Embed Size (px)
Citation preview

Retículo Endoplasmático
Aparelho de Golgi
Lisossoma
Núcleo
Peroxissoma
Mitocôndria
Tráfego intracelular de proteínas
Compartimentos intracelulares e
Tráfego de proteínas

Compartimentos intracelulares
Organelos da célula eucariótica

Núcleo
Compartimentos intracelulares e
Tráfego de proteínas

NÚCLEO
Ultraestrutura
O núcleo é o organelo mais volumoso da célula e serve de abrigo ao genoma tornando‐se assim o centro de controlo da célula. No núcleo ocorre a replicação do DNA, a transcrição e o processamento do RNA, enquanto que a fase final da expressão genética (i.e.tradução) ocorre no citoplasma.
RE
filamentosintermédios
poronuclear
memb. externa
memb. internainvólucronuclear
microtúbulo
centrossoma
nucléolo
cromatina
Lamina nuclear
invólucronuclear
membranacitoplasmática
NÚCLEO
CITOSOL
DNA
RNAtranscrição
splicing
transportedo RNA
ribossomastradução
proteínas

Invólucro nuclear
O invólucro nuclear separa o nucleoplasma (substância amorfa de naturezaproteica que preencha a matriz nuclear) do citoplasma.
É constituído por duas biomembranas (membrana externa e membrana interna) que delimitam um espaço perinuclear que varia com o estado fisiológico da célula.
À membrana nuclear externa estão associados ribossomas que originam umacerta continuidade entre esta membrana e o RER, o que serve de base paraadmitir que as membranas do RE se originam a partir da membrana nuclearexterna.
O invólucro nuclear não é uma estrutura contínua uma vez que há pontos dedescontinuidade (poros nucleares) que correspondem a zonas de contactoentre as duas membranas nucleares.
Ultraestrutura
NÚCLEO

Invólucro nuclear
invólucronuclear
membrananuclear externa
membrananuclearexterna
membrana do RE
lúmen do RE
laminanuclearporo
nuclearespaço
perinuclear
Ultraestrutura
NÚCLEO

Invólucro nuclear e lâmina nuclear
Associada à membrana nuclear interna, existe uma estrutura fibrosa densa:lâmina nuclear que serve de suporte mecânico ao núcleo.
Lâmina nuclear observada em TEM
Ultraestrutura
NÚCLEO

Organização interna
O núcleo tem uma estrutura interna que organiza o material genético e localiza as funções nucleares em zonas discretas.
São exemplos desta organização: o nucléolo (local de transcrição dos genes de rRNAs e de síntese de ribossomas), a organização dos cromossomas, a localização em domínios distintos das funções de replicação do DNA e de processamento do pré‐mRNA.
invólucronuclear
poronuclear
lâminanuclear
heterocromatina
eucromatina
Ultraestrutura
NÚCLEO

No núcleo dos eucariotas, o DNA encontra‐se associado a proteínas (histonas),constituindo a cromatina.
Durante a interfase, a cromatina é dispersa e não condensada chamando‐se eucromatina. Há no entanto uma parte da cromatina que se mantém condensada chamando‐se heterocromatina. Esta encontra‐se inactiva em termos de transcrição.
Ultraestrutura
Organização interna
NÚCLEO
invólucronuclear
poronuclear
lâminanuclear
heterocromatina
eucromatina

Ultraestrutura
Complexo poro nuclear (NPC)
O complexo poro nuclear é constituído por canais de 10 nm de diâmetro que permitem o transito de pequenas moléculas, iões e macromoléculas (proteínas e RNAs) entre o núcleo e o citoplasma.
Os poros nucleares não são estruturas estáticas, estando o seu número e a sua localização directamente relacionados com a actividade fisiológica da célula: eles abrem e fecham onde e quando são ou não são necessários.
Técnica de crifractura
NÚCLEO
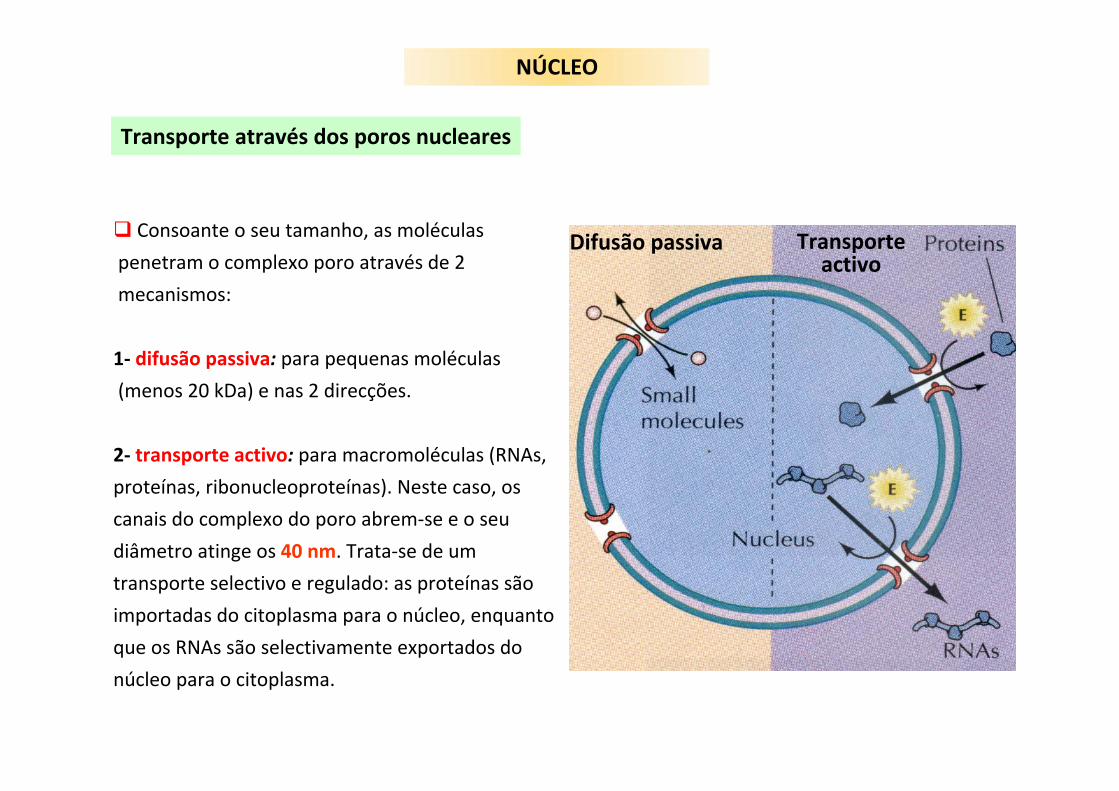
Transporte através dos poros nucleares
Consoante o seu tamanho, as moléculas
penetram o complexo poro através de 2
mecanismos:
1‐ difusão passiva: para pequenas moléculas
(menos 20 kDa) e nas 2 direcções.
2‐ transporte activo: para macromoléculas (RNAs,
proteínas, ribonucleoproteínas). Neste caso, os
canais do complexo do poro abrem‐se e o seu
diâmetro atinge os 40 nm. Trata‐se de um
transporte selectivo e regulado: as proteínas são
importadas do citoplasma para o núcleo, enquanto
que os RNAs são selectivamente exportados do
núcleo para o citoplasma.
Difusão passiva Transporteactivo
NÚCLEO

Nucléolo
é a estrutura mais proeminente do núcleo
é o local de transcrição e processamento dos rRNAs e de montagem dos ribossomas
muda de tamanho consoante a actividade de síntese proteica da célula
O ribossoma é formado por rRNAs associados a proteínas ribossomais
Os ribossomas dos eucariotas contêm 4 tipos de RNAs: rRNAs 5S, 5.8 S, 18S e 28S.
Os ribossomas catalisam a síntese proteica nos procariotas e eucariotas.
NÚCLEO

Os ribossomas dos procariotas (70S) e dos eucariotas (80S) consistem numa subunidade maior e numa subunidade menor, ambas formadas por rRNAs e proteínas ribossomais.
70S 80S
50S 30S 60S 40S
Nucléolo
NÚCLEO

Retículo Endoplasmático
REL RER ribossomasmitocôndria
Compartimentos intracelulares e
Tráfego de proteínas

RETÍCULO ENDOPLASMÁTICO
Ultraestrutura e funções
O retículo endoplasmático (RE) é um complexo membranar de cisternas, canais, vesículas, vacúolos, geralmente anastomosados.
O RE no seu conjunto é rodeado por uma extensíssima membrana contínua que delimita um interior comum: o lúmen.
A membrana do RE está em continuidade com a membrana externa do núcleo.
Ultraestruturalmente, distingue‐se dois tipos funcionais de RE:
‐ Retículo Endoplasmático Rugoso (RER) apresenta ribossomas aderentes à sua facecitosólica e é implicado na síntese e processamento das proteínas.
‐ Retículo Endoplasmático Liso (REL) não apresenta ribossomas à sua superfície e éimplicado na síntese dos lípidos.
No entanto, existe uma continuidade física e funcional entre o RER e REL.

RE rugoso RE liso
lúmenAspecto ultraestrutural do RER
O tipo de retículo e a sua quantidade por célula variam entre os diferentes tipos celulares e de acordo com a actividade de síntese da célula.
RETÍCULO ENDOPLASMÁTICO
Ultraestrutura e funções
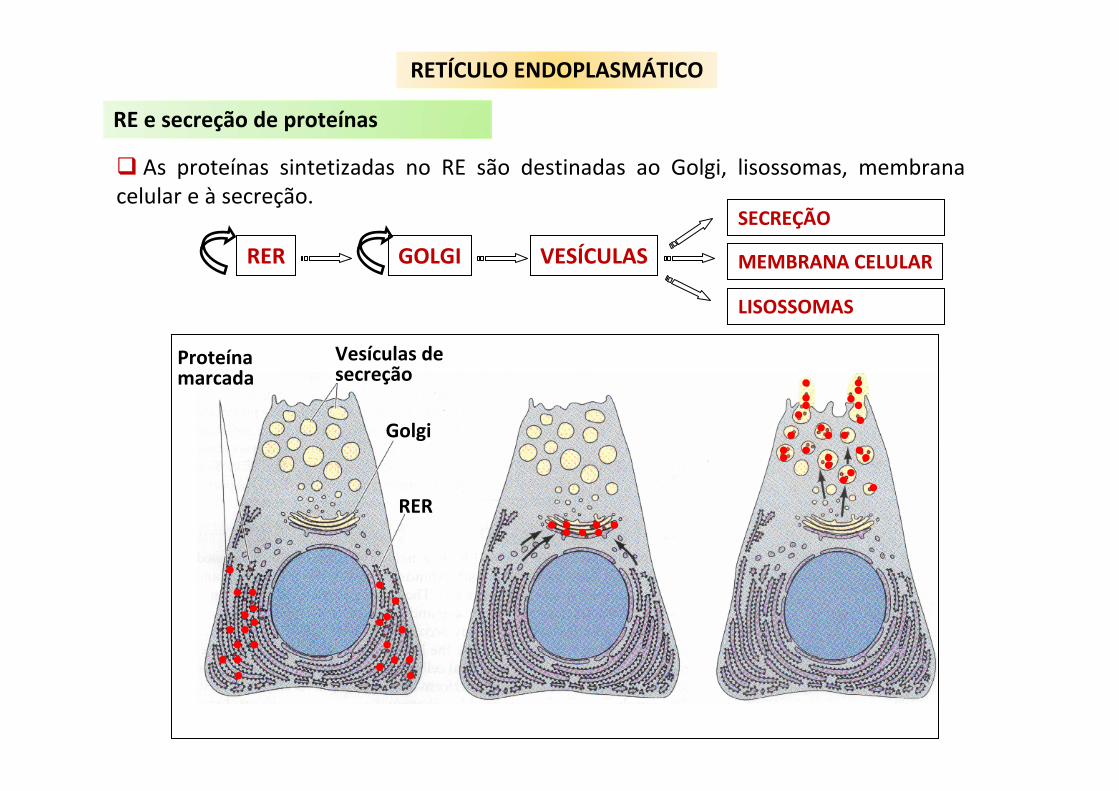
As proteínas sintetizadas no RE são destinadas ao Golgi, lisossomas, membrana celular e à secreção.
Proteínamarcada
Vesículas desecreção
Golgi
RER
RER GOLGI VESÍCULAS
SECREÇÃO
MEMBRANA CELULAR
LISOSSOMAS
RETÍCULO ENDOPLASMÁTICO
RE e secreção de proteínas

Muitas proteínas sofrem alterações estruturais antes de adquirir a sua função.
Estas modificações dependem da localização final da proteína activa.
As proteínas destinadas ao RE, lisossomas ou à secreção sofrem modificaçõesespecíficas:
aquisição da estrutura tridimensional (Folding)
formação de complexos multiméricos
formação de pontes dissulfureto
glicosilação
RETÍCULO ENDOPLASMÁTICO
Modificações das proteínas no RE

Depois de incorporadas no lúmen do RE,
as proteínas sofrem a adição de um
oligossacarídeo.
Os oligossacarídeos mais comuns são
constituídos por:
2 resíduos de N‐acetilglucosamina (GlcNAc),
9 de manose
3 de glucose
Estes oligossacarídeos ligam‐se, através de
GlcNAc, ao grupo NH2 de um resíduo de
asparagina da proteína.
N‐acetilgluco‐samina
Manose
Glucose
(asparagina)
Glicosilação de proteínas no RE
RETÍCULO ENDOPLASMÁTICO

A maioria dos lípidos de membrana (fosfolípidos e colesterol) são sintetizados no REL.
O fosfolípido mais abundante é a fosfatidilcolina.
A síntese de fosfolípidos derivados do glicerol faz‐se em 3 etapas partindo‐se de2 moléculas de ácidos gordos, 1 de glicerofosfato e outra de cabeça polar (exemplo: colina):
As enzimas destas reacções encontram‐se na face citosólica do RE e utilizam precursoresdados pelo citoplasma.
Retículo Endoplasmático liso e síntese de lípidos
RETÍCULO ENDOPLASMÁTICO

(ácido fosfatídico) (diacilglicerol) (fosfatidilcolina)
RETÍCULO ENDOPLASMÁTICO
1 ‐ a acil transferase associa os 2 ácidos gordos ao glicerofosfato originando o ácidofosfatídico. 2 ‐ este serve de substrato à fosfatase, formando‐se 1 molécula dediacilglicerol. 3 ‐ esta associa‐se finalmente a 1 molécula de fosfocolina, sob aacção de colina fosfotransferase, formando‐se 1 molécula de fosfatidilcolina.
Retículo Endoplasmático liso e síntese de lípidos

Translocadores de
fosfolípidos (flipases)
transportam lípidos de
uma camada lipídica
para outra.
FlipaseFlipase
RETÍCULO ENDOPLASMÁTICO
Retículo Endoplasmático liso e síntese de lípidos
Tráfego intracelular de fosfolípidos
Proteínamarcada
Vesículas desecreção
Golgi
RER
RETÍCULO ENDOPLASMÁTICO
Retículo Endoplasmático liso e síntese de lípidos

Proteínas de troca de fosfolípidos ajudam a transportar fosfolípidos do RE para as membranas da mitocôndrias e peroxissomas.
RETÍCULO ENDOPLASMÁTICO
Retículo Endoplasmático liso e síntese de lípidos

Complexo de Golgi
Compartimentos intracelulares e
Tráfego de proteínas

Ultraestrutura e funções
O complexo de Golgi (ou dictiossoma) é constituído por um conjunto de sáculos achatados dispostos paralelamente uns aos outros formando uma espécie de “pilha de pratos”. No interior delimitam uma cavidade: o lúmen.
O Golgi está intimamente associado ao retículo endoplasmático.
invólucronuclear
RER
face cisde Golgi
Aspecto ultraestrutural de Golgi
vacúolos do Golgi
COMPLEXO DE GOLGI
vesículasdo Golgi

Na periferia das cisternas existem vesículas ou vacúolos golgianos, que se difundem pelo citoplasma transportando substâncias sintetizadas/modificadas no Golgi para os lisossomas, membrana celular ou para a secreção.
Ultraestrutura e funções
COMPLEXO DE GOLGI
invólucronuclear
RER
face cisde Golgi
Aspecto ultraestrutural de Golgi
vacúolos do Golgi
vesículasdo Golgi

Compartimentação morfológica e funcional
O Golgi é uma estrutura polarizada. As proteínas do RE entram no Golgi pela face cis,transitam no compartimento mediano e saem pela face trans.
face cis
face trans
Golgi cis
Golgimediano
Golgitrans
Vesículagolgiana
Vesículade secreção
COMPLEXO DE GOLGI

À medida que atravessam estes compartimentos, as proteínas vão sendo modificadas paradepois serem transportadas para os seus destinos na célula.
Assim, a face cis é a face de formação de novas cisternas por fusão das vesículas do RE.A face trans é a face de maturação onde as cisternas se desintegram em vesículas.
face cis
face trans
Golgi cis
Golgimediano
Golgitrans
Vesículagolgiana
Vesículade secreção
Compartimentação morfológica e funcional
COMPLEXO DE GOLGI

Funções do complexo de Golgi
COMPLEXO DE GOLGI
• Glicosilação de proteínas e de lípidos (formação de glicoproteínas e glicolípidos)
• Secreção celular
• Formação dos lisossomas
• Síntese da porção glucídica dos proteoglicanos
• Participação no metabolismo dos lípidos
• Sulfatação de várias biomoléculas
• Biogénese e recuperação de membranas• Regulação do teor hídrico da célula (por formação de vesículas que acumulam água e a expelem para o exterior da célula).
• Nas células vegetais: síntese dos componentes da matriz da parede celular

Glicosilação de proteínas no lúmen de Golgi
A glicosilação de proteínas iniciada no RER continua no lúmen de Golgi. A glicosilação destas proteínas é diferente consoante o seu destino final na célula:
1. Se forem para a membrana citoplasmática ou para a secreção, as glicoproteínas são transformadas em oligossacarídeos complexos.
2. Se forem para os lisossomas, as glicoproteínas adquirem apenas o grupo fosfato. O terminal manose 6‐fosfato que se forma nestas proteínas é extremamente importante para o seu destino para os lisossomas.
Oligossacarídeo tipo complexo
1 2
COMPLEXO DE GOLGI

COMPLEXO DE GOLGI
Transporte de proteínas a partir do complexo de Golgi
As proteínas são transportadas da região trans de Golgi, dentro de vesículas para o seu desIno final:1. Na ausência de sinais específicos internos, as proteínas são transportadas para a superfície celular (membrana celular ou matriz extracelular) através da secreção constitutiva.Exemplos: formação de glicocálice, produção de anticorpos pelos plasmócitos
2. Outras proteínas são destinadas aos lisossomas ou à secreção regulada.Exemplos: produção de hormonas pelas células endócrinas ou de neurotransmissores pelos neurónios
Mistura de proteínas Triagem
Receptor demanose 6‐fosfato
Secreçãoconstitutiva
Secreção regulada
Lisossomas
Fluxo para oEspaço extracelular

Lisossoma
Compartimentos intracelulares e
Tráfego de proteínas

LISOSSOMA
Ultraestrutura e função
Os lisossomas são definidos como vesículas
delimitadas por uma membrana, e implicados
no processo de digestão intracelular.
Os lisossomas contêm várias enzimas hidrolíticas
(hidrolases ácidas) que são activas em meio ácido
(pH ~ 5) e não a pH neutro do citosol.
Estas enzimas são capazes de hirolisar proteínas,
DNA, RNA, polissacarídeos e lípidos.
O pH da matriz lisossomal é mantido ácido pela
bomba de protões que utiliza a energia do ATP
para bombear os H+ do citosol para dentro.

LISOSSOMA
Ultraestrutura e função
Ultraestruturalmente, os lisossomas são organelos heterogéneos, podendo apresentar uma diversidade morfológica que reflecte as variações na quan;a e natureza do material ingerido.
É possível distinguir 3 tipos de lisossomas:1. Lisossomas primários: de aspecto homogéneo, são lisossomas recem‐formados contendo as
hidrolases mas ainda sem material ingerido2. Lisossomas secundários: heterogéneos, com hidrolases e material em degradação3. Lisossomas secundários com corpos residuais: acumulam material incompletamente degradado
Lisossomas em MET Lisossomas de macrófago

Biogénese
A formação dos lisossomas implica a via de
secreção e a via de endocitose:
1. O material extracelular é interiorizado por
vesículas endocíticas que se formam a partir da
membrana e fundem com os endossomas
precoces.
2. A maturação dos endossomas precoces em
endossomas tardios passa pela diminuição do
pH interno (3 a 6), o que permite a aquisição
das hidrolases ácidas lisossomais a partir da
rede trans de Golgi e a subsequente maturação
dos endossomas tardios em lisossomas
secundários.
LISOSSOMA
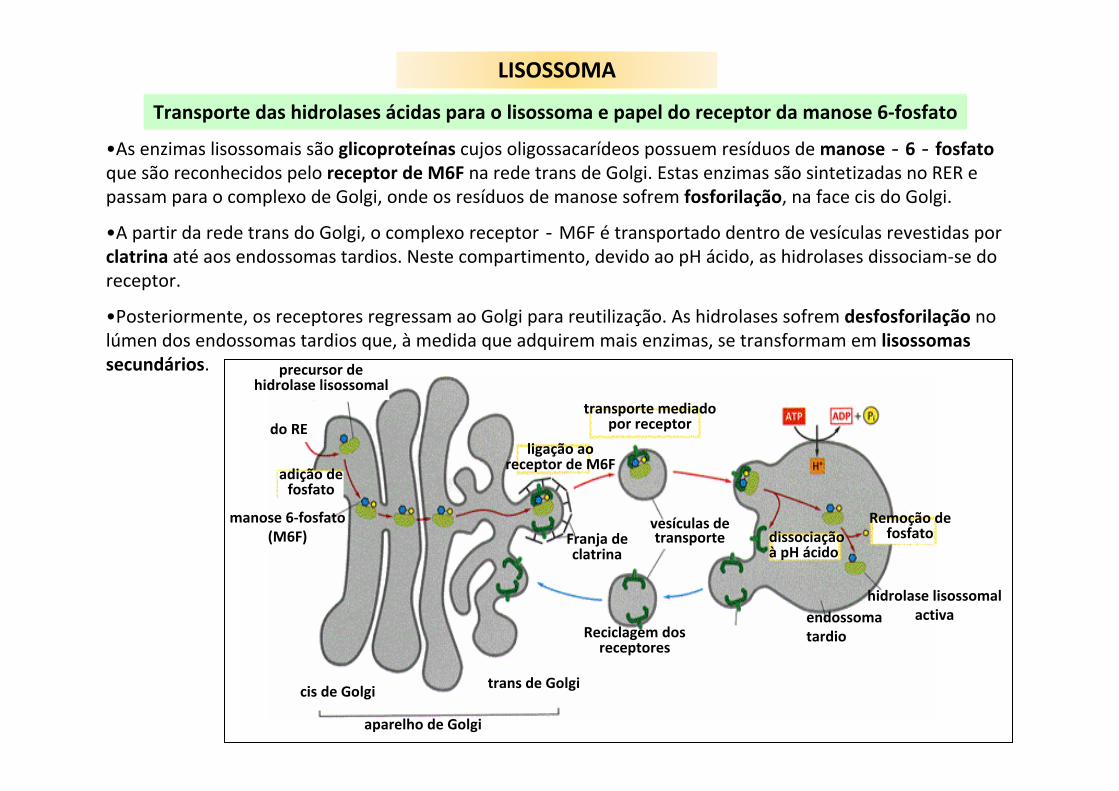
LISOSSOMA
Transporte das hidrolases ácidas para o lisossoma e papel do receptor da manose 6‐fosfato
adição defosfato
manose 6‐fosfato(M6F)
do RE
precursor dehidrolase lisossomal
cis de Golgi trans de Golgi
aparelho de Golgi
Reciclagem dosreceptores
vesículas detransporteFranja de
clatrina
ligação aoreceptor de M6F
transporte mediadopor receptor
dissociaçãoà pH ácido
Remoção defosfato
hidrolase lisossomalactivaendossoma
tardio
•As enzimas lisossomais são glicoproteínas cujos oligossacarídeos possuem resíduos de manose‐6‐fosfato que são reconhecidos pelo receptor de M6F na rede trans de Golgi. Estas enzimas são sintetizadas no RER e passam para o complexo de Golgi, onde os resíduos de manose sofrem fosforilação, na face cis do Golgi.
•A partir da rede trans do Golgi, o complexo receptor‐M6F é transportado dentro de vesículas revestidas por clatrina até aos endossomas tardios. Neste compartimento, devido ao pH ácido, as hidrolases dissociam‐se do receptor.
•Posteriormente, os receptores regressam ao Golgi para reutilização. As hidrolases sofrem desfosforilação no lúmen dos endossomas tardios que, à medida que adquirem mais enzimas, se transformam em lisossomas secundários.

Digestão intracelular
Além da degradação das moléculas interiorizadas por endocitose, os lisossomas digerem material
das vias de fagocitose e de autofagia:
1. Fagocitose: incorporação e digestão de grandes partículas ou células (bactérias, células
envelhecidas, etc.) provenientes do exterior da célula, originando um heterolisossoma. As
células humanas especializadas nesta função são os macrófagos e os neutrófilos.
2. Autofagia: digestão intracelular de organelos e estruturas da própria célula. Neste caso o
organelo (p. e. mitocôndria) é rodeado por uma membrana derivada do retículo
endoplasmático e a vesícula resultante (autofagossoma) funde‐se com o lisossoma
(autolisossoma). O conteúdo do autolisossoma é, então, digerido pelas enzimas lisossomais.
LISOSSOMA

FAGOCITOSE
ENDOCITOSE
AUTOFAGIA
Endosoma precoce
Endolisossomatardio
lisossoma
autofagossoma
Membranacitoplasmática
fagossomabactéria
As 3 vias de digestão intracelular pelos lisossomas
LISOSSOMA
Digestão intracelular

Peroxissoma
Compartimentos intracelulares e
Tráfego de proteínas

Ultraestrutura
Os peroxissomas são
estruturas esféricas ou ovais
(diâmetro 0,5‐1,5 μm) rodeadas
de uma membrana, que engloba
uma matriz peroxissomal
electronodensa, onde por vezes
se observa (dependente do
plano do corte) o nucleóide com
aspecto cristalino (cristalóide)
ou amorfo.
Aspecto ultraestrutural deperoxissomas do fígado de rato.
PEROXISSOMA

Os peroxissomas contêm várias enzimas:
1. Oxidases: intervêm em processos oxidativos utilizando o oxigéniopara remover o hidrogénio de determinados substratos:
RH2 + O2 R + H2O2
2. Catalase: desdobra o H2O2 (peróxido de hidrogénio) em água eoxigénio, o H2O2 seria tóxico para a célula se acumulado:
2H2O2 2H2O + O2
R´H2 + H2O2 R’ + 2H2O
Nas células animais, os peroxissomas são particularmente activos no fígado e rinsonde as reacções oxidativas permitem a eliminação de substâncias tóxicas.Exemplo: é através dos peroxissomas que o álcool que ingerimos é facilmente excretado sob a forma de acetaldeído.
CH3–CH2–OH + H202 CH3–CHO + 2H20
Funções
PEROXISSOMA

Oxidação dos ácidos gordos, acompanhada de produção de H2O2 a partir de O2, o H2O2 é posteriormente degradado pela catalase
Oxidação de aminoácidos e do ácido úrico
Biossíntese de colesterol, ácidos biliares, plasmalogénios
Nas plantas: ciclo glioxílico (síntese de carbohidratos a partir dos ácidos gordos) e fotorespiração
Funções
PEROXISSOMA
Degradado pela catalase

Os peroxissomas são estruturas semi‐autónomas. Embora não tenham DNA nemribossomas, derivam de peroxissomas pré‐existentes, por um processo semelhante ao das mitocôndrias e plastídeos.
As proteínas dos peroxissomas são sintetizadasno citosol e importadas para os peroxissomaspré‐existentes. Esta importação de proteínasresulta no crescimento dos peroxissomas pré‐existentes que se dividem em novos peroxissomas.
peroxissoma
proteínas específicasde importação de
proteínas
crescimento por importação deproteínas específicas do citosol
fissão
novos peroxissomas
Biogénese dos peroxissomas
PEROXISSOMA
As proteínas destinadas aos peroxissomaspossuem um peptídeo sinal (PTS1 ou PTS2) que se liga a seu receptor na membrana do organelo, permitindo a sua translocação para a matriz peroxissomal.

Mitocôndria
Compartimentos intracelulares e
Tráfego de proteínas

Ultraestrutura e função
Em média tem 1‐2 μm de comprimento e0,5‐1 μm de largura
É rodeada por 2 membranas, uma internae outra externa separadas pelo espaçointermembranar.
A membrana interna projecta vários pregosou cristas para o interior da mitocôndria oumatriz mitocondrial. Na matriz, encontra‐seDNA, ribossomas e pequenos grânulos.
De ponto de vista funcional, a membranainterna e a matriz são os compartimentosmais importantes da mitocôndria.
MITOCÔNDRIA
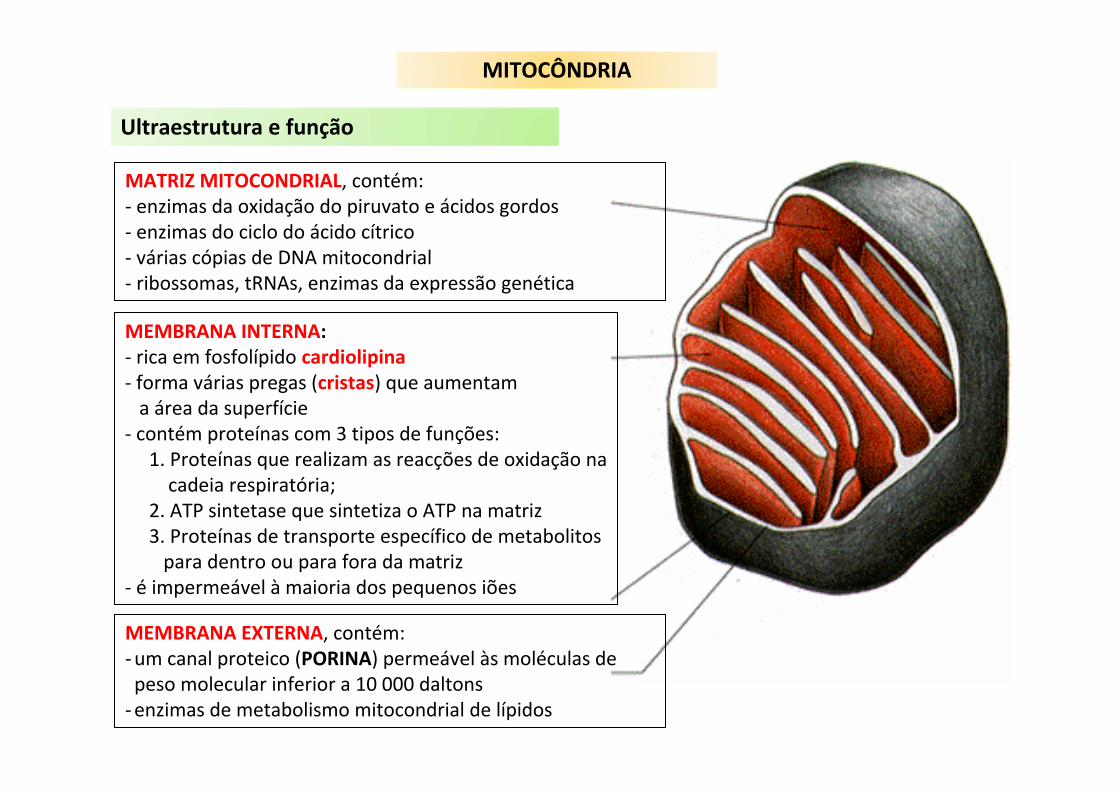
MATRIZ MITOCONDRIAL, contém:‐ enzimas da oxidação do piruvato e ácidos gordos‐ enzimas do ciclo do ácido cítrico‐ várias cópias de DNA mitocondrial‐ ribossomas, tRNAs, enzimas da expressão genética
MEMBRANA INTERNA:‐ rica em fosfolípido cardiolipina‐ forma várias pregas (cristas) que aumentama área da superfície
‐ contém proteínas com 3 tipos de funções:1. Proteínas que realizam as reacções de oxidação nacadeia respiratória;
2. ATP sintetase que sintetiza o ATP na matriz3. Proteínas de transporte específico de metabolitospara dentro ou para fora da matriz
‐ é impermeável à maioria dos pequenos iões
MEMBRANA EXTERNA, contém:‐um canal proteico (PORINA) permeável às moléculas de peso molecular inferior a 10 000 daltons‐enzimas de metabolismo mitocondrial de lípidos
Ultraestrutura e função
MITOCÔNDRIA

É a principal fonte de energia – sob a forma de ATP ‐ das células eucarióticas através do processo de fosforilação oxidativa.
Importa a maioria das suas proteínas do citosol.
Possui o seu próprio DNA que codifica para tRNAs, rRNAs e proteínas mitocondriais.
A sua biogénese depende de proteínas codificadas por seu genoma e de proteínas codificadas pelo genoma nuclear.
MITOCÔNDRIA
Ultraestrutura e função

Biogénese
À medida que crescem e aumentam de tamanho, as mitocôndrias dividem‐se por fissão de modo semelhante à divisão das bactérias.
As mitocôndrias não são sintetizadas de novo; os seus constituintes de origem citosólica são acrescentados durante o processo de crescimento e divisão.
Replicação de DNA
MITOCÔNDRIA

TRÁFEGO INTRACELULAR DE PROTEÍNAS
Cytosol
Os ribossomas livres
no citoplasma
sintetizam proteínas
destinadas a:
núcleo,
mitocôndria,
cloroplastos e
peroxissomas.
Os ribossomas
associados à
membrana do RE
sintetizam proteínas
destinadas a:
lisossomas,
membrana
citoplasmática e
secreção.

Peptídeo Sinal
O destino final das proteínas produzidas no citosol ou pelo RER é determinado
por uma sequência presente nestas proteínas chamada peptídeo sinal.
NH2 COOH
peptídeo sinal
Outras proteínas sofrem modificações que determinam o seu destino.
Por exemplo: manose 6 fosfato nas proteínas destinadas aos lisossomas.
Tráfego intracelular de proteínas

Peptídeo Sinal
Tráfego intracelular de proteínas





























