S.; VERVOORT, S.; SEAGER, S.W.J. Rectal probe electrostimulation in the treatment of anejaculatory spinal cord injured men. Paraplegia, v.25, p.120-129, 1987. HAMMERSTEDT, R. H.; GRAHAM, J. K.; NOLAN, J. P. Cryopreservation of mammalian sperm: what we ask them to survive. Journal of Andrology, v.11, p.73-88, 1990.

49
HARAYAMA, H.; MIYAKI, M. A cyclic adenosine 3’,5’-monophosphate dependent protein kinase C activation is involved in the hyperactivation of boar spermatozoa. Molecular reproduction and development, v.73, p. 1169-1178, 2006. HERNÁNDEZ-LÓPEZ, L.; CERDA-MOLINA, A.L.; MONDRAGÓN-CEBALLOS, R. Seasonal emission of seminal coagulum and in vivo sperm dynamics in the Black-handed Spider Monkey (Ateles geoffroyi). Theriogenology, v.69, p. 466-472, 2008. HERNÁNDEZ-LÓPEZ, L. et al. The seminal coagulum favours passage of fast-moving sperm into the uterus in the black-handed spider monkey. Reproduction, v.136, p. 411-421, 2008. HERNÁNDEZ-LÓPEZ, L. et al. Digestion by trypsin enhances assessment of sperm parameters in the black-handed spider monkey (Ateles geoffroyi). Laboratory Primate Newsletter, v.41, p. 4-6, 2002. HOLT, W.V. Basic aspects of frozen storage of semen. Animal of Reproduction Science, v.62, p.3-22, 2000. IBAMA. Portaria nº 1.522, 19 dez. 1.989, Diário Oficial da União, 23 fev. 1989. IUCN. Disponível em. <http://cms.iucn.org>. Acesso em: 16 maio 2008. KRAEMER, D.C.; VERA CRUZ, N.C. Collection, gross characteristics and freezing of baboon semen. Journal of Reproduction and Fertility, v.20, p.345-348, 1969. KUEDERLING, I.; MORREL, J.M.; NAYUDU, P.L. Collection of semen from marmoset monkeys (Callithrix jacchus) for experimental use by vaginal washing. Laboratory Animals, v.30, p.260-266, 1996. KUEDERLING, I. et al. Non-invasive collection of ejaculates from the common marmoset (Callithrix jacchus) using penile vibrostimulation. American Journal of Primatology, v.52, n.3, p.149-154, 2000. KUSUNOKI, H. et al. Birth of a Chimpanzee (Pan troglodytes) after artificial insemination with cryopreserved epididymal spermatozoa collected postmortem. Zoo Biology, v.20, p. 135-143, 2001.

50
LAMBERT, H. et al. Penetration of zona-free hamster oocytes by ejaculated cryopreserved gorilla spermatozoa. Fertility and Sterility, v.56, p.1201-1203, 1991. LANG, C.M. A Technique for the Collection of Semen from Squirrel Monkeys (Saimiri
Sciureus) by Electro-ejaculation. Laboratory Animal Care, v.17, n.2, p. 218-221, 1967. LANZENDORF, S.E. et al. In vitro fertilization and gamete micromanipulation in the lowland gorilla. Journal of Assisted Reproduction and Genetic, v.9, p.358–364, 1992. LEÃO, Danuza Leite et al. Quantitative analysis of spermatogenesis in capuchin monkey (Cebus apella). In: CONGRESSO BRASILEIRO DE ANATOMIA, 23. CONGRESSO LUSO BRASILEIRO DE ANATOMIA, 11. CONGRESSO DO CONE SUL, 9. CONGRESSO CHILENO DE ANATOMIA, 28., 2008, Belém - PA. Anais... Belém, 2008. LI, X.; GAO, D.; JI, W. Cryopreservation of sperm of an endangered species-Assamese macaque. Cell Preservation Technology, v.2, p.29-33, 2004. LI, Y. et al. Effect of Amino Acids on Cryopreservation of Cynomolgus Monkey (Macaca
fascicularis) Sperm. American Journal of Primatology, v.59, p.159-165, 2003. LI, Y.H. et al. Cryopreservation of cynomolgus monkey (Macaca fascicularis) spermatozoa in a chemical defined extender. Asian Journal of Andrology, v.7, n.2, p.139-144, 2005b. LI, Y.H. et al. Effects of various extenders and permeating cryoprotectants on cryopreservation of Cynomolgus monkey (Macaca fascicularis) Spermatozoa. Journal of Andrology, v.26, n.3, p.387-395, 2005a. LOVELOCK, J.E.; POLGE, C. The immobilization of spermatozoa by freezing and thawing and the protective action of glycerol. Byochemical Journal, v.58, p.318-322, 1954. MAHONE, J.P.; DUKELOW, W.R. Semen Preservation in Macaca fascicularis. Laboratory of Animal Science, v.28, n.5, p.556-561, 1978. MAHONY, M.C. et al. Effect of caffeine and dbcAMP on zona pellucida interactions by epididymal spermatozoa of cynomolgus monkeys (Macaca fascicularis). Molecular Reproduction and Development, v.43, p.530-535, 1996. MCKINNON, A.O. Artificial insemination of cooled, transported and frozen semen. Australian Equine Veterinary Journal, v.14, p.156-175, 1996.

51
MCPHAIL, D.B.; GOODMAN, B.A. TRIS buffer – a case for caution in its use for cooper containing systems. Biochemical Journal, v.221, p.559-560, 1984. MIES FILHO, A. Reprodução dos animais domésticos e inseminação artificial. Porto Alegre: Sulina, 1987. MONTEZUMA JÚNIOR, P. A.; VIANA NETO, R; NUNES, J.F. Utilização da água de coco “in natura”, com adição de água gema de ovo, como diluente de congelação do sêmen canino, em paillets de 0,5ml. In: CONGRESSO BRASILEIRO DE MEDICINA VETERINÁRIA, 23., 1994, Olinda-PE. Anais... Olinda, 1994. p 535. MORELAND, R.B. et al. Characterizing the reproductive physiology of the male Southern Black Howler Monkey, Alouatta caraya. Journal of Andrology, v.22, n.3, p.395-403, 2001. MORRELL, J.M. Cryopreservation of marmoset sperm (Callithrix jacchus). Cryo-letters, v.18, p. 45-54, 1997. MORRELL, J.M.; HODGES, J.K. Cryopreservation of non-human primate sperm: priorities for future research. Animal Reproduction Science, v.53, p.43-63, 1998. MORRELL, J.M. et al. Birth of offspring following artificial insemination in the common marmoset, Callithrix jacchus. American Journal of Primatology, v.41, n.1, p.37-43, 1997. MORRELL, J.M. et al. Artificial insemination in Callithrix jacchus using fresh or cryopreserved sperm. Animal Reproduction Science, v.52, p.165–174, 1998. NAGLE, C. A.; DENARI, J.H. The Cebus Monkey (Cebus apella). In: HEARN, John P. Reproduction of new world primates. Lancaster: MTP Press, 1983, p. 39-67. NAGLE, C.A. et al. The plasma pattern of ovarian steroids during the menstrual cycle in capuchin monkeys (Cebus apella). Biology of Reproduction, v.21, p.979-983, 1979. NAPIER, J R; NAPIER, P. H.. Structure and Function. In:________.The Natural history of the primates. Cambridge: The MITT Press, 1986. p. 30-59. NOWAK, R. M. Primates. In: ________. Walker’s mammals of the world. Baltimore: The Johns Hopkins University Press, 1991. p.400-514.

52
NUNES, J. F. A inseminação artificial como método alternativo para o melhoramento da caprinocultura leiteira. In: SIMPÓSIO DE CAPRINOCULTURA DO ESTADO DO RIO, 1986, Niterói-RJ. Anais... Niterói, 1986. NUNES, J.F. Utilização da água de coco como diluidor do sêmen de caprinos e ovinos. Ciência Animal, v.7, p.62-69, 1995. NUNES, J.F. Utilização da água de coco como diluidor do sêmen de animais domésticos e do homem. Revista Brasileira de Reprodução Animal, v.22, p.109-112, 1998. NUNES, J.F; COMBARNOUS, Y. Utilização da água de coco e suas frações ativas como diluidor de sêmen dos mamíferos domésticos. Ciência Animal, v.5, p.15-21, 1995. O’BRIEN, J.K. et al. Flow cytometric sorting of frozen-thawed spermatozoa in sheep and non-human primates. Reproduction, Fertility and Development, v.15, p. 367-375, 2003. OHL, D.A.; MCCABE, M.; SØNKSEN, J. Management of infertility in spinal cord injury. Topics Spinal Cord Injury Rehabilitation, v.1, p. 65-75, 1996. OHL, D.A.; SØNKSEN, J.; BOLLING, R. New stimulation pattern for electroejaculation based on physiological studies. Journal of Urology, v.163 (Suppl.), p.15-22, 2000. OLIVEIRA, K.G. et al. Conservação do sêmen e liquefação do coágulo seminal de macaco-prego (Cebus apella) em água de coco em pó (ACP-118®), em diferentes temperaturas. Ciência Rural, v.40, n.3, p. 617-621, 2010.
PAZ, R.C.R. et al. O efeito das enzimas hialuronidase e tripsina na liquefação do sêmen de macacos pregos (Cebus apella). Brazilian Journal Veterinary Animal Science, v.43, p.196-201, 2006. RODRIGUES, B. A. Efeito do diluidor à base de albumina sérica bovina (BSA) sobre a viabilidade in vitro do sêmen canino criopreservado. 1997. 176 f. Dissertação (Mestrado em Medicina Veterinária) - Universidade Federal do Rio Grande do Sul, 1997 RODRIGUES, J. L. Aspectos da congelação de embriões bovinos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE TRANSFERÊNCIA DE EMBRIÕES, 1992, Jaboticabal-SP. Anais… Jaboticabal, 1992. p 55-79.

53
ROTA, A. Studies on Preservation capacitation and fertility of dog spermatozoa. 1998. Tese (Doutorado) - Swedish University of Agricultural Sciences, 1998. ROUSSEL, J.D.; AUSTIN, C.R. Preservation of primate spermatozoa by freezing. Journal of Reproduction and Fertility, v.13, p.333-335, 1967. ROWE, N. The pictorial guide to the living primates. East Hampton: Pogonias Press, 1996. 263p. SADLEIR, R.M.F.S. The preservation of mammalian spermatozoa by freezing. Laboratory Practice, v.15, p.413, 1966. SALLES, M.G.F. Água de coco (Cocus nucifera L.) ‘’in natura’’ e sob a forma de gel e estabilizadas como diluidor de sêmen caprino. 1989. Dissertação (Mestrado em Medicina Veterinária) - Universidade Federal do Rio Grande do Sul, 1989. SANCHEZ-PARTIDA, L.G. et al. Live rhesus offspring by artificial insemination using fresh sperm and cryopreserved sperm. Biology Reproduction, v.63, p. 1092–1097, 2000. SANKAI, T. et al. Cryopreservation of spermatozoa from Cynomolgus monkeys (Macaca
fascicularis). Journal of Reproduction and Fertility, n.101, p.273-278, 1994. SANKAI, T. et al. In vitro fertilization of follicular oocytes by frozen-thawed spermatozoa in Japanese monkeys (Macaca fuscata). Laboratory Animal Science, v.47, n.1, p.58-62, 1997. SCHNEIDERS, A.; SONKSEN, J.; HODGES, J.K. Penile vibratory stimulation in the marmoset monkey: a practical alternative to electro-ejaculation, yielding ejaculates of enhanced quality. Journal of Medical Primatology, v.33, n.2, p.98-104, 2004. SEAGER, S.W.J. et al. Semen collection and evaluation in Gorilla gorilla gorilla. American Journal of Primatology, v.3, n.1, p.13, 1982. SEIER, J.V. et al. Cryopreservation of vervet monkey semen and recovery of progressively motile spermatozoa. Journal of Medical Primatology, v.22, p.355-359, 1993. SI, W. et al. Cryopreservation of rhesus macaque (Macaca mulatta) spermatozoa and their functional assessment by in vitro fertilization. Cryobiology, v.41, p.232-240, 2000.

54
SILVA, A.R. Criopreservação do sêmen canino diluído em Tris: Avaliação morfológica, funcional e de suas interações com oócitos homólogos. 2005a. 165 f. Tese (Doutorado em Ciências Veterinárias) – Universidade Estadual do Ceará, Faculdade de Veterinária, 2005a. SILVA, A.R. et al. Quality of canine semen submitted to single or fractionated glycerol addition during the freezing process. Theriogenology, v.59, p.821-829, 2003. SILVA, J.R.V. et al. Effect of coconut water and Braun–Collins solutions at different temperatures and incubation times on the morphology of caprine preantral follicles preserved in situ. Theriogenology, v. 54, p. 809–822, 2000. SILVA, K.S.M. Avaliação andrológica e criopreservação de sêmen de Ateles (macaco-aranha) mantidos em cativeiro. 2005b. 83 f. Dissertação (Mestrado em Ciência Animal) - Universidade Federal do Pará, 2005b. SØNKSEN, J; OHL, D. Penile vibratory stimulation and electroejaculation in the treatment of ejaculatory dysfunction. International Journal of Andrology, v.25, p.324-332, 2002. SØNKSEN, J.; OHL, D.A.; WEDEMEYER, G. Sphincteric events during penile vibratory ejaculation and electroejaculation in men with spinal cord injuries. Journal of Urology, v.165, p. 426-429, 2001. TAUBER, P.F. et al. A new technique to measure the liquefaction rate of human semen: the bag method. Fertility and Sterility, v.33, p. 567-570, 1980. TEIXEIRA, D.G. Estudo anatômico descritivo dos órgãos genitais masculinos do macaco-prego (Cebus apella Linnaeus, 1758). 2005. 193 f. Tese (Doutorado em Ciências) - Universidade de São Paulo, 2005. THOMSON, J.A. et al. Collection and fertilization potential of sperm from the sulawesi crested black macaque (Macaca nigra). American Journal of Primatology, v.28, n.4, p.289-297, 1992. TOLLNER, T.L. et al. Cryopreservation of Spermatozoa from cynomolgus monkeys (Macaca
fascicularis). Journal of Reproduction and Fertility, v.90, p.347-352, 1990. TONIOLLI, R. Estudo das características “in vitro” do sêmen caprino de raças nativas do nordeste brasileiro diluído em água de coco sob a forma ‘’in natura’’, estabilizada e em gel. Revista Brasileira de Reprodução Animal, v.13, p.209-220, 1989a.

55
TONIOLLI, Ricardo. Conservação do sêmen suíno em água de coco. In: CONGRESSO BRASILEIRO DE REPRODUÇÃO ANIMAL, 8, 1989b, Belo Horizonte-MG. Anais... Belo Horizonte, 1989b. p 138-142. TONIOLLI, R. et al. Effect of indole-3-acetic acid (plant auxin) on the preservation at 15 °C of boar semen for artificial insemination. Reproduction, Nutrition Development, v.36, p. 503-511, 1996. VALE, W.G. et al. Inseminação artificial em búfalos com sêmen congelado em diluente a base de água de coco (Cocos nucifera). Revista Brasileira de Reprodução Animal, v.23, p.354-356, 1999. VALLE, R.R. Características físicas e morfológicas do sêmen de Alouatta caraya
(Humboldt, 1812) mantidos em cativeiro. 2002. 66 f. Dissertação (Mestrado em Reprodução Animal) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, 2002. VALLE, R.R. et al. Collection and evaluation of semen from captive howler monkeys (Alouatta caraya). Theriogenology, v.62, p.131-138, 2004. VALTONEN-ANDRÉ, C. et al. Ejaculates from the common marmoset (Callithrix jacchus) contain semenogelin and beta-microseminoprotein but not prostate-specific antigen. Molecular Reproduction and Development, v.71, p. 247-255, 2005. VANDEVOORT, C.A. High quality sperm for nonhuman primate ART: production and assessment. Reproductive Biology and Endocrinology, v.2, p.33, 2004. VIDAL, F.D. et al. Coleta de sêmen em mico-leão-de-cara-dourada (Leontopithecus
chrysomelas) (Kuhl, 1820) através da eletroejaculação Callitrichidae-primates. Revista Brasileira de Ciência Veterinária, v.14, p. 67-71, 2007. WATSON, P.F. The causes of reduced fertility with cryopreserved semen. Animal Reproduction Science, v.60- 61, p.481-492, 2000. WATSON, P.F. The preservation of semen in mammals. Oxford Reviews of Reproductive Biology, v.1, p.183-350, 1979. WILSON, D.E.; REEDER, D.A.M. Mammal species of the world: A taxonomic and geographic reference. 3. ed. Baltimore: Johns Hopkins University Press, 2005.

56
WOLF, D.P. et al. Use of assisted reproductive technologies in the propagation of Rhesus macaque offspring. Biology of Reproduction, v.71, p.486-493, 2004. WRIGHT, E.M.; BUSH, D.E. The reproductive cycle of the Capuchin (Cebus apella). Laboratory of Animal Science, v.27, p.651-654, 1977. YEOMAN, R.R. et al. Low temperature storage of rhesus monkey spermatozoa and fertility evaluation by intracytoplasmic injection. Theriogenology, v.63, p. 2356-2371, 2005. YEOMAN, R.R. et al. Penile vibratory stimulation yields increased spermatozoa and accessory gland production compared with rectal electroejaculation in a neurologically intact primate (Saimiri boliviensis). Human Reproduction, v.13, p.2527-2531, 1998. YOUNG, L.G. et al. Sodium, potassium and protein concentrations and 2D-SDS-PAGE of epididymal luminal and ejaculated seminal fluids of the adult chimpanzee (Pan troglodytes). American Journal of Primatology, v.34, p.249-259, 1994. YOUNG, L.G.; SMITHWICK, E.B.; GOULD, K.G. Characteristics of Chipanzee (Pan
troglodytes) ejaculates collected by rectal probe electrostimulation and by artificial vagina. American Journal of Primatology, v.35, n.4, p.293-304, 1995.