Embed Size (px)
Citation preview


Disponível no Site www.lisina.com.br

Disponível no Site www.lisina.com.br 3
Há muito que ser celebrado: estima-se que um em cada três leitões no Brasil recebam rações com a suplementação de AminoGut e a melhoria de desempenho nas fases iniciais dos animais resulta em uma contribuição de quase 2% do volume total de toda a carne suína produzida no Brasil!
O início de sua história é anterior a 2005: foram anos de pesquisa que contou com a participação de cientistas e nu-tricionistas do mundo todo, que puderam suprir as exigências dos leitões em fase de desmame com um aminoácido sabidamente essencial, usado anteriormente apenas em situações especificas de nutrição clínica de humanos.
Ainda hoje, a suplementação de AminoGut, uma combinação de Glutamina e Ácido Glutâmico (aminoácidos es-senciais para leitões em desmame), é muito estudada: são mecanismos complexos do metabolismo que equipes de cientistas no mundo todo estão dedicados a compreender e explicar seus efeitos na melhoria do desempenho e saúde dos animais que receberam AminoGut nas dietas de pré e pós desmame.
É com muito orgulho que podemos afirmar que o sucesso do AminoGut deve-se a participação de muitos profissio-nais que contribuíram para aplicar na prática o que a ciência comprovou nos últimos anos: a importância de uma nutrição precisa.
A essencialidade da suplementação de aminoácidos funcionais para tornar a produção animal mais econômica e responsável é responsabilidade de todos nós, pois ela será imperativa para alimentar os nove bilhões de habitantes no mundo, que nas próximas três décadas, consumirá em média, mais proteína do que no passado.
Com o objetivo de reunir as mais recentes informações sobre a importância do AminoGut na nutrição de leitões este material contou com a participação de renomados cientistas que vivenciaram em seus trabalhos os efeitos da suplementação do AminoGut, e ou L-Glutamina e L- Ácido Glutâmico.
Mais uma vez agradecemos a todos os pesquisadores e profissionais, das pequenas e grandes empresas às instituições de pesquisa, que comemoram conosco e fazem parte desses 10 anos de conquistas com o desenvolvimento e pro-dução do AminoGut.
Boa Leitura!
AminoGut 10 AnosAjinomoto do Brasil - Ajinomoto Animal Nutrition GroupMarianne Kutschenko, Eduardo Nogueira, Gabriel Pessôa e Luciana Lima
Suplementação de Glutamina e Glutamato na Nutrição AnimalProf. Dr. Malcolm Watford, D.Phil.
A L-Glutamina é Essencial na Dieta para Maximizar o Crescimento e a Eficiência Alimentar em Suínos e Aves Prof. Dr. Guoyao Wu, Ph.D.
Em 2015, o AminoGut completa 10 anos do início de sua produção industrial.

Disponível no Site www.lisina.com.br
Figura 1. Progressão da maturação gastrointestinal de três distintas espécies. Em leitões, essa maturação ocorre em
até 12 semanas de idade (Adaptado de Sangild, 2000).
AminoGut 10 Anos
A importância da nutrição do leitão ao desmame
Os períodos críticos na produção de leitões são a fase neonatal (os primeiros dias após o nascimento), o período de desmame (as duas semanas posteriores ao desmame) e as primeiras semanas após a transferência para as insta-lações das fases de crescimento e terminação. As taxas glo-bais de mortalidade na suinocultura são consideravelmente mais altas para suínos que para outros animais de produção (Knudsen et. al., 2008).
De acordo com esses autores, os principais problemas enfrentados pelos leitões ao desmame são únicos e não experimentados em outras fases de seu crescimento:
A alimentação muda: do leite líquido da fêmea suína para um alimento sólido e estruturalmente complexo;
Os leitões são misturados e transferidos, incluindo mudanças na temperatura do ambiente;
Há mudanças marcantes estruturais e funcionais do intestino delgado que ocorrem em até 24 horas após o desmame;
Frequentemente há surtos de diarreias devido à proliferação de bactérias enterotoxigênicas (principalmente Escherichia coli) no intestino delgado e/ou fermentação de nutrientes menos digestíveis no intestino grosso (Pluke et. al., 1995, 1997; Hamson et al., 1999, citados por Knudsen et al., 2008).
Dos problemas citados, três tem relação direta com a nu-trição. Dessa maneira, mesmo que todos os cuidados sejam tomados para que o novo ambiente seja favorável às mu-danças necessárias ao desmame, ferramentas e estratégias nutricionais econômicas devem ser tomadas para minimizar ou reverter o impacto do desmame na fisiologia e, conse-quentemente, na produção de leitões. Segundo Machado (2008), o desempenho dos leitões na primeira semana após o desmame é um excelente preditor do desempenho geral dos animais nas fases de recria e terminação: quanto maior o ganho de peso ao 7º dia após o desmame, menos dias são necessários para que o peso ao abate seja atingido pelo suíno.
Marianne Kutschenko, Eduardo Nogueira, Gabriel Pessôa e Luciana LimaAjinomoto do Brasil - Ajinomoto Animal Nutrition Group
Disponível no Site www.lisina.com.br 5
A primeira ingestão de alimento é feita por somente meta-de dos leitões nas primeiras 24 horas pós-desmame: os leitões conseguem ingerir os requerimentos de energia
Alterações intestinais frequentemente observadas durante o período pós-desmame incluem mudanças na morfolo-gia vilos e cripta e na atividade das enzimas da borda em escova (Lallès et. al., 2004). O intestino delgado perde de 20 a 30% de seu peso relativo durante as duas semanas pós-desmame e sua regeneração completa demora de 5 a 10 dias, sendo que os vilos intestinais atrofiam para 45 a 70% comparativamente aos valores anteriores ao desma-me (Spreeuwenbergh 2002, citado por Lallès et. al., 2004).
O intestino dos leitões, assim como em muitos animais, é o maior site de oxidação (glutamina, ácido glutâmico, asparagina), síntese (prolina, alanina, tirosina, arginina, ornitina, citrulina) e utilização para síntese de proteína (treonina, lisina, fenilalanina, aminoácidos de cadeia
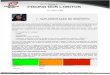
para manutenção apenas três dias após o desmame, e le-vam de 8 a 14 dias para recuperar os níveis de energia ingerida antes de serem desmamados, conforme Tabela 1 (Lallès et. al., 2004).
ramificada e metionina) dos aminoácidos (Burrin e Stoll, 2000, citado por Lallès et. al., 2004).
Alguns aminoácidos são essenciais e indispensáveis para proteção e defesa do intestino: treonina para mucina, cisteína para glutationa, triptofano e histidina para 5-HT e histamina, metionina para poliaminas, arginina para óxido nítrico, etc. Em especifico, o metabolismo daglutamina, arginina e citrulina, está particularmente aumentado nos enterócitos na fase pós-desmame, comparado à fase pré-desmame e a suplementação oral de glutamina ou ácido glutâmico previne a atrofia das vilosidades intestinais (Ewtushick et al. 2000; Wu et al. 1996; citados por Lallès et. al., 2004).
Tabela 1. Mudanças em funções arquiteturais e funcionais de parâmetros do intestino
delgado de leitões desmamados aos 21 dias de idade (valores expressos em porcentagem
dos valores pré-desmame) (Lallès et. al., 2004).
1Ingestão de alimentos de 9, 61 e 80g por peso corporal0,75 por dia, nos dias 2, 8 e 15
pós-desmame, respectivamente.

Disponível no Site www.lisina.com.br
Algumas estratégias podem ser adotadas para reduzir os impactos do desmame na estrutura intestinal dos leitões, entre elas: fornecer rações compostas por alimentos de ener-gia e proteínas de alta digestibilidade (Spreeuwenbergh 2001, citado por Lallès et. al., 2004); aumentar os níveis de lactose reduzindo a proteína das rações; e suplementar glutamina (1-4%) e ácido glutâmico (6,5%) para melhorar a arquitetura intestinal e eficiência alimentar, pós-desma-me (Lallès et al., 2004).
A conversão da proteína da ração para proteína animal requer complexos processos bioquímicos e fisiológicos, os quais envolvem a digestão, a absorção, o metabolismo (incluindo microrganismos do lúmen intestinal, leitos esplâncnicos -fígado, trato gastrintestinal, pâncreas-, e outros órgãos) e a regulação desses eventos via múltiplos sinalizadores (Wu, 2010a).
De acordo com o mesmo autor, o metabolismo intestinal dos aminoácidos tem profundos impactos na nutrição e saúde:
O catabolismo de glutamina, glutamato e aspartato provem a maior parte do ATP para manter a integridade e funções intestinais;
Elevados níveis de glutamina, glutamato e aspartato no plasma sanguíneo exercem efeitos neurotóxicos, e seu ca-tabolismo extensivo pelo intestino delgado é essencial para a sobrevivência do organismo;
As transformações dos aminoácidos pelo intestino de-sempenha um papel importante na regulação da síntese endógena dos aminoácidos não essenciais (como a citruli-na, arginina, prolina e alanina) modulando a disponibili-dade de aminoácidos para os tecidos extra intestinais;
As relações com a lisina da maioria dos aminoácidos nas dietas (proteína ideal) diferem substancialmente daquelas que entram na veia portal do lúmen do intestino delgado ou aparecem no plasma e nas proteínas corporais. As dis-crepâncias entre os padrões de aminoácidos entre a proteí-na da dieta e a corporal são particularmente maiores para a arginina, histidina, metionina, prolina, glutamina, glicina e serina.
Aminoácidos condicionalmente essenciais
Dos mais de 700 aminoácidos encontrados na natureza, somente 20 são utilizados para a formação de proteínas, sendo estes, classificados como essenciais (ou não sin-tetizáveis), condicionalmente essenciais ou não essenciais (ou sintetizáveis).
Sua classificação pode variar de acordo com a espécie animal e também com o estado fisiológico dos indivíduos (Tabela 2). De acordo com Wu (2013), a Glutamina é um aminoácido essencial para neonatos (incluindo o huma-no e o suíno), adultos sob estresse (por exemplo, estresse térmico, queimaduras e infecções) e animais reprodutores (machos e fêmeas).
Tabela 2. Classificação dos aminoácidos como Essenciais, Não
Essenciais e Condicionalmente Essenciais para Mamíferos de
acordo com Wu, 2013.
*Condições para que esses aminoácidos se tornem essenciais:
ser neonato, macho ou fêmea destinados à reprodução, e estresse.

Disponível no Site www.lisina.com.br 7
Como os animais não podem sintetizar os aminoácidos essenciais para satisfazer suas exigências, eles devem ser obrigatoriamente fornecidos por intermédio da dieta, já que sua produção endógena não é capaz de atender às suas necessidades nutricionais para determinada fase ou maior desafio sanitário. É possível que para os aminoáci-dos condicionalmente essenciais não se possa estabelecer uma exigência nutricional fixa, porque ela pode variar de acordo com a intensidade dos fatores que influenciam sua demanda.
Além de distintas outras funções biológicas, os aminoácidos sintetizáveis também são importantes e devem ser estudadospor serem precursores dos aminoácidos essenciais. Por exemplo, (a arginina é considerada um aminoácido essencial para quase todos os neonatos mamíferos, mas é sintetizada por adultos. As exceções são os mamíferos es-tritamente carnívoros, como os gatos e os ferretes) a Glu-tamina, que desempenha mais de 60 funções biológicas distintas descritas na literatura, entre elas a prevenção da atrofia de vilosidades intestinais, comum durante o des-mame de leitões. A habilidade de manter a saúde, por exemplo, significa que alguns aminoácidos são sintetizáveis em um corpo saudável e se tornam essenciais em certas condições fisiológicas ou patológicas. Assim, esses amino-ácidos são considerados condicionalmente essenciais (Wat-ford, 2011).
Historicamente, as rações de leitões foram inicialmente formuladas utilizando exclusivamente um nível mínimo de proteína bruta, e posteriormente formuladas com base em aminoácidos totais, exclusivamente em lisina e metionina. Nos últimos anos as rações se tornaram muito mais preci-sas e, por meio da proteína ideal, prove-se sem deficiências
ou excessos, as necessidades absolutas de todos os amino-ácidos essenciais requeridos para mantença e crescimen-to corporal. Esse ajuste de nutrientes e a formulação sem excessos são cruciais para a saúde dos leitões: a transição do leite da fêmea suína para a alimentação composta por ingredientes vegetais sólidos mais a imaturidade do sistema digestivo, que produz quantidade insuficiente de ácido clo-rídrico e enzimas para digestão de proteínas, reduzem a ca-pacidade de digestão pelos leitões. O aumento de proteína não digerida eleva o pH intestinal e fornece substrato para a proliferação de microrganismos patogênicos no intestino e, ainda, altera o balanço hidroeletrolítico intestinal, que podem levar à diarreia e prejudicar o estado sanitário do rebanho (Nogueira, 2005).
De acordo com Watford (2011), em estado de saúde, o maior sítio de síntese endógena e estocagem de glutamina no corpo é o musculo esquelético (Figura 2), onde outros aminoácidos (especialmente os aminoácidos de cadeia ramificada: valina, leucina e isoleucina) são metabolizados para a síntese de glutamina endógena. Outros sítios de síntese de glutamina incluem o tecido adiposo, pulmões e fígado. Quantitativamente, o maior sítio de utilização de glutamina são os enterócitos do intestino delgado e as células do sistema imune (macrófagos, timócitos e lin-fócitos), onde a glutamina é utilizada como o combustível respiratório principal para a síntese de ATP. Os rins, por sua vez, utilizam quantidade importante de glutamina como substrato para a produção de amônia, requerida para a manutenção do equilíbrio ácido-básico (com o uso da glutamina, o carbono é recuperado como glucose), e em alguns momentos, o fígado também pode utilizar glutami-na sendo a glucose e a ureia os principais produtos gerados.
Figura 2. Metabolismo da glutamina em estados de saúde
(Watford et al., 2011)

Disponível no Site www.lisina.com.br
Figura 3. Metabolismo da glutamina em estados catabólicos
(Watford et al., 2011)
O organismo saudável é capaz de sintetizar quantidades consideráveis de glutamina, e em adultos saudáveis não há evidências de qualquer escassez desse aminoácido. Mas há duas décadas, tornou-se apropriado concluir que em certos momentos, o organismo tem suas necessidades de glutamina aumentadas acima do que poderia ser suprido pela síntese endógena. Essa descoberta tem sido mais ex-tensivamente estudada em condições clínicas humanas, e a glutamina passa a ser considerada um aminoácido condi-
A glutamina é um dos aminoácidos mais versáteis no me-tabolismo celular e fisiologia. E embora esteja presente abundantemente nas proteínas dos tecidos vivos vegetais e animais na forma ligada, a suplementação de glutamina livre (L-Glutamina) previne a atrofia intestinal, melhora a função imune e o desempenho de leitões recém-desmama-dos (Wu, 2008).
A glutamina é o mais importante substrato de energia para células em divisão rápida, como as do sistema imune en-terócitos e linfócitos, e para outros tipos de células, como os macrófagos e células renais, regula a proliferação de linfócitos-T: a glutamina fornece ATP para o turnover da proteína intracelular e síntese protéica, é o maior com-bustível celular para a produção das citocinas, ativação de macrófagos, inibição da apoptose, transporte de nutrientes através da membrana plasmática, crescimento e migração celular, e manutenção da integridade da célula (Li et al., 2007).
Por ser o aminoácido mais abundante encontrado no plasma sanguíneo, a glutamina é o maior transportador de nitrogênio de sítios de sínteses de glutamina (múscu-lo esquelético, fígado e pulmões) para sítios de utilização
cionalmente essencial para humanos também, o que sig-nifica que quando o organismo não é capaz de produzi-la em quantidade suficiente para suprir seu requerimento, ela deve ser suplementada na dieta. Durante períodos de traumas severos ou infecção, os referi-dos estados supercatabólicos, aumentam os requerimen-tos de glutamina pelos tecidos como as células do sistema imune, assim como rins e fígado (Watford et al., 2011), como mostra a Figura 3.
(rins, intestinos, neurônios, células do sistema imune e fígado). Ela age como precursora chave para a formação de nucleotídeos, e em muitas circunstâncias fisiológicas a glutamina e o glu-tamato estão sensivelmente relacionados com a função de requerimento da célula. Além disso, ambos não podem ser substituídos por outros inputs metabólicos (caso os existentes falhem), portanto o metabolismo da glutamina e do glutamato deve ser considerado tão importante como o metabolismo da glucose (Newlsrohme, 2003).
O desmame de leitões é um dos principais responsáveis por perdas econômicas observadas em criações comerciais de suínos. Seus efeitos ocorrem em cascata no metabolismo do leitão: ele altera sua fisiologia gastrintestinal e por con-sequência seu aporte de glutamina, com isso é diminuída a capacidade digestiva e absortiva de nutrientes, o que final-mente compromete o desempenho de todas as fases sub-sequentes da criação.
A suplementação de todos os aminoácidos essenciais, incluindo a glutamina, no período que antecede, duran-te e pós-desmame, reduz as perdas produtivas e permite suporte para o desempenho adequado dos animais.
Disponível no Site www.lisina.com.br 9
Metabolismo da Glutamina e do Ácido Glutâmico no intestino delgado de leitões
O desmame de leitões agride a mucosa intestinal direta e indiretamente: de forma direta com a mudança da alimen-tação líquida (leite) para sólida (ração), granulometria da ração, introdução de matérias-primas de origem vegetal, e indiretamente pela liberação de hormônios relacionados ao estresse (cortisol), pela mudança no ambiente sanitário, térmico e social (mistura de leitegadas), o que resulta em redução de consumo e aumento do catabolismo e da de-manda por glutamina e por glutamato.
Quase todo o glutamato e aspartato, aproximadamente 70% da glutamina e 30 a 40% da prolina na dieta en-teral são catabolizados pelo intestino do neonato, leitões desmamados e fêmeas suínas gestantes. Enquanto que so-mente 5% do glutamato e aspartato, 30 a 33% da gluta-mina e 60 a 65% da prolina da dieta entram na circulação portal. Em humanos e suínos, 40% da arginina da dieta é catabolizada pelo intestino delgado na primeira passagem.
Considerando todos os aminoácidos da dieta, o glu-tamato é o aminoácido com as maiores taxas de de-gradação no intestino delgado, seguido pela glutami-na, aspartato e prolina. Porém, é importante notar que o intestino delgado retira uma quantidade grande de glutamina do sangue arterial, e consequentemente, a taxa total de utilização de glutamina pelo intestino é maior que a de glutamato.
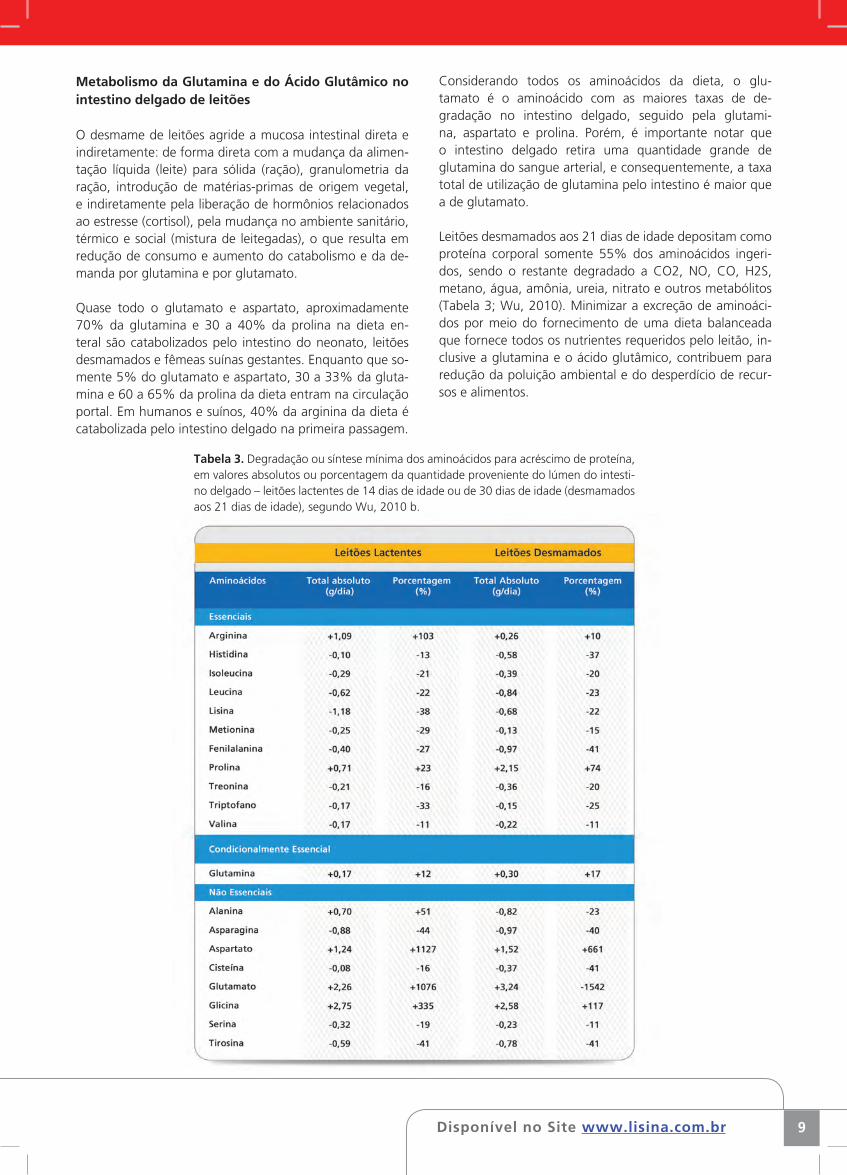
Leitões desmamados aos 21 dias de idade depositam como proteína corporal somente 55% dos aminoácidos ingeri-dos, sendo o restante degradado a CO2, NO, CO, H2S, metano, água, amônia, ureia, nitrato e outros metabólitos (Tabela 3; Wu, 2010). Minimizar a excreção de aminoáci-dos por meio do fornecimento de uma dieta balanceada que fornece todos os nutrientes requeridos pelo leitão, in-clusive a glutamina e o ácido glutâmico, contribuem para redução da poluição ambiental e do desperdício de recur-sos e alimentos.
Tabela 3. Degradação ou síntese mínima dos aminoácidos para acréscimo de proteína, em valores absolutos ou porcentagem da quantidade proveniente do lúmen do intesti-no delgado – leitões lactentes de 14 dias de idade ou de 30 dias de idade (desmamados aos 21 dias de idade), segundo Wu, 2010 b.

Disponível no Site www.lisina.com.br
A dieta perfeita
Tradicionalmente acreditava-se que o leite materno suí-no provia quantidades adequadas de aminoácidos para os leitões. Resultados recentes, porém, indicaram que o leite da fêmea suína não provê quantidades adequadas de glutamina para síntese protéica necessária aos tecidos não intestinais de leitões, pois as taxas de síntese de novo são muito mais altas no leitão lactente (pelo menos 0,88 g/kg de peso vivo ao dia). Adicionalmente, o leite materno suíno satisfaz no máximo 23%, 66%, 23% e 42% das exigências de glicina, alanina, aspartato + asparagina e glutamato + glutamina, respectivamente. Similarmente, uma ração típica a base de milho e farelo de soja não pode prover suficientes quantidades de arginina, prolina, aspar-
Wu (2013) publicou que o requerimento de Glutamina pelo leitão é de no mínimo 1.949 mg/kg de peso corporal/dia. Esse requerimento dietético dos aminoácidos essenciais, não essenciais e condicionalmente essenciais por todo o corpo ou tecido (por exemplo, o intestino delgado), pode ser estimado pelo autor como base fatorial de análise, como a soma do nitrogênio fecal e urinário em resposta a uma dieta livre de proteína (mantença) mais os aminoácidos depositados no corpo e os aminoácidos excretados pelos produtos animais (leite, ovos, lã e crescimento fetal). Para certos aminoácidos, o método fatorial pode ainda ter como base a soma das necessidades de aminoácidos pelas vias metabólicas e perdas obrigatórias via secreções corporais.
Nos primeiros estudos realizados, a suplementação deglutamina indicava que se poderia aumentar o ganho de peso de leitões à saída de creche, como consequência da prevenção da atrofia das vilosidades intestinais, e se melhorar a conversão alimentar de leitões desmamados em condições comerciais. A diminuição da ocorrência de diarreia também foi observada com a suplementação de Glutamina, pois a melhor digestão e absorção de nutrientes
tato, glutamato, glutamina ou glicina para o acréscimo de proteína corporal requerido pelo suíno em crescimento em fase de pós desmame (Wu, 2010).
O leite da fêmea suína apresenta considerável aumento da concentração de Glutamina livre principalmente a partir do 14º dia de lactação (Figura 4) e praticamente dobra e triplica de concentração aos 21 e 28 dias de lactação, respectivamente. O leitão das criações comerciais é restringido do recebimento da glutamina livre proveniente do leite materno por causa do desmame por volta dos 21 dias de idade. Ele passa a receber um alimento sólido, a base de vegetais, e com conteúdo mínimo de glutamina total e nulo de glutamina livre.
reduz o substrato para proliferação de microrganismosindesejáveis (patogênicos). A melhoria de desempenho na fase de creche tem como consequência melhor desempenhonas fases subsequentes de recria e terminação, o que resulta em suínos mais pesados ou abatidos mais precoce-mente ao termino do ciclo de criação.
Embora não seja considerado um aminoácido condicional-mente essencial, o glutamato também desempenha muitas funções no metabolismo, como a produção de ATP, sín-tese de arginina e glutationa no epitélio celular do intestino delgado. Além disso, já é sabido que o glutamato inibe a degradação da glutamina pela glutaminase mitocondrial fosfato-dependente nos tecidos extra-hepáticos e células (Curthoys e Watford, 1995; citado por Wu, 2008).
Entre as funções da Glutamina e do Ácido Glutâmico, destacam-se:
Precursores de Nucleotídeos, fundamentais para síntese de DNA/RNA (purina e pirimidina) e multiplicação celular;
Figura 4. Concentração de Glutamina e Ácido Glutâmico livres no
leite da fêmea suína de acordo com os dias de lactação

Disponível no Site www.lisina.com.br 11
Precursores de Poliaminas que participam da proliferação celular e reparo das células intestinais (putrescina, espermi-dina e espermina);
Precursores da síntese de mucina, por intermédio da N-acetilglucosamina e N-acetilgalactosamina. A mucina é a primeira barreira de proteção intestinal contra agressões intestinais;
Estimulam a ação do m-TOR, substância que estimula a síntese protéica e a multiplicação celular;
Fonte de energia para a renovação (turnover) da mucosa, por intermédio do ATP produzido a partir do ciclo de Krebs;
A história da L-Glutamina e do L-Ácido Glutâmico
O Ácido Glutâmico (como ácido alfa-aminoglutárico) foi isolado pela primeira vez em 1866 a partir do hidroli-sado de glúten de trigo (gliadina). Sete anos depois, o glutamato foi obtido a partir da caseína, a primeira proteína de origem animal a mostrar conteúdo em aminoácidos e em 1890 o Glutamato foi sintetizado quimicamente do ácido levulínico (Wu, 2013). Mas, foi em 1908, que o professor e pesquisador japonês da Universidadede Tóquio, Kikunae Ikeda, conseguiu isolar o Ácido Glutâmico, como uma substância presente no Kombu, um tipo de alga Japonesa, o qual promovia o “umami” reconhecido quinto gosto básico, distinto dos outros gostos (amargo, salgado, azedo e doce). Após processado, o ácido glutâmico deu origem ao glutamato monossódico, que passou a ser fabricado industrialmente em 1909 e utilizado naalimentação humana como o intensificador de sabor chamado “AJI-NO-MOTO”, ou “Essência do Sabor” em japonês (AJINOMOTO, 2013). Dois derivativos do Glutamato (N-Acetilglutamato e fosfoglutamato) estão primariamente presentes nos animais como aminoácidos livres (Wu, 2013).
A L-Glutamina (ácido alfa-aminoglutarâmico) por sua vez, foi obtida pela primeira vez na forma de aminoácido livre do suco da beterraba em 1883. Em 1904 foi reconhecida por E. Fischer como a maior fonte de amônia produzida a partir da hidrólise de proteínas. E depois de muito re-finamento dos procedimentos laboratoriais, a glutamina foi isolada por hidrólise enzimática a partir da gliadina em 1932. Sendo que na década de 60, foi considerado um nu-triente necessário em dietas animais, mas recebeu pouca ou nenhuma atenção nos livros clássicos de nutrição. Foi a partir de 1965, que E.M.Neptune descobriu que o intestino delgado (íleo) de varias espécies animais (como macacos, coelhos e ratos) tem altas taxas de oxidação de glutamina, e houve o inicio de uma nova era das pesquisas e resultados bioquímicos e nutricionais da importância da Glutamina
Fonte de nitrogênio para síntese de aminoácidos e outros compostos nitrogenados;
Precursores da glutationa, um antioxidante intracelular que auxilia na manutenção da integridade intestinal.
A glutamina é utilizada para a síntese de várias molécu-las com importantes funções metabólicas relacionadas à homeostase e ao desenvolvimento animal. Sua utilização resulta em melhora do desempenho de leitões desmama-dos e pode ser incorporada às rações na forma do produto comercial AminoGut (Abreu e Donzele, 2008).
para humanos e outros animais (Wu, 2013). É essencial ajudar o trato intestinal a recuperar sua for-ma da atrofia causada pelo desmame o mais rápido pos-sível. Os aminoácidos funcionais como a Glutamina e o Ácido Glutâmico desempenham um papel crucial nessa recuperação e desenvolvimento, por serem precursores de replicação e desenvolvimento celular e por melhorar o estado imunológico do leitão desmamado.
Como surgiu o AminoGut
Com base nas descrições da literatura sobre sua essencialidade para a fase de desmame dos leitões, a importância dos aminoácidos funcionais e os efeitos de sua suplementação para animais de produção, a Glutamina e o Ácido Glutâmico têm sido estudados desde a década de 2000 pela Ajinomoto.
Os primeiros estudos foram conduzidos no Japão e na Uni-versidade do Missouri na década de 2000. Inicialmente, a suplementação de Glutamina e Ácido Glutâmico foi feita dos aminoácidos isolados e em fases tenras do pós desma-me.
Em 2000 um dos primeiros desses estudos foi conduzido nos Estados Unidos, na Universidade do Missouri (Touchette et al., 2000). Cento e vinte e cinco leitões desmamados aos 17 dias de idade (5,09 kg) receberam por 14 dias, dietas adicionadas de L-Arginina ou L-Glutamina. Os leitões que haviam recebido L-Glutamina apresentaram aos 24 dias de idade, maior consumo de ração diário, aos 31 dias de idade, maior profundidade de cripta e, aos 45 dias de idade possuíram peso corporal superior que os animais que receberam a ração controle. Os autores concluíram que 1,20% de suplementação de L-Glutamina resultou em efeitos positivos sobre o desempenho e a morfologia intestinal dos leitões. Em 2002, o total de 210 leitões com 17 dias de idade e

Disponível no Site www.lisina.com.br
4,8 kg de peso vivo médio, receberam rações a base de milho, soja, lactose e farinha de peixe com e sem plasma (controles negativo e positivo) suplementadas, individual-mente ou em combinação (sete tratamentos), com L-Glu-tamina, L-Ácido Glutâmico ou nucleotídeos. Os animais de cada baia tiveram contato “nariz com nariz” com leitões mais velhos (22 kg de peso vivo) para aumentar a carga de patógenos endêmicos. Os resultados dessa avaliação mostraram que a combinação do L-Ácido Glutâmico, Glu-tamina e nucleotídeo melhorou o consumo diário de ração e altura das vilosidades intestinais (jejuno) dos leitões na primeira semana pós desmame (Yi et al., 2002).
Yi et al. (2005) estudaram o efeito da suplementação de L-Glutamina e plasma sobre o desempenho de leitões (5,32 kg de peso vivo aos 17 dias de idade) desafiados com Escherichia coli K88+ aos 12 dias após o desmame. Os autores concluíram que a suplementação de Glutamina teve efeitos benéficos em aliviar a depressão que a E. coli K88+ causou no crescimento dos animais desafiados, prin-cipalmente mantendo a morfologia e função intestinal, e/ou possivelmente via modulação dos eixos somatotrópicos.
Após a compilação e análise crítica dos dados encontra-dos, concluiu-se naquele momento que a suplementação de Ácido L-Glutâmico promovia efeitos semelhantes no desempenho de leitões no período pós-desmame àqueles que haviam sido suplementados com a L-Glutamina. Mas até então, a suplementação de ambos os aminoácidos em um mesmo tratamento não havia sido feita para notar-se efeito sinérgico.
Então, em um estudo conduzido pela Universidade do Missouri, leitões com peso inicial de 4,76kg e 17 dias de idade foram divididos em sete tratamentos, os quais eram compostos por suplementação de L-Glutamina, ou L-ÁcidoGlutâmico, Nucleotídeos, Plasma ou uma combinação de L-Glu-
tamina + L-Ácido Glutâmico. As dietas experimentais foram for-necidas por duas semanas após o desmame. Os resultados desse trabalho impressionaram pelo efeito positivo do tratamento com a mistura de L-Glutamina e L-Ácido Glutâmico que gerou desempenho superior e melhor estrutura intestinal dos leitões. Concluiu-se com esse trabalho que a combinação dos aminoácidos promovia melhores resultados de desempenho dos animais que quando suplementados isoladamente. E que esse feito sinérgico era especialmente pronunciado na melhoria da altura das vilosidades intestinais.
Os resultados de desempenho superior com a suplemen-tação combinada de Glutamina e Ácido Glutâmico são coerentes com o metabolismo da Glutamina e do Glutamato no organismo animal.
A Glutamina é sintetizada a partir do Glutamato e da amônia pela ação da enzima glutamina sintetase e é hidrolisada no-vamente à Glutamato e amônia pela glutaminase (Figura 5). A maioria dos tecidos expressa ambas as enzimas, mas em geral uma delas predomina, dependendo se o tecido cor-poral é sítio de produção ou de utilização da Glutamina. A glutamina é substrato para inúmeras reações biosintéticas, principalmente as amidotransferases, que resultam na sín-tese de glucosamina, pirimidinas, purinas e NAD. Esses substratos são importantes para todas as células, particu-larmente naquelas de divisão rápida. Mas a glutamina pro-veniente da ação da glutaminase corresponde a menos de 5% da glutamina total do metabolismo, enquanto que o glutamato produzido da glutamina, é o principal substrato para a gluconeogênese e síntese de ATP, além de ser im-portante precursor para a síntese de glutationa e inúmeros outros aminoácidos, como a prolina, ornitina e a arginina (Watford, 2009).
Figura 5. A Glutamina é sintetizada a partir do glutamato e amônia
pela ação da enzima glutamina sintetase e a reação reversa pela enzima
glutaminase (Watford, 2009).
Disponível no Site www.lisina.com.br 13
Nos primeiros anos desse milênio várias pesquisas demonstraram que a suplementação de glutamina e ácido glutâmico nas rações de leitões pode melhorar as condições intestinais e os parâmetros de desempenho desses animais (Kutschenko et al., 2008). No trato gastrintestinal, em es-pecial, a Glutamina tem um efeito sinérgico com o Ácido Glutâmico, os quais desempenham funções primordiais para a manutenção da estrutura funcional.
Concomitantemente aos estudos realizados no Japão e Estados Unidos, o Brasil foi escolhido para a continuidade
das pesquisas e aplicação prática dos resultados científicos encontrados.
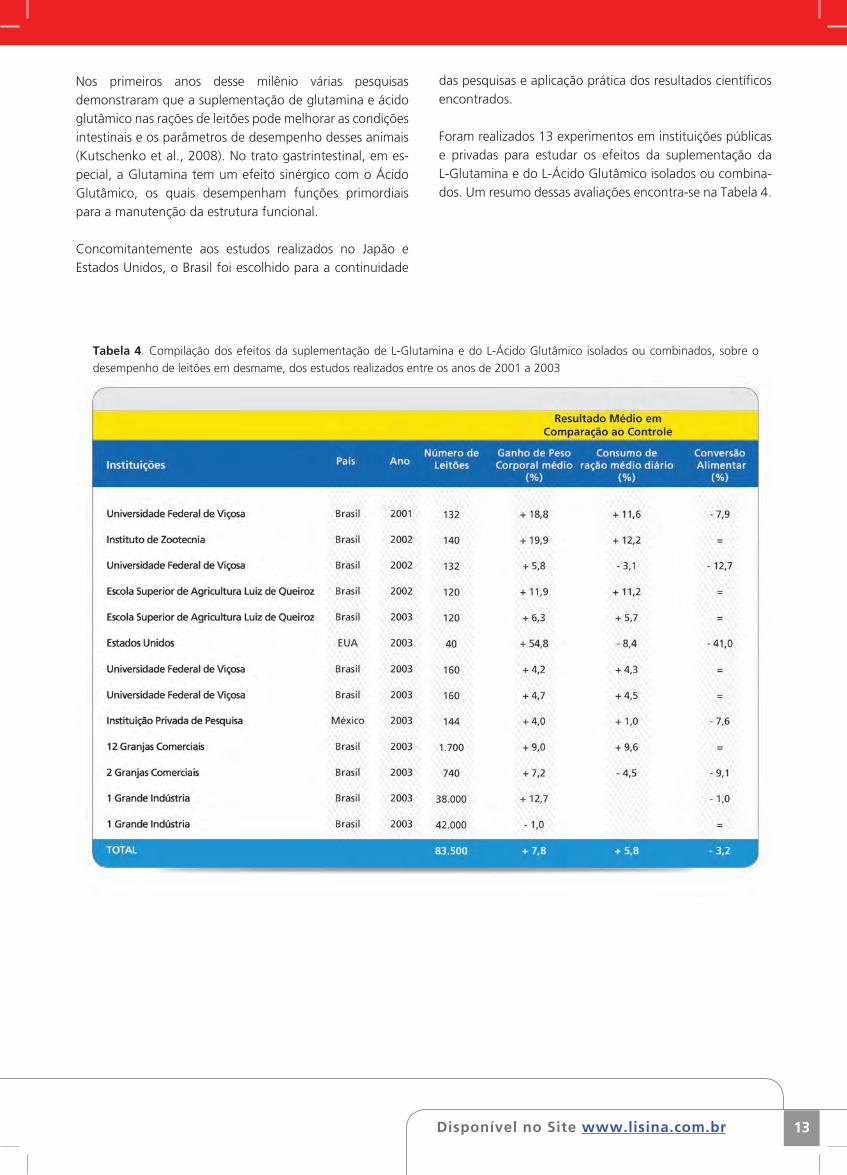
Foram realizados 13 experimentos em instituições públicas e privadas para estudar os efeitos da suplementação da L-Glutamina e do L-Ácido Glutâmico isolados ou combina-dos. Um resumo dessas avaliações encontra-se na Tabela 4.
Tabela 4. Compilação dos efeitos da suplementação de L-Glutamina e do L-Ácido Glutâmico isolados ou combinados, sobre o
desempenho de leitões em desmame, dos estudos realizados entre os anos de 2001 a 2003
Disponível no Site www.lisina.com.br
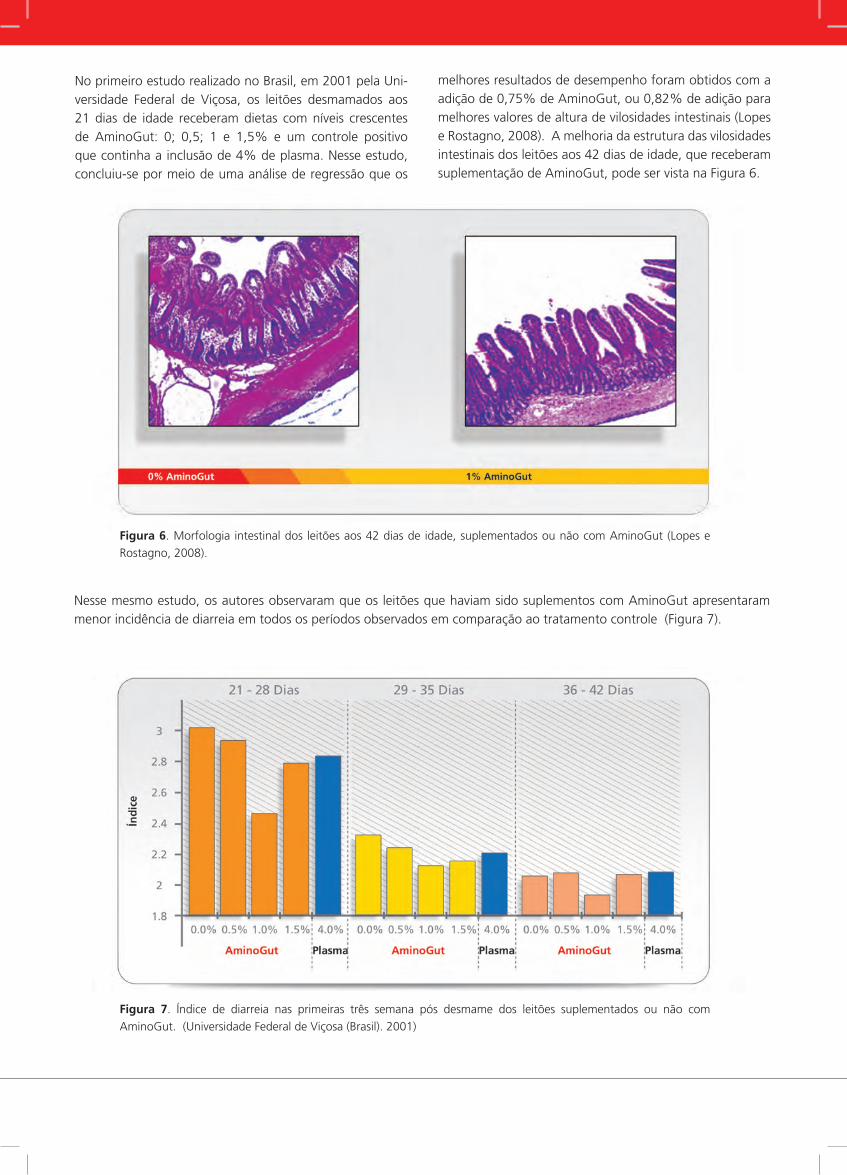
No primeiro estudo realizado no Brasil, em 2001 pela Uni-versidade Federal de Viçosa, os leitões desmamados aos 21 dias de idade receberam dietas com níveis crescentes de AminoGut: 0; 0,5; 1 e 1,5% e um controle positivo que continha a inclusão de 4% de plasma. Nesse estudo, concluiu-se por meio de uma análise de regressão que os
melhores resultados de desempenho foram obtidos com a adição de 0,75% de AminoGut, ou 0,82% de adição para melhores valores de altura de vilosidades intestinais (Lopes e Rostagno, 2008). A melhoria da estrutura das vilosidades intestinais dos leitões aos 42 dias de idade, que receberam suplementação de AminoGut, pode ser vista na Figura 6.
Figura 6. Morfologia intestinal dos leitões aos 42 dias de idade, suplementados ou não com AminoGut (Lopes e
Rostagno, 2008).
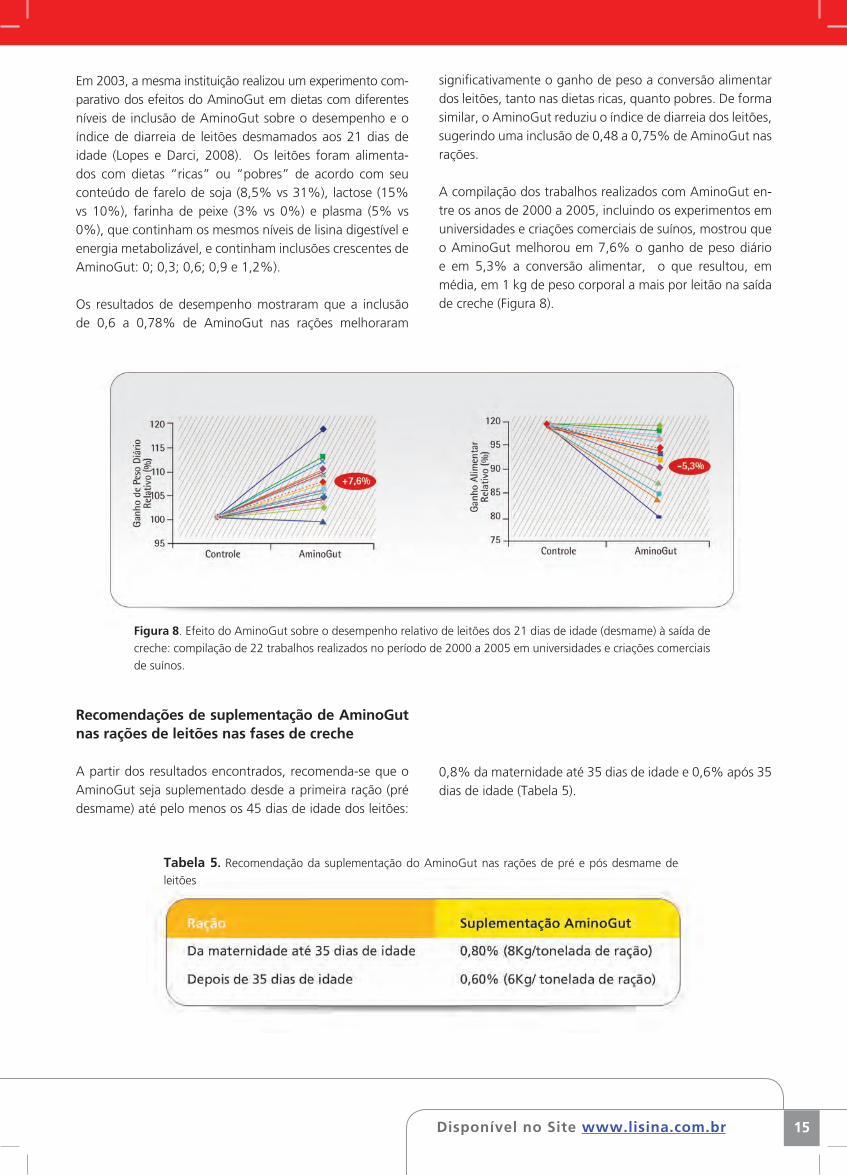
Figura 7. Índice de diarreia nas primeiras três semana pós desmame dos leitões suplementados ou não com
AminoGut. (Universidade Federal de Viçosa (Brasil). 2001)
Nesse mesmo estudo, os autores observaram que os leitões que haviam sido suplementos com AminoGut apresentaram menor incidência de diarreia em todos os períodos observados em comparação ao tratamento controle (Figura 7).
Disponível no Site www.lisina.com.br 15
Em 2003, a mesma instituição realizou um experimento com-parativo dos efeitos do AminoGut em dietas com diferentes níveis de inclusão de AminoGut sobre o desempenho e o índice de diarreia de leitões desmamados aos 21 dias de idade (Lopes e Darci, 2008). Os leitões foram alimenta-dos com dietas “ricas” ou “pobres” de acordo com seu conteúdo de farelo de soja (8,5% vs 31%), lactose (15% vs 10%), farinha de peixe (3% vs 0%) e plasma (5% vs 0%), que continham os mesmos níveis de lisina digestível e energia metabolizável, e continham inclusões crescentes de AminoGut: 0; 0,3; 0,6; 0,9 e 1,2%).
Os resultados de desempenho mostraram que a inclusão de 0,6 a 0,78% de AminoGut nas rações melhoraram
Recomendações de suplementação de AminoGut nas rações de leitões nas fases de creche
A partir dos resultados encontrados, recomenda-se que o AminoGut seja suplementado desde a primeira ração (pré desmame) até pelo menos os 45 dias de idade dos leitões:
significativamente o ganho de peso a conversão alimentar dos leitões, tanto nas dietas ricas, quanto pobres. De forma similar, o AminoGut reduziu o índice de diarreia dos leitões, sugerindo uma inclusão de 0,48 a 0,75% de AminoGut nas rações.
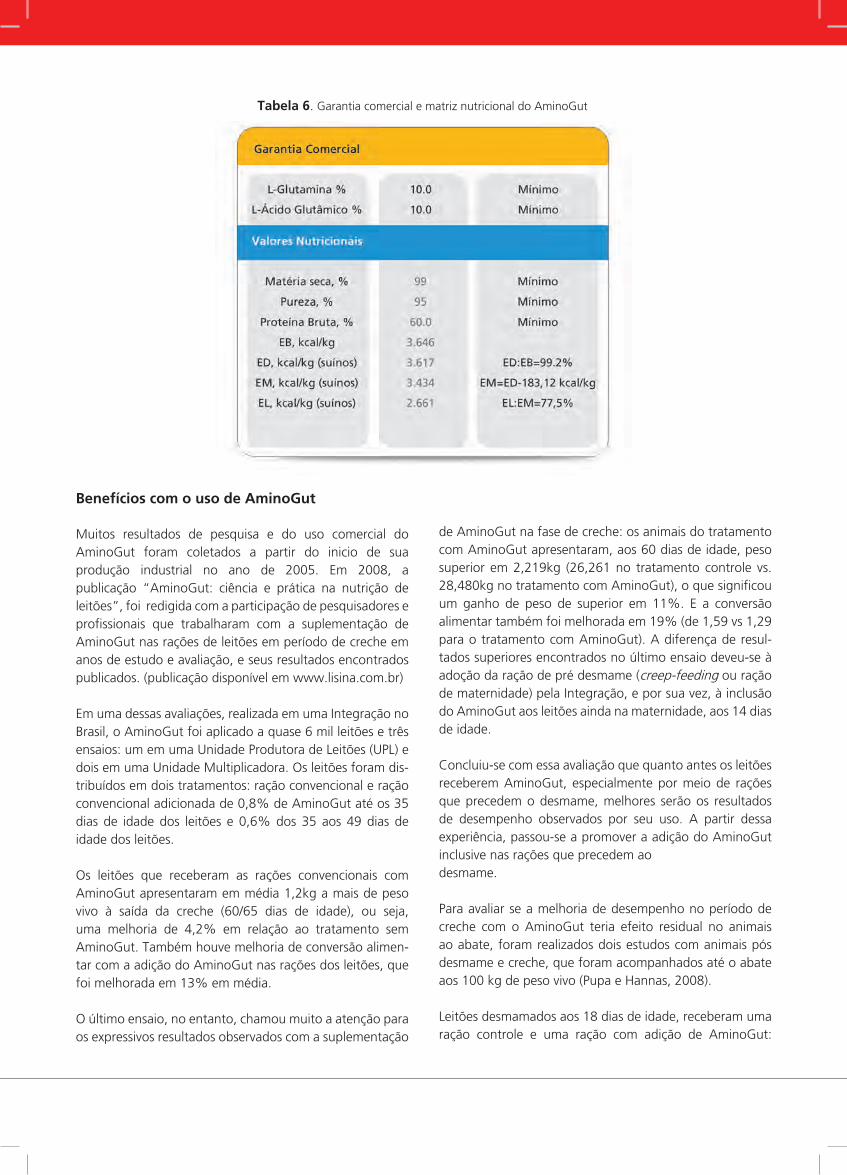
A compilação dos trabalhos realizados com AminoGut en-tre os anos de 2000 a 2005, incluindo os experimentos em universidades e criações comerciais de suínos, mostrou que o AminoGut melhorou em 7,6% o ganho de peso diário e em 5,3% a conversão alimentar, o que resultou, em média, em 1 kg de peso corporal a mais por leitão na saída de creche (Figura 8).
0,8% da maternidade até 35 dias de idade e 0,6% após 35 dias de idade (Tabela 5).
Figura 8. Efeito do AminoGut sobre o desempenho relativo de leitões dos 21 dias de idade (desmame) à saída de
creche: compilação de 22 trabalhos realizados no período de 2000 a 2005 em universidades e criações comerciais
de suínos.
Tabela 5. Recomendação da suplementação do AminoGut nas rações de pré e pós desmame de
leitões
Disponível no Site www.lisina.com.br
Benefícios com o uso de AminoGut
Muitos resultados de pesquisa e do uso comercial do AminoGut foram coletados a partir do inicio de sua produção industrial no ano de 2005. Em 2008, a publicação “AminoGut: ciência e prática na nutrição de leitões”, foi redigida com a participação de pesquisadores e profissionais que trabalharam com a suplementação de AminoGut nas rações de leitões em período de creche em anos de estudo e avaliação, e seus resultados encontrados publicados. (publicação disponível em www.lisina.com.br)
Em uma dessas avaliações, realizada em uma Integração no Brasil, o AminoGut foi aplicado a quase 6 mil leitões e três ensaios: um em uma Unidade Produtora de Leitões (UPL) e dois em uma Unidade Multiplicadora. Os leitões foram dis-tribuídos em dois tratamentos: ração convencional e ração convencional adicionada de 0,8% de AminoGut até os 35 dias de idade dos leitões e 0,6% dos 35 aos 49 dias de idade dos leitões.
Os leitões que receberam as rações convencionais com AminoGut apresentaram em média 1,2kg a mais de peso vivo à saída da creche (60/65 dias de idade), ou seja, uma melhoria de 4,2% em relação ao tratamento sem AminoGut. Também houve melhoria de conversão alimen-tar com a adição do AminoGut nas rações dos leitões, que foi melhorada em 13% em média.
O último ensaio, no entanto, chamou muito a atenção para os expressivos resultados observados com a suplementação
de AminoGut na fase de creche: os animais do tratamento com AminoGut apresentaram, aos 60 dias de idade, peso superior em 2,219kg (26,261 no tratamento controle vs. 28,480kg no tratamento com AminoGut), o que significou um ganho de peso de superior em 11%. E a conversão alimentar também foi melhorada em 19% (de 1,59 vs 1,29 para o tratamento com AminoGut). A diferença de resul-tados superiores encontrados no último ensaio deveu-se à adoção da ração de pré desmame (creep-feeding ou ração de maternidade) pela Integração, e por sua vez, à inclusão do AminoGut aos leitões ainda na maternidade, aos 14 dias de idade.
Concluiu-se com essa avaliação que quanto antes os leitões receberem AminoGut, especialmente por meio de rações que precedem o desmame, melhores serão os resultados de desempenho observados por seu uso. A partir dessa experiência, passou-se a promover a adição do AminoGut inclusive nas rações que precedem ao desmame.
Para avaliar se a melhoria de desempenho no período de creche com o AminoGut teria efeito residual no animais ao abate, foram realizados dois estudos com animais pós desmame e creche, que foram acompanhados até o abate aos 100 kg de peso vivo (Pupa e Hannas, 2008).
Leitões desmamados aos 18 dias de idade, receberam uma ração controle e uma ração com adição de AminoGut:
Tabela 6. Garantia comercial e matriz nutricional do AminoGut

Disponível no Site www.lisina.com.br 17
0,8% nas rações fornecidas aos animais dos 18 aos 35 dias de idade e 0,6% de inclusão nas rações para os animais de 36 a 45 dias de idade.
Os leitões, que haviam recebido AminoGut, de ambos os ensaios apresentaram melhoria (P<0,05) de 8% e 12% no peso corporal dos leites aos 45 dias de idade, o que cor-respondeu a mais 0,997 e 1,423kg de peso corporal por leitão, nos ensaios um e dois, respectivamente.
Os animais do experimento um foram acompanhados até o abate. Os animais que haviam recebido AminoGut nas rações iniciais, atingiram o peso vivo de 100 kg cinco dias antes dos animais que não haviam consumido AminoGut no período de creche, ou seja, quase 4 kg de peso vivo a mais ao abate por animal. Considerando-se a estimativa do volume de produção de carne suína brasileira projetada pelo Ministério da Agricultura em 2015, estima-se que o AminoGut contribuirá com um incremento de 1,7% do to-tal de carne produzida no Brasil, como efeito subsequente de seu uso na fase de creche
Em 2010, avaliações práticas e resultados de campo foram publicadas pela Ajinomoto, nomeado de “AminoGut na Prática”, a chamada de Cartilha AminoGut, trouxe com uma visão simplificada experiências comerciais resumidas e seus resultados (publicação disponível em www.lisina.com.br)
Esse material contém resultados de campo encontrados em granjas comerciais brasileiras (nomeadas de Granja A, B, C, D e E, para manter a confidencialidade dos produtores) que suplementaram AminoGut nas rações de leitões em pré e pós desmame e comparam seu efeitos com suas rações convencionais.
Em todos os resultados encontrados (P<0,05) com a suple-mentação do AminoGut percebe-se sempre a mesma tendência: maior resultado de peso corporal e ganho de peso médio diário, melhoria da conversão alimentar e re-dução das taxas de mortalidade e dos índices de diarreia dos animais.
Concluiu-se, dessa compilação de resultados encontrados em condições comerciais e produtores de médio e pequeno porte, que em média o AminoGut aumentou o peso corpo-ral dos leitões em 1kg à saída de creche.
Estudos e resultados recentes
No trabalho de dose-resposta, Teixeira et al. (2011) con-cluíram que associação de Glutamina e Ácido Glutâmico melhorou o desempenho e a morfo-fisiologia gastrintesti-nal de leitões desmamados aos 21 dias de idade. O menor
índice de diarréia foi observado com a inclusão de 1,0% de AminoGut, e os melhores resultados de altura de vilosidade intestinal e relação vilosidade:cripta foram obtidos com a inclusão de 0,82% de AminoGut.
Os mesmos autores (Teixeira et al., 2011b) estudando leitões de 21 a 28 dias de idade, concluíram que os animais que haviam consumido a dieta contendo 1,0% de AminoGut tiveram maior (P<0,05) ganho de peso diário (GPD) e consumo médio diário de dieta (CMD) quando comparados com a dieta controle. Enquanto que no perío-do de 21 a 42 dias de idade, animais alimentados com as dietas contendo 0,5 e 1,0% de AminoGut tiveram melhor CA em comparação com a dieta controle. Estes resulta-dos corroboram o estudo de Lackeyram et al. (2001), que relataram que a suplementação de 0,8% de glutamina em dietas a base de milho e farelo de soja foi eficaz para aumentar o GPD de leitões após o desmame. No período de 21 a 28 dias de idade, os animais que consumiram a dieta contendo 1,0% de AminoGut tiveram menos (P<0,05) diarreia.
Da mesma maneira, Lima et al. (2012) estudando alimen-tos de alto valor nutricional para leitões, concluíram que a suplementação de glutamina melhora o desempenho de leitões desmamados em período de creche, quando com-parados ao desempenho dos animais do grupo controle.
Todos os resultados encontrados com o uso do AminoGut em dietas de leitões estão em acordo com os primeiros es-tudos que foram realizados desde antes dos anos 2000. No entanto, as rações de leitões, seus requerimentos nutricionais e os ingredientes utilizados, mudaram mui-to nessa ultima década. Dessa maneira, dois estudos de dose-resposta foram realizados pela Universidade Federal de Viçosa em 2012 (Lescano et al., 2013), para reafirmar as recomendações dos níveis de inclusão dos AminoGut nas rações de leitões em pré e pós desmame.
No experimento 1, o total de 44 leitões desmamados, de 18 aos 46 dias de idade, receberam rações a base de milho e farelo de soja, milho pré-cozido, lácteos e plasma sanguí-neo, isoenergéticas e isoaminoacídicas, exceto pela inclusão de níveis crescentes de AminoGut: 0, 0.4%; 0.8%; 1.2%. A adição de AminoGut melhorou linearmente (P<0,05) o con-sumo diário de ração e a conversão alimentar dos leitões. Houve ainda efeito linear (P<0,01) e quadrático (P<0,04) sob o peso corporal final, ganho de peso total e ganho de peso diário dos leitões (Tabela 7). Concluiu-se que a adição de 0,8% de AminoGut promoveu os melhores resultados de desempenho para leitões de 18 a 46 dias de idade.
Disponível no Site www.lisina.com.br
Tabela 7. Desempenho de leitões alimentados com AminoGut de 18 a 46 dias de idade
Tabela 8. Parâmetros histológicos intestinais médios de leitões aos 25 dias de idade
No experimento dois (Lescano et al., 2013b) foram obser-vados os efeitos da suplementação crescente de AminoGut nas rações de leitões (0, 0.4%; 0.8%; 1.2%) sobre os parâmetros de morfologia intestinal. O total de 24 leitões foram eutanasiados 7 dias após o desmame para coleta de amostras histológicas de intestino delgado. Os autores con-cluíram que a adição de AminoGut melhorou linearmente (P<0,05) a altura das vilosidades intestinais das porções do
duodeno e do íleo, e a relação altura de cripta: profundi-dade de vilos no duodeno, jejuno e íleo. Houve também efeitos linear e quadrático (P<0,05) da profundidade de criptas da porção do duodeno (Tabela 8). Concluiu-se que a suplementação de 1,2% de AminoGut nas rações de leitões é benéfica para amenizar os efeitos do desmame sob os parâmetros morfológicos intestinais do intestino del-gado de leitões de 18 a 25 dias de idade.
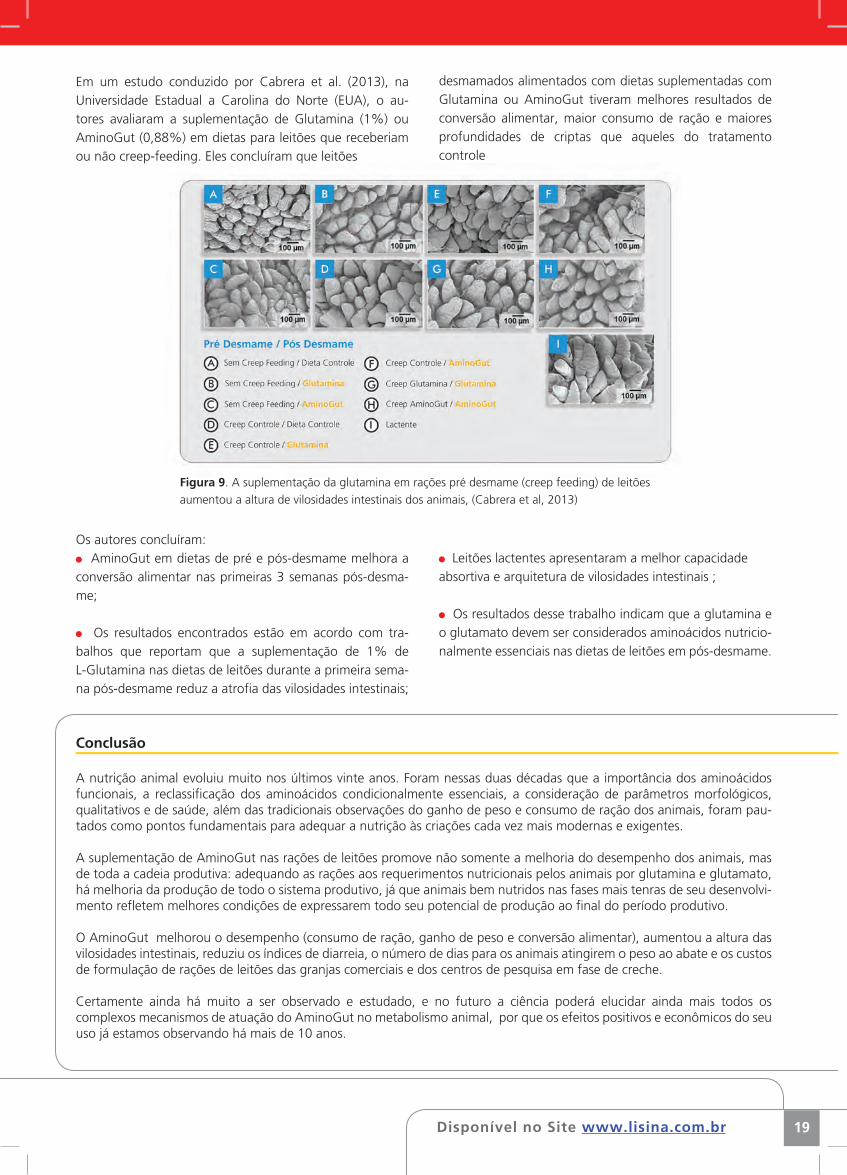
Disponível no Site www.lisina.com.br 19
Figura 9. A suplementação da glutamina em rações pré desmame (creep feeding) de leitões
aumentou a altura de vilosidades intestinais dos animais, (Cabrera et al, 2013)
Em um estudo conduzido por Cabrera et al. (2013), na Universidade Estadual a Carolina do Norte (EUA), o au-tores avaliaram a suplementação de Glutamina (1%) ou AminoGut (0,88%) em dietas para leitões que receberiam ou não creep-feeding. Eles concluíram que leitões
Os autores concluíram: AminoGut em dietas de pré e pós-desmame melhora a
conversão alimentar nas primeiras 3 semanas pós-desma-me;
Os resultados encontrados estão em acordo com tra-balhos que reportam que a suplementação de 1% de L-Glutamina nas dietas de leitões durante a primeira sema-na pós-desmame reduz a atrofia das vilosidades intestinais;
desmamados alimentados com dietas suplementadas com Glutamina ou AminoGut tiveram melhores resultados de conversão alimentar, maior consumo de ração e maiores profundidades de criptas que aqueles do tratamento controle
Conclusão
A nutrição animal evoluiu muito nos últimos vinte anos. Foram nessas duas décadas que a importância dos aminoácidos funcionais, a reclassificação dos aminoácidos condicionalmente essenciais, a consideração de parâmetros morfológicos, qualitativos e de saúde, além das tradicionais observações do ganho de peso e consumo de ração dos animais, foram pau-tados como pontos fundamentais para adequar a nutrição às criações cada vez mais modernas e exigentes.
A suplementação de AminoGut nas rações de leitões promove não somente a melhoria do desempenho dos animais, mas de toda a cadeia produtiva: adequando as rações aos requerimentos nutricionais pelos animais por glutamina e glutamato, há melhoria da produção de todo o sistema produtivo, já que animais bem nutridos nas fases mais tenras de seu desenvolvi-mento refletem melhores condições de expressarem todo seu potencial de produção ao final do período produtivo.
O AminoGut melhorou o desempenho (consumo de ração, ganho de peso e conversão alimentar), aumentou a altura das vilosidades intestinais, reduziu os índices de diarreia, o número de dias para os animais atingirem o peso ao abate e os custos de formulação de rações de leitões das granjas comerciais e dos centros de pesquisa em fase de creche.
Certamente ainda há muito a ser observado e estudado, e no futuro a ciência poderá elucidar ainda mais todos os complexos mecanismos de atuação do AminoGut no metabolismo animal, por que os efeitos positivos e econômicos do seu uso já estamos observando há mais de 10 anos.
Leitões lactentes apresentaram a melhor capacidade absortiva e arquitetura de vilosidades intestinais ;
Os resultados desse trabalho indicam que a glutamina e o glutamato devem ser considerados aminoácidos nutricio-nalmente essenciais nas dietas de leitões em pós-desmame.

Disponível no Site www.lisina.com.br
Referências Bibliográficas
AJINOMOTO. Encyclopedia of amino acids: production processes.
[S.1., 2015]. Disponivel em: <http://www.ajinomoto.com/amino/
eng/product_print.html>. Acesso em 10 set. 2015.
Cabrera R, Usry J, Arrellano C, Nogueira E, Kutschenko M, Moeser
A, Odle J: Effects of creep feeding and supplemental glutamine or
glutamine plus glutamate (AminoGut) on pre- and post-weaning
growth performance and intestinal health of piglets. J. Anim. Sci.
Biotechnology 2013, 4:29.
Knudsen KEB, Laerke HN, Hedemann MS: The Hole of fibre in
piglet gut health. In: Gut efficiency; the key ingredient in pig and
poultry production. Elevating animal performance and healthy.
Taylor-Pickard J.A.; Spring P. 2008. ISBN 978-90-8686-060-9
Kutschenko M, Nogueira ET, Haese D, Lima L: AminoGut:
ciência e prática na nutrição de leitões. Kutschenko M, Nogueira
ET, Haese D, Lima L. 2008. Disponível em: <http://www.lisina.com.
br/publicacoes_detalhes.aspx?id=2174> Acesso em 30/09/2015.
Lallès JP, Boudry G, Favier C, Le Floc´H N, Luron I, Montagne L,
Oswald I. P, Pié S, Pile C, Sève B: Gut function and dysfuncion in
young pigs: physiology. Anim. Res. 53. P. 301-316. 2004. DOI
10.1051/Animal Research: 2004018
Lescano D, Albin L, Hannas M, Salguero S, Kutschenko M, Noguei-
ra E, Rostagno H: Evaluation of dietary glutamic acid plus gluta-
mine levels on the growth performance of piglets. 2013. J. Anim.
Sci. Vol. 91, E-Suppl. 2/J. Dairy Sci. Vol. 96, E-Suppl. 1
Lescano D, Albin L, Hannas M, Salguero S, Kutschenko M, Noguei-
ra E, Rostagno H: Effect of glutamic acid plus glutamine levels on
the intestinal morphology of piglets. 2013b. J. Anim. Sci. Vol. 91,
E-Suppl. 2/J. Dairy Sci. Vol. 96, E-Suppl. 1
Li P.; Yin Y. L.; Li d. F.; Kim S. W.; Wu G. Amino acids and immune
function. 2007. British Journal Nutrition 98, 237-252.
Lima GJMM, Kutschenko M, Nogueira ET: Effects of steam pro-
cessed rice, natural vitamin E and glutamine in diets for weaning
piglets. J. Anim. Sci. 2012. Vol. 90, Suppl. 3/J. Dairy Sci. Vol. 95,
Suppl. 2.
Lopes D, Rostagno HS: Desempenho e morfologia intestinal de
leitões alimentados com AminoGut e plasma. In: Kutschenko M,
Nogueira ET, Haese D, Lima L. AminoGut: ciência e prática na
nutrição de leitões. 2008. Disponível em: <http://www.lisina.com.
br/publicacoes_detalhes.aspx?id=2174> Acesso em 30/09/2015.
Lopes D, Rostagno HS: Utilização do AminoGut em dietas
ricas e pobres em leitões desmamados aos 21 dias de idade.
In: Kutschenko M, Nogueira ET, Haese D, Lima L. AminoGut:
ciência e prática na nutrição de leitões. 2008. Disponível em:
<http://www.lisina.com.br/publicacoes_detalhes.aspx?id=2174>
Acesso em 30/09/2015.
Machado G. S. O presente e o futuro dos desafios relacionados
ao desmame e à nutrição na fase pós-desmame. In: Kutschenko
M, Nogueira ET, Haese D, Lima L. AminoGut: ciência e prática na
nutrição de leitões. 2008. Disponível em: <http://www.lisina.com.
br/publicacoes_detalhes.aspx?id=2174> Acesso em 30/09/2015.
Newsholme P, Lima MMR, Procopio J, Pithon-Curi TC, Doi SQ,
Bassotes, RB, Curi R: Glutamine an glutamate as vital metabolites.
Brazilian Journal of Medical and Biological Research (2003) 36:
153-163.
Nogueira ET: Aminoácidos: essenciais para Suínos. Suinocultura
Industrial, Edição 191, v. 08, p. 26 - 28. 2005
Pupa JMR, Hannas MI: AminoGut: desempenho e retorno
econômico em granjas comerciais de suínos. In: Kutschenko M,
Nogueira ET, Haese D, Lima L. AminoGut: ciência e prática na
nutrição de leitões. 2008. Disponível em: <http://www.lisina.com.
br/publicacoes_detalhes.aspx?id=2174> Acesso em 30/09/2015.
Sangilt P. T. Symposium on Digestive Physiology in Pigs. 8th
Edition. 2000, Uppsale, Sweden. Digestive physiology of pigs:
proceedings of the 8th Symposium. Lindberg J.E.; Ogle B. ISBN
0-85199-517-9
Teixeira AO, Nogueira ET, Kutschenko M, Rostagno HS, Lopes DC:
Inclusão de glutamina associada ao ácido glutâmico em dietas
para leitões ao desmame dos 21 aos 42 dias de idade. In: XV Con-
gresso da ABRAVES, 2011, Fortaleza. Anais do Congresso, 2011.
Teixeira AO, Nogueira ET, Kutschenko M, Rostagno HS, Lopes DC:
Avaliação da inclusão de glutamina associada ao ácido glutâmico
em dietas sobre a morfológica de leitões ao desmame dos 21 aos
42 dias de idade. In: XV Congresso da ABRAVES, 2011, Fortaleza.
Anais do Congresso, 2011b.
Touchette J, Allee G.L, Watanabe K, Toride Y, Shinzato I, Usry
JL: The effect of arginine and glutamine on post-weaning perfor-

Disponível no Site www.lisina.com.br 21
mance and intestinal morphology of pigs. J. Anim. Sci. Vol. 78,
Suppl. 1/J.Dairy Sci. Vol.83, Suppl.1/2000.
Watford M.; Kutschenko M.; Nogueira E. T. Optimal dietary gluta-
mine for growth and development. R. Bras. Zootec., v.40, p.384-
390, 2011 (supl. especial). 2011.
Watford M: Functional amino acids and intestinal immune func-
tion in neonates. VI Congresso Latino-Americano de Nutrição An-
imal, CBNA, 2014.
Watford M: Glutamine metabolism and function in relation to
proline synthesis and the safety of glutamine and proline supple-
mentation. Journal of Nutrition, v.134, p.2003S- 2007S, 2008.
Wu G: Amino Acids: Biochemistry and Nutrition. Boca Raton,
Florida: CRC Press; 2013.
Wu G: Functional amino acids in growth, reproduction and health.
American Society for Nutrition. Adv. Nutr. 1: 31-37. 2010a. DOI
10.3945/an.110.1008.
Wu G: Biochemical and physiological limitations to efficiency of
amino acid utilization for animal growth. In: Energy and protein
metabolism and nutrition. 3rd EAAP International Symposium on
Energy and Protein Metabolism and Nutrition, 2010b. ISBN 978-
90-8686-153-8.
Wu G: Papéis importantes da glutamina na nutrição e produção
animal. In: Kutschenko M, Nogueira ET, Haese D, Lima L. Amino-
Gut: ciência e prática na nutrição de leitões. 2008. Disponível em:
<http://www.lisina.com.br/publicacoes_detalhes.aspx?id=2174>
Acesso em 30/09/2015.
Wu G, Meier SA, Knabe DA: Dietary glutamine supplementation
prevents jejunal atrophy in weaned pigs. Journal of Nutrition 126:
2578-2584. 1996.
YI GF, Carrol JA, Allee GL, Gaines AM, Kendall DC, Usry JL, Toride
Y, Izuru S: Effect of glutamine and spray-dried plasma on growth
performance, small intestinal morphology, and immune responses
of Escherichia coli K88+ - challenged weaned pigs. J. Anim. Sci.
2005. 83:634-643.
YI GF, Allee GL, Toride Y, Usry JL, Gaines AM: Impact of gluta-
mine, glutamate, and nucleotides on the growth performance
and intestinal morphology of weaned piglets. J. 2002. Anim. Sci.
Vol. 80, Suppl. 1/ J. Dairy Sci. Vol 85, Suppl. 1.

Disponível no Site www.lisina.com.br Disponível no Site www.lisina.com.br
Malcolm Watford, [email protected]
O Prof. Dr. Malcolm Watford tem trabalhado com o metabolismo da glutamina
por mais de 40 anos: por meio do estudo do metabolismo intermediário e espe-
cificamente, a relação da gluconeogênese e a excreção de nitrogênio.
Na década de 70 foi orientado pelo Dr. Hans Krebs na Trent Polytechnic em Bio-
logia Aplicada, quando recebeu a honra de bolsa de pesquisa de pós-graduação
do Medical Research Council da Grã-Bretanha para estudar na Universidade de
Oxford na Grã Bretanha (1974-77) a Regulação Metabólica, do fígado, rins e
intestinos. Em seu pós-doutorado, na Universidade de Montreal (1977-79), inves-
tigou os papeis dos músculos e rins no metabolismo da glutamina, e a regulação
da gluconeogênese pelos aminoácidos no Departamento de Bioquímica da Esco-
la da Medicina da Case Western Reserve University (1979-1980).
Como cientista independente, encontrou que a isoenzima glutaminase dos
diferentes tecidos são reguladas diferentemente em resposta à diabetes insuli-
na-dependente e acidose metabólica. Essa descoberta foi seguida pela primeira
purificação da glutaminase encontrada no fígado (hoje conhecida como GLS2),
pela identificação do primeiro cDNA e do DNA genômico dessa enzima, e a
caracterização da regulação da expressão hormonal usando o promotor isolado
para esse gene. Ele continuou estudando o metabolismo da glutamina, desco-
brindo que os enterócitos conservam o esqueleto de carbono da glutamina como
alanina (ou seja, gluconeogênese).
Mais recentemente, seu trabalho está centrado no estudo do papel do tecido
adiposo no metabolismo da glutamina, primeiro descobrindo que o tecido adi-
poso in vivo é um produtor importante de glutamina, e depois, demonstrando
que a glutamina é requerida pelos adipócitos tanto no processo de diferenciação,
quanto durante o processo de maturação de adipócitos funcionais. Além disso,
o Prof. Dr. Watford tem conduzido extensas pesquisas com a suplementação de
glutamina e AminoGut em animais vivos (suínos e ratos) durante a lactação e
tem encontrado que a glutamina pode evitar a perda de massa magra corporal
das fêmeas e aumentar a concentração de glutamina e lipídios no leite para o
neonato.
Atualmente o Prof. Dr. Malcolm Watford é Professor Associado da Universidade
de Rutgers (desde 1990), é Diretor do Programa de Honras a Estudiosos da School
of Environmental & Biological Sciences na mesma instituição, e Pesquisador
Adjunto do Centro de Biotecnologia Animal e Terapia Gênica (CBATEG) da
Universidade Autônoma de Barcelona, além de seu trabalho como Editor
Associado das principais revistas e publicações científicas do mundo.

Disponível no Site www.lisina.com.br 23 Disponível no Site www.lisina.com.br
Suplementação de Glutamina e Glutamato na Nutrição Animal
Introdução
A necessidade de proteína dietética está tradicionalmente relacionada às exigências de aminoácidos para síntese de proteína. Há mais de 100 anos, desde a época de Osborne e Mendel [1] e depois de Rose [2-4], sabemos que os ami-noácidos da dieta são indispensáveis (essenciais) e outros são dispensáveis (não-essenciais), com referência à ca-pacidade do organismo sintetizar aminoácidos específicos em quantidades suficientes para sustentar o crescimento ideal ou manter o equilíbrio de nitrogênio. Na produção industrial de animais domésticos, qualquer aminoácido in-dispensável limitante – como a lisina em dietas à base de grãos – é rotineiramente suplementado na forma de ami-noácido purificado, sem a necessidade de aumentar muito o teor proteico da dieta. Está se tornando cada vez mais importante manter os níveis de proteína da dieta baixos por causa do custo financeiro e do impacto ambiental da excreção de nitrogênio. Este artigo destaca uma outra con-sideração: alguns aminoácidos comumente chamados de dispensáveis, como a glutamina e o glutamato, também podem ser limitantes nestas dietas. Recentemente, uma mistura de glutamina e glutamato (AminoGut) se tornou disponível como aditivo para a alimentação animal e os primeiros estudos demonstraram que, quando adicionado à dieta a 0,8-1,0%, este suplemento traz benefícios tanto para leitões em crescimento como para porcas em lactação.
Aminoácidos funcionais
Como destacado por Rose [2-4] há mais de 90 anos, a de-terminação do que é – e do que não é – um aminoácido indispensável é puramente uma questão de definição. Ao longo dos anos, a definição de aminoácido indispensável limitou-se à exigência para crescimento máximo ou para a manutenção do balanço de nitrogênio [5]. Em parte, esta definição se devia às limitações técnicas à época em que
os aminoácidos individuais foram isolados pela primeira vez e usados em estudos de alimentação. Na medida em que passamos a conhecer outros papéis importantes dos aminoácidos além de substratos para a síntese proteica, se tornou evidente que fornecer aminoácidos em níveis que sustentem o crescimento e o balanço de nitrogênio não é suficiente para outras funções [5,6]. Portanto, as exigências de aminoácidos estão hoje sendo cada vez mais avaliadas em relação aos seus papéis não-proteicos e alguns aminoácidos dispensáveis estão sendo reclassificados como aminoácidos funcionais e muitos são agora considerados como condicionalmente essenciais. De fato, até mesmo Rose [4] reconheceu este potencial ao escrever que “não se pode excluir a possibilidade de que os aminoácidos que não são necessários para o balanço de nitrogênio em indivíduos normais possam se tornar indispensáveis no caso de doenças ou para funções especiais, como reprodução e lactação”.
Glutamina e Glutamato
A glutamina e o glutamato são os aminoácidos mais abundantes no organismo, mas ambos são considerados dispensáveis porque são prontamente sintetizados pelo organismo [7-10]. Geralmente, estes dois aminoácidos são expressos simplesmente como “glutamato” porque a hidrólise ácida das proteínas, usada para determinar o teor de aminoácidos específicos, de fato hidrolisa toda a glutamina em glutamato e, portanto, a estimativa final de “glutamato” nos ingredientes de ração usualmente se refere à soma de glutamato e glutamina. Isto tem poucas consequências, uma vez que o glutamato e a glutamina da dieta são amplamente catabolizados dentro da muco-sa do intestino delgado e apresentam apenas pequenas diferenças nos produtos finais.
A glutamina é o alfa-aminoácido mais abundante na maio-ria das espécies e está presente em grandes quantidades
Prof. Dr. Malcolm Watford, D.Phil.Departamento de Ciências Nutricionais - Escola de Ciências Ambientais e BiológicasUniversidade Rutgers- EUA
23

Disponível no Site www.lisina.com.br
(>90%) nas células do músculo esquelético. Da mesma for-ma, a glutamina é o alfa-aminoácido mais abundante no plasma sanguíneo da maioria dos mamíferos e este pool é rapidamente renovado (na ordem de 60-80g por dia em pessoas adultas). Por outro lado, embora as concentrações de glutamato sejam relativamente altas na maioria das cé-lulas, a concentração de glutamato livre no plasma é muito mais baixa [10]. Portanto, a glutamina é uma das principais formas de transporte de nitrogênio, carbono e energia en-tre os tecidos, enquanto que o glutamato tem um papel intracelular importante nos metabolismos da energia e de aminoácidos.
A glutamina e o glutamato também são muito abundan-tes na maioria dos alimentos e, juntos, são responsáveis por 5-20% dos aminoácidos das proteínas [11,12], mas em nível de consumo normal, não há absorção líquida na circulação portal. Depois do consumo de alimento, os enterócitos do intestino delgado utilizam a glutamina e o glutamato derivados da dieta, assim como a glutamina cir-culante, como principal substrato respiratório. Portanto, o grande pool corporal de glutamina é sintetizado de novo através da ação da enzima glutamina sintetase [9]. O prin-cipal sítio de síntese de glutamina é o músculo esquelético, com pequenas contribuições do fígado, pulmões, tecido adiposo e, possivelmente, dos rins em espécies herbívoras. Em indivíduos saudáveis, não-gestantes e não-lactantes, os principais sítios de utilização de glutamina são os enteróci-tos do intestino delgado e as células do sistema imune, onde a glutamina é o principal combustível respiratório [9,13]. Há alguma absorção de glutamina pelo fígado, para gliconeogênese e síntese de ureia, e pelos rins, para a pro-dução de amônia para o equilíbrio ácido-base, sendo o car-bono da glutamina recuperado através da gliconeogênese renal [7-10].
O nitrogênio da amida da glutamina é utilizado para a síntese de purinas, pirimidinas, NAD, NADP, glicosamina e as paragina através da ação de diversasamidotransferases. No entanto, a maior parte da degradação da glutamina (>97%) ocorre via hidrólise a glutamato por enzimas glutaminases [7]. O glutamato pode formar glutationa, prolina, ornitina, citrulina e ar-ginina, mas, novamente, a maior parte do catabolismo do glutamato passa através do alfa-cetoglutarato, en-tra no ciclo de Krebs e é usado para a produção de ATP (enterócitos, células imunes) ou gliconeogênese (fígado, rins). Além disso, o metabolismo da glutamina e do glutamato nos enterócitos também forma citrulina (e, portanto, arginina), alanina, prolina, lactato e glutationa. Foi observado recen-temente que, além destes papéis metabólicos, a glutami-na é uma importante molécula reguladora da sinalização
do anabolismo (geralmente via estimulação do mTOR), aumentando a síntese proteica e a proliferação e diferen-ciação celular e, ao mesmo tempo, reduzindo a proteólise e a apoptose [8]. Portanto, a glutamina e o glutamato têm papeis essenciais e quantitativamente importantes no me-tabolismo e na função de diversas células e tecidos, espe-cialmente durante o crescimento e a diferenciação.
Metabolismo da glutamina em estadoscatabólicos
Há 30 anos, foi observado que, em paciente hipercatabóli-cos, as exigências de glutamina aumentavam além da ca-pacidade de síntese de glutamina pelo organismo. Nestas condições, há aumento da necessidade de glutamina para o sistema imune, correção da acidose, síntese de proteínas de fase aguda e gliconeogênese. Uma das primeiras respos-tas à injúria é a rápida liberação de glutamina do pool in-tramuscular, resultando em uma nova e mais baixa concen-tração de glutamina intramuscular e taxas altas e contínuas de liberação de glutamina, que são mantidas pelo aumento da síntese de glutamina [7]. Os únicos substratos disponíveis para esta síntese são os aminoácidos liberados pela degra-dação das proteínas musculares e, portanto, estes quadros são acompanhados de perda excessiva de massa corporal magra [14]. Estados catabólicos são sempre acompanhados de baixos níveis musculares e plasmáticos de glutamina e têm sido feitas diversas tentativas de suplementar glutami-na para tentar reduzir a necessidade de síntese muscular de glutamina, limitando assim a proteólise líquida de músculo. Em diversos quadros, como transplante de medula óssea e sepse grave, têm sido obtidos bons resultados com glu-tamina intravenosa [15], enquanto que a glutamina oral parece ter efeitos benéficos sobre a manutenção da saúde intestinal [16,17]. Recentemente, foram relatados proble-mas com o fornecimento de glutamina a pacientes com falência múltipla dos órgãos [18,19], mas exceto nestes ca-sos extremos, a suplementação de glutamina é considera-da segura. Além disso, suplementos orais de glutamina são amplamente usados na área de esportes e halterofilismo e doses de 20-40g por dia são bem toleradas.
A glutamina e o glutamato na produção animal
Na produção animal, o tratamento de quadros catabóli-cos não é uma preocupação, mas a pergunta importante é: “o glutamato e a glutamina são condicionalmente essenciais em outros momentos”? O papel da glutami-na na proliferação, diferenciação e crescimento celular é claramente importante no neonato em desenvolvimento, mas o metabolismo interórgãos da glutamina e do gluta-

Disponível no Site www.lisina.com.br 25
mato também é alterado durante a prenhez e a lactação. Durante a gestação, há grande absorção de glutamina cir-culante através da placenta que, junto com a síntese de glutamina pela placenta, fornece grandes quantidades de glutamina para o crescimento fetal [20]. Além disso, du-rante a lactação, o glutamato e a glutamina livres e ligados à proteína, derivados da circulação e da síntese na glândula mamária, perfazem 20% dos aminoácidos do leite (tanto como aminoácidos livres, como os incorporados à proteína) [21,22]. Os altos níveis de glutamina e glutamato no leite estão relacionados ao fato de que estes aminoácidos são essenciais para o crescimento e diferenciação das células epiteliais intestinais e para a manutenção da altura das vi-losidades intestinais e da integridade intestinal, que são da maior importância para o neonato. Além disso, a glutamina é exigida pelo tecido linfoide associado ao intestino (GALT), o maior órgão imune do organismo e essencial para o de-senvolvimento da imunidade do neonato [23]. Portanto, a manutenção da homeostase do glutamato e da glutamina é importante para o bem-estar de diversos tecidos, espe-cialmente durante a gestação, a lactação e o crescimento. Acreditava-se que os substratos para esta síntese adicional de glutamina fossem os aminoácidos em excesso consu-midos na dieta materna. Entretanto, há alguns anos, foi observado que a lactação em equinos estava associada com um leve estado catabólico, indicado pela perda de massa corporal magra e redução dos níveis plasmático e muscular de glutamina [24]. Achados semelhantes foram relatados em outras espécies [25], como o suíno [26,27], sugerindo que as reservas de proteína materna são usadas, em parte, para fornecer glutamina. Portanto, é extremamente impor-tante manter a homeostase do glutamato e da glutamina durante a gestação e a lactação e sugere-se que a suple-mentação de glutamina e/ou glutamato na dieta materna possa ser útil.
O trato gastrointestinal e o tecido linfoide associado ao intestino
Ao nascimento e ao desmame, ocorrem mudanças im-portantes no desenvolvimento e no metabolismo do trato gastrointestinal (TGI) e no GALT [28-30]. Ao nascimento, o TGI é imaturo e o intestino permanece “aberto” por alguns dias para a transferência da imunidade do colostro para o neonato. Mamar fornece ao leitão uma dieta láctea alta-mente palatável, rica em gordura, de fácil digestão, mas que também representa a introdução inicial de micróbios no TGI. No desmame, o leite é substituído por uma die-ta com níveis relativamente altos de carboidratos, mais complexa e de mais difícil digestão. O desmame também está associado com mudanças importantes no número e na variedade de micróbios ingeridos. Da mesma forma, ao
nascimento, o GALT ainda está pouco desenvolvido; alguns aspectos da função do GALT podem levar até seis semanas para amadurecer. Na natureza, os leitões são gradualmente desmamados em 10-12 semanas, mas na suinocultura moderna, o desmame é abrupto e ocorre muito antes (3-5 semanas), causando considerável estresse ao TGI e ao GALT [28-32]. O desmame precoce está associado com redução do consumo de ração, perda de peso corporal, diarreia e baixa eficiência alimentar. O desmame também causa uma redução marcante do ganho de peso, do peso e do compri-mento do intestino delgado e da altura das vilosidades, re-sultando em aumento da permeabilidade intestinal e maior relação de glutamina oxidada para glutationa. Isto é acom-panhado por aumento da expressão de genes envolvidos no estresse oxidativo e na ativação imune, mas também à redução da expressão de genes relacionados ao metabo-lismo de macronutrientes e à proliferação celular, além de maior atrofia das vilosidades no intestino delgado. Foram propostas várias medidas para lidar com os problemas do desmame precoce e muitas se baseiam na adição de suple-mentos à dieta, como probióticos, prebióticos, pequenos compostos individuais como aminoácidos, diversas prepa-rações de plasma e outras proteínas e ingredientes mais complexos [31,32]. Alguns estudos avaliaram a suplemen-tação de aminoácidos tradicionalmente indispensáveis em uma dieta de baixa proteína, e demonstraram que permite crescimento adequado e efetivamente diminui a incidên-cia de diarreia em leitões desmamados de forma abrupta [33]. Como tanto a glutamina, quanto o glutamato são necessários para o desenvolvimento e a manutenção das funções do TGI e do GALT, o uso da suplementação de glutamina e glutamato começou a ser avaliado na suino-cultura.
Suplementação de glutamina e glutamato para leitões desmamados
Os primeiros estudos com a suplementação de glutamina foram realizados em leitões desmamados há 10 anos. A suplementação de 1% de glutamina para leitões por 14 dias pós-desmame preservou a altura das vilosidades do jejuno e melhorou o desempenho (eficiência alimentar) em 25% [34]. Recentemente, Zou et al. [35] confirmaram es-tes achados, com 28% maior taxa de crescimento, melhor conversão alimentar e redução da incidência de diarreia em leitões desmamados suplementados com 1% de glutami-na. Hanczakowska e Niwinska [36] revisaram o uso de glu-tamina em nutrição de suínos e citam seu próprio trabalho, não publicado, em que obtiveram aumento do ganho depeso (8,7%) em leitões alimentados com 2% de glutamina. Recentemente, estes autores relataram que a suplementação de 1% de glutamina teve pouco efeito

Disponível no Site www.lisina.com.br
quando fornecida individualmente, mas aumentou os efei-tos da suplementação de butirato sobre a estrutura do epi-télio intestinal. No entanto, em sua revisão, destacam que nem todos os estudos demonstram alterações no ganho de peso de leitões. Isto pode estar relacionado ao fato de que muitos experimentos terminaram apenas alguns dias de-pois do desmame, antes que os benefícios para os leitões, como maior ganho de peso ou menor incidência de diar-reia, ficassem evidentes.
Foi relatado que um alto nível de glutamina (4,4%) preve-niu a queda do nível de glutamina no músculo esquelético de leitões recém-desmamados, além de melhorar a função imune dos linfonodos mesentéricos e normalizar a função dos linfócitos em leitões infectados com E. coli [38,39]. Este alto nível de glutamina suplementar (4,4%) também atenuou o catabolismo, suprimiu a inflamação intestinal e a liberação de citocinas e manteve a expressão da proteína das junções firmes e a função de barreira epitelial em leitões infectados com E. coli. Em outros estudos com leitões infec-tados com E. coli e suplementados com glutamina (2%), a depressão do crescimento induzida pela E. coli foi aliviada, em parte devido à manutenção da morfologia intestinal e da função imune [40,41]. Também foi demonstrado que níveis muito mais baixos de suplementação de glutamina (0,5%) beneficiaram o intestino aumentando a altura das vilosidades, reduzindo o número de células apoptóticas e melhorando o estado imunológico de leitões desmamados [42,43]. Estes resultados se devem, em parte, ao efeito positivo sobre a função de barreira intestinal, aumentan-do o número de células epiteliais e de células mitóticas no epitélio. Manzke et al. [44] avaliaram a suplementação de leitões desmamados com 2g de glutamina, glutamato ou AminoGut por sete dias. Verificaram que a imunidade celu-lar melhorou e que houve melhor resposta ao teste cutâneo com Phaseolus vulgaris (que avalia a função imune) depois da suplementação. Da mesma forma, a suplementação de glutamina (1%) a leitões desmamados alimentados com a micotoxina desoxinivalenol aliviou o efeito da toxina sobre o consumo de ração, a resposta imune e a expressão de transportadores intestinais de aminoácidos [45]. Jiang et al. [46] avaliaram leitões desmamados muito precocemente, aos 14 dias, que foram repetidamente (dias 7, 14 e 21) de-safiados com uma endotoxina e suplementados com uma glicil-glutamina. Verificaram que o suplemento melhorou a taxa de crescimento e a morfologia intestinal tanto nos leitões-controle, como nos desafiados com a endotoxi-na. Um suplemento de N-acetil-L-glutamina protegeu os leitões contra alterações do peso corporal e da imunidade intestinal após o desmame [47].
A suplementação de AminoGut (0,4-1,2%) a leitões do
desmame (dia 18) ao dia 46 resultou em aumento linear do consumo de ração, da eficiência alimentar e do peso corporal, assim como aumentou a altura das vilosidades intestinais. Estes resultados indicam que 0,8% AminoGut parece ser o nível ideal de suplementação para leitões des-mamados [48,49].
Os mecanismos moleculares responsáveis pelos efeitos protetores da glutamina no desmame precoce foram es-tudados usando microarranjos intestinais de leitões des-mamados aos 21 dias e alimentados com dietas com e sem glutamina e amostrados aos 28 dias e de leitões que permaneceram mamando até os 28 dias [50]. A suple-mentação da dieta pós-desmame com 1% de glutami-na aliviou parcialmente os efeitos do desmame sobre o ganho de peso, peso intestinal, altura das vilosidades, níveis de glutationa e melhorou a capacidade de defesa antioxi-dante. Além disso, foi verificada maior expressão intestinal de genes envolvidos no crescimento celular e na remoção de oxidantes e menor expressão de genes que codificam fatores que promovem o estresse oxidativo e a ativaçãoimune nos leitões suplementados com glutamina. Zhong et al. [51,52] investigaram os mecanismos da ação da glu-tamina em leitões desmamados aos 21 dias e alimentados com 1g de glutamina por kg de peso corporal em intervalos de 12 horas por 14 dias. O tratamento com glutamina au-mentou o ganho de peso médio diário, reduziu a incidên-cia de diarreia, aumentou o peso do intestino delgado e a altura das vilosidades e aumentou a expressão do mRNA da proteína de choque térmico 70 (HSP70) e desta proteína. Acredita-se que a HSP70 esteja envolvida na manutenção da estrutura e da função celular durante o estresse e foi demonstrado que tem papel citoprotetor do epitélio. Relatos de alterações metabolômicas no intestino do leitão em resposta à suplementação pós-desmame com glutamina (1%) indicam alterações importantes na digestão luminal de ingredientes da ração e no metabolismo intermediário dos enterócitos [53-55]. Em cultura de células intestinais suínas, a glutamina é necessária para manter a sinalização mTOR e MAPK e para reduzir a autofagia [56]. In vivo, isto se reflete no fato de que 1% de glutamina pode prevenir a atrofia das vilosidades e manter a altura das vilosidades intestinais e a barreira à permeabilidade no pós-desmame, em parte por aumentar a expressão de proteínas-chave das junções firmes [41,57] e, ao mesmo tempo, reduzir a expressão de CRF, um importante hormônio relacionado ao estresse que é ativado pelo estresse de desmame precoce [57].
Os efeitos da suplementação de glutamato no pós-desma-me ainda não foram muito estudados. Ewtushik et al. [58] verificaram que o glutamato (6,51%) evitou a atrofia das vilosidades induzida pelo desmame e promoveu o cresci-

Disponível no Site www.lisina.com.br 27
mento total e da mucosa do intestino delgado. Renazi et al. [59] forneceram glutamato (até 4%) por três semanas a leitões submetidos a desmame precoce e verificarammelhor eficiência alimentar, redução da diarreia, melhor morfologia intestinal e aumento da altura das vilosidades, do teor de DNA e da capacidade antioxidante. Wu et al. [60] também observaram que o glutamato (1%) melhorou a altura das vilosidades intestinais, assim como a proliferação e a morfologia epiteliais. Portanto, o glutama-to parece ter pelo menos parte do potencial da glutamina.
Suplementação de glutamina e glutamatodurante a amamentação
Embora as concentrações de glutamina e glutamato sejam altas no leite, isto não significa que sejam ideais para o crescimento dos leitões. Haynes e colaboradores [61] for-neceram glutamina ou alanil-glutamina por via oral duas vezes por dia por 7 dias (a partir de 7 dias de idade) a leitões que ainda estavam mamando na porca. Os leitões então receberam uma injeção de endotoxina de E. coli e foram amostrados 24 e 48 horas depois. Os autores verificaram que a glutamina extra reduziu a expressão do receptor tipo Toll 4, caspase 3 ativa e NF-kapa b (todos aumentados em resposta à endotoxina), com uma melhora geral das lesões intestinais, redução da temperatura retal e melhor desem-penho. Estes estudos indicam um papel potencial para a suplementação de glutamina antes do desmame, mas não é prático tratar leitões individuais duas vezes por dia por via oral na suinocultura comercial. Cabrera e colaboradores [62], usando um método alternativo e menos invasivo, for-neceram ração aos leitões com e sem glutamina antes do desmame. Verificaram que o fornecimento de glutamina (1%) ou uma mistura de glutamina mais glutamato (0,88% AminoGut) na ração dos leitões dos dias 14 a 21 e na dieta pós-desmame aumentou a altura das vilosidades intesti-nais e melhorou a conversão alimentar até seis semanas de idade.
Suplementação para porcas em lactação
Como descrito acima, nós e outros pesquisadores verifi-camos que a lactação em suínos e em outras espécies é acompanhada de um estado catabólico leve, demonstrado pela perda de massa corporal magra e redução do teor de glutamina intramuscular [24]. Portanto, outra abordagem seria suplementar glutamina e glutamato diretamente para a porca para evitar perda de massa corporal magra e aumentar o fornecimento de glutamina e de glutama-to no leite. Kitt et al. [63] verificaram que o fornecimento de 2,5% de glutamina a porcas aumentou o teor de glu-
tamina no leite e nós observamos resultados semelhantes com glutamina (2,5%) ou AminoGut (2,5%). Em ambos os estudos, não houve alteração do desempenho dos leitões. No entanto, verificamos que houve manutenção do teor de glutamina muscular no músculo esquelético das porcas suplementadas com glutamina ou AminoGut, indicando certa limitação da resposta catabólica à lactação. Em um estudo mais recente, confirmamos o aumento da glutami-na no leite com a suplementação de AminoGut e vimos que a contagem de células somáticas no leite também aumen-tou com a suplementação [65]. Recentemente, observamos que porcas suplementadas com AminoGut retornaram ao cio mais cedo que as que consumiram dietas não-suple-mentadas (resultados não-publicados).
Portanto, a suplementação de glutamina e/ou glutama-to para leitões recém-desmamados ou em amamentação (ração para leitões) e para porcas em lactação pode melhorar a saúde intestinal e a função imune dos neonatos, aliviando assim parte do estresse do desmame precoce. Além disso, a suplementação da dieta materna também alivia a perda de massa corporal magra durante a lactação.
Suplementação de glutamina e glutamato para outras espécies domésticas
Meijer propôs que a disponibilidade de glutamina é limitante no leite bovino [66,67]. Usando a glutamina protegida no (rúmen bypass), Caroprese e colaboradores verificaram que a glutamina (160 ou 320 g/d) melhora a resposta imune e aumenta o teor de gordura no leite de vacas holandesas adultas [68,69].
Há evidências de que o glutamato (ou a glutamina) é necessário para frangos de corte porque o seu organismo não consegue sintetizar quantidades suficientes para o crescimento [70-71].Yi et al. [73] observaram que a suple-mentação de frangos de corte em estresse por calor com glutamina (1% na água e na ração) melhorou a função imune, o ganho de peso e a eficiência alimentar e reduziu a mortalidade. Da mesma forma, Bartell e Batal [74] re-lataram maior crescimento e melhor morfologia intestinal e função imune, além de melhor qualidade de carne, em frangos de corte estressados por calor e suplementados com 0,5 e 1% de glutamina. Suplementando glutamina a 5g/kg, Dai et al. [75] relataram melhor qualidade de carne em frangos de corte sob estresse por calor. Recentemente, os autores confirmaram estes achados e relataram que a glutamina reduziu os marcadores de resposta ao estresse nestes frangos [76]. Outros autores também observaram melhor desempenho e estado imune com a suplementação de glutamina para frangos de corte. A suplementação de

Disponível no Site www.lisina.com.br
Aminogut (0,5%) foi avaliada em frangos mantidos em diferentes densidades em condições tropicais quentes e úmidas e, embora não tenha havido diferenças em peso corporal e outros parâmetros de desempenho, a mortali-dade foi reduzida (5,6% e 9,9% no grupo que recebeu AG e no controle, respectivamente).
Glutamina e glutamato como aminoácidosindispensáveis
Estas melhorias promovidas com a suplementação de glu-tamina e glutamato sugerem que estes aminoácidos de-vem ser considerados condicionalmente essenciais. Rose [2-4] verificou que animais alimentados com uma mistura de aminoácidos essenciais (para suprir as exigências míni-mas) não cresceram tão bem quanto os alimentados com
Conclusões
O conhecimento dos importantes papéis não-proteicos dos aminoácidos significa que devemos reclassificar alguns dos aminoácidos tradicionalmente não-essenciais como “funcionais”. Foi comprovado que a glutamina e o glutamato ajudam a manter as funções do TGI e do GALT. São necessários na dieta em quantidades suficientes para obter o desempenho ideal de leitões desmamados precocemente e possivelmente para a manutenção da massa corporal magra da porca durante a lactação. Questões importantes e ainda não elucidadas são a quantidade ideal do suplemento, se os suplementos devem ser fornecidos individualmente ou combinados, qual o papel da dieta basal, qual o animal (a porca ou os leitões) a ser suple-mentado e o momento e duração do tratamento. O ganho de peso e a eficiência alimentar são claramente importantes no longo prazo, mas os resultados de estudos experimentais são essenciais para identificar os benefícios e mecanismos de ação destes suplementos.
estes aminoácidos mais aminoácidos dispensáveis e mes-mo fontes de nitrogênio não-amino. Estes e outros estudos [80] ilustram claramente que é necessária certa quantidade de aminoácidos não-essenciais na dieta e que o glutamato pode ser a fonte ideal. Outros estudos com aves mostraram que o glutamato dietético é essencial para o crescimento desta espécie [70-72]. A glutamina não era avaliada nos primeiros estudos porque era difícil sua determinação, mas devido ao glutamato provavelmente não ser limitante em dietas tradicionais, este tema recebeu pouca atenção nos últimos 30-40 anos. A tendência de usar dietas com baixa proteína, geralmente à base de ingredientes que contêm baixos teores de glutamina, e a disponibilidade de novas fontes comerciais de glutamato e de glutamina (AminoGut) renovou o interesse no uso destes aminoácidos na forma suplementar.

Disponível no Site www.lisina.com.br 29
Referências Bibliográficas
1. Osbourne TB, Mendel LB (1914) Amino-acids in nutrition
and growth. J Biol Chem 17:325-349
2. Rose WC, Oesterling, MJ, Womack, M. (1948) Compar-
ative growth on diets containing ten and 19 amino acids, with
further observations upon the role of glutamic and aspartic acids.
J Biol Chem 176:753-62.
3. Rose WC, Smith, LC, Womack, M, Shane, MJ. (1949)
The utilization of the nitrogen of ammonium salts, urea, and cer-
tain other compounds in the synthesis of non-essential amino ac-
ids in vivo. J Biol Chem. 181:307-16.
4. Rose WC (1957) The amino acid requirements of adult
man. Nutr Abst Rev 27:631-647
5. Wu G (2010) Functional amino acids in growth, repro-
duction and health. Adv Nutr 1:31-37
6. Wu G (2014) Dietary requirements of synthesizable ami-
no acids by animals: a paradigm shift in protein nutrition. J Anim
Sci Biotech Doi:10.1186/2049-1891-5-34
7. Curthoys NP, Watford M. (1995) Regulation of glu-
taminase activity and glutamine metabolism. Annu Rev Nutr
1995:15:133-59.
8. Curi R, Newsholme P, Procopio J, Lagranha C, Gorjão
R, Pithon-Curi TC. (2007) Glutamine, gene expression, and cell
function. Front Biosci 12:344-57
9. Watford M. (2008) Glutamine metabolism and function
in relation to proline synthesis and the safety of glutamine and
proline supplementation. J Nutr 138:2003S-2007S
10. Watford M, Reeds, PJ (2003) Glutamate metabolism in
the gut. Forum Nutr 56:81-82
11. Lenders CM, Liu S, Wilmore DW, Sampson L, Dougherty
LW, Spiegelman D, Willett WC. (2009) Evaluation of a novel food
composition database that includes glutamine and other amino
acids derived from gene sequencing data. Eur J Clin Nutr 63:1433-
9. doi: 10.1038/ejcn.2009.110.
12. Li X, Rezaei R, Li P, Wu G (2011) Composition of amino
acids in feed ingredients for animal diets. Amino Acids 40:1159-
68. doi: 10.1007/s00726-010-0740-y.
13. Kim H (2011) Glutamine as an immunonutrient. Yonsei
Med J 52:892-897
14. Watford M (1989) Does glutamine regulate skeletal
muscle turnover? Trends Biochem Sci 14:1-2
15. Wernerman J (2008) Clinical use of glutamine supple-
mentation. J Nutr 138:2040S-2044S
16. Gleeson, M (2008) Dosing and efficacy of glutamine
supplementation in human exercise and sport training. J Nutr
138:2045S-2049S
17. Wang Y, Jiang ZM, Nolan MT, Jiang H, Han HR, Yu K, Li
HL, Jie B, Liang XK (2010) The impact of glutamine dipeptide-sup-
plemented parenteral nutrition on outcomes of surgical patients:
a meta-analysis of randomized clinical trials. J Parenter Enteral
Nutr 34:521-9. doi: 10.1177/0148607110362587
18. Heyland DK, Elke G, Cook D, et al. (2015) Glutamine
and antioxidants in the critically ill patient: A post hoc analysis of
a large scale randomized trial. J Parenter Enter Nutr 39:401-409
19. Prieser JC, van Zanten ARH, Berger, MM, et al. (2015)
Metabolic and nutritional support of critically ill patients: consen-
sus and controversies. Critical Care 19:35 doi 10.1186/s13054-
015-0737-8
20. Wu X, Xie C, Zhang Y, Fan Z, Yin Y, Blachier F (2015)
Glutamine-glutamate cycle and exchange in the placenta-fetus
unit during late pregnancy. Amino Acids 47: 45-53
21. Wu G, Knabe DA (1994) Free and protein-bound amino
acids in sow’s colostrum and milk. J Nutr 124:415-24.
22. Davis TA, Nguyen HV, Garcia-Bravo R, Fiorotto ML, Jack-
son EM, Reeds PJ (1994) Amino acid composition of the milk of
some mammalian species changes with stage of lactation. Br J
Nutr 72:845-53.
23. Ruth MR, Field CJ (2013) The immune modifying effects
of amino acids on gut-associated lymphoid tissue. J Animal Sci
Biotech 4.27 doi10.1186/2049-1891-4-27
24. Manso Filho HC, McKeever KH, Gordon ME, Costa HE,
Lagakos WS, Watford M. (2008) Changes in glutamine metabo-
lism indicate a mild catabolic state in the transition mare. J Anim
Sci 86:3424-31. doi: 10.2527/jas.2008-1054.
25. Pine AP, Jessop NS, Allan GF, Oldham JD. (1994) Ma-
ternal protein reserves and their influence on lactational perfor-
mance in rats. 4. Tissue protein synthesis and turnover associated
with mobilization of maternal protein. Br J Nutr 72:831-44.

Disponível no Site www.lisina.com.br
26. Clowes EJ, Williams IH, Baracos VE, et al. (1998) Feeding
lactating pimiparous sows to establish three divergent metabolic
states: II. Effect on nitrogen partitioning and skeletal muscle com-
position. J Anim Sci 76:1154-1164
27. Clowes EJ, Aherne FX, Baracos VE. (2005) Skeletal mus-
cle protein mobilization during the progression of lactation. Am J
Physiol Endocrinol Metab 288:E564-72.
28. Baile M, Haverson K, Inman C, Harris P, Jones P, Corfield
G, Miller B, Stokes C (2005) The development of the mucosal im-
mune system pre- and post-weaning: balancing regulatory and
effector function. Proc Nutr Soc 64:451-457
29. Lalles JP, Bosi P, Smidt H, Stokes CR (2007) Nutritional
management of gut health in pigs around weaning. ProcNutr Soc
66:260-268
30. Campbell JM, Crenshaw JD, Polo J (2013) The biologi-
cal stress of early weaning piglets. J Animal Sci Biotech 4:19 doi:
10.1186/2049-1891-4-19
31. Pluske JR (2013) Feed- and feed additives-related as-
pects of gut health and developments in weanling pigs. JAnim Sci
Biotech 4:1 doi: 10.1186/2049-1891-4-1
32. Jacobi SK,Odle J (2012) Nutritional factors influencing
intestinal health of the neonate. Adv Nutr 3:687-696
33. Lordelo MM, Gaspar AM, Le Bellego L, Freire JPB (2008)
Isoleucine and valine supplementation of a low-protein corn-
wheat-soybean meal-based diet for piglets: Growth performance
and nitrogen balance. J Anim Sci86:2936-2941
34. Wu G, Meier SA, Knabe DA. (1996) Dietary glutamine
supplementation prevents jejunal atrophy in weaned pigs. J Nutr
126:2578-84.
35. Zou XT, Zheng GH, Fang XJ, Jiang JF (2006) Effects of
glutamine on growth performance of weanling piglets. Czech J
Anim Sci 51:444-448
36. Hanczakowska E, Niwinska B (2013) Glutamine as a feed
supplement for piglets: A review. Ann Anim Sci 13:5-15
37. Hanczakoska E, Niwinska B (2014) Effect of dietary glu-
tamine, glucose and/or sodium butyrate on piglet growth, intes-
tinal environment, subsequent fattner performance, and meat
quality. Czech J Anim Sci 59:460-470
38. Yi GF, Carroll JA, Allee GL, Gaines AM, Kendall DC, Usry
JL, Toride Y, Izuru S. (2005) Effect of glutamine and spray-dried
plasma on growth performance, small intestinal morphology, and
immune responses of Escherichia coli K88+-challenged weaned
pigs. J Anim Sci 83:634-43.
39. Johnson IR, Ball RO, Baracos VE, Field CJ (2006) Glu-
tamine supplementation influences immune development in the
newly weaned piglet. Develop. Comp. Immunol. 30:1191-1201
40. Yoo SS, Field CJ, McBurney MI. (1997) Glutamine sup-
plementation maintains intramuscular glutamine concentrations
and normalizes lymphocyte function in infected early weaned
pigs. J Nutr 127:2253-9.
41. Ewaschuk, JB, Murdoch, GK, Johnson, IR, Madsen, KL,
Field, CJ (2011) Glutamine supplementation improves intestinal
barrier function in a weaned piglet model of Escherichia coli infec-
tion. Br J Nutr 206:870-877
42. Domeneghini C, Di Giancamillo A, Savoini G, Paratte R,
Bontempo V, Dell’Orto V (2004) Structural patterns of swine ileal
mucosa following L-glutamine and nucleotide administration dur-
ing the weaning period. An histochemical and histometrical study.
Histo Histopathol 19:49-58
43. Domeneghini C, Di Giancamillo A, Bosi G, Arrighi S
(2006) Can nutraceuticals after the structure of intestinal mus-
cosa? Qualitative and quantitative microanatomy in L-glutamine
diet-supplemented weaning piglets. Vet Res Comm 30:331-342
44. Manzke, NE, Scapini, LB, Loyola, W, et al. (2013) Oral
administration of amino acids as energy sources for newborn pig-
lets. J Anim Sci 91: E-Suppl. 1, 107 (abstract)
45. Wu J, Wang W, Yao K, et al. (2013) Effects of dietary
arginine and glutamine on alleviating the impairment induced by
deoxynivalenol stress and immune relevant cytokines in growing
pigs. PLOS ONE 8:e69502.doi:10.1371/journal.pone.0069502
46. Jiang ZY, Sun LH, Lin YC, Ma XY, Zheng CT, Zhou GL,
Chen F, Zou ST. 2009) Effects of dietary glycyl-glutamine on
growth performance, small intestinal integrity, and immune re-
sponses of weaning piglets challenged with lipopolysaccharide. J
Anim Sci 87:4050-6.
47. López-Pedrosa JM, Manzano M, Baxter JH, Rueda R.
(2007) N-acetyl-L-glutamine, a liquid-stable source of glutamine,
partially prevents changes in body weight and on intestinal im-
munity induced by protein energy malnutrition in pigs. Dig Dis Sci
52:650-8.
48. Lescano D, Albino L, Hannas M, Salguero S, Kutschenko
M, Nogueira E, Rostagno H (2013) Evaluation of dietary glutamic
acid plus glutamine levels on the growth performance of piglets. J
Anim Sci 91: E-Suppl. 1, 341 (abstract)

Disponível no Site www.lisina.com.br 31
49. Lescano D, Albino L, Hannas M, Salguero S, Kutschenko
M, Nogueira E, Rostagno H (2013) Effect of glutamic acid and
glutamine on the intestinal morphology of piglets. J Anim Sci 91:
E-Suppl. 1, 107 (abstract)
50. Wang J, Chen L, Li P, Li X, Zhou H, Wang F, Li D, Yin
Y, Wu G. (2008) Gene expression is altered in piglet small intes-
tine by weaning and dietary glutamine supplementation. J Nutr
138:1025-1032
51. Zhong X, Zhang XH, Li XM, Zhou YM, Li W, Huang XX,
Zhang LL, Wang T. (2011) Intestinal growth and morphology is as-
sociated with the increase in heat shock protein 70 expression in
weaning piglets through supplementation with glutamine. J Anim
Sci 89:3634-3642
52. Zhong X, Li W, Wang Y, Zhang L, Zhou Y, Hussain A,
Wang T. (2012) Effects of glutamine supplementation on the im-
mune status in weaning piglets with intrauterine growth retarda-
tion. Arch Anim Nutr 66:347-356
53. Xiao Y, Wu T, Hong Q, Sun J, Chen A, Yang C, Li X.
(2012) Response to weaning and dietary L-glutamine supplemen-
tation metabolomics analysis in piglets by gas chromatography/
mass spectrometry. J. Zhejiang Univ Sci B (Biomed and Biotechnol)
13:567-578
54. Xiao YP, Wu TX, Sun JM, Yang L, Hong QH, Chen AG,
Yang CM (2012) Response to dietary L-glutamine supplementa-
tion in weaned piglets: a serum metabolomics comparison and
hepatic metabolic regulation analysis. J Animal Sci 90:4421-4430
55. Xiao YP, Li XY, Wu TX, Yang L, Hong QH, Yang CM,
Chen AC (2012) Effects of dietary glutamine supplementation on
nutrient absorption and activity of enzymes involved in glutamine
metabolism and energy production in the jejunum of weaned pig-
lets. J Anim Vet Adv 11:1441-1449
56. Zhu Y, Lin G, Dai Z, et al. (2014) L-Glutamine depriva-
tion induces autophagy and alters the mTOR and MAPK signaling
pathways in porcine intestinal epithelial cells. Amino Acids DOI
10.1007/s00726-014-1785-0
57. Wang H, Zhang C, Wu G, Sun Y, Wang B, He B, Dai Z,
Wu Z. (2015) Glutamine enhances tight junction protein expres-
sion and modulates corticotropin releasing factor signaling in the
jejunum of weanling piglets. J Nutr 245:25-31
58. Ewtushik AL, Bertolo RFP, Ball RO (2000) Intestinal de-
velopment of early-weaned piglets receiving diets supplemented
with selected amino acids or polyamines. Can J Anim Sci 80:653-
662
59. Renzaei R, Knabe DA, Tekwe CD, Dahanayaka S, Ficken
MD, Fielder SE, Eide SJ, Lovering SL, Wu G (2013) Dietary supple-
mentation with monosodium glutamate is safe improves growth
performance in postweaning pigs. Amino Acids 44:911-923
60. Wu X, Zhang Y, Liu Z, Li, TJ, Yin YL (2012) Effects of oral
supplementation with glutamate or a combination of glutamate
and N-carbamylglutamate mucosa morphology and epithelium
cell proliferation in weanling piglets. J. Anim. Sci. 90:337-339
61. Haynes TE, Li P, Li X, Shimotori K, Sato H, Flynn NE,
Wang J, Knabe DA, Wu G. (2009) L-Glutamine or L-alanyl-L-glu-
tamine prevents oxidant- or endotoxin-induced death of neonatal
enterocytes. Amino Acids 37:131-42. doi: 10.1007/s00726-009-
0243-x.
62. Cabrera RA, Usry, J., Arrellano C, Nogueira ET, Kutschen-
ko M, Moeser AJ, Odle, J (2013) Effects of creep feeding and sup-
plemental glutamine or glutamine plus glutamate (Aminogut) on
pre- and post-weaning growth performance and intestinal health
of piglets. J. Anim. Sci. Biotech. 4:29 doi:10 1186/2046-1891-4-
29
63. Kitt SJ, Miller PS, Fischer RL (2004) Effect of sow dietary
glutamine supplementation on sow and litter performance, sub-
sequent weanling pig performance and intestinal development
after an immune challenge. Nebraska Swine Report 14-17
64. Manso HE, Manso Filho HC, de Carvalho LE, Kutschen-
ko M, Nogueira ET, Watford M (2012) Glutamine and glutamate
supplementation raise milk glutamine concentrations in lactating
gilts. J Anim Sci Biotechnol 3:2. doi: 10.1186/2049-1891-3-2.
65. Santos de Aquino R, Dutra WM, MansoHECC, Manso
Filho HC, Kutschenko M, Nogueira ET, Watford M (2014) Glu-
tamine and glutamate (AminoGut) supplementation influences
sow colostrum and mature milk composition. LiveStock Science
2014;169:112-117 http://dx.doi.org/10.1016/j.livsci.2014.07.009
1871-1413
66. Meijer GAL, van der Meulen J, van Vuures AM (1993)
Glutamine is a potentially limiting amino acid for milk production
in dairy cow: a hypothesis. Metabolism 42:358-364
67. Meijer GAL, van der Meulen J, Bakker JGM, van der Koe-
len CJ, van Vuuren AM (1995) Free amino acids in plasma and
muscle of high yielding dairy cows in early lactation. J. Dairy Sci.
78: 1502-1508
68. Caropese M, Albenzio M, Marino R, Santillo A, Sevi A
(2012) Immune response and milk production of dairy cows fed
graded levels of rumen-protected glutamine. Res. Vet. Sci. 93:
202-208

Disponível no Site www.lisina.com.br Disponível no Site www.lisina.com.br
69. Caropese M, Albenzio M, Marino R, Santillo A, Sevi A
(2013) Dietary glutamine enhances immune responses of dairy
cows under high ambient temperature. J. Dairy Sci. 96: 3002-
3011
70. Featherston WR, Bird HR, Harper AE (1962) Effectiveness
of urea and ammonium nitrogen for the synthesis of dispensable
amino acids by the chick. J Nutr 78:198-206.
71. Maruyama K, Sunde ML, Harper AE (1976) Is L-glutamic
acid nutritionally a dispensable amino acid for the young chick?
Poult Sci 55:45-60.
72. Maruyama K, Harper AE, Sunde ML. (1975) Effects of
D-, DL-and L-glutamic acid on chicks. J Nutr 105:1012-9.
73. Yi, GF, Allee, GL, Knight, CD, Dibner, JJ (2004) Impact
of glutamine and Oasis Hatchling Supplement on growth per-
formance, small intestinal morphology and immune response of
broilers vaccinated and challenged with Eimeria maxima. Poultry
Sci 84:283-293
74. Bartel SM, Batal AB (2007) The effect of supplemental
glutamine on growth performance, development of the gastroin-
testinal tract, and humoral immune response of broilers. Poulty Sci
86:1940-1947
75. Dai SF, Wang LK, Wen AY, Wang LX, Jin GM (2009) Di-
etary glutamine supplementation improves growth performance,
meat quality and colour stability of broilers under heat stress. Brit
Poultry Sci 50:333-340
76. Dai SF, Gao F, Xu XL, Zhang, WH, Song SX, Zhou GH
(2012) Effects of dietary glutamine and gamma-aminobutyric acid
on meat colour, pH, composition, and water holding characteris-
tic in broilers under cyclic heat stress. Br Poultry Sci 53:471-481
77. Szabo J, Andrasofszky E, Tuboly T, Bersenyi A, Weisz A,
Hetenyi N, Hullar I (2014) Effect of arginine or glutamine sup-
plementation on production, organ weights, interferon gamma,
interleukin 6 and antibody titre of broilers. Acta Vet Hung 62:348
78. Hu H, Bai X, Shah AA, Wen AY, Hua JL, Che CY, He SJ,
Jiang JP, Ca ZH, Dai SE (2015) Dietary supplementation with glu-
tamine and gamma aminobutyric acid improves growth perfor-
mance and serum parameters in 22 to 35 day old broilers exposed
to hot environment. J Anim Physiol Anim Nutr doi: 10.1111/
jpn.12346
79. Shakeri M, Zulkifli I, Soleimani AF, O’Reilly EL, Eckersall
PD, Anna AA, Kumari S, Abdullah FFJ. (2014) Response to die-
tary supplementation of L-glutamine and L-glutamate in broiler
chickens reared at different stocking densities under hot, humid
tropical conditions. Poultry Sci 93:2700-2708
80. Rechcigl M, Loosli JK, Williams HH (1957) The net utiliza-
tion of non-specific nitrogen sources for the synthesis of non-es-
sential amino acids. J Nutr 63:177-192

Disponível no Site www.lisina.com.br Disponível no Site www.lisina.com.br
Guoyao Wu, Ph.D.Department of Animal Science, Texas A&M University
O Prof. Dr. Guoyao Wu é Professor Emérito da Universidade Texas A&M.
Graduou-se em Zootecnia pela Universidade de Agricultura do Sul da China e
recebeu grau de mestre em Nutrição Animal pela Universidade de Agricultura de
Pequim. Fez mestrado e doutorado em Bioquímica Animal na Universidade de
Alberta, Canadá.
Depois de realizar estágio pós-doutoral em Bioquímica e Nutrição na
Universidade McGill e na Universidade Memorial de Newfoundland, no Canadá,
o Dr. Wu entrou para o corpo docente da Texas A&M em 1991.
Realiza pesquisas em nutrição de proteínas e aminoácidos em nível molecular,
celular e do organismo. O Dr. Wu publicou 487 artigos em periódicos científicos,
56 capítulos de livros e um livro: “Aminoácidos: Bioquímica e Nutrição” (CRC
Press 2013). É um dos cientistas mais citados no SCI.
O Dr. Wu recebeu vários prêmios importantes da profissão, como o prêmio de
Pesquisador Destacado da Associação Americana do Coração, Pesquisa em Nu-
trição de Não-Ruminantes da Sociedade Americana de Zootecnia, Novas Frontei-
ras na Pesquisa em Nutrição Animal FASS-AFIA da Federação de Sociedades de
Zootecnia e da Associação Americana da Indústria de Rações (o maior prêmio em
pesquisa em nutrição animal dos EUA), além do prêmio de Pesquisa Destacada
Sigma Xi. É membro eleito da Associação Americana para o Avanço da Ciência.
33

Disponível no Site www.lisina.com.br
Resumo: Na natureza, a L-glutamina (Gln) é um aminoácido neutro. A falta de capacidade de analisar a Gln emingredientes de ração e na proteína animal resultou na visão tradicionalmente errônea de que a Gln não era necessária para os animais. Há cada vez mais evidências que a síntese endógena de Gln é inadequada para maximizar o crescimento, a lactação e a reprodução de mamíferos (inclusive suínos) e para maximizar o desempenho zootécnico de aves. Isto ocorre porque são necessárias grandes quantidades de Gln para muitas vias metabólicas essenciais para a síntese proteica, homeostase do organismo, saúde e bem-estar. Além disso, a Gln regula a expressão gênica e vias de sinalização celular por melhorar a sobrevivência, a deposição de tecido magro e a adaptação ao estresse. Portanto, espera-se que a disponibilidade de Gln (como o AminoGut) para rações de baixo custo melhore a eficiência alimentar e o retorno econômico da produção animal em todo o mundo.
Palavras-chave: Glutamina, nutrição, animais de produção, suínos, aves
A L-Glutamina é essencial na dieta para maximizar o crescimento e a eficiência alimentar em suínos e aves
Introdução
Na natureza, a L-glutamina (Gln) é um aminoácido neutro. Na sua forma cristalina, a Gln é estável em temperatura ambiente (p. ex., 20-25ºC) e é insípida (não tem gosto) (Wu, 2013). É um excelente transportador de nitrogênio e carbono para o metabolismo entre órgãos de aminoácidos em animais, e há interconversão entre a Gln e o glutamato via reações enzimáticas. A concentração de Gln no plasma sanguíneo (0,5 a 1 mM) é aproximadamente 10 vezes maior que a de glutamato e a Gln é o aminoácido livre mais abun-dante no organismo (Curthoys e Watford, 1995). Embora a Gln não seja tradicionalmente descrita como componente da proteína ou mesmo mencionada em livros-texto clássi-cos de nutrição animal (Bondi, 1987; Maynard et al., 1979), cada vez mais pesquisas demonstram que a Gln dietética é essencial para maximizar o desempenho de crescimento, lactação e reprodução, assim como a saúde e o bem-estar de animais, especialmente em condições de estresse (Wu et al., 2011). O principal objetivo deste artigo é dar uma perspectiva histórica das pesquisas sobre a bioquímica, fisiologia e nutrição da Gln relevantes para a produçãoanimal.
Perspectivas históricas das pesquisas sobre a química, bioquímica e fisiologia da Gln
As pesquisas sobre a química da Gln datam do século 19 (ver revisão de Wu, 2013). Foi isolada pela primeira vez como aminoácido livre do suco de beterraba em 1883 e, em 1904, foi reconhecida como uma das principais fontes de amônia produzida pela hidrólise ácida de proteínas. De-pois de avanços nos métodos analíticos, a Gln foi obtida de hidrolisados enzimáticos da gliadina (a proteína mais abun-dante no trigo) em 1932. Um ano depois, este aminoácido foi sintetizado quimicamente pela primeira vez a partir do ácido anidro carbobenzoxi-L-glutâmico.
A disponibilidade de Gln cristalina na forma quimicamente pura facilitou muito as pesquisas sobre sua bioquímica e fisiologia. Uma série de estudos fundamentais, realizados por Krebs na década de 1930, revelou o catabolismo da Gln pela glutaminase ativada por fosfato no fígado e nos rins de ratos (ver a revisão de Brosnan, 2001). Na déca-da de 1940 e 1950, foram realizados estudos clínicos que demonstraram que a Gln é a principal fonte de amônia para a remoção renal de H+ em resposta à acidose metabólica. Em 1965, Neptune verificou que preparações do intestino
Guoyao Wu, Ph.D.Department of Animal Science, Texas A&M UniversityE-mail: [email protected]

Disponível no Site www.lisina.com.br 35
delgado de diversas espécies animais (cobaias, porquinhos da Índia, hamsters, macacos, coelhos e ratos) têm altas taxas de oxidação de Gln. Posteriormente, Windmueller e Spaeth (1975) relataram que: (a) o intestino delgado de ra-tos utiliza grandes quantidades de Gln dietética e arterial como principal combustível metabólico e (b) a maior parte da Gln enteral não entra na circulação do sistema porta. Em suínos, apenas 33% da Gln dietética está disponível para a utilização de tecidos extra-intestinais. Estes dados indicam que o intestino é possivelmente o órgão-alvo das ações fisiológicas da Gln em mamíferos.
Os principais sítios da síntese de Gln em animais só foram conhecidos depois que Goldberg e Chang (1978) desco-briram que o músculo esquelético de ratos sintetiza glu-tamina a partir de aminoácidos de cadeia ramificada (leu-cina, isoleucina e valina, doadores do grupo amino na Gln) e alfa-cetoglutarato (esqueleto de carbono da Gln). A sín-tese de Gln nas células da mucosa do intestino mamífe-ro é muito limitada devido à baixa atividade da glutamina sintetase. Com sua massa relativamente grande (40-45% do peso corporal), o músculo esquelético é a principal fon-te de Gln endógena em animais. Maclennan et al. (1987)
demonstraram que a Gln estimula a síntese proteica e inibe a degradação de proteína no músculo esquelético de ratos. Wu e colaboradores relataram na década de 1990 (a) a presença do ciclo glutamina-glutamato no múscu-lo esquelético de frangos de corte e ratos para regular a liberação de glutamina do tecido (Wu et al., 1991) e (b) um efeito anabólico direto da glutamina sobre a deposição de proteína no músculo esquelético de frangos (Wu e Thompson, 1990a). O mecanismo subjacente envolve a ativação do alvo mecanicista da via de sinalização celular da rapamicina para promover a iniciação da formação de polipeptídeos e a supressão da autofagia (uma das principais vias de pro-teólise intracelular) nos tecidos (Wu, 2013).
Versatilidade metabólica da Gln em animais
As taxas de turnover de Gln são altas em suínos recém-nascidos, desmamados, em gestação e em lactação, sendo a meia vida da Gln no sangue de 0,65 horas (Curi et al., 2005; Wu et al., 2010). Isto corrobora a participação ati-vada Gln em várias vias metabólicas (Figura 1).
Figura 1. Diferentes vias metabólicas da utilização de Gln em animais. Observe que a
síntese de ornitina, citrulina e arginina a partir da Gln ocorre exclusivamente no intesti-
no delgado. Outros metabólitos da Gln são o piruvato, a asparagina, a ureia e, possivel-
mente, a glicina. O asterisco indica que a reação não pode ser substituída por nenhum
outro AA nas células. AA = aminoácidos; Cir = citrulina; Lact = lactato; Orn = ornitina.
Fonte: Wu et al. (2011).

Disponível no Site www.lisina.com.br
Em primeiro lugar, como observado anteriormente, a Gln é um dos principais substratos energéticos para células em divisão rápida, como enterócitos e linfócitos submeti-dos a desafio imune, e outros tipos de células (p. ex., rins, durante a privação de alimento, e macrófagos ativados), fornecendo ATP para turnover da proteína intracelular, transporte de nutrientes através da membrana plasmática, crescimento e migração celular e manutenção da integri-dade e função das células. Em segundo lugar, a formação de amônia e bicarbonato pelo metabolismo da Gln nos rins é vital para a regulação do equilíbrio ácido-base em animais, especialmente em condições acidóticas. Em ter-ceiro lugar, a Gln serve como precursor para a síntese dos nucleotídeos purina e pirimidina, que são essenciais para a proliferação celular, inclusive das células do embrião, trofoblastos, linfócitos, células epiteliais mamárias, miócitos e enterócitos. Em quarto lugar, a Gln contribui consideravelmente para o fornecimento endógeno de arginina na maioria dos mamíferos, como suínos, bovinos e ovinos. Esta via de síntese compensa: 1) a deficiência de arginina (aminoácido nutricionalmente essencial para
suínos jovens) no leite durante o período de amamentação e 2) o extenso catabolismo da arginina da dieta pelo intes-tino delgado de animais pós-desmame. Em quinto lugar, a Gln é convertida em alanina nos enterócitos através da via glutaminolítica, que compensa o teor relativamente baixo de alanina no leite para leitões em amamentação ou em dietas para porcas em gestação e em lactação. Em sexto lugar, a Gln é necessária para a formação de N-acetilglu-cosamina-6-fosfato, um substrato comum para a síntese de glicoproteínas, que são especialmente abundantes nas células da mucosa, membrana plasmática e embriões em desenvolvimento. Em sétimo lugar, como precursora do glutamato, a Gln atua na síntese de glutationa, o antiox-idante de baixo peso molecular mais abundante nas célu-las. Isto é especialmente importante para células como as hemácias, que não absorvem glutamato. Finalmente, além de seu papel como substrato em diversas vias metabólicas, as concentrações fisiológicas de Gln regulam a expressão gênica e a transdução de sinais e, portanto, o metabolismo celular
Figura 2. Funções regulatórias da Gln em animais. A L-Glutamina pode regular di-
versos processos fisiológicos em diferentes tipos de células, como a expressão gênica,
sinalização celular e turnover de proteína. Estes efeitos da Gln são específicos de células
e tecidos, indicando que é necessária a cooperação inter-órgãos para que a Gln exerça
seus efeitos benéficos em todo o organismo. Fonte: Wu et al. (2011).

Disponível no Site www.lisina.com.br 37
Figura 3. Utilização da Gln circulante pelos principais órgãos
de leitões. Os valores na figura estão expressos em mg de Gln/
kg de peso corporal/dia. O leitão (7,92 kg PV) consome 789
mg de Gln/kg PV/dia da dieta e ganha 293 g de PV/dia. O fluxo
líquido de Gln no sangue arterial para o intestino delgado é
de 438 mg/kg PV/dia. A taxa de extração de Gln pelas vísceras
drenadas pelo sistema porta,
Claramente, a Gln é um aminoácido funcional que pro-move a homeostase e a eficiência da utilização da ração em animais.
Exceto na placenta, glândula mamária e intestino delgado, ocorre um ciclo nutricionalmente significativo de Gln-gluta-mato em diversos tipos de célula (Wu, 2013). Portanto, o glutamato pode substituir parcialmente a Gln em diversas vias, como a produção de ATP e síntese de arginina, alani-na, prolina e aspartato, se houver amônia disponível para estas vias. Além disso, o glutamato inibe o catabolismo da Gln pela glutaminase em tecidos extra-hepáticos e células, poupando a Gln para uso como combustível metabólico e substrato para processos de síntese. Deve ser lembrado que
algumas funções da Gln (p.ex., síntese de Gln-tRNA, aminoaçúcares, NAD(P) e de purinas e pirimidinas; amoniogênese renal e regulação da expressão da ornitina descarboxilase para a geração de poliaminas) não podem ser supridas pelo glutamato. Além disso, embora a Gln e o glutamato da dieta sejam amplamente catabolizados no intestino delgado, este órgão absorve Gln, mas não glu-tamato, a partir do sangue arterial. Finalmente, devido à complexa compartimentalização do metabolismo celular, o glutamato extracelular pode ser canalizado preferencial-mente para o citoplasma em vez das mitocôndrias e, por-tanto, ter efeitos diferentes do glutamato gerado a partir da Gln nas mitocôndrias.
Reconhecimento da Gln como nutrienteessencial em dietas de suínos
A Gln não é tradicionalmente considerada um nutriente necessário em dietas de qualquer espécie animal e não é destacada em livros-texto clássicos de nutrição. Isto mudou fundamentalmente quando Wu e Knabe (1994a) relata-ram pela primeira vez o alto teor de glutamina e glutamato no leite da porca (4 e 1 mM, respectivamente). Isto levanta uma importante questão quanto à significância nutricional destes aminoácidos para o crescimento e desenvolvimento
neonatal. Estes autores verificaram que a Gln é utilizada pelos enterócitos (células epiteliais absortivas do intestino delgado) de suínos para sintetizar citrulina e arginina (Wu et al., 1994b) de forma a compensar parcialmente o forne-cimento inadequado de arginina no leite e em dietas à base de milho e farelo de soja. Posteriormente, os autores relata-ram que: (a) os rins têm um papel importante na conversão de citrulina em arginina (um aminoácido nutricionalmente essencial para mamíferos jovens) em leitões pós-desmame (Wu e Knabe, 1995) e (b) a suplementação da dieta com glutamina efetivamente aumenta a disponibilidade de Gln para utilização no intestino delgado, prevenindo assim a

Disponível no Site www.lisina.com.br
atrofia intestinal e promovendo o desempenho zootécnico de leitões desmamados precocemente (Wu et al., 1996). Este estudo fundamental levou ao desenvolvimento e à disponibilidade comercial da Gln para alimentação animal (AminoGut) para a suinocultura. Cada vez mais evidências mostram que a Gln dietética é essencial para a integridade, saúde e função do intestino delgado de mamíferos, aves e animais aquáticos, especialmente em condições de estresse, como no desmame, lactação, gestação, altas temperaturas ambientais e infecções (Andersen et al., 2016; Bartell e Batal, 2007; Wu et al., 2011). Além disso, há relatos que: (a) a suplementação Gln na dieta de marrãs em gestação reduz o número de leitões com baixo peso ao nascer e a mortalidade de leitões pré-desmame e (b) a suplementação de Gln na dieta de porcas em lactação aumenta a disponi-bilidade de glutamina no leite e promove o crescimento e a sobrevivência pré-desmame dos leitões (Wu et al., 2011; Manso et al., 2012). Portanto, os suínos não conseguem sintetizar Gln em quantidade suficiente e isto também se aplica a aves após a eclosão. Com base nestes acha-dos, a edição atual do National Research Council (2012) reconhece a Gln como aminoácido condicionalmente es-sencial para suínos.
Reconhecimento da Gln como nutriente dietético para maximizar o desempenho zootécnico de aves
A concentração de Gln no plasma de galinhas é o dobro daquela de mamíferos após o nascimento (Watford e Wu, 2005), indicando possivelmente maior demanda metabólica de Gln em aves. Como a absorção da glicose pelo múscu-lo esquelético de galinhas é limitado devido à ausência do transportador de glicose 4, a Gln facilita a oxidação de áci-dos graxos e de corpos cetônicos neste tecido, fornecendo oxaloacetato (Wu e Thompson, 1990b). Embora o músculo esquelético das aves possa produzir glutamato e Gln (Wu et al., 1989), esta via de síntese de novo não é suficiente para maximizar o crescimento das aves. Corroborando esta visão, foi verificado que a suplementação dietética de 0,5 a 1% de glutamato aumentou a altura das vilosidades do intestino delgado, o ganho de peso e a eficiência alimentar e reduziu a mortalidade pós-eclosão de frangos de corte de
1 a 42 dias de idade (Olubodun et al., 2015). Também foi relatado que a suplementação dietética de 1% ou 0,7% de glutamina aumentou o ganho de peso e a eficiência ali-mentar de frangos de corte de 1 a 21 dias de idade (Bartell e Batal, 2007) e de perus de 21 a 42 dias (Salmanzadeh e Shahryar, 2013), respectivamente. As necessidades de Gln são maiores quando as aves são alimentadas com dietas com baixos níveis de proteína.
Segurança da suplementação de Gln parasuínos e aves
A L-Glutamina em si não é tóxica para as células, uma vez que grandes quantidades de Gln (4 mM ou aproximada-mente 8 a 10 vezes a concentração fisiológica no plasma) são geralmente incluídas em meios de cultura para todos os tipos de células. Além disso, a Gln está presente em al-tas concentrações nos fluidos fetais de espécies de animais de produção (p.ex., 25 nM no fluido alantoide ovino) no início e na metade da gestação (Kwon et al., 2003; Gao et al. 2009). A Gln administrada oralmente também de-saparece quase completamente da circulação em 4 horas. Portanto, a suplementação de Gln por curto ou longo prazo em doses adequadas (p. ex., até 1% da dieta na matéria natural) é segura para suínos recém-nascidos, em gestação e em lactação, assim como para frangos em crescimento. Por exemplo, suínos recém-nascidos, em gestação e em lactação toleram altas quantidades de Gln suplementar (da-dos fornecidos pela Ajinomoto Inc., Tóquio, Japão) por pelo menos 3 semanas a pelo menos 1,0; 0,20 e 0,32 g de Gln/kg de peso corporal/dia, respectivamente, acima da Gln na dieta basal (1,2; 0,24 e 0,55 g de Gln/kg de peso corpo-ral/dia, respectivamente), sem qualquer efeito adverso (Wu et al., 2011). Também verificamos que a suplementação dietética de até 1% de Gln (na matéria natural) por 6 se-manas não causou redução do consumo de ração, doenças ou mortalidade em suínos e aves. Não foram observados efeitos colaterais da suplementação de até 1% de Gln na dieta (na matéria natural) em leitões pós-desmame por pelo menos 3 meses depois do encerramento de períodos de suplementação de 2 semanas, 1 mês e 3 meses e, em fran-gos de corte, em pelo menos 4 semana depois do final de períodos de suplementação de 2 e 6 semanas.

Disponível no Site www.lisina.com.br 39
Conclusões
Nos últimos 25 anos, pesquisas amplas e sistemáticas sobre a nutrição de Gln em animais de produção, originadas da observação original da abundância de Gln no leite da porca, transformaram a Gln de uma substância não-descrita em livros-texto clássicos de nutrição animal em um dos principais nutrientes que comprovadamente melhoram a saúde, a sobrevivência, o crescimento, o desenvolvimento e a reprodução de suínos. Da mesma forma, a Gln foi identificada como suplemento dietético eficaz para melhorar o desempenho zootécnico de aves. No início da década de 1990, o preço da Gln era extremamente alto, tornando seu uso prático na produção animal proibitivo. Felizmente, com o desenvolvimento de no-vas técnicas para a produção de Gln e Glu através da fermentação microbiana, a Ajinomoto Co., Inc. (Tóquio, Japão) lançou com sucesso em 2005 a Gln para alimentação animal de baixo custo (AminoGut) para dietas de suínos e aves. Este avanço significativo na nutrição animal tem contribuído de forma significativa para melhorar a saúde intestinal, o crescimento de tecido magro e a eficiência alimentar dos animais de produção na última década.

Disponível no Site www.lisina.com.br
Referências Bibliográficas
Andersen, S.M., Waagbø, R., and Espe, M. 2016. Functional ami-no acids in fish nutrition, health and welfare. Front Biosci. 8:E143-169.
Bartell, S.M., and Batal AB. 2007. The effect of supplemental glu-tamine on growth performance, development of the gastrointes-tinal tract, and humoral immune response of broilers. Poult Sci. 86:1940-1947.
Bondi, A. A. 1987. Animal Nutrition. John Wiley & Sons. New York .
Brosnan, J.T. 2001. Amino acids, then and now--a reflection on Sir Hans Krebs’ contribution to nitrogen metabolism. IUBMB Life. 52:265-270.
Curi, R., C. J. Lagranha, S. Q. Doi, D. F. Sellitti, J. Procopio, T. C. Pithon-Curi, M. Corless, and P. Newsholme. 2005. Molecular mechanisms of glutamine action. J. Cell. Physiol. 204:392-401.
Curthoys, N. P., and M. Watford. 1995. Regulation of glutaminase activity and glutamine metabolism. Annu. Rev. Nutr. 15:133-159.
Gao, H. J., G. Wu, T. E. Spencer, G. A. Johnson, X. L. Li, and F. W. Bazer. 2009. Select nutrients in the ovine uterine lumen: I. Amino acids, glucose and ions in uterine lumenal flushings of cyclic and pregnant ewes. Biol. Reprod. 80:86-93.
Goldberg, A.L. and T.W. Chang. 1978. Regulation and signif-icance of amino acid metabolism in skeletal muscle. Fed. Proc. 37:2301-2307.
Kwon, H., T. E. Spencer, F. W. Bazer, and G. Wu. 2003. Develop-mental changes of amino acids in ovine fetal fluids. Biol. Reprod. 68:1813-1820.
Maclennan, P. A, R. A. Brown, and M. J. Rennie. 1987. A positive relationship between protein synthetic rate and intracellular glu-tamine concentration in perfused rat skeletal muscle. FEBS Lett. 215:187-191.
Manso HE, Filho HC, de Carvalho LE, Kutschenko M, Nogueira ET, Watford M. 2012. Glutamine and glutamate supplementation raise milk glutamine concentrations in lactating gilts. J Anim Sci Biotechnol. 3(1):2.
Maynard, L. A., J. K. Loosli, H. F. Hintz, and R. G. Warner. 1979. Animal Nutrition. 7th ed., McGraw-Hill Book Company, New York.
National Research Council. 2012. Nutrient Requirements of Swine. National Academy Press, Washington, D.C.
Olubodun JO, Zulkifli I, Farjam AS, Hair-Bejo M, Kasim A. 2015. Glutamine and glutamic acid supplementation enhances perfor-mance of broiler chickens under the hot and humid tropical con-dition. Italian J Anim Sci. 14 (1): Article Number 3263.
Salmanzadeh M, Shahryar HA. Effects of dietary supplementation with glutamine on growth performance, small intestinal morphol-ogy and carcass traits in turkey poults under heat stress. Revue Med Vet. 2013;164:476-480.
Watford, M., and G. Wu. 2005. Glutamine metabolism in uric-otelic species: variation in skeletal muscle glutamine synthetase, glutaminase, glutamine levels and rates of protein synthesis. Comp. Biochem. Physiol. B. 140:607-614.
Windmueller, H.G., and Spaeth, A.E. 1975. Intestinal metabolism of glutamine and glutamate from the lumen as compared to glu-tamine from blood. Arch Biochem Biophys. 171:662-672.
Wu, G. 2013. Amino Acids: Biochemistry and Nutrition, CRC Press, Boca Raton, Florida.
Wu, G., and D.A. Knabe. 1995. Arginine synthesis in enterocytes of neonatal pigs. Am. J. Physiol. 269:R621-R629.
Wu, G., and J.R. Thompson. 1990a. The effect of glutamine on protein turnover in chick skeletal muscle in vitro. Biochem. J. 265:593-598.
Wu, G., and J.R. Thompson. 1990b. The effect of ketone bodies on protein turnover in isolated skeletal muscle from the fed and fasted chick. Int. J. Biochem. 22:263-268.
Wu, G., J.R. Thompson, G. Sedgwick, and M. Drury. 1989. Forma-tion of alanine and glutamine in chick (Gallus domesticus) skeletal muscle. Comp. Biochem. Physiol. 93B: 609-613.
Wu, G., J.R. Thompson, and V.E. Baracos. 1991. Glutamine metabolism in skeletal muscle from the broiler chick (Gallus do-mesticus) and the laboratory rat (Rattus norvegicus). Biochem. J. 274:769-774.
Wu, G., F.W. Bazer, G.A. Johnson, D.A. Knabe, R.C. Burghardt, T.E. Spencer, X.L. Li, and J.J. Wang. 2011. Important roles for L-glutamine in swine nutrition and production. J. Anim. Sci. 89:2017-2030.
Wu, G., F.W. Bazer, R.C. Burghardt, G.A. Johnson, S.W. Kim, D.A. Knabe, X.L. Li, M.C. Satterfield, S.B. Smith, and T.E. Spencer. 2010. Functional amino acids in swine nutrition and production. In: Dynamics in Animal Nutrition (J. Doppenberg and P. van der Aar, ed.), Wageningen Academic Publishers, The Netherlands, pp. 69-98.
Wu, G. and D. A. Knabe. 1994a. Free and protein-bound amino acids in sow’s colostrum and milk. J Nutr. 124:415-24.Wu, G., D. A. Knabe, and N. E. Flynn. 1994b. Synthesis of citrul-line from glutamine in pig enterocytes. Biochem. J. 299:115-121.
Wu, G., S. A. Meier, and D. A. Knabe. 1996. Dietary glutamine supplementation prevents jejunal atrophy in weaned pigs. J. Nutr. 126:2578-2584.
Wu, G., and J. R. Thompson. 1990. The effect of glutamine on protein turnover in chick skeletal muscle in vitro. Biochem. J.
265:593-598.

Disponível no Site www.lisina.com.br 41

Disponível no Site www.lisina.com.br