Embed Size (px)
Citation preview

Efeito do exercício físico nas adaptações
musculares esqueléticas induzidas pela
obesidade
Dissertação apresentada com vista à
obtenção do 2º ciclo em Atividade Física
e Saúde, da Faculdade de Desporto da
Universidade do Porto ao abrigo do
Decreto de Lei nº.74/2006 de 24 de
Março
Orientadores: Prof. Doutor José Magalhães e Prof. Doutora Inês Gonçalves
Autora: Inês Ferreira Moreira Marinho
Porto, outubro de 2015

Ficha de Catalogação
Marinho, I. (2015). Efeito do exercício físico nas adaptações
musculares esqueléticas induzidas pela obesidade. Porto: I.
Marinho. Dissertação de Mestrado em Atividade Física e Saúde
apresentada à Faculdade de Deporto da Universidade do Porto.
Palavras-Chave: MÚSCULO-ESQUELÉTICO, EXERCÍCIO
FÍSICO, OBESIDADE, BIOGÉNESE, STRESS OXIDATIVO

I
Agradecimentos
Este trabalho não seria possível sem a preciosa ajuda e compreensão dos
meus orientadores, Professor Doutor José Magalhães e Professora Doutora
Inês Gonçalves, por todas as horas dedicadas ao meu estudo.
Quero também agradecer a toda a equipa do laboratório, Sílvia Rodrigues,
Telma Bernardo, Estela Alves, pela disponibilidade demonstrada em ensinar-
me e ajudar-me sempre que necessário.
Às amigas de sempre pelo apoio incondicional, Márcia, Catarina, Andreia,
Sofia, Joana, Sara, Maria, Nita. Às minhas amigas e amigos que a faculdade
me deu e que guardo para sempre, Marta, Rai, Jé, Rita, Caças, João, Manel.
Às minhas atletas pela paciência e compreensão.
Por ultimo, mas mais importante, à minha família pelos valores que me
transmitiram e pelo apoio incondicional, seja perto ou longe. Em especial ao
meu pai, à minha mãe pelas noitadas, por todas as horas de pesquisa e leitura,
pela paciência e amor permanentes, à minha irmã que apesar de longe está
sempre perto. E por fim, à minha avó Tété, pela lutadora que é, por ser um
exemplo para mim.

II

III
Índice Geral
Índice Geral III
Índice de Figuras V
Índice de Tabelas VI
Resumo VII
Abstract VIII
Abreviaturas IX
1. Introdução 1
2. Revisão de literatura
2.1. Obesidade: Conceito, prevalência e características 5
2.2. Modelos de dieta animal para indução de obesidade 7
2.3. Obesidade e tecido adiposo 8
2.4. Obesidade em outros tecidos: coração, fígado, cérebro 10
2.5. Obesidade e Musculo: stress oxidativo, biogénese mitocondrial 13
2.6. Papel do exercício físico na obesidade: stress oxidativo,
biogénese mitocondrial 16
3. Objetivos 19
3.1. Objetivo Geral 19
3.1.1. Objetivos específicos 19

IV
4. Metodologia 21
4.1. Caracterização da amostra 21
4.2. Programa de exercício 22
4.3. Western Blotting 22
4.5. Procedimentos Estatísticos 23
5. Resultados 25
6. Discussão 29
7. Conclusões 33
8. Referências bibliográficas 35

V
Índice de Figuras
Figura 1. Distância percorrida no tapete por semana 25
Figura 2. Efeito do exercício físico e da dieta nos marcadores de stress
oxidativo (SH, MDA, Sirt3 e P66Shc) 26
Figura 3. Efeito do exercício físico e da dieta nos marcadores de
biogénese mitocondrial (Tfam e PGC1a) e no conteúdo dos complexos
mitocondriais (OXPHOS) 27

VI
Índice de Tabelas
Tabela 1. Características anatómicas e consumo calórico após 17
semanas de tratamento 25

VII
Resumo
A obesidade é uma doença global, associada a um conjunto de co-
morbilidades metabólicas, como a diabetes, a esteatose hepática entre outras.
Apesar da obesidade ter uma etiologia multifactorial, o sedentarismo é uma das
causas primárias, sendo por conseguinte o aumento da atividade física uma
das mais importantes estratégias preventivas e terapêuticas para esta
patologia. Atendendo ao envolvimento do músculo-esquelético no exercício
físico e ao seu papel no dispêndio energético, o objetivo do presente estudo foi
analisar os mecanismos celulares (biogénese e stress oxidativo) subjacentes
ao potencial efeito protetor do exercício contra as alterações metabólicas
induzidas pela obesidade. Para tal, vinte e quatro ratos Sprague-Dawley foram
divididos em dois grupos: um grupo dieta-controlo (CS, n=11) e um grupo dieta-
gorda sedentários (GS, n=13). Após 9 semanas de dieta, metade dos animais
do grupo CS e do grupo GS, iniciaram um programa de exercício em tapete
rolante durante 8 semanas, 5 dias/semana e 1h/dia (CT, n=5 e GT, n=7).
Foram avaliados marcadores associados à lesão (SH e MDA) ou indução de
stress oxidativo (Sirt 3 e P66shc), assim como marcadores de biogénese
mitocondrial (PGC-1α e Tfam). Para além disto, foi ainda analisado o conteúdo
proteico das subunidades dos complexos mitocondriais. Nos animais do grupo
GS, observou-se um aumento dos grupos tióis e do conteúdo de Sirt3, PGC-1α
e complexo III. O exercício crónico induziu uma redução dos grupos tióis e do
conteúdo proteico do complexo III, enquanto a expressão do complexo IV
aumentou significativamente.
Em conclusão, a dieta rica em gordura induziu um aumento da biogénese,
provavelmente na tentativa de reduzir a produção de espécies reativas de
oxigénio, através do aumento da oxidação de ácidos gordos. O aumento da
expressão do complexo IV nos animais do grupo GT, poderá contribuir para
uma melhoria da capacidade oxidativa mitocondrial e assim atenuar o ambiente
pro-oxidante associado a esta patologia.
Palavras-Chave: MÚSCULO-ESQUELÉTICO, EXERCÍCIO FÍSICO,
OBESIDADE, BIOGÉNESE, STRESS OXIDATIVO

VIII
Abstract
Obesity as been described as a global epidemic, with several associated
metabolic comorbidities, such as diabetes, hepatic steatosis and others.
Although obesity etiology is multifactorial, sedentary behaviors are accepted as
a primary cause for this disorder and increased physical activity one of the first
line therapies for the prevention and treatment of this pathology. Considering
the involvement of skeletal muscle in exercise and its role in energy
expenditure, the aim of this study was to analyze the cellular mechanism
(biogenesis and oxidative stress) behind the potential protective effect of
exercise against metabolic alterations-induced by obesity. For that, twenty-four
Sprague-Dawley rats were divided in two groups: a standard diet (SS, n=11)
and high-fat diet sedentary group (HS, n=13). After 9 weeks of diet treatment,
half of SS and HS group were engaged in an exercise program on treadmill for
8 weeks, 5days/week and 1h/day (ST, n=5 and HT, n=7). Oxidative damage
(SH, MDA) and pro-oxidant signaling (Sirt3 and P66shc), biogenesis markers
(Tfam, PGC-1α), as well as mitochondrial complex content were assessed. HS
showed an increase of thiol groups and Sirt 3, PGC-1α and complex III content.
Chronic exercise induced a decrease of thiol groups and complex III content,
whereas complex IV was significantly increased.
In conclusion, high-fat diet feeding increased biogenesis probably as an attempt
to reduce reactive oxygen production and increased fatty acid oxidation, which
will ultimately contribute to ameliorate metabolic conditions. The increased
expression of complex IV-induced by chronic exercise in obese animals, might
had contributed to increase mitochondrial oxidation capacity and therefore
attenuate the pro-oxidant environment.
Keywords: SKELETAL MUSCULE, PHYSICAL EXERCISE, OBESITY,
BIOGENESIS AND OXIDATIVE STRESS

IX
Abreviaturas
ADN Ácido desoxirribonucleico
ADNmt Ácido desoxirribonucleico mitocondrial
ATP Adenosina trifosfato
AVC Acidente vascular cerebral
BSA Albumina do soro de boi
CS Dieta controlo sedentário
CT Dieta controlo treinado
CTE Cadeia transportadora de eletrões
DC Dieta de cafetaria
EROs Espécies reativas de oxigénio
GS Dieta gorda sedentário
GT Dieta gorda treinado
MDA Malondialdeído
Mg Miligramas
Min Minuto
Ml Mililitro
mM Milimolar
NAFLD Doença hepática não alcoólica
NASH Esteatohepatite não alcoólica
OMS Organização Mundial de Saúde

X
OXPHOS do inglês, oxidative phosphorylation
PGC-1α do inglês, peroxisome proliferator-activated receptor gamma
coactivator 1alpha
SDS/PAGE do inglês, polyacrylamide gel electrophoresis
SH Grupo sulfidrilo
Sirt3 Sirtuina 3
TBST Tris buffered saline with tween
Tfam do inglês, mitochondrial transcription factor A
VO2 Volume de oxigénio
WHO World Health Organization
α Alfa

1
1. Introdução
A atividade física é fundamental na melhoria da aptidão cardiorrespiratória bem
como na saúde mental dos indivíduos. Os seus benefícios estão também
relacionados com uma maior interação e envolvimento social das populações
(Cavill, Kahlmeier, Racioppi, 2008). O exercício desencadeia ainda alterações
metabólicas, fisiológicas e hormonais em diferentes órgãos, nomeadamente
adaptações musculares esqueléticas. Por outro lado, a predominância do estilo
de vida sedentário e, por consequência, a falta de exercício físico, tem levado
ao aumento significativo de indivíduos com obesidade desde idades precoces
(WHO, 2015). Há uma necessidade urgente em combater esta doença crónica
que eleva o risco de desenvolvimento de outras doenças como a diabetes,
doenças cardiovasculares, cancro, entre outras, sendo um importante fator de
morbilidade e mortalidade (Beja, Ferrinho & Craveiro, 2014; Hariri, & Thibault,
2010; Peters, 2014; Sarzani, Bordicchia, Spannella, Dessi-Fulgheri,
Fedecostante, 2014; Schmidt, 2012).
O músculo-esquelético adapta a sua capacidade oxidativa às necessidades
bioenergéticas que, por sua vez, são largamente determinadas pelos hábitos
de atividade física ou comportamento sedentário. Na obesidade, a capacidade
oxidativa do músculo é reduzida, característica relacionada com a resistência à
insulina (Menshikova et al., 2007). Parte da energia gerada pela degradação de
substratos (catabolismo) é utilizada para a produção de adenosina trifosfato
(ATP), esta molécula de elevada capacidade energética é importante para a
contração muscular. A fosforilação oxidativa é o processo de síntese de ATP a
partir de diferentes nutrientes. Este processo ocorre na mitocôndria sendo o
mais rentável e eficiente na produção de energia. Em situações de exercício,
há uma necessidade acrescida do consumo de mais substratos energéticos
para poder levar o oxigénio às células, dar resposta às necessidades
fisiológicas e manter o equilíbrio metabólico.
.

2
A atividade física provoca adaptações musculares esqueléticas específicas
relativas ao tipo, intensidade e duração do treino. Com o exercício físico, há um
aumento do consumo de oxigénio com consequente situação favorável à
produção de espécies reativas de oxigénio (EROs) e aparente aumento do
stress oxidativo a nível celular, tecidual e orgânico, avaliado por marcadores
tais como Sirtuina3 (Sirt3), grupo sulfidrilo (SH), proteína oxidante P66Shc. As
EROs são assim indicativas de stress oxidativo durante o exercício
encontrando-se também associadas a danos celulares (Ascensão et al., 2003;
Castrogiovanni & Imbesi 2012). Na realidade, durante o exercício observa-se
um aumento de radicais livres que retiram os eletrões da membrana lipídica
produzindo uma cadeia de reações que conduz à peroxidação lipídica (MDA)
assim como danos no ADN mitocondrial e nas proteínas (Ascensão et al.,
2003; Castrogiovanni & Imbesi, 2012).
Por seu lado, a adaptação do músculo ao treino está associada a mudanças na
expressão génica, regulação dos mecanismos de proteção celular e
remodelação da própria estrutura muscular bem como a biogénese
mitocondrial. Este processo de biogênese mitocondrial exprime-se não apenas
na formação de novas mitocôndrias, mas também na melhoria da expressão da
sua enzimática (Silva, 2014). Esta resulta da ativação do coativador 1α do
receptor gama ativado pelo proliferador de peroxisoma - PGC-1α, proteína
reguladora de uma variedade de processos metabólicos, incluindo a formação
de novas mitocôndrias nos músculos esqueléticos e no coração, bem como o
controle da β-oxidação mitocondrial (Lin et al., 2005; Silveira et al., 2006) e a
resposta celular ao stress oxidativo (St-pierre et al., 2006). Assim sendo, de
uma forma geral, a PGC-1α estimula a biogénese mitocondrial e promove a
remodelação do tecido muscular para uma composição que é mais oxidativa e
menos glicolítica. A sua relevância na modulação do metabolismo lipídico
torna-se um fator chave na intervenção na obesidade e diabetes através do
exercício (Kang & Ji, 2012; Liang & Ward, 2006).

3
À semelhança de outros estudos, neste trabalho foi utilizado o método de
Western Blotting de semi-quantificação de proteínas para perceber de que
forma, comparativamente à condição de sedentário, as proteínas PGC1α, T-
Fam, envolvidas na biogénese mitocondrial, se alteram com o exercício em
animais obesos e não obesos sujeitos a exercício. Adicionalmente, foram ainda
determinados marcadores de stress oxidativo, Sirt3, MDA, SH, p66Shc, bem
como o conteúdo proteico das subunidades dos complexos mitocondriais I, II,
III e IV.

4

5
2. Revisão de literatura
2.1. Obesidade: Conceito, prevalência e características
O excesso de peso e a obesidade constituem um problema de saúde cada vez
mais grave e atual. A obesidade é definida pela World Health Organization
(WHO, 2015) como uma acumulação anormal de gordura que se poderá tornar
um fator de risco para a saúde do individuo.
Em 2014, mais de 1,9 milhões de adultos (> 18 anos) tinham excesso de peso
e, destes, 600 milhões eram obesos (WHO, 2014). Tendo em conta estes
dados e comparando-os com os dados relativos a 1980, a prevalência de
obesidade a nível mundial mais do que duplicou. Os fatores genéticos poderão
ter alguma influência na propensão para obesidade, no entanto, esta condição
parece dever-se sobretudo a um desequilíbrio energético resultante de um
aporte calórico muito superior às necessidades orgânicas e a uma diminuição
significativa da atividade física, muito relacionada com o sedentarismo (Beja,
Ferrinho & Craveiro, 2014; Hariri, & Thibault, 2010; Peters, 2014; Sarzani,
Bordicchia, Spannella, Dessi-Fulgheri, Fedecostante, 2014; Schmidt, 2012).
Este estilo de vida tem efeitos negativos não só na saúde física mas
igualmente na saúde mental (Peters, 2014), constituindo-se como um fator de
risco para dificuldades de aprendizagem, baixa autoestima, distúrbios mentais
(depressão, perturbações de ansiedade) e, na idade adulta, doenças
cardiovasculares, diabetes, distúrbios músculo-esqueléticos (ex: osteoartrite),
cancro (ex: endométrio, mama e cólon). Atendendo ao aumento global da
obesidade e consequentemente das comorbilidades associadas, vários estudos
têm sido realizados nesta área (Beja et al, 2014; Buettner, Scholmerich,
Bollheimer, 2007; Freeman, Haley-Zitlin, Rosenberger, Granholm, 2014; Galic,
Oakhill, Steinberg, 2010; Lindinger et al, 2015; Romacho et al, 2014;
WHO/Europe, 2007).
Na Europa, em 2008, mais de 50% dos indivíduos com idade superior a 20
anos apresentavam excesso de peso e mais de 20% apresentavam obesidade.
No grupo de obesidade a prevalência por género era, aproximadamente, 20%

6
homens e 23% mulheres. Nos adolescentes, dados do inquérito Health
Behaviour in School-aged Children (2009-2010) revelam uma elevada
prevalência de sobrepeso em vários países da Europa com Portugal a registar
32% neste grupo (WHO/Europe, 2013). Entre 1990 e 2008 verificou-se que
cerca de 60% das crianças que apresentavam sobrepeso antes da puberdade
tinham elevada probabilidade de o manter na idade adulta (Beja et al., 2014;
Hariri, & Thibault, 2010; Lehnert et al., 2013; WHO/Europe, 2007).
Dados do Instituto Nacional Ricardo Jorge no âmbito do Childhood Obesity
Surveillance Iniciative - COSI Portugal 2010 (Rito, 2012) mostram que 14,3%
das crianças com idades compreendidas entre os 6-8 anos são obesas e
30,2% encontram-se em excesso de peso sendo a prevalência mais elevada
na região da grande Lisboa e no Arquipélago da Madeira.
Com o aumento do número de obesos numa população de determinado país,
os valores monetários investidos nas despesas de saúde relacionadas com a
obesidade e as doenças associadas, são maiores (Kershaw & Flier, 2004;
Lehnert et al., 2013). Na Europa estima-se que os custos diretos e indiretos
sejam de 0,09% a 0,61% da receita bruta total anual nos países Ocidentais.
Estes custos indiretos estão associados com a redução de produtividade,
problemas a nível dos sistemas de saúde, empregadores e com a economia do
país em geral (Lehnert et al., 2013; Von Lengerke & Krauth, 2011). Assim,
parece evidente que é imperativo um forte investimento nas estratégias de
prevenção, a fim de limitar o crescimento das despesas de saúde imputáveis à
obesidade em toda a Europa (Von Lengerken & Krauth, 2011).
No sentido de prevenir os aspetos negativos acima referidos, a investigação
tem procurado perceber de que forma se poderá combater esta epidemia e
controlar os custos económicos a ela associados. Desta forma têm sido
registadas uma panóplia de intervenções preventivas e terapêuticas,
atendendo à relação custo-eficácia sendo privilegiadas intervenções mais
económicas e, simultaneamente, com elevado grau de eficácia. A intervenção
centrada na prevenção consiste na sensibilização junto das populações mais

7
jovens para uma alimentação saudável, variada e livre de gorduras, bem como
o incentivo à prática de exercício físico (Faeh, 2012; Lehnert et al., 2013).
De facto, as intervenções não farmacológicas propostas pela União Europeia
com vista a combater a obesidade, nomeadamente a atividade física regular e
uma alimentação nutricionalmente equilibrada, são prioridades a serem
atendidas nos vários estados membros nomeadamente nas instituições
educativas (Faeh, 2012). No mesmo sentido a OMS realça estes dois pontos
no seu plano de ação elaborado para 2013-2020 (WHO, 2013).
2.2. Modelos de dieta animal para indução de obesidade
O modelo animal tem sido utilizado frequentemente para mimetizar a
obesidade, todavia algumas variáveis potencialmente confundidoras deverão
ser controladas nomeadamente a espécie, a idade, o nível de gordura da dieta
e ainda o tipo de dieta de controlo (Crescenzo et al., 2013; Farias et al., 2013;
Gajda, 2015; Hariri & Thibault, 2010; Rosini, Silva, Moraes, 2012).
Na literatura, têm sido referidos vários modelos nutricionais de indução da
obesidade nomeadamente: Modelo de cafeteria, Modelo hipercalórico, Modelo
de LiberDeCarli. O modelo nutricional para indução da obesidade, designado
por dieta de cafeteria baseia-se na ingestão ad libitum biscoitos, doces, queijos
e carnes processadas (Gajda, 2015; Hariri & Thibault, 2010). A vantagem
principal desta dieta está na semelhança com a dieta humana ocidental que é
rica em sal, açúcar e gorduras, sem grande definição dos componentes
nutritivos e não nutritivos. Por outro lado, uma das desvantagens desta dieta é
a escolha aleatória de alimentos, pois o animal pode fazer uma seleção de
alimentos diferente em cada dia. Este tipo de dieta, foi utilizado no estudo de
Casanova et al. (2014), para induzir obesidade, tendo o peso corporal
aumentado significativamente nos animais sujeitos a DC comparativamente
com os alimentados por uma dieta controlo (CS).

8
O modelo hipercalórico utilizado continha níveis elevados de gordura animal de
banha de porco (> 30% de gordura) (Farias et al., 2013; Rosini, Silva, Moraes,
2012). Os resultados mostram que a obesidade provoca grandes alterações
relacionadas com diversos parâmetros bioquímicos e moleculares no tecido
adiposo, bem como valores mais elevados do peso corporal (Hariri & Thibault,
2010). Os níveis de glicemia, plasma sanguíneo, triglicerídeos, bem como os
níveis de glicose no sangue, também demonstraram aumentos significativos
(Tinkov, Popova, Polyakova, Skalny, Nikonorov, 2014; Yokota et al., 2009).
Umas das desvantagens das dietas hipercalóricas é o seu tempo de indução –
2 a 3 meses.
Outro modelo de dieta utilizado é a dieta líquida (isocalórica) - modelo de
LiberDeCarli. Este tipo de dieta tem a vantagem de mostrar resultados em
apenas 3 semanas, sendo mais significativos após 5/6 semanas. Estes
resultados traduzem-se no aumento de peso, alterações ao nível da cadeia
respiratória e desenvolvimento de várias características-chave para doenças
associadas à obesidade (Abdelmegeed et al., 2011). Outra das vantagens
deste tipo de dieta é o facto de evitar uma deficiência nutricional. Esta dieta foi
apresentada pela primeira vez no estudo realizado por Lieber et al. (2004) com
ratos Sprague-Dawley machos que utilizou uma dieta líquida rica em gordura
(71% de energia proveniente de gordura, 11% de hidratos de carbono, 18% de
proteínas) e uma dieta controlo (35% de energia proveniente de gordura, 47%
de hidratos de carbono, 18% de proteína). Os autores puderam constatar que a
dieta com elevado teor de gordura causou anomalias mitocondriais,
inflamações mononucleares e aumento do stress oxidativo (Abdelmegeed et
al., 2011; Wang et al., 2009). A desvantagem desta dieta prende-se com a
exigência do procedimento de alimentação pois deverá ser diário e feito
individualmente.
2.3. Obesidade e tecido adiposo
O tecido adiposo é um órgão endócrino complexo e com uma elevada atividade
metabólica, fonte de energia e de secreção de proteínas com uma importante

9
função endócrina e anti-inflamatória. Este tecido é composto por adipócitos e
subdivide-se em: tecido adiposo branco – armazenamento de lípidos – e tecido
adiposo castanho que tem como principal e crucial função a termorregulação
(Lagowska & Jeszka, 2011). Na sua constituição encontra-se ainda tecido
nervoso, células imunes, não sendo considerado apenas um local de
armazenamento de lípidos e triglicerídeos (fontes de energia) e de proteção
(contra a perda de calor) (Galic, Oakhill, Steinberg, 2010; Kershaw & Flier,
2004; Lindinger et al, 2015; Paglialunga, Ludzki, Root-McCaig, Holloway,
2015).
Gollish et al. (2009) mostraram que uma alimentação rica em gorduras
aumentava drasticamente a quantidade total de tecido adiposo. Este estudo
tinha como objetivo estudar os efeitos do treino no tecido adiposo subcutâneo e
visceral em ratos que foram alimentados com uma dieta de ração controlo
(13% gordura) ou tornados resistentes à insulina por uma dieta rica em
gorduras (60%). Já outro estudo realizado por Crescenzo et al, (2013) com
ratos machos Sprague–Dawley, revelou que a dieta rica em frutose utilizada
induz o desenvolvimento de obesidade tendo por base o aumento dos níveis de
gordura em todo o corpo mais especificamente gordura intra-abdominal.
Estudos mostram uma associação entre a síndrome metabólica e a obesidade
abdominal que parece estar altamente dependente da distribuição da gordura
corporal, principalmente no que se refere à gordura periférica (Romancho et al.,
2014).
No que se refere à função endócrina e secretora, o tecido adiposo produz
citocinas (ou adipocinas), proteínas de sinalização intercelulares durante a
resposta imunológica ou inflamatória. Estas podem atuar localmente no tecido
adiposo mas também chegam a outros órgãos através da circulação sistémica
onde intervêm ao nível da regulação da ingestão alimentar e peso corporal,
sensibilidade à insulina, inflamação, coagulação ou função vascular. Esta
função endócrina é potenciada pelas consequências metabólicas adversas
resultantes tanto do excesso de tecido adiposo como da sua redução (Kershaw
& Flier, 2004; Romancho et al., 2014). Os locais de deposição do tecido

10
adiposo afetam a função endócrina uma vez que as hormonas endócrinas
originárias do tecido adiposo visceral são segregadas no sistema portal tendo
assim acesso direto ao fígado (Kershaw & Flier, 2004).
Relativamente à função anti-inflamatória do tecido adiposo, na obesidade os
fatores que fazem desencadear a produção de macrófagos, principal fonte de
citocinas inflamatórias, são elevados e surgem com maior frequência levando a
uma resposta inflamatória crónica, que tem sido associada ao desenvolvimento
da resistência à insulina e aos diabetes (Lagowska & Jeszka, 2011; Romancho
et al., 2014). Alguns estudos referem especificamente os efeitos do tecido
adiposo na resistência à insulina (Lagowska & Jeszka, 2011; Romancho et al.,
2014). Amano et al. (2014) verificaram que o tecido adiposo acumula células
imunes que irão libertar citocinas que poderão desencadear a resistência à
insulina. Os macrófagos existentes no tecido adiposo sofreram uma divisão
celular inesperada o que mostrou um aumento da proliferação na obesidade.
As desordens metabólicas do tecido adiposo estão assim intimamente ligadas
à inflamação que acompanha o recrutamento e infiltração de células imunes o
que inclui macrófagos do tecido adiposo (Cai, Wang, Ji, Meyer, van der
Westhuyzen, 2012). Em síntese, uma dieta rica em gorduras num curto período
de tempo induz um aumento da resistência à insulina e hipóxia em sujeitos
obesos (Furukawa & et al, 2004; Lagowska & Jeszka, 2011), o que poderá ter
consequências noutros tecidos.
2.4. Obesidade em outros tecidos: fígado, coração, cérebro
A obesidade pode manifestar-se por alterações ao nível dos principais órgãos
do ser humano, alterações essas que por sua vez serão fatores de morbilidade
(Wang et al., 2015). De facto, a obesidade é a causa primária de várias
doenças metabólicas, como a diabetes e a doença hepática não alcoólica
(NAFLD). A NAFLD é considerada um síndrome com um amplo espetro de
lesões hepáticas, desde a esteatose simples, à esteatohepatite não alcoólica
(vulgarmente designada por NASH) e, finalmente, fibrose ou cirrose não

11
alcoólica. A NASH, para além da acumulação de gotículas lipídicas nos
hepatócitos caracteriza-se pela apresentação de células inflamatórias (Wang et
al., 2008). Apesar da esteatose ser designada como benigna, a possibilidade
desenvolvimento e progressão da doença é evidente (Abdelmegeed et al,
2011; Lieber et al, 2004; Woo et al., 2014). Machado et al. (2012) concluíram
que a obesidade constitui um dos principais riscos para o desenvolvimento da
esteatose hepática não-alcoólica devido à capacidade de armazenamento do
tecido adiposo estar saturada, levando a uma acumulação de gordura na parte
exterior dos órgãos. Esta acumulação pode desencadear a ativação dos
macrófagos e, consequente, libertação de citocinas pró-inflamatórias. A
inflamação, por sua vez, conduz ao desenvolvimento de fibrose, cirrose e
doença hepática crónica (Negrin et al., 2014). Mais tarde Larter et al. (2013)
demonstrou que ratos (Alms1 mutant) alimentados com dieta gorda
desenvolvem obesidade hiperfágica, diabetes, síndrome metabólica e
esteatose hepática não-alcoólica (Negrin et al., 2014).
Para além destes problemas, a obesidade está associada ao agravamento da
doença inflamatória intestinal sendo o excesso de ingestão de gorduras
saturadas um fator de risco para esta doença (Gruber et al., 2013). Kredel e
Siegmund (2014) demonstraram que a deposição de tecido adiposo está
associada a um aumento da resposta inflamatória no intestino com
consequências sistémicas. O estado de inflamação crónica daí resultante pode
contribuir para o aparecimento da doença de Crohn, cancro do colon, entre
outras. De facto, estudos recentes têm sugerido uma associação entre
alterações na flora microbiana intestinal e várias doenças metabólicas,
nomeadamente a obesidade e a diabetes tipo 2 (Gruber et al., 2013; Ojeda et
al., 2015). Por outro lado, estudos evidenciam que dietas ricas em gordura
induzem hiperinsulinemia e acumulação lipídica que, por sua vez, está
associada a um aumento do stress oxidativo e alterações morfológicas levando
a lesões em tecidos como o fígado, rins e coração (Muthulakshmi & Saravanan,
2013).
Para além de patologias hepáticas e gastrointestinais, outros estudos têm
evidenciado o risco de doença cardíaca associada à obesidade, principalmente

12
se a obesidade estiver presente desde as idades mais jovens podendo refletir-
se no batimento cardíaco e problemas associados à vasodilatação dos
capilares do músculo cardíaco (Lacerda-Miranda et al., 2012; Sarzani,
Bordicchia, Spannella, Dessi-Fulgheri, Fedecostante, 2014). Os mesmos
resultados têm sido evidenciados em humanos (Mogoi, Ilie, Paul, Velea, 2014).
Um estudo realizado por Sarzani et al, (2014), na Alemanha e na Suíça com
260,000 crianças obesas ou com excesso de peso, revelou que 35% tinham
hipertensão com aumento do ventrículo ou rigidez arterial. Com efeito, os
autores concluíram que crianças que mantêm um padrão de obesidade até à
idade adulta estão propensas a desenvolver hipertensão, diabetes,
aterosclerose (Mogoi, Ilie, Paul, Velea, 2014).
As alterações metabólicas induzidas pela obesidade estão também associadas
a alterações no cérebro, tanto ao nível estrutural como funcional (Bilbo &
Tsang, 2010). De facto, tem sido sugerido que a hiperglicémia provoca danos
graves no tecido cerebral levando ao desenvolvimento de doenças neuro
degenerativas como a doença de Alzheimer e Parkinson (Ashrafian, Harling,
Darzi, Athanasiou, 2013; Davidson et al., 2013; Jayaraman, Lent-Schochet,
Pike, 2014; Sheikhpour, & Khoradmehr, 2014; Wang et al., 2015). Para além
destas patologias, a demência surge também associada a uma deficiente
alimentação, principalmente à base de carnes vermelhas, açucares e grãos
refinados e alimentos com elevado teor de gordura, como também, alimentação
rica em ácidos gordos saturados está associada a uma pior performance
cognitiva (Davidson et al., 2013; Ashrafian, Harling, Darzi, Athanasiou, 2013;
Freeman, Haley-Zitlin, Rosenberger, Granholm, 2014). A demência define-se
por uma deterioração de uma ou mais partes do cérebro levando a uma
diminuição da performance cognitiva. Estudos comprovam que a obesidade e o
consumo de uma dieta com elevado teor de gordura aumenta o risco de
desenvolvimento de demência, estando associada a uma diminuição da
integridade dendrítica e ativação das células microglias no hipocampo
(Davidson et al., 2013; Freeman, Haley-Zitlin, Rosenberger, Granholm, 2014).
Assim a relação entre a obesidade e a diminuição da capacidade cognitiva é

13
cada vez mais evidente. (Jayaraman, Lent-Schochet, Pike, 2014; Sheikhpour, &
Khoradmehr, 2014).
Para além dos efeitos da obesidade ao nível dos órgãos referidos
anteriormente, existem igualmente consequências nefastas ao nível muscular.
2.5. Obesidade e Músculo: stress oxidativo, biogénese mitocondrial
Num estado de obesidade, a condição física do individuo torna-se limitada, o
que por sua vez limita a sua capacidade de trabalho, nomeadamente através
da restrição de determinados movimentos e da diminuição da agilidade e força
gerada pelo músculo.
Com o incremento da obesidade, a prevalência dos efeitos negativos no
músculo-esquelético aumentaram. Neste sentido, os estudos nesta área têm-se
tornado relevantes a nível clinico. Lafortuna, Tresoldi, Rizzo, (2014)
demonstraram que a mudança de peso de normal para obeso estava
associado a um aumento progressivo do volume muscular do membro inferior e
da área de secção transversal do músculo-esquelético. Isto poderá ser visto
como uma resposta adaptativa muscular do aumento do peso corporal. Para
além disto, vários estudos têm reportado a alteração do metabolismo
mitocondrial no músculo-esquelético, não estando contudo esclarecido quais as
vias celulares envolvidas nestas alterações e quais as suas consequências
(Baker II et al., 2015).
O músculo-esquelético é um tecido altamente especializado com excelente
plasticidade em resposta a estímulos externos, como o exercício físico. As
contrações musculares realizadas durante o treino desencadeiam uma
variedade de características fenotípicas e respostas fisiológicas incluindo a
ativação da biogénese mitocondrial, transformação do tipo de fibra, fibra mais
oxidativa e menos glicolítica, e angiogénese. Em conjunto, estas respostas
aumentam a capacidade do metabolismo aeróbico do músculo e a sua
resistência à fadiga (Liang & Ward, 2006; Steinbacher & Eckl, 2015).

14
A obesidade, por seu lado, provoca vários efeitos nefastos no músculo-
esquelético. Por exemplo, ao nível da sua vasculatura podemos observar um
comprometimento da função endotelial, aumento da vulnerabilidade dos sinais
vasoconstritores e resistência vascular à insulina. Apesar disto, várias
evidências propõem que estas consequências não são exatamente iguais para
todos os indivíduos pois nem todos os sistemas vasculares são igualmente
afetados. Jenkins et al. (2014) relataram recentemente que as artérias dos
músculos gastrocnemius e solear apresentam diversas vulnerabilidades na
disfunção endotelial e diferenciam-se nas ações dos vasodilatadores endotélio-
dependente. Outros estudos mostram que a ingestão de uma dieta altamente
calórica resulta num estado de hiperglicemia e de níveis elevados de ácidos
gordos na circulação sanguínea, levando a um estado de stress oxidativo. Com
efeito, o aumento do stress oxidativo no músculo-esquelético pode prejudicar a
função mitocondrial e limitar a capacidade de exercício físico. O dano oxidativo
pode atingir primariamente as mitocôndrias o que poderá fazer com que o ADN
mitocondrial afete a biogénese mitocondrial e, por consequência, induzir
disfunção mitocondrial (Farias et al., 2012).
A este respeito, Boudina et al. (2012) salientam que as adaptações
mitocondriais no músculo-esquelético de animais submetidos a uma dieta rica
em gorduras são complexas e específicas. No entanto, não parece provável
que o stress oxidativo tenha um impacto direto na diminuição da absorção da
glicose mediada pela insulina que caracteriza os estádios inicias da resistência
à insulina. Da mesma forma, os antioxidantes parecem ter um papel limitado na
prevenção primária da resistência à insulina. Por sua vez, a redução da beta-
oxidação mitocondrial é responsável pelo desenvolvimento da resistência à
insulina no músculo-esquelético. Esta resistência à insulina está ainda
associada a uma diminuição da expressão genética dos componentes que
medeiam a fosforilação oxidativa (Boudina et al., 2012; Buchner et al., 2011).
Neste sentido, sendo o músculo-esquelético um tecido heterogéneo constituído
por diferentes tipos de fibras contráteis, é particularmente importante estudar a
fosforilação mitocondrial oxidativa e a glicólise na produção de energia. Os

15
músculos com características mais glicolíticos geram energia por processos
metabólicos essencialmente anaeróbicos, enquanto os músculos mais
oxidativos estão implicados na resistência à fadiga e obtenção de energia a
partir do processo metabólico oxidativo, cuja capacidade é principalmente
determinada pela função mitocondrial e pela biogénese (Gómez-Pérez et al.,
2012). Skovbro, Boushel, Hansen, Helge, (2011) concluíram que o grupo de
ratos alimentado com dieta isocalórica sofreu uma inibição do estado 3 da
resposta respiratória mitocondrial ao nível muscular na presença dos
substratos glicolíticos visto que estes não estavam presentes após estimulação
com ácidos gordos.
Os resultados do estudo de Crescenzo et al. (2015) mostraram que uma
alimentação com alto teor de gordura induz alterações no equilíbrio energético,
fazendo com que haja uma diminuição do gasto de energia despendida e um
aumento de lípidos. Este tipo de alimentação poderá levar a um aumento do
fluxo de lípidos e da eficiência energética no músculo-esquelético, tendo como
consequência principal a redução da oxidação de ácidos gordos e consequente
acumulação de lípidos que poderão desencadear resistência à insulina no
músculo e mais tarde progredir para diabetes tipo 2 (Crescenzo et al., 2015;
Standford & Goodyear, 2014). Desta forma, quando o músculo se encontra em
repouso, as mudanças no número de organelos e/ou na atividade do músculo
podem não ter efeitos negativos na bioenergética celular, enquanto
modificações na eficiência energética mitocondrial farão variar a energia
oxidada pela célula mesmo que o ATP (turnover) não varie (Crescenzo et al.,
2015).
A biogénese mitocondrial pode ser entendida como uma adaptação
vocacionada para neutralizar a quantidade elevada de substrato disponível. De
facto, estudos mostram que uma alimentação rica em gordura induz a
biogénese mitocondrial através do aumento da PGC-1α, principal regulador da
energia metabólica. O aumento dos níveis desta proteína implica um reforço da
capacidade oxidativa do músculo enquanto os níveis de ADN mitocondrial
(ADNmt) apontam para um aumento do conteúdo mitocondrial (Gómez-Pérez et

16
al., 2012; Kang & Ji, 2012; Tadaish et al., 2011). Estas alterações estão
altamente relacionadas com a prática de atividade física, ou a falta desta, pelo
que se torna relevante compreender de que forma o exercício físico pode ser
um fator de prevenção ou de terapia na obesidade.
2.6. Papel do exercício físico na obesidade: stress oxidativo, biogénese
mitocondrial
O exercício físico tem um papel crucial no combate à obesidade, quando
realizado de forma regular, estando associado a uma redução da gordura
abdominal (Ross, Hudson, Stotz, Lam, 2015). Segundo a European Society of
Lifestyle Medicine (2015), a prática semanal de atividade física moderada
contribui para a redução do risco de doença cardíaca isquémica (em 30%),
diabetes (27%), de cancro da mama ou colon (21-25%) reduzindo ainda o risco
de AVC, depressão e hipertensão.
Decorrente da prática regular de exercício, há um aumento do consumo de
oxigénio, tornando-se numa situação favorável à produção de EROs e aparente
aumento do stress oxidativo. Esta produção elevada de EROs é por vezes
associada a efeitos secundários negativos como a redução da capacidade de
gerir força e um aumento da atrofia muscular. De facto, este aumento dos
radicais livres pode causar peroxidação lipídica assim como danos no ADNmt e
em proteínas. No entanto, estudos recentes têm vindo a comprovar que as
EROs também produzem efeitos positivos pois o aumento de EROs vai
influenciar os processos celulares que se relacionam com o aumento da
expressão dos antioxidantes, os quais neutralizam os efeitos dos radicais livres
estando envolvidos na adaptação muscular induzida pelo exercício
(Castrogiovanni & Imbesi, 2012; Steinbacher & Eckl, 2015). Para perceber de
que forma o stress oxidativo aumenta com o exercício, existem marcadores
que sinalizam este aumento e que identificam ocorrência de peroxidação
lipídica e modificação de proteínas. Um dos marcadores utilizados é o MDA
que tem origem na peroxidação dos lípidos e é normalmente medido através da

17
sua reação com o ácido tiobarbitúrico (Stanković & Radovanović, 2012). A Sirt3
é outro dos marcadores utilizados como indicador da ocorrência de stress
oxidativo. Esta proteína tem uma função protetora em diferentes tecidos
nomeadamente no tecido muscular e no tecido cardíaco. Os seus valores
aumentam em situação de exercício de modo a proteger a mitocôndria dos
danos provocados pelas EROs (Bause & Haigis, 2013; Palacios, et al., 2009;
Sundaresan, et al. 2008)
Assim, o aumento do metabolismo aeróbio durante o exercício é
potencialmente uma fonte de stress oxidativo. Uma vez que os benefícios do
exercício na saúde são bem conhecidos, a possibilidade de redução do stress
oxidativo causado pela adaptação à atividade física implica o aumento da
capacidade antioxidante e a redução da formação de radicais durante a
fosforilação oxidativa (Stanković & Radovanović, 2012).
A obesidade aponta para níveis elevados de hiperglicemia o que leva à
produção de EROs na mitocôndria. Quando o gradiente de protões excede o
limiar, a transferência de eletrões no complexo III da cadeia respiratória é
bloqueada, dando-se a formação do ião superóxido. Há uma redução da
capacidade oxidativa e mitocondrial no músculo-esquelético o que está
relacionado com o ganho de peso e a resistência à insulina. Estudos mostram
que o exercício aumenta a atividade oxidativa enzimática bem como um
aumento do número das mitocôndrias, aumento dos antioxidantes e da via
lipolítica (Farias et al., 2012; Menshikova et al., 2007). Hariri e Thibault (2010)
realizaram um estudo com ratos submetidos a dieta gorda e a exercício até à
exaustão. Os resultados mostraram que, em comparação com os ratos com
dieta controlo, atingiram níveis de exaustão mais rapidamente tendo sido ainda
observado um decréscimo do pico de VO2. Os autores mostraram também que
o exercício físico reequilibra estas alterações o que induz um menor ganho de
peso.
A PGC-1α possui um papel fundamental na regulação do metabolismo
energético e está intimamente relacionada com o aumento da biogénese
mitocondrial. Vários estudos comprovam que o treino de endurance está

18
relacionado com o aumento de conteúdo mitocondrial e capacidade respiratória
do músculo-esquelético resultando numa taxa mais lenta de utilização de
glicogénio no músculo e glicose no sangue e, por outro lado, uma maior
dependência da oxidação de gordura e menor acumulação de lactato durante o
exercício (Kang & Ji, 2012; Skovbro et al., 2011). O exercício físico é um fator
fundamental para a regulação da expressão génica da PGC-1α e,
consequente, regulação positiva da biogénese mitocondrial (Kang & Ji, 2012;
Steinbacher & Eckl, 2015). Outros estudos mostram ainda que a PGC-1α
aumenta com o exercício e promove a biogénese mitocondrial e a angiogénese
no músculo, tem o papel de ativar simultaneamente os fatores de transcrição
na resposta às solicitações energéticas resultando na ativação dos genes
nucleares envolvidos na biogénese mitocondrial, nomeadamente na transcrição
mitocondrial da TFAM. No entanto, curiosamente, uma dieta rica em gordura
induz a biogénese mitocondrial músculo-esquelética observando-se um
aumento da PGC-1α e dos níveis da TFAM. (Gómez-Pérez et al., 2012;
Steinbacher & Eckl, 2015; Tadaish et al., 2011).

19
3. Objetivos
3.1. Objetivo Geral
O objetivo fundamental deste estudo foi compreender os mecanismos celulares
(stress oxidativo e biogénese) associados ao potencial efeito protetor do
exercício físico no músculo-esquelético em situação de obesidade.
3.1.1. Objetivos específicos
Pretendeu-se analisar as alterações induzidas pelo exercício crónico realizado
concomitantemente com uma dieta rica em gordura:
- em marcadores de stress oxidativo (SH, MDA, Sirt3, p66Shc);
- em marcadores de biogénese mitocondrial (Tfam e PGC-1α);
- no conteúdo proteico das subunidades dos complexos da CTE.

20

21
4. Metodologia
4.1. Caraterização da Amostra
A amostra foi constituída por 24 ratos Sprague-Dawley macho com 6 semanas
de idade e um peso entre 125-150g. No decorrer do protocolo, os animais
foram mantidos em gaiolas individuais, em biotério e numa atmosfera
controlada (21-22ºC; 50-60% humidade), com comida e água ad libitum, e num
ciclo de 12 horas dia/noite.
Durante as 17 semanas do protocolo experimental, os animais foram
submetidos a dietas isocalóricas com diferentes quantidades de lípidos: dieta
controlo (35% das Kcal derivadas dos lípidos) ou dieta gorda (71% das Kcal
derivadas dos lípidos).
Para se adaptarem à consistência da dieta, todos os animais foram
alimentados com a dieta de controlo liquida durante a primeira semana.
Posteriormente, os animais foram distribuídos aleatoriamente por quatro
grupos:
1) Dieta controlo sedentário (CS, n=6),
2) Dieta controlo e treino aeróbio (CT, n=5),
3) Dieta gorda sedentário (GS, n=6),
4) Dieta gorda e treino aeróbio (GT, n=7)
Após 9 semanas, os grupos CT e GT iniciaram o programa de exercício,
mantendo-se o protocolo das dietas. Semanalmente, procedeu-se à
quantificação do volume de dieta ingerida pelos animais. O consumo
energético por semana foi calculado da seguinte forma: 1 mL =1 Kcal para
ambas as dietas.

22
4.2. Programa de exercício
Os animais submetidos ao protocolo de treino de endurance (CT e GT) foram
adaptados durante a primeira semana à passadeira rolante, 30 minutos a uma
velocidade de 15 m/min-1 e sem inclinação. Posteriormente, os animais
correram 60 minutos a uma velocidade inicial de 15 m/min-1 aumentando
progressivamente até 25 m/min-1, 5 dias por semana, durante 8 semanas.
Durante o protocolo de treino, os animais sedentários (CS e GS) foram
colocados numa passadeira rolante imóvel de forma a expor os animais às
mesmas condições ambientais e de manuseamento (Gonçalves et al., 2014;
Passos et al., 2015).
4.3. Western Blotting
A expressão de proteínas (PGC1α, T-FAM, Sirt3 e P66shc) foi determinada por
Western-blot, utilizando-se quantidades iguais de proteína (50µg) que foram
posteriormente desnaturadas em tampão de lise e separadas por eletroforese
em gel de dodecil sulfato de poliacrilamida (SDS/PAGE) como descrito por
Laemmli (1970), seguida de uma transferência para uma membrana de
nitrocelulose (Hybond-ECL, Amersham Biosciences). Após a secagem, as
membranas foram bloqueadas com uma solução de 5% (w/v) de leite em pó ou
de BSA em TBST-T. As membranas foram incubadas com o anticorpo primário
(diluição 1:1000) em 1% de leite em pó ou de BSA durante 2 horas à
temperatura ambiente (22-25ºC). Após o período de incubação, as membranas
foram lavadas três vezes e incubadas durante 2 horas com o anticorpo
secundário. A visualização das bandas foi feita através de reagentes
quimioluminescentes ChemiDoc (BioRad Laboratories) o software Image Lab
(BioRad Laboratories) para análise das mesmas.

23
4.4. Procedimentos Estatísticos
O tratamento estatístico dos dados foi realizado recorrendo ao programa
Statistical Package for the Social Sciences (SPSS Inc. version 21.0 para
Windows).
A média e o erro padrão da média foi calculado para todas as variáveis. Na
comparação de médias foi utilizado o teste paramétrico, a ANOVA a dois
fatores de medidas independentes seguida pelo teste à post hoc de Bonferroni.
Em ambos os testes o nível de significância foi estabelecido em 5 %.

24
25
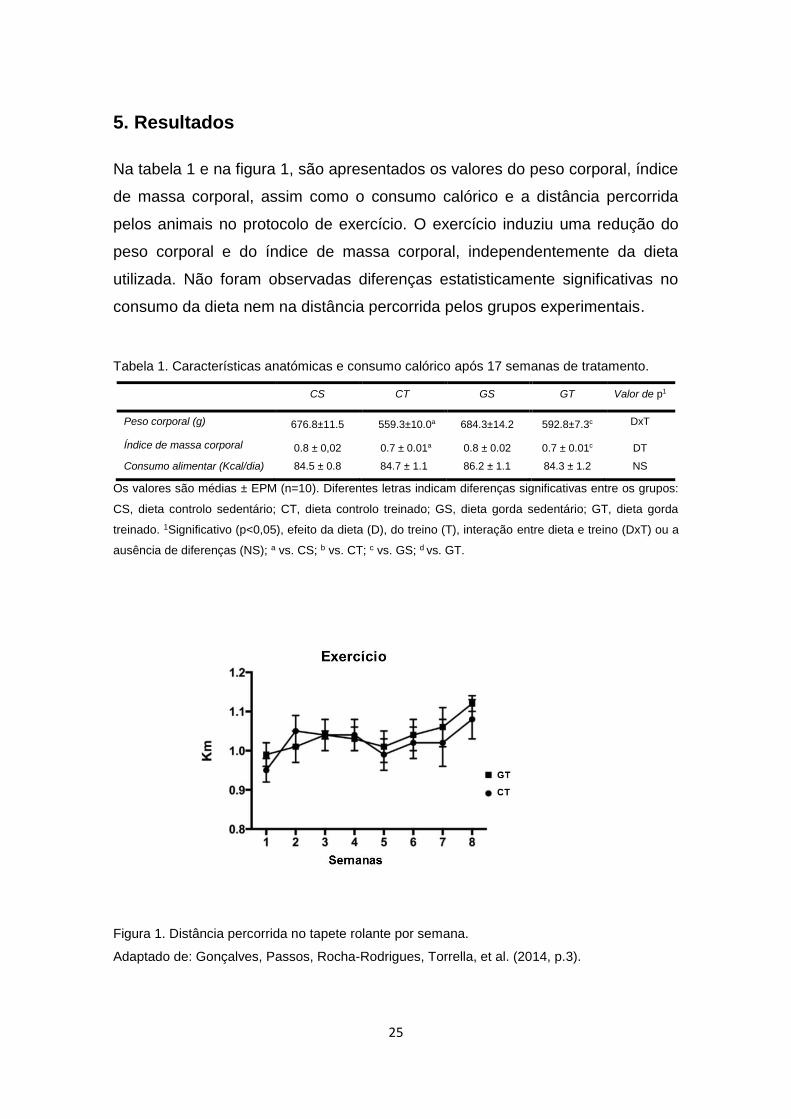
5. Resultados
Na tabela 1 e na figura 1, são apresentados os valores do peso corporal, índice
de massa corporal, assim como o consumo calórico e a distância percorrida
pelos animais no protocolo de exercício. O exercício induziu uma redução do
peso corporal e do índice de massa corporal, independentemente da dieta
utilizada. Não foram observadas diferenças estatisticamente significativas no
consumo da dieta nem na distância percorrida pelos grupos experimentais.
Tabela 1. Características anatómicas e consumo calórico após 17 semanas de tratamento.
CS CT GS GT Valor de p1
Peso corporal (g) 676.8±11.5 559.3±10.0a
684.3±14.2 592.8±7.3c DxT
Índice de massa corporal 0.8 ± 0,02 0.7 ± 0.01a
0.8 ± 0.02
0.7 ± 0.01c DT
Consumo alimentar (Kcal/dia) 84.5 ± 0.8 84.7 ± 1.1 86.2 ± 1.1 84.3 ± 1.2 NS
Os valores são médias ± EPM (n=10). Diferentes letras indicam diferenças significativas entre os grupos:
CS, dieta controlo sedentário; CT, dieta controlo treinado; GS, dieta gorda sedentário; GT, dieta gorda
treinado. 1Significativo (p<0,05), efeito da dieta (D), do treino (T), interação entre dieta e treino (DxT) ou a
ausência de diferenças (NS); a vs. CS; b vs. CT; c vs. GS; d vs. GT.
Figura 1. Distância percorrida no tapete rolante por semana.
Adaptado de: Gonçalves, Passos, Rocha-Rodrigues, Torrella, et al. (2014, p.3).

26
Após 17 semanas, os animais sedentários alimentados com dieta gorda
apresentaram um aumento do conteúdo de Sirt 3, assim como dos grupos tióis
(figura 2). Todavia, o treino de endurance apenas reverteu as alterações
induzidas na Sirt 3 (GS vs. GT). Não foram observadas diferenças no marcador
de peroxidação lipídica (MDA) nem na expressão da P66Shc.

27
Na figura 3, encontram-se representados alguns marcadores de biogénese
mitocondrial (PGC-1α e Tfam) e a expressão dos complexos mitocondriais.
Como é possível observar, a dieta rica em gordura aumentou o conteúdo da
proteína PGC-1α, assim como a subunidade do complexo III mitocondrial nos
animais sedentários (GS vs. CS). O exercício por um lado, aumentou a
expressão do complexo IV mitocondrial e, por outro lado, diminuiu a expressão
do complexo III nos animais obesos (GT vs. GS).

28

29
6. Discussão
O estilo de vida ocidental, caracterizado por dietas hipercalóricas ricas em
gordura e sedentarismo, tem sido descrito como a causa primária para o
desenvolvimento de várias doenças metabólicas (Willett et al., 2006). Com
efeito, alterações destes fatores de risco são aceites como as principais
estratégias na prevenção de patologias crónicas como a obesidade, diabetes e
cancro (Willett et al., 2006). Em particular, estudos têm demonstrado que o
exercício físico possui um papel fundamental na modelação de mecanismos
intracelulares e sistémicos (Gonçalves et al., 2014; Gonçalves et al., 2015)
nomeadamente a nível da funcionalidade mitocondrial (Gonçalves et al., 2014)
e da resistência à insulina (Ross et al., 2004). Considerando o papel do
músculo-esquelético na regulação da glucose sanguínea e o envolvimento da
mitocôndria na utilização e produção energética, procuramos neste estudo
compreender o efeito do exercício nas alterações celulares (stress oxidativo e
biogénese) do músculo-esquelético induzidas pela obesidade. Os resultados do
presente estudo mostraram que a obesidade estimulou alguns mecanismos
associados a um aumento da proteção contra o stress oxidativo e a um maior
consumo energético (PGC-1α, CIII). Por outro lado, o exercício crónico parece
ter contribuído para uma melhor funcionalidade tecidual observada pelo
aumento da expressão da subunidade do complexo IV mitocondrial.
Stress oxidativo e Biogénese na “obesidade muscular”
O stress oxidativo é descrito como um desequilibro entre a produção de
espécies reativas de oxigénio (EROs) e as defesas antioxidantes a favor da
primeira (Halliwell et al., 1994). Esta alteração da homeostasia redox poderá
ocorrer devido ao aumento do fluxo de eletrões ou a um comprometimento da
cadeia transportadora, com consequências deletérias para alguns dos
componentes celulares (proteínas, lípidos e ácido nucleicos) (Begriche et al.,
2006). Na obesidade, estudos em humanos e modelos animais têm
demonstrado um aumento dos marcadores de stress oxidativo (Banse et al.,

30
2015; Bonnard et al., 2008; SamJoo et al., 2013) e uma diminuição das defesas
antioxidantes no músculo-esquelético (Banse et al., 2015 & Pimenta et al.,
2015). Todavia, os resultados do presente estudo parecem contrariar os dados
da literatura, na medida em que observamos um aumento dos grupos tióis e do
conteúdo de Sirtuina 3, ambos compatíveis com a melhoria da antioxidante nos
animais obesos (GS vs. CS). De facto, a expressão da Sirtuina 3 no músculo
tem sido descrita como determinante no controlo da produção de ROS e
consequentemente da energia (Jing et al., 2011; Lin et al., 2014). Num estudo
desenvolvido com ratinhos knockout para Sirt3, Jing et al. (2011) observaram
um aumento do stress oxidativo e uma diminuição do consumo de oxigénio
mitocondrial nestes animais. Adicionalmente, Lin et al. (2014) utilizando um
modelo genético com expressão muscular da isoforma Sirt3 (SIRT3M3),
verificaram que estes animais apresentavam um aumento do dispêndio
energético comparativamente com os controlos. Com efeito, o fenótipo protetor
aparentemente observado nos animais obesos poderá constituir uma estratégia
defensiva/adaptativa, no sentido de procurar aumentar o dispêndio energético,
reduzindo-se o “leakage” de eletrões na CTE e, por conseguinte, a produção de
EROs nesta patologia.
Segundo alguns autores o exercício crónico contribui para a tolerância
cruzada em relação a estímulos indutores de stress oxidativo adicional, como
por exemplo a obesidade e diabetes (Nojima et al., 2008; Oh et al., 2013).
Neste sentido, seria expectável que o exercício induzisse adaptações positivas
nos marcadores de oxidação proteica (SH), peroxidação lipídica (MDA) ou de
algumas proteínas pro-oxidantes (Sirt3 e P66Shc). Os resultados do presente
estudo parecem, contudo, contrariar esta premissa verificando-se uma maior
oxidação dos grupos tiol e uma redução da expressão da proteína Sirt3 no
grupo GT. Todavia, estudos anteriores em tecidos não contrateis têm
igualmente demonstrado o aumento de marcadores de stress oxidativo com o
exercício regular (Ascensão et al., 2012; Gonçalves et al., 2014), tendo sido
sugerido que outros mecanismos anti-pro-oxidantes poderão ser estimulados
pelo exercício, nomeadamente aqueles associados à glutationa (Navarro et al.,
2004; Sun et al., 2010). Neste sentido, outros marcadores deverão ser

31
analisados em trabalhos futuros, para melhor descrever o papel do exercício na
modelação do stress oxidativo muscular. Recentemente, Strobel et al. (2014)
demonstraram ainda que a depleção da glutationa muscular, conjuntamente
com o exercício agudo (indutor de stress) parece ter uma importante função na
regulação da PGC-1α e, consequentemente, na biogénese mitocondrial. No
presente estudo observamos que os animais obesos apresentavam um
aumento da PGC-1α, o que parece sugerir uma estimulação da biogénese
mitocondrial. Este resultado é corroborado por Gomez-Perez et al. (2012),
segundo os quais a PGC-1α muscular poderá aumentar em situações de
obesidade, como uma estratégia compensatória da disfunção mitocondrial.
Para além disto, um aumento do número de mitocôndrias poderá estimular a
produção de energia com recurso aos ácidos gordos, promovendo um aumento
do dispêndio energético. No entanto, importa referir que outros estudos têm
sugerido uma diminuição do conteúdo de mitocôndrias musculares na
obesidade por comprometimento da síntese de novos organelos (Andersen et
al., 2005; Crunkhorn et al., 2007; Sparks et al., 2005). Apesar das alterações
observadas na PGC-1α e considerando a importância da biogénese
mitocondrial muscular no controlo energético, não foram observadas diferenças
no conteúdo de Tfam. Este resultado parece estar de acordo com o estudo de
Goto et al. (2001) que sugere a ocorrência da transcrição mitocondrial sem
uma intervenção significativa da Tfam. Para além de uma função reguladora na
biogénese, a PGC-1α apresenta igualmente um papel central no controlo da
formação de EROS (St-Pierre et al., 2003; St-Pierre et al., 2006; Vale et al.,
2005), uma vez que um aumento do volume mitocondrial permitirá dissipar a
força protomotriz, reduzindo a probabilidade de formação de ROS. De acordo
com St-Pierre et al. (2003), os animais knockout para a PGC-1α apresentam
uma maior suscetibilidade para estímulos indutores de stress. Deste modo, a
diminuição da oxidação de grupos tióis no grupo alimentado com dieta gorda
(GS), poderá ser também o resultado do aumento da expressão da PGC-1α.
Apesar do exercício crónico ser descrito como um indutor de biogénese
muscular (Hood et al., 2009; Hood et al., 2011), observamos uma diminuição
significativa no conteúdo da PGC-1α. Todavia, este resultado não é

32
completamente surpreendente, uma vez que alguns autores (Leick et al., 2008)
sugerem que a PGC-1α poderá não ser essencial para a síntese de
determinados genes/proteínas mitocondriais (citocromo c e citocromo oxidase).
Este facto, poderá justificar o aumento do conteúdo da subunidade do
complexo IV mitocondrial na ausência de um aumento simultâneo da PGC-1α
no grupo GT. O aumento da expressão dos complexos mitocondriais (CIII) foi
igualmente observado nos animais obesos sedentários, o que parece estar de
acordo com o estudo de Jain et al. (2014), no qual é demostrado que a
obesidade de origem genética ou nutricional induz um aumento dos complexos
da cadeia respiratória, com concomitante aumento da biogénese mitocondrial,
provavelmente numa tentativa de aumentar a oxidação de ácidos gordos e, por
outro lado, de reduzir a produção de ROS.
Em suma, o aumento da biogénese mitocondrial observado nos animais
obesos (GS), poderá ser interpretado como estratégia protetora no sentido de
atenuar uma condição pro-oxidante através da utilização dos ácidos gordos
como substrato energético. O exercício parece ter um efeito positivo na
modelação do conteúdo do complexo IV mitocondrial, uma vez que o aumento
da sua expressão está associado a uma melhoria da capacidade oxidativa
mitocondrial (Larsen et al., 2012).

33
7. Conclusões
Após a análise do efeito do exercício crónico realizado concomitantemente com
uma dieta rica em gordura em marcadores de stress oxidativo e biogénese
mitocondrial parece-nos possível retirar as seguintes conclusões:
I) A obesidade induziu um aumento da biogénese mitocondrial (PGC-
1α), o que se traduziu em melhorias no ambiente pro-oxidante
(grupos tiol e Sirt3).
II) A obesidade induziu um aumento do conteúdo do complexo III
mitocondrial o que poderá ter contribuído igualmente para uma
redução da pressão redox (tiol e Sirt3).
III) O exercício crónico induziu um aumento do conteúdo proteico das
subunidades do complexo IV mitocondrial, contribuindo para uma
melhoria da oxidação mitocondrial dos substratos energéticos e
assim para uma redução da pressão redox induzida pelo tempo de
redução dos complexos mitocondriais.

34

35
8. Referências Bibliográficas
Abdelmegeed, M. A., Yoo, S-H., Henderson, L. E., Gonzalez, F. J., Woodcroft,
K. J., Song, B.J. (2011). PPARa Expression protects male mice from
high fat–induced nonalcoholic fatty liver. The Journal of Nutrition:
Nutrition and Disease, 603-610.
Alexey A. T., Elizaveta V. P., Valentina S. P., Anatoly V. S., & Alexandra N.
(2014). Effect of high fat diet on macroelement content in hair and
adipose tissue of Wistar rats. Trace Elements and Electrolytes, 31 (4),
156-159. Doi: 110.5414/TEX01351
Amano S., Cohen J., Vangala P., Tencerova M., Nicoloro S., Yawe J., Shen Y.,
Czech M., Aouadi M. (2014). Local Proliferation of Macrophages
Contributes to Obesity-Associated Adipose Tissue Inflammation. Cell
Metabolism 19 (1), 162-171. Doi:10.1016/j.cmet.2013.11.017
Andersin, G., Wegner, L., Yanagisawa, K., Rose, C. S., Lin, J., Glumer, C.,
Drivsholm, T., Broch-Johnsen, K., Jorgensen, T., Hansen, T.,
Spiegelman, B. M., Pedersen, O. (2005). Evidence of an association
between genetic variation of the coactivator PGC-1beta and obesity.
Jornal of Medical Genetics 42 (5), 402-407. Doi:
10.1136/jmg.2004.026278
Ascensão, A., Gonçalves, I. O., Lumini-Oliveira, J., Marques-Aleixo, I., Dos
Passos, E., Rocha-Rodrigues, S., Machado, N. G., Moreira, A. C.,
Oliveira, P. J., Torrella, J. R., Magalhães, J. (2012). Endurance training
and chronic intermittent hypoxia modulate in vitro salicylate-induced
hepatic mitochondrial dysfunction. Mitochondrion 12 (6), 607-16. Doi:
10.1016/j.mito.2012.10.007
Ascensão, A., Magalhães, J., Soares, J., Oliveira, J., Duarte, J. A. (2003).
Exercício e Stress Oxidativo Cardíaco. Revista Portuguesa Cardiologia
22 (5), 651-678.

36
Ashrafian, H., Harling, L., Darzi, A., Athanasiou, T. (2013). Neurodegenerative
disease and obesity: what is the role of weight loss and bariatric
interventions? Metabolic Brain Disease 28 (3), 341-353. Doi:
10.1007/s11011-013-9412-4
Baker II, P. R., Boyle, K. E., Koves, T. R., Ilkayeva, O. R., Muoio, D. M.,
Houmard, J. A., Friedman, J. E. (2015). Metabolomic analysis reveals
altered skeletal muscle amino acid and fatty acid handling in obese
humans. Obesity Biology And Integrated Physiology 23 (5), 981 – 988.
Doi:10.1002/oby.21046
Banse, H. E., Frank, N., Kwong, G. P., McFarlane, D. (2015). Relationship of
oxidative stress in skeletal muscle with obesity and obesity-associated
hyperinsulinemia in horses. Canadian Journal Veterinary Research 79
(4), 329-338.
Bause, A. & Haigis, M. C. (2013). SIRT3 regulation of mitochondrial oxidative
stress. Experimental Gerontology 48, 634 - 639.
Doi.org/10.1016/j.exger.2012.08.007
Begriche, K., Igoudjil, A., Pessayre, D., Fromenty, B. (2006). Mitochondrial
dysfunction in NASH: causes, consequences and possible means to
prevent it. Mitochondrion 6 (1), 1-28. Doi: 10.1016/j.mito.2005.10.004
Beja, A., Ferrinho, P., Craveiro, I. (2014). Evolução da prevenção e combate à
obesidade de crianças e jovens em Portugal ao nível do planeamento
estratégico. Revista Portuguesa de Saúde Pública 32 (1), 10 – 17.
Bilbo, S. D. & Tsang, V. (2010). Enduring consequences of maternal obesity for
brain inflammation and behavior of offspring. The FASEB Journal, 24 (6),
2104-15. Doi: 10.1096/fj.09-144014
Bonnard, C., Durand, A., Peyrol, S., Chanseaume, E., Chauvin, M. A., Morio,
B., Vidal, H., Rieusset, J. (2008). Mitochondrial dysfunction results from
oxidative stress in the skeletal muscle of diet-induced insulin-resistant

37
mice. Journal of Clinical Investigation 118 (2), 789-800.
Doi:10.1172/jci32601
Boudina, S., Sena, S., Sloan, C., Tebbi, A., Han, Y. H., O’Neill, B. T., Cooksey,
R. C., Jones, D., Holland, W. L., McClain, D. A., Abe, E. D. (2012). Early
mitochondrial adaptations in skeletal muscle to diet-induced obesity are
strain dependent and determine oxidative stress and energy expenditure
but not insulin sensitivity. Endocrinology 153 (6), 2677 – 2688. Doi:
10.1210/en.2011-2147
Buchner, D. A., Yazbek, S. N., Solinas, P., Burrage, L. C., Morgan, M. G.,
Hoppel, C. L., Nadeau, J. H. (2011). Increased mitochondrial oxidative
phosphorylation in the liver is associated with obesity and insulin
resistance. Obesity 19 (5), 917 – 924. Doi:10.1038/oby.2010.214
Buettner, R., Scho¨lmerich, J., Bollheimer, L. C. (2007). High-fat diets: modeling
the metabolic disorders of human obesity in rodents. OBESITY 15 (4),
798-808.
Cai, L., Wang, Z., Ji, A., Meyer, J. M., van der Westhuyzen, D. R. (2012).
Scavenger receptor cd36 expression contributes to adipose tissue
inflammation and cell death in diet-induced obesity. Plos ONE 7 (5). Doi:
10.1371/journal.pone.0036785
Casanova E., Baselga-Escudero L., Ribas-Latre A., Cedó L., Arola-Arnal A.,
Pinent M., Bladé C., Arola L., Salvadó M. J. (2014). Chronic intake of
proanthocyanidins and docosahexaenoic acid improves skeletal muscle
oxidative capacity in diet-obese rats. Journal of Nutritional Biochemistry
25 (10), 1003-1010. Doi: 10.1016/j.jnutbio.2014.05.003
Castrogiovanni, P. & Imbesi, R. (2012). Oxidative stress and skeletal muscle in
exercise. Italian Journal of Anatomy and Embryology 117 (2), 107-116.
Cavill, N., Kahlmeier, S., Racioppi, F. (2008). Actividade Física e Saúde na
Europa. Evidências para a acção. Centro de Investigação em Actividade

38
Física, Saúde e Lazer. Porto, Faculdade de Desporto da Universidade
do Porto
Crescenzo R., Bianco F., Coppola P., Mazzoli A., Cigliano, L., Liverini, G.,
Lossa, S. (2015). The effect of high-fat–high-fructose diet on skeletal
muscle mitochondrial energetics in adult rats. European Journal of
Nutrition 54 (2), 183-192. Doi: 10.1007/s00394-014-0699-7
Crescenzo R., Bianco F., Coppola P., Mazzoli A., Valiante S., Liverini G., Lossa
S. (2013). Adipose tissue remodeling in rats exhibiting fructose-induced
obesity. European Journal of Nutrition 53 (2), 413-420. Doi:
10.1007/s00394-013-0538-2
Crunkhorn, S., Dearie, F., Mantzoros, C., Gami, H., da Silva, W. S., Espinoza,
D., Faucette, R., Barry, K., Bianco, A. C., Patti, M. E. (2007). Peroxisome
proliferator activator receptor gamma coactivator-1 expression is reduced
in obesity: potential pathogenic role of saturated fatty acids and p38
mitogen-activated protein kinase activation. Journal of Biological
Chemistry 282 (21), 15439-15550. Doi:10.1074/jbc.M611214200
Davidson, T. L., Hargrave, S. L., Swithers, S. E., Sample, C. H., Kinzig, K. P.,
Zheng, W. (2013). Inter-relationships among diet, obesity and
Hippocampal-dependent cognitive function. Neuroscience 253, 110 –
122. Doi: 10.1016/j.neuroscience.2013.08.044
European Society of Lifestyle Medicine (2015). Obesity and overweight are
leading risk factors for chronic diseases. Disponível em: http://eu-
lifestylemedicine.org/lifestyle-medicine-obesity-overweight/
Faeh, A. (2012). Obesity in Europe: The Strategy of the European Union from a
Public Health Law Perspective. European Journal of Health Law 19, 69-
86. Doi: 10.1163/157180912X615149
Farias, J. M., Bom, K. F., Tromm, C. B., Luciano, T. F., Marques, S. O., Tuon,
T., Silva, L. A.,. Lira, F. S. Souza, C. T., Pinho, R. A. (2013). Effect of
Physical Training on the Adipose Tissue of Diet-induced Obesity Mice:

39
Interaction Between Reactive Oxygen Species and Lipolysis. Hormone
Metabolic Research 45, 190 – 196. Doi: http://dx.doi.org/10.1055/s-0032-
1323740
Freeman, L. R., Haley-Zitlin, V., Rosenberger, D. S., Granholm, A. (2014).
Damaging effects of a high-fat diet to the brain and cognition: A review of
proposed mechanisms. Nutritional Neuroscience 17 (6), 241-251. Doi:
10.1179/1476830513Y.0000000092
Furukawa, S., Fujita, T., Shimabukuro, M., Iwaki,M., Yamada, Y., Nakajima, Y.,
Nakayama, O., Makishima, M., Matsuda, M., Shimomura, I. (2004).
Increased oxidative stress in obesity and its impact on metabolic
syndrome. The Journal of Clinical Investigation 114 (12), 1752-1761.
Gajda, A. M. (2015). Brief Scientific Literature Review- High Fat Diets for Diet-
Induced Obesity Models. Research Diets, Inc. Disponível em:
http://www.researchdiets.com/opensource-diets/diet-induced-disease-
models/obesity
Galic, S., Oakhill, J. S., Steinberg, G. R. (2010). Adipose tissue as an endocrine
organ. Molecular and Cellular Endocrinology, 316 (2), 129 – 139.
Doi:10.1016/j.mce.2009.08.018
Gollisch K., Brandauer J., Jessen N., Toyoda T., Nayer A., Hirshman M.,
Goodyear L. (2009). Effects of exercise training on subcutaneous and
visceral adipose tissue in normal - and high-fat diet-fed rats. American
Journal Of Physiology Endocrinology And Metabolism 297 (2), 495–504
Doi: 10.1152/ajpendo.90424.2008
Gómez-Pérez, T., Capllonch-Amer, G., Gianotti, M., Lladó, I., Proenza, A. M.
(2012). Long-term high-fat-diet feeding induces skeletal muscle
mitochondrial biogenesis in rats in a sexdependent and muscle-type
specific manner. Nutrition & Metabolism 9 (15), 1-10. Doi:10.1186/1743-
7075-9-15

40
Gonçalves, I. O., Maciel, E., Passos, E., Torrella, J. R., Rizo, D., Viscor, G.,
Rocha-Rodrigues, S., Santos-Alves, E., Domingues, M. R., Oliveira, P.,
Ascensão, A., Magalhães, J. (2014). Exercise alters liver mitochondria
phospholipidomic profile and mitochondrial activity in non-alcoholic
steatohepatitis. The International Journal of Biochemistry & Cell Biology
54, 163 – 173. Doi:10.1016/j.biocel.2014.07.011
Gonçalves, I. O., Passos, E., Rocha-Rodrigues, S., Diogo, C. V., Torrella, J. R.,
Rizo, D., Viscor, G., Santos-Alves, E., Marques-Aleixo, I., Oliveira, P. J.,
Ascensão, A., Magalhães, J. (2014). Physical exercise prevents and
mitigates non-alcoholic steatohepatitis-induced liver mitochondrial
structural and bioenergetics impairments. Mitochondrion 15, 40-51.
Doi:10.1016/j.mito.2014.03.012
Gonçalves, I. O., Passos, E., Rocha-Rodrigues, S., Torrella, J. R., Rizo, D.,
Santos-Alves, E., Portincasa, P., Martins, M. J., Ascensão, A.,
Magalhães, J. (2015). Physical exercise antagonizes clinical and
anatomical features characterizing Lieber-DeCarli diet-induced obesity
and related metabolic disorders. Clinical Nutrition 34 (2), 241-247.
Doi:10.1016/j.clnu.2014.03.010
Goto, A., Matsushima, Y., Kadowaki, T., andKitagawa, Y. (2001). Drosophila
mitochondrial transcription factor A (d-TFAM) is dispensable for the
transcription of mitochondrial DNA in Kc167 cells. Biocheminal Journal
354 (2), 243-248.
Gruber, L., Kisling, S., Lichti, P., Martin, F-P., May, S., Klingenspor, M.,
Lichtenegger, M., Rychlik, M., Haller, D. (2013). High fat diet accelerates
pathogenesis of murine crohn’s disease-like ileitis independently of
obesity. PLoS ONE 8 (8), 1-13. Doi:10.1371/journal.pone.0071661
Halliwell, B. & Cross, C. E. (1994). Oxygen-derived species: their relation to
human disease and environmental stress. Environmental Health
Perspectives 102 (10), 5-12.

41
Hariri, N. & Thibault, L. (2010). High-fat diet-induced obesity in animal models.
Nutrition Research Reviews 23, 270–299. Doi:
10.1017/S0954422410000168
Hood, D. A. (2009). Mechanisms of exercise-induced mitochondrial biogenesis
in skeletal muscle. Applied Physiology, Nutrition, and Metabolism 34 (3),
465-72. Doi:10.1139/h09-045
Hood, D. A., Uguccioni, G., Vainshtein, A., D'Souza, D. (2011). Mechanisms of
exercise-induced mitochondrial biogenesis in skeletal muscle:
implications for health and disease. Comprehensive Physiology 1 (3),
1119-34. Doi:10.1002/cphy.c100074
Hood, D.A., Irrcher, I., Ljubicic, V., Joseph A.M. (2006) Coordination of
metabolic plasticity in skeletal muscle. The Journal of Experimental
Biology 209 (12), 2265-2275.
Jain, S. S., Paglialunga, S., Vigna, C., Ludzki, A., Herbst, E. A., Lally, J. S.,
Schrauwen, P., Hoeks, J., Tupling, A. R., Bonen, A., Holloway, G. P.
(2014). High-fat diet-induced mitochondrial biogenesis is regulated by
mitochondrial-derived reactive oxygen species activation of CaMKII.
Diabetes 63 (6), 1907-1913. Doi:10.2337/db13-0816
Jayaraman, A., Lent-Schochet, D., Pike, C. (2014). Diet-induced obesity and
low testosterone increase neuroinflammation and impair neural function.
Journal of Neuroinflammation 11 (1), 1-32. Doi:10.1186/s12974-014-
0162-y
Jenkins, N. T., Padilla, J., Thorne, P. K., Martin, S. J., Rector, R. S., Davis, J.
W., Laughlin, M. H. (2014). Transcriptome-wide RNA sequencing
analysis of rat skeletal muscle feed arteries. I. Impact of obesity. Journal
of Applied Physiology, 116 (7-8), 1017-1032. Doi:
10.1152/japplphysiol.01233.2013

42
Jing, E., O'Neill, B. T., Rardin, M. J., Kleinridders, A., Ilkeyeva, O. R., Ussar, S.,
Bain, J. R., Lee, K. Y., Verdin, E. M., Newgard, C. B., Gibson, B. W.,
Kahn, C. R. (2013). Sirt3 regulates metabolic flexibility of skeletal muscle
through reversible enzymatic deacetylation. Diabetes 62 (10), 3404-
3417. Doi:10.2337/db12-1650
Kang, C. & Ji, L. L. (2012). Role of PGC-1α signaling in skeletal muscle health
and disease. New York Academy of Science 1271, 110-117. Doi:
10.1111/j.1749-6632.2012.06738.x
Kershaw, E. E. & Flier, J. S. (2004). Adipose Tissue as an Endocrine Organ.
The Journal of Clinical Endocrinology & Metabolism, 89 (6), 2548 – 2556.
Doi: 10.1210/jc.2004-0395
Kredel, L. I. & Siegmund, B. (2014). Adipose-tissue and intestinal inflammation
– visceral obesity and creeping fat. Frontiers in Immunology, 5 (462), 1-
12. Doi: 10.3389/fimmu.2014.00462
Lacerda-Miranda, G., Soares, V. M., Veira, A. K. G., Lessa, J. G., Rodrigues-
Cunha, A. C. S., Cortez, E., Garcia-Sousa, E. P., Moura, A. S. (2012).
Ghrelin signaling in heart remodeling of adult obese mice. Peptides, 35
(1), 65 – 73. Doi:10.1016/j.peptides.2012.02.025
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of
the head of bacteriophage T4. Nature, 227, 680-685.
Lafortuna, C. L., Tresoldi, D., Rizzo, G. (2014). Influence of body adiposity on
structural characteristics of skeletal muscle in men and women. Clinical
Physiology & Functional Imaging, 34 (1), 47-56. Doi: 10.1111/cpf.12062
Lagowska, K. & Jeszka, J. (2011). Adipose tissue as an endocrine organ.
Medicina Sportiva,15 (3), 140-146. Doi: 10.2478/v10036-011-0024-y
Larsen, S., Nielsen, J., Hansen, C. N., Nielsen, L. B., Wibrand, F., Stride, N.,
Schroder, H. D., Boushel, R., Helge, J. W., Dela, F., Hey-Mogensen, M.
(2012). Biomarkers of mitochondrial content in skeletal muscle of healthy

43
young human subjects. Journal of Physiology 590 (14), 3349-60.
Doi:10.1113/jphysiol.2012.230185
Larter, C. Z., C. Z., Yeh, M. M., Haigh, W. G., Van Rooyen, D. M., Brooling, J.,
Heydet D., Nolan, C. J., Teoh, N. C., Farrell, G. C. (2013). Dietary
modification dampens liver inflammation and fibrosis in obesity-related
fatty liver disease. Obesity, 21 (6), 1189-1199. Doi: 10.1002/oby.20123
Lehnert, T., Sonntag, D., Konnopka, A., Riedel-Heller, S., König, H-H. (2013).
Economic costs of overweight and obesity. Best Practice & Research
Clinical Endocrinology & Metabolism, 27 (2), 105-
115. http://dx.doi.org/10.1016/j.beem.2013.01.002
Leick, L., Wojtaszewski, J.F., Johansen, S.T., Kiilerich, K., Comes, G., Hellsten,
Y., Hidalgo, J., andPilegaard, H. 2008. PGC-1alpha is not mandatory for
exercise- and training-induced adaptive gene responses in mouse
skeletal muscle. American Journal of Physiology Endocrinology
Metabolism 294 (2), 463-474. Doi:10.1152/ajpendo.00666.2007
Liang, H. & Ward, W. F. (2006). PGC-1α: a key regulator of energy metabolism.
Advances in Physiology Education 30, 145 – 151.
Doi:10.1152/advan.00052.2006.
Lieber, C. S., Leo, M. A., Mak, K. M., Xu, Y., Cao, Q., Ren, C., Ponomarenko,
A., DeCarli, L. M. (2004). Model of nonalcoholic steatohepatitis.
American Journal of Clinical Nutrition, 79, 502 – 509.
Lin, J., Handschin, C., Spiegelman, B. M. (2005). Metabolic control through the
PGC- 1 family of transcription coactivators. Cell Metabolism 1 (6), 361-
370
Lin, L., Chen, K., Abdel Khalek, W., Ward, J. L., 3rd, Yang, H., Chabi, B.,
Wrutniak-Cabello, C., Tong, Q. (2014). Regulation of skeletal muscle
oxidative capacity and muscle mass by SIRT3. PLoS One 9 (1), e85636.
Doi:10.1371/journal.pone.0085636

44
Lindinger, P. W., Christe, M., Eberle, A. N., Kern, B., Peterli, R., Peters, T.,
Jayawardene, K. J. I., Fearnley, I. M., Walker, J. E. (2015). Important
mitochondrial proteins in human omental adipose tissue show reduced
expression in obesity. Jornal of Proteomics. Doi:
10.1016/j.jprot.2015.03.037
Lumini-Oliveira, J., Magalhaes, J., Pereira, C. V., Moreira, A. C., Oliveira, P. J.,
Ascensao, A. (2011). Endurance training reverts heart mitochondrial
dysfunction, permeability transition and apoptotic signaling in long-term
severe hyperglycemia. Mitochondrion 11 (1), 54-63. Doi:
10.1016/j.mito.2010.07.005
Machado, M. V., Duarte M. S. Ferreira, D. M. S., Castro, R. E., Silvestre, A. R.,
Evangelista, T., Coutinho, J., Carepa, F., Costa, A., Rodrigues, C. M. P.,
Pinto, H. C. (2012). Liver and muscle in morbid obesity: the interplay of
fatty liver and insulin resistance. PLoS ONE, 7 (2).
Doi:10.1371/journal.pone.0031738
Menshikova, E. V., Ritov, V. B., Ferrell, R. E., Azuma K., Goodpaster, B. H.,
Kelley, D. E. (2007). Characteristics of skeletal muscle mitochondrial
biogenesis induced by moderate-intensity exercise and weight loss in
obesity. Journal of Applied Physiology, 103, 21-27.
Doi:10.1152/japplphysiol.01228.2006
Mogoi, M., Ilie, C., Paul, C., Velea J. P. (2014). The evaluation of obesity-
related cardiometabolic diseases: a simple anthropometric tool for a
complicated matter. Romanian Journal of Pediatrics, 63 (3), 259-325.
Muthulakshmi, S. & Saravanan, S. (2013). Protective effects of azelaic acid
against high-fat diet-induced oxidative stress in liver, kidney and heart of
C57BL/6J mice. Molecular and Cellular Biochemistry, 377 (1-2), 23-33.
Doi: 10.1007/s11010-013-1566-1
Navarro, A., Gomez, C., Lopez-Cepero, J. M., Boveris, A. (2004). Beneficial
effects of moderate exercise on mice aging: survival, behavior, oxidative

45
stress, and mitochondrial electron transfer. American Journal of
Physiology Regulatory Integrative and Comparative Physiology 286 (3),
R505-511. Doi: 10.1152/ajpregu.00208.2003
Negrin, K. A., Flach, R. J. R., DiStefano, M. T., Matevossian, A., Friedline, R.
H., Jung, DaeY., Kim, J. K., Czech, M. P. (2014). IL-1 signaling in
obesity-induced hepatic lipogenesis and steatosis. PLoS ONE, 9 (9), 1-
15. Doi:10.1371/journal.pone.0107265
Ngo, H. T., Hetland, R. B., Nygaard, U. C., Steffensen, I-L. (2015). Genetic and
diet-induced obesity increased intestinal tumorigenesis in the double
mutant mouse model multiple intestinal neoplasia x obese via disturbed
glucose regulation and inflammation. Journal of Obesity, 2015, 1-21.
Doi:10.1155/2015/343479
Nojima, H., Watanabe, H., Yamane, K., Kitahara, Y., Sekikawa, K., Yamamoto,
H., Yokoyama, A., Inamizu, T., Asahara, T., Kohno, N. (2008). Effect of
aerobic exercise training on oxidative stress in patients with type 2
diabetes mellitus. Metabolism 57 (2), 170-176.
Doi:10.1016/j.metabol.2007.08.021
Oh, S., Tanaka, K., Warabi, E., Shoda, J.(2013). Exercise reduces inflammation
and oxidative stress in obesity-related liver diseases. Medicine & Science
in Sports & Exercise 45 (12), 2214-2222.
Doi:10.1249/MSS.0b013e31829afc33
Ojeda, P., Bobe, A., Dolan, K., Leone, V., Martinez, K. (2015). Nutritional
modulation of gut microbiota — the impact on metabolic disease
pathophysiology. Journal of Nutritional Biochemistry,
Doi.org/10.1016/j.jnutbio.2015.08.013
Paglialunga, S., Ludzki, A., Root-McCaig, J., Holloway G. P. (2015). In adipose
tissue, increased mitochondrial emission of reactive oxygen species is
important for short-term high-fat diet-induced insulin resistance in mice.
Diabetologia, 58, 1071 – 1080. Doi: 10.1007/s00125-015-3531-x

46
Palacios, O. M., Carmona, J. J., Michan, S., Chen, K. Y., Manabe, Y., Ward III,
J. L., Goodyear, L. J., Tong, Q. (2009). Diet and exercise signals regulate
SIRT3 and activate AMPK and PGC‐1α in skeletal muscle. Aging 1 (9),
771-783.
Passos, E., Pereira, C.D., Gonçalves, I. O., Rocha-Rodrigues, S., Silva, N.,
Guimarães, J. T., Neves, D., Ascensão, A., Magalhães, J., Martins, M. J.
(2015). Role of physical exercise on hepatic insulin, glucocorticoid and
inflammatory signaling pathways in an animal model of non-alcoholic
steatohepatitis. Life Sciences, 123, 51-60. Doi:10.1016/j.lfs.2014.12.013
Peters, R. (2014). Case Studies of Schools Exemplifying Wellness and Lifetime
Fitness Models. International Journal of Sport & Society, 4 (3), 55-69.
Pimenta, M., Bringhenti, I., Souza-Mello, V., Dos Santos Mendes, I. K., Aguila,
M. B., Mandarim-de-Lacerda, C. A. (2015). High-intensity interval training
beneficial effects on body mass, blood pressure, and oxidative stress in
diet-induced obesity in ovariectomized mice. Life Science 139, 75-82.
Doi:10.1016/j.lfs.2015.08.004
Rito, A. (2012) Childhood Obesity Surveillance Iniciative - COSI Portugal 2010.
Boletim Epidemiológico do Instituto de Nacional de Saúde Doutor
Ricardo Jorge, 1, 2ª série, 6.
Romacho, T., Elsen, M., Rohrborn, D., Eckel, J. (2014). Adipose tissue and its
role in organ crosstalk. Acta Physiologica, 210 (4), 733-753.
Rosini, T., Silva, A., Moraes, C. (2012). Obesidade induzida por consumo de
dieta: modelo em roedores para o estudo dos distúrbios relacionados
com a obesidade. Revista da Associação Médica Brasileira; 58 (3), 383-
387.
Ross, R., Hudson, R., Stotz, P. J., Lam, M. (2015). Effects of exercise amount
and intensity on abdominal obesity and glucose tolerance in obese
adults. Annals of internal medicine, 162 (5), 325-334. Doi: 10.7326/M14-
1189

47
Ross, R., Janssen, I., Dawson, J., Kungl, A. M., Kuk, J. L., Wong, S. L.,
Nguyen-Duy, T. B., Lee, S., Kilpatrick, K., Hudson, R. (2004). Exercise-
induced reduction in obesity and insulin resistance in women: a
randomized controlled trial. Obesity Research 12 (5), 789-798.
Doi:10.1038/oby.2004.9
Samjoo, I. A., Safdar, A., Hamadeh, M. J., Raha, S., Tarnopolsky, M. A. (2013).
The effect of endurance exercise on both skeletal muscle and systemic
oxidative stress in previously sedentary obese men. Nutrition & Diabetes
3, e88. Doi:10.1038/nutd.2013.30
Sarzani, R., Bordicchia, M., Spannella, F., Dessi-Fulgheri, P., Fedecostante, M.
(2014). Hypertensive Heart Disease and Obesity: A complex interaction
between hemodynamic and not hemodynamic factors. High Blood
Pressure & Cardiovascular Prevention, 21 (2), 81-87. Doi:
10.1007/s40292-014-0054-3
Schmidt, S. (2012). Obesity and Exercise. American College of Sports
Medicine. Disponível em: https://www.acsm.org/access-public-
information/articles/2012/01/19/obesity-and-exercise
Sheikhpour, R. & Khoradmehr, A. (2014). New insights into the effect of
diabetes and obesity in alzheimer’s disease. Iranian Journal of Diabetes
and Obesity, 6 (1), 41-45.
Silva, C. (2014). Biogênese mitocondrial e marcadores de estresse oxidativo
em ratos treinados e suplementados com larginina. Dissertação de
Mestrado em Ciências Da Motricidade – Biodinâmica Da Motricidade
Humana, UNESP.
Silveira, L.R., Pilegaard, H., Kusuhara, K., Curi, R., Hellsten, Y. (2006). The
contraction induced increase in gene expression of peroxisome
proliferator-activated receptor (PPAR)-gamma coactivator 1alpha (PGC-
1alpha), mitochondrial uncoupling protein 3 (UCP3) and hexokinase II

48
(HKII) in primary rat skeletal muscle cells is dependent on reactive
oxygen species. Biochimica et Biophysica Acta 1763 (9), 969-976.
Skovbro M., Boushel R., Hansen C. H., Helge J. W. (2011). High-fat feeding
inhibits exercise-induced increase in mitochondrial respiratory flux in
skeletal muscle. Journal of Applied Physiology, 110, 1607-1614 Doi:
10.1152/japplphysiol.01341.2010
Sparks, L. M., Xie, H., Koza, R. A., Mynatt, R., Hulver, M. W., Bray, G. A.,
Smith, S. R. (2005). A high-fat diet coordinately downregulates genes
required for mitochondrial oxidative phosphorylation in skeletal muscle.
Diabetes 54 (7), 1926-1933.
Standford, K. I. & Goodyear, L. J. (2014). Exercise and type 2 diabetes:
molecular mechanisms regulating glucose uptake in skeletal muscle.
Advances in Physiology Education 38, 308-314.
Doi:10.1152/advan.00080.2014
Stanković, M. & Radovanović, D. (2012). Oxidative stress and physical activity.
SportLogia 8 (1), 1 – 11. Doi: 10.5550/sgia.120801.en.001S
Steinbacher, P. & Eckl, P. (2015). Impact of oxidative stress on exercising
skeletal muscle. Biomolecules 5, 356-377. Doi:10.3390/biom5020356
St-Pierre, J., Drori, S., Uldry, M., Silvaggi, J. M., Rhee, J., Jager, S., Handschin,
C., Zheng, K., Lin, J., Yang, W., Simon, D. K., Bachoo, R., Spiegelman,
B. M. (2006). Suppression of reactive oxygen species and
neurodegeneration by the PGC-1 transcriptional coactivators. Cell 127
(2), 397-408. Doi:10.1016/j.cell.2006.09.024
St-Pierre, J., Lin, J., Krauss, S., Tarr, P. T., Yang, R., Newgard, C. B.,
Spiegelman, B. M. (2003). Bioenergetic analysis of peroxisome
proliferator-activated receptor gamma coactivators 1alpha and 1beta
(PGC-1alpha and PGC-1beta) in muscle cells. Journal of Biological
Chemistry 278 (29), 26597-26603. Doi:10.1074/jbc.M301850200

49
Strobel, N. A., Matsumoto, A., Peake, J. M., Marsh, S. A., Peternelj, T. T.,
Briskey, D., Fassett, R. G., Coombes, J. S., Wadley, G. D. (2014).
Altering the redox state of skeletal muscle by glutathione depletion
increases the exercise-activation of PGC-1alpha. Physiological Reports 2
(12), e12224. Doi:10.14814/phy2.12224
Sun, L., Shen, W., Liu, Z., Guan, S., Liu, J., Ding, S. (2010). Endurance
exercise causes mitochondrial and oxidative stress in rat liver: effects of
a combination of mitochondrial targeting nutrients. Life Sciences 86 (1-2),
39-44. Doi: 10.1016/j.lfs.2009.11.003
Sundaresan, R. N., Samant S. A., Pillai, V. B., Rajamohan, S. B., Gupta, M. P.
(2008). Sirt3 is a stress-responsive deacetylase in cardiomyocytes that
protects cells from stress-mediated cell death by deacetylation of ku70.
Molecular And Cellular Biology 28 (20), 6384 – 6401.
Doi:10.1128/MCB.00426-08
Tadaish, M., Miura, S., Ka, Y., Kano, Y., Oish, Y., Ezaki, O. (2011). Skeletal
muscle-specific expression of pgc-1a-b, na exercise-responsive isoform,
increases exercise capacity and peak oxygen uptake. PLoS ONE, 6 (12),
e28290. Doi:10.1371/journal.pone.0028290
Valle, I., Alvarez-Barrientos, A., Arza, E., Lamas, S., Monsalve, M. (2005).
PGC-1alpha regulates the mitochondrial antioxidant defense system in
vascular endothelial cells. Cardiovascular Research 66 (3), 562-573.
Doi:10.1016/j.cardiores.2005.01.026
Von Lengerke, T. & Krauth, C. (2011). Economic costs of adult obesity: A
review of recent European studies with a focus on subgroup-specific
costs. Maturitas, 69, 220 – 229.
Wang, J., Freire, D., Knable. L., Zhao, W., Gong, B., Mazzola, P., Ho, L.,
Levine, S., Pasinetti, G. M. (2015). Childhood and adolescent obesity
and long-term cognitive consequences during aging. The Journal of
Comparative Neurology, 523 (5), 757-68. Doi: 10.1002/cne.23708

50
Wang, Y., Ausman, L. M., Russell, R. M., Greenberg, A. S., Wang, X-D. (2009).
Increased apoptosis in high-fat diet–induced nonalcoholic steatohepatitis
in rats is associated with c-jun nh2-terminal kinase activation and
elevated proapoptotic bax. Journal of Nutrition, 138 (10), 1866 – 1871.
WHO (2015). Obesity and overweight. Disponível em:
http://www.who.int/mediacentre/factsheets/fs311/en/
WHO/Europe (2007). Childhood obesity surveillance in the WHO European
Region, 5. Disponível em: http://www.euro.who.int/en/health-
topics/noncommunicable-diseases/obesity/publications/pre-2009/fact-
sheet-5-childhood-obesity-surveillance-in-the-who-european-region
WHO/Europe (2013). Country profiles on nutrition, physical activity and obesity
in the 53 WHO European Region Member States. WHO Regional Office
for Europe, disponível em: http://www.euro.who.int/pubrequest
Willett, W.C., Koplan, J. P., Nugent, R., Dusenbury, C., Puska, P., Gaziano, T.
A. (2006). Prevention of chronic disease by means of diet and lifestyle
changes. Disease Control Priorities in Developing Countries 44, 833-850.
Woo, S-L., Xu, H., Li, H., Zhao, Y., Hu, X., Zhao, J., Guo, X., Guo, T., Botchlett,
R., Qi, T., Pei, Y., Zheng, J., Xu, Y., Na, X., Chen, L., Chen, L., Li, Q.,
Xiao, X., Huo, Y., Wu, C. (2014). Metformin ameliorates hepatic steatosis
and inflammation without altering adipose phenotype in diet-induced
obesity. PLoS ONE, 9 (3), 1-10. Doi:10.1371/journal.pone.0091111
World Health Organization – WHO (2013). Global action plan for the prevention
and control of noncommunicable diseases 2013-2020. Geneve,
Switzerland: WHO.
Yim, E. H. & Yoo, H. K. (2015). Early life obesity and chronic kidney disease in
later life. Pediatric Nephrology, 30, 1255 – 1263. Doi: 10.1007/s00467-
014-2922-4

51
Yokota, T., Kinugawa, S., Hirabayashi, K., Matsushima, S., Inoue, N., Ohta, Y.,
Hamaguchi, S., Sobirin, M. A., Ono, T., Suga, T., Kuroda, S., Tanaka, S.,
Terasaki, F., Okita, K., Tsutsui, H. (2009). Oxidative stress in skeletal
muscle impairs mitochondrial respiration and limits exercise capacity in
type 2 diabetic mice. American Journal Physiology: Heart and Circulation
Physiology, 297, 1069–1077. Doi:10.1152/ajpheart.00267.2009





























