Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL PAULISTA - UNESP
CAMPUS DE JABOTICABAL
HUANGLONGBING E Diaphorina citri: ESTUDOS DAS
RELAÇÕES PATÓGENO-VETOR-HOSPEDEIRO
Juan Camilo Cifuentes Arenas
Engenheiro Agrônomo
2017

UNIVERSIDADE ESTADUAL PAULISTA - UNESP
CAMPUS DE JABOTICABAL
HUANGLONGBING E Diaphorina citri: ESTUDOS DAS
RELAÇÕES PATÓGENO-VETOR-HOSPEDEIRO
Juan Camilo Cifuentes Arenas
Orientador: Dr. Silvio Aparecido Lopes
Tese apresentada à Faculdade de Ciências Agrárias e Veterinárias – Unesp, Câmpus de Jaboticabal, como parte das exigências para a obtenção do título de Doutor em Agronomia (Produção Vegetal)
2017

Arenas, Juan Camilo Cifuentes
A681h Huanglongbing e Diaphorina citri: estudos das relações patógeno-vetor-hospedeiro / Juan Camilo Cifuentes Arenas. – – Jaboticabal, 2017
xii, 133 p. : il. ; 29 cm Tese (doutorado) - Universidade Estadual Paulista, Faculdade de
Ciências Agrárias e Veterinárias, 2017 Orientador: Silvio Aparecido Lopes
Banca examinadora: António de Góes, Glauco de Souza Rolim, Renato Beozzo Bassanezi, Alécio Souza Moreira
Bibliografia 1. Citrus sinensis. 2. Fenologia. 3. Porta-enxertos. I. Título. II.
Jaboticabal-Faculdade de Ciências Agrárias e Veterinárias.
CDU 581.2:634.31
Ficha catalográfica elaborada pela Seção Técnica de Aquisição e Tratamento da Informação – Diretoria
Técnica de Biblioteca e Documentação - UNESP, Câmpus de Jaboticabal.


DADOS CURRICULARES DO AUTOR
JUAN CAMILO CIFUENTES ARENAS – nascido em 09 de setembro de 1985, no
município de Turbo – Antioquia (Colômbia). Iniciou o curso de graduação em
Engenharia Agronômica, em agosto de 2003, na Universidad Nacional de Colombia na
cidade de Medellín – Antioquia, concluindo-o em março de 2012. Durante a graduação
atuou durante 4 anos como Estudante Monitor Auxiliar no Centro de Produção
Audiovisual da mesma Universidade, dando assistência como cinegrafista e editor de
conteudos audiovisuais emitidos no “Canal Universitário de Antioquia – Canal U –“.
Seu trabalho de conclusão de curso intitulado “Efecto del uso de biofertilizantes en la
fisiologia del cultivo del frijol (Phaseolus vulgaris)” recebeu reconhecimento à
excelência no concurso “Mejores Trabajos de Grado versión XXI”. Em julho de 2012
começou trabalhar como Auxiliar de pesquisa no projeto “Identificación de la dinámica
poblacional de Diaphorina citri (Hemiptera: Psyllidae) en los cultivos de cítricos de
Colombia: una herramienta para implementar un sistema piloto de seguimiento de
poblaciones del insecto vector del HLB”, um projeto pioneiro que foi desenvolvido
durante um ano. Já em agosto de 2013 ingressou no curso de mestrado junto ao
Programa de Pós-Graduação em Agronomia (Produção Vegetal) na Faculdade de
Ciências Agrárias e Veterinárias (UNESP - Jaboticabal), sendo bolsista CNPq, mas em
agosto de 2015 foi aprovada a sua transferência para o Doutorado Direto na mesma
instituição, sendo bolsista CAPES.

AGRADECIMENTOS
À minha família que sempre esteve presente nos momentos de alegrias e
tristezas, mesmo a milhares de quilômetros, sendo o meu suporte.
Ao Brasil pela oportunidade que tive de realizar meus estudos graças ao grande
investimento que fazem em educação e pesquisa, estarei eternamente agradecido.
Ao meu orientador Prof. Dr. Silvio Aparecido Lopes, pela confiança depositada,
pelos ensinamentos recebidos, pelo incentivo e amizade.
À UNESP/Jaboticabal, ao atual coordenador Prof. Dr. Rouverson Pereira da
Silva e ao ex-coordenador Prof. Dr. Arthur Bernardes e demais professores do
Programa de Pós-Graduação em Agronomia (Produção Vegetal) pelo aperfeiçoamento
e formação acadêmica.
Aos funcionários da Seção de Pós-Graduação pela disponibilidade no
atendimento e informações prestadas.
Aos meus amigos Laudecir, Fernanda Queiróz, Priscila Alves, Hermes Teixeira,
Everton e Luís, por me ajudarem na realização deste trabalho de doutorado e pela
companhia e amizade... muito obrigado.
À CAPES e ao CNPq pela concessão das bolsas.
Ao FUNDECITRUS pelo apoio financeiro e técnico, fundamentais ao
desenvolvimento da pesquisa.
Muito obrigado a todos.

vi
SUMÁRIO
RESUMO..................................................................................................................... ix
ABSTRACT ................................................................................................................. xi
CAPÍTULO 1 – Considerações gerais ....................................................................... 1
1. REVISÃO DE LITERATURA .................................................................................. 1
1.1. Os citros e a citricultura brasileira ................................................................... 1
1.2. Huanglongbing (HLB) ...................................................................................... 3
1.2.1. Os patógenos associados ........................................................................ 3
1.2.2. Processo infeccioso, efeitos na planta e manejo da doença .................... 5
1.3. Bioecologia de Diaphorina ciri Kuwayama ...................................................... 7
1.4. Ecofisiologia dos citros e modelos de crescimento ......................................... 8
2. REFERÊNCIAS ................................................................................................... 11
CAPÍTULO 2 – Potencial de Swinglea glutinosa como criadouro de Diaphorina
citri e fonte de Candidatus Liberibacter asiaticus ................................................. 21
RESUMO.................................................................................................................... 21
ABSTRACT ................................................................................................................ 22
1. INTRODUÇÃO ..................................................................................................... 23
2. MATERIAL E MÉTODOS .................................................................................... 25
2.1. Material vegetal e criação do inseto .............................................................. 25
2.2. Extração e quantificação de DNA de material vegetal e de inseto ................ 26
2.3. Experimento 1: Potencial de Swinglea glutinosa e de algumas variedades de
laranjeira doce e de limoeiro ‘Siciliano’ em atuar como hospedeiro de Diaphorina citri
...................................................................................................................... 28
2.4. Experimento 2: Potencial de Swinglea glutinosa, Murraya exotica e laranjeira
‘Valência’ em multiplicar ‘Candidatus Liberibacter asiaticus’ .................................. 29
2.5. Experimento 3: Potencial de Swinglea glutinosa, Murraya exotica e laranjeira
‘Valência’ em atuar como fonte de inóculo .............................................................. 30
2.6. Avaliações e análise dos dados .................................................................... 30
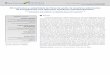
3. RESULTADOS .................................................................................................... 32
4. DISCUSSÃO ........................................................................................................ 40
5. REFERÊNCIAS ................................................................................................... 44

vii
CHAPTER 3 – Citrus Flush Shoot Ontogeny Modulates Biotic Potential of
Diaphorina citri ......................................................................................................... 50
ABSTRACT ................................................................................................................ 50
1. INTRODUCTION ................................................................................................. 51
2. MATERIAL AND METHODS ............................................................................... 53
3. RESULTS ............................................................................................................ 57
4. DISCUSSION ...................................................................................................... 65
5. REFERENCES .................................................................................................... 69
RESUMO.................................................................................................................... 77
CAPÍTULO 4 – Degree-day model for estimating flush shoot ontogeny in Valencia
sweet orange and scheduling insecticide sprays to control Diaphorina citri ..... 79
ABSTRACT ................................................................................................................ 79
1. INTRODUÇÃO ..................................................................................................... 81
2. MATERIAL E MÉTODOS .................................................................................... 84
2.1. Material vegetal, adequação das plantas e descrição dos ambientes .......... 84
2.2. Experimento 1: desenvolvimento do broto em câmaras de crescimento ...... 85
2.3. Experimento 2: desenvolvimento do broto em plantas no campo e influência
do déficit hídrico ...................................................................................................... 86
2.4. Graus dia de desenvolvimento (GDD)........................................................... 86
2.5. Validação dos resultados .............................................................................. 87
2.6. Proposta de modelo geral aplicação de medidas de proteção dos brotos .... 87
2.7. Análise dos dados ......................................................................................... 89
3. RESULTADOS .................................................................................................... 90
4. DISCUSSÃO ...................................................................................................... 100
5. REFERÊNCIAS ................................................................................................. 103
CAPÍTULO 5 – Limitada influência do porta-enxerto sobre a biologia de
Diaphorina citri em citros ...................................................................................... 108
RESUMO.................................................................................................................. 108
ABSTRACT .............................................................................................................. 110
1. INTRODUÇÃO ................................................................................................... 112
2. MATERIAL E MÉTODOS .................................................................................. 114
2.1. Material vegetal e criação de Diaphorina citri .............................................. 114

viii
2.2. Descrição dos experimentos ....................................................................... 115
2.3. Análises de dados ....................................................................................... 119
3. RESULTADOS .................................................................................................. 119
4. DISCUSSÃO ...................................................................................................... 127
5. REFERÊNCIAS ................................................................................................. 130

ix
HUANGLONGBING E Diaphorina citri: ESTUDOS DAS RELAÇÕES PATÓGENO-
VETOR-HOSPEDEIRO
RESUMO
RESUMO. O huanglongbing (HLB), associada a bactérias de floema (Candidatus
Liberibacter spp) transmitidas por insetos (Diaphorina citri), é a doença mais destrutiva
dos citros e muitos têm sido os esforços visando reduzir sua disseminação nos
pomares, porém, nem sempre bem-sucedidos. Somente no cinturão citrícola paulista
mais de 45 milhões de plantas já foram erradicadas por causa do HLB. O objetivo deste
trabalho foi investigar diferentes aspectos associados ao patossistema HLB, visando
trazer informações que ajudem na contenção da doença. Foram estudados: 1)
hospedeiros alternativos do inseto vetor e do patógeno; 2) ontogenia do broto
vegetativo de citros e seu potencial em multiplicar D. citri; 3) requerimentos térmicos
para o desenvolvimento do broto; e 4) influência do porta-enxerto na biologia de D. citri.
No primeiro capítulo é apresentada revisão de literatura sobre os aspectos mais
relevantes da citricultura e do patossistema em estudo e, nos demais, os resultados
das pesquisas. No segundo é apresentada a importância Swinglea glutinosa (rutácea
muito comum em certas regiões citrícolas) como fonte de Ca. Liberibacter asiaticus e
criadouro de D. citri. A bactéria do HLB multiplicou na planta, porém atingiu populações
consideravelmente menores do que em laranjeira ‘Valência’. Apresenta, portanto, baixo
potencial como fonte de inóculo para os citros. Por outro lado, D. citri reproduziu
abundantemente em S. glutinosa, tanto quanto nas principais variedades de laranjeiras
doces e limoeiros do grupo ‘Siciliano’ cultivados no Brasil. No terceiro capítulo são
apresentados detalhes do desenvolvimento do broto em laranjeira ‘Valência’ e como
os diferentes estádios influenciam o potencial biótico de D. citri. O inseto completou
seu ciclo de vida em todos os estádios, mas em diferentes intensidades, sendo maiores
nos estádios iniciais. Com os dados criou-se um índice de favorabilidade à reprodução
do inseto, que leva em consideração o peso relativo de cada estádio do broto na
biologia do inseto. No quarto capítulo é apresentado o impacto da temperatura do ar
sobre o broto, o que permitiu determinar os graus-dia necessários ao seu
desenvolvimento (GDD) e, com base no GDD, estabelecer um modelo geral inovador
de previsão de pulverizações. No quinto capítulo é demonstrado que o porta-enxerto

x
(limoeiro Cravo, tangerineira Sunki e citrumeleiro Swingle), por mecanismos de
antibiose e/ou antixenose, afetam D. citri. Mas esses fenômenos ocorrem somente na
fase de seedlings, e nas copas neles enxertadas somente enquanto a planta é jovem
(muda). Em plantas adultas, de maior porte, tais fenômenos não foram detectados.
Podem ter sido mascarados por influência da variedade copa (laranjeira).
Palavras-chave: Citrus sinensis, murta, hospedeiros alternativos, qPCR, Swinglea
glutinosa.

xi
HUANGLONGBING AND Diaphorina citri: STUDIES ON PATHOGEN-HOST-
VECTOR INTERACTIONS
ABSTRACT
ABSTRACT. Huanglongbing (HLB), associated with insect-borne phloem bacteria
(Candidatus Liberibacter spp), is the most destructive disease of citrus and many efforts
have been made to reduce its spread in orchards, but not always well-succeeded. Only
in the citrus belt of São Paulo state (Brazil), more than 45 million plants have already
been eradicated because of HLB. The objective of this work was to investigate different
aspects associated to the HLB pathosystem, aiming to bring information that helps in
the management of the disease. We studied: 1) alternative vector insect and pathogen
hosts; 2) ontogeny of the citrus vegetative shoot and its potential to multiply D. citri; 3)
thermal requirements for shoot development; and 4) influence of the rootstock on the
biology of D. citri. In the first chapter, a literature review on the most relevant aspects
of citriculture and pathosystem in study is presented and, in the others, the results of
the experiments. In the second one the importance of Swinglea glutinosa (a Rutaceae
very common in certain citrus regions) as a source of Ca. Liberibacter asiaticus and
host for D. citri is presented. The HLB bacteria multiplied in the plant, but reached
populations considerably smaller than in 'Valencia' sweet orange. Therefore, it presents
low potential as a source of inoculum for citrus. On the other hand, D. citri reproduced
abundantly in S. glutinosa, as well as in the main sweet orange and lemon varieties of
the 'Sicilian' group cultivated in Brazil. In the third chapter, it is presented details of the
development of the 'Valencia' orange new shoot and how the different stages influence
the biotic potential of D. citri. The insect completed its life cycle at all stages of new
shoots, but at different intensities, being larger in the early stages. With the data, an
insect reproduction index was created, which takes into account the relative weight of
each shoot stage in the insect biology. In the fourth chapter, it is presented the impact
of the air temperature on the shoot, which allowed to determine the degree-days
required for its development (GDD) and, based on the GDD, to establish an innovative
general model of spray forecasting. In the fifth chapter it is demonstrated that the
rootstock (‘Rangpur’ lime, ‘Sunki’ mandarin and ‘Swingle’ citrumelo), by mechanisms of
antibiosis and / or antixenosis, affect D. citri. But these phenomena occur only in the

xii
stage of seedlings, and in the scions grafted on them only while the plant is young
(nursery tree). In larger adult plants, such phenomena were not detected. They may
have been masked by the influence of the scion (orange) variety.
Key words: Citrus sinensis, orange jasmine, alternative host, qPCR, Swinglea
glutinosa.

1
CAPÍTULO 1 – Considerações gerais
1. REVISÃO DE LITERATURA
1.1. Os citros e a citricultura brasileira
A maioria das espécies cítricas cultivadas hoje em dia tem como progenitores
Citrus maxima (Burm.) Merrill e Citrus reticulata Blanco (GMITTER; HU, 1990; WU et
al., 2014), com origem no sudeste asiático (província de Yunnan, China), com exceção
do pomelo (Citrus paradisi Macf.), um híbrido interespecífico entre Citrus grandis (L.)
Osb. e Citrus sinensis (L.) Osb., de origem no Caribe, no início do século XVIII
(GMITTER, 1995). Os citros espalharam-se pelo mundo seguindo rotas de comércio
fluvial e terrestre. A grande preferência das pessoas por este grupo de frutas é
indicada pelo tamanho da área em produção. Em 2014, por exemplo, os citros eram
plantados em aproximadamente 7,6 milhões de hectares no mundo, sendo 51,5, 30,3,
14,0 e 4,2% ocupados por laranjeiras, tangerineiras, limoeiros/limeiras e pomelos,
respectivamente (FAO, 2014).
Os citros foram trazidos para o Brasil pelos portugueses no início do século
XVI. Aqui se expandiu por todo o país, em particular nos estados de São Paulo e Rio
de Janeiro (HASSE, 1987 apud DONADIO; MOURÃO FILHO; MOREIRA, 2005),
atingindo o ápice durante a década de 1990 e início dos anos 2000 (BOTEON;
NEVES, 2005). Significativa redução da área cultivada ocorreu desde então,
possivelmente como consequência da ocorrência de importantes problemas
fitossanitários. Em 2016 a área citrícola foi estimada em 758,5 mil hectares, ocupadas
principalmente com laranjeiras (87,9%) (AGRIANUAL, 2017). A maior parte desta área
está concentrada no cinturão citrícola, que engloba diversas regiões do estado de São
Paulo e a região do Triângulo Mineiro em Minas Gerais, com 402,6 mil ha e estimados
364,5 milhões de caixas de 40,8 kg para a safra de 2017/2018. Este volume de fruta
resulta do cultivo de 174,8 milhões de árvores com mais de três anos de idade
(FUNDECITRUS, 2017a).

2
A citricultura brasileira está estabelecida no uso de diversas variedades de
copa e de porta-enxertos. As variedades de copa diferem entre si principalmente pela
qualidade de seus frutos e épocas de floração e colheita (PIO et al., 2005). No último
inventário de árvores produtivas feito pelo Fundecitrus, o parque citrícola estava
composto por 19,9% de variedades precoces (Hamlin, Westin, Rubi, Valência
Americana, e outras), 38,2% de variedade meia estação (Pêra Rio), e 44,9% de
variedades tardias (Valência, Valência Folha Murcha, e Natal) (FUNDECITRUS,
2017a).
No caso dos porta-enxertos, as diferenças mais marcantes observadas entre
eles têm a ver com o porte que conferem às copas, e com sua grande variação em
relação à tolerância a estresse hídrico e a agentes patogênicos. Os mais importantes
são o limoeiro Cravo, o citrumeleiro Swingle e as tangerineiras Sunki e Cleópatra (IAC,
2014; POMPEU JUNIOR, 2005). O Cravo já atingiu no passado mais de 70% da área
plantada, sendo ainda o mais preferido, presente em 50% de toda a área plantada. A
influência dos porta-enxertos se verifica não só sobre a produtividade, a qual é ditada
principalmente por variações hormonais (LIU et al., 2015; TAN et al., 2015), mas
também sobre o vigor das copas (SAEED; DODD; SOHAIL, 2010), tolerância a
diversas condições de estresse abiótico, especialmente excesso ou déficit hídrico
(ALLARIO et al., 2013; ARGAMASILLA; GÓMEZ-CADENAS; ARBONA, 2014;
TRAMONTINI et al., 2013; TREEBY et al., 2007), e resistência ou tolerância a pragas
e doenças, especialmente as que afetam as raízes (AGUT et al., 2014; LAPOINTE;
BOWMAN, 2002).
Apesar do pouco uso como porta-enxerto, Swinglea glutinosa (Blanco) Merr.,
conhecida também como “Tabog”, “Tabog of Luzon” e “Phillippine Tabog” (Ásia),
“limón swingle” (Colômbia), e “limoncillo” (Venezuela) (SWINGLE; REECE, 1967), é
muito utilizada na Colômbia como cerca viva para delimitação de condomínios,
parques, lotes e fazendas agrícolas (EBRATT-RAVELO et al., 2011; KING, 2012;
LEÓN et al., 2008). No Brasil, sabe-se de sua presença e uso como ornamental no
estado de Goiás. Esta espécie parece apresentar algumas características desejáveis
como resistência a seca, resistência à gomose e redução do porte da copa, resultando
em aumento da eficiência produtiva (RESTREPO; TORRES; SALAZAR, 1987;
RONDÓN et al., 1993; VENNING, 1957).

3
1.2. Huanglongbing (HLB)
1.2.1. Os patógenos associados
Quando relatada pela primeira vez no sul da China (REINKING, 1919), a
doença, hoje conhecida como Huanglongbing (HLB), foi descrita inicialmente como
uma deficiência nutricional (TIRTAWIDJAJA, 1988). Na África do Sul, plantas com
sintomas similares (ramos amarelos) eram conhecidos desde 1928 na província de
Transvaal, pais no qual a doença recebeu o nome de “Greening” (OBERHOLZER;
VON STADEN; BASSON, 1965). Apesar de, no início, acreditou-se tratar de problema
de toxidez mineral (VAN DER MERWE; ANDERSEN, 1937), este ocorria
frequentemente associado à presença de um inseto, acreditando ser ele o potencial
transmissor de um possível agente causal (SCHWARZ, 1964). A transmissão do
agente (ainda desconhecido) foi conseguido por meio de enxertia e por exposição das
plantas ao inseto naturalmente estabelecido em pomares afetados (MCCLEAN;
OBERHOLZER, 1965; TIRTAWIDJAJA; HADIWIDJAJA; LASHEEN, 1965). Levantou-
se, então, a hipótese de que o agente causador do problema se tratava de um vírus.
Na Índia, onde a doença já ocorria muito provavelmente há décadas, o mal recebeu o
nome de “Citrus Vein-phloem Degeneration” - CVPD. Em 1970, com a observação de
imagens sugestivas de procariotos no floema, acreditou-se que a doença era causada
por micoplasmas (LAFLECHE; BOVÉ, 1970 apud van LELYVELD; van VUUREN,
1988). No entanto, a presença nesses organismos, de parede celular semelhante à
que ocorre em bactérias Gram-negativas, levou à conclusão de que se tratava de uma
bactéria (GARNIER; BOVÉ, 1977; MOLL; MARTIN, 1974). Hoje sabe-se que HLB está
associado às α-proteobactérias, Gram-negativas e restritas ao floema, Candidatus
Liberibacter spp. São conhecidas três espécies: ‘Ca. L. americanus’ (CLam), ‘Ca. L.
africanus’ (CLaf) e ‘Ca. L. asiaticus’ (CLas) (JAGOUEIX; BOVE; GARNIER, 1994;
TEIXEIRA et al., 2005a, 2005c). Em 2013 e 2014, estudos moleculares revelaram a
presença de uma nova variante de Liberibacter em amostras de DNA do inseto
Diaphorina citri coletados na Colômbia, a qual foi denominada ‘Ca. L. caribbeanus’,

4
(MANJUNATH et al., 2015). Sobre esta suposta nova espécie nada se sabe sobre sua
ocorrência em citros.
CLaf é intolerante a altas temperaturas (>32ºC) e está restrita ao continente
africano, a parte da península arábica e a algumas ilhas do Oceano Índico
(JAGOUEIX; BOVÉ; GARNIER, 1996; PHAHLADIRA; VILJOEN; PIETERSEN, 2012;
PIETERSEN et al., 2010). Em 2000 foi reportada a presença de uma variante de CLaf
afetando uma rutácea ornamental nativa na África do Sul (Calodendrum capensis), a
qual foi denominada de ‘Ca. Liberibacter africanus subsp. capensis’ (GARNIER et al.,
2000). CLaf também foi detectada em Clausena anisata, Vepris lanceolata, e
Zanthoxylum capense), sendo denominada ‘LafCl’, ‘LafV’ e ‘LafZ’, respectivamente
(ROBERTS; STEENKAMP; PIETERSEN, 2015). Aparentemente essas variantes não
ocorrem naturalmente nos pomares de Citrus sp. (PHAHLADIRA; VILJOEN;
PIETERSEN, 2012). Em contraste, dois haplótipos de CLaf descritos recentemente
(‘LafA’ e ‘LafB’) ocorrem em pomares comerciais na mesma região (NELSON et al.,
2015).
CLam foi reportada pela primeira vez em amostras foliares coletadas na região
de Araraquara – SP (Brasil) em 2004. Nessa época CLam a espécie mais amplamente
detectada nos pomares paulistas (TEIXEIRA et al., 2005a, 2005b, 2005c). Contudo,
hoje em dia CLam é raramente detectada nos pomares (menos de 0,5% das amostras
com Liberibacter), ao contrário de CLas (LOPES et al., 2013). CLam está restrita ao
Brasil, apesar de recentemente ter sido reportada a presença de uma variante dessa
espécie no Texas (DA GRAÇA; KUNTA, 2015). Além de CLam ser intolerante a altas
temperaturas (>30°C), sua concentração nos tecidos vegetais é geralmente menor do
que a de CLas, mas causa maior dano à planta cítrica (LOPES et al, 2009). Esses
aspectos podem limitar a sua dispersão em regiões mais quentes (GASPAROTO et
al., 2012; LOPES et al., 2009), resultando em declínio em sua incidência ao longo do
tempo (LOPES et al., 2017b).
Apesar de mais tolerante, CLas também é afetada por altas temperaturas,
levando a redução na sua concentração nas plantas e nos insetos que nelas sugaram
(LOPES et al., 2013). Recentemente observou-se que além das altas temperaturas, o
déficit hídrico (menor quantidade de chuvas acumuladas nas semanas anteriores à
coleta de brotos novos) também afeta negativamente a população de CLas nos brotos

5
(LOPES et al., 2017b) a qual foi menor em plantas da região norte (Triângulo Mineiro)
do que em plantas da região central (Analândia) do parque citrícola, e que pode
contribuindo para as menores taxas de disseminação da doença observadas no norte.
CLas também é a espécie de Liberibacter mais amplamente distribuída no
mundo (BOVÉ, 2006, 2014). Destaca-se sua rápida expansão nos últimos anos para
áreas produtoras de citros no continente americano, como no Brasil em 2004
(TEIXEIRA et al., 2005c), Estados Unidos em 2005 (WANG; TRIVEDI, 2013), Cuba
em 2007 (LUIS et al., 2009), República Dominicana em 2008 (MATOS; HILF;
CAMEJO, 2009), México, Belize, Jamaica e Porto Rico em 2009 (HALL et al., 2013),
Costa Rica em 2011 (SFE, 2011), Argentina em 2012 (SENASA, 2013), Paraguai em
2013 (SENAVE, 2013), e Colômbia no final de 2015 (ICA, 2015).
1.2.2. Processo infeccioso, efeitos na planta e manejo da doença
Nesta revisão limitar-se-á a apresentar dados sobre a espécie CLas. Isto
porque é ela a mais disseminada no mundo e praticamente a única hoje presente no
Brasil, e sobre a qual maior volume de informação existe. CLas pode ser transmitida
artificialmente por meio de enxertia de material vegetal infectado, e naturalmente por
meio de D. citri. O processo de aquisição e de inoculação pelo inseto pode ocorrer
após poucos minutos de alimentação (BONANI et al., 2010). A aquisição é mais
eficiente quando inseto se encontra no estágio de ninfa (INOUE et al., 2009), porém
taxas de aquisição por ninfas similares às obtidas por adultos foram observadas
quando a população da bactéria nos brotos é relativamente ou muito baixa ou muito
alta (< 103 ou > 105 células g-1 de tecido) (LOPES et al., 2017a).
Por insetos adultos, a inoculação só é bem sucedida depois de terem
passados 10 a 18 dias entre o momento de sua alimentação na planta doente e o
momento de alimentação na planta sadia (AMMAR et al., 2016; CANALE et al., 2017;
INOUE et al., 2009). Logo após a infecção, CLas se movimenta mais rapidamente
para as partes que estão servindo de dreno dos fotoassimilados, ou seja, partes em
desenvolvimento nas extremidades da raiz ou ramos (RAIOL JR et al., 2016). A

6
velocidade de movimentação, no sentido descendente, pode chegar a estimados 6,9
cm dia-1 (RAIOL-JÚNIOR; CIFUENTES-ARENAS; LOPES, 2017).
Ainda na fase pré-sintomática, a planta responde à infecção ativando genes
de defesa, relacionados ao transporte de minerais e à biossíntese de amido, e
desativando genes relacionados ao funcionamento do floema, metabolismo da parede
celular, lipídeos e nucleotídeos, entre muitos outros (BOAVA; CRISTOFANI-YALY;
MACHADO, 2017; FOLIMONOVA; ACHOR, 2010; NWUGO et al., 2013). Mais tarde,
já na fase sintomática, ocorre acúmulo de calose nos tubos crivados, levando a
redução do tamanho dos poros e diminuição do fluxo simplástico de fotoassimilados
das células companheiras para o floema. Como consequência ocorre acúmulo de
amido nas folhas (FAN et al., 2010; KOH et al., 2012). Com o bloqueio no transporte
de açúcares, os fluxos vegetativos tornam-se amarelos, as folhas manifestam clorose
assimétrica característica (“mosqueado”), os frutos crescem de forma irregular, com
menor produção de etileno, maiores níveis de ácido abscísico e indol-acético e
apresentam maturação invertida (iniciando-se do pedúnculo) (DA GRAÇA et al., 2016;
ROSALES; BURNS, 2011). Além disso, há progressiva perda de massa de raízes, o
que limita a capacidade de absorção de nutrientes das plantas (GRAHAM et al., 2013;
JOHNSON et al., 2014).
Os diferentes genótipos de Citrus reagem diferentemente à infecção por CLas.
Apesar de a bactéria conseguir se estabelecer e atingir altas concentrações, em
alguns genótipos os sintomas foliares são menos intensos (FOLIMONOVA et al.,
2009). Maior ou menor nível de expressão de genes de defesa (ALBRECHT;
BOWMAN, 2012) e maior ou menor rapidez com que as mudanças anatômicas
acontecem após a infecção, são alguns dos aspectos que diferenciam genótipos
tolerantes de suscetíveis (FAN et al., 2013). Neste sentido, de particular interesse
estão, na atualidade, alguns de Poncirus trifoliata e seus híbridos (ALBRECHT;
BOWMAN, 2012; BOAVA; CRISTOFANI-YALY; MACHADO, 2017), o limoeiro
‘Rugoso’ (Citrus jambhiri) (FAN et al., 2012), o ‘Carrizo citrange’ (CEVALLOS-
CEVALLOS et al., 2012) e a lima ácida ‘Tahiti’ (Citrus limettioides) (BOSCARIOL-
CAMARGO et al., 2010).
Os prejuízos diretos causados pelo HLB estão associados à rápida
progressão da incidência tanto de sintomas na copa como de plantas infectadas no

7
pomar, o que diminui notoriamente a quantidade e qualidade dos frutos (BALDWIN et
al., 2010; BASSANEZI et al., 2011). Os prejuízos indiretos estão associados aos altos
custos envolvidos no manejo da doença, já que leva a um aumento considerável no
custo total de produção (BOVÉ, 2006; DA GRAÇA, 1991; GOTTWALD, 2010).
O plantio de mudas produzidas em viveiros protegidos com malhas a prova
de insetos, a eliminação das plantas sintomáticas para reduzir as fontes de inóculo, e
o monitoramento e controle do inseto vetor são os pilares fundamentais do manejo da
doença (BASSANEZI et al., 2013; BELASQUE JÚNIOR et al., 2009, 2010). Apesar de
todos esses esforços, o HLB tem sido o principal responsável pela erradicação de
mais de 46 milhões de plantas somente no estado de São Paulo. No ano de 2017 a
doença foi estimada estar presente em 16,7% dos 191,7 milhões de plantas do
cinturão citrícola, sendo as maiores incidências encontradas nas regiões Centro e Sul
e nas propriedades de menor tamanho (< 300 mil plantas) (FUNDECITRUS, 2017b).
Uma das principais causas do insucesso do manejo do HLB tem a ver com a
dificuldade de se proteger os brotos do acesso de psilídeos criados em plantas
doentes em pomares abandonados ou em fundos de quintal, onde não são alvo de
controle. Para minimizar este problema, muitos produtores vêm fazendo um trabalho
forte de conscientização de seus vizinhos, negociando a erradicação das plantas
sintomáticas, ou solicitando autorização para pulverizá-las regularmente. Quando não
permitem a eliminação dessas plantas ou a aplicação de produtos químicos, solicitam
autorização para liberar Tamarixia radiata, uma pequena vespa que parasita as ninfas
de D. citri. Essas ações em conjunto (controle do vetor e eliminação das plantas
doentes) têm reduzido a pressão de inóculo externo e facilitado o controle da doença
dentro das propriedades comerciais (MICHIGAMI, 2015; MONTEIRO, 2013).
1.3. Bioecologia de Diaphorina ciri Kuwayama
Diaphorina citri é um inseto de 2,5 a 3,5 mm de comprimento que, depois da
eclosão do ovo, passa por cinco estádios ninfais antes de virar adulto. Altamente
prolífico na presença de brotações e sob condições ambientais favoráveis

8
(temperatura média diária entre 24 a 29 °C, e umidade relativa entre 50 e 75%), se
amplamente disseminado nos pomares cítricos e áreas urbanas nos países onde está
presente (AMMAR et al., 2011; LIU; TSAI, 2000; NAVA et al., 2007). Os adultos recém
emergidos são brancos com as asas cristalinas e enrugadas e caminham sobre a folha
procurando permanecer, preferencialmente, na parte abaxial das mesmas,
provavelmente por ser esta a região da folha de mais fácil acesso aos vasos do floema
(GEORGE et al., 2017). Em questão de aproximadamente uma hora, o inseto vai
adquirindo suas cores características – marrom escura no corpo e manchas brancas
e cinzas nas asas. Aparentemente, machos e fêmeas emergem simultaneamente,
sem evidência de protandria ou protogenia (WENNINGER; HALL, 2007). Os adultos
conseguem se acasalar logo após a emergência e as fêmeas a colocar ovos depois
de 8h do acasalamento (PANDE, 1971). Porém, esse acasalamento precoce nem
sempre resulta em ovos férteis. O macho precisa de mais tempo para estar
sexualmente maduro (WENNINGER; HALL, 2007). A oviposição normalmente ocorre
em brotos novos e tenros com folhas em desenvolvimento e entre as margens de
folhas ainda fechadas, próximas ao meristema. Existem relatos de que uma fêmea
pode colocar até 1900 ovos durante sua vida (CATLING, 1970; HUANG, 1990).
1.4. Ecofisiologia dos citros e modelos de crescimento
As espécies cítricas se caracterizam por emitir fluxos de brotação em
intervalos mais ou menos definidos em resposta principalmente a fatores ambientais.
Este padrão é também conhecido como crescimento simpodial (SWINGLE; REECE,
1967). Esses fluxos, dependendo da época do ano, podem ser exclusivamente
vegetativos (responsáveis pelo crescimento da planta), reprodutivos (responsáveis
pela reprodução e frutificação), ou mistos (SPIEGEL-ROY; GOLDSCHMIDT, 1996).
No grupo das laranjeiras esses ciclos são mais bem definidos e também mais afetados
pela latitude. Dessa forma, em regiões de clima temperado e parte da região
subtropical, ocorrem em laranjeiras, usualmente dois fluxos anuais, enquanto que na
região tropical, ciclos adicionais. Já no grupo dos limoeiros e limeiras ácidas não existe

9
separação tão definida dos ciclos de brotação. Nestas plantas, até em clima
subtropical os fluxos são contínuos (SPIEGEL-ROY; GOLDSCHMIDT, 1996).
Os brotos originam-se de gemas (2 a 3) localizadas nas inserções das folhas
com o ramo, ou no final do ramo. Eles crescem emitindo folhas novas que vão se
expandindo até que o broto amadurece e sofre abscisão do ponteiro (SPIEGEL-ROY;
GOLDSCHMIDT, 1996). O início da divisão das células nas gemas é influenciado pela
temperatura e disponibilidade hídrica. Por sua vez, a diferenciação das gemas em
brotos reprodutivos ou vegetativos é determinada pelo balanço hormonal e acúmulo
de horas de frio ou de dias sem chuva, o que vai determinar a intensidade de flores
(ABBOTT, 1935; GARCIA-LUIS et al., 1992; RANDHAWA; DINSA, 1947;
STATHAKOPOULOS; ERICKSON, 1966). Os ciclos de crescimento na parte aérea e
na raiz normalmente se alternam e são definidos pela dinâmica de acúmulo de
fotoassimilados, que irão servir de reserva para o seguinte ciclo de crescimento, o
qual diminui em intensidade de brotos à medida em que a planta cresce (BEVINGTON;
CASTLE, 1985; GIRTON, 1927; REED, 1921; REED; MCDOUGAL, 1937).
Embora para Reed e McDougal (1937) a emissão dos fluxos obedece mais a
fatores genéticos, temperatura e disponibilidade hídrica têm papéis fundamentais
nesse comportamento. Trabalhando com seedlings e durante um curto período (1 ou
2 semanas), Girton (1927) determinou que existe diferença entre parte aérea e raiz
quanto a temperatura base inferior de desenvolvimento (Tb) – aquela abaixo da qual
o crescimento é praticamente imperceptível. De acordo com o autor, para o
crescimento de raízes de laranjeira doce e azeda a Tb está entre 12 e 13 °C, e para
pomelo entre 10 e 11 °C. Neste estudo, o crescimento da parte aérea de laranja doce
foi, no período avaliado, pequeno em qualquer temperatura, consequência
possivelmente da maior influência da raiz (BEVINGTON; CASTLE, 1985). As
temperaturas de 12 a 13 °C são, portanto, as Tb aceitas para uso em estudos que
envolvem modelos de crescimento vegetativo ou de acúmulo de graus dia para Citrus
spp. (AURAMBOUT et al., 2009; MENDEL, 1969). No entanto, essas temperaturas de
referência podem não ser as mais certas, pois foram definidas com base em um curto
período de crescimento e para seedlings que, por suas características, podem ser
especialmente sensíveis a variações ambientais. Provavelmente a forma mais correta

10
de se definir a Tb nos citros seja considerar ciclos completos de desenvolvimento,
desde o estímulo da brotação até a emissão do seguinte fluxo.
Em citros, os estudos estão voltados principalmente em estimar o impacto das
condições ambientais de temperatura e disponibilidade hídrica sobre o
desenvolvimento reprodutivo e produção de frutos (ALBRIGO; SAÚCO, 2004). Várias
metodologias podem ser usadas, como o acúmulo de graus dia (unidades de calor ou
de frio) sendo uma das mais aceitas. Basta contabilizar o número de horas dentro de
uma faixa de temperatura (ALLEN, 1976; WILSON; BARNETT, 1983). Por exemplo,
segundo Stathakopoulos e Erickson (1966) são necessários aproximadamente 3000
graus-hora após transferência das plantas para casa de vegetação para as gemas de
Poncirus trifoliata brotarem, e Moss (MOSS, 1969, 1976) determinou que entre maior
o número de horas abaixo de 18 °C maior é a proporção de brotos florais em laranja
doce. No caso da produção de frutos, o porta-enxerto afetou a o acúmulo de energia
necessário desde a antese até a maturação fiosiológica do fruto da laranjeira Folha
Murcha, necessitando em média de 4462 a 5090 graus dia (STENZEL et al., 2006).
No entanto, quanto ao crescimento dos brotos individuais não existem
modelos que permitam fazer uma modelagem das brotações no campo. Os modelos
existentes relacionados foram realizados há bastante tempo e descrevem o
crescimento das plantas como um todo, na sua altura e volume principalmente ou são
realizados com base em medições feitas a intervalos de tempo muito espaçados
(DUBEY; YADAV, 2004; TURRELL, 1961; TURRELL et al., 1969). Pelo contrário, são
abundantes os modelos de requerimentos térmicos de diversas pragas. Por exemplo,
Auramblout et al. (2009) desenvolveram um modelo para estimar o potencial de
distribuição de Diaphorina citri na Austrália diante de diversos cenários de mudanças
climáticas. No modelo, os autores consideraram a presença dos brotos e o ciclo de
vida de Diaphorina citri, porém desconsideraram o diferencial no potencial biótico que
tem o broto durante seus diferentes estádios de desenvolvimento, nem os
requerimentos para o surgimento dos brotos de ciclo a ciclo de brotação.

11
2. REFERÊNCIAS
ABBOTT, C. E. Blossom-bud differentiation in citrus trees. American Journal of Botany, v. 22, n. 4, p. 476, 1935.
AGRIANUAL. Citros. In: Anuário Estatístico da Agricultura Brasileira. São Paulo: FNP, Consultoria e Comércio, 2017. p. 225–253.
AGUT, B. et al. Different metabolic and genetic responses in citrus may explain relative susceptibility to Tetranychus urticae. Pest Management Science, v. 70, n. 11, p. 1728–1741, 2014.
ALBRECHT, U.; BOWMAN, K. D. Transcriptional response of susceptible and tolerant citrus to infection with Candidatus Liberibacter asiaticus. Plant Science, v. 185–186, p. 118–130, 2012.
ALBRIGO, L. G.; SAÚCO, V. G. Flower bud induction, flowering and fruit-set of some tropical and subtropical fruit tree crops with special reference to citrus. Acta Horticulturae, v. 632, p. 81–90, 2004.
ALLARIO, T. et al. Tetraploid Rangpur lime rootstock increases drought tolerance via enhanced constitutive root abscisic acid production. Plant, Cell and Environment, v. 36, n. 4, p. 856–868, 2013.
ALLEN, J. C. A modified sine wave method for calculating degree days. Environmental Entomology, v. 5, n. 3, p. 388–396, 1976.
AMMAR, E.-D. et al. Detection and relative titer of Candidatus Liberibacter asiaticus in the salivary glands and alimentary canal of Diaphorina citri (Hemiptera: Psyllidae) vector of citrus huanglongbing disease. Annals of the Entomological Society of America, v. 104, n. 3, p. 526–533, 2011.
AMMAR, E.-D. et al. Acquisition, replication and inoculation of Candidatus Liberibacter asiaticus following various acquisition periods on Huanglongbing-infected citrus by nymphs and adults of the asian citrus psyllid. PLoS ONE, v. 11, n. 7, p. e0159594, 2016.
ARGAMASILLA, R.; GÓMEZ-CADENAS, A.; ARBONA, V. Metabolic and regulatory responses in citrus rootstocks in response to adverse environmental conditions. Journal of Plant Growth Regulation, v. 33, n. 2, p. 169–180, 2014.
AURAMBOUT, J. P. et al. A concept model to estimate the potential distribution of the Asiatic citrus psyllid (Diaphorina citri Kuwayama) in Australia under climate change-A means for assessing biosecurity risk. Ecological Modelling, v. 220, n. 19, p. 2512–2524, 2009.

12
BALDWIN, E. et al. Effect of Liberibacter infection (Huanglongbing disease) of citrus on orange fruit physiology and fruit/fruit juice quality: chemical and physical analyses. Journal of Agricultural and Food Chemistry, v. 58, n. 2, p. 1247–62, 2010.
BASSANEZI, R. B. et al. Yield loss caused by huanglongbing in different sweet orange cultivars in São Paulo, Brazil. European Journal of Plant Pathology, v. 130, n. 4, p. 577–586, 2011.
BASSANEZI, R. B. et al. Efficacy of area-wide inoculum reduction and vector control on temporal progress of Huanglongbing in young sweet orange plantings. Plant Disease, v. 97, n. 6, p. 789–796, 2013.
BELASQUE JÚNIOR, J. et al. Base científica para a erradicação de plantas sintomáticas e assintomáticas de Huanglongbing (HLB, Greening) visando o controle efetivo da doença. Tropical Plant Pathology, v. 34, n. 3, p. 137–145, 2009.
BELASQUE JÚNIOR, J. et al. Controle do huanglongbing no estado de São Paulo, Brasil. Citrus Research & Technology, v. 31, n. 1, p. 53–64, 2010.
BEVINGTON, K. B.; CASTLE, W. S. Annual root growth pattern of young citrus trees in relation to shoot growth, soil temperature, and soil water content. Journal of the American Society for Horticultural Science, v. 110, n. 6, p. 840–845, 1985.
BOAVA, L. P.; CRISTOFANI-YALY, M.; MACHADO, M. A. Physiologic, anatomic, and gene expression changes in Citrus sunki, Poncirus trifoliata, and their hybrids after “Candidatus Liberibacter asiaticus” infection. Phytopathology, v. 107, n. 5, p. 590–599, 2017.
BONANI, J. P. et al. Characterization of electrical penetration graphs of the asian citrus psyllid, Diaphorina citri, in sweet orange seedlings. Entomologia Experimentalis et Applicata, v. 134, n. 1, p. 35–49, 2010.
BOSCARIOL-CAMARGO, R. L. et al. Avaliação de diferentes genótipos de citros à infecção por Candidatus Liberibacter asiaticus. Citrus Research & Technology, v. 31, n. 1, p. 85–90, 2010.
BOTEON, M.; NEVES, E. M. Citricultura brasileira: aspectos econômicos. In: MATTOS JÚNIOR, D. DE et al. (Eds.). . Citros. Campinas: Instituto Agronômico e Fundag, 2005. p. 19–36.
BOVÉ, J. M. Huanglongbing: a destructive, newly-emerging, century-old disease of citrus. Journal of Plant Pathology, v. 88, n. 1, p. 7–37, 2006.
BOVÉ, J. M. Heat-tolerant Asian HLB meets heat-sensitive African HLB in the Arabian Peninsula. Why? Journal of Citrus Pathology, v. 1, n. 1, p. 1–78, 2014.
CANALE, M. C. et al. Latency and persistence of “Candidatus Liberibacter asiaticus” in its psyllid vector, Diaphorina citri (Hemiptera: Liviidae). Phytopathology, v. 107, n. 3, p. 264–272, 2017.

13
CATLING, H. D. Distribution of the psyllid vectors of citrus greening disease, with notes on the biology and bionomics of Diaphorina citri. FAO Plant Protection Bulletin, v. 18, n. 1, p. 8–15, 1970.
CEVALLOS-CEVALLOS, J. M. et al. GC–MS metabolomic differentiation of selected citrus varieties with different sensitivity to citrus huanglongbing. Plant Physiology and Biochemistry, v. 53, p. 69–76, 2012.
DA GRAÇA, J. V.; KUNTA, M. Detection of Ca. Liberibacter americanus variant in Asian citrus psyllids, Diaphorina citri, in Texas. Proceedings of the APS Annual Meeting. Anais...Pasadena, California: 2015
DA GRAÇA, J. V. Citrus Greening Disease. Annual Review of Phytopathology, v. 29, n. 1, p. 109–136, 1991.
DA GRAÇA, J. V et al. Huanglongbing: An overview of a complex pathosystem ravaging the world’s citrus. Journal of Integrative Plant Biology, v. 58, n. 4, p. 373–387, 2016.
DONADIO, L. C.; MOURÃO FILHO, F. DE A. A.; MOREIRA, C. S. Centros de origem, distribuição geográfica das plantas cítricas e histórico da citricultura no Brasil. In: MATTOS JÚNIOR, D. DE et al. (Eds.). . Citros. Campinas: Instituto Agronômico e Fundag, 2005. p. 1–18.
DUBEY, A. K.; YADAV, D. S. Studies on growth pattern of different citrus species during the spring flush. Indian Journal of Agricultural Research, v. 38, n. 1, p. 50–54, 2004.
EBRATT-RAVELO, E. E. et al. Diaphorina citri (Kuwayama , 1907) and Tamarixia radiata (Waterson, 1922) in citrus crops of Cundinamarca, Colombia. Agronomía Colombiana, v. 29, n. 3, p. 487–493, 2011.
FAN, J. et al. Changes in carbohydrate metabolism in Citrus sinensis infected with “Candidatus Liberibacter asiaticus”. Plant Pathology, v. 59, n. 6, p. 1037–1043, 2010.
FAN, J. et al. Comparative transcriptional and anatomical analyses of tolerant Rough lemon and susceptible Sweet orange, in response to “Candidatus Liberibacter, Asiaticus” infection. Molecular Plant-Microbe Interactions, v. 25, n. 11, p. 1396–1407, 2012.
FAN, J. et al. Differential anatomical responses of tolerant and susceptible citrus species to the infection of “Candidatus Liberibacter asiaticus”. Physiological and Molecular Plant Pathology, v. 83, p. 69–74, jul. 2013.
FAO. FAOSTAT. Disponível em: <http://www.fao.org/faostat/en/#data>. Acesso em: 1 abr. 2017.
FOLIMONOVA, S. Y. et al. Examination of the responses of different genotypes of citrus to huanglongbing (citrus greening) under different conditions. Phytopathology, v. 99, n. 12, p. 1346–1354, dez. 2009.

14
FOLIMONOVA, S. Y.; ACHOR, D. S. Early events of citrus greening (Huanglongbing) disease development at the ultrastructural level. Phytopathology, v. 100, n. 9, p. 949–58, 2010.
FUNDECITRUS. Tree inventory and orange production forecast for the 2017-2018 season of the São Paulo and west-southwest of Minas Gerais citrus belt. Araraquara - SP, Brazil: Fundecitrus, 2017a. Disponível em: <http://www.fundecitrus.com.br>.
FUNDECITRUS. Mesmo estabilizada, insidência de greening permanece alta no cinturão citrícola de São Paulo e Triângulo/Sudoeste Mineiro. Araraquara - SP, Brazil: Fundecitrus, 2017b. Disponível em: <http://www.fundecitrus.com.br>.
GARCIA-LUIS, A. et al. Low temperature influence on flowering in Citrus. The separation of inductive and bud dormancy releasing effects. Physiologia Plantarum, v. 86, n. 4, p. 648–652, 1992.
GARNIER, M. et al. Genomic characterization of a Liberibacter present in an ornamental rutaceous tree, Calodendrum capense, in the Western Cape province of South Africa. Proposal of “Candidatus Liberibacter africanus subsp. capensis”. International Journal of Systematic and Evolutionary Microbiology, v. 50, n. 6, p. 2119–2125, 2000.
GARNIER, M.; BOVÉ, J. M. Structure trimellaire des deux membranes qui entoment les organisms Prokaryotes associes a la maladie du “Greening” des Agrumes. Fruits, v. 32, p. 749–752, 1977.
GASPAROTO, M. C. G. et al. Influence of temperature on infection and establishment of “Candidatus Liberibacter americanus” and “Candidatus Liberibacter asiaticus” in citrus plants. Plant Pathology, v. 61, n. 4, p. 658–664, 2012.
GEORGE, J. et al. Sclerenchymatous ring as a barrier to phloem feeding by Asian citrus psyllid: Evidence from electrical penetration graph and visualization of stylet pathways. PLoS ONE, v. 12, n. 3, p. e0173520, 2017.
GIRTON, R. E. The growth of citrus seedlings as influenced by environmental factors. University of California Publications in Agricultural Sciences, v. 5, p. 83–117, 1927.
GMITTER. Origin, evolution and breeding of the Grapefruit. In: JANICK, J. (Ed.). . Plant Breeding Reviews. New York: John Wiley & Sons, Inc., 1995. v. 13p. 345–363.
GMITTER, F. G. J.; HU, X. The possible role of Yunnan, China, in the origin of contemporary Citrus species (Rutaceae). Economic Botany, v. 44, n. 2, p. 267–277, 1990.
GOTTWALD, T. R. Current epidemiological understanding of citrus Huanglongbing. Annual Review of Phytopathology, v. 48, p. 119–139, 2010.
GRAHAM, J. H. et al. Presymptomatic fibrous root decline in citrus trees caused by

15
huanglongbing and potential interaction with Phytophthora spp. Plant Disease, v. 97, n. 9, p. 1195–1199, 2013.
HALL, D. G. et al. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomologia Experimentalis et Applicata, v. 146, n. 2, p. 207–223, 2013.
HUANG, B. Citrus psyllid. In: WU, Z. F. et al. (Eds.). . Chinese Agricultural Encyclopedia, Insect Volume Insect. Beijing, China: Agriculture Press, 1990. p. 119–120.
IAC. Instituto Biológico de Campinas, 2014. Disponível em: <http://www.iac.sp.gov.br/>. Acesso em: 2 abr. 2017
ICA. Resolución No. 00002390. Bogotá, Colombia: Instituto Colombiano Agropecuario, 2015. Disponível em: <http://www.ica.gov.co/getattachment/814e6823-aa26-49ee-a2c0-a814e2e61967/2015R2390.aspx>.
INOUE, H. et al. Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Annals of Applied Biology, v. 155, n. 1, p. 29–36, 2009.
JAGOUEIX, S.; BOVE, J.-M.; GARNIER, M. The phloem-limited bacterium of greening disease of citrus is a member of the α subdivision of the proteobacteria. International Journal of Systematic Bacteriology, v. 44, n. 3, p. 379–386, 1994.
JAGOUEIX, S.; BOVÉ, J. M.; GARNIER, M. PCR detection of the two “Candidatus” Liberobacter species associated with greening disease of citrus. Molecular and Cellular Probes, v. 10, n. 1, p. 43–50, 1996.
JOHNSON, E. G. et al. Association of “Candidatus Liberibacter asiaticus” root infection, but not phloem plugging with root loss on huanglongbing-affected trees prior to appearance of foliar symptoms. Plant Pathology, v. 63, n. 2, p. 290–298, 2014.
KING, C. W. H. Dispersión de Diaphorina citri (Hemiptera: Psyllidae) en el departamento del Tolima (Colombia). Revista Tumbaga, v. 2, n. 7, p. 51–60, 2012.
KOH, E.-J. et al. Callose deposition in the phloem plasmodesmata and inhibition of phloem transport in citrus leaves infected with “Candidatus Liberibacter asiaticus”. Protoplasma, v. 249, n. 3, p. 687–97, 2012.
LAPOINTE, S.; BOWMAN, K. D. Is there meaningful plant resistance to Diaprepes abbreviatus (Coleoptera: Curculionidae) in citrus rootstock germplasm? Journal of Economic Entomology, v. 95, n. 5, p. 1059–1065, 2002.
LEÓN, M. G. et al. Natural Infection of Swinglea glutinosa by the Citrus leprosis virus Cytoplasmic Type (CiLV-C) in Colombia. Plant Disease, v. 92, n. 9, p. 1364, 2008.
LIU, X. et al. Mechanisms for the influence of citrus rootstocks on fruit size. Journal of Agricultural and Food Chemistry, v. 63, n. 10, p. 2618–2627, 2015.

16
LIU, Y. H.; TSAI, J. H. Effects of temperature on biology and life table parameters of the asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Annals of Applied Biology, v. 137, n. 3, p. 201–206, 2000.
LOPES, S. et al. Additional evidence that the environment may be impacting HLB spread in Brazil. Journal of Citrus Pathology, v. 4, n. 1, p. 25, 2017a.
LOPES, S. A. et al. Liberibacters associated with citrus huanglongbing in Brazil: “Candidatus Liberibacter asiaticus” is heat tolerant, “Ca. L. americanus” is heat sensitive. Plant Disease, v. 93, n. 3, p. 257–262, 2009.
LOPES, S. A. et al. “Candidatus Liberibacter asiaticus” titers in citrus and acquisition rates by Diaphorina citri are decreased by higher temperature. Plant Disease, v. 97, n. 12, p. 1563–1570, 2013.
LOPES, S. A. et al. Seasonal variation of “Candidatus Liberibacter asiaticus” titers in new shoots of citrus in distinct environments. Plant Disease, v. 101, n. 4, p. 583–590, 2017b.
LUIS, M. et al. Occurrence of citrus Huanglongbing in Cuba and association of the disease with Candidatus Liberibacter asiaticus. Journal of Plant Pathology, v. 91, n. 3, p. 709–712, 2009.
MANJUNATH, K. L. et al. Candidatus Liberibacter caribbeanus, a new citrus associated Liberibacter from Colombia, South America. Journal of Citrus Pathology, v. 2, n. 1, p. 21, 2015.
MATOS, L.; HILF, M. E.; CAMEJO, J. First report of “Candidatus Liberibacter asiaticus” associated with citrus huanglongbing in the Dominican Republic. Plant Disease, v. 93, n. 6, p. 668–668, 2009.
MCCLEAN, A. P. D.; OBERHOLZER, P. C. J. Greening disease of the sweet orange: evidence that it is caused by a transmissible virus. South African Journal of Agricultural Sciences, v. 8, p. 253–276, 1965.
MENDEL, K. The influence of temperature and light on the vegetative development of citrus trees. Proceedings of the 1st International Citrus Symposium. Anais...1969
MICHIGAMI, F. A. B. Benefícios das medidas de manejo adotadas interna e externamente a uma propriedade comercial de citros na redução do progresso do Huanglongbing. Araraquara - SP, Brazil: Fundo de Defesa da Citricultura, 2015.
MOLL, J. N.; MARTIN, M. M. Comparison of the organism causing greening disease with several plant pathogenic Gram-negative bacteria, Rickettsia-like organisms and mycoplasma-like organisms. Inserm, v. 33, p. 87–96, 1974.
MONTEIRO, A. B. Efeito de um pomar sem manejo de Huanglongbing sobre a ocorrência de Diaphorina citri e incidência de plantas doentes em uma área vizinha com manejo da doença. Araraquara - SP, Brasil: Fundo de Defesa da

17
Citricultura, 2013.
MOSS, G. Influence of temperature and photoperiod on flower induction and inflorescence development in sweet orange (Citrus sinensis L. Osbeck). Journal of Horticultural Science, v. 44, n. 4, p. 311–320, 1969.
MOSS, G. Temperature effects on flower initiation in sweet orange (Citrus sinensis). Australian Journal of Agricultural Research, v. 27, n. 3, p. 399, 1976.
NAVA, D. E. et al. Biology of Diaphorina citri (Hem., Psyllidae) on different hosts and at different temperatures. Journal of Applied Entomology, v. 131, n. 9–10, p. 709–715, 2007.
NELSON, W. et al. Cryptic haplotypes of “Candidatus Liberibacter africanus”. Journal of Plant Pathology, v. 97, p. 291–295, 2015.
NWUGO, C. C. et al. The effect of “Candidatus Liberibacter asiaticus” infection on the proteomic profiles and nutritional status of pre-symptomatic and symptomatic grapefruit (Citrus paradisi) plants. BMC Plant Biology, v. 13, n. 1, p. 59, 2013.
OBERHOLZER, P. C. J.; VON STADEN, D. F. A.; BASSON, W. J. Greening disease of sweet orange in South Africa. Proceedings of the 3rd Conference of the IOCV. Anais...South Africa: 1965
PANDE, Y. D. Biology of citrus psylla, Diaphorina citri Kuw. (Hemiptera: Psyllidae). Israel Journal of Entomology, v. 6, n. 2, p. 307–311, 1971.
PHAHLADIRA, M. N. B.; VILJOEN, R.; PIETERSEN, G. Widespread occurrence of “Candidatus liberibacter africanus subspecies capensis” in Calodendrum capense in South Africa. European Journal of Plant Pathology, v. 134, n. 1, p. 39–47, 2012.
PIETERSEN, G. et al. A survey for “Candidatus Liberibacter” species in South Africa confirms the presence of only “Ca. L. africanus” in commercial citrus. Plant Disease, v. 94, n. 2, p. 244–249, 2010.
PIO, R. M. et al. Variedades copas. In: MATTOS JÚNIOR, D. DE et al. (Eds.). . Citros. Campinas: Instituto Agronômico e Fundag, 2005. p. 37–60.
POMPEU JUNIOR, J. Porta-enxertos. In: MATTOS JÚNIOR, D. DE et al. (Eds.). . Citros. Campinas: Instituto Agronômico e Fundag, 2005. p. 61–104.
RAIOL-JÚNIOR, L. L.; CIFUENTES-ARENAS, J. C.; LOPES, S. A. Speed of Candidatus Liberibacter asiaticus movement in citrus plants. Journal of Citrus Pathology, v. 4, n. 1, p. 45, 2017.
RAIOL JR, L. L. et al. Liberibacter movement in citrus scion and rootstock. (D. Mattos Jr. et al., Eds.). International Citrus Congress. Anais...Foz do Iguaçu, PR: 2016
RANDHAWA, G. S.; DINSA, H. S. Time of blossom-bud differentiation in Citrus. Proceedings of the American Society of Horticultural Science, v. 50, p. 165–171,

18
1947.
REED, H. S. Growth and sap concentration. Journal of Agricultural Research, v. 21, n. 1, p. 81–98, 1921.
REED, H. S.; MCDOUGAL, D. T. Periodicity in the growth of the orange tree. Growth, v. 1, p. 371–373, 1937.
REINKING, O. A. Diseases of economic plants in southern China. Philippine Agriculturist, v. 8, p. 109–135, 1919.
RESTREPO, O. C.; TORRES, R. M.; SALAZAR, R. C. Comparación del “Tabog” (Swinglea glutinosa Merr) con el limón “Rugoso” (Citrus jambhiri Swingle) y la mandarina “Cleópatra” (C. reticulata Swingle) como porta-injerto para la lima ácida “Tahiti” (C. aurantifolia Swingle). Acta Agronómica, v. 37, n. 3, p. 43–50, 1987.
ROBERTS, R.; STEENKAMP, E. T.; PIETERSEN, G. Three novel lineages of “Candidatus Liberibacter africanus” associated with native rutaceous hosts of Trioza erytreae in South Africa. International Journal of Systematic and Evolutionary Microbiology, v. 65, n. 2, p. 723–731, 2015.
RONDÓN, A. et al. Reacción de patrones cítricos a Phytophthora nicotianae B. de Haan. var. parasitica (Dastur) Waterh, en condiciones de umbráculo. Agronomía Tropical, v. 43, n. 3–4, p. 117–125, 1993.
ROSALES, R.; BURNS, J. K. Phytohormone changes and carbohydrate status in sweet orange fruit from huanglongbing-infected trees. Journal of Plant Growth Regulation, v. 30, n. 3, p. 312–321, 2011.
SAEED, M.; DODD, P. B.; SOHAIL, L. Anatomical studies of stems, roots and leaves of selected rootsock varieties in relation to their vigour. Journal of Horticulture and Forestry, v. 2, n. 4, p. 87–94, 2010.
SCHWARZ, R. E. An insect-transmissible virus, affecting sweet orange seedlings, trapped in orchards where greening disease is common. South African Journal of Agricultural Sciences, v. 7, p. 885–890, 1964.
SENASA. Resolución 959/2009: Declara la emergencia fitosanitaria con respecto al Huanglongbing (HLB) en todo el Territorio Nacional. Argentina: [s.n.]. Disponível em: <http://www.senasa.gov.ar/contenido.php?to=n&in=1402&io=11771>.
SENAVE. Resolución No 80 de 2013: “Por la cual se declara la emergência fitossanitária por la presencia de la plaga denominada huanglongbing (HLB) de los cítricos, en todo el território nacional”. Paraguay: [s.n.]. Disponível em: <http://www.senave.gov.py/docs/resoluciones/senave/Res80-13.pdf>.
SFE. Autoridades del SFE confirman presencia del “Dragón Amarillo” en árboles de la Zona Norte. Costa Rica: [s.n.]. Disponível em: <http://www.sfe.go.cr/documentos/comunicados/2011/21_02_2011_SFE_confirma_presencia_HLB_en_zona_norte.pdf>.

19
SPIEGEL-ROY, P.; GOLDSCHMIDT, E. E. Biology of Citrus. Cambridge: Cambridge University Press, 1996.
STATHAKOPOULOS, N.; ERICKSON, L. The effect of temperature on bud break in Poncirus trifoliata. Proceedings of the American Society of Horticultural Science, v. 89, p. 223–225, 1966.
STENZEL, N. M. C. et al. Maturation curves and degree-days accumulation for fruits of “Folha Murcha” orange trees. Scientia Agricola, v. 63, n. 3, p. 219–225, 2006.
SWINGLE, W. T.; REECE, P. C. The botany of Citrus and its wild relatives. In: REUTHER, W.; WEBBER, H. J.; BATCHELOR, L. D. (Eds.). . The citrus industry. 2. ed. Berkeley, California: University of California, 1967. v. 1p. 190–430.
TAN, F. Q. et al. Comparative metabolic and transcriptional analysis of a doubled diploid and its diploid citrus rootstock (C. junos cv. Ziyang xiangcheng) suggests its potential value for stress resistance improvement. BMC Plant Biology, v. 15, n. 1, p. 1–14, 2015.
TEIXEIRA, D. C. et al. “Candidatus Liberibacter americanus”, associated with citrus huanglongbing (greening disease) in Sao Paulo State, Brazil. International Journal of Systematic and Evolutionary Microbiology, v. 55, n. 5, p. 1857–1862, 2005a.
TEIXEIRA, D. C. et al. First report of a huanglongbing-like disease of Citrus in Sao Paulo State, Brazil and association of a new Liberibacter species, “Candidatus Liberibacter americanus”, with the disease. Plant Disease, v. 89, n. 1, p. 107, 2005b.
TEIXEIRA, D. DO C. et al. Citrus huanglongbing in São Paulo State, Brazil: PCR detection of the “Candidatus” Liberibacter species associated with the disease. Molecular and Cellular Probes, v. 19, n. 3, p. 173–9, 2005c.
TIRTAWIDJAJA, S. Citrus virus research in Indonesia. (E.C. Calavan, S. M. Garnsey, L. W. Timmer, Eds.)Proceedings of the 8th Conference of the IOCV. Anais...Riverside: IOCV, 1988
TIRTAWIDJAJA, S.; HADIWIDJAJA, T.; LASHEEN, A. M. Citrus vein phloem degeneration, a possible cause of citrus chlorosis in Java. Proceedings of the American Society of Horticultural Science, v. 86, p. 235–243, 1965.
TRAMONTINI, S. et al. Rootstock control of scion response to water stress in grapevine. Environmental and Experimental Botany, v. 93, p. 20–26, 2013.
TREEBY, M. T. et al. Irrigation management and rootstock effects on navel orange [Citrus sinensis (L.) Osbeck] fruit quality. Agricultural Water Management, v. 91, n. 1–3, p. 24–32, 2007.
TURRELL, F. M. Growth of the photosynthetic area of citrus. Botanical Gazette, v. 122, n. 4, p. 284–298, 1961.
TURRELL, F. M. et al. Growth equations and curves for citrus trees. Hilgardia, v. 39,

20
n. 16, p. 429–445, 1969.
VAN DER MERWE, A. J.; ANDERSEN, F. G. Chromium and manganese toxicity. Is it important in Transvaal citrus greening? Farming South Africa, v. 4, p. 439–440, 1937.
VAN LELYVELD, L. J.; VAN VUUREN, S. P. Peroxidase activity as a marker in Greening disease of citrus for assessment of tolerance and susceptibility. Journal of Phytopathology, v. 121, n. 4, p. 357–362, 1988.
VENNING, F. D. Trials with Swinglea glutinosa (Blanco) Merr. as a rootstock for citrus. Proceedings of the Florida Horticultural Society. Anais...Florida: 1957
WANG, N.; TRIVEDI, P. Citrus huanglongbing: a newly relevant disease presents unprecedented challenges. Phytopathology, v. 103, n. 7, p. 652–65, 2013.
WENNINGER, E. J.; HALL, D. G. Daily timing of mating and age at reproductive maturity in Diaphorina citri (Hemiptera: Psyllidae). Florida Entomologist, v. 90, n. 4, p. 715–722, 2007.
WILSON, L. T.; BARNETT, W. W. Degree-days: An aid in crop and pest management. California Agriculture, v. 37, n. 1, p. 4–7, 1983.
WU, G. A. et al. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nature Biotechnology, v. 32, n. 7, p. 656–662, 2014.

21
CAPÍTULO 2 – Potencial de Swinglea glutinosa como criadouro de Diaphorina
citri e fonte de Candidatus Liberibacter asiaticus
RESUMO
RESUMO. O huanglongbing é a doença mais destrutiva dos citros e Diaphorina citri o
inseto vetor das bactérias associadas à doença. O risco de uma rápida expansão da
doença para novas áreas pode ser incrementado se espécies hospedeiras alternativas
tanto das bactérias como do vetor estão presentes. O objetivo deste trabalho foi
investigar a importância relativa de Swinglea glutinosa como criadouro de D. citri e
como fonte de Candidatus Liberibacter asiaticus. Esta espécie vegetal pertence à
família Rutáceae, é compatível fisiologicamente com os citros comerciais, e usada
como cerca viva em certas regiões ou países, como a Colômbia, por exemplo. No
estudo sobre biologia do inseto, comparou-se S. glutinosa com as laranjeiras doces
‘Valência’, ‘Hamlin’, ‘Rubi’, ‘Pera’, ‘Valência Americana’, ‘Folha Murcha’ e ‘Natal’, e
com os limoeiros Siciliano ‘AB’, ‘Femminelo’, e ‘Limonera’. A taxa de reprodução do
inseto foi maior nas laranjeiras e menor nos limoeiros, com S. glutinosa ficando na
posição intermediária. Quanto a multiplicação de Liberibacter, comparou-se S.
glutinosa com a ornamental murta (Murraya exotica) e com a laranjeira ‘Valência’,
tanto em inoculação por inseto quanto por garfagem de topo. O título bacteriano foi
maior em ‘Valência’ do que em S. glutinosa ou murta sem diferença entre os métodos
de inoculação. ‘Valência’ representou de 3 a 20 vezes maior risco de servir como fonte
de inóculo do que S. glutinosa. Nenhum dos insetos que se alimentaram de plantas
de murta infectadas por Liberibacter adquiriram a bactéria. Este é o primeiro relato de
Swinglea glutinosa infectada por Ca. L. asiaticus. Apesar de os títulos da bactéria e
as taxas de sua aquisição por D. citri serem baixos em S. glutinosa, recomenda-se
sua erradicação em áreas próximas a pomares cítricos comerciais, pois ainda assim
pode atuar como reservatórios tanto da bactéria do HLB como do seu inseto vetor e
atuarem como fontes de inóculo.
Palavras-chave: Huanglongbing, Tabog, Candidatus Liberibacter asiaticus,
hospedeiros alternativos, Citrus sinensis.

22
CHAPTER 2 – Potential of Swinglea glutinosa as host for Diaphorina citri and
as inoculum source of Candidatus Liberibacter asiaticus
ABSTRACT
ABSTRACT. Huanglongbing is the most destructive disease of citrus, and Diaphorina
citri is the vector of the bacteria associated with the disease. The risk of a rapid spread
of the disease to new areas may be increased where alternative hosts for both bacteria
and vector are present. The objective of this work was to investigate the relative
importance of Swinglea glutinosa, a Rutaceae physiologically compatible with
commercial citrus and widely used in Colombia as a fence, for D. citri reproduction and
as a source of Candidatus Liberibacter asiaticus. For the insect biology studies,
Swinglea was compared to the sweet oranges Valencia, Hamlin, Rubi, Pera, Valencia
Americana, Folha Murcha, Natal, and the Sicilian lemons AB, Femminelo, and
Limonera. The sweet orange trees were the most suitable for reproduction of the
insect, while the lemon trees the least. S. glutinosa was rated as intermediate. In the
studies involving Ca. Liberibacter asiaticus, S. glutinosa was compared to orange
jasmine (Murraya exotica) and to Valencia, inoculated by both insect and top grafting.
The bacterial titer was higher in Valencia than in S. glutinosa or orange jasmine, with
no difference between inoculation methods. The risk of serving as a source of inoculum
was 3 to 20 times higher for Valencia than for S. glutinosa. None of the insects that fed
on Las-positive orange jasmine acquired the bacteria. This is the first report of S.
glutinosa infected by Ca. L. asiaticus confirmed by PCR. Despite the low titer of the
bacterium and its low acquisition rates by D. citri, S. glutinosa could still be a reservoir
of the HLB bacterium and of its vector D. citri. Therefore, should be eliminated if
growing near commercial citrus orchards.
Keywords: Huanglongbing, Tabog, swingle lemon, , Diaphorina citri, alternative
hosts, Citrus sinensis.

23
1. INTRODUÇÃO
O Huanglongbing (HLB) está associado às α-proteobactérias gram-negativas,
e restritas ao floema, Candidatus Liberibacter americanus (Lam), Ca. L. asiaticus (Las)
e Ca. L. africanus (Laf) (BOVÉ, 2006; JAGOUEIX; BOVE; GARNIER, 1994; TEIXEIRA
et al., 2005a). É considerada a doença mais destrutiva dos citros, com Las sendo a
espécie mais amplamente distribuída no mundo (BOVÉ, 2014). A expansão mais
relevante foi a que ocorreu nos últimos anos, para as principais áreas produtoras e
exportadoras de sucos de laranja do mundo - Brasil e Estados Unidos. A doença foi
relatada no Brasil em 2004 (COLETTA-FILHO et al., 2004), e em pouco mais de uma
década já estava presente na maioria dos países americanos, indo do sul da Califórnia
e toda a Flórida nos Estados Unidos até o norte da Argentina. Exclui-se nesta grande
area o centro-norte do Brasil e alguns países da américa latina . Os prejuízos do HLB
estão associados à rápida progressão tanto da incidência de plantas afetadas quanto
da severidade dos sintomas, provocando perdas significativas em produtividade
(BALDWIN et al., 2010; BASSANEZI et al., 2011; GOTTWALD, 2010).
Um aspecto que dificulta o controle do HLB é a não existência de variedades
cítricas resistentes à doença. Por isso, intensos esforços têm sido feitos na busca de
fontes de resistência dentro da família Rutaceae. Busca-se materiais que possam
servir como doadores de material genético em programas de melhoramento ou, caso
sejam fisiologicamente compatíveis com as variedades cítricas plantadas, materiais
que possam ser usados diretamente como porta-enxertos. Estudos indicam que
quanto mais filogeneticamente distantes dos citros for a rutácea maior é a
probabilidade desta conter genes para resistência ou tolerância à bactéria. Dentro do
gênero Citrus só algumas espécies como a cidra (Citrus medica L.) e parentes
próximos (MILES et al., 2017; RAMADUGU et al., 2016) o limoeiro ‘Rugoso’ (Citrus
jambhiri) (FAN et al., 2012), o citrange ‘Carrizo’ (CEVALLOS-CEVALLOS et al., 2012)
e a ‘lima da Pérsia’ (Citrus limettioides) (BOSCARIOL-CAMARGO et al., 2010)
mostraram certa tolerância. Fora dos Citrus tem se o Poncirus trifoliata e seus híbridos
(ALBRECHT; BOWMAN, 2012; BOAVA; CRISTOFANI-YALY; MACHADO, 2017).

24
O segundo aspecto que pode estar dificultando o seu controle tem a com a
possibilidade de a bactéria e seu vetor se hospedar em outras espécies vegetais, com
elas atuando como reservatórios de ambos os organismos. Sabe-se, por exemplo,
que em murta (Murraya exotica L.) e em ‘curry’ (Bergera koenigii L.), duas espécies
amplamente usadas como ornamentais, Diaphorina citri é altamente prolífico (HALL
et al., 2013). Porém estas espécies não são consideradas boas hospedeiras para Las.
Nelas a bactéria atinge baixos títulos (log 3 a 4 do número de células por grama de
tecido) quando comparados com os citros (log 6 a 7) (DAMSTEEGT et al., 2010;
LOPES et al., 2010). Contudo, o fato de mudas desses materiais serem produzidas
em áreas abertas, diferentemente do que acontece no caso das mudas cítricas no
estado de São Paulo, e também por não haver qualquer controle de seu comércio e
distribuição, é possível que possam estar contribuindo, inadvertidamente, para
dispersão de Las e do inseto para áreas ainda indenes.
Existe ainda uma terceira espécie vegetal sobre a qual pouco se sabe quanto
a hospedar ou não a bactéria do HLB e seu vetor. Trata-se da Swinglea glutinosa
(Blanco) Merr. Também conhecida como “Tabog” na Ásia, S. glutinosa é uma árvore
trifoliada de porte pequeno ou médio com frutos duros, sem suco e com muitas
sementes (SWINGLE; REECE, 1967). Na Colômbia, onde o HLB foi relatado no final
de 2015 (ICA, 2015), S. glutinosa é muito usada como cerca viva em condomínios,
parques, lotes e fazendas agrícolas (EBRATT-RAVELO et al., 2011; KING, 2012). É
fisiologicamente compatível com laranjeiras doces, pomeleiros, tangerineiras e com a
lima Tahiti. Apesar de não ser muito usada como porta-enxerto, parece apresentar
algumas características desejáveis como resistência a seca, resistência à gomose e
ao vírus da tristeza dos citros, e também promover redução do porte de copas com
consequente aumento da eficiência produtiva (DEL VALLE; RODRÍGUEZ;
RODRÍGUEZ, 2004; RESTREPO; TORRES; SALAZAR, 1987; RONDÓN et al., 1993;
VENNING, 1957).
No que diz respeito ao seu potencial em disseminar Las, nenhuma informação
consistente existe. O único registro encontrado é o trabalho de Tirtawidjaja (1981) na
Indonésia, que observou sintomas em três de 15 plantas aparentemente inoculadas
por inseto. Naquela época, naquele país, o HLB não tinha ainda sua etiologia
conhecida como acontece hoje. Atribuía-se como causa da doença, conhecida como

25
Citrus Phloem Vein Degeneration um vírus ou complexo viral. O não uso da técnica
PCR, a qual seria desenvolvida mais tarde, impede maior assertividade sobre o
achado.
Considerando-se o potencial impacto que a S. glutinosa poderia ter como
porta- enxerto para copas de citros explorados comercialmente, e a dúvida sobre o
quanto esta espécie poderia estar contribuindo para o aumento da população de D.
citri e Las e, portanto, para a epidemia do HLB, foi conduzido este trabalho. Teve como
objetivos: (i) determinar o potencial de S. glutinosa como criadouro de D. citri em
relação a algumas variedades de laranja doce e de limão verdadeiro (Siciliano), e (ii)
avaliar seu potencial em multiplicar Las, em relação a um material altamente favorável
(laranja Valência), e outro sabidamente desfavorável (murta).
2. MATERIAL E MÉTODOS
2.1. Material vegetal e criação do inseto
Foram usadas plantas de 2 anos de idade de S. glutinosa e de murta (Murraya
exotica) em vaso de 2 L, e mudas de 2,5 anos de idade das laranjeiras doces
‘Valência’, ‘Valência americana’, ‘Hamlin’, ‘Folha murcha’, ‘Natal’, ‘Pêra’, ‘Rubi’, e dos
limoeiros ‘Siciliano’ ‘AB’, ‘Femminello’ e ‘Limonera’, todas enxertadas em limão ‘Cravo’
e plantadas em vasos de 4.7L, com substrato comercial Plantmax Citrus®. Todas as
plantas foram produzidas em viveiro protegido. As plantas foram irrigadas de acordo
às necessidades e ferti-irrigadas quinzenalmente com 100 mL por vaso contendo uma
solução nutritiva de Ca(NO3)2, MAP, MgSO4, Cu, Zn, Mn-EDTA, (NH4)2MoO4, e Fe-
EDTA, às concentrações de 1,35, 0,111, 0,4, 0,015, 0,01, 7,5x10-3, 4,5x10-4, 0.075
g/L, respectivamente.
Insetos livres de CLas foram criados em plantas de murta dentro de uma sala
climatizada a partir de uma colônia matriz mantida no Fundecitrus em condições de
telado e livre da presença de qualquer planta com HLB. Plantas de murta foram

26
podadas para emissão de brotos novos e 50 insetos foram confinados por 5 dias em
cada planta, com auxílio de um voal, para acasalamento e oviposição. Após o período
de desenvolvimento, os adultos foram separados por sexo a cada 24 h, assegurando
indivíduos não acasalados para os experimentos (WENNINGER; HALL, 2007). Os
adultos permaneceram na sala por mais 15 dias antes de serem usados nos
experimentos.
Foram usadas duas fontes de Las. A primeira foi inseto infectivo criado em
condições de telado usando mudas de Valência positivas para Las, com 3 anos de
idade, enxertada em Swingle. As plantas foram ligeiramente podadas e, após início
da nova brotação, 20-30 insetos foram confinados em cada planta, por oito dias, e
depois retirados. O desenvolvimento do inseto em plantas doentes permite longo
período de aquisição da bactéria e assegura produção de adulto já altamente
infectivos logo após sua emergência (INOUE et al., 2009; PELZ-STELINSKI et al.,
2010). Os adultos emergidos foram deixados nas mesmas plantas até que a maioria
completasse ao menos 20 dias de idade (CANALE et al., 2017; INOUE et al., 2009).
Logo depois, os adultos de todas as plantas foram transferidos para as três plantas
mais sintomáticas, as quais foram levadas para a sala climatizada no Departamento
de Fitossanidade da Faculdade de Ciências Agrárias e Veterinárias da UNESP em
Jaboticabal, mantidas na sala por mais 8 dias de aclimatação, antes de serem usadas
no experimento.
A segunda fonte de Las o foi segmento apical de ramo (garfo) de 7 a 10 cm
de comprimento retirados de plantas em vaso de Valência/Swingle doentes, no
mesmo dia da inoculação. As inoculações tanto por inseto como por enxertia foram
feitas entre os dias 20/11 e 20/12//2014.
2.2. Extração e quantificação de DNA de material vegetal e de inseto
As amostras consistiram de folhas maduras sintomáticas emitidas em fluxos
vegetativos posteriores às inoculações ou, no caso de não haver folhas sintomáticas,
em folhas assintomáticas próximas ao ponto de inoculação. Para extração de DNA

27
total das folhas, alíquota de 0,5 g da nervura central de cada amostra foi finamente
picada sobre papel toalha e colocada em sacos plásticos, nos quais foram adicionados
2 mL de solução de extração (MURRAY; THOMPSON, 1980). O material foi em
seguida macerado com homogeneizador Homex 6. Para extração de DNA total de
cascas e raízes fibrosas, foram picadas sobre papel toalha 0,3 g de material vegetal
de cada amostra e colocadas em tubo eppendorf de 2 mL. Foi adicionado 1,5 mL de
solução de extração. O material vegetal (caule ou raiz) foi incubado em banho maria
a 65ºC por duas horas e homogeneizado por inversão dos tubos a cada 30 min. A
extração do DNA de folhas, casca e raízes foi conduzida como descrito por Teixeira
et al. (2005). O DNA extraído foi quantificado, ajustado à concentração de 100 ng µL-
1 e analisado por PCR em tempo real ou quantitativo (qPCR), em volume final de 12
µL contendo 6 µL de master mix (Invitrogen, Carlsbad, CA). Para análise do DNA de
folhas, casca de caule e raízes, o volume final continha 3 µL de DNA e 150 nM dos
primers e sonda HLBaspr (LI et al., 2006). A PCR foi realizada no termociclador
StepOne Plus (Applied Biosystems). Os valores de Cq (quantification cycle) foram
ajustados automaticamente usando o software StepOne. Somente amostras com CT
≤ 34 foram consideradas positivas. Os valores de CT foram convertidos a log10 do
número de cópias de amplicons por grama de tecido, usando a equação y = –0,2998x
+ 11,042 (LOPES et al., 2013).
De D. citri o DNA foi extraído de insetos indivídualizados ou de lotes de cinco
adultos. Os insetos foram macerados com pistilo em tubos eppendoff de 1,5 mL
contendo 400 µL de tampão de extração CTAB (MURRAY; THOMPSON, 1980). As
amostras foram incubadas por 30 min em banho maria a 65ºC e centrifugadas a 956
x g por 5 min. A extração foi conduzida como descrito (TEIXEIRA et al., 2005b). O
DNA foi desidratado por 6 min no speed vácuo e re-suspenso em 25 µL de água Milli-
Q autoclavada e deixado overnight a 4ºC. As amostras foram analisadas por qPCR
utilizando o equipamento StepOne Plus thermocycler (Applied Biosystems). Para a
reação de qPCR foi utilizado o kit ‘TaqMan qPCR Master Mix’ (Ambricon®). O volume
final da reação foi de 12 µL contendo 3 µL de DNA total, 150 nM dos primers, sonda
HLBaspr, que tem como alvo o 16S rDNA de Las (LI; HARTUNG; LEVY, 2006), e 150
nM dos primers e sondas da sequência parcial do gene wingless (Wg) de D. citri
(THAO et al., 2000). Os valores de CT ≤ 34 foram considerados como positivos e foram

28
convertidos em títulos de CLas usando a equação y = – 0.2904x + 11.085 , como
descrito (LOPES et al., 2013).
2.3. Experimento 1: Potencial de Swinglea glutinosa e de algumas variedades
de laranjeira doce e de limoeiro ‘Siciliano’ em atuar como hospedeiro de
Diaphorina citri
Este experimento foi conduzido no departamento de Fitossanidade da FCAV
– UNESP, Jaboticabal (SP, Brasil). As mudas (n = 5 a 13) de ‘Valência’, ‘Valência
americana’, ‘Hamlin’, ‘Folha Murcha’, ‘Natal’, ‘Pêra’, ‘Rubi’, ‘Siciliano’ ‘AB’,
‘Femminello’ e ‘Limonera’, todas enxertadas em ‘Cravo’ foram podadas e desfolhadas
15 – 20 cm acima da linha de enxertia, e as de Swinglea a 25 cm acima do nível do
substrato. A poda foi feita para estimular a emissão de novos brotos e favorecer a
reprodução e sobrevivência dos insetos. Foi selecionado um broto no estádio V2 por
planta o qual foi envolvido dentro de saco de malha e confinados durante 72h dois
casais virgens de 15 dias de idade. Como em S. glutinosa o desenvolvimento do broto
é diferente ao dos brotos em laranjeiras e limoeiros, e mais parecido com o do
citrumeleiro ‘Swingle’ ou do Poncirus trifoliata, foi selecionado um broto com tecidos
tenros com pelo menos 1 trifolio em expansão e outro apenas se desenrolando mas
sem os folíolos claramente definidos; normalmente os brotos de S. glutinosa estão
cobertos por abundante pilosidade. Após esse período os adultos foram retirados e
foram realizadas contagens de ovos e ninfas eclodidas (com auxílio de lupa manual
com aumento 30X) além de adultos emergidos. Foi estimada a viabilidade de ovos e
ninfas e viabilidade total, assim como a razão sexual e duração do ciclo de ovo a
adulto. Esse experimento foi repetido duas vezes, a primeira em sala climatizada (26
± 2 °C, 70 ± 5% UR, 12 h fotoperiodo PAR = 60 µmol m-2 s-1) e a segunda em telado
(23ºC ±10, 60% ±17 UR).

29
2.4. Experimento 2: Potencial de Swinglea glutinosa, Murraya exotica e
laranjeira ‘Valência’ em multiplicar ‘Candidatus Liberibacter asiaticus’
O experimento foi conduzido em sala climatizada e telado no departamento
de Fitossanidade da FCAV – UNESP, Jaboticabal (SP, Brasil), e em estufa no
Fundecitrus, Araraquara (SP, Brasil). Foram usados dois métodos de inoculação (por
inseto e por enxertia). O experimento foi instalado entre 30/11/2014 e 10/12/2014.
A inoculação por inseto foi feita em sala climatizada. Para estimular brotação,
as plantas de S. glutinosa (n = 44), ‘Valência’/ ‘Swingle’ (n = 39), e de murta (n = 42)
foram podadas cortando-se 10-15 cm do topo e retirando as folhas mais próximas do
ponto de poda. Após o surgimento de novos brotos, foi selecionado um por planta,
destacando-se os demais. O broto de S. glutinosa e de murta tinham uma a duas
folhas compostas em expansão e outra apenas começando a se desenrolar (pleno
desenvolvimento). No caso da ‘Valência’, o broto selecionado estava no estádio V3
(em pleno desenvolvimento). Em cada broto foram confinados 5 a 7 insetos CLas+ por
um período de aceso à inoculação (PAI) de 8 dias. Após o término desse período os
insetos foram retirados e guardados (separados por planta) em congelador a -20 °C.
As plantas foram pulverizadas com inseticida e após 15 dias dentro da sala climatizada
foram transferidas para o telado.
A inoculação por enxertia foi feita usando o método tipo “wedge”
(SHOKROLLAH et al., 2009), também chamado de ‘cunha’ ou ‘garfagem de topo em
fenda’. Foi usada uma única planta fonte (‘Valência’) para cada planta inoculada
(murta: n = 92; S. glutinosa: n = 40; ‘Valência’; n = 40). As plantas-fonte apresentavam
sintomas característicos de HLB, sendo confirmadas positivas para CLas por qPCR.
Dos ramos das plantas fontes foram destacados os 7,5 cm ± 1,5 finais (“garfos”)
contendo 5 a 7 folhas, as quais foram reduzidas a ¼ - ½ do seu tamanho. O diâmetro
dos garfos foi similar ao diâmetro da planta que ia ser enxertada. O ponto de união
entre o garfo e a planta inoculada foi protegido usando Parafilm© M. As plantas
permaneceram em casa de vegetação.

30
2.5. Experimento 3: Potencial de Swinglea glutinosa, Murraya exotica e
laranjeira ‘Valência’ em atuar como fonte de inóculo
Este experimento foi conduzido logo após a terceira amostragem folhas feita
em 06/12/2015. Independentemente do método de inoculação, foram selecionadas
plantas de S. glutinosa, murta e ‘Valência’ positivas para CLas. As plantas foram
ligeiramente podadas para emissão de novos brotos e depois confinados 18 a 23
insetos CLas-negativos em cada planta e mantidos em câmara de crescimento
(CMP4030, Conviron, Winnipeg, Canada) a 26 °C ± 1, 70% ± 10 UR, e fotoperíodo de
12 h (PAR = 100 µmol m-2 s-1). Havia sido feita amostragem aleatória de insetos da
mesma colônia fonte usada no confinamento (equivalente a ≈ 5% da quantidade a ser
utilizada no experimento) e confirmada a ausência de CLas nos mesmos. Os primeiros
15 insetos confinados foram retirados a cada 24, 48, e 72 h. Nas plantas que
presentaram oviposição, os indivíduos foram permitidos a se desenvolverem e assim
que se tornaram adultos foram coletados para posterior análise. Todos os insetos
coletados foram armazenados em freezer (-20 °C) por aproximadamente um mês até
serem processados e analisados individualmente por qPCR. O último grupo de insetos
foi subdividido após PAI de 144 h e transferido para mudas de ‘Valência’ CLas-
negativas (8 plantas para os insetos vindos de Swinglea e 8 plantas para os vindos de
murta), confinando-os por PAA = 8 dias e retirando-os para análise individual por
qPCR. As plantas inoculadas foram analisadas para presença de sintomas ou da
bactéria 12 meses depois.
2.6. Avaliações e análise dos dados
No que se refere à biologia do inseto, os ambientes foram analisados
separadamente. Os dados de oviposição, duração do ciclo ovo – adulto, e de
viabilidade foram analisados para normalidade e igualdade das variâncias. Quando
necessário foram transformados para logaritmo ou arcoseno, e em seguida foi

31
realizada uma análise de variância com separação de médias pelo teste Scott-Knott
(SCOTT; KNOTT, 1974). A razão sexual foi analisada pelo teste Chi-quadrado.
Para determinar se existia relação entre a variação na população de estádios
imaturos do inseto no broto e sobrevivência total (quantos indivíduos tornam-se
adultos) foi realizada análise de regressão linear simples (y = β0+ β1*x), com a
viabilidade total sendo a variável dependente e o número de ovos ou número máximo
de ninfas observado as variáveis independentes. A hipótese levantada foi a de que
quanto maior fosse a população do inseto no broto maior seria a mortalidade de
indivíduos (possivelmente por competição) e, consequentemente, menor seria o
número de adultos emergidos. A significância da regressão foi determinada por meio
de análise de variância e os parâmetros da equação pelo teste t de Student.
No que se refere aos experimentos 2 e 3, os dados de porcentagem de
infecção e de sucesso na transmissão de CLas por garfagem foram comparados pelo
teste Chi-quadrado ou, no caso de frequências < 5, pelo teste exato de Fisher, na
primeira avaliação, feita quatro meses depois das inoculações. Em ambos os métodos
de inoculação, foi determinada a presença, características e intensidade de sintomas
típicos de clorose assimétrica associada ao HLB ou a deficiência mineral. Foi
determinado também o título de CLas em folhas das plantas onde a garfagem bem
sucedida e em todas as plantas inoculadas por inseto. Sucessivas avaliações foram
feitas a cada 4 – 6 meses para estudar as variações de título bacteriano nas folhas.
Adicionalmente, após a primeira avaliação de folias foram selecionadas 5 plantas de
cada espécie na inoculação por enxertia e 9 a 10 plantas na inoculação por inseto
para determinar o título bacteriano em folhas, caule e raiz, totalizando três amostras
por planta
Na primeira amostragem de folhas para estimar a presença e título de Las foi
realizada análise de variância para determinar o efeito da espécie (Swinglea, murta,
‘Valência’) e do método de inoculação (enxertia de topo ou inseto) no título da bactéria.
Em seguida foram selecionadas algumas plantas de cada espécie e amostrados seus
caules, raízes e novamente folhas. Visou-se com isso determinar o efeito da espécie
e da parte da planta amostrada (folha, raiz e caule) na multiplicação de CLas.

32
O nível de 5% de probabilidade foi usado em todas as análises, realizadas
com os programas Statgraphics Centurion VXII (Statpoint Technologies Inc.) e R (R
CORE TEAM, 2014).
3. RESULTADOS
Experimento 1: Na sala climatizada houve influência significativa do
hospedeiro no número de ovos, sem clara separação entre laranjeiras, limoeiros e S.
glutinosa (Tabela 1). No grupo formado por ‘Natal’, ‘Pera’, ‘AB’ e ‘Hamlin’ foram
encontrados média de 50,3 ovos por broto, enquanto que no grupo formado por
‘Valência Americana’, ‘Feminello’, ‘Rubi’, ‘Limonera’, S. glutinosa, ‘Valência’ e ‘Folha
Murcha’ a média foi 23,9 ovos. Tanto a viabilidade de ovos, ninfas e total foi
influenciada pelo hospedeiro. No caso da viabilidade de ovos foram claramente
diferenciados 2 grupos: o primeiro formado pelas laranjeiras doces ‘Natal’, ‘Valência
Americana’, ‘Hamlin’, ‘Rubi’ e ‘Valência’ (média = 84,64%) e o segundo pelos
limoeiros, S. glutinosa, e as laranjeiras ‘Folha Murcha’ e ‘Pera’ (média = 72,85%)
(Tabela 1). No caso da viabilidade de ninfas e total, foram claramente separados 3
grupos, sendo que S. glutinosa se destacou como o pior hospedeiro e os limoeiros
significativamente inferiores a todas as laranjeiras doces avaliadas (Tabela 1). Nem a
duração do ciclo ovo-adulto (média = 14,9 dias) nem a razão sexual (média = 0,53) foi
influenciada pelo hospedeiro.
Contrário ao acontecido na sala climatizada, em condições de telado houve
clara inferioridade dos limoeiros (média = 54,17 ovos) em relação às laranjeiras doces
(média = 123,70 ovos) (Tabela 1), com apenas S. glutinosa e ‘Rubi’ classificadas
como intermédias (média = 90,80 ovos). No caso da viabilidade de ovos, S. glutinosa
ficou agrupada com as laranjeiras doces ‘Valencia Americana’, ‘Pera’ e ‘Natal’ (média
= 86,40%), enquanto que os limoeiros e as laranjeiras doces ‘Hamlin’, ‘Valencia’,
‘Folha Murcha’ e ‘Rubi’ foram significativamente inferiores (média = 78,59%).
Similar ao acontecido na sala climatizada, a viabilidade de ninfas e total
mostrou clara separação entre os limoeiros e a maioria das laranjeiras doces, sendo
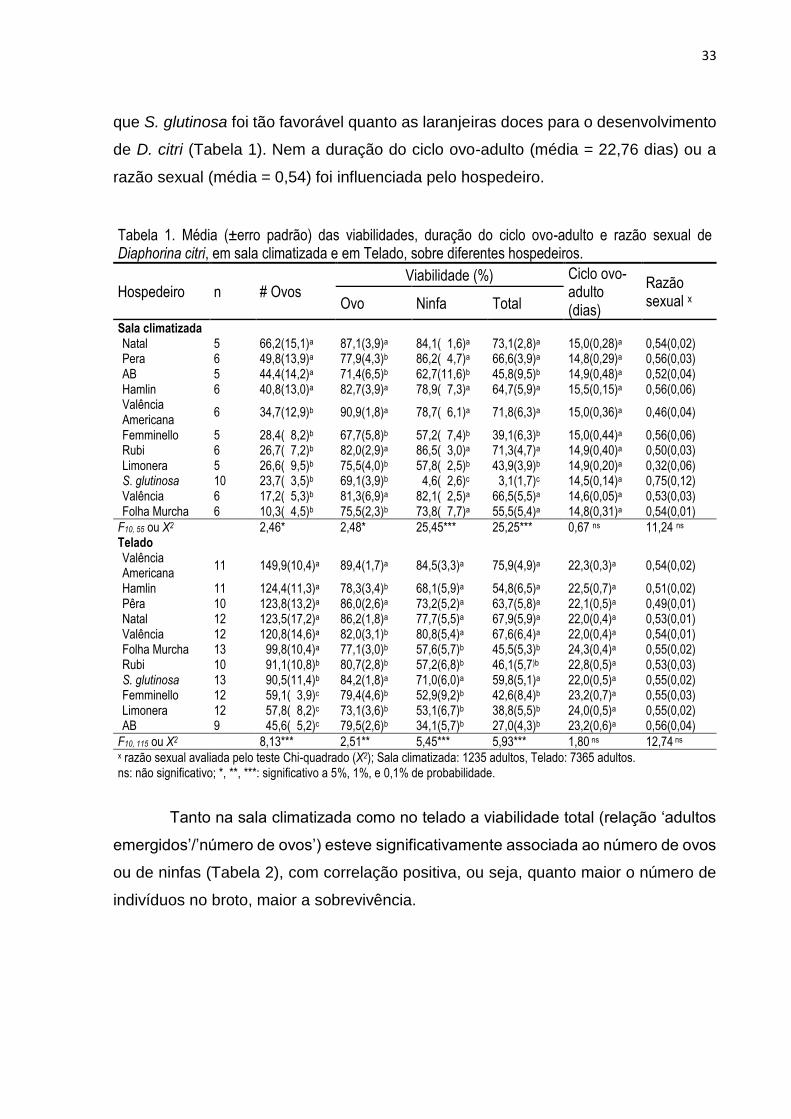
33
que S. glutinosa foi tão favorável quanto as laranjeiras doces para o desenvolvimento
de D. citri (Tabela 1). Nem a duração do ciclo ovo-adulto (média = 22,76 dias) ou a
razão sexual (média = 0,54) foi influenciada pelo hospedeiro.
Tabela 1. Média (±erro padrão) das viabilidades, duração do ciclo ovo-adulto e razão sexual de Diaphorina citri, em sala climatizada e em Telado, sobre diferentes hospedeiros.
Hospedeiro n # Ovos Viabilidade (%) Ciclo ovo-
adulto (dias)
Razão sexual x Ovo Ninfa Total
Sala climatizada Natal 5 66,2(15,1)a 87,1(3,9)a 84,1(11,6)a 73,1(2,8)a 15,0(0,28)a 0,54(0,02) Pera 6 49,8(13,9)a 77,9(4,3)b 86,2(14,7)a 66,6(3,9)a 14,8(0,29)a 0,56(0,03) AB 5 44,4(14,2)a 71,4(6,5)b 62,7(11,6)b 45,8(9,5)b 14,9(0,48)a 0,52(0,04) Hamlin 6 40,8(13,0)a 82,7(3,9)a 78,9(17,3)a 64,7(5,9)a 15,5(0,15)a 0,56(0,06) Valência Americana
6 34,7(12,9)b 90,9(1,8)a 78,7(16,1)a 71,8(6,3)a 15,0(0,36)a 0,46(0,04)
Femminello 5 28,4(18,2)b 67,7(5,8)b 57,2(17,4)b 39,1(6,3)b 15,0(0,44)a 0,56(0,06) Rubi 6 26,7(17,2)b 82,0(2,9)a 86,5(13,0)a 71,3(4,7)a 14,9(0,40)a 0,50(0,03) Limonera 5 26,6(19,5)b 75,5(4,0)b 57,8(12,5)b 43,9(3,9)b 14,9(0,20)a 0,32(0,06) S. glutinosa 10 23,7(13,5)b 69,1(3,9)b 74,6(12,6)c 73,1(1,7)c 14,5(0,14)a 0,75(0,12) Valência 6 17,2(15,3)b 81,3(6,9)a 82,1(12,5)a 66,5(5,5)a 14,6(0,05)a 0,53(0,03) Folha Murcha 6 10,3(14,5)b 75,5(2,3)b 73,8(17,7)a 55,5(5,4)a 14,8(0,31)a 0,54(0,01)
F10, 55 ou X2 2,46* 2,48* 25,45*** 25,25*** 0,67 ns 11,24 ns Telado Valência Americana
11 149,9(10,4)a 89,4(1,7)a 84,5(3,3)a 75,9(4,9)a 22,3(0,3)a 0,54(0,02)
Hamlin 11 124,4(11,3)a 78,3(3,4)b 68,1(5,9)a 54,8(6,5)a 22,5(0,7)a 0,51(0,02) Pêra 10 123,8(13,2)a 86,0(2,6)a 73,2(5,2)a 63,7(5,8)a 22,1(0,5)a 0,49(0,01) Natal 12 123,5(17,2)a 86,2(1,8)a 77,7(5,5)a 67,9(5,9)a 22,0(0,4)a 0,53(0,01) Valência 12 120,8(14,6)a 82,0(3,1)b 80,8(5,4)a 67,6(6,4)a 22,0(0,4)a 0,54(0,01) Folha Murcha 13 199,8(10,4)a 77,1(3,0)b 57,6(5,7)b 45,5(5,3)b 24,3(0,4)a 0,55(0,02) Rubi 10 191,1(10,8)b 80,7(2,8)b 57,2(6,8)b 46,1(5,7)b 22,8(0,5)a 0,53(0,03) S. glutinosa 13 190,5(11,4)b 84,2(1,8)a 71,0(6,0)a 59,8(5,1)a 22,0(0,5)a 0,55(0,02) Femminello 12 159,1(13,9)c 79,4(4,6)b 52,9(9,2)b 42,6(8,4)b 23,2(0,7)a 0,55(0,03) Limonera 12 157,8(18,2)c 73,1(3,6)b 53,1(6,7)b 38,8(5,5)b 24,0(0,5)a 0,55(0,02) AB 9 145,6(15,2)c 79,5(2,6)b 34,1(5,7)b 27,0(4,3)b 23,2(0,6)a 0,56(0,04)
F10, 115 ou X2 8,13*** 2,51** 5,45*** 5,93*** 1,80 ns 12,74 ns x razão sexual avaliada pelo teste Chi-quadrado (X2); Sala climatizada: 1235 adultos, Telado: 7365 adultos. ns: não significativo; *, **, ***: significativo a 5%, 1%, e 0,1% de probabilidade.
Tanto na sala climatizada como no telado a viabilidade total (relação ‘adultos
emergidos’/’número de ovos’) esteve significativamente associada ao número de ovos
ou de ninfas (Tabela 2), com correlação positiva, ou seja, quanto maior o número de
indivíduos no broto, maior a sobrevivência.

34
Tabela 2. Coeficientes da regressão linear simples e estatísticas do modelo para predizer a viabilidade total em função do número total de ovos ou de ninfas no broto como população inicial.
Variável independente β0 ± EP t P β1 ± EP t P R2Ajustado
Sala climatizada Número de ovos 35,69 ± 5,64 6,33 <0,0001 0,41 ± 0,13 3,19 0,0023 13,16% Número de ninfas 34,80 ± 4,99 6,98 <0,0001 0,53 ± 0,13 4,00 0,0002 20,55% Telado z Número de ovos 33,40 ± 4,35 7,68 <0,0001 0,21 ± 0,04 5,27 <0,0001 17,62% Número de ninfas 29,82 ± 3,74 7,97 <0,0001 0,29 ± 0,04 7,33 <0,0001 29,69%
Experimento 2: A frequência de sobrevivência dos ‘garfos’ vindos de plantas
CLas-negativas usados como controle foi independente da espécie enxertada (P =
0,5324), ou seja, a compatibilidade dos garfos foi boa quando enxertados em S.
glutinosa, ‘Valência’ ou murta. Porém, a sobrevivência dos garfos vindos de plantas
CLas-positivas variou significativamente dependendo da espécie (X2 = 18,852, GL =
2, P < 0,001) (Tabela 3), com maiores índices de sobrevivência na murta (77,17%) do
que na S. glutinosa (52,5%) ou ‘Valência’ (40%), na primeira avaliação 4 meses após
inoculação. Porém, a sobrevivência dos garfos até o final do experimento foi maior em
Valência e Swinglea do que em murta. A frequência de infecção foi independente da
espécie, com 61,90% em Valência, 66,67% em Swinglea e 61,97% em murta (X2 =
1,876, GL = 2, P = 0,3913).
Tabela 3. Frequências e título de ‘Candidatus Liberibacter asiaticus’ em folhas de murta, Swinglea e ‘Valência’ após 4 meses
de inoculação por enxertia e insetox
Inoculação por enxertia Inoculação por inseto
Espécie Controle y CLas+ y Log ± EP z CLas+ y Log ± EP z
Murta 90% 63,4% (77,2%) 2,77 ± 0,06 aA 29,5% 2,76 ± 0,26 aA
Swinglea 90% 66,7% (52,6%) 3,74 ± 0,09 bA 20,5% 3,58 ± 0,22 aA
Valência 100% 65,0% (50,0%) 5,14 ± 0,43 cA 87,2% 5,45 ± 0,25 bA x As médias foram calculadas sobre as amostras qPCR positivas; médias com diferentes letras dentro da mesma coluna
(letras minúsculas) ou entre as colunas (letras maiúsculas) diferem significativamente (P < 0,05). y Controle: sobrevivência dos garfos controle (n = 20); CLas+: porcentagem de infecção considerando só plantas com garfos
vivos (porcentagem sucesso na garfagem) ou plantas inoculadas por inseto. z Log10 de genomas de ‘Ca. L. asiaticus’ por grama de tecido (y = -0,2998x+11,042; (LOPES et al., 2013)), ± erro padrão.
Todos os lotes de insetos usados na inoculação continham CLas, que
transmitiram a bactéria para apenas 9,52% e 20,45% das plantas de murta e Swinglea,
respectivamente, mas para 87,18% das plantas de ‘Valência’, (X2 = 60,484, GL = 2, P
< 0,001) (Tabela 3). Os métodos de inoculação não influenciaram no título de CLas
em folhas em nenhuma das espécies estudadas (P > 0,05), maiores na laranjeira
35
‘Valência’ do que em murta ou Swinglea (P < 0,05), que se manteve sem grandes
variações em todas as avaliações (Figura 1).
Figura 1. Variação sazonal do título de ‘Candidatus Liberibacter asiaticus’ em folhas de murta (Murraya exotica), Swinglea glutinosa e laranja Valência inoculadas por inseto (a) e por enxertia no topo de segmentos de ramos doentes ou ‘garfos’ (b).
Com exceção da Swinglea, a bactéria foi detectada em mais amostras de folha
do que em caules ou raízes, resultado de provável distribuição irregular dentro da
planta, mesmo depois de quatro meses de terem sido inoculadas. Na Swinglea
inoculada por enxertia a bactéria não foi detectada na raiz. Não houve diferenças
estatísticas significativas (P > 0,05) entre as partes da planta quanto ao título
bacteriano. Porém, os valores foram significativamente menores em murta e Swinglea
do que em ‘Valência’ (Tabela 4).
Nas plantas inoculadas por enxertia houve presença de sintomas
característicos de HLB em todas as espécies, mas com diferentes intensidades. Em
murta, 51% das plantas apresentaram clorose assimétrica associada ao HLB muito
leve e 45% clorose associada a deficiência mineral. Na Swinglea a porcentagem de
plantas com clorose associada ao HLB ou a deficiência mineral foi de 33% e 40%,
respectivamente, em contraste 14% e 46% em ‘Valência’. Geralmente os sintomas de
HLB estiveram presentes em folhas novas completamente expandidas e próximas ao
ponto de garfagem. A maioria dos garfos sobreviventes sofreu abscisão de folhas, e
nos poucos que conservaram as folhas a clorose associada ao HLB, presente no
momento da inoculação, não progrediu. Além disso, alguns poucos garfos
apresentaram brotos novos com aparência sadia mas de tamanho menor que nos
garfos sadios.
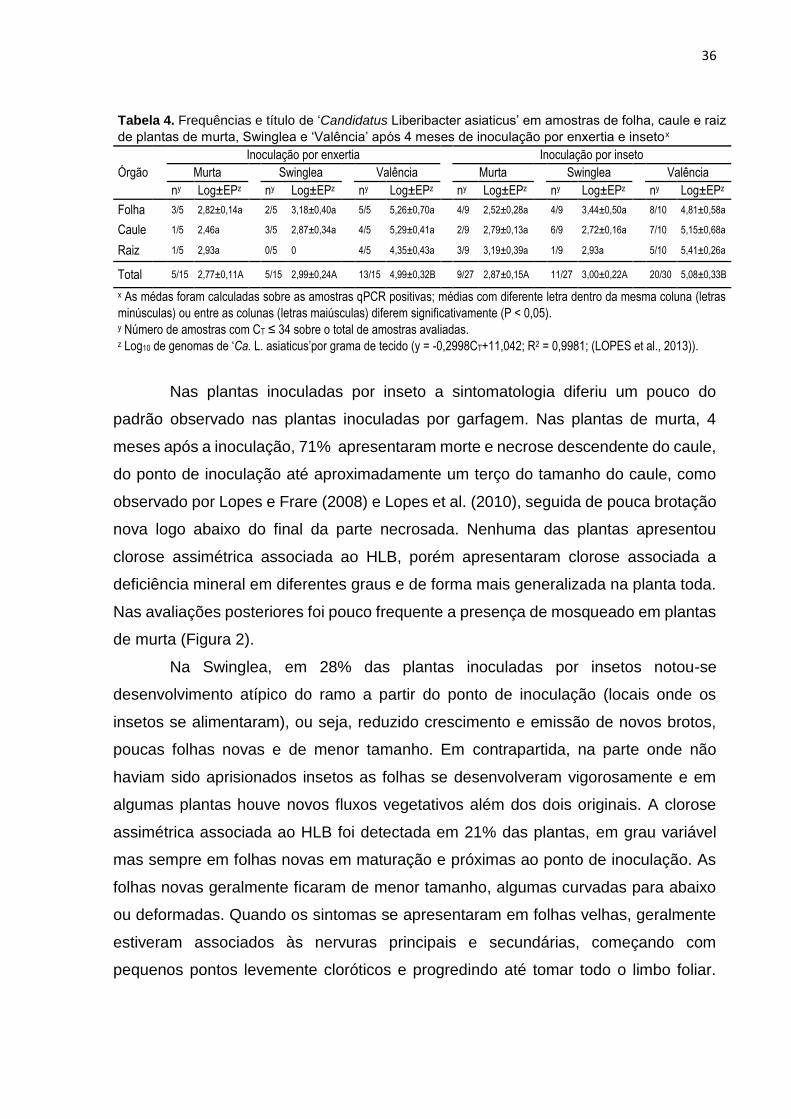
36
Tabela 4. Frequências e título de ‘Candidatus Liberibacter asiaticus’ em amostras de folha, caule e raiz
de plantas de murta, Swinglea e ‘Valência’ após 4 meses de inoculação por enxertia e insetox
Órgão
Inoculação por enxertia Inoculação por inseto Murta Swinglea Valência Murta Swinglea Valência ny Log±EPz ny Log±EPz ny Log±EPz ny Log±EPz ny Log±EPz ny Log±EPz
Folha 3/5 2,82±0,14a 2/5 3,18±0,40a 5/5 5,26±0,70a 4/9 2,52±0,28a 4/9 3,44±0,50a 8/10 4,81±0,58a
Caule 1/5 2,46a 3/5 2,87±0,34a 4/5 5,29±0,41a 2/9 2,79±0,13a 6/9 2,72±0,16a 7/10 5,15±0,68a
Raiz 1/5 2,93a 0/5 0 4/5 4,35±0,43a 3/9 3,19±0,39a 1/9 2,93a 5/10 5,41±0,26a
Total 5/15 2,77±0,11A 5/15 2,99±0,24A 13/15 4,99±0,32B 9/27 2,87±0,15A 11/27 3,00±0,22A 20/30 5,08±0,33B
x As médas foram calculadas sobre as amostras qPCR positivas; médias com diferente letra dentro da mesma coluna (letras
minúsculas) ou entre as colunas (letras maiúsculas) diferem significativamente (P < 0,05). y Número de amostras com CT ≤ 34 sobre o total de amostras avaliadas. z Log10 de genomas de ‘Ca. L. asiaticus’por grama de tecido (y = -0,2998CT+11,042; R2 = 0,9981; (LOPES et al., 2013)).
Nas plantas inoculadas por inseto a sintomatologia diferiu um pouco do
padrão observado nas plantas inoculadas por garfagem. Nas plantas de murta, 4
meses após a inoculação, 71% apresentaram morte e necrose descendente do caule,
do ponto de inoculação até aproximadamente um terço do tamanho do caule, como
observado por Lopes e Frare (2008) e Lopes et al. (2010), seguida de pouca brotação
nova logo abaixo do final da parte necrosada. Nenhuma das plantas apresentou
clorose assimétrica associada ao HLB, porém apresentaram clorose associada a
deficiência mineral em diferentes graus e de forma mais generalizada na planta toda.
Nas avaliações posteriores foi pouco frequente a presença de mosqueado em plantas
de murta (Figura 2).
Na Swinglea, em 28% das plantas inoculadas por insetos notou-se
desenvolvimento atípico do ramo a partir do ponto de inoculação (locais onde os
insetos se alimentaram), ou seja, reduzido crescimento e emissão de novos brotos,
poucas folhas novas e de menor tamanho. Em contrapartida, na parte onde não
haviam sido aprisionados insetos as folhas se desenvolveram vigorosamente e em
algumas plantas houve novos fluxos vegetativos além dos dois originais. A clorose
assimétrica associada ao HLB foi detectada em 21% das plantas, em grau variável
mas sempre em folhas novas em maturação e próximas ao ponto de inoculação. As
folhas novas geralmente ficaram de menor tamanho, algumas curvadas para abaixo
ou deformadas. Quando os sintomas se apresentaram em folhas velhas, geralmente
estiveram associados às nervuras principais e secundárias, começando com
pequenos pontos levemente cloróticos e progredindo até tomar todo o limbo foliar.

37
Quanto à clorose associada a deficiência mineral, estes foram de leves a médios
(Figura 2) em 82% das plantas.
As plantas de ‘Valência’ inoculadas por inseto também apresentaram
sintomas típicos de HLB (Figura 2), com 23% das plantas desenvolvendo clorose
assimétrica e 56% clorose típica de deficiência mineral. Os sintomas apareceram
principalmente em folhas novas em processo de maturação, as quais ficaram de
menor tamanho e geralmente curvadas para baixo.
Quanto aos títulos bacterianos, houve relativa constância nos valores ao longo
das avaliações para os dois métodos de inoculação (Figura 1), porém, para murta,
Swinglea e ‘Valência’ houve perda progressiva de plantas, não sendo sempre as que
tinham sido confirmadas CLas-positivas. Na inoculação por inseto houve intermitência
na detecção de plantas positivas, especialmente de em murta e Swinglea (Tabela 5),
maior número de amostras positivas foi encontrado na primeira avaliação, diminuindo
na segunda, aumentando na 3ª e 4ª, e finalizando com menos na última. Em
‘Valência’, apesar da perda de plantas (Tabela 5), a frequência foi mais uniforme. No
lote de plantas de murta e Swinglea inoculado por meio de garfos, houve diminuição
gradativa na frequência de plantas positivas, mesmo depois do inóculo ter sido
removido, o que não se observou na ‘Valência’.

38
Figura 2. Sintomas foliares de huanglongbing em folhas de Swinglea glutinosa controle sadio (a) e inoculada (b, c, d, e), de Valência sadia (f) e inocuada (g, h), e de murta inoculada (i, j, k, l).
Procurou se determinar se a planta inoculada, por variar em sua
suscetibilidade a CLas, estaria influenciando a bactéria no garfo usado como inóculo.
Numa seleção aleatória de plantas com garfagem foi determinado o título bacteriano
tanto no garfo quanto na planta. Os resultados indicam que no caso de Valência a
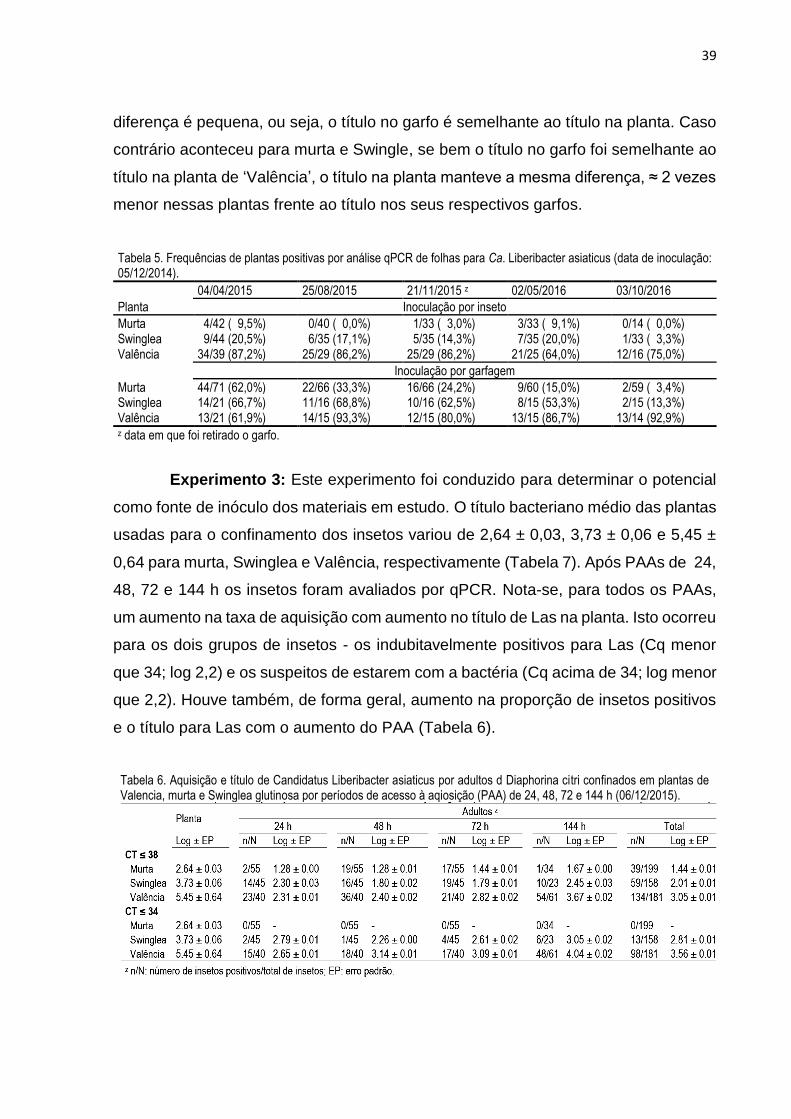
39
diferença é pequena, ou seja, o título no garfo é semelhante ao título na planta. Caso
contrário aconteceu para murta e Swingle, se bem o título no garfo foi semelhante ao
título na planta de ‘Valência’, o título na planta manteve a mesma diferença, ≈ 2 vezes
menor nessas plantas frente ao título nos seus respectivos garfos.
Tabela 5. Frequências de plantas positivas por análise qPCR de folhas para Ca. Liberibacter asiaticus (data de inoculação: 05/12/2014).
04/04/2015 25/08/2015 21/11/2015 z 02/05/2016 03/10/2016
Planta Inoculação por inseto
Murta 4/42 ( 9,5%) 0/40 ( 0,0%) 1/33 ( 3,0%) 3/33 ( 9,1%) 0/14 ( 0,0%) Swinglea 9/44 (20,5%) 6/35 (17,1%) 5/35 (14,3%) 7/35 (20,0%) 1/33 ( 3,3%) Valência 34/39 (87,2%) 25/29 (86,2%) 25/29 (86,2%) 21/25 (64,0%) 12/16 (75,0%)
Inoculação por garfagem
Murta 44/71 (62,0%) 22/66 (33,3%) 16/66 (24,2%) 9/60 (15,0%) 2/59 ( 3,4%) Swinglea 14/21 (66,7%) 11/16 (68,8%) 10/16 (62,5%) 8/15 (53,3%) 2/15 (13,3%) Valência 13/21 (61,9%) 14/15 (93,3%) 12/15 (80,0%) 13/15 (86,7%) 13/14 (92,9%) z data em que foi retirado o garfo.
Experimento 3: Este experimento foi conduzido para determinar o potencial
como fonte de inóculo dos materiais em estudo. O título bacteriano médio das plantas
usadas para o confinamento dos insetos variou de 2,64 ± 0,03, 3,73 ± 0,06 e 5,45 ±
0,64 para murta, Swinglea e Valência, respectivamente (Tabela 7). Após PAAs de 24,
48, 72 e 144 h os insetos foram avaliados por qPCR. Nota-se, para todos os PAAs,
um aumento na taxa de aquisição com aumento no título de Las na planta. Isto ocorreu
para os dois grupos de insetos - os indubitavelmente positivos para Las (Cq menor
que 34; log 2,2) e os suspeitos de estarem com a bactéria (Cq acima de 34; log menor
que 2,2). Houve também, de forma geral, aumento na proporção de insetos positivos
e o título para Las com o aumento do PAA (Tabela 6).
Tabela 6. Aquisição e título de Candidatus Liberibacter asiaticus por adultos d Diaphorina citri confinados em plantas de Valencia, murta e Swinglea glutinosa por períodos de acesso à aqiosição (PAA) de 24, 48, 72 e 144 h (06/12/2015).

40
Em murta CLas-positiva, nenhum dos insetos que se alimentaram em seus
tecidos, mesmo por período de 144 h, conseguiram adquirir CLas (CT ≤ 34) (Tabela
6). Quando comparadas aos insetos que se alimentaram de Valência (15/40, 18/40,
17/40, 48/61), de Swinglea, as proporções de insetos que adquiriram Las foram
sempre baixas (2/45, 1/45, 4/45, 6/24). As taxas de aquisição também estiveram
quase sempre associadas positivamente aos títulos de CLas nos insetos. Assim,
Valência apresentou entre 3,02 e 20,25 vezes (média final 8,04 vezes) maior potencial
de atuar como fonte de CLas para D. citri do que Swinglea glutinosa. Somente 2/8
plantas de Valência que receberam inseto de planta fonte de Swinglea em dezembro
de 2015 resultaram positivas 10 meses mais tarde (outubro de 2016) com título
bacteriano médio de 2,68 log de células de CLas por grama de folha, mas sem
expressar os sintomas característicos de HLB.
4. DISCUSSÃO
O objetivo deste trabalho foi comparar a importância relativa de S. glutinosa
como criadouro de D. citri e multiplicadora e fonte de inóculo de Ca. L. asiaticus, em
relação aos citros e à murta. S. glutinosa apresentou potencial de criar D. citri similar
ao das laranjas e superior ao dos limões em condições de telado. Porém, nas
laranjeiras doces foi constante a maior favorabilidade à reprodução deste inseto
quando comparadas aos limoeiros. Nelas a viabilidade de ovos e ninfas foi maior tanto
na sala climatizada como no telado.
O ambiente mais estável em termos de temperatura (sala climatizada)
favoreceu o desenvolvimento do inseto. O ciclo ovo-adulto foi mais curto, tendência
que já relatada por diversos autores (LIU; TSAI, 2000; NAVA et al., 2007, 2010). Outra
provável causa das diferenças entre ambientes do comportamento de D. citri nos
hospedeiros avaliados, especialmente na S. glutinosa, é a qualidade da fonte de luz
utilizada (lâmpadas fluorescentes e incandescentes com menor radiação
fotossintéticamente ativa (PAR) na sala, e radiação solar natural com maior PAR no
telado). Insetos sugadores como D. citri se veem diretamente influenciados pela fonte

41
de luz (PARIS et al., 2015), e indiretamente pelas alterações no fluxo da seiva,
condutância estomática, transpiração (COHEN et al., 1997; ORTUÑO et al., 2006), ou
bioquímica nas folhas e seiva do floema (DÁDER et al., 2014; HU; ZHAO; THIEME,
2013), todos decorrentes da qualidade da luz.
O fato da laranjeira ‘Folha Murcha’ e limoeiro ‘Limonera’ terem se destacado
como os hospedeiros onde o ciclo ovo-adulto foi maior, contrasta com o observado
em vários estudos, onde o hospedeiro não afetou este parâmetro (ALVES; DINIZ;
PARRA, 2014; NAVA et al., 2007; TSAI; LIU, 2000), mas corrobora outros. Na
tangerina ‘Cleópatra’ a duração do ciclo foi maior do que na laranja azeda
(TSAGKARAKIS; ROGERS, 2010). Quanto à preferência de D. citri por limoeiros,
Nehru, Bhagat e Koul (2004) encontraram que o limão ‘Rugoso’ (Citrus jambheri) e a
limeira ‘Kagzi’ (Citrus aurantifolia) foram menos favoráveis para o desenvolvimento do
psilídeo quando comparados com a laranja doce, e Patt e Sétamou (2010)
encontraram que os adultos responderam preferencialmente aos voláteis emanados
por brotos de Murraya paniculata e de limoeiro ‘Eureka’ (Citrus lemon) sobre aqueles
da laranjeira doce (Citrus sinensis). Porém, somente a presença dos adultos sobre os
brotos ou a oviposição nas diferentes espécies não são bons indicativos de
preferência. Estes parâmetros nem sempre se correlacionam com alimentação (YANG
et al., 2013), que pode estar mais relacionado a características morfológicas dos
tecidos que definem o comportamento alimentar (AMMAR et al., 2014; GEORGE et
al., 2017).
Associações entre viabilidade total e número de ovos ou ninfas (Tabela 3)
foram positivas, mas os coeficientes de determinação baixos. Isto pode ser o resultado
do peso que um pequeno número de insetos mortos tem sobre a viabilidade total
dentro de uma população com baixo número de indivíduos. Um indivíduo representa
5% em uma população de 20, mas apenas 1% numa de 100. Portanto, é plausível
considerar que, pelo menos dentro da faixa de tamanho populacional do inseto no
broto observada neste experimento (a qual deve ter minimizado qualquer competição
por sítios de alimentação), e com os brotos nas características especificadas, as
variações na população inicial não devem ter influenciado a viabilidade total. Há que
se considerar também o rápido crescimento do broto, com incremento em sua
superfície à medida que passa pelos vários estádios de desenvolvimento

42
(CIFUENTES-ARENAS et al., 2016; CIFUENTES-ARENAS; RAIOL JÚNIOR; LOPES,
2017). A metodologia empregada na estimação da sobrevivência dos indivíduos foi
portanto adequada, tornando desnecessária transferências de ninfas como conduzido
em outros estudos (LIU; TSAI, 2000).
O sucesso do uso da garfagem na inoculação de Las dependeu da
compatibilidade entre os materiais (fonte do garfo e planta sendo inoculada) e do
status sanitário do garfo, conforme demonstrado neste estudo. Garfos sadios
apresentaram maior sobrevivência que garfos doentes. A manifestação dos sintomas
de HLB, geralmente observada após meses de incubação, é precedida por uma série
de eventos e processos que começa logo após a infecção da bactéria. Existem
mudanças anatômicas acompanhadas por acúmulo de calose nos elementos crivados
do floema, obstruindo-os e desencadeando acúmulo de outras substâncias
(FOLIMONOVA; ACHOR, 2010). Isto pode ter influenciado a baixa porcentagem de
sucesso na garfagem em ‘Valência’. Por se tratar de garfos oriundos de plantas
doentes, a concentração bacteriana neles pode ter impedido a conexão e fluxo
adequado de seiva entre os dois tecidos.
A transmissão de Las varia muito dependendo do método de inoculação.
Neste estudo a porcentagem de plantas de ‘Valência’ que se infectaram por ambos os
métodos de inoculação foi maior do que a relatada por Lopes et al. (2009). No entanto,
o tempo de ocorrência e a intensidade de sintomas de HLB coincidiram nos dois
estudos, ou seja, quatro meses após inoculação algumas plantas já manifestavam
sintomas leves. A variação na frequência de infecção nos dois estudos pode ser
devido à variação no ambiente em que os estudos foram conduzidos e ao tamanho
do tecido sendo usado como inóculo, como demonstrado por Lopes e Frare (2008).
A mensuração da importância que têm hospedeiros alternativos tanto do
inseto vetor como da bactéria é de vital importância para o entendimento da
progressão do HLB no campo e para tomadas de decisão sobre o seu controle. A
transmissão e expressão de sintomas de HLB em murta obtidas nesta pesquisa
contradizem observações sobre a imunidade da murta feitas por Chen e Liao (1982),
e os resultados obtidos por Hungm, Wu e Su, (2000), ao inocular, por meio de
borbulhas oriundas de laranjeira ‘Valência’ doente, plantas de Murraya paniculata
(muito provavelmente sinônimo de M. exotica) e outras três espécies rutáceas não

43
cítricas. Segundo eles, a compatibilidade entre os materiais foi baixa. Para aqueles
bem-sucedidos não houve transmissão da bactéria. Alta incompatibilidade entre murta
e citros também foi relatada por Lopes et al. (2010).
Ao se comparar o título bacteriano entre murta, Swinglea e ‘Valência’ nota-se
evidente estratificação, com ‘Valência’ como a espécie de maior título, Swinglea
intermediária e a murta como a espécie com menor concentração bacteriana. A
inferioridade da murta como reservatório de Las é relatada na literatura, inicialmente
por Lopes et al (2010) e depois por Damsteegt et al. (2010) e Walter, Duan e Hall
(2012). Esta estratificação também parece refletir o nível de parentesco filogenético
entre as espécies vegetais e sugere que Las tenha co-evoluido com Citrus. Murraya
paniculata pertence a um grupo (cluster) distante de Citrus e Swinglea se encontra
entre os dois grupos (MORTON; GRANT; BLACKMORE, 2003).
O fato da frequência de plantas de Swinglea e murta positivas ir diminuindo
com tempo pode ser devido à inviabilidade de Las para sobreviver na planta por longos
períodos ou, o que pode ser mais provável, ao fato dela não conseguir acompanhar o
crescimento da planta acabando com ter um efeito de “diluição” da chance de
detecção da bactéria a medida que a planta cresce. A distribuição de Las e Lam em
plantas cítricas é bastante irregular especialmente na parte aérea (LOUZADA et al.,
2016; TEIXEIRA et al., 2008). Além disso, evidências existem de que a bactéria
acompanha passivamente o fluxo da seiva do floema seguindo sempre a sazonalidade
dos fluxos de crescimento tanto na parte aérea como na de raiz (RAIOL JÚNIOR,
2017).
Em nenhum outro estudo o potencial da Swinglea glutinosa como criatório de
D. citri e multiplicação de Las havia sido estudada no nível de detalhes aqui
apresentado e em relação a diversas variedade e espécies de citros. Demonstrou-se
que Las consegue se estabelecer nas plantas, mas em baixos títulos, e que a planta
recupera seu crescimento normal após alguns meses da infecção. Quanto a D. citri,
foi estabelecido que, caso se alimente em plantas infectadas podem adquirir a bactéria
e inocular mudas de laranja, porém em baixas proporções quando comparadas aos
citros. A baixa população de Las em Swinglea pode torná-la forte candidata a ser
explorada como porta-enxerto para citros. Há intensa busca por porta-enxerto
ananicante para copas de laranjeira, de tal forma a facilitar colheita e permitir

44
adensamento da cultura (FUNDECITRUS, 2017), e S. glutinosa tem esse potencial.
Há que determinar se algum componente da Swinglea responsável pela baixa
população bacteriana se transfere para a copa ou ao menos limite seu movimento
dentro da planta. Apesar do potencial de uso desta espécie como porta-enxerto,
recomenda-se a sua eliminação nos locais onde ocorre naturalmente, como é o caso
da Colômbia. Naquele país esta espécie é usada como cerca viva em propriedades
rurais, cidades e chácaras. Para evitar crescimento exagerado as plantas são ainda
submetidas a podas frequentes o que provoca constante oferta de brotos, ambiente
altamente favorável à alimentação e reprodução de D. citri. Por não ser ser submetida
a qualquer tratamento fitossanitário, esta espécie vegetal poderá vir a se constituir em
importantes reservatórios de psilídeo e, se infectadas, importantes fontes de inóculo
para as plantas cítricas.
5. REFERÊNCIAS
ALBRECHT, U.; BOWMAN, K. D. Transcriptional response of susceptible and tolerant citrus to infection with Candidatus Liberibacter asiaticus. Plant Science, v. 185–186, p. 118–130, 2012.
ALVES, G. R.; DINIZ, A.; PARRA, J. Biology of the huanglongbing vector Diaphorina citri (Hemiptera: Liviidae) on different host plants. Journal of Economic Entomology, v. 107, n. 2, p. 691–696, 2014.
AMMAR, E.-D. et al. Differences in stylet sheath occurrence and the fibrous ring (sclerenchyma) between xCitroncirus plants relatively resistant or susceptible to adults of the asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae). PLoS ONE, v. 9, n. 10, p. e110919, 2014.
BALDWIN, E. et al. Effect of Liberibacter infection (Huanglongbing disease) of citrus on orange fruit physiology and fruit/fruit juice quality: chemical and physical analyses. Journal of Agricultural and Food Chemistry, v. 58, n. 2, p. 1247–62, 2010.
BASSANEZI, R. B. et al. Yield loss caused by huanglongbing in different sweet orange cultivars in São Paulo, Brazil. European Journal of Plant Pathology, v. 130, n. 4, p. 577–586, 2011.
BOAVA, L. P.; CRISTOFANI-YALY, M.; MACHADO, M. A. Physiologic, anatomic, and gene expression changes in Citrus sunki, Poncirus trifoliata, and their hybrids after

45
“Candidatus Liberibacter asiaticus” infection. Phytopathology, v. 107, n. 5, p. 590–599, 2017.
BOSCARIOL-CAMARGO, R. L. et al. Avaliação de diferentes genótipos de citros à infecção por Candidatus Liberibacter asiaticus. Citrus Research & Technology, v. 31, n. 1, p. 85–90, 2010.
BOVÉ, J. M. Huanglongbing: a destructive, newly-emerging, century-old disease of citrus. Journal of Plant Pathology, v. 88, n. 1, p. 7–37, 2006.
BOVÉ, J. M. Huanglongbing or yellow shoot, a disease of Gondwanan origin: Will it destroy citrus worldwide? Phytoparasitica, v. 42, n. 5, p. 579–583, dez. 2014.
CANALE, M. C. et al. Latency and persistence of “Candidatus Liberibacter asiaticus” in its psyllid vector, Diaphorina citri (Hemiptera: Liviidae). Phytopathology, v. 107, n. 3, p. 264–272, 2017.
CEVALLOS-CEVALLOS, J. M. et al. GC–MS metabolomic differentiation of selected citrus varieties with different sensitivity to citrus huanglongbing. Plant Physiology and Biochemistry, v. 53, p. 69–76, 2012.
CHEN, X. Y.; LIAO, C. Q. Observation of biological characteristics of citrus psyllid, Diaphorina citri and its relationship with citrus huanglongbin (in Chinese). China Citrus, v. 4, p. 14–15, 1982.
CIFUENTES-ARENAS, J. C. et al. Monitoring citrus flush shoot ontogeny as a potential strategy for HLB and psyllid management. Journal of Citrus Pathology, v. 3, n. 1, p. 4, 2016.
CIFUENTES-ARENAS, J. C.; RAIOL JÚNIOR, L. L.; LOPES, S. A growing degree-day model for assessing flush shoot ontogeny in citrus. Journal of Citrus Pathology, v. 4, n. 1, p. 7, 2017.
COHEN, S. et al. Response of citrus trees to modified radiation regime in semi-arid conditions. Journal of Experimental Botany, v. 48, n. 1, p. 35–44, 1997.
COLETTA-FILHO, H. D. et al. First report of the causal agent of huanglongbing (“Candidatus Liberibacter asiaticus”) in Brazil. Plant Disease, v. 88, n. 12, p. 1382, dez. 2004.
DÁDER, B. et al. Impact of UV-A radiation on the performance of aphids and whiteflies and on the leaf chemistry of their host plants. Journal of Photochemistry and Photobiology B: Biology, v. 138, p. 307–316, set. 2014.
DAMSTEEGT, V. D. et al. Murraya paniculata and related species as potential hosts and inoculum reservoirs of “Candidatus Liberibacter asiaticus”, causal agent of huanglongbing. Plant Disease, v. 94, n. 5, p. 528–533, maio 2010.
DEL VALLE, N.; RODRÍGUEZ, R.; RODRÍGUEZ, K. Comportamiento del tangelo Orlando injertado sobre patrones tolerantes a la tristeza de los cítricos. Centro

46
Agrícola, v. 31, n. 3–4, p. 38–42, 2004.
EBRATT-RAVELO, E. E. et al. Diaphorina citri (Kuwayama , 1907) and Tamarixia radiata (Waterson, 1922) in citrus crops of Cundinamarca, Colombia. Agronomía Colombiana, v. 29, n. 3, p. 487–493, 2011.
FAN, J. et al. Comparative transcriptional and anatomical analyses of tolerant Rough lemon and susceptible Sweet orange, in response to “Candidatus Liberibacter, Asiaticus” infection. Molecular Plant-Microbe Interactions, v. 25, n. 11, p. 1396–1407, 2012.
FOLIMONOVA, S. Y.; ACHOR, D. S. Early events of citrus greening (Huanglongbing) disease development at the ultrastructural level. Phytopathology, v. 100, n. 9, p. 949–58, 2010.
FUNDECITRUS. Plantio e manejo diferenciados nas bordas ajudam a manter o psilídeo fora do pomar. Revista Citricultor, v. 8, n. 40, p. 8–11, 2017.
GEORGE, J. et al. Sclerenchymatous ring as a barrier to phloem feeding by Asian citrus psyllid: Evidence from electrical penetration graph and visualization of stylet pathways. PLoS ONE, v. 12, n. 3, p. e0173520, 2017.
GOTTWALD, T. R. Current epidemiological understanding of citrus Huanglongbing. Annual Review of Phytopathology, v. 48, p. 119–139, 2010.
HALL, D. G. et al. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomologia Experimentalis et Applicata, v. 146, n. 2, p. 207–223, 2013.
HU, Z.-Q.; ZHAO, H.-Y.; THIEME, T. The effects of enhanced ultraviolet-B radiation on the biology of green and brown morphs of Sitobion avenae (Hemiptera: Aphididae). Environmental Entomology, v. 42, n. 3, p. 578–585, 1 jun. 2013.
HUNG, T. H.; WU, M. L.; SU, H. J. Identification of alternative hosts of the fastidious bacterium causing citrus greening disease. Journal of Phytopathology, v. 148, n. 6, p. 321–326, 2000.
ICA. Resolución No. 00002390. Bogotá, Colombia: Instituto Colombiano Agropecuario, 2015. Disponível em: <http://www.ica.gov.co/getattachment/814e6823-aa26-49ee-a2c0-a814e2e61967/2015R2390.aspx>.
INOUE, H. et al. Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Annals of Applied Biology, v. 155, n. 1, p. 29–36, 2009.
JAGOUEIX, S.; BOVE, J.-M.; GARNIER, M. The phloem-limited bacterium of greening disease of citrus is a member of the α subdivision of the proteobacteria. International Journal of Systematic Bacteriology, v. 44, n. 3, p. 379–386, 1994.
KING, C. W. H. Dispersión de Diaphorina citri (Hemiptera: Psyllidae) en el departamento del Tolima (Colombia). Revista Tumbaga, v. 2, n. 7, p. 51–60, 2012.

47
LI, W.; HARTUNG, J. S.; LEVY, L. Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus huanglongbing. Journal of Microbiological Methods, v. 66, n. 1, p. 104–115, 2006.
LIU, Y. H.; TSAI, J. H. Effects of temperature on biology and life table parameters of the asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Annals of Applied Biology, v. 137, n. 3, p. 201–206, 2000.
LOPES, S. A. et al. Graft transmission efficiencies and multiplication of “Candidatus Liberibacter americanus” and “Ca. Liberibacter asiaticus” in citrus plants. Phytopathology, v. 99, n. 3, p. 301–306, 2009.
LOPES, S. A. et al. Liberibacters associated with orange jasmine in Brazil: incidence in urban areas and relatedness to citrus liberibacters. Plant Pathology, v. 59, n. 6, p. 1044–1053, 2010.
LOPES, S. A. et al. “Candidatus Liberibacter asiaticus” titers in citrus and acquisition rates by Diaphorina citri are decreased by higher temperature. Plant Disease, v. 97, n. 12, p. 1563–1570, 2013.
LOPES, S. A.; FRARE, G. F. Graft transmission and cultivar reaction of citrus to “Candidatus Liberibacter americanus”. Plant Disease, v. 92, n. 1, p. 21–24, 2008.
LOUZADA, E. S. et al. Distribution of “Candidatus Liberibacter asiaticus” above and below ground in Texas citrus. Phytopathology, v. 106, n. 7, p. 702–709, 2016.
MILES, G. P. et al. Apparent tolerance to huanglongbing in Citrus and Citrus-related germplasm. HortScience, v. 52, n. 1, p. 31–39, 2017.
MORTON, C. M.; GRANT, M.; BLACKMORE, S. Phylogenetic relationships of the Aurantioideae inferred from chloroplast DNA sequence data. American Journal of Botany, v. 90, n. 10, p. 1463–1469, 2003.
MURRAY, M. G.; THOMPSON, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research, v. 8, n. 19, p. 4321–4326, 1980.
NAVA, D. E. et al. Biology of Diaphorina citri (Hem., Psyllidae) on different hosts and at different temperatures. Journal of Applied Entomology, v. 131, n. 9–10, p. 709–715, 2007.
NAVA, D. E. et al. The effects of host, geographic origin, and gender on the thermal requirements of Diaphorina citri (Hemiptera: Psyllidae). Environmental Entomology, v. 39, n. 2, p. 678–684, 2010.
NEHRU, R. K.; BHAGAT, K. C.; KOUL, V. K. Influence of citrus species on the development of Diaphorina citri. Annals of Plant Protection Sciences, v. 12, n. 2, p. 425–438, 2004.
ORTUÑO, M. F. et al. Relationships between climatic variables and sap flow, stem water potential and maximum daily trunk shrinkage in lemon trees. Plant and Soil, v.

48
279, n. 1–2, p. 229–242, 2006.
PARIS, T. M. et al. Temporal response and attraction of Diaphorina citri to visual stimuli. Entomologia Experimentalis et Applicata, v. 155, n. 2, p. 137–147, 2015.
PATT, J. M.; SÉTAMOU, M. Responses of the asian citrus psyllid to volatiles emitted by the flushing shoots of its rutaceous host plants. Environmental Entomology, v. 39, n. 2, p. 618–24, 2010.
PELZ-STELINSKI, K. S. et al. Transmission parameters for Candidatus liberibacter asiaticus by Asian citrus psyllid (Hemiptera: Psyllidae). Journal of Economic Entomology, v. 103, n. 5, p. 1531–1541, 2010.
R CORE TEAM. R: A language and environment for statistical computingViena, AustriaR Fundation for Statistical Computing, , 2014. Disponível em: <http://www.r-project.org/>
RAIOL JÚNIOR, L. L. Translocação de Candidatus Liberibacter asiaticus em citros. [s.l.] Universidade Estadual Paulista “Júlio de Mesquita Filho”, 2017.
RAMADUGU, C. et al. Long-term field evaluation reveals Huanglongbing resistance in Citrus relatives. Plant Disease, v. 100, n. 9, p. 1858–1869, 2016.
RESTREPO, O. C.; TORRES, R. M.; SALAZAR, R. C. Comparación del “Tabog” (Swinglea glutinosa Merr) con el limón “Rugoso” (Citrus jambhiri Swingle) y la mandarina “Cleópatra” (C. reticulata Swingle) como porta-injerto para la lima ácida “Tahiti” (C. aurantifolia Swingle). Acta Agronómica, v. 37, n. 3, p. 43–50, 1987.
RONDÓN, A. et al. Reacción de patrones cítricos a Phytophthora nicotianae B. de Haan. var. parasitica (Dastur) Waterh, en condiciones de umbráculo. Agronomía Tropical, v. 43, n. 3–4, p. 117–125, 1993.
SCOTT, A. A. J.; KNOTT, M. A closter analysis method for grouping means in the analysis of variance. Biometrics, v. 30, n. 3, p. 507–512, 1974.
SHOKROLLAH, H. et al. Determination of the Presence of Huanglongbing in Seeds and Movement of the Pathogen in Citrus reticulata. American Journal of Applied Sciences, v. 6, n. 6, p. 1180–1185, 2009.
SWINGLE, W. T.; REECE, P. C. The botany of Citrus and its wild relatives. In: REUTHER, W.; WEBBER, H. J.; BATCHELOR, L. D. (Eds.). . The citrus industry. 2. ed. Berkeley, California: University of California, 1967. v. 1p. 190–430.
TEIXEIRA, D. C. et al. “Candidatus Liberibacter americanus”, associated with citrus huanglongbing (greening disease) in Sao Paulo State, Brazil. International Journal of Systematic and Evolutionary Microbiology, v. 55, n. 5, p. 1857–1862, 2005a.
TEIXEIRA, D. C. et al. Distribution and quantification of Candidatus Liberibacter americanus, agent of huanglongbing disease of citrus in São Paulo State, Brasil, in leaves of an affected sweet orange tree as determined by PCR. Molecular and

49
Cellular Probes, v. 22, n. 3, p. 139–150, jun. 2008.
TEIXEIRA, D. DO C. et al. Citrus huanglongbing in São Paulo State, Brazil: PCR detection of the “Candidatus” Liberibacter species associated with the disease. Molecular and Cellular Probes, v. 19, n. 3, p. 173–9, 2005b.
THAO, M. L. et al. Cospeciation of psyllids and their primary prokaryotic endosymbionts. Applied and Environmental Microbiology, v. 66, n. 7, p. 2898–2905, 2000.
TIRTAWIDJAJA, S. Insect, dodder, and seed transmissions of citrus vein phloem degeneration (CVPD). Proceedings of the 4th Internacional Citrus Congress. Anais...Tokyo: International Society of Citriculture, 1981
TSAGKARAKIS, A. E.; ROGERS, M. E. Suitability of “Cleopatra” mandarin as a host plant for Diaphorina citri (Hemiptera: Psyllidae). Florida Entomologist, v. 93, n. 3, p. 451–453, set. 2010.
TSAI, J. H.; LIU, Y. H. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. Journal of Economic Entomology, v. 93, n. 6, p. 1721–1725, 2000.
VENNING, F. D. Trials with Swinglea glutinosa (Blanco) Merr. as a rootstock for citrus. Proceedings of the Florida Horticultural Society. Anais...Florida: 1957
WALTER, A. J.; DUAN, Y.; HALL, D. G. Titers of “Ca. Liberibacter asiaticus” in Murraya paniculata and Murraya-reared Diaphorina citri Are Much Lower than in Citrus and Citrus-reared Psyllids. HortScience, v. 47, n. 10, p. 1449–1452, 2012.
WENNINGER, E. J.; HALL, D. G. Daily timing of mating and age at reproductive maturity in Diaphorina citri (Hemiptera: Psyllidae). Florida Entomologist, v. 90, n. 4, p. 715–722, 2007.
YANG, Y. et al. Influences of leaf age and type, non-host volatiles, and mineral oil deposits on the incidence, distribution, and form of stylet tracks of Diaphorina citri. Entomologia Experimentalis et Applicata, v. 147, n. 1, p. 33–49, 2013.

50
CHAPTER 3 – Citrus Flush Shoot Ontogeny Modulates Biotic Potential of
Diaphorina citri
ABSTRACT
ABSTRACT
The biology and behaviour of the psyllid Diaphorina citri Kuwayama (Hemiptera:
Sternorrhyncha: Liviidae), the major insect vector of bacteria associated with
huanglongbing, have been extensively studied with respect to host preferences,
thermal requirements, and responses to visual and chemical volatile stimuli. However,
development of the psyllid in relation to the ontogeny of immature citrus flush growth
has not been clearly defined or illustrated. Such information is important for
determining the timing and frequency of measures used to minimize populations of the
psyllid in orchards and spread of HLB. Our objective was to study how flush ontogeny
influences the biotic potential of the psyllid. We divided citrus flush growth into six
stages within four developmental phases: emergence (V1), development (V2 and V3),
maturation (V4 and V5), and dormancy (V6). Diaphorina citri oviposition and nymph
development were assessed on all flush stages in a temperature-controlled room, and
in a screen-house in which ambient temperatures varied. Our results show that biotic
potential of Diaphorina citri is not a matter of the size or the age of the flushes (days
after budbreak), but the developmental stage within its ontogeny. Females laid eggs
on flush V1 to V5 only, with the time needed to commence oviposition increasing with
the increasing in flush age. Stages V1, V2 and V3 were most suitable for oviposition,
nymph survival and development, and adult emergence, which showed evidence of
protandry. Flush shoots at emerging and developmental phases should be the focus
of any chemical or biological control strategy to reduce the biotic potential of D. citri, to
protect citrus tree from Liberibacter infection and to minimize HLB dissemination.
Kay words: Citrus sinensis, huanglongbing, Asian citrus psyllid, protandry.

51
1. INTRODUCTION
The Asiatic citrus psyllid Diaphorina citri Kuwayama (Hemiptera:
Sternorrhyncha: Liviidae) is the only known vector of the fastidious phloem restricted
α-Proteobacteria, 'Candidatus Liberibacter asiaticus' and 'Ca. L. americanus',
associated in Brazil with huanglongbing (HLB), the most devastating disease of citrus
(JAGOUEIX; BOVE; GARNIER, 1994; TEIXEIRA et al., 2005). It was first recorded in
Brazil in 1942 (COSTA LIMA, 1942) and subsequently spread throughout the main
citrus producing regions of the country (BELOTI et al., 2013) but did not become
economically important until after the first report of HLB in citrus orchards in São Paulo
State (SPS) in 2004 (COLETTA-FILHO et al., 2004). Planting of certified nursery citrus
trees produced in insect-proof screen-houses, elimination of symptomatic trees to
reduce inoculum sources, and monitoring and insecticide applications (BASSANEZI et
al., 2013; BELASQUE JÚNIOR et al., 2009, 2010) were subsequently implemented in
order to minimize the spread of the disease. Insecticide applications during the citrus
dormant seasons (late autumn and winter), when adult insects survive by feeding on
mature leaves, have also been recommended (MONZO; QURESHI; STANSLY, 2014;
QURESHI; KOSTYK; STANSLY, 2014; QURESHI; STANSLY, 2008, 2010; ROGERS,
2008). Despite these efforts the disease spread and 46,2 millions of trees had been
removed by 2017. Currently, the disease was estimated to be present in 16,73% of the
191.7 million trees in the citrus belt of SPS and Triângulo Mineiro region
(FUNDECITRUS, 2017a).
The difficulty in successfully managing HLB is related in part to the criteria
used to determine the most appropriate timing and frequency of insecticide
applications. In SPS these criteria are based mainly on the presence of adult psyllids
captured on yellow sticky traps (YST). However, the number of adults per YST may
not reflect the extent of psyllid populations (HALL, 2009), as they are influenced by
trap density and location within orchards, positioning within tree canopies, population
densities of the psyllid, weather conditions, and the nature and abundance of new flush
growth (HALL, 2009; HALL; HENTZ; CIOMPERLIK, 2007; HALL; SÉTAMOU; MIZELL,
2010; LEWIS-ROSENBLUM et al., 2015; MONZO et al., 2015; TOMASETO;

52
KRUGNER; LOPES, 2016; WU et al., 2015). Moreover, the impact of flush growth
ontogeny on populations of D. citri is not clearly understood.
In SPS, which has a temperate/subtropical climate, citrus trees usually
produce two main flushes of new growth annually in relatively well-defined cycles, one
related to plant growth in summer-autumn, and another related to flowering and fruiting
in spring. The flushes show sympodial growth as they pass through predicable and
recognizable stages of development (termed ontogeny) (BARTON; KORICHEVA,
2010) that are genetically and environmentally governed with temperature,
photoperiod, solar radiation and rainfall playing important roles (CHANG et al., 2014;
FENNER, 1998; HORVATH et al., 2003; MOSS, 1969, 1976; OLESEN; SMITH;
MULDOON, 2013; ORDUZ-RODRÍGUEZ; FISCHER, 2007).
Although eggs, nymphs or adults of D. citri can be found on reproductive
flushes, vegetative flushes are apparently the main sites for psyllid development. This
explains the strong association between vegetative flush phenology and the dynamics
of D. citri populations in citrus groves (CHAVAN; SUMMANWAR, 1993; PLUKE;
QURESHI; STANSLY, 2008; TSAI; WANG; LIU, 2002; YAMAMOTO; PAIVA;
GRAVENA, 2001), and the rate of spread of HLB (MONTEIRO, 2013). The psyllid is
attracted to flushes that offer optimal conditions for feeding and oviposition, especially
those at the very initial stage of development, by a combination of chemical volatiles
and visual stimuli (AMMAR; HALL; SHATTERS, 2013; KARLEY; DOUGLAS;
PARKER, 2002; PATT et al., 2014; PATT; SÉTAMOU, 2010; WENNINGER;
STELINSKI; HALL, 2009; YANG et al., 2013).
It has been known since Husain and Nath (HUSAIN; NATH, 1927) published
the first major study on D. citri in 1927 that oviposition occurs almost exclusively on
very young shoots. They wrote “the period of greatest activity of the insects and most
rapid increase in their numbers corresponds with that of the sprouting of new shoots
and the appearance of new leaves” (HUSAIN; NATH, 1927). Other authors have
reported that females lay their eggs in the growing tips of young host plants, preferring
flush growth < 6 mm in length (AZAM et al., 2013; CHEN; LIAO, 1982; LIN; KE; TAO,
1973; XU; XIA; KE, 1994) and that most eggs are laid within 14 days of the
commencing of new growth (LIN; KE; TAO, 1973). Initiation of oogenesis, and
subsequent maturation of eggs within ovaries, is closely related to the presence of

53
buds (DAI et al., 1982; HUANG, 1990; HUANG et al., 1999). ‘Feather flush’ has been
cited as the growth stage on which eggs are laid (CATLING, 1970; GRAFTON-
CARDWELL; STELINSKI; STANSLY, 2013; HALBERT; MANJUNATH, 2004; PLUKE;
QURESHI; STANSLY, 2008) but this term, incorrectly ascribed to Chavan and
Summanwar (1993) by Halbert and Manjunath (2004), appears to be the first of five
stages related to monitoring of the black citrus aphid Toxoptera citricida (Kirkaldy)
(Hemiptera: Sternorrhyncha: Aphididae) (MICHAUD; BROWNING, 1999) and a stage
of growth with ‘leaves still folded’.
Although an overall description of flush development is available in the
literature (HALL; ALBRIGO, 2007), no detailed phenotypical illustration of each stage
during shoot ontogeny exist, as well as the risk that each growth stage of shoots
represents to the development and increasing of D. citri population. The goal of this
study was (i) to define, clearly describe and illustrate the stages during the ontogeny
of flush growth in citrus, and (ii) to determine the relative importance of each stage on
insect oviposition, nymph survival and adult emergence.
2. MATERIAL AND METHODS
There was was carried out two experiments at the Plant Production
Department of the São Paulo State University, in Jaboticabal, SPS, Brazil (21.2522°
S, 48.3257° W), in two different environments: (i) a 3 × 3 × 3 m temperature-controlled
environmental room (CER) (average 26 ± 2 °C, 70 ± 5% RH, 12 h photoperiod) with
artificial light provided by 16 fluorescent bulbs (32 W each) and 5 incandescent lamps
(250 W each) yielding approximately 60 µmol m-2 s-1, and (ii) a 8 × 12 × 5 m screen-
house (SH) under ambient temperatures (average 25 ºC, max. 42ºC, min. 12ºC) and
relative humidity (average 53%, max. 93%, min. 15%). Data on temperature and
relative humidity were recorded with a HT-500 data logger (Instrutherm, São Paulo,
SP). The experiments were carried out from March to April 2014 (autumn) inside CER,
and from September to October 2014 (spring) inside SH.

54
Plant material. Two-year-old nursery trees of 'Valencia' sweet orange (Citrus
sinensis (L.) Osbeck.) grafted on 'Swingle' citrumelo (Citrus paradisi MacFaden ×
Poncirus trifoliata (L.) Raf. – based on (SWINGLE; REECE, 1967)) were maintained in
4.7 L plastic pots containing substrate composed of 80% Pinus sp. bark, 15%
vermiculite and 5% charcoal (Multiplant citrus®; Terra do Paraiso, Holambra, SP).
Before starting the experiments, the plants were kept in greenhouse irrigated four times
a week and ferti-irrigated biweekly with 100 mL pot-1 of a nutritive solution containing
Ca(NO3)2, MAP, MgSO4, Cu, Zn, Mn-EDTA, (NH4)2MoO4, and Fe-EDTA, at
concentrations of 1.35, 0.111, 0.4, 0.015, 0.01, 0.0075, 0.00045, 0.075 g L-1,
respectively.
Insect rearing. Fifteen to twenty D. citri adults of each sex from a colony on
healthy orange jasmine plants (Murraya exotica L.) at Fundecitrus (Araraquara, SP)
were confined for oviposition in sleeve cages for 5 days in the CER, on immature
growth flushes of previously uninfested orange jasmine plants, and then removed to
allow eggs laid to hatch and nymphs to develop to a new generation of adults. After
eclosion, the adults were separated daily and placed individually in 50 mL glass test
tubes for sexing (AUBERT, 1987). The insects were then transferred to new non-
infested orange jasmine plants, grouping by sex. Daily insect separation of emerging
adults ensured collection of unmated adults (WENNINGER; HALL, 2007). A manual
aspirator, made of a mesh-covered plastic Pasteur pipette inserted into a plastic tube,
was used to collect the adults, and a 30× magnification hand lens was used to
distinguish males from females. Given that adult emergence fits Gaussian curve, the
few adults that emerged during the first three days were discarded. Only those at the
top of the bell shape emergence growth curve (days 4 to 7) were used. These
procedures allowed the use of a relatively uniform group of insects that were further
maintained on orange jasmine in the CER for additional 15 days before use in the
experiments. This methodology was adapted from Skelley & Hoy (2004).
Ontogenesis of flush shoots. Previous observations of potted plants in
greenhouses and mature trees in orchards led us to divide the growth of citrus flush
shoots into six discrete stages of development. To describe the stages and their

55
durations, four healthy two-year-old potted plants of ‘Valência’ on ‘Swingle’ citrumelo
rootstocks were pruned 20 cm above the bud union and fully defoliated. When the 3rd
or 4th bud below the pruning site started swelling, its length was measured with a
Vernier calliper (Mitutoyo 530 Series, Suzano, SP, Brazil). Any subsequent new shoots
that appeared were detached (Figure 1A to C).
Figure 1. General schematic representation of treatments of the mounting process. (A) Intact plant. (B) Pruned and defoliated branch. (B) Newly developed shoots on the defoliated branch. (C) Selected shoot at the beginning of stage V2. (D) Adult Diaphorina citri individuals positioned on the defoliated branch with selected new shoot.
Influence of flush ontogeny on the biotic potential of Diaphorina citri. To
evaluate the influence of flush ontogeny on D. citri multiplication, 100 potted ‘Valencia’
plants were divided into five groups, each comprising 20 plants. The plants were then
sequentially pruned and defoliated weekly over a 5-week interval (Figure 1A to C).
Using a sleeve cage (Figure 1D), two unmated 15-day-old D. citri couples were
confined for mating and oviposition on each flush stage. In the CER the insects were
confined for 72 h. In the SH, the flushes were observed for the presence of eggs every
24 h and, when present, the adults remained confined for additional 48 h to allow 72 h

56
for oviposition. Eggs and nymphs were then counted, and emerging adults collected
daily in SH or every four days in CER. All adults were stored at -20 °C in 5 × 20 cm
transparent plastic bags for further sex identification using a stereo-microscope. Egg,
nymph, and total viability (Ve = [nymph/eggs]*100, Vn = [adults/nymphs] *100, and Vt =
[adults/eggs] *100, respectively), as well as sex ratio (SR = % female) and duration of
egg to adult cycle, were determined for both ambient, whereas time to oviposition and
synchrony of male/female emergence were determined only for SH. Egg to adult cycle
was estimated in replicate for each flush shoot, summarizing the period from the
second oviposition day (24 to 48 h) until the day previous to the appearance of the first
adult, plus the number of days between the appearance of first adult up to the
appearance of 50% of the adults emerged. This value was estimated through nonlinear
regression analysis of the data. All variables studied was used to determine the relative
weight of each shoot stage on the reproduction of the insect. This was proceeded using
a 0 to 1 scale, with 1 the most and 0 the less suitable stage.
Hypothetical scenarios. The model for the relative weight of the new shoot
on D. citri development was applied in two hypothetical situations involving the
presence of new shoots in two groves of Valencia sweet orange assessed in two
different dates. For both situations the existence of a heterogeneous population of flush
shoots (NSraw), equivalent to 100 new shoots tree-1, was assumed. For both groves
and assessing dates the composition of the new shoot population per tree, in terms of
Index of Relative Favourability for psyllid reproduction (IRF), was determined by the
equation 1:
.
𝐼𝑅𝐹 = ∑ 𝑁𝑆𝑉 × 𝑤𝑉
6
𝑉=1
....…………………….. Eq. (1)
where NSV is the number of new shoots in stages V1 to V6, and wV is the
relative weight of each new shoots in stage V1 to V6 (values from 0 to 1), determined
in the results section as the product of the relative importance of the shoot stage on
the percentage of shoots with eggs, time for the female to begin oviposition, number
of eggs and total survival.

57
Statistical analysis. To describe flush shoot elongation, several non-linear
regression analyses were carried out. The final model was chosen based on the Akaike
Information Criteria (AIC) (AKAIKE, 1998), for which the simplest and smaller model
that maximizes the goodness-of-fit should the best for explaining the data (HILBE,
2011). Prior to statistical analysis, data on number of eggs and the percentages were
log (y’ = Log10 (y + 1)) and arcsine (y’ = arcsine(sqrt(y/100))) transformed, respectively
(ZAR, 2010). The effect of the different flush stages on psyllid reproduction were
analysed using the general linear model (GLM) procedure. Each ambient was
analysed separately. When significant differences were detected, the means were
compared by Fisher’s Least Significance Difference (LSD) test. Sex ratio and the
percentage of oviposition were analysed by the Chi-square test, or by Fisher’s Exact
test, when the frequencies were less than 5 (ZAR, 2010). To evaluate the patterns of
adult emergence distribution, two procedures were applied: (i) a linear regression of
the quantiles (1, 5, 10, 25, 50, 75, 90, 95, 99) (for the female emergence as a function
of male emergence) (WENNINGER; HALL, 2007), and (ii) a Linear Trend test
complemented with analysis of the adjusted residuals (AGRESTI, 2002). Adjusted
residuals was interpreted as indicated by Haberman (HABERMAN, 1973), for which
absolute values greater than │1.96│ (P = 0.05) or │1.65│ (P = 0.10) indicate lack of
independence or that the observed frequency (Xij) is more variable than should be. The
Statgraphics Centurion XVII software (Statpoint Technologies Inc.) was used, with P <
0.05 for all analyses.
3. RESULTS
Flush shoot ontogenesis
Flush shoot elongation in the SH fitted a sigmoidal logistic curve (Figure 2)
spanning four developmental phases over an average 35 days. Phase 1 or
emergence comprised a single stage, V1 (Vegetative 1), commenced with bud
swelling followed by opening of the protective scales (Figure 3 A and B). In this phase,
elongation rates reached a maximum of ≈ 2.5 mm day-1 over an interval of 3 days
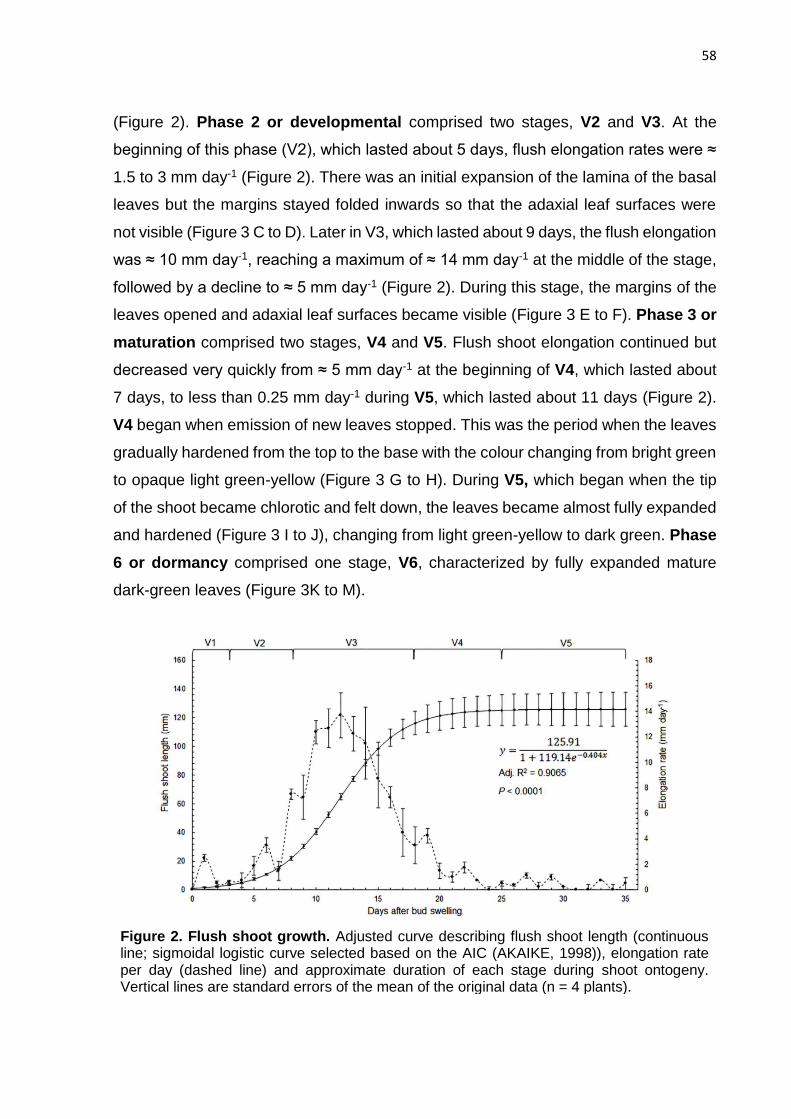
58
(Figure 2). Phase 2 or developmental comprised two stages, V2 and V3. At the
beginning of this phase (V2), which lasted about 5 days, flush elongation rates were ≈
1.5 to 3 mm day-1 (Figure 2). There was an initial expansion of the lamina of the basal
leaves but the margins stayed folded inwards so that the adaxial leaf surfaces were
not visible (Figure 3 C to D). Later in V3, which lasted about 9 days, the flush elongation
was ≈ 10 mm day-1, reaching a maximum of ≈ 14 mm day-1 at the middle of the stage,
followed by a decline to ≈ 5 mm day-1 (Figure 2). During this stage, the margins of the
leaves opened and adaxial leaf surfaces became visible (Figure 3 E to F). Phase 3 or
maturation comprised two stages, V4 and V5. Flush shoot elongation continued but
decreased very quickly from ≈ 5 mm day-1 at the beginning of V4, which lasted about
7 days, to less than 0.25 mm day-1 during V5, which lasted about 11 days (Figure 2).
V4 began when emission of new leaves stopped. This was the period when the leaves
gradually hardened from the top to the base with the colour changing from bright green
to opaque light green-yellow (Figure 3 G to H). During V5, which began when the tip
of the shoot became chlorotic and felt down, the leaves became almost fully expanded
and hardened (Figure 3 I to J), changing from light green-yellow to dark green. Phase
6 or dormancy comprised one stage, V6, characterized by fully expanded mature
dark-green leaves (Figure 3K to M).
Figure 2. Flush shoot growth. Adjusted curve describing flush shoot length (continuous line; sigmoidal logistic curve selected based on the AIC (AKAIKE, 1998)), elongation rate per day (dashed line) and approximate duration of each stage during shoot ontogeny. Vertical lines are standard errors of the mean of the original data (n = 4 plants).

59
Figure 3. Ontogeny of flush shoot of ‘Valencia’ sweet orange plants.
Stage V1: from bud swelling (A) to shoot emergence (B) (white square bracket indicates flush stem with less than 2 mm length). Stage V2: from initial stem elongation to clear separation between basal petioles and leaves (C) with all leaves remaining closed (D). Stage V3: from initial leaf blade expansion and separation from the shoot axis (E) and opening of the margins of lower leaves (F) until new the emission of leaves stops. Stage V4: from the unfolding of all leaf (G) and definition of final leaf number (H) to shoot tip chlorosis. Stage V5: from the time all leaves are fully expanded, green-light yellow coloured, and gradually hardened from the top to the base (I) to shoot tip chlorosis and/or abscission (J). Stage V6: the vegetative shoot is fully mature with the leaves hardened and green-dark (K) and the presence of dormant buds (L). A comparison of a fully expanded, partially hardened and green-light-yellow leaf from a V5 shoot with one fully expanded, hardened and final green-dark leaf from a V6 flush is also shown (M). For relative sizes and duration of each stage see Figure 2.
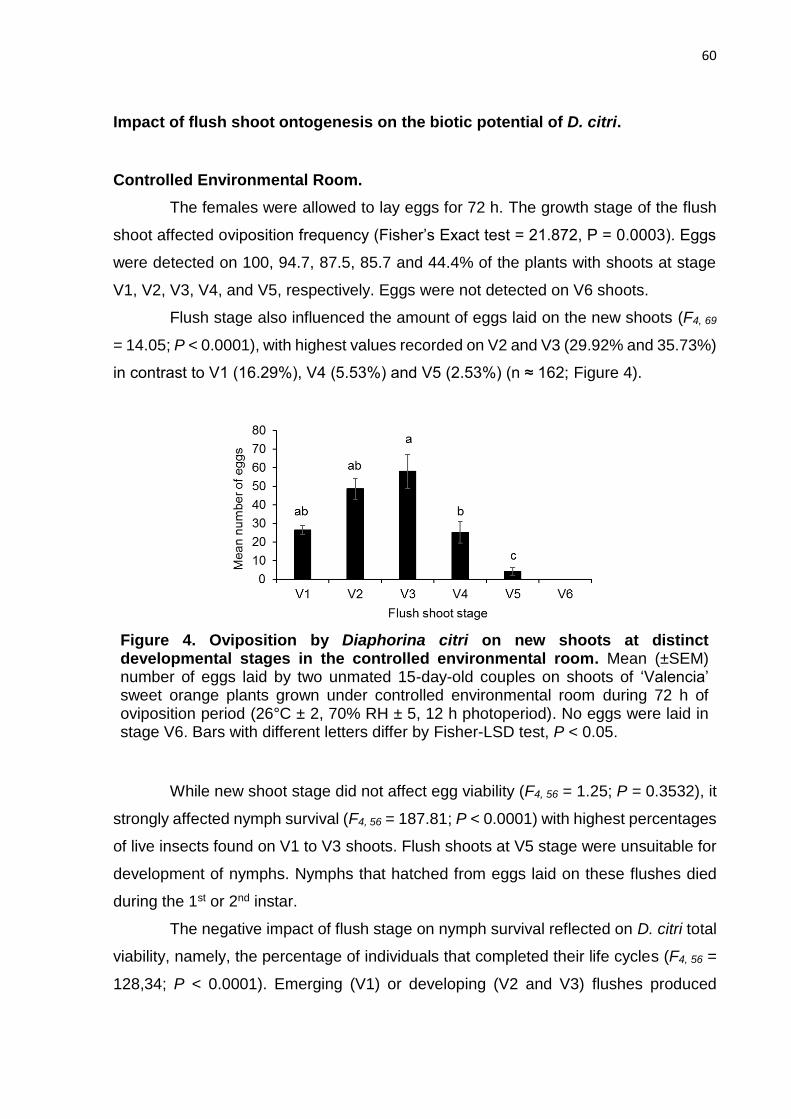
60
Impact of flush shoot ontogenesis on the biotic potential of D. citri.
Controlled Environmental Room.
The females were allowed to lay eggs for 72 h. The growth stage of the flush
shoot affected oviposition frequency (Fisher’s Exact test = 21.872, P = 0.0003). Eggs
were detected on 100, 94.7, 87.5, 85.7 and 44.4% of the plants with shoots at stage
V1, V2, V3, V4, and V5, respectively. Eggs were not detected on V6 shoots.
Flush stage also influenced the amount of eggs laid on the new shoots (F4, 69
= 14.05; P < 0.0001), with highest values recorded on V2 and V3 (29.92% and 35.73%)
in contrast to V1 (16.29%), V4 (5.53%) and V5 (2.53%) (n ≈ 162; Figure 4).
Figure 4. Oviposition by Diaphorina citri on new shoots at distinct developmental stages in the controlled environmental room. Mean (±SEM) number of eggs laid by two unmated 15-day-old couples on shoots of ‘Valencia’ sweet orange plants grown under controlled environmental room during 72 h of oviposition period (26°C ± 2, 70% RH ± 5, 12 h photoperiod). No eggs were laid in stage V6. Bars with different letters differ by Fisher-LSD test, P < 0.05.
While new shoot stage did not affect egg viability (F4, 56 = 1.25; P = 0.3532), it
strongly affected nymph survival (F4, 56 = 187.81; P < 0.0001) with highest percentages
of live insects found on V1 to V3 shoots. Flush shoots at V5 stage were unsuitable for
development of nymphs. Nymphs that hatched from eggs laid on these flushes died
during the 1st or 2nd instar.
The negative impact of flush stage on nymph survival reflected on D. citri total
viability, namely, the percentage of individuals that completed their life cycles (F4, 56 =
128,34; P < 0.0001). Emerging (V1) or developing (V2 and V3) flushes produced
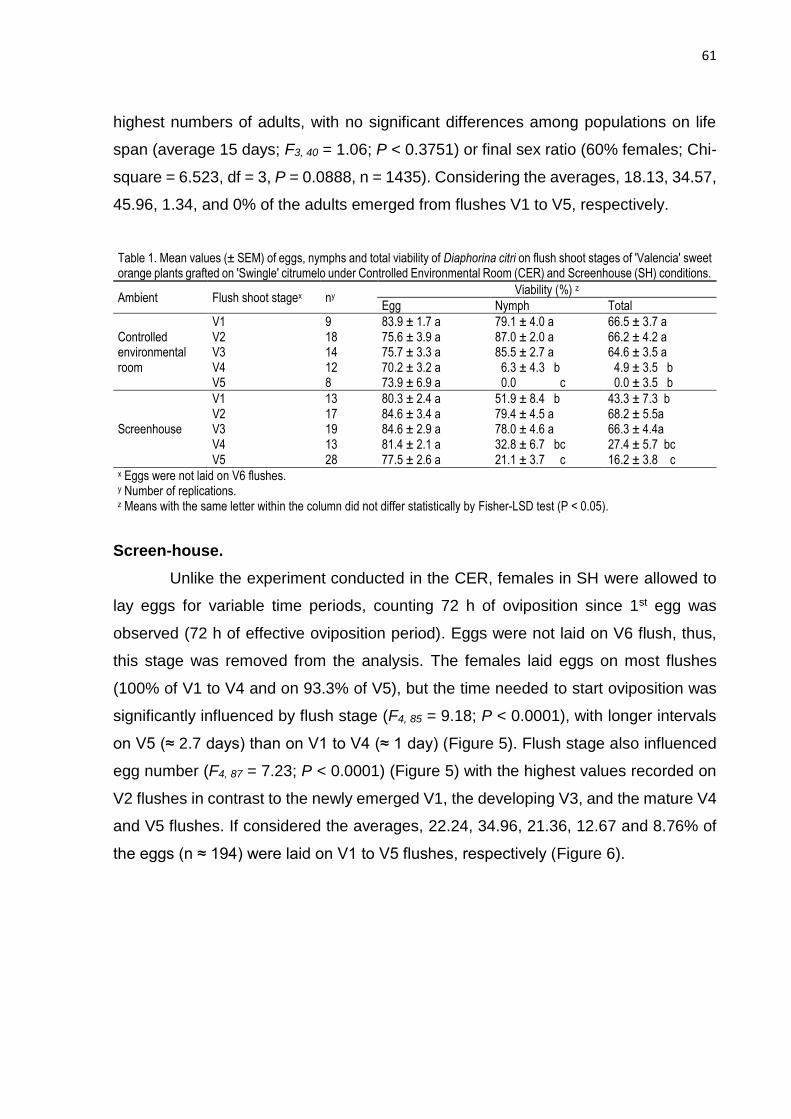
61
highest numbers of adults, with no significant differences among populations on life
span (average 15 days; F3, 40 = 1.06; P < 0.3751) or final sex ratio (60% females; Chi-
square = 6.523, df = 3, P = 0.0888, n = 1435). Considering the averages, 18.13, 34.57,
45.96, 1.34, and 0% of the adults emerged from flushes V1 to V5, respectively.
Table 1. Mean values (± SEM) of eggs, nymphs and total viability of Diaphorina citri on flush shoot stages of 'Valencia' sweet orange plants grafted on 'Swingle' citrumelo under Controlled Environmental Room (CER) and Screenhouse (SH) conditions.
Ambient Flush shoot stagex ny Viability (%) z
Egg Nymph Total
Controlled environmental room
V1 9 83.9 ± 1.7 a 79.1 ± 4.0 a 66.5 ± 3.7 a V2 18 75.6 ± 3.9 a 87.0 ± 2.0 a 66.2 ± 4.2 a V3 14 75.7 ± 3.3 a 85.5 ± 2.7 a 64.6 ± 3.5 a V4 12 70.2 ± 3.2 a 86.3 ± 4.3 b 64.9 ± 3.5 b V5 8 73.9 ± 6.9 a 60.0 ± 2.7 c 60.0 ± 3.5 b
Screenhouse
V1 13 80.3 ± 2.4 a 51.9 ± 8.4 b 43.3 ± 7.3 b V2 17 84.6 ± 3.4 a 79.4 ± 4.5 a 68.2 ± 5.5a V3 19 84.6 ± 2.9 a 78.0 ± 4.6 a 66.3 ± 4.4a V4 13 81.4 ± 2.1 a 32.8 ± 6.7 bc 27.4 ± 5.7 bc V5 28 77.5 ± 2.6 a 21.1 ± 3.7 c 16.2 ± 3.8 c
x Eggs were not laid on V6 flushes. y Number of replications. z Means with the same letter within the column did not differ statistically by Fisher-LSD test (P < 0.05).
Screen-house.
Unlike the experiment conducted in the CER, females in SH were allowed to
lay eggs for variable time periods, counting 72 h of oviposition since 1st egg was
observed (72 h of effective oviposition period). Eggs were not laid on V6 flush, thus,
this stage was removed from the analysis. The females laid eggs on most flushes
(100% of V1 to V4 and on 93.3% of V5), but the time needed to start oviposition was
significantly influenced by flush stage (F4, 85 = 9.18; P < 0.0001), with longer intervals
on V5 (≈ 2.7 days) than on V1 to V4 (≈ 1 day) (Figure 5). Flush stage also influenced
egg number (F4, 87 = 7.23; P < 0.0001) (Figure 5) with the highest values recorded on
V2 flushes in contrast to the newly emerged V1, the developing V3, and the mature V4
and V5 flushes. If considered the averages, 22.24, 34.96, 21.36, 12.67 and 8.76% of
the eggs (n ≈ 194) were laid on V1 to V5 flushes, respectively (Figure 6).
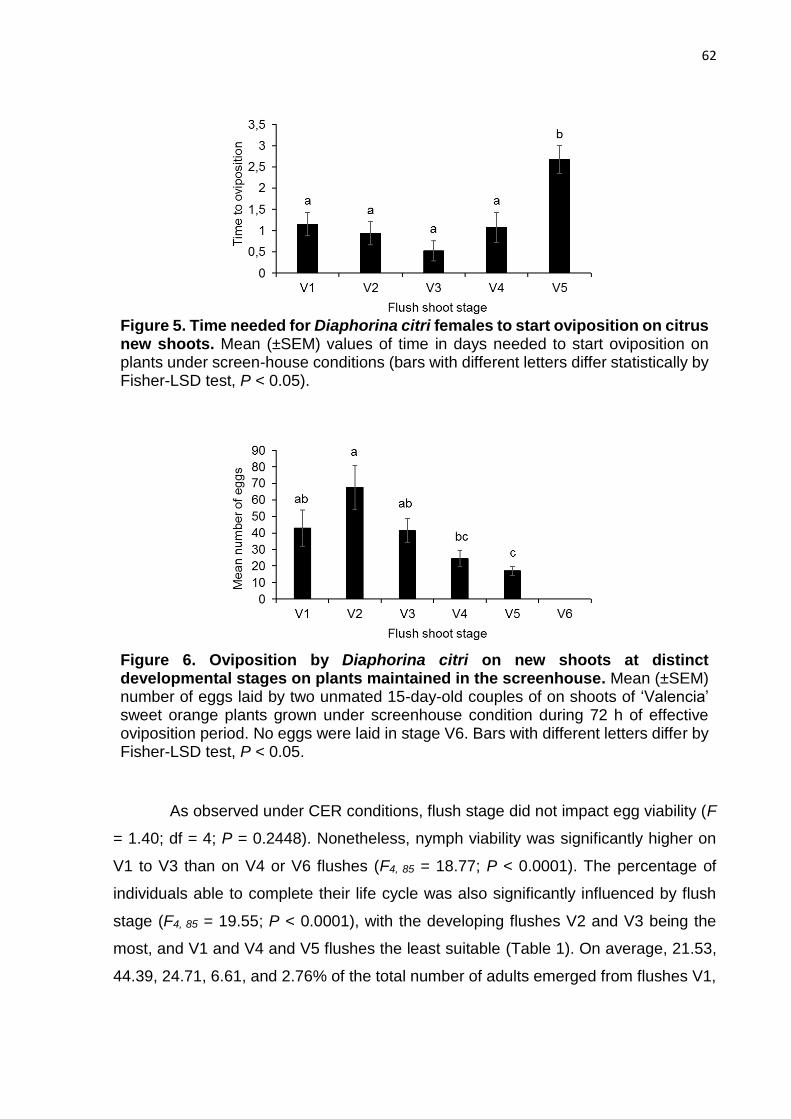
62
Figure 5. Time needed for Diaphorina citri females to start oviposition on citrus new shoots. Mean (±SEM) values of time in days needed to start oviposition on plants under screen-house conditions (bars with different letters differ statistically by Fisher-LSD test, P < 0.05).
Figure 6. Oviposition by Diaphorina citri on new shoots at distinct developmental stages on plants maintained in the screenhouse. Mean (±SEM) number of eggs laid by two unmated 15-day-old couples of on shoots of ‘Valencia’ sweet orange plants grown under screenhouse condition during 72 h of effective oviposition period. No eggs were laid in stage V6. Bars with different letters differ by Fisher-LSD test, P < 0.05.
As observed under CER conditions, flush stage did not impact egg viability (F
= 1.40; df = 4; P = 0.2448). Nonetheless, nymph viability was significantly higher on
V1 to V3 than on V4 or V6 flushes (F4, 85 = 18.77; P < 0.0001). The percentage of
individuals able to complete their life cycle was also significantly influenced by flush
stage (F4, 85 = 19.55; P < 0.0001), with the developing flushes V2 and V3 being the
most, and V1 and V4 and V5 flushes the least suitable (Table 1). On average, 21.53,
44.39, 24.71, 6.61, and 2.76% of the total number of adults emerged from flushes V1,
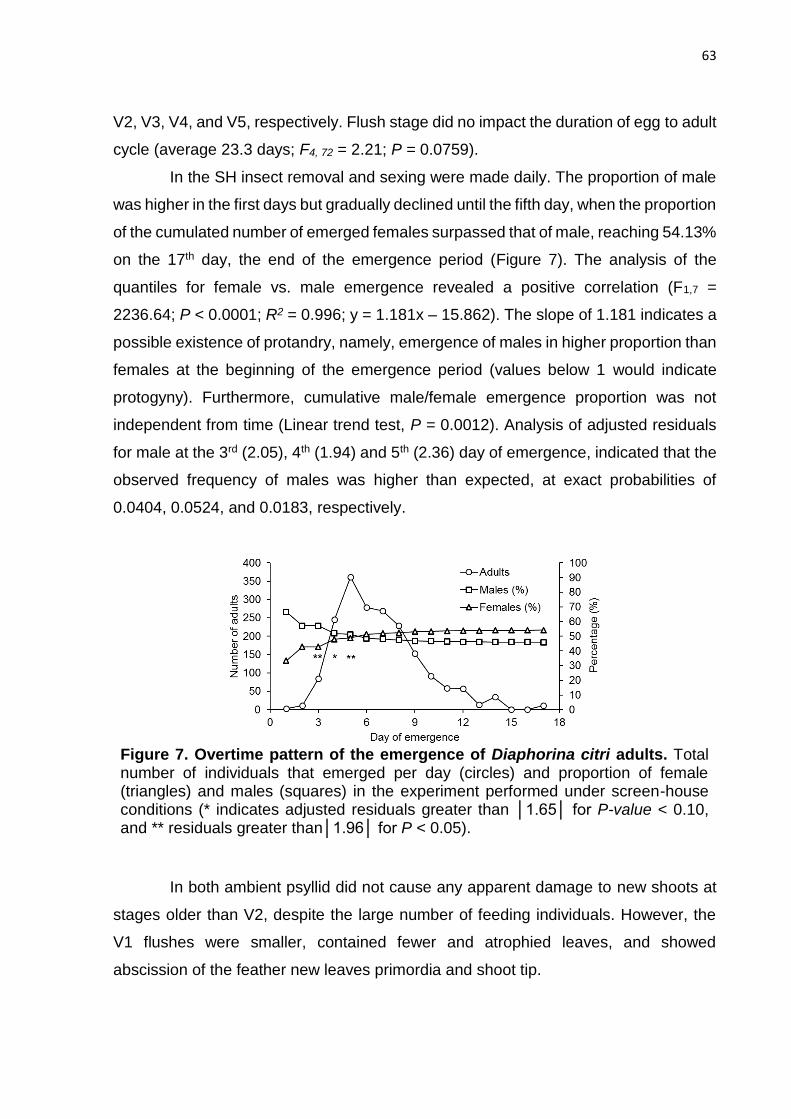
63
V2, V3, V4, and V5, respectively. Flush stage did no impact the duration of egg to adult
cycle (average 23.3 days; F4, 72 = 2.21; P = 0.0759).
In the SH insect removal and sexing were made daily. The proportion of male
was higher in the first days but gradually declined until the fifth day, when the proportion
of the cumulated number of emerged females surpassed that of male, reaching 54.13%
on the 17th day, the end of the emergence period (Figure 7). The analysis of the
quantiles for female vs. male emergence revealed a positive correlation (F1,7 =
2236.64; P < 0.0001; R2 = 0.996; y = 1.181x – 15.862). The slope of 1.181 indicates a
possible existence of protandry, namely, emergence of males in higher proportion than
females at the beginning of the emergence period (values below 1 would indicate
protogyny). Furthermore, cumulative male/female emergence proportion was not
independent from time (Linear trend test, P = 0.0012). Analysis of adjusted residuals
for male at the 3rd (2.05), 4th (1.94) and 5th (2.36) day of emergence, indicated that the
observed frequency of males was higher than expected, at exact probabilities of
0.0404, 0.0524, and 0.0183, respectively.
Figure 7. Overtime pattern of the emergence of Diaphorina citri adults. Total number of individuals that emerged per day (circles) and proportion of female (triangles) and males (squares) in the experiment performed under screen-house conditions (* indicates adjusted residuals greater than │1.65│ for P-value < 0.10, and ** residuals greater than│1.96│ for P < 0.05).
In both ambient psyllid did not cause any apparent damage to new shoots at
stages older than V2, despite the large number of feeding individuals. However, the
V1 flushes were smaller, contained fewer and atrophied leaves, and showed
abscission of the feather new leaves primordia and shoot tip.
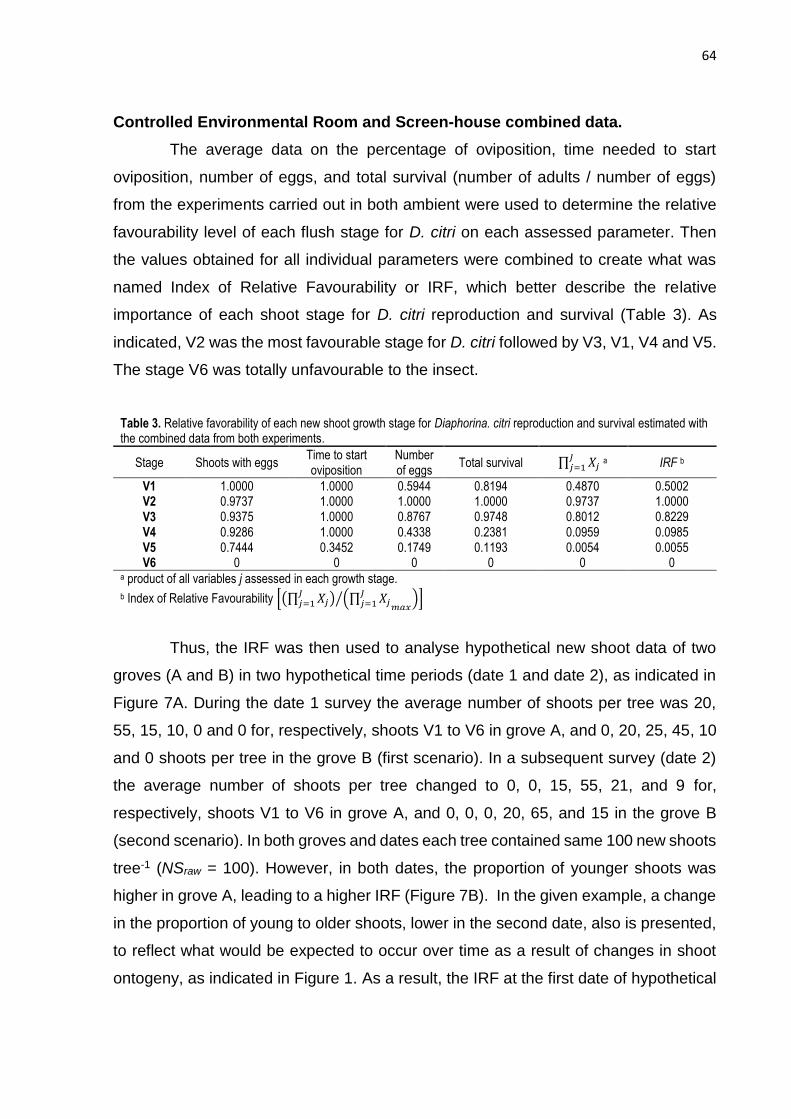
64
Controlled Environmental Room and Screen-house combined data.
The average data on the percentage of oviposition, time needed to start
oviposition, number of eggs, and total survival (number of adults / number of eggs)
from the experiments carried out in both ambient were used to determine the relative
favourability level of each flush stage for D. citri on each assessed parameter. Then
the values obtained for all individual parameters were combined to create what was
named Index of Relative Favourability or IRF, which better describe the relative
importance of each shoot stage for D. citri reproduction and survival (Table 3). As
indicated, V2 was the most favourable stage for D. citri followed by V3, V1, V4 and V5.
The stage V6 was totally unfavourable to the insect.
Table 3. Relative favorability of each new shoot growth stage for Diaphorina. citri reproduction and survival estimated with the combined data from both experiments.
Stage Shoots with eggs Time to start oviposition
Number of eggs
Total survival ∏ 𝑋𝑗𝐽𝑗=1 a IRF b
V1 1.0000 1.0000 0.5944 0.8194 0.4870 0.5002 V2 0.9737 1.0000 1.0000 1.0000 0.9737 1.0000 V3 0.9375 1.0000 0.8767 0.9748 0.8012 0.8229 V4 0.9286 1.0000 0.4338 0.2381 0.0959 0.0985 V5 0.7444 0.3452 0.1749 0.1193 0.0054 0.0055 V6 0 0 0 0 0 0
a product of all variables j assessed in each growth stage.
b Index of Relative Favourability [(∏ 𝑋𝑗𝐽𝑗=1 ) (∏ 𝑋𝑗
𝐽𝑗=1 𝑚𝑎𝑥
)⁄ ]
Thus, the IRF was then used to analyse hypothetical new shoot data of two
groves (A and B) in two hypothetical time periods (date 1 and date 2), as indicated in
Figure 7A. During the date 1 survey the average number of shoots per tree was 20,
55, 15, 10, 0 and 0 for, respectively, shoots V1 to V6 in grove A, and 0, 20, 25, 45, 10
and 0 shoots per tree in the grove B (first scenario). In a subsequent survey (date 2)
the average number of shoots per tree changed to 0, 0, 15, 55, 21, and 9 for,
respectively, shoots V1 to V6 in grove A, and 0, 0, 0, 20, 65, and 15 in the grove B
(second scenario). In both groves and dates each tree contained same 100 new shoots
tree-1 (NSraw = 100). However, in both dates, the proportion of younger shoots was
higher in grove A, leading to a higher IRF (Figure 7B). In the given example, a change
in the proportion of young to older shoots, lower in the second date, also is presented,
to reflect what would be expected to occur over time as a result of changes in shoot
ontogeny, as indicated in Figure 1. As a result, the IRF at the first date of hypothetical
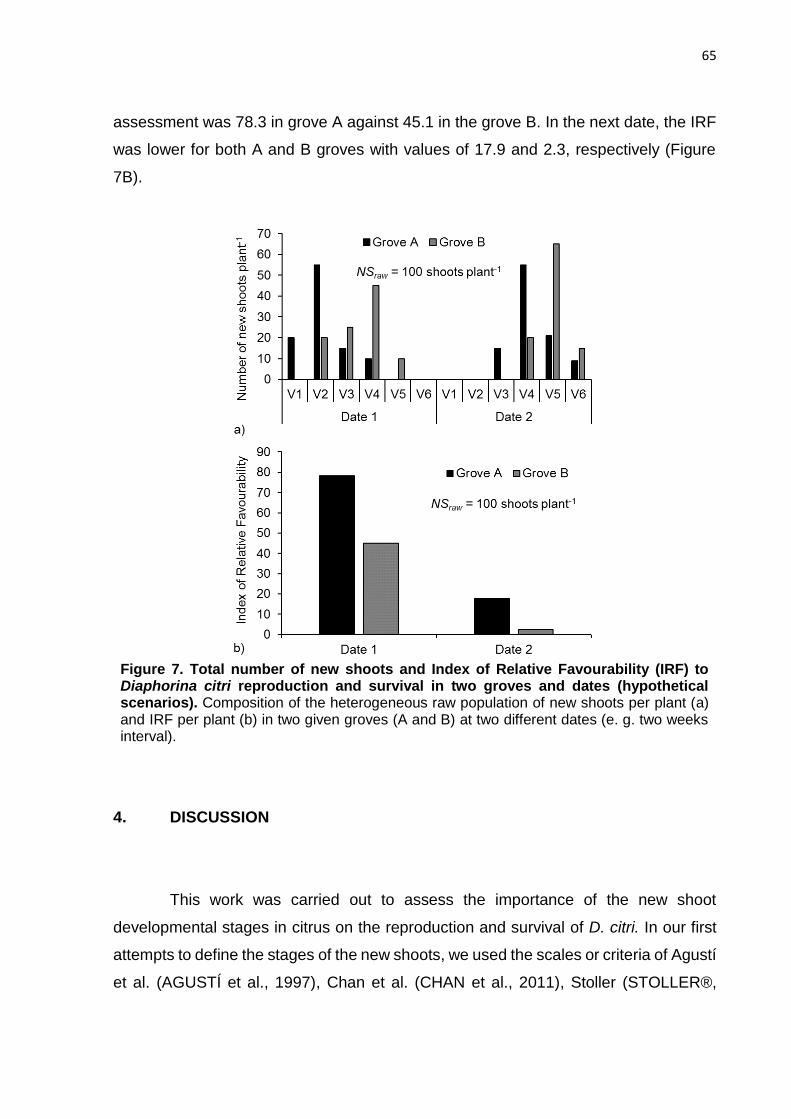
65
assessment was 78.3 in grove A against 45.1 in the grove B. In the next date, the IRF
was lower for both A and B groves with values of 17.9 and 2.3, respectively (Figure
7B).
Figure 7. Total number of new shoots and Index of Relative Favourability (IRF) to Diaphorina citri reproduction and survival in two groves and dates (hypothetical scenarios). Composition of the heterogeneous raw population of new shoots per plant (a) and IRF per plant (b) in two given groves (A and B) at two different dates (e. g. two weeks interval).
4. DISCUSSION
This work was carried out to assess the importance of the new shoot
developmental stages in citrus on the reproduction and survival of D. citri. In our first
attempts to define the stages of the new shoots, we used the scales or criteria of Agustí
et al. (AGUSTÍ et al., 1997), Chan et al. (CHAN et al., 2011), Stoller (STOLLER®,

66
2013), and Yang et al. (YANG et al., 2013), which were developed based on flush size
and colour, or leaf expansion. The simultaneous occurrence of all those variables
limited their use. In greenhouses and orchards, shoots of distinct sizes but with similar
phenotypic characteristics and apparent similar levels of tissue softness, have
frequently been observed. This led us to develop a new scale composed of six stages
(V1-V6) of growth within four phases of flush development. It proved useful to
distinguish the stages not only in potted plants but also in young and adult trees
growing in the field (data not shown). The favourability of each stages to D. citri
reproduction was then determined.
Diaphorina citri females laid eggs on all shoot stages with the exception of
mature V6. Oviposition was higher on stages V2 and V3, and gradually declined as the
shoots matured. Flush ontogeny also influenced the time required for females to
commence oviposition, which took longer on harder tissues of stage V5 (≈3 days) than
on the younger tissues of V1 to V4 (about 1 day). Younger shoots are softer and
nutritionally richer (AMMAR; HALL; SHATTERS, 2013; AZAM et al., 2013;
GOLDSCHMIDT, 1976; SÉTAMOU et al., 2016) which may stimulate the females to
lay eggs on sites more suitable for the progeny to develop. Similar behaviour is
exhibited by Toxoptera citricida (MICHAUD; BROWNING, 1999), Trioza erytreae Del
Guercio (Triozidae) (MORAN; BUCHAN, 1975), and Phyllocnistis citrella (Lepidoptera:
Gracilariidae) (VERCHER et al., 2008).
Flush ontogeny also impacted nymph viability, which was lower on stages V4
to V5. The few live young nymphs observed on older tissues moved actively (which is
in opposition to the suggestions that younger nymphs walk less, apparently to reduce
the risk of dehydration (CHHETRY; GUPTA; TARA, 2012), in contrast to those found
on younger less lignified tissues. Since younger nymphs have shorter and weaker
stylets (AMMAR; HALL; SHATTERS, 2013), the relatively thicker leaf cuticles (HAUKE;
SCHREIBER, 1998) and larger and deeper vascular vessels, characteristic of mature
leaves (AMMAR et al., 2014; AMMAR; HALL; SHATTERS, 2013), may have affected
nymph probing and survival. Nutritional changes could be an additional factor. It is
already known that the mineral composition of citrus leaves varies from organogenesis
to maturation and senescence, with calcium remaining immobilized and nitrogen and
potassium moving from older to younger tissues (FISCHER, 2007; HAUKESFORD et

67
al., 2012). Nutritional variation in plant tissues may cause changes in tissue palatability
and phloem sap quality, and consequently affect the activity of chewing and sucking
insects (DOUGLAS, 2006; MATTSON, WILLIAM, J., 1980; MULDER; BAKEMA, 1956;
POSCHENRIEDER; TOLRÀ; BARCELÓ, 2006). For instance, changes in essential
amino acid composition changed the performance of Manduca sexta Linnaeus (CHEN
et al., 2005), and the sucking insects Bemisia tabaci Gennadius (CRAFTS-
BRANDNER, 2002; ISAACS; BYRNE; HENDRIX, 1998) and aphids (KARLEY;
DOUGLAS; PARKER, 2002). Changes in calcium supply also have shown to affect the
integrity of plant cell wall and tissue hardness, leading to decreased damages caused
by bacterial or fungal infections (NANAYAKKARA; UDDIN; DATNOFF, 2008;
SUGIMOTO et al., 2010) and damages caused by feeding insects. Increased rigidity
of leaf tissues in response to calcium supply caused attrition to the mouthparts of the
chewing insects Spodoptera exigua Hübner, Eldana saccharina Walker, and
Deroceras reticulatum Müller (GRIFFIN; HOGAN; SCHMIDT, 2015; KORTH et al.,
2006; KVEDARAS et al., 2009), and might impede D. citri stylet penetration. Non
conclusive association between physicochemical characteristics of the shoots and
psyllid populations in the field was recently reported (SÉTAMOU et al., 2016).
The early emergence of males in proportion higher than that of females at the
beginning of the emergence period (Figure 6), suggests protandry, a phenomenon that
represents a reproductive advantage for insects with high fecundity rates (ALVES;
DINIZ; PARRA, 2014), as seems to be the case of D. citri. Protandry had been
described in Cardiaspina densitexta Taylor (WHITE, 1970) and Cacopsylla pyri L.
(Psyllidae) (BURTS; FISHER, 1967) but not in D. citri (WENNINGER; HALL, 2007).
Regarding D. citri, in previous studies relatively low numbers of adults, as compared
to those in the present work, were observed, which may have influenced the results
and conclusions. The existence of protandry would benefit D. citri reproduction as
males require more time to reach sexual maturity (WENNINGER; HALL, 2007) and
have shorter life spans (NAVA et al., 2007; RUSSELL et al., 2014) than females.
Currently in SPS, effective measures to minimise the impact of HLB have been
based on removal of symptomatic trees and area-wide suppression of D. citri
populations (BASSANEZI et al., 2013). Vector suppression has benefited from
information generated through the “phytosanitary alert system”, which involves

68
fortnightly inspection of some 20,000 yellow sticky traps distributed within the major
citrus growing areas of the state (FUNDECITRUS, 2017b). The presence of new
shoots on trees is also monitored. Regional and coordinated applications of
insecticides start when numbers of adult psyllids per trap exceed set intervention
thresholds (< 0.5 psyllid per trap, varying with the region) and when the new shoots
indicates favourable conditions for rapid increase in psyllid populations. Yet, psyllid
records on YST would reflect a process that begun a few weeks before. The shoot
development criteria on which decisions are made are almost exclusively based on
shoot size, not on shoot ontogeny, which was shown here a better indicator of
favourability to insect reproduction and survival.
As shown in this study, shoot ontogeny provides a better evidence for
phenotypic characterization and duration of the flush stages that mostly influence D.
citri reproduction. Combined to an estimation of flush density using, for example, the
methodology proposed by Hall and Albrigo (2007), it may be possible to determine at
a given moment the potential risk for psyllid multiplication in a particular orchard, and
the best moment for psyllid control based on the proportion of most suitable flushes.
Published models to estimate favourability to psyllid reproduction consider mainly the
temperature (AURAMBOUT et al., 2009). Addition of the IFR of new shoots, as
described in this work, could improve the certainty of the model. As shown in Figure
2, in the ambient conditions the study was conducted, the period of time most suitable
to D. citri spanned around 20 days, from bud-break to the end of V3 flushes, when D.
citri chemical control practices should be intensified. Outside that period, when leaf
emission and expansion rate decrease quickly or when there are not new shoots, it
would be probably more reasonable to alternate insecticide or mineral oil applications
with massive release of bio control agents (HUANG et al., 2005; KEATHLEY;
STELINSKI; LAPOINTE, 2016; LEONG et al., 2012; PATT et al., 2015; QURESHI;
STANSLY, 2010; RAE et al., 2003, 2006; TANSEY et al., 2015; YANG et al., 2013).
This work focused on the impact of shoot ontogeny on D. citri reproduction and
survival. As D. citri, other insects that prefer younger tissues for feeding and
reproduction also affect citrus, directly as parasite or indirectly as a vector of citrus
diseases. This include aphids, sharpshooters, and leaf miner, which, as D. citri, require
insecticide applications to reduce insect populations. We envisage that the control of

69
these insects may benefit from the information generated in this work. Furthermore,
since a citrus orchard should be seen as a heterogeneous population of flush shoots,
D. citri control also could be benefited from the establishment of a ‘reproduction’ or
‘potential risk’ threshold value based on IFR at a given moment. It is understandable
that direct extrapolation of the growth stages or the values of wv for other varieties of
sweet oranges are not completely recommendable, but it is plausible that processes
that define the transitions between growth stages (see Figure 3) are also present in
the development of vegetative shoots of these other citrus, e.g., shoot emergence,
initial elongation, leaf emission period, leaf expansion, shoot tip chlorosis and
abscission, and complete hardening of the leaves.
5. REFERENCES
AGRESTI, A. Categorical data analysis. 2. ed. Hoboken - NJ: Wiley, 2002.
AGUSTÍ, M. et al. Adaptation de l’échelle BBCH à la description des stades phénologiques des agrumes du genre Citrus. Fruits, Paris, v. 52, n. 5, p. 287–295, 1997.
AKAIKE, H. Information theory and an extension of the maximum likelihood principle. In: PARZEN, E.; TANABE, K.; KITAGAWA, G. (Eds.). . Selected Papers of Hirotugu Akaike. [s.l.] Springer New York, 1998. p. 199–213.
ALVES, G. R.; DINIZ, A.; PARRA, J. Biology of the huanglongbing vector Diaphorina citri (Hemiptera: Liviidae) on different host plants. Journal of Economic Entomology, v. 107, n. 2, p. 691–696, 2014.
AMMAR, E.-D. et al. Differences in stylet sheath occurrence and the fibrous ring (sclerenchyma) between xCitroncirus plants relatively resistant or susceptible to adults of the asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae). PLoS ONE, v. 9, n. 10, p. e110919, 2014.
AMMAR, E.-D.; HALL, D. G.; SHATTERS, R. G. Stylet morphometrics and citrus leaf vein structure in relation to feeding behavior of the asian citrus psyllid Diaphorina citri, vector of citrus huanglongbing bacterium. PLoS ONE, v. 8, n. 3, p. e59914, 2013.
AUBERT, B. Le psylle asiatique des agrumes (Diaphorina citri KUWAYAMA) au Brésil. Situation actuelle et perspectives de lutte. Fruits, Paris, v. 42, n. 4, p. 225–229, 1987.
AURAMBOUT, J. P. et al. A concept model to estimate the potential distribution of the

70
Asiatic citrus psyllid (Diaphorina citri Kuwayama) in Australia under climate change-A means for assessing biosecurity risk. Ecological Modelling, v. 220, n. 19, p. 2512–2524, 2009.
AZAM, M. et al. Citrus leaf volatiles as affected by developmental stage and genetic type. International Journal of Molecular Sciences, v. 14, n. 9, p. 17744–17766, 2013.
BARTON, K. E.; KORICHEVA, J. The Ontogeny of plant defense and herbivory: characterizing general patterns using meta-analysis. The American Naturalist, v. 175, n. 4, p. 481–493, 2010.
BASSANEZI, R. B. et al. Efficacy of area-wide inoculum reduction and vector control on temporal progress of Huanglongbing in young sweet orange plantings. Plant Disease, v. 97, n. 6, p. 789–796, 2013.
BELASQUE JÚNIOR, J. et al. Base científica para a erradicação de plantas sintomáticas e assintomáticas de Huanglongbing (HLB, Greening) visando o controle efetivo da doença. Tropical Plant Pathology, v. 34, n. 3, p. 137–145, 2009.
BELASQUE JÚNIOR, J. et al. Controle do huanglongbing no estado de São Paulo, Brasil. Citrus Research & Technology, v. 31, n. 1, p. 53–64, 2010.
BELOTI, V. H. et al. Population dynamics of Diaphorina citri Kuwayama (Hemiptera: Liviidae) in orchards of “Valencia” orange, “Ponkan” mandarin and “Murcott” tangor trees. Florida Entomologist, v. 96, n. 1, p. 173–179, 2013.
BURTS, E. C.; FISHER, W. R. Mating behaviour, egg production and egg fertility in the pear psylla. Journal of Economic Entomology, v. 60, p. 1297–1300, 1967.
CATLING, H. D. Distribution of the psyllid vectors of citrus greening disease, with notes on the biology and bionomics of Diaphorina citri. FAO Plant Protection Bulletin, v. 18, n. 1, p. 8–15, 1970.
CHAN, S. L. T. et al. Influence of host plant species and flush growth stage on the Asian Citrus Psyllid, Diaphorina citri Kuwayama. American Journal of Agricultural and Biological Sciences, v. 6, n. 4, p. 536–543, 2011.
CHANG, Y. et al. Temperature effects on shoot growth and flowering of kumquat trees. Korean Journal of Horticultural Science and Technology, v. 32, n. 1, p. 1–9, 2014.
CHAVAN, V. M.; SUMMANWAR, A. S. Population dynamics and aspects of the biology of citrus psylla, Diaphorina citri Kuw., in Maharashtra. Proceedings of the 12th Conference of the IOCV. Anais...1993
CHEN, H. et al. Jasmonate-inducible plant enzymes degrade essential amino acids in the herbivore midgut. Proceedings of the National Academy of Sciences, v. 102, n. 52, p. 19237–19242, 2005.
CHEN, X.; LIAO, C. Studies on the biology of Diaphorina citri and its relationship with

71
HLB. China Citrus, v. 4, p. 4–17, 1982.
CHHETRY, M.; GUPTA, R.; TARA, J. S. Bionomics of Diaphorina citri Kuwayama (Hemiptera: Psyllidae), on Citrus sinensis in Jammy region of J & K state. Munis Entomology & Zoology, v. 7, n. 1, p. 304–308, 2012.
COLETTA-FILHO, H. D. et al. First report of the causal agent of huanglongbing (“Candidatus Liberibacter asiaticus”) in Brazil. Plant Disease, v. 88, n. 12, p. 1382, dez. 2004.
COSTA LIMA, A. M. Insetos do Brasil, Homoptera. [s.l.] Escola Nacional de Agronomia, 1942.
CRAFTS-BRANDNER, S. J. Plant nitrogen status rapidly alters amino acid metabolism and excretion in Bemisia tabaci. Journal of Insect Physiology, v. 48, p. 33–41, 2002.
DAI, Y. et al. Studies on the vector of citrus huanglongbing citrus psylla Diaphorina citri Kuw. China Citrus, v. 3, p. 1–2, 1982.
DOUGLAS, A. E. Phloem-sap feeding by animals: problems and solutions. Journal of Experimental Botany, v. 57, n. 4, p. 747–754, 2006.
FENNER, M. The phenology of growth and reproduction in plants. Perspectives in Plant Ecology, Evolution and Systematics, v. 1, n. 1, p. 78–91, 1998.
FISCHER, A. M. Nutrient remobilization during leaf senescence. In: GAN, S. (Ed.). . Senescence Processes in Plants. Ithaca, NY, USA: Blackwell Publishing, 2007. p. 87–107.
FUNDECITRUS. Mesmo estabilizada, insidência de greening permanece alta no cinturão citrícola de São Paulo e Triângulo/Sudoeste Mineiro. Araraquara - SP, Brazil: Fundecitrus, 2017a. Disponível em: <http://www.fundecitrus.com.br>.
FUNDECITRUS. Alerta Fitossanitário. Disponível em: <http://www.fundecitrus.com.br/alerta-fitossanitario>.
GOLDSCHMIDT, E. E. Endogenous growth substances of citrus tissues. HortScience, v. 11, n. 2, p. 95–99, 1976.
GRAFTON-CARDWELL, E. E.; STELINSKI, L. L.; STANSLY, P. A. Biology and management of asian citrus psyllid, vector of the huanglongbing pathogens. Annual Review of Entomology, v. 58, n. 1, p. 413–432, 2013.
GRIFFIN, M.; HOGAN, B.; SCHMIDT, O. Silicon reduces slug feeding on wheat seedlings. Journal of Pest Science, v. 88, p. 17–24, 2015.
HABERMAN, S. J. The analysis of residuals in cross-classified tables. Biometrics, v. 29, n. 1, p. 205, 1973.
HALBERT, S. E.; MANJUNATH, K. L. Asian citrus psyllids (Sternorrhyncha: Psyllidae)

72
and greening disease of citrus: A literature review and assessment of risk in Florida. Florida Entomologist, v. 87, n. 3, p. 330–353, 2004.
HALL, D. G. An Assessment of Yellow Sticky Card Traps as Indicators of the Abundance of Adult Diaphorina citri (Hemiptera: Psyllidae in citrus. Journal of Economic Entomology, v. 102, n. 1, p. 446–452, 2009.
HALL, D. G.; ALBRIGO, L. G. Estimating the relative abundance of flush shoots in citrus with implications on monitoring insects associated with flush. HortScience, v. 42, n. 2, p. 364–368, 2007.
HALL, D. G.; HENTZ, M. G.; CIOMPERLIK, M. A. A comparison of traps and stem tap sampling for monitoring adult asian citrus psyllid (Hemiptera: Psyllidae) in citrus. Florida Entomologist, v. 90, n. 2, p. 327–334, 2007.
HALL, D. G.; SÉTAMOU, M.; MIZELL, R. F. A comparison of sticky traps for monitoring Asian citrus psyllid (Diaphorina citri Kuwayama). Crop Protection, v. 29, n. 11, p. 1341–1346, 2010.
HAUKE, V.; SCHREIBER, L. Ontogenetic and seasonal development of wax composition and cuticular transpiration of ivy (Hedera helix L.) sun and shade leaves. Planta, v. 207, n. 1, p. 67–75, 1998.
HAUKESFORD, M. et al. Function of macronutrients. In: MARSCHNER, H. (Ed.). . Marschner’s Mineral Nutrition of Higher Plants. 3rd. ed. Waltham, MA, USA: Elsevier Academic Press, 2012. p. 125–189.
HILBE, J. M. Negative Binomial Regression. 2nd. ed. New York: Cambridge University Press, 2011.
HORVATH, D. P. et al. Knowing when to grow: signals regulating bud dormancy. Trends in Plant Science, v. 8, n. 11, p. 534–540, 2003.
HUANG, B. Citrus psyllid. In: WU, Z. F. et al. (Eds.). . Chinese Agricultural Encyclopedia, Insect Volume Insect. Beijing, China: Agriculture Press, 1990. p. 119–120.
HUANG, J. et al. Studies on Citrus Psylla, Diaphorina citri Kuwayama. Entomological Journal of East China, v. 81, p. 26–34, 1999.
HUANG, M. et al. Green Technology for Citrus Pest and Disease Control. Guangzhou, China: Guangdong Science & Technology Press, 2005.
HUSAIN, M. A.; NATH, D. The citrus psylla (Diaphorina citri, Kuw.) (Psyllidae: Homoptera). Memoirs of the Department of Agriculture India, v. 10, p. 1–27, 1927.
ISAACS, R.; BYRNE, D. N.; HENDRIX, D. L. Feeding rates and carbohydrate metabolism by Bemisia tabaci (Homoptera: Aleyrodidae) on different quality phloem saps. Physiological Entomology, v. 23, n. 3, p. 241–248, 1998.

73
JAGOUEIX, S.; BOVE, J.-M.; GARNIER, M. The phloem-limited bacterium of greening disease of citrus is a member of the α subdivision of the proteobacteria. International Journal of Systematic Bacteriology, v. 44, n. 3, p. 379–386, 1994.
KARLEY, A. . J.; DOUGLAS, A. E.; PARKER, W. E. Amino acid composition and nutritional quality of potato leaf phloem sap for aphids. The Journal of Experimental Biology, v. 205, n. 19, p. 3009–18, 2002.
KEATHLEY, C. P.; STELINSKI, L. L.; LAPOINTE, S. L. Investigating dormant-season application of pheromone in citrus to control overwintering and spring populations of Phyllocnistis citrella (Lepidoptera: Gracillariidae). Pest Management Science, v. 72, n. 7, p. 1405–1410, 2016.
KORTH, K. L. et al. Medicago truncatula mutants demonstrate the role of plant calcium oxalate crystals as an effective defense against chewing insects. Plant Physiology, v. 141, n. 1, p. 188–195, 2006.
KVEDARAS, O. L. et al. Influence of plant silicon and sugarcane cultivar on mandibular wear in the stalk borer Eldana saccharina. Agricultural and Forest Entomology, v. 11, p. 301–306, 2009.
LEONG, S. C. T. et al. Impacts of horticultural mineral oils and two insecticide practices on population fluctuation of Diaphorina citri and spread of huanglongbing in a citrus orchard in Sarawak. The Scientific World Journal, v. 2012, p. 1–7, 2012.
LEWIS-ROSENBLUM, H. et al. Seasonal movement patterns and long-range dispersal of Asian Citrus Psyllid in florida citrus. Journal of Economic Entomology, v. 18, n. 1, p. 3–10, 2015.
LIN, S.; KE, Y.; TAO, C. C. Bionomics observation and integrated control of citrus psylla, Diaphorina citri Kuwayama. Journal of the Horticultural Society of China, v. 19, n. 4, p. 234–242, 1973.
MATTSON, WILLIAM, J., J. Herbivory in relation to plant nitrogen content. Annual Review of Ecology and Systematics, v. 11, n. 1980, p. 119–161, 1980.
MICHAUD, J. P.; BROWNING, H. W. Seasonal abundance of the Brown Citrus Aphid, Toxoptera citricida, (Homoptera: Aphididae) and its natural enemies in Puerto Rico. Florida Entomologist, v. 82, n. 3, p. 424–447, 1999.
MONTEIRO, A. B. Efeito de um pomar sem manejo de Huanglongbing sobre a ocorrência de Diaphorina citri e incidência de plantas doentes em uma área vizinha com manejo da doença. Araraquara - SP, Brasil: Fundo de Defesa da Citricultura, 2013.
MONZO, C. et al. Sampling methods for detection and monitoring of the asian citrus psyllid (Hemiptera: Psyllidae). Environmental Entomology, v. 44, n. 3, p. 780–788, 2015.
MONZO, C.; QURESHI, J. A.; STANSLY, P. A. Insecticide sprays, natural enemy

74
assemblages and predation on Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Bulletin of Entomological Research, v. 104, n. 5, p. 576–585, 2014.
MORAN, V. C.; BUCHAN, P. R. Oviposition by the citrus psylla, Trioza erytreae (Homoptera: Psyllidae), in relation to leaf hardness. Entomologia Experimentalis et Applicata, v. 18, n. 1, p. 96–104, 1975.
MOSS, G. Influence of temperature and photoperiod on flower induction and inflorescence development in sweet orange (Citrus sinensis L. Osbeck). Journal of Horticultural Science, v. 44, n. 4, p. 311–320, 1969.
MOSS, G. Temperature effects on flower initiation in sweet orange (Citrus sinensis). Australian Journal of Agricultural Research, v. 27, n. 3, p. 399, 1976.
MULDER, E. G.; BAKEMA, K. Effect of the nitrogen, phosphorus, potassium and magnesium nutrition of potato plants on the content of free amino-acids and on the amino-acid composition of the protein of the tubers. Plant and Soil, v. 7, n. 2, p. 135–166, 1956.
NANAYAKKARA, U. N.; UDDIN, W.; DATNOFF, L. E. Effects of soil type, source of silicon, and rate of silicon source on the development of gray leaf spot of perennial ryegrass turf. Plant Disease, v. 92, n. 6, p. 870–877, 2008.
NAVA, D. E. et al. Biology of Diaphorina citri (Hem., Psyllidae) on different hosts and at different temperatures. Journal of Applied Entomology, v. 131, n. 9–10, p. 709–715, 2007.
OLESEN, T.; SMITH, G.; MULDOON, S. J. Flush development in Tahitian lime. Australian Journal of Botany, v. 61, n. 5, p. 358–364, 2013.
ORDUZ-RODRÍGUEZ, J. O.; FISCHER, G. Balance hídrico e influencia del estrés hídrico en la inducción y desarrollo floral de la mandarina “Arrayana” en el piedemonte llanero de Colombia. Agronomía Colombiana, v. 25, n. 2, p. 255–263, 2007.
PATT, J. et al. Innate and conditioned responses to chemosensory and visual cues in asian citrus psyllid, Diaphorina citri (Hemiptera: Liviidae), vector of huanglongbing pathogens. Insects, v. 5, n. 4, p. 921–941, 2014.
PATT, J. M. et al. Efficacy of an autodisseminator of an entomopathogenic fungus, Isaria fumosorosea, to suppress Asian citrus psyllid, Diaphorina citri, under greenhouse conditions. Biological Control, v. 88, p. 37–45, set. 2015.
PATT, J. M.; SÉTAMOU, M. Responses of the asian citrus psyllid to volatiles emitted by the flushing shoots of its rutaceous host plants. Environmental Entomology, v. 39, n. 2, p. 618–24, 2010.
PLUKE, R.; QURESHI, J.; STANSLY, P. Citrus flushing patterns, Diaphorina citri (Hemiptera: Psyllidae) populations and parasitism by Tamarixia radiata (Hymenoptera: Eulophidae) in Puerto Rico. Florida Entomologist, v. 91, n. 1, p. 36–42, 2008.

75
POSCHENRIEDER, C.; TOLRÀ, R.; BARCELÓ, J. Can metals defend plants against biotic stress? Trends in Plant Science, v. 11, n. 6, p. 288–95, 2006.
QURESHI, J. A.; KOSTYK, B. C.; STANSLY, P. A. Insecticidal suppression of asian citrus psyllid Diaphorina citri (Hemiptera: Liviidae) vector of huanglongbing pathogens. PLoS ONE, v. 9, n. 12, p. e112331, 2014.
QURESHI, J. A.; STANSLY, P. A. Rate, placement and timing of aldicarb applications to control Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), in oranges. Pest Management Science, v. 64, p. 1159–1169, 2008.
QURESHI, J. A.; STANSLY, P. A. Dormant season foliar sprays of broad-spectrum insecticides: An effective component of integrated management for Diaphorina citri (Hemiptera: Psyllidae) in citrus orchards. Crop Protection, v. 29, n. 8, p. 860–866, 2010.
RAE, D. et al. Use of Horticultural and Agricultural Mineral Oils in Citrus IPDM. Ha Noi, Việt Nam.: Ha Noi Agricultural Press, 2003.
RAE, D. et al. Mineral oils and their use: sustainable pest management and green agriculture. Guangzhou, Guangdong, People’s Republic of China.: Guangdong Science & Technology Press, 2006.
ROGERS, M. E. General Pest Management Considerations. Citrus Industry, v. 89, n. 3, 2008.
RUSSELL, D. N. et al. Host suitability of Citrus and Zanthoxylum</i> spp. for Leuronota fagarae Burckhardt and Diaphorina citri Kuwayama (Hemiptera: Psylloidea). Florida Entomologist, v. 97, n. 4, p. 1481–1492, 2014.
SÉTAMOU, M. et al. Quality Matters: Influences of Citrus Flush Physicochemical Characteristics on Population Dynamics of the Asian Citrus Psyllid (Hemiptera: Liviidae). PLoS ONE, v. 11, n. 12, p. e0168997, 28 dez. 2016.
SKELLEY, L. H.; HOY, M. A. A synchronous rearing method for the asian citrus psyllid and its parasitoids in quarantine. Biological Control, v. 29, n. 1, p. 14–23, 2004.
STOLLER®. Guia de fases de desenvolvimento - CitrosCampinas - SPStoller do Brasil, , 2013. Disponível em: <http://www.stoller.com.br/guias/guia-de-fases>
SUGIMOTO, T. et al. Field application of calcium to reduce Phytophthora Stem Root of soybean, and calcium distribution in plants. Plant Disease, v. 94, n. 7, p. 812–819, 2010.
SWINGLE, W. T.; REECE, P. C. The botany of Citrus and its wild relatives. In: REUTHER, W.; WEBBER, H. J.; BATCHELOR, L. D. (Eds.). . The citrus industry. 2. ed. Berkeley, California: University of California, 1967. v. 1p. 190–430.
TANSEY, J. A. et al. Costs and benefits of frequent low-volume applications of horticultural mineral oil for management of Asian citrus psyllid, Diaphorina citri

76
Kuwayama (Hemiptera: Psyllidae). Crop Protection, v. 76, p. 59–67, 2015.
TEIXEIRA, D. C. et al. First report of a huanglongbing-like disease of Citrus in Sao Paulo State, Brazil and association of a new Liberibacter species, “Candidatus Liberibacter americanus”, with the disease. Plant Disease, v. 89, n. 1, p. 107, 2005.
TOMASETO, A. F.; KRUGNER, R.; LOPES, J. R. S. Effect of plant barriers and citrus leaf age on dispersal of Diaphorina citri (Hemiptera: Liviidae). Journal of Applied Entomology, v. 140, n. 1–2, p. 91–102, 2016.
TSAI, J. H.; WANG, J.-J.; LIU, Y.-H. Seasonal abundance of the asian citrus psyllid, Diaphorina citri (Homoptera: Psyllidae) in Southern Florida. Florida Entomologist, v. 85, n. 3, p. 446–451, 2002.
VERCHER, R. et al. Factors influencing adult female oviposition in the citrus leafminer Phyllocnistis citrella. Agricultural and Forest Entomology, v. 10, n. 1, p. 45–51, 2008.
WENNINGER, E. J.; HALL, D. G. Daily timing of mating and age at reproductive maturity in Diaphorina citri (Hemiptera: Psyllidae). Florida Entomologist, v. 90, n. 4, p. 715–722, 2007.
WENNINGER, E. J.; STELINSKI, L. L.; HALL, D. G. Roles of olfactory cues, visual cues, and mating status in orientation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) to four different host plants. Environmental Entomology, v. 38, n. 1, p. 225–234, 2009.
WHITE, T. Some aspects of the life history, host selection, dispersal and ovipostion of adult Cardiaspina densitexta (Homoptera : Psyllidae). Australian Journal of Zoology, v. 18, n. 1, p. 105, 1970.
WU, F. et al. Movement of Diaphorina citri (Hemiptera: Liviidae) adults between huanglongbing-infected and healthy citrus. Florida Entomologist, v. 98, n. 2, p. 410–416, 2015.
XU, C. F.; XIA, Y. H.; KE, C. Study on the biology and control of the citrus psylla. Acta Phytophylacica Sinica, v. 21, n. 1, p. 53–56, 1994.
YAMAMOTO, P. T.; PAIVA, P. E. B.; GRAVENA, S. Flutuação populacional de Diaphorina citri Kuwayama (Hemiptera: Psyllidae) em pomares de citros na região Norte do Estado de São Paulo. Neotropical Entomology, v. 30, n. 1, p. 165–170, 2001.
YANG, Y. et al. Influences of leaf age and type, non-host volatiles, and mineral oil deposits on the incidence, distribution, and form of stylet tracks of Diaphorina citri. Entomologia Experimentalis et Applicata, v. 147, n. 1, p. 33–49, 2013.
ZAR, J. H. Biostatistical Analysis. 5th. ed. Upper Saddle River, New Jersey: Pearson, 2010.

77
CAPÍTULO 4 – Modelo de graus dia para estimação da ontogenia do broto de
laranjeira Valência e definição de intervalos de pulverização de inseticidas para
controle de Diaphorina citri
RESUMO
RESUMO. Os brotos vegetativos de citros vêm sendo alvo de intensa atenção por
serem os locais de alimentação e reprodução preferidos pelo inseto Diaphorina citri,
vetor das bactérias associadas ao huanglongbing, doença mais destrutiva dessa
cultura. Para o controle do inseto tem sido feitas pulverizações de inseticidas de forma
indiscriminada, com a frequência obedecendo calendários fixos, não se importando,
de forma geral, no momento da pulverização, nem com a presença do broto na planta
nem com o estádio de seu desenvolvimento, fatores que podem ser críticos para o
sucesso desta medida. Ao se desenvolver os brotos adicionam novos tecidos em sua
extremidade distal que, se não protegidos, servirão de locais de alimentação do inseto
vetor e porta de entrada de Liberibacter na planta. Dessa forma, entender como os
brotos se desenvolvem e quais os requerimentos térmicos para tal poderão ser de
grande valia para se estabelecer melhores estratégias de proteção contra o inseto e,
consequentemente, de infecções por Liberibacter. Portanto, o objetivo deste trabalho
foi (i) estimar o número de graus-dia necessário para o desenvolvimento (GDD) do
broto, desde o momento em que foi estimulado a surgir por meio da poda até a
emissão do fluxo seguinte, (ii) comparar crescimento em câmaras de crescimento e
campo, em condição de sequeiro e irrigado, e (iii) propor, baseado nos GDDs, um
modelo teórico de pulverizações. Para o experimento em câmaras de crescimento
foram usadas mudas de ‘Valência’/’Swingle’ de três anos de idade. Para o
experimento em campo foram usadas plantas de 3,5 anos de idade de
‘Valência’/’Swingle’, em área com e sem irrigação, estudando-se a brotação natural
depois de um período de chuvas. Desde o momento da poda até a emissão do fluxo
vegetativo seguinte foram necessários o acúmulo de aproximadamente 970,03 GDD,
sendo os estádios V4, V3 e V5 os mais demorados. Os brotos desenvolvidos na área
com irrigação foram maiores (tanto em comprimento quanto em área foliar) do que os
da área sem irrigação, o que influenciou os GDDs necessários. Os dados das câmaras
de crescimento foram mais similares aos do campo na área com irrigação. O modelo

78
teórico de frequência de pulverização permitiu visualizar o que pode ocorrer no campo
em termos de acúmulo de área foliar desprotegida do acesso de D. citri (ausência de
cobertura inseticida), em função de um crescimento mais ou menos rápido do broto e
de ocorrência de chuvas, por exemplo. Estes resultados poderão ser úteis na tomada
de decisão no controle do psilídeo e, consequentemente, no manejo do HLB.
Palavras-chave: huanglongbing, graus dia de desenvolvimento, frequência de
pulverizações, Citrus sinensis, Liberibacter.

79
CAPÍTULO 4 – Degree-day model for estimating flush shoot ontogeny in Valencia
sweet orange and scheduling insecticide sprays to control Diaphorina citri
ABSTRACT
ABSTRACT. Studies on vegetative shoots in citrus have gained attention because
they are the preferred sites for feeding and reproduction of Diaphorina citri, the vector
of bacteria associated with huanglongbing, the most destructive citrus disease.
Insecticide sprays to control of the vector have generally been made based on calendar
with fixed intervals between applications. Details of shoot appearance and growth,
important for the success of control, have not been taken into consideration. The rate
of shoot growth should be considered because, as it grows, new unprotected tissue is
added to the top of the shoot providing opportunities for the coming insect to feed and
eventually inoculated the HLB pathogen. Understanding shoot appearance and growth
and the thermal requirements for these processes is therefore important. The objective
of this work was then (i) to estimate the growing-degree days (GDD) required for the
new shoot to develop, from the pruning date until it fully matures, (ii) to compare shoot
growth and GDD requirements for shoots developed in growth chambers and field in
irrigated and non-irrigated conditions, and (iii) to propose a theoretical model of
scheduling sprays for D. citri based on GDD. For the experiment in growth chambers,
we used 3-year-old potted plants of ‘Valencia’/’Swingle’, and for the field experiment,
3,5-year-old plants of ‘Valencia’/’Swingle’. In growth chamber, we assessed new
shoots that developed on pruned branches and in the field new shoots that appeared
naturally after a short rainy season. Approximately 970,03 GDDs were needed for
shhot to develop from the moment of pruning to the emission of the following vegetative
flow. The developmental stages V4, V3 and V5 required the highest GDDs. In the field,
the shoots on the irrigated trees were larger (both in length and in leaf area) than those
in on the non-irrigated trees, which influenced the required GDD. GDDs from the
growth chambers were similar to those of the irrigated area. The theoretical model for
scheduling spray frequencies allowed to visualize what could happen in terms of
accumulation of new tissues unprotected from the access of D. citri (not covered by
insecticide) as a function of rate of shoot growth and rain occurrence, for example. The

80
model can help the grower to decide when and how frequent actions against D. citri
should be taken.
Key words: huanglongbing, day-degrees of development, frequency of spraying,
Citrus sinensis, Liberibacter.

81
1. INTRODUÇÃO
O crescimento das plantas obedece à atividade metabólica dos pontos de
crescimento (meristemas) e envolve intrincados processos biológicos que definem a
dinâmica das relações fonte-dreno (FOURCAUD et al., 2008). Esse crescimento pode
ser de dois tipos: indeterminado, caracterizado por crescimento contínuo e ausência
de morte e abscisão do ponto apical, como no caso de algumas variedades de tomate
ou de feijão (DAVIS; GARCIA, 1983; HURD; GAY; MOUNTIFIELD, 1979), ou do tipo
determinado no qual, após ativação do meristema, segue um período de crescimento
até a morte e abscisão do ponteiro, um processo que, nos citros, parece estar
governado por genes relacionados à morte celular programada (ZHANG et al., 2014).
Esse tipo de crescimento é repetitivo nos citros e faz com que as plantas emitam fluxos
vegetativos, reprodutivos ou mistos em eventos de brotação que delimitam ciclos mais
ou menos definidos em resposta principalmente a fatores ambientais, também
conhecido como crescimento simpodial (SPIEGEL-ROY; GOLDSCHMIDT, 1996;
SWINGLE; REECE, 1967). No grupo das laranjeiras esses ciclos são mais bem
definidos e mais afetados pela latitude, contrário do que ocorre no grupo dos limoeiros
e limeiras ácidas nas quais não ocorre separação tão definida com as brotações
ocorrendo em fluxos contínuos (SPIEGEL-ROY; GOLDSCHMIDT, 1996).
Localizados nas inserções das folhas com o ramo ou no final deste, os brotos
originam-se de duas as três gemas e crescem emitindo folhas novas até seu completo
amadurecimento (SPIEGEL-ROY; GOLDSCHMIDT, 1996). O início da brotação exige
condições favoráveis de temperatura e disponibilidade hídrica. As gemas podem se
diferenciar em brotos reprodutivos ou vegetativos dependendo do balanço hormonal
e do acúmulo de horas de frio nas semanas que antecedem seu desenvolvimento
(ABBOTT, 1935; GARCIA-LUIS et al., 1992; RANDHAWA; DINSA, 1947;
STATHAKOPOULOS; ERICKSON, 1966). Os brotos vegetativos passam por seis
estádios que compreendem 4 fases: emergência (estádio V1); desenvolvimento
(estádios V2 e V3); maturação (estádios V4 e V5); e dormência (estádio V6)
(CIFUENTES-ARENAS et al., 2016). Geralmente o crescimento da parte aérea se
alterna com o da raiz, dinâmica definida pela dinâmica dos fluxos de fotoassimilados

82
(BEVINGTON; CASTLE, 1985; GIRTON, 1927; REED, 1921; REED; MCDOUGAL,
1937).
As épocas de brotação das plantas cítricas vêm geralmente acompanhadas
de surtos populacionais de diversas pragas como pulgões, psilídeos ou larvas
minadoras que, além de ver facilitados seus ciclos biológicos, podem servir como
veículo ou facilitadores da entrada na planta de diversos patógenos (CHAGAS et al.,
2001; MICHAUD; BROWNING, 1999; PLUKE; QURESHI; STANSLY, 2008;
YAMAMOTO; PAIVA; GRAVENA, 2001), ou seja, os brotos podem ser considerados
facilitadores das relações patógeno-vetor-hospedeiro. No caso de Diaphorina citri
Kuwayama, principal vetor das bactérias associadas à doença mais destrutiva dos
citros, o huanglongbing (BOVÉ, 2006), essa relação tem sido amplamente estudada.
Sabe-se que, por uma série de estímulos visuais e olfatórios, o inseto é mais atraído
pelos brotos novos, os quais são constituídos de tecidos tenros (SÉTAMOU et al.,
2014; SULE et al., 2012). Sabe-se também que na presença de brotos o inseto tende
a se manter na planta e não se dispersar (TOMASETO; KRUGNER; LOPES, 2016),
que a taxa aquisição de ‘Candidatus Liberibacter asiaticus’ é diretamente proporcional
aos títulos da bactéria no broto (LOPES et al., 2013, 2017), e que a presença dos
brotos pode facilitar tanto o processo de aquisição como o de inoculação (HALL;
ALBRECHT; BOWMAN, 2016; SÉTAMOU et al., 2016). Como consequência de todos
os fatores que contribuem para as atividades do inseto, além daqueles que contribuem
para a multiplicação da bactéria nos brotos, a incidência de plantas com HLB
aumentou muito desde o momento em que foi detectado no centro do estado de São
Paulo em 2004, levando à erradicação de mais de 46 milhões de plantas somente
neste Estado. No ano de 2017, 16,73% de um total aproximado de 191,7 milhões de
árvores produtivas manifestavam sintomas de HLB (FUNDECITRUS, 2017).
Uma das principais causas do insucesso no controle do HLB tem a ver com a
dificuldade de se proteger os brotos do acesso de psilídeos infectivos, o que leva os
produtores a fazer pulverizações geralmente em esquema tipo calendário fixo
(MONTEIRO, 2013), desconsiderando a presença ou não de brotos. Com isto,
períodos de desenvolvimento críticos à alimentação e reprodução do inseto podem
não estar sendo adequadamente protegidos. O conhecimento detalhado da dinâmica
de brotação e do desenvolvimento dos brotos em função das variáveis ambientais

83
traria, portanto, grandes benefícios para o controle de D. citri e, consequentemente,
para o manejo da doença. Em citros, os estudos nesta área têm focado em estimar o
impacto da temperatura e disponibilidade hídrica sobre o desenvolvimento reprodutivo
e a produção (ALBRIGO; SAÚCO, 2004) com poucos voltados a brotos vegetativos.
Para o estudo do desenvolvimento reprodutivo e produção várias
metodologias têm sido usadas com o acúmulo de graus dia (unidades de calor ou de
frio), por sua simplicidade, sendo uma das mais aceitas. Consiste em contabilizar,
dentro de uma faixa de temperatura, o número de horas necessário para que um
evento ocorra (ALLEN, 1976; WILSON; BARNETT, 1983). Por exemplo, são
necessários aproximadamente 3000 graus-hora para que as gemas de Poncirus
trifoliata brotem (STATHAKOPOULOS; ERICKSON, 1966). Também foi demonstrado
que quanto maior o número de horas abaixo de 18°C maior é a proporção de brotos
florais em laranja doce (MOSS, 1969, 1976). Em lichia (Litchi chinensis Sonn.), por
sua vez, foi necessário o acúmulo de 7373 graus-hora de teemperatura abaixo de
23,43 °C, desde o momento de maturação das folhas, para que houvesse indução
floral, e adicionais 2397 graus-hora de calor da indução floral para que ocorresse a
antese (CHEN et al., 2016). Na laranjeira Folha Murcha foram necessários em média
de 4462 a 5090 graus-dia para que o fruto amadurecesse, dependendo do porta-
enxerto sobre o qual a copa estava enxertada (STENZEL et al., 2006).
Para estudo de brotos vegetativos, os modelos existentes ou foram
desenvolvidos para descrever o crescimento das plantas como um todo, em altura e
volume principalmente, ou se basearam em medições feitas a intervalos de tempo
muito espaçados, sem se considerar os diferentes estádios fenológicos (DUBEY;
YADAV, 2004; TURRELL, 1961; TURRELL et al., 1969). Por causa disto podem não
ser suficientemente precisos. Se modelos para desenvolvimento vegetativo são
escassos, são abundantes os modelos de requerimentos térmicos para diversas
pragas (MILLER; LANIER; BRANDT, 2001; PRUESS, 1983). Um estudo mais
detalhado sobre os requerimentos termicos para brotos poderia tornar mais completo
e preditivo, por exemplo, o modelo proposto por Aurambout et al. (2009), usado para
se estimar o potencial de distribuição de D. citri na Austrália frente a diversos cenários
de mudanças climáticas.

84
Em vista do exposto, este trabalho teve por objetivos (i) determinar os
requerimentos térmicos dos diferentes estádios de desenvolvimento brotos
vegetativos na laranjeira doce ‘Valência’, (ii) determinar a influência do regime de
irrigação no desenvolvimento do broto e em seus requerimentos térmicos, e (iii) propor
um modelo geral, baseado em graus-dia, de aplicação de medidas de proteção dos
brotos em desenvolvimento.
2. MATERIAL E MÉTODOS
2.1. Material vegetal, adequação das plantas e descrição dos ambientes
Foram realizados dois experimentos em quatro ambientes (3 câmaras de
crescimento e campo) e dois locais (Fundecitrus, Araraquara, SP e pomar comercial
em fazenda em Matão, SP), entre setembro de 2015 e dezembro de 2016. Para o
experimento em câmaras de crescimento (CMP4030, Conviron, Winnipeg, Canadá)
foram usadas mudas de 3 anos de idade de laranja ‘Valência’ enxertada em citrumelo
‘Swingle’ e plantadas em citrovaso de 4,7 L com substrato orgânico comercial
(Plantmax Citrus®). As plantas foram irrigadas 1 a 3 vezes por semana segundo as
suas necessidades e fertirrigadas quinzenalmente com 100 mL vaso-1 de uma solução
nutritiva composta por Ca(NO3)2, MAP, MgSO4, Cu, Zn, Mn-EDTA, (NH4)2MoO4, e Fe-
EDTA, em comcentrações de 1,35, 0,111, 0,4, 0,015, 0,01, 0,0075, 0,00045, 0,075 g
L-1, respectivamente. Para o experimento em campo foram usadas plantas de 3.5 anos
de idade de Valência enxertada em ‘Swingle’ distribuídas em duas áreas, uma sem
irrigação e outra com irrigação. A irrigação consistiu em aplicação de lâminas de água
de aproximadamente 2 mm dia-1 para mitigação do estresse hídrico da planta por
déficit após estiagem muito prolongada, mas que insuficiente em estimular brotação
ou florada. A adubação dessas plantas foi feita de acordo com o manejo normal da
fazenda. Os dados de temperatura e umidade foram registrados em data loggers
LogBox-RHT-LCD (NOVUS Produtos Eletrônicos Ltda, Porto Alegre, RS, Brasil), a
intervalos de um minuto nas câmaras de crescimento e de uma hora no campo.

85
A descrição dos regimes de temperatura usados nas câmaras e a variação da
temperatura durante o tempo em que o experimento foi conduzido no campo são
apresentados na Tabela 1.
Tabela 1. Regimes de temperatura [°C] (umidade relativa [%]) e duração dos diferentes períodos do dia e noite nos três ambientes das câmaras de crescimento e no campo durante o período em que os experimentos foram conduzidos.
Regime Dia (≈ 300 µmol m-2 s-1)
Noite �̅�
°C (% UR) 3h 6h 3h 12h
Câmaras de crescimento
Frio 14 (79,61) 20 (77,75) 14 (76,56) 10 (84,51) 14,83 (79,85)
Médio 24 (78,28) 30 (74,25) 24 (75,64) 18 (80,06) 24,38 (76,15)
Quente 30 (64,00) 38 (54,48) 30 (66,94) 24 (74,23) 30,30 (64,45)
Campo 26,10 (67,24) 33,79 (45,45) 30,29 (54,36) 22,81 (73,95) 26,74 (63,64)
2.2. Experimento 1: desenvolvimento do broto em câmaras de crescimento
As plantas em vaso foram podadas e desfolhadas 15 – 20 cm da linha de
enxertia e colocadas no ambiente quente (Tabela 1) para estimular brotação durante
5 dias. Quando o inchamento da gema e a abertura de suas escamas protetoras eram
evidentes as plantas foram divididas em três grupos de 13 a 15 plantas cada, e
distribuídas nos três ambientes usados (Tabela 1). A partir desse momento foram
feitas medições diárias (exceto finais de semana) do comprimento do broto,
contabilizada a emissão de folhas e anotados os momentos de transição entre as
fases e os estádios de desenvolvimento do broto, de acordo com Cifuentes-Arenas et
al. (2016). Uma breve descrição das fases é dada na Tabela 2.O incremento na área
folhar foi determinado por método não destrutivo. Foram selecionadas as três folhas
basais (as primeiras em se separar do eixo central) de 8/13, 6/14 e 6/15 das plantas
do ambiente frio, médio e quente, respectivamente. Uma escala de referência foi
colocada entre a folha e o eixo do broto e foram tomadas fotografias com câmera
digital. Posteriormente as imagens foram processadas para determinar a área foliar
usando ImageJ 1,50 (Image Processing and Analysis in Java), um programa livre
desenvolvido pelo National Institutes of Health (NIH) (SCHNEIDER; RASBAND;
ELICEIRI, 2012).

86
Tabela 2. Descrição das fases e estádios de desenvolvimento dos brotos em laranjeira Valência (CIFUENTES-ARENAS et al., 2016) Fase Estádios Características
Emergência V1 Inchamento da gema, abertura de escamas protetoras e aparecimento dos primórdios foliares, tecidos moles.
Desenvolvimento
V2 Eixo do broto visível com > 2 mm, nenhuma das folhas se separando do eixo, tecidos moles.
V3 Primeiras folhas basais se separando do eixo e iniciando expansão, rápido crescimento em comprimento, emissão de todas as folhas, tecidos moles.
Maturação V4
Número de folhas já definido, pouco crescimento em comprimento, expansão foliar, tecidos das folhas basais começando a endurecer, mas as partes medianas e apicais ainda moles.
V5 Clorose e morte do ponteiro, pouca expansão foliar, uniformidade na firmeza e transição da cor verde clara para verde amarelada.
Dormência V6 Folhas firmes com cor verde escura, gemas dormentes.
2.3. Experimento 2: desenvolvimento do broto em plantas no campo e
influência do déficit hídrico
Este experimento foi desenvolvido de setembro a outubro de 2015 em fazenda
comercial no município de Matão (SP, Brazil: 21°38’31” S, 48°30’02” O) em duas
áreas: uma com irrigação por gotejamento e outra sem irrigação. Em cada área foram
selecionadas 5 plantas da laranjeira ‘Valência’ enxertada em ‘Swingle’ com 3,5 anos
de idade, as quais, após um período curto de chuvas no começo de setembro, foram
monitoradas regularmente para determinar o momento de emissão natural de brotos.
No momento da instalação do experimento (22/09/2015), todos os brotos estavam no
final do estádio V2. A partir desse momento foram marcados 3 a 5 brotos por planta.
Foram feitas medições diárias do comprimento e monitorada a transição entre as fases
e estádios de desenvolvimento (Tabela 2). No final do experimento, os brotos foram
destacados e levados para o laboratório para determinar o número de nós (gemas)
(devido a que, provávelmente por injúria ou abscisão, alguns brotos careciam de
várias folhas), número de folhas, e área foliar (AF). As folhas foram secas em estufa
a 60 °C até peso constante, e determinado massa foliar específico (massa/AF).
2.4. Graus dia de desenvolvimento (GDD)
A determinação dos graus dia de desenvolvimento, tanto para cada fase e
estádio de desenvolvimento como para o ciclo completo, desde a poda até a emissão
do seguinte fluxo vegetativo, foi determinada como 1 𝛽1⁄ derivada da regressão linear
simples (y = β1x – β0) entre a taxa de desenvolvimento (inverso da duração das fases,
estádios ou ciclo completo, dias-1) e a temperatura média dos ambientes nas câmaras

87
de crescimento. A temperatura base de desenvolvimento (Tb) foi determinada como
𝛽0 𝛽1⁄ . A temperatura limite superior (Tsup), a partir da qual houve um efeito inibitório
no tamanho do broto e insignificante encurtamento do tempo de desenvolvimento, foi
determinada por como |𝛽1 𝛽2⁄ | 2⁄ derivados da regressão polinomial de segunda
ordem do tamanho do broto (mm) em função da temperatura (°C) (y = –β2x2 + β1x +
β0).
2.5. Validação dos resultados
A validação dos resultados foi feita comparando os dados obtidos nas
câmaras de crescimento com os obtidos no campo (experimento 2). Visto que no
experimento 2 não foi possível determinar com exatidão o momento em que as plantas
receberam o estímulo para iniciar brotação nem o início da emissão dos seguintes
fluxos vegetativos, foi calculado o acúmulo de GDD desde o último período de chuvas
no começo de setembro até o final do estádio V2, e comparado com os GDDs desde
a poda até atingir o estádio V2 nas câmaras (experimento 1). A partir deste momento
foi estimado o acúmulo de GDD necessários para completar os estádios V3 e atingir
V5 (inclusive) no experimento 2 e comparado com o mesmo período do experimento
1.
2.6. Proposta de modelo geral aplicação de medidas de proteção dos brotos
Com base nos resultados obtidos neste estudo, foi gerada proposta de
aplicação de medidas de proteção (pulverizações) baseado nos GDDs. A proposta
considera que durante o ano as plantas cítricas de um pomar emitem fluxos
vegetativos em eventos de brotação que delimitam ciclos mais ou menos definidos.
Em cada evento de brotação a área foliar nova que precisa ser protegida (NLA) vai se
incrementando a uma taxa σ, que por sua vez depende tanto da expansão de cada
folha como da taxa π em que cada folha é emitida no broto, até completar um número
definido de folhas por broto (LS) (Fig. 1). Por sua vez, em cada planta os brotos vão
sendo emitidos a uma taxa β durante um período definido de tempo até completarem
a quantidade total de brotos (ST) destinados a saírem desse evento específico de
brotação. Já em escala geográfica de quadra ou pomar, o número árvores que
começam a emitir brotos novos (Tws) irão aparecendo a uma taxa α durante um período

88
de tempo definido. Outros parâmetros que influenciam a dinâmica do modelo são: a
taxa de perda de plantas no pomar ou quadra (ω) que diminui o Tws (mas a taxa de
replanta (γ) o incrementa); a taxa de perda de brotos (τ), que pode acontecer por dano
mecânico ou por “aborto” do broto, que diminui ST; a taxa de perda de folhas (δ), que
pode acontecer por herbivoria ou dano mecânico, que reduz o LS; a aplicação de
medidas de proteção (ξ) com determinada eficiência, que reduz a NLA, enquanto que
os eventos ou processos como precipitação ou radiação solar (ψ) reduzem a eficiência
das ξ causando um incremento na proporção da NLA (Figura 1). O resumo dos
parâmetros e suas unidades são apresentadas na Tabela 3.
Figura 1. Diagrama de fluxo que representa o modelo geral de incremento na área foliar nova desprotegida (NLA) num determinado pomar e tempo. Os compartimentos representam o número de folhas por broto (Ls), o número de brotos por planta (ST) e o número de plantas com brotos (Tws) (ver Tabela 3).
Tabela 3. Definição dos parâmetros usados no modelo geral de aplicação de medidas de proteção num pomar considerando os graus-dia durante um evento de brotação.
Parâmetros Definição Unidades
Tws Número de plantas com brotos novos no pomar unidade
ST Número de brotos na planta unidade
LS Número de folhas no broto unidade
NLA Área foliar nova desprotegida proporção
α Taxa de surgimento plantas com brotos novos dentro do pomar GDD-1
ω Taxa de perda de plantas no pomar GDD-1
β Taxa de surgimento de brotos dentro de cada planta GDD-1
τ Taxa de perda de brotos por planta GDD-1
π Taxa de emissão de folhas por broto GDD -1
δ Taxa de perda de folhas por broto GDD -1
σ Taxa de incremento da área foliar GDD -1
ξ Redução da NLA por medidas de proteção %
ψ Incremento da NLA por eventos que reduzem a eficiência das medidas de proteção %
Com os parâmetros da Tabela 3 foi feita simulação em três cenários. Nestes,
fatores adicionais que poderiam afetar a eficácia das medidas de controle não foram
considerados:
γ

89
1) Incremento da proporção da área foliar nova sem pulverizações (C1).
2) Incremento da proporção da área foliar nova com pulverizações a cada
120 GDD (eficiência de 80%) (C2).
3) Cenário 2 + um evento de chuva a 50 GDD depois da primeira pulverização
(perda da eficiência em 80%) (C3).
2.7. Análise dos dados
O crescimento em comprimento e a expansão foliar (proporção) nas câmaras
de crescimento foram submetidas a regressão não linear ajustando os dados aos
modelos sigmoide Logístico [𝑦 = 1 (1 + 𝑒(−𝑘 ∗ (𝑑𝑖𝑎−𝑇50)))⁄ ] ou sigmoide Gompertz
[𝑦 = 𝑒(−𝑒−𝑘 ∗ (𝑑𝑖𝑎−𝑇50))], comparando-os por meio do critério AIC (AKAIKE, 1998), onde
k é a proporção inicial e T50 o tempo necessário para atingir a metade do tamanho
final.
T50 entre os ambientes foi comparado por meio de análise de variância, com
separação das médias pelo teste Tukey (P < 0,05). A relação entre T50 e tamanho final
do broto ou a área foliar total foi analisada por meio de uma regressão linear simples.
A taxa de emissão foliar foi determinada por regressão linear em função do tempo.
Para o experimento em câmaras de crescimento foi realizada regressão linear múltipla
do número de folhas em função da temperatura e do tamanho do broto. Para
determinar se a posição da folha no broto influenciou a área foliar ou o T50, foi feita
análise de covariância considerando a temperatura como a covariável e a posição da
folha como a variável independente. Para o experimento em campo, o tamanho do
broto, a área foliar média por broto, a área foliar específica (g cm-2) e os GDDs entre
a área com e sem irrigação foram comparadas por meio do teste t. Para todas as
análises foi usado o programa Statgraphics Centurion XVII (Statpoint Technologies
Inc.).
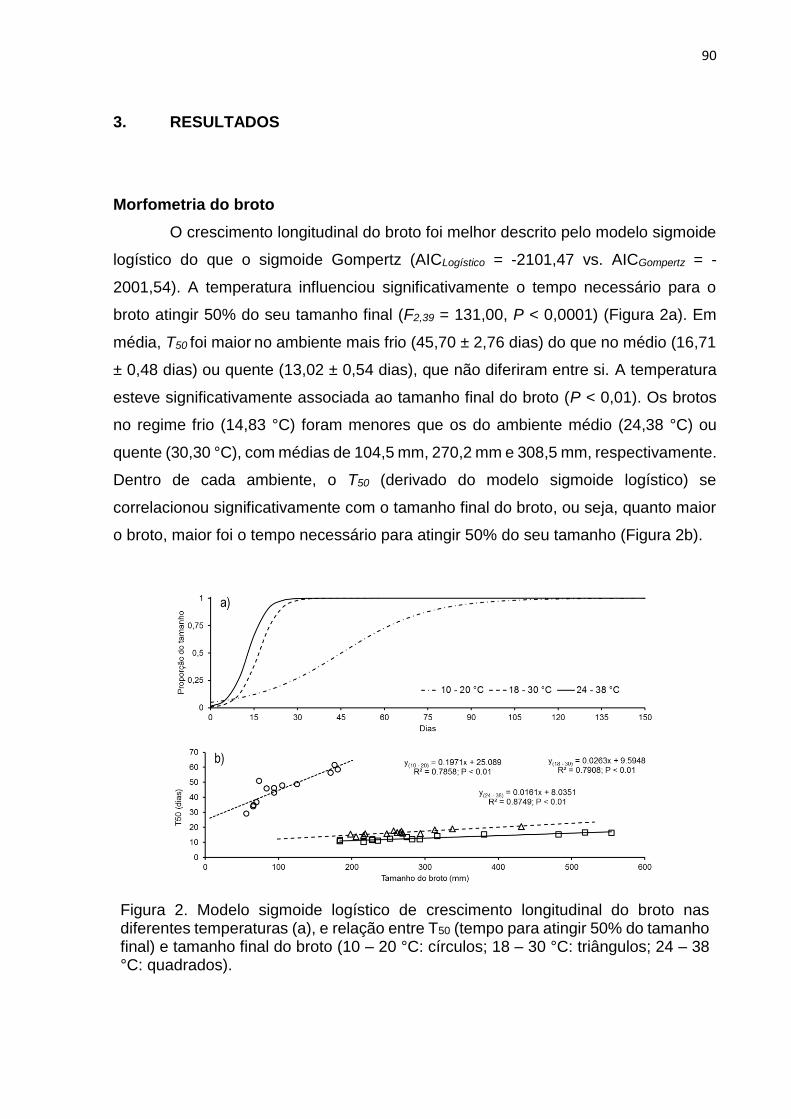
90
3. RESULTADOS
Morfometria do broto
O crescimento longitudinal do broto foi melhor descrito pelo modelo sigmoide
logístico do que o sigmoide Gompertz (AICLogístico = -2101,47 vs. AICGompertz = -
2001,54). A temperatura influenciou significativamente o tempo necessário para o
broto atingir 50% do seu tamanho final (F2,39 = 131,00, P < 0,0001) (Figura 2a). Em
média, T50 foi maior no ambiente mais frio (45,70 ± 2,76 dias) do que no médio (16,71
± 0,48 dias) ou quente (13,02 ± 0,54 dias), que não diferiram entre si. A temperatura
esteve significativamente associada ao tamanho final do broto (P < 0,01). Os brotos
no regime frio (14,83 °C) foram menores que os do ambiente médio (24,38 °C) ou
quente (30,30 °C), com médias de 104,5 mm, 270,2 mm e 308,5 mm, respectivamente.
Dentro de cada ambiente, o T50 (derivado do modelo sigmoide logístico) se
correlacionou significativamente com o tamanho final do broto, ou seja, quanto maior
o broto, maior foi o tempo necessário para atingir 50% do seu tamanho (Figura 2b).
Figura 2. Modelo sigmoide logístico de crescimento longitudinal do broto nas diferentes temperaturas (a), e relação entre T50 (tempo para atingir 50% do tamanho final) e tamanho final do broto (10 – 20 °C: círculos; 18 – 30 °C: triângulos; 24 – 38 °C: quadrados).
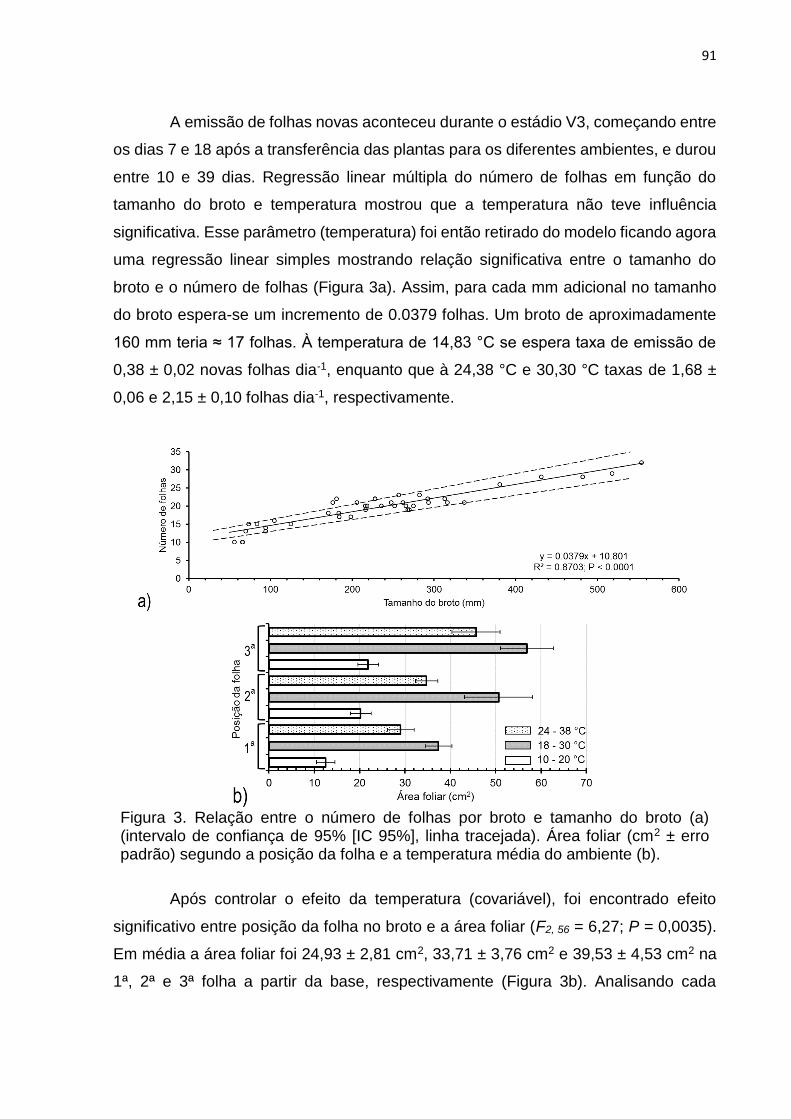
91
A emissão de folhas novas aconteceu durante o estádio V3, começando entre
os dias 7 e 18 após a transferência das plantas para os diferentes ambientes, e durou
entre 10 e 39 dias. Regressão linear múltipla do número de folhas em função do
tamanho do broto e temperatura mostrou que a temperatura não teve influência
significativa. Esse parâmetro (temperatura) foi então retirado do modelo ficando agora
uma regressão linear simples mostrando relação significativa entre o tamanho do
broto e o número de folhas (Figura 3a). Assim, para cada mm adicional no tamanho
do broto espera-se um incremento de 0.0379 folhas. Um broto de aproximadamente
160 mm teria ≈ 17 folhas. À temperatura de 14,83 °C se espera taxa de emissão de
0,38 ± 0,02 novas folhas dia-1, enquanto que à 24,38 °C e 30,30 °C taxas de 1,68 ±
0,06 e 2,15 ± 0,10 folhas dia-1, respectivamente.
Figura 3. Relação entre o número de folhas por broto e tamanho do broto (a) (intervalo de confiança de 95% [IC 95%], linha tracejada). Área foliar (cm2 ± erro padrão) segundo a posição da folha e a temperatura média do ambiente (b).
Após controlar o efeito da temperatura (covariável), foi encontrado efeito
significativo entre posição da folha no broto e a área foliar (F2, 56 = 6,27; P = 0,0035).
Em média a área foliar foi 24,93 ± 2,81 cm2, 33,71 ± 3,76 cm2 e 39,53 ± 4,53 cm2 na
1ª, 2ª e 3ª folha a partir da base, respectivamente (Figura 3b). Analisando cada

92
posição da folha, observa-se relação quadrática entre temperatura e área folhar (y = -
β2x2 + β1x - β0, P < 0.01; Figura 3b), sendo que para desenvolvimento da folha a
temperatura ótima variou de 23,8 °C a 24,7 °C, com média de 24,34 °C, determinado
da mesma forma que a temperatura limite superior para crescimento em comprimento
do broto. O modelo sigmoide logístico foi o que melhor descreveu o incremento na
área foliar só no ambiente frio (AICLogístico = -138,81 vs. AICGompertz = -121,85) em
contraste com o médio (AICLogístico = -116,43 vs. AICGompertz = -127,73) e o quente
(AICLogístico = -83,01 vs. AICGompertz = -89,33), cujos tempos médios necessários para
atingir 50% da área foliar foram 27,20 ± 0,75 dias, 11,02 ± 0,26 dias e 6,69 ± 0,21
dias para os três ambientes, respectivamente. A posição da folha não influenciou
significativamente o T50 em nenhum dos ambientes (P > 0,05) (Figura 4a).
Contrário ao que aconteceu com o comprimento do broto, na maioria dos
casos o T50 das folhas não se correlacionou com a sua área final e, quando a
correlação foi significativa, houve geralmente baixíssima representação da
variabilidade dos dados no modelo de regressão linear (baixo R2), indicando que
quanto maior o tamanho da folha, mais rápido ela cresce sem precisar de mais tempo
(Figura 4b, c, d).
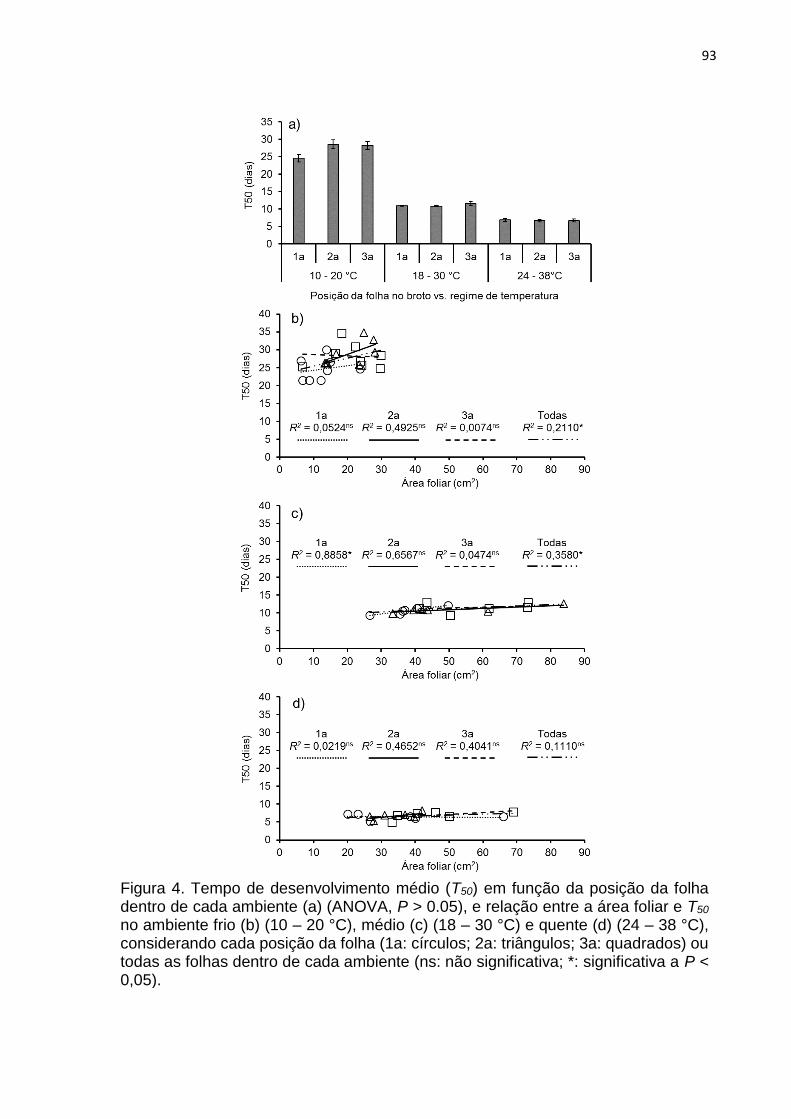
93
Figura 4. Tempo de desenvolvimento médio (T50) em função da posição da folha dentro de cada ambiente (a) (ANOVA, P > 0.05), e relação entre a área foliar e T50 no ambiente frio (b) (10 – 20 °C), médio (c) (18 – 30 °C) e quente (d) (24 – 38 °C), considerando cada posição da folha (1a: círculos; 2a: triângulos; 3a: quadrados) ou todas as folhas dentro de cada ambiente (ns: não significativa; *: significativa a P < 0,05).

94
Graus dia de desenvolvimento (GDD)
A temperatura influenciou significativamente cada uma das fases de
desenvolvimento do broto assim como a duração total do ciclo (do estádio V1 ao fim
do V6), quando teve início o seguinte fluxo vegetativo. Em média, os estádios que
mais duraram nas três temperaturas foram V5, V6, e V3, em contraste com V4, V2 e
V1 (Tabela 4). Foram necessários de 41 a 162 dias para que o broto completasse seu
desenvolvimento (V1 à emissão do fluxo seguinte) (Tabela 4), o que resultaria
potencialmente entre 8,9 e 2,3 ciclos vegetativos por ano.
As temperaturas limites superior (Tsup) e a temperatura base (Tb) foram de
31,95 °C (Figura 4a) e 9,00 °C (Figura 5b). Com essas duas temperaturas foi
determinado os GDDs durante o período entre a poda e o dia em que as plantas foram
distribuídas para cada um dos ambientes, integrando área abaixo da curva e variação
da temperatura. Os GDDs durante esse período foram 136,70 (Tabela 5, Fig. 6).
Tabela 4. Tempo médio de desenvolvimento (dias ± erro padrão) de cada um dos estádios ontogênicos do broto de laranja Valência em diferentes regimes de temperatura. Temperatura (°C) V1 V2 V3 z V4 V5 V6 V1-V6
30,30 2,67 ± 0,23 03,93 ± 0,15 09,93 ± 0,61 08,80 ± 0,46 08,53 ± 0,36 07,33 ± 0,46 141,40 ± 0,71 24,38 3,07 ± 0,20 04,14 ± 0,14 12,43 ± 0,52 07,64 ± 0,72 15,71 ± 0,69 14,64 ± 1,42 157,64 ± 2,19 14,83 6,23 ± 0,59 11,92 ± 0,69 38,92 ± 2,55 16,62 ± 0,99 45,08 ± 2,76 43,54 ± 5,61 162,31 ± 8,12
Média
F1, 40 40,64*** 122,30*** 138,17*** 43,26*** 225,84*** 66,91*** 221,07***
ns: não significativo; *, **, ***: significativo a α = 0.05, 0.01, 0,001, respectivamente. z período de emissão de folhas.
Para cada um dos estádios de desenvolvimento do broto (V1 – V6) assim
como para as fases (emergência [V1], desenvolvimento [V2, V3], maturação [V4, V5],
e dormência [V6]) foram determinados os GDDs como o inverso da taxa de
desenvolvimento (1/𝛽1) derivada da regressão linear (Tabela 5). Os coeficientes de
correlação mais altos foram obtidos quando consideradas as fases ao invés dos
estádios de desenvolvimento individuais. A Tb variou de 2,68 até 12,01°C (Tabela 5).
Do momento da poda até o início da emissão do fluxo vegetativo seguinte foram
necessários aproximadamente 970,03 GDD (Tabela 5). Considerando a Tb para o ciclo
completo de brotação (da poda até início do fluxo vegetativo seguinte) e a Tsup, foi
determinada a oferta térmica de três locais (municípios) com os registros de
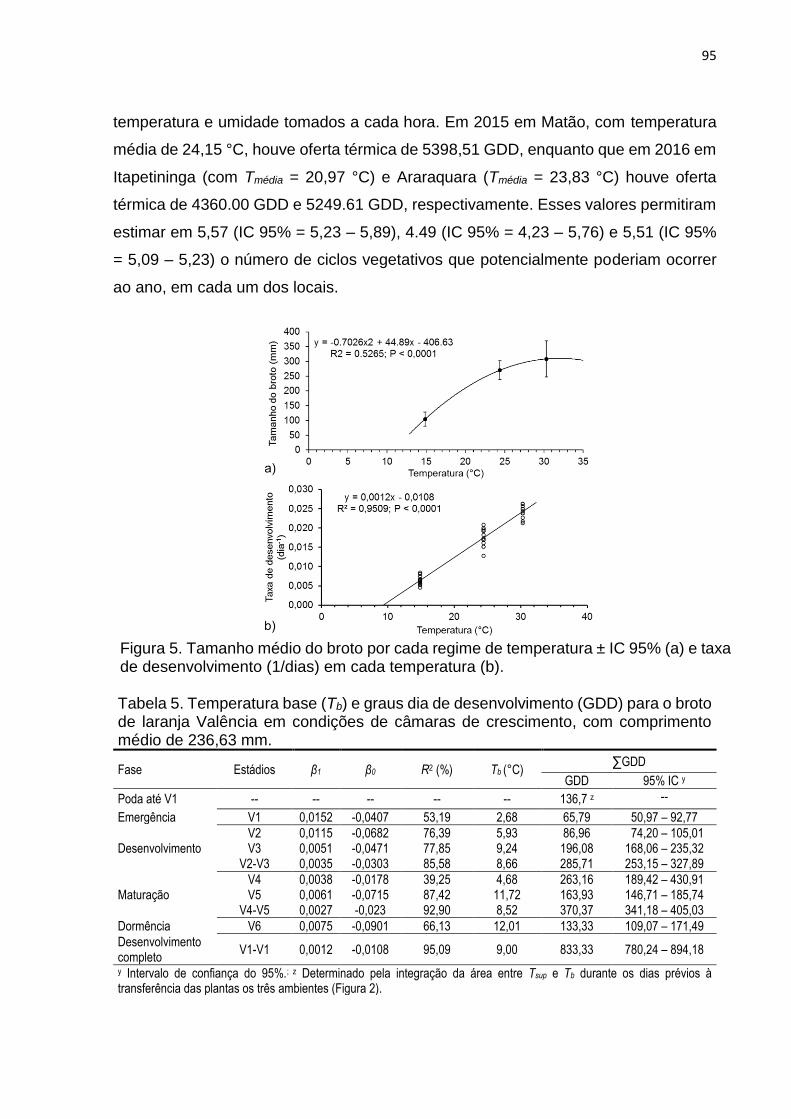
95
temperatura e umidade tomados a cada hora. Em 2015 em Matão, com temperatura
média de 24,15 °C, houve oferta térmica de 5398,51 GDD, enquanto que em 2016 em
Itapetininga (com Tmédia = 20,97 °C) e Araraquara (Tmédia = 23,83 °C) houve oferta
térmica de 4360.00 GDD e 5249.61 GDD, respectivamente. Esses valores permitiram
estimar em 5,57 (IC 95% = 5,23 – 5,89), 4.49 (IC 95% = 4,23 – 5,76) e 5,51 (IC 95%
= 5,09 – 5,23) o número de ciclos vegetativos que potencialmente poderiam ocorrer
ao ano, em cada um dos locais.
Figura 5. Tamanho médio do broto por cada regime de temperatura ± IC 95% (a) e taxa de desenvolvimento (1/dias) em cada temperatura (b).
Tabela 5. Temperatura base (Tb) e graus dia de desenvolvimento (GDD) para o broto de laranja Valência em condições de câmaras de crescimento, com comprimento médio de 236,63 mm.
Fase Estádios β1 β0 R2 (%) Tb (°C) ∑GDD
GDD 95% IC y
Poda até V1 -- -- -- -- -- 136,7 z --
Emergência V1 0,0152 -0,0407 53,19 2,68 65,79 50,97 – 92,77
Desenvolvimento V2 0,0115 -0,0682 76,39 5,93 86,96 174,20 – 105,01 V3 0,0051 -0,0471 77,85 9,24 196,08 168,06 – 235,32
V2-V3 0,0035 -0,0303 85,58 8,66 285,71 253,15 – 327,89
Maturação V4 0,0038 -0,0178 39,25 4,68 263,16 189,42 – 430,91 V5 0,0061 -0,0715 87,42 11,72 163,93 146,71 – 185,74
V4-V5 0,0027 -0,023 92,90 8,52 370,37 341,18 – 405,03
Dormência V6 0,0075 -0,0901 66,13 12,01 133,33 109,07 – 171,49
Desenvolvimento completo
V1-V1 0,0012 -0,0108 95,09 9,00 833,33 780,24 – 894,18
y Intervalo de confiança do 95%.; z Determinado pela integração da área entre Tsup e Tb durante os dias prévios à transferência das plantas os três ambientes (Figura 2).

96
Experimento em campo
O tamanho do broto foi menor na área sem irrigação do que na área com
irrigação (t = 3,76, P = 0,0010), com média de 69,00 mm ± 8,26 e 164,16 mm ± 21,37
respectivamente. A relação entre o número de nós e o comprimento do broto foi
significativa tanto na área irrigada (F1, 12 = 75,21, P < 0,01, R2 = 86,24%; β1 = 0,032, t
= 8,67, P < 0,01; β0 = 6,25, t = 8,67, P < 0,01) quanto na não irrigada (F1, 9 = 10,40, P
= 0,01, R2 = 53,60%; β1 = 0,040, t = 3,22, P = 0,01; β0 = 4,77, t = 5,17, P < 0,01) assim
como para o conjunto dos dois locais (F1, 23 = 161,99, P < 0,01, R2 = 87,57%; β1 =
0,037, t = 12,73, P < 0,01; β0 = 5,32, t = 12,83, P < 0,01) (Figura 7). Os coeficientes β1
ou β0 da regressão não diferiram entre área irrigada e não irrigada, (Pβ0 = 0,1282 e
Pβ1 = 0,5537). Quando comparados os dados conjuntos do experimento em câmaras
de crescimento com os dados obtidos em campo, os valores de β1 não foram
diferentes (P = 0,2221) mas sim β0 (P < 0,01), ou seja, o número de folhas por cada
mm de comprimento do broto foi igual nos brotos desenvolvidos nas câmaras de
crescimento e no campo, mas o tamanho médio do broto foi maior nas câmaras de
crescimento. As folhas dos brotos desenvolvidos na área não irrigada foram menores
em área foliar do que na irrigada (t = 6,82, P < 0,0001), com 14,55 cm2 ± 3,25 vs.
33,58 cm2 ± 9,14 respectivamente. A massa foliar específica também foi menor na
área não irrigada (68,79 mg cm-2) do que na irrigada (91,77 mg cm-2) (t = 0,34, P =
0,0286).
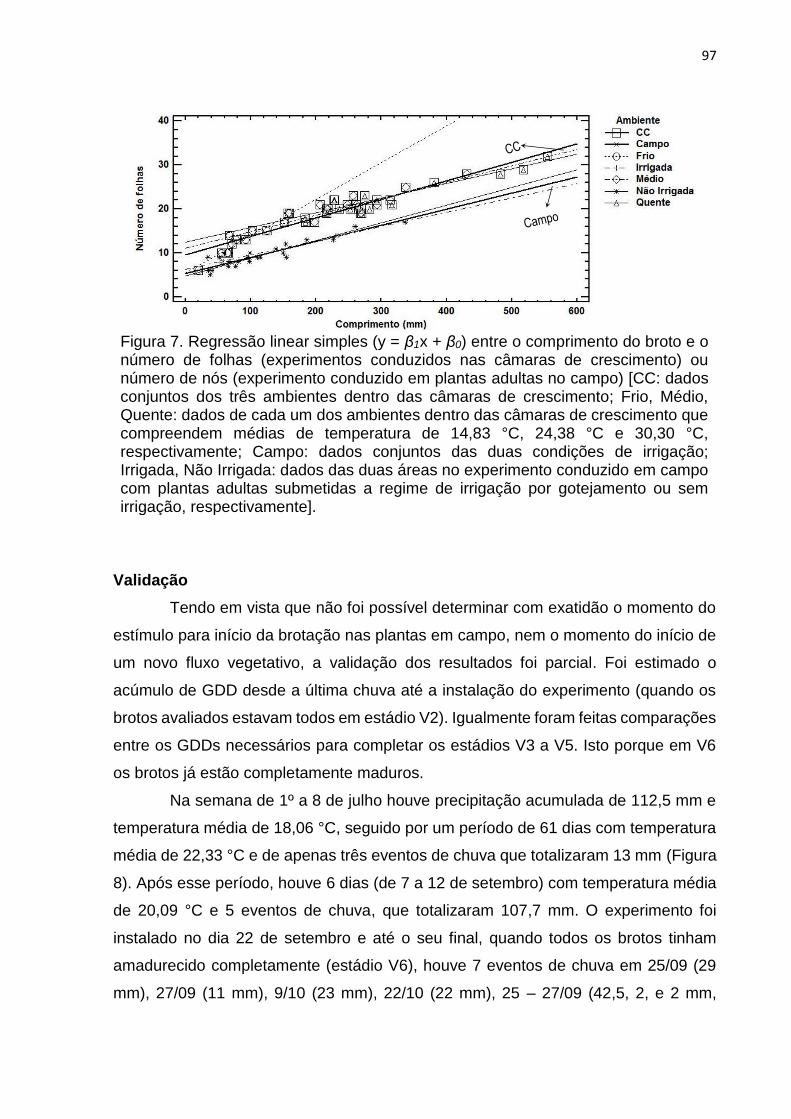
97
Figura 7. Regressão linear simples (y = β1x + β0) entre o comprimento do broto e o número de folhas (experimentos conduzidos nas câmaras de crescimento) ou número de nós (experimento conduzido em plantas adultas no campo) [CC: dados conjuntos dos três ambientes dentro das câmaras de crescimento; Frio, Médio, Quente: dados de cada um dos ambientes dentro das câmaras de crescimento que compreendem médias de temperatura de 14,83 °C, 24,38 °C e 30,30 °C, respectivamente; Campo: dados conjuntos das duas condições de irrigação; Irrigada, Não Irrigada: dados das duas áreas no experimento conduzido em campo com plantas adultas submetidas a regime de irrigação por gotejamento ou sem irrigação, respectivamente].
Validação
Tendo em vista que não foi possível determinar com exatidão o momento do
estímulo para início da brotação nas plantas em campo, nem o momento do início de
um novo fluxo vegetativo, a validação dos resultados foi parcial. Foi estimado o
acúmulo de GDD desde a última chuva até a instalação do experimento (quando os
brotos avaliados estavam todos em estádio V2). Igualmente foram feitas comparações
entre os GDDs necessários para completar os estádios V3 a V5. Isto porque em V6
os brotos já estão completamente maduros.
Na semana de 1º a 8 de julho houve precipitação acumulada de 112,5 mm e
temperatura média de 18,06 °C, seguido por um período de 61 dias com temperatura
média de 22,33 °C e de apenas três eventos de chuva que totalizaram 13 mm (Figura
8). Após esse período, houve 6 dias (de 7 a 12 de setembro) com temperatura média
de 20,09 °C e 5 eventos de chuva, que totalizaram 107,7 mm. O experimento foi
instalado no dia 22 de setembro e até o seu final, quando todos os brotos tinham
amadurecido completamente (estádio V6), houve 7 eventos de chuva em 25/09 (29
mm), 27/09 (11 mm), 9/10 (23 mm), 22/10 (22 mm), 25 – 27/09 (42,5, 2, e 2 mm,
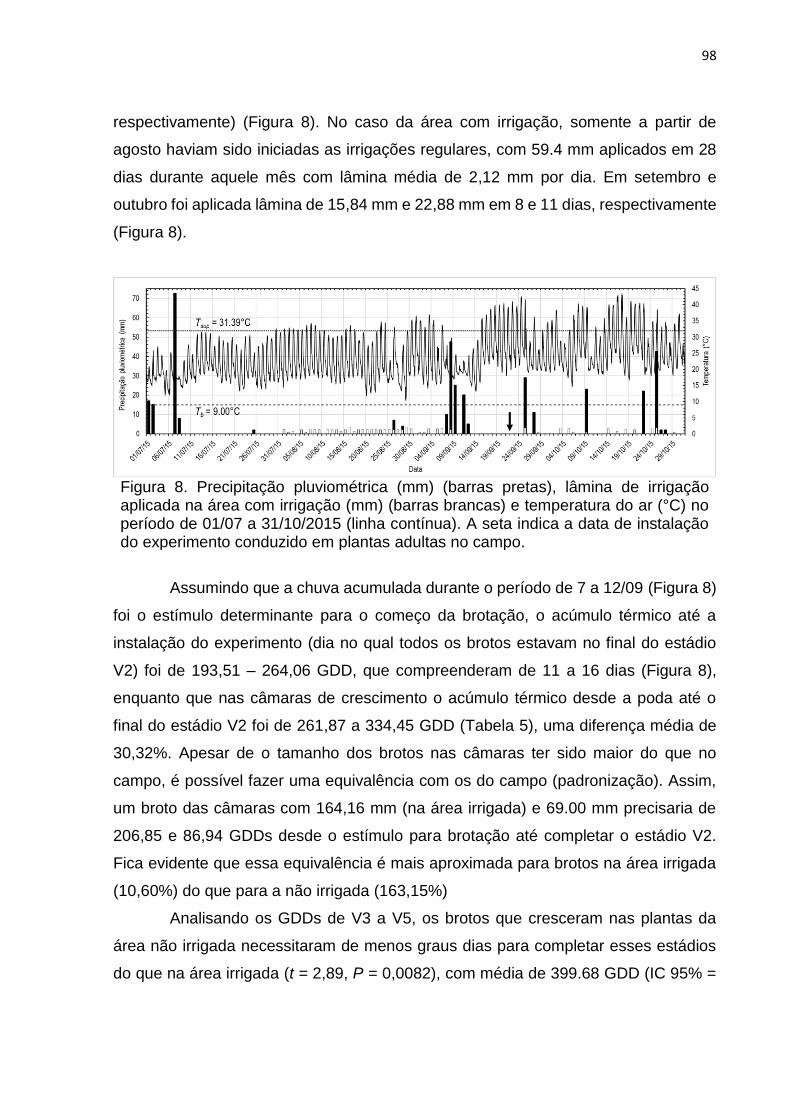
98
respectivamente) (Figura 8). No caso da área com irrigação, somente a partir de
agosto haviam sido iniciadas as irrigações regulares, com 59.4 mm aplicados em 28
dias durante aquele mês com lâmina média de 2,12 mm por dia. Em setembro e
outubro foi aplicada lâmina de 15,84 mm e 22,88 mm em 8 e 11 dias, respectivamente
(Figura 8).
Figura 8. Precipitação pluviométrica (mm) (barras pretas), lâmina de irrigação aplicada na área com irrigação (mm) (barras brancas) e temperatura do ar (°C) no período de 01/07 a 31/10/2015 (linha contínua). A seta indica a data de instalação do experimento conduzido em plantas adultas no campo.
Assumindo que a chuva acumulada durante o período de 7 a 12/09 (Figura 8)
foi o estímulo determinante para o começo da brotação, o acúmulo térmico até a
instalação do experimento (dia no qual todos os brotos estavam no final do estádio
V2) foi de 193,51 – 264,06 GDD, que compreenderam de 11 a 16 dias (Figura 8),
enquanto que nas câmaras de crescimento o acúmulo térmico desde a poda até o
final do estádio V2 foi de 261,87 a 334,45 GDD (Tabela 5), uma diferença média de
30,32%. Apesar de o tamanho dos brotos nas câmaras ter sido maior do que no
campo, é possível fazer uma equivalência com os do campo (padronização). Assim,
um broto das câmaras com 164,16 mm (na área irrigada) e 69.00 mm precisaria de
206,85 e 86,94 GDDs desde o estímulo para brotação até completar o estádio V2.
Fica evidente que essa equivalência é mais aproximada para brotos na área irrigada
(10,60%) do que para a não irrigada (163,15%)
Analisando os GDDs de V3 a V5, os brotos que cresceram nas plantas da
área não irrigada necessitaram de menos graus dias para completar esses estádios
do que na área irrigada (t = 2,89, P = 0,0082), com média de 399.68 GDD (IC 95% =

99
345,83 – 387,83) vs. 455,46 GDD (IC 95% = 425,69 – 485,23), respectivamente,
enquanto que os brotos que se desenvolveram nas câmaras de crescimento
completaram os mesmos estádios em 566,45 GDD (IC 95% = 509,24 – 640,38)
(Tabela 5), uma diferença de 41,76% a 24,37%. No mesmo sentido que a comparação
dos GDD desde a chuva até V2, a diferença entre os resultados das câmaras de
crescimento e campo podem ser reduzidas se se considerar o tamanho médio do
broto. Assim, o acúmulo de GDD de um broto das câmaras com 164,16 mm e de 69.00
mm seria de 392.97 (IC 95% = 292,66 – 493,18) e 165,15 (IC 95% = 126,40 – 203,90).
Torna-se evidente também que esta padronização foi mais apropriada para os brotos
na área irrigada do que na não irrigada, com diferença média de 15,90% 142,01%,
respectivamente.
Proposta de modelo geral de aplicação de medidas de proteção do broto de
acordo com os GDD
O modelo teórico proposto mostrou que a proporção da área foliar nova
desprotegida (NLA) cresce rapidamente atingindo uma proporção > 0,9 em
aproximadamente 210 GDD (C1). Com pulverizações a cada 120 GDD, a NLA chega
até 0,3 no momento da primeira pulverização no C2, quando é reduzida até ≈ 0,06
(Figura 9). No entanto, ao se tratar de um período de rápido crescimento e expansão
foliar, há um rápido aumento da NLA até atingir ≈ 0,65 na segunda pulverização,
momento no qual a NLA é reduzida novamente e fica em torno de 0,12. A partir deste
momento o incremento da área foliar é muito reduzido e o incremento em NLA é quase
imperceptível (Fig. 9). Já depois da terceira e última pulverização a NLA é reduzida
até menos de 0,05. Considerando uma chuva ocorrida 50 GDD após a primeira
pulverização, reduzindo a eficácia da pulverização um 80% (C3), a NLA foi
incrementada e atingiu ≈ 0,9 no momento da segunda pulverização, quando foi
reduzida para ≈ 0,2. Já na terceira pulverização, a NLA ficou bem próxima da do C2,
≈ 0,05 (Figura 9).
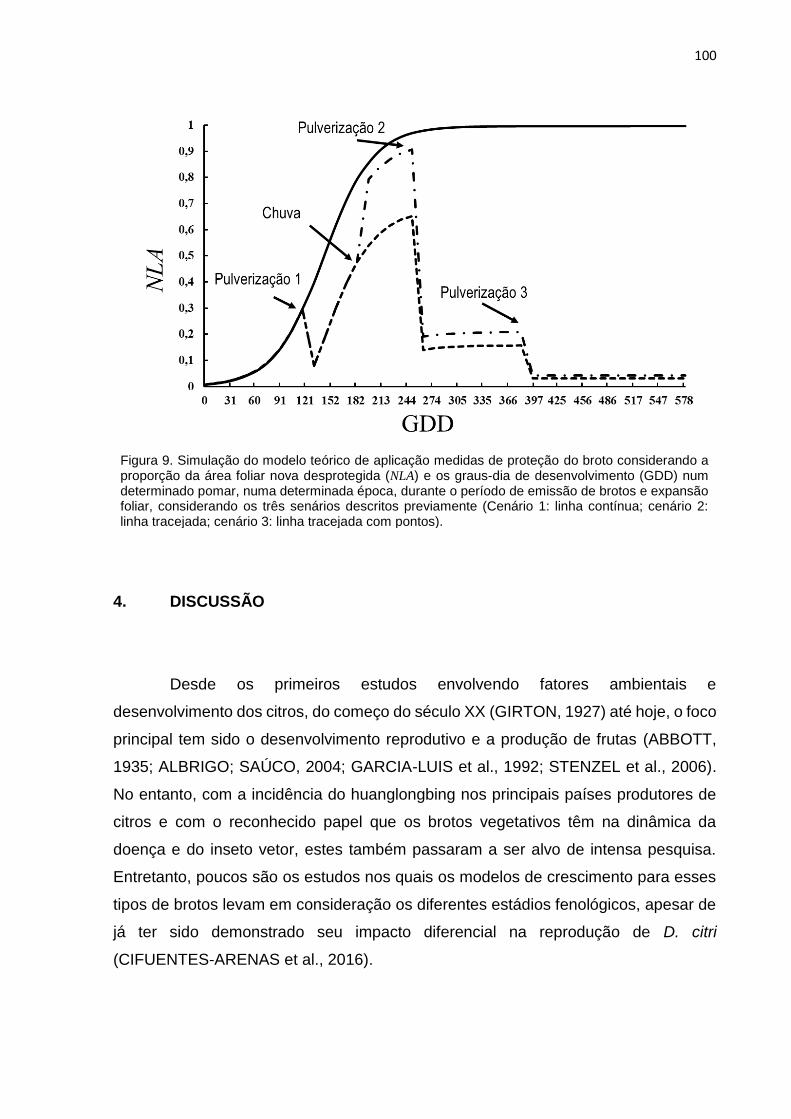
100
Figura 9. Simulação do modelo teórico de aplicação medidas de proteção do broto considerando a proporção da área foliar nova desprotegida (NLA) e os graus-dia de desenvolvimento (GDD) num determinado pomar, numa determinada época, durante o período de emissão de brotos e expansão foliar, considerando os três senários descritos previamente (Cenário 1: linha contínua; cenário 2: linha tracejada; cenário 3: linha tracejada com pontos).
4. DISCUSSÃO
Desde os primeiros estudos envolvendo fatores ambientais e
desenvolvimento dos citros, do começo do século XX (GIRTON, 1927) até hoje, o foco
principal tem sido o desenvolvimento reprodutivo e a produção de frutas (ABBOTT,
1935; ALBRIGO; SAÚCO, 2004; GARCIA-LUIS et al., 1992; STENZEL et al., 2006).
No entanto, com a incidência do huanglongbing nos principais países produtores de
citros e com o reconhecido papel que os brotos vegetativos têm na dinâmica da
doença e do inseto vetor, estes também passaram a ser alvo de intensa pesquisa.
Entretanto, poucos são os estudos nos quais os modelos de crescimento para esses
tipos de brotos levam em consideração os diferentes estádios fenológicos, apesar de
já ter sido demonstrado seu impacto diferencial na reprodução de D. citri
(CIFUENTES-ARENAS et al., 2016).

101
Estudos comparativos dos dados obtidos dentro de ambientes controlados
(câmaras de crescimento) com os obtidos em condições de campo têm mostrado que,
mesmo que a equivalência não seja direta (HENNESSEY, 1976), os resultados são
muito úteis para estudar uma grande diversidade de condições que, no campo, levaria
um grande esforço e gasto de recursos. Neste experimento, os resultados obtidos nas
câmaras se aproximaram dos obtidos no campo em área irrigada, indicando que
outros fatores devem ser considerados antes de se extrapolar os resultados para
plantas estabelecidas em condições de sequeiro.
Observação interessante foi a de que no ambiente controlado o broto foi quase
10 cm maior do que em condições de campo (quando consideradas a média geral dos
dois ambientes), mas o número de folhas por broto (nas câmaras), ou de nós por broto
(no campo) foi igual para os dois ambientes. Caso particular aconteceu no ambiente
frio, pois o número de folhas por broto foi menor, o que pode estar relacionado à
dificuldade de absorção, translocação ou metabolização de nutrientes pelas plantas
cítricas a baixas temperaturas (SCHOLBERG et al., 2002). Isto pode indicar que,
possivelmente, o número de folhas por cada cm de broto seja uma informação própria
do genótipo (porta-enxerto, copa, ou combinação dos dois), enquanto que o tamanho
do broto ou da área foliar decorrente da adaptação das plantas às diversas condições
ambientais (NICOTRA et al., 2010; REWALD et al., 2012).
O déficit hídrico, avaliado em condições de campo, diminuiu os valores de
todas as variáveis avaliadas (comprimento dos brotos, a área foliar e a massa foliar
específica) e levou à necessidade de um menor valor de graus-dias para que o broto
se desenvolvesse por completo. Esses resultados estão de acordo com os relatados
na literatura. Para a tangerineira ‘Clementina de Nules’, González-Altozano e Castel
(2000) encontraram que o déficit hídrico equivalente a 25% da evapotranspiração
(medida por lisímetro), aplicado desde o início da brotação, foi o que mais
drasticamente reduziu o tamanho dos brotos. Em outro estudo, plantas de laranjeira
‘Valência’ submetidas a déficit hídrico apresentaram menor condutância estomática,
menor potencial hídrico nas folhas e menor taxa de assimilação de CO2, quando
enxertadas em citrumeleiro ‘Swingle’ (sabidamente mais exigente em água) do que
em ‘Cravo’ (PEDROSO et al. (2014),. Os mecanismos envolvidos na redução do
tamanho dos brotos desenvolvidos em condição de déficit hídrico podem estar

102
relacionados à interação entre as rotas da autofagia seletiva e dos brassinoesteróides
[BRs] (fitohormônios promotores de crescimento), na qual os BRs são degradados
limitando o crescimento da planta, assegurando sua sobrevivência durante épocas de
déficit hídrico (NOLAN et al., 2017). Os BRs também têm sido reportados como
redutores das populações de Ca. L. asiaticus nas plantas afetadas por HLB
(CANALES et al., 2016).
Este trabalho foi o primeiro a estimar os requerimentos térmicos da brotação
em citros, desde sua indução até a maturação e a emissão natural do seguinte fluxo
vegetativo, considerando-se os brotos como unidade individual de estudo. Outros
aspectos como a área foliar, foi função do número total de brotos que, por sua vez,
surgem com frequências e intervalos definidos. Por isso, futuros estudos deverão
considerar os fatores determinantes para emissão de fluxos vegetativos, no campo,
assim como sua relação com as variáveis climáticas. Aqui, apesar de no campo não
ter sido possível determinar o momento exato em que os brotos avaliados receberam
o estímulo necessário para emergir, a metodologia de GDD permitiu estimar que esse
estímulo provavelmente ocorreu após curto período de tempo com ocorrência de
chuvas de mais de 100 mm que seguiu um período de estiagem de cerca de dois
meses. Para outras situações há necessidade de mais estudos. Com base nos
resultados das câmaras de crescimento (Tabela 4) estima-se que, no campo, sob
condições equivalentes haveria, por ano, em torno de 4,5 a quase 6 fluxos vegetativos
em regiões subtropicais (que abrange a região citrícola de São Paulo) contra 7,1 ciclos
em regiões tropical, como, por exemplo, Aracataca na Colômbia (MENDEL, 1969).
Não se considera aqui a intensidade de brotação que pode ser variável.
Com base nas informações geradas neste estudo foi desenvolvido um modelo
geral de suporte a tomada de decisões sobre o melhor momento de início e frequência
de pulverizações das plantas cítricas visando o controle de insetos e pragas que
afetam os brotos. O modelo considera que, em cada evento de brotação, o número
de brotos com determinado número de folhas irá aparecer a taxas definidas. Haverá,
consequentemente, uma área foliar nova que irá crescer rapidamente e precisa ser
protegida. Na simulação, o intervalo entre pulverizações foi definido em função de um
número fixo de graus dia. No entanto, isto pode ser definido também por uma
proporção aceitável de área foliar nova desprotegida (NLA), ou seja, pela proporção

103
da área foliar desprotegida que se está disposto a tolerar, assumindo-se que as
pulverizações anteriores não acompanham a expansão dos tecidos. Considerando-se
que durante o período de brotação a NLA cresce constantemente, poderia se definir,
empiricamente, a necessidade de pulverizações quando a NLA atingisse 0,5, ou 0,3.
Os limites da NLA não são conhecidos. Requerem estudos adicionais para seu
estabelecimento. Irão depender, obviamente, entre outros fatores, da dinâmica das
populações do agente alvo do controle (vetor ou praga) e do seu potencial de dano.
Especial atenção deve ser dada a D. citri, responsável pela disseminação do patógeno
que mais danos tem causado nas principais citriculturas mundiais. Fatores adicionais
importantes a serem considerados na definição dos limites de NLA são (i) proximidade
do pomar comercial que se pretende proteger de áreas abandonadas afetadas por
HLB promovendo alta pressão de inóculo (MONTEIRO, 2013), (ii) flutuação nas
populações de psilídeos portadores de Liberibacter (SASSI et al., 2017), e (iii) variação
nos títulos de CLas nos brotos em função da época do ano, que, por sua vez,
influencia na habilidade do psilídeo em adquirir a bactéria (LOPES et al., 2017).
Este estudo permitiu estabelecer os requerimentos térmicos para o
desenvolvimento do broto vegetativo em copas de laranjeira ‘Valência’, desde o
inchamento das gemas até a maturação do broto, separando os requerimentos para
cada um dos estádios de desenvolvimento durante a ontogenia. Estudos adicionais
precisam ser feitos para se determinar quais fatores ambientais impactam a
frequência e intensidade de brotos, incluindo-se o déficit hídrico, que, conforme
demonstrado neste e em outros estudos, tem papel relevante em todo o processo. A
condição fitossanitária da planta também deveria ser estudada pois as plantas
doentes aparentemente possuem padrão de emissão de brotos totalmente distinto do
de plantas sadias (MONTESINO, 2011). O padrão de desenvolvimento dos mesmos
também parece ser diferente (observação pessoal), assunto sob investigação.
5. REFERÊNCIAS
ABBOTT, C. E. Blossom-bud differentiation in citrus trees. American Journal of Botany, v. 22, n. 4, p. 476, 1935.

104
AKAIKE, H. Information theory and an extension of the maximum likelihood principle. In: PARZEN, E.; TANABE, K.; KITAGAWA, G. (Eds.). . Selected Papers of Hirotugu Akaike. [s.l.] Springer New York, 1998. p. 199–213.
ALBRIGO, L. G.; SAÚCO, V. G. Flower bud induction, flowering and fruit-set of some tropical and subtropical fruit tree crops with special reference to citrus. Acta Horticulturae, v. 632, p. 81–90, 2004.
ALLEN, J. C. A modified sine wave method for calculating degree days. Environmental Entomology, v. 5, n. 3, p. 388–396, 1976.
AURAMBOUT, J. P. et al. A concept model to estimate the potential distribution of the Asiatic citrus psyllid (Diaphorina citri Kuwayama) in Australia under climate change-A means for assessing biosecurity risk. Ecological Modelling, v. 220, n. 19, p. 2512–2524, 2009.
BEVINGTON, K. B.; CASTLE, W. S. Annual root growth pattern of young citrus trees in relation to shoot growth, soil temperature, and soil water content. Journal of the American Society for Horticultural Science, v. 110, n. 6, p. 840–845, 1985.
BOVÉ, J. M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. Journal of Plant Pathology, v. 88, n. 1, p. 7–37, 2006.
CANALES, E. et al. “Candidatus Liberibacter asiaticus”, causal agent of citrus huanglongbing, is reduced by treatment with brassinosteroids. PLoS ONE, v. 11, n. 1, p. e0146223, 2016.
CHAGAS, M. C. M. et al. Phyllocnistis citrella Stainton (Lepidoptera: Gracillariidae) and its relationship with the citrus canker bacterium Xanthomonas axonopodis pv citri in Brazil. Neotropical Entomology, v. 30, n. 1, p. 55–59, mar. 2001.
CHEN, P.-A. et al. Temperature model of litchi flowering—From induction to anthesis. Scientia Horticulturae, v. 205, p. 106–111, 2016.
CIFUENTES-ARENAS, J. C. et al. Monitoring citrus flush shoot ontogeny as a potential strategy for HLB and psyllid management. Journal of Citrus Pathology, v. 3, n. 1, p. 4, 2016.
DAVIS, J. H. C.; GARCIA, S. Competitive ability and growth habit of indeterminate beans and maize for intercropping. Field Crops Research, v. 6, n. C, p. 59–75, 1983.
DUBEY, A. K.; YADAV, D. S. Studies on growth pattern of different citrus species during the spring flush. Indian Journal of Agricultural Research, v. 38, n. 1, p. 50–54, 2004.
FOURCAUD, T. et al. Plant growth modelling and applications: the increasing importance of plant architecture in growth models. Annals of Botany, v. 101, n. 8, p. 1053–1063, 2008.
FUNDECITRUS. Mesmo estabilizada, insidência de greening permanece alta no

105
cinturão citrícola de São Paulo e Triângulo/Sudoeste Mineiro. Araraquara - SP, Brazil: Fundecitrus, 2017. Disponível em: <http://www.fundecitrus.com.br>.
GARCIA-LUIS, A. et al. Low temperature influence on flowering in Citrus. The separation of inductive and bud dormancy releasing effects. Physiologia Plantarum, v. 86, n. 4, p. 648–652, 1992.
GIRTON, R. E. The growth of citrus seedlings as influenced by environmental factors. University of California Publications in Agricultural Sciences, v. 5, p. 83–117, 1927.
GONZÁLEZ-ALTOZANO, P.; CASTEL, J. R. Regulated deficit irrigation in “Clementina de Nules” citrus trees. II: Vegetative growth. Journal of Horticultural Science and Biotechnology, v. 75, n. 4, p. 388–392, 2000.
HALL, D. G.; ALBRECHT, U.; BOWMAN, K. D. Transmission rates of “Ca. Liberibacter asiaticus” by asian citrus psyllid are enhanced by the presence and developmental stage of citrus flush. Journal of Economic Entomology, v. 109, n. 2, p. 558–563, 2016.
HENNESSEY, T. C. A comparison of field and growth chamber productivity of three poplar clones. [s.l.] Iowa State University - Ames, Iowa, 1976.
HURD, R. G.; GAY, A. P.; MOUNTIFIELD, A. C. The effect of partial flower removal on the relation betwen root, shootand fruti growth in the indeterminate tomato. Annals of Applied Biology, v. 93, n. 1, p. 77–89, 1979.
LOPES, S. A. et al. “Candidatus Liberibacter asiaticus” titers in citrus and acquisition rates by Diaphorina citri are decreased by higher temperature. Plant Disease, v. 97, n. 12, p. 1563–1570, 2013.
LOPES, S. A. et al. Seasonal variation of “Candidatus Liberibacter asiaticus” titers in new shoots of citrus in distinct environments. Plant Disease, v. 101, n. 4, p. 583–590, 2017.
MENDEL, K. The influence of temperature and light on the vegetative development of citrus trees. Proceedings of the 1st International Citrus Symposium. Anais...1969
MICHAUD, J. P.; BROWNING, H. W. Seasonal abundance of the Brown Citrus Aphid, Toxoptera citricida, (Homoptera: Aphididae) and its natural enemies in Puerto Rico. Florida Entomologist, v. 82, n. 3, p. 424–447, 1999.
MILLER, P.; LANIER, W.; BRANDT, S. Using growing degree days to predict plant stages. Bonzeman, MO: [s.n.].
MONTEIRO, A. B. Efeito de um pomar sem manejo de Huanglongbing sobre a ocorrência de Diaphorina citri e incidência de plantas doentes em uma área vizinha com manejo da doença. Araraquara - SP, Brasil: Fundo de Defesa da Citricultura, 2013.

106
MONTESINO, L. H. Evolução dos sintomas de Huanglongbing em laranjeiras jovens: relação com época do ano, fenologia das plantas, flutuação populacional de Diaphorina citri Kuwayama (Hemiptera: Psyllidae) e medidas de controle do vetor. [s.l.] Fundo de Defesa da Citricultura, 2011.
MOSS, G. Influence of temperature and photoperiod on flower induction and inflorescence development in sweet orange (Citrus sinensis L. Osbeck). Journal of Horticultural Science, v. 44, n. 4, p. 311–320, 1969.
MOSS, G. Temperature effects on flower initiation in sweet orange (Citrus sinensis). Australian Journal of Agricultural Research, v. 27, n. 3, p. 399, 1976.
NICOTRA, A. B. et al. Plant phenotypic plasticity in a changing climate. Trends in Plant Science, v. 15, n. 12, p. 684–692, 2010.
NOLAN, T. M. et al. Selective autophagy of BES1 mediated by DSK2 balances plant growth and survival. Developmental Cell, v. 41, n. 1, p. 33–46, 2017.
PEDROSO, F. K. J. V. et al. Drought tolerance in citrus trees is enhanced by rootstock-dependent changes in root growth and carbohydrate availability. Environmental and Experimental Botany, v. 101, p. 26–35, 2014.
PLUKE, R.; QURESHI, J.; STANSLY, P. Citrus flushing patterns, Diaphorina citri (Hemiptera: Psyllidae) populations and parasitism by Tamarixia radiata (Hymenoptera: Eulophidae) in Puerto Rico. Florida Entomologist, v. 91, n. 1, p. 36–42, 2008.
PRUESS, K. P. Day-degree methods for pest management. Environmental Entomology, v. 12, n. 3, p. 613–619, 1983.
RANDHAWA, G. S.; DINSA, H. S. Time of blossom-bud differentiation in Citrus. Proceedings of the American Society of Horticultural Science, v. 50, p. 165–171, 1947.
REED, H. S. Growth and sap concentration. Journal of Agricultural Research, v. 21, n. 1, p. 81–98, 1921.
REED, H. S.; MCDOUGAL, D. T. Periodicity in the growth of the orange tree. Growth, v. 1, p. 371–373, 1937.
REWALD, B. et al. Phenotypic plasticity and water flux rates of Citrus root orders under salinity. Journal of Experimental Botany, v. 63, n. 7, p. 2717–2727, 2012.
SASSI, R. S. et al. Incidence and distribution o Diaphorina citri carrying Candidatus Liberibacter asiaticus. 5th International Research Conference on Huanglongbing. Anais...Orlando: USDA-ARS, University of Florida, 2017
SCHNEIDER, C. A; RASBAND, W. S.; ELICEIRI, K. W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods, v. 9, n. 7, p. 671–675, 2012.
SCHOLBERG, J. M. S. et al. Soil temperature, nitrogen concentration, and residence

107
time affect nitrogen uptake efficiency in Citrus. Journal of Environment Quality, v. 31, n. 3, p. 759, 2002.
SÉTAMOU, M. et al. Visual responses of adult asian citrus psyllid (Hemiptera: Liviidae) to colored sticky traps on citrus trees. Journal of Insect Behavior, v. 27, n. 4, p. 540–553, 2014.
SÉTAMOU, M. et al. Enhanced acquisition rates of “Candidatus Liberibacter asiaticus” by the asian citrus psyllid (Hemiptera: Liviidae) in the presence of vegetative flush growth in citrus. Journal of Economic Entomology, v. 109, n. 5, p. 1973–1978, 2016.
SPIEGEL-ROY, P.; GOLDSCHMIDT, E. E. Biology of Citrus. Cambridge: Cambridge University Press, 1996.
STATHAKOPOULOS, N.; ERICKSON, L. The effect of temperature on bud break in Poncirus trifoliata. Proceedings of the American Society of Horticultural Science, v. 89, p. 223–225, 1966.
STENZEL, N. M. C. et al. Maturation curves and degree-days accumulation for fruits of “Folha Murcha” orange trees. Scientia Agricola, v. 63, n. 3, p. 219–225, 2006.
SULE, H. et al. Response of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) to volatiles emitted from leaves of two rutaceous plants. Journal of Agricultural Science, v. 4, n. 6, p. 152–159, 2012.
SWINGLE, W. T.; REECE, P. C. The botany of Citrus and its wild relatives. In: REUTHER, W.; WEBBER, H. J.; BATCHELOR, L. D. (Eds.). . The citrus industry. 2. ed. Berkeley, California: University of California, 1967. v. 1p. 190–430.
TOMASETO, A. F.; KRUGNER, R.; LOPES, J. R. S. Effect of plant barriers and citrus leaf age on dispersal of Diaphorina citri (Hemiptera: Liviidae). Journal of Applied Entomology, v. 140, n. 1–2, p. 91–102, 2016.
TURRELL, F. M. Growth of the photosynthetic area of citrus. Botanical Gazette, v. 122, n. 4, p. 284–298, 1961.
TURRELL, F. M. et al. Growth equations and curves for citrus trees. Hilgardia, v. 39, n. 16, p. 429–445, 1969.
WILSON, L. T.; BARNETT, W. W. Degree-days: An aid in crop and pest management. California Agriculture, v. 37, n. 1, p. 4–7, 1983.
YAMAMOTO, P. T.; PAIVA, P. E. B.; GRAVENA, S. Flutuação populacional de Diaphorina citri Kuwayama (Hemiptera: Psyllidae) em pomares de citros na região Norte do Estado de São Paulo. Neotropical Entomology, v. 30, n. 1, p. 165–170, 2001.
ZHANG, J.-Z. et al. Involvements of PCD and changes in gene expression profile during self-pruning of spring shoots in sweet orange (Citrus sinensis). BMC Genomics, v. 15, n. 1, p. 892, 2014.

108
CAPÍTULO 5 – Limitada influência do porta-enxerto sobre a biologia de
Diaphorina citri em citros
RESUMO
RESUMO. Diaphorina citri é o vetor das bactérias associadas ao HLB, a doença mais
destrutiva dos citros. Os processos de aquisição e infecção da bactéria pelo inseto
ocorrem principalmente nos brotos jovens da planta. Estudos já conduzidos indicam
que existem diversos genótipos que emitem brotos poucos favoráveis ao inseto,
porém, a maioria dos estudos incluiu seedlings, ou seja, plantas oriundas de
sementes. Esta situação é muito diferente da situação real, onde o seedling atua como
porta-enxerto sobre o qual irá desenvolver a variedade copa. Estudos como os
conduzidos não permitiram, portanto, determinar o quanto uma possível
antibiose/antixenose poderia se transferir do porta-enxerto para a copa. Neste estudo
procurou-se corrigir este problema avaliando-se a influência de diferentes porta-
enxertos de citros na biologia de Diaphorina citri em copas de laranjeiras, com as
brotações que ocorreram a diferentes distâncias (alturas) do porta-enxerto. Em casa
de vegetação e câmaras de crescimento a preferência dos insetos pelos brotos foi
testada com e sem chance de escolha usando-se seedlings de citrumeleiro ‘Swingle’,
tangerineira ‘Sunki’, Poncyrus trifoliata, e dos híbridos 75AB, 79AC, 61AA3, 81G220
e 81G513, e mudas de laranjeira ‘Valência’ sobre o porta-enxerto limoeiro ‘Cravo’. Os
insetos também foram avaliados sobre mudas de ‘Valência’, ‘Hamlin’ e ‘Folha Murcha’
enxertadas em ‘Cravo’, ‘Swingle’ e ‘Sunki’ de 3 anos de idade, em casa de vegetação
e campo. Foi corroborada a baixa preferência de D. citri por ‘Sunki’, P. trifoliata e
‘Swingle’, também observada nas copas de mudas formadas sobre ‘Sunki’ e ‘Swingle’.
Adicionalmente foi estudada a oviposição, sem chance de escolha, em plantas adultas
de aproximadamente 2,5 m de altura estabelecidas em fazenda comercial. No campo,
o efeito do porta-enxerto já não foi mais significativo, talvez em decorrência da
“diluição” da(s) substância(s) do porta-enxerto durante seu trajeto até a extremidade
das copas de plantas de maior porte. Para testar esta hipótese, foram usadas mudas
de ‘Valência’ enxertadas em ‘Cravo’ e ‘Sunki’ (os mais contrastantes nos testes
anteriores), com a poda efetuada a duas alturas (15 e 150 cm acima da linha de
enxertia). Confirmou-se a menor preferência de D. citri por brotos localizados mais

109
próximos da linha de enxertia. Demostrou-se com este estudo a impropriedade de se
extrapolar resultados de testes feitos unicamente com seedlings ou mudas pequenas
para a variedade copa de laranjeiras, quando o objetivo é identificar porta-enxertos
com potencial impacto negativo sobre o inseto D. citri. A pequena ou ausência de
antibiose/antixenose dos porta-enxertos testados também explica a não observação
de qualquer efeito desses porta-enxertos na incidência de HLB no campo, sobre os
quais as copas devem estar atuando de forma similar na atração, alimentação e
reprodução de D. citri.
Palavras chave: Citrus sinensis, interação copa/porta-enxerto, HLB, antibiose,
antixenose, brotação.

110
CHAPTER 5 – Limited influence of the rootstock on the biology of Diaphorina
citri in citrus
ABSTRACT
ABSTRACT. Diaphorina citri is the vector of the bacteria associated with HLB, the
most destructive citrus disease. The acquisition or inoculation process occurs mainly
through the new shoots that develop at the end of branches. Several plant genotypes
has been reported as being less preferred by the insect. However, most studies used
young seedlings not allowing to determine if any of the antibiosis/antixenosis observed
is transferred to the scion of larger plants. The objective of this study was then to
evaluate the influence of different rootstocks on the biology of D. citri using seedlings
and young and adult trees of various rootstock/scion combinations. The insect
preference was assessed in several free- and no-choice tests on potted seedlings of
'Swingle', 'Sunki', Poncyrus trifoliata, and the hybrids ‘75AB’, ‘79AC’, ‘61AA3’, ‘81G220’
and 81G513, and young trees of ‘Valencia’/’Cravo’. Also, 3-year old potted plants of
‘Valencia’, ‘Hamlin’, ‘Folha Murcha’, all grafted in ‘Cravo’, ‘Swingle’ and ‘Sunki’, and
plants of the same combinations growing in the field were used. The low preference of
‘Sunki’, P. trifoliata and ‘Swingle’ in the biology of D. citri observed in other studies was
confirmed. The effect was transferred from seedlings to the scion in potted plants, with
lower insect preference for the scions grafted in ‘Sunki’ and ‘Swingle’. After these tests,
a no-choice oviposition test was carried out on 2,5 m height adult plants growing in a
commercial farm. Here, the effect of the rootstock was no longer significant. It was
hypothesized that the putative substance(s) present in the rootstock became less
concentrated as it moved through the scion of larger trees. In order to test this
hypothesis, nursery potted plants of ‘Valencia’ grafted on ‘Cravo’ and ‘Sunki’ (the most
contrasting rootstock in the previous tests), pruned at two distances above the grafting
line (15 and 150 cm) were used. It was confirmed the lower preference of D. citri for
new shoots emitted near of the rootstock, while the most distant were most suitable.
Overall this study showed how inappropriate it could be to extrapolate to adult citrus
trees the significance of results of tests carried out using young seedlings, in studies
carried out with the objective of finding rootstock useful to produce trees repellant or
not suitable to D. citri. The study also explains the lack of any influence of rootstock on

111
HLB incidence observed in the field, on which the scions might have similar
attractiveness to the insect.
Key words: Citrus sinensis, rootstock scion communication, huanglongbing,
antibiosis, antixenosis, flush shoots.

112
1. INTRODUÇÃO
O psilídeo Diaphorina citri Kuwayama (Hemiptera: Liviidae) é o inseto vetor
das bactérias associadas ao huanglongbing ‘Candidatus Liberibacter asiaticus’ (Las)
e ‘C. L. americanus’ (Lam), a doença mais destrutiva dos citros (DA GRAÇA et al.,
2016). Do primeiro relato do inseto no Brasil em 1942 (COSTA-LIMA, 1942) até o ano
de 2004, o psilídeo passava invariavelmente despercebido nos pomares. Mas, com o
relato da ocorrência de Liberibacter nos pomares paulistas em 2004 e 2005
(COLETTA-FILHO et al., 2004; TEIXEIRA et al., 2005), o inseto passou a ser alvo de
controle químico, que se intensificou ao longo do tempo. Esta medida faz parte de
pacote que inclui também o plantio de mudas produzidas em ambientes protegidos e
certificados e redução das fontes de inóculo (erradicação de plantas) (BASSANEZI et
al., 2013; BASSANEZI; BELASQUE; MONTESINO, 2013; BOINA; BLOOMQUIST,
2015). Infelizmente, apesar de todos esses esforços, a doença continuou avançando,
levando à erradicação de aproximadamente 46,2 milhões de plantas somente no
cinturão citrícola de São Paulo e Triângulo Mineiro. Em 2017 a doença havia sido
detectada em 16,73% dos 191,7 milhões de árvores plantadas no mencionado
cinturão (FUNDECITRUS, 2017).
Para o controle do HLB duas estratégias potencialmente promissoras se
destacam, mas ainda sob investigação. A primeira consiste na incorporação nos citros
de genes que confiram à planta resistência ou tolerância à multiplicação da bactéria
do HLB, o que só é possível por meio de técnicas biotecnológicas e, para fontes dos
genes, empregando espécies filogeneticamente distantes do gênero Citrus (MILES et
al., 2017; RAMADUGU et al., 2016). A segunda consiste no uso de variedades/
espécies de plantas pouco atrativas ou inapropriadas à alimentação e/ou reprodução
do psilídeo. Contrário do que acontece em relação à bactéria, já foram identificadas
em Citrus e parentes próximos espécies mais ou menos resistentes ao inseto
envolvendo os fenômenos de antixenose e/ou antibiose (BORGONI et al., 2014; NAVA
et al., 2007). Outras foram adequadas à sobrevivência de D. citri mas não à sua
reprodução (RUSSELL et al., 2014; SÉTAMOU; DA GRAÇA; SANDOVAL, 2016). De
especial interesse têm sido o porta-enxerto Poncirus trifoliata e seus híbridos que,

113
além de apresentar relativa tolerância à infecção por Las (ALBRECHT; BOWMAN,
2012a, 2012b; BOAVA; CRISTOFANI-YALY; MACHADO, 2017), também repele o
psilídeo (BORGONI et al., 2014; HALL; GEORGE; LAPOINTE, 2015; RICHARDSON;
HALL, 2013). No Brasil este porta-enxerto é pouco empregado, contrário do que
acontece com o limoeiro ‘Cravo’ (Citrus limonia Osbeck), o citrumeleiro ‘Swingle’ (P.
trifoliata (L.) Raf. × Citrus paradisi Macfad.) e a tangerineira ‘Sunki’ (Citrus sunki)
(POMPEU JUNIOR, 2005). Em estudos já conduzidos, os dois últimos foram
relativamente menos apropriados para o desenvolvimento do inseto que o ‘Cravo’
(BORGONI et al., 2014; NAVA et al., 2007).
Na maioria dos trabalhos em que foi estudada a biologia de D. citri em
hospedeiros cítricos e não cítricos foram empregados seedlings (ALVES; DINIZ;
PARRA, 2014; NAVA et al., 2007; TSAI; LIU, 2000), ou seja, plantas oriundas de
sementes de poucas semanas ou meses de idade. Efeitos desfavoráveis a D. citri
foram verificados (NAVA et al., 2007), mas os resultados não condizem com o que se
observa no campo. A razão pode estar no uso de seedlings, possivelmente
inadequados a tais estudos. Em primeiro lugar, nos seedlings os tecidos são
ontogeneticamente juvenis, contrário do que ocorre com as copas, todas constituídas
por tecido maduro. Evidências existem de que a biologia de certos insetos se vê
afetada pela idade fisiológica da planta (KEARSLEY; WHITHAM, 1989). Além disso,
há que se considerar que uma planta cítrica explorada comercialmente consiste numa
combinação de duas espécies, com a que serve de suporte (o porta-enxerto) podendo
influenciar inúmeros aspectos da fisiologia da copa nele enxertada (YONEMOTO et
al., 2004). Vale lembrar que são as extremidades da copa os locais onde D. citri irá se
desenvolver, e essas extremidades podem estar a diferentes distâncias do porta-
enxerto, dependendo da altura da planta. Assim, surge a dúvida: até que ponto a
influência de um porta-enxerto, identificado em plantas jovens e pé franco (seedlings),
irá se refletir na extremidade da copa de uma planta adulta?
Evidência da influência do porta-enxerto sobre a variedade copa vem do
trabalho de Bruessow et al. (2010). Enquanto que a laranjeira azeda usada como pé
franco afetou diversos parâmetros da biologia do ácaro Tetranychus urticae, quando
usada como porta-enxerto, só alguns parâmetros foram afetados negativamente na
copa da tangerineira Satsuma cv. ‘Hashimoto’. Sugeriu-se que os efeitos sobre o

114
ácaro deviam-se a incrementos induzidos na atividade das rotas metabólicas do ácido
jasmônico e de flavonoides (AGUT et al., 2014), enquanto que poucos efeitos
sistêmicos verificados na copa foram mediados por aminoácidos (AGUT et al., 2016).
Dada a ausência de informação sobre o real impacto de porta-enxertos na
performance de D. citri na copa de plantas adultas, foi proposto este trabalho para
determinar o alcance relativo da influência dos porta-enxertos sobre a biologia de
Diaphorina citri.
2. MATERIAL E MÉTODOS
2.1. Material vegetal e criação de Diaphorina citri
Foram utilizados três grupos de plantas. O primeiro, usado nos experimentos
1 e 2, foi constituído por mudas de laranjeira ‘Valência’ enxertadas sobre limoeiro
‘Cravo’, e seedlings pé-franco de citrumelo ‘Swingle’, tangerina ‘Sunki’, P. trifoliata, e
dos híbridos 75AB (Citrus paradisi x Poncirus trifoliata), ‘79AC’ (Citrus reshni x
(Poncirus trifoliata x Citrus paradisi)), ‘61AA3’ (Citrus reshni x Poncirus trifoliata),
‘81G220’ e ‘81G513’ (Citrus volkameriana x Citrus reshni). Também foram incluídos
seedlings pé-franco de murta (Murraya exotica). Todos os materiais foram plantados
em vaso de 1,7 L, apresentando um ano de idade quando os experimentos iniciaram.
As sementes dos cinco últimos híbridos foram importadas da Estação Experimental
Obispo Colombres, Argentina (Processo MAPA: 21052.003390/2014 – 48). O
segundo grupo de plantas, usado nos experimentos 3 e 5, foi constituído por mudas
de 2,5 anos de idade das laranjeiras doces ‘Valência’, ‘Hamlin’ e ‘Folha Murcha’
enxertadas em Cravo, Swingle e Sunki plantadas em vasos de 4.7 L (ECOVASO®,
SP). O substrato usado para esses dois grupos foi Plantmax Citrus®, com irrigação
feita de acordo com a necessidade e fertirrigação quinzenal. O terceiro grupo de
plantas (experimento 4) foi constituído pelas mesmas variedades do grupo 2,
plantadas em fazenda comercial e com aproximadamente 3 anos de idade.

115
A criação dos adultos foi feita usando como base uma colônia estabelecida no
Fundo de Defesa da Citricultura (Fundecitrus, Araraquara – SP). Antes da realização
de cada experimento, 10 a 15 murtas foram podadas no topo para estimular a
brotação. Cada planta foi totalmente envolvida por um saco de tecido de tunil medindo
30 x 40 cm. Trinta a quarenta adultos foram confinados em cada planta de murta
mantida em sala climatizada (28 ºC ± 2, 75% UR ± 6 e fotoperíodo 12h), metodologia
adaptada de Skelley e Hoy (2004). Após eclosão e desenvolvimento ninfal, os adultos
emergidos foram coletados a cada 24 h quando é maior a chance de ainda serem
imaturos sexualmente e não terem se acasalado (WENNINGER; HALL, 2007). Foram
em seguida sexados com lupa manual de 30X, observando-se a genitália no extremo
abdominal (AUBERT, 1987), e colocados separadamente machos de fêmeas em
novas plantas de murtas com brotos em desenvolvimento, mantidos aí até serem
utilizados em cada experimento (15 ± 1 dia). Cada lote de insetos foi constituído por
adultos coletados em todas as plantas no pico gaussiano de emergência, ou seja,
entre o terceiro e o quinto dia após início da emergência.
2.2. Descrição dos experimentos
Experimento 1: Dez plantas de cada seedling (murta, ‘Cravo’, ‘Swingle’,
‘Sunki’, ‘Trifoliata’, ‘81G220’, ‘81G513’, ‘61AA3’, ‘75AB’, ‘79AC’) e dez plantas de
laranjeira ‘Valência’ enxertada em limoeiro ‘Cravo’ foram podadas e totalmente
desfolhadas 25 cm acima do nível do substrato (seedling) e 15 cm acima da linha de
enxertia (muda). Quando os brotos em estádio V2 (CIFUENTES-ARENAS et al., 2016)
estavam presente, selecionou-se um broto (destacando os subsequentes que saíam)
e sobre ele foram confinados para acasalamento e oviposição em cada planta, durante
72 h, dois casais de D. citri. Esse tempo passou a ser contado somente a partir do
momento em que foi confirmada a presença de ovos em observações feitas a cada
24 h. Foi assim que se determinou o tempo necessário para começo da oviposição
(TCO).

116
Durante o período de confinamento (TCO + 72h) foi registrado a cada 24 h se
os insetos se encontravam posicionados sobre o broto ou no saco de tunil. Isto
permitiu determinar a média ponderada do número de insetos encontrados pousados
sobre o broto (PP) como 𝑃𝑃 = [(∑ 𝐴𝐷𝐵𝑟𝑖
𝑛𝑖=1 ) (𝐴𝐷𝑇×𝑛)⁄ ]×100, onde ADBr.i: número de
adultos pousados no broto na avaliação i, ADT: número total de adultos confinados por
broto, e n: número total de avaliações. O número de ovos e ninfas emergidas foi
contabilizado com auxílio de uma lupa manual de 30X e estimada a viabilidade de
ovos. A sobrevivência de ninfas foi estimada transferindo-se, com auxílio de uma lupa
hipodérmica de 5 mL, 20 ninfas de primeiro instar com menos de 6 h de idade, de
brotos de murta para brotos em estádios V2 de plantas dos onze materiais (n = 8),
apresentando todas as mesmas características acima descritas. Após o
desenvolvimento ninfal, os adultos emergidos foram congelados e sexados com
auxílio de estereoscópio para determinar a razão sexual (fêmeas/(adultos
emergidos)).
Este experimento foi desenvolvido no Fundo de Defesa da Citricultura
(Fundecitrus, Araraquara – SP) dentro de câmara de crescimento (modelo CMP4030,
Conviron Inc., Winnipeg, Canada) onde a temperatura e umidade variaram tentando
simular condições aproximadas de um dia “normal” em uma região de clima mediano,
ou seja, fotoperíodo de 12h D:N com radiação fotossinteticamente ativa (PAR) de 300
μmol m−2 s−1. A temperatura e a umidade foram de 17,84 °C ± 0,05 e 86,84% ± 0,38
(12h, período noturno), seguidos de 25,43 °C ± 0,05 e 69.93% ± 0.72 (3h, as primeiras
horas do dia), de 30,83 °C ± 0,04 e 72,22% ± 0,69 (6h, período mais quente do dia),
e finalmente 24,82 °C ± 0,05 e 77,09% ± 0,85 (3h, final de tarde e início da noite). Os
registros de temperatura e umidade relativa em todos os ambientes onde foram
desenvolvidos os diferentes experimentos foram coletados com registradores LogBox-
AA ou LogBox-RHT-LCD (NOVUS Produtos Eletrônicos Ltda, Porto Alegre, RS, Brasil)
em intervalos de uma h.
Experimento 2: Foram usadas plantas com as mesmas características que
as descritas para as usadas no Exp. 1. Quando dois a três brotos em estádio V2
estavam presentes, as plantas foram colocadas individualmente dentro de uma gaiola
de 45 cm x 45 cm x 71 cm feita com tela antiafídeos transparente (0,87 mm x 0,30

117
mm) e com estrutura em tubos de ½” de PVC. No centro de cada gaiola foram
liberados e mantidos durante 7 dias 100 adultos em proporção de 1:1 macho:fêmea
para acasalamento e oviposição. Após o período de confinamento, os adultos foram
retirados registrando-se o lugar onde estavam pousados (planta ou paredes da
gaiola). Os brotos foram destacados e examinados sob microscópio estereoscópio
para contagem de ovos e estimativa da percentagem de brotos com ovos. Foram
usadas 9 gaiolas. A murta não foi incluída nesse teste devido a sua possível
interferência em D. citri na livre escolha de hospedeiro ao ser esta a planta usada na
criação e conservação do inseto. Este estudo foi conduzido dentro de estufa com
controle de temperatura e irrigação no Fundecitrus. Durante esse experimento a
temperatura (°C) e a umidade relativa (%) dentro da estufa oscilaram da seguinte
forma: 20,59 °C ± 0,93 e 89,93% ± 1,48 entre 06:00h e 08:59h, 32,50 °C ± 0,71 e
59,61% ± 4,18 entre 09:00h e 14:59h, 31,60 °C ± 0,37 e 61,91% ± 3,80 entre 15:00h
e 17:59h, e 19,86 °C ± 0,81 e 89,75% ± 1,44 entre 18:00h e 05:59h.
Experimento 3: Após o primeiro ‘screening’ dos materiais nos primeiros dois
experimentos, foram selecionados os três porta-enxertos mais plantados no Brasil
(POMPEU JUNIOR, 2005), para tentar estabelecer se as diferenças observadas na
fase de pé franco seriam transmitidas para a copa. Neste caso foram usadas mudas
de laranjeiras doces das variedades ‘Valência’, ‘Hamlin’ e ‘Folha Murcha’ todas
enxertadas em limoeiro ‘Cravo’, citrumelo ‘Swingle’ e tangerineira ‘Sunki’ em esquema
fatorial 33. As plantas foram podadas e desfolhadas 20 cm acima da linha de enxertia
e quando um broto no estádio V2 estava presente (destacando os emergidos
subsequentes) dois casais de D. citri foram confinados nesse único broto por planta
(repetição) durante 72 horas para acasalamento e oviposição. Foi determinada a
preferência de pouso, a frequência de oviposição e o número de ovos em dois
ambientes. O primeiro foi conduzido em sala climatizada com fotoperíodo de 12 h,
iluminação fornecida por tubos fluorescentes e lâmpadas incandescentes (60 μmol
m−2 s−1 PAR) e temperatura e umidade médias de 28,25 °C ± 0,79 e 79,15% °C ± 1,20,
respectivamente. O segundo foi conduzido em condições de telado onde, durante o
estudo, a temperatura e umidade foram 22,37 °C ± 0,41 e 62,95% ± 1.81 entre 06:00h
e 08:59h, 32,33 °C ± 0,54 e 36,93% ± 2,08 entre 09:00h e 14:59h, 32,13 °C ± 0,51 e

118
35,49% ± 2,14 entre 15:00h e 17:59h, e 21,16 ± 0,33 e 62,55% ± 1,78 entre 18:00h e
05:59h.
Experimento 4: esse experimento consistiu de um teste de oviposição sem
chance de escolha usando plantas de 3 anos de idade estabelecidas em fazenda
comercial no município de Onda Verde, SP, das mesmas variedades usadas no
experimento 3. Os ramos secundários de 5 plantas de cada combinação foram
podados e desfolhados para estimular emissão de novos brotos. Quando a maioria
dos brotos estavam em estádio V2 ou começo do V3 (≈ 1 mês depois da poda), 3 a 5
brotos por planta foram selecionados para confinamento, durante 72 h, de dois casais
virgens em cada um. Após o período de confinamento foi registrada a presença dos
insetos sobre ou fora do broto e uma porção do ramo (aproximadamente 5 cm) no
qual o saco de tunil foi prendido e cortado mantendo tanto o broto como os insetos
ainda dentro para evitar que escapassem ao pomar. Os sacos de tunil foram
colocados cuidadosamente dentro de caixas de isopor refrigeradas e transportados
rapidamente para o laboratório (< 4 h) onde os adultos foram retirados e os brotos
guardados em câmara fria (4 °C) até a contagem de ovos. Cada broto foi considerado
como uma repetição. Foi determinada a preferência de pouso, a percentagem de
brotos com ovos, e a quantidade de ovos. Durante o período de brotação das plantas
a temperatura e a umidade relativa oscilou da seguinte forma: 17,15 °C ± 0,46 e
96,68% ± 0,17 entre 06:00h 08:59h, 26,03 °C ± 0,33 e 77,24% ± 1,15 entre 09:00h e
14:59h, 30,93 °C ± 0,43 e 60,09% ± 2,15 entre 15:00h e 17:59h, e 20,99 °C ± 0,31 e
89,49% ± 0,61 entre 18:00h e 05:59h. Durante o período de confinamento de 72 h a
variação foi: 14,42 °C ± 1,21 e 96,47% ± 0,18 entre 06:00h – 08:59h, 23,49 °C ± 0,77
e 77,81% ± 1,36 entre 09:00h e 14:59h, 28,94 °C ± 0,45 e 63,39% ± 5,16 entre 15:00h
e 17:59h, e 18,88 °C ± 0,87 e 92,16% ± 1,16 18:00h – 05:59h.
Experimento 5: o objetivo deste experimento foi determinar a influência da
altura da planta (distância entre o broto e o porta-enxerto) na possível “diluição” do
efeito do porta-enxerto no comportamento de D. citri sobre os brotos emitidos na copa.
Foram usadas mudas de laranjeira ‘Valência’ enxertadas em limoeiro ‘Cravo’ e
tangerina ‘Sunki’ com características semelhantes às usadas no experimento 3,

119
descrito acima. As plantas foram divididas em dois grupos: no primeiro, foi feita uma
poda e desfolha 15 cm acima da linha de enxertia; no segundo, a poda foi realizada
nos ponteiros dos ramos mais distantes da linha de enxertia (130 a 150 cm). O
confinamento dos dois casais virgens e os parâmetros avaliados (preferência de
pouso, tempo para começo da oviposição, número de ovos) foram feitos conforme já
descrito. O desenho experimental foi o fatorial 22 (duas alturas e dois porta-enxertos).
2.3. Análises de dados
Os dados de preferência de pouso, porcentagem de brotos com ovos, e
viabilidade de ovos e ninfas foram transformados (y’ = arcseno(sqrt(y/100))) quando
necessário e submetidos à análise de variância usando o Modelo Linear Generalizado
(GLM), com o teste Tukey-HSD para separação de médias, ou análise de residuais
padronizados para determinar frequências que fogem do esperado. A preferência de
pouso no Exp. 2 foi analisada pelo teste Friedman para múltiplas amostras
dependentes, e as médias foram separadas por meio do teste de Nemenyi. O tempo
para começo da oviposição e o número de ovos foi submetida à análise de variância
por meio de GLM. No experimento 6 houve interação significativa, motivo pelo qual a
natureza dessa interação foi analisada por meio do teste t Student para amostras
independentes. Todas as análises foram realizadas nos programas “R” (R CORE
TEAM, 2014), pacotes glm, multcomp e PMCMR, e Statgraphics Centurion XVII
(Statpoint Tecnologies Inc.) considerando α = 0,05.
3. RESULTADOS
Preferência para pouso
A preferência de D. citri por permanecer pousado sobre o broto no Exp. 1
(Figura 1a) foi influenciada significativamente pelo hospedeiro (F10, 99 = 5,35; P <
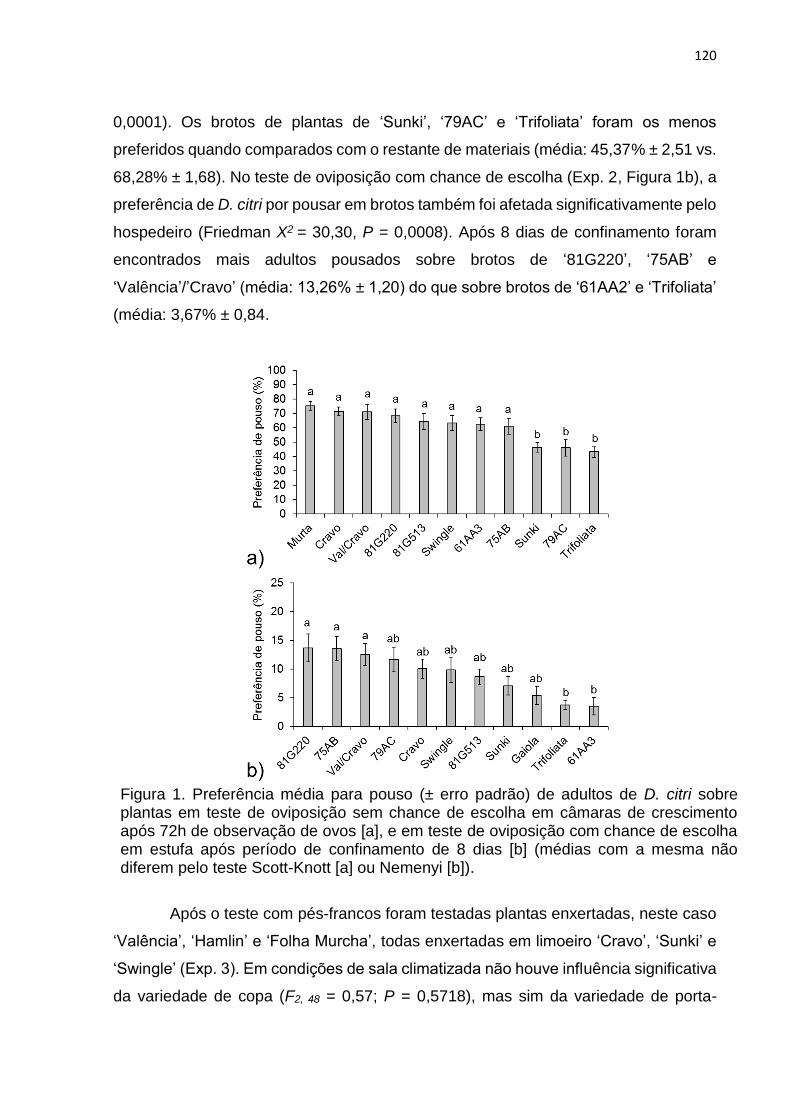
120
0,0001). Os brotos de plantas de ‘Sunki’, ‘79AC’ e ‘Trifoliata’ foram os menos
preferidos quando comparados com o restante de materiais (média: 45,37% ± 2,51 vs.
68,28% ± 1,68). No teste de oviposição com chance de escolha (Exp. 2, Figura 1b), a
preferência de D. citri por pousar em brotos também foi afetada significativamente pelo
hospedeiro (Friedman X2 = 30,30, P = 0,0008). Após 8 dias de confinamento foram
encontrados mais adultos pousados sobre brotos de ‘81G220’, ‘75AB’ e
‘Valência’/’Cravo’ (média: 13,26% ± 1,20) do que sobre brotos de ‘61AA2’ e ‘Trifoliata’
(média: 3,67% ± 0,84.
Figura 1. Preferência média para pouso (± erro padrão) de adultos de D. citri sobre plantas em teste de oviposição sem chance de escolha em câmaras de crescimento após 72h de observação de ovos [a], e em teste de oviposição com chance de escolha em estufa após período de confinamento de 8 dias [b] (médias com a mesma não diferem pelo teste Scott-Knott [a] ou Nemenyi [b]).
Após o teste com pés-francos foram testadas plantas enxertadas, neste caso
‘Valência’, ‘Hamlin’ e ‘Folha Murcha’, todas enxertadas em limoeiro ‘Cravo’, ‘Sunki’ e
‘Swingle’ (Exp. 3). Em condições de sala climatizada não houve influência significativa
da variedade de copa (F2, 48 = 0,57; P = 0,5718), mas sim da variedade de porta-

121
enxerto (F2, 48 = 39,53; P < 0.0001). A interação Copa*Porta-enxerto foi insignificante
(F4, 48 = 1,63; P = 0,1844). Os insetos preferiram, em média, 52,94% ± 8,79, 56,28%
± 8,70 e das copas de ‘Valência’, das de ‘Folha Murcha’ e 62,50% ± 7,65 das de
‘Hamlin’. Já para os porta-enxertos o ‘Cravo’ diferiu significativamente do ‘Swingle’ e
do ‘Sunki’ (teste Tukey, P < 0,0001), com 93,06% ± 2,64, 35,00% ± 6,15 e 37,50%
±6,61, respectivamente.
Similar à sala climatizada, no telado a variedade de copa não mostrou
influência significativa (F2, 102 = 0,70; P = 0,4975), porém a variedade de porta-enxerto
mostrou-se altamente determinante na preferência de pouso (F2, 102 = 16,01; P <
0,0001); a interação entre os fatores também não foi significativa (F4, 102 = 1,88; P =
0,1202). A preferência de pouso foi de 53,57% ± 6,16, 59,46% ± 7,24 e 65,32% ± 6,62
para ‘Valência’, ‘Folha Murcha’ e ‘Hamlin’, respectivamente, enquanto que ‘Cravo’
diferiu significativamente de ‘Sunki’ e ‘Swingle’ (teste Tukey, P < 0,01), com 84,72% ±
4,74, 34,38 ± 6,32 e 55,71 ± 6,32, respectivamente.
Depois de ter testado esses materiais em sala climatizada (condições
controladas) e em telado (condições semi-controladas), foi realizado um teste de
oviposição sem chance de escolha em fazenda comercial com as mesmas
combinações (Exp. 4). Desta vez não foi evidenciada influência significativa da
variedade de Copa ou de Porta-enxerto na percentagem de adultos encontrados sobre
o broto no momento de sua remoção (Copa: X2 = 1,93, GL = 2, P = 0,3797; P. Enx.:
X2 = 1,80, GL = 2, P = 0,4075; CxP: X2 = 3,95, GL = 4, P = 0,4131; n = 219 brotos e
876 adultos). Em média 82,21% ± 1,20 dos adultos foram encontrados sobre os
brotos.
Preferência para oviposição
A preferência de oviposição foi estudada considerando-se o tempo para
começo da oviposição no Experimentos 1 e 5 e a percentagem de brotos nos quais D.
citri colocou ovos nos experimentos 2, 3 e 4. O tempo necessário para começo da
oviposição não foi influenciado pelo material vegetal no Experimento 1 (F10, 109 = 0,92;
P = 0,5181; �̅� = 5,30 ± 0,32 dias) (Figura 2a). No teste de oviposição com chance de
escolha (Exp. 2) a proporção de brotos com ovos foi influenciada pela planta (X2 =
41,58; GL = 9; P < 0,0001), maior em ‘Valência’/’Cravo’, ‘Sunki’ e ‘79AC’ (+16,0%,
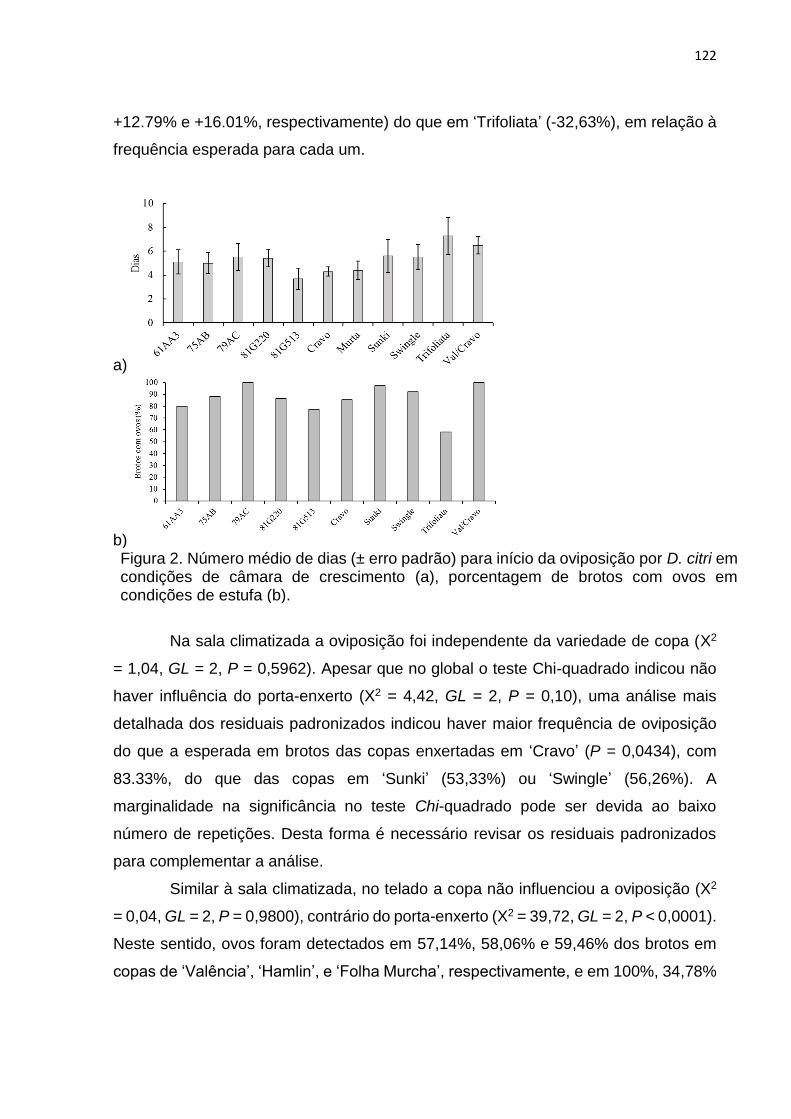
122
+12.79% e +16.01%, respectivamente) do que em ‘Trifoliata’ (-32,63%), em relação à
frequência esperada para cada um.
a)
b) Figura 2. Número médio de dias (± erro padrão) para início da oviposição por D. citri em condições de câmara de crescimento (a), porcentagem de brotos com ovos em condições de estufa (b).
Na sala climatizada a oviposição foi independente da variedade de copa (X2
= 1,04, GL = 2, P = 0,5962). Apesar que no global o teste Chi-quadrado indicou não
haver influência do porta-enxerto (X2 = 4,42, GL = 2, P = 0,10), uma análise mais
detalhada dos residuais padronizados indicou haver maior frequência de oviposição
do que a esperada em brotos das copas enxertadas em ‘Cravo’ (P = 0,0434), com
83.33%, do que das copas em ‘Sunki’ (53,33%) ou ‘Swingle’ (56,26%). A
marginalidade na significância no teste Chi-quadrado pode ser devida ao baixo
número de repetições. Desta forma é necessário revisar os residuais padronizados
para complementar a análise.
Similar à sala climatizada, no telado a copa não influenciou a oviposição (X2
= 0,04, GL = 2, P = 0,9800), contrário do porta-enxerto (X2 = 39,72, GL = 2, P < 0,0001).
Neste sentido, ovos foram detectados em 57,14%, 58,06% e 59,46% dos brotos em
copas de ‘Valência’, ‘Hamlin’, e ‘Folha Murcha’, respectivamente, e em 100%, 34,78%

123
e 37,14% dos brotos de copas enxertadas em ‘Cravo’, ‘Sunki’, e ‘Swingle’,
respectivamente.
No teste de oviposição em campo, não houve efeito do porta-enxerto na
preferência por oviposição (X2 = 3,50, GL = 2, P = 0,1742), mas sim o fator Copa (X2
= 6,88, GL = 2, P = 0,0321); a interação copa*porta-enxerto foi significativa (X2 = 11,14,
GL = 4, P = 0,0251), indicando que o efeito da copa depende do porta-enxerto.
Analisando a natureza da interação, observou-se maior frequência de ovos nas copas
‘Hamlin’ e ‘Valência’ sobre ‘Cravo’ e ‘Sunki’ do que na copa de Hamlin sobre Sunki
(Tabela 1).
Tabela 1. Percentagem de brotos com ovos no teste de oviposição sem chance de escolha após confinamento por 72 h em brotos de plantas de três anos de idade cultivadas em fazenda comercial no município de Onda Verde – SP.
Cravo +* Sunki +* Swingle +* Total copas +*
Valência 61,11 +* 75,00 +* 78,57 +* 53.66+*
Hamlin 88,89 +* 36,36 -* 60,00 +* 68.25+*
Folha Murcha 60,00 +* 46,43 +* 54,17 +* 72.97+*
Total porta-enxertos 70.67+* 56.72+* 64.94+* 64.38+* +* Combinação com frequência significativamente maior à esperada (P < 0.05). -* Combinação com frequência significativamente inferior à esperada (P < 0.05).
Número e viabilidade de ovos e ninfas
No Exp. 1 o material vegetal influenciou a oviposição (F10, 99 = 16,57; P <
0,0001) e a viabilidade de ninfas (F10, 77 = 4,53; P < 0,0001) mas não influenciou a
viabilidade de ovos (F10, 99 = 1,80; P = 0,0689), com média geral de 69,05% ±1,43,
nem a razão sexual (X2 = 6,17, GL = 10, P = 0,8004), com uma proporção de f[emeas
de 0,545 ± 0,025. Menor número de ovos foi encontrado em ‘Trifoliata’, ‘Swingle’ e
‘Sunki’, com destaque para ‘Trifoliata’ (Tabela 2). ‘Murta’, ‘Cravo’, ‘81G220’ e
‘Valência’/’Cravo’ foram os mais favoráveis. Os materiais argentinos ‘75AB’, ‘79AC’,
‘81G513’ e ‘61AA3’ foram medianamente favoráveis para a oviposição. As ninfas
sobreviveram em maiores proporções em ‘Valência’/’Cravo’, ‘Murta’ e ‘Cravo’ do que
em ‘Sunki’, ‘61AA3’, ‘75AB’, ‘81G220’ e ‘Trifoliata’.
No Exp. 2 foi avaliada somente a oviposição que, similar ao Exp. 1, também
foi afetada pelo hospedeiro (F9, 89 = 13,87; P < 0,0001), com ‘Trifoliata’ sendo o
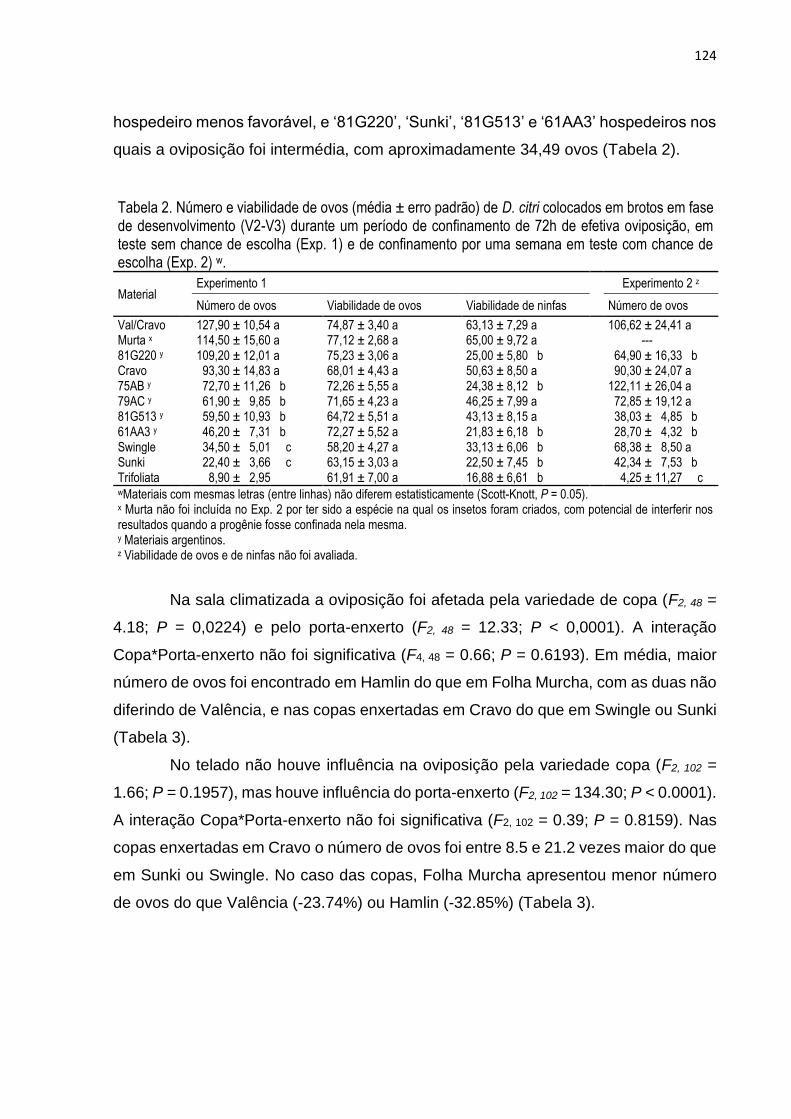
124
hospedeiro menos favorável, e ‘81G220’, ‘Sunki’, ‘81G513’ e ‘61AA3’ hospedeiros nos
quais a oviposição foi intermédia, com aproximadamente 34,49 ovos (Tabela 2).
Tabela 2. Número e viabilidade de ovos (média ± erro padrão) de D. citri colocados em brotos em fase de desenvolvimento (V2-V3) durante um período de confinamento de 72h de efetiva oviposição, em teste sem chance de escolha (Exp. 1) e de confinamento por uma semana em teste com chance de escolha (Exp. 2) w.
Material
Experimento 1 Experimento 2 z
Número de ovos Viabilidade de ovos Viabilidade de ninfas Número de ovos
Val/Cravo 127,90 ± 10,54 a 74,87 ± 3,40 a 63,13 ± 7,29 a 106,62 ± 24,41 a Murta x 114,50 ± 15,60 a 77,12 ± 2,68 a 65,00 ± 9,72 a --- 81G220 y 109,20 ± 12,01 a 75,23 ± 3,06 a 25,00 ± 5,80 abcde 164,90 ± 16,33 b Cravo 193,30 ± 14,83 a 68,01 ± 4,43 a 50,63 ± 8,50 a 190,30 ± 24,07 a 75AB y 172,70 ± 11,26 b 72,26 ± 5,55 a 24,38 ± 8,12 abcde 122,11 ± 26,04 a 79AC y 161,90 ± 19,85 b 71,65 ± 4,23 a 46,25 ± 7,99 a 172,85 ± 19,12 a 81G513 y 159,50 ± 10,93 b 64,72 ± 5,51 a 43,13 ± 8,15 a 138,03 ± 4,85 b 61AA3 y 146,20 ± 17,31 ab 72,27 ± 5,52 a 21,83 ± 6,18 abcdef 128,70 ± 4,32 b Swingle 134,50 ± 15,01 c 58,20 ± 4,27 a 33,13 ± 6,06 b 168,38 ± 8,50 a Sunki 122,40 ± 13,66 c 63,15 ± 3,03 a 22,50 ± 7,45 abcdef 142,34 ± 7,53 b Trifoliata 118,90 ± 12,95 c 61,91 ± 7,00 a 16,88 ± 6,61 abcdef 114,25 ± 11,27 c wMateriais com mesmas letras (entre linhas) não diferem estatisticamente (Scott-Knott, P = 0.05). x Murta não foi incluída no Exp. 2 por ter sido a espécie na qual os insetos foram criados, com potencial de interferir nos resultados quando a progênie fosse confinada nela mesma. y Materiais argentinos. z Viabilidade de ovos e de ninfas não foi avaliada.
Na sala climatizada a oviposição foi afetada pela variedade de copa (F2, 48 =
4.18; P = 0,0224) e pelo porta-enxerto (F2, 48 = 12.33; P < 0,0001). A interação
Copa*Porta-enxerto não foi significativa (F4, 48 = 0.66; P = 0.6193). Em média, maior
número de ovos foi encontrado em Hamlin do que em Folha Murcha, com as duas não
diferindo de Valência, e nas copas enxertadas em Cravo do que em Swingle ou Sunki
(Tabela 3).
No telado não houve influência na oviposição pela variedade copa (F2, 102 =
1.66; P = 0.1957), mas houve influência do porta-enxerto (F2, 102 = 134.30; P < 0.0001).
A interação Copa*Porta-enxerto não foi significativa (F2, 102 = 0.39; P = 0.8159). Nas
copas enxertadas em Cravo o número de ovos foi entre 8.5 e 21.2 vezes maior do que
em Sunki ou Swingle. No caso das copas, Folha Murcha apresentou menor número
de ovos do que Valência (-23.74%) ou Hamlin (-32.85%) (Tabela 3).
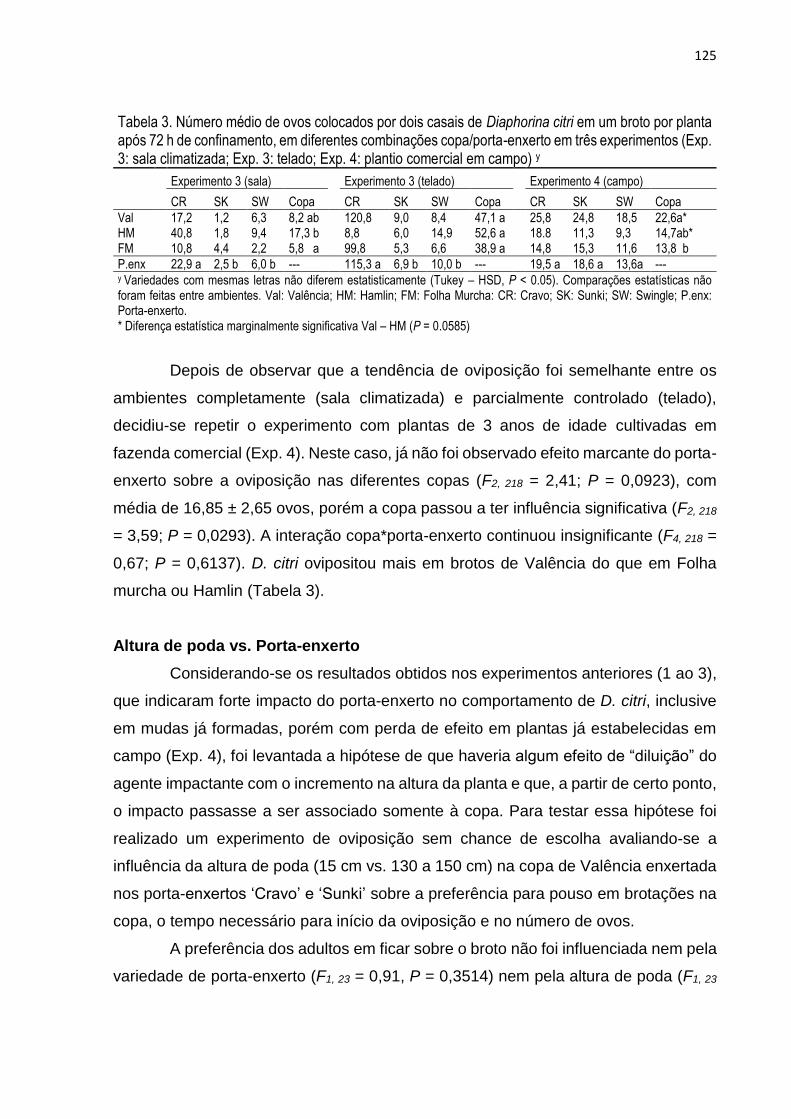
125
Tabela 3. Número médio de ovos colocados por dois casais de Diaphorina citri em um broto por planta após 72 h de confinamento, em diferentes combinações copa/porta-enxerto em três experimentos (Exp. 3: sala climatizada; Exp. 3: telado; Exp. 4: plantio comercial em campo) y
Experimento 3 (sala) Experimento 3 (telado) Experimento 4 (campo)
CR SK SW Copa CR SK SW Copa CR SK SW Copa
Val 17,2 1,2 6,3 8,2 ab 120,8 9,0 8,4 47,1 a 25,8 24,8 18,5 22,6a* HM 40,8 1,8 9,4 17,3 b 8,8 6,0 14,9 52,6 a 18.8 11,3 9,3 14,7ab* FM 10,8 4,4 2,2 5,8 a 99,8 5,3 6,6 38,9 a 14,8 15,3 11,6 13,8 b
P.enx 22,9 a 2,5 b 6,0 b --- 115,3 a 6,9 b 10,0 b --- 19,5 a 18,6 a 13,6a --- y Variedades com mesmas letras não diferem estatisticamente (Tukey – HSD, P < 0.05). Comparações estatísticas não foram feitas entre ambientes. Val: Valência; HM: Hamlin; FM: Folha Murcha: CR: Cravo; SK: Sunki; SW: Swingle; P.enx: Porta-enxerto. * Diferença estatística marginalmente significativa Val – HM (P = 0.0585)
Depois de observar que a tendência de oviposição foi semelhante entre os
ambientes completamente (sala climatizada) e parcialmente controlado (telado),
decidiu-se repetir o experimento com plantas de 3 anos de idade cultivadas em
fazenda comercial (Exp. 4). Neste caso, já não foi observado efeito marcante do porta-
enxerto sobre a oviposição nas diferentes copas (F2, 218 = 2,41; P = 0,0923), com
média de 16,85 ± 2,65 ovos, porém a copa passou a ter influência significativa (F2, 218
= 3,59; P = 0,0293). A interação copa*porta-enxerto continuou insignificante (F4, 218 =
0,67; P = 0,6137). D. citri ovipositou mais em brotos de Valência do que em Folha
murcha ou Hamlin (Tabela 3).
Altura de poda vs. Porta-enxerto
Considerando-se os resultados obtidos nos experimentos anteriores (1 ao 3),
que indicaram forte impacto do porta-enxerto no comportamento de D. citri, inclusive
em mudas já formadas, porém com perda de efeito em plantas já estabelecidas em
campo (Exp. 4), foi levantada a hipótese de que haveria algum efeito de “diluição” do
agente impactante com o incremento na altura da planta e que, a partir de certo ponto,
o impacto passasse a ser associado somente à copa. Para testar essa hipótese foi
realizado um experimento de oviposição sem chance de escolha avaliando-se a
influência da altura de poda (15 cm vs. 130 a 150 cm) na copa de Valência enxertada
nos porta-enxertos ‘Cravo’ e ‘Sunki’ sobre a preferência para pouso em brotações na
copa, o tempo necessário para início da oviposição e no número de ovos.
A preferência dos adultos em ficar sobre o broto não foi influenciada nem pela
variedade de porta-enxerto (F1, 23 = 0,91, P = 0,3514) nem pela altura de poda (F1, 23
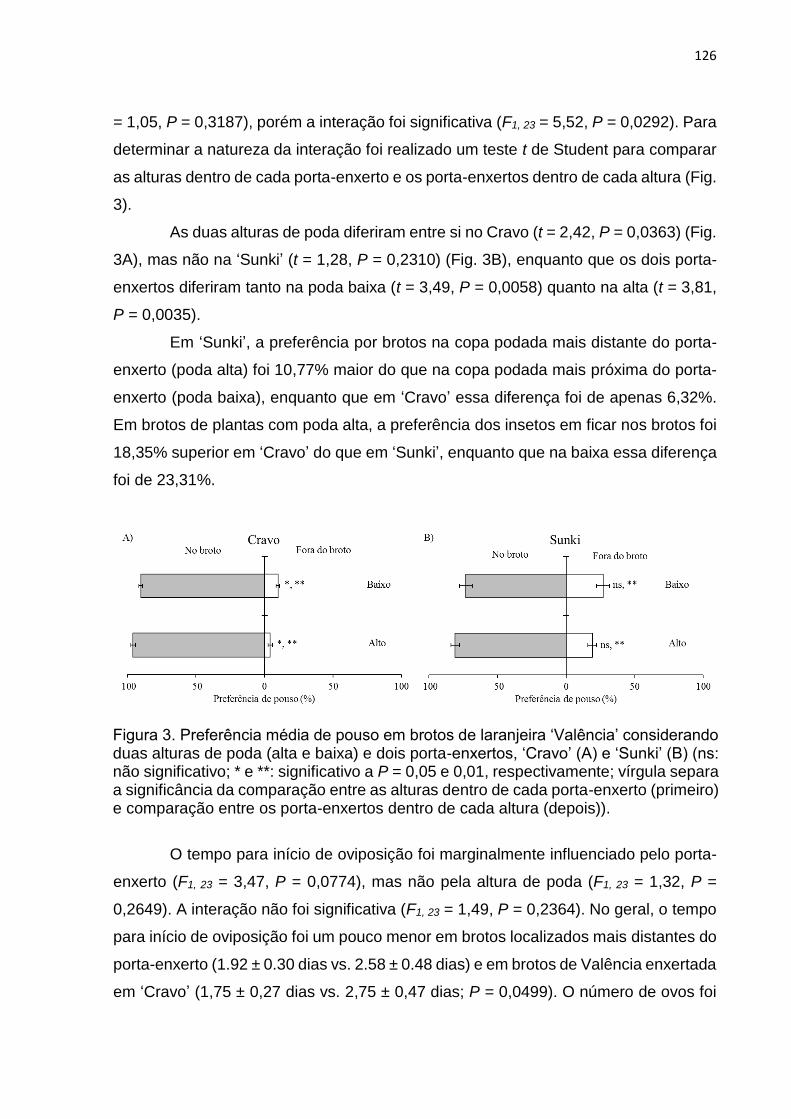
126
= 1,05, P = 0,3187), porém a interação foi significativa (F1, 23 = 5,52, P = 0,0292). Para
determinar a natureza da interação foi realizado um teste t de Student para comparar
as alturas dentro de cada porta-enxerto e os porta-enxertos dentro de cada altura (Fig.
3).
As duas alturas de poda diferiram entre si no Cravo (t = 2,42, P = 0,0363) (Fig.
3A), mas não na ‘Sunki’ (t = 1,28, P = 0,2310) (Fig. 3B), enquanto que os dois porta-
enxertos diferiram tanto na poda baixa (t = 3,49, P = 0,0058) quanto na alta (t = 3,81,
P = 0,0035).
Em ‘Sunki’, a preferência por brotos na copa podada mais distante do porta-
enxerto (poda alta) foi 10,77% maior do que na copa podada mais próxima do porta-
enxerto (poda baixa), enquanto que em ‘Cravo’ essa diferença foi de apenas 6,32%.
Em brotos de plantas com poda alta, a preferência dos insetos em ficar nos brotos foi
18,35% superior em ‘Cravo’ do que em ‘Sunki’, enquanto que na baixa essa diferença
foi de 23,31%.
Figura 3. Preferência média de pouso em brotos de laranjeira ‘Valência’ considerando duas alturas de poda (alta e baixa) e dois porta-enxertos, ‘Cravo’ (A) e ‘Sunki’ (B) (ns: não significativo; * e **: significativo a P = 0,05 e 0,01, respectivamente; vírgula separa a significância da comparação entre as alturas dentro de cada porta-enxerto (primeiro) e comparação entre os porta-enxertos dentro de cada altura (depois)).
O tempo para início de oviposição foi marginalmente influenciado pelo porta-
enxerto (F1, 23 = 3,47, P = 0,0774), mas não pela altura de poda (F1, 23 = 1,32, P =
0,2649). A interação não foi significativa (F1, 23 = 1,49, P = 0,2364). No geral, o tempo
para início de oviposição foi um pouco menor em brotos localizados mais distantes do
porta-enxerto (1.92 ± 0.30 dias vs. 2.58 ± 0.48 dias) e em brotos de Valência enxertada
em ‘Cravo’ (1,75 ± 0,27 dias vs. 2,75 ± 0,47 dias; P = 0,0499). O número de ovos foi
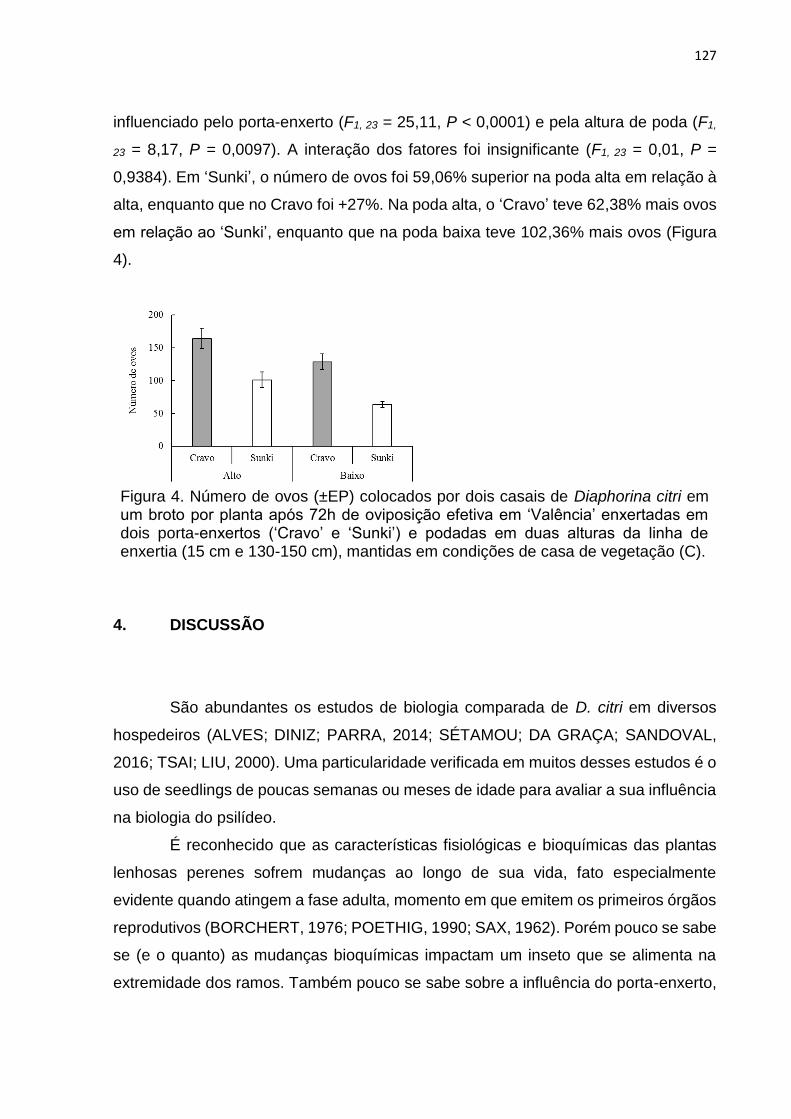
127
influenciado pelo porta-enxerto (F1, 23 = 25,11, P < 0,0001) e pela altura de poda (F1,
23 = 8,17, P = 0,0097). A interação dos fatores foi insignificante (F1, 23 = 0,01, P =
0,9384). Em ‘Sunki’, o número de ovos foi 59,06% superior na poda alta em relação à
alta, enquanto que no Cravo foi +27%. Na poda alta, o ‘Cravo’ teve 62,38% mais ovos
em relação ao ‘Sunki’, enquanto que na poda baixa teve 102,36% mais ovos (Figura
4).
Figura 4. Número de ovos (±EP) colocados por dois casais de Diaphorina citri em um broto por planta após 72h de oviposição efetiva em ‘Valência’ enxertadas em dois porta-enxertos (‘Cravo’ e ‘Sunki’) e podadas em duas alturas da linha de enxertia (15 cm e 130-150 cm), mantidas em condições de casa de vegetação (C).
4. DISCUSSÃO
São abundantes os estudos de biologia comparada de D. citri em diversos
hospedeiros (ALVES; DINIZ; PARRA, 2014; SÉTAMOU; DA GRAÇA; SANDOVAL,
2016; TSAI; LIU, 2000). Uma particularidade verificada em muitos desses estudos é o
uso de seedlings de poucas semanas ou meses de idade para avaliar a sua influência
na biologia do psilídeo.
É reconhecido que as características fisiológicas e bioquímicas das plantas
lenhosas perenes sofrem mudanças ao longo de sua vida, fato especialmente
evidente quando atingem a fase adulta, momento em que emitem os primeiros órgãos
reprodutivos (BORCHERT, 1976; POETHIG, 1990; SAX, 1962). Porém pouco se sabe
se (e o quanto) as mudanças bioquímicas impactam um inseto que se alimenta na
extremidade dos ramos. Também pouco se sabe sobre a influência do porta-enxerto,

128
que pode estar posicionado próximo ou distante das brotações, o que vai depender
da altura da planta. Em citros, vários estudos já relataram a influência do porta-enxerto
sobre D. citri mas em nenhum a distância do porta-enxerto às brotações foi
considerado. A única referência sobre este assunto encontrada se refere ao ácaro
vermelho Tetranychus urticae e a resistência a este agente oferecida pelo porta-
enxerto laranja azeda, resistência esta transferida para a copa de outra variedade nele
enxertada (AGUT et al., 2014, 2016; BRUESSOW et al., 2010).
Visando verificar se o mesmo fenômeno se aplica a D. citri uma série de
experimentos foi conduzida, envolvendo seedlings de porta-enxertos comerciais,
mudas cítricas já formadas, mas de pequeno porte (≈ 60 cm) e ainda mantida em
vaso, e plantas adultas (≈ 2.5 m de altura) cultivadas em campo. Os resultados com
seedlings e mudas pequenas obtidos no presente estudo corroboraram resultados já
publicados (ALVES; DINIZ; PARRA, 2014; BORGONI et al., 2014; RICHARDSON;
HALL, 2013), que indicam que os porta-enxertos ‘Sunki’ e ‘Swingle’ são desfavoráveis
a D. citri. Observou-se também que a variedade ou híbrido preferido para oviposição
não foi necessariamente o mais adequado ao desenvolvimento do inseto, conforme já
registrado (MAYHEW, 2001; WISE et al., 2008). Isto aconteceu com os híbridos
81G220 e 75AB, que se apresentaram mais adequados à oviposição, mas com baixas
taxas de viabilidade de ninfa. Este fenômeno pode ser explicado pelas diferenças em
compostos, voláteis ou não, presentes nas plantas, e pelas diferentes necessidades
e ou estímulos de ninfas e adultos, afetando seu comportamento de escolha para
alimentação e oviposição (antixenose) e desenvolvimento (antibiose) (WESTBROOK
et al., 2011).
Os importantes contrastes de níveis de antibiose ou antixenose a D. citri
verificados neste estudo entre porta-enxertos, nas fases de seedlings e muda jovem,
levaram a conduzir experimento de oviposição, sem chance de escolha, em plantas
adultas de diferentes combinações copa (‘Valência’, ‘Folha Murcha’ e ‘Hamlin’) e
porta-enxerto (‘Cravo’, ‘Swingle’ e ‘Sunki’), em condições de campo. As diferenças
observadas entre os porta-enxertos na fase de seedlings ou muda pequena já não
foram tão evidentes no campo.
Se diversas características dos porta-enxertos como produtividade (BREMER
NETO et al., 2013; CANTUARIAS-AVILÉS et al., 2011, 2012; LIU et al., 2015; TAN et

129
al., 2015), vigor (SAEED; DODD; SOHAIL, 2010) e tolerância a excesso ou déficit
hídrico (ALLARIO et al., 2013; ARGAMASILLA; GÓMEZ-CADENAS; ARBONA, 2014;
TRAMONTINI et al., 2013; TREEBY et al., 2007) são transferidas às copas de plantas
adultas, por que o mesmo não ocorreu em relação às características de
antibiose/antixenose contra D. citri? Duas hipóteses surgem para explicar os
resultados desta pesquisa. Uma está relacionada à idade fisiológica dos tecidos. Se
bem que em sentido inverso, um exemplo deste fenômeno é reportado por Kearsley
e Whitham (1989) para o afídeo Pemphigus betae em álamo (Populus angustifolia). A
performance do inseto foi maior em plantas em estádio juvenil do que em estádio
maduro. A segunda hipótese está relacionada à comunicação porta-enxerto / copa e
à interação entre metabólitos de rotas específicas, como por exemplo as do ácido
salicílico (AS, sintetizado principalmente em folhas) ou do ácido jasmônico (AJ,
sintetizado principalmente em raízes), ambos os fitohormônios reportados em citros
(DE OLLAS et al., 2013; OLIVEIRA et al., 2015), e reconhecidos como mediadores
nas respostas induzidas de plantas a condições de estreses bióticos e abióticos
(ARGAMASILLA; GÓMEZ-CADENAS; ARBONA, 2014; GLAZEBROOK, 2005;
HOWE; SCHALLER, 2008). Porém, os efeitos desses dois fitohormônios sobre a
biologia de insetos são maiores isoladamente do que juntos (BOSTOCK, 2005;
WALTERS; HEIL, 2007; SCHWEIGER et al., 2014).
Este trabalho apresenta a primeira evidência dos limites que estudos
conduzidos em condições de telado, casa de vegetação ou câmaras de crescimento,
com plantas jovens mantidas em vasos, podem ter no entendimento das relações
planta-inseto, em particular se as informações geradas têm como objetivo aprimorar
práticas de controle direto ou indireto de pragas e doenças em citros. Ficou aqui
demonstrado que a antibiose/antixenose de citros à D. citri é transferida do porta-
enxerto para a copa, mas somente em plantas de pequeno porte. Em plantas adultas,
onde os brotos (locais de alimentação e reprodução do inseto) se encontram mais
distantes do porta-enxerto, independentemente do diâmetro do tronco deste, este
efeito se perde. A razão desta perda requer novos estudos. É razoável supor, no
entanto, que tal fenômeno esteja associado a influência do metabolismo da variedade
usada como copa, tanto na dinâmica de fotoassimilados como na dinâmica de
metabólitos secundários, com impacto direto sobre o inseto.

130
5. REFERÊNCIAS
AGUT, B. et al. Different metabolic and genetic responses in citrus may explain relative susceptibility to Tetranychus urticae. Pest Management Science, v. 70, n. 11, p. 1728–1741, 2014.
AGUT, B. et al. Systemic resistance in citrus to Tetranychus urticae induced by conspecifics is transmitted by grafting and mediated by mobile amino acids. Journal of Experimental Botany, v. 67, n. 19, p. erw335, 2016.
ALBRECHT, U.; BOWMAN, K. D. Transcriptional response of susceptible and tolerant citrus to infection with Candidatus Liberibacter asiaticus. Plant Science, v. 185–186, p. 118–130, 2012a.
ALBRECHT, U.; BOWMAN, K. D. Tolerance of trifoliate citrus rootstock hybrids to Candidatus Liberibacter asiaticus. Scientia Horticulturae, v. 147, p. 71–80, nov. 2012b.
ALLARIO, T. et al. Tetraploid Rangpur lime rootstock increases drought tolerance via enhanced constitutive root abscisic acid production. Plant, Cell and Environment, v. 36, n. 4, p. 856–868, 2013.
ALVES, G. R.; DINIZ, A.; PARRA, J. Biology of the huanglongbing vector Diaphorina citri (Hemiptera: Liviidae) on different host plants. Journal of Economic Entomology, v. 107, n. 2, p. 691–696, 2014.
ARGAMASILLA, R.; GÓMEZ-CADENAS, A.; ARBONA, V. Metabolic and regulatory responses in citrus rootstocks in response to adverse environmental conditions. Journal of Plant Growth Regulation, v. 33, n. 2, p. 169–180, 2014.
BASSANEZI, R. B. et al. Efficacy of area-wide inoculum reduction and vector control on temporal progress of Huanglongbing in young sweet orange plantings. Plant Disease, v. 97, n. 6, p. 789–796, 2013.
BASSANEZI, R. B.; BELASQUE, J.; MONTESINO, L. H. Frequency of symptomatic trees removal in small citrus blocks on citrus huanglongbing epidemics. Crop Protection, v. 52, p. 72–77, out. 2013.
BOAVA, L. P.; CRISTOFANI-YALY, M.; MACHADO, M. A. Physiologic, anatomic, and gene expression changes in Citrus sunki, Poncirus trifoliata, and their hybrids after “Candidatus Liberibacter asiaticus” infection. Phytopathology, v. 107, n. 5, p. 590–599, 2017.
BOINA, D. R.; BLOOMQUIST, J. R. Chemical control of the Asian citrus psyllid and of huanglongbing disease in citrus. Pest Management Science, v. 71, n. 6, p. 808–823, 2015.

131
BORCHERT, R. The concept of juvenility on woody plants. Acta Horticulturae, v. 56, n. 56, p. 21–36, 1976.
BORGONI, P. C. et al. Resistance of Citrus and related genera to Diaphorina citri Kuwayama (Hemiptera: Liviidae). Neotropical Entomology, v. 43, n. 5, p. 465–469, 2014.
BOSTOCK, R. M. Signal crosstalk and induced resistance: straddling the line between cost and benefit. Annual Review of Phytopathology, v. 43, n. 1, p. 545–580, 2005.
BREMER NETO, H. et al. The horticultural performance of five “Tahiti” lime selections grafted onto “Swingle” citrumelo under irrigated and non-irrigated conditions. Scientia Horticulturae, v. 150, p. 181–186, 2013.
BRUESSOW, F. et al. Replacement of CTV-susceptible sour orange rootstock by CTV-tolerant ones may have triggered outbreaks of Tetranychus urticae in Spanish citrus. Agriculture, Ecosystems & Environment, v. 137, n. 1–2, p. 93–98, 2010.
CANTUARIAS-AVILÉS, T. et al. Horticultural performance of “Folha Murcha” sweet orange onto twelve rootstocks. Scientia Horticulturae, v. 129, n. 2, p. 259–265, 2011.
CANTUARIAS-AVILÉS, T. et al. Rootstocks for high fruit yield and quality of “Tahiti” lime under rain-fed conditions. Scientia Horticulturae, v. 142, p. 105–111, 2012.
CIFUENTES-ARENAS, J. C. et al. Monitoring citrus flush shoot ontogeny as a potential strategy for HLB and psyllid management. Journal of Citrus Pathology, v. 3, n. 1, p. 4, 2016.
COLETTA-FILHO, H. D. et al. First report of the causal agent of huanglongbing (“Candidatus Liberibacter asiaticus”) in Brazil. Plant Disease, v. 88, n. 12, p. 1382, 2004.
COSTA-LIMA, Â. M. DA. Homópteros. In: Insetos do Brasil. Rio de Janeiro: Escola Nacional de Agronomia, 1942. v. 3,p. 327.
DA GRAÇA, J. V et al. Huanglongbing: An overview of a complex pathosystem ravaging the world’s citrus. Journal of Integrative Plant Biology, v. 58, n. 4, p. 373–387, 2016.
DE OLLAS, C. et al. Jasmonic acid transient accumulation is needed for abscisic acid increase in citrus roots under drought stress conditions. Physiologia Plantarum, v. 147, n. 3, p. 296–306, 2013.
FUNDECITRUS. Mesmo estabilizada, insidência de greening permanece alta no cinturão citrícola de São Paulo e Triângulo/Sudoeste Mineiro. Araraquara - SP, Brazil: Fundecitrus, 2017. Disponível em: <http://www.fundecitrus.com.br>.
GLAZEBROOK, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annual Review of Phytopathology, v. 43, n. 1, p. 205–227, 2005.

132
HALL, D. G.; GEORGE, J.; LAPOINTE, S. L. Further investigations on colonization of Poncirus trifoliata by the asian citrus psyllid. Crop Protection, v. 72, p. 112–118, 2015.
HOWE, G. A.; SCHALLER, A. Direct defenses in plants and their induction by wounding and insect herbivores. In: Induced Plant Resistance to Herbivory. Dordrecht: Springer Netherlands, 2008. p. 7–29.
KEARSLEY, M. J. C.; WHITHAM, T. G. Developmental changes in resistance to herbivory: implications for individuals and populations. Ecology, v. 70, n. 2, p. 422–434, 1989.
LIU, X. et al. Mechanisms for the influence of citrus rootstocks on fruit size. Journal of Agricultural and Food Chemistry, v. 63, n. 10, p. 2618–2627, 2015.
MAYHEW, P. J. Herbivore host choice and optimal bad motherhood. Trends in Ecology & Evolution, v. 16, n. 4, p. 165–167, 2001.
MILES, G. P. et al. Apparent tolerance to huanglongbing in Citrus and Citrus-related germplasm. HortScience, v. 52, n. 1, p. 31–39, 2017.
NAVA, D. E. et al. Biology of Diaphorina citri (Hem., Psyllidae) on different hosts and at different temperatures. Journal of Applied Entomology, v. 131, n. 9–10, p. 709–715, 2007.
OLIVEIRA, T. M. et al. Comparative study of the protein profiles of Sunki mandarin and Rangpur lime plants in response to water deficit. BMC Plant Biology, v. 15, n. 1, p. 69, 2015.
POETHIG, R. S. Phase change and the regulation of shoot morphogenesis in plants. Science, v. 250, n. 4983, p. 923–930, 1990.
POMPEU JUNIOR, J. Porta-enxertos. In: MATTOS JÚNIOR, D. DE et al. (Eds.). . Citros. Campinas: Instituto Agronômico e Fundag, 2005. p. 61–104.
R CORE TEAM. R: A language and environment for statistical computingViena, AustriaR Fundation for Statistical Computing, , 2014. Disponível em: <http://www.r-project.org/>
RAMADUGU, C. et al. Long-term field evaluation reveals Huanglongbing resistance in Citrus relatives. Plant Disease, v. 100, n. 9, p. 1858–1869, 2016.
RICHARDSON, M. L.; HALL, D. G. Resistance of Poncirus and Citrus × <I>Poncirus</i> germplasm to the asian citrus psyllid. Crop Science, v. 53, n. 1, p. 183, 2013.
RUSSELL, D. N. et al. Host suitability of Citrus and Zanthoxylum</i> spp. for Leuronota fagarae Burckhardt and Diaphorina citri Kuwayama (Hemiptera: Psylloidea). Florida Entomologist, v. 97, n. 4, p. 1481–1492, 2014.
SAEED, M.; DODD, P. B.; SOHAIL, L. Anatomical studies of stems, roots and leaves

133
of selected rootsock varieties in relation to their vigour. Journal of Horticulture and Forestry, v. 2, n. 4, p. 87–94, 2010.
SAX, K. Aspects of aging in plants. Annual Review of Plant Physiology, v. 13, n. 1, p. 489–506, 1962.
SÉTAMOU, M.; DA GRAÇA, J. V.; SANDOVAL, J. L. Suitability of native North American Rutaceae to serve as host plants for the Asian citrus psyllid (Hemiptera: Liviidae). Journal of Applied Entomology, v. 140, n. 9, p. 645–654, 2016.
SKELLEY, L. H.; HOY, M. A. A synchronous rearing method for the asian citrus psyllid and its parasitoids in quarantine. Biological Control, v. 29, n. 1, p. 14–23, 2004.
TAN, F. Q. et al. Comparative metabolic and transcriptional analysis of a doubled diploid and its diploid citrus rootstock (C. junos cv. Ziyang xiangcheng) suggests its potential value for stress resistance improvement. BMC Plant Biology, v. 15, n. 1, p. 1–14, 2015.
TEIXEIRA, D. C. et al. First report of a huanglongbing-like disease of Citrus in Sao Paulo State, Brazil and association of a new Liberibacter species, “Candidatus Liberibacter americanus”, with the disease. Plant Disease, v. 89, n. 1, p. 107, 2005.
TRAMONTINI, S. et al. Rootstock control of scion response to water stress in grapevine. Environmental and Experimental Botany, v. 93, p. 20–26, 2013.
TREEBY, M. T. et al. Irrigation management and rootstock effects on navel orange [Citrus sinensis (L.) Osbeck] fruit quality. Agricultural Water Management, v. 91, n. 1–3, p. 24–32, 2007.
TSAI, J. H.; LIU, Y. H. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. Journal of Economic Entomology, v. 93, n. 6, p. 1721–1725, 2000.
WALTERS, D.; HEIL, M. Costs and trade-offs associated with induced resistance. Physiological and Molecular Plant Pathology, v. 71, n. 1–3, p. 3–17, jul. 2007.
WENNINGER, E. J.; HALL, D. G. Daily timing of mating and age at reproductive maturity in Diaphorina citri (Hemiptera: Psyllidae). Florida Entomologist, v. 90, n. 4, p. 715–722, 2007.
WESTBROOK, C. J. et al. Colonization of Citrus and Citrus-related Germplasm by Diaphorina citri (Hemiptera: Psyllidae). Hortscience, v. 46, n. 7, p. 997–1005, 2011.
WISE, M. J. et al. Good mothers, bad mothers, and the nature of resistance to herbivory in Solidago altissima. Oecologia, v. 155, n. 2, p. 257–266, 2008.
YONEMOTO, Y. et al. Effects of rootstock and crop load on sap flow rate in branches of “Shirakawa Satsuma” mandarin (Citrus unshiu Marc.). Scientia Horticulturae, v. 102, n. 3, p. 295–300, 2004.