Embed Size (px)
Citation preview

ESTABILIDADE DE PRODUÇÃO E DA CAPACIDADE DE COMBINAÇÃO EM
HÍBRIDOS DE MILHO
JUAREZ CAMPOLINA MACHADO
2007

Ficha Catalográfica Preparada pela Divisão de Processos Técnicos da Biblioteca Central da UFLA
Machado, Juarez Campolina Estabilidade de produção e da capacidade de combinação em híbridos de milho / Juarez Campolina Machado. – Lavras : UFLA, 2007.
68 p. : il.
Orientador: João Cândido de Souza. Dissertação (Mestrado) – UFLA. Bibliografia.
1. Milho. 2. Produção. 3. Capacidade de combinação. 4. Híbrido. I. Universidade Federal de Lavras. II. Título.
CDD-633.1523

JUAREZ CAMPOLINA MACHADO
ESTABILIDADE DE PRODUÇÃO E DA CAPACIDADE DE COMBINAÇÃO EM HÍBRIDOS DE MILHO
Dissertação apresentada à Universidade Federal de Lavras como parte das exigências do Programa de Pós-graduação em Agronomia, área de concentração em Genética e Melhoramento de Plantas, para a obtenção do título de "Mestre".
Orientador
Prof. Dr. João Cândido de Souza
LAVRAS
MINAS GERAIS - BRASIL
2007

JUAREZ CAMPOLINA MACHADO
ESTABILIDADE DE PRODUÇÃO E DA CAPACIDADE DE COMBINAÇÃO EM HÍBRIDOS DE MILHO
Dissertação apresentada à Universidade Federal de Lavras como parte das exigências do Programa de Pós-graduação em Agronomia, área de concentração em Genética e Melhoramento de Plantas, para a obtenção do título de "Mestre".
APROVADA em 26 de fevereiro de 2007.
Dr. Antônio Carlos de Oliveira EMBRAPA/CNPMS
Dr. Paulo Evaristo de Oliveira Guimarães EMBRAPA/CNPMS
Prof. Dr. João Cândido de Souza
UFLA
(Orientador)
LAVRAS
MINAS GERAIS – BRASIL

A todos que me incitam na busca pelo conhecimento
DEDICO
“Jamais se poderá expressar em frias letras a ternura de um filho ao compreender os sacrifícios de seus pais."
(Raumsol)

AGRADECIMENTOS
A Deus.
À Universidade Federal de Lavras, pela oportunidade concedida.
Ao CNPq, pelo auxílio financeiro e pela concessão da bolsa de estudos.
Aos meus pais, Quélvia e Machado, pelo apoio constante, em todos os
momentos de minha vida.
Ao meu avô Machado, pelo carinho.
Ao professor, amigo e orientador João Cândido, pela prazerosa
convivência, ensinamentos transmitidos e confiança em todos os momentos da
pós-graduação.
Ao professor Magno, pela dedicada co-orientação, estímulo, exemplo
profissional e pelo incansável esforço em transmitir seus conhecimentos.
Ao professor Daniel Furtado Ferreira, pelas valiosas sugestões e pela
disponibilidade nas etapas finais da realização deste trabalho.
À pesquisadora Ângela de Fátima Barbosa Abreu (“tia” Ângela), pela
prazerosa convivência ao longo do curso.
Ao professor Moacil, pela orientação durante a graduação e pelo
incentivo profissional.
Aos membros da banca, pela disponibilidade e valiosas sugestões
apresentadas para a melhoria do presente trabalho.
Ao amigo Lauro, pelas sugestões na redação.
Aos colegas do “milho” e “feijão”, pela colaboração nos trabalhos e
amigável convivência.
Aos demais colegas do GEN, pela possibilidade de aprendermos juntos,
em especial aos amigos Alex, Aisy, Matheus, Vanessa, Bruzi, Ricardo, Dheyne,

Aninha, Marcus, Admilson, César, Marciane, Mônica, Cris, Diego, Márcio,
Flávio, Wila, Helton, Marcelo, Lívia, Francine, Josi e Fabrício.
A querida amiga Flavinha, pelo carinho, incentivo, exemplo e pelos
valiosos conselhos ao longo da pós-graduação.
Aos grandes amigos “irmãos”, José Ângelo e José Luís, pela amizade
cultivada, confiança e prazerosa convivência.
Aos professores César, João Bosco, Elaine, Flávia Avelar, Samuel e
Renzo, pelos ensinamentos.
Aos funcionários de campo, Leo e Lindolfo, pelo auxílio na condução
dos experimentos e pelo companheirismo.
Aos funcionários do DBI, Zélia, Rafaela e Irondina, pela convivência
amigável.
À grande companheira Elaine, pela amizade e ajuda em todos os
momentos.
Aos produtores rurais, pela disponibilidade em fornecer as áreas para
condução dos experimentos, em especial aos Srs. Adelino, Fausto, Pedro
Milanez e Bráulio.
Enfim, a todos aqueles que, direta ou indiretamente, estiveram
envolvidos na realização deste trabalho, meu MUITO OBRIGADO!!!

SUMÁRIO
Página
RESUMO.......................................................................................................... i ABSTRACT..................................................................................................... ii 1 INTRODUÇÃO ............................................................................................ 1 2 REFERENCIAL TEÓRICO .......................................................................... 3 2.1 Heterose no melhoramento do milho........................................................... 3 2.2 Análise dialélica ......................................................................................... 7 2.3 Interação genótipos x ambientes................................................................ 13 2.4 Estatísticas não-paramétricas .................................................................... 17 3 MATERIAL E MÉTODOS ......................................................................... 19 3.1 Material genético ...................................................................................... 19 3.2 Condução dos experimentos...................................................................... 20 3.3 Análise estatística dos dados ..................................................................... 23 3.3.1 Adaptabilidade e estabilidade................................................................. 23 3.3.2 Análise dialélica .................................................................................... 25 3.3.3 Estabilidade dos efeitos da capacidade de combinação ........................... 27 4 RESULTADOS E DISCUSSÃO ................................................................. 29 4.1 Adaptabilidade e estabilidade de híbridos simples e duplos de milho......... 29 4.2 Análise dialélica de híbridos comerciais de milho ..................................... 36 4.3 Estabilidade dos efeitos da capacidade de combinação .............................. 40 5 CONCLUSÕES........................................................................................... 43 REFERÊNCIAS BIBLIOGRÁFICAS ............................................................ 44
ANEXOS ....................................................................................................... 53

i
RESUMO
MACHADO, Juarez Campolina. Estabilidade de produção e da capacidade de combinação em híbridos de milho. 2007. 68 p. Dissertação (Mestrado em Genética e Melhoramento de Plantas) – Universidade Federal de Lavras, MG.* O presente trabalho foi conduzido com os objetivos de estudar a adaptabilidade e a estabilidade de híbridos simples e duplos de milho, estimar os efeitos da capacidade geral e específica de combinação, e avaliar a estabilidade das estimativas desses parâmetros. Foram avaliados 55 tratamentos, sendo dez híbridos simples comerciais (DKB 199, 30F90, A2555, DKB 333B, DOW 657, AG 8060, 30F87, DOW 8420, 30K75, AG 7000) utilizados como parentais e 45 híbridos duplos resultantes de um dialelo completo. As sementes dos híbridos duplos foram obtidas na área experimental do Departamento de Biologia da Universidade Federal de Lavras (DBI/UFLA). Os experimentos foram conduzidos em 15 ambientes, no ano agrícola de 2005/06, em propriedades de agricultores e estações experimentais. Os tratos culturais foram os normalmente utilizados pelos agricultores para a cultura do milho. Os tratamentos foram avaliados utilizando-se o delineamento de blocos casualizados. O caráter avaliado foi a produtividade de espigas despalhadas (kg/ha), corrigida para 13% de umidade. Para o estudo da adaptabilidade e da estabilidade dos híbridos, adotou-se a estatística não-paramétrica. Obteve-se a contribuição de cada híbrido para a interação genótipos x ambientes e o desvio em relação ao desempenho máximo em cada ambiente. Procedeu-se à análise dialélica conjunta envolvendo todos os ambientes e realizou-se o estudo da estabilidade dos efeitos da capacidade geral e específica de combinação. Adotando-se a estatística não-paramétrica obteve-se também a contribuição de cada efeito para a interação genótipos x ambientes. Os híbridos duplos foram, em média, mais estáveis, contudo, identificaram-se híbridos simples tão estáveis quanto os duplos, como, por exemplo, o AG 8060. Os efeitos não aditivos foram mais importantes que os efeitos aditivos para a variação dos híbridos. Os genitores com as maiores estimativas do efeito da CGC foram AG 8060 e 30F90. O genitor AG 8060 associa alta estimativa do efeito da CGC à estabilidade desse parâmetro. *Comitê de orientação: João Cândido de Souza – UFLA (Orientador); Magno Antonio Patto Ramalho – UFLA (Co-orientador)

ii
ABSTRACT
MACHADO, Juarez Campolina. Yield and combining ability stabilities in maize hybrids. 2007. 68 p. Dissertation (Master’s degree in Genetics and Plant Breeding) – Universidade Federal de Lavras, Minas Gerais, Brazil.* The objectives of the present work were to study the adaptability and stability of single and double-cross maize hybrids, estimate the effects of general and specific combining abilities and to evaluate the stability of the estimates of these parameters. The commercial single-cross hybrids DKB 199, 30F90, A2555, DKB 333B, DOW 657, AG 8060, 30F87, DOW 8420, 30K75, AG 7000 and all possible double-cross hybrids, obtained from a complete diallel, were the treatments evaluated. Seeds of the double-cross hybrids were obtained on an experimental area of the Biology Department at Universidade Federal de Lavras (DBI/UFLA). The experiments were conducted in 15 environments in the 2005/06 growing season, on farms and in research stations. The cultural practices were the ones normally used by maize farmers. Treatments were evaluated in randomized complete blocks design. The trait under study was husked ears yield (kg/ha), corrected to 13% moisture content. Nonparametric statistics were used to study hybrids adaptability and stability. The contribution of each hybrid to the genotype-by-environment interaction and the deviation in relation to the maximum performance in each environment was determined. The joint diallel analysis involving all environments was performed and the stability of general and specific combining ability effects were investigated. The underlying nonparametric statistics evaluated the contribution of each effect to the genotype-by-environment interaction. The double-cross hybrids were on average more stable, although some single-cross hybrids were identified as stable as the double-cross hybrids, as for instance, the AG 8060. The non-additive effects were more important than the additive effects for hybrid variation. The parents with the highest GCA effect estimates were AG 8060 and 30F90. AG 8060 associated a high GCA effect with stability of the parameter. *Advisor committee: João Cândido de Souza – UFLA (Advisor); Magno Antonio Patto Ramalho – UFLA (Co-advisor)

1
1 INTRODUÇÃO
A cultura do milho alcança altos níveis de produtividade, em ambientes
apropriados e com técnicas de cultivo adequadas (Duvick, 2005). Em um
ambiente, a manifestação fenotípica é o resultado da ação do genótipo sob
influência do meio. Entretanto, quando se considera uma série de ambientes,
detecta-se, além dos efeitos genéticos e ambientais, um efeito adicional,
proporcionado pela interação desses fatores. A interação é um complicador no
momento de recomendar cultivares superiores, pois, quando significativa, pode
ocorrer alteração na classificação relativa dos genótipos avaliados nos diferentes
ambientes, dificultando a identificação de cultivares estáveis e exigindo a
adoção de medidas que controlem ou minimizem seus efeitos (Cruz et al., 2004;
Ramalho et al., 1993).
Um dos fatores que exercem grande influência na adaptabilidade e na
estabilidade de uma cultivar é sua estrutura genética. Na literatura, há relato de
que os híbridos duplos de milho são mais estáveis que os híbridos simples para a
produtividade de grãos (Guillen-Portal et al., 2003), entretanto, podem ser
identificados híbridos simples tão estáveis quanto os duplos para o mesmo
caráter (Carvalho et al., 2005). Nas condições brasileiras, a comparação entre
híbridos com diferentes estruturas genéticas é particularmente importante,
devido às variadas condições ambientais nas quais o milho é cultivado e,
principalmente, porque, no Brasil, os diferentes tipos de híbridos disponíveis são
ainda bastante utilizados pelos agricultores.
Em programas de melhoramento de milho, as estimativas da capacidade
de combinação auxiliam na escolha de genitores e no entendimento dos efeitos
genéticos envolvidos na determinação dos caracteres (Cruz et al., 2004). Essas
estimativas podem ser obtidas por meio de análises dialélicas e têm sido

2
amplamente empregadas (Melani & Carena, 2005; Rodrigues et al., 2006;
Welcker et al., 2005). Contudo, são escassas as informações a respeito da
estabilidade dos parâmetros da capacidade de combinação. Pacheco et al. (1999)
estudaram a estabilidade desses parâmetros por meio do desdobramento dos
desvios de regressão de Eberhart & Russel, em efeitos genéticos aditivos ( ig e
jg ) e de dominância ( ijs ), utilizando variedades de polinização aberta.
Entretanto, não foram encontrados relatos, na literatura, envolvendo híbridos
comerciais de milho.
Diante do exposto, os objetivos do presente trabalho foram estudar a
adaptabilidade e a estabilidade de híbridos simples e duplos de milho, estimar os
efeitos da capacidade geral e específica de combinação, e avaliar a estabilidade
das estimativas desses parâmetros.

3
2 REFERENCIAL TEÓRICO
2.1 Heterose no melhoramento do milho
O milho híbrido foi um dos primeiros exemplos de sucesso na aplicação
da teoria genética à produção de alimentos (Duvick, 2001). A heterose, definida
há quase um século por Shull e aplicada à produção de híbridos, foi uma das
maiores contribuições práticas da genética à agricultura mundial.
O procedimento para a obtenção de híbridos de milho foi proposto por
Shull e East independentemente, ao constatarem que, quando plantas eram
autofecundadas, ocorria perda de vigor. No entanto, quando cruzadas, o
desempenho do híbrido poderia ser superior à média da população original
(Troyer, 2006).
Uma grande limitação à aplicabilidade dessa técnica era o alto custo, o
que inviabilizava a produção comercial das sementes híbridas, sobretudo devido
à baixa produtividade das linhagens disponíveis. Esse problema foi contornado
após o trabalho de Jones (1918), que sugeriu a produção de híbridos duplos.
Nesse caso, como a produção de sementes era realizada a partir de um híbrido
simples, o processo tornou-se comercialmente viável (Paterniani, 2001).
A utilização de híbridos possui enormes vantagens, tais como: associar
fenótipos de interesse que se encontram separados nos genitores, utilizar
interações alélicas na geração híbrida e obter cultivares superiores e uniformes
em um prazo relativamente curto. Existem diferentes tipos de híbridos que
podem ser produzidos: a) híbrido Top cross; b) híbrido simples; c) híbrido
simples modificado; d) híbrido triplo; e) híbrido triplo modificado; f) híbrido
duplo e g) híbrido intervarietal. Detalhes de cada tipo podem ser encontrados em
Miranda Filho & Viégas (1987).

4
Nos Estados Unidos, os primeiros híbridos duplos foram lançados entre
os anos de 1920 e 1930 e superavam as variedades de polinização aberta. A
partir de então, a produtividade, que estava estagnada, aumentou
consideravelmente. Por volta dos anos de 1960, quando os híbridos simples
foram disponibilizados aos agricultores, verificaram-se incrementos ainda
maiores, evidenciando o êxito com a utilização de híbridos de milho na
agricultura norte-americana (Figura 1). Atualmente, a produtividade média de
milho nos Estados Unidos é da ordem de 10.000 kg/ha, ou seja, 400% superior à
produtividade média no início do século XX.
variedadesb= 0.88
híbrido duplob = 83.97
híbrido simplesb= 188.43
0
2000
4000
6000
8000
10000
12000
1885 1895 1905 1915 1925 1935 1945 1955 1965 1975 1985 1995 2005
Ano
Prod
utiv
idad
e m
édia
de
milh
o (k
g/ha
)
FIGURA 1. Produtividade média de milho e os principais tipos de cultivares
utilizadas nos Estados Unidos, de 1885 a 2006. Valores de “b” (kg/ha) indicando o ganho de produtividade por ano (USDA-NASS, 2006).

5
Esse enorme sucesso tem sido atribuído, em grande parte, ao
melhoramento genético. Alguns pesquisadores quantificaram que mais de 50%
dos ganhos em produtividade observados se devem à inclusão das cultivares
híbridas no sistema produtivo (Duvick, 2005; Troyer, 1999). No Brasil, o êxito
dos programas de melhoramento de milho também foi expressivo. Os resultados
apresentados na Tabela 1, envolvendo estimativas de progresso genético,
evidenciam esse fato.
Apesar de muito bem utilizada na produção de alimentos, a heterose é
um fenômeno pouco compreendido (Troyer, 2006). As principais hipóteses
explicativas são: da dominância, da sobredominância e da epistasia. As
evidências empíricas indicam que a heterose é função do papel da dominância
parcial e ou dominância completa, porém, a conclusão sobre as causas genéticas
da heterose é difícil, pois os genes quantitativos envolvidos no controle dos
caracteres são ainda pouco conhecidos (Bernardo, 2002).
TABELA 1. Estimativas do progresso genético com a cultura do milho no Brasil, relatadas na literatura.
Anos Aumento de produtividade (kg/ha/ano) Fonte
1964-1983 72-109 (PE)1/ Vencovsky et al. (1988)
1946-1986 60 (PG) Paterniani (1990)
1970-1990 31-51 (PG) Araújo (1995)
1964-1993 123 (PE) Fernandes & Frazon (1997)
1998-2002 175 (PG) Storck et al. (2005) 1/ PE - peso de espigas; PG - peso de grãos.

6
Surpreendentemente, aumentos na heterose não têm sido os principais
responsáveis pelas produtividades alcançadas. Alguns trabalhos têm
demonstrado que a heterose se manteve constante ao longo dos anos. Dessa
forma, o desenvolvimento de híbridos superiores tem sido atribuído ao progresso
genético com o melhoramento das linhagens endogâmicas (Duvick, 2001;
Troyer, 2006).
Em um programa de melhoramento visando à obtenção de híbridos de
milho, as etapas envolvidas são: a escolha das populações, a obtenção das
linhagens, a avaliação das capacidades de combinação das mesmas e o teste
intensivo dos híbridos obtidos (Paterniani & Campos, 1999).
Entre essas etapas, a escolha das populações é de fundamental
importância, pois todo o sucesso dependerá dela. As populações com maior
potencial poderão ser utilizadas para extrair linhagens e ou para iniciar um
programa de seleção recorrente recíproca. Nesse intuito, os melhoristas de milho
têm preferido utilizar populações de base genética estreita (Koutsika-Sotiriou &
Karagounis, 2005; Troyer, 1999). A utilização de híbridos comerciais é uma das
alternativas. Os híbridos comerciais são adaptados, possuem produtividade
média alta e, provavelmente, concentram alta freqüência de alelos favoráveis,
devido ao longo processo de melhoramento e à pressão de seleção durante as
etapas de desenvolvimento. Como existem inúmeras opções de híbridos
comerciais, é importante identificar populações, derivadas desses híbridos, com
o máximo potencial de originar cultivares superiores.
Dessa forma, para se ter segurança na escolha das populações mais
promissoras, é essencial que a avaliação seja realizada no maior número possível
de ambientes representativos da região alvo do programa de melhoramento.

7
2.2 Análise dialélica
O milho, entre as espécies vegetais, foi o que mais contribuiu para o
conhecimento dos princípios teóricos do melhoramento de plantas e para o
desenvolvimento de delineamentos genéticos para diversos tipos de
cruzamentos. Nesse contexto, os cruzamentos dialélicos se destacaram no que
diz respeito à sua importância e abrangência de utilização (Miranda Filho &
Gorgulho, 2001).
Por definição, cruzamentos dialélicos são delineamentos genéticos
estatísticos em que n parentais são cruzados dois a dois (Cruz et al., 2004;
Ramalho et al., 1993). Os procedimentos de análise dialélica têm por finalidade
a estimação de parâmetros úteis na seleção de genitores e no entendimento dos
efeitos genéticos envolvidos na determinação dos caracteres (Cruz et al., 2004).
Entre os métodos mais utilizados, citam-se os propostos por Griffing
(1956), pelos quais são estimados os efeitos da capacidade geral e específica de
combinação; o método proposto por Gardner & Eberhart (1966), no qual são
avaliados os efeitos de variedades e heterose varietal e o proposto por Hayman
(1954), que fornece informações sobre os mecanismos básicos de herança do
caráter em estudo, dos valores genéticos e do limite de seleção.
A origem dos procedimentos para análises dialélicas é atribuída a
Schmidt, por volta dos anos de 1920 (Ghosh & Biswas, 2003). Porém, os
primeiros a definirem os termos capacidade geral de combinação (CGC) e
capacidade específica de combinação (CEC) foram Sprague & Tatum (1942).
Segundo esses autores, a CGC refere-se ao comportamento médio de cada
genitor em cruzamento com os demais do grupo estudado e a CEC é interpretada
como um efeito na expressão do híbrido que não é explicado pelos efeitos da
CGC dos genitores.

8
Nos vários métodos de análise dialélica que foram propostos, as
combinações híbridas são avaliadas em experimentos com repetições e as
médias dos tratamentos são utilizadas para formar uma tabela dialélica n x n. As
n2 combinações possíveis podem ser avaliadas, incluindo os genitores e seus
híbridos recíprocos. Dessa forma, Griffing (1956) idealizou quatro possíveis
métodos de análise (Tabela 2). No método 1 são analisados todos os
cruzamentos possíveis, incluindo os genitores e os recíprocos; no método 2 são
incluídos somente os genitores e os híbridos; no método 3 são incluídos apenas
os híbridos e seus recíprocos e no método 4 somente os híbridos são utilizados
na análise.
Os métodos apresentados por Griffing (1956) podem ser aplicados a um
conjunto fixo ou aleatório de genótipos, dependendo de os tratamentos
experimentais fornecerem conclusões que lhes são particulares ou que possam
ser extrapoladas para uma população na qual os tratamentos representam uma
amostra (Hallauer & Miranda Filho, 1988). Apesar de terem sido desenvolvidos
para linhagens, os métodos de Griffing podem ser utilizados para outros tipos de
populações com qualquer grau de endogamia.
TABELA 2. Métodos de análise dialélica propostos por Griffing (1956).
Método Inclusão dos genitores
Inclusão dos recíprocos
Números de genótipos avaliados
1 sim sim 2n
2 sim não 2
)1( +nn
3 não sim )1( −nn
4 não não 2
)1( −nn
Fonte: Adaptado de Wricke & Weber, 1986.

9
Além dos métodos propostos por Griffing (1956), outros também foram
sugeridos. O modelo de análise dialélica proposto por Gardner & Eberhart
(1966) considera um conjunto fixo de variedades e seus híbridos, tendo como
base os efeitos de variedade ( iv ) e de heterose ( 'iih ), sendo este último
subdividido nos componentes: heterose média ( h ), heterose de variedades ( ih ) e
heterose específica ( 'jjs ).
Apesar das diferenças existentes entre os procedimentos de análise
dialélica de Griffing e Gardner & Eberhart, em um trabalho visando comparar os
dois métodos, Cruz & Vencovsky (1989) demonstraram como os vários
parâmetros se inter-relacionam e concluíram que os dois métodos são
basicamente equivalentes na quantidade de informações genéticas que fornecem.
Concluíram também que a estimativa de ig (efeito da capacidade geral de
combinação do genitor i) ainda é o melhor critério para a seleção de genitores.
Nesse mesmo trabalho, os autores interpretaram o parâmetro iis (capacidade
específica de combinação de um genitor com ele mesmo) e demonstraram que
esse parâmetro é um importante indicador da existência de dominância
unidirecional e de heterose varietal.
É oportuno destacar o método proposto por Hayman (1954), o qual
exige que o conjunto de genótipos avaliados seja aleatório, isto é, representem
uma amostra aleatória da população para a qual as informações serão
generalizadas. Por meio desse procedimento, os dados da tabela dialélica podem
ser utilizados para testar a presença dos efeitos genéticos aditivos e dominantes,
além de possibilitar a estimação de componentes da variância genética e
parâmetros de importância prática, como herdabilidade e grau médio de
dominância (Mather & Jinks, 1971). Os dados da tabela dialélica permitem,
ainda, efetuar uma análise gráfica que classifica os genótipos quanto à proporção
de alelos dominantes e recessivos e estimar os limites seletivos das populações
avaliadas (Hayman, 1954; Santos, 1984).

10
Algumas variações metodológicas para a análise de dialelos foram
sugeridas. Kempthorne & Curnow (1961) propuseram o método de cruzamentos
dialélicos parciais, os quais envolvem dois grupos diferentes de genitores e seus
respectivos cruzamentos. Esse procedimento apresenta, como principal
vantagem, uma maior flexibilidade aos cruzamentos dialélicos, possibilitando o
emprego de um grande número de genitores, de forma a maximizar as
informações sobre os genótipos estudados (Miranda Filho & Geraldi, 1984;
Ramalho et al., 1993).
Outros métodos de análise dialélica que podem também ser utilizados
são: dialelos circulantes, dialelos incompletos e dialelos desbalanceados.
Detalhes desses métodos poderão ser encontrados em Cruz et al. (2004) e
Vencovsky & Barriga (1992).
Na literatura, são encontrados vários exemplos de aplicação dos métodos
de análise dialélica. A relação de alguns trabalhos que utilizaram modelos
dialélicos na cultura do milho é apresentada na Tabela 3. Pode-se observar que
diferentes métodos foram adotados para os mais diversos caracteres.
Apesar de amplamente empregados, são escassas as informações em
relação à estabilidade dos parâmetros da análise dialélica. Pacheco (1997)
desdobraram os desvios de regressão de Eberhart & Russel em efeitos genéticos
aditivos ( ig e jg ) e de dominância ( ijs ) e concluíram que os programas de
melhoramento teriam sua eficiência melhorada se fossem selecionados genitores
que apresentassem estabilidade dos efeitos da capacidade geral e específica de
combinação.

11
TABELA 3. Aplicação de métodos de análise dialélica na cultura do milho.
Método empregado Número de genitores Tipo de genitor Caráter1/ Fonte
Griffing (1956) - Método I 4 Linhagens
MS da parte aérea MS da raiz Conteúdo de clorofila Altura de plantas Comprimento da raiz
Silva et al., 2006
Griffing (1956) - Método II 7 Linhagens Reação à Phaeospheria
maydis Pegoraro et al., 2002
Griffing (1956) - Método II 10 Linhagens
Produtividade de espigas Produtividade de espigas despalhadas Rendimento de espigas
Lemos et al., 2002
Griffing (1956) - Método IV 10 Híbridos simples e
triplos Produtividade de espigas despalhadas Melo et al., 2001
Griffing (1956) - Método IV 11 Variedades
Vigor Altura de plantas Dias para o florescimento ASI Espigas por planta Produção de grãos
Welcker et al., 2005
Griffing (1956) - Método IV 17 Linhagens Produção de grãos Bétran et al., 2003
“...continua...”
11

12
“TABELA 3. Cont.”
Método empregado Número de genitores Tipo de genitor Caráter1/ Fonte
Griffing (1956) - Método IV 14 Linhagens
Concentração de Fe em farelo Concentração de Zn em farelo Concentração de Fe no grão Concentração de Zn no grão
Long et al., 2004
Griffing (1956) - Métodos IV;
Gardner & Eberhart (1966)
10 Variedades
Produção de grãos Umidade de grãos % plantas acamadas % plantas quebradas
Melani & Carena, 2005
Dialelo parcial adaptado por Geraldi
& Miranda Filho (1984)
16 Variedades
% plantas acamadas % plantas quebradas Altura de plantas Florescimento
Rodrigues et al., 2006
Hayman (1954) 6 Linhagens
Fileiras por espiga Grãos por fileira Peso de 100 grãos Produção de grãos por planta
Saeed et al., 2000
1/ MS - matéria seca; ASI - assincronia entre o florescimento masculino e feminino
12

13
2.3 Interação genótipos x ambientes
É de amplo conhecimento dos melhoristas que, ao se avaliar cultivares
em vários ambientes, não se nota um comportamento consistente das mesmas.
Esta resposta diferenciada dos genótipos à variação do ambiente é denominada
interação genótipos x ambientes. Isso significa que os efeitos genéticos e
ambientais não são independentes, uma vez que as respostas dos genótipos
podem diferir com as variações ambientais (Souza Júnior & Vencovsky, 1989).
A existência da interação está associada a fatores fisiológicos e
bioquímicos de cada cultivar. Em termos genéticos, a interação ocorre quando a
contribuição dos alelos dos diferentes genes que controlam o caráter ou o nível
de expressão dos mesmos não é coincidente entre os ambientes. Isso porque a
expressão dos genes é influenciada e ou regulada pelo ambiente (Kang & Gauch
Jr., 1996).
Como o principal objetivo de um programa de melhoramento é
selecionar genótipos de consistente e elevada produtividade, nos mais diversos
ambientes, a interação desses fatores pode representar um problema para os
melhoristas, por reduzir a precisão na seleção de um ambiente para outro. A
presença da interação genótipos x ambientes provoca um aumento no desvio
padrão fenotípico, reduz a herdabilidade ao longo dos ambientes e,
conseqüentemente, diminui os ganhos genéticos potenciais (Matheson &
Raymond, 1986).
Pela importância dessa interação, cabe ao melhorista avaliar sua
magnitude e significância, quantificar seus efeitos e fornecer subsídios que
possibilitem adotar procedimentos para a sua minimização e ou o seu
aproveitamento (Cruz et al., 2004). Detectada a interação, é possível a seleção de
genótipos de adaptação ampla ou específica, escolher locais de seleção,
identificar o nível de estresse nos ambientes escolhidos para as fases iniciais de

14
seleção e determinar o número ideal de ambientes e de genótipos a serem
avaliados em cada fase (Fox et al., 1997). Nesse contexto, os métodos para
estudos de adaptabilidade e estabilidade merecem destaque. Na literatura, são
encontradas diferentes definições para esses termos.
Adaptabilidade refere-se à capacidade dos genótipos em assimilarem
vantajosamente o estímulo ambiental, uma vantagem do ponto de vista do
rendimento agrícola, isto é, a adaptabilidade é avaliada pelo desempenho médio
do genótipo (Mariotti et al., 1976).
A estabilidade, para Mariotti et al. (1976), é definida como a capacidade
dos genótipos de exibirem um desempenho o mais constante possível, em
função das variações da qualidade do ambiente.
Lin et al. (1986) sugeriram três conceitos: a) estabilidade do tipo 1, a
cultivar será considerada estável se sua variância entre ambientes for pequena;
b) tipo 2, em que a cultivar será estável se sua resposta aos ambientes for
paralela ao desempenho médio de todas cultivares avaliadas e c) tipo 3, será
estável a cultivar que apresentar o quadrado médio do desvio da regressão baixo,
próximo a zero, ou seja, alta confiabilidade na resposta estimada.
Os três conceitos apresentados por Lin et al. (1986) não levam em
consideração as variações imprevisíveis do ambiente. Nesse sentido, Lin &
Binns (1988a) sugeriram o tipo 4, no qual a cultivar com maior estabilidade será
aquela que apresentar menor quadrado médio da interação genótipos x anos, ou
seja, indivíduos estáveis frente às variações imprevisíveis.
Becker & Léon (1988) dividem a estabilidade em dois tipos: estática,
associando aqueles genótipos que apresentam desempenho constante com as
variações ambientais, e dinâmico, associando os genótipos que apresentam um
comportamento previsível dentro das variações ambientais.
Vários estudos têm sido realizados, no intuito de verificar o controle
genético da estabilidade. Torres (1988) constatou que o grau de determinação

15
genético e a repetibilidade da produtividade de grãos foram maiores que os da
estabilidade e, assim, é de esperar maiores dificuldades em selecionar para esse
último caráter. Lee et al. (2003), estudando os componentes genéticos da
estabilidade, concluíram que esse caráter é herdável, preditível e com
predominância de efeitos gênicos aditivos. Entretanto, não há nenhum resultado
conclusivo em relação ao número de locos que o controlam, mas acredita-se que
seja um número grande e de herança complexa, o que dificultaria a seleção
(Rosse, 1999).
São muitas as metodologias de análise de adaptabilidade e estabilidade
destinada a um grupo de cultivares testadas em vários ambientes. A diferença
entre elas origina-se nos próprios conceitos de estabilidade e nos procedimentos
biométricos de quantificar a interação entre cultivares e ambientes (Vencovsky
& Barriga, 1992). Existem métodos baseados em variância da interação
cultivares x ambientes, regressão linear, regressão linear bissegmentada,
regressão não-linear, métodos não-paramétricos, métodos multivariados e
aqueles que integram métodos univariados e multivariados.
Esses métodos deverão ser empregados quando ocorrerem interações
entre cultivares e ambientes significativas, sendo complementares às análises de
variância, individuais e conjunta, realizadas em uma série de ambientes.
Revisões sobre esses procedimentos são comuns na literatura (Cruz et al., 2004;
Langer et al., 1979; Lin et al., 1986; Oliveira, 1976; Veronesi, 1995).
Estudos têm sido realizados com o objetivo de verificar se a estrutura
genética de uma população lhe confere maior estabilidade ou adaptação. No caso
do milho, uma variedade de polinização aberta apresenta maior heterogeneidade
que uma linhagem ou híbrido simples, entretanto, sua heterozigosidade pode ser
menor. Dessa forma, um híbrido simples deve possuir maior homeostase
individual e uma variedade maior homeostase populacional (Allard & Bradshaw,
1964).

16
Existem relatos na literatura que comparam a magnitude relativa da
interação genótipos x ambientes, utilizando populações com diferentes estruturas
genéticas (Tabela 4). Observe que a estrutura genética das populações influencia
a magnitude da interação genótipos x ambientes. Em híbridos de milho, a
estabilidade está intensamente relacionada com o nível de heterozigosidade de
cada grupo, porém, heterozigosidade intrínseca não é suficiente para explicar a
estabilidade nesta espécie (Adams & Shank, 1959).
Realizando uma comparação entre genótipos de milho com diferentes
estruturas genéticas, Schnell & Becker (1986) constataram que os híbridos
duplos foram mais estáveis que os simples para produtividade de grãos,
concluindo que a estrutura genética exerce grande influência na estabilidade de
uma população. Entretanto, podem ser identificados híbridos simples tão
estáveis quanto os duplos para o mesmo caráter (Carvalho et al., 2005). Dessa
forma, estratégias utilizadas para o desenvolvimento de cultivares de melhor
adaptação às diferentes condições ambientais requerem cuidadosa escolha da
estrutura genética das populações a serem recomendadas aos agricultores.
TABELA 4. Magnitude relativa do quadrado médio da interação genótipos x ambientes (QMGA), em híbridos simples (HS) e duplos (HD) de milho.
QMGA Fonte
HS HD
Eberhart & Russel (1969) 100 68
Weatherspoon (1970) 100 45
Schnell & Becker (1986) 100 57

17
2.4 Estatísticas não-paramétricas
Estatística não-paramétrica pode ser considerada como sendo os
procedimentos estatísticos para os quais não são relacionados parâmetros
específicos. Algumas razões para a sua utilização foram apresentadas por
Campos (1983): a) menor exigência nas pressuposições a serem atendidas; b)
obtenção de probabilidades nos testes com exatidão; c) independência quanto à
forma da população da qual a amostra foi obtida; d) maior facilidade de
aplicação; e) menor volume de cálculos e f) maior eficiência quando os dados
não apresentam distribuição normal.
No melhoramento de plantas, os métodos não-paramétricos podem ser
utilizados para o estudo da interação genótipos x ambientes e para a avaliação da
estabilidade fenotípica de cultivares. Alguns métodos foram propostos com esse
objetivo. Huehn (1990) sugeriu a avaliação de forma não-paramétrica da
estabilidade fenotípica baseada na classificação dos genótipos em cada
ambiente, utilizando o princípio da homeostase para caracterizar os genótipos.
No método em questão, um genótipo é considerado estável se a classificação
apresentada pelo efeito da interação genótipos x ambientes é similar. Nesse caso,
os parâmetros que medem a estabilidade (S1, S2 e S3) são iguais a zero.
Apesar da simplicidade de obtenção das estatísticas que avaliam a
estabilidade, o método de Huehn (1990) tem recebido algumas críticas, inerentes
ao processo de classificação. A primeira é a de não levar em consideração a
magnitude dos valores obtidos; outro aspecto é que as estatísticas S1, S2 e S3
contemplam a estabilidade, independente da classificação ser boa ou ruim. Dessa
forma, as estatísticas só serão úteis se consideradas simultaneamente à média
dos genótipos avaliados (Cruz & Carneiro, 2003).
Além do método sugerido por Huehn (1990), outros procedimentos
também foram propostos. O método de Kang (1988), no qual se faz uma

18
classificação para a média do caráter (posto um para o maior valor) e para o
parâmetro de estabilidade de Shukla (1972) (posto um para o menor valor). A
soma dos dois postos provê um índice final, sendo o valor mais baixo relativo ao
genótipo desejável.
O método de Fox et al. (1990) utiliza uma estratificação da posição da
cultivar em três grupos (superior, médio, inferior). Um genótipo que concentra-
se, principalmente, no grupo superior é considerado como o desejável.
Outro método que também pode ser utilizado é o de Thennasaru (1993),
citado por Sabaghnia et al. (2006). Nesse método, as medidas de estabilidade
NPi(1), NPi(2), NPi(3) e NPi(4), baseadas em postos ajustados para cada ambiente,
são utilizadas no estudo. O genótipo mais estável é aquele cuja posição em
relação aos outros permanece inalterada nos ambientes avaliados.
Uma alternativa que pode ser implementada para o estudo da
estabilidade fenotípica de forma não-paramétrica é a efetuação dos testes via
reamostragem. Desse modo, podem-se adaptar alguns métodos clássicos de
estudos da estabilidade ao enfoque não-paramétrico.
Um método de reamostragem que vem sendo cada vez mais utilizado é o
“bootstrap”. Nesse método, os dados são utilizados em sua própria análise.
Dessa forma, todo resultado “bootstrap” depende diretamente da amostra
observada, isto é, os resultados são consistentes para a amostra original. A noção
básica é que os dados em si, vistos como distribuições de freqüência,
representam a melhor imagem disponível da distribuição da qual eles são
amostrados (Crowley, 1992).
Nos casos não-paramétricos, o método reamostra os dados, com
reposição, de acordo com uma distribuição empírica estimada, tendo em vista
que, em geral, não se conhece a distribuição subjacente aos dados (Lavoranti,
2003).

19
3 MATERIAL E MÉTODOS
3.1 Material genético
Foram avaliados 55 tratamentos, sendo dez híbridos simples comerciais
utilizados como parentais (Tabela 5) e 45 híbridos duplos resultantes de um
dialelo completo. As sementes dos 45 híbridos duplos foram obtidas na área
experimental do Departamento de Biologia da Universidade Federal de Lavras
(DBI/UFLA), no ano agrícola de 2004/05. Para cada cruzamento, as espigas
foram debulhadas em conjunto e as sementes misturadas, de forma a representar
uma amostra única para o preparo dos experimentos de avaliação.
TABELA 5. Descrição dos dez híbridos simples comerciais.
Híbrido Nome Empresa Textura dos grãos Ciclo1/
1 DKB 199 Monsanto Semiduro SMP
2 30F90 Pioneer Duro SMP
3 A 2555 Nidera Sementes Duro SMP
4 DKB 333B Monsanto Semiduro SMP
5 DOW 657 Dow AgroScience Semiduro P
6 AG 8060 Monsanto Semiduro P
7 30F87 Pioneer Duro SMP
8 DOW 8420 Dow AgroScience Duro P
9 30K75 Pioneer Semiduro SMP
10 AG 7000 Monsanto Semiduro SMP 1/ P- ciclo precoce; SMP- ciclo semiprecoce

20
3.2 Condução dos experimentos
Os experimentos foram conduzidos em 15 ambientes (Tabela 6), no ano
agrícola de 2005/06, em propriedades de agricultores e estações experimentais.
Os tratamentos foram avaliados utilizando-se o delineamento de blocos
casualizados. O caráter avaliado foi a produtividade de espigas despalhadas
(kg/ha), corrigida para 13% de umidade.
Em todos os ambientes, procedeu-se à correção da produtividade de
espigas utilizando o estande como covariável, conforme Schmildt (2000). Por
ocasião da semeadura, foram distribuídas 50 sementes por parcela, sendo
realizado o desbaste quando as plantas apresentavam em torno de 4 a 5 folhas
completamente expandidas. Em todos os ambientes as parcelas foram
constituídas de duas fileiras, empregando-se uma densidade populacional
equivalente a 55.000 plantas por hectare. Os demais tratos culturais foram os
normalmente utilizados pelos agricultores para a cultura do milho. Detalhes
experimentais dos ambientes de condução dos experimentos são apresentados na
Tabela 7.

21
TABELA 6. Características dos ambientes de condução dos experimentos.
Ambiente Local Município Latitude Longitude Altitude (m)
1 Área Experimental DBI/UFLA Lavras, MG 21º13’S 44º58’W 910
2 Área Experimental Geneze Guarda-Mor, MG 17º34’S 47º08’W 1010
3 Área Experimental Bionacional Barreiras, BA 12º08’S 45º00’W 452
4 Área Experimental Prezzotto Jussara, GO 23º35’S 52º28’W 250
5 Fazenda Experimental Vitorinha Lavras, MG 21º12’S 44º58’W 951
6 Área Experimental Coopadap São Gotardo, MG 19º18’S 46º03’W 1058
7 Fazenda Faepe Ijaci, MG 21º09’S 44º56’W 859
8 Fazenda Faepe Ijaci, MG 21º09’S 44º56’W 859
9 Fazenda Mato Dentro Lavras, MG 21º13’S 45º03’W 918
10 Fazenda Morro do Guerra Ribeirão Vermelho, MG 21º10’S 45º04’W 884
11 Fazenda Candeias Candeias, MG 20º46’S 45º19’W 967
12 Fazenda Chimarrão Paracatu, MG 17º13’S 46º39’W 580
13 Fazenda Milanez Carrancas, MG 21º24’S 44º38’W 1005
14 Fazenda dos Coelhos Itutinga, MG 21º23’S 44º46’W 958
15 Fazenda da Pedra Ingaí, MG 21º22’S 44º45’W 980
21

22
TABELA 7. Detalhes experimentais dos ambientes de condução dos experimentos.
Ambiente Data de plantio Repetições Tamanho da parcela (m2)
Espaçamento entre linhas (m)
Produtividade média de espigas (kg/ha)
1 08/11/2005 3 4,0 0,50 10803
2 21/12/2005 3 4,8 0,80 6212
3 13/12/2005 3 4,8 0,80 4549
4 05/04/2006 3 4,8 0,80 5152
5 12/11/2005 3 4,8 0,60 6246
6 07/11/2005 3 4,8 0,80 8085
7 13/10/2005 3 4,0 0,50 13192
8 31/01/2006 3 4,8 0,60 8896
9 28/11/2005 3 4,8 0,80 8685
10 03/11/2005 3 4,0 0,50 8737
11 13/11/2005 3 4,8 0,80 9876
12 20/02/2006 3 4,8 0,80 7768
13 21/12/2005 3 4,8 0,80 7353
14 06/12/2005 3 4,8 0,80 8041
15 17/11/2005 2 4,8 0,80 9166
22

23
3.3 Análise estatística dos dados
3.3.1 Adaptabilidade e estabilidade
As análises individuais por ambientes, tendo como referência a
produtividade de espigas despalhadas (kg/ha), foram realizadas no PROC GLM
do SAS (SAS, 2000), adotando-se o modelo estatístico:
ijjiij erhY +++ = µ
em que:
ijY : observação referente ao híbrido i na repetição j;
µ : média geral;
ih : efeito fixo do híbrido i , i=1, 2, 3,...,55;
jr : efeito aleatório da repetição j , j=1, 2, 3;
ije : erro experimental )N(0, 2∩ σije .
O efeito de híbridos foi decomposto ortogonalmente em efeitos de
híbridos simples, híbridos duplos e o contraste entre os dois tipos de híbridos.
Posteriormente, procedeu-se à análise conjunta envolvendo todos os ambientes,
também no PROC GLM do SAS (SAS, 2000), adotando-se o seguinte modelo
estatístico:
ijlilljliijl eharahY +++++= )()(µ
em que:
ijlY : observação referente ao híbrido i na repetição j no ambiente l;

24
µ : média geral;
ih : efeito fixo do híbrido i , i= 1, 2, 3,..., 55;
la : efeito fixo do ambiente l , l= 1, 2, 3,..., 15;
)(ljr : efeito aleatório da repetição j no ambiente l;
ilha)( : efeito fixo da interação híbridos x ambientes;
ijle : erro experimental )N(0, 2∩ σijle .
Para o estudo da adaptabilidade e da estabilidade dos híbridos em
avaliação, adotou-se a estatística não-paramétrica.
Primeiramente, o estudo da estabilidade foi realizado utilizando-se a
contribuição de cada híbrido para a interação genótipos x ambientes. A partir das
médias dos híbridos, foi realizada a ordenação dos dados em uma tabela de
dupla entrada entre híbridos (dispostos nas linhas) e ambientes (dispostos nas
colunas), tendo o híbrido de menor média em cada ambiente recebido o posto 1
e o de maior média, o posto 55. A partir dessa classificação, foi obtida a
estimativa da ecovalência (Wricke, 1965), utilizando-se a seguinte expressão:
2
.... )( RRRRW jiiji +−−= ∑
em que:
iW : ecovalência;
ijR : posto referente à observação do híbrido i no ambiente j;
.iR : posto médio do híbrido i;
jR . : posto médio do ambiente j;
..R : média geral.

25
Estimou-se também o desvio de cada híbrido em relação ao desempenho
máximo em cada ambiente. Nesse caso, realizou-se a ordenação de todos os
dados em uma tabela de dupla entrada entre híbridos (dispostos nas linhas) e
ambientes (dispostos nas colunas), tendo o híbrido de menor média recebido o
posto 1 e o de maior média, o posto 825. A partir dessa classificação, foi obtida
a estimativa do parâmetro de adaptabilidade e estabilidade de Lin e Binns
(1988b), com os testes realizados via “bootstrap”, utilizando-se a seguinte
expressão:
∑=
−=n
j
jiji nMRP1
2 2/)(
em que:
iP : índice de adaptabilidade e estabilidade do híbrido i;
ijR : posto referente à observação do híbrido i no ambiente j;
jM : posto máximo entre todos os híbridos no ambiente j;
n : número de ambientes.
As análises de adaptabilidade e estabilidade foram realizadas com o
auxílio do aplicativo computacional R (R CORE TEAM, 2006).
3.3.2 Análise dialélica
Para as análises dialélicas individuais utilizou-se o modelo I (modelo
fixo), proposto por Griffing (1956) – Método II, no qual são avaliados todos os
cruzamentos possíveis, incluindo os genitores. O modelo estatístico adotado foi:
ijkijjikijk e s g g r Y +++++= µ

26
em que:
ijkY : observação referente à combinação híbrida ij ou ao genitor (i=j), na
repetição k;
µ : média geral;
kr : efeito aleatório da repetição k , k=1, 2, 3;
g g ji , : efeito fixo da capacidade geral de combinação do i-ésimo ou j-ésimo
genitor (i , j= 1, 2, 3,..., 10), respectivamente;
ijs : efeito fixo da capacidade específica de combinação para os cruzamentos
entre genitores de ordem i e j;
ijke : erro experimental
A análise dialélica conjunta foi realizada conforme Zhang et al. (2005),
de acordo com o seguinte modelo estatístico:
ijklijljlilijjilklijkl easagag s g gra Y +++++++++= )()()()(µ
em que:
ijklY : observação referente à combinação híbrida ij ou ao genitor (i=j), na
repetição k no ambiente l;
µ : média geral;
la : efeito fixo do ambiente l , l= 1, 2, 3,..., 15;
)(lkr : efeito aleatório da repetição k no ambiente l;
ig , jg : efeito fixo da capacidade geral de combinação do i-ésimo ou j-ésimo
genitor (i , j= 1, 2 , 3,...,10), respectivamente;
ijs : efeito fixo da capacidade específica de combinação para os cruzamentos
entre genitores de ordem i e j;

27
ilag)( , jlag)( : efeito fixo da interação capacidades gerais de combinação x
ambientes;
ijlas)( : efeito fixo da interação capacidades específicas de combinação x
ambientes;
ijkle : erro experimental.
Foram estimados os componentes quadráticos que expressam a
variabilidade dos genitores estudados, referentes à capacidade geral de
combinação ( gφ ) e específica de combinação ( sφ ), conforme Cruz et al. (2004).
Para verificar se as estimativas das capacidades gerais e específicas de
combinação são diferentes de zero, foi realizado o teste de significância t de
Student (Zhang et al., 2005).
3.3.3 Estabilidade dos efeitos da capacidade de combinação
A estabilidade das estimativas dos parâmetros da análise dialélica de
Griffing foi obtida de forma não-paramétrica, utilizando-se a contribuição de
cada estimativa para a interação genótipos x ambientes.
Os efeitos estimados das capacidades gerais de combinação, obtidos na
análise dialélica, foram ordenados em uma tabela de dupla entrada entre efeitos
(dispostas nas linhas) e ambientes (dispostos nas colunas). A menor estimativa
em cada ambiente recebeu o posto 1 e a maior recebeu o posto 10. O mesmo
procedimento foi adotado para os efeitos das capacidades específicas de
combinação, porém, a maior estimativa em cada ambiente recebeu o posto 55.
A partir dessa classificação, foi obtida a estimativa da ecovalência
(Wricke, 1965), utilizando-se a seguinte expressão:

28
2.... )( RRRRW jiiji +−−= ∑
em que:
iW : ecovalência;
ijR : posto referente à observação da estimativa do parâmetro i no ambiente j;
.iR : posto médio da estimativa do parâmetro i;
jR . : posto médio do ambiente j;
..R : média geral.
As análises de estabilidade foram realizadas com o auxílio do aplicativo
computacional R (R CORE TEAM, 2006).

29
4 RESULTADOS E DISCUSSÃO
4.1 Adaptabilidade e estabilidade de híbridos simples e duplos de milho
As análises de variância individuais evidenciaram que a precisão
experimental avaliada pelo coeficiente de variação (CV) foi relativamente boa,
com estimativas inferiores a 21% em todos os experimentos. Esse fato também é
comprovado pela proporção da variação explicada pelo modelo (R2), que foi de
magnitude elevada na maioria dos casos e sempre superior a 44% (Tabela 1A).
Foram constatadas diferenças significativas (P≤0,05) para a fonte de
variação híbridos, em 93% dos experimentos. A média dos híbridos simples
(HS) foi superior à média dos híbridos duplos (HD), na maioria dos
experimentos, tendo o contraste entre os dois tipos de híbridos sido significativo
(P≤0,05) nos ambientes 1, 4, 6, 8, 9, 10, 12 e 13 (Tabela 1A).
Na análise de variância conjunta, envolvendo os 15 ambientes,
verificou-se que o CV foi de 13,9%. Ressalta-se que o modelo explicou 87,6%
da variação total, confirmando que os híbridos foram avaliados sob boas
condições experimentais (Tabela 8). É oportuno comentar que, dos 15
experimentos de avaliação, nove foram conduzidos em propriedades de
agricultores, utilizando-se os tratos culturais normalmente empregados pelos
mesmos. Apesar das dificuldades de ordem prática, a representação desses
ambientes aumenta a probabilidade de sucesso no desenvolvimento de cultivares
superiores, uma vez que é a condição para a qual se pretende recomendar
(Ceccarelli et al., 2006).
Foi constatado, na análise de variância conjunta, que a fonte de variação
ambientes foi significativa pelo teste de F (P≤0,01) (Tabela 8), com
produtividade média variando de 4549 kg/ha, no ambiente 3 a 13192 kg/ha, no

30
ambiente 7. Esse fato era esperado, uma vez que as condições ambientais
diferiram bastante em relação a alguns fatores, como a fertilidade do solo, o
espaçamento entre plantas e a época de semeadura.
As demais fontes de variação também apresentaram significância pelo
teste de F (P≤0,01). Infere-se, portanto, que há diferença entre os híbridos
avaliados e que o desempenho desses híbridos não foi coincidente nos diferentes
ambientes. Verificou-se também que a média dos híbridos simples diferiu da
média dos híbridos duplos, tendo o desempenho desses diferentes tipos de
híbridos variado em função do ambiente (Tabela 8).
A superioridade dos híbridos simples em relação aos híbridos duplos foi,
em média, de 6,9%, considerando todos os ambientes. O híbrido simples mais
produtivo (AG 8060) foi 3,3% superior ao híbrido duplo de melhor desempenho
(AG 8060 x 30F87) (Tabelas 8 e 9), contudo, formaram um único grupo pelo
teste de Scott & Knott (1974). Resultados semelhantes foram relatados por
Wricke & Weber (1986) em um estudo teórico de comparação de diferentes
tipos de híbridos. Estes autores consideraram que, se os valores genotípicos de
híbridos simples seguem uma distribuição normal, a média dos diferentes tipos
de híbridos pode ser predita. No exemplo apresentado por Wricke & Weber
(1986), o híbrido simples mais produtivo foi 3% superior ao híbrido duplo de
melhor desempenho, valor muito próximo ao obtido no presente trabalho.
No Brasil, tem-se constatado uma rápida substituição dos híbridos
duplos pelos triplos e simples. Em conseqüência, nota-se uma tendência de
elevação no custo das sementes híbridas, o que inviabiliza a sua adoção por um
grande contingente de agricultores (Cruz & Pereira Filho, 2006). Dessa forma, é
importante levar em consideração, no momento da recomendação da cultivar, se
o maior potencial produtivo dos híbridos simples compensaria o maior custo das
sementes.

31
TABELA 8. Resumo da análise de variância conjunta para produtividade de espigas despalhadas (kg/ha) de dez híbridos simples (HS) e 45 híbridos duplos (HD) de milho, avaliados em 15 ambientes, no ano agrícola de 2005/2006.
F.V. G.L. QM AMBIENTES (A) 14 794873096** REPETIÇÃO/AMBIENTE 29 6585709** HÍBRIDOS (H) 54 18097540** HS 9 19467194** HD 44 15885586** HS vs HD 1 103096622** H X A 756 2554880** HS x A 126 2928380** HD x A 616 2396571** HS vs HD x A 14 6158962** ERRO 1566 1289652 CV (%) 13,9 R2 (%) 87,6 MÉDIA (kg/ha) 8162 MÉDIA HS (kg/ha) 8643 MÉDIA HD (kg/ha) 8082 ** Significativo, pelo teste de F, a 1% de probabilidade.
Para o estudo da estabilidade dos híbridos, adotou-se o método de
Wricke (1965). Verificou-se, pelas estimativas de ecovalência (Wi), que os
híbridos duplos (HD) contribuíram, em média, com 1,73% para a interação
genótipos x ambientes, e os híbridos simples (HS), com 2,20% (Tabela 9). Essa
diferença pode parecer pequena, contudo, correspondem a estimativas de Wi,
21% inferiores para os híbridos duplos, em relação às obtidas pelos híbridos
simples.

32
TABELA 9. Produtividade média de espigas despalhadas (PED), em kg/ha e estimativas da contribuição para a interação genótipos x ambientes (Wi) obtidas na avaliação de híbridos de milho, envolvendo 15 ambientes, no ano agrícola de 2005/2006.
HÍBRIDO PED (kg/ha) Wi (%) HÍBRIDO PED
(kg/ha) Wi (%)
DKB 199 9013b1/ 2,24 A 2555 x DOW 657 6531g 0,07
30F90 9054b 2,54 A 2555 x AG 8060 7907e 1,88
A 2555 7923e 2,33 A 2555 x 30F87 8007e 2,41
DKB 333B 9187b 2,59 A 2555 x DOW 8420 8242d 2,08
DOW 657 7475e 2,21 A 2555 x 30K75 7783e 1,99
AG 8060 9656a 0,55 A 2555 x AG 7000 8060d 2,46
30F87 8870c 1,92 DKB 333B x DOW 657 7971e 3,59
DOW 8420 8187d 2,34 DKB 333B x AG 8060 8388c 2,96
30K75 8225d 3,00 DKB 333B x 30F87 7163f 0,29
AG 7000 8839c 2,30 DKB 333B x DOW 8420 7718e 0,96
DKB 199 x 30F90 8973b 1,56 DKB 333B x 30K75 8497c 2,25
DKB 199 x A 2555 8243d 1,27 DKB 333B x AG 7000 7004f 0,75
DKB 199 x DKB 333B 6678g 1,17 DOW 657 x AG 8060 8397c 1,86
DKB 199 x DOW 657 7929e 1,45 DOW 657 x 30F87 8137d 2,74
DKB 199 x AG 8060 9036b 1,16 DOW 657 x DOW 8420 7723e 1,47
DKB 199 x 30F87 7958e 1,45 DOW 657 x 30K75 7956e 2,28
DKB 199 x DOW 8420 8655c 1,88 DOW 657 x AG 7000 8177d 1,41
DKB 199 x 30K75 8264d 2,06 AG 8060 x 30F87 9344a 0,32
DKB 199 x AG 7000 6635g 0,36 AG 8060 x DOW 8420 8272d 2,05
30F90 x A 2555 8341d 1,83 AG 8060 x 30K75 8320d 1,68
30F90 x DKB 333B 8875c 1,80 AG 8060 x AG 7000 8868c 1,02
30F90 x DOW 657 7971e 2,49 30F87 x DOW 8420 7538e 0,98
30F90 x AG 8060 8618c 1,85 30F87 x 30K75 8019e 1,25
30F90 x 30F87 8738c 1,58 30F87 x AG 7000 8101d 3,49
30F90 x DOW 8420 8217d 2,12 DOW 8420 x 30K75 7885e 2,14
30F90 x 30K75 8008e 1,63 DOW 8420 x AG 7000 8157d 1,58
30F90 x AG 7000 8597c 2,91 30K75 x AG 7000 8004e 1,36
A 2555 x DKB 333B 7790e 2,11 1/ Médias seguidas pela mesma letra pertencem ao mesmo grupo, pelo teste de Scott & Knott (1974), a 5% de probabilidade.

33
Na literatura, são encontrados estudos com o objetivo de verificar se a
estrutura genética de uma população confere-lhe maior estabilidade. No caso do
milho, um híbrido duplo apresenta maior heterogeneidade que um híbrido
simples; conseqüentemente, deve possuir maior homeostase populacional
(Allard & Bradshaw, 1964).
A maior estabilidade dos híbridos duplos de milho tem sido relatada na
literatura (Guillen-Portal et al., 2003; Schnell & Becker, 1986), indicando que a
homeostase populacional é importante para a obtenção de cultivares estáveis.
Entretanto, podem ser identificados híbridos simples tão estáveis quanto os
duplos para o mesmo caráter (Carvalho et al., 2005). No presente trabalho,
identificaram-se híbridos simples com estimativas de Wi equivalentes às dos
híbridos duplos, como o híbrido AG 8060, que contribuiu com 0,55% para a
interação genótipos x ambientes (Tabela 9).
O híbrido duplo A 2555 x DOW 657 foi o mais estável, apresentando
menor contribuição para a interação genótipos x ambientes (Tabela 9),
entretanto, está entre os de menor produtividade média de espigas despalhadas.
Esse tipo de estabilidade é referido, por Lin et al. (1986), como estabilidade
biológica. A utilização de híbridos que apresentam estabilidade do tipo biológica
não é desejável, pois, normalmente, está associada a baixas produtividades (Cruz
et al., 2004).
Na Tabela 10 está apresentada a estimativa do parâmetro de
adaptabilidade e estabilidade (Pi) de Lin & Binns (1988b). O híbrido AG 8060
(HS) apresentou o melhor desempenho, seguido do híbrido DKB 333B (HS) e
do híbrido AG 8060 x 30F87 (HD). Constatou-se que, na média, os híbridos
simples obtiveram menores estimativas de Pi, em comparação aos híbridos
duplos, ou seja, apresentaram menores desvios, em relação ao desempenho
máximo em cada ambiente.

34
Considerando os critérios produtividade média de espigas despalhadas,
contribuição para a interação genótipos x ambientes e a estimativa do desvio em
relação ao máximo em cada ambiente, os híbridos que se destacaram foram o
AG 8060 (HS) e AG 8060 x 30F87 (HD), associando alta produtividade média,
baixa estimativa de Pi e estabilidade de produção nos ambientes avaliados
(Tabela 9 e 10).
Na comparação dos híbridos simples e duplos de milho, verificou-se que
podem ser selecionados híbridos estáveis e produtivos, independente da estrutura
genética. Desse modo, para os agricultores que utilizam altos níveis de
tecnologia, objetivando máxima produtividade e uniformidade, a utilização de
híbridos simples é justificável. Contudo, a utilização dos híbridos duplos pode
ser vantajosa para condições de baixo investimento (Ribeiro, 1998),
principalmente com o emprego da geração F2 ou das populações em equilíbrio,
para a produção de sementes híbridas (Souza Sobrinho, 2001).

35
TABELA 10. Estimativas do parâmetro de adaptabilidade e estabilidade de Lin & Binns (1988b), obtidas na avaliação de híbridos de milho, envolvendo 15 ambientes no ano agrícola de 2005/2006.
HÍBRIDO Pi Valor-p1/ HÍBRIDO Pi Valor-p1/ DKB 199 19539 0,50 A 2555 x DOW 657 93828 0,00
30F90 19982 0,46 A 2555 x AG 8060 47787 0,00
A 2555 40605 0,00 A 2555 x 30F87 41234 0,00
DKB 333B 10133 0,99 A 2555 x DOW 8420 31469 0,01
DOW 657 58442 0,00 A 2555 x 30K75 46067 0,00
AG 8060 7874 1,00 A 2555 x AG 7000 43484 0,00
30F87 19926 0,46 DKB 333B x DOW 657 44192 0,00
DOW 8420 38785 0,00 DKB 333B x AG 8060 30317 0,01
30K75 36079 0,00 DKB 333B x 30F87 63821 0,00
AG 7000 22521 0,27 DKB 333B x DOW 8420 47703 0,00
DKB 199 x 30F90 13528 0,91 DKB 333B x 30K75 22229 0,29
DKB 199 x A 2555 29072 0,03 DKB 333B x AG 7000 72590 0,00
DKB 199 x DKB 333B 80714 0,00 DOW 657 x AG 8060 30651 0,01
DKB 199 x DOW 657 38932 0,00 DOW 657 x 30F87 41148 0,00
DKB 199 x AG 8060 14848 0,85 DOW 657 x DOW 8420 48874 0,00
DKB 199 x 30F87 38877 0,00 DOW 657 x 30K75 39108 0,00
DKB 199 x DOW 8420 24865 0,14 DOW 657 x AG 7000 31073 0,01
DKB 199 x 30K75 26147 0,09 AG 8060 x 30F87 11553 0,97
DKB 199 x AG 7000 84312 0,00 AG 8060 x DOW 8420 34819 0,00
30F90 x A 2555 30072 0,02 AG 8060 x 30K75 29478 0,02
30F90 x DKB 333B 17848 0,65 AG 8060 x AG 7000 19040 0,54
30F90 x DOW 657 41365 0,00 30F87 x DOW 8420 55476 0,00
30F90 x AG 8060 23748 0,19 30F87 x 30K75 38093 0,00
30F90 x 30F87 20013 0,46 30F87 x AG 7000 40311 0,00
30F90 x DOW 8420 40917 0,00 DOW 8420 x 30K75 48121 0,00
30F90 x 30K75 39461 0,00 DOW 8420 x AG 7000 38053 0,00
30F90 x AG 7000 26672 0,07 30K75 x AG 7000 44314 0,00
A 2555 x DKB 333B 46095 0,00 1/ Nível crítico para testar a hipótese de que um genótipo não desvia do desempenho máximo.

36
4.2 Análise dialélica de híbridos comerciais de milho
Os resumos das análises de variância individuais com a decomposição
do efeito de híbridos em capacidade geral de combinação (CGC) e capacidade
específica de combinação (CEC) são apresentados na Tabela 2A. Foram
constatadas diferenças significativas (P≤0,05) em 13 dos 15 experimentos, para
as fontes de variação CGC e CEC (Tabela 2A).
Na análise de variância conjunta, envolvendo os 15 ambientes foram
detectadas diferenças significativas (P≤0,01) para todas as fontes de variação.
Infere-se, portanto, que os genitores em avaliação diferem nas suas freqüências
alélicas médias, contribuindo diferentemente para os cruzamentos em que eles
estão envolvidos e que as combinações híbridas apresentaram desempenhos
diferentes dos esperados somente pelos efeitos das CGC. Os efeitos
significativos das fontes de variação CGC x A e CEC x A indicam que tanto os
efeitos das CGC quanto os da CEC não foram coincidentes nos ambientes
avaliados (Tabela 11).
O componente quadrático referente à CEC ( sφ ), foi 4,1 vezes superior
ao componente quadrático referente à CGC ( gφ ), indicando que os efeitos não
aditivos foram mais importantes que os efeitos aditivos. É importante comentar
que as estimativas das capacidades gerais de combinação são funções das
diferenças genéticas dos genitores e do efeito médio de uma substituição alélica,
])21()[( dtappCGC ii −+−= ou ])21()[( dtappCGC jj −+−= , e estão
associadas aos efeitos aditivos (Hallauer & Miranda Filho, 1988; Vencovsky,
1987). As capacidades específicas de combinação, por sua vez, são funções dos
efeitos de dominância e do produto das diferenças de freqüências alélicas dos
genitores, )])([(2 ii ttppdCEC −−= , fazendo com que as mesmas estejam
relacionadas aos efeitos de dominância e epistáticos (Hallauer & Miranda Filho,

37
1988; Vencovsky, 1987). A predominância dos efeitos não aditivos, assim como
constatado no presente trabalho, tem sido relatada na literatura (Fuzatto et al.,
2002; Machado et al., 2006).
As estimativas das capacidades gerais de combinação ( ig ) dos híbridos
avaliados variaram de -358,1 a 540,4. As maiores estimativas foram obtidas
pelos híbridos AG 8060 e 30F90, indicando que esses híbridos possuem alta
freqüência de alelos favoráveis para a produtividade de espigas (Tabela 12). É
conveniente salientar que esses dois genitores, que obtiveram as maiores
estimativas de ig , também foram os que apresentaram as maiores
produtividades.
Em relação às estimativas de capacidades específicas de combinação,
verificou-se que 15% foram significativamente diferentes de zero (P≤0,05). A
porcentagem de estimativas positivas foi de 38% e negativas de 62%, variando
de -1532,7 a 1619,4 (Tabela 12).
Constatou-se, ainda, que a estimativa da capacidade específica de
combinação de um genitor com ele mesmo ( iis ) foi positiva para todos os casos
(Tabela 12). Segundo Cruz & Vencovsky (1989), isso é um indicativo de
dominância unidirecional no sentido de diminuir a expressão do caráter. Esse
resultado era esperado, uma vez que os genitores em avaliação eram híbridos
simples, com alto potencial produtivo, logo os híbridos duplos oriundos do
cruzamento desses híbridos deveriam apresentar menores produtividades, como
constatado no presente trabalho.
Foram identificados genitores que, além de possuir alta estimativa de
ig , produzem combinações híbridas superiores. Dentre essas, se destacaram as
combinações AG8060 x 30F87, DKB 199 x 30F90, DKB 199 x AG 8060 e
30F90 x AG 8060, que apresentaram alta produtividade, alta freqüência de
alelos favoráveis dos seus genitores e possibilidade de exploração da heterose de
forma satisfatória (Tabela 12).

38
TABELA 11. Resumo da análise de variância conjunta da produtividade de espigas despalhadas (kg/ha), com decomposição do efeito de híbridos na capacidade geral (CGC) e específica (CEC) de combinação, obtida na avaliação do cruzamento dialélico entre dez genitores de milho envolvendo 15 ambientes, no ano agrícola de 2005/2006.
F.V. G.L. QM AMBIENTES (A) 14 794873096** REPETIÇÃO/AMBIENTE 29 6585709** HÍBRIDOS (H) 54 18097540** CGC 9 39020099** CEC 45 14193119** H X A 756 2554880** CGC x A 126 5159456** CEC x A 630 2033964** ERRO 1566 1289652 CV (%) 13,9
R2 (%) 87,6 MÉDIA (kg/ha) 8162 ** Significativo, pelo teste de F, a 1% de probabilidade.

39
TABELA 12. Efeitos das capacidades gerais ( ig ) e específicas de combinação ( ijs ) para a produtividade de espigas despalhadas (kg/ha), nos 15 ambientes de condução dos experimentos, no ano agrícola de 2005/2006.
GENITOR DKB 199 30F90 A 2555 DKB 333B DOW 657 AG 8060 30F87 DOW 8420 30K75 AG 7000 ig
DKB 199 789,4* 365,1ns 302,9ns -1376,2** 79,5ns 237,4ns -325,6ns 550,3ns 120,6ns -1532,7** 22,6ns
30F90 129,0ns 75,0ns 451,1ns -218,8ns -440,3ns 140,4ns -236,3ns -471,7ns 77,5ns 359,4**
A 2555 319,0ns -26,2ns -1002,9** -525,3ns 34,0ns 431,3ns -96,0ns 169,2ns -275,7**
DKB 333B 1268,6** 250,5ns -178,9ns -965,0** -226,1ns 542,9ns -1009,3** -123,3ns
DOW 657 18,3ns 2,7ns 278,9ns -8,6ns 176,7ns 405,5ns -358,1**
AG 8060 424,7ns 532,0ns -334,2ns -358,7ns 216,2ns 540,4**
30F87 573,8ns -630,1ns -150,5ns -61,8ns 69,6ns
DOW 8420 212,6ns -124,6ns 153,2ns -97,8ns
30K75 199,4ns -37,3ns -78,5ns
AG 7000 1619,4* -58,6ns * e ** Significativamente diferente de zero, pelo teste t, a 5% e a 1 % de probabilidade, respectivamente. ns Não significativo.
39

40
4.3 Estabilidade dos efeitos da capacidade de combinação
Admitindo que a estabilidade esteja sob controle genético, os
melhoristas deveriam incluir, como critério de escolha de genitores para a
formação de uma população base para os programas de melhoramento, aqueles
que possuam estabilidade de parâmetros que auxiliem em tal escolha, como os
parâmetros da análise dialélica. No estudo da estabilidade dos parâmetros da
análise dialélica do método II de Griffing constatou-se que tanto os efeitos da
capacidade geral de combinação ( ig ) quanto os efeitos da capacidade específica
de combinação ( ijs ) variaram em função do ambiente, contribuindo de forma
diferenciada para a interação.
Na Tabela 13, estão apresentados os valores da ecovalência (Wi), para os
efeitos da capacidade geral de combinação ( ig ), bem como suas estimativas. Os
valores de Wi para os efeitos de ig variaram de 3% a 13,8%. Constatou-se que o
híbrido AG 8060 associa estabilidade do efeito de capacidade geral de
combinação a uma estimativa de alta magnitude, sendo promissor para a
formação de populações base para a extração de linhagens que gerem híbridos
estáveis e produtivos.

41
TABELA 13. Efeitos das capacidades gerais de combinação ( ig ) e respectivas estimativas da ecovalência (Wi), obtidas na avaliação do cruzamento dialélico entre dez genitores de milho, envolvendo 15 ambientes, no ano agrícola de 2005/2006.
Genitor ig Wi (%)
DKB 199 22,6 9,5
30F90 359,4 9,9
A 2555 -275,7 10,1
DKB 333B -123,3 13,8
DOW 657 -358,1 8,2
AG 8060 540,4 3,0
30F87 69,6 12,6
DOW 8420 -97,8 12,3
30K75 -78,5 7,0
AG 7000 -58,6 13,6
Para o efeito da capacidade específica de combinação ( ijs ), os valores de
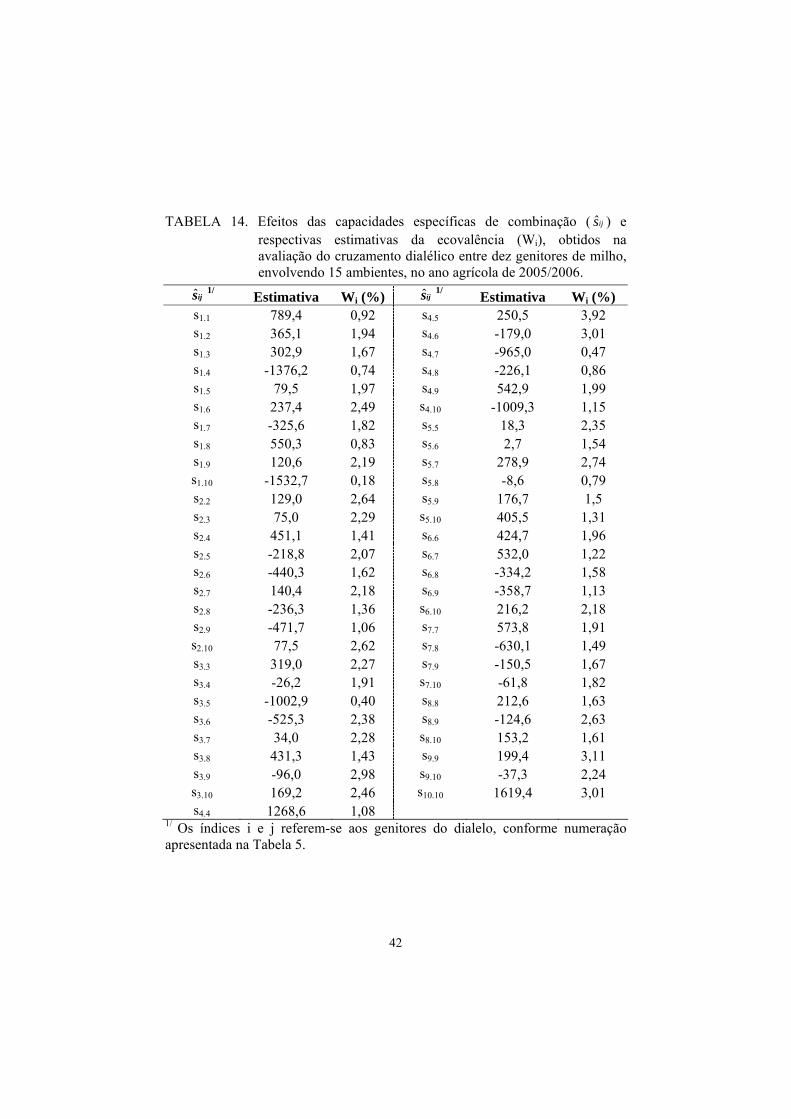
Wi variaram de 0,18% a 3,92%. Foi verificado que os efeitos de ijs com as
menores contribuições para a interação normalmente estão associados a
estimativas de baixa magnitude, como no caso dos efeitos s1.10, s3.5 e s4.7, que
contribuíram com menos de 0,50% para a interação, porém, apresentaram
estimativas negativas de ijs iguais a -1532,7, -1002,9 e -965,0, respectivamente
(Tabela 14).
42
TABELA 14. Efeitos das capacidades específicas de combinação ( ijs ) e respectivas estimativas da ecovalência (Wi), obtidos na avaliação do cruzamento dialélico entre dez genitores de milho, envolvendo 15 ambientes, no ano agrícola de 2005/2006.
ijs 1/ Estimativa Wi (%) ijs 1/ Estimativa Wi (%) s1.1 789,4 0,92 s4.5 250,5 3,92 s1.2 365,1 1,94 s4.6 -179,0 3,01 s1.3 302,9 1,67 s4.7 -965,0 0,47 s1.4 -1376,2 0,74 s4.8 -226,1 0,86 s1.5 79,5 1,97 s4.9 542,9 1,99 s1.6 237,4 2,49 s4.10 -1009,3 1,15 s1.7 -325,6 1,82 s5.5 18,3 2,35 s1.8 550,3 0,83 s5.6 2,7 1,54 s1.9 120,6 2,19 s5.7 278,9 2,74 s1.10 -1532,7 0,18 s5.8 -8,6 0,79 s2.2 129,0 2,64 s5.9 176,7 1,5 s2.3 75,0 2,29 s5.10 405,5 1,31 s2.4 451,1 1,41 s6.6 424,7 1,96 s2.5 -218,8 2,07 s6.7 532,0 1,22 s2.6 -440,3 1,62 s6.8 -334,2 1,58 s2.7 140,4 2,18 s6.9 -358,7 1,13 s2.8 -236,3 1,36 s6.10 216,2 2,18 s2.9 -471,7 1,06 s7.7 573,8 1,91 s2.10 77,5 2,62 s7.8 -630,1 1,49 s3.3 319,0 2,27 s7.9 -150,5 1,67 s3.4 -26,2 1,91 s7.10 -61,8 1,82 s3.5 -1002,9 0,40 s8.8 212,6 1,63 s3.6 -525,3 2,38 s8.9 -124,6 2,63 s3.7 34,0 2,28 s8.10 153,2 1,61 s3.8 431,3 1,43 s9.9 199,4 3,11 s3.9 -96,0 2,98 s9.10 -37,3 2,24 s3.10 169,2 2,46 s10.10 1619,4 3,01 s4.4 1268,6 1,08
1/ Os índices i e j referem-se aos genitores do dialelo, conforme numeração apresentada na Tabela 5.

43
5 CONCLUSÕES
Os híbridos duplos foram, em média, mais estáveis, contudo,
identificaram-se híbridos simples tão estáveis quanto os duplos, como, por
exemplo, o híbrido AG 8060.
Os efeitos não aditivos foram mais importantes que os efeitos aditivos
para a variação dos híbridos. Os genitores com as maiores estimativas do efeito
da capacidade geral de combinação (CGC) foram AG 8060 e 30F90.
O genitor AG 8060 associa alta estimativa do efeito da CGC à
estabilidade desse parâmetro.

44
REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, M. W.; SHANK, D. B. The relationship of heterozygosity to homeostasis in maize hybrids. Genetics, Baltimore, v. 44, n. 5, p. 777-786, 1959. ALLARD, R. W.; BRADSHAW, A. D. Implications of genotype-environment interactions in applied plant breeding. Crop Science, Madison, v. 4, n. 5, p. 503-508, July/Aug. 1964. ARAÚJO, J. S. Ganhos genéticos obtidos em híbridos e variedades de milho representativos de três décadas de melhoramento no Brasil. 1995. 64 p. Dissertação (Genética e Melhoramento de Plantas) - Universidade Federal de Lavras, Lavras, MG.
BECKER, H. C.; LÉON, J. Stability analysis in plant breeding. Plant Breeding, Berlin, v. 101, n. 1, p. 1/23. Aug. 1988.
BERNARDO, R. Breeding for quantitative traits in plants. Woodbury, Minnesota, 2002. 368 p.
BÉTRAN, F. J.; BECK, D.; BANZIGER, M.; EDMEADS, G. O. Genetic analysis of inbred and hybrid grain yield under stress and nonstress environments in tropical maize. Crop Science, Madison, v. 43, n. 3, p. 807-817. May/June 2003.
CAMPOS, H. Estatística experimental não paramétrica. Piracicaba: ESALQ, 1983. 349 p.
CARVALHO, H. W. L. de; CARDOSO, M. J.; LEAL, M. de L. da S.; SANTOS, M. X. dos; TABOSA, J. N.; SOUZA, E. M. Adaptabilidade e estabilidade de cultivares de milho no Nordeste brasileiro. Pesquisa Agropecuária Brasileira, Brasília, v. 40, n. 5, p. 471-477, maio 2005.
CECCARELLI, S.; GRANDO, S.; BAUM, M. Plant breeding for dry areas In: Anais do X Simpósio Sobre Atualização em Genética e Melhoramento de Plantas. Lavras: UFLA, 2006. p. 29-57.
CROWLEY, P. H. Resampling methods for computation-intensive data analysis in ecology and evolution. Annual Review of Ecology and Systematics, Palo Alto, v. 23, p. 405-447, 1992.

45
CRUZ, C. D.; CARNEIRO, P. C. de S. Modelos biométricos aplicados ao melhoramento genético. Viçosa: UFV, 2003, v. 2. 585 p.
CRUZ, C. D.; REGAZZI, A. J.; CARNEIRO, P. C. de S. Modelos biométricos aplicados ao melhoramento genético. 3. ed. Viçosa: UFV, 2004. v. 1, 480 p.
CRUZ, C. D.; VENCOVSKY, R. Comparação de alguns métodos de análise dialélica. Revista Brasileira de Genética, Ribeirão Preto, v. 12, n. 2, p. 425-438, jun. 1989.
CRUZ, J. C.; PEREIRA FILHO, I. A. Cultivares de milho disponíveis no mercado de sementes do Brasil para a safra 2006/07. 2006. Disponível em: <http://www.cnpms.embrapa.br/milho/cultivares/index.php>. Acesso em: 31 jan. 2007.
DUVICK, D. N. Biotechnology in the 1930’s: the development of hybrid maize. Nature Reviews Genetics, Hampshire, v. 2, n. 1, p. 69-74, Jan. 2001.
DUVICK, D. N. The contribution of breeding to yield advances in maize (Zea mays L.), Advances in Agronomy, San Diego, v. 86, p. 83-145, 2005.
EBERHART, S. A.; RUSSEL, W. A. Yield stability for a 10-line diallel of single-cross and double-cross maize hybrids. Crop Science, Madison, v. 9, n. 3, p. 357-361, May/June 1969.
FERNANDES, J. S. C.; FRAZON, J. F. Thirthy years of genetic progress in maize (Zea mays L.) in a tropical environment. Maydica, Bergamo, v. 42, n. 1, p. 21-27, May/June 1997.
FOX, P. N.; CROSSA, J.; ROMAGOSA, I. Multi-environment testing and genotype x environment interaction. In: KENPTON R. A.; FOX, P. N. [Eds.] Statistical methods for plant variety evaluation. London: Chapman and Hall, 1997. p. 117-138.
FOX, P. N.; SKOVMAND, B.; THOMPSON, B. K.; BRAUN, H. J.; CORMIER, R. Yield and adaptation of hexaploid spring triticale. Euphytica, Dordrecht, v. 47, n. 1, p. 57-64, Apr. 1990.
FUZATTO, S. R.; FERREIRA, D. F.; RAMALHO, M. A. P.; RIBEIRO, P. H. E. Divergência genética e sua relação com os cruzamentos dialélicos na cultura do milho. Ciência e Agrotecnologia, Lavras, v. 26, n. 1, p. 22-32, jan./fev. 2002.

46
GARDNER, C. O.; EBERHART, S. A. Analysis and interpretation of the variety cross diallel and related population. Biometrics, Alexandria, v.22, n. 3, p. 439-452, Sept. 1966.
GHOSH, D. K.; BISWAS, P. C. Complete diallel crosses plans through balanced incomplete block designs. Journal of Applied Statistics, Hants, v. 30, n. 6, p. 697-708, July 2003.
GRIFFING, B. A. Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal of Biological Sciences, East Melbourne, v. 9, p. 463-493, 1956.
GUILLEN-PORTAL, F. R.; RUSSELL, W. K.; BALTENSPERGER, D. D.; ESKRIDGE, K. M.; D'CROZ-MASON, N. E.; NELSON, L. A. Best types of maize hybrids for the western high plains of the USA Crop Science, Madison, v. 43, n. 6, p. 2065-2070, Nov./Dec.2003.
HALLAUER, A. R.; MIRANDA FILHO, J. B. de Quantitative genetics in maize breeding. Ames: Iowa State University Press, 1988. 468 p.
HAYMAN, B. I. The theory and analysis of diallel crosses. Genetics, Baltimore, v. 39, n. 6, p. 789-809, 1954.
HUEHN, M. Nonparametric measures of phenotypic stability. Part I: Theory. Euphytica, Dordrecht, v. 47, n. 3, p. 189-194, June 1990.
JONES, D. F. The effects of inbreeding and crossbreeding upon development. Connecticut Agricultural Experimental Station Bulletin, New Haven, p. 5-100, 1918.
KANG, M. S. A rank-sum method for selecting high-yielding, stable corn genotypes. Cereal Research Communications, Szeged, v. 16, n. 1/2, p. 113-115, 1988.
KANG M. S.; GAUCH Jr., H. G. Genotype by environment interaction. New York: CRC Press, 1996. 416 p.
KEMPTHORNE, O.; CURNOW, R. N. The partial diallel cross. Biometrics, Alexandria, v. 17, n. 2, p. 229-250, 1961.
KOUTSIKA-SOTIRIOU, M. S.; KARAGOUNIS, C. A. Assessment of maize hybrids. Maydica, Bergamo, v. 50, n. 1, p. 63-70, 2005.

47
LAVORANTI, O. J. Estabilidade e adaptabilidade fenotípica através da reamostragem "Bootstrap" no modelo AMMI. 2003. 166 p. Tese (Doutorado em Genética e Melhoramento) - Escola Superior de Agricultura "Luiz de Queiroz", Piracicaba, SP.
LANGER, I; FREY, K. J.; BAILEY, T. Associations among productivity, production response and stability indexes in oat varieties. Euphytica, Dordrecht, v. 28, n. 1, p. 17-24, 1979.
LONG, J. K.; BANZIGER M.; SMITH M. E. Diallel analysis of grain iron and zinc density in southern African-adapted maize inbreds. Crop Science, Madison v. 44, n. 6, p. 2019-2026, Nov./Dec. 2004.
LEE, E. A.; DOERKSEN, T. K.; KANNENBERG, L. W. Genetic components of yield stability in maize breeding populations. Crop Science, Madison, v. 43, n. 6, p. 2018-2027, Nov./Dec. 2003.
LEMOS, M. A.; GAMA, E. E. G. e; MENEZES, D.; SANTOS, V. F. dos; TABOSA, J. N. Avaliação de dez linhagens e seus híbridos de milho superdoce em um dialelo completo. Horticultura Brasileira, Brasília, v. 20, n. 2, p. 167-170, jun. 2002.
LIN, C. S.; BINNS M. R. A method of analyzing cultivar x location x year experiments: a new stability parameter. Theoretical and Applied Genetics, Berlin, v. 76, n. 3, p. 425-430, 1988a.
LIN, C. S.; BINNS, M. R.; LEFKOVITCH, L. P. Stability analysis: where do we stand? Crop Science, Madison, v. 26, n. 5, p. 894-900, Sept./Oct. 1986.
LIN, C. S.; BINNS M. R. A superiority measure of cultivar performance for cultivar x location data. Canadian Journal of Plant Science, Ottawa, v. 68, n. 3, n. 1, p. 193-198, Jan. 1988b.
MACHADO, J. C.; SOUZA, J. C. de; LIMA, J. L.; REIS, M. C. dos; Capacidade de combinação entre híbridos de milho no sul de Minas Gerais. In: CONGRESSO NACIONAL DE MILHO E SORGO, 26., 2006, Belo Horizonte, Resumos... Sete Lagoas: ABMS, 2006. 1CD-ROM.
MARIOTTI, J. A.; OYARZABAL, E. S.; OSA, J. M.; BULACIO, A. N. R.; ALMADA, G. H. Análisis de estabilidad y adaptabilidad de genotipos de caña de azúcar I: Interacciones dentro de una localidad experimental. Revista Agronómica del Noroeste Argentino, Tucumán, v. 13, n. 14, p. 105-127, 1976.

48
MATHER, K.; JINKS, J. L. Biometrical genetics. Ithaca: Cornell University Press, 1971. 382p.
MATHESON, A. C.; RAYMOND, C. A. A review of provenance x environment interaction: its pratical importance and use with particular reference to the tropics. Commonwealth Forestry Review, Oxford, v. 65, n. 4, p. 283-302, 1986.
MELANI, M. D.; CARENA M. J. Alternative maize heterotic patterns for the northern Corn Belt. Crop Science, Madison, v. 45, n. 6, p. 2186-2194, Nov./Dec. 2005.
MELO, W. M. C.; VON PINHO, R. G.; FERREIRA, D. F. Capacidade combinatória e divergência genética em híbridos comerciais de milho. Ciência e Agrotecnologia, Lavras, v. 25, n. 4, p. 821-830, jul./ago. 2001.
MIRANDA FILHO, J. B. de.; GERALDI, I. O. An adapted model for the analysis of partial diallel crosses. Revista Brasileira de Genética, Ribeirão Preto, v. 7, n. 4, p. 677-688, Dec. 1984.
MIRANDA FILHO, J. B. de; GORGULHO, E. P. Cruzamentos com testadores e dialelos. In: NASS, L. L.; VALOIS, A. C. C.; MELO, I. S. de; VALADARES-INGLIS, M. C. (Ed.). Recursos genéticos e melhoramento: plantas. Rondonópolis: Fundação MT, 2001, p. 649-671.
MIRANDA FILHO, J. B. de; VIÉGAS, G. P. Milho híbrido. In: PATERNIANI, E.; VIÉGAS, G. P. [Ed.]. Melhoramento e produção de milho. Campinas: Fundação Cargill, 1987. v. 1, p. 275-340.
OLIVEIRA, A. C. Comparação de alguns métodos de determinação da estabilidade em plantas cultivadas. 1976. 64 p. Dissertação (Mestrado em Ciências Agrárias) – Universidade de Brasília, Brasília, DF. PACHECO, C. A. P. Associação das metodologias de análise dialélica de Griffing e de análise de adaptabilidade e de estabilidade de Eberhart e Russell. 1997. 118 p. Tese (Doutorado em Genética e Melhoramento) - Universidade Federal de Viçosa, Viçosa, MG. PACHECO, C. A. P.; CRUZ, C. D.; SANTOS, M. X. dos. Association between Griffing's diallel and the adaptability and stability of Eberhart and Russel. Genetics and Molecular Biology, São Paulo , v. 22, n. 3, p. 451-456, Sept. 1999.
PATERNIANI, E. Maize breeding in the tropics. Critical Review in Plant Sciences, Philadelphia, v. 9, n. 2, p. 125-154, 1990.

49
PATERNIANI, E.; CAMPOS, M. S. Melhoramento do milho. In: BORÉM, A. [Ed.]. Melhoramento de espécies cultivadas. Viçosa: UFV, 1999. p. 429-485.
PATERNIANI, M. E. A. G. Z. Use of heterosis in maize breeding: history, methods and perspectives: A review. Crop Breeding and Applied Biotechnology, Viçosa, v. 1, n. 2, p. 159-178, Apr./June 2001.
PEGORARO, D. G.; BARBOSA NETO, J. F.; DAL SOGLIO, F. K.; VACARO, E.; NUSS, C. N. ; CONCEIÇÃO, L. D. H. Herança da resistência à mancha foliar de feosféria em milho. Pesquisa Agropecuária Brasileira, Brasília, v. 37, n. 3, p. 329-336, mar. 2002.
R DEVELOPMENT CORE TEAM. R: A language and environment for statistical computing. Vienna, Áustria: R Foundation for Statistical Computing, 2006.
RAMALHO, M. A. P.; SANTOS, J. B. dos; ZIMMERMANN, M. J. de O. Genética quantitativa em plantas autógamas: aplicações ao melhoramento do feijoeiro. Goiânia: UFG, 1993. 271 p.
RIBEIRO, P. H. E. Adaptabilidade e estabilidade de cultivares de milho em diferentes épocas de semeadura, níveis de adubação e locais do estado de Minas Gerais. 1998. 126 p. Tese (Doutorado em Genética e Melhoramento de Plantas) - Universidade Federal de Lavras, Lavras, MG.
RODRIGUES, M. C.; CHAVES, L. J.; PACHECO, C. A. P. Heterosis in crosses among white grain maize populations with high quality protein. Pesquisa Agropecuária Brasileira, Brasília, v. 41, n. 1, p. 59-66, jan. 2006.
ROSSE, L. N. Modelo de regressão não-linear aplicado na avaliação da estabilidade fenotípica em plantas. 1999. 179 p. Tese (Doutorado em Genética e Melhoramento) - Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, SP.
SABAGHNIA, N.; DEHGHANI, H.; SABAGHPOUR, S. H. Nonparametric methods for interpreting genotype x environment interaction of lentil genotypes. Crop Science, Madison, v. 46, n. 3, p. 1100-1106, May/June 2006.
SAEED, M. T.; SALEEM, M.; AFZAL, M. Genetic analysis of yield and its components in maize diallel crosses (Zea mays L.). International Journal of Agriculture & Biology, Hamilton, v. 2, n. 4, p. 376-378. 2000.
SANTOS, J. B. dos. Controle genético de caracteres agronômicos e potencialidades de cultivares de feijão (Phaseolus vulgaris L.) para o

50
melhoramento genético. 1984. 223 p. Tese (Doutorado em Genética e Melhoramento) - Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, SP.
SAS INSTITUTE. SAS/STAT User’s Guide, Version 8. Cary, 2000.
SCHMILDT, E. R. Correção de rendimento de parcelas, estratificação ambiental e adaptabilidade e estabilidade de cultivares de milho. 2000. 110 p. Tese (Doutorado em Genética e Melhoramento) - Universidade Federal de Viçosa, Viçosa, MG.
SCHNELL, F. W.; BECKER, H. C. Yield and yield stability in a balanced system of widely differing population structures in Zea mays L.. Plant Breeding, Berlin, v. 97, n. 1, p. 30-38, 1986.
SCOTT, A. J.; KNOTT, M. A. A cluster analysis method for grouping means in the analysis of variance. Biometrics, Raleigh, v. 30, n. 3, p. 507-512, Sept. 1974.
SHUKLA, G. K. Some aspects of partitioning genotype-environmental components of variability. Heredity, London, v. 28, p. 237-245, Oct. 1972.
SILVA, S. D. A. e; SERENO, M. J. C. M.; SILVA, C. F. L; BARBOSA NETO, J. F. Capacidade combinatória de genótipos de milho para tolerância ao encharcamento do solo. Ciência Rural, Santa Maria, v. 36, n. 2, p. 391-396, mar./abr. 2006.
SOUZA JUNIOR, C. L.; VENCOVSKY, R. Covariância entre parentes na presença da interação genótipos x ambientes. In: SIMPÓSIO DE ESTATÍSTICA APLICADA A EXPERIMENTAÇÃO AGRONÔMICA, 3., Resumos... Lavras: ESAL, 1989. p. 50-51.
SOUZA SOBRINHO, F. de. Divergência genética de híbridos simples e alternativa para obtenção de híbridos duplos de milho, 2001. 96 p. Tese (Doutorado em Genética e Melhoramento de Plantas) - Universidade Federal de Lavras, Lavras, MG.
SPRAGUE, G. F.; TATUM, L. A. General vs. specific combining ability in single crosses of corn. Journal of the American Society of Agronomy, Madison, v. 34, n. 10, p. 923-932, Oct. 1942.
STORCK, L.; BISOGNIN, D. A.; CARGNELUTTI FILHO, A. Ganho genético decorrente da substituição anual de cultivares de milho. Pesquisa Agropecuária Brasileira, Brasília, v. 40, n. 9, p. 881-886, set. 2005.

51
TORRES, R. A. de A. Estudo do controle genético da estabilidade fenotípica de cultivares de milho (Zea mays L.). 1988. 133 p. Tese (Doutorado em Genética e Melhoramento) - Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, SP.
TROYER, A. F. Adaptedness and heterosis in corn and mule hybrids. Crop Science, Madison. v. 46, n. 2, p. 528-543. Mar./Apr. 2006.
TROYER, A. F. Background of U. S. hybrid corn. Crop Science, Madison, v. 39, n. 3, p. 601-626, May/June 1999. USDA-NASS. USDA National Agricultural Statistics Service. 2006. Disponível em: <http://www.nass.usda.gov/QuickStats/PullData_US.jsp>. Acesso em: 29 nov. 2006. VENCOVSKY, R. Herança quantitativa. In: PATERNIANI, E., VIÉGAS, G.P. [Ed.]. Melhoramento e produção do milho. 2. ed. Campinas : Fundação Cargill, 1987. p. 137-214.
VENCOVSKY, R.; BARRIGA, P. Genética biométrica no fitomelhoramento. Ribeirão Preto: Sociedade Brasileira de Genética, 1992. 486 p. VENCOVSKY, R.; MORAIS, A. R.; GARCIA, J. C.; TEIXEIRA, N. M. Avaliação do progresso genético devido ao melhoramento de milho no Brasil. In: CONGRESSO NACIONAL DE MILHO E SORGO, 16., 1986, Belo Horizonte. Anais... Sete Lagoas: EMBRAPA-CNPMS, 1988. VERONESI, J. A. Comparação de métodos e avaliação da adaptabilidade e estabilidade de comportamento de vinte genótipos de milho (Zea mays L.) em dez ambientes do estado de Minas Gerais. 1995. 90 p. Dissertação (Mestrado em Genética e Melhoramento) - Universidade Federal de Viçosa. Viçosa, MG.
WEATHERSPOON, J. H. Comparative yields of single, three-way, and doublé crosses of maize. Crop Science, Madison, v. 10, n. 2, p. 157-159. Mar./Apr. 1970.
WELCKER, C.; THE, C.; ANDREAU, B.; DE LEON, C.; PARENTONI, S. N.; BERNAL, J.; FÉLICITÉ, J.; ZONKENG, C.; SALAZAR, F.; NARRO, L.; CHARCOSSET, A.; HORST, W. J. Heterosis and combining ability for maize adaptation to tropical acid soils: implications for future breeding strategies. Crop Science, Madison, v. 45, n. 6, p. 2405-2413, Nov./Dec. 2005.
WRICKE, G. Zur berechning der okovalenz bei sommerweizen und hafer. Zertschrift fur Pflanzenzuchtung, Hamburg, v. 52, n. 2, p. 127-138, 1965.

52
WRICKE, G.; WEBER, W. E. Quantitative genetics and selection in plant breeding. New York: W. de Gruyter, 1986. 406 p.
ZHANG, Y.; KANG, M. S.; LAMKEY, K. R. Diallel-SAS05: a comprehensive program for griffing’s and gardner-eberhart analyses. Agronomy Journal, Madison, v. 97, p. 1097-1106, 2005.

53
ANEXOS
TABELA 1A. Resumos das análises individuais, por ambientes, para a
produtividade de espigas despalhadas (kg/ha) obtidas na
avaliação de dez híbridos simples (HS) e 45 híbridos duplos
(HD) de milho, no ano agrícola de 2005/2006.......................54
TABELA 2A. Resumos das análises individuais, para a produtividade de
espigas despalhadas (kg/ha) com decomposição do efeito de
híbridos na capacidade geral (CGC) e específica (CEC) de
combinação, obtidas na avaliação do cruzamento dialélico
entre dez genitores de milho, no ano agrícola de
2005/2006...............................................................................57
ROTINA 1. Rotina utilizada para a obtenção da estimativa da ecovalência
(Wi), de Wricke (1965), utilizando estatística não-
paramétrica, com o auxílio do aplicativo computacional
R.............................................................................................58
ROTINA 2. Rotina utilizada para a obtenção dos parâmetros de
adaptabilidade e estabilidade de Lin & Binns (1988b),
utilizando estatística não-paramétrica, com o auxílio do
aplicativo computacional R....................................................63

54
TABELA 1A. Resumos das análises individuais, por ambientes, para a produtividade de espigas despalhadas (kg/ha) obtidas na avaliação de dez híbridos simples (HS) e 45 híbridos duplos (HD) de milho, no ano agrícola de 2005/2006.
AMBIENTES F.V. 1 2 3 4 5 HÍBRIDOS 10676229** 2384802** 924703** 1247119** 3074601** HS 4440473ns 3660722** 1292575** 676938ns 4398337** HD 10614596** 2170705** 860476** 947837* 2870591* HS vs HD 69509889** 321759ns 439859ns 19547130** 137412ns ERRO 2333345 699880 296402 416969 1692794 CV (%) 14,1 13,5 12,0 12,5 20,8 R2 (%) 69,7 63,0 61,8 60,1 49,1 MÉDIA (kg/ha) 10803 6212 4549 5152 6246 MÉDIA HS (kg/ha) 12180 6306 4440 5882 6185 MÉDIA HD (kg/ha) 10497 6191 4573 4990 6260 “...continua...”
54

55
“TABELA 1A, Cont.”
AMBIENTES F.V. 6 7 8 9 10 HÍBRIDOS 4337997** 6218107** 3014583** 2489157* 4671759** HS 4504569** 5433782** 5866474** 1772356ns 8835568** HD 4007224** 6392960** 2334399** 2360923ns 3037492ns HS vs HD 17392843** 5583476ns 7275678** 14582653** 39105204** ERRO 835100 1791855 912827 1606169 2243216 CV (%) 11,3 10,1 10,7 14,6 17,1 R2 (%) 72,7 64,3 65,0 44,2 51,3 MÉDIA (kg/ha) 8085 13192 8896 8685 8737 MÉDIA HS (kg/ha) 8773 13582 9341 9316 9770 MÉDIA HD (kg/ha) 7932 13105 8797 8545 8508 “...continua...”
55

56
“TABELA 1A, Cont.”
AMBIENTES F.V. 11 12 13 14 15 HÍBRIDOS 1860229* 4205038** 2745087** 3372170** 2877687ns HS 1221894ns 7402960** 4989321** 4459572* 1922185ns HD 1963835* 2964374** 2260901** 3109595* 3110114ns HS vs HD 3046575ns 30012939** 3851158* 5138814ns 1250424ns ERRO 1136266 871661 879601 1904152 2159436 CV (%) 10,8 12,0 12,8 17,2 16,0 R2 (%) 51,0 75,0 61,3 48,7 61,5
MÉDIA (kg/ha) 9876 7768 7353 8041 9166 MÉDIA HS (kg/ha) 10164 8672 7677 8415 8940 MÉDIA HD (kg/ha) 9812 7567 7281 7958 9217 * , ** Significativo, a 5% e 1% de probabilidade, pelo teste de F. ns Não significativo
56

57
TABELA 2A. Resumos das análises individuais para a produtividade de espigas despalhadas (kg/ha) com decomposição do efeito de híbridos na capacidade geral (CGC) e específica (CEC) de combinação, obtidas na avaliação do cruzamento dialélico entre dez genitores de milho, no ano agrícola de 2005/2006.
AMBIENTES F.V. 1 2 3 4 5 HÍBRIDOS 10676229** 2384802** 924703** 1247119** 3074601** CGC 19519047** 7065954** 2754199** 877389* 3391938* CEC 8907665** 1448571** 558804** 1321065** 3011133** ERRO 2333345 699880 296402 416969 1692794
6 7 8 9 10 HÍBRIDOS 4337997** 6218107** 3014583** 2489157* 4671759** CGC 13565878** 16588791** 6051963** 760723.5ns 8622292** CEC 2492420** 4143970** 2407107** 2834844** 3881652* ERRO 835100 1791855 912827 1606169 2243216
11 12 13 14 15 HÍBRIDOS 1860229* 4205038** 2745087** 3372170** 2877687ns CGC 4529294** 11848405** 6227269** 5164352** 4284991ns CEC 1326416ns 2676364** 2048651** 3013733* 2596226ns ERRO 1136266 871661 879601 1904152 2159436 * , ** Significativo, a 5% e 1% de probabilidade, pelo teste de F. ns Não significativo
57

58
ROTINA 1. Rotina utilizada para a obtenção da estimativa da ecovalência (Wi), de Wricke (1965), utilizando estatística não-paramétrica, com o auxílio do aplicativo computacional R.
# programa para obtenção do Wi não-paramétrico de Wricke (1965) # (Ecovalência) # será utilizado um exemplo com 55 genótipos de milho e 15 ambientes # Leitura dos dados dados = read.table("milhofim.txt",header=TRUE) dados = as.matrix(dados) p = 55; q = 15; # transformação em postos Rij <- rank(dados[,3]) # Acrescentando a coluna de postos dados = cbind(dados,Rij) # transformando a matriz de dados em uma matriz G # genotipos nas linhas e ambientes nas colunas GRij = function(X,g,a,col) { cont = 1 G=matrix(0,g,a) for (i in 1:a) { for (j in 1:g) { G[X[cont,2],X[cont,1]] = X[cont,col] cont = cont + 1 } } G } # função que transforma em postos os dados dos genótipos (1 a p) # em cada ambiente e substitui o conjunto de dados na 4 col pelos Rij's y1_a_p_em_dij = function(dad,g,a) { G = GRij(dad,g,a,3)

59
for (j in 1:a) { G[,j] = rank(G[,j]) } GA = matrix(0,1,g*a) cont = 1 for (i in 1:a) { for (j in 1:g) { GA[cont] = G[dad[cont,2],dad[cont,1]] cont = cont + 1 } } dad[,4] = GA dad } # função das médias genótipos, ambientes, geral e máximo em cada # ambiente(Mj)-entrar com matriz de postos p x q (G), p=g, q=a Gmedia = function(G,g,a) { # médias dos ambientes um = matrix(1,g,1) ambbar = (t(G) %*% um)/g # médias dos genótipos um = matrix(1,a,1) genbar = (G %*% um)/a # média geral xbargeral = mean(G) # Máximo Mj Mj = matrix(0,a) for (i in 1:a) { Mj[i] = max(G[,i]) } # QMGA

60
QMGA = 0 for (i in 1:g) { QMGA = QMGA + sum((G[i,]- ambbar - matrix(genbar[i],a)+ matrix(xbargeral,a))^2) } QMGA = QMGA/((a-1)*(g-1)) list(Rip=genbar, Rpj=ambbar,Mj = Mj, Rpp=xbargeral,QMGA = QMGA) } # Transformar os Yij em dij (efeitos de interação) e retorna conjunto de dados # com a 4 coluna com os efeitos dos postos da interação - deve ser chamado # como alternativa a primeira geração de Postos yij_em_dij = function(dad,g,a) { G = GRij(dad,g,a,3) GAsum = Gmedia(G,g,a) GA = matrix(0,g,a) for (i in 1:a) { for (j in 1:g) { GA[j,i] = G[j,i]-GAsum$Rpj[i]-GAsum$Rip[j]+GAsum$Rpp } } G = matrix(0,1,g*a) cont = 1; for (i in 1:a) { for (j in 1:g) { G[cont] = GA[dad[cont,2],dad[cont,1]] cont = cont + 1 } } Rij = rank(G) dad[,4] = Rij dad }

61
# Transformar os Yij em dij (dij=Yij-Yip+Ypp) e retorna conjunto de # dados com a 4 coluna com os efeitos dos postos da interação - deve # ser chamado como alternativa na geração de postos (5a opção) yijhuhn_em_dij = function(dad,g,a) { G = GRij(dad,g,a,3) GAsum = Gmedia(G,g,a) GA = matrix(0,g,a) for (i in 1:a) { for (j in 1:g) { GA[j,i] = G[j,i]-GAsum$Rip[j]+GAsum$Rpp } } for (j in 1:a) { GA[,j] = rank(GA[,j]) } G = matrix(0,1,g*a) cont = 1 for (i in 1:a) { for (j in 1:g) { G[cont] = GA[dad[cont,2],dad[cont,1]] cont = cont + 1 } } dad[,4] = GA dad } # função para obter a ecovalência Wi a partir dos postos # para o i-ésimo genótipo retorna uma matriz com: # 1 col: Rank médio, col 2: Desvio padrão Rij, col 3: Wi # op = 1: utilizar dados originais, op = 2: postos de todos os Yij, # op = 3: postos das interações, op = 4: postos dos dados por ambiente # op = 5: tranforma dados em dij=Yij-Yip+Ypp (Huhn) e os postos/ambiente

62
Wi_ecov = function(dad,g,a,op) { if (op == 3) dad = yij_em_dij(dad,g,a) if (op == 4) dad = y1_a_p_em_dij(dad,g,a) if (op == 5) dad = yijhuhn_em_dij(dad,g,a) if (op == 1) { G = GRij(dad,g,a,3) } else G = GRij(dad,g,a,4) IR = matrix(0,g,3) # médias dos genótipos um = matrix(1,a,1) IR[,1] = (G %*% um)/a # desvio padrão IR[,2] = sqrt(diag(var(t(G)))) # médias dos ambientes um = matrix(1,g,1) Mj = (t(G) %*% um)/g # Ecovalência MG = mean(G) for (i in 1:g) { IR[i,3] = sum((G[i,] - IR[i,1]-Mj+MG)^2) } IR } # op = 1: utilizar dados originais, op = 2: postos de todos os Yij, # op = 3: postos das interações, op = 4: postos dos dados por ambiente # op = 5: tranforma dados em dij=Yij-Yip+Ypp (Huhn) e os postos/ambiente op = 4 Wi_ecov(dados,p,q,op)

63
ROTINA 2. Rotina utilizada para obtenção dos parâmetros de adaptabilidade e estabilidade de Lin & Binns (1988b), utilizando estatística não-paramétrica, com o auxílio do aplicativo computacional R.
# programa para obtenção do Pi não-paramétrico de Lin e Binns (1988b) # cujos testes são efetuados via bootstrap ou via teste da permutação # será utilizado um exemplo com 55 genótipos de milho e 15 ambientes # Leitura dos dados dados = read.table("milhofim.txt",header=TRUE) dados = as.matrix(dados) p = 55; q = 15; # transformação em postos Rij <- rank(dados[,3]) # Acrescentando a coluna de postos dados = cbind(dados,Rij) # transformando a matriz de dados em uma matriz G # genotipos nas linhas e ambientes nas colunas GRij = function(X,g,a,col) { cont = 1 G=matrix(0,g,a) for (i in 1:a) { for (j in 1:g) { G[X[cont,2],X[cont,1]] = X[cont,col] cont = cont + 1 } } G } # função das médias genótipos, ambientes, geral e máximo em cada ambiente(Mj) # entrar com matriz de postos p x q (G), p=g, q=a Gmedia = function(G,g,a)

64
{ # médias dos ambientes um = matrix(1,g,1) ambbar = (t(G) %*% um)/g # médias dos genótipos um = matrix(1,a,1) genbar = (G %*% um)/a # média geral xbargeral = mean(G) # Máximo Mj Mj = matrix(0,a) for (i in 1:a) { Mj[i] = max(G[,i]) } # QMGA QMGA = 0 for (i in 1:g) { QMGA = QMGA + sum((G[i,]- ambbar - matrix(genbar[i],a)+ matrix(xbargeral,a))^2) } QMGA = QMGA/((a-1)*(g-1)) list(Rip=genbar, Rpj=ambbar,Mj = Mj, Rpp=xbargeral,QMGA = QMGA) } # Transformar os Yij em dij (efeitos de interação) e retorna conjunto de dados # com a 4 coluna com os efeitos dos postos da interação - deve ser chamado # como alternativa a primeira geração de Postos yij_em_dij = function(dad,g,a) { G = GRij(dad,g,a,3) GAsum = Gmedia(G,g,a) GA = matrix(0,g,a) for (i in 1:a) {

65
for (j in 1:g) { GA[j,i] = G[j,i]-GAsum$Rpj[i]-GAsum$Rip[j]+GAsum$Rpp } } G = matrix(0,1,g*a) cont = 1; for (i in 1:a) { for (j in 1:g) { G[cont] = GA[dad[cont,2],dad[cont,1]] cont = cont +1 } } Rij = rank(G) dad[,4] = Rij dad } # função para obter os Pi's e sua decomposição em efeito genético # e contribuição para a interação. Os resultados serão armazenados em # um matriz G.result com: col 1: Pi, col 2: efeito genético, col 3: contr # para interação, col 4: % explicação, col 5: estatística F e col 6: valor-p GPi = function(G,g,a,Gmedia) { G.Result = matrix(0,g,6) for (i in 1:g) { G.Result[i,1] = sum((G[i,]-Gmedia$Mj)^2)/(2*a) G.Result[i,2] = (Gmedia$Rip[i]- mean(Gmedia$Mj))^2/2 G.Result[i,3] = sum((G[i,]- Gmedia$Mj-matrix(Gmedia$Rip[i],a)+ matrix(mean(Gmedia$Mj),a))^2)/(2*a) } G.Result[,4] = G.Result[,3]/(sum(G.Result[,3]))*100 G.Result[,5] = G.Result[,1]/Gmedia$QMGA G.Result } # função para permutar a matriz dos postos

66
Gpermut = function(G,g,a) { np = g*a Gp = G while (np >= 2) { ip = trunc(runif(1)*np)+1 # gera numero aleatório entre 1 e np # desmonta o índice do número gerado em ip e ia ig = ip %/% a ia = ip - ig*a if (ia!=0) ig = ig + 1 if (ia==0) ia = a # desmonta o índice do maior elemento np de Gp em ipp e iaa igg = np %/% a iaa = np - igg*a if (iaa!=0) igg = igg + 1 if (iaa==0) iaa = a # permuta o elemento Gp[ig,ia] com Gp[igg,iaa] aux = Gp[igg,iaa]; Gp[igg,iaa] = Gp[ig,ia] Gp[ig,ia] = aux # np é atualizado para np-1 np = np - 1 } Gp } # função para reamostrar (bootstrap) a matriz dos postos Gboot = function(G,g,a) { Gb = matrix(0,g,a) for (ii in 1:g) { for (jj in 1:a) { ig = trunc(runif(1)*g)+1 # gera numero aleatório entre 1 e g ia = trunc(runif(1)*a)+1 # gera numero aleatório entre 1 e a Gb[ii,jj] = G[ig,ia] } } Gb

67
} # Fazer análise de variancia, obter Fi = Pi/QMGA, fazer permutação e # confrontar Pi com Pib, para obter p-valores H0: Pi != Máximo # op = 1: utilizar dados originais, op = 2: postos de todos os Yij, # op = 3: postos das interações Pi.permut = function(dad,g,a,op,nsim) { if (op == 3) dad = yij_em_dij(dad,g,a) if (op == 1) { G = GRij(dad,g,a,3) } else G = GRij(dad,g,a,4) GA.sumario = Gmedia(G,g,a) Pi = GPi(G,g,a,GA.sumario) for (i in 1:nsim) { Gp = Gpermut(G,g,a) GA.sumariop = Gmedia(Gp,g,a) Pip = GPi(Gp,g,a,GA.sumariop) for (ii in 1:g) { if (abs(Pip[ii,5])>=abs(Pi[ii,5])) { Pi[ii,6] = Pi[ii,6] + 1/nsim; } } } Pi } # Fazer análise de variancia, obter Fi = Pi/QMGA, fazer bootstrap e # confrontar Pi com Pib, para obter p-valores H0: Pi != Máximo # op = 1: utilizar dados originais, op = 2: postos de todos os Yij, # op = 3: postos das interações Pi.boot = function(dad,g,a,op,nsim) { if (op == 3) dad=yij_em_dij(dad,g,a) if (op == 1) { G = GRij(dad,g,a,3) } else G = GRij(dad,g,a,4) GA.sumario = Gmedia(G,g,a) Pi = GPi(G,g,a,GA.sumario)

68
for (i in 1:nsim) { Gb = Gpermut(G,g,a) GA.sumariob = Gmedia(Gb,g,a) Pib = GPi(Gb,g,a,GA.sumariob) for (ii in 1:g) { if (abs(Pib[ii,5])>=abs(Pi[ii,5])) { Pi[ii,6] = Pi[ii,6] + 1/nsim; } } } Pi } # op = 1: dados, op = 2: postos dados, op = 3: postos interação # neste caso - não faz sentido fazer em cima de postos de interação # não usar op = 3 op = 2 nsim=10000 Pi = Pi.permut(dados,p,q,op,nsim) Pi Pi = Pi.boot(dados,p,q,op,nsim)