Embed Size (px)
Citation preview

1
Montes Claros
Minas Gerais – Brasil
Maio - 2016
UNIVERSIDADE ESTADUAL DE MONTES CLAROS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
BIOLÓGICAS - PPGCB
Letícia Fernanda Ramos Leite
Estrutura da comunidade de galhas em uma planta super-hospedeira:
testando a hipótese da competição em habitats xéricos

2
UNIVERSIDADE ESTADUAL DE MONTES CLAROS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
BIOLÓGICAS - PPGCB
Letícia Fernanda Ramos Leite
Estrutura da comunidade de galhas em uma planta super-hospedeira:
testando a hipótese da competição em habitats xéricos
Orientador: Dr. Marcílio Fagundes
Dissertação apresentada pela
mestranda, Letícia Fernanda
Ramos Leite, para obtenção do
título de Mestre, pelo Programa
de Pós-graduação em Ciências
Biológicas – PPGCB -
UNIMONTES

3
L533e
Leite, Letícia Fernanda Ramos.
Estrutura da comunidade de galhas em uma planta super-hospedeira
[manuscrito] : testando a hipótese da competição em habitats xéricos / Letícia
Fernanda Ramos Leite. – 2016.
38. : il.
Bibliografia: f. 33-38.
Dissertação (mestrado) - Universidade Estadual de Montes Claros - Unimontes,
Programa de Pós-Graduação em Ciências Biológicas/PPGCB, 2016.
Orientador: Prof. Dr. Marcílio Fagundes.
1. Índice de coocorrência - C-score. 2. Insetos galhadores. 3. Copaifera
langsdorffii - Espécie arbórea tropical. 4. Competição. I. Fagundes, Marcílio. II.
Universidade Estadual de Montes Claros. III. Título. IV. Título: Testando a
hipótese da competição em habitats xéricos.
Catalogação Biblioteca Central Professor Antônio Jorge
Catalogação Biblioteca Central Professor Antônio Jorge

4

5
Agradecimentos
A realização deste trabalho contou com a colaboração direta de algumas pessoas
que não posso deixar de agradecê-las:
Primeiramente ao programa de mestrado PPGCB e a Unimontes pela
infraestrutura e por liberar carro e motorista para as minhas coletas. A Capes pela
concessão de bolsa. E colegas do mestrado pelas conversas, troca de ideias e resenhas
bacanas, principalmente a Mariana, Bráulio e Maiara.
Ao meu orientador Marcílio Fagundes, que mostrou confiança ao propor a ideia
do projeto e sempre dispôs a me ajudar. Soube chamar a atenção quando preciso, apoiou
nos momentos de desespero e neste tempo de convívio tornou um bom amigo, saiba que
você é à base do meu conhecimento.
Ao nosso amigo “Pós-doc” por curto tempo, Ricardo Solar, por dar um Up nas
minhas perspectivas, por ter sido um ótimo professor com ideias claras e diferentes! E
principalmente, pela ajuda no delineamento do projeto, ajudando a ordenas as ideias e a
sanar as dúvidas, valeu demais RR Solar.
Aos amigos professos do programa que pelos bons momentos de discussão e
pelo empenho em tirar dúvidas e ajudar. Ao Ronaldo, pela paciência com o R, ao
Maurício por sempre colocar grilos na minha cabeça, principalmente ao falar de
competição. Aos demais pelos bons momentos de diversão nas festas da bio e
principalmente nos botecos, risada sempre garantida!
A galera do LBC, que mesmo barulhando demais são bacanas. Agradeço pela
disponibilidade em ajudar, principalmente ao Henrique (Peppa), a Rita e a Lara por irem
às coletas e ajudar nas infinitas triagens, ralamos muito, mais foi gratificante. A Ani por

6
ajudar na triagem também (Santo ImageJ!!!) e Siqueira por dar uma ajuda inicial na id
das galhas e pelas discussões ecológicas. A Karen, Pri, Renatinha, e Jú por torcerem pra
dar certo! Ao Serapas pelos momentos de discussão, ajuda nas coletas, ajuda no
trabalho e por sempre mandar artigos bacanas. Obrigada galera, vocês brilharam!!
Aos amigos mestres e os aspiras a mestre dessa biologia, Kátia, Mary, Paulinho,
Gui, Igão, Cléo, Karenzinha por sempre garantir o riso, pelas palhaçadas sem fim, pelas
festinhas e fugidas pros botecos, e claro, pelas discussões produtivas também. Ao
Lucketinho que mesmo resmungando me ajudava com o R e sempre tinha um palpite
bacana sobre o meu trabalho. Ao amigo Arleu, que mesmo longe, eu uma força na
plotagem das minhas figuras, obrigada.
A Huguinho, que me deu uma força inicial no EcoSim, pelas longas discursões
ecológicas, pelas críticas e sugestões, e mais ainda, pelo apoio emocional nos momentos
difíceis que passei (...) não tenho como te agradecer, você me ajudou a ter forças
quando o desespero era forte demais, obrigada gato.
Aos meus pais, pela oportunidade e incentivo, sei que estão orgulhosos. Aos
meus irmãos por serem companheiro, Mickael pela “cobertura” (rsrsr) e paciência,
Robson pelo incentivo. As cunhadas Suzy e Letícia pela amizade e apoio e por me
darem esses sobrinhos lindos que me enche de orgulho e felicidade, é muito gostoso um
abraço e um beijo de uma criança, é uma força pra ir mais longe.
Em especial a minha mãe, que teve um problema sério de saúde e eu a
acompanhava nas internações e consultas, e via de perto o sofrimento, mas a sua força
sempre me admirava, eu via seu esforço pra não demostrar o quanto estava sofrendo,
que mesmo sentindo dor, com a respiração ofegante sempre me sorria e falava “pode ir

7
fazer a suas coisas, não se preocupe comigo, não quero te atrapalhar”. Eu a admiro
muito! Muito obrigada por me apoiar e me encorajar.
Resumo
As forças bottom-up, top-down e as interações dentro do mesmo nível trófico
determinam a organização das comunidades de galhas nos diferentes ambientes. No
entanto, fatores de estresse (elevada radiação, estresse hídrico e menor disponibilidade
de recurso) são barreiras para sobrevivência desses indivíduos, favorecendo uma
elevada abundância de galhas nos ambientes xéricos. Nos ambientes com elevada
abundância de indivíduos e o recurso (sítios para oviposição) é limitado, é possível que
exista uma força competitiva capaz de atuar nestas comunidades. Neste sentido, estamos
propondo que a competição interespecífica é uma força atuante na estruturação da
comunidade de galhas em ambientes xéricos. Foram avaliadas sete populações de C.
langsdorffii com características peculiaridades que formam um gradiente de estresse.
Em cada população foram amostradas características vegetativas das plantas
(indicadoras de estresse ambiental) e avaliado a coexistência das galhas dentro de um
mesmo folíolo. O índice de coocorrência (C-score) observado foi comparado com o
modelo nulo para aceite ou rejeição da hipótese biológica, sendo uma análise para cada
população. Os valores de C-score observados em todos os habitats foram comparados
com os parâmetros vegetativos indicadores de estresse. As galhas encontradas nos
ambientes úmidos coocorrem livremente dentro do folíolo, já nos ambientes xéricos as
galhas coocorreram menos do que o esperado pelo acaso, sustentando a nossa hipótese.
Desta forma, assumimos que a competição é uma força estruturadora das comunidades
de galhas nos ambientes xéricos.

8
Palavras-chave: Coocorrência; C-score; Insetos galhadores; Copaifera langsdorffii;
Competição.
Introdução
A organização das comunidades dos insetos galhadores pode ser moldada por
fatores ambientais (Blanche 2000, Price et al. 2002, Cuevas-Reyes et al. 2006, Craig et
al. 2007, Fagundes & Fernandes 2011, Butterrill e Novotny 2015), forças top-down
(Price et al. 1997, Fagundes et al. 2005), bottom-up (Hunter e Price 1992, Espírito-
Santo 2007, Egan & Ott 2007, Malinga et at 2014) e pelas interações que ocorrem
dentro do mesmo nível trófico (Fagundes & Fernandes 2011, Cornelissen et al. 2013). A
hipótese do estresse ambiental (Fernandes & Price 1988) sugere que a ação dos
parasitas e predadores (terceiro nível trófico) é mais efetiva na regulação da diversidade
de galhas em habitats mésicos. Diversos estudos conduzidos em habitats tropicais (Lara
et al. 2002, Fernandes et al. 2005, Jesus et al. 2012, Julião et al. 2014) e temperados
(Fernandes & Price 1988, Fernandes & Price 1992, Fernandes et al. 2003) corroboram
esta hipótese. Além disto, plantas que ocorrem em habitat ricos em nutrientes
apresentam maior capacidade para manifestar respostas induzidas ao ataque de
galhadores (Fernandes 1990, Fernandes e Negreiros 2001, Höglund et al. 2005).
Portanto, plantas que ocorrem em ambientes estressados devem apresentar maior
abundância de insetos galhadores (Price et al. 1987, Fernandes e Price 1992). Neste
cenário, onde forças top-down são mais efetivas em habitats mésicos e as populações de
galhadores alcançam maior densidade em habitat xéricos, seria razoável admitir que
outras interações que ocorrem dentro do mesmo nível trófico (e.g. competição) são mais
Catalogação Biblioteca Central Professor Antônio Jorge
Catalogação Biblioteca Central Professor Antônio Jorge

9
importantes para a organização das comunidades de insetos galhadores em habitats
xéricos do que em habitas mésicos.
Características ambientais adversas como extremo de temperatura, baixa
disponibilidade de agua, luminosidade, fertilidade do solo são fatores de estresse que
afetam diretamente o desenvolvimento das plantas (Pennington & Collins 2007,
Sapijanskas 2014; Lázaro-Nogal et al. 2015). Usualmente estes fatores atuam
sinergicamente no ambiente, desta forma, torna-se mais fácil avaliar a resposta da planta
aos fatores de estresse. Estratégias adaptativas como ajustes morfológicos e fisiológicos
nas plantas foram os fatores chaves que determinam colonização e sobrevivência deste
grupo nos diferentes ambiente (Sultan 2001). Variações nas características das folhas,
no crescimento dos ramos, têm sido correlacionadas com a eficiência de uso dos
recursos e com a plasticidade fenotípica das plantas em resposta às variações abióticas
(Chaturvedi 2014, Lázaro-Nogal et al. 2015). Neste sentido, espécies amplamente
distribuídas devem sofrer variação no seu desenvolvimento e estender este efeito para os
níveis tróficos superiores.
A teoria da competição prediz que a disputa por recursos entre dois organismos
é mais intensa quando a densidade, a similaridade ecológica e a coocorrência espaço/
temporal aumentam (Diamond 1975, Stone e Roberts 1990). As espécies competidoras
raramente conseguem coexistir no mesmo recurso, a menos que haja uma diferenciação
mínima no habitat, o que culmina na redução da fecundidade, sobrevivência e
crescimento dos organismos envolvidos (Begon 2005). Algumas características da
interação insetos galhadores e sua planta hospedeira em ambientes xéricos poderiam
eleger a competição como força organizadora da comunidade destes herbívoros nestes
ambientes. Por exemplo, os galhadores são altamente específicos com a sua planta

10
hospedeira e com o órgão alvo que ovipositam (Joy e Crespi 2007, Carneiro et al.
2009). As fêmeas dos galhadores apresentam elevada sincronia com o surgimento dos
órgãos alvo da planta que são limitados no tempo (Yukawa 2000). Minutos após a
oviposição a larva do galhador induz rápidas mudanças morfológicas e fisiológicas no
tecido da planta drenando o recurso para o seu desenvolvimento (Ollerstam et al. 2002,
Höglund 2014), que potencialmente interfere no comportamento de outros herbívoros
(Cornelissen et al. 2013).
A prevalência da competição como força organizadora das comunidades pode
ser testada através de experimentos de exclusão (Diamond 1986, Connell 1961), ou
testando a hipótese biológica com modelos nulos (Gotelli & Entsminger 2001). Modelos
nulos são modelos de comunidades que possuem características das suas equivalentes
reais, mas mantém a distribuição das espécies ao acaso, especificamente excluindo os
efeitos das interações biológicas (Gotelli & Graves 1996). Estes modelos avaliam se as
espécies de uma comunidade natural coocorrem significativamente mais ou menos que
o esperado pelo acaso (Gotelli & Graves 1996). Exemplo de uso de modelos nulos para
testar a prevalência da competição pode ser observados para plantas (Sanders et al.
2003), comunidade de artrópodes (Ellwood et al. 2009), minadores de folhas
(Cornelissen & Stiling 2008), formigas (Ribas e Schoereder 2002) liquens (Browker &
Maestre 2012) e insetos galhadores (Cornelissem et al. 2013).
Plantas super-hospedeiras (plantas que possuem elevada diversidade de galhas)
de insetos galhadores, que apresentam ampla distribuição geográfica e ocorrem em
diferentes habitats, constituem sistemas adequados para avaliar os mecanismos e
processos que regulam a organização da comunidade de galhas entre diferentes
ambientes (Fagundes 2014). Copaifera langsdorffii (Fabaceae) é uma espécie

11
amplamente distribuída nas diversas formações vegetais do Brasil e comporta-se como
uma super-hospedeira de insetos galhadores (Fagundes 2014). Assim, este sistema é um
bom modelo para avaliar os efeitos bottom-up na comunidade de insetos galhadores.
Usando este sistema, estamos propondo a hipótese da Competição em Habitat Xéricos,
com a predição que a competição interespecífica é uma força estruturadora da
comunidade de insetos galhadores em habitat xéricos melhor do que em mésicos.
Materiais e Métodos
1- Sistema de estudo
Copaifera langsdorffii Desf. (Fabaceae: Caesalpinioideae), é uma espécie arbórea
tropical que varia de 8 a 25 m de altura, apresenta ampla distribuição geográfica,
ocorrendo na Argentina, Bolívia e em todo o Cerrado brasileiro (Carvalho 2003). A
variabilidade do tamanho das sementes e a alta plasticidade fenotípica favorecem a
vasta distribuição geográfica desta espécie (Souza e Fagundes 2014). Esta planta
apresenta deciduidade marcante nos meses de julho a agosto, com emissão de folhas
logo em seguida (Souza e Fagundes 2015). Foram descritos 23 morfoespécies de galhas
em C. langsdorffii (Costa et al. 2010), o que leva a classificação desta espécies como
um super-hospedeiro de insetos galhadores (Fagundes 2014). A ampla distribuição
geográfica, a assincronia fenológica intra e inter-anual e a arquitetura da planta são
fatores que favorecem diversificação dos insetos galhadores nesta espécie (Costa et al.
2011, Fagundes 2014)
2- Áreas de estudo

12
Este estudo foi desenvolvido em sete populações de C. lagsdorffii localizadas em
diferentes formações vegetais (Canga, Campo Rupestre, Cerrado sentido restrito,
Cerradão, Mata seca, Mata Atlântica e Mata ciliar (Fig. 1 Tab. 1). Estas formações
foram escolhidas por simular um gradiente de variação de características ambientais do
habitat. Por exemplo, a Canga é caracterizada pelos solos ferruginosos que formam uma
carapaça dificultando a infiltração da água e a penetração das raízes. As plantas são
esclerófilas adaptadas às variações bruscas de temperaturas, ventos fortes, e solos
pobres em nutrientes (Giulietti et al. 1987, Oliveira-Filho & Ratter 2002). O Campo
rupestre é uma formação vegetal com predomínio de espécies arbóreo-arbustivo que
crescem entre os afloramentos rochosos. Possui vegetação esclerofilia adaptada a
condições extremas tais como elevada radiação, variações diária de temperatura, solos
rasos, arenosos com baixa umidade e nutricionalmente pobres (Giulietti et al. 1987).
O Cerrado sentido restrito apresenta solos ácidos, pobres em nutrientes. Esta região
possui clima semiárido com estações secas e chuvosas bem definidas, a vegetação
lenhosa apresenta folhas coriáceas como adaptação às condições de seca (Rizzini 1997,
Oliveira 1998). O cerradão é uma formação vegetal que apresenta dossel contínuo,
principalmente na estação chuvosa, a vegetação possui porte elevado com
características esclerófilas e xeromórfica. Os solos são profundos, ligeiramente ácidos
com médio teor de matéria orgânica proveniente da queda das folhas na estação seca
(Rizzini 1997, Sano 2008).
As Florestas estacionais deciduais (matas secas) apresentam vegetação com porte
elevado (que podem ultrapassar 25 metros) e a abscisão foliar na estação seca é a
principal característica deste ambiente. Os solos são alcalinos, ricos em nutrientes e a
queda das folhas contribui para fertilidade dos solos (Sano 2008, Fagundes et al. 2011).
A Mata atlântica apresenta vegetação com porte elevado formando um dossel contínuo

13
com elevada umidade. Os solos são úmidos, rasos e pobres em nutrientes minerais,
contudo, a decomposição da matéria orgânica é um importante fator na disponibilidade
de nutrientes para as plantas (Sano 2008). A Mata Ciliar é composta pela vegetação que
forma corredores acompanhando o leito do rio, apresenta variação na sua estrutura e
composição florística, elevada umidade relativa com solos férteis muito visados para
agricultura (Oliveira-Filho & Ratter 2002).
Provavelmente, variações na disponibilidade hídrica, qualidade dos solos e
topografia são alguns fatores que determinam as variações fisionômicas nas diferentes
formações vegetais (Oliveira-Filho & Ratter 2002). Portanto, podemos traçar um
gradiente de estresse nestes ambientes, partindo do ambiente mais xérico como Canga,
Campo rupestre e Cerrado sentido restrito, Cerradão, Mata secas a ambientes mais
úmidos como, Mata Atlântica e Mata ciliar.
3- Coleta de dados
Nos meses de abril e maio de 2015, período que antecede a queda das folhas e que
todas as galhas já haviam estabelecido nas plantas, foram amostradas galhas em 15
plantas adultas de C. langsdorffii de cada população. As plantas selecionadas
apresentam distribuição irregular no ambiente, no entanto, mantivemos uma distância
mínima de 10 m entre cada amostragem para garantir independência das amostras. Em
cada indivíduo foram coletados 10 ramos terminais de aproximadamente 30 cm de
comprimento para avaliar os parâmetros biológicos (tamanho dos ramos, massa foliar e
área foliar). Os ramos foram coletados ao entorno da copa para evitar os efeitos do
micro-habitat no desenvolvimento dos ramos e na comunidade de galhas (Costa et al.
2010). Em cada ramo foi medido o último investimento vegetativo (corresponde ao
último investimento em crescimento anual que pode ser identificado através da cicatriz

14
visível no ramo), a massa e a área dos folíolos (esclerofilia) e quantificado as galhas dos
folíolos.
Para avaliar os parâmetros vegetativos (crescimento do ramo, massa foliar e área
foliar) e o ataque de insetos galhadores foram analisadas as três últimas folhas,
completamente expandidas e saudáveis, do ramo. O crescimento de cada ramo foi
determinado medindo a distância entre cada inserção das folhas no ramo. A massa foliar
específica foi medida utilizando discos foliares (0,38 cm²) do segundo par de folíolos
que foram secos em estufa a 40° C por 92 horas e pesados em balança analítica. Foram
coletados 20 folíolos por população para medir a área foliar, os folíolos foram
digitalizados e usado o software ImageJ (Maloof et al. 2013). Para quantificar as galhas
foi avaliado cada folíolo separadamente e criado uma matriz de presença e ausência de
galhas, padronizamos usar 137 folhas para cada população. As galhas foram
morfoespeciadas segundo a sua cor, textura e tamanho (Costa et al. 2011, Fagundes et
al. 2014).
4- Análises de dados
As diferenças dos parâmetros vegetativos (crescimento dos ramos, massa foliar
específica e área foliar) entre os diferentes habitats foram testadas usando Modelos
Lineares Generalizados (GLM) seguidos por Análise de Deviance (ANODEV). Assim,
os habitats foram usados como variáveis explicativas e os parâmetros vegetativos
usados como variáveis respostas. Os modelos foram testados utilizando o teste “F” com
distribuição de erros Gaussian (dados contínuos).
Para quantificar os efeitos dos habitats, dos parâmetros vegetativos (crescimento
de ramos, massa foliar e área foliar) na riqueza e abundância de galhas, foram criados

15
GLM’s seguido pela ANCOVA. Os parâmetros vegetativos, os ambientes e a interação
entre todas as variáveis foram usados como variáveis explicativas, a riqueza e
abundância foram usadas como variáveis respostas. Utilizamos distribuição de erros
Poisson (dados de contagem), com função de ligação logarítmica. Para eliminar o efeito
da ordem de entrada das variáveis no modelo foi realizada uma seleção de modelos por
Stepwise (KamilBartoń 2015). A seleção do modelo mais parcimonioso neste caso é
feita com base nos valores do critério de informação de Akaike (AICc). Finalmente, o
modelo mais parcimonioso foi submetido à análise de resíduos para verificar a
adequabilidade do modelo em relação aos seus requisitos (Crawley 2007).
Para avaliar se a composição da fauna de insetos galhadores variou entre as
populações analisadas, foi usado um teste de permutação Anosim. O índice de
similaridade de Jaccard, calculado com base na matriz de presença e ausência de
morfoespécies de galhas, foi usado nos testes acima.
Modelos nulos foram desenvolvidos para comparar os padrões observados e
aleatórios na ocorrência de galhas em cada população analisada. A hipótese nula prediz
que a presença de uma galha no folíolo não interfere na presença de outra no mesmo
folíolo. O índice de coocorrência é dado pela média dos possíveis pares de interações.
Se o índice de coocorrência da matriz original encontrar dentro de 95% da frequência de
distribuição da matriz randomizada, a hipótese nula é aceita, ou seja, não há evidência
de mecanismos biológicos agindo na estruturação desta comunidade. No entanto, se o
índice da matriz original estiver fora dos 95% da matriz simulada, a hipótese nula é
rejeitada e podemos inferir que existe uma força biológica moldando a ocorrência das
espécies nesta comunidade (Ribas & Schoereder 2002).
Obtivemos a matriz de análises de ocorrência de galhas usando o programa
EcoSim (Gotelli e Entsminger 2001). Este software testa o padrão de ocorrência dos

16
organismos randomizando a matriz original com o número pré-especificado de
aleatorizações. Usamos o índice de C-score (Stone e Roberts, 1990) cuja métrica de
quantificação dos padrões de coocorrência ou “checkerboard units (CU)” entre os
possíveis pares de galhas é dada pela fórmula: CU = (ri– S) (rj– S), onde S é o número
de espécies compartilhadas no folíolo e o ri e rj é o número total de espécies de galhas (i
e j). O C-score é a média de todos os possíveis pares de espécies que interagem pelo
menos uma vez na matriz. Usamos um algoritmo “fixo-fixo” com 5000 mil
randomizações onde as linhas e colunas da matriz original foram preservadas. Isto
significa que o número de ocorrência de cada espécie e o número de espécies no folíolo
dos modelos nulos é igual aos da matriz original (Gotelli & Entsminger 2001). As
análises foram conduzidas separadamente para cada ambiente analisado.
Para validarmos a hipótese que a competição atua nos ambientes xéricos, nós
relacionamos os resultados da coocorrência de galhas entre os habitats com os
parâmetros vegetativos das plantas (indicadores de estresse) de todos os habitats. Foi
construído GLM onde a média da massa foliar específica das plantas de cada ambiente
foi usada como variável explicativa e os índices da coocorrência de galhas (C-score)
usados como variável resposta. Construímos outro modelo (GLM) utilizando a média de
crescimento dos ramos das plantas de todos os habitats (variável explicativa) com os
valores de C-score (variável resposta). Utilizamos a distribuição de erros Gaussian
(dados contínuos) com Identidade sendo a função de ligação. Todos os modelos foram
submetidos à análise de resíduos para verificar sua adequabilidade a distribuição de
erros assumida (Crawley 2007). Todos os GLM’s, o ANOSIM e a seleção de modelos
(pacote MuMIn) foram realizados no software R (R Core Team, 2015).
Resultados

17
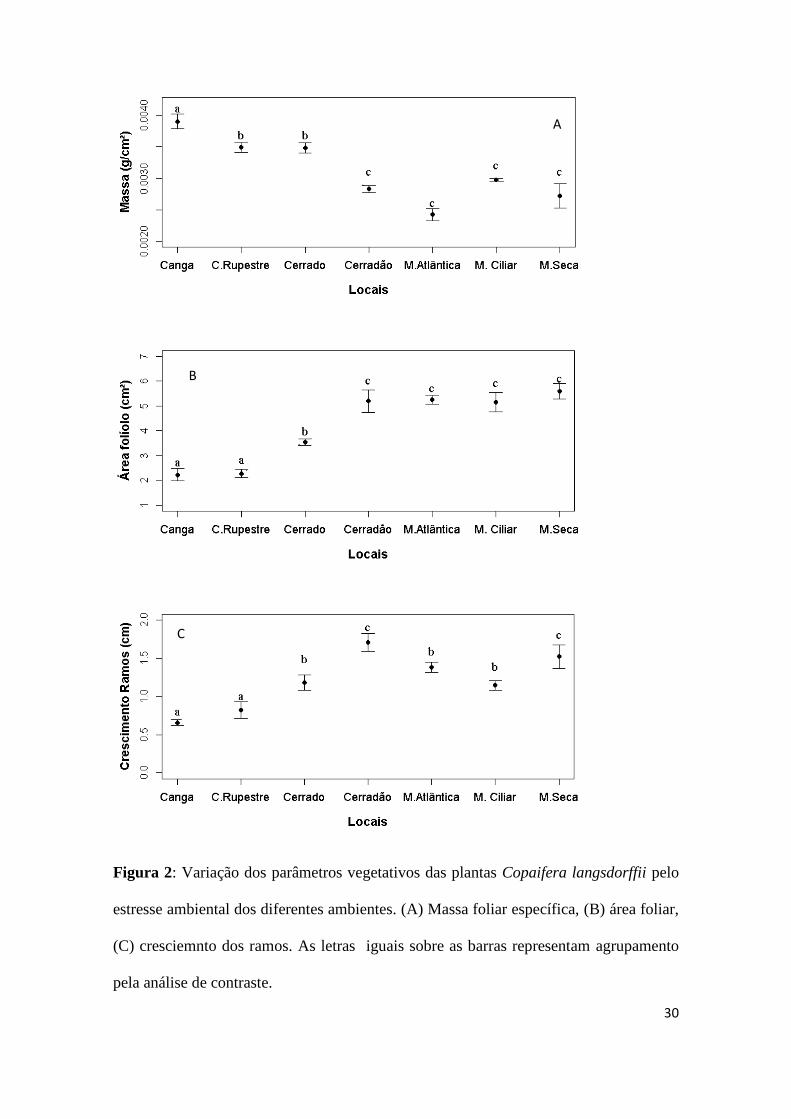
A massa foliar, o tamanho dos folíolos e o crescimento dos ramos variaram entre
os ambientes avaliados (Tab. 2). Nos ambientes xéricos (Canga, Campo Rupestre,
Cerrado sentido restrito) as plantas são mais esclerófilas (maior massa foliar e menor
área dos folíolos) e apresentam menor crescimento dos ramos quando comparado com
os ambientes menos estressados (Mata Seca, Cerradão, Mata Atlântica e Mata Ciliar)
(Fig 2A, 2B, 2C).
Foram amostradas 2550 galhas distribuídas em 25 morfoespécies nos diferentes
ambientes (Tab. 3). Os morfotipos M3, M13 e M21 foram os mais abundantes e
estavam presentes em quase todos os ambientes avaliados, estes morfotipos juntos
responderam por mais de 40% do total de galhas amostradas. Alguns morfotipos são
específicos dos ambientes secos, como por exemplo, o M23 e o M25 que foram
encontrados somente nas regiões de Canga e Campo rupestre.
Dentre as variáveis explicativas testadas, apenas a massa foliar teve influência
significativa na riqueza das galhas (Tab. 4). Há uma relação positiva entre a riqueza de
galhas e massa foliar, sugerindo que as morfoespécies de galhas atacam
preferencialmente plantas mais esclerófilas (Fig. 3). Entretanto, a abundância de galhas
foi afetada somente pelos ambientes avaliados (Tab. 4), sendo significativamente
elevada nos ambientes xéricos, como a Canga e o Campo rupestre (Fig. 4).
A composição da comunidade de galhas variou entre os ambientes analisados
(Tab. 6 Fig. 5). Somente a composição dos galhadores do Campo Rupestre e da Canga é
similar, mas diferem dos demais ambientes. A composição do Cerrado sentido restrito,
Mata Atlântica, Mata Ciliar, Mata Seca e do Cerradão diferiram entre si. Este resultado
sugere a formação de um grupo de galhas especializada a ambientes xéricos.

18
Os valores de C-score foram maiores que o esperado pelo acaso (Tab. 7 Fig. 5)
nas comunidades de Canga, Campo Rupestre e Cerrado sentido estrito, portanto, a
hipótese biológica foi aceita. No Cerradão, Mata seca, Mata atlântica e Mata ciliar a
coocorrência das espécies estavam dentro do limite de 95% da frequência de
distribuição das matrizes randomizadas (Tab. 7 Fig. 5), deste modo, a hipótese nula foi
aceita.
Os índices de C-score apresentaram correlação significativa com os parâmetros
vegetativos das plantas (massa foliar e crescimento dos ramos) (Fig. 6A e 6B Tab. 5).
As plantas que tiveram menor crescimento e maior massa foliar apresentaram valores
significativos de C-score (Vide Fig. 7 Tab. 7). De fato, a competição pode ser
importante para organização de galhas nas vegetações mais esclerófilas.
Discussão
Variações nas respostas plásticas das plantas determinam a gama de condições
em que as espécies podem sobreviver e reproduzir com sucesso (Sultan 2001). Estresse
ambiental inclui os fenômenos que reduz a produção fotossintética das plantas, por
exemplo, oferta limitada dos recursos e fatores ambientais (nutrientes, água, luz,
temperaturas extremas e elevada radiação) que estão diretamente ligadas à fotossíntese
(Brooker e Callaghan 1998). Variações ambientais causam alterações no crescimento,
área foliar e massa foliar das plantas (Chaturvedi 2014, Lázaro-Nogal et al. 2015). A
esclerofilia foliar (massa foliar específica) é uma resposta da planta aos fatores de
estresse, por exemplo, as folhas apresentam maiores tricomas, cutícula mais espessa,
maior densidade estomática e maior espessura (resultado do maior número de estratos
de células do parênquima paliçádico) (Wilians e Blackc 1993, Reich et al. 2003, Bussoti
et al. 2015). Assim, variações nas características foliares são interpretadas como

19
resposta a variação do ambiente e permitem que as plantas aumentem a sua capacidade
de distribuição. Neste estudo nós observamos que plantas presentes nos ambientes
xéricos apresentaram maior massa foliar específica, menor área foliar e menor
crescimento dos ramos quando comparado aos ambientes menos estressados. É
importante ressaltar que de fato existe um gradiente de estresse nos ambientes avaliados
e que este se manifesta através das diferenças estruturais encontra nas folhas nos
diferentes habitats. Estas variações nos parâmetros vegetativos de C. langsdorffii podem
causar alterações nas interações planta-galhadores e nos efeitos dos níveis tróficos
superiores (Egan & Ott 2007, Craig et al. 2007).
Diversos estudos mostram que a comunidade de galhadores varia em função do
estresse ambiental. Por exemplo, a ação de parasitas e patógenos é maior em habitats
mésicos, favorecendo o aumento da diversidade de galhas em habitats xéricos
(Fernandes e Price 1992, Price et al. 2002, Lara et al. 2002). Além disto, plantas
presentes em habitas xéricos geralmente possuem maior disponibilidade de aminoácidos
e compostos de nitrogênio orgânico livre e são mais nutritivas (White 1969, Price
1991). Estas plantas possuem menor capacidade para apresentar defesas induzidas que
são menos eficazes nos ambientes com temperaturas elevadas e com limitações de água
(Fernandes 1990, Fernandes et al. 2000, Fernandes e Negreiros 2001, Höglund et al.
2005), favorecendo o ataque de galhadores nos ambientes estressados (Barbosa e
Fernandes 2014).
Muitos fatores abióticos alteram a composição das espécies de galhas nos
ambientes, algumas espécies respondem positivamente ao estresse sendo classificadas
como bioindicadoras de estresse ambiental (Fernandes et al. 1995, Julião et al. 2005,
Toma et al. 2014). Nossos resultados mostram que composição das espécies de galhas

20
foi semelhante entre os habitats com maior estresse (Canga e Campo rupestre) e diferiu
entre os demais. Possivelmente há uma fauna de galhadores especializados em viver
nestes ambientes que possuem características semelhantes, por exemplo, elevada
radiação, solos com baixa retenção de água (Lara et al. 2002). De fato, as morfoespécies
M23 e M25 (Tab. 2) amostradas são exclusivas de ambientes estressados, estas
morfoespécies de galhas poderiam ser utilizadas como bioindicadores de estresse (Toma
et al. 2014). Flutuações nas características de umidade e nutrientes do solo podem
afetar o desempenho dos galhadores nos ambientes úmidos, desta forma, características
intrínsecas destes habitats podem selecionar algumas morfoespéceis específicas de
galhadores.
Variações da qualidade do habitat causam importantes modificações nas
comunidades de galhas e alteram os mecanismos de controle destas comunidades.
Estudos recentes tem mostrado que a competição entre insetos herbívoros parece ser
bem comum (Reitz & Trumble 2002, Kaplan & Denno 2007, Cornelissen et al. 2013) e
potencialmente atuam na organização das comunidades. Nossos resultados mostraram
que a há um padrão de organização das galhas nos habitas xéricos. Vários fatores
podem condicionar a distribuição não aleatória nestes ambientes, por exemplo, a
elevada abundância de galhas, a menor disponibilidade de recursos (espaço) e menor
ação dos inimigos naturais. Outro fator que opera na escala de folíolos e potencialmente
favorece um padrão de distribuição das espécies de galhas é a competição por
interferência de recurso. Organismos que possuem hábito de vida séssil são susceptíveis
a influências competitivas, pois, depois de estabelecidos, eles não podem escapar dos
vizinhos e os resultados desta interação resultam em seu padrão de distribuição. Por
exemplo, Sanders e colaboradores (2003) em um trabalho envolvendo plantas invasoras
mostraram que a competição por interferência também molda o padrão de distribuição

21
destas espécies (Sanders et al. 2003). Apesar dos insetos galhadores avaliados ocorrem
em uma pequena ilha de recursos (representado pelo folíolo) observamos que nos
ambientes que os folíolos eram menores (ambientes xéricos) as galhas ocorriam
geralmente uma única morfoespécies por folíolo. Resultados semelhantes foram
encontrados para outros sistemas (Connor e Simberloff 1983, Sanders et al. 2003,
Ellwood et al. 2009, Cornelissen et al. 2013) e a competição foi sugerida como fonte
dominante de organização.
O uso dos modelos nulos para detectar padrões de distribuição das espécies nas
comunidades podem prever a aleatoriedade (Boschilia et al. 2008), a agregação
(Krasnov et al. 2006) e a segregação das espécies (Connor & Simberloff 1983, Sanders
et al. 2003, Ellwood et al. 2009). Organizações não aleatórias de comunidades sugerem
que exista pelo menos um fator de estruturação, como interações biológicas específicas
atuando na distribuição dos organismos (Gotelli 2000, Boschilia et al. 2008). Os efeitos
da remoção de uma galha sobre as demais são difíceis de serem mensurados em campo,
desta forma, a observação da distribuição da comunidade completa e a avaliação com
modelos nulos tornam-se mais eficaz nos teste de competição (Cornelissen et al. 2013).
Observamos relação significativa entre os índices de C-score e os parâmetros
indicadores de estresse (massa foliar especifica e crescimento dos ramos), assumimos
que a competição é uma força de estruturação em ambientes com elevado estresse
abióticos. São descritos nove algoritmos para testes dos modelos nulos, sendo a escolha
do mais apropriado depende da estrutura da matriz original (Gotteli 2000). Os índices
C-score (Stone e Roberts 1990) mede o padrão de exclusão das espécies que reflete as
interações competitivas. No entanto, este índice não incorpora a abundância das
espécies e são sensíveis a variações da riqueza de espécies (Gotelli e Graves 1996), por
outro lado, é menos propenso aos Erros do Tipo I (rejeitar a hipótese nula sendo ela

22
verdadeira) e Erros do Tipo II (aceitar a hipótese nula quando ela é falsa) (Gotelli 2000).
Alguns autores ressaltam que ao analisar a distribuição das espécies com o C-score o
índice mostra padrões de segregação, no entanto as espécies não interagem pelo simples
fato de haver uma diferenciação no uso dos recursos (Diamond e Gilpin 1982, Gotelli e
Rohde 2002). Contudo, neste trabalho as espécies avaliadas utilizam o mesmo recurso
(folíolo). Portanto, as interações bióticas diferem entre os habitas avaliados e a hipótese
nula no favorece inferir que a um padrão de organização de galhas nos ambientes
xéricos, e que possivelmente a competição é uma força importante nestes ambientes.
Conclusão
Os diferentes ambientes causam modificações nas plantas hospedeiras com impacto
direto na organização das comunidades associadas. Nos ambientes mésicos não há um
padrão de organização das comunidades de galhas associadas a C. langsdorffii, já nos
ambientes xéricos é notado certo padrão de organização destas comunidades e elegemos
a competição como uma possível força estruturadora neste ambiente.

23
Tabelas e Figuras
Figura 1: Mapa das regiões de amostragem. Foram amostradas sete regiões com
diferentes tipos de vegetação no estado de Minas Gerais – Brasil. (Detalhes de cada
região vide Tabela 1).

24
Tabela 1: Caracterização climática histórica das variáveis ambientais das regiões de
estudo. Temperatura máxima média °C (T. max.), Temperatura mínima média °C (T.
mim.) e Evaporação de Piche (mm) (Evapo). Dados referentes a 55 anos obtidos do site
do INMET em agosto de 2015. Os dados correspondiam às estações meteorológicas
mais próximas das áreas de estudo.
Local Ambiente Coordenadas Altitude Insolação T.max. T.mim Evapo.
Ibirité Canga 20°04’S,
43°59’W
1423 185.21 28.12 14.65 89.43
Serra do
Cipó
Campo
Rupestre
19°16’S,
43°35’W
1200 168.73 28.05 15.39 87.71
Montes
Claros
Cerrado
Sentido
Restrito
16o40’S,
43o48’W
652 222.86 29.73 17.54 138.52
Paraopeba Cerradão 19°20’S
44°24’W
732 221.39 28.51 15.89 88.66
Japonvar Mata
Seca
15º58’S,
44º16’W
826 240.36 31.44 18.39 143.51
Belo
Horizonte
Mata
Atlântica
19°53'S
43°58'W
915 208.05 27.13 17.26 116.92
Jequitaí Mata
Ciliar
56°25’S,
80°96’W
480 229.28 31.03 18.30 125.62

25
Tabela 2: Analise de deviance dos modelos mínimos adequados que mostra os efeitos
dos diferentes habitats sobre os parâmetros vegetativos de Copaifera langsdorffii.
Variáveis
respostas
Variáveis
explicativas
Deviance Residual
Deviance
DF F p
Cresc. ramos Local 10.792 9.2088 78 15.235 < 0.001
Massa foliar Local 1.9315e-05 9.3463e-06 78 26.866 < 0.001
Área Folíolo Local 163.72 95.951 78 22.182 < 0.001
Tabela 3: Lista de Morfosespécies de galhas associados à Copaifera langsdorffii nos
ambientes de Campo Rupestre, Canga, Cerrado sentido restrito, Cerradão, Mata
Atlântica, Mata Seca e Mata ciliar. (Morfosespécies segundo Costa et al. 2010 e
Fagundes 2014)
Campo
Rupestre
Canga Cerrado Cerradão Mata
Atlântica
Mata
Seca
Mata
Ciliar
Total
M1 0 0 0 0 0 0 47 47
M2 12 19 62 20 4 49 58 224
M3 22 44 35 5 123 150 81 460
M4 19 22 9 22 41 72 56 241
M5 0 0 4 1 5 4 4 18
M6 0 0 0 0 1 0 1 2

26
... Continuação Tab. 3
Campo
Rupestre
Canga Cerrado Cerradão Mata
Atlântica
Mata
Seca
Mata
Ciliar
Total
M7 1 18 26 0 2 8 8 63
M8 16 23 13 16 2 17 23 110
M9 0 1 5 0 0 5 3 14
M10 0 0 1 0 0 0 0 1
M11 0 9 19 1 0 8 17 54
M12 0 8 1 0 0 1 0 10
M13 21 19 157 36 96 0 28 357
M14 0 1 0 0 0 0 0 1
M15 88 58 7 0 36 39 17 245
M16 0 1 53 1 0 0 7 62
M18 0 0 0 0 0 2 0 2
M19 28 11 0 19 17 12 0 87
M20 0 5 14 2 1 2 5 29
M21 24 52 87 11 19 54 62 309
M23 56 118 0 0 0 0 0 174

27
... Continuação Tab. 3
Campo
Rupestre
Canga Cerrado Cerradão Mata
Atlântica
Mata
Seca
Mata
Ciliar
Total
M24 1 1 2 0 0 0 0 4
M25 3 33 0 0 0 0 0 36
Riq 12 18 16 11 14 15 16
Abun. 291 443 495 134 347 423 417 2550
Tabela 4: Analise de deviance dos modelos mínimos adequados para avaliar os efeitos
dos diferentes ambinetess e os parâmetros vegetativos de Copaifera langsdorffii sobre a
riqueza e abundância de galhas.
Variáveis
resposta
Variáveis
explicativas
Deviance Residual
Deviance
DF F p
Riqueza Massa foliar 16.241 65.940 78 23.1 < 0.001
Abundância Local 27.297 93.162 78 < 0,001
Tabela 5: Análise de deviance dos modelos mínimos adequados para avaliar os efeitos dos
parâmetros vegetativos (Variáveis explicativas) sobre os índices de coocorrência de galhas
associadas a C. langsdorffii em sete ambientes diferentes.
Variáveis
resposta
Variáveis
explicativas
Deviance Residual
Deviance
DF F p

28
C-score Massa 1331 627.9 6 10.59 0.01
C-score Cresc. ramo 118.8 840.9 6 6.659 0.04
Tabela 6: Análise de similaridade (ANOSIM) comparando a composição das
morfoespécies de galhas nos ambientes de Canga, Campo Rupestre, Cerrado sentido
estrito, Mata Seca, Mata Atlântica e Mata ciliar. (Valor superior a 0.05 representa
composição semelhante)
Canga Campo
Rupestre
Cerrado Cerradão Mata
Seca
Mata
Atlântica
Mata
Ciliar
Canga 0.62 0.0001 0.0001 0.0001 0.0008 0.0001
Campo R 0.0001 0.0001 0.0001 0.0219 0.0001
Cerrado 0.0001 0.0005 0.0001 0.0001
Cerradão 0.0002 0.0037 0.0001
Mata Seca 0.001 0.0128
Mata A. 0.0002
Mata C.

29
Tabela 7: Índices de C-score da ocorrênica de galhas presente em Copaifera
langsdorffii nas diferentes populações analisadas. Os índices máximo e mínimo foram
calculados a partir de 5000 randomizações da matriz original. Os valores de P foram
obtidos pelo teste bi-caudal, representando a probabilidade do índice observado ser
maior, menor ou igual ao esperado pelas matrizes randomizadas. Observado (obs),
esperado (esp)
Local Índices da matriz randomizada Índice obs. Valores de P
Mínimo Máximo Obs.>=esp. Obs.=<esp.
Campo R. 120.30 123.84 123.25 0.001 0.98
Canga 74.83 76.85 76.81 0.0002 0.9998
Cerrado 114.82 121.92 121.66 0.003 0.9970
Cerradão 128.66 132.98 130.78 0.552 0.456
Mata S. 115.62 118.62 117.89 0.252 0.757
Mata C. 99.84 103.24 101.92 0.15 0.846
Mata A. 122.45 127.21 123.40 0.89 0.11
30
Figura 2: Variação dos parâmetros vegetativos das plantas Copaifera langsdorffii pelo
estresse ambiental dos diferentes ambientes. (A) Massa foliar específica, (B) área foliar,
(C) cresciemnto dos ramos. As letras iguais sobre as barras representam agrupamento
pela análise de contraste.
A
C
B

31
Figura 3: Relação entre a riqueza de galhas e a massa foliar específica das plantas de
C. langsdorffii em sete ambientes diferentes.
Figura 4: Relação entre a abundância média de galhas associada a C. langsdorffii em
sete ambientes. As letras sobre as barras representam a difrença dos grupos

32
Figura 5: Variação na composição da comunidade de insetos galhadores entre plantas
de Copaifera langsdorffii em sete ambientes analisados. Existe uma semelhança na
comunidade de galhas entre a área de Canga e o Campo Rupestre (p = 0.62), a
composição de galhas dos demais ambientes diferiu entre si.

33
Figura 6: Histograma das frequências de simulações dos C-scores simulados usando o
modelo fixo-fixo para morfotipos de galhas nos folíolos de Copaifera langsdorffii em
sete locais. As setas mostram os índices observados a partir de 5000 randomizações.
Veja a tabela 7 com os valores máximos e mínimos das simulações.
Mata ciliar Índice obs.
Cerrado
Índice obs.
Canga
índice obs.
Mata
Atlântica índice
obs.
Cerradão Índice obs.
.
Mata seca
Índice obs.
Campo R.
Índice obs.

34
Figura 7: Variação da coocorrência (C-score) das morfoespécies de galhas associadas a
Copaifera langsdorffii pelos parâmetros vegetativos (indicadores de estresse da planta).
(A) Massa foliar, (B) crescimento dos ramos. Valores baixos do índice de C-score
indicam baixa co-ocorrência de galhas (valores de significância dos índices descritos na
Tab. 7).
A
B

35
Referências Bibliográficas
Barbosa, M. and Fernades G. W. 2014. Bottom-Up Effects on Gall Distribution. - In:
Fernandes, G. W & Santos, J. C (ed.), Neotropical Insect Galls. Springer pp: 99-114 (In
press).
Barton K. 2015. MuMIn: Multi-Model Inference. R Package Version 1.15.1. Available
from URL: http://CRAN.R-project. org/package=MuMIn
Begon, M. et al. 2007. Ecology From Individuals and Ecosystemas. – Blackwell
publishing.
Blanche, K. R. 2000. Diversity of insect galls along a temperature-rainfall gradient in
the tropical savannah region of the northern territory Australia. Austral Ecolo. 25: 311-
318.
Boschilia, S. M. et al. 2008. Do aquatic macrophytes co-occur randomly? An analysis of
null models in a tropical floodplain. – Oecologia 156: 203–214.
Browker, M. A. and Maestre, F. T. 2012. Inferring local competition intensity from
patch size distributions: a test using biological soil crusts. Oikos 121: 1914–1922.
Brooker, R. W. and Callaghan, T. V. 1998. The balance between positive and negative
plant interactions and its relationship to environmental gradients: a model. – Oikos 81: 1
Bussotti, F. et al. 2015 Functional traits and adaptive capacity of European forests to
climate change. - Environmental and Experimental Botany 111: 91-113.
Butterill, P and Novotny, V. 2015. Gall-forming insects in a lowland tropical rainforest:
low species diversity in an extremely specialised guild. - Ecological Entomology
Carneiro, M. A. A. et al. 2009. Are gall midge species (Diptera, Cecidomyiidae) host-
plant specialists? - Rev. Bras. entomol. 53: 365-378.
Carvalho, P. E. R. (2003). Espécies arbóreas brasileiras. Brasília, Colombo - Embrapa
Florestas.
Chaturvedi, R. K. et al. 2014. Relative effects of different leaf attributes on sapling
growth in tropical dry forest. - Journal of Plant Ecology 7: 544–558.
Connell, J. H. 1961. The Influence of Interspecific Competition and Other Factors on
the Distribution of the Barnacle Chthamalus Stellatus. - Ecology 42: 710-723.
Connor, E. F. and D. Simberloff. 1983. Interspecific competition and species co-
occurrence patterns on islands: null models and the evaluation of evidence. - Oikos 41:
455-465.
Cornelissen, T. and Stiling, P. 2008. Clumped distribution of oak leaf-miners between
and within plants. - Basic Appl Ecol 9: 67-77.

36
Cornelissen, T. et al. 2013. Interspecific competition influences the organization of a
diverse sessile insect community. - Acta Oecologica 52: 15-18.
Costa, F. V et al. 2010. Arquitetura da planta e diversidade de galhas associadas à
Copaifera langsdorffii (Fabaceae). – Ecol. Aust. 20: 9-17.
Costa, F. V et al. 2011. Relationship between plant development, tannin concentration
and insects associated with Copaifera langsdorffii (Fabaceae). – Arthropod Plant
Interact 5: 9-18.
Craig, T. P et al. 2007. Host plant genotype influences survival of hybrids between
Eurosta solidaginis host races. - Evolution 61: 2607–2613
Crawley, M. J. 2007. The R Book. New York: John Wiley and Sons.
Cuevas-Reyes, P et al. 2006. Abundance and Leaf Damage Caused by Gall-Inducing
Insects in a Mexican Tropical Dry Forest. - Biotropica 38: 107–115.
Diamond, J. M. 1973. Distributional ecology of New Guinea birds. - Science, N.Y 179:
759-69.
Diamond, J. M and Gilpin, M. E. 1982. Examination of the “null” model of connor
and simberloff for species co-occurrences on Islands. - Oecologia 52: 64-74.
Diamond, J. M. 1986. Overview: laboratory experiments, field experiments and natural
experiments. - In: Diamond, J. and Case, T.J. (ed.), Community Ecology.Harper & Row
pp: 3-22.
Egan S. P and Ott J. R. 2007. Host plant quality and local adaptation determine the
distribution of a gall-forming herbivore. - Ecology 88: 2868-2879.
Ellwood, M. D. F. et al. 2009. Stochastic and deterministic processes jointly structure
tropical arthropod communities. - Ecology Letters 12: 277–284.
Espírito-Santo M. M, et al. 2007. Plant architecture and meristem dynamics as the
mechanisms determining the diversity of gall-inducing insects. - Oecologia 153: 353-
364.
Fagundes M. 2014. Gall community associated with Copaifera langsdorffii (Fabaceae):
The role of inter and intra-annual plant phenology. - In: Fernandes, G. W & Santos, J. C
(ed.), Neotropical Insect Galls. Springer pp: 163-177 (In press).
Fagundes M. and Fernandes G, W. 2011 Insect herbivores associated with Baccharis
dracunculifolia (Asteraceae): responses of gall-forming and free-feeding insect
herbivores to latitudinal variation. Revista de Biología Tropical.
Fagundes, M. 2005. Direct and indirect interactions involving ants, insect herbivores,
parasitoids, and the host plant Baccharis dracunculifolia (Asteraceae). Ecological
Entomology 30: 28-35.

37
Fernandes, G. W . et al. 2000. Plant fight gall formation: hipersensitivity. – Cienc. Cult
52: 49-54.
Fernandes, G. W. et al. 1995. Distribuição diferencial de insetos galhadores entre
habitats e seu possível uso como bioindicadores. - Vida Silvestre Neotropical 4: 133-
139
Fernandes, G. W, and Price, P. W. 1988. Biogeographical gradients in galling species
richness: tests of hypotheses. - Oecologia 76:161-167.
Fernandes, G. W. 1990. Hypersensitivity: a neglected plant resistance mechanism
against insect herbivores. - Environmental Entomology 19: 1173-1182
Fernandes, G. W. and Price, P. W. 1992. The adaptative significance of insect gall
distribution: survivorship of species in xeric and mesic habitats. - Oecologia 90:14-20.
Fernandes, G. W and Negreiros D. 2001. The occurrence and effectiveness of
hypersensitive reaction against galling herbivores across host taxa. - Ecological
Entomology 26: 46–55.
Fernandes, G. W. et al. 2005. Habitat-driven effects on the diversity of gall-inducing
insects in the Brazilian cerrado. pp 693-708. In, A. Raman, C. W. Schaefer, and T. M.
Withers (Editors). Biology, ecology and evolution of gall-Inducing arthropods. Science
Pub - lishers, New Hampshire. 817 pp.
Fernandes, G. W. et al. 2003. Hypersensitivity of Fagus sylvatica L. against leaf galling
insects. Trees - 17: 407- 411.
Giulietti, A. M. et al. 1987. Flora da Serra do Cipó, MG: caracterização e lista das
espécies. - Boletim de Botânica da Universidade de São Paulo 9: 1-151
Gotelli, N. J and Entsminger, G. L. 2001. EcoSim: null models software for ecology,
version 6.21. Acquired Intelligence, Kesey-Bear, http://homepages.together.net/-
gentsmin/ecosim.html
Gotelli, N. J. 2000. Null model analysis of species co-occurrence patterns. – Ecology
81: 2606–2621
Gotelli, N. J. and Graves, G. R. 1996. Null Models in Ecology. Smithsonian Institute
Press, Washington.
Gotelli, N. J. and Rohde, K. 2002. Co-occurrence of ectoparasites of marine fishes: a
null model analysis. - Ecology Letters, 5, 86–94.
Höglund, S. 2014. Timing of growth determines fitness and performance of a galling
insect on willow. - Ecological Entomology 39: 159-67.
Höglund, S. et al. 2005. Both hypersensitive and non-hypersensitive responses are
associated with resistance in Salix viminalis against the gall midge Dasineura
marginemtorquens. - Journal of Experimental Botany 56: 3215–3222

38
Hunter, M. D. and Price, P. W. 1992. Playing chutes and ladders: heterogeneity and the
relative roles of bottom-up and top-down forces in natural communities. - Ecology 73:
724-732.
Jesus, F.M. et al. 2012. Differential Female Attack and Larval Performance of a Galling
Cecidomyiid on the Host, Astronium fraxinifolium (Anacardiaceae), in Contrasting
Habitats. - Entomological News 122: 10-21
Joy, J. B and Crespi, B. J .2007. Adaptive radiation of gall-inducing insects within a
single host-plant species. - Evolution 61-4: 784–795.
Julião, G. J. et al. 2005. Insetos galhadores associados a duas espécies de plantas
invasoras de áreas urbanas e peri-urbanas.- Revista Brasileira de Entomologia 49: 97-
106.
Julião, G. J. et al. 2014. Galling Insects in the Pantanal Wetland and Amazonian
Rainforest. The role of inter and intra-annual plant phenology. - In: Fernandes, G. W &
Santos, J. C (ed.), Neotropical Insect Galls. Springer pp: 377-403 (In press).
Kaplan, I. and Denno, R. F. 2007. Interspecific interactions in phytophagous insects
revisited: a quantitative assessment of competition theory. - Ecol. Lett. 10: 977-994.
Krasnov, B. R. et al. 2006. Are ectoparasite communities structured? Species co-
occurrence, temporal variation and null models. - Journal of Animal Ecology 75: 1330–
1339
Lara, A. C. F. et al. 2002. Tests of hypotheses on patterns of gall distribution along an
alti- tudinal gradient. -Tropical Zoology 15: 219-232.
Lázaro-Nogal, A. et al. 2015. Environmental heterogeneity leads to higher plasticity in
dry-edge populations of a semi-arid Chilean shrub: insights into climate change
responses. - Journal of Ecology 103: 338–350.
Malinga, M. G. et al. 2014. Bottom‑up impact on the cecidomyiid leaf galler and its
parasitism in a tropical rainforest. – Oecologia 176:511–520.
Maloof, J. N. et al. 2013. Leaf J: An ImageJ Plugin for Semi-automated Leaf Shape
Measurement. - J. Vis. Exp. 71.
Matthew, A. Bowker, M. A. and Maestre, F. T. 2012. Inferring local competition
intensity from patch size distributions: a test using biological soil crusts. - Oikos 121:
1914–1922.
Oliveira-Filho, A. T. and Ratter, J. A. 1995. A study of the origin of central Brazilian
forests by the analysis of plant species distribution patterns. -Edinburg Journal of
Botany 52: 141-194.

39
Oliveira P. E. 1998. Fenologia e biologia reprodutiva das espécies de cerrado. In: Sano
S.M, Almeida SP (eds) Cerrado: ambiente e flora. Embrapa-CPAC, Planaltina, Brazil,
pp 169–192.
Oliveira-Filho, A. T. and Ratter, J. A. 2002. Vegetation physiognomies and woody flora
of the Cerrado Biome. - In: Oliveira, P. S. and Marquis, R. J. (ed.), The Cerrados of
Brazil: ecology and natural history of a Neotropical savanna. Columbia University Press
pp: 91-120.
Ollerstam, O. et al. 2002. A rapid hypersensitive response associated with resistance in
the willow Salix viminalis against the gall midge Dasineura marginemtorquens. -
Entomologia Experimentalis et Applicata 102: 153–162.
Pedroni, F. et al. 2002. Fenologia da copaíba (Copaifera langsdorffii Desf. -
Leguminosae, Caesalpinioideae) em uma floresta semidecídua no sudeste do Brasil. -
Revista Brasileira de Botanica 25: 183-194.
Pennington, D. D. and Collins, S. L. 2007. Response of an aridland ecosystem to
interannual climate variability and prolonged drought. - Landscape Ecol 22:897–910.
Price, P. W. 1990. Evaluating the role of natural enemies in latent and eruptive species:
new approaches in life table construction. - In: A. D. Watt, S. R. Leather, M. D. Hunter,
and N. A. C. Kidd (ed), Population dynamics of forest insects pp. 221-232.
Price, P. W. 1991. The plant vigor hypothesis and herbivore attack. - Oikos 62: 244-
251.
Price, P. W. 1997. Insect Ecology, 3rd edn. Wiley. New York
Price, P. W. 2002. Resource-driven terrestrial interactions webs. – Ecological Research
17: 241-247.
Price, P. W. et al. 1987. Adaptive nature of insect galls. – Environ. Entomol. 16: 15-24.
R Core Team 2015. R: A language and environment for statistical computing.R
Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-
project.org/.
Reich, P. B. et al. 2003. The evolution of plant functional variation: traits, spectra, and
strategies.- International Journal of Plant Sciences 164: 143–164
Reitz, S. T. and Trumble, J. T. 2002. competitive displacement among insects and
arachnids. - Annu. Rev. Entomol. 47:435–65.
Ribas, C. R. and Schoereder, J. H. 2002. Are all ant mosaics caused by competition? -
Oecologia 131: 606-611.
Rizzini, C. T. 1997. Tratado de fitogeografia do Brasil: aspectos ecológicos,
sociológicos e florísticos. Rio de Janeiro: Âmbito Cultural.

40
Sanders, N. J. et al. 2003. Community disassembly by an invasive species. – PNAS 100:
2474-2477.
Sano, S. M et al. 2008. Cerrado: ecologia e flora vol. 1. 1. ed. Brasília: Embrapa, 2008.
v.2
Sapijanskas, J. et al. 2014. Tropical tree diversity enhances light capture through crown
plasticity and spatial and temporal niche differences. – Ecology 95: 2479–2492.
Souza, M. L et al. 2015. Reproductive strategy of Copaifera langsdorffii (Fabaceae):
more seeds or better seeds? - Revista de Biología Tropical 63:1161-1184.
Souza, M. L and Fagundes, M. 2014. Seed Size as Key Factor in Germination and
Seedling Development of Copaifera langsdorffii (Fabaceae).- American Journal of Plant
Sciences 5: 2566-2573
Stone, L. and Roberts, A. 1990. The checkerboard score and species distributions. -
Oecologia 85: 74-79.
Sultan, S.E. 2001. Phenotypic plasticity for fitness components in polygonum species of
contrasting ecological breadth. - Ecology 82: 328–343.
Toma, T.S. et al. 2014. Galling Insects as Indicators of Habitat Quality. - In: Fernandes,
G. W & Santos, J. C (ed.), Neotropical Insect Galls. Springer pp: 143-150 (In press).
White, T.C.R. 1969. An Index to Measure Weather-Induced Stress of Trees Associated
With Outbreaks of Psyllids in Australia. - Ecology 50: 905 - 909.
Williams, D.G. and Blackc, R.A. 1993. Phenotypic variation in contrasting temperature
environments: growth and photosynthesis in Pennisetum setaceum from different
altitudes on Hawaii. - Functional Ecology 7: 623-633.
Yukawa, J. 2000. Synchronization of gallers with host plant phenology. Population
Ecology 42: 105-113





























