Embed Size (px)
Citation preview

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 1/46
ÍNDICE……………………………………………………………………………………………….....................................1
1. INTRODUÇÃO……………………………………………………………………………………….................................3
2. EUTROFIZAÇÃO – UM POUCO DA SUA HISTÓRIA……………………………………………........……………….5
2.1. AS ALTERAÇÕES DA PRODUTIVIDADE DO LITORAL E A EUTROFIZAÇÃO…………………………………. 7
3. O CICLO DO FÓSFORO GLOBAL…………………………..………………………………………………………..…….9
3.1. O FÓSFORO NA ÁGUA DOCE……………………….........................................................................9
3.2. A DISTRIBUIÇÃO DO FÓSFORO ORGÂNICO E INORGÂNICO NOS LAGOS ………………………10 3.3. O FÓSFORO E SUA INTERACÇÃO COM OS SEDIMENTOS……………………………………………..13
3.4. CONTEÚDO DE OXIGÉNIO DA MICROZONA……………………………………………………………..14
3.5. LIBERTAÇÃO DE FÓSFORO PELOS SEDIMENTOS………………………………………………………16 3.6. INTERACÇÃO COM ANGIOSPÉRMICAS AQUÁTICAS E ORGANISMOS BENTÓNICOS……….…17 3.7. A FLORA LITORAL E OS LOCAIS DE FLUXO DE FÓSFORO…………………………………………….19 3.8. OS INVERTEBRADOS BENTÓNICOS E O TRANSPORTE DE FÓSFORO……………………………..20 3.9. CICLO DO FÓSFORO DENTRO DO EPILIMNION…………………………………………………………21 3.10. RECICLAGEM DO FÓSFORO E DO AZOTO PELO ZOOPLANCTON………………………………….25 3.11. AS NECESSIDADES DE FÓSFORO DAS ALGAS…………………………………………………………26 3.12. INFLUÊNCIA DO PH E DA LUZ SOBRE A ABSORÇÃO DE FOSFATO…………………………….....27 3.13. CONCENTRAÇÕES DE FÓSFORO NECESSÁRIAS PARA O DESENVOLVIMENTO……………..…27 3.14. ABSORÇÃO DE FÓSFORO PELAS ALGAS VS. BACTÉRIAS……………………………………………29
4. O HOMEM E O CICLO DO FÓSFORO NOS LAGOS……………………………………………………………………29
4.1. ORIGEM DO FÓSFORO…………………………………………………………………………………………29
4.1.1. PRECIPITAÇÃO……………………………………………………………………………………...29
4.1.2. A ÁGUA SUBTERRÂNEA……………………………………………………………………………30
4.1.3. ESCORRÊNCIA TERRESTRE E ÁGUAS CORRENTES…………………………………………30
4.2. EFEITO DA CONCENTRAÇÃO DE FÓSFORO SOBRE A PRODUTIVIDADE DOS LAGOS………….33 4.3. A SOBRECARGA DE FÓSFORO E AZOTO E A PRODUTIVIDADE DAS ALGAS……………………...36
4.4. SOBRECARGA DE NUTRIENTES……………………………………………………………………………..36
4.5. RESPOSTAS BIOLÓGICAS À SOBRECARGA DE NUTRIENTES………………………………………..38

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 2/46
4.6. APLICAÇÃO DOS MODELOS DE PREVISÃO E A REALIDADE DOS DADOS………………………...39
5. POSSIVEIS MEDIDAS DE MINIMIZAÇÃO E REGRESSÃO TRÓFICA (RECUPERAÇÃO)…………………..…41
5.1. CONTROLO DA EUTROFIZAÇÃO E REGRESSÃO TRÓFICA………………………………………….…41
5.2. DIAGNÓSTICO…………………………………………………………………………………………………..41 5.3. CONTROLO EXTERNO DAS FONTES DE FÓSFORO………………………………………………………42 5.4. CONTROLO INTERNO DO FÓSFORO……………………………………………………………………….43
6. CONSIDERAÇÕES FINAIS………………..……………………………………………………………………………….44
REFERÊNCIAS BIBLIOGRÁFICAS……………..………………………………………………………….....................46

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 3/46
B I O G E O Q U Í M I C A D O F Ó S F O R O E E U T R O F I Z A Ç Ã O –
E C O S S I S T E M A S L Ê N T I C O S E M P A R T I C U L A R
U N I V E R S I D A D E D E É V O R A
Ciências do Ambiente, ramo de Qualidade do Ambiente
Autor: Ferreira, C. J. n.º 15193 E-mail: [email protected]
1. INTRODUÇÃO
É da maior importância que compreendamos a estrutura e o modo de funcionamento dos sistemas de água doce. A quantidade de água doce da Terra é muito pequena em comparação com a água dos oceanos mas tem, por outro lado, tempos de renovação muito mais rápidos,
sendo um factor importante a considerar quando se faz a análise dos seus recursos. Também tem de se ter em conta que tanto o desenvolvimento das populações humanas como a utilização da água doce se fazem segundo uma curva exponencial.
O Homem é um componente destes ecossistemas, e os seus efeitos sobre eles continuarão
a ser fortemente acelerados até que se consiga estabilizar o desenvolvimento demofórico. O
factor que poderá tornar-se o mais sério de todos os originados pelo crescimento demofórico é a degradação severa da qualidade da água. A água doce representa um recurso limitado que pode ser aumentado apenas ligeiramente, como é o caso via dessalinização, o que actualmente
acarreta um grande dispêndio de energia. O emocionalismo e as reacções alarmistas ao momentum da exploração dos limites da biosfera pelo sistema tecnológico não levam muito longe e, como tem vindo a ser repetidamente demonstrado, são por vezes antagónicos ao
melhoramento.
É necessário compreender as respostas metabólicas dos ecossistemas aquáticos para que se lhes possa fazer face a eliminar os efeitos dessas alterações, de modo a maximizar a gestão significativa dos recursos de água doce. É necessária uma integração do crescimento demofórico
humano com o metabolismo das águas doces, para conseguir reduzir ao mínimo as alterações prejudiciais.
Um efeito bem documentado do impacto do Homem sobre os ecossistemas aquáticos é a eutrofização, um termo multifacetado, em geral associado ao aumento da produtividade, à simplificação estrutural dos componentes bióticos e à redução da habilidade metabólica que os
organismos possuem para se adaptarem às alterações impostas (redução da estabilidade). Nesta situação de eutrofização as entradas excessivas parecem em geral ultrapassar a capacidade de equilíbrio do ecossistema (Wetzel, 1983). Contudo, o que acontece na realidade é que estes
sistemas estão desequilibrados apenas no que diz respeito às características químicas e bióticas da água doce que o Homem deseja para fins determinados. Para que haja esperança de integrar efectivamente o Homem como um componente dos ecossistemas lacustres e de registar a sua
utilização deste tipo de recursos, é imperioso que conheçamos com algum pormenor as propriedades funcionais das águas doces. Só então se poderá avaliar, com precisão razoável, a
influência que as actividades humanas terão sobre as características metabólicas destes sistemas.

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 4/46
Ao estudo das relações funcionais e da produtividade das comunidades de água doce e do efeito
nelas causado pelo seu ambiente físico, químico e biológico denomina-se Limnologia (Wetzel, 1983).
Mais recentemente, o Homem tem vindo a criar um grande número de albufeiras e pequenas represas até ao ponto de estas formarem sistemas lacustres importantes. Embora as
albufeiras sejam em muitos aspectos fundamentalmente diferentes dos lagos naturais, consegue-se a partir de conhecimentos bem cimentados sobre a dinâmica dos lagos, fazer uma transição com relativa facilidade para o estudo das características mais variáveis e individualizadas das
albufeiras. Na sua maioria os lagos, tanto naturais como artificiais, são relativamente pouco profundos, em geral com uma profundidade inferior a 20m. Assim, visto serem pouco profundos e possuírem grandes áreas são propícias ao desenvolvimento de macrófitas. Este tipo de cobertura
vegetal modifica radicalmente a produtividade do sistema. Além disso, as pequenas albufeiras recebem em geral entradas de nutrientes que são elevadas se considerarmos o seu volume, e isto vai aumentar ainda mais a sua capacidade produtiva. A maior parte das albufeiras existe durante
um período de tempo relativamente curto devido à enorme carga de sedimentos trazida pelos afluentes. Um exemplo típico deste caso encontra-se nas albufeiras do Alentejo. Todas as características morfométricas atrás descritas faz com que uma proporção cada vez maior do
volume dos lagos está exposta a, e interactua com, os processos químicos e metabólicos do solo e dos sedimentos.
A morfometria e os substratos geológicos das bacias dos lagos têm uma importância fundamental na determinação das interacções sedimento-água e da produtividade daí resultante,
bem como na significância da produtividade do litoral em relação à produtividade total de um ecossistema lêntico (águas “paradas”). Os lagos pouco profundos, que tem uma área de sedimento maior por unidade de volume de água, são de um modo geral mais produtivos que os
lagos profundos, sendo as comunidades fixas do litoral que contribuem, nestes lagos, com a proporção maior para a produtividade total.
Mas para uma melhor avaliação da produtividade dos ecossistemas lênticos temos que tratar da química básica de nutrientes dos ecossistemas aquáticos. Os biota estão ligados de forma inseparável à dinâmica de muitos elementos químicos, em particular à distribuição dos
nutrientes e à sua regulação biogeoquímica. Os elementos químicos, incluindo todos os elementos essenciais do protoplasma, tendem a circular na biosfera, por vias características, do ambiente para os organismos e destes novamente para o ambiente. Estas vias mais ou menos circulares
são conhecidas por ciclos biogeoquímicos (Odum, 1971). Do ponto de vista da biosfera como um todo, os ciclos biogeoquímicos classificam-se em
dois grupos básicos que são o de tipo gasosos, nos quais o depósito está na atmosfera ou na hidrosfera (oceanos) e os de tipo sedimentar, nos quais o depósito está na crosta terrestre que envolve o elemento fósforo. Os de tipo sedimentar tendem a ser menos perfeitos e mais
facilmente rompíveis por perturbações locais (Carrapeto, 1994), dado que a grande massa de material se encontra num depósito relativamente inactivo e imóvel na crosta terrestre.
Consequentemente, uma certa porção do material de troca tende a «perder-se» por longos períodos de tempo quando o movimento «encosta abaixo» é mais rápido do que o retorno «encosta acima». Os mecanismos de retorno ou de «reciclagem» são em muitos casos
principalmente bióticos (Odum, 1971).

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 5/46
«Bio» refere-se a organismos vivos e «Geo» às rochas, ao ar e à água da Terra. A
geoquímica é uma ciência relacionada com a composição química da terra e com a troca de elementos entre os diversos pontos da crosta terrestre e dos oceanos, rios e outras massas de água como é o caso dos sistemas lênticos. A biogeoquímica tornou-se, assim, o estudo da troca
ou de fluxo (isto é, do movimento de um lados para o outro) dos materiais, entre os componentes vivos e não vivos da biosfera (Odum, 1971)
Para o caso de um ecossistema lêntico chegou-se à conclusão, depois de estudos ao longo
dos tempos, que as “análises-dentro-do-lago” compreendem apenas uma parte do ecossistema.
Os nutrientes vindos da bacia de drenagem são bastante importantes na regulação do metabolismo dos lagos embora a sua importância ainda esteja grandemente limitada à sobrecarga de nutrientes inorgânicos.
A influência que os biota terrestres da zona húmida e do litoral exercem sobre a
quantidade e a qualidade da sobrecarga orgânica e inorgânica que entra num lago pode ser
profunda. A água carregada de substâncias orgânicas e inorgânicas escorre das regiões de altitude para a bacia do lago receptor, através do subsolo e dos cursos de água da superfície. Dão-se no percurso reacções químicas e biológicas que vão alterando de maneira selectiva a
qualidade e a quantidade de nutrientes que entram no lago. Os cursos de água à superfície atravessam muitas vezes o complexo zona húmida-litoral, podendo ainda perder ou ganhar compostos orgânicos e inorgânicos, também de modo selectivo, antes de atingirem a água do
lago (Wetzel, 1983). As actividades industriais e agrícolas do Homem levam ao lançamento de uma sobrecarga apreciável de substancias orgânicas e inorgânicas para o solo, atmosfera, água
acabando por ir parar à bacia de drenagem e ao próprio lago por precipitação seca ou húmida sendo por vezes muito significativa.
Tendo feita uma abordagem geral ao tema proposto com alguns conceitos que são a base da discussão passar-se-à para uma análise mais profunda do assunto. Visto ter diversas vertentes de estudo o problema será dividido em secções e que ao longo da monografia se tentarão
relacionar entre si. A Monografia tem como objectivo desenvolver o tema BioGeoQuímica do Fósforo e
relacioná-lo com os problemas associados aos ecossistemas lênticos e semi-lênticos (caso das albufeiras), como é o caso específico da eutrofização.
2. EUTROFIZAÇÃO – UM POUCO DA SUA HISTÓRIA
O termo eutrofia (derivado do adjectivo alemão eutrophe) refere-se em geral à riqueza de nutrientes. Foi Naumann que em 1919 criou os conceitos gerais de oligotrofia e eutrofia,
distinguindo-os, segundo as populações de fitoplâncton. Nos lagos oligotróficos havia poucas algas planctónicas, encontrando-se estes apenas nas zonas em que as rochas primitivas são dominantes. Nos lagos eutróficos havia mais fitoplâncton, sendo estes lagos frequentes nas
regiões mais baixas, naturalmente férteis, em que a actividade humana fornece uma dose crescente de nutrientes. Embora naquela altura a metodologia química fosse grosseira ou mesmo não existisse para certas substâncias, Naumann chamou, a atenção para factores químicos como
o fósforo, o azoto combinado e o cálcio, considerando-os determinantes do estado dos lagos. As provas limnológicas que se têm vindo a acumular têm revelado a existência de uma relação muito

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 6/46
forte entre a dinâmica biológica de um lago e o seu nível nutricional (concentração de nutrientes)
(Wetzel, 1983). Pouco depois da viragem deste século, verificou-se que nos lagos alpinos e subalpinos as
larvas de dípteros, em grande parte do género Tanytarsus, eram características dos lagos pouco produtivos e fundos em que a água hipolimnética perde muito pouco oxigénio durante a
estratificação de verão. Os lagos eutróficos eram menos profundos, mais ricos em plâncton e a água pouco oxigenada do hipolimnion era dominada por fauna que pode tolerar concentrações muito baixas de oxigénio, como o díptero Chíronomus. Foram os estudos exaustivos e os
conceitos que estes primeiros investigadores criaram que constituíram os fundamentos sobre os quais assenta toda a limnologia regional dos anos 30.
Depois destes existe uma superabundância de estudos limnológicos orientados para a avaliação das características dos diferentes tipos de lagos. Foi na verdade fenomenal a terminologia que para eles surgiu nas décadas que se seguiram. Muitos dos tipos de lagos
distinguiam-se com base na existência de espécies indicadoras. Os critérios para muitas destas classificações descritivas baseavam-se no controlo de variáveis físicas e químicas, enquanto que para outras classificações isso não interessava. A terminologia e as classificações dicotómicas que
se sucederam, embora instrutivas, eram demasiadas. Os lagos foram classificados de acordo com quase todos os aspectos limnológicos - geomorfológico, físico, químico - e ainda segundo as espécies indicadoras ou agregados de organismos de quase todos os grupos, desde as bactérias
até aos peixes. Até foram estudados os tipos extremos de lagos baseados nas aves aquáticas que com eles estão relacionadas. À medida que se estudavam mais lagos encontravam-se excepções
que conduziram a outras divisões e deram origem ao aparecimento de mais nomes. Os estudos exaustivos, principalmente de Naumann foram sendo menos usados porque a terminologia se tornava mais complexa. Os lagos possuem características tão únicas que, se se abusar dos
pormenores da classificação, em breve serão necessárias chaves taxonómicas para os classificar, o que já sucedeu.
Ilustração 1 – A eutrofização do Lago Medical, nos EUA, no seu esplendor.
O “bloom” das algas assim o confirma. (Botkin & Keller, 1995)
Os lagos devem ser agrupados segundo um espectro de estados de metabolismo entre
oligotrofia e eutrofia. A história demonstra que em muitos lagos pequenos das zonas temperada e glaciária é frequente registar-se uma sucessão de sedimentos predominantemente inorgânicos, com fósseis indicadores de oligotrofia, para sedimentos mais orgânicos, com fósseis indicadores
de um estado eutrófico (Hutchinson, 1975). As provas indicam que, uma vez estabelecida a sedimentação orgânica após a fase inicial oligotrófica, se verifica o estabelecimento de um tipo de equilíbrio trófico ou de um razoável equilíbrio dinâmico sazonal. Isto implica que as entradas de

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 7/46
nutrientes vindos da bacia de drenagem sejam relativamente constantes durante longos períodos
de tempo, sofrendo apenas alterações menores com as oscilações climatéricas e as entradas provenientes da cobertura vegetal e da erosão. Os sistemas podem, e acontece muitas vezes, sofrer uma regressão na produtividade à medida que a camada superficial do solo vai sendo
lixiviada e contém menos nutrientes. De um modo inverso, a produtividade pode ser grandemente acelerada pelo aumento das entradas de nutrientes (Goldman & Horne, 1995).
2.1. AS ALTERAÇÕES DA PRODUTIVIDADE DO LITORAL E A EUTROFIZAÇÃO
Como a água dos lagos recebe sobrecargas cada vez maiores de nutrientes, há uma enorme tendência para o fitoplâncton se desenvolver até à capacidade máxima dentro das limitações impostas pela temperatura e pela luz disponível. É no entanto imperativo que a
eutrofização dos sistemas aquáticos não seja encarada no sentido restrito da produtividade do fitoplâncton. Dentro das óbvias restrições geomorfológicas que se impõem ao desenvolvimento no litoral, é frequente a produtividade do litoral desempenhar um papel fundamental nos estados
inicial e final da fertilidade crescente do sistema de lago visto como um todo. Há com certeza excepções, mas esta condição de uma produção litoral relevante afecta nitidamente uma grande percentagem dos lagos do mundo.
As baixas de produtividade dos lagos oligotróficos são muitas vezes determinadas por
entrada de nutrientes inorgânicos em pequenas quantidades, vindos de fontes externas. Certas
características morfométricas como dimensões e profundidade relativamente grandes que estão na origem de razões elevadas de volume do hipolimnion para volume do epilimnion são em geral
típicas de lagos oligotróficos e afectam os ciclos de nutrientes. A baixa produção de matéria orgânica, as baixas taxas de decomposição daí resultantes e as condições hipolimnéticas favoráveis à oxidação têm como consequência taxas relativamente baixas de libertação de
nutrientes dos sedimentos num sistema causal cíclico. As baixas concentrações de substratos orgânicos dissolvidos de decomposição relativamente rápida tanto de origem planctónica como oriundos do litoral fazem com que existam populações pequenas de bactérias e taxas baixas de
metabolismo microbiano. A síntese de micronutrientes orgânicos, essenciais à maior parte das algas planctónicas, será correspondentemente limitada. A formação de complexos de micronutrientes orgânicos essenciais com compostos orgânicos dissolvidos, através da qual a
solubilidade e a disponibilidade fisiológica podiam ser parcialmente mantidas em condições de oxidação, será menos eficaz em oligotrofia com pouca síntese de matéria orgânica e taxas elevadas de degradação dos substratos disponíveis. A importância da limitação pelo fósforo e pelo
azoto para a manutenção de uma produtividade baixa numa grande percentagem de lagos do mundo.
As circunstâncias verificadas nos Lagos Shield, no Canadá, são ideais para demonstrar a importância da limitação pelo fósforo e a rápida recuperação a partir de condições de eutrofia após o controlo das entradas. Os exemplos do Lago Washington e do Lago Swiss (mais à frente
falar-se-à mais pormenorizadamente) representam casos análogos de sucesso no combate à eutrofização que é possível quando se tomam medidas de controlo do fósforo. Apesar das
cautelas que obviamente devem ser tomadas quando se fazem generalizações a respeito de sistemas tão complexos e variados como são os lagos, é importante salientar uma vez mais que a redução do fósforo não faz voltar todos os lagos às condições anteriores ao enriquecimento. A
reciclagem do fósforo a partir de fontes internas nos sedimentos pode manter taxas de produção

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 8/46
elevadas durante muitos anos após a redução da sobrecarga de nutrientes. A importância da
necessidade de fósforo e do seu fornecimento para o desenvolvimento vegetal é tal que a primeira coisa a fazer é reduzir as entradas para as águas doces, sendo provável que este trata-mento produza resultados positivos na maioria dos casos. Além disso, para que as entradas de
fósforo para as águas superficiais sejam reduzidas de modo mais efectivo, as fontes pontuais devem ser eliminadas o mais rapidamente possível. Os milhares de toneladas de fosfato que são
actualmente introduzidos nos sistemas de água doce pelos detergentes sintéticos podem ser eliminados de modo relativamente fácil: já existe tecnologia para a remoção do fosfato durante o tratamento das águas residuais.
Esta resolução chama a atenção para o papel crítico desempenhado pelo fósforo na
eutrofização rápida das águas interiores e para a necessidade de controlar as adições deste
elemento a todas elas por todos os meios disponíveis. Além do tratamento secundário dos esgotos, os métodos de controlo incluem:
Restrições ao uso de produtos de limpeza que contenham fosfatos ou outras
substâncias ecologicamente perigosas; Remoção do fosfato nas estações de tratamento de esgotos que libertem efluentes
para as águas interiores;
Controlo da drenagem proveniente de campos de engorda de animais, zonas agrícolas, tanques sépicos e outras fontes difusas de fósforo.
Devem também ser adoptadas medidas de controlo para o azoto nas bacias hidrográficas em que se torna evidente a sua necessidade. A implementação de tais medidas de controlo é
socialmente complexa e politicamente controversa, mas é acessível em termos tecnológicos.
Em condições de eutrofia, as taxas de sobrecarga de fósforo e de azoto, bem como as de
outros nutrientes cuja necessidade não é tão aguda, são relativamente elevadas. À medida que a taxa de produtividade fotossintética aumenta durante a eutrofização, as interacções cíclicas relacionadas com a regeneração de nutrientes inorgânicos a partir dos compostos orgânicos
aumentam de intensidade. A produtividade do plâncton sofre um aumento acentuado, tendo como resultado uma compressão do limite de intensidade luminosa da zona trofogénica. Esta redução da profundidade da zona trofogénica continua com o agravamento da eutrofização. Mais
tarde, o auto-ensombramento pelas populações de fitoplâncton atinge um grau a partir do qual não é possível haver mais aumentos da radiação solar incidente nas condições naturais. A taxa de produção fitoplanctónica atinge então uma plataforma, seguida de progressão gradual para a
extinção posterior através da sedimentação da matéria orgânica que se acumula mais rapidamente do que é removida por decomposição. Em certas condições de morfometria, meteorologia e produtividade, não é raro, em lagos mesotróficos ou eutrófico
Os percursos determinísticos mais importantes para governar a ontogenia e a eutrofização
progressiva dos lagos têm por base os mecanismos que sinergisticamente regulam o metabolismo
autotrófico. A discussão é aqui orientada primariamente para a maioria dos lagos temperados de origem glaciária, tectónica e vulcânica, de dimensões médias. Certamente existem excepções. Em
muitos casos, a discussão que se segue é aplicável a certos lagos desde a sua formação; noutros casos, os percursos descritos aplicam-se somente aos tempos mais recentes, e.g. desde as últimas perturbações provocadas pelo homem ou àquelas alterações que podiam ser previstas a
par de modificações induzidas pelas actividades humanas que afectam as taxas de sobrecarga de

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 9/46
nutrientes e de matéria orgânica.
3. O CICLO DO FÓSFORO GLOBAL
Antes de tudo há que referir que o fósforo é um nutriente. Segundo Mackson & Jackson (1996) o nutriente é uma espécie química em actividade e que é aproveitada por um organismo e
usado para manter as suas funções metabólicas. Os nutrientes são os elementos químicos necessários para um normal crescimento de um organismo, sendo assim denominados de elementos essenciais ou bioelementos. O fósforo é um dos nove bioelementos que é necessário
em maiores quantidades do que os restantes. O fósforo desempenha um papel central no metabolismo e na reprodução dos organismos.
O ciclo do fósforo é apresentado de uma forma simplificada na Ilustração 2. Em essência é um ciclo geoquímico que segue o curso do ciclo das rochas de erosão, transporte, sedimentação e uplift, seguido de um renovar da erosão, e assim sucessivamente. O maior reservatório deste ciclo
é a crosta. Isto acontece porque naturalmente os compostos contendo fósforo são de menor solubilidade e muito baixa volatilidade. Assim, comparativamente com outros elementos, é baixa a concentração de fósforo em sistemas aquáticos, mais especificamente em ecossistemas lênticos e
virtualmente nenhum é encontrado na atmosfera. O fluxo entre reservatórios resulta em maior parte do movimento de partículas sólidas quer por transporte aéreo quer em suspensão na água. A magnitude destes fluxos não é totalmente conhecido visto que o processo de transporte varia
com o tempo e varia de local para local (Mackson & Jackson, 1996).
A baixa solubilidade do fósforo contendo minerais significa que muito do fósforo nos solos e sedimentos debaixo dos corpos de água (como os lagos) é inviável para plantas e outros organismos. Assim, até em ambientes onde o fosfato mineral é relativamente abundante, a baixa
concentração de fósforo em solução poderá limitar o crescimento. Isto verifica-se especialmente em organismos que são capazes de fixar azoto sendo que nestas espécies o
crescimento não é limitada pelo fornecimento de azoto. Por conseguinte, a adição de fertilizante com fósforo a um
solo suportando leguminosas irá aumentar a proporção de azoto fixado. A importância do fósforo como nutriente
significa que tem feito um trabalho importantíssimo no comportamento deste elemento no ambiente do solo (Butcher e t al., 1992). Ilustração 2 – O Ciclo do Fósforo. (Mackson & Jackson, 1996)
3.1. O FÓSFORO NA ÁGUA DOCE
Não existe outro elemento na água doce que tenha sido tão estudado como o fósforo. Há

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 10/46
imensos dados quantitativos sobre a distribuição sazonal do fósforo em lagos e as taxas de
sobrecarga deste elemento a partir das bacias de drenagem respectivas. O interesse do fósforo em ecologia vem do papel principal que ele desempenha no metabolismo biológico relativamente à pequena quantidade que está à disposição na hidrosfera. Quando comparado com a abundância
natural de outros componentes nutricional e estruturalmente importantes para os biota (carbono, hidrogénio, azoto, oxigénio, enxofre), o fósforo é o menos abundante de todos, sendo portanto o
que mais vezes limita a produtividade biológica.
3.2. A DISTRIBUIÇÃO DO FÓSFORO ORGÂNICO E INORGÂNICO NOS LAGOS
Contrastando com as numerosas formas de azoto que têm importância nos sistemas de lago, a forma mais significativa de fósforo inorgânico é o ortofosfato (PO4
3-). Uma grande proporção, maior do que 90%, do fósforo na água doce existe sob a forma de fosfatos orgânicos
e constituintes celulares dos biota adsorvidos a partículas inorgânicas e à matéria orgânica particulada morta. É melhor discutir em primeiro lugar os aspectos gerais das formas de fósforo e da sua distribuição na água doce, antes de passar à dinâmica das trocas entre os diversos
compartimentos (Wetzel, 1983). O fósforo total orgânico e inorgânico tem sido objecto de várias classificações com base em
análises químicas; estas fracções assim obtidas têm por vezes muito pouco a ver com o metabolismo do fósforo. Talvez a determinação mais importante seja a do conteúdo de fósforo total na água não filtrada, que compreende o fósforo da fase particulada e o da fase «dissolvida».
Ambos os compartimentos consistem em vários componentes. O fósforo particulado divide-se em: Fósforo nos seres vivos sob a forma de:
ácidos nucleicos, DNA e RNA, relativamente estáveis, e fosfoproteínas, que
não estão envolvidas no ciclo rápido do fósforo; ésteres de enzimas de baixo peso molecular, vitaminas;
fosfatos de nucleotidos, como o difosfato de adenosina (ADP) e o 5-trifosfato de adenosina (ATP) que é utilizado nos percursos bioquímicos da respiração e na assimilação do CO2;
Fases minerais das rochas e do solo, como a hidroxiapatite, em que o fósforo se encontra adsorvido a complexos inorgânicos como argilas, carbonatos e hidróxidos de ferro;
Fósforo adsorvido à matéria orgânica particulada morta ou constituindo agregados macroorgânicos.
Contrastando com o fósforo da matéria particulada, o fósforo dissolvido é constituído por: ortofosfato (P04
3-); polifosfatos, muitas vezes provenientes de detergentes sintéticos;
colóides orgânicos ou fósforo combinado com colóides por adsorção e; ésteres fosfóricos de baixo peso molecular.
Em virtude da importância fundamental do fósforo como nutriente e por este ser um dos constituintes celulares principais, tem-se dado muita atenção à sua determinação analítica. As análises químicas do fósforo giram à volta da sua reactividade com o molibdato e de alterações
verificadas nessa reactividade durante a hidrólise enzimática e ácida das formas complexas de compostos de fósforo à medida que eles são convertidos em ortofosfato. As determinações mais

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 11/46
pormenorizadas diferenciam oito formas de fósforo (Gower, 1980), que se distinguem pelo modo
como reagem com o molibdato, pela facilidade com que são hidrolisadas e pelas dimensões das partículas (Strickland e Parsons, 1972 in Wetzel, 1983). Por consequência temos quatro classes operacionais:
P solúvel reactivo; P solúvel não reactivo;
P particulado reactivo e; P particulado não reactivo.
Estes métodos operacionais não correspondem necessariamente, no entanto, nem às espécies químicas do fósforo nem aos seus papéis no ciclo biótico do fósforo.
A maior parte dos dados de fósforo referentes às águas doces representam o fósforo total e o fósforo inorgânico solúvel (ortofosfato), embora se tenham identificado quatro fracções gerais em estudos mais pormenorizados (Hutchinson, 1975). Estas quatro fracções assemelham-se aos
quatro grupos operacionais já citados: fosfato solúvel; fósforo em suspensão (sestónico) solúvel em ácido, principalmente fosfato férrico e
fosfato de cálcio; fósforo orgânico solúvel e fósforo coloidal e; fósforo orgânico em suspensão (sestónico).
As concentrações de fosfato total encontradas nas águas naturais não poluídas abrangem
uma gama muito larga, desde menos de 1 µg/l até mais do que 200 mg/l em alguns lagos halinos fechados. A concentração de fósforo total da maior parte das águas de superfície não contaminadas varia entre 10 e 50 µg P/l. A variação é no entanto muito grande, podendo estar
relacionada com características geológicas regionais. Os níveis mais baixos de fósforo das águas doces encontram-se geralmente nas regiões montanhosas em que a geomorfologia do estrato de rocha é cristalina, aumentando nas águas de terras baixas que derivam de depósitos de rochas
sedimentares. Os lagos com muita matéria orgânica, como os lagos ácidos, possuem normalmente concentrações elevadas de fósforo total. Algumas zonas costeiras sedimentares, como as do sudeste dos Estados Unidos, contêm muitas rochas fosfatadas. Os lagos cuja
drenagem se faz através destes depósitos contêm níveis anormalmente elevados de fósforo. Num trabalho extremamente pormenorizado em que relaciona o fósforo e o azoto com a
produtividade dos lagos, Vollenweider (1968 in Wetzel, 1983) demonstrou, por meio de vários critérios, que a quantidade de fósforo total aumenta geralmente com a produtividade dos lagos (Ilustração 3/Ilustração 4). Embora haja excepções a esta regra, ela constitui um princípio geral
muito útil para a resolução de questões aplicadas de eutrofização.
Ilustração 3 – Relação geral da produtividade dos lagos com a concentração de fósforo total.
NÍVEL GERAL DA PRODUTIVIDADE DO LAGO
FÓSFORO TOTAL (µG/L-1)
ULTRA-OLIGOTRÓFICO <5
OLIGO-MESOTRÓFICO 5 – 10
MESO-EUTRÓFICO 10 – 30
EUTRÓFICO 30 – 100
HIPEREUTRÓFICO >100

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 12/46
Além da apresentação em tabela é possível representar tais valores de forma gráfica (Ilustração 4) que se apresenta em seguida.
Ilustração 4 – Probabilidade de ocorrência dos diversos estados tróficos em consequência da maior ou menor concentração de fósforo. (Freedman, 1995
in Vollendeiwer & Kerekes, 1982)
O fósforo solúvel inorgânico é sempre muito pouco, constituindo apenas uma pequena percentagem do fósforo total, e circula muito rapidamente nas zonas de utilização. A razão do fósforo solúvel inorgânico para as outras formas de fósforo é aproximadamente 1: 20, ou inferior
a 5%, pois o fosfato inorgânico é surpreendentemente constante numa grande variedade de lagos da zona temperada.
É provável que a percentagem do fósforo total que existe como verdadeiro ortofosfato
iónico seja consideravelmente menor que 5% na maior parte das águas naturais (Prepas e Rigler, 1982 in Wetzel, 1983).
De uma maneira geral as observações que se fazem nas zonas trofogénicas dos lagos dizem respeito à distribuição do fósforo pelas formas acabadas de discutir. O fosfato, o pirofosfato, o trifosfato e os aniões polifosfato de maior valência também formam complexos,
quelatos e sais insolúveis com numerosos iões metálicos (Stumm e Morgan, 1981 in Wetzel, 1983). O grau de complexação e de formação de quelatos entre os vários fosfatos e os iões metálicos que existem nas águas naturais depende das concentrações relativas dos fosfatos e dos
iões metálicos, do pH e da presença de outros ligandos (sulfato, carbonato, fluoreto e espécies orgânicas) (Gower, 1980).
Em virtude de as concentrações de fosfato serem de um modo geral baixas, a formação de complexos que envolve estes catiões principais e vários aniões fosfato terá forçosamente muito
pouco efeito sobre a distribuição dos iões metálicos, podendo no entanto ter um efeito acentuado sobre a distribuição do fosfato (Wetzel, 1983). Os iões metálicos, como o ferro férrico, o manganésio manganoso, o zinco, o cobre, entre outros, estão presentes em concentrações
semelhantes ou ainda mais baixas que as dos fosfatos. Para estes iões, a formação de complexos pode afectar de modo significativo a distribuição do ião metálico, dos fosfatos, ou de ambos. Por exemplo, a solubilidade do fosfato de alumínio (AlP04) é mínima a pH 6, aumentando para valores
de pH superiores e inferiores àquele. O fosfato férrico (FeP04) comporta-se de maneira semelhante mas é um pouco mais solúvel que o AlP04. A concentração de cálcio interfere com a formação de hidroxilapatite [Ca5(OH)(P04)3] (LeCren & Lowe-McConell,????). Numa solução
aquosa em que não haja outros compostos, a concentração de cálcio de 40 mg/l limita a solubilidade do fosfato a aproximadamente 10 µg /l ao pH 7. Se o nível de cálcio for 100 mg/l o equilíbrio máximo do fosfato baixa para 1 µg/l . O aumento do pH das águas que contêm
concentrações normais de cálcio deveria conduzir à formação de apatite. Além disso o aumento

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 13/46
do pH leva à formação de carbonato de cálcio, o qual coprecipita o fosfato com os carbonatos
(Otsuki e Wetzel, 1972 in Wetzel, 1983). É bem conhecida a adsorção de fosfatos e polifosfatos às superfícies, particularmente às de minerais de argila (Stumm e Morgan, 1981 in Wetzel, 1983), por ligação química dos aniões às arestas com cargas positivas das partículas de argila e por
substituição de silicatos por fosfatos na estrutura da argila. Em geral os valores baixos de pH (entre 5 e 6 aproximadamente) favorecem a adsorção elevada de fosfato às argilas.
Ilustração 5 – Distribuição vertical generalizada do
fósforo solúvel (Ps) e total (Pt) nos lagos estratificados com produtividade muito baixa e com produtividade elevada.
Na sequência destas interacções e da distribuição do fósforo pelas fracções orgânica e
inorgânica, a tendência geral é para que os lagos pouco produtivos que apresentam curvas ortogradas de oxigénio tenham um conteúdo de fósforo relativamente constante com a
profundidade (Ilustração 5). Do mesmo modo, durante os períodos de circulação do Outono e da Primavera, a distribuição vertical do fósforo é mais ou menos uniforme. Os metais oxidados, como o ferro, e os catiões principais, em particular o cálcio, podem induzir a precipitação do fósforo.
Nos lagos que durante os períodos de estratificação apresentam curvas clinogradas de
oxigénio, contudo, a distribuição vertical do fósforo é muito mais variável. Observa-se
normalmente um aumento acentuado do conteúdo de fósforo no hipolimnion inferior, especialmente nas fases mais adiantadas da estratificação térmica (Ilustração 5). A maior parte deste aumento hipolimnético corresponde a fósforo solúvel, junto dos sedimentos. A fracção de
fósforo do seston varia muito com a profundidade. O fósforo do seston no epilimnion apresenta grandes flutuações, de acordo com as oscilações das populações de plâncton. O fósforo do seston do metalimnion e do hipolimnion varia com a sedimentação do plâncton, as taxas de
decomposição que por sua vez dependem da profundidade, e com o desenvolvimento de populações bacterianas e outro plancton que habitam em profundidade (caso das euglenofíceas).
3.3. O FÓSFORO E SUA INTERACÇÃO COM OS SEDIMENTOS
A troca de fósforo entre os sedimentos e a água suprajacente é um dos componentes mais
importantes do ciclo do fósforo nas águas naturais. Aparentemente existe na maioria dos lagos um movimento líquido de fósforo para dentro dos sedimentos. A eficácia deste fluxo líquido de
fósforo e a rapidez com que se dão os processos que regeneram o fósforo, voltando a trazê-lo para a água, dependem de uma série de factores físicos, químicos e metabólicos. Existe uma correlação fraca entre a quantidade de fósforo nos sedimentos e a profundidade da água supraja-
cente, podendo o conteúdo de fósforo nos sedimentos ser várias ordens de grandeza superior ao da água. Os factores mais importantes a considerar são:
A capacidade de retenção de fósforo pelos sedimentos;
As condições da água suprajacente e; Os biota que habitam os sedimentos e alteram os equilíbrios de troca, efectuando o
transporte do fósforo novamente para a água.

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 14/46
As trocas efectuadas através da interface dos sedimentos são reguladas por mecanismos
relacionados com os equilíbrios entre os minerais e a água, processos de adsorção (principalmente troca iónica), interacções do tipo redox, dependentes do oxigénio, e a actividade de bactérias, fungos, plâncton e invertebrados. As taxas de troca são função dos coeficientes
locais de difusão e do controlo ambiental entre o orgânico e o inorgânico, isto é, reacções enzimáticas. A interface sedimentos-água efectua a separação entre dois domínios muito
diferentes. Em apenas poucos milímetros da camada superior dos sedimentos, dá-se a regulação das trocas por movimentos à escala molecular, com taxas de difusão correspondentemente baixas. Na água as trocas são reguladas por taxas muito mais elevadas e muito mais variáveis de
difusão por turbulência (Mortimer, 1971 in Wetzel, 1983). 3.4. CONTEÚDO DE OXIGÉNIO DA MICROZONA
As características reguladoras mais conspícuas da fronteira dos sedimentos são a interface sedimentos-água e o seu conteúdo de oxigénio. O conteúdo de oxigénio desta microzona é
afectado principalmente pelo metabolismo de bactérias, fungos, invertebrados planctónicos que migram para a interface e invertebrados bentónicos sésseis. O processo que consome mais
oxigénio nas zonas de água profunda dos lagos é a degradação microbiana da matéria orgânica particulada morta, que sedimenta para o hipolimnion e se deposita nos sedimentos. A taxa de esgotamento do oxigénio está condicionada pelas taxas de sobrecarga orgânica para o
hipolimnion e pela morfologia do lago ou da albufeira. A decomposição das fracções de matéria orgânica mais frágil, em grande parte de origem vegetal, dá-se durante o percurso para os sedimentos e, conforme as taxas de entrada e de deposição, os sedimentos recebem muitas
vezes apenas resíduos orgânicos relativamente resistentes a decomposição posterior. Os sedimentos têm uma necessidade absoluta e imprescindível de oxigénio, a qual é
governada pela intensidade do metabolismo microbiano e respiratório, pela lentidão das taxas de difusão e pela acumulação de elementos inorgânicos na forma reduzida, como o Fe2+, que são libertados para os sedimentos durante a decomposição dos biota. O transporte é regulado por
difusão e faz-se essencialmente ao nível molecular nos sedimentos, a menos que a superfície dos mesmos seja perturbada por turbulência da água suprajacente. O oxigénio da água bem arejada suprajacente, como a dos lagos oligotróficos ou a dos lagos mais produtivos durante os períodos
de circulação completa, penetra apenas poucos centímetros nos sedimentos, por difusão. A entrada de oxigénio é regulada pela taxa de fornecimento deste gás aos sedimentos, pela mistura
por turbulência dos sedimentos superficiais, se ela existir, e pela necessidade de oxigénio por unidade de volume dos sedimentos. Mortimer (1971 in Wetzel, 1983) demonstrou a importância da existência de uma microzona oxidada para que se efectuem as trocas químicas, especialmente
as que dizem respeito ao fósforo, nos sedimentos. À superfície dos sedimentos o factor crítico na regulação das trocas entre os sedimentos e a água é uma diferença de poucos milímetros na penetração do oxigénio. São exemplos destas relações dois lagos com sedimentos orgânicos: no
primeiro lago a concentração de oxigénio na interface dos sedimentos manteve-se acima de 8 mg/l durante a estratificação do verão, enquanto que no segundo lago o nível de oxigénio na interface desceu para menos de 1mg/l.
No primeiro caso, nos estudos de Mortimer no Lago Windermere, no Reino Unido, a
concentração de oxigénio à superfície dos sedimentos nunca foi inferior a 1 ou 2 mg/l. Os
potenciais redox, que se aproximam dos potenciais redox compostos da água suprajacente

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 15/46
oxigenada e dos sedimentos superficiais até à profundidade de cerca de 5 mm, foram sempre
elevados (de + 200 a + 300 mv). Abaixo de 40 ou 50 mm nos sedimentos registaram-se poten-ciais uniformemente baixos (aproximadamente - 200 mv), indicadores de condições redutoras extremas e anoxia total (Ilustração 6). Os sedimentos mantiveram-se oxidados até à profundidade
de perto de 5 mm durante todo o período de estratificação de verão. Foram observadas diferenças na profundidade a que se dava a transição de potencial elevado para baixo nos
sedimentos, mas a zona de baixo potencial não chegou nunca a atingir a água. Depois de cinco meses de estratificação o ponto de zero mv deslocou-se para a superfície dos sedimentos, para -5 mm, partindo de aproximadamante -12 mm no tempo da circulação da primavera, e deslocou-se
novamente para baixo, para -10 mm, durante a circulação do Outono. A integridade da microzona oxidada foi sempre mantida, numa camada fina mas operacionalmente muito significativa, durante os períodos de estratificação. Depois disso a microzona oxidada foi mantida por difusão e
pelo deslocamento por turbulência dos sedimentos localizados mais à superfície para a água suprajacente durante os períodos de circulação (Gorham, 1958 in Wetzel, 1983). A eficácia da microzona oxidada na prevenção da libertação de quantidades significativas de componentes
solúveis da água intersticial dos sedimentos para a água suprajacente foi demonstrada em câmaras experimentais. O fósforo, principalmente, foi impedido de migrar para cima.
Ilustração 6 – Diagrama do perfil do potencial redox
composto, não corrigido para as variações do pH, através da
interface sedimentos-água em tarolos não perturbados da
porção mais funda do Lago Windermere, antes, durante e
depois da estratificação. (Baseado em dados de Mortimer,
1971 in Wetzel, 1983)
A capacidade dos sedimentos para reterem fósforo abaixo de uma microzona oxidada na interface está relacionada com vários factores que actuam de um modo sinergístico. A maior parte do fósforo orgânico que chega aos sedimentos por deposição é decomposta e hidrolisad. Quase
todo o fósforo dos sedimentos é inorgânico, como a apatite por exemplo, derivado da bacia de drenagem e adsorvido a argilas e hidróxidos de ferro. Além disso o fósforo coprecipita com o ferro, o manganésio e os carbonatos (Wetzel, 1972 in Wetzel, 1983). O trabalho realizado sobre
os sedimentos do Lago Wisconsin e dos Grandes Lagos forneceu indicações de que o fósforo dos sedimentos está predominantemente sob as formas de apatite, fósforo orgânico e iões ortofosfato ligados por covalência a óxidos de ferro hidratados (Shukla, 1971 in Wetzel, 1983). Nos
sedimentos calcários dos lagos de águas duras com uma percentagem de CaCO3 compreendida entre 30 e 60% do peso, não foi possível estabelecer uma relação directa entre o nível de CaCO3 e o fósforo inorgânico ou o fósforo total. Verificou-se existir nestes sedimentos uma capacidade
de adsorção do fósforo inorgânico inferior à dos sedimentos não calcários. Deduz-se destas observações que a adsorção do CaCO3 não tem tanta importância como os complexos de ferro e fosfato para a regulação da concentração de fósforo nos sedimentos (Frevert, 1980 in Wetzel,
1983). A camada oxidada constitui uma armadilha eficaz para o ferro e o manganésio, assim como para o fosfato, reduzindo, desta maneira, enormemente o transporte de materiais para a
água e fazendo nesta uma depuração de substâncias como o fosfato.
A barreira constituída pela microzona oxidada enfraquece à medida que o conteúdo de

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 16/46
oxigénio junto da interface dos sedimentos diminui. Segundo Mortimer (1971 in Wetzel, 1983), a
libertação de fósforo, ferro e manganésio aumentou imenso quando diminuiu o potencial redox. Com a redução dos hidróxidos de ferro e outros complexos, o ferro ferroso e o fosfato a ele adsorvido foram mobilizados e apareceram na água. Observaram-se as mesmas reacções gerais
na água do hipolimnion mesmo justaposta aos sedimentos de um lago eutrófico, segundo um padrão repetidamente observado nos lagos dimícticos produtivos. Quando a linha isovolt + 0,20
(E7 = + 200 mv) emerge acima da superfície da interface, dá-se a libertação súbita do ferro ferroso e do fosfato para a água. Este acontecimento é precedido de redução do nitrato e libertação lenta de bases (alcalinidade), CO2 e amónia. O manganésio é reduzido, e portanto
mobilizado, a um potencial redox mais elevado que o ferro. A introdução de oxigénio no hipolimnion durante a circulação do Outono provoca a
oxidação do ferro ferroso e a redução simultânea do fosfato, em parte como fosfato férrico, menos solúvel que o hidróxido férrico, e em parte por adsorção ao hidróxido férrico e ao CaCO3. O manganésio, embora seja oxidado mais lentamente que o ferro, está mesmo assim
completamente precipitado quando chega o momento da circulação. O ferro ferroso libertado dos sedimentos é sempre em maior quantidade que o fosfato, de modo que quando é oxidado precipita a maior parte deste. Parte do fosfato férrico particulado pode sofrer hidrolise muito lenta
e devolver algum fosfato à água nos sedimentos das zonas superiores e da zona litoral do lago (Hutchinson, 1957 in Wetzel, 1983). No entanto, quase todo o fosfato retoma posteriormente aos sedimentos.
Nos lagos muito produtivos onde a decomposição da matéria orgânica que sofre deposição
no hipolimnion origina condições de anoxia e produz sulfureto de hidrogénio, dá-se a precipitação de sulfureto de ferro (FeS). O sulfureto de ferro, tal como muitos outros sulfuretos de metais, é extremamente insolúvel e forma-se a um potencial redox de cerca de +100 mv. Se houver
precipitação de grandes quantidades de FeS, pode retirar-se da água ferro suficiente para permitir que algum do fosfato acumulado no hipolimnion permaneça em solução durante a circulação do Outono. Foi sugerida, como um método de fertilização de lagos através da regeneração de fosfato
dos sedimentos, a adição de sulfato aos lagos de modo a fazer aumentar a produção de sulfureto de hidrogénio pelas bactérias para acelerar a perda de ferro (Hasler e Einsele, 1948 in Wetzel, 1983).
3.5. LIBERTAÇÃO DE FÓSFORO PELOS SEDIMENTOS
Devido à importância óbvia do fósforo como um dos nutrientes que muito frequentemente fazem aumentar a produtividade das águas doces, tem sido dedicado muito interesse ao conteúdo deste elemento nos sedimentos e ao seu movimento para a água suprajacente. A concentração
de fósforo nos sedimentos dos lagos é muito mais elevada que na água (Schlesinger, 1991). Em condições de aerobiose, os equilíbrios das trocas são geralmente unidireccionais, para os sedimentos. Quando existe anaerobiose, no entanto, as trocas inorgânicas que se dão na
interface sedimentos-água são fortemente influenciadas pelas condições redox (Frevert, 1979 in Wetzel, 1983). A profundidade dos sedimentos que participam na migração activa do fósforo para
a água é considerável. Verificou-se que em sedimentos anóxicos não perturbados, dado tempo suficiente (2 a 3 meses), o fósforo se deslocou sem dificuldade para cima, de uma profundidade de pelo menos -10 cm para a água suprajacente, independentemente de os sedimentos serem
lama eutrófica calcária ou de natureza turfosa e ácida (Hynes e Greib, 1970 in Wetzel, 1983). A

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 17/46
comparação deste movimento de deslocação em sedimentos esterilizados e sedimentos contendo
bactérias anaeróbias não apresentou diferenças significativas; portanto, a difusão é o processo predominante.
Verificou-se que as bactérias mobilizadoras de fósforo, especialmente as dos géneros Pseudomonas, Bacterium e Chromobacterium, existiam até pelo menos 15 cm nos sedimentos de
albufeiras. A abundância variava com o tipo de sedimentos, bem como a sua distribuição vertical. Havia poucas em sedimentos arenosos com pequenas quantidades de siltes, concentrando-se as bactérias junto da interface. O seu número aumentou e tornou-se mais uniforme com a
profundidade em sedimentos arenosos que possuíam quantidades moderadas de matéria orgânica e de siltes. O maior número de bactérias foi encontrado em siltes ricos em matéria orgânica.
Se por um lado as bactérias são de uma importância fundamental na dinâmica do ciclo do fósforo na água, como será discutido na secção que se segue, por outro lado o seu papel na aceleração das trocas de fósforo através da interface dos sedimentos é relativamente
insignificante quando comparado com o dos processos químicos de equilíbrio. A importância da microflora dos sedimentos reside no seu papel em fazer aumentar a concentração de fósforo dissolvido na água intersticial (Fleischer, 1978 in Wetzel, 1983). O metabolismo das bactérias da
interface, no entanto, possui relativamente pouco efeito sobre a fixação biogénica e a remoção de fosfato da água suprajacente. Descobriu-se, com o auxílio de sedimentos naturais esterilizados, que a fixação microbiana e o transporte de fósforo da água para os sedimentos correspondia a
menos do que 5% do movimento total em condições redutoras de anaerobiose. Em aerobiose as bactérias da interface aumentaram de modo significativo o transporte microbiano de fósforo para
os sedimentos, tendo esta perda sido relacionada com a quantidade de fósforo microbiano que se depositava na interface (Frevert, 1979 in Wetzel, 1983).
A taxa de libertação de fósforo dos sedimentos dos lagos aumenta bastante (quase duplica) se houver agitação devida a turbulência (Zicker et al., 1965 in Wetzel, 1983). Se os sedimentos anaeróbios forem cobertos com areia ou com toalhas de polietileno, consegue-se
impedir em grande parte a perda de oxigénio pela água suprajacente, diminuindo a libertação de fósforo, ferro e amónio pelos sedimentos (Hynes & Greib, 1970 in Wetzel, 1983).
As algas que se desenvolvem sobre os sedimentos são capazes de utilizar efectivamente o fósforo neles contido (Freedman, 1995). Além disso, as algas em suspensão na água que continha vários compostos inorgânicos particulados de solubilidade extremamente baixa foram capazes de
extrair fósforo suficiente para o seu desenvolvimento; em condições experimentais, suprimidas as fontes de fósforo dos sedimentos, o desenvolvimento das algas foi limitado pelo conteúdo de fósforo da água. A presença ou a ausência de bactérias teve pouco efeito sobre a utilização do
fósforo pelas algas. Estes resultados salientam a importância dos fosfatos susceptíveis de ser extraídos dos sedimentos se estes forem agitados na coluna de água, como acontece nos lagos pouco profundos, mesmo quando a solubilidade dos compostos de fósforo é muito baixa.
3.6. INTERACÇÃO COM ANGIOSPÉRMICAS AQUÁTICAS E ORGANISMOS BENTÓNICOS
A importância das angiospérmicas submersas, de folhas flutuantes e emergentes, para a
dinâmica do ciclo do fósforo foi já demonstrada por vários estudos. Embora se conheçam
bastante bem os fenómenos de absorção de fósforo pelo sistema radicular das plantas e o seu

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 18/46
retorno para seguir o ciclo (principalmente através da decomposição da matéria orgânica) no que
respeita às plantas terrestres, acreditou-se durante muitos anos que este ciclo não existia, ou pelo menos não era relevante, nos habitats aquáticos. O sistema vascular e os tecidos estruturais das angiospérmicas aquáticas são muito reduzidos. Além disso, a morfologia das folhas destas plantas
foi simplificada pela redução ou perda completa da cutícula das paredes das células da epiderme. A constatação destes factos, juntamente com numerosos estudos fisiológicos que demonstraram
absorção activa de nutrientes por folhas submersas, levou à generalização de que os nutrientes eram absorvidos principalmente através das folhas, servindo as raízes destas angiospérmicas apenas para as fixar ao sedimento. A absorção de nutrientes, quer por adsorção à folhagem quer
através do sistema raízes-rizomas, varia com a espécie nas plantas aquáticas. A importância da vegetação litoral tem sido repetidamente salientada desde as primeiras
tentativas de estudo da dinâmica do ciclo do fósforo nos lagos pela adição de fósforo radioactivo à água da superfície (Hayes et al., 1951 in Wetzel, 1983). Estes estudos demonstraram claramente haver absorção rápida de fósforo pela vegetação litoral, assim como pelo fitoplâncton,
sempre que aquele nutriente era adicionado à água. Para além disso observou-se também a libertação lenta de fósforo pelas plantas e a libertação um pouco mais rápida pelas algas epífitas nessas mesmas plantas. Postulou-se ainda a libertação de fósforo com a senescência das
macrófitas anuais no fim do verão. A libertação de fósforo pelas folhas e raízes das macrófitas emergentes, flutuantes e
submersas, após a morte, demonstra a importância da macrovegetação como fonte de fósforo em muitos sistemas aquáticos. A lixiviação do fósforo de macrófitas mortas, em condições de
esterilidade, foi rápida e teve como consequência a perda de 20 a 50% do conteúdo de fósforo total em poucas horas, tendo sido entre 65 e 85% a perda em períodos mais longos. A lixiviação deu-se mais rapidamente a partir de plantas secas do que das plantas húmidas, mas as taxas não
foram praticamente afectadas pela fragmentação da matéria vegetal ou pela variação da temperatura dentro da gama limnológica normal. As taxas de lixiviação das raízes foram superiores às das folhas. Pelo facto de o conteúdo de fósforo que constitui compostos orgânicos
nos tecidos vegetais variar durante o período vegetativo, diminuindo geralmente com a idade, a quantidade de fósforo lixiviado para a água varia sazonalmente, dependendo do conteúdo de fósforo total das plantas. O fósforo assim rapidamente libertado (numa questão de dias) das
macrófitas senescentes depressa é mineralizado, podendo ser utilizado pelo metabolismo das bactérias e das algas ou ser perdido através da microzona oxidada dos sedimentos (Solski, 1962 in Wetzel, 1983).
Verificou-se também que todas as partes das plantas absorviam 32P igualmente e que
aproximadamente uma proporção igual à do fósforo adsorvido era translocada dos ápices ou dos
rizóides para outras partes da planta (Hayes, 1955 in Wetzel, 1983). Os estudos de McRoy et al. (1972 in Wetzel, 1983) contribuíram muito para esclarecer o
papel das macrófitas aquáticas no ciclo do fósforo. Descobriu-se que as taxas de absorção e excreção do fósforo pelas raízes e folhas de Zostera marina L. dependiam da concentração de
ortofosfato do meio. A concentração do fósforo inorgânico intersticial era cerca de duas ordens de grandeza maior que a da água. As plantas absorveram 108 mg P/m2 dia-1 dos sedimentos, assimilaram 45 mg na produção de material novo e excretaram 62 mg para a água (Ilustração 7).

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 19/46
Ilustração 7 – Fluxo de fósforo num coberto da macrófita Zostera marina L. À esquerda: Gradiente de fósforo, unidades em µg P(g planta)-1dia -1. À direita: Movimento diário liquído do fósforo (mg m-2). A quantidade de fósforo (mg m-2) de cada compartimento está dentro de parêntesis. (McRoy et al., 1972 in Wetzel, 1983)
Uma quantidade equivalente a 41% do fósforo reactivo inorgânico foi exportada dos tapetes de plantas para a água ao largo. Embora esta trajectória do bombeamento de fósforo
reactivo dos sedimentos para a água pelas plantas vasculares aquáticas tenha sido demonstrada numa zona litoral marinha, não há razão para pensar que seja quantitativamente menos
importante nos sistemas de água doce. Na realidade pode explicar-se muita da informação que existe sobre o ciclo quantitativo na água ao largo dos lagos com base na exportação contínua de fósforo dos sedimentos, conjugada com a rapidez com que o ciclo do fósforo se faz no
fitoplâncton. 3.7. A FLORA LITORAL E OS LOCAIS DE FLUXO DE FÓSFORO
Uma maneira de distinguir os principais locais de fluxo de fósforo é dividindo-os em três
compartimentos:
A água ao largo e os seres vivos do epilimnion; Os organismos do litoral e; O hipolimnion e os sedimentos (Ilustração 8).
O lago comunica com a bacia de drenagem através do epilimnion, entrando o fósforo com
a água que flui para dentro do lago e saindo com a água que flui para fora e também por
sedimentação. Nesta divisão a flora litoral, que constitui um dos componentes mais importantes em muitos se não na maioria dos lagos, considera-se incluída no epilimnion e na zona trofogénica,
pois ambos se estendem virtualmente até à mesma profundidade em muitos lagos pequenos. A aplicação da análise compartimental aos resultados de experiências com indicadores
demonstrou que o fósforo no epilimnion é extremamente móvel. O tempo de renovação do fósforo no epilimnion (isto é, o tempo necessário para que uma quantidade de fósforo equivalente à quantidade total deste elemento num compartimento deixe esse compartimento e uma
quantidade semelhante de fósforo entre nele) foi determinado por Rigler (1964 in Wetzel, 1983) como sendo de 3,6 dias. Vinte minutos após ter entrado, mais de 95% do fósforo adicionado à água tinham sido absorvidos pelo plâncton; o tempo de renovação foi inferior a 20 minutos. Num
pequeno lago ácido com uma zona litoral bastante ampla em que se desenvolvem plantas aquáticas com raízes, a zona litoral foi o contribuinte mais importante para a renovação do fósforo no epilimnion; a perda de fósforo para esse compartimento fez-se 10 vezes mais rapidamente do
que para o hipolimnion e os sedimentos, e 50 vezes mais rapidamente do que a perda através do efluente. A devolução de fósforo à água feita na zona litoral durante o verão foi cerca de 20% superior à perda. Assim, torna-se evidente que a flora litoral pode desempenhar um papel

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 20/46
fundamental na dinâmica do ciclo do fósforo em que plantas vasculares fazem a assimilação e o
transporte de fósforo a partir dos sedimentos.
Ilustração 8 – Os três compartimentos principais do
ciclo do fósforo num lago. Os números representam os
fluxos relativos de fósforo entre os compartimentos no
lago Toussaint, Ontário, em meados do Verão.
(Segundo Rigler, 1964 in Wetzel, 1983)
Embora não se possam derivar as constantes das taxas das perdas de fósforo do epilimnion a partir dos dados existentes, Rigler (1973 in Wetzel, 1983) voltou a analisar os resultados de
vários estudos. Verificou que o tempo de renovação do fósforo no epilimnion, que variava entre 20 e 45 dias, apresentava uma correlação negativa com as áreas dos lagos e as estimativas de desenvolvimento da vegetação litoral.
Algumas conclusões aparentemente conflituosas sobre a importância da flora litoral para a
circulação do fósforo nos lagos, baseadas em experiências de adição à água durante o verão de fósforo indicador radioactivo, podem ser esclarecidas por meio de análise das diferenças entre as comunidades dos lagos estudados. Em sistemas de tanques experimentais com regulação do
fósforo que entra e sai do epilimnion de modo a conservar um equilíbrio estável, verificou-se que as algas filamentosas da zona litoral simulada eram um dos locais referenciais de assimilação de fósforo antes de este elemento atingir o epilimnion. O fósforo era libertado para a água ao largo
quando se dava a senescência das algas filamentosas. Numa relação deste tipo, o sistema de equilíbrio de trocas entre a entrada de fósforo para ou a libertação de fósforo da flora litoral e de ou para a água do epilimnion varia com a quantidade de fósforo que chega da bacia de
drenagem. Do mesmo modo, a assimilação e a libertação de fósforo pelas macrófitas do litoral e algas a elas fixas irá sofrer variações de acordo com as limitações físicas impostas ao desenvolvimento da flora litoral e determinadas pela morfologia da bacia. Além disso, esta relação
ficará sujeita a viragens sazonais, proporcionais às alterações verificadas no desenvolvimento activo da flora litoral e senescência subsequente das plantas anuais no fim do verão. Quando as plantas vasculares submersas são perenes, ou nos lagos tropicais onde o desenvolvimento no
litoral é mais ou menos contínuo, a regra geral é haver uma troca constante de fósforo entre a zona litoral e o epilimnion.
3.8. OS INVERTEBRADOS BENTÓNICOS E O TRANSPORTE DE FÓSFORO
Não está ainda completamente esclarecido o efeito que os invertebrados bentónicos exercem sobre a dinâmica do ciclo do fósforo entre os sedimentos e a água. Parece que a acção de escavar, característica de muitos deles, pode alterar de forma significativa a zona oxidada
superficial, através da penetração física dos animais. Não se sabe ainda quais os efeitos que uma disrupção da superfície do tipo da efectuada por densas populações de fauna bentónica ou peixes que se alimentam no fundo (como a carpa ou a perca) possa ter sobre as trocas de fósforo em
resultado da manipulação física da interface, mas é natural que sejam bastante pequenos em comparação com a regulação química-microbiana dominante. Durante o desenvolvimento das

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 21/46
populações de invertebrados bentónicos, o fósforo é incorporado na fauna a partir da matéria
orgânica de que eles se alimentam nos sedimentos. A adsorção, bem como a assimilação directa, de fósforo inorgânico, não são quantitativamente significativas, pelo menos no que respeita aos micro-crustáceos (Rigler, 1964 in Wetzel, 1983). Quando os invertebrados bentónicos se
transformam em adultos podem emigrar dos sedimentos, fazendo deste modo o transporte do fósforo para outros compartimentos do sistema. Nos cursos de água, por exemplo, tem-se
verificado uma migração significativa de fósforo em direcção a montante, efectuada por peixes e invertebrados (Ball et al., 1963 in Wetzel). No entanto, esta deslocação de fósforo pelos invertebrados desempenha apenas um pequeno papel na totalidade do ciclo quantitativo do
fósforo no sistema do lago. O papel desempenhado no transporte de fósforo para a água pela actividade dos micro-
invertebrados na interface dos sedimentos também ainda não foi esclarecido. Os ciliados que vivem nos sedimentos são capazes de hidrolisar ácidos orgânicos dissolvidos e libertar fosfato inorgânico para a água (Hooper & Elliot, 1953 in Wetzel, 1983). As concentrações baixas de
oxigénio, no entanto, não só produzem um ambiente pouco favorável para os ciliados como também inibem a libertação de fosfato pelas suas células. Supõe-se que os cladóceros do zooplancton, que possuem fototactismo negativo e que migram para a interface dos sedimentos
durante as horas do dia, se alimentam de forma muito activa da rica microflora que aí existe. Não se sabe até que ponto o transporte de fósforo para o epilimnion aquando da sua subsequente migração nocturna é importante, assunto este certamente digno de ser investigado.
Em estudos realizados com macroinvertebrados bentónicos demonstrou-se o potencial que
estes organismos possuem para perturbar a interface sedimentos-água através da sua actividade escavadora e promover as trocas de fósforo. As espécies de maiores dimensões provocam efeitos maiores do que os mais pequenos. A libertação de fósforo aumentou de forma aproximadamente
linear (de 0,3 a 9,4 mg P m-2 dia-1). Quase todo o fósforo libertado para a água suprajacente estava na forma de ortofosfato, que é prontamente assimilado pelas bactérias ou pelas algas se a intensidade luminosa for suficiente. Supõe-se que o fósforo foi directamente excretado pelos
invertebrados, a julgar pelo que já foi verificado para o zooplancton. Quando as larvas de inver-tebrados atingem densidades extremas (Lindegaard & Jónasson, 1979 in Wetzel, 1983), a sua actividade pode afectar o transporte de fósforo através da interface sedimentos-água.
3.9. CICLO DO FÓSFORO DENTRO DO EPILIMNION
Os estudos clássicos de Einsele (1941 in Wetzel, 1983) e muitas análises teóricas e aplicadas que se lhes seguiram sobre a circulação e o destino do fósforo na água ao largo do epi-limnion demonstraram que o fósforo é incorporado muito rapidamente nas algas fitoplanctónicas
e nas bactérias. Os estudos mais recentes têm focado as taxas do movimento do fósforo entre as formas biologicamente importantes na água dos lagos. O resultado foi o aparecimento de duas categorias gerais de investigação:
Mecanismos de transferência de fósforo em que os organismos vivos são inter-mediários e;
Reacções abióticas de complexação.
Da primeira categoria fazem parte os estudos sobre a transferência de fósforo entre o
seston e as várias formas de fósforo dissolvido, incluindo a formação de fósforo coloidal com

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 22/46
intervenção dos biota e a utilização facilitada por enzimas de ésteres de fósforo dissolvidos. A
última categoria consiste em estudos sobre a adsorção e a libertação de fósforo aos e dos compostos húmicos dissolvidos, carbonato de cálcio coloidal e outras partículas.
Lean (1973 in Wetzel, 1983) propôs um modelo quantitativo, para um estado de equilíbrio de trocas de fósforo na água do epilimnion durante o verão, com a seguinte composição
(Ilustração 9): fósforo particulado, ou seja a fracção retirada por filtração da água através de um
filtro com 0,5 µm de poro, que contém a maior parte do fósforo;
ortofosfato solúvel (P043-), inorgânico, reactivo, que possui um tempo de renovação
extremamente curto; um composto orgânico de fósforo (XP), de baixo peso molecular (aproximadamente
250) e; fósforo coloidal solúvel, macromolecular, de peso molecular superior a 5000000.
Ilustração 9 – Movimento do fósforo dentro da zona de
água ao largo do epilimnion do lago Heart, Ontário,
mostrando o mecanismo de troca entre o fosfato e as
fracções particuladas (Wetzel, 1983 in Lean, 1973)
O mecanismo predominante é a troca entre o fosfato inorgânico e a fracção particulada, mas os microorganismos excretam algum fósforo sob a forma de composto de baixo peso mole-
cular (XP). Os compostos coloidais de elevado peso molecular provêm da policondensação dos compostos de baixo peso molecular (XP). Ambas as fracções, mas principalmente a última, libertam fosfato inorgânico solúvel, o qual fica à disposição do plancton.
As constantes de equilíbrio da Ilustração 9 atestam a rapidez com se faz a dinâmica do fósforo
neste compartimento misto de algas e bactérias num lago hipotético, a meio do verão:
K1, assimilação de P04 = 0,9 unidades de massa relativas min-1 ou 0,002 µg/l min-1 de fósforo biologicamente activo total.
K2, libertação de XP, e K3, ligação de XP por condensação ao P coloidal = 0,022 min-1. A
excreção de compostos de fósforo orgânico extracelular é um fenómeno bem conhecido nas algas marinhas e de água doce durante os períodos de desenvolvimento intenso e de senescência.
K4, hidrólise de P coloidal para P04 = 0,0017 min-1. Depois da rápida formação de P coloidal via XP, em menos de dois minutos, a adição de XP separa o P do colóide, ficando
novamente a forma solúvel inorgânica P04 disponível para assimilação.
K5, libertação de P04 inorgânico pelo seston, que se verificou ser muito superior à excreção
de fósforo orgânico solúvel (XP). Os cálculos de equilíbrio de massa demonstraram que K5
é cerca de 70 vezes maior que K2
K6, hidrólise directa de XP para P04 que não foi significativa quando comparada com a libertação na forma coloidal.
K7, constante de equilíbrio para a perda de fósforo coloidal, considerada pequena em

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 23/46
relação às outras constantes de equilíbrio. É preciso notar, no entanto, que a agregação
posterior da fracção coloidal para levar à forma particulada. ou a adsorção à matéria parti-culada, seguidas de sedimentação, representam um depósito de fósforo formado a partir do epilimnion. Semelhantemente, a sedimentação do fósforo particulado representa uma
perda lenta, mas contínua, do epilimnion. Sendo assim, durante o desenvolvimento activo de algas e bactérias heterotróficas, deve haver uma substituição contínua de fósforo por
meio de entradas dos afluentes ou da zona litoral, ou ainda por reciclagem através da ingestão pelo zooplancton e subsequente excreção. Nos sistemas experimentais a maior parte da fracção coloidal sofre deposição, deixando de estar biologicamente disponível
passados um a cinco dias.
Partindo destas primeiras luzes e dos estudos que se seguiram sobre o ciclo do fósforo na
água ao largo, tornou-se evidente que o fósforo orgânico sestónico teria de ser dividido em pelo menos duas fracções:
uma fracção de circulação rápida, que vai sendo trocada entre as formas solúveis;
nesta fracção o fosfato é rapidamente transferido através da fase particulada para compostos de baixo peso molecular;
uma fracção de fósforo sestónico que é libertada mais lentamente.
A transferência de fósforo coloidal do fitoplancton para a água faz-se minutos depois de se
ter, dado a assimilação de fosfato solúvel pelas bactérias e algas (Paerl e Lean, 1976 in Wetzel,
1983). As bactérias e as algas excretam quantidades significativas de compostos de fósforo orgânico dissolvido. Como ficou dito atrás, a assimilação de fosfato pelas bactérias é superior à
feita pelas algas, mas como a biomassa das algas é superior à das bactérias, a incorporação de fosfato pelas algas normalmente domina a assimilação e a libertação de fósforo no epilimnion.
A cinética da assimilação e da renovação do fósforo foi estudada em numerosos lagos de produtividade tanto baixa como elevada e em diferentes estações do ano (Ilustração 10).
Ilustração 10 – Tempo de renovação do fosfato de água doce.(Wetzel, 1983)
A renovação do fósforo é extremamente rápida nos períodos de verão em que se verificam grandes exigências e as entradas são relativamente baixas, mas torna-se muito mais lenta, cerca de duas ordens de grandeza, durante os períodos de Inverno. A renovação do fósforo é mais
rápida em ambientes oligotróficos onde é maior a deficiência deste elemento (Peters, 1979 in Wetzel, 1983).
Fazendo parte da constituição do fitoplâncton existem muitos ésteres complexos de fósforo (açúcares fosfatados, fosfonucleótidos, polifosfatos). No modelo de Lean a hidrólise enzimática
LAGO TEMPO DE RENOVAÇÃO (MIN)
Lago Maggiore, Itália – epilimnion, verão 10-200
Lago Maggiore, Itália – epilimnion, Inverno 200-10000
Lago Maggiore, Itália – hipolimnion 1000-100000
15 Lagos europeus (epilimnia) 4-74400
Lagos do Sul de Ontário (epilimnia) – verão 2-8
Lagos do Sul de Ontário (epilimnia) – Inverno 10-10000
Lagos da África Oriental 1-1000
Lago 227, Ontário – verão 0,4-15
Lago 227, Ontário – Inverno 120-11700

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 24/46
directa das fracções XP de baixo peso molecular é tida como negligenciável. Na realidade é
provável que existam mais mecanismos de regeneração do fósforo complexo. A hidrólise enzimática dos ésteres de fosfato que ocorrem nos ambientes naturais é um dos
mecanismos mais plausíveis para a regeneração do fósforo no epilimnion. A quebra enzimática directa de compostos de fósforo inorgânico dissolvido e de polifosfatos para dar fósforo inorgânico
dissolvido, feita por uma actividade fosfatásica relativamente pouco específica (fosfomonoesterase) tem sido objecto de estudos intensivos. Há numerosas investigações que demonstram que a actividade fosfatásica incorporada na membrana das algas aumenta quando a
deficiência em fósforo se torna aguda. Os aumentos verificados na actividade da fosfatase alcalina (APA: «alkaline phosphatase activity») em condições de limitação pelo fosfato, podem melhorar a capacidade competitiva das algas, fornecendo-lhes um meio que lhes permite utilizar
organofosfatos ou polifosfatos inorgânicos como substrato, constituindo fontes alternativas de fósforo. Embora na forma livre a fosfatase alcalina dissolvida tenha uma duração curta (Pettersson, 1980 in Wetzel, 1983) podem encontrar-se quantidades substanciais de APA (muitas
vezes ultrapassando 50% do total) na fase dissolvida (Pettersson, 1980 in Wetzel, 1983). A importância da actividade fosfatásica no ciclo do fósforo nos lagos complica-se porque há produção de fosfatase alcalina pelas bactérias nos ambientes pelágicos, mas esta não é induzida
estritamente pela limitação pelo fosfato (Wilkins, 1972 in Wetzel, 1983). Observa-se uma grande variação sazonal e espacial da APA solúvel e da APA associada às
algas e a outra matéria particulada (Wetzel, 1983). A APA do epilimnion é normalmente elevada durante a primavera e os períodos de verão em que a necessidade de fósforo é máxima. Nos
anos em que a circulação da primavera é incompleta ou quando um lago sofre meromixia tem-porária, encontram-se valores extremamente altos de APA concomitantes com a limitação do fitoplancton pelo fósforo durante os meses de primavera e verão (Wetzel, 1983).
Foram encontrados na água dos lagos quantidades substanciais de fosfomonoésteres
(PME: «phosphomonoesters» (55 µg PME-P/l nos lagos Twin, eutróficos, em Ohio; Heath &
Cooke, 1975 in Wetzel, 1983). Verificou-se que havia uma relação inversa entre as concentrações de PME e a APA durante a estratificação de verão. Foram determinadas taxas potenciais de hidrólise dos PME pela APA superiores e 0,05 µmoles por hora. Estes valores são semelhantes aos
encontrados nos sistemas marinhos quando se usou a fosfatase alcalina das algas como enzima hidrolítico (Rivkin & Swift, 1980 in Wetzel, 1983). Estes resultados, juntamente com análises químicas muito diferentes do reservatório de fósforo complexo na água dos lagos (Lean, 1973 in Wetzel, 1983) sugerem que a composição do reservatório de fósforo solúvel varia bastante com os diferentes tipos de lagos. Enquanto alguns lagos se aproximam do modelo do ciclo do fósforo apresentado na Ilustração 9, noutros este ciclo pode ser dominado pela produção e pela hidrólise
de PME feita por intermédio dos biota. Alguns processos físico-químicos podem afectar o ciclo do fósforo e, por consequência,
influenciar a produtividade primária. Certos complexos húmicos, de baixo e de elevado peso molecular, contêm fósforo. O ortofosfato adsorvido aos compostos húmicos de ferro pode ser
libertado por fotorredução, induzida pela luz UV, do ferro férrico. Os compostos húmicos de baixo peso molecular podem libertar ortofosfato após serem expostos à fosfatase alcalina.
Actualmente ainda não se dá o devido valor à importância que têm os complexos húmicos

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 25/46
de fósforo para o fitoplancton. Por exemplo, as taxas de assimilação do carbono pelo fitoplâncton
diminuíram quando as algas foram expostas a baixas concentrações de matéria húmiça dissolvida originária do litoral e das zonas húmidas (Stewart & Wetzel, 1982 in Wetzel, 1983). Observou-se selectividade do efeito em relação à espécie, sendo o maior efeito observado referente ao
fitoplâncton de menores dimensões (de 1 a 5µm) cultivado em condições de baixa intensidade luminosa. A actividade da fosfatase alcalina foi altamente estimulada pela presença de compostos
húmicos dissolvidos de baixo peso molecular em concentrações baixas. As provas obtidas indicam que estes materiais húmicos podem desempenhar o papel de agentes sequestradores dos compostos orgânicos de fósforo, reduzindo assim a disponibilidade deste elemento para o
fitoplâncton. Estas relações podem ser importantes tanto para o ciclo do fósforo como para a produtividade do fitoplâncton, pois as chuvas abundantes podem introduzir quantidades substanciais de fósforo e compostos húmicos dissolvidos nas zonas pelágicas dos lagos pequenos
e de tamanho médio. 3.10. RECICLAGEM DO FÓSFORO E DO AZOTO PELO ZOOPLANCTON
A assimilação de nutrientes, especialmente fósforo e azoto, através da ingestão de
partículas alimentares pelo zooplancton herbívoro, pode em certas alturas do ano ser
considerável. Durante a dejecção dá-se a libertação de nutrientes sob a forma de iões fosfato solúveis, e alguns compostos orgânicos de fósforo e amónia (Lehman, 1980 in Wetzel, 1983). Parte do fósforo e do azoto permanece nas partículas fecais que se depositam, saindo da zona
trofogénica.
Os nutrientes que o zooplâncton liberta voltam rapidamente a ser assimilados pelo fitoplâncton. Em certas circunstâncias, quando a concentração de fósforo na zona trofogénica é baixa, o fósforo e o azoto regenerados pelo zooplâncton podem constituir uma fracção
significativa dos requisitos das algas (Lehman, 1980 in Wetzel, 1983). Por exemplo, com base em determinações in situ e nos cômputos de nutrientes, calcula-se que o zooplâncton do epilimnion do Lago Washington fornece a esta camada superficial em circulação dez vezes mais fósforo e
três vezes mais azoto durante os meses de verão (de Junho a Setembro) do que a quantidade destes elementos que provém de todas as fontes externas combinadas (Edmonson, 19??).
O grau de assimilação de nutrientes e de reciclagem é função do estado de nutrição das células das algas (Lehman, 1980 in Wetzel, 1983). Quando se fizeram comparações com as células cultivadas em quantidades suficientes de nutrientes, verificou-se que as células das algas
com deficiência de nutrientes assimilaram fosfato e amónia mais rapidamente e apresentaram taxas de remineralização mais baixas.
A herbivoria praticada pelo zooplâncton acelera as taxas de regeneração de fósforo e de azoto. É provável também que tanto as algas planctónicas como as bactérias assimilem o fósforo logo que ele é libertado. Mesmo sendo baixa a concentração média de fósforo (e muitas vezes a
do azoto complexado) no epilimnion, a libertação de nutrientes pelo zooplâncton pode provocar uma distribuição micro-heterogénea de concentração de nutrientes, pois a distribuição do
zooplâncton é muito mais agregada que a do fitoplâncton. Considerando o período anual, é menos óbvia a importância que a reciclagem de nutrientes
tem para as taxas gerais de produtividade das algas. A remoção de algas e bactérias pelo

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 26/46
zooplâncton herbívoro sofre grandes flutuações com as estações do ano e a distribuição espacial.
Além disso, a herbivoria praticada sobre as algas é muitas vezes selectiva em relação às dimensões. Esta selectividade pode fazer aumentar a disponibilidade de nutrientes em relação às algas menos procuradas pelos herbívoros. As relações complicam-se ainda mais devido aos
padrões sazonais de predação sobre o zooplâncton pelos peixes planctívoros. Uma predação deste tipo faz reduzir a eficácia da regeneração de nutrientes pelo zooplâncton. Bartell & Kitchell (1978 in Wetzel, 1983), por exemplo, calcularam que a redução nas taxas de libertação de fósforo pelo zooplancton pelágico fosse de uma ordem de grandeza durante os meses de Junho e Julho, quando a predação pelos peixes atingiu o máximo sazonal e aos fornecimentos alóctones
correspondia o período baixo da estação.
Finalmente, deve recordar-se que quase todo o fósforo está ligado à matéria particulada:
algas, bactérias e detritos. As taxas de regeneração de nutrientes a partir destas fontes à medida que o fitoplâncton envelhece e morre não estão ainda bem estudadas. Certamente que estes organismos com tempos de geração curtos reciclam os nutrientes rapidamente na zona
trofogénica antes de afundarem para profundidades maiores. A grandeza relativa da reciclagem que eles fazem, em relação à do zooplâncton, é desconhecida, mas deve variar tanto espacial como sazonalmente. Actualmente pode-se apenas concluir que a herbivoria realizada pelo zoo-
plâncton e a reciclagem concomitante de nutrientes podem fazer aumentar as taxas de produtividade primária das algas durante os períodos em que as elevadas exigências de nutrientes excedem as entradas da sobrecarga de fontes externas para a zona trofogénica.
3.11. AS NECESSIDADES DE FÓSFORO DAS ALGAS
Os compostos que contêm fósforo desempenham papéis importantes em praticamente
todas as fases do metabolismo, em particular nas transformações de energia das reacções de
fosforilação da fotossíntese. O fósforo é necessário para a síntese de nucleótidos, fosfolípidos, açúcares fosfatados e outros compostos intermediários fosforilados. Além disso, o fosfato liga-se, geralmente como éster, a um grande número de enzimas e vitaminas de baixo peso molecular
que são essenciais ao metabolismo das algas. A importância do fósforo para a fisiologia das algas tem um interesse especial para o
limnologista por este ser o menos abundante de todos os elementos principais necessários para o desenvolvimento das algas na grande maioria dos ambientes de água doce. Além disso, a forma de fósforo mais importante para a nutrição vegetal é o P04 inorgânico ionizado. Embora as algas
do plâncton possam utilizar ésteres orgânicos de fósforo, como os glicerofosfatos, esta capacidade varia com a espécie, tal como a capacidade de obtenção de grupos fosfato enzimaticamente ou através da libertação de exoenzimas para a água a fim de provocar
dissociação catalítica (Krauss, 1958 in Wetzel, 1983). Quase todo o fósforo libertado para a água durante o desenvolvimento activo das algas é fosfato solúvel inorgânico e ésteres orgânicos, os quais são, por sua vez, reciclados muito rapidamente. Durante a lise e a decomposição das
células, a maior parte do fósforo libertado das algas está sob a forma orgânica e sofre degradação bacteriana (Krauss, 1964 in Wetzel, 1983).

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 27/46
3.12. INFLUÊNCIA DO PH E DA LUZ SOBRE A ABSORÇÃO DE FOSFATO
Para muitas espécies de algas o desenvolvimento e a absorção máxima de fósforo dão-se
dentro de uma gama distinta de pH do meio. O pH pode provocar a alteração da taxa de absorção
de fosfato por meio de efeitos directos sobre a actividade das enzimas, sobre a permeabilidade da membrana celular ou através da modificação do grau de ionização do fosfato. Por exemplo, a
absorção de fósforo pela diatomácea Asterionella é máxima para valores de pH compreendidos entre 6 e 7 (Mackereth, 1953 in Wetzel, 1983). Observou-se uma correlação positiva entre as taxas de absorção de fósforo e a presença na água de numerosos compostos iões, como o
potássio, a disponibilidade de micronutrientes e os compostos orgânicos. Os mecanismos que explicam este fenómeno não estão ainda completamente esclarecidos.
Para muitas das algas estudadas em cultura, a absorção inicial do fosfato e também a assimilação posterior são superiores em presença da luz, especialmente se houver limitação pelo CO2. Em resultado disto faz-se uma acumulação reversível de fosfato na célula, muito do qual
pode ser libertado para o meio. As populações naturais de algas apresentam os ciclos de produção e de crescimento mais ou menos sincronizados, predominando a síntese de proteínas durante a parte inicial do ciclo diurno de luz. Na alga verde Scenedesmus verificou-se que a liber-
tação de fosfato pelas células era máxima durante a fase final do período de luz, antes e durante a divisão celular às escuras (Overbeck, 1962 in Wetzel, 1983).
A absorção de fosfato por células em presença da luz depende em geral da concentração de fósforo no meio, dentro de uma gama que é bastante específica para uma dada espécie de
algas. A assimilação às escuras pelas células que apresentam deficiência em fósforo não depende da concentração de fosfato. Quando existe excesso de fosfato, o conteúdo de fósforo por célula permanece praticamente constante; a quantidade em excesso em relação às necessidades fisio-
lógicas é muitas vezes incorporada e armazenada sob a forma de polifosfatos (Baird, 1995). As taxas de assimilação de fósforo são bastante específicas para os diversos grupos de algas e muitas vezes mesmo para as espécies, dentro do mesmo género. Se, por outro lado, a absorção
de nitrato é independente da concentração de fosfato, o desenvolvimento óptimo de muitas algas dá-se em presença de concentrações mais elevadas de fosfato quando o nitrato, de preferência a amónia, é a fonte de azoto (Wetzel, 1983).
3.13. CONCENTRAÇÕES DE FÓSFORO NECESSÁRIAS PARA O DESENVOLVIMENTO
Do ponto de vista ecológico, o desenvolvimento das algas, tanto em ambientes naturais como nas culturas de laboratório, apresenta dependência em relação à quantidade de fósforo disponível e também à taxa a que este elemento circula. Investigações exaustivas sobre as
concentrações mínima e máxima de fósforo, realizadas principalmente por Rodhe (1948 in Wetzel, 1983), levaram ao agrupamento das algas de água doce em categorias consoante a sua gama de tolerância ficava abaixo de, à volta de, ou acima de 20 µg P04-P/l:
Espécies cujo desenvolvimento óptimo e limite superior de tolerância estão abaixo de 20µg P04-P l-1, como a Uroglena e algumas espécies da macroalga Chara.
Espécies cujo desenvolvimento óptimo se faz abaixo de 20 µg P04-P/l, mas cujo limite de tolerância está bastante acima deste nível, como a Asterionella e outras diatomáceas.

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 28/46
Espécies cujo desenvolvimento óptimo e limite superior de tolerância estão acima de
20 µg P04-P /l, por exemplo algas verdes como Scenedesmus, Ankistrodesmus e muitas outras.
Em todos aqueles estudos, as concentrações de fósforo dos meios de cultura necessárias para um desenvolvimento óptimo eram quase sempre superiores às encontradas na água dos
habitats naturais onde essas algas vivem. Foram sugeridas muitas explicações para esta diferença, entre elas que factores orgânicos desconhecidos estariam presentes no lago mas não nos meios artificiais das culturas assépticas. Sabe-se actualmente que a massa química de fósforo
inorgânico na água apresenta uma relação muito fraca com a cinética do crescimento. De importância primordial é a rapidez com que o fósforo percorre o ciclo e é permutado entre o fósforo particulado e as fases solúveis orgânica e inorgânica, como já foi discutido
pormenorizadamente. Em segundo lugar tem grande importância ecológica o facto de muitas algas, quando se lhes apresenta um fornecimento suficiente de fósforo, poderem absorvê-lo em quantidades muito superiores às necessidades reais (Kuhl, 1974 in Wetzel, 1983). Parte deste
fósforo é gasto no desenvolvimento activo normal sob a forma de compostos orgânicos e inorgânicos, sendo reciclado rapidamente, pelo menos em parte. Mas uma grande maioria (mais de 95%) do fósforo faz parte da fase particulada das algas e do seston orgânico morto. A
assimilação de quantidades excessivas de fósforo pelas algas, muitas vezes referida como consumo «de luxo», pode fornecer fósforo suficiente para manter o desenvolvimento das algas no epilimnion mesmo quando a concentração externa é muito baixa ou está esgotada.
É claro que esta reciclagem e utilização das reservas de fósforo armazenado não pode
durar muito. Estão continuamente a dar-se perdas através da componente de fósforo coloidal e também por sedimentação da componente de fósforo particulado. São necessárias entradas constantes de fósforo para o sistema, ou da zona litoral, ou por transferência dos estratos
inferiores devida à turbulência, ou ainda de origem externa, de modo a manter o desenvolvimento por períodos longos. A quantidade de fósforo que chega ao sistema, quer de fontes internas quer externas, torna-se assim importante para a manutenção do desenvolvimento das algas, sendo um
dos muitos factores que influenciam o tipo de algas que se desenvolvem num determinado lago ou num certo período do ano.
É portanto mais relevante para a questão do aumento da produtividade das algas encarar as concentrações de fósforo em termos de fósforo total, uma vez que quase todo ele está ligado à componente particulada numa altura ou noutra. Dos poucos estudos que existem em que se
analisaram as espécies mais frequentes de algas em relação às necessidades de fósforo, em lagos de diferentes produtividades, pode avaliar-se a quantidade mínima de fósforo necessária por volume das células (Ilustração 11).
Ilustração 11 – Requisitos mínimos de fósforo por
unidade de volume das células de várias algas frequentes
em lagos de produtividade progressivamente crescente.
ALGAS REQUISITO MÍNIMO DE P,
EM µG MM-3 VOLUME CÉL.
Asterionella <0,2
Fragilaria 0,2-0,35
Tabellaria 0,45-0,6
Scenedesmus >0.5
Oscillatoria >0,5
Microcytis >0,5

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 29/46
A diatomácea colonial Asterionella formosa encontra-se frequentemente em águas
oligotróficas e necessita de pequenas quantidades de fósforo, podendo atingir a densidade máxima com concentrações muito baixas deste elemento. A quantidade de fitoplâncton, expressa em volume de células, que pode ser produzida com 1 µg P /l é entre 2 e 5 mm3 P/l (Vollenweider,
1968 in Wetzel, 1983). Esta é uma quantidade de algas que se encontra frequentemente nos lagos de baixa produtividade. As diatomáceas Tabellaria e Fragilaria atingem densidades
populacioais máximas a concentrações de fósforo de aproximadamente 45 µPO4-P/l, ao passo que Scenedesmus necessita de concentrações mais elevadas, de cerca de 500 µg-1.
3.14. ABSORÇÃO DE FÓSFORO PELAS ALGAS VS. BACTÉRIAS
Foram comparados, para duas diatomáceas e três bactérias, o conteúdo mínimo de fósforo,
as taxas de crescimento quando o fósforo não era limitante, a constante de semi-saturação de Michaelis para a absorção e a taxa máxima de absorção de fósforo. Estas descobertas particularmente brilhantes de Fuhs e seus colaboradores têm implicações de alcance muito
grande. O gama de concentrações para a limitação da taxa de crescimento pelo fosfato inorgânico é inferior a 3 µg P/l, desde que não haja esgotamento à superfície da célula. Verificou-se que as taxas máximas de crescimento variavam imenso, dependendo no entanto o conteúdo de fósforo
do volume do protoplasma. As taxas máximas de absorção de ortofosfato por unidade de área da superfície celular foram semelhantes para todas as espécies. As bactérias apresentaram uma menor afinidade para o ortofosfato do que as algas, mas podiam potencialmente desenvolver-se
melhor devido a uma razão área superficial/volume mais favorável.
Com os estudos sobre competição pelo fosfato entre a alga Scenedesmus e a bactéria Pseudomonas provou-se que o desenvolvimento das algas sofre uma limitação aguda em presença das bactérias, sendo o desenvolvimento destas pouco afectadas pelas algas (Rhee, 1972 in Wetzel, 1983). Considerando como primeiro sintoma de competição a cessação do desenvolvimento exponencial das algas, pode dizer-se que ela foi sempre observada algum tempo depois de a concentração externa de fosfato se ter esgotado. A concentração de polifosfatos
armazenados internamente nas algas diminuiu para um valor crítico de cerca de zero na altura em que o seu desenvolvimento foi suspenso e se deu uma progressão muito mais rápida das bactérias.
4. O HOMEM E O CICLO DO FÓSFORO NOS LAGOS
4.1. ORIGEM DO FÓSFORO 4.1.1. PRECIPITAÇÃO
O conteúdo de fósforo na precipitação e na deposição seca de matéria particulada varia
muito, tendo como resultado, portanto, uma sobrecarga também variável de fósforo para os lagos
(Chapin & Uttormark, 1973 in Wetzel, 1983). Em geral, a precipitação contribui com menos fósforo que azoto. Nas zonas agrícolas onde se emprega muito a fertilização, o conteúdo de
fósforo da precipitação é muito mais elevado durante a estação activa de crescimento do que no Inverno. Os ventos dominantes e a água das tempestades que se dirigem do mar para a terra transportam normalmente pouco fósforo, podendo conter maiores quantidades nas zonas
costeiras (Graham et al, 1979 in Wetzel, 1983). A fonte principal de fósforo na precipitação é a

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 30/46
poeira originada sobre a terra, proveniente da erosão do solo e também da contaminação urbana
e industrial da atmosfera. O conteúdo de fósforo da precipitação é geralmente baixo, sendo normalmente inferior a
30 µg P/l nas regiões não povoadas, sobre a terra. Este conteúdo aumenta consideravelmente, para bastante mais que 100 µg/l, perto dos aglomerados urbanos e industriais. Com base em
relativamente poucos dados, sabe-se que a contribuição em fósforo por parte da atmosfera é aproximadamente de 0,01 até 0,1 g m-2 ano-1 (0,1-1,0 Kg ha-1 ano-1), estando a maior parte dos valores actualmente situados na porção mais baixa daquela gama. Esta entrada de fósforo vindo
da atmosfera é, no entanto, mais significativa do que geralmente se pensa, a julgar pelo valor 0,07 g m-2 ano-1 que Vollenweider (1968 in Wetzel, 1983) estimou como sendo, em geral, a taxa de sobrecarga permissível de fósforo para os lagos. Valores como 0,13 g m-2 ano-1 ou mais são
considerados perigosos do ponto de vista de controlo da eutrofização nos lagos com profundidade média inferior a 5 m.
Foi já feito e continua a fazer-se um grande número de análises químicas das precipitações húmidas e seca provenientes da atmosfera, a nível internacional, devido ao enorme potencial de sobrecarga vinda da poluição atmosférica. Há imensos relatórios publicados sobre a sobrecarga
atmosférica que afecta bacias de drenagem inteiras. A sobrecarga de origem atmosférica varia imenso, mas na maioria dos casos as quantidades de fósforo e azoto com esta proveniência constituem uma parte significativa da sobrecarga total, não podendo ser ignoradas. De notar
também que os nutrientes se acumulam nos montes de neve e no gelo durante o Inverno, sendo libertados rapidamente em grandes quantidades durante o degelo da primavera (English, 1978 in Wetzel, 1983). 4.1.2. A ÁGUA SUBTERRÂNEA
O conteúdo de fósforo da água subterrânea é normalmente baixo; a concentração média é
de cerca de 20 µg P/l, mesmo nas regiões em que o solo contém quantidades de fósforo
relativamente grandes. Este baixo conteúdo de fósforo é uma consequência da natureza relativamente insolúvel dos minerais que o contêm e da depuração do fosfato que se encontra à superfície do solo feita pelos biota escavadores.
4.1.3. ESCORRÊNCIA TERRESTRE E ÁGUAS CORRENTES
Em geral as características químicas regionais das águas de superfície estão bastante relacionadas com as características do solo das respectivas bacias de drenagem (Vollenweider, 1968 in Wetzel, 1983). Os solos são o reflexo da regionalidade geológica e climática. A drenagem
superficial contribui, muitas vezes de forma importante, com fósforo para os cursos de água e para os lagos. As quantidades de fósforo que são adicionadas a esta drenagem superficial variam com a quantidade do elemento existente no solo, a topografia, a cobertura vegetal, ao material
da rocha-mãe a partir do qual o solo se forma por meteorização possui um conteúdo muito variável de fósforo, aumentando esta variabilidade com a espessura e a heterogeneidade das
camadas estratificadas de solo. As rochas ígneas básicas contêm relativamente pouco fósforo, sob a forma de apatite; as percentagens de fósforo das outras rochas são as seguintes: arenito, aproximadamente 0,02, gneisse 0,04, loess não meteorizado 0,07, andesite 0,16 e diabase 0,03.
Os calcários que contêm aproximadamente 1,3% de P e os poucos depósitos de fosfato rocha

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 31/46
(com 10 a 15% de P) em que se concentraram acumulações de origem biótica, quer fossem os
próprios organismos quer o guano, são em grande parte de origem sedimentar (Hutchinson, 1950 in Wetzel, 1983). As camadas superficiais do solo são relativamente ricas em fósforo orgânico proveniente dos detritos vegetais em vários estados de decomposição pelos fungos e bactérias do
solo. A capacidade de troca dos solos em relação ao fósforo depende da composição destes e aumenta com a presença de quantidades crescentes de colóides orgânicos e inorgânicos. O
fósforo está mais disponível e é mais facilmente lixiviado de solos com pH compreendido entre 6 e 7. Para valores mais baixos de pH o fósforo combina-se rapidamente com o alumínio, o ferro e o manganésio, enquanto que para valores de pH 6 e superiores há quantidades progressivamente
maiores de fosfato que se combinam com o cálcio para formar apatites e fosfatos de cálcio.
O relevo da bacia de drenagem determina o grau de erosão e a exportação subsequente
de nutrientes. As terras planas onde a escorrência é pouca e as taxas de infiltração são relativamente elevadas contribuem com uma carga menor de nutrientes para a escorrência do que as terras de características semelhantes com gradientes mais inclinados (Ilustração 12).
Ilustração 12 – Concentração de fósforo na escorrência
de Miami Silt Loam com diferentes inclinações (Wetzel,
1983).
Além disso a erosão relativa é grandemente afectada pelo tipo de vegetação e pela
aplicação que se faz da terra. Foi feita uma tentativa para classificar as regiões naturais e agrícolas com base nas quantidades de fósforo contidas na escorrência (Ilustração 13). A aplicação de fertilizantes e os processos de gestão das terras, tanto na agricultura como nas
florestas, irão modificar e fazer aumentar em geral estes valores de uma forma considerável.
EXPORTAÇÃO / G M-2 ANO-1
Produtividade Geral do Solo Fósforo PO4-P Fósforo Total
Baixa <0,01 <0,02
Média 0,01 – 0,025 0,02-0,05
Elevada >0,025 >0,05
Ilustração 13 – Agrupamentos de solos de regiões naturais e agrícolas com base na sua
produtividade geral e na exportação de fósforo.
A urbanização tem como consequência o aumento das quantidades de fósforo lançadas nas águas de superfície em proporção aproximadamente directa com a densidade populacional
(Botkin & Keller, 1995). O fósforo proveniente de fertilização residencial intensa, da drenagem de esgotos de tempestades e das folhas (Baird, 1995) pode provocar o aumento das entradas de fósforo para a drenagem superficial. No Lago Gull, por exemplo, que é o maior lago do sudoeste
de Michigan, verificou-se que aproximadamente 24% do fósforo total que entra neste lago em processo rápido de eutrofização era proveniente da fertilização dos relvados a ele adjacentes. O
solo de 75% dos relvados estava, contudo, saturado de fósforo, o que significa que os fertilizantes não eram necessários (Moss, 1972 in Wetzel, 1983). A escorrência superficial recebe grandes quantidades de fósforo dos esgotos domésticos. É evidente que a sobrecarga que vem
nestas águas varia grandemente com a densidade populacional, com o tratamento que se faz aos esgotos para remoção de nutrientes e com os locais de descarga dos efluentes. Os valores médios são apresentados na Ilustração 14. As entradas de origem industrial, especialmente as
INCLINAÇÃO (%) FÓSFORO / G M-2 ANO-1
8 0,06
20 0,2
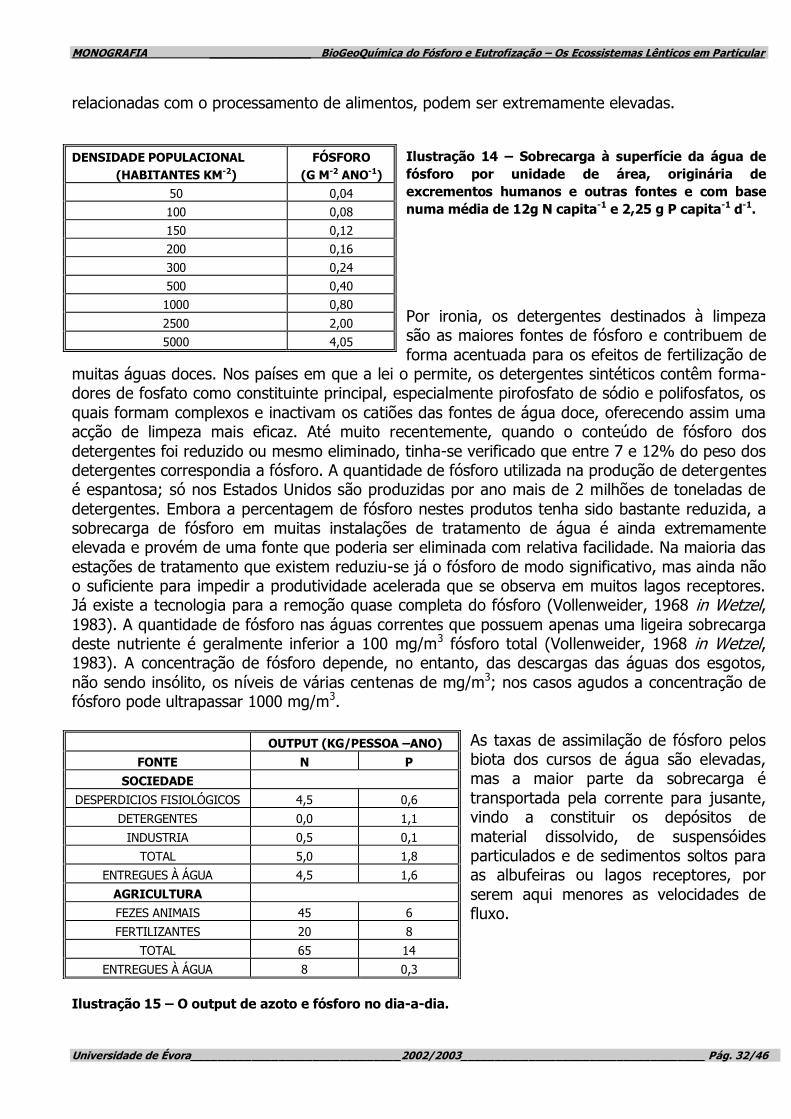
MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 32/46
relacionadas com o processamento de alimentos, podem ser extremamente elevadas.
Ilustração 14 – Sobrecarga à superfície da água de
fósforo por unidade de área, originária de
excrementos humanos e outras fontes e com base
numa média de 12g N capita-1 e 2,25 g P capita-1 d-1.
Por ironia, os detergentes destinados à limpeza são as maiores fontes de fósforo e contribuem de
forma acentuada para os efeitos de fertilização de muitas águas doces. Nos países em que a lei o permite, os detergentes sintéticos contêm forma-dores de fosfato como constituinte principal, especialmente pirofosfato de sódio e polifosfatos, os
quais formam complexos e inactivam os catiões das fontes de água doce, oferecendo assim uma acção de limpeza mais eficaz. Até muito recentemente, quando o conteúdo de fósforo dos
detergentes foi reduzido ou mesmo eliminado, tinha-se verificado que entre 7 e 12% do peso dos detergentes correspondia a fósforo. A quantidade de fósforo utilizada na produção de detergentes é espantosa; só nos Estados Unidos são produzidas por ano mais de 2 milhões de toneladas de
detergentes. Embora a percentagem de fósforo nestes produtos tenha sido bastante reduzida, a sobrecarga de fósforo em muitas instalações de tratamento de água é ainda extremamente elevada e provém de uma fonte que poderia ser eliminada com relativa facilidade. Na maioria das
estações de tratamento que existem reduziu-se já o fósforo de modo significativo, mas ainda não o suficiente para impedir a produtividade acelerada que se observa em muitos lagos receptores. Já existe a tecnologia para a remoção quase completa do fósforo (Vollenweider, 1968 in Wetzel, 1983). A quantidade de fósforo nas águas correntes que possuem apenas uma ligeira sobrecarga deste nutriente é geralmente inferior a 100 mg/m3 fósforo total (Vollenweider, 1968 in Wetzel, 1983). A concentração de fósforo depende, no entanto, das descargas das águas dos esgotos,
não sendo insólito, os níveis de várias centenas de mg/m3; nos casos agudos a concentração de fósforo pode ultrapassar 1000 mg/m3.
As taxas de assimilação de fósforo pelos biota dos cursos de água são elevadas, mas a maior parte da sobrecarga é
transportada pela corrente para jusante, vindo a constituir os depósitos de
material dissolvido, de suspensóides particulados e de sedimentos soltos para as albufeiras ou lagos receptores, por
serem aqui menores as velocidades de fluxo.
Ilustração 15 – O output de azoto e fósforo no dia-a-dia.
DENSIDADE POPULACIONAL
(HABITANTES KM-2)
FÓSFORO
(G M-2 ANO-1)
50 0,04
100 0,08
150 0,12
200 0,16
300 0,24
500 0,40
1000 0,80
2500 2,00
5000 4,05
OUTPUT (KG/PESSOA –ANO)
FONTE N P
SOCIEDADE
DESPERDICIOS FISIOLÓGICOS 4,5 0,6
DETERGENTES 0,0 1,1
INDUSTRIA 0,5 0,1
TOTAL 5,0 1,8
ENTREGUES À ÁGUA 4,5 1,6
AGRICULTURA
FEZES ANIMAIS 45 6
FERTILIZANTES 20 8
TOTAL 65 14
ENTREGUES À ÁGUA 8 0,3

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 33/46
Ilustração 16 – Comparação de valores da concentração de
fósforo e azoto em diferentes domínios. (Fonte: Council of
Environmental Quality, 1978 in Botkin & Keller, 1995)
4.2. EFEITO DA CONCENTRAÇÃO DE FÓSFORO SOBRE A PRODUTIVIDADE DOS LAGOS
Embora existam muitas definições de eutrofização dos lagos, baseadas numa série de condições sinergísticas relacionadas com a produtividade crescente, o consenso entre os limnologistas é de que o termo eutrofização e sinónimo de aumento nas taxas de crescimento dos
biota dos lagos, e de que a taxa de aumento da produtividade é acelerada em relação à que existiria se o sistema não tivesse sido perturbado. A variável mais conspícua, mais básica e mais fácil de medir, para averiguar o aumento da produtividade, é o aumento da quantidade de
carbono assimilado pelas algas e pelas plantas vasculares numa dada área (Levins, 1968). Na grande maioria das circunstâncias os factores mais importantes a considerar na viragem
de um estado menos produtivo para outro mais produtivo são o fósforo e o azoto. A matéria orgânica tipicamente vegetal das algas e das macrófitas aquáticas contém fósforo, azoto e carbono nas seguintes proporções aproximadas (Vallentyne, 1974 in Wetzel, 1983):
1P: 7N: 40C por 100 de peso seco, ou
1P: 7N: 40C por 500 de peso húmido.
Se um destes três elementos for limitante e todos os outros estiverem presentes em
excesso em relação às necessidades físicas, o fósforo pode teoricamente traduzir-se em 500 vezes o seu peso de algas vivas, o azoto em 71 (500:7) vezes e o carbono em 12 (500:40) vezes.
A comparação das quantidades relativas dos diferentes elementos necessários ao desenvolvimento das algas com as quantidades dos mesmos à disposição na água doce ilustra bem a importância geral do fósforo e do azoto. Foram demonstradas razões de proporcionalidade
semelhantes para a comparação entre as exigências das plantas terrestres e a quantidade dos elementos acessíveis na litosfera (Hutchinson, 1975). Embora possa acontecer que haja variações das condições de solubilidade ou de disponibilidade que por vezes transformem elementos muito
abundantes (como a sílica, ferro e certos micronutrientes) em quase impossíveis de obter, o fósforo, e logo a seguir o azoto, são geralmente os primeiros a impor limitações ao sistema. Esta relação torna-se mais nítida quando se compara a razão necessidade/fornecimento médios no fim
do Inverno, antes do pico de algas que aparece nos lagos das regiões temperadas na Primavera, e também durante os meses intermédios do Verão, em que a produtividade é mantida nos valores

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 34/46
máximos (Ilustração 17).
RAZÃO DA PROCURA PARA A DISPONIBILIDADE
ELEMENTO FIM DO INVERNO MEADOS DO VERÃO
FOSFORO 80000 ATÉ 800000
AZOTO 30000 ATÉ 300000
CARBONO 5000 ATÉ 6000
FERRO, SILICIO GERALMENTE BAIXA E VARIAVEL
TODOS OS OUTROS ELEMENTOS <1000 <1000
Ilustração 17 – Comparação das razões das concentrações necessárias de nutrientes inorgânicos para
as concentrações médias disponíveis em água doce.
Os lagos oligotróficos são frequentemente limitados pelo fósforo e possuem azoto em
excesso. O primeiro efeito que se verifica à medida que estes lagos se tornam mais produtivos é um aumento da sobrecarga de fósforo. Como foi dito anteriormente, as concentrações
instantâneas de fósforo normalmente diminuem e variam bastante, mas a taxa de renovação aumenta muito. Aumentam as taxas de perda mas a elevada produtividade das algas exige uma sobrecarga contínua de fósforo para o sistema, proveniente de fontes alóctones e do litoral.
Ilustração 18 – A posição de importantes lagos
relativamente à recepção anual de fósforo e sua
profundidade, diferenciando, assim, lagos
eutróficos de lagos oligotróficos. Para lagos que
tem elevadas descargas poluentes, verificou-se a
passagem da condição (O) para condição ().
(Fonte: Vollendeiwe, 1968 in The Functioning of
Freshwater Ecosystems (LE CREN & LOWE-
McCONELL, ????).
O azoto combinado, embora se encontre em quantidades avultadas na litosfera, não está em grande parte à disposição das plantas. O fornecimento de azoto às águas doces é aumentado mais facilmente a partir do exterior do que o do fósforo. Em circunstâncias de extrema eutrofia a
utilização pelo plâncton do azoto combinado pode ultrapassar as entradas e de facto esgotar a reserva existente na zona trofogénica. Quando isto acontece pode dar-se um aumento das entradas de azoto através da fixação de azoto molecular pelas cianobactérias possuidoras de
heterocistos, embora isso seja conseguido à custa de um elevado preço metabólico. O potencial que o carbono inorgânico possui para se tornar limitante da eutrofização tem
sido objecto de muita discussão. Em algumas circunstâncias de extrema produtividade, como acontece nas lagoas formadas por águas de esgotos, em que o fósforo e o azoto estão à disposição em quantidades muito superiores às necessárias, o carbono pode tornar-se o elemento
limitante do desenvolvimento das algas (Kerr et al., 1972 in Wetzel, 1983). Embora haja algumas provas de que o carbono inorgânico é o elemento limitante em lagos de águas extremamente
macias (Allen, 1972 in Wetzel, 1983), a difusão do CO2 atmosférico é em geral suficiente para sustentar as necessidades de carbono das populações fitoplanctónicas. É extremamente improvável que a limitação pelo carbono seja significativa em relação ao desenvolvimento das

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 35/46
populações de algas na maior parte das águas duras em que se verifica a existência de elevadas
densidades destes organismos em condições de eutrofia. A importância do fósforo em comparação com o azoto e o carbono ficou particularmente
bem ilustrada pelas experiências de fertilização em grande escala (Schindler, 1974 in Wetzel, 1983). O Lago 226, localizado a noroeste de Ontário sobre rocha do Escudo Precambriano, é
química e biologicamente semelhante a mais de 50% das águas que drenam os Grandes Lagos Laurentinos. Tirando partido de uma constrição existente na bacia, a zona trofogénica do Lago 226 foi dividida em dois lagos (Ilustração 19). Uma das bacias resultantes foi fertilizada com
fósforo, azoto e carbono e a outra com concentrações equivalentes de azoto e carbono (adicionou-se o equivalente a 3,16g de NO3-N e 6,05g de sacarose-C por ano a ambas as bacias, em 20 incrementos semanais iguais. A bacia de nordeste recebeu ainda 0,6g m-2 ano-1 de PO4-P.
As razões N/P e C/P eram superiores às dos esgotos tratados, mas a quantidade de P adicionado à água não foi excepcionalmente elevada se se comparar com lagos que recebem normalmente poluição de origem doméstica). A bacia enriquecida com fosfato depressa se tornou altamente
eutrófica, enquanto que a bacia que recebia apenas azoto e carbono permaneceu nas condições que apresentava anteriormente à fertilização (Ilustração 19).
Nesta bacia enriquecida com fósforo, bem como em vários outros lagos sujeitos a tratamento análogo durante um período de vários anos, a biomassa das algas aumentou duas ordens de grandeza em relação à dos lagos que receberam apenas enriquecimento em azoto e
carbono. A recuperação e o regresso a perto dos níveis de pré-fertilização foram muito rápidos, uma vez cortado o fornecimento de fosfato.
É tão grande a quantidade de provas deste tipo de relações, tanto da investigação
experimental como da aplicada, que é difícil acreditar que se tenha gerado no Canadá, nos fins
dos anos 60, uma controvérsia tão acesa sobre a maior importância de carbono em relação ao fósforo como elemento limitante na água doce. Em consequência do empolamento que se deu a alguns resultados obtidos com culturas de cianobactérias e de uns poucos, raros, casos de
descoberta de pouca disponibilidade de carbono em habitats naturais em que existiam quantidades excessivas de azoto e fósforo, este último foi apontado como de menor importância que o carbono para a aceleração do processo de eutrofização.
Esta interpretação errónea dos resultados feita por alguns cientistas foi tão convincente, e
tão eficazmente foi a situação explorada pela indústria de detergentes e por jornalistas
irresponsáveis, que se tornou extremamente urgente legislar no sentido de reduzir a sobrecarga de fósforo nos efluentes que eram lançados nas águas de superfície. O assunto foi finalmente encerrado depois de ter sido refutado por Vallentyne (1970 in Wetzel, 1983) e numerosas
investigações subsequentes, acrescidas de um simpósio nacional sobre o assunto (Likens, 1972 in Wetzel, 1983). Continuam no entanto a sentir-se efeitos da controvérsia tanto na investigação que se faz nesta área como nos esforços que são feitos a fim de reduzir a sobrecarga das águas
doces em milhões de toneladas de fósforo por ano para impedir que os lagos não se transformem em taças cheias de algas.

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 36/46
Ilustração 19 - O Lago 226 do noroeste de Ontário, que foi dividido na constrição da bacia, em 4 de Setembro de 1973. A bacia situada a nordeste, mais afastada, fertilizada com fósforo, azoto e carbono, ficou coberta por um “bloom” denso de algas num período de dois meses. Não
foram observados aumentos nas populações de algas nem alterações da composição específica na bacia mais próxima, que recebeu quantidades semelhantes de azoto e carbono mas não fósforo. (Schindler, D.W. in Freedman, 1995)
4.3. A SOBRECARGA DE FÓSFORO E AZOTO E A PRODUTIVIDADE DAS ALGAS
Quando se adiciona fósforo de uma forma intermitente aos lagos ou tanques pouco produtivos, quer experimentalmente para efectuar uma fertilização intencional quer através de efluentes que resultam da actividade do homem, a resposta do sistema é geralmente um
aumento muito rápido da produtividade das algas (Vollenweider, 1968 in Wetzel, 1983). Esta produtividade crescente não pode, no entanto, ser mantida de forma contínua, diminuindo
rapidamente em poucas semanas ou meses para níveis próximos dos anteriores à adição do nutriente. Dão-se perdas contínuas de fósforo na zona trofogénica, tanto na forma coloidal como por sedimentação do fósforo particulado. As entradas para o sistema têm de ser mantidas, ou de
forma contínua ou intermitentemente, de modo a conservar o aumento da produtividade. Por outras palavras, considera-se um factor crítico para a manutenção de uma produtividade crescente na maioria dos lagos com produtividade biológica baixa ou média que haja uma
sobrecarga de fósforo administrada continuamente ao sistema. Pelo contrário, para reduzir a produtividade de um lago que está a receber uma sobrecarga
contínua de nutrientes, obtém-se em geral mais eficazmente a diminuição do desenvolvimento das algas com uma redução das entradas de fósforo para valores abaixo da quantidade de fósforo perdido pelo sistema. O objectivo é a redução do fósforo total da água, pois este é o nutriente
principal que é necessário em quantidades muito superiores à oferta. Por outro lado o fósforo reage quimicamente, é tecnologicamente mais fácil de remover da água que o azoto e não possui reservatórios importantes na atmosfera.
4.4. SOBRECARGA DE NUTRIENTES
O conceito de sobrecarga de nutrientes implica uma relação entre a quantidade de um

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 37/46
nutriente que entra numa massa de água e a resposta desta a essa entrada. Os efeitos desta
relação podem ser expressos por um índice quantificável da produtividade ou por uma variável relacionada com a qualidade da água (como a concentração da clorofila, transparência da água).
Foi reconhecido há já algum tempo que o conceito de sobrecarga era aplicável às alte-rações a que ficam sujeitos os lagos em resposta a um enriquecimento com fósforo e azoto
(Edmondson, 19??). Sawyer (1947 in Wetzel, 1983) sugeriu que, se os níveis críticos de azoto inorgânico dissolvido (300 µg N/l) e fósforo inorgânico dissolvido (10 µg P/l) medidos por altura da circulação da primavera nos lagos de Wisconsin fossem ultrapassados, então dar-se-ia
provavelmente o desenvolvimento exagerado de populações de fitoplâncton durante a estação de crescimento. Vollenweider (1968 in Wetzel, 1983) foi o primeiro investigador a formular critérios quantitativos definidos de sobrecarga para o fósforo e o azoto bem como as condições tróficas
esperadas das massas de água. Definiu fronteiras entre os lagos oligotróficos e eutróficos por meio de relações entre as sobrecargas de nutrientes e a profundidade média (tomada como uma medida do volume dos lagos), e mais tarde aperfeiçoou estas relações (Vollenweider, 1975 in Wetzel, 1983). Os critérios de sobrecarga e os modelos com eles relacionados preocupam-se sobretudo com o fósforo porque este é, com muita frequência, o nutriente que primeiro se torna limitante para as algas, como já foi discutido, e porque os muitos processos que constituem o
ciclo do azoto (nitrificação, desnitrificação, fixação de azoto) tornam complicadas as medições rigorosas da sobrecarga de azoto para os lagos.
As relações de sobrecarga baseiam-se todas na equação de equilíbrio de massas de uma substância M entre as suas fontes e as suas perdas (depósitos) (Vollenweider et al., 1980):
ΔM//Δt = I-O-(S-R) (1)
onde ΔM/Δt = ganho ou perda da reserva do nutriente M durante o tempo Δt I = sobrecarga do nutriente vinda do exterior O = perda do nutriente por efluente
S = perda do nutriente para os sedimentos R = regeneração do nutriente a partir dos sedimentos (sobrecarga interna).
Enquanto que I e O podem ser medidos rigorosamente e de uma forma directa, (S-R) é mais difícil de medir, obtendo-se muitas vezes por diferença. Em condições de estado estacionário, ΔM/Δt = 0. Embora este estado na realidade não exista num lago, pode observar-se
naqueles que oscilam em torno de sobrecargas de nutrientes relativamente constantes com períodos de vários anos, que apresentam uma série de características tróficas, podendo considerar-se como se estivessem num estado estacionário repetitivo. Por vezes este é definido
como o conteúdo de reserva de nutrientes medido durante a circulação da primavera ou como o conteúdo anual da reserva. Nos modelos que se baseiam no equilíbrio de massas supõe-se ainda que a sobrecarga do nutriente se mistura instantânea e completamente na água do lago, uma
suposição que para a maior parte dos lagos é verdadeira apenas em parte. Estes modelos destinam-se, por consequência, a ser aplicados a uma grande população de lagos e não à
descrição do comportamento específico de um lago em particular. Supondo condições de estado estacionário, a equação simples do equilíbrio de massas tem
em conta a sobrecarga de fósforo por volume, assim como as perdas por descarga e por

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 38/46
sedimentação (Vollenweider e Kerekes, 1980 in Wetzel, 1983).
Quando qualquer destes modelos é aplicado e com os dados se constrói um gráfico para
mostrar o grau de previsão da concentração de fósforo proveniente da sobrecarga na circulação
da primavera, o que se obtém é uma descrição do comportamento estatístico médio dos lagos em termos da relação entre a sobrecarga de fósforo e a acumulação do elemento dentro do lago
(Vollenweider, 1976 in Wetzel, 1983). Nos lagos cujo comportamento se afasta do modelo pode provar-se que existem mecanismos que conduzem à sobrestimativa e à subestimativa da sobrecarga (como as taxas de sedimentação elevadas, regeneração de muito fósforo a partir dos
sedimentos).
4.5. RESPOSTAS BIOLÓGICAS À SOBRECARGA DE NUTRIENTES
Com base nestas dependências da sobrecarga de nutrientes podem relacionar-se as
sobrecargas para um determinado lago com as respostas biológicas como a biomassa do
fitoplancton (como a concentração de clorofila) e a produtividade. As primeiras previsões dos níveis de clorofila feitas a partir das concentrações de fósforo nos lagos durante a circulação da primavera ou na estação de crescimento (Schindler, 1978 in Wetzel, 1983), foram mais tarde
alargadas a maiores conjuntos de dados relativos a todo o mundo (Vollenweider, 1976 in Wetzel, 1983). Dentro de certos limites, a regressão entre a sobrecarga de fósforo e a concentração média anual de clorofila obtém-se aproximadamente pela fórmula:
76,0
) ))1/()((55,0).( wPachl
Foram verificadas relações para muitos lagos, apenas com desvios de menor importância. A taxa anual da produtividade primária das algas foi também relacionada com a con-
centração prevista de fósforo (Ilustração 20). O gráfico log-log não é linear devido aos efeitos de redução da luz por ensombramento das populações muito densas de algas (biomassa elevada)
quando os níveis de produtividade são elevados. Esta relação pode representar-se por:
gC( m-2 ano-1) = 7
76,0
76,0
1
)1/()(011,03,0
))1/()((
wi
w
P
P
que funciona de modo semelhante ao integral da fotossíntese diária e se baseia nas concen-trações médias de clorofila e na extinção da luz em consequência da turbidez e da presença de substâncias orgânicas dissolvidas (Vollenweider e Kerekes, 1980 in Wetzel, 1983).
Ilustração 20 – Produtividade primária anual (g C m-2 ano-1) expressa como função da concentração prevista de fósforo total (Vollenweider, 1979 in Wetzel, 1983)
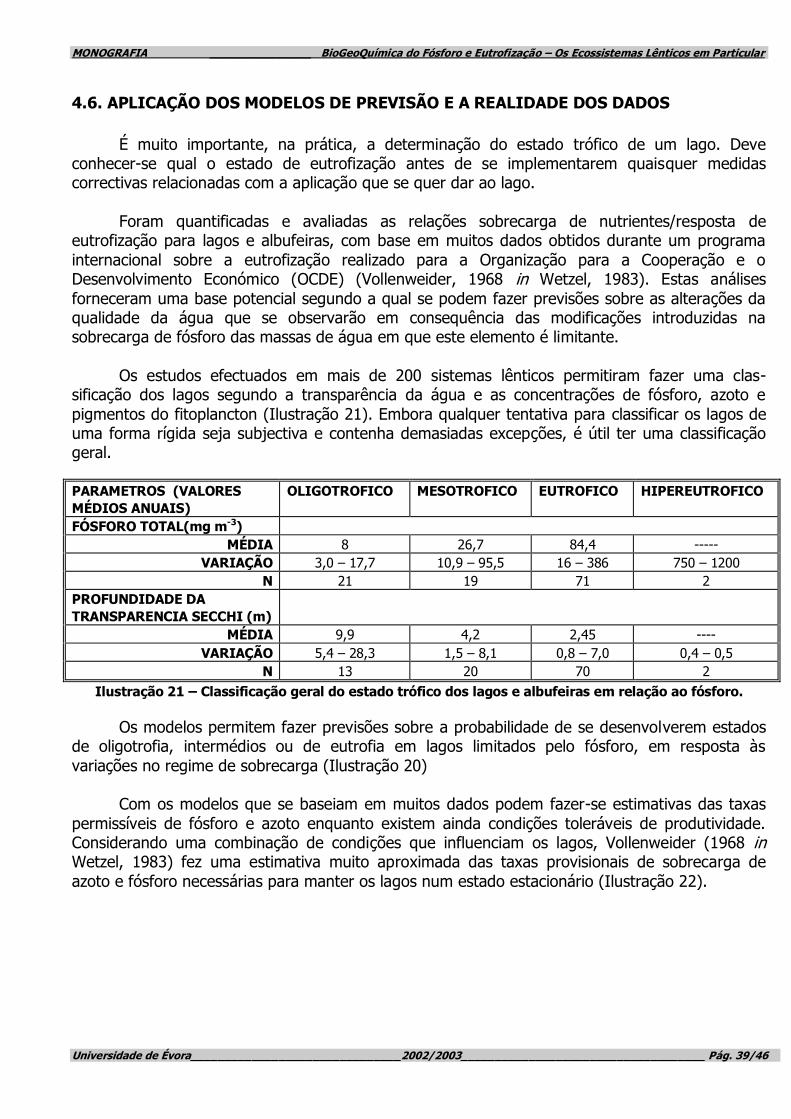
MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 39/46
4.6. APLICAÇÃO DOS MODELOS DE PREVISÃO E A REALIDADE DOS DADOS
É muito importante, na prática, a determinação do estado trófico de um lago. Deve conhecer-se qual o estado de eutrofização antes de se implementarem quaisquer medidas correctivas relacionadas com a aplicação que se quer dar ao lago.
Foram quantificadas e avaliadas as relações sobrecarga de nutrientes/resposta de
eutrofização para lagos e albufeiras, com base em muitos dados obtidos durante um programa
internacional sobre a eutrofização realizado para a Organização para a Cooperação e o Desenvolvimento Económico (OCDE) (Vollenweider, 1968 in Wetzel, 1983). Estas análises
forneceram uma base potencial segundo a qual se podem fazer previsões sobre as alterações da qualidade da água que se observarão em consequência das modificações introduzidas na sobrecarga de fósforo das massas de água em que este elemento é limitante.
Os estudos efectuados em mais de 200 sistemas lênticos permitiram fazer uma clas-
sificação dos lagos segundo a transparência da água e as concentrações de fósforo, azoto e
pigmentos do fitoplancton (Ilustração 21). Embora qualquer tentativa para classificar os lagos de uma forma rígida seja subjectiva e contenha demasiadas excepções, é útil ter uma classificação geral.
PARAMETROS (VALORES
MÉDIOS ANUAIS)
OLIGOTROFICO MESOTROFICO EUTROFICO HIPEREUTROFICO
FÓSFORO TOTAL(mg m-3)
MÉDIA 8 26,7 84,4 -----
VARIAÇÃO 3,0 – 17,7 10,9 – 95,5 16 – 386 750 – 1200
N 21 19 71 2
PROFUNDIDADE DA
TRANSPARENCIA SECCHI (m)
MÉDIA 9,9 4,2 2,45 ----
VARIAÇÃO 5,4 – 28,3 1,5 – 8,1 0,8 – 7,0 0,4 – 0,5
N 13 20 70 2
Ilustração 21 – Classificação geral do estado trófico dos lagos e albufeiras em relação ao fósforo.
Os modelos permitem fazer previsões sobre a probabilidade de se desenvolverem estados de oligotrofia, intermédios ou de eutrofia em lagos limitados pelo fósforo, em resposta às
variações no regime de sobrecarga (Ilustração 20) Com os modelos que se baseiam em muitos dados podem fazer-se estimativas das taxas
permissíveis de fósforo e azoto enquanto existem ainda condições toleráveis de produtividade. Considerando uma combinação de condições que influenciam os lagos, Vollenweider (1968 in Wetzel, 1983) fez uma estimativa muito aproximada das taxas provisionais de sobrecarga de
azoto e fósforo necessárias para manter os lagos num estado estacionário (Ilustração 22).

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 40/46
Quando se aumenta a sobrecarga é de esperar que haja um aumento rápido da eutrofização. Embora estes modelos previsivos tenham sido
muito melhorados desde o seu aparecimento, o nível de sobrecarga tolerável de 10 mg P/ m3
mantém-se sem alteração, enquanto que o nível de sobrecarga excessiva foi ligeiramente aumentado para 25 mg P/m3 (Vollenweider e
Kerekes, 1980 in Wetzel, 1983).
Ilustração 22 – Níveis permitidos de
sobrecarga para o fósforo totais
(bioquimicamente activos) em g m-2 ano-1.
Podem conseguir-se muito bons resultados na redução da produtividade dos lagos diminuindo a sobrecarga de fósforo. Entre os exemplos mais frequentemente discutidos está o do
Lago Washington, em Seattle, Washington (Freedman, 1995). O Lago Washington foi enriquecido com volumes crescentes de um efluente de estações de tratamento secundário de esgotos desde
1941 até 1953. A produção aumentou enormemente e as algas tornaram-se mais abundantes. A concentração de fosfato aumentou também proporcionalmente, muito mais que as de nitrato e de dióxido de carbono. Em 1963 o efluente foi desviado por volta de 1969, o fosfato tinha diminuído
para 28 por cento do seu valor em 1963. A concentração de clorofila no verão diminuiu praticamente na mesma proporção, mas o nitrato e o CO2 total flutuavam de um ano para o outro à volta de valores relativamente elevados. A redução do fitoplâncton e do fósforo continua a
verificar-se (Edmondson, 19??). A comparação de dois lagos, o Char Lake e o Meretta Lake, no árctico vem também
confirmar que os lagos com maior concentração de fósforo e azoto são os que estão mais eutrofizados.
Ilustração 23 – Comparação de 2 lagos High Arctic Lakes a 75ºN na Ilha Cornawallis, Canada. Char
Lake é um típico lago polar ultraoligotrofico enquanto o Meretta Lake é eutrofico. (Freedman, 1995)
A taxa a que se dá a regressão da produtividade de um lago para as condições existentes antes de se ter aumentado a sobrecarga varia muito e depende da morfologia da bacia, das
características químicas da água e da natureza das fontes de fósforo (difuso ou concentrado em fontes pontuais). A sobrecarga de azoto e fósforo para os lagos é ainda influenciada pela razão da área da superfície da bacia de drenagem para a área da superfície do lago (Freedman, 1995).
Dependendo das perdas percentuais sofridas pela terra circundante, que pode em parte ser terra cultivada, as entradas aumentam grosso modo em proporção à razão do aumento verificado nos
PROFUNDIDA
DE MÉDIA (M)
SOBRECARGA
PERMITIDA
(P)
SOBRECARG
A
PERIGOSA
(P)
5 0,07 0,13
10 0,10 0,20
50 0,25 0,50
100 0,40 0,80
150 0,50 1,00
200 0,60 1,20

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 41/46
«arredores».
Depois da redução das taxas de sobrecarga para as anteriores ao aumento das entradas ou
para ainda menos que estas, são necessários entre 2 a 10 anos para que os lagos de dimensões e
tempo de reenchimento hidrológico médios possam recuperar. A taxa a que se faz a recuperação, no entanto, é muito mais lenta nos lagos menos profundos onde a «sobrecarga interna» de
fósforo vindo dos sedimentos é elevada. Por exemplo no lago Shagawa, em Minnesota, que é eutrófico, a sobrecarga externa de fósforo foi reduzida em mais de 70% e mesmo assim não se atingiram os níveis baixos de equilíbrio que tinham sido previstos (os níveis da concentração de
fósforo observados depois da diversão ultrapassavam em mais de 100% aquilo que tinha sido previsto) (Larsen et al., 1975 in Wetzel, 1983). O Lago Shagawa é pouco profundo, possui um hipolimnion anaeróbio e uma zona litoral muito extensa e coberta de macrófitas submersas,
contribuindo todos estes factores para fazer aumentar a libertação de fósforo dos sedimentos. A sobrecarga interna derivada dos sedimentos é típica de numerosos outros lagos pouco profundos (Cooke et al., 1977 in Wetzel, 1983), o que faz com que a recuperação dos lagos por intermédio
da redução da sobrecarga alóctone de fósforo seja um processo muito mais lento. Na maior parte dos lagos, no entanto, a contribuição da «sobrecarga interna» constitui uma porção relativamente pequena em relação à sobrecarga externa de fósforo (Edmondson, 19??), de modo que o controlo
das taxas de sobrecarga produz efeito na maior parte das vezes quando é aplicado na gestão do estado trófico dos lagos.
5. POSSIVEIS MEDIDAS DE MINIMIZAÇÃO E REGRESSÃO TRÓFICA (RECUPERAÇÃO)
5.1. CONTROLO DA EUTROFIZAÇÃO E REGRESSÃO TRÓFICA
Associando um pouco a cadeira de Métodos de Controlo e Monitorização a este trabalho,
poder-se-à também tentar levar à recuperação dos lagos eutrofizados. É uma área científica relativamente recente que só nos anos setenta se implantou, à medida que a gravidade e extensão das situações de eutrofização eram evidentes um pouco por toda a zona norte
temperada. Há cerca de 15 a 20 técnicas diferentes que podem ser utilizadas para melhorar ou manter o estado trófico de uma massa de água lêntica, podendo ser classificadas em formas de controlo externo e interno. Ambas se encontram dedicadas quase em exclusivo ao controlo do
fósforo, porque a resposta do azoto é em geral menos conhecida, menos evidente e menos previsível (Welch, 1992 in Ferreira, ????). Como formas de controlo externo, têm-se: desvio de fontes poluidoras pontuais, tratamento secundário e terciário de efluentes e retenção, desvio ou
tratamento de águas de drenagem. As formas de controlo interno incluem: inactivação do fósforo, diluição das águas enriquecidas, dragagem de sedimentos, circulação artificial da coluna de água, arejamento hipolimnêtico, extracção de água hipolimnética, adição de azoto, biomanipulação (por
introdução de espécies planctívoras, ou seja, comedoras de plâncton), colheita mecânica ou biológica de plantas (por peixes herbivoros) e impermeabilização dos fundos. As acções conducentes à manutenção ou regressão do estado trófico podem ser sistematizadas da seguinte
forma: diagnóstico, controlo externo e controlo interno, das fontes de fósforo, controlo da biomassa vegetal e biomanipulação.
5.2. DIAGNÓSTICO
Verificados os sintomas de eutrofização, tais como concentrações elevadas de fósforo,

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 42/46
diminuição da transparência da água, determinada composição e biomassa de algas e presença
de espumas e aglomerados biológicos nas margens, é necessário definir a situação de referência inicial, antes de ser iniciado um programa de recuperação. Esta situação de referência permite o estabelecimento de um quadro diagnosticante suficientemente pormenorizado para delinear o tipo
e forma de intervenção (ou decidir, pelo contrário, que ainda não é altura de intervir) e também avaliar a eficácia e o custo-benefício das medidas de controlo que venham a ser tomadas. Entre
os elementos mais importantes que devem ser recolhidos para este diagnóstico, incluem-se: Localização, volume, carga nutritiva e periodicidade (por impulsos, continuas, por
arrastamento de chuvas) das fontes pontuais de nutrientes que entram na
albufeira; e avaliação da carga (ou pelo menos da susceptibilidade) de entrada proveniente de fontes difusas de nutrientes que entram na albufeira (por escoamento, drenagem ou mais raramente a partir do nível freático);
Caracterização do regime de variação do volume armazenado na albufeira, ou seja, o balanço das entradas e saidas de água e tempos de retenção hidráulica;
Análise das características da massa de água e diagnóstico pormenorizado do seu
estado trófico, incluindo: concentrações de nutrientes, perfis de temperatura, D.O., pH e condutividade ao longo da coluna de água, transparência de Secchi, biomassa clorofilina, composição e abundância quantitativa do fitoplâncton, num número de
estações de amostragem representativo da heterogeneidade da albufeira, com recolhas de periodicidade, no mínimo, mensal, por um período de um ano.
Este diagnóstico fino é necessário porque a resposta biológica à eutrofização varia ao longo do ano em função de variáveis ambientais como o volume da água armazenada, a temperatura e
a insolação, e registam-se frequentemente manifestações de eutrofização temporalmente isoladas, por exemplo, ao diminuir acentuada e rapidamente o volume da água armazenada. Por vezes este diagnóstico pormenorizado já está realizado quando se verifica a decisão de
recuperação da massa de água. Frequentemente este diagnóstico pormenorizado é utilizado no desenvolvimento e calibração de modelos preditivos da evolução da eutrofização.
5.3. CONTROLO EXTERNO DAS FONTES DE FÓSFORO
O desvio ou o tratamento de efluentes têm sido as técnicas mais utilizadas no controlo da
eutrofização. O tratamento de efluentes deve incluir a retirada dos nutrientes e nomeadamente do fósforo, o que frequentemente é realizado por via química na estação de tratamento (normalmente precipitando o fósforo e matéria orgânica da água residual tratada com sulfato de
alumínio, carbonato de cálcio ou cloreto de ferro) embora daí resulte, em geral, largo volume de lamas mais ou menos tóxicas. Uma alternativa ecologicamente mais interessante consiste em conduzir esta água tratada, mas muito rica e mineralizada, para bacias de recepção (lagoas) onde
é realizada a retirada de nutrientes por incorporação biológica (são utilizados no crescimento de algas e plantas, por sua vez consumidas por invertebrados e peixes, sendo esta biomassa periodicamente removida e rentabilizada). Outra alternativa muito utilizada em pequenos lagos
europeus é a abertura de valas paralelas à margem para onde a água tratada acede, sendo os nutrientes absorvidos por plantas aquáticas e estas utilizadas por fauna selvagem, normalmente
patos (Ferreira, 19??). A recuperação da massa de água após desvio e tratamento dos efluentes pode não ser evidente ou completa, uma vez que as fontes nutritivas internas de fósforo existentes nos sedimentos podem ser suficientes para manter o estado trófico existente (o fósforo
é libertado dos sedimentos em condições anaeróbias, que ocorrem durante parte do ano). Os

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 43/46
lagos mais pequenos e com quociente epilímnio/ hipolímnio elevado são os mais difíceis de
recuperar desta forma. Na maior parte dos casos, ocorre de facto alguma recuperação após desvio de efluentes, dependendo a magnitude desta, também de outros factores, como o tempo de retenção da massa de água.
As fontes difusas de nutrientes são por vezes muito importantes, sobretudo em bacias
muito agricultadas. A remoção destes nutrientes é bem mais difícil e, de uma forma geral, é realizada pela passagem desta água de escoamento em pequenas bacias de retenção e zonas semi-artificiais de arrelvados ou de alagados, interpostos entre as zonas de máximo escoamento e
a massa de água, sendo a água enriquecida aí retida por um período variável. Quanto maior a retenção, maior o efeito de absorção de nutrientes e sua incorporação na biomassa vegetal. A implementação de algumas práticas agrícolas e de uso da bacia hidrográfica é também uma ajuda
no controlo de eutrofização, como a criação de socalcos em curvas de nível, a criação de terraços, o reflorestamento de declives íngremes, a manutenção de uma faixa de vegetação marginal ou a sementeira de corredores de macrófitos (Ferreira, ????).
As concentrações de nutrientes na água podem também ser diminuidas por introdução de
volumes de água com menores concentrações destes, em geral com um aumento associado do
fluxo da água para fora do lago. Esta técnica depende da existência de volumes adequados de água com baixo teor em nutrientes e dos custos envolvidos na operação. Tem sido utilizada com sucesso em Espanha, junto aos Pirinéus, sendo a água introduzida oriunda de uma albufeira
oligotrófica a montante e o transporte efectuado por gravidade. Uma outra técnica consiste simplesmente em diminuir o tempo de retenção da massa de água e aumentar o fluxo de
passagem da água, embora tal exija grandes quantidades de água disponível e apresente fracos resultados se a própria água adicionada estiver eutrofizada (Ferreira, ????)
5.4. CONTROLO INTERNO DO FÓSFORO
As cinco técnicas mais utilizadas neste controlo são a inactivação do fósforo, a técnica
"riplox", o arejamento hipolimnético, a extracção de água hipolimnética e as dragagens. O sulfato de alumínio, quando adicionado à água, normalmente em conjunto com argila bentonítica, forma flocos que precipitam sobre os fundos anóxicos. Estes flocos cobrem os sedimentos evitando a
libertação de fósforo. Contudo, os residuos que permanecem na massa de água são bastante tóxicos. A técnica "riplox" reduz o fósforo solúvel intersticial dos sedimentos por oxidação da matéria orgânica. Esta oxidação é obtida pela desnitrificação resultante da adição de Ca(NO)2 em
condições alcalinas, sendo adicionado ferro quando este está presente em níveis baixos ou se os niveis de enxofre são elevados. O objectivo é manter o ferro sob a forma oxidada de compostos férricos, para que o fósforo se mantenha complexado nestes (Cooke et ai., 1986 in Ferreira, 19??)
embora bastante eficazes, trata-se de métodos ecologicamente pouco interessantes, sobretudo o primeiro, visto que alteram profunda e bruscamente a química do ecossistema.
O arejamento hipolimnético é uma técnica bastante praticada em lagos relativamente profundos embora em geral com resultados pouco permanentes e muito localizados (Welch, 1992
in Ferreira, 19??). A libertação de fósforo dos sedimentos pára enquanto se processa o arejamento, mas recomeça de imediato após o tratamento. Os arejadores apresentam um raio de acção limitado. Na albufeira do Vilar esta técnica vem sendo utilizada para melhorar a qualidade
da água na zona junto à captação, com bons resultados. A extracção de água hipolimnética

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 44/46
apresentou grande sucesso em vários casos, não só diminuindo o fósforo total hipolimnético,
como também mostrando-se correlacionada a diminuição do fósforo epilimnético com a magnitude da extracção da água hipolimnética para fora do lago. Contudo, este método não é eficaz quando a carga externa de nutrientes é elevada e se mantém. A água extraída não deve
ser lançada no rio a jusante, sendo normalmente utilizada para irrigação.
A dragagem, embora de custos elevados, é um dos métodos mais eficazes para promover a regressão trófica, ao extrair as camadas superficiais dos sedimentos ricas em fósforo, normalmente até 1 m de espessura. Um dos exemplos mais conhecidos e bem sucedidos é o do
lago Trummen, mas outros exemplos de massas de água apresentaram melhorias evidentes, nomeadamente portugueses, como a albufeira da Lagoa Azul (Sintra), dragada nos anos oitenta.
6. CONSIDERAÇÕES FINAIS
Muitas definições foram atribuídas ao termo eutrofização entendida aqui como antrópica, para a distinguir do processo de envelhecimento natural das massas de água de lagos naturais. O envelhecimento natural realiza-se um período bastante alargado e consiste na diminuição
progressiva do volume de água por acumulação de sedimentos orgânicos, aumento da concentração de nutrientes e da importância das camadas profundas como fonte destes, avanço da vegetação litoral até ao centro da bacia e incorporação final no meio terrestre. A eutrofização é
um processo que se realiza à escala histórica e é definida como o enriquecimento nutritivo do meio aquático, causado directa ou indirectamente por actividades humanas e provocando o
aumento da produtividade biológica e alterações gerais do metabolismo aquático. E é aqui que entra o fósforo.
O fósforo desempenha um papel muito importante no metabolismo. Em comparação com outros macronutrientes necessários aos biota, o fósforo é o menos abundante de todos. O aumento desmesurado do fluxo de fósforo para a água é devido à crescente erosão dos solos
devido à moderna tecnologia agrícola, o uso de fertilizantes e também devido à prática da desflorestação (Mackson & Jackson, 1996). Os detergentes também desempenham um papel fundamental na eutrofização.
Hoje em dia é globalmente aceite que a eutrofização dos sistemas resulta de uma elevada
produtividade biológica, consequência das descargas de azoto e fósforo existindo uma relação
entre o seu teor e o estado trófico das massas de água. Esta relação é particularmente evidente para o fósforo, considerado elemento-chave com papel limitante na eutrofização, isto é, na produtividade biológica (Fonseca, 2002). A eutrofização causa a diminuição do valor biológico, pa-
trimonial e estético das massas de água e provoca a diminuição da sua potabilidade e a sua insalubridade.
As taxas de renovação do fósforo são geralmente mais rápidos em condições de maior oligotrofia e maior deficiência de fósforo. Quando se adiciona fósforo a águas doces não
produtivos, quer seja experimentalmente quer em resultado das actividades do homem, a resposta é, em geral, um rápido aumento da produtividade das algas. Uma grande quantidade, por vezes superior a 95%, do fósforo, está ligada à fase particulada dos biota, especialmente as
algas.

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 45/46
Foram criados numerosos modelos de equilíbrio de mossas para prever, com base na sobrecarga de fósforo e nos tempos de retenção, as respostas antecipados da biomassa e da produtividade das algas. Estes modelos fazem uma previsão razoavelmente rigoroso do
sobrecarga de fósforo permissível necessário poro atingir um determinado nível de produtividade, se for reduzido a sobrecarga.
Com tudo isto podemos afirmar que cabe a nós tentar minimizar a presença elevada de fósforo nas águas que nos circundam deixando que a Natureza faça o seu trabalho, se possível
sem ser “importunada”.

MONOGRAFIA _______________ BioGeoQuímica do Fósforo e Eutrofização – Os Ecossistemas Lênticos em Particular
Universidade de Évora_______________________________2002/2003____________________________________ Pág. 46/46
REFERÊNCIAS BIBLIOGRÁFICAS BAIRD, C., 1995. Environmental Chemistry. W.H.Freeman and Company, New York;
BOTKIN, D. & E.KELLER, 1995. Environmental Science – Earth as a Living Planet. Jonh Wiley & Sons, Inc; BUTCHER, S.S. & R.J. CARLSON & G.H. ORIANS & G.V. WOLFE, 1992. Global Biogeochemical Cycles. In:
Academic Press; CARRAPETO, C., 1994. Ecologia: Princípios e Conceitos. Universidade Aberta, Lisboa;
EDMONSON, W.T., 1973. Water Quality Manegement and Lake Eutrophication – The Lake Washington Case. Nº XI 139-178
FERREIRA, M.T., ????. Curso de Exploração e Segurança de Barragens. ?????? FONSECA, R.M.F., 2002. Impactos Ambientais Associados a Barragens e a Albufeiras. Estratégia de Re-
Aproveitamento dos Sedimentos Depositados. Dams: Impacts and Hazards. Centro de Geofísica de Évora,
Universidade de Évora FREEDMAN, B., 1995. Environmental Ecology – The Ecological Effects of Pollution, Disturbance and other Stresses.
2ndEdition, Academic Press, Inc.; GOLDMAN, C.R. & A.J.HORNE, 1995. Limnolgy. Internacional Student Edition. McGraw-Hill International Book
Company; GOWER, A., 1980. Water Quality in Catchement Ecosystems. John Wiley & Sons, New York; HUTCHINSON, G.E., 1975. A Treatise on Limnology - Chemistry of Lakes. Volume I - Part 2. John Wiley & Sons,
New York; LE CREN, E.D. & R.H.LOWE-McCONELL, ????. The Functioning of Freshwater Ecosystems. In: Internacional
Biological Programme 22, Cambridge University Press; LEVINS, R., 1968. Evolution in Changing Environments. Princeton University Press. Princeton, New Jersey; MACKSON, A.R.W. & J.M. JACKSON, 1996. Environmental Science. The Natural Environment and Human Impact.
Longman; ODUM, E.P.,1997. Fundamentos de Ecologia. 5ª Edição, Fundação Calouste Gulbenkian, Lisboa; SCHLESINGER, W.H., 1991. Biogeochemistry – An Analyses of Global Change. 2nd Edition, Academic Press, Inc.;
WETZEL, R.G., 1983. Limnologia. 2ª Edição, Fundação Calouste Gulbenkian, Lisboa;





























