Embed Size (px)
Citation preview

Homem, Ambiente e Ecologia no Estado de Rot Barbosa R. I., Ferreira E. J. G. & Castellón E. G.
INPA -
Flutu ações do lim lite
Thierry Desjardins (I); Arnaldo Carneiro Filho (‘I) e Armand Chauve1 (Il1)
Estudos palinológicos e geomorfológicos mostram que nas Nplanícies” da América do Sul, mudanças de vegetação ocorreram durante o período final do Quaternário (Servant et al., 1993). Esses estudos sugerem que as superfícies ocupadas por florestas e cerrados evoluiram muito durante esse período, sendo essa ,evolução principal mente controlada por variações paleocl i máticas (Servant et al., 1981; van der Hammen, 1991, Bush et al., 1990). Alguns trabalhos mostram qpe, na bacia amazônica, ocorreram regressões regionais da floresta, associadas à modificações da composição florística em consequência de períodos climáticos mais secos (Absy & van der Hammen, 1976; Liu & Colinvaux, 1988; Absy et al., 1991).
Atualmente, uma parte d o Estado do Roraima, assim como outras partes da zona tropical da América do Sul está coberta por um mosaico de floresta e cerrado. A distribuição dessas formações vegetais não esta sempre relacionada à fatores geomorfológicos, hidrológicos e edáficos (Carneiro Filho, 1993). A hipótese mais plausível para explicar a origem e a distribuição atual desse mosaico esta relacionada a mudanças paleoclimáticas, embora poucos dados estejam disponíveis para Roraima.
Pelo estudo da composição isotópica da matéria orgânica d o solo, a diferença natural da composição em isótopos d e carbono
aima (ed4 1997
I I ) Institut Français de Recherche Scientifique pour le Dévelopement en Coopération (ORSTOM), Instituto Nacional de Pesquisas da Amazônia, Caixa Postal 478, 6901 1-970 ManaudAmazonas. Instituto Nacional de Pesquisas da Amazônia, CoordenaÇä0 de Pesquisas em Ecologia, Caixa Postal 478,6901 1-970 Manaus/Amazonas.
1111) Institut Français de Recherche Scientifique pour fe Dévelopement en Coopération (ORSTOM), 32
i 1 Fonds Documentalre zo

Homem, Ambiente e Ecologia no Estado de Roraima
308 estável das plantas com ciclo fotossintético c3 e c 4 (Bender, 1971) fornece uma oportunidade de avaliar as mudanças de vegetação em regiões onde florestas ( ~ 3 ) e gramineas tropicais de cerrado ( ~ 4 ) coexistem'. O fracionamento isotópico do carbono associado à fotossintese é menor nas plantas ~4 do que nas plantas ~ 3 : os valores de ¿Y3C2 das plantas em C3 variam de -23 a -34% (média de -260/00) enquanto que os valores de 613C das plantas em ~4 variam de -9 a -17%0 (média de -12%~). A composição isotópica da matéria orgânica do solo reflete aquela da vegetacão crescendo sobre esse solo; com exceção de um ligeiro aumento de alguns (%.) com a profundidade. Esse enriquecimento em cl 3 com a profundidade pode ser explicado por: (I) uma alteração com o tempo da composição isotópica da vegetação em consequência de recentes variações do conteúdo em C13 do CO, atmosférico; (2) uma possível decomposição diferencial dos compostos bioquimicos das plantas, isotopicamente heterogêneos e (3) um fracionamento isotópico durante a mineralização da matéria orgânica (ver Deines, 1980; Balesdent et al., 1987; Mariotti, I 991 ; Mariotti & Peterschmitt, 1994). Esse enriquecimento, sempre inferior a 40/,, não 6 suficiente para esconder a diferença de 14%~ existente entre as liteiras das plantas ~3 e ~ 4 . Portanto, é possível determinar, uti l izando o conteúdo em C13 da matéria orgânica do solo, se uma mudança na composição ~ 3 / c 4 de uma communidade de plantas ocorreu no passado (Dzurec et al., 1985; Delaune, 1986; Schwartz et al., 1986; Guillet et al., 1988; Mariotti & Balesdent, 1990; Ambrose & Sikes, 1991; M c Pherson et al., 1993; Wang et a/., 1993; Mariot t i & Peterschmitt, 1994).
Objetivo
O objetivo do estudo apresentado nesse artigo é de fornecer, através de medidas da abundância isotópica em carbono 13 da
Todas as plantas tem a capacidade de utilizar o CO, do ar para sintetizar compostos orgânicos. O conjunto de reações químicas envolvidas neste processo recebe o nome de ciclo fotossintético. Pela diferença nos compostos orgânicos formados, podemos encontrar três tipos de ciclos na natureza : ~3 (plantas com maior tolerância à pouca luminosidade, ex: maioria dos vegetais superiores); C4 (plantas com alta eficiência luminosa, ex: maioria das gramheas) e CAM (plantas com respostas intermediárias à luminosidade).
O elemento carbono possui dois isótopos estáveis (lzC e lac) que se diferem pelo peso atômico. A relação isotópica 13R=13C/12C 6 da ordem de 1,1%. Como na natureza a variação natural dessa relação é pequena, costuma-se usar a unidade relativa (definida na metodologia).
k
1

971) I Ç ã 0
'(C4) ìo à
a de .iam téria esse m a lade ) da Lntes uma t das ?n to i nes, mitt, ente das
lo o a na J no
i kes, ti &
: os
1986;
ecer, 3 da
cos. o tético. los na 2getais llantas
jmico.
~
Flutuações do limite floresta-cerrado durante o Holoceno em Roraima
matéria orgânica do solo, evidências geoquímicas da evolução d o 309 limite floresta-cerrado e m Roraima durante o Holoceno.
Material e métodos
Local d e estudo e amostragem
O estudo foi desenvolvido na Fazenda Paraviana (02' 52' norte, 60' 32' oeste), vinte quilômetros ao norte de Boa Vista (Fig. 1). O clima é caracterizado por uma temperatura anual média de 26,5'c e uma pluviosidade anual média de 1500 mm, 80 o/o dessa chuva ocorrendo entre abril e outubro (Nimer, 1989). A zona de estudo esta localizada n u m plano recoberto por um mosaico de floresta e cerrado. No cerrado, regionalmente conhecido como lavrado, a
l
Figura I . Mapa de localização dos sítios estudados (ABC = Agua Boa de Cima; c = 'Confiança; FSB = três transectos no limite floresta-cerrado).

Homem, Ambiente e Ecologia no Estado de Roraima ?
31 O cobertura herbácea é contínua e dominada pelos gêneros Arsitida, Andropogon e Trachypogon (Silva, I 993), enquanto q u e nos estratos arbustivos e arbóreos predominam as espécies Curatella americana, Byrsonimia crassifolia e Bowdichia virgiloides. Nas margens do cerrado desenvolve-se uma floresta tropical semi-decidua, geralmente considerada como mata de transição entre a floresta sempre-verde e o cerrado (RADAMBRASIL, 1975).
Os solos são originários dos sedimentos continentais da Formação Boa Vista (ver Ab'Saber neste volume). São geralmente latossolos de textura média, Cujas principais características estão apresentadas na Tab. I . Elas indicam que debaixo dos dois tipos de vegetação, a parte inferior dos solos é quase idêntica, devido aos processos de Iatossolização. A parte superficial dos solos (0-40 cm) apresenta algumas diferenças: (I) os solos sob o cerrado são mais arenosos na superfície que os solos sob floresta; (2) a capacidade de troca catiônica (CTC) e a soma das bases trocáveis são levemente mais elevadas nos solos sob floresta. Essas diferenças nas camadas superficiais parecem estar ligadas ao tipo de vegetação.
Em maio de 1992 e 1993, três transectos (Tr I, Tr II, Tr I I I ) foram traçados perpendicularmente ao limite floresta-cerrado. Os transectos (FSB na Fig. I), distantes 60 m um dos outros, mediam 140 m de comprimento, sendo 70 m na floresta e 70 m no cerrado. Para cada transecto, seis tradagens foram feitas: 3 na floresta a 3, 15 e 70 m do limite, e três no cerrado, a 3, 15 e 70 m do limite. Os solos foram amostrados usando u m trado de 6cm d e diâmetro. A profundidade da tradagem variou de 120 a 460 cm. As amostras de
Tabela I. Algumas caracteristicas dos solos estudados.
" Profundidade Argila PH CTC Bases trocáveis (meq 1OOg-') (cm) P I O ) (H,O) (meq 1OOg')
Solo sob floresta 0-1 o 20-30 70-80
160-1 70 250-260
Solo sob savana 0-1 o 20-30 70-80
160-1 70 250-260
19.1 20.3 34.1 32.7 25.7
8 .O 15.3 37.5 30.6 25.7
4.05 4.08 4.53 4.40 4.77
4.55 4.34 4.37 4.74 4.81
2.1 o 1.98 1.74 0.94 0.96
1.56 1.96 1.68 0.86 0.80
0.84 0.1 6 0.1 6 0.08 0.10
0.57 0.24 0.12 0.05 0.05
1
! I
I I
i
l l
I t 1 I
1
I
l I
1
I
I
i
1
I i

Flutuações do limite floresta-cerrado durante o Holoceno em Roraima
solo foram “secas” ao ar, peneiradas .em crivo de 2mm, homo- 31 1
geneizadas e moídas até obtenção de partículas < 200 pm. A liteira da floresta assim como folhas frescas das espécies arbóreas e herbáceas da cerrado foram amostradas, secas a .5OoC e moídas. Para amostrar carvões, três trincheiras de 150 cm de comprimento e 120cm de profundidade foram cavadas no primeiro transecto: duas no cerrado e uma na floresta.
Dois outros solos’ foram amostrados como referência: um perfil sob floresta sempre-verde (Confiança, C na Fig. I), localizada 50km ao sul do limite floresta-cerrado estudado (02’ 25’ norte, 60’ 40’ oeste) e um outro perfil numa zona de cerrado (Agua Boa de Cima, ABC na Fig. I), 5 kni ao noroeste do limite estudado (02’ 55’ norte, 60’ 34’ oeste). Os dois solos foram amostrados até a profundidade de 450 cm.
Métodos analíticos
O carbono orgânico e o nitrogênio total foram determinados por combustão por via seca, utilizando um analisador elementar cHN “Carlo Erba 1.500”. As relações isotópicas da matéria orgânica do solo foram medidas com um espectrômetro de massa ”Fisons SlRA IO” acoplado a um analisador elementar (Girardin & Mariotti, 1991). A abundância natural em carbono 13 fo i expressa em unidades 6, tendo como referência o padrão internacional PDB (Craig, 1957), segundo a seguinte equação:
Medidas repetidas de 613C de u m solo de referência finamente moído e homogeneizado atingiram uma precisão de 0,060/00. Para minimizar os efeitos da heterogeneidade das amostras, elas foram moídas e peneiradas até 200 pm. Amostras de carvão encontradas nos solos foram datados com o carbono 14.
Resultados Distribui@o do carbono orgânico
Os solos de cerrado e floresta apresentam nítidas diferenças no cooteúdo de carbono total (Fig. 2). N o solo de floresta, a

Homem, Ambiente e Ecologia no Estado de Roraima
I I
31 2
al U
U
L? m 200' o 1
.2 303- 2 : a !
3
Cerrado /-, Cerrado d=15m ,+ d=3m
400 ; i Cerrado
.iM) i d=70m ! I
Cerrado I i
- :
I .io0 j
Floresta d = 3 m
Floresta d=15m
p' 2O Carbono (mg. 9")
LO *'* _._.........._.__I 1 .. ......~+.....-.g.,- (9" 6
CI c
I
Floresta d=70m
Figura 2. Conteúdo de carbono orgânico do solo nos perll= estudados nos três transectos, Tr I,Tr I I , Tr I I I . (d) representa a distância, e m metros, entre um dado perfil de solo e o limite floresta-cerrado.
concentração d e carbono nos primeiros centímetros varia d e 14 m g g ' a 20 mg.g-l. A seguir o teor d e carbono diminui rapidamente e d e maneira contínua, variando d e 5,3 até 9,3 mg.g-l na camada 30-40Cm. Abaixo desse nível, o conteúdo d e carbono diminui lentamente, mas d e maneira contínua. N a floresta perto do limite com o cerrado (d = 3 m), a concentração d e carbono é superior a 30 mg.g-l e diminui até 9 o u I O m g g l na camada 30-40 cm. N o solo d e cerrado, a 15 e 70 m do limite, o conteúdo d e carbono dos 50Cm superficiais é nitidamente menor que no solo d e floresta. O teor d e carbono na superfície varia d e 5,2 até. I 1,9 mg.g-l e a 35cm d e profundidade atinge 5-6 mg.g-l. Próximo d o limite com a floresta (d = 3 m), o teor d e carbono da camada superior do solo d e cerrado é quase igual àquele d o solo de floresta.
i 1
i I I
l 1
1 i
I
i l
i l I b
J
U+
ì'
! i i i
lono !T')
)s nos ietros,
le 14 2ente nada n i n u i imite Gor a ) solo 50Cm or d e m d e sta (d
.rrado
I Flutuaçdes do limite floresta-cerrado durante o Holoceno em Roraima
¿Y3C das plantas e da liteira
Os valores de 6'3C d e uma amostra da liteira da floresta variaram d e -30 ,2%~ para as folhas inteiras a té - ~ 8 , 3 % ~ para os fragmentos d e folhas. Os galhos apresentaram um valor inter- mediário d e -29,1%~. Esses valores são típicos de plantas em ~3 e similares à outros resultados obtidos para a liteira de floresta na Amazônia (Desjardins et al., 1994). O valor médio obtido para as gramíneas do cerrado foi de - 1 4 ~ 2 % ~ ~ típico d e plantas c4, enquanto que as folhas das árvores do cerrado apresentaram um valor d e - 29,6~,~, típico de plantas C3.
Distribuiqão do isótopo C13
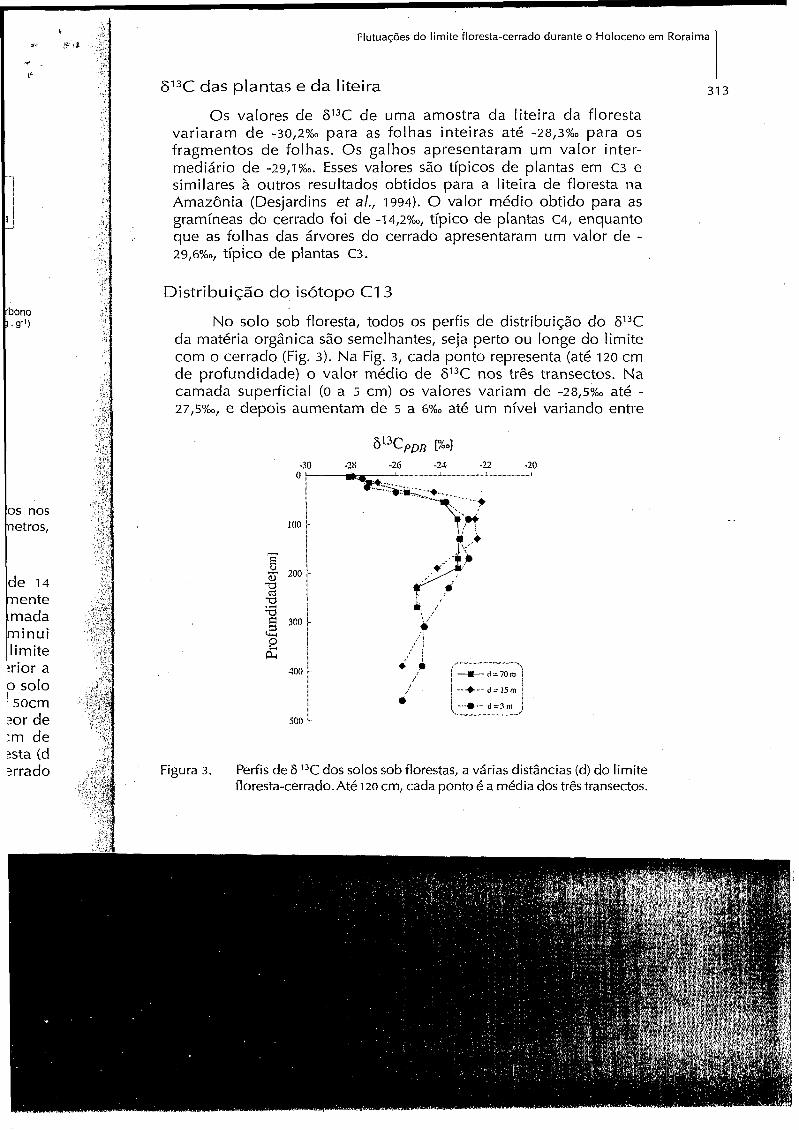
No solo sob floresta, todos os perfis de distribuição do 613C da matéria orgânica são semelhantes, seja perto ou longe do limite com o cerrado (Fig. 3). Na Fig. 3, cada ponto representa (até 120 cm d e profundidade) o valor médio de 613C nos três transectos. Na camada superficial (O a 5 cm) os valores variam de - 2 ~ ~ ~ 5 % ~ até - 2 7 , ~ % ~ , e depois aumentam de 5 a 6%0 até um nivel variando entre
S'.3Cp** @o]
-30 -28 -26 -23 -22 0
JO0
_1
E u 5 200
2 2
2
-U 3 300 %I
k 400
500 ................. '
Figura 3. Perfis de 6 13C dos solos sob florestas, a várias distâncias (d) do limite floresta-cerrado. Até 120 cm, cada ponto é a média dos três transectos.
I 31 3
I Homem, Ambiente e Ecologia no Estado de Roraima
I
314 60 e gocm, segundo o perfil. O valor máximo observado varia de - 2 3 ~ até -22,b0. Esses valores ficam relativamente constantes até 2
metros de profundidade. Na parte inferior dos perfis, os valores de 613C diminuem ligeiramente, passando de -23,7 até -25,8%0 entre 2 e 4 metros de profundidade. Para uma dada profundidade, as diferenças de valores de 613C entre os transectos são geralmente pequenas, mais ou menos uma unidade 6.
No solo localizado sob floresta sempre-verde (Confiança, C na Fig. I), o perfil de distribuição do 613C (Fig. 5) é semelhante com os perfis ,dos solos sob floresta semi-decidua (Fig. 3). Da superfície até 60Cm de profundidade, observa-se um aumento de -28,9%,, até - 23,9%0, seguido por um decréscimo regular com a profundidade, atingindo -26,5% até 4m. Todavia, a não ser para os primeiros centímetros, os valores de 613C são sempre u m pouco mais negativos que nos perfis sob floresta semi-decidua.
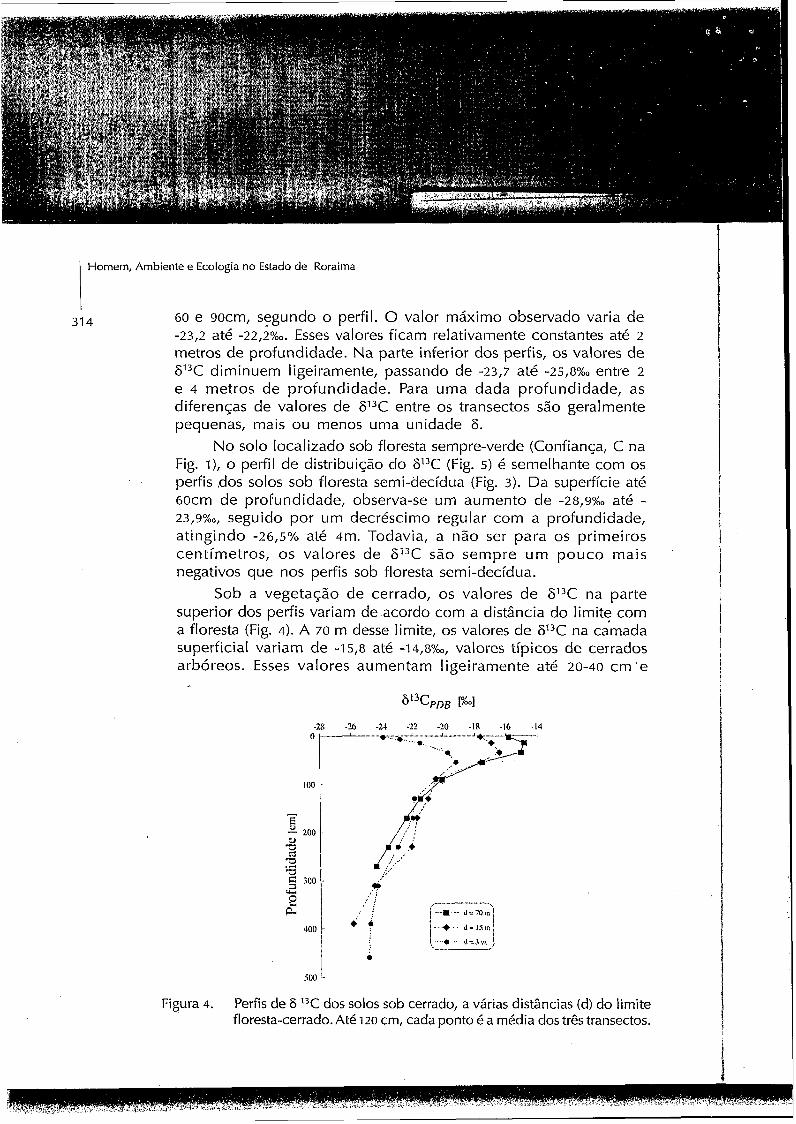
Sob a vegetação de cerrado, os valores de 613C na parte superior dos perfis variam de acordo com a distância do limite com a floresta (Fig. 4). A 70 m desse limite, os valores de 613C na camada superficial variam de -15,8 até -14,8%,,, valores típicos de cerrados arbóreos. Esses valores aumentam ligeiramente até 20-40 cm 'e
-28 -26 -23. -22 -20 -18 -16 .I4
Figura 4. Perfis de 6 I3C dos solos sob cerrado, a várias distâncias (d) do limite floresta-cerrado. Até 120 cm, cada ponto é a média dos três transectos.
......... .
a€ . ' e . v?
ia d e até 2 es d e itre 2 e, as iente
C na jm os ¡e até a té - jade, eiros mais
parte I com mada rados cm e
limite ;ectos.
Flutuaçöes do limite floresta-cerrado durante o Holoceno em Roraima
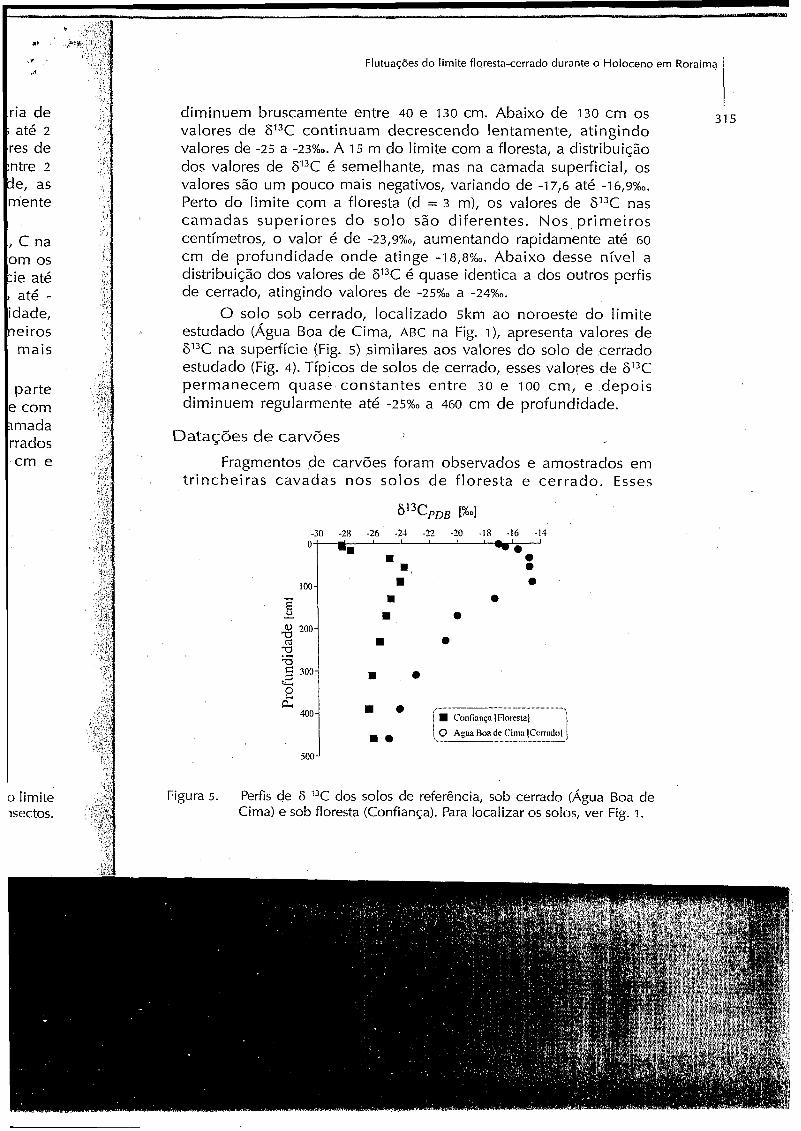
31 5 diminuem bruscamente entre 40 e 130 cm. Abaixo d e 130 cm os valores d e 613C continuam decrescendo lentamente, atingindo valores de -25 a - ~ 3 % ~ . A 15 m do limite com a floresta, a distribuição dos valores de 613C é semelhante, mas na camada superficial, os valores são um pouco mais negativos, variando de -I 7,G até -1 6,9%o. Perto do limite com a floresta (d = 3 m), os valores de 613C nas c a m a d a s superiores d o solo são diferentes. Nos, primeiros centímetros, o valor é d e - ~ , 9 % ~ , aumentando rapidamente até GO cm d e profundidade o n d e atinge -18,8%~. Abaixo desse nível a distribuição dos valores de 6I3C é quase identica a dos outros perfis d e cerrado, atingindo valores d e
O solo sob cerrado, localizado 5km a o noroeste d o limite estudado (Agua Boa de Cima, ABC na Fig. I ) , apresenta valores d e 6'3C na superficie (Fig. 5) similares aos valores d o solo d e cerrado estudado (Fig. 4). Típicos d e solos d e cerrado, esses valores d e 613C permanecem quase constantes entre 30 e 100 c m , e depois diminuem regularmente até -25%,, a 460 cm d e profundidade.
a - 2 4 ~ ~ .
DataGaes de carvões
Fragmentos d e carvões foram observados e amostrados em tr incheiras cavadas nos solos d e floresta e cer rado . Esses
wo
son
S'3CPDR ["ho]
-28 -26 -24 -22 -20 -18 -16 -14 I 9 I 9 1 - 1 I
- 0 . e
O H * O
H O
O
e . O
'i 0 .... .......... " ...... ~
O Agua ßoa de (hi:: (Ccrradol 1 D O ___l.."l" I
i C o n h p \Florestal
Figura 5. Perfis de F 13C dos solos de referência, sob cerrado (Agua Boa de Cima) e sob floresta (Confiança). Para localizar os solos, ver Fig. I.

Homem, Ambiente e Ecologia no Estado de Roraima ,
31 6 fragmentos foram principalmente observados nos solos de cerrado. Alguns desses fragmentos foram datados usando o carbono 14 (Tab. 2) e, separados e m dois grupos: um grupo com idades variando de I .mo a 3.230 anos A.P. (antes do presente) e outro grupo com idades variando de 6.540 a 7.630 anos A.P.
Discussão
O conteúdo e a distribuição do carbono observados no solo de floresta são comparáveis aos resultados obtidos para vários latossolos da Amazônia (Volkoff & Cerri, 1988). O conteúdo de carbono no solo de cerrado se encontra na mesma faixa de valores mencionados por Askew et a/. (1970), Eden (1974) e Ross et a/. (1992) para outros solos d e cerrado na Amazônia. O menor conteúdo de carbono nas camadas superficias do solo de cerrado pode estar relacionado com a menor produção de liteira e com o menor teor de argila nesta camada. Na maioria dos latossolos, o conteúdo de matéria orgânica está fortemente relacionado a textura do solo (Feller et al., 1991).
Os teores mais elevados de carbono observados nos solos sob cerrado pr6ximos do contato com a floresta, podem estar ligados em parte a características intrínsecas do solo, como o teor mai5 elevado de argila e um aporte de folhas vindas da floresta
Tabela 2. Origem dos carvões e datações 14C corrigidas (a determinação da origem dos carvões foi feita por R. Dechamps, do Musée Royal de l'Afrique Centrale, Tervuren, Belgique)
I
Profundidade Especies classificadas Idade (B. P.) I
(cm) a partir dos carvões I
Floresta 20 Guatteria schomburgkiana (flo.) I
35 Copaifera langsdorfii (flo.) t
! 52 Connarus sp. (flo. ou sav.) 2750 2 60 f
Savana trincheira 1 30 Bauhinia sp. (flo.) 30 Terminalia amazonica (flo.) 1790 2 60
6540 2 30 62 7630 C 70 78
Connarus sp. (fio. ou sav.) Wtx sp. (flo. ou sav.)
Savana trincheira 2 15 Aspidosperma spruceanum (flo.1 1
I
I 40 Terminalia amazonica (flo.) 31 60 ? 50 !
67 Wtex sp. Mo. ou sav.) I
34 Terminalia amazonica (flo.) 34
45 Peltogyne densiflora (flo.) 3230 2 60 67 Terminalia amazonica (fio.) 2780 2 60
Cassia sp. (flo. ou sav.)
!
I
!
e*
* r3
jo. 2)
d e ies
d e I os 310 lor los ias 3m sta Ica
los Sar eor !sta
da de
I Flutuaçöes do limite floresta-cerrado durante o Holoceno em Roraima
I
31 7 vizinha. Estes resultados estão de acordo com os dados d e Sanchez et al. (1982) que mostraram que, nas camadas superficiais, os solos sob florestas tropicais possui geralmente teores de carbono superiores aos solos sob cerrado.
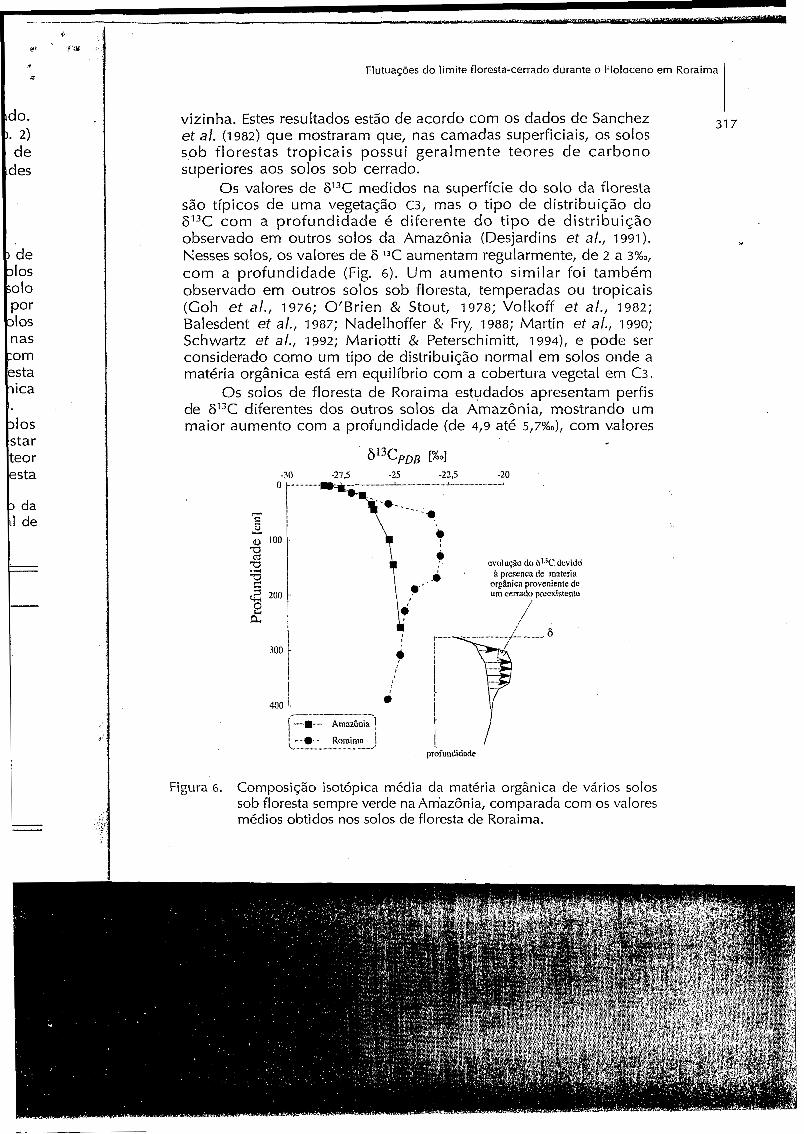
Os valores d e 613C medidos n a superfície do solo da floresta são típicos d e uma vegetação c3, mas o tipo de distribuição do 613C com a profundidade é diferente d o t ipo d e distribuição observado em outros solos da Amazônia (Desjardins et al., 1991). Nesses solos, os valores d e 6 13C aumentam regularmente, de z a 3%0, com a profundidade (Fig. 6). Um aumento similar foi também observado em outros solos sob floresta, temperadas ou tropicais (Goh et al., 1976; O’Brien & Stout, 1978; Volkoff et al., 1982; Balesdent et al., 1987; Nadelhoffer & Fry, 1988; Martin et al., 1990; Schwartz et al., 1992; Mariotti & Peterschimitt, 1 9 9 4 ) ~ e pode ser considerado como um tipo de distribuição normal em solos onde a matéria orgânica está em equilíbrio com a cobertura vegetal em C3.
Os solos d e floresta d e Roraima estudados apresentam perfis de 613C diferentes dos outros solos da Amazônia, mostrando um maior aumento com a profundidade (de 4,9 até 5,7%0), com valores
6’3C,/j [“Ao]
-30 -27.5 -25 -22.5 -20 () ........................ d I I 7 ’ .‘.e-. ..... . . * .
evolu@l do h’3C devido Q pmsenca de rnateila
org3iiica proveniente de um cerrado preexisten\c:
i
/i
....... L. 6
i f
. I.-.--I. ~ .......... \ ( --.-- Amazbnia \ I- I L - -O.- ....... ~ ........................ Roraima I 1 I ..................... _<’
Profundidade
Figura 6 . Composição isotópica média da matéria orgânica de vários solos sob floresta sempre verde na Am’azônia, comparada com os valores médios obtidos nos solos de floresta de Roraima.
J

1 Homem, Ambiente e Ecologia no Estado de Roraima
de 613C entre 50 e 200 cm de profundidade “menos” negativos do que aqueles geralmente observados. Entretanto na parte inferior dos perfis estudados, os valores dos perfis de 613C voltam a ser mais negativos, semelhante aos valores observados nos horizontes de profundidade de outros solos de floresta na Amazônia (Fig. 6).
Os solos estudados são latossolos bem drenados, com uma distribuição do carbono similar àquela da maioria dos latossolos e podzólicos da Amazônia (Desjardins et al., 1991). Pode-se considerar que os processos de decomposição e humificação da matéria orgânica são parecidos nesses solos, e portanto não podem ser responsáveis pelas diferenças observadas nos perfis de 613C. Além disto, os teores de carbono nestes solos de Roraima decrescem regularmente com a profundidade, sem mostrar nenhuma evidência de paleo-horizonte orgânico.
A hipótese mais provável para explicar esses perfis de P3C, seria a presença, no passado, de uma vegetação de cerrado mais expandida do que hoje, com os valores de 613C observados entre 50 e 200cm devido à presenca de uma pequena quantidade de matéria orgânica derivada dessa antiga vegetação de cerrado, resistente à biodegradação.
Na camada superior do solo de floresta, os valores de 613C sãd típicos de vegetação em ~ 3 , tanto nos lugares afastados do limite com o cerrado quanto às margens do limite. Isso indica que atualmente, a floresta não cstá invadindo o cerrado. Mariotti & Peterschmitt (1 994) mostraram que quando uma vegetação C3 está invadindo uma vegetação ~ 4 , os valores de 6I3C do solo da floresta próxima do limite com a savana (cerrado) são intermediários entre os valores de 613C da matéiia orgânica derivada de plantas em C3 e os valores de 613C da matéria orgânica derivada de plantas em ~ 4 . Isto é devido à persistência de matéria orgânica derivada do cerrado. O limite floresta-cerrado estudado indica estar estável atualmente. A atividade antrópica pode ser responsável por esta estabilidade, em particular a queima anual do cerrado para favorecer a criação de gado.
Os valores de 613C da camada superior do solo do cerrado são típicos de uma vegetação dominada por plantas em C4. Perto do limite com a floresta, a influência da liteira das árvores é nítida, os 613C sendo intermediários entre os valores de vegetação ~3 e ~ 4 . Nos solos de savana, uma pequena diminução dos valores de 613C com a profundidade é geralmente observada (Martin et al., 1990; Mariotti & Peterschimitt, 1994). Nos solos de cerrado estudados em Roraima, essa diminução é maior, com valores de 6I3C. encontrados
I 31 8

,D
a-
do *¡or ser \tes
6 ) . ma 1s e -se da ' em 3c. ma :rar
'3C, lais i tre de
do,
i'3C do
que i & ?Stá ?sta itre c3
em do
ive I ?sta )ara
ado erto ;i da, ? c4. F13C 990; em
idos
Flutuaçóes do limite floresta-cerrado durante o Holoceno em Roraima
I nos horizontes de profundidade, sendo similares aos valores 31 9 observados sob floresta. Um perfil similar foi descrito em um latossolo de cerrado no Congo (África) por Schwartz et al. (1992). Os baixos valores de 613C encontrados em maiores profundidades são o resultado da conservação de uma antiga matéria orgânica proveniente de uma vegetação de floresta anterior ao cerrado atual. A presença frequente de carvões nestes solos indica que essa região foi submetida ao fogo no passado. A maioria dos carvões encontrados nos solos atualmente sob cerrado são provenientes de espécies arbóreas de floresta, o que confirma os resultados isotópicos. A distribuição da idade dos carvões com a profundidade não mostra um modelo bem definido. Possivelmente, a atividade biológica provocou uma redistribuição dos fragmentos de carvão. Em solos do norte-oeste e do leste da Amazônia, carvões datados do Holoceno médio e tardio, não diretamente relacionados com a atividade humana, já foram encontrados (Saldarriaga & West, 1986; Soubiès, 1980).
Uma grande extensão de floresta tropical, na zona tropical sul da América do Sul, ocorreu entre 10.000-9.500 e 8.000 anos A.P. (Absy et al., 1991; van der Hammen, 1991; Servant et al., 1993). Apesar da falta de datações, parece provável que os valores de 613C da matéria orgânica das camadas profundas do solo de cerrado sejam a marca dessa extensão máxima da floresta. Estudos palinilÓgicos e paleolimnológicos no Brasil Central e na Amazônia Oriental têm mostrado que os cerrados apareceram, com o desenvolvimento de um clima mais seco, a partir de 8.000 A.P. Esse período seco atingiu seu máximo em 6.000-5.000 anos A.P. (Ledru, 1992; Absy et al., 1991). Os cerrados atuais expandiram-se provavelmente durante esse periodo. A existência, nos solos de cerrado, de carvões derivados de espécies de árvores da floresta, datados de 6.000 a 7.500 anos A.P. conforta essa hipótese. Parece igualmente possível que durante esse periodo, os cerrados tenham coberto áreas maiores do que atualmente. Isso poderia explicar os valores de 613C observados entre 50 e 200 cm nos solos de floresta (Fig. 6). Durante a transição, entre o Holoceno médio e a época atual, o clima tornou-se mais úmido. Entretanto, uma grande variabilidade climática parece ter atuado, provocando a aparição regular de fogos na América do Sul (Servant et al., 1993). A presenca de vários carvões, datados entre 3.230 e 1.790 anos A.P., confirma o papel desses fogos na dinâmica do limite floresta- cerrado durante o Holoceno.
.
.
Os dados apresentados para Roraima confirmam as fl utuações do limite floresta-cerrado ocorridas no norte da Amazônia

Homem, Ambiente e Ecologia no Estado de Roraima
3 20 brasileira no Holoceno. Esses resultados são coerentes com os dados obtidos em outras regiões da Bacia Amazônica. Entretanto novas investigações, em outros limites floresta-cerrado são necessárias para confirmar esses resultados. U m esforço na ampliação dos estudos, além de favorecer um melhor entendimento deste processo na Amazônia, pode fornecer respostas sobre a dinâmica da biodiversidade regional e a fragilidade dos ecossistemas locais. Estes parâmetros fazem parte da base potencial de informações necessárias para o desenvolvimento e o aproveitamento dos recursos naturais de Roraima de forma sustentada e racional.
Agradecimentos
Esse trabalho foi financiado pela ORSTOM e CNRS (França) no quadro do programa internacional ECOFlT (Ecologie et paléoécologie des forêts intertropicales - Ecologia e paleoecologia das florestas intertropicais). Os autores agradecem a Edileuza L.S. Silva e Reinaldo I . Barbosa pela ajuda no campo e pelas discussões estim u I antes.
Bibliografia
Absy,M.L. & van der Hammen,T. (I 976). Some paleoecological data from Rondonia, southern part of the Amazon Basin. Acta Amazonica 6: 293-299.
Absy,M.L.; Cleef,A.; Fournier,M.; Martin,L.; Servant,M.; Sifeddine’A.; da Silva,M.F.; Soubiès,F.; Suguio,K.; Turcq,B. & van der Hammen‘T. (1 991). Mise en évidence de quatre phases d‘ouverture de la forêt dense dans le sud-est de l’Amazonie au cours des 60.000 dernières années. Premières comparaisons avec d’autres forêts tropicales. C.R. Acad. Sci. Paris ll(312): 673-678.
Ambr0se)S.H. & Sikes,N.E. (I 991). Soil carbon isotope evidence for Holocene habitat change in the Kenya Rift Valley. Science 253: 1402-1405.
Askew,G.P.; Moffatt,D.J.; Montgomery,R.F. & Sear1 P.L. (1970). Soil landscapes in north-eastern Mato Grosso. Geographical lourna/ 136: 21 1-227.
BalesdentJ.; Mariotti,A. & Guillet B. (1987). Natural 13C abundance as a tracer for soil organic matter dynamics. studies. Soil Bio/. Biochem. 19: 25-30.
Bender,M. (1971). Variations in the 13CP2C ratios of plants in relation to the pathway of photosynthetic carbon dioxide fixation. Phytochemistry 1 O: 1239- 1244.
Bush,M.B.; Colinvaux,P.A.; Wiemann,M.C.; Pipern0,D.R. & Liu,K.B. (1990). Late Pleistocene temperature depression and vegetation change in Ecuadorian Amazonia. Quat. Res. 34: 330-345.
Carneiro Filho,A. (1 993). Cerrados amazônicos: fósseis vivos? Algumas reflexões. Rev. IC/USP 14: 63-68.
I
1
I I I
l
6
l
i
l
I i I
, I
l
I
l
I
l
I
I
I
I
l
I
\
I
1

I
a P * I .y9
6
#*
3m os etanto o são ;o na mento lbre a ,temas al de nento :ional.
;a) no ologie irestas ilva e issões
Indon i a,
,A.; da ). Mise ì-est de raisons
habitat
apes in
acer for
to the 1: 1239-
o). Late kdorian
Flutuaçóes do limite floresta-cerrado durante o Holoceno em Roraima
321 Craig,H. (1 957). lsotopics standards for carbon and oxygen and correction factors
for mass spectrometric analysis of carbon dioxide. Geochim. Cosmochim. Acta 12: 133-149.
Deines,P. (1980). The isotopic composition of reduced organic carbon. In: P. Fritz & J. C. Fontes (eds.) Handbook of Environmental lsotope Geochemistry - The Terrestrial Environment, vol 1. Elsevier, Amsterdam. pp. 329-346.
DeLaune,R.D. (1986). The use of 6 ”C signature of C, and C, plants in determining past depositional environments in rapidly accreting marshes of the Mississippi river deltaic plain, Louisiana, USA. Chem. Geol. (Isot, Geosc. Sect.) 59: 315-320.
Desjardins,T.; Volkoff,B; Andreux,F. & Cerri;C.C. (1 991). Distribution du carbone total et de l’isotope 13C dans des sols ferrallitiques du Brésil. Science du Sol
Desjardins,T.; Andreux’F.; Volkoff,B & Cerri,C.C. (1 994). Organic carbon and 13C contents in soils and soil size-fractions, and their changes due to deforestation and pasture installation in eastern Amazonia. Geoderma 61 : 103-1 18.
Dzurec,R.S.; Boutton,%W.; Caldwel1,M.M. & Smith,B.N. (I 985). Carbon isotope ratios of soil organic matter and their use in assessing community composition change in Curlew Valley, Utah. Oecologia 66: 17-24.
Eden,M.J. (7974). Paleoclimatic influences and the development of savanna in southern Venezuela. lourna/ of Biogeography I : 95-1 09.
Feller,C.; Fritsch,E.; Poss,R. & Valentin,C. (1991). Effet de la texture sur le stockage et la dynamique des matières organiques dans quelques sols ferrugineux et ferrallitiques (Afrique de l‘Ouest, .en particulier). Cah. Orstom (sér. Pédol.)
29: 175-1 87.,
XXVI: 25-36. Girardin,C. & Mariotti,A. (1991). Analyse isotopique du 13C en abondance
naturelle dans le carbone organique: un système automatique avec robot préparateur. Cab. Orstom (sér. Pédol.) XXVI: 371 -380.
Goh,K.M.; Rafter,T.A.; Stout,J.D.& Wa1ker)T.W. (1 976). The accumulation of soil organic matter and its carbon isotope content in a chronosequence of soils developped on aeolian sand in New Zealand. /. Soil Science 27: 89-100.
Cuillet,B.; Faivre,P.; Mariotti,A. & Khobzi,J. (1988). The evidence of open landscapes in valleys adjacent to the “Sabana de Bogota” during Holocene. Paleogeography, Edeoclimatolog~ Paleoecology 65: 51 -58.
Ledru,M.P. (1992). Modifications de la végétation du Brésil Central entre la dernière époque glaciaire et l’interglaciaire actuel. C.R.Ac.Sci. Paris II (31 4): 117-123.
Liu,K.B. & Colinvaux,P.A. (1988). A 5200-year history of Amazon rain forest. /. Biogeogr. 15: 23 1-248.
Mariotti’A. (1 991). Le carbone-13 en abondance naturelle, traceur de la dynamique de la matière organique des sols et de I’évolution des paléoenvironnments continentaux. Cah. Orstom (sér. Pédol.) XXVI: 299-31 3.
Mariotti,A. & BalesdentJ. (1990). - 13C natural abundance as a tracer of soil organic matter turnover and paleoenvirbnment dynamics. Chem. Geol. 84: 31 7-31 9.
Mariotti,A. & Peterschmitt,E. (1 994). Forest savanna ecotone dynamics in India as revealed by carbon isotope ratios of soil organic matter. Oecologia 97: 475-480.

I Homem, Ambiente e Ecologia no Estado de Roraima
Martin,A.; Mariotti,A.; BalesdentJ.; Lavelle,P. & Vuattoux R. (1 990). Estimate of organic matter turnover rate in a savanna soil by 13C natural abundance measurements. Soil Biol. Biochem. 22: 51 7-523.
Mc Pherson,G.R.; Boutton,T.W. & Midwood A.J. (1 993). Stable carbon isotope analysis of soil organic matter illustrates vegetation change at the grassland/ woodland boundary in southeastern Arizona, USA. Oecologia 93: 95-1 01.
Nadelhoffer,K.J. & Fry,B. (1 988). Controls on natural nitrogen-I 5 and carbon-13 abundances in forest soil organic matter. Soil Sci. Soc. Am. /. 52: 1633-1 640.
Nimer,E. (1989). Climatologia do Brasil. IBGE, Rio de Janeiro, 421pp.
0’Brien’B.J. & Stout,J.D. (1978). Movement and turnover of soil organic matter as indicated by carbon isotope measurements. Soil Bio/. Biochem. 1 O: 309-31 7.
Radambrasil (1 975). Projeto Radambrasil. Levantamento de Recursos Naturais, vol. 8, Folha NB.20, Boa Vista. Ministério das Minas e Energia, Brasil.
Ross,S.M.; Luizã0,F.J. & Luizã0,R.C.C. (1 992). Soil conditions and soi l biology in different habitats across a forest-savanna boundary on Maracá Island, Roraima, Brazil. In: P. A. Furley; J. Proctor & J. A. Ratter (eds.) Nature and Dynamics of Forest-Savanna Boundaries. Chapman et al., London, pp. 145-1 70.
Sa1darriagaJ.G. & West,D.C. (1 986). Holocene fires in the northern Amazon Basin. Quaternary Research 26: 358-366.
Sanchez,P.A.; Gichuru,M.P. & Kat2,L.B. (1 982). Organic matter in major soils of the tropical and temperate regions. In: Non symbiotic nitrogen fixation and organic matter in the tropics. 12th ICSS, New Delhi. pp. 99-114.
Schwartz,D.; Mariotti,A.; Lanfranchi,R. & Cuillet,B. (1 986). 13C/1zC ratios of soi l organic matter as indicators of ecosystem changes in tropical regions. Geoderma 39: 97-1 03.
Schwartz,D.; Mariotti,A.; Trouvé,C.; Van den Borg,K. & Guillet,B. (1 992). Etude des profils isotopiques 13C et 14C d’un sol ferrallitique sableux du littoral congolais. Implications sur la dynamique de la matière organique et l’histoire de la vegétation. C.R.Acad.Sci. Paris II (31 5): 141 1-1 41 7.
Servant,M.; Fontes,J.C.; Rieu,M. & Saliège,J.F. (1 981). Phases climatiques arides holocènes dans le sud-ouest de l’Amazonie (Bolivie). C.R. Acad. Sci. Paris
Servant,M.; Maley,J.; Turcq,B.; Absy,M.L.; Bennac,P.; Fournier,M. & Ledru,M.P. (1993). Tropical forest changes during the Late Quaternary in African and South American lowlands. Global and Planetary Change 7: 25-40.
Silva,E.L.S. (1 993). Inventário preliminar das espécies arbóreas das florestas dos arredores de Boa Vista (Roraima) - Uma abordagem fitossociológica. Dissertação de Mestrado, INPA/Fundação Universidade do Amazonas, Manaus, 194 pp.
Soubiès,F. (1 980). Existence d’une phase sèche en Amazonie brésilienne datée par la présence de charbons dans les sols (6000-3000 ans B.P.). Cah. Orstom (sér.
van der Hammen,T. (1 991). Palaeoecological background : neotropics. Clim. Change 19: 37-47.
Volkoff,B. & Cerri,C.C. (1988). L’humus des sols du Brésil. Nature et relations avec l’environnement. Cah. ORSJOM (sér. Pédol.) XXIV: 83-95.
I 322
II(292): 1295-1 297.
Géol.) XI: 133-148.

' C/im.
s avec
Flutuaçöes do limite floresta-cerrado durante o Holoceno em Roraima I
I
323 Volkoff,B.; Matsui,E. & Cerri,C.C. (1 982). Discriminação isotópica do carbono nos humus de latossolos e podzol na região amazônica do Brasil. In: Proc. ofthe Regional Colloquium on Soi/ Organic Matter, CENA-PROMOCET, Piracicaba, Brasil. pp. 147-1 53.
Wang,Y.; Cerling,T.E. & Effland,W.R. (1 993). Stable isotope ratios of soil carbonate and soil organic matter as indicators of forest invasion of prairie near Ames, Iowa. Oecologia 95: 365-369.
Thierry Desjardins é PhD e m pedologia pela Universidade d e Nancy, França. Atualmente exerce a função de pesquisador d a ORSTOM resi- dente e m Manaus. Seu campo de interesse é a ecologia d o solo c o m ênfase no comportamento da matéria orgânica diante das alterações antrópicas e naturais e m sistemas tropicais.
Arnaldo Carneiro Filho é Mestre pelo International Institute for Aerospace Survey and Earth, Holanda, especialista e m ecologia d e paisagem e geoprocessamento. É pesquisador d o INPA e seu campo d e interesse são as transformações da paisagem na Amazônia em função d a atividade humana na regiso.
Armand Chauve1 é PhD em pedologia pela Universidade d e Strasbourg, França. É pesquisador d a ORSTOM e seu c a m p o de interesse é a pedogênese e a ecologia do solo.

1