Embed Size (px)
Citation preview

I - INTRODUÇÃO
A lactação representa uma das fases de maior demanda energética do ciclo da vida
em fêmeas de mamíferos.1 Numerosas adaptações fisiológicas, metabólicas e
comportamentais ocorrem no organismo materno nesta fase, para garantir o crescimento,
desenvolvimento e nutrição do neonato, bem como a recuperação da mãe no pós-parto.2 O
aumento das necessidades de energia é suprido primariamente pelo consumo alimentar.3
Adicionalmente, ocorrem adaptações que melhoram a eficiência metabólica
favorecendo a utilização de nutrientes pela glândula mamária.3 Em roedores, ocorre
diminuição da termogênese no tecido adiposo marrom para preservar combustíveis
metabólicos,4 redução na lipogênese no tecido adiposo branco, hipotireoidismo e
hipoinsulinemia.5,6 Apesar desse esforço, em estudos realizados em roedores os animais
apresentam balanço energético negativo, com mobilização de cerca de 1g de tecido
adiposo por dia para suprir as suas necessidades nutricionais.7
As alterações metabólicas desencadeadas no organismo materno durante a lactação
podem ser compreendidas a partir do estudo dos substratos requeridos pela glândula
mamária para a produção de leite. Em condições de suprimento dietético normal, todo o
aporte de lactose e caseína e, aproximadamente 50% dos lipídios, são sintetizados dentro
da própria glândula. Assim, no pico da lactação, a captação de glicose pela glândula
mamária passa a ser ao redor de 5,4g/24h, sendo uma parte usada para a síntese de lactose
e o restante para a lipogênese,8 ocorrendo um aumento acentuado na taxa de síntese de
lipídios nesse tecido.9 Os requerimentos necessários para completar a demanda de lipídios
para o leite são obtidos a partir dos graxos livres (AGL) e dos triglicerídeos (TG) captados
do sangue.10 E os requerimentos de glicose são obtidos a partir de mudanças no
metabolismo de outros tecidos7.

2
O tecido adiposo tem um papel fundamental no suprimento de energia durante a
lactação. No inicio desta fase, a síntese de lipídios fica reduzida nesse tecido, sendo muito
aumentada na glândula mamária.11 Assim, a taxa de lipogênese in vivo e in vitro e a
atividade de algumas enzimas chave nesse processo são deprimidas, com similar
diminuição na atividade da enzima lípase lipoprotéica (LPL) ao final da prenhez,
permanecendo reduzida durante toda a lactação.12,13,14,15,16,17 A lipólise, ao contrario,
encontra-se aumentada no tecido adiposo e como resultado, maior quantidade de ácidos
graxos livres e glicerol são liberados para a corrente sangüínea.7,18 Em ratos e
camundongos foi observada a utilização de, aproximadamente, 60-70% das reservas do
tecido adiposo durante a lactação.6
O papel do fígado no metabolismo lipídico é mais complexo do que o do tecido
adiposo ou da glândula mamária,19,20 sendo o maior regulador do fluxo de metabólitos do
corpo. A dieta é a principal fonte de substratos metabólicos para o fígado, sendo que tanto
carboidratos, proteínas e lipídios, podem fornecer substratos para a síntese de ácidos
graxos nesse tecido.21 Essas substâncias provêm do intestino via veia porta, da circulação
geral pela artéria hepática e pelo sistema linfático, sendo captados pelos hepatócitos em
quantidade muito maior que a necessária para atender as suas necessidades, podendo isso
explicar a ocorrência de esteatose, principalmente pela captação excessiva de lipídios.21
Os lipídios alcançam o fígado sob a forma de ácidos graxos livres e partículas de
lipoproteínas. No hepatócito os AGL podem ser convertidos em TG ou oxidados. A β-
oxidação mitocondrial irá resultar tanto em energia para a célula como também na
produção de corpos cetônicos (acetoacetato e β-hidroxibutirato), os quais são liberados na
circulação. Os AGL convertidos em TG podem ser utilizados na produção de lipoproteínas
de muito baixa densidade (VLDL) que são exportadas para o sangue, sendo o excesso de
TG armazenado no fígado.17,19,21

3
Na lactação, o metabolismo hepático sofre profundas alterações.17,20 Vários
mecanismos intracelulares promovem a oxidação de ácidos graxos durante esse período, o
que é mediado pela redução na insulinemia, associada à elevação do glucagon e de AGL.22
Apesar da reduzida razão sérica insulina/glucagon e de outros mecanismos que promovem
a oxidação de ácidos graxos, a esterificação desses ácidos é também aumentada nessa fase,
devido ao aumento da captação de AGL associada à elevação de algumas enzimas de
esterificação.23
Em contraste, a secreção de VLDL não se modifica, podendo mesmo diminuir,
possivelmente devido à redução na produção de apoproteína B.20 A apoproteína B100
(Apo B100) é a maior proteína componente das lipoproteínas plasmáticas e é requerida
para a síntese e secreção das lipoproteínas ricas em TG circulantes, como as VLDL.24,25
Em conseqüência, freqüentemente a produção excede a secreção de VLDL e, assim, ocorre
um aumento no conteúdo de lipídios nas células hepáticas. Parece curioso que o fígado não
aumenta a liberação de VLDL, porém por ação da LPL, libera ácidos graxos de seus TG,
direcionando-os para serem utilizados pela glândula mamária.18,21
No início da lactação é comum ocorrer tanto aumento na massa como também
acúmulo de TG no fígado.8,18 Porém, o propósito fisiológico desse processo ainda não está
totalmente compreendido, podendo ser devido a um defeito no sistema de captação de
ácidos graxos livres pelo fígado, que é determinado mais pelo suprimento do que pela
necessidade, conforme descrito anteriormente, refletindo o papel desse órgão na regulação
da composição de nutrientes do sangue.26
As mudanças comportamentais e metabólicas que acompanham a lactação estão
associadas com alterações hormonais. A insulina é o mais importante fator hormonal na
regulação do metabolismo na glândula mamária.8 Entretanto, as suas concentrações
apresentam-se reduzidas durante a lactação, possivelmente refletindo a elevada utilização

4
de glicose pela glândula nesse período, o que favorece a maior disponibilidade dos
nutrientes armazenados para a síntese de leite.27,28
O glucagon é um potencial modulador do metabolismo no tecido adiposo e a razão
insulina/glucagon pode ser determinada pelo balanço entre a lipólise e a lipogênese nesse
tecido. Durante a lactação essa razão passa a ser reduzida na circulação em,
aproximadamente 50%, e isso favorece a maior liberação de substratos para serem
destinados principalmente para a glândula mamária.29
A prolactina é um hormônio que tem função tanto no crescimento celular, como na
diferenciação, secreção e na involução da glândula mamária. Ela age como um sensor
fisiológico durante a lactação, respondendo às demandas para a produção de leite e
direcionando os nutrientes do tecido adiposo para a glândula.30 Esse hormônio interfere na
síntese dos principais constituintes do leite: aumento da síntese de proteínas; de lactose,
por aumento da captação de glicose; e de lipídeos, por aumento coordenado das enzimas
chave nesse processo.31 O efeito estimulatório da prolactina sobre o consumo alimentar
pode contribuir para o aumento na ingestão tanto no período da gestação, como na
lactação.1
Contudo, a avaliação das ações metabólicas desse hormônio é dificultada por
algumas limitações, como: 1- o processo de iniciação e manutenção da lactação é
complexo e exige a ação de vários hormônios; 2- o lactogênio placentário, a prolactina e o
hormônio do crescimento têm funções similares ou sobrepostas; 3- a maior parte dos
efeitos conhecidos da prolactina é obtida mediante estudos de supressão in vivo desse
hormônio por bromocriptina ou através da remoção das crias.32
As concentrações de leptina em ratas encontram-se deprimidas durante a lactação,
sendo este um fator que contribui para a hiperfagia observada nessa fase.33 Este hormônio
encontra-se envolvido na redução da lipogênese34,35 e esse efeito é obtido através da

5
inibição de genes relacionados com a síntese de ácidos graxos e de TG,35 sendo mediado
por proteínas ligadoras dos elementos regulados por esteróis (SREBP1).36,37
Durante a lactação, a ingestão inadequada de proteínas em quantidade ou qualidade
tem sido associada com a redução na ingestão alimentar e a diminuição da performance
lactacional em ratas.38,39,40,41 A concentração de gordura na carcaça parece não se
modificar em função do teor de proteínas na dieta, mas a massa corpórea magra é
catabolizada na restrição protéica associada à lactação.38,42 Adicionalmente, ocorre redução
na taxa de metabolismo basal43, na termogênese no tecido adiposo marrom4 e na
lipogênese no tecido adiposo branco,44,6 visando atender a alta demanda energética para a
produção de leite.
Poucos relatos na literatura avaliaram os efeitos da restrição protéica materna sobre
a concentração de lipídios em tecidos específicos durante a lactação, e os estudos
avaliados, foram desenvolvidos na fase inicial, que corresponde ao período entre o 1º ao
14º dia, considerada como o pico da lactação.45,46,47
Em nosso laboratório, Ferreira et al. (2007)48 demonstraram que ratas lactantes
submetidas à restrição protéica exibiram, no 14º dia da lactação, aumento na massa do
tecido adiposo gonadal e maior deposição de gordura na carcaça, comparadas a ratas
controles. Porém, nesse estudo não foram avaliados parâmetros relacionados à deposição
de lipídios no fígado.
Um estudo recente em ratas, descrito por Erhuma et al. (2007),49 abordando a
restrição protéica materna durante a prenhez, observou na prole adulta (180 dias), elevação
da trigliceridemia, resistência à insulina e evidência histológica de esteatose hepática.
Assim, considerou-se de fundamental relevância avaliar alguns aspectos
metabólicos, na fase tardia da lactação, em ratas submetidas à restrição protéica.

6
II – OBJETIVOS
2.1 – Geral
Ø Avaliar alguns aspectos metabólicos na lactação tardia em ratas submetidas à
restrição protéica.
2.2 - Específicos
Ø Monitorar o consumo alimentar e a massa corporal durante o período experimental;
Ø Verificar a glicemia, a proteinemia e as concentrações hormonais de insulina,
leptina, glucagon e prolactina, no 18º dia de lactação;
Ø Avaliar no fígado: a massa, o conteúdo de glicogênio, lipídios e a atividade das
enzimas: glicose-6-fosfato desidrogenase, málica e ATP- citrato liase;
Ø Verificar a relação lipídio/glicogênio e a correlação entre a prolactina sérica e as
enzimas glicose-6-fosfato desidrogenase e enzima málica no fígado;
Ø Verificar a massa e o conteúdo de lipídios no tecido adiposo branco gonadal, na
glândula mamária e na carcaça;
Ø Verificar o efeito da restrição protéica sobre a performance lactacional materna e o
desenvolvimento da massa corporal das crias.

7
III - MATERIAIS E MÉTODOS
Este estudo foi desenvolvido no Laboratório de Avaliação Biológica de Alimentos
(LABA), do Departamento de Alimentos e Nutrição (DAN) da Faculdade de
Nutrição/FANUT/UFMT, Cuiabá–MT, Brasil.
3.1 Animais e dietas
Todos os procedimentos experimentais foram realizados de acordo com as
recomendações do Colégio Brasileiro de Experimentação Animal (COBEA),50 adotado
pela UFMT.
Foram utilizadas ratas fêmeas da linhagem Wistar - Rattus Norvergicus, variedade
Albinus, ordem Rodentia Mammalia, família Muridae, com aproximadamente 90 dias,
fornecidas pelo Biotério Central da UFMT. O acasalamento foi realizado abrigando 4
fêmeas e 1 macho, em gaiolas coletivas de polipropileno durante a noite e a prenhez foi
confirmada por esfregaço vaginal e constatação da presença de espermatozóides. As ratas
foram separadas aleatoriamente nos seguintes grupos: ratas controles não-prenhes e
prenhes, tratadas com dieta controle (17% de proteína) e ratas submetidas à restrição
protéica não-prenhes e prenhes, tratadas com dieta hipoprotéica (6% de proteína), durante
o período correspondente à prenhez. Os dois tipos de dieta eram isocalóricas (15,74 kJ/g),
recomendadas para roedores em fases de crescimento, gravidez e lactação, segundo as
recomendações do American Institute of Nutrition (AIN 93-G)51 (Tabela 1).
O nascimento dos filhotes ocorreu no 22º dia de prenhez (considerado como
primeiro dia de lactação) e, no terceiro dia, as crias foram reduzidas a oito filhotes por
grupo, assegurando assim, um número padrão de crias para cada mãe.

8
De acordo com o estado fisiológico e o tipo de dieta oferecida, os grupos de ratas
passaram a ser assim designados:
Ø Controle não-lactante (CNL): ratas não-lactantes tratadas com dieta controle;
Ø Controle lactante (CL): ratas lactantes tratadas com dieta controle;
Ø Hipoproteica não-lactante (HPNL): ratas não-lactantes tratadas com dieta hipoprotéica;
Ø Hipoproteica lactante (HPL): ratas lactantes tratadas com dieta hipoprotéica.
As ratas foram mantidas em gaiolas individuais (uma fêmea e suas crias ou uma
fêmea), sob temperatura constante (24±1º C) e ciclo claro/escuro de 12 em 12 horas, tendo
livre acesso à água e à dieta, até o décimo oitavo dia (18°) de lactação.
Todas as dietas foram elaboradas na forma de pó, sendo os ingredientes secos
pesados em balança marca Marte (modelo A500), peneirados (malha 20) e
homogeneizados. Preparou-se quantidade suficiente de dieta para atender toda a fase
experimental, sendo acondicionadas em porções, em recipientes de vidro com tampa e
armazenadas a 5° C, para serem oferecidas aos animais gradualmente.

9
Tabela 1. Composição das dietas normoprotéica e hipoprotéica, utilizadas durante 18 dias
de tratamento de ratas não-lactantes e lactantes dos grupos controle (CNL, CL) e
hipoprotéico (HPNL e HPL).
Ingredientes Dieta Controle
(17% proteína)
Dieta Hipoprotéica
(6% proteína)
g /Kg
Caseína (84% proteína) 202,0 71,5
Amido de milho1 397,0 480,0
Amido de milho dextrinizado1 130,5 159,0
Sacarose 100,0 121,0
Óleo de soja 70,0 70,0
Fibra 50,0 50,0
Mistura de minerais (AIN-93G)* 35,0 35,0
Mistura de vitaminas (AIN-93G)* 10,0 10,0
L-cistina 3,0 1,0
Bitartarato de colina 2,5 2,5
*Para composição detalhada, em Reeves et al (1993) 1Fornecidos por Corn Products of Brazil
3.2 Procedimentos Experimentais
3.2.1 Determinação do consumo alimentar, evolução da massa corporal e ingestão
protéica
O consumo de dietas e a evolução da massa corporal das mães e das crias foram
registrados três vezes por semana, durante os 18 dias de lactação. A ingestão alimentar
absoluta foi calculada pela diferença entre a quantidade de ração oferecida e a dieta

10
rejeitada após 48h, sendo as sobras peneiradas antes da sua pesagem. Para calcular o
consumo relativo, considerou-se o consumo absoluto relacionado ao peso final de cada
rata, em percentual. A avaliação da massa corporal das mães foi realizada sempre pela
manhã e efetuada em balança semi-analítica, previamente tarada e com precisão de 0,01 g,
sendo determinada a massa corporal e o consumo alimentar total (g) e proporcional à
massa corporal (g/100g). Nas crias avaliou-se a massa corporal inicial e final (g) e a
evolução da massa corporal durante o período experimental (18 dias). Para o cálculo da
proteína ingerida, em valores absolutos (g) e relativos (g/100g), calculou-se o percentual de
proteínas do total de dieta consumido por cada animal.
3.2.2 Produção de leite
Foi adaptada da técnica inicialmente descrita por Russel52 e modificada por Tavares
do Carmo,53 estimando-se a produção de leite entre o 11º e o 12º dias de lactação. Separou-
se as crias em quatro grupos:
Ø Crias amamentadas controle (CCA);
Ø Crias amamentadas restritas em proteína (CHPA), que foram mantidas em
amamentação regular;
Ø Crias controle jejum (CCJ);
Ø Crias de mães submetidas à restrição protéica (CHPJ), submetidas a jejum de 12 horas
(das 18h do 11º dia às 06h do 12° dia de lactação).
O cálculo da produção de leite foi realizado com base na seguinte fórmula:
L= Pa2 - Pa1 x (1-k), onde:
L= produção de leite (g/dia);
Pa1= peso das proles amamentadas no início do período de 12h;
Pa2= peso das proles amamentadas no final do período de 12h;

11
K= perda relativa de peso das proles jejuadas (média).
K= (Pj1-Pj2)/Pj1; sendo:
Pj1= peso inicial do período de 12h;
Pj2= peso final do período de 12h.
O fator K foi calculado para cada prole jejuada.
3.2.3 Coleta de amostras de sangue e de tecidos
Todos os procedimentos tiveram inicio às 7:00h e se estenderam por toda a manhã.
As coletas de sangue para as determinações bioquímicas e hormonais, bem como a coleta
de tecidos e carcaças para as análises foram realizadas com os animais no estado
alimentado, no 18° dia de lactação. Através de uma pequena incisão na extremidade
inferior da cauda dos animais, foram retiradas amostras de sangue para a determinação de
glicemia. Após o sacrifício das ratas, coletaram-se novas amostras de sangue. O soro foi
separado em alíquotas e armazenado a -80º C para posteriores determinações bioquímicas
(albumina e proteínas totais) e hormonais (insulina, glucagon, prolactina e leptina).
Após laparotomia mediana foram retirados o tecido adiposo branco gonadal
(GON), a glândula mamária (GMA) e o fígado (FIG). Esses tecidos e a carcaça (CARC)
foram pesados imediatamente para a obtenção do peso fresco em valor absoluto (g) e
relativo (g/100g de massa corporal), sendo armazenados em freezer à -80° C para análises
posteriores.

12
3.3 Determinações Bioquímicas e Hormonais
3.3.1 Determinações Bioquímicas
3.3.1.1 Glicose
Foi determinada utilizando monitor com tiras de teste para determinação da
glicemia (glicosímetro) por fotômetro de refletância da marca Accu-Chek®, Laboratório
Roche Diagnósticos, Alemanha. Os resultados foram expressos em mg/dL.
3.3.1.2 Proteínas Totais
A determinação de proteínas séricas totais foi realizada e seguindo o método do
reativo de biureto modificado.54 O ensaio baseia–se na reação de íons cobre com as
ligações peptídicas das proteínas séricas. A concentração de proteínas totais foi avaliada
em espectofotômetro, em comprimento de onda de 545 nm e os resultados foram expressos
em g/dL.
3.3.1.3 Albumina
A concentração de albumina sérica foi determinada pelo método colorimétrico do
verde de bromocresol55 tamponado em pH 3,8, empregando Kit comercialmente disponível
(Labtest). A concentração de albumina foi medida em espectrofotômetro com
comprimento de onda de 630 nm, sendo os resultados expressos em g/dL.
3.3.2 Determinações Hormonais
As concentrações séricas dos hormônios foram determinadas por ELISA, utilizando
kits comercialmente disponíveis: Insulina (Rat Insulin Elisa Kit – Linco Research - USA),
sendo os resultados expressos em pg/mL; Prolactina (Rat Prolactin EIA – ALPCO

13
Diagnostics - USA) e os resultados foram expressos em ng/mL; Leptina (Antigenix
América’s Leptin Elisa Consctruction Kit, Inc. USA), e os resultados foram expressos em
pg/mL; Glucagon (Rat Glucagon Elisa Kit Wako – USA), com os resultados foram
expressos em pg/mL.
3.4 Determinação de glicogênio hepático
O conteúdo de glicogênio no fígado (FIG) foi determinado segundo o método
descrito por Sjögren et al. (1938).56 Foram colocados 0,5g do respectivo tecido em tubos
de centrífuga graduados, contendo 2,0mL de KOH a 30%. Os tubos foram então
transferidos para um banho-maria à temperatura de 100º C durante 1 hora para a fase de
digestão e adicionadas cinco gotas de Na2SO4 e álcool etanólico PA, de tal maneira que a
solução apresentasse uma concentração suficiente para a extração do glicogênio. A
dosagem foi então realizada mediante a adição do reagente de fenol. Tomando-se como
referência uma curva padrão de glicose determinou-se o teor de glicose obtido nas
amostras. Calculou-se, posteriormente, a concentração de glicogênio e os valores foram
expressos em mg de glicogênio/100mg de peso fresco de tecido hepático.
3.5 Determinação do conteúdo de lipídios nos tecidos e na carcaça
A extração de lipídios do tecido adiposo branco gonadal (GON), glândula mamária
(GMA) e fígado (FIG), foi realizada pelo método descrito por Folch et al. (1957).57 Os
tecidos foram colocados em clorofórmio:metanol (2:1), homogeneizados e o volume
completado para 10mL de solução. As amostras eram deixadas “overnight” para a extração
dos lipídios, sendo posteriormente filtradas. A uma alíquota de 8mL do filtrado eram
adicionados 1,6mL de salina. Após separação da fase clorofórmica e aquosa, o material era
centrifugado a 2500rpm durante 5 minutos. Uma alíquota da fase clorofórmica e aquosa,

14
do material foi transferida para tubos previamente pesados e evaporada à temperatura
ambiente. A quantidade de gordura em gramas foi obtida pela diferença entre a pesagem
do tubo após evaporação e a pesagem inicial (tubo vazio). Os valores foram expressos em
mg de lipídio/100mg de peso fresco dos tecidos.
O conteúdo de lipídios na carcaça foi determinado por método gravimétrico,
conforme descrito por Oller do Nascimento e Williamson (1986).58 As carcaças após serem
evisceradas, foram pesadas e submetidas a calor úmido sob pressão por aproximadamente
40 minutos em água (1:2). A seguir, liquidificou-se e alíquotas de 3,0 g do homogeneizado
foram submetidas a banho-maria à 70º C por 2 horas na presença de KOH a 30 % e etanol
absoluto. Após acidificação com H2SO4 6M, os ácidos graxos totais foram extraídos
mediante três lavagens sucessivas com éter de petróleo. Em cada lavagem, as amostras
foram agitadas por 10 minutos e centrifugadas a 1500rpm. O volume total da extração foi
evaporado em capela à temperatura ambiente e o teor de gordura foi determinado
gravimetricamente. O ensaio foi realizado em triplicata e os resultados obtidos foram
expressos em gramas de lipídios por 100g de carcaça.
3.6 Determinação da atividade de enzimas no fígado
Um fragmento de fígado foi extraído, imediatamente armazenado em nitrogênio
líquido e mantido em freezer a -80° C até sua utilização. Todos os procedimentos após a
retirada do tecido foram realizadas em gelo e para o ensaio das enzimas glicose-6-P-
desidrogenase (G6PDH), enzima málica (EM) e ATP-citrato liase (ATP-cit) foram
utilizados os mesmos homogeneizados. Cada 0,5g de FIG foi homogeneizado em 5 ml de
tampão Tris 10mM, pH 7,4; contendo sacarose 0,32M, EDTA 2mM e 2-β-mercaptoetanol
5mM e centrifugado por 10 minutos a 11000rpm. A camada superior foi descartada e a
fração sobrenadante centrifugada a 11000rpm à temperatura de 4° C por duas horas, para

15
obtenção da fração citosólica. As atividades das enzimas avaliadas foram expressas em
nmol. mg de proteína-1.min-1. O conteúdo protéico do homogenado foi determinado pelo
método de Lowry et al. (1951).59
3.6.1 Glicose- 6-P desidrogenase (G6PDH)
A atividade da glicose-6-P desidrogenase foi medida em espectrofotômetro a 340
nm, de acordo com o método de Lee (1982),60 que se baseia na redução de NADP. A
mistura de reação continha Tris 0,1 M, pH 8,0, NADP+ 1mM e D-glicose-6-fosfato 1mM,
à qual se adicionava a proteína citosólica. A reação foi iniciada pela adição de glicose-6-
fosfato sendo omitida no branco.
3.6.2 Enzima málica (EM)
A atividade da enzima málica foi avaliada medindo-se a formação de NADPH a
partir de L-malato e NADP+. O método utilizado foi descrito por Ochoa (1955),61 e
modificado por Hsu e Lardy (1969).62 A formação do NADPH foi monitorada em
espectrofotômetro a 340nm, e o ritmo de sua formação, foi proporcional à concentração da
enzima. O sistema de ensaio continha tampão trietanolamina 67mM, L-malato 5mM,
MnCl2. 4 H2O 4mM, NADP+ 0,2mM. A reação foi iniciada com L-malato, sendo omitida
no branco.
3.6.3 ATP-citrato liase (ATP-cit)
O ensaio da ATP-citrato liase foi realizado de acordo com o método descrito por
Srere (1959),63 acompanhando a oxidação do NADH, em espectrofotômetro a 340nm por 3
minutos. A mistura da reação continha tampão trietanolamina 50mM pH 7,7, ATP-Mg 7
mM, citrato de potássio 10mM, NADH 0,1mM, 2-β-mercaptoetanol 10mM, coenzima A

16
0,24mM, NADH-malato desidrogenase 1U/ml. A reação foi iniciada pela adição de citrato
de potássio e o NADH sendo omitido no branco.
3.7 Análise Estatística
Os resultados foram apresentados em média e erro padrão da média (EPM) com o
número de animais indicados entre parênteses. Os dados foram submetidos à análise de
variância (ANOVA) a duas vias e precedidas pelo teste de Bartlett para verificar a
homogeneidade das variâncias. Os resultados com variâncias heterogêneas e sem
distribuição normal foram inicialmente transformados (logarítmica - Log ou Ln da
variável). Quando necessário essas análises foram complementadas por testes de
comparações múltiplas de médias (Tukey HSD para grupos com n iguais e diferentes).
Quanto comparados apenas dois grupos, os valores obtidos foram avaliados pelo test-t de
Student para amostras independentes, após as variâncias homogêneas e distribuição
normal. O coeficiente de correlação foi usado para verificar a relação entre a concentração
de prolactina sérica e a atividade das enzimas G6PDH e EM, pelo método de regressão
linear simples. Em todas as análises estabeleceu-se um nível de significância para rejeição
da hipótese de nulidade de 5%. Para análise dos resultados utilizou-se o programa Statistic
for Windows, versão 4.3 (StatSoft, Tulsa, OK, USA).

17
IV – RESULTADOS
O consumo alimentar absoluto e relativo e a ingestão protéica total e relativa foram
modificados em função do estado nutricional e do fisiológico, com interação entre esses
fatores (Tabela 2). O consumo alimentar absoluto nos grupos HPL, HPNL e CNL foi
similar e significativamente menor comparado ao grupo CL. Entretanto, em proporção à
massa corporal, o consumo alimentar nos grupos lactantes foi mais elevado, sendo que no
grupo HPL foi menor em comparação ao CL (p<0,0002).
A ingestão protéica absoluta e relativa foi reduzida nos grupos HPL e HPNL
quando comparados aos grupos controles.
A massa corporal inicial foi maior nos grupos lactantes em relação aos não
lactantes (F1,25=140,96; p<0,001) e menor nos grupos submetidos à restrição protéica em
comparação aos grupos que receberam dieta controle (F1,25=5,21; p<0,003).
O estado nutricional e a interação entre o estado nutricional e fisiológico
(F1,25=31,95; p<0,000 e F1,25=19,45; p<0,000), contribuíram para a alteração da massa
corporal ao final do período experimental, sendo esta menor no grupo HPL do que no
grupo HPNL e maior no grupo CL, em relação ao CNL. O grupo HPL mostrou massa
corporal significativamente menor que os demais grupos ao final do estudo (Tabela 2).

18
Tabela 2. Consumo alimentar e ingestão protéica absoluta e relativa e massa corporal
inicial e final de ratas não-lactantes e lactantes tratadas com dieta controle (CNL e CL) ou
hipoprotéica (HPNL e HPL) durante 18 dias de lactação.
GRUPOS
CNL (7)
CL (7)
HPNL (7)
HPL (8)
Consumo alimentar absoluto (g) 293±18 b 573±26 a 276±11 b 308±9,0 b
Consumo alimentar relativo(g/100g) 103±7 c 175±10 a 103±3 c 129±5 b
Ingestão protéica absoluta (g) 49,9±8,2 b 97,4±4,3 a 16,5±0,6 c 19,3±0,9 c
Ingestão protéica relativa (g/100g) 17,4±1,2 b 29,7±1,8 a 6,2±0,2 d 7,7±0,2 c
Massa corporal inicial (g) 268±4 404±10* 255±2 § 362±16 § *
Massa corporal final (g) 300±7 b 330±8 a 287±5 b 249±10 c
Valores representam média ± EPM do número de ratas entre parênteses.
§ Diferença estatística com relação às ratas controles (ANOVA a dois fatores; p<0,05)
* Diferença estatística com relação às ratas não-lactantes (ANOVA a dois fatores; p<0,05)
Letras diferentes indicam diferenças estatísticas (Teste de Tukey HSD; p<0,05)
As concentrações séricas de proteínas totais foram alteradas pelo estado nutricional
e houve interação entre o estado nutricional e o fisiológico (F1,24=4,74; p<0,05 e
F1,24=4,32; p<0,05). No grupo HPL as concentrações de proteínas foram significantemente
menores que nos demais grupos. A restrição protéica reduziu as concentrações de albumina
(F1,24=19,00; p<0,000) e elevou a glicemia (F1,24=4,94; p<0,04).
O conteúdo de glicogênio no fígado não diferiu entre os grupos (Tabela 3).

19
Tabela 3. Concentrações séricas de proteínas totais, albumina, glicose e glicogênio hepático
em ratas não-lactantes e lactantes tratadas com dietas controle (CNL e CL) ou hipoprotéica
(HPNL e HPL) durante 18 dias de lactação.
GRUPOS
CNL CL HPNL HPL
Proteínas totais (g/dL) 4,9±0,4 a
(7) 5,2±0,4 a
(7) 4,9±0,6 a
(7) 3,4±0,3 b
(7) Albumina (g/dL) 3,1±0,1
(7) 2,7±0,2
(7) 2,2±0,3 §
(7) 1,8±0,2 §
(7) Glicose (mg/dL) 109±1,6
(7) 108±5,8
(7) 124±3,6 §
(7) 112±4,3 §
(8) Glicogênio hepático (mg/100mg) 3,8±0,3
(7) 5,1±0,5
(7) 5,2±0,4
(7) 4,7±0,8
(8) Os valores representam média ± EPM do número de ratas entre parênteses.
§ Diferença estatística com relação às ratas controles (ANOVA a dois fatores; p<0,05)
Letras diferentes indicam diferenças estatísticas (Teste de Tukey HSD; p<0,05)
As concentrações séricas de insulina e de glucagon foram modificadas em função
da restrição protéica (F1,23=8,53; p<0,008 e F1,23=5,88; p<0,02) e da lactação (F1,23=21,36;
p<0,00 e F1,23=4,70; p<0,04). A insulinemia e a glucagonemia foram reduzidas nos grupos
lactantes em relação aos não-lactantes e nos grupos restritos em proteína, comparados aos
grupos controles. A razão insulina/glicose foi influenciada tanto pelo estado nutricional
como pelo estado fisiológico (F1,23=10,24; p<0,004; F1,23=18,60; p<0,0002). A razão
insulina/glicose foi menor nos grupos lactantes em relação aos não-lactantes. A razão
insulina/glucagon foi influenciada pelo estado fisiológico.
A prolactina foi modificada tanto pelo estado nutricional como pelo fisiológico
(F1,18=28,37; p<0,000 e (F1,18=126,16; p<0,000), havendo interação entre essas condições
(F1,18=20,85; p<0,000). Apesar dos grupos lactantes terem maior concentração desse
hormônio, no grupo HPL esta foi 50% menor que no CL. Nas concentrações de leptina
apenas a lactação interferiu reduzindo esse parâmetro (F1,20=22,60; p<0,000) (Tabela 4).

20
Tabela 4. Concentrações séricas de insulina, glucagon, leptina, prolactina, razão
insulina/glicose e insulina/glucagon, em ratas não-lactantes e lactantes tratadas com dieta
controle (CNL e CL) ou hipoprotéica (HPNL e HPL) durante 18 dias de lactação.
GRUPOS
CNL (7)
CL (7)
HPNL (6)
HPL (7)
Insulina (pg/mL) 3490±800
1660±500 *
1810±200 §
520±80 * §
Glucagon (pg/mL) 1130±180
620±100 *
590±200 §
510±150 * §
Prolactina (ng/mL) 26,5±4,3 c
130±3,9 a
21,6±8 c
65±6 b
Leptina (pg/mL) 2458±680
1136±220 *
3691±557
1055±256 *
Razão insulina/glicose (pmol/L:mmol/L)
102±22
53±18 *
47±3 §
15±2 * §
Razão insulina/glucagon (pg/mL)
2,28±0,6
1,76±0,4 *
2,43±0,4
0,77±0,2 *
Os valores expressam média ± EPM de animais entre parênteses
§ Diferença estatística com relação às ratas controles (ANOVA a dois fatores; p<0,05)
* Diferença estatística com relação às ratas não-lactantes (ANOVA a dois fatores; p<0,05)
Letras diferentes indicam diferenças estatísticas (Teste de Tukey HSD; p<0,05)
No FIG, a massa absoluta foi modificada pelos estados nutricional (F1,25=5,22;
p<0,03) e fisiológico (F1,25=61,33 p<0,000). Assim, a lactação aumentou a massa desse
tecido, mas a restrição protéica reduziu esse parâmetro. Independente do estado nutricional
as ratas lactantes mostraram massa relativa do fígado maior que as não-lactantes
(F1,25=95,22; p<0,000) (Tabela 5).
Na avaliação da massa da GMA ocorreu interação entre o estado fisiológico e o
nutricional, tanto em termos absolutos como relativos (F1,25=23,16; p<0,000 e F1,25=15,93;
p<0,000). O grupo HPL teve massa absoluta da GMA similar ao grupo HPNL e o grupo
CL maior, em relação ao CNL (Tabela 5).

21
O estado fisiológico determinou redução significativa na massa do GON (total e
proporcional) (F1,25=9,79; p<0,004 e F1,25=14,71; p<0,000). Assim, ratas lactantes
mostraram redução ao redor de 25% na massa absoluta e relativa desse tecido adiposo,
comparadas às não-lactantes (Tabela 5).
A massa absoluta da CARC foi influenciada pelo estado nutricional (F1,25=34,71;
p<0,000) e este interagiu com o fisiológico (F1,25=20,02; p<0,000). Ratas do grupo HPL
tiveram massa absoluta da carcaça menor em relação ao grupo HPNL, enquanto as ratas do
grupo CL tiveram essa variável maior em relação à CNL. Em termos relativos houve
influência do estado fisiológico, sendo que as ratas lactantes apresentaram redução nesse
parâmetro, comparadas às não-lactantes (F1,25=11,40; p<0,002) (Tabela 5).
22
Tabela 5. Massas do fígado (FIG), glândula mamária (GMA), tecido adiposo branco
gonadal (GON) e carcaça (CARC) absoluta e relativa de ratas não-lactantes e lactantes
tratadas com dieta controle (CNL e CL) ou hipoprotéica (HPNL e HPL) durante 18 dias de
lactação.
GRUPOS
CNL (7)
CL (7)
HPNL (7)
HPL (8)
g
FIG 9,4±0,3 14,7±0,8 * 8,9±0,3 § 12,4±0,7 § *
GMA 6,1±0,6 b 10,5±0,6 a 8,6±0,9 b 6,1±0,5 b
GON 16,2±1,4 14,1±1,2 * 18,0 ±1,6 11,1±1,5 *
CARC 215,4±4,2 b 228,3±4,4 a 208,5±2,3 b 177,7±6,7 c
g/100g
FIG 3,2±0,09 4,5±0,2 * 3,1±0,09 5,0±0,2 *
GMA 2,0±0,2 c 3,2±0,2 a 3,0±0,3 b 2,5±0,2 c
GON 5,4±0,3 4,2±0,3 * 6,2±0,5 4,4±0,4 *
CARC 72,6±0,8 69,2±0,5 * 72,4±0,8 71,4±0,5 *
Os valores expressam média ± EPM do número de ratas entre parênteses.
§ Diferença estatística com relação às ratas controles (ANOVA a dois fatores; p<0,05)
* Diferença estatística com relação às ratas não-lactantes (ANOVA a dois fatores; p<0,05)
Letras diferentes indicam diferenças estatísticas (Teste de Tukey HSD; p<0,05)
A Figura 1 apresenta o conteúdo proporcional de lipídios nos tecidos: FIG, GON,
GMA e na CARC. Em função do estado fisiológico houve aumento de depósito de lipídios
no FIG e redução no GON e na GMA (F1,25=41,93; p<0,001; F1,25=11,62; p<0,002 e
F1,25=78,78; p<0,001, respectivamente). Porém, nas carcaças esse parâmetro não diferiu
entre os grupos.

23
CNL CL HPNL HPL0
5
10
15
20
A
**
Con
teúd
o de
lip
ídeo
s no
FIG
(m
g/10
0mg)
CNL CL HPNL HPL0
25
50
75
100
B
* *
Con
teúd
o de
lipí
dios
no
GO
N(m
g/10
0mg)
CNL CL HPNL HPL0
25
50
75
100
*
*
C
Con
teúd
o de
lip
ídeo
s na
GM
A (
mg/
100m
g)
CNL CL HPNL HPL0
5
10
15
20
D
Con
teúd
o de
lip
ídeo
s na
CA
RC
(m
g/10
0mg)
CNL CL HPNL HPL0
1
2
3
4
c
b
c
a
E
Raz
ão l
ipíd
io:g
lico
gêni
ono
FIG
(m
g/10
0mg)
Figura 1. Conteúdo de lipídios no fígado (FIG) (A), tecido adiposo branco gonadal (GON)
(B), glândula mamária (GMA) (C), carcaça (CARC) (D) e razão lipídio/glicogênio (E) no
fígado de ratas não-lactantes e lactantes, tratadas com dieta controle (CNL e CL) ou
hipoprotéica (HPNL e HPL) durante 18 dias de lactação. As colunas representam o
conteúdo de lipídios expressa em média ± EPM de 7 a 8 ratas.* Diferença estatística com
relação às ratas não-lactantes (ANOVA a dois fatores; p<0,05). Letras diferentes indicam
diferenças estatísticas (Teste de Tukey HSD; p<0,05).

24
A razão lipídio/glicogênio no fígado foi maior no grupo HPL em relação aos
demais grupos (F1,24=6,32; p<0,02).
A lactação aumentou a atividade das enzimas avaliadas no FIG (Figura 2). Com
relação a G6PDH, as lactantes apresentaram maior atividade que as não-lactantes, porém o
grupo HPL apresentou atividade aumentada em, aproximadamente 3,5 vezes, comparado
ao HPNL, enquanto que no grupo CL a atividade desta enzima foi 1,5 vezes maior em
comparação ao CNL (F1,16=;10,06; p<0,001). A atividade da enzima ATP-cit foi maior em
ratas lactantes em relação às não-lactantes (F1,16=17,22; p<0,000), enquanto que a da
enzima málica foi significativamente mais elevada no grupo HPL, em comparação aos
demais grupos.
Em adição, a Figura 3 mostra uma correlação positiva entre a concentração sérica
de prolactina e a atividade das enzimas G6PDH e EM no fígado de ratas lactantes restritas
em proteína (HPL) (r=0,97; p<0,05).

25
CNL CL HPNL HPL0
100
200
300
A
b
aa
cGlic
ose-
6-fo
sfat
ode
sidr
ogen
ase
(nm
ol.m
g de
pro
t-1.m
in-1
)
CNL CL HPNL HPL0
100
200
300
B
*
*
AT
P c
itrat
o lia
se(n
mol
.mg
de p
rot-1
.min
-1)
CNL CL HPNL HPL0
50
100
150
200
C
bc b
c
a
Enz
ima
mál
ica
(nm
ol.m
g de
pro
t-1.m
in-1
)
Figura 2. Atividade das enzimas glicose-6-fosfato desidrogenase (A), ATP-citrato liase (B)
enzima málica (C) no fígado de ratas não-lactantes e lactantes, tratadas com dieta controle
(CNL e CL) ou hipoprotéica (HPNL e HPL) durante 18 dias de lactação. As colunas
representam a atividade das enzimas expressa em média ± EPM de 5 ratas. *Diferença
estatística com relação às ratas não-lactantes. Letras diferentes indicam diferenças
estatísticas pela ANOVA a dois fatores seguidos de Teste de Tukey HSD (p<0,05).

26
0 50 100 150 2000
100
200
300
400CL r=0,28 p=0,64HPL r=0,97 p<0,05A
Prolactina(ng/mL)
Glic
ose-
6-fo
sfat
ode
sidr
ogen
ase
(nm
ol.m
g de
pro
t-1.m
in-1
)
0 50 100 150 2000
50
100
150
200CL r= -0,54 p=0,35HPL r=0,97 p<0,05B
Prolactina(ng/mL)
Enz
ima
mál
ica
(nm
ol.m
g de
pro
t-1.m
in-1
)
Figura 3. Correlação entre a concentração sérica de prolactina e a atividade da enzima
glicose-6-fosfato desidrogenase (A) e enzima málica (B) no figado, de ratas lactantes
tratadas com dieta controle (CL) ou hipoprotéica (HPL), durante 18 dias de lactação.
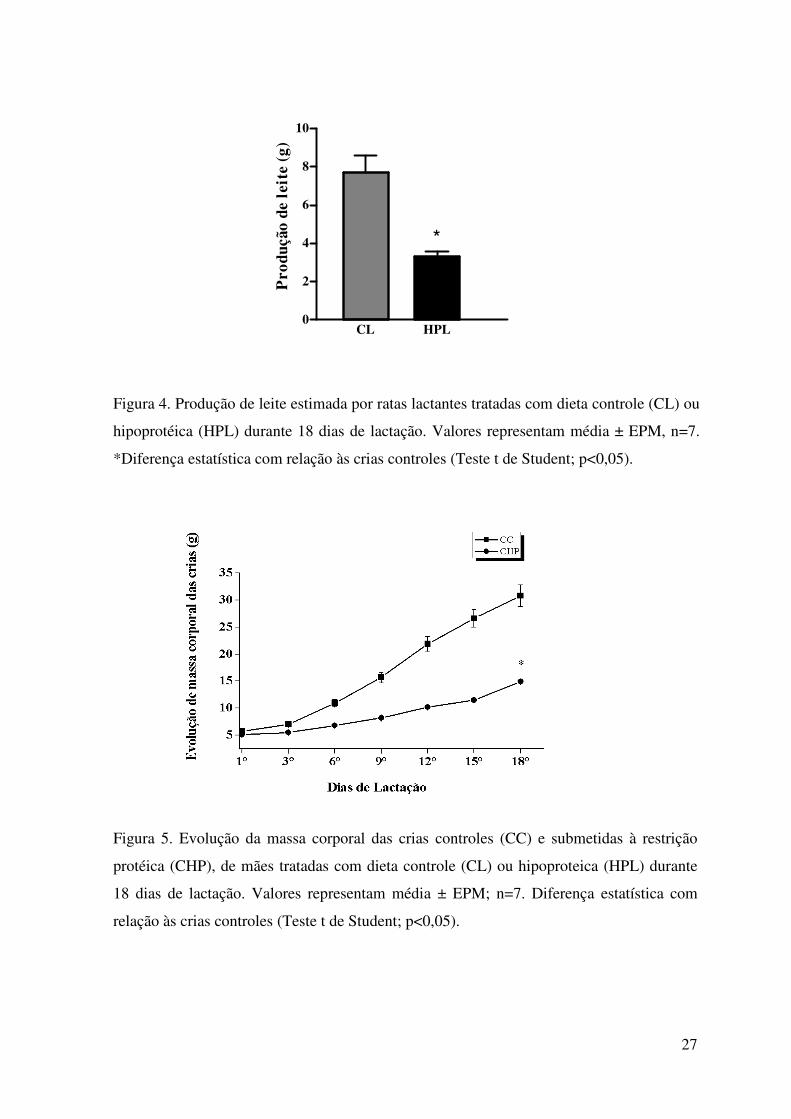
O estado nutricional reduziu a produção de leite nas ratas HPL em cerca de 50%,
em comparação às ratas CL (p<0,000) (Figura 4).
Observa-se que a massa das crias dos dois grupos avaliados, durante o período
experimental, sofreu elevação gradativa. Entretanto, desde o nascimento essa elevação foi
estatisticamente mais acentuada no grupo de crias controle (CC) comparada às crias de
ratas submetidas à restrição protéica (CHP) (Figura 5).
27
CL HPL0
2
4
6
8
10
*
Pro
duçã
o de
lei
te (
g)
Figura 4. Produção de leite estimada por ratas lactantes tratadas com dieta controle (CL) ou
hipoprotéica (HPL) durante 18 dias de lactação. Valores representam média ± EPM, n=7.
*Diferença estatística com relação às crias controles (Teste t de Student; p<0,05).
Figura 5. Evolução da massa corporal das crias controles (CC) e submetidas à restrição
protéica (CHP), de mães tratadas com dieta controle (CL) ou hipoproteica (HPL) durante
18 dias de lactação. Valores representam média ± EPM; n=7. Diferença estatística com
relação às crias controles (Teste t de Student; p<0,05).

28
V - DISCUSSÃO
Durante a lactação, o fígado, o tecido adiposo e a glândula mamária, são os
principais sítios do metabolismo de lipídios, podendo sintetizar ácidos graxos de novo e
esterificá-los em triglicerídeos.18
No presente estudo foi observado no 18º dia de lactação, que a massa e o conteúdo
de lipídios no fígado encontravam-se elevados de forma similar nos grupos lactantes (CL e
HPL), comparados aos não-lactantes (CNL e HPNL), havendo concomitante redução na
massa e no conteúdo de lipídios no tecido adiposo gonadal nos mesmos grupos. O
metabolismo no tecido adiposo encontra-se normalmente em estado catabólico durante a
lactação, devido ao aumento na mobilização dos substratos desse tecido gorduroso.16,17
Por outro lado, o mecanismo fisiológico do acúmulo de TG no fígado ainda não
está totalmente esclarecido, podendo ser devido à elevada concentração de ácidos graxos
livres e/ou ésteres de coenzima A na célula,64 ou pelo aumento na atividade de algumas
enzimas de esterificação.23 Pode ser ainda em decorrência de uma reduzida produção de
apoproteína B, com menor secreção de VLDL20 e, conseqüentemente, maior deposição de
lipídios nos hepatócitos.
Tem sido descrito que na lactação inicial, um moderado acúmulo de TG no fígado é
usual e fisiológico, sem prejuízo para as suas funções. Porém, à medida que a lactação
progride para a fase tardia, é esperado que esse acúmulo decresça gradativamente.18
Embora o conteúdo de glicogênio no fígado não tenha sido diferente entre os
grupos, um fator importante a ser considerado é a concentração de lipídios em relação ao
conteúdo de glicogênio, ou razão lipídio/glicogênio.65 Assim, preconiza-se como normal
uma razão que não ultrapasse valores acima de 2/1, o que pode indicar acúmulo de lipídios
nos hepatócitos e desencadear efeitos deletérios, como a ocorrência de esteatose.20 No

29
grupo HPL, a razão lipídio/glicogênio foi de 3/1, e isso sugere que a restrição protéica
associada à lactação, tenha favorecido a maior concentração de lipídios no fígado.
Observou-se, também, que a lactação aumentou a atividade das enzimas
lipogênicas avaliadas no fígado e que a restrição protéica associada à lactação, acentuou a
atividade das enzimas G6PDH e EM.
Prévios estudos em ratos, avaliando o efeito de dietas com altas concentrações de
carboidratos e baixas concentrações de lipídios ofertadas ad libitum, sobre a atividade de
enzimas lipogênicas no fígado, mostraram que quanto mais elevado o aporte energético
consumido, maior o estímulo para o aumento de atividade dessas enzimas.66
A lactação diminuiu as concentrações de insulina e leptina. A restrição protéica
associada à lactação (HPL) reduziu em aproximadamente dois terços a insulinemia, quando
comparado à CL. De acordo com Sipols et al. (1995)67animais com baixas concentrações
de insulina são hiperfágicos, porém esse resultado não ocorreu no presente estudo.
A hipoleptinemia parece ser o achado mais consistente na lactação e pode resultar
principalmente do balanço energético negativo1, contribuindo para o aumento do apetite.19
Por outro lado, a hipoleptinemia pode promover o acúmulo de TG no fígado durante a
lactação.18
As baixas concentrações de insulina e leptina durante a lactação associada à
restrição protéica podem ser resultantes do processo catabólico, que favorece a liberação
de ácidos graxos do tecido adiposo, direcionando-os para o tecido hepático.21,25
Ao considerar a razão insulina/glicose, observou-se que as ratas lactantes foram
mais sensíveis à insulina, comparadas às não-lactantes e que a restrição protéica aumentou
essa sensibilidade. Além do mais, a razão insulina/glucagon nas lactantes foi 46% menor
que nas não-lactantes, e isso pode ter favorecido o processo de catabolismo observado no
tecido adiposo.

30
Com relação à prolactina, embora os grupos lactantes tenham apresentado
concentrações mais elevadas, foi observado que a restrição protéica associada à lactação
contribuiu para a redução nas concentrações desse hormônio em 50% nas ratas HPL,
comparadas às CL. Por outro lado, no mesmo grupo de ratas, ocorreu correlação positiva
entre as concentrações séricas de prolactina e as enzimas G6PDH e EM avaliadas no
fígado.
Pesquisas recentes demonstram que a prolactina é altamente expressa no fígado,
onde tem um papel central na homeostase metabólica. Em adição, este hormônio regula a
atividade de enzimas e transportadores que estão associados com o metabolismo de glicose
e de lipídios em outros órgãos alvo.68
Nesse período, a utilização preferencial dos precursores lipogênicos é a glândula
mamária visando a produção de leite6 e tornando essa glândula, o sítio mais ativo da
síntese de lipídios in vivo, com atividade lipogênica aumentada em cerca de cinco vezes,
comparada à do fígado.8 Na glândula mamária, foi observada redução proporcional da
massa no grupo HPL, enquanto que no grupo CL ocorreu aumento. Por outro lado, o teor
de lipídios na GMA foi reduzido de forma concomitante nos grupos lactantes.
Apesar do aumento no consumo alimentar relativo, ratas HPL apresentaram
características típicas de desnutrição protéica, como redução da massa corporal e
hipoproteinemia. Esses achados associados à redução na massa da glândula mamária,
possivelmente, contribuíram para a menor produção de leite e, conseqüentemente,
interferiram no desenvolvimento somático das crias nesse grupo. De acordo com Dewey
(1997),69 um aporte adequado de proteínas na dieta materna eleva a produção láctea, além
de aumentar positivamente o balanço de proteínas do leite.
O conjunto dos resultados apresentados demonstra que na lactação tardia ocorreu
maior concentração de lipídios no fígado e que a restrição protéica nesse período favoreceu

31
esse processo, evidenciado pelo aumento na atividade da enzima málica e na razão lipídio/
glicogênio.

32
VI – REFERÊNCIAS 1. Woodside B. Prolactin and the hyperfhagia of lactation. Physiol. Behav. In press.
2007.
2. Leite MS, Azevedo VB, Carmo MG, Boaventura GT. Utilização da multimistura
durante a lactação e seus efeitos na produção e composição do leite materno de ratas.
Rev. Nut. Campinas. 2002; 15(2):211-21.
3. Denis RGP, Williams G, Vernon RG. Regulation of serum leptin and its role in the
hyperphagia of lactation in the rat. J. Endocrinol. 2003; 176:193-203.
4. Trayhurn P. Thermogenesis and the energetics of pregnancy and lactation. Can. J.
Physiol. Pharmacol. 1989; 67:370-5.
5. Williamson DH, Lund P. Cellular mechanisms for the regulation of adipose tissue
lipid metabolisms in pregnancy and lactation. Adv. Exp. Med. Biol.1994; 352:45-70.
6. Vernon RG, Pond CM. Adaptations of maternal adipose tissue to lactation. J.
Mammary Gland Biol. Neoplasia. 1997; 2(3):231-41.
7. Barber MC, Clegg RA, Travers MT, Vernon RG. Lipid metabolism in the lactating
mammary gland. Biochem. Biophys. Acta. 1997; 1347:101-26.

33
8. Williamson DH. Integration of metabolism in tissues of the lactating rat. Febs
Letters. 1980; 117: K93-K105.
9. Robinson AM, Girard JR, Williamson DH. Evidence or a role of insulin regulation of
lipogenesis in lactanting rat mammary gland. Measurements of lipogenesis in vivo
and plasma hormone concentrations in response to starvation and refeeding.
Biochem. J. 1978; 170:609-13.
10. Hawkins RA, Williamson DH. Measurements of substrate uptake by mammary gland
of the rat. Biochem. J. 1972; 129:1171-3.
11. Flint DJ, et al. Effects of growth hormone and prolactin on adipose tissue
development and function. Pituitary. 2003; 6:97-102.
12. Flint DJ, Sinnett-Smith PA, Clegg RA, Vernon RG. Role of insulin receptors in the
changing metabolism of adipose tissue during pregnancy and lactation in the rat.
Biochem. J. 1979; 182(2):421-7.
13. Smith RW. The effects of pregnancy, lactation and involution on the metabolism of
glucose by rat parametrial adipose tissue. J. Dairy. Res. 1973; 40(3):353-60.
14. Otaway S, Robinson DS. The significance of changes in tissue clearing-factor lipase
activity in relation to the lipaemia of pregnancy. Biochem. J. 1968; 106(3):677-82.

34
15. Vernon RG, Flint DJ. Control of fatty acid synthesis in lactation. Proc. Nutr. Soc.
1983; 42:315-31.
16. Bauman DE. Regulation of nutrient partitioning during lactation: homeostasis and
homeorhesis revisited. In: Cronjé PB. Ruminant Physiology: Digestion, Metabolism,
Growth and reproduction, Wallingford: CABI. 2000; (18):311-28.
17.
Vernon RG, Denis RGP, Sorensen A, Williams G. Leptin and the adaptations of
lactation in rodents and ruminants. Horm. Metab. Res. 2002; 34:678-85.
18. Vernon RG. Lipid metabolism during lactation: a review of adipose tissue-liver
interactions and the development of fatty liver. J. Dairy Res. 2005; 72:460-9.
19. Zammit VA. Ketogenesis in the liver of ruminants – adaptations to a challenge. J.
Agric. Sci. 1990; 115:155-62.
20. Drackley JK, Overton TR, Douglas GN. Adaptations of glucose and long-chain fatty
acid metabolism in liver of dairy cows during periparturient period. J. Dairy Sci.
2001; 84:E100-E12
21. Bradbury MW. Lipid metabolism and liver inflammation. I Hepatic fatty acid uptake:
possible role in steatosis. A. J. Physiol. Gastroint. Liver. Physiol. 2006; 290:6194-8.

35
22. Dann HM, Douglas GN, Overton TR, Drackley JK. Carnitine palmitoyltransferase
activity in liver of periparturient dairy cows. J. Dairy Sci. 2000; 83(S1):251.
23. Van den Top AM, Wensing T, Geelen MJH, Wentink GH, Van’t Klooster AT,
Beynen AC. Time trends of plasma lipids and enzymes synthesizing hepatic
triacylglycerol during postpartum development of fatty liver in dairy cows. J. Dairy
Sci. 1995; 78:2208-20.
24. Avramglu RK, Qiu W, adeli K. Mechanisms of metabolic in insulin resistant states:
desregulation of hepatic and intestinal lipoprotein secretion. Front. Biosci. 2003; 8:
D464-D76.
25.
Canbay A, Bechmann L, Gerken G. Lipid metabolism in the liver. Z. Gastroenterol.
2007; 45:35-41.
26. Vernon RG. Adipose tissue: an expanding role in the regulation of energy
metabolism. In: Souffrant WB and Metges CC, editors. Proceedings of the EAAP-
symposium on energy and protein metabolism and nutrition; 2003 Sep 13-18;
Rostock-Warnemünde, Germany. EAAP publication No. 109. Wageningen:
Wageningen Academic Publishers; 2003. p. 451-64.

36
27. Oller do Nascimento CM, Ilic V,Williamson DH. Re-examination of the putative roles
of insulin and prolactin in the regulation of lipid deposition and lipogenesis in vivo in
mammary gland and white and brown adipose tissue of lactating rats and litter-
removed rats. Biochem. J. 1989; 258:273-8.
28. Neville MC, McFadden TB, Forsyth I. Hormonal regulation of mammary
differentiation and milk secretion. J. Mammary. Gland Biol. Neoplasia. 2002;
7(1):49-66.
29. Robinson AM, Girard JR, Williamson DH. Evidence for a role of insulin in the
regulation of lipogenesis in lactating rat mammary gland. Measurements of
lipogenesis in vivo and plasma hormone concentrations in response to starvation and
refeeding. Biochem. J. 1978; 176(1):343-6.
30. Flint DJ, Knight CH. Interactions of prolaction and growth hormone (GH) in the
regulation of mammary gland function and epithelial cell survival. J. Mammary
Gland Biol. Neoplasia 1997; 2:41-8.
31. Groner, B. Transcription factor regulation in mammary epithelial cells. Domest.
Anim. Endocrinol. 2002; 23:25-32.
32. Neville MC. et al. Hormonal regulation of mammary differentiation and milk
secretion. J. Mammary Gland Biol. Neoplasia. 2002; 7:49-66.

37
33. Woodside B, Abizaid A, Walker C. Changes in leptin levels during lactation:
implications for lactational hyperphagia and anovulation. Horm. Behav. 2000; 37(4):
353-65.
34. Bai Y, Zhang S, Kim KS, Lee JK, Kim KH. Obese gene expression alters the ability
of 30A5 preadipocytes to respond to lipogenic hormones. J. Biol. Chem. 1996;
271:13939-42.
35. Wang MY, Lee Y, Unger RH. Novel form of lipolysis induced by leptin. J. Biol.
Chem. 1999; 274:17541-4.
36. Soukas A, Cohen P, Socci ND, Friedman JM. Leptin-specific patterns of gene
expression in white adipose tissue. Genes Dev. 2000; 14:963-80.
37. Kakuma T, Lee Y, Higa M, Wang ZW, Pan W, Shimomura I, et al. Leptin,
troglitazone, and the expression of sterol regulatory element binding proteins in liver
and pancreatic islets. Proc. Natl. Acad. Sci. 2000; 97:8536-41.
38. Naismith DJ, Richardson DP and Pritchard AE. The utilization of protein and energy
during lactation in the rat, with particular regard to the use of fat accumulated in
pregnancy. J. Nutr. 1982; 48:433-41.
39. Jansen GR, Hunsaker H. Effect of dietary protein and energy on protein synthesis
during lactation in rats. J. Nutr. 1986; 116:957-68.

38
40. Friggens NC. The effecs of feed composition and level on lactational performance in
rats and dairy cows: a basic approach to feed description. PhD thesis, University of
Edinburgh, London. 1990.
41. Cambraia RP, Vannucchi H, De-Oliveira LM. Food intake and weight of lactating
rats maintained on different protein-calorie diets, and pup growth. Braz. J. Med. Biol.
Res. 1997; 30(8):985-88.
42. Naismith DJ, Robinson SM. Adaptations in protein metabolism during lactation in
the rat. Br. J. Nutr. 1987; 58:533-38.
43. Shetty PS. Physiological mechanisms in the adaptive response of metabolic rates to
energy restriction. Nutr. Res. Rev. 1990; 3:49-74.
44. McNamara JP Role and regulation of metabolism in adipose tissue during lactation.
Nutr. Biochem. 1995; 6:120-9.
45. Sainz RD, Calvert CC, Baldwin RL. Relationship among dietary protein, feed intake
and changes in body and tissue composition of lactating rats. J. Nutr. 1986;
116:1529-39.
46. Taylor JB, Calvert CC, Baldwin RL, Sainz RD. Effects of dietary protein, fat and
restriction on body composition and energy balance in lactating rats. J. Nutr. 1986;
116:1519-28.

39
47. Grigor MR, Allan JE, Carrigton JM, Carne A, Geursen A, Young D, et al. Effect of
dietary protein and food restriction, maternal tissues and enzymes in lactating rats. J.
Nutr. 1987; 117:1247-58.
48. Ferreira CLP, Macêdo GM, Latorraca MQ, Arantes VC, Veloso RV, Carneiro AC, et
al. Serun leptin and insulin levels in lactating protein-restricted rats: implications for
energy balance. Br. J. Nutr. 2007; 97:27-34.
49. Erhuma A, Salter AM, Sculley DV, Langley-Evans SC, Bennett AJ. Prenatal
exposure to a low-protein diet programs disordered regulation of lipid metabolism in
the aging rat. Am. J. Physiol. Endocrinol. Metab. 2007; 292(6):E1702-E14.
50. Manual para técnicos de laboratorios. Comissão de Ensino do Colégio Brasileiro de
Experimentação Animal (COBEA). 1996; 2ª ed.
51. Reeves PG, Nielsen FH, Fahey GC. AIN-93 Purified diets for laboratory rodents:
final report of the American Institute of Nutrition ad hoc writing committee on
reformulation of the AIN-76A rodent diet. J. Nutr. 1993; 123(6):1939-51.
52. Russel JA. Milk Yield, suckling behavior and ejection in the lactating rat nursing
litters of different sizes. J. Physiol. 1980; 303:403-15.

40
53. Tavares do Carmo MG, Oller do Nascimento CM, Martin A, Herrera E. Ethanol
Intake during lactation impairs milk production in rats and affects growth and
metabolism of suckling pups. Alcohol. 1999; 18(1):71-6.
54. Wolfson WQ, Cohn C, Calvary F, Ichiba F. Studies in serum proteins 5. A rapid
procedure for the estimation of total protein, true albumin, total globulin, alpha
globulin, and gamma globulin in 1.0ml of serum. Am. J. Clin. Pathol. 1948; 18:723-
30.
55. Doumas BT, Watson W A, Biggs HG. Albumin standards and the measurements of
serum albumin with bromocresol green. Clin. Chim. Acta. 1971; 31:87-96.
56. Sjögren B, Koning WA, Holmgeen H, E Möllersöm J. Betrag zur Kennynies der
leberrthytlmik (glycogen, phosphor und calcium in des kaninchenleber). Pflüger
Arch. Ges. Physiol. 1938; 240:427-36.
57. Folch J, Lees M, Sloane-Stanley GH. A simple method for the isolation and
purification of total lipids from animal tissues. J. Biol. Chem. 1957; 226(34):497-
509.
58. Oller do Nascimento CM, Williamson DH. Tissue-specific effects of starvations and
refeeding on the disposal of oral (1-14C) triolen in the rat during lactation and on
removal of litter. Biochem. J. 1986; 254:539-46.

41
59. Lowry OH. Protein measurement with of the folin phenol reagent. J. Biol. Chem.
1951; 193:265-8.
60. Lee CY. Glucose-6-phosphate dehydrogenase from mouse. Methods Enzymol. 1982;
89:252-7.
61. Ochoa S. Malic enzyme. Methods Enzymol. 1955; 1:739-53.
62. Hsu RY, Lardy HA. Malic enzyme. Methods Enzymol. 1969; 13:230.
63. Srere PA. The citrate cleavage enzyme. I. Distribution and purification. J. Biol.
Chem. 1959; 234: 2544-47.
64. McGarry JD. Dysregulation of fatty acid metabolism in the etiology of type 2
diabetes. Diabetes. 2002;517-18.
65. Grummer RR. Etiology of lipid-related metabolic disorders in periparturient dairy
cows. J. Dairy Sci. 1993; 76:3882-96.
66. Iritani N, Katsurada A, Fukuda H. Regulation of hepatic lipogenic enzyme gene
expression by diet quantity in rats fed a fat-free, high carbohydrate diet. J. Nutr.
1992; 122:28-36.

42
67. Sipols AJ, Baskin DG, Schwartz MW. Effect of intracerebroventricular insulin
infusion on diabetic hyperphagia and hypothalamic neuropeptide gene expression.
Diabetes. 1995; 44:147-51.
68. Ben-Jonathan N, Hugo ER, Brandebourg TD, LaPensee C. Focus on prolaction as a
metabolic hormone. Trends Endocrinol. Metab. 2006; 17(3); 110-6.
69. Dewey KG. Energy and protein requirements during lactation. Ann. Rev. Nutr. 1997;
17:19-36.