Embed Size (px)
Citation preview

1
INTRODUÇÃO GERAL
A busca por uma vida saudável e de corpo e mente em harmonia, vem
se tornando uma preocupação de pessoas do mundo inteiro nos últimos anos.
Neste contexto, os consumidores se deparam com a falta do consumo regular
de produtos na forma in natura. As frutas estão, cada vez mais, sendo grandes
aliados na prevenção de doenças, em razão de suas propriedades
antioxidantes atribuídas à presença de compostos fenólicos, dentre eles os
subgrupos antocianinas e flavonóis.
A escassez de trabalhos sobre as frutas nativas da Região Sul da
América do Sul, a crescente demanda de informações, o apelo mercadológico
dos consumidores ávidos por novos produtos e a possibilidade de utilização
das frutas nas indústrias, são parâmetros que impulsionam a pesquisa e
viabilidade de aumento da produção e do consumo dessas frutas.
Vários estudos associam o consumo de frutas com a redução de
diversas doenças crônicas (Kritchevski, 1999; Nicoli, Anese e Parpinel, 1999).
Mansouri et al. (2005) e Vayalil (2002) enfatizam que as frutas são ricas em
compostos solúveis em água, dentre eles, metabólitos secundários (compostos
fenólicos e carotenóides) que possuem ação antioxidante frente a radicais
livres no organismo humano. Os radicais livres, quando em grandes
quantidades e em condições específicas, são responsáveis pelo
desenvolvimento de doenças crônicas degenerativas, tais como doenças
cardiovasculares e câncer.
Os compostos fenólicos também apresentam possíveis ações
antiproliferativas frente a linhagens de células cancerígenas, impedindo a

2
multiplicação dessas células no organismo humano, combatendo assim o
surgimento e proliferação de diversos tipos de câncer (Fang, Yang e Wu, 2002;
Adhami et al., 2007 e Nooman, Benelli e Albini, 2007 ).
O objetivo principal desse estudo foi quantificar os teores totais de
compostos fenólicos, de antocianinas, de carotenóides e do ácido L-ascórbico
em frutas nativas da Região Sul do Brasil, além de avaliar a atividade
antioxidante e antiproliferativa destas frutas frente a células tumorais.

3
CAPÍTULO I – QUANTIFICAÇÃO DE COMPOSTOS BIOATIVOS EM
FRUTAS NATIVAS DA REGIÃO SUL DA AMÉRICA DO SUL
INTRODUÇÃO
O Brasil possui a flora mais complexa do planeta, com espécies de
frutas nativas adaptados às diferentes condições climáticas, incluindo em clima
equatorial, tropical, semi-árido e temperado. Vários estudos vêm sendo
realizados com plantas (Silva et al., 2005) e frutas tropicais (Lim et al., 2004,
Kukoski et al., 2005), mas são escassos os dados sobre espécies frutíferas
nativas de clima temperado. Os benefícios à saúde, atribuídos aos alimentos
ricos em compostos fenólicos e outros antioxidantes naturais (como ácido
ascórbico e carotenóides) na sua composição química, têm elevado a procura
por novas espécies botânicas que possuam, além dessa propriedade, uma
atividade biológica complementar relevante (Céspedes et al., 2008). Dentre
essas propriedades, destacam-se aquelas relacionadas à proteção contra
doenças crônicas e processos infecciosos ou imunodepressivos (Prior et al.,
1998; Weisburger, 1999; Ross e Kasum, 2002; Adhami et al., 2007). Acredita-
se que as pequenas frutas nativas de clima temperado, com destaque para as
mirtáceas (gênero Myrtaceae), como pitanga (Eugenia uniflora), araçá (Psidium
cateleianum) e guabiroba (Campomanesia xanthocarpa), possam ser
portadoras de algumas dessas propriedades. Essas espécies estão

4
amplamente distribuídas nos estados do Sul do Brasil, mas são também
encontradas em outras regiões do País e da América do Sul.
A Embrapa Clima Temperado vem estruturando acervos de acessos de
pitanga e de araçá, buscando desenvolver seleções que contemplem aspectos
agronômicos como porte e arquitetura, produção e qualidade (tamanho,
aparência e sabor) das frutas. A expressão comercial ainda é limitada, devido,
sobretudo, à insuficiente disponibilidade de material propagativo e também pelo
desconhecimento da população sobre essas frutas, tendo em vista que o
consumo ainda é restrito às microrregiões de produção e à população do meio
rural, a qual, pelo conhecimento local, valoriza esses produtos. Além disso, há
materiais ainda não coletados e espécies pouco estudadas, como é o caso da
guabiroba, cereja-do-mato e uvaia. Porém, os efeitos benéficos ao trato
digestivo e respiratório, proteção a infecções, além de “bem estar geral”, são
amplamente divulgados pelos consumidores desses frutos.
As frutas nativas do Hemisfério Norte têm sido estudadas e avaliadas
quanto a sua composição química e propriedades funcionais. Sabe-se que
frutas como o mirtilo, framboesa e amora são fontes importantes de
substâncias com propriedades bioativas, potencializando o mercado de
produção, pela popularização dos benefícios que proporcionam (Pool-Zobel et
al., 1999; Smith et al., 2000).
Ervas, como o chá verde (Camellia Sinensis L. Kuntz), uma bebida
consumida nos países asiáticos e, nas últimas décadas, popularizada em todo
planeta, contém compostos com forte capacidade antioxidante. Essa planta
possui diversos polifenóis importantes na sua composição química, tais como a
(-) epicatequina e (-) epigalocatequina, que têm atraído a atenção da área da
saúde por ser um agente biologicamente ativo com efeitos anticancerígenos,
antidiabéticos, antiviral, cardioprotetor, hepatoprotetor e neuroprotetor
(Anderson e Polanski, 2002; Adhami et al., 2007; Nooman, Benelli e Albini,
2007).

5
Nesse estudo, pretendeu-se verificar se as frutas nativas da Região Sul
do Brasil apresentam uma elevada concentração de compostos bioativos
oriundos do metabolismo secundário das plantas, e sua influencia sobre a
atividade antioxidante das frutas.
Para esta finalidade, o objetivo desse estudo foi avaliar a composição
química de espécies nativas da Região Sul do Brasil, quanto aos seus teores
de compostos fenólicos totais, antocianinas totais, carotenóides totais e
conteúdo em ácido L-ascórbico, e avaliar a atividade antioxidante das frutas.

6
REVISÃO DE LITERATURA
2.1 Frutas Nativas
A fruticultura brasileira ocupa posição de destaque no cenário mundial,
devido a sua elevada produção. Apesar do crescimento da área de plantio e
produção de algumas frutíferas, as espécies nativas ainda não recebem a
devida importância econômica, apesar de seu grande potencial de exploração
(D’eeckenbrugge et al., 1998), tanto para o mercado interno como para o
externo (Bezerra et al., 2003), sendo ainda pouco exploradas (Ferreira, 1999;
Harder et al., 2004).
Entre as diversas plantas nativas, destacam-se as da familia Myrtaceae.
Essa família possui cerca de 140 gêneros e mais de 3.000 espécies
distribuídas em dois centros de dispersão, na América e na Austrália. Estes
gêneros incluem frutas comestíveis e de sabor agradável, como o araçá,
cereja, uvaia, guabiroba, pitanga e jabuticaba (Silva et al., 2001).
A pitangueira (Eugenia uniflora) é uma planta originária do Brasil.
Encontra-se por todo o país, desde o Nordeste até o Rio Grande do Sul,
ultrapassando fronteiras inclusive de algumas regiões do Uruguai e Argentina.
É uma árvore que nas regiões subtropicais alcança 2 a 4m de altura, mas,
vegetando sob condições ótimas de clima e de solo, pode alcançar até 10m
quando adulta (Pereira, 2006). A pitanga consiste em uma baga globosa, com
sete a dez sulcos longitudinais de 1,5 a 5,0cm de diâmetro, coroado com
sépalas persistentes que possui aroma característico intenso e sabor doce e
ácido. No processo de maturação, o epicarpo passa de verde para amarelo,

7
alaranjado, vermelho, vermelho escuro, podendo chegar até quase negro (Bezerra,
Silva Júnior e Lerdeman, 2000).
O araçá (Psidium cateleianum) é uma das frutíferas nativas do Brasil que está
sendo muito estudada devido às suas características de alta qualidade da fruta e
alta produção da planta. Segundo Donadio (2000), este arbusto tem porte pequeno a
médio, atingindo de 1 a 10m de altura. A fruta é de coloração amarelo a amarelo-
esverdeado, além de possuir frutas de coloração roxo-avermelhada. Possui sabor
adocicado e é muito consumida em feiras livres, cuja safra ocorre nos meses de
fevereiro a abril (Manica, 2000).
A jabuticabeira (Myrciaria cauliflora) é uma espécie nativa da Mata Atlântica
brasileira, que foi domesticada e incorporada pelos indígenas tupis (Mattos, 1983). A
fruta consiste de uma baga, subglobosa, negra lisa com 1,2 a 2,2cm de diâmetro,
contendo 1 a 4 sementes (Donadio, Môro e Servidone, 2002). Suas frutas são
consumidas in natura ou processadas na forma de geléias, licores e vinhos.
A cereja-do-mato (Eugenia involucrata), também conhecida como cereja-da-
terra ou cereja-do-rio-grande, também é uma espécie nativa do Brasil. A planta é
rústica e ornamental, com boa formação de copa, podendo ser utilizada para a
arborização urbana. Suas frutas podem ser utilizadas na fabricação de doces,
geléias, licores e na produção de mel (Sanchotene, 1989; Lorenzi, 1998).
A uvaia (Eugenia uvalha), conhecida também por uvalha, é uma árvore de 6 a
13m de altura, dotada de copa arredondada, tronco geralmente ereto, com 30 a
50cm de diâmetro. A época de florescência ocorre entre os meses de agosto a
setembro, com início de maturação dos frutos em setembro, prolongando-se até
novembro. As frutas são druplas globosas de cor amarela e carnosa, contendo de
uma a três sementes. A fruta é muito apreciada na forma de sucos (Lorenzi, 1998;
Andrade e Ferreira, 2000).
A guabirobeira (Campomanesia xanthocarpa) também é uma planta nativa da
Região Sul do Brasil, Uruguai, Argentina e Paraguai; apresenta um grande porte,
atingindo mais de 10m de altura, quando adulta. A guabiroba é uma fruta de
coloração amarelo-esverdeada, consumida in natura ou utilizada na elaboração de
sucos, licores e doces (Biavatti et al., 2004).

8
2.2 Radicais Livres e Atividade Antioxidante
Os consumidores estão cada vez mais preocupados com a qualidade de vida
e em possuir bons hábitos alimentares, como consumir alimentos que ajudem a
manter a boa saúde. As frutas são alimentos excelentes, pois além de possuírem
baixas calorias, contêm quantidades significativas de substâncias antioxidantes, as
quais são capazes de prevenir uma grande variedade de patologias, como câncer,
doenças cardiovasculares, derrames e isquemias (Tavarini et al., 2008). Por esta
razão, frutas e hortaliças representam dentro da pirâmide alimentar, a maior fonte de
antioxidantes naturais na dieta (Tavarini et al., 2008). O crescimento no consumo de
frutas, associado às contínuas melhorias que estão sendo introduzidas na qualidade
dos alimentos, indicam que as frutas e outros vegetais devem ganhar, cada vez
mais, espaço no mercado consumidor (Agostini-Costa, 2003).
Os compostos antioxidantes, presentes nas frutas e hortaliças atuam nas
espécies reativas de oxigênio, que incluem radicais livres reativos, tais como
superóxidos, hidroxilas e peroxila, e também com não radicais, como peróxido de
hidrogênio. Estas substâncias atuam na etapa de iniciação e propagação dos
radicais livres ou supressão da formação dos mesmos (Lim, Lim e Tee, 2007).
Os radicais livres são átomos ou moléculas produzidas continuamente
durante os processos metabólicos e atuam como mediadores para a transferência
de elétrons em várias funções relevantes no metabolismo. As principais fontes de
radicais livres são as organelas citoplasmáticas que metabolizam o oxigênio, o
nitrogênio e o cloro, gerando grande quantidade de metabólitos (Shami e Moreira,
2004). Porém, fontes exógenas também contribuem na formação de radicais livres
no organismo humano, como por exemplo, o tabagismo e a poluição (Elsayed, 2001;
Lachance et al., 2001).
Quimicamente, os radicais livres são quaisquer átomos, moléculas ou
fragmentos de molécula contendo um ou mais elétrons desemparelhados nas suas
camadas de valência (Halliwell e Gutteridge, 1990). Essa configuração faz dos
radicais livres moléculas altamente instáveis, com curto tempo de meia-vida e
quimicamente muito reativas (Pompella, 1997). Das espécies reativas de oxigênio, o
radical hidroxil (OH•) e o ânion superóxido (O2•) são os que têm maior importância

9
biológica, porque são formados durante o processo normal ou exarcebado de
redução de O2 no interior das mitocôndrias (Benzi, 1993).
Diversos estudos têm demonstrado que uma série de doenças entre as
quais, câncer, aterosclerose, diabetes, artrite, malária, AIDS e doenças
cardiovasculares podem estar ligadas aos danos causados por formas de oxigênio
extremamente reativas (Brenna e Pagliarini, 2001; Yldrim, Mavi e Kara, 2002). Os
radicais livres podem atacar vários substratos no corpo humano e contribuir para o
desenvolvimento de doenças crônicas, como por exemplo, oxidação da fração LDL
causando doenças cardio-vasculares (Touyz, 2004). À oxidação do DNA pode-se
atribuir um importante papel no desenvolvimento de diversos tipos de câncer (Lim et
al., 2004).
As lesões causadas pelos radicais livres nas células podem ser prevenidas ou
reduzidas por meio da atividade de antioxidantes, sendo estes encontrados em
muitos alimentos. Os antioxidantes são capazes de interceptar os radicais livres
gerados pelo metabolismo celular ou por fontes exógenas, impedindo o ataque
sobre os lipídeos, nos aminoácidos das proteínas, nas duplas ligações dos ácidos
graxos polinssaturados e as bases de DNA, evitando a formação de lesões e a
perda de integridade celular (Bianchi e Antunes, 1999).
Vários estudos têm quantificado a atividade antioxidante em diversos
alimentos, especialmente em produtos de origem vegetal, tais como grãos, vegetais
e frutas (Connor et al., 2002; Karl e John, 2002; Imeh e Khokhar, 2002; Ou et al.,
2002; Sun et al., 2002). O interesse crescente em consumir frutas deve-se ao seu
conteúdo de compostos antioxidantes, além de ser de fácil consumo, já que podem
ser ingeridas frescas, na forma de sucos ou em outros tipos de alimentos
processados. Especial atenção é dada às frutas, fontes ricas de compostos
fenólicos (Kalt et al., 1999; Robards et al., 1999; Wang e Lin, 2000).
2.3 Compostos Bioativos
De modo geral, as frutas e hortaliças contêm significantes teores de
compostos bioativos com funções fisiológicas e bioquímicas, conferindo benefícios à

10
saúde humana. Nos últimos tempos, os alimentos que contêm grandes teores de
compostos fenólicos, dentre eles as antocianinas, carotenóides e vitaminas, como a
E e a C na sua composição, assumiram o status de “alimentos funcionais”, pois além
de satisfazerem as necessidades nutricionais, oferecem benefícios fisiológicos, tais
como a prevenção de importantes doenças crônicas (Tavarini et al., 2008).
Os compostos fenólicos (Figura 1) compreendem uma grande variedade de
substâncias divididas em várias classes, assim como os ácidos fenólicos (derivados
do ácido hidrobenzóico e ácido cinâmico) e flavonóides (flavonas, flavonoóis,
flavanonas e isoflavonas).
Figura 1. Estrutura química geral de compostos fenólicos.
Esses compostos estão presentes na maioria dos vegetais, frutas, sementes,
chás, vinhos e sucos (Manach et al., 2004; Lu e Foo, 2001; Murthy, Singh e
Jayaprakasha, 2002). Recentemente, os compostos fenólicos têm recebido atenção
desde que estudos epidemiológicos sugeriram que o consumo de alimentos e
bebidas ricos em polifenóis está associado com a redução no risco de doenças
cardiovasculares, derrame cerebral e alguns tipos de câncer. Esses efeitos
protetores têm sido atribuídos às propriedades antioxidantes especialmente dos
flavonóides (Figura 2) (Kaur e Kapoor, 2001). Pesquisa recente conduzida por Lau,
Shukitt-Hale e Joseph (2005) demonstraram que antioxidantes naturais, presentes
em mirtilo (Vaccinium spp.) podem reverter declínios fisiológicos decorrentes da

11
idade avançada, como a perda da função motora. Kraft et al. (2005) relatam que
espécies de mirtilo contêm cadeias de compostos como os fitosteróis, ácidos
fenólicos, flavanas-3-óis, antocianinas e proantocianidinas oligoméricas que
protegem contra os estádios de iniciação, promoção e progressão de
carcinogenesis.
Figura 2. Estrutura química dos principais flavonóides.
As antocianinas (Figura 3), uma subclasse pertencente aos flavonóides, são
amplamente encontrada em frutas, flores e vegetais, em geral, sendo responsáveis
pelas cores laranja, vermelho e azul. Além disso, além da função do pigmento
colorido nos alimentos as antocianinas estão relacionadas com a atividade
antioxidante (Kim, Jeong e Lee, 2003; Philpott, et al., 2004), promovendo efeitos
benéficos à saúde, reduzindo a incidência de doenças crônicas, além de ser um
importante agente antiinflamatório (Wang, Cao e Prior, 1996; Wang e Jiao, 2000).
Estudos em células humanas e em ratos demonstraram que em dietas ricas
destes compostos, as antocianinas podem exercer ação benéfica contra o início e

12
desenvolvimento de doenças cardiovasculares. As antocianinas e os outros
subgrupos de compostos fenólicos, devido à presença de vários radicais fenólicos,
evitam a oxidação da fração LDL e a deposição de gorduras nas artérias (Negre-
Salvagyre, Salvagyre, 1992). Elas também podem agir na redução de doenças
neurológicas (Andriambelosom et al. 1998; Tsuda, Horio e Osawa, 1998).
Figura 3. Estrutura química das principais moléculas de antocianinas.
Os carotenóides (Figura 4) exercem ações relacionadas à redução do risco de
doenças degenerativas, prevenção da formação de catarata, redução da
degeneração macular relacionada ao envelhecimento e redução do risco de doenças
coronárias (Krinski, 2001). Além disso, os carotenóides desempenham um papel
fundamental como pigmento acessório na fotossíntese, agindo como coletor de
energia e protetor contra foto-oxidação (Krinski, 2001; Silva e Mercadante, 2002).
Fraser e Bramley (2004) associam a alta ingestão de alimentos ricos em β-caroteno
com a redução da incidência de câncer de mama. A luteína está inversamente
relacionada à incidência de câncer de cólon tanto em homens como em mulheres
(Slaterry et al., 2000). Segundo Olson (1999), os carotenóides seqüestram o
oxigênio singlete, removem os radicais peróxidos, modulam o metabolismo
carcinogênico, inibem a proliferação celular, estimulam a comunicação entre células
(junções gap), e elevam a resposta imune.

13
Figura 4. Estrutura química das principais moléculas de carotenóides.
Vários estudos têm investigado o efeito de vitamina C (Figura 5) de frutas e
vegetais e o desenvolvimento de câncer. Estudos epidemiológicos sugerem que
vitaminas antioxidantes, incluindo a vitamina C, em concentrações suficientes inibem
doenças no coração e câncer (Flagg, Coates e Greenberg, 1995). Alguns estudos
recentes têm mostrado uma redução significante no risco de câncer de pulmão pelo
aumento de vitamina C na dieta (Ocké et al., 1997).
Figura 5. Estrutura química do ácido L-ascórbico.

14
Devido às espécies nativas de clima temperado terem co-evoluido em
ecossistema com significativas variações de temperatura, oscilações hídricas, assim
como de luminosidade, acredita-se que essas espécies tenham desenvolvido um
complexo metabolismo secundário, resultando, no mínimo, em frutas ricas nesses
compostos, que além de serem mecanismos de defesa, exercem algum efeito
fisiológico de propriedade terapêutica. Por este motivo, é importante conhecer a
composição química dessas frutas e verificar suas potencialidades como agentes
antioxidantes.

15
MATERIAL E MÉTODOS
3.1 Material Vegetal
Foram utilizadas frutas das espécies Psidium cateleianum (araçá), Eugenia
involucra (cereja-do-mato), Campomanesia xanthocarpa (guabiroba), Myrciaria
cauliflora (jabuticaba), Eugenia uniflora (pitanga), Eugenia uvalha (uvaia), todos da
familia Myrtaceae, colhidos nos anos 2005 e 2008 (Tabela 1). Dos 48 acessos
estudados de frutas de araçá, cereja-do-mato, guabiroba, jabuticaba, pitanga e
uvaia, 44 genótipos foram provenientes do Banco de Germoplasma da Embrapa -
Clima Temperado, Pelotas/RS. As demais frutas foram coletadas em propriedades
rurais da Região Sul do RS. A colheita das frutas foi realizada no estádio de
maturação característica de cada espécie, pela coloração externa da casca. Após a
colheita, as frutas foram imediatamente congeladas a -80ºC até o momento das
avaliações. Todas as avaliações foram feitas com extratos das frutas com casca,
pois essa é a forma como são comumente consumidos.
Para efeito comparativo, utilizaram-se duas espécies frutíferas, Rubus sp.
(amora-preta) e Vaccinium myrtillus (mirtilo) não nativas da Região Sul, mas que se
adaptaram ao solo e ao clima da região (Tabela 2). Ambas as espécies são
amplamente conhecidas como fontes de compostos biologicamente ativos e de alta
atividade antioxidante. Essas frutas foram colhidas também no Banco de
Germoplasma da Embrapa Clima Temperado, Município de Pelotas/RS.

16
TABELA 1. Frutas nativas da Região Sul da América do Sul utilizadas.
Fruta Nome Científico Coloração Codificação
Amarela
AR012, AR031, AR052,
AR271, AR462, AR472,
AR482, AR492, AR572,
AR582, AR592, AR602,
AR642, AR722, AR732,
AR742, AR752 Araçá Psidium cateleianum
Roxa
AR082, AR092, AR101,
AR112, AR182, AR192,
AR282, AR292, AR422,
AR442, AR942, AR1002,
AR1012
Cereja-do-
Mato Eugenia involucra Roxa CER3
Guabiroba Campomanesia
xanthocarpa Laranja Gua011, Gua023, Gua033
Jabuticaba Myrciaria cauliflora Roxa JAB3
Laranja Pit151, Pit711
Roxa PitH1, Pit401
Pitanga Eugenia uniflora
Vermelha
PitU1, Pit411, Pit492, Pit502,
Pit532, Pit552, Pit612,
Pit622
Uvaia Eugenia uvalha Laranja UVI3
1 Colhidos Safra 2005-2006; 2 Colhidos Safra 2006-2007; 3 Colhidos Safra-2007-2008.
TABELA 2. Frutas não-nativas da Região Sul da América do Sul estudados.
Fruta Nome Científico Coloração Codificação
Amora-Preta Rubus sp. Roxa AMR1
Mirtilo Vaccinium myrtillus Roxa MIR1
1 Colhidos Safra 2007-2008.

17
3.2 Avaliações
3.2.1 Determinação de Sólidos Solúveis Totais (SST), pH, Acidez Total
Titulável (ATT) e Relação SST/ATT
Foram realizadas as avaliações de sólidos solúveis totais (SST), pH e acidez
total titulável (ATT) segundo método de Manzino, Silvestrini e Rearte (1987). A
determinação de SST foi feita por meio de um refratômetro manual, com
compensação de temperatura, sendo os resultados expressos em graus Brix. O pH
foi determinado, por via direta, e, potenciômetro digital. A ATT foi realizada por
titulometria de neutralização com hidróxido de sódio 0,1N, sendo os resultados
expressos em mol (m/v) de equivalentes de ácido cítrico.
3.2.2 Determinação de Antocianinas Totais
O teor de antocianinas totais foi determinado por espectrofotometria de
acordo com Lees e Francis (1972). Dois gramas de fruta foram macerados e, em
seguida, 50mL de etanol pH 1,0 foi adicionado para a extração das antocianinas. A
solução foi homogeneizada a cada cinco minutos, em um intervalo de uma hora.
Após esse período, filtrou-se a solução para um balão volumétrico de 100mL e
completou-se o volume. A leitura foi realizada a 520nm, utilizando etanol pH 1,0
como referência. O cálculo da concentração das antocianinas totais foi baseado na
Lei de Beer e os resultados obtidos foram expressos em miligramas de cianidina-3,5-
glicosídeo por 100 gramas de peso fresco.
3.2.3. Determinação de Compostos Fenólicos Totais
O teor de compostos fenólicos totais foi determinado pelo método modificado
de Folin-Ciocalteu (Singleton e Rossi, 1965). Um grama de polpa do fruto foi
macerado e, em seguida, adicionou-se 60mL de água Milli-Q e 5mL do Reagente
de Folin-Ciocalteu. Seguiu-se a neutralização com 20mL de solução saturada de

18
carbonato de sódio. A absorbância da solução resultante foi medida a 720nm após
um período de duas horas de reação. Como ensaio controle, realizou-se todas as
etapas citadas anteriormente, excetuando-se a adição de um grama de polpa, que
foi substituído por 1mL de água Milli-Q. Ácido gálico foi usado para construir uma
curva padrão (0,00-120,00 mg.100g-1; y = 0,0112 x; R2 = 0,9978). Os resultados
foram expressos em equivalente de ácido gálico (miligramas por 100 gramas de
peso fresco).
3.2.4 Determinação de Carotenóides Totais
O teor de carotenóides totais foi determinado pelo método proposto por
Rodriguez-Amaya (1999). Cinco gramas de amostras foram macerados e adicionou-
se 2g de celite. Acrescentou-se 20mL de acetona na amostra e homogenizou-se por
10 minutos, e filtrou-se a vácuo. O resíduo foi lavado com acetona até que o mesmo
ficasse incolor. Então, tranferiu-se essa solução para um funil de separação e
acrescentou-se 200mL de éter de petróleo. A amostra foi lavada com água destilada
até remoção completa da acetona. Após a remoção completa do solvente transferiu-
se a amostra para um balão volumétrico de 250mL e completou-se o volume com
éter de petróleo. A leitura foi realizada a 450nm, utilizando éter de petróleo como
controle, e os resultados foram expressos em microgramas de β-caroteno por grama
de peso-fresco, seguindo a fórmula abaixo:
Carotenóides Totais = Absorbância X Volume Extrato (mL) X 106
2500 X 100 X peso amostra (g)
3.2.5 Determinação do Ácido L-Ascórbico
O método de extração foi realizado segundo Vinci, Rot e Mele (1995). Dez
gramas de amostra e solução de ácido metafosfórico a 4,5% foram deixados em
repouso por uma hora, em frasco protegido da luz. Transferiu-se a amostra para um
balão de 50mL e completou-se o volume com água Milli-Q, filtrando a amostra

19
posteriormente. Centrifugou-se o filtrado a 7000rpm por 10minutos. O sobrenadante
dessa centrifugação injetou-se diretamente no cromatógrafo.
A análise cromatográfica foi realizada em cromatógrafo Shimadzu, equipado
com módulo de mistura de solventes LC-10ATVP, desgaseificador FCV-10AVP,
bomba reodine DGU-14A, sistema de controle SCL-10AVP, forno de coluna CTO-
10ASVP e amostrador automático SIL-10AF. Foi utilizada uma coluna de separação
analítica de fase reversa, Nova-Pak C18 (3,9cm x 150mm x 4µm). Como fase móvel
utilizou-se dois solventes: (A) solução de ácido acético 0,1% e (B) metanol 100%
(Tabela 3). Injetou-se 10µL de amostra, com fluxo de 0,8mL.min-1, e temperatura de
coluna mantida a 25°C. O tempo de eluição da amostra foi de 12 minutos. A
identificação do composto foi realizada através de informações obtidas dos
espectros de absorção no UV-visível, modelo SPD-10AVVP em um comprimento de
onda de 254nm. Os dados obtidos foram adquiridos e processados com o uso do
software Class-VP, por meio de uma curva de calibração externa quadrática,
preparada com ácido L-ascórbico (reagente padrão com 99,97% de pureza), em
concentrações que variaram entre 10-100mg.100mL-1, utilizando fase móvel e
gradiente de eluição igual ao utilizado. Na determinação de ácido L-ascórbico, com
tempo de eluição de 10 minutos e fluxo de 0,8mL.min-1. Os resultados foram
expressos em miligramas de ácido ascórbico por 100 gramas de peso fresco.

20
Tabela 3. Gradiente de eluição dos solventes A e B na determinação de ácido L-
ascórbico.
Tempo (minutos) Solvente A (%)1 Solvente B (%)2
0 100 0
5 98 2
7 98 2
10 100 0
1Solvente A: solução de água Milli-Q:ácido acético p.a. (99,9:0,1% v/v); 2Solvente B: metanol p.a.
100%.
3.2.6 Determinação da Atividade Antioxidante
Em extratos de fruto, foi quantificado a atividade antioxidante segundo método
adaptado de Brand-Willians, Cuvelier e Berset (1995), que se baseia na redução do
radical estável 2,2-difenil-1-picrylhidrazil (DPPH). Para o preparo dos extratos
utilizaram-se 5 g de polpa de fruta trituradas em Ultraturrax®, aos quais foi
adicionado 20 mL de metanol. Essa mistura foi mantida durante 24 h a 4ºC, após o
qual se centrifugou a 12000g a 4ºC durante 20 minutos. Usou-se 0,1mL de extrato
em 3,9mL de DPPH a 0,1mM, e realizou-se a leitura após 30 minutos de incubação
a 25ºC. Porém, em ensaios preliminares, verificou-se que nessas concentrações
havia a redução total do radical DPPH antes dos 30 minutos preconizados para
algumas frutas. Por isso, para araçás, cereja-do-mato, pitangas e uvaia padronizou-
se o uso de 0,01 mL de extrato metanólico, e para a guabiroba e a jabuticaba
utilizou-se 0,001 mL. A leitura para estes extratos foi realizada após 1 minuto ao
invés de 30 minutos.
Todas as operações foram realizadas na ausência de incidência direta de luz.
No ensaio controle, ao invés da adição do extrato metanólico, acrescentou-se
apenas metanol. A concentração do radical DPPH remanescente foi lida a 517nm.
Os resultados foram expressos em mM de Trolox (6-hidroxi-2,5,7,8-
tetrametilcroman-2-ácido carboxílico), que consiste de um antioxidante sintético de

21
referência. Para a quantificação, foi realizada uma curva-padrão de Trolox, que
variou entre 0,20 a 1,40mM (y = 0,032 x; R2 = 0,9643).
3.3 Delineamento Experimental
O delineamento experimental foi inteiramente casualizado, com três
repetições para cada unidade de experimental. A unidade experimental consistiu em
500g de cada uma das seleções colhidas. Os dados foram expressos em médias
aritméticas. O teste de Duncan a 5% de probabilidade foi aplicado para determinar
as diferenças das médias. As análises estatísticas foram realizadas com o auxílio do
programa de computação STATISTICA 6.0.

22
RESULTADOS E DISCUSSÃO
4.1 Teor de Sólidos Solúveis Totais, pH, Acidez Total Titulável e
Relação SST/ATT
As frutas de guabiroba foram os que apresentaram os maiores teores de
sólidos solúveis totais (SST), que em média foi de 14,3°Brix. A seleção de pitanga-
vermelha Pit53 foi a que apresentou menor teor de SST de todas as frutas avaliados
neste trabalho, com valor médio de 3,27. Dentro da mesma espécie também houve
diferenças significativas. No caso dos acessos de araçá-amarelo houve variação
entre 5,4 e 10,8°Brix, no araçá-roxo entre 6,4 e 13,6°Brix, na pitanga-laranja entre
7,0 e 9,8°Brix, na pitanga-roxa entre 8,2 e 11,4°Brix e na pitanga-vermelha entre 4,0
e 8,8°Brix. Os teores de SST para as frutas de cereja-do-mato, jabuticaba e uvaia
foram, respectivamente, 6,6°Brix, 12,4ºBrix e 6,0°Brix (Tabela 4).
A diferença significativa entre os acessos de mesma espécie pode ser devido
ao estádio de maturação, pois todas as frutas foram colhidas quando estavam
maduras do ponto de vista de consumo. Observaram-se diferenças entre a
quantidade de frutas por planta e época de floração entre os acessos, o que pode
ser uma das causas dessas diferenças. Os teores de SST podem variar, de acordo,
com o genótipo e fatores climáticos, tais como, a pluviosidade. O excesso de chuvas
ou até mesmo irrigação excessiva diminui o teor de SST nas frutas, em virtude da
diluição do suco celular (Gerhardt et al., 1997; Nogueira et al., 2002).
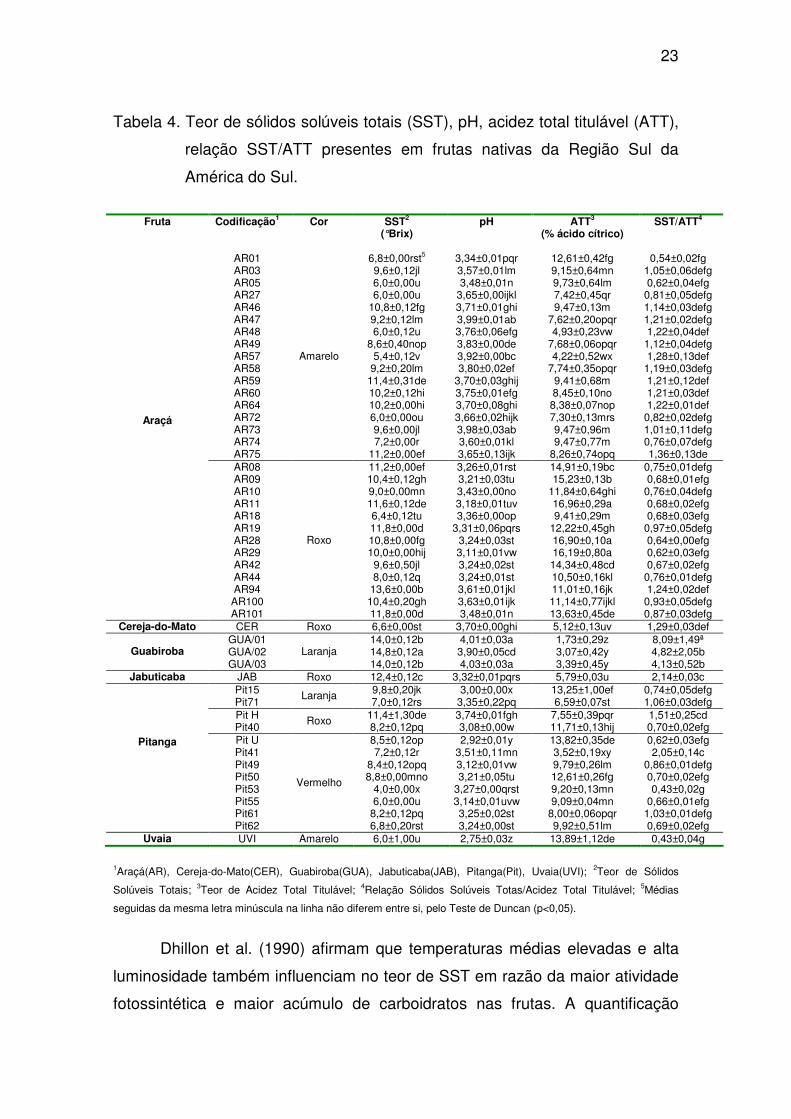
23
Tabela 4. Teor de sólidos solúveis totais (SST), pH, acidez total titulável (ATT),
relação SST/ATT presentes em frutas nativas da Região Sul da
América do Sul.
Fruta Codificação1 Cor SST2 pH ATT3 SST/ATT4
(°Brix) (% ácido cítrico)
AR01 6,8±0,00rst5 3,34±0,01pqr 12,61±0,42fg 0,54±0,02fg AR03 9,6±0,12jl 3,57±0,01lm 9,15±0,64mn 1,05±0,06defg AR05 6,0±0,00u 3,48±0,01n 9,73±0,64lm 0,62±0,04efg AR27 6,0±0,00u 3,65±0,00ijkl 7,42±0,45qr 0,81±0,05defg AR46 10,8±0,12fg 3,71±0,01ghi 9,47±0,13m 1,14±0,03defg AR47 9,2±0,12lm 3,99±0,01ab 7,62±0,20opqr 1,21±0,02defg AR48 6,0±0,12u 3,76±0,06efg 4,93±0,23vw 1,22±0,04def AR49 8,6±0,40nop 3,83±0,00de 7,68±0,06opqr 1,12±0,04defg AR57 5,4±0,12v 3,92±0,00bc 4,22±0,52wx 1,28±0,13def AR58 9,2±0,20lm 3,80±0,02ef 7,74±0,35opqr 1,19±0,03defg AR59 11,4±0,31de 3,70±0,03ghij 9,41±0,68m 1,21±0,12def AR60 10,2±0,12hi 3,75±0,01efg 8,45±0,10no 1,21±0,03def AR64 10,2±0,00hi 3,70±0,08ghi 8,38±0,07nop 1,22±0,01def AR72 6,0±0,00ou 3,66±0,02hijk 7,30±0,13mrs 0,82±0,02defg AR73 9,6±0,00jl 3,98±0,03ab 9,47±0,96m 1,01±0,11defg AR74 7,2±0,00r 3,60±0,01kl 9,47±0,77m 0,76±0,07defg AR75
Amarelo
11,2±0,00ef 3,65±0,13ijk 8,26±0,74opq 1,36±0,13de AR08 11,2±0,00ef 3,26±0,01rst 14,91±0,19bc 0,75±0,01defg AR09 10,4±0,12gh 3,21±0,03tu 15,23±0,13b 0,68±0,01efg AR10 9,0±0,00mn 3,43±0,00no 11,84±0,64ghi 0,76±0,04defg AR11 11,6±0,12de 3,18±0,01tuv 16,96±0,29a 0,68±0,02efg AR18 6,4±0,12tu 3,36±0,00op 9,41±0,29m 0,68±0,03efg AR19 11,8±0,00d 3,31±0,06pqrs 12,22±0,45gh 0,97±0,05defg AR28 10,8±0,00fg 3,24±0,03st 16,90±0,10a 0,64±0,00efg AR29 10,0±0,00hij 3,11±0,01vw 16,19±0,80a 0,62±0,03efg AR42 9,6±0,50jl 3,24±0,02st 14,34±0,48cd 0,67±0,02efg AR44 8,0±0,12q 3,24±0,01st 10,50±0,16kl 0,76±0,01defg AR94 13,6±0,00b 3,61±0,01jkl 11,01±0,16jk 1,24±0,02def AR100 10,4±0,20gh 3,63±0,01ijk 11,14±0,77ijkl 0,93±0,05defg
Araçá
AR101
Roxo
11,8±0,00d 3,48±0,01n 13,63±0,45de 0,87±0,03defg Cereja-do-Mato CER Roxo 6,6±0,00st 3,70±0,00ghi 5,12±0,13uv 1,29±0,03def
GUA/01 14,0±0,12b 4,01±0,03a 1,73±0,29z 8,09±1,49ª GUA/02 14,8±0,12a 3,90±0,05cd 3,07±0,42y 4,82±2,05b Guabiroba GUA/03
Laranja 14,0±0,12b 4,03±0,03a 3,39±0,45y 4,13±0,52b
Jabuticaba JAB Roxo 12,4±0,12c 3,32±0,01pqrs 5,79±0,03u 2,14±0,03c Pit15 9,8±0,20jk 3,00±0,00x 13,25±1,00ef 0,74±0,05defg Pit71
Laranja 7,0±0,12rs 3,35±0,22pq 6,59±0,07st 1,06±0,03defg
Pit H 11,4±1,30de 3,74±0,01fgh 7,55±0,39pqr 1,51±0,25cd Pit40
Roxo 8,2±0,12pq 3,08±0,00w 11,71±0,13hij 0,70±0,02efg
Pit U 8,5±0,12op 2,92±0,01y 13,82±0,35de 0,62±0,03efg Pit41 7,2±0,12r 3,51±0,11mn 3,52±0,19xy 2,05±0,14c Pit49 8,4±0,12opq 3,12±0,01vw 9,79±0,26lm 0,86±0,01defg Pit50 8,8±0,00mno 3,21±0,05tu 12,61±0,26fg 0,70±0,02efg Pit53 4,0±0,00x 3,27±0,00qrst 9,20±0,13mn 0,43±0,02g Pit55 6,0±0,00u 3,14±0,01uvw 9,09±0,04mn 0,66±0,01efg Pit61 8,2±0,12pq 3,25±0,02st 8,00±0,06opqr 1,03±0,01defg
Pitanga
Pit62
Vermelho
6,8±0,20rst 3,24±0,00st 9,92±0,51lm 0,69±0,02efg Uvaia UVI Amarelo 6,0±1,00u 2,75±0,03z 13,89±1,12de 0,43±0,04g
1Araçá(AR), Cereja-do-Mato(CER), Guabiroba(GUA), Jabuticaba(JAB), Pitanga(Pit), Uvaia(UVI); 2Teor de Sólidos
Solúveis Totais; 3Teor de Acidez Total Titulável; 4Relação Sólidos Solúveis Totas/Acidez Total Titulável; 5Médias
seguidas da mesma letra minúscula na linha não diferem entre si, pelo Teste de Duncan (p<0,05).
Dhillon et al. (1990) afirmam que temperaturas médias elevadas e alta
luminosidade também influenciam no teor de SST em razão da maior atividade
fotossintética e maior acúmulo de carboidratos nas frutas. A quantificação

24
dessa variável é importante, pois frutas com teores maiores de SST são mais
indicados para a elaboração de produtos, tais como geléias, doces em massa,
sucos e polpas, obtendo-se produtos característicos sensoriais mais
agradáveis e com maior rendimento industrial (Pinto et al., 2003).
Dentre os 48 acessos de frutas avaliados, houve diferença significativa
quanto aos valores de pH, que se situaram entre 3,34 e 3,99 para os acessos
de araçá-amarelo, 3,11 e 3,99 para araçá-roxo, 3,70 para cereja-do-mato, 3,90
e 4,03 para guabiroba, 3,32 para jabuticaba, 3,00 e 3,35 para pitanga-laranja,
3,98 e 3,74 para pitanga-roxa. 2,92 e 3,74 para pitanga-vermelha e 2,75 para
uvaia.
O fato de as frutas apresentarem valores de pH entre 2,75 e 4,03, os
caracterizam como ácidos, constituindo-se numa característica importante no
que tange a potenciais aspectos de processamento, pois é sabido que produtos
com pH inferior a 4,5 exigem tratamentos térmicos menos drásticos para a
conservação e ou são mais facilmente conserváveis pela combinação de
métodos, além de não ser necessário a adição de ácido cítrico nas formulações
(Semensato e Pereira, 2000). Isso é importante pensando-se na utilização
dessas frutas, seja na forma de produtos liofilizados, desidratados, sucos,
polpas, congelados, apertizados e outros.
Os teores de acidez total titulável (ATT) também apresentaram
diferenças significativas entre as frutas de mesma espécie e entre espécies
diferentes. Em média, os acessos de araçá-roxo foram os que apresentaram os
maiores de ATT, expressos em % de ácido cítrico equivalente, entre os 48
genótipos avaliados. A seleção AR11 (araçá-roxo) o fruto que apresentou teor
de ATT superior, 16,90% seguido pelo araçá-roxo AR28, com 16,19%. A uvaia
também apresentou alto teor de ATT com 13,89% de ácido cítrico. As frutas de
araçá-amarelo apresentaram valores de ATT que variaram entre 4,22 e 12,61%
de ácido cítrico. As frutas de guabiroba destacaram por não serem frutas com
um alto teor de ATT. Observou-se mesmo comportamento nas frutas de
jabuticaba.

25
Embora tenha havido marcantes variações nos teores de ATT entre as
frutas avaliadas, as variações de pH foram menores. Esse comportamento
sugere que os principais ácidos orgânicos das frutas não sejam de caráter
forte. As células buscam manter uma condição tamponante associando ou
dissociando os ácidos presentes de modo a manter o pH citosólico próximo a
6,8. Por isso que o pH vascular varia fortemente com a finalidade de manter o
pH do citosol.
Do ponto de vista tecnológico, a relação SST/ATT consiste em um
parâmetro de palatibilidade nos alimentos, que indica um equilíbrio entre os
sabores doce e ácido presentes no alimento. Quanto maior a razão entre SST
e ATT, mais “doce” o fruto é mais apreciado pelos consumidores e mais
indicado para ser destinado à indústria (Teixeira et al., 2001). As frutas de
guabiroba foram os que apresentaram maior relação SST/ATT (5,68), sendo
esses mais indicados como matéria-prima na indústria.
4.2 Teor de Compostos Fenólicos Totais
Dentre os 48 genótipos de frutas avaliados, houve diferença significativa
entre as distintas espécies e acessos estudados (Tabela 5). As frutas de
guabiroba do acesso GUA/01 foram os que apresentaram maior teor de
compostos fenólicos (1062,03 mg ácido gálico equivalente por 100g de peso
fresco). Os demais acessos de guabiroba apresentaram 269,88mg.100g-1
(GUA/02) e 277,46mg.100g-1 (GUA/02). Houve diferença significativa elevada
entre os teores de compostos fenólicos totais entre os acessos de mesma
espécie, devido provavelmente ao local de produção e/ou diferenças
genotípicas. O acesso que apresentou maior teor de compostos fenólicos totais
(GUA/01) foi em uma safra que a Região Sul do Estado do Rio Grande do Sul,
Brasil sofreu uma grande seca, havendo um período de escassez de chuvas na
época de floração e colheita (dados não apresentados). Provavelmente, as
condições climáticas proporcionaram um estresse à planta que,
conseqüentemente, produziu e acumulou uma concentração maior de
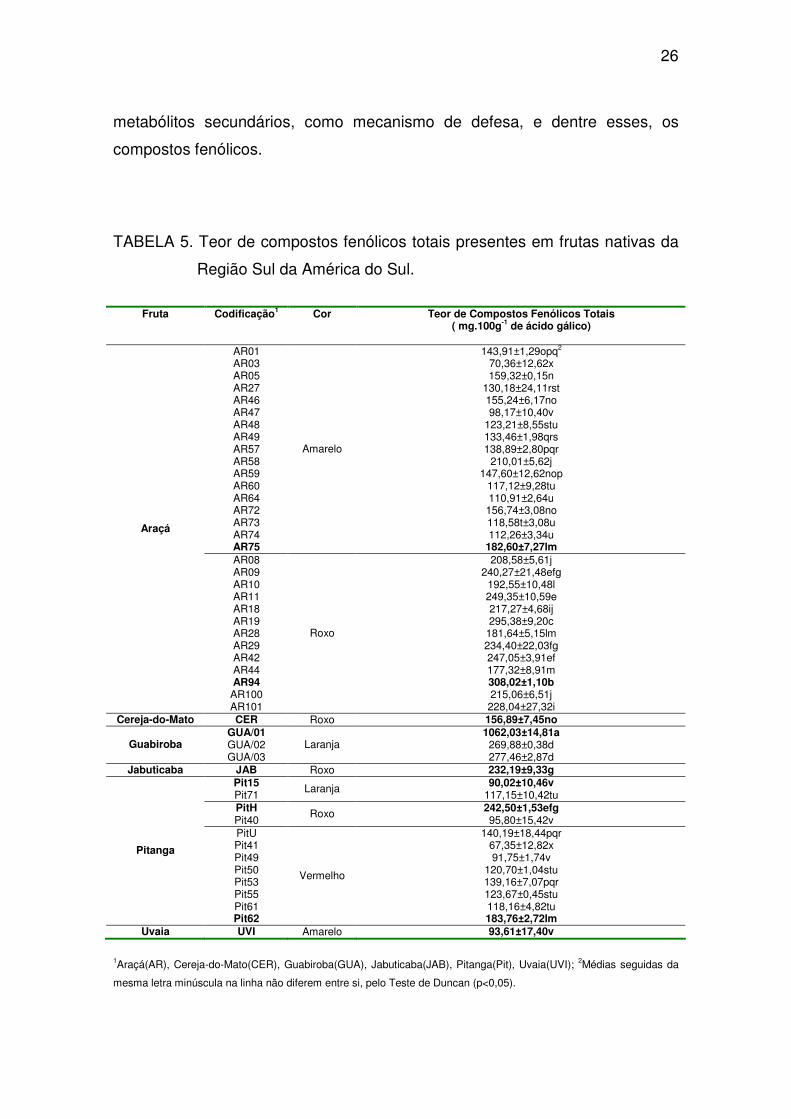
26
metabólitos secundários, como mecanismo de defesa, e dentre esses, os
compostos fenólicos.
TABELA 5. Teor de compostos fenólicos totais presentes em frutas nativas da
Região Sul da América do Sul.
Fruta Codificação1 Cor Teor de Compostos Fenólicos Totais ( mg.100g-1 de ácido gálico)
AR01 143,91±1,29opq2 AR03 70,36±12,62x AR05 159,32±0,15n AR27 130,18±24,11rst AR46 155,24±6,17no AR47 98,17±10,40v AR48 123,21±8,55stu AR49 133,46±1,98qrs AR57 138,89±2,80pqr AR58 210,01±5,62j AR59 147,60±12,62nop AR60 117,12±9,28tu AR64 110,91±2,64u AR72 156,74±3,08no AR73 118,58t±3,08u AR74 112,26±3,34u AR75
Amarelo
182,60±7,27lm AR08 208,58±5,61j AR09 240,27±21,48efg AR10 192,55±10,48l AR11 249,35±10,59e AR18 217,27±4,68ij AR19 295,38±9,20c AR28 181,64±5,15lm AR29 234,40±22,03fg AR42 247,05±3,91ef AR44 177,32±8,91m AR94 308,02±1,10b AR100 215,06±6,51j
Araçá
AR101
Roxo
228,04±27,32i Cereja-do-Mato CER Roxo 156,89±7,45no
GUA/01 1062,03±14,81a GUA/02 269,88±0,38d Guabiroba GUA/03
Laranja 277,46±2,87d
Jabuticaba JAB Roxo 232,19±9,33g Pit15 90,02±10,46v Pit71
Laranja 117,15±10,42tu
PitH 242,50±1,53efg Pit40
Roxo 95,80±15,42v
PitU 140,19±18,44pqr Pit41 67,35±12,82x Pit49 91,75±1,74v Pit50 120,70±1,04stu Pit53 139,16±7,07pqr Pit55 123,67±0,45stu Pit61 118,16±4,82tu
Pitanga
Pit62
Vermelho
183,76±2,72lm Uvaia UVI Amarelo 93,61±17,40v
1Araçá(AR), Cereja-do-Mato(CER), Guabiroba(GUA), Jabuticaba(JAB), Pitanga(Pit), Uvaia(UVI); 2Médias seguidas da
mesma letra minúscula na linha não diferem entre si, pelo Teste de Duncan (p<0,05).

27
Além do acesso de GUA/01, o araçá-roxo AR94 (308,02 mg.100g-1)
também mostrou-se rico em compostos fenólicos totais. Pode-se observar que
a maioria dos acessos de araçá-roxo avaliados possui teores de compostos
fenólicos totais superiores que os acessos de pitanga-vermelha, pitanga-
laranja, cereja-do-mato e uvaia. Isso sugere que as frutas da espécie Psidium
cateleianum (araçá) de coloração roxa possuem essa característica genética
de produzir uma concentração maior de metabólitos secundários que os outras
frutas avaliados nesse estudo, apesar de todos as frutas avaliados fazerem
parte da mesma família botânica, a Myrtaceae. Estudos posteriores, de
expressão de genes ou da via de biossíntese de compostos fenólicos e as
enzimas envolvidas são necessários para testar essa hipótese.
Observaram-se diferenças nos teores de compostos fenólicos totais em
frutas de guabiroba. As frutas de GUA/01 apresentaram um alto teor de
compostos fenólicos totais quando comparado ao teor dos outros dois acessos
(GUA/02 e GUA/03) avaliados e o restante das frutas. Esse comportamento foi
observado em frutas colhidas em safras diferentes, o que sugere que as
condições edafoclimáticas, época de floração e de colheita podem diferir na
composição química dos mesmos. Robards et al. (1999) afirmam que a
composição dos compostos fenólicos é em decorrência de fatores genéticos e
ambientais.
Dentre as pitangas, a pitanga-roxa (PitH) apresentou o maior teor de
compostos fenólicos totais (242,50 mg.100g-1 de ácido gálico). Outro fruto
nativa que apresentou teor de compostos fenólicos superior aos araçás de
coloração amarela e pitangas de coloração vermelha e laranja, foi a jabuticaba
com 232,19 mg.100g-1. As frutas que apresentaram os teores mais baixos
foram os genótipos de araçá-amarelo, pitanga-laranja e uvaia.
Quando se compara o teor de compostos fenólicos das frutas nativas da
Região Sul da América do Sul com as frutas não nativas estudadas nesse
trabalho como tratamento controle, se observa que vários acessos avaliados
possuem teor de compostos fenólicos totais superior ao da amora-preta (AMR)
e do mirtilo (MIR), conhecidos por possuírem, na sua composição química,

28
elevados teores desses compostos. Dos genótipos que apresentaram os
maiores teores, dentre as diversas seleções e suas diferentes colorações, o
teor total de compostos fenólicos das frutas não nativas foi superior apenas aos
teores apresentados pela pitanga-laranja e a uvaia. Os demais acessos
avaliados apresentaram concentrações superiores as da amora-preta e do
mirtilo (Figura 6). Nesse caso, era esperado que os acessos de amora-preta e
mirtilo colhidos na Região Sul do Estado do Rio Grande do Sul do Brasil
apresentassem teores de compostos fenólicos totais superiores, pois são frutas
conhecidos cientificamente por serem ricos em compostos fenólicos (Ehlenfeld
e Prior, 2001). Esse fato põe em evidência o fato que as espécies nativas
possuem um grande potencial, inclusive superando espécies tidas como
padrões de qualidade quando avaliadas sob esse aspecto.
Pelo método utilizado para a determinação de compostos fenólicos
totais, extraem-se todos os compostos fenólicos presente na amostra, desde as
antocianinas, flavonols, flavonas a compostos mais complexos, como por
exemplo, taninos não condensáveis. Esses compostos são produzidos em
função da característica genotípica de cada espécie e também como respostas
a estresses bioticos e abioticos. É possível que pelas características
edafoclimaticas (variações de temperatura, luminosidade, luz ultravioleta (UV),
ventos, aporte hídrico e nutricional) e fitopatias (ataque de patógenos e pragas)
nos quais esses acessos são cultivados sejam fatores estimuladores da síntese
desses metabólitos típicos do metabolismo secundário (Simões et al., 2007).
Essa pode ser a razão dos elevados teores desses compostos em algumas
frutas nativas estudados, que um acesso apresentou um alto teor de
compostos fenólicos totais e os outros não.
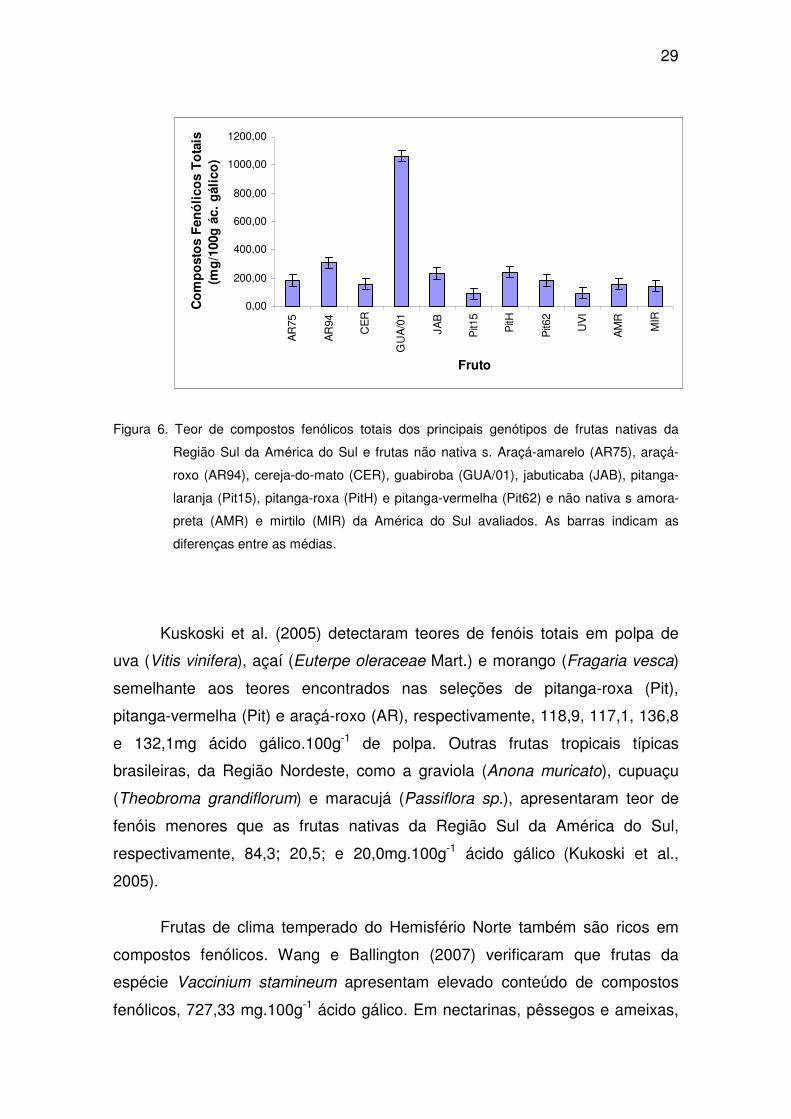
29
0,00
200,00
400,00
600,00
800,00
1000,00
1200,00
AR
75
AR
94
CE
R
GU
A/0
1
JAB
Pit1
5
PitH
Pit6
2
UV
I
AM
R
MIR
Fruto
Co
mp
ost
os
Fen
óli
cos
To
tais
(m
g/1
00g
ác.
gál
ico
)
Figura 6. Teor de compostos fenólicos totais dos principais genótipos de frutas nativas da
Região Sul da América do Sul e frutas não nativa s. Araçá-amarelo (AR75), araçá-
roxo (AR94), cereja-do-mato (CER), guabiroba (GUA/01), jabuticaba (JAB), pitanga-
laranja (Pit15), pitanga-roxa (PitH) e pitanga-vermelha (Pit62) e não nativa s amora-
preta (AMR) e mirtilo (MIR) da América do Sul avaliados. As barras indicam as
diferenças entre as médias.
Kuskoski et al. (2005) detectaram teores de fenóis totais em polpa de
uva (Vitis vinifera), açaí (Euterpe oleraceae Mart.) e morango (Fragaria vesca)
semelhante aos teores encontrados nas seleções de pitanga-roxa (Pit),
pitanga-vermelha (Pit) e araçá-roxo (AR), respectivamente, 118,9, 117,1, 136,8
e 132,1mg ácido gálico.100g-1 de polpa. Outras frutas tropicais típicas
brasileiras, da Região Nordeste, como a graviola (Anona muricato), cupuaçu
(Theobroma grandiflorum) e maracujá (Passiflora sp.), apresentaram teor de
fenóis menores que as frutas nativas da Região Sul da América do Sul,
respectivamente, 84,3; 20,5; e 20,0mg.100g-1 ácido gálico (Kukoski et al.,
2005).
Frutas de clima temperado do Hemisfério Norte também são ricos em
compostos fenólicos. Wang e Ballington (2007) verificaram que frutas da
espécie Vaccinium stamineum apresentam elevado conteúdo de compostos
fenólicos, 727,33 mg.100g-1 ácido gálico. Em nectarinas, pêssegos e ameixas,

30
produzidos no Estado da Califórnia, EUA, o conteúdo de compostos fenólicos
totais (mg.100g-1 ácido gálico) oscilaram entre 14-102, 21-111 e 42-109,
respectivamente (Gil et al. 2002), semelhante ao que foi detectado nas frutas
de araçá-amarelo, pitanga-laranja e pitanga-vermelha. Luximon-Ramma et al.
(2003) observaram teor semelhante em carambola (142mg.100g-1), aos
encontrados na maioria dos araçá-amarelo, pitanga-vermelha, pitanga-laranja e
uvaia avaliados neste estudo.
Ehlenfeldt e Prior (2001) analisaram 87 cultivares de mirtilo e detectaram
teor de compostos fenólicos totais de ate (95,0 mg.100g-1), portanto inferiores
aos observados na maioria das frutas nativas da Região Sul da América do Sul
(araçá-amarelo, araçá-roxo, cereja-do-mato, guabiroba, jabuticaba, pitanga-
roxa e pitanga-vermelha). O teor encontrado nas frutas de mirtilo foi
semelhante aos teores encontrados nos acesso de pitanga-laranja (Pit15 e
Pit71) e uvaia.
Sun et al. (2002) também quantificaram o teor de compostos fenólicos
totais em frutas de grande consumo, como no caso de morango, banana,
abacaxi, pêssego, pêra, maçã e laranja. Esses autores verificaram em maçãs
um alto teor de compostos fenólicos totais e semelhante aos encontrados nos
acessos de guabiroba (GUA/02 e GUA/03) e em alguns genótipos de araçá-
roxo (AR09, AR94, AR100 e AR01), em torno de 272,1mg.100g-1 de ácido
gálico. O teor de compostos fenólicos totais em morangos (147,8mg.100g-1)
foram semelhantes aos encontrados nas frutas de pitanga-laranja (Pit15 e
Pit71). Em frutos, tais como, abacaxi (40,4mg.100g-1), banana (56,1mg.100g-1),
pêra (53,6mg.100g-1) e pêssego (65,3 mg.100g-1), os teores de compostos
fenólicos totais foram inferiores aos 48 frutas avaliados nesse estudo.
Patthamakanokporn et al. (2008) encontraram valores inferiores de compostos
fenólicos totais para frutas de goiaba (Psidium guajava) e mamão (Carcinia
papaya) aos acessos de guabiroba, araçá-amarelo, araçá-roxo, cereja-do-mato
e jabuticaba, respectivamente 148,0mg.100g-1 e 54,0mg.100g-1.
Vasco, Ruales e Kamal-Eldin (2008) encontraram valores semelhantes,
aos encontrado na guabiroba, ao avaliar frutas nativas do Equador. Esses

31
autores encontraram teores de compostos fenólicos totais superiores aos
encontrados nas frutas da Região do Sul do Brasil, com 2167 mg.100g-1 de
ácido gálico equivalente para andean blackberry (Rubus, Berth), 1494 mg.100g-
1 de ácido gálico equivalente para capulí cherry (Prunus serotina, var. Capulí) e
1010 mg.100g-1 de ácido gálico equivalente para banana passion (Passiflora
mollissima L.). Esses autores também encontraram teores semelhantes aas
frutas da Região Sul da América do Sul, que variaram entre 250 e 400
mg.100g-1 de ácido gálico equivalente para ameixa (Prunus salicina), goiaba
(Psidium guajava L.) e cherimóia (Annona cherimólia) e também teores
inferiores a 100 mg.100g-1 de ácido gálico equivalente, semelhante aos
encontrados em araçá-amarelo, cereja-do-mato, pitanga-laranja, pitanga-
vermelha e uvaia, nas frutas naranjilla (Solarum quitoense), physalis (Physalis
peruviana), tomate (Lycopersicon esculentum) e maracujá (Passiflora edulis,
var. flavicarpa).
Liu et al. (2008) avaliaram frutas nativas da Ásia, em seis regiões
chinesas (Phyllanthus emblica L.) e também encontraram teores semelhantes,
entre 81,5 e 120,9 mg.100g-1 de ácido gálico , às frutas nativas da Região Sul
da América do Sul avaliados neste estudo, tais como uvaia, maioria dos 13
acessos de araçá-amarelo, pitanga-laranja e pitanga-vermelha) . Liu et al.
(2008) verificaram que as diferentes condições de crescimento das frutas
podem contribuir no teor de compostos fenólicos totais e de outros metabólitos.
Ercisli e Orhan (2007) verificaram altos teores de compostos fenólicos
totais em frutas de mulberrie vermelha (Morus rubra) e mulberrie negra (Morus
nigra), respectivamente, 1035 e 1422 mg.100g-1 equivalente a ácido gálico.
Esses teores são semelhantes ao encontrado em um dos acessos de
guabiroba avaliados nesse estudo.
4.3 Teor de Antocianinas Totais
A cereja-do-mato foi o fruto que apresentou o teor mais elevado de
antocianinas totais dentre os 48 avaliados, com 89,00 mg.100g-1 em

32
equivalente de cianidina-3,5-glicosídeo, seguido da pitanga-roxa (PitH) com
76,73 mg.100g-1 e jabuticaba com 67,39 mg.100g-1 (Tabela 6). Dentre os 13
genótipos de araçá-roxo, o AR94 foi o que apresentou maior teor com 10,54
mg.100g-1. Os dois acessos de pitanga-laranja (Pit15 e Pit71) apresentaram
teores de antocianinas totais superiores aos de araçá-roxo, entre 8,53 e 9,02
mg.100g-1. Dentre os acessos de araçá-roxo avaliados, houve diferença
significativa entre os teores de antocianinas totais que variaram entre 0,89 e
10,54 mg.100g-1. Nos acessos de pitanga-vermelha também observou-se uma
grande diferença entre os teores encontrados nas frutas de mesma espécie e
mesma coloração, que variaram entre 5,11 e 20,65 mg.100g-1. Entre os
genótipos de araçá-amarelo, o teor de antocianinas totais variou entre 0,35 e
8,01 mg.100g-‘. Porém observou-se que essas frutas apresentaram teores bem
inferiores que as cultivares avaliadas de pitanga, guabiroba, cereja-do-mato,
jabuticaba e uvaia.
Ehlenfeldt e Prior (2001) analisaram 87 cultivares de mirtilo e
encontraram teor de antocianinas totais superior (179,0 mg.100g-1) aos
detectados nas frutas nativas da Região Sul da América do Sul. Porém
observou-se que nos mirtilos colhidos na Região Sul da América do Sul, o teor
de antocianinas totais foi inferior (54,85mg.100g-1) ao observado por aqueles
autores com o mesmo material, mas cultivado no Estado de New Jersey, EUA.
Esse comportamento sugere que como as frutas analisados foram no seu
centro de origem, nessas frutas ocorreu sua expressão na máxima capacidade.
Reynertson et al. (2008) estudaram frutas da família Myrtaceae, que consiste
do mesmo gênero as frutas nativas da Região Sul da América do Sul e
observaram teores de antocianinas totais semelhantes aos encontrados nos
acessos de araçá-amarelo, araçá-roxo e guabiroba, ao avaliarem frutas das
espécies Eugenia brasilienses, Eugenia aggregata, Myricia cauliflora.

33
Tabela 6. Teor de antocianinas totais presentes em frutas nativas da Região
Sul da América do Sul.
Fruta Codificação1 Cor Teor de Antocianinas Totais ( mg.100g-1 de cianidina 3,5-glicosídeo)
AR01 1,04±0,17q2 AR03 1,25±0,32q AR05 0,90±0,74q AR27 8,01±0,32hijlmno AR46 2,55±0,16opq AR47 0,35±0,10q AR48 0,54±0,17q AR49 0,99±0,16q AR57 1,75±0,20pq AR58 2,33±0,46opq AR59 1,41±0,12pq AR60 0,63±0,17q AR64 1,94±0,27pq AR72 1,28±0,02q AR73 1,27±0,02q AR74 1,67±0,14pq AR75
amarelo
1,08±0,03q AR08 3,52±0,14mnopq AR09 2,05±1,11pq AR10 0,89±0,26q AR11 6,09±0,09jlmnopq AR18 4,05±0,21mnopq AR19 6,29±0,08ijlmnopq AR28 11,82±0,36fghi AR29 4,82±0,30lmnopq AR42 3,10±0,24nopq AR44 3,93±0,04mnopq AR94 10,54±0,41ghijl AR100 1,18±0,31pq
Araçá
AR101
roxo
2,15±0,14opq Cereja-do-mato CER roxo 89,00±22,51ª
GUA/01 4,39±0,39mnopq GUA/02 5,28±2,22lmnopq Guabiroba GUA/03
laranja 7,36±0,57ijlmnop
Jabuticaba JAB roxo 67,39±18,45c Pit15 8,53±0,28hijlmn Pit71
laranja 9,02±1,59ghijlm
PitH 76,73±0,46b Pit40
roxo 27,15±2,55d
PitU 11,60±0,89fghi Pit41 13,28±0,37fgh Pit49 16,65±1,58ef Pit50 11,98±0,86fghi Pit53 6,08±0,11jlmnopq Pit55 5,11±0,11lmnopq Pit61 11,63±0,96fghij
Pitanga
Pit62
vermelho
20,65±0,73e Uvaia UVI amarelo 14,51±0,27fg
1Araçá(AR), Cereja-do-Mato(CER), Guabiroba(GUA), Jabuticaba(JAB), Pitanga(Pit), Uvaia(UVI); 2Médias seguidas da
mesma letra minúscula na linha não diferem entre si, pelo Teste de Duncan (p<0,05).
Quando se comparam as frutas nativas da Região Sul da América do Sul
com as frutas não nativas da América do Sul, quanto ao conteúdo de
antocianinas totais (Figura 7), verifica-se que a maioria das frutas nativas
analisados apresentou teores de antocianinas totais inferiores aos obtidos a um
das frutas não nativa s avaliado (amora-preta), conhecido cientificamente por

34
possuir na sua composição química, elevados teores de antocianinas (Dugo et
al., 2001; Mota, 2006).
-20,00
0,00
20,00
40,00
60,00
80,00
100,00
120,00
140,00
AR
75
AR
94
CE
R
GU
A/0
1
JAB
Pit1
5
PitH
Pit6
2
UV
I
AM
R
MIR
Fruto
An
toci
anin
as T
ota
is (
mg
/100
g
cian
idin
a 3,
5-g
lico
sila
da)
Figura 7. Teor de antocianinas totais dos principais genótipos de frutas nativas da Região Sul
da América do Sul e frutas não nativa s. Araçá-amarelo (AR75), araçá-roxo (AR94),
cereja-do-mato (CER), guabiroba (GUA/01), jabuticaba (JAB), pitanga-laranja (Pit15),
pitanga-roxa (PitH) e pitanga-vermelha (Pit62) e não nativa s amora-preta (AMR) e
mirtilo (MIR) da América do Sul avaliados. As barras indicam as diferenças entre as
médias.
Os três genótipos que mais se aproximaram dos valores de antocianinas
totais na amora-preta (AMR) (115,98 mg.100g-1 cianidina 3,5-glicosídeo) foram
a cereja-do-mato, a pitanga-roxa (PitH) e a jabuticaba, respectivamente, com
89,00 mg.100g-1, 76,73 mg.100g-1 e 67,39 mg.100g-1. Esses três genótipos
apresentaram teor de antocianinas totais superiores ao conteúdo presente no
fruto não nativa mirtilo (54,85 mg.100g-1). O mirtilo é uma fruta nativa do
Hemisfério Norte conhecido cientificamente como rico em compostos
antociânicos, porém o acesso colhido na Região Sul da América do Sul
apresentou um valor abaixo do que outros detectam em mirtilo colhidos em
outras regiões (Ehlenfeldt e Prior, 2001; Katsube et al., 2003; Pantelidis et al.,
2007). Esse dado sugere que o centro de origem propicia a máxima expressão
do potencial genético, ou seja, os mirtilo, embora se adaptem ao clima e solo

35
da Região Sul da América do Sul, são frutas típicas do Hemisfério Norte e
nessas regiões as plantas apresentam o máximo de seu potencial genético no
que concerne ao metabolismo secundário.
4.4 Teor de Carotenóides Totais
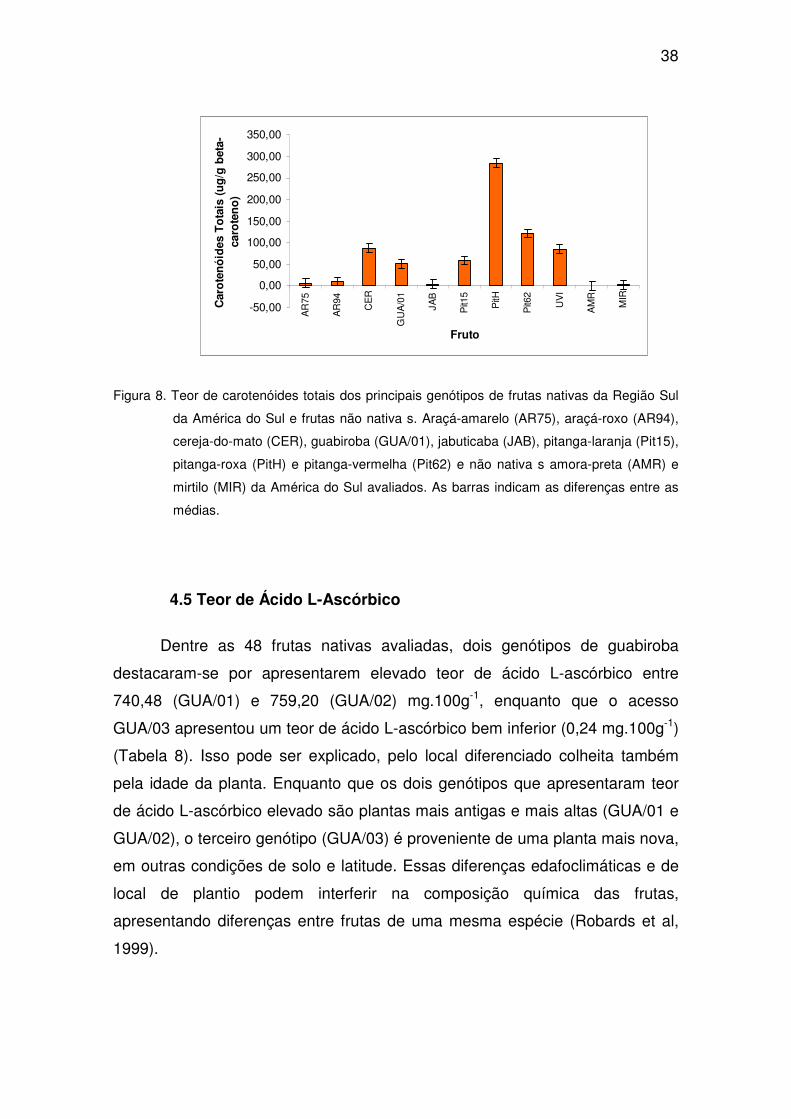
Os acessos de araçá, tanto os de coloração roxa como a amarela,
apresentaram baixo teores de carotenóides totais, não ultrapassando 11,34 e
10,84 µg.g-1 de β-caroteno, respectivamente, quando comparados com os
teores de carotenóides em pitangas-roxas e pitangas-vermelhas (Tabela 7).
Dos 48 genótipos avaliados, os dois acessos de pitanga-roxa (PitH e Pit40)
apresentaram alto teor de carotenóides totais na sua composição, entre 117,17
e 285,14 µg.g-1 de β-caroteno. Entre os oito genótipos de pitanga-vermelha
analisados, dois apresentaram os maiores teores de carotenóides totais, entre
213,25 e 231,71 µg.g-1 de β-caroteno. Das seis espécies avaliadas (araçá,
cereja-do-mato, guabiroba, jabuticaba, pitanga e uvaia, os genótipos de
pitanga-roxa e pitanga-vermelha foram os que apresentaram os teores de
carotenóides totais mais elevados, seguidos pela cereja-do-mato (88,82 µg.g-1
de β-caroteno) e uvaia (85,61 µg.g-1 de β-caroteno). Os três acessos de
guabiroba (GUA/01, GUA/02 e GUA/03) apresentaram valores intermediários,
variando entre 32,51 e 52,26 µg.g-1 de β-caroteno. Dos 17 acessos de araçá-
amarelo e 13 de araçá-roxo, todos apresentaram os menores teores de
carotenóides totais, que variou entre 3,15 e 10,84 µg.g-1 de β-caroteno para as
frutas de araçá de coloração amarela e entre 3,02 e 11,34 µg.g-1 de β-caroteno
para as frutas de coloração roxa (Tabela 7).
Lima, Melo e Lima (2002) avaliaram o teor de carotenóides em pitangas,
de coloração roxa e vermelha, (Eugenia uniflora L.) encontradas na Região
Nordeste. Em todas as cultivares analisadas, o teor de carotenóides foi inferior
aos encontrados nos acessos de pitanga-roxa e pitanga-vermelha nativa s
avaliados neste estudo, em média 111,00µg.g-1 para as cultivares de cor roxa e
104,00µg.g-1 para as de cor vermelha. Essa diferença na composição química,
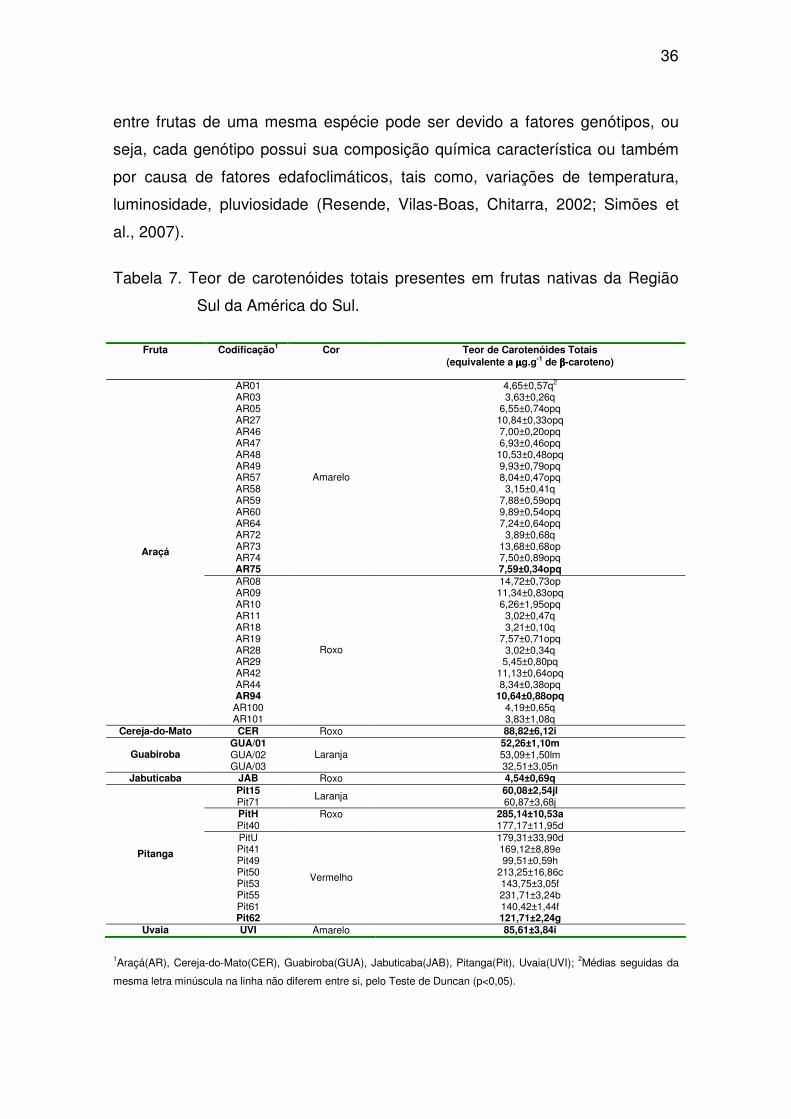
36
entre frutas de uma mesma espécie pode ser devido a fatores genótipos, ou
seja, cada genótipo possui sua composição química característica ou também
por causa de fatores edafoclimáticos, tais como, variações de temperatura,
luminosidade, pluviosidade (Resende, Vilas-Boas, Chitarra, 2002; Simões et
al., 2007).
Tabela 7. Teor de carotenóides totais presentes em frutas nativas da Região
Sul da América do Sul.
Fruta Codificação1 Cor Teor de Carotenóides Totais (equivalente a µµµµg.g-1 de ββββ-caroteno)
AR01 4,65±0,57q2 AR03 3,63±0,26q AR05 6,55±0,74opq AR27 10,84±0,33opq AR46 7,00±0,20opq AR47 6,93±0,46opq AR48 10,53±0,48opq AR49 9,93±0,79opq AR57 8,04±0,47opq AR58 3,15±0,41q AR59 7,88±0,59opq AR60 9,89±0,54opq AR64 7,24±0,64opq AR72 3,89±0,68q AR73 13,68±0,68op AR74 7,50±0,89opq AR75
Amarelo
7,59±0,34opq AR08 14,72±0,73op AR09 11,34±0,83opq AR10 6,26±1,95opq AR11 3,02±0,47q AR18 3,21±0,10q AR19 7,57±0,71opq AR28 3,02±0,34q AR29 5,45±0,80pq AR42 11,13±0,64opq AR44 8,34±0,38opq AR94 10,64±0,88opq AR100 4,19±0,65q
Araçá
AR101
Roxo
3,83±1,08q Cereja-do-Mato CER Roxo 88,82±6,12i
GUA/01 52,26±1,10m GUA/02 53,09±1,50lm Guabiroba GUA/03
Laranja 32,51±3,05n
Jabuticaba JAB Roxo 4,54±0,69q Pit15 60,08±2,54jl Pit71
Laranja 60,87±3,68j
PitH 285,14±10,53a
Pit40 Roxo
177,17±11,95d PitU 179,31±33,90d Pit41 169,12±8,89e Pit49 99,51±0,59h Pit50 213,25±16,86c Pit53 143,75±3,05f Pit55 231,71±3,24b Pit61 140,42±1,44f
Pitanga
Pit62
Vermelho
121,71±2,24g Uvaia UVI Amarelo 85,61±3,84i
1Araçá(AR), Cereja-do-Mato(CER), Guabiroba(GUA), Jabuticaba(JAB), Pitanga(Pit), Uvaia(UVI); 2Médias seguidas da
mesma letra minúscula na linha não diferem entre si, pelo Teste de Duncan (p<0,05).

37
Marinova e Ribarona (2007) também detectaram teores de carotenóides
totais semelhantes aos da pitanga-roxa (PitH) e pitanga-vermelha (Pit55) ao
analisarem amora (Rubus fruticosus L.) e framboesa (Rubus ideaus L.)
oriundos da Europa. Também verificaram teores de carotenóides superiores,
aos encontrados nos 48 frutas nativas da Região Sul da América do Sul
avaliado, com 440µg.g-1 e 370µg.g-1 para morango Fragaria vesca L.) e
framboesa (Rubus ideaus), respectivamente.
Zanatta e Mercadante (2007) analisaram o teor de carotenóides em
frutas de camu-camu (Myrciaria dúbia), fruta nativa da Região Amazônica e da
mesma família botânica que as 48 frutas avaliados nesse estudo (família
Myrtaceae), verificaram que essas frutas possuem teor de carotenóides totais
superior (354,8 e 1095,3 µg.g-1 β-caroteno) a todos as frutas nativas da Região
Sul da América do Sul avaliados nesse estudo. Como as frutas de camu-camu
também foram avaliadas nos seus centros de origem, o fato de possuírem o
teor de carotenóides total mais elevado que as frutas nativas da Região Sul da
América do Sul é devido a características genotípicas, próprias da espécie.
As frutas não nativas avaliadas nesse estudo, amora-preta (AMR) e
mirtilo (MIR), apresentam valores baixos de teores de carotenóides, quando
comparados com as frutas nativas da Região Sul, com 1,48 e 2,93 µg.g de β-
caroteno, respectivamente. Os 48 genótipos de frutas nativas avaliados
apresentaram valores superiores, destacando-se a pitanga-roxa (PitH) e
pitanga-vermelha (Pit62) (Figura 8).
38
-50,00
0,00
50,00
100,00
150,00
200,00
250,00
300,00
350,00
AR
75
AR
94
CE
R
GU
A/0
1
JAB
Pit1
5
PitH
Pit6
2
UV
I
AM
R
MIR
Fruto
Car
ote
nó
ides
To
tais
(u
g/g
bet
a-ca
rote
no
)
Figura 8. Teor de carotenóides totais dos principais genótipos de frutas nativas da Região Sul
da América do Sul e frutas não nativa s. Araçá-amarelo (AR75), araçá-roxo (AR94),
cereja-do-mato (CER), guabiroba (GUA/01), jabuticaba (JAB), pitanga-laranja (Pit15),
pitanga-roxa (PitH) e pitanga-vermelha (Pit62) e não nativa s amora-preta (AMR) e
mirtilo (MIR) da América do Sul avaliados. As barras indicam as diferenças entre as
médias.
4.5 Teor de Ácido L-Ascórbico
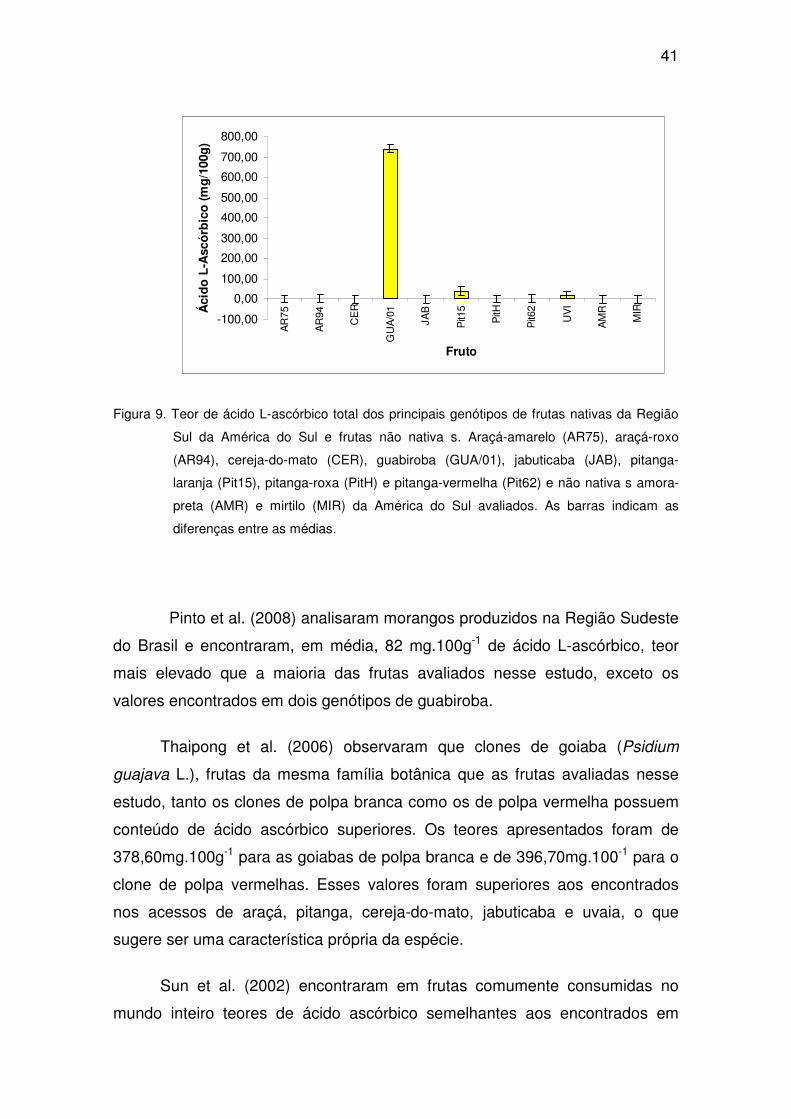
Dentre as 48 frutas nativas avaliadas, dois genótipos de guabiroba
destacaram-se por apresentarem elevado teor de ácido L-ascórbico entre
740,48 (GUA/01) e 759,20 (GUA/02) mg.100g-1, enquanto que o acesso
GUA/03 apresentou um teor de ácido L-ascórbico bem inferior (0,24 mg.100g-1)
(Tabela 8). Isso pode ser explicado, pelo local diferenciado colheita também
pela idade da planta. Enquanto que os dois genótipos que apresentaram teor
de ácido L-ascórbico elevado são plantas mais antigas e mais altas (GUA/01 e
GUA/02), o terceiro genótipo (GUA/03) é proveniente de uma planta mais nova,
em outras condições de solo e latitude. Essas diferenças edafoclimáticas e de
local de plantio podem interferir na composição química das frutas,
apresentando diferenças entre frutas de uma mesma espécie (Robards et al,
1999).

39
Tabela 8. Teor de ácido L-ascórbico presente em frutas nativas da Região Sul
da América do Sul.
Fruta Codificação1 Cor Teor de Ácido L-Ascórbico Total (mg.100g-1)
AR01 1,30±0,07vhi2 AR03 0,00±0,00i AR05 2,81±0,57hi AR27 0,00±0,00i AR46 7,20±0,98ghi AR47 4,97±0,74hi AR48 6,68±1,16ghi AR49 6,73±1,72ghi AR57 0,75±0,09i AR58 3,37±0,02hi AR59 3,80±0,15hi AR60 4,17±0,10hi AR64 8,53±0,32gh AR72 4,72±0,04hi AR73 0,10±0,04i AR74 0,13±0,07i AR75
amarelo
1,59±0,07hi AR08 2,50±0,35hi AR09 0,00±0,00i AR10 0,00±0,00i AR11 2,22±0,17hi AR18 0,00±0,00i AR19 0,45±0,22i AR28 0,00±0,00i AR29 0,00±0,00i AR42 0,38±0,16i AR44 12,89±0,22g AR94 3,16±0,25hi AR100 1,46±0,13hi
Araçá
AR101
roxo
2,38±0,09hi Cereja-do-Mato CER roxo 0,00±0,00i
GUA/01 740,68±33,56b GUA/02 759,20±9,59a Guabiroba GUA/03
laranja 0,24±0,00i
Jabuticaba JAB roxo 0,71±0,11i Pit15 40,95±3,26c Pit71
laranja 27,75±1,88e
PitH 2,50±0,22hi Pit40
roxo 34,10±3,22d
PitU 35,53±1,81cd Pit41 4,20±0,63hi Pit49 2,31±1,35hi Pit50 3,97±0,78hi Pit53 3,22±0,20hi Pit55 2,56±0,46hi Pit61 4,89±0,06hi
Pitanga
Pit62
vermelho
4,38±0,31hi Uvaia UVI amarelo 19,86±0,68f
1Araçá(AR), Cereja-do-Mato(CER), Guabiroba(GUA), Jabuticaba(JAB), Pitanga(Pit), Uvaia(UVI); 2Médias seguidas da
mesma letra minúscula na linha não diferem entre si, pelo Teste de Duncan (p<0,05).
Dos oito genótipos de pitanga-vermelha avaliados, apenas uma
apresentou teor de ácido L-ascórbico alto (PitU), com 35,53 mg.100g-1; nos
outros sete frutas o teor variou entre 2,31 e 4,89 mg.100g-1 de ácido L-
ascórbico. Entre os dois acessos de pitanga-roxa (PitH e Pit40), um apresentou
teor de ácido L-ascórbico bem superior (PitH) e outro não (Pit40), o que sugere

40
que a concentração desse nutriente no fruto não é uma característica da
espécie, mas sim de cada genótipo. A grande maioria das frutas de araçá-roxo
e a cereja-do-mato não apresentou quantidades significativas de ácido L-
ascórbico (Tabela 8).
Os 17 genótipos de araçá-amarelo apresentaram teores menores de
ácido L-ascórbico, os quais foram inferiores a 8,53 mg.100g-1, quando
comparados aos teores encontrados nos dois acessos de guabiroba (GUA/01 e
GUA/02).
Assim como foi observado na avaliação de carotenóides totais, as
frutas não nativas apresentaram valores relativamente menores de ácido L-
ascórbico: 0,26 mg.100g-1 para a amora-preta e 0,12 mg.100g-1 para o mirtilo,
no entanto, o teor desse componentes encontrado na guabiroba (GUA/01) foi
muito superior ao encontrados nas frutas não nativas, obtendo-se valores de
até 759,20mg.100g-1 (Figura 9).
A acerola, fruta nativa da Região Nordeste do Brasil, conhecida
cientificamente como fonte rica de vitamina C, apresenta teor de ácido
ascórbico entre 695 e 4827 mg de ácido ascórbico por 100g de fruta (Santos et
al., 1999; Gomes et al., 2000). Das 48 frutas analisados, apenas dois acessos
de guabiroba (GUA/01 e GUA/02) apresentaram teor semelhante (740,68 e
759,20mg.100g-1). O restante das frutas avaliadas apresentou teores de ácido
L-ascórbico inferiores aos detectados na acerola, apesar de ambos as frutas
terem sido avaliados nos seus respectivos centros de origem, o que significa
que o teor de vitamina C presente nessas frutas também é uma característica
genotípica de cada espécie e susceptível a fatores ambientais (Simões et al.,
2007).
41
-100,00
0,00
100,00
200,00
300,00
400,00
500,00
600,00
700,00
800,00
AR
75
AR
94
CE
R
GU
A/0
1
JAB
Pit1
5
PitH
Pit6
2
UV
I
AM
R
MIR
Fruto
Áci
do
L-A
scó
rbic
o (
mg
/100
g)
Figura 9. Teor de ácido L-ascórbico total dos principais genótipos de frutas nativas da Região
Sul da América do Sul e frutas não nativa s. Araçá-amarelo (AR75), araçá-roxo
(AR94), cereja-do-mato (CER), guabiroba (GUA/01), jabuticaba (JAB), pitanga-
laranja (Pit15), pitanga-roxa (PitH) e pitanga-vermelha (Pit62) e não nativa s amora-
preta (AMR) e mirtilo (MIR) da América do Sul avaliados. As barras indicam as
diferenças entre as médias.
Pinto et al. (2008) analisaram morangos produzidos na Região Sudeste
do Brasil e encontraram, em média, 82 mg.100g-1 de ácido L-ascórbico, teor
mais elevado que a maioria das frutas avaliados nesse estudo, exceto os
valores encontrados em dois genótipos de guabiroba.
Thaipong et al. (2006) observaram que clones de goiaba (Psidium
guajava L.), frutas da mesma família botânica que as frutas avaliadas nesse
estudo, tanto os clones de polpa branca como os de polpa vermelha possuem
conteúdo de ácido ascórbico superiores. Os teores apresentados foram de
378,60mg.100g-1 para as goiabas de polpa branca e de 396,70mg.100-1 para o
clone de polpa vermelhas. Esses valores foram superiores aos encontrados
nos acessos de araçá, pitanga, cereja-do-mato, jabuticaba e uvaia, o que
sugere ser uma característica própria da espécie.
Sun et al. (2002) encontraram em frutas comumente consumidas no
mundo inteiro teores de ácido ascórbico semelhantes aos encontrados em

42
frutas nativas do Sul da América do Sul. O teor de vitamina C avaliados em
pêssegos (6,6mg.100g-1) são similares aos encontrados nos acessos de araçá-
amarelo (AR46, AR47, AR48 e AR49). Os teores de ácido L-ascórbico em pêra
(4,0 mg.100g-1) e em laranja (4,5 mg.100g-1) foram semelhantes ao teor de
ácido L-ascórbico encontrados nos acessos de pitanga-vermelha. O teor de
ácido L-ascórbico em abacaxi (15,4 mg.100g-1) foi similar ao da uvaia. Esses
autores também não observaram teores elevados de ácido L-ascórbico, em
frutas de cramberrie, que são da mesma família botânica do mirtilo.
Ercisli e Orhan (2007) avaliaram frutas de mulberry de três colorações
diferentes, branca (Morus alba), vermelha (Morus rubra) e negra (Morus nigra).
O teor de ácido L-ascórbico encontrado nas cultivares branca, vermelha e
negra foi, respectivamente de 22,4, 19,4 e 21,8 mg.100g-1. Esses teores são
superiores aos encontrados nas frutas de araçá-amarelo, araçá-roxo e pitanga-
vermelha avaliados nesse estudo, e semelhantes ao encontrado na uvaia.
Pantelidis et al. (2007) verificaram um teor elevado de ácido L-ascórbico
em uma fruta nativa do Hemisfério Norte, Cornelian cherry (Cornus mas cv.
Vermio). Esses autores encontraram, nessa fruta, 103,30 mg.100g-1 de ácido L-
ascórbico. Esse valor não foi superior, quando comparado com as frutas da
Região Sul da América do Sul, aos encontrados nos dois acessos de guabiroba
(GUA/01 e GUA/02).
Medina, Rodríguez e Romero (2007) avaliaram a composição química
de frutas nativas da América Central (Opontia dillenii e Opuntia fícus indica).
Esses autores verificaram que, respectivamente, esses dois frutas
apresentaram 29,7 e 17,2 mg.100g-1 ácido L-ascórbico. Esses valores são
semelhantes aos teores encontrados na uvaia, um acesso de pitanga-laranja
(Pit15) e um acesso de pitanga-roxa (Pit40).

43
4.6 Atividade Antioxidante
Tabela 9. Atividade Antioxidante, expressa em mM de Trolox, em frutas nativas
da Região Sul da América do Sul.
Fruta Codificação1 Cor Atividade Antioxidante (mm Trolox)
AR01 5,32±0,69e2 AR03 9,67±0,66e AR05 4,13±0,24e AR27 11,27±0,40e AR46 4,21±0,32e AR47 4,41±1,30e AR48 5,78±1,18e AR49 4,29±0,78e AR57 4,57±0,49e AR58 4,58±0,44e AR59 4,12±0,75e AR60 4,21±0,29e AR64 4,58±0,37e AR72 6,06±0,70e AR73 4,81±0,70e AR74 5,24±0,29e AR75
Amarelo
5,81±0,28e AR08 7,85±0,98e AR09 6,73±1,56e AR10 9,71±0,67e AR11 7,33±0,91e AR18 8,71±0,74e AR19 7,36±1,33e AR28 6,93±0,75e AR29 8,97±0,27e AR42 7,29±0,44e AR44 4,21±1,41e AR94 7,45±2,04e AR100 8,52±0,98e
Araçá
AR101
Roxo
8,32±1,08e Cereja-do-Mato CER Roxo 8,59±1,18e
GUA/01 1695,59±131,31d GUA/02 2654,00±839,74c Guabiroba GUA/03
Laranja 3519,85±932,07a
Jabuticaba JAB Roxo 3361,26±676,55b Pit15 4,10±0,69e Pit71
Laranja 3,45±0,25e
PitH 9,91±0,28e Pit40
Roxo 6,63±0,60e
PitU 4,65±0,17e Pit41 4,78±0,44e Pit49 4,40±0,50e Pit50 3,92±0,78e Pit53 3,42±0,22e Pit55 4,07±0,09e Pit61 2,78±0,20e
Pitanga
Pit62
Vermelho
3,88±0,34e Uvaia UVI Amarelo 5,64±1,09e
1Araçá(AR), Cereja-do-Mato(CER), Guabiroba(GUA), Jabuticaba(JAB), Pitanga(Pit), Uvaia(UVI); 2Médias seguidas da
mesma letra minúscula na linha não diferem entre si, pelo Teste de Duncan (p<0,05).
Os três genótipos de guabiroba (GUA/01, GUA/02 e GUA/03) e o de
jabuticaba apresentaram os maiores valores de atividade antioxidante
comparados com os demais avaliados nesse estudo. Não houve diferença

44
significativa entre os resultados obtidos no araçá-amarelo, araçá-roxo, pitanga-
roxa, pitanga-vermelha, pitanga-laranja, cereja-do-mato e uvaia (Tabela 9).
Dentre as seleções de pitangas, observou-se que a cultivar roxa, colhida
na propriedade rural (PitH), foi a que apresentou maior atividade antioxidante, e
ao mesmo tempo, os maiores teores de compostos fenólicos totais e de, além
de uma grande concentração de carotenóides totais. Presume-se que, os altos
teores destes compostos contribuíram para a maior atividade antioxidante
dessa fruta. Os teores encontrados no acesso de pitanga-roxa (PiH) sugere
que, por ter sido colhido no seu centro de origem, a expressão gênica foi
máxima. Bao et al. (2005), quando analisaram a atividade antioxidante de
quatro “chinese berry” (Myrica rubra), verificaram que as frutas mais ricas em
compostos antociânicos foram também as que apresentaram maior atividade
antioxidante. As seleções de araçá-amarelo, araçá-roxo e guabiroba
apresentaram atividade antioxidante superior às seleções de pitanga, porém,
apresentaram teores inferiores de antocianinas totais, os quais são os outros
compostos que supostamente conferem a estas frutas, o potencial antioxidante.
A atividade antioxidante das duas frutas não nativas avaliadas não
diferiu significativamente aos encontrados nos genótipos de frutas nativas
(Figura 10). Apenas as frutas de guabiroba e jabuticaba diferiram
significativamente entre os demais acessos avaliados.
Os três acessos de guabiroba (GUA/01, GUA/02 e GUA/03) não
apresentaram correlação entre as variáveis analisadas (compostos fenólicos
totais, antocianinas totais, carotenóides totais e ácido L-ascórbico) e a
respectiva atividade antioxidante. A jabuticaba (JAB) apresentou correlação
positiva (R2 = 0,5437) somente quando se correlacionou o teor de antocianinas
totais com sua atividade antioxidante (Tabela 10).

45
-500,00
0,00
500,00
1000,00
1500,00
2000,00
2500,00
3000,00
3500,00
4000,00
AR
75
AR
94
CE
R
GU
A/0
1
JAB
Pit1
5
PitH
Pit6
2
UV
I
AM
R
MIR
Fruto
Ati
vid
ade
An
tio
xid
ante
(m
M
Tro
lox)
Figura 10. Atividade Antioxidante dos principais frutas nativas da Região Sul da América do Sul
e frutas não nativa s. Araçá-amarelo (AR75), araçá-roxo (AR94), cereja-do-mato
(CER), guabiroba (GUA/01), jabuticaba (JAB), pitanga-laranja (Pit15), pitanga-roxa
(PitH) e pitanga-vermelha (Pit62) e não nativa s amora-preta (AMR) e mirtilo (MIR) da
América do Sul avaliados. As barras indicam as diferenças entre as médias.
Quatro acessos de araçá-amarelo (AR47, AR57, AR64 e AR73), dois
de araçá-roxo (AR11 e AR28), dois de pitanga-roxa (PitH e Pit40) e um de
pitanga-vermelha (Pit41) obtiveram resultados significativos quando
correlacionou-se seus respectivos teores de compostos fenólicos totais e sua
atividade antioxidante (Tabela 10).
Poucas frutas apresentaram boa correlação entre o teor de
antocianinas totais e a atividade antioxidante (araçá-amarelo AR05, araçá-roxo
AR11, araçá-roxo AR94, jabuticaba e pitanga-vermelha Pit50). O restante das
frutas avaliadas apresentou correlações negativas (Tabela 10).
Dois acessos de araçá-amarelo (AR58 e AR60) apresentaram alta
correlação, R2 = 0,9412 e R2 = 0,9728, respectivamente, quando analisou o
teor de carotenóides totais e suas respectivas atividades antioxidantes. Um
único fruto (PitU) apresentou alta correlação (R2 = 0,7794) entre o teor de
ácido L-ascórbico e a atividade antioxidante (Tabela 10).

46
Porém, verificou-se que as frutas que apresentaram boas correlações
não foram as frutas que obtiveram atividade antioxidante superiores; com isso
não se observou uma alta correlação entre os compostos biologicamente ativos
e a atividade antioxidante. Estes dados sugerem que a mistura dos compostos
presentes nas frutas e seu sinergismo são responsáveis pela atividade
antioxidante e não apenas uma classe específica de compostos.
Gil et al. (2002) também verificaram que não houve uma relação nítida
entre o conteúdo de compostos fenólicos totais e a capacidade antioxidante,
quando analisou cinco cultivares de nectarinas e de pêssegos. Não é possível
generalizar esse tipo de relação, o que se presume que a atividade
antioxidante é mais relacionada com cada cultivar (Gil et al., 2002). Porém,
vários autores correlacionam o alto teor de compostos fenólicos e antociânicos
com uma alta atividade antioxidante (Imeh e Khokhar, 2002; Pinelo et al., 2004;
Roberts e Gordon, 2003; Sun et al., 2002).
Há estudos que correlacionam o conteúdo total de compostos fenólicos
com a atividade antioxidante em frutas, vegetais, ervas e especiarias
(Ehlenfeldt e Prior, 2001; Jimenez-Escrig et al., 2001; Scalzo et al., 2005;
Velioglu et al., 1998; Vinson et al., 2001). Gil et al. (2002) também obtiveram
correlações similares quando analisaram compostos fenólicos totais e atividade
antioxidante (R2 entre 0,93 e 0,96) em nectarinas, pêssegos e ameixas, porém
para estas mesmas frutas não obtiveram boa correlação ao comparar a
atividade antioxidante com os teores de ácido ascórbico. Sun et al. (2002)
encontraram correlação entre o teor de compostos fenólicos e a atividade
antioxidante nos extratos de frutos, tais como, maçã, cramberrie, morango e
laranja (R2 = 0,9788).
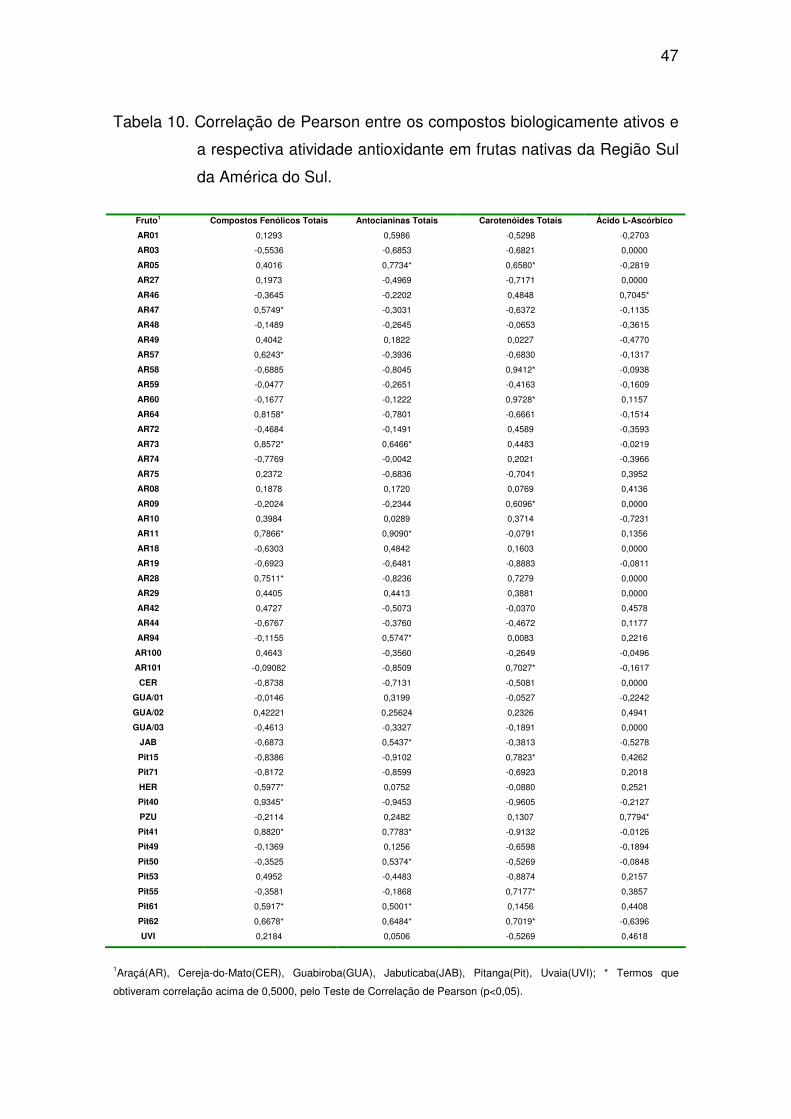
47
Tabela 10. Correlação de Pearson entre os compostos biologicamente ativos e
a respectiva atividade antioxidante em frutas nativas da Região Sul
da América do Sul.
Fruto1 Compostos Fenólicos Totais Antocianinas Totais Carotenóides Totais Ácido L-Ascórbico
AR01 0,1293 0,5986 -0,5298 -0,2703
AR03 -0,5536 -0,6853 -0,6821 0,0000
AR05 0,4016 0,7734* 0,6580* -0,2819
AR27 0,1973 -0,4969 -0,7171 0,0000
AR46 -0,3645 -0,2202 0,4848 0,7045*
AR47 0,5749* -0,3031 -0,6372 -0,1135
AR48 -0,1489 -0,2645 -0,0653 -0,3615
AR49 0,4042 0,1822 0,0227 -0,4770
AR57 0,6243* -0,3936 -0,6830 -0,1317
AR58 -0,6885 -0,8045 0,9412* -0,0938
AR59 -0,0477 -0,2651 -0,4163 -0,1609
AR60 -0,1677 -0,1222 0,9728* 0,1157
AR64 0,8158* -0,7801 -0,6661 -0,1514
AR72 -0,4684 -0,1491 0,4589 -0,3593
AR73 0,8572* 0,6466* 0,4483 -0,0219
AR74 -0,7769 -0,0042 0,2021 -0,3966
AR75 0,2372 -0,6836 -0,7041 0,3952
AR08 0,1878 0,1720 0,0769 0,4136
AR09 -0,2024 -0,2344 0,6096* 0,0000
AR10 0,3984 0,0289 0,3714 -0,7231
AR11 0,7866* 0,9090* -0,0791 0,1356
AR18 -0,6303 0,4842 0,1603 0,0000
AR19 -0,6923 -0,6481 -0,8883 -0,0811
AR28 0,7511* -0,8236 0,7279 0,0000
AR29 0,4405 0,4413 0,3881 0,0000
AR42 0,4727 -0,5073 -0,0370 0,4578
AR44 -0,6767 -0,3760 -0,4672 0,1177
AR94 -0,1155 0,5747* 0,0083 0,2216
AR100 0,4643 -0,3560 -0,2649 -0,0496
AR101 -0,09082 -0,8509 0,7027* -0,1617
CER -0,8738 -0,7131 -0,5081 0,0000
GUA/01 -0,0146 0,3199 -0,0527 -0,2242
GUA/02 0,42221 0,25624 0,2326 0,4941
GUA/03 -0,4613 -0,3327 -0,1891 0,0000
JAB -0,6873 0,5437* -0,3813 -0,5278
Pit15 -0,8386 -0,9102 0,7823* 0,4262
Pit71 -0,8172 -0,8599 -0,6923 0,2018
HER 0,5977* 0,0752 -0,0880 0,2521
Pit40 0,9345* -0,9453 -0,9605 -0,2127
PZU -0,2114 0,2482 0,1307 0,7794*
Pit41 0,8820* 0,7783* -0,9132 -0,0126
Pit49 -0,1369 0,1256 -0,6598 -0,1894
Pit50 -0,3525 0,5374* -0,5269 -0,0848
Pit53 0,4952 -0,4483 -0,8874 0,2157
Pit55 -0,3581 -0,1868 0,7177* 0,3857
Pit61 0,5917* 0,5001* 0,1456 0,4408
Pit62 0,6678* 0,6484* 0,7019* -0,6396
UVI 0,2184 0,0506 -0,5269 0,4618
1Araçá(AR), Cereja-do-Mato(CER), Guabiroba(GUA), Jabuticaba(JAB), Pitanga(Pit), Uvaia(UVI); * Termos que
obtiveram correlação acima de 0,5000, pelo Teste de Correlação de Pearson (p<0,05).

48
CONCLUSÕES
Considerando-se os dados obtidos neste estudo, pode-se concluir que:
1. Há significativa variabilidade na composição química entre os
acessos e espécies, e há acessos produtores de frutas com altos
teores de compostos potencialmente bioativos, com destaque para
compostos fenólicos;
2. Frutas de guabirobeira e jabuticabeira são aqueles que
apresentaram a maior atividade antioxidante;
3. Pitanga, uvaia e guabiroba são fontes ricas de carotenóides;
4. Não houve correlação positiva na maioria dos acessos com o
conteúdo de compostos fenólicos totais, antocianinas totais,
carotenóides totais e ácido L-ascórbico com a atividade antioxidante
apresentada pela fruta.

49
CAPÍTULO II – QUANTIFICAÇÃO DE COMPOSTOS FENÓLICOS
INDIVIDUAIS EM FRUTAS NATIVAS DA REGIÂO SUL DA AMÉRICA DO
SUL E AVALIAÇÃO DA ATIVIDADE ANTIPROLIFERATIVA FRENTE A
CÉLULAS TUMORAIS
INTRODUÇÃO
A Região Sul da América do Sul possui espécies vegetais nativas
produtoras de frutas ricas em compostos considerados como biologicamente
ativos do ponto de vista de alimentos funcionais, como é o caso da pitanga e
do araçá. Essas frutas contêm altos teores de compostos fenólicos, dentre
eles, as subclasses dos ácidos fenólicos e de flavonóides.
Esses compostos possuem ação antioxidante, combatendo os radicais
livres que provocam o estresse oxidativo nas células, interrompendo a cadeia
de reações de oxidação em diversas moléculas, como lipídeos, proteínas e
ácidos nucléicos, atuando na inibição das etapas de iniciação e propagação,
retardando o processo oxidativo. Outros benefícios à saúde são atribuídos a
esses compostos, dentre os quais atividade anticancerígena, antiproliferativa,
antimicrobiana, antiinflamatórica e antiterarogênica (Ross e Kasum, 2002).
O câncer é uma das doenças de maior morbidez na atualidade, tendo
em vista que a expectativa média de vida aumentou e a suscetibilidade a
carcinogenese aumenta com o envelhecimento da população. Há a
necessidade de desenvolver novas drogas, combinação de moléculas e novos
tratamentos utilizando compostos sintéticos, biológicos ou produtos naturais

50
para a prevenção ou cura dessas doenças. Os diversos compostos presentes
naturalmente nas espécies vegetais podem auxiliar a combater o avanço
dessas doenças no organismo humano. Diversos estudos epidemiológicos
associam dietas contendo alimentos ricos em compostos fenólicos com uma
baixa incidência de câncer (Block, Patterson e Subar, 1992; Willet, 1994; Fang,
Yang e Wu, 2002).
Na primeira etapa desse estudo comprovou-se a hipótese de que as
frutas da família das mirtáceas, mais especificamente a guabiroba
(Campomanesia xanthocarpa) e a jabuticaba (Mircyaria cauliflora), possuem
alta atividade antioxidante e altos teores de compostos fenólicos totais.
Compostos fenólicos e derivados, como a quercetina, luteolina e catequinas,
são considerados melhores antioxidantes que a vitamina C, vitamina E e β-
caroteno, em uma mesma concentração molar (Gao et al., 1999). Acredita-se
que esses compostos apresentem potencial como agentes antiproliferativos de
células tumorais humanas com linhagens de células de carcinoma cervical
(Narayanan et al., 1999), câncer de colon (Jayaprakasha et al., 2007) e câncer
de mama (Saleem et al., 2002). Estudos demonstram que compostos dos
grupos fenólicos, dentre eles o ácido elágico e compostos do grupo dos
carotenóides, dentre eles a luteolina, podem apresentar essa propriedade.
Acredita-se que as frutas nativas possuem uma composição de
compostos fenólicos individuais característica da espécie e que esses
compostos são importantes agentes antiproliferativos frente a células tumorais.
Nesse contexto, com o objetivo de estudar o potencial dos extratos das
frutas ricas em compostos fenólicos, obtidos a partir das frutas nativas da
Região Sul do Brasil e da América do Sul, buscou-se quantificar os compostos
fenólicos individuais presentes e avaliar a atividade antiproliferativa dos
extratos frente à linhagem células cancerígenas.

51
REVISÃO DE LITERATURA
2.1 Compostos Bioativos em Frutas e Ação Terapêutica
Em geral, os alimentos de origem vegetal são ricos em compostos
biologicamente ativos, principalmente compostos fenólicos, e dentre eles as
antocianinas, flavonóis, flavononas e ácidos fenólicos (Lu e Foo, 2001; Murthy,
Singh e Jayaprakasha, 2002; Manach et al., 2004).
Especial atenção tem sido dada a estes compostos devido à correlação
com alta atividade antioxidante (Kalt et al., 1999; Wang e Lin, 2000). Os
compostos fenólicos, especialmente os flavonóides, são conhecidos pela
elevada capacidade de captar radicais livres causadores de estresse oxidativo,
atribuindo um efeito preventivo de doenças crônicas (Tsuda et al., 1994; Wang
et al., 2002; Katsube et al., 2003; Lapidot et al., 2004). Além disso, possuem
propriedades anticancerígenas, antiinflamatórias, antimicrobianas e
antiprolierativas (Satué-Gracia, Heinonen e Frankel, 1997; Prior et al., 1998;
Sanchéz-Moreno, Larrauri e Saura-Calixto, 1998; Ross e Kasum, 2002).
Compostos fenólicos tais como, ácido elágico, catequinas e ácidos clorogênico,
caféico e ferúlico, podem atuar como potentes agentes antimutagenicos e
anticarcinogênico (Ayrton et al., 1992; Edenharden e Tang, 1997).

52
2.2 Câncer e Ação Antiproliferativa
O câncer surge devido a mutações genéticas, espontâneas ou induzidas
por agentes bióticos ou abióticos. Os radicais livres que promovem desordens
no ciclo celular podem aumentar a taxa de proliferação de células anormais e/
ou induzir deficiências nas taxas de morte celular, culminando com a formação
de agrupamentos de clones de células neoplásicas, os tumores (Ferrari e
Torres, 2002).
Os mecanismos de ação dos fitoquímicos na prevenção do câncer
superam a atividade antioxidante frente a radicais livres. Eles se
complementam e sobrepõem mecanismos de ação, incluindo atividade
antioxidante frente aos radicais livres; regulação da expressão genética em
células em proliferação, diferenciação celular, oncogenes e genes supressores
de tumor; indução de apoptose; modulação da atividade enzimática na
detoxicação, oxidação e redução; estimulação do sistema imune; regulação do
metabolismo de hormônios; e efeitos antibacterianos e antivirais (Liu, 2004).
Estudos epidemiológicos revelam que dietas contendo alimentos ricos
em compostos fenólicos estão associadas a uma menor incidência de câncer
(Fang, Yang e Wu, 2002). Adhami et al., 2007 e Nooman, Benelli e Albini, 2007
observaram que polifenóis, tais como a epicatequina, presente no chá verde
(Camellia sinensis), são importantes agentes biológicos com potencial
anticancerígeno e antiproliferativo. Yang and Wang (1993) e Stroner e Mukhtar
(1995) verificaram que polifenóis presente em pequenas frutas possuem
importante ação em células cancerígenas e no processo de metástase, se
destacando o ácido elágico. Narayanan et al. (1999) verificaram o efeito do
ácido elágico na inibição da proliferação de células e na indução da apoptose
em linhagens de células tumorais cervicais. Yu et al. (2005), por meio de
ensaios in vivo, observaram que o ácido elágico atua na redução do estresse
oxidativo, o qual é responsável pela formação de diversas doenças no
organismo humano, tais como os diferentes tipos de câncer. Também foi
comprovado em testes in vitro e in vivo que os outros compostos fenólicos

53
induzem respostas consistentes através de efeito protetor em condições
degenerativas (Chung et al., 2002; Manach, Mazur e Scalburt, 2005).
Bomser et al., (1996), e Prochaska, (1994) ao estudarem extratos
obtidos a partir de espécies de Vaccinium, verificaram que composto presentes
tem potencial anticarcinogênico e que uma fração específica isolada do extrato
tem atividade indutora da quinona redutase, enzima essencial na detoxicação
de xenobióticos e carcinogênicos. Cheng et al. (2006) sugerem que as
antocianinas presentes em mulberrie (Morus alba L.) poderiam diminuir a
proliferação de células cancerígenas A549 (câncer humano de pulmão).
Atividade antiproliferativa foi testada usando frutos, tais como cranberrie
(Vaccinium macrocarpon) (Yan et al., 2002; Sun e Liu, 2006), mulberrie (Morus
alba) (Cheng et al., 2006), litchi (Litchi chinensis) (Wang et al., 2006) e citros
(Citrus sinensis) (Camarda et al., 2007), obtendo resultados satisfatórios, ou
seja, inibição da proliferação de células tumorais. Além desses autores, outros
realizam estudos com diversos tipos de extratos vegetais e linhagens de
células tumorais obtendo resultados significativos (Lu et al., 2005; Cheng et al.,
2006; Veeriah et al., 2006; Sestili et al., 2007).
Ao avaliar a atividade antiproliferativa do extrato bruto das folhas da L.
divaricata frente à linhagens de células tumorais humanas, Tanaka et al. (2005)
observaram o efeito citostático sem seletividade de extrato bruto das folhas, na
concentração de 250µg.mL-1, inibindo em aproximadamente 85% o
crescimento celular das linhagens de UACC.62, MCF-7 e C786.0; além de um
efeito citotóxico com seletividade nesta mesma concentração para as linhagens
OVCAR.3 (20% de morte), K.562 (30% de morte) e NCL.ADR (50% de morte).
Cheng et al. (2006) isolaram antocianinas presentes em extratos de
mulberry (Morus alba L.) e verificaram que cianidina 3-rutinosidase e cianidina
3-glicosidase possuem forte efeito anticancerígeno frente a células de A549
(linhagens de células de câncer de pulmão). Shih, Yeh e Yen (2005)
analisaram malvinidina, um exemplo de antocianina, frente em linhagens de

54
células de adenocarcinoma gástrico e verificaram eficácia no apoptose dessas
células.
Eberhardt et al. (2000) e Veeriah et al. (2006) verificaram que compostos
fenólicos presentes em maçãs podem inibir células de câncer de cólon in vitro.
Veeriah et al. (2006) encontraram que uma mistura de flavonóides obtidos a
partir de maçãs inibe a proliferação de células de HT29 em concentrações sub-
citotóxicas e induzem a expressão de vários genes.

55
MATERIAL E MÉTODOS
3.1 Material Vegetal
Foram utilizadas frutas nativas da família Myrtaceae (pitanga araçá,
guabiroba, jabuticaba, cereja-do-mato e uvaia) colhidos entre 2005 e 2008
(Tabela 1). Dentre todos os 48 acessos de frutas nativas estudadas no Capítulo
I, apenas nove destas foram selecionadas para a segunda parte desse estudo,
uma de cada seleção de pitanga-roxa (PitH), pitanga-vermelha (Pit62), pitanga-
laranja (Pit15), araçá-roxo (AR94), araçá-amarelo (AR75), jabuticaba (JAB),
cereja-do-mato (CER), guabiroba (GUA/01) e uvaia (UVI). A escolha desses
genótipos foi feita em função da sua composição em relação aos seus teores
totais de compostos fenólicos. A colheita das frutas foi realizada no estádio de
fruta madura, pela cor externa da casca característica de cada fruta. Após a
colheita, cada genótipo foi imediatamente estocado a -80ºC até o momento das
avaliações. Todas as avaliações foram feitas com extratos com as frutas non-
peeled, pois essa é a forma como são comumente consumidas. Dentre as
frutas selecionadas, a pitanga-roxa (PitH) e a guabiroba (GUA/01) foram
colhidas em propriedades rurais localizadas no Município de Pelotas/RS, e as
demais foram colhidas no Banco de Germoplasma da Embrapa Clima
Temperado, também localizado no Município de Pelotas/RS.

56
3.2. Identificação e Quantificação de Compostos Fenólicos
Individuais
A extração e quantificação dos compostos fenólicos individuais, por
cromatografia líquida de alta eficiência foi segundo método adaptado de
Hänikken e Törrönen (2000) e Zambiazi (1997).
3.2.1 Padrões Cromatográficos
Ácido elágico, ácido gálico, ácido caféico, ácido hidroxobenzóico, ácido
p-cumárico, ácido ferúlico, epicatequina, catequina, miricetina, kaempferol e
quercetina. (Sigma Chemical Co. St. Louis, USA e Fluka Milwaukee, WI, USA).
3.2.2 Método de Extração
As frutas (5g) foram maceradas e adicionou-se 4,9 mL de HCL p.a. com
o objetivo de realizar uma hidrólise ácida e 30mL de metanol p.a.. As amostras
foram mantidas em banho-maria a 35°C por 24 horas, após o qual foram
filtradas em algodão e lavadas com duas porções de 10mL de metanol p.a.
Após a lavagem, completou-se o volume de 50mL com metanol p.a. e
concentrado em rota-evaporador a 40°C até o volume final de 5mL. Completou-
se o volume do extrato para 5mL e centrifugou-se o concentrado a 7000rpm
por 10 minutos. O sobrenadante foi usado para análise cromatógrafica.
3.2.3 Análise Cromatográfica
A análise cromatográfica foi realizada em cromatógrafo Shimadzu,
equipado com módulo de mistura de solventes LC-10ATVP, desgaseificador
FCV-10AVP, bomba reodine DGU-14A, sistema de controle SCL-10AVP, forno de
coluna CTO-10ASVP e amostrador automático SIL-10AF. Foi utilizada uma

57
coluna de separação analítica de fase reversa, Nova-Pak C18 (3,9cm x 150mm
x 4µm). Como fase móvel utilizou-se dois solventes: (A) solução de metanol (B)
solução de ácido acético 1% (Tabela 11). Injetou-se 20µL de amostra, com
fluxo de 0,8mL.min-1, e temperatura de coluna mantida entre 25°C. O tempo de
eluição da amostra foi de 45 minutos. A identificação do composto foi realizada
através de informações obtidas dos espectros de absorção no UV-visível,
modelo SPD-10AVVP em um comprimento de onda de 280nm.
Tabela 11. Gradiente de eluição dos solventes A e B na determinação de
compostos fenólicos individuais.
Tempo (minutos) Solvente A (%)1 Solvente B (%)2
0 100 0
25 60 40
27 60 40
37 95 5
42 95 5
45 100 0
1Solvente A: solução de água Milli-Q:ácido acético p.a. (99:1% v/v); 2Solvente B: metanol p.a.
100%.
Os dados obtidos foram adquiridos e processados com o uso do
software Class-VP. Os compostos fenólicos individuais foram quantificados
com base de curva de calibração externa, cujos padrões foram dissolvidos em
metanol p.a. A concentração das soluções, obtidas a partir dos padrões
cromatográficos, variou entre 0,125-12,5µg.25µL-1 para os padrões de ácido p-
coumárico, ácido caféico, quercetina, ácido ferúlico, epicatequina, ácido p-
hidroxibenzóico, ácido gálico e ácido elágico; 1,25-87,5µg.25µL-1 para a
catequina e 0,125-6,25µg.25µL-1 para os padrões miricetina e kaempferol. Os
resultados foram expressos em mg.100g-1 de peso fresco.

58
3.3 Atividade Antiproliferativa frente a Células Tumorais
3.3.1 Preparo do Extrato
As frutas foram maceradas e, posteriormente, centrifugadas, sob
refrigeração a 4°C e 4000 g por 30 minutos. Após esse período, o
sobrenadante foi armazenado a -80°C até o momento das avaliações.
3.3.2 Linhagens de Células e Cultivo
Linhagens de células tumorais humanas MCF-7 (adenocarcinoma de
mama) adquiridas no Banco Celular da Universidade do Rio de Janeiro/RJ.
Como controle foi usado linhagens de células 3T3 (fibroblastos embrionários de
ratos). As células foram estocadas em nitrogênio líquido e descongeladas em
banho-maria. Para o cultivo celular (5 x 105 mL/células) foi adicionado meio
suplementar com 20% de soro fetal bovino. O meio de cultivo foi trocado a
cada dois ou três dias e o acompanhamento do crescimento celular se deu a
cada 24 horas por meio de um microscópio óptico invertido. As células foram
mantidas a 37°C, em atmosfera úmida contendo 95% de O2 e 5% de CO2.
3.3.3 Determinação da Atividade Antiproliferativa
A atividade antiproliferativa foi avaliada nas linhagens de células
tumorais e não tumorais já citadas. Essas linhagens celulares foram
microtituladas na mesma densidade e pré-incubadas por 24 horas.
As células foram colocadas em cultivo monocamada, segundo Monks et
al., 1991. As placas, contendo as células aderidas, durante 24 horas a 37°C e
100% de UR (umidade relativa) foram expostas a cada um dos extratos
vegetais, em três diluições diferentes, por 48 horas e comparadas a um grupo

59
controle. Foram realizadas três repetições para cada diluição de extrato. Após
o período de incubação, nas condições de 37°C, 5% de CO2 e 100% de UR, as
células foram fixadas mediante a adição de TCA (ácido tricloroacético) e
incubadas em refrigerador a 4°C durante uma hora. O ensaio colorimétrico foi
realizado mediante a adição de solução de sulforodamina B 0,4% p/v (com
ácido acético a 1%) em cada uma das cavidades da placa de cultivo. Essa
placa permaneceu em repouso, em temperatura ambiente por 30 minutos, e
posteriormente, a solução não fixada foi desprezada mediante lavagens com
ácido acético a 1%. O corante fixado nas proteínas celulares foi ressolubilizado
com tampão Tris Base 10 mM (pH 10,0) sob agitação giratória, em uma
velocidade de 50 rpm, à temperatura ambiente, por um tempo de cinco minutos
(Rubinstein et al., 1990; Skehan et al., 1990). A leitura da densidade óptica foi
realizada em espectrofotômetro leitor de placa ELISA, em um comprimento de
onda de 492 e 560 nm (Ithrat et al., 2004; Skehan et al., 1990).
A medida da viabilidade e sobrevivência celular, segundo Monks et al.,
1991, foi expressa em %T/C [(densidade óptica das células tratadas /
densidade óptica do controle celular) x 100]. Por meio destas medidas, a
resposta celular foi calculada nos termos de estimulação do crescimento,
ausência do efeito extrato/fração, inibição do crescimento, morte celular e
proliferação celular.
3.4 Delineamento Experimental
O delineamento foi inteiramente casualizado, com três repetições para
cada tratamento. Os resultados foram expressos como médias. As diferenças
das variáveis durante o armazenamento foram avaliadas pelo Teste de
Duncan, em nível de 5%. As análises estatísticas foram realizadas com o
auxílio do programa de computação STATISTICA 6.0.
Os resultados da análise de atividade antiproliferativa e atividade
antioxidante foram tabulados e analisados em software estatístico GraphPad
Prism, versão 4.00 para Windows (GraphPad Software, San Diego,
Califórnia, EUA, ‘www.graphpad.com’, 2003). Com base nos resultados, além

60
do crescimento celular, foi avaliada a taxa de proliferação celular ao longo dos
dias de teste.

61
RESULTADOS E DISCUSSÃO
4.1 Compostos Fenólicos Individuais
Nas nove frutas avaliadas, nessa etapa do trabalho, verificou-se que,
dentre os compostos fenólicos, dois compostos apresentaram picos
representativos em todos os acessos avaliados, o ácido gálico (ácido fenólico)
e a catequina (flavanol). Em menor concentração, o ácido caféico, ácido
ferúlico, ácido elágico (ácidos fenólicos) e o kaempferol (flavonol) foram
identificados em frutas de cereja-do-mato (CER), pitanga-vermelha (Pit62),
uvaia (UVI), pitanga-roxa (PitH), jabuticaba (JAB) e guabiroba (GUA/01)
(Tabela 12).
A soma do conteúdo de todos os compostos fenólicos individuais
quantificados, através da determinação por cromtografia líquida de alta
eficiência, perfaz de 40 a 94,62% do total de compostos fenólicos determinados
por espectrofotometria (Tabela 5, Capítulo I). A diferença no conteúdo dos
compostos fenólicos se deve pela presença de outros compostos presentes em
seus subgrupos, os quais não foram quantificados como ácido fenólico ou
flavanol, mas que são quantificados na determinação de compostos fenólicos
totais.
Os frutos de guabiroba (GUA/01) apresentaram os maiores teores de
ácido gálico (305mg.100g-1), entre as nove frutas avaliadas, seguido dos frutos
de jabuticaba (JAB) com 137,64mg.100g-1 e de araçá-roxo (AR94) com

62
131,64mg.100g-1. O menor conteúdo deste composto foi identificado no fruto
de pitanga-laranja (Pit15), 19,57mg.100g-1.
Tabela 12. Teor de compostos fenólicos individuais em frutos nativos da
Região Sul da América do Sul.
Fruta Ácido Gálico1 Catequina Ácido Caféico
Ácido Ferúlico
Ácido Elágico
Kaempferol Total
Araçá
Amarelo (AR75)
56,25±0,50d2 19,45±0,84d nd3 nd nd nd 75,70
Araçá Roxo
(AR94)
131,64±11,16b 95,14±22,45b nd nd nd nd 226,78
Cereja do
Mato (CER)
28,10±5,70ef 64,27±2,92bc 8,80±1,57b 1,06±0,09a nd nd 102,23
Guabiroba (GUA/01) 305,08±36,37a 576,04±84,17a nd nd 7,29±3,08b nd 888,41
Jabuticaba (JAB) 137,64±12,52b 23,99±5,75d 7,44±0,51b nd 1,99±1,42c 23,17±0,63a 194,23
Pitanga Laranja (Pit15)
19,57±1,50f 27,19±3,78cd nd nd nd nd 46,76
Pitanga Roxa (PitH)
22,64±0,79f 38,24±3,21cd 70,44±14,29a nd 15,88±0,48a nd 147,20
Pitanga Vermelha
(Pit62) 86,48±0,99c 23,41±0,29d 1,85±0,42c nd nd nd 111,74
Uvaia (UVI) 45,51±27,67de 42,42±6,78cd 0,54±0,04c nd nd nd 88,47
1Resultados expressos em mg.100g-1; 2 Médias seguidas da mesma letra minúscula na linha não diferem entre si, pelo
Teste de Duncan (p<0,05); 3Não detectado.
Em relação ao outro composto fenólico majoritário (catequina),
novamente a guabiroba (GUA/01) foi o acesso que apresentou o maior teor,
com 576,04mg.100g-1. A catequina foi identificada como composto majoritário
nos frutos de cereja-do-mato (CER), guabiroba (GUA/01) e pitanga-laranja
(Pit15), com 64,27mg.100g-1, 576,04mg.100g-1 e 27,19mg.100g-1,

63
respectivamente. O menor conteúdo foi identificado no fruto de araçá-amarelo
AR75 (19,45mg.100g-1).
O ácido caféico foi identificado em cinco frutas nativas, apresentando-
se em quantidades superiores apenas nos frutos de pitanga-roxa (PitH) quando
comparado com as demais frutas avaliadas. Este composto fenólico também foi
o composto majoritário presente apenas nos frutos de pitanga-roxa (PitH).
O ácido ferúlico foi identificado em apenas um dos acessos de frutas
nativas avaliadas nesse estudo, cereja-do-mato CER (1,06mg.100g-1), mesmo
assim em pequenas quantidades. O ácido elágico foi identificado em três frutas
nativas, guabiroba (GUA/01), jabuticaba (JAB) e pitanga-roxa (PitH), em todos
eles também em pequenas quantidades. O kaempferol foi identificado somente
em um das frutas avaliadas, na jabuticaba (JAB), com 23,17mg.100g-1.
A guabiroba (GUA/01) foi o acesso que apresentou a maior
concentração de ácido gálico e de catequina, entre os nove acessos avaliados
nesse estudo. A atividade antioxidante destes frutos também foi elevada
(1.695,59mM Trolox) quando comparada com a atividade das outras 47 frutas
avaliadas na primeira etapa, além de apresentar alto teor de compostos
fenólicos totais e de ácido L-ascórbico (Tabela 5, 8 e 9, Capítulo I). Embora o
conteúdo de antocianinas e de carotenóides totais seja intermediário, quando
comparado com o conteúdo das demais frutas. Esses dados sugerem que a
atividade antioxidante pode estar relacionada ao teor de algum composto
fenólico individual presente nas frutas. A combinação destes compostos e o
sinergismo com outros, podem ser os principais fatores que conferem a
atividade antioxidante as frutas. Estes dados estão em desacordo com outros
autores, que afirmam que a atividade antioxidante está diretamente relacionada
aos altos teores de compostos fenólicos totais e de antociânicos total presentes
nos frutos (Imeh e Khokhar, 2002; Sun et al., 2002; Roberts e Gordon, 2003;
Pinelo et al., 2004).
A pitanga-vermelha (Pit62) foi a fruta que apresentou a menor atividade
antioxidante (3,88mM Trolox) entre as nove frutas estudadas nessa etapa.

64
Porém, apresentou um conteúdo de ácido gálico e de catequina superiores aos
acessos de araçá-amarelo (AR75). O acesso de araçá-amarelo (AR75)
apresentou teor inferior de ácido gálico e de catequina, com respectivamente,
46,54mg.100g-1 e 19,40mg.100g-1, mas apresentou uma atividade antioxidante
superior ao acesso de pitanga-vermelha (Pit62).
A pitanga-roxa (PitH) foi uma das frutas, entre os 48 acessos avaliados
na primeira etapa, que apresentou a maior atividade antioxidante (Tabela 9,
Capítulo I), diferindo significativamente apenas dos três acessos de guabiroba
(GUA/01, GUA/02 e GUA/03) e de jabuticaba (JAB). Na determinação de
compostos fenólicos individuais, na pitanga-roxa (PitH), observou-se um teor
inferior de ácido gálico e de catequina, em relação aos acessos de uvaia (UVI)
e araçá-roxo (AR94), mas estes dois acessos apresentaram atividade
antioxidante inferior ao da pitanga-roxa (Tabela 9, Capítulo I). Porém, a
pitanga-roxa (PitH) apresentou teor de ácido caféico elevado, em relação aos
outros compostos avaliados (Tabela 12), o que sugere que não é apenas o
conteúdo de compostos fenólicos totais, mas talvez a presença de alguns
ácidos fenólicos, ou o sinergismo e/ou combinação de um ou mais compostos
que são responsáveis pela atividade antioxidante das frutas.
Com isso pode-se sugerir que a composição química de compostos
fenólicos individuais não está linearmente correlacionada com a respectiva
atividade antioxidante. A combinação de compostos ou de uma subclasse de
compostos e seus sinergismos podem ser os responsáveis por conferir à
atividade antioxidante às frutas.
Frutas de outros centros de origem (Hemisfério Norte), tais como, amora
e mirtilo apresentaram uma composição de compostos fenólicos individuais
distintos. Zadernowski, Naczk e Nesterowicz (2005) avaliaram diversas
pequenas frutas e verificaram concentrações de ácido gálico inferior as frutas
nativas da Região Sul da América do Sul. Como foi sugerido no capítulo
anterior, a composição química é uma característica genotípica e está sujeita a
fatores pré-colheita climáticos e ambientais (Robards et al., 1999). Esses
autores verificaram que frutos de mirtilo (Vaccinium myrtillus) possuem na sua

65
composição química, além do ácido gálico (9,36mg.100g-1) e do caféico
(11,72mg.100g-1), presentes nas frutas nativas da Região Sul da América do
Sul, outros ácidos derivados do ácido p-hidroxibenzóico, tais como, ácido
siríngico (4,16mg.100g-1) e ácido vanílico (11,7mg.100g-1), além de ácido p-
coumarico (76,18mg.100g-1) e hidroxicaféico (72,38mg.100g-1). Chiou et al.
(2007), ao avaliarem frutos de Vitis vinifera var. Vostizza, verificaram teor de
ácidos fenólicos e flavonols individuais distinto aos teores identificados nas
frutas nativas da Região Sul da América do Sul, em média, 0,52mg.100g-1 para
ácido gálico, 0,55 mg.100g-1 para ácido caféico, 0,05 mg.100g-1 para
kaempferol é de 0,37 mg.100g-1 para a quercetina.
Reynertson et al. (2008) também analisaram frutas da família
Myrtaceae e verificaram baixos teores de flavonols (quercetina, kaempferol e
miricetina) inferior a 24mg.100g-1, em média.
Das frutas nativas avaliadas neste estudo, apenas a jabuticaba (JAB)
apresentou teor de kaempferol, o que sugere que seja uma característica da
família botânica a baixa incidência dessa classe de compostos fenólicos na
composição química das frutas nativas da Região Sul da América do Sul.
4.2. Atividade Antiproliferativa
O efeito inibitório dos extratos vegetais na proliferação das linhagens de
células de adenocarcinoma de mama em humanos (MCF-7) foi investigado
nesse estudo, através de ensaio com duas linhagens de células, uma linhagem
controle de fibroblastos embrionários de ratos (3T3) e outra de adenocarcinoma
de mama em humanos (MCF-7); e três concentrações de extratos (6µL.200mL-
1, 4µL.200mL-1 e 2µL.200mL-1).
Verificou-se que, dentre as nove frutas avaliadas, apenas os extratos de
pitanga-vermelha (AR62) e de araçá-roxo (AR94) apresentaram inibição de
proliferação de células tumorais (Figuras 12 e 14).

66
Nos extratos obtidos a partir do acesso de araçá-roxo (AR94), a taxa de
inibição da proliferação das células de adenocarcinoma de mama em humanos
(MCF-7) foi entre 40 e 50%, enquanto que nos extratos obtidos a partir dos
frutos de pitanga-vermelha (Pit62), na concentração de 6µL.200µL-1, a inibição
foi de quase 100%. Em relação às células controle de fibroblastos (linhagem de
células 3T3), em ambos os extratos houve morte celular, porém observou-se
uma inibição de proliferação celular inferior a proliferação observada nas
células de MCF-7 tratadas com os extratos.
Dos nove extratos de frutas avaliadas, apenas duas apresentaram
atividade antiproliferativa frente a células tumorais, pitanga-vermelha (Pit62) e
araçá-roxo (AR94). Os outros sete extratos, araçá-amarelo (AR75), cereja-do-
mato (CER), guabiroba (GUA/01), jabuticaba (JAB), pitanga-laranja (Pit15),
pitanga-roxa (PitH) e uvaia (UVI) não apresentaram efeito inibitório da
proliferação celular. Esses dados sugerem que os extratos de araçá-roxo
(AR94) e de pitanga-vermelha (Pit62) possuem na sua composição química,
algum composto e/ou de um subgrupo de compostos que possuam ação
antiproliferativa frente às células tumorais de MCF-7.
0.0 0.2 0.4 0.6 0.8 1.0-50
-40
-30
-20
-10
log10[Araçá-Roxo - AR94]
% P
rolif
eraç
ão c
elu
lar
Figura 11. Atividade citotóxica do extrato de araçá-roxo AR94 sobre a linhagem de células 3T3.

67
0.0 0.2 0.4 0.6 0.8 1.0-50
-40
-30
-20
-10
log10[Araçá-Roxo - AR94]
% P
rolif
eraã
o c
elu
lar
Figura 12. Atividade citotóxica do extrato de araçá-roxo AR94 sobre a linhagem de células
MCF-7.
0.00 0.25 0.50 0.75 1.00-40
-30
-20
-10
0
log10[Pitanga-Vermelha Pit62]
% P
roli
fera
ção
cel
ula
r
Figura 13. Atividade citotóxica do extrato de pitanga-vermelha Pit62 sobre a linhagem de
células 3T3.

68
0.2 0.4 0.6 0.8 1.0
-100
-50
0
50
100
log10[Pitanga-Vermelha Pit62]
% P
rolif
eraç
ão c
elu
lar
Figura 14. Atividade citotóxica do extrato de pitanga-vermelha Pit62 sobre a linhagem de
células MCF-7.
Vários autores relacionam o teor de compostos fenólicos com a atividade
antiproliferativa (García-Alonso, Ros e Periago, 2006; Kaur e Singh, 2008;
Saleem et al., 2002; Singh e Kaur, 2008). Porém nesse estudo, verificou-se
que, dentre as frutas avaliadas, o acesso (GUA/01) que apresentou o maior
teor de compostos fenólicos totais (1.062,03mg.100g-1) e individuais
(888,41mg.100g-1), não apresentou atividade antiproliferativa frente as células
tumorais MCF-7.
A alta capacidade antioxidante não demonstra estar lineamente
relacionada à atividade antiproliferativa (Sun et al., 2002). Essa afirmação
coincide, com o estudo realizado, que indicou que as frutas com maior
atividade antioxidante, guabiroba (GUA/01) e jabuticaba (JAB) não
apresentaram atividade antiproliferativa frente a células tumorais de
adenocarcinoma de mama (MCF-7). Dentre as nove frutas avaliadas, a
pitanga-vermelha (Pit62) foi o acesso que apresentou atividade antioxidante
inferior (3,88mM Trolox), um teor de compostos fenólicos totais inferior aos de
acesso de araçá-roxo (AR94), jabuticaba (JAB), guabiroba (GUA) e pitanga-
roxa (PitH), além de não ser o acesso com os teores mais elevados, dentre as
48 frutas estudadas, de antocianinas totais, carotenóides totais e ácido L-
ascórbico (Tabelas 1, 2, 3 e 4, Capítulo I); porém, apresentou atividade
antiproliferativa frente a linhagem de células MCF-7, com taxa de inibição de
quase 100% (Figura 14). Em relação à determinação de compostos fenólicos

69
individuais, a pitanga-vermelha (Pit62) apresentou o maior conteúdo de ácido
gálico em relação aos outros dois acessos de pitangas avaliados. No entanto,
além da pitanga-vermelha (Pit62), os acessos de araçá-amarelo (AR75),
guabiroba (GUA/01), jabuticaba (JAB) e uvaia (UVI), apresentam o ácido gálico
como o composto fenólico individual majoriário; porém, não apresentaram
atividade antiproliferativa frente a células tumorais de MCF-7.
Esses dados sugerem que nenhum dos compostos específicos
avaliados neste estudo presente na composição química dos frutos de pitanga-
vermelha (Pit62) possui essa ação antiproliferativa frente às células tumorais
de adenocarcinoma de mama. Sun et al. (2002), ao avaliarem a atividade
antiproliferativa em frutas, tais como, maçã, morango, abacaxi, banana,
pêssego, laranja e pêra, sugerem que seja algum composto fenólico específico
ou um subgrupo de compostos fenólicos, o(s) responsável por essa atividade.

70
CONCLUSÕES
Considerando-se os dados obtidos neste estudo, pode-se concluir que:
1. Os principais compostos fenólicos identificados nas frutas nativas da
Região Sul da América do Sul são o ácido gálico e a catequina,
sendo a guabiroba, a fruta que apresentou os maiores teores desses
compostos;
2. As frutas de pitanga-vermelha (Pit62) e de araçá-roxo (AR94)
apresentaram efeito antiproliferativo e citotóxico nas linhagens
tumorais humanas de MCF-7;
3. Não se observou uma relação direta entre o conteúdo dos
compostos fenólicos individuais com a atividade antioxidante e a
atividade antiproliferativa frente a linhagem de células tumorais de
MCF-7.

71
REFERÊNCIAS BIBLIOGRÁFICAS
ADHAMI, V.M.; MALIK, A.; ZAMAN, N.; SARFARAZ, S.; SIDDIQUI, I.A.; SYED,
D.N.; et al. Combined inhibitory effects of green tea polyphenols and selective
cyclooxygenase-2 inhibitors on the growth of human prostate cancer cells both
in vitro and in vivo. Clinical Cancer Research, v.13, p.1611-1619, 2007.
AGOSTINI-COSTA, T.S. et al. Efeito do Congelamento e do Tempo de
Estocagem da Polpa de Acerola sobre o Teor de Carotenóides. Revista
Brasileira de Fruticultura, v.25, n.1, abr.2003. p. 56-58.
ANDERSON, R.A.; POLANSKI, M.M. Tea enhances insulin activity. Journal of
Agriculture and Food Chemistry, v.50, p.7182-7186, 2002.
ANDRADE, R.N.B.; FERREIRA, A.G. Germinação e armazenamento de
sementes de uvaia (Eugenia pyriformis Camb.) – Myrtaceae. Revista
Brasileira de Sementes, v.12, n.2, p.66-73, 1990.
ANDRIAMBELOSOM, E.; MAGNIER, C.; HAAN-ARCHIPOFF, G.; LOBSTEIN,
A.; ANTON, R.; BERETZ, A.; STOCLET, J.; ANDRIANTSITOHAIMA, R. Natural
dietary polyphenolic compounds cause endothelium-dependent vasorelaxation
in rat thoracic aorta. Journal of Nutrition, v.128, p.2324-2333, 1998.

72
AYRTON, A.D.; LEWIS, D.F.V.; WALKER, R.; LOANNIDES, C.
Antimutagenicity of ellargic acid towards the food mutagen IQ: investigation
intopossible mechanisms of action. Food and Chemical Toxicology, v.30,
p.289-295, 1992.
BAO, J.; CAI, Y.; SUN, M.; WANG, G.; CORKE, H. Anthocyanins, Flavonols,
and Free Radical Scavenging Activity of Chinese Bayberry (Myrica rubra)
Extracts and Their Color Properties and Stability. Journal of Agricultural and
Food Chemistry, v.53. p;2327-2332, 2005.
BENZI, G. Aerobic performance and oxygen free-radicals. The Journal of
Sports Medical Physical Fitness, v.33, p.205-222, 1993.
BEZERRA, J.E.F.; SILVA JÚNIOR, J.F.; LEDERMAN, I.E. Pitanga (Eugenia
uniflora L.). Jaboticabal: FUNEP, 2000. 308p. (Série Frutas Nativas 1).
BEZERRA, M.A.; ALVES, J.D.; OLIVEIRA, L.E.M.; PRISCO, J.T.
Caracterização morfológica e mobilização de reservas durante os estádios
iniciais de desenvolvimento de plântulas de Vigna unguiculata (L.) Walp.
Revista Ciência Agronômica. V.34, n.2, p.253-259, 2003.
BIANCHI, M.L.P.; ANTUN ES, L.M.G. Radicais livres e os principais
antioxidantes da dieta. Revista de Nutrição, v.12, n.2, p.123-130, 1999.
BIAVATTI, M.W.; FARIAS, C.; CURTIUS, F.; BRASIL, L.M.; HORT, S.;
SCHUSTH, L.; LEITE, S.N.; PRADO, S.R.T. Prelimnary studies on
Campomanesia xanthocarpa (Berg.) and Cuphea carthagenensis (Jacq.) J.F.
Macbr. Aqueous extract: weigh control and biochemical parameters. Journal of
Ethnopharmacology, v.93, p.385-389, 2004.
BLOCK, G.; PATTERSON, B.; SUBAR, A. Fruit, vegetables, and cancer
prevention – a review of the epidemiologic evicence. Nutrition of Cancer
International Journal, v.18, p.1-29, 1992.

73
BLOT, W.J.; LI, J.Y.; TAYLOR, P.R. Nutrition intervention trials in Linxian,
China: supplementation with specific vitamin/mineral combinations, cancer
incidence, and disease-specific mortality in the general population. Journal of
National Cancer Institute, v.85, p.1483-1492, 1993.
BOMSER, J.; MADHAVI, D.L.; SINGLETARY, K.; SMITH, M.A. In vitro
anticancer activity of fruit extracts from Vaccinium species. Planta Medica,
v.62, p.212-216, 1996.
BOWLES, B.L.; JUNEJA, V.K. Inhibition of foodborne bacterial pathogens by
naturally occurring food additives. Journal of Food Safety, v.18, p.101-112,
1998.
BRAND-WILLIANS, W.; CUVELIER, M.E.; BERSET, C. use of a free radical
method to evaluate antioxidant activity. Lebensmittel-Wissenschaft und
Technologie/Food Science and Technology, v.28, p.25-30, 1995.
BURNS, A.E. et al. Antioxidant properties of evening primrose seed extracts.
Journal of Agricultural and Food Chemistry, v.49, p.4502-4507, 2001.
CAMARDA, L.; STEFANO, V.D.; DEL BOSCO, S.F.; SCHILLACI, D.
Antiproliferative activity of Citrus juices an HPLC evaluation of their flavonoid
composition. Fitoterapia, v.78, p.426-429, 2007.
CÉSPEDES, C.L.; EL-HAFIDI, M.; PAVON, N.; ALARCON, J. Antioxidant and
cardioprotective activities of phenolic extracts from fruits of Chilean blackberry
Aristotelia chilensis (Elaeocarpaceae), Maqui. Food Chemistry, v.107, p.820-
829, 2008.
CHENG, P.; CHU, S.; CHIOU, H.; KUO, W.; CHIANG, C.; HSIEH, Y. Mulberry
anthocyanins, cyaniding 3-rutinoside and cyaniding 3-glucoside, exhibited an
inhibitory effect on the migration and invasion of human lung cancer cell line.
Cancer Letters, v.235, p.248-259, 2006.

74
CHIOU, A.; KARATHANOS, V.T.; MYLONA, A.; SALTA, F.N.; PREVENTI, F.;
ANDRIKOPOULOS, N.K. Currant (Vitis vinifera) content of simple phenolic and
antioxidant activity. Food Chemistry, b.102, p.516-522, 2007.
CHUNG, Y.C.; CHANG, C.T.; CHAO, W.W.; LIN, C.F.; CHOU, S.T.
Antioxidantive activity and safety of the 50% ethanolic extract from red bean
fermented by Bacillus subtilis IMR-NKI. Journal of Agricultural and Food
Chemistry, v.50, p.2454-2458, 2002.
CONNOR, A.M.; LUBY, J.J.; HANCOCK, J.F.; BERKHEIMER, S.; HANSON,
E.J. Changes in fruit antioxidant activity among blueberry cultivars during cold-
temperature storage. Journal of Agricultural and Food Chemistry, v.50,
p.893-898, 2002.
D’EECKENBRUGGE, G.C.; FERLA, D.L.; FERREIRA, F.R. Diversidade e
potencial das fruteiras neotropicais. In: Congresso Brasileiro de Fruticultura.
Anais... Poços de Calda, 1998. p.19-47.
DHILLON, B.S.; SINGH, S.N.; KUNDAL, G.S. Studies on the developmental
physiology of guavaa fruit (Psodium guajava L.) II. Biochemical characteres.
Pujab Horticultural Journal, v.27, n.3-4, p.212-221, 1990.
DONADIO, L.C. Jabuticaba (Myrciaria jaboticaba (Verg.) Berg. Jabotical:
FUNEP, 2000. 55p. (Série Frutas Nativas 3).
DONADIO, L.C.; MÔRO, F.V.; SEVIDONE, A.A. Frutas Brasileiras. Jabotical:
FUNEP, 2002. 288p.
DUAN, X.; JIANG, Y.; SU, X .; ZHANG, Z.; SHI, J. Antioxidant properties of
anthocyanins extracted from litchi (Litchi chinenesis Sonn.). Food Chemistry,
v.101, p.1365-1371, 2007.
DUGO, P.; MONDELLO, L.; ERRANTE, G.; ZAPPIA, G.; DUGO, G.
Identification of anthocyanins in berries by narrow-bore high-performance liquid
chromatography with electrospray ionization detection. Journal of Agricultural
and Food Chemistry, v.49, p.3987-3992, 2001.

75
EBERHARDT, M.V.; LEE, C.Y.; LIU, R.H. Antioxidant activity of fresh apples.
Nature, v.405, p.903-904, 2000.
EDENHARDER, R.; TANG, X. Inhibition of the mutagenicity of 2-nitrofluorene,
3-nitrofluoranthene and 1-nitropyrene by flavonoids, coumarines, quinines and
other phenolic compounds. Food and Chemical Toxicology, v.35, p.357-372,
1997.
EHLENFELDT, M.K.; PRIOR, R.L. Oxygen radical absorbance capacity
(ORAC) and phenolic and anthocyanin concentrations in fruit and leaf tissues of
highbush blueberry. Journal of Agricultural and Food Chemistry, v.49,
p.222-227, 2001.
ELSAYED, N.M. Antioxidant mobilization in response to oxidative stress: a
dynamic environmental nutrition intervention. Nutrition, v.17, n.828-834, 2001.
ERCISLI, S.; ORHAN, E. Chemical composition of white (Morus alba), red
(Morus rubra) and black (Morus nigra) mulberry fruits. Food Chemistry, v.103,
p.1380-1384, 2007.
FANG, Y.Z.; YANG, S.; WU, G. Free radicals, antioxidants, and nutrition.
Nutrition, v.18, p.872-879, 2002.
FERRARI, C.K.B.; TORRES, E.A.F.S. Novos compostos dietéticos com
propriedades anticarcinogênicas. Revista Brasileira de Cancerologia, v.48,
n.3, p.375-382, 2002.
FERREIRA, F.R. Recursos Genéticos de Espécies Frutíferas no Brasil.
Brasília: Embrapa Recursos Genéticos e Biotecnologia, 1999. 190p.
FLAGG, E.W.; COATES, R.J.; GREENBERG, R.S. Epidemiologic studies of
antioxidants and cancer in humans. Journal of American Collage Nutrition.
v.14, p. 419., 1995.
FRASER, P.D.; BRAMLEY, P.M. The biosynthesis and nutritional uses of
carotenoids. Progress in Lipid Research, v.43, p.228-265, 2004.

76
GAO, Z.; HUANG, K.; YANG, X.; XU, H. Free radical scavenging and
antioxidant activities of flavonoids extractes from the radix of Scutellaria
baicalensis Georgi. Biochima et Biophysica Acta, v.1472, p.643-650, 1999.
GARCÍA-ALONSO, J.; ROS, G.; PERIAGO, M.J. Antiproliferative and
cytoprotective activities of a phenolic-rich juice in HepG2 cells. Food Research
International, v.39, p.982-991, 2006.
GERHARDT, L.B.A..; MANICA, I.; KIST, H.; SIELER, R.L.. Características
físico-químicas das frutas de quatro cultivares e três clones de goiabeira em
Porto Lucena, RS. Pesquisa Agropecuária Brasileira, v.32, n.2, p.185-192,
1997.
GRAPHPAD SOFTWARE INC. – Prism 4 for Windows. La Joella: GraphPad,
2003, software version 4.0.
GREENBERG, E.R.; BARON, J.A.; STUCKEL, T.A.; STEVENS, M.M.;
MANDEL, J.S. A clinical trial of β-carotene to prevent basal cell and squamous
cell cancer of the skin. The New England Journal of Medicine, v.323, p.789-
795, 1990.
GIL, M.I.; TOMÁS-BARBERÁN, F.A.; HESS-PIERCE, B.; KADER, A.A.
Antioxidant capacities, phenolic compounds, carotenoids, and vitamin C
contents of nectarine, peach, and plum cultivars from California. Journal of
Agricultural and Food Chemistry, v.50, p.4976-4982, 2002.
GOMES, E.; DILERMANDO, P.; MARTINS, A. B. G.; FERRAUDO, A. S.
Análise de grupamentos e de componentes principais no processo seletivo em
genótipos de aceroleira (Malpighia emarginata DC). Revista Brasileira de
Fruticultura, v.22, n.1, p.36-39, 2000.
HALLIWELL, B.; GUTTERIDGE, J.M.C. Role of free radicals and catalytic metal
ions in human disease: and overview. Methods in Enzymology, v.186, p.1-85,
1990.

77
HÄKKINEN, S.H.; TÖRRÖNEN, R. Content of flavonols and selected phenolics
acids in strawberries and Vaccinium species: influence of cultivar, cultivation
site and technique. Food Research International, v.33, p.517-524, 2000.
HARDER, I.C.F.; TEIXEIRA, L.A.J.; POMMER, C.V.; GALLO, P.B.
Desenvolvimento inicial de plantas de espécies frutíferas nativas e exóticas em
Mococa, Sp. In: Congresso Brasileiro de Fruticultura. Anais... Florianópolis,
2004. p.16-22.
HENNEKENS, C.H.; BURING, J.E.; MANSON, J.E.; STAMPFER, M.;
ROSNER, B.; COOK, N.R.; et al. Lack of effect of long-term supplementation
with beta carotene on the incidence og malignant neoplasms and
cardiovascular disease. The New England Journal of Medicine, v.334,
p.1145-1149, 1996.
IMEH, U.; KHOKHAR, S. Distribution of conjugates and free phenols in fruits:
antioxidant and cultivar variations. Journal of Agricultural and Food
Chemistry, v.50, p. 6301-6306, 2002.
ITHARAT, A.; HOUGHTON, P.J.; ENO-AMOOQUAYE, E.; BURKE, P.J.;
SAMPSON, J.H.; RAMAN, A. In vitro cytotoxic activity of Thai medicinal plants
used traditionally to treat cancer. Journal of Ethnopharmacology, n.90, p.33-
38, 2004.
JAYAPRAKASHA, G.K.; MANDADI, K.K.; POULOSE, S.M.; JADEGOUD, Y.;
GOWDA, G.A.N.; PATIL, B. Inhibition of colon cancer cell growth and
antioxidant activity of bioactive compounds from Poncirus trifoliate (L.) Raf.
Bioorganic & Medicinal Chemistry, v.15, p.4923-4932, 2007.
JYMENEZ,-ESCRIG, A.; RINCON, M.; PULIDO, R.; SAURA-CALIXTO, F.
Guava fruit (Psidium guajava L.) as a new source of antiixidant dietary fiber.
Journal of Agricultural and Food Chemistry, v.49, p.5489-5493, 2001.

78
KAHKONEN, M.P.; HOPIA, A.I.; VUORELA, H.J.; RAUCHA, J.P.; PIHLAJA, K.;
KUJALA, T.S. et al. Antioxidant capacity of plants extracts containg phenolic
compounds. Journal of Agricultural and Food Chemistry, v.47, p.3954-3962,
1999.
KALT, W.; FORNEY, C.F.; MARTIN, A.; PRIOR, R. Antioxidant capacity,
vitamin C, phenolics and anthocyanin after fresh storage of small fruits. Journal
of Agricultural and Food Chemistry, v.47, p.4638-4644, 1999.
KANDIL, F.E.; SMITH, M.A.L..; ROGERS, R.B.; PEPIN, M.F.; SONG, L.L.;
PEZZUTO, J.M.; et al. Composition of a chemoppreventive proanthocyanidin-
rich fraction from cranberry fruits responsible for the inhibition of 12-O-
tetradecanoyl phorbol-13-acetate (TPA)-induced ornithine decarboxylase (ODC)
activity. Journal of Agricultural and Food Chemistry, v.1063-1069, 2002.
KARL, J.H.; JOHN, M.F. The antioxidant activity and composition of fresh,
frozen, jarred and canned vegetables. Innovate Food Science and Emerging
Technologies, v.3, p.399-406, 2002.
KATSUBE, N.; KEIKO, I.; TSUSHIDA, T.; YAMAKI, K.; KOBORI, M. Induction of
apoptosis in cancer cells by bilberry (Vaccinium mirtillus ) and the anthocyanins.
Journal of Agricultural and Food Chemistry, v.51, p.68-75, 2003.
KAUR, C.; KAPOOR, H.C. Antioxidants in fruits and vegetables – the
millennium’s health. International Journal of Food Science and Technology,
v.36, p.703-725, 2001.
KAUR, G.; SINGH, R.P. Antibacterial and membrane damaging activity
Livistona chinensis fruit extract. Food Chemistry Toxicology, 2008.
doi:10.1016/j.fct.2008.03.026.
KIM, D.; JEONG, J.; LEE, C.Y. Antioxidant capacity of phenolics
phytochemicals from various cultivars of plums. Food Chemistry, v.81, p.321-
326, 2003.

79
KIM, D.; LEE, K.W.; LEE, H.J.; LEE, C.Y. Vitamin C Equivalent Antioxidant
Capacity (VEAC) of Phenolic Phytochemicals. Journal of Agricultural and
Food Chemistry, v.50, p.3713-3717, 2002.
KRAFT, T.F.B.; SCHMIDT, B.M.; YOUSEF, G.G.; KNIGHT, C.T.G. CUENDT,
M.; KANG, Y.H.; et al. Chemopreventive potential of wild, lowbush blueberry
fruits in multiple stages of carcinogenesis. Journal of Food Science, v.70,
p.S159-S166, 2005.
KRINSKI, N.I. Carotenoids as antioxidants. Nutrition, v.17, p.815-817, 2001.
KRITCHEVSKI, B.S. β-Carotene, carotenoids and the prevention of coronary
heart disease. Journal of Nutrition, v.128, p.5-8, 1999.
KUKOSKI, E.M.; ASUERO, A.G.; TRONCOSO, A.M.; MANCINI-FILHO, J.;
FETT, R. Aplicación de diversos métodos químicos para determinar actividad
antioxidante em pulpa de frutos. Ciência e Tecnologia de Alimentos, v.25,
n.4, p.726-732, 2005.
LACHANCE, P.A.; NAKAT, Z.; JEONG, W. Antioxidants: an integrative
approach. Nutrition, v.17, p.835-838, 2001.
LAPIDOT, T.; HAREL, S.; AKIRI, B.; GRANIT, R.; KANNER, J. pH-dependent
forms of red wine anthocyanins as antioxidants. Journal of Agricultural and
Food Chemistry, v.47, p.67-70, 2004.
LAU, F.C.; SHUKITT-HALE, B.; JOSEPH, J.A. The beneficial effects of fruit
polyphenols on brain aging. Neurobiology of Aging, v.26, p.128-132, 2005.
LEES, D.H.; FRANCIS, F.J. Stardardization of pigment analyses in cranberries.
Hortscience, v.7, n.1, p.83-84, 1972.
LIM, P.; WUENSCHELL, G.E.; HOLLAND, V.; LEE, D.; PFEIFER, G.P.;
RODRIGUEZ, H.; TERMINI, J. Peroxyl radical mediated oxidative DNA base
damage: implications for lipid peroxidation induced mutagenesis.
Biochemistry, v.43, p.15339-15348, 2004.

80
LIM, Y.Y.; LIM, T.T.; TEE, J.J. Antioxidant properties of several tropical fruits: A
comparative study. Food Chemistry, v.103, p.1003-1008, 2007.
LIMA, V.L.A.G.; MÉLO, E.A.; LIMA, D.E.S. Fenólicos e carotenóides totais em
pitanga. Scientia Agrícola, v.59, p.447-450, 2002.
LIU, R. Potencial synergy of phytochemicals in cancer prevention: mechanism
of action. Journal of Nutrition, v.134, p.3479S-3485S, 2004.
LIU, X.; ZHAO, M.; WANG, J.; YANG, B.; JIANG, Y. Antioxidant activity of
methanolic extract of emblica fruit (Phyllanthus emblica L.) from six regions in
China. Journal of Food Composition and Analysys, v.21, p.219-228, 2008.
LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de
plantas árboreas nativas do Brasil. Nova Odessa: Ed. Plantarum, 1998.
352p.
LU, Q.; ARTEAGA, J.R.; ZHANG, Q.; HUERTA, S.; GO, V.L.W.; HEBER, D.
Inhibition of prostate cancer growth by an avocado extracts: role of lipid-soluble
bioactive substances. Journal of Nutritional Biochemistry, v.16, p.23-30,
2005., n.5, p.496-502, 2005.
LU, Y.; FOO, Y.L. Antioxidant activities of polyphenols from sage (Salvia
officinalis). Food Chemistry, v.75, p.197-202, 2001.
LUXIMON-RAMMA, A.; BAHORUN, T.; CROZIER, A. Antioxidant actions and
phenolic and vitamin C contents of common Mauritian exotic fruits. Journal of
the Science of Food and Agricultura, v.83, p.496-502, 2003.
MANACH, C.; MAZUR, A.; SCALBERT, A. Polyphenols and prevention of
cardiovascular diseases. Current Opinion in Lipidology, v.16, p.77-84, 2005.
MANACH, C.; SCALBERT, A.; MORAND, C.; REMESY, C.; JIMENEZ, L.
Polyphenols: food sources and bioavailability. American Journal of Clinical
Nutrition, v.79, p.727-747, 2004.

81
MANSOURI, A.; EMBAREK, G.; KOKKALOU, E.; KEFALAS, P. Phenolic profile
and antioxidant activity of the Algerian ripe date palm fruit (Phoenix dactylifera).
Food Chemistry, v.89, p.411-420, 2005.
MANICA, I. Frutas nativas, silvestres e exóticas 1 - técnicas de produção e
mercado: abili, amora-preta, araçá, bacuri, biribá, carambola, cereja-do-
rio-grande, jabuticaba. Porto Alegre: Cinco Continentes, 2000. 327p.
MANZINO, M.B.; SILVESTRINI, M.P.; REARTE, A.E. Identidad y calidad de los
alimentos frutihortícolas industrilizados. Mendoza, v.8, p.4-5, 1987.
MARINOVA, D.; RIBAROVA, F. HPLC determination of carotenoids in
Bulgarian berries. Journal of Food Composition and Analysis, v.20, p.370-
374, 2007.
MATTOS, J.R. Frutas Nativas do Brasil: Jabuticabeira. Porto Alegre, 1983.
92p.
MEDINA, E.M.D.; RODRÍGUEZ, E.M.R.; ROMERO, C.D. Chemical
characterization of Opuntia dilleni and Opuntia ficus indica fruits. Food
Chemistry, v.103, p.38-45, 2007.
MELO, E.A.; GUERRA, N.B. Ação antioxidante de compostos fenólicos
naturalmente presentes em alimentos. Boletim SBCTA, v.36, n.1, p.1-11,
2002.
MIDDLETON, E.; KANDASWAMI, C.; THEOHARIDES, T.C. The effects of plant
flavonoids on mammalian cells: implications for inflammation, heart disease,
and cancer. Pharmacology Review, v.52, p.673-751, 2000.
MONKS, A.; SCUDIERO, D.; SKEHAN, P.; SHOEMAKER, R.; PAULL, K.;
VISTICA, D.; HOSE, C.; LANGLEY, J.; CRONISE, P.; VAIGRO-WOLFF, A.; et
al. Feasibility of a high-flux anticancer drug screen using a diverse panel of
cultured human tumor cell lines. Journal of National Cancer Institute, v.83,
p.757-766, 1991.

82
MOTA, R.V. Caracterização do suco de amora-preta elaborado em extrator
caseiro. Ciência e Tecnologia de Alimentos, v.26, n.12, p.303-308, 2006.
MÜLLER, J.B. Avaliação das atividades antimicrobiana, antioxidante e
antinociceptiva das folhas Liehea divaricata Martius. 2006. 99p.
Dissertação (Mestrado em Ciências Farmacêuticas), Universidade Federal de
Santa Maria, Santa Maria.
MURTHY, K.N.C.; SINGH, R.P.; JAYAPRAKASHA, G.K. Antioxidant capacity of
grape (Vitis vinifera) pomace extracts. Journal of Agricultural and Food
Chemistry, v.50, p.5909-5912, 2002.
NAIDU, A.S. Natural food antimicrobial systems. Boca Raton, CRC Press.
2000. 818p.
NARAYANAN, B.A.; GEOFFROY, O.; WILLINGHAM, M.C.; RE, G.G.; NIXON,
D.W. p53/p21 (WAF1/CIPI1) expression and its possible role in G1 arrest and
apoptosis in ellagic acid treated cancer cells. Cancer Letters, v.136, p.215-221,
1999.
NEGRE-SALVAGYRE, A.; SALVAGYRE, R. Quercetin prevents the cytoxicity
of oxidized low-density lipoproteins by macrophages. Free Radical Biology
Medicine, v.12, p.101-106, 1992.
NICOLI, MC.; ANESE, M; PARPINEL, M. Influence of processing on the
antioxidant properties of fruit and vegetables. Trends in Food Science and
Technology, v.10, p.94-100, 1999.
NOGUEIRA, R.J.M.C.; MORAES, J.A.P.V. BURITY, H.A. JUNIOR, J.F.S. Efeito
do estádio de maturação das frutas nas características físico-químicas de
acerola. Pesquisa Agropecuária Brasileira, v.37, n.4, p.463-470, 2002.
NOONAN, D.M.; BENELLI, R.; ALBINI, A. Angiogenesis and cancer prevention:
a vision. Recent Results. Cancer Research, v.174, p.219-224, 2007.

83
OCKÉ, M.C.; BUENO-DE-MESQUITA, H.B.; FESKENS, E.J.M.; VAN
STAVEREN, W.A.; KROMHOUT, D. Repeat measurements of vegetables,
fruits, β-carotene and vitamins C and E in relation to lung cancer. American
Journal of Epidemiology, p. 145, 358, 1997.
OLSON, J.A. Carotenoids and human health. Arch Latinoamerican Nutrition,
v.49, p.183-198, 1999.
OU, B.; HUANG, D.; MAUREEN, H.W.; FLANAGAN, J.A.; DEEMERI, E.K.
Analysis of antioxidant activities of common vegetable employing oxygen
radical absorbance capacity (ORAC) and ferric reducing antioxidsnt power
(FRAP) assays: a comparative study. Journal of Agricultural and Food
Chemistry, v.50, p.3122-3128, 2002.
PANTELIDIS, G.E.; VASILAKAKIS, M.; MANGANARIS, G.A.; DIAMANTIDIS,
G. Antioxidant capacity, phenol, anthocyanin and ascorbic acid contents in
raspberries, blackberries, red currants, gooseberries and Cornelian cherries.
Food Chemistry, v.102, p.777-783, 2007.
PATTHAMAKANOKPORN, O.; PUWASTIEN, P.; NITITHAMYOUNG, A.;
SIRICHAKWAL, P.P Changes of antioxidant activity and total phenolic
compounds during storage of selected fruits. Journal of Food Composition
and Analysis, v.21, p.241-248, 2008.
PEREIRA, J.F.M. Propagação e manejo da pitangueira. In: III Simpósio
Nacional do Morango e II Encontro sobre Pequenas Frutas Nativas do
Mercosul. Palestras... Pelotas, 2006. p.113-115.
PHILPOTT, M.; GOULD, K.S.; LIM, C.; FERGUSON, L.R. In situ and in vitro
antioxidant capacity of sweet potato anthocyanins. Journal of Agricultural and
Food Chemistry, v.52, p.1511-1513, 2004.
PINTO, M.S.; LAJOLO, F.M.; GENOVESE, M.I. Bioactive compounds and
quantification of total ellagic acid in strawberries (Fragaria x ananassa Duch.).
Food Chemistry, v.107, p.1629-1635, 2008.

84
PINTO, W.S.; DANTAS, A.C.V.L.; FONSECA, A.A.O.; LEDO, C.A.S.; JESUS,
S.C.; CALAFANGE, P.L.P.; ANDRADE, E.M. Caracterização física, físico-
química e química de frutas de genótipos de cajazeiras. Pesquisa
Agropecuária Brasileira, v.38, n.9, p.1059-1066, 2003.
PINELO, M.; MANZOCCO, L.; NUÑEZ, M.J.; NICOLI, M.C. Interaction among
phenols in food fortification: negative synergism on antioxidant capacity.
Journal of Agricultural and Food Chemistry, v.52, p.1177-1180, 2004.
POMPELLA, A. Biochemistry and histochemistry of oxidant stress and lipid
peroxidation. International Journal of Vitamin and Nutrition Research, v.67,
n.5, p.289-297, 1997.
POOL-ZOBEL, B.L.; SCHRÖDER, N.; RECHKEMMER, G. Anthocyanins are
potent antioxidants in model systems but do not reduce endogenous oxidative
DNA damage in human colon cells. European Journal of Nutrition, v.38,
p.227-234, 1999.
PRIOR, R.L.; CAO, G.; MARTIN, A.; SOFIC, E.; MCEWAN, J.; O’BRIEN, C.;
LISCHNER, N.; EHLENFELDT, M.; KALT, W.; KREWER, G.; MAINLAND, C.M.
Antioxidant capacity as influenced by total phenolic and anthocyanin content,
maturity and variety of Vaccinium specie. Journal of Food Chemistry, v.46,
p.2686-2693, 1998.
PROCHASKA, H.J. Screening strategies for the detection of anticarcinogenic
enzyme inducers. Journal of Nutrition Biochemistry, v.5, p.360-368, 1994.
RESENDE, J.M.; VILAS BOAS, E.V.B.; CHITARRA, M.I.F. Uso de atmosfera
modificada na conservação pós-colheita do maracujá-amarelo. Ciência
Agrotécnica, v.25, n.1, p.159-168, 2001.
REYNERTSON, K.A.; YANG, H.; JIANG, B.; BASILE, M.J.; KENNELLY, E.J.
Quantitative analysis of antiradical phenolic constituents from fourteen edible
Myrtaceae fruits. Food Chemistry, v.109, p.883-890, 2008.

85
ROBARDS, K.; PRENZLER, P.D.; TUCKER, G.; SWATSITANG, P.; GLOVER,
W. Phenolic compounds and their role in oxidative processes in fruits. Food
Chemistry, v.66, p. 401-436, 1999.
ROBERTS, G.; GORDON, M.H. Determination of the antioxidant activity of
fruits and vegetables by a liposome assay. Journal of Agricultural and Food
Chemistry, v.51, p.1486-1493, 2003.
RODRIGUEZ-AMAYA, D.B. A guide to carotenoids analysis in foods. ILSI
Press: Washington, 1999. 64p.
ROSS, J.A.; KASUM, C.M. Dietary flavonoids: bioavailability , metallic effects
and safety. Annual Review and Nutrition, v.22, p.19-34, 2002.
RUBINSTEIN, L.V.; SHOEMAKER, R.H.; PAULL, K.D.; SIMON, R.M.; TOSINI,
S.; SKEHAN, P.; SCUDIERO, D.A.; MONKS, A.; BOYD, M.R. Comparison of in
vitro anticancer-drug-screening data generated with a tetrazolium assay versus
a protein assay against a diverse panel of human tumor cell lines. Journal of
National Cancer Institute, v.82, 1113-118, 1990.
SALEEM, A.; HUSHEEM, M.; HÄRKÖNEN, P.; PIHLAJA, K. Inhibition of cancer
growth by crude extract and the phenolics of Terminalia chebula retz. Fruit.
Journal of Ethnopharmacology, v.81, p.327-336, 2002.
SALONEN, J.T.; NYYSONEN, K.; SALONEN, R. Antioxidant supplementation
in atherosclerosis prevention (ASAP) study: a randomized trial of the effect of
vitamins E and C on 3-year progression of carotid atherosclerosis. Journal of
Internal Medicine, v.248, p.377-386, 2000.
SÁNCHEZ-MORENO, C.; LARRAURI, J.A.; SAURA-CALIXTO, F.A. Procedure
to measure the antiradical efficiency of polyphenols. Journal of the Science of
Food and Agriculture, v.76, p.270-276, 1998.
SANCHOTENE, M.C.C. Frutíferas nativas úteis à fauna na arborização
urbana. Porto Alegre: FEPLAN, 1989. 304p.

86
SANTOS, A. R. L.; REINHARDT, D. H.; SILVEIRA, W. R.; OLIVEIRA, J. R. P.;
CALDAS, R. C. Qualidade pós-colheita de acerola para processamento, em
função de estádios de maturação e condições de armazenamento. Revista
Brasileira de Fruticultura, Jaboticabal, v.21, n.3, p.365-371, 1999.
SANTOS, R.I. Metabolismo básico e origen dos metabólitos secundários. In:
SIMÕES, C.M.O., et al., Farmagnosia: da planta ao medicamento, Porto
Alegre: Editora da Universidade Federal do Rio Grande do Sul, 2000. p.323-
354.
SATUÉ-GRACIA, M.T.; HEINONEN, M; FRANKEL, E.N. Anthocyanin as
antioxidants on human low-density lipoprotein and lecithin-liposome systems.
Journal of Agricultural and Food Chemistry, v.45, p.3362-3367, 1997.
SCALZO, J.; POLITI, A.; PELLEGRINI, N.; MEZZETTI, N.; BATTINO, M. Plant
genotype affects total antioxidant capacity and phenolic contents in fruit.
Nutrition, v.21, p.207-213, 2005.
SEMENSATO, L.R.; PEREIRA, A.S. Características de frutas de genótipos de
aceroleira cultivados sob elevada altitude. Pesquisa Agropecuária Brasileira,
v.35, n.12, p.2529-2536, 2000.
SESTILI, P.; MARTINELLI, C.; RICCI, D.; FRATERNALE, D.; BUCCHINI, A.;
GIAMPERI, L.; CURCIO, R.; PICCOLI, G.; STOCCHI, V. Cytoprotective effect
of preparations from various parts of Punica granatum L. fruits in oxidatively
injured mammalian, cells in comparison with their antioxidant capacity in cell
free systems. Pharmacological Research, v.56, p.18-26, 2007.
SHAMI, N.L.I.E; MOREIRA, E.A.M. Licopeno como agente antioxidante.
Revista da Nutrição. Campinas, v.17, n.2, p.227-236, 2004.
SHIH, P.; YEH, C.; YEN, G. Effects of anthocyanin on the inhibition of
proliferation and induction of apoptosis in human gastric adenicarcinoma cells.
Food and Chemical Toxicology, v.43, p.1557-1566, 2005.

87
SKEHAN, P.; STORENG, R.; SCUDIERO, D.; MONKS, A.; MCMAHON, J.;
VISTICA, D.; WARREN, J.T.; BOKESH, H; KENNEY, S.; BOYD, M.R. New
colorimetric cytotoxic assay for anticancer-drug screening. Journal of National
Cancer Institute, v.82, p.1107-1112, 1990.
SILVA, C.G.; HERDEIRO, R.S.; MATHIAS, C.J.; PANEK, A.D.; SILVEIRA, C.S.;
RODRIGUES, V.P.; RENNÓ, M.N.; FALCÃO, D.Q.; CERQUEIRA, D.M.;
MINTO A.B.M.; NOGUEIRA, F.L.P.; QUARESMA, C.H.; SILVA, J.F.M.;
MENEZES, F.S.; ELEUTHERIO, E.C.A. Evaluation of antioxidant activity of
Brazilian plants. Pharmacological Research, v.52, p.229-233, 2005.
SILVA, R.P.; PEIXOTO, J.R.; JUNQUEIRA, N.T.V. Influência de diversos
substratos no desenvolvimento de muda de maracujazeiro-azedo (Passiflora
edulis Sims. f. flavicarpa Deg.). Revista Brasileira de Fruticultura, v.23, n.2,
p.377-381, 2001.
SILVA, S.R.; MERCADANTE, A.Z. Composição de carotenóides de maracujá-
amarelo (Passiflora edulis flavicarpa) in natura. Ciência e Tecnologia de
Alimentos, v.22, n.3, p.254-258, 2002.
SIMÕES, C.M.O. et al. Farmacognosia da planta ao medicamento. Porto
Alegre: Editora da UFRGS; Florianópolis: Editora da UFSC, 2007. 1102p.
SINGH, R.P.; KAUR, G. Hemolitic activity of aqueous extract of Livistona
chinensis fruits. Food and Chemical Toxicology, v.46, p.553-556, 2008.
SLATTERY, M.L.; BENSON, J.; CURTIN, K.; MA, K.N.; SCHAEFFER, D.;
POTTER, J.D. Carotenoids and colon cancer. American Journal of Clinical
Nutrition, v.71, p.575-582, 2000.
SMITH, M.A.L.; MARLEY, K.A.; SEIGLER, D.S.; SINGLETARY, K.W.; MELINE,
B. Bioactive properties of wild blueberry fruits. Journal of Food Science, v.65,
p.352-256, 2000.

88
SINGLETON, V.L.; ROSSI, J.A. Colorimetry of Total Phenolics with
Phosphomolybdic-Phosphotungstic Acid Reagents. American Journal of
Enology Viticulture, v.20, p.144-158, 1965.
SOFOS, J.N.; BEUCHAUT, L.R.; DAVISDON, P.M.; JOHNSON, E.A. Naturally
occurring antimicrobials in food. Regulatory Toxicology and Pharmacology,
v.28, p.71-72, 1998.
STATSOFT INC. – Statistica for Windows. Tulsa: Statsoft, 2000, software
version 6.0.
STRONER, G.D.; MUKHTAR, H. Polyphenols as cancer chemopreventive
agents. Journal of Cellular Biochemistry, v.22, p.169-180, 1995.
SUN, J.; CHU, Y.F.; WU, X.; LIU, R.H. Antioxidant and antiproliferative activities
of common fruits. Journal of Agricultural and Food Chemistry, v.50, p.7449-
7454, 2002.
SUN, J.; LIU, R.H. Cranberry phytochemical extracts induce cell cycle arrest
and apoptosis in human MCF-7 breast cancer cells. Cancer Letters, v.241,
p.124-134, 2006.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 3ed. Porto Alegre, Artmed, 2004.
719p.
TANAKA, J.C.A.; et al. Constituintes químicos de Lyehea divaricara Marticus
(Tiliaceae). Química Nova, v.28, n.5, p.834-837, 2005.
TAVARINI, S.; DEGL’INNOCENTI, E.; REMORINI, D.; MASSAI, R.; GUIDI, L.
Antioxidant capacity, ascorbic acid, total phenols and carotenoids changes
during harvest and after storage of Hayward kiwifruit. Foof Chemistry, v.107,
p.282-288, 2008.
TEIXEIRA, G.H.A.; DURIGAN, J.F.; DONADIO, L.C.; SILVA, J.A.A.
Caracterização pós-colheita de seis cultivares de carambola (Averrhoa
carambola L.). Revista Brasileira de Fruticultura, v.23, n.3, p.546-550, 2001.

89
THAIPONG, K; BOONPRAKOB, U.; CROSBY, K.; CISNEROS-ZEVALLOS, L.;
BYRNE, D.H. Comparison of ABTS, DPPH, FRAP and ORAC assays fos
estimating antioxidant activity from guava fruit extracts. Journal of Food
Composition and Analysis, v.19, p.669-675, 2006.
TSUDA, T.; HORIO, F.; OSAWA, T. Dietary cyanindin3-O-β-D-glucoside
increases ex vivo oxidation resistance of serum in rats. Lipids, v.33, p.583-588,
1998.
TSUDA, T.; WATANABE, M.; OHSHIMA, K.; NORINOBU, S.; CHOI, S.W.;
KAWAKISHI, S.; OSAWA, T. Antioxidative activity of the anthocyanin pigments
cyaniding 3-O-β-D glucoside and cyaniding. Journal of Agricultural and Food
Chemistry, v.42, p.2407-2410, 1994.
TOUYZ, R.M. Reactive oxygen species, vascular oxidative stress, and redox
signaling in hypertension. What is the clinical significance? Hipertension, v.44,
p.248, 2004.
VASCO, C.; RUALES, J.; KAMAL-ELDIN, A. Total phenolic compounds and
antioxidant capacities of major fruits from Ecuador. Food Chemistry, 2008. doi:
10.1016/j.foodchem.2008.04.054.
VAYALIL, P.K. Antioxidant and antimutagenic properties of aqueous extract of
date fruit (Phoenix dactylifera L. arecaceae). Journal of Agriculture and Food
Chemistry, v.50, p.610-617, 2002.
VEERIAH, S.; KAUTENBURGER, T.; HABERMANN, N.; SAUER, J.;
DIETRICH, H.; WILL, F.; POOL-ZOBEL, B.L. Apple flavonoids inhibit growth of
HT29 human colon cancer cells and modulate expression of genes involved in
the biotransformation of xenobiotics. Molecular Carcinogenesis, v.45, p.164-
174, 2006.
VELIOGLU, V.S.; MAZZA, G.; GAO, I.; OOMAH, B.D. Antioxidative capacity
and total phenolics in selected fruits, vegetables and grain products. Journal of
Agricultural and Food Chemistry, v.46, p.4113-4117, 1998.

90
VINCI, G.; ROT, F.; MELE, G. Ascorbic acid in fruits, a liquid chromatographic
investigation. Food Chemistry, v.53, p.4183-4189, 1995.
VINSON, J.A; SU, X.; ZUBIK, L.; BOSE, P. Phenol antioxidant quantity and
quality in foods: Fruits. Journal of Agricultural and Food Chemistry, v.49,
p.5315-5321, 2001.
WALRANT, P.V.F. Como detector ações farmacológicas de extratos vegetais
em modelos experimentais “in vivo”. XI Simpósio de Plantas Medicinais do
Brasil. João Pessoa (PB): Programa de Resumos, 1990.
WANG. H.; CAO, G.; PRIOR, R.L. Total antioxidant capacity of fruits. Journal
of Agricultural and Food Chemistry, v.44, p.701-705, 1996.
WANG, H.; JIAO, H. Scanvenging capacity of berry crops on superoxide
radicals, hydrogen peroxide, hydroxyl radicals, and singlet oxygen. Journal of
Agricultural and Food Chemistry, v.48, p.5677-5684, 2000.
WANG, J.; MAZZA, G.; GAO, I.; OOMAH, B.D. Antioxidative activity and total
phenolics in selected fruits, vegetables and grain prpducts. Journal of
Agricultural and Food Chemistry, v.50, p.4183-4189, 2002.
WANG, S.Y.; BALLINGTON, J.R. Free radical scavenging capacity and
antioxidant enzyme activity in deerberry (Vaccinium Stamineum L.). LWT –
Food Science and Technology, v.40, n.8, p.1352-1361, 2007.
WANG, S.Y.; LIN, H.S. Antioxidant activity in fruits and leaves of blackberry,
raspberry, and strawberry varies with cultivar and development stage. Journal
of Agricultural and Food Chemistry, v.48, p.140-146, 2000.
WANG, X.; WEI, Y.; YUAN, S.; LIU, G.; ZHANG, Y.L.J.; WANG, W. Potential
anticancer activity of litchi fruit pericarp extract against hepatocellular carcinoma
in vitro and in vivo. Cancer Letters, v.239, p.144-150, 2006.

91
WEISBURGER, J.H. Mechanisms of action of antioxidants as exemplified in
vegetables, tomatoes and tea. Food Chemistry Toxicology, v.37, p.104-113,
1999.
WHEELER, G.L.; JONES, M.A.; SMIRNOFF, N. The biosynthetic pathway of
vitamin C in higher plants. Nature, v.393, p.365-369, 1998.
WILLET, W.C. Diet and health: what should we eat? Science, v.264, p.532-
537, 1994.
YAN, X.; MURPHY, B.T.; HAMMOND, G.B.; VINSON, J.A.; NETO, C.C.
Antioxidant activities and antitumor screening of extracts from cranberry fruit
(Vaccinium macrocarpon). Journal of Agricultural and Food Chemistry, v.50,
p.5844-5849, 2002.
YANG, C.S.; WANG, Z.Y. Tea and cancer. Journal of National Cancer
Institute, v.85, p. 1038-1049, 1993.
YU, Y.; CHANG, W.; WU, C.; CHIANG, S. Reduction of oxidation stress and
apoptosis in hyperlipidemic rabbits by ellagic acid. Journal of Nutritional
Biochemistry, v.16, p.675-681, 2005.
ZADERNOWSKI, R.; NACZK, M.; NESTEROWICZS, J. Phenolic acid profiles in
some small berries. Journal of the Agricltural and Food Chemistry, v.53,
p.2118-2124, 2005.
ZAMBIAZI, R.C. The role of endogenou lipi components on vegetable oil
stability. Tese de Doutorado. Winnipeg, University of Manitoba, Canadá. 304p.
ZANATTA, C.F.; MERCADANTE, A.Z. Carotenoid composition from the
Brazilian tropical fruit camu-camu (Myrciaria dubia). Food Chemistry, v.101,
p.1526-1532, 2007.





























