Embed Size (px)
Citation preview

DESAFIOS DA TOMATICULTURA EM REGIÕES QUENTES E ÚMIDAS:
ADAPTAÇÃO GENÉTICA OU ARTIFICIALIZAÇÃO DOS AMBIENTES
DE CULTIVO?
Hiroshi Noda1
Introdução
O processo de domesticação de plantas teve inicio há pouco mais de dez milênios e
ocorreu, simultaneamente, em diferentes regiões do planeta. Assim, no mesmo espaço de tempo,
nas terras baixas da América Tropical, plantas alimentares como a mandioca, batata-doce, ariá,
cará, taioba, entre outras, foram domesticadas. A domesticação de plantas, a dispersão das
espécies agrícolas e o desenvolvimento da agricultura possibilitaram o surgimento de sociedades
humanas complexas.
O tomateiro é originário dos países andinos, desde o norte do Chile até a Colômbia e sua
domesticação ocorreu no México. Originalmente, era uma espécie geneticamente adaptada aos
ambientes de clima temperado. O seu cultivo nas regiões tropicais úmidas é um desafio
agronômico, pois, além das condições climáticas adversas ao desenvolvimento vegetativo e
reprodutivo da planta devem ser considerados outros fatores que podem prejudicar a
produtividade e qualidade da produção: doenças causadas por bactérias e fungos. Nessas
ocorrências, as perdas podem ser totais, pois, as condições ambientais prevalecentes (altas
temperaturas e umidade excessiva) atuam como fatores estressantes ao desenvolvimento do
tomateiro e, ao mesmo tempo, aceleram os ciclos reprodutivos e a dispersão dos patógenos.
O uso de tecnologias de controle ambiental associada à aplicação de agrotóxicos para o
controle de pragas e doenças tem sido uma alternativa para viabilizar o cultivo do tomateiro em
ambientes adversos. Nessas condições, as variedades geneticamente não adaptadas ao ambiente
natural podem completar satisfatoriamente o ciclo vegetativo e reprodutivo. Outra alternativa é
promover a melhoria genética do tomateiro visando sua adaptação aos fatores ambientais
adversos. A história da pesquisa agronômica brasileira é extremamente rica em exemplos
exitosos nos seus programas de melhoramento genético do tomateiro para cultivo nos
1 Pesquisador Titular aposentado do Instituto Nacional de Pesquisas da Amazônia/INPA.
Professor dos Programas de Pós-Graduação Ciências do Ambiente e Sustentabilidade na Amazônia/Universidade
Federal do Amazonas e Agricultura no Trópico Úmido/INPA.

diversificados ecossistemas do território nacional. Em um importante e histórico artigo, publicado
em 2009, Paulo Cesar Tavares de Melo, Arlete Marchi Tavares de Melo e Fernando Antonio de
Souza Aragão relatam os resultados obtidos por professores e pesquisadores de instituições
públicas e privadas relacionados aos estudos e criação de cultivares de tomate adaptadas para o
cultivo em ambientes sub-tropicais e tropicais úmidos e semi-áridos.
O tomateiro e o contexto ambiental no Trópico Úmido
Uma densa floresta tropical úmida constitui, ainda hoje, a paisagem predominante da
porção centro-ocidental da Amazônia brasileira. Aproximadamente 80% dessa floresta ocorre
sobre solos Latossolos e Argissolos de terra firme (áreas não inundáveis nos períodos de
enchentes dos rios). São solos frágeis e a floresta é mantida em equilíbrio por meio de uma
eficiente ciclagem de nutrientes envolvendo uma complexa rede de interações entre plantas,
animais e microrganismos (Alfaia et al., 2013). A transformação das florestas de terra firme em
áreas agrícolas exige o uso de técnicas adequadas uma vez que o estoque de nutrientes encontra-
se na fitomassa aérea e a retirada da floresta, interrompendo a dinâmica de reciclagem, pode ter,
como consequência, a exaustão do solo. Portanto, o manejo inadequado desse recurso natural
pode comprometer a sustentabilidade da atividade agrícola, principalmente em se tratando de
cultivo de espécies de ciclo anual, como no caso da maioria das hortaliças. Os solos dos
ecossistemas de várzea (áreas inundáveis) dos rios de água branca apresentam alta fertilidade
natural e são utilizadas para cultivo de espécies anuais, principalmente hortaliças. Entretanto, são
utilizáveis apenas no período das vazantes.
As características meteorológicas prevalecentes no trópico úmido são fatores que podem
afetam negativamente desenvolvimento vegetativo e reprodutivo do tomateiro. As temperaturas
diurna e noturna são consideradas elevadas e o curto período de insolação, devido a presença
constante de nuvens no inverno (época chuvosa) afeta o desenvolvimento vegetativo.
Temperaturas elevadas afetam a fisiologia da reprodução prejudicando a qualidade do pólen e
provocam o abortamento das flores e, consequentemente, a frutificação.
No final da década de setenta e início de oitenta do século passado olericultores das áreas
de produção próximas de Manaus já utilizavam técnicas para o controle da “murcha bacteriana”,
causada pela bactéria Ralstonia solanacearum, um fator limitante ao cultivo do tomateiro no
trópico úmido. Por exemplo, a enxertia de tomateiros suscetíveis em porta enxertos resistentes e
tratamento do solo para eliminação do patógeno. No primeiro caso, a baixa compatibilidade do
porta-enxerto em relação ao tomateiro não propiciava uma produtividade capaz viabilizar

economicamente o uso da técnica. No mesmo período, a Escola Agrotécnica Adventista, no
município de Rio Preto da Eva, AM, utilizou a tecnologia de cultivo de tomateiro em caixas com
solo esterilizado com brometo de metila em galpões com cobertura de plástico. As variedades
utilizadas foram Tropic e Floradel, de origem norte-americana. Durante alguns anos o uso desse
sistema de produção garantiu o abastecimento de significativa parte da demanda de Manaus.
Entretanto, o custo da manutenção do sistema devido ao processo de re-infecção do solo e a
ocorrência de epidemias da doença “mancha alvo”, causada por Corynespora cassiicola,
inviabilizou a continuidade do uso da tecnologia.
Melhoramento genético do tomateiro para cultivo no trópico úmido
O Programa de Melhoramento Genético de Hortaliças do INPA tem como objetivo gerar
conhecimentos e produtos destinados aos agricultores familiares da região do trópico úmido e,
assim, contribuir para uma produção diversificada de alimentos de qualidade obtida a partir da
observância aos preceitos agroecológicos e como isso garantir a segurança alimentar e nutricional
das populações humanas na Amazônia. As espécies convencionais mais conhecidas e consumidas
na região amazônica são as mesmas de outras regiões brasileiras. O tomate é um ingrediente
muito utilizado na culinária regional e, por isso, o tomateiro é cultivado mesmo nas remotas
comunidades rurais da Amazônia. O consumo, per capita, na Região Norte é 3,30 kg, ou seja,
cerca da 64 % da média brasileira ou, ainda, 68 % da média da Região Sul (IBGE, 2004). O
tomate é a mais importante hortaliça convencional que vem sendo estudada pelo INPA e a
“murcha bacteriana” é um fator limitante ao cultivo do tomateiro. A bactéria R. solanacearum,
agente causal da doença, é parte da biodiversidade da região tropical úmida de baixa altitude.
Assim, as pesquisas têm sido direcionadas no sentido do melhoramento para resistência do
tomateiro ao patógeno R. solanacearum e tolerância ao cultivo sob condições de temperaturas
elevadas.
Variabilidade genética e dinâmica de populações em R. solanacearum
R. solanacearum é o agente causal da “murcha bacteriana” em mais de 200 espécies
hospedeiras. É encontrada em solos ainda não cultivados e possui aptidão para sobreviver no solo
ou na rizosfera de seus hospedeiros e de outras espécies vegetais. Boher et al. (2013), em
levantamentos realizados em vinte e dois municípios abrangendo todas as microrregiões do
Estado do Amazonas encontraram isolados de R. solanacearum classificados como biovar 1, N2,

também denominada biovar 2 tropical, e biovar 3 presentes em doze espécies e seis famílias
(Figura 1). Em tomateiro, a biovar mais frequentemente encontrada foi o 1. Em pimentão e
pimenta-de-cheiro foram encontrados as biovares 1 e 3, sendo, a biovar 3, mais frequente. Os
isolados da biovar 1 foram encontrados em bananeira e de Heliconia sp., provocando murcha em
bananeiras e apresentando níveis de agressividade variáveis em solanáceas.
Figura 1. Ocorrência e variabilidade de R. solanacearum no Estado do Amazonas. Fonte: Boher
et al., 2013.
Boher (2013) demonstrou, em experimentos realizados em ambiente de terra firme e
várzea, a alta variabilidade genética do patógeno mesmo em áreas restritas. Numa mesma parcela
(5m2) foram isoladas de plantas infectadas com biovares 1 e 3 de R. solanacearum (Figura 2).
Nas parcelas com a testemunha suscetível Santa Cruz Kada foi detectada a presença das biovares
1 e 3 em plantas diferentes ou ambas biovares na mesma planta. Nas parcelas com a testemunha
resistente Yoshimatsu foi detectada somente a presença da biovar 1 nas plantas infectadas.
Interação hospedeiro x patógeno x ambiente
Noda et al. (1986), avaliando genótipos de tomateiro em experimento de campo, em solos
naturalmente infestados por R. solanacearum, já haviam verificado que a evolução do processo

da doença “murcha bacteriana” apresentava características epidemiológicas bem definidas, onde
níveis elevados de resistência, conferida por um genótipo, eram correlacionados aos baixos níveis
de velocidade do progresso da doença na população hospedeira. Na figura 3 é apresentado a
distribuição espacial das parcelas e os blocos de um experimento, realizado no ano de 2010 em
área de várzea, para avaliação da resistência ao patógeno R. solanacearum. Nas parcelas bz, cw,
fy e Hz, ocupadas pela cultivar suscetível Santa Cruz Kada é possível observar o padrão da dinâmica
epidemiológica da doença. O início do aparecimento dos sintomas ocorreu a partir de dezenove dias após
o transplante das mudas no campo (parcela bz) e a seguir o processo epidemiológico expande-se por toda
a parcela ocupada pela cultivar. Por outro lado, nas parcelas ocupadas pela variedade resistente
Yoshimatsu 4-11 (ax, dy, fx, hx) não houve ocorrência da doença ou, quando ocorreu, a planta infectada
expressou o sintoma da doença após um período mais longo em relação à data do transplante e o processo
epidemiológico não se expandiu por toda a parcela. Em relação ao total de plantas infeccionadas foi
observado que o genótipo suscetível apresentou 92% de ocorrência, ao passo que no genótipo resistente a
ocorrência foi de 7%. Os resultados mostram que a ocorrência total de 17% e 8% de plantas infectadas,
respectivamente, nas parcelas ocupadas pelas cultivares Caraiba e Yoshimatsu 4 – 11 concordam com os
obtidos por Prior et al. (1996) que utilizaram como critério para registro de ocorrência da doença a
presença do patógeno nos tecidos vasculares das plantas infectadas. Esses autores alcançaram resultados
mostrando que a severidade da doença é maior na estação mais quente.
Metodologia para incorporação de resistência genética
Uma vez que nas condições de cultivo, a expressão da resistência genética do tomateiro é
resultante da sua interação com populações de R. solanaceraum, presume-se que sua estabilidade,
no tempo e espaço, possa ser estimada a partir de estudos que levem em conta a alta
variabilidade genética do patógeno e a capacidade do hospedeiro em completar um ciclo
reprodutivo quando cultivado em ambientes desfavoráveis e heterogêneos (temperatura e
umidades elevadas) e na presença de solos naturalmente infestados pelo patógeno. A resistência

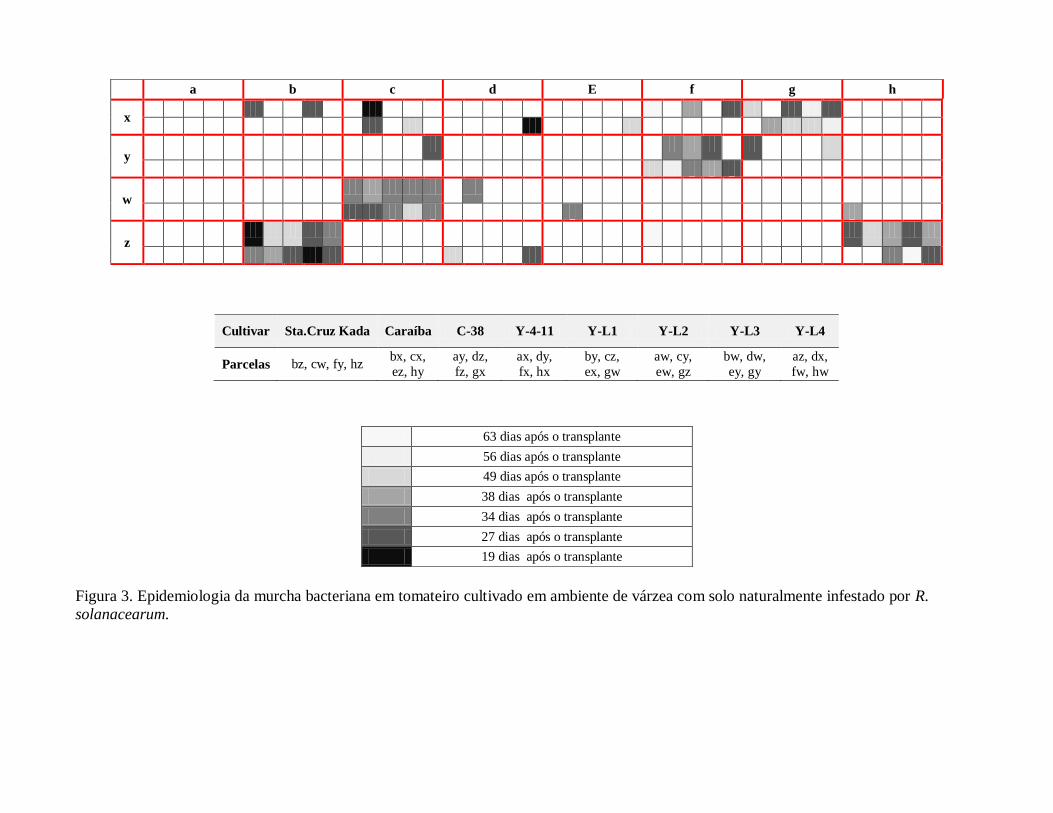
Figura 3. Epidemiologia da murcha bacteriana em tomateiro cultivado em ambiente de várzea com solo naturalmente infestado por R.
solanacearum.
a b c d E f g h
x
y
w
z
Cultivar Sta.Cruz Kada Caraíba C-38 Y-4-11 Y-L1 Y-L2 Y-L3 Y-L4
Parcelas bz, cw, fy, hz bx, cx,
ez, hy
ay, dz,
fz, gx
ax, dy,
fx, hx
by, cz,
ex, gw
aw, cy,
ew, gz
bw, dw,
ey, gy
az, dx,
fw, hw
63 dias após o transplante
56 dias após o transplante
49 dias após o transplante
38 dias após o transplante
34 dias após o transplante
27 dias após o transplante
19 dias após o transplante

genética do hospedeiro ao patógeno deve permanecer estável seja em relação ao tempo, aos
diferentes espaços de cultivo frente à extrema variabilidade genética do patógeno e aos estresses
ambientais que podem interferir negativamente na expressão do caráter. Dessa forma, foram
adotados métodos que, levando em conta todos os fatores intervenientes nas complexas
interações hospedeiro x patógeno x ambiente, permitissem avaliações confiáveis dos níveis de
resistência das progênies selecionadas.
Até o final da década de oitenta, do século passado, havia duas tendências distintas no que
tange à interpretação dos mecanismos controladores da resistência genética do tomateiro em
relação ao patógeno R. solanacearum. Alguns autores encontraram evidências de controle
oligogênico na reação de resistência (Acosta et al., 1964; Digat & Derieux, 1968). Entretanto, as
cultivares americanas Saturn e Venus, consideradas resistentes segundo esta interpretação,
apresentaram reação de susceptibilidade à R. solanacearum quando avaliadas, no Brasil, em casas
de vegetação, com temperaturas máximas (acima de 30oC), consideradas elevadas para o
tomateiro (Couto et al., 1979 e Martins et al., l988). Por outro lado, Russel (1978) e Villareal,
(1980), afirmam, respectivamente, que a herança é complexa e o controle da expressão de
resistência está fortemente correlacionado com as condições ambientais.
Os ensaios para discriminação das progênies resistentes frente à alta variabilidade do
patógeno foram realizados em ambientes contrastantes da várzea (áreas inundáveis) e terra firme
(áreas não inundáveis), naturalmente infestados pelo patógeno e, para estimar os níveis de
resistência das progênies, foi utilizado o parâmetro epidemiológico para doenças monocíclicas
Taxa de Infecção (QR), segundo recomendação de Planck (1963).
Os padrões constantes de referência (testemunhas) de reação do hospedeiro ao patógeno
são as cultivares Santa Cruz Kada, com reação de suscetibilidade e Caraiba, com reação de
resistência (Noda et al., 1986; Noda & Machado, 1993; Noda, 1997 e Noda et al., 1997). A
presença da cultivar Santa Cruz Kada em todos os ensaios tem, também, a finalidade de certificar
a presença do patógeno e o potencial de inoculo.
Nas avaliações de resistência ao patógeno é utilizada uma variação do método descrito por
Kuriyama (1975), no qual as plantas com ausência ou pouco sintomas da doença no final do ciclo
são consideradas resistentes. A presença da bactéria nos feixes vasculares é constatada, segundo
método descrito por Király et al., (1970) e consiste na exudação de células bacterianas na forma
de um líquido de coloração leitosa quando pequenos pedaços de tecido afetado pelo patógeno são
colocados sobre uma placa de vidro com água. As leituras de ocorrência de murcha bacteriana

são iniciadas após o transplante das mudas no campo e, em intervalos semanais, faz-se o registro
das plantas afetadas pela doença, anotando-se, também, a posição das plantas nas parcelas.
Avaliação continuada no tempo usando testemunhas constantes
Durante o processo de avaliação e seleção, no período de 1983/92, observou-se,
consistentemente, a ocorrência de contrastes significativos para os caracteres de resistência à R.
solanacearum e de produção de frutos sob condição de cultivo em solos infestados pelo
patógeno. As progênies selecionadas para as etapas subsequentes apresentaram maiores níveis de
resistência ao patógeno e maior capacidade de produção em solos infestados quando comparadas
com a testemunha susceptível (grupo Santa Cruz) e a testemunha resistente (Caraiba). Noda et
al., (1986), observaram que o caráter Taxa de Infecção (QR), por se tratar de um parâmetro
epidemiológico, consegue detectar um maior número de contrastes significativos quando
comparado com outros parâmetros que não consideram a resistência do hospedeiro como
resultante de um processo cuja expressão varia em função do tempo.
Com base nos dados obtidos em experimentos realizados no período de 1983 até 2010 foi
observado a estabilidade nos níveis de resistência das progênies e linhagens obtidas derivadas do
cruzamento HT – 16, que deu origem à cultivar Yoshimatsu, quando comparadas com os
referenciais constantes (Tabela 1). Quando ocorreu ausência completa da doença nas progênies,
como foi observado em alguns experimentos (gerações F8, em 1988; F10, em 1992 e linhagem
Yoshimatsu L-3, período 2005 - 2010), supõe-se que tenha sido devida aos menores níveis do
potencial de inoculo do solo, ocorridos naqueles experimentos. Os dados sobre os níveis de
resistência obtidos em 1988, pela testemunha resistente Caraíba (QR = 0,0079), em 1992 (QR =
0,0039) e em 2005 (QR = 0,0079) que foram os mais elevados, considerando-se todo o período
de 1986 a 2005, reforçam a sugestão da ocorrência de níveis mais baixos de potencial de inóculo
nos locais onde se realizaram os experimentos de 1988 e 2005. Por outro lado, nos ensaios
realizados em 1987 e 2001, para avaliação das progênies F7, F9 e F13 (Linhagem Yoshimatsu L-3),
respectivamente, observaram-se os maiores valores da Taxa de Infecção para a testemunha
Caraiba. Mesmo naquelas condições, as resistências estimadas das progênies selecionadas na
geração F7 foram 17 vezes mais elevadas, em relação à cultivar Caraiba, 29 vezes superior na
geração F9 e 17 vezes superior, na geração F13. Assim, o uso de testemunhas susceptível e
resistente constantes mostrou ser muito recomendável para a avaliação do nível de contaminação
do solo com o patógeno, em cada experimento e esse procedimento é um recurso muito valioso
para se poder estimar o ganho genético ocorrido no processo de seleção das progênies.
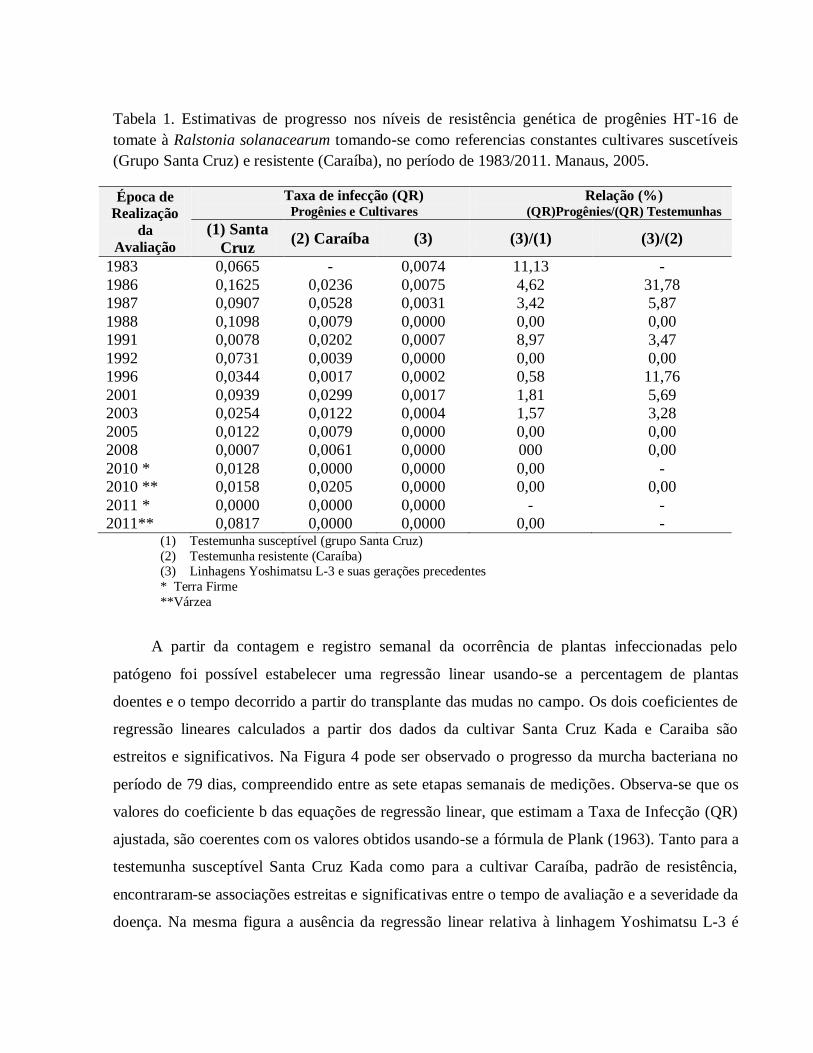
Tabela 1. Estimativas de progresso nos níveis de resistência genética de progênies HT-16 de
tomate à Ralstonia solanacearum tomando-se como referencias constantes cultivares suscetíveis
(Grupo Santa Cruz) e resistente (Caraíba), no período de 1983/2011. Manaus, 2005.
Época de
Realização
da
Avaliação
Taxa de infecção (QR) Progênies e Cultivares
Relação (%) (QR)Progênies/(QR) Testemunhas
(1) Santa
Cruz (2) Caraíba (3) (3)/(1) (3)/(2)
1983 0,0665 - 0,0074 11,13 -
1986 0,1625 0,0236 0,0075 4,62 31,78
1987 0,0907 0,0528 0,0031 3,42 5,87
1988 0,1098 0,0079 0,0000 0,00 0,00
1991 0,0078 0,0202 0,0007 8,97 3,47
1992 0,0731 0,0039 0,0000 0,00 0,00
1996 0,0344 0,0017 0,0002 0,58 11,76
2001 0,0939 0,0299 0,0017 1,81 5,69
2003 0,0254 0,0122 0,0004 1,57 3,28
2005
2008
2010 *
2010 **
2011 *
2011**
0,0122
0,0007
0,0128
0,0158
0,0000
0,0817
0,0079
0,0061
0,0000
0,0205
0,0000
0,0000
0,0000
0,0000
0,0000
0,0000
0,0000
0,0000
0,00
000
0,00
0,00
-
0,00
0,00
0,00
-
0,00
-
- (1) Testemunha susceptível (grupo Santa Cruz)
(2) Testemunha resistente (Caraíba) (3) Linhagens Yoshimatsu L-3 e suas gerações precedentes
* Terra Firme
**Várzea
A partir da contagem e registro semanal da ocorrência de plantas infeccionadas pelo
patógeno foi possível estabelecer uma regressão linear usando-se a percentagem de plantas
doentes e o tempo decorrido a partir do transplante das mudas no campo. Os dois coeficientes de
regressão lineares calculados a partir dos dados da cultivar Santa Cruz Kada e Caraiba são
estreitos e significativos. Na Figura 4 pode ser observado o progresso da murcha bacteriana no
período de 79 dias, compreendido entre as sete etapas semanais de medições. Observa-se que os
valores do coeficiente b das equações de regressão linear, que estimam a Taxa de Infecção (QR)
ajustada, são coerentes com os valores obtidos usando-se a fórmula de Plank (1963). Tanto para a
testemunha susceptível Santa Cruz Kada como para a cultivar Caraíba, padrão de resistência,
encontraram-se associações estreitas e significativas entre o tempo de avaliação e a severidade da
doença. Na mesma figura a ausência da regressão linear relativa à linhagem Yoshimatsu L-3 é

explicada pela ausência de ocorrência de plantas infectadas durante todo o período do
experimento (QR = zero).
ID: Índice de Doença
**: Significativo ao nível de 1% de probabilidade pelo teste t.
Figura 4. Progresso da murcha bacteriana, causada por Ralstonia solanacearum, em cultivares e
linhagem de tomate cultivadas em Manaus, em 2005 em solo naturalmente infestado pelo
patógeno.
Caraíba
y = 0,0632x - 0,1532
r = 0,9038 **
Santa Cruz
y = 0,1306x - 0,2189
r = 0,9746 **
-0.2
0
0.2
0.4
0.6
0.8
1
30 37 44 51 58 65 71 79
log e
[ 1
/(1
-ID
)]
Dias após o transplante
Cultivar ou
Linhagem QR
Coeficiente
b r
Santa Cruz 0,0122 0,1306 0,9746
Caraíba 0,0079 0,0632 0,9038
Yoshimatsu L-3 0

Herança da resistência do tomateiro ao patógeno Ralstonia solanacearum
Um aspecto importante a se considerar no processo de incorporação da resistência genética
no tomateiro está relacionado à dinâmica de populações do patógeno no processo de interação
com o hospedeiro. Resultados obtidos por Lopes et al (1994), em experimento em ambiente
controlado e inoculação artificial, mostraram que a cultivares resistentes de modo geral
apresentam alta resistência em relação às biovares do grupo 3 e resistência parcial às biovares do
grupo 1. Esses resultados coincidem com aqueles obtidos por Boher et al (2013) nos ensaios em
áreas naturalmente infestados. O fato das cultivares de tomateiro com maiores níveis de
resistência serem infectadas somente por isolados da biovar 1 reforça a hipótese do nível mais
elevado de agressividade dessa biovar em relação à biovar 3. Ou seja, na expressão da resistência
genética, não ocorre interação específica entre o patógeno e o hospedeiro, mas diferentes níveis
de agressividade do patógeno, expressos sob a forma de diferentes níveis de resistência do
hospedeiro.
Levando-se em conta os resultados obtidos em experimentos e os conceitos enunciados por
Plank (1975) presume-se que a capacidade patogênica dentro da variabilidade genética de R.
solanacearum expressa-se em níveis de agressividade e não em virulência. Nesta circunstância
considera-se que a resistência genética do tomateiro ao patógeno, baseada na relação gene a gene,
resistência do hospedeiro e virulência do patógeno, não seja efetiva ao longo do tempo. Por outro
lado, considera-se que a modalidade de resistência expressa pela variedade Yoshimatsu, é do tipo
horizontal ou poligênico. Assim, pode ser afirmado que esta variedade apresentará variação nos
níveis de resistência em função da agressividade da população de patógeno ou do potencial de
inoculo do solo infestado, mas não ocorrerá risco de quebra da resistência incorporada.
Evidências adicionais de que o controle envolvido no mecanismo de resistência da variedade
Yoshimatsu é poligênico têm sido obtidas por meio de diferentes modalidades de ensaios, em
diferentes locais, inclusive fora dos trópicos úmidos.
No exterior, experimentos realizados por Prior et al. (1996) confirmaram os resultados
obtidos no Brasil. Menezes (1998), utilizando a metodologia de triagem de genótipos resistentes
em solos naturalmente infestadas pelo patógeno, no Estado de Pernambuco, demonstrou de que a
herança envolvida no controle da expressão de resistência na variedade Yoshimatsu é governada
por mais de um gene ou bloco gênico, exibindo dominância, além de efeitos aditivos no aumento
do caráter. Este autor encontrou, também, efeitos positivos na capacidade geral e específica de
combinação para o caráter pegamento de frutos, produção de frutos, número de frutos, peso

médio de frutos, número de lóculos e sobrevivência. Lopes et al. (1994), efetuando inoculações
artificiais com isolados de diversas regiões do Brasil e Campos et al. (1998), no Estado de
Tocantins, realizando inoculações com isolados locais, confirmaram a resistência da variedade
Yoshimatsu em relação à R. solanacearum. Oliveira et al., (1998) inocularam a cultivar
Yoshimatsu 4-11 com dois isolados, um classificado como biovar 1 e outro como biovar 3 e
chegaram à conclusão que a herança da resistência é de natureza quantitativa com dominância
parcial, sendo observado a presença de efeito aditivo significativo.
As maiores magnitudes dos coeficientes de variação experimentais têm ocorrido, de
maneira geral, para o caráter produtivo quando comparados com aqueles observados para o
parâmetro Taxa de Infecção, que expressa níveis de resistência. Esses dados sugerem que, apesar
de tratar-se de um caráter de herança complexa, a magnitude da herdabilidade da resistência do
tomateiro ao patógeno R. solanacearum é superior às características relacionadas à produção.
Mesmo porque, quando o cultivo é realizado sob condição de solo infestado pelo patógeno, a
resistência genética é um componente que interfere decisivamente no ciclo de vida e capacidade
produtiva do hospedeiro.
Seleção para tolerância ao calor
O tomateiro quando cultivado em locais com temperaturas elevadas apresenta a tendência
de abortamento das flores comprometendo a frutificação e, por conseguinte, a produção. Por isso,
os genótipos selecionados para resistência à R. solanacearum foram avaliados, também, para
tolerância ao calor. A temperatura noturna ótima para o pegamento de frutos variam de 15 a 20oC
que são temperaturas incomuns no ambiente tropical. Em altas temperaturas a polinização do
tomateiro é prejudicada porque o estilete alonga-se ultrapassando o cone de anteras da flor
(Villareal, 1980). O pólen não atinge o estigma receptivo, impedindo a polinização e os demais
processos seguintes, fecundação e formação do fruto. Noda e Machado (1992) realizaram três
experimentos no período de maio em 1988 nos quais progênies F-7 do cruzamento HT-16 foram
cultivadas no interior de casa de vegetação com cobertura de plástico e envolta com tela de
plástico. A amplitude de variação da temperatura do ar foi de 190 C, registrado por volta de 06:00
horas até 430
C, registrado próximo às 14:00 horas. No experimento foi possível detectar a
variabilidade genética, ainda disponível, dentro das progênies (Tabela 2). A partir dos resultados
obtidos naquele experimento foi possível selecionar progênies com níveis mais elevados de
tolerância ao cultivo em condições de temperaturas elevadas, confirmada por Silva et al. (2010)
em experimento realizado na Universidade Federal Rural de Pernambuco.

Tabela 2. Médias de percentagem de pegamento de frutos entre plantas dentro
de progênies e médias das progênies quando cultivadas sob condições de
temperatura elevadas. Manaus. (Fonte: Noda & Machado (1992)).
Plantas
dentro
de Progênies
PROGÊNIES
HT-16-9-2-7-5-1 HT-16-9-2-7-5-3 H6-16-9-4-12-2-5
Percentagem de pegamento de frutos
1 97,87 a 55,06 abcd 45,83 d
2 80,04 b 39,05 d 73,66 bc
3 91,87 ab 65,21 abcd 46,87 d
4 87,38 ab 76,13 a 73,23 bc
5 94,37 ab 74,37 ab 89,46 ab
6 100,00 a 44,58 bcd 87,50 ab
7 93,75 ab 70,30 abc 100,00 a
8 90,97 ab 82,29 a 75,00 bc
9 100,00 a 70,16 abc 67,45 cd
Médias 92,59 A 64,13 B 73,22 AB Nas colunas as médias seguidas das mesmas letras minúsculas não diferem entre si pelo teste de
Duncan ao nível de 5% de probabilidade.
Na linha as médias seguidas pelas mesmas letras maiúsculas não diferem entre si pelo teste de
Duncan ao nível de 5% de probabilidade.
Situação atual e próxima estapa das pesquisas
Os trabalhos posteriores realizados no período de 1997 a 2009 apontaram para a linhagem
Y-95-3 (Yoshimatsu), como promissora para resistência à murcha bacteriana, com produtividade
(38,40 t/ha) e qualidade do fruto (Figura 5). Atualmente, a prioridade das pesquisas é selecionar,
dentro do cruzamento HT-16, genótipos capazes de produzir frutos de melhor qualidade, como a
resistência à podridão estilar e menor índice de rachadura.
No processo de seleção para o desenvolvimento da cultivar Yoshimatsu foram levados em
conta, também, a adaptabilidade e estabilidade dos genótipos perante ambientes heterogêneos.
Para isso, foram realizados ensaios multilocais para estimar os níveis de adaptabilidade genética e
estabilidade fenotípica das linhagens selecionadas, segundo procedimento proposto por Eberhart
e Russel (1966). Em 2002 um conjunto de quatro experimentos em ambientes contrastantes de
terra firme e várzea mostrou que as linhagens avançadas do cruzamento HT-16 (Yoshimatsu L-1
e Yoshimatsu L-2) são adaptadas para o cultivo em ambientes de terra firme e várzea. As

Figura 5. Cultivo em solos infestados por R. solanacearum. Protegido (esquerda) e campo aberto
(direita). Estação Experimental de Hortaliças “Dr. Alejo Von der Pahlen”. Manaus.
progênies avançadas L-3-2002 e L-4-2002 foram consideradas estáveis, rústicas e produtivas em
qualquer ambiente e pouco responsivas à melhora ambiental, quanto aos caracteres produtivos
(Noda et al., 2013; Pena et al. 2010). Em outro conjunto de cinco experimentos realizados em
terra firme e várzea entre 2008 a 2011 os resultados obtidos confirmaram os níveis de resistência
da cultivar Yoshimatsu 4-11 e das linhagens Yoshimatsu L-1, Yoshimatsu L-2, Yoshimatsu L-3 e
Yoshimatsu L-4 e superiores em relação à cultivar Caraiba (Tabela 3). Em relação à
produtividade em massa Yoshimatsu 4-11 e as linhagens apresentaram médias semelhantes à
Caraiba exceto a linhagem Yoshimatsu L-3 que apresentou média superior. As estimativas dos
coeficientes de regressão linear (β) de todos os genótipos avaliados apresentaram valores não
significativos revelando boa adaptabilidade aos ambientes onde ocorreram os experimentos.
Neste aspecto, para o caráter Taxa de Infecção (QR), a cultivar Caraíba e Santa Cruz Kada
apresentaram valores de β > 1 indicando adaptação apenas aos ambientes favoráveis. Por outro
lado, os genótipos Santa Cruz Kada, Caraíba e Yoshimatsu L-3 apresentaram valores dos desvios
de regressão σ2d significativos e R
2 inferior a 60% indicando baixa previsibilidade de
comportamento dos genótipos. Em relação caráter Produção de Massa (PTM) o genótipo Santa
Cruz Kada, apresentou valor de σ2d significativo indicando, também, baixa previsibilidade. Em
relação ao mesmo caráter, C-38 e Yoshimatsu L-4 apresentaram σ2d significativos mas, os R
2
estimados foram superiores a 60%, indicando ajustamento à regressão devido ao componente
genético em detrimento do ambiental, segundo Vencovsky e Barriga (1992).
As pesquisas devem seguir a orientação inicial da concepção do material geneticamente
melhorado. Isso significa continuar avaliando as progênies avançadas do material desenvolvido,

quanto à adaptabilidade genética e estabilidade fenotípica, resistência, produção e qualidade do
fruto. Outro ponto importante é o compromisso social de continuar na perspectiva do
melhoramento genético vegetal onde o seu produto possa ser usado de forma generalizada pelas
populações locais. Ou seja, a sustentabilidade da agricultura familiar praticada na região
amazônica em base agroecológica, mais independente de insumos comerciais e contribuir para o
aumento da produção e disponibilidade de cultivos agrícolas indispensáveis para a alimentação
humana.

Tabela 3. Valores médios de Taxa de Infecção (QR), rendimento em frutos expresso em produção total de massa (PTM) em tomateiros (Solanum
lycopersicum L.) cultivados em cinco ambientes de várzea e terra firme, no período de 2008, 2009, 2010 e 2011, com solos naturalmente
infestados por Ralstonia solanacearum e estimativas de parâmetros de adaptabilidade genética e estabilidade fenotípica.
(1)Dados não transformados;
Taxa de Infecção (QR);
Produção Total de Massa (PTM) expressa em g 0,5 m-2;
QML = Quadrado Médio dos Desvios de Regressão Linear; β = Coeficiente de Regressão Linear; σ2
d = Variância dos Desvios da Regressão; R2
=
Coeficiente de Determinação; Na coluna QR as médias seguidas pelas mesmas letras não diferem estatisticamente entre si pelo teste Scott e Knott a nível de 5% de probabilidade;
Na coluna PTM as médias seguidas pelas mesmas letras não diferem estatisticamente entre si pelo teste Tukey a nível de 5% de probabilidade;
Nas colunas QML e σ2d pelo teste F e na Coluna β pelo Teste t;
** Significativo a 1% de probabilidade; * Significativo a 5% de probabilidade; ns - Não Significativo estatisticamente; (i) Valor calculado menor do que zero.
Genótipos Médias
(1) QML β σ
2d R
2
QR PTM QR PTM QR PTM QR PTM QR PTM
Caraíba 0,0060 b
527,40 bc 9,0306 ns 55,8653 ns 1,5792 ns 1,0193 ns 1,8286** 8,4233 ns 34,8628 92,8471
C-38 0,0042 a 814,60 abc
0,6795 ns 192,3839 ns 1,1770 ns 1,0990 ns 00000(i) ns 42,5530** 79,8037 81,4192
Y-4-11 0,0003 a 982,20 ab 0,5412 ns 49,9458 ns 0,4838 ns 1,0049 ns 00000(i) ns 6,9434 ns 45,5980 93,3831
Y-L1 0,0024 a
972,00 ab 1,4738 ns 94,5350 ns 1,1130 ns 0,9999 ns 0,0000(i) ns 8,1907 ns 61,9613 92,7017
Y-L2 0,0003 a 946,40 ab 1,8723 ns 45,1039 ns 0,8999 ns 1,0205 ns 0,0390 ns 5,7330 ns 45,5980 94,1579
Y-L3 0,0006 a 1.115,80 a 5,7651* 27,2976 ns 0,9790 ns 0,9030 ns 1,0122* 1,2814 ns 24,3687 95,4227
Y-L4 0,0024 a 963,60 ab 1,1859 ns 139,0374** 1,3033 ns 1,0706 ns 0,0000(i) ns 29,2163** 73,5179 85,1946
Santa Cruz Kada 0,0205 b 408,20 c 13,0361** 163,4515** 0,4648 ns 0,8828 ns 2,8299** 35,3199** 3,1113 76,8937

Referências Bibliográficas
ACOSTA, J. C.; GILBERT, J. C. ; QUINON, V.L. 1964. Heritability of bacterial wilt resistance
in tomato. Proceedings of American Society for Horticultural Science, Geneva, 84: 455 –
462.
ALFAIA, S. S.; RODRIGUES, M. R. L.; UGUEN, K.; AYRES, M. I. C. Manejo do nitrogênio
em agrossistemas de terra firme e várzea na Amazônia. In: Noda, H.; Souza, L. A. G.; Silva
Filho, D. F. Pesquisas Agronômicas para a Agricultura Sustentável na Amazônia Central.
Manaus : Wega. p: 201 -217. 2013.
BOHER, B.; COELHO NETTO, R. A.; NODA, H. Caracterização de Ralstonia solanacearum no
Estado do Amazonas. . In: Noda, H.; Souza, L. A. G.; Silva Filho, D. F. Pesquisas
Agronômicas para a Agricultura Sustentável na Amazônia Central. Manaus : Wega. p: 73 -
86. 2013.
CAMPOS, G.A.; SILVEIRA, M.A.; AZEVEDO, S.M.; MALUF, W.R.; RESEND, J.T.V. 1998.
Resistência de linhagens de tomateiro à murcha bacteriana no Estado do Tocantins.
Horticultura Brasileira, Brasília, 16 (1): (Resumo n.º 046).
COUTO, F. A. A.; MIZUBUTI, A.; MATSUOKA, K.; CAMPOS, J. P. 1979. Avaliação do grau
de resistência à Pseudomonas solanacearum de cinco cultivares de tomateiro e das
progênies resultantes do cruzamento entre eles. Revista de Olericultura, XVII: 48-58
DIGAT, B.; DERIEUX, M. 1968. A study of the varietal resistance of tomato to bacterial wilt II.
The practical value of F1 hybrids ans their contribuition to the genetic basis of resistance. In:
Proceedings of the Annual Meeting Caribbean Food Crops Society. St. Augustine, p: 85 –
100.
EBERHART, S. A,; RUSSEL, W. A. 1966. Stability parameters for comparing varieties. Crop
Science, v.6: 36 – 40.
IBGE. 2004. Pesquisa de orçamentos familiares 2002-2003: aquisição alimentar domiciliar per
capita: Brasil e grandes regiões. IBGE, Coordenação de Índices de Preços. Rio de
Janeiro:IBGE. 280 p.
KURIYAMA, T. 1975. Testing methods for breeding disease-resistant vegetables in Japan.
Japan. Agriculture Research Quaterly, 9:96-100.
LOPES, C.A., QUEZADO-SOARES, A.M.; MELO, P.E. 1994. Differential resistance of tomato
cultigens to biovares I e III of Pseudomonas solanacearum. Plant Disease, 78: 1091-1094.
MARTINS, O. M.; REIFSCHNEIDER, F. J. B.; TAKATSU, A.; PESSOA, H. B. S. V. 1988.
Fonte de resistência em tomateiro à Pseudomonas solacearum. Horticultura Brasileira, 6
(2): 17-19.
MELO, P. C. T.; MELO,A. M. T.; ARAGÃO, F. A. S. Melhoramento genético de hortaliças no
Brasil: Retrospectiva e perspectiva. In: Vidal Neto, F. C. et al. I Simpósio Nordestino de
Genética e Melhoramento de Plantas. Anais. Fortaleza : Sociedade Brasileira de
Melhoramento de Plantas, 2009. p. 60 - 82.

MENEZES, D. 1998. Análise Genética de um Cruzamento Dialelico em Tomateiro
(Lycopersicon esculentum Mill). Universidade Federal Rural de Pernambuco. Recife. 95 p.
(Tese de Doutorado).
NODA, H. & MACHADO, F. M. 1992. Avaliação de progênies de tomate (Lycopersicon
esculentum) para cultivo sob temperaturas elevadas. Acta Amazonica, Manaus, 22 (2): 183 –
190.
NODA, H. 1997. Tomato breeding for disease resistence in humid tropical conditions.
Proceedings of the 1st International Conference on the Processing Tomato and 1st
International Symposium on Tropical Tomato Diseases. ASHS Press, Alexandria: 133 - 138.
NODA, H.; MACHADO, F.M.; COELHO NETTO, R. A.; SILVA FILHO, D. F. 2013.
Estabilidade da resistência genética da cultivar Yoshimatsu ao agente patogênico da
“murcha bacteriana” do tomateiro. In: Noda, H.; Souza, L. A. G.; Silva Filho, D. F.
Pesquisas Agronômicas para a Agricultura Sustentável na Amazônia Central. Manaus :
Wega. p: 43 – 60. 2013.
NODA, H.; PAHLEN, A. Von Der; SILVA FILHO, D.F. 1986. Avaliação da resistência de
progênies de tomate à murcha bacteriana em solo naturalmente infestado por Pseudomonas
solanacearum. Revista Brasileira de Genética, IX (1): 55 – 66.
NODA, H.; PAIVA, W. O.; SILVA FILHO, D. F. & MACHADO, F. M. 1997. Melhoramento de
hortaliças convencionais para cultivo no Trópico Úmido Brasileiro. In: Noda, H.; Souza, L.
A. G. e Fonseca, O. J. M. Duas Décadas de Contribuições do INPA à Pesquisa Agronômica
no Trópico Úmido. Instituto Nacional de Pesquisas da Amazônia. Ministério da Ciência e
Tecnologia. Manaus. P: 59 - 87.
Nordestino de Genética e Melhoramento de Plantas. Fortaleza : Embrapa Agroindústria
Tropical. 2009. P: 60 – 82.
OLIVEIRA, W.F.; GIORDANO, L. B.; LOPES, C. A. 1998. Herança da resistência em tomateiro
à murcha-bacteriana. Fitopatologia Brasileira, 24(1): 49 – 53.
PENA, M. A. A.; NODA, H.; MACHADO, F.M.; PAIVA, M. S. S. 2010. Adaptabilidade e
estabilidade de genótipos de tomateiro sob cultivo em solos de terra firme e várzea da
Amazônia infestados por Ralstonia solanacearum. Bragantia, Campinas, v.69, n.1. p: 27-37.
PLANK, J. E. van der. 1963. Plant Disease: Epidemics and Control. Academic Press. New York.
349 p.
PLANK, J. E. van der. 1975. Principles of Plant Infection. Academic Press. New York York. 216
p.
PRIOR, P.; BART, S.; LECLERCQ, S.; DARRASSE, A.; ANAIS, G. 1996. Resistance to
bacterial wilt in tomato as discerned by spread of Pseudomonas (Burholderia) solacearum in
the stem tissues. Plant Disease, 45: 720 - 726.
RUSSEL, G. E. 1978. Plant Breeding for Pest and Disease Resistance. Butterworths. London
and Boston. 485 p.
SCOTT, A.J.; KNOTT, M.A. 1974. Cluster analyses methods for grouping means in the analyses
of variance. Biometrics. 30:507-512.

SILVA, E. B.; MENEZES, D.; ARAUJO, A. L. R.; A;VES, R. N.; CARVALHO FILHO, J. L.
Avaliação de híbridos de tomateiro e seus genitores. X Jornada de Ensino, Pesquisa e
extensão. Universidade Federal Rural de Pernambuco. Recife.
VENCOVSKY, R..; BARRIGA, P. Genética Biométrica no Fitomelhoramento. Ribeirão Preto :
Sociedade Brasileira de Ganética. 1992. 496 p.
VILLAREAL, R. L. 1980. Tomatoes in the Tropics. Westview, Boulder. 174 p.





























