Embed Size (px)
Citation preview

Ardeola MI), 1987. 37-59
COMUNIDADES DE PASSERIFORMES EN BOSQUES MIXTOS DE ALTITUDES MEDIAS
DE LA SIERRA DE CAZORLA
Las comunidades de psseriformes son el resultado de componentes espacia- les y temporales. Los espaciales suponen la localizacibn geográfica a escala amplia y el habitat concreto en que se sitúa la comunidad. Mientras que por otra parte, el componente temporal implica la situación de la comunidad en una escala de tiempo determinada, por contemplarla como un ente dinámico en el que cada composición estructural instanthnea es consecuencia de la situacibn anterior y va a condicionar las sucesivas. Esta visión de la comunidad como resultado de un proceso cambiante espacio-temporal, que se traduce en reajustes en la composición y abundancia de las especies, es una visión relativamente moderna que se inició con los trabajos de ~ L L (1972) y HERRERA (1980a, 1980b, 1981). En los inicios del estudio de las comunidades de aves se hizo hincapié en las llamadas comunidades nidificantes, pero en la actualidad cada vez son más numerosos los trabajos que contemplan los cambios estacionales en el seno de la comunidad (HOGSTAD 1967, BLONDEL 1969, FROCHOT 1971, EYBERT 1973, PURROY 1975, HOLMES y SURGES 1975, HERRERA 1980a, ZAMORA y CAMACHO 1984 a y b), si bien se dista mucho de tener información completa para un buen número de habitats. Por ello, describiré aquí dos comunidades de passeriformes de bosque mixto mediterráneo de montaña para las que aún no se dispone de idormación.
El área de estudio se localiza en un enclave denominado Roblehondo, situado en Ia Sierra del Pozo, dentro del Parque Natural de las Sierras de Cazorla y Segura, en el Este de la provincia de Jaén (02-48 0, 37-56 N). Está situado a 1.300-1.400 m a.s.n.m. y tiene un clima de montaña: la media anual de precipitación es de 1.527 mm, concentrándose en otoño-invierno y primavera, las temperaturas medias estimadas para el mes más frío y más cálido son 2,9 y 22,5"C. La vegetación esta constituida por un bosque de Pims nigra con Querrus ilex y sotobosque disperso de Juniperus sp. y Crataegus sp. Define una interfase wtonat entre el matorral esclerafdo mediterráneo de montaña (bosques de Phillyrea lar#iilia, Q. ilex, Arbutus unedo y J . oxycedrus) y el bosque
Estación Biológica Doñana. Apariado 1056, 41013 Sevilla.

38 ARDEOLA 34(1), 1987
de las partes altas de la sierra (P. nigra con sotobosque de J. communis y B. hispanica).
En esta zona se han escogido dos parcelas de estudio RH y CO; la primera constituida por un bosque de ladera (con cobertura arbórea y de matorral del 60 % aproximadamente) y la segunda por un bosque más clareado (cobertura del 30 %) en el nivel cacuminal del valle, a 1 km de la primera. En el Apéndice 1 indicamos la cobertura de las especies leñosas más importantes.
Para el análisis de la comunidad de Passeriformes se realizaron censos periódicos en ambas parcelas, empleando el método lineal descrito por EMLEN (1971). Los censos se llevaron a cabo a lo largo de la línea de base de dos transectos de 1.000 m en RH y CO. Se realizaron 2-8 veces por mes en ambas parcelas y dentro de lo posible se procuraron realizar con buen tiempo y a una velocidad media de 1,75 km/h. Durante el verano se hicieron dentro de las dos primeras horas después de la salida del sol y en invierno cuando el sol comenzaba a dar en las parcelas de estudio, ya que era entonces cuando podía apreciarse mayor actividad en los pájaros. Siguiendo la línea del transecto se establecieron 10 bandas paralelas de 5 m-de anchura cada una (cinco a cada lado de la línea base). Con los contactos obtenidos en cada banda para cada especie se calcularon los coeficientes de detectabilidad (C.D.) según indica EMLEN (1971). De este modo, la densidad de cada especie i (Di) vendrá dada por la expresión:
Di = KCi S/CDi (d)L2d, donde
Ci=número de contactos para la especie i. S = superficie total censada. d= anchura de la banda base. CD(i)(d)=coeficiente de detectabilidad para la especie i y banda base dada. L = longitud del transecto. K = constante de detectabilidad basal.
He empleado como constante de detectabilidad basal, K, los valores 1,50 (para primavera-verano) y 1,15 (para invierno) (EMLEN, 197 1 ; SHIELDS, 1979).
El estudio ha sido realizado durante 17 meses (junio 1983-octubre 1984) y los resultados se exponen mensualmente excepto para el período de 1983 (agosto- noviembre) en el que se recogió la información de forma más detallada y se exponen los resultados quincenales.
Análisis cualitativo
Composición especifica.-En los Apéndices 11 y 111 se detallan las densidades medias obtenidas para los diferentes meses de estudio.

En la parcela de RH se registraron 46 especies diferentes du;ante todo el período de estudio, mientras que en CO fueron 47; de eflas 41 son comunes a ambos sitios. Las exclusivas de CO son especies con tendencia a ocupar terrenos más abiertos y no propias de bosques: L. arborea, L. excubifor, P. pyrrhocorax, C. corax, 0. oewnihe y C. eannabina. Es de destacar, sin embargo, la presencia de L. arborea en CO como vidente y en densidades apreciables, wnstituyendo la diferencia más relevante y en relación con la cobertura del medio.
Según la clasificación de Voo (1960), en la composición de especies 7 predominan los elementos Paleártico y Europeos, siendo escasos los Europeo- turquestániws y los Mediterráneos. Es de destacar la presencia de un par de taxones correspondientes a una fauna de tipo paleoxeromontano (P. rupesrris y Ph. ochruros). La importancia a nivel cuantitativo recae sobre los dos primeros tipos señalados y esta composición específica caracteriza una fauna de passeri- formes típicamente eurosiberiana.
He comparado la composición de las comunidades estudiadas con la información disponible para la avifauna de otras localidades tomando datos de otras quince comunidades de bosque para las que se dispone información bas- tante completa sobre el ciclo anual. Para eliminar el abultamiento en número de especies que pueden introducir las especies migrantes en paso en aquellas comunidades para las que se dispone de información del ciclo anual completo no se han tenido en cuenta estas especies. Sobre la base de los datos de presencia-ausencia de las diferentes especies de pájaros se construyó la matriz de similitud cualitativa mediante el índice de Czechanovsky (MARGALEF, 1977). Como es lógico esperar, RH y CO se parecen más entre sí que a ningún otro lugar (el valor de similitud calculado es de 0 9 ) y la localidad con la que muestran mayor afinidad es con el bosque mixto de quejigos, pinos y hayas de Navarra (similitud de 0,74 w n RH y 0,67 con CO); le siguen el robledal de Ancares (0,72) y los bosques de altitudes medias en la Sima de Cazorla (encinar con Phillyrea y Arbutus) (0,70) para RH. El CO, después del bosque mixto navarro, se parece más a los encinares leoneses de montaña (0,64) y al robledal de Ancares (0.63). Las comunidades que tienen menos parecido serían los pinares de Doñana y los de Teruel para RH y CO respectivamente.
El tipo de avifauna que está más asociado al de RH es la del bosque mixto de I m (Navarra), situado a 1.000 m de altitud y de wmposición estructural muy similar. Las diferencias básicas serían la presencia de hirundínidos en Roblehon- do que están ausentes en el bosque de 1x0 (estas especies no están asociadas a bosques sino a roquedos, de ahí su presencia en nuestro área de estudio) y la ausencia en RH de elementos típicamente centroeuropeos cuya área de distribu- ción cae fuera de nuestro área de estudio y sí están presentes en Izco (P. palusfris, A. trivialis, P. pyrrhula y F. montifrhgilla).
Hemos rea l ido un anhlisis de «clusten> mediante un programa BMDPIM (DEON, 1983) a partir de la matriz de similitud calculada (véase fig. 1). Con ello pretendemos agrupar las diferentes localidades según su grado de asociación siguiendo un método aglomerativo, diferenciándose agrupaciones con una

Fio. l . -hdcgam L c W i n &.k 15 40- cuya campo&& cuaiitativa se ha o o m p a r a d o u m ~ n i i a r t r a h d c ~ o . P.?ác&uhba ' delamatrizdcaimilandad ~ t m n ~ n a n ~ b a i u l . . a i a t r r d c p r s r n a n ~ ~ ~ m ~ ~ i 7 m r n ~ d s a e r : 11 R o M c d a l d o S m r a N c v a d a ~ y C u u ~ , 1984b) , 2 )Enanard tSmNevada (ZAMo~~y CAMAMO. 19&4a)*, 3) Rnares da Doflana LCosr~, l.)*, 4) Pipnreci de Brctnh (EYWXT, 1973), 5) J h q u e mixto (Navarra) (PURROY, 1975) , 6) Eaanar dt Sierra Morcaa (Hwlva) (HBRReRA, 1980 7) Jhhar montano (Lbbn) GARNIcA, 1978), 8) R o b W (Borgoña ( F R m , 1971), 9 R o b h dila-) (Fh~or, 1977). \ O) PAu nXgo m. P. p k u w (T-0 bw. 1918). 111 P. nigm (& puro ( O o W , 197% 12 Bcqm de P"" , PhyIS7ea Y Arbum WWJM, iw, 13) R O d dc Airuo -1 /M. Iw*. 19 m. 19 m. 16) «NO)
Ymi lP6? *, 17) Piaar montano (tebn) (ALVAREZ, 1931).
[ C h f i r o n o 17 pseruld conatunifk b d an ecv~ comparltbn (premtce-obsace). M- b tW.7
relación geogdf~ca bastante estrecha. La composici6n de la avifam de nuestro 8rea de estudio se agrupa con la de los bosques de las partes medias de la Sierra y los bosques de Sierra Nevada, aicimm y robledales. Este grupo se une pmmiomnte con bosques del Suroeste de Espaíia (encinares de Sierra Morena y pinares de Doñana) y enciníues de la monta&+ l eona. A pesar de la escasa similitud encontrada entre nuestro sistema y las dos
localidades del Suroeste de Eqafía, todas ellas se asocian formando un .grupo. Por otra parte, se asociarían los pinarcB montanos de León y el bosque mixto de Lzco con robledales del norte de Eqmih y robfedales y pinares franceses, f do se este conjunto como una tendencia de tipo atlántico centroeuropeo. Estos resultados demuestran las conexiones existentes entre las formaciones boscosas montanas del sur de España, que tienen una composición faunística diferenciada dentro del marco mediterráneo.
Evoluci6n mlitotiva: aspectos genetc~ies.-La composición eqedica de nuesaas comunidades varía a lo largo del periodo de estudio tan@ en el número como en la identidad de las especies preaentq. Sobre una situación media en la que es lmtante estable el número;& especies se tienen incremgtos co&&endo con la mi@6n, postgenerativa y valores mfnimos para. las condiciones

temporales más duras (mano 1984) (wmo se verá más adelante, tabla 1). Las especies ((wnstantew representan el 39,O % en RH y el 373 %en CO. Dentro de este grupo están aquellas especies propias de bosque (Páridos, S. europaea, C. brachydactyla, G. glandorius, F. coelebs, T. viscivorus) y algunas otras que ocupan mayor diversidad de hábitats (T. merula, E. rubenrla, E. cia). L. nrrvirostra y P. cristatus aparecen como especies características de pinares. El grupo de especies «constantes» tiene entre sus componentes diversos grados de sedentarismo (según se revela mediante los datos obtenidos por anillamiento en el área de estudio desde 1979). Existe toda una gradación desde especies wmo C. brachydactyla que parecen ser muy sedentarios, hasta algunos pándos (por ej., P. ater y P. caeruleus) que renuevan sus poblaciones en diferentes épocas del año, aunque no se sabe nada de la magnitud de sus desplazamientos. Las especies ((temporales)), a pesar de elevado número, tienen menos importancia cuantitativa en la wmunidad y serían grupos o «bloques» de especies que discurrirían sobre la base establecida por las «constantes».
Evolución cualitativa: reemplazamiento espec~jico y ordenacióri temporal.-Una forma adecuada de medir la intensidad de estos cambios es mediante el re- emplazamiento especifico (HERRERA, 1980a) que se cuantiticó wmo la di- ferencia a la unidad del coeficiente de similitud de Czechanovsky entre pares de meses consecutivos (¡-Si). En la figura 2 se muestra la evolución de este reemplazamiento especifiw mensual y en ella es relevante el fenómeno antenor- mente comentado de que en algunos periodos (septiembre-octubre para los dos años) los valores son destacadamente superiores al resto de los meses. El reemplazamiento especifico puede dar una idea del grado de estabilidad cualitativa del sistema y sobre esta base podemos caWcar el período otoñal o de migración postgenerativa wmo el más inestable o en el que se suceden los cambios con mayor rapidez y el de principios de verano o época de cría wmo una época muy estable. Del mismo modo habría que clasificar al wrto período invernal 12.83-01.84 para la parcela de RH.
Una v a que se ha hecho referencia a los reemplazamientos especificas
FIG. 2.-Evolución temporal del rsemplazamiento cualitativo entre psres de m- oonssutivos, (1-Sij)/mes. RH (drcuioa nllnos). (drculos vados). .
[Seasonal chmrges in !he quollitatiw replocement, or dwmilwity. betweenpairs o/ consecutive months. (1-Sijl/monrh. See rexl for defmirion. RH (filed circles), CO (open circles).]

Evolucibn temporal del número de cspcics (S). d ivmidd tspedúca (H3, domiDaocia (ID), equjlcibilided (Y) y dcaaidad de m o r m e s @) (expmda en número de individuos - IWi Ha) calculadas para las das psratas.
f.Soarwirrl chrmges Ur p x & s richness (S). dlwrsiry (H. ) , &nnbimrce ( I P ) , evomesz (J') adpprvrU1P aburdonce (DI f e pc o/ WnlduoLr.. 10 Ha-]) for RH ami CO plors) .j
RH co Mu S H- ID J- D S H- ID * J- D
06 16 2.44 38.83 101.2 18 . 2.59 , 33.76 0.89 1282 104.6 07-1 - 15 2.39 40.04 0.00 18 . 2.63 M36 0.91 138.6
19 2.78 20.57 0.95 1 S8.5 16 2.56 30.65 - 0.93 76.1 18 2.73 25.51 0.94 96.6, 18 272 25.95 -0.94 75.6
I 18 2.77 18.72 O.% 117.5 18 2.75 16.69" 0.95 i 8 2.73 22.03 0.95 113.1 2 1 3.02 24.04 11.51 '. y 16 2.58 26.58 0.93 97.9 2o 2.77 23x2 ,093 o.m 19.11 19 2.64 31.70 0.89 85.6 19. 2.78 22.43: 0.94 79.4 . 19 2.58 30.09 0.88 M3 18 - 258 34.80 0.89 723 16 2.48 33.88 0.89 71.7 . 13 2.33 36.96 0.91 38.3
a 12 11 2.14 4125 0.89 40.3 9 2.M 39.95 0.92 31.6 O1 1 O 1 .W 53.36 0.86 50.1 9 2.12 32.69 0.97 40.5 02 11 1.81 6215 0.75 48.3 .lo 2.15 3627 0.W 40.1 03 9 1 .n 65.42 0.78 25.6 8 1.82 56.30 0.00 15.1 04 19 2.57 36.40 0.67 825 8 1334 . 50.74 0.89 39,s O5 16 2.53 31.19 0.91 80.8 . 13 . 220 45.35 0.86 545 06 17 2.46 32.19 0.87 74.5 15 2.33 39.64 0.86 61.6 W 1 S 2.41 44.22 0.84 59.1 18 238 36.77 0.62 52.5 08 1 S 2,SI 25.80 0.93 51.9 15 . 2.54 28.78 56.4 09 18 258 24.'17 0.89 75.1 23 2.67 . 34.49 O.* 0.85 5 3 2 10 '19 2.63 27.89 0.89 66.1 14 238 38.62 0.90 63.7 11 1 O 2.lS 37.05 0.94 M.9 14 2-38, 38.62 -0.90 27.4

intermensuales debemos tratar de ver si éstos son similares para los mismos meses de diferentes años, y por otra parte clasificar los diferentes meses en base a su composición específica en períodos cualitativamente homogéneos. Para ello se ha utilizado una vez más el índice de Czechanovsky pata calcular las matrices de similitud cualitativa intermensual como fase previa y a partir de ellas se realizó un análisis de clasificación (fíg. 3), tomando como base las matrices de similaridad cualitativas binaria. Un resultado claro a primera vista es la unión de los meses idénticos para los dos aÍíos, lo que prueba la recurrencia interanual (dentro de la breve d a de tiempo considerada) y la asociación en muchos casos de meses consecutivos de forma ordenada y jerárquica. Esto iiftimo sería indicativo de la existencia de periodos homogéneos cualitativamente y determi- na momentos clave en los que los cambios son más bruscos. En CO los meses de junio, julio, agosto y septiembre se asocian de forma ordenada secuencia1 y jerárquica, produciéndose sucesivamente mayores diferencias, de manera que septiembre constituiría un período algo más separado del resto que puede interpretarse como una transición a la fase siguiente. A partir de octubre hasta marzo, los meses no se presentan de forma tan ordenada pero constituyen claramente un grupo aparte. Los resultados que se tienen a la vista de los dendrogramas engranan con la evolucibn del reemplazamiento especifico intermensual al coincidir el paso de septiembre a octubre como el de mayor reemplazamiento con la separación de los dos racimos más importantes dentro
ho. 3.-Dendrogamas que agrupan los di fmtes períodos de muestreo en función de la cibn cualitativa de las comunidades de RH y CO.
~ C l u r ~ t i o n of m ~ m ~ ~ i o d s h e d on rhe quaiiiotire rompositim o/ ,he puse& rommuniry.]

44 ARDEOLA 34(1), 1987
de los dendrogramas. El cambio que se tiene de invierno a la situación de «verano» (marzo a abril-junio) no es tan apreciable en los reemplazamientos calculados.
Análisis cuantitativo
La densidad total de passeriformes en cada mes se muestra en la fig. 4. Las tendencias estacionales son muy similares en RH y CO (r= 0,785, p< 0,001, g.l.=21), pero las densidades son significativamente diferentes en ambas parcelas (test de W n c o x o ~ , T=42, p<0,01). La evolución numérica muestra marcadas diferencias entre 1983 y 1984. El primer año parece haber sido excepcionalmente bueno para la reproducción y las densidades de pájaros fueron muy elevadas. El número de individuos desciende rápidamente al aproximarse y avanzar el invierno, que es cuando se censan las densidades más bajas de todo el período estudiado, a la par que se producen las condiciones ambientales más adversas. En primavera se producen incrementos que son más atenuados en CO y durante la época de cría del segundo año estudiado no se aprecia ningún aumento de la población. El aumento en la densidad que se aprecia en abril se debe a la afluencia de migrantes, la mayoría probablemente de áreas cercanas como las partes bajas de la sierra. Con el avance del verano se aprecia un nuevo descenso y con la migración postgenerativa una subida para llegar otra vez a las reducidas densidades de noviembre. La disminución de aves al avanzar el verano, después de tener una densidad elevada tras el período de cría, el aumento con la migración otoñal, también se observan en CO en 1983, pero en RH queda enmascarado por el elevado número de pájaros censados al iniciarse agosto.
J J A S O N D E F M A M J J A S O N
FIG. 4.-Evolución estaciona1 de la densidad total de passeriformes en las dos parcelas: RH (círculos rellenos) y CO (círculos vacíos).
[Seasonal variation of passerine abundance in both study plots: RH (filled circles) and CO (open circles) . J

Comparacibn de las densidades en épocas de cría e invernada .4n el fin de realizar comparaciones de densidad durante la época de cría, he decidi- do adoptar un valor medio de los dos años y parcelas para el mes de junio -(prácticamente el único mes de cría en Roblehondo): 91,O indiv./lO Ha. Nuestro área de estudio se encuadra entre los bosques maduros de Pinus sylvesrris, que presentan densidades similares (PURROY, 1974) o superiores (ALVAREZ, 1981), 1 16 indiv./Ha., 1981) y los de P. pinea (COSTA, 1984) que quedan por debajo (80 individuos/lO Ha.). Las densidades en estos pinares están muy por encima de las que se encuentran en plantaciones jovenes de P. sylvestris (entre 12 y 50 pájaros/lO Ha., SANTOS Y SUAREZ, 198 1).
Las densidades de formaciones de aestilignosa, a excepción de los robledales, muestran en general en nuestra Península valores inferiores a los del bosque de Roblehondo; entre 25 y 76 indiv./lO Ha. para hayedos y bosques de castaños del norte de Espafia (~LIRRoY, 1977b, SUAW y SANTOS, 1981, BAS et al., 1978, en GvrnA~, 1984). Y ya por encima de nuestros valores se sitúan los robledales (por ej., PURROY, 1977a, ZAMORA y CAMACHO, 1984a).
La dedsidad invernal de passeiiformes, cerca de 50 indiv./lO Ha. en RW y aproximadamente 40 en CO, está próxima a la del bosque mixto de Lzco (59,l indiv./lO Ha.) y por encima de otros resultados obtenidos para medios similares (por ej., GONZALEZ, 1975; TELLER~A, 1983; S m s y SUAREZ, 1981).
Comparación de la evolución anual de la densiahí.-Hemos recogido informa- ción sobre trece comunidades además de nuestra área de estudio (las referencias son: BLONDEL, 1969; FROCHOT, 1973; EYBERT, 1973; ALATALO, 1978, ALVA- REZ, 1981; GARNICA en prensa y las marcadas con * en el pie de la fig. l), y se han comparado porcentajes (respecto al mes en que tiene lugar el valor rnaxirno), que serán mejores indicadores de las tendencias estacionales que los valores absolutos. Se ba tomado siempre un ciclo anual, los 12 primeros meses para RH y CO.
Con toda la información se calculó la matriz de correlación entre todas las comunidades, encontrando que las mas parecidas en su evolución estacional (coeficientes de correlación más elevados) a la parcela de RH son el bosque de coniferas Finlandés (ALATALO, 1978) y el pinar de Lillo, León (ALVAREZ, 1981). La característica común más destacable de estas tres comunidades es la epoca estival representativa de las mayores densidades de pájaros y el considerable descenso en invierno, que constituyen las épocas más favorable y desfavorable, respectivamente. Las correlaciones más negativas que presenta nuestro área de estudio con otras localidades se dan con el pinar de Doñana y la garriga Francesa, que tienen la particularidad de aumentar la densidad durante la invernada.
Evolución estaciona1 & lar dferentes categorías de pasetifoms.-Se ha calculado la variación del porcentaje de abundancia numérica de especies constantes y temporales respecto al total (fig. 5). En ambas parcelas de estudio se aprecia una escasa importancia cuantitativa de las especies temporales (inferior al 10 % de media en M), indicando que la casi totalidad de la va-

ARDEOLA 34(1), 1987
01 . . . . . . . . . . . # . . . . 1 J J A S O N D E F M A M J J A S O N
SPP-
FIG. 5.-Cambios estacionales de la importancia numérica relativa de las especies «constantes» y «temporales», para RH (línea continua) y CO (línea de puntos).
[Seasonal changes in the relative abundance of ((resident)) and ((temporal)) species for RH (solid line) and CO (dashed line) .]
..-. . ---id-- -
riación cuantitativa estaciona1 es atribuible a cambios en las abundancias de las especies constantes. La ocupación de la comunidad a nivel cuantitativo por las especies constantes deja un espacio muy reducido a la introducción de las temporales, que lo hacen en baja abundancia en relación al número de especies que suponen. Este efecto es notorio sobre todo en invierno, ya que por producirse la desaparición de algunos recursos (muchos frutos y semillas) y al disminuir y hacerse escasamente renovables otros (artrópodos en general), las propias especies constantes estructuran su número reduciéndolo considerable- mente. Esto se manifiesta claramente en CO, donde no hay especies temporales de enero a abril y en RH, donde desde diciembre a abril alcanzan niveles mínimos de abundancia.
La densidad media de especies temporales es significativamente mayor en CO que en RH (t= 3,395; p<0,01, g.l.=21). Aunque en los meses más duros del invierno no hay especies temporales, en el resto del año se encuentran en mayor número que en RH; algunas especies que usan más frecuentemente espacios abiertos (E. cirlus, Ph. ochruros y C. corone) son responsables de las diferencias encontradas.
Otra forma en la que se suele dividir el espectro de especies presentes en una comunidad es según sus categorías tróficas. He considerado la evolución temporal en porcentaje de frugívoros, granívoros y un grupo formado por el resto que van desde insectívoros puros hasta polífagos (fig. 6). La categoría más importante numéricamente en la comunidad durante todo el año es la de los polífagos (en valores medios 74 % en RH, 67 % en CO), dentro de la que se encuadran la mayoría de las especies constantes. Unicamente algunas de ellas (T. viscivorus, T. merula y E. rubecula) son fmgívoros durante períodos más o menos prolongados. Esta última categoría representa el 13,8 % en RH y el 18,4 % en CO como situación media.
temporales
La abundancia de frugívoros no está relacionada de manera simple con la disponibilidad de fmtos en el área de Roblehondo (sólo abundantes de agosto a diciembre, observ. pers.), al igual que se ha puesto de manifiesto en otras

O J J A S O N D E F M A M J J A S D N
RG. 6.-Importancia estaciona1 relativa de las diferentes cateponas tr6ficas que se msideran en el texto: Polifagos (en blanw), Frugivoros (sombreado) y Gnuúvoros (punteado).
[ S e a m ~ I voririiion m rhe relairve imporiance o/ dflrerenr irophic caregorres: polyphngow (whire), f iug~vorm (black) Mdgrmtiwrour (dorred).]
ocasiones (JMOMPSON y WILLSON, 1979; BAIRD, 1980; JORDANO, 1982; HERRE- RA, 1982) e incluso puede verse en otras zonas de la Sierra de Cazorla. Aunque en RH este grupo muestra una tendencia a ser más abundante durante agosto- octubre, muestran mayor abundancia proporcional (respecto al total de espe- cies) en primavera (cuando ya no hay frutos) y en CO la homogeneidad estaciona1 es aún mayor. Los granivoros representan la fracción más reducida dentro de las categonas consideradas (medias de 11,s % en RH y 14,3 % en CO). Se incluyen aquí L. curvirosira (como consumidor de semillas de pino) y E. cia dentro de las especies constantes, y E. cirlus junto con algunos fringílidos de presencia más o menos irregular dentro de las temporales. Tampoco es sencillo aquí asociar su evolución temporal con la abundancia de semillas porque si bien tienen tendencia a presentar un número más elevado en verano, se producen irrupciones de fringílidos en otoño avanzado.
Evolución temporal de la diversidad, dominancia y equitabi1idad.-El número de especies presentes en CO varía entre 15 (marzo 1984) y 36 (septiembre 1984) (25,7+ 5,44, para el primer ciclo anual), mientras que en RH oscila entre 16 y 31, para los mismos meses (24,ll k 4.09).
El índice de diversidad (H') utilizado se ha calculado con la expresión de Shannon (por ej.: PIELOU, 1966; MAY, 1975). El cálculo de la equitabilidad se ha efwtuado mediante la expresión: J=H'/H máx., donde H máx.=log S, siendo S la riqueza específica (PIELOU, 1975). También se calculó un índice inverso de diversidad, o dominancia, mediante la expresión de MCNAUGHTON y WOLF (1970): ID=(D,+D,) 100/Dt, donde DI y D2 son las densidades de las dos especies más abundantes y Q la densidad total. Estos índices fueron obtenidos a partir de los apéndices 11 y III de densidades de especies para todo el período de estudio y los resultados se representan en la tabla l.

48 ARDEOLA 34(1), 1987
La evolución temporal de H' sigue una tendencia parecida en RH y CO (r=0,712, g.l.=21, p<0,01), con máximos en otoño (septiembre de 1983 y octubre en 1984) y un mínimo en invierno (marzo). Durante la época de cría y verano los valores se mantienen más o menos estables y relativamente altos, bastante por encima de la situación que se produce en la estación desfavorable. La dominancia (ID), como es de esperar, sigue una tendencia opuesta a la diversidad (r= -0,942, g.l.=2l, p<0,01), obteniéndose máximos en marzo y mínimos en septiembre y octubre. La situación media está en 34,9 % en RH y 33,9 % en CO. Unicamente en el período de diciembre a abril la dominancia sobrepasa el 40 %, pero la situación general es de un elevado valor de este índice, que en invierno es producido por P. ater y P. major, mientras que F. coelebs y S. europaea lo producen en verano de 1983 y F. coelebs y P. ater en 1984.
Se han obtenido para RH las curvas de importancia mensuales (WHITTA- KER, 1972; MAY, 1975), que son el resultado de ordenar las especies según su importancia numérica relativa en una escala logarítmica (fig. 7). A pesar de que los valores de dominancia no son excesivamente altos, las curvas que se han obtenido tienen mucha tendencia a la verticalidad, indicando diferencias acusadas de importancia entre las especies más abundantes y las más escasas. Las curvas más verticales corresponden al período de enero a marzo y únicamente algunos meses como agosto y septiembre de 1983 tienen pendientes poco pronunciadas; esta tendencia apenas se repite en 1984. Aun a riesgo de que la separación anterior sea subjetiva y de que en ningún caso las diferencias sean muy importantes, podemos asignar las curvas a dos grupos diferentes: diciem- bre a marzo con tendencia a una serie geométrica y resto de los meses (junio a noviembre en 1983 y abril a noviembre en 1984) donde se vería disminuida esta tendencia. Normalmente, las curvas de importancia suelen situarse entre dos tipos de distribuciones: una serie geométrica y la distribución lognormal. El
Secuencia de especies FIG. 7 .4urvas de importancia relativa de las especies de la comunidad para la parcela de RH.
[Relative importance curves for passerine species in the RH plot.]

primer caso seria típico de wmunidades controladas por un solo factor, estableciéndose una gradación hasta las wmunidades controladas por muchos factores y con recursos equilibrados (WHITTAKER, 1970; MAY, 1975, 1981). En nuestro caso vemos cómo el periodo invernal tendria tendencia a estar controlado por un único factor que seria la adversidad del medio, mientras que durante el resto del año habría una mayor heterogeneidad.
Les especies ((wnstantesw del área de estudio corresponden a los tipos Holártiw, Pal&iw y Europeo de Voous (1960), constituyendo en definitiva un tipo de fauna europea típica. Como se comentó anteriormente, la identidad de estas especies es altamente coincidente con la de los bosques mixtos de wniferas y frondosas del Pirineo Navarro que se sitúan en la misma altitud WRROY, 1975). A nivel global, ras wmunidades estudiadas se asocian con otras localidades forestales del sur de España (fig. l), pero una característica muy importante de nuestro área de estudio es la ausencia de elementos mediterráneos entre las especies «constantes», siendo muy raros cuando se consideran todas las especies y que siempre están presentes en otras comunida- des forestales del área mediterránea (HERRERA, 1980a; ZAMORA y CAMACHO, 1984 a y b). También es destacable la presencia de la práctica totalidad de los elementos europeos característicos de este tipo de habitat por lo que no es observable el empobrecimiento de esta fauna comentado por HERRERA (1980a) como consecuencia de la progresiva mediterraneización del clima. La diferencia- ción en la composición fadstica del sistema estudiado puede apreciarse cuando se compara con las partes más bajas de la Sierra: en el piso inmediatamente inferior (1.100 m) hacen su aparición varios elementos mediterráneos (p. ej., Sylvia cantillans, Sylvia melanocephnla) (HERRERA, 1984).
En cuanto a la evolución temporal de las densidades de Passeriformes, G U ~ N (1984), al comparar los perfiles anuales de ocho wmunidades, llega a la conclusión de que éstos vienen determinados por la influencia de factores como el perfrl anual de temperaturas y los sistemas de desplazamientos wmunitanos, bien a escala local o a una escala más amplia. Nuestras correlaciones pueden encuadrarse muy bien dentro de esta explicación porque en los climas nórdicos y de montaña las densidades mayores se tienen en verano (encinares y robledales de Sierra Nevada, Sierra de Ancares, Montaña Leonesa y Finlandia) y las duras temperaturas invernales hacen disminuir mucho la densidad de los pájaros. Los inviernos suaves (p. ej., pinares de Doñana y gamga Francesa), por el contrario, permitirían una buena invernada de passenfonnes provenientes de otros medios. Algunos medios de montaña como el bosque mixto de Izco alcanzan su máximo anual en otoño, wmo wnsecuen- cia de la afluencia otoñal que casi con toda probabilidad se enmarca dentro de los grandes movimientos migratorios paleártico-africanos que deben atravesar

los PSimos. Un fai6mmo similar debe pducirse en la Borgoña Franasa y en el sur de No-.
El bosque de Roblehondo tiene un porcentaje de especKJ «COnstantnu, muy superior al encontrado en otras comunidrrdea de nuestras latitudes. En la garriga francca es iud i i por BLONDBL (1969), la abundanQa de especies temporales es más importante que la de constantes durante la invernada. En bosques del sur de España los valores medios de la abundancia de temporales también sod mayores que en el área de estudio, tanto en encinares (HERRERA, 1980a; ZAMORA y CAMAao, 19848) como en pinares (CosrA, 1984) y en algunos casos se asocian los fuertes contingentes de temporales con las épocas de las grandes migraciones (H~RERA, 1980a; -A, 1984). La abundancia de especies temporales también se hace muy patente en abctales boreales dunuite la época de cría (HOO~~AD, 1967). Situaciones más similares a la nuestra deben arperarsc en zonas de montaña y de hecho asi micede en los robledates de Sierra Nevada. No es así sin embargo en enoinares de la misma Sierra Nevada (ZAMoRA y Ch%%QIO, 1984a) ni en la Sierra de los AM.ares (Gvrrvw, 1984). donde encinas y acebos, respectivamente, parecen ejerar un efecto de tampbn durante las situaciones adversas, pudiendo aim mantenerse las especies tempo- rales también durante el invierno.
En cuanto a b diversidad espMf~ca, loa valores que tenemos en RH durante la época de reproducción en los dos años (544 y 2,46) son del todo coincidentes con los que expone TRAMER (1969) para los bosques de coníferas norteamerica- nos, al igual que los valores de equitabüidad (0,88 y 0,87 para los dos añoa considerados) y de riqueza especifica (23 y 26 iqmtivamente). Para CO se tienen resultados similares. La diversidad invernal tiene valores comprendidos entre 1,7 y 2,1 según los meses y parcelas, que son muy pancidos a los que describen A u m y TOMOFF (1978) para diversas localidades y tipos de vegetación de Atizona: Quereus-Jmipm (1,89), Pinus-Quercus (2,02), Pinus (1.65) y Abies-Pk (1,83); entre los que únicamente la comunidad de Pinm Q m c u s tiene valores claramente sup%iorai. Los mismos autores caldaron los valores estacionales de estos p r h e t r o s en Nevada para comunidades de Pinus- Jmiprmr, PUrus-Cerc~~nrrpus y Pinus-Abies encontrando unas pautas para H' muy pancidas en las tres: máximo en verano u otoiío y mínimo en invierno.
Los valores de diversidad a cualquier nivel trófico pueden depender de la diversidad a niveles inferiores y a un balance entre la tendencia a la generaliza- ciónola- . . 'ón dentro de are nivel (PAINE, 1971; A u W y TOMOFF, 1978). En aianto a la equitabiüdad, J', se ha discutido bastante su validez como indice desnipíivo @e DE~mi?Drcri.s, 1973; H ~ L , 1973) y en algunos casos se utilizan sus valores modiIicados ( H ~ L , 1973; ALATALO y AJATALO, 1980). TRAMER (1969) ha sugerido que las poblaciones no nidificantes tendrían valores más bajos al comparar su abundancia relativa debido al efecto amortiguador que ejemrían los ienitorios en Cpoca de d a . Pero A u ~ y TOMOFF (1978) aportan muchos datos sobre la constnncia de ese índice en todas las estaciones del año y lo mismo demuestran R ~ E R R Y , et al. (1979). y ALATALO y

COMUNIDADES DE PASSERlFORMES W CA20üJ.A 51
ALATALO (1980). En el área de estudio, la equitabilidad calculada resulta ser muy poco variable a lo largo del periodo observado y se mantiene en valores muy altos, quizás con la única excepción de dos meses de invierno en RH. Los valores medios, 0,88 para R H y 0,90 para CO entran dentro del rango de valores que se dan para varios tipos de bosques norteamericanos en TRAMER (1969) y la homogeneidad estacional de J' está de acuerdo con las referencias citadas anteriormente.
Los recursos tr0ficos que ofrece el medio deben ir reemplazándose en sucesivos «bloqu~» de diferente amplitud y período en la escala temporal, como consecuencia de la estacionalidad del medio. Por su parte, el grupo de pájaros que forman las especies constantes constituyen un gran «bloque» de importancia numérica estacional variable, sobre el que transcurren pequeños «paquetes» de especies temporales que alteran la estructura de la comunidad, pero presumiblemente en muy pequeño grado debido a su escasa importancia cuantitativa. Las especies temporales se insertan en la comunidad en cualquier época del año, aunque su importancia cualitativa y cuantitativa es mayor en otoño (septiembre-octubre).
Como se ha demostrado en numerosas ocasiones, aunque no faltan excepcio- nes @. ej., WIENS y ROTENBERRY, 1981), la diversidad especifica de las aves está directamente relacionada con la complejidad estructural del medio, particular- mente con su estratificación vertical (MACARTHUR y MACARTHUR, 1961; MACARTHWR, 19&1; RECHER, 1969; KARR y R o n , 1971; CAROTHERS, el al., 1974; TOMOFF, 1974). Nuestra área de estudio presenta en época de cría valores de H' muy similares a otros medios de vegetación parecida, pero su variación estacional probablemente no puede ser explicada en su totalidad en base a esta similitud de la vegetación. En la bajada invernal de los valores de H' también debe influir en alto grado la disminución de la diversificación y abundancia de los recursos. Este último factor, además de aiectar a las densidades de pájaros puede producir reestmcturaciones en el nicho que ocupa cada especie y en general, provoca la disminución de la diverstdad. La llegada del verano, por el contrario, trae consigo un aumento de diversificación de recursos que provoca un incremento del numero de posibilidades efectivas de explotación que pueden ser desempeñadas por las especies. Y de hecho así sucede, tanto para las especies constantes que podrían ampliar sus nichos como por las temporales afluentes en esa época. El aumento en el número de especies que ocurre en verano con respecto al periodo precedente es un indicativo de este «llenado» de las sucesivas posibilidades.
La mejor situación en cuanto a recursos, dentro de esta época favorable, pone un limite a la densidad y diversidad de aves que puede mantener un medio dado. El aumento de recursos que tiene lugar entonces puede interpretarse gráficamente según el modelo de HERRERA (1981), aunque en este caso las poblaciones residentes aumentan de tamaño cíclicamente. El número de pájaros que pueden usar el medio es mayor durante el tiempo que duran los recursos temporales. Aunque en este modelo la nueva situación de abundancia se

52 ARDEOLA 34(1), 1987
ocuparía por especies temporales; en nuestro caso, la mayor parte es utilizada también por las especies constantes que incrementan su número. El dominio de la situación temporal precedente y su importancia numérica hace que ocupen el habitat dejando muy poco margen a la inserción de las especies temporales, que como se ha demostrado no deben tener mucha incidencia en la comunidad.
Pero esto no significa que en cada estación deba desembocarse en situaciones similares y completamente prefijadas. Si consideramos las comunidades de aves como sistemas de evolución constante (HERRERA, 1981) no podemos ver a la comunidad en un momento dado como un patrón fijo que ha de repetirse cíclicamente. Podemos determinar algunas situaciones que sí pueden repetirse, pero el grado de predecibilidad va a ser reducido desde el momento en que la comunidad se encuentra inmersa en un contexto temporal y cada etapa del ciclo va a ser consecuencia de las situaciones precedentes por las que ha atravesado. Esto puede apreciarse muy bien en el área de estudio en la primavera-verano de 1984; las nevadas y fuertes lluvias que se produjeron hasta finales de mayo provocaron un retraso general en la fenología con respecto al año anterior y afectaron en gran medida a la reproducción de los passeriformes. Muchos abandonaron el área después de haberse instalado para nidificar y comenzar a hacerlo. La situación meteorológica no permitió establecerse un número de parejas suIiciente para producir un «llenado» del habitat (de acuerdo con su productividad estival) y además la cría fue deficiente. Como consecuencia de ello, en primavera-verano de 1984 tenemos densidades de passeriformes que suponen la mitad de las que se tenían en el año precedente en las mismas fechas. Es de esperar por tanto, que en verano de 1984 el medio tendría capacidad para soportar muchos más pájaros de los que realmente tenía y esos «espacios» o «huecos» libres no han sido utilizados.
AGRADECIMIENTOS
Francisco Salas, del antiguo I.C.O.N.A., me proporcionó alojamiento en la Sierra de Cazorla. Carlos Herrera y Juan Amat me ayudaron con diversos aspectos de este manuscrito. Durante el período de realización de este trabajo fui becario del Plan de Formación del Personal Investigador.
RESUMEN Se ha analizado la evolución temporal de una comunidad de Passeriformes en bosques de Pinus
nigra de altitudes medias de las Sierra de Cazorla, empleando transectos lineales como método de muestreo.
La composición de la avifauna es similar a la de bosques mixtos de latitudes templadas y los reemplazamientos específicos más acusados se producen en otoño. La evolución cuantitativa de la comunidad tiene valores máximos en verano (después de la época de cría) y mínimos en invierno (coincidiendo con las condiciones ambientales más desfavorables).
Las especies «constantes» en la comunidad acaparan la mayor parte de la composición cuantitativa durante todo el período de estudio.
El índice de diversidad presenta máximos en otoño y valores mínimos en invierno, mientras que la equitabilidad presenta escasas variaciones estacionales.
PALABRAS CLAVE: Passeriformes, ciclo anual, comunidad, bosque mixto, Sierra de Cazorla.

SUMMARY Passerine communiries in rhe mLxed woodlondr oj Cazorla Mouniains, SE Spain.
Seasonal variations in a &ne communily w m studied in the mixed woodlands (pine-oalrs) of Sierra of Cazorla, al middle altitudes. Line uansea was vsed as sampling methcd.
The mmposition of this wmmunity is similar to that of temperate, mixed woodland. The qualitative variation &es a maximum in autumn. Numefical fluctuations showed that thm is a peak in summer ( d e r b rd ing season), the lowest numbers being recorded in winter (when the habitats conditions w m most unfavourable).
Resident species are the most imponant during the whok study pcricd. The diversity index presents the highest value in autumn and the lowest in winter. while evenness shows slight annual variation.
KEY WORDS: Passmformes, annual cycle. wmmuniiy, mixed fomt, Sierra of Cazorla.
BIBLlOGRAFlA ALATALO, R. V. (1978). Bird communities energetics in a boreal comferow fomt. Holarfic. Eeol .. 1:
?67.?76 -. . - . y AUTAW, R. H. (1980). Seasonal vadation in evenness in foresi bird wmmunities. Ornis
Scandi~vica, 1 1: 217-222. ALVAREZ, A. (1981). Estudio estnictural de la comunidad odtica del Pinar de LiUo W n ) . Estudio
del ciclo anual. Tesina de Licenciatura. Univ. de León. A v m ~ , O. T. y TOMOF'F, C. S. (1978). Relative abundante in b id populations. Am. Nat., 112: 695-
699. BAIRD, J. W. (1980). The seleaion and u% of fmit by buds in an eastem fomt. W i h n Bull., 92: 63-
7'4 BLON&L, J. (1969). Sedentsnté el migration des oiseaux dans une gmigue mediterranécnne. Teme
er Vie. 23: 269-314. CAROTHERS, S. W., JOHNSON, R. R., y AIM~MSON, S. W. (1974). Population stniclure and sonal
organuation of southwstcm npwian birds. Am. Zd., 14: 97-108. CGSTA, L (1984). Composición de la comunidad de aves en pinares del Parque Nacional de Doíiana
(suroeste de España) Doñana Acr. Verf., 11: 151-183. DE DEBENEDIC~IS, P. A (1973). On the oomlation between artain d i w i t y indiocs. Am. Nof., 107:
295-302. DIXON. W. J. (1983). BMDP staliSIicnl software. Univ. Califomia Pnui, Bcrkeley. EMLEN, J. T. (1971). Population densities of b i d ddved fmm franssl m t s . A& 88: 323-342. EYBERT. M. C. (1973) Lc ncle anual des oisaux daas trois stades evolutifes d'une pinede de
Bretagae. Terre VE, 27: 507422. FRsnueLL, S. D. (1972). Populatwns m a seasonal mviromnent. Princcton Univ. Rss, Riaaton.
217 p. FROCHOT, B. (1971). Ecologie &S oiseaux forestiers de Bourgogne e: du Jura. Tesis Doctoral, Univ.
Dijon. GARNICA, R. (1978). Comunidades de avea ni los encinam lwneses. Nat. HW.. 13, 1.CO.N.A. - (en prni&a). &lo anual de la omitocenosis del encinar de llanura m la pmvincia de León:
priodos y gnipos. S d i o Oecol. G~NZ~LE% J. M. (1975). Descri~5Ón de la fa& de vertebrados de la zona de Mora de Rubielor
(T-1). Bol. list. ~ e n r . E&, 4: 63-78. G u ~ A N , J. (1984); Eeologin de w a comunia'ad & pauerfomres en w bosyue monfano de lu
Cordillera Cantabrico occúkntal. Tesis Doctoral. Universidad de Santiago. Heaauu. C. M. 11980a). CommsiciOn v eslrvctura de dos comunidades meditedneas de ~ .~.. .-~ -~-, -
PassrriFomes. Doñono Acta ~ k . , 7: 1-%O. - (1980b). Seasonal patterns in bird community orpanization. Local asd global approachcs. Acr. XVII Cong. Inr. Om., Berlin., pp.: 1082-1087.

54 ARDEOLA 34(1), 1987
- (1981). Organización temporal en las comunidades de aves. Doñana Acta Vert., 8: 79-102. - (1982). Seasonal variation in the quality of fruits and diffuse coevolution between plants and
avian dispersers. Ecology, 63: 773-785. - (1984). A study of avian frugivores, bird-dispersed plants and their interaction in Mediterra-
nean shrublands. Ecol. Monogr., 54: 1-23. HILL, M. 0 . (1973). Diversity and evenness: A unifying notation and its consequences. Ecology, 54:
427-432. HOGSTAD, 0 . (1967). Seasonal fluctuation in bird populations within a forest area near Oslo
(Southern Norway). Nytt. Mag. Zool., 15: 81-96. HOLMES, R. T. y STURGES, F. W. (1975). Bird community dynamics and energetics in a northern
hardwoods ecosystem. J. Anim. Ecol., 44: 175-200. JORDANO, P. (1982). Migrant birds are the main seed dispersers of blackberries in Southern Spain.
Oikos, 38: 183-193. - (1984). Relaciones entre plantas y aves frugivoras en el matorral mediterráneo del área de
Doñana. Tesis Doctoral. Universidad de Sevilla. KARR, J. R. y ROTH, R. R. (1971). Vegetation structure and avian diversity in severa1 New World
areas. Am. Nat., 105: 423-435. MACARTHUR, R. H. (1964). Environmental factors affecting bird species diversity. Am. Nat., 98: 387-
397. - y MACARTHUR, J. W. (1961). On bird species diversity. Ecology, 42: 594-598. MARGALEF, R. (1977). Ecologia. Omega, Barcelona. MAY, R. M. (1975). Patterns of species abundance and diversity. (Ed. por M. L. Cody y J. M.
Diamond). Ecology and evolution of communities, págs. 81-120. Belknap Press, Cambridge. - (1981). Patterns in multi-species communities. Theoretical ecology, principies and applications.
(Ed. por R. M. May), págs. 197-227. Blackwell Sci. mibl., Oxford. MCNAUGHTON, S. J. y WOLF, L. L. (1970). Dominante and the niche in ecological systems. Science,
167: 131-139. PAINE, R. T. (1971). A short-term experimental investigation of resource partitioning in New Zeland
rocky intertidal habitat. Ecology, 52: 1096- 1 106. PIELOU, E. C. (1966). The measurement of diversity in different types of biological collections. J.
Theor. Biol., 13: 131-144. PURROY, F. J. (1974). Contribución al estudio ornítico de los pinares pirenaicos. Ardeola, 20: 245-
261. - (1975). Evolución anual de la avifauna de un bosque mixto de coníferas y frondosas en
Navarra. Ardeola, 21: 669-697. (1977a). Avifauna nidificante e invernante del robledal atlántico de Quercus sessilzj7ora.
Ardeola, 22: 87-95. - (1977b). Avifauna nidificante en hayedos, quejigales y encinares del Pirineo. Bol. Est. Cent.
Ecol., 11: 93-103. RECHER, H. F. (1969). Bird species diversity and habitat diversity in Australia and North America.
Am. Nat., 103: 75-80. ROTENBERRY, J. T., FITZNER, R. 1. y RICKARD, W. H. (1979). Seasonal variation in avian community
structure: Differences in mechanisms regulating density. Auk, 96: 499-505. SANTOS, T. y SUÁREZ, F.(1981). The bird community of the heathlands of Palencia. The effects of
coniferous plantations. VII Int. Conf. Bird Census Work, León. SHIELDS, W. M. (1979). Avian census techniques: an analytical review. The role of insectivorous birds
in forest ecosystems. (Ed. por J. G. Dickson, R. N. Conner, R. R.,Fleet, J. A. Jackson y J. C. Kroll. págs. 23-51. Academic Press, New York.
TELLER~A, J. L. (1983). La distribución invernal de las aves en el país Vasco atlántico. Munibe, 35: 101-108.
~ O M P S O N , J. N. y WILLSON, M. F. (1979). Evolution of temperate fruitlbird interactions: phenological strategies. Evol~ion, 33: 973-982.
TOMOFF, C. S. (1974). Avian species diversity in desert shcrub. Ecology, 55: 396-403.

TRAUER, E. J. (1969). Bird s p d a divasity: componeni3 of Shannon's formula Eeology. 50: 927- 929.
VWUS, K. (1960). Atlas of wopem bir&. Nelson. WHITTAKBR, R. H. (1970). Communifit's and ecosysrents. MacMillsn. - (1972). Evohition and measuremmt of diversity. Taxon: 213-250. W m . J. A. y ROIINBERRY. J. T. (1981). Habitat assoOstions and eommunity stnidun ofbirds in
shnibstepp eowonments. Eeol. Monogr.. 51: 21-41. ZAMORA, R Y Cuiu\cwo, I.(19&la). Evolución estscronal de la comunidad de avfs de un encinas de
Sierra Nevada. Doi7Mn Acto Vert.. 11: 25-43. - y CAMALHO, 1.(1984b). Evolución estaciona1 de la comunidad de ava de un robledal de Sicm
Nevada. Doñuna Acto Vert.. 11: 129-1%. [Recibido: 11.3.86/
-DI= 1 Cobemrra media de las ePpaies a r M m y ahustivas detuladas en 20 nansectos l i n d a de 100 m
para cada parcela. a) Parala de RH. b) Pamla de CO. IMem cow of ~hrUbS and trees species in M line t r m c r s of 100 m. o) RHplor. b) CO plor.1
3
especie Cobertura medi0id.e. C.V.
PaDlsnigra .................. 36,68+17,51 47,37 QWCLU ilex.. ............... 12.0426.W 50,16 $.f"U"" .................. 1,202 2.W 170,oO
raroegw monogyrur. .......... 3,14+4,06 129.29 C. kzcmiata ................. 0,17+0,63 3 7 0 3 ñerberishipunica ............. 1,33+2.69 202.25 Junipenrs oxyctdnu ........... 4,W+ 5.57 1 13.67 3. phoenicea ................. 0,38+ 1.29 339.47 J. convnunts. ................. 0,28+0,77 275.00 Ro sosp ..................... 3,06c2,85 93,14 Rubw sp.. .................. 2,06+2,87 139,32 Lonicera spIend& ............ 0,1120,49 445.45 P m i 0 broteroi. .... : ........ 0,36+0,67 186,ll Dqhne h e o l a .............. 0,22+ 0,41 186,36 Hedera helix. ................ 0,42+ 1,42 338.09 P m sp ................... 0.06~0,17 283,33 Acer sp ..................... 0,1320,54 415.38 Pistacia ferebmtlrus. ........... 0,212 0,83 395.23 Lonicera elnwnr. ............. 0,022 0,M 303.00
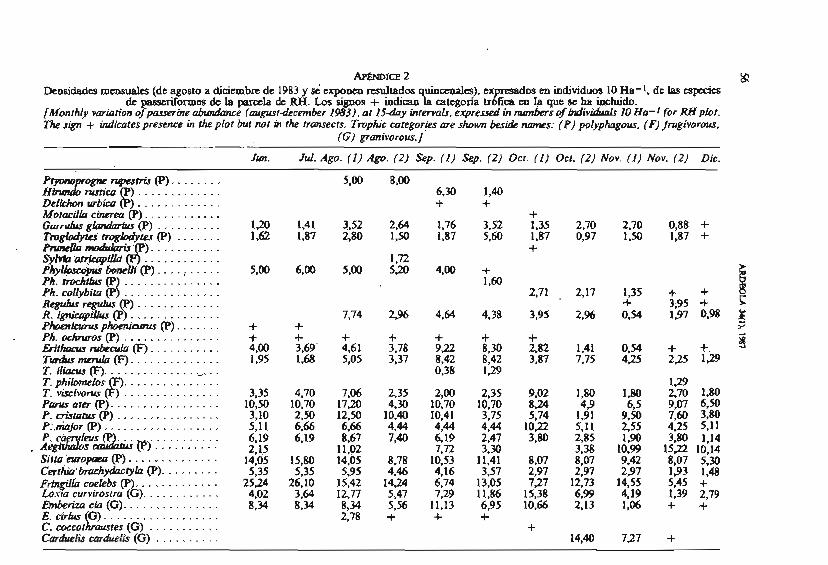
APwOlce 2 tQ Densidades mensuales (de agosto a diciembre de 1983 y sc' c q m n m resulisdos uinecioales). ex m m individuos 10 Ha- 1, de las cspecics
de +ormm ~i praia m. ím signos + indica98 aitcgoxia u P fia m I* p e r h. iahiido. (Monthlv vcwioh'on of pusserhe abumhce (aupst-december 1983). o! 15-day inrervub. expreued in mPnbcrs of b d i v & h í s IO Ha- 1 for RH pbr. m -+ indkaiespreme in hpplor bur no1 hi ik hawecfs. Tmphic caregor.ies me shown besi& mmes: (P) polyphagou~. (F) Jnigivorou.
(C) gzmiivorous. /
Jm. J d . Ago. (1 ) A@. (2) Sep. (1) Sep. (2) 00. ( 1 ) 0 c r . (2) Nov. (1)=(2) Dic.
. . . . . . . . . . . . . . . . . . . . Morailúr c w m (P)
Gamdus g* (P) .......... ~~ hagh&h3 @') .......
... . . . . . Ph. ~~ (P . . . . - - - - . . . . . . . Ph. col!ybiiui (P! . . . . . . . . . . . . . . . ~r~ ) ............. R. (Py. ............. Phoenrcvncr p h 4 e n i n m (P) ....... Ph. W o s (P) ............... Eritt~acw Nbrcvla 0. . . . . . . . . . . TimlLFmmrúr(F). . . . . . . . . . . . . . T. Iif'aclu 0. ...... . - - . . . 4 . '
.............. T. philwmIosCp T. virclwrur ) ............... Ppnrr arer (P). ................ P. aiir&aais CP) ................ . , P: . thbr (P) . . . . . . . . . . . . . . . . . . P ca Ieu~ Cp . . . . . . . . . . . A pj. .... , . . . . Sirm 'éurop~ca (P) ..............
........ Cerrhio'Wy&cryla (P). F&gzlI)r c ~ k b s (P). . . . . . . . . - . . . Lo.rro curvrrosrra (G). ........... EmbPrlln c& (O). . . . . . . . . . . . . . . E. cirbrr O). . . . . . . . . . . . . . . . . . L C. cmcoi mstes (G) ........... Cmúuelb cm&ii& (G) . . . . . . . . . .

Ene. Feb. Mar. Abril Mayo Jun. Jul. Agos. Sep. Ocr. Nov.
Pryono rogne rupesrris. .......... ~Yun$ rrrsfktl. . . . . . . . . . . . . . . . . Delichon wbica . . . . . . . . . . . . . . . .
.............. M o ~ i l l r r cinereu. ............. G d u glmrdarhrr
. . . . . . . . . . . . . . C o n w corone (P Troglodytes rro odyres . . . . . . . . . . H @ ~ O ~ L ~ pa/lu% (P) . . . . . . . . . . . . Syivia atricapilla . . . . . . . . . . . . . . . P h y l b s c o p boneIIi. . . . . . . . . . . . . Ph. ~rochilus. . . . . . . . . . . . . . . . . . Ph. collybita . . . . . . . . . . . . . . . . . . R . i g n w i l b ................. Fke& hypofeucn (P). . . . . . . . . . . Muwicapa srriato (P). . . . . . . . . . . . Oenmrrhe hispanica (P) . . . . . . . . . . Phoenirrtrus p h w n i ~ u s . . . . . . . . . . Ph. odvuros.. ................ Erithacur rubecuia. .............
................ Twdw rnerula. T. phibmelos . . . . . . . . . . . . . . . . . Twdw viscivow. . . . . . . . . . . . . . . P m arer. . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . P. crirrartu.. P. m j o r . . . . . . . . . . . . . . . . . . . . . P. Cnerule~p . . . . . . . . . . . . . . . . . . Aegirhalos Em<datiu. ............ Sitra twopaea. . . . . . . . . . . . . . . . . Cerfhia brachydoctyla . . . . . . . . . . . Fringiíía cwlebs . . . . . . . . . . . . . . .
.............. h x i n muosfra . . . . . . . . . . . . . . . . . . . Emberizac io
. . . . . . . . . . . . . . . . . . . . E . c u ~ . Coccorhrausihes coccothraustes. . . . .
. . . . . . . . . . . . . Cmduelis cardtcelk. Serima serinrc~ (P). .............

APBNDlCz 3 Deosidades en indiv./lO Ha. para la parala CO durante el período junio de 1983 a noviembre de 1984.
[Monthly varíotiun of passerine a h d z m e e x p r e d in manbers o/ individuals. 10 ha-! for CO plof; sep Appettdix 2.1
Jan. Jul. Ago. (1) Ago. (2) Sep. (1) Sep. (2) Oct. (1) Ocr. (2) Nov. (1) Nov. (2) Dic.
L.rnboreo . . . . . . . . . . . . . . . . . . . 8.18 + + 4- 1 0 3 1227 6,13 6,13 + . . . . . . P. npeshLI.. . . . . . . . . . . : + + H.rustica ..................... 2.00 f f.:e&tw .................. 0.66 G . g & d d ~ ................. 1.71 3,42 1.71 1,71 2.57 3.42 1.31 + 0.98 4- P. p y n h r r x ................ + + C.cw~~se ................... 0.85 + 0.85 + 1.71 0,85 0,85 + 0.85 + + T. t r0g Idp-s : . ............... 1 ,B7 1.43 (47 1
.................. Ph.bonelr 2,00 7.50 6,00 4,00 + Ph. c s l ~ a . ................. 2.00 1.53 2.29 3.06 -t R.igníccprUhrs ................. 5.15 3.86 321 1,93 2 3 1.60 2.50 1,60 S.*m . . . . . . . . . . . . . . . . . . . . S. iorqwta.
&o6 .................. + Ph. phmdamu. ............... 4,M 5,M + Ph: ikbumt. ................. + + + 6,W [,O1 E.Nbeailp ................... 4.68 3.90 4,68 1,56 456 4,68 4.06 5,03 3.12 + 8 T. &.. .................. 8.75 14.00 3,50 2,62 8,75 525 8,05 6,03 4,02 3,35 2,68 Y T. iHacus.. . . . . . . . . . e . . . . . . . .
T. vlsriwKlcr 466
9.67 5.64 2,90 3.22 6,45 4.19 3,70 1,85 1.R 2,47 2.47- - f" ' ..................
P. ater ...................... 10.25 1 8 3 11.39 7.97 6.83 6,83 10,48 6,11 3,14 3,49 3,49 .. P.crirtuhu ................... 5.86 8,08 3,03 4.04 6.06 6,M 3.09 5.42 3,09 532 1,54
P. mqbr'. ..................... 7.46 5.80 4,W 3,31 6,63 7,% 2,54 4.45 5,08 2,s 127 P.coenrlevp .................. + 3,OO + + 6,OO &O0 + 1,38 0,92 A.'iud2tw.. ................. + 11.22 + 7,48 + 5.61 2,86 5.73 + + 5,01 S.ewopaea ................... 4,óO. 7.50 3,75 5.62 6,37 9.37 4,31 7,W 7.18 515 1.43
. . . . . . . . . . . . . . . C.brrrdiydBcty& 2,66 4.09 2,04 1.84 504 2,66 0.94 1 , s 1 , s 0,85 F. c w h . . . . , .. . . . . . . . . . . . . . 27.72 23,87 11,93 5.96 7.95 13.92 6,lO 9.91 14.48 10,67 7.62 L . m m ................. 15.51 12,M 8,62 3.44 12,93 13,79 6,60 3,30 528 2,64 + E . & ....................... 3,33 2,49 3,33 11,M 8,33 6,66 6,38 2,SS 2.55 + + E. cfrbu.. ................... 6,66 3,33 4.16 3,33 8,33 2,49 0,83 549 127 555 C. cocurthruuszes. .............. + + + * C. .&lb: .................. + 3.05 1,52 6,10 C. JIP" .................... + 10.67 3.05

Ene. Feb. Marzo Abril Mayo Junw u Agos. Sepr. Oci. Nov.
L. arboreu . . . . . . . . . . . . . . . . . . . 7,84 + 6,13 3,M + 9 2 7 27,222 1,02 P. rupesirir. . . . . . . . . . . . . . . . . . . + + H. wtica . . . . . . . . . . . . . . . . . . . . + + L. excubitor . . . . . . . . . . . . . . . . . . 0.30 G. glandarius . . . . . . . . . . . . . . . . . + 1.71 + 2,14 1,71 1,71 + + P.pyrrhocorax . . . . . . . . . . . . . . . . + 1 ,O2 C. corone.. . . . . . . . . . . . . . . . . . . f + + 0,85 1,70 0,51 0,68 0,51
. . . . . . . . . . . . . . . . . . . C. corax.. + T. ~roglodytes. . . . . . . . . . . . . . . . . 0.35 0,71 1.86 0.57 + Ph. trochilw . . . . . . . . . . . . . . . . . . 1.32 Ph. bomlli . . . . . . . . . . . . . . . . . . . 8,OO 2,W 9,OO 8,50 4,W 0160 Ph. colíybira . . . . . . . . . . . . . . . . . . + 0.61 R. ignicapilius. . . . . . . . . . . . . . . . . 2,SO 2.50 0,96 2,41 0.80 + 0,96 3.21 1.60
M. sfriafa. . . . . . . . . . . . . . . . . . . . O. mnmrrhe . . . . . . . . . . . . . . . . . . O. hupanica . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . S. torquara Ph. phoeninuur, . . . . . . . . . . . . . . . Ph. ochruros. . . . . . . . . . . . . . . . . . E. rubecuh. . . . . . . . . . . . . . . . . . . T. menda. . . . . . . . . . . . . . . . . . . . 4,02 T. iliacrcs . . . . . . . . . . . . . . . . . . a .
T. viscivorus . . . . . . . . . . . . . . . . . . T. lorquarus . . . . . . . . . . . . . . . . . . P. ater . . . . . . . . . . . . . . . . . . . . . . P . crislaiw. . . . . . . . . . . . . . . . . . . P. mojor. . . . . . . . . . . . . . . . . . . . . P. caeruleur . . . . . . . . . . . . . . . . . . A. &rus. . . . . . . . . . . . . . . . . . . S. europaea. . . . . . . . . . . . . . . . . . . C. brach ydocryla . . . . . . . . . . . . . . . F. coelebs . . . . . . . . . . . . . . . . . . . . L. curvirosfra . . . . . . . . . . . . . . . . . E.c ln . . . . . . . . . . . . . . . . . . . . . . . E. cirlus. . . . . . . . . . . . . . . . . . . . . S. serinus. . . . . . . . . . . . . . . . . . .





























