Embed Size (px)
Citation preview

1
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E
ZOOTECNIA
Programa de Pós-graduação em Agricultura Tropical
EPIDEMIOLOGIA, CONTROLE QUÍMICO E
DISTRIBUIÇÃO ESPACIAL DE Sclerotinia sclerotiorum EM
MATO GROSSO
ROSELI MUNIZ GIACHINI
CUIABÁ-MT
2013

2
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E
ZOOTECNIA
Programa de Pós-graduação em Agricultura Tropical
EPIDEMIOLOGIA, CONTROLE QUÍMICO E
DISTRIBUIÇÃO ESPACIAL DE Sclerotinia sclerotiorum EM
MATO GROSSO
ROSELI MUNIZ GIACHINI
Engenheira Agrônoma
Orientadora: Profª. Drª. MARIA CRISTINA DE F. E ALBUQUERQUE
Co-Orientadores: Prof. Dr. DANIEL CASSETARI NETO
Prof. Dr. MARCELO DE CARVALHO ALVES
CUIABÁ-MT
2013
Tese apresentada à Faculdade de Agronomia, Medicina Veterinária e Zootecnia da Universidade Federal de Mato Grosso para obtenção do título de Doutora em Agricultura Tropical.

3
FICHA CATALOGRÁFICA

4

5
A meu esposo Vilmar
Aos meus filhos Fernando e Fabiola
Aos meus pais Ladir Lau e Leonor
pelo apoio e força nos momentos difíceis
Dedico...........

6
AGRADECIMENTOS
À Universidade Federal de Mato Grosso (UFMT), pela oportunidade da
realização do curso.
À Faculdade de Agronomia, Medicina Veterinária e Zootecnia (FAMEVZ)
pelas condições oferecidas na realização do curso.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
pelo apoio financeiro na concessão da bolsa.
À professora Maria Cristina de Figueiredo e Albuquerque, minha
orientadora, pela dedicação, orientação e amizade.
Aos professores Daniel Cassetari Neto e Marcelo de Carvalho Alves, meus
co-orientadores, pela amizade, sugestões e apoio na realização deste trabalho.
A todos os colegas do curso de Pós-graduação em Agricultura Tropical,
pela amizade e alegria no período de convivência.
Aos amigos inseparáveis do Laboratório em especial a Diele, Carmen, Zé
Luis, Marcela, Claudinéia, Rafael, e estagiários, pela amizade, companheirismo e
ajuda na realização dos testes.
Aos funcionários e amigos do Laboratório, Sidnéa (amiga de toda hora) e
Sr. João, pela colaboração e constante amizade.
Aos meus familiares, que nos momentos mais difíceis sempre me apoiaram
para que eu não desistisse.
Ao meu esposo Vilmar e meus filhos Fernando e Fabiola pela paciência,
dedicação, companheirismo, apoio e compreensão nos momentos em que estive
ausente.
À Deus acima de tudo, por ter me guiado até aqui, concebendo a realização
de mais um de meus objetivos.
...Obrigado

7
SUMÁRIO
Página
1 INTRODUÇÃO ....................................................................................... 11
2 REVISÃO DE LITERATURA ............................................................. 14
2.1 Podridão na Haste ou Mofo Branco na Cultura da Soja causada por
Sclerotinia sclerotiorum ............................................................................. 14
2.2 Taxonomia............................................................................................ 16
2.3 Aspectos Biológicos ............................................................................ 16
2.3.1 Produção de escleródios .................................................................. 16
2.3.2 Germinação dos escleródios ............................................................ 18
2.4 Mecanismo de Patogenicidade: Produção de Ácido Oxálico .............. 21
2.5 Sintomas e Epidemiologia.................................................................... 22
2.6 Variabilidade do Patógeno .................................................................. 26
2.7 Métodos de Detecção de Sclerotinia sclerotiorum............................... 27
2.8 Controle de Sclerotinia sclerotiorum..................................................... 30
2.9 Geoestatística....................................................................................... 33
2.10 Ocorrência de S. sclerotiorum em Mato Grosso ............................... 34
2.11 Referências Bibliográficas.................................................................. 36
3 CARACTERÍSTICAS MORFOLÓGICAS E FISIOLÓGICAS DE
Sclerotinia sclerotiorum ISOLADO DA CULTURA DA SOJA E
USO DA RESTRIÇÃO HÍDRICA ........................................................ 46
Resumo..................................................................................................... 46
Abstract...................................................................................................... 47
3.1 Introdução............................................................................................ 48
3.2 Material e Métodos............................................................................... 50
3.3 Resultados e Discussão....................................................................... 56
3.4 Conclusões........................................................................................... 70
3.5 Referências Bibliográficas.................................................................... 71
4 EFICIÊNCIA DE FUNGICIDAS NO CONTROLE DE MOFO BRANCO
DA CULTURA DA SOJA E NA GERMINAÇÃO DE ESCLERÓDIOS
DE Sclerotinia sclerotiorum............................................................ 75

8
Resumo...................................................................................................... 75
Abstract...................................................................................................... 76
4.1 Introdução............................................................................................ 77
4.2 Material e Métodos............................................................................... 79
4.3 Resultados e Discussão....................................................................... 83
4.4 Conclusões........................................................................................... 91
4.5 Referências Bibliográficas.................................................................... 92
5 EFEITO DA TEMPERATURA E UMIDADE EM Sclerotinia
sclerotiorum DA SOJA ...................................................................... 95
Resumo...................................................................................................... 95
Abstract....................................................................................................... 96
5.1 Introdução ............................................................................................ 97
5.2 Material e Métodos .............................................................................. 99
5.3 Resultados e Discussão ...................................................................... 102
5.4 Conclusões .......................................................................................... 113
5.5 Referências Bibliográficas................................................................... 114
6 DISTRIBUIÇÃO ESPACIAL DE Sclerotinia sclerotiorum EM
SOJA..................................................................................................... 117
Resumo...................................................................................................... 117
Abstratc...................................................................................................... 118
6.1 Introdução ........................................................................................... 119
6.2 Material e Métodos .............................................................................. 121
6.3 Resultados e Discussão ...................................................................... 124
6.4 Conclusões ......................................................................................... 134
6.5 Referências Bibliográficas.................................................................. 135

9
EPIMEMIOLOGIA, CONTROLE QUÍMICO E DISTRIBUIÇÃO ESPACIAL
DE Sclerotinia sclerotiorum EM MATO GROSSO
RESUMO - Na cultura da soja, o mofo branco, causado pelo fungo por S.
sclerotiorum, que sempre foi de ocorrência esporádica em áreas de cultivo
no estado de Mato Grosso, não havia causado problemas até a safra
2008/09, pois ocorria em pequenas reboleiras, com poucas plantas
infectadas por hectare. Uma série de fatores como monocultivo, sucessão
de culturas com espécies altamente suscetíveis e o plantio de sementes
infectadas, fizeram com que a doença se tornasse um dos principais
problemas, com perdas significativas na produtividade de soja. Assim,
objetivou-se, neste trabalho, estudar o fungo S. sclerotiorum e os fatores
que influenciam no seu crescimento. Dessa forma, foram verificados (1) a
biometria e germinação de escleródios; (2) inoculação e detecção do
patógeno em sementes; (3) a sensibilidade de escleródios à fungicidas; (4)
o crescimento e desenvolvimento do patógeno em função das temperaturas
e disponibilidade hídrica; e (5) a distribuição espacial de S. sclerotiorum. O
comprimento, a largura e a espessura dos escleródios em soja variam de
2,41 a 13,62 mm; 2,06 a 3,46 mm e 1,58 a 2,38 mm, respectivamente. O
meio mais adequado para inoculação em sementes de soja é o Batata-
Dextrose-Agar sem restrição hídrica. O período mais eficiente para a
inoculação é de 72 horas. O método rolo de papel permite a detecção mais
rápida e eficiente do patógeno em lotes de sementes de soja com 5% de
contaminação. Os fungicidas apresentam eficiência na redução do mofo
branco, mesmo com apenas uma aplicação. O efeito da aplicação de
fungicida na cultura da soja sobre a germinação de escleródios e formação
de apotécios ocorre até os 50 dias de incubação de escleródios. A umidade
do solo afeta a germinação carpogênica, o número de apotécios produzidos
por escleródios, a massa, comprimento e diâmetro dos apotécios. A
variação espacial de S. sclerotiorum ocorre em reboleiras.
Palavras-chave: mofo branco, escleródios, apotécios.

10
EPIDEMIOLOGY, CHEMICAL CONTROL AND SPATIAL DRISTRIBUTION
OF Sclerotinia sclerotiorum IN MATO GROSSO
ABSTRACT - In soybean, the white mold, caused by S. sclerotiorum, which
has always been sporadic in areas of soybean cultivation in the state of
Mato Grosso, there were problems caused by the 2008/09 as occurred in
small foci, with few infected plants per hectare. A number of factors such as
monoculture, crop succession with highly susceptible species and planting of
infected seed, caused the disease became a major problem, with significant
losses in soybean yield. Thus, the aim of this work was to study S.
sclerotiorum and the factors that influence its growth. Thus, it was verified
(1) biometrics and germination of sclerotia, (2) and detection of the pathogen
inoculation of seeds, (3) the sensitivity of the sclerotia fungicides, (4) growth
and development of the pathogen as a function of temperature and water
availability, and (5) the spatial distribution of S. sclerotiorum. The length,
width and thickness of sclerotia in soybean varies from 2.41 to 13.62 mm,
2.06 to 3.46 mm 1.58 to 2.38 mm, respectively. The most appropriate means
of inoculation in soybean seeds is the Potato Dextrose Agar without fluid
restriction. The most effective period for inoculation is 72 hours. The paper
roll method allowed the detection faster and more efficient pathogen in
batches of seeds with 5% contamination. Fungicides have efficiency in
reducing white mold, even with just one application. The effect of fungicide
application on soybean on germination of sclerotia and apothecia formation
occurs up to 50 days of incubation of sclerotia. Soil moisture affects the
germination carpogenic, the number of apothecia produced by sclerotia,
mass, length and diameter of the apothecia. The spatial variation of S.
sclerotiorum occurred in foci.
Keywords: white mold, sclerotia, apothecia.

11
1 INTRODUCÃO
A soja (Glycine max L. Merrill) é a principal espécie cultivada no país.
Na região Centro-Oeste, o Mato Grosso continua líder no ranking de
produção nacional, com estimativa de safra em 21,5 milhões de toneladas
obtidas em 2011/12, com área plantada de 7,89 milhões de hectares e
produção estimada de 24,13 milhões de toneladas para a safra 2012/13
(Imea, 2012). Dentre os fatores que limitam a produtividade, a lucratividade
e o sucesso de produção desta leguminosa destacam-se as doenças
(Juliatti et al., 2004).
Na cultura da soja, o mofo branco, causado pelo fungo Sclerotinia
sclerotiorum (Lib.) de Bary, que sempre foi de ocorrência esporádica em
áreas de cultivo no estado de Mato Grosso, não havia causado problemas
até a safra 2008/09, pois ocorria em pequenas reboleiras, com poucas
plantas infectadas por hectare, na região sul do Estado e em pivôs centrais.
Dessa forma, não havia perdas significativas que justificassem a
preocupação dos agricultores (Silva et al., 2010). Em função de uma série
de fatores como monocultivo, sucessão de culturas com espécies altamente
suscetíveis tais como feijão, girassol, algodão, e o plantio de sementes
infectadas, fizeram com que a doença se tornasse um dos principais
problemas no sul do estado de Mato Grosso na safra 2009/10. Na safra
2009/10, foram constatadas perdas significativas na produtividade de soja
na região.

12
O fungo S. sclerotiorum é considerado um dos fitopatógenos mais
importantes no mundo e está distribuído em todas as regiões produtoras de
soja, sejam elas de regiões temperadas, subtropicais ou tropicais (Leite,
2005). Em regiões de altitudes elevadas, normalmente acima de 800 m,
onde as temperaturas noturnas são amenas e ocorre ampla formação de
orvalho, tem sido comum observar áreas com incidência da doença superior
a 50%.
O controle do mofo branco é considerado difícil em função da
ausência de cultivares resistentes, sobrevivência do fungo no solo por
longos períodos, ampla gama de hospedeiros, grande número de
ascósporos produzidos pelo apotécio e sua rápida disseminação a distância
relativamente longa a partir da fonte produtora e, sobrevivência em
sementes na forma de micélio dormente ou escleródios aderidos ou em
misturas às mesmas.
O fato de o controle ser considerado difícil, como exposto acima,
torna-se necessário que várias medidas sejam adotadas. Entretanto,
segundo McDonald (1997), os sistemas agrícolas impõem um forte
direcionamento de seleção em populações de patógenos para adaptar
constantemente as mudanças em seu ambiente, garantindo sua
sobrevivência. Estas imposições podem ser em decorrência de mudanças
climáticas, aplicações de fungicidas, rotação de culturas, entre outras. No
Brasil, fungicidas para controle de mofo branco em soja só foram liberados
em 2011, pelo Ministério da Agricultura, Pecuária e Abastecimento (MAPA),
com os princípios ativos Fluazinam e Procimidona, com custo elevado para
a cultura da soja.
De modo geral, os trabalhos com mofo branco na cultura da soja são
necessários devido a importância do patógeno, por isso, é primordial que
pesquisas nessa área de conhecimento continuem acontecendo, como
estudar o desenvolvimento do patógeno a partir da sua estrutura de
resistência, avaliar métodos de detecção do patógeno na semente para
quantificar o percentual de contaminação ou infecção e estudar o efeito de

13
fungicidas sobre escleródios de S. sclerotiorum para melhor direcionamento
de medidas integradas de controle.
Diante disso, o objetivo neste trabalho foi estudar o fungo S.
sclerotiorum e os fatores que influenciam no seu crescimento como: (1) a
biometria e germinação de escleródios; (2) restrição, inoculação e detecção
do patógeno em sementes de soja; (3) a sensibilidade de escleródios à
fungicidas; (4) o crescimento e desenvolvimento do patógeno em função
das temperaturas de incubação e disponibilidade hídrica e (5) a distribuição
espacial de S. sclerotiorum.

14
2 REVISÃO DE LITERATURA
2. 1 Podridão na Haste ou Mofo Branco na Cultura de Soja causada
por Sclerotinia sclerotiorum
O fungo Sclerotinia sclerotiorum é considerado um dos patógenos
mais importantes no mundo e está distribuído em todas as regiões
produtoras, sejam temperadas, subtropicais ou tropicais. O mofo branco
causado pelo fungo S. sclerotiorum (Lib.) de Bary é conhecido e estudado
desde 1837 (Bolton et al., 2006). Entretanto, no Brasil, o primeiro relato da
doença ocorreu em 1921, por Saccá, que diagnosticou o fungo em plantas
de batata (Solanum tuberosum L.), no Estado de São Paulo. Nos anos
seguintes o patógeno foi constatado em diferentes hospedeiros em outros
estados do país (Chaves, 1964).
O fungo afeta várias espécies de plantas, sendo 408 espécies
suscetíveis, pertencentes a 278 gêneros e 75 famílias (Boland e Hall, 1994).
Algumas culturas como a batata (Solanum tuberosum L.), tomate (Solanum
lycopersicum Mill.), ervilha (Pisum sativum L.), amendoim (Arachis hypogea
L.), girassol (Helianthus annuus L.), alface (Lactuca sativa L.) (Purdy, 1979),
e espécies de plantas daninhas como picão (Bidens pilosa), carrapicho
(Acanthospermun hispidum), caruru (Amaranthus spp) e mentrasto
(Ageratum conyzoides) são suscetíveis à S. sclerotiorum (Paula Júnior et
al., 2006).
Com a expansão da fronteira agrícola na região dos cerrados, e com
a agricultura praticada no período frio do ano e sob sistemas irrigados, esta
doença se destacou na cultura do feijão e da ervilha, pois o patógeno
encontrou temperaturas amenas e umidade alta para seu estabelecimento e
desenvolvimento (Café-Filho, 1985; Santos et al., 1990). A doença na
cultura da soja só foi constatada no Brasil em 1975, no Estado do Paraná,
causando perdas de até 70% em lavouras destinadas a produção de
sementes (Ferreira et al., 1981). Neste período, as perdas da produção de
soja devido a doença em lavouras comerciais nos municípios de Castro,

15
Ponta Grossa, Palmeira e Guarapuava, no estado do Paraná, variaram de
70 a 92% do rendimento de grãos, com incidência variando de 20 a 90%
(Homechin, 1982).
No início da década de 1980, foram registradas perdas de até 30%
em lavouras de soja nos municípios de São Gotardo, Rio Parnaíba, e
Carmo do Parnaíba, em Minas Gerais, devido a grande ocorrência de S.
sclerotiorum (Nasser et al., 1984). Em Goiás, os primeiros relatos da doença
aconteceram na safra de 2001/02. Na safra de 2005/06 algumas lavouras
apresentaram altas incidências chegando a ocorrer perdas de até 33%; na
safra de 2007/08, foi observada presença da doença em quase todas as
áreas de cultivo da leguminosa (Görgen, 2009). A doença aumentou
consideravelmente, afetando cerca de 45% da área cultivada na safra de
2009/10 (Campos et al., 2010). De modo geral, nas regiões sudoeste, leste
de Goiás e entorno do Distrito Federal, as perdas já alcançaram 60% nas
produtividades (Nunes Júnior et al., 2009). Perdas de até 60% da
produtividade, tanto em áreas experimentais, quanto dos próprios
agricultores têm sido constatadas no Sudoeste Goiano.
O mofo branco vem, comprovadamente, trazendo prejuízos
significativos aos sojicultores de vários estados produtores, como Minas
Gerais, Goiás, São Paulo, Paraná, Mato Grosso do Sul e Bahia (Furlan,
2008).
No estado de Minas Gerais, a doença que proporcionou principal
atenção na safra de 2007/08 foi o mofo branco, devido a sua alta incidência
nas áreas acima de 900 m de altitude (Zanetti, 2009). Na região de
Chapadão do Sul (MS), o cultivo do girassol safrinha causou grande
disseminação do patógeno comprometendo outras espécies suscetíveis,
dentre elas a soja (Pitol, 2009).
No estado de Mato Grosso na safra 2009/10, principalmente nos
municípios de Campo Verde e Jaciara (região sul do estado), foi constatada
a presença do mofo branco, o qual causou perdas significativas na
produtividade da cultura da soja.

16
2.2 Taxonomia
O patógeno, Sclerotinia sclerotiorum, pertence ao Reino Fungi, filo
Ascomycota, Classe Ascomycetes, ordem Helotiales e família
Sclerotiniaceae (Bolton et al., 2006; Kirk et al., 2008). Do gênero Sclerotinia,
são conhecidas as espécies Sclerotinia sclerotiorum, S. minor, S. borealis,
S. babulbarium, S. glacidis, S. homeocarpa, S. nivalis, S. pirolae, S.
spermophila, S. subarctica, S. tetraspora, S. trifoliorum e S. veratri (NCBI,
2013). A característica mais marcante da Ordem Helotiales é a formação de
escleródios bem desenvolvidos, já a família Sclerotiniaceae caracteriza-se
pela produção, no ciclo sexual, de apotécios com estipe a partir da
germinação dos escleródios. O micélio desse grupo de fungos é constituído
por hifas hialinas, septadas, multinucleadas e ramificadas (Bolton et al.,
2006).
O filo Ascomycotina constitui o grupo mais numeroso de fungos,
apresentando como característica básica a formação, após a meiose, de
esporos sexuais, os ascósporos, dentro de uma estrutura em forma de
saco, o asco (Massola e Krugner, 2011). O fungo produz estruturas de
resistência denominadas de escleródios, que podem sobreviver no solo por
até oito anos (Adams e Ayers, 1979).
2.3 Aspectos Biológicos
2.3.1 Produção de escleródios
Os escleródios são constituídos de agregados de hifas e são
formados em resposta às mudanças na disponibilidade de nutrientes
(LeTourneau, 1979). Estas estruturas apresentam formato irregular,
variando de um a vários milímetros de diâmetro e comprimento, a princípio
de coloração branca e, posteriormente, tornam-se negras e duras (Agrios,
1997). As hifas vegetativas geralmente se desenvolvem de forma
distanciada uma da outra; no início há uma atração entre elas de modo que
ocorre a fusão das mesmas, e com uma mudança na disponibilidade dos

17
nutrientes ocorre a formação dos escleródios. A formação de escleródios
envolve mudanças celulares, mobilização e deposição de muitas
substâncias, sendo que alguns isolados perdem a habilidade de produzir
escleródios após repicagens consecutivas em meio de cultura; isto pode ser
devido à inabilidade do fungo sintetizar compostos específicos exigidos para
a sua formação (Le Tourneau, 1979).
Os estágios de desenvolvimento para formação dos escleródios
podem ser caracterizados em três fases: iniciação, que é a agregação de
hifas para formar uma massa branca chamada de escleródio inicial;
desenvolvimento, caracterizada pelo crescimento das hifas e agregação
para aumentar de tamanho; e a maturação de limitação da superfície,
depósito de melanina e consolidação interna (Bolton et al., 2006).
Os escleródios são compostos por três camadas distintas: uma
parede grossa rica em melanina, responsável pela coloração negra, uma
parede fina (córtex) e a medula branca, que é o micélio dormente do fungo.
Metabólitos presente em Sclerotinia sp., conhecido como “sclerin”, em
combinação com outros compostos, como fenóis, estão envolvidos na
melanogênese e na formação de agregados de hifas (Le Tourneau, 1979).
A melanina confere resistência aos escleródios, às condições adversas do
solo, fazendo com que esses permaneçam viáveis por vários anos, mesmo
na ausência de plantas hospedeiras (Rocha, 2007).
O fungo produz ß-caroteno em baixos e altos níveis durante o estágio
de não diferenciação e diferenciação, respectivamente, em isolados que
produzem escleródios. A produção de ß-caroteno depende das condições
de luz durante os estágios de desenvolvimento, enquanto que em isolados
que não produzem escleródios, a produção de ß-caroteno é muito baixa e
independe de luz e do período de crescimento (Georgiou et al., 2001).
Os escleródios são considerados como estrutura de resistência, de
consistência firme, que desempenham papel importante na sobrevivência
do fungo durante a entressafra, conservando intacto seu poder patogênico.
Esse processo se dá pela constituição física dura, devido à presença da
proteção exterior negra formada pela concentração de melanina, altamente

18
resistente à degradação microbiana, envolvendo a parte interna formada
pelo micélio do fungo, composto por uma matriz fibrosa composta de
carboidratos, quitina e ß-glucanos (Le Tourneau, 1979). Os escleródios
produzidos nas plantas infectadas retornam ao solo pela queda natural dos
mesmos ou após a colheita com os resíduos da cultura, garantindo a
perpetuação do fungo no solo (Schwartz e Steadman, 1978; Leite, 2005).
Os escleródios não são capazes de germinar sem antes maturar
fisiologicamente na mesma safra (Abawi e Grogan, 1979), sugerindo ser o
mofo branco uma doença monocíclica (Souza, 1999). Steadman (1983)
relatou que somente os escleródios localizados nos 5 cm superficiais do
solo são capazes de produzir apotécios. Mas alguns escleródios localizados
até 10 cm também podem produzir apotécios, como citado anteriormente
por Cook et al. (1975). O apotécio tem 3 mm de diâmetro e eleva-se de 3 a
6 mm da superfície do solo (Schwartz e Steadman, 1989).
Segundo Huang (1983), o fungo também pode produzir escleródios
diferenciados em plantas de girassol. O primeiro relato foi em Manitoba, nos
anos de 1977/79, sendo chamados escleródios anormais aqueles que
apresentaram a superfície enrugada quando comparados com escleródios
normais, cuja superfície é lisa. Segundo esse mesmo autor, os escleródios
anormais perderam a viabilidade mais rapidamente, comparando com os
normais, e a redução na longevidade foi proporcional ao grau da má
formação. A viabilidade dos escleródios, na faixa de temperatura de 0,5 a
30 ºC, diminui com o aumento da temperatura e, dentro de cada
temperatura, a viabilidade reduz com o aumento do período de
armazenamento e grau de anomalia do escleródios (Huang e Kozub, 1994).
2.3.2 Germinação dos escleródios
Vários fatores influenciam a germinação do escleródio, tais como: os
nutrientes do substrato no qual este é formado, a sua idade, os fatores
ambientais como umidade, temperatura, luz, pH do solo e aeração, e a
profundidade em que se encontra no solo (Willetts e Wong, 1980; Philips,
1987).

19
As condições favoráveis ao desenvolvimento da doença são
temperaturas amenas, abaixo de 20 °C, e alta umidade relativa, acima de
70%. Nestas condições, os apotécios liberam os ascósporos, que são
responsáveis pela infecção da parte aérea (Leite, 2005). Outro fator
importante para que os escleródios sejam formados é a umidade do solo.
Em geral, umidade do solo correspondendo a tensão acima de 5 bar
favorecem a formação de apotécios (Boland e Hall, 1987a).
Os escleródios podem germinar na forma miceliogênica e
carpogênica. Na forma miceliogênica ocorre a produção de micélio hialino e
septado (Purdy, 1979), e não há formação de esporos. Na forma
carpogênica ocorre a formação dos apotécios (Schwartz e Steadman, 1989;
Napoleão, 2001; Napoleão et al., 2005). O escleródio pode produzir um ou
mais apotécios, nos quais estão os ascos contendo os ascósporos
(Schwartz e Steadman, 1989).
Venette (1998) observou que, em situações de limitações de
nutrientes, pode ser desencadeada a germinação carpogênica; já com a
disponibilidade de nutrientes, os escleródios podem germinar formando um
novo micélio.
A germinação carpogênica inicia-se ativando o desenvolvimento do
fungo na região do córtex esclerodial ou medular; assim as células fúngicas
crescem formando primórdios que rompem a capa do escleródio e
continuam o crescimento como ramificações em forma de tubo,
denominados estipes (Bolton et al., 2006). Esse processo ocorre sob
condições de alta umidade, temperatura amena entre 18 a 22 °C e
luminosidade moderada (Boland e Hall, 1987b). As estipes sob o efeito da
luz, especialmente a ultravioleta (< 390 nm), diferenciam os apotécios, ou
seja, a ponta da estipe expande para formar uma superfície superior
(himênio), dando origem ao aparecimento do apotécio, uma estrutura em
forma de taça (Bolton et al., 2006). Logo serão produzidos dentro das
ascas, os ascósporos (esporos sexuais) que são a principal fonte de inóculo
de S. sclerotiorum (Boland e Hall, 1987b).

20
Os apotécios são a maior fonte de inóculo do fungo, pois produzem
grande quantidade de ascósporos que, ejetados, são facilmente
transportados pelo vento e podem infectar plantas em um raio de 50 m a
100 m da fonte produtora (Steadman, 1983). Cada apotécio contém
centenas de ascos de forma cilíndrica. Em cada asco, há oito ascósporos,
que são ovóides (Schwartz e Steadman, 1989).
O apotécio libera ascósporos continuamente por 2 a 17 dias, com
média de 9 dias. A produção máxima de ascósporos ocorre em intervalo de
2 a 3 dias entre o quarto e nono dia de vida ativa do apotécio (Schwartz e
Steadman, 1978). Um apotécio descarrega, simultaneamente, 10.000 a
30.000 ascósporos maduros, e que podem produzir, em cinco a dez dias de
vida funcional, mais de 2.000.000 novos esporos (Steadman, 1983).
A influência do tamanho dos escleródios na germinação carpogênica
apresenta resultados contraditórios. Para Budge e Whipps (1991), maior
produção de apotécios foi obtida em escleródios com massa menor que 10
mg e 2 mm de diâmetro. Em escleródios maiores a produção de apotécios
também é maior, sendo que um escleródio de 13 x 5 mm, dá origem a 15
apotécios (Bedi, 1963). Dillard et al. (1995) observaram que quanto maiores
os escleródios maior a porcentagem de escleródios germinados e maior o
número de apotécios produzidos.
Os escleródios produzidos em plantas infectadas vão retornar ao solo
após a colheita agregados aos resíduos das culturas, podendo então
garantir a perpetuação do fungo nos próximos cultivos (Schwartz e
Steadman, 1978). Quando ocorre estresse hídrico, logo após a iniciação da
germinação dos escleródios, pode ocorrer o retardamento da formação dos
apotécios (Abawi e Grogan, 1975).
Quando os ascósporos estão maduros dentro dos ascos, um ligeiro
decréscimo da umidade relativa do ar provoca a liberação por ejeção (Natti,
1978). No apotécio também são formadas numerosas ascas estéreis,
denominadas de paráfises, que podem servir como estruturas de suporte.
Dentro das ascas, um vacúolo que é responsável pelo aumento da pressão
hidrostática forma-se abaixo dos ascósporos e à medida que a pressão

21
aumenta, a asca se expande, porém a sua pressão lateral é limitada pela
paráfise e ascas vizinhas. Entretanto a expansão continua relativamente
além da paráfise e, em algum momento, a pressão excedente torna-se
insuportável pela parede dilatada, então as ascas se rompem e liberam os
ascósporos para o ambiente (Venette, 1998).
2.4 Mecanismo de Patogenicidade: Produção de Ácido Oxálico
A patogenicidade pode ser facilitada em seus hospedeiros pela
produção de uma ampla gama de enzimas degradantes da parede celular,
que incluem pectinases, ß-1,3-glucanases, glicosidases, celulases,
xilanases e cutinases, dando grande flexibilidade ao patógeno para a
penetração e colonização do hospedeiro (Bolton et al., 2006).
A capacidade de sintetizar o ácido oxálico tem sido um fator
determinante para a patogenicidade; plantas de feijão e girassol tratadas
com ácido oxálico sintético e um filtrado de S. sclerotiorum apresentaram os
mesmos sintomas exibidos por plantas naturalmente infectadas em campo
(Noyes e Hancoock, 1981).
O ácido oxálico é um composto que pode ser encontrado na forma de
ácido livre, em forma solúvel, como oxalato de sódio e potássio e, insolúvel,
como oxalato de cálcio, que é frequentemente associado a desordens
metabólicas e doenças infecciosas (Guimarães e Stolz, 2004). Um grande
número de fungos fitopatogênicos secreta o ácido oxálico incluindo as
espécies S. sclerotiorum e S. minor.
O ácido oxálico e as enzimas pectolíticas estão associados com o
desenvolvimento do mofo branco causado por S. sclerotiorum. O ácido
oxálico secretado pelo patógeno penetra no tecido ao redor da lesão,
reduzindo o pH de aproximadamente 6,8 para 4,0, fornecendo um pH ótimo
para a ação da enzima pectolítica (Echandi e Walker, 1957; Maxwell e
Lumsden, 1970; Lumsden, 1979). As diferenças na tolerância ao ácido
oxálico e/ou resistência à sua difusão no tecido do hospedeiro podem
resultar em regiões de encharcamento variáveis ao redor das lesões (Tu,

22
1980; Beversdorf, 1982). Espécies de Sclerotinia podem tolerar ampla faixa
de pH, mas se adaptam melhor ao substrato ácido. O ácido oxálico é
produzido por S. sclerotiorum no tecido do hospedeiro e em cultura,
contribuindo para o decréscimo do pH de substratos alcalinos (Willetts e
Wong, 1980). A produção do ácido durante a infecção altera o
funcionamento da célula guarda, causando a abertura dos estômatos e
avanço das hifas no tecido infectado (Guimarães e Stolz, 2004).
2.5 Sintomas e Epidemiologia
A doença conhecida como mofo branco recebe esse nome em
função dos sintomas e sinais externos causados na planta. Na cultura do
feijoeiro, assim como na soja, o sintoma inicial é a murcha de algumas
plantas, resultado do apodrecimento do caule causado pelo fungo.
Posteriormente, folhas, hastes e vagens apresentam manchas
encharcadas, de coloração parda e consistência mole, seguidas de
crescimento de micélio branco de aspecto cotonoso, cobrindo porções dos
tecidos (Leite, 2005; Paula Júnior et al., 2006; Paula Júnior et al., 2008).
Com o progresso da doença, em poucos dias, o micélio transforma-
se em escleródio, facilmente visível, podendo ser formado tanto na
superfície, como no interior da haste e das vagens infectadas (Almeida et
al., 2005; Paula Júnior et al., 2006; Paula Júnior et al., 2008). Desta forma,
os tecidos tornam-se secos, leves e quebradiços. As sementes infectadas
são pequenas, sem brilho, descoloridas, enrugadas, ou não apresentam
sintomas (Paula Júnior et al., 2008). Na soja, os sintomas ocorrem
geralmente no terço médio das plantas, atingindo a haste principal,
pecíolos, folhas e vagens (Leite, 2005).
Os sintomas atingem outros órgãos da planta e pode envolver toda a
haste, impedindo o fluxo de água e nutrientes, levando a planta à morte
(Grau, 1989).
A fase mais vulnerável da cultura da soja vai do estádio da floração
plena (R2) ao início da formação das vagens (R3/R4). O fungo é capaz de

23
infectar qualquer parte da planta, porém, as infecções iniciam-se com mais
frequência a partir das inflorescências e das axilas das folhas e dos ramos
laterais (Almeida et al., 2005), cujos sintomas, geralmente, ocorrem no terço
médio das plantas (Leite, 2005), momento em que o patógeno encontra
fonte exógena de energia nas flores senescentes para os ascósporos
germinarem (Vieira, 1994). Nesta fase, o microclima é mais favorável ao
patógeno, devido ao maior índice de área foliar. Além disso, a maior
cobertura foliar durante o fechamento da cultura permite que plantas
doentes entrem em contato com plantas sadias, aumentando os focos da
doença e/ou a sua disseminação radial (Garcia, 2008).
Tecidos vegetais jovens e, principalmente flores e fragmentos de
flores, órgãos que possuem altas concentrações de α-celulose, são a base
nutritiva para o início da colonização do fungo (Sutton e Deveral, 1983).
Os ascósporos liberados pelos apotécios constituem a fonte primária
de infecção de plantas (Abawi e Grogan, 1979). Os ascósporos colonizam
as flores em dois a três dias, e o micélio do fungo infecta tecidos de folhas,
caule e vagens que entra em contato. Geralmente, numerosos escleródios
são produzidos na superfície do micélio, de sete a dez dias após a
colonização (Abawi e Grogan, 1979). Caso os ascósporos sejam liberados
antes que a planta floresça, estes podem sobreviver por cerca de duas
semanas na superfície da planta ou do solo. Uma vez colonizadas as flores,
o micélio permanece viável por até um mês (Steadman, 1983). Além do
vento, as abelhas também podem servir como agente de disseminação dos
ascósporos.
Uma substância mucilaginosa é liberada junto com os ascósporos, o
que auxilia na sua adesão aos tecidos dos hospedeiros ou a outros
obstáculos encontrados durante o seu percurso aéreo. A maioria dos
ascósporos fica retido dentro do dossel das plantas, possibilitando alto
potencial de infecção local. A liberação dos ascósporos ocorre tanto durante
o dia quanto à noite, no entanto, a duração entre cada liberação pode variar
de 36 a 168 horas, havendo declínio quando a umidade relativa do ar atinge
valores entre 65 a 75% (Clarkson et al., 2003). A germinação constante de

24
escleródios e a liberação contínua dos ascósporos de cada apotécio
asseguram adequado potencial de infecção, superior a duas semanas
(Vieira, 1994).
Após os ascósporos serem depositados sobre os tecidos florais
senescentes, germinam, infectam e formam o micélio que invadem também
outros tecidos sadios do hospedeiro (Lu, 2003). Essa colonização também
poderá ocorrer por escleródios que germinam na forma miceliogênica no
solo, produzindo hifas e colonizando diretamente a matéria orgânica em
decomposição, formando um micélio que, em contanto com a planta
hospedeira, provoca a infecção. No entanto, isso ocorre devido à produção
de alguns compostos voláteis produzidos pela decomposição, estimulando a
germinação dos escleródios (Punja, 1985).
A infecção do tecido sadio do hospedeiro depende da formação do
apressório, que pode ser simples ou complexo, dependendo da estrutura da
superfície do hospedeiro. Na maioria dos casos, a penetração ocorre
diretamente através da cutícula e não através dos estômatos. O apressório
desenvolve-se da ramificação dicotômica das hifas que crescem na
superfície do hospedeiro e consiste de um aglomerado mucilaginoso de
hifas largas, multi-septadas e curtas. Embora a penetração da cutícula seja
um processo puramente mecânico, a degradação enzimática da cutícula
auxilia no processo de penetração, afetando a lamela média das células
(Lumsden, 1979; Hegedus e Rimmer, 2005). A Figura 1 resume de forma
esquemática o ciclo de vida do mofo branco da soja causado por Sclerotinia
sclerotiorum, incluindo os processos de infecção via germinação
miceliogênica e carpogênica.

25
FIGURA 1. Esquema do ciclo de vida do fungo Sclerotinia
sclerotiorum, agente causal do mofo branco da soja.
Adaptado de Wharton e Kirk (2007).
Quanto aos escleródios, estes podem ser disseminados pelo
transporte de solo contaminado, água de irrigação, e misturados às
sementes e fezes de animais que se alimentaram de plantas doentes (Tu,
1998).
Em áreas livres do patógeno, quando o inóculo não está presente no
solo, uma epidemia da doença pode ser iniciada por meio de sementes
contaminadas internamente pelo micélio dormente do fungo, ou com
escleródios associados ao lote de sementes. O micélio dormente, presente
na testa e nos cotilédones das sementes, pode manter sua viabilidade por
mais de três anos; quando semeadas, e sob condições favoráveis de
umidade e temperatura, o micélio se desenvolve e dá início à infecção.
Muitas das sementes contaminadas podem não germinar, mas o patógeno
associado pode produzir micélio e escleródios. Cada semente infectada
pode produzir de três a seis escleródios, aumentando a quantidade de
inóculo no solo (Tu, 1998).

26
Os primeiros sintomas nas folhas são manchas de anasarca que
evoluem para coloração castanho-clara (Yorinori, 1997). Sobre as áreas
afetadas, desenvolvem abundante formação de micélio branco e de aspecto
cotonoso. Esses são formados tanto na superfície, como no interior da
haste e das vagens infectadas (Almeida et al., 2005). Os escleródios, por
sua vez, podem germinar produzindo micélio ou apotécios e atacar grande
número de plantas.
Todavia, a transmissão por escleródios pode ser reduzida, uma vez
que, durante o beneficiamento de sementes, grande parte destes pode ser
removida. Ainda que a taxa de transmissão por micélio dormente seja
bastante baixa em um lote de sementes, a sua importância reside na
possibilidade da introdução do inóculo em novas áreas de cultivo (Henning,
2004).
2.6 Variabilidade do Patógeno
O fungo S. sclerotiorum foi estudado por mais de 150 anos e muitas
informações foram geradas sobre sua biologia, sintomatologia,
patogenicidade e morfologia. Entretanto, ainda se sabe pouco sobre seus
aspectos genéticos (Arbaqui et al., 2008). Assim, a variabilidade quanto à
agressividade entre isolados de S. sclerotiorum em soja tem sido muito
demandado em estudos de melhoramento visando resistência ao patógeno
(Kull et al., 2004).
Estudos em população de S. sclerotiorum demonstram a diversidade
genética e fornecem evidência para a reprodução clonal e sexual.
Geralmente, a reprodução clonal predomina em regiões de clima temperado
(Cubeta et al., 1997; Hambleton et al., 2002), enquanto que a recombinação
sexual ocorre em regiões de temperaturas amenas (Atallah et al., 2004;
Malvárez et al., 2007). Kohn et al. (1990), estudando a compatibilidade
micelial entre 35 isolados de S. sclerotiorum, de diferentes localidades,
verificaram que 23 isolados foram incompatíveis com todos os outros, sendo

27
que 12 isolados formaram cinco grupos de compatibilidade, com dois a três
isolados por grupo, denotando heterogeneidade genética dentro da espécie.
Kohli et al. (1992) verificaram que, em um único campo, pode haver
mais de um clone de S. sclerotiorum, e que em diferentes campos e
distâncias geográficas relativamente longas pode-se encontrar um único
clone. No Brasil, entre 40 isolados coletados em diferentes localidades
brasileiras, foi verificada a formação de cinco grupos de compatibilidade
micelial, sugerindo elevada variabilidade genética e recombinação sexual
entre os isolados estudados (Litholdo Júnior et al., 2011).
Outros estudos também demonstraram variabilidade genética em
populações de S. sclerotiorum. Meinhardt et al. (2002) observaram dois
grupos de compatibilidade entre 23 isolados, sendo 21 isolados de feijoeiro,
um isolado de batata e um isolado de pimenta, além de polimorfismo entre
os isolados dentro de um mesmo grupo de compatibilidade micelial.
Diversidade genética em 79 isolados coletados em áreas de feijoeiro,
no Cerrado brasileiro, por meio de marcadores microssatélites também foi
verificada por Gomes et al. (2011).
Kull et al. (2004) constataram 42 grupos de compatibilidade micelial
entre 299 isolados provenientes de Diverse, DeKalb, Watseka Sets e
Argentina. Estes grupos de compatibilidade apresentaram diferenças quanto
à agressividade em plantas de soja. Otto-Hanson et al. (2011) também
verificaram a existência de 64 grupos de compatibilidade micelial em uma
amostra de 156 isolados coletados em diferentes regiões, dos 64 grupos, 36
grupos foram formados de um único isolado.
2.7 Métodos de Detecção de Sclerotinia sclerotiorum
O conhecimento prévio das formas de interação do inóculo de
patógenos com sementes é importante em patologia de sementes, dentre
vários aspectos, para a escolha de métodos de detecção destes agentes.
De modo geral, o transporte de microrganismos por sementes em um dado
lote pode se dar de três maneiras. No primeiro caso, o microrganismo,

28
separado ou não, encontra-se em mistura com as sementes, fazendo parte
da fração impura do lote. Fazem parte desta fração: fragmentos vegetais,
sementes de plantas invasoras e partículas do solo que podem, todos, ser
portadoras de micélio dormente, corpos frutíferos e esporos de fungos,
cistos ou galhas de nematóides, células bacterianas e partículas de vírus,
escleródios ou estromas fúngicos. Uma segunda maneira pela qual certos
patógenos podem se associar e ser(em) transportados pelas sementes é
por adesão passiva à superfície destas. A terceira forma de associação de
microrganismos com sementes é a presença do inóculo nos tecidos das
sementes, seja em estruturas superficiais ou mais interno no embrião. Essa
é a forma de interação e transporte mais comum entre os agentes
transmitidos por sementes. É preciso salientar que, apesar da distinção que
se faz entre esses três tipos de interação de inóculo com as sementes, um
mesmo patógeno pode estar presente em um lote, sob uma ou mais dessas
formas de associação (Brasil, 2009b).
O objetivo dos métodos é determinar o estado sanitário de uma
amostra de sementes e, consequentemente, do lote, obtendo-se
informações confiáveis para comparar a qualidade de diferentes lotes,
fornecendo informações para programas de certificação, serviços de
vigilância vegetal, tratamento de sementes, melhoramento de plantas e
outros (Henning, 1994; Machado, 2000).
Há três métodos mais utilizados, citados em Brasil (2009b) e por
Nasser et al. (1995). O primeiro é a Incubação em Substrato de Papel ou
método do Papel de Filtro (“blotter test”), onde as sementes dispostas
individualmente sobre camada de papel de filtro umedecido em recipientes
como placas de Petri, gerboxes ou equivalentes, por sete a oito dias (Brasil,
2009b). Para reduzir o processo de germinação das sementes de espécies
de dicotiledôneas, durante o período de incubação, o substrato de papel
pode ser umedecido em solução de sal de 2,4-D (2,4-diclorofenoxiacetato
de sódio) a 5 ppm de concentração. Em alguns casos é necessário
prolongar o período de incubação, por exemplo, para detectar Sclerotinia
sclerotiorum em sementes de soja e feijão, por mais uma semana a

29
temperatura de 10-15 ºC, com umedecimento adicional do substrato (Brasil,
2009b).
O segundo método é a Incubação em rolo de papel, que é utilizado
para detecção de S. sclerotiorum tanto para sementes de soja quanto de
feijão. São usadas folhas de papel de germinação, em forma de rolo, por 14
dias. A desinfestação superficial das sementes é recomendada por meio de
solução 1% de hipoclorito de sódio por três minutos. Para a avaliação das
sementes, deve-se observar a presença de micélio tipicamente branco do
fungo, com formação de escleródios negros, os quais apresentam forma
esférica, sendo irregulares e de tamanho de 2 mm a 10 mm, ao redor das
sementes infectadas (Brasil, 2009b).
O terceiro método Incubação em meio Agar-Bromofenol (Neon),
proposto por Nasser et al. (1995), utiliza um meio de cultura denominado
meio neon, para detecção da viabilidade de escleródios de S. sclerotiorum.
Esse meio é composto de 1,0 L de BDA (batata-dextrose-ágar) autoclavado
+ 150 mg de azul de bromofenol + 150 mg de penicilina G + 150 mg de
estreptomicina, pH ajustado com HCl para 4,7. O princípio de sua utilização
baseia-se na mudança de cor do meio, do azul para o amarelo, devido ao
ácido oxálico produzido pelo patógeno.
Nasser et al. (1999) fizeram alterações em sua composição (1,0 L de
BDA autoclavado + 75 mg de azul de bromofenol + 75 mg de cloranfenicol +
75 mg de ácido 2,4 diclorofenoxiacético (2,4-D), sendo o pH ajustado com
HCl para 4,7). Napoleão et al. (2006) modificaram o meio neon e o
denominaram de neon-S. Os autores procuravam um antibiótico de amplo
espectro e uma forma do ácido 2,4 D que suportassem a autoclavagem
para serem incorporados ao novo meio. Desta maneira, o meio neon-S
constitui-se de 1,0 L de BDA + 50 mg de azul de bromofenol + 50 mg de
cloranfenicol + 50 mg de ácido livre 2,4-D. Faz-se a autoclavagem à 121°C
por 20 minutos, a incubação das sementes à 19°C ±1°C no escuro, sendo a
leitura realizada de sete a doze dias. Todos os novos ingredientes são
adicionados antes da autoclavagem e o ajuste de pH é dispensado. As
Regras para Análise de Sementes (RAS) recomendam a incubação em

30
meio ágar-bromofenol (meio neon), no qual algumas modificações foram
feitas com base nos meios anteriormente descritos (Brasil, 2009b). A partir
do terceiro dia de incubação das sementes, as observações começam a ser
realizadas para verificar a formação de halos amarelo avermelhados ao
redor das sementes. A visualização de micélio superficial sobre o meio na
zona do halo amarelado, partindo das sementes, confirma a presença do
fungo em foco. Este método é específico para detecção de S. sclerotiorum,
porém algumas espécies de Aspergillus, Rhizopus e Sclerotium podem
também provocar alteração da cor do meio. Nestes casos, a formação de
frutificações típicas dessas espécies pode ser facilmente distinguida de
Sclerotinia (Brasil, 2009b).
2.8 Controle de Sclerotinia sclerotiorum
O controle do mofo branco é considerado difícil devido à ausência de
cultivares resistentes, produção de estruturas de resistência do fungo,
sobrevivência do mesmo no solo por vários anos na ausência de
hospedeiros ou sob condições desfavoráveis, ampla gama de hospedeiros,
elevado número de ascósporos produzidos por apotécio e sua rápida e
longa disseminação a partir da fonte produtora, sobrevivência nas sementes
sob a forma de micélio dormente ou escleródios aderidos às mesmas e
devido à dificuldade de se atingir os sítios de infecção através do controle
químico. Desta forma, o controle mais efetivo baseia-se em um programa
integrado de diversas práticas culturais (Leite, 2005).
Uma medida de controle refere-se ao beneficiamento de sementes, o
qual pode ajudar a reduzir a quantidade de inóculo que poderia retornar à
área de plantio, tanto na forma de sementes infectadas, como de
escleródios (Napoleão, 2001; Napoleão et al., 2005). Para sementes de
soja, no beneficiamento pode-se fazer a separação dos escleródios pelo
uso do separador espiral seguido da mesa de gravidade (Henning, 2004;
Leite, 2005). Considerando-se que o mofo branco é uma doença
monocíclica, ou seja, apenas o inóculo inicial é responsável pela quantidade

31
de doença, o beneficiamento pode representar um grande passo para a
redução deste inóculo e, consequentemente, da doença.
Yang et al. (1999) verificaram que a incidência de S. sclerotiorum em
cultivares de soja está relacionada com grupos de maturação. Segundo
esses autores, cultivares de ciclo longo são mais suscetíveis, devido o
período de florescimento ser maior, período no qual se tem bastante
infecção, em função da liberação dos ascósporos.
A utilização do controle químico, integrado a outras práticas culturais,
pode prevenir o aparecimento da doença, visando proteger as flores da
infecção pelos ascósporos produzidos em apotécios. Entretanto, na prática,
o controle químico para essa doença tem sido extremamente difícil, seja
pela ausência de informações ou pela própria natureza do fungo, que pode
sobreviver até 11 anos no solo (Farias Neto, 2012).
Segundo Furlan (2008), o fungicida a ser aplicado, seja de ação de
contato ou sistêmico, deve ser posicionado no alvo no momento correto e
de forma adequada para se obter controle econômico e racional da doença.
As pulverizações devem ser realizadas uniformemente, com boa
distribuição nos tecidos da planta e, se possível, alcançando a superfície do
solo, onde surgem os apotécios e desenvolvem os micélios. A primeira
pulverização deve ser feita preventivamente, na abertura das primeiras
flores.
A sensibilidade de S. sclerotiorum à determinados fungicidas também
tem sido relatada na literatura (Gindrat, 1993; Mueller et al., 2002). Menten
et al. (1995) avaliaram a eficiência in vitro de alguns fungicidas no controle
de S. sclerotiorum do feijoeiro comum. Esses autores verificaram para os
fungicidas clorotalonil, tebuconazole, procimidone, iprodione, tiofanato
metílico + clorotalonil, captan, vinclozolin, tiofanato metílico e benomyl que a
DL50 foi menor que 1 μg.mL-1 de ingrediente ativo, portanto, considerados
altamente eficientes, enquanto que para os fungicidas fentin-acetato,
mancozeb e óxido cuproso as DL50 foram respectivamente, 1-10 μg.mL-1,
10-50 μg.mL-1 e maior que 50 μg.mL-1 de ingrediente ativo, considerados
moderadamente eficiente, pouco eficiente e ineficiente.

32
Campos et al. (2008) demonstraram que a eficácia do controle
químico depende do princípio ativo utilizado, do número de aplicações, do
momento da aplicação e da tecnologia utilizada. De acordo com os
resultados de pesquisa já obtidos, melhor controle da doença tem sido
verificado com fungicidas pertencentes aos grupos químicos
dicarboximidas, benzimidazóis e fenilpirimidinamida. Porém, vale chamar a
atenção para o manejo de resistência de fungos a fungicidas, pois
Sclerotinia é um fungo altamente adaptável, e a utilização de um manejo
inadequado dos ingredientes ativos no controle da doença, poderá induzir a
seleção de isolados para a resistência a esses produtos. Para isso,
aconselha-se a associação de medida de controle, como por exemplo:
controle cultural, controle químico e controle biológico concomitantemente,
além de realizar uma alternância entre os ingredientes ativos utilizados
(Fundação Bahia, 2011).
Atualmente no mercado não existem variedades de soja resistentes
ao mofo branco, porém existem características morfológicas e fisiológicas
da planta que ajudam a reduzir a incidência da mesma, como arquitetura
ereta, plantas precoces, resistentes ao acamamento, entre outros (Campos
et al., 2010). Como o patógeno pode atacar todas as partes da planta, um
genótipo que apresenta resistência a uma forma de ataque à planta pode
apresentar suscetibilidade a outro mecanismo de ataque (Tourvieille e Vear,
1990).
O controle genético também é uma ferramenta útil a ser utilizada
para o controle de mofo branco. Diferenças entre cultivares de soja quanto à
reação à S. sclerotiorum têm sido avaliadas em condições de campo, casa
de vegetação e laboratório, sendo observadas respostas variadas ao
patógeno.
Zito et al. (2005) verificaram em condições de campo, em
Sacramento, MG, que as cultivares BR97-11548 e Potenza foram mais
suscetíveis ao mofo branco do que BRSMG Garantia, Monarca, MG/BR 46
(Conquista) e MGBR99-4656. Boland e Hall (1987b) verificaram diferença
de suscetibilidade entre 42 cultivares de soja em condições de campo. Os

33
autores observaram que a incidência da doença foi correlacionada com a
altura da cultivar, maturidade e número de apotécios sob o dossel,
indicando que o escape à doença é um mecanismo importante que afeta
algumas cultivares de soja.
2.9 Geoestatística
Existe forte estrutura de dependência espacial entre plantas durante
o progresso espacial de doenças, em função da origem e da disseminação
do inóculo, principalmente quando existem reboleiras (Campbell e Madden,
1990; Alves et al., 2006). Portanto, cada valor carrega consigo forte
interferência dos valores de sua vizinhança, de maneira a ilustrar a
continuidade espacial (Isaaks e Srivastava, 1989). Havendo dependência
espacial, podem-se estimar valores da variável em estudo para os locais
não amostrados, sem tendenciosidade e com variância mínima, por meio do
método da krigagem (Vieira, 2000).
Como ao longo do espaço existe correlação espacial entre plantas no
progresso de epidemias, a geoestatística tem sido aplicada para modelar
padrões espaço-temporais e gerar hipóteses sobre aspectos ecológicos e
epidemiológicos de doenças de plantas (Chellemi et al., 1988; Larkin et al.,
1995; Nelson et al., 1999; Wallace e Hawkins, 1994; Jaime-Garcia et al.,
2001; Charest et al., 2002; Alves et al., 2006). A geoestatística tem se
destacado no estudo de patógenos presentes no solo, principalmente para a
confecção de mapas de gradiente e também para determinar o padrão de
distribuição da doença, devido ao caráter agregado do inóculo e da doença
(Alves et al., 2006).
Issaks e Srivastava (1989) afirmaram que na teoria fundamental da
geoestatística espera-se que, na média, as amostras próximas no tempo e
no espaço sejam mais similares entre si do que as que estiverem distantes.
As técnicas geoestatísticas podem ser usadas para descrever e modelizar
padrões espaciais (variografia), para predizer valores em locais não
amostrados (krigagem), para obter a incerteza associada a um valor

34
estimado em locais não amostrados (variância de krigagem) e para otimizar
malhas de amostragem (Andriotti, 2003).
O semivariograma é a técnica mais usada para descrever a
dependência espacial na geoestatistica, o qual descreve a associação
espacial dos pontos amostrais em função da distância entre eles. É uma
técnica exploratória que auxilia os métodos geoestatisticos, contribuindo na
modelagem da dependência espacial, determinando os parâmetros básicos
do modelo: alcance, efeito pepita e patamar (Ribeiro Jr e Diggle, 2001).
Entre os diversos modelos de semivariogramas os mais utilizados são o
esférico, o exponencial e o gaussiano.
O padrão espacial de doença é definido como arranjo ou
posicionamento das plantas doentes, umas em relação às outras (Gilligan,
1983) e depende não só da forma de dispersão do patógeno, como também
da arquitetura das plantas hospedeiras e do arranjo destas no campo
(Gilligan,1982). Assim, é possível relacionar, indiretamente, o padrão
espacial encontrado às práticas de manejo adotadas nos locais estudados
por meio de técnicas estatísticas (Laranjeira, 2002). Basicamente a
distribuição espacial de uma doença no campo pode apresentar três tipos
de padrões: regular, casualizado (aleatório) e agregado (agrupado)
(Bergamin Filho et al., 2002).
2.10 Ocorrência de S. sclerotiorum em Mato Grosso
Na safra 2007/08 havia informação de ocorrência de mofo
branco em Campo Verde-MT, entretanto somente na safra 2008/09
observou-se maior incidência nas avaliações onde ocorreu quantificação da
doença, no final do ciclo da cultura da soja. Verificou-se incidência de até
65,4% em talhão com a cultivar Tabarana. A partir da safra 2009/10
algumas lavouras apresentaram altas incidências de mofo branco, o qual
causou perdas significativas na produtividade da cultura da soja,
principalmente em Campo Verde e Jaciara. Na região Norte do estado em

35
2008, contatou-se a presença do patógeno somente em áreas de feijão
irrigado com pivô central no período de entressafra da soja.

36
2.11 Referências Bibliográficas
ABAWI, G.S.; GROGAN, R.G. Epidemiology of disease caused by Sclerotinia species. Phytopathology, v. 63, n. 8, p. 899-903, 1979. ABAWI, G. S.; GROGAN, R. G. Source of primary inoculum and effects of temperature and moisture on infection of beans by Whetzelinia sclerotiorum. Phytopathology, v. 65, n.3, p. 300-309, 1975. ADAMS, P. B.; AYERS, W. A. Ecology of Sclerotinia species. Phytopathology, v. 69, n. 8, p.896-898, 1979. AGRIOS, G. Plant pathology. 4 ed. San Diego: Academic Press, 1997. 635 p. ALMEIDA, A. M. R.; PEREIRA, L. P.; YORINORI, J. T.; SILVA, J. F. V.; HENNING, C. V.; GODOY, L. M.; COSTAMILAN, L. M.; MEYER, M. C. Doenças da soja. In: KIMATI, H.; AMORIM, L.; REZENDE, J. A. M.; BERGAMIN FILHO, A.; CAMARGO, L. E. A. (Ed.). Manual de fitopatologia: doenças das plantas cultivadas. 4 ed. São Paulo: Agronômica Ceres, 2005. v. 2, p. 569-588. ALVES, M. C.; POZZA, E. A.; MACHADO, J. C.; ARAÚJO, D. V.; TALAMINI, V. ; OLIVEIRA, M. S. Geoestatística como metodologia para estudar a dinâmica espaço-temporal de doenças associadas a Colletotrichum spp. transmitidos por sementes. Fitopatologia Brasileira, v. 31, p. 557-563, 2006. ANDRIOTTI, J. L.S. Fundamentos de estatística e geoestatistica. São Leopoldo,RS: Unisinos, 2003. 165 p. ARBAQUI, M.; KRAIC, J.; HUSZÁR, J. Genetic variation of Sclerotinia sclerotiorum isolates from different conditions. Agriculture (Pol’nohospodárstvo), v.54, p. 36-39, 2008. ATALLAH, Z. K.; LARGET, B.; CHEN, X. JOHNSON, D. A. High genetic diversity, phenotypic uniformity, and evidence of outcrossing in Sclerotinia sclerotiorum in the Columbia basin of Washington State. Phytopathology, v. 94, p. 737-742, 2004. BEDI, K.S; The age of sclerotia of Sclerotinia sclerotiorum (Lib.) de Bary in relation to the formation of aphotecia. Jornal of the Indian Botanical Society, v.42, p.204-207, 1963. BERGAMIN FILHO, A. HAU, B.; AMORIM, L. et al. Análise espacial de epidemias. Revisão Anual de Patologia de Plantas, v.10, 2002, p.155-218.

37
BEVERSDORF, W. D. Tolerance to White mold ( Sclerotinia sclerotiorum. Phytopathology, n. 70. v. 7, p. 670-674, 1982. BOLAND, G. J.; HALL, R. Index of plants of hostes Sclerotinia sclerotiorum. Canadian Journal of Plant Pathology, v. 16, n.1, p. 93–108, 1994. BOLAND, G. J.; HALL, R. Epidemiology of white bean in Ontario. Canadian Journal of Plant Pathology, v. 9, n.2, p.218-224, 1987a. BOLAND, G. J.; HALL, R. Evaluating soybean cultivars for resistance to Sclerotinia sclerotiorum under field conditions. Plant Disease, v. 71, p.934-936,1987b. BOLTON, M. D.; THOMMA, B. P. H. J.; NELSON, B. D. Sclerotinia sclerotiorum (Lib.) de Bary: biology and molecular traits of a cosmopolitan pathogen. Molecular Plant Pathology, v.11, n. 7, p.1-16, 2006. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária/MAPA/ACS, 2009a. 395 p. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Manual de análise sanitária de sementes. Brasília: Secretaria de Defesa Agropecuária/MAPA/ACS, 2009b. 200 p. BUDGE, S.P.; WHIPPS, J.M. Effec of sucrose concentration on sclerotia production and subsequent apothecial formation by Sclerotinia sclerotiorum. Mycological Research, v. 95, n.2, p. 195-198, 1991. CAFÉ-FILHO, A.C. Alerta aos produtores de ervilha: Podridão de Sclerotinia. Horticultura Brasileira, v.3, p. 57, 1985. CAMPBELL, C. L.; MADDEN, L. V. Introduction to plant disease epidemiology. New York: J. Wiley & Sons, 1990. 532 p. CAMPOS, H. D. ; SILVA, L.H.C.P. ; SILVA, J.R.C. ; MONTEIRO, F. Effect of fungicides in the development of Colletotrichum truncatum in controlled conditions. In: 4 Top Ciência, 2008, Heideberg - Germany. Top Ciência Basf. São Paulo: BASF The Chemical Company, 2008. v. 5. p. 1-2. CAMPOS, H.D.; SILVA,L.H.C.P.; MEYER,M.C.; SILVA,J.R.C.; JUNIOR, J.N. Mofo branco na cultura da soja e os desafios da pesquisa no Brasil. Tropical Plant Pathology, n. 35, p. 99-100, 2010. CESSNA, S.G.; SEARS, V.E.; DICKMAN, M.B.; LOW, P.S. Oxalic acid, a pathogenicity factor for Sclerotinia sclerotiorum, suppresses the oxidative burst of the host plant, The Plant Cell, v.12, n.11, p. 2191–2199, 2000.

38
CHAREST, J.; DEWDNEY, M.; PAULITZ, T.; PHILION, V.; CARISSE, O. Spatial Distribution of Venturia inaequalis Airborne Ascospores in Orchards. Phytopathology, v.92, p. 769-779, 2002. CHAVES, G.M. Estudos sobre S. sclerotiorum (Lib.) de Bary. Experientiae, v.4, n.2, p. 69-133, 1964. CHELLEMI, D. O.; ROHRBACH, K. G.; YOST, R. S.; SONODA, R. M. Analysis of the spatial pattern of plant pathogens on diseased plants using geoestatistics. Phytopathology, v. 78, p. 221-226, 1988. CLARKSON, J.P.; STAVELEY, J. PHELPS, K.; YOUNG, C.S.; WHIPPS, J.M. Ascorpore release and survival in Sclerotinia sclerotiorum. Mycological Research, v.107, p. 213-222, 2003. COOK, G.E.; STEADMAN, J.R.; BOOSALIS, M.G. Surviral of Wetzelinia sclerotiorum and initial infection of dry edible beans in Western Nebraska. Phytophatology, v.65, n. 3, p. 250-255, 1975. CUBETA, M.; CODY, B.; KOHLI, Y.; KOHN L.M. Clonality in S. sclerotiorum on infected cabbage in eastern North Carolina. Phytopathology, v.87, n.10, p. 1000–1004, 1997. DILLARD, H.R.; LUDWIG, J.W.; HUNTER, J.E. Conditioning sclerotia os Sclerotinia sclerotiorum for carpogenic germination. Plant Disease, v.4, n.79, p.411-415, 1995. ECHANDI, E.; WALKER, J.C. Pectolytic enzymes produced by Sclerotinia sclerotiorum. Phytopathology, v. 47, p.303-306, 1957. FARIAS NETO, A.L. Embrapa estuda manejo para controle do mofo branco. Disponível em: <http://www.reporternews.com.br/noticia.php?cod=268553> Acesso em: out. de 2012. FERREIRA, L.P; LEHMAN, P.S; ALMEIDA, A.M.R. Moléstias e seu controle. In: MIYASAKA, S.J; MEDINA, J.C. (eds.) A soja no Brasil. 1981. p.603-639. FUNDAÇÃO BAHIA. Mofo branco e seu manejo no oeste Baiano. Disponível em: <http://www.fundacaoba.com.br/index.php?p=mofo branco>. Acesso em: maio de 2011. FURLAN, S. H. Importância e manejo do mofo branco na cultura da soja. Revista Plantio Direto, Passo Fundo, edição 107, p. 28-31, 2008. GARCIA, R. A. Produção de inóculo, efeitos de extratos vegetais e de fungicidas e a reação de genótipos de soja a Sclerotinia sclerotiorum.

39
2008.154f. Dissertação (Mestrado em Fitopatologia) - Universidade Federal de Uberlândia, Uberlândia, 2008. GILLIGAN, N.A. Statistical analysis of the spatial pattern of Botrytis fabae on Vicia faba: a methological study. Transactions of the British Mycology Society, v.79, p.193-200, 1982. GILLIGAN, C.A. Modeling of soilborne pathogens. Annual Review of Phytopathology, v.21, p.45-64, 1983. GINDRAT, D. La sclérotiniose: sensibilité du Sclerotinia sclerotiorum au Carbendazime et à la vinclozoline. Revue Suisse D’Agriculture, v. 25, n. 2, p. 115-119, 1993. GEORGIOU, D. C.; TAIRIS, N.; POLYCRATIS, A. Production of B-caroteno by Sclerotinia sclerotiorum and its role in sclerotium differentiation. Mycological Research, v.9, n.105, p 1110-1115, 2001. GOMES, E. V.; NASCIMENTO, L. B. do; FREITAS, M. A. de; NASSER, L. C. B.; PETROFEZA, S. Microsatellite markers reveal genetic variation within Sclerotinia sclerotiorum populations in irrigated dry bean crops in Brazil. Journal of Phytopathology, v. 159, p. 94-99, 2011. GÖRGEN, C. A. Manejo do mofo branco da soja com palhada de Brachiaria ruziziensis e Trichoderma harzianum‘1306’. 2009. 72f. Dissertação ( Mestrado em Agronomia) - Universidade Federal de Goiás, Campus Jataí, 2009. GRAU, C. R. Sclerotinia stem rot. In: Compendium of soybean diseases. Saint. Paul, APS Press. 1989, p. 47-48. GUIMARÃES, R.L.; STOTZ, H.U. Oxalate production by Sclerotinia sclerotiorum deregulates guard cells during infection. Plant Physiology, v.136, p. 3703-3711, 2004. HAMBLETON S., WALKER C., KOHN L.M. Clonal lineages of Sclerotinia sclerotiorum previously known from other crops predominate in 1999–2000 samples from Ontario and Quebec soybean. Canadian Journal Plant Pathology, v. 3, n. 24, p. 309–315, 2002. HANCOCK, J. G. Degradation of pectic substances associated with pathogenesis by Sclerotinia sclerotiorum in sunflower and tomato stems. Phytopathology, v. 56, p. 975-979, 1966. HEGEDUS, D.D.; RIMMER, S.R. Sclerotinia sclerotiorum: When “to be or not to be” a pathogen? Fems Microbiology Letters, v. 251, p. 177-184, 2005.

40
HENNING, A. A. Patologia de sementes. Londrina: EMBRAPA-CNPSo, 1994. 43 p. (Documentos, 90) HENNING, A. A. Patologia e tratamento de sementes: noções gerais. Londrina: Embrapa Soja, 2004. 51 p. (Documentos 235). HOMECHIN, M. Determinação de perdas da produção de soja, devido a incidência do fungo Sclerotinia sclerotiorum. In: CONGRESSO BRASILEIRO DE FITOPATOLOGIA, 15. 1982, São Paulo. Anais... São Paulo: Sociedade Brasileira de Fitopatologia. p 476. HUANG, H.C; KOZUB, G.C. Longevity of normal and abnormal scleortia of Sclerotinia sclerotiorum. Plant Disease, v.78, n.12, p. 1164-1166, 1994. HUANG, H.C; Morphologically abnormal sclerotia of Sclerotinia sclerotiorum. Canadian Journal of Microbiology, v.28, p.87-91, 1983. IMEA. Instituto Mato-Grossense de Economia Agropecuária. Disponível em:< http://www.imea.com.br>. Acesso em: 29 out. 2012. ISAAKS, E. H.; SRIVASTAVA, R. M. Applied geostatistics. New York: Oxford University Press, 1989. 561p. JAIME-GARCIA, R.; ORUM, T.V.; FELIX-GASTELUM, R.; TRINIDAD-CORREA, R.; VANETTEN, H.D.; NELSON, M.R. Spatial analysis of Phytophthora infestans genotypes and late blight severity on tomato and potato in the del fuerte valley using geostatistics and geographic information systems. Phytopathology, v. 91, p.156-165, 2001. JULIATTI, F.; POLIZEL, A. C.; JULIATTI, F. C. Manejo integrado de doenças na cultura da soja. Uberlândia: Composer, 2004. 327 p. KIRK, P.M.; CANNON, P.F.; MINTER D.W. ; STALPERS, J.A. Ainsworth and Bisby’s Dictionary of The Fungi. 10th edition. Wallingford, UK: CAB International, 2008. 640 p. KOHN L.M., CARBONE I., ANDERSON J.B. Mycelial interactions in Sclerotinia sclerotioum. Experimental Mycology, v. 2, n.14, p. 255–267, 1990. KOHLI, Y.; MORRAL, A.A.; ANDERSON, J. B.; KOHN, L. C. Local and transcanadian clonal distribution of Sclerotinia sclerotiorum on Canola. Phytopathology, v. 82, p. 875-880, 1992. KULL L.S., PEDERSEN W.L., PALMQUIST D., HARTMAN G.L. Mycelial compatibility grouping and aggressiveness of Sclerotinia sclerotiorum. Plant Disease, v.4, n. 88, p. 325– 332, 2004.

41
LARANJEIRA, F. F. Epidemiologia da clorose variegada dos citros no Estado de São Paulo. 2002. 158f. Tese (Doutorado em Fitopatologia), Escola Superior de Agricultura Luiz de Queiroz, Piracicaba – SP, 2002. LARKIN, R. P.; GUMPERTZ, M. L.; RISTAINO, J. B. Geostatistical analysis of Phytophthora epidemic development in commercial bell pepper fields. Phytopathology, v. 85, n.2, p. 191-203, 1995. LEITE, R.M.V.B.C. Ocorrência de doenças causadas por Sclerotinia sclerotiorum em girassol e soja. Londrina: Embrapa Soja, 2005. 3p. (Embrapa Soja. Circular Técnica, 76). LE TOURNEAU, D. Morphology, cytology and physiology of Sclerotinia species in culture. Phytopathology, v. 69, p. 887–890, 1979. LITHOLDO JÚNIOR, C. G.; GOMES, E. V.; LOBO JÚNIOR, M.; NASSER, L. C. B.; PETROFEZA, S. Genetic diversity and mycelial compatibility groups of the plant-pathogenic fungus Sclerotinia sclerotiorum in Brazil. Genetics and Molecular Research, v. 10, n. 2, p. 868-877, 2011. LU, G. Engineering Sclerotinia sclerotiorum resistance in oilseed crops. African Journal of Biotechnology, v. 2, n 12, p. 509-516, 2003. LUMSDEN, R.D. Histology and physiology of pathogenesis in plant disease caused by Sclerotinia species. Phytopathology, v. 69, n. 8, p. 890-896, 1979. MACHADO, J. C. Tratamento de sementes no controle de doenças. Lavras:LAPS/UFLA/FAEPE, 2000. 138 p. MALVÁREZ, G.; CARBONE, I.; GRUNWALD, N. J.; SUBBARAO, K. V.; SCHAFER, M.; KOHN, L. M. New populations of Sclerotinia sclerotiorum from Lettuce in California and Peas and Lentils in Washington. Phytopathology, v. 97, p. 470-483, 2007. MASSOLA JR, N. S.; KRUGNER, T. L. Fungos fitopatogênicos. In: AMORIN, L.; REZENDE, J.A.M.; BERGAMIN FILHO, A. Manual de fitopatologia: princípios e conceitos. São Paulo: Ceres, 2011. Cap.8, p. 149-206. MAXWELL, D. P.; LUMSDEN, R.D. Oxalic acid production by Sclerotinia sclerotiorum in infected bean and in culture. Phytopathology, v. 60, p. 1395-1398, 1970. MCDONALD, B. A. The populations genetics of fungi: tools and techniques. Phytopathology, v. 87, n. 4, p. 448-452, 1997.

42
MEINHARDT, L.W.; NELSON, A.W.; BELLATO, C.M.; TSAI, S.M. Telomere and microsatellite primers reveal diversity among Sclerotinia sclerotiorum isolates from Brazil. Fitopatologia Brasileira, v. 27, n. 2, p. 211-215, 2002. MENTEN, J. O. M.; PARADELA, A.; GALLI, M. A.; GONELLA, L. G. R. Eficiência in vitro de diversos fungicidas sobre Sclerotinia sclerotiorum do feijoeiro. In: CONGRESSO BRASILEIRO DE FITOPATOLOGIA, 28. 1995, Ilhéus. Anais... Ilhéus: Sociedade Brasileira Fitopatologia. P. 320. MUELLER, D. S.; DORRANCE, A. E.; DERKSEN, R. C.; OZKAN, E.; KURLE, J. E.; GRAU, C. R.; GASKA, J. M.; HARTMAN, G. L.; BRADLEY, C. A.; PEDERSEN, W. L. Efficacy of fungicides on Sclerotinia sclerotiorum and their potencial for control of Sclerotinia stem rot on soybean. Plant Disease, v. 86, n. 1, p. 26-31, 2002.
NAPOLEÃO, R.; CAFÉ FILHO, A. C.; NASSER, L. C. B.; LOPES, C. A.; SILVA , H. R. Intensidade do mofo branco do feijoeiro em plantio convencional e direto sob diferentes lâminas d'água. Fitopatololgia Brasileira, v. 30, n. 4, p.374-379, 2005. NAPOLEÃO, R.; NASSER, L. C. B.; LOPES, C. A.; CAFÉ FILHO, A. C. Neon-S, um novo meio de detecção de Sclerotinia sclerotiorum em sementes. Summa Phytopathologica, v. 32, n. 2, p. 180-182, 2006. NAPOLEÃO, R. L. Efeito do sistema de plantio, da irrigação e do espaçamento sobre o mofo branco do feijoeiro, causado por Sclerotinia sclerotiorum. 2001. 72f. Tese (Doutorado em Fitopatologia) - UNB, Brasília, 2001. NASSER, L.C.B.; ANJOS, J.R.N.; PERES, J.R.R.; MEDEIROS, A.C.S.; SPEHAR, C.R.; UIBEN FILHO, G.; SOUSA, P.I.M. Fungicidas para tratamento de sementes de soja. Planaltina; Embrapa cerrados, 1984, 6p. (Comunicado técnico 40). NASSER, L. C. B.; BOLAND, G. J.; SUTTON, J. C. Meio semi-seletivo para detecção da viabilidade de escleródios de Sclerotinia sclerotiorum. Fitopatologia Brasileira, v.20, supl, p.376, 1995. NASSER, L. C. B.; ARANCIBIA, R. C.; NAPOLEÃO, R. Uso do meio Neon modificado para determinação de Sclerotinia sclerotiorum em sementes de feijão produzidas em áreas irrigadas do cerrado. Fitopatologia Brasileira, v. 24, suplem., p.309, 1999. NCBI, The NCBI Taxonomy Homepage. Disponível em: http://www.ncbi.nlm.nih.gov/Taxonomy/taxonomyhome.html/. Acesso em: 25 de jan. de 2013.

43
NATTI, J.J. Epidemiology and control of bean white mold. Phytopathology, v.61, p-669-674, 1978. NELSON, M. R.; ORUM, T. V.; JAIME-GARCIA, R.; NADEEM, A. Applications of geographic information systems and geostatistics in plant disease epidemiology and management. Plant Disease, v. 83, n. 4, p. 308-319, 1999. NOYES, R,D.; HANCOCK, J,G. Role of oxalic acid in the Sclerotinia sclerotiorum wilt of sunflower Helianthus annuus. Physiol Plant Pathology, v.18, p. 123–132, 1981. NUNES JÚNIOR, J.; PIMENTA, C. B.; NUNES SOBRINHO, J. B.; FERREIRA, L. C.; COSTA, N. B.; ANDRADE, P. J. M.; MEYER, M. C. Avaliação da eficácia de fungicidas no controle de mofo branco (Sclerotinia sclerotiorum) em soja, no Estado de Goiás. Tropical Plant Pathology, v. 34, p. 102, 2009. OTTO-HANSON, L.; STEADMAN, J. R.; HIGGINS, R.; ESKRIDGE, K. M. Variation in Sclerotinia sclerotiorum bean isolates from multisite resistance screening locations. Plant Disease, v. 95, p. 1370-1377, 2011. PAULA JÚNIOR, T. J.; VIEIRA, R. F.; TEIXEIRA, H.; CARNEIRO, J. E. S. Manejo do mofo branco do feijoeiro. Belo Horizonte: Epamig, mar. 2008, 4 p. (Circular Técnica n. 13). PAULA JÚNIOR, T. J.; VIEIRA, R. F.; LOBO JÚNIOR, M.; MORANDI, M. A. B.;CARNEIRO, J. E. S.; ZAMBOLIM, L. Manejo integrado do mofo branco do feijoeiro. Viçosa: Epamig, 2006. 48 p. PITOL,C. Relatos por estado sobre o comportamento da cultura de soja na safra 2007/2008: Mato Grosso do Sul. In: REUNIÃO DE PESQUISA DA SOJA DA REGIÃO CENTRAL DO BRASIL, 30., 2009. Anais... Londrina, EMBRAPA-Soja, 2009. Sessao 2.3, p.24. Londrina: Embrapa Soja, 2009. 350 p. PHILIPS, A. J. L. Carpogenic germination of sclerotia of Sclerotinia sclerotiorum. Phytophylactica, v. 19, n. 3, p.279-283, 1987. PUNJA, Z. K. The biology, ecology and control of Sclerotium rolfsii. Annual. Review of Phytopathology, v. 23, p.97-127, 1985. PURDY, L.H. Sclerotinia sclerotiorum: history, diseases and symptomatology, host range, geographic distribution, and impact. Phytopathology, v. 69, p. 875- 880, 1979.

44
REIS, E. M.; TOMAZINI, S. L. Viabilidade de escleródios de Sclerotinia sclerotiorum, no campo, em duas profundidades do solo. Summa Phytopathologica, v.31, n. 1, p.97-99, 2005. RIBEIRO, J. R. AND DIGGLE, P. J. GeoR: A package for geostatistical analysis. R-News, v.1. n. 2, 2001. Disponivel em: http://www.r-project.org/doc/Rnews/Rnews_2001-2.pdf. Acesso em: 20 de jun.de 2013. ROCHA, R.P. Manejo da podridão de sclerotinia (Sclerotinia sclerotiorum) e míldio (Bremia lactucae) na cultura da alface. 2007. Dissertação (Mestrado em Agronomia) – Universidade Estadual de Ponta Grossa, Paraná. 2007. SANTOS, J. R. M.; CHARCHAR, M. J.; NASSER, L. C. B. Levantamento de patógenos que afetam ervilha irrigada no Distrito Federal. Fitopatologia Brasileira, v. 15, p. 98-99, 1990. SCHWARTZ, H.F.; STEADMAN, J.R. Factors affecting sclerotium populations of, and apothecium production by Sclerotinia sclerotiorum, Phytopathology, v. 68, n. 383-388, 1978. SCHWARTZ, H.F.; STEADMAN, J.R. White mold. In: SCHWARTZ, H.F.; PASTOR-CORSALES, M.A. (ed.). Bean production problems in the tropics. 2. ed. Cali: CIAT, 1989. p. 736. SILVA, L. H. C. P.; CAMPOS, H. D.; SILVA, J. R. C. Manejo do mofo branco da soja. In: SILVA, L. H. C. P.; CAMPOS, H. D.; SILVA, J. R. C. (Ed.). Manejo fitossanitário de cultivos agroenergéticos. Lavras: UFLA, 2010. p. 205-214. SOUZA, E.D.T. Métodos de Inoculação para avaliação e seleção de germoplasma de Phaseolus spp. quanto à resistência a Sclerotinia sclerotiorum (Lib.) de Bary. 1999. 82f. Dissertação (Mestrado em Agronomia) – Universidade Federal de Goiás, Goiânia, 1999. SUTTON, D.C.; DEVERAL, B.J. Studies on enfection of bean (Phaseolus vulgaris) and soybean (Glycine max) by ascósporos of Sclerotinia sclerotiorum. Plant Pathology, v.32, p. 251-261, 1983. STEADMAN, J.R. White mold – a serius yield-limiting desease of bean. Plant Disease, v. 67, n.4, p. 346-350, 1983. TOURVIEILLE, D., VEAR, F. Heredity of resistance to Sclerotinia sclerotiorum in sunflower. Study of reaction to artificial infection of roots and cotyledons. Agronomie, v.10, p. 323–330, 1990.

45
TU, C. The role of mold-infected white bean (Phaseolus vulgaris L.) seeds in the dissemination of Sclerotinia slcerotiorum de Bary. Journal of Phytopathology, v. 121, n. 1, p.40-50, 1998. TU , J. C. Gliocadium virens, a destructive mycoparasite of Sclerotinia sclerotiorum. Phytopathology, v. 70, n. 7, p. 670-674, 1980. VIEIRA, R. O mofo branco do feijoeiro – feijão no inverno. Informe Agropecuário, v. 17, n. 178. p. 54-63, 1994. VIEIRA, S. R. Geoestatística aplicada a agricultura de precisão. IN: BORÉM, A.; GIÚDICE, M. P.; QUEIROZ, D. M.; MANTOVANI, E. C.; FERREIRA, L. R.; VALLE, F. X. R.; GOMIDE, R. L. Agricultura de Precisão, 2000. p. 93-108. VENETTE, J. Sclerotinia spore formation, transport, and infection. In: Proceedings of the sclerotinia. Workshop. North Dakota State University. 21 de janeiro de 1998. Fargo North Dakota. USA. WALLACE, M. K.; HAWKINS, D. M. Applications of geoestatistics in plant nematology. Journal of Nematology, n. 26, n.4S, p. 626-634. 1994. WHARTON, P; KIRK, W. White mold Michigan potato diseases series; MSU Extension bulletin E-2989, 2007. WILLETTS, H.J.; WONG, J.A.L. The biology of Sclerotinia sclerotiorum, S. trifoliorum, and S. minor with emphasis on specific nomenclature. The Botanical Review, v. 46, p.101-165, 1980. YANG, X. B.; LUNDEEN, P.; UPHOFF, D. Soybean varietal response and yield caused by Sclerotinia sclerotiorum. Plant Disease, v. 83, n. 5, p.456-461, 1999. YORINORI, J. T. Soja. In: VALE, F. X. R. do; ZAMBOLIM, L. Controle de doenças de plantas. Viçosa: UFV, 1997. p. 973. ZANETTI, A. L. Relatos por Estado sobre o comportamento da cultura de soja na safra 2007/2008: Minas Gerais. Reunião de Pesquisa da Soja da Região Central do Brasil, 30., 2009. Anais... Londrina, Embrapa-Soja, 2009. Sessão 2.3, p. 24. ZITO, R. K.; WRUCK, D. S. M.; FRONZA, V.; ARANTES, N. E. Reação de genótipos de soja a Sclerotinia sclerotiorum. In: REUNIÃO DE PESQUISA DE SOJA DA REGIÃO CENTRAL DO BRASIL, 27, 2005, Cornélio Procópio. Anais... Embrapa Soja, p. 362.

46
3 CARACTERÍSTICAS MORFOLÓGICAS E FISIOLÓGICAS DE
Sclerotinia sclerotiorum ISOLADO DA CULTURA DA SOJA E USO
DA RESTRIÇÃO HÍDRICA
RESUMO - Os objetivos neste trabalho foram caracterizar, fisicamente,
escleródios de S. sclerotiorum em soja, pela avaliação do tamanho e da
massa de escleródios; estudar a germinação de escleródios e formação de
apotécios em relação ao tamanho do escleródio; determinar o potencial
osmótico e o período adequado para inoculação artificial de S. sclerotiorum
em sementes e verificar a viabilidade dos métodos de detecção “blotter
test”, neon e rolo de papel para detecção do patógeno em lotes de
sementes de soja contaminadas. A partir de amostras de escleródios, os
mesmos foram separados visualmente em quatro tamanhos. Após foram
determinados o comprimento, a largura e a espessura. Para determinar o
potencial osmótico foi utilizado o soluto NaCl com BDA, nos potenciais: 0,0;
-0,4; -0,6; -0,8; -1,0; e -1,2 MPa. As variáveis analisadas foram índice de
velocidade de crescimento micelial, quantificação e massa de escleródios.
Para determinação do tempo adequado de inoculação, os tratamentos
usados foram cinco períodos: 24, 48, 72, 96 e 120 h, mais o controle O
comprimento, a largura e a espessura dos escleródios de S. sclerotiorum
em soja variam de 2,41 a 13,62 mm; 2,06 a 3,46 mm e 1,58 a 2,38 mm,
respectivamente. Com o aumento do tamanho dos escleródios, o número e
comprimento de apotécios produzidos por escleródio também aumenta. O
meio mais adequado para inoculação de S. sclerotiorum em sementes de
soja é o BDA sem potencial osmótico. O período mais eficiente para a
inoculação é 72 horas e o método rolo de papel detecta de forma eficiente o
patógeno em lotes com 5% de contaminação.
Palavras-chave: Glycine max, mofo branco, germinação carpogênica.

47
MORPHOLOGICAL AND PHYSIOLOGICAL CHARACTERISTICS OF
ISOLATED Sclerotinia sclerotiorum SOYBEAN CROP AND USE OF
WATER RESTRICTION
ABSTRACT - The objectives of this study were to characterize physically
sclerotia of S. sclerotiorum in soybean by evaluating the size and mass of
sclerotia ; study the germination and the formation of sclerotia apothecia in
relation to the size of sclerodia; osmotic potential and determine the suitable
period for artificial inoculation S. sclerotiorum in seeds and verify the
feasibility of the detection methods " blotter test " , neon and scroll to detect
the pathogen in seed lots contaminated soybeans. From samples of
sclerotia, they were separated visually in four sizes. After were determined
length, width and thickness. To determine the osmotic potential was used
with NaCl solute PDA, the potentials of 0.0, -0.4, -0.6 , -0.8, -1.0 , and -1.2
MPa . The variables were speed index mycelial growth, quantification and
mass of sclerotia. For determining the timing of inoculation , treatments used
were five periods : 24, 48 , 72, 96 and 120 h , most control the length, width
and thickness of the sclerotia of S. sclerotiorum in soybean varies from 2.41
to 13.62 mm, 2.06 to 3.46 mm 1.58 to 2.38 mm, respectively . With the
increase in size of sclerotia the number and length of apothecia produced by
sclerotia also increases. The most appropriate means of inoculation of S.
sclerotiorum in soybean seeds is the PDA without osmotic potential. The
most effective period for inoculation is 72 hours and the paper roll method to
efficiently detect the pathogen batch with 5% contamination.
Keywords: Glycine max, white mold, carpogenic germination.

48
3.1 Introdução
O mofo branco, causado pelo fungo Sclerotinia sclerotiorum, é hoje
no Brasil um dos principais problemas nas áreas produtoras de soja,
principalmente no sul e no centro do Brasil. Nesses locais o fungo encontra
condições ótimas de crescimento em temperaturas mais amenas de 15 a 25
°C, e ambientes com alta umidade, condições essas favoráveis para o
desenvolvimento da doença (Purdy, 1979).
O escleródio é um agregado de hifas com o exterior preto formado
por melanina, que desempenha papel importante no ciclo da doença, sendo
a principal estrutura de sobrevivência do patógeno a longo prazo (Willetts e
Wong, 1980). Segundo Reis e Tomazini (2005), o período de sobrevivência
dos escleródios no solo, em condições de campo, varia com a sua
profundidade. Escleródios apresentam maior viabilidade quando enterrados
a 10 cm do solo e a mantém por até 35 meses e, quando mantidos na
superfície do solo, por 13 meses. Alguns autores, como Adams e Ayres
(1979), citaram a persistência dos escleródios no solo por mais de oito
anos.
Apesar de sua importância no ciclo da doença, não há referências
sobre as características biométricas dos escleródios de S. sclerotiorum em
soja e de informações sobre a germinação e formação de apotécios.
A utilização da restrição hídrica, por meio dos solutos Manitol, NaCl e
KCl, revela-se como um procedimento promissor para impedir ou reduzir a
germinação de sementes de soja por ocasião do teste de incubação em
substrato de papel visando a detecção dos principais fungos associados às
sementes desta espécie (Machado et al., 2003). Os testes de sanidade
aplicados para detectar S. sclerotiorum em sementes baseiam-se na
presença ou ausência de escleródio em uma amostra de sementes, sem
considerar a presença do micélio do patógeno no interior ou na superfície
das sementes, sendo esta a principal forma de trasmissão da doença
(Peres, 1996). Prejuízos causados por este fungo à cultura da soja têm
exigido estudos sobre os métodos de inoculação artificial que representem

49
com maior eficiência tanto a detecção do patógeno como a inoculação
natural. Em lotes de sementes infectadas naturalmente, ou seja, oriundas
de lotes provenientes de áreas infectadas com S. sclerotiorum, geralmente
o patógeno não é facilmente detectado (Nasser et al., 1999; Henning et al.,
2009), havendo necessidade de aumentar o período de incubação de sete
dias para trinta dias (Brasil, 2009), quando se utiliza o método de detecção
“blotter test”.
Machado et al. (2007) desenvolveram uma metodologia de
inoculação de sementes de soja com os fungos Colletotrichum truncatum,
Phomopsis sojae e Sclerotinia sclerotiorum por meio da técnica de potencial
osmótico, de modo a obter sementes infectadas por estes patógenos.
Napoleão et al. (2006) e Parisi et al. (2006) evidenciaram a necessidade de
modificação dos testes de detecção, neon e rolo de papel, para efetivar a
identificação do patógeno e reduzir o tempo de detecção em sementes
inoculadas com 100% de infecção.
Em condições naturais de campo, lotes de semente de soja
contaminados com S. sclerotiorum apresentam baixa incidência do
patógeno na semente, em torno de 2%, quando submetidos a testes de
sanidade (Peres, 1996). Para Hennenberg et al. (2011), é necessária a
realização de estudos envolvendo métodos de detecção mais sensíveis de
S. sclerotiorum em sementes de soja. Portanto, os testes de detecção
devem ser sensíveis, reprodutíveis e viáveis economicamente. Dessa
forma, tornam-se importantes pesquisas sobre a capacidade de
recuperação de S. sclerotiorum por meio dos testes de detecção em lotes
de sementes de soja com baixos níveis de contaminação.
Este estudo objetivou caracterizar morfologicamente escleródios de
S. sclerotiorum em soja provenientes da região de Jaciara-MT, verificar a
germinação e formação de apotécios em relação ao tamanho do escleródio,
determinar o potencial osmótico e o período adequado para inoculação
artificial de S. sclerotiorum em sementes e verificar a viabilidade dos
métodos de detecção “blotter test”, neon e rolo de papel para detecção do
patógeno em lotes de sementes de soja.

50
3.2 Material e Métodos
3.2.1 Escleródios – biometria, germinação e formação de apotécios
Foram avaliados 5.000 escleródios provenientes de área
naturalmente infectada com mofo branco, na Fazenda São Jorge, município
de Jaciara-MT, cujas coordenadas geográficas são: 15º 48’ 51,05”S e 55º
12’ 86,96”W e altitude média de 750 m. Os escleródios foram coletados no
dia da colheita de soja da variedade P 98Y11, em um talhão com alta
incidência e severidade da doença na safra 2011/12. Os escleródios foram
coletados nas plantas e na massa de grãos na hora da colheita. Nessa
área, durante a condução da cultura da soja foram realizadas duas
aplicações de fungicida (Azoxtrobin+Ciproconazol) para controle da
ferrugem asiática (Phakopsora pachyrhizi).
No Laboratório de Sementes da Faculdade de Agronomia, Medicina
Veterinária e Zootecnia (FAMEVZ) da Universidade Federal de Mato Grosso
(UFMT), Cuiabá – MT, os escleródios coletados foram separados
visualmente em quatro tamanhos (Figura 2): muito pequeno (MP), pequeno
(P), médio (M) e grande (G). Após foram determinados o comprimento
(mm), a largura (mm), a espessura (mm), todos com uso de paquímetro
digital de precisão de 0,01, e a massa de escleródios (g) com o uso de
balança digital, de precisão de 0,0001 g. As médias dos dados biométricos
dos escleródios foram calculadas com 100 unidades por tamanho.

51
FIGURA 2. Separação dos escleródios muito pequeno (MP), pequeno (P),
médio (M) e grande (G) de S. sclerotiorum, coletados em
lavoura de soja. UFMT, Cuiabá-MT, 2012.
Foi utilizado o delineamento experimental inteiramente casualizado
com quatro tratamentos (tamanhos de escleródios) e cinco repetições.
Para o teste de germinação de escleródios, para cada tamanho (MP,
P, M e G), foram utilizadas 100 unidades em cinco subamostras de 20
escleródios. Estes foram colocados para germinar em solo tipo Latossolo
Vermelho textura argilosa coletado na região da fazenda, mas sem
infestação de escleródios. O solo foi esterelizado e, após, distribuído em
caixas de plástico tipo “gerbox”. Cada parcela foi composta por uma caixa
contendo 250 g de solo e sobre esse substrato, os escleródios (20 por
caixa) foram distribuídos equidistantemente. O solo foi umedecido com
100% da capacidade de campo com base na massa (Brasil, 2009) e
reumedecido semanalmente. Essas caixas foram incubadas à temperatura
de 20 °C ± 2 °C com fotoperíodo de 12 horas luz, por 100 dias.
Foram realizadas avaliações do número de escleródios germinados,
número de estipes e número de apotécios por escleródio e tamanho de
apotécios aos 27, 40, 51, 70 e 100 dias após o início da incubação (DAI). A
MP P M G

52
massa (g) de apotécios dos escleródios foi avaliada com o uso de balança
digital, de precisão de 0,0001 g e o comprimento com paquímetro digital de
precisão de 0,01, aos 51 DAI.
Os resultados foram analisados pelo teste F e as médias comparadas
pelo teste de Tukey a 5% de probabilidade, utilizando-se o programa
computacional SISVAR 4.2 (Ferreira, 2003).
3.2.2 Determinação do potencial osmótico, do período adequado para
inoculação artificial de S. sclerotiorum em sementes de soja e da
viabilidade dos métodos “blotter test”, neon e rolo de papel para
detecção do patógeno em lotes de sementes de soja contaminados
Para esses experimentos foram utilizadas sementes de soja da
variedade SYN 9070 RR, nos Laboratórios de Sementes e de Fitopatologia
da FAMEVZ/ UFMT, em Cuiabá - MT.
3.2.2.1 Determinação do potencial osmótico para o desenvolvimento
de S. sclerotiorum
O experimento foi conduzido em delineamento inteiramente
casualizado com seis tratamentos (potenciais osmóticos) e 20 repetições.
O fungo S. sclerotiorum foi isolado de sementes de soja submetidas a
análise de rotina (blotter test), no laboratório de Fitopatologia, e colocado
em meio de cultura BDA (extrato de 200 g de batata, 20 g de dextrose e 20
g de ágar), ao qual foi adicionado o soluto NaCl nos potenciais osmóticos:
0,0 (controle); -0,4; -0,6; -0,8; -1,0 e -1,2 MPa .
O meio BDA+NaCl, nos referidos potenciais, foi distribuído em placas
de Petri de 15 cm de diâmetro, na proporção de 50 mL por placa, sendo em
seguida transferido, para cada meio, um disco de 0,6 cm de diâmetro da
colônia pura do fungo. As placas foram colocadas para incubação a 20 ± 2
°C e fotoperíodo de 12 horas por 10 dias.
As avaliações foram realizadas por meio do índice de velocidade de
crescimento micelial (IVCM), quantificação e massa de escleródios. Para o

53
IVCM, as avaliações foram feitas diariamente durante o período de
incubação, medindo-se o diâmetro médio das colônias e aplicando a
fórmula utilizada por Oliveira (1991):
IVCM = C1/N1+ C2/N2 + ... + Cn/Nn (1)
Em que:
IVCM = índice de velocidade de crescimento micelial;
C1, C2 e Cn = crescimento micelial na primeira, segunda e n-ésima
avaliação;
N1, N2 e Nn = número de dias de avaliação.
A quantificação foi realizada pelo número de escleródios formados
por repetição e a massa por meio de balança com precisão de 0,0001 g.
3.2.2.2 Período adequado de inoculação de S. sclerotiorum em
sementes de soja e viabilidade dos métodos de detecção “blotter test”,
neon e rolo de papel para recuperação do patógeno em lotes de
sementes de soja
Logo após a determinação do potencial osmótico para inoculação, foi
verificado o período de inoculação de S. sclerotiorum em sementes de soja,
em delineamento inteiramente casualizado com seis tratamentos e quatro
repetições. Os tratamentos foram cinco períodos de inoculação (24, 48, 72,
96 e 120 h) mais a testemunha em quatro repetições, compondo o total de
24 parcelas.
Trabalhou-se com o mesmo isolado de S. sclerotiorum com meio
BDA sem restrição em placas de Petri de 15 cm de diâmetro. Após o
período de desenvolvimento do fungo no meio, as sementes de soja foram
inoculadas pelo período estabelecido nos tratamentos. Cada placa continha
150 sementes distribuídas sobre a colônia do fungo, em camada única, e
levemente prensadas com os dedos, sobre o meio.
As placas de Petri contendo o fungo com as sementes de soja foram
vedadas com filme de polipropileno e distribuídas em BOD com temperatura
de 20 °C ± 2 ºC pelo período estabelecido nos tratamentos, sob escuro
contínuo.

54
Ao término de cada período de inoculação, as sementes foram
retiradas das placas, secadas em temperatura ambiente por 24 horas,
colocadas diretamente em sacos de papel e misturadas às sementes
sadias, tendo-se obtido seis lotes de sementes com 5% de contaminação,
na proporção de cinquenta sementes para mil sementes, para cada período
de inoculação, com finalidade de representar a contaminação natural em
campo. Os lotes de sementes contaminados artificialmente foram
armazenados em câmara refrigerada à temperatura de 18 ± 2 ºC e 75 ± 4%
de umidade relativa. Após 30 dias, foram realizados os testes de sanidade
“blotter test” e Neon, para detecção do patógeno e confirmação da
inoculação. Foi realizado o teste padrão de germinação (Brasil, 2009a) dos
lotes de sementes de soja inoculados e avaliada a influência do período de
inoculação sobre a porcentagem de germinação após 30 dias de
inoculação.
Para verificar a viabilidade dos métodos “blotter test”, neon e rolo de
papel para detecção do patógeno em lotes de sementes de soja
contaminados, foi realizado um experimento em delineamento inteiramente
casualizado, com dezoito tratamentos e quatro repetições. Os tratamentos
foram seis periodos de inoculação de sementes com 5% de sementes
contaminadas, cada lote representando um período de inoculação (24, 48,
72, 96 e 120 h) mais a testemunha, submetidos a três testes de detecção de
patógenos em sementes em quatro repetições, compondo o total de 72
parcelas.
Blotter test: Duzentas sementes de soja inoculadas com S.
sclerotiorum de cada tratamento (50 sementes/sub-amostra) foram
distribuídas em placas de Petri (15 cm) contendo três folhas de papel de
filtro qualitativo previamente umedecidas com solução de NaCl a - 0,8 MPa
acrescido de ágar (0,2 g de ágar diluído em 1.000 mL de água e 10,3 g de
NaCl) e incubadas por sete dias à 22 oC, sob fotoperíodo de 12 horas
(lâmpadas fluorescentes tipo “luz do dia” e negra “NUV”) conforme Brasil
(2009a). Após o período de incubação, foi avaliado sob microscópio

55
estereoscópico a presença e ausência do fungo nas sementes, sendo os
resultados expressos em porcentagem.
Incubação em rolo de papel: seguindo as mesmas recomendações
do teste de germinação conforme Brasil (2009a), porém sendo mantidos em
câmara de incubação por 14 dias a 15 ºC, em posição vertical, na ausência
de luz, e acondicionados em recipientes com atmosfera próxima a
saturação (Brasil, 2009a).
Incubação em meio ágar-bromofenol (Neon-S): Foi utilizado o
meio BDA (extrato de 200 g de batata, 20 g de dextrose e 20 g de ágar)
fundido (50 °C) após autoclavagem sendo adicionado 150 mg/L de azul de
bromofenol, 50 g de clorafenicol e pH final ajustado para 4,7 com HCl.
Posteriormente foram colocados 20 mL do meio por placa de Petri de 9 cm
de diâmetro e depois as sementes foram distribuídas sobre o meio. As
placas ficaram expostas a luz negra, com 12 horas de fotoperíodo a 20 ºC
por 8 dias (Brasil, 2009a). A partir do terceiro dia de incubação, foram
observadas a formação de halos amarelo-avermelhados ao redor das
sementes. Para a confirmação da presença de S. sclerotiorum, foi feito a
visualização do micélio superficial sobre o meio de cultura na zona do halo
amarelado, partindo das sementes.
Os resultados foram previamente submetidos aos testes de
normalidade (Lillefors) e de homogeneidade (Cochran), e posteriormente
realizada a análise de variância. A análise dos dados foi feita pelo teste F e
as médias comparadas pelo teste de Scott-Knott a 5% de probabilidade.
Quando necessário os dados de porcentagem foram transformados para
.
Para a análise foi usado o programa estatístico SISVAR 4.2 (Ferreira,
2003).

56
3.3 Resultados e Discussão
3.3.1 Escleródios – biometria, germinação e formação de apotécios
Nas amostras onde foram separados visualmente os escleródios em
quatro tamanhos (Figura 2), observou-se microescleródios que não foram
medidos por serem menores que um milimetro de comprimento e espessura
(Figura 3), constatando-se grande quantidade desses microescleródios nas
amostras originadas da lavoura de soja. Por serem muito pequenos esses
microescleródios podem estar infestando lotes de sementes sem serem
detectados, aderidos às sementes, e não são separados das mesmas, em
áreas onde produtores usam sementes “salvas”, sem beneficiamento.
FIGURA 3. Amostra contendo escleródios muito pequeno, pequeno, médio,
grande e microescleródios (no círculo) de S. sclerotiorum,
coletados em lavoura de soja. UFMT, Jaciara-MT, 2012.
Outro aspecto importante relacionado aos microescleródios é a sua
dispersão dentro do talhão e para outros locais da propriedade favorecido
pela pequena massa e reduzido tamanho. Como são muito pequenos
podem ser levados pela chuva, vento, implementos e máquinas agrícolas,
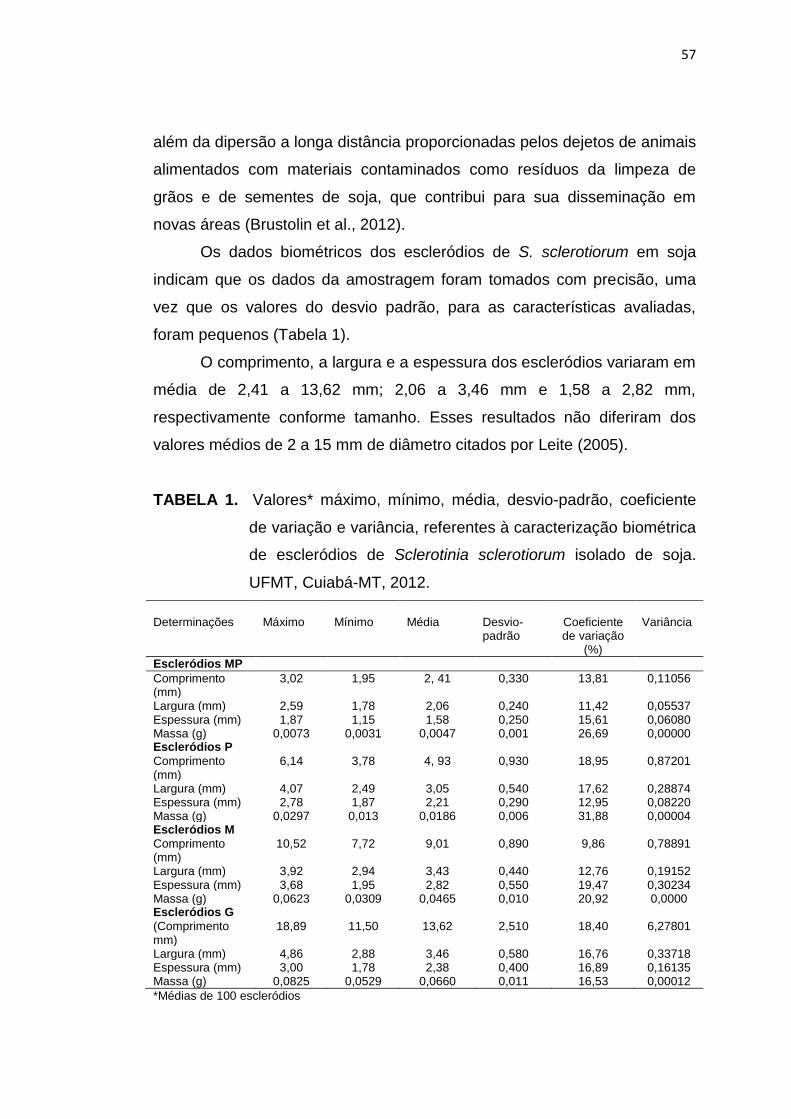
57
além da dipersão a longa distância proporcionadas pelos dejetos de animais
alimentados com materiais contaminados como resíduos da limpeza de
grãos e de sementes de soja, que contribui para sua disseminação em
novas áreas (Brustolin et al., 2012).
Os dados biométricos dos escleródios de S. sclerotiorum em soja
indicam que os dados da amostragem foram tomados com precisão, uma
vez que os valores do desvio padrão, para as características avaliadas,
foram pequenos (Tabela 1).
O comprimento, a largura e a espessura dos escleródios variaram em
média de 2,41 a 13,62 mm; 2,06 a 3,46 mm e 1,58 a 2,82 mm,
respectivamente conforme tamanho. Esses resultados não diferiram dos
valores médios de 2 a 15 mm de diâmetro citados por Leite (2005).
TABELA 1. Valores* máximo, mínimo, média, desvio-padrão, coeficiente
de variação e variância, referentes à caracterização biométrica
de escleródios de Sclerotinia sclerotiorum isolado de soja.
UFMT, Cuiabá-MT, 2012.
Determinações
Máximo
Mínimo
Média
Desvio-padrão
Coeficiente de variação
(%)
Variância
Escleródios MP
Comprimento (mm)
3,02 1,95 2, 41 0,330 13,81 0,11056
Largura (mm) 2,59 1,78 2,06 0,240 11,42 0,05537 Espessura (mm) 1,87 1,15 1,58 0,250 15,61 0,06080 Massa (g) 0,0073 0,0031 0,0047 0,001 26,69 0,00000 Escleródios P Comprimento (mm)
6,14 3,78 4, 93 0,930 18,95 0,87201
Largura (mm) 4,07 2,49 3,05 0,540 17,62 0,28874 Espessura (mm) 2,78 1,87 2,21 0,290 12,95 0,08220 Massa (g) 0,0297 0,013 0,0186 0,006 31,88 0,00004 Escleródios M Comprimento (mm)
10,52 7,72 9,01 0,890 9,86 0,78891
Largura (mm) 3,92 2,94 3,43 0,440 12,76 0,19152 Espessura (mm) 3,68 1,95 2,82 0,550 19,47 0,30234 Massa (g) 0,0623 0,0309 0,0465 0,010 20,92 0,0000 Escleródios G (Comprimento mm)
18,89 11,50 13,62 2,510 18,40 6,27801
Largura (mm) 4,86 2,88 3,46 0,580 16,76 0,33718 Espessura (mm) 3,00 1,78 2,38 0,400 16,89 0,16135 Massa (g) 0,0825 0,0529 0,0660 0,011 16,53 0,00012
*Médias de 100 escleródios

58
Para a massa dos esclerodios, foi constatada uma grande variação
entre tamanhos, 0,0047 a 0,0660 g, ainda que o desvio padrão tenha
expressado os menores valores. Os maiores valores de desvio padrão
indicam que há maior variância amostral para os comprimentos de
escleródios, em relação às demais características avaliadas (Tabela 1).
As formas e dimensões dos escleródios variam em função do
hospedeiro onde eles são produzidos. Na cultura do girassol, por exemplo,
os escleródios podem variar de 1 a 35 cm de diâmetro, podendo tomar
dimensões semelhantes às de um capítulo floral. Na cultura do feijão
observaram-se escleródios globosos variando de 2 a 10 mm de diâmetro
(Bolton et al., 2006).
No teste de germinação verificou-se que aproximadamente 100% dos
escleródios germinaram. Observou-se que não houve diferença entre os
tamanhos de escleródios quanto à germinação carpogênica, pois ambos os
tamanhos germinaram, entretanto os escleródios de tamanho grande
produziram estipes em maior quantidade em relação aos tamanhos
menores (Tabela 2).
TABELA 2. Números de escleródios, de estipes e de apotecios nos
tamanhos muito pequeno (MP), pequeno (P), médio (M) e
grande (G) de S. sclerotiorum. UFMT, Cuiabá-MT, 2012.
DAI (dias após incubação). Médias seguidas da mesma letra, na coluna, não diferem estatitiscamente
pelo teste Tukey a 5% de probabilidade.
Houve diferença entre tamanho de escleródios na formação de
estipes (p<0,05). Decorridos 27 dias após a incubação, observou-se que
Tamanho de
escleródios
Escleródios germinados
(27 DAI)
Escleródios germinados
(51 DAI)
Estipes (27 DAI)
Estipes (40 DAI)
Estipes (51 DAI)
Estipes (70 DAI)
Estipes (100DAI)
estipes/ escleródio (27 DAI)
MP 19,4ns
20ns
96,6 b 105,6 b 114,4 c 99,8 c 89,0 c 5,0 b P 20,0 20 107,6 b 136,0 a 154,0 b 143,6 a 123,8 a 5,9 ab
M 19,0 20 101,6 b 138,0 a 148,6 b 130,2 b 99,6 b 5,3 ab
G 20,0 20 133,0 a 145,4 a 159,8 a 141,6 a 121,0 a 6,7 a
CV % 3,42 - 6,69 4,01 2,98 2,69 5,38 12,9

59
havia grande quantidade de estipes e todos os escleródios haviam
germinado, entretanto sem formação de apotécio. A proporção de estipes
por escleródios foi superior nos escleródios de tamanho grande, com média
de 6,7 estipes por escleródios (Tabela 2), porém não diferiram dos
tamanhos P e M.
Na avaliação aos 51 dias, os escleródios grandes, de tamanho G,
formaram apotécios com média de 12,7 mm de comprimento, e diferenciou
do número de apotécios formados dos demais tamanhos (Tabela 3).
Embora não diferenciem do tamanho M em comprimento médio de
apotécios, os escleródios grande, diferenciaram na velocidade de formação
da estrutura de reprodução que formam os ascósporos. Não houve
regressão de estipes e sim transformaram-se em apotécios.
TABELA 3. Número de apotécios produzidos de S. sclerotiorum a partir de
escleródios, muito pequenos (PP), escleródios pequenos (P),
médios (M) e grandes (G). UFMT, Cuiabá-MT, 2012.
Tamanho de escleródios
Apotécios* (40 DAI)
Apotécios* (51 DAI)
Apotécios* (70 DAI)
Apotécios* (100 DA)
Comprimento apotécios (mm)
(51 DAI)
MP 1,0 c 7,4 c 7,6 c 8,8 c 9,3 b P 1,0 c 32,6 b 16,8 b 22,0 b 10,6 b M 1,8 b 35,2 b 15,4 b 43,6 a 13,2 a G 3,0 a 62,0 a 19,8 a 24,8 b 12,7 a
CV % 10,62 19,6 8,56 29,52 12,99 DAI (dias após incubação). *número de apotécios por escleródio. Médias seguidas da mesma letra, na coluna, não diferem estatisticamente pelo teste de Tukey a 5% de probabilidade.
Houve diferença no número total de apotécios formados (p<0,5). Os
escleródios tamanho G formaram maior número e comprimento de
apotécios, importantes no processo de infecção na planta e esses tinham
estipes mais longas. Schwartz e Steadman (1989) verificaram que o
apotécio com 3 mm de diâmetro eleva-se de 3 a 6 mm da superfície do solo.
Neste trabalho verificou-se que os escleródios com comprimento acima de 3
mm elevaram-se, em média a valores superiores a 9 mm. Os escleródios de
tamanho MP e P formaram apotécios pequenos, diferenciando
significativamente dos tamanhos M e G (Tabela 3). Assim, esses
escleródios com tamanho reduzido têm menor capacidade de se elevar se

60
situados em camadas mais profundas no solo, ou se estiverem recobertos
por palhada, devido ao comprimento da estipe ser menor.
A massa média dos escleródios G foi 66 mg (Tabela 1) e os mesmos
produziram apotécios com massa média de 0,0234 g (dados não
apresentados), discordando de Budge e Whipps (1991), que obtiveram
maior produção de apotécios em escleródios com peso menor que 10 mg e
2 mm de diâmetro.
O número de apotécios produzidos pelos escleródios foi
significativamente maior para os escleródios grandes, exceto na avaliação
aos 100 DAI, e decresceu com o tamanho de escleródios. Escleródios muito
pequenos produziram, aos 51 DAI, cinco vezes menos apotécios que os
escleródios pequenos e médios e oito vezes menos que escleródios
grandes (Tabela 3). Resultados semelhantes foram observados por Dillard
et al. (1995), em que os escleródios pequenos produziram cinco vezes
menos apotécios que escleródios grandes. O número de apotécios
produzidos por escleródio, citado por esses autores, foi significativamente
maior em escleródios grandes e decresceram com o tamanho. Essa
diferença entre tamanho de escleródios do trabalho citado e os tamanhos
dos escleródios utilizados neste estudo (Tabela 1), possivelmente se deve a
produção dos mesmos em ambiente diferentes e diferença de isolados.
Neste estudo, os apotécios mediram de 9,3 mm a 13,2 mm e
diferiram entre apotécios formados de escleródios muito pequenos em
relação aos escleródios médios e grandes. Escleródios que variaram de
1,95 mm a 6,14 mm formaram apotécios menores que escleródios médios e
grandes (Tabela 3). De forma semelhante, Bedi (1963) observou que em
escleródios maiores a produção de apotécios também é maior, sendo que
um escleródio de 13 mm de comprimento por 5 mm de largura, originou 15
apotécios. Na Figura 4 verifica-se a diferença visual entre a produção de
apotécios de escleródios pequenos com média de 2,4 mm e grandes com
média de 13,6 mm após 51 dias de incubação.

61
FIGURA 4. Escleródios de Sclerotinia sclerotiorum de tamanho muito
pequeno (esquerda) e grande (direita) com produção de
estipes e apotécios aos 51 dias após incubação. UFMT,
Cuiabá-MT, 2012.
Com o aumento do tamanho dos escleródios, o número de estipes
formadas e apotécios produzidos por escleródio também aumentou
(Tabelas 2 e 3). Esses resultados estão em acordo com os de Ben-Yephet
et al. (1993), que observaram que houve aumento significativo no número
de apotécios produzido por escleródios com o aumento do peso.
Em estudos com escleródios, Görgen et al. (2010) verificaram, em
condições naturais, que o escleródio pode ser formado e atingir sua
maturidade funcional em diferentes períodos de tempo, influenciado pelas
condições ambientais. Em avaliações em campo, esses autores observaram
escleródios menores do que 2 mm que apresentavam apotécios. Em outro
estudo, Hao et al. (2003) relataram a produção de apotécios em escleródios
grandes (>4,75 mm), médios (2,0 a 4,75 mm), e pequenos (<2.0 mm) e
verificaram, ainda, que o número de apotécios produzidos é proporcional ao
tamanho do escleródio de S. sclerotiorum. Embora a biometria dos
escleródios se diferenciem dos encontrados no presente estudo, as
respostas são semelhantes em relação ao tamanho dos escleródios.

62
Resultados semelhantes foram obtidos por Huang e Kozub (1994),
em estudo com escleródios, onde verificaram que o número de estipes
produzidos por S. sclerotiorum foi altamente correlacionado com o tamanho
dos escleródios. Os resultados sugeriram que existe um limite de tamanho
de escleródios abaixo do qual os apotécios não são produzidos. Houve
germinação de 100% para escleródios maiores e de 30% para escleródios
menores.
Como a produção de apotécios foi diretamente proporcional ao
tamanho de escleródios, o uso de medidas de controle que reduzam a
germinação e o tamanho de escleródios podem reduzir apotécios formados.
Menor número de apotécios formados, por sua vez, produziriam menor
quantidade de ascósporos (Abawi e Grogan, 1979), porque cada apotécio
pode produzir de 2 a 30 milhões de ascósporos durante um período de até
10 dias (Clarkson et al., 2003). A cobertura do solo pode funcionar como
barreira física, reduzindo a entrada de luz necessária à formação dos
apotécios (Görgen et al., 2009; Paula Júnior et al., 2009) e
consequentemente a liberação de ascósporos, reduzindo a germinação e a
elevação de apotécios originados de escleródios pequenos.
Em estudo comparando a germinação de escleródios na presença e
ausência de Brachiaria ruziziensis, Görgen et al. (2009) também verificaram
redução no número de apotécios formados pela utilização da cobertura
vegetal. Para Lobo Junior et al. (2009), a redução do número de apotécios
formados sobre o solo, devido à camada de palha, é o efeito mais notável e
imediato capaz de reduzir a severidade do mofo branco.
Com exceção do cártamo, constata-se que o uso de cobertura
vegetal do solo reduz o número de estipes e apotécios de S. sclerotiorum.
As coberturas com braquiária, girassol e nabo forrageiro aumentam o
período de tempo até a formação do apotécio, porém, somente braquiária
reduz a porcentagem de apotécios formados (Venturoso, 2013). Entretanto
é importante considerar que o número de estipes por escleródio de S.
sclerotiorum não é alterado pelos sistemas de manejo (Gorgen et al., 2010).

63
Os resultados deste estudo estabeleceram claramente que o
tamanho de escleródios é um dos principais determinantes para a formação
de estipes e apotécios. Mesmo que os escleródios menores sejam menos
eficazes em produzir apotécios (Huang e Kozub, 1994; Dillard et al. 1995), a
produção de apotécios, varia com o tamanho dos escleródios.
Este é o primeiro estudo que identifica a importância do escleródio
proveniente de lavoura de soja, como um fator fundamental para a produção
de apotécios de S. sclerotiorum, embora os fragmentos menores
(microescleródios) possam não possuir reserva suficiente para a produção
de apotécios (Hao et al., 2003). Em vez disso, pode ocorrer a germinação
miceliogênica quando as condições de solo são apropriadas (Abawi e
Grogan, 1979).
O tamanho de escleródios é um dos fatores que pode ter papel
importante na indução da produção de apotécios. Fatores que afetam a
germinação de escleródios e a sobrevivência são também importantes para
a infecção subsequente de S. sclerotiorum. Após a germinação
carpogênica, vários fatores afetam como a liberação de ascósporos,
propagação, germinação e infecção (Abawi et al., 1979; Hao et al., 2003).
Assim em estudos de epidemiologia de S. sclerotiorum devem ser evitados
o uso de pequenos escleródios (<3mm) para a produção de ascósporos
(Dillard et al., 1995), por terem reduzida produção de apotécios.
3.3.2 Determinação do potencial osmótico para o desenvolvimento de
S. sclerotiorum
Verificou-se que não houve diferença entre os tratamentos de
restrição hídrica para o número de escleródios produzidos, mas houve
redução na massa de escleródios em relação à testemunha com BDA,
evidenciando o efeito da restrição hídrica com NaCl. Pelos resultados,
observou-se que o índice de velocidade de crescimento micelial (IVCM)
reduziu para as restrições a partir de -1,0 MPa de NaCl (Tabela 4).
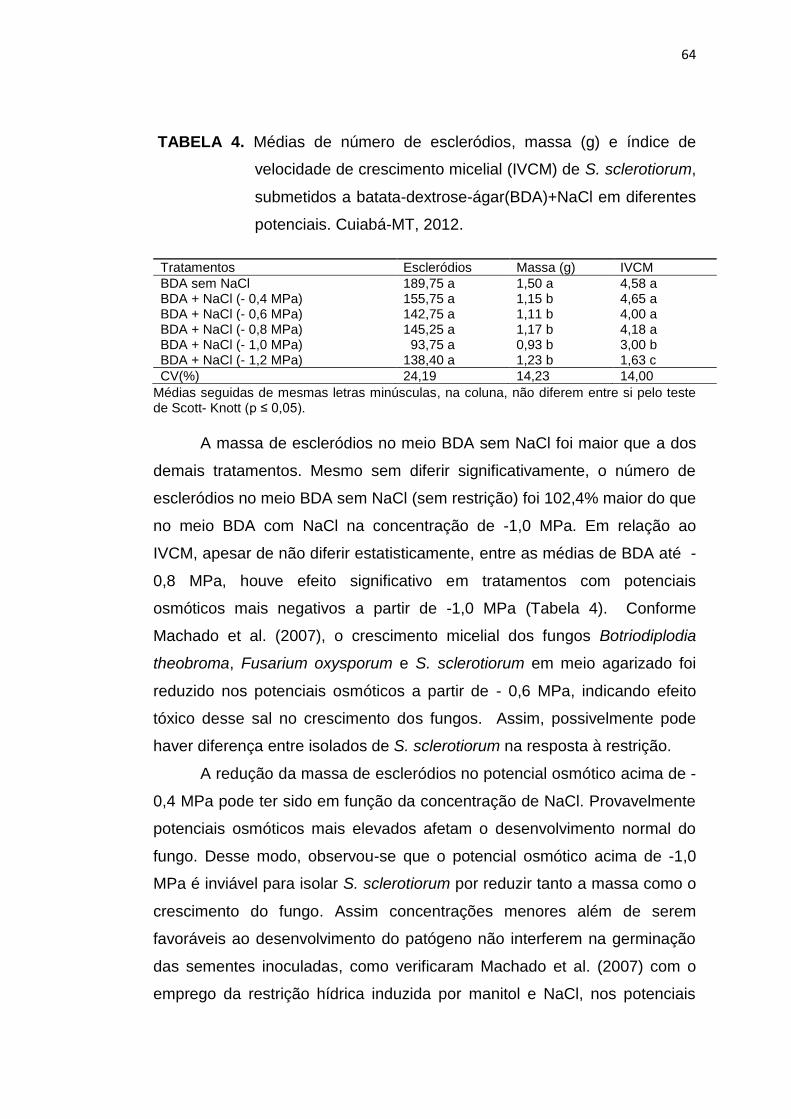
64
TABELA 4. Médias de número de escleródios, massa (g) e índice de
velocidade de crescimento micelial (IVCM) de S. sclerotiorum,
submetidos a batata-dextrose-ágar(BDA)+NaCl em diferentes
potenciais. Cuiabá-MT, 2012.
Médias seguidas de mesmas letras minúsculas, na coluna, não diferem entre si pelo teste de Scott- Knott (p ≤ 0,05).
A massa de escleródios no meio BDA sem NaCl foi maior que a dos
demais tratamentos. Mesmo sem diferir significativamente, o número de
escleródios no meio BDA sem NaCl (sem restrição) foi 102,4% maior do que
no meio BDA com NaCl na concentração de -1,0 MPa. Em relação ao
IVCM, apesar de não diferir estatisticamente, entre as médias de BDA até -
0,8 MPa, houve efeito significativo em tratamentos com potenciais
osmóticos mais negativos a partir de -1,0 MPa (Tabela 4). Conforme
Machado et al. (2007), o crescimento micelial dos fungos Botriodiplodia
theobroma, Fusarium oxysporum e S. sclerotiorum em meio agarizado foi
reduzido nos potenciais osmóticos a partir de - 0,6 MPa, indicando efeito
tóxico desse sal no crescimento dos fungos. Assim, possivelmente pode
haver diferença entre isolados de S. sclerotiorum na resposta à restrição.
A redução da massa de escleródios no potencial osmótico acima de -
0,4 MPa pode ter sido em função da concentração de NaCl. Provavelmente
potenciais osmóticos mais elevados afetam o desenvolvimento normal do
fungo. Desse modo, observou-se que o potencial osmótico acima de -1,0
MPa é inviável para isolar S. sclerotiorum por reduzir tanto a massa como o
crescimento do fungo. Assim concentrações menores além de serem
favoráveis ao desenvolvimento do patógeno não interferem na germinação
das sementes inoculadas, como verificaram Machado et al. (2007) com o
emprego da restrição hídrica induzida por manitol e NaCl, nos potenciais
Tratamentos Escleródios Massa (g) IVCM
BDA sem NaCl 189,75 a 1,50 a 4,58 a BDA + NaCl (- 0,4 MPa) 155,75 a 1,15 b 4,65 a BDA + NaCl (- 0,6 MPa) 142,75 a 1,11 b 4,00 a BDA + NaCl (- 0,8 MPa) 145,25 a 1,17 b 4,18 a BDA + NaCl (- 1,0 MPa) 93,75 a 0,93 b 3,00 b BDA + NaCl (- 1,2 MPa) 138,40 a 1,23 b 1,63 c
CV(%) 24,19 14,23 14,00

65
osmóticos -0,6 MPa, -0,8 MPa, -1,0 MPa e -1,2 MPa, que reduziram o
processo de germinação de sementes de algodoeiro.
Diferenças entre os solutos quanto à eficiência em reduzir e/ou
retardar a germinação de sementes foram relatadas por Heydecker et al.
(1975), e a redução acentuada da germinação de sementes em maiores
concentrações de soluções osmóticas tem sido atribuída à redução da
quantidade de água absorvida pelas sementes em meio salino, em
decorrência da redução do potencial osmótico das soluções (Braccini et al.,
1996).
Em relação a Sclerotinia sclerotiorum, Machado et al. (2001)
observaram que houve comportamento semelhante, ou seja, as sementes
que ficaram menos tempo em contato com o inóculo do fungo apresentaram
germinação abaixo de 20% e nos potenciais de -0,6; -0,8 e -1,0 MPa
praticamente não houve germinação, com mais de 90% de sementes
mortas.
Assim, de acordo com os resultados encontrados, para índice de
velocidade de crescimento micelial, massa e número de escleródios em
meio BDA, não se recomenda o uso de NaCl nos referidos potenciais
osmóticos para inoculação de S. sclerotiorum em sementes de soja. É mais
prático a utilização do método sem a adição de NaCl, sem necessidade de
custos adicionais e é uma alternativa economicamente viável, além de ser
um meio eficiente para o desenvolvimento do fungo por proporcionar maior
massa de escleródios (g) em sete dias de incubação.
3.3.3 Período adequado de inoculação de S. sclerotiorum em sementes
de soja e viabilidade dos métodos de detecção “blotter test”, neon e
rolo de papel para recuperação do patógeno em sementes de soja
inoculadas
Por meio do método rolo de papel foi possível detectar o fungo S.
sclerotiorum nas sementes de soja inoculadas artificialmente com o
patógeno em menor período de inoculação, após 24 horas, enquanto que a
identificação do patógeno por meio do “blotter test” e teste neon só foram
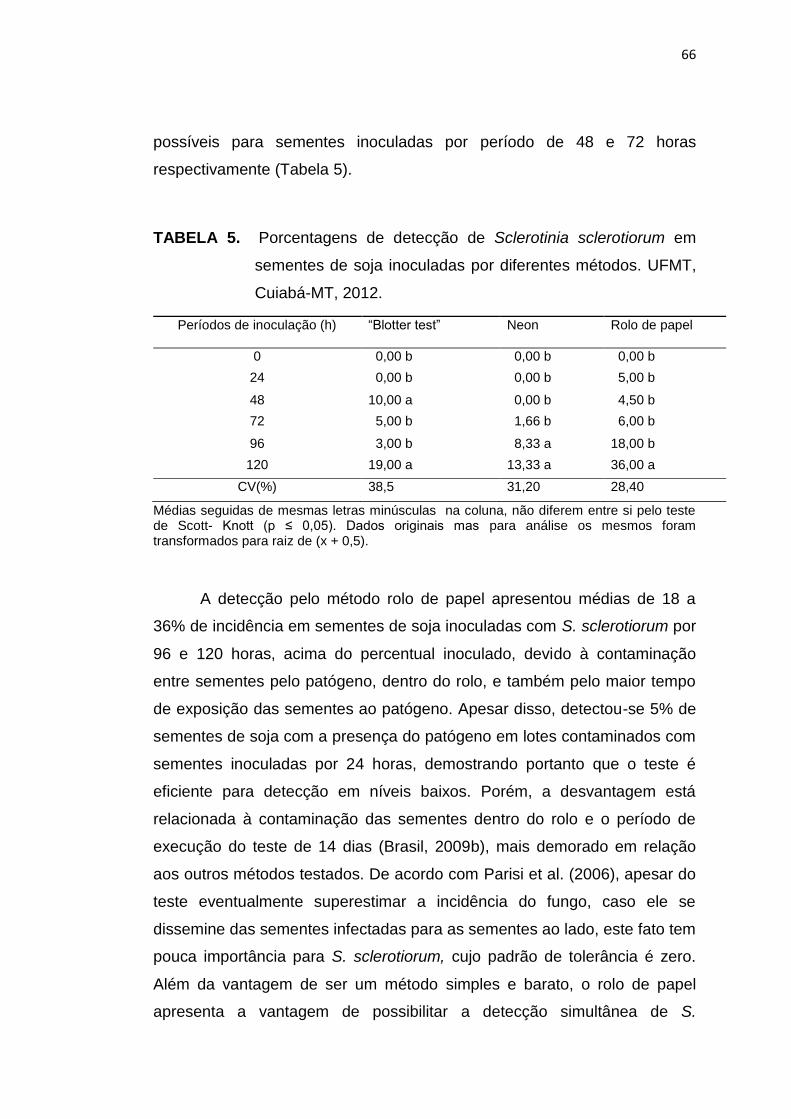
66
possíveis para sementes inoculadas por período de 48 e 72 horas
respectivamente (Tabela 5).
TABELA 5. Porcentagens de detecção de Sclerotinia sclerotiorum em
sementes de soja inoculadas por diferentes métodos. UFMT,
Cuiabá-MT, 2012.
Médias seguidas de mesmas letras minúsculas na coluna, não diferem entre si pelo teste de Scott- Knott (p ≤ 0,05). Dados originais mas para análise os mesmos foram transformados para raiz de (x + 0,5).
A detecção pelo método rolo de papel apresentou médias de 18 a
36% de incidência em sementes de soja inoculadas com S. sclerotiorum por
96 e 120 horas, acima do percentual inoculado, devido à contaminação
entre sementes pelo patógeno, dentro do rolo, e também pelo maior tempo
de exposição das sementes ao patógeno. Apesar disso, detectou-se 5% de
sementes de soja com a presença do patógeno em lotes contaminados com
sementes inoculadas por 24 horas, demostrando portanto que o teste é
eficiente para detecção em níveis baixos. Porém, a desvantagem está
relacionada à contaminação das sementes dentro do rolo e o período de
execução do teste de 14 dias (Brasil, 2009b), mais demorado em relação
aos outros métodos testados. De acordo com Parisi et al. (2006), apesar do
teste eventualmente superestimar a incidência do fungo, caso ele se
dissemine das sementes infectadas para as sementes ao lado, este fato tem
pouca importância para S. sclerotiorum, cujo padrão de tolerância é zero.
Além da vantagem de ser um método simples e barato, o rolo de papel
apresenta a vantagem de possibilitar a detecção simultânea de S.
Períodos de inoculação (h) “Blotter test”
Neon Rolo de papel
0 0,00 b 0,00 b 0,00 b
24 0,00 b 0,00 b 5,00 b
48 10,00 a 0,00 b 4,50 b
72 5,00 b 1,66 b 6,00 b
96 3,00 b 8,33 a 18,00 b
120 19,00 a 13,33 a 36,00 a
CV(%) 38,5 31,20 28,40

67
sclerotiorum e de outros importantes patógenos transmitidos por sementes
como C. lindemuthianum, Macrophomina phaseolina e Rhizoctonia solani
(Parisi et al., 2006).
Os resultados estão de acordo com os obtidos na literatura, em que a
inoculação artificial da semente foi utilizada para avaliar a sensibilidade dos
métodos rolo, blotter e neon em detectar o fungo (Peres et al., 2002;
Napoleão et al., 2006; Henneberg et al., 2012).
A detecção pelo método neon foi facilitada pela liberação de ácido
oxálico no meio de cultura pelo S. sclerotiorum, formando um halo
amarelado. Porém, a presença de Aspergillus sp. pode prejudicar a
avaliação, sendo esta uma desvantagem do teste. Em estudo realizado por
Peres et al. (2002), a coloração produzida por isolados de S. sclerotiorum
iniciou a partir de 24 horas de incubação por este método. Entretanto, os
isolados de Penicillium sp. e Aspergillus sp. apresentaram coloração média,
e a coloração produzida de S. sclerotiorum foi mais intensa.
As altas porcentagens de S. sclerotiorum, detectados pelos métodos
neon e “blotter test”, ocorreram em sementes de soja inoculadas com o
patógeno por períodos de 96 e 120 horas, podendo estar relacionado ao
maior tempo de exposição das sementes ao patógeno que pode ter
favorecido a contaminação de sementes na placa.
O período mais eficiente, pelos testes de detecção “blotter test” e
neon, para a inoculação de S. sclerotiorum em sementes de soja, foi 72
horas (Tabela 5). Nesse período, observou-se a incidência do patógeno
pelos três métodos de detecção e, ainda em percentuais semelhantes ao
percentual inoculado, exceto o método de detecção neon, que detectou o
patógeno em menores percentuais (1,66%). Também nesse período de
inoculação não houve perdas significativas no potencial germinativo das
sementes de soja (Figura 5). Em 72 horas de inoculação, o “blotter test” foi
o mais preciso, recuperando 5% de incidência do patógeno nos lotes de
sementes de soja com 5% de contaminação.
O teste neon, no período de 72 horas de inoculação, permitiu a
detecção de 1,66% de incidência de S. sclerotiorum após oito dias de
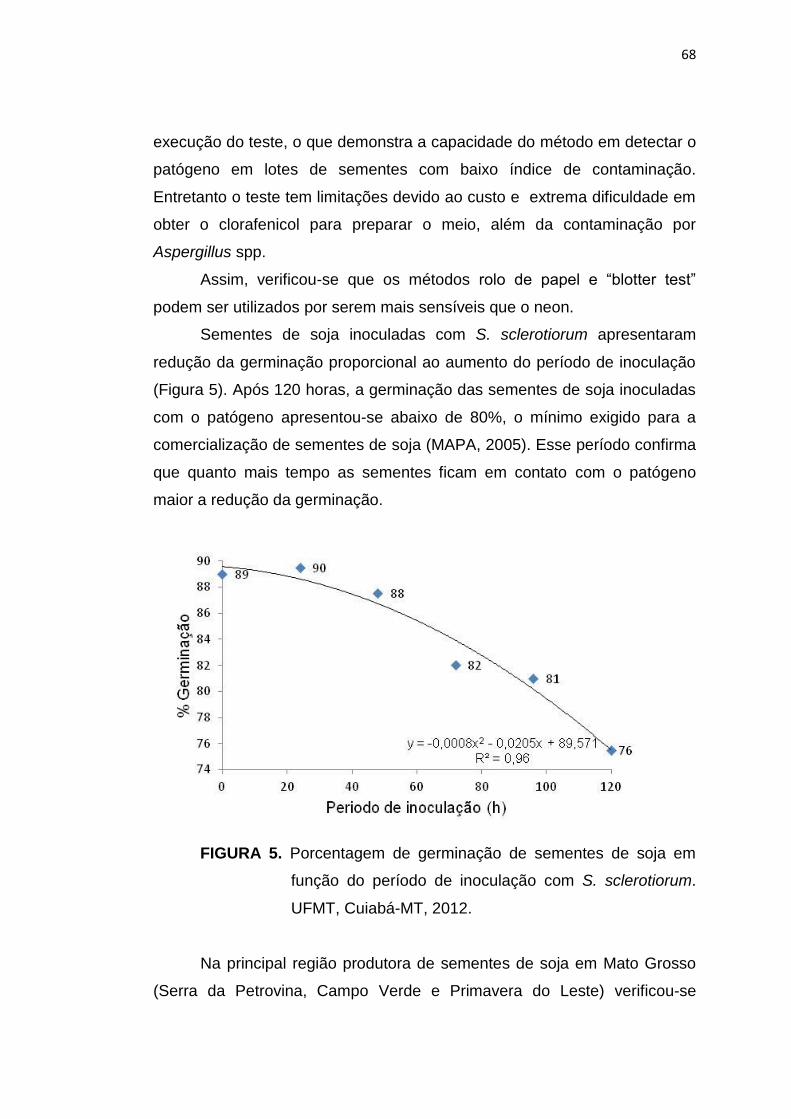
68
execução do teste, o que demonstra a capacidade do método em detectar o
patógeno em lotes de sementes com baixo índice de contaminação.
Entretanto o teste tem limitações devido ao custo e extrema dificuldade em
obter o clorafenicol para preparar o meio, além da contaminação por
Aspergillus spp.
Assim, verificou-se que os métodos rolo de papel e “blotter test”
podem ser utilizados por serem mais sensíveis que o neon.
Sementes de soja inoculadas com S. sclerotiorum apresentaram
redução da germinação proporcional ao aumento do período de inoculação
(Figura 5). Após 120 horas, a germinação das sementes de soja inoculadas
com o patógeno apresentou-se abaixo de 80%, o mínimo exigido para a
comercialização de sementes de soja (MAPA, 2005). Esse período confirma
que quanto mais tempo as sementes ficam em contato com o patógeno
maior a redução da germinação.
FIGURA 5. Porcentagem de germinação de sementes de soja em
função do período de inoculação com S. sclerotiorum.
UFMT, Cuiabá-MT, 2012.
Na principal região produtora de sementes de soja em Mato Grosso
(Serra da Petrovina, Campo Verde e Primavera do Leste) verificou-se

69
aumento da área com incidência de mofo branco, contaminadas tanto por
sementes como por escleródios. Isto ocorreu mesmo com a Portaria no 47,
de 26 de fevereiro de 2009, do Ministério da Agricultura, Pecuária e
Abastecimento, que estabeleceu padrão zero para Sclerotinia
sclerotiorum em sementes de feijão, soja e girassol, não podendo haver
nenhum escleródio no exame da amostra de sementes secas. Também
ocorre a detecção de sementes infectadas pelo fungo, que apesar de baixa
pode vir a infestar áreas isentas.
A contaminação em novas áreas isentas do patógeno pode ser
evitada, sendo necessário o aprimoramento de testes de sanidade para
detecção mais rápida e eficiente do patógeno. Esses testes são ferramentas
úteis no controle de disseminação da doença em novas áreas, quando
efetuados nos lotes de sementes originados dessa região.
Neste estudo comprovou-se que o método rolo de papel é sensível,
simples e de baixo custo, podendo ser utilizado nas análises de rotina,
como medida preventiva da disseminação do patógeno e também poderá
ser utilizado para a detecção de S. sclerotiorum em sementes de outras
culturas (Parisi et al., 2006). Entretanto Henneberg et al. (2012a) verificaram
que os métodos papel de filtro “blotter test”, rolo de papel e neon, não foram
eficientes em detectar o patógeno S. sclerotiorum em sementes de soja
naturalmente contaminadas em níveis até 1% e que não houve relação
entre a incidência do patógeno no campo e a incidência verificada nas
sementes de soja colhidas. Portanto, níveis de incidência do patógeno em
sementes de soja interferem na sensibilidade dos métodos de detecção.
Em outro estudo, Hennerberg et al. (2012b) observaram que os
métodos estudados não apresentaram repetibilidade, sendo pouco
sensíveis para a detecção do patógeno em sementes provenientes de áreas
com incidência da doença; porém, foram eficientes para a detecção em
sementes inoculadas artificialmente.
Assim, a eficiência dos métodos na detecção de S.sclerotiorum, em
amostras com inoculação artificial, pode ser atribuída ao fato de o patógeno
apresentar maior potencial de infecção com esse tipo de inoculação, o que

70
teria facilitado o seu desenvolvimento nas condições de incubação
utilizadas nos testes de sanidade, pois o fungo depende somente do
ambiente favorável para se desenvolver (Sousa et al., 2008).
Esses resultados representam um avanço nos estudos com esse
patógeno associado às sementes, por ser de difícil detecção e apresentar
baixa transmissibilidade da planta para a semente (Henning et al., 2009). A
técnica da restrição hídrica possibilitou a obtenção de sementes
efetivamente infectadas pelo patógeno, as quais poderão ser utilizadas em
estudos, tais como transmissibilidade do patógeno da semente para a
planta, considerando os diferentes níveis de inóculo que a técnica pode
proporcionar.
3.4 Conclusões
O tamanho de escleródios não afeta na germinação carpogênica,
mas influencia no número e comprimento de apotécios.
O meio de cultura adequado para inoculação de S. sclerotiorum em
sementes de soja é o BDA (Batata-Dextrose e Agar) sem restrição hídrica
de NaCl.
Os métodos de detecção, “blotter test”, neon e rolo de papel são
eficientes na detecção de S. sclerotiorum quando sementes de soja foram
inoculadas artificialmente por um período de 72 horas.
O método rolo de papel permite a detecção de forma eficiente de S.
sclerotiorum em lotes de sementes de soja com 5% de incidência.

71
3.5 Referências Bibliográficas
ABAWI, G.S.; GROGAN, R.G. Epidemiology of disease caused by Sclerotinia species. Phytopathology, v. 63, n. 8, p. 899-903, 1979. ADAMS, P. B.; AYERS, W. A. Ecology of Sclerotinia species. Phytopathology, v. 69, n. 8, p.896-898, 1979. BEN-YEPHET, Y.; GENIZI, A. ; SITI, E. Sclerotial survival and apothecial production by Sclerotinia sclerotiorum following out-breaks of lettuce drop. Phytopathology, v. 83, p. 509-513, 1993. BEDI, K.S; The age of sclerotia of Sclerotinia sclerotiorum (Lib.) de Bary in relation to the formation of aphotecia. Jornal of the Indian Botanical Society, v.42, p.204-207, 1963. BOLTON, M. D.; THOMMA, B. P. H. J.; NELSON, B. D. Sclerotinia sclerotiorum (Lib.) de Bary: biology and molecular traits of a cosmopolitan pathogen. Molecular Plant Pathology, v.11,n.7, p.1-16, 2006. BRACCINI, A. L. Relação entre potencial hídrico, condicionamento osmótico e qualidade fisiológica de sementes de soja (Glycine max (L.) Merrill). 1996. 135 f. Tese (Doutorado em Fitotecnia) – Universidade Federal de Viçosa, Viçosa-MG, 1996. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária/MAPA/ACS, 2009a. 395 p. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Manual de análise sanitária de sementes. Brasília: Secretaria de Defesa Agropecuária/MAPA/ACS, 2009b. 200 p. BRUSTOLIN, R.; ROSSI, L.D.; REIS, E.M. Mofo branco. In: REIS, E.M.; CASA, R.T. (Eds). Doenças da soja. Passo Fundo: Berthier. 2012. p. 217-232. BUDGE, S.P.; WHIPPS, J.M. Effec of sucrose concentration on sclerotia production and subsequent apothecial formation by Sclerotinia sclerotiorum. Mycological Research, v. 95, n.2, p. 195-198. 1991. CESSNA, S.G.; SEARS, V.E.; DICKMAN, M.B.; LOW, P.S. Oxalic acid, a pathogenicity factor for Sclerotinia sclerotiorum, suppresses the oxidative burst of the host plant. The Plant Cell, v. 12, n. 11, p. 2191–2199, 2000.

72
CLARKSON, J.P.; STAVELEY, J. PHELPS, K.; YOUNG, C.S.; WHIPPS, J.M. Ascorpore release and survival in Sclerotinia sclerotiorum. Mycological Research, v.107, p. 213-222, 2003. DILLARD, H.R.; LUDWIG, J.W.; HUNTER, J.E. Conditioning sclerotia os Sclerotinia sclerotiorum for carpogenic germination. Plant Disease, v. 79, v.4, p.411-415, 1995. FERREIRA, D.F. Sisvar: versão 4.2. Lavras: UFLA, 2003. GÖRGEN, C. A.; SILVEIRA NETO, A. N.; CARNEIRO, L. C.; RAGAGNIN, V.; LOBO JUNIOR, M. Controle do mofo branco com palhada e Trichoderma harzianum 1306 em soja. Pesquisa Agropecuária Brasileira, v. 44, n. 12, p. 1583-1590, 2009. GÖRGEN, C.A.; CIVARDI, E.A; RAGAGNIM, V.A.; SILVEIRA NETO, A.N. da.; CARNEIRO, L.C.; LOBO JUNIOR, M. Redução do inóculo inicial de Sclerotinia sclerotiorum em soja cultivada após uso do sistema Santa Fé. Pesquisa Agropecuária Brasileira, v.45, p.1102-1108, 2010. HAO, J.J.; SUBBARAO, K.V.; DUNIWAY, J.M. Germination of Sclerotinia minor and S. sclerotiorum sclerotia under various soil moisture and temperature combinations. Phytopathology, v.93, p.443-450, 2003.
HEYDECKER, W.; HIGGINS, J.; TURNER, Y.J. Invigoration of seeds? Seed Science and Technology, v.3, n.3, p.881- 888, 1975. HENNENBERG, L.; JACCOUB FILHO, D. S.; GRZYBOWSKI, C. R. S.PANOBIANCO, M. Importância da detecção de Sclerotinia sclerotiorum em sementes de soja. Informativo Abrates, v. 21, n. 3, 2011. HENNEBERG, L.; GRABICOSKI, E. M. G.; JACCOUD-FILHO, D. S.; PANOBIANCO, M. Incidência de Sclerotinia sclerotiorum em sementes de soja e sensibilidade dos testes de detecção. Pesquisa Agropecuária Brasileira,[online], v.47, n.6, p. 763-768, 2012a. HENNEBERG, L.; JACCOUD FILHO, D. de S.; RUARO, L.; PANOBIANCO, M. Efficiency of methods to detect Sclerotinia sclerotiorum in commercial soybean seed lots. Revista Brasileira de Sementes, v.34,n.1 p.61-69, 2012b. HENNING, A.A.; PAULA, F.Y.H.; MOMTEMEZZO, C.A.O.; BOSSE, E.J.; BERGONSI, J.S.S. Avaliação de princípios ativos para o controle químico de mofo branco (Sclerotinia sclerotiorum) em soja – safra 2008/2009. Informativo ABRATES, v.19, n.1, p.29-31, 2009. http://www. abrates.org.br/portal/images/stories/informativos/v19n1/artigo01.pdf HUANG, H. C.; KOZUB, G. C. Longevity of normal and abnormal

73
sclerotia of Sclerotinia sclerotiorum. Plant Disease, v.78, p.1164-1166, 1994. LOBO JUNIOR, M.; BRANDÃO, R. S.; CORRÊA, C. A.; GÖRGEN, C. A.; CIVARDI, E. A.; OLIVEIRA, P. Santo Antônio de Goiás: Embrapa Arroz e Feijão, 2009. 8 p. (Comunicado técnico, 183). MACHADO, J. C., OLIVEIRA, J. A., VIEIRA, M. G.G.C. et al. Inoculação artificial de sementes de soja por fungos, utilizando a solução de manitol, Revista Brasileira de Sementes, v. 23, n. 2, 95 – 101, 2001. MACHADO, J.C; OLIVEIRA, J.A; VIEIRA, M.G.G.C; ALVES, M.C. Controle da germinação de sementes de soja em testes de sanidade pelo uso da restrição hídrica. Revista Brasileira de Sementes, v.25, n.2, 2003. MACHADO, A. Q., MACHADO, J. C., VIEIRA, M. D.G.G.C. et al. Potencial do Uso da Restrição Hídrica em Testes de Sanidade de Sementes de Algodoeiro, Fitopatologia Brasileira, v. 32, n. 5, p. 408 – 414, 2007. MAPA.MINISTÉRIO DE AGRICULTURA, PECUARIA E ABASTECIMENTO. 2005. Disponível em: <http://www.agricultura.pr.gov.br/arquivos/File/PDF/padroes_soja.pdf. Acesso em: 24 de fev. 2013. NAPOLEÃO, R.; NASSER, L.; LOPES, C.; FILHO, A. Neon – S, novo meio para detecção de Sclerotinia sclerotiorum em sementes, Summa Phytopathologica, v. 32, n. 2, p. 180–182, 2006. NASSER, L.C.B.; NAPOLEÃO, R.; CARVAJAL, R.A. Mofo branco - cuidado com a semente. Revista Cultivar Grandes Culturas, n.4, 1999. OLIVEIRA, J. A. Efeito do tratamento fungicida em sementes no controle de tombamento de plântulas de pepino (Cucumis sativus L.) e pimentão (Capsicum annum L.). 1991. 111 f. Dissertação (Mestrado em Fitossanidade) – Universidade Federal de Lavras, Lavras, MG. PAULA JÚNIOR, T.J. ; VIEIRA, R.F.; ROCHA, P.R.R.; BERNARDES, A.; COSTA, E.L.; CARNEIRO, J.E.S.; RIBEIRO DO VALE, F.X.; ZAMBOLIM, L. White mold intensity on common bean in response to plant density, irrigation frequency, grass mulching, Trichoderma spp., and fungicide. Summa Phytopathologica, v. 35, n.1, p. 44-48, 2009. PARISI, J. J. D.; PATRÍCIO, F. R. A; OLIVEIRA, S. H. F. Método do rolo de papel toalha modificado para a detecção de Sclerotinia sclerotiorum em sementes de feijão, Summa Phytopathologica, v. 32, n. 3, p. 288-290, 2006.

74
PERES, A. P. Detecção de Sclerotinia sclerotiorum (Lib.) de Bary em sementes de feijão (Phaseolus vulgaris L.) e soja (Glycine max (L.) Merrill). 1996. 51 f. Dissertação (Mestrado em Fitopatologia) – Universidade Federal de Lavras. PERES, A. P.; NASSER, L. C. B.; MACHADO, J. C. Use os semi-seletive media for detection of Sclerotinia sclerotiorum on bean and soybean seeds. Fitopatologia Brasileira, v. 27, n. 2, p. 123 – 127, 2002. PURDY, L.H. Sclerotinia sclerotiorum: history, diseases and symptomatology, host range, geographic distribution, and impact. Phytopathology, v. 69, p. 875- 880, 1979. REIS, E. M.; TOMAZINI, S. L. Viabilidade de escleródios de Sclerotinia sclerotiorum, no campo, em duas profundidades do solo. Summa Phytopathologica, v.31, n. 1, p.97-99, 2005. SCHWARTZ, H.F.; STEADMAN, J.R. White mold. In: SCHWARTZ, H.F.; PASTOR-CORSALES, M.A. (ed.). Bean production problems in the tropics. 2. ed. Cali: CIAT, 1989. p. 736. SOUSA, M.V.; MACHADO, J.C.; PFENNING, L.H.; KAWASAKI, V.H.; ARAÚJO, D.V.; SILVA, A.A.; MARTINI NETO, A. Métodos de inoculação e efeitos de Fusarium oxysporum f. sp. vasinfectum em sementes de algodoeiro. Tropical Plant Pathology, v.33, p.41-48, 2008. SILVA, F. DE A. S. E.; AZEVEDO, C. A. V. DE. Principal components analysis in the software assistat-statistical attendance. In: WORLD CONGRESS ON COMPUTERS IN AGRICULTURE, 7, Reno-NV-USA: American Society of Agricultural and Biological Engineers, 2009. VENTUROSO, L. R.; BACCHI, L.M.A.; GAVASSONI, W.L.; VENTUROSO, L. A. C. et al. Produção de soja e germinação carpogênica de Sclerotinia sclerotiorum sob diferentes coberturas de solo. Semina: Ciências Agrárias, v. 34, n. 2, p. 615-626, 2013. WILLETTS, H. J.; WONG, J. A. The biology of Sclerotinia sclerotiorum, S. trifoliorum, and S. minor with emphasis on specific nomenclature. The Botanical Review, v.46, p. 101-165,1980.

75
4 EFICIÊNCIA DE FUNGICIDAS NO CONTROLE DE MOFO BRANCO NA
CULTURA DA SOJA E NA GERMINAÇÃO DE ESCLERÓDIOS DE
Sclerotinia sclerotiorum
RESUMO - A incidência e a severidade do mofo branco (Sclerotinia
sclerotiorum) na região sul do estado de Mato Grosso têm aumentado
devido às condições de ambiente favoráveis ao desenvolvimento da
doença. Assim, o objetivo neste trabalho foi avaliar o efeito do controle
químico na incidência e severidade de mofo branco em soja, e verificar o
efeito do tratamento químico com fungicidas sobre a viabilidade dos
escleródios formados. O delineamento foi em blocos ao acaso com quatro
tratamentos, mais a testemunha, e quatro repetições. Os tratamentos foram:
Fluazinam 1,0 L.ha-1, aplicados em R1; duas aplicações de Fluazinam (1,0
L.ha-1) – R1; R1+ 10 dias; duas aplicações de Procimidona (1,0 kg.ha-1) -
R1; R1+10 dias; Fluazinam (1,0 L.ha-1) + Carbendazim (1,0 L.ha-1) em R1 e
segunda aplicação de Procimidona (1,0 kg.ha-1) + Carbendazin (1,0 L.ha-1),
10 dias após a primeira aplicação) e a testemunha sem controle químico.
Para a germinação dos escleródios, foram colocados 250 g de solo em
caixas de plástico tipo “gerbox”, e, posteriormente, foi efetuado o enterrio de
25 escleródios coletados nas parcelas onde foram aplicados os fungicidas
na cultura da soja. Os fungicidas reduziram o crescimento de S.
sclerotiorum, diminuindo a incidência e severidade em relação à
testemunha. O controle de mofo branco foi superior com o uso de
Fluazinam em relação à testemunha e à Procimidona. O efeito da aplicação
de fungicida na cultura da soja sobre a germinação de escleródios e
formação de apotécios ocorreu antes dos 50 dias de incubação. Os
fungicidas aplicados reduzem a incidência e severidade de mofo branco na
soja.
Palavras-chave: mofo branco, incidência, controle químico.

76
FUNGICIDES EFFICIENCY IN CONTROL OF MOLD IN WHITE AND
SOYBEAN CROP OF sclerotial germination Sclerotinia sclerotiorum
ABSTRACT - The incidence and severity of white mold ( Sclerotinia
sclerotiorum ) in the southern state of Mato Grosso have increased due to
environmental conditions favorable for disease development . The objective
of this study was to evaluate the effect of chemical control on the incidence
and severity of white mold in soybeans , and verify the effect of chemical
treatment with fungicides on the viability of sclerotia formed . The
experimental design was a randomized block design with four treatments ,
plus the control , and four replications . The treatments were : 1.0 Fluazinam
L.ha - 1 , applied in R1 , two applications Fluazinam (1.0 L.ha- 1 ) - R1 , R1 +
10 days , two applications Procymidone (1.0 kg ha-1) - R1 , R1 +10 days;
Fluazinam (1.0 L.ha-1 ) + Carbendazim (1.0 L.ha-1) R1 and the second
application Procymidone (1.0 kg ha-1 ) + Carbendazin (1.0 L.ha- 1 ) , 10 days
after the first application ) and control without chemical control . For
germination of sclerotia were placed 250 g of soil in plastic type " gerbox " ,
and was later performed the burial of sclerotia collected in 25 plots where
fungicides were applied on soybean . Fungicides reduced the growth of S.
sclerotiorum , reducing the incidence and severity compared to the control .
The control of white mold was higher with the use of Fluazinam compared to
control and Procymidone . The effect of fungicide application on soybean on
germination of sclerotia and apothecia formation occurred before 50 days of
incubation . Fungicides applied to reduce the incidence and severity of white
mold in soybeans
Keywords: white mold, incidence, chemical control.

77
4.1 Introdução
O mofo branco ou podridão branca da haste, doença causada pelo
fungo Sclerotinia sclerotiorum (Lib.) de Bary, tem se destacado nas
principais regiões produtoras de soja em função dos danos proporcionados
à lavoura. O fungo em condições favoráveis ao seu desenvolvimento, pode
causar perdas expressivas como as relatadas por Almeida et al. (2005) e
Leite (2005).
A partir da safra 2004/05, em lavouras do Sudoeste de Goiás com
alta pressão de inóculo (escleródios) associada às condições climáticas
favoráveis, ocorreram perdas na produção de grãos de soja que
ultrapassaram a 40% (Campos et al., 2008; Meyer e Campos, 2009). No
estado de Mato Grosso, na safra 2009/10, ocorreram várias áreas afetadas
com mofo branco na região Sul do Estado como nos municípios de Campo
Verde e Jaciara, levando a perdas significativas na produtividade da cultura
da soja.
O controle de S. sclerotiorum é considerado difícil devido à ausência
de cultivares de soja resistentes, à produção de estruturas de resistência
que permitem a sobrevivência do fungo no solo por vários anos na ausência
de hospedeiros e/ou sob condições desfavoráveis, a ampla gama de
hospedeiros, ao elevado número de ascósporos produzidos nas
frutificações sexuais (apotécio) e sua rápida e longa disseminação a partir
da fonte produtora, à sobrevivência nas sementes sob a forma de micélio
dormente ou escleródios aderidos às mesmas e devido à dificuldade de se
atingir os sítios de infecção através das pulverizações com fungicidas.
Desta forma, o controle mais efetivo baseia-se em um programa integrado
de diversas práticas (Leite, 2005).
Atualmente, não há disponibilidade de cultivares de soja resistentes
ao mofo branco, com isso, a maioria das medidas é preventiva, devendo-se
adotar sementes certificadas, fazer rotação de culturas com gramíneas,
espaçamento e população de plantas adequadas, uso de fungicidas em
tratamentos de semente e parte aérea.

78
O controle químico dessa doença apresentou diferenças de eficiência
e algumas vezes resultados contraditórios (Gasparotto, 1980; Vieira, 1994).
O uso de fungicida na parte aérea deve ser associado a outras estratégias
de manejo e tem sido necessário em áreas já com a presença do inóculo,
ou seja, com histórico da doença.
Portanto, o conhecimento da atividade dos fungicidas sobre o agente
causal da doença é de fundamental importância para o seu manejo. A
eficiência desses produtos depende de vários fatores, como a densidade de
inóculo no solo, o estágio da doença, o grau de cobertura das plantas pelo
fungicida, o número de pulverizações, a sua fungitoxidade, dose, época,
volume e equipamento de aplicação, espaçamento de plantas, incidência e
severidade da doença (Vieira, 1994).
Até o momento, apenas os ingredientes ativos tiofanato metílico,
Fluazinam, Procimidona e Fluazinam + Tiofanato metílico apresentam
registro junto ao Ministério da Agricultura, Pecuária e Abastecimento
(MAPA, 2012) para o controle do mofo branco na cultura da soja. Essa
limitação de produtos pode impossibilitar a recomendação dentro de
programas de aplicações mais efetivos, além de proporcionar riscos da
perda de sensibilidade do fungo aos fungicidas. Esses princípios ativos são
recomendados em outras culturas visando o controle do mesmo patógeno
(Vieira, 1994; Reis et al., 2010). Em outros países, a menor sensibilidade de
S. sclerotiorum a diferentes fungicidas foi evidenciada, conforme
demonstrado para os ingredientes ativos Carbendazim, Iprodione e
Boscalida (Liu et al., 2009; Ma et al., 2009) e Benomyl (Attanayake et al.,
2009). Além de poucos produtos registrados para a cultura da soja, esses,
têm custo elevado e não apresentam controle satisfatório.
Assim sendo, os objetivos neste trabalho foram avaliar o controle
químico na incidência e severidade de mofo branco em soja, e verificar o
efeito do tratamento químico de fungicidas sobre a viabilidade dos
escleródios formados.

79
4.2 Material e Métodos
Os experimentos foram conduzidos na Fazenda Planalto, no
município de Jaciara-MT, cujas coordenadas geográficas são 15º 47’ 25,90”
S e 55º 12’ 46,90” W, em área com histórico de infestação por S.
sclerotiorum, com população acima de 15 escleródios/m2, e no Laboratório
de Fitopatologia da Faculdade de Agronomia, Medicina Veterinária e
Zootecnia (FAMEVZ) da Universidade Federal de Mato Grosso (UFMT),
campus Cuiabá-MT, no período de outubro de 2010 a outubro de 2012.
4.2.1 Experimento 1: Avaliação do controle de mofo branco na cultura
da soja com o uso de fungicidas
O delineamento experimental foi em blocos ao acaso, com cinco
tratamentos e quatro repetições, sendo as parcelas de 4,0 x 6,0 m. A
parcela foi constituída de oito linhas espaçadas de 0,50 m com seis metros
de comprimento e a área útil de 12 m2. A variedade semeada foi P98Y11,
grupo de maturação 8.1, com população de 300 mil plantas.hectare-1.
Os tratamentos utilizados foram três ingredientes ativos de fungicidas
em doses distintas e uma testemunha sem fungicida: Fluazinam 1,0 L.ha-1,
aplicados em R1 (início de florescimento); duas aplicações de Fluazinam
(1,0 L.ha-1) – R1; R1+ 10 dias; duas aplicações de Procimidona (1,0 kg.ha-1)
- R1; R1+10 dias; Fluazinam (1,0 L.ha-1) + Carbendazim (1,0 L.ha-1) em R1
e segunda aplicação de Procimidona (1,0 kg.ha-1) + Carbendazin (1,0 L.ha-
1), 10 dias após a primeira aplicação) (Tabela 1).
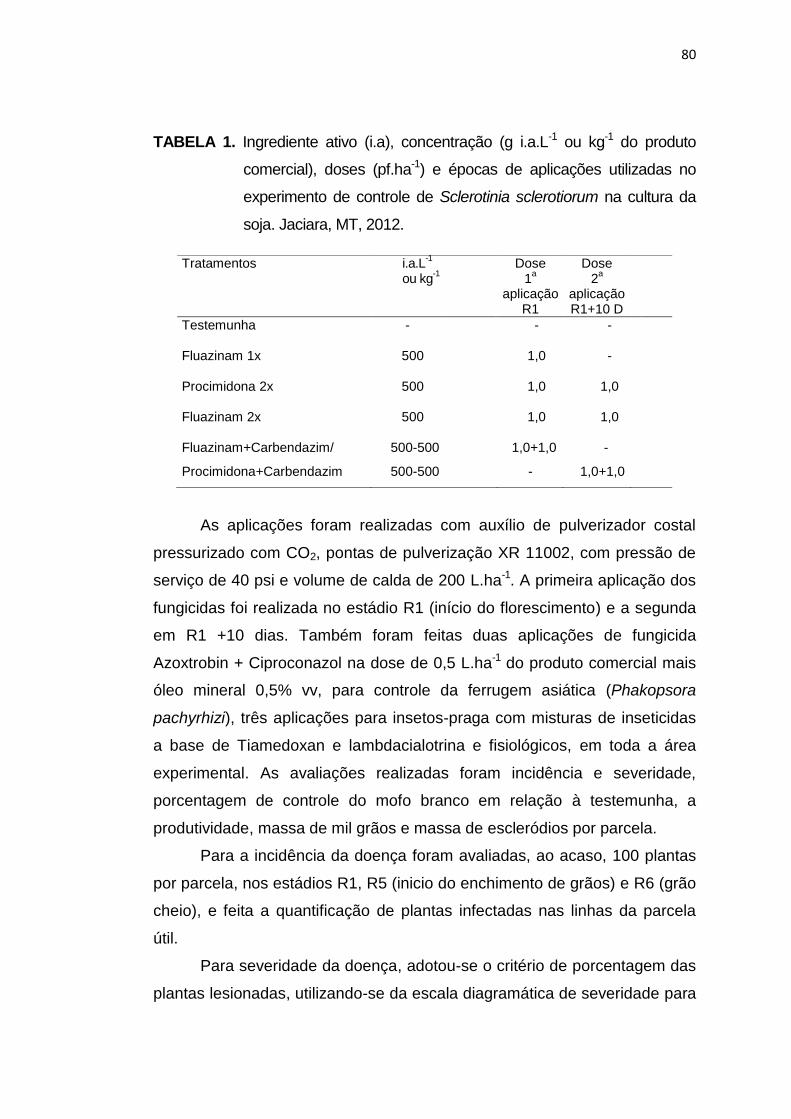
80
TABELA 1. Ingrediente ativo (i.a), concentração (g i.a.L-1 ou kg-1 do produto
comercial), doses (pf.ha-1) e épocas de aplicações utilizadas no
experimento de controle de Sclerotinia sclerotiorum na cultura da
soja. Jaciara, MT, 2012.
As aplicações foram realizadas com auxílio de pulverizador costal
pressurizado com CO2, pontas de pulverização XR 11002, com pressão de
serviço de 40 psi e volume de calda de 200 L.ha-1. A primeira aplicação dos
fungicidas foi realizada no estádio R1 (início do florescimento) e a segunda
em R1 +10 dias. Também foram feitas duas aplicações de fungicida
Azoxtrobin + Ciproconazol na dose de 0,5 L.ha-1 do produto comercial mais
óleo mineral 0,5% vv, para controle da ferrugem asiática (Phakopsora
pachyrhizi), três aplicações para insetos-praga com misturas de inseticidas
a base de Tiamedoxan e lambdacialotrina e fisiológicos, em toda a área
experimental. As avaliações realizadas foram incidência e severidade,
porcentagem de controle do mofo branco em relação à testemunha, a
produtividade, massa de mil grãos e massa de escleródios por parcela.
Para a incidência da doença foram avaliadas, ao acaso, 100 plantas
por parcela, nos estádios R1, R5 (inicio do enchimento de grãos) e R6 (grão
cheio), e feita a quantificação de plantas infectadas nas linhas da parcela
útil.
Para severidade da doença, adotou-se o critério de porcentagem das
plantas lesionadas, utilizando-se da escala diagramática de severidade para
Tratamentos i.a.L-1
ou kg-1
Dose 1
a
aplicação R1
Dose 2
a
aplicação R1+10 D
Testemunha
- - -
Fluazinam 1x
500 1,0 -
Procimidona 2x
500 1,0 1,0
Fluazinam 2x
500 1,0 1,0
Fluazinam+Carbendazim/ 500-500 1,0+1,0 -
Procimidona+Carbendazim 500-500 - 1,0+1,0

81
mofo branco, adaptada de Azevedo (1998), com as notas: 0- ausência de
sintomas; 1- presença de micélio circundando a base da haste; 2- presença
de micélio e escleródios; 3- micélio aéreo, escleródios, e folhas/ramos
murchos; 4- planta seca. Foram avaliadas ao acaso, 40 plantas por parcela
no estádio R6. O critério de análise percentual de controle da doença em
relação à testemunha foi considerado pela relação entre incidência da
doença na parcela tratada no estádio R6 e a média da incidência na
testemunha.
A colheita foi manual e a produtividade da soja foi dada pelas massas
de grãos obtidas em cada parcela útil, transformadas em kg.ha-1 com ajuste
do teor de água para 13%. Posteriormente, determinou-se a massa de mil
grãos (Brasil, 2009a), com oito subamostras de 100 grãos. A massa (g) de
escleródios foi obtida apenas dos escleródios recolhidos após a trilha das
plantas colhidas em cada parcela.
Os resultados foram previamente submetidos aos testes de
normalidade (Lillefors) e de homogeneidade (Cochran), posteriormente foi
realizada análise de variância com o teste de F, e as médias, comparadas
pelo teste de Scott-Knott a 5%. O processamento estatístico foi realizado
pelo programa computacional ASSISTAT 7.2 versão beta (Silva e Azevedo,
2009).
4.2.2 Experimento 2: Efeito dos fungicidas aplicados na cultura da soja
sobre a germinação carpogênica dos escleródios no solo
O experimento foi montado em 03/05/2012, com amostras de
escleródios obtidas, aleatoriamente, de plantas colonizadas pelo patógeno
nas parcelas de soja infestadas em cada repetição/tratamento do
Experimento 1. A amostra de escleródios foi composta de escleródios
formados nas hastes das plantas, formados externamente e escleródios
obtidos da trilha das parcelas com tratamento de fungicidas. Os escleródios
foram acondicionados em recipientes de vidro, lacrados, identificados e
acondicionados em temperatura de 25 ºC no Laboratório de Fitopatologia da
FAMEVZ (UFMT).

82
O delineamento experimental foi inteiramente casualizado com cinco
tratamentos e quatro repetições. Os tratamentos foram os mesmos do
Experimento 1 com os escleródios originados de cada parcela tratada com
ingredientes ativos dos fungicidas utilizados no Experimento 1: Fluazinam
1,0 L.ha-1, aplicados em R1; duas aplicações de Fluazinam (1,0 L.ha-1) – R1,
R1+ 10 dias; duas aplicações de Procimidona (1,0 kg.ha-1) - R1, R1+10 dias;
Fluazinam (1,0 L.ha-1) + Carbendazim (1,0 L.ha-1) em R1 e segunda
aplicação de Procimidona (1,0 kg.ha-1) + Carbendazin (1,0 L.ha-1) - 10 dias
após a primeira aplicação) e a testemunha.
Os escleródios foram colocados para incubar, em solo cultivado e
isento de infestação de S. sclerotiorum. O solo foi esterilizado e distribuído
em caixas de plástico tipo “gerbox”. Cada parcela foi composta por uma
caixa contendo camada de 250 g de solo e 25 escleródios de tamanhos
variados equidistantemente distribuídos sobre esse substrato. O substrato
foi com 100% da capacidade de campo, com base na massa (Brasil,
2009a). As caixas foram colocadas em câmara tipo BOD à temperatura de
20 °C ± 2 com fotoperíodo de 12 horas luz, por um período de 100 dias.
As variáveis analisadas foram: percentual de escleródios germinados,
emissão de estipes, número de estipes por escleródios e formação de
apotécios. As avaliações foram efetuadas contando-se o número de estipes
e apotécios formados em cada caixa. As avaliações foram aos 27, 40, 51,
70 e 100 dias após a incubação dos escleródios.
Para análise dos dados, foi realizada análise de variância pelo teste
F, e as médias comparadas pelo teste de Scott-Knott e Tukey a 5% de
probabilidade, utilizando o sistema de análise estatística ASSISTAT, versão
7.2 beta (Silva e Azevedo, 2009).

83
4.3 Resultados e Discussão
4.3.1 Avaliação do controle de mofo branco na cultura da soja com o
uso de fungicidas
Não houve incidência do mofo branco, na primeira aplicação de
fungicida (R1). Na segunda avaliação (R5), vinte dias após a primeira
aplicação dos fungicidas, a incidência atingiu 27% na testemunha,
confirmando a pressão da doença na área (Tabela 2). A testemunha, sem
aplicação de fungicida, apresentou as maiores porcentagens de incidência
quando comparada aos demais tratamentos.
TABELA 2. Incidência (%) de mofo branco em soja, variedade P98Y11,
nos estádios R1, R5 e R6, após controle químico. Jaciara-MT,
2011/12.
*Médias seguidas das mesmas letras, nas colunas, não diferem entre si pelo teste de Scott-Knott a 5% de probabilidade.
Após as aplicações dos fungicidas, verificou-se que os tratamentos
com controle químico apresentaram diferenças significativas na incidência
da doença em relação à testemunha, embora não tenham ocorrido
diferenças significativas entre os fungicidas avaliados (Tabela 2).
No estádio R5, a incidência variou de 3,25% no tratamento com
Fluazinam com duas aplicações a 6% no tratamento com Procimidona em
duas aplicações. Observou-se que o controle químico com fungicidas
Tratamentos (i.a) Incidência R1
Incidência* R5
Incidência R6
Testemunha
- 27,0 a 42,75 a
Fluazinam 1x - 3,42 b 8,00 b
Procimidona 2x - 6,00 b 10,50 b
Fluazinam 2x - 3,25 b 5,75 b
Fluazinam+Carbendazim/ Procimidona+Carbendazim
- 4,25 b 6,75 b
CV (%) - 46,65 29,49

84
reduziu o crescimento micelial e inibiu a formação de escleródios de S.
sclerotiorum em relação à testemunha.
Após a avaliação de incidência em R6, verificou-se que a testemunha
apresentava maior incidência da doença mostrando evolução da doença de
27% para 42,7%. Verificou-se nas parcelas sem aplicação de fungicidas
(testemunha), alta disseminação da doença, provavelmente devido às
condições ambientais favoráveis, proporcionando que o micélio do fungo se
multiplicasse indefinidamente (Brustolin et al., 2012).
Resultados de pesquisa sobre a eficiência de fungicidas indicam que
também o Fluazinam, além de Benomyl, Iprodione, Fluopyram,
Procimidona, a mistura Dimoxystrobina + Boscalid e Vinclozolin, foram mais
eficientes no controle de S. sclerotiorum (Vieira et al., 2001; Costa e Costa,
2004; Henning et al., 2009; Paula Júnior et al., 2009).
Em relação à severidade de mofo branco na cultura da soja, todos os
fungicidas avaliados proporcionaram menor severidade da doença em
relação à testemunha. A maior eficiência no controle da doença foi
observada nas parcelas dos tratamentos contendo Fluazinam (Figura 1). A
aplicação preventiva pode ter influenciado na eficiência de controle. Como
uma das principais formas de infecção ocorre pelos ascósporos por meio
das flores das plantas de soja, estas plantas devem estar protegidas por
fungicidas de forma preventiva, principalmente entre os estádios R1 e R4
(Meyer, 2009; Machado e Cassetari Neto, 2010).
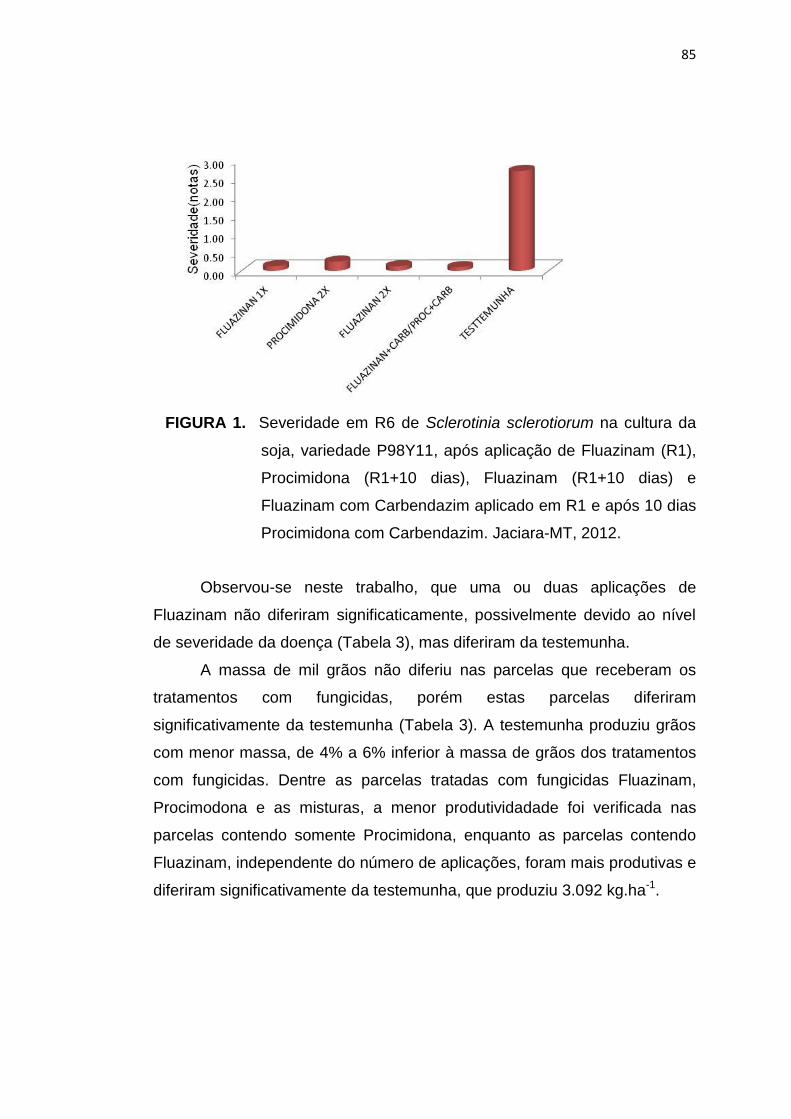
85
FIGURA 1. Severidade em R6 de Sclerotinia sclerotiorum na cultura da
soja, variedade P98Y11, após aplicação de Fluazinam (R1),
Procimidona (R1+10 dias), Fluazinam (R1+10 dias) e
Fluazinam com Carbendazim aplicado em R1 e após 10 dias
Procimidona com Carbendazim. Jaciara-MT, 2012.
Observou-se neste trabalho, que uma ou duas aplicações de
Fluazinam não diferiram significaticamente, possivelmente devido ao nível
de severidade da doença (Tabela 3), mas diferiram da testemunha.
A massa de mil grãos não diferiu nas parcelas que receberam os
tratamentos com fungicidas, porém estas parcelas diferiram
significativamente da testemunha (Tabela 3). A testemunha produziu grãos
com menor massa, de 4% a 6% inferior à massa de grãos dos tratamentos
com fungicidas. Dentre as parcelas tratadas com fungicidas Fluazinam,
Procimodona e as misturas, a menor produtividadade foi verificada nas
parcelas contendo somente Procimidona, enquanto as parcelas contendo
Fluazinam, independente do número de aplicações, foram mais produtivas e
diferiram significativamente da testemunha, que produziu 3.092 kg.ha-1.
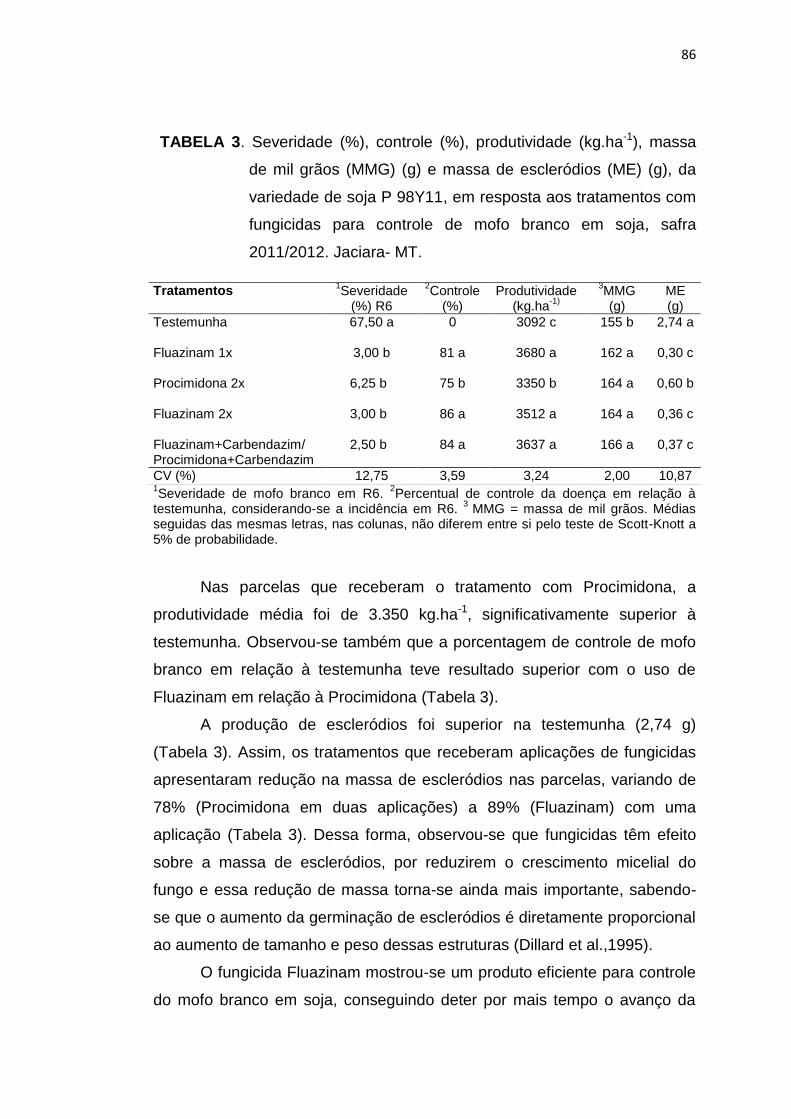
86
TABELA 3. Severidade (%), controle (%), produtividade (kg.ha-1), massa
de mil grãos (MMG) (g) e massa de escleródios (ME) (g), da
variedade de soja P 98Y11, em resposta aos tratamentos com
fungicidas para controle de mofo branco em soja, safra
2011/2012. Jaciara- MT.
1Severidade de mofo branco em R6.
2Percentual de controle da doença em relação à
testemunha, considerando-se a incidência em R6. 3
MMG = massa de mil grãos. Médias seguidas das mesmas letras, nas colunas, não diferem entre si pelo teste de Scott-Knott a 5% de probabilidade.
Nas parcelas que receberam o tratamento com Procimidona, a
produtividade média foi de 3.350 kg.ha-1, significativamente superior à
testemunha. Observou-se também que a porcentagem de controle de mofo
branco em relação à testemunha teve resultado superior com o uso de
Fluazinam em relação à Procimidona (Tabela 3).
A produção de escleródios foi superior na testemunha (2,74 g)
(Tabela 3). Assim, os tratamentos que receberam aplicações de fungicidas
apresentaram redução na massa de escleródios nas parcelas, variando de
78% (Procimidona em duas aplicações) a 89% (Fluazinam) com uma
aplicação (Tabela 3). Dessa forma, observou-se que fungicidas têm efeito
sobre a massa de escleródios, por reduzirem o crescimento micelial do
fungo e essa redução de massa torna-se ainda mais importante, sabendo-
se que o aumento da germinação de escleródios é diretamente proporcional
ao aumento de tamanho e peso dessas estruturas (Dillard et al.,1995).
O fungicida Fluazinam mostrou-se um produto eficiente para controle
do mofo branco em soja, conseguindo deter por mais tempo o avanço da
Tratamentos 1Severidade
(%) R6
2Controle
(%) Produtividade
(kg.ha-1)
3MMG (g)
ME (g)
Testemunha 67,50 a 0 3092 c 155 b 2,74 a
Fluazinam 1x
3,00 b 81 a 3680 a 162 a 0,30 c
Procimidona 2x
6,25 b 75 b 3350 b 164 a 0,60 b
Fluazinam 2x
3,00 b 86 a 3512 a 164 a 0,36 c
Fluazinam+Carbendazim/ Procimidona+Carbendazim
2,50 b 84 a 3637 a 166 a 0,37 c
CV (%) 12,75 3,59 3,24 2,00 10,87

87
incidência e severidade da doença na cultura da soja, reduzindo a massa de
escleródios, principal fonte de inóculo de S. sclerotiorum (Abawi e Grogan,
1979) para a safra seguinte. A eficiência de Fluazinam, também foi
verificada por Vieira et al. (2001) e Vieira et al. (2010) em feijoeiro e por
Henning et al. (2009) em soja. Assim, com o registro desses produtos para
uso na cultura da soja, o produtor deverá avaliar os custos e optar pela
aplicação que tenha eficiência no controle com menor custo.
Com base nesse estudo, o Fluzinam mesmo com uma aplicação foi
superior a Procimidona e às misturas com Carbendazim no controle de
mofo branco em soja (Tabela 3). Em estudo com mofo branco em feijoeiro,
Paula Júnior et al. (2009), verificaram que o fungicida Fluazinam foi mais
eficiente que Procimidona em reduzir o crescimento micelial do patógeno, e
incrementaram a produtividade entre 32 e 41%.
Os resultados satisfatórios da aplicação de Fluazinam observados
por Vieira et al. (2001), na cultura do feijoeiro, foram devido à aplicação do
fungicida na água de irrigação por aspersão. Os autores verificaram que o
Fluazinam sobre o solo mostrou-se eficiente na redução da incidência do
mofo branco em plantas, talvez por inibir a formação de apotécios. Segundo
Furlan (2008), as pulverizações devem ser realizadas uniformemente, com
boa distribuição nos tecidos da planta e, se possível, alcançando a
superfície do solo, onde surgem os apotécios e desenvolvem os micélios.
Assim o controle químico deve atingir os escleródios no solo por terem
efeito na germinação e formação de apotécios. Portanto, a eficiência do
fungicida depende da técnica de aplicação (Venegas e Saad, 2010).
O fungicida Fluazinam apresenta pequena atividade curativa e
sistêmica, mas bom efeito residual e seu mecanismo de ação atua no
sentido de inibir os processos de infecção, tais como a germinação de
esporos e a formação de apressórios, penetração e crescimento de hifas.
Por outro lado, o Procimidona, possui ação sistêmica moderada e ação
curativa e preventiva e seu modo de ação baseia-se na paralisação do
crescimento de hifas (Picinini e Goulart, 2002). Portanto, é necessário que
ocorra uma boa aplicação, pois os fungicidas tornam-se mais eficientes

88
quando há uma boa cobertura ou proteção das flores (Steadman, 1983).
Além disso, é importante que a técnica de aplicação atinja o solo em que se
encontram os escleródios do fungo, alvo indireto de controle da doença.
Como estes fungicidas foram aplicados de forma preventiva, e com baixa
incidência em R1 (27%), os resultados obtidos neste estudo podem ser
alterados se a porcentagem de incidência da doença na lavoura for
diferente da apresentada neste estudo.
4.3.2 Efeito dos fungicidas aplicados na cultura da soja, sobre a
germinacao carpogênica dos escleródios no solo.
Não houve diferença estatística entre os tratamentos de controle
químico com Fluazinam, Procimidona e misturas com Carbendazim, para
mofo branco aplicados na cultura da soja para porcentagem de escleródios
germinados, número de estipes, nem na relação do número de estipes por
número de escleródios germinados (Tabela 4).
Até a segunda avaliação, aos 40 dias após incubação do escleródio,
o Fluazinam em duas aplicações havia sido o melhor tratamento,
apresentando eficiência de 97% na inibição da formação de apotécios em
relação à testemunha (Tabela 4). Houve 100% de formação de estipes, mas
permitiu apenas a formação de estipes inviáveis, resultando em baixa
formação de apotécios. Os outros tratamentos foram menos eficientes na
inibição da germinação carpogênica dos escleródios, mas diferiram
significaticamente da testemunha. Entretanto, o efeito do controle químico
com fungicidas na inibição inicial de formação de estipes, produção de
estipes e apotécios inviáveis foi superado após 50 dias de incubação dos
escleródios.
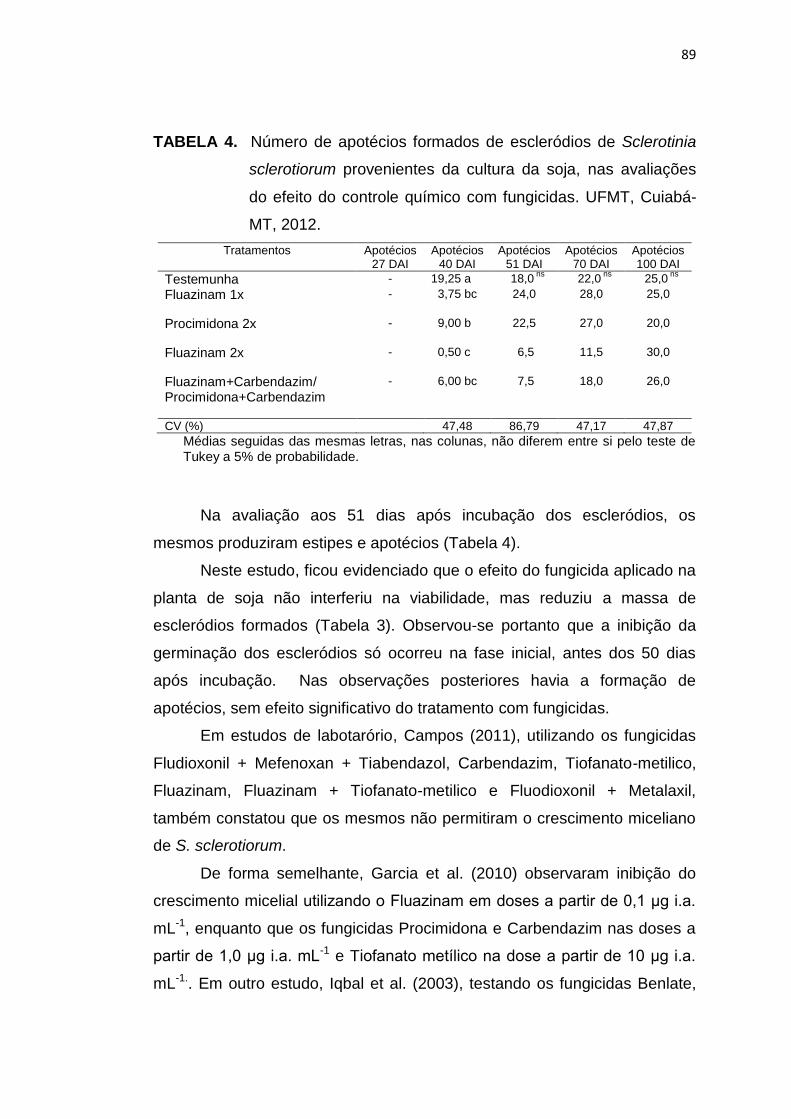
89
TABELA 4. Número de apotécios formados de escleródios de Sclerotinia
sclerotiorum provenientes da cultura da soja, nas avaliações
do efeito do controle químico com fungicidas. UFMT, Cuiabá-
MT, 2012.
Tratamentos Apotécios 27 DAI
Apotécios 40 DAI
Apotécios 51 DAI
Apotécios 70 DAI
Apotécios 100 DAI
Testemunha - 19,25 a 18,0 ns
22,0 ns
25,0 ns
Fluazinam 1x
- 3,75 bc 24,0 28,0 25,0
Procimidona 2x
- 9,00 b 22,5 27,0 20,0
Fluazinam 2x
- 0,50 c 6,5 11,5 30,0
Fluazinam+Carbendazim/ Procimidona+Carbendazim
- 6,00 bc 7,5 18,0 26,0
CV (%) 47,48 86,79 47,17 47,87
Médias seguidas das mesmas letras, nas colunas, não diferem entre si pelo teste de Tukey a 5% de probabilidade.
Na avaliação aos 51 dias após incubação dos escleródios, os
mesmos produziram estipes e apotécios (Tabela 4).
Neste estudo, ficou evidenciado que o efeito do fungicida aplicado na
planta de soja não interferiu na viabilidade, mas reduziu a massa de
escleródios formados (Tabela 3). Observou-se portanto que a inibição da
germinação dos escleródios só ocorreu na fase inicial, antes dos 50 dias
após incubação. Nas observações posteriores havia a formação de
apotécios, sem efeito significativo do tratamento com fungicidas.
Em estudos de labotarório, Campos (2011), utilizando os fungicidas
Fludioxonil + Mefenoxan + Tiabendazol, Carbendazim, Tiofanato-metilico,
Fluazinam, Fluazinam + Tiofanato-metilico e Fluodioxonil + Metalaxil,
também constatou que os mesmos não permitiram o crescimento miceliano
de S. sclerotiorum.
De forma semelhante, Garcia et al. (2010) observaram inibição do
crescimento micelial utilizando o Fluazinam em doses a partir de 0,1 μg i.a.
mL-1, enquanto que os fungicidas Procimidona e Carbendazim nas doses a
partir de 1,0 μg i.a. mL-1 e Tiofanato metílico na dose a partir de 10 μg i.a.
mL-1.. Em outro estudo, Iqbal et al. (2003), testando os fungicidas Benlate,

90
Ridomil, Tecto 60 e Topsin – M, observaram inibição completa do
crescimento e da germinação dos escleródios nas doses de 50 e 100 ppm.
No entanto, Costa e Costa (2004) observaram a inibição de 100% na
formação de estipes e apotécios de S. sclerotiorum utilizando o fungicida
Vinclozolin. O fungicida Fluazinam permitiu apenas a formação de estipes
inviáveis, não formando apotécios, entretanto para a germinação
miceliogênica, o Tiofanato metílico reduziu 75%, seguido dos fungicidas
Procimidona e Vinclozolin. Os fungicidas Benomyl, Fluazinam e Iprodione
foram menos eficientes.
Neste esperimento, somente ocorreu o efeito inibitório dos fungicidas
sobre a germinação e produção de apotécios na fase inicial da incubação,
possivelmente pelo efeito residual, sendo o efeito dos fungicidas superado
durante o período de avaliação (Figura 3). Portanto seria importante o uso
desses fungicidas na fase mais vulnerável da cultura da soja (R1), com
intervalos de aplicação visando a inibição da germinação dos escleródios e
a liberação de ascósporos pelos apotécios. Verificou-se que o controle
químico com fungicida reduz a formação de escleródios e a emissão de
apotécios, interferindo no progresso da doença, já que cada escleródio pode
produzir de um a 20 apotécios, e, durante o período funcional (de cinco a
dez dias), cada apotécio pode liberar mais de dois milhões de ascósporos
(Schwartz e Steadman, 1978; Steadman 1983). É fundamental utilizar o
controle químico com fungicidas eficientes que reduzam a produção do
escleródio como forma de reduzir o inóculo na área de soja.
A severidade do mofo‑branco em diferentes hospedeiros de S.
sclerotiorum é, em geral, proporcional à densidade de inóculo do patógeno
no solo. Portanto, a redução da população de escleródios é essencial para
o controle efetivo do mofo‑branco, que além disso também deve bloquear a
formação de apotécios e a ejeção de ascósporos e minimizar a produção de
novos escleródios pelo controle preventivo da doença na parte aérea das
plantas (Görgen et al., 2010).
.

91
FIGURA 3. Formação de apotécios de escleródios de S. sclerotiorum
provenientes da cultura da soja, após o controle químico com
fungicidas aos 40, 51, 70 e 100 dias após incubação de
escleródios. Jaciara-MT, 2012. A figura 3 e a tabela 4 informam
a mesma coisa. Decidir somente por uma forma de
apresentação dos dados. Figura ilustra melhor, veja se é
possível colocar letras na mesma.
4.4 Conclusões
Os fungicidas Fluazinam, Procimidona e em misturas com
Carbendazim reduzem a incidência e severidade de mofo branco na cultura
da soja.
A aplicação do fungicida Fluazinam na cultura da soja inibe a
germinação dos escleródios e formação de apotécios somente na fase
inicial de incubação destes escleródios, ou seja, anterior aos 50 dias após o
início da incubação.

92
4.5 Referências Bibliográficas
ABAWI, G.S.; GROGAN, R.G. Epidemiology of disease caused by Sclerotinia species. Phytopathology, v. 63, n. 8, p. 899-903, 1979. ALMEIDA, A. M. R.; PEREIRA, L. P.; YORINORI, J. T.; SILVA, J. F. V.; HENNING, C. V.; GODOY, L. M.; COSTAMILAN, L. M.; MEYER, M. C. Doenças da Soja. In: KIMATI, H.; AMORIM, L.; REZENDE, J. A. M.; BERGAMIN FILHO, A.; CAMARGO, L. E. A. (Ed.). Manual de Fitopatologia: doenças das plantas cultivadas. 4 ed. São Paulo: Agronômica Ceres, v. 2, 2005. p. 569-588. ATTANAYAKE, R.N.; GLAWE,D.A.; DUGAN, F.M.; CHEN, W. (2009) Erysiphe trifolii causing powdery mildew of lentil (Lens culinaris). Plant Disease, v. 93, p. 797–803, 2009. AZEVEDO, L. A. S. Manual de quantificação de doenças de plantas. São Paulo: Grupo Quattro Digital Media, 1998. 114 p. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária/MAPA/ACS, 2009. 395 p. BRUSTOLIN, R.; DE ROSSI, L.; REIS, E. M. Mofo branco. In: REIS, E. M.; CASA, R. T. (Org.). Doenças da soja: etiologia, sintomatologia, diagnose e manejo integrado. 1ed. Passo Fundo-RS: Berthier, 2012. v. 1, p. 217-232. CAMPOS, H. D. ; SILVA, L.H.C.P. ; SILVA, J.R.C. ; MONTEIRO, F. . Effect of fungicides in the development of Colletotrichum truncatum in controlled conditions. In: Top Ciência, 4. 2008, Heideberg - Germany. Top Ciência Basf. São Paulo : BASF The Chemical Company, 2008. v. 5. p. 1-2. CAMPOS, B. A.. Efeito de fungicidas no crescimento miceliano de Sclerotinia sclerotiorum. 2011. 33f. Monografia (Graduação em Agronomia) – Fesurv, Universidade de Rio Verde, 2011. COSTA, G. R.; COSTA, L.S.J. . Efeito da aplicação de fungicidas no solo sobre a germinação carpogênica e miceliogênica de escleródios de Sclerotinia sclerotiorum. Pesquisa Agropecuária Tropical, n. 34, v.3, p.133-138, 2004. DILLARD, H.R.; LUDWIG, J.W.; HUNTER, J.E. Conditioning sclerotia os Sclerotinia sclerotiorum for carpogenic germination. Plant Disease, v. 79, v.4, p.411-415, 1995. FURLAN, S. H. Importância e manejo do mofo branco na cultura da soja. Revista Plantio Direto, edição 107, p.28-31, 2008.

93
GARCIA, R.A.;SANTOS, R.C.; LOBO JÚNIOR, M.; MEYER, M.C.; OLIVEIRA, M.;CUNHA, M.G. Sensibilidade de isolados de Sclerotinia sclerotiorum à fungicidas. REUNIÃO DE PESQUISA DE SOJA DA REGIÃO CENTRAL DO BRASIL, 31. Resumos... Brasília, DF, 2010. GASPAROTTO, L. Sobrevivência de Sclerotinia sclerotiorum em solos cultivados com gramíneas e controle químico da podridão de alface. 1980. 42f. Dissertação (Mestrado em Agronomia) - Universidade Federal de Viçosa. Viçosa-MG, 1980. GÖRGEN, C.A.; CIVARDI, E.A; RAGAGNIM, V.A.; SILVEIRA NETO, A.N. da.; CARNEIRO, L.C.; LOBO JUNIOR, M. Redução do inóculo inicial de Sclerotinia sclerotiorum em soja cultivada após uso do sistema Santa Fé. Pesquisa Agropecuária Brasileira, v.45, p.1102-1108, 2010. HENNING, A.A.; PAULA, F.Y.H.; MOMTEMEZZO, C.A.O.; BOSSE, E.J.; BERGONSI, J.S.S. Avaliação de princípios ativos para o controle químico de mofo branco (Sclerotinia sclerotiorum) em soja – safra 2008/2009. Informativo ABRATES, v.19, n.1, p.29-31, 2009. http://www. abrates.org.br/portal/images/stories/informativos/v19n1/artigo01.pdf IQBAL, S.M.; GHAFOOR, A.; AHMAD, Z.; HAQQANI, A.M. Pathogenicity and Fungicidal Efficacy for Sclerotinia Rot of Brinjal. International Journal of Agriculture and Biology, v. 5, n. 4, p.199-206, 2003. LEITE, R.M.V.B.C. Ocorrência de doenças causadas por Sclerotinia sclerotiorum em girassol e soja. Londrina: Embrapa Soja, 2005. 3p. (Embrapa Soja. Circular Técnica, 76). LIU, XIN et al. Sensitivity to iprodione and Boscalid of Sclerotinia sclerotiorum isolates collected from rapeseed in China. Pesticide Biochemistry and Physiology, v.95, n.2, p. 106-112, 2009. MA, H; CHEN, Y.;WANG,J.;YU, W; TANG, Z.;CHEN, C.; ZHOU, M. Activity of Carbendazim, dimethachlon, iprodione, procymidone and Boscalid against Sclerotinia stem rot in Jiangsu Province of China. Phytoparasitica, v.37, n.5, p. 421-429, 2009. MACHADO, A.Q.; CASSETARI NETO, D. Epidemia branca. Cultivar Grandes Culturas, n. 130, ano 12, p. 20-23. 2010. MAPA. Ministério da Agricultura Pecuária e Abastecimento. Coordenação Geral de Agrotóxicos e Afins/DFIA/DAS. Disponível em: <http://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons> Acesso em: 20 de dezembro de 2012.. MEYER, M.C.; CAMPOS, H.D. Guerra ao mofo. Cultivar Grandes Culturas. Ano 11, n. 120, p.16-18, 2009.

94
PAULA JÚNIOR, T.J. ; VIEIRA, R.F.; ROCHA, P.R.R.; BERNARDES, A.; COSTA, E.L.; CARNEIRO, J.E.S.; RIBEIRO DO VALE, F.X.; ZAMBOLIM, L. White mold intensity on common bean in response to plant density, irrigation frequency, grass mulching, Trichoderma spp., and fungicide. Summa Phytopathologica, v.35, n.1, p.44-48, 2009. PICININI,E.C.; GOULART, A.C.P. Novos fungicidas para tratamento de sementes. Reunião Anual de Patologia de Plantas, p. 33-63, 2002. REIS, E.M.; REIS, A.C.; CARMONA, M.A. Manual de fungicidas: guia para controle químico de doenças de plantas. 6. ed. Passo Fundo: Ed. Universidade de Passo Fundo, 2010. 226 p. SCHWARTZ, H.F.; STEADMAN, J.R. Factors affecting sclerotium populations of, and apothecium production by Sclerotinia sclerotiorum, Phytopathology, v. 68, n. 383-388, 1978. SILVA, F. DE A. S. E. ; AZEVEDO, C. A. V. DE. Principal Components Analysis in the Software Assistat-Statistical Attendance. In:WORLD CONGRESS ON COMPUTERS IN AGRICULTURE, 7, Reno-NV-USA: American Society of Agricultural and Biological Engineers, 2009. STEADMAN, J.R. White mold – a serius yield-limiting desease of bean. Plant Disease, v. 67, n.4, p. 346-350, 1983. VENEGAS, F; SAAD, J.C.C. Fungigação no controle de mofo branco e produtividade do feijoeiro em condições do Cerrado. Irriga, v.15, n.2, p.159-172, 2010. VIEIRA, R. O mofo branco do feijoeiro – feijão no inverno. Informe Agropecuario, v. 17, n. 178, p. 54-63, 1994. VIEIRA, R. F.; PAULA JÚNIOR, T. J.; TEIXEIRA, H.; CARNEIRO, J. E. White mold management in common bean by increasing within-row distance between plants. Plant Disease, v. 94, n. 3, 2010. VIEIRA, R.F., PAULA JÚNIOR, T.J. de, PERES, A.P. ; MACHADO, J. da C. Fungicidas aplicados via água de irrigação no controle do mofo branco no feijoeiro e incidência do patógeno na semente. Fitopatologia Brasileira, v. 26, p. 770-773, 2001.

95
5 EFEITO DA TEMPERATURA E UMIDADE EM Sclerotinia sclerotiorum
DA SOJA
RESUMO - Os objetivos neste trabalho foram avaliar diferentes
temperaturas e níveis de disponibilidade de água no solo sobre a
germinação carpogênica de S. sclerotiorum e verificar o crescimento do
patógeno isolado da cultura da soja, na presença de restritores hídricos em
diferentes temperaturas. Foram feitos dois ensaios e, no primeiro,
escleródios de S. sclerotiorum, oriundos de área de produção de soja, foram
submetidos em câmara de crescimento (BOD), a diferentes temperaturas
(30, 25 e 20 °C ± 2 °C) e umidades do solo (100, 80, 60, 40 e 20% da
capacidade de campo), com fotoperíodo de 12 horas. Os escleródios foram
avaliados quanto à germinação carpogênica, número de estipes, número de
apotécios por escleródio, comprimento, diâmetro e massa de apotécios nos
diferentes níveis de disponibilidades de água e temperatura. No segundo
ensaio, o crescimento de S. sclerotiorum foi avaliado na presença de
restritores hídricos em diferentes temperaturas. O fungo S. sclerotiorum foi
colocado para desenvolver em meio de cultura BDA com o soluto NaCl nos
potenciais osmóticos: 0,0; - 0,4; - 0,6; - 0,8; - 1,0 e – 1,2 MPa em
germinadores com temperaturas controladas de 5 a 40 °C, por 10 dias e
fotoperíodo de 12 horas. As avaliações foram realizadas por meio do índice
de velocidade de crescimento micelial, quantificação e massa de
escleródios. A quantidade de água disponível no solo afeta a germinação
carpogênica, o número de apotécios produzidos por escleródios, a massa, o
comprimento e diâmetro dos apotécios. A temperatura de 20 °C favorece o
índice de velocidade de crescimento micelial, produção de escleródios e
massa.
Palavras-chave: restrição, escleródios, germinação carpogênica.

96
EFFECT OF TEMPERATURE AND HUMIDITY IN SOY Sclerotinia
sclerotiorum
ABSTRACT- The objective of this study was to evaluate the temperature
variation and different levels of soil water availability on germination of S.
carpogenic sclerotiorum and check the growth of the pathogen isolated from
soybean in the presence of water restriction at different temperatures. Two
experiments were done. In the first, sclerotia of S. sclerotiorum, coming from
the area of soybean production, were submitted in a growth chamber (BOD),
at different temperatures (30, 25 and 20 ° C ± 2 ° C) and soil moisture (100,
80, 60, 40 and 20 % of field capacity), with a photoperiod of 12 hours. The
sclerotia were evaluated for germination carpogenic, number of stems,
number of apothecia per escleródio, length, diameter and mass of apothecia
at different levels of water availability and temperature. In the second
experiment, the growth of S. sclerotiorum was evaluated restrictors in the
presence of water at different temperatures. The fungus S. sclerotiorum was
isolated from soybean seeds from sanity was put in to develop PDA culture
medium with NaCl solute osmotic potentials: 0.0, - 0.4, - 0.6, - 0.8, - and 1.0
- 1.2 MPa in temperature controlled germination of 5, 10,15, 20, 25, 30, 35
and 40 ° C for 10 days and 12 hours photoperiod. The evaluations were
performed using the index mycelial growth rates, quantification and mass of
sclerotia. The amount of water available in the soil affect the germination
carpogenic, the number of apothecia produced by sclerotia, mass, length
and diameter of the apothecia. A temperature of 20 ° C favored the speed
index mycelial growth, sclerotia production and mass.
Keywords: restraint, sclerotia, carpogenic germination.

97
5.1 Introdução
O mofo branco, causado pelo fungo Sclerotinia sclerotiorum (Lib.) de
Bary, é uma doença que atinge várias culturas de importância econômica
(Boland e Hall, 1994). Os ascósporos são a principal fonte de inóculo desse
patógeno, os quais são produzidos nos apotécios, resultante da germinação
de escleródios, em um processo denominado de germinação carpogênica
(Boland e Hall, 1987).
Para o controle do mofo branco da soja é requerida a integração de
várias medidas, inclusive o manejo cultural em função das condições
ambientes favoráveis à germinação carpogênica, visando o esgotamento do
potencial de inóculo (Meyer e Campos, 2009). A germinação carpogênica
de escleródios é altamente dependente da disponibilidade de água no solo
e pode ocorrer em condições que vão desde solo saturado até
relativamente seco, com tensão de água variando de zero a 750 kPa
(Ferraz et al., 1999). Não só a umidade do solo é importante, mas também o
tempo que o solo se mantém na umidade adequada à germinação
carpogênica (Phillips, 1987).
Em cultivos irrigados, o intervalo entre regas é muito importante, pois
regas mais frequentes, mesmo com volumes menores de água, favorecem
a maior formação de apotécios (Napoleão et al., 2007). Em zonas de clima
temperado, as condições ideais partem de uma combinação de eventos
úmidos e secos, baixas temperaturas e tempo (Bardin e Huang, 2001). Em
contraste, isolados oriundos dos trópicos não requerem frio no processo de
germinação carpogênica. Portanto, a origem geográfica dos isolados é de
fundamental importância em estudos com S. sclerotiorum (Bolton et al.,
2006). Estudos anteriores mostraram que temperatura e umidade do solo
têm influência direta na germinação carpogênica dos escleródios (Huang e
Kozub, 1991; Clarkson et al., 2004).
Os objetivos neste trabalho foram avaliar diferentes temperaturas e
níveis de disponibilidade de água no solo sobre a germinação carpogênica
de S. sclerotiorum e verificar o crescimento do patógeno isolado da cultura

98
da soja da região sul do estado de Mato Grosso, na presença de restritores
hídricos em diferentes temperaturas.

99
5.2 Material e Métodos
O experimento foi conduzido nos Laboratórios de Sementes e de
Fitopatologia da Faculdade de Agronomia, Medicina Veterinária e Zootecnia
(FAMEVZ) da Universidade Federal de Mato Grosso (UFMT), em Cuiabá –
MT, em dois ensaios:
5.2.1 Ensaio 1 - Efeito de diferentes temperaturas e níveis de
disponibilidade de água no solo sobre a germinação carpogênica de S.
sclerotiorum
Escleródios de S. sclerotiorum, oriundos de área de produção de soja
em Campo Verde-MT, foram submetidos em câmara de crescimento (BOD),
a diferentes temperaturas (20, 25 e 30 °C ± 2 °C) e umidade do solo (100,
80, 60, 40 e 20% da capacidade de campo, CC) com fotoperíodo de 12
horas. A capacidade de campo foi calculada segundo Brasil (2009).
Foi utilizado o delineamento experimental inteiramente casualizado,
em esquema fatorial 5x3, sendo cinco tratamentos de disponibilidade hídrica
e três temperaturas, com quatro repetições. As parcelas foram compostas
por caixas de plástico, tipo gerbox (11 x 11 x 3,5 cm) com solo cultivado,
retirado da camada de 0-5 cm de profundidade, livre de escleródios. O solo
foi esterilizado, e após, 25 escleródios foram dispostos sobre o substrato,
em caixas com 250 g de solo, equidistantemente distribuídos sobre o
mesmo. A quantidade de água foi determinada pelo método das pesagens.
O solo foi umedecido e mantido semanalmente com as referidas
porcentagens da capacidade de campo, com base na massa.
As avaliações ocorreram até os 78 dias após a incubação. Os
escleródios foram avaliados quanto à germinação carpogênica, número de
estipes, número de apotécios por escleródio, comprimento, diâmetro e
massa de apotécios nos diferentes níveis de disponibilidades de água e
temperatura.
Os dados das variáveis avaliadas foram submetidos à análise de
variância e aquelas afetadas pelo fator temperatura x umidade foram
analisadas por regressão linear. Para descrever o efeito da temperatura e

100
da umidade sobre a germinação de escleródios foi utilizada a análise com
modelo linear com cálculo de superfícies de tendência e equações
polinomiais de superfície de terceira ordem: eq.(1).
Z*= a+bX+cY+dX2+eXY+fY2+gX3+hX2Y+iXY2+jY3 (1)
Em que:
Z*= valor estimado de Z*i para o nó da célula (variável dependente);
X e Y= coordenadas
X= capacidade de campo em porcentagem (CC)
Y = temperatura em graus Celsius (T)
Xi e Yi = variáveis independentes
a....j= coeficientes que proporcionam o melhor ajuste aos dados amostrados
R2 = porcentagem de ajuste da superfície
5.2.2 Ensaio 2 - Crescimento de S. sclerotiorum isolado da cultura da
soja da região sul do estado de Mato Grosso, na presença de
restritores hídricos em diferentes temperaturas
O delineamento foi inteiramente casualizado em esquema fatorial
6x8, sendo seis concentrações de restritores hídricos e oito temperaturas
com três repetições. O fungo S. sclerotiorum foi isolado de sementes de
soja a partir do teste de sanidade e colocado em meio de cultura BDA
(extrato de 200 g de batata, 20 g de dextrose e 20 g de ágar), segundo
Machado et al. (2001), modificado com o uso de potencial osmótico
(Machado et al., 2007), ao qual foi adicionado o soluto NaCl nos potenciais
osmóticos: 0,0; - 0,4; - 0,6; - 0,8; - 1,0 e - 1,2 MPa.
O meio BDA+NaCl, nos referidos potenciais, foi distribuído em placas
de Petri de 9 cm de diâmetro, na proporção de 20 mL por placa, sendo em
seguida, transferido um disco de 0,6 cm de diâmetro da colônia pura do
fungo. Após as placas foram colocadas em germinadores com temperaturas
controladas de 5, 10, 15, 20, 25, 30, 35 e 40 °C, por 10 dias e fotoperíodo
de 12 horas.

101
As avaliações foram realizadas por meio do índice de velocidade de
crescimento micelial (IVCM), quantificação e massa de escleródios. Para o
IVCM, as avaliações foram realizadas diariamente durante o período de
incubação, medindo-se o diâmetro médio das colônias e aplicando a
fórmula utilizada por Oliveira (1991): form. (1).
IVCM = C1/N1+ C2/N2 + ... + Cn/Nn (1)
Em que:
IVCM = índice de velocidade de crescimento micelial;
C1, C2 e Cn = crescimento micelial na primeira, segunda e n-ésima
avaliação;
N1, N2 e Nn = número de dias de avaliação.
O índice de crescimento micelial foi avaliado diariamente até que a
placa fosse tomada pelo micélio. A quantificação foi realizada pelo número
de escleródios formados por repetição e a massa em balança com precisão
de 0,0001 g. Os dados das variáveis avaliadas foram submetidos à análise
de variância e aquelas afetadas pelo fator temperatura x potenciais
osmóticos foram analisadas por regressão linear. Para descrever o efeito da
temperatura e da restrição hídrica sobre o crescimento do patógeno foi
utilizada a análise com modelo linear com cálculo de superfícies de
tendência e equações polinomiais de superfície de terceira ordem: eq. (2).
Z*= a+bX+cY+dX2+eXY+fY2+gX3+hX2Y+iXY2+jY3 ( 2)
Em que:
Z*= valor estimado de Z*i para o nó da célula (variável dependente)
X e Y= coordenadas
X= temperatura em graus Celcius (T)
Y = potencial osmótico (P)
Xi e Yi = variáveis independentes
a....j = coeficientes que proporcionam o melhor ajuste aos dados
amostrados
R2 = porcentagem de ajuste da superfície
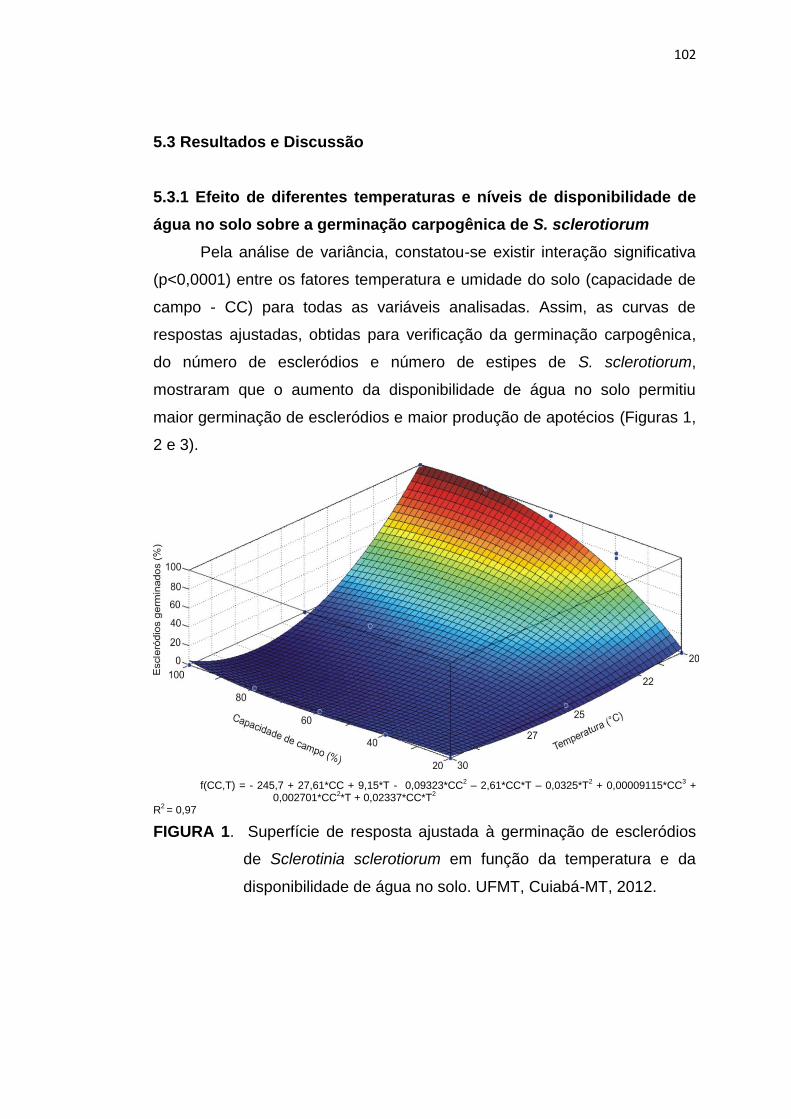
102
5.3 Resultados e Discussão
5.3.1 Efeito de diferentes temperaturas e níveis de disponibilidade de
água no solo sobre a germinação carpogênica de S. sclerotiorum
Pela análise de variância, constatou-se existir interação significativa
(p<0,0001) entre os fatores temperatura e umidade do solo (capacidade de
campo - CC) para todas as variáveis analisadas. Assim, as curvas de
respostas ajustadas, obtidas para verificação da germinação carpogênica,
do número de escleródios e número de estipes de S. sclerotiorum,
mostraram que o aumento da disponibilidade de água no solo permitiu
maior germinação de escleródios e maior produção de apotécios (Figuras 1,
2 e 3).
f(CC,T) = - 245,7 + 27,61*CC + 9,15*T - 0,09323*CC2 – 2,61*CC*T – 0,0325*T
2 + 0,00009115*CC
3 +
0,002701*CC2*T + 0,02337*CC*T
2
R2 = 0,97
FIGURA 1. Superfície de resposta ajustada à germinação de escleródios
de Sclerotinia sclerotiorum em função da temperatura e da
disponibilidade de água no solo. UFMT, Cuiabá-MT, 2012.
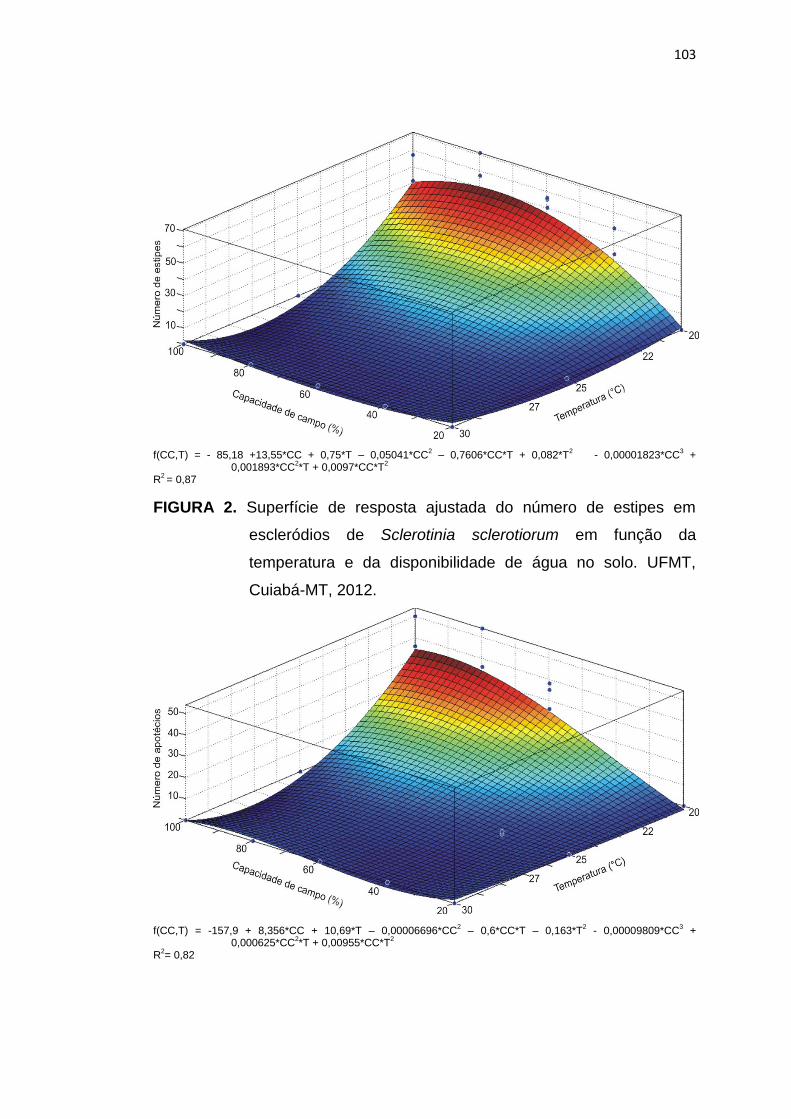
103
f(CC,T) = - 85,18 +13,55*CC + 0,75*T – 0,05041*CC2 – 0,7606*CC*T + 0,082*T
2 - 0,00001823*CC
3 +
0,001893*CC2*T + 0,0097*CC*T
2
R2 = 0,87
FIGURA 2. Superfície de resposta ajustada do número de estipes em
escleródios de Sclerotinia sclerotiorum em função da
temperatura e da disponibilidade de água no solo. UFMT,
Cuiabá-MT, 2012.
f(CC,T) = -157,9 + 8,356*CC + 10,69*T – 0,00006696*CC2 – 0,6*CC*T – 0,163*T
2 - 0,00009809*CC
3 +
0,000625*CC2*T + 0,00955*CC*T
2
R2= 0,82

104
FIGURA 3. Superfície de resposta ajustada para número de apotécios em
escleródios de Sclerotinia sclerotiorum em função da
temperatura e da umidade do solo. UFMT, Cuiabá-MT, 2012.
Não houve germinação de escleródios na temperatura de 30 °C como
também na capacidade de campo de 20% (Figura 1). Assim, verificou-se
que a umidade exerceu um efeito importante, mesmo quando da incubação
na temperatura de 20 °C; quanto maior a disponibilidade de água, maior a
formação de estipes e apotécios (Figuras 2 e 3). Constatou-se, portanto,
que o efeito da umidade do substrato tende a estimular a germinação dos
escleródios, o que não foi verificado no solo mais seco, onde não houve
sinalização para a germinação dos escleródios nas temperaturas incubadas.
Também, Abawi e Grogan (1975) e Steadman (1983) haviam citado que a
umidade desempenha um papel fundamental no desenvolvimento de S.
scleorotiorum.
A temperatura de 20 °C favoreceu a germinação do maior número de
escleródios, produção de estipes e apotécios (Figuras 1 a 3), corroborando
com o que foi observado por Boland e Hall (1987), que relataram o
desenvolvimento do mofo branco de 15 a 25 °C, sendo mais rapidamente a
20 °C; a 30 °C o mesmo não se desenvolveu.
A partir de 60% CC, a 20 °C, não foram observadas diferenças
significativas no número de escleródios germinados e, a 25 °C, verificou-se
produção esporádica de apotécios. Na umidade do solo com 40% CC, os
escleródios germinaram e formaram apotécios somente a 20 °C (Figura 3).
Possivelmente a temperatura seja fator determinante para que ocorra a
germinação carpogênica, pois mesmo em alta disponibilidade de água no
substrato (100% CC), não ocorreu germinação acima de 25 °C. Observou-
se que as estipes emitidas na temperatura de 25 °C não se desenvolveram
e se desidrataram rapidamente.
O número de apotécios atingiu o seu valor máximo entre 48 e 78
dias, exceto na menor disponibilidade de água (20% CC) (Figura 3). Este
período coincide com o estádio de floração média da maioria das cultivares

105
de soja (cerca de 50 dias), época de maior suscetibilidade da cultura. Na
condição a 40% da capacidade de campo, houve produção de apotécios
durante todo o ensaio, entretanto somente a 20 °C, evidenciando que a
umidade do solo onde estavam os escleródios não foi fator tão determinante
para a germinação dos escleródios, sendo que a temperatura teve efeito
mais direto. Entretanto, Tokeshi et al. (1995) concluíram ser a umidade da
superfície um dos principais fatores no controle da produção de apotécios.
Nos tratamentos incubados a 20 °C, onde o solo foi regado com 60 a
100% da capacidade de campo, 95% dos escleródios germinaram, contra
60% de germinação a 40% CC (Figuras 1, 2 e 3). A mesma tendência foi
observada em relação ao número de estipes e número total de apotécios
(Figuras 2 e 3), onde se verificou que acima de 40% CC, houve aumento na
produção de apotécios. Isso possivelmente explica a capacidade de S.
sclerotiorum para produzir apotécios em solos com níveis relativamente
baixos de umidade. Quando ocorre estresse hídrico, logo após a iniciação
da germinação dos escleródios, pode ocorrer o retardamento da formação
dos apotécios (Abawi e Grogan, 1975). Outros estudos também
confirmaram a correlação positiva existente entre temperatura e umidade
para a germinação carpogênica de escleródios de S. sclerotiorum
(Schuwartz e Steadman, 1978; Tu, 1989; Tokeshi et al.,1995; Mila e Yang,
2008; Wu e Subbaro, 2008).
Foi ajustado um modelo linear (Figuras 1 a 3) para estimar a
germinação de escleródios, produção de estipes e apotécios em função da
umidade e da temperatura do solo. A qualidade do ajuste foi satisfatória
pelas correlações entre os valores observados e estimados pelo modelo,
pois o mesmo apresentou qualidade de ajuste regular (Figura 3). Segundo
esse modelo, condições de temperatura próximas de 20 °C e umidades
acima de 60% CC são mais favoráveis à germinação carpogênica de
escleródios, em condições de ambiente controlado.
Os dados evidenciaram que, embora a disponibilidade de água
aplicada no solo tenha sido importante, a temperatura foi altamente
limitante. Em áreas com alta infestação de escleródios, que tenham

106
disponibilidade hídrica e temperatura em torno de 20 °C haveria condições
necessárias para a germinação carpogênica de escleródios, o que
consequentemente, implicaria na produção de apotécios e aumentaria a
quantidade de inóculo na forma de ascósporos (Abawi et al., 1975). Assim,
o monitoramento da temperatura é fator decisivo para auxiliar no controle
integrado da germinação de escleródios e produção de apotécios. Embora a
temperatura de 20 °C seja a mais favorável para germinação carpogênica
de escleródios, como também verificado por Boland e Hall (1987) e Sun e
Yang (2000), a germinação também ocorre em temperaturas próximas à
temperatura ótima (20 °C).
Na análise de regressão do tamanho de apotécios de S. sclerotiorum
(Figura 4), verificou-se que o comprimento, diâmetro e massa de apotécios
variaram nos tratamentos com umidade do solo. A disponibilidade de água
no solo influenciou significativamente no tamanho do apotécio. O
comprimento, diâmetro e massa foram menores no tratamento 40% CC.
Verificou-se que as respostas do tamanho de apotécios podem ser
estimados pelas equações de regressão. As respostas são proporcionais à
disponibilidade de água até 100% CC.
O comprimento, diâmetro e massa de apotécios aumentaram com a
disponibilidade hídrica. Observou-se que a 100% da capacidade de campo,
quando ocorreu germinação de somente um apotécio por escleródio, esta
estrutura teve diâmetro maior que apotécios formados em escleródios que
continham mais apotécios. Nesta mesma disponibilidade hídrica, verificou-
se maior quantidade formada de estipes, maior número de apotécios, com
diâmetros maiores, do que os formados em 80% CC; entretanto esta
umidade (80% CC) favoreceu a formação de apotécios com comprimento e
diâmetro maiores que os formados a 60% CC.
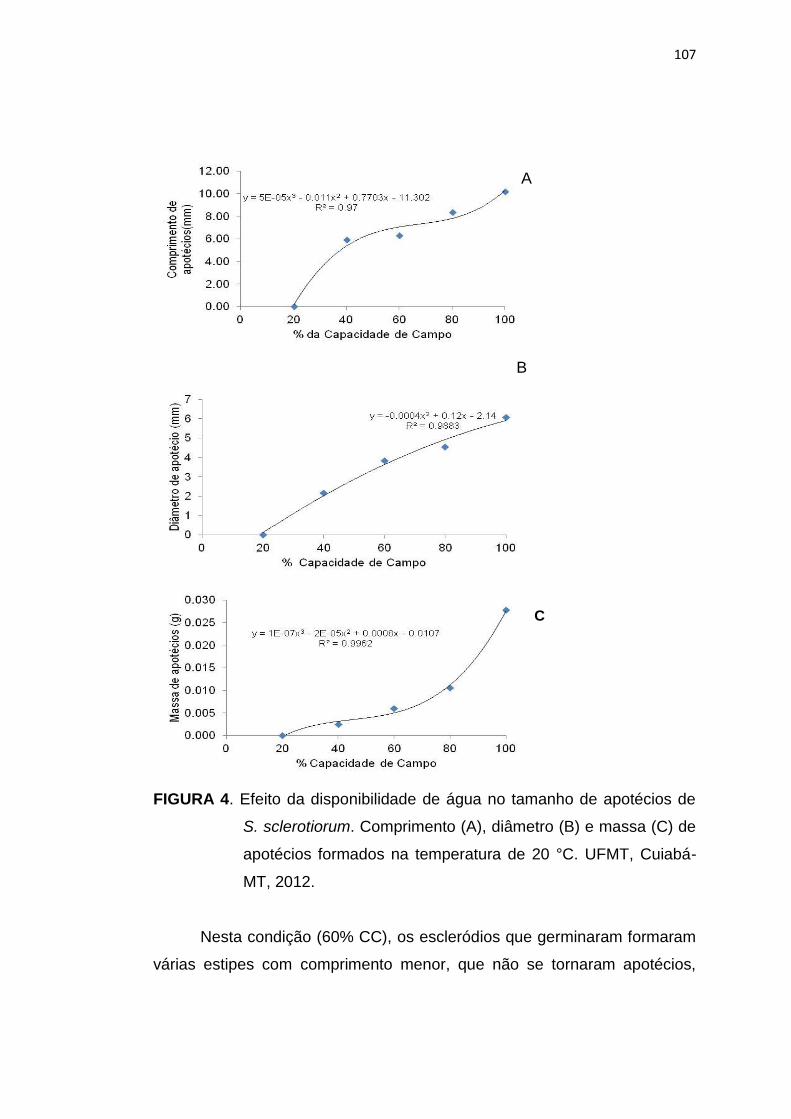
107
FIGURA 4. Efeito da disponibilidade de água no tamanho de apotécios de
S. sclerotiorum. Comprimento (A), diâmetro (B) e massa (C) de
apotécios formados na temperatura de 20 °C. UFMT, Cuiabá-
MT, 2012.
Nesta condição (60% CC), os escleródios que germinaram formaram
várias estipes com comprimento menor, que não se tornaram apotécios,
B
C
A

108
devido à restrição de água; dessa forma ocorreu a formação de novas
estipes e parte delas se tornou apotécios.
Alguns fatores como temperatura, umidade, aeração e intensidade
luminosa influenciam a germinação carpogênica. Estudos realizados por
Hao et al. (2003) obtiveram germinação carpogênica de 5 a 25 °C com
ótima em 15 °C, enquanto nos estudos de Sun e Yang (2000) a produção
dependeu da intensidade luminosa, além do efeito na produção de
apotécios. Ascósporos somente germinaram em umidade livre por mais de
72 horas (Abawi et al., 1975; Purdy, 1979). A temperatura ótima para
germinação do ascósporo liberado pelo apotécio foi 25 °C, a germinação
boa a 20 °C e fraca a 10 e 30 °C (Abawi e Grogan, 1975). Em ambiente
controlado a doença desenvolveu-se de 15 a 25 °C, mas rapidamente a 20
°C, mas não a 30 °C (Bolland e Hall, 1987). Portanto, como ocorreu neste
trabalho a temperatura de 20 °C foi mais favorável a germinação
carpogênica favorecendo o desenvolvimento do fungo.
O efeito da temperatura na germinação carpogênica varia com o
isolado, principalmente com sua origem geográfica (Huang e Kozub, 1993;
Huang e Kozub, 1991). Assim as respostas obtidas neste trabalho, para os
fatores ambientais estudados, podem estar relacionadas à variabilidade
genética do isolado de S. sclerotiorum dessa região, podendo ser realizadas
novas pesquisas para confirmar a viabilidade dos ascósporos produzidos
com restrição de umidade, pois a produção de apotécios não significa que
todos tenham ascósporos viáveis.
5.3.2 Crescimento de S. sclerotiorum isolado da cultura da soja da
região sul do estado de Mato Grosso, na presença de restritores
hídricos em diferentes temperaturas
Na análise do crescimento de S. sclerotiorum isolado da cultura da
soja da região sul do estado de Mato Grosso, na presença de restritores
hídricos em diferentes temperaturas, observou-se que houve interação
significativa entre temperatura e restrição hídrica para crescimento micelial
(Figuras 5 e 6). O índice de velocidade de crescimento micelial variou de
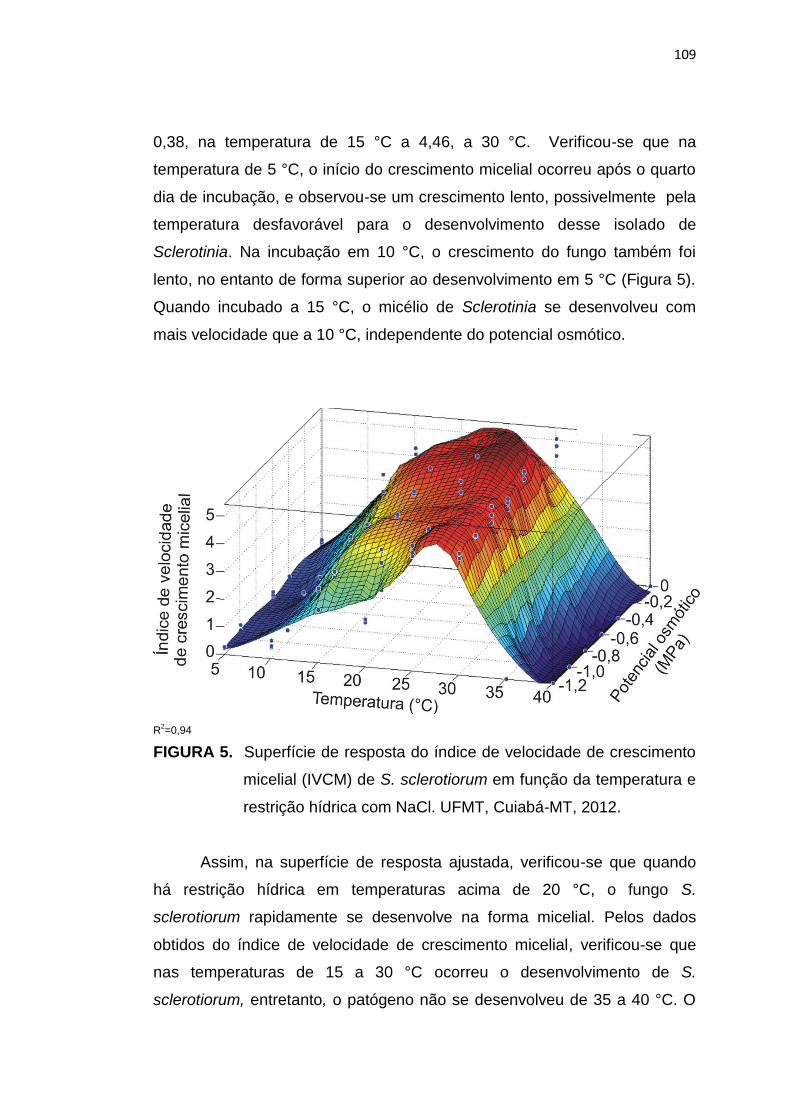
109
0,38, na temperatura de 15 °C a 4,46, a 30 °C. Verificou-se que na
temperatura de 5 °C, o início do crescimento micelial ocorreu após o quarto
dia de incubação, e observou-se um crescimento lento, possivelmente pela
temperatura desfavorável para o desenvolvimento desse isolado de
Sclerotinia. Na incubação em 10 °C, o crescimento do fungo também foi
lento, no entanto de forma superior ao desenvolvimento em 5 °C (Figura 5).
Quando incubado a 15 °C, o micélio de Sclerotinia se desenvolveu com
mais velocidade que a 10 °C, independente do potencial osmótico.
R2=0,94
FIGURA 5. Superfície de resposta do índice de velocidade de crescimento
micelial (IVCM) de S. sclerotiorum em função da temperatura e
restrição hídrica com NaCl. UFMT, Cuiabá-MT, 2012.
Assim, na superfície de resposta ajustada, verificou-se que quando
há restrição hídrica em temperaturas acima de 20 °C, o fungo S.
sclerotiorum rapidamente se desenvolve na forma micelial. Pelos dados
obtidos do índice de velocidade de crescimento micelial, verificou-se que
nas temperaturas de 15 a 30 °C ocorreu o desenvolvimento de S.
sclerotiorum, entretanto, o patógeno não se desenvolveu de 35 a 40 °C. O
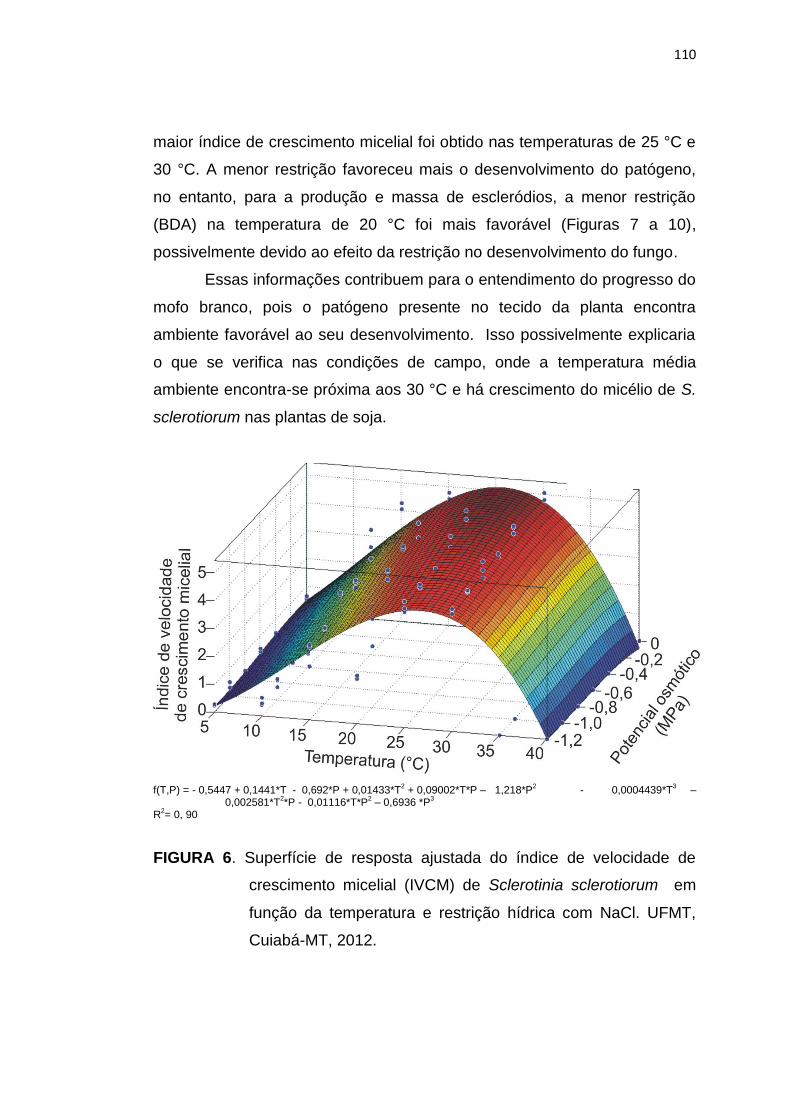
110
maior índice de crescimento micelial foi obtido nas temperaturas de 25 °C e
30 °C. A menor restrição favoreceu mais o desenvolvimento do patógeno,
no entanto, para a produção e massa de escleródios, a menor restrição
(BDA) na temperatura de 20 °C foi mais favorável (Figuras 7 a 10),
possivelmente devido ao efeito da restrição no desenvolvimento do fungo.
Essas informações contribuem para o entendimento do progresso do
mofo branco, pois o patógeno presente no tecido da planta encontra
ambiente favorável ao seu desenvolvimento. Isso possivelmente explicaria
o que se verifica nas condições de campo, onde a temperatura média
ambiente encontra-se próxima aos 30 °C e há crescimento do micélio de S.
sclerotiorum nas plantas de soja.
f(T,P) = - 0,5447 + 0,1441*T - 0,692*P + 0,01433*T2 + 0,09002*T*P – 1,218*P
2 - 0,0004439*T
3 –
0,002581*T2*P - 0,01116*T*P
2 – 0,6936 *P
3
R2= 0, 90
FIGURA 6. Superfície de resposta ajustada do índice de velocidade de
crescimento micelial (IVCM) de Sclerotinia sclerotiorum em
função da temperatura e restrição hídrica com NaCl. UFMT,
Cuiabá-MT, 2012.
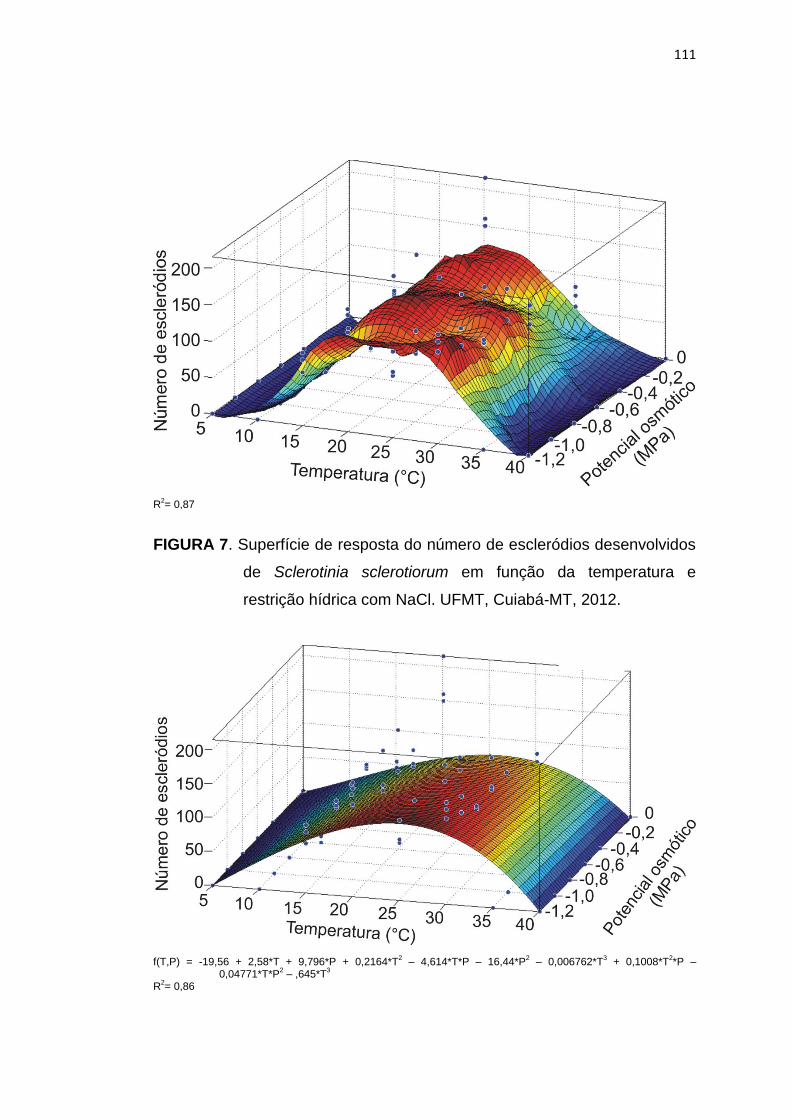
111
R2= 0,87
FIGURA 7. Superfície de resposta do número de escleródios desenvolvidos
de Sclerotinia sclerotiorum em função da temperatura e
restrição hídrica com NaCl. UFMT, Cuiabá-MT, 2012.
f(T,P) = -19,56 + 2,58*T + 9,796*P + 0,2164*T2 – 4,614*T*P – 16,44*P
2 – 0,006762*T
3 + 0,1008*T
2*P –
0,04771*T*P2 – ,645*T
3
R2= 0,86
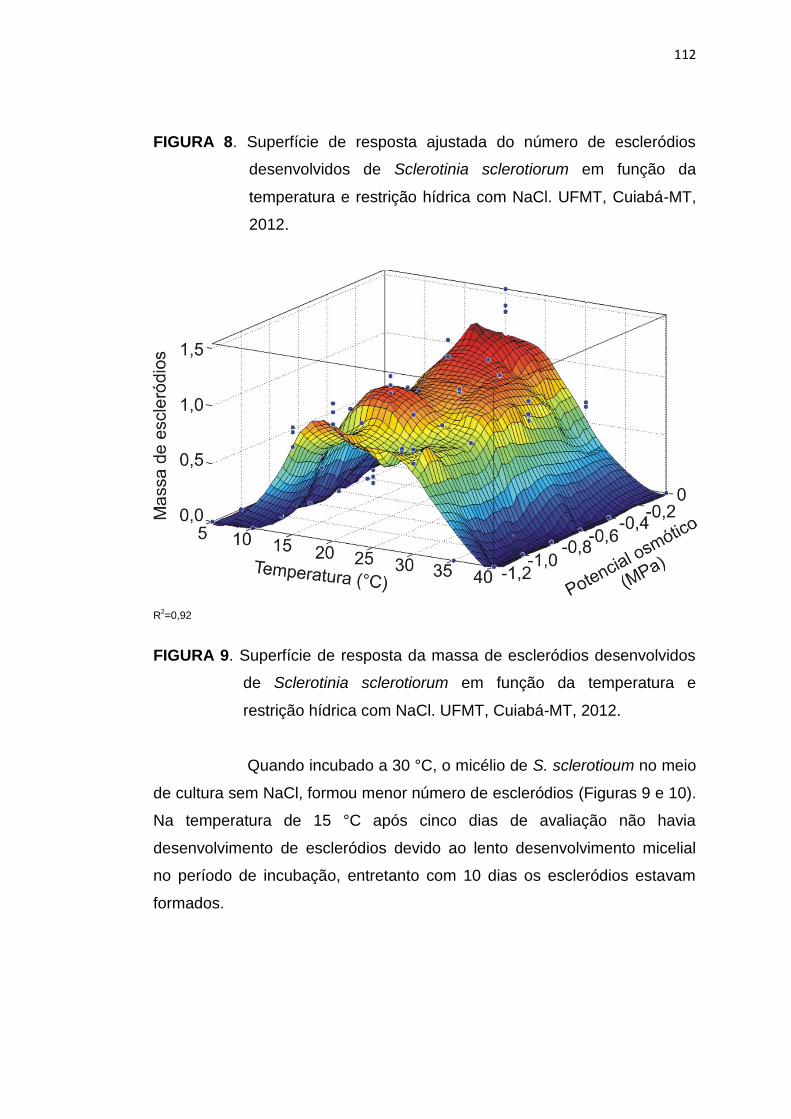
112
FIGURA 8. Superfície de resposta ajustada do número de escleródios
desenvolvidos de Sclerotinia sclerotiorum em função da
temperatura e restrição hídrica com NaCl. UFMT, Cuiabá-MT,
2012.
R2=0,92
FIGURA 9. Superfície de resposta da massa de escleródios desenvolvidos
de Sclerotinia sclerotiorum em função da temperatura e
restrição hídrica com NaCl. UFMT, Cuiabá-MT, 2012.
Quando incubado a 30 °C, o micélio de S. sclerotioum no meio
de cultura sem NaCl, formou menor número de escleródios (Figuras 9 e 10).
Na temperatura de 15 °C após cinco dias de avaliação não havia
desenvolvimento de escleródios devido ao lento desenvolvimento micelial
no período de incubação, entretanto com 10 dias os escleródios estavam
formados.
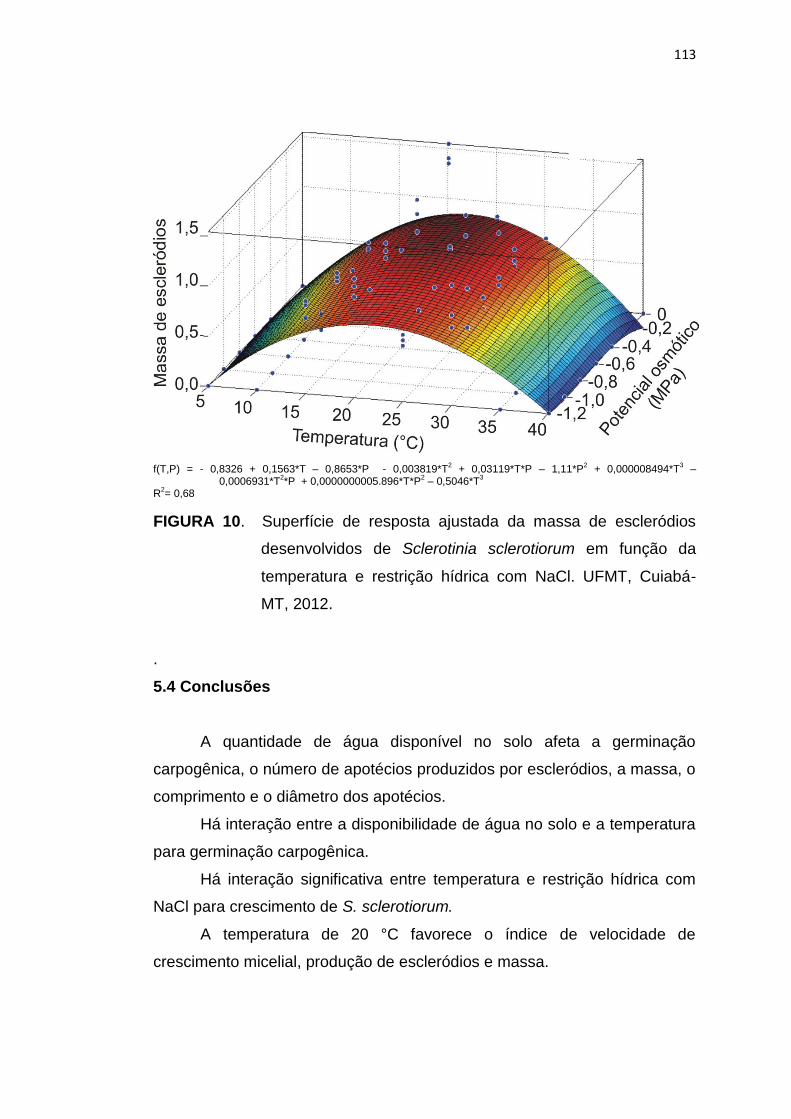
113
f(T,P) = - 0,8326 + 0,1563*T – 0,8653*P - 0,003819*T2 + 0,03119*T*P – 1,11*P
2 + 0,000008494*T
3 –
0,0006931*T2*P + 0,0000000005.896*T*P
2 – 0,5046*T
3
R2= 0,68
FIGURA 10. Superfície de resposta ajustada da massa de escleródios
desenvolvidos de Sclerotinia sclerotiorum em função da
temperatura e restrição hídrica com NaCl. UFMT, Cuiabá-
MT, 2012.
.
5.4 Conclusões
A quantidade de água disponível no solo afeta a germinação
carpogênica, o número de apotécios produzidos por escleródios, a massa, o
comprimento e o diâmetro dos apotécios.
Há interação entre a disponibilidade de água no solo e a temperatura
para germinação carpogênica.
Há interação significativa entre temperatura e restrição hídrica com
NaCl para crescimento de S. sclerotiorum.
A temperatura de 20 °C favorece o índice de velocidade de
crescimento micelial, produção de escleródios e massa.

114
Há desenvolvimento micelial do fungo S. sclerotiorum em
temperaturas até 30 °C com produção de escleródios.
5.5 Referências Bibliográficas
ABAWI, G. S.; GROGAN, R. G. Source of primary inoculum and effects of temperature and moisture on infection of beans by Whetzelinia sclerotiorum. Phytopathology, v. 65, n.3, p. 300-309, 1975. ABAWI, G.,S.; POLACH, F. J.; MOLIN, W. T. Infection of bean by ascospores of Whetzelinia sclerotiorum. Phytopathology, v. 65, p. 673-678, 1975. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária/MAPA/ACS, 2009. 395 p. BOLAND, G. J.; HALL, R. Epidemiology of white bean in Ontario. Canadian Journal of Plant Pathology, v. 9, n.2, p. 218-224, 1987. BOLAND, G. J.; HALL, R. Index of plants of hostes Sclerotinia sclerotiorum. Canadian Journal of Plant Pathology, v. 16, n.1, p. 93-108, 1994. BARDIN, S. D;HUANG, H. C. Research on biology and control of Sclerotinia diseases in Canada. Canadian Journal Plant Pathology, v. 23, n. 1, p. 88–98, 2001. CLARKSON, J. P.; PHELPS, K., WHIPPS, J. M.; YOUNG, C. S.; SMITH, J. A.; WATLING, M. Forecasting sclerotinia disease on lettuce: a predictive model for carpogenic germination of Sclerotinia sclerotiorum sclerotia. Phytopathology, v. 97, p. 621-631, 2007. CLARKSON, J. P.; PHELPS, K., WHIPPS, J. M.; YOUNG, C. S.; SMITH, J. A.; WATLING, M. Forecasting sclerotinia disease on lettuce: Toward developing a prediction model for carpogenic germination of sclerotia. Phytopathology, v. 94, p. 268-279, 2004. FERRAZ, L. C. L.; CAFÉ-FILHO, A. C.; NASSER, L. C. B.; AZEVEDO, J. Effects of soil moisture, organic matter and grass mulching on the carpogenic germination of sclerotia and infection of bean by Sclerotinia sclerotiorum. Plant Pathology, v. 48, n. 1, p.77- 82, 1999. GROGAN, R. G.; ABAWI, G. S. Influence of water potential on growth and survival of Whetzelina sclerotiorum. Phytopathology, v. 65, p.122-128, 1975.

115
HAO, J. J.; SUBBARAO, K. V.; DUNIWAY, J. M. Germination of Sclerotinia minor and S. sclerotiorum sclerotia under various soil moisture and temperature combinations. Phytopathology, v. 93, n. 4, 2003. HUANG, H. C.; KOZUB, G. C. Temperature requirements for carpogenic germination of Sclerotinia sclerotiorum isolates of different geographic origins. Botanical Bulletin of Academia, v. 32, p. 279-286, 1991. HUANG, H. C.; KOZUB, G. C. Influence of inoculum production temperature on carpogenic germination of sclerotia of Sclerotinia sclerotiorum. Canadian Journal of Microbiology, v. 39, n. 5, p. 548-550, 1993. MACHADO, J. C., OLIVEIRA, J. A., VIEIRA, M. G.G.C. et al. Inoculação artificial de sementes de soja por fungos, utilizando a solução de manitol, Revista Brasileira de Sementes, v. 23, n. 2, 95 – 101, 2001. MACHADO, A. Q., MACHADO, J. C., VIEIRA, M. D.G.G.C. et al. Potencial do Uso da Restrição Hídrica em Testes de Sanidade de Sementes de Algodoeiro, Fitopatologia Brasileira, v. 32, n. 5, p. 408 – 414, 2007. MEYER, M. C; CAMPOS, H. D. Guerra ao mofo. Cultivar grandes culturas. Ano 11, n. 120, p.16-18, 2009. MILA, A. L.; YANG, X.B. Effects of Fluctuating Soil Temperature and Water Potential on Sclerotia Germination and Apothecial Production of Sclerotinia sclerotiorum. Plant Disease, v. 92, n. 1, p. 78-82, 2008. NAPOLEÃO, R.; CAFÉ-FILHO, A.C.; LOPES, C.A.; NASSER, L.C.B.; MAROUELLI, W.A. Efeito da freqüência de rega e da umidade do solo sobre a germinação carpogênica de Sclerotinia sclerotiorum. Summa Phytopathologica, v. 33, n. 1, p. 80-82, 2007. NEPA, A. ; MENDOZA, L. E. D. R. Effect of sclerotial water content on carpogenic germination of Sclerotinia sclerotiorum. Plant Disease, v. 96, n. 9, p. 1315-1322, 2012. OLIVEIRA, J. A. Efeito do tratamento fungicida em sementes no controle de tombamento de plântulas de pepino (Cucumis sativus L.) e pimentão (Capsicum annum L.). 1991. 111 f. Dissertação (Mestrado em Fitossanidade) – Universidade Federal de Lavras, Lavras, MG. PHILLIPS, A. J. L. Carpogenic germination of sclerotia of Sclerotinia sclerotiorum: a review. Phytophylactica, v.19, n. 2, p. 279-283, 1987. PURDY, L. H. Sclerotinia sclerotiorum: history, diseases and symptomatology, host range, geographic distribution, and impact. Phytopathology, v. 69, p. 875- 880, 1979.

116
SCHUWARTZ, H. F.,;STEADMAN, J. R. Factors affecting sclerotium populations of, and apothecium production by, Sclerotinia sclerotiorum. Phytopathology, v. 68, n. 3 , p. 383-388, 1978. STEADMAN, J. R. White mold – a serius yield-limiting desease of bean. Plant Disease, v. 67, n. 4, p. 346-350, 1983. SUN, P.; YANG, X. B. Light, temperature, and moisture effects on apothecium production of Sclerotinia sclerotiorum. Plant Disease, v. 84, n. 12, p. 1287-1293, 2000. TOKESHI, H.; ALVES, M. C.; SANCHES, A. B.; HARADA, D. Y. In: International Coference on Kyusei Nature Farming, Fourth, 1995, Paris. Effective Microorganisms for Controlling the Phytoipathogenic Fungus Sclerotinia sclerotiorum in Lettuce. Washington: 1995. p. 131-139. TU, J. C. Management of white mold of white beans in Ontario. Plant Disease, v. 73, n. 3, p. 281-285, 1989. WU, B. M; PENG, Y.L. ; QIN, Q. M; SUBBARAO, K. V. Incubation of excised apothecia enhances ascus maturation of Sclerotinia sclerotiorum. Mycologia, v. 99, n. 1, p. 33–41, 2007. WU, B. M.; SUBBARAO, K. V. Effects of soil temperature, moisture, and burial depths on carpogenic germination of Sclerotinia sclerotiorum and S. minor. Phytopathology, v. 98, n. 10, p. 1144-1152, 2008.

117
6 DISTRIBUIÇÃO ESPACIAL DE S. sclerotiorum EM SOJA
RESUMO - Neste trabalho o objetivo foi avaliar a variabilidade espacial e
mapear a incidência S. sclerotiorum na cultura da soja, e sua relação com a
textura do solo em Jaciara-MT, por meio da geoestatística. As amostragens
foram realizadas quando a variedade de soja TMG 132 RR estava no
estádio R.9 (ponto de colheita). Os pontos amostrais das áreas foram
demarcados com o auxílio de um GPS de posicionamento absoluto Marca
Garmin 12. Na área da Fazenda São Jorge, as dimensões da grade
amostral foi constituída de pontos distanciados de 20 a 25 m, totalizando 90
pontos. Na Fazenda Planalto, foi realizado um caminhamento aleatório e no
ponto amostral foi realizada a contagem de escleródios na lavoura, em 30
pontos aleatórios em 0,50 m2. Para quantificar, em cada ponto, a incidência
de S. sclerotiorum nas sementes, foram coletadas e debulhadas vagens de
soja para realização dos testes de sanidade das sementes, pelo método de
incubação em rolo de papel e “blotter test”. Nos mesmos pontos amostrais,
foram coletadas amostras de solo, na profundidade de 0 a 20 cm, com trado
holandês, para análise textural. A análise da distribuição espacial foi
realizada através do semivariograma no programa computacional ArcGis,
que definiu o modelo da variabilidade. Com a análise geoestatística é
possível quantificar o grau de dependência espacial e mapear a
variabilidade espacial de escleródios e da incidência de S. sclerotiorum em
soja. A variação espacial de S. sclerotiorum ocorre em reboleiras. Há
correlação positiva da presença de escleródios com a textura argilosa.
Palavras-chave: padrão espacial, escleródios, semivariograma.

118
SPATIAL DISTRIBUTION OF S. sclerotiorum IN SOYBEAN
ABSTRACT - In this study we evaluated the spatial variability and mapping
the incidence S. sclerotiorum in soybean, and its relationship with soil
texture in Jaciara-MT through geostatistics. The study was conducted in
2011/2012 season, in two areas of commercial production of soybeans,
Farm São Jorge and Farm Planalto. Samples were taken when the soybean
variety TMG 132 RR were stage R.9 (point of harvest). The sampling points
were demarcated areas with the aid of a GPS absolute positioning Brand
Garmin 12. In the area of Farm São Jorge, the dimensions of the sampling
grid consisted of points spaced 20-25 m, totaling 90 points. In Farm Planalto,
we performed a random pathway and the sample point were counted
sclerotia in the field, 30 random points at 0.50 m2. To quantify the incidence
at each point S. sclerotiorum in the seeds were collected and threshed
soybean pods for carrying out the tests for seed, by the method of incubation
in paper roll and "blotter test". In the sampling points, soil samples were
collected at a depth 0-20 cm, with Dutch auger for textural analysis. A spatial
distribution analysis was performed using the computer program ArcGIS
semivariogram, which defined the model variability. With geostatistical
analysis is to quantify the degree of spatial dependence and map the spatial
variability of sclerotia and the incidence of S. sclerotiorum in soybean. The
spatial variation of S. sclerotiorum occurs in foci. There is a positive
correlation of the presence of sclerotia with clayey.
Keywords: spatial pattern, sclerotia, semivariogram.

119
6.1 Introdução
As sementes são uma das principais fontes de inóculo, disseminação
e sobrevivência de microrganismos fitopatogênicos, e podem provocar
redução na germinação, vigor e originar focos primários de infecção. Goulart
(2005) afirmou que a maioria das doenças de importância econômica que
ocorrem na cultura da soja são causadas por patógenos disseminados
pelas sementes. Entre esses, Bolton et al. (2006) ressaltaram o patógeno
Sclerotinia sclerotiorum (Lib) de Barry causador do mofo branco ou podridão
branca da haste, que sobrevive no solo por meio de estruturas de
resistência, escleródios, e ser hospedeiro de mais de 400 espécies de
plantas, destacando-se soja, feijão, algodão e girassol. Por causa disto, o
índice de tolerância para este patógeno em sementes é zero, de acordo
com programas de certificação de sementes no Brasil (MAPA, 2009).
De acordo com Furlan (2010), levantamentos de áreas de cultura de
soja contaminadas por S. sclerotiorum no Brasil indicam extensões
superiores a 500 mil hectares a partir da safra 2007/08. Essa grande
extensão de área afetada pode ter ocorrido pela forma de disseminação do
fungo, ou pelas condições favoráveis à manutenção das estruturas de
resistência do patógeno e pela dificuldade de controle da doença.
Em decorrência da importância dessa doença, alguns trabalhos
necessitam ser desenvolvidos, abordando principalmente a incidência do
patógeno no campo, porém estudos voltados à avaliação da dependência
espacial de S. sclerotiorum, nas lavouras de soja, são escassos. Para
estudar a dependência espacial, a geoestatística é a ferramenta mais
adequada, pois considera a posição no espaço da variável em estudo
(Liebhold et al., 1993). Com o uso da geoestatística, é possível determinar a
dependência espacial da doença, através da elaboração de
semivariogramas que são ajustados a um modelo que fornece o raio de
agregação de plantas doentes ou escleródios no solo e, a partir desse
semivariograma, é feita a elaboração de mapas que mostram como ocorre a
expansão da doença na área.

120
A análise do padrão de incidência no campo é precursor da
investigação de estratégias amostrais e da análise dos dados que elas
geram (Hughes e Gottwald, 1998). Estudos de estimativas mais precisas
sobre o crescimento da doença, no tempo e no espaço, atentando para a
localização e tamanho dos focos, podem viabilizar o controle da doença nas
áreas de localização desses focos da doença ao invés da ação em área
total. O estudo do padrão espacial de plantas doentes gera informações
sobre a importância do inóculo primário, os mecanismos de dispersão do
patógeno, a direção e a distância de propagação da doença e sobre o efeito
de fatores ambientais na epidemia (Wu et al., 2001).
A geoestatística é uma importante ferramenta de análise da
distribuição espacial de doenças (Alves et al., 2006). O processo de
georreferenciamento e a utilização de SIG (Sistema de Informação
Geográfica) permitem o armazenamento, processamento e análise dos
dados relativos à variabilidade espacial e auxiliam na modelagem e
confecção de mapas de variabilidade espacial de doenças. Nesse contexto,
o conhecimento da dinâmica espacial de doenças de plantas, bem como
dos fatores ambientais que podem influenciar a epidemia, é de grande
importância para a definição de estratégias de manejo eficiente e
sustentável. Sendo assim, objetivou-se avaliar a variabilidade espacial e
mapear a incidência S. sclerotiorum na cultura da soja, e sua relação com a
textura do solo em Jaciara-MT, por meio da geoestatística.

121
6.2 Material e Métodos
O estudo foi realizado na safra 2011/2012, em duas áreas de
produção comercial de soja, na Fazenda São Jorge, coordenadas
geográficas 15º 48’ 51,05” S e 55º 12’ 86,96” W, e Fazenda Planalto,
coordenadas 15º 47’ 25,90”S e 55º 12’ 46,90” W, localizadas no município
de Jaciara, Mato Grosso. O solo das áreas foi classificado como Latossolo
Vermelho distrófico, de textura muito argilosa. As áreas vêm sendo
manejadas há muitos anos sob plantio direto, e cultivadas com soja, milho
ou algodão com histórico de incidência de mofo branco. Na Fazenda
Planalto houve duas aplicações de Fluazinam para controle de S.
sclerotiorum e, na Fazenda São Jorge, não houve aplicação visando
controle de mofo branco. As amostragens foram realizadas quando a
variedade de soja TMG 132 RR estava no estádio R.9 (ponto de colheita).
Os pontos amostrais das áreas foram demarcados com o auxílio de
um GPS de posicionamento absoluto Marca Garmin 12. Na área da
Fazenda São Jorge, as dimensões da grade amostral foi constituída de
pontos distanciados de 20 a 25 m e o número de pontos totalizou 90 pontos
amostrais (Figura 1). A amostragem foi realizada no dia 11/02/2012 e para
quantificar em cada ponto a incidência de S. sclerotiorum nas sementes,
foram coletadas e debulhadas vagens de soja para realização dos testes de
sanidade das sementes, pelo método de incubação em rolo de papel e
“blotter test”. Nos mesmos pontos amostrais, foram coletadas amostras de
solo, na profundidade de 0 a 20 cm, com trado holandês, para análise
textural.
Para a realização do experimento e coleta das amostras da Fazenda
Planalto, foi realizado um caminhamento aleatório e no ponto amostral foi
realizada a contagem de escleródios na lavoura, em 30 pontos aleatórios
em 0,50m2 (Figura 1). Nos mesmos pontos, também foram coletadas
amostras de solo, na profundidade de 0 a 20 cm, com trado holandês, para
análise textural e coletadas e debulhadas vagens de soja para teste de
sanidade das sementes, pelo método de incubação em rolo de papel.
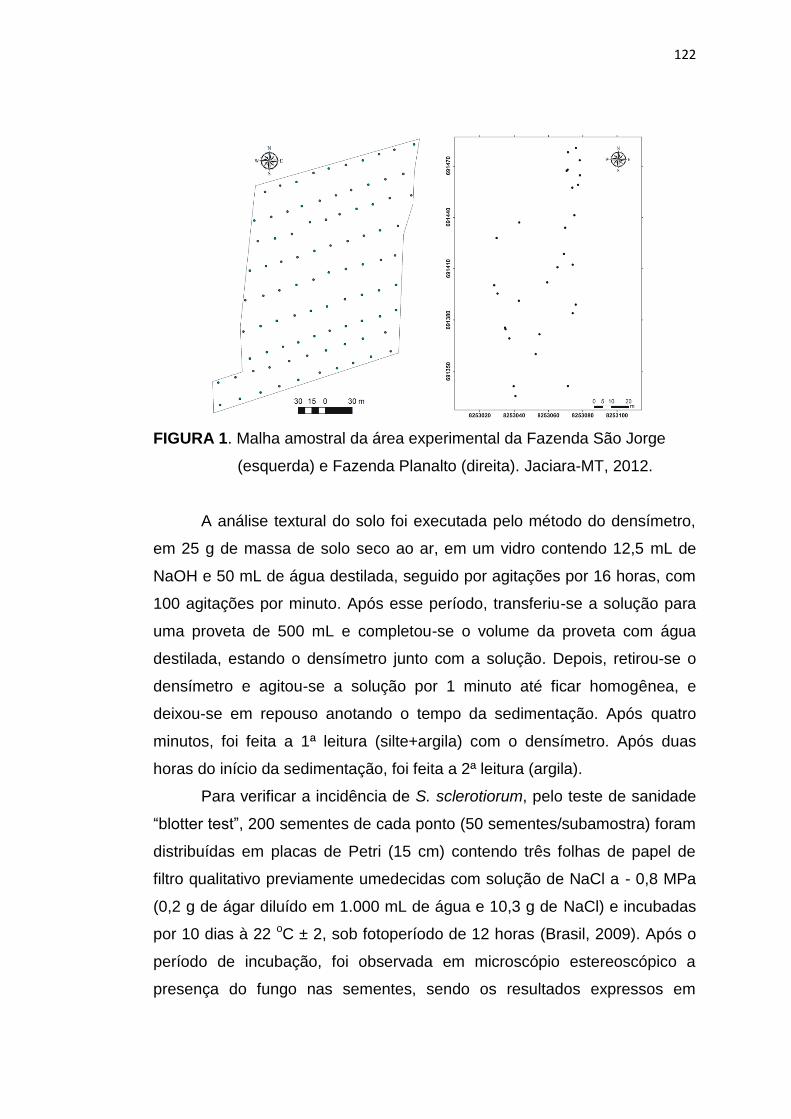
122
FIGURA 1. Malha amostral da área experimental da Fazenda São Jorge
(esquerda) e Fazenda Planalto (direita). Jaciara-MT, 2012.
A análise textural do solo foi executada pelo método do densímetro,
em 25 g de massa de solo seco ao ar, em um vidro contendo 12,5 mL de
NaOH e 50 mL de água destilada, seguido por agitações por 16 horas, com
100 agitações por minuto. Após esse período, transferiu-se a solução para
uma proveta de 500 mL e completou-se o volume da proveta com água
destilada, estando o densímetro junto com a solução. Depois, retirou-se o
densímetro e agitou-se a solução por 1 minuto até ficar homogênea, e
deixou-se em repouso anotando o tempo da sedimentação. Após quatro
minutos, foi feita a 1ª leitura (silte+argila) com o densímetro. Após duas
horas do início da sedimentação, foi feita a 2ª leitura (argila).
Para verificar a incidência de S. sclerotiorum, pelo teste de sanidade
“blotter test”, 200 sementes de cada ponto (50 sementes/subamostra) foram
distribuídas em placas de Petri (15 cm) contendo três folhas de papel de
filtro qualitativo previamente umedecidas com solução de NaCl a - 0,8 MPa
(0,2 g de ágar diluído em 1.000 mL de água e 10,3 g de NaCl) e incubadas
por 10 dias à 22 oC ± 2, sob fotoperíodo de 12 horas (Brasil, 2009). Após o
período de incubação, foi observada em microscópio estereoscópico a
presença do fungo nas sementes, sendo os resultados expressos em

123
porcentagem de incidência do patógeno. Para o método de incubação em
rolo de papel foram usadas as recomendações do teste de germinação para
sementes de soja (Brasil, 2009), porém sendo mantidos em câmara de
incubação por 14 dias a 15 ºC ± 2, em posição vertical e na ausência de luz,
e acondicionados em recipientes com atmosfera próxima a saturação
(Brasil, 2009).
As coordenadas geográficas dos pontos amostrais foram locadas por
meio de sistema de posicionamento global por satélite (GPS). Com os
dados efetuou-se a estatística descritiva, onde se calculou média, mínimo,
máximo, desvio padrão e coeficiente de variação. Com base nos valores, foi
realizada a análise de correlação (correlação de Pearson) entre os dados,
por meio do programa Statistic 7.0.
Posteriormente, de posse dos dados de análise textural de solos,
testes de sanidade e número de escleródios, elaborou-se uma tabela
contendo as coordenadas geográficas (x e y) dos pontos amostrais e os
valores correspondentes a cada variável estudada. Após, para a análise
geoestatística, onde foi ajustado o semivariograma esférico e exponencial
procedeu-se à verificação da existência de dependência espacial, por meio
do semivariograma, estimado conforme a eq.(1).
2)(
1)()(
)(2
1)(
hN
i ii hxZxZhN
hy (1)
Em que: N(h) representa o número de pares de valores medidos [Z(xi), Z(xi
+ h)] separados por um vetor h. Para o estudo em questão, os valores de Z
foram as variáveis estudadas, enquanto os valores de xi e xi + h foram
definidos de acordo com as posições das amostras no campo.
A análise da dependência espacial é baseada na suposição de que
medições separadas por distâncias pequenas são mais semelhantes umas
às outras, que aquelas separadas por distâncias maiores. A distância limite
de dependência espacial, denominada alcance (A), indica que amostras
localizadas a distâncias menores que o alcance são correlacionadas umas

124
com as outras. O alcance depende do tamanho da área amostrada e da
escala de observação realizada, sendo tanto maior quanto maior for o
intervalo entre medidas (Trangmar et al., 1985).
Os parâmetros dos semivariogramas foram representados nos
gráficos pelo efeito pepita (Co), patamar (Co+C) e alcance (A). Para a
análise do grau de dependência espacial das variáveis, foi utilizada a
classificação de Cambardella et al. (1994), em que é considerado forte
dependência espacial o semivariograma que tem efeito pepita 25% do
patamar, moderada quando está entre 25 e 75% e de fraca quando for
>75%.
A análise da distribuição espacial foi realizada por meio do
semivariograma no programa computacional ArcGis, que definiu o modelo
da variabilidade.
6.3 Resultados e Discussão
Após análise dos dados da estatística descritiva das variáveis
(média, mínimo, máximo, desvio padrão e coeficiente de variação) foi
realizada a análise dos variogramas. A média da incidência de S.
sclerotirum detectada pelo método rolo de papel, na Fazenda Planalto, foi
de 5,33%, variando de zero ao máximo de 11% de incidência. Maior
variação ocorreu na presença de escleródios nos pontos amostrais, com
amostras variando de zero a 206 escleródios e média de 72 escleródios por
ponto amostral. O coeficiente de variação obtido foi alto, para as variáveis
avaliadas, e essa alta variação de incidência deve-se possivelmente ao fato
dos métodos de detecção não apresentarem os mesmos resultados nas
repetições, sendo pouco sensíveis para a detecção do patógeno em
sementes provenientes de áreas com incidência da doença (Henneberg et
al., 2012) e da dispersão das estruturas de resistência do fungo por
máquinas e implementos em pontos aleatórios.
Verificou-se que na Fazenda São Jorge, a média de incidência de
mofo branco nas sementes de soja variou de 5,2% detectado pelo método
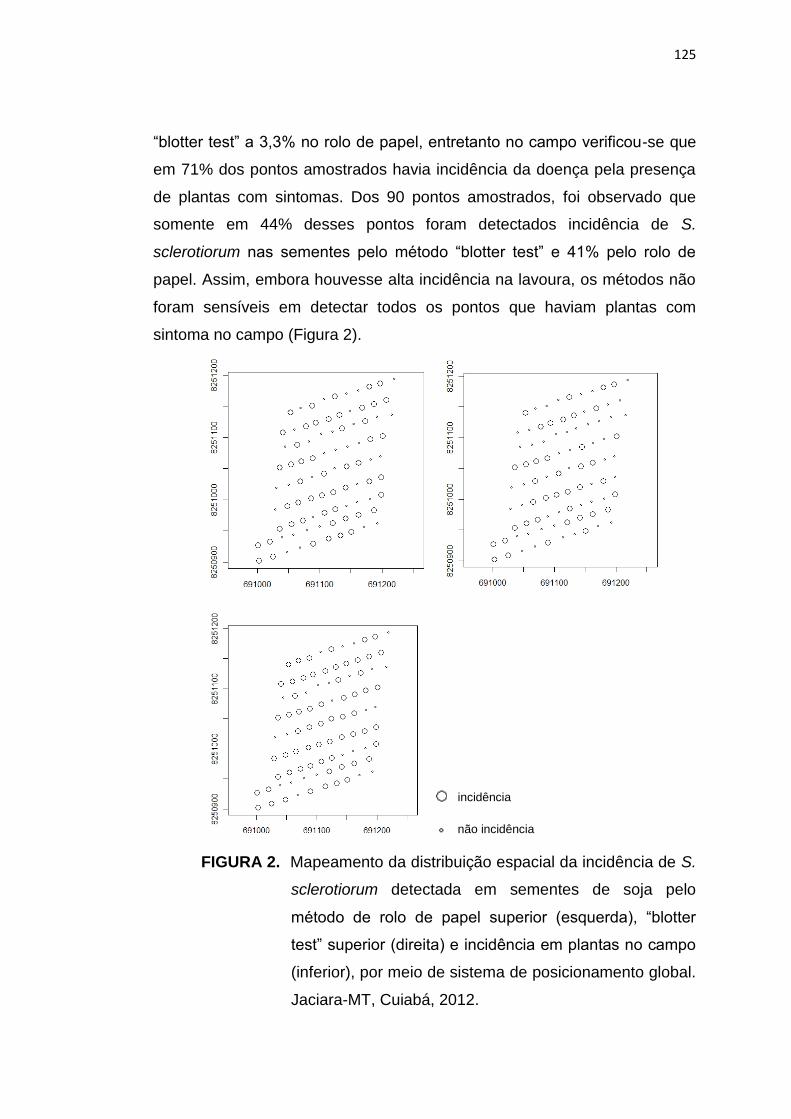
125
“blotter test” a 3,3% no rolo de papel, entretanto no campo verificou-se que
em 71% dos pontos amostrados havia incidência da doença pela presença
de plantas com sintomas. Dos 90 pontos amostrados, foi observado que
somente em 44% desses pontos foram detectados incidência de S.
sclerotiorum nas sementes pelo método “blotter test” e 41% pelo rolo de
papel. Assim, embora houvesse alta incidência na lavoura, os métodos não
foram sensíveis em detectar todos os pontos que haviam plantas com
sintoma no campo (Figura 2).
FIGURA 2. Mapeamento da distribuição espacial da incidência de S.
sclerotiorum detectada em sementes de soja pelo
método de rolo de papel superior (esquerda), “blotter
test” superior (direita) e incidência em plantas no campo
(inferior), por meio de sistema de posicionamento global.
Jaciara-MT, Cuiabá, 2012.
incidência
não incidência
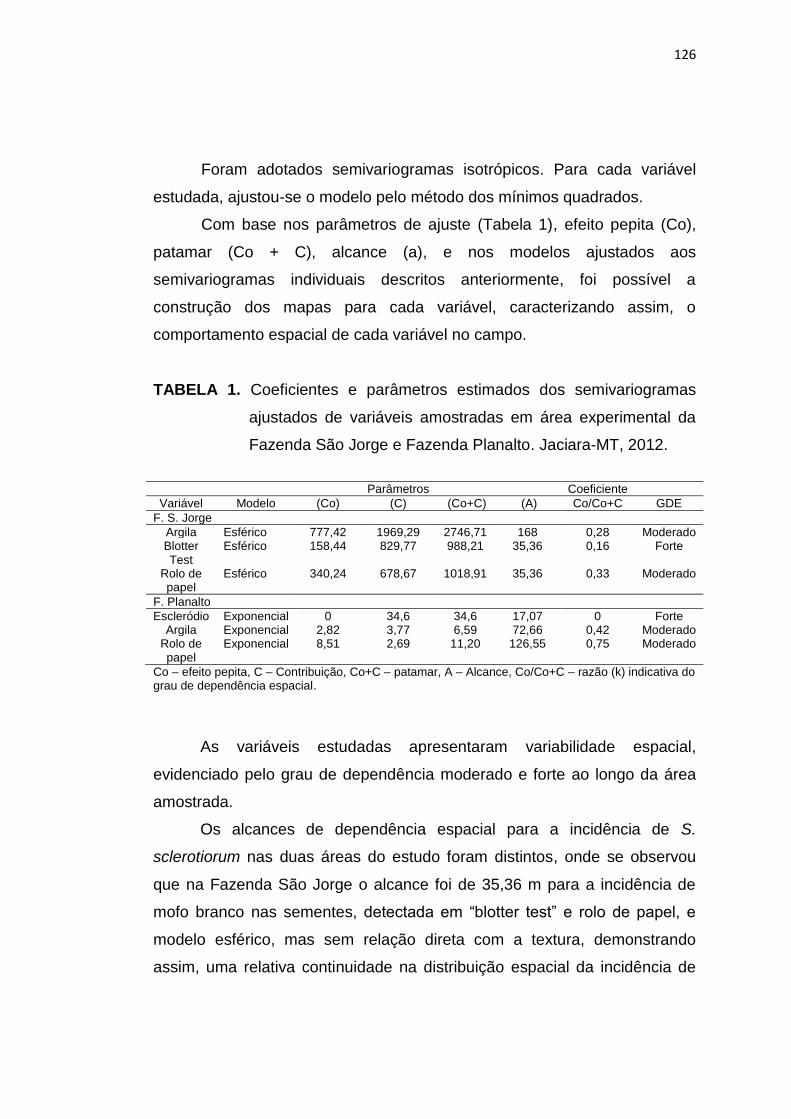
126
Foram adotados semivariogramas isotrópicos. Para cada variável
estudada, ajustou-se o modelo pelo método dos mínimos quadrados.
Com base nos parâmetros de ajuste (Tabela 1), efeito pepita (Co),
patamar (Co + C), alcance (a), e nos modelos ajustados aos
semivariogramas individuais descritos anteriormente, foi possível a
construção dos mapas para cada variável, caracterizando assim, o
comportamento espacial de cada variável no campo.
TABELA 1. Coeficientes e parâmetros estimados dos semivariogramas
ajustados de variáveis amostradas em área experimental da
Fazenda São Jorge e Fazenda Planalto. Jaciara-MT, 2012.
Parâmetros Coeficiente
Variável Modelo (Co) (C) (Co+C) (A) Co/Co+C GDE
F. S. Jorge
Argila Esférico 777,42 1969,29 2746,71 168 0,28 Moderado Blotter Test
Esférico 158,44 829,77 988,21 35,36 0,16 Forte
Rolo de papel
Esférico 340,24 678,67 1018,91 35,36 0,33 Moderado
F. Planalto
Escleródio Exponencial 0 34,6 34,6 17,07 0 Forte Argila Exponencial 2,82 3,77 6,59 72,66 0,42 Moderado
Rolo de papel
Exponencial 8,51 2,69 11,20 126,55 0,75 Moderado
Co – efeito pepita, C – Contribuição, Co+C – patamar, A – Alcance, Co/Co+C – razão (k) indicativa do grau de dependência espacial.
As variáveis estudadas apresentaram variabilidade espacial,
evidenciado pelo grau de dependência moderado e forte ao longo da área
amostrada.
Os alcances de dependência espacial para a incidência de S.
sclerotiorum nas duas áreas do estudo foram distintos, onde se observou
que na Fazenda São Jorge o alcance foi de 35,36 m para a incidência de
mofo branco nas sementes, detectada em “blotter test” e rolo de papel, e
modelo esférico, mas sem relação direta com a textura, demonstrando
assim, uma relativa continuidade na distribuição espacial da incidência de

127
mofo branco, possivelmente em função da dispersão da doença de forma
diferenciada em função do manejo adotado de controle da doença.
Para a variável argila, o ajuste de dados ao modelo esférico refletiu-
se num maior alcance para a dependência espacial, que foi de 168 m.
Na área experimental da Fazenda Planalto, foi constatada
dependência espacial entre os dados de incidência de mofo branco
detectado pelo método rolo de papel. O padrão agregado de dispersão foi
constatado por meio do ajuste de modelo de semivariograma aos dados,
pois o valor absoluto da diferença entre duas amostras observadas cresceu
com o aumento da distância entre elas, até um valor no qual os efeitos
locais não apresentam mais influência (Andriotti, 2003; Alves et al., 2006).
O semivariograma isotrópico esférico foi ajustado para incidência de
S. sclerotiorum, com alta dependência espacial e alcance de 126,55 m para
a Fazenda Planalto (Figura 3). Observou-se que o alcance para a incidência
de escleródio foi de 17,07 m e forte grau de dependência espacial, com
modelo exponencial. Nessa área experimental, o alcance da incidência de
esclerotinia detectada pelo rolo de papel foi maior que na área da Fazenda
São Jorge, com modelo exponencial.
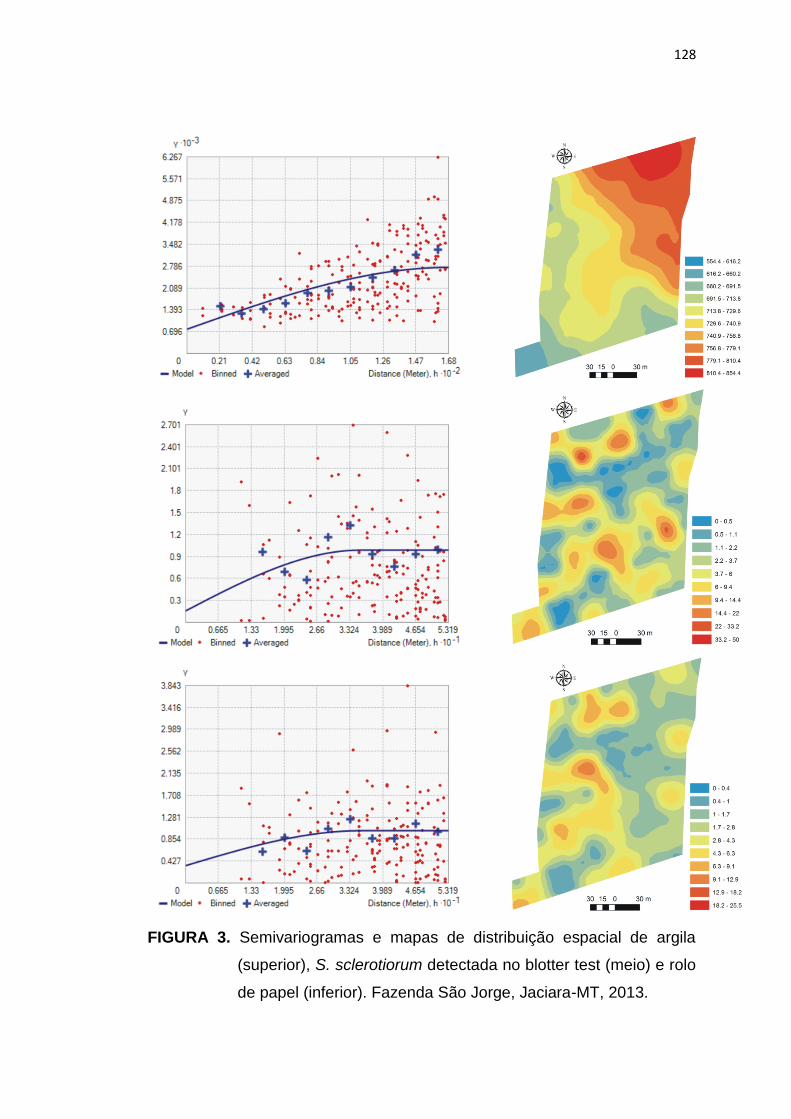
128
FIGURA 3. Semivariogramas e mapas de distribuição espacial de argila
(superior), S. sclerotiorum detectada no blotter test (meio) e rolo
de papel (inferior). Fazenda São Jorge, Jaciara-MT, 2013.

129
Com relação à incidência de S. sclerotiorum, pelos testes de
sanidade de sementes nas áreas experimentais, constatou-se padrão de
variabilidade espacial semelhante entre os métodos. Não houve correlação
com a textura do solo, possivelmente devido ao alto teor de argila das áreas
(Tabela 2).
TABELA 2. Coeficientes de correlação (r) de Pearson entre argila,
escleródios e incidência de S. sclerotiorum pelo testes de
sanidade rolo de papel e “blotter test”. Cuiabá-MT, 2012.
Fazenda Variável Escleródios Rolo de papel “Blotter test”
Planalto Argila 0,54* 0,21 -
Areia -0,58* -0,34 -
São Jorge Argila - 0,21 0,10
Areia - -0,34 0,05
P* significativo a 1%.
O estudo da variabilidade espacial dos escleródios no solo
proporcionou o mapeamento da área de estudo na Fazenda Planalto, onde
se verificou que há dependência espacial e correlação positiva com teor de
argila, possivelmente devido a maior retenção de água (Figuras 4 e 5),
favorecendo a germinação carpogênica de escleródios e a incidência da
doença.
Em solo com textura franco-argilosa, a variância de sobrevivência de
escleródios e apotécios foram explicadas principalmente pela estrutura
espacial de atributos físicos e químicos do solo, enquanto o índice de
severidade da doença não distribuiu esta estrutura espacial, e foi em grande
parte explicado pelos efeitos da rotação com milho (Rousseau et al., 2006).
De acordo com os mapas de krigagem, gerados após o ajuste dos
semivariogramas isotrópicos experimentais, pode-se observar o padrão
agregado da incidência do patógeno, mas não houve correlação com a
textura do solo nas duas áreas experimentais.
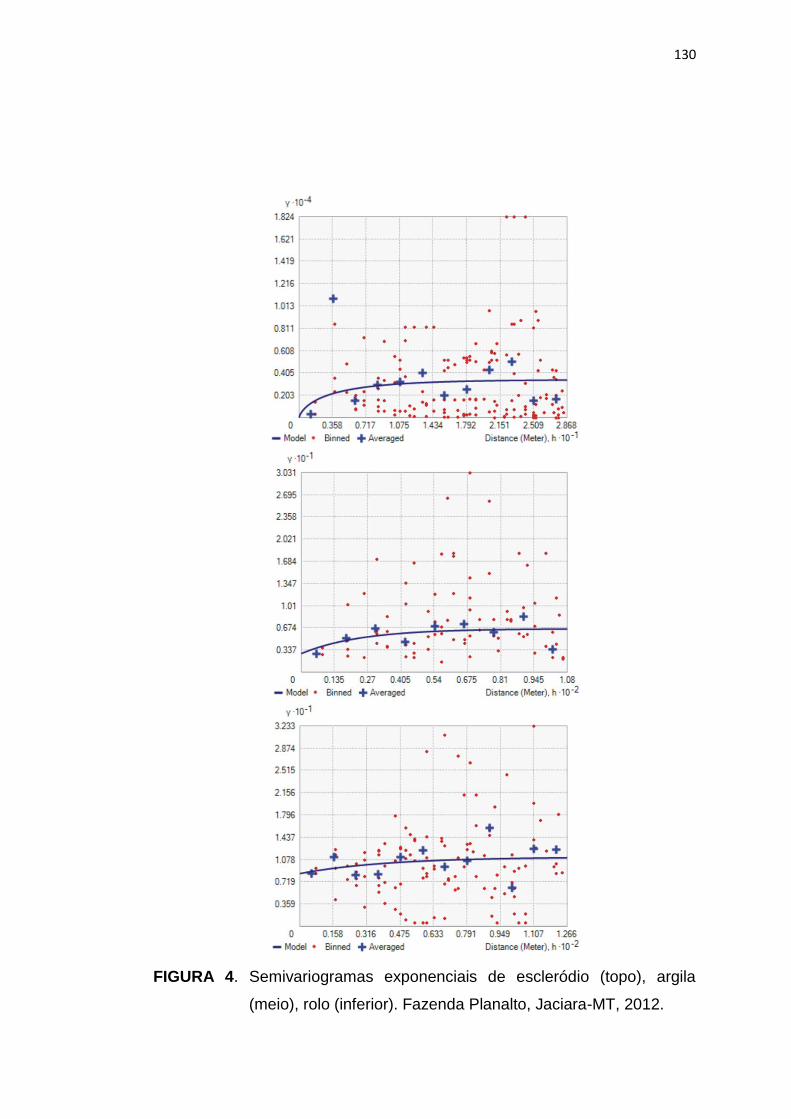
130
FIGURA 4. Semivariogramas exponenciais de escleródio (topo), argila
(meio), rolo (inferior). Fazenda Planalto, Jaciara-MT, 2012.
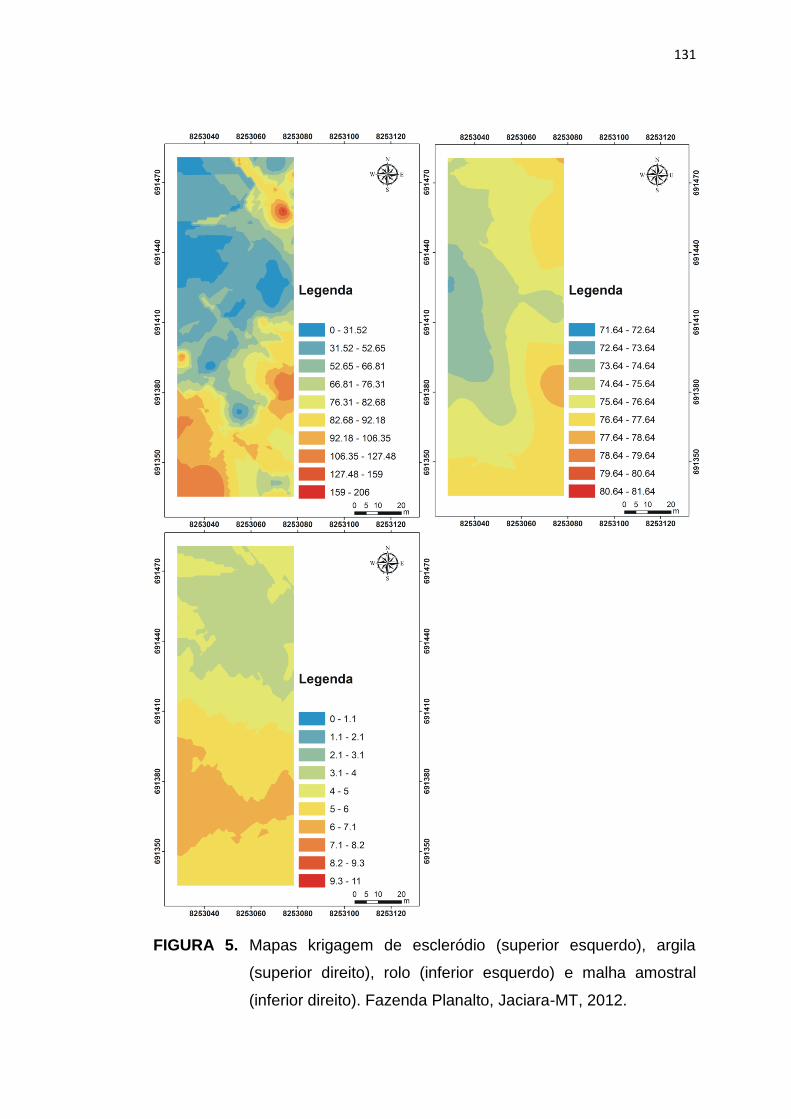
131
FIGURA 5. Mapas krigagem de escleródio (superior esquerdo), argila
(superior direito), rolo (inferior esquerdo) e malha amostral
(inferior direito). Fazenda Planalto, Jaciara-MT, 2012.

132
Embora as áreas sejam próximas e possuam textura muito argilosa,
o manejo adotado para controle da doença foi diferente. Na Fazenda São
Jorge não houve aplicação de fungicidas visando ao controle do mofo
branco, pois o produtor não tinha conhecimento da ocorrência da doença na
área. Na Fazenda Planalto houve controle preventivo, pois já havia sido
observada a ocorrência da doença na área.
A distribuição de escleródios normalmente não ocorre de modo
uniforme nas áreas infestadas e sua variabilidade espacial e localização
puderam ser determinadas por análises geoestatísticas. Mesmo assim,
houve dispersão dos escleródios ao longo da linha de plantio pelas
máquinas e implementos, alterando a dispersão do fungo e o seu padrão
agregado. Nesse estudo, observou-se a ocorrência da incidência de
escleródios ao longo da linha de plantio, provavelmente devido à dispersão
pelas máquinas na semeadura, na aplicação de defensivos e colheita.
O efeito pepita foi um parâmetro importante na análise do
semivariograma e indicou variabilidade não explicada, considerando a
distância de amostragem utilizada (McBratney e Webster, 1986). Esse
parâmetro foi expresso como porcentagem do patamar, objetivando facilitar
a comparação do grau de dependência espacial das variáveis em estudo
(Trangmar et al., 1985).
Para análise do grau de dependência espacial dessas variáveis,
utilizou-se a classificação de Cambardella et al. (1994). O alcance foi uma
medida importante para o planejamento e avaliação experimental, já que
pôde auxiliar na definição de procedimento de amostragem (Burgess et al.,
1981; Webster, 1985). Esse parâmetro representou a distância, em metros,
em que os pontos amostrais foram correlacionados entre si (Journel e
Huijbregts, 1991).
Conforme Cambardella et al. (1994), as variáveis que apresentaram
forte dependência espacial são mais influenciadas pela dispersão do
patógeno e pelo padrão agregado da doença, como número de escleródios.
Por outro lado, as variáveis com fraca dependência foram mais
influenciadas por fatores externos, tais como: presença ou não do patógeno

133
da semente, transmissibilidade e ainda a sensibilidade do teste para
detectar o fungo na semente, ou seja, pelas condições controladas do teste
de sanidade. A partir dos parâmetros dos modelos de semivariogramas
ajustados para cada variável, foram estimados os valores em locais não
amostrados por meio do interpolador geoestatístico krigagem. A magnitude
baixa do efeito pepita foi fundamental nesse processo, pois, quanto maior
for a diferença do efeito pepita em relação ao patamar do semivariograma,
maior será a continuidade do fenômeno e menor a variância da estimativa,
ou seja, maior a confiança que se pode ter na estimativa gerada (Isaaks e
Srivastava, 1989). Com os valores estimados por krigagem, foram gerados
os mapas com a espacialização do número de escleródios, incidência de
sclerotinia e teor de argila (Figuras 2 e 4).
A análise geoestatística foi utilizada para estudar a dependência
espacial de outros patógenos, causadores de doenças, como antracnose.
Alves et al. (2006) estudaram a dinâmica espaço-temporal da antracnose
em feijoeiro (Colletotrichum lindemuthianum) e da ramulose do algodão (C.
gossypii (South) var. cephalosporioides), e, também encontraram padrão
agregado dessas doenças da parte aérea veiculadas por sementes. Outros
patógenos também foram estudados, como Fusarium oxysporum em
tomateiro (Rekah et al., 1999), Phytophthora infestans em tomateiro e
batateira (Jaime-Garcia et al., 2001), Venturia inaequalis em macieira
(Charest et al., 2002) e Greening em citrus (Leal et al., 2010).
Em outros estudos com dependência espacial de plantas doentes,
Amaral Mello (2007) constatou o fitoplasma associado ao enfezamento do
repolho e, utilizando sequências ordinárias, concluiu que o padrão espacial
da doença foi agregado. Noetzold (2011) também verificou por meio dos
mapas de distribuição de C. truncatum, padrão espacial agregado na
primeira avaliação de incidência dos folíolos de soja para os inóculos
testados.
A caracterização do padrão espacial da doença e do patógeno pode
gerar informações essenciais na elaboração de estratégias de manejo do
mofo branco da soja. Como a variação espacial de S. sclerotiorum ocorreu

134
em reboleiras (focos), padrão agregado, poderia se utilizar controle químico
com fungicida em taxa variável e avaliar a eficiência de controle baseado na
incidência de plantas com sintomas nas zonas de manejo, o que evitaria
aplicação de fungicida para controle da doença em área total, reduzindo o
custo de aplicação.
Embora não tenha sido avaliada a produtividade da cultura da soja, a
ocorrência de S. sclerotiorum em lavoura de soja com alta incidência pode
afetar a produtividade (Lee et al., 2000; Kurle et al., 2001; Mueller et al.,
2004; Lee et al., 2005) pelo efeito do patógeno nas plantas com incidência,
além do aumento do inóculo para a próxima safra. Após a colheita, a
dispersão de escleródios na área pelas máquinas e implementos pode
aumentar os focos de incidência para a safra seguinte, dependendo das
condições climáticas, manejo da cultura e do potencial de inóculo na área. A
associação entre o padrão espacial da incidência de plantas infectadas por
S. sclerotiorum e da incidência de sementes com o patógeno, levantadas na
mesma oportunidade, não leva à relação infecção-infestação devido à baixa
transmissibilidade da planta para a semente, mas a relação infestação de
escleródios e incidência no campo é diretamente proporcional.
6.4 Conclusões
Com a análise geoestatística é possível quantificar o grau de
dependência espacial e mapear a variabilidade espacial de escleródios e da
incidência de S. sclerotiorum em soja.
A variação espacial de S. sclerotiorum ocorre em reboleiras. O
padrão da incidência da doença é agregado.
Não há relação da incidência da doença pelos testes de sanidade
blotter test e rolo de papel com a textura do solo.
Há correlação positiva da presença de escleródios de S. sclerotiorum
com a textura argilosa da área.

135
6.5 Referências Bibliográficas
ALVES, M. C.; POZZA, E. A.; MACHADO, J. C.; ARAÚJO, D. V.; TALAMINI, V.; OLIVEIRA, M. S. Geoestatística como metodologia para estudar a dinâmica espaço-temporal de doenças associadas a Colletotrichum spp. transmitidos por sementes. Fitopatologia Brasileira, v. 31, n. 6, p.557-563, 2006. AMARAL MELLO, A. P. O. Identificação molecular de fitoplasmas associados ao enfezamento do repolho e análise epidemiológica da doença. 2007. 64f. Tese (Doutorado em Fitopatologia) - Escola Superior de Agricultura Luiz de Queiroz, Piracicaba – SP, 2007. ANDRIOTTI, J. L. S. Fundamentos de estatística e geoestatistica. São Leopoldo,RS:Unisinos, 2003. 165 p. BOLTON, M. D.; THOMMA, B. P. H. J.; NELSON, B. D. Sclerotinia sclerotiorum (Lib.) de Bary: biology and molecular traits of a cosmopolitan pathogen. Molecular Plant Pathology, v.11, n. 7, p. 1-16, 2006. BURGESS, T. M.; WEBSTER, R.; MCBRATNEY, A. B. Optimal interpolation and isarithmic mapping of soil properties. IV.Sampling strategy. Journal Soil Science, v. 32, n. 4, p. 643-659, 1981. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília: Secretaria de Defesa Agropecuária/MAPA/ACS, 2009. 395p. CAMBARDELLA, C. A.; MOORMAN, T. B.; NOVAK, J. M.; PARKIN, T. B.; KARLEN, D. L.; TURCO, R. F.; KONOPKA, A. E. Field-scale variability of soil properties in Central Iowa Soils. Soil Science Society of America Journal, v. 58, n. 2, p. 1501-1511, 1994. CHAREST, J.; DEWDNEY, M.; PAULITZ, T.; PHILION, V.; CARISSE, O. Spatial distribution of Venturia inaequalis airborne ascospores in orchards. Phytopathology, v. 92, n.7, p. 769-779, 2002. FURLAN, S.H. Importância do tratamento de sementes no manejo de Sclerotinia sclerotiorum. In: III WORKSHOP BRASILEIRO DE CONTROLE DE QUALIDADE DE SEMENTES: Desafios Tecnológicos dos Atributos Genéticos e Sanitários de Sementes. Informativo Abrates... Uberlândia: Abrates, 2010. p. 58. GOULART, A. C. P. Fungos em sementes de soja: detecção, importância e controle. Dourados: Embrapa Agropecuária Oeste, 2005. 72 p.

136
HENNEBERG, L.; JACCOUD FILHO, D. de S.; RUARO, L.; PANOBIANCO, M. Efficiency of methods to detect Sclerotinia sclerotiorum in commercial soybean seed lots. Revista Brasileira de Sementes, v. 34, n.1 p.61-69, 2012. HUGHES, G.; GOTTWALD, T. R. Survey methods for assessment of citrus tristeza virus incidence. Phytopathology, v. 88, n. 7, p. 715-723, 1998. ISAAKS, E. H.; SRIVASTAVA, R. M. An introduction to applied geostatistics. New York: Oxford University Press, 1989. 561 p. JAIME-GARCIA, R.; ORUM, T. V.; FELIX-GASTELUM, R.; TRINIDAD-CORREA, R.; VANETTEN, H. D.; NELSON, M. R. Spatial analysis of Phytophthora infestans genotypes and late blight severity on tomato and potato in the del fuerte valley using geostatistics and geographic information systems. Phytopathology, v. 91, p.156-165, 2001. JOURNEL, A. G.; HUIJBREGTS, C. J. Mining geostatistics. London: Academic Press, 1991. 600 p. KURLE, J. E. ; GRAU, C. R. ; OPLINGER, E. S. ; MENGISTU, A. Tillage, crop sequence, and cultivar effects on sclerotinia stem rot incidence and yield in soybean. Agronomy Journal, v. 93, n. 5, p. 973-982, 2001. LEAL, R. M. ; BARBOSA, J. C.; COSTA, M. G. ; BELASQUE JUNIOR,J. ; YAMAMOTO, P. T.; DRAGONE, J. Distribuição espacial de huanglongbing (greening) em citros utilizando a geoestatística. Revista Brasileira de Fruticultura, v. 32, n. 3, p. 808-818, 2010. LEE, C. D.; RENNER, K. A.; PENNER, D.; HAMMERSCHMIDT, R.; KELLY, J.D. Glyphosate-resistant soybean management system effect on sclerotinia stem rot. Weed Technology, v. 19, n. 3, p. 580-588, 2005. LEE, C. D.; PENNER, D.; HAMMERSCHMIDT, R. Influence of formulated glyphosate and activator adjuvants on Sclerotinia sclerotiorum in glyphosate-resistant and-susceptible Glycine max. Weed Science, v. 48, n.6, p. 710-715, 2000. LIEBHOLD, A. M.; ROSSI, R. E.; KEMP, W. P. Geostatistic and geographic information system in applied insect ecology. Annual Review of Entomo-logy, v. 38, n.1, p. 303-327, 1993. MAPA. Ministério da Agricultura, Pecuária e Abastecimento. Diário Oficial da União. Portaria n. 47, 2009. Disponível em: http://extranet.agricultura.gov.br/sislegis-consulta/consultarLegislacao. Acesso em: 04 de jun.de 2013.

137
MCBRATNEY, A. B.; WEBSTER, R. Choosing functions for semi-variograms of soil properties and fitting them to sampling estimates. Journal Soil Science, v. 3, n. 3, p. 617-639, 1986. MUELLER, D. S.; BRADLEY, C. A.; GRAU, C. R.; GASKA, J. M.; KURLE, J. E.; PEDERSEN, W. L. Application of thiophanate-methyl at different host growth stages for management of sclerotinia stem rot in soybean. Crop Protection, v. 23, n. 10, p. 983-988, 2004. NOETZOLD , R. Avaliação de metodologias de análise estatística e geoestatística para estudar o progresso espacial de Colletotrichum truncatum em sementes de soja. 2011. 94f. Dissertacão (Mestrado em Agricultura Tropical) - Universidade Federal de Mato Grosso, Cuiabá-MT, 2011. REKAH, Y.; SHTIENBERG, D.; KATAN, J. Spatial distribution and temporal development of Fusarium crown and root rot of tomato and pathogen dissemination in field soil. Phytopathology, v. 89, n. 9, p. 831-839, 1999. ROUSSEAU, G. X.; RIOUX, S.; DOSTALER, D. Partitioning the spatial and environmental variation of Sclerotinia stem rot on soybean. Soil Biology & Biochemistry, v. 38, n. 12, p. 3343-3358, 2006. TRANGMAR, B. B.; YOST, R. S.; UEHARA, G. Application of geostatistics to spatial estudies of soil properties. Advances Agronomic, v. 38, n.1, p. 45-93, 1985. WEBSTER, R. Quantitative spatial analysis of soil in field. Advances in Soil Science, v. 3, n. 1, p. 2-56, 1985. WU, B. M; Van BRUGGEN, A. H. C.; SUBBARO, K. V.; PENNINGS, G. G. H. Spatial analysis of lettuce downy mildew using geostatistics and geographic information systems. Phytopathology, v. 91, n. 2, p.134-142, 2001.





























