Embed Size (px)
Citation preview

Ilhéus, Bahia
2007
Universidade Estadual de Santa Cruz Programa Regional de Pós-graduação em Desenvolvimento e Meio Ambiente
Mestrado em Desenvolvimento Regional e Meio Ambiente
IDENTIFICAÇÃO DE BIOINDICADORES E
PLANEJAMENTO DE MINI-CORREDORES ECOLÓGICOS
NA ÁREA DE PROTEÇÃO AMBIENTAL
COSTA DE ITACARÉ/SERRA GRANDE, BAHIA
HILDA SUSELE RODRIGUES ALVES

HILDA SUSELE RODRIGUES ALVES
Dissertação apresentada ao Programa Regional de Pós-graduação em Desenvolvimento Regional e Meio Ambiente, Sub-programa Universidade Estadual de Santa Cruz, como parte dos requisitos para obtenção do título de Mestre em Desenvolvimento Regional e Meio Ambiente, Sub-área de concentração: Planejamento e Gestão Ambiental no Trópico Úmido.
Orientador: Profº Dr. Jacques H. C. Delabie
Co-orientador: Profº Dr. Maurício Moreau
Ilhéus, Bahia
2007
IDENTIFICAÇÃO DE BIOINDICADORES E
PLANEJAMENTO DE MINI-CORREDORES ECOLÓGICOS
NA ÁREA DE PROTEÇÃO AMBIENTAL
COSTA DE ITACARÉ/SERRA GRANDE, BAHIA

. . . . ALVES, Hilda Susele Rodrigues Criação de Instrumentos visando a conservação e recuperação ambientais na Área de Proteção Ambiental Costa de Itacaré/Serra Grande, Bahia. Hilda Susele Rodrigues Alves. – Ilhéus, BA: UESC/PRODEMA, 2007. XXXfl. : il. Orientador: Jacques H. C. Delabie e Maurício Moreau Dissertação (mestrado) – Universidade Estadual de Santa Cruz. Pós-graduação em Desenvolvimento Regional e Meio Ambiente. Bibliografia: X. [xx] - xx 1. Bioindicadores 2. Corredor Ecológico 3. Formigas 4. Mata
Atlântica 5. SIG. I. Título. CCD xxx.x

HILDA SUSELE RODRIGUES ALVES
Aprovada em 20 de setembro de 2007
Comissão Examinadora:
Profº Dr. Jacques Hubert Charles Delabie (Orientador) UESC – Universidade Estadual de santa Cruz
Profº Drª. Raquel Maria de Oliveira UESC – Universidade Estadual de santa Cruz
Profº Dr. Ivan Cardoso Nascimento CEPEC/CEPLAC
IDENTIFICAÇÃO DE BIOINDICADORES E
PLANEJAMENTO DE MINI-CORREDORES ECOLÓGICOS
NA ÁREA DE PROTEÇÃO AMBIENTAL
COSTA DE ITACARÉ/SERRA GRANDE, BAHIA

DEDICATÓRIA
Ao Profº Jacques Delabie (Orientador) e Profº Maurício Moreau (Co-orientador),
pelos ensinamentos valiosos, orientação e amizade demonstrada
durante a elaboração desse estudo, dedico.

AGRADECIMENTOS
O meu eterno agradecimento a Deus, que me concede o dom da vida.
A Universidade Estadual de Santa Cruz e ao Programa Regional de Pós-graduação em
Desenvolvimento Regional e Meio Ambiente, pela oportunidade de realização do curso.
A Fapesb, pelo apoio financeiro.
A Jacques Delabie e Maurício Moreau, pela orientação e dedicação.
A Profª Drª Raquel, pela amizade e palavras de autoconfiança.
Ao Profº Drº Mariano e Profº Drº Ivan Nascimento, pelas correções e sugestões na
qualificação.
Ao Conselho Gestor da APA (em especial Marco Aurélio e Mônica), pelo suporte à
participação nas reuniões do Conselho e acesso a documentos de interesse desse estudo.
Aos professores do curso, pelos ensinamentos recebidos.
A minha família, meus pais Rita e Jovino (in memorian), Helena (minha segunda
mãe), minhas irmãs Adriana, Saadi e Samile, meus sobrinhos Matheus, Malthus e Nathália,
meu companheiro e sempre amigo Tadeu, Tia Cidinha, Tio Hemilson e Tio Halemil.
Agradeço pelo amor, carinho e apoio recebido.
À equipe do Laboratório de Mirmecologia, pelo apoio e amizade.
Aos amigos Benoit Jahyny, Luiza Carla e Yana Teixeira pela contribuição com o
inglês e com estatística.
Aos colegas de turma, pela amizade, momentos polêmicos e também momentos de
descontração.
Serei eternamente grata a todos vocês que de alguma forma contribuíram para que este
estudo fosse possível.

SUMÁRIO
Resumo........................................................................................................................... xi
Abstract.......................................................................................................................... xii
1 INTRODUÇÃO............................................................................................................. 1
2 A FRAGMENTAÇÃO FLORESTAL DA MATA ATLÂNTICA........................... 4
3 CORREDOR ECOLÓGICO....................................................................................... 11
3.1 Corredor Central da Mata Atlântica – CCMA.......................................................... 15
4 INSTRUMENTOS LEGAIS QUE APÓIAM AS AÇÕES NO
ESTABELECIMENTO DE CORREDORES ECOLÓGICOS................................
19
5 FORMICIDAE COMO BIOINDICADORES............................................................ 23
6 SISTEMAS DE INFORMAÇÃO GEOGRÁFICA NO ESTUDO DE ÁREAS
PRIORITÁRIAS À FORMAÇÃO DE CORREDORES ECOLÓGICOS.............. 26
7 METODOLOGIA......................................................................................................... 28
7.1 Caracterização da área de estudo.................................................................................... 28
7.2 Procedimentos metodológicos........................................................................................ 37
7.2.1 Identificação de Áreas de Preservação Permanente........................................................ 37
7.2.2 Identificação dos mini-corredores ecológicos................................................................. 37
7.2.3 Análise de Formicidae..................................................................................................... 39
7.2.4 Análises Faunísticas........................................................................................................ 42
8 RESULTADOS E DISCUSSÃO.................................................................................. 44
8.1 Áreas de Preservação Permanente................................................................................... 44
8.2 Formigas como bioindicadores....................................................................................... 53
8.2.1 Composição da comunidade de Formicidae.................................................................... 53
8.2.1.2 Análise da diversidade das áreas de florestas, cacauais e pastagens amostradas em
1996/1998........................................................................................................................ 62
8.2.1.3 Índice de Similaridade..................................................................................................... 66
8.2.1.4 Gêneros bioindicadores 69
8.3 Identificação de mini-corredores ecológicos 75
9 CONSIDERAÇÕES FINAIS....................................................................................... 96
10 REFERÊNCIAS BIBLIOGRÁFICAS........................................................................ 99

LISTA DE SIGLAS
APA Área de Proteção Ambiental
CAR Companhia de Desenvolvimento e Ação Regional
CCMA Corredor Central da Mata Atlântica
CEPLAC Comissão Executiva do Plano da Lavoura Cacaueira
CI Conservation International
CONAMA Conselho Nacional do Meio Ambiente
EUA Estados Unidos da América
GPS Sistemas de Posicionamento Global
IBAMA Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis
IBGE Instituto Brasileiro de Geografia e Estatística
IESB Instituto de Estudos Sócio-Ambientais do Sul da Bahia
MMA Ministério do Meio Ambiente
MVSP Multi-Variate Statistical Package
ONG’s Organizações não Governamentais
PPG7 Programa Piloto para a Proteção das Florestas Tropicais do Brasil
SEI Superintendência de Estudos Econômicos e Sociais da Bahia
SIG Sistemas de Informações Geográficas
SNUC Sistema Nacional de Unidades de Conservação
WWF Fundo Mundial para Conservação da Vida Silvestre

LISTA DE FIGURAS
1 Representação esquemática dos corredores lineares e de paisagem........................ 14
2 Proposta para criação de corredores ecológicos no Brasil. Projeto Parques e
Reservas (BRASIL-MMA, 1998)............................................................................ 16
3 Corredor Central da Mata Atlântica......................................................................... 18
4 Municípios onde se localiza a APA Costa de Itacaré/Serra Grande, Bahia............. 30
5 Área de Proteção Ambiental Costa de Itacaré/Serra Grande, Bahia........................ 31
6 Hidrografia da área de estudo - APA Costa de Itacaré/Serra Grande, Bahia........... 33
7 Solos da área de estudo - APA Costa de Itacaré/Serra Grande, Bahia..................... 34
8 Classes de Altitude (Relevo) da área de estudo - APA Costa de Itacaré/Serra
Grande, Bahia........................................................................................................... 35
9 Uso do Solo da área de estudo - APA Costa de Itacaré/Serra Grande, Bahia.......... 36
10 Fluxograma com os dados e comandos utilizados no Idrisi 2.0 para elaboração de
cada corredor ecológico........................................................................................... 38
11 Pontos de coletas na área de estudo e áreas próximas da APA Costa de
Itacaré/Serra Grande, Bahia..................................................................................... 41
12 Área de Preservação Permanente da área de estudo (Vegetação) - APA Costa de
Itacaré/Serra Grande, Bahia..................................................................................... 45
13 Área de Preservação Permanente da área de estudo (Hidrografia e Nascentes) -
APA Costa de Itacaré/Serra Grande, Bahia............................................................. 52
14 Riqueza de espécies de formigas por subfamílias.................................................... 55
15 Riqueza de espécies de formigas por gêneros.......................................................... 56
16 Curva de Riqueza Observada nas oito áreas de coletas........................................... 63
17 Similaridade entre as oito áreas de coletas. Índice de Jaccard, APA Costa de
Itacaré/Serra Grande, Bahia..................................................................................... 67
18 Relação entre distância e similaridade nas áreas de floresta na APA Costa de
Itacaré/Serra Grande, Bahia..................................................................................... 68
19 Relação entre distância e similaridade nas áreas de floresta e cacauais na APA
Costa de Itacaré/Serra Grande, Bahia...................................................................... 69
20 Número de ocorrências dos gêneros bioindicadores................................................ 73
21 Razão das ocorrências de Pheidole/Wasmannia...................................................... 74
22 Proposta do mini-corredor 1 na APA Itacaré/Serra Grande, Bahia......................... 77

23 Proposta de mini-corredor 2 na APA Itacaré/Serra Grande, Bahia.......................... 78
24 Proposta de mini-corredor 3 na APA Itacaré/Serra Grande, Bahia.......................... 80
25 Proposta de mini-corredor 4 na APA Itacaré/Serra Grande, Bahia.......................... 81
26 Proposta de mini-corredor 5 na APA Itacaré/Serra Grande, Bahia.......................... 83
27 Proposta de mini-corredor 6 na APA Itacaré/Serra Grande, Bahia.......................... 84
28 Proposta de mini-corredor 7 na APA Itacaré/Serra Grande, Bahia.......................... 86
29 Proposta de mini-corredor 8 na APA Itacaré/Serra Grande, Bahia.......................... 87
30 Proposta do mini-corredor 9 na APA Itacaré/Serra Grande, Bahia......................... 89
31 Proposta do mini-corredor 10 na APA Itacaré/Serra Grande, Bahia....................... 90
32 Proposta de mini-corredores na APA Costa de Itacaré/Serra Grande, Bahia.......... 95

LISTA DE TABELAS
1 Níveis de dificuldade para estabelecimento do corredor ecológico conforme
percepção do autor................................................................................................... 39
2 Siglas que serão utilizadas no texto para indicar as áreas de estudo........................ 53
3 Composição de formigas realizadas em oito áreas na APA Itacaré/Serra Grande,
Bahia, Brasil, (1996/1998)....................................................................................... 53
4 Formicidae coletados com extrator de Winkler em oito áreas na APA Costa de
Itacaré/Serra Grande, Bahia, Brasil (1996/1998)..................................................... 56
5 Riquezas Observadas e Estimadas, e Índice de diversidade das espécies coletadas 62
6 Representatividade das ocorrências dos gêneros indicadores em relação ao total
de ocorrências........................................................................................................... 73

xi
IDENTIFICAÇÃO DE BIOINDICADORES E PLANEJAMENTO DE MINI-CORREDORES ECOLÓGICOS NA APA COSTA DE
ITACARÉ/SERRA GRANDE, BAHIA
RESUMO
A Área de Proteção Ambiental (APA) Costa de Itacaré/Serra Grande foi criada em 1993 pelo Governo do Estado da Bahia, com o objetivo de preservar remanescentes de Mata Atlântica, proteger recursos hídricos e paisagens naturais. Em 2003, a APA foi ampliada, e hoje possui 62.960 ha. Está inserida no “Corredor Central da Mata Atlântica”, e abrange parte dos municípios de Uruçuca e Itacaré. Este estudo tem como objetivo identificar gêneros de formiga que podem ser utilizados como indicadores biológicos e planejar uma rede de mini-corredores ecológicos na APA. Para atingir estes objetivos, utilizou-se: o software ArcView 9.1 para elaboração do banco de dados cartográficos (localização da área de estudo; hidrografia; pedologia; geomorfologia; uso do solo e Áreas de Preservação Permanente); análises estatísticas com base no material biológico conservado na coleção do Laboratório de Mirmecologia, a fim de identificar formigas como bioindicadores de áreas antropizadas ou áreas conservadas; elaboração da rede de mini-corredores ecológicos da APA, utilizando os softwares ArcView 9.1 e Idrisi 2.0, visando sua incorporação no Corredor Central da Mata Atlântica. Os resultados obtidos evidenciaram que os fragmentos de floresta primária sofrem de um forte efeito de borda, pois observa que na sua vizinhança predominam pastagens, rodovias, estradas e cultivos agrícolas; sessenta e nove por cento da área é considerada Área de Preservação Permanente de acordo com a Lei Federal Nº 4.771; os ambientes de florestas e cacauais apresentaram diversidade de espécies de formigas superior à das áreas de pastagens; as áreas de floresta primária apresentam uma similaridade com as áreas de cabrucas em relação à composição de comunidades; existe uma correlação positiva entre similaridade e distância entre áreas de floresta primária; dos seis gêneros selecionados a priori como bioindicadores, dois (Pheidole e Wasmannia) foram confirmados, sendo que a diversidade de Pheidole é indicativa de áreas conservadas e a ocorrência de Wasmannia auropunctata é indicativa de áreas degradadas; a Floresta Secundária em Estágio Avançado de Regeneração/Cabruca é o elemento que predomina na matriz (35,84%); o SIG mostrou-se ser eficiente para interligar os fragmentos florestais de floresta primária; foram identificados dez mini-corredores na área, com indicações sobre as áreas propícias à implementação desses, recomendações para políticas públicas e ONGs que deverão assim beneficiar a qualidade de vida dos organismos nativos.
Palavras-chave: Bioindicadores, Corredor Ecológico, Formigas, Mata Atlântica, SIG.

xii
IDENTIFICATION OF BIOINDICATORS AND PLANIFICATION OF MINI-ECOLOGICAL CORRIDORS IN THE “APA” COSTA DE
ITACARÉ/SERRA GRANDE, BAHIA, BRAZIL
ABSTRACT
The Area of Environmental Protection (APA) Costa de Itacaré-Serra Grande was created in 1993 by the Bahia State government aiming the preservation of remnants of the Atlantic coastal forest of Brazil that, to protect water resources and native landscapes. In 2003, the APA was augmented, and has today 62.960 ha. It is inserted in the “Central Corridor Central of the Brazilian Atlantic Forest”, enclosing part of the counties of Ilhéus, Uruçuca and Itacaré. This study aims to identify ant genus as biological indicators and to planned web of mini-ecological corridors in the APA. With these objectives, we used: the software ArcView 9.1 to elaborate the bank of cartographic data (location of the study areas; soils, geomorphology; land use and permanent protection areas); statistics were performed using biological material of the Myrmecology Laboratory, aiming to identify ant species as biological indicators of anthropized or preserved areas; and the net of mini-ecological corridors of the APA was elaborated using ArcView 9.1 and Idrisi 2.0 softwares, aiming their integration in the Central Corridor Central of the Brazilian Atlantic Forest. Our results made evident that the pristine forest remnants suffer a strong border effect, as it was observed in the neighboring predominate pastures, roads, and crops; sixty nine percent of the area is considered as permanent preservation land, according the Federal Law Nº 4.771; forest and cocoa plantation environments showed an ant diversity upper to that of pastures; The comunities' composition is similar between pristine forest and cabruca area; it exists a positive correlation between similarity and distance between pristine forest areas; between the six genera that were selected a priori, two of them (Pheidole and Wasmannia) were confirmed, showing that the Pheidole diversity is indicative of conserved lands and the Wasmannia auropunctata occurrence indicates degraded land; the secondary forest in advanced stage of the regeneration/Cabruca contributes the most to the matrix (35,84%); the use of GIS was efficient to interlink pristine forest remnants; ten mini-ecological corridors were identified, with indications on the more propitious areas to the implantation of these ones, recommendations for public polities and ONGs that will benefit the life quality of native organisms.
Key-Words: Biological indicators, ecological corridor, ants, Atlantic coastal forest of Brazil, GIS.

1
1 INTRODUÇÃO
A fragmentação da Mata Atlântica sempre esteve associada aos ciclos econômicos
brasileiros e à expansão urbana (DEAN, 2006). O processo de ocupação do Brasil ocorreu
baseado na destruição dos recursos naturais, exploração social e em arranjos espaciais
desordenados. A Região Sul da Bahia por ser uma região litorânea teve seu processo de
ocupação iniciado na época da colonização do Brasil (COMPANHIA DE
DESENVOLVIMENTO E AÇÃO REGIONAL – CAR, 1995) e também foi submetida a um
processo de ocupação desordenado.
Desde o século XVI, inicio da colonização até o presente, o sul da Bahia passou por
transformações na sua estrutura econômica e social, com reflexos na paisagem, em
conseqüência do papel que lhe foi atribuída pelos seus colonizadores. Nesse processo, grandes
extensões de terra foram desmatadas e posteriormente ocupadas pela agricultura e pecuária
(Companhia de Desenvolvimento e Ação Regional - CAR, 1995). Ainda segundo a CAR, os
ciclos econômicos como a extração do pau-brasil e a cultura da cana-de-açúcar fizeram parte
desse processo. No final do século XVIII, foram introduzidos na região os cultivos do
algodão, café e cacau. No início do século XIX, a cacauicultura se consolidava na região e na
passagem para o século XX, a importância econômica do cacau torna-se definitiva para o
Estado da Bahia. A expansão da pecuária é posterior à introdução da cacauicultura e as duas
atividades passam a coexistir nas propriedades (CAR, 1995; LEITE et al., 1976). O processo

2
de ocupação dos Municípios costeiros do Estado da Bahia só se deu a partir do final do século
XIX, com o desenvolvimento das cidades e consequentemente da população que passou a
ocupar as cidades litorâneas (ANDRADE, 1996). A partir dos anos 60 com a construção do
Porto do Malhado em Ilhéus e com a ampliação da malha rodoviária (construção da BA 001
que liga Ilhéus a Canavieiras no sentido sul e liga Ilhéus a Itacaré no sentido norte) os
ecossistemas litorâneos passam a sofrer maior intervenção humana (ANDRADE, 1996).
No ano de 1987, a decadência da cacauicultura levou muitos agricultores a usar as
árvores nativas, vendendo a madeira e as substituírem por espécies de importância econômica,
ou substituir as plantações de cacaueiros por pastagens ou outros tipos de cultivos (CAR,
1995; REID; BLANES, 1997). Hoje o que se ver é a Mata Atlântica da Região Sul da Bahia
relativamente isolada do restante da Mata Atlântica e bastante fragmentada devido ao
processo histórico de ocupação ao qual foi submetido.
Algumas medidas estão sendo tomadas para minimizar os impactos causados pela
fragmentação. Uma delas é a criação de unidades de conservação que são áreas protegidas por
lei com específicas restrições de usos. Um exemplo é a criação da Área de Proteção
Ambiental (APA) Costa de Itacaré/Serra Grande em 1993 localizada na Região Sul da Bahia
(BRASIL-MMA, 2004).
A criação de unidades mantidas como fragmento isolado não é suficiente para a
conservação dos recursos naturais, tornando imprescindível o manejo sustentável nas
chamadas Zonas Tampão. Nessa perspectiva, o conceito de corredor ecológico foi
consolidado para que a proteção ambiental não fique restrita a áreas protegidas isoladas entre
si (IESB, 1997).
A justificativa central para a criação de corredores é que apenas as zonas tampão ou
áreas de entorno das unidades de conservação por si só não propiciam a proteção da fauna e

3
flora. Além disso, o objetivo primordial do corredor é manter e restaurar a conectividade entre
áreas que contribuem à preservação (IESB, 1997; ARRUDA, 2004; AYRES et al., 2005).
A APA Costa de Itacaré/Serra Grande é uma área de extrema importância biológica e
está inserida nos limites do Corredor Central da Mata Atlântica. No entanto, não existe
nenhum estudo identificando as áreas prioritárias para a implementação de mini-corredores
ecológicos da APA.
Outro método utilizado para minimizar os impactos causados pela fragmentação é o
monitoramento ambiental, utilizando-se como indicadores biológicos plantas ou animais. As
formigas (DELABIE, 1999; ANDERSEN, 1997; AGOSTI et al, 2000) são um bom exemplo
de indicadores biológicos. Vários estudos utilizam a fauna de formigas para análise qualitativa
de diferentes ambientes, pois a estrutura de comunidade de formigas é potencialmente
indicadora de mudanças ambientais (DELABIE, 1999). Formigas são utilizadas como
bioindicadores porque ocorre em quase todos os ambientes e por meio das espécies consegue
caracterizar o grau de perturbação ambiental de uma determinada área (DELABIE, 1999).
Sendo assim, este estudo tem como objetivo identificar gêneros de formigas que
podem ser utilizados como indicadores biológicos e fazer o planejamento de mini-corredores
ecológicos na APA Costa de Itacaré-Serra Grande, Bahia. Esses instrumentos visam a
conservação e recuperação ambiental nesta APA. Para isto propõe-se:
a) Elaborar mapas com as Áreas de Preservação Permanente;
b) Identificar a ocorrência de gêneros bioindicadores (Formicidae) na área em estudo;
c) Planejar a matriz de mini-corredores ecológicos da APA utilizando Sistemas de
Informações Geográficas, para serem incorporadas na política do Corredor Central da Mata
Atlântica.

4
2 A FRAGMENTAÇÃO FLORESTAL DA MATA ATLÂNTICA
O processo de fragmentação florestal da Mata Atlântica é um assunto bastante
discutido, de grande importância ambiental, social e econômica e que necessita de medidas
conservacionistas complexas, que leve em consideração sua extensão territorial e as
diferenças culturais das populações que reside nos seus domínios.
O conceito de Mata Atlântica e sua delimitação tem sido objeto de diversos debates. O
fato de existir vários sistemas de classificação da vegetação baseados em diferentes
parâmetros (abióticos e fisionômicos) favorece a falta de consenso.
Em janeiro de 1996 foi realizado em Belo Horizonte um Workshop sobre a Mata
Atlântica que teve como um dos resultados o estabelecimento de um conceito. Este conceito
tomou como base a área territorial do mapa de vegetação do IBGE de 1988, que abrange a
Floresta Ombrófila Densa, Ombrófila com Araucária, Floresta Estacional Decidual e Semi-
Decidual, não acatando a nomenclatura específica adotada pelo IBGE e incluindo
ecossistemas associados como ilhas oceânicas, restingas, manguezais, florestas costeiras,
campos de altitude e encraves de campos rupestres e cerrados no Sudeste do Brasil (PINTO et
al., 1996).
A utilização do conceito sensu stricto da Mata Atlântica, que a considera sinônimo de
uma única tipologia, representada apenas pela faixa estreita próxima ao litoral, reconhecida
como Floresta Ombrófila Densa para fins de conservação implica em não reconhecê-la como

5
um mosaico de tipologias vegetacionais integradas. Ao se conceituar Mata Atlântica para
efeito de conservação deve-se buscar uma visão global que norteie a prática da conservação,
principalmente em virtude do alto índice de degradação a que tem sido submetida (PINTO et
al., 1996).
A conceituação sensu lato exprime a idéia de continuidade do bioma, isto é, a Mata
Atlântica é composta de uma série de tipologias ou unidades fitogeográficas, constituindo um
mosaico vegetacional que proporciona a grande biodiversidade reconhecida para o bioma.
Uma delas é a Floresta Ombrófila Densa, que, em alguns trabalhos, tem sido citada como
sinônimo da Mata Atlântica como um todo, apesar de ser apenas um dos seus ecossistemas
significativos. Sendo assim, não se devem confundir tipologias com biomas. Enquanto só
existe um único bioma Mata Atlântica, as Florestas Ombrófilas Densas são encontradas
também na Amazônia, nos países andinos, na mesoamérica e em outros continentes, como a
África e a Ásia (PINTO et al., 1996).
O Decreto 750/93, considera adequada a definição de Mata Atlântica sensu lato
(BRASÍLIA, 1993). Já Oliveira (2003), discorda da abrangência geográfica da Mata Atlântica
segundo o conceito sensu lato. Para este autor, esses limites são maiores daqueles
estabelecidos nos documentos técnicos oficiais que tratam do assunto. Segundo ele, a
verdadeira e única Mata Atlântica é a vegetação Floresta Ombrófila Densa.
A Lei da Mata Atlântica nº 11.428, de 22 de dezembro de 2006, também considera
integrantes do Bioma Mata Atlântica as formações florestais nativas e ecossistemas
associados, com as respectivas delimitações estabelecidas no mapa do IBGE (BRASÍLIA,
2006). A Lei da Mata Atlântica reforça o conceito definido pelo workshop. Neste estudo será
considerado o conceito sensu lato com vista à conservação.
A Mata Atlântica abrange 14 Estados brasileiros das Regiões Nordeste, Sudeste e Sul
do país. Estima-se que mais de 60% da população brasileira viva na área de abrangência

6
original da Mata Atlântica, onde se concentram os principais pólos de urbanização, assim
como o desenvolvimento econômico desde o início de nossa história (FUNDAÇÃO SOS
MATA ATLÂNTICA; INSTITUTO NACIONAL DE PESQUISAS ESPACIAIS, 2001;
GUIMARÃES, 2003; JESUS; ROLIM, 2003; RINALDI; LIMA, 2003). Este fato faz com que
a Mata Atlântica apresente uma grave situação de fragmentação e seja um dos biomas mais
ameaçados do Planeta. Estima-se que restam somente cerca de 7% a 8% de sua área original
(MYERS et al., 2000; FUNDAÇÃO SOS MATA ATLÂNTICA, 2007), mesmo assim a Mata
Atlântica preserva uma grande diversidade biológica com altíssimo grau de endemismo
(MYERS et al., 2000; RINALDI; LIMA, 2003; JESUS; ROLIM, 2003).
A maior parte da diversidade biológica dos ecossistemas primários nordestinos
provavelmente foi destruída nos 300 anos de colonização da região, exceto, em parte, as
matas do sudeste baiano, que passaram a ser mais visadas há cerca de 60 anos, apesar de ter
tido anteriormente intensa atividade madeireira seletiva (COIMBRA-FILHO; CÂMARA,
1996).
O processo de fragmentação da Mata Atlântica iniciou com a derrubada das matas
ainda pelos indígenas para construções de habitações, estradas e utilização da madeira. Mas
foi a partir da colonização européia em 1500 que o processo de degradação da mata se
intensificou. Atividades econômicas como extração de madeira (pau-brasil, jacarandá),
cultivo de cana-de-açúcar, pecuária ou mineração contribuíram significadamente para a atual
situação de fragmentação da Mata Atlântica (COIMBRA-FILHO; CÂMARA, 1996; DEAN,
1996; YOUNG, 2005). A fragmentação afetou consideravelmente a Mata Atlântica, tanto no
que se refere à fitofisionomia quanto à desestabilização das condições ecológicas
(COIMBRA-FILHO; CÂMARA, 1996).
O desmatamento da Mata Atlântica é o resultado inevitável das políticas sociais e
econômicas recentes que são levadas a cabo em nome do desenvolvimento. Muitas

7
instituições de desenvolvimento e grupos políticos consideram a pressão da população como o
fator principal da destruição da floresta. Esta convicção é usada por muitos governos para
insinuar que pouco ou nada pode ser feito a respeito do problema do desmatamento. Na
realidade, as grandes companhias e as injustiças do comércio internacional são as reais causas
de destruição das florestas tropicais (REVINGTON, 1992). Segundo Young (2005), outra
falsa convicção é a afirmação da ligação entre desmatamento e geração de empregos já que o
aumento do desmatamento vem sendo acompanhado pela diminuição de oportunidades de
trabalho nas áreas da Mata Atlântica convertidas para uso agrícola. Para este autor, o
desmatamento da Mata Atlântica é o outro lado do processo de exclusão social e econômica
que caracterizou a formação da nação brasileira, onde a maioria dos trabalhadores rurais está
limitada em pequenas áreas e poucos grandes proprietários dominam extensas faixas de terras
mais produtivas.
Cerqueira et al. (2003) definem fragmentação como sendo a separação em partes de
uma dada unidade do ambiente que passa a ter condições ambientais diferentes em sua
vizinhança. Complementam que a fragmentação ocorre quando um hábitat contínuo (terrestre
ou aquático) é dividido em manchas mais ou menos isoladas. Quando a continuidade é
destruída, os fragmentos restantes ficam isolados uns dos outros por uma paisagem
modificada ou degradada (PRIMACK; RODRIGUES, 2001).
A fragmentação pode ocorrer por causas naturais - flutuações climáticas,
heterogeneidade de solos, processos de sedimentação e hidrodinâmica, processos
hidrogeológicos (CONSTANTINO et al., 2003), ou ainda por causas antrópicas - atividades
socioeconômicas, ocupação de áreas de florestas para implantação de infra-estrutura urbana
(FISZON et al., 2003). A grande questão é que o processo de fragmentação causada pelo
homem ocorre num período de tempo mais curto que a fragmentação natural (RODRIGUES;
NASCIMENTO, 2006).

8
A importância no estudo da fragmentação florestal é que a maior parte da
biodiversidade vive nos fragmentos florestais remanescentes que são pouco estudados e
somente um pequeno número destes fragmentos é protegido por lei, ficando o restante nas
mãos de proprietários particulares.
Os principais efeitos ligados à fragmentação de habitats que influenciam a dinâmica
biológica da floresta são: o grau de isolamento; o tamanho e a forma do fragmento; o tipo de
matriz circundante e o efeito de borda (CERQUEIRA et al., 2003). A qualidade conservativa
dos fragmentos varia ainda em relação ao processo histórico de fragmentação destes.
Grau de isolamento – O grau de isolamento afeta o fluxo gênico entre os fragmentos
florestais e a sustentabilidade de populações naturais (VIANA; PINHEIRO, 1998), levando à
perda e diminuição da riqueza em espécies. Se a paisagem possuir uma fração considerável de
habitat original, a distância entre os fragmentos tem chances de ser pequeno e um número
maior de espécies será capaz de atravessar a matriz e colonizar outros fragmentos (VIEIRA et
al., 2003).
Tamanho e forma do fragmento (mancha) – Várias perguntas a cerca do tamanho e
forma do fragmento costumam ser levantadas: Que extensão deve ter um fragmento para
melhor manutenção das espécies? Qual é a forma ideal de um fragmento? O ideal seria ter um
fragmento grande ou vários fragmentos pequenos?
O tamanho e a forma dos fragmentos diferem do habitat original de duas formas: 1) o
tamanho da borda por área de habitat nos fragmentos aumenta; 2) o centro de cada fragmento
de habitat fica mais próximo da borda (CERQUEIRA et al., 2003; PRIMACK; RODRIGUES,
2001). Isso leva a crer que manchas menores sejam compostas quase que exclusivamente por
ambientes de margem e as manchas maiores possuirão mais espécies do que as manchas
menores (SOARES FILHO, 1998; SCARIOT et al, 2003).

9
A forma do fragmento em relação à distribuição da borda tem um significado. Por
exemplo, um fragmento no formato de um círculo ou um quadrado contém mais áreas
interiores do que a borda, um retângulo com a mesma área tem proporcionalmente maior
relação borda/interior e um fragmento estreito de mesma área pode ser composto inteiramente
pela sua borda (FORMAN; GODRON, 1986 citado por SOARES FILHO, 1998; PRIMACK;
RODRIGUES, 2001; SCARIOT et al., 2003).
Matriz circundante – Forman e Godron (1986), citado por Soares Filho (1998),
definem matriz como o elemento mais extensivo e conectado da paisagem que possuiu um
importante papel no seu funcionamento. Complementando, se nenhum tipo de elemento da
paisagem for preponderante, o maior grau de conectividade indicará qual elemento
corresponde à matriz, ou seja, o papel dominante na dinâmica da paisagem determinará a
matriz (SOARES FILHO, 1998).
A matriz determina a possibilidade de deslocamento das espécies entre os fragmentos,
determina a probabilidade de invasões biológicas nos fragmentos e afeta também a
sobrevivência das populações (SCARIOT et al., 2003).
Efeito de borda – Os fragmentos de mata são fortemente influenciados por efeitos de
borda, conceituado por Laurance (1991), como várias mudanças ecológicas associadas às
abruptas bordas artificiais dos fragmentos de floresta. A radiação solar e os ventos quentes e
secos penetram à floresta alterando o microclima. Os fragmentos circundados por pastagens
são mais propícios a efeito de borda do que fragmentos circundados por floresta secundária
(VIANA; PINHEIRO, 1998).
De acordo com Rodrigues (1993) citado por Queiroga e Rodrigues (2001), os efeitos
de borda são divididos em dois tipos: abióticos e bióticos. Os abióticos envolvem mudanças
nos fatores climáticos ambientais (umidade, radiação solar e vento). Os bióticos podem ser
diretos e indiretos. Os diretos envolvem mudanças na abundância e na distribuição de

10
espécies provocados pelos fatores abióticos e os indiretos envolvem mudanças na interação
entre as espécies (predação, parasitismo, herbivoria, dispersão de sementes, competição e
polinização). Há vários estudos mostrando que esses impactos não se restringem à borda, eles
atingem também porções teoricamente mais protegidas, distante da borda (VIANA;
PINHEIRO, 1998; SCARIOT et al., 2003; UEHARA-PRADO, 2003; PACIENCIA; PRADO,
2004; TABARELLI et al., 2004; BALDISSERA; GANADE, 2005; NASCIMENTO;
LAURANCE, 2006; RODRIGUES; NASCIMENTO, 2006).
Os fragmentos florestais não são auto-sustentáveis, a degradação destes é resultado da
complexa interação entre fatores inerentes ao processo de fragmentação, como redução do
habitat, maior exposição ao efeito de borda, isolamento e constante pressão antrópica
(VIANA; PINHEIRO, 1998).

11
3 CORREDOR ECOLÓGICO
A maior parte dos remanescentes florestais de Mata Atlântica da Região Sul da Bahia
encontra-se isolada e fragmentada. Mesmo assim, a região tem experimentado um aumento
das taxas de desmatamento que piorou o grau de fragmentação florestal, dinâmica que se
acelerou devido à crise da lavoura cacaueira (ROCHA, 2002). A Conservation International -
CI e Instituto de Estudos Sócio-Ambientais do Sul da Bahia - IESB (2000) ressaltam que
ainda restam extensões significativas de florestas nessa área com níveis de conectividade
florísticas e faunísticas que sugerem a existência da viabilidade ecológica em longo prazo.
Em resposta a fragmentação florestal nasce o conceito de corredor ecológico ou
corredor biológico (tratados neste estudo como sinônimos). Em 1936, Simpson utiliza pela
primeira vez o termo “corredores” com um sentido biológico em estudo de dispersão entre
continentes (CREEL, 1999). No entanto, a teoria dos corredores idealizados atualmente foi
criada por volta dos anos de 1960 e 1970 por Preston, onde recomendava o estabelecimento
de corredores entre reservas com a finalidade de aumentar as possibilidades de sobrevivência
das espécies (CREEL, 1999; DAMSCHEN, 2006).
Segundo Forman e Godron (1986), os corredores podem ser divididos em três tipos,
conforme a função e utilização dada por diferentes organismos, plantas, animais e humanos:
1- Linhas-Corredores: estradas, trilhas, cercas, canais e outros (forma linear); 2- Faixas-

12
Corredores: com presença de vegetação e mais largas do que as linhas; e 3- Cursos-d’água-
Corredores: considerando a vegetação que margeia os mesmos.
Anderson et al. (2003), identificam dois grandes tipos de corredores ecológicos:
a) Corredor Linear - consiste em uma ligação estreita e unidirecional entre dois blocos
maiores de habitat nativo estendendo-se a distâncias de até 10 km. São adequados para
realizar objetivos específicos e possivelmente eficazes em circunstância onde:
• uma grande parte da paisagem é alterada ficando inabitável às espécies nativas;
• as espécies críticas são condicionadas a habitats sem distúrbio;
• e/ou o ecossistema desejado pode ser restabelecido ou mantido em corredores lineares,
como ao longo de córregos (matas ciliares).
b) Corredor de paisagem - Esta proposta está associada à conservação em larga escala, como
uma unidade de planejamento regional e podem estender-se sobre áreas de milhares de km2.
Consiste na ligação multidirecional e compreende uma rede de unidades de conservação e
outras áreas de usos menos intensivo, que são gerenciadas de maneira integrada para garantir
a sobrevivência do maior número possível de espécies de uma região.
A definição de corredores passou por uma evolução no decorrer do tempo. Atualmente
alguns conceitos de corredores ecológicos são discutidos:
a) Corredor Linear:
“Faixa de cobertura vegetal existente entre remanescente de vegetação primária, em estágio médio e avançado de regeneração, capaz de propiciar habitat ou servir de área de trânsito para a fauna residente nos remanescentes. Os corredores entre remanescentes constituem-se de faixas de cobertura vegetal existentes, nas quais seja possível a interligação de remanescentes, em especial às unidades de conservação e áreas de preservação permanentes” e define as matas ciliares incluindo as áreas de preservação permanente como passíveis a formação de corredores ecológicos (BRASIL. RESOLUÇÃO CONAMA nº 9/96).
“Áreas homogêneas (numa determinada escala) de uma unidade de paisagem, que se distinguem das unidades vizinhas e que apresentam disposição espacial linear. Em estudos de fragmentação, consideram-se corredor apenas os elementos anteriormente conectados” (METZGER, 2001).

13
Rede de ligação através de estradas e/ou trilhas (sistema viário) e de córregos e/ou canais (sistema de drenagem), sendo que a arranjo espacial do mosaico de manchas e de redes compõe o padrão paisagístico que permiti a conectividade entre os organismos existentes (BUREL; BAUDRY, 2002).
b) Corredor de Paisagem:
“Grandes extensões de ecossistemas florestais biologicamente prioritários na Amazônia e na Mata Atlântica, delimitados em grande parte por conjuntos de unidades de conservação (existentes ou propostas) e pelas comunidades ecológicas que contêm. O conceito de corredores ecológicos permite ainda o incremento do grau de conectividade entre as áreas naturais remanescentes, sob diferentes categorias de proteção e manejo, através de estratégias de fortalecimento e expansão do número de unidades de conservação, incluindo-se aqui as RPPNs, além da recuperação de ambientes degradados, quando considerado compatível” (BRASIL. Ministério do Meio Ambiente - MMA, Projetos Corredores Ecológicos, 1998).
“Porções de ecossistemas naturais ou semi-naturais, ligando unidades de conservação, que possibilitam entre elas o fluxo de genes e o movimento da biota, facilitando a dispersão de espécies e a recolonização de áreas degradadas, bem como a manutenção de populações que demandam para sua sobrevivência áreas com extensão maior do que aquela das unidades individuais” (BRASIL. Sistema Nacional de Unidades de Conservação - SNUC, 2000).
“São ecossistemas naturais ou semi-naturais que garantem a manutenção das populações biológicas e a ligação entre as áreas protegidas. São geridos como unidades de planejamento visando a conservação da biodiversidade, o uso sustentável dos recursos naturais e a repartição eqüitativa das riquezas para as presentes e futuras gerações” (BRASIL. IBAMA; WWF, 2000).
“Um mosaico de usos da terra que conectam fragmentos de floresta natural através da
paisagem... O planejamento de corredores é feito em escala regional” (CI; IESB, 2000).
Analisando esses conceitos é possível perceber que existem dois entendimentos sobre
os corredores (ANDERSON et al., 2003) (Figura 1). A área da Biologia da conservação
considera corredores ecológicos como uma faixa de vegetação que conectam fragmentos –
corredor linear. Na biologia da conservação, os corredores têm a função primordial de
proporcionar vias de intercâmbio e incrementar as possibilidades de movimento de indivíduos
pertencentes a populações que se encontram isolados (AYRES et al., 2005). Porém a CI,
IBAMA, SNUC, MMA e WWF consideram os corredores ecológicos como uma unidade de
planejamento regional, composta por um mosaico de unidades de conservação e áreas com
diferentes usos da terra – corredor de paisagem.

14
Para Brasil-MMA (2006), os corredores ecológicos não são unidades políticas ou
administrativas, são extensas áreas geográficas definidas a partir de critérios biológicos para
fins de planejamento e conservação. Os cordões de vegetação nativa que conectam
fragmentos são um dos componentes dos corredores, mas não o único. Ayres et al. (2005)
acrescentam que as áreas de corredores não constituem figuras legais de preservação ou
categorias de unidades de conservação e dentro dessas áreas é possível a existência de
iniciativas conservacionistas e desenvolvimentistas. Por esse motivo os corredores ecológicos
abrangem grandes cidades e pólos tradicionais de desenvolvimento regional.
Fragmentos Corredor Linear Corredor dePaisagem
Figura 1 - Representação esquemática dos corredores lineares e de paisagem. Fonte: Conservation International 2000.
De acordo com Anderson et al. (2003), os corredores de paisagem podem ser inseridos
para cumprir um vasto conjunto de metas do planejamento ambiental, envolvendo a
conservação da biodiversidade, serviços ambientais, usos de recursos naturais, lazer e estética.
São possivelmente eficientes em situações onde:
• uma amostra significativa da paisagem está intacta;
• e/ou espécies alvo exigem grandes áreas de habitat (Bennett 1999, apud Anderson et al.
2003).

15
As estratégias de planejamento e implementação desses dois tipos de corredores se
diferenciam, mas ambos ajudam a maximizar a conectividade entre habitats e ecossistemas e,
como resultado, minimiza os efeitos danosos da fragmentação florestal.
Pesquisas orientadas por Damschen (2006), demonstram que os corredores são
ferramentas práticas para a preservação da biodiversidade, pois auxiliam a diversidade de
plantas ao melhorar a dispersão das sementes e a polinização e alterar a competição entre as
plantas. O experimento consiste em dividir uma vasta área de floresta de pinus na Carolina do
Sul (EUA) em seis lotes de 500 m². Eles verificaram que as manchas de habitats que foram
conectadas a outras manchas através de corredores apresentaram um crescimento de cerca de
20% no número de espécies de plantas em cada lote, comparando com as manchas isoladas.
Este estudo é a confirmação de que os corredores funcionam e que mesmo corredores
estreitos podem ser eficazes (neste caso, as espécies estudadas foram às plantas).
Alguns pesquisadores especulam sobre os riscos que os corredores podem vir a trazer,
como facilitar o trânsito de doenças e espécies daninhas, que poderia se espalhar nas áreas de
reservas naturais antes isoladas e causar extinções de espécies raras. Outro risco seria que os
animais que utilizam os corredores poderiam ser expostos a caçadores. Embora essas
possibilidades sejam aceitáveis não existem dados que as comprovem (PRIMACK;
RODRIGUES, 2001).
3.1 Corredor Central da Mata Atlântica - CCMA
A partir de 1997, o Governo Federal do Brasil através do Ministério do Meio
Ambiente desenvolve uma estratégia geopolítica de trabalho que visa equalizar o problema da

16
fragmentação, através da criação e manutenção dos corredores ecológicos para a Amazônia
Brasileira e para a Mata Atlântica (BRASIL-MMA, 1998). Este projeto propõe a adição ao
paradigma das “ilhas biológicas”, o paradigma dos “corredores ecológicos”, conectando áreas
protegidas dentro dos biomas regionais, desenvolvendo modelos inovadores de manejo que
incorporem as comunidades tradicionais e outros atores da sociedade (AYRES et al., 2005).
Foram indicados sete corredores: cinco na Amazônia e dois na Mata Atlântica (Figura
2).
Figura 2 – Proposta para criação de corredores ecológicos no Brasil. Projeto Parques e Reservas (BRASIL-MMA, 1998). Fonte: AYRES et al., 2005.

17
Baseados em critérios de riqueza de espécies, diversidade de comunidades e
ecossistemas, grau de conectividade, integridade dos blocos de paisagem natural e riqueza de
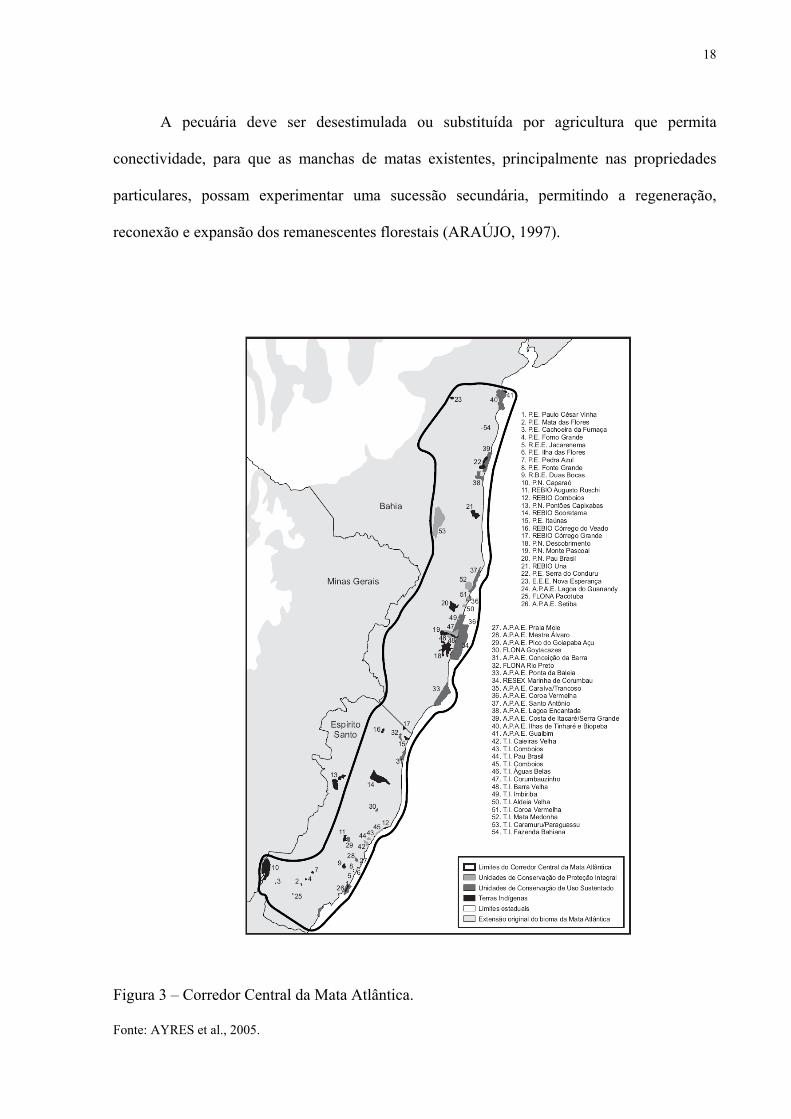
espécies endêmicas, ficou definido o Corredor Central da Mata Atlântica – CCMA (Figura 3)
que abrange a faixa litorânea compreendida desde o sul do Recôncavo Baiano até o centro-sul
do Espírito Santo. Sua distância média da costa para o interior é de 300 km. Dos 8.635.900 ha
que abrange o CCMA, 314.562 ha encontram-se teoricamente protegidos em unidades de
conservação, 30.543 ha por instituições de pesquisas ou entidades do setor privado e 52.973
ha são áreas indígenas sob jurisdição da FUNAI (AYRES et al., 2005), ou seja, somente 4,6
% da área encontra-se protegida. Os limites do CCMA não são definitivos e podem ser
aperfeiçoados se necessário (www.corredores.org.br, 2006; FONSECA, et al., 2004).
O CCMA é definido como uma linha imaginária que agrupa grande diversidade
biológica e pode ser comparada a um mosaico de diferentes usos da terra
(www.corredores.org.br). O CCMA abrange centros de endemismo, grande riqueza biológica
e abriga espécies ameaçadas de extinção e possui 83 unidades de conservação. As principais
atividades econômicas baseiam-se no cultivo do cacau, do eucalipto e café, na pecuária e na
exploração do turismo (BRASIL-MMA, 2006).
A criação dos corredores ecológicos nos domínios da Mata Atlântica, de acordo com o
IESB (1997), tem a função primordial de proporcionar vias de intercâmbio e incrementar as
possibilidades de movimentação de indivíduos pertencentes às populações que se encontram,
em maior ou menor grau, isolados em áreas de habitat mais propício à sua sobrevivência.
Ressalta ainda que mesmo paisagens fragmentadas oferecem oportunidades de movimentação
de organismos. Sob este aspecto o sistema agroflorestal cabruca no qual raleia-se a floresta e
introduz-se o cacau sob a sombra das árvores nativas (LOBÃO, 1997), deverá merecer
incentivos para a sua recuperação principalmente quanto ao seu papel de incrementar o grau
de conectividade entre os remanescentes florestais.
18
A pecuária deve ser desestimulada ou substituída por agricultura que permita
conectividade, para que as manchas de matas existentes, principalmente nas propriedades
particulares, possam experimentar uma sucessão secundária, permitindo a regeneração,
reconexão e expansão dos remanescentes florestais (ARAÚJO, 1997).
Figura 3 – Corredor Central da Mata Atlântica. Fonte: AYRES et al., 2005.

19
4 INSTRUMENTOS LEGAIS QUE APÓIAM AS AÇÕES NO ESTABELECIMENTO DE CORREDORES ECOLÓGICOS
Diante de tal quadro de degradação a sociedade hoje recorre ao Direito para salvar o
que resta dos recursos naturais. O Direito Ambiental surgiu nos EUA e na Europa nas décadas
60/70 influenciado pela crise ambiental. No Brasil, o Direito Ambiental nasceu em 1981 da
preocupação com o meio ambiente como uma tentativa de conter os impactos ambientais
(Rocha, com Pess.). Serrano (1998), conceitua Direito Ambiental como “[...] sistema de
normas, princípios, instituições, práticas operativas e ideologias jurídicas que regulam as
relações entre os sistemas sociais e seus contornos naturais”. No Brasil, a Constituição de
1988 foi a primeira a tratar da questão do meio ambiente, atribuindo-lhe a feição de direito
fundamental de terceira dimensão, que se refere aos direitos e deveres de solidariedade de
toda humanidade (Rocha, com. Pess.). Atualmente a legislação brasileira abarca grande parte
das questões ambientais.
No Brasil, a primeira criação de uma área protegida foi o Parque Nacional de Itatiaia
situado na Serra da Mantiqueira, na divisa dos estados do Rio de Janeiro e de Minas Gerais
em 1937 (FUNDAÇÃO BRASILEIRA PARA O DESENVOLVIMENTO SUSTENTÁVEL,
2000). Sob o aspecto legal, só com a promulgação da Constituição de 1988, cinqüenta e um
anos depois da criação do primeiro parque, ocorreram no país avanços no que diz respeito às
nossas áreas protegidas. Ficou então estabelecido que seja obrigação do Estado a proteção da
biodiversidade brasileira, e que essa proteção se dará através da criação de áreas protegidas ao

20
longo do território nacional, entre outras estratégias. Diz ainda a Constituição que essas áreas
naturais somente poderão ser modificadas através de lei (BRASIL-Constituição da República
Federativa do Brasil, 1988).
Uma iniciativa relevante é o projeto de Lei nº 9.985 de 2000 que instituiu o Sistema
Nacional de Unidades de Conservação (SNUC), como estratégia para reverter o quadro de
fragmentação florestal. O SNUC estabelece critérios e normas para a criação, implantação e
gestão das unidades de conservação. Traz também alguns conceitos como o de unidade de
conservação, diversidade biológica, uso indireto, uso direto, uso sustentável, corredores
ecológicos, entre outros. O SNUC contempla dois grupos de categorias de unidades de
conservação: Unidades de Proteção Integral que são definidas como áreas destinadas a
manutenção dos ecossistemas livres de alterações causadas por interferência humana,
admitindo-se apenas o uso indireto de seus atributos naturais (Estação Ecológica, Reserva
Biológica, Parque Nacional, Monumento Natural e Refúgio de Vida Silvestre) e Unidades de
Uso Sustentável definidas como áreas destinadas a exploração do ambiente de maneira a
garantir a perenidade dos recursos ambientais renováveis e dos processos ecológicos,
mantendo a biodiversidade e os demais atributos ecológicos de forma socialmente justa e
economicamente viável (Área de Proteção Ambiental, Área de Relevante Interesse Ecológico,
Floresta Nacional, Reserva Extrativista, Reserva de Fauna, Reserva de Desenvolvimento
Sustentável e Reserva Particular do Patrimônio Natural) (BRASIL-SNUC, 2000).
Santilli (2005) faz uma crítica ao SNUC por priorizar, claramente, as unidades de
proteção integral, que não admitem presença humana. Apesar de criar categoria de
desenvolvimento sustentável, estas são concebidas como de importância secundária em
relação às unidades de proteção integral. Segundo Dourojeanni (1997) e Milano (2000) apud
Cabral e Souza (2002), esse modelo brasileiro de unidades de conservação não é cópia do
modelo norte-americano apresentando características próprias, pois se baseou em diferentes

21
níveis e de diferentes maneiras, em convenções internacionais. O SNUC foi resultado de
quase uma década de debate nacional.
No “IV Congresso Internacional de Áreas Protegidas” em Caracas, 1992, o Ministério
do Meio Ambiente chegou à conclusão de que o total de áreas protegidas por bioma é
insuficiente para a conservação da biodiversidade e que as áreas já criadas ainda não
atingiram os objetivos que motivaram sua criação (União Mundial para Conservação da
Natureza, 1992).
Uma nova estratégia de conservação que é a criação de corredores ecológicos
unificando fragmentos isolados dos principais biomas ameaçados está prevista por lei. Dentre
as várias abordagens possíveis, o estabelecimento da política dos corredores ecológicos pelo
governo representa uma das mais novas estratégias para um planejamento regional eficaz.
Assim, a proteção dos fragmentos florestais não fica restrita a unidades isoladas. O SNUC é
mais detalhista e complementa as demais leis quanto aos critérios para a formação e uso em
áreas de corredores ecológicos.
O Decreto 750/93 no seu Artigo 7º é o primeiro instrumento legal que proíbe a
exploração de vegetação que tenha a função de formar corredores entre remanescentes de
vegetação primária ou em estágio avançado e médio de regeneração (BRASÍLIA, Decreto
750/93).
A Resolução CONAMA 09 de 1996, introduz o conceito de Corredores Ecológicos:
"faixa de cobertura vegetal existente entre remanescentes de vegetação primária em estágio
médio e avançado de regeneração, capaz de propiciar habitat ou servir de área de trânsito para
a fauna residente nos remanescentes" e define as matas ciliares incluindo as áreas de
preservação permanente como passíveis a formação de corredores ecológicos. O Artigo 3º
fixa a largura para os corredores que deverá ser de 10% (dez por cento) do seu comprimento

22
total, sendo que a largura mínima será de 100 metros. Essa medida diz respeito ao modelo de
corredor linear.
A Resolução CONAMA 11 de 1987 passa a considerar os corredores ecológicos como
Unidades de Conservação.
O Brasil tem hoje um grande aparato de leis de proteção ambiental, mas apesar do
endurecimento da legislação ambiental, a degradação dos ecossistemas avança sem dar sinais
de retrocesso ou de desaceleração.

23
5 FORMICIDAE COMO BIOINDICADORES
Por causa de crescentes problemas ambientais, estão sendo desenvolvidos métodos
voltados para o monitoramento ambiental, utilizando-se como indicadores biológicos plantas
ou animais (TOLMASQUIM, 2001; ELLENBERG et al. citado por MAZZONI-VIVEIROS;
TRUFEM, 2004). Os bioindicadores passam a expressar reações características quando
expostos aos diferentes tipos de degradação ambiental, fornecendo informações sobre a
situação do ambiente (MAZZONI-VIVEIROS; TRUFEM, 2004).
Segundo Hawksworth (1992), são considerados bioindicadores os organismos ou
comunidades que apresentam sintomas específicos ou respostas que indiquem mudanças em
algum controle ambiental, geralmente de forma qualitativa. Para Wolterbeek et al. (1995), os
bioindicadores mostram informações sobre a qualidade ou modificações do ambiente, mas
não possibilita quantificar tais informações, isso sendo possível só com o biomonitoramento.
Um indicador é um meio encontrado para reduzir uma ampla quantidade de dados à
sua forma mais simples, retendo o significado essencial do que esta sendo perguntado sobre o
dado (OTT, 1978 apud TOLMASQUIM, 2001). O conceito de bioindicador se refere a um
parâmetro biológico de comportamento que surge diante da alteração de determinada
condição externa ou de estresse do ambiente (ACCACIO et al. 2003).
Um bom bioindicador deve ser capaz de responder de forma rápida e objetiva por
intermédio da análise de seu estado a condição em que se encontra um sistema biológico

24
(SCHECK, 1990 apud ACCACIO et al. 2003), deve ser notável e facilmente amostrável
(BROWN, 1991 apud ACCACIO et al. 2003) e deve estar associado aos grandes processos do
ecossistema, ser sensível a variações no manejo e no clima (DORAN; PARKIN, 1994).
A fauna de solo é sensível às práticas de manejo do solo, por isso é um potencial
indicador biológico da qualidade ambiental (CORREIA, 2002). Atualmente estudos que
discutem a importância dos insetos nos ecossistemas terrestres como bioindicadores de
qualidade ambiental é crescente, pois os insetos desempenham papel chave nos ecossistemas
terrestres (ANTONINI et al., 2003). Alguns estudiosos aceitam a idéia de que algumas
espécies são mais importantes para a comunidade do que outras, pois essas espécies chave
exercem interações ecológicas com plantas, animais e microorganismos, decomposição,
ciclagem de nutrientes, dispersão e predação de sementes, fluxo de energia, polinização, entre
outros (DIDHAM, et al. 1996; BROWN, 1997; WINK, 2005).
A escolha de um bioindicador vai depender do problema científico a ser discutido.
Como neste estudo, o objetivo é identificar gêneros que podem ser utilizados como
indicadores biológicos, é necessário, para tanto, avaliar qualitativamente os fragmentos
remanescentes da área e outros ambientes como pastagens e cacauais. Sendo assim, optou-se
pelo uso da mirmecofauna de solo como bioindicador. Vários estudos, tanto no exterior como
no Brasil, utilizam a fauna de formigas para análise qualitativa de diferentes ambientes, pois a
estrutura de comunidade de formigas é potencialmente indicadora de mudanças ambientais
(DELABIE, 1990; ANDERSEN; SPARLING, 1997; DELABIE, 1999; CORREIA, 2002;
UNDERWOOD; FICHER, 2006; DELABIE et al. 2007).
As formigas serão utilizadas aqui para indicar, através de sua ocorrência áreas
degradadas ou conservadas (DELABIE, 1999; DELABIE et al. 2000). Elas são componentes
fundamentais dos ecossistemas tropicais, sua biomassa representa cerca de um quarto da
biomassa animal de invertebrados, enquanto elas interagem com todos os outros segmentos da

25
fauna e da flora, por predação e transporte de sementes (HÖLLDOBLER; WILSON, 1990).
As formigas que se propõe amostrar são utilizadas como ferramenta para estimativa da
diversidade local, pois são geralmente consideradas organismos representativos dos demais
invertebrados. As principais vantagens de usar formigas como bioindicadores é porque elas
são abundantes e facilmente amostradas, e podem ser identificadas até gênero ou espécie
(diferentemente de outros insetos) (ANDERSEN 1997; AGOSTI et al, 2000).

26
6 SISTEMAS DE INFORMAÇÃO GEOGRÁFICA NO ESTUDO DE ÁREAS PRIORITÁRIAS À FORMAÇÃO DE CORREDORES ECOLÓGICOS
“O termo Geoprocessamento denota a disciplina do conhecimento que utiliza técnicas
matemáticas e computacionais para o tratamento de informação geográfica” (CÂMARA;
MEDEIROS, 1998). Os Sistemas de Informação Geográfica (SIG) são ferramentas para o
Geoprocessamento. Este, por sua vez, é um termo amplo que engloba várias tecnologias de
tratamento e manipulação de dados geográficos, através de programas computacionais, tais
como: sensoriamento remoto, digitalização de dados, automação de tarefas cartográficas,
utilização de Sistemas de Posicionamento Global (GPS) e os SIGs. O SIG pode englobar
todas as demais técnicas do Geoprocessamento, mas nem todo Geoprocessamento é um SIG
(CARVALHO, 2000).
SIG foi definido por Teixeira et al. (1995), como “conjunto de programas,
equipamentos, metodologias, dados e pessoas (usuário), perfeitamente integrados, de forma a
tornar possível a coleta, o armazenamento, o processamento e a análise de dados
georreferenciados, bem como a produção de informação derivada de sua aplicação”.
Outra definição considera os SIG como sistemas computacionais (hardware, software)
dados geográficos, usados para a investigação de fenômenos que ocorrem no espaço
geográfico (geologia, bacias hidrográficas, pedologia, engenharia urbana, vegetação, etc). São
capazes de capturar, armazenar, atualizar, manipular, analisar e apresentar a possibilidade de

27
visualizar as informações referenciadas geograficamente (CARVALHO, 2000; CALIJURI;
RÖHM, 1995).
As propriedades de um SIG consistem: na capacidade de apresentação cartográfica de
informações complexas; numa sofisticada base integrada de objetos espaciais e de seus
atributos ou dados; e um engenho analítico formado por um conjunto de procedimentos e
ferramentas de análise espacial (MAGUIRRE et al., citado por CARVALHO, 2000).
Gulinck et al., (1991), utilizaram imagem de sensoriamento remoto (imagens SPOT)
para visualização de corredores e concluíram que esta é uma ferramenta útil para o
mapeamento de elementos da paisagem e modelos que tem um possível papel como corredor
ecológico.
Szmuchrowski e Martins (2001) realizaram um estudo utilizando geoprocessamento
para interligar fragmentos florestais a áreas de proteção ambiental no Município de Palmas,
no Tocantins. Eles concluíram que a aplicação do SIG mostrou-se uma ferramenta eficiente
para traçar corredores ecológicos interligando os fragmentos florestais.
Nunes et al. (2005) também utilizaram SIG como ferramenta para identificar
corredores ecológicos na Sub-Bacia Hidrográfica do Rio Vacacaí-Mirim (RS). Concluíram
que a metodologia utilizada para o traçado de corredores ecológicos mostrou ser eficiente
devendo ser escolhidos os maiores fragmentos, e mais distantes entre si, atribuindo menores
graus de atrito às áreas remanescentes para traçar os corredores. Acrescentam que a utilização
de SIG para diagnosticar os fragmentos florestais e também as outras classes de usos da terra
pode representar uma evolução para o planejamento espacial.
Mesmo o SIG sendo uma excelente ferramenta para escolhas de áreas prioritárias
(entre outras utilizações) deve-se checar em campo os resultados obtidos em laboratório. Os
SIG´s auxiliam na escolha dessas áreas, mas não deve ser considerado como o fim, e sim,
como um meio para obter resultados.

28
7 METODOLOGIA
7.1 Caracterização da área de estudo
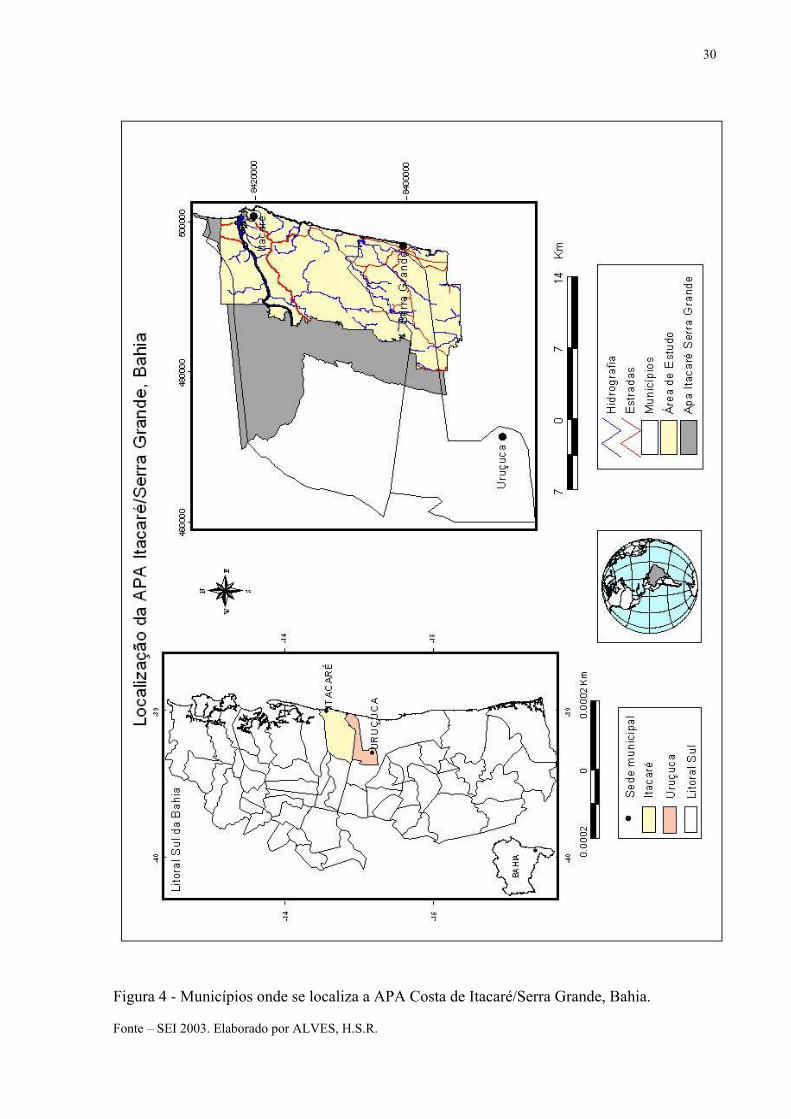
A Área de Proteção Ambiental Costa de Itacaré/Serra Grande foi criada em 1993, pelo
Decreto Estadual nº. 2.186. Dez anos mais tarde, teve sua área ampliada conforme Decreto nº.
8.649 para um total de 62.960 ha. A APA localiza-se na Região Sul do Estado da Bahia, entre
as coordenadas 14º16’2” e 14º30’3” de Latitude Sul e 39º5’18“ e 38º58’38” de Longitude
Oeste. Ocupa uma faixa litorânea de 28 km de extensão, abrangendo os municípios de
Uruçuca e Itacaré (Figura 4 e 5) (BRASIL-MMA, 1996; BRASIL-MMA, 2004; MURICY,
2004).
O Plano de Manejo e Zoneamento econômico da APA foi realizado em 1996 e
revisado em 2004 pelo IESB em parceria com a Conservação Internacional do Brasil. Este
plano de manejo engloba apenas a primeira delimitação da APA antes da sua ampliação
(BRASIL-MMA, 1996; BRASIL-MMA, 2004).
O Conselho Gestor da APA foi criado em maio do ano 2000. Fazem parte do Conselho
Gestor representantes do poder público, sociedade civil de Itacaré e de Serra Grande,
Organizações Não Governamentais (ONG’s) e comunidades rurais da nova área que foi
ampliada em 2003 (BRASIL-MMA, 2004).

29
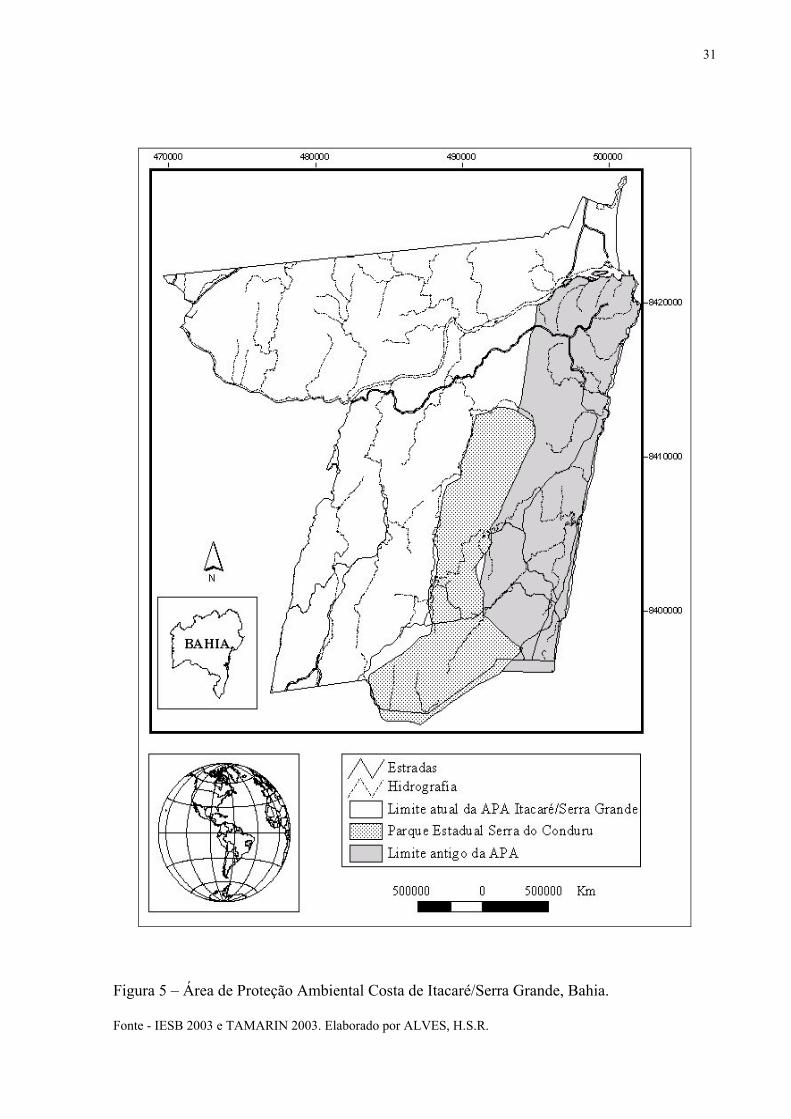
Não será incluso nesse estudo a área total da APA devido à insuficiência de dados que
a abrangem. Sendo assim, a área que será aqui considerada é de 40.178,11 ha (Figura 6). Esta
área necessita uma maior urgência de ações conservacionistas por ser uma área da APA que
sofre maior pressão demográfica. O que não significa que o restante da APA (parte oeste) não
seja prioritário. Por ter sido incluída apenas em 2003, visitas realizadas às propriedades
mostraram que alguns moradores desta região (pequenos produtores) ainda desconhecem o
significado de uma APA.
A região da APA Itacaré/Serra Grande tem clima Tropical Super Úmido (Köffen), sem
estação seca acentuada e temperatura média anual de 21º C a 25º C. A região apresenta
precipitação pluviométrica regular e uma pluviosidade de 1.750 mm a 2.000 mm anuais
(BRASIL-MMA, 1996).
30
Figura 4 - Municípios onde se localiza a APA Costa de Itacaré/Serra Grande, Bahia. Fonte – SEI 2003. Elaborado por ALVES, H.S.R.
31
Figura 5 – Área de Proteção Ambiental Costa de Itacaré/Serra Grande, Bahia. Fonte - IESB 2003 e TAMARIN 2003. Elaborado por ALVES, H.S.R.

32
Os principais cursos d´água que banham a APA são: o Rio de Contas, Riacho Capitão,
Rio Aderno, Rio Jeribucaçu, Rio Burundanga, Rio Itacarezinho, Rio Tijuípe, Rio Tijuipinho,
Rio Sargi, (Figura 6). A Bacia do Rio de Contas é impactada pela falta de saneamento básico
nas cidades que o margeiam. Nos demais cursos d´água da APA, não se constatam fontes
poluidoras significativas, exceto os pequenos cursos d´água no perímetro urbanos de Itacaré,
onde a falta de saneamento básico traz problemas diretos ao meio ambiente e a saúde dos
moradores (BRASIL-MMA, 1996).
A geologia é predominantemente formada por rochas pré-cambrianas que formam o
substrato cristalino local, recoberto pelos sedimentos tércio-quaternários (BRASIL-MMA,
1996). A área apresenta diversos tipos de solos: Argissolo Vermelho-Amarelo Eutrófico;
Latossolo Amarelo-Distrófico; Latossolo Vermelho-Amarelo Distrófico; Neossolo
Quartzarênico e Tipos de Terreno (Figura 7) (SEI, 2003).
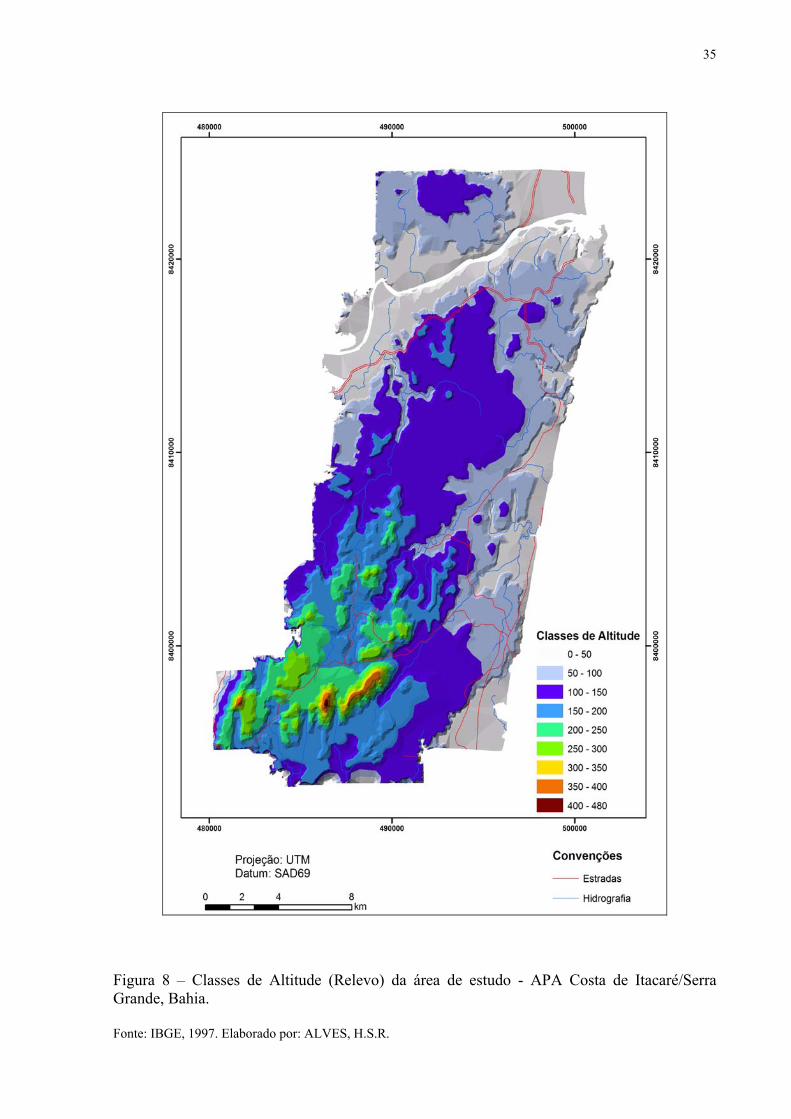
Sua geomorfologia é composta por complexos formados de praias, aluviais e
estuarinos, tabuleiros costeiros e tabuleiros pré-litorâneos (BRASIL-MMA, 1996). Na Figura
8 é possível verificar as classes de altitude que atinge o máximo de 480 m (IBGE, 1997).
A APA possui ainda ecossistemas de grande importância: Floresta Ombrófila Densa
em estágio inicial, médio e avançado de regeneração; Restinga; Manguezal; uma área de
Vegetação Relictual denominada localmente de “campo cheiroso” (Figura 9) (IESB, 2002). A
Floresta Ombrófila Densa caracteriza-se pela marcada predominância de árvores de grande
porte, associadas às várias outras formas biológicas, com temperaturas relativamente elevadas
e ausência de período seco, com precipitação abundante e bem distribuída o ano todo
(SILVA, 2002).

33
Figura 6 – Hidrografia da área de estudo - APA Costa de Itacaré/Serra Grande, Bahia. Fonte: SEI, 2003. Elaborado por: ALVES, H.S.R.

34
Figura 7 – Solos da área de estudo - APA Costa de Itacaré/Serra Grande, Bahia. Fonte: SEI, 2003. Elaborado por: ALVES, H.S.R.
35
Figura 8 – Classes de Altitude (Relevo) da área de estudo - APA Costa de Itacaré/Serra Grande, Bahia. Fonte: IBGE, 1997. Elaborado por: ALVES, H.S.R.
36
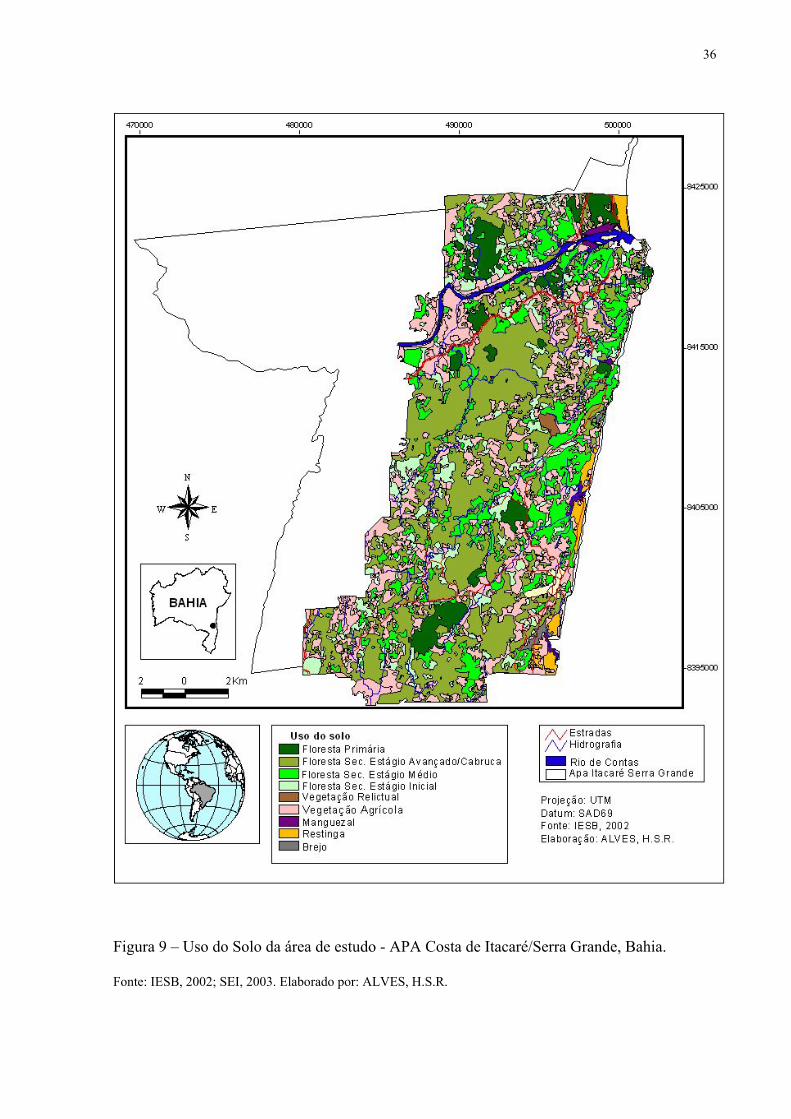
Figura 9 – Uso do Solo da área de estudo - APA Costa de Itacaré/Serra Grande, Bahia. Fonte: IESB, 2002; SEI, 2003. Elaborado por: ALVES, H.S.R.

37
7.2 Procedimentos metodológicos
7.2.1 Identificação de Áreas de Preservação Permanente
Para elaboração dos mapas de Áreas de Preservação Permanente utilizou-se o
programa ArcView 9.1, adotando as restrições conforme o Código Florestal (BRASÍLIA, Lei
nº 4.771).
7.2.2 Identificação dos mini-corredores ecológicos
Para delimitação dos Corredores Ecológicos utilizou-se um mapa de vegetação e uso
da terra de 2002 elaborado pelo setor de geoprocessamento do Instituto de Estudos Sócio-
Ambientais do Sul da Bahia – IESB, no formato shape do sistema de informações geográficas
ArcView 9.1.
A partir deste mapa base foram selecionados os fragmentos florestais acima de 50 ha
os quais foram identificados para conexão através de corredores ecológicos. Em seguida
exportou-se o mapa de uso da terra de 2002 e os polígonos dos fragmentos florestais
selecionados para o sistema de informações geográficas Idrisi 2.0 onde se aplicou a
metodologia de elaboração de corredores ecológicos adaptada da metodologia de distâncias de
custo e caminhos de menor custo de Eastmam (1995).
38
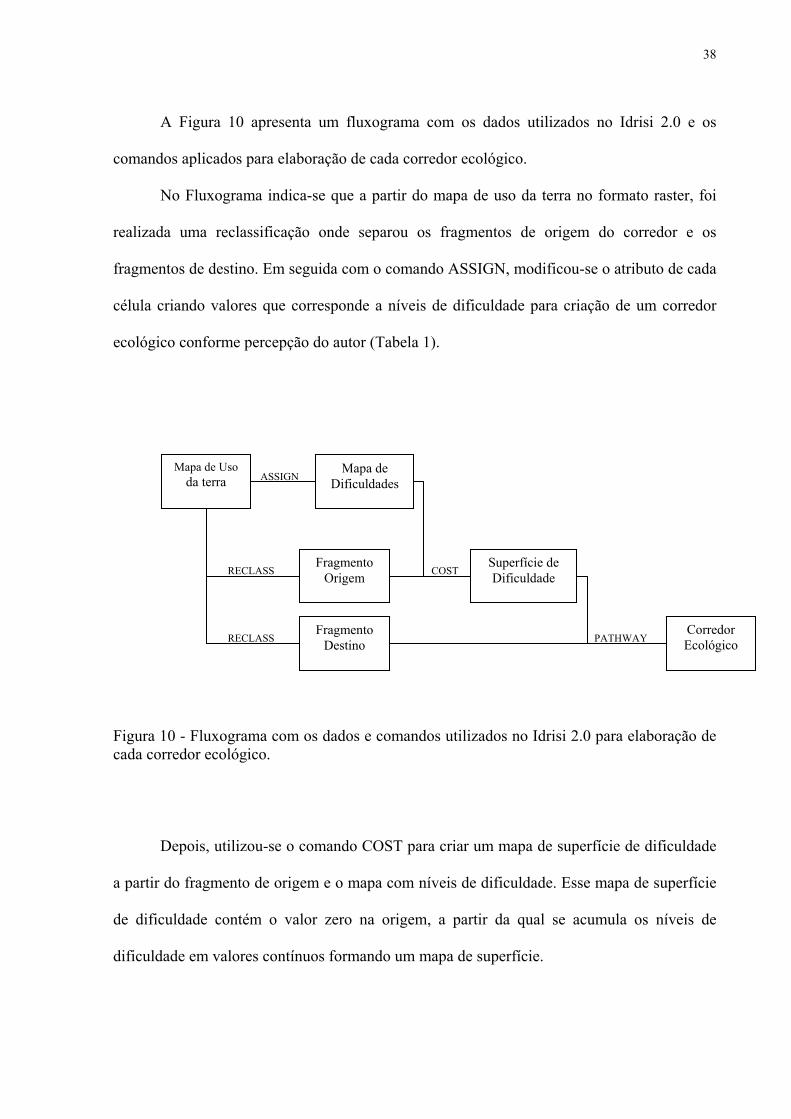
A Figura 10 apresenta um fluxograma com os dados utilizados no Idrisi 2.0 e os
comandos aplicados para elaboração de cada corredor ecológico.
No Fluxograma indica-se que a partir do mapa de uso da terra no formato raster, foi
realizada uma reclassificação onde separou os fragmentos de origem do corredor e os
fragmentos de destino. Em seguida com o comando ASSIGN, modificou-se o atributo de cada
célula criando valores que corresponde a níveis de dificuldade para criação de um corredor
ecológico conforme percepção do autor (Tabela 1).
Figura 10 - Fluxograma com os dados e comandos utilizados no Idrisi 2.0 para elaboração de cada corredor ecológico.
Depois, utilizou-se o comando COST para criar um mapa de superfície de dificuldade
a partir do fragmento de origem e o mapa com níveis de dificuldade. Esse mapa de superfície
de dificuldade contém o valor zero na origem, a partir da qual se acumula os níveis de
dificuldade em valores contínuos formando um mapa de superfície.
Corredor Ecológico
Fragmento Origem
Fragmento Destino
Mapa de Dificuldades
Mapa de Uso da terra
Superfície de Dificuldade COST
PATHWAY
ASSIGN
RECLASS
RECLASS

39
Tabela 1 - Níveis de dificuldade para estabelecimento do corredor ecológico conforme percepção do autor.
Uso da Terra Nível de Dificuldade Descrição Mata Primária 1 Sem dificuldade para estabelecimento do corredor Mata Séc. Estágio Avançado 2 Fácil estabelecimento do corredor Mata Séc. Estágio Médio 3 Fácil estabelecimento do corredor Mata Séc. Estágio Inicial 4 Fácil estabelecimento do corredor Vegetação Relictual 1 Sem dificuldade para estabelecimento do corredor Restinga 1 Sem dificuldade para estabelecimento do corredor Manguezal 1 Sem dificuldade para estabelecimento do corredor Brejo 5 Fácil estabelecimento do corredor Vegetação Agrícola 500 Recuperação para estabelecimento do corredor Rios 500 Barreira física para muitas espécies de fauna Área Urbana 1000 Área impossibilitada de ser um corredor
Como último procedimento para obtenção de cada corredor, utiliza-se o comando
PATHWAY, que calcula o caminho mais indicado para implementação de um corredor
ecológico entre fragmentos, tomando por base o mapa de superfície de dificuldade e o
fragmento de destino. O corredor é indicado como arquivo raster entre o fragmento que para
elaboração dos mapas temáticos foi transformado em arquivo vetorial do Idrisi e em seguida
exportado no formato shapefile para o ArcView 9.1.
7.2.3 Análise de Formicidae
Foi realizada uma consulta na coleção do Laboratório de Mirmecologia da CEPLAC
para verificar as coletas existentes na APA ou próxima a ela. Já existiam dados oriundos de
oito pontos de coletas (Figura 11). Estas coletas foram realizadas entre 1996 e 1998 em três
ambientes diferentes: florestas (4 áreas), cacauais (2) e pastagens (2) (Tabela 2). O objetivo
desta análise foi verificar o estado de conservação das áreas e verificar as diferenças na

40
composição da mirmecofauna nos diferentes tipos vegetacionais (florestas, cacauais e
pastagens).
O método de coleta utilizado é conhecido como "Winkler litter sifting method"
(BESTELMEYER et al., 2000). Este método é o mais indicado para amostrar formigas de
serapilheira. Ele consiste em recolher 1m² de serapilheira, peneirá-la, colocá-la num saco e
levá-la ao laboratório. Em seguida coloca-se o substrato em sacos de malha de rede que ficam
dependurados dentro de um saco de pano fechado por três dias, onde as formigas caem num
copo plástico contendo álcool ao fugir de um meio cada vez mais seco. São coletados 50
pontos de amostra a intervalos de 50 m entre os pontos (DELABIE, et al. 1999). As formigas
coletadas foram triadas, montadas e identificadas pela equipe do Laboratório de
Mirmecologia da Ceplac e posteriormente depositadas na coleção do referido laboratório.

41
Figura 11 – Pontos de coletas na área de estudo e áreas próximas da APA Costa de Itacaré/Serra Grande, Bahia. Fonte: Elaborado por: ALVES, H.S.R.

42
7.2.4 Análises Faunísticas
Para analisar os dados utilizou os programas Excel, EstimateS (Statistical Estimation
of Species Richness and Share Species from Samples) versão 7.5 (COLWELL, 1997) e o
programa MVSP (Multi-Variate Statistical Package) versão 3.1. Esses programas permitiram
gerar análises faunísticas das espécies em cada área separadamente e a similaridade entre as
áreas.
Riqueza observada - A riqueza observada é o número absoluto de morfo-espécies
encontradas nas armadilhas (MAGURRAN, 1983).
Índice de riqueza estimada (Chao 2) - A riqueza de uma comunidade deve ser estimada por
que um senso completo de espécies (principalmente de formigas) em condições naturais é
impossível na prática. A riqueza estimada foi obtida pelo índice de Chao 2 por meio do
programa EstimateS que usa os dados de freqüência das espécies. Serve para visualizar se os
levantamentos dos táxons foram completos ou se haveria um acréscimo no número de
espécies com o aumento do esforço de coleta (MAGURRAN, 1983).
Índice de diversidade (H’) - A fim de comparar as comunidades por um único valor, usou-se
o índice de diversidade de Shannon-Wiener (H’) calculado com o auxílio do programa
MVSP. Esse índice usa o número de espécies (riqueza) e a distribuição dos indivíduos entre
as espécies (equitatividade) para representar a diversidade de indivíduos de uma área
independente do tamanho da amostra (MAGURRAN, 1983).

43
Similaridade entre áreas - Para determinar o Índice de Similaridade entre os ambientes
estudados usou-se o índice de Jaccard calculado pelo programa MVSP. Este índice é
apropriado para indicar semelhanças entre comunidades, com relação à composição de seus
indivíduos (MAGURRAN, 1983). Espera-se, que quanto menor a distância entre as
localidades, maior a similaridade encontrada.

44
8 RESULTADOS E DISCUSSÃO
8.1 Áreas de Preservação Permanente
A fragmentação e o isolamento dos remanescentes florestais trazem grandes
preocupações. Na APA Itacaré/Serra Grande, as manchas de florestas primárias sofrem com
problemas de fragmentação no seu entorno, com seus limites sofrendo efeito de borda, pois
observou-se na sua vizinhança pastagens, rodovias, estradas, cultivos agrícolas ou áreas
degradadas. Os fragmentos, quando estão isolados, podem diminuir sua biodiversidade com o
tempo, pois há redução na troca de material genético entre populações e os processos
ecológicos (polinização, predação e comportamento territorial) são afetados (METZGER,
2003).
O tamanho total da área considerada neste estudo é de 40.178,11 ha. Deste total,
28.053,25 ha é considerado Área de Preservação Permanente (APP) de acordo com a Lei
Federal Nº 4.771 (Figura 12), ou seja, (69,82%) de toda a área. Esta lei que institui o novo
Código Florestal considera às florestas e demais formas de vegetação natural área de
preservação permanente com algumas restrições de usos.
A porcentagem de (69,82%) da área de estudo considerada de preservação permanente
evidencia o potencial preservacionista da APA. Esta é uma área considerada como prioritária
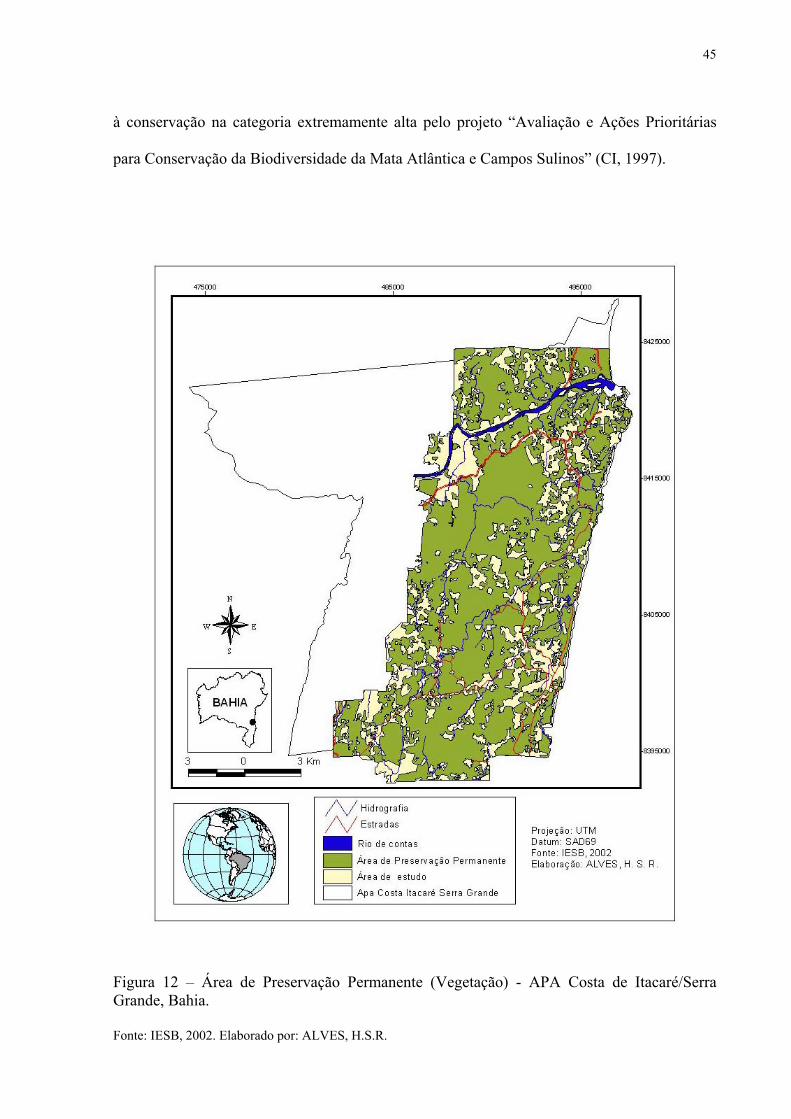
45
à conservação na categoria extremamente alta pelo projeto “Avaliação e Ações Prioritárias
para Conservação da Biodiversidade da Mata Atlântica e Campos Sulinos” (CI, 1997).
Figura 12 – Área de Preservação Permanente (Vegetação) - APA Costa de Itacaré/Serra Grande, Bahia. Fonte: IESB, 2002. Elaborado por: ALVES, H.S.R.

Na APA Itacaré/Serra Grande as áreas que são consideradas de preservação
permanente são as restingas e os brejos, os manguezais e os fragmentos florestais primários e
secundários (estes em estágio inicial, médio e avançado de regeneração).
As restingas e os brejos correspondem juntos a 806,58 ha da área de estudo (2%). As
restingas são ambientes geradas pela deposição de sedimentos marinhos e continentais em
áreas costeiras (ALBERTONI; ESTEVES, 1999), e nas restingas o solo não constitui a
principal fonte de nutrientes da vegetação (ARAÚJO; LACERDA, 1992). As restingas
exercem papel importante no fluxo migratório de numerosas espécies de ecossistemas
associados e na manutenção de numerosas espécies endêmicas (ODUM, 1988) tais como os
insetos, aves, mamíferos, anfíbios e répteis (CÂMARA, 1991). Áreas de restinga são
exploradas com pastagens e plantações de dendê (Elaeis guineensis), piaçava (Attalea
funífera) e coco da Bahia (Cocos nucifera). As áreas de brejos são dominadas por plantas
higrófilas, com solos ricos em matéria orgânica, espécies das famílias Gramineae, Ciperaceae
e Tifaceae (ARAÚJO; LACERDA, 1992).
Durante reconhecimento da área, foi possível observar o processo de antropização nas
restingas com a urbanização desordenada devido à especulação imobiliária e construção da
rodovia BA-001 que favorece o acesso ao longo da costa, provocando alterações no ambiente
como introdução de espécies exóticas (animal e vegetal), remoção de areia de praias e
desmatamento da vegetação. Apesar de estarem protegidas por lei, as áreas de restinga
encontram-se bastante degradadas. A recuperação e manutenção das restingas são essenciais,
pois além de oferecer proteção às espécies de fauna e à Floresta Ombrófila Densa, podem ser
considerada como um importante corredor biológico.
Os manguezais correspondem a 289,85 ha da área de estudo (0,72 %). Os manguezais
possibilitam a manutenção de diversos habitats propícios à proteção contra predadores,
reprodução, desova e crescimento de diversas espécies marinhas ou de água doce. Dois terços

47
das espécies de peixes marinhos dependem desse ecossistema (GUSMÃO, 2002). O
manguezal apresenta uma rica e abundante fauna, composta por aves, crustáceos, moluscos,
mamíferos e peixes. As espécies animais do manguezal não são exclusivas desse ambiente
(LACERDA, 1999). Setenta por cento (70%) dos organismos pescados são originários de
estuários ou de ecossistemas estuarino-lagunares (SCHAEFFER-NOVELLI et al., 2001).
Apesar da preservação dos manguezais terem grande importância econômica, estes são
aterrados para construções de casas, grandes hotéis, lançamento de esgoto. Os manguezais
também são protegidos por lei, mas continuam sendo ocupados das mais diferentes formas
pela população.
Outra Área de Preservação Permanente é o “Campo Cheiroso” que se caracteriza por
apresentar formações florísticas e fatores edáficos diferenciados do restante da região, sendo
chamado pelos especialistas de comunidade relíquia. A vegetação do Campo Cheiroso está
associado a Restinga e Campo Rupestre, com solos principalmente arenosos, apresentando
afloramento rochoso (lajedo), possibilitando o transporte superficial da água nessas áreas
(BRASIL-MMA, 2004). Segundo Jardim (2003), a vegetação nesse ambiente é semelhante à
encontrada nos campos de restinga e em áreas de “Mussununga” herbácea-arbustiva
encontradas nas matas de tabuleiro na região Extremo Sul da Bahia. O Zoneamento
Ecológico-Econômico (ZEE) da APA sugere a inclusão dessa área no Parque Estadual Serra
do Conduru que é uma Unidade de Conservação com categoria mais restritiva que a APA
(BRASIL-MMA, 2004).
A vegetação primária e secundária (em estágios inicial, médio e avançado de
regeneração) também é considerada área de preservação permanente e está sujeito a restrições
de uso. A resolução nº 5, de 04 de maio de 1994 do Conselho Nacional do Meio Ambiente, a
fim de orientar os procedimentos de licenciamento de atividades florestais no Estado da Bahia
define vegetação primária e secundária (BRASIL-Resolução CONAMA, 1994).

48
O artigo 1º considera a Vegetação primária “aquela de máxima expressão local, com
grande diversidade biológica, sendo os efeitos das ações antrópicas mínimos, a ponto de não
afetar significativamente suas características originas de estrutura e de espécies”.
O artigo 2º considera vegetação secundária ou em regeneração “aquela resultante dos
processos naturais de sucessão, após supressão total ou parcial da vegetação primária por
ações antrópicas ou causas naturais, podendo ocorrer árvores remanescentes da vegetação
primária”.
O artigo 3º caracteriza os estágios de regeneração da vegetação secundária levando em
consideração: sua fisionomia; DAP das espécies lenhosas; existência de epífitas e trepadeiras;
espessura da serapilheira; diversidade biológica; cobertura arbórea ou ausência de subosque;
espécies pioneiras abundantes ou espécies emergentes ocorrendo com diferentes graus de
intensidade, entre outros.
Vale ressaltar que, dos 40.178,11 ha da área de estudo, somente 2.326,12 ha (5,8%)
são de áreas de vegetação primária, e 14.403,79 ha (35,84%) correspondem à floresta
secundária em estágio avançado/cabruca. Mesmo utilizando fotografia aérea, não foi possível
separar a floresta secundária em estágio avançado das cabrucas devido à forma de seu plantio.
No sistema cabruca raleia-se a floresta e introduz-se o cacau sob a sombra das árvores nativas
(LOBÃO, 1997), por isso a dificuldade em separar a floresta da cabruca. Existem poucas
áreas sob cultivo de cacau na região da APA, sendo a maioria localizada nos vales de rios a
exemplo das margens do Rio de Contas no limite norte do polígono (BRASIL-MMA, 2004),
ou seja, existe mais vegetação secundária em estágio avançado de regeneração do que
Cabruca.
Esse mosaico formado pelas florestas e pelas cabrucas tem importância estratégica
para a conservação da biodiversidade da região, pois funcionam como corredores biológicos
entre os fragmentos (RICE; GREENBERG, 2000). No entanto, segundo Faria (2002), as

49
cabrucas contribuem para a conservação dependendo do contexto da paisagem em que se
insere. Comparando a utilização das cabrucas em Una e Uruçuca, a autora constatou que em
Una, a paisagem é predominantemente florestal, com fragmentos de floresta íntegros,
enquanto em Uruçuca, existem muitas cabrucas e poucos fragmentos. Os resultados
demonstraram que cabrucas próximas a fragmentos florestais ou unidades de conservação
contribuem para a conservação da biodiversidade, mas não acontece o mesmo em locais onde
a proporção de cabrucas é maior do que a floresta. O corredor neste caso não terá a mesma
funcionalidade. Analisando tudo isso, percebe-se que para a cabruca funcionar como corredor,
é preciso ter mais florestas protegidas como APP, Reserva Legal, Unidades de Conservação.
A proporção maior de área de vegetação secundária em estágio avançado de
regeneração na APA pode ser resultante da recuperação de estágios sucessionais
intermediários ou da antropização da floresta primária. A APA possui mais de (70%) do seu
território com florestas e sistemas agroflorestais associados ao cacau (BRASIL-MMA, 2004).
O desenvolvimento da cacauicultura foi, em grande parte responsável pela degradação
da Mata Atlântica no Sul da Bahia, mesmo quando implantada sob o sistema agroflorestal
cabruca, pois a maior parte da área ocupada anteriormente por floresta encontra-se atualmente
ocupada por cacauais (FARIA, 2002). A policultura e a pecuária intensificaram ainda mais o
desmatamento. Se a redução e a fragmentação dos remanescentes continuarem, poderá ocorrer
a redução da diversidade e riqueza de espécies, uma vez que o tamanho do fragmento e seu
isolamento interferem nestes índices (MACRTHUR; WILSON, 1967).
A atual política de criação de unidades de conservação não irá garantir por si só a
sobrevivência dos ecossistemas. Diante desta problemática, faz-se necessário que se proíba
implantação de novas áreas de cultivos ou pastagens dentro das áreas ainda ocupadas por
fragmentos de floresta primária e secundária (nos diferentes estágios de regeneração), para

50
que seja contido o processo de isolamento das ilhas remanescentes através da política dos
corredores ecológicos.
Foi gerado também o mapa de APPs considerando a hidrografia na área de estudo
(Figura 13). De acordo com o Código Florestal, toda vegetação natural ao longo dos rios ou
de qualquer curso d'água também é considerada de preservação permanente. A largura da
faixa de mata ciliar que deve ser preservada está relacionada com a largura do curso d’água.
Neste estudo, é considerada de preservação permanente a vegetação ao longo dos rios cuja
largura mínima de 50 (cinqüenta) metros para os cursos d'água que tenham de 10 (dez) a 50
(cinqüenta) metros de largura; de 100 (cem) metros para os cursos d'água que tenham de 50
(cinqüenta) a 200 (duzentos) metros de largura; ao redor das nascentes, ainda que
intermitentes e nos chamados "olhos d'água", qualquer que seja a sua situação topográfica,
num raio mínimo de 50 (cinqüenta) metros de largura.
O desmatamento das matas ciliares traz como conseqüência o assoreamento dos rios.
A ausência da mata ciliar deixa o solo desprotegido, ficando sujeito a erosões. Com a chuva, a
terra é lixiviada para o rio, o qual fica assoreado, com tendência a ficar cada vez mais raso.
Isso também diminui a qualidade da água, afetando os ecossistemas que habitam o rio
(GORGÔNIO, 1998; CARDOSO-LEITE et al., 2004; PRIMO; VAZ, 2006).
É cada vez mais aceito que existe uma relação de interdependência entre a floresta e o
ecossistema aquático, e que a degradação ou escassez de um perturba profundamente a
existência e a qualidade do outro. A floresta, além de tantas outras funções, desempenha
também funções hidrológicas, pois é fundamental na conservação dos ecossistemas aquáticos,
inclusive da biota a eles associada. As florestas influenciam sobre processos hidrológicos
como: atenuação dos picos de vazão, influência da qualidade da água, ciclagem de nutrientes,
proteção dos corpos d’água e geração dos serviços ambientais (LINO; DIAS, 2003).

51
Garantir a integridade das Áreas de Preservação Permanente tem sido um grande
desafio. As APPs, como já foi dito, são áreas protegidas com base no Código Florestal como
sendo áreas públicas ou particulares, nas quais a supressão total ou parcial da vegetação
natural só é permitida mediante previa autorização do Poder Executivo Federal, quando
necessária à execução de obras, planos, atividades ou projetos de utilidade pública ou de
interesse social.

52
Figura 13 – Área de Preservação Permanente (Hidrografia e Nascentes) - APA Costa de Itacaré/Serra Grande, Bahia. Fonte: SEI, 2003. Elaborado por: ALVES, H.S.R.

53
8.2 Formigas como bioindicadores
8.2.1 Composição da comunidade de Formicidae
Nas oito áreas estudadas (Tabela 2) foram coletadas 234 espécies de formigas
pertencendo a 55 gêneros, 27 tribos e 10 subfamílias (Tabela 3).
Tabela 2 – Siglas que serão utilizadas no texto para indicar as áreas de estudo.
ÁREAS LOCALIDADE AMBIENTE SIGLA ÁREAS LOCALIDADE AMBIENTE SIGLA Área 1 Taboquinhas Floresta (A1)T-F Área 5 Itacaré Cacaual (A5)I-C Área 2 Uruçuca Floresta (A2)U-F Área 6 Serra Grande Cacaual (A6)SG-C Área 3 Itacaré Floresta (A3)I-F Área 7 Itacaré Pastagem (A7)I-P Área 4 Serra Grande Floresta (A4)SG-F Área 8 Serra Grande Pastagem (A8)SG-P
Tabela 3 – Composição de formigas realizadas em oito áreas na APA Itacaré/Serra Grande, Bahia, Brasil, (1996/1998).
FLORESTA CACAUAL PASTAGEM
(A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-P Total Nº Subfamílias 7 7 8 7 8 8 6 8 10Nº Tribos 16 15 19 16 23 19 16 15 27
Nº Gêneros 32 28 29 29 35 33 25 22 55Nº de Espécies 109 83 89 86 87 81 52 43 234
Obs.: (A1)T-F = Taboquinhas Floresta; (A2)U-F = Uruçuca Floresta; (A3)I-F = Itacaré Floresta; (A4)SG-F = Serra Grande Floresta; (A5)I-C = Itacaré Cacau; (A6)SG-C = Serra Grande Cacau; (A7)I-P = Itacaré Pastagem e (A8)SG-P = Serra Grande Pastagem.

54
A subfamília mais diversificada foi Myrmicinae com 14 tribos, 30 gêneros e 134
espécies correspondendo a 57% das espécies encontradas. Duas subfamílias apresentaram
apenas uma tribo, um gênero e uma espécie o que corresponde a 0,5% das espécies
encontradas Cerapachyinae e Proceratinae (Figura 14).
Bolton (2003) registrou 14 subfamílias na região Neotropical. Então o fato de
encontrar dez subfamílias nas coletas pode ser considerado um número alto e também permite
afirmar que essas áreas possuem alta diversidade. A subfamília Myrmicinae apresentou o
maior número de táxons corroborando os resultados de Feitosa e Ribeiro (2005) e muitos
outros em geral para a Região Neotropical. Mesmo utilizando outros métodos de coletas esta
subfamília geralmente apresenta maior diversidade. Segundo Fowler et al. (1991), as formigas
dessa subfamília são as mais diversificadas quando se trata de hábitos alimentares e
nidificação.
As subfamílias Cerapachyinae e Proceratinae apresentaram somente uma tribo, um
gênero e uma espécie cada uma. A primeira provavelmente foi encontrada uma só espécie
porque ocorre ocasionalmente na serrapilheira predando ovos de outras formigas (DELABIE
et al., 2000).
Em relação aos gêneros, dos 55 observados, os mais ricos em espécies foram Pheidole
(36 espécies), Hypoponera e Solenopsis (16), Paratrechina (12), Brachymyrmex (11),
Crematogaster, Pachycondyla e Pyramica (10) (Figura 15 e Tabela 4).
Pheidole, Hypoponera e Solenopsis estão entre os gêneros mais diversificados do
mundo (BOLTON, 1995). Nos estudos de Ramos et al. (2003), Pheidole também foi o gênero
predominante. Este gênero apresenta mais de 600 espécies na região Neotropical (WILSON,
2003), e segundo Fowler (1993) é considerado um dos mais competitivos, pois domina seus
recursos alimentares conseguindo de forma altamente eficiente a eliminação dos seus
concorrentes. Nos estudos de Silvestre (2000), em ambiente de Cerrado, o gênero mais rico
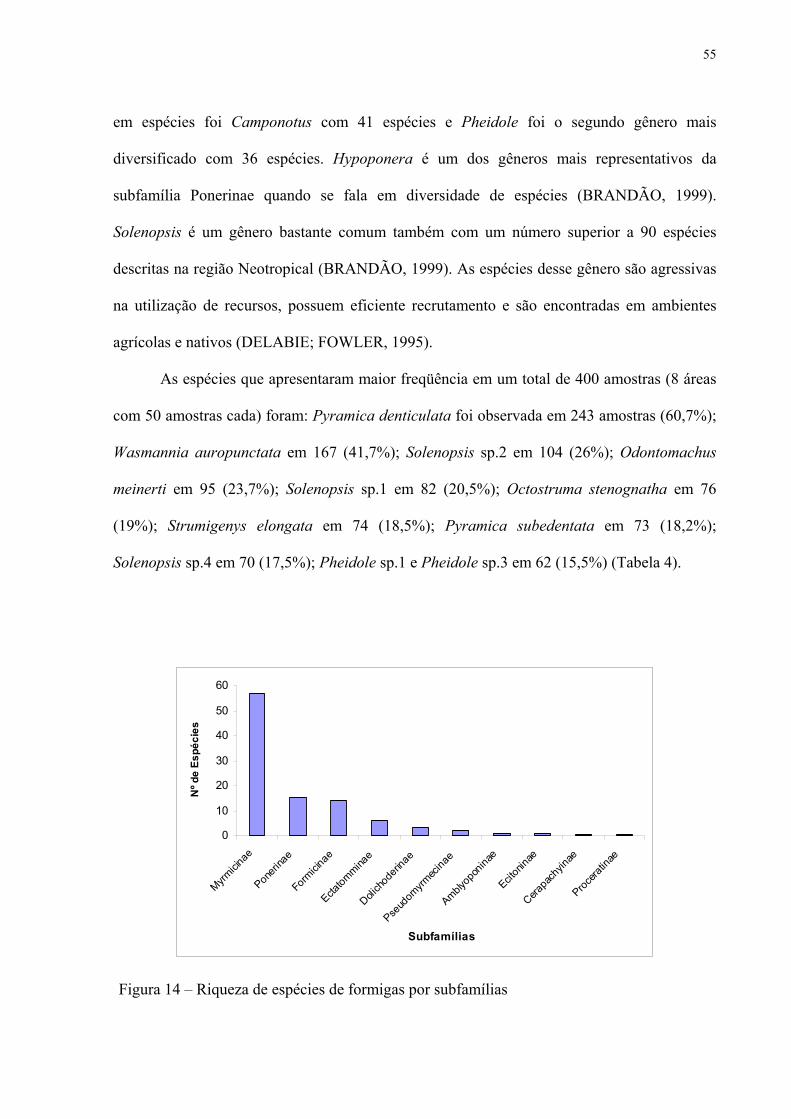
55
em espécies foi Camponotus com 41 espécies e Pheidole foi o segundo gênero mais
diversificado com 36 espécies. Hypoponera é um dos gêneros mais representativos da
subfamília Ponerinae quando se fala em diversidade de espécies (BRANDÃO, 1999).
Solenopsis é um gênero bastante comum também com um número superior a 90 espécies
descritas na região Neotropical (BRANDÃO, 1999). As espécies desse gênero são agressivas
na utilização de recursos, possuem eficiente recrutamento e são encontradas em ambientes
agrícolas e nativos (DELABIE; FOWLER, 1995).
As espécies que apresentaram maior freqüência em um total de 400 amostras (8 áreas
com 50 amostras cada) foram: Pyramica denticulata foi observada em 243 amostras (60,7%);
Wasmannia auropunctata em 167 (41,7%); Solenopsis sp.2 em 104 (26%); Odontomachus
meinerti em 95 (23,7%); Solenopsis sp.1 em 82 (20,5%); Octostruma stenognatha em 76
(19%); Strumigenys elongata em 74 (18,5%); Pyramica subedentata em 73 (18,2%);
Solenopsis sp.4 em 70 (17,5%); Pheidole sp.1 e Pheidole sp.3 em 62 (15,5%) (Tabela 4).
0
10
20
30
40
50
60
Myrmicin
ae
Poneri
nae
Form
icinae
Ectatom
minae
Dolich
oderi
nae
Pseud
omyrm
ecina
e
Amblyop
onina
e
Eciton
inae
Cerapa
chyin
ae
Procera
tinae
Subfamílias
Nº d
e Es
péci
es
Figura 14 – Riqueza de espécies de formigas por subfamílias

56
0
5
10
15
20
25
30
35
40
Pheidole
Hypopo
nera
Solenop
sis
Paratre
china
Brachy
myrmex
Cremato
gaster
Pachycon
dyla
Pyramica
Gnampto
genys
Strumigeny
s
Apterost
igma
Campo
notus
Sericom
yrmex
Acropy
ga
Octostr
uma
Gêneros
Nº d
e E
spéc
ies
Figura 15 – Riqueza de espécies de formigas por gêneros.
Tabela 4 – Formicidae coletados com extrator de Winkler em oito áreas na APA Costa de Itacaré/Serra Grande, Bahia, Brasil (1996/1998).
FLORESTA CACAU PASTAGEM ESPÉCIES (A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-P
Subfamília Amblyoponinae Tribo Amblyoponini
Amblyopone armigera Mayr, 1897 1 Prionopelta antillana Forel, 1909 7 5 5 16 1 1
Subfamília Cerapachyinae Tribo Cerapachyini
Cerapachys splendens Borgmeier, 1957 1 Subfamília Dolichoderinae
Tribo Dolichoderini Azteca sp.1 3 Dolichoderus attelaboides (Fabricius, 1775) 1 Dolichoderus imitator Emery, 1894 13 6 9 18 2 4 Dolichoderus lamellosus (Mayr, 1870) 1 Dolichoderus lutosus (Fr. Smith, 1858) 1 Dorymyrmex thoracicus Gallardo, 1916 1 Linepithema humile (Mayr, 1866) 1 1 1
Subfamília Ecitoninae Tribo Ecitonini
Labidus coecus (Latreille, 1802) 2 3 Neivamyrmex sp.1 2
“... continua...”
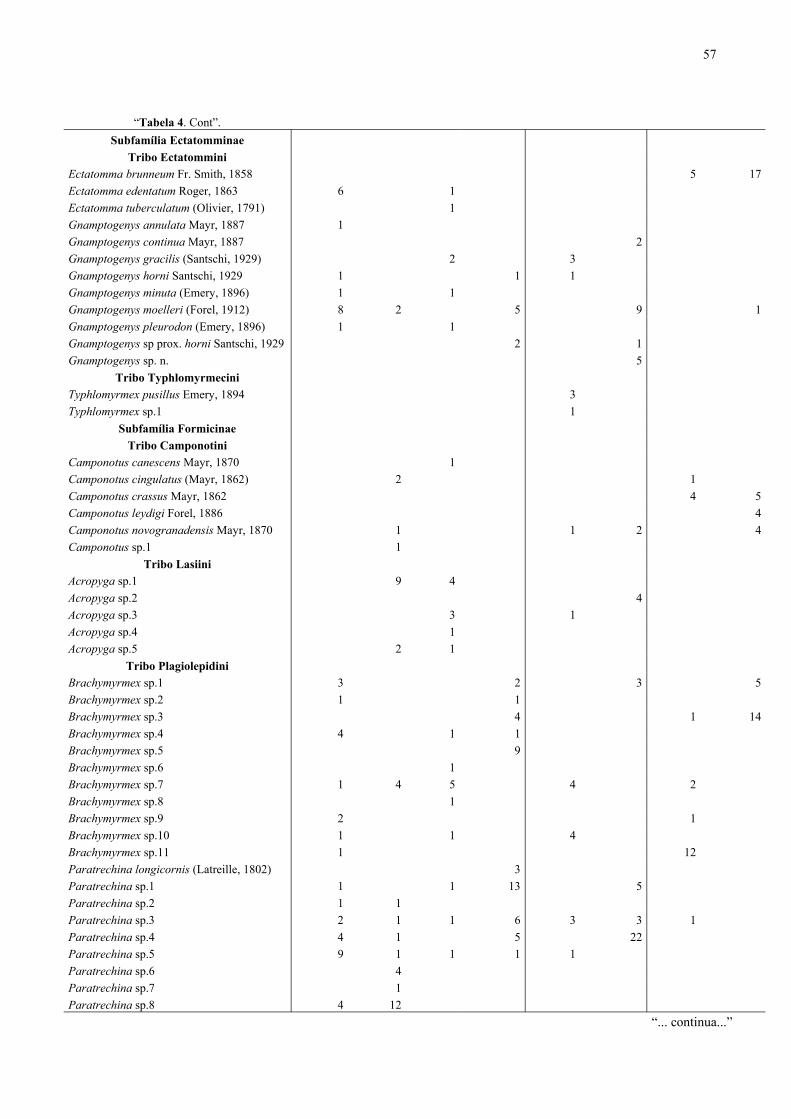
57
“Tabela 4. Cont”. Subfamília Ectatomminae
Tribo Ectatommini Ectatomma brunneum Fr. Smith, 1858 5 17Ectatomma edentatum Roger, 1863 6 1 Ectatomma tuberculatum (Olivier, 1791) 1 Gnamptogenys annulata Mayr, 1887 1 Gnamptogenys continua Mayr, 1887 2 Gnamptogenys gracilis (Santschi, 1929) 2 3 Gnamptogenys horni Santschi, 1929 1 1 1 Gnamptogenys minuta (Emery, 1896) 1 1 Gnamptogenys moelleri (Forel, 1912) 8 2 5 9 1Gnamptogenys pleurodon (Emery, 1896) 1 1 Gnamptogenys sp prox. horni Santschi, 1929 2 1 Gnamptogenys sp. n. 5
Tribo Typhlomyrmecini Typhlomyrmex pusillus Emery, 1894 3 Typhlomyrmex sp.1 1
Subfamília Formicinae Tribo Camponotini
Camponotus canescens Mayr, 1870 1 Camponotus cingulatus (Mayr, 1862) 2 1 Camponotus crassus Mayr, 1862 4 5Camponotus leydigi Forel, 1886 4Camponotus novogranadensis Mayr, 1870 1 1 2 4Camponotus sp.1 1
Tribo Lasiini Acropyga sp.1 9 4 Acropyga sp.2 4 Acropyga sp.3 3 1 Acropyga sp.4 1 Acropyga sp.5 2 1
Tribo Plagiolepidini Brachymyrmex sp.1 3 2 3 5Brachymyrmex sp.2 1 1 Brachymyrmex sp.3 4 1 14Brachymyrmex sp.4 4 1 1 Brachymyrmex sp.5 9 Brachymyrmex sp.6 1 Brachymyrmex sp.7 1 4 5 4 2 Brachymyrmex sp.8 1 Brachymyrmex sp.9 2 1 Brachymyrmex sp.10 1 1 4 Brachymyrmex sp.11 1 12 Paratrechina longicornis (Latreille, 1802) 3 Paratrechina sp.1 1 1 13 5 Paratrechina sp.2 1 1 Paratrechina sp.3 2 1 1 6 3 3 1 Paratrechina sp.4 4 1 5 22 Paratrechina sp.5 9 1 1 1 1 Paratrechina sp.6 4 Paratrechina sp.7 1 Paratrechina sp.8 4 12 “... continua...”

58
“Tabela 4. Cont”. FLORESTA CACAU PASTAGEM
ESPÉCIES (A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-PParatrechina sp.9 1 1 Paratrechina sp.10 1 8 11 Paratrechina sp.11 2 10 1 1 1
Subfamília Myrmicinae Tribo Attini
Acromyrmex balzani (Emery, 1890) 1Acromyrmex rugosus (Fr. Smith, 1858) 2 7Apterostigma acre Lattke, sp.n. 1 1 2 Apterostigma auriculatum Wheeler, 1925 3 1 4 1 1 Apterostigma ierense Weber, 1937 2 1 Apterostigma madidiense Weber, 1938 1 1 1 Apterostigma sp complexo pilosum 1 3 Apterostigma sp.1 1 2 5 Atta sexdens rubropilosa (Forel, 1908) 1 Cyphomyrmex peltatus Kempf, 1965 4 4 4 1 4 1 Cyphomyrmex rimosus (Spinola, 1853) 5 11 4 1 Cyphomyrmex transversus Emery, 1894 7 10 4 4 18 1 7Mycocepurus smithi Forel, 1893 7 5 7 5 9 4 1 Myrmicocrypta sp. prox. buenzlii 1 Sericomyrmex bondari Borgmeier, 1937 5 6 3 4 8 19 Sericomyrmex sp.1 1 Sericomyrmex sp.2 1 Sericomyrmex sp.3 1 2 Sericomyrmex sp.4 7 7 24 Sericomyrmex sp.5 3 6 1 Trachymyrmex cornetzi Forel, 1912 18 2 7 5 8 Trachymyrmex relictus Borgmeier, 1934 1 2 Trachymyrmex sp.1 2
Tribo Adelomyrmicini Cryptomyrmex boltoni Fernández, 2003 1
Tribo Basicerotini Octostruma balzani (Emery, 1894) 1 Octostruma jheringhi (Emery, 1887) 1 4 1 Octostruma petiolata (Mayr, 1887) 1 4 16 2 Octostruma rugifera (Mayr, 1887) 5 7 5 Octostruma stenognatha Brown & kempf, 1960 21 13 13 11 4 14
Tribo Blepharidattini Blepharidatta sp.1 2 Wasmannia auropunctata (Roger, 1863) 13 20 22 31 37 28 16Wasmannia sp.1 1 13Wasmannia sp.2 8 Wasmannia sp.3 1
Tribo Cephalotini Procryptocerus pictipes Emery, 1896 1
Tribo Crematogastrini Crematogaster acuta (Fabricius, 1804) 1 1 Crematogaster sp.1 1 7 1 Crematogaster sp.2 6 4 Crematogaster sp.3 1 2 9 Crematogaster sp.4 1 3 3
“... continua...”

59
“Tabela 4. Cont”. FLORESTA CACAU PASTAGEM
ESPÉCIES (A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-PCrematogaster sp.5 3 1 30Crematogaster sp.6 2 Crematogaster sp.7 1 Crematogaster sp.8 1 Crematogaster sp.9 14 17 7
Tribo Dacetonini Acanthognathus ocellatus Mayr, 1887 1 Pyramica alberti (Forel, 1893) 1 Pyramica appretiatus (Borgmeier, 1954) 1 1 Pyramica crassicornis (Mayr, 1887) 1 Pyramica denticulata (Mayr,1887) 43 32 46 26 45 37 14 Pyramica eggersi (Emery,1890) 1 2 3 11 2Pyramica rugithorax (Kempf,1959) 5 1 6 9 23 Pyramica schulzi (Emery, 1894) 1 1 1 9 15 4Pyramica sp.1 1 Pyramica sp.2 17 1 3 6 15 7 Pyramica subedentata (Mayr,1887) 10 6 10 7 22 18 Strumigenys carinithorax Borgmeier, 1934 2 13 2 1 2 1 Strumigenys dolichognatha Weber, 1934 1 Strumigenys elongata Roger, 1863 7 17 8 3 18 19 2 Strumigenys louisianae Roger, 1863 1 5 5Strumigenys perparva Brown, 1957 1 2 Strumigenys sp.1 2 Strumigenys subedentata Mayr 1887 1
Tribo Formicoxenini Cardiocondyla emeryi Forel, 1881 1 Nesomyrmex tristani (Emery, 1896) 1 1 1
Tribo Myrmicini Hylomyrma balzani (Emery, 1894) 4 1 Hylomyrma immanis Kempf, 1973 2 3 6 3 6 2 Hylomyrma sagax Kempf, 1973 5 4 1 3 1
Tribo Pheidolini Pheidole fimbriata Roger, 1863 6 2 8 6 Pheidole sp.1 20 9 18 13 2 Pheidole sp.2 25 3 3 Pheidole sp.3 9 1 16 19 16 1Pheidole sp.4 22 6 7 5 9 5 Pheidole sp.5 1 Pheidole sp.6 6 3 14 2 1 4Pheidole sp.7 1 1 Pheidole sp.8 1 1 3 Pheidole sp.9 4 4 3 1 4Pheidole sp.10 2 1 2 4Pheidole sp.11 2 1 20 24 Pheidole sp.12 1 Pheidole sp.13 9 7 6 3 Pheidole sp.14 1 1 Pheidole sp.15 6 1 1 2 7 3Pheidole sp.16 13 4Pheidole sp.17 1
“... continua...”

60
“Tabela 4. Cont”. FLORESTA CACAU PASTAGEM
ESPÉCIES (A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-P Pheidole sp.18 1 4 1Pheidole sp.19 1 Pheidole sp.20 1Pheidole sp.21 1 1 Pheidole sp.22 1 Pheidole sp.23 5 Pheidole sp.24 3 Pheidole sp.25 1 1 Pheidole sp.26 1 1 1Pheidole sp.27 1 2 Pheidole sp.28 3 5 Pheidole sp.29 6 Pheidole sp.30 3 Pheidole sp.31 1 6 Pheidole sp.32 3 1 Pheidole sp.33 3 Pheidole sp.34 1 5 Pheidole sp.35 1
Tribo Pheidologetonini Carebara panamensis (Wheeler, 1925) 2 2 14 1 1Carebara urichi (Wheeler, 1922) 3 1
Tribo Solenopsidini Carebarella bicolor Emery, 1905 3 Carebarella sp.1 12 Megalomyrmex drifti Kempf, 1961 2 Megalomyrmex goeldii Forel, 1912 7 1 2 Megalomyrmex silvestrii Wheeler, 1909 1 1Monomorium floricola (Jerdon, 1852) 1 1 3 5 5Monomorium pharaonis (Linnaeus, 1758) 23 Monomorium sp.1 3 1 2 Solenopsis geminata (Fabricius, 1804) 1 4 1 5 Solenopsis interrupta Santschi, 1916 2 Solenopsis sp.1 13 17 23 17 11 1Solenopsis sp.2 16 11 16 10 36 10 5Solenopsis sp.3 4 6 3 16 7 3 Solenopsis sp.4 4 28 6 9 7 16 Solenopsis sp.5 1 2 4 Solenopsis sp.6 1 3 2 1 Solenopsis sp.7 1 1 1 2 12Solenopsis sp.8 5 1 1 Solenopsis sp.9 2 16 3 12 11 Solenopsis sp.10 1 Solenopsis sp.11 10 Solenopsis sp.12 1 Solenopsis sp.13 1 Solenopsis virulens (Fr. Smith, 1858) 3 15 4 15 2
Tribo Stegomyrmecini Stegomyrmex vizottoi Diniz, 1990 1 3
“... continua...”
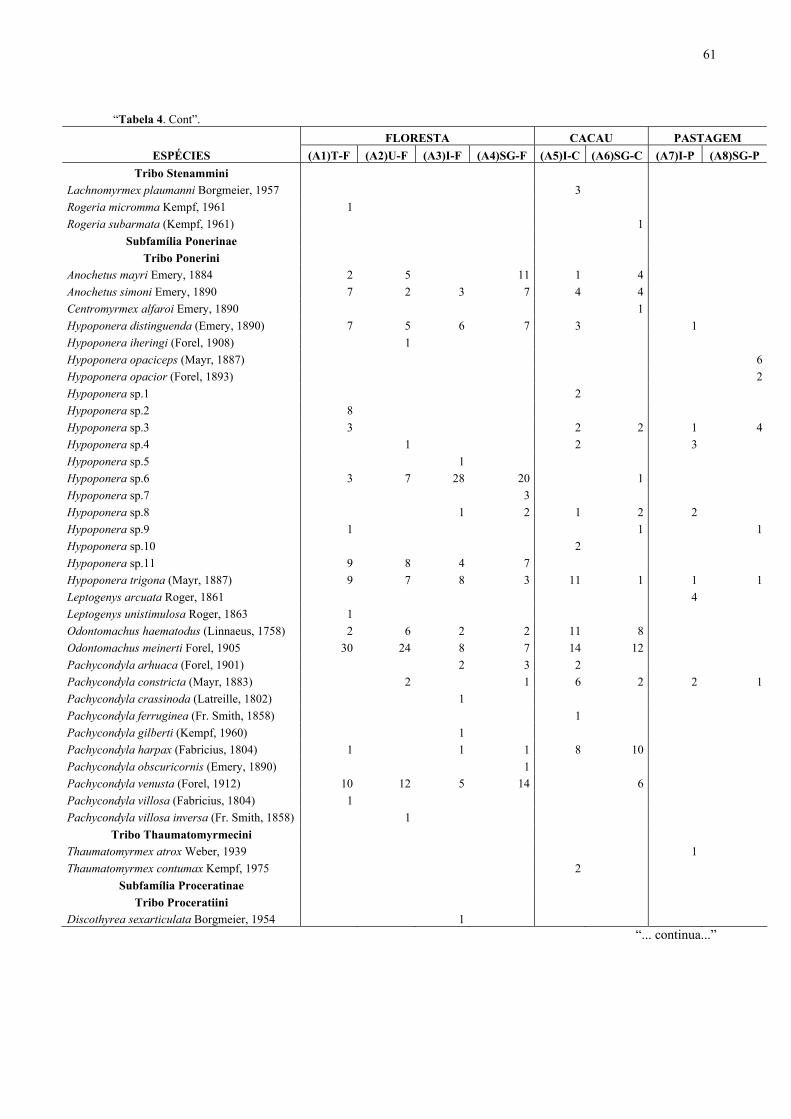
61
“Tabela 4. Cont”. FLORESTA CACAU PASTAGEM
ESPÉCIES (A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-P Tribo Stenammini
Lachnomyrmex plaumanni Borgmeier, 1957 3 Rogeria micromma Kempf, 1961 1 Rogeria subarmata (Kempf, 1961) 1
Subfamília Ponerinae Tribo Ponerini
Anochetus mayri Emery, 1884 2 5 11 1 4 Anochetus simoni Emery, 1890 7 2 3 7 4 4 Centromyrmex alfaroi Emery, 1890 1 Hypoponera distinguenda (Emery, 1890) 7 5 6 7 3 1 Hypoponera iheringi (Forel, 1908) 1 Hypoponera opaciceps (Mayr, 1887) 6Hypoponera opacior (Forel, 1893) 2Hypoponera sp.1 2 Hypoponera sp.2 8 Hypoponera sp.3 3 2 2 1 4Hypoponera sp.4 1 2 3 Hypoponera sp.5 1 Hypoponera sp.6 3 7 28 20 1 Hypoponera sp.7 3 Hypoponera sp.8 1 2 1 2 2 Hypoponera sp.9 1 1 1Hypoponera sp.10 2 Hypoponera sp.11 9 8 4 7 Hypoponera trigona (Mayr, 1887) 9 7 8 3 11 1 1 1Leptogenys arcuata Roger, 1861 4 Leptogenys unistimulosa Roger, 1863 1 Odontomachus haematodus (Linnaeus, 1758) 2 6 2 2 11 8 Odontomachus meinerti Forel, 1905 30 24 8 7 14 12 Pachycondyla arhuaca (Forel, 1901) 2 3 2 Pachycondyla constricta (Mayr, 1883) 2 1 6 2 2 1Pachycondyla crassinoda (Latreille, 1802) 1 Pachycondyla ferruginea (Fr. Smith, 1858) 1 Pachycondyla gilberti (Kempf, 1960) 1 Pachycondyla harpax (Fabricius, 1804) 1 1 1 8 10 Pachycondyla obscuricornis (Emery, 1890) 1 Pachycondyla venusta (Forel, 1912) 10 12 5 14 6 Pachycondyla villosa (Fabricius, 1804) 1 Pachycondyla villosa inversa (Fr. Smith, 1858) 1
Tribo Thaumatomyrmecini Thaumatomyrmex atrox Weber, 1939 1 Thaumatomyrmex contumax Kempf, 1975 2
Subfamília Proceratinae Tribo Proceratiini
Discothyrea sexarticulata Borgmeier, 1954 1 “... continua...”

62
“Tabela 4. Cont”. FLORESTA CACAU PASTAGEM
ESPÉCIES (A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-P Subfamília Pseudomyrmecinae
Tribo Pseudomyrmecini Pseudomyrmex sp.1 1 Pseudomyrmex sp.2 2 1 7Pseudomyrmex tenuis (Fabricius, 1804) 1 1 1 1 Pseudomyrmex termitarius (Fr. Smith, 1855) 5 2Total de ocorrência 556 409 444 495 504 555 201 214
Obs.: (A1)T-F = Taboquinhas Floresta; (A2)U-F = Uruçuca Floresta; (A3)I-F = Itacaré Floresta; (A4)SG-F = Serra Grande Floresta; (A5)I-C = Itacaré Cacau; (A6)SG-C = Serra Grande Cacau; (A7)I-P = Itacaré Pastagem e (A8)SG-P = Serra Grande Pastagem.
8.2.1.2 Análise da diversidade das áreas de florestas, cacauais e pastagens amostradas em 1996/1998
Na tabela 5, estão informados a riqueza observada, riqueza estimada (Chao 2) e índice
de diversidade (H’) de cada área separadamente.
Tabela 5 - Riquezas Observadas e Estimadas, e Índice de diversidade das espécies coletadas.
FLORESTA CACAUAIS PASTAGEM
(A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-P Total
Riqueza observada de spp 109 83 89 86 87 81 52 43 234
Riqueza estimada (Chao 2) 209.49 118.53 191.9 114.67 110.15 90.86 87.42 60.84 -
Índice de diversidade (H’) 4,69 4,41 4,48 4,45 4,46 4,39 3,95 3,76 - Obs.: (A1)T-F = Taboquinhas Floresta; (A2)U-F = Uruçuca Floresta; (A3)I-F = Itacaré Floresta; (A4)SG-F = Serra Grande Floresta; (A5)I-C = Itacaré Cacau; (A6)SG-C = Serra Grande Cacau; (A7)I-P = Itacaré Pastagem e (A8)SG-P = Serra Grande Pastagem.
Nas áreas estudadas foram observadas entre 109 e 43 espécies de formigas (Tabela 5).
Na área de floresta (A1)T-F foi observado o maior número de espécies equivalente a 47% de
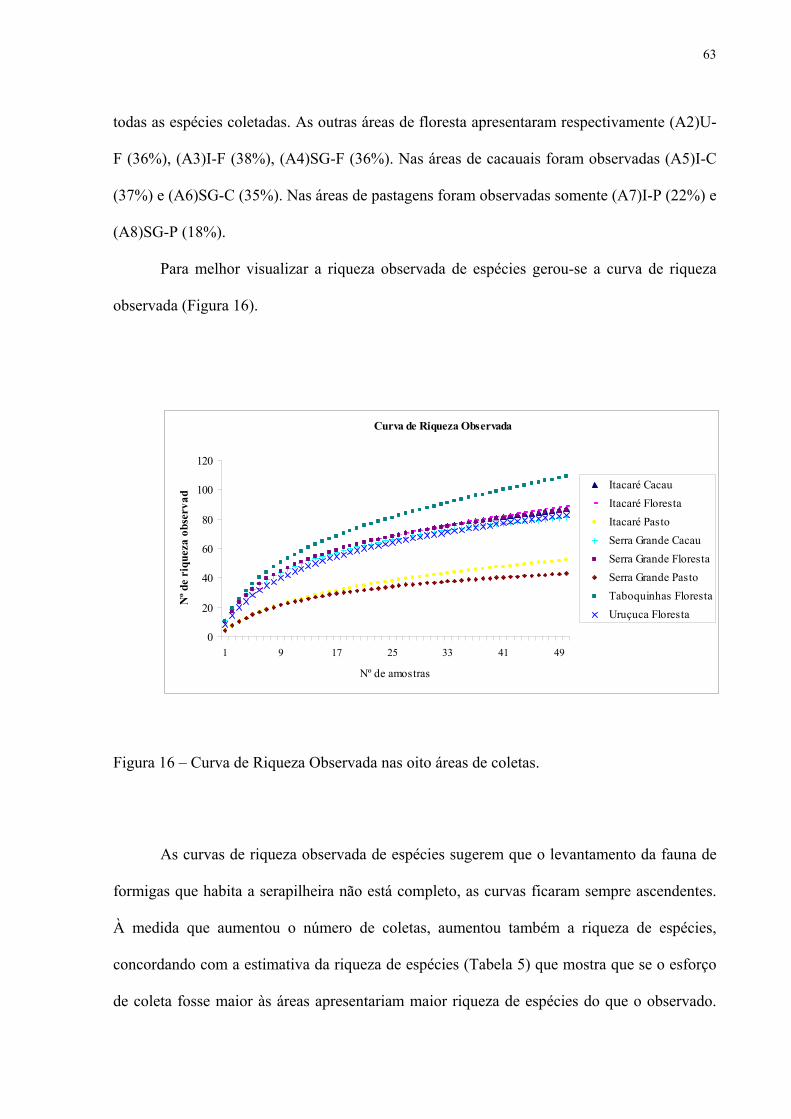
63
todas as espécies coletadas. As outras áreas de floresta apresentaram respectivamente (A2)U-
F (36%), (A3)I-F (38%), (A4)SG-F (36%). Nas áreas de cacauais foram observadas (A5)I-C
(37%) e (A6)SG-C (35%). Nas áreas de pastagens foram observadas somente (A7)I-P (22%) e
(A8)SG-P (18%).
Para melhor visualizar a riqueza observada de espécies gerou-se a curva de riqueza
observada (Figura 16).
Curva de Riqueza Observada
0
20
40
60
80
100
120
1 9 17 25 33 41 49
Nº de amostras
Nº d
e ri
quez
a ob
serv
ad
Itacaré CacauItacaré FlorestaItacaré PastoSerra Grande CacauSerra Grande FlorestaSerra Grande PastoTaboquinhas FlorestaUruçuca Floresta
Figura 16 – Curva de Riqueza Observada nas oito áreas de coletas.
As curvas de riqueza observada de espécies sugerem que o levantamento da fauna de
formigas que habita a serapilheira não está completo, as curvas ficaram sempre ascendentes.
À medida que aumentou o número de coletas, aumentou também a riqueza de espécies,
concordando com a estimativa da riqueza de espécies (Tabela 5) que mostra que se o esforço
de coleta fosse maior às áreas apresentariam maior riqueza de espécies do que o observado.

64
Segundo Delabie (com. pess.), em ambientes de florestas tropicais é muito difícil a curva de
riqueza observada estabilizar. A extrapolação da estimativa de riqueza só é possível se a curva
de espécies observadas resulta de uma comunidade homogênea e relativamente estável.
Em geral, os ambientes de floresta e cacauais apresentaram diversidade de espécies
bem superiores ao número de espécies encontradas nas áreas de pastagens que apresentaram a
metade do número de espécies dos outros dois ambientes. Isso acontece devido ao fato dessas
áreas possuírem mais recursos tróficos do que as áreas de pastagens, disponibilidade de locais
de nidificação e microclimas mais ameno. Os habitats mais complexos criam melhores
oportunidades para um maior número de espécies sobreviverem (OLIVEIRA et al., 1995;
BRUHL et al., 1998). A vegetação arbórea da cabruca apesar da baixa densidade de
indivíduos e forte influência antrópica, apresenta uma alta diversidade de espécies, refletindo
assim a alta diversidade biológica natural na região (SAMBUICHI, 2002).
Belshaw e Bolton (1993) realizaram estudos em áreas de floresta primária, floresta
secundária e cacauais e verificaram que, em relação à composição e riqueza de espécies
nessas áreas, não há diferenças. Segundo estes autores a manutenção de floresta primária
próxima às áreas alteradas funciona como fonte, contribuindo para a recolonização das áreas e
a serapilheira é um habitat semelhante nesses ambientes. Esta afirmação comprova o fato de
que a área de cacaual (A5)I-C apresentou maior diversidade do que duas áreas de floresta
(A2)U-F e (A4)SG-F.
Nas áreas estudadas foram observadas entre 109 e 43 espécies e estima-se que nessas
áreas poderiam ser encontradas entre 209,49 e 60,84 espécies (Tabela 5). De acordo com os
dados de riqueza estimada, nas áreas (A1)T-F e (A3)I-F poderia ser encontrado a mais
respectivamente 100 e 103 espécies. Esta disparidade de valores entre a riqueza observada e
estimada de espécies pode indicar uma provável diversificação de ambientes, aumentando as

65
vantagens de colonização para as espécies exigentes quanto ao local de nidificação e
instalação de outros grupos.
Na área de cacaual (A6)SG-C segundo a riqueza estimada Chao 2, somente 10
espécies a mais poderiam ser encontradas se o esforço de coleta fosse aumentado. Na área de
pastagem (A8)SG-P poderia ser amostradas 18 espécies a mais. Nas áreas de florestas (A2)U-
F e (A4)SG-F poderiam ser encontrado respectivamente 36 e 29 espécies a mais. Segundo
Santos (2004), quanto maior a heterogeneidade na distribuição espacial das espécies, maior
será a distância entre a riqueza observada e a riqueza estimada por Chao 2.
Diante desse resultado pode-se inferir que as áreas de floresta (A1)T-F e (A3)I-F estão
mais conservada e possuem alta riqueza de espécies, pois o número estimado de espécies é o
dobro do que foi observado, mostrando que ambientes mais complexos favorecem alta
diversidade.
Nos ambientes mais complexos, as oportunidades de instalação e sobrevivência das
espécies são maiores, pois a capacidade de suporte do meio favorece a permanência das
mesmas. Outros fatores importantes são as condições de umidade, temperatura e insolação
que estão distribuídas de forma diferente nas áreas com cobertura florestal (neste caso floresta
e cacauais) que das áreas de pastagens (ANDOW, 1991). Isso explica o fato das áreas de
floresta e cacauais apresentarem maior riqueza observada e maior riqueza estimada de
espécies.
Talvez se nas áreas de pastagens que é um ambiente relativamente homogêneo, tivesse
sido adicionado ao winkler outro tipo de armadilha teria apresentado um número maior de
espécies e a curva de riqueza observada teria estabilizado.
A partir da comparação de valores, a área que apresentou o maior índice de
diversidade de Shannon foi a área (A1)T-F com 4,69. As outras áreas de floresta e as áreas de
cacauais variam entre 4,39 a 4,48. Embora a região em estudo tenha passado por um histórico

66
de perturbação, os valores de H’ foram mais elevados nas áreas de florestas e cacauais, isso
ocorre porque a estrutura de vegetação e estratificação de plantações de cabruca são
semelhante, embora menos complexo que, o de florestas naturais (SAATCHI et al. 2001), e o
sistema de cabruca, protege um número grande de espécies de formigas (DELABIE, 2007).
Nas pastagens, os valores de H’ foram mais baixos (3,76 e 3,95).
8.2.1.3 Índice de Similaridade
Analisando o dendograma (Figura 17) observou-se que as áreas (A1)T-F e (A3)I-F e
as áreas (A4)SG-F e (A6)SG-C foram as mais similares respectivamente 44% e 42%. As
primeiras localizam-se dentro do município de Itacaré, distantes 16 km uma da outra e das
áreas de floresta estudadas estas são as mais próximas (figura 11). As áreas (A4)SG-F e
(A6)SG-C localizam-se em Serra Grande (distrito de Uruçuca) são ambientes diferentes
(floresta e cabruca), mas são áreas vizinhas.
Provavelmente as espécies de floresta colonizaram a cabruca corroborando Belshaw e
Bolton (1993) que hipotetizam que a manutenção de floresta próxima às áreas antropizadas
funciona como fonte de espécies, contribuindo para a recolonização.

67
Figura 17 - Similaridade entre as oito áreas de coletas. Índice de Jaccard, APA Costa de Itacaré/Serra Grande, Bahia. Obs.: (A1)T-F = Taboquinhas Floresta; (A2)U-F = Uruçuca Floresta; (A3)I-F = Itacaré Floresta; (A4)SG-F = Serra Grande Floresta; (A5)I-C = Itacaré Cacau; (A6)SG-C = Serra Grande Cacau; (A7)I-P = Itacaré Pastagem e (A8)SG-P = Serra Grande Pastagem.
As localidades de floresta (A2)U-F e de cacaual (A5)I-C não possuem similaridade
direta uma com a outra, porém são mais similares em torno de (31%) e (34%)
respectivamente das outras áreas de floresta e cacauais. A área de cacaual (A5)I-C está
distante da outra área de cacaual (A6)SG-C 14 km e estão distante 10 km e 5 km das áreas de
florestas (A1)T-F e (A3)I-F respectivamente. A área de floresta (A2)U-F é a mais distante de
todas as outras áreas, localiza-se próximo à sede do município de Uruçuca e está fora dos
domínios da APA. Foi inclusa neste estudo para uma comparação já que este fragmento de
floresta está no entorno da APA e o objetivo é encontrar bioindicadores para a região.
Jaccard's Coefficient
(A5)I-C
(A3)I-F
(A1)T-F
(A6)SG-C
(A4)SG-F
(A2)U-F
(A7)I-P
(A8)SG-P
0,04 0,2 0,36 0,52 0,68 0,84 1
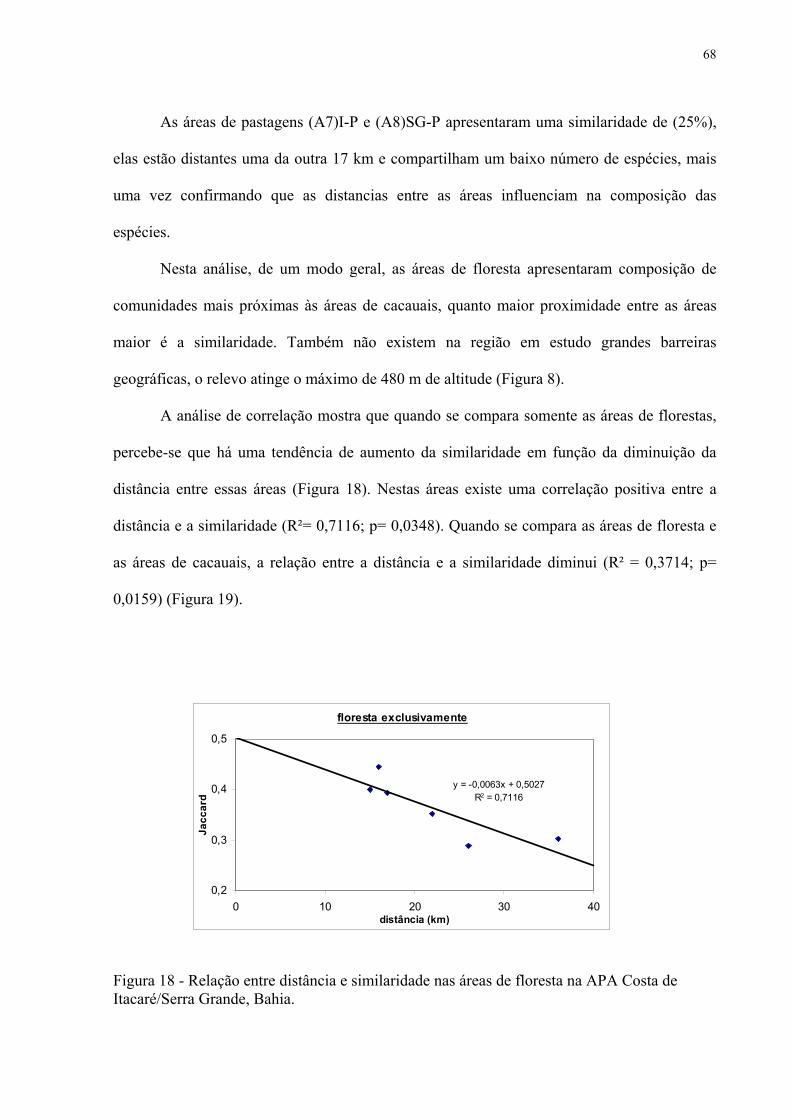
68
As áreas de pastagens (A7)I-P e (A8)SG-P apresentaram uma similaridade de (25%),
elas estão distantes uma da outra 17 km e compartilham um baixo número de espécies, mais
uma vez confirmando que as distancias entre as áreas influenciam na composição das
espécies.
Nesta análise, de um modo geral, as áreas de floresta apresentaram composição de
comunidades mais próximas às áreas de cacauais, quanto maior proximidade entre as áreas
maior é a similaridade. Também não existem na região em estudo grandes barreiras
geográficas, o relevo atinge o máximo de 480 m de altitude (Figura 8).
A análise de correlação mostra que quando se compara somente as áreas de florestas,
percebe-se que há uma tendência de aumento da similaridade em função da diminuição da
distância entre essas áreas (Figura 18). Nestas áreas existe uma correlação positiva entre a
distância e a similaridade (R²= 0,7116; p= 0,0348). Quando se compara as áreas de floresta e
as áreas de cacauais, a relação entre a distância e a similaridade diminui (R² = 0,3714; p=
0,0159) (Figura 19).
floresta exclusivamente
y = -0,0063x + 0,5027R2 = 0,7116
0,2
0,3
0,4
0,5
0 10 20 30 40distância (km)
Jacc
ard
Figura 18 - Relação entre distância e similaridade nas áreas de floresta na APA Costa de Itacaré/Serra Grande, Bahia.

69
floresta + cacauais
y = -0,0034x + 0,4152R2 = 0,3714
0,2
0,3
0,4
0,5
0 10 20 30 40distância (km)
Jacc
ard
Figura 19 - Relação entre distância e similaridade nas áreas de floresta e cacauais na APA Costa de Itacaré/Serra Grande, Bahia.
8.2.1.4 Gêneros bioindicadores
Do total de 55 gêneros coletadas, foram selecionados seis para serem utilizados como
bioindicadores: Apterostigma (8 espécies), Camponotus (6), Ectatomma (3), Pachycondyla
(10), Pheidole (36) e Wasmannia (4), num total de 67 espécies. Foram escolhidos estes
gêneros por serem reconhecidamente aceitos como indicadores de áreas conservadas ou
degradadas (DELABIE, ref. pess.).
Como já era de se esperar, o gênero que apresentou maior número de espécies em
todos ambientes estudados foi Pheidole com 36 espécies, sendo mais presentes nas áreas de
floresta, seguida das áreas de cacauais e pastagens (Figura 20). Pertence à tribo Pheidolini, é
um gênero que apresenta hiperdiversidade e é freqüentemente amostrado em levantamentos
de áreas tropicais (WILSON, 2003). A diversidade elevada de Pheidole numa área é um
indicativo de área conservada. Pheidole é um dos gêneros dessa subfamília de maior

70
relevância, pois apresenta uma ampla distribuição e expansão, o que indica uma área em bom
estado de conservação.
O segundo gênero com maior número de espécies foi Pachycondyla com 10 espécies.
Pertence a tribo Ponerini e é constituído principalmente de espécies que podem forragear a
longas distâncias. As espécies deste gênero são geralmente relevantes predadoras generalistas
muito importantes no manejo dos ecossistemas florestais (MARIANO, 2004). Isso justifica
que o maior número de espécies, tenha sido coletado em áreas com características ambientais
mais conservadas, tais como nas florestas e nos cacauais (Figura 19). Somente uma espécie
desse gênero ocorreu nas pastagens: P. constricta. Essa espécie ocorreu em 6 áreas estudadas,
exceto nas duas áreas de floresta mais conservadas (A1)T-F e (A3)I-F, o que nos leva a
deduzir que pode se tratar de uma espécie mais adaptada a diferentes graus de degradação. As
espécies do gênero Pachycondyla são mais raras no pasto e são características de ambientes
conservados.
O gênero Camponotus, da tribo Camponotini, apresentou seis espécies distribuídas em
todas as áreas, sendo mais freqüente nas áreas de pastagens (Figura 20). O gênero
Camponotus é considerado o segundo gênero mais diversificado em espécies do mundo
(WILSON, 1976). É um gênero cosmopolita, pois possui ampla distribuição geográfica e
domina numericamente os ecossistemas terrestres das regiões tropicais. Nidifica dentro de
cavidades variadas: galhos, troncos, epífitas, montes de matéria orgânica e embaixo de pedras.
Apresenta grande abundância em determinado ambiente e é característico de ambientes
degradados (MARIANO, 2004). As espécies desse gênero são na maioria oportunistas e
generalistas em termo de dieta e local de nidificação (SILVESTRE, 2000).
O gênero Apterostigma pertence à tribo Attini e é composto por espécies cultivadoras
de fungo que vivem na serapilheira. Neste estudo está representado por oito morfo-espécies.
Ocorreu nas áreas de florestas, seguida das áreas de cacauais, não sendo amostradas nas

71
pastagens (Figura 20). Sua abundância pode ser explicada pelo microclima úmido no interior
dos fragmentos que proporciona acúmulo de serapilheira e maior disponibilidade de matéria
orgânica. São encontrados em locais mais fechados da floresta, com um comportamento
críptico (LATTKE, 1997; SILVESTRE, 2000). Quando uma área apresenta uma riqueza
elevada em espécies deste gênero, isto pode indicar riqueza da serapilheira em demais
espécies de formigas (CAMPIOLO et al., 2001). É um gênero característico de ambientes
conservados.
O gênero Wasmannia pertence à tribo Blepharidattini e está representado por quatro
morfo-espécies neste estudo. Foi observado em todos os ambientes, floresta, cacaual e
pastagem, mas não foi observado em uma área de floresta (Figura 20). O gênero Wasmannia
tem como espécie mais comum a W. auropunctata (DELABIE, 1988). Espécie nativa da
região neotropical que possui grande capacidade de adaptação e de multiplicação (ULLOA-
CHACÓN; CHERIX, 1990). Segundo Young citado por Delabie (1988), geralmente dominam
o meio onde se instala influenciando na composição da mirmecofauna local. Wasmannia
auropunctata, conhecida popularmente na região como ‘pixixica’, é um conhecido indicador
de áreas degradadas.
O gênero Ectatomma pertence à tribo Ectatommini e está representado neste estudo
por três espécies. Ocorreu em dois ambientes, florestas e pastagens, ficando ausente nas áreas
de cacauais (Figura 20). Provavelmente isso aconteceu por um artefato da coleta: o Winkler
não é apropriado para amostrar espécies de tamanho como as deste gênero, melhor coletado
com pit-fall, (DELABIE, 1990). O gênero Ectatomma apresenta espécies que possuem ampla
distribuição geográfica e nidificam no solo de diferentes ambientes, inclusive áreas urbanas
(FERNÁNDEZ, 1991). Ectatomma edentatum é uma espécie que ocorre na Mata Atlântica,
principalmente nas áreas mais úmidas perto da costa. Vive em áreas de climas quentes e
úmidos com ou sem estação seca. Ectatomma tuberculatum vive em habitats arbóreos (é a

72
única do gênero que é arborícola). Esta espécie é extremamente comum, principalmente nos
cacauais do sudeste da Bahia e nas capoeiras em toda a região estudada. Ectatomma
brunneum nidifica em áreas de restinga não arbórea, na caatinga e todas as formações vegetais
secundárias dos demais ecossistemas com ocorrência de gramíneas e ciperáceas (DELABIE et
al. no prelo). É um gênero característico de ambientes degradados.
Para comparar a dominância e freqüência dos gêneros bioindicadores, analisou-se a
freqüência desses gêneros por área (Figura 20 e Tabela 6). Verifica-se que os gêneros
Pheidole e Wasmannia tiveram a maior freqüência. Pheidole foi o gênero mais freqüente e
dominante nas áreas de florestas e cacauais, somente não dominou nas pastagens
apresentando freqüência bastante reduzida nestes ambientes. Wasmannia apesar de aparecer
nas áreas de florestas e cacauais, pois são espécies nativas, só dominou nas pastagens, áreas
de ambiente mais homogêneo e bastante degradado. Além disso, o gênero Pheidole possui
recrutamento eficiente, domina os seus recursos alimentares e isso permite excluir seus
competidores (FOWLER, 1993).
A espécie W. auropunctata a mais representativa desse gênero tem populações
maiores em ambientes degradados, mas está presente em qualquer ambiente desta região com
freqüência parecida (DELABIE et al., 2007). De acordo com a figura 20, Wasmannia
realmente só dominou nas pastagens, mas apresenta freqüência semelhante nas áreas de
pastagens e cacauais e menor freqüência nas áreas de florestas. Provavelmente isso ocorre
porque o ambiente de pastagem é mais homogêneo impedindo que espécies mais
especializadas nidifiquem sendo assim dominadas por espécies generalistas.

73
Nº de ocorrência por gêneros bioindicadores
0
20
40
60
80
100
120
140
(A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-P
Áreas de coletas
Nº d
e oc
orrê
ncia
Pheidole
Wasmannia
Pachycondyla
Apterostigma
Ectatomma
Camponotus
Figura 20 – Número de ocorrências dos gêneros bioindicadores. Obs.: (A1)T-F = Taboquinhas Floresta; (A2)U-F = Uruçuca Floresta; (A3)I-F = Itacaré Floresta; (A4)SG-F = Serra Grande Floresta; (A5)I-C = Itacaré Cacau; (A6)SG-C = Serra Grande Cacau; (A7)I-P = Itacaré Pastagem e (A8)SG-P = Serra Grande Pastagem.
Tabela 6 - Representatividade das ocorrências dos gêneros indicadores em relação ao total de ocorrências.
FLORESTA CACAUAL PASTAGEM % (A1)T-F (A2)U-F (A3)I-F (A4)SG-F (A5)I-C (A6)SG-C (A7)I-P (A8)SG-P
Nº de ocorrências
Ectatomma 3,8 - 1,7 - - - 6,1 20,4 30
Camponotus - 5,7 0,8 - 1,0 1,5 6,1 15,6 26
Apterostigma 4,5 7,2 5,2 3,3 - 7,5 - - 32
Wasmannia 8,3 - 17,5 20,1 33,6 28,0 44,4 34,9 190
Pheidole 75,4 65,2 65,7 59,6 46,7 49,2 40,7 27,7 472
Pachycondyla 7,7 21,7 8,7 16,8 18,4 13,6 2,4 1,2 95
Total 155 69 114 119 92 132 81 83 Obs.: (A1)T-F = Taboquinhas Floresta; (A2)U-F = Uruçuca Floresta; (A3)I-F = Itacaré Floresta; (A4)SG-F = Serra Grande Floresta; (A5)I-C = Itacaré Cacau; (A6)SG-C = Serra Grande Cacau; (A7)I-P = Itacaré Pastagem e (A8)SG-P = Serra Grande Pastagem.
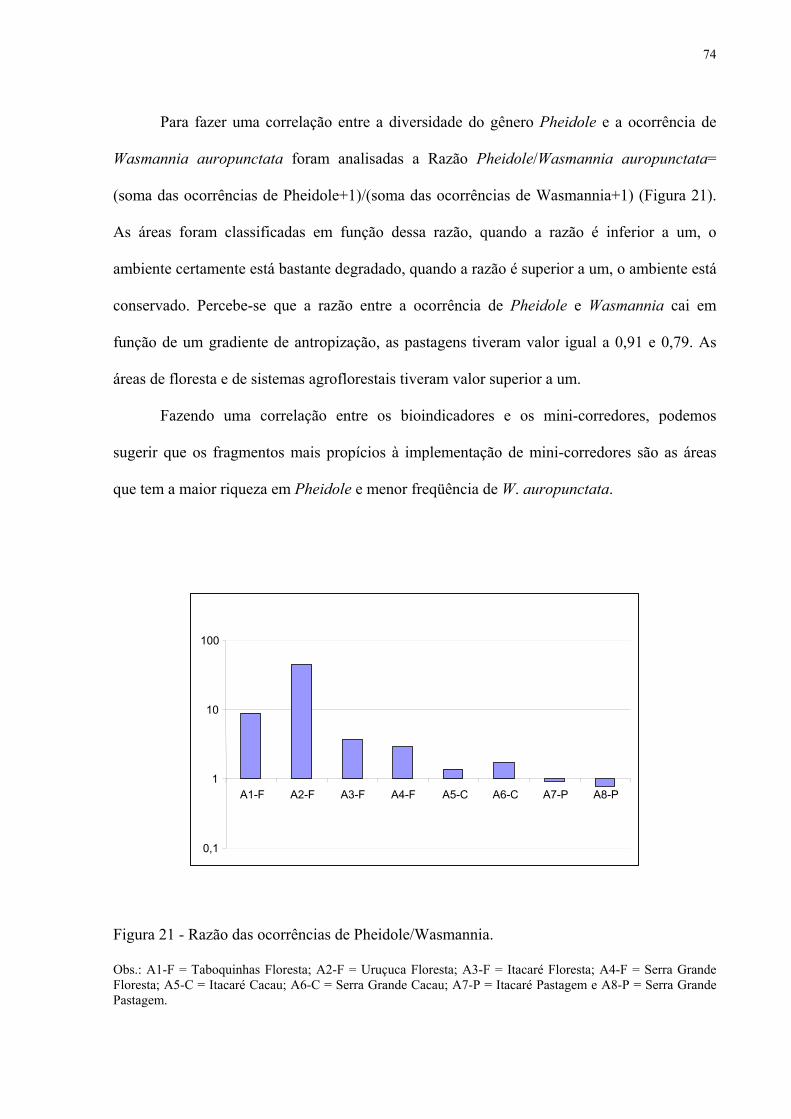
74
Para fazer uma correlação entre a diversidade do gênero Pheidole e a ocorrência de
Wasmannia auropunctata foram analisadas a Razão Pheidole/Wasmannia auropunctata=
(soma das ocorrências de Pheidole+1)/(soma das ocorrências de Wasmannia+1) (Figura 21).
As áreas foram classificadas em função dessa razão, quando a razão é inferior a um, o
ambiente certamente está bastante degradado, quando a razão é superior a um, o ambiente está
conservado. Percebe-se que a razão entre a ocorrência de Pheidole e Wasmannia cai em
função de um gradiente de antropização, as pastagens tiveram valor igual a 0,91 e 0,79. As
áreas de floresta e de sistemas agroflorestais tiveram valor superior a um.
Fazendo uma correlação entre os bioindicadores e os mini-corredores, podemos
sugerir que os fragmentos mais propícios à implementação de mini-corredores são as áreas
que tem a maior riqueza em Pheidole e menor freqüência de W. auropunctata.
Figura 21 - Razão das ocorrências de Pheidole/Wasmannia. Obs.: A1-F = Taboquinhas Floresta; A2-F = Uruçuca Floresta; A3-F = Itacaré Floresta; A4-F = Serra Grande Floresta; A5-C = Itacaré Cacau; A6-C = Serra Grande Cacau; A7-P = Itacaré Pastagem e A8-P = Serra Grande Pastagem.
0,1
1
10
100
A1-F A2-F A3-F A4-F A5-C A6-C A7-P A8-P

75
8.3 Identificação de mini-corredores ecológicos
De acordo com o mapa base de vegetação (IESB, 2002) a área em estudo possui 18
fragmentos de floresta primária dos mais variados tamanhos (de 5 ha 544 ha).
Como o número de fragmentos de floresta primária é pouco, para aplicar a
metodologia de Eastmam (1995) foram utilizados quase todos os fragmentos florestais, exceto
dois fragmentos pequenos de 5,31 ha e 7,17 ha. Através dessa metodologia foi possível
identificar o caminho de menor custo para o estabelecimento de mini-corredores ecológicos
entre os fragmentos, onde o nível 1 é o mais indicado e o nível 1000 é o menos indicado
(Tabela 1).
Com base na corrente de corredor linear foram criados 10 mini-corredores. A corrente
de corredor linear utiliza o termo “corredor” em relação a faixas de vegetação ligando
fragmentos de habitat nativo. Para uma melhor análise os corredores foram analisados
separadamente (Figuras 22 a 31).
A resolução do CONAMA nº 9/96 fixa a largura para os corredores que deverá ser de
10% do seu comprimento total, sendo que a largura mínima será de 100 m. Os mini-
corredores propostos neste estudo possuem entre 654 m a 6,4 km de comprimento. De acordo
com Forman (1995), o nível de resistência da paisagem ao movimento de organismos é
relativo, em situações de conectividade reduzida, as aves podem movimentar-se entre os
fragmentos, pequenos mamíferos demonstram padrões intermediários e grandes mamíferos
encontram maior resistência. Então a largura do corredor vai depender do objetivo do
corredor, em particular da população de organismos que se pretende beneficiar com a
conexão.

76
Mini-corredor ecológico 1
O mini-corredor 1 possui 1,4 km de comprimento e 140 m de largura. Este corredor
está conectando um fragmento de floresta primária de 544,15 ha a outro de 61,89 ha. A matriz
em que o corredor está inserido inclui uma larga parte de floresta secundária em estágio
avançado de regeneração/cabruca e uma pequena parte de vegetação agrícola/pastagens. No
local onde o corredor é interrompido por vegetação de tipo agrícola ou pastagens, destacada
no mapa por um círculo vermelho, o ideal é que seja realizado um manejo adequado para que
a área seja restaurada e assim facilite a dispersão das espécies.
Mini-corredor ecológico 2
O mini-corredor 2 possui 3,5 km de comprimento e 350 m de largura. Está conectando
um fragmento de floresta primária de 61,89 ha a outro fragmento de 210,25 ha. Este corredor
passa na sua maior parte por uma matriz de floresta secundária em estágio avançado de
regeneração/cabruca, uma pequena parte em floresta secundária em estágio médio de
regeneração, e chegando ao fragmento final, passa por uma pequena área de floresta
secundária em estágio inicial de regeneração. Para implantação deste corredor, a vegetação
secundária em estágio inicial de regeneração (capoeira), por onde o corredor passa, deve ser
adequadamente manejada.

77
Figura 22 - Proposta do mini-corredor 1 na APA Itacaré/Serra Grande, Bahia.

78
Figura 23 - Proposta do mini-corredor 2 na APA Itacaré/Serra Grande, Bahia.

79
Mini-corredor ecológico 3
O mini-corredor 3 possui 3,5 km de comprimento e 350 m de largura. Está conectando
um fragmento de floresta primária de 22,65 ha a outro fragmento de 55,41 ha. A matriz a qual
este corredor está inserido passa por vegetação secundária em estágio avançado de
regeneração/cabruca e estágio inicial de regeneração. A grande dificuldade para
implementação deste corredor é o fato dele atravessar uma grande parte de vegetação agrícola
ou pastagens. Deve ser realizado um manejo adequado nas áreas de vegetação secundária em
estágio inicial de regeneração e nas áreas de vegetação para facilitar a dispersão das espécies.
Mini-corredor ecológico 4
O mini-corredor 4 possui 2,7 km de comprimento e 270 m de largura. Está conectando
um fragmento de floresta primária de 50,14 ha a outro fragmento de 22,65 ha. Este corredor
passa na sua maior parte por uma vegetação secundária em estágio avançado de
regeneração/cabruca. Para implementação deste corredor, por conseqüente, não será
necessário reflorestar.
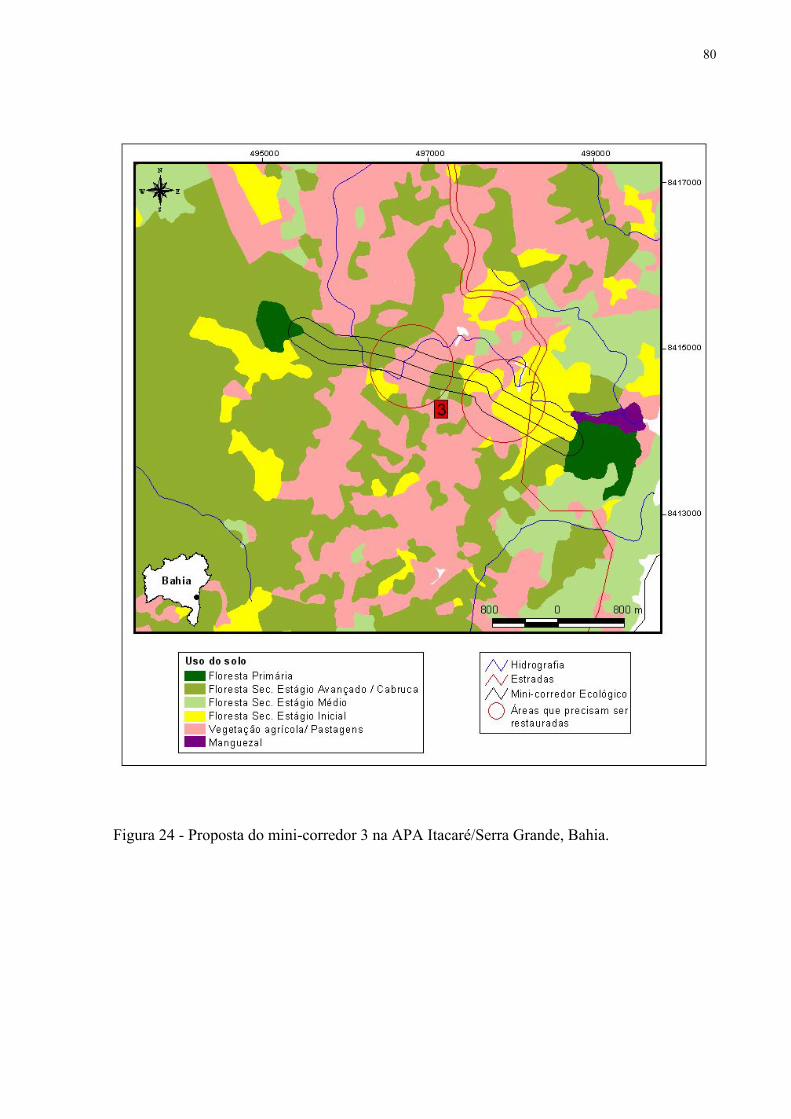
80
Figura 24 - Proposta do mini-corredor 3 na APA Itacaré/Serra Grande, Bahia.

81
Figura 25 - Proposta do mini-corredor 4 na APA Itacaré/Serra Grande, Bahia.

82
Mini-corredor ecológico 5
O mini-corredor 5 possui 1,3 km de comprimento e 130 m de largura. Está conectando
um fragmento de floresta primária de 50,14 ha a outro fragmento de 126,75 ha. A matriz em
que o corredor está inserido é de floresta secundária em estágio avançado de regeneração e
cabruca. Para implementação deste corredor não será necessário reflorestar. Deverá ser
previsto uma estratégia particular quando o corredor cruza a estrada que liga o município de
Itacaré a Taboquinhas.
Mini-corredor ecológico 6
O mini-corredor 6 possuiu 1,8 km de comprimento e 180 m de largura. Está
conectando um fragmento de floresta primária de 73,05 ha a outro fragmento de 50,14 ha. A
matriz em que este corredor está inserido é na sua maior parte de floresta secundária em
estágio avançado de regeneração/cabruca, e uma pequena parte de vegetação secundária em
estágio médio de regeneração.
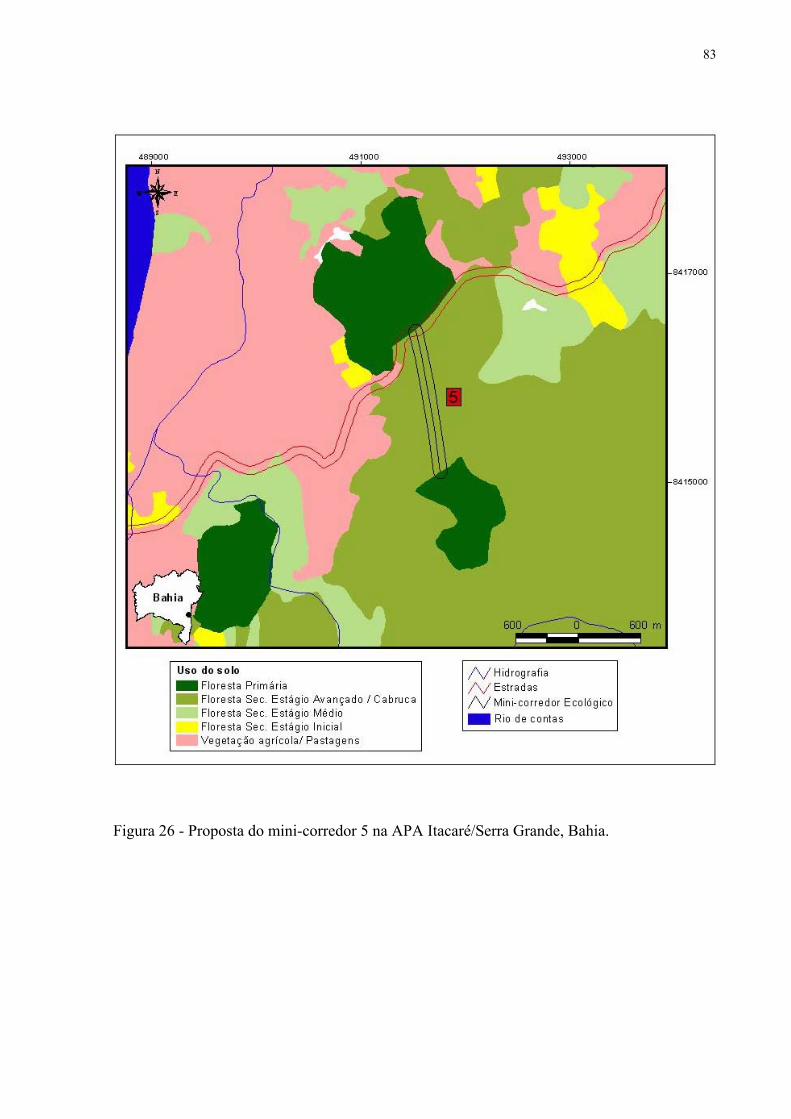
83
Figura 26 - Proposta do mini-corredor 5 na APA Itacaré/Serra Grande, Bahia.

84
Figura 27 - Proposta do mini-corredor 6 na APA Itacaré/Serra Grande, Bahia.

85
Mini-corredor ecológico 7
O mini-corredor 7 possui 3,4 km de comprimento e 340 m de largura. Está conectando
um fragmento de floresta primária de 126,75 ha a outro fragmento de 149,82 ha. A matriz em
que este corredor está inserido é na sua maior parte de floresta secundária em estágio
avançado de regeneração/cabruca. Também passa por floresta secundária em estágio médio e
inicial de regeneração e perto de brejo. Os círculos vermelhos no mapa indicam as áreas que
necessitam de maiores cuidados, como reflorestamento, manejo, entre outros.
Mini-corredor ecológico 8
O mini-corredor 8 possui 654 m de comprimento e 100 m de largura. É o menor
corredor proposto para a APA. Está conectando um fragmento de floresta primária de 14,95
ha a outro fragmento de 28,95 ha. A matriz em que o corredor está inserido é constituída
praticamente de floresta secundária em estágio avançado de regeneração/cabruca e uma
pequena parte de floresta secundária em estágio médio de regeneração. Este corredor está bem
próximo à área urbana do município de Itacaré e próximo da costa.
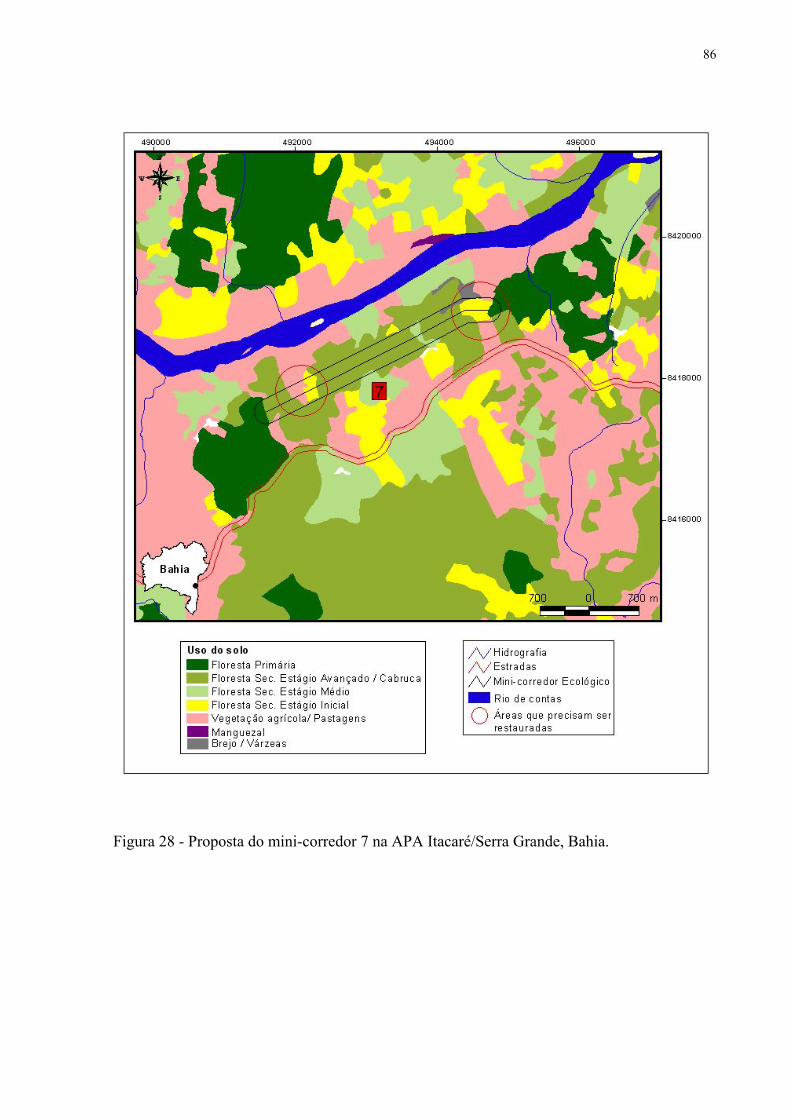
86
Figura 28 - Proposta do mini-corredor 7 na APA Itacaré/Serra Grande, Bahia.

87
Figura 29 - Proposta do mini-corredor 8 na APA Itacaré/Serra Grande, Bahia.

88
Mini-corredor ecológico 9
O mini-corredor 9 possui 1,7 km de comprimento e 170 m de largura. Está conectando
um fragmento de floresta primária de 126,75 ha a outro fragmento de 516,16 ha. A matriz em
que o corredor está inserido é na sua maior parte constituída por vegetação agrícola ou
pastagens e de floresta secundária em estágio avançado de regeneração/cabruca. Este corredor
corta o Rio de Contas ligando os fragmentos que estão ao sul do rio aos fragmentos que estão
ao norte. Este corredor está sendo sugerido para facilitar a dispersão de médios e grandes
mamíferos que conseguem atravessar o rio. A complexidade deste corredor é que ele passa
por uma grande área de vegetação agrícola.
Mini-corredor ecológico 10
O mini-corredor 10 possui 6,4 km de comprimento e 640 de largura. Este corredor está
conectando três fragmentos de floresta primária de tamanhos variados 516,16 ha, 79,44 ha e
284,34 ha. A matriz em que o corredor está inserido é na sua maior parte constituída de
floresta secundária em estágio avançado de regeneração/cabruca e estágio médio de
regeneração. Este corredor passa também pela floresta secundária em estágio inicial de
regeneração e pela vegetação agrícola/pastagens. A sugestão é que seja realizado um manejo
apropriado nas áreas de floresta secundária em estágio médio e inicial de regeneração
principalmente na área onde está demarcada pelo círculo vermelho. Dois fragmentos menores

89
que estão a oeste do maior fragmento não foram conectados pelo corredor por estarem muito
próximos podendo funcionar como stepping stones para a fauna.
Figura 30 - Proposta do mini-corredor 9 na APA Itacaré/Serra Grande, Bahia.
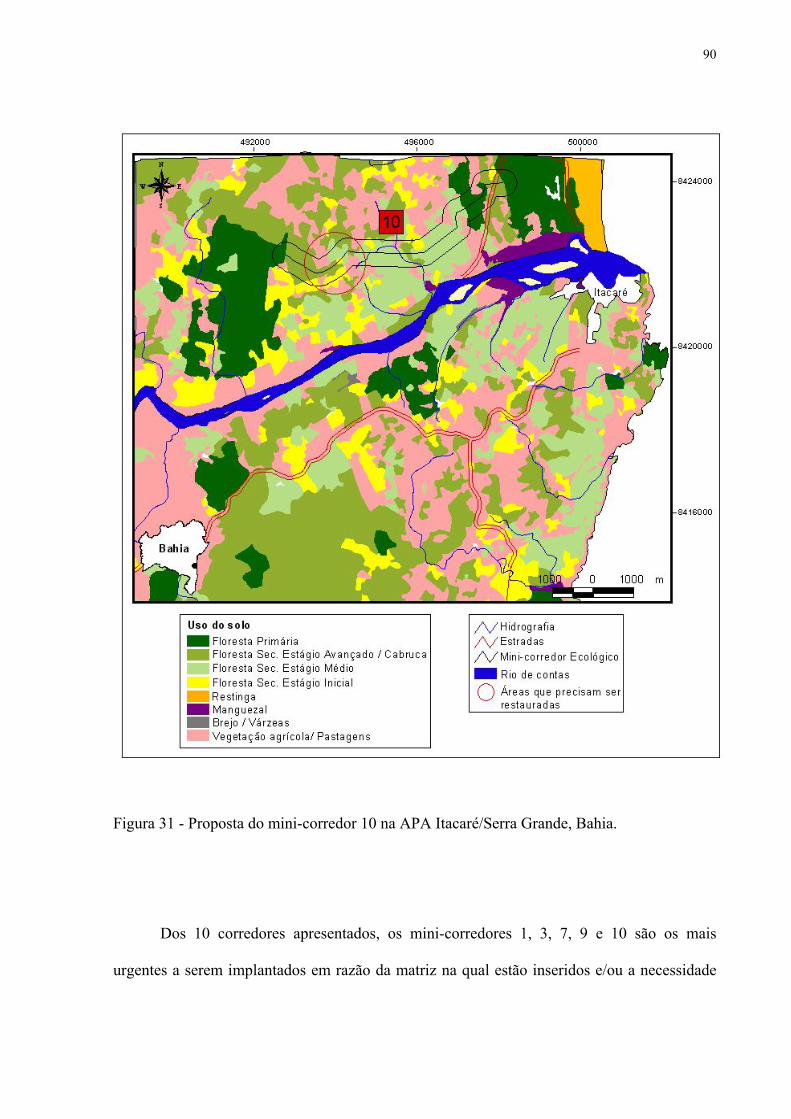
90
Figura 31 - Proposta do mini-corredor 10 na APA Itacaré/Serra Grande, Bahia.
Dos 10 corredores apresentados, os mini-corredores 1, 3, 7, 9 e 10 são os mais
urgentes a serem implantados em razão da matriz na qual estão inseridos e/ou a necessidade

91
de reflorestamento. Para reflorestamento, podem ser desenvolvidos projetos em parcerias com
empresas privadas utilizando espécies de interesse econômico e espécies nativas.
A ausência de zonas de contato físico entre fragmentos não significa que estes sejam
impermeáveis ao intercâmbio. Fragmentos isolados, porém próximos, podem proporcionar
vias de acesso, funcionando como stepping stones (trapolins) (FORMAN, 1995). Onde a
conexão física dos corredores não for possível, Arruda (2004) sugere que sejam plantados
bosques de árvores frutíferas entre os fragmentos para aumentar a heterogeneidade na
paisagem estimulando movimentos saltitantes de dispersão para muitas espécies.
A matriz na qual os fragmentos estão inseridos é de fundamental importância para
diminuir os custos de implementação dos corredores. No caso da área em estudo, a
implementação física desses mini-corredores não teria um custo financeiro muito elevado,
pois, apesar de só ter 18 fragmentos de floresta primária, totalizando 2.326,12 ha (5,8%), a
principal matriz é de floresta secundária em estágio avançado de regeneração/cabruca
totalizando 14.403,79 ha (35,84%), evitando assim a necessidade de criação de grandes áreas
artificiais de floresta para interligar os fragmentos. Apesar da floresta dominante na área da
APA ser praticamente toda secundária, de onde foi extraída grande parte das madeiras nativas
(BRASIL-MMA, 2004), esta matriz é bem melhor do que se fosse constituída
majoritariamente por agricultura ou pastagens. A matriz que circunda o fragmento é
fundamental, pois os sistemas ecológicos estão abertos e sujeitos a troca de energia com a
matriz que os cerca (ARRUDA, 2004).
A proposta de criação dos corredores sugerida pelo SIG mostra o caminho de menor
custo para unir os fragmentos, mas o desenho definitivo do corredor deve ser bem analisado
para determinar a melhor forma de desenhá-los. Neste caso, a criação dos corredores tem por
objetivo facilitar o movimento de organismos pelo meio de “ligações” entre fragmentos
próximos, o caminho ideal sendo aquele que passa por paisagens menos fragmentadas ou que

92
interliga o maior número possível de fragmentos. A proposta de desenho dos corredores
apresentada resulta de uma análise científica, mas deve ainda ser discutida no conselho gestor
da APA, pelo governo local e proprietários de terras onde os corredores serão criados. É
preciso convencer os proprietários que a criação de corredor não significa mais uma restrição
de uso do solo nas suas propriedades, mas que, pelo contrário, terão suas terras melhoradas e
consequentemente valorizadas. Implantados estrategicamente, os corredores podem mudar
fundamentalmente o papel ecológico das áreas protegidas (ARRUDA, 2004) e a relação das
populações com o meio ambiente.
A proposta apresentada aqui aponta que as duas correntes que definem o conceito de
corredores ecológicos (corredor Linear e Corredor de Paisagem) são atreladas. A área em
estudo pode ser considerada como um corredor natural, pois os fragmentos de floresta
primária analisados neste estudo estão imersos numa matriz de floresta secundária em estágio
avançado de regeneração/cabruca que é bastante permeável. Segundo Metzger (1995), o
isolamento depende não só da distância entre os fragmentos de mesmo tipo, mas também das
características da matriz. Neste caso, a estratégia é garantir a conectividade entre as áreas por
meio de ações que visem à manutenção e conservação. No local onde a conexão está
interrompida, a estratégia é assegurar a proteção dos remanescentes florestais mais
importantes e incrementar o grau de ligação entre os fragmentos. Para atingir o objetivo da
criação de corredores ecológicos, é necessária a introdução de estratégias adequadas de uso da
terra, assim como a restauração de alguns trechos degradados nas áreas que une os
fragmentos. Nessas áreas-chave podem ser criadas faixas de vegetação para completar a
conexão entre fragmentos de floresta primária e assim contribuir para otimizar os benefícios
ecológicos das florestas e conservarem os recursos genéticos.
As faixas de corredores indicadas com o auxílio do SIG podem também ser
consideradas como a zona núcleo do corredor. Após análise, algumas faixas foram reajustadas

93
levando em consideração a matriz mais adequada para desenhar o corredor. A proposta final
para criação de corredores na área em estudo é apresentada na Figura 32. Os círculos
vermelhos no mapa indicam as áreas que precisam com urgência de um investimento para
serem restauradas e apropriadamente manejadas, pois se tratam de locais onde domina a
vegetação de tipo agrícola ou pastagens e vegetação secundária em estágio inicial de
regeneração (capoeira). Os limites dos mini-corredores não são definitivos, eles são
reajustáveis em função de futuros estudos científicos.
Precisa deixar claro que somente o estabelecimento de corredores não assegura que
fragmentos cumpram seu papel na preservação das espécies, mas estes elevam as
probabilidades de sobrevivência do conjunto de populações outrora isoladas de cada espécies.
Para completar a estratégia de criação do corredor, deve ainda ser feito: restauração e manejo
de algumas áreas florestais situadas estrategicamente; pesquisas biológicas e sócio-
econômicas; incentivo à agricultura e pecuária familiar que utilize técnicas conservacionistas
e harmonize os objetivos econômicos com os ambientais; combate à caça ilegal; criação de
novas RPPN’s; estabelecimento de “zonas-tampão”, entre outras.
A inserção da comunidade local em todas as fases de implantação dos corredores para
garantir a sustentabilidade em longo prazo dessa estratégia é fundamental. O corredor
ecológico é mais do que uma simples conexão entre fragmentos, o envolvimento da
comunidade local é decisivo para o êxito do projeto (BRASIL-MMA, 2006). A APA
Itacaré/Serra Grande, como outras unidades de conservação no Brasil, foi criada sem o
conhecimento da população local e isso gerou inicialmente um problema de aceitação e
entendimento por parte dela. Por isso, a fim que a estratégia de criação de corredores seja
aceita e entendida pela população, se faz necessário um trabalho prévio e aprofundado de
informação. Sem a mobilização e o efetivo envolvimento da população local, o corredor

94
ecológico terá baixíssima chance de sucesso e permanecerá apenas no papel (GALINKIN et
al., 2004).
Vale lembrar que, apesar dos corredores serem considerados como unidades de
conservação, não são tratados como unidades políticas ou administrativas, e sim como
unidades geopolíticas ambientais, prioritárias à conservação por apresentar atributos
ecológicos, biológicos, econômicos e por isso devem ser utilizados e conservados de forma
extremamente planejada. A lei do SNUC não determina que sejam criados corredores
ecológicos nas APAs, esta criação fica a critério dos gestores da unidade.
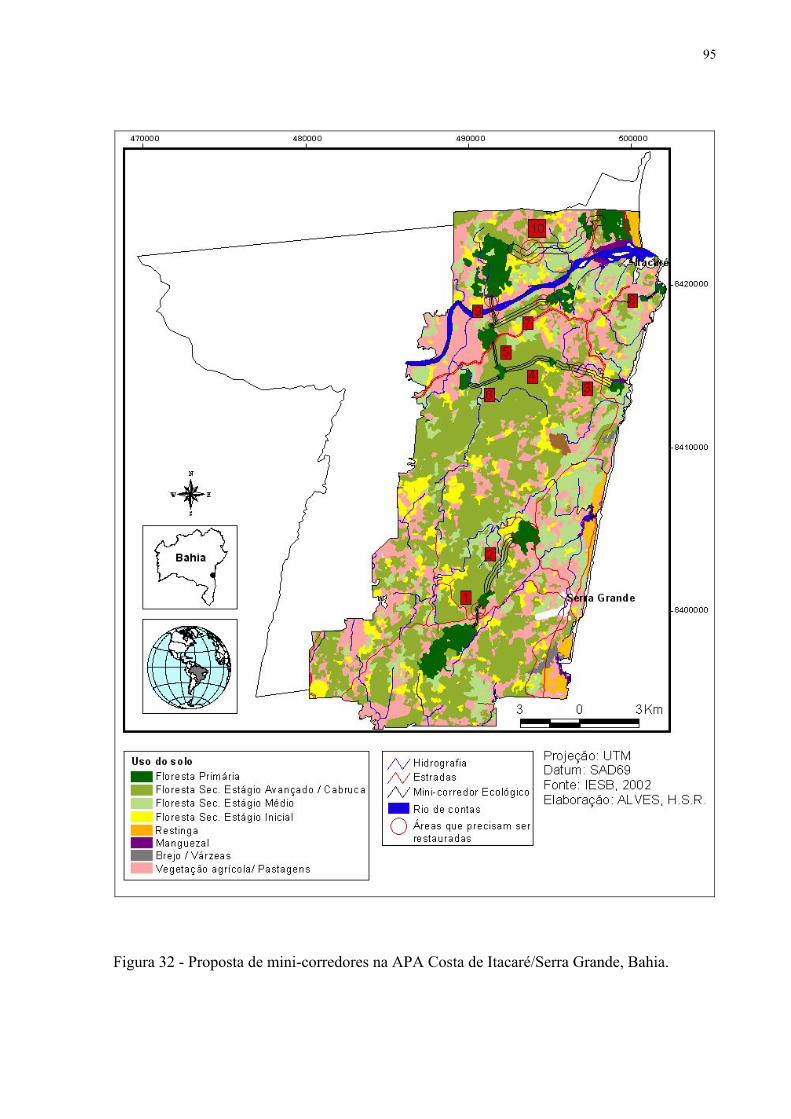
95
Figura 32 - Proposta de mini-corredores na APA Costa de Itacaré/Serra Grande, Bahia.

96
9 CONSIDERAÇÕES FINAIS
- Os resultados obtidos evidenciam que as manchas de florestas primárias podem estar
sofrendo com problemas de fragmentação como por exemplo, efeito de borda, pois observou-
se na sua vizinhança, floresta secundária em diferentes estágios de regeneração, pastagens,
rodovias, estradas e cultivos agrícolas;
- Sessenta e nove por cento de toda a área em estudo é considerada Área de Preservação
Permanente de acordo com a Lei Federal Nº 4.771;
- As áreas de preservação permanente em uso antrópico, devem ser detalhadamente estudadas
para que possam subsidiar a elaboração de projetos de recuperação de áreas degradadas;
- Os inventários de insetos nos ecossistemas são importantes a fim de que se identifiquem
grupos de bioindicadores potencias, pois à medida que ocorre o equilíbrio ambiental os
insetos respondem também em diversidade e densidade, cumprindo a sua função indicadora;
- As áreas de florestas e cacauais apresentaram proporcionalmente maior diversidade de
espécies de formigas do que as áreas de pastagens, isso significa que estas áreas permitem
manter uma melhor fração da biodiversidade nativa;
- As áreas de cabrucas apresentam uma certa similaridade com as áreas de florestas;
- Existe uma correlação positiva entre similaridade e distâncias entre as áreas de floresta
primária;

97
- Dos seis gêneros selecionados como bioindicadores, dois (Pheidole e wasmannia) foram
representativos e confirmam a hipótese de que a diversidade de Pheidole é indicativa de áreas
conservadas e a ocorrência de Wasmannia auropunctata é indicativa de áreas degradadas;
- Os fragmentos mais propícios à implementação de mini-corredores são as áreas que tem a
maior riqueza em Pheidole e menor freqüência de W. auropunctata.
- A principal matriz da área em estudo é a Floresta Secundária em Estágio Avançado de
Regeneração/Cabruca (35,84%);
- Foram identificados 10 mini-corredores na área em estudo, a fim de dar suporte à escolha
das áreas propícias à implementação desses corredores, recomendar políticas públicas e
orientar ONG’s que irão beneficiar a melhoria da qualidade ambiental;
- Este estudo científico só realizou a primeira fase de implementação dos corredores, falta
ainda convencer conselho gestor da APA, sociedade civil local e ONG’s e em seguida
implementar o corredor criando projetos com ênfase na sustentabilidade;
- Se faz necessário identificar as áreas de floresta do lado oeste da APA que são prioritárias
para formação de mini-corredores (áreas que ficaram de fora deste estudo), e incluí-las na
proposta de criação de corredores ecológicos;
- Serão necessárias visitas in loco nas áreas consideradas ‘chave’ para a conexão dos
fragmentos de floresta primária;
- A metodologia utilizando o SIG mostrou-se ser eficiente, pois interligou os fragmentos de
floresta primária considerando os pesos de menores custos sendo estes na sua maioria a
floresta secundária em estágio avançado de regeneração;
- A restauração de Áreas de Preservação Permanente e criação de Reservas Legais podem
contribuir para a conexão entre os fragmentos florestais;
- O desenho de corredores ecológicos deve ser incorporado no Zoneamento Ecológico-
Econômico da APA;

98
- A APA Costa de Itacaré-Serra Grande pode ser considerada neste estudo como um corredor
ecológico natural que deve ser monitorado para que sua função de proteger a biodiversidade
possa de fato servir a este propósito.

99
10 REFERÊNCIAS BIBLIOGRÁFICAS
ACCACIO, G. de M. et al. Ferramentas biológicas para avaliação e monitoramento de habitats naturais fragmentados. In: Fragmentação de ecossistemas: Causas, Efeitos sobre a Biodiversidade e Recomendações de Políticas Públicas. Denise Marçal Rambaldi, Daniela América Suárez de Oliveira (orgs.), Brasília: MMA/SBF, 2003.
AGOSTI, D.; MAJER, J.D.; ALONSO, L.E.; SCHULTZ, T.R. Ants: standard methods for measuring and monitoring biodiversity. Smithsonian Institution Press, Washington, 2000.
ALBERTONI, E. F.; ESTEVES, F. A. Jurubatiba, uma restinga peculiar. Revista Ciência Hoje. Rio de Janeiro, abril de 1999.
ANDERSEN, A. N. Functional groups and patterns of organization in North American ant communities: a comparison with Australia. Journal of Biogeography, 1997. 24: 433-460.
ANDERSEN, A. N; SPARLING, G. P. Ants as Indicators of Restoration Success: Relationship with Soil Microbial Biomass in the Australian Seasonal Tropics. Restoration Ecology. Vol. 5, Nº 2, 1997. pp. 109-114.
ANDERSON, A. B.; JENKINS, C.; MAY, P. H. Cordão de Mata: Planejamento e implementação de um corredor biológico na Mata Atlântica do Rio de Janeiro. In: Fundação CIDE (Centro de Informações e Dados do RJ). (Org.). Índice de Qualidade dos Municípios (IQM Verde). 2 ed. Rio de Janeiro: Fundação CIDE (Governo do Estado do Rio de Janeiro), 2003. p. 133-140.
ANDOW, D. A. Vegetacional diversity and arthropod population responses. Annual Review of Entomology, Palo Alto, v. 36, 1991. p. 561-586.
ANDRADE, M. P. Ilhéus: passado e presente. Salvador, BDA-Bahia, 1996.

100
ANTONINI, Y. et al. Insetos. In: Fragmentação de ecossistemas: Causas, Efeitos sobre a Biodiversidade e Recomendações de Políticas Públicas. Denise Marçal Rambaldi, Daniela América Suárez de Oliveira (orgs.) Brasília: MMA/SBF, 2003.
ARAÚJO, M. Conservação da Mata Atlântica na Região Cacaueira da Bahia. In: Anais do V Seminário da reserva da Biosfera da Mata Atlântica. AL, 10-11 de Nov, 1997. 16 p.
ARAÚJO, S. D. A.; LACERDA, L. D. A natureza das restingas. Revista Ciência Hoje. Eco Brasil, Edição Especial, Rio de Janeiro, 1992.
ARRUDA, M. B. Corredores Ecológicos no Brasil-Gestão Integrada de ecossistemas. In: Corredores ecológicos: Uma abordagem integradora de ecossistemas no Brasil. IBAMA, Brasília-DF. p. 11-46, 2004.
AYRES, J. M. et al. Os Corredores Ecológicos das Florestas Tropicais do Brasil. Belém, PA: Sociedade Civil Mamirauá – SCM, 2005.
BALDISSERA, R.; GANADE, G. Predação de sementes ao longo de uma borda de Floresta Ombrófila Mista e pastagem. Acta Botânica Brasileira. 19(1): 161-165. 2005.
BELSHAW, R.; BOLTON, B. The effect of forest disturbance on the leaf litter ant fauna in Ghana. Biodiversity and Conservation. v. 2, p. 656-666, 1993.
BESTELMEYER, B.T. et al. Field techniques for the study of ground-dwelling ants. In: Agosti, D.; Majer, J.D.; Tennant de Alonso, L.; Schultz, T. (Eds.). Measuring and monitoring biological diversity: standard methods for ground living ants. Smithsonian Institution Press, Washington, p.122-144, 2000.
BOLTON, B. Synopysis and classificatin of Formicidae. Memoirs of the America Entomological Institute. v. 71, p.1-370, 2003.
BOLTON, B. Taxonomic and zoogeographical census of the extant ant taxa (Hymenoptera: Formicidae). Journal of Natural History. v. 29, p. 1037-1056, 1995.
BRANDÃO, C. R. F. Reino Animalia: Formicidae. In: JOLY, C. A.; CANCELLO, E, M. (Ed.). Invertebrados terrestres. São Paulo: FAPESB, p. 58-63, 1999.

101
BRASIL. Resolução CONAMA 09 de 1996. Define corredores entre remanescentes e estabelece parâmetros e procedimentos para a sua identificação e proteção. Brasília, DF: Publicação DOU: 07/11/1996.
______. Resolução CONAMA 05 de 1994. Define vegetação primária e os estágios sucessionais de Mata Atlântica no Estado da Bahia. Brasília, DF: Publicação DOU: 04/05/1994.
______. Resolução CONAMA 11 de 1987. Dispõe sobre a declaração da Unidades de Conservação, várias categorias e sítios ecológicos de relevância cultural. Brasília, DF: Publicação DOU: 18/03/1988.
BRASIL. Constituição da República Federativa do Brasil. Brasília: Congresso Nacional, 1988.
BRASIL. Instituto Brasileiro de Meio Ambiente dos Recursos Naturais Renováveis; Fundo Mundial para Conservação da Vida Silvestre - WWF. Ecorregiões e estudo de representatividade ecológica nos biomas brasileiros. In: ANAIS do SBPC. Brasília, 2000.
BRASIL. Sistema Nacional de Unidade de Conservação. Lei 9.985. 2000.
______. Ministério do Meio Ambiente. O corredor central da Mata Atlântica: uma nova escala de conservação da biodiversidade. Brasília: Ministério do Meio Ambiente; Conservação Internacional, 2006.
______. Ministério do Meio Ambiente. Corredor Central da Mata Atlântica: Ecologia e Conservação. Projeto Corredores Ecológicos. Disponível em: http://www.corredores.org.br. Acesso em: 9 jun. 2006.
______. Ministério do Meio Ambiente. Revisão do Zoneamento Ecológico-Econômico: APA Costa de Itacaré-Serra Grande. Instituto de Estudos Sócio-Ambientais do Sul da Bahia. 2004.
______. Ministério do Meio Ambiente, dos Recursos Hídricos e da Amazônia Legal. Programa Piloto para Proteção das Florestas Tropicais do Brasil (PPG7). Projeto Corredores Ecológicos. Brasília: Secretaria de Coordenação da Amazônia. Instituto Brasileiro de Meio Ambiente e Recursos Naturais Renováveis, 1998.
______. Ministério do Meio Ambiente. Zoneamento Ecológico-econômico: APA Costa de Itacaré-Serra Grande. Instituto de Estudos Sócio-Ambientais do Sul da Bahia. 1996.

102
BRASÍLIA. DECRETO Nº 750, de 10.02.1993. Publicado no DOU de 11.02.93.
BRASÍLIA. LEI Nº 11.428, de 22.12.2006. Publicada no DOU de 26.12.2006. Retificado no DOU de 9.1.2007.
______. LEI Nº 4.771, de 15.09.1965. Publicado no DOU de 16.09.1965.
BROWN, K. S. Insetos como rápidos e sensíveis indicadores de uso sustentável de recursos naturais. In: MARTOS, H. L.; MAIA, N. B. Indicadores ambientais. 1º ed. Sorocaba: s.n., p. 143-151, 1997.
BRUHL, C. A.; MOHAMED, M.; LINSENMAISR, K. E. Altitudinal distribution of leaf litter ants along a transect in primary forest in Mount Kinabalu, Sabah, Maysia. Journal of Tropical Ecology, Cambridge, v. 15, n. 3, p. 265-277, 1998.
BUREL, F.; BAUDRY, J. Ecologia del paisaje: conceptos, métodos y aplicaciones. Madrid: Mundi-Prensa, 2002.
CABRAL, N. R. A. J.; SOUZA, M. P. de. Área de Proteção Ambiental: planejamento e gestão de paisagens protegidas. São Carlos: RiMa, 2002.
CALIJURI, M. L.; RÖHM, A. S. Sistemas de Informações Geográficas. Universidade Federal de Viçosa. Imprensa Universitária, Viçosa, Minas Gerais, 1995.
CÂMARA, I. G. Plano de ação para a Mata Atlântica. São Paulo: Interação Ltda, 1991.
CÂMERA, G.; MEDEIROS, J. S. de. Princípios Básicos em Geoprocessamento. In: Sistema de Informação Geográfica: Aplicações na agricultura. Eduardo Delgado Assad; Edson Eyjisano (Edts). 2 ed., rev. e ampl. – Brasília: Embrapa – SPI/ Embrapa- CPAC, 1998.
CAMPIOLO, S.; et al. Relação riqueza genérica e específica: um instrumento de apoio para inventários rápidos em áreas de Mata Atlântica. In: Encontro de Mirmecologia, 15, 2001, Londrina. Anais… Londrina: IAPAR, p. 301-303, 2001.
CARDOSO-LEITE, E., COVRE, T. B., OMETTO, R. G., CAVALCANTI, D. C., PAGANI, M. I. Fitossociologia e caracterização sucessional de um fragmento de mata ciliar, em RioClaro/SP, como subsídio à recuperação da área. Revista Instituto Florestal, São Paulo, v. 16, n. 1,p.31-41,jun. 2004.

103
CARVALHO, M. S. (Org.). Conceitos básicos de Sistemas de Informações Geográficas e Cartográficas aplicados à Saúde. Brasília: Organização Panamericana da Saúde/ Ministério da Saúde, 2000.
CERQUEIRA, R. et al. Fragmentação: alguns conceitos. In: Fragmentação de ecossistemas: Causas, Efeitos sobre a Biodiversidade e Recomendações de Políticas Públicas. Denise Marçal Rambaldi, Daniela América Suárez de Oliveira (orgs.) Brasília: MMA/SBF, 2003.
COIMBRA-FILHO, A. F.; CÂMARA, I. de G. Os Limites Originais do Bioma Mata Atlântica na Região Nordeste do Brasil. Fundação Brasileira para a Conservação da Natureza – FBCN, 1996.
COLWELL, R. K. Estimates: Statistical estimation of species richness and shared apecies from sample. 1997. User’s guide ande application published at: http://purl.oclc.org/estimates.
COMPANHIA DE DESENVOLVIMENTO E AÇÃO REGIONAL - CAR. Sul da Bahia: Perfil Regional. Programa de Desenvolvimento Regional Sustentável – PDRS. Salvador, out. 1995.
CONSERVATION INTERNATIONAL (CI). The Economics of Biodiversity Conservation in the Brazilian Atlantic Forest. Washington, D.C., 1997.
CONSERVATION INTERNATIONAL (CI); INSTITUTO DE ESTUDOS SÓCIO-AMBIENTAIS DO SUL DA BAHIA (IESB). Designing Sustainable Landscapes. Center for Applied Biodiversity Science at Conservation International and Institute for Social and Envirommental Studies of Southern Bahia. Washigton, D.C., 2000.
CONSTANTINO, R. et al. Causas da Fragmentação: Causas Naturais. In: Fragmentação de ecossistemas: Causas, Efeitos sobre a Biodiversidade e Recomendações de Políticas Públicas. Denise Marçal Rambaldi, Daniela América Suárez de Oliveira (orgs.) Brasília: MMA/SBF, 2003.
CORREIA, M. E. F. Potencial de utilização dos atributos das comunidades de fauna de solo e de grupos chave de invertebrados como bioindicadores do manejo de ecossistemas. Seropédica: Embrapa Agrobiologia, dez. 2002.
CREEL, J. E. B. ¿QUÉ ES UN CORREDOR BIOLÓGICO? Disponível em: <http://wbln0018.worldbank.org/mesoam/malib.nsf/11ab7b3e4d19f4328525660c0077aefb/26e78c2a754c954d85256810004b6365. 1999. Acesso em: 10 jul. 2006.

104
DAMSCHEN, E. I.; HADDAD, N. M.; ORROCK, J. L.; TEWKSBURY, J. J.; LEVEY, D. J. 2006. Corridors Increase Plant Species Richness at Large Scales. Science 313 (5197): 1284-1286.
DEAN, W. A ferro e fogo: A História e a devastação da Mata Atlântica. São Paulo: companhia das Letras, 1996.
DELABIE, J. H. C. et al. Contribution of cocoa plantations to the conservation of native ants (Insecta: Hymenoptera: Formicidae) with a special emphasis on the Atlantic Forest fauna of southern Bahia, Brazil. Biodivers Conserv. 16:2359-2384, 2007.
DELABIE, J. H. C. Comunidades de formigas (Hymenoptera; Formicidae): Métodos de estudo e estudos de casos na Mata Atlântica. In: XII ENCONTRO DE ZOOLOGIA DO NORDESTE, 1999, Feira de Santana. Resumos. Feira de Santana: UEFS, p. 58-68, 1999.
DELABIE, J. H. C. The Ant Problems of Cocoa Farms in Brazil. In: Robert K. V. Meer, Klaus Jaffe, and Aragua Cedeno. Applied Myrmecology: a World Perspective. p. 555-569, 1990.
DELABIE, J. H. C. Ocorrência de Wasmannia auropunctata (Roger, 1863) (Hymenoptera, Formicidae, Myrmicinae) em cacauais na Bahia, Brasil. Revista Theobroma 18 (1):29-37. Centro de Pesquisa do Cacau Ilhéus, Bahia, Brasil. 1988.
DELABIE, J. H. C.; FISHER, B. L.; MAJER, J. D.; WRIGHT, I. W. Sampling effort and choice of methods. In: D. Agosti, J. D. Majer, L. Tennant de Alonso e T. Schultz (eds.) Ants: Standard methods for measuring and monitoring biodiversity. Washington: Smithsonian Institution Press, 2000. p. 145-154.
DELABIE, J. H. C.; FOWLER, H. G. Soil and litter cryptic ant assemblages of Bahia cocoa plantations. Pedobiologia. v. 39, n. 5, p. 423-433, 1995.
DELABIE, J.; NASCIMENTO, I.; REIS, Y.; CAMPIOLO, S. O gênero como unidade de análise em estudos usando os Formicidae como indicadores biológicos. In: VI Seminário de Pesquisa da UESC, 1999, Ilhéus. Resumos. Ilhéus: Editus, p. 37, 1999.
DIDHAM, R. K., et al. Insects in fragmented forests: a functional approach. Tree 11(6): 255-260, 1996.

105
DORAN, J. W.; PARKIN, T. B. Defining and asswssing soil quality. In: DORAN, J. W. et al. Defining soil quality for a sustainable environment. Madison: Soil Society of America, 1994.
EASTMAN, J. R. Idrisi for Windows User´s Guide. Worcester, Clark University, 1995.
FARIA, D. M. de (Coord.). Projeto de Pesquisa: Remanescentes de Floresta da Região de Una. Ministério do Meio Ambiente (Auxílio financeiro). Universidade Estadual de Santa Cruz. 1999-2002.
FEITOSA, R. dos S. M.; RIBEIRO, A. S. Mirmecofauna (Hymenoptera, Formicidae) de serapilheira de uma área de Floresta Atlântica no Parque Estadual da Cantareira – São Paulo, Brasil. Biotemas, 18 (2): 51 - 71, 2005.
FERNÁNDES, F. Las hormigas cazadoras del género Ectatomma (Formicidae: Ponerinae) en Colombia. Caldasia, 16: p. 551-564, 1991.
FISZON, J. T. et al. Causas da Fragmentação: Causas Antrópicas. In: Fragmentação de ecossistemas: Causas, Efeitos sobre a Biodiversidade e Recomendações de Políticas Públicas. Denise Marçal Rambaldi, Daniela América Suárez de Oliveira (orgs.) Brasília: MMA/SBF, 2003.
FONSECA, G. A. B. et al. Corredores de Biodiversidade: o Corredor Central da Mata Atlântica. In: Corredores ecológicos: uma abordagem integradora de ecossistemas no Brasil. Moacir B. Arruda, Luís F. S. N. de Sá (orgs). Brasília: Ibama, 2004.
FORMAN, R. T. T. Land Mosaics – the ecology of landscapes and regions. Cambridge: Cambridge University Press, 1995.
FORMAN, R.T. T.; GODRON, M. Landscape Ecology. Wiley: New York. 1986.
FOWLER, H. G. Relative representation of Pheidole (Hymenoptera: Formicidae) in local ground ant assemblages of the Américas. Annales de Biologia. v. 19, p. 29-37. 1993.
FOWLER, H. G. et al. Ecologia Nutricional de formigas. In: Panizzi, A. R.; PARRA, J. R. P. (Ed.). Ecologia nutricional de insetos e suas implicações no manejo de pragas. São Paulo: Manole, 1991.

106
FUNDAÇÃO BRASILEIRA PARA O DESENVOLVIMENTO SUSTENTÁVEL. O Parque Nacional do Itatiaia. Cadernos para o Desenvolvimento Sustentável. Ângelo A. dos Santos (Ed.) Ponto de Vista Comunicação Ltda. 2000.
FUNDAÇÃO SOS MATA ATLÂNTICA. MATA ATLÂNTICA. Disponível em: http://www.sosmatatlantica.org.br/index.php?section=info&action=mata. Acesso em: 10 jul. 2007.
FUNDAÇÃO SOS MATA ATLÂNTICA; INSTITUTO NACIONAL DE PESQUISAS ESPACIAIS. Atlas dos remanescentes florestais da Mata Atlântica e ecossistemas associados no período de 1995-2000. Fundação SOS Mata Atlântica e INPE, São Paulo, 2001.
GALINKIN, M. et al. Projeto Corredor Ecológico Araguaia-Bananal. In: Corredores ecológicos: Uma abordagem integradora de ecossistemas no Brasil. ARRUDA, M. B.; SÁ, L. F. S. N. (Orgs.) IBAMA, Brasília-DF. p.81-131, 2004.
GORGÔNIO, A. de S. Estudo ambiental de alterações antrópicas nas matas de galeria da bacia hidrográfica do Ribeirão Taboca. Brasília: Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis, 1998.
GUIMARÃES, A. L. Instituto BioAtlântica – IBIO. Revista Ação Ambiental: Experiências de preservação, conservação e recuperação da Mata Atlântica. Universidade Federal de Viçosa. Ano VI, n. 25. Maio/Junho/2003.
GULINCK, H. et al. The visualização of corridors in the landscape using SPOT data. In: SAUNDERS, D. A.; HOBBS, R. J. (Edts). Nature Conservation 2: The Role of Corridors. Surrey Beatty; Sons Pty Limited. Austrália. 1991.
GUSMÃO, L. Manguezais: Fonte de vida e renda. Jornal Agora, Itabuna, 07-13 set. Caderno Meio Ambiente. p. 6, 2002.
HAWKSWORTH, D.L. Litmus tests for ecosystem helth: the potential of bioindicator in the monitoring of biodiversity. IN: SWAMINATHAN, M. S.; JANA, S. (eds.) Biodiversity. Implications for global food security. Madras: Macmillan Índia, 1992. 17: 184-204.
HÖLLDOBLER, B.; WILSON, E. O. The Ants. Massachusetts: Harvard University Press. 1990. 732p

107
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Geociências: Cartografia. (1997). Disponível em: http://www.ibge.gov.br/home/geociencias/cartografia/default.shtm. Acesso em: 3 mar. 2006.
INSTITUTO DE ESTUDOS SÓCIO-AMBIENTAIS DO SUL DA BAHIA - IESB. Abordagens Ecológicas e Instrumentos Econômicos para o Estabelecimento do Corredor do Descobrimento: uma estratégia para reverter à fragmentação florestal na Mata Atlântica do Sul da Bahia: Proposta submetida ao Probio – Edital 01/97. Ilhéus, 1997.
______. Mapa de Vegetação. 2002.
JARDIM, J.G. Uma cracterização parcial da vegetação na região sul da Bahia, Brasil. In Prado P.I., Landau, E.C., Moura R.T., Pinto, L.P.S., Fonseca, G.A.B., Alger K. (orgs.) Corredor de Biodiversiade da Mata Atlântica do Sul da Bahia. Publicação em CD-ROM, Ilhéus, IESB/CI/CABS/UFMG/UNICAMP. 2003.
JESUS, R. M.; ROLIM, S. G. Pesquisa em Conservação na Mata Atlântica na reserva natural do Vale do Rio Doce, Linhares (ES). Revista Ação Ambiental: Experiências de preservação, conservação e recuperação da Mata Atlântica. Universidade Federal de Viçosa. Ano VI, n. 25. Maio/Junho/2003.
LACERDA, L. D. Os Manguezais do Brasil. In: VANNUCCI, M. Os manguezais e nós. 1ª ed. São Paulo: EDUSP. 1999.
LATTKE, J. E. Revisión del Género Apterostigma Mayr (Hymenoptera: Formicidae). Arquivos de Zoologia. v. 34, n. 5. Museu de Zoologia da Universidade de São Paulo, 1997. p. 121-221.
LAURANCE, W. F. Edge effects in tropical forest fragments: Application of a model for the design of nature reserves. Biological Conservation 57: 205-219. 1991.
LEITE, J. de O.; SANTOS, J. E. L. L. dos; MENDONÇA, J. R. Dinâmica do Uso da Terra: diagnóstico Sócio Econômico da Região Cacaueira. Ilhéus, CEPLAC/IICA, v.3. 1976.
LINO, C. F.; DIAS, H. Águas e Florestas da Mata Atlântica: por uma Gestão Integrada. São Paulo: Conselho Nacional da Reserva da Biosfera da Mata Atlântica, 2003.
LOBÃO, D. E. Cacau-Cabruca, um modelo sustentável de agricultura tropical. Revista Indícios Veemente. Ilhéus, BA. Ano III. 1997. 10-24 p.

108
MACARTHUR, R. H.; WILSON, E. O. The Theory of Island Biogeography. Princeton NJ. University Press 1967.
MAGURRAN, A. E. Ecological Diversity and its Measurement. London: Chapmann e Hall. 1983.
MARIANO, C. dos S. F. Evolução cariotípica em diferentes grupos de Formicidae. Doutorado em Entomologia. Universidade Federal de Viçosa, UFV, Brasil. Tese. 2004.
MAZZONI-VIVEIROS, S. C.; TRUFEM, S. F.B. Effects of air and soil pollution on the root system of the Tibouchina pulchra Cogn. (Melastomataceae): arbuscular mycorrhizal associations and morphology in Atlantic Forest area. Rev. bras. Bot. Apr./June v. 27, n.2, p.337-348, 2004.
METZGER, J. P. O que é Ecologia de Paisagens? Biota Neotropica v. 1, números 1 e 2, 2001. Disponível em: http://www.biotaneotropica.org.br/v1n12/pt/abstract?thematic-review+BN00701122001. Acesso em: 09 abr. 2006.
METZGER, J. P. Estrutura da Paisagem: o uso adequado de métricas. In: JUNIOR, L. C.; PÁDUA, C. V.; RUDRAN, R. Métodos de estudos em biologia da conservação e manejo da vida silvestre. Curitiba, PR. Ed. da UFPR. Fundação O Boticário de Proteção à Natureza, 2003.
METZGER, J. P. Structure du paysage et diversité des peuplements ligneux fragmentés du rio Jacaré-Pepira (sud-est du Brésil). Tese (Doutorado). Universidade Paul Sabatier, Toulouse. 1995.
MURICY, L. A ampliação da APA Costa de Itacaré/Serra Grande. Jornal da APA. Gráfica Agora, Itacaré-BA, ano III, n. 31, mar 2004.
MYERS, N.; MITTERMEIER, R. A.; FONSECA, G. A. B.; KENT, J. Biodiversity hotspots for conservation priorities. Nature. 403: 845-853. 2000.
NASCIMENTO, H. E. M.; LAURANCE, W. F. Efeitos de área e de borda sobre a estrutura florestal em fragmentos de floresta de terra-firme após 13-17 anos de isolamento. Acta Amazônica. v. 36 (2), p. 183-192, 2006.

109
NUNES, G. M. et al. Sistemas de Informações Geográficas aplicadas na implantação de corredores ecológicos na Sub-Bacia Hidrográfica do Rio Vacacaí-Mirim (RS). In: XII Simpósio Brasileiro de Sensoriamento Remoto. Anais. Goiânia, Brasil, 16-21 abril 2005, INPE, p. 3183-3189. 2005.
ODUM, E. P. Ecologia. Guanabara: Rio de Janeiro. 1988.
OLIVEIRA, C. A. S. Mata Atlântica, desmatamento zero: o que deve ser esclarecido. Revista Ação Ambiental: Experiências de preservação, conservação e recuperação da Mata Atlântica. Universidade Federal de Viçosa. Ano VI, n. 25. Maio/Junho/2003.
OLIVEIRA, M. A.; DELLA-LUCIA, T. M. C.; ARAÚJO, M. S.; CRUZ, A. P. A fauna de formigas em povoamentos de eucalipto e mata nativa no Estado do Amapá. Acta Amazônica, 25(1): 117-126, 1995.
PACIÊNCIA, M. L. B.; PRADO, J. Efeitos de borda sobre a comunidade de pteridófitas na Mata Atlântica da região de Una, sul da Bahia, Brasil. Revista Brasileira de Botânica. v. 27, n. 4, p. 641-653, out.-dez. 2004.
PINTO, L. P. de S. et al. (Eds). Mata Atlântica: Ciência, Conservação e Políticas. Workshop Científico sobre a Mata Atlântica. Belo Horizonte, 22-23 de janeiro de 1996. Disponível em: http://www.aliancamataatlantica.org.br/limites.html. Acesso em: 31 jan de 2007.
PRIMACK, R. B.; RODRIGUES, E. Biologia da Conservação. Londrina: E. Rodrigues, 2001.
PRIMO, D. C.; VAZ, L. M. S. Degradação e Perturbação Ambiental em Matas Ciliares: Estudo de Caso do Rio Itapicuru-Açu em Ponto Novo e Filadélfia Bahia. Diálogos & Ciência – Revista Eletrônica da Faculdade de Tecnologia e Ciências. Ano IV, n. 7, jun. 2006. Disponível em: http://www.ftc.br/revistafsa. Acesso em: 9 dez. 2006.
QUEIROGA, J. L. de; RODRIGUES, E. Efeito de borda em fragmentos de Cerrado em áreas de agricultura do Maranhão, Brasil. Dissertação de Mestrado. Universidade Estadual de Londrina, Paraná, Brasil. 2001.
RAMOS, L. S.; MARINHO, C. G. S.; ZANETTI, R.; DELABIE, J. H. C.; SCHLINDWEIN, M. N. Impacto de iscas formicidas granuladas sobre a mirmecofauna não-alvo em eucaliptais segundo duas formas de aplicação. Neotropical Entomology. v. 32, n. 2, p. 231-237, 2003.

110
REID, J.; BLANES, J. A pecuária extensiva, da forma como é realizada na região cacaueira, não é rentável. Resumos de Pesquisa. 3ª ed. Ilhéus, BA: Instituto de Estudos Sócios-Ambientais do Sul da Bahia. p. 22-25, 1997.
REVINGTON, J. The Causes of Tropical Deforestation. New Renaissance Magazine. v. 3, n. 2. 1992. Disponível em: http:/www.ru.org/32defore.html. Acesso em 20 ago. 2005.
RICE, R. A.; GREENBERG, R. Cacao Cultivation and the Conservation of Biological Diversity. Ambio, 29(3): 167- 173, 2000.
RINALDI, R. R.; LIMA, G. S. Unidades de Conservação: estratégia de proteção da Mata Atlântica. Revista Ação Ambiental: Experiências de preservação, conservação e recuperação da Mata Atlântica. Universidade Federal de Viçosa. Ano VI, n. 25. Maio/Junho/2003.
ROCHA, R. B. O Futuro da Mata Atlântica: um olhar sobre o sul da Bahia. UMA. Salvador, World Watch, v. 14, n. 6, p. 40-46, 2002.
RODRIGUES, P. J. F. P.; NASCIMENTO, M. T. Fragmentação Florestal: Breves considerações teóricas sobre efeito de borda. Rodriguésia 57 (1): 63-74. 2006.
SAATCHI, S.; AGOSTI, D.; ALGER, K.; DELABIE, J.; MUSINSKI, J. Examining fragmentation and loss of primary forest in the southern Bahian Atlantic Forest of Brazil with radar imagery. Conserv Biol 15(4):867–875. 2001.
SAMBUICHI, R. H. R. Fitossociologia e diversidade de espécies arbóreas em cabruca (Mata Atlântica raleada sobre plantação de cacau) na Região Sul da Bahia, Brasil. Acta bot. Bras. 16(1): 89-101, 2002.
SANTILLI, J. A Lei 9.985/2000, que instituiu o Sistema Nacional de Unidades de Conservação da natureza (SNUC): uma abordagem socioambiental. Revista de Direito Ambiental. Ed. Revista doa Tribunais, São Paulo, SP, Brasil. Ano 10, n. 40, outubro-dezembro de 2005.
SANTOS, M. S. Estrutura da comunidade de formigas (Hymenoptera: Formicidae) em fragmentos de floresta semidecídua. Lavras, 2004. Dissertação (Mestrado). Universidade Federal de Lavras.

111
SCARIOT, A. et al. Efeitos da fragmentação sobre a biodiversidade: vegetação e flora. In: Fragmentação de ecossistemas: Causas, Efeitos sobre a Biodiversidade e Recomendações de Políticas Públicas. Denise Marçal Rambaldi, Daniela América Suárez de Oliveira (orgs.) Brasília: MMA/SBF, 2003.
SCHAEFFER-NOVELLI, Y.; JUNIOR, C. C.; TOGNELLA-DE-ROSA, M. Manguezais. São Paulo: Ática, 1. ed. 2001.
SERRANO, José-Luis. Concepto, formación y autonomia Del Derecho Ambiental. In: Valella, M. D.; Borges, M. R. C. B. (orgs.). O novo em Direito Ambiental. Belo Horizonte: Del Rey, p. 33-49, 1998.
SILVA, S.M. Considerações Fitogeográficas e Conservacionistas sobre a Floresta Atlântica no Brasil. 2002. Disponível no site http://www.conservation.org.br/ma/rp_flora.htm.
SILVESTRE, R. Estrutura de comunidades de formigas do Cerrado. Ribeirão Preto, 2000. Tese (Doutorado). Universidade de São Paulo.
SOARES FILHO, B. S. Modelagem da dinâmica de uma região de fronteira de colonização amazônica. Tese de Doutorado. Escola Politécnica da Universidade de São Paulo. São Paulo, 1998.
SUPERINTENDÊNCIA DE ESTUDOS ECONÔMICOS E SOCIAIS DA BAHIA - SEI. Base Cartográfica Digital. 2003.
SZMUCHROWSKI, M. A. e MARTINS, I. C. de M. Geoprocessamento para a Indicação de corredores ecológicos Interligando os fragmentos de florestais e áreas de proteção ambiental no Município de Palmas – TO. Anais. X SBSR, Foz do Iguaçu,21-26 abril, INPE, p. 675-681, 2001.
TABARELLI, M.; SILVA, J. M. C. da; GASCON, C. Forest fragmentation, synergisms and the impoverishment of neotropical forests. Biodiverty and Conservation 13: 1419-1425, 2004.
TEIXEIRA, A. et al. Qual a Melhor Definição de SIG? Fator GIS 11 – A Revista do Geoprocessamento. 10/12: p. 20-23, 1995.

112
TOLMASQUIM, M. T. Estrutura conceitual para a elaboração de indicadores de sustentabilidade ambiental para o Brasil. In: Garay, I. E. G.; Dias, Braulio F. S. Conservação da Biodiversidade em ecossistemas tropicais: avanços conceituais e revisão de novas metodologias de avaliação e monitoramento. Petrópolis: Editora Vozes, 2001.
UEHARA-PRADO, M. Efeitos de fragmentação florestal na guilda de borboletas frugívoras do Planalto Atlântico Paulista. Dissertação de mestrado. Universidade Estadual de Campinas, SP, 2003.
ULLOA-CHACÓN, P.; CHERIX, D. The Little Fire Ant Wasmannia auropunctata R. (Hymenoptera: Formicidae). In R. K Vander Meer, K. Jaffee, and A. Cedenõ [eds.], Applied Myrmecology: a world perspective. Westview Press, Boulder, Colorado. p. 281-289, 1990.
UNDERWOOD, E. C.; FISHER, B. L. The role of ants in conservation monitoring: If, when, and how. Biological Conservation. 166-182, 2006.
UNIÃO MUNDIAL PARA CONSERVAÇÃO DA NATUREZA - UICN. Proposta e Projetos. In IV Congresso Mundial de Áreas Protegidas, Caracas. 1992.
VIANA, V. M.; PINHEIRO, L. A. F. V. Conservação da biodiversidade em fragmentos florestais. Série Técnica IPEF, ESALQ/USP, v. 12, n. 32, p. 25-42, 1998.
VIEIRA, M. V. et al. Efeitos da fragmentação sobre a biodiversidade: mamíferos. In: Fragmentação de ecossistemas: Causas, Efeitos sobre a Biodiversidade e Recomendações de Políticas Públicas. Denise Marçal Rambaldi, Daniela América Suárez de Oliveira (orgs.) Brasília: MMA/SBF, 2003.
YOUNG, C. E. F. Causas socioeconômicas do desmatamento da Mata Atlântica brasileira. In: Carlos Galindo-Leal; Ibsen de Gusmão Câmara (Edts). Mata Atlântica: biodiversidade, ameaças e perspectivas. Belo Horizonte e São Paulo, 2005.
WILSON, E. O. Pheidole in the New World, a dominant, hyperdiverse ant genus. Massachusetts: Harvard University Press, 2003.
WILSON, E. O. Which are the most prevalent ant genera? Studia Entomológica. Petrópolis, v. 19, n. 1/4, p. 187-200, 1976.
WINK, C. et al. Insetos edáficos como indicadores da qualidade ambiental. Revista de Ciências Agroveterinárias, Lages, v. 4, n. 1, p. 60-71, 2005.

113
WOLTERBEEK, H. T. H.; KUIK, P.; VERBURG, T. G.; HERPIN, U.; MARKERT, B.; THÖNI, L. Moss interspecies comparisons in trace element concentrations. Environmental Monitoring and Assessment, 35:263-286. 1995. Disponível em: http://biomonitor.ist.utl.pt/biomonitor - Portal Biomonitor Powered by Mambo Generated. Acesso em 2 set. 2006.
WWW.CORREDORES.ORG.BR. O que é o Corredor Central da Mata Atlântica. Disponível em: http://www.corredores.org.br. Acesso em: 8 mar. 2006.

A474 Alves, Hilda Susele Rodrigues.
Identificação de bioindicadores e planejamento de mi- ni-corredores ecolológicos na área de proteção ambiental costa de Itacaré/ Serra Grande, Bahia / Hilda Susele Rodrigues Alves. – Ilhéus, BA: UESC, 2008.
xii, 113f. : il.
Orientador: Jacques H. C. Delabie. Co-orientador: Maurício Moreau. Dissertação (Mestrado) – Universidade Estadual de Santa Cruz. Programa de Pós-graduação em Desenvol- vimento Regional e Meio Ambiente.
Bibliografia: f. 99 -113. 1. Proteção ambiental. 2. Mata Atlântica – Conserva- ção. 3. Formiga. 4. Sistema de informação geográfica. 5. Área de proteção ambiental – Itacaré (BA). I. Título. CDD 333.73





























