Embed Size (px)
Citation preview

ESTUDOS ECOLÓGICOS SOBRE MOSQUITOS CULICIDAE NOSISTEMA DA SERRA DO MAR, BRASIL.
1 — Observações no ambiente extradomiciliar.*
Oswaldo Paulo Forat t ini**Almério de Castro Gomes**Eunice Aparecida Bianchi Galati **Ernesto Xavier Rabello **Lygia Busch Iversson**
RSPUB9/418
FORATTINI, O. P. et al. Estudos ecológicos sobre mosquitos Culicidae no sistemada Serra do Mar, Brasil, 1 — Observações no ambiente extradomiciliar. Rev.Saúde públ., S. Paulo, 12:297-325, 1978.
RESUMO : Apresentam-se os resultados das coletas extradomiciliares demosquitos Culicidae no Vale do Ribeira, Estado de São Paulo, Brasil. Asobservações foram levadas a efeito visando obter informações que esclareçamo mecanismo de transmissão de encefalite por vírus ocorrida em caráter epidê-mico na região. Com o emprego de várias técnicas, foram feitas coletas emáreas com aspectos variáveis em relação à conservação do ambiente. Funda-mentalmente, a diferenciação baseou-se em áreas alteradas, com matas residuaise ocorrência de casos de encefalite, e áreas conservadas, com matas primáriase sem a presença da doença, até o momento. Foram obtidos dados que sugerempossível associação entre a ocorrência da virose e o comportamento de Aedesserratus, Aedes scapularis e Culex (Melanoconion) sp. Admite-se também apossível participação de representantes de Limatus e Psorophora, embora prova-velmente de maneira variável e menos intensa do que a dos anteriores. Apossível ocorrência de ciclos prolongados de baixa densidade daqueles mosquitosna zona florestal da escarpa serrana, poderá explicar o possível papel debarreira que a Serra de Paranapiacaba e do Mar desempenham para a propa-gação da epidemia em direção ao planalto do Estado de São Paulo.
UNITERMOS: Culicidae, Vale do Ribeira, SP, Brasil. Arboviroses. Encefaliteepidêmica, S. Paulo, Brasil. Ecologia. Aedes serratus. Aedes scapularis. Culex(Melanoconion) sp.
* Trabalho do Centro Brasileiro de Estudos Entomológicos em Epidemiologia (CENTEP) erealizado com auxílio financeiro do Grupo de Avaliação de Projetos de Pesquisa (GAP) doMinistério da Saúde.
** Do Departamento de Epidemiologia da Faculdade de Saúde Pública da Universidade de SãoPaulo — Avenida Dr. Arnaldo, 715 — 01255 — São Paulo, SP — Brasil.
INTRODUÇÃO
A partir dos primeiros meses de 1975registrou-se a ocorrência de surtos de ence-falite na região sul do Estado de São Paulo,
Brasil. Os estudos iniciais visaram os aspec-tos clínicos e, no decurso da epidemia, asmedidas de controle orientaram-se no com-

bate a culicídeos. E isso face à suposiçãode se tratar de arbovirose veiculada por esseshematófagos (Tiriba19 1975; SUCEN 17
1975; Tiriba e col. 20 1976). O isolamentode agente identificado como um Flavivirusdeu razão a essa conduta. Por sua vez, oestudo da casuística ocorrida no período1975-1976 permitiu conhecer alguns aspectosdo quadro epidemiológico, descrevendo-se adistribuição cronológica e geográfica doscasos observados, além de algumas dascaracterísticas dessa população atingida(Tiriba e col.20 1976; Iversson9 1977).
Tais circunstâncias levaram à necessida-de de realização de estudos, com o objetivode esclarecer os possíveis mecanismos locaisde transmissão da doença. Como foi men-cionado, desde o início as evidências suge-riram tratar-se de arbovirose. Por sua vez,a riqueza faunística regional em dípteroshematófagos, principalmente Culicidae, fezcom que as atenções se voltassem paraesses mosquitos. Dessa maneira, em 1976foi iniciada a execução de projeto, de longoprazo, visando obter informações sobre anatureza e comportamento da fauna culici-diana local. E isso tanto no ambienteextradomiciliar como no domiciliar. Nopresente trabalho são apresentados dadosiniciais relativos ao primeiro.
REGIÃO ESTUDADA
Inicialmente, a epidemia mostrou seusmaiores coeficientes de morbidade, no pri-meiro semestre de 1975, na zona da Baixa-da Santista. Posteriormnte, e no mesmoperíodo do ano seguinte, o fenômeno repe-tiu-se na região conhecida como o Vale doRio Ribeira de Iguape (Iversson9 , 1977).Em vista disso, esta última foi escolhidapara sede das investigações atuais.
A região aqui referida como Vale doRibeira compreende, além das vertentesdesse curso de água, o extremo meridional
da zona litorânea do Estado de São Paulo,Brasil (Fig. 1). Sob o ponto de vista admi-nistrativo nela estão incluídos 16 municí-pios. Na verdade, porém, esse sistema hi-drográfico engloba porções dos territóriosde outras quatro áreas municipais paulis-tas, além de unidades análogas do vizinhoEstado do Paraná. Contudo, nestas pesqui-sas foi considerada a área correspondenteà parte daquela bacia hidrográfica e à zonalitorânea adjacente, constituída pelo con-junto territorial dos supracitados municí-pios15. Encontra-se, aproximadamente, entre24° e 25° 16' de latitude sul e de 46° 50'e 49° 20' de longitude oeste. É percorrida,em praticamente toda sua extensão nordes-te -sudoeste, pela estrada de rodagem BR116 que une as capitais dos dois Estados,representadas pelos centros urbanos deSão Paulo e Curitiba. A rede rodoviárialocal inclui ramais que comunicam entresi os vários municípios e com o portomarítimo de Santos, passando pela cidadede Peruibe. O maior núcleo urbano é re-presentado por Registro, de onde tambémse origina a via que, atingindo a sede deSete Barras, alcança São Miguel Arcanjosituada no planalto.
Como é entendido nestas pesquisas, osistema da Serra do Mar inclui-se no cha-mado domínio tropical atlântico de acordocom Ab'Saber (1977)*, e constitui o centrohomônimo de dispersão de vertebrados, naconceituação de Müller12,13 (1972, 1973).Todavia, em relação aos domínios morfo-estruturais da região Sudeste do Brasil, oVale do Ribeira situa-se nas unidades doLitoral e da Serra do Mar. A primeiracompreendendo o aspecto caracterizado porpraias e baixadas limitadas pela segunda,enquanto esta participa do sistema de es-carpas que forma a borda ocidental doplanalto cristalino interiorano. Em sua totalextensão de mais de 1000 km, e que seprolonga desde o Estado de Santa Catarinaaté o do Rio de Janeiro, a Serra do Mar
* Cominação pessoal
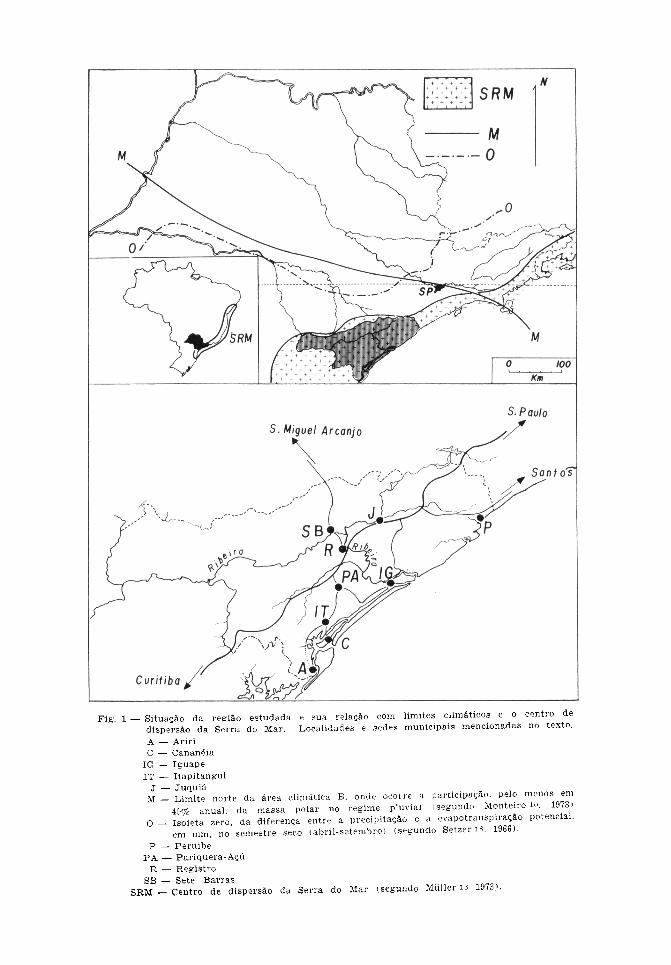

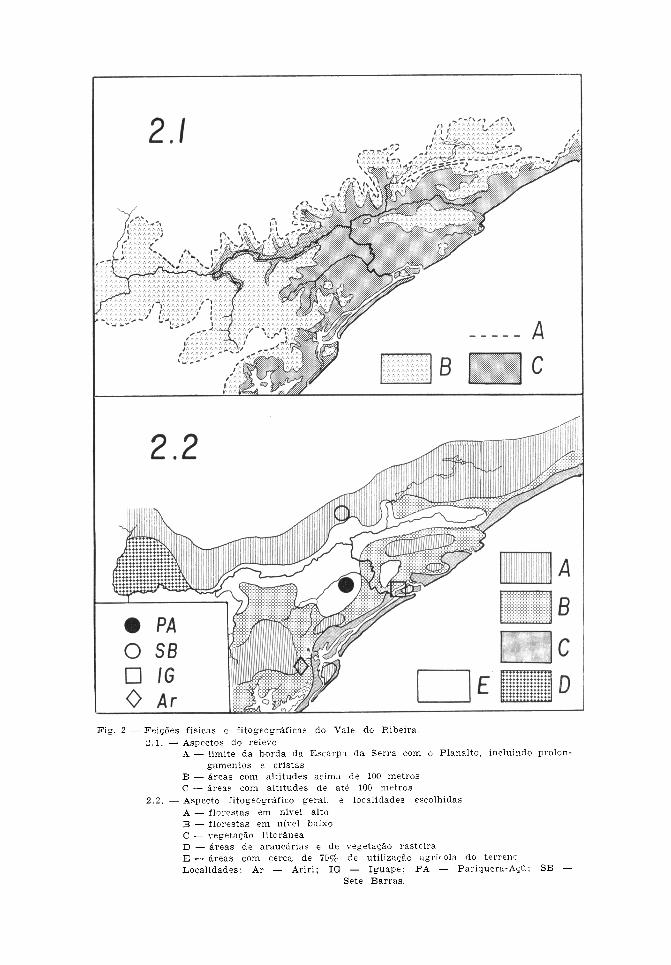
apresenta altitudes variáveis entre 800 e1.800 m, chegando a ultrapassar os 2.000m em alguns pontos. Assim sendo, a regiãoaqui estudada do Vale do Ribeira participada chamada Província Costeira do Estadode São Paulo. Nela, o paredão montanhosorepresenta a zona denominada de SerraniaCosteira, e o terreno que se estende do seusopé até a orla marítima constitui a Baixa-da Litorânea. Aquela apresenta-se acentua-damente interiorizada, recebendo aqui onome de Serra de Paranapiacaba, e dotadade alguns cimos que atingem os 1.200 a1.300 m. Tal recuo, que chega até umacentena de quilômetros do mar, permite queo traçado dos cursos de água, em boa par-te, se faça em sentido paralelo à costa epercorrendo grandes extensões em planurade quotas variáveis abaixo dos 100 m (Fig.2). Considerando-se, pois, tais aspectos dorelevo, esta região pode ser subdividida emtrês sub-regiões. A primeira é a menciona-da Baixada (Fig. 3), com cerca de 2.400km2, de formação predominantemente sedi-mentaria constituindo uma das maioresplanícies litorâneas tropicais e onde geral-mente se situam os núcleos urbanos locais.A segunda é a também referida Escarpada Serra (Fig. 4), com extensão aproxima-da de 8.600 km2, orientando-se no sentidonordeste-sudoeste e aproximando-se dacosta, tanto ao norte como ao sul. Assimsendo, esse paredão mostra, a grosso modo,percurso com aspecto geral de arco deconcavidade voltada para o mar e emoldu-rando toda a extensão do Vale do Ribeira,aqui considerada. Finalmente, nos limitesentre essas duas pode-se reconhecer a sub-região Pré-Serrana (Fig. 5), de relevoondulado e com feições transicionais entreaquelas (Camargo e col.5 , 1972; Almeida2 ,1974; Moreira e Camalier 11, 1977).
Considerando-se os elementos climáticose pedológicos, esta área participa do Com-plexo Cristalino e da Baixada Litorânea, nosistema de divisão ecológica de Setzer18
(1966) para o Estado de São Paulo. Demaneira geral, as cotas superiores (acima
de 350 m) da Escarpa da Serra situam-sena sub-região fria da primeira (CCf),enquanto os sopés dessa encosta e asmenores altitudes participam, respectiva-mente, das duas sub-regiões, fr ia e quente,da segunda (BLf e BLq). Por sua vez,levando-se em conta critérios fenológicos,o Vale do Ribeira representa a primeirae constitui parte da quarta, das regiõesecológicas no sistema de Troppmair2 1
(1975). Em relação àquela, designada deLitoral Sul, inclui a já descrita planície,enquanto que para a última, denominadade Planalto Cristalino, contribui com asáreas de maiores altitudes encontradas naparte da escarpa serrana. Dessa maneira,os caracteres climáticos da região fazemcom que se considere dois aspectos. Corres-pondem, respectivamente, às partes maisaltas e à planície. Para ambas as áreaspode-se adotar a classificação fundamen-tada no Sistema de Koeppen, de acordocom Setzer18 (1966) e Nimer14 (1977) eno de Monteiro10 (1973). Aquela baseia-senos dados pluviométricos e de temperaturasmédias, enquanto a última utiliza o critérioda atuação das massas de ar envolvidas nacirculação regional e que se refletem naprodução de chuvas. Assim sendo, o pri-meiro inc lu i as zonas altas no tipo Cfbcorrespondente a clima temperado, sub-tropical ou mesotérmico brando superúmido,caracterizando-se pela ausência de estaçãoseca e com menos de 22°C como tempera-tura média do mês mais quente (janeiro).Quanto à planície, o mesmo critério classi-fica-a como Cfa, com as mesmas caracterís-ticas da anterior porém com mais de 22°Cde temperatura média no mês mais quente(janeiro). Acrescente-se, contudo, que emrelação à baixada litorânea propriamentedita, a classificação corresponde ao tipoAf, ou seja, de clima tropical úmido ousubseqüente superúmido, sem estiagem,com mais de 60 mm de chuva no mês maisseco ( ju lho) ; e com temperatura médiasuperior a 22°C para o mês mais quente(janeiro). No que concerne à atuação dasmassas de ar sobre a pluviosidade, o Vale


do Ribeira situa-se ao sul do limite quedivide o Estado de São Paulo, de acordocom a participação anual da massa polar.Assim, nessa parte meridional do territóriopaulista essa atividade não é inferior a40%, podendo chegar a 75%. Em vistadisso, dá lugar a clima permanentementeúmido, caracterizado pela ausência de pe-ríodos secos. Neste sistema, a região aquiestudada encontra-se incluída no tipo B,correspondente a esse comportamento damassa polar (Fig. 1), e na feição climáticaIIa que se estende até a borda planaltiva.Assim sendo, a participação polar relevantesobrepuja com freqüência a das massastropicais, resultando em maior significadodas chuvas frontais com certa variabilidadedas precipitações. Por sua vez, as caracte-rísticas já mencionadas do relevo (Fig. 2),com o acentuado afastamento da Serra deParanapiacaba da orla marítima e a pre-sença de elevações isoladas, permite maiorpenetração das massas polares e passagensfrontais. Daí a influência da topografiana distribuição quantitativa das precipita-ções atmosféricas. Pode-se pois compre-ender que, o elevado grau de pluviosidadedurante o ano resulta da atuação alternadadas massas de ar polar e tropical, sejaprovocando chuvas orográficas na escarpada serra e elevações isoladas, seja causan-do precipitações frontais pelo encontro dasduas massas, além da atuação da convec-ção térmica no período estival. De qualquermaneira, as características climáticas destaparte paulista do Vale do Ribeira corres-pondem ao tipo subtropical, no interior, eao tropical, na costa. Ambos com feiçãosuperúmida, apresentando média anual deprecipitações equivalentes a 1.800 mm,atingindo 2.000 a 3.000 mm na zona daescarpa da Serra, e distribuídas sobre 175dias por ano. As temperaturas, sofrendo ainfluência marítima, são de valor médiosuperior a 22°C no mês mais quente e naplanície onde, em virtude da atuação da
massa tropical atlântica, pode alcançarvalores de 35°C a 40°C. Quanto aos valo-res mínimos, a média do mês mais friocorresponde a 13°C e a atividade da massapolar pode determinar valores de até 3°Cna planície e ao redor de 0°C na escarpa.(Camargo e col.5 1972, Monteiro10 1973,Troppmair21 1975, Nimer14 1977).
Considerando-se a precipitação efetivacomo índice de umidade climática, o terri-tório do Estado de São Paulo pode serdividido por isoietas indicativas das dife-renças entre a precipitação e a evapotrans-piração potencial no semestre seco (abril-setembro). Dessa forma, a linha corres-pondente ao valor zero representa, demaneira geral, o limite demarcatório entreas áreas com e sem estiagem. A regiãodo Vale do Ribeira encontra-se ao suldessa isoieta e o excedente acusa valor de800 a 1.200 mm, não ocorrendo deficiênciade água no solo mesmo no período menoschuvoso (Fig. 1). Assim sendo, à ausênciade estiagem correspondem feições edáficasque, permitindo a retenção de água, condi-cionam maior persistência de umidade(Setzer l8 1966, Camargo4 1971). A carac-terística primitiva da fitogeografia localconsiste no aspecto predominantementeflorestal do chamado domínio tropicalatlântico, acrescido de alguma participação,na parte sul desta região, do correspon-dente ao do planalto de araucárias(Ab'Saber*, 1977). Como exceção a esseaspecto geral, deve-se considerar a faixacosteira com a vegetação litorânea repre-sentada pela rasteira das praias, pelo tipopantanoso marinho dos mangues e pelacobertura arbórea-arbustiva com predomi-nância de indivíduos de porte médio, conhe-cida como "restinga" ou "jundu" (Fig. 6).Acrescente-se, em virtude do sistema hidro-gráfico da planície, a existência de forma-ções hidrófitas situadas nas áreas alagadas.Estas apresentam extensões consideráveisnas zonas da baixada e pré-serrana,
* Comunicação pessoal


acompanhando os numerosos cursos de água.Contudo, como foi mencionado a predomi-nância marcante pertence ao aspecto flo-restal que corresponde àquele da mataperenifólia higrófila cuja distribuiçãocorresponde à ocorrência da elevada umi-dade que caracteriza a região. Essa forma-ção pode ser considerada como climax eé conhecida também pelas denominações defloresta latifoliada tropical úmida da en-costa, e floresta perenifólia latifoliadahigrófila costeira. A sua fisionomia érepresentada por indivíduos arbóreos deporte elevado, podendo alcançar 30 a 40 mde altura, de aspecto denso e comelevado grau de heterogeneidade (Fig. 7).Tal cobertura é encontrada tanto naspartes elevadas da escarpa como na pla-nície e pode assim ser dividida em florestaem níveis altos e em níveis baixos. Aprimeira ocorre nas áreas pré-serranas enas da encosta da serra, além de naselevações isoladas da região, caracterizan-do-se por desenvolver maior número devegetais distribuídos, em linhas gerais, emtrês andares. A segunda situa-se na baixadae os indivíduos distribuem-se em doisandares. A maior luminosidade dos níveisaltos, como conseqüência da inclinação doterreno, faz com que a diminuição da luzno interior da mata seja de apenas 5 vezesem relação ao exterior, contra a reduçãode 8 a 10 vezes que se observa na baixada.Em ambos os tipos ressalta, como aspectopaisagístico marcante, a riqueza de brome-liáceas em suas situações epífitas, terrestresou rupestres, e invadindo não apenas asáreas florestais, como também a restingae os terrenos abertos da orla marítima(Fig. 8). As matas em níveis baixos,freqüentemente sofrem a ação da estagna-ção dos cursos de água, tornando alagadiçoo seu solo. A atuação humana sobre essacobertura florestal tem sido intensa e datada época do descobrimento do Brasil. Tendoem vista a utilização do terreno para fina-lidades agrícolas e pastoris, a devastaçãoatingiu, em maior grau, as matas da planície.
Face à sua menor acessibilidade, as da en-costa sofreram alterações menos intensas.Todavia, o caráter primitivo essencial en-contra-se atualmente limitado a áreas res-tritas. Assim sendo, ao se utilizar os termosde florestas primárias e secundárias, não sepretende significar ausência ou presença demodificações antrópicas. Na verdade, oaspecto primário indica que essa atividadefoi menos marcante ou se limitou a açõesextrativas pouco intensas, de maneira que amata conservou características de primitivis-mo. Por sua vez, no tipo secundário, aatuação humana implicou a ocorrência detransformações profundas nas característicasiniciais. O mapa constante da Fig. 2, forneceidéia da atual situação fitogeográfica nestaregião. Pode-se verificar que, em sua maiorextensão, a cobertura florestal atinge valoresapreciáveis constituindo ainda aspecto domi-nante na paisagem geográfica local (Ca-margo e col.5 1972; SUDELPA16 1974;Alonso3 1977).
Finalmente, deve-se considerar que o Valedo Ribeira encontra-se incluído em regiãoconsiderada como centro de dispersão devertebrados; nos conceitos de Müller 12,13
(1972, 1973), participa do assim chamadoCentro da Serra do Mar que compreendetodo o sistema da escarpa atlântica e a zonalitorânea. Situa-se pois no bioma das citadasflorestas perenifólias higrófilas que se esten-dem ao longo da costa brasileira, desde oEstado de Santa Catarina até os Estadosnordestinos, sofrendo apenas alguma inter-rupção ao nível das regiões limítrofesentre os Estados do Rio de Janeiro e doEspírito Santo (Fig. 1). A parte que seestende ao sul desse limite é consideradacomo o subcentro Paulista e os estudospaleoclimáticos sugerem que o sistemaflorestal ali existente, nos períodos glaciaisquaternários estaria limitado a áreas derefúgio constituídas por manchas de matastropicais descontínuas. Estas estariamlocalizadas na testada superior das escarpasda Serra do Mar mais sujeitas à umidadee circundadas por áreas de semi-aridez


(Ab'Saber1 , 1977). Tais aspectos, junta-mente com observações relativas à fauna devertebrados, permitem considerar essa regiãocomo detentora de elevado caráter deendemicidade com apreciável riqueza deespécies (Müller 13, 1973).
Como foi mencionado, a realização destaspesquisas tencionou a abordagem dospossíveis mecanismos de transmissão deinfecções, ali ocorrentes, mediante a atuaçãohematófaga de dípteros culicídeos. Dessamaneira, o programa de observações a seremefetuadas teve de levar em conta as váriascaracterísticas regionais supradescritas.Assim, de início tornou-se necessária aescolha de várias áreas considerando-se,não apenas as condições fitogeográficasprimárias, como também as resultantes daalteração antrópica da vegetação e do solo.Na circunstância particular da ocorrênciade surto regional de encefalite, a focalizaçãoteve de ser feita visando, tanto os ambientesextradomiciliares, primário e modificado,como os artificiais representados pelosmeios domiciliares, rural e urbano. Assimsendo, procedeu-se à escolha de localidadesem que figurassem essas várias feições e,no presente trabalho, estão especificadas asrepresentantes do ambiente extradomiciliar.
Localidades escolhidas — Em relação aosistema florestal, uma das orientaçõesconsistiu na escolha de área modificadasituada na planície, com a presença dematas residuais e a ocorrência de casoshumanos de encefalite. Tal aspecto foiencontrado no município de Pariquera-Açú,e a estação foi instalada na FazendaExperimental do Instituto Agronômico deCampinas. Ali o terreno é intensamenteutilizado para agricultura e pecuária, con-tando-se com a existência de manchas deflorestas do tipo já descrito para os níveisbaixos. A outra escolha norteou-se parazona pouco modificada, coberta de mataprimária, e onde não tivesse sido assinaladaa ocorrência de casos humanos da epidemia.Para tanto, selecionou-se localidade nomunicípio de Sete Barras correspondenteà Reserva Florestal da Secretaria da
Agricultura do Estado de São Paulo, situadana encosta da Serra de Paranapiacaba. Alias condições são de mata primária emnível alto, com o terreno conservado em suagrande extensão. Finalmente, e em relaçãoainda ao meio extradomiciliar, algumasobservações foram realizadas em áreas in-tensamente moditicadas, e em outras cominício de transformação, em ambos oscasos acusando a ocorrência de casoshumanos de encefalite. O aspecto corres-pondente às primeiras foi encontrado nosarredores do centro urbano de Iguape oqual, por sua vez, foi também escolhidocomo seae de investigações focalizando oambiente domiciliar. Tais localidades fo-ram representadas pelo sítio Embu e peloterreno da Escola Agrícola da Secretariada Agricultura, ali existente. Em ambas,à alteração ambiental soma-se a presençade mata primária em níveis baixos e altos.Quanto às zonas em início de modificação,a escolha recaiu na extremidade da estradaem construção de Itapitangui para Ariri, nomunicípio de Cananéia. O acampamentoali existente abrigou a estação que, pelaprópria mecânica da construção da rodovia,tornou-se móvel, atravessando áreas comflorestas tanto em níveis baixos como altos.
Em resumo, de acordo com os critériosmencionados, foram selecionadas quatrolocalidades onde procedeu-se a instalaçãodas seguintes estações (Figs. 1 e 2):
A (Pariquera-Açú) — Ambiente acentuada-mente alterado, com a presença de ma-tas residuais em níveis baixos e aocorrência de casos de encefalite.Município — Pariquera-Açú.Localidade — Fazenda Experimental de
Pariquera-Açú, do Instituto Agronô-mico de Campinas (Secretaria daAgricultura do Estado de São Paulo).
B (Sete Barras) — Floresta primária emníveis altos, e sem ocorrência de casosde encefalite, pelo menos que tenhamsido detectados até fevereiro de 1978.Município — Sete Barras.

Localidade — Reserva Florestal deSete Barras, na Serra de Parana-piacaba (Secretaria da Agriculturado Estado de São Paulo).
C (Iguape) — Ambiente alterado, nas pro-ximidades da costa marítima, com apresença de matas, primária e residual,em níveis baixos e a ocorrência decasos de encefalite.
Município — Iguape.
Localidades — Sítio Embu.
Escola Agrícola (Secretaria da Edu-cação do Estado de São Paulo).
Centro urbano.
D (Ariri) — Ambiente pouco modificado,em início de colonização, com a pre-sença de matas primárias, em níveisbaixos e altos, e a ocorrência de casosesparsos de encefalite.Município — Cananéia.
Localidade — Ariri (estrada em cons-trução, de Itapitangui a Ariri).
MATERIAL E MÉTODOS
A atividade culicidiana no ambiente ex-tradomiciliar foi observada mediante aorganização de coletas levadas a efeito demaneira regular, ao lado de outras execu-tadas isoladamente ou em reduzido númerode vezes. Nelas empregou-se técnicasbaseadas na utilização de isca humana,armadilha tipo Shannon e armadilha lumi-nosa automática tipo New Jersey.
As observações regulares foram realiza-das nas matas das estações de Pariquera-Açú e de Sete Barras. Em ambas, osambientes focalizados foram o intra e oextraflorestal. No primeiro utilizou-se iscahumana para capturas executadas, de ma-neira simultânea, em dois níveis e quecorrespondem ao solo e à copa arbórea.Para isso, estabeleceu-se posto de coletarepresentado por uma árvore, adrede esco-
lhida, em cujas ramificações superioresprocedeu-se à instalação de plataforma aces-sível mediante escada construída ao longode seu tronco (Fig. 9). Cada estaçãocontou com um posto operando semanal-mente com o emprego de indivíduos pre-viamente treinados. Trabalhando, de ma-neira concomitante, na plataforma e nosolo, a captura atingiu todos os mosquitosque pousavam ou se aproximavam parasugar. Assim sendo, cada coletor desem-penhou também o papel de isca humana.Em dia pré-estabelecido da semana, taisatividades foram realizadas em três etapas,correspondentes aos horários das 9:00 às11:00, das 16:00 às 18:00 e das 20:00 às22:00 horas. Em algumas ocasiões, esseritmo semanal teve de ser interrompido facea ocorrência de circunstâncias diversas,principalmente a queda de chuvas prolon-gadas que dificultaram ou mesmo impossi-bilitaram a realização das capturas. Toda-via tais eventos não chegaram a compro-meter sensivelmente a regularidade pro-gramada.
Em relação ao meio extraflorestal, asobservações foram realizadas em locaisabertos, previamente escolhidos, correspon-dentes à margem da mata e a terrenos uti-lizados e modificados para lavouras ouformação de pastagens. Ali empregou-sea armadilha tipo Shannon, em ritmo mensalde captura e obedecendo aos horáriossupracitados. Em Pariquera-Açú, além delocal na imediata vizinhança da bordaflorestal, foi escolhido outro em plantaçãode árvores frutíferas situado a cerca de200 m, em linha reta, da mata residualonde foram levadas a efeito as outrascoletas. Em Sete Barras, face às caracte-rísticas locais, foi maior a proximidade doambiente florestal. Assim sendo, a maiordistância que o separou do local dascapturas, não ultrapassou os 50 m, emlinha reta. As Figs. 10 e 11 fornecemidéia dessa situação e dos aspectos paisa-gísticos desses locais.

Ainda no que concerne ao ambiente flo-restal, levou-se a efeito coletas com a utili-zação de armadilha tipo New Jersey. Foramrealizadas nas estações de Ariri e de Iguape.Em relação à primeira, os dados de suaexecução e resultados serão apresentadosem trabalho posterior. Quanto à segunda,as coletas deixaram de ser efetuadas emritmo regular e nelas foram empregadas,de maneira variável, as três técnicas men-cionadas, tendo a armadilha automáticafuncionado no horário das 18:00 às 22:00horas. Os locais em que foram realizadas
são representados pelas matas do SítioEmbu e da Escola Agrícola (Fig. 12).
Nas capturas levadas a efeito com ritmoregular, procurou-se obter informaçõessobre a atividade hematófaga mediante ocálculo das médias de Williams ([]w),sempre que os dados obtidos o permitissem(Haddow7 , 8 1954, 1960). Para tanto epara a representação gráfica, as médiasforam transformadas em percentagens dasoma total. Com o objetivo de facilitar amanipulação, os valores obtidos forammultiplicados por 100,0.


No que concerne à identificação dosespécimens coletados, deve-se assinalar queapresentou as dif iculdades conhecidas, paravários gêneros, em relação às fêmeas, Nestaetapa inicial dos trabalhos não foi possívela realização de estudos baseados emcriação de formas imaturas, motivo peloqual vários grupos foram determinadosapenas até gênero. Assim é que, paraCulex, somente algumas espécies do sub-gênero Culex foram identificadas com boamargem de segurança, ao passo que paraMelanoconion não houve nem mesmo essapossibilidade. Assinale-se contudo que, emrelação a este último, alguns machos cole-tados permitiram assinalar a presença deCulex taeniopus. Porém, a sua real pro-porção na amostra permaneceu desconhe-cida. Acrescente-se a presença tambémde indivíduos masculinos dos subgênerosAedinus e Eubonnea. Face a isso incluimoscomo Culex (Melanoconion) sp., fêmeas que
possivelmente pertencessem a esses grupos,sem mencionar Mochlostyrax, cuja posiçãosubgenérica é questionável. Fato análogoocorreu com as fêmeas de Psorophora albi-pes e Ps.lutzii, que não foi possível separar.Todavia deve-se levar em conta que se tratade espécies muito próximas, não se afastandoa hipótese de sinonímia. Finalmente, resul-tado análogo teve de ser aceito em relaçãoa alguns Limatus e à maioria de represen-tantes de Phoniomyia, Trichoprosopon eWyeomyia. Tratam-se de grupos de siste-mática ainda não bem estabelecida, o queno estudo atual dos conhecimentos tornaproblemática a determinação de indivíduosfemininos .
Com as ressalvas feitas acima, adotou-sea orientação de considerar somente as es-pécies ou gêneros que, de maneira indivi-dual , compareceram com percentagenssuperiores a 1,0% do total geral. As demais

constam dos resultados obtidos, apenaspara o registro de sua presença na com-posição faunística local. Tal conduta foiaplicada a todos os tipos de coleta e, nocaso das realizadas nas duas alturas flo-restais, teve sua aplicação prioritária nosresultados obtidos ao nível do solo. Com-preende-se esta última atitude uma vez quese considere os objetivos dos atuais estudose que residem, fundamentalmente, na verifi-cação de culicídeos que se relacionam coma população humana. Tal relacionamentoforçosamente se fará a baixas alturas,interessando saber se tais mosquitos pode-riam ou não freqüentar também a copadas árvores, onde encontrariam possíveisreservatórios naturais da virose. Essa regrageral, aparentemente deixou de ser seguidano caso de Phoniomyia sp. Contudo, éde se considerar a presença altamente fre-qüente de Phoniomyia davisi e os indíciosque indicaram a ocorrência de grandenúmero de representantes naquele grupo.Tal fato levou a supor que a participaçãoindividual de cada um não fosse possivel-mente comparável com a daquela espécie.Por sua vez, os aspectos observados comPh. davisi, e até que ulteriores estudostragam maiores esclarecimentos, podem sergenericamente aplicados a Phoniomyia sp.como um todo. Finalmente, a inclusão dosdados de Anopheles (Kerteszia) prende-seà reconhecida importância desses anofelinosno relacionamento humano já mencionado.
RESULTADOS
Os dados apresentados a seguir resultamde observações levadas a efeito no períodode julho de 1976 a setembro de 1977. Comofoi mencionado, os que se referem às esta-ções de Pariquera-Açú e Sete Barrasrefletem regularidade em seu ritmo deexecução durante aquele espaço de tempo.Por outro lado, nas coletas realizadas naestação de Iguape, não foi possível conse-guir seqüência regular. Em vista disso, osresultados ali obtidos serão apresentadosapenas em seus aspectos globais.
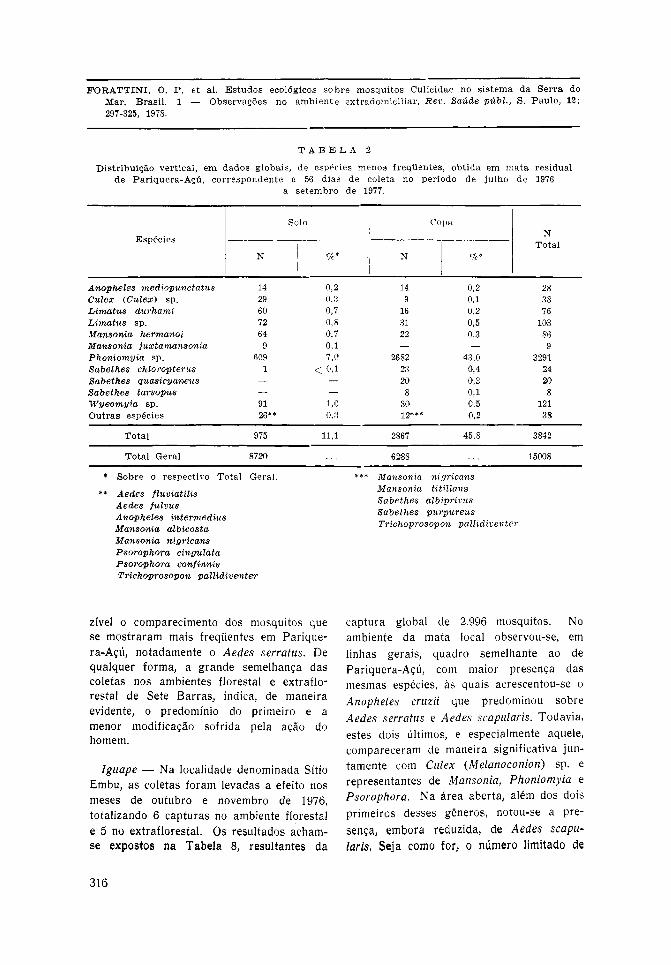
Pariquera-Açú — No ambiente da mataresidual levou-se a eleito o total de 56 diasde coleta, obtendo-se o número global de15.008 mosquitos. As tabelas 1 e 2 apre-sentam a distribuição específica bem comoa referente aos dois níveis de captura, ouseja, o solo e a copa. A primeira incluios representantes que ocorreram com maiorfreqüência, para os quais se calculou amédia por coleta ([]w). Pode-se notar opapel preponderante que, nesse sentido, oAedes serratus desempenhou ao nível dosolo florestal, com média de 56,87 porcoleta e constituindo 46,3% dos mosquitoscapturados. Na copa, essa primazia coubea Phoniomyia davisi com 20,84 e 25,6%para esses valores. Contudo, note-se queaquele mosquito, mesmo a esse nível,compareceu com a média de 6,28, enquantoque este, no solo, participou com 7,58. Asdemais espécies mostraram menor atividade,podendo-se assinalar alguma relevânciapara o Aedes scapularis ao nível do solo,com 5,36 por coleta. No que concerneà segunda dessas tabelas, estão ali indi-cados os representantes que, em conjunto,formaram pouco mais de 11,0% dos culicí-deos coletados. Entre eles destaca-se osincluídos em Phoniomyia sp. os quais, comojá foi mencionado, são representados pornumerosas espécies que, de maneira isolada,torna-se lícito admitir que provavelmentenão tivessem desempenhado papel compa-rável ao apresentado por Ph. davisi.
Levando-se em conta o número de 11.166mosquitos, que se constituíram em 74,4%do material coletado (Tabela 1), pôde-secalcular as médias correspondentes a cadauma das espécies participantes desse grupo.Isso foi feito para cada nível de capturana mata, transformando-as em percentuaissobre o total correspondente às []w.Considerando-se os que participaram comvalores iguais ou superiores a 5,0%, pelomenos em um dos níveis, obteve-se osresultados representados no gráfico da Fig.13. Pode-se, assim, verificar a distribuiçãovertical desses mosquitos, com ampla pre-
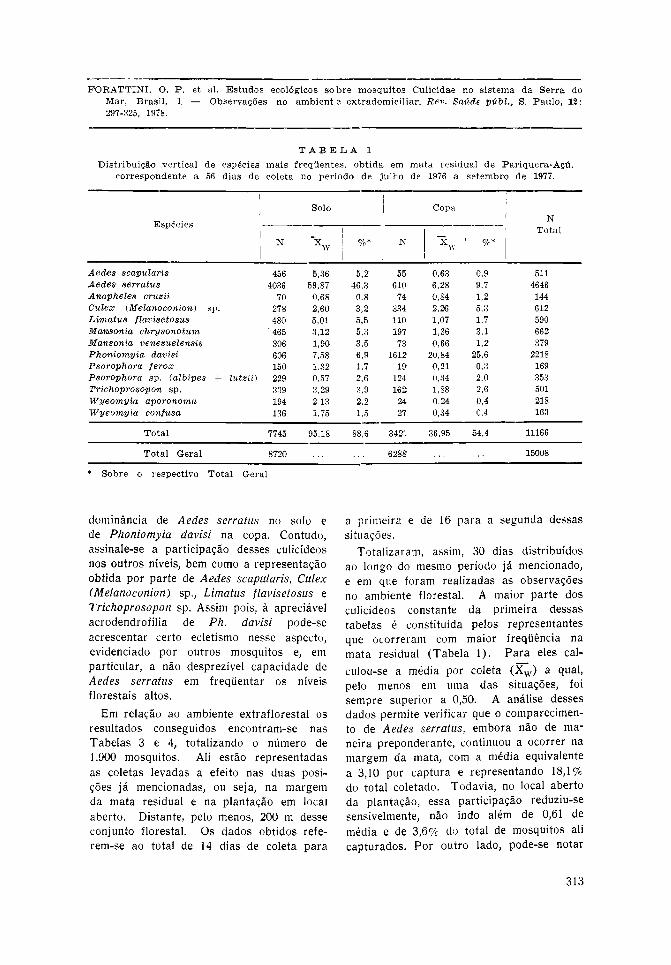
dominância de Aedes serratus no solo ede Phoniomyia davisi na copa. Contudo,assinale-se a participação desses culicídeosnos outros níveis, bem como a representaçãoobtida por parte de Aedes scapularis, Culex(Melanoconion) sp., Limatus flavisetosus eTrichoprosopon sp. Assim pois, à apreciávelacrodendrofilia de Ph. davisi pode-seacrescentar certo ecletismo nesse aspecto,evidenciado por outros mosquitos e, emparticular, a não desprezível capacidade deAedes serratus em freqüentar os níveisflorestais altos.
Em relação ao ambiente extraflorestal osresultados conseguidos encontram-se nasTabelas 3 e 4, totalizando o número de1.900 mosquitos. Ali estão representadasas coletas levadas a efeito nas duas posi-ções já mencionadas, ou seja, na margemda mata residual e na plantação em localaberto. Distante, pelo menos, 200 m desseconjunto florestal. Os dados obtidos refe-rem-se ao total de 14 dias de coleta para
a pr imeira e de 16 para a segunda dessassituações.
Totalizaram, assim, 30 dias distribuídosao longo do mesmo período já mencionado,e em que foram realizadas as observaçõesno ambiente florestal. A maior parte dosculicídeos constante da primeira dessastabelas é constituída pelos representantesque ocorreram com maior freqüência namata residual (Tabela 1). Para eles cal-culou-se a média por coleta ([]w) a qual,pelo menos em uma das situações, foisempre superior a 0,50. A análise dessesdados permite verificar que o comparecimen-to de Aedes serratus, embora não de ma-neira preponderante, continuou a ocorrer namargem da mata, com a média equivalentea 3,10 por captura e representando 18,1%do total coletado. Todavia, no local abertoda plantação, essa participação reduziu-sesensivelmente, não indo além de 0,61 demédia e de 3,6% do total de mosquitos alicapturados. Por outro lado, pode-se notar
que o Aedes scapularis manteve-se presentede maneira constante, com valores maioresdo que os obtidos na copa arbórea damata. Na área aberta, sua presença foimais acentuada, com média de 1,22 e consti-tuindo 11,1% dos espécimens, sendo ultra-passado ali somente pela Mansonia chry-sonotum com 36,2% embora este mosquitotenha apresentado média pouco superior eequivalente a 1,95. Além dessas espécies,chamou a atenção na margem da mata apresença de Culex (Melanoconion) sp.,Limatus f íavisetosus; Mansonia venezue-lensis, Phoniomyia davisi e Wyeomyiaconfusa. Nesse particular, convém assinalarque somente o primeiro, constituído pelo
conjunto subgenérico de Melanoconion,mostrou sensível aumento, quando compa-rado com o observado no ambiente flores-ta l . Com efeito, as médias ali obtidas fo-ram de 2,60 e 2,26 para os dois níveis,enquan to que nesta margem da mata resi-dual , esse valor foi de 4,68. Os demais,ou d iminu i ram a sua presença, ou a aumen-ta ram de maneira pouco acentuada. Emrelação ao local aberto situado na planta-ção distante da mata, observou-se diminui-ção geral das médias, comparecendo comvalores ao redor de 1,0 ou superior, apenasos representantes do gênero Mansonia,Aedes scapularis e Culex (Melanoconion) sp.
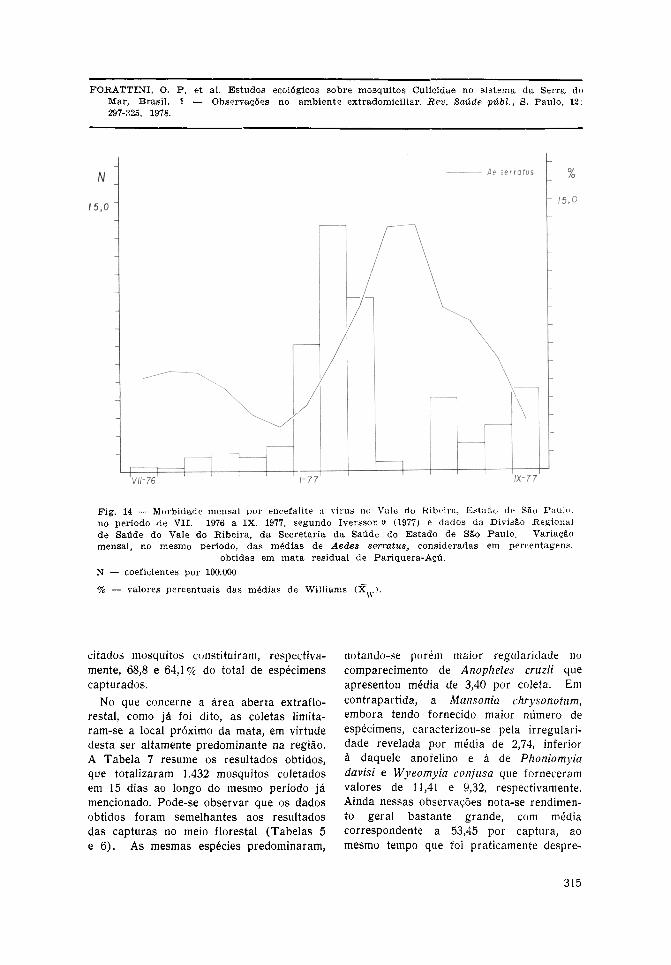
Considerando-se Aedes serratus como aespécie que mostrou maior atividade noambiente f lores ta l , com presença signif icantetambém fora da mata, procedeu-se ao cál-culo das médias mensais no espaço detempo já citado ( ju lho de 1976 a setembrode 1977). Transformadas em percentuaissobre o total da soma dos valores mensais,fo ram colocados em gráfico representadopela Fig. 14. A essa curva mensal adicio-nou-se os dados referentes aos casos deencefalite, ocorridos no mesmo período, paratodo o Vale do Ribeira. Pode-se observarcerta concordância das incidências, emboraas informações sobre o supracitado mos-quito se re f i r am a uma localidade apenas.
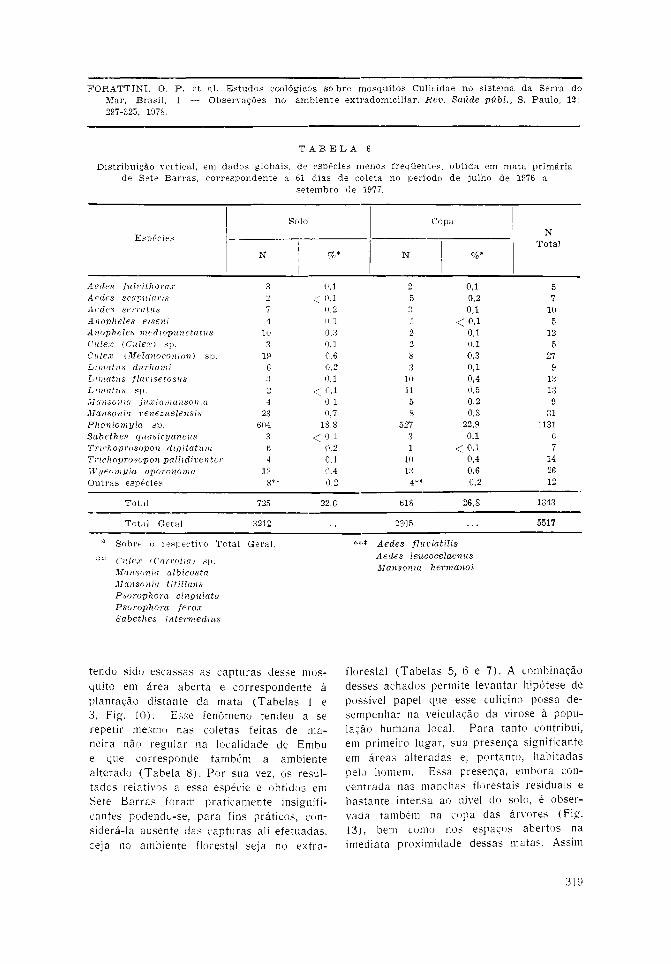
Sete Barras — As observações efetuadasna floresta p r imár ia , num total de 61 diasde coletas, resultaram na obtenção de 5.517mosquitos. As distribuições, específica evertical, encontram-se nas Tabelas 5 e 6.Na pr imeira estão incluídos os represen-tantes mais freqüentes, para os quais foram
calculadas as médias por captura ([]w).Ver i f i cou-se o predomínio de Phoniomyiadavisi, Wyeomyia confusa e Mansonia chry-sonotum sem, contudo, terem ocorridodiferenças sensíveis entre os dois níveis.Pode-se notar a reduz ida presença de espé-cies que se revelaram como as mais fre-qüentes na mata residual de Pariquera-Açú.Assim, no solo e na copa, os três supra-
citados mosquitos constituíram, respectiva-mente, 68,8 e 64,1% do total de espécimenscapturados.
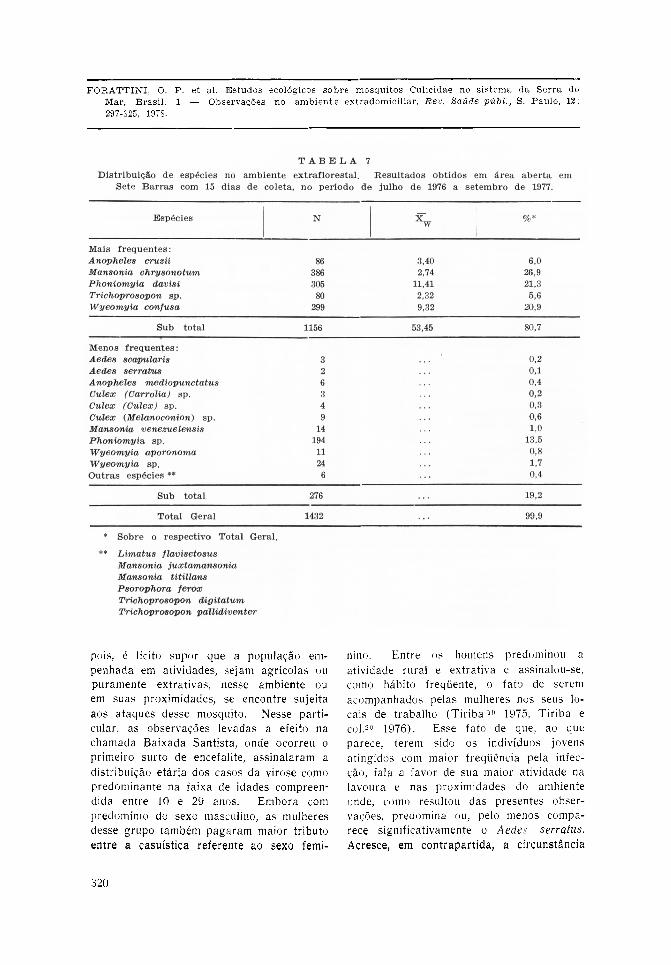
No que concerne a área aberta extraflo-restal, como já foi dito, as coletas limita-ram-se a local próximo da mata, em virtudedesta ser altamente predominante na região.A Tabela 7 resume os resultados obtidos,que totalizaram 1.432 mosquitos coletadosem 15 dias ao longo do mesmo período jámencionado. Pode-se observar que os dadosobtidos foram semelhantes aos resultadosdas capturas no meio florestal (Tabelas 5e 6). As mesmas espécies predominaram,
notando-se porém maior regularidade nocomparecimento de Anopheles cruzii queapresentou média de 3,40 por coleta. Emcontrapartida, a Mansonia chrysonotum,embora tendo fornecido maior número deespécimens, caracterizou-se pela irregulari-dade revelada por média de 2,74, inferiorà daquele anofelino e à de Phoniomyiadavisi e Wyeomyia confusa que forneceramvalores de 11,41 e 9,32, respectivamente.Ainda nessas observações nota-se rendimen-to geral bastante grande, com médiacorrespondente a 53,45 por captura, aomesmo tempo que foi praticamente despre-
zível o comparecimento dos mosquitos quese mostraram mais freqüentes em Parique-ra-Açú, notadamente o Aedes serratus. Dequalquer forma, a grande semelhança dascoletas nos ambientes florestal e extraflo-restal de Sete Barras, indica, de maneiraevidente, o predomínio do primeiro e amenor modificação sofrida pela ação dohomem.
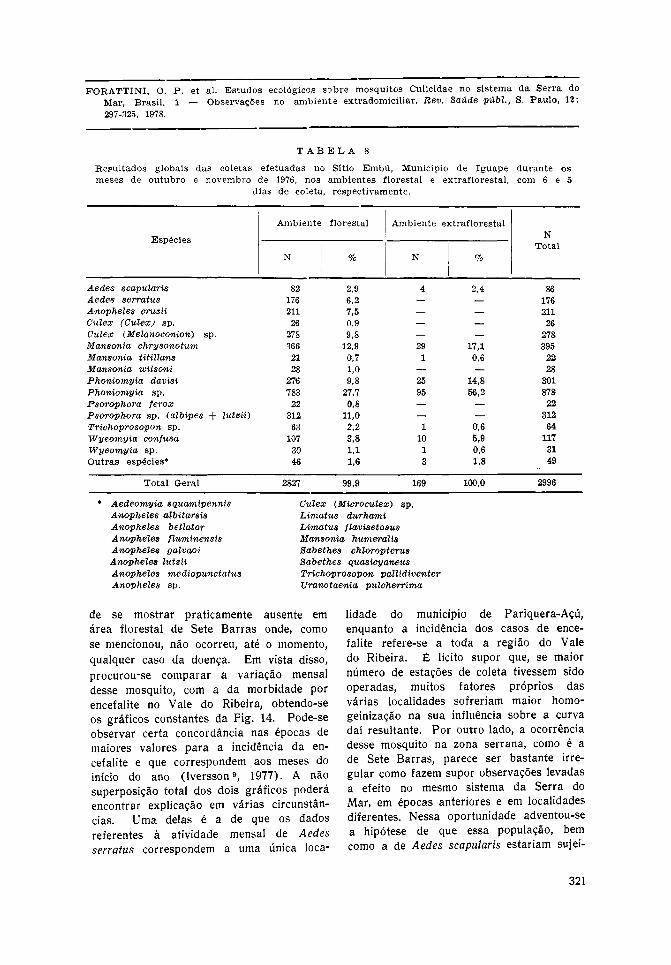
Iguape — Na localidade denominada SítioEmbu, as coletas foram levadas a efeito nosmeses de outubro e novembro de 1976,totalizando 6 capturas no ambiente florestale 5 no extraflorestal. Os resultados acham-se expostos na Tabela 8, resultantes da
captura global de 2.996 mosquitos. Noambiente da mata local observou-se, emlinhas gerais, quadro semelhante ao dePariquera-Açú, com maior presença dasmesmas espécies, às quais acrescentou-se oAnopheles cruzii que predominou sobreAedes serratus e Aedes scapularis. Todavia,estes dois últimos, e especialmente aquele,compareceram de maneira significativa jun-tamente com Culex (Metanoconion) sp. erepresentantes de Mansonia, Phoniomyia ePsorophora. Na área aberta, além dos doisprimeiros desses gêneros, notou-se a pre-sença, embora reduzida, de Aedes scapu-laris. Seja como for, o número limitado de
observações não permitiu deduzir quadrolocal mais preciso.
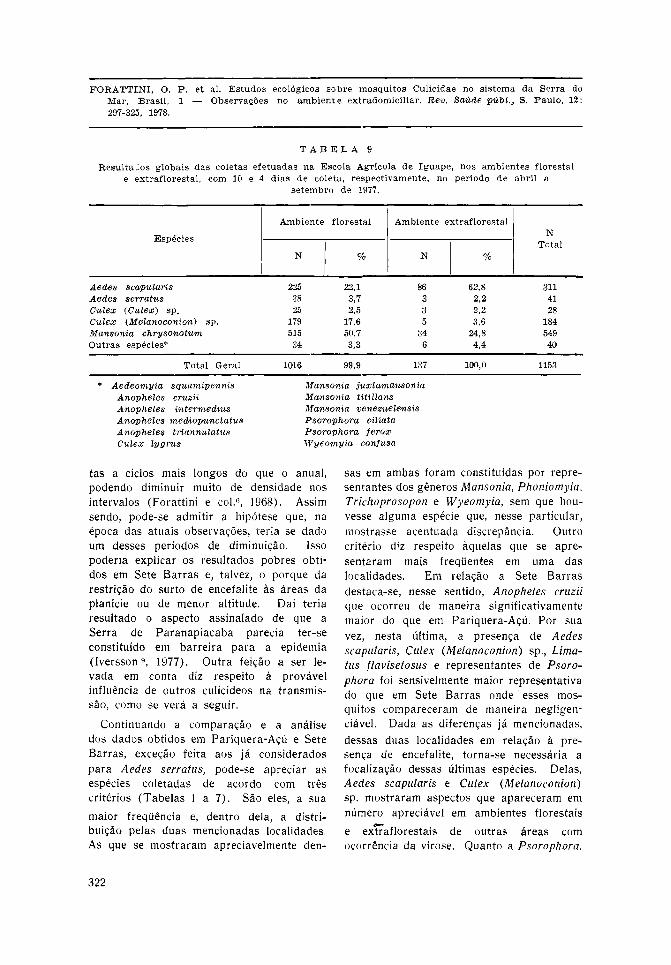
Quanto às coletas levadas a efe i to naárea da Escola Agrícola de Iguape, os
resultados encontram-se na Tabela 9, ecorrespondem aos totais obtidos em 10 diasde capturas no meio florestal e 4 no extra-
f loresta l . Pode-se observar presença mar-cante de Aedes scapularis nos dois ambien-
tes, ao lado de Culex (Melanoconion) sp.e Mansonia chrysonotum, ao lado da ocor-rência de Aedes serratus, embora de maneiradiscreta. Cabe aqui considerar as mesmascircunstâncias mencionadas no parágrafoanterior, ou seja, o reduzido número e afalta de regular idade no ritmo dessasobservações. Tais aspectos constituíram-seem fatores limitantes, para possível aprecia-
ção do quadro conseguido.
COMENTÁRIOS
Os dados obtidos com estas observaçõesque focal izaram o ambiente extradomici l iarpermitem chamar atenção sobre algunsaspectos. De início considere-se que asestações de Pariquera-Açú e de Sete Barrasrepresentam, respectivamente, áreas deterreno alterado e preservado, aliado à pre-sença e ausência de casos de encefalite.Assim sendo, os resultados podem sercomparados à luz dessas diferenças, umavez que as coletas foram conduzidas regu-larmente e de maneira comparável, nomesmo período compreendido entre ju lho de1976 e setembro de 1977.
A composição cul icidiana obtida em Pari-quera-Açú revelou nítido predomínio deAedes serratus no solo de mata residual,mantendo signif icat iva presença na copa enas proximidades desse conjunto florestal.Por outro lado, sua ocorrência parecedecrescer sensivelmente com a distância,
tendo sido escassas as capturas desse mos-quito em área aberta e correspondente àplantação distante da mata (Tabelas 1 e3, Fig. 10). Esse fenômeno tendeu a serepetir mesmo nas coletas feitas de ma-neira não regular na local idade de Embue que corresponde também a ambientealterado (Tabela 8). Por sua vez, os resul-tados relativos a essa espécie e obt idos emSete Barras fo ram pra t icamente in s ign i f i -cantes podendo-se, para f i n s práticos, con-siderá-la ausente das capturas ali efetuadas,ceja no ambiente f lorestal seja no extra-
florestal (Tabelas 5, 6 e 7). A combinaçãodesses achados permite levantar hipótese depossível papel que esse cu l i c ino possa de-sempenhar na veiculação da virose à popu-lação humana local. Para tanto contr ibui ,em pr imei ro lugar , sua presença s igni f ican teem áreas alteradas e, portanto, habi tadaspelo homem. Essa presença, embora con-centrada nas manchas florestais residuais ebastante intensa ao nível do solo, é obser-vada também na copa das árvores (Fig.13), bem como nos espaços abertos naimediata proximidade dessas matas. Assim
pois, é lícito supor que a população em-penhada em atividades, sejam agrícolas oupuramente extrativas, nesse ambiente ouem suas proximidades, se encontre sujeitaaos ataques desse mosquito. Nesse parti-cular, as observações levadas a efeito nachamada Baixada Santista, onde ocorreu oprimeiro surto de encefalite, assinalaram adistr ibuição etária dos casos da virose comopredominante na faixa de idades compreen-dida entre 10 e 29 anos. Embora compredomínio do sexo masculino, as mulheresdesse grupo também pagaram maior tributoentre a casuística referente ao sexo femi-
nino. Entre os homens predominou aatividade rural e extrativa e assinalou-se,como hábito freqüente, o fato de seremacompanhados pelas mulheres nos seus lo-cais de trabalho (Tiriba1 9 1975, Tiriba ecol.20 1976). Esse fato de que, ao queparece, terem sido os indivíduos jovensatingidos com maior f reqüência pela infec-ção, fala a favor de sua maior atividade nalavoura e nas proximidades do ambienteonde, como resultou das presentes obser-vações, predomina ou, pelo menos compa-rece significativamente o Aedes serratus.Acresce, em contrapartida, a circunstância
de se mostrar praticamente ausente emárea florestal de Sete Barras onde, comose mencionou, não ocorreu, até o momento,qualquer caso da doença. Em vista disso,procurou-se comparar a variação mensaldesse mosquito, com a da morbidade porencefalite no Vale do Ribeira, obtendo-seos gráficos constantes da Fig. 14. Pode-seobservar certa concordância nas épocas demaiores valores para a incidência da en-cefalite e que correspondem aos meses doinício do ano (Iversson9, 1977). A nãosuperposição total dos dois gráficos poderáencontrar explicação em várias circunstân-cias. Uma delas é a de que os dadosreferentes à atividade mensal de Aedesserratus correspondem a uma única loca-
lidade do município de Pariquera-Açú,enquanto a incidência dos casos de ence-falite refere-se a toda a região do Valedo Ribeira. É lícito supor que, se maiornúmero de estações de coleta tivessem sidooperadas, muitos fatores próprios dasvárias localidades sofreriam maior homo-geinização na sua influência sobre a curvadaí resultante. Por outro lado, a ocorrênciadesse mosquito na zona serrana, como é ade Sete Barras, parece ser bastante irre-gular como fazem supor observações levadasa efeito no mesmo sistema da Serra doMar, em épocas anteriores e em localidadesdiferentes. Nessa oportunidade adventou-sea hipótese de que essa população, bemcomo a de Aedes scapularis estariam sujei-
tas a ciclos mais longos do que o anual,podendo diminuir muito de densidade nosintervalos (Forattini e col.6, 1968). Assimsendo, pode-se admitir a hipótese que, naépoca das atuais observações, teria se dadoum desses períodos de diminuição. Issopoderia explicar os resultados pobres obti-dos em Sete Barras e, talvez, o porque darestrição do surto de encefalite às áreas daplanície ou de menor altitude. Daí teriaresultado o aspecto assinalado de que aSerra de Paranapiacaba parecia ter-seconstituído em barreira para a epidemia(Iversson9 , 1977). Outra feição a ser le-vada em conta diz respeito à provávelinfluência de outros culícideos na transmis-são, como se verá a seguir.
Continuando a comparação e a análisedos dados obtidos em Pariquera-Açú e SeteBarras, exceção feita aos já consideradospara Aedes serratus, pode-se apreciar asespécies coletadas de acordo com trêscritérios (Tabelas 1 a 7). São eles, a sua
maior freqüência e, dentro dela, a distri-buição pelas duas mencionadas localidades.As que se mostraram apreciavelmente den-
sas em ambas foram constituídas por repre-sentantes dos gêneros Mansonia, Phoniomyia,Trichoprosopon e Wyeomyia, sem que hou-vesse alguma espécie que, nesse particular,mostrasse acentuada discrepância. Outrocritério diz respeito àquelas que se apre-sentaram mais freqüentes em uma daslocalidades. Em relação a Sete Barrasdestaca-se, nesse sentido, Anopheles cruziique ocorreu de maneira significativamentemaior do que em Pariquera-Açú. Por suavez, nesta última, a presença de Aedesscapularis, Culex (Melanoconion) sp., Lima-tus flavisetosus e representantes de Psoro-phora foi sensivelmente maior representativado que em Sete Barras onde esses mos-quitos compareceram de maneira negligen-ciável. Dada as diferenças já mencionadas,dessas duas localidades em relação à pre-sença de encefalite, torna-se necessária afocalização dessas últimas espécies. Delas,Aedes scapalaris e Culex (Melanoconion)sp. mostraram aspectos que apareceram emnúmero apreciável em ambientes florestais
e extraflorestais de outras áreas comocorrência da virose. Quanto a Psorophora,

embora sem essa constância, pôde-se obser-var aspectos semelhantes (Tabelas 8 e 9).Assim pois, é de se admit i r a hipótese deque essas espécies também contribuam paraa transmissão, devendo-se chamar a atençãopara a feição apresentada por Aedes sca-pularis e Culex (Melanoconion) sp. com suafreqüência aos locais abertos do meio extra-florestal. Esse aspecto poderia torná-losaptos a atingir a população que trabalhanas plantações, bem como as habitaçõesconstruídas nesses locais. Quanto a Limatuse Psorophora, a sua presença limitada àfloresta e de maneira não tão constantequanto à apresentada por Aedes serratus,poderia reduzir a sua responsabilidade decontato com a população humana.
Finalmente, deve-se considerar o grandenúmero de espécies coletadas, indicativasda riqueza faunística regional. A extrati-ficação observada na mata residual dePariquera-Açú (Tabelas 1 e 2, Fig. 13)não foi verificada em Sete Barras ondenão se notou evidências desse fenômeno.É possível que, para explicar essa discre-pância, tenha influído a diferença já men-cionada, de iluminação para o solo e acopa, entre as florestas em nível alto ebaixo (Camargo e col.5, 1972). Sendo estassensivelmente mais escuras no solo do queaquelas, a extratificação dos mosquitospoderá tender a se tornar mais evidente.
Em resumo, as observações levadas aefeito em várias situações, sugerem que,do grande número de espécies de mosquitosencontradas, algumas apresentam maiorprobabilidade de contatos freqüentes com apopulação humana. Nesse sentido, desta-cam-se Aedes serratus, Aedes scapularis eCulex (Melanoconion) sp., sem contudodeixar de admitir a possibilidade de atuaçãode outros culicídeos.
CONCLUSÕES
1 — Admitindo-se a hipótese de que o surtode encefalite ocorrido no Vale do Ri-beira e na Baixada Santista, tenhasido causado por uma arbovirose, deve
ser considerada a responsabilidade ve-tora de mosquitos Culicidae.
2 — A fauna culicidiana regional é extre-mamente rica, apresentando variações,no que concerne à freqüência das váriasespécies, em relação às característicaslocais.
3 — Em áreas de ambiente alterado, commatas residuais e ocorrência de casosda doença, os representantes mais sig-nificantes foram Aedes serratus, Aedesscapularis e Culex (Melanoconion) sp.
4 — A significância desses culicídeos baseia-se na sua freqüência em várias situa-ções, e na sua ausência ou presençanegligenciável em locais onde nãoocorreram casos da epidemia.
5 — A presença desses culicídeos apresentacaracterísticas que permite associá-laaos hábitos da população local, rela-tivos à ocupação e ao trabalho noambiente extradomiciliar.
6 — Em que pesem essas evidências, nãose exclui a participação, de caráterprovavelmente variável, de outras es-pécies de mosquitos, como dos gênerosLimatus e Psorophora.
7— A baixa freqüência de Aedes serratus,Aedes scapularis e Culex (Melano-conion) sp. na encosta da Serra deParanapiacaba, revelada pelas obser-vações em Sete Barras, poderá obedecera fenômeno cíclico de baixa densidade.Esse fato poderá contribuir para expli-car o aparente papel de barreira quea Serra de Paranapiacaba e a Serra doMar propriamente dita desempenharampara a propagação da epidemia emdireção ao planalto.
AGRADECIMENTO
Ao Dr. Rui Ribeiro dos Santos, Diretorda Fazenda Experimental de Pariquera-Açú,da Secretaria da Agricultura do Estado deSão Paulo, pelas facilidades proporcionadasque tornaram possível a realização destasobservações.

RSPUB9/418
FORRATTINI, O. P. et al. [Ecologic studies on Culicidae mosquitoes in the Serra doMar System, Brazil. 1 — Observations in the outdoor environment] Rev.Saúde públ., S. Paulo, 12:297-325, 1978.ABSTRACT: This paper presents the results of out door catches of Culicidae
mosquitoes performed in the Ribeira Valley, State of S. Paulo, Brazil, where anoutbreak of encephalits occurred in 1976 and 1977. These entomologicalinvestigations were made using several techniques for collecting mosquitoes,including human bait. Several places were chosen considering the environmentalaspects related to human activities. In regions where high agriculturalactivities predominate and the primary forest was reduced to remaining spots,large numbers of Aedes serratus, Aedes scapularis and Culex (Melanoconion) sp.were caught. These mosquitoes were scarcely found in primary forest regionswhere no human cases of encephalitis occurred. These aspects suggest theirparticipation in virus transmission. Beside this, other species, mainly Limatusand Psorophora, may possibly act as vectors, although with less epidemiologicalsignificance. It was noted that the scarcity of Aedes in the forest of the "Serrado Mar" system may be a cyclic phenomenon, as was pointed out in otherinvestigations. This circunstance may explain the apparent barrier role to theepidemic that this mountain system seems to fulfill.
UNITERMS: Culex, Vale do Ribeira, SP, Brazil. Arboviruses. Encephalitis,epidemic, S. Paulo, Brazil. Ecology. Aedes serratus. Aedes scapularis. Culex (Mela-noconion) sp.
REFERÊNCIAS BIBLIOGRÁFICAS
1. AB'SABER, A. N. Espaços ocupados pelaexpansão dos climas secos na Américado Sul, por ocasião dos períodos gla-ciais quaternários. São Paulo, Institutode Geografia da USP, 1977. (SériePaleoclimas, 3).
2. ALMEIDA, F. F. M. de Fundamentos geo-lógicos do relevo paulista. São Paulo,Instituto de Geografia da USP, 1974.(Série Teses e Monografias, 14).
3. ALONSO, M. T. A. Vegetação. In: Funda-ção IBGE. Geografia do Brasil: regiãosudeste. Rio de Janeiro, 1977. v. 3,p. 91-118.
4. CAMARGO, A. P. de Balanço hídrico noEstado de São Paulo. 3a. ed. Cam-pinas, Instituto Agronômico, 1971.(Boletim, 116).
5. CAMARGO, J. C. G. et al. Estudo f i togeo-gráfico e ecológico da bacia hidro-gráfica paulista do Rio Ribeira. SãoPaulo, Instituto de Geografia da USP,1972. (Série Biogeografia, 5).
6. FORATTINI, O. P. et al. Investigaçõessobre o comportamento de formasadultas de mosquitos silvestres noEstado de São Paulo, Brasil. Rev.Saúde públ., S. Paulo, 2:11-73, 1968.
7. HADDOW, A. J. Studies of the biting-habits of African mosquitos. Anappraisal of methods employed, withspecial reference to the twenty-four-hour catch. Bull. ent. Res., 45:199-242,1954.
8. HADDOW, A. J. Studies on the bitinghabits and medical importance of eastAfrican mosquitos in the genus Aedes.I — Sbgenera Aëdimorphus, Banksi-nella and Dunnius. Bull. ent. Res,50:759-79, 1960.
9. IVERSSON, L. B. Epidemia de encefalitepor arbovírus na região sul do Estadode São Paulo, Brasil, em 1975 e 1976:aspectos da distribuição cronológica egeográfica dos casos. Rev. Saúdepúbl., S. Paulo, 11:375-88, 1977.
10. MONTEIRO, C. A. de F. A dinâmica cli-mática e as chuvas no Estado de SãoPaulo. São Paulo, Instituto de Geo-grafia da USP, 1973.

11. MOREIRA, A. A. N. & CAMELIER, C.Relevo. In: Fundação IBGE. Geografiado Brasil: região sudeste. Rio deJaneiro, 1977. v. 3, p. 1-50.
12. MÜLLER, P. Centres of dispersal andevolution in the neotropical region.Stud. neotrop. Fauna, 7:173-85, 1972.
13. MÜLLER, P. The dispersal centres ofterrestrial vertebrates in the neotro-pical realm. The Hague, W. Junk B.V. Publ., 1973.
14. NIMER, E. Clima. In: Fundação IBGE.Geografia do Brasil: região sudeste.Rio de Janeiro, 1977. v. 3, p. 51-89.
15. SECRETARIA DE ECONOMIA E PLANE-JAMENTO. Conheça seu município:região do Vale do Ribeira. São Paulo,1974. v. 1.
16. SECRETARIA DE ECONOMIA E PLANE-JAMENTO. SUDELPA: área de atua-ção; caracterização físico-econômico-social. São Paulo 1974.
17. SUPERINTENDÊNCIA DE CONTROLE DEENDEMIAS (SUCEN). Combate a
vetores em municípios do Estado deSão Paulo atingidos por encefalite.[Apresentado ao 8o Congresso Brasi-
leiro de Engenharia Sanitária, Rio deJaneiro, 1975].
18. SETZER, J. Atlas climático e ecológico doEstado de São Paulo. São Paulo,Comissão Interestadual da Bacia Pa-raná-Uruguai e Centrais Elétricas deSão Paulo, 1966.
19. TIRIBA, A. da C. Epidemia de encefaliteatribuída a arbovírus ocorrida nolitoral sul de São Paulo em 1975:contribuição para o estudo clínico. SãoPaulo, 1975. [Tese de livre docência— Escola Paulista de Medicina].
20. TIRIBA, A. da C. et al. Encefalite humanaprimária epidêmica por arbovírusobservada no litoral sul do Estado deSão Paulo: estudo clínico efetuado emhospital de emergência. Rev. Ass. med.bras., 22:415-20, 1976.
21. TROPPMAIR, H. Regiões ecológicas doEstado de São Paulo. São Paulo,Instituto de Geografia da USP, 1975.(Série Biogeografia, 10).
Recebido para publicação em 05/04/1978Aprovado para publicação em 13/04/1978





























