Embed Size (px)
Citation preview

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
1
IMUNIDADE ADAPTATIVA: LINFÓCITOS B
IMUNOLOGIA

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
2
IMUNIDADE ADAPTATIVA: LINFÓCITOS B
CONTEÚDO: HIASMIN ACOSTA ALVES CURADORIA: FABRÍCIO MONTALVÃO

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
3
SUMÁRIO
IMUNIDADE HUMORAL .................................................................................. 4
ATIVAÇÃO DA CÉLULA B .............................................................................. 5
RESPOSTAS DE ANTICORPOS T DEPENDENTES A ANTÍGENOS PROTEICOS ........................................................................... 6
Troca de isotipo da cadeia pesada ................................................................. 8
Maturação da afinidade ................................................................................... 10
Plasmócitos secretores de anticorpos ............................................................ 11
Células B de memória .......................................................................................... 12
RESPOSTAS DE ANTICORPOS A ANTÍGENOS T INDEPENDENTES ........... 13
INIBIÇÃO DA PRODUÇÃO DE ANTICORPOS ............................................. 14
REFERÊNCIAS ............................................................................................... 15

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
4
IMUNIDADE HUMORAL
A imunidade humoral é mediada por an-ticorpos secretados por linfócitos B. Os linfócitos B maduros, capazes de res-ponder ao antígeno, se desenvolvem a partir de células precursoras da medula óssea e migram para os órgãos linfoides periféricos, onde ocorre a interação dos linfócitos com os antígenos.
As respostas imunes humorais se desen-volvem a partir do reconhecimento de antígenos por linfócitos B específicos. O antígeno se liga ao BCR (receptor de cé-lulas B) que se localiza na membrana das células B virgens maduras, ativando-as. Nos linfócitos B maduros que saem da medula óssea, o BCR é uma molécula de Imunoglobulina (Ig) M inserida na mem-brana. Também existem moléculas de IgD que funcionam como BCR, mas o pa-pel desta classe de Ig não é completa-mente conhecido.
A ativação acarreta na proliferação de células específicas para o antígeno e na diferenciação, gerando plasmócitos (cé-lulas B diferenciadas) secretores de anti-corpos e células de memória.
Algumas células B ativadas podem pro-duzir outros tipos de anticorpos além da IgM e IgD, o que configura a troca de iso-tipo (classe) de cadeia pesada. De acordo com o desenvolvimento da res-posta imune humoral, as células B ativa-das se ligam a antígenos com afinidade crescente, processo chamado de matu-ração da afinidade.
O tipo e a quantidade de anticorpos produzidos dependem do tipo de antí-geno que está induzindo a resposta imune, do envolvimento de células T, da exposição prévia ao antígeno e do local em que ocorre a ativação.
As respostas de anticorpos a antígenos proteicos necessitam que o antígeno seja internalizado por células B específi-cas e que seus peptídeos sejam proces-sados e apresentados aos linfócitos Th CD4+ para que estes ativem as células B.
Devido à necessidade de ativação pelos linfócitos T, as proteínas são classifica-das como antígenos T dependentes. Além disso, os linfócitos T são chamados também de auxiliares por desempenhar a função de estimular os linfócitos B a produzir anticorpos.
Um tipo especializado de célula T auxi-liar, denominada célula T auxiliar folicu-lar, facilita a formação dos centros ger-minativos nos órgãos linfoides, onde acontecem as respostas imunes humo-rais T-dependentes.
As respostas de anticorpos para antíge-nos como polissacarídeos, alguns lipídios e ácidos nucleicos, não necessitam da indução dos linfócitos T CD4+ antígeno-específicos, ou seja, são antígenos T in-dependentes. No caso desta ativação, é necessário que estes carboidratos/lipí-deos possuam repetições na sua estru-tura molecular, de modo a engajar múl-tiplos BCRs na superfície do linfócito B.

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
5
Somente assim, é possível a ativação T-independente.
As células B ativadas se diferenciam em plasmócitos e algumas dessas células B ativadas de forma T dependente se di-ferenciam em células de memória. As cé-lulas B de memória ficam em um estado de repouso, ou seja, sem secretar anti-corpos, porém, quando necessário, elas montam respostas rápidas para o mesmo antígeno que estimulou sua for-mação.
Processos como a troca de isotipo e ma-turação da afinidade ocorrem em res-postas imunes humorais T dependentes quando há presença de antígenos pro-teicos. As respostas humorais primárias e secundárias contra antígenos proteicos diferem qualitativamente e quantitati-vamente.
As respostas primárias são resultado da ativação de células B imaturas e as res-postas secundárias ocorrem somente após a estimulação das células B de me-mória. Logo, a resposta secundária é mais rápida que a primária e as quanti-dades de anticorpos produzidos são maiores.
Subpopulações de células B respondem a diferentes tipos de antígenos. Células B foliculares em órgãos linfoides periféri-cos respondem principalmente à antíge-nos proteicos que necessitam das célu-las T CD4+.
Já as células B localizadas na zona mar-ginal no baço reconhecem antígenos multivalentes e suas respostas são em maioria T independentes. As células B-1, localizadas em tecidos de mucosa e em cavidades (peritônio, pleura) também possuem respostas principalmente T in-dependentes.
ATIVAÇÃO DA CÉLULA B
A maioria dos linfócitos B virgens madu-ros são células B foliculares que recircu-lam no sangue e vão de um órgão lin-foide secundário para outro na busca de antígenos. As células B foliculares entram nos tecidos linfoides secundários através dos vasos sanguíneos e migram para os folículos, que são as zonas de célula B. O movimento para os folículos linfoides é induzido pela quimiocina CXCL13, secre-tada por células dendríticas foliculares e por outras células estromais.
A maioria dos antígenos nos tecidos são transportados para os linfonodos atra-vés dos vasos linfáticos aferentes que drenam para o seio subcapsular dos lin-fonodos. Assim, os antígenos solúveis podem chegar até a zona de células B.
Os macrófagos presentes no seio sub-capsular capturam grandes microrga-nismos e complexos antígeno-anticorpo e os apresenta nos folículos. Antígenos relativamente grandes que entram a partir dos vasos linfáticos aferentes po-dem ser capturados por células

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
6
dendríticas da zona medular e transpor-tados até as células B.
Antígenos de imunocomplexos podem se ligar a receptores do complemento em células B da zona marginal que os trans-portam para as células B foliculares. Os imunocomplexos também podem se li-gar ao receptor do complemento CR2 na superfície das células dendríticas folicu-lares para serem apresentados as célu-las B antígeno-específicas. O antígeno que é apresentado as células B está, ge-ralmente, em sua conformação nativa e não é processado por células apresen-tadoras de antígeno.
O antígeno e as citocinas desempenham um papel importante na sobrevivência das células B imaturas. A sobrevivência das células B foliculares depende dos si-nais do BCR (receptor de células B) e também da citocina BAFF (fator ativador de célula B), que fornece sinais de matu-ração e de sobrevivência.
A ativação dos linfócitos B antígeno-es-pecíficos se dá pela ligação do antígeno as moléculas de Ig de membrana, que, em conjunto com a Igα e Igβ, formam o receptor de antígeno de células B madu-ras (BCR).
O reconhecimento do antígeno pode desencadear respostas de células B, mas geralmente ele é insuficiente para estimular de forma significativa a prolife-ração e a diferenciação das células B.
A ativação da célula B é facilitada pelo correceptor CR2/CD21, que reconhece fragmentos de complemento ligados ao antígeno ou que fazem parte dos imu-nocomplexos contendo o antígeno.
Os produtos microbianos se acoplam aos receptores do tipo toll nas células B, o que também aumenta a ativação dessa célula. A ligação cruzada no BCR com o antígeno gera diferentes reações de acordo com o tipo de antígeno: antí-genos multivalentes iniciam a prolifera-ção e a diferenciação da célula B e os antígenos proteicos preparam as células B para interagir com as células T CD4+.
RESPOSTAS DE ANTICORPOS T DEPENDENTES A ANTÍGENOS PROTEICOS
Antígenos proteicos são reconhecidos por linfócitos B e T específicos nos órgãos linfoides periféricos. As células ativadas migram para esses órgãos, onde iniciam as respostas imunes humorais.
A ativação de células B e T específicas para o mesmo antígeno é muito impor-tante para a interação funcional entre elas. As células Th CD4 diminuem a ex-pressão do receptor de quimiocina CCR7 e aumentam a expressão de CXCR5, dessa forma, deixam a zona da célula T e migram para o folículo.
O ligante para CXCR5, o CXCL13, é se-cretado pelas células dendríticas

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
7
foliculares e por células do estroma foli-cular e atrai as células T CD4+ ativadas para o folículo. As células B respondem a esses antígenos reduzindo a expressão do CXCR5 de suas superfícies celulares e aumentam a expressão do receptor CCR7. O que as atrai para a zona de lin-fócitos T.
Dessa forma, as células B ativadas mi-gram para a zona da célula T por ação de um gradiente de CCL19 e de CCL21, os ligantes para CCR7. As células B ativa-das por antígenos proteicos também podem expressar CD69, causando re-tenção de células B ativadas nos linfono-dos.
Antígenos proteicos que se ligam em re-ceptores da célula B são endocitados e processados para gerar peptídeos que se ligam a moléculas de MHC de classe II e são apresentados as células T CD4.
Durante a ativação, as células T CD4+ expressam o ligante de CD40 (CD40L), o qual se acopla ao seu receptor, CD40, nas células B estimuladas pelo antígeno, gerando a proliferação e diferenciação da célula B.
Mutações no gene do CD40L causam uma doença chamada síndrome da hi-per-IgM ligada ao X, em que o indivíduo possui defeitos na produção de anticor-pos, troca de isotipo, maturação da afi-nidade e na geração de células B de me-mória em resposta a antígenos protei-cos.
A ativação extrafolicular da célula B gera uma resposta precoce de anticor-pos a
antígenos proteicos e se ajusta à forma-ção da resposta do centro germinativo, desenvolvida mais lentamente.
A ativação T dependente de células B nos focos extrafoliculares produz anti-corpos de baixa afinidade que possuem a capacidade de circular e limitar uma infecção. Além disso, a resposta extrafo-licular também auxilia na formação de células T CD4+ foliculares que se trans-portam para o folículo e são necessárias para a formação do centro germinativo.
As respostas humorais dependentes das células T CD4+, como a maturação da afinidade, a troca de isotipo e a geração de plasmócitos e células B de memória, ocorrem principalmente nos centros ger-minativos dentro de folículos linfoides.
Para que ocorra a reação no centro ger-minativo é necessária a presença de cé-lulas dendríticas foliculares (FDCs). As FDCs são encontradas somente em folí-culos linfoides e expressam receptores do complemento (CR1, CR2 e CR3) e re-ceptores de Fc, que estão envolvidos na retenção do antígeno para a seleção de células B do centro germinativo.
Depois de 4 a 7 dias da exposição ao an-tígeno, as células B ativadas específicas induzem a diferenciação de algumas cé-lulas Tfh, que expressam altos níveis do receptor CXCR5, que se liga à quimiocina

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
8
CXCL13, e são direcionadas para os folí-culos linfoides.
As células Tfh também expressam ICOS (coestimulador indutível), PD-1 (morte programada-1), IL-21 e o fator de trans-crição Bcl-6.
Troca de isotipo da cadeia pesada
Após realizar a interação com o linfócito T, algumas células B ativadas que ex-pressam IgM e IgD realizam a troca de isotipo (classe). As diferentes classes de Igs representam diferentes regiões cons-tantes da cadeia pesada.
Essa característica das células B de pro-duzirem diferentes isotipos de anticorpos é muito importante para as respostas imunes humorais, já que geram anticor-pos com funções efetoras diferentes para atuarem na defesa contra diferen-tes tipos de agentes infecciosos.
A troca de isotipo é regulada por citoci-nas produzidas pelas células T CD4+ ati-vadas. O IFN-γ (interferon-gama) esti-mula a troca para a classe IgG em célu-las B e a IL-4 induz a troca para IgE. A IgG atua principalmente em resposta a vírus e bactérias intracelulares, bloqueando a entrada dos microrganismos nas células hospedeiras e também promovendo a fagocitose por macrófagos.
Vírus e bactérias que ativam células T CD4+, principalmente Th1, e as células Tfh, estimulam a produção de IFN-γ. Já
a IgE atua contra parasitas helmintos, auxiliando sua eliminação mediada por eosinófilos e mastócitos. Além disso, os anticorpos IgE também controlam as re-ações de hipersensibilidade imediata (alergias).
A IgA é a principal imunoglobulina secre-tada por plasmócitos presentes em teci-dos de mucosa, pois pode ser facilmente transportada dos epitélios para as se-creções mucosas com o objetivo de rea-lizar a defesa contra microrganismos que tentam invadir os epitélios.
A mudança de isotipo para IgA é indu-zida pelo fator transformador de cresci-mento-β (TGF-β), produzido por célu-las T CD4+, pela mucosa e por outros te-cidos. As citocinas da família do TNF (fa-tor de necrose tumoral), BAFF e APRIL (li-gante indutor de proliferação) também estimulam a mudança para IgA. Além das citocinas citadas, os sinais do CD40 também são relevantes na indução da troca de isotipo.
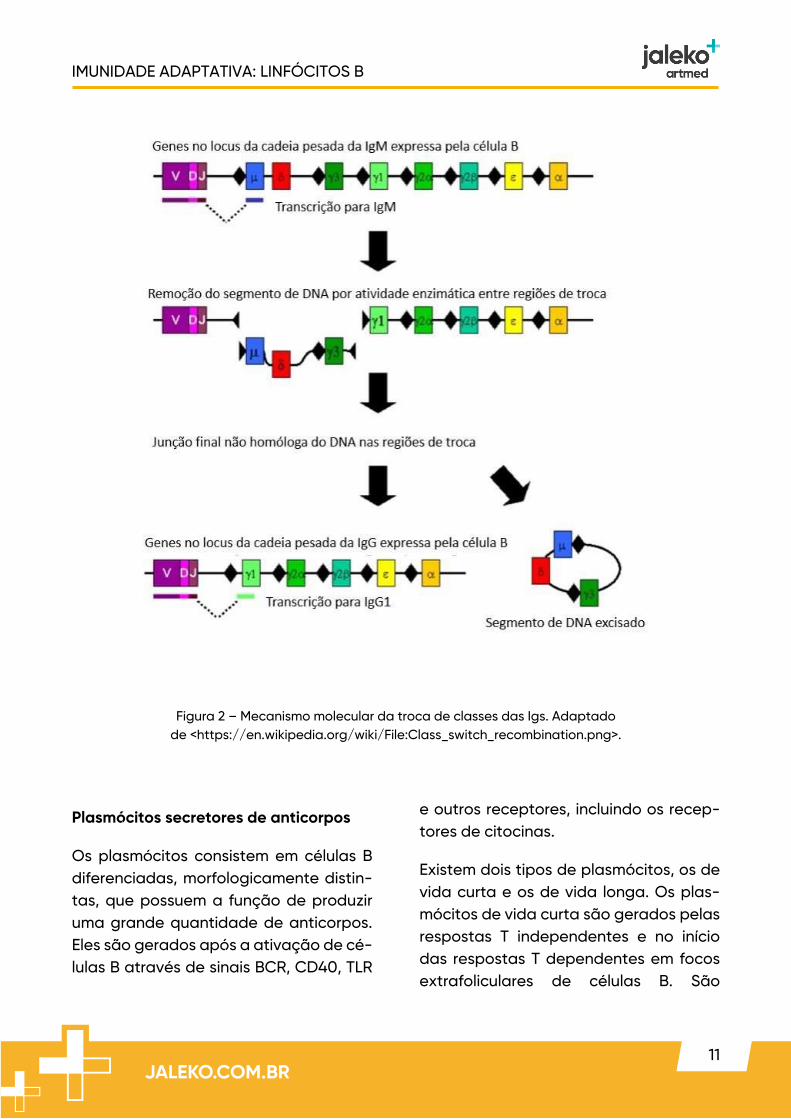
A ligação do CD40 com seu ligante esti-mula a enzima deaminase induzida por ativação (AID), muito importante tanto para troca de isotipo como da matura-ção de afinidade. O mecanismo molecu-lar que origina a troca de isotipo é cha-mado de recombinação de troca.
Na recombinação de troca, o DNA da cadeia pesada de Ig em células B é cor-tado e recombinado de modo que um éxon VDJ já formado, que codifica o do-mínio V, é posto em uma posição

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
9
adjacente a uma região C subsequente e o DNA intercalado entre essas regiões é excluído.
Esse processo envolve sequências de nucleotídeos chamadas de regiões de troca, que se localizam nos íntrons entre os segmentos J e C, nas extremidades 5’ de cada lócus CH.
As regiões de troca são compostas por numerosas repetições de sequências de DNA e
são encontradas na porção inicial dos genes da cadeia pesada. A porção ini-cial de cada região de troca é um pe-queno éxon chamado de éxon I (iniciador de transcrição), precedido por uma re-gião I, promotora.
Os sinais das citocinas e do CD40 indu-zem a transcrição de uma determinada região I promotora lendo um éxon I, a re-gião de troca e éxons CH adjacentes. Es-tes transcritos são conhecidos como transcritos da linha germinativa.
Os transcritos da linha germinativa são encontradas no lócus μ e no lócus pos-terior da cadeia pesada para a qual uma célula B ativada está sendo induzida a troca. Em cada região de troca, os
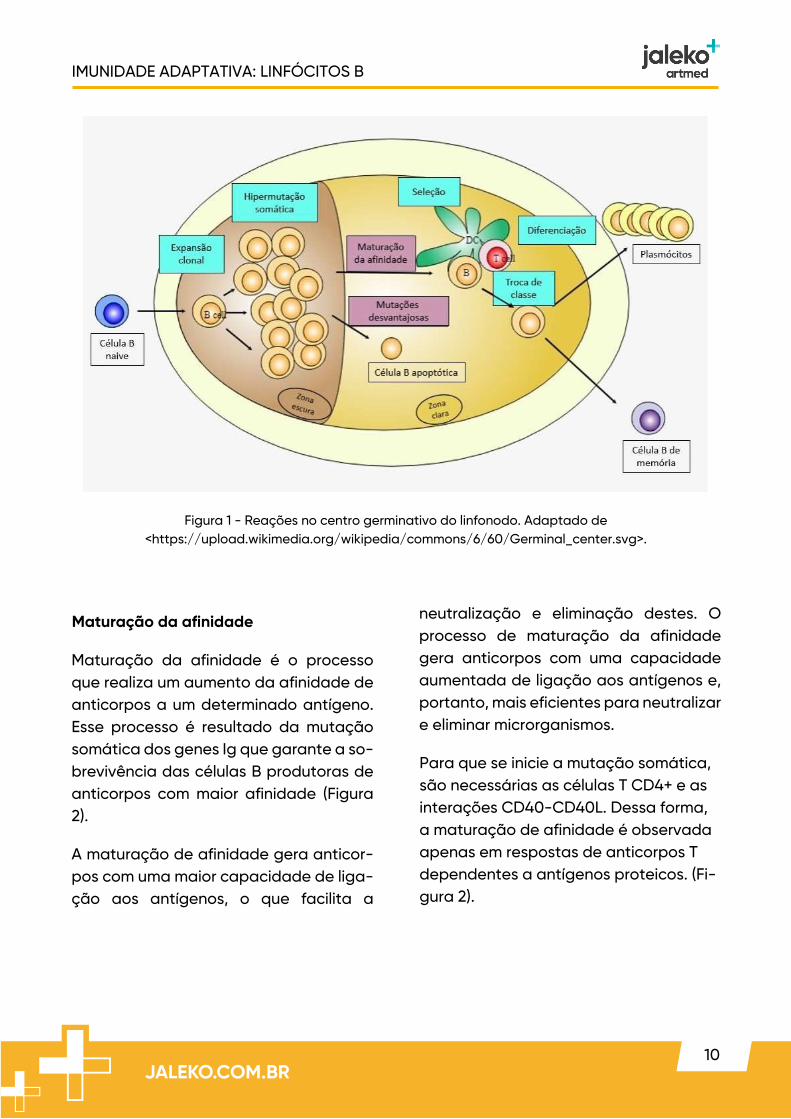
transcritos da linha germinativa facilitam a geração de quebras no DNA de cadeia dupla (Figura 1).
A quebra de DNA na região inicial de troca (μ) se une as quebras na região de troca posterior selecionada. Assim, o éxon VDJ rearranjado mais próximo à porção inicial da região de troca μ na célula B produtora de IgM é recombi-nado com o gene da cadeia pesada de Ig localizado imediatamente após a re-gião de troca posterior.
As citocinas determinam qual região CH irá sofrer transcrição da linha germina-tiva. Como já mencionada, a enzima chave necessária para a troca de isotipo é a AID, que é ativada principalmente por sinais de CD40 de células Tfh.
A AID desamina citosinas em moldes de DNA de fita simples, convertendo resí-duos de citosina (C) em resíduos de ura-cila (U), o que causa uma aberração no DNA, já que o DNA não possui uracilas. Após um longo processo, o resultado é que o DNA entre as duas regiões de troca é eliminado e o resultado final é que a região V rearranjada original torna-se adjacente à uma nova região constante (Figura 1).
IMUNIDADE ADAPTATIVA: LINFÓCITOS B
10
Figura 1 - Reações no centro germinativo do linfonodo. Adaptado de <https://upload.wikimedia.org/wikipedia/commons/6/60/Germinal_center.svg>.
Maturação da afinidade
Maturação da afinidade é o processo que realiza um aumento da afinidade de anticorpos a um determinado antígeno. Esse processo é resultado da mutação somática dos genes Ig que garante a so-brevivência das células B produtoras de anticorpos com maior afinidade (Figura 2).
A maturação de afinidade gera anticor-pos com uma maior capacidade de liga-ção aos antígenos, o que facilita a
neutralização e eliminação destes. O processo de maturação da afinidade gera anticorpos com uma capacidade aumentada de ligação aos antígenos e, portanto, mais eficientes para neutralizar e eliminar microrganismos.
Para que se inicie a mutação somática, são necessárias as células T CD4+ e as interações CD40-CD40L. Dessa forma, a maturação de afinidade é observada apenas em respostas de anticorpos T dependentes a antígenos proteicos. (Fi-gura 2).
IMUNIDADE ADAPTATIVA: LINFÓCITOS B
11
Figura 2 – Mecanismo molecular da troca de classes das Igs. Adaptado de <https://en.wikipedia.org/wiki/File:Class_switch_recombination.png>.
Plasmócitos secretores de anticorpos
Os plasmócitos consistem em células B diferenciadas, morfologicamente distin-tas, que possuem a função de produzir uma grande quantidade de anticorpos. Eles são gerados após a ativação de cé-lulas B através de sinais BCR, CD40, TLR
e outros receptores, incluindo os recep-tores de citocinas.
Existem dois tipos de plasmócitos, os de vida curta e os de vida longa. Os plas-mócitos de vida curta são gerados pelas respostas T independentes e no início das respostas T dependentes em focos extrafoliculares de células B. São

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
12
geralmente encontrados em órgãos lin-foides secundários e em tecidos não lin-foides periféricos.
Os plasmócitos de vida longa são gera-dos em respostas T dependentes a antí-genos proteicos nos centros germinati-vos. Os sinais do BCR e da IL-21 atuam juntos na diferenciação de plasmócitos e de seus precursores, chamados plasma-blastos.
Os plasmablastos são encontrados em sua maioria na circulação e são identifi-cados por serem células secretoras de anticorpo que não expressam CD20, um marcador de células B maduras.
Os plasmablastos são direcionados para a medula óssea e lá se diferenciam em plasmócitos de vida longa. Assim, a me-dula óssea se torna o local principal de produção de anticorpos aproximada-mente 2 a 3 semanas após a imunização com um antígeno T dependente.
A diferenciação das células B em plas-mócitos necessita de adaptações na es-trutura do retículo endoplasmático, um aumento da produção de Ig e uma alte-ração da forma ligada à membrana para a forma secretada dos anticorpos.
Células B de memória
As células B de memória são geradas durante a reação no centro germinativo e são capazes de gerar respostas rápi-das ao antígeno. As células B presentes no centro germinativo em proliferação sofrem mutações pontuais nos genes V
de Ig. O ponto mais importante destas mutações é que estão relacionadas ao aumento da afinidade dos anticorpos ao antígeno que induziu a resposta. As cé-lulas B que se ligam a antígenos com alta afinidade são selecionadas para sobre-viver nos centros germinativos.
Inicialmente, o reconhecimento do antí-geno já induz a expressão de proteínas antiapoptóticas da família Bcl-2. Além disso, as células B de alta afinidade te-rão preferência na endocitose e apre-sentação do antígeno e, consequente-mente, terão prioridade durante a inte-ração com o limitado número de células Tfh no centro germinativo, que estimu-lam a sobrevivência das células B. A mu-tação somática ocorre na zona escura basal dos centros germinativos, em célu-las B que possuem AID nuclear, chama-das de centroblastos.
Essas células com mutações se diferen-ciam em células chamadas centrócitos. Assim, os centrócitos de alta afinidade podem ser selecionados na zona clara pelo antígeno com o auxílio de células Tfh e podem sofrer troca de isotipo adi-cional.
As células selecionadas se diferenciam em células B de memória ou em precur-sores de plasmócitos secretores de anti-corpos de alta afinidade que migram do centro germinativo.
A diferenciação das células B é regulada pela indução e ativação de diferentes fatores de transcrição, como o Bcl-6, o

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
13
Blimp-1 e o IRF4. O Blc-6 consiste em um repressor transcricional que mantém a reação no centro germinativo, principal-mente a geração de células B.
O Bcl-6 é produzido pelas células B partir dos sinais gerados pelo CD40 e pelo re-ceptor da IL-21. Ele reprime a expressão de inibidores da quinase dependente de ciclina, estimulando ativadores transcri-cionais, como c-Myb, que medeia a en-trada das células B no centro germina-tivo. Ele também reprime p53, um fator de transcrição que acarreta a morte ce-lular por apoptose após lesão do DNA.
O Blimp-1 é um repressor de transcrição e o IRF4 é um ativador de transcrição que são induzidos em algumas das células B ativadas, não permitindo sua diferencia-ção. Blimp1 também suprime Bcl-6 e o Pax5, necessários para a manutenção de células B maduras.
Os fatores de transcrição utilizados no desenvolvimento da célula B de memória ainda não foram identificados. Vacinas contra microrganismos e toxinas micro-bianas devem induzir a maturação de afinidade e a formação de células B de memória a partir da ativação das células T CD4+.
RESPOSTAS DE ANTICORPOS A ANTÍGENOS T INDEPENDENTES
Antígenos T independentes (TI) são antí-genos não proteicos que estimulam a produção de anticorpos sem necessitar
das células T CD4+. Há muitas diferenças entre as respostas T independentes e T dependentes aos antígenos proteicos. Os anticorpos que são produzidos a par-tir de respostas T independentes são, geralmente, de baixa afinidade e são principalmente IgM.
Assim, a zona marginal e as subpopula-ções B-1 de células B são importantes para as respostas de anticorpos aos an-tígenos TI. Após sua ativação, essas cé-lulas se diferenciam em plasmócitos de vida curta que secretam principalmente IgM.
As respostas de anticorpos TI podem co-meçar no baço, medula óssea, cavidade peritoneal e mucosas. Os principais antí-genos TI são polissacarídeos, glicolipí-dios e ácidos nucleicos, pois possuem a capacidade de estimular a produção de anticorpos específicos sem necessitar de células T.
Isso ocorre porque não é possível que es-tes antígenos sejam processados e apresentados em associação a molécu-las de MHC à células T CD4+ CD4+. A maior parte dos antígenos TI são multi-valentes e conseguem estimular uma li-gação cruzada máxima do complexo BCR em células B específicas, acarre-tando à ativação sem a necessidade de uma célula T auxiliar.
Além do mais, muitos polissacarídeos ativam o sistema complemento pela via alternativa, gerando C3d, que se liga ao

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
14
antígeno e é reconhecido por CR2, au-mentando assim a ativação de células B.
INIBIÇÃO DA PRODUÇÃO DE ANTICORPOS
Os anticorpos que são secretados ini-bem a continuação da ativação das cé-lulas B por meio da formação de comple-xos entre o antígeno e o anticorpo, que se ligam em receptores dos antígenos e receptores Fcγ inibitórios nas células B específicas para aquele antígeno.
Esse processo é chamado de retroali-mentação de anticorpos, ou seja, a re-gulação negativa da produção de anti-corpos pelos próprios anticorpos IgG se-cretados. Os complexos antígeno-anti-corpo se ligam a um receptor de células B para as porções Fc de IgG, chamado de receptor II Fcγ ou Fcγ RIIB. A cauda ci-toplasmática do Fcγ RIIB contém um mo-tivo de inibição de imunorreceptor base-ado em tirosina (ITIM).
Quando o receptor Fcγ de células B é acoplado, o ITIM na cauda citoplasmá-tica do receptor é fosforilado nos resí-duos tirosina e forma um local de anco-ramento para a fosfatase-5 inositol (SHIP). A SHIP recrutada hidrolisa um fos-fato no lipídio de sinalização intermediá-ria fosfatidilinositol trifosfato (PIP3) e o inativa.
Esse processo de feedback negativo da produção de anticorpos mediada pelo receptor de FcRIIb constitui um meca-nismo fisiológico de controle das respos-tas imunes humorais, pois é desencade-ada pelo anticorpo secretado e também bloqueia a produção de anticorpos de modo a garantir a homeostase e evitar a produção descontrolada de anticorpos.
Estudos apontam que um polimorfismo no gene Fcγ RIIB pode estar associado à suscetibilidade ao desenvolvimento da doença autoimune conhecida como lú-pus eritematoso sistêmico (LES) em hu-manos.

IMUNIDADE ADAPTATIVA: LINFÓCITOS B
15
@jalekoacademicos Jaleko Acadêmicos @grupoJaleko
REFERÊNCIAS
ABBAS, Abul K.; LICHTMAN, Andrew H.; PILLAI, Shiv. Imunologia celular e molecular. 8ed. Rio de Janeiro: Elsevier, 2015.
VISITE NOSSAS REDES SOCIAIS



![351lulas imune nat e adaptativa [Modo de Compatibilidade]) · fungos, fosfolipideos de parasitas, envelope viral, proteínas de choque térmico do proprio ... Imunidade adaptativa:](https://img.document.onl/doc/110x75/5be5e14a09d3f28a428ce150/351lulas-imune-nat-e-adaptativa-modo-de-compatibilidade-fungos-fosfolipideos.jpg)






![aula indicadores imunol gicos 2013 [Modo de compatibilidad]) · organismo: vírus, bactérias, parasitas ou ... Imunidade específica Resposta adaptativa Reconhece invasores, destrói](https://img.document.onl/doc/110x75/5be5e14a09d3f28a428ce14a/aula-indicadores-imunol-gicos-2013-modo-de-compatibilidad-organismo-virus.jpg)