Embed Size (px)
Citation preview

UNIVERSIDADE DE SÃO PAULO
INSTITUTO DE QUÍMICA
Construção e análise de mutantes fluorescentes
da troponina I
Deodoro Camargo Silva Gonçalves de Oliveira
Tese de doutorado submetida ao Departamento de Bioquímica do Instituto de
Química da Universidade de São Paulo.
Orientador: Prof. Dr. Fernando de Castro Reinach
São Paulo
2001

1
Para a minha irmã Carolina (em memória).

2
AGRADECIMENTOS
Ao Fernando Reinach pela orientação.
Ao Chuck Farah que me guiou nos primeiros passos.
Ao colega Fernando Fortes (DF) que muito colaborou no meu
desenvolvimento científico.
Aos muitos amigos do Instituto de Química que me acompanharam
nestes anos.
À Maria Isabel (Bezinha) pela correção ortográfica.
Ao CNPq e a FAPESP pelas bolsas concedidas.
Ao CNPq, a FAPESP e a Howard Hughes Medical Institute pelo suporte
financeiro.

3
ÍNDICE
AGRADECIMENTOS 2
ÍNDICE 3
LISTA DE FIGURAS E TABELAS 6
ABREVIATURAS 8
RESUMO 10
SUMMARY 11
INTRODUÇÃO 12
O mecanismo da contração muscular 12
A regulação da contração muscular 15
A organização do complexo troponina 20
Propriedades de ligação do cálcio 21
Proteínas recombinantes e mutação sítio-dirigida 23
As propriedades fluorescentes dos aminoácidos 24
Troponinas I recombinantes contendo 5-hidroxitriptofano 26
OBJETIVOS 29
MATERIAIS E MÉTODOS 30
Técnicas básicas de biologia molecular 30
Construção dos mutantes de TnI usando o fago M13 31
Construção dos mutantes de TnI por PCR 33

4
Indução de proteínas recombinantes 35
Indução de proteínas recombinantes contendo 5-hidroxitriptofano 35
Purificação de TnI recombinante 36
Purificação de TnC recombinante 37
Purificação de TnT recombinante 38
Reconstituição do complexo troponina 39
Eletroforese em géis de uréia-poliacrilamida (uréia / PAGE) 39
Experimentos de fluorescência 40
RESULTADOS 42
Novos mutantes de TnI 42
Regiões da TnI sensíveis à ligação de Ca2+ na TnC 46
Identificação do domínio da TnC que promove a mudança
conformacional percebida pelas sondas na TnI 55
DISCUSSÃO 60
O mecanismo induzido por cálcio 61
O efeito modulador da TnI na afinidade da TnC por Ca2+ 64
Hipótese dos sítios de maior afinidade no complexo troponina
serem os sítios regulatórios I e II no N-domínio da TnC 66
ANEXO 1 70
Caracterização dos sítios de alta e baixa afinidade no complexo
troponina por diálise de fluxo 70
Reconstituição do complexo troponina para diálise de fluxo 71

5
Diálise de fluxo 72
Resultados das diálises de fluxo 73
REFERÊNCIAS BIBLIOGRÁFICAS 77
CURRICULUM VITAE 86

6
LISTA DE FIGURAS E TABELAS
Figura 1: O sarcômero e a teoria do deslizamento dos filamentos. 13
Figura 2: Modelo da molécula de miosina e esquema do ciclo da
ATPase da actomiosina. 14
Figura 3: O filamento fino do músculo esquelético. 17
Figura 4: Estruturas da troponina C. 19
Figura 5: Modelo de uma EF-hand. 22
Figura 6: Fluorescência dos aminoácidos. 25
Figura 7: Modelo esquemático da TnC e TnI. 27
Figura 8: Esquema das mutações por PCR. 34
Figura 9: Complexos troponina reconstituídos. 43
Figura 10: Espectros de excitação. 44
Figura 11: Formação do complexo TnC-TnI em uréia / PAGE. 45
Figura 12: Complexos troponina binários reconstituídos com mutantes
fluorescentes de TnI sensíveis à ligação de Ca2+. 46
Figura 13: Complexos troponina ternários reconstituídos com mutantes
fluorescentes de TnI sensíveis à ligação de Ca2+. 48
Figura 14: Titulação com cálcio dos complexos troponina fluorescentes. 51
Figura 15: Complexo troponina ternário reconstituído com a TnCF29HW. 52
Figura 16: Titulação com cálcio dos complexos troponina ternários
contendo as TnIs fluorescentes e TnCwt, TnCD30A, TnCD66A,
TnCD106A ou TnCD142A. 56

7
Figura 17: Titulação com cálcio dos complexos troponina binários
contendo a TnIF106W e TnCwt, TnCD30A, TnCD66A, TnCD106A ou
TnCD142A. 58
Figura 18: Estrutura modelo da interação regulatória TnC / TnI. 61
Figura 19: Modelo da mudança da estrutura da interação TnC / TnI
promovido por Ca2+. 63
Figura 20: Experimentos de diálise de fluxo. 74
Tabela 1: Variações na intensidade da fluorescência dos complexos
troponina reconstituídos com as TnIs mutantes provocadas pela ligação
de Ca2+. 47
Tabela 2: Parâmetros das curvas de emissão de fluorescência das
titulações com Ca2+. 54
Tabela 3: Parâmetros das curvas de diálise de fluxo. 73

8
ABREVIATURAS
ADP - adenosina bifosfato
ATP - adenosina trifosfato
BSA - albumina de soro bovino
cDNA - DNA complementar
CM - carboxi-metil
DEAE - dietilaminoetil
DO - densidade óptica
DTT - ditiotreitol
EDTA - ácido etileno-diamino-tetraacético
EGTA - ácido etileno-glicol-bis(β-aminoetil-éter)-n,n,n',n'-tetraacético
IPTG - isopropil β-D-tiogalactopiranosídeo
kDa - quilodalton
Kd - constante de dissociação
MOPS - ácido 3-(N-morfolino) propanosulfônico
n - constante de Hill
PAGE - eletroforese em gel de poliacrilamida
PEG - polietilenoglicol
pCa - -log [Ca2+]
Pi - fosfato inorgânico

9
PCR - reação em cadeia da polimerase
SDS - dodecil sulfato de sódio
TCA - ácido tricloroacético
Tn - complexo troponina
TnC - troponina C
TnI - troponina I
TnT - troponina T
Tris - tris-(hidroximetil)-aminometano

10
RESUMO
A troponina (Tn) regula a contração do músculo estriado esquelético de
vertebrados. Ela é composta de três subunidades: troponina I (TnI), troponina C
(TnC) e troponina T (TnT). A TnI tem a função inibitória que é neutralizada pela
ligação de Ca2+ nos sítios regulatórios do N-domínio da TnC, e a TnT posiciona
o complexo no filamento fino.
Para monitorar o sinal do Ca2+ sendo transmitido da TnC para a TnI as
propriedades espectrais únicas do 5-hidroxitriptofano (5HW) foram utilizadas. O
5HW foi incorporado em mutantes pontuais de TnI com um único códon para
triptofano. Foram identificadas duas sondas espectrais intrínsecas na TnI
capazes de detectar a ligação de Ca2+ na Tn: as TnIs com 5HW nas posições
100 e 121. Complexos troponina reconstituídos com estes mutantes
fluorescentes de TnI, Tn-TnIF100HW e Tn-TnIM121HW, apresentaram
respectivamente 12 e 70 % de aumento na intensidade do espectro de emissão
devido à ligação de Ca2+ na TnC. Nos complexos binários (TnC-TnI) as TnIs com
5HW nas posições 106 e 121 também captam a ligação do Ca2+ na TnC.
A análise da fluorescência destas sondas demonstrou que: 1) as regiões
da TnI que respondem ao N-domínio regulatório da TnC ocupado com Ca2+ são
a região inibitória da TnI, resíduos 96 até 116, e a região vizinha que inclui a
posição 121 da TnI; 2) mutações pontuais e a incorporação de 5HW na TnI
podem afetar tanto a afinidade como a cooperatividade da ligação de Ca2+ na
TnC, confirmando o papel da TnI em modular a afinidade da TnC por Ca2+; 3) as
constantes de dissociação de Ca2+ surpreendentemente altas, Kd ~ 10-8 M,
calculadas a partir dos sinais das sondas na região inibitória da TnI, sugerem a
possibilidade de que os sítios do domínio N-terminal da TnC sejam os sítios de
ligação de Ca2+ de maior afinidade no complexo troponina.

11
SUMMARY
Vertebrate striated muscle contraction is regulated by troponin (Tn). Tn is
composed of three subunits: troponin I (TnI), troponin C (TnC) and troponin T
(TnT). TnI has an inhibitory role that is neutralized by calcium binding to the
regulatory sites in the N-domain of TnC, and TnT positions the troponin complex
on the thin filament.
In order to follow the Ca2+ induced conformational change that is
transmitted from TnC to TnI, the unique spectral properties of
5-hydroxytryptophan (5HW) incorporated as point-mutants of TnI were used. It
was possible to identify two new TnI intrinsic spectral probes sensitive to Ca2+
binding to Tn: TnI with single 5HW at positions 100 and 121. Trimeric troponin
complexes reconstituted with two fluorescent mutants of TnI, Tn-TnIF100HW and
Tn-TnIM121HW, showed respectively 12 and 70 % increase in the emission
spectra when Ca2+ bound to TnC. In the binary complexes (TnC-TnI) two TnIs
with 5HW at positions 106 and 121 were also sensitive to Ca2+ binding to TnC.
Fluorescence analysis of these probes showed: 1) the regions in TnI that
respond to Ca2+ binding to the regulatory N-domain of TnC are the inhibitory
region of TnI (residues 96 to 116), and a neighbor region that includes position
121; 2) point mutations and incorporation of 5HW in TnI can affect both the
affinity and the cooperativity of Ca2+ binding to TnC, confirming the role of TnI as
a modulator of the Ca2+ affinity of TnC; 3) the high dissociation constant for sites
in the N-terminal domain of TnC (Kd ~ 10-8 M), derived from data using probes in
the inhibitory region of TnI suggested the possibility that these sites are the high
affinity Ca2+ binding sites in the troponin complex.

12
INTRODUÇÃO
Esta tese descreve a construção e análise de mutantes fluorescentes de
troponina I (TnI). A TnI é uma das três subunidades polipeptídicas que compõe o
complexo troponina. O complexo troponina está envolvido na regulação da
contração muscular. Nesta introdução apresentamos uma breve revisão sobre o
músculo estriado de vertebrados e sobre as proteínas envolvidas. Também
incluímos os fundamentos da estratégia empregada para investigar o papel
regulatório da TnI, a mutação sítio-dirigida em proteínas recombinantes e as
propriedades fluorescentes dos aminoácidos.
O mecanismo da contração muscular
Existem dois tipos de músculos em vertebrados, lisos e estriados. Os
músculos lisos estão sob controle involuntário do sistema nervoso central. Eles
estão presentes em órgãos internos e vasos sanguíneos. Os músculos
estriados, cardíaco ou esquelético, apresentam uma aparência estriada em
microscópico óptico. Os músculos estriados esqueléticos estão sob controle
nervoso voluntário, conectados aos ossos e estão envolvidos em postura
corporal e movimento (para introdução na área ver Squire, 1981 e Bagshaw,
1993, para revisões recentes ver Squire e Morris, 1998 e Gordon et al, 2000).
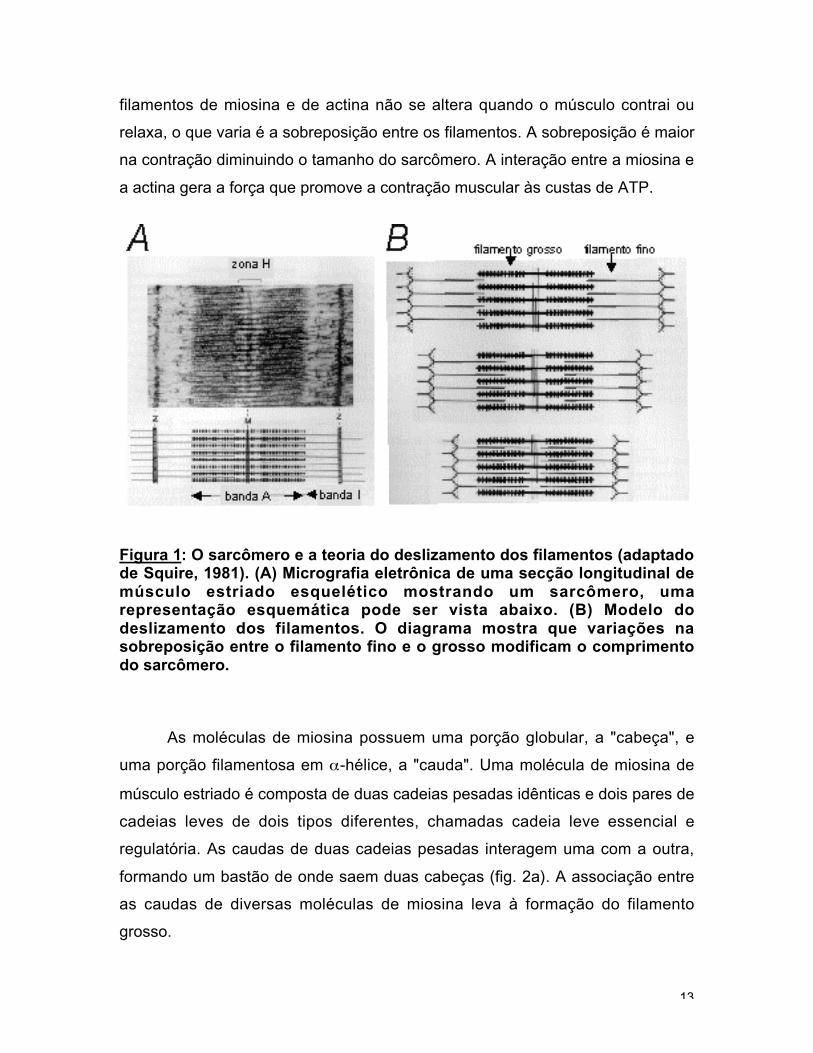
A contração do músculo estriado esquelético de vertebrados se deve a
interação entre os miofilamentos que estão organizados paralelamente no
sarcômero (fig. 1a). A aparência estriada é dada pela repetição de sarcômeros
adjacentes na célula muscular, alternando bandas claras (banda I) e escuras
(banda A). Os miofilamentos são de dois tipos, o filamento grosso e o filamento
fino, ambos são polímeros formados respectivamente de monômeros de miosina
e actina. O modelo mais aceito estabelece que a contração ocorre pelo
deslizamento entre o filamento fino e o grosso (fig. 1b). O comprimento dos
13
filamentos de miosina e de actina não se altera quando o músculo contrai ou
relaxa, o que varia é a sobreposição entre os filamentos. A sobreposição é maior
na contração diminuindo o tamanho do sarcômero. A interação entre a miosina e
a actina gera a força que promove a contração muscular às custas de ATP.
Figura 1: O sarcômero e a teoria do deslizamento dos filamentos (adaptadode Squire, 1981). (A) Micrografia eletrônica de uma secção longitudinal demúsculo estriado esquelético mostrando um sarcômero, umarepresentação esquemática pode ser vista abaixo. (B) Modelo dodeslizamento dos filamentos. O diagrama mostra que variações nasobreposição entre o filamento fino e o grosso modificam o comprimentodo sarcômero.
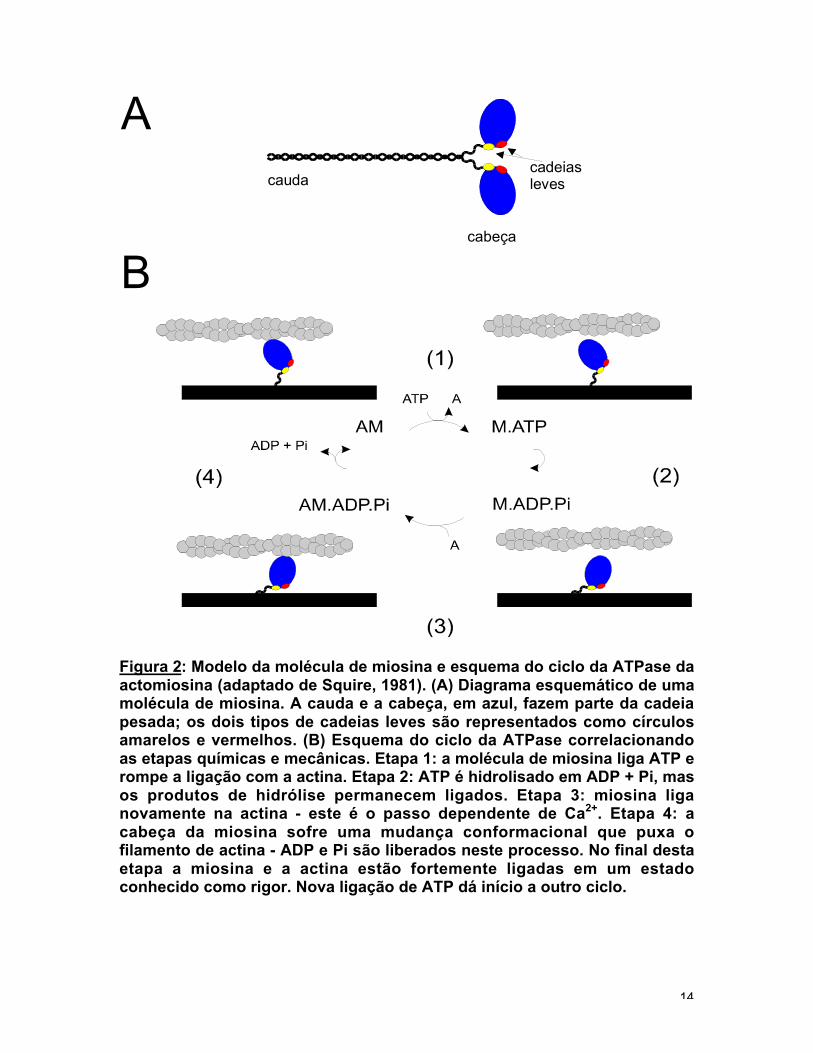
As moléculas de miosina possuem uma porção globular, a "cabeça", e
uma porção filamentosa em α-hélice, a "cauda". Uma molécula de miosina de
músculo estriado é composta de duas cadeias pesadas idênticas e dois pares de
cadeias leves de dois tipos diferentes, chamadas cadeia leve essencial e
regulatória. As caudas de duas cadeias pesadas interagem uma com a outra,
formando um bastão de onde saem duas cabeças (fig. 2a). A associação entre
as caudas de diversas moléculas de miosina leva à formação do filamento
grosso.
14
Figura 2: Modelo da molécula de miosina e esquema do ciclo da ATPase daactomiosina (adaptado de Squire, 1981). (A) Diagrama esquemático de umamolécula de miosina. A cauda e a cabeça, em azul, fazem parte da cadeiapesada; os dois tipos de cadeias leves são representados como círculosamarelos e vermelhos. (B) Esquema do ciclo da ATPase correlacionandoas etapas químicas e mecânicas. Etapa 1: a molécula de miosina liga ATP erompe a ligação com a actina. Etapa 2: ATP é hidrolisado em ADP + Pi, masos produtos de hidrólise permanecem ligados. Etapa 3: miosina liganovamente na actina - este é o passo dependente de Ca2+. Etapa 4: acabeça da miosina sofre uma mudança conformacional que puxa ofilamento de actina - ADP e Pi são liberados neste processo. No final destaetapa a miosina e a actina estão fortemente ligadas em um estadoconhecido como rigor. Nova ligação de ATP dá início a outro ciclo.
cauda
cabeça
cadeiasleves
A
B

15
A molécula de miosina pode ser clivada por digestão pela protease
papaína. A miosina é dividida em cauda e cabeça. A cabeça da miosina isolada
é chamada S1 e retém a habilidade de ligar actina, hidrolisar o ATP e mantém as
duas cadeias leves ligadas. O fragmento S1 de músculo esquelético de galinha
foi cristalizado (Rayment et al., 1993). O sítio de ligação do ATP está na cabeça
da miosina, a miosina é portanto uma ATPase. As cabeças da molécula de
miosina se projetam do filamento grosso para interagir com o filamento de actina
de uma maneira cíclica. O filamento de actina ativa a ATPase. A energia liberada
pela hidrólise do ATP em ADP e fosfato inorgânico é de alguma forma convertida
em trabalho e promove a contração. A figura 2b apresenta uma representação
esquemática do ciclo mecanoquímico da ATPase da actomiosina.
A actina é uma proteína globular presente em células musculares e não
musculares. Todas as células eucarióticas contêm actina em associação com
diversas proteínas ligantes de actina, formando o esqueleto celular. Todas as
isoformas de actina são bastante conservadas, apresentam propriedades
semelhantes e peso molecular ao redor de 42 kDa. A actina monomérica é
chamada G-actina e polimeriza formando filamentos de F-actina. A F-actina
possui uma estrutura de dupla hélice quando observada em microscópio
eletrônico. A estrutura do monômero de actina foi resolvida por cristalografia de
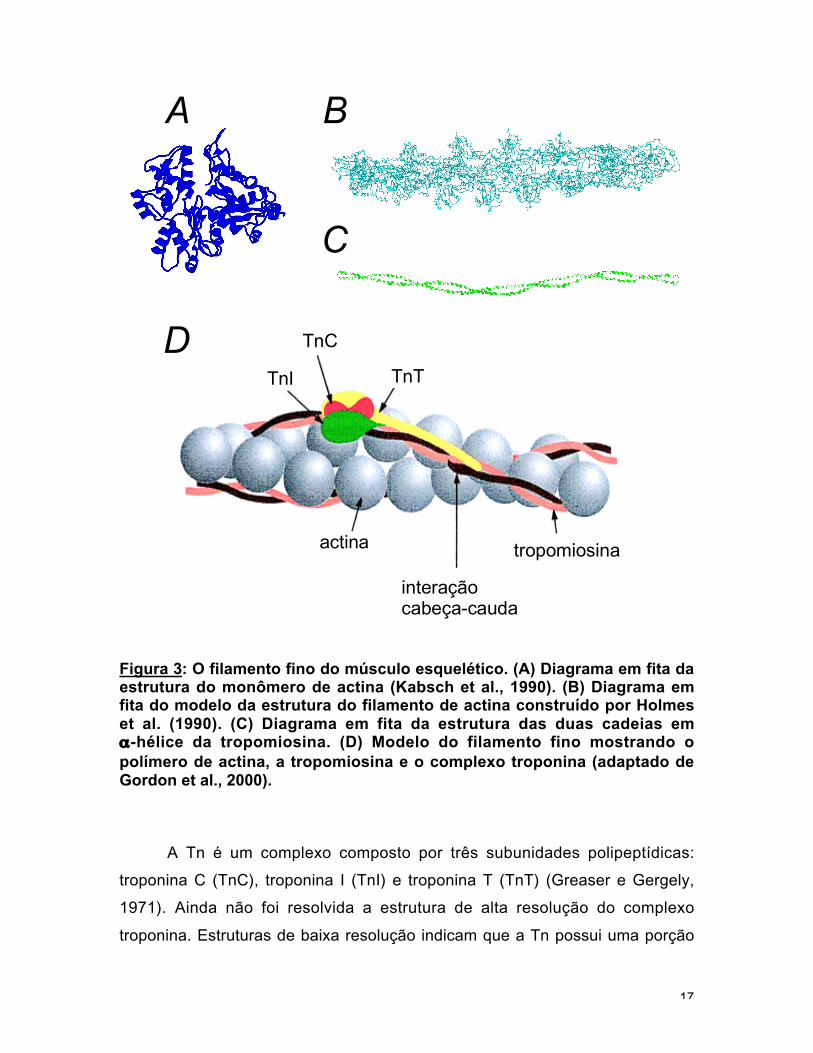
raios-X formando um complexo com a DNAse I (fig. 3a, Kabsch et al., 1990).
Assumindo que a conformação do monômero seria a mesma no filamento de
actina Holmes et al. (1990) propuseram um modelo estrutural para o filamento
(fig 3b).
A regulação da contração muscular
No músculo estriado esquelético o impulso nervoso resulta na liberação
de acetilcolina na sinapse neuromuscular, o que por sua vez provoca a
despolarização da membrana plasmática da célula muscular. A despolarização é
propagada pela superfície da célula e penetra nos túbulos T, invaginações da

16
membrana plasmática. Como consequência o Ca2+ armazenado no retículo
sarcoplasmático é liberado no interior da célula muscular. Isso provoca um
aumento na concentração intracelular de Ca2+ de 0,1 µM para 10 µM. Em baixa
concentração intracelular de Ca2+ o músculo está em repouso, o sinal nervoso
promove um aumento da concentração de Ca2+ e a ativação da contração
muscular. Bombas de cálcio na membrana do retículo sarcoplasmático retiram o
Ca2+ e o músculo relaxa. A importância dos íons Ca2+ como mensageiros
químicos intracelulares fica evidenciada no músculo (Squire, 1981 e Bagshaw,
1993).
A regulação da contração dos músculos estriados de vertebrados é
efetuada por uma proteína associada com a actina no filamento fino, a troponina
(Tn) (Ebashi et al., 1969). A Tn funciona como um sensor da concentração
intracelular de cálcio (Farah e Reinach, 1995). A Tn regula a interação entre a
actina e a miosina. A ligação de cálcio na troponina é transmitida para sete
monômeros de actina via tropomiosina (Smillie, 1979). Ou seja, a unidade do
sistema regulatório é formada por uma troponina, uma tropomiosina e sete
actinas (fig. 3d).
O modelo do bloqueamento estérico propõe que na presença de Ca2+ a
tropomiosina muda de posição desbloqueando o sítio de interação da actina com
a miosina o que conseqüentemente possibilita a contração muscular (Huxley,
1972). Neste modelo a interação entre actina e miosina é fisicamente impedida
pela tropomiosina em baixas concentrações de cálcio.
A tropomiosina é um dímero "coiled coil" (fig. 3c). Cada tropomiosina é
uma proteína alongada que se conecta com a tropomiosina seguinte, em uma
interação conhecida como "cabeça-cauda", e com isso forma uma fita contínua
ao longo do filamento de actina (fig. 3d). As suas principais funções parecem ser
a de posicionar a Tn e amplificar a informação proveniente de um complexo
troponina, permitindo regular sete monômeros de actina.
17
Figura 3: O filamento fino do músculo esquelético. (A) Diagrama em fita daestrutura do monômero de actina (Kabsch et al., 1990). (B) Diagrama emfita do modelo da estrutura do filamento de actina construído por Holmeset al. (1990). (C) Diagrama em fita da estrutura das duas cadeias emα-hélice da tropomiosina. (D) Modelo do filamento fino mostrando opolímero de actina, a tropomiosina e o complexo troponina (adaptado deGordon et al., 2000).
A Tn é um complexo composto por três subunidades polipeptídicas:
troponina C (TnC), troponina I (TnI) e troponina T (TnT) (Greaser e Gergely,
1971). Ainda não foi resolvida a estrutura de alta resolução do complexo
troponina. Estruturas de baixa resolução indicam que a Tn possui uma porção
TnC
TnTTnI
actina tropomiosina
interaçãocabeça-cauda
A B
C
D

18
globular e uma cauda (Ohtsuki, 1974). A região globular seria formada pela TnC,
TnI e região C-terminal da TnT (região conhecida como T2, Ohtsuki, 1979,
Heeley et al., 1987). A região N-terminal da TnT formaria a cauda (T1). A
troponina se posiciona no terço mais C-terminal da molécula de tropomiosina
(fig. 3d).
A troponina T é o componente que se liga a actina-tropomiosina e ancora
o complexo no filamento fino (Ebashi et al., 1972). A regulação sensível ao cálcio
assim como a ativação da ATPase não são verificadas na ausência da TnT
(Farah et al., 1994). A região T1 (aminoácidos 1 até 158) interage com a
tropomiosina e a região globular T2 (aminoácidos 159 até 258) com a
tropomiosina, a TnC e a TnI (Heeley et al., 1987). Malnic et al. (1998)
demonstraram que a porção N-terminal da TnT possui as propriedades
ativatórias da ATPase da actomiosina. Recentemente Oliveira et al., (2000)
demonstraram que um fragmento da região dos resíduos 77 até 191 continha as
propriedades ativatórias da TnT.
A troponina C tem os sítios de ligação do Ca2+, o cálcio ligando na TnC
remove a inibição promovida pela TnI para a contração do músculo. A estrutura
da TnC foi resolvida (Herzberg e James, 1985). A TnC possuí dois domínios
globulares conectados por uma α-hélice (fig 4a). Cada domínio possuí duas
EF-hands, um motivo estrutural que é um sítio responsável pela ligação de
metais divalentes. As EF-hands da TnC são numeradas de I a IV pela ordem que
aparecem na seqüência primária (fig. 7). Os sítios I e II localizados na
extremidade amino, N-domínio, ligam Ca2+ e tem a função regulatória. Na
extremidade carboxi, C-domínio, os sítios III e IV também ligam Ca2+, porém
parecem ter uma função principalmente estrutural.
A subunidade capaz de inibir a atividade ATPásica da actomiosina é a
troponina I. A TnI esquelética possui aproximadamente 180 aminoácidos e peso
molecular de 21 kDa. A TnI é capaz de ligar actina, tropomiosina, TnC e TnT.
Syska et al. (1976) identificaram um fragmento obtido pela digestão com
brometo cianogênico (resíduos 96 até 116) que é capaz de inibir a atividade

19
ATPásica da actomiosina. Esta região passou a ser conhecida como região
inibitória. Foi demonstrado posteriormente que a região entre os resíduos 104
até 115 era a seqüência mínima capaz de manter a característica inibitória da
TnI (Talbot e Hodges, 1981). A TnI forma complexo com a TnC de maneira
Ca2+ independente, mas o cálcio aumenta a afinidade dessa interação (Head e
Perry, 1974). A região N-terminal da TnI se liga estruturalmente com o
C-domínio da TnC e com a porção mais C-terminal da TnT, estabilizando o
complexo (Hitchcock-DeGregory, 1982, Farah et al., 1994, Malnic et al., 1998).
Figura 4: Estruturas da troponina C. (A) Na esquerda a representação emfita da estrutura cristalina da TnC com 2 Ca2+ ligados no C-domínio(Herzberg e James, 1985). Na direita a representação em fita da TnC com 4Ca2+ ligados (Houdusse et al., 1997). O íon Ca2+ é representado como umaesfera preta. Cada uma das hélices está identificada. Note o movimentorelativo das hélices B e C no N-domínio provocado pela ligação de Ca2+
(adaptado de Gordon et al., 2000). (B) Diagrama em fita da estrutura daTnC / TnI1-47 (adaptado de Vassylyev et al., 1998). O peptídeo TnI1-47 queforma uma única α-hélice ligada no bolso hidrofóbico do C-domínio da TnCestá em azul claro. A hélice central está dobrada, em azul, sugerindo que aTnC pode assumir uma conformação mais compacta no complexotroponina.
Apesar da TnC possuir os sítios de ligação do cálcio, ela não interage
diretamente com o filamento de actina, portanto, a informação tem que
A B

20
necessariamente ser transmitida através dos demais componentes do complexo
troponina, TnI e TnT. Ou seja, a ligação de cálcio na TnC inicia uma série de
eventos moleculares que resultam na contração do músculo esquelético. As
mudanças conformacionais provocadas pelo cálcio são transmitidas via TnI e
TnT para a actina-tropomiosina.
A organização do complexo troponina
Até agora apenas estruturas de alta resolução da TnC isolada são
conhecidas (fig 4, Herzberg e James, 1985, Gagné et al., 1995, Houdusse et al.,
1997). Como as estruturas dos complexos binários (TnC-TnI) e ternários (TnC-
TnI-TnT) não foram resolvidas, ainda não sabemos precisamente como essas
proteínas interagem. E principalmente como o sinal da ligação do Ca2+ na TnC é
transmitido via TnI e TnT possibilitando a regulação da ATPase da actomiosina.
Uma série de investigações bioquímicas tem procurado desvendar as
interações entre as subunidades do complexo troponina e deste com a
tropomiosina e a actina. Como resultado, dois tipos de interação têm sido
propostos: as estruturais ou Ca2+ insensíveis, e as regulatórias ou
Ca2+ sensíveis. Acredita-se que parte das interações seriam apenas estruturais,
e ocorrem para manter a integridade e a organização do complexo troponina e
do filamento fino. Este tipo de interação seria Ca2+ independente. O outro tipo
corresponde às interações que são formadas ou rompidas em resposta às
variações na concentração do cálcio, ou Ca2+ dependentes. São estas
transições conformacionais sofridas pelo complexo troponina que possibilitam a
regulação da contração muscular.
Utilizando mutantes de deleção Farah et al., 1994 demonstraram que a
associação entre as cadeias polipeptídicas da TnI e da TnC é antiparalela. As
interações estruturais envolvem a ligação do domínio C-terminal da TnC com a
região N-terminal da TnI (Farah et al., 1994); a ligação da região N-terminal da
TnI, resíduos 40 a 96, com a região C-terminal da TnT (Hitchock-De Gregori,

21
1982); e a ancoragem do complexo troponina no filamento fino dada pela porção
N-terminal da TnT (fragmento tríptico conhecido como T1 que tem grande
afinidade por actina-tropomiosina - Ohtsuki, 1979). Vassylyev et al. (1998)
obtiveram a estrutura da TnC ligada a um fragmento N-terminal de TnI, TnI1-47,
demonstrando a interação estrutural TnC / TnI (fig. 4b). Nesta estrutura a TnC
tem uma dobra na hélice central o que possibilitaria a interação entre os
domínios da TnC.
A ligação de Ca2+ no N-domínio dispara os rearranjos conformacionais
Ca2+ dependentes. O N-domínio com Ca2+ ligado tem uma conformação aberta
que expõe uma área hidrofóbica que é supostamente um sítio de ligação com a
TnI (fig. 4a). A abertura do N-domínio foi inicialmente proposta como um modelo
por Herzberg et al., (1986). Posteriormente estruturas da TnC com Ca2+ ligado
no N-domínio confirmaram a hipótese (Gagné et al., 1995, Houdusse et al.,
1997). Ainda não está claro qual região da TnI está diretamente envolvida na
interação com o domínio regulatório da TnC. Mas é evidente que com a ligação
de cálcio a região inibitória da TnI se afasta da actina, possibilitando que a actina
e a miosina interajam produzindo a força muscular (Syska et al., 1976). Este
sinal é espalhado para sete monômeros de actina através do deslocamento da
tropomiosina da posição "off" para a posição "on" possibilitando a interação da
cabeça da miosina com a actina.
Propriedades de ligação do cálcio
As propriedades de ligação de Ca2+ da TnC isolada são bem conhecidas
(Potter e Gergely, 1975, Leavis et al., 1978). Os sítios do C-domínio ligam Ca2+
com maior afinidade, constante de dissociação (Kd) ~ 10-7 M. Os sítios
regulatórios do N-domínio ligam Ca2+ com menor afinidade, Kd ~ 10-5 M. Os
sítios C-terminais também ligam magnésio em concentrações fisiológicas. Por
isso considera-se que estejam sempre ocupados por Mg2+ no músculo. Nos
complexos binários e ternários duas classes de sítios foram caracterizadas pelos

22
seus Kds, ~ 10-8 M e ~ 10-6 M (Potter e Gergely, 1975). Porém até agora não
existem dados que permitam associar inequivocamente cada uma destas
classes de sítio a um domínio estrutural da TnC. Claramente a TnI influencia a
afinidade da TnC por Ca2+. A maneira como isto ocorre é um fenômeno pouco
compreendido.
No N-domínio da TnC existem cinco hélices, N, A, B, C e D. No C-domínio
existem mais quatro hélices, E, F, G, H. Cada um dos quatro sítios de ligação do
Ca2+ é formado por um par destas hélices separadas por uma alça com doze
resíduos. Por exemplo, o sítio I é formado pela hélice A - alça - hélice B (fig. 4,
fig. 5 e fig. 6).
Figura 5: Modelo de uma EF-hand (adaptado de Kendrick-Jones et al.,1991). A cadeia principal da proteína é representada por uma fita. Apenasas cadeias laterais dos resíduos envolvidos na coordenação do íon estãorepresentadas. As seis posições de coordenação da alça de doze resíduosestão indicadas.
posição: 1 2 3 4 5 6 7 8 9 10 11 12X Y Z -X-Y -Z

23
O motivo estrutural EF-hand está envolvido na coordenação de metais
divalentes e é comum em várias proteínas que ligam Ca2+ (Moncrief et al., 1990).
Eles participam em mudanças conformacionais importantes e possuem grande
diversidade de afinidade e especificidade por íons divalentes. Cada EF-hand
possui uma estrutura secundária característica em forma de hélice-alça-hélice
(fig. 5). A primeira posição (posição X) de coordenação do Ca2+ é sempre um
aspartato (Asp). Tem sido demonstrado que a substituição do Asp por alanina
(Ala) reduz fortemente a capacidade da ligação do cálcio (Reinach et al., 1986,
Farah et al., 1994, Sorenson et al., 1995).
Proteínas recombinantes e mutação sítio-dirigida
Nosso laboratório clonou e padronizou o processo de purificação de todas
as proteínas regulatórias: TnC, TnI, TnT e tropomiosina (Quaggio et al., 1993,
Farah et al., 1994, Monteiro et al., 1994). O cDNA para todas estas proteínas
foram clonados no vetor de expressão pET 3a (Studier et al., 1990).
O sistema pET emprega a cepa de Escherichia coli BL21 (DE3) pLys S ou
pLys E. A BL21 é uma linhagem deficiente nas proteases lon e ompT o que
facilita o processo de purificação da proteína recombinante. O cDNA da proteína
está clonado em pET sob controle de um promotor específico para T7 RNA
polimerase (RNA polimerase do fago T7) - um promotor forte e que não ocorre
em Escherichia coli. A T7 RNA polimerase é fornecida pelo fago lambda DE3. O
gene para T7 RNA polimerase está clonado neste fago sobre controle do
promotor lacUVC. Desta maneira o sistema pET permite que a expressão da
proteína seja induzida em bactéria transformada com o vetor desejado pela
adição de IPTG. A bactéria transformada com pET pode ser selecionada pela
resistência para ampicilina / carbenicilina. Quando o produto do DNA clonado é
tóxico mesmo um nível de indução basal pode impedir a manutenção do vetor
nas células. A T7 lisozima é um inibidor natural da T7 RNA polimerase. Os
plasmídeos pLys S e pLys E possuem o gene da T7 lisozima clonado,

24
respectivamente sob controle de um promotor mais fraco e mais forte. Estes
plasmídeos são mantidos na BL21 (DE3) por seleção para resistência a
cloranfenicol e inibem a produção basal, não induzida, da proteína
recombinante.
O uso de mutagênese sítio-dirigida tem sido empregado com sucesso no
estudo de estrutura e função de proteínas. A proteína recombinante pode ser
redesenhada de várias formas: produção de fragmentos da proteína por inserção
de códons de iniciação ou terminação (ex. Farah et al., 1994); construção de
proteínas quiméricas com a união de partes de duas ou mais proteínas
diferentes (ex. da Silva et al., 1992) e substituições pontuais de aminoácidos
com base nas propriedades específicas das cadeias laterais (ex. Pearlstone et
al., 1992). Uma estratégia muito empregada é a introdução de códons para
cisteína (Cys). A Cys é muito reativa e possibilita que moléculas sejam ligadas
covalentemente. Resíduos de Cys portanto permitem que se acople alguma
sonda extrínseca fluorescente ou capaz de fazer "cross-link" com resíduos
próximos por foto-ativação, permitindo assim mapear regiões de interação
proteína-proteína (ex. Luo et al., 1999). Mutações para aminoácidos
fluorescentes são uma possibilidade muito interessante já que funcionam como
uma sonda intrínseca e permitem monitorar o comportamento da proteína.
As propriedades fluorescentes dos aminoácidos
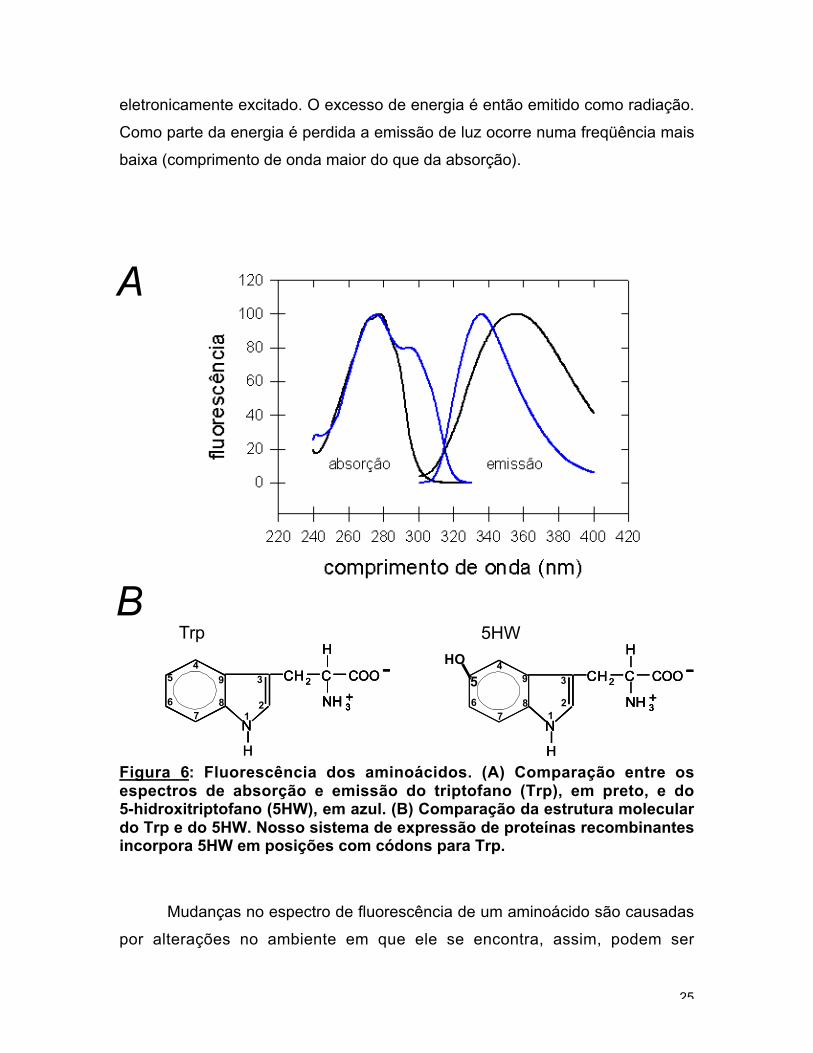
Os aminoácidos com cadeias aromáticas apresentam propriedades
fluorescentes, a fenilalanina (Phe), a tirosina (Try) e principalmente o triptofano
(Trp) (para introdução a fluorescência ver Lakowicz, 1983).
Fluorescência é um processo de decaimento radiativo no qual a molécula
descarta a energia de excitação na forma de um fóton. A absorção da luz leva a
molécula para um estado eletronicamente excitado. A molécula excitada perde
energia de forma não radiativa por colisão com as moléculas vizinhas (ou seja,
calor). Isso ocorre até atingir o nível vibracional mais baixo do estado
25
eletronicamente excitado. O excesso de energia é então emitido como radiação.
Como parte da energia é perdida a emissão de luz ocorre numa freqüência mais
baixa (comprimento de onda maior do que da absorção).
Figura 6: Fluorescência dos aminoácidos. (A) Comparação entre osespectros de absorção e emissão do triptofano (Trp), em preto, e do5-hidroxitriptofano (5HW), em azul. (B) Comparação da estrutura moleculardo Trp e do 5HW. Nosso sistema de expressão de proteínas recombinantesincorpora 5HW em posições com códons para Trp.
Mudanças no espectro de fluorescência de um aminoácido são causadas
por alterações no ambiente em que ele se encontra, assim, podem ser
B
H
N
H
CH2 C COO
3NH +
-4
8
5
67
9
12
3
H
N
H
CH2 C COO
3NH +
-
N
H
CH2 C COO
3NH 3NH +
-
Trp
N
H
CH2 C COO
3NH
H
+
-
12
3
4
867
9
HO
N
H
CH2 C COO
3NH
H
+
-
N
H
CH2 C COO
3NH 3NH
H
+
-
5HW
5
A

26
correlacionadas com modificações na conformação das proteínas. Quando uma
proteína sofre uma alteração estrutural, um aminoácido pode ter a área de
exposição ao solvente alterada, ou pode ter a interação com outros aminoácidos
modificada. Estas mudanças podem ser refletidas no espectro de fluorescência.
Vários estudos utilizam variações nas propriedades de fluorescência do
Trp ou da Tyr para monitorar alterações conformacionais na proteína. Um bom
exemplo é um mutante de TnC onde a Phe da posição 29 foi mutada para Trp,
TnCF29W (Pearlstone et al., 1992). A posição 29 é vizinha ao sítio I do
N-domínio da TnC. Este mutante possibilita acompanhar a ligação de Ca2+ no
domínio regulatório da TnC e o aumento de três vezes na intensidade da
fluorescência está correlacionado com as mudanças conformacionais
provocadas pela ligação de Ca2+.
A fluorescência de vários Trp se somam e são observadas como um
único espectro. Quando uma proteína ou complexo protéico possui diversos Trp
a análise se torna complicada. O 5-hidroxitriptofano (5HW), um análogo do Trp
pode ser incorporado em proteínas recombinantes com uma cepa de
Escherichia coli auxotrófica para Trp (Hogue et al., 1992, Farah e Reinach,
1999). A vantagem do 5HW é que o único 5HW pode ser seletivamente excitado
na presença de vários Trp, entre 310 e 320 nm. Como cada proteína
recombinante mutante pode ter um único 5HW em uma posição de interesse,
isso permite que suas propriedades fluorescentes funcionem como uma sonda
sítio específica para rearranjos conformacionais (fig. 6).
Troponinas I recombinantes contendo 5-hidroxitriptofano
Esta tese descreve a construção por mutagênese sítio-dirigida de um
grupo de mutantes fluorescentes de TnI. Utilizamos mutantes pontuais de TnI
nos quais foi introduzido um códon para Trp. Como descrito acima, as
propriedades fluorescentes do 5HW e do Trp possibilitam que estes aminoácidos
sejam utilizados como sondas intrínsecas na molécula de proteína (fig. 6).
27
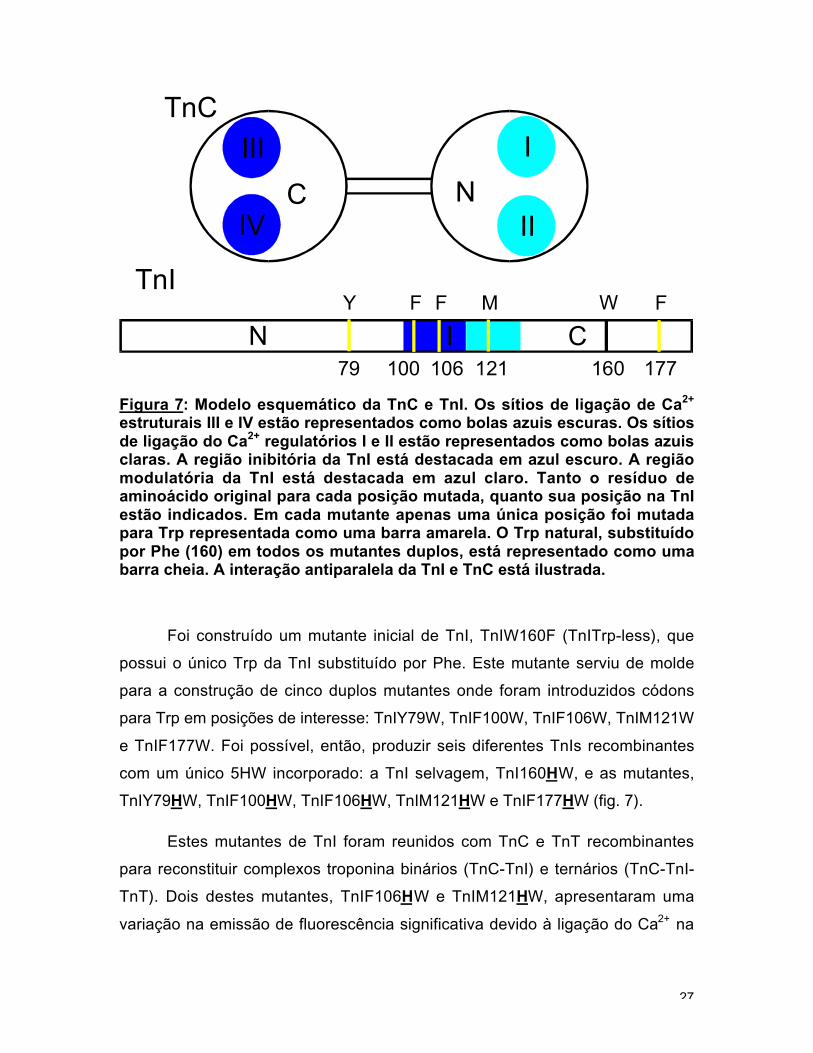
Figura 7: Modelo esquemático da TnC e TnI. Os sítios de ligação de Ca2+
estruturais III e IV estão representados como bolas azuis escuras. Os sítiosde ligação do Ca2+ regulatórios I e II estão representados como bolas azuisclaras. A região inibitória da TnI está destacada em azul escuro. A regiãomodulatória da TnI está destacada em azul claro. Tanto o resíduo deaminoácido original para cada posição mutada, quanto sua posição na TnIestão indicados. Em cada mutante apenas uma única posição foi mutadapara Trp representada como uma barra amarela. O Trp natural, substituídopor Phe (160) em todos os mutantes duplos, está representado como umabarra cheia. A interação antiparalela da TnI e TnC está ilustrada.
Foi construído um mutante inicial de TnI, TnIW160F (TnITrp-less), que
possui o único Trp da TnI substituído por Phe. Este mutante serviu de molde
para a construção de cinco duplos mutantes onde foram introduzidos códons
para Trp em posições de interesse: TnIY79W, TnIF100W, TnIF106W, TnIM121W
e TnIF177W. Foi possível, então, produzir seis diferentes TnIs recombinantes
com um único 5HW incorporado: a TnI selvagem, TnI160HW, e as mutantes,
TnIY79HW, TnIF100HW, TnIF106HW, TnIM121HW e TnIF177HW (fig. 7).
Estes mutantes de TnI foram reunidos com TnC e TnT recombinantes
para reconstituir complexos troponina binários (TnC-TnI) e ternários (TnC-TnI-
TnT). Dois destes mutantes, TnIF106HW e TnIM121HW, apresentaram uma
variação na emissão de fluorescência significativa devido à ligação do Ca2+ na
C N
TnC
TnI
106 160 17779 121100
F F FWMY
CIN
I
II
III
IV

28
TnC, nos complexos binários TnC-TnI. Quando a Tn completa foi reconstituída,
TnC-TnI-TnT, os mutantes de TnI, TnIF100HW e TnIM121HW, demonstraram
sensibilidade ao mecanismo induzido por Ca2+.
A conclusão é que as regiões da TnI que sofrem mudanças
conformacionais disparadas pelo Ca2+ são a região inibitória e uma região
adjacente que inclui a posição 121 da TnI. Esta última região é provavelmente o
sítio de ligação para o bolso hidrofóbico no N-domínio da TnC (Herzberg at al.,
1986, Vassylyev et al., 1998). Também foi investigado o papel da TnI em
modular a afinidade da TnC por Ca2+. A determinação de uma afinidade por
cálcio relacionada com os sítios I e II da TnC surpreendentemente alta, levou à
proposição de uma hipótese onde a TnI inverteria a afinidade por Ca2+ dos
domínios da TnC no complexo troponina. Neste modelo quando a TnC está
ligada na TnI o N-domínio conteria os sítios da maior afinidade pelo Ca2+ e o
C-domínio conteria os sítios de menor afinidade pelo Ca2+.

29
OBJETIVOS
Este trabalho teve como objetivos:
• Construir mutantes fluorescentes de troponina I sensíveis às interações
com os demais componentes do filamento fino.
• Utilizar estes mutantes de troponina I para melhor compreender algumas
das interações estruturais e regulatórias da troponina I. Entre elas
investigar a transmissão do sinal do Ca2+ através do complexo troponina e
como a troponina I influencia a afinidade dos domínios C-terminal e
N-terminal da troponina C por cálcio.

30
MATERIAIS E MÉTODOS
Técnicas básicas de biologia molecular
As técnicas básicas de biologia molecular utilizadas tais como preparação
e purificação de plasmídeos ou eletroforese em géis de agarose estão descritas
em Sambrook et al. (1989).
As enzimas de restrição, a T4 DNA ligase, a polinucleotídeo quinase e
seus respectivos tampões foram adquiridos da New England Biolabs e utilizados
como recomendado pelo fabricante.
As reações de PCR foram realizadas com a enzima Platinum Taq DNA
Polimerase, Gibco BRL, de acordo com as recomendações do fabricante.
O seqüenciamento de DNA foi feito usando “Sequenase Version 2.0 DNA
Sequencing Kit” e de acordo com o protocolo sugerido pelo fabricante
(Amershan Life Science). Parte do seqüenciamento de DNA utilizou o kit para
seqüenciador automático "DNA sequencing kit: BigDyetm terminator cycle
sequencing ready reaction", Applied Biosystems, como recomendado.
As bactérias foram transformadas de acordo com o método de alta
eficiência de Hanahan (1985). As características das cepas de Escherichia coli
utilizadas podem ser encontradas em Sambrook et al. (1989).
A eletroforese em gel de poliacrilamida, SDS / PAGE, foi feita de acordo
com Laemmli (1970).
A concentração das proteínas foi determinada com a técnica descrita por
Hartree (1972) usando como padrão 1 mg / mL BSA.

31
Construção dos mutantes de TnI usando o fago M13
O mutante TnIW160F (TnITrp-less) foi construído através da substituição
do único códon para Trp no cDNA de músculo esquelético rápido de galinha
(clone M13R-75, Quaggio et al., 1993) por Phe, criando o fago M13-TnIW160F.
Dois duplos mutantes foram construídos usando o fago M13-TnIW160F como
molde. Eles tiveram os códons para Phe-106, TnIF106W, ou a Phe-177,
TnIF177W, mutados para Trp (fig. 7).
As mutagêneses sítio-dirigidas mediadas por oligonucleotídeo foram
realizadas pelo método descrito por Carter et al. (1985) com as modificações
sugeridas por Kunkel et al. (1987). Como se segue:
Uma placa isolada do fago M13 contendo o cDNA da TnI foi crescido na
cepa de Escherichia coli CJ236 em 50 mL meio 2xTY com 200 µg / mL
cloranfenicol succinato acrescido de 25 µg / mL uridina por 6 horas. A cultura foi
centrifugada 1.800 xg, 4 oC, por 30 min. O fago simples fita contendo o DNA
uridinilado foi transferido, 30 mL de sobrenadante, para um tubo de centrifuga
(para o ciclo de vida do bacteriófago M13 ver Sambrook et al, 1989).
2xTY: 16 g / L triptona; 10 g / L extrato de levedura; 5 g / L NaCl; pH 7,4.
Para isolar o DNA do capsídeo protéico do vírus M13 foi adicionado
7,5 mL de NaCl / PEG 15 % (1 / 4 do volume) ao sobrenadante. A solução foi
agitada levemente e deixada 1 hora no gelo. Depois, foi centrifugada 1.800 xg,
4 oC, por 20 min. O sobrenadante foi descartado. O precipitado foi
ressuspendido em 4 mL de TE e transferido para tubo de vidro. O precipitado foi
lavado com mais 2 mL de TE. O volume reunido foi agitado vigorosamente por
30 seg; deixado no gelo por 1 hora e novamente agitado vigorosamente por mais
30 seg. A solução foi centrifugada 1.800 xg, 4 oC, por 20 min. O sobrenadante foi
transferido para um tubo de polipropileno. Para purificar o DNA, os
contaminantes foram extraídos duas vezes com igual volume de fenol
equilibrado, e uma vez com igual volume de clorofórmio. A fase aquosa foi

32
transferida para novo tubo onde se acrescentou 1 / 10 volume de 3 M acetato de
sódio pH 5,2 e 2 volumes de etanol absoluto a 0 oC para precipitar o DNA. A
mistura foi deixada no gelo por 30 min. O DNA foi recuperado por centrifugação
1.800 xg, 4 oC, por 20 min. Cuidadosamente o sobrenadante foi removido. O
DNA foi lavado com 10 mL de etanol 70 % a temperatura ambiente e
centrifugado 1.800 xg, 4 oC, por 20 mim. O tubo foi seco e o DNA foi
ressuspendido em 200 µL de TE.
NaCl / PEG 15 %: 15 % (p / v) PEG; 2,5 M NaCl.
TE: 10 mM Tris-HCl pH 8,0; 0,1 mM EDTA.
A reação de mutagênese consistiu em reunir o oligonucleotídeo
mutagênico fosforilado pela polinucleotídeo quinase com o DNA do bacteriófago
M13 simples fita uridinilado: 5 µL de DNA M13 simples fita uridinilado (0,5 pmol
ou 1 µg), 2 µL de oligonucleotídeo mutagênico fosforilado (10 pmol),
1 µL 10 x PE1, 2 µL de H20. A mistura foi aquecida por 5 min 20 oC acima da TM,
e depois foi deixada esfriar até a temperatura ambiente para o oligonucleotídeo
anelar na posição desejada. A seguir foi adicionada a solução de extensão /
ligação: 10 µL de 10 x PE3 gelado. A mistura de extensão / ligação foi incubada
por 5 min no gelo, 5 min na temperatura ambiente, 2 horas à 37 oC. O produto foi
utilizado para transformar a cepa de Escherichia coli JM101. O seqüenciamento
parcial das placas de vírus isoladas permitiu selecionar os clones mutantes. O
inserto de TnI pode ser liberado da forma replicativa do M13 (DNA dupla fita)
pela digestão com Nde I / Bam HI. O inserto contendo o cDNA da TnI mutado foi
subclonado em pET 3a (Studier et al., 1990) digerido com as mesmas enzimas e
foram obtidos assim os vetores de expressão.
PE1: 200 mM Tris-HCl pH 7,5; 100 mM MgCl2; 500 mM NaCl; 10 mM DTT.
PE2: 200 mM Tris-HCl pH 7,5; 100 mM MgCl2.

33
PE3: 1 µl 10x PE2, 1 µL 10 mM rATP, 2 µL 5 mM dNTPs, 1 µL 100 mM
DTT, 2 µL H2O, 2 µL seqüenase diluída 1:8 em “Enzime Diluition Buffer”, 1 µL T4
DNA ligase.
Os primers mutagênicos foram:
W160F 5'-TGGGTGACTTCAGGAAGAACA-3'
F106W 5'-GGGCAAGTGGAAGAGGCCA-3'
F177W 5'-GAAGAAGATGTGGGAGGCCGG-3'
Os códons modificados estão sublinhados.
Construção dos mutantes de TnI por PCR
Os duplos mutantes TnIY79W, TnIF100W e TnIM121W foram produzidos
por PCR. Eles tiveram respectivamente a Tyr-79, Phe-100 e Met-121
substituídos por Trp (fig. 7a).
A técnica de mutagênese por PCR foi descrita por Higuchi (1990). As
mutações foram feitas usando como molde o vetor pET-TnIW160F (TnITrp-less).
Os oligonucleotídeos mutagênicos foram:
Y79W 5’-GGATGAGGAAAGGTGGGACACAGAG-3’
Y79W (rev) 5'-TCACCTCTGTGTCCCACCTTTCCTC-3’
F100W 5’-GAGCCAGAAGCTGTGGGACCTGAG-3’
F100W (rev) 5'-GCCCCTCAGGTCCCACAGCTTCTG-3'
M121W 5’-GTCTGCTGATGCCTGGCTGCGTG-3’
M121W (rev) 5'-CAGGGCACGCAGCCAGGCATCAG-3’
Os códons para Trp estão sublinhados.

34
T7 promoter e T7 terminator são oligonucleotídeos complementares com
a seqüência do pET 3a e externos aos sítios de restrição Nde I e Bam HI:
T7 promoter 5’-TACGACTCACTATAGGGAGACCAC-3’
T7 terminator 5'-TAGTTATTGCTCAGCGGTGGCAGC-3’
Duas reações iniciais de PCR, uma combinando os oligonucleotídeos T7
promoter e um mutagênico (rev) e outra um mutagênico e o T7 terminator,
geram os produtos primários que são complementares na região da mutação
(fig. 8). Os produtos são reunidos e amplificados em uma segunda reação de
PCR com os primers T7 promoter e T7 terminator produzindo um fragmento de
DNA que contém a seqüência de TnI mutada. A digestão com Nde I / Bam HI
libera o cDNA completo da TnI possibilitando a subclonagem em pET 3a digerido
com as mesmas enzimas (Studier et al., 1990). Todas as mutações foram
confirmadas através de seqüenciamento do DNA (Sanger et al., 1977).
Figura 8: Esquema das mutações por PCR (Higuchi, 1989). Duas reaçõesde PCR primárias utilizando a combinação dos oligonucleotídeos T7promoter e mutagênico (rev) ou dos oligonucleotídeos mutagênico e T7terminator geram os produtos primários que são complementares naregião da mutação. Os dois produtos primários foram reunidos em umanova reação de PCR e amplificados com os oligonucleotídeos externos T7promoter e T7 terminator gerando um fragmento de DNA como acima,contendo a mutação. A digestão com Nde I / Bam HI libera o cDNAcompleto da TnI.

35
Indução de proteínas recombinantes
Todas as proteínas recombinantes (mutantes ou não) possuem o
respectivo cDNA clonado em pET-3a (Studier et al., 1990). Elas foram
produzidas em larga escala.
A expressão das proteínas foi induzida em Escherichia Coli BL21 (DE3)
Plys S. Colônias isoladas da bactéria transformada com o vetor desejado foram
selecionadas em placa TYE contendo 200 µg / L carbenicilina e 200 µg / L
cloranfenicol succinato. A cultura da placa foi utilizada para inocular 50 mL de
2xTY contendo 200 µg / L carbenicilina e 200 µg / L cloranfenicol succinato e
crescida por 3 h, 37 oC. Este pré-inóculo foi utilizado para inocular 4 L do mesmo
meio. A cultura foi crescida até DO600 ~ 0,8 quando IPTG foi adicionado para
0,4 mM. A cultura foi incubada por mais 3 horas e as bactérias coletadas por
centrifugação. A purificação continuou como descrito para TnC, TnI ou TnT
recombinante (vide abaixo).
TYE: 10 g / L triptona; 5 g / L extrato de levedura; 8 g / L NaCl; 15 g / L
agar.
Indução de proteínas recombinantes contendo 5-hidroxitriptofano
Foi desenvolvida uma linhagem para a incorporação de 5HW em
proteínas recombinantes (Quaggio, 1994, Farah e Reinach, 1999), a partir de
uma cepa de Escherichia coli auxotrófica para Trp (Drapeau et al., 1968). Esta
linhagem, CY (DE3) Plys S, permite a utilização do sistema pET (Studier et al.,
1990).
Uma colônia transformada com o vetor desejado foi selecionada em TYE
contendo 200 µg / L carbenicilina e 200 µg / L cloranfenicol succinato e foi
crescida em 50 mL de meio mínimo acrescido de 50 µg / L L-triptofano,
200 µg / L carbenicilina e 200 µg / L cloranfenicol succinato, 37 oC. Esta cultura

36
foi usada para inocular 4 L do mesmo meio. Quando a DO600 da cultura atingiu
0,8 - 1,0 as bactérias foram recuperadas por centrifugação 3.000 xg, 4 oC, 15
min e o sobrenadante foi descartado. As bactérias foram ressuspendidas no
mesmo meio sem L-triptofano. A cultura foi posta para crescer novamente e
IPTG foi adicionado para 0,4 mM. Após 15 min 100 µg / L L-5-hidroxitriptofano
foi adicionado. A cultura de bactéria foi incubada por 3 h e coletada por
centrifugação. A purificação continuou como descrito para TnC, TnI ou TnT
recombinante (vide abaixo).
meio mínimo: 17 g / L Na2HPO4.12H2O; 3 g / L KH2PO4; 0,5 g / L NaCl;
1 g / L NH4Cl; 0,5 g / L MgCl2.6H2O; 0,05 g / L CaCl2.2H2O; 4 g / L glicose;
1 mg / L vitamina B1 (tiamina).
Purificação de TnI recombinante
A TnI recombinante, TnITrp-less, TnIF106W, assim como TnIY79HW,
TnIF100HW, TnIF106HW, TnIM121HW, TnI160HW e TnIF177HW foram
purificadas como descrito (Quaggio et al. 1993 e Farah et al. 1994).
A cultura de bactéria (4 L, vide acima) foi ressuspendida em 100 mL de
Tampão - A. A cultura de bactéria foi lisada com três passagens na "French
Press", 16.000 psi. A mistura foi ultracentrifugada, 100.000 xg, 1 hora, 4 oC, e o
sobrenadante descartado. O precipitado foi ressuspendido em 80 mL de Tampão
- B e homogeneizado manualmente. A preparação ressuspendida foi aplicada
em uma coluna CM Sepharose F. F. equilibrada com Tampão - B. A amostra foi
fracionada com um gradiente de 0 até 250 mM NaCl em Tampão - B. As frações
contendo a TnI purificada foram reunidas e dialisadas contra o Tampão - C. A
TnI pura foi armazenada a -20 oC.
Tampão - A: 50 mM Tris-HCl pH 8,0; 1 mM EDTA; 14 mM
2-mercaptoetanol.

37
Tampão - B: 50 mM Tris-HCl pH 8,0; 8 M uréia; 1 mM EDTA; 14 mM
2-mercaptoetanol.
Tampão - C: 50 mM Tris-HCl pH 7,0; 1 M KCl; 1 mM EDTA; 0,01 % NaN3;
14 mM 2-mercaptoetanol.
Purificação de TnC recombinante
O vetor para expressão de TnC recombinante (Quaggio et al., 1993) e a
construção dos mutantes TnCD30A, TnCD66A, TnCD106A e TnCD142A (Farah
et al., 1994) já foram descritos. O mutante TnCF29W também foi previamente
descrito (Pearlstone et al., 1992).
Os mutantes de TnC foram preparados pelo mesmo método utilizado para
TnC recombinante (Fujimori et al., 1990).
A cultura de bactéria (4 L, vide acima) foi ressuspendida em 120 mL de
Tampão - A. A cultura de bactéria foi lisada com três passagens na "French
Press", 16.000 psi. A mistura foi ultracentrifugada 50.000 xg, 30 min, 4 oC, e o
sobrenadante descartado. O volume da solução foi aumentado para 200 mL
usando Tampão - A. Na solução foi adicionado TCA para 5 %, em um banho de
gelo com agitação constante. A solução foi deixada no gelo por 20 min. A
mistura foi centrifugada 5.000 xg, 30 min, 4 oC e o sobrenadante descartado. O
precipitado foi ressuspendido em 200 mL de Tampão - B. O TCA foi removido
por diálise contra o Tampão - B. Após a diálise o material insolúvel foi removido
por centrifugação 25.000 xg, 30 min 4 oC. Foi adicionada uréia ao sobrenadante
para 8 M. O sobrenadante foi aplicado em uma coluna DEAE Sepharose F. F.
equilibrada com Tampão - C. A amostra foi fracionada com um gradiente de
0 até 400 mM NaCl em Tampão - C. As frações contendo TnC foram agrupadas
e dialisadas contra Tampão - D. A TnC pura foi armazenada a -20 oC.
Tampão - A: 50 mM Tris-HCl pH 8,0; 25 % Sacarose; 1 mM EDTA.

38
Tampão - B: 25 mM Tris-HCl pH 8,0; 1 mM MgCl2; 1,4 mM
2-mercaptoetanol.
Tampão - C: 25 mM Tris-HCl pH 8,0; 8 M uréia; 1 mM MgCl2; 1,4 mM
2-mercaptoetanol.
Tampão - D: 50 mM Tris-HCl pH 8,0; 100 mM KCl; 1 mM EDTA; 1 mM
DTT.
Purificação de TnT recombinante
A preparação da TnT recombinante seguiu o procedimento descrito por
Malnic et al. (1998).
A cultura de bactéria (4 L, vide acima) foi ressuspendida em 100 mL de
Tampão - A. A cultura de bactéria foi lisada com três passagens na "French
Press", 16.000 psi. A mistura foi ultracentrifugada 50.000 xg, 40 min, 4 oC, e o
precipitado descartado. O sobrenadante foi aplicado em uma coluna DEAE
Sepharose F. F. equilibrada com Tampão - A. A amostra foi fracionada com um
gradiente de 0 até 600 mM NaCl em Tampão - A. As frações contendo TnT
foram agrupadas e dialisadas contra Tampão - B. O dialisado foi aplicado numa
coluna CM Sepharose F. F. equilibrada com Tampão - B. A amostra foi
fracionada com um gradiente de 0 até 600 mM NaCl em Tampão - B. As frações
contendo a TnT purificada foram reunidas e dialisadas contra o Tampão - C. A
TnT pura foi armazenada a -20 oC.
Tampão - A: 50 mM Tris-HCl pH 8,0; 6 M uréia; 1 mM EDTA; 1,4 mM
2-mercaptoetanol.
Tampão - B: 50 mM NaAc pH 5,0; 6 M uréia; 1 mM EDTA; 1,4 mM
2-mercaptoetanol.
Tampão - C: 50 mM Tris-HCl pH 8,0; 1 M KCl; 1 mM EDTA; 0,01 % NaN3;
1 mM DTT.

39
Reconstituição do complexo troponina
Os complexos binários (TnC-TnI) e ternários (TnC-TnI-TnT, fig. 9c) foram
reconstituídos como descrito em Farah et al. (1994) com algumas modificações.
Quantidades equimolares de proteína foram reunidas e dialisadas uma vez
contra os seguintes tampões:
a) 50 mM Tris-HCl pH 8,0; 4,6 M uréia; 1 M KCl; 50 µM CaCl2; 0,01 %
NaN3; 10 mM 2-mercaptoetanol.
b) 50 mM Tris-HCl pH 8,0; 2 M uréia; 1 M KCl; 50 µM CaCl2; 0,01 %
NaN3; 10 mM 2-mercaptoetanol.
c) 50 mM MOPS pH 7,0; 1 M KCl; 5 µM CaCl2; 0,01 % NaN3; 10 mM
2-mercaptoetanol.
E finalmente três vezes contra o tampão de fluorescência:
d) 50 mM MOPS pH 7,0; 100 mM KCl; 1 mM EGTA; 0,01 % NaN3; 10 mM
2-mercaptoetanol.
Após a diálise os complexos troponina foram centrifugados 10.000 xg, 15
min, 4 oC para remover as proteínas precipitadas.
Eletroforese em géis de uréia-poliacrilamida (uréia / PAGE)
A habilidade da TnC em formar complexos estáveis com cada mutante de
TnI foi visualizada por uréia / PAGE (ver Farah et al., 1994, Head e Perry, 1974):
gel: 6 M uréia; 8 % poliacrilamida; 25 mM Tris; 80 mM glicina; pH 8,6; na
presença de 0,5 mM EDTA, 10 mM MgCl2 / 1 mM EGTA ou 0,5 mM CaCl2.
O tampão de corrida foi o mesmo do gel, sem uréia e poliacrilamida.

40
As amostras foram diluídas para uma concentração final de tampão: 6 M
uréia; 25 mM Tris; 80 mM glicina; pH 8,6; mais 0,5 mM EDTA, 10 mM MgCl2 /
1 mM EGTA ou 0,5 mM CaCl2. A concentração das proteínas na alíquota (20 µL)
aplicada no gel era de 5 µM.
Experimentos de fluorescência
Os ensaios de fluorescência foram realizados em espectrofluorímetro
F-4500 Hitachi, em cubeta de quartzo Hellma 101-QS.
As amostras foram diluídas em tampão de fluorescência (50 mM MOPS
pH 7,0; 100 mM KCl; 1 mM EGTA; 0,01 % NaN3; 10 mM 2-mercaptoetanol) para
concentração de 2 µM, volume final 1,5 mL. As proteínas equilibraram por
20 min a 25 oC antes do início dos experimentos. Para os espectros de excitação
a emissão foi registrada em 340 nm. Para os espectros de emissão a excitação
foi a 315 nm para proteínas com 5HW e 295 nm para proteínas com Trp. As
fendas de emissão e excitação foram sempre de 5 nm. O branco foi subtraído e
os espectros corrigidos para a diluição. Tampões de fluorescência mais 5 mM ou
50 mM CaCl2 foram usados nos experimentos de titulação. A área total do
espectro de emissão entre 325 e 345 nm retirado para cada adição de Ca2+ foi
usada em cada ponto da curva de titulação.
A concentração de Ca2+ livre foi calculada com o programa "Sliders" de
Chris Patton (www.stanford.edu/~cpatton/maxc.html).
Os dados da emissão de fluorescência nas titulações com Ca2+ foram
ajustados para a equação:
ΔF = (ΔFmax x [Ca2+]n) / (Kdn + [Ca2+]n)

41
Onde ΔF é a variação na fluorescência; ΔFmax é a variação máxima da
fluorescência; Kd é a constante de dissociação aparente do Ca2+ e n é o
coeficiente de Hill.
Apenas para TnC-TnIF106HW (e TnC-TnIF106W) foi utilizada uma
equação que descreve uma curva bifásica:
ΔF = (ΔFmax1 x [Ca2+]n1) / (Kd1n1 + [Ca2+]n1) + + (ΔFmax2 x [Ca2+]n2) / (Kd2
n2 +
[Ca2+]n2)
Onde ΔF é a variação na fluorescência; ΔFmax1 é a variação máxima na
fluorescência; Kd1 é a constante de dissociação aparente do Ca2+ e n 1 é o
coeficiente para a primeira parte da curva; ΔFmax2 é a variação máxima na
fluorescência; Kd2 é a constante de dissociação aparente do Ca2+ e n 2 é o
coeficiente de Hill para a segunda parte da curva.

42
RESULTADOS
Novos mutantes de TnI
O objetivo inicial foi identificar mutantes de TnI que se comportassem
como sondas fluorescentes sensíveis às interações com os outros componentes
do filamento fino. Pretendeu-se usar as propriedades espectrais únicas do 5HW.
Proteínas recombinantes puderam ser produzidas em um sistema que permitiu a
incorporação do 5HW em posições que codificam para Trp (Hogue et al., 1992,
Farah e Reinach, 1999). O 5HW difere do Trp apenas devido à presença de uma
hidroxila na posição 5 do anel indólico (fig. 6b). Um único 5HW pode ser
seletivamente excitado entre 310 - 320 nm na presença de diversos Trp. A
fluorescência do 5HW depende do ambiente ao redor do resíduo. O intuito era
que o 5HW funcionasse como um sonda sítio-específica para rearranjos
conformacionais na molécula de TnI em experimentos de fluorescência.
Cinco mutantes duplos de TnI foram construídos. A TnI de músculo
esquelético rápido de galinha tem um único Trp na posição 160 (Quaggio et al.,
1993). O Trp natural foi substituído por Phe (TnITrp-less) e este mutante foi
utilizado como molde para adicionar códons para Trp em posições que se
pretendia investigar. O 5HW foi incorporado em seis posições da TnI: na tipo-
selvagem TnI160HW e nos mutantes duplos TnIY79HW, TnIF100HW,
TnIF106HW, TnIM121HW e TnIF177HW (fig. 7). Os mutantes de TnI purificados
demonstraram que a incorporação do 5HW foi bem sucedida (ver Materiais e
Métodos e fig. 10).
Todas as TnIs mutantes se comportaram como a TnIwt nas etapas de
purificação (dados não apresentados) e apresentaram a mesma mobilidade em
eletroforese em géis de poliacrilamida (fig. 9). A quantidade de TnI contendo
5HW purificada foi entre 5 a 10 mg / L de cultura. A taxa de incorporação de
43
5HW para este método foi estimada ser maior do que 90 % (Farah e Reinach,
1999).
A reconstituição dos complexos binários (TnC-TnI) e ternários (TnC-TnI-
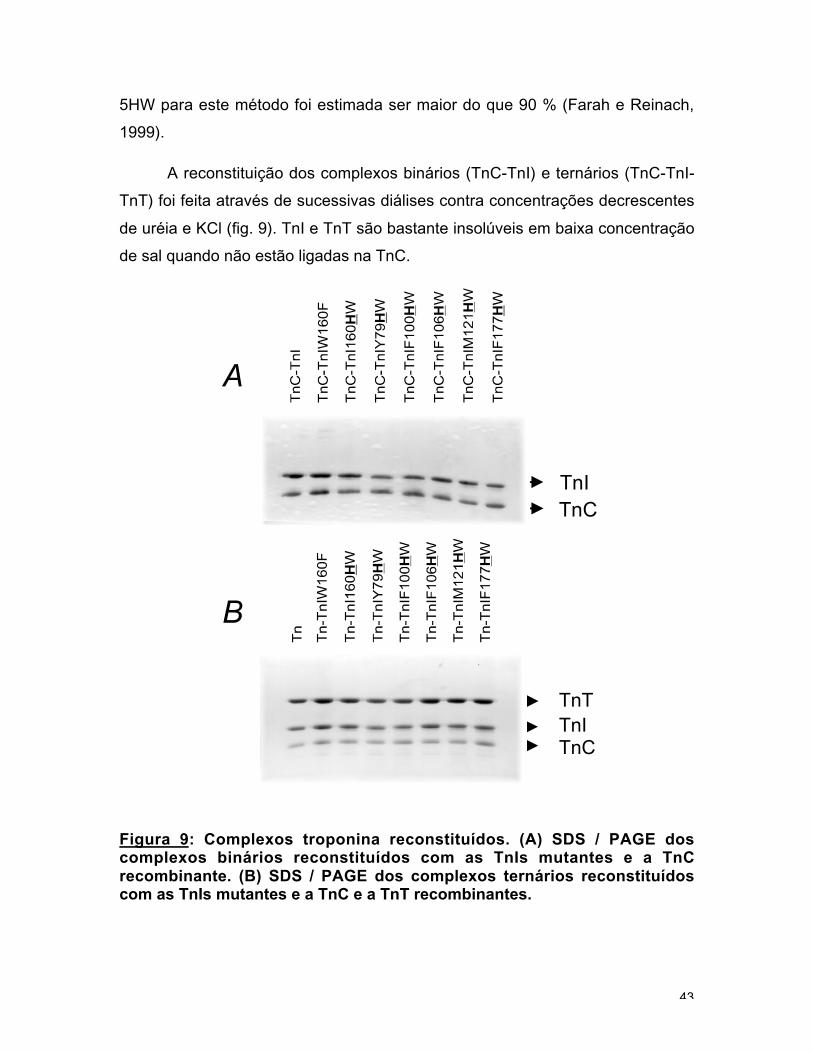
TnT) foi feita através de sucessivas diálises contra concentrações decrescentes
de uréia e KCl (fig. 9). TnI e TnT são bastante insolúveis em baixa concentração
de sal quando não estão ligadas na TnC.
Figura 9: Complexos troponina reconstituídos. (A) SDS / PAGE doscomplexos binários reconstituídos com as TnIs mutantes e a TnCrecombinante. (B) SDS / PAGE dos complexos ternários reconstituídoscom as TnIs mutantes e a TnC e a TnT recombinantes.
B
TnITnT
TnC
TnCTnI
A
44
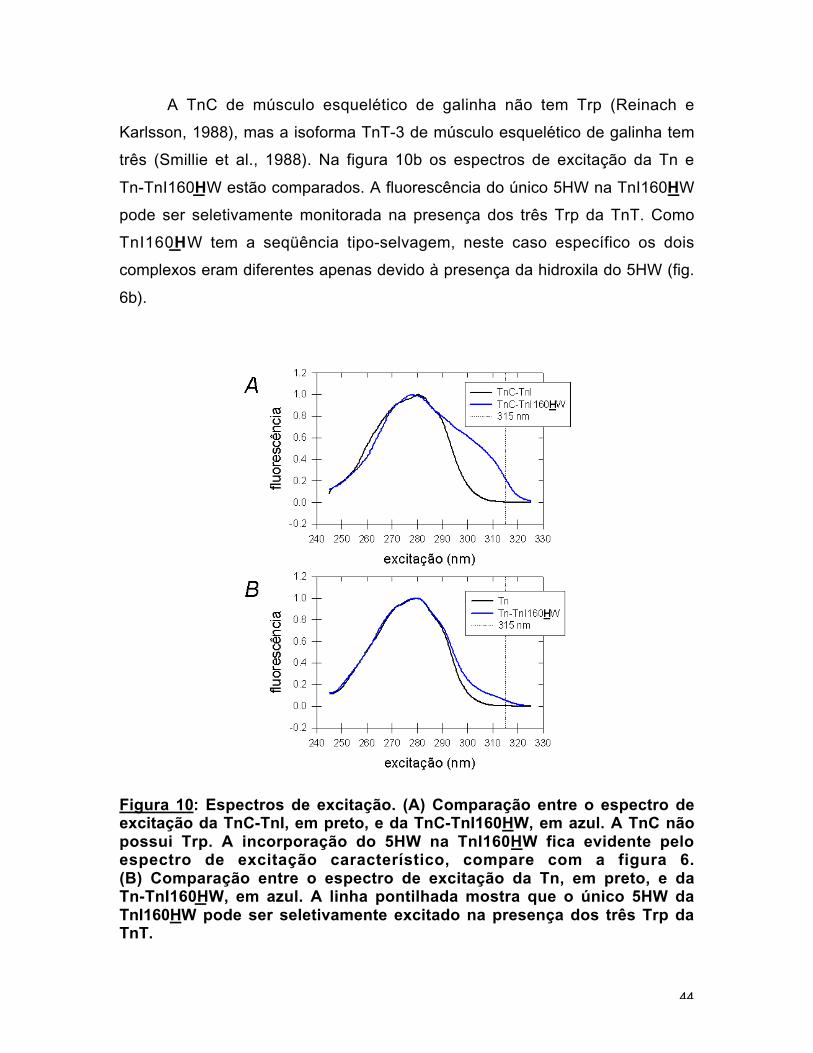
A TnC de músculo esquelético de galinha não tem Trp (Reinach e
Karlsson, 1988), mas a isoforma TnT-3 de músculo esquelético de galinha tem
três (Smillie et al., 1988). Na figura 10b os espectros de excitação da Tn e
Tn-TnI160HW estão comparados. A fluorescência do único 5HW na TnI160HW
pode ser seletivamente monitorada na presença dos três Trp da TnT. Como
TnI160HW tem a seqüência tipo-selvagem, neste caso específico os dois
complexos eram diferentes apenas devido à presença da hidroxila do 5HW (fig.
6b).
Figura 10: Espectros de excitação. (A) Comparação entre o espectro deexcitação da TnC-TnI, em preto, e da TnC-TnI160HW, em azul. A TnC nãopossui Trp. A incorporação do 5HW na TnI160HW fica evidente peloespectro de excitação característico, compare com a figura 6.(B) Comparação entre o espectro de excitação da Tn, em preto, e daTn-TnI160HW, em azul. A linha pontilhada mostra que o único 5HW daTnI160HW pode ser seletivamente excitado na presença dos três Trp daTnT.
45
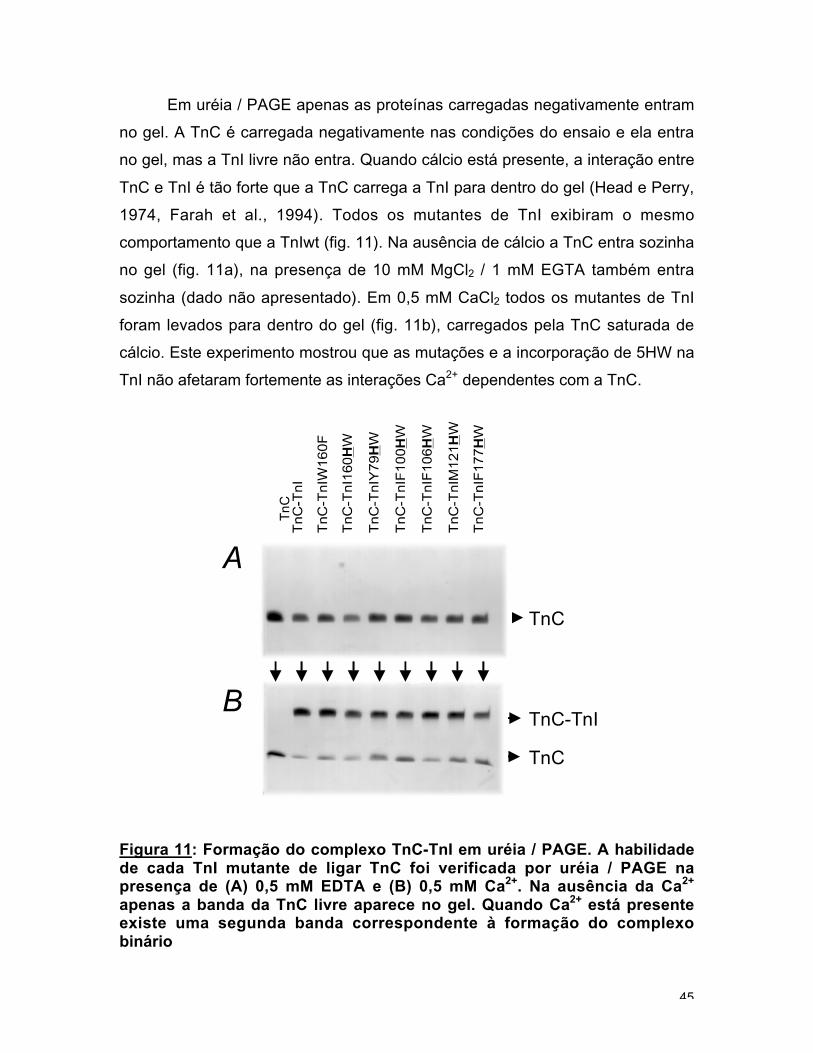
Em uréia / PAGE apenas as proteínas carregadas negativamente entram
no gel. A TnC é carregada negativamente nas condições do ensaio e ela entra
no gel, mas a TnI livre não entra. Quando cálcio está presente, a interação entre
TnC e TnI é tão forte que a TnC carrega a TnI para dentro do gel (Head e Perry,
1974, Farah et al., 1994). Todos os mutantes de TnI exibiram o mesmo
comportamento que a TnIwt (fig. 11). Na ausência de cálcio a TnC entra sozinha
no gel (fig. 11a), na presença de 10 mM MgCl2 / 1 mM EGTA também entra
sozinha (dado não apresentado). Em 0,5 mM CaCl2 todos os mutantes de TnI
foram levados para dentro do gel (fig. 11b), carregados pela TnC saturada de
cálcio. Este experimento mostrou que as mutações e a incorporação de 5HW na
TnI não afetaram fortemente as interações Ca2+ dependentes com a TnC.
Figura 11: Formação do complexo TnC-TnI em uréia / PAGE. A habilidadede cada TnI mutante de ligar TnC foi verificada por uréia / PAGE napresença de (A) 0,5 mM EDTA e (B) 0,5 mM Ca2+. Na ausência da Ca2+
apenas a banda da TnC livre aparece no gel. Quando Ca2+ está presenteexiste uma segunda banda correspondente à formação do complexobinário
TnC
TnC-TnI
TnC
A
B
TnC
46
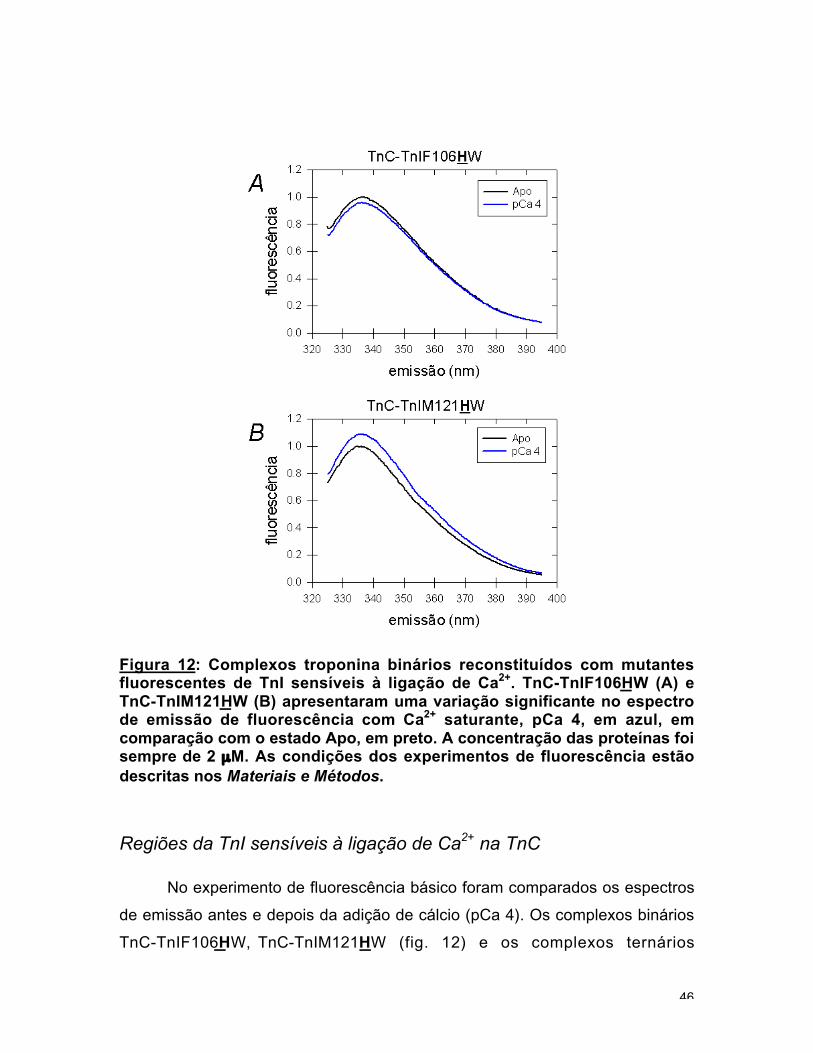
Figura 12: Complexos troponina binários reconstituídos com mutantesfluorescentes de TnI sensíveis à ligação de Ca2+. TnC-TnIF106HW (A) eTnC-TnIM121HW (B) apresentaram uma variação significante no espectrode emissão de fluorescência com Ca2+ saturante, pCa 4, em azul, emcomparação com o estado Apo, em preto. A concentração das proteínas foisempre de 2 µM. As condições dos experimentos de fluorescência estãodescritas nos Materiais e Métodos.
Regiões da TnI sensíveis à ligação de Ca2+ na TnC
No experimento de fluorescência básico foram comparados os espectros
de emissão antes e depois da adição de cálcio (pCa 4). Os complexos binários
TnC-TnIF106HW, TnC-TnIM121HW (fig. 12) e os complexos ternários

47
Tn-TnIF100HW e Tn-TnIM121HW (fig. 13) apresentaram variação significante na
emissão de fluorescência no estado Ca2+ saturado em comparação com o
estado Apo. Todos os outros complexos não foram sensíveis ao Ca2+ ligado (as
variações na intensidade da fluorescência foram menores que 3 %, tabela 1).
Tabela 1: Variações nas intensidades da fluorescência dos complexostroponina reconstituídos com as TnIs mutantes provocadas pela ligação deCa2+.
A tabela resume a variação relativa na intensidade da fluorescência paracada um dos mutantes de TnI reconstituindo os complexos troponina. Osvalores descrevem a diferença na intensidade do espectro de emissão noestado Ca2+ saturado, pCa 4, em comparação com o estado Apo. * Narealidade uma curva bifásica, vide tabela 2 e figura 14. Os valores paraTnCF29HW também são apresentados. NV significa que a variação naintensidade da fluorescência foi inferior a 3 %.
Para todos os complexos, o comprimento de onda máximo de emissão foi
ao redor de 335 nm e nenhum deslocamento significativo foi observado após a
adição de Ca2+.
complexos binários(TnC-TnI)
complexos ternáriosTnC-TnI-TnT
TnIY79HW NV NV
TnIF100HW NV + 12 %
TnIF106HW - 4 %* NV
TnIM121HW + 10 % + 70 %
TnI160HW NV NV
TnIF177HW NV NV
TnCF29HW + 500 % + 450 %
48
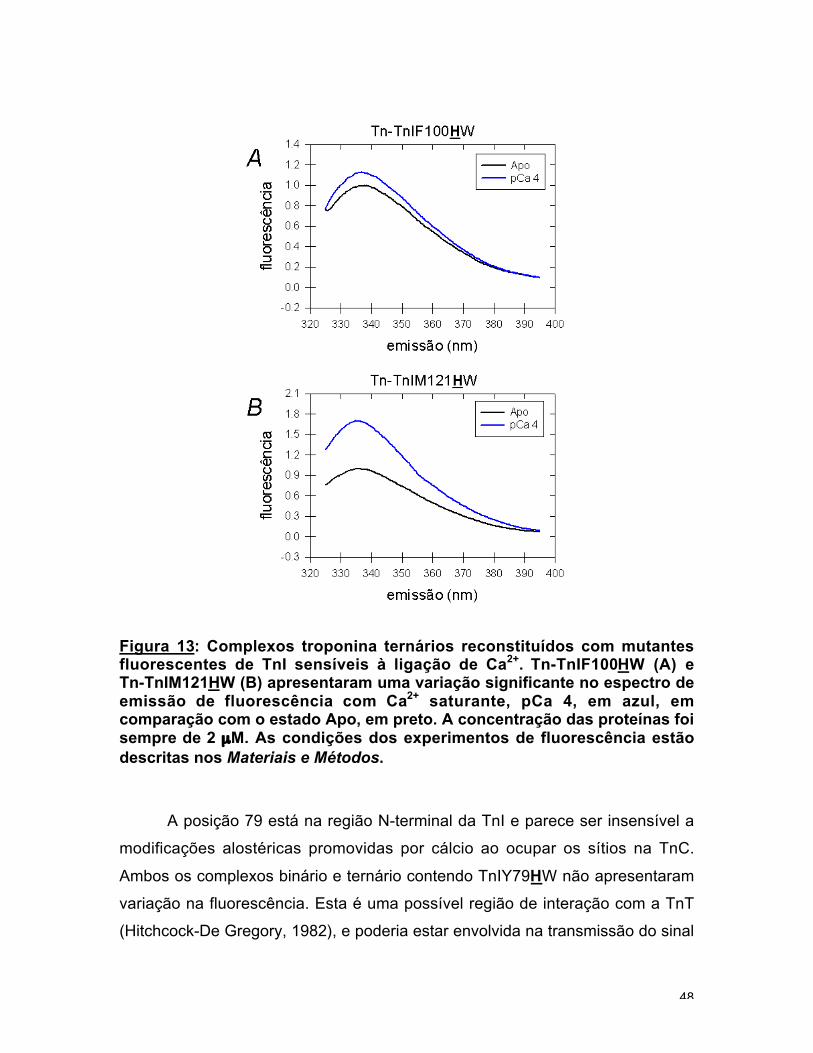
Figura 13: Complexos troponina ternários reconstituídos com mutantesfluorescentes de TnI sensíveis à ligação de Ca2+. Tn-TnIF100HW (A) eTn-TnIM121HW (B) apresentaram uma variação significante no espectro deemissão de fluorescência com Ca2+ saturante, pCa 4, em azul, emcomparação com o estado Apo, em preto. A concentração das proteínas foisempre de 2 µM. As condições dos experimentos de fluorescência estãodescritas nos Materiais e Métodos.
A posição 79 está na região N-terminal da TnI e parece ser insensível a
modificações alostéricas promovidas por cálcio ao ocupar os sítios na TnC.
Ambos os complexos binário e ternário contendo TnIY79HW não apresentaram
variação na fluorescência. Esta é uma possível região de interação com a TnT
(Hitchcock-De Gregory, 1982), e poderia estar envolvida na transmissão do sinal

49
de Ca2+ para a TnT. Mutantes de deleção da TnI que não têm a região
N-terminal inteira demonstraram propriedades regulatórias da ATPase da
actomiosina similares a TnI completa (Farah et al., 1994, Tripet et al., 1997, Van
Eyk et al., 1997, Ramos, 1999). A região N-terminal da TnI interage com a
porção globular C-terminal da TnT de uma forma Ca2+ independente para manter
a estrutura do complexo troponina (Farah et al., 1994, Malnic et al., 1998).
O complexo Tn-TnIF100HW apresentou um aumento de 12 % no
espectro de emissão (fig. 13b). Nos experimentos de titulação com Ca2+ ficou
evidente que o aumento na fluorescência era causado pelo preenchimento dos
sítios de alta afinidade na TnC, Kd = 3,1 x 10-8 M (fig. 14a). Nenhuma
cooperatividade foi observada, n = 1,0. A TnIF100HW no complexo binário se
mostrou praticamente insensível à presença de cálcio. A Phe na posição 100 é
parte da região inibitória, resíduos 96 até 116 (Syska et al., 1976), e pode estar
interagindo com o C-domínio da TnC (Vassylyev et al., 1998). Uma análise mais
detalhada desta região concluiu que a posição 100 não faz parte da seqüência
inibitória mínima, resíduos 104 até 114 (Talbot e Hodges, 1981). Mutantes de
deleção N-terminais que não possuem esta posição, TnI103-182 (Farah et al.,
1994) ou TnI103-156 (Ramos, 1999), continuaram capazes de regular a ATPase da
actomiosina. Isto significa que a Phe-100 não é um aminoácido central no
mecanismo regulatório e que o 5HW nesta posição pode funcionar bem como
um sonda pouco invasiva para a transmissão do sinal de Ca2+ na Tn.
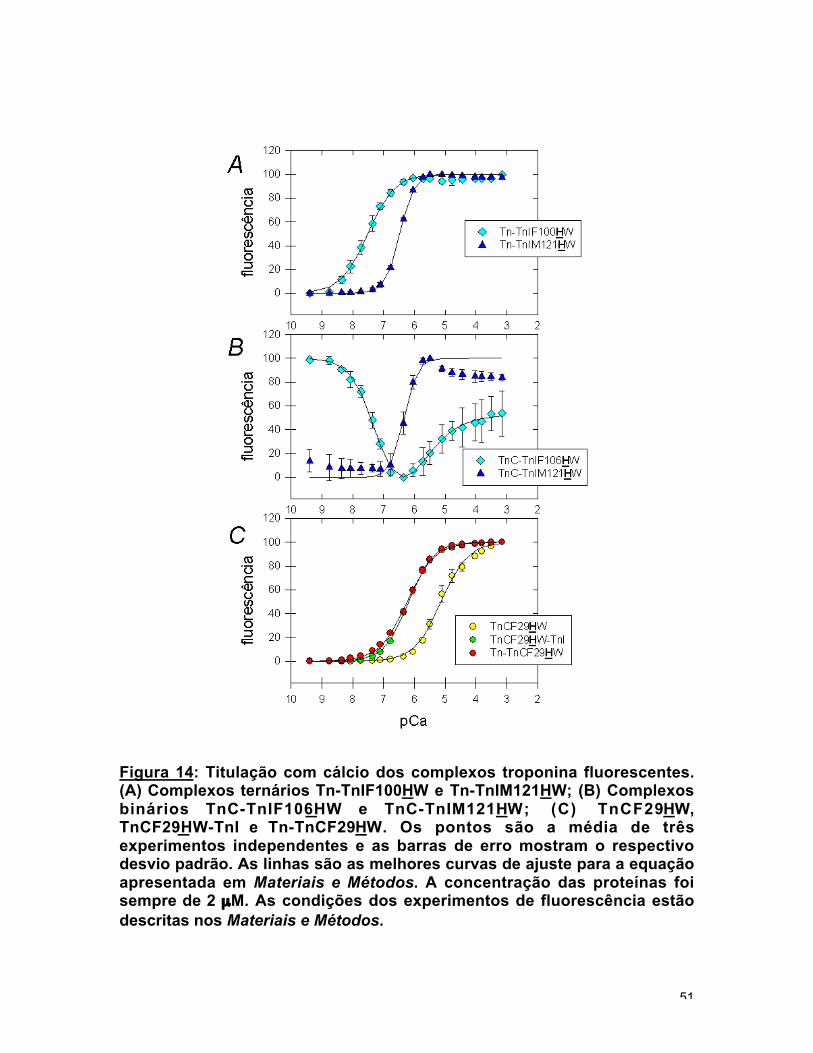
Os dados para o complexo binário TnC-TnIF106HW apresentaram uma
pequena e complicada variação na intensidade da emissão de fluorescência. No
experimento de titulação com cálcio uma curva de duas fases foi obtida: primeiro
houve uma queda na fluorescência (-6 %), Kd1 = 4,5 x 10-8 M e em seguida
houve um aumento (3 %), Kd2 = 2,8 x 10-6 M (fig. 14b). A posição 106 está no
meio da região inibitória (Syska et al., 1976). Não está claro com qual domínio
da TnC a região inibitória interage. Tem sido investigado se ela interage com o
C-domínio estrutural, com o N-domínio regulatório ou com os dois (Swenson e
Fredricksen, 1992, Farah et al., 1994, Chandra et al., 1994a, Pearlstone e

50
Smillie, 1995). Se a segunda parte da curva for considerada consistente, a
TnIF106HW seria uma sonda para ligação de cálcio em ambos os domínios da
TnC. A presença da TnT aboliu o sinal de fluorescência.
A maior variação encontrada foi um aumento de 70 % na fluorescência de
Tn-TnIM121HW promovida pela adição de cálcio (fig. 13b). TnC-TnIM121HW
exibiu um espectro de emissão 10 % mais alto quando Ca2+ estava ligado em
comparação com o espectro na ausência de Ca2+ (fig. 12b) Apesar de ter sido
analisado como uma curva de uma fase, este complexo binário apresentou uma
queda na curva de titulação em pCa baixos (fig. 14b). Esta queda poderia ser
também uma indicação de Ca2+ ligando em uma diferente classe de sítios.
Novamente a presença de TnT afetou de alguma maneira o fenômeno; ela não
alterou significativamente o Kd nem o n, mas aumentou a variação na
fluorescência (tabela 2). A alta cooperatividade, n ~ 2, significa que os dois
sítios ligam Ca2+ simultaneamente. A Met na posição 121 da TnI tem sido
considerada um resíduo chave no mecanismo regulatório (Tripet et al., 1997,
Vassylyev et al., 1998), e foi incluída na região da TnI que tem um papel em
modular a afinidade por Ca2+ do N-domínio da TnC (Ramos, 1999).
A TnI160HW apresentou pouca sensibilidade a modificações estruturais
induzidas por ligação de Ca2+ na TnC. Também a posição 177 da TnI não tem
sensibilidade ao Ca2+ ligando na TnC. Estas posições C-terminais parecem estar
envolvidas principalmente em estabilizar a interação da TnI com o filamento de
actina-tropomiosina (Ramos, 1999). Além do mais, vários autores têm
demonstrado com fragmentos de TnI truncados que apenas a primeira metade
da porção C-terminal da molécula de TnI é necessária para a regulação in vitro
da ATPase (Farah et al., 1994, Tripet et al., 1997, Van Eyk et al., 1997, Ramos,
1999).
51
Figura 14: Titulação com cálcio dos complexos troponina fluorescentes.(A) Complexos ternários Tn-TnIF100HW e Tn-TnIM121HW; (B) Complexosbinários TnC-TnIF106HW e TnC-TnIM121HW; (C) TnCF29HW,TnCF29HW-TnI e Tn-TnCF29HW. Os pontos são a média de trêsexperimentos independentes e as barras de erro mostram o respectivodesvio padrão. As linhas são as melhores curvas de ajuste para a equaçãoapresentada em Materiais e Métodos. A concentração das proteínas foisempre de 2 µM. As condições dos experimentos de fluorescência estãodescritas nos Materiais e Métodos.
52
Juntos, os dados dos mutantes fluorescentes de TnI apontam que as
partes da TnI que respondem a ligação de Ca2+ na TnC são a região inibitória
mais uma região vizinha que incluí a posição 121 (fig. 7). Sugerimos que a
região da TnI do resíduo 117 até o resíduo 127 seja chamada região
modulatória. O segmento regulatório da TnI, resíduos 96 até 127, cobriria as
regiões inibitória e modulatória (ver Discussão).
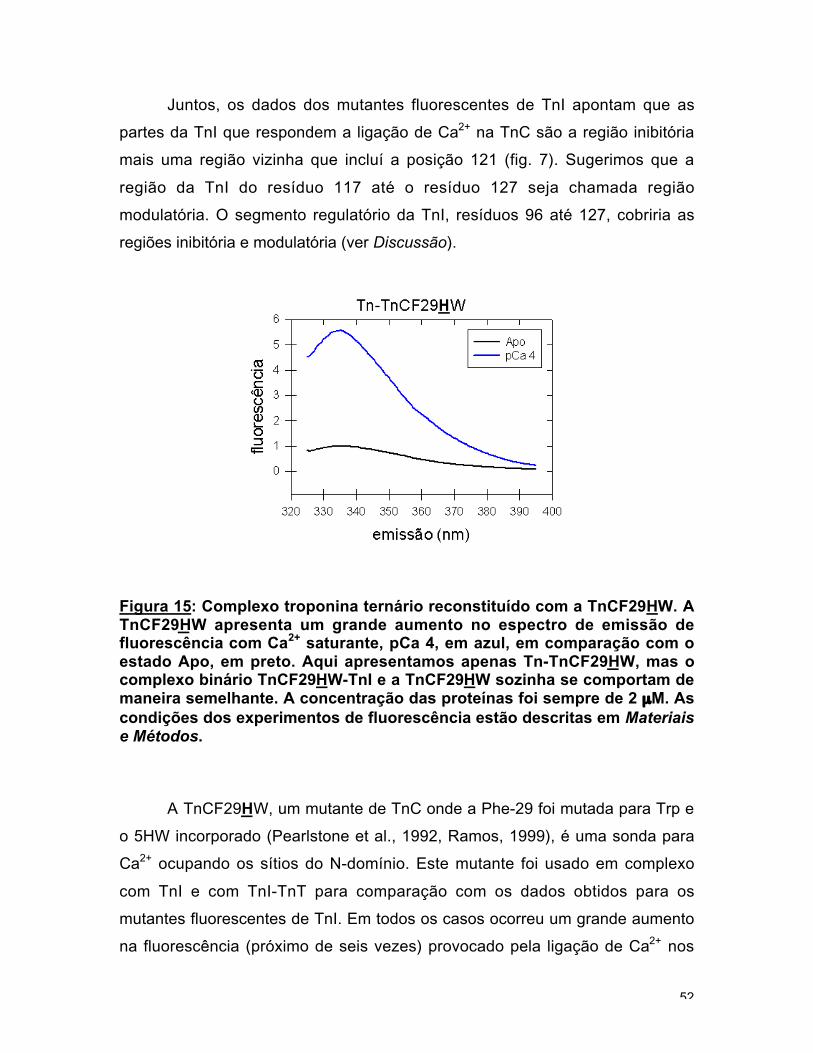
Figura 15: Complexo troponina ternário reconstituído com a TnCF29HW. ATnCF29HW apresenta um grande aumento no espectro de emissão defluorescência com Ca2+ saturante, pCa 4, em azul, em comparação com oestado Apo, em preto. Aqui apresentamos apenas Tn-TnCF29HW, mas ocomplexo binário TnCF29HW-TnI e a TnCF29HW sozinha se comportam demaneira semelhante. A concentração das proteínas foi sempre de 2 µM. Ascondições dos experimentos de fluorescência estão descritas em Materiaise Métodos.
A TnCF29HW, um mutante de TnC onde a Phe-29 foi mutada para Trp e
o 5HW incorporado (Pearlstone et al., 1992, Ramos, 1999), é uma sonda para
Ca2+ ocupando os sítios do N-domínio. Este mutante foi usado em complexo
com TnI e com TnI-TnT para comparação com os dados obtidos para os
mutantes fluorescentes de TnI. Em todos os casos ocorreu um grande aumento
na fluorescência (próximo de seis vezes) provocado pela ligação de Ca2+ nos

53
sítios regulatórios (fig. 15). A presença de TnI aumentou o Kd uma ordem de
magnitude. A TnT praticamente não afetou as propriedades de ligação de cálcio
(fig. 14c e Tabela 2, Potter e Gergely, 1975). Titulações com TnCF29HW e
TnCF29HW-TnI foram apresentadas anteriormente e os dados estão em boa
concordância (Ramos, 1999). As pequenas diferenças nos valores de Kd
ocorreram, provavelmente, por causa da força iônica diferente no tampão de
fluorescência, 100 mM KCl ao invés dos 6,5 mM KCl usado por ele. Aqui a
titulação deste mutante de TnC reconstituindo a troponina inteira,
Tn-TnCF29HW, é apresentada pela primeira vez.
A presença da TnT teve um efeito mínimo nos parâmetros de ligação de
Ca2+, Kd e n, para os respectivos complexos binários TnC-TnI (Potter e Gergely,
1975). Por outro lado, a TnT alterou a intensidade de emissão de fluorescência
para todas as sondas na TnI, como explicado acima. Apesar de TnC-TnI ligar
Ca2+ da mesma maneira que a troponina inteira, o complexo binário não regula a
ATPase da actomiosina na ausência de TnT (Farah et al., 1994, Malnic et al.,
1998). Os resultados aqui apresentados mostram que a TnT provoca alterações
no ambiente ao redor das regiões da TnI envolvidas no processo regulatório.
A tabela 2 resume os dados de fluorescência obtidos. Os parâmetros para
Tn-TnIF100HW estão em concordância com a primeira parte da curva de
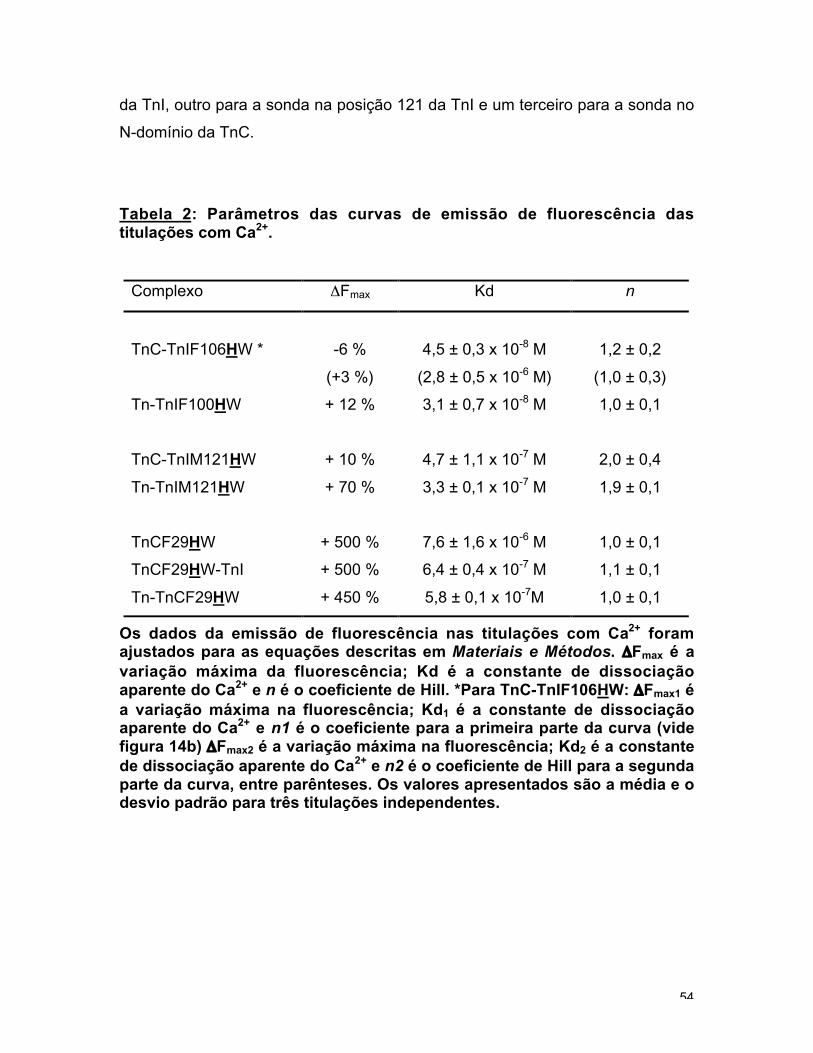
TnC-TnIF106HW, para ambos Kd e n (fig. 14a e 14b). Posições 100 e 106 fazem
parte da região inibitória e responderam ao mesmo evento: Ca2+ ligando na
classe de sítios de alta afinidade da TnC. A sonda na posição 121 da TnI nos
complexos binário e ternário apresentou um Kd que poderia ser consistente com
a ocupação dos sítios de ligação ao Ca2+ de baixa afinidade da TnC com uma
cooperatividade muito alta. Os valores de Kd para os complexos TnCF29HW-TnI
e Tn-TnCF29HW são apenas um pouco diferentes dos respectivos complexos
com TnIM121HW e poderiam ser considerados compatíveis. Porém, a alta
cooperatividade não foi observada nas titulações com TnCF29HW. Conclui-se
que há três diferentes grupos de dados: um para as sondas na região inibitória
54
da TnI, outro para a sonda na posição 121 da TnI e um terceiro para a sonda no
N-domínio da TnC.
Tabela 2: Parâmetros das curvas de emissão de fluorescência dastitulações com Ca2+.
Complexo ΔFmax Kd n
TnC-TnIF106HW * -6 %
(+3 %)
4,5 ± 0,3 x 10-8 M
(2,8 ± 0,5 x 10-6 M)
1,2 ± 0,2
(1,0 ± 0,3)
Tn-TnIF100HW + 12 % 3,1 ± 0,7 x 10-8 M 1,0 ± 0,1
TnC-TnIM121HW + 10 % 4,7 ± 1,1 x 10-7 M 2,0 ± 0,4
Tn-TnIM121HW + 70 % 3,3 ± 0,1 x 10-7 M 1,9 ± 0,1
TnCF29HW + 500 % 7,6 ± 1,6 x 10-6 M 1,0 ± 0,1
TnCF29HW-TnI + 500 % 6,4 ± 0,4 x 10-7 M 1,1 ± 0,1
Tn-TnCF29HW + 450 % 5,8 ± 0,1 x 10-7M 1,0 ± 0,1
Os dados da emissão de fluorescência nas titulações com Ca2+ foramajustados para as equações descritas em Materiais e Métodos. ΔFmax é avariação máxima da fluorescência; Kd é a constante de dissociaçãoaparente do Ca2+ e n é o coeficiente de Hill. *Para TnC-TnIF106HW: ΔFmax1 éa variação máxima na fluorescência; Kd1 é a constante de dissociaçãoaparente do Ca2+ e n1 é o coeficiente para a primeira parte da curva (videfigura 14b) ΔFmax2 é a variação máxima na fluorescência; Kd2 é a constantede dissociação aparente do Ca2+ e n2 é o coeficiente de Hill para a segundaparte da curva, entre parênteses. Os valores apresentados são a média e odesvio padrão para três titulações independentes.

55
Identificação do domínio da TnC que promove a mudança
conformacional percebida pelas sondas na TnI.
O modelo regulatório mais aceito sugere que quando o N-dominio é
preenchido por cálcio ele expõe uma superfície hidrofóbica. Esta superfície seria
um novo sítio de interação com a TnI. A região inibitória da TnI, resíduos 96 até
116, e a região vizinha C-terminal, que inclui a posição 121 e deve estar
diretamente envolvida na interação, seriam atraídas em direção à TnC. Como
resultado, a região inibitória se desligaria da actina permitindo a contração
(Syska et al., 1976, Herzberg et al., 1986, Tripet et al., 1997, Vassylyev et al.,
1998). Isto significa que seria esperado que as sondas nas posições 29 da TnC
e posições 100, 106 e 121 da TnI respondessem ao mesmo evento: Ca2+ ligando
nos sítios I e II no N-domínio regulatório da TnC (fig. 7).
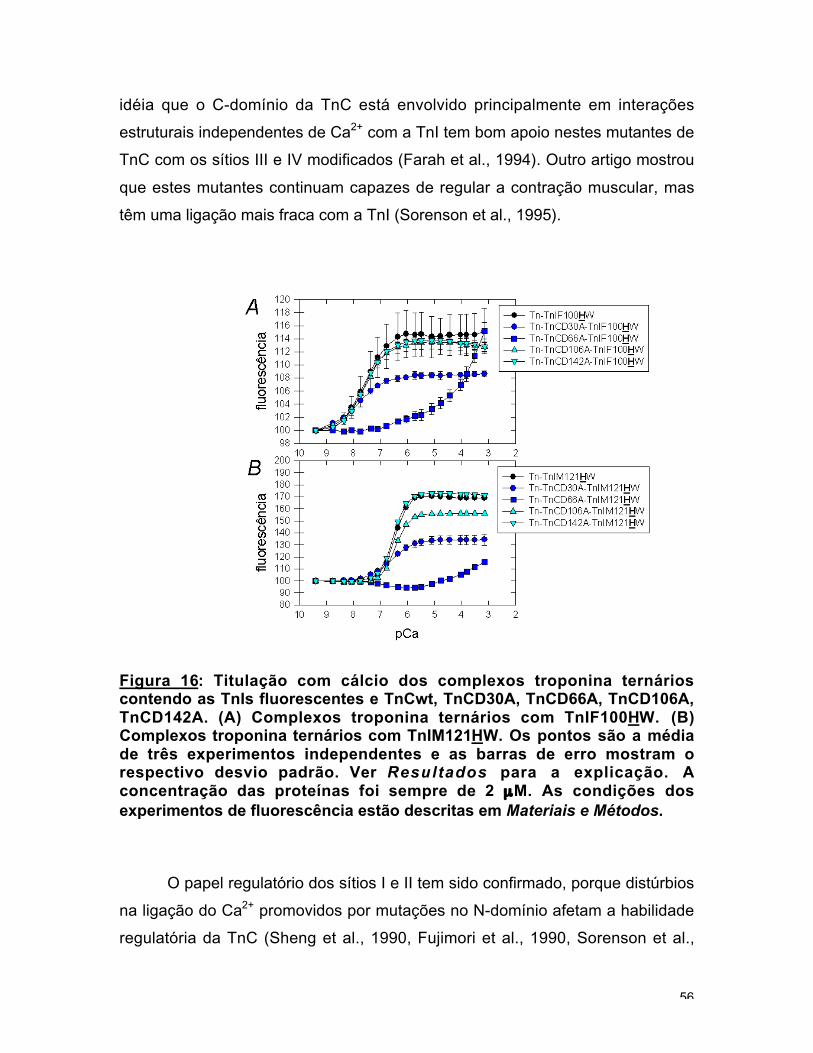
Anteriormente foi descrita a construção de quatro mutantes de TnC que
tinham cada um dos sítios de ligação de Ca2+ alterado por uma substituição de
um Asp por uma Ala (Farah et al., 1994, Sorenson et al., 1995): TnCD30A
(sítio I), TnCD66A (sítio II), TnCD106A (sítio III), TnCD142A (sítio IV). Com o
objetivo de verificar se as variações em Kd e n eram um efeito das mutações ou
fenômenos diferentes, foram reconstituídos complexos com estes mutantes de
TnC e os mutantes fluorescentes de TnI, TnIF100HW e TnIM121HW.
Ambos os complexos ternários com o sítio IV da TnC destruído,
TnCD142A, apresentaram a mesma curva de titulação que os respectivos
complexos com TnCwt. De maneira similar, a mutação que bloqueia a ligação de
Ca2+ no sítio III da TnC, TnCD106A, não teve efeito na curva de titulação da
TnIF100HW e apenas uma pequena diminuição na intensidade da variação para
TnIM121HW. Nem a afinidade por Ca2+ nem a cooperatividade apresentada pela
TnC com os quatro sítios funcionais no complexo ternário com estas TnIs
fluorescentes foram afetadas pelas mutações nos sítios III e IV da TnC.
Conclui-se que as sondas nas posições 100 e 121 da TnI não são sensíveis à
ligação de Ca2+ nos sítios estruturais III e IV do C-domínio da TnC (fig. 16). A
56
idéia que o C-domínio da TnC está envolvido principalmente em interações
estruturais independentes de Ca2+ com a TnI tem bom apoio nestes mutantes de
TnC com os sítios III e IV modificados (Farah et al., 1994). Outro artigo mostrou
que estes mutantes continuam capazes de regular a contração muscular, mas
têm uma ligação mais fraca com a TnI (Sorenson et al., 1995).
Figura 16: Titulação com cálcio dos complexos troponina ternárioscontendo as TnIs fluorescentes e TnCwt, TnCD30A, TnCD66A, TnCD106A,TnCD142A. (A) Complexos troponina ternários com TnIF100HW. (B)Complexos troponina ternários com TnIM121HW. Os pontos são a médiade três experimentos independentes e as barras de erro mostram orespectivo desvio padrão. Ver Resultados para a explicação. Aconcentração das proteínas foi sempre de 2 µM. As condições dosexperimentos de fluorescência estão descritas em Materiais e Métodos.
O papel regulatório dos sítios I e II tem sido confirmado, porque distúrbios
na ligação do Ca2+ promovidos por mutações no N-domínio afetam a habilidade
regulatória da TnC (Sheng et al., 1990, Fujimori et al., 1990, Sorenson et al.,

57
1995). Em concordância com isto pode ser claramente visto nas figuras 16a e
16b que as curvas de titulação com Ca2+ da fluorescência dos complexos Tn
foram fortemente perturbadas pela substituição do Asp-66 da TnC por Ala,
TnCD66A. Dados recentes do nosso laboratório confirmam que esta mutação
diminuiu severamente a afinidade por Ca2+ do domínio regulatório da TnC,
afetando não apenas o sítio II, mas também o sítio I (Valencia e Reinach
comunicação pessoal). Com os dados dos complexos Tn-TnCD66A-TnIF100HW
e Tn-TnCD66A-TnIM121HW demostrou-se que a região inibitória e a região
mudulatória da TnI são sensíveis ao sinal disparado por Ca2+ vindo do N-
domínio da TnC.
Interessante notar que a TnC cardíaca não tem o sítio I funcional (Sia et
al., 1997), então a TnCD30A pode ser considerada "tipo-cardiaca". Os
complexos ternários com essas TnCs tiveram a amplitude da mudança da
emissão de fluorescência diminuída, mas as constantes de afinidade não foram
afetadas. Para todos os complexos com TnCs com os dois sítios do N-domínio
funcionais na presença da TnIM121HW mostraram forte cooperatividade, perto
de 2 (tabela 2, fig. 15 e fig. 16b). TnCD30A tem apenas um sítio funcional no
domínio regulatório e a cooperatividade seria impossível. De fato, TnCD30A
elegantemente derrubou o valor de n para 1 (fig. 16b). Isto implica que a
presença de um 5HW na posição 121 da TnI de alguma maneira induziu
cooperatividade entre os sítios regulatórios I e II da TnC.
Complexos binários com este grupo de TnCDAs e a TnIF106HW foram
formados e submetidos a análises de fluorescência (dados não apresentados). A
TnIF106W também foi investigada, o mesmo mutante de TnI com Trp ao invés
de 5HW na posição 106, em experimentos similares. Como a TnC não tem Trp
(Reinach e Karlsson, 1998), a fluorescência do único Trp da TnIF106W pode ser
seletivamente excitado a 295 nm no complexo binário. A TnIF106W se comporta
como a TnIF106HW, mas a variação no sinal da fluorescência foi um pouco
maior, uma queda de 10 % na primeira parte da curva e de 4 % de aumento na
segunda parte (fig. 17). A obstrução dos sítios I e II, principalmente sítio II,
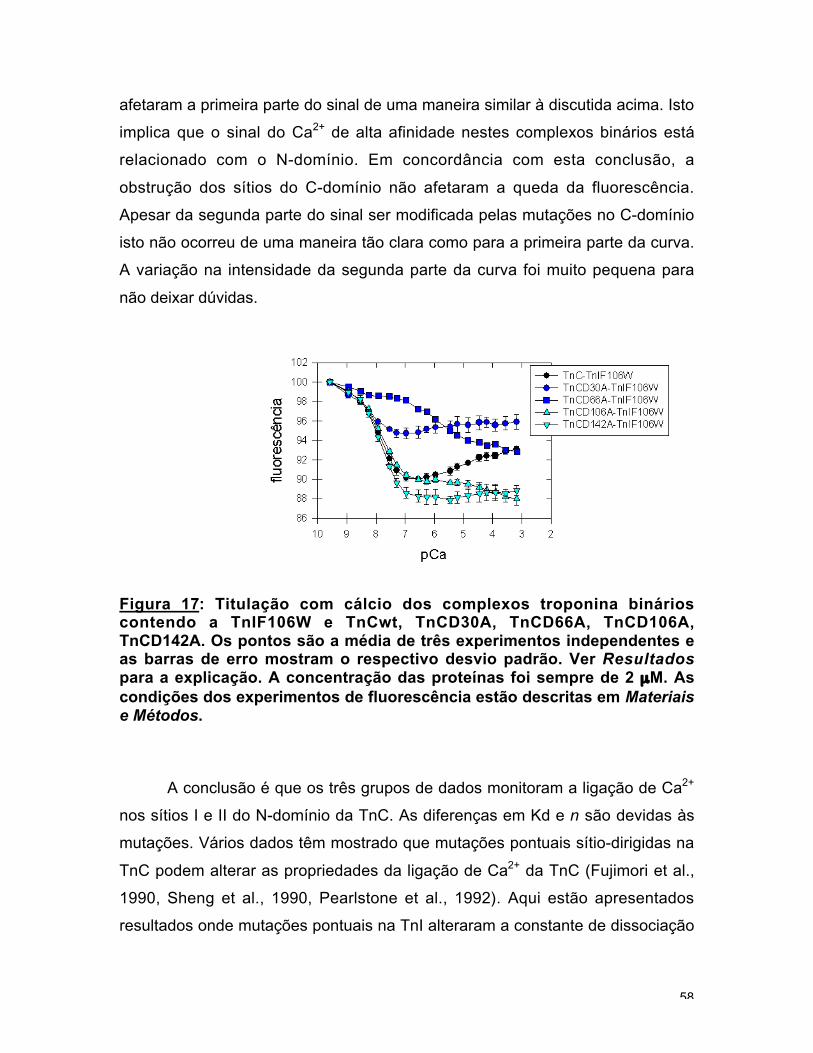
58
afetaram a primeira parte do sinal de uma maneira similar à discutida acima. Isto
implica que o sinal do Ca2+ de alta afinidade nestes complexos binários está
relacionado com o N-domínio. Em concordância com esta conclusão, a
obstrução dos sítios do C-domínio não afetaram a queda da fluorescência.
Apesar da segunda parte do sinal ser modificada pelas mutações no C-domínio
isto não ocorreu de uma maneira tão clara como para a primeira parte da curva.
A variação na intensidade da segunda parte da curva foi muito pequena para
não deixar dúvidas.
Figura 17: Titulação com cálcio dos complexos troponina binárioscontendo a TnIF106W e TnCwt, TnCD30A, TnCD66A, TnCD106A,TnCD142A. Os pontos são a média de três experimentos independentes eas barras de erro mostram o respectivo desvio padrão. Ver Resultadospara a explicação. A concentração das proteínas foi sempre de 2 µM. Ascondições dos experimentos de fluorescência estão descritas em Materiaise Métodos.
A conclusão é que os três grupos de dados monitoram a ligação de Ca2+
nos sítios I e II do N-domínio da TnC. As diferenças em Kd e n são devidas às
mutações. Vários dados têm mostrado que mutações pontuais sítio-dirigidas na
TnC podem alterar as propriedades da ligação de Ca2+ da TnC (Fujimori et al.,
1990, Sheng et al., 1990, Pearlstone et al., 1992). Aqui estão apresentados
resultados onde mutações pontuais na TnI alteraram a constante de dissociação

59
e a cooperatividade da ligação de Ca2+ na TnC, o que evidencia ainda mais o
papel modulador da TnI na afinidade da TnC por Ca2+. Os resultados com as
TnCDAs associaram altos valores de constante de dissociação obtidos com
Tn-TnIF100HW e TnC-TnIF106HW com a ligação de cálcio no N-domínio da
TnC (tabela 2, fig. 16 e fig. 17). O que abriu uma possibilidade nunca antes
considerada, de que de fato a TnI aumenta a afinidade do domínio regulatório da
TnC por Ca2+ mais de cem vezes. Neste modelo os sítios I e II da TnC seriam os
sítios de alta afinidade nos complexos troponina binário e ternário.

60
DISCUSSÃO
Um dos objetivos mais almejados na pesquisa bioquímica é a capacidade
de monitorar rearranjos conformacionais nas proteínas. A troponina é um
biosensor da concentração de Ca2+ e rearranjos alostéricos complicados são
esperados para promover controle na interação geradora de força entre a actina
e a miosina. Muitos estudos têm descrito o uso de aminoácidos fluorescentes
que ocorrem naturalmente Tyr e Trp, ou proteínas marcadas com sondas
extrínsecas fixadas covalentemente para analisar interações com ligantes,
interações proteína-proteína e caminhos de enovelamento (Leavis et al., 1978,
Rosenfeld e Taylor, 1985, Grebarek et al., 1986, Pearlstone et al., 1992,
Swenson e Fredricksen, 1992, Chandra et al., 1994a, Pearlstone e Smillie, 1995,
Pearlstone et al., 1997). Entretanto o uso de aminoácidos que ocorrem
naturalmente é limitado porque se mais de um está presentes a interpretação
dos dados fica difícil. O uso de sondas extrínsecas fixadas covalentemente pode
ser prejudicial devido ao tamanho e ao potencial para formar ou romper
interações, o que pode levar a alterações estruturais na proteína. A incorporação
do 5HW e de outros análogos de aminoácidos que não ocorrem naturalmente
parece ser uma boa alternativa. Eles podem ser utilizados como sondas sítio-
específicas, com um dano conformacional esperado menor (Hogue et al., 1992,
Farah e Reinach, 1999, Ramos, 1999).
Esta tese demonstra a possibilidade de se construir mutantes
fluorescentes de TnI que tiveram seus espectros de emissão afetados por Ca2+
ligando na TnC, uma cadeia polipeptídica diferente (fig. 12 e fig. 13). E que se
pode utilizar a fluorescência destes mutantes de TnI para investigar o sinal da
ligação do Ca2+ nos sítios regulatórios da TnC sendo transmitido para a região
inibitória da TnI e analisar o efeito modulador da TnI nas propriedades de ligação
de Ca2+ da TnC.
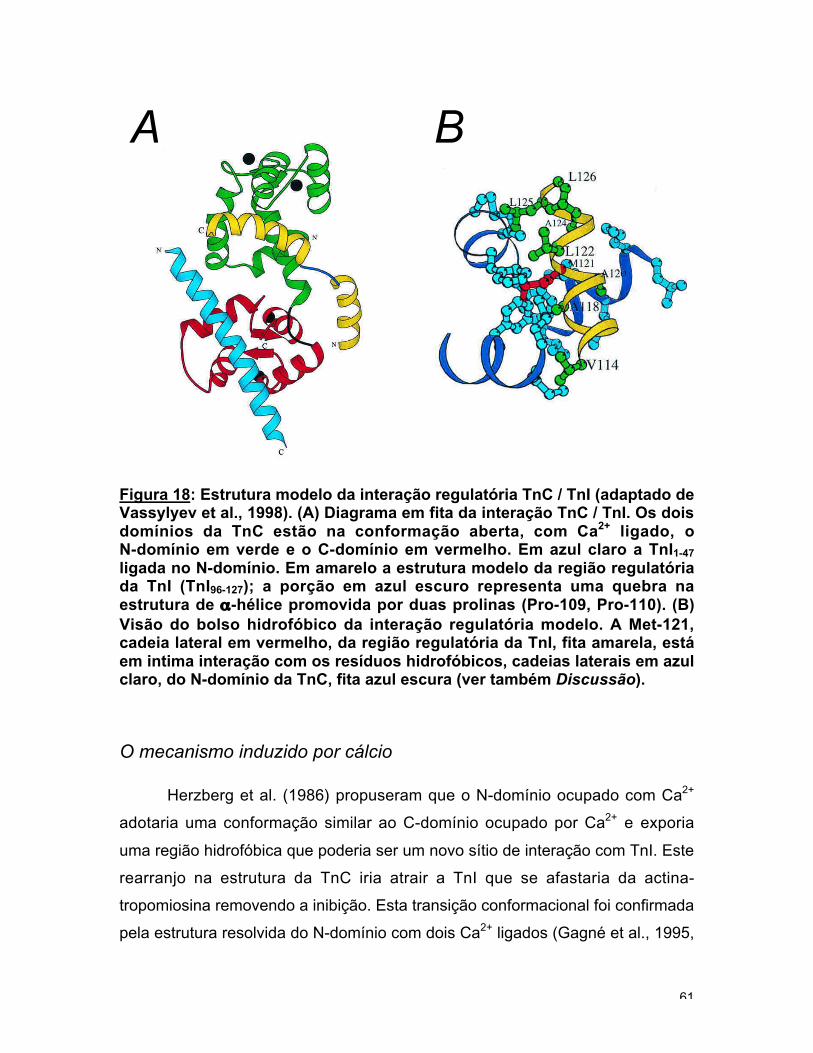
61
Figura 18: Estrutura modelo da interação regulatória TnC / TnI (adaptado deVassylyev et al., 1998). (A) Diagrama em fita da interação TnC / TnI. Os doisdomínios da TnC estão na conformação aberta, com Ca2+ ligado, oN-domínio em verde e o C-domínio em vermelho. Em azul claro a TnI1-47
ligada no N-domínio. Em amarelo a estrutura modelo da região regulatóriada TnI (TnI96-127); a porção em azul escuro representa uma quebra naestrutura de α-hélice promovida por duas prolinas (Pro-109, Pro-110). (B)Visão do bolso hidrofóbico da interação regulatória modelo. A Met-121,cadeia lateral em vermelho, da região regulatória da TnI, fita amarela, estáem intima interação com os resíduos hidrofóbicos, cadeias laterais em azulclaro, do N-domínio da TnC, fita azul escura (ver também Discussão).
O mecanismo induzido por cálcio
Herzberg et al. (1986) propuseram que o N-domínio ocupado com Ca2+
adotaria uma conformação similar ao C-domínio ocupado por Ca2+ e exporia
uma região hidrofóbica que poderia ser um novo sítio de interação com TnI. Este
rearranjo na estrutura da TnC iria atrair a TnI que se afastaria da actina-
tropomiosina removendo a inibição. Esta transição conformacional foi confirmada
pela estrutura resolvida do N-domínio com dois Ca2+ ligados (Gagné et al., 1995,
A B

62
Houdusse et al., 1997). A região precisa da TnI envolvida nesta interação ainda
é tentativa. Recentemente Vassylyev et al. (1998) obtiveram a estrutura da
porção N-terminal da TnI ligada ao C-domínio estrutural da TnC. Eles usaram
uma abordagem semelhante à de Herzberg e colegas para propor um modelo
para a interação regulatória entre TnC-TnI (fig. 18). Neste modelo a região
inibitória, 96-116, mais um prolongamento até o resíduo 127, chamado de TnIreg,
está ligado na superfície hidrofóbica do N-domínio. A Met-121 da TnI seria um
resíduo fundamental nesta interação. Portanto Met-121 e os resíduos vizinhos
seriam a região diretamente em contato com o bolso hidrofóbico, e não a região
inibitória. As mudanças na fluorescência do 5HW na posição 121 da TnI
promovidas por Ca2+ estão em concordância com esta idéia (fig. 16).
Muitos outros estudos têm apontado que a região da TnI que realmente
interage com o bolso hidrofóbico do N-domínio é a região adjacente ao lado
carboxi da região inibitória. Aplicando proteólise limitada na Tn, Takeda et al.
(1997) demonstraram que a proteção do segmento 116 até 134 (ou 140) ocorre
na presença de Ca2+ e não ocorre na ausência. A ligação do peptídeo da TnI,
TnI115-131, na superfície hidrofóbica do N-domínio saturado com Ca2+ foi
monitorada por espectroscopia multidimensional de NMR (McKay et al., 1997).
Um papel modulador na afinidade por Ca2+ do N-domínio da TnC foi associado à
região entre os resíduos 96 até 129 por Ramos (1999). A interação preferencial
dos resíduos 117 até 148 da TnI com o N-domínio da TnC também foi apontada
por Pearlstone et al. (1997). Resíduos do N-domínio foram ligados por
"photocrosslink" com a Met-121 da TnI (Luo et al., 1999). Tripet et al. (1997)
propuseram que este segmento vai do aminoácido 116 até o 131 e chamaram
de "segundo sítio de ligação na TnC". Conseqüentemente a região inibitória
deve ligar-se em algum outro lugar que não diretamente no bolso hidrofóbico.
Ainda não está claro se a região inibitória da TnI interage com o C, o N ou os
dois domínios da TnC (Swenson e Fredricksen, 1992, Farah et al., 1994,
Chandra et al., 1994a, Pearlstone e Smillie, 1995). Apesar dos dados aqui
apresentados não resolverem esta questão, os complexos TnC-TnIF106HW e
Tn-TnIF100HW apresentaram sensibilidade à ligação de Ca2+ no domínio

63
regulatório da TnC (fig. 16 e fig. 17), o que demonstra que a ocupação com
cálcio dos sítios no N-domínio promoveu rearranjos conformacionais que foram
transmitidos para a região inibitória da TnI.
Figura 19: Modelo da mudança da estrutura da interação TnC / TnIpromovido por Ca2+ (adaptado de Luo et al., 2000). (A) Na presença de Ca2+
os resíduos ao redor da posição 121 estão interagindo com o bolso doN-domínio. (B) Na ausência de Ca2+ os resíduos ao redor da posição 121deixam de ocupar o bolso do N-domínio e conjuntamente a região 89-104se afasta da TnC para interagir com a actina.
Um mapa das interações da TnI pode ser desenhado. A região N-terminal
da TnI, posição 1 até 96, parece ter principalmente uma função estrutural de
manter a organização da Tn (Farah et al., 1994, Malnic et al., 1998). A primeira
metade, resíduos 1 até 47, se ligaria com o C-domínio (Vassylyev et al., 1998). A
parte seguinte, aminoácidos 48 até 95, se ligaria à região C-terminal da TnT
(Hitchcock-De Gregory, 1982). O meio da molécula da TnI, uma região que vai
da posição 96 até 131, retém a maior parte da função regulatória. Os dados
mostraram que ela responde ao sinal regulatório do Ca2+. De acordo com estes
dados sugerimos que ela deva ser chamada de "região regulatória". A região
inibitória, aminoácidos 96 até 116, tem sido conhecida há bastante tempo e é
A B

64
bem caracterizada (Syska et al., 1976). As posições seguintes na seqüência
primária até aproximadamente o resíduo 131 é provavelmente a região da TnI
que é atraída para a TnC em resposta ao mecanismo regulatório promovido por
Ca2+ (ver acima). Esta parte da molécula de TnI modula a afinidade da TnI por
actina ou TnC e também parece modular a afinidade do domínio regulatório por
Ca2+ (Ramos, 1999). Então sugerimos que ela passe a ser conhecida como
"região modulatória". A função da parte C-terminal da TnI é menos
compreendida. Um mapeamento das interações da TnI com as outras proteínas
do filamento fino obtido por estudos de “photocrosslink”, realizado por Luo et al.
(2000), está em boa conformidade com este esquema (fig. 19).
O efeito modulador da TnI na afinidade da TnC por Ca2+
Desde os experimentos originais da ligação de Ca2+ feitos por Potter e
Gergely (1975) ficou claro que a TnI modula a afinidade da TnC por Ca2+,
enquanto a TnT não provocava um efeito maior. Tem sido considerado que a TnI
aumenta a afinidade dos dois domínios uma ordem de magnitude - Kd para os
dois sítios de alta afinidade de ~ 10-7 M para ~ 10-8 M e Kd para os dois sítios de
baixa afinidade de ~ 10-5 M para ~ 10-6 M. Os sítios de alta afinidade por Ca2+
também ligariam Mg2+, enquanto os sítios de baixa afinidade seriam específicos
para o Ca2+. No mesmo artigo foi concluído que os dois sítios de baixa afinidade
seriam responsáveis pela regulação da ATPase da actina-miosina. Não havia a
identificação de quais seriam os dois sítios de alta e os dois sítios de baixa
afinidade, nem para TnC sozinha, nem para o complexo troponina. Naquela
época a estrutura da TnC e a relativa independência do N e do C-domínios eram
desconhecidas. Quando Leavis et al., 1978 usando fragmentos proteolíticos de
TnC identificaram os sítios de alta afinidade, III e IV, e por exclusão os sítios de
baixa afinidade, I e II, para TnC livre, foi assumido que também era o caso para
TnC-TnI e TnC-TnI-TnT. Muitos dados têm dado suporte à conclusão para TnC
isolada (Fujimori et al., 1990, Pearlstone et al., 1992, Chandra et al., 1994a,
Fredricksen e Swenson, 1996, Pearlstone et al., 1997, Mercier et al., 2000). Não

65
conhecemos nenhum dado que ligue diretamente a afinidade com um domínio
específico da TnC para proteína inteiras nos complexos binários e ternários,
além dos dados com TnCF29HW formando estes complexos (esta tese e Ramos
1999). Dados de sondas extrínsecas fixadas geralmente na Cys-98 da TnC são
sensíveis ao Ca2+ ligando nas duas classes de sítios. Apesar dos autores
normalmente interpretarem que os sítios de alta afinidade estão no C-domínio e
que os sítios de baixa afinidade no N-domínio da TnC uma associação sem
dúvidas não poderia ser feita (Rosenfeld e Taylor, 1985. Grebarek et al., 1986 e
referências citadas). A investigação mais compreensiva que vincula a afinidade
por cálcio para cada um dos sítios de ligação na TnC nos complexo troponina foi
apresentada aqui.
A Tn-TnCF29HW e a TnCF29HW-TnI apresentaram um aumento de uma
ordem de magnitude na afinidade por cálcio dos sítios regulatórios em
comparação com a TnCF29HW sozinha (tabela 2 e fig. 14c, Ramos, 1999). O
que está em muito boa concordância com a literatura (ver acima). O grande
aumento na emissão de fluorescência, ao redor de cinco vezes, nos convida a
acreditar nisto. Isto implicaria que as mutações na região inibitória da TnI,
substituição das Phe-100 e Phe 106 por 5HW, causaram um aumento extra na
afinidade dos sítios I e II por Ca2+, ou alguma outra justificativa mais intrincada.
Os experimentos com as TnCDAs claramente assinalam que estes mutantes de
TnI são sondas para o Ca2+ ligando no N-domínio da TnC (fig. 16). Uma
explicação alternativa consideraria que os valores de Kd, ~ 3 x 10-8 M, obtidos
com as sondas na região inibitória da TnI estão de fato corretos. Nunca valores
de afinidade por Ca2+ tão altos tinham sido relacionados com os sítios I e II da
TnC. O valor de Kd por ele mesmo poder não dar uma informação segura já que
ele pode variar dependendo das condições experimentais. A comparação da
afinidade por Ca2+ do N-domínio na TnC sozinha, TnCF29HW, e no complexo
troponina, Tn-TnIF100HW, obtidos em condições experimentais idênticas, dá um
aumento de aproximadamente 250 vezes da constante de dissociação (tabela 2
e fig. 14). Para assumir esta proposta é preciso argumentar que a incorporação

66
do 5HW na posição 29 da TnC e na posição 121 da TnI reduziram a afinidade
dos sítios regulatórios por Ca2+.
Hipótese dos sítios de maior afinidade no complexo troponina serem
os sítios regulatórios I e II no N-domínio da TnC
Existe um bom suporte inicial pelo modelo estrutural proposto por
Vassylyev et al. (1998). Eles modelaram a região ao redor da Met-121 da TnI em
íntima interação com a superfície hidrofóbica do N-domínio da TnC. Phe-29 da
TnC é parte desta superfície hidrofóbica. Esta seria a interação regulatória entre
TnC e TnI (fig. 18). Se este modelo estiver correto não seria surpreendente que
a substituição dos dois Met-121 da TnI ou a Phe-29 da TnC por 5HW poderia
potencialmente afetar a interação TnC-TnI dependente de Ca2+, diminuindo o
efeito modulador da TnI na afinidade por Ca2+ do N-domínio regulatório da TnC.
Tripet et al. (1997) descreveram que alterações na Met-121 ou na região ao
redor reduziram a interação Ca2+ dependente com a TnC. Ramos (1999) indicou
a importância dos resíduos 117 até 129 no aumento da afinidade por Ca2+ dos
sítios do N-domínio, e pode-se esperar que alterações nesta região devam afetar
o efeito modulador total. Apesar dos dados regulatórios para TnCF29HW não
estarem disponíveis, sua contraparte TnCF29W é um dos mutantes de TnC mais
estudados. TnCF29W é ainda capaz de regular a ATPase da actomiosina,
porém não retém toda a propriedade reguladora (Chandra et al., 1994b). Uma
análise comparativa entre a TnC e a TnCF29W mostrou que o Trp na posição 29
desestabiliza o N-domínio (Yu et al., 1999), portanto um efeito comparável pode
ser predito para TnCF29HW. Valencia e Reinach, comunicação pessoal,
compararam as propriedades de ligação de Ca2+ da TnCF29W e da TnCF29HW
livres em solução. Eles demonstram que TnCF29W tem afinidade por Ca2+ mais
alta que TnCF29HW, oito vezes mais, diretamente relacionado com os sítios do
N-domínio. O que implica que 5HW ou Trp na posição 29 da TnC tem efeitos
diferentes na capacidade de ligação Ca2+.

67
Esta proposta implica que os sítios III e IV no C-domínio da TnC teriam
menor afinidade por Ca2+ que os sítios I e II quando no complexo troponina. E
também implica que o efeito da TnI nos sítios estruturais seria de reduzir a
afinidade por Ca2+, ou pelo menos não aumentar. Contra nossa hipótese estão
dados de titulação com Ca2+ de mutantes de TnC com um único Trp no
C-domínio, TnCF105W e TnCF154W (Chandra et al., 1994a, Pearlstone e
Smillie, 1995). Estas duas proteínas apresentaram aumento, apesar de menor
que dez vezes, na afinidade por Ca2+ quando na presença de um excesso molar
do peptídeo inibitório. Swenson e Fredricksen (1992) reportaram resultados
semelhantes. É importante notar que peptídeos com a seqüência inibitória
podem ligar no C-domínio, mas esta deve ser uma interação não fisiológica. A
principal região de ligação da TnI com este domínio deve ser a região dos
primeiros 40 aminoácidos, TnI1-40 (Mercier et al., 2000 e Discussão acima).
Quando livre em solução o C-domínio da TnC está desordenado
enquanto o N-domínio está enovelado (Mercier et al., 2000). Então parece que
para a TnC isolada o enovelamento do C-domínio só é conseguido com a ajuda
de um metal divalente, e por isso a forma com Ca2+ ligado seria energeticamente
favorável. Quando o complexo está montado o C-domínio está estruturalmente
interagindo com a região N-terminal da TnI (Vassylyev et al., 1998). Nesta
condição ele não está mais desordenado, e a superfície hidrofóbica também não
está exposta. Pode não haver razão para a afinidade por Ca2+ aumentar, nesta
situação a necessidade por um metal ligado pode até diminuir. Portanto pode ser
razoável que o requerimento por Ca2+ ligado no C-dominio não aumente quando
a TnI está presente. O N-domínio já está estruturado sozinho em solução,
portanto ele não apresenta o mesmo requerimento por metal que o C-domínio na
forma livre da TnC. A ligação de Ca2+ iria causar a abertura da superfície
hidrofóbica, o que é energeticamente adverso na ausência da TnI. Quando o
complexo troponina está montado na presença de Ca2+ o equilíbrio entre as
formas aberta e fechada do N-domínio seria altamente favorável à forma aberta.
Prevalecendo a forte interação TnC-TnI dependente de Ca2+. Uma análise da

68
relação entre a afinidade por Ca2+ e estabilidade conformacional dos domínios
da TnC foi feita por Fredricksen e Swenson (1996).
Como são eventos energeticamente acoplados, a TnI deve aumentar a
afinidade da TnC por Ca2+ na mesma potência que o Ca2+ aumenta a afinidade
da TnC por TnI. Isto pode ser uma importante questão regulatória, quando o
Ca2+ ocupa o domínio regulatório da TnC a afinidade pela região modulatória da
TnI aumenta, não apenas dez, mas mais de cem vezes. O que deve ser
necessário para desfazer a interação TnI-actina. Nos experimentos de ATPase
as condições do ensaio são diferentes comparadas com os nossos experimentos
de fluorescência. A região inibitória da TnI está ligada na actina. Como a actina
desfaz a interação regulatória TnC-TnI é provável que reduza a constante de
dissociação aparente do domínio regulatório da TnC. Então o estado fechado,
Ca2+ não ligado (Kd ~ 10-5 M) e o estado aberto, Ca2+ ligado (Kd ~10-8 M), do
domínio regulatório da TnC deveriam ser vistos em experimentos de titulação
com Ca2+ do filamento fino (Farah e Reinach, 1999) ou em experimentos de
ATPase (Farah et al., 1994, Van Eyk et al., 1997) com uma constante de
dissociação aparente entre 10-6 M e 10-7 M, o que de fato ocorre.
Neste trabalho não foram investigadas as propriedades de ligação de
Mg2+ do complexo troponina. A possibilidade dos sítios I e II da TnC serem os
sítios de alta afinidade teria que levar em consideração que eles também ligam
magnésio (Potter e Gergelly, 1975). Normalmente existem quantidades
milimolares de Mg2+ em experimentos de ATPase e isso também pode contribuir
para a diminuição do ponto médio de transição do Ca2+ neste tipo de
experimento. Um debate bem quente apareceu recentemente na literatura em
razão da crença de que o Mg2+ está sempre ligado nos sítios estruturais do C-
domínio da TnC na Tn no sarcômero (Finley et al., 2000, Harada et al., 2001).
Parece que as propriedades de ligação de metais na troponina completa livre ou
no filamento fino precisam de suporte experimental suplementar.
Acima foi apresentada uma hipótese baseada na possibilidade de que os
sítios regulatórios I e II da TnC são os sítios de maior afinidade por Ca2+ no

69
complexo troponina. Apesar de nós não resistirmos a propor isto, estamos
convencidos de que apenas dados experimentais muito bem planejados usando
a troponina completa e uma assinatura direta para cada sítio de ligação de Ca2+
irá resolver a questão. Nossos dados mostraram que pequenas modificações na
TnI como mutações pontuais e uma sonda pouco invasiva podem afetar a
afinidade e a cooperatividade dos sítios de ligação de Ca2+ da TnC. Portanto é
necessário cuidado ao considerar dados que venham de peptídeos, fragmentos
proteolíticos, mutantes de deleção e de sondas extrínsecas fixadas. Além do
mais deve-se ter consciência de que assim como as propriedades da TnC livre
não são iguais as da TnC no complexo troponina, do mesmo jeito, conclusões
tiradas da troponina sozinha podem não representar as condições no filamento
fino onde a troponina é, provavelmente, fortemente afetada pela interação com
actina-tropomiosina.

70
ANEXO 1
Caracterização dos sítios de alta e baixa afinidade no complexo
troponina por diálise de fluxo
Os experimentos realizados utilizando-se os complexos com as TnIs
fluorescentes e com os mutantes de TnC, com cada um dos sítios de ligação de
cálcio destruídos, TnCD30A, TnCD66A, TnCD106A e TnCD142A (Farah et al.,
1994 e Sorenson et al., 1995) indicaram que as mutações no N-domínio da TnC
afetavam a curva relacionada com sítios de alta afinidade (ver Resultados e
Discussão). Estes dados são surpreendentes, pois contrariam a literatura. Na
TnC isolada está bem caracterizado que os sítios de alta e baixa afinidade estão
localizados respectivamente nos domínios C e N-terminais. Sabia-se que a TnI
influenciava a afinidade da TnC por Ca2+. Porém o que se pensava era que a TnI
aumentava a afinidade de ambos os domínios por Ca2+ uma ordem de
magnitude - constante de dissociação dos sítios C-terminais ~ 10-7 M para
~ 10-8 M e constante de dissociação dos sítios N-terminais ~ 10-5 M para
~ 10-6 M (Potter e Gergely, 1975 e Leavis et al., 1978). Nossos dados indicam
que a TnI na verdade inverteria a afinidade dos sítios.
Em nosso laboratório o mutante TnCF29H W isolado foi
caracterizado com uma combinação das técnicas de diálise de fluxo (Womack e
Colowick, 1973) e fluorescência (Valencia e Reinach, comunicação pessoal). A
diálise de fluxo é um ensaio direto de ligação de Ca2+ na TnC, enquanto a
fluorescência é um ensaio indireto que permite a análise de mudanças
conformacionais em regiões específicas na proteína. O mutante TnCF29HW é
uma excelente sonda de ligação de Ca2+ nos sítios N-terminais da TnC - ele
apresenta um aumento ao redor de seis vezes na fluorescência devido à ligação
de Ca2+ na TnC isolada (Valencia e Reinach, comunicação pessoal), e como
contém 5HW pode ser analisado complexado com TnI, que contém um Trp e

71
com a TnT que possui três Trp (fig. 14, fig.15, Ramos, 1999). Quando a
TnCF29HW é investigada claramente o aumento na fluorescência acompanha a
ligação de Ca2+ nos sítios de menor afinidade (fig 20a). Pretendeu-se utilizar
esta abordagem para verificar se a variação da fluorescência nos complexos
TnCF29HW-TnIwt e Tn-TnCF29HW acompanhava a ligação de Ca2+ nos sítios
de maior afinidade determinada por diálise de fluxo, o que daria suporte ao
modelo de inversão da afinidade dos sítios. Estes experimentos foram iniciados
por Anna Victoria Rotberg, que realizou um estágio de seis meses em nosso
laboratório.
Reconstituição do complexo troponina para diálise de fluxo
Os complexos binários (TnC-TnI) e ternários (TnC-TnI-TnT, fig. 2c) foram
reconstituídos como descrito em Farah et al. (1994) com algumas modificações.
Quantidades equimolares de proteína foram reunidas e dialisadas uma vez
contra os seguintes tampões:
a) 50 mM Tris-HCl pH 8,0; 4,6 M uréia; 1 M KCl; 50 µM CaCl2; 0,01 %
NaN3; 10 mM 2-mercaptoetanol.
b) 50 mM Tris-HCl pH 8,0; 2 M uréia; 1 M KCl; 50 µM CaCl2; 0,01 %
NaN3; 10 mM 2-mercaptoetanol.
c) 50 mM MOPS pH 7,0; 1 M KCl; 5 µM CaCl2; 0,01 % NaN3; 10 mM
2-mercaptoetanol.
d) 50 mM MOPS pH 7,0; 100 mM KCl; 1 mM EDTA; 0,01 % NaN3; 10 mM
2-mercaptoetanol.
E de quatro até seis trocas contra o tampão de diálise de fluxo:
e) 50 mM MOPS pH 7,0; 100 mM KCl; 0,01 % NaN3; 5 mM DTT.

72
Os complexos foram centrifugados 100.000 xg, 15 min, 4 oC para retirada
das proteínas precipitadas.
Diálise de fluxo.
Os experimentos de diálise de fluxo foram realizados como descrito (da
Silva et al., 1993), com algumas modificações: a concentração final das
proteínas foi de 12,5 µM e o tampão de diálise de fluxo: 50 mM MOPS pH 7,0;
100 mM KCl; 0,01 % NaN3; 5 mM DTT. A concentração do estoque de TnC (e
TnCF29HW) para diálise de fluxo foi determinada por análise de aminoácidos. E
a concentração dos complexos foi determinada através da técnica de Hartree
(1972) utilizando como padrão uma TnC com concentração determinada por
análise de aminoácidos.
O método de diálise de fluxo utiliza um aparato composto de duas
câmaras separadas por uma membrana de diálise. A proteína foi colocada na
câmara superior e pela câmara inferior passou um fluxo constante de tampão. O
ligante marcado (no caso 45Ca2+) foi adicionado na câmara superior. Adicionou-
se posteriormente, em intervalos regulares, concentrações crescentes de ligante
frio. Frações do tampão em fluxo foram coletadas e a contagem é proporcional à
concentração de ligante livre (não ligado na proteína). Como as concentrações
de proteína e ligante eram conhecidas foi possível calcular a quantidade ligada
na proteína. Os valores obtidos foram analisados com a equação:
[Ca2+ligado] / [TnC] = ((2*[Ca2+
livre]n1) / (Kd1n1 + [Ca2+
livre]n1 )) + ((2*[Ca2+
livre]n2) / (Kd2n2 + [Ca2+
livre]n2 ))
onde Kd1, Kd2, n1 e n 2 são respectivamente as constantes de dissociação
aparente e o coeficiente de Hill para os sítios de alta e baixa afinidade.

73
Experimentos de fluorescência foram feitos de forma a reproduzir as
condições dos ensaios de diálise de fluxo, de maneira que cada adição de Ca2+
simulasse a adição de Ca2+ na diálise de fluxo. Desta forma a emissão de
fluorescência em cada ponto foi correlacionada com o valor da pCa obtido na
diálise de fluxo.
Tabela 3: Parâmetros das curvas de diálise de fluxo.
proteína Kd1 n1 Kd2 n2
TnC
diálise de fluxo
5,2 ± 0,1 x 10-7 M 1,0 ± 0,1 2,1 ± 0,2 x 10-5 M 1,5 ± 0,1
TnCF29HW
diálise de fluxo
5,7 ± 1,1 x 10-7 M 1,0 ± 0,1 2,0 ± 0,4 x 10-5 M 1,3 ± 0,1
TnCF29HW
fluorescência
____ ____ 2,5 ± 0,1 x 10-5 M 1,1 ± 0,1
Os dados de diálise de fluxo foram ajustados para a equação descrita emDiálise de Fluxo. Os dados de fluorescência foram ajustados para aequação descrita em Materiais e Métodos. Kd1 é a constante dedissociação aparente do Ca2+ e n1 é o coeficiente para os sítios de maiorafinidade (vide figura 20a) Kd2 é a constante de dissociação aparente doCa2+ e n2 é o coeficiente de Hill para os sítios de menor afinidade. Osvalores apresentados são a média e o desvio padrão para trêsexperimentos independentes.
Resultados das diálises de fluxo
Os experimentos de diálise de fluxo foram iniciados com a TnCwt e
TnCF29HW isoladas. Os resultados obtidos (fig. 20) e os valores das constantes
de dissociação obtidos por diálise de fluxo estão em acordo com o esperado
(tabela 3). Os parâmetros para TnCF29HW são ligeiramente diferentes daqueles
obtidos por fluorescência (tabela 2), esta discrepância foi notada anteriormente,
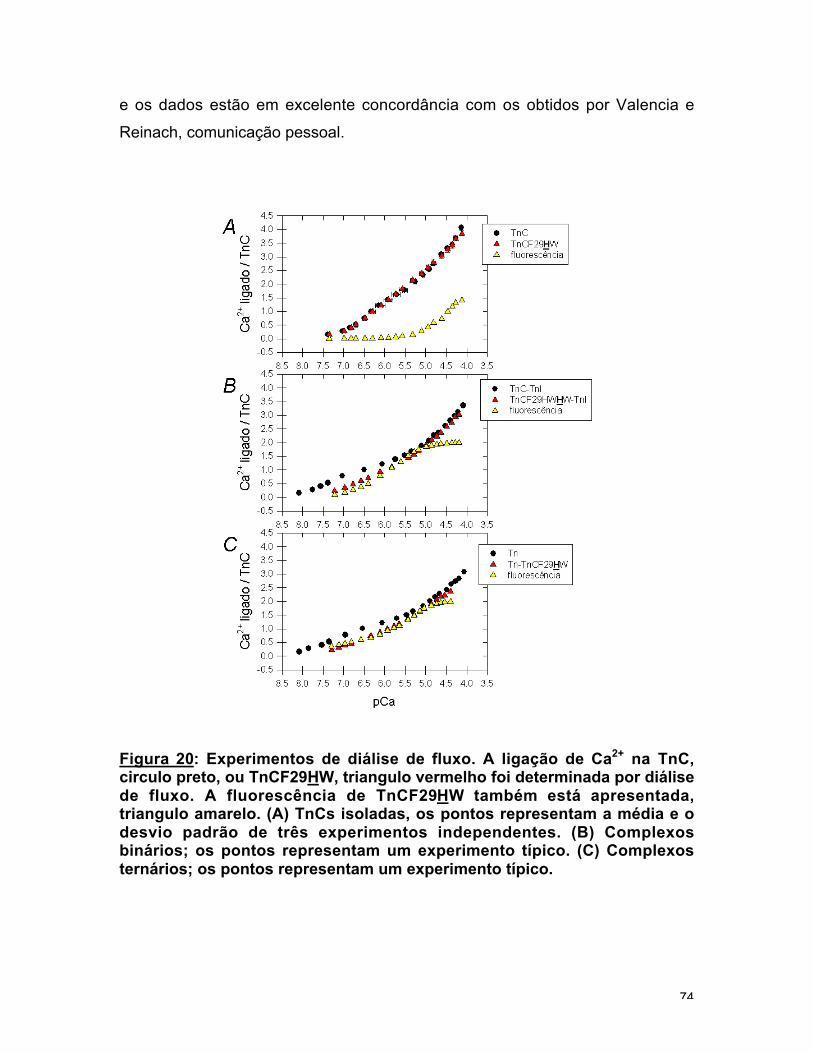
74
e os dados estão em excelente concordância com os obtidos por Valencia e
Reinach, comunicação pessoal.
Figura 20: Experimentos de diálise de fluxo. A ligação de Ca2+ na TnC,circulo preto, ou TnCF29HW, triangulo vermelho foi determinada por diálisede fluxo. A fluorescência de TnCF29HW também está apresentada,triangulo amarelo. (A) TnCs isoladas, os pontos representam a média e odesvio padrão de três experimentos independentes. (B) Complexosbinários; os pontos representam um experimento típico. (C) Complexosternários; os pontos representam um experimento típico.

75
Os complexos binários TnC-TnI e TnCF29HW-TnI e ternários Tn e Tn-
TnCF29HW foram preparados para experimentos de diálise de fluxo. Entretanto
a qualidade dos experimentos para os complexos ficou abaixo da desejada.
Importante notar que TnC e TnCF29HW preparadas em paralelo apresentam a
curva bifásica mostrando claramente a ligação de 2 Ca2+ nos sítios de alta
afinidade e 2 Ca2+ nos sítios de baixa afinidade (fig. 20). No caso da TnCF29HW
isolada, a curva obtida por fluorescência acompanha a ligação de Ca2+ nos sítios
de baixa afinidade da TnC, N-terminais. O objetivo era demonstrar que nos
complexos binário e ternário este fenômeno seria invertido, a curva de
fluorescência acompanharia a ligação de Ca2+ nos sítios de alta afinidade. Nas
diálises de fluxo para os complexos não foi possível caracterizar claramente a
presença de duas classes de sítios. Porém, duas coisas suportam o modelo da
inversão: a fluorescência de TnCF29HW-TnI e de Tn-TnCF29HW acompanhou o
início da curva de ligação de Ca2+ na diálise de fluxo, e a ligação de Ca2+ nos
complexos com TnCF29HW sempre começou em pCas mais baixos do que nos
com TnC. Esta última indica que a presença de um 5HW na posição 29 diminui a
afinidade dos sítios de maior afinidade, como discutido (ver Discussão).
Os experimentos de diálise de fluxo são muito sensíveis a erros na
determinação das concentrações, tanto da proteína, como do Ca2+. Uma das
dificuldades para melhorar a qualidade dos dados para as diálises de fluxo dos
complexos deve ser a imprecisão maior na determinação na concentração de
proteína. Os complexos também foram submetidos a uma diálise bastante longa,
chegando a seis dias. A TnC é muito mais solúvel que a TnI e a TnT. A TnI
forma dímeros através de interligações dissulfeto por resíduos de cistina
intercadeias. A TnT é comumente degradada por proteases contaminantes.
Portanto, é provável que exista um excesso de TnC, e que parte dos complexos
não esteja funcional, tudo isso leva a uma imprecisão na determinação da
concentração dos complexos. Outra dificuldade é a retirada do Ca2+ das
preparações dos complexos troponina. Como os complexos possuem uma
afinidade por cálcio maior do que a TnC sozinha, nas mesmas condições é
esperado que exista mais Ca2+ ligado nos complexos (vide equação em Diálise

76
de Fluxo). Isto implicaria em uma situação onde os sítios de alta afinidade dos
complexos já estariam parcialmente ocupados por Ca2+ no início do experimento
de diálise da fluxo. Dados preliminares de absorção atômica confirmam que os
complexos binários e ternários possuem realmente mais do que um Ca2+ ligado
por TnC (não apresentado), mesmo após diálise mais longa. O que eleva a
concentração de Ca2+ contaminando a preparação para a mesma concentração
do complexo - ao redor de 10-5 M contra 10-6 M das preparações de TnC
isoladas. Isso quer dizer que para a realização das diálises de fluxo com TnC-
TnI ou Tn seria necessário modificar o tampão utilizado no ensaio.

77
REFERÊNCIAS BIBLIOGRÁFICAS
1. Bagshaw, C. R. (1993). Muscle contraction, second edition. Chapman &
Hall, London.
2. Chandra, M., McCubbin, W. D., Oikawa, K., Kay, C. M. e Smillie, L. B.
(1994). Ca2+, Mg2+, and troponin I inhibitory peptide binding to a Phe-154
to Trp mutant of chicken muscle troponin C. Biochemistry 33, 2961-2969
3. Chandra, M., da Silva, E. F., Sorenson, M. M., Ferro, J. A., Pearlstone, J.
R., Nash, B. E., Borgford, T., Kay, C. M. e Smillie, L. B. (1994). The effects
of N helix deletion and mutant F29W on the Ca2+ binding and functional
properties of chicken skeletal muscle troponin. J. Biol. Chem. 269, 14988-
14994
4. Carter, P., Bedouelle, H. e Winter, G. (1985). Improved oligonucleotide
site-directed mutagenesis using M13 vectors. Nucleic Acids Res. 13,
4431-4443
5. da Silva, A. C., Kendrick-Jones, J. e Reinach, F. C. (1992). Hybrid myosin
light chains containing a calcium-specific site from troponin C. Eur. J.
Biochem. 15, 85-91
6. da Silva, A. C., de Araujo, A. H., Herzberg, O., Moult, J., Sorenson, M. e
Reinach, F. C. (1993). Troponin-C mutants with increased calcium affinity.
Eur. J. Biochem. 213, 599-604
7. Drapeau, G. R., Brammar, W. J. e Yanofsky, C. (1968). Amino acid
replacements of the glutamic acid residue at position 48 in tryptophan
synthetase A of Escherichia coli. J. Mol. Biol. 35, 357-367
8. Ebashi, S., Endo, M. e Ohtsuki, I. (1969). Control of muscle contraction.
Quart. Rev. Biophys. 2, 351-384

78
9. Ebashi, S., Ohtsuki, I. e Mihashi, K. (1972). Regulatory proteins of muscle
with special reference to troponin. Cold Spring Harbor Symp. Quant. Biol.
37, 215-223
10. Farah, C. S., Miyamoto, C. A., Ramos, C. H. I., Silva, A. C. R., Quaggio,
R. B., Fujimori, K., Smillie, L. B. e Reinach, F. C. (1994). Structural and
regulatory functions of the NH2- and COOH-terminal regions of skeletal
muscle Troponin I. J. Biol. Chem. 269, 5230-5240
11. Farah, C. S. e Reinach, F. C. (1995). The troponin complex and regulation
of muscle contraction. FASEB J. 9, 755-767
12. Farah, C. S. e Reinach, F. C. (1999). Regulatory properties of
recombinant tropomyosins containing 5-hydroxytryptophan: Ca2+-binding
to troponin results in a conformational change in a region of tropomyosin
outside the troponin binding site. Biochemistry 38, 10543-10551
13. Finley, N., Dvoretsky, A. e Rosevear, P. R. (2000). Magnesium-calcium
exchange in cardiac troponin C bound to cardiac troponin I. J. Mol. Cell.
Cardiol. 32, 1439-1446
14. Fredricksen, R. S. e Swenson, C. A. (1996). Relationship between stability
and function for isolated domains of troponin C. Biochemistry 35, 14012-
14026
15. Fujimori, K., Sorenson, M., Herzberg, O., Moult, J. e Reinach, F. C.
(1990). Probing the calcium-induced conformational transition of troponin
C with site-directed mutants. Nature 345, 182-184
16. Gagné, S. M., Tsuda, S., Li, M. X., Smillie, L. B. e Sykes, B. D. (1995).
Structures of the troponin C regulatory domains in the apo and calcium-
saturated states. Nat. Struct. Biol. 2, 784-789
17. Gordon, A. M., Homsher, E. e Regnier, M. (2000). Regulation of
contraction in striated muscle. Physiol. Rev. 80, 853-924

79
18. Greaser, M. L. e Gergely, J. (1971). Reconstitution of troponin activity from
three proteins components. J. Biol. Chem. 246, 4226-4233
19. Grebarek, Z., Leavis, P. C. e Gergely, J. (1986). Calcium binding to the
low affinity sites in troponin C induces conformational changes in high
affinity domain: A possible route of information transfer in activation of
muscle contraction. J. Biol. Chem. 261, 608-613
20. Hanahan, D. (1985). Techniques for transformation in E. coli. In: DNA
cloning: a practical approach. Glover, D. M. IRL Press, Oxford.
21. Harada, K., Arana, C. e Potter, J. D. (2001). Magnesium-calcium
exchange with the high affinity Ca2+-Mg2+ binding sites of cardiac troponin.
J. Mol. Cell. Cardiol. 33, 593-596
22. Hartree, E. F. (1972). Determination of protein: a modification of Lowry
method that gives a linear photometric response. Anal. Biochem. 4, 422-
427
23. Head, J. F. e Perry, S. V. (1974). The interaction of the calcium binding
troponin (troponin C) with bivalent cations and the inhibitory protein
(troponin I). Biochem. J. 137, 145-154
24. Heeley, D. H., Golosinska, K. e Smillie, L. B. (1987). The effects of
troponin T fragments T1 and T2 on the binding of nonpolymerizable
tropomyosin to F-actin in the presence and absence of troponin I and
troponin C. J. Biol. Chem. 263, 9971-9978
25. Herzberg, O. e James, M. N. G. (1985). Structure of the calcium regulatory
muscle protein troponin-C at 2.8 Å resolution. Nature 313, 653-659
26. Herzberg, O., Moult, J. e James, M. N. G. (1986). A model for the Ca2+-
induced conformational transition of troponin C. A trigger for muscle
contraction. J. Biol. Chem. 261, 2638-2644

80
27. Higuchi, R. (1990). PCR Protocols: A Guide to Methods and Applications.
Academic Press, London, United Kingdom.
28. Hitchcock-De Gregori, S. E. (1982). Study of the structure of troponin I by
measuring the relative reactivities of lysines with acetic anhydride. J. Biol.
Chem. 257, 7372-7380
29. Hogue, C. W. V., Rasquinha, I., Szabo, A. G. e MacManus, J. P. (1992). A
new intrinsic fluorescent probe for proteins. FEBS 310, 269-272
30. Holmes, K. C., Popp, D., Gebhard, W. e Kabsch, W. (1990). Atomic model
of the actin filament. Nature 347, 44-49
31. Houdusse, A., Love, M. L., Dominguez, R., Grabarek, Z. e Cohen, C.
(1997). Structures of four Ca2+ bound troponin C at 2.0 Å resolution:
further insights into the Ca2+ switch in the calmodulin superfamily.
Structure 5, 1695-1711
32. Huxley, H. E. (1972). Structural changes in the actin- and myosin-
containing filaments during contraction. Cold Spring Harbor Symp. Quant.
Biol. 37, 361-376
33. Kabsch, W., Mannherz, H. G., Suck, D., Pai, E. F. e Holmes K. C. (1990).
Atomic structure of the actin: DNase I complex. Nature 347, 37-44
34. Kendrick-Jones, J., Rasera da Silva, A. C., Reinach, F. C., Messer, N.,
Rowe, T. e McLaughlin, P. (1991). Recombinant DNA approaches to study
the role of the regulatory light chains (RLC) using scallop myosin as a test
system. J. Cell. Sci. Suppl. 14, 55-58
35. Kunkel, T. A., Roberts, J. D. e Zakour, R. A. (1987). Rapid and efficient
site-specific mutagenesis without phenotypic selection. Methods. Enzymol.
154, 367-382
36. Laemmli, U. K. (1970). Cleavage of structural proteins during assembly of
head of the bacteriophage T4. Nature 227, 680-685

81
37. Lakowicz, J. R. (1983). Principles of fluorescence spectroscopy. Plenum
Press, New York.
38. Leavis, P. C., Rosenfeld, S. S., Gergely, J., Grabarek, Z. e Drabikowski,
W. (1978). Proteolytic fragments of troponin C: Localization of high and
low affinity Ca2+ binding sites and interactions with troponin I and troponin
T. J. Biol. Chem. 253, 5452-5459
39. Luo, Y., Wu, J. L., Li, B., Langsetmo, K., Gergely, J. e Tao, T. (2000).
Photocrosslinking of benzophenone-labeled single cysteine troponin I
mutants to other thin filament proteins. J. Mol. Biol. 296, 899-910
40. Luo, Y., Leszyk, J., Qian, Y., Gergely, J. e Tao, T. (1999). Residues 48
and 82 at the N-terminal hydrophobic pocket of rabbit skeletal muscle
troponin-C photo-cross-link to Met121 of troponin-I. Biochemistry 38,
6678-6688
41. McKay, R. T., Tripet, B. P., Hodges, R. S. e Sykes, B. D. (1997).
Interaction of the second binding region of troponin I with the regulatory
domain of skeletal muscle troponin C as determined by NMR
spectroscopy. J. Biol. Chem. 272, 28494-28500
42. Malnic, B., Farah, C. S. e Reinach, F. C. (1998). Regulatory properties of
the NH2- and COOH-terminal domains of troponin T: ATPase activation
and binding to troponin I and troponin C. J. Biol. Chem. 273, 10594-10601
43. Mercier, P., Li, M. X. e Sykes, B. D. (2000). Role of the structural domain
of troponin C in muscle regulation: NMR studies of Ca2+ binding and
subsequent interactions with regions 1-40 and 96-115 of troponin I.
Biochemistry 39, 2902-2911
44. Moncrief, N. D., Kretsinger, R. H. e Goodman, M. (1990). Evolution of EF-
hand calcium-modulated proteins. I - Relationships based on amino acid
sequences. J. Mol. Evol. 30, 522-562

82
45. Monteiro, P. B., Lataro, R. C., Ferro, J. A. e Reinach, F. C. (1994).
Functional α-tropomyosin produced in Escherichia coli. A dipeptide
extension can substitute the amino-terminal acetyl group. J. Biol. Chem.
269, 10461-10466
46. Ohtsuki, I. (1974). Localization of troponin in the filament and tropomyosin
paracrystal. J. Biochem. (Tokyo) 75, 753-765
47. Ohtsuki, I. (1979). Molecular arrangement of troponin-T in the thin
filament. J. Biochem. 86, 491-497
48. Oliveira, D. M., Nakaie, C. R., Sousa, A. D., Farah, C. S. e Reinach, F. C.
(2000). Mapping the domain of troponin T responsible for the activation of
actomyosin ATPase activity. Identification of residues involved in binding
to actin. J. Biol. Chem. 276, 27513-27519
49. Pearlstone, J. R., Borgford, T., Chandra, M., Oikawa, K., Kay, C. M.,
Herzberg, O., Moult, J., Herklotz, A., Reinach, F. C. e Smillie, L. B. (1992).
Construction and characterization of a spectral probe mutant of TnC:
Applications to analyses of mutants with increased Ca2+ affinity.
Biochemistry 31, 6545-6553
50. Pearlstone, J. R. e Smillie, L. B. (1995). Evidence for two-site binding of
troponin I inhibitory peptides to the N and C Domains of TnC. Biochemistry
34, 6932-6940
51. Pearlstone, J. R., Sykes, B. D. e Smillie, L. B. (1997). Interactions of
structural C and regulatory N domains of troponin C with repeated
sequence motifs in troponin I. Biochemistry 36, 7601-7606
52. Potter, J. D. e Gergely, J. (1975). The calcium and magnesium binding
sites on troponin and their role in the regulation of myofibrilar adenosine
triphosphatase. J. Biol. Chem. 250, 4628-4633

83
53. Quaggio, R. B., Ferro, J. A., Monteiro, P. B. e Reinach, F. C. (1993).
Cloning and expression of chicken skeletal muscle troponin I in
Escherichia coli: The role of rare codons on the expression level. Protein
Sci. 2, 1053-1056
54. Quaggio, R. B. (1994). Estudo da atividade inibitória da troponina I através
de mutações sítio-dirigidas. Tese de doutorado apresentada ao
Departamento de Bioquímica do Instituto de Química, USP.
55. Ramos, C. H. I. (1999). Mapping subdomains in the C-terminal region of
troponin I involved in its binding to troponin C and thin filament. J. Biol.
Chem. 274, 18189-18195
56. Reyment, I., Rypniewski, W. R., Schmidt-Bäse, K., Smith, R., Tomchick,
D. R., Benning, M. M., Winkelmann, D. A., Wesenberg, G. e Holden, H. M.
(1993). Three-dimensional structure of myosin subfragment-1: a molecular
motor. Science 261, 50-58
57. Reinach, F. C., Nagai, K. e Kendrick-Jones, J. (1986). Site-directed
mutagenesis of the regulatory light-chain Ca2+ / Mg2+ binding site and its
role in hybrid myosins. Nature 322, 80-83
58. Reinach, F. C. e Karlsson, R. (1988). Cloning, expression and site-
directed mutagenesis of chicken skeletal muscle troponin C. J. Biol.
Chem. 263, 2371-2376
59. Rosenfeld, S. S. e Taylor, E. W. (1985). Kinetic studies of calcium and
magnesium binding to troponin C. J. Biol. Chem. 260, 242-251
60. Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989). Molecular cloning: a
laboratory manual. Second ed., Cold Spring Harbor Laboratory Press,
Cold Spring Harbor, NY.
61. Sanger, F.; Nicklen, S. e Coulson, A. R. (1977). DNA sequencing with
chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74, 5463-5467

84
62. Sia, S. K., Li, M. X., Spyracopoulos, L., Gagné, S. M., Liu, W., Putkey, J.
A. e Sykes, B. D. (1997). Structure of cardiac muscle troponin C
unexpectedly reveals a closed regulatory domain. J. Biol. Chem.
272,18216-18221
63. Sheng, Z., Strauss, W. L., François, J. M. e Potter, J. D. (1990). Evidence
that both Ca(2+)-specific sites of skeletal muscle TnC are required for full
activity. J. Biol. Chem. 265, 21554-21660
64. Smillie, L. B. (1979). Structure and functions of tropomyosins from muscle
and non-muscle sources. Trends Biochem. Sci. 4, 151-155
65. Smillie, L. B., Golosinska, K. e Reinach, F. C. (1988). Sequences of
complete cDNAs encoding four variants of chicken skeletal muscle
troponin T. J. Biol. Chem. 263, 18816-18820
66. Sorenson, M. M., da Silva, A. C. R., Gouveia, C. S., Sousa, V. P., Oshima,
W., Ferro, J. A. e Reinach, F. C. (1995). Concerted action of the high
affinity calcium binding sites in skeletal muscle troponin C. J. Biol. Chem.
270, 9770-9777
67. Squire J. (1981). The structural basis of muscular contraction. Plenum
Press. New York, NY.
68. Squire, J. M. e Morris, E. P. (1998). A new look at thin filament regulation
in vertebrate skeletal muscle. FASEB J. 12, 761-771
69. Studier, F. W., Rosenberg, A. H., Dunn, J. J. e Dubendorff, J. W. (1990).
Use of T7 RNA polymerase to direct expression of cloned genes. Methods
Enzymol. 185, 60-89
70. Syska, H., Wilkinson, J. M., Grand, R. J. A. e Perry, S. V. (1976). The
relationship between biological activity and primary structure of troponin I
from white skeletal muscle of the rabbit. Bichem. J. 153, 375-387

85
71. Swenson, C. A. e Fredricksen, R. S. (1992). Interaction of troponin C and
troponin C fragments with troponin I and the troponin I inhibitory peptide.
Biochemistry 31, 3420-3429
72. Talbot, J. A. e Hodges, R. S. (1981). Synthetic studies on the inhibitory
region of rabbit skeletal troponin I. Relationship of amino acid sequence to
biological activity. J. Biol. Chem. 256, 2798-2802
73. Tripet, B., Van Eyk, J. E. e Hodges, R. S. (1997). Mapping of a second
actin-tropomyosin and a second troponin C binding sites within the C
terminus of troponin I, and their importance in the Ca2+-dependent
regulation of muscle contraction. J. Mol. Biol. 271, 728-750
74. Van Eyk, J. E., Thomas, L. T., Tripet, B., Wiesner, R. J., Pearlstone, J. R.,
Farah, C. S., Reinach, F. C. e Hodges, R. S. (1997). Distinct regions of
troponin I regulate Ca2+-dependent activation and Ca2+ sensivity of the
Acto-S1-TM ATPase activity of the thin filament. J. Biol. Chem. 272,
10529-10537
75. Vassylyev, D. G., Takeda, S., Wakatsuki, S., Maeda, K. e Maeda, Y.
(1998). Crystal structure of troponin C in complex with troponin I fragment
at 2.3-Å resolution. Proc. Natl. Acad. Sci. 95, 4847-4852
76. Womack, F. C. e Colowick, S. P. (1973). Rapid measurement of binding of
ligands by rate dialysis. Meth. Enzymol. 25, 464-471
77. Yu, A., Ballard, L., Smillie, L., Pearlstone, J., Foguel, D., Silva, J., Jonas,
A. e Jonas, J. (1999). Effects of high pressure and temperature on the
wild-type and F29W mutant forms of the N-domain of avian troponin C.
Biochim. Biophys. Acta. 1431, 53-56

86
CURRICULUM VITAE
Deodoro Camargo Silva Gonçalves de Oliveira
Brasileiro, nascido em 3 de Agosto de 1969
Educação
2o grau no Logos Escola de 2o Grau, 1986.
Bacharel em Ciências Biológicas no Instituto de Biociências, Universidade de
São Paulo, 1992.
Licenciado em Ciências Biológicas no Instituto de Biociências / Faculdade de
Educação, Universidade de São Paulo, 1994.
Doutorado em Biologia Molecular no Instituto de Química; Universidade de São
Paulo, até o presente.
Bolsas
Bolsista de Mestrado do CNPq, de Março de 1996 até Julho de 1998.
Bolsista de Doutorado da FAPESP, de Agosto de 1998 até Fevereiro de 2001.
Publicações
Oliveira, D. C. S. G. e Reinach, F. C., "The Calcium Induced Switch in the
Troponin Complex Probed by Fluorescent Mutants of Troponin I", in press.



















![Contração muscular-2010 [Modo de Compatibilidade]](https://img.document.onl/doc/110x75/5870be151a28ab30658b7e18/contracao-muscular-2010-modo-de-compatibilidade.jpg)









