Embed Size (px)
Citation preview

Universidade de Aveiro 2007
Departamento de Biologia
Carla Sofia dos Santos Ferreira
Estudos in vitro e in vivo em peixes da Ria de Aveiro
Dissertação apresentada à Universidade de Aveiro para cumprimento dos requisitos necessários à obtenção do grau de Mestre em Toxicologia e Ecotoxicologia, realizada sob a orientação científica da Professora Doutora Maria Ana Dias Monteiro Santos, Professora Catedrática do Departamento de Biologia da Universidade de Aveiro

o júri
presidente Prof. Dr. Fernando José Mendes Gonçalves Professor Associado com Agregação do Departamento de Biologia da Universidade de Aveiro
Prof. Dra. Maria Ana Dias Monteiro Santos Professora Catedrática do Departamento de Biologia da Universidade de Aveiro
Prof. Dr. Mário Guilherme Garcês Pacheco Professor Auxiliar do Departamento de Biologia da Universidade de Aveiro
Doutora Vera Lúcia de Almeida Maria Investigadora Auxiliar do Centro de Investigação Marinha e Ambiental (CIMA) da Universidade do Algarve

agradecimentos
Quero expressar o meu profundo agradecimento à minha orientadora Prof. Dra. Maria Ana Santos, não só pelo seu apoio científico e pessoal, mas principalmente pela sua amizade e companheirismo para com os alunos. O seu incentivo e acompanhamento permanente foram decisivos durante a realização deste trabalho científico. O seu ensinamento foi extremamente gratificante e enriquecedor, uma vez que me permitiu crescer e amadurecer cientificamente. Ao Prof. Doutor Mário Pacheco agradeço o facto de ter aceite a co-orientação da minha tese de Mestrado. Agradeço aos meus colegas de laboratório, nomeadamente ao Mestre Miguel Oliveira e aos Doutores Iqbal Ahmad e Vera Maria, a pronta colaboração, bem como a disponibilidade e apoio dispensados. Ao Mestre Miguel Oliveira quero ainda agradecer em especial, a sua amizade, incentivo e apoio científico, os quais foram extremamente importantes para mim durante a realização deste trabalho. Agradeço aos meus pais e avó tudo o que sou e tenho. Sem eles nada disto seria possível. À minha mãe, em particular, quero agradecer a força que sempre me transmitiu para ir mais além. O seu estímulo e a sua presença sempre foram, são e serão fundamentais para mim. Agradeço ao meu namorado, Ricardo, a sua paciência, compreensão e carinho ao longo deste ano. Sei que não foi fácil mas foi por uma boa razão. O seu apoio e ajuda foram fundamentais para levar a bom porto este objectivo pessoal.

palavras-chave
Contaminantes ambientais, biotransformação, biomarcadores, EROD, anomalias nucleares eritrocíticas, genotoxicidade, hormonas esteróides, hidrocarbonetos aromáticos policíclicos.
resumo
Dado a importância e complementaridade existente entre estudos de campo e estudos de laboratório, o presente trabalho engloba ambos os tipos de estudo. Os peixes desempenham um papel ecológico bastante relevante ao nível das cadeias tróficas aquáticas, pelo que são frequentemente usados como organismos sentinela em estudos de biomonitorização. Assim, para o presente trabalho, foram adoptadas duas espécies representativas da ictiofauna da Ria de Aveiro: a tainha garrento (Liza aurata) e a enguia europeia (Anguilla anguilla L.). O estudo de campo consistiu na avaliação genotóxica sazonal de uma laguna poluída (Ria de Aveiro), através da detecção de anomalias nucleares eritrocíticas (ANEs) em espécimes juvenis de L. aurata, colhidos em seis locais da Ria. A investigação revelou um local crítico – Vagos – onde foi possível observar elevadas frequências de ANEs na Primavera, no Verão e no Inverno. Neste contexto, o teste de ANEs revelou ser um teste genotóxico relativamente rápido, fácil de executar e sensível. Para a realização da componente laboratorial e in vitro da presente tese, adoptou-se como modelo biológico A. anguilla L., a qual foi injectada intraperitonealmente com β-naftoflavona (4mg/kg). A β-naftoflavona é um composto flavonóide sintético reconhecido como um forte indutor das monooxigenases hepáticas, como por exemplo a etoxiresorufina O-desetilase (EROD). A EROD tem demonstrado ser muito sensível à indução causada por vários contaminantes, pelo que a sua actividade é frequentemente usada como biomarcador in vivo de exposição a poluentes químicos, nomeadamente hidrocarbonetos aromáticos policíclicos (HAPs) e compostos estruturalmente semelhantes. Apesar deste efeito causado, ao nível do fígado, pelos HAPs ou por compostos estruturalmente semelhantes, também pode ocorrer inibição sob diversas circunstâncias, tais como a presença de elevadas concentrações de HAPs ou de outros compostos orgânicos ou de metais pesados. Assim, os microsomas hepáticos foram isolados a partir do fígado de A. anguilla L. e posteriormente usados nos estudos in vitro da actividade EROD efectuados, Os HAPs são compostos toxicologicamente relevantes devido à sua dispersão e persistência nos meios aquáticos, bem como ao seu potencial genotóxico. Assim, procedeu-se ao estudo in vitro dos efeitos de dois HAPs -benzo[a]pireno (B[a]P) e fluoranteno (FL) - sobre a actividade EROD microsomal hepática. Uma vez que o sistema endócrino desempenha um papel extremamente relevante nos mecanismos de stress dos peixes, avaliou-se ainda o efeito de duas hormonas esteróides - cortisol (C) e 17β-estradiol (E2) sobre a referida actividade enzimática, isoladamente e em combinação com o B[a]P ou o FL. A concentração adoptada para as duas hormonas esteróides foi de 22.02 nM. O B[a]P revelou ser um forte inibidor in vitro da actividade hepática de EROD. O FL 0.1 e 0.3 µM induziu significativamente a actividade EROD microsomal hepática, enquanto que as concentrações maiselevadas (0.9 e 2.7 µM) levaram a uma forte inibição desta actividade enzimática, contudo inferior à induzida pelas mesmas concentrações de B[a]P. A exposição dos microsomas hepáticos de A. anguilla L. ao C causou uma indução significativa da actividade EROD hepática. A exposição in vitro ao E2
também levou ao aumento desta actividade enzimática.

O C e o E2, exerceram um efeito protector sobre a actividade EROD hepática, uma vez que, na sua presença, verificou-se uma significativa redução dos efeitos inibitórios causados pelo B[a]P e pelo FL 0.9 e 2.7 µM. Os resultados da avaliação in vitro da actividade EROD hepática obtidos, sob as condições em cima referidas, confirmam a importância da estrutura dos HAPs e respectiva concentração molar, assim como das condições fisiológicas do indivíduo, como por exemplo diferentes status hormonais para a indução in vivo da actividade EROD, i. e. em estudos de laboratório e em estudos de campo.

keywords
Environmental contaminants, biotransformation, biomarkers, EROD, erythrocityc nuclear abnormalities, genotoxicity, steroid hormones, polycyclic aromatic hydrocarbons.
abstract
The present research work has been focused on the importance and complementarity of field and laboratory in vivo and in vitro studies. Fish are often used as sentinel organisms in biomonitorization studies because they play a major ecological role in the aquatic food-webs. Thus, two representative species of the Aveiro Lagoon’s ichthyo fauna were adopted in the present work: the gold grey mullet (Liza aurata) and the european eel (Anguilla anguilla L.). The field study consisted in the seasonal genotoxic assessment of a polluted lagoon (Ria de Aveiro), using erythrocytic nuclear abnormalities (ENAs) of L. aurata juvenile specimens, collected at six stations of Ria de Aveiro. The ENAs survey revealed a critical site – Vagos – where an elevated ENA frequency was observed in Spring, Summer and Winter. In this context, the ENA assay revealed to be a relatively rapid, easy to perform and sensitive genotoxicity test. The laboratorial and in vitro component of the present thesis was carried out using A. anguilla L. as a biological model, which was intraperitoneally injected with β-naphthoflavone (4mg/kg). β-naphthoflavone is a synthetic flavonoid compound known as a strong hepatic monooxigenases inducer such as ethoxyresorufin O-deethylase (EROD). Liver EROD activity induction showed to be very sensitive to several contaminants, thus her activity is often used as an in vivo biomarker of exposure to chemical pollutants, namely polycyclic aromatic hydrocarbons (PAHs) and structural similar compounds. Despite this effect caused by PAHs or PAH like substances upon the liver, inhibition may also occur under different circumstances such as the presence of high PAH concentrations or other organic compounds or heavy metals. Therefore, A. anguilla L. hepatic microsomes were isolated and then used in EROD in vitro activity studies where those PAHs that are toxicologically relevant due to their dispersion and persistence in the aquatic environments were assayed. Thus, the in vitro effects of two PAHs - benzo[a]pyrene (B[a]P) and fluoranthene (FL) - on liver microsomal EROD activity were assessed. Since the endocrine system plays a major role in fish stress mechanisms, the effect of two steroid hormones such as cortisol (C) and 17β-estradiol (E2) - on liver microsomal EROD activity was also studied, individually and in combination with B[a]P or FL. The same concentration (22.02 nM) was adopted for both steroid hormones. B[a]P revealed to be a strong in vitro inhibitor of liver EROD activity. FL 0.1 e 0.3 µM significantly induced liver microsomal EROD activity, whereas the highest concentrations (0.9 e 2.7) µM caused a strong inhibition of the mentioned enzymatic activity, though lower than the inhibition induced by the same B[a]P concentrations. A. anguilla L. liver microsomes in vitro exposure to C significantly induced EROD activity. E2 in vitro exposure also caused an elevation of this enzymatic activity. Both C and E2 exerted a protective effect in liver EROD activity, since a significant reduction of the inhibitory effects caused by B[a]P and FL 0.9 e 2.7 µM, was observed in their presence. The data concerning the assessment of in vitro hepatic EROD activity under the above conditions confirm the importance of PAHs structure and molar concentration as well as the organism’s physiological conditions such as different hormonal status for EROD activity in vivo induction i.e. either laboratory or field surveys.

ÍNDICE
CAPÍTULO I – Introdução geral ……………………………………………….
1. Poluição aquática........................................................................................
1.1. Hidrocarbonetos aromáticos policíclicos......................................
2. Biomarcadores............................................................................................
2.1. Organismos sentinela e biomarcadores de peixes......................
3. Respostas biológicas a poluentes..............................................................
3.1. Biotransformação……………………………………………………...
3.2. Etoxiresorufina O-desetilase.........................................................
3.3. Sistema endócrino…………………………………………………….
4. Avaliação da genotoxicidade......................................................................
5. Orientações e objectivos da tese…………………………………………….
6. Referências bibliográficas……………………………………………………...
CAPÍTULO II – Avaliação genotóxica sazonal de uma laguna poluída
(Ria de Aveiro, Portugal) através do estudo das anomalias nucleares
eritrocíticas de Liza aurata.............................................................................
CAPÍTULO III – Estudo in vitro do efeito do benzo[a]pireno, cortisol e
17ß-estradiol sobre a actividade microsomal hepática de EROD de
Anguilla anguilla L..........................................................................................
CAPÍTULO IV – Estudo in vitro do efeito do fluoranteno, cortisol e 17ß-
estradiol sobre a actividade EROD microsomal hepática, em Anguilla
anguilla L.. ………………………………………………………………………….
CAPÍTULO V – Discussão geral…………………………………………………
1. Avaliação genotóxica da Ria de Aveiro ……………………………………..
2. Efeitos in vitro de HAPs e esteróides, isoladamente ou em conjunto ….
2.1. Exposição in vitro ao benzo[a]pireno e ao fluoranteno…………
9
10
12
13
17
18
18
22
23
25
26
28
36
51
72
92
93
94
94

2.2. Exposição in vitro ao cortisol e 17β-estradiol……………………
2.3. Exposição in vitro ao benzo[a]pireno e ao fluoranteno em
combinação com o cortisol ou 17β-estradiol…………………………………..
3. Considerações finais e perspectivas futuras.............................................
4. Referências bibliográficas..........................................................................
96
99
100
101

CAPÍTULO I
Introdução geral

Introdução geral
10
1. POLUIÇÃO AQUÁTICA
A libertação acidental e/ou deliberada de efluentes, contendo compostos
químicos nocivos para o meio ambiente, potencia distúrbios na estrutura e no
funcionamento dos ecossistemas (Cavas e Ergene-Gözükara, 2005a). Desde o
início dos anos 60 que a humanidade tem vindo a tomar consciência dos
potenciais efeitos adversos a longo prazo destes compostos e do seu potencial
risco para os ecossistemas aquáticos e terrestres (Van der Oost et al., 2003).
As indústrias erguem-se nas margens dos cursos de água, atraídas pela água
necessária aos processos industriais, descarregando frequentemente os
diversos tipos de efluentes directamente na massa de água (rios, lagos),
através da canalização dos esgotos. O desenvolvimento da tecnologia
industrial tem originado fontes de descarga industrial mais complexas, levando
a que a água usada nesses processos também apresente uma maior e mais
complexa toxicidade (Cavas e Ergene-Gözükara, 2005a). As cidades, também
por se localizarem nas margens dos cursos de água, resolvem o problema dos
resíduos por despejo simples dos esgotos nos rios, na maior parte dos casos,
sem qualquer tipo de tratamento prévio. Uma vez que o ambiente aquático é o
último receptor dos poluentes contidos nos efluentes industriais, a sua
acumulação e persistência neste ambiente constitui uma ameaça à vida
(Fleeger et al., 2003). Assim, existe a necessidade de se proceder a uma
avaliação efectiva do risco ecológico associado. A avaliação do risco ecológico
ou ambiental é definida como sendo o procedimento através do qual os efeitos
adversos prováveis ou reais dos poluentes e de outras actividades
antropogénicas, são estimados através de métodos científicos nos
ecossistemas e nos seus constituintes, com um certo grau de certeza
(Depledge e Fossi, 1994). A avaliação do risco ecológico apresenta várias
características vantajosas no que diz respeito a tomadas de decisão no
contexto ambiental, fornecendo uma base quantitativa para a comparação e o
estabelecimento de prioridades ao nível dos riscos existentes, assim como
formas sistemáticas de melhorar a compreensão e conhecimento dos riscos,
permitindo também estimar, de forma clara e consistente, quais os valores
ambientais a proteger (Suter, 1993).

Capítulo I
11
A Ria de Aveiro é um sistema lagunar costeiro que se encontra
permanentemente ligado ao mar, localizada no noroeste de Portugal, rodeada
de vários agregados urbanos e unidades industriais (Pacheco et al., 2005).
Este sistema lagunar tem sido, ao longo de vários anos, o principal receptor de
descargas antropogénicas, resultantes da actividade industrial, bem como de
efluentes domésticos e municipais que nem sempre são sujeitos a tratamento
nas estações de águas residuais, o que leva ao aumento dos níveis de
contaminação orgânica e inorgânica. A intensa actividade portuária também
tem contribuído para o aumento da poluição, uma vez que produtos
resultantes desta actividade, tal como restos de combustíveis e tintas usadas
para pintar pequenas embarcações e barcos de grande porte, acabam
frequentemente nas águas da ria. Atendendo aos factos anteriormente
referidos e à persistência de alguns contaminantes, existem algumas zonas
críticas onde é imperativo um programa efectivo de biomonitorização (Pacheco
et al., 2005). A presença destes compostos na água pode interferir com os
sistemas biológicos aí existentes. A absorção desses contaminantes pode
ocorrer a partir do sedimento, da coluna de água e/ou da fonte de alimento.
No entanto, as principais vias de absorção irão depender da dieta de cada
organismo em particular e da sua ecologia (Van Veld, 1990; Livingstone,
1998). Sendo virtualmente impossível monitorizar todos os contaminantes de
origem antropogénica e natural que constituem uma potencial ameaça à
qualidade ambiental, tem sido usada uma abordagem promissora que consiste
no uso de biomarcadores para se efectuar a avaliação do estado de saúde dos
organismos e para se obter sinais antecipados de aviso da existência de riscos
ambientais (Van der Oost et al., 2003). Neste contexto, os peixes são
frequentemente usados como organismos sentinela uma vez que intervêm a
vários níveis na cadeia trófica, acumulando substâncias tóxicas e respondendo
a baixas concentrações de mutagénicos (Cavas e Ergene-Gözükara, 2005). A
diversidade de espécies de peixes existente, aliada ao facto de ocuparem
diferentes tipos de habitats, torna-os particularmente úteis em estudos de
ecotoxicologia, permitindo avaliar os efeitos de vários tipos de contaminantes
sob variadas condições de exposição.

Introdução geral
12
1.1. Hidrocarbonetos aromáticos policíclicos
Os hidrocarbonetos aromáticos policíclicos (HAPs) são um grupo de
compostos ubíquos no meio ambiente, tanto sob a forma de produtos naturais
(como resultado, por exemplo, da actividade vulcânica) bem como
contaminantes ambientais (Oliveira et al., 2007) e que possuem, no mínimo,
dois anéis aromáticos fundidos (Menzie et al., 1992). Estes compostos
ocorrem na maioria das substâncias petrolíferas encontrando-se fortemente
dispersos nos ambientes aquáticos, principalmente como resultado de
descargas petroquímicas, derrames acidentais, resíduos domésticos (Connell,
1974), assim como da actividade portuária (Pacheco e Santos, 1997). Eisler
afirmou em 1987 que a nível mundial entravam anualmente no meio aquático,
cerca de duzentas e trinta mil toneladas de HAPs (Eisler, 1987). Desde então
tem-se assistido a um crescente aumento da industrialização e urbanização,
pelo que é expectável que estes valores tenham sofrido alterações no sentido
de um aumento significativo. A contaminação por este tipo de compostos
constitui uma preocupação generalizada devido à sua forte capacidade de
bioconcentração (Connell, 1990), à sua persistência no ambiente, potencial
mutagénico e cancerígeno anteriormente demonstrado em peixes (Gravato e
Santos, 2002; Pacheco e Santos, 2002; Maria et al., 2002; Busetti et al.,
2006). A relativa insolubilidade dos HAPs na água e a sua forte capacidade de
adsorção a partículas faz com que se acumulem facilmente nos sedimentos
(Seruto et al., 2005). Por outro lado a sua lipofilicidade faz com que
atravessem facilmente as membranas biológicas, podendo bioacumular-se em
diferentes tecidos (Billiard et al., 2002). Os organismos aquáticos,
nomeadamente peixes, que vivem em ambientes contaminados por HAPs
podem, por isso, absorver estes compostos, sobretudo através da água que
entra pela boca e atravessa as guelras, assim como pela ingestão de
sedimentos ou comida contaminados (Varasani et al., 1989). Contudo, a
concentração de HAPs nos tecidos dos peixes, pode ser significativamente
reduzida através do metabolismo e excreção (Billiard et al., 2002).

Capítulo I
13
Figura 1 – Representação da estrutura molecular de alguns HAPs. O composto
sublinhado é carcinogénico (modificado de Fent e Bätscher, 2000).
2. BIOMARCADORES
Os efeitos adversos nas populações resultantes da exposição a
contaminantes são frequentemente difíceis de detectar, uma vez que muitos
destes efeitos tendem a manifestar-se apenas a longo prazo (Van der Oost et
al., 2003). Quando o efeito adverso se torna finalmente visível, o processo
destrutivo pode já ter ido além do ponto onde ainda era possível reverter o
processo anteriormente mencionado, através de acções remediadoras ou
redução do risco (Van der Oost et al., 2003) (figura 2). Este tipo de cenário
levou a investigação a estabelecer sinais antecipados de aviso ou
biomarcadores, que reflectissem as respostas biológicas adversas perante a
existência de contaminação ambiental (Bucheli e Fent, 1995). Os
biomarcadores representam mudanças nos sistemas biológicos induzidas por
contaminantes, podendo servir como ligação entre a existência de
contaminação ambiental (causa) e os seus efeitos, fornecendo deste modo,
informação relevante sobre a saúde do ecossistema (Pacheco et al., 2005).
Naftaleno
Fenantreno
Fluoranteno
Benzo[a]pireno
Benzeno
Fluoreno
Pireno
Criseno
Antraceno

Introdução geral
14
Figura 2 – Representação esquemática da ordem sequencial de respostas de
um sistema biológico ao stress causado por exposição a um poluente (Adaptado de
Van der Oost et al., 2003 e modificado de Bayne et al., 1985).
As respostas dos biomarcadores são de extrema importância e utilidade
porque integram uma larga variedade de factores ambientais, toxicológicos e
ecológicos que controlam e modulam a exposição a xenobióticos e seus
efeitos, podendo mesmo ser idênticas numa larga variedade de organismos
(Van der Oost et al., 2003). Assim, os biomarcadores podem fornecer
informação sobre os efeitos biológicos dos poluentes para além da mera
quantificação dos seus níveis no ambiente (Van der Oost et al., 2003). Van
Gestel e Van Brummelen (1994) redefiniram os termos “biomarcador” e
“bioindicador” associando-os a diferentes níveis de organização biológica.
Nesta perspectiva, um biomarcador pode ser definido como qualquer resposta
biológica a um composto ambiental ao nível sub-individual, medida dentro do
organismo ou nos seus produtos (urina, fezes, pêlo, penas, etc.), indicando
um desvio do estado normal que não pode ser detectado no organismo como
um todo. Um bioindicador é definido como sendo um organismo que fornece
informação sobre as condições ambientais do seu habitat, através da sua

Capítulo I
15
presença ou ausência ou através do seu comportamento. Efeitos nos níveis
hierárquicos mais elevados são sempre precedidos por mudanças prévias nos
processos biológicos, pelo que, foi reconhecido que a resposta bioquímica
dever-se-ia tornar no primeiro aviso, sendo sinónimo da capacidade
integrativa dos seres vivos de responder a mudanças físicas e químicas da
qualidade do ambiente (Bayne et al., 1985). No contexto ambiental, os
biomarcadores apresentam-se como uma ferramenta promissora, já que são
indicadores sensíveis, demonstrando que os tóxicos que entram nos
organismos, se distribuem pelos tecidos e induzem efeitos tóxicos em alvos
críticos (McCarthy e Shugart, 1990).
Normalmente, uma situação de stress causada por exposição a
poluentes despoleta uma cascata de respostas biológicas, cada uma das quais,
em teoria, serve como biomarcador (McCarthy et al., 1991). A aplicação ou
interpretação imprópria das respostas de biomarcadores pode, contudo, levar
a falsas conclusões acerca do stress causado pelos poluentes ou contaminação
ambiental. Determinadas respostas estabelecidas para uma espécie não são
necessariamente válidas para outra espécie (Van der Oost et al., 2003). A
aplicação de biomarcadores em ambos os estudos de laboratório e de campo,
pode fornecer uma importante ligação entre a toxicidade observada no
laboratório e a avaliação no campo (Van der Oost et al., 2003).
De acordo com o Conselho Nacional de Investigação (NRC, 1987) e com
a Organização Mundial de Saúde (WHO, 1993), os biomarcadores podem ser
subdivididos em três classes:
- biomarcadores de exposição: englobam a detecção e quantificação de
uma substância exógena, ou do seu metabolito, ou o produto da interacção
entre um agente xenobiótico e uma determinada molécula ou célula alvo, que
é medida dentro do organismo;
- biomarcadores de efeito: incluem alterações bioquímicas, fisiológicas
ou outro tipo de alterações mensuráveis ao nível dos tecidos ou fluidos
corporais de um organismo que pode ser reconhecido como estando associado
com uma possível ou já estabelecida doença ou alteração do estado de saúde;
- biomarcadores de susceptibilidade: indicam a capacidade inerente ou
adquirida de um organismo responder ao desafio da exposição a uma

Introdução geral
16
substância xenobiótica específica, incluindo factores genéticos e alterações em
receptores que alteram a susceptibilidade de um organismo a essa exposição
(Van der Oost et al., 2003).
Com o propósito de avaliar, de forma objectiva a fiabilidade dos
biomarcadores de peixes, foi proposto por Van der Oost et al. (2003), com
base nos critérios formulados por Stegeman et al. (1992), seis critérios que
compreendem a informação mais importante que deve estar disponível ou que
tem de ser estabelecida para cada candidato a biomarcador:
1 - O ensaio usado para quantificar o biomarcador deve ser fiável (com
certificação de qualidade), relativamente barato e fácil de efectuar;
2 - A resposta do biomarcador deve ser sensível à exposição a
poluentes e/ou aos seus efeitos, de forma a poder ser usada como uma
informação antecipada;
3 - A actividade basal do biomarcador deve ser bem definida de forma a
se poder distinguir entre variabilidade natural (ruído de fundo) e o stress
induzido pela exposição a contaminantes (sinal);
4 - O impacto de factores que possam interferir com a resposta do
biomarcador devem ser bem estabelecidos;
5 - O mecanismo subjacente às relações existentes entre a resposta do
biomarcador e a exposição a poluentes (dosagem e tempo) devem igualmente
ser estabelecidas;
6 - A significância ecológica do biomarcador, isto é, as relações entre a
sua resposta e o impacto (a longo prazo) para o organismo, devem ser
estabelecidas.
Além destes critérios tem também sido sugerido que os biomarcadores
devem, preferencialmente, ser não-invasivos ou não destrutivos, de forma a
permitir ou a facilitar a monitorização ambiental dos efeitos de poluentes em
espécies protegidas ou em perigo de extinção (Fossi e Marsili, 1997).
Uma implementação bem sucedida dos biomarcadores em programas de
monitorização ambiental requer, portanto, uma boa compreensão dos
mecanismos subjacentes às respostas dos mesmos. Um dos maiores desafios
do desenvolvimento e aplicação dos biomarcadores, no que diz respeito à

Capítulo I
17
avaliação do risco ecológico, é definir a significância das respostas dos
biomarcadores em termos de efeitos ecológicos dos poluentes (Suter, 1990).
2.1. Organismos sentinela e biomarcadores de peixes
O uso de organismos sentinela para a monitorização biológica pode
constituir uma abordagem sensível na estimativa dos efeitos potenciais dos
poluentes (Cavas et al., 2005b). Os peixes podem ser, virtualmente,
encontrados em qualquer lado no ambiente aquático e desempenham um
papel ecológico bastante relevante ao nível das cadeias alimentares devido à
sua função de transportadores de energia dos níveis tróficos mais baixos para
os mais elevados (Beyer, 1996). O conhecimento da via de entrada do tóxico,
assim como do comportamento e respostas dos peixes, pode ter uma elevada
relevância ecológica. Os biomarcadores que têm sido mais extensivamente
estudados são as enzimas envolvidas na desintoxicação dos xenobióticos e
seus metabolitos (enzimas envolvidas na biotransformação) (Van der Oost et
al., 2003). Vários parâmetros bioquímicos têm sido testados, nos peixes, no
que diz respeito às suas respostas a substâncias tóxicas e à sua potencial
utilização como biomarcadores de exposição ou efeito. A maior parte dos
critérios gerais de biomarcadores parecem ser directamente aplicáveis a
determinados biomarcadores de peixes (Stegeman et al., 1992). Apesar das
suas limitações, como por exemplo o facto de terem uma mobilidade
relativamente elevada, os peixes são, geralmente, considerados os
organismos mais adequados para monitorizar a poluição nos sistemas
aquáticos (Van der Oost et al., 2003). Portanto, o uso de biomarcadores de
peixes como indicadores dos efeitos causados pela poluição, é de crescente
importância, podendo permitir a detecção antecipada de problemas nos
ecossistemas aquáticos (Van der Oost et al., 2003).
Teoricamente, a espécie ideal de peixe a ser usada na avaliação da
toxicidade de um determinado tipo de poluente deve obedecer aos seguintes
critérios: (1) disperso por diferentes ecossistemas, cosmopolita se possível, de
forma a poder ser usada em estudos in situ; (2) suficientemente sensível na
detecção da genotoxicidade de uma larga variedade de poluentes em doses
baixas; (3) adequada para condições de cultura de forma a se poder efectuar

Introdução geral
18
experiências laboratoriais (in vitro) (4) a sua abundância na natureza e a
existência de populações numerosas de forma a permitir a captura de
indivíduos sem que isso ponha em risco a sua conservação (Sanchez-Galan et
al., 1999).
3. RESPOSTAS BIOLÓGICAS A POLUENTES
3.1. Biotransformação
Vários xenobióticos são encontrados em ambientes aquáticos
antropogenicamente alterados, encontrando-se frequentemente disponíveis a
serem absorvidos pelos organismos aquáticos. Uma das questões mais
importantes da toxicologia aquática é a capacidade de prever a velocidade e a
extensão dessa absorção pelos organismos, bem como o destino e os efeitos
dos compostos absorvidos (James et al., 1994).
Várias vias de exposição são possíveis. Os compostos dissolvidos ou
suspensos na água podem entrar através das guelras, da pele ou do sistema
gastrointestinal. Os compostos presentes no sedimento podem ser absorvidos
por contacto directo com a derme e por ingestão, enquanto que os compostos
presentes nas plantas e em organismos pertencentes a baixos níveis tróficos,
podem ser ingeridos e absorvidos através do sistema gastrointestinal. A
importância relativa de cada via de entrada varia com as propriedades
químicas intrínsecas de cada xenobiótico e também com a possibilidade de o
composto em questão ser degradado no ambiente aquático ou
biotransformado por organismos de baixos níveis tróficos (James et al., 1994).
Os xenobióticos parecem ser mais rapidamente absorvidos pelos
organismos quando presentes na água. Este facto é, em grande parte, devido
às propriedades da água como solvente universal. Contudo, compostos que
são virtualmente insolúveis em água, são frequentemente “solúveis” na
matéria orgânica suspensa na coluna de água (James et al., 1994). As
propriedades físico-químicas do xenobiótico, como por exemplo lipofilicidade,
hidrofilicidade, presença de grupos acídicos ou básicos e o seu pKa, assim
como o tamanho da molécula, influenciarão separada e conjuntamente, o
processo de absorção dos compostos ao nível do sistema gastrointestinal. Uma

Capítulo I
19
vez que a difusão passiva é o processo mais comum e importante através do
qual estes compostos entram nos organismos, a solubilidade do xenobiótico
em água e nos lípidos é de grande importância (James et al., 1994).
Contudo, os organismos têm duas grandes formas de eliminar um
composto: ou é excretado na sua forma original ou é biotransformado pelo
organismo. A biotransformação leva, geralmente, à formação de um composto
mais hidrofílico, mais facilmente excretável que o composto original
(Vermeulen, 1996). O fígado é o órgão que mais activamente se encontra
envolvido na biotransformação de compostos estranhos ao organismo, devido
à sua função, posição e circulação sanguínea (Van der Oost et al., 2003). No
entanto, existem outros órgãos com capacidade biotransformadora, tais como
o rim, as guelras e o intestino, embora com menor capacidade que a do
fígado. A biotransformação também pode alterar a toxicidade de um
composto, o que pode ser benéfico ou prejudicial para o organismo (Van der
Oost et al., 2003). Caso haja uma reacção de desintoxicação, a toxicidade do
composto é reduzida enquanto a excreção é geralmente elevada (Van der Oost
et al., 2003). No caso de haver bioactivação o composto é, contudo,
transformado num metabolito reactivo que é mais tóxico que o composto
original (Van der Oost et al., 2003). Os efeitos tóxicos podem-se manifestar
quando o composto original ou os seus metabolitos se ligam a macromoléculas
celulares, o que pode, em última instância, levar à ruptura membranar, lesão
celular e/ou efeitos genotóxicos que, por sua vez, podem levar ao
desenvolvimento e progressão de doenças (por exemplo cancro) (Van der
Oost et al., 2003) (figura 3).

Introdução geral
20
Metabolito tóxico
(reactivo)
XENOBIÓTICO
Ligação a moléculas celulares
Metabolito estável
Metabolito estável
Efeito Tóxico
(lesão celular, ruptura
membranar, cancro)
Excreção
1 2 3 4
5 6
A B C
Figura 3 – Possíveis vias de desintoxicação e activação dos xenobióticos: (1)
efeito tóxico directo (A); (2) activação metabólica; (3) formação de um metabolito
estável passível de causar efeito tóxico (C); (4) desintoxicação. O metabolito reactivo
formado por bioactivação (2) pode causar efeito tóxico (B) através da reacção com
alvos críticos (5) ou ser eliminado através da reacção com um agente protector (6)
(Adaptado de Van der Oost et al., 2003).
A biotransformação é, portanto, determinante no que respeita à
actividade de um determinado composto, duração dessa mesma actividade,
envolvendo, frequentemente, enzimas com um elevado grau de especificidade
por um determinado substrato. A biotransformação dos xenobióticos no fígado
pode levar, como já foi referido, à desintoxicação ou bioactivação dos
mesmos, podendo-se subdividir este processo em três fases: Fase I, Fase II e
Fase III. A fase I consiste numa alteração não sintética (oxidação, redução, ou
hidrólise) da molécula original, que pode ser subsequentemente conjugada na
fase II e catabolizada na Fase III (Commandeur et al., 1995).

Capítulo I
21
Geralmente, os biomarcadores mais sensíveis de efeito são alterações
nos níveis e na actividade de enzimas envolvidas no processo de
biotransformação. Contudo, a actividade destas enzimas pode ser induzida ou
inibida em peixes, após exposição a xenobióticos (Bucheli e Fent, 1995). A
indução enzimática consiste num aumento da actividade das enzimas em
questão, enquanto que em caso de inibição, a actividade enzimática é
bloqueada, possivelmente devido a forte ligação ou formação de um complexo
entre a enzima e os inibidores. Duas grandes classes de enzimas estão
envolvidas na biotransformação de xenobióticos: as enzimas da fase I e as
enzimas e cofactores da fase II (Van der Oost et al., 2003). No que respeita, à
grande maioria dos compostos xenobióticos, as reacções da fase I são
catalizadas por monooxigenases (MO), existentes ao nível dos microsomas,
também designadas por oxidases multi-funções (MFOs), como por exemplo as
monooxigenases associadas ao complexo citocromo P450. Os citocromos P450
são proteínas membranares que estão, predominantemente, localizadas no
retículo endoplasmático do fígado, e que compreendem uma vasta, e ainda em
expansão, família de proteínas heme (Stegeman et al., 1992; Bucheli e Fent,
1995), as quais foram também encontradas noutros organelos e tecidos de
peixes mas em menor quantidade (Celander, 1993). A maioria das enzimas da
fase II catalizam reacções sintéticas de conjugação, facilitando, portanto, a
excreção dos compostos através da adição de grupos mais polares, podendo,
por isso, desempenhar um importante papel não só na manutenção da
homeostasia como também na desintoxicação e eliminação de muitos
compostos (Commandeur et al., 1995; Van der Oost et al., 2003). As reacções
catalisadas pelas MFOs resultam na inserção ou adição de um átomo de
oxigénio numa molécula substrato. Assim, a hidrossolubilidade e/ou
capacidade de conjugação da molécula substrato aumenta, o que facilita a sua
excreção (Masfaraud et al., 1990). Assim, a característica mais importante do
sistema MFO é a sua capacidade de facilitar a excreção de certos compostos,
na fase I de biotransformação, transformando compostos lipofilicos em
compostos mais hidrofílicos (Bucheli e Fent, 1995). Uma vez que o sistema
MFO é sensível a determinados poluentes ambientais, a sua actividade pode

Introdução geral
22
ser usada na monitorização biológica na exposição a determinadas classes de
xenobióticos (Bucheli e Fent, 1995).
3.2. Etoxiresorufina O-desetilase (EROD)
A etoxiresorufina O-desetilase (EROD) é uma MFO que tem
demonstrado ser muito sensível à indução causada por vários compostos
químicos, nomeadamente hidrocarbonetos aromáticos policíclicos (HAPs).
Assim, a avaliação da actividade da EROD em peixes tem sido fortemente
usada como um biomarcador in vivo de exposição a HAPs, como por exemplo
o benzo[a]pireno (B[a]P), e a compostos estruturalmente semelhantes tal
como a ß-naftoflavona (BNF) (Buchelli e Fent, 1995). O mecanismo de indução
de CYP1A por alguns HAPs, parece estar relacionado com a sua capacidade em
se ligar ao receptor citosólico hidrocarboneto arilo (RAh) e activar a
transcrição do gene que codifica para CYP1A, iniciando-se assim a sua síntese.
Os HAPs são facilmente metabolizados pelas enzimas da fase I da
biotransformação (MFOs) em produtos mais hidrofílicos. Peixes apanhados em
águas predominantemente contaminadas por compostos orgânicos,
nomeadamente HAPs, ou por misturas complexas (Kosmala et al., 1998)
apresentam indução da actividade da EROD ao nível hepático (Pacheco et al.,
2005). A utilização da EROD como biomarcador aumentou nos últimos anos
devido, essencialmente à optimização de protocolos para a medição da sua
actividade catalítica de uma forma rápida e relativamente pouco dispendiosa
(Burke e Mayer, 1974). A actividade da EROD é medida através da
monitorização do aumento de fluorescência como consequência da formação
do produto da reacção – resorufina (Burke e Mayer, 1974).
Em várias espécies de peixe, os níveis de proteína de CYP1A no fígado
demonstraram ser um biomarcador muito sensível à exposição a HAPs, o que
certamente se enquadra nos procedimentos necessários a uma correcta
avaliação do risco ecológico (Van der Oost et al., 2003). A avaliação de CYP1A
pode ser usada em várias fases do processo de avaliação do risco ecológico,
nomeadamente, ao nível da quantificação do impacto e exposição a vários
poluentes orgânicos, monitorização ambiental da “sáude” dos organismos e
dos ecossistemas, identificação de efeitos tóxicos primários subtis,

Capítulo I
23
investigação dos mecanismos de toxicidade dos xenobióticos e identificação da
exposição a compostos específicos (Stegeman et al., 1992).
3.3. Sistema endócrino
O sistema endócrino desempenha um papel extremamente relevante
nos mecanismos de stress dos peixes, pelo que, alterações em funções
hormonais específicas e consequentes efeitos bioquímicos podem constituir
importantes biomarcadores de stress (Pacheco e Santos, 2001a), sendo ainda
particularmente importante devido à sua função crucial de manter a
homeostasia dos peixes. Neste contexto, o cortisol desempenha um papel
extremamente relevante, como produto final do eixo hipotálamo-pituitária-
tecido interrenal (HPI), em resposta a diferentes agentes causadores de
stress. O papel do cortisol no metabolismo intermediário é vital, mobilizando
reservas, como por exemplo aminoácidos, e convertendo-as em glucose e
lípidos, exercendo, portanto, efeitos directos e indirectos no referido
metabolismo (Teles et al., 2004a). Os níveis de cortisol no plasma são o
indicador de stress mais vulgarmente usado nos peixes. O aumento da
concentração de cortisol no plasma de peixes foi demonstrado após exposições
de curto prazo a uma vasta gama de contaminantes, desde metais pesados
(De Boeck et al., 2003), ácidos resínicos (Teles et al., 2003), HAPs (Pacheco e
Santos, 2001a) e pesticidas (Waring e Moore, 2004), até hormonas “naturais”
como o 17β-estradiol (E2) (Pottinger et al., 1996). A própria manipulação dos
animais, condicionamento dos mesmos, bem como a fraca qualidade da água,
também constituem factores de stress, podendo igualmente provocar uma
rápida elevação do cortisol plasmático. (Wendelaar-Bonga, 1997).
Vários estudos, maioritariamente in vivo, têm procedido à avaliação dos
níveis de cortisol plasmático em diferentes espécies de peixes, tais como
Anguilla anguilla L. (Pacheco e Santos, 2001a e 2001b; Teles et al., 2003,
2004b), Sparus aurata L. (Teles et al., 2005), Liza aurata (Oliveira et al.,
2007) e Dicentrarchus labrax L. (Teles et al., 2004a), sob diversas condições
ambientais. Todos eles demonstraram a inquestionável relevância do cortisol
como indicador biológico de stress nos peixes.

Introdução geral
24
De acordo com Hontela et al. (1997), peixes colhidos em locais poluídos
revelaram incapacidade de aumentar os seus níveis plasmáticos de cortisol
como consequência de stress agudo e longa exposição a poluentes.
Níveis reduzidos de esteróides sexuais, assim como uma performance
sexual diminuída foram igualmente observados em peixes provenientes de
áreas nos Grandes Lagos, afectadas pelo impacto dos efluentes da pasta de
papel (Hontela et al., 1997). As hormonas esteróides sexuais são produzidas
em grande quantidade nas gónadas, embora a sua síntese também ocorra no
córtex adrenal. Essas hormonas são produzidas a partir do colesterol, que
numa primeira fase é convertido em progesterona, a qual é em seguida
transformada em androgénios. Estes últimos são então convertidos em
estrogénios, dos quais o E2 é o mais potente, desempenhando um importante
papel em vários aspectos do crescimento, desenvolvimento e diferenciação
morfológica, assim como no desenvolvimento e regulação dos
comportamentos e ciclos reprodutivos e sexuais de ambos sexos (Randall et
al., 2001). Os efeitos do E2 nos peixes têm sido relatados sobretudo no que diz
respeito a aspectos endócrinos, nomeadamente sobre os seus efeitos na
reprodução. Contudo, há evidências recentes de que o E2 também pode
exercer disrupção sobre eventos endócrinos não-reprodutivos, nomeadamente
sobre respostas associadas ao stress, medidas através dos níveis plasmáticos
de cortisol (Hontela, 1997, Teles et al., 2005).
A detecção relativamente recente de concentrações significativas de E2
em efluentes domésticos, aliado à sua capacidade de causar alterações
endócrinas nos organismos, levou a que fosse considerado como contaminante
ambiental, e revelou a necessidade de uma avaliação efectiva da toxicidade
deste estrogénio natural como potencial contaminante do meio aquático
(Kramer et al., 1998). Assim, durante as últimas décadas tem-se verificado
um aumento significativo das concentrações de E2 no meio aquático,
principalmente em zonas urbanas, sendo os seus níveis na água na ordem das
ng/L, podendo atingir 200 ng/L (Bowman et al., 2000). Teles et al. (2007)
num estudo de campo in situ realizado numa lagoa de água doce (Pateira de
Fermentelos) sujeita a vários tipos de contaminantes antropogénicos,
relataram elevados níveis de E2 (na ordem das 150/180 ng/L) em dois dos

Capítulo I
25
cinco locais de exposição seleccionados para o referido estudo. Os elevados
níveis de E2 foram coincidentes com a medição de níveis elevados de nitritos,
os quais constituem um indício da existência de decomposição parcial de
matéria orgânica resultante de descargas antropogénicas.
Ao contrário das enzimas envolvidas no processo de biotransformação,
cuja resposta à exposição a xenobióticos orgânicos é bem conhecida, a ligação
entre este tipo de resposta e outras funções biológicas, nomeadamente a
regulação endócrina, é ainda um desafio para a toxicologia ambiental (Teles et
al., 2005).
4. AVALIAÇÃO DA GENOTOXICIDADE
A indução do CYP1A em peixes, associada com a existência de danos ao
nível do ADN, tem vindo a ser considerada como uma abordagem sensível e
adequada na avaliação da genotoxicidade causada por exposição a poluentes
(Maria et al., 2002).
Os intermediários reactivos dos HAPs têm potencial para, ao nível
celular, interagir com qualquer local nucleofílico, incluindo os que existem nas
proteínas e no DNA, induzindo lesões genéticas (Stegeman e Lech, 1991). O
estudo de danos no DNA, ao nível do cromossoma, é uma parte essencial da
toxicologia genética, uma vez que a mutação é um evento importante no
processo cancerígeno (Fenech, 2000). A constatação de que danos ao nível do
cromossoma podem ser causados por exposição a radiação ionizante ou
químicos carcinogénicos esteve entre as primeiras evidências plausíveis das
alterações relevantes que determinados agentes físicos e químicos podem
causar ao nível do material genético, em células eucarióticas (Evans, 1977).
Em 1970, Miller estabeleceu a relação entre a alteração do material genético e
a cancerogénese, defendendo a necessidade do desenvolvimento de um teste
que possibilitasse obter resultados a curto prazo, que servisse como um
indicador simples na avaliação do risco de cancro. Neste contexto, testes
genotóxicos como a detecção de anomalias nucleares em eritrócitos aliados ao
estudo das enzimas envolvidas na biotransformação, quando feitos em
espécimes juvenis, merecem especial atenção de um ponto de vista

Introdução geral
26
ecotoxicológico (Pacheco et al., 1997; Gravato e Santos, 2003). A prova que
compostos provenientes da actividade industrial podem induzir efeitos
genotóxicos, vem reforçar a urgente necessidade de se efectuar testes
sensíveis de forma a proceder à avaliação do potencial genotóxico destes
mesmos compostos (Kohlpoth et al., 1999).
Os micronúcleos são massas citoplasmáticas de cromatina com a
aparência de pequenos núcleos que têm origem em fragmentos de
cromossomas ou até em cromossomas inteiros que não são sujeitos à fase
anafásica da divisão celular. A presença de micronúcleos em células é reflexo
de aberrações (estruturais ou numéricas) cromossómicas que emergem
durante a mitose (Fenech et al., 1999). Nos últimos anos, vários estudos têm
descrito a presença de anomalias nucleares, para além dos micronúcleos, em
células de peixes expostos a substâncias genotóxicas (Cavas e Ergene-
Gözükara, 2003). Em geral, estas anomalias são consideradas indicadores de
danos genotóxicos e, como tal, podem complementar as contagens de
micronúcleos em estudos de genotoxicidade (Cavas e Ergene-Gözükara,
2005a). O teste de anomalias nucleares eritrocíticas, baseado na detecção de
micronúcleos e outras anomalias nucleares, é visto como sendo um teste
sensível, simples, rápido, de baixo custo, aplicado com sucesso em diferentes
espécies de peixes, tais como Anguilla anguilla (Pacheco e Santos, 1997,
1998, 1999, 2001a e b), Dicentrarchus labrax (Gravato e Santos, 2002),
Oncorhynchus mykiss (Ayllón and Garcia-Vasquez, 2001) e Liza aurata
(Oliveira et al., 2007), expostas a várias classes de genotoxinas, sendo por
isso, frequentemente usado como biomarcador de genotoxicidade (Pacheco et
al., 2005).
5. ORIENTAÇÕES E OBJECTIVOS DA TESE
Os testes toxicológicos in vitro têm vantagens sobre os sistemas
tradicionais in vivo pelo sacrifício de um menor número de organismos, o
melhor controlo das variáveis ambientais, a possibilidade de uma amostragem
simultânea ou repetida ao longo do tempo, o uso de menor quantidade de
compostos químicos (xenobióticos), permitindo estudar uma reacção de

Capítulo I
27
biotransformação específica sob condições estritamente controladas (Moore e
Simpson, 1992). Contudo, uma vez que tanto a sub-estimação como a sobre-
estimação dos efeitos pode acontecer, é importante validar os estudos
laboratoriais (nomeadamente in vitro) de biomarcadores com estudos de
campo (Van der Oost et al., 2003). Neste contexto, e atendendo à relevância
de ambos os tipos de estudo (in vivo e in vitro), a presente tese encontra-se
subdividida em cinco capítulos. No capítulo II foi efectuada a avaliação sazonal
da frequência de anomalias nucleares eritrocíticas (ANEs) numa espécie
de tainha - Liza aurata – colhida em diferentes locais de amostragem ao longo
de uma laguna poluída – Ria de Aveiro. Esta espécie foi escolhida como bio
indicadora neste estudo, tendo em conta a sua relevância ecológica,
abundância e distribuição geográfica, assim como a fácil captura e
manipulação.
Nos capítulos III e IV procedeu-se ao estudo dos efeitos in vitro de dois
hidrocarbonetos aromáticos policíclicos - B[a]P e fluoranteno (FL) - e de
duas hormonas esteróides - cortisol e 17ß-estradiol -, separadamente e em
conjunto, sobre a actividade EROD microsomal hepática de Anguilla anguilla
L., previamente induzida in vivo por ß-naftoflavona (BNF). Uma das
metodologias frequentemente usada nos estudos in vitro em peixes é a
utilização de microsomas, já que a obtenção de um stock de microsomas
hepáticos com características homogéneas se torna fácil, a partir de um único
animal de grandes dimensões, por indução prévia através da injecção
intraperitoneal ou exposição em água contendo BNF. Esta metodologia, para
além de não ser dispendiosa, permite a preservação do stock microsomal
durante meses a -80 ºC tendo demonstrado a sua fiabilidade, no que respeita
à sensibilidade e reprodutibilidade da actividade da EROD (Yamazaki et al.,
1997). A escolha da enguia deve-se ao facto de ter sido anteriormente
comprovada a sua sensibilidade aos HAPs e BNF, após a sua administração por
via intraperitoneal ou exposição na água, através da elevada indução de EROD
hepática o que permitiu a obtenção de um stock concentrado de microsomas a
partir de um único animal, com uma elevada actividade enzimática. No
capítulo V realizou-se uma discussão geral dos resultados obtidos nos
capítulos II, III e IV.

Introdução geral
28
6. REFERÊNCIAS BIBLIOGRÁFICAS
Ayllón F. e Garcia-Vazquez E. (2001). Micronuclei and other nuclear lesions as
genotoxicity indicators in rainbow trout Oncorhynchus mykiss. Ecotoxicol.
Environ. Saf. 49:221-225.
Bayne, B. L., Brown, D. A., Burns, K., Dixon, D. R., Ivanovici, A., Livingstone, D. A.,
Lowe, D. M., Moore M. N., Stebbing, A. R. D. e Widdings, J. (1985). The effects
of stress and pollution on marine animals. Praeger, New York, USA.
Beyer, J. (1996). Fish biomarkers in marine pollution monitoring; evaluation and
validation in laboratory and field studies. Em: Van der Oost R., Beyer J.,
Vermeulen, N. P. E. (2003). Fish bioaccumulation and biomarkers in
environmental risk assessment: a review. Environ. Toxicol. Pharm. 13:57-149.
Billiard, S. M., Hahn, M E., Franks, D. G., Peterson, R. E., Bols, N C. e Hodson, P. V.
(2002).Binding of polycyclic aromatic hydrocarbons (PAHs) to teleost aryl
hydrocarbon receptors (AHRs). Comp. Biochem. Physiol. B: Biochem. Mol. Biol.
133:55-68.
Bowman, C. J., Kroll, K. J., Hemmer, M. J., Folmar, L. C. e Denslow, N. D. (2000).
Estrogen-induced vitellogenin mRNA and protein in sheepshead minnow
(Cyprinodon variegatus). Gen. Comp. Endocrinol. 120:300-313.
Bucheli, T. D., e Fent, K. (1995). Induction of cytocrome P450 as a biomarker for
environmental contamination in aquatic ecosystems. Crit. Rev. Environ. Sci.
Technol. 25:201-268.
Burke, M. D. e Mayer, M. T. (1974). Ethoxyresorufin: direct fluorometric assay of a
microsomal O-dealkylation which is preferentially inducible by 3-
methylcholanthrene. Drug. Metab. Dispos. 2:583-588.
Busetti, F., Heitz, A., Cuomob, M., Badoer, S., e Traverso, P. (2006) Determination of
sixteen polycyclic aromatic hydrocarbons in aqueous and solid samples from an
Italian wastewater treatment plant. J. Chromatogr. A. 1102:104-115.

Capítulo I
29
Cavas, T. e Ergene-Gözükara, S. (2003). Micronuclei, nuclear lesions and interphase
silver-stained nucleolar organizer regions (AgNORs) as cyto-genotoxicity
indicators in Oreochromis niloticus exposed to textile mill effluent. Mut.
Res./Gen. Toxicol. Environ. Mutagen. 538:81-91.
Cavas, T. e Ergene-Gözükara, S. (2005a). Induction of micronuclei and nuclear
abnormalities in Oreochromis niloticus following exposure to petroleum refinery
and chromium processing plant effluents. Aquat. Toxicol. 74:264-271.
Cavas T., Garanko, N. N. e Arkhipchuck, V. V. (2005b). Induction of micronuclei and
binuclei in blood, gill and liver cells of fishes subchronically exposed to chloride
and copper sulphate. Food Chem. Toxicol. 43:569-574
Celander, M., Leaver, M. J., George, S. G. e Förlin, L. (1993). Induction of cytocrome
P450 1A1 and conjugating enzymes in rainbow trout (Oncorhyncus mykiss) liver:
a time course study. Comp. Biochem. Physiol. 106C:343-349.
Commandeur, J. N. M., Stijntjes, G. J. e Vermeulen, N. P. E. (1995). Enzymes and
transport systems involved in the formation and disposition of glutathione S-
conjugates. Role in bioactivation and detoxication mechanisms of xenobiotics.
Pharmacol. Rev. 47:271-330.
Connell, D. W. (1974). A kerosene-like taint in the sea mullet - Mugil cephalus L. –
composition and environmental occurrence of the tainting substance. Aust. J.
Mar. Freshwater Res. 25:7-24.
Connell, D. W. (1990). Bioaccumulation of xenobiotic compounds. CRC Press, Boca-
Raton, FL.
Depledge, M. H. e Fossi, M. C. (1994). The role of biomarkers in environmental
assessment (2). Ecotoxicol. 3:161-172.
De Boeck, G., De Wachter, B., Vlaeminck, A. e Blust, R. (2003). Effect of cortisol
treatment and/or sublethal copper exposure on copper uptake and heat shock
protein levels in common carp, Cyprinus carpio. Environ. Toxicol. Chem. 22:122-
1126.

Introdução geral
30
Eisler R. (1987). Polycyclic aromatic hydrocarbons hazards to fish, wildlife and
invertebrates: a synoptic review. Fish Wildl. Serv. Biol. Rep. (US) Washington
DC. 85(1.11).
Evans, H. J. (1977). Molecular mechanism in the induction of chromosome
aberrations. Em: Fenech, M., (2000). The in vitro micronucleus technique.
Mutat. Res. 455:81-95.
Fenech, M., Holland, N., Chang, W. P., Zeiger, E. e Bonassi, S. (1999). The human
micronucleus project – an international collaborative study on the use of
micronucleus technique for measuring DNA damages in humans. Mutat. Res.
428:271-283.
Fenech M. (2000). The in vitro micronucleus technique. Mutat. Res. 455:81-95.
Fent K. e Bätscher, R. (2000). Cytocrome P4501A induction potencies of polycyclic
aromatic hydrocarbons in a fish hepatoma cell line: demonstration of additive
interactions. Environ. Toxicol. Chem. vol. 19, nº 8, 2047-2058.
Flegger, J. W., Carman, K. R. e Nisbet, R. M. (2003). Indirect effects of contaminants
in aquatic ecosystems. Sci. Total Environ. 317 (1-3):207-233.
Fossi, M. C. e Marsili, L. (1997). The use of non-destructive biomarkers in the study of
marine mammals. Em: Van der Oost R., Beyer J., Vermeulen, N. P. E. (2003).
Fish bioaccumulation and biomarkers in environmental risk assessment: a
review. Environ. Toxicol. Pharm. 13:57-149.
Gravato C. e Santos, M. A. (2002). Juvenile sea bass liver P450 EROD induction and
erythrocytic genotoxic responses to PAH and PAH-like compounds. Ecotoxicol.
Environ. Saf. 51:115-127.
Gravato C. e Santos, M. A. (2003). Genotoxicity biomarkers’ association with B(a)P
biotransformation in Dicentrarchus labrax L.. Ecotoxicol. Environ. Saf. 55:352-
358.
Hontela, A. (1997). Endocrine and physiological responses of fish to xenobiotics: role
of glucocorticosteroid hormones. Rev. Toxicol. 1:1-46.

Capítulo I
31
James, M. O e Kleinow, K. M. (1994). Trophic transfer of chemicals in the aquatic
environment. In Aquatic Toxicology - Molecular, Biochemical, and Cellular
Perspectives (Mallins, D. C. e Ostrander, G. K., Eds.) pp. 1-35, CRC Press, Boca
Raton, FL.
Kohlpoth, M., Rusche, B. e Nüsse, M. (1999). Flow cytometric measurement of
micronuclei induced in a permanent fish cell line as a possible screening test for
genotoxicity of industrial waste waters. Mutagenesis. 14 (4): 397-402.
Kosmala, A., Migeon, B., Flammarion, P. e Garric, J. (1988). Impact assessment of a
wastewater treatment plant effluent using the fish biomarker ethoxyresorufin-O-
deethylase: field and on-site experiments. Ecotoxicol. Environ. Saf. 41:19-28.
Kramer, V. J., Miles-Richardson, S., Pierens, S. L., Giesy, J. P. (1998). Reproductive
impairment and induction of alkaline-labile phosphate, a biomarker of estrogen
exposure, in fathead minnows (Pimephales Promelas) exposed to water-borne
17β-estradiol. Aquat. Toxicol. 40:335-360.
Livingstone, D. R. (1998). The fate of organic xenobiotics in aquatic ecosystems:
quantitative and qualitative differences in biotransformation by invertebrates and
fish. Em: Livingstone, D. R. (2001). Contaminant-stimulated reactive oxygen
species production and oxidative damage in aquatic organisms. Mar. Pollut.
Bulletin. vol. 42, nº 8, 656-666.
Maria, V. L., Correia, A. C., Santos, M. A. (2002). Anguilla anguilla L. biochemical and
genotoxic responses to benzo[a]pyrene. Ecotoxicol. Environ. Saf. 53:86-92.
Masfaraud, J. F., Monod, G. e Devaux, A. (1990). Use of the fish cytocrome P450-
dependent 7-ethylresorufin O-deethylase activity as a biochemical indicator of
water pollution. Study of the liver and the kidney of male and female nase
(Chondrostoma nasus) from the river Rhône. Sci. Total Environ. 97/98:729-
738.
McCarthy, J. F. e Shugart, L. R. (1990). Biological markers of environmental
contamination. Em: Van der Oost R., Beyer J. e Vermeulen, N. P. E. (2003). Fish
bioaccumulation and biomarkers in environmental risk assessment: a review.
Environ. Toxicol. Pharm. 13:57-149.

Introdução geral
32
McCarthy, J. F., Halbrook, R. S. e Shugart, L. R. (1991). Conceptual strategy for
design, implementation and validation of a biormarker-based biomonitoring
capability. TM-11783. Oak Ridge, TN:Oak Ridge National Laboratory.
Menzie, C. A., Potocki, B. B. e Santodonato, J. (1992). Exposure to carcinogenic PAHs
in the environment. Environ. Sci. Techno. 26:1278-1284.
Moore, M. N. e Simpson, M. G. (1992). Molecular and cellular pathology in
environmental impact assessment. Aquat. Toxicol. 22:313-322.
NRC: Committee on biological markers of the Nacional Research Council (1987).
Biological markers in environmental health research. Environ. Health. Perspect.
74:3-9.
Oliveira M., Santos, M. A., e Pacheco M. (2005). Heavy metal inhibition of Anguilla
anguilla L. liver microsomal EROD activity and thiol protection. Fresen. Environ.
Bulletin. vol. 14, nº 1, 59-64.
Oliveira M., Pacheco M. e Santos, M. A. (2007). Cytochrome P4501A, genotoxic and
stress responses in golden grey mullet (Liza aurata) following short-term
exposure to phenanthrene. Chemosphere. 66:1284-1291.
Pacheco M. e Santos, M. A. (1997). Induction of EROD activity and genotoxic effects
by polycyclic aromatic hydrocarbons and resin acids on the juvenile eel (Anguilla
anguilla L.). Ecotoxicol. Environ. Saf. 38:252-259.
Pacheco M. e Santos M. A. (1998). Induction of liver EROD and erythrocytic nuclear
abnormalities by cyclophosphamide and PAHs in Anguilla anguilla L.. Ecotoxicol.
Environ. Saf. 40:71-76.
Pacheco M. e Santos, M. A. (1999). Biochemical and genotoxic responses of adult eel
(Anguilla anguilla L.) to resin acids and pulp mill effluent: laboratoty and field
experiments. Ecotoxicol. Environ. Saf. 42:81-93.
Pacheco, M. e Santos, M. A. (2001a). Biotransformation, endocrine and genetic
responses of Anguilla anguilla to petroleum distillate products and environmental
contaminated waters. Ecotoxicol. Environ. Saf. 49:64-75.

Capítulo I
33
Pacheco, M. e Santos, M. A. (2001b). Tissue distribution and temperature-dependence
of Anguilla anguilla L. EROD activity following exposure to model inducers and
relationship with plasma cortisol, lactate and glucose levels. Environ. Int.
26:149-155.
Pacheco, M. e Santos, M. A. (2002). Naphthalene and ß-naphthoflavone toxicity to
Anguilla anguilla L.: Hepatic metabolism and erythrocytic nuclear abnormalities.
Environ. Int. 28:285-293.
Pacheco M., Santos, M. A., Teles M., Oliveira M., Rebelo, J. E. e Pombo L. (2005).
Biotransformation and genotoxicity biomarkers in mullet species (Liza sp.) from
a contaminated coastal lagoon (Ria de Aveiro, Portugal). Environ. Monit. Assess.
107:133-153.
Pottinger, T. G., Carrick, T. R., Hughes, S. E. e Balm, P. H. (1996). Testosterone, 11-
ketotestosterone, and estradiol-17 beta modify baseline and stress-induced
interrenal and corticotropic activity in trout. Gen. Comp. Endocrinol. 104:284-
295; 1996.
Randall, D., Warren Burggren, W. e French, K. (2001). Glands and Hormones –
Physiological effects of hormones. Em: Animal Physiology – mechanisms and
adaptations (5ª edição). Cap 9, pp. 338-347. W. H. Freeman and Company, New
York.
Sanchez-Galan S., Linde, A. R., e Garcia-Vazquez E. (1999). Brown trout and
European minnow as target species for genotoxicity tests: Differential sensitivity
to heavy metals. Ecotoxicol. Environ. Saf. 43:301-304.
Seruto, C., Sapozhnikova, Y. e Schlenk, D. (2005). Evaluation of the relationships
between biochemical endpoints of PAH exposure and physiological endpoints of
reproduction in male California Halibut (Paralichthys californicus) exposed to
sediments from a natural oil seep. Mar. Environ. Res. 60:454-465.
Stegeman, J. J. e Lech, J. J. (1991). Cytocrome P-450 monooxygenase systems in
aquatic sprecies – carcinogen metabolism and biomarkers for carcinogen and
pollutant exposure. Environ. Health Perspect. 90:101-109.

Introdução geral
34
Stegeman, J. J., Brouwer, M., Richard, T. D. G., Förlin, L., Fowler, B. A., Sanders, B.
M. e Van Veld, P. A. (1992). Molecular responses to environmental
contamination enzyme and protein systems as indicators of chemical exposure
and effect. Em: Van der Oost R., Beyer J., Vermeulen, N. P. E. (2003). Fish
bioaccumulation and biomarkers in environmental risk assessment: a review.
Environ. Toxicol. Pharm. 13:57-149.
Suter, G. W., II (1990). Use of biomarkers in ecological risk assessment. Em: Van der
Oost R., Beyer J., Vermeulen, N. P. E. (2003). Fish bioaccumulation and
biomarkers in environmental risk assessment: a review. Environ. Toxicol. Pharm.
13:57-149.
Suter, G. W., II (1993). Ecological risk assessement. Lewis Publishers, Boca Raton, FL,
USA, pp 538.
Teles M., Maria, V. L., Pacheco M. e Santos M. A. (2003). Anguilla anguilla L. plasma
cortisol, lactate and glucose responses to abietic acid, dehydroabietic acid and
retene. Environ. Int. 29:995-1000.
Teles M., Santos M. A., e Pacheco M. (2004b). Responses of European eel (Anguilla
anguilla L.) in two polluted environments: in situ experiments. Ecotoxicol.
Environ. Saf. 58:373-378.
Teles M., Santos M. A., e Pacheco M. (2004b). Responses of European eel (Anguila
anguilla L.) in two polluted environments: in situ experiments. Ecotoxicol.
Environ. Saf. 58:373-378.
Teles, M., Pacheco, M. e Santos, M. A. (2005). Sparus aurata L. liver EROD and GST
activities, plasma cortisol, lactate, glucose and erythrocytic nuclear anomalies
after short-term exposure either to 17ß-estradiol (E2) or E2 combined with 4-
nonylphenol. Sci. Total Environ. 336:57-69.
Teles M., Pacheco M. e Santos M. A. (2007). Endocrine and metabolic responses of A.
anguilla L. caged in a freshwater-wetland (Pateira de Fermentelos - Portugal).
Sci. Total Environ. 372:562-570.

Capítulo I
35
Van der Oost R., Beyer J. e Vermeulen, N. P. E. (2003). Fish bioaccumulation and
biomarkers in environmental risk assessment: a review. Environ. Toxicol. Pharm.
13:57-149.
Van Gestel, C. A. M. e Van Brummelen, T. C. (1990). Incorporation of the biomarker
concept in ecotoxicology calls for a redefinition of terms. Ecotoxicol. 5:217-225.
Van Veld, P. A. (1990). Absorption and metabolism of dietary xenobiotics by the
intestine of fish. Em: Livingstone, D. R. (2001). Contaminant-stimulated reactive
oxygen species production and oxidative damage in aquatic organisms. Mar.
Pollut. Bulletin. vol. 42, nº 8, 656-666.
Varanasi, U., Stein, J. E. e Nishimoto, M. (1989). Biotransformation and disposition of
PAH in fish. Em: Oliveira, M., Pacheco, M. e Santos, M. A. (2007). Cytocrome
P4501A genotoxic and stress responses in golden grey mullet (Liza aurata)
following short-term exposure to phenantrene. Chemosphere. 66:1284-1291.
Vermeulen, N. P. E. (1996). Role of metabolism in chemical toxicity. Em: Van der Oost
R., Beyer J., Vermeulen, N. P. E. (2003). Fish bioaccumulation and biomarkers in
environmental risk assessment: a review. Environ. Toxicol. Pharm. 13:57-149.
Waring, C. P. e Moore, A. (2004). The effect of atrazine on Atlantic salmon (Salmo
salar) smolts in fresh water and after sea water transfer. Aquat. Toxicol. 66:93-
104; 2004.
Wendelaar-Bonga, S. E. W. (1997). The stress response in fish. Physiol. Rev. 77:591-
625.
WHO: International Programme on Chemical Safety (IPCS) (1993). Biomarkers and
risk assessment: concepts and principles. Environ. Health Criteria 155. World
Health Organization, Geneva.
Yamazaki, H., Inoue, K., Turvy, C. G., Guengerich, F. P. e Shimada, T. (1997). Effects
of freezing, thawing, and storage of human liver samples on the microsomal
contents and activities of cytochrome P450 enzymes. Drug Metab. Dispos. 25:
168-174.

CAPÍTULO II
Avaliação genotóxica sazonal de uma laguna poluída (Ria de
Aveiro) através do estudo das anomalias nucleares eritrocíticas de
Liza aurata

Capítulo II
37
RESUMO
A Ria de Aveiro é um sistema lagunar costeiro que se encontra
permanentemente ligado ao mar, tendo sido ao longo de vários anos, o
principal receptor de efluentes industriais e domésticos. Este estudo teve
como objectivo avaliar os efeitos genotóxicos dos poluentes existentes na Ria
de Aveiro, pelo que se usou Liza aurata como espécie bioindicadora. Liza
aurata é uma espécie de tainha bastante abundante na Ria de Aveiro,
apresentando uma larga distribuição geográfica e ocorrendo regularmente ao
longo do ano. A frequência de anomalias nucleares eritrocíticas (ANEs) foi
usada como biomarcador de genotoxicidade e avaliada em Liza aurata. Os
espécimes foram colhidos sazonalmente entre Maio de 2006 e Fevereiro de
2007, em seis locais distintos da Ria de Aveiro. A investigação revelou um
local crítico – Vagos – onde foi possível verificar uma elevada frequência de
ANEs na Primavera, Verão e Inverno, quando comparada com a referência
(Torreira) e com os restantes locais de amostragem. Estes resultados revelam
a existência de substâncias genotóxicas na Ria, muito provavelmente,
hidrocarbonetos aromáticos policíclicos (HAPs), e confirmam os resultados
obtidos em anteriores estudos de campo efectuados nesta mesma espécie e
na referida área. Liza aurata revelou ser um bom bioindicador, uma vez que
demonstrou ser uma espécie sensível permitindo assim detectar diferenças
entre os diferentes locais de amostragem.
INTRODUÇÃO
A Ria de Aveiro é um sistema lagunar, com 47 Km2 de superfície,
permanente ligada ao mar, localizada na região noroeste de Portugal (Pacheco
et al., 2005). A existência de unidades industriais junto à Ria, bem como o
intenso tráfego naval, têm contribuído para o aumento da poluição, uma vez
que os produtos resultantes destas mesmas actividades acabam,
frequentemente, nas águas da Ria. Além dos efluentes industriais, há ainda os
efluentes de origem doméstica e agrícola, que também atingem a Ria e nem
sempre estão sujeitos a tratamento, o que leva ao aumento dos níveis de
contaminação orgânica. Em consequência dos factos anteriormente descritos e

Avaliação genotóxica sazonal da Ria de Aveiro
38
também devido à persistência de alguns desses contaminantes, existe a
necessidade urgente de um programa efectivo de biomonitorização (Pacheco
et al., 2005), o qual permita, por exemplo, avaliar os efeitos genotóxicos daí
resultantes. Os peixes são frequentemente usados como organismos teste em
estudos de genotoxicidade (Arkhipchuk e Garanko, 2005), uma vez que
intervêm a vários níveis na cadeia trófica, acumulando substâncias tóxicas e
respondendo a baixas concentrações de agentes mutagénicos (Cavas e
Ergene-Gözüraka, 2005). Liza aurata tem demonstrado ser uma boa espécie
bioindicadora na monitorização de águas contaminadas por xenobióticos
orgânicos (Pacheco et al., 2005; Oliveira et al., 2007). Num estudo de campo
efectuado por Pacheco et al. (2005), na mencionada Ria de Aveiro, foram
adoptadas três espécies de tainha - Liza ramada, Liza aurata e Liza saliens. L.
aurata revelou ser a mais abundante, apresentando uma maior distribuição
geográfica e ocorrendo mais regularmente ao longo do ano. Assim,
demonstrou ser a espécie mais apropriada para a biomonitorização de toda a
ria, revelando ser uma boa candidata como espécie bioindicadora (Pacheco et
al., 2005). Apesar das eventuais migrações sazonais, esta espécie de tainha
apresenta um raio de mobilidade reduzido, o que também constitui uma
característica favorável na sua escolha como bioindicadora, encontrando-se
bem adaptada a diferentes salinidades que vão desde 0 até 35 (‰), atributo
muito importante quando se pretende biomonitorizar ecossistemas costeiros
onde exista uma grande variação da salinidade da água (Pacheco et al.,
2005).
Os micronúcleos são usados como biomarcadores de danos estruturais
ao nível dos cromossomas (Arkhipchuk e Garanko, 2005). Durante os últimos
anos, vários estudos têm descrito a presença de anomalias nucleares, para
além dos referidos micronúcleos, em eritrócitos de peixes expostos a
substâncias genotóxicas (Ayllón e Garcia-Vazquez 2000; Cavas e Ergene-
Gözüraka, 2003b). Os micronúcleos têm origem em fragmentos de
cromossomas ou cromossomas inteiros que não foram sujeitos à fase
anafásica durante a divisão nuclear (Fenech e Crott, 2002). O teste das
anomalias nucleares (AN), que tem sido aplicado com sucesso em peixes e
usado como uma medida de genotoxicidade (Pacheco e Santos, 1997, 1998,

Capítulo II
39
1999, 2001a e 2001b; Oliveira et al., 2007), baseia-se na detecção de
micronúcleos e/ou estruturas nucleares anormais em eritrócitos maduros
(Pacheco et al., 1997). Este teste tem demonstrado ser um teste com um
elevado grau de sensibilidade, de fácil execução e grande eficiência, tanto em
estudos laboratoriais como em estudos de campo (Pacheco et al., 1999).
Este trabalho teve como objectivo avaliar, sazonalmente, a frequência
de anomalias nucleares eritrocíticas em espécimes de L. aurata. Os espécimes
foram colhidos em locais pré seleccionados da Ria de Aveiro, sujeitos a
diferentes tipos e fontes de contaminação, com o intuito de avaliar e comparar
a genotoxicidade existente nos referidos locais.
MATERIAL E MÉTODOS
Amostragem – animais e locais de colheita
A amostragem decorreu, sazonalmente, entre Maio de 2006 e Fevereiro
de 2007, durante a maré baixa, tendo sido usada para o efeito uma rede
tradicional designada “chincha”. Foram colhidos espécimes juvenis de uma
espécie de tainha vulgarmente designada de tainha garrento (L. aurata).
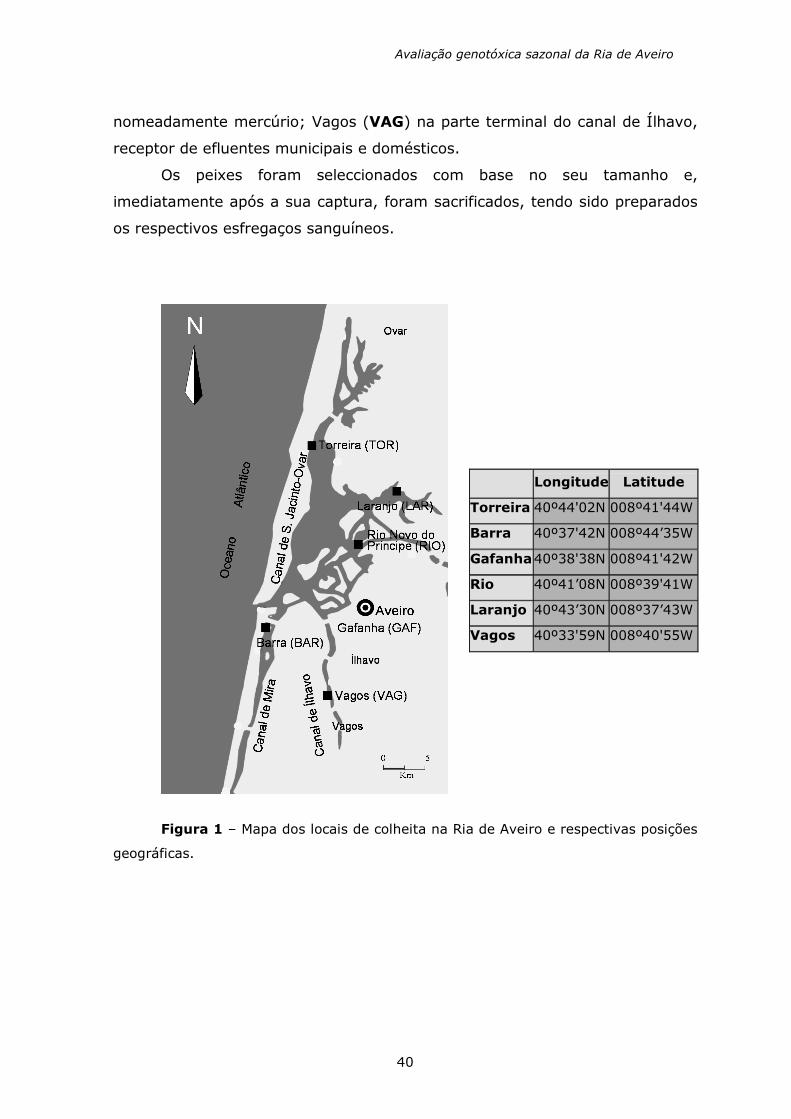
Os locais de amostragem, na Ria de Aveiro (figura 1), foram
seleccionados com base numa distribuição geográfica ao longo dos canais
principais e na entrada da ria, tendo em consideração os vários tipos e fontes
de contaminação, bem como a selecção (teórica) de um ponto de referência
não poluído. Os locais de amostragem seleccionados foram os seguintes:
Torreira (TOR) como ponto de referência, uma vez que não se encontra
próximo das principais fontes de poluição; Barra (BAR) situado na parte inicial
do Canal de Mira, próximo da entrada da ria que dá para o mar e sujeita a um
tráfico naval considerável; Gafanha (GAF) localizado na proximidade de uma
zona portuária; Rio Novo do Príncipe (RIO) situado na parte terminal do rio
Vouga e principal fonte de água doce, a 6 Km de distância de uma fábrica de
pasta de papel; Laranjo (LAR) próximo de uma fábrica de soda cáustica, a
qual constitui uma importante fonte de contaminação de metais pesados,
Avaliação genotóxica sazonal da Ria de Aveiro
40
nomeadamente mercúrio; Vagos (VAG) na parte terminal do canal de Ílhavo,
receptor de efluentes municipais e domésticos.
Os peixes foram seleccionados com base no seu tamanho e,
imediatamente após a sua captura, foram sacrificados, tendo sido preparados
os respectivos esfregaços sanguíneos.
Figura 1 – Mapa dos locais de colheita na Ria de Aveiro e respectivas posições
geográficas.
Longitude Latitude
Torreira 40º44'02N 008º41'44W
Barra 40º37'42N 008º44’35W
Gafanha 40º38'38N 008º41'42W
Rio 40º41’08N 008º39'41W
Laranjo 40º43’30N 008º37’43W
Vagos 40º33'59N 008º40'55W

Capítulo II
41
Contagem das Anomalias Nucleares Eritrocíticas
Os esfregaços sanguíneos foram fixados com metanol durante 10
minutos, corados com Giemsa (5%) durante 30 minutos e sujeitos a secagem
à temperatura ambiente, de forma a serem usados no teste das anomalias
nucleares eritrocíticas. A contagem das anomalias nucleares teve lugar em
seguida, numa amostra de 1000 eritrócitos maduros por peixe, de acordo com
os critérios de Schmid (1976), Carrasco et al. (1990) e Smith (1990),
adaptado por Pacheco e Santos (1996), com o objectivo de avaliar a
genotoxicidade. De acordo com estes autores, as lesões nucleares são
incluídas e contadas numa das seguintes categorias: micronúcleo, núcleo
lobulado, núcleo segmentado e núcleo reniforme. O resultado final foi
expresso como o valor médio (‰) da soma de todas a lesões observadas em
cada indivíduo.
Análise Estatística
Os resultados estão expressos em médias ± erro padrão,
correspondentes a grupos de cinco peixes capturados por cada local de
amostragem (n=5). O software SIGMASTAT 2.03 foi usado para efectuar a
análise estatística. Os dados experimentais foram inicialmente testados para
normalidade e homogeneidade da variância, de forma a obedecer a requisitos
estatísticos e, posteriormente, com o teste de Tukey (Zar, 1999). As
diferenças entre médias foram consideradas estatisticamente significativas
quando p <0.05.
Avaliação genotóxica sazonal da Ria de Aveiro
42
RESULTADOS
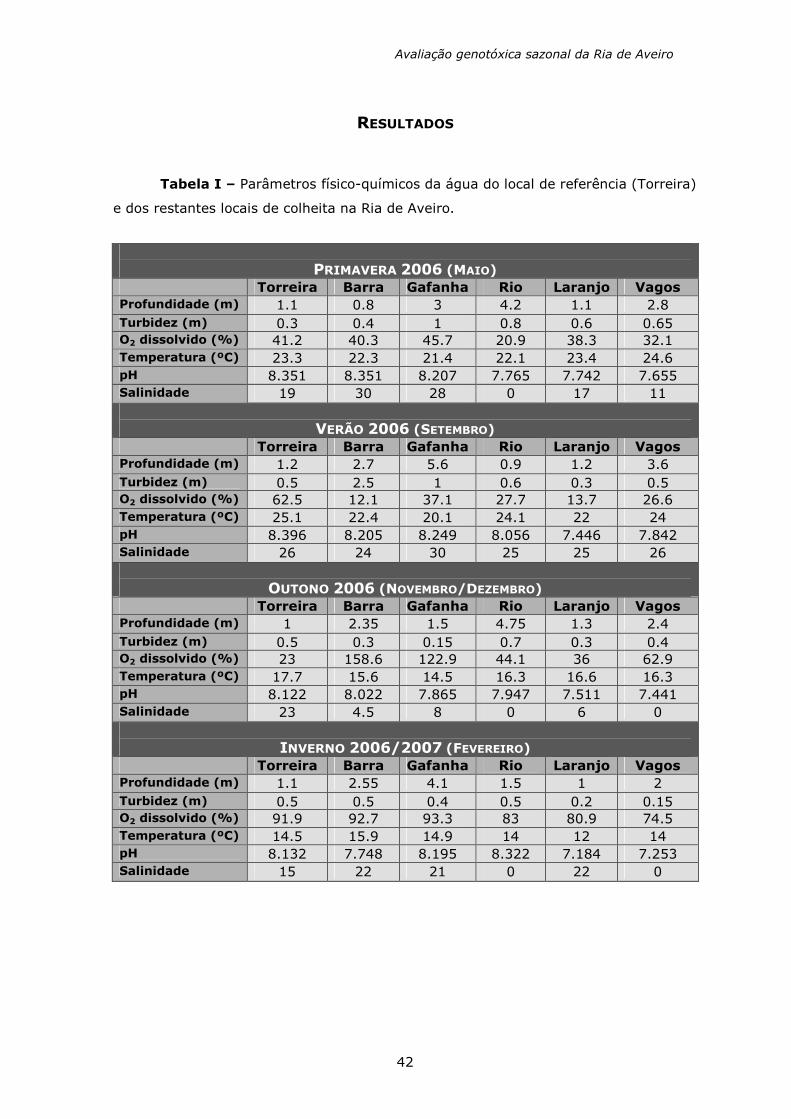
Tabela I – Parâmetros físico-químicos da água do local de referência (Torreira)
e dos restantes locais de colheita na Ria de Aveiro.
PRIMAVERA 2006 (MAIO) Torreira Barra Gafanha Rio Laranjo Vagos Profundidade (m) 1.1 0.8 3 4.2 1.1 2.8 Turbidez (m) 0.3 0.4 1 0.8 0.6 0.65 O2 dissolvido (%) 41.2 40.3 45.7 20.9 38.3 32.1 Temperatura (ºC) 23.3 22.3 21.4 22.1 23.4 24.6 pH 8.351 8.351 8.207 7.765 7.742 7.655 Salinidade 19 30 28 0 17 11
VERÃO 2006 (SETEMBRO)
Torreira Barra Gafanha Rio Laranjo Vagos Profundidade (m) 1.2 2.7 5.6 0.9 1.2 3.6 Turbidez (m) 0.5 2.5 1 0.6 0.3 0.5 O2 dissolvido (%) 62.5 12.1 37.1 27.7 13.7 26.6 Temperatura (ºC) 25.1 22.4 20.1 24.1 22 24 pH 8.396 8.205 8.249 8.056 7.446 7.842 Salinidade 26 24 30 25 25 26
OUTONO 2006 (NOVEMBRO/DEZEMBRO)
Torreira Barra Gafanha Rio Laranjo Vagos Profundidade (m) 1 2.35 1.5 4.75 1.3 2.4 Turbidez (m) 0.5 0.3 0.15 0.7 0.3 0.4 O2 dissolvido (%) 23 158.6 122.9 44.1 36 62.9 Temperatura (ºC) 17.7 15.6 14.5 16.3 16.6 16.3 pH 8.122 8.022 7.865 7.947 7.511 7.441 Salinidade 23 4.5 8 0 6 0
INVERNO 2006/2007 (FEVEREIRO)
Torreira Barra Gafanha Rio Laranjo Vagos Profundidade (m) 1.1 2.55 4.1 1.5 1 2 Turbidez (m) 0.5 0.5 0.4 0.5 0.2 0.15 O2 dissolvido (%) 91.9 92.7 93.3 83 80.9 74.5 Temperatura (ºC) 14.5 15.9 14.9 14 12 14 pH 8.132 7.748 8.195 8.322 7.184 7.253 Salinidade 15 22 21 0 22 0

Capítulo II
43
Primavera
Figura 2 – Frequência de anomalias nucleares eritrocíticas (ANEs) em Liza
aurata, obtida para os diferentes locais da Ria de Aveiro, na Primavera. Os valores
encontram-se expressos em médias ± erro padrão (n=5). As diferenças significativas
(p<0.05) são: T vs Torreira (local de referência); B vs Barra; G vs Gafanha; R vs Rio.
A frequência de ANEs foi significativamente mais elevada em Vagos
quando comparada com o local de referência (Torreira) e com os restantes
locais de amostragem, com excepção do Laranjo. Também foi possível
observar uma forte elevação da frequência de ANEs no Laranjo, a qual é
significativa quando comparada com a Torreira e com o Rio. A frequência mais
baixa de ANEs ocorreu no Rio e foi significativamente mais baixa que a
frequência obtida para a Gafanha.
0
100
200
300
400
TOR BAR GAF RIO LAR VAG
Locais de Amostragem
Fre
quên
cia
de A
NE
(‰)
0
100
200
300
TOR BAR GAF LAR RIO VAG
T B G R T
R
G
400

Avaliação genotóxica sazonal da Ria de Aveiro
44
Verão
Figura 3 – Frequência de anomalias nucleares eritrocíticas (ANEs) em Liza
aurata, obtida para os diferentes locais da Ria de Aveiro, no Verão. Os valores
encontram-se expressos em médias ± erro padrão (n=5). As diferenças significativas
(p<0.05) são: T vs Torreira (local de referência); B vs Barra; G vs Gafanha; R vs Rio.
A frequência de ANEs foi significativamente mais elevada em Vagos,
quando comparada com o local de referência (Torreira) e com a Barra, a
Gafanha e o Rio. A frequência mais baixa de ANEs foi obtida para a Barra e foi
significativamente diferente do local de referência (Torreira).
0
100
200
300
400
TOR BAR GAF RIO LAR VAG
0
100
200
300
400
TOR BAR GAF LAR RIO VAG
Fre
quên
cia
de A
NE
(‰
)
T B G R
T
Locais de Amostragem

Capítulo II
45
Outono
Figura 4 – Frequência de anomalias nucleares eritrocíticas (ANEs) em Liza
aurata, obtida para os diferentes locais da Ria de Aveiro, no Outono. Os valores
encontram-se expressos em médias ± erro padrão (n=5). As diferenças significativas
(p<0.05) são: B vs Barra; G vs Gafanha; L vs Laranjo.
A frequência mais elevada de ANEs ocorreu na Gafanha, sendo
significativamente diferente da Barra, Rio e Laranjo. A frequência mais baixa
de ANEs foi obtida para o Rio e para o Laranjo.
A frequência de ANEs observada em Vagos foi significativamente mais
elevada que aquela obtida para o Laranjo.
0
100
200
300
400
1 2 3 4 5 6
0
100
200
300
400
TOR BAR GAF LAR RIO VAG
Fre
quên
cia
de A
NE
(‰
)
Locais de Amostragem
L B
G G

Avaliação genotóxica sazonal da Ria de Aveiro
46
Inverno
Figura 5 – Frequência de anomalias nucleares eritrocíticas (ANEs) em Liza
aurata, obtida para os diferentes locais da Ria de Aveiro, no Inverno. Os valores
encontram-se expressos em médias ± erro padrão (n=5). As diferenças significativas
(p<0.05) são: T vs Torreira (local de referência); B vs Barra; G vs Gafanha; R vs Rio;
L vs Laranjo.
A frequência de ANEs foi significativamente mais elevada em Vagos,
quando comparada, não só com o local de referência (Torreira), mas também
com os restantes locais de amostragem (Barra, Gafanha, Rio e Laranjo). Não
se observaram diferenças significativas entre a Barra, Gafanha Rio e Laranjo e
entre estes últimos e o local de referência.
DISCUSSÃO
De uma forma geral, e tendo em conta os resultados obtidos para as
quatro estações do ano, importa realçar que a presença de ANEs foi notória e
bastante expressiva num local – Vagos, o que é concordante com um estudo
de campo anteriormente feito nas mesmas condições (Pacheco et al., 2005).
Nesse mesmo estudo foram detectados elevados níveis de HAPs,
0
100
200
300
400
1 2 3 4 5 6
0
100
200
300
400
TOR BAR GAF LAR RIO VAG
Fre
quên
cia
de A
NE
(‰
)
Locais de Amostragem
T B G R L

Capítulo II
47
nomeadamente B[a]P, em Vagos, na água, os quais foram apontados como
uma causa provável para a indução de ANEs. Este facto apoia os resultados
obtidos neste trabalho, uma vez que Vagos continua a ser um receptor de
efluentes domésticos e municipais. Alguns estudos têm demonstrado o
potencial genotóxico dos HAPs, nomeadamente através da sua capacidade de
indução de ANEs (Pacheco e Santos, 1997; Pacheco e Santos, 2001a; Oliveira
et al., 2007), já que as reacções envolvidas na biotransformação destes
compostos, podem levar à formação de metabolitos reactivos que, por sua
vez, podem ser mais tóxicos que o composto original e, como tal, interagir
com o ADN, e em última instância causar danos a este nível.
Contudo, de acordo com Pacheco e Santos (2002) o estudo de ANEs
pode revelar-se inadequado em situações de intensa contaminação, já que,
nestas situações, podem ocorrer interferências ao nível da expressão
genotóxica, como resultado do balanço entre o aumento do catabolismo
eritrocítico no baço e um decréscimo da velocidade da eritropoiese. De acordo
com Das e Nanda (1986), a indução de lesões nucleares poderá por vezes ser
mascarada através de uma acção citotóxica, havendo morte dos eritrócitos em
vez de ocorrer formação de lesões nucleares não-letais. Para além disso, as
células portadoras de danos/anomalias tendem a ser removidas do organismo
mais rapidamente que as normais (Das e Nanda, 1986). Estes factos poderão
explicar a ausência de expressão genotóxica, nomeadamente em Vagos, no
Outono. Parâmetros físico-químicos, tais como temperatura, pluviosidade, pH
e oxigénio dissolvido, influenciam a biodisponibilidade dos contaminantes,
constituindo por isso uma importante variável que também tem de ser tida em
conta. O Outono de 2006 (ao contrário das restantes estações) foi
extremamente chuvoso (Instituto de Meteorologia de Portugal, 2006) pelo
que, a pluviosidade intensa poderá ter originado um efeito de diluição das
substâncias genotóxicas presentes na água, o que poderá ter contribuído para
uma alteração dos níveis de contaminação. Contudo, os resultados obtidos
para cada estação do ano devem ser interpretados com precaução, atendendo
a que, como não se procedeu à análise da água e do sedimento, desconhece-
se qual a natureza e concentração dos compostos presentes nos mesmos à
data das amostragens.

Avaliação genotóxica sazonal da Ria de Aveiro
48
A espécie adoptada neste trabalho – L. aurata – demonstrou, uma vez
mais, a sua utilidade como espécie bioindicadora, uma vez que permitiu
detectar diferenças entre os diferentes locais de amostragem.
CONCLUSÕES
O presente estudo revelou a existência de um local problemático –
Vagos - onde se verificou a existência de uma elevada frequência de ANEs em
todas as estações do ano, com excepção do Outono. Estes resultados indiciam
a existência de substâncias genotóxicas na água, muito provavelmente,
hidrocarbonetos aromáticos policíclicos (HAPs), confirmando assim os
resultados obtidos em anteriores estudos de campo efectuados na referida
área (Pacheco et al., 2005).
L. aurata demonstrou a sua utilidade, enquanto espécie bioindicadora,
em estudos de monitorização dos efeitos biológicos de contaminantes
antropogénicos, nomeadamente na Ria de Aveiro.
REFERÊNCIAS BIBLIOGRÁFICAS
Arkhipchuck, V. V. e Garanko, N. N. (2005). Using the nucleolar biomarker an the
micronucleus test on in vivo fish fin cells. Ecotoxicol. Environ. Saf. 62:42-52.
Ayllón F. e Garcia-Vazquez E. (2000). Induction of micronuclei and other nuclear
abnormalities in european minnow Phoxinus phoxinus and mollie Poecilia
latipinna: as assessment of the fish micronucleus test. Mut. Res./Gen. Toxicol.
Environ. Mutagen. 467:177-186.
Carrasco, K. R., Tilbury, K. L. e Myers, M. S. (2005). Assessment of the piscine
micronuclei test as an in situ biological indicator of chemical contaminant effects.
Can. J. Fish. Aquat. Sci. 47:2123-2136.

Capítulo II
49
Cavas, T. e Ergene-Gözüraka, S. (2003b). Micronuclei, nuclear lesions and interphase
silver-stained nucleolar organizer regions (AgNORs) as cyto-genotoxicity
indicators in Oreochromis niloticus exposed to textile mill effluent. Mutat. Res.
538(1-2): 81-91.
Cavas, T. e Ergene-Gözüraka, S. (2005). Induction of micronuclei and nuclear
abnormalities in Oreochromis niloticus following exposure to petroleum refinery
and chromium processing plant effluents. Aquat. Toxicol. 74:264-271.
Das, R. K. e Nanda, R. K. (1986). Induction of micronuclei in peripheral erythrocytes
of the fish, Heteropneustes fossilis, by mitomycin C and paper mill effluent.
Mutat. Res. 175: 67-71.
Fenech, M. e Crott. J. W. (2002). Micronuclei nucleoplasmatic bridges and nuclear
buds induced in folic acid deficient human lymphocytes – evidence for breakage-
fusion-bridge cycles in the cytokinesis-block micronucleus assay. Mutat. Res.
504: 131-136.
Instituto de Meteorologia de Portugal (2006). Relatório climático anual de 2006.
http://www.meteo.pt/resources/im/pdfs/clim_ap_00_06.pdf.
Oliveira M., Pacheco M., Santos, M. A. (2007). Cytochrome P4501A, genotoxic and
stress responses in golden grey mullet (Liza aurata) following short-term
exposure to phenanthrene. Chemosphere. 66:1284-1291.
Pacheco M. e Santos, M. A (1996). Induction of micronuclei and nuclear abnormalities
in the erythrocytes of Anguilla anguilla L. exposed either cyclophosphamide or to
bleached kraft pulp mill effluent. Fresen. Environ. Bulletin. 5: 746-751.
Pacheco M. e Santos, M. A. (1997). Induction of EROD activity and genotoxic effects
by polycyclic aromatic hydrocarbons and resin acids on the juvenile eel (Anguilla
anguilla L.). Ecotoxicol. Environ. Saf. 38:252-259.
Pacheco M. e Santos, M. A. (1998). Induction of liver EROD and erythrocytic nuclear
abnormalities by cyclophosphamide and PAHs in Anguilla anguilla L.. Ecotoxicol.
Environ. Saf. 40:71-76.

Avaliação genotóxica sazonal da Ria de Aveiro
50
Pacheco M. e Santos, M. A. (1999). Biochemical and genotoxic responses of adult eel
(Anguilla anguilla L.) to resin acids and pulp mill effluent: laboratoty and field
experiments. Ecotoxicol. Environ. Saf. 42:81-93.
Pacheco M. e Santos, M. A. (2001a). Biotransformation, endocrine, and genetic
responses of Anguilla anguilla L. to petroleum distillate products and
environmentally contaminated waters. Ecotoxicol. Environ. Saf. 49:64-75.
Pacheco, M. e Santos, M. A. (2001b). Tissue distribution and temperature-dependence
of Anguilla anguilla L. EROD activity following exposure to model inducers and
relationship with plasma cortisol, lactate and glucose levels. Environ. Int.
26:149-155.
Pacheco M. e Santos, M. A. (2002). Biotransformation, genotoxic and histopathological
effects of environmental contaminants in European eel (Anguilla anguilla L.).
Ecotoxicol. Environ. Saf. 53:331-347.
Pacheco M., Santos, M. A., Teles M., Oliveira M., Rebelo, J. E. e Pombo L. (2005).
Biotransformation and genotoxicity biomarkers in mullet species (Liza sp.) from
a contaminated coastal lagoon (Ria de Aveiro, Portugal). Environ. Monit. Assess.
107: 133-153.
Schmid, W. (1976) The micronucleus test for cytogenetic analysis, in A. Hollaender
(ed.), Chemical Mutagen. vol. 4, 31-53, Plenum Press, New York.
Smith, I. R. (1990). Erythrocytic micronuclei in wild fish from Lakes Superior and
Ontario that have pollution-associated neopasia. J. Great Lakes Res. 16: 139-
142.

CAPÍTULO III
Estudo in vitro do efeito do benzo[a]pireno, cortisol e 17ß-
estradiol sobre a actividade microsomal hepática de EROD de Anguilla
anguilla L..

Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
52
RESUMO
Uma enguia adulta foi injectada intraperitonealmente com ß-
naftoflavona (BNF) 4mg/Kg. Após 24 horas de exposição, o respectivo fígado
foi extraído, tendo-se procedido ao isolamento dos seus microsomas. Os
microsomas hepáticos foram expostos numa cuvette – in vitro - a
benzo[a]pireno (B[a]P) 0.1, 0.3, 0.9 e 2.7 µM. O efeito protector de cortisol
(C) e 17ß-estradiol (E2) 22,02 nM sobre actividade microsomal hepática de
EROD, foi testado através da sua inclusão no tampão substrato previamente à
exposição às diferentes concentrações de B[a]P testadas. A exposição prévia
dos microsomas hepáticos ao tampão-substrato contendo C (22.02 nM) levou
a uma elevação acentuada da actividade de EROD. A exposição dos
microsomas hepáticos a B[a]P (0.1, 0.3, 0.9 e 2.7 µM) causou uma forte
inibição da actividade de EROD. Ambas as hormonas demonstraram um efeito
protector ao nível da inibição causada pelo B[a]P (0.1, 0.3, 0.9 e 2.7 µM),
efeito esse que se traduziu num aumento da actividade da EROD.
INTRODUÇÃO
A poluição dos recursos hídricos é um problema sério e crescente.
Apesar da existência de legislação relevante, a poluição do meio aquático por
poluentes químicos tóxicos continua a ocorrer, sendo os efluentes de origem
doméstica e industrial as principais fontes responsáveis pela contaminação do
meio aquático (Matsumoto et al., 2006). De entre os diferentes tipos de
poluentes tipicamente atribuíveis às actividades humanas, os produtos
derivados do petróleo, como por exemplo os hidrocarbonetos aromáticos
policíclicos (HAPs), são os mais ecotoxicologicamente relevantes (Pacheco e
Santos, 2001). O sistema endócrino dos peixes encontra-se, por isso, exposto
a uma larga variedade de compostos naturais e antropogénicos, os quais
podem causar danos ao nível do sistema reprodutor, bem como outros efeitos
adversos, incluindo alterações das enzimas envolvidas na metabolização
desses mesmos compostos (Vaccaro et al., 2005)

Capítulo III
53
Após absorção, o destino e efeitos da grande maioria dos xenobióticos,
incluindo os HAPs, são fortemente governados pela actividade catalítica de
várias enzimas envolvidas na metabolização dos xenobióticos. Estas enzimas
desempenham um papel central na biotransformação deste tipo de compostos
havendo, neste contexto, um interesse significativo na avaliação da actividade
da etoxiresorufina-O-desetilase (Pacheco et al., 2001), a qual é usada como
um biomarcador in vivo de exposição aos referidos HAPs, como por exemplo o
benzo[a]pireno (B[a]P), e a compostos estruturalmente semelhantes tal como
a ß-naftoflavona (BNF) (Buchelli e Fent, 1995). O BNF é reconhecido como um
forte indutor da actividade hepática da EROD (Pacheco e Santos, 1998). Neste
contexto, os bioensaios in vitro de rápida execução podem facilitar a avaliação
do risco, devido à facilidade que proporcionam a análise e identificação da
presença de compostos potencialmente tóxicos pertencentes à família dos
HAPs, individualmente e/ou em misturas complexas (Billiard, 2004). Contudo,
na natureza nem sempre existe uma relação directa entre a concentração de
um composto químico indutor e a resposta do sistema CYP1A, devido à
existência de uma série de factores que podem alterar a resposta indutora
(Sarasquete et al., 2000). A existência de parâmetros exógenos não químicos,
tais como a temperatura ambiental ou a estação do ano, e factores
endógenos, tais como os níveis hormonais ou a condição fisiológica do peixe,
podem de forma análoga, modular a resposta do sistema CYP1A (Sarasquete
et al., 2000). O fígado é, geralmente, referido como sendo o órgão mais
importante, envolvido na metabolização dos xenobióticos (Pacheco et al.,
2001).
Os indutores típicos do citocromo P4501A (CYP1A), tais como os HAPs,
são reconhecidos por exercerem disrupção endócrina nos peixes, avaliada
através dos baixos níveis de E2 no plasma (Casillas et al., 1991), maturação
sexual precoce (Collier et al., 1998) e fertilidade reduzida (Cheek et al.,
2001). O E2 é o mais potente dos estrogénios, desempenhando um importante
papel em ambos os sexos, em vários aspectos do crescimento,
desenvolvimento e diferenciação morfológica, assim como no desenvolvimento
e regulação dos comportamentos e ciclos reprodutivos e sexuais (Randall et
al., 2001). Vários estudos (maioritariamente de campo) têm demonstrado que

Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
54
E2 afecta a actividade hepática de CYP1A nos peixes (Navas e Segner, 2001;
Vaccaro et al., 2005; Teles et al., 2005) levando, geralmente, ao decréscimo
da mesma.
O cortisol (C) é exemplo de outro esteróide, cuja síntese ocorre em
células do tecido interrenal, sendo o principal corticoesteróide em peixes assim
como o mais abundante e activo. Os principais órgãos-alvo do cortisol são as
brânquias, fígado e intestino, sendo também importante a sua acção sobre o
músculo esquelético (Hontela, 1997). O aumento da concentração de cortisol
no plasma, foi observado em peixes após exposição de curto prazo a uma
vasta gama de contaminantes, tais como metais pesados (De Boeck et al.,
2003), ácidos resínicos (Teles et al., 2003), HAPs (Pacheco e Santos, 2001) e
pesticidas (Waring e Moore, 2004). Assim, o nível de cortisol no plasma é
usado como um indicador de rotina da magnitude e duração da resposta ao
stress causado pela exposição a contaminantes (Ruane et al., 2001).
Apesar da existência de bastante informação científica acerca da
importância do sistema endócrino em peixes, nomeadamente a resposta de
hormonas esteróides a situações de stress, pouco se conhece sobre os níveis
fisiológicos celulares destas hormonas e da sua influência na actividade
microsomal hepática de EROD, in vitro. Este trabalho pretendeu avaliar in
vitro, o efeito de B[a]P isoladamente e também com exposição prévia a C e
E2, de forma a estudar o efeito protector destas duas hormonas sobre a
actividade EROD microsomal hepática de A. anguilla L.
MATERIAL E MÉTODOS
Compostos químicos
β-naftoflavona (BNF), dimetil sulfóxido (DMSO), resorufina e
benzo[a]pireno (B[a]P) (Sigma Chemical Co, EUA); 7-etoxiresorufina e NADPH
(Roche); 17ß-estradiol (Sigma-Aldrich, Alemanha); cortisol (Merck Sharp &
Dohme).

Capítulo III
55
Animais
Uma enguia adulta – Anguilla anguilla L. – com cerca de 500 gramas,
adquirida num mercado de peixe local, foi mantida no laboratório, num tanque
de água salobra com arejamento durante um período de aclimatização de 48
horas. A enguia foi injectada intraperitonealmente com ß-naftoflavona
4mg/Kg, após o referido período de aclimatização, e sacrificada após 24 horas.
O fígado foi então retirado, sendo imediatamente congelado em azoto líquido,
e armazenado a -80ºC até se proceder à respectiva homogeneização. O
animal não foi alimentado quer durante o período de recobro quer durante o
período experimental.
Isolamento dos microsomas
Procedeu-se à homogeneização do fígado previamente extraído em
tampão TRIS-HCL 0.1 M, pH 7.4 + KCL (0.15M) + Glicerol (20%). Os
microsomas hepáticos foram obtidos de acordo com os métodos de Lange et
al. (1993), Monod e Vindimian (1991), adaptados por Pacheco e Santos
(1998).
Actividade da EROD
A actividade da EROD foi determinada de acordo com o método de
Burke e Meyer (1974), a 25ºC, através do aumento progressivo de
fluorescência, resultante da formação de resorufina, durante 3 minutos
(comprimento de onda de excitação 530nm; comprimento de onda de emissão
585nm), num espectrofluorímetro Jasco FP 750, e expressa em
picomoles/min/mg de proteína. A reacção foi iniciada com a adição de 10 µl de
NADPH (10 mM).
Determinação da concentração de proteína
A concentração microsomal de proteína foi determinada de acordo com
o método de Biureto (Gornall et al., 1949), usando a albumina (proteína
bovina) como padrão.

Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
56
Tabela 1 – Tampões, solventes e soluções usados durante o protocolo
experimental
Tampões
TS 0.5 µM etoxiresorufina em Tris-HCl 0,1 M pH 7.4 com KCl 0,15M e 20%
T Tris-HCl 0,1 M pH 7.4 com KCl 0,15M e 20% glicerol
TSC TS com cortisol (concentração final na cuvette = 22.02nM)
TSE2 TS com E2 (concentração final na cuvette = 22.02nM)
Solventes
DMSO Dimetil sulfóxido
Soluções
B[a]P Benzo[a]pireno (0.1, 0.3, 0.9 e 2.7µM) em DMSO
Tabela 2 – Procedimento experimental para avaliação da actividade
microsomal hepática de EROD. A numeração (1, 2, 3 e 4) indica a ordem pela qual os
compostos intervenientes na reacção foram adicionados na cuvette.
Ensaio
5 µ
l m
icro
som
as
1090 µ
l T
S
1090 µ
l T
SC
1090 µ
l T
SE
2
5 µ
l T
5 µ
l D
MS
O
5 µ
l B
[a]P
(em
D
MSO
)
10 µ
l N
AD
PH
Controlo 1 2 3 4
Controlo DMSO 1 2 3 4 Efeitos do C in vitro
Controlo DMSO+C 1 2 3 4
Controlo 1 2 3 4
Controlo DMSO 1 2 3 4 Efeitos do E2 in vitro
Controlo DMSO+E2 1 2 3 4
Controlo 1 2 3 4
Controlo DMSO 1 2 3 4 Efeitos do B[a]P in vitro B[a]P (0.1, 0.3, 0.9 e
2.7 µM) 1 2 3 4
Controlo DMSO 1 2 3 4
Controlo DMSO+C 1 2 3 4 Efeitos in vitro do
B[a]P após exposição a C (22.02 nM) C+B[a]P (0.1, 0.3, 0.9 e
2.7 µM) 1 2 3 4
Controlo DMSO 1 2 3 4
Controlo DMSO+E2 1 2 3 4 Efeitos in vitro do
B[a]P após exposição a E2 (22.02 nM) E2+B[a]P (0.1, 0.3, 0.9
e 2.7 µM) 1 2 3 4

Capítulo III
57
Os microsomas hepáticos foram expostos a B[a]P, C e E2, de acordo
com as seguintes condições: a uma suspensão de 5 µl de microsomas
hepáticos foi adicionado
(1) 1090 µl de solução tampão com substrato (TS) e 5 µl de solução
tampão sem substrato (T), e designado como Controlo;
(2) 1090 µl TS e 5 µl de dimetil sulfóxido (DMSO), e designado como
Controlo DMSO;
(3) 1090 µl TS e 5 µl de B[a]P 2.7, 0.9, 0.3 e 0.1 µM, diluídos em
DMSO, e designado como B[a]P;
(4) 1090 µl TS contendo 22,02 nM de C (TSC) e 5 µl de DMSO, e
designado como Controlo DMSO+C;
(5) 1090 µl de TSC mais 5 µl de B[a]P, e designado como C+B[a]P;
(6) 1090 µl TS contendo 22.02 nM de E2 (TSE2) e 5 µl de DMSO, e
desigando como Controlo DMSO+E2;
(7) 1090 µl de TSE2 mais 5 µl de B[a]P, e designado como E2+B[a]P;
A concentração final respectiva de C e E2, na cuvette, foi de 22.02 nM.
Análise Estatística
Os resultados estão expressos em médias ± erro padrão,
correspondentes a quatro réplicas de cada ensaio efectuado (n=4). O software
SIGMASTAT 2.03 foi usado para efectuar a análise estatística. Os dados
experimentais foram inicialmente testados para normalidade e homogeneidade
da variância, de forma a obedecer a requisitos estatísticos e, posteriormente,
com o teste de Tukey (Zar, 1999). As diferenças entre médias foram
consideradas estatisticamente significativas quando p <0.05.

Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
58
RESULTADOS
Figura 1 – Efeitos in vitro de B[a]P 0.1, 0.3, 0.9, 2.7 µM. Os valores
encontram-se expressos em médias ± erro padrão (n=4). As diferenças significativas
(p<0.05) são: * vs Controlo; # vs Controlo DMSO; 0.1 vs B[a]P 0.1; 0.3 vs
B[a]P 0.3; 0.9 vs B[a]P 0.9.
Após exposição intraperitoneal durante 24 horas a BNF 4mg/Kg, a
actividade microsomal hepática de EROD de Anguilla anguilla L., foi de 9.65
pmol.min-1.mg de proteína-1. A adição de 5 µl de DMSO ao Controlo induziu
significativamente a actividade de EROD quando comparado com o Controlo
sem DMSO.
A inibição significativa da actividade microsomal hepática de EROD foi
observada após exposição ao B[a]P 0.1, 0.3, 0.9 e 2.7 µM quando comparada
com Controlo DMSO. A anteriormente referida inibição da actividade EROD
foi proporcional ao aumento da concentração de B[a]P.
Controlo Controlo DMSO
B[a]P 0.1 µM
B[a]P 0.3 µM
B[a]P 0.9 µM
B[a]P 2.7 µM
0,00
5,00
10,00
15,00
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
*
#
0.1 0.3 0.9 #
0.1 #
0.1 #

Capítulo III
59
Figura 2 – Efeitos in vitro de B[a]P 0.1, 0.3, 0.9, 2.7 µM, após exposição a C
(22.02 nM). Os valores encontram-se expressos em médias ± erro padrão (n=4). As
diferenças significativas (p<0.05) são: * vs Controlo; ** vs Controlo DMSO; ***
vs Controlo DMSO+C; 0.1 vs C+B[a]P 0.1; 0.3 vs C+B[a]P 0.3; 0.9 vs C+B[a]P
0.9.
Na figura 2 observa-se que actividade EROD hepática é
significativamente mais elevada no Controlo contendo DMSO quando
comparado com o Controlo sem DMSO.
A exposição prévia dos microsomas hepáticos ao C (22.02 nM) levou a
uma forte indução da actividade de EROD, quando comparada com Controlo
DMSO.
Após exposição prévia ao C, o efeito inibitório de B[a]P 0.1, 0.3, 0.9 e
2.7 µM sobre a actividade microsomal hepática de EROD foi significativo,
quando comparado com Controlo DMSO+C, pelo que, o C não foi capaz de
impedir a inibição da actividade EROD hepática pelo B[a]P.
Apesar da presença do C, o efeito inibitório das concentrações 0.1, 0.3,
0.9 e 2.7 µM de B[a]P sobre a actividade de EROD, foi também proporcional
ao aumento das referidas concentrações de B[a]P.
0,00
5,00
10,00
15,00
20,00
25,00 A
ctiv
idad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
Controlo Cont. DMSO
C+B[a]P 0.1 µM
C+B[a]P 0.3 µM
C+B[a]P 0.9 µM
C+B[a]P 2.7 µM
Cont. DMSO+C
**
*
***
0.1 0.3 0.9 ***
0.1 0.3 ***
*
0.1 ***
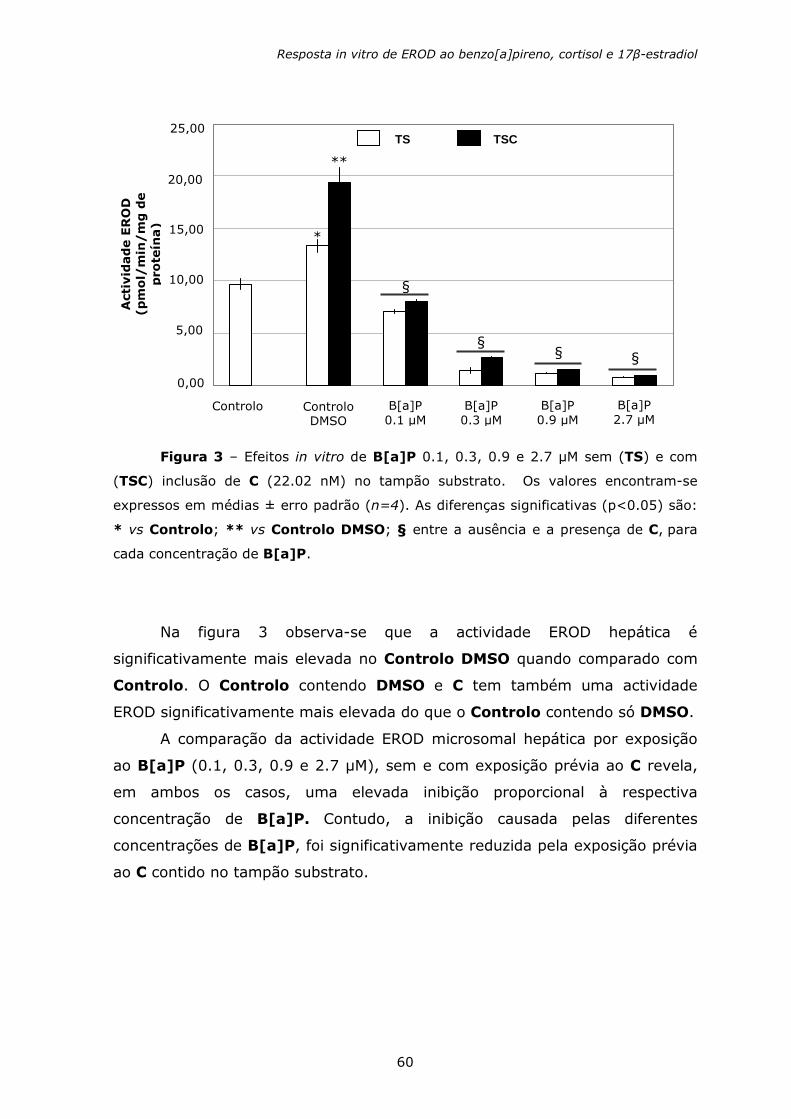
Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
60
Figura 3 – Efeitos in vitro de B[a]P 0.1, 0.3, 0.9 e 2.7 µM sem (TS) e com
(TSC) inclusão de C (22.02 nM) no tampão substrato. Os valores encontram-se
expressos em médias ± erro padrão (n=4). As diferenças significativas (p<0.05) são:
* vs Controlo; ** vs Controlo DMSO; § entre a ausência e a presença de C, para
cada concentração de B[a]P.
Na figura 3 observa-se que a actividade EROD hepática é
significativamente mais elevada no Controlo DMSO quando comparado com
Controlo. O Controlo contendo DMSO e C tem também uma actividade
EROD significativamente mais elevada do que o Controlo contendo só DMSO.
A comparação da actividade EROD microsomal hepática por exposição
ao B[a]P (0.1, 0.3, 0.9 e 2.7 µM), sem e com exposição prévia ao C revela,
em ambos os casos, uma elevada inibição proporcional à respectiva
concentração de B[a]P. Contudo, a inibição causada pelas diferentes
concentrações de B[a]P, foi significativamente reduzida pela exposição prévia
ao C contido no tampão substrato.
0,00
5,00
10,00
15,00
20,00
25,00
Controlo Controlo DMSO
B[a]P 0.1 µM
B[a]P 0.3 µM
B[a]P 0.9 µM
B[a]P 2.7 µM
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
TS TSC
§
§ § §
*
**

Capítulo III
61
Figura 4 – Efeitos in vitro de B[a]P 0.1, 0.3, 0.9, 2.7 µM. Os valores
encontram-se expressos em médias ± erro padrão (n=4). As diferenças significativas
(p<0.05) são: # vs Controlo DMSO; 0.1 vs B[a]P 0.1; 0.3 vs B[a]P 0.3; 0.9 vs
B[a]P 0.9.
A actividade EROD microsomal hepática de Anguilla anguilla L., após
exposição intraperitoneal durante 24 horas a BNF 4mg/Kg, foi de 13.36
pmol.min-1.mg de proteína-1. A adição de 5 µl de DMSO ao Controlo não
afectou significativamente a actividade de EROD, quando comparado com o
Controlo sem DMSO.
A exposição às diferentes concentrações de B[a]P testadas (0.1, 0.3,
0.9 e 2.7 µM) inibiu de forma significativa a actividade EROD microsomal
hepática, quando comparado com Controlo DMSO.
0,00
4,00
8,00
16,00 A
ctiv
idad
e E
RO
D
(pm
ol/
min
/m
g
de p
rote
ína)
12,00
Controlo Controlo DMSO
B[a]P 0.1 µM
B[a]P 0.3 µM
B[a]P 0.9 µM
B[a]P 2.7 µM
# 0.1 0.3 #
0.1 #
0.1 0.3 #
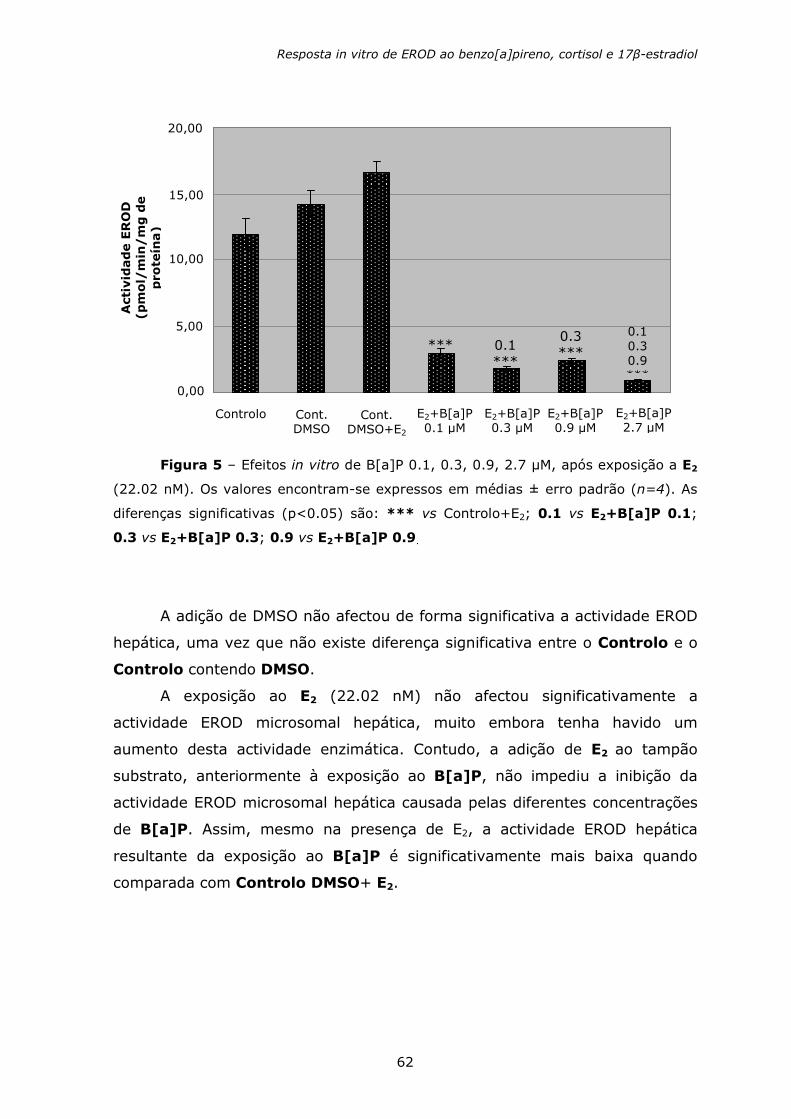
Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
62
Figura 5 – Efeitos in vitro de B[a]P 0.1, 0.3, 0.9, 2.7 µM, após exposição a E2
(22.02 nM). Os valores encontram-se expressos em médias ± erro padrão (n=4). As
diferenças significativas (p<0.05) são: *** vs Controlo+E2; 0.1 vs E2+B[a]P 0.1;
0.3 vs E2+B[a]P 0.3; 0.9 vs E2+B[a]P 0.9.
A adição de DMSO não afectou de forma significativa a actividade EROD
hepática, uma vez que não existe diferença significativa entre o Controlo e o
Controlo contendo DMSO.
A exposição ao E2 (22.02 nM) não afectou significativamente a
actividade EROD microsomal hepática, muito embora tenha havido um
aumento desta actividade enzimática. Contudo, a adição de E2 ao tampão
substrato, anteriormente à exposição ao B[a]P, não impediu a inibição da
actividade EROD microsomal hepática causada pelas diferentes concentrações
de B[a]P. Assim, mesmo na presença de E2, a actividade EROD hepática
resultante da exposição ao B[a]P é significativamente mais baixa quando
comparada com Controlo DMSO+ E2.
0,00
5,00
10,00
15,00
20,00
Controlo Cont. DMSO
E2+B[a]P 0.1 µM
E2+B[a]P 0.3 µM
E2+B[a]P 0.9 µM
E2+B[a]P 2.7 µM
Cont. DMSO+E2
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
*** 0.1 0.3 0.9 ***
0.3 *** 0.1
***

Capítulo III
63
Figura 6 – Efeitos in vitro de B[a]P 0.1, 0.3, 0.9 e 2.7 µM, sem (TS) e com
(TSE2) inclusão de E2 (22.02 nM) no tampão substrato. Os valores encontram-se
expressos em médias ± erro padrão (n=4). As diferenças significativas (p<0.05) são:
§ entre a ausência e a presença de E2, para cada concentração de B[a]P.
Na figura 6 observa-se que a actividade EROD hepática não é
significativamente afectada pela adição de 5 µl de DMSO ao Controlo, uma
vez que não existe diferença significativa entre o Controlo contendo DMSO e
o Controlo sem DMSO. A adição de E2 ao Controlo contendo DMSO também
não afectou a actividade EROD significativamente uma vez que não existe
diferença significativa entre o Controlo DMSO contendo E2 e o Controlo só
com DMSO.
A comparação da actividade EROD microsomal hepática por exposição
ao B[a]P (0.1, 0.3, 0.9 e 2.7 µM), sem e com exposição prévia ao E2 revela,
em ambos os casos, uma inibição significativa. Contudo, a inibição causada
pelas diferentes concentrações de B[a]P, foi significativamente reduzida pela
exposição prévia ao E2 contido no tampão substrato.
0,00
5,00
10,00
15,00
20,00
25,00
Controlo Controlo DMSO
B[a]P 0.1 µM
B[a]P 0.3 µM
B[a]P 0.9 µM
B[a]P 2.7 µM
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
TS TSE2
§ §
§
§

Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
64
DISCUSSÃO
O BNF é reconhecido como um forte indutor da actividade EROD
hepática em peixes, pelo que foi usado neste estudo, de modo a obter in vivo
um controlo positivo com elevada actividade de EROD (Pacheco e Santos,
1996; Gravato e Santos, 2002; Oliveira e Santos, 2003a; Oliveira et al.,
2003b; Oliveira et al., 2005; Santos e Maria, 2005) com o intuito de observar
a inibição (provavelmente) causada pelas diferentes concentrações de B[a]P
a serem testadas. As diferentes concentrações de B[a]P testadas (0.1, 0.3,
0.9 e 2.7 µM) demonstraram in vitro a inibição, de forma significativa, da
actividade EROD microsomal hepática, sendo estes resultados concordantes
com aqueles obtidos in vitro em Dicentrarchus labrax por Gravato e Santos
(2002), onde foi sugerido a possibilidade de tais resultados se deverem à
ocorrência de inibição enzimática.
A concentração de E2 usada baseou-se nos valores plasmáticos de E2
obtidos por Teles et al. (2005) em Sparus aurata L. após 4 horas deste
esteróide ter sido adicionado à água dos aquários que continham os
espécimes. A mesma concentração molar foi adoptada para o cortisol de forma
a permitir a comparação entre o efeito do mesmo número de moléculas das
duas hormonas, e de forma a assegurar a uniformidade e coerência das
condições testadas. Ao contrário dos estudos in vivo que têm vindo a ser feitos
neste âmbito (Teles et al., 2004a, 2005, 2006 e 2007; Vaccaro et al., 2005),
são escassos os estudos in vitro que relatam o papel de E2 no que diz respeito
ao seu efeito sobre a actividade EROD.
A exposição in vitro dos microsomas hepáticos a E2, não afectou
significativamente a actividade EROD microsomal hepática, o que é
concordante com os resultados obtidos em estudos anteriormente feitos, os
quais relatam inibição ou inalteração dessa mesma actividade (Teles et al.,
2004a; 2005; 2006; Vaccaro et al., 2005). O mecanismo subjacente à inibição
da actividade da EROD na presença de compostos estrogénicos não é
completamente conhecido (Andersson et al., 2007). Contudo existem estudos
que relatam que após exposição a E2, os níveis proteicos e mRNA associados a
CYP1A, são reduzidos, o que indicia que, de alguma forma, o E2 interfere ao
nível do gene (Navas e Segner, 2001). A exposição a E2 em combinação com

Capítulo III
65
um antagonista dos receptores de estrogénio, anulou o efeito inibitório de E2
na actividade de EROD em hepatócitos de Oncorhynchus mykiss (Navas e
Segner, 2001), o que sugere o envolvimento dos receptores de estrogénio
(Andersson et al., 2007).
No que diz respeito ao cortisol o cenário é idêntico; são muito poucos os
estudos in vitro realizados neste contexto. A maioria dos estudos de campo
efectuados tem demonstrado a elevação do nível plasmático de cortisol em
resposta à curta exposição a poluentes (Pacheco e Santos, 2001; Teles et al.,
2004b e 2007), nomeadamente HAPs (Oliveira et al., 2007). No presente
trabalho, verificou-se o aumento da actividade de EROD após exposição in
vitro dos microsomas hepáticos ao C, bem como a redução dos efeitos
inibitórios de B[a]P 0.1, 0.3, 0.9 e 2.7 µM após exposição prévia dos
microsomas hepáticos à referida hormona. Santos et al. (2000), num estudo
de cultura de orgãos em A. anguilla L., também observaram um aumento da
actividade hepática de EROD, quando o C foi adicionado ao meio de cultura.
Contudo, de acordo com o estudo efectuado por Hontela et al. (1997), peixes
colhidos numa área sujeita a contaminação por efluentes da pasta de papel,
demonstraram uma reduzida capacidade de elevar o nível plasmático de C,
como consequência de stress agudo e longa exposição aos referidos efluentes.
Por outro lado os peixes colhidos no local de referência revelaram possuir a
expectável e normal capacidade de elevar o C plasmático. Os resultados
obtidos in vitro, no presente estudo, demonstram que o C exerce um efeito
protector sobre a actividade microsomal hepática de EROD quando exposta a
contaminantes inibidores.
Verificou-se ainda que tanto na presença do C como do E2 (22.02 nM) a
inibição causada pelo B[a]P foi significativamente reduzida, revelando o efeito
protector destas duas hormonas esteróides sobre a actividade EROD
microsomal hepática. Gravato e Santos (2002), obtiveram resultados
semelhantes com naftaleno 2.7 µM; na presença deste composto, a inibição
causada por B[a]P 0.1 e 0.01 µM na actividade microsomal hepática de EROD
foi reduzida, indiciando que de alguma forma o naftaleno (tal como o C e o E2)
também atenua o efeito inibitório de B[a]P.

Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
66
O efeito exercido pelas duas hormonas (C e E2), isoladamente ou em
combinação com o B[a]P, sobre a actividade EROD hepática foi idêntico.
Contudo, nem o C nem o E2 demonstraram um efeito preventivo no que diz
respeito à inibição causada pelo B[a]P, o que sugere que, muito
provavelmente, a concentração de C e E2 usada (22.02 nM) não foi a
adequada para que tal efeito fosse observável. Neste contexto seria, por isso,
interessante testar o efeito de outras concentrações de C e E2 sobre a
actividade EROD microsomal hepática, para as condições estabelecidas neste
trabalho.
CONCLUSÕES
- A exposição in vitro dos microsomas hepáticos de A. anguilla L. a
benzo(a)pireno, previamente induzidos pela β-naftoflavona, resultou numa
forte inibição da actividade de EROD.
- A exposição in vitro ao cortisol induziu significativamente a actividade
EROD microsomal hepática, enquanto que a exposição in vitro ao 17β-
estradiol resultou num aumento, embora não significativo, desta actividade
enzimática.
- Tanto na presença do cortisol como do 17β-estradiol (22.02 nM),
observou-se uma redução significativa dos efeitos inibitórios causados pela
exposição in vitro ao benzo(a)pireno, sobre a actividade EROD microsomal
hepática.
- O efeito exercido pelo cortisol e pelo 17β-estradiol sobre a actividade
EROD hepática, isoladamente ou em combinação com o B[a]P, foi idêntico.

Capítulo III
67
REFERÊNCIAS BIBLIOGRÁFICAS
Andersson, C., Katsiadaki, I., Lundstedt-Enkel, K. e Örberg, J. (2007). Effects of 17α-
ethynylestradiol on EROD activity, spiggin and vitellogenin in three-spined
stickleback (Gasterosteus aculeatus). Aquat. Toxicol. 83:33-42.
Billiard, S. M., Bols, N C. e Hodson, P. V. (2004). In vitro and in vivo comparisons of
fish-specific CYP1A induction relative potency factors for selected polycyclic
aromatic hydrocarbons. Ecotoxicol. Environ. Saf. 59:292-299.
Bucheli, T. D., e Fent, K. (1995). Induction of cytocrome P450 as a biomarker for
environmental contamination in aquatic ecosystems. Crit. Rev. Environ. Sci.
Technol. 25:201-268.
Casillas, E., Misitano, D., Johnson, L. L., Rhodes, L. D., Collier, T. K., Stein, J. E.,
McCain, B. B. e Varanasi, U. (1991). Inducibility of spawning and reproductive
success of female english sole (Parophrys vetulus) from urban and non urban
areas of Puget sound, Washington. Mar. Environ. Res. 31:99-122.
Cheek, A. O., Brouwer, T. H., Carroll, S., Manning, S., McLachlan, J. A. e Brouwer, M.
(2001). Experimental evaluation of vitellogenin as a predictive biomarker for
reproductive disruption. Environ. Health Perspect. 109:681-690.
Collier, T. K., Johnson, L. L., Stehr, C. M., Myers, M. S. e Stein, J. E. (1998). A
comprehensive assessment of the impacts of contaminants on fish from an urban
waterway. Mar. Environ. Res. 46:243-247.
De Boeck, G. De Wachter, B., Vlaeminck, A. e Blust, R. (2003). Effect of cortisol
treatment and/or sublethal copper exposure on copper uptake and heat shock
protein levels in common carp, Cyprinus carpio. Environ. Toxicol. Chem. 22:122-
1126.
Gornall, A. C., Bardawill, C. J. e David, M. M. (1949). Determination of serum proteins
by means of biuret reaction. J. Biol. Chem. 177(2):751-766.

Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
68
Gravato C. e Santos, M. A. (2002). Juvenile sea bass liver P450 EROD induction and
erythrocytic genotoxic responses to PAH and PAH-like compounds. Ecotoxicol.
Environ. Saf. 51:115-127.
Hontela, A., Daniel, C. e Rasmussen, J. B. (1997). Structural and functional
impairment of the hypothalamo-pituitary-interrenal axis in fish exposed to
bleached kraft mill effluent in the St Maurice River, Québec. Ecotoxicol. 6:1-12.
Lange, U., Danischewski, D. e Siebers, D. (1993). Regional variability and sexual
differences in ethoxyresorufin-O-deethylase activities and cytochrome P450
concentration in the liver of mature dab (Limanda limanda L.) in the German
Bight. Em: Variability of EROD and Cytochrome P450 in North Sea Dab
(Braumbeck, T., Hanke, W. and Segner, H., Eds.) VCH, Verlag Chemie, New
York.
Matsumoto, S. T., Mantovani, S. M., Malaguttii, M. I. A., Dias, A. L., Fonseca, I. C., e
Marin-Morales, M. A. (2006). Genotoxicity and mutagenicity of water
contaminated with tannery effluents, as evaluated by the micronucleus test and
comet assay using the fish Oreochromis niloticus and chromossome aberrations
on onion root-tips. Gen. Mol. Biol. 29 (1):148-158.
Monod, G. e Vindimian, E. (1991). Effect of storage conditions and subcellular
fractionation of fish and cytochrome P-450-dependent enzymatic activities used
for the monitoring of water pollution. Water Res. 25:173-177.
Navas, J. M. e Segner, H. (2001). Estrogen-mediated suppression of cytocrome
P4501A (CYP1A) in rainbow trout hepatocytes: role of estrogen receptor. Chem.
Biol. Interact. 138: 285-298
Oliveira M. e Santos, M. A. (2003a). In vitro Dicentrarchus labrax L. liver microsomal
EROD activity under different thiol concentrations and its protective role against
chromium, iron and zinc. Fresen. Environ. Bulletin. vol. 12, nº 12, 1476-1482.
Oliveira M., Santos, M. A., Gravato, C. e Pacheco M. (2003b). Chromium effects on
Anguilla anguilla liver organ culture. Fresen. Environ. Bulletin. vol. 12, nº 4,
349-352.

Capítulo III
69
Oliveira M., Santos, M. A., e Pacheco M. (2005). Heavy metal inhibition of Anguilla
anguilla L. liver microsomal EROD activity and thiol protection. Fresen. Environ.
Bulletin. vol. 14, nº 1, 59-64.
Oliveira M., Pacheco M. e Santos, M. A. (2007). Cytochrome P4501A, genotoxic and
stress responses in golden grey mullet (Liza aurata) following short-term
exposure to phenanthrene. Chemosphere. 66:1284-1291.
Pacheco, M. e Santos, M. A. (1998). Induction of liver EROD and erythrocytic nuclear
abnormalities by cyclophosphamide and PAH’s in Anguilla anguilla L.. Ecotoxicol.
Environ. Saf. 40:71- 76.
Pacheco M. e Santos, M. A. (1999). Anguilla anguilla L. liver organ culture as a
toxicological model: in vivo and in vitro EROD induction by β-naphthoflavone.
Fres. Environ. Bulletin. 9: 527-534
Pacheco, M. e Santos, M. A. (2001). Tissue distribution and temperature-dependence
of Anguilla anguilla L. EROD activity following exposure to model inducers and
relationship with plasma cortisol, lactate and glucose levels. Environ. Int.
26:149-155.
Randall, D., Warren Burggren, W. e French, K. (2001). Glands and Hormones –
Physiological effects of hormones. Em: Animal Physiology – mechanisms and
adaptations (5ª edição). Cap 9, pp. 338-347. W. H. Freeman and Company, New
York.
Ruane, N. M., Huisman, E. A. e Komen, J. (2001). Plasma cortisol and metabolite level
profiles in two isogenic strains of common carp during confinement. J. Fish Biol.
59:1-12.
Santos, M. A., e Maria V. L. (2005). Abietic, Dehydroabietic acids and retene in vitro
effets on Anguilla anguilla L. liver microsomal EROD activity. Fresen. Environ.
Bulletin. vol. 14, nº 8, 698-702.
Sarasquete, C. e Segner, H. (2000). Los citocromos P-450. Distribución tisular e
inducción xenobiótica de CYP1A en la biota acuática. Patologia, fisiologia e

Resposta in vitro de EROD ao benzo[a]pireno, cortisol e 17β-estradiol
70
biotoxicología en acuicultura (en prensa). Madrid: Servicio Publicaciones, CSIC,
2000, Spain.
Teles M., Gravato, C., Pacheco M. e Santos M. A. (2004a). Juvenile sea bass
biotransformation, genotoxic and endocrine responses to β-naphthoflanove, 4-
nonylphenol and 17ß-estradiol individual and combined exposures.
Chemosphere. 57:147-158.
Teles M., Santos M. A., e Pacheco M. (2004b). Responses of European eel (Anguilla
anguilla L.) in two polluted environments: in situ experiments. Ecotoxicol.
Environ. Saf. 58:373-378.
Teles, M., Pacheco, M. e Santos, M. A. (2005). Sparus aurata L. liver EROD and GST
activities, plasma cortisol, lactate, glucose and erythrocytic nuclear anomalies
after short-term exposure either to 17ß-estradiol (E2) or E2 combined with 4-
nonylphenol. Sci. Total Environ. 336:57-69.
Teles, M., Santos, M. A. e Pacheco, M. (2006). Biotransformation, stress and genotoxic
effects of 17β-estradiol in juvenile sea bass (Dicentrarchus labrax L.). Environ.
Int. 32:470-477.
Teles M., Pacheco M. e Santos M. A. (2007). Endocrine and metabolic responses of A.
anguilla L. caged in a freshwater-wetland (Pateira de Fermentelos - Portugal).
Sci. Total Environ. 372:562-570.
Vaccaro, E., Meucci, V., Intorre, L., Soldani, G., Di Bello, D., Longo, V., Gervasi, P. G.
e Pretti, C. (2005). Effects of 17ß-estradiol, 4-nonylphenol and PCB 126 on the
estrogenic activity and phase I and 2 biotransformation enzymes in male sea
bass (Dicentrarchus labrax). Aquat. Toxicol. 75:293-305.
Waring, C. P., Moore, A. (2004). The effect of atrazine on Atlantic salmon (Salmo
salar) smolts in fresh water and after sea water transfer. Aquat. Toxicol. 66:93-
104.

CAPÍTULO IV
Estudo in vitro do efeito do fluoranteno, cortisol e
17ß-estradiol sobre a actividade EROD microsomal hepática, em
Anguilla anguilla L..

Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
72
RESUMO
Uma enguia adulta foi intraperitonealmente injectada com ß-
naftoflavona (BNF) 4mg/Kg. Após 24 horas de exposição, o respectivo fígado
foi extraído, tendo-se procedido ao isolamento dos seus microsomas. Os
microsomas hepáticos foram expostos numa cuvette - in vitro - ao fluoranteno
(FL) 0.1, 0.3, 0.9 e 2.7 µM. O efeito protector de cortisol (C) e 17β-estradiol
(E2) 22,02 nM sobre a actividade microsomal hepática de EROD, foi testado
através da sua adição ao tampão-substrato, previamente à exposição a
diferentes concentrações de FL anteriormente seleccionadas. A exposição
prévia dos microsomas hepáticos ao tampão-substrato contendo C e E2 levou a
uma indução significativa da actividade de EROD. A exposição dos microsomas
hepáticos de A. anguilla L. ao FL 0.1 e 0.3 µM, resultou numa indução
significativa da actividade de EROD, enquanto que a exposição a
concentrações mais elevadas do referido composto (0.9 e 2.7 µM) levou a uma
forte inibição da mesma. As hormonas demonstraram um efeito protector no
que respeita à inibição desta actividade enzimática causada pelas
concentrações mais elevadas de FL (0.9 e 2.7 µM).
INTRODUÇÃO
Os ambientes aquáticos têm sido usados, durante décadas, como o
maior depósito de descargas antropogénicas (Pacheco e Santos, 2001a), pelo
que são continuamente sobrecarregados com compostos químicos estranhos
(xenobióticos) resultantes da actividade das comunidades urbanas e
industriais (Van der Oost et al., 2003). Assim, durante o século XX foram
produzidos, e em parte libertados para o meio ambiente, milhares de
poluentes orgânicos tais como policlorobifenis (PCBs), hidrocarbonetos
aromáticos policíclicos (HAPs) e pesticidas organoclorados (Van der Oost et al.,
2003).
A indústria mundial da pasta de papel tem vindo a sofrer, durante os
últimos anos, uma rápida expansão no que diz respeito à utilização de
sofisticados processos mecânicos e químico-mecânicos (O´Connor et al.,
1992). Estes últimos produzem elevadas quantidades de efluente que são
libertadas para o ambiente aquático, causando sérios efeitos ecológicos e

Capítulo IV
73
toxicológicos nos ecossistemas receptores (Pacheco e Santos, 1999). Estes
efluentes são compostos por misturas complexas de substâncias
ambientalmente activas, que contêm cerca de 300 químicos actualmente
conhecidos (Nestmann et al., 1980), que pertencem a várias classes de
compostos orgânicos, nomeadamente ácidos gordos insaturados, ácidos
resínicos, HAPs, entre outros (Pacheco e Santos, 1999). De entre os diferentes
efeitos tóxicos induzidos por estes compostos, há um interesse significativo na
indução das monooxigenases associadas ao citocromo P450. A relação entre a
actividade da etoxiresorufina O-desetilase (EROD) hepática (Pacheco e Santos,
1998) e os níveis plasmáticos e celulares de determinadas hormonas
esteróides, tais como cortisol (C) e 17ß-estradiol (E2) em peixes é pouco
conhecida. Contudo, os níveis de cortisol plasmático foram amplamente
estudados in vivo em Anguilla anguilla L. (Santos e Pacheco, 1996; Pacheco e
Santos, 2001a e 2001b; Teles et al., 2004b), Sparus aurata L. (Teles et al.,
2005), Liza aurata (Oliveira et al., 2007) e em Dicentrarchus labrax L. (Teles
et al., 2004a) sob diversas condições ambientais, e in vitro (Santos et al.,
2000) mas de forma menos exaustiva. O nível plasmático de C é o indicador
de stress mais vulgarmente usado em peixes devido à rápida elevação que
ocorre em resposta a vários agentes stressantes, nomeadamente
contaminantes (Wendelar-Bonga, 1997). O E2 é o mais potente dos
estrogénios, desempenhando um papel preponderante em vários aspectos do
crescimento, desenvolvimento e diferenciação morfológica (Randall et al.,
2001). No entanto, este estrogénio natural também pode ser considerado um
contaminante aquático. A existência de elevados níveis de E2 na água,
principalmente como consequência de efluentes domésticos e industriais, pode
causar efeitos adversos nos peixes, nomeadamente indução de
intersexualidade em machos (Metcalfe et al., 2001). Vários estudos têm sido
realizados in vivo, neste âmbito, em Sparus aurata L. (Teles et al., 2005) e
Dicentrarchus labrax L. (Teles et al., 2004a; Teles et al., 2006).
O fígado é, geralmente, o órgão onde a maioria dos xenobióticos
absorvidos são sujeitos a um processo crítico de activação metabólica
(oxidação), mediado pelas referidas monooxigenases, e posterior inactivação
mediada pelas conjugases, permitindo a excreção e eliminação dos mesmos

Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
74
(Buhler e Williams, 1988). A EROD tem demonstrado ser muito sensível à
indução causada por vários poluentes químicos, nomeadamente HAPs
(Pacheco e Santos, 1998). O fluoranteno (FL) é um HAP vulgarmente
encontrado no sedimento resultante dos efluentes da pasta de papel, sendo
constituído por quatro anéis, e com actividade mutagénica (Kinae et al., 1988)
e cancerígena relativamente baixa em mamíferos (Willett et al., 2001). A
informação acerca da toxicidade do FL em peixes é limitada, uma vez que
pouco se sabe acerca das consequências da exposição a este composto,
nomeadamente, efeitos ao nível da actividade de CYP1A, em particular na
actividade de EROD, e demais respostas de stress. Fent e Bätscher (2000),
num estudo in vitro, não obtiveram indução significativa da actividade EROD
microsomal hepática na presença de FL, enquanto que Willett et al., (2001),
relatou a inibição in vitro e in vivo em Fundulus heteroclitus da actividade de
EROD.
A escassez de informação científica, nos peixes, no que diz respeito aos
efeitos in vitro do FL, C e E2 isolados ou em conjunto, sobre a actividade
microsomal hepática de EROD, serviu de estímulo ao presente trabalho. Deste
modo, procedeu-se ao estudo dos efeitos in vitro de diferentes concentrações
de FL com e sem exposição prévia ao C e E2, de forma a tentar compreender
o papel destas duas hormonas esteróides sobre a actividade EROD microsomal
hepática de A. anguilla L..
MATERIAL E MÉTODOS
Compostos químicos
β-naftoflavona, dimetil sulfóxido (DMSO) e resorufina (Sigma Chemical
Co, EUA) 7-etoxiresorufina e NADPH (Roche); 17ß-estradiol e fluoranteno
(Sigma-Aldrich - Alemanha); cortisol (Merck Sharp & Dohme)
Animais
Uma enguia adulta – Anguilla anguilla L. – com cerca de 650 gramas,
adquirida num mercado de peixe local, foi mantida no laboratório, num tanque

Capítulo IV
75
de água salobra com arejamento durante um período de aclimatização de 48
horas. A enguia foi injectada intraperitonealmente com ß-naftoflavona
4mg/Kg, após o referido período de aclimatização, e sacrificada após 24 horas.
O fígado foi então retirado, sendo imediatamente congelado em azoto líquido,
e armazenado a -80ºC até se proceder à sua homogeneização. O animal não
foi alimentado nem durante o período de recobro nem durante o período
experimental.
Isolamento dos microsomas
Procedeu-se à homogeneização do fígado previamente extraído em
tampão TRIS-HCL 0.1 M pH 7.4 + KCL (0.15M) + Glicerol (20%). Os
microsomas hepáticos foram obtidos de acordo com os métodos de Lange et
al. (1993), Monod e Vindimian (1991), adaptados por Pacheco e Santos
(1998).
Actividade da EROD
A actividade da EROD foi determinada de acordo com o método de
Burke e Meyer (1974), a 25ºC, através do aumento progressivo de
fluorescência, resultante da formação de resorufina, durante 3 minutos
(comprimento de onda de excitação 530nm; comprimento de onda de emissão
585nm), num espectrofluorímetro Jasco FP 750, e expressa em
picomoles/min/mg de proteína. A reacção foi iniciada com a adição de 10 µl de
NADPH (10 mM).
Determinação da concentração de proteína
A concentração microsomal de proteína foi determinada de acordo com
o método de Biureto (Gornall et al., 1949), usando a albumina (proteína
bovina) como padrão.
Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
76
Tabela 1 – Tampões, solventes e soluções usados durante o protocolo
experimental
Tampões
TS 0.5 µM etoxiresorufina em Tris-HCl 0,1 M pH 7.4 com KCl 0,15M e 20%
T Tris-HCl 0,1 M pH 7.4 com KCl 0,15M e 20% glicerol
TSC TS com cortisol (concentração final na cuvette = 22.02nM)
TSE2 TS com E2 (concentração final na cuvette = 22.02nM)
Solventes
DMSO Dimetil sulfóxido
Soluções
B[a]P Benzo[a]pireno (0.1, 0.3, 0.9 e 2.7µM) em DMSO
Tabela 2 – Procedimento experimental para avaliação da actividade
microsomal hepática de EROD. A numeração (1, 2, 3 e 4) indica ordem pela qual os
intervenientes na reacção foram adicionados na cuvette.
Ensaio
5 µ
l m
icro
som
as
1090 µ
l T
S
1090 µ
l T
SC
1090 µ
l T
SE
2
5 µ
l T
5 µ
l D
MS
O
5 µ
l FL (
em D
MS
O)
10 µ
l N
AD
PH
Controlo 1 2 3 4
Controlo DMSO 1 2 3 4 Efeitos do C in vitro
Controlo DMSO+C 1 2 3 4
Controlo 1 2 3 4
Controlo DMSO 1 2 3 4 Efeitos de E2 in vitro
Controlo DMSO+E2 1 2 3 4
Controlo 1 2 3 4
Controlo DMSO 1 2 3 4 Efeitos do FL in vitro FL (0.1, 0.3, 0.9 e 2.7 µM) 1 2 3 4
Controlo DMSO 1 2 3 4
Controlo DMSO+C 1 2 3 4 Efeitos in vitro do FL após exposição a C
(22.02 nM) C+FL (0.1, 0.3, 0.9 e 2.7 µM) 1 2 3 4
Controlo DMSO 1 2 3 4
Controlo DMSO+E2 1 2 3 4 Efeitos in vitro do FL após exposição a E2
(22.02 nM) E2+FL (0.1, 0.3, 0.9 e 2.7 µM) 1 2 3 4

Capítulo IV
77
Os microsomas hepáticos foram expostos durante 3 minutos a FL, C e
E2, de acordo com as seguintes condições: a uma suspensão de 5 µl de
microsomas hepáticos foi adicionado
(1) 1090 µl de solução tampão com substrato (TS) e 5 µl de solução
tampão sem substrato (T), e designado como Controlo;
(2) 1090 µl TS e 5 µl de dimetil sulfóxido (DMSO), e designado como
Controlo DMSO;
(3) 1090 µl TS e 5 µl de FL 2.7, 0.9, 0.3 e 0.1 µM, diluídos em DMSO, e
designado como FL;
(4) 1090 µl TS contendo 22,02 nM de C (TSC) e 5 µl de DMSO, e
designado como Controlo DMSO+C;
(5) 1090 µl de TSC mais 5 µl de FL, e designado como C+FL;
(6) 1090 µl TS contendo 22.02 nM de E2 (TSE2) e 5 µl de DMSO, e
desigando como Controlo DMSO+E2;
(7) 1090 µl de TSE2 mais 5 µl de FL, e designado como E2+FL;
A concentração final respectiva de C e E2, na cuvette, foi de 22.02 nM.
Análise Estatística
Os resultados estão expressos em médias ± erro padrão,
correspondentes a quatro réplicas de cada ensaio efectuado (n=4). O software
SIGMASTAT 2.03 foi usado para efectuar a análise estatística. Os dados
experimentais foram inicialmente testados para normalidade e homogeneidade
da variância, de forma a obedecer a requisitos estatísticos e, posteriormente,
com o teste de Tukey (Zar, 1999). As diferenças entre médias foram
consideradas estatisticamente significativas quando p <0.05.

Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
78
RESULTADOS
Figura 1 – Efeitos in vitro de FL 0.1, 0.3, 0.9, 2.7 µM. Os valores encontram-
se expressos em médias ± erro padrão (n=4). As diferenças significativas (p<0.05)
são: # vs Controlo DMSO; 0.1 vs FL 0.1; 0.3 vs FL 0.3; 0.9 vs FL 0.9.
A actividade microsomal hepática de EROD de Anguilla anguilla L.
determinada, após exposição intraperitoneal ao BNF 4mg/Kg, foi de 7.46
pmol.min-1.mg de proteína-1. A actividade da EROD não foi significativamente
afectada pela adição de 5 µl de DMSO ao respectivo meio de reacção.
A exposição in vitro dos microsomas hepáticos de A. anguilla L. às
concentrações mais baixas de FL (0.1 e 0.3 µM) levou a uma forte indução da
actividade de EROD. No entanto, a exposição às concentrações mais elevadas
do referido composto (0.9 e 2.7 µM), resultou na inibição significativa desta
actividade enzimática, quando comparada com Controlo DMSO.
Controlo Controlo DMSO
FL 0.1 µM FL 0.3 µM FL 0.9 µM FL 2.7 µM
5,00
10,00
15,00
0,00
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
#
0.1 0.3 0.9 #
0.1 0.3 #
0.1 #

Capítulo IV
79
Figura 2 – Efeitos in vitro de FL 0.1, 0.3, 0.9, 2.7 µM, após exposição a C
(22.02 nM). Os valores encontram-se expressos em médias ± erro padrão (n=4). As
diferenças significativas (p<0.05) são: ** vs Controlo DMSO; *** vs Controlo
DMSO+C; 0.1 vs C+FL 0.1; 0.3 vs C+FL 0.3; 0.9 vs C+FL 0.9.
A exposição ao C (22.02 nM), na presença de DMSO, induziu
significativamente a actividade microsomal hepática de EROD, quando
comparado com Controlo DMSO.
A presença de C aumentou de forma significativa a actividade EROD
microsomal hepática no caso da exposição à concentração FL 0.1 µM, em
relação ao Controlo DMSO+C.
A adição de C ao tampão substrato previamente à exposição ao FL 0.3 e
0.9 µM, levou a uma actividade EROD hepática idêntica à que se obteve para
Controlo DMSO+C. Contudo, no que diz respeito à exposição à concentração
mais elevada de FL testada (2.7 µM), a adição de C não evitou a inibição
causada pela referida exposição.
0,00
4,00
8,00
12,00
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
Controlo Cont. DMSO
C+FL 0.1 µM
C+FL 0.3 µM
C+FL 0.9 µM
C+FL 2.7 µM
Cont. DMSO+C
**
***
0.1 0.3 0.9 ***
0.1

Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
80
Figura 3 – Efeitos in vitro de FL 0.1, 0.3, 0.9 e 2.7 µM sem (TS) e com (TSC)
inclusão de C (22.02 nM) no tampão substrato. Os valores encontram-se expressos
em médias ± erro padrão (n=4). As diferenças significativas (p<0.05) são: ** vs
Controlo DMSO; § entre a ausência e a presença de C, para cada concentração de
FL.
Na figura 3 observa-se que a actividade EROD hepática é
significativamente mais elevada no Controlo contendo DMSO e C quando
comparado com o Controlo contendo só DMSO. Assim, apesar do aumento
significativo da actividade EROD microsomal hepática induzido pelo FL,
verifica-se que esse aumento é significativamente maior para as
concentrações 0.1 e 0.3 de FL sempre que o C é previamente adicionado. A
actividade EROD microsomal hepática começa a ser inibida pela concentração
0.9 de FL a qual se estende à concentração 2.7 de FL. Contudo, o C contido no
TS consegue impedir a acção inibidora dessa concentração 0.9 de FL, o qual é
comprovado pelo facto de não existirem diferenças significativas entre C+FL
0.9 e C+FL 0.3. Apesar da concentração 2.7 de FL inibir substancialmente a
actividade EROD, a acção protectora do C é ainda manifesta mas não tão
potente quanto para a concentração 0.9 de FL.
A medição da actividade EROD microsomal hepática, com (TSC) e sem
(TS) a adição de C ao referido tampão, após exposição ao FL 0.1, 0.3, 0.9 e
5,00
10,00
15,00
20,00
25,00
0,00 Controlo Controlo
DMSO FL 0.1 µM FL 0.3 µM FL 0.9 µM FL 2.7 µM
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
TS TSC
§
§
§
§
**

Capítulo IV
81
2.7 µM, demonstra que a inclusão de C no tampão-substrato estimula e
impede significativamente o decréscimo dessa mesma actividade.
Figura 4 – Efeitos in vitro de FL 0.1, 0.3, 0.9, 2.7 µM. Os valores encontram-
se expressos em médias ± erro padrão (n=4). As diferenças significativas (p<0.05)
são: # vs Controlo DMSO; 0.1 vs FL 0.1; 0.3 vs FL 0.3; 0.9 vs FL 0.9.
A actividade microsomal hepática de EROD determinada em Anguilla
anguilla L., foi de 6.85 pmol.min-1.mg de proteína-1, após exposição
intraperitoneal a BNF 4mg/Kg durante 24 horas. A adição de 5 µl de DMSO à
reacção não afectou significativamente a actividade EROD hepática.
A exposição ao FL 0.1 e 0.3 µM, levou a uma indução significativa da
actividade microsomal hepática de EROD quando comparada com Controlo
DMSO. Contudo, a inibição significativa da actividade microsomal hepática de
EROD foi observada após a exposição às concentrações mais elevadas de FL
(0.9 e 2.7 µM), quando comparada com Controlo DMSO.
Controlo Controlo DMSO
FL 0.1 µM FL 0.3 µM FL 0.9 µM FL 2.7 µM
5,00
10,00
15,00
0,00
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
#
0.1 0.3 0.9 #
0.1 0.3 #
0.1 #
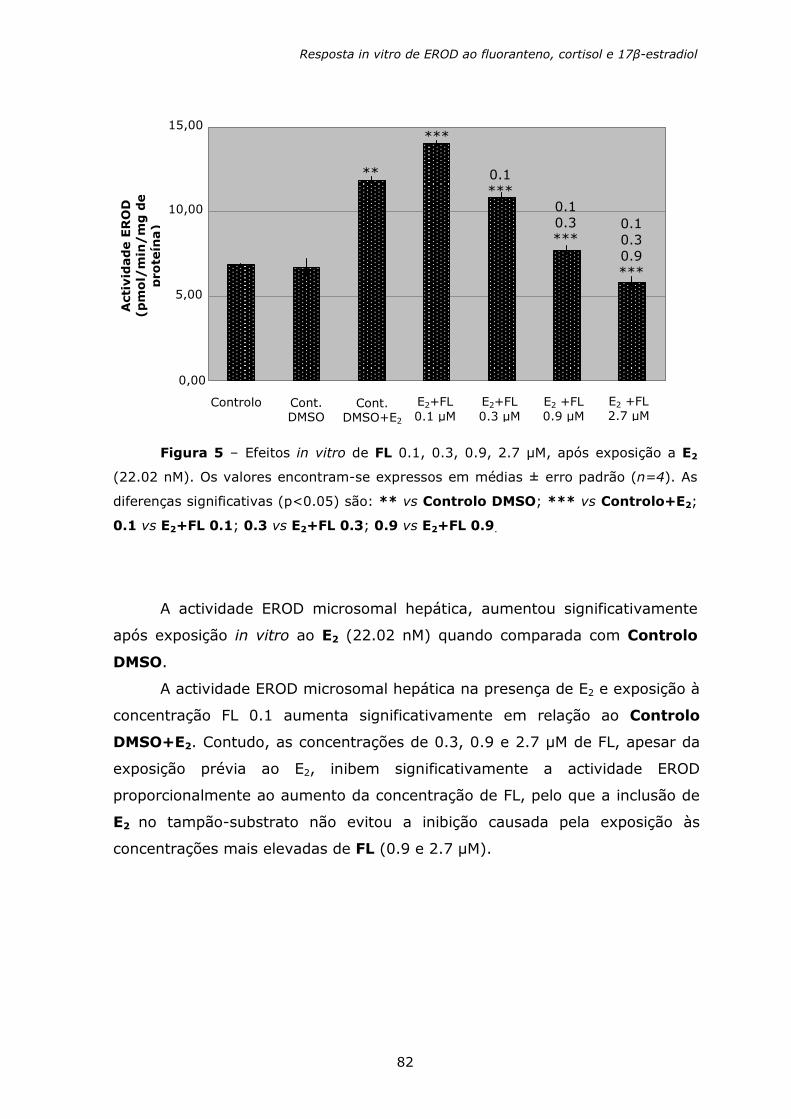
Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
82
Figura 5 – Efeitos in vitro de FL 0.1, 0.3, 0.9, 2.7 µM, após exposição a E2
(22.02 nM). Os valores encontram-se expressos em médias ± erro padrão (n=4). As
diferenças significativas (p<0.05) são: ** vs Controlo DMSO; *** vs Controlo+E2;
0.1 vs E2+FL 0.1; 0.3 vs E2+FL 0.3; 0.9 vs E2+FL 0.9.
A actividade EROD microsomal hepática, aumentou significativamente
após exposição in vitro ao E2 (22.02 nM) quando comparada com Controlo
DMSO.
A actividade EROD microsomal hepática na presença de E2 e exposição à
concentração FL 0.1 aumenta significativamente em relação ao Controlo
DMSO+E2. Contudo, as concentrações de 0.3, 0.9 e 2.7 µM de FL, apesar da
exposição prévia ao E2, inibem significativamente a actividade EROD
proporcionalmente ao aumento da concentração de FL, pelo que a inclusão de
E2 no tampão-substrato não evitou a inibição causada pela exposição às
concentrações mais elevadas de FL (0.9 e 2.7 µM).
5,00
10,00
15,00
0,00
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
Controlo Cont. DMSO
E2+FL 0.1 µM
E2+FL 0.3 µM
E2 +FL 0.9 µM
E2 +FL 2.7 µM
Cont. DMSO+E2
**
***
0.1 0.3 0.9 ***
0.1 0.3 ***
0.1 ***

Capítulo IV
83
Figura 6 – Efeitos in vitro de FL 0.1, 0.3, 0.9 e 2.7 µM sem (TS) e com (TSE2)
inclusão de E2 (22,02 nM) no tampão substrato. Os valores encontram-se expressos
em médias ± erro padrão (n=4). As diferenças significativas (p<0.05) são: ** vs
Controlo DMSO; § entre a ausência e a presença de E2, para cada concentração de
FL.
A actividade EROD aumentou significativamente nos microsomas
hepáticos expostos a DMSO+E2 quando comparado com o Controlo DMSO. A
presença de E2 não afectou de forma significativa a actividade EROD
microsomal hepática após exposição ao FL 0.1 e 0.3 µM. No entanto, a inibição
causada por FL 0.9 e 2.7 µM foi significativamente reduzida sempre que
previamente se administrou E2 à solução tampão-substrato, para uma
concentração final (na cuvette) de 22.02 nM. A inclusão de E2 no tampão-
substrato (TSE2) aumentou significativamente a actividade EROD microsomal
hepática quando comparada com a exposição apenas ao FL 0.9 e 2.7 µM, isto
é, sem a adição prévia de E2 ao referido tampão (TS).
Controlo Controlo DMSO
FL 0.1 µM FL 0.3 µM FL 0.9 µM FL 2.7 µM
5,00
10,00
15,00
20,00
25,00
0,00
Act
ivid
ad
e E
RO
D
(pm
ol/
min
/m
g d
e
pro
teín
a)
TS TSE2
§ §
**

Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
84
DISCUSSÃO
Os resultados obtidos neste trabalho mostram que a actividade EROD
microsomal hepática vai aumentando à medida que as concentrações de FL
diminuem. A exposição às concentrações mais baixas (0.1 e 0.3 µM) causou
uma forte indução da actividade EROD microsomal hepática, enquanto que a
exposição às concentrações mais elevadas de FL (0.9 e 2.7 µM) teve o efeito
oposto, levando a uma forte inibição desta actividade enzimática.
Fent e Bätscher (2000), num estudo in vitro, não obtiveram indução
significativa da actividade da EROD na presença de FL, enquanto que Willett
et al., (2001), relatou a inibição in vitro e in vivo em Fundulus heteroclitus da
actividade de EROD, tendo sido sugerido por estes autores que o mecanismo
in vitro de acção parece ocorrer através de inibição enzimática não-
competitiva, enquanto que os resultados in vivo sugerem incapacidade do FL
em se ligar ao RAh e iniciar a transcrição. Já os resultados obtidos por Oliveira
et al. (2007), demonstram que a actividade da EROD foi induzida, em Liza
aurata, após rápida exposição dos microsomas hepáticos a fenantreno 0.3, 0.9
e 2.7 µM, cuja estrutura e peso molecular é similar ao FL. Importa referir que
os dados disponíveis acerca da indução de CYP1A nos peixes, por HAPs de
pequeno tamanho, com 4 ou menos anéis aromáticos fundidos,
disponibilizaram resultados não consensuais, sendo essas respostas
dependentes da espécie testada (Shailaja e D’Silva, 2003). Contudo, é sabido
que vários indutores da síntese de CYP1A (sub família do gene P450) também
podem inibir a actividade catalítica de EROD (Stegeman e Hahn, 1994),
quando presentes em elevadas doses (Haasch et al., 1993).
A concentração de E2 usada baseou-se nos valores plasmáticos de E2
obtidos por Teles et al. (2005) em Sparus aurata L. após 4 horas desta ter
sido adicionada à água dos aquários que continham os espécimes. A mesma
concentração molar foi adoptada para o cortisol de forma a permitir a
comparação entre o efeito do mesmo número de moléculas das duas
hormonas, e de forma a assegurar a uniformidade e coerência das condições
testadas. É reconhecido que as hormonas esteróides fazem parte dos factores
endógenos que têm a capacidade de regular a expressão de CYP1A, contudo
os mecanismos de interacção subjacentes não estão completamente

Capítulo IV
85
estabelecidos (Navas e Segner, 2000). Vários estudos têm demonstrado, in
vivo um efeito inibidor do E2 sobre a actividade de EROD (Vaccaro et al.,
2005; Teles et al., 2005), o que poderá ser explicado pela interferência do E2
ao nível do gene, tendo-se observado da redução da quantidade de proteína e
mRNA associados a CYP1A, após exposição ao referido esteróide (Navas e
Segner, 2001). Contudo, o mecanismo subjacente à inibição da actividade da
EROD na presença de compostos estrogénicos não é completamente conhecido
(Andersson et al., 2007). O aumento significativo da actividade da EROD, após
exposição in vitro dos microsomas hepáticos ao E2, não é pois concordante
com os resultados obtidos em estudos in vivo anteriormente feitos, o que
justifica a necessidade de estudos comparativos adicionais, de forma a se
obter uma melhor compreensão dos efeitos de E2 na expressão de CYP1A,
mais particularmente ao nível da actividade de EROD.
No que diz respeito ao C, a maioria dos estudos tem relatado a rápida
elevação, do nível plasmático de cortisol após breve exposição in vivo a
diferentes contaminantes (Pacheco e Santos, 2001b; Teles et al., 2004),
nomeadamente HAPs (Oliveira et al., 2007), pelo que o nível plasmático de
cortisol pode ser usado como um indicador antecipado da existência de
contaminação aquática (Pacheco e Santos, 2001b). O aumento significativo da
actividade da EROD, após exposição in vitro dos microsomas hepáticos ao C, é
concordante com os resultados anteriormente obtidos em estudos in vivo.
Uma vez que não se obtiveram diferenças significativas entre o Controlo
DMSO e o Controlo sem DMSO, os resultados obtidos in vitro neste
trabalho, sugerem uma acção estimulante/estabilizadora no que se refere ao
papel do C e do E2 no aumento significativo da actividade EROD hepática.
Verificou-se ainda que tanto na presença do C como do E2 (22.02 nM) a
inibição causada pelo FL 0.9 e 2.7 µM, foi significativamente reduzida,
revelando o efeito protector destas duas hormonas esteróides ao nível da
actividade de EROD. Contudo, nem o C nem o E2 demonstraram um efeito
preventivo no que diz respeito à inibição causada pelo FL o que sugere que,
muito provavelmente, a concentração de C e E2 usada (22.02 nM) não foi
suficiente para que tal efeito fosse observável.

Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
86
CONCLUSÕES
- A exposição in vitro dos microsomas às concentrações mais baixas de
fluoranteno - 0.1 e 0.3 µM - levou à indução significativa da actividade EROD
hepática, enquanto que a exposição às concentrações mais elevadas de
fluoranteno - 0.9 e 2.7 µM -, resultou numa forte inibição desta actividade
enzimática. Os resultados sugerem que o fluoranteno apesar de
eventualmente ser um indutor da actividade EROD hepática, quando presente
em elevadas doses, também pode exercer um efeito inibidor sobre esta
actividade enzimática.
- A exposição in vitro ao cortisol e ao 17β-estradiol induziu
significativamente a actividade EROD microsomal hepática, pelo que ambas as
hormonas tiveram um efeito idêntico sobre esta actividade enzimática.
- Tanto na presença do cortisol como do 17β-estradiol (22.02 nM),
verificou-se uma redução significativa dos efeitos inibitórios causados pela
exposição ao fluoranteno 0.9 e 2.7 µM. O cortisol evitou a inibição causada
pelo fluoranteno 0.9 µM. Contudo esta hormona não foi capaz de evitar a
inibição causada pelo fluoranteno 2.7 µM.
- O efeito exercido pelas duas hormonas quando em combinação com o
FL, sobre a actividade EROD hepática, foi distinto. A exposição prévia ao C
aumentou significativamente a actividade EROD microsomal hepática após
exposição ao fluoranteno 0.1, 0.3, 0.9 e 2.7 µM, enquanto que o E2 só exerceu
efeito idêntico para as concentrações de FL inibidoras (0.9 e 2.7 µM).

Capítulo IV
87
REFERÊNCIAS BIBLIOGRÁFICAS
Andersson, C., Katsiadaki, I., Lundstedt-Enkel, K. e Örberg, J. (2007). Effects of 17α-
ethynylestradiol on EROD activity, spiggin and vitellogenin in three-spined
stickleback (Gasterosteus aculeatus). Aquat. Toxicol. 83:33-42.
Billiard, S. M., Hahn, M E., Franks, D. G., Peterson, R. E., Bols, N C. e Hodson, P. V.
(2002).Binding of polycyclic aromatic hydrocarbons (PAHs) to teleost aryl
hydrocarbon receptors (AHRs). Comp. Biochem. Physiol. B: Biochem. Mol. Biol.
133:55-68.
Buhler, D. R. e Williams D. E. (1988). The role of biotransformation in the toxicity of
chemicals. Aquat. Toxicol. 11:19-28.
Connell, D. W. (1974). A kerosene-like taint in the sea mullet - Mugil cephalus L. –
composition and environmental occurrence of the tainting substance. Aust. J.
Mar. Freshwater Res. 25:7-24.
Fent K. e Bätscher, R. (2000). Cytocrome P4501A induction potencies of polycyclic
aromatic hydrocarbons in a fish hepatoma cell line: demonstration of additive
interactions. Environ. Toxicol. Chem. vol. 19, nº 8, 2047-2058.
Gornall, A. C., Bardawill, C. J. e David, M. M. (1949). Determination of serum proteins
by means of biuret reaction. J. Biol. Chem. 177(2):751-766.
Gravato C. e Santos, M. A. (2002). Juvenile sea bass liver P450 EROD induction and
erythrocytic genotoxic responses to PAH and PAH-like compounds. Ecotoxicol.
Environ. Saf. 51:115-127
Haasch, M. L., Quardokus, E. M., Sutherland, L. A., Goodrich, M. S. e Lech, J. J.
(1993). Hepatic CYP1A induction in rainbow trout by continuous follow-through
exposure to β-naphthoflavone. Toxicol. Sci. 20:72 – 82.
Kinae N., Hashizume, T., Makita, T., Tomita, I., Kimura, I. e Karamori, H. (1981).
Studies on the toxicity of pulp and paper mill effluents-I. Mutagenicity of the
sediment samples derived from kraft paper mills. Water Res. 15:17-24.

Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
88
Lange, U., Danischewski, D. e Siebers, D. (1993). Regional variability and sexual
differences in ethoxyresorufin-O-deethylase activities and cytochrome P450
concentration in the liver of mature dab (Limanda limanda L.) in the German
Bight. Em: Variability of EROD and Cytochrome P450 in North Sea Dab
(Braumbeck, T., Hanke, W. and Segner, H., Eds.) VCH, Verlag Chemie, New
York.
Metcalfe, C. D., Metcalfe, T. L., Kiparissis, Y., Koenig, B. G., Khan, C., Hughes, R. J.,
Croley, T. R., March, R. E. e Potter, T. (2001) Estrogenic potency of chemicals
detected in sewage treatment plant effluents as determined by in vivo assays
with Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem. 20:297-308.
Monod, G. e Vindimian, E. (1991). Effect of storage conditions and subcellular
fractionation of fish and cytochrome P-450-dependent enzymatic activities used
for the monitoring of water pollution. Water Res. 25:173-177.
Navas, J. M. e Segner, H. (2000). Modulation of trout 7-ethoxyresorufin-O-deethylase
(EROD) activity by estradiol and octylphenol. Mar. Environ. Res. 50:157-162.
Navas, J. M. e Segner, H. (2001). Estrogen-mediated suppression of cytocrome
P4501A (CYP1A) in rainbow trout hepatocytes: role of estrogen receptor. Chem.
Biol. Interact. 138: 285-298
Nestmann, E. R., Lee, E. G. H., Matula, T. I., Douglas, G. R. e Mueller, J. C. (1980).
Mutagenicity of constituents identified in pulp and paper mill effluent using the
Salmonella/Mammalia-microsome assay. Mutat. Res. 79:203-212.
O’Connor, B. I., Kovacs, T. G. e Voss R. H. (1992). The effect of wood species
composition on the toxicity of simulated mechanical pulping effluents. Environ.
Toxicol. Chem. 11:1259-1270.
Oliveira M., Santos, M. A., Gravato C., Pacheco M. (2003). Chromium effects on
Anguilla anguilla liver organ culture. Fresen. Environ. Bulletin. vol. 12, nº 4,
349-352.

Capítulo IV
89
Oliveira M., Pacheco M. e Santos, M. A. (2007). Cytochrome P4501A, genotoxic and
stress responses in golden grey mullet (Liza aurata) following short-term
exposure to phenanthrene. Chemosphere. 66:1284-1291.
Pacheco M. e Santos M. A. (1998). Induction of liver EROD and erythrocytic nuclear
abnormalities by cyclophosphamide and PAHs in Anguilla anguilla L.. Ecotoxicol.
Environ. Saf. 40:71-76.
Pacheco, M. e Santos, M. A. (1999). Biochemical and genotoxic responses of adult eel
(Anguilla anguilla L.) to resin acids and pulp mill effluent: laboratory and field
experiments. Ecotoxicol. Environ. Saf. 42:81-93.
Pacheco, M. e Santos, M. A. (2001a). Biotransformation, endocrine and genetic
responses of Anguilla anguilla L. to petroleum distillate products and
environmentally contaminated waters. Ecotoxicol. Environ. Saf. 49:64-75.
Pacheco, M. e Santos, M. A. (2001b). Tissue distribution and temperature-dependence
of Anguilla anguilla L. EROD activity following exposure to model inducers and
relationship with plasma cortisol, lactate and glucose levels. Environ. Int.
26:149-155.
Pacheco, M. e Santos, M. A. (2002). Naphthalene and ß-naphthoflavone toxicity to
Anguilla anguilla L.: Hepatic metabolism and erythrocytic nuclear abnormalities.
Environ. Int. 28:285-293.
Randall, D., Warren Burggren, W. e French, K. (2001). Glands and Hormones –
Physiological effects of hormones. Em: Animal Physiology – mechanisms and
adaptations (5ª edição). Cap 9, pp. 338-347. W. H. Freeman and Company, New
York.
Santos, M. A. e Pacheco, M. (1996). Anguilla anguilla L. stress biomarkers recovery in
clean water and secondary treated pulp mill effluent. Ecotoxicol. Environ. Saf.
35: 96-100.
Santos, M. A., Pacheco, M. e Magalhães, I. (2000). Anguilla anguilla L. liver organ
culture as a toxicological model: in vivo and in vitro EROD induction by β-
naphthoflavone. Fres. Environ. Bulletin. 9: 527-534.

Resposta in vitro de EROD ao fluoranteno, cortisol e 17β-estradiol
90
Shailaja, M. S. e D’Silva, C. (2003). Evaluation of impact of PAH on a tropical fish,
Oreochromis mossambicus using multiple biomarkers. Chemosphere. 53:835-
841.
Stegeman, J. J. e Hahn, M. E. (1994). Biochemestry and molecular biology of
monooxigenases. Em: Pacheco M. e Santos, M. A. (1999). Biochemical and
genotoxic responses of adult eel (Anguilla anguilla L.) to resin acids and pulp mill
effluent: laboratoty and field experiments. Ecotoxicol. Environ. Saf. 42:81-93.
Teles M., Gravato, C., Pacheco M. e Santos M. A. (2004a). Juvenile sea bass
biotransformation, genotoxic and endocrine responses to β-naphthoflanove, 4-
nonylphenol and 17ß-estradiol individual and combined exposures.
Chemosphere. 57:147-158.
Teles M., Santos M. A., e Pacheco M. (2004b). Responses of European eel (Anguila
anguilla L.) in two polluted environments: in situ experiments. Ecotoxicol.
Environ. Saf. 58:373-378.
Teles M., Pacheco M. e Santos, M. A. (2005). Sparus aurata L. liver EROD and GST
activities, plasma cortisol, lactate, glucose and erythrocytic nuclear anomalies
following short-term exposure either to 17β-estradiol (E2) or E2 combined with4-
nonylphenol. Sci. Total Environ. 336:57-69.
Teles, M., Santos, M. A. e Pacheco, M. (2006). Biotransformation, stress and genotoxic
effects of 17β-estradiol in juvenile sea bass (Dicentrarchus labrax L.). Environ.
Int. 32:470-477.
Vaccaro, E., Meucci, V., Intorre, L., Soldani, G., Di Bello, D., Longo, V., Gervasi, P. G.
e Pretti, C. (2005). Effects of 17ß-estradiol, 4-nonylphenol and PCB 126 on the
estrogenic activity and phase I and 2 biotransformation enzymes in male sea
bass (Dicentrarchus labrax). Aquat. Toxicol. 75:293-305.
Van der Oost R., Beyer J. e Vermeulen, N. P. E. (2003). Fish bioaccumulation and
biomarkers in environmental risk assessment: a review. Environ. Toxicol. Pharm.
13:57-149.

Capítulo IV
91
Varanasi, U., Stein, J. E. e Nishimoto, M. (1989). Biotransformation and disposition of
PAH in fish. Em: Oliveira, M., Pacheco, M. e Santos, M. A. (2007). Cytocrome
P4501A genotoxic and stress responses in golden grey mullet (Liza aurata)
following short-term exposure to phenantrene. Chemosphere. 66:1284-1291.
Wendelaar-Bonga, S. E. W. (1997). The stress response in fish. Physiol. Rev. 77:591-
625.
Willett, K. L., Wassenberg, D., Lienesch, L., Reichert, W. e Di Giulio, R. T. (2001). In
vivo and in vitro inhibition of CYP1A-dependent activity in Fundulus heteroclitus
by the polynuclear aromatic hydrocarbon fluoranthene. Toxicol. Apllied Pharm.
177:264-271.

CAPÍTULO V
Discussão geral

Discussão geral
93
A discussão geral pretende ser uma análise crítica global da tese, uma
vez que cada capítulo inclui uma discussão específica do seu conteúdo.
1. Avaliação genotóxica da Ria de Aveiro
Verificou-se uma elevada frequência de ANEs em espécimes colhidos em
Vagos em todas as estações do ano, à excepção do Outono. De acordo com
um estudo efectuado na mesma área e nas mesmas condições (Pacheco et al.,
2005), foram obtidos resultados similares nas colheitas então efectuadas em
Julho e Setembro de 2000, os quais foram correlacionados com a existência de
elevados níveis de HAPs existentes na água do referido local. A
biotransformação da maior parte dos compostos químicos actua como um
mecanismo de desintoxicação. Contudo, a metabolização dos HAPs origina,
frequentemente, intermediários reactivos altamente tóxicos, mutagénicos, ou
cancerígenos para os peixes (Van der Oost et al., 1994). Assim, após
absorção, os HAPs poderão ser biotransformados no fígado, tornando-se
genotóxicos, o que justificaria o aparecimento de ANEs (Pacheco e Santos,
2001b), como resultado das lesões causadas ao nível do DNA.
A ausência de expressão genotóxica em Vagos no Outono, poderá estar
relacionada com a existência de acções inibitórias resultantes de elevados
níveis de contaminação. Resultados similares foram obtidos por Pacheco et al.
(2005) no mês de Maio, no qual, apesar da existência de elevadas
concentrações de HAPs na água, em Vagos, não se verificou expressão
genotóxica, ao contrário do que seria previsível. A indução de lesões nucleares
pode ser mascarada através de uma acção citotóxica, havendo morte dos
eritrócitos em vez de ocorrer formação de lesões nucleares não-letais (Das e
Nanda, 1986). Das e Nanda (1986) relataram um decréscimo gradual da
frequência de micronúcleos em função do aumento da concentração do
xenobiótico, como consequência, provavelmente, de um efeito inibitório ao
nível da divisão celular, o qual, por sua vez, poderá resultar de uma inibição
da síntese de DNA ou inibição directa da eritropoiese. Para além disso, as
células portadoras de danos/anomalias tendem a ser removidas do organismo
mais rapidamente que as normais (Das e Nanda, 1986).

Capítulo V
94
No seguimento do que anteriormente foi exposto e, com o objectivo de
evitar interpretações erróneas, principalmente resultantes de acções inibitórias
da eritropoiese, recomenda-se fortemente a contagem do número de
eritrócitos adultos e imaturos, assim como a medição da hemoglobina no
sangue por cada eritrócito e a análise complementar dos contaminantes
presentes na água, sedimento e tecidos dos peixes (Pacheco et al., 2005).
Portanto, muito embora os resultados obtidos e o estudo anteriormente
efectuado por Pacheco et al. (2005) sugiram uma relação entre a existência de
HAPs e a elevada expressão genotóxica obtida em Vagos, há que fazer uma
interpretação cautelosa, uma vez que não se efectuou análise à água e ao
sedimento (em 2006/2007).
2. Efeitos in vitro de HAPs e esteróides, isoladamente ou em
conjunto
O BNF é reconhecido como um forte indutor da EROD hepática (Pacheco
e Santos, 1996), pelo que foi usado, neste estudo, como controlo in vivo
positivo da actividade de EROD. A exposição ao referido composto foi feita por
injecção intraperitoneal. Pretendeu-se assim obter uma elevada actividade
EROD microsomal hepática, de forma a observar in vitro possíveis alterações
desta actividade enzimática quando exposta a:
1) diferentes concentrações de benzo[a]pireno (B[a]P) e fluoranteno
(FL);
2) cortisol (C) e 17β-estradiol (E2) (22.02 nM);
3) diferentes concentrações de B[a]P e FL após exposição prévia ao C
ou ao E2.
Verificou-se que o contacto dos microsomas, na cuvette, com os
referidos HAPs e esteróides, isoladamente ou em combinação, pode resultar
em alterações significativas da actividade de EROD.
2.1. Exposição in vitro ao benzo[a]pireno e ao fluoranteno
As concentrações dos HAPs testados in vitro (0.1, 0.3, 0.9 e 2.7 µM)
podem ser considerados, de uma forma geral, ecotoxicologicamente

Discussão geral
95
relevantes, atendendo às concentrações que podem ser encontradas em
ecossistemas poluídos (Anyakora et al., 2005).
A exposição in vitro dos microsomas de A. anguilla L. ao B[a]P 0.1, 0.3,
0.9 e 2.7 µM causou uma forte inibição da actividade EROD hepática, o que é
concordante com os resultados obtidos in vitro por Gravato e Santos (2002)
em Dicentrarchus labrax L., tendo sido sugerido por estes autores a
possibilidade de tais resultados se deverem à ocorrência de inibição
enzimática. A inibição da actividade EROD microsomal hepática, em A. anguilla
L., também foi demonstrada após exposição in vitro ao reteno e ácido abiético
(Santos e Maria, 2005), assim como a metais, nomeadamente cobre, zinco e
crómio (Oliveira et al., 2005).
A exposição in vitro ao FL 0.1 e 0.3 µM induziu de forma significativa a
actividade EROD microsomal hepática. Vários estudos têm relatado a indução
da actividade EROD hepática em A. anguilla L., após exposição in vivo a outros
HAPs, tais como reteno (Maria et al., 2002 e 2005) e naftaleno (Maria et al.,
2002; Pacheco e Santos, 2002) assim como ao BNF (Maria et al., 2002;
Pacheco e Santos, 2002) e ácidos resínicos (Maria et al., 2004a e 2004 b;
Pacheco e Santos, 1997). Oliveira et al. (2007) demonstraram igualmente a
indução da actividade EROD hepática, em L. aurata, após exposição in vivo ao
fenantreno.
Santos e Maria (2005) também descreveram a indução da actividade
EROD após exposição in vitro dos microsomas hepáticos de A. anguilla L. ao
ácido desidroabiético.
A exposição in vitro dos microsomas ao FL 0.9 e 2.7 µM inibiu
significativamente a actividade EROD hepática. Contudo, a inibição causada
pelo FL 0.9 e 2.7 µM foi inferior à inibição causada pelas mesmas
concentrações de B[a]P. É sabido que vários indutores da síntese de CYP1A
(sub família do gene P450) também podem inibir a actividade catalítica de
EROD, dependendo essa inibição da concentração utilizada na exposição
(Stegeman e Hahn, 1994). Este tipo de inibição é evidente através da

Capítulo V
96
diminuição da actividade catalítica de CYP1A em microsomas hepáticos de
peixes ou células de peixe expostas a elevadas doses de xenobióticos, tais
como os HAPs (Haasch et al., 1993). Assim, os factos anteriormente
mencionados poderão explicar a indução de EROD observada para as
concentrações mais baixas de FL (0.1 e 0.3 µM) e a inibição verificada para as
concentrações mais elevadas (0.9 e 2.7 µM).
Os resultados obtidos por Fent e Bätscher (2000) demonstraram que a
resposta indutora da EROD mostrou ser fortemente dependente da estrutura
do composto ao qual é exposta. Os resultados obtidos, na presente
dissertação, para o B[a]P e o FL também evidenciam esta dependência. Assim,
a exposição microsomal hepática ao B[a]P e ao FL, não se traduziu no mesmo
tipo de resposta no que respeita à actividade EROD. Os dados disponíveis
referentes à indução de CYP1A em peixes, por HAPs de pequeno tamanho,
com 4 ou menos anéis aromáticos, correspondem a resultados não
consensuais, sendo essas respostas dependentes da espécie animal testada
(Shailaja e D’Silva, 2003). Como tal, e tendo em consideração que os referidos
resultados se referem a estudos in vitro com a Anguilla anguilla L., torna-se
necessário estudos adicionais com outras espécies de forma a validar os
mesmos.
No caso do FL os resultados obtidos incentivam a exploração, em
estudos futuros, de concentrações compreendidas entre 0.3 e 0.9 µM, uma
vez que para 0.9 µM observa-se uma inibição significativa da actividade de
EROD enquanto que para 0.3 µM se observa o oposto, ou seja, um forte
indução.
2.2. Exposição in vitro ao cortisol e 17β-estradiol
É sabido que o sistema endócrino desempenha um papel extremamente
relevante no mecanismo de stress dos peixes, havendo bastante informação
científica acerca da resposta de hormonas esteróides, nomeadamente cortisol
e 17β-estradiol, a situações de stress em diferentes espécies de peixes tais
como Anguilla anguilla L. (Pacheco e Santos, 2001a e 2001b; Santos e
Pacheco, 1996; Santos et al., 2000; Teles et al., 2004b), Dicentrarchus labrax
L. (Teles et al., 2004a e 2006), Sparus aurata L. (Teles et al., 2005) e Liza

Discussão geral
97
aurata (Oliveira et al., 2007). Contudo, pouco é sabido acerca da ligação entre
a resposta de enzimas envolvidas no processo de biotransformação (como por
exemplo a EROD) e a influência deste tipo de hormonas sobre esta actividade
enzimática, in vivo e in vitro. De forma análoga, pouco é sabido acerca dos
níveis fisiológicos celulares destas hormonas e de que forma a sua
biodisponibilidade interfere/influencia a resposta/actividade deste tipo de
enzimas.
A concentração adoptada durante os ensaios in vitro para as duas
hormonas esteróides foi de 22.02 nM e baseou-se no nível plasmático de E2
obtido por Teles et al. (2005) em Sparus aurata L., 4 horas após a adição
deste esteróide à água dos aquários que continham os espécimes. Assim, este
valor foi usado como referência do nível fisiológico celular provável desta
hormona, tendo-se adoptado a mesma concentração para o C de forma a
assegurar a uniformidade e coerência das concentrações testadas e a
possibilitar comparações entre os efeitos produzidos pelos dois esteróides.
A exposição in vitro dos microsomas hepáticos da enguia ao C 22.02 nM
induziu significativamente a actividade de EROD. A maioria dos estudos
laboratoriais (Santos e Pacheco, 1996) e de campo (Pacheco e Santos, 2001b;
Teles et al., 2004, Oliveira et al., 2007) tem descrito a elevação do nível de C
plasmático como uma resposta característica à exposição a compostos
químicos stressantes isolados ou em misturas complexas (Teles et al., 2004),
incluindo HAPs e efluentes da fábrica de pasta de papel, razão pela qual o C é
frequentemente usado como indicador de stress. Os resultados obtidos
sugerem que o C é capaz de regular a expressão de CYP1A. É reconhecido que
as hormonas esteróides fazem parte dos factores endógenos que têm a
capacidade de regular a expressão de CYP1A, contudo os mecanismos de
interacção subjacentes não estão completamente estabelecidos (Navas e
Segner, 2000). Santos et al. (2000), num estudo de cultura de orgãos em A.
anguilla L., também observaram um aumento da actividade hepática de EROD,
quando o cortisol foi adicionado ao meio de cultura.
De acordo com Hontela (1997) nos estudos in vivo, os níveis
plasmáticos de C podem ser afectados por algumas variáveis bióticas (sexo,

Capítulo V
98
predação, confinação, e manuseamento) e abióticas (fotoperíodo, temperatura
e saturação de oxigénio na água). Assim, nos presentes estudos in vitro, as
condições de controlo laboratoriais adoptadas foram cuidadosamente definidas
de forma a evitar interferências significativas nos resultados obtidos.
A exposição in vitro dos microsomas hepáticos de A. anguilla L. ao E2
também se traduziu num aumento da actividade EROD hepática. Vários
estudos têm demonstrado que a exposição in vivo ao E2 afecta a actividade
hepática de CYP1A nos peixes (Navas e Segner, 2001; Vaccaro et al., 2005;
Teles et al., 2005). Contudo, Vaccaro et al. (2005) e Teles et al. (2005)
demonstraram, respectivamente, em Dicentrarchus labrax L. e Sparus aurata
L, in vivo um efeito inibidor do E2 sobre a actividade EROD hepática. De
acordo com Navas e Segner (2001) este efeito inibidor poderá estar
relacionado com a interferência do E2 ao nível do gene, uma vez que estes
autores observaram uma redução da quantidade de proteína e mRNA
associados ao CYP1A, após exposição de hepatócitos de Oncorhynchus mykiss
ao referido esteróide (Navas e Segner, 2001). No entanto, o mecanismo
subjacente ao alegado efeito inbitório de compostos esteróides sobre a
actividade da EROD não é completamente conhecido (Andersson et al., 2007;
Navas e Segner, 2001).
Contudo, este estrogénio natural (E2) pode também, ser considerado um
contaminante aquático. Durante as últimas décadas tem-se verificado um
aumento significativo das concentrações de E2 no meio aquático,
principalmente em zonas urbanas, sendo os seus níveis na água na ordem das
ng/L, podendo atingir 200 ng/L (Bowman et al., 2000). Este facto aliado à sua
capacidade de causar alterações endócrinas nos organismos, revelou a
necessidade de uma avaliação efectiva da toxicidade deste estrogénio natural
como potencial contaminante do meio aquático (Kramer et al., 1998). Assim,
vários estudos têm sido realizados in vivo, neste âmbito, em Sparus aurata L.
(Teles et al., 2005) e Dicentrarchus labrax L. (Teles et al., 2004a; Teles et al.,
2006). Teles et al. (2007) num estudo in situ realizado numa lagoa de água
doce (Pateira de Fermentelos) sujeita a vários tipos de contaminantes
antropogénicos, encontrou elevados níveis de E2 (na ordem das 150/180 ng/L)
no plasma de Anguilla anguilla L., em dois dos cinco locais de exposição

Discussão geral
99
seleccionados para o referido estudo. Apesar de não ter sido levada a cabo a
determinação de E2 na água dos locais de exposição de Anguilla anguilla L. na
Pateira, os elevados níveis de E2 encontrados no plasma desta espécie, foram
coincidentes com a medição de níveis elevados de nitritos, os quais constituem
um indício da existência de decomposição parcial de matéria orgânica
resultante de descargas antropogénicas.
A exposição in vitro dos microsomas hepáticos da enguia ao C e ao E2
levou, portanto, a um aumento da actividade EROD microsomal hepática, pelo
que o efeito dos dois esteróides sobre esta actividade enzimática foi idêntico.
2.3. Exposição in vitro ao benzo[a]pireno e ao fluoranteno em
combinação com o cortisol ou 17β-estradiol.
A exposição in vitro ao C e ao E2, quando combinados com B[a]P,
levou ao aumento significativo da actividade EROD microsomal hepática
quando comparada com a exposição in vitro apenas ao B[a]P. Assim, foi
possível verificar uma redução significativa dos efeitos inibitórios causados
pelo B[a]P como consequência da exposição prévia, na cuvette, dos
microsomas hepáticos aos referidos esteróides. O efeito sequencial exercido
pelo C e/ou E2 com o B[a]P, sobre a actividade EROD hepática, foi idêntico.
Contudo, nem o C nem o E2 foram capazes de evitar a inibição causada pelas
diferentes concentrações de B[a]P.
O contacto dos microsomas hepáticos com o C, previamente à exposição
ao FL 0.1, 0.3, 0.9 e 2.7 µM, levou à elevação significativa da actividade
EROD quando comparado com a exposição apenas ao FL. No caso do E2
apenas se verificou o aumento significativo da actividade EROD microsomal
hepática aquando da exposição in vitro ao referido esteróide em combinação
com FL 0.9 e 2.7 µM, uma vez que presença de E2 não afectou de forma
significativa a actividade EROD microsomal hepática após exposição ao FL 0.1
e 0.3 µM. Assim, o efeito exercido pelo C e pelo E2 quando em combinação
com o FL, sobre a actividade EROD hepática, foi distinto. No entanto, foi

Capítulo V
100
possível observar uma redução significativa dos efeitos inibitórios causados
pelo FL 0.9 e 2.7 µM, como consequência da exposição prévia, na cuvette, dos
microsomas hepáticos aos referidos esteróides. Os dois esteróides (C e E2)
demonstraram, portanto, exercer um efeito protector sobre a actividade EROD
hepática, o que demonstra a capacidade que os mesmos têm de regular a
expressão de CYP1A, nomeadamente a sua actividade catalítica. Contudo, os
mecanismos subjacentes a esta interacção ainda não foram completamente
estabelecidos (Navas e Segner, 2000).
O C foi mesmo capaz de evitar a inibição da actividade EROD hepática
causada pelo FL 0.9 µM ao contrário do E2, que não evitou a inibição desta
actividade enzimática quando exposta ao FL 0.9 e 2.7 µM.
Gravato e Santos (2002), num estudo in vitro obtiveram resultados
semelhantes com naftaleno 2.7 µM, uma vez que, na presença deste
composto, a inibição causada por B[a]P 0.1 e 0.01 µM na actividade
microsomal hepática de EROD também decresceu, o que indicia que de
alguma forma o naftaleno tal como o C e o E2, também atenua o efeito
inibitório de B[a]P. O naftaleno, tal como o C foi mesmo capaz de evitar a
inibição causada pelo B[a]P 0.01 µM.
3. Considerações finais e perspectivas futuras
Os ensaios efectuados com base nos microsomas hepáticos isolados a
partir do fígado de Anguilla anguilla L. demonstraram grande utilidade, no que
respeita ao fornecimento de informação relevante sobre o efeito in vitro de
dois HAPs ubíquos nos ambientes aquáticos sobre a actividade EROD hepática,
assim como acerca da protecção desta actividade enzimática conferida pelo
cortisol e pelo 17β-estradiol. Os resultados da avaliação da actividade EROD
hepática obtidos confirmam a importância da estrutura dos HAPs, da
respectiva concentração molar e ainda das condições fisiológicas do indivíduo
ao longo da sua vida e/ou das condições do respectivo ensaio, vindo ainda
reforçar a necessidade de estudos adicionais in vitro que possibilitem uma
melhor compreensão acerca da influência e efeito protector que as hormonas
esteróides poderão exercer sobre este tipo de enzimas, de forma a permitir

Discussão geral
101
estabelecer uma ligação entre a regulação endócrina e a resposta de
determinadas enzimas envolvidas no processo de biotransformação,
nomeadamente a EROD.
Os resultados obtidos incentivam ainda à realização de estudos adicionais
que avaliem a influência do C e do E2, sobre a actividade EROD microsomal
hepática, quando exposta a outros HAPs (individualmente e/ou em misturas) e
a metais pesados. Para as mesmas condições estabelecidas e descritas nos
capítulos III e IV, é igualmente interessante e pertinente proceder a estudos
adicionais que contemplem outras concentrações de C e de E2.
REFERÊNCIAS BIBLIOGRÁFICAS
Anyakora, C., Ogbeche, A., Palmer, P. e Cocker, H. (2005). Determination of
polynuclear aromatic hydrocarbons on marine samples of Siokolo Fishing
Settlement. J. Chromatogr. A 1073: 323-330.
Das, R. K. e Nanda, R. K. (1986). Induction of micronuclei in peripheral erythrocytes
of the fish, Heteropneustes fossilis, by mitomycin C and paper mill effluent.
Mutat. Res. 175: 67-71.
Fent K. e Bätscher, R. (2000). Cytocrome P4501A induction potencies of polycyclic
aromatic hydrocarbons in a fish hepatoma cell line: demonstration of additive
interactions. Environ. Toxicol. Chem. vol. 19, nº 8, 2047-2058.
Gravato C. e Santos, M. A. (2002). Juvenile sea bass liver P450 EROD induction and
erythrocytic genotoxic responses to PAH and PAH-like compounds. Ecotoxicol.
Environ. Saf. 51:115-127.
Haasch, M. L., Quardokus, E. M., Sutherland, L. A., Goodrich, M. S. e Lech, J. J.
(1993). Hepatic CYP1A induction in rainbow trout by continuous follow-through
exposure to β-naphthoflavone. Toxicol. Sci. 20:72 - 82.
Hontela, A. (1997). Endocrine and physiological responses of fish to xenobiotics: role
of glucocorticosteroid hormones. Rev Toxicol. 1:1-46

Capítulo V
102
Maria, V. L., Gravato, C., Correia, A. C. e Santos, M. A. (2002). Biotransformation and
genotoxic responses to PAHs in two teleost species. Fres. Environ. Bulletin. vol.
9a, nº 11, 609-615.
Maria, V. L., Correia, A. C. e Santos, M. A. (2004a). Anguilla anguilla L. genotoxic and
liver biotransformation responses to abietic acid exposure. Ecotoxicol Environ.
Saf. 58:202-210.
Maria, V. L., Correia, A. C. e Santos, M. A. (2004b). Genotoxic and liver
biotransformation responses in Anguilla anguilla L. after exposure desidroabietic
acid. Fres. Environ. Bulletin. vol. 13, nº 1, 56-65.
Maria, V. L., Correia, A. C. e Santos, M. A. (2005). Anguilla anguilla L. liver EROD
induction and genotoxic responses after retene exposure. Ecotoxicol Environ.
Saf. 61:230-238.
Navas, J. M. e Segner, H. (2000). Modulation of trout 7-ethoxyresorufin-O-deethylase
(EROD) activity by estradiol and octylphenol. Mar. Environ Res. 50:157-162.
Oliveira M., Santos, M. A., e Pacheco M. (2005). Heavy metal inhibition of Anguilla
anguilla L. liver microsomal EROD activity and thiol protection. Fresen. Environ.
Bulletin. vol. 14, nº 1, 59-64.
Oliveira M., Pacheco M. e Santos, M. A. (2007). Cytochrome P4501A, genotoxic and
stress responses in golden grey mullet (Liza aurata) following short-term
exposure to phenanthrene. Chemosphere. 66:1284-1291.
Pacheco M. e Santos, M. A. (1997). Induction of EROD activity and genotoxic effects
by polycyclic aromatic hydrocarbons and resin acids on the juvenile eel (Anguilla
anguilla L.). Ecotoxicol. Environ. Saf. 38:252-259.
Pacheco, M. e Santos, M. A. (2001a). Biotransformation, endocrine and genetic
responses of Anguilla anguilla to petroleum distillate products and environmental
contaminated waters. Ecotoxicol. Environ. Saf. 49:64-75.
Pacheco, M. e Santos, M. A. (2001b). Tissue distribution and temperature-dependence
of Anguilla anguilla L. EROD activity following exposure to model inducers and

Discussão geral
103
relationship with plasma cortisol, lactate and glucose levels. Environ. Intl.
26:149-155.
Pacheco, M. e Santos, M. A. (2002). Naphthalene and ß-naphthoflavone toxicity to
Anguilla anguilla L.: Hepatic metabolism and erythrocytic nuclear abnormalities.
Environ. Int. 28:285-293.
Santos, M. A. e Pacheco, M. (1996). Anguilla anguilla L. stress biomarkers recovery in
clean water and secondary treated pulp mill effluent. Ecotoxicol. Environ. Saf.
35: 96-100.
Santos, M. A., Pacheco, M. e Magalhães, I. (2000). Anguilla anguilla L. liver organ
culture as a toxicological model: in vivo and in vitro EROD induction by β-
naphthoflavone. Fres. Environ. Bulletin. 9: 527-534.
Shailaja, M. S. e D’Silva, C. (2003). Evaluation of impact of PAH on a tropical fish,
Oreochromis mossambicus using multiple biomarkers. Chemosphere. 53:835-
841.
Stegeman, J. J. e Hahn, M. E. (1994). Biochemestry and molecular biology of
monooxigenases. Em: Pacheco M. e Santos, M. A. (1999). Biochemical and
genotoxic responses of adult eel (Anguilla anguilla L.) to resin acids and pulp mill
effluent: laboratoty and field experiments. Ecotoxicol. Environ. Saf. 42:81-93.
Teles M., Gravato, C., Pacheco M. e Santos M. A. (2004a). Juvenile sea bass
biotransformation, genotoxic and endocrine responses to β-naphthoflanove, 4-
nonylphenol and 17ß-estradiol individual and combined exposures.
Chemosphere. 57:147-158.
Teles M., Santos M. A., e Pacheco M. (2004b). Responses of European eel (Anguila
anguilla L.) in two polluted environments: in situ experiments. Ecotoxicol.
Environ. Saf. 58:373-378.
Teles, M., Pacheco, M. e Santos, M. A. (2005). Sparus aurata L. liver EROD and GST
activities, plasma cortisol, lactate, glucose and erythrocytic nuclear anomalies
after short-term exposure either to 17ß-estradiol (E2) or E2 combined with 4-
nonylphenol. Sci. Total Environ. 336:57-69.

Capítulo V
104
Teles, M., Santos, M. A. e Pacheco, M. (2006). Biotransformation, stress and genotoxic
effects of 17β-estradiol in juvenile sea bass (Dicentrarchus labrax L.). Environ.
Int. 32:470-477.
Teles M., Pacheco M. e Santos M. A. (2007). Endocrine and metabolic responses of A.
anguilla L. caged in a freshwater-wetland (Pateira de Fermentelos - Portugal).
Sci. Total Environ. 372:562-570.