Embed Size (px)
Citation preview

Emílio López Couto
Estudo da Catepsina B e da Heparanase em Carcinoma Espinocelular de Cabeça e Pescoço
MOGI DAS CRUZES 2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

- 1 -
UNIVERSIDADE DE MOGI DAS CRUZES EMÍLIO LÓPEZ COUTO
ESTUDO DA CATEPSINA B E DA HEPARANASE EM CARCINOMA ESPINOCELULAR DE CABEÇA E PESCOÇO
Dissertação Apresentada à Universidade de
Mogi das Cruzes para a Obtenção de Título de
Mestre em Biotecnologia
Prof Orientador: Dr. Ivarne Luis dos Santos Tersariol Profa a Co-Orientadora: Dr Maria Aparecida da Silva Pinhal
MOGI DAS CRUZES, SP 2006

- 2 -

- 3 -

- 4 -
Dedicatória
Terminando uma nova fase da vida acadêmica, vislumbramos novos desafios, e
relembramos o caminho percorrido. O prazer que advém do estudo gera a ansiedade de mais
querer saber, e humildemente nos restringimos à nossa incapacidade de tudo aprender.
Foram tantas as pessoas que participaram da minha formação, familiares, professores,
amigos...Com certeza se fosse descrever o nome de todos, cometeria o erro imperdoável de
esquecer alguém. Meus pais que me deram as primeiras lições, minha tia Martha e família,
com eles, pela primeira vez, falei corretamente frases longas (já tinha cinco anos), minha
primeira professora, primeira escola, primeiras provas, trabalhos.
Fiquei cerca de seis anos afastado dos bancos escolares, resolvi ganhar a vida, mas
sempre sentia falta das discussões e das descobertas. Sentia que faltava algo, exercitar a
mente... Difícil foi a volta, mas com encontro do meu orientador, pesquisador de invejável
conhecimento, sempre entusiasmado, nos estimulando a procurar o saber, foi mais fácil.
Dedico a cada um deles a participação neste trabalho, e os convido a continuar na
minha jornada

- 5 -
Agradecimentos
Aos meus pais, meus primeiros mestres, sempre me estimularam ao estudo e à leitura, e com
seus exemplos de esforço, dedicação e superação das dificuldades, ainda são o norte em
minha vida.
À minha esposa, Elizabete Nunes Giarola, quanta paciência precisou ter para me ajudar nos
momentos de estresse. E nas discussões sobre o trabalho e a pesquisa, sem ela com certeza
seria muito mais difícil a realização do projeto.
Ao meu orientador Prof Dr Ivarne Luis dos Santos Tersariol, grande pesquisador, mesmo
quando os resultados não eram o esperado consegue retirar conhecimento e vislumbrar uma
nova linha de pesquisa. Nunca desiste.
À minha co – orientadora Profa a Dr Maria Aparecida da Silva Pinhal, abriu seu laboratório
e nos forneceu um inestimável auxílio em recursos humanos para a realização de nossos
estudos imunoistoquímicos.
Ao colega de mestrado Luís Gabriel Nunes, e à colega Thais Peretti inestimável ajuda na
realização dos ensaios de atividade enzimática e de imunoistoquímica
Aos amigos e cirurgiões que me auxiliaram nos procedimentos e coleta de material César
Augusto Simões, Francisco Amorim, Sérgio Uchoa, Marcos Loretto, e mais recentemente
Samara Novak
a aÀ Prof Dr Cláudia Bincoletto Trindade e à Profa a Dr Regina L. B. C. Oliveira que
participaram da banca de qualificação, pelas observações e orientações.
Ao meu irmão Arthur López Couto, pudera eu escrever com a clareza e precisão que ele
tem.

- 6 -
Ao meu primo Lincoln Gonçalves Couto, desde a adolescência um grande parceiro em
discussões científicas e no xadrez.
A todos aqueles que direta ou indiretamente participaram da elaboração e realização deste
projeto

- 7 -
“O caminho da vida, o mais agradável e o mais inofensivo, passa pelas avenidas da ciência e
do saber; e quem quer que possa remover quaisquer obstáculos desta via ou abrir uma nova
perspectiva, deve ser considerado um benfeitor da humanidade. Embora estas pesquisas
possam parecer árduas e fatigantes, ocorre aqui como com certos espíritos e corpos que, por
estarem dotados de grande vitalidade, necessitam de exercícios severos e colhem prazer
daquilo que, para a maioria dos homens, parece penoso e laborioso. A obscuridade é, de fato
penosa tanto para o espírito como para os olhos, todavia, trazer luz da obscuridade, por mais
trabalhoso que seja, deve ser agradável e regozijado.”
(David Hume)

- 8 -
Resumo Metástase e invasão tumoral das células malignas envolvem interação célula a célula e célula-matriz, as quais são reguladas pela expressão e localização de proteinases e glicosidases nas células tumorais. O objetivo deste estudo é elucidar relações entre a expressão de Heparanase e a Catepsina B (Cat B) nos carcinomas espinocelulares primários de cabeça e pescoço. As amostras de tecidos foram tiradas da cavidade oral, orofaringe, laringe e hipofaringe de pacientes com carcinoma espinocelulares de cabeça e pescoço. A expressão imunoistoquímica da heparanase e da cat B foram analisadas em 20 amostras de carcinoma espinocelulares. O método de coloração padrão de imunoperoxidase avidina-biotina foi realizado nas lâminas de tecidos fixadas em parafina de 4 µm com anticorpos murinos de anti-heparanase e anticorpos monoclonais anti-catepsina. Também foram realizadas dosagens da atividade enzimática da catepsina B com adição do substrato fluorogênico Z-F-R-MCA em fragmentos de tecidos coletados a fresco. Níveis elevados de catepsina B apresentaram uma significativa correlação com o estadiamento avançado dos tumores (p<0,05), os níveis de catepsina B aumentam com o avançar do estadiamento tumoral, 3,4 ± 0,5 UAF/min/μg em lesões benignas, 15 ± 2 UAF/min/μg para tumores T1-3 e 25 ± 5 UAF/min/μg nos T4. Foi estatisticamente significativa a associação da expressão de catepsina B nos tumores primários de orofaringe e o comprometimento linfonodal (p<0,05), os níveis de catepsina B na presença de metástase (26 ± 5 UAF/min/μg) foram maiores do que na ausência de metástase (15 ± 2 UAF/min/μg). Também os níveis elevados de catepsina B observados em cânceres de laringe invasivos estavam fortemente associados a fenotipos mais agressivos tumorais. Entretanto, não observamos correlação da catepsina B, nos tumores de laringe, com o comprometimento linfonodal. Aumento da expressão da catepsina B correlaciona-se significativamente com a presença de metástase, invasão local tumoral, piores graus histológicos de malignidade e altas taxas de proliferação celular. Esses achados indicam uma forte relação causa/efeito entre a expressão da catepsina B em carcinomas espinocelulares de cabeça e pescoço e sua invasibilidade local e metastatização. Diferentemente da catepsina B, a heparanase, em tecidos normais (não displásicos), como em adenomas de baixa displasia , apresentou uma reatividade grande com o anticorpo anti-heparanase e até maior em focos de moderada displasia. Contudo, interessantemente, essa reatividade com o anticorpo anti-heparanase foi marcadamente reduzida em displasias intensas e em carcinomas espinocelulares. Esses resultados indicam que a heparanase é um bom marcador da diferenciação celular. Esses resultados, em conjunto, indicam que a transformação epitélio-mesênquimal de desdiferenciação do carcinoma espinocelular aumenta a expressão da catepsina B e diminui a expressão da heparanase. Palavras-chaves: Catepsina B, heparanase, carcinoma espinocelular, cabeça e pescoço

- 9 -
Abstract Metastasis and local invasion of malignant tumor cells involve cell-cell and cell-matrix interactions, which is regulated by the expression and localization of proteinases and glycosidases secreted by tumoral cells. The aim of this study was to clarify the relationship between the expression of heparanase and Cathepsin B (CathB) at the main tumor mass of head and neck squamous cell carcinoma. The tissues samples were taken from oral cavity, oropharynxes, larynxes and hypopharynxes of patients with squamous cell carcinoma. The immunohistochemical expressions of heparanase and CathB were studied in 20 specimens of squamous cell carcinoma. A standard avidin-biotin immunoperoxidase staining method was performed on 4-micrometer paraffin-embedded tissue sections with a mouse anti-human heparanase and cathepsin B monoclonal antibody. Also, we determined the enzymatic activity of cathepsin B with the aid of fluorogenic substrate Z-F-R-MCA in biopsies of fresh tumor tissues. Increased level of cathepsin B in oral carcinomas was significantly associated with advanced tumor stage (p < 0.05), the amount of cathepsin B increases with advanced tumor stage, 3.4 ± 0.5 AFU/min/μg for benign lesions, 15 ± 2 AFU/min/μg for T1-3 stage and 25 ± 5 AFU/min/μg for T4 stage. A statistically significant association was found between the expressions of cathepsin B in the primary tumor of oropharynxes cavity and lymph node involvement (p<0.05), the amount of cathepsin B in the presence of metastasis (26 ± 5 AFU/min/μg) was higher than without metastasis (15 ± 2 AFU/min/μg). Also, the high levels of cathepsin B observed in larynx cancer were strongly associated with more aggressive tumor phenotype. However, cathepsin B in larynx tumors was not associated with lymph node involvement. Increased expression of cathepsin B correlated significantly with the presence of metastasis, local tumoral invasion, poor histologic malignancy grade and high proliferation rate. These findings indicate that there is a strong cause/effect relationship between the expression levels of cathepsin B in squamous cell head and neck cancers and their local invasive and metastatic growth patterns. Different from cathepsin B, heparanase in normal epithelium (nondysplastic), as well as areas of mild dysplasia in adenomas, exhibited a strong reactivity with antibody anti-heparanase that became even stronger in foci of moderate dysplasia. Interestingly, the reactivity with antibody anti-heparanase was markedly reduced in severe dysplasia and in squamous cell carcinoma. These results indicate that heparanase is a good marker of squamous cell differentiation. Taken together, these results show that the epithelial to mesenchymal transformation process in squamous cells carcinoma increases cathepsin B expression and decreases heparanase expression. Key words: cathepsin B, heparanase, squamous cell carcinoma

- 10 -
Lista de Tabelas
Tabela 1 Catepsina B em Tumores Humanos............................................ ..............................17
Tabela 2 Localização Celular Alterada da Catepsina B em Tumores..... ..............................21
Tabela 3 Estadiamento conforme AJCC e UICC, 2002............................ ..............................32
Tabela 4 Patologia, Estadio, Sexo e Idade dos Pacientes.......................... ..............................69
Tabela 5 Atividade da Catepsina B, Estadiamento e Patologia................ ..............................70
Tabela 6 Atividade da Catepsina B nas Lesões Benignas e nos
diferentes Estadiamentos dos Cânceres...................................... ..............................71
Tabela 7 Expressão Enzimática da Catepsina B e a Classificação do
Tumor...........................................................................................................................74
Tabela 8 Expressão Enzimática da Catepsina B nas Diferentes Regiões
Anatômicas.................................................................................... ..............................75
Tabela 9 Expressão Quantitativa de Heparanase, Estadiamento e
Patologia dos Pacientes................................................................. ..............................76
Tabela 10 Classificação quantitativa da Heparanase por Índice de
Positividade.................................................................................... ..............................77

- 11 -
LISTA DE ILUSTRAÇÕES
Figura 1 Representação Esquemática da Estrutura
Tridimensional da Catepsina B...................................... ..................................15
Estadiamento Linfonodal..........................................................Figura 2 .................................28
Estadiamento de Tumores de Laringe.....................................Figura 3 .................................29
Estadiamento dos Tumores de Cavidade Oral....................... Figura 4 .................................30
Figura 5 Atividade da Catepsina B em Função da Idade dos
Pacientes..................................................................................... .................................31
Figura 6 Atividade Enzimática da Catepsina B em Carcinomas
Espinocelulares e Hiperplasias Benignas................................................................39
Figura 7 Representação Gráfica dos Valores Médios da Atividade
da Catepsina B...............................................................................................40
Figura 8 Detecção de Catepsina B por Reações de Imunoistoquímica
em Carcinomas Espinocelulares e Hiperplasias
Benignas..................................................................................... .................................41
Atividade da Catepsina B e Classificação do Tumor.............Figura 9 .................................43
Atividade de Catepsina B em Tumores Espinocelulares....... Figura 10 .................................44
Figura 11 Quantidade de Catepsina B em Relação ao Estadiamento
dos Tumores de Laringe........................................................... .................................45
Atividade de Catepsina B em Tumores de Laringe............... Figura 12 .................................46
Figura 13 Atividade de Catepsina B em Tumores da Cavidade
Orofaríngea................................................................................ ..................................47
Figura 14 Reações de Imunoistoquímica de Heparanase em
Carcinomas Espinocelulares.................................................... .................................48

- 12 -
Lista de Abreviaturas e Siglas
AJCC American Joint Comittee on Cancer
AMP Adenosina Monofosfato
AMPc AMP cíclico
Arg Arginina
Asp Ácido aspártico
ATP Adenosina Trifosfato
Cat B Catepsina B
Cys Cisteína
DNA Ácido Desoxirribonucléico
cDNA DNA complementar
EDTA Ácido Etilenodiaminotetracético
Glu Glutamina
HEMC Hospital Estadual Mário Covas – Santo André – SP
His Histidina
HPV Papiloma Vírus Humano
INCA Instituto Nacional do Cancer
IP Índice de positividade
MS Ministério da Saúde
PBS Tampão Fosfato de Sódio
RNA Ácido Ribonucléico
RNAm RNA mensageiro
TNF Fator de Necrose Tumoral
TNM Sistema de Estadiamento do Câncer sendo, T: tumor;
N: linfonodos regionais comprometidos; M: metástase
à distância
TSH Hormônio Estimulante da Tireóide
UAF Unidade de Atividade Formadora
UICC União Internacional Contra o Câncer
UMC Universidade de Mogi das Cruzes
Z-F-R-MCA Carbobenzoxil-L-fenilalanil-L-arginina-7-amido-4-
metilcoumarina

- 13 -
12-S-HETE Ácido 12-S-hidroxitetraeônico

- 14 -
SUMÁRIO
1 INTRODUÇÃO: ESTADO DA ARTE...............................................................................................14
1.1 CatepsinaB.........................................................................................................................14
1.1.1 Estrutura, Função e Localização celular da catepsina B............... ............................14
1.1.2 Participação de Catepsina B na Metástase Tumoral...................................................16
1.1.3 Controle da Expressão Gênica e da Secreção de Catepsina B.................... ............................19
1.1.4 Papel da Catepsina B e dos Lisossomos na Apoptose de Células
Tumorais....................................................................................................... ............................22
1.2 Heparanase em Células Tumorais e Metástases............................................. ............................24
1.3 Câncer de Cabeça e Pescoço...........................................................................................25
2 MATERIAIS......................................................................................................................................31
2.1 Obtenção do tecido tumoral.............................................................................. ............................31
2.2 Detecção das Enzimas em Cortes Histológicos................................................ ............................33
2.3 Obtenção do Extrato Bruto Tecidual............................................................... ............................33
2.4 Dosagem da Atividade Enzimática da Catepsina B nos Tecidos................. ............................34
2.5 Quantificação e Interpretação de Heparanase nas Reações de
Imunoistoquímica.................................................................................................... ............................34
2.6 Análise Estatística da Atividade Enzimática................................................... ............................35
3 Resultados.............................................................................................................. ............................36
3.1 Atividade da Catepsina B.................................................................................. ............................36
3.2 Classificação TNM e Região Anatômica.......................................................... ............................42
3.3 Atividade da Heparanase.................................................................................. ............................48
4 Discussão.............................................................................................................. ............................49
5 Conclusões e Sugestões........................................................................................ ............................54
REFERÊNCIAS....................................................................................................... ............................55
Anexo – Comitê de Ética........................................................................................ ............................64 Apêndices.................................................................................................................. ............................66

- 15 -
1 INTRODUÇÃO: ESTADO DA ARTE
1.1 CATEPSINA B
1.1.1 ESTRUTURA, FUNÇÃO E LOCALIZAÇÃO CELULAR DA CATEPSINA B
A catepsina B é uma cisteíno-proteinase lisossomal que possui tanto atividade
endopeptidásica quanto atividade carboxidipeptidil-peptidásica, este fato a distingue das
demais enzimas da família da papaína (BARRET, 1981; NÄGLER et al., 1997). A principal
característica estrutural que difere a catepsina B das demais cisteíno-peptidases é a inserção
de uma alça estrutural, denominada alça de oclusão (Fig.1), que ocupa os subsítios S2’ ’– S3 da
enzima (MUSIL et al., 1991). O sítio catalítico da catepsina B é formado pelos resíduos Cys29
e His199. Os dois resíduos de His110 e His111, presentes na alça de oclusão, determinam a
atividade carboxidipeptidil-peptidase da catepsina B por ancorar a carboxila C-teminal dos
substratos peptídicos (MUSIL et al., 1991). O par iônico Cys29-His199 também é determinante
para a estabilidade da α-hélice central da enzima, bem como outras pontes salinas tais como:
Asp22-His110 e Arg116-Asp224, localizadas na alça de oclusão. Ainda, ligações interdomínios,
Asp40-Arg202 e Arg41-Glu163, são responsáveis pela estabilização desta hélice. A ruptura do
par iônico Cys29-His199, pela desprotonação do imidazol em meio alcalino, catalisa a
desnaturação da enzima por desorganizar a estrutura de sua α hélice central (TURK et al.,
1994). A pré-pro-catepsina B é sintetizada por apenas um gene, com localização no 8p22,
que pode transcrever mais de um tipo de RNA mensageiro, que variam em tamanho
dependendo da extensão da região não traduzida. O RNA mensageiro é traduzido ligado à
membrana dos ribossomos. A pró-enzima recém sintetizada no retíulo endoplasmático rugoso
passa para o aparelho de Golgi, onde recebera, na sua estrutura primária, oligossacarídeos do
tipo alta-manose, que contém manose-6-fosfato. A glicoproteína fosforilada se liga aos
receptores de manose-6-fosfato na membrana do Golgi e é direcionada ao lisossomo. Durante
este processo, as vesículas de transporte são acidificadas, fato este que permite a dissociação
do receptor, e também desencadeia o autoprocessamento da pró-enzima, pela liberação
proteolítica do seu pró-peptídeo. O processamento intermolecular, feito pela catepsina B ou
outras peptidases, pode ser um mecanismo alternativo de ativação destas enzimas (ROWAN
et al., 1991).

- 16 -
FIGURA 1. Representação Esquemática da Estrutura Tridimensional da Catepsina B. A figura mostra o diagrama da estrutura cristalina da enzima catepsina B lisossomal humana. A molécula, que possui aproximadamente 28 kDa, é orientada por dois domínios globulares, direito e esquerdo (por convenção), e o sítio ativo se encontra inserido entre estes dois domínios. Os resíduos de aminoácidos que formam o sítio ativo estão representados na figura (Cys29 199/His ). As estruturas de α hélice estão representadas em vermelho e as estruturas β pregueadas estão representadas em azul. A estrutura representada em verde recobrindo o sítio ativo é a alça de oclusão, que através dos resíduos de histidina (His110 e His111), determina a atividade de carboxidipeptidil-peptidase da catepsina B.
Além da função de inibir a enzima proteolítica durante a sua biossíntese, o pró-peptídeo
da catepsina B também é responsável por estabilizar a enzima em pH neutro. Em condições
normais de ativação, o pró-peptídeo é degradado após ser clivado da pró-enzima. Entretanto,
em algumas circunstâncias, a catepsina B pode ser secretada da célula como um complexo
não covalente formado pelo pró-peptídeo e a enzima madura. Esta forma pode representar um
potencial reservatório extracelular de enzimas proteolíticas (MORT; BUTTLE, 1997).
Devido ao ambiente pouco favorável dos lisossomos, a meia-vida da maioria das
proteínas lisossomais fica restrita em torno de 24 h. A prolina localizada na porção N-
terminal é uma característica marcante das cisteíno-peptidases lisossomais. A presença de

- 17 -
prolina protege a enzima de possíveis ataques de amino-peptidases (MORT; BUTTLE,
1997).
1.1.2 Participação de Catepsina B na Metástase Tumoral
O estudo da catepsina B e dos componentes da matriz extracelular tem sido de
relevante interesse nos processos fisiopatológicos que envolvem a degradação de
componentes da matriz. Entre esses processos incluem-se a angiogênese (MIGNATTI;
RIFKIN, 1996), o “turnover” da matriz extracelular, bem como processos invasivos em geral
(MIGNATTI; RIFKIN, 1993). As enzimas lisossomais, entre elas, a catepsina B, tem sido
alvo constante de estudos que visam um melhor entendimento dos processos de remodelação
tecidual, evolução tumoral e metástases (MIGNATTI; RIFKIN, 1993).
O alto potencial destrutivo das cisteíno-peptidases lisossomais é regulado por
inibidores protéicos endógenos, tais como: cistatinas, cininogênio e tireoglobulinas do tipo I
(TURK et al., 1997). As cisteíno-peptidases lisossomais também são controladas pelo pH do
meio, estas enzimas sofrem desnaturação em pH 7,4 e alcalino (TURK et al., 1994). Desta
forma, inibidores protéicos, juntamente com o pH fisiológico, são responsáveis por manter o
“balanço” proteolítico desta classe de enzima no tecido. Alterações do conteúdo enzimático
ou dos inibidores podem resultar em diversas patologias que envolvem estados de
remodelação tecidual.
A elevação da atividade da catepsina B está particularmente bem documentada em
carcinomas de cólon, pâncreas e cérebro (SLOANE et al., 1986). Além do lisossomo, a
catepsina B tem sido localizada na membrana plasmática de células tumorais (GUINEC et al.,
1992; MOIN et al., 1998). Esta distribuição permite a enzima degradar os componentes da
matriz extracelular e da membrana basal (SHEAHAN et al., 1989; BUCK et al., 1992).
Estudos sobre a progressão do câncer correlacionados aos níveis de catepsina B, têm
mostrado que essa enzima freqüentemente aparece elevada na maioria dos tumores em
estádios invasivos (Tabela 1). A elevação dos níveis de catepsina B, em carcinoma de cólon
humano, tem sido notada nos estadios precoces em relação aos estadiamentos avançados.
Ainda, neste tumor não foi notado um decréscimo da quantidade de inibidores endógenos
presentes, quando comparados aos níveis da mucosa normal (IACOBUZIO-DONAHUE et
al., 1997).
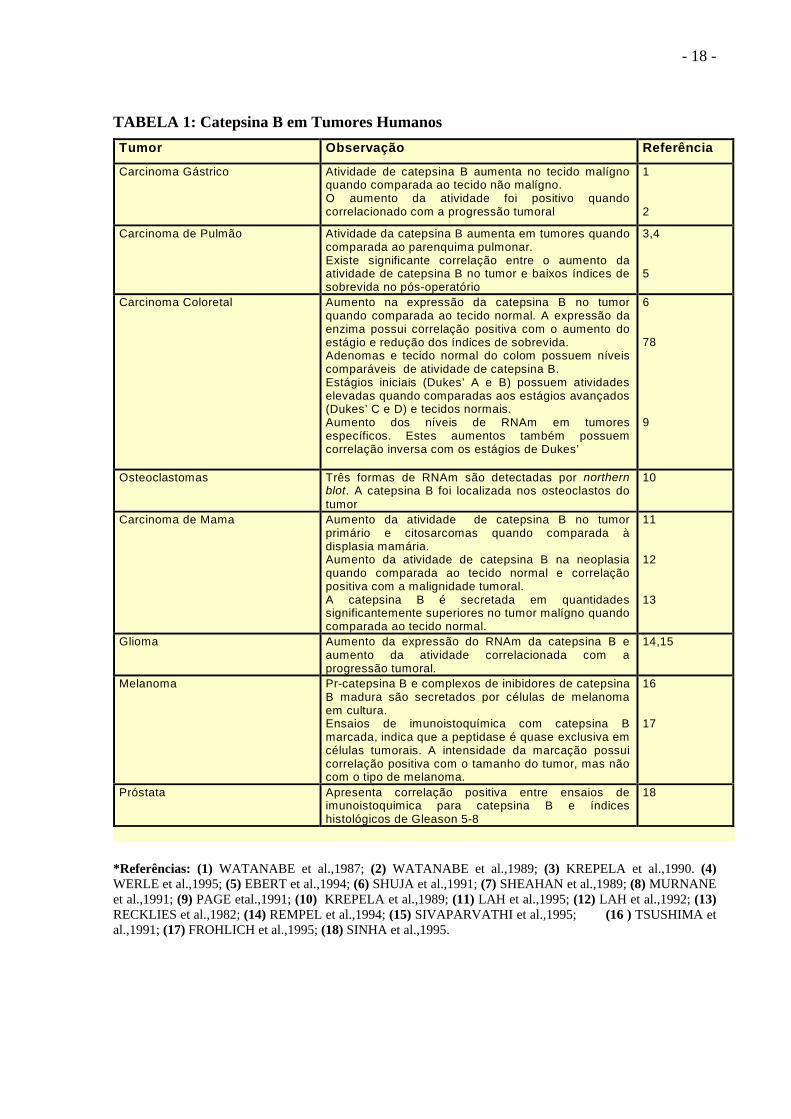
- 18 -
TABELA 1: Catepsina B em Tumores Humanos Tumor Observação Referência
Carcinoma Gástrico Atividade de catepsina B aumenta no tecido malígnoquando comparada ao tecido não malígno.O aumento da atividade foi positivo quandocorrelacionado com a progressão tumoral
1
2
Carcinoma de Pulmão Atividade da catepsina B aumenta em tumores quandocomparada ao parenquima pulmonar.Existe significante correlação entre o aumento daatividade de catepsina B no tumor e baixos índices desobrevida no pós-operatório
3,4
5
Carcinoma Coloretal Aumento na expressão da catepsina B no tumorquando comparada ao tecido normal. A expressão daenzima possui correlação positiva com o aumento doestágio e redução dos índices de sobrevida.Adenomas e tecido normal do colom possuem níveiscomparáveis de atividade de catepsina B.Estágios iniciais (Dukes’ A e B) possuem atividadeselevadas quando comparadas aos estágios avançados(Dukes’ C e D) e tecidos normais.Aumento dos níveis de RNAm em tumoresespecíficos. Estes aumentos também possuemcorrelação inversa com os estágios de Dukes’
6
78
9
Osteoclastomas Três formas de RNAm são detectadas por northernblot. A catepsina B foi localizada nos osteoclastos dotumor
10
Carcinoma de Mama Aumento da atividade de catepsina B no tumorprimário e citosarcomas quando comparada àdisplasia mamária.Aumento da atividade de catepsina B na neoplasiaquando comparada ao tecido normal e correlaçãopositiva com a malignidade tumoral.A catepsina B é secretada em quantidadessignificantemente superiores no tumor malígno quandocomparada ao tecido normal.
11
12
13
Glioma Aumento da expressão do RNAm da catepsina B eaumento da atividade correlacionada com aprogressão tumoral.
14,15
Melanoma Pr-catepsina B e complexos de inibidores de catepsinaB madura são secretados por células de melanomaem cultura.Ensaios de imunoistoquímica com catepsina Bmarcada, indica que a peptidase é quase exclusiva emcélulas tumorais. A intensidade da marcação possuicorrelação positiva com o tamanho do tumor, mas nãocom o tipo de melanoma.
16
17
Próstata Apresenta correlação positiva entre ensaios deimunoistoquimica para catepsina B e índiceshistológicos de Gleason 5-8
18
*Referências: (1) WATANABE et al.,1987; (2) WATANABE et al.,1989; (3) KREPELA et al.,1990. (4) WERLE et al.,1995; (5) EBERT et al.,1994; (6) SHUJA et al.,1991; (7) SHEAHAN et al.,1989; (8) MURNANE et al.,1991; (9) PAGE etal.,1991; (10) KREPELA et al.,1989; (11) LAH et al.,1995; (12) LAH et al.,1992; (13) RECKLIES et al.,1982; (14) REMPEL et al.,1994; (15) SIVAPARVATHI et al.,1995; (16 ) TSUSHIMA et al.,1991; (17) FROHLICH et al.,1995; (18) SINHA et al.,1995.

- 19 -
O tráfego celular e o direcionamento de enzimas lisossomais são controlados,
classicamente, por vias que envolvem o reconhecimento de receptores de manose-6-fosfato
(BROWN et al.,1986). No entanto, vários trabalhos mostram que esta classe de receptores não
é suficiente para o direcionamento de várias enzimas lisossomais, seja por uma mudança na
expressão destes receptores (KASPER et al., 1996), seja por alterações no padrão de
glicosilação de enzimas lisossomais, tal como observado para a catepsina B em células
tumorais (IACOBUZIO-DONAHUE et al., 1997). De fato, o direcionamento lisossomal de
catepsina B, em células normais, independente do receptor de manose-6-fosfato tem sido
proposto (HANEWINKEL et al., 1987), bem como para a catepsina B de células de
carcinoma de cólon humano HT-29 (DE STEFANIS et al., 1997). Altos níveis de catepsina B,
além de modificações qualitativas na sua expressão protéica, o que inclui alteração do padrão
normal de glicosilação da enzima, são determinantes na manutenção do fenótipo de
malignidade de células tumorais (IACOBUZIO-DONAHUE et al., 1997). Alterações na
expressão, no processamento e na localização celular de catepsina B têm sido observados em
vários tecidos tumorais; investigações clínicas têm mostrado que a catepsina B é um indicador
altamente confiável para o diagnóstico e prognóstico de câncer (KOS; LAH, 1998).
Proteínas da matriz extracelular (HUMPHRIES et al., 1983; BUCK et al., 1992) e de
membrana basal (BARICOS et al., 1988; BRIOZZO et al., 1988) podem ser hidrolisadas pela
catepsina B. Estudos de imunoistoquímica realizados em vários tipos de carcinomas humanos:
carcinomas de bexiga (VISSHER et al., 1994), cólon (KHAN et al., 1998a), de estômago
(KHAN et al., 1998b) e pulmão (SUKOH et al. 1994), mostraram que a intensidade de
marcação para catepsina B e colágeno tipo IV ou laminina foram inversamente proporcionais
entre si, ou seja, quanto maior a quantidade de catepsina B detectada, menor foi a quantidade
de colágeno e lamina detectada nestes tumores.
Também foi demonstrado que as células de estroma, vizinhas ao tumor de mama, além
das células tumorais, também apresentam aumento na expressão de catepsina B
(CASTIGLIONI et al., 1994). Joensuu et al. (1995) obtiveram resultados similares para
catepsina D em carcinomas de mama.

- 20 -
1.1.3 Controle da Expressão Gênica e da Secreção de Catepsina B
A capacidade de componentes da matriz extracelular de mediar o controle da
expressão de cisteíno e aspartil-peptidases ainda não está totalmente elucidada. A hipótese
que os elementos da matriz extracelular das células de estroma possam modular a expressão
de catepsina B e D tumoral se torna muito importante quando analisada pelo ponto de vista
que estas enzimas possam estar contribuindo para a atividade degradativa tumoral
(KOBLINSKI et al., 2002). Foi observado que os fibroblastos adjacentes ao tecido tumoral
maligno apresentam elevada produção de serino-peptidases, metalo-peptidases e de catepsinas
B e D, em relação aos fibroblastos encontrados em estroma de neoplasia benigna (BASSET et
al., 1990). Interessantemente, foi demonstrado, que o colágeno tipo I é capaz de promover a
expressão e secreção da catepsina B em fibroblastos, via sua interação com as integrinas ·1·1 e
·2·2. Por outro lado, esta interação não foi capaz de estimular a síntese e secreção da catepsina
D. Este diferencial na indução de síntese e secreção de enzimas lisossomais distintas, mediado
por uma proteína da matriz, pode indicar diferentes papeis destas enzimas na fisiopatologia
dos processos tumorais (KOBLINSKI et al., 2002).
As interações entre integrinas com colágeno do tipo I, não são só responsáveis pelo
aumento da expressão de catepsina B, como também são responsáveis pelo aumento da
secreção de pró-catepsina B, sugerindo que a enzima secretada por esta via ainda não tenha
alcançado o endossomo ou o compartimento lisossomal. A quantidade de catepsina B ativa
que é secretada em resposta ao colágeno tipo I (KOBLINSKI et al., 2002), pode estar
refletindo o processamento da pró-catepsina B extracelularmente ou a exocitose da catepsina
B ativa dos lisossomos/endossomos, como já foi mostrado em uma grande variedade de
células (ANDREWS et al., 2000).
A secreção de catepsina B é regulada pela via do ras (SLOANE et al., 1994). Uma
grande variedade de agentes e tratamentos podem desencadear a exocitose dos lisossomos
dependendo do tipo celular (GUO et al., 2002). Exposição às partículas de zimosan em
macrófagos alveolares, agentes que inibem a H+-ATPase do vacúolo ou agentes que elevam o
pH do lisossomo, são situações que provocam a exocitose lisossomal (TAPPER et al., 1990;
SUNDLER et al., 1997). O metabólito de ácido araquidônico, o ácido12-S-hidroxi-
eicosatetraeonico (12-S-HETE) induz a exocitose de lisossomos em uma grande variedade de
células em cultura (HONN et al., 1994; ULBRICHT et al., 1996). Finalmente, condições que
rapidamente elevam as concentrações citosólicas de Ca2+ também induzem a exocitose dos
lisossomos em várias linhagens celulares. Recentes estudos têm identificado agentes
potencializadores, mas não iniciadores do processo de exocitose lisossomal, tais como o

- 21 -
AMPc e agentes que aumentam os níveis de AMPc, potencializam a ação do Ca2+,
desencadeando a exocitose do lisossomo (RODRIGUEZ et al., 1999). Igualmente, o TPA e
outros ésteres de forbol são agentes potencializadores, mas não desencadeadores da exocitose
lisossomal (TAPPER et al., 1995).
Ainda não se sabe o motivo porque são encontradas populações distintas de
lisossomos exocitados, ora apresentando teor de catepsina L, ora apresentando catepsina L
com catepsina B e vice-versa. As condições capazes de desencadear a exocitose lisossomal
seletiva de catepsina L, enzima da mesma família da catepsina B, ainda não foram
identificadas. Estudos recentes mostram que a catepsina L é exocitada por populações
lisossomais que são sensíveis à elevação de concentração do Ca2+ citosólico (MARTINEZ et
al., 2000). A literatura tem mostrado que o TPA induz a exocitose de lisossomos contendo
apenas catepsina B, mas não promove a secreção de catepsina L e D. As similaridades das
ações do TPA e 12-S-HETE são mais que apenas coincidências, a ênfase na natureza
heterogênea dos lisossomos/endossomos e a demonstração que estas organelas diferem em
suas susceptibilidades no processo de sinalização e no processo de desencadeamento da sua
exocitose, é um ponto crítico no estudo da secreção celular da catepsina B. Os recentes
estudos sugerem a existência de vários sinalizadores e mecanismos envolvidos na exocitose
lisossomal (GUO et al., 2002). Dados da literatura mostram que fisiologicamente as células
epiteliais de tireóide secretam catepsina B ativa por exocitose lisossomal estimulada pelo
hormônio TSH. A secreção da catepsina B ativa promove a liberação de tiroxina (T4) pela
degradação extracelular da tireoglobulina (LINKE, et al., 2002).
As secreções das formas ativas e da pró-catepsina B têm sido descritas em vários
tecidos neoplásicos (Tabela 2). A saturação da via da manose-6-fosfato pode provocar o
redirecionamento das enzimas lisossomais, fazendo com que estas enzimas sejam secretadas
na suas formas inativas (pró-catepsina B). A secreção da enzima madura é muito mais
complexa. Aparentemente, a secreção da enzima madura pode indicar que ela esteja sendo
processada intracelularmente em excesso, passando diretamente pelo tráfego lisossomal ou
também pode estar sendo ativada extracelularmente (FROSCH et al., 1999). Tooze at al.
(1991) mostrou que a catepsina B madura é secretada pelo pâncreas exócrino de rato pela
fusão de lisossomos com grânulos de reserva citoplasmático. Estes grânulos podem se
apresentar de dois modos: corpos crinofágicos, os quais degradam intracelularmente material
secretório, ou como grânulos secretórios, responsáveis por exocitar produtos de degradação
catabólica (HASHIZUME et al., 1993).

- 22 -
Os múltiplos caminhos relacionados no processo de secreção celular da catepsina B,
tanto da forma ativa como da pró-enzima, sugerem que a secreção de catepsina B pode ter
conseqüências funcionais determinantes para a degradação dos componentes da matriz
extracelular (KOBLINSKI et al., 2002).
TABELA 2: Localização Celular Alterada da Catepsina B em Tumores
*Referências. (1) FROHLICH et al.,1995; (2) TERADA et al., 1995; (3) SINHA et al.,1995; (4) RECKLIES et al.,1982; (5) POOLE et al.,1978; (6) VISSCHER et al.,1994; (7 ) SLOANE et al.,1986.
Tecido Tumoral Observação Referência*Melanoma Catepsina B foi encontrada em estruturas vesiculáres e
espaços extracelulares. 1
Colângio- e hepatocelular carcinomas
Catepsina B foi encontrada em todos os tipos celulares tumorais. Em tecidos normais, apenas em hapatócitos e raramente em fibroblastos, em ducto intra-hepático bilial as células epiteliais apresentaram marcação.
2
Próstata Catepsina B foi localizada nos compartimentos citoplasmático e nuclear das células tumorais.
3
Câncer de mama Tumores de mama humanos secretam pró-catepsina B. Quando comparado ao fibrossarcoma benigno, câncer de mama secreta maior quantidade da enzima.
4,5
Câncer de bexiga Catepsina B foi encontrada em tumores em estágios iniciais colocalizada aos lisossomos. Tumores em estágios avançados de evolução apresentaram marcação difusa por todo citoplasma.
6
Melanoma murino Fracionamento subcelular e ensaios de atividade indicam a ocorrência de uma mudança da atividade enzimática para a membrana plasmática. Este dado possui correlação positiva com o aumento do potencial metastático.
7
Nosso grupo demonstrou que a atividade da catepsina B é regulada pela presença de
glicosaminoglicanos “heparin-like” (ALMEIDA et al., 2001). Foi observado que a heparina e
o heparam sulfato são capazes de interagir com a His111 da alça de oclusão da catepsina B e
promover a estabilização da α-hélice central do sítio ativo da enzima em pH 7,4, a
estabilização conformacional favorece a atividade endopeptidásica da enzima por aumentar
em cinco vezes a meia-vida desta enzima em pH 7,4. A catepsina B tem sido envolvida nos
processos de invasão tumoral pela degradação de componentes da matriz extracelular (BUCK
et al., 1992). A degradação dos componentes da matriz pode ocorrer intracelularmente, pela
intensa atividade proteolítica da célula tumoral, ou extracelularmente, pela catepsina B
presente na superfície celular associada à membrana plasmática (SLOANE et al. , 1996). A
presença de catepsina B na membrana plasmática resulta em dissolução focal de proteínas da
matriz extracelular e permite a invasão de células tumorais (KOBLINSKI; SLOANE, 1997).
Nossos resultados sugerem que o heparam sulfato pode ancorar a catepsina B na superfície
celular, e esta interação favorece a atividade endopeptidásica da enzima por aumentar em
cinco vezes sua meia-vida em pH fisiológico (ALMEIDA et al., 2001), o que está de acordo

- 23 -
com a observação de que formas de catepsina B ligadas à membrana são muito resistentes à
inativação em pH neutro (SLOANE et al., 1988).
Os proteoglicanos de heparam sulfato da superfície celular estão em constante
renovação como resultado de sua secreção e endocitose contínua. Foi demonstrado que
algumas proteínas ligadas à cadeias de heparam sulfato são endocitadas juntamente com
proteoglicanos, como por exemplo: o fator de crescimento de fibroblastos (REILAND;
RAPRAEGER, 1993) e lipases lipoprotéicas (JACKSON et al., 1991). É interessante observar
que em vesículas lisossomais existe uma alta concentração de catepsina B (KOS; LAH, 1998)
e que heparam sulfato também está presente neste compartimento durante seu tráfego
intracelular (BIENKOWISKI; CONRAD, 1984). Estas observações sugerem que o
mecanismo de inserção da catepsina B na membrana plasmática, bem como seu tráfego
celular, possa ser dependente de proteoglicanos de heparam sulfato.
1.1.4 Papel da Catepsina B e dos Lisossomos na Apoptose de Células Tumorais
Além de sua função de degradar proteínas da matriz extracelular e de garantir
capacidade de invasão de células tumorais, a catepsina B tem sido implicada no controle de
apoptose. A catepsina B possui atividade de protease ativadora de morte celular dependente e
independente de caspases, disparando os mecanismos de apoptose em células tumorais.(FU et
al, 1998). Foi demonstrado que a ativação de apoptose em células infectadas por HPV,
induzidas pelo complexo inibitório da oncoproteína E7 de HPV e p21, está associada à
ativação da catepsina B, e este poderia ser um mecanismo alternativo para garantir a morte
celular das células infectadas. Também foi observado que a ativação da catepsina B ocorre em
células tumorais com mutações de caspases, bem como superexpressão de proteínas Bcl-2 e
p21 (KAZNELSON et al, 2004). Drogas antitumorais que induzem a permeabilização da
membrana lisossomal liberam catepsinas B e D para o citoplasma que, por sua vez, podem
induzir apoptose dependente de Bax (ERDAL et al.,2005). Interessantemente, compostos que
promovem a translocação de enzimas lisossomais para o citoplasma induzem morte celular
independentemente de p53 (ERDAL et al.,2005). Tem sido observado que TNF, via receptor-
1 (TNF-R1), além de ativar caspases, também promove apoptose mediada pela mobilização
de enzimas lisossomais para o citoplasma. Antagonicamente, NF-kappaB é capaz de bloquear
o efeito pró-apoptótico do TNF pela indução de uma proteína inibidora da atividade de
catepsina B, denominada Spi2A (LIU et al., 2003). Dados na literatura têm mostrado que a
catepsina B é capaz de ativar a proteína Bid na sua forma pró-apoptótica tBid (CIRMAN et
al., 2004) e degradar a enzima SK-1 que é inibidora de apoptose (TAHA et al.,2005). Desta

- 24 -
forma, as enzimas lisossomais, em especial a catepsina B, tem sido diretamente implicadas no
controle da apoptose.
As células tumorais possuem vários defeitos na suas vias de morte celular, tais como:
exacerbação da expressão gênica de proteínas antiapoptóticas, Bcl-2, Bcl-xL, Akt/PKB e
inibidores, bem como, apresentam mutações em proteínas pró-apoptóticas, com: Bax, Apaf-1
e p53. Essas alterações favorecem o desenvolvimento tumoral por bloquear a interferir
negativamente na ativação das caspases e, dessa forma, bloquear a apoptose pelas vias
clássicas. Por outro lado, durante os estágios iniciais da tumorigênese, muitas células
transformadas podem ser sensibilizadas por vários estímulos de morte e, freqüentemente,
muitas delas podem entrar em apoptose espontânea (JAATTELA, 2004).
Alterações marcantes observadas no compartimento lisossomal das células tumorais e o
direto envolvimento das enzimas lisossomais no controle da apoptose tumoral, sugerem que a
via lisossomal de disparo da morte celular pode contribuir, significantemente, para a
sensibilização do programa de morte em células tumorais. Dentre essas alterações lisossomais
promovida pela imortalização celular podemos destacar: 1) a elevação da sensibilidade da
membrana lisossomal a agentes lisossomotrópicos; 2) o aumento do volume dos lisossomos e
3) a elevação da sensibilidade da morte celular pela via lisossomal. Também, podem-se notar
as alterações lisossomais promovidas pela transformação celular: 1) elevação na expressão
gênica das catepsinas lisossomais; 2) alteração do tráfego celular dos lisossomos; 3) elevação
na taxa de secreção das catepsinas e 4) aparecimento da “heat shock protein” Hsp70 na
membrana lisossomal induzindo maior estabilidade da membrana lisossomal para as células
tumorais (FEHRENBACHER & JAATTELA, 2005).
Tem sido mostrado que a imortalização e a transformação, em fibroblastos embrionários
murinos, sensibilizam essas células para a indução de morte celular pela via lisossomal
disparada por TNF. Por si mesmo, a espontânea imortalização observada nessas células,
gerada pela inativação do seu sistema de p53, foi suficiente para sensibilizar em mais de 1000
vezes os fibroblastos embrionários murinos à ação apoptótica induzida pelo TNF. Essa
sensibilização foi altamente dependente da atividade da enzima lisossomal catepsina B.
Interessantemente, foi mostrado que o nocaute do gene da catepsina B nessas células de
fibroblasto embrionárias imortalizadas geram células resistentes à indução de apoptose pelo
TNF, de modo semelhante à ação do TNF nas células selvagens não imortalizadas. A
reintrodução da catepsina B nessas células mutantes defectivas de catepsina B foi suficiente
para promover a ressensibilização ao TNF, enquanto que a inibição farmacológica da
catepsina B ou a depressão da expressão gênica dessa enzima por RNA de interferência

- 25 -
significativamente protegeram as células imortalizadas contra a apoptose induzida por TNF.
Ainda, a transformação dos fibroblastos embrionários murinos imortalizados induzida por v-
Ha-ras ou c-Src promoveu uma grande elevação na taxa de expressão gênica das catepsinas B
e L (FEHRENBACHER et al., 2004). Dessa forma, os autores especularam que a
sensibilização destas células para o estímulo apoptótico disparado pelo TNF se deve à
exacerbação da via lisossomal presentes em células transformadas (FEHRENBACHER &
JAATTELA, 2005).
1.2 Heparanase em Células Tumorais e Metástases.
Em 1975, Hook descreveu a atividade de uma endoglicosidase de degradação do
heparam sulfato, primeiro relato da heparanase. Somente em 1983 relacionaram com a
invasibilidade e metastatização tumoral. A purificação da heparanase foi dificultada por sua
limitada disponibilidade e sua instabilidade (SIMIZU et al., 2004).
Os glicosaminoglicanos “heparin-like” estão amplamente distribuídos em organismos
multicelulares (Nader & Dietrich, 1989). Particularmente, a heparina e heparam sulfato,
dentre os demais glicosaminoglicanos, apresentam elevada capacidade de interação com
variado número de proteínas. Os glicosaminoglicanos “heparin-like” exercem um complexo
papel na regulação da matriz extracelular, controlando uma grande variedade de processos
biológicos, tais como: hemostase, inflamação, angiogênese, ação de fatores de crescimento,
adesão celular, entre outros (Conrad et al., 1998). Heparina e heparam sulfato são reguladores
da atividade proteolítica de algumas proteinases, bem como de seus inibidores (TERSARIOL
et al., 2002).
A heparanase é uma endo-β-glucuronidase que quebra ligações glicosídicas
intrassacarídicas do heparam sulfato. O gene da heparanase determina um polipeptídio de 543
aminoácidos com peso molecular de 61,2 kDa, entretanto a heparanase purificada apresenta
uma diferença de 156 aminoácidos a menos, esses resultados sugerem a existência de
proteólise pós-traducional para a enzima madura encontrada nos tecidos ( SIMIZU et
al.,2004). Essa enzima foi recentemente clonada, sendo que o gene da heparanase humana
apresenta um cDNA de 3.7 kb e encontra-se distribuído em cerca de 50 kb, contendo 14 éxons
e 13 íntrons, este gene está localizado no cromossomo humano 4q22 (DONG et al., 2000).
A heparanase promove a degradação de cadeias de heparam sulfato dos proteoglicanos
que são componentes da matriz extracelular, de membrana basal e da superfície celular. Em
tumores, a heparanase pode facilitar a invasão de células tumorais e metástases pela

- 26 -
degradação da membrana basal vascular e matriz extracelular (Nurcombe et al., 2000;
Vlodavski et al., 1999).
Pacientes que apresentavam altos níveis de expressão de heparanase em carcinoma
espinocelular de esôfago apresentaram pior prognóstico. Células endoteliais de novos vasos e
de vasos formados na periferia do tumor apresentavam uma intensa expressão de heparanase.
A heparanase facilita a angiogênese por liberar para o meio extracelular os fatores de
crescimentos angiogênicos ligados aos proteoglicanos de heparam sulfato. A elevação da
densidade de microvasos está correlacionada com a expressão de heparanase. (Takaomi,2004)
Há uma correlação entre pior prognóstico e a expressão aumentada da heparanase e o
carcinoma ductal de mama in situ. Sendo que em tecidos normais não foi encontrada dosagem
de heparanase ( Maxhimer et al., 2005). Também se identificou uma marcação intensa de
heparanase em áreas de invasão tumoral em cânceres ductais mamários ( ZCHARIA et
al.,2001).
Nos adenocarcinomas colorretais foi descrito a presença da heparanase na superfície
celular e no citoplasma (FRIEDMAN et al.,2000).
O conjunto destas observações sugere que o mecanismo de inserção da catepsina B na
membrana plasmática, bem como seu tráfego celular, possam ser dependente de
proteoglicanos de heparam sulfato e a liberação de catepsina B da superfície celular pode
estar relacionada com a atividade da heparanase. Assim, as enzimas catepsina B e a
heparanase são moléculas alvo para o desenvolvimento de agentes antitumorais e também
podem servir como marcadores moleculares para o diagnóstico e prognóstico de tumores.
1.3 Câncer de Cabeça e Pescoço
Os cânceres de cabeça e pescoço são os tumores malignos que acometem as estruturas
anatômicas do lábio, cavidade oral (mucosa bucal, gengiva, palato duro, língua oral, assoalho
da boca), orofaringe (amídalas, base da língua, palato mole), nasofaringe, fossa nasal e seios
paranasais, laringe, hipofaringe, glândulas salivares e tireóide. Excluímos no presente trabalho
os cânceres de tiróide e de glândulas salivares, pois são histologicamente diferentes.
A incidência de casos novos diagnosticados nos EUA, excluindo os cânceres de pele, é
estimada em 70.000 casos anualmente. Representa 5% de todos os cânceres diagnosticados.
Predominantemente são espinocelulares (90%). E são responsáveis por cerca de 8100
óbitos/ano (cânceres de cavidade oral e faringe) e 4200 óbitos/ano em câncer de laringe
(American Cancer Society,1997). Nas Estimativas de Incidência e Mortalidade por Câncer no
Brasil em 2003, as neoplasias são responsáveis por cerca de 12,7% dos óbitos no Brasil sendo

- 27 -
destes 2,84% cânceres de cavidade oral e 2,09% cânceres de laringe. As estimativas de casos
novos de câncer de cavidade oral para 2003 eram de 10.600 caso com 3200 óbitos
(INCA/MS/2003)
Há uma forte associação entre os cânceres do trato aerodigestivo alto e o uso do
tabaco. Estudos epidemiológicos mostram um aumento do risco em 20 a 40 vezes para os
fumantes. O consumo de álcool também é um dos maiores fatores de risco, acrescentando um
risco de 5 a 11 vezes de ter a doença em relação ao só fumante ou ao não fumante
(TALAMINI et al., 2002).
O paciente que apresenta um tumor de cavidade oral, em geral, se queixa de uma
úlcera na boca que não cicatriza, perda de dentes, prótese dentária que não se adapta mais,
trismo ou perda de peso. Também, podem reclamar de disfagia, odinofagia, sangramento ou
otalgia. É importante para o médico ter um alto nível de suspeita principalmente em fumantes
e etilistas. Cerca 30 % dos pacientes com neoplasia de cabeça e pescoço tem como primeira
queixa uma metástase em linfonodo cervical (SHAH et al, 1990)
A localização do linfonodo metastático ajuda na procura do primário, pois apresenta
uma correlação positiva do primário e seu primeiro sítio de metastatização.
Os estudos de imagens são fundamentais no estadiamento, principalmente a
Tomografia Computadorizada, ajudando na avaliação da extensão do tumor, sua infiltração de
estruturas anexas, e aumenta de 70 a 93 % a efetividade na detecção de metástase cervical (
STEVENS et al, 1985 ).
O sistema TNM para o estadiamento dos cânceres de cabeça e pescoço permite ao
médico comparar resultados, discutir prognóstico, e decidir o tratamento adequado. A
classificação T se refere ao tamanho e invasão de estruturas anexas da lesão primária, N para
a situação dos linfonodos cervicais e o M para a presença ou não de metástase à distância, nas
figuras 2, 3 e 4 temos modelos de classificação linfonodal, laríngea e de cavidade oral,
respectivamente (AMERICAN JOINT COMMITTEE ON CANCER E UICC, 2002 ).
A cirurgia se mantém como padrão de tratamento para os carcinomas espinocelulares
de cabeça e pescoço. Tendo como objetivo englobar o tratamento da lesão primária e o
pescoço envolvido. Principalmente nos estadios avançados (estadio III e IV). A radioterapia
pode ser usada como tratamento adjuvante pré- ou pós-cirúrgico. Usualmente é administrada
pós-operatoriamente. Está particularmente indicada nos casos com linfonodos positivos.

- 28 -
Primariamente está indicada nos casos precoces de câncer de laringe (estadio I e II) e nos
carcinomas de nasofaringe.
A quimioterapia tem sido intensamente estudada, estando presente em vários
protocolos de tratamento neoadjuvante. Muito interesse vem apresentado os modelos de
preservação de órgãos nos casos de câncer de laringe (WOLF,et al, 1993)
Como nova terapêutica, mostrando ser promissora, está a terapia genética onde a
introdução, nas células somáticas do paciente, de genes para fabricarem proteínas com efeito
terapêutico (LIU , et al, 1994).

- 29 -
Figura 2. Estadiamento Linfonodal. N1, linfonodo ipsilateral da lesão com até 3 cm. N2a, linfonodo ipsilateral da lesão de 3 a 6 cm. N2b, mais de um linfonodo ipsilateral da lesão menores de 6 cm. N2c, presença de linfonodo contralateral da lesão menor de 6 cm. N3, linfonodo de tamanho igual ou maior de 6 cm (American Joint Committee on Cancer Staging Manual, 6th ed., 2002)

- 30 -
Figura 3. Estadiamento de tumores de laringe. T1, limitado a uma subregião da supraglote, ou limitado a prega vocal com motilidade preservada (T1a, uma prega e T1b, ambas as pregas), ou limitado a subglote. T2, mucosa de mais que uma sub-região da supraglote ou glote ou região adjacente, sem fixação da prega vocal; ou nos tumores glóticos extensão para supra ou subglote, ou diminuição da mobilidade de prega vocal; ou nos de subglote extensão para prega vocal com mobilidade normal ou diminuída. T3, fixação de prega vocal ou invasão da área pós-cricóide, pré-epiglótica, espaço paraglótico ou erosão da cartilagem tireóidea. T4a, extravasamento da laringe e T4b, invasão do espaço pré-vertebral, mediastino ou artéria carótida (American Joint Committee on Cancer Staging Manual, 6th ed., 2002).
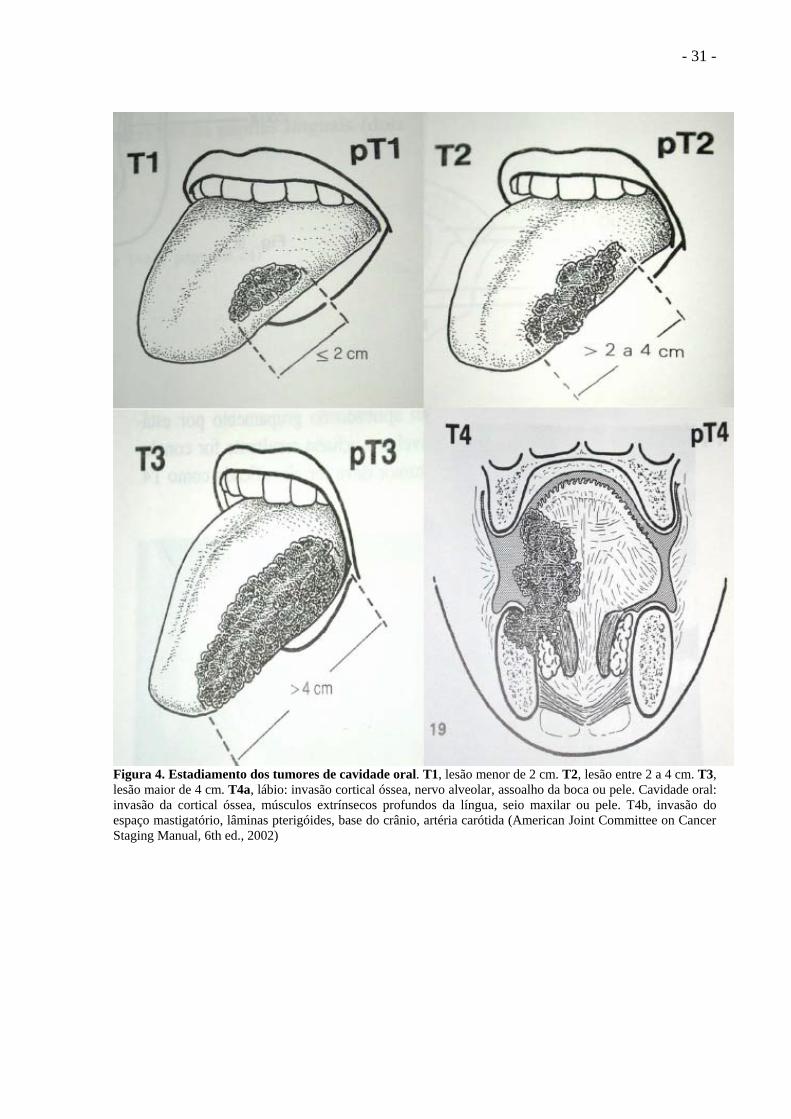
- 31 -
Figura 4. Estadiamento dos tumores de cavidade oral. T1, lesão menor de 2 cm. T2, lesão entre 2 a 4 cm. T3, lesão maior de 4 cm. T4a, lábio: invasão cortical óssea, nervo alveolar, assoalho da boca ou pele. Cavidade oral: invasão da cortical óssea, músculos extrínsecos profundos da língua, seio maxilar ou pele. T4b, invasão do espaço mastigatório, lâminas pterigóides, base do crânio, artéria carótida (American Joint Committee on Cancer Staging Manual, 6th ed., 2002)

- 32 -
2 MATERIAIS
2.1 Obtenção do tecido tumoral
Foram separados fragmentos de tecidos provenientes de biópsias, previamente indicados
para exame anátomo-patológico, de pacientes encaminhados ao Departamento de Cirurgia de
Cabeça e Pescoço do Hospital Estadual Mário Covas – Santo André (HEMC), SP. Nos casos
de câncer, no ato do exame clínico do paciente foi feito o estadiamento clínico da lesão, pelo
do sistema TNM (T, extensão do tumor primário; N, linfonodos regionais; M, metástase à
distância), proposto pela American Joint Comittee of Cancer de 1988 e União Internacional
de Combate ao Câncer, revisada de 2002 (Tabela 3). Os procedimentos semiológicos,
cirúrgicos e triagem dos pacientes são da responsabilidade do cirurgião Emílio López Couto
da equipe de cirurgia de cabeça e pescoço do HEMC. Partes dos fragmentos de tecidos
removidos de forma rotineira por biópsias para análise histopatológica foram imediatamente
congelados e enviados ao laboratório do CIIB-UMC, para o devido processamento, num total
de 9 lesões benignas.
Conforme o anexo A obtivemos aprovação da comissão de ética em pesquisa da
faculdade de medicina ABC utilizando o modelo de autorização do anexo B.
Foram divididos pela localização anatômica em Cavidade Oral, Orofaringe, Laringe e
Hipofaringe. As lesões de lábio da Cavidade Oral foram excluídas por terem outro fator
desencadeante (exposição solar). Foram analisados 20 pacientes no período de julho de 2004
a fevereiro de 2006. Sendo 18 do sexo masculino e 2 femininos. Divididos por região
anatômica, obtivemos: 3 de cavidade oral, 6 de orofaringe, 9 de laringe e 2 de hipofaringe.
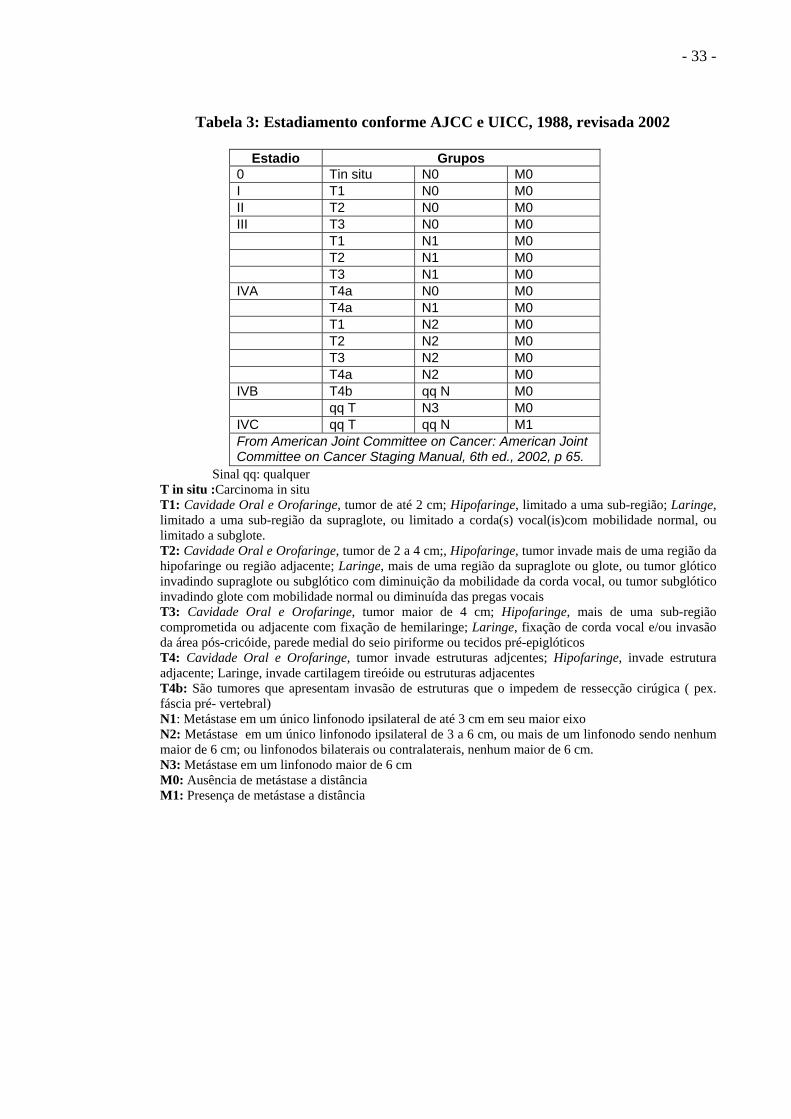
- 33 -
Tabela 3: Estadiamento conforme AJCC e UICC, 1988, revisada 2002
Estadio Grupos 0 Tin situ N0 M0 I T1 N0 M0 II T2 N0 M0 III T3 N0 M0 T1 N1 M0 T2 N1 M0 T3 N1 M0 IVA T4a N0 M0 T4a N1 M0 T1 N2 M0 T2 N2 M0 T3 N2 M0 T4a N2 M0 IVB T4b qq N M0 qq T N3 M0 IVC qq T qq N M1 From American Joint Committee on Cancer: American Joint Committee on Cancer Staging Manual, 6th ed., 2002, p 65.
Sinal qq: qualquer T in situ :Carcinoma in situ T1: Cavidade Oral e Orofaringe, tumor de até 2 cm; Hipofaringe, limitado a uma sub-região; Laringe, limitado a uma sub-região da supraglote, ou limitado a corda(s) vocal(is)com mobilidade normal, ou limitado a subglote. T2: Cavidade Oral e Orofaringe, tumor de 2 a 4 cm;, Hipofaringe, tumor invade mais de uma região da hipofaringe ou região adjacente; Laringe, mais de uma região da supraglote ou glote, ou tumor glótico invadindo supraglote ou subglótico com diminuição da mobilidade da corda vocal, ou tumor subglótico invadindo glote com mobilidade normal ou diminuída das pregas vocais T3: Cavidade Oral e Orofaringe, tumor maior de 4 cm; Hipofaringe, mais de uma sub-região comprometida ou adjacente com fixação de hemilaringe; Laringe, fixação de corda vocal e/ou invasão da área pós-cricóide, parede medial do seio piriforme ou tecidos pré-epiglóticos T4: Cavidade Oral e Orofaringe, tumor invade estruturas adjcentes; Hipofaringe, invade estrutura adjacente; Laringe, invade cartilagem tireóide ou estruturas adjacentes T4b: São tumores que apresentam invasão de estruturas que o impedem de ressecção cirúgica ( pex. fáscia pré- vertebral) N1: Metástase em um único linfonodo ipsilateral de até 3 cm em seu maior eixo N2: Metástase em um único linfonodo ipsilateral de 3 a 6 cm, ou mais de um linfonodo sendo nenhum maior de 6 cm; ou linfonodos bilaterais ou contralaterais, nenhum maior de 6 cm. N3: Metástase em um linfonodo maior de 6 cm M0: Ausência de metástase a distância M1: Presença de metástase a distância

- 34 -
2.2 Detecção das Enzimas em Cortes Histológicos
As biópsias de mucosa orofaríngea foram fixadas em formol tamponado e incluídas em
parafina. Para as reações de imunoistoquímica os cortes histológicos de 4 μm foram
colocados em lâminas previamente silanizadas com organosilano 4 % em acetona (3-
aminopropril trietoxisilano – SIGMA). As lâminas foram submetidas a banhos consecutivos
em baterias de xilol e álcool para a completa desparafinização. A atividade da peroxidase
endógena foi bloqueada tratando as lâminas com 2 banhos consecutivos de peróxido de
hidrogênio 3% em metanol (v/v), por 15 minutos, à temperatura ambiente. A seguir, para a
recuperação antigênica, os cortes foram submetidos a fervura em panela à vapor do tipo
steam-cuisine, imersos em tampão citrato de sódio 10mM, pH 6,0 durante 70 minutos, a
95°C. Após o resfriamento, as lâminas foram lavadas 3 vezes com PBS, por 5 minutos cada
lavagem. A seguir, para o bloqueio de reações inespecíficas, as lâminas foram incubadas com
leite desnatado 1% em PBS por 40 minutos. A seguir, as lâminas foram, novamente, lavadas
em PBS e adicionado sobre os cortes histológicos o anticorpo primário monoclonal
anticatepsina B e anti-heparanase (Sigma), as lâminas foram incubadas à temperatura
ambiente, overnight em câmara úmida. Decorrido o período de incubação, as lâminas foram
submetidas à lavagem com PBS e incubadas com o anticorpo secundário marcado com biotina
(LSAB, DAKO) em câmara úmida por 30 minutos à 37ºC. A seguir, as lâminas foram
novamente lavadas, 3 vezes em PBS, por 5 minutos cada lavagem, e incubadas com
complexo Streptavidina-Biotina-Peroxidase (LSAB, DAKO), em câmara úmida por 30
minutos, à 37ºC. Em seguida, os cortes foram lavados em PBS, e a reação revelada com
incubação dos cortes em solução de diaminobenzidina e peróxido de hidrogênio à 37ºC, por
cinco minutos. Após a incubação, as lâminas foram lavadas em água corrente, e finalmente
coradas com hematoxilina de Carrazi por 60 segundos, lavados novamente em água corrente,
desidratadas em baterias de álcool e xilol e montadas em Permount (Fisher Scientific).
2.3 Obtenção do Extrato Bruto Tecidual
Fragmentos de tecidos humanos, provenientes das biopsias da mucosa orofaríngea,
foram homogeneizados em Polter, utilizando pestilo revestido de teflon, no tampão acetato de
sódio 0,05M, pH 5,0 a 4°C. Para remover fragmentos de tecidos insolúveis da preparação, o
homogenato foi centrifugado por 5 mim 10.000 x g a 4°C, o sobrenadante foi recuperado
obtendo-se, desta forma,l o extrato bruto tecidual. O extrato bruto foi estocado a -20°C e
dosado quanto a quantidade de proteína presente, pelo método de Comassie Blue (Spector) e
quanto ao teor da enzima catepsina B.

- 35 -
2.4 Dosagem da Atividade Enzimática da Catepsina B nos Tecidos
A atividade de catepsina B presente no extrato bruto foi monitorada pela hidrólise de
substrato fluorogênico Z-F-R-MCA a 37 ºC, utilizando espectrofluorímetro Hitachi F-2500. O
comprimento de onda do fluorímetro foi ajustado para, λ = 380 nm e λexcitação emissão= 460nm,
respectivamente, fenda 10/10 nm. Foram utilizadas cubetas de caminho óptico de 10 mm,
volume final de 2 ml. As soluções para reação foram mantidas no compartimento
termostatizado durante 5 minutos antes da adição da enzima. Antes do ensaio, as cisteíno-
proteases presentes no extrato bruto foram ativadas com DTT 2 mM, 5 min. a 37 ºC em
tampão fosfato de sódio 50 mM, pH 6.0, NaCl 200 mM e EDTA 1 mM. A seguir, foi
adicionada à preparação enzimática 5 μM do substrato fluorogênico Z-F-R-MCA e a
fluorescência liberada decorrente da hidrólise do substrato foi monitorada em tempo real. A
atividade endopeptidásica da catepsina B presente no extrato bruto tecidual foi certificada
pela adição de 5 μM de seu inibidor irreversível específico CA-074 à preparação. O resultado
da quantidade enzimática foi expresso em unidades de enzima por miligramas de proteína do
tecido.
2.5 Quantificação e Interpretação de Heparanase nas Reações de Imunoistoquímica.
Cerca de 8 a 10 campos com células marcadas foram selecionados de cada lâmina para a
análise quantitativa da heparanase. As células positivas para heparanase foram quantificadas
em 100 células/biópsia. Assim, a porcentagem de expressão imunoistoquímica foi
determinada para a heparanase no presente estudo. A expressão quantitativa de heparanase foi
classificada em índices de positividade (IP): negativo IP0 (0 a 5%); IP1 (> 5% a 30% de
positividade), IP2 (> 30% a 60%) e IP3 (>60% a 83%), de acordo com MILLON et al. (2001).
Ainda, a expressão de todas as proteínas foi avaliada segundo a intensidade e distribuição
tecidual da coloração. Para análise estatística das lâminas de imunoistoquímica, a
significância foi determinada pelo teste de Chi-quadrado (χ2). Foi utilizado o software de
análise de imagem: Sistema de Processamento e Análise de Imagem Imagelab ® (Softium
Informática).
As lâminas foram analisadas em microscópio óptico Nikon Eclipse® TS100, sendo a
área mais representativa determinada em aumento 40 e 100x, e para análise da
imunoexpressão 400x, foram realizadas captações de imagens com a câmera digital Nikon
Coolpix® 4300, com zoom óptico máximo, abertura F13.4. As imagens foram transferidas
para um computador Pentium 4 ®

- 36 -
2.6 Análise Estatística da Atividade Enzimática.
Os dados foram analisados estatisticamente pelo teste t de student, com o auxílio do Programa
computacional Excell da Microsoft. Os dados foram considerados estatisticamente
significantes com p < 0,05, ou seja, 95% de confiabilidade Exceto a idade, o tamanho dos
tumores (T),e a presença de metástase em tumores da laringe em função do teor de catepsina
B, todos os outros dados foram estatisticamente significantes.

- 37 -
3 Resultados
3.1 Atividade da Catepsina B
Avaliamos a atividade da Catepsina B em 20 amostras de carcinomas espinocelulares
de cabeça e pescoço, excluindo lesões de pele e de lábio. Realizando a análise sob diversos
parâmetros procurando padrões e diferenças entre os grupos.
No período de Julho de 2004 a Fevereiro de 2006 coletamos amostras de 20 pacientes
que foram submetidos à ressecções cirúrgicas de carcinomas espinocelulares de cavidade oral,
orofaringe, laringe e hipofaringe. Dos 20 pacientes, 18 eram do sexo masculino e 2 do
feminino. Com idade variando de 42 anos a 71 anos. Em 3 pacientes tivemos lesões de
cavidade oral, 6 pacientes com acometimento da orofaringe, 9 pacientes encontramos tumores
de laringe e 2 com lesões de hipofaringe. O estadiamento variou entre Estadio II a IV, não
sendo obtidas amostras de pacientes Estadio I. No Estadio II foi somente 1 paciente, Estadio
III foram 4 pacientes e Estadio IV foram 15 pacientes. Conforme a revisão de 2002 da UICC
nos Estadio IV encontramos 1 Estadio IVc, 1 Estadio IVb e 13 Estadio IVa (Tabela IV).
Dividimos em dois grupos por idade, sendo o primeiro grupo de 12 pacientes com 40 a
55 anos de idade e um segundo grupo de 8 pacientes com idade de 56 a 75 anos. Ao
analisarmos a atividade média e o erro padrão da atividade da Catepsina B por grupo, não
observamos diferença significativa. No primeiro grupo (40 – 55 anos, N=12) os valores foram
de 24 ± 5 UAF/min.μg e no segundo grupo (56 – 75 anos, N=8) os valores foram de 23 ± 6
UAF/min.μg (Figura 5).
Na Tabela 5 foram colocados os valores individualizados da atividade da Catepsina B
encontradas nos pacientes portadores de carcinoma espinocelular, com suas diferentes
apresentações patológicas e o estadiamento conforme a American Joint Comittee of Cancer de
2002. Como todos os pacientes foram passíveis de ressecção cirúrgica não temos representado
T4b, que são tumores avançados com impossibilidade de ressecção cirúrgica. Apresenta um
paciente estadio IVc, pois apresentava metástase à distância. A atividade específica da enzima
em carcinoma espinocelulares variou de 3,4 – 62,1 UAF/min.μg de proteína e para as lesões
benignas variou de 1,8 – 6,2 UAF/min.μg de proteína.
Os resultados da atividade da Catepsina B também foram distribuídos pelo
estadiamento comparativamente com os resultados das amostras de hiperplasia benignas de
mucosa oral. Não obtivemos amostra estadio I. Nos estadios II tivemos um paciente com
atividade enzimática de 14,1 UAF/min/µg de proteína. Obtivemos quatro pacientes estadio III

- 38 -
com variação da atividade enzimática de 3,4 a 26,6 UAF/min/µg. O maior grupo foi o do
estadio IV com 15 pacientes com valores de catepsina B variando de 9,0 a 62,1 UAF/min/µg.
As lesões benignas em número de nove tiveram a atividade da catepsina B variando de 1,8 a
6,2 UAF/min/µg (Tabela 6).
Realizamos um gráfico de dispersão com os resultados da atividade da Catepsina B
nos Estadiamentos conforme AJCC 2002 e com as lesões benignas, observando o padrão de
dispersão dos resultados. Onde nas lesões benignas a atividade se concentrava na base do
gráfico e no estadio IV se dispersava em valores mais elevados (Figura 6).
O valor médio da atividade da Catepsina B e o erro padrão determinado nas lesões
cancerosas foi de 23,5 ± 3,6 UAF/min.μg de proteína e nas lesões benignas o valor médio e
erro padrão foi de 3,4 ± 0,5 UAF/min.μg de proteína. Os dados apresentados mostram que as
lesões cancerosas apresentam cerca de 8 vezes mais atividade enzimática do que as lesões
benignas (Figura 7). O gráfico representa o elevado teor de atividade da Catepsina B nas
lesões malignas comparativamente com as lesões benignas.
A Figura 7 mostra a distribuição de Catepsina B em amostras teciduais provenientes
de lesões benignas (Figs. 8A e 8C) e lesões malignas (Figs. 8B e 8D). A Figura 8 apresenta
cortes teciduais provenientes da mucosa orofaríngea revelados por reações de
imunoistoquímica para Catepsina B em marrom e núcleo celular em azul. As Figs. 8A e 8C,
representativas das lesões benignas, mostram bordas bem delimitadas e em intensa
proliferação epitelial, bem como áreas de tecido conjuntivo íntegro adjacente à hiperplasia. Foi
verificada que a Catepsina B, nas lesões benignas, aparece fracamente marcada em marrom de
localização exclusivamente intracelular em grânulos ao redor do núcleo, sugerindo uma
localização lisossomal perinuclear. As Figs 8B e 8D são representativas de carcinomas
espinocelulares, claramente pode-se observar a presença de tecido epitelial invadindo a matriz
extracelular adjacente ao epitélio tumoral. Ao contrário do padrão de quantidade e localização
celular de catepsina B observada para as lesões benignas, foi verificado que a Catepsina B em
carcinoma espinocelular aparece fortemente marcada em marrom e com localização celular
anômala, pois foi detectada a presença dessa enzima na matriz extracelular. Também as Figs.
8B e 8D mostram que a Catepsina B aparece particularmente concentrada na porção infiltrante
tumoral, região de intensa remodelação do tecido conjuntivo adjacente.

- 39 -
0
5
10
15
20
25
30
1
Cate
psin
aB(
UAF
/min
*ug)
40 - 55 56 - 75
Figura 5. Atividade da Catepsina B em Função da Idade dos Pacientes. Grupo de paciente de 45 a 55 anos apresentou atividade específica de catepsina B igual a 24 ± 5 UAF/min/μg (média ± erro padrão, N=12) e o grupo de 56 aos 75 anos apresentou 23 ± 6 UAF/min/μg (média ± erro padrão, N=08).

- 40 -
0
5
10
15
20
25
30
35
40
45
50
55
60
65
0 1 2 3 4 5 E I E II E III E IV Benig
Cat
epsi
na B
(UA
F/m
in*u
g)
Figura 6. Atividade Enzimática da Catepsina B em Carcinomas Espinocelulares e Hiperplasias Benignas. A atividade enzimática da catepsina B nos tecidos provenientes da mucosa orofaríngea humana sendo:E I= Estadio I ( Coluna 1, N=0 ), E II= Estadio II (Coluna 2, N = 1), E III= Estadio III ( Coluna 3 , N = 4 ), E IV= Estádio IV ( Coluna 4, N = 15 ) e hiperplasias benignas (Coluna 5, N = 09) foi determinada espectrofluorimetricamente utilizando o substrato fluorigênico Z-FR-MCA. Cerca de 10 μl do homogenato tecidual foram incubados a 37ºC com 5 μM do substrato Z-FR-MCA no tampão fosfato de sódio 50 mM, pH 6,3 contendo 200 mM NaCl, 1 mM EDTA e 2 mM DTT, na presença ou na ausência de 2μM do inibidor irreversível específico da catepsina B, CA-074.
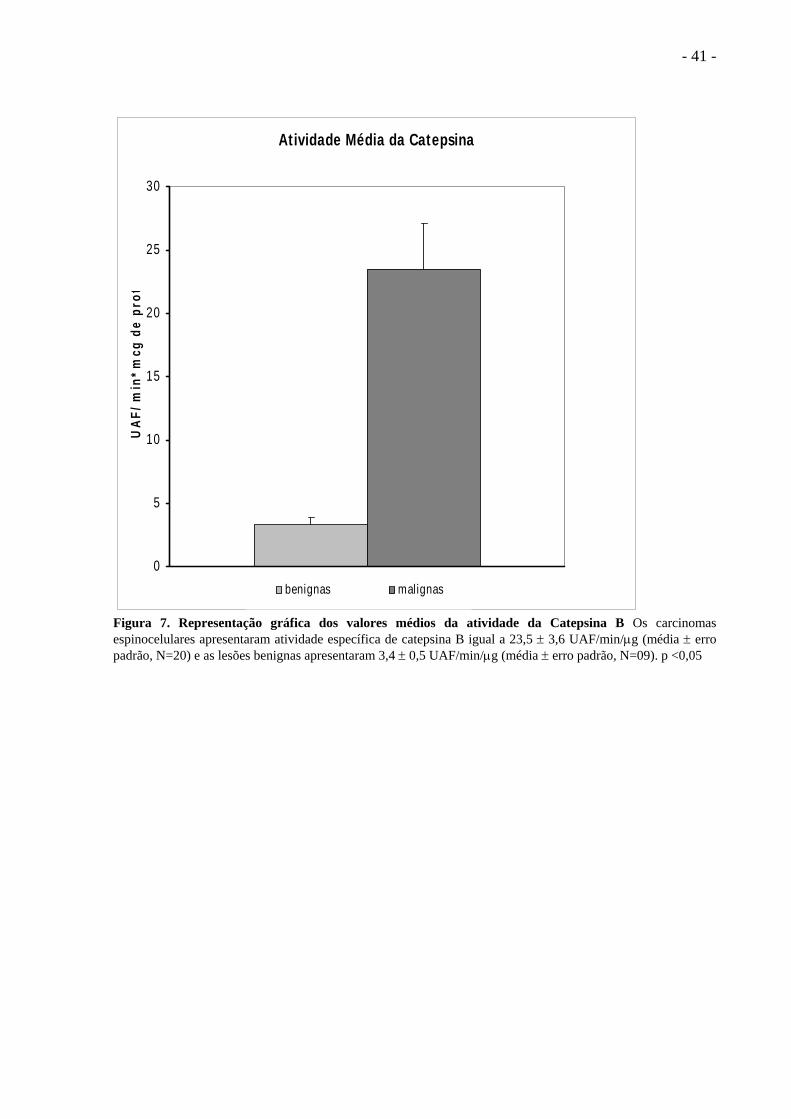
- 41 -
Atividade Média da Catepsina
0
5
10
15
20
25
30
1
UAF
/min
*mcg
de
prot
benignas malignas
Figura 7. Representação gráfica dos valores médios da atividade da Catepsina B Os carcinomas espinocelulares apresentaram atividade específica de catepsina B igual a 23,5 ± 3,6 UAF/min/μg (média ± erro padrão, N=20) e as lesões benignas apresentaram 3,4 ± 0,5 UAF/min/μg (média ± erro padrão, N=09). p <0,05

- 42 -
A B
C D
AA BB
CC DD
Figura 8. Detecção de Catepsina B por Reações de Imunoistoquímica em Carcinomas Espinocelulares e Hiperplasias Benignas. Cortes histológicos de 4 μm de tecidos embebidos em parafina provenientes da mucosa orofaríngea humana foram pré-tratados para recuperação antigênica, conforme descrito em Métodos, e incubados com anticorpo anticatepsina B humano na diluição 1:80. A ligação do anticorpo primário anticatepsina B ao tecido foi monitorada pela adição de anticorpo secundário conjugado com peroxidase. Após reação de peroxidase, as lâminas foram contra-coradas por hematoxilina. As Figs. 8A e 8C representam amostras de hiperplasias benignas, as Figs. 8B e 8D representam amostras de carcinoma espinocelular. Símbolos: a seta indica o epitélio espinocelular, a cruz indica o tecido conjuntivo adjacente e a barra representa a escala de 50 μm. (fonte do Autor)

- 43 -
3.2 Classificação TNM e Região Anatômica
Observando os valores elevados da atividade da Catepsina B nos tumores,
desdobramos os resultados por classificação do Tumor, por presença de Linfonodos
comprometidos, presença de Metástase e localização Anatômica do tumor. Tentamos observar
um padrão de maior ou menor expressão enzimática, o que foi prejudicado pelo N pequeno.
Na Tabela 7 comparamos a classificação T do estadiamento TNM com as lesões
benignas. Nos casos benignos N=9 tivemos uma variação da atividade da Catepsina B de 1,8
a 6,2 UAF/min/μg , obtivemos um paciente T1 com atividade de 15,57 UAF/min/μg , na
classificação T2 tivemos 4 pacientes com atividade enzimática de 3,39 a 48,83 UAF/min/μg
, pacientes T3 em número de 3 com Catepsina B de 5,39 a 26,62 e no grupo classificado com
T4 com N=12 e atividade enzimática de 9,03 a 62,12 UAF/min/μg .
Dividimos em três grupos, grupo A com lesões benignas em número de 9 e atividade
da Catepsina B e erro padrão de 3,4 ± 0,5 UAF/min/μg com Grupo B pacientes com tumores
mais precoces T1, T2 e T3, obtivemos valores de atividade enzimática e erro padrão de 19,8 ±
5,6 UAF/min/μg, N=8 e no Grupo C os tumores avançados T4 com atividade de 25,4 ± 4,9
UAF/min/μg , N=12 (Figura 9).
Há uma forte correlação entre a progressão da doença para uma atividade
metastatizante e o aumento dos valores da atividade da catepsina B. Em lesões benignas a
atividade foi de 3,4 ± 0,5 UAF/min/µg, N = 9. Nos tumores malignos sem metástase os
valores foram de 15 ± 2 UAF/min/µg, N = 8 e nos Tumores com metástase a atividade
enzimática foi de 26 ± 5 UAF/min/µg, N = 9 (Figura 10).
Em relação à metástase à distância, somente tivemos um paciente submetido à
ressecção cirúrgica com presença de metástase à distância. O paciente com a metástase
apresentava um valor alto de atividade da Catepsina B de 48,8 UAF/min/µg.
Na avaliação dos tumores laríngeos dividimos em dois grupos, o primeiro com
pacientes estadio II e III com atidade da Catepsina e erro padrão de 11 ± 4 UAF/min/ug, N=4,
nas lesões mais invasivas estadio IV obtivemos uma atividade de 31 ± 10 UAF/min/ug, N=5
(Figura 11).
Continuando na avaliação das lesões laríngeas dividimos em dois grupos o primeiro
sem metástase e o segundo com, obtivemos resultados sem diferença significativa da
atividade da Catepsina B nos dois grupos. No grupo sem metástase a atividade foi de 16 ± 4
UAF/min/ug, e no grupo com metástase 16 ± 3 UAF/min/ug (Figura 12).

- 44 -
A presença de metástase nas lesões de orofaringe está positivamente correlacionada
com a atividade enzimática da Catepsina B conforme a avaliação dos dois grupos (Figura
13). No grupo sem metástase a atividade foi de 15 ± 3 UAF/min/ug e no grupo com metástase
31 ± 6 UAF/min/ug. Apresentando uma atividade duas vezes maior nas lesões com metástase.
Na Tabela 8 dividimos por regiões anatômicas, onde encontramos uma tendência de
termos valores mais elevados da atividade enzimática nas lesões da Cavidade Oral, sendo na
laringe encontrado o menor valor, porém pertence a um paciente com doença localmente
precoce, e o maior valor em um paciente com doença localmente
invasiva.
0
5
10
15
20
25
30
35
1
Cate
psin
aB(
UAF
/min
*ug)
A B C
Figura 9. Atividade da Catepsina B e Classificação do Tumor. Grupo A, atividade específica de catepsina B nas lesões benignas 3,4 ± 0,5 UAF/min/μg ( N=9). Grupo B, teor de catepsina B nos pacientes com tumores mais precoces T1, T2 e T3 igual a 19,8 ± 5,6 UAF/min/μg ( N=8) e no Grupo C os tumores avançados T4 apresentam 25,4 ± 4,9 UAF/min/μg ( N=12). p <0,05

- 45 -
0
5
10
15
20
25
30
35
A B C
Cate
psin
a B
(UAF
/min
*ug)
Figura 10. Atividade de Catepsina B em Tumores Espinocelulares. Os valores representam a média e o erro padrão da atividade específica de catepsina B presente em tumores do epitélio espinocelular.(A) tumores benignos, hiperqueratose (3,4 ± 0,5 UAF/min.ug, N = 9); (B) tumores sem metástase (15 ± 2 UAF/min/ug, N = 8) e (C) tumores com metástase (26 ± 5 UAF/min/ug, N = 9). p <0,05

- 46 -
0
5
10
15
20
25
30
35
40
45
A B
Cate
psin
a B
(UAF
/min
*ug)
Figura 11. Quantidade de Catepsina B em Relação ao Estadiamento dos Tumores de Laringe. Os valores representam a média e o erro padrão da atividade específica de catepsina B presente em tumores de laringe.(A) tumores em estadios II e III (11 ± 4 UAF/min/ug, N = 4) e (C) tumores em estadio IV (31 ± 10 UAF/min/ug, N = 5). p <0,05

- 47 -
0
5
10
15
20
25
A B
Cate
psin
a B
(UAF
/min
*ug)
Figura 12. Atividade de Catepsina B em Tumores de Laringe. Os valores representam a média e o erro padrão da atividade específica de catepsina B presente nas lesões cancerosas de laringe.(A) tumores sem metástase (16 ± 4 UAF/min/ug, N=4) e (B) tumores com metástase (16 ± 3 UAF/min/ug, N=5).

- 48 -
0
5
10
15
20
25
30
35
40
A B
Cate
psin
a B
(UAF
/min
*ug)
Figura 13. Atividade de Catepsina B em Tumores da Cavidade Orofaríngea. Os valores representam a média e o erro padrão da atividade específica de catepsina B presente nas lesões cancerosas da cavidade orofaríngea.(A) tumores sem metástase (15 ± 3 UAF/min/ug, N=5) e (B) tumores com metástase (31 ± 6 UAF/min/ug, N=6). p <0,05
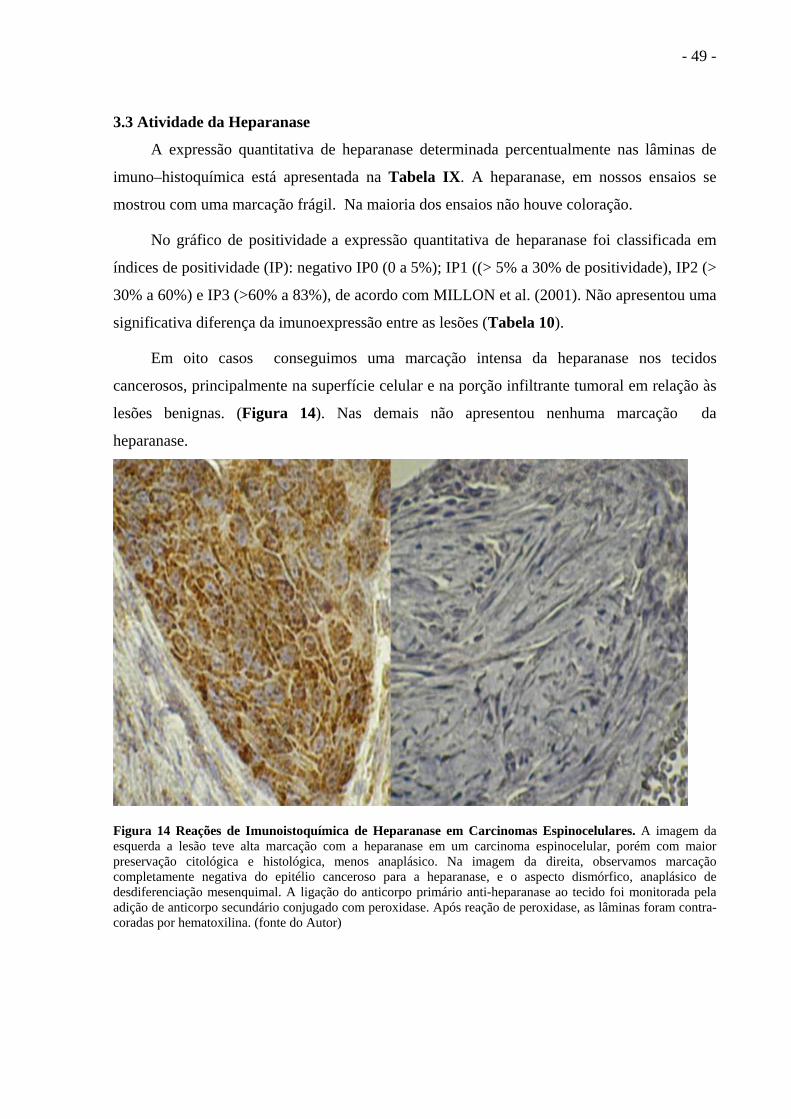
- 49 -
3.3 Atividade da Heparanase
A expressão quantitativa de heparanase determinada percentualmente nas lâminas de
imuno–histoquímica está apresentada na Tabela IX. A heparanase, em nossos ensaios se
mostrou com uma marcação frágil. Na maioria dos ensaios não houve coloração.
No gráfico de positividade a expressão quantitativa de heparanase foi classificada em
índices de positividade (IP): negativo IP0 (0 a 5%); IP1 ((> 5% a 30% de positividade), IP2 (>
30% a 60%) e IP3 (>60% a 83%), de acordo com MILLON et al. (2001). Não apresentou uma
significativa diferença da imunoexpressão entre as lesões (Tabela 10).
Em oito casos conseguimos uma marcação intensa da heparanase nos tecidos
cancerosos, principalmente na superfície celular e na porção infiltrante tumoral em relação às
lesões benignas. (Figura 14). Nas demais não apresentou nenhuma marcação da
heparanase.
Figura 14 Reações de Imunoistoquímica de Heparanase em Carcinomas Espinocelulares. A imagem da esquerda a lesão teve alta marcação com a heparanase em um carcinoma espinocelular, porém com maior preservação citológica e histológica, menos anaplásico. Na imagem da direita, observamos marcação completamente negativa do epitélio canceroso para a heparanase, e o aspecto dismórfico, anaplásico de desdiferenciação mesenquimal. A ligação do anticorpo primário anti-heparanase ao tecido foi monitorada pela adição de anticorpo secundário conjugado com peroxidase. Após reação de peroxidase, as lâminas foram contra-coradas por hematoxilina. (fonte do Autor)

- 50 -
4 Discussão
O carcinoma espinocelular se origina nas camadas mais profunda do epitélio, e
dependendo de seu grau de diferenciação histológica, cresce rapidamente, invade e destrói os
tecidos adjacentes, segundo os planos de menor resistência, podendo emitir metástase para os
linfonodos da região, ou mais raramente, acometer outros órgãos utilizando-se da via
hematogênica. Conforme mencionados anteriormente, estudos sobre a progressão do câncer
correlacionados aos níveis de catepsina B, têm mostrado que essa enzima freqüentemente
aparece elevada em tumores em estadios invasivos.
Câncer de cabeça e pescoço é uma doença séria, na maioria das vezes sequelante e
incapacitante do indivíduo, que para o ano de 2003 o Instituto Nacional do Câncer estimou
em 10.600 casos novos, somente de Cavidade Oral, com 3200 óbitos. Nos Estados Unidos a
estimativa total de Cânceres de Cabeça e Pescoço para 2003 são de 36500 com 11000 óbitos
(JORNAL A., et al 2003). No nosso grupo observamos uma predominância do sexo
masculino de nove vezes o número de mulheres, acima da média que é de cerca de duas a três
vezes o número de homens a mais do que o de mulheres (SEER,1999). A idade média em
torno dos 55 anos está dentro do esperado, apesar de que vem aumentando a incidência em
pacientes mais novos. Obtivemos 45 % de cânceres de laringe sendo a média em torno de 40
% (Tabela 4). Porém, dentro do estadiamento geral, os lábios entram como lesões de
cavidade oral. Nós não incluímos estes pacientes, pois o mecanismo de cancerização do lábio,
em geral, decorre da exposição solar, diferentemente das lesões de língua e assoalho de boca,
onde o tabagismo e etilismo são os principais causadores. A maior parte dos pacientes foram
operados no estadio IV (Tabela 5), pois normalmente chegam ao serviço público de
referência com tumores avançados, por erros diagnósticos e atraso nas condutas (COLLINS,
et al, 1995).
O Estadiamento pela Classificação TNM (AJCC,2002) apresenta uma grande
vantagem na sua simplicidade, sua universalidade (em qualquer serviço é capaz de realizá-lo)
e garante uma padronização da informação. Porém em suas vantagens encontramos suas
limitações, pois não se levam em conta aspectos histológicos, biológicos do tumor e do
paciente. Podendo ter dois pacientes do mesmo estadiamento com evoluções distintas. Outros
modelos de estadiamentos têm sido propostos, dentre eles os que agregam aspectos biológicos
do tumor. Os pacientes analisados todos foram passíveis de ressecção cirúrgica, não sendo

- 51 -
representados pacientes estadio IVb, que são tumores com critérios locais de inoperabilidade
(pex. invasão de fáscia pré-vertebral).
Em relação à metástase à distância, somente tivemos um paciente submetido à
ressecção cirúrgica com presença de metástase à distância (Classificação IVc). Pacientes com
metástase à distância, são classificados como doença sistêmica, sem controle regional com
cirurgia e radioterapia, não sendo indicada cirurgia pelo grupo, com exceções (por exemplo:
metástase única no pulmão, onde a equipe de tórax irá realizar a ressecção da lesão). Neste
caso em particular, a metástase foi em região inguinal e encontrada no dia da cirurgia, sendo
inicialmente classificada como tumor a se esclarecer.
Quando avaliada a presença de linfonodos comprometidos, observamos uma
apresentação inversa no número de pacientes em relação aos tumores avançados, sendo
encontrados mais pacientes N 0 (N=11) e menos N 3 (N=1), e nos tumores encontramos mais
pacientes T 4 (N=12) do que T 1 (N=1). Isto é decorrente que pacientes com linfonodos N3
(lesões maiores de 6 cm), são menos passíveis de ressecção cirúrgica, porém, normalmente os
pacientes chegam até os serviços públicos de referência com tumores localmente avançados
(COLLINS, et al, 1995).
Encontramos uma larga literatura relacionando a atividade das Cisteíno
proteases e seu envolvimento na metastatização e evolução tumoral, em diversos sítios
conforme mostrado na Tabela 1. Porém foi pouco estudada em Cânceres de Cabeça e
Pescoço (SIEWISKI, et al 1992: RUSSO, et al 1995). Contudo, apesar de poucos, estes
trabalhos observam uma correlação positiva entre a gravidade das lesões com estadiamentos
avançados e a atividade aumentada da catepsina. Os resultados obtidos por nós confirmam
uma correlação positiva entre o estadiamento avançado com maior positividade da atividade
da Catepsina B em comparação com as lesões benignas (Figura 7).
Dividimos em dois grupos em relação à idade dos pacientes, pois há uma impressão
entre os cirurgiões que em pacientes mais jovens os tumores são mais agressivos (não tendo
base estatística), não encontramos diferenças entre os dois grupos (Figura 5).
Observamos que na presença de metástases há um aumento significativo da catepsina.
Como o mecanismo de metastatização ocorre com invasão das estruturas adjacentes, e a
catepsina é uma enzima proteolítica relacionada a invasibilidade tumoral, o seu aumento é um
fator de marcação da capacidade metastatizante do tumor. O mesmo foi observado por
Watanabe em carcinoma de cólon. Poderíamos argumentar que ela só se eleva quando já
ocorreu a metástase, porem o material foi colhido da lesão tumoral primária, sendo uma
característica do tumor e não da metástase(Figura 10).

- 52 -
Sabendo-se que o principal fator prognóstico do tumor é sua metastatizaçao, drogas
inibidoras da catepsina poderiam diminuir esta capacidade facilitando o controle local do
tumor.
Dividimos em dois grupos os tumores laríngeos, um grupo estadio II e estadio III, e
outro estadio IV. Observa-se que as lesões avançadas apresentam atividade aumentada em
relação a lesões menos invasivas de cerca de três vezes (Figura 11). A catepsina é
participante dos processos invasivos, e os resultados confirmam o esperado mostrando seu
significativo aumento neste tipo de tumores, participando do processo remodelação e invasão
tecidual que foi confirmada pela imunoistoquímica onde observamos a Catepsina B em
carcinoma espinocelular fortemente marcada em marrom e com localização celular anômala,
pois foi detectada a presença dessa enzima na matriz extracelular (Figura 8).
Em contraste com as avaliações gerais de atividade de catepsina B, quando analisamos
especificamente a presença de metástase ou não em tumores de laringe não observamos uma
diferença na atividade enzimática (Figura 12). Tumores de laringe apresentam uma
característica menos metastatizante que das outras localizações, porém apresentam uma
agressividade local grande. Isto decorre da anatomia laríngea, que na região glótica apresenta
uma menor rede linfática, e o cone elástico que funciona com uma barreira para a invasão
tumoral. Em contrapartida há um espaço muito pequeno separando a mucosa da cartilagem.
Tumores glóticos de comissura anterior podem evoluir rapidamente de um T2 para T4 com
invasão cartilaginosa (WELSH LW., et al, 1984). Conforme já avaliamos há uma correlação
positiva da atividade enzimática e as progressões locais do tumor, que nos nossos dados
sugerem ser mais importante na secreção enzimática na laringe do que a capacidade de
metastatização.
Diferentemente das lesões laríngeas, nas lesões de orofaringe e cavidade oral há um
significativo aumento da atividade da Catepsina B nas lesões metastáticas em relação às sem
metástase linfonodal (Figura 13). Conforme o descrito acima acredito no aspecto anatômico
possa estar influenciando, contudo não posso descartar que aspectos biomoleculares do tumor
laríngeo levem a uma produção semelhante de Catepsina B nos tumores com e sem
metástase. E nas demais regiões aumento nas lesões metastatizadas (CLOSE LG ,1987).
Pode-se especular que a secreção aumentada da catepsina é uma conseqüência da
progressiva desdiferenciação tumoral e sua capacidade de invasão.
Como informado anteriormente a heparanase promove a degradação de cadeias de
heparam sulfato dos proteoglicanos que são componentes da matriz extracelular e da
membrana basal. Em tumores, a heparanase pode facilitar a invasão de células tumorais e

- 53 -
metástases pela degradação da membrana basal vascular e matriz extracelular (NURCOMBE
., 2000; VLODAVSKI., 1999). O mecanismo de inserção da catepsina B na membrana
plasmática, bem como seu tráfego celular, devem ser dependente de proteoglicanos de
heparam sulfato e a liberação de catepsina B da superfície celular pode estar relacionada com
a atividade da heparanase
As imagens de imunoistoquímica apresentam uma grande marcação para
Catepsina B, não só intralisossomal perinuclear, mas principalmente aparece concentrada na
porção infiltrante tumoral, região de intensa remodelação do tecido conjuntivo adjacente.
Esses dados sugerem que a Catepsina B pode estar participando do processo de degradação da
matriz extracelular e facilitando a infiltração do tumor epitelial no mesênquima (Figura 8).
Em relação a Heparanase não encontramos uma marcação intensa, sendo em apenas
oito casos positivos. Em todos demais casos a marcação no tecido tumoral foi totalmente
negativa (Figura 14).
O índice de positividade foi baixo, não apresentando diferença significativa entre as
lesões benignas e malignas (Tabela 10). Mesmo após repetições dos ensaios os resultados não
se alteraram. O número de casos pequeno poderia ser o fator determinante, se não fosse a
grande maioria dos casos negativa. Encontramos em alguns casos tecidos glandulares
positivos, podendo ser a heparanase um marcador melhor para tumores de origem glandular
do que de linhagem epitelial. Porém, a literatura já apresenta trabalhos correlacionando
evolução tumoral de carcinoma espinocelulares e níveis de heparanase.
A Heparanase, em nosso estudo, não se mostrou um marcador das lesões cancerosas
ou da desdiferenciação celular. Na verdade a heparanase foi detectada nas células tumorais
que ainda preservavam as características das células epiteliais e não nas células tumorais mais
desdiferenciadas. Recentemente (DOVINER et al., 2006), foi demonstrando que durante o
processo de malignização das células epiteliais de cólon, ocorre uma grande alteração do
padrão de expressão da heparanase ativa (50 kDa), de modo geral foi verificado que as células
menos displásicas dos tumores, bem como as células normais apresentavam elevados teores
da forma ativa da enzima, enquanto que as células tumorais mais displásicas e mais
desdiferenciadas do tumor apresentavam baixíssimas quantidade da enzima ativa e elevada
quantidade da enzima inativa, a pró-heparanase (65 kDa). Esses resultados sugerem que a
transformação epitélio-mesenquimal (DECLERCK et al, 2004), observada no processo de
malignização das células espinocelulares leva ao decréscimo da quantidade de heparanase
ativa nos tecidos tumorais. Esses dados indicam que a heparanase pode ser um marcador da
diferenciação celular nas células epiteliais. As células embrionárias apresentam elevados

- 54 -
teores dos glicosaminoglicanos condroitim-sulfato e ácido hialurônico durante o período
embrionário, e baixos teores desses glicosaminoglicanos durante a fase de diferenciação
celular. A presença do proteoglicano de heparam sulfato só ocorre na fase embrionária
durante a diferenciação celular (SAMPAIO et al, 1981). Interessantemente, os tumores
perdem os proteoglicanos de heparam sulfato (INKI et JALKANEN, 1996; SAMPAIO et al,
1977) e ganham os de condroitim sulfato, bem como, o ácido hialurônico (DIETRICH et al,
1993). Esse fato parece ser característico da transformação epitélio-mesenquimal observado
durante o processo de malignização. Assim sendo, parece que o decréscimo da quantidade de
heparanase observada na transformação epitélio-mesenquimal correlaciona com a diminuição
da quantidade do proteoglicano de heparam sulfato observado nos tecidos tumorias. Esses
resultados sugerem que os proteoglicanos de heparam sulfato controlam positivamente a
biossíntese da heparanase. A perda de heparam sulfato observada nos tecidos tumorais
aparentemente não está ligada ao aumento da enzima heparanase, mais sim a alterações de
enzimas envolvidas na biossíntese desses glicosaminoglicanos durante o processo de
desdiferenciação celular observado nos tumores. Outro fato importante da perda de heparam
sulfato da superfície celular é a disseminação de catepsina B para a matriz extracelular,
conforme mencionado previamente o heparam sulfato pode ancorar a catepsina B na
superfície celular e promover sua endocitose. Assim sendo, a deficiência de proteoglicanos da
superfície celular das células tumorais facilita a difusão de catepsina B para a matriz
extracelular e sua subseqüente degradação. Também, é conhecido que a enzima catepsina B
quando secretada para o meio extracelular é capaz de degradar várias proteínas da matriz
extracelular, inclusive o proteoglicano de heparam sulfato. Assim a presença dessa proteinase
na matriz pode modular a biossíntese da enzima heparanase, pela degradação do
proteoglicano de heparam sulfato. Como já observado anteriormente, o processo de
desenvolvimento tumoral, a desdiferenciação epitélio-mesenquimal, possui características
similares ao processo da embriogênese.

- 55 -
5 Conclusões e Sugestões
A Catepsina B é um bom marcador da malignidade e da evolução dos carcinomas
espinocelulares de cabeça e pescoço. Apresenta uma correlação positiva entre os estadios
avançados e os níveis de atividade enzimática.
Especificamente nas lesões laríngeas, a atividade da Catepsina B é mais acentuada e
determinada no processo de invasibilidade local do que nos processos de metastatização.
Nas lesões de orofaringe a característica metastatizante parece ser mais determinante
do que a invasibilidade local nas elevações mais acentuadas da atividade enzimática.
A Catepsina B está correlacionada com a degradação da matriz celular, com os dados
obtidos concluímos que ela está participando do processo de invasibilidade local do câncer e
de sua metastatização. E, sua produção e atividade são progressivamente maiores com a
desdiferenciação celular, ou seja, na transformação epitélio-mesenquimal observadas nas
células tumorais durante o processo de malignização (DECLERCK et al, 2004). O
desenvolvimento de inibidores específicos da Catepsina B pode ser importante para a
diminuição da invasibilidade e da metastatização do câncer. Sendo esta, em geral, o principal
fator prognóstico da doença, o seu controle seria um fator determinante na sobrevida dos
pacientes.
No nosso estudo, diferentemente da literatura ou do esperado na elaboração do
projeto, a heparanase não foi um marcador significativo das lesões malignas, ou da progressão
tumoral. A desdiferenciação tumoral com sua transformação epitélio – mesenquimal, com
perda de proteoglicanos de heparam sulfato, promovem uma diminuição da biossíntese da
heparanase. Desta forma a heparanase se mostrou um marcador de células diferenciadas.
Estudos subseqüentes e a continuação das análises da heparanase em carcinomas
espinocelulares de cabeça e pescoço poderiam confirmar ou alterar nosso trabalho.
Os dados obtidos em conjunto mostraram que a desdiferenciação tumoral aumenta a
atividade e expressão da Catepsina B e diminui a expressão da Heparanase.

- 56 -
REFERÊNCIAS
AMERICAN CANCER SOCIETY: Cancer Facts and Figures. Washington, DC, American Cancer Society, 1997
AMERICAN JOINT COMMITTEE FOR CANCER STAGING AND END-RESULTS REPORTING.: American Joint Committee on Cancer Staging Manual, 6th ed., 2002.
ANDREWS, N. W. (2000) Regulated secretion of conventional lysosomes. Trends. Cell. Biol. 10, 361-321. ALMEIDA, P. C., NANTES, I. L, CHAGAS, J. R., RIZZI, C.C.A., FALJONI-ALÁRIO, A., CARMONA, E., JULIANO, L., NADER, H. B. & TERSARIOL, I. L. S. (2001). Cathepsin B activity regulation. Heparin-like glycosaminoglycans protect human cathepsin B from alkaline pH-induced inactivation. J. Biol. Chem. 276, 944-951. BARICOS, W. H., ZHOU, Y., MASON, R. W. & BARRET, A. J. (1988) Human kidney cathepsins B and L. Characterization and potential role in degradation of glomerular basement membrane. Biochem. J. 252, 301-304. BARRETT, A. J. & RAWLINGS, N.D. (1996) Perspectives in Drug Discovery and Design. 6, 1-11. BARRETT, A. J. & KIRSCHKE, H. (1981). Cathepsin B, Cathepsin H, and Cathepsin L. Methods Enzymol. 80, 535-561. BASSET, P., BELLOCQ, J. P., WOLF, C., STOLL, I., HUTIN, P., LIMACHER, J. M., PODHAJCER, O. L., CHENARD, M. P., RIO, M. C. & CHAMBON, P. (1990) A novel metalloproteinase gene specifically expressed in stromal cells of breast carcinomas. Nature. 348, 699-704. BIENKOWISKI, M. J. & CONRAD, H. E. (1984) Kinetics of proteoheparan sulfate synthesis, secretion, endocytosis, and catabolism by a hepatocyte cell line. J. Biol. Chem. 259, 12989-12996. BRIOZZO, P., MORISSET, M., CAPONY, F., ROUGEOT, C. & ROCHEFORT, H. (1988) In vitro degradation of extracellular matrix with Mr 52,000 cathepsin D secreted by breast cancer cells. Cancer Res. 48, 3688-3692. BUCK, M. R., KARUSTIS, D. G., DAY, N. A., HONN, K. V., & SLOANE, B. F. (1992) Degradation of extracellular matrix proteins by human cathepsin B from normal and tumour tissues. Biochem J. 282, 273-278. CASTIGLIONI, T., MERINO, M. J., ELSNER, B., LAH, T. T., SLOANE, B. F. & EMMERT-BUCK, M. R. (1994) Immunohistochemical analysis of cathepsins D, B, and L in human breast cancer. Hum. Pathol. 25, 857-862. CIRMAN, T., ORESIC, K., MAZOVEC, G.D., TURK, V., REED, J. C., MYERS, R. M., SALVESEN, G. S. & TURK, B. (2004) Selective disruption of lysosomes in HeLa cells triggers apoptosis

- 57 -
mediated by cleavage of Bid by multiple papain-like lysosomal cathepsins. J. Biol. Chem. 279, 3578-3587. CLOSE LG, BURNS D.K., REISCH J., SCHAEFER S. D., (1987). Microvascular invasion in cancer of the oral cavity and oropharynx. Arch Otolaryngol Head Neck Surg 113, 1191–1195. COLLINS SL. Avoiding delay and misdiagnosis of head and neck cancer: Rare tumors with common symptoms. Compr Ther 21, 59–67. DE STEFANIS, D., DEMOZ, M., DRAGONETTI, A., HOURI, J. J., ORGIER-DENIS, E., CODOGNO, P., BACCINO, F. M. & ISIDORO, C. (1997) Differentiation-induced changes in the content, secretion, and subcellular distribution of lysosomal cathepsins in the human colon cancer HT-29 cell line. Cell Tissue Res. 289, 109-117. DIETRICH, C.P., MARTINS, J.R., SAMPAIO, L.O. & NADER, H.B. (1993). Anomalous structure of urinary chondroitin sulfate from cancer patients. A potential new marker for diagnosis of neoplasias. Lab. Ivest. 68, 439-445. DECLERCK, Y. A., MERCURIO, A.M., STACK, M.S.,CHAPMAN, H.A., ZUTTER, M.M., MUSCHEL, R.J., RAZ, A., MATRISIAN, L.M., SLOANE, B. F., NOEL, A., HENDRIX, M.J., COUSSENS, L., PADARATHSINGH, M. (2004). Proteases, extracellular matrix, and cancer. Am. J. Pathol. 164, 1131-1139. DOVINER, V., MALY, B., KAPLAN, V., GINGIS-VELITSKI, S., ILAN, N., VLODAVSKY, I. & SHERMAN, Y. (2006) Spatial and temporal heparanase expression in colon mucosa throughout the adenoma-carcinoma sequence. Mod Pathol. 19, 878-888. EBERT, W., KNOCH, H., WERLE, B., TREFZ, G., MULEY, T. & SPIESS, E. (1994) Prognostic value of increased lung tumor tissue cathepsin B. Anticancer Res. 14, 895-900. ERDAL, H., BERNDTSSON, M., CASTRO, J., BRUNK, U., SHOSHAN, M.C., & LINDER, S. (2005) Introduction of lysosomal membrane permeabilization by compounds that activate p53-independent apoptosis. Proc. Natl. Acad. Sci USA 102, 192-197. FEHRENBACHER, N. & JAATTELA, M. (2005) Lysosomes as targets for cancer therapy. Cancer Res. 65, 2993-2995. FRIEDMAN, Y., VLODAVSKY, I., AIGORN, H., AVIV, A., PERETZ, T., PECKER, I., PAPOO,O. (2000) Expression of Heparanase in normal, dysplastic, and neoplastic human colonic mucosa and stroma. Evidence for its role in colonic tumorigenesis. Am. J. Pathol. 157(4), 1167-1175 FROHLICH, E., SCHAUMBURG-LEVER, S. & KLESSEN, C. (1995) Immunocytochemical and immunoelectron microscopic demosnstration of cathepsin B in human malignant melanoma. Br. J. Dermatol. 132, 867-875.

- 58 -
FROCH, B. A., BERQUIN, I., EMMERT-BUCK, M. R., MOIN, K. & SLOANE, B. F. (1999) Molecular regulation, membrane association and secretion of tumor cathepsin B. APMIS 107, 28-37. FU, Y.H., NISHINAKA, T., YOKOYAMA, K., & CHIU, R. (1998) A retinoblastoma susceptibility gene product RB, targeting protease is regulated through the cell cycle. FEBS Letters. 421, 89-93. GUINEC, N.; DALET-FUMERON, V. & PAGANO, M. (1992) Quantitative study of the binding of cysteine proteinases to basement membranes. FEBS lett. 24, 305-308. GUO, M., MATHIEU, P. A., LINEBAUGH, B., SLOANE, B. F. & REINERS, J. J. (2002) Phorbol ester activation of a proteolitic cascade capable of activating latent TGF-β: A process initiated by the exocitoses of cathepsin B. J. Biol. Chem. 26, 14829-14837. HASHIZUME, Y., WAGURI, S., WATANABE, T., KOMINAMI, E. & UCHIYAMA, Y. (1993) Cysteine proteinases in rat parathyroid cells with special reference to their correlation with parathyroid hormone in storage granules. J. Histochem. Cytochem. 41, 855-864. HANEWINKEL, H., GLOSSL, J. & KRESSE, H. (1987) Biosynthesis of cathepsin B in cultured normal and I-cell fibroblasts. J. Biol. Chem. 262, 12351-12355. HONN, K. V., TIMAR, J., ROZHIN, J., BAZAZ, R., SAMENI, M., ZIEGLER, G. & SLOANE, B. F. (1994) A lipoxygenase metabolite, 12-(S)-HETE, stimulates protein kinase C-mediated release of cathepsin B from malignant cells. Exp Cell Res. 214,120-130. HUMPHRIES, M. J. & AYAD, S. R. (1983) Stimulation of DNA synthesis by cathepsin D digests of fibronectin. Nature 305, 811-813. IACOBUZIO-DONAHUE, C. A., SHUJA, S., CAI, J., PENG, P. & MURNAME, M. J. (1997) Elevations in cathepsin B protein content and enzyme activity occur independently of glycosylation during colorectal tumor progression. J. Biol. Chem. 272, 29290-29199. IMBERTI, R.; NIEMINEN, A-L.; HERMAN, B.; LEMASTERS, J.J (1993). Mitochondrial and glycolytic dysfunction in lethal injury to hepatocytes by t-butylhydroperoxide: protection by fructose, cyclosporin A and trifluoperazine. J. Pharmacol. Exp. Ther. 265, 392-400. INKE, P. & JALKANEN, M. (1996). The role of syndecan-1 in malignancies. Ann. Med. 28, 63-67. JAATTELA, M. (2004) Multiple cell death pathways as regulators of tumour initiation and progression. Oncogene 23, 2746-2756. JACKSON, R. L., BUSCH, S. J. & CARDIN, A. D. (1991) Glycosaminoglycans: molecular properties, protein interactions, and role in physiological processes. Physiol. Rev. 71,481-539. JOENSUU, H., TOIKKANEN, S. & ISOLA, J. (1995) Stromal cell cathepsin D expression and long-term survival in breast cancer. Br. J. Cancer 71, 155-159.

- 59 -
JORNAL, A., MURRAY, T., THOMAS, A, et al (2003) Cancer statistic, CA Cancer J Clin 2003, 53: 5 -26 KAGEDAL, K., ZHAO, M., SVENSSON, I., & BRUNK, U.T. (2001) Sphingosine-induced apoptosis is dependent on lysosomal proteases. Biochem J. 359, 335-343 KASPER, D., DITTMER, F., VON FIGURA, K. & POHLMANN, R. (1996) Neither type of mannose 6-phosphate receptor is sufficient for targeting of lysosomal enzymes along intracellular routes. J. Cell. Biol. 134, 615-623. KAZNELSON D.W., BRUNN S., MONRAD A., GJERLOV S., BIRK J., RÖPKE C. & NORRILD B.(2004) Simultaneous human papilloma virus type 16 E7 and CDK inhibitor p21 expression induces apoptosis and cathepsin B activation. Virology. 320, 301-312. KHAN, A., KRISHNA, M., BAKER, S. P. & BANNER, B. F. (1998) Cathepsin B and tumor-associated laminin expression in the progression of colorectal adenoma to carcinoma. Mod. Pathol. 11, 704-708. KHAN, A., KRISHNA, M., BAKER, S. P., MALHOTRA, R. & BANNER, B. F. (1998) Cathepsin B and tumor-associated laminin expression in the progression of colorectal adenoma to carcinoma. Mod Pathol 8, 704-708. . KOBLINSKI, J. E., DOSESCUT, J., SAMENI, M., MOIN, K., CLARK, K. & SLOANE, B. F. (2002) Interaction of Human Fibroblasts with collagen I Increases Secretion of Procathepsin B. J. Biol. Chem. 277, 32220-32227. KOBLINSKI, J. E. & SLOANE, B. F. (1997). Medical aspects of Protease and Protease Inhibitors. IOS Press, Amsterdam, Netherlands, pp. 185-193. KOS, J. & LAH, T. T. (1998) Cysteine proteinases and their endogenous inhibitors: Target proteins for prognosis, diagnosis and therapy in câncer (Review). Oncol. Rep. 5, 1349-1361. KREPELA, E., VICAR, J. & CERNOCH, V. (1989) Cathepsin B in human breast tumor tissue and cancer cells. Neoplasma 36, 41-52. KREPELA, E., KASAFIREK, E., NOVAK, K. & VIKLICKY, J. (1990) Increased cathepsin B activity in human lung tumors. Neoplasma 37, 61-70. LAH, T. T., CALEF, G., KALMAN, E., SHINDE, B. G., RUSSO, J., JAROSZ, D., ZABREKY, J., SOMERS, R. & DASKAL, I. (1995) Cathepsins D, B and L in breast carcinoma and in transformed human breast epithelial cells (HBEC). Biol. Chem. Hoppe-Seyler 376, 357-363. LAH, T. T., KOKALJ-KUNOVAR, M., STRUKELJ, B., PUNGERCAR, J., BARLIC-MAGANJA, D., DROBNIC-KOSOROK, M., KASTELIC, L., BABINIK, J., GOLOUH, R. & TURK, V. (1992) Stefins and lysosomal cathepsins B, L and D in human breast carcinoma. Int. J. Cancer 50, 36-44.

- 60 -
LINKE, M., HERZOG, V. & BRIX, K. (2002) Trafficking of lysosomal cathepsin B-green fluorescent protein to the surface of thyroid epithelial cells involves the endosomal/lysosomal compartment. J. Cell Sci. 115, 4877-4899. LIU, N.,RAJA, S.M. ZAAZZERONI, F. METKAR, S. S., SHAH, R., ZHANG, M., WANG, Y., BROMME, D., RUSSIN, W.A., LEE, J. C., PETER, M.E., FROELICH, J.C., FRANZOSO, G. & ASHTON-RICKARDT, P. G. (2003) NF-kappaB protects from the lysosomal pathway of cell death. EMBO J. 22, 5313-5322. MARTINEZ, I., CHAKRABARTI, S., HELLEVICK, T., MOREHEAD, J., FOWLER, K. & ANDREWS, N. W. (2000) Synaptotagmin VII regulates Ca(2+)-dependent exocytosis of lysosomes in fibroblasts. J. Cell. Biol 148,1141-1149. MIGNATTI, P. & RIFKIN, D. B. (1996) Plasminogen activators and matrix metalloproteinases in angiogenesis. Enzyme Protein. 49, 117-137. MIGNATTI, P. & RIFKIN, D. B. (1993) Biology and biochemistry of proteinases in tumor invasion. Physiological Reviews. 73, 161-195. MINISTÉRIO DA SAÚDE. INSTITUTO NACIONAL DE CÂNCER – INCA, Estimativas da incidência e mortalidade por câncer. Rio de Janeiro: INCA, 2003 MOIN, K.; CAO, L. DAY, N. A.; KOBLISKI, J. E. & SLOANE, B F. (1998) Tumor cell membrane cathepsin B. Biol. Chem. 379, 1093-1099. MONTASER, M.; LALMANACH, G. & MACH, L. (2002) CA-074, but not its methyl ester CA-074Me, is a selective inhibitor of cathepsin B within living cells. Biol. Chem. 387, 1305-1308. MORT, J. S. & BUTTLE, D. J. (1997) Molecules in focus. Cathepsin B. Int. J. Biochem. Cell. Biol. 29, 715-720. MURNANE, M. F., SHEAHAN, K., OZDEMIRLI, M. & SHUJA, S. (1991) Stage-specific increases in cathepsin B messenger RNA competent in human colorectal carcinoma. Cancer Res. 51, 1137-1142. MUSIL, D., ZUCIC, D., TURK, D., ENGH, R. A., MAYR, I., HUBER, R., POPOVIC, T., TURK, V., TOWATARI, T., KATUNUMA, N., & BODE, W. (1991) The refined 2.15 A X-ray crystal structure of human liver cathepsin B: the structural basis for its specificity. EMBO J. 10, 2321-2330. NAGLER, D.K., STORER, A.C., PORTARO, F.C.V., CARMONA, E., JULIANO, L & MÉNARD, R. (1997) Major increase in endopeptidase activity of human cathepsin B upon removal of occluding loop contacts. Biochemistry 36, 12608-12615. PAGE, A. E., WARBURTON, M. J., CHAMBERS, T. J., PRINGLE, J. A. S. & HAYMAN, A. R. (1991) Human osteoclastomas contain multiple forms of cathepsin B. Biochm. Biophys. Acta 1116, 57-66.

- 61 -
PERCIVAL, M. D., OUELLET, M., CAMPAGNOLO, C., CLAVEAU, D. & LI, C. (1999). Inhibition of cathepsin K by nitric oxide donors: evidence for the formation of mixed disulfides and a sulfenic acid. Biochemistry 38, 13574-13583. POOLE, A. R., TILTMAN, J. K., RECKELIES, A. D. & STOCKER, T. A. M. (1978) Differences in secretion of the proteinase cathepsin B at the edges of human breast carcinomas and fibroadenomas. Nature 273, 545-547. RECKLIES, A. D., POOLE, A. R. & MORT, J. S. (1982) A cysteine proteinase secreted from human breast tumours is immunologically related to cathepsin B. Biochem. J. 207, 633-636. REILAND, J. & RAPRAEGER, A.C. (1993) Heparan sulfate proteoglycan and FGF receptor target basic FGF to different intracellular destinations. J. Cell Sci. 105,1085-1093. REMPEL, S. A., ROSENBLUM, M. L., MIKKELSEN, T., YAN, P. S., ELLIS, K. D. GOLEMBIESKI, W. A., SAMENI, M., ROZHIN, J., ZIEGLER, G. & SLOANE, B. F. (1994) Cathepsin B expression and localization in glioma progression and invasion. Cancer Res. 54, 6027-6031. RODRIGUEZ, A., MARTINEZ, I., CHUNG, A., BERLOT, C. H. & ANDREWS, N. W. (1999) cAMP regulates Ca2+-dependent exocytosis of lysosomes and lysosome-mediated cell invasion by trypanosomes. J Biol Chem. 274,16754-16759. ROWAN, A. D., MASON, P., MACH, L. & MORT, J. S. (1991) Rat procathepsin B. Proteolitic processing to the mature form in vitro. J. Biol. Chem. 267, 15993-15999. RUSSO A., BAZAN V., GEBBIA N., PIZZOLANTI G., TUMMINELLO F. M., DARDANONI G., INGRIA F., RESTIVO S., TOMASINO R. M., LETO G. (1995) Flow cytometric DNA analysis and lysosomal cathepsins B and L in locally advanced laryngeal cancer. Cancer (Phila.), 76: 1757-1764. SAMPAIO, L.O. & DIETRICH, C.P. (1981). Changes in sulfated mucopolysaccharide composition during fetal development. J. Biol. Chem. 256, 757-765. SAMPAIO, L.O., DIETRICH, C.P., & GIANOTTI, F. O.(1977) Changes in sulfated mucopolysaccharide compositionof mammalian tissues during growth and cancer tissues. Biochem. Biophys. Acta 498, 123-130. SHAH JP, CANDELA FC, PODDAR AK. (1990) The patterns of cervical lymph node metastasis from squamous carcinoma of the oral cavity. Cancer 66:109-113 SHEAHAN, K.; SHUJA, S. & MURNANE, M. J (1989). Cysteine protease activities and tumor development in human colorectal carcinomas. Cancer Res. 49, 3809-3814. SHUJA, S., SHEAHAN, K. & MURNANE, M. J. (1991) Cysteine endopeptidases activity levels in normal human tissues, colorectal adenomas and carcinomas. Int. J. Cancer 49, 341-346.

- 62 -
SIEWISKI M., KRCICKIT., JARMUOWICZ J., BERDOWSKA I (1992). Cysteine proteinase inhibitors in serum of patients with head and neck cancer. Diagn. Oncol., ,: 323-326 SIMIZU S,. ISHIDA K., OSADA H (2004). Heparanase as a molecular target of cancer chemoterapy. Cancer Sci 95, 7:553 - 558 SINHA, A. A., WILSON, M. J., GLEASON, D. F., REDDY, P. K., SAMENI, M. & SLOANE, B. F. (1995) Immunohistochemical localization of cathepsin B in neoplastic human prostate. Prostate 26, 171-178. SIVAPARVATHI, M., SAWAYA, R., WANG, S. W., RAYFOR, A., YAMAMOTO, M., LIOTTA, L. A., NICOLSON, G. L. & RAO, J. S. (1995) Overexpression and localization of cathepsin B during the progression of human gliomas. Clin. Exp. Metast. 13, 49-56. SLOANE, B. F. (1996) Suicidal tumor proteases. Nat. Biotech. 14, 826-827. SLOANE, B. F., ROZHIN, J., JOHNSON, K., TAYLOR, H., CRISSMAN, J. D., & HONN, K. V. (1986) Cathepsin B: association with plasma membrane in metastatic tumours. Proc. Natl. Acad. Sci. 83, 2483-2487. SLOANE, B. F., ROZHIN, J., LAH, T. T., DAY, N. A., BUCK, M., RYAN, R. E., CRISSMAN, J. D., & HONN, K. V. (1988) Tumor cathepsin B and its endogenous inhibitors in metastasis. Adv. Exp. Med. Biol. 233, 259-268. SLOANE, B. F., MOIN. K., SAMENI, M. TAIT, L. R., ROZHIN, J., & ZIEGLER, G (1994) Membrane association of cathepsin B can be induced by transfection of human breast epithelial cells with c-Ha-ras oncogene. J. Cell. Sci. 107, 373-384.
STEVENS MH, HARNSBERGER HR, MANCUSO AA (1985), et al: Computed tomography of cervical lymph nodes. Arch Otolaryngol 111:735
SUKOH, N., ABE, S., OGURA, S., ISOBE, H., TAKEKAWA, H., INOUE, K., & KAWAKAMI, Y. (1994) Immunohistochemical study of cathepsin B. Prognostic significance in human lung cancer. Cancer 74, 46-51. SUNDLER, R. (1997) Lysosomal and cytosolic pH as regulators of exocytosis in mouse macrophages. Acta Physiol Scand.161, 553-556. TAHA, T. A., KITATANI, K., BIELAWSKI, J., CHO, W., HANNUN, Y. A. OBEID, L. M. (2005) TNF induces the loss of sphingosine kinase-1 by a cathepsin B dependent mechanism. J. Biol. Chem. TALAMINI R (2002), et al: Combined effect of tobacco and alcohol on laryngeal cancer risk: A case-control study. Cancer Causes Control;13(10):957–964. TAKAOMI O., YOSHIO N., MUNENORI T., TETSUJI N., KAZUHIRO N., TAKAYUKI M., TOSHIHIRO M., HIROKAZU U., MASAHIKO K., YASUHIRO S., TOMOKI Y., NAGAHIDE M., JUNJI M., MINORU H., MEHMET G., HIDETSUGU T., HITOSHI N.,

- 63 -
MASAO H. MOTOWO N., NORIAKI T.(2004), Localization of heparanase in esophageal cancer cells: respective roles in prognosis and differentiation. Lab Invest., 84(10): 1289 - 304 TAPPER, H. & SUNDLER, R. (1990) Role of lysosomal and cytosolic pH in the regulation of macrophage lysosomal enzyme secretion. Biochem. J . 272, 407-414. TAPPER, H. & SUNDLER, R. (1995) Protein kinase C and intracellular pH regulate zymosan-induced lysosomal enzyme secretion in macrophages. J Leukoc Biol. 58, 485-494. TERADA, T., OHTA, T., MINAMOTO, H. & NAKAMURA, Y. (1995) Expression of pancreatic trypsinogen/trypsin and cathepsin B in human cholangiocarcinomas and hepatocellular carcinomas. Hum. Pathol. 26, 746-752. TOOZE, J. HOLLINSHEAD. M., HENSEL, G., KERN, H. F. & HOFLACK, B. (1991) Regulated secretion of mature cathepsin B from rat exocrine pancreatic cells. Eur. J. Cell. Biol. 56, 187-200. TSUSHIMA, H., YOSHIDA, H. E., UEKI, Y. & HOPSU-HAVU, V, K. (1991) Inactive cathepsin B-like enzyme in human melanoma culture medium. Melanoma Res. 1, 341-347. TURK, B., DOLENC, I., ZEROVNIK, E., TURK, D., GUBENSEK, F. & TURK, V. (1994) Human cathepsin B is a metastable enzyme stabilized by specific ionic interactions associated with the active site. Biochemistry 33, 14800-14806. TURK, B.; TURK, V. & TURK, D. (1997) Structural and functional aspects of papain-like cysteine proteinases and their protein inhibitors. Biol. Chem. 378, 141-150. ULBRICHT, B., HAGMANN, W., EBERT, W. & SPIESS, E. (1996) Differential secretion of cathepsins B and L from normal and tumor human lung cells stimulated by12(S)-hydroxy-eicosatetraenoic acid. Exp Cell Res.1,255-263. VISSHER, D. W., SLOANE, B. F., SAMENI, M., BABIARZ, J. W., JACOBSON, J. & CRISSMAN, J. D. (1994) Clinicopathologic significance of cathepsin B immunostaining in transitional neoplasia. Mod. Phatol. 7, 76-81. WATANABE, M., HIGASHI, T., HASHIMOTO, M., TOMODA, I., TOMINAGA, S., HASHIMOTO, M., MORIMOTO, S., YAMAUCHI, Y., NAKATSUKASA, H., KOBAYASHI, M., WATANABE, A. & HAGASHIMA, H. (1997) Elevation of tissue cathepsin B and L activities in gastric cancer. Hepato-gastroenterol 4, 120-122. WATANABE, M., HIGASHI, T., WATANABE, A., OSAWA, T., SATO, Y., KIMURA, Y., TOMINAGA, S., HASHIMOTO, M., YOSHIDA, Y., MORIMOTO, S., SHIOTA, T., HASINOTO, M., KOBAIASHI, M., TOMODA, J. & TSUJI, T. (1989) Cathepsin B and L activities in gastric cancer tissue: correlation with histological findings. Bio. Chem., Med. Metabol. Biol. 42 WELSH LW, WELSH JJ, RIZZO TA: Laryngeal lymphatics: current anatomic concepts, Ann Otol Rhinol Laryngol 93:19, 1984.

- 64 -
ZCHARIA, E., METZGER, S., CHAJEK-SHAUL, T., ELKIM, M., FRIEDMAN, Y., PECKER, I., PERETZ, T., VLODAVSKY, I. (2001), Molecular properties and involvment of heparanase in cancer progression and mammary gland morphogenesis. J. Mammary Gland. Biol. Neoplasia 6(3), 311-322

- 65 -
Anexo – Comitê de Ética

- 66 -

- 67 -
Apêndice A - TERMO DE LIVRE CONSENTIMENTO ESCLARECIDO

- 68 -
Prezado Paciente O projeto de pesquisa intitulado: “Estudo da Catepsina B e da Heparanase em Carcinoma Espinocelular de Cabeça e Pescoço” dos autores Emílio López Couto, Profª Drª Maria A. da Silva Pinhal e Prof. Dr . Ivarne L. S. Tersariol tem por objetivo auxiliar o diagnóstico de tumores de cabeça e pescoço, bem como avaliar o grau de gravidade da doença. Se obtivermos resultados favoráveis, poderemos contribuir para detecção precoce do câncer e conhecer melhor como estas doenças evoluem no nosso organismo. Baseado nestas informações poderemos melhorar o modo de tratamento destes tumores. Desta forma, necessitamos obter acesso às amostras de sangue e de tecidos tumorais provenientes de Cirurgias de Cabeça e Pescoço, que serão realizadas no Hospital Estadual Mário Covas – santo André, SP. Bem como, precisamos também obter informações contidas no seu prontuário médico. Informamos que a coleta de amostras de tecidos tumorais não acarretará em danos maiores do que aqueles promovidos pelo procedimento cirúrgico de remoção do tumor. O paciente submetido à cirurgia será acompanhado pela equipe de Cirurgia de Cabeça e Pescoço do Hospital Estadual Mário Covas – Santo André. Todas as informações sobre a doença e possíveis seqüelas cirúrgicas serão fornecidas pela equipe cirúrgica. Comprometemos manter o sigilo das informações coletadas e o anonimato das pessoas cujos materiais biológicos e documentos médicos forem consultados, também garantimos que as informações colhidas serão utilizadas exclusivamente para o cumprimento do propósito deste projeto de pesquisa. O paciente em qualquer momento da pesquisa poderá retirar o livre consentimento e automaticamente seus dados serão retirados do trabalho em questão, sem que isto lhe acarrete ônus de qualquer espécie. Eu,_____________________________________________ Identidade________________ R:_________________________, nº__________, CEP_____________________________ Cidade_______________________________________Tel__________________________ fui claramente informado sobre os propósitos deste projeto de pesquisa e voluntariamente aceito a participar desta pesquisa. _________________________________________________________________________
Emílio López Couto, Cirurgião de Cabeça e Pescoço – Hospital Estadual Mário Covas – Fundação ABC, Santo André, SP

- 69 -
Apêndice B - Tabelas

- 70 -
Tabela 4: Patologia, Estadio, Sexo e Idade dos Pacientes
Amostra Patologia Estadio Sexo Idade 1 T4aN2BM0 Cavidade Oral IVa masc 42
2 T4aN0M0 Larringe IVa masc
60 3 T3N1M0 Laringe III masc
60
4 T4aN2bM0 Hipofaringe IVa masc
54 5 T4aN0M0 Laringe IVa masc 54 6 T2N0M1 de Cavidade Oral IVc masc 49 7 T3N1M0 Orofaringe III masc 50
8 T4aN0M0 Cavidade Oral IVa masc
66 9 T4aN0M0 Laringe IVa masc 71
10 T4aN0M0 Orofaringe IVa masc
55 11 T4aN0M0 Orofaringe IVa masc 51 12 T4aN2cM0 Hipofaringe IVa masc 70 13 T1N3M0 Laringe IVb masc 62
14 T4aN2bM0 Laringe IVa masc
60 15 T2N2M0 Orofaringe IVa masc
47
16 T4aN0M0 Laringe IVa fem
51 17 T2N1M0 Laringe III masc 64 18 T2N0M0 Orofaringe II masc 55 19 T4aN0M0 Laringe IVa masc 45
20 T3N0M0 Orofaringe III fem
52
As amostras foram coletadas no período Julho de 2004 a Fevereiro de 2006. Estadiamento conforme a padronização da American Joint Comittee of Cancer de 1988 e União Internacional de Combate ao Câncer, revisada de 2002

- 71 -
Tabela 5: Atividade da Catepsina B, Estadiamento e Patologia
Resultados da atividade enzimática correlacionando com a patologia e o estadiamento dos pacientes portadores de carcinoma espinocelular. Os valores estão expressos em atividade específica de catepsina B (UAF/min/µg de proteína )
Amostra Patologia Estadio Atividade Catepsina
UAF/min*µg
1 T4aN2BM0 Cavidade Oral IVa 46,51
2 T4aN0M0 Laringe IVa 48,57 3 T3N1M0 Laringe III 20,69 4 T4aN2bM0 Hipofaringe IVa 19,72 5 T4aN0M0 Laringe IVa 16,71 6 T2N0M1 de Cavidade Oral IVc 48,83 7 T3N1M0 Orofaringe III 26,62 8 T4aN0M0 Cavidade Oral IVa 17,14 9 T4aN0M0 Laringe IVa 62,12
10 T4aN0M0 Orofaringe IVa 23,32 11 T4aN0M0 Orofaringe IVa 14,84 12 T4aN2cM0 Hipofaringe IVa 12,13 13 T1N3M0 Laringe IVb 15,57 14 T4aN2bM0 Laringe IVa 11,83 15 T2N2M0 Orofaringe IVa 30,63 16 T4aN0M0 Laringe IVa 23
17 T2N1M0 Laringe III 3,39 18 T2N0M0 Orofaringe II 14,1 19 T4aN0M0 Laringe IVa 9,03
20 T3N0M0 Orofaringe III 5,39
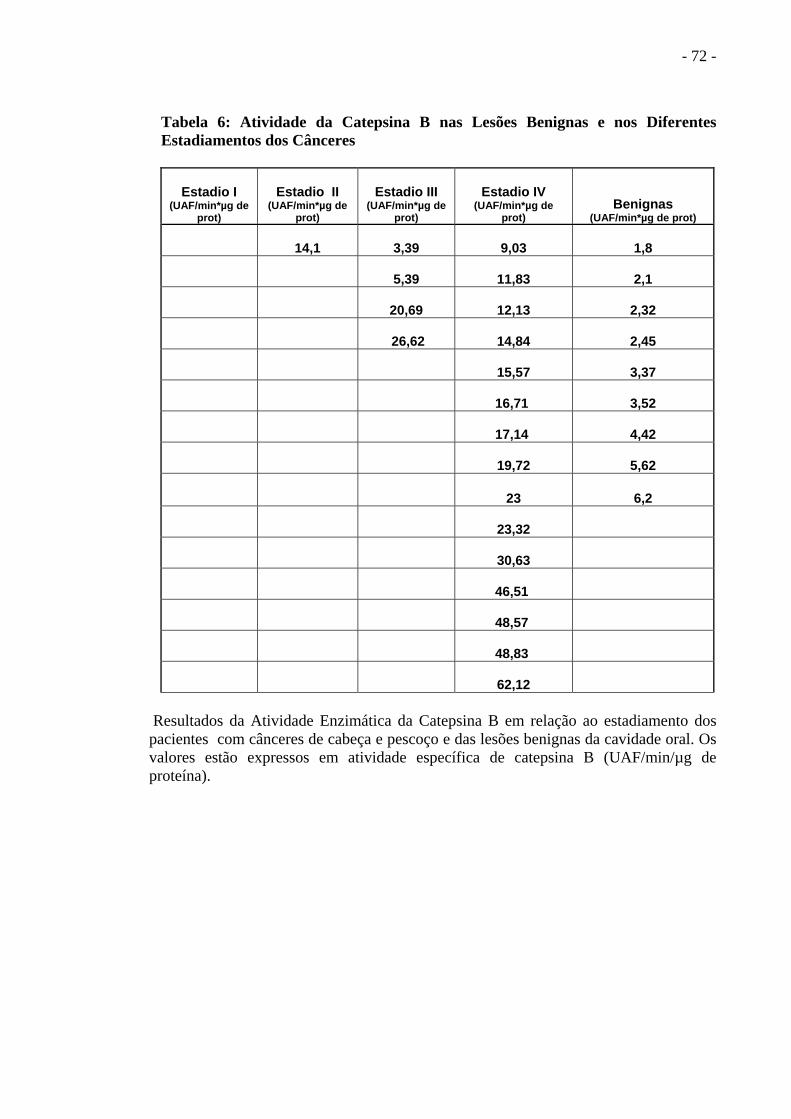
- 72 -
Tabela 6: Atividade da Catepsina B nas Lesões Benignas e nos Diferentes Estadiamentos dos Cânceres
Estadio II Estadio III Estadio IV Estadio I
Benignas (UAF/min*µg de prot)
(UAF/min*µg de prot)
(UAF/min*µg de prot)
(UAF/min*µg de prot) (UAF/min*µg de prot)
14,1 3,39 9,03 1,8
5,39 11,83 2,1
20,69 12,13 2,32
26,62 14,84 2,45
15,57 3,37
16,71 3,52
17,14 4,42
19,72 5,62
23 6,2
23,32
30,63
46,51
48,57
48,83
62,12
Resultados da Atividade Enzimática da Catepsina B em relação ao estadiamento dos pacientes com cânceres de cabeça e pescoço e das lesões benignas da cavidade oral. Os valores estão expressos em atividade específica de catepsina B (UAF/min/µg de proteína).

- 73 -
Tabela 7: Expressão Enzimática da Catepsina B e a Classificação do Tumor.
Benignas T1 T2 T3 T4 (UAF/min*µg de
prot) (UAF/min*µg de
prot) (UAF/min*µg de
prot) (UAF/min*µg de
prot) (UAF/min*µg de
prot)
1,8 15,57 3,39 5,39 9,03
2,1 14,1 20,69 11,83
2,32 30,63 26,62 12,13
2,45 48,83 14,84
3,37 16,71
2,52 17,14
4,42 19,72
5,62 23
6,2 23,32
46,51
48,57
62,12
Utilizou – se o Estadiamento conforme a padronização da American Joint Comittee of Cancer de 1988 e União Internacional de Contra o Câncer, revisada de 2002 . Os valores estão expressos em atividade específica de catepsina B (UAF/min/µg de proteína).
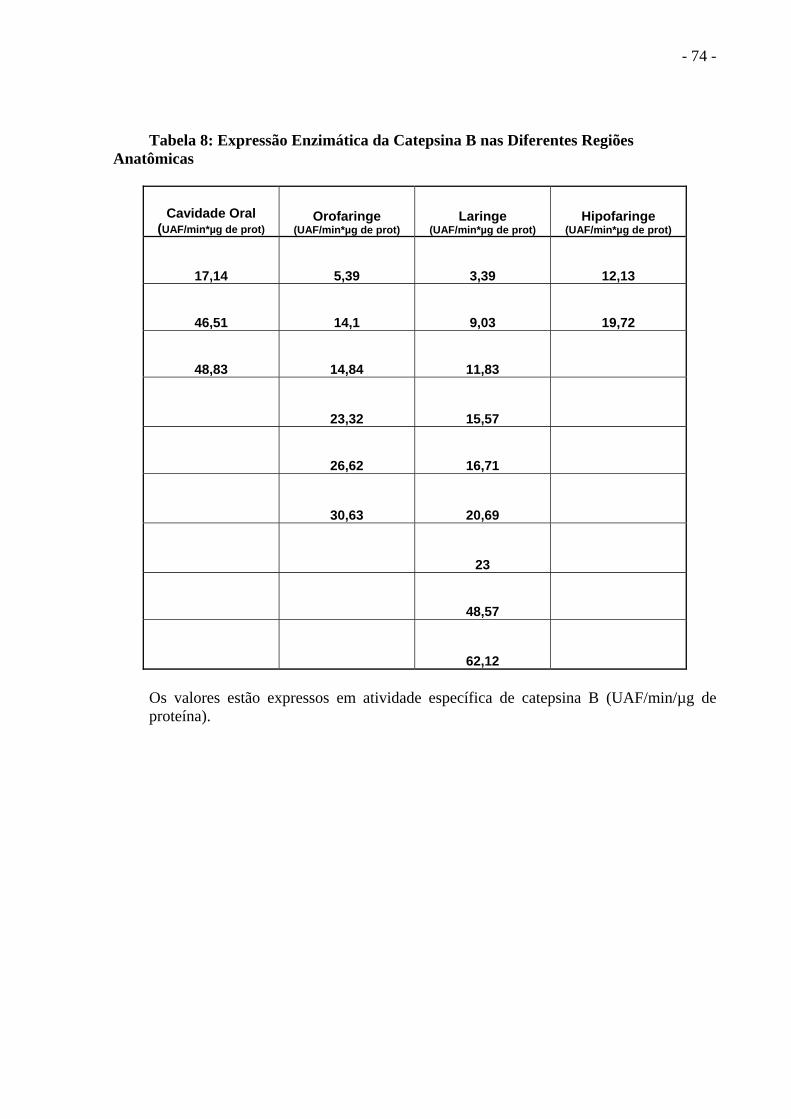
- 74 -
Tabela 8: Expressão Enzimática da Catepsina B nas Diferentes Regiões
Anatômicas
Cavidade Oral Laringe Hipofaringe Orofaringe (UAF/min*µg de prot) (UAF/min*µg de prot) (UAF/min*µg de prot) (UAF/min*µg de prot)
17,14 5,39 3,39 12,13
46,51 14,1 9,03 19,72
48,83 14,84 11,83
23,32 15,57
26,62 16,71
30,63 20,69
23
48,57
62,12
Os valores estão expressos em atividade específica de catepsina B (UAF/min/µg de proteína).
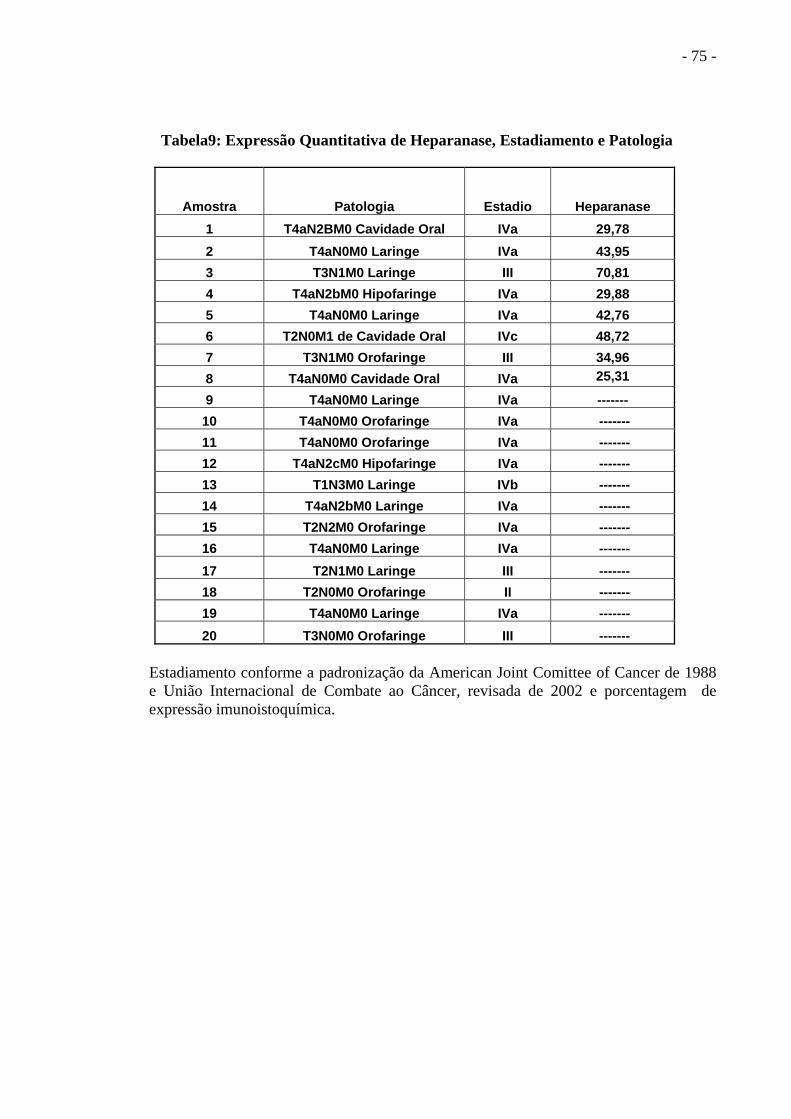
- 75 -
Tabela9: Expressão Quantitativa de Heparanase, Estadiamento e Patologia
Estadiamento conforme a padronização da American Joint Comittee of Cancer de 1988 e União Internacional de Combate ao Câncer, revisada de 2002 e porcentagem de expressão imunoistoquímica.
Amostra Patologia Estadio Heparanase 1 T4aN2BM0 Cavidade Oral IVa 29,78 2 T4aN0M0 Laringe IVa 43,95 3 T3N1M0 Laringe III 70,81 4 T4aN2bM0 Hipofaringe IVa 29,88 5 T4aN0M0 Laringe IVa 42,76 6 T2N0M1 de Cavidade Oral IVc 48,72 7 T3N1M0 Orofaringe III 34,96
25,31 8 T4aN0M0 Cavidade Oral IVa 9 T4aN0M0 Laringe IVa -------
10 T4aN0M0 Orofaringe IVa ------- 11 T4aN0M0 Orofaringe IVa ------- 12 T4aN2cM0 Hipofaringe IVa ------- 13 T1N3M0 Laringe IVb ------- 14 T4aN2bM0 Laringe IVa ------- 15 T2N2M0 Orofaringe IVa ------- 16 T4aN0M0 Laringe IVa ------- 17 T2N1M0 Laringe III ------- 18 T2N0M0 Orofaringe II ------- 19 T4aN0M0 Laringe IVa ------- 20 T3N0M0 Orofaringe III -------

- 76 -
Tabela 10: Classificação quantitativa da Heparanase por Índice de Positividade
Heparanase Total IP0 IP1 IP2 IP3
Lesões Benignas
6 3 0 0 9
Carcinoma Espinocelular
12 3 4 1 20
IP0 (0 a 5%); IP1 ((> 5% a 30% de positividade), IP2 (> 30% a 60%) e IP3 (>60% a 83%)
A expressão quantitativa de heparanase foi classificada em índices de positividade (IP): negativo IP0 (0 a 5%); IP1 ((> 5% a 30% de positividade), IP2 (> 30% a 60%) e IP3 (>60% a 83%), de acordo com MILLON et al. (2001) Oral Oncol.. 37, 620-631. Para análise estatísticas, a significância foi verificada pelo teste de Chi-quadrado (χ2).

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo













