Embed Size (px)
DESCRIPTION
Testes bioquímicas utilizados na identificação bacteriana.
Citation preview

BACTERIOLOGIA DE PEIXES
Identificação bacteriana com base em métodos bioquímicos, serológicos e moleculares
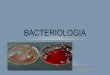
Cultura pura
Sub-culturas

IDENTIFICAÇÃO BACTERIANA
Existem vários métodos de identificação bacteriana:
Métodos bioquímicos:
Métodos clássicos;
Sistemas multiprova.
Métodos serológicos;
Métodos moleculares.

IDENTIFICAÇÃO BIOQUÍMICA TRADICIONAL

IDENTIFICAÇÃO BACTERIANA
A utilização de métodos de identificação mais avançados, exige quase sempre uma identificação bioquímica prévia;
A identificação bacteriana clássica inicia-se com:
Reacção de Gram, baseada na estrutura da parede celular;
Morfologia e mobilidade;
Oxidase;
Metabolismo oxidativo/fermentativo;
Descarboxilação de aminoácidos;

IDENTIFICAÇÃO BIOQUÍMICA
São vários os testes bioquímicos utilizados na identificação bacteriana:
Catalase;
Produção de ácido e/ou gás a partir de vários açúcares;
Redução de nitratos;
Hidrólise da ureia (produção da enzima urease);
Utilização de citrato;
Hidrólise da gelatina;
Teste do Vermelho de metilo (MR);
Teste de Voges-Proskauer (VP);

IDENTIFICAÇÃO BIOQUÍMICA
Testes bioquímicos:
Produção de indol;
Produção de sulfureto de hidrogénio (H2S);
β - Galactosidase (ONPG);
Crescimento a várias concentrações de sal (NaCl);
Crescimento a várias temperaturas;
Sensibilidade a antibióticos.

TESTES BIOQUÍMICOS

GRAM
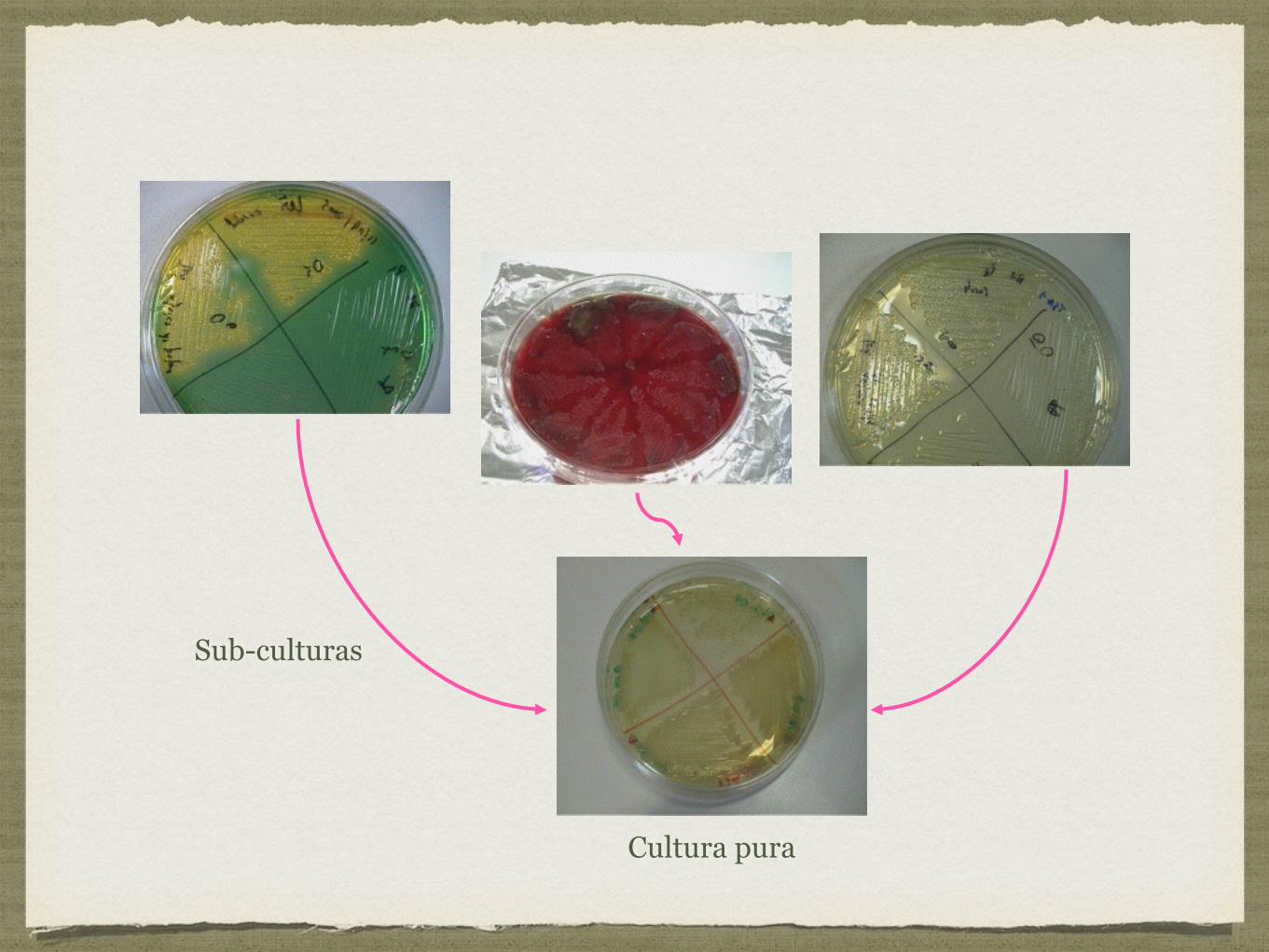
Teste de Gram:
Primeiro teste a ser efectuado quando se pretende identificar uma bactéria;
Permite dividir as bactérias em 2 grandes grupos: Gram positivas e Gram negativas;
Este teste baseia-se na constituição da parede bacteriana.

COMPOSIÇÃO DA PAREDE BACTERIANA
Constituição da parede de uma bactéria Gram-positiva

Constituição da parede de uma bactéria Gram-negativa

MÉTODOS
Pode ser realizado por dois métodos distintos:
Coloração de Gram tradicional;
Método de Ryu (1938), em que se utiliza uma solução de KOH a 3%.

COLORAÇÃO DE GRAM
Na coloração de Gram, realizam-se em 4 passos:
Cobrir o esfregaço com uma solução de violeta de cristal durante 30 segundos;
Lavar com água e aplicar uma solução de lugol durante 30 segundos;
Lavar com água e aplicar uma solução de acetona;
Lavar com água e aplicar uma solução de safranina durante 30 segundos e voltar a lavar com água.

COLORAÇÃO DE GRAM
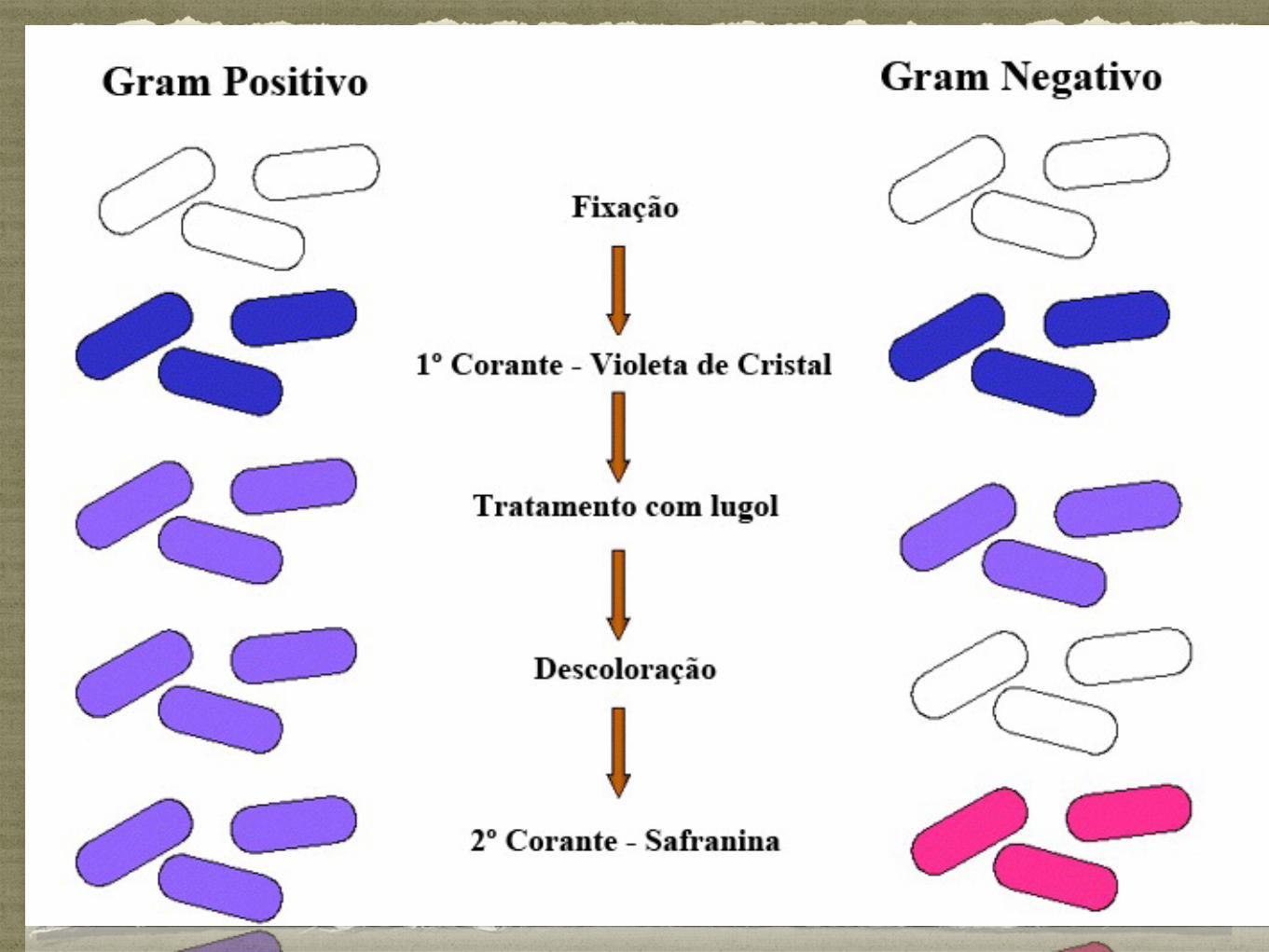
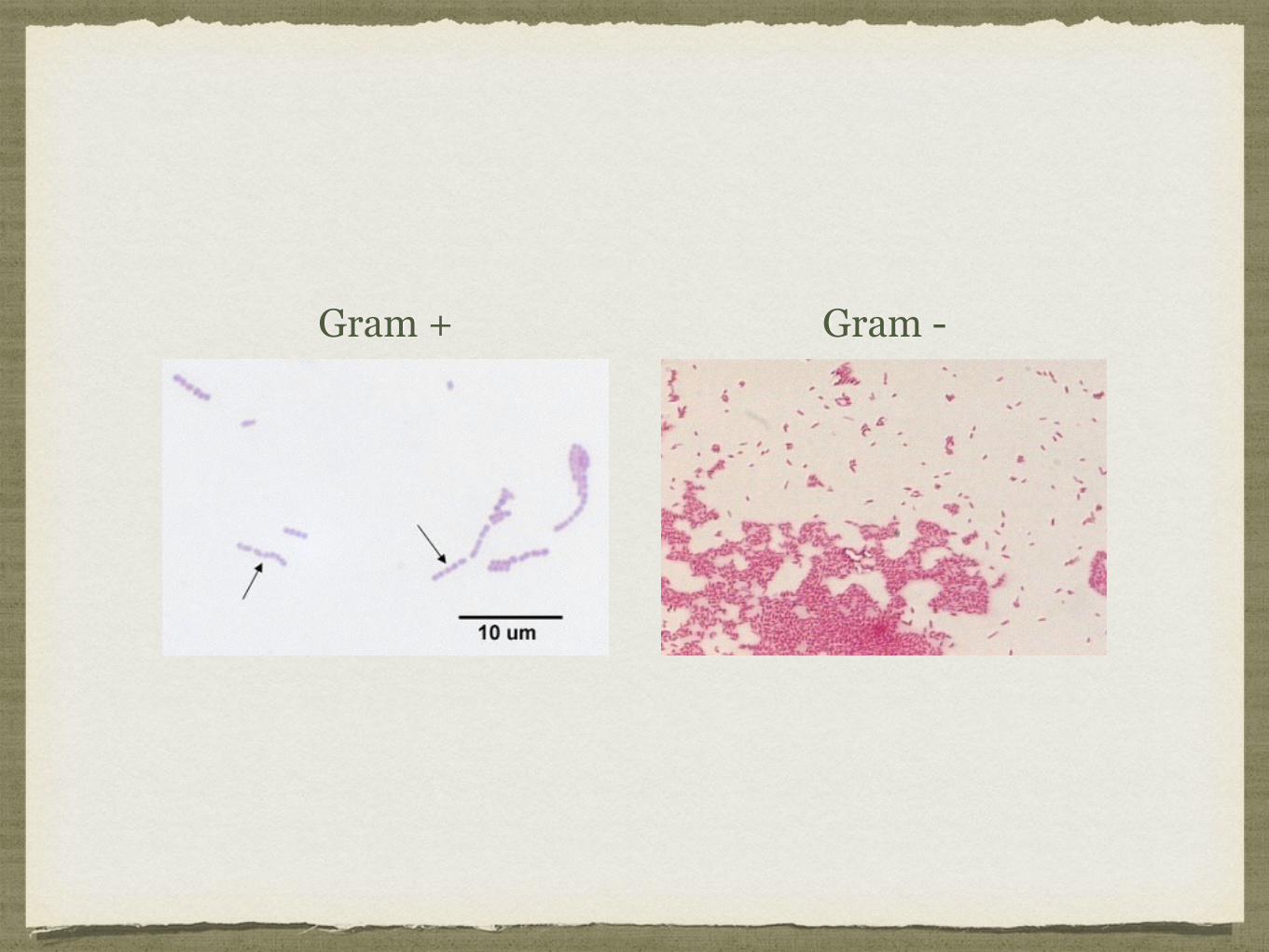
Bactérias Gram positivas, possuem um peptidoglicano espesso:
Coram de azul/violeta, já que retêm o violeta de cristal;
Bactérias Gram negativas, possuem um peptidoglicano fino:
Coram de vermelho porque não são capazes de reter o violeta de cristal durante o processo de descoloração, corando com a safranina ;
Gram + Gram -

FORMA
As bactérias têm 3 formas características:
Cocos;
Bacilos;
Espiroquetas - forma de saca-rolhas.

MOBILIDADE
Quanto à mobilidade, as bactérias podem ser:
Móveis:
Devido aos flagelos, que são estruturas proteicas que se estendem para fora da membrana celular de algumas bactérias Gram positivas e negativas;
Imóveis:
Não confundir com os movimentos Brownianos.

CITOCROMO OXIDASE
As bactérias que contêm o sistema citrocromo, produzem uma enzima oxidativa que cataliza a oxidação do citocromo c;
Os organismos que possuem no seu sistema respiratório o citocromo c são oxidase positivos, dando uma cor azul/purpura com o reagente N, N, N’, N’-tetrametil-p-fenilenodiamina;
Os organismos que não possuem citocromo c, são oxidase negativas e não vão oxidar o reagente mantendo a cor original.
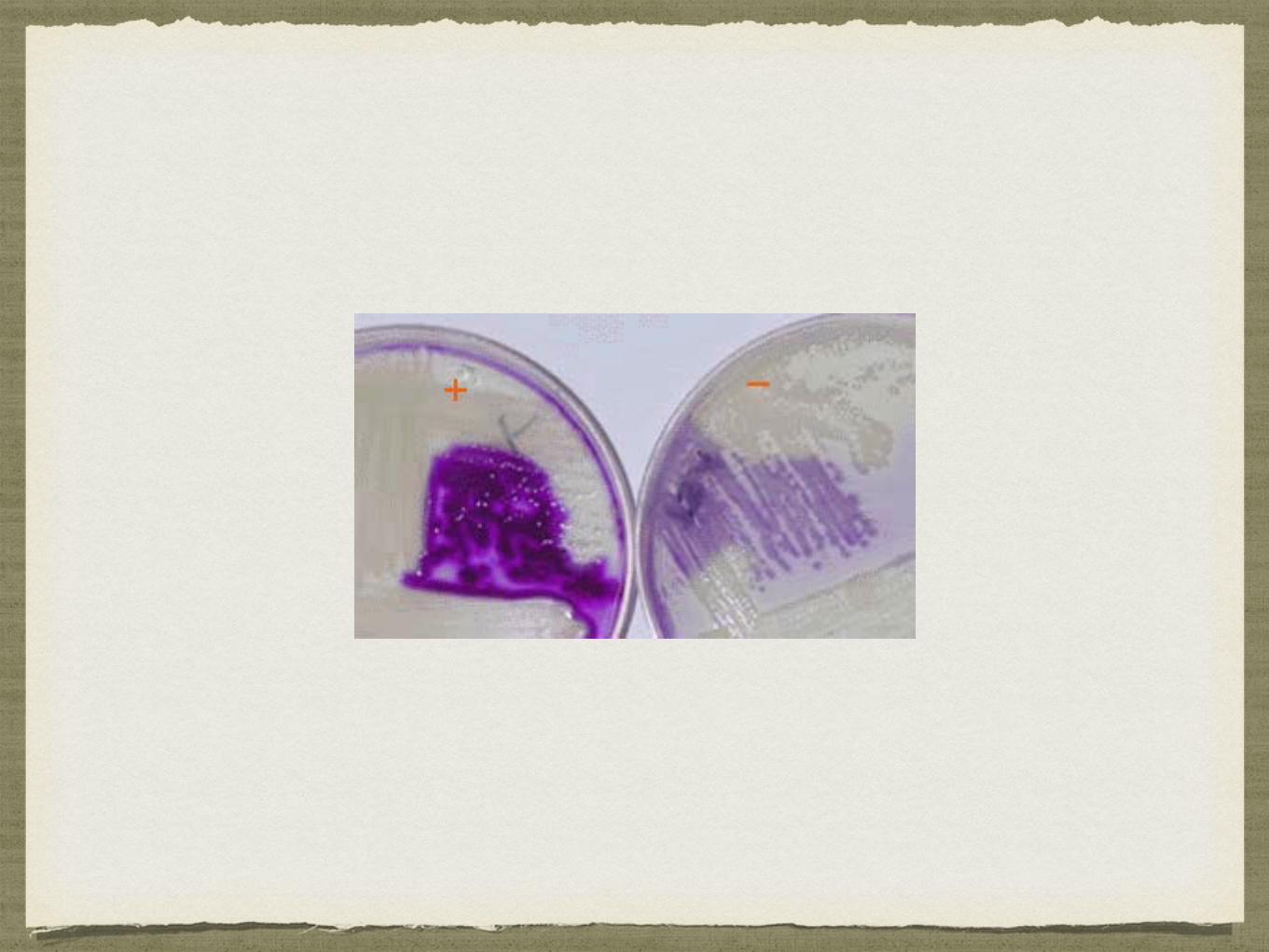

CATALASE
Durante a respiração aeróbia, as bactérias produzem peróxido de hidrogénio e em alguns casos ião superóxido:
Estes compostos são moléculas altamente reactivas;
A sua acumulação vai resultar na morte das bactérias, se estas não forem capazes de os degradar enzimaticamente.
A enzima catalase é a responsável pela degradação destes compostos;

CATALASE
A produção da enzima catalase pode ser determinada pela adição de peróxido de hidrogénio a uma cultura bacteriana:
As bactérias capazes de produzirem esta enzima, degradam rapidamente o peróxido de hidrogénio, levando à produção de água e oxigénio, o que corresponde a uma reacção positiva;
A ausência de bolhas de oxigénio corresponde a uma reacção negativa.

Reacção catalase positiva

METABOLISMO DA GLUCOSE
As bactérias podem utilizar duas vias metabólicas distintas para obterem energia a partir da glucose:
Respiração: na presença de oxigénio, as moléculas de glucose são completamente oxidadas em dióxido de carbono e água;
Fermentação: se o oxigénio não está presente, a glucose é convertida em ácido pirúvico e outros produtos terminais como etanol, ácido acético e butírico, levando a uma acidificação do meio onde se encontram.

METABOLISMO DA GLUCOSE
Metodologia:
O/F Basal Medium, segundo Hugh & Leifson, suplementado com 1% de glucose;
Utilizam-se 2 tubos, um dos quais é coberto com parafina líquida estéril;
Incubação até 7 dias;
A mudança da cor do meio para amarelo é considerado um resultado positivo.

METABOLISMO DA GLUCOSE
Bactérias de água salgada:
Suplementar o O/F Basal Medium com 1% de NaCl;
Utilizar ZOF (Lemos et al., 1985).

O/F segundo Hugh & Leifson
A B C
ZOF
A B C
A - Meio não inoculado; B - Reacção negativa; C - Reacção positiva.
A - Meio não inoculado; B - Reacção negativa; C - Reacção positiva.

PROVA DE INDOL
Baseia-se na capacidade das bactérias desdobrarem a molécula de triptofano levando à produção de indol;
O triptofano é um aminoácido essencial;
A sua oxidação origina ácido pirúvico e amoníaco:
As várias enzimas que intervêm neste processo recebem o nome colectivo de triptofanase.
A capacidade de hidrolisar o triptofano, com a produção de indol, não é uma característica de todas as bactérias.

PROVA DE INDOL
A triptofanase catalisa uma reacção de desaminação:
A hidrólise da molécula de triptofano ocorre apenas na sua cadeia lateral;
O anel aromático fica intacto sob a forma de indol.
A presença de indol é detectada pela adição do reagente de Kovacs, que é constituído por p-dimetilamino-benzaldeído, butanol e ácido clorídrico;

PROVA DE INDOL
O indol presente no meio é extraído para a camada de reagente de Kovacs pelo butanol acidificado e nesta, vai reagir com o p-dimetilamino-benzaldeído, adquirindo uma cor vermelho cereja.

Depois da adição do reagente de Kovacs: A - Camada amarela - bactéria indol negativa;
B - Camada vermelho cereja - bactéria indol positiva.
A B

SULFURETO DE HIDROGÉNIO (H2S)
Determinar se ocorre a libertação de sulfureto de hidrogénio a partir de aminoácidos que possuem enxofre na sua constituição;
Aminoácidos que contêm enxofre:
Metionina;
Cisteína.

SULFURETO DE HIDROGÉNIO (H2S)
Na presença de qualquer destes aminoácidos, o enxofre é reduzido por hidrogenação, levando à produção de sulfureto de hidrogénio sob a forma gasosa:
Gás incolor:
É necessário um indicador para a sua detecção;
São vários os compostos que podem funcionar como indicadores:
Ferro, sulfato ferroso, sulfato de amónio ferroso ou férrico, tiosulfato de sódio ou sulfito de bismuto.

SULFURETO DE HIDROGÉNIO (H2S)
Meio SIM, possui como indicadores o sulfato de amónia ferroso e o tiosulfato de sódio;
É necessário a presença dos dois indicadores, já que a detecção do sulfureto de hidrogénio só é possível ao fim de duas etapas:
A bactéria vai reagir com o tiosulfato de sódio, dando origem a sulfato ou sulfito de sódio e sulfureto de hidrogénio;
O sulfureto de hidrogénio vai reagir com o sulfato de amónia ferroso, produzindo um precipitado negro insolúvel, o sulfato de ferro.

Reacção de sulfureto de hidrogénio: A - Reacção positiva; B - Reacção negativa.
A B

UTILIZAÇÃO DO CITRATO
Pretende-se determinar se uma bactéria é capaz de utilizar o citrato como única fonte de carbono;
O citrato é um intermediário do ciclo de Krebs;
As bactérias que possuem a enzima citrato-permease vão transportar o citrato para o interior da célula e transforma-lo em piruvato.

UTILIZAÇÃO DO CITRATO
O agar citrato de Simmons possui na sua constituição citrato de sódio como única fonte de carbono e fosfato de amónia como única fonte de azoto;
As bactérias capazes de utilizar o citrato como única fonte de carbono vão degradar o fosfato de amónia em amoníaco ou hidróxido de amónia, levando à alcalinização do meio (passa a azul).

CITRATO DE SIMMONS
A - Resultado negativo B - Resultado positivo
A B