Embed Size (px)
Citation preview

0
LUIZ RODRIGO SALDANHA GAZZANEO
Indução de Mutação e Seleção em Feijão-
Caupi [Vigna unguiculata (L.) Walp.] Visando
Tolerância à Salinidade
Recife, PE Novembro 2007
Dissertação apresentada ao Programa de Pós-
Graduação em Genética da Universidade Federal
de Pernambuco, como parte dos requisitos
necessários para a obtenção do grau de Mestre em
Genética.
Orientador: Prof. Dr. Éderson Akio Kido,
Depto. de Genética, Centro de Ciências
Biológicas, UFPE.
Co-Orientador: Profa. Dra. Laureen M.
Houllou-Kido, Lab. de Genoma e Cultura de
Tecido, IPA.

1
Gazzaneo, Luiz Rodrigo Saldanha Indução de mutação e seleção em Feijão-Caupi [Vigna unguiculata (L.) Walp.] visando tolerância à salinidade / Luiz Rodrigo Saldanha Gazzaneo . – Recife: O Autor, 2007. 100 folhas : il., fig., tab.
Dissertação (mestrado) – Universidade Federal de Pernambuco. CCB. Departamento de Genética, 2007.
Inclui bibliografia e anexo.
1. Biologia de Fungos 2. Myxomycetes 3. Herbário Dárdano de Andrade Lima (IPA) 4. Herbário padre Camille Torrend (URM) Título. 531.52 CDU (2.ed.) UFPE 631.52 CDD (22.ed.) CCB – 2007-178

2
UNIVERSIDADE FEDERAL DE PERNAMBUCO PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA
PARECER DA COMISSÃO EXAMINADORA DA DEFESA DE DISSERTAÇÃO DE MESTRADO DE
LUIZ RODRIGO SALDANHA GAZZANEO
“Indução de Mutação e Seleção em Feijão-Caupi [Vigna unguiculata (L.) Walp.] Visando Tolerância à Salinidade”
Área de concentração: BIOLOGIA MOLECULAR
A comissão examinadora, composta pelos professores abaixo, sob a presidência do primeiro, considera o candidato
LUIZ RODRIGO SALDANHA GAZZANEO como
Recife, 30 de Outubro de 2007.
________________________________________________________ Orientador: Dr. Éderson Akio Kido (Genética-UFPE)
______________________________________________________ Co-Orientador: Dra. Laureen M. Houllou-Kido (Cultura de Tecido-IPA)
________________________________________________________ Prof. Dr. Mauro Guida dos Santos (Botânica-UFPE)
________________________________________________________ Prof. Dr. Péricles de Albuquerque Melo Filho (Agronomia-UFRPE)
________________________________________________________ Profa. Dr. Ana Maria Benko-Iseppon (Genética-UFPE)
________________________________________________________ Coordenador do Programa: Dr. Marcos Antonio de Morais Júnior

3
Dedico esta dissertação a todos aqueles que de uma forma ou de outra me apoiaram nessa nova etapa vencida.

4
AGRADECIMENTOS
Ao Programa de Pós-graduação em Genética por me possibilitar o ambiente e condições para aprimorar o meu até então parco conhecimento na área.
Ao CNPq por fazer o indispensável papel de fomentador de bolsas e projetos, sem os quais teria sido muito difícil completar o presente trabalho.
Ao Professor Éderson Kido e Professora Laureen Houllou-Kido que me acolheram em seu laboratório e me orientaram durante todo esse longo processo.
Ao IPA-Empresa Pernambucana de Pesquisa Agropécuária por fornecer o material biológico usado durante todo o curso dos experimentos.
Ao Diretor do Centro de Ciências biológicas da UFPE, Professor Gilberto Dias, por disponibilizar gentilmente suas instalações para desenvolvimento dos experimentos in vivo.
Ao Professor Waldeciro Colaço (DEN-Departamento de Energia Nuclear-UFPE), por permitir que as irradiações fossem realizadas nas dependências de seu departamento.
À Professora Ana Maria Benko-Iseppon, por me possibilitar vivenciar a experiência de participar, ora como monitor, ora como organizador dos diversos cursos e Workshops e com isso ter a plena idéia do que é interdiciplinaridade e network.
À Michely Diniz, pois sem sua presença (ora me apoiando, ora permitindo que a apoiasse, ora me alegrando, ora sorrindo com minhas brincadeiras) essa experiência não seria completa. Conquistou minha admiração incondicional no campo pessoal, e mostrou que podem existir amizades verdadeiras e livres de inveja no campo profissional. À Pedranne Barbosa que junto com Bárbara, seu alter-ego, foi muito presente com seu alto astral de menina e doces prejuízos. À D. Zizi, que sempre me ajudou e aliviou minhas manhãs e tardes de trabalho. Muito mais do que uma simples funcionária, uma voluntária disposta ajudar a todos (coisa que está em falta nesse país) e dar puxões de orelhas nos “maleducados”. Ao Professor Paulo Paes de Andrade, um exemplo a ser seguido (só lembrem de pedir o script antes, pois seu mainframe não são todos que entendem).
À trupe da LGBV, Mário, Hayanna, Kênia, Diego, Geyner, Ana Maria, Isaac. Que muitas outras excursões, congressos, viagens (mesmo que astrais ou ego-trips) surjam e nos unam sempre.
Ao Velho Gazza e D. Maria que apesar da pressão alta tê-los mantidos afastados da minha defesa, tenho certeza que se orgulham de mim.
À George Modesto pelo apoio emocional e logístico durante todo o período da produção da dissertação e andamento dos experimentos.

SUMÁRIO
Página
Lista de Figuras i
Lista de Tabelas iii
Lista de Abreviações V
Abstract 11
Resumo 12
1. Introdução 13
2. Objetivos 15
3. Revisão bibliográfica: 16
3.1. Feijão-caupi [Vigna unguiculata (L.) Walp.] 16
3.1.1. Programas de melhoramento 18
3.2. Salinização 19
3.3. Cultivo de vegetais in vitro 24
3.4. Indução de Mutação 26
3.5. Seleção in vitro de Plantas 32
4. Referências Bibliográficas 35
5. Manuscrito
5.1. Manuscrito 1
47
48
7. Conclusões 75
8. Anexos 76
7.1. Dados complementares da seleção in vitro 77
7.2. Dados complementares da seleção in vivo 85
7.3. Instruções para autores – Genetics and Molecular Biology 96

7
LISTA DE FIGURAS
Página
Manuscrito 1
Figura 1. Cultivo in vitro de Vigna unguiculata (L.) Walp cv. IPA 206. 68
Figura 2. Efeito da concentração de NaCl na freqüência de eventos do processo de
regeneração in vitro de eixos embrionários de feijão-caupi cv. IPA206 (mudança de cor,
início de regeneração, inibição do desenvolvimento e necrose). 69
Figura 3. Efeito da concentração de NaCl na freqüência de organogênese direta e
indireta de eixos embrionários de IPA 206. 69
Figura 4. Efeito de diferentes doses de radiação gama na freqüência de organogênese
direta de cotilédones e eixos embrionários de feijão-caupi cv. IPA 206. 70
Figura 5. Efeito de diferentes doses de radiação gama na freqüência da organogênese
indireta em eixos embrionários de IPA 206. 70
Figura 6. Efeito de diferentes doses de radiação gama na freqüência da organogênese
total (direta e indireta) de eixos embrionários de feijão-caupi cv. IPA 206. 71
Figura 7. Cultivo in vivo de Vigna unguiculata (L.) Walp. cv. IPA 206. 72
Anexo 2. Dados complementares in vivo 85
Figura 1. Plantas com melhores resultados para precocidade após seleção in vivo para
tolerância a NaCl em cv. IPA 206 88
Figura 2. Efeito da radiação gama no peso de 100 sementes de Vigna unguiculata (L.)
Walp cv. IPA 206 após seleção in vivo. 89
Figura 3. Plantas com melhores resultados para “peso de 100 sementes” após seleção in
vivo para tolerância a NaCl em cv. IPA 206. 90
Figura 4. Efeito da radiação gama no comprimento de vagens de Vigna unguiculata
(L.) Walp cv. IPA 206 após seleção in vivo. 90
Figura 5. Plantas com melhores resultados para comprimento de vagem após seleção
in vivo para tolerância a NaCl em cv. IPA 206. 91
Figura 6. Efeito da radiação gama no número de sementes por vagens de Vigna
unguiculata (L.) Walp cv. IPA 206 após seleção in vivo 92
i

8
Figura 7. Plantas com melhores resultados para número de sementes por vagem após
seleção in vivo para tolerância a NaCl em cv. IPA 206. 92
ii

9
LISTA DE TABELAS
Página
Revisão da Literatura
Tabela 1. Número de plantas de diferentes tipos agronômicos geradas a partir de
indução de mutação via radiação gama, em diferentes regiões agrícolas (Fonte:
FAO/IAEA, 2007). 19
Tabela 2. Cultivares de V. unguiculata obtidos pelo uso de radiação gama (Fonte:
FAO/IAEA, 2007). 21
Manuscrito 1
Tabela 1. Valores médios das características agronômicas (tamanho da vagem, número
de sementes por vagem, peso de 100 sementes, dias até colheita) de feijão-caupi cv. IPA
206 oriundas de sementes irradiadas (SI) após seleção (OS) in vivo para tolerânica ao
NaCl (60 mM). 73
Tabela 2. Melhores resultados (número de vagens, comprimento das vagens, peso de
100 sementes, número de sementes por vagem e dias para a colheita) observados após
seleção in vivo para tolerância ao NaCl (60 mM) em feijão-caupi cv. IPA 206. 74
Anexo 1
Tabela 1 Resultados do teste in vitro de sensibilidade ao NaCl em eixos embrionários
de feijão-caupi Vigna unguiculata (L.) Walp cv. IPA 206. 78
Tabela 2. Efeito de diferentes doses de radiação gama na organogêse in vitro de
cotilédones de feijão-caupi Vigna unguiculata (L.) Walp cv. IPA 206. 79
Tabela 3. Efeito da radiação gama na organogênese in vitro de eixos embrionários de
feijão-caupi Vigna unguiculata (L.) Walp cv. IPA 206. 83
Anexo 2
Tabela 1. Dados coletados das plantas durante seleção in vivo. 86
Teste F
Tabela 1. Análise da variância da freqüência média de organogênese indireta de eixos
embrionários de Vigna unguiculata (L.) Walp. cultivados em diferentes doses de NaCl 93
iii

10
Tabela 2. Análise da variância da freqüência média de organogênese direta de
cotilédones de Vigna unguiculata (L.) Walp. expostos a diferentes doses de radiação
gama. 93
Tabela 3. Análise da variância da freqüência média de organogênese direta de eixos
embrionários de Vigna unguiculata (L.) Walp. expostos a diferentes doses de radiação
gama. 93
Tabela 4. Análise da variância da freqüência média de organogênese total (direta e
indireta) de eixos embrionários de Vigna unguiculata (L.) Walp. expostos a diferentes
doses de radiação gama. 94
Tabela 5. Análise da variância do número médio de dias para a colheita (precocidade)
de vagens de plantas de Vigna unguiculata (L.) Walp. irradiadas em diferentes doses de
radiação gama. 94
Tabela 6. Análise da variância do tamanho médio de vagens de plantas de Vigna
unguiculata (L.) Walp. irradiadas em diferentes doses de radiação gama. 94
Tabela 7. Análise da variância da produção de sementes por vagens de plantas de Vigna
unguiculata (L.) Walp. irradiadas em diferentes doses de radiação gama. 95
iv

11
LISTA DE ABREVIAÇÕES
AGL Agriculture Land and Water Development Division
BAP 6-Benzilaminopurina
CONAB Companhia Nacional de Abastecimento
DL100 Dose letal de 100%
DMS Dimetil sulfato
EMS Ethylmethane Sulphonate
FAO Food and Agriculture Organization of the United Nations
GA3 Ácido giberélico
IAEA International Atomic Energy Agency
IITA International Institute of Tropical Agriculture
InR Início de regeneração dos explantes
ISRA Instituto Senegalense de Pesquisa Agrícola
MS Murashigue e Skoog
Nec Necrose
Din Desenvolvimento inibido dos explantes
CSS Controle sem sal
CCS Controle com sal
IMc Intumescimento e mudança de cor dos explantes
SOS Salt overly sensitive
UNESCO Organização das Nações Unidas para a Educação, a Ciência e a Cultura
VMA Vírus do mosáico amarelo.
v

12
Abstract
Cowpea [Vigna unguiculata (L.) Walp.] is an important culture for the economy
and nutrition of developing countries peoples, especially to the northeast Brazilian
population. Altought its easy cultivation under diverse soil and weather conditions
(rusticity), its culture suffers with large productivity loss due to damage cause by biotic
and abiotic factors. Soil salinization, originated by incorrect soil and water management,
grows and affects mainly the arid and semiarid zones of the Globe, being a major
limitation to cowpea cultivation area expansion. Because of its high production potencial,
nutrition and genetic variability, the cowpea presents itself as a promising breeding
program incoming material. New tools and non-conventional approaches on plant breeding
(molecular markers, mutagenesis, etc.) may help the process of obtaining new cultivars.
The present study evaluated the gamma radiation induced mutation and the in vivo and in
vitro selection of V. unguiculata cv. IPA 206, targeting salt tolerance (NaCl). Results
showed that a 258 mM NaCl concentration dose proved to be ideal for in vitro selection,
allowing the identification of 12 tolerant explants among the 3000 inoculated. On the in
vivo selection it was possible to identify four tolerant plants under NaCl 60 mM. The
results lead to the conclusion that induced mutation, specially the utilization of gamma
rays, together with in vivo or in vitro cultivation and selection techniques can produce
cowpea plants with new agronomic features of great interest.
Keywords: Cowpea, mutagenesis, gamma rays, tissue culture, saline stress.

13
RESUMO
O feijão-caupi [Vigna unguiculata (L.) Walp.] é uma importante cultura para a
economia e nutrição dos povos de países em desenvolvimento, em especial para a
população do Nordeste brasileiro. Apesar da sua capacidade de cultivo nas mais diversas
condições de solo e clima (rusticidade), é uma cultura que ainda sofre com grandes perdas
de produção devido a danos causados por fatores bióticos e abióticos. A salinização de
solos, fenômeno originado em parte pelo manejo incorreto do solo e irrigação, cresce e
afeta, principalmente, as zonas áridas e semi-áridas do globo, sendo um dos maiores
limitantes da expansão do cultivo do feijão-caupi nessas áreas. Devido a seu alto potencial
produtivo, nutricional e variabilidade genética, se apresenta como promissor material para
programas de melhoramento genético. Novas ferramentas e abordagens não-convencionais
no melhoramento vegetal (marcadores moleculares, mutagêneses, etc.) podem auxiliar o
processo de obtenção de novas cultivares. O presente trabalho avaliou indução de mutação
via radiação gama e a seleção in vivo e in vitro em V. unguiculata cv. IPA 206, visando
tolerância à salinidade (NaCl). Os resultados do trabalho mostraram que a dose de 258 mM
foi identificada como ideal para a seleção in vitro, permitindo selecionar 12 plantas
tolerantes em 3000 sementes inoculadas. Na seleção in vivo foi possível identificar quatro
plantas tolerantes à concentração de 60 mM de NaCl. Tais resultados atestam que a
indução de mutação, em especial a utilização de raios gama, aliada às técnicas de cultivo e
seleção in vitro ou in vivo podem gerar plantas com novas características agronômicas de
grande interesse em feijão-caupi.
Palavras-Chave: Feijão macassar, mutagênese, radiação gama, cultura de tecidos, estresse
salino.

14
1. INTRODUÇÃO
Um dos principais objetivos mundiais é garantir o acesso a uma alimentação básica
por toda a população, mas o mesmo está longe de ser alcançado, em parte pela
insuficiência na produção agrícola e, principalmente, pela falta de poder econômico da
maioria pobre. Não obstante a questão sócio-econômica, a diminuição das áreas cultivadas
devido ao empobrecimento do solo, salinização e o aumento progressivo da população
global afetam de maneira direta a disponibilidade de recursos agrícolas. Tal realidade
demanda estudos para a formulação de estratégias de manejo e remediação mais eficientes.
O fenômeno da salinização do solo já afeta 25% das áreas irrigadas das regiões
áridas e semi-áridas ou cerca de 45 dos 230 milhões de hectares irrigados do globo
(http://www.fao.org/ag/agl/agll/spush/intro.htm). As altas concentrações de sais no solo
tornam-se tóxicas para as plantas causando alterações no crescimento ou mesmo
impedindo seu desenvolvimento. Porém, plantas tolerantes ou mesmo resistentes podem
apresentar um crescimento até normal em certas condições salinas. A resistência pode se
dar por diversos mecanismos, dentre eles o seqüestro de íons pelos vacúolos, cuja redução
do potencial osmótico permite a entrada de água pelas raízes. Esse ajuste osmótico é
necessário, pois um dos efeitos iniciais do estresse é a restrição da absorção de água pelas
raízes, que evoluem para o acúmulo de íons provocando a toxidez, o que acarreta
distúrbios nutricionais e metabólicos. Assim, plantas de interesse comercial ou nutricional
tolerantes à salinidade seriam de extremo valor como recurso agronômico e alimentício
para populações carentes localizadas em áreas afetadas pela salinização.
Dentre as plantas cultivadas, o feijão-caupi [Vigna unguiculata (L.) Walp.],
também conhecido como feijão de corda, fradinho, macassar ou feijão-verde, devido à sua
versatilidade e plasticidade fenotípica, que lhe permitem crescer em condições
desfavoráveis, é uma cultura muito importante para as regiões Norte-Nordeste do Brasil.
Sua utilização se dá na alimentação humana, tendo propriedades nutricionais e
digestibilidade superiores ao feijão comum (Phaseolus vulgaris L.), na alimentação animal
como forrageira, além de servir de fonte de genes para o melhoramento de diversas
leguminosas. Apresenta também potencial para uso em bioremediação e fertilização de
solos pobres. Em geral, a produtividade do feijão-caupi é menor que a do feijão comum
(Phaseolus spp.). Isso se dá em decorrência do baixo nível tecnológico empregado e/ou de
cultivo em áreas adversas (pouco férteis e secas ou salinas). Por outro lado, a obtenção de

15
genótipos tolerantes à salinidade através de melhoramento genético clássico não tem
mostrado os resultados desejados. Assim, a adoção de outras tecnologias, como a indução
de mutação e as técnicas de cultura de tecidos vegetais, poderia auxiliar nos programas de
melhoramento. A indução de mutação via radiação gama permitiria a geração de
variabilidade genética, necessária para que o processo de seleção seja realizado, enquanto
que a cultura de tecidos vegetais permitiria que um grande número de explantes fosse
avaliado no processo seletivo, o que poderia culminar no desenvolvimento de novas
cultivares ou de materiais importantes para cruzamentos com os materiais comerciais. Esse
trabalho teve como objetivo a indução de mutação via radiação gama em feijão-caupi para
seleção visando à obtenção de tolerância à salinidade.

16
2. OBJETIVOS
2.1 Objetivo geral
Obtenção de variantes de feijão-caupi V. unguiculata tolerantes à salinidade.
2.2 Objetivos específicos
* Avaliar o efeito da radiação gama e do NaCl sobre o desenvolvimento “in vitro” e “in
vivo” de feijão-caupi V. unguiculata cv. IPA 206
* Selecionar in vitro e in vivo variantes promissores de feijão-caupi V. unguiculata cv. IPA
206 com tolerância ao NaCl (mínimo de 60 mM).

17
3. REVISÃO BIBLIOGRÁFICA
3.1. Feijão-caupi [V. unguiculata (L.) Walp.]
O feijão-caupi é uma planta dicotiledônea que pertence à ordem Fabales, família
Fabaceae, subfamília Faboideae, tribo Phaseoleae, subtribo Phaseolinae, gênero Vigna,
secção Catiang e espécie Vigna unguiculata (L.) Walp. (National Plant Germplasm
System; http://www.ars-grin.gov/cgi-bin/npgs/html/taxon.pl?41647).
Esta espécie é cultivada desde áreas tropicais e subtropicais de baixa altitude até
altitudes acima de 1300 m, chegando a 1600 m no Kênia e Camarões (Freire-Filho et al.,
1997). Sua produção e consumo estão espalhados por áreas do oeste da África, Índia, sul e
sudeste da Ásia, Austrália e sul da Europa, além de sudeste e sudoeste da América do
Norte e em vastas áreas das Américas Central e do Sul. Na América do Sul, a região
Nordeste do Brasil é sua maior produtora (Ehlers e Hall, 1997). Acredita-se que o feijão-
caupi tenha sido introduzido na América Latina no século XVI pelos colonizadores
espanhóis e portugueses primeiramente nas colônias espanholas, e em seguida no Brasil,
provavelmente pelo Estado da Bahia (Freire-Filho, 1988). A partir da Bahia, o feijão-caupi
foi levado pelos colonizadores para outras áreas da região Nordeste e para as outras regiões
do país. No Estado da Paraíba, o feijão-caupi é cultivado em quase todas as microrregiões,
detendo 75% das áreas de cultivo com feijão, exercendo sua efetiva participação na dieta
alimentar da população humana (Silva e Oliveira et al. 2002; IBGE, 1996). Estatísticas
atuais de produção em nível nacional não estão disponíveis pelo fato de serem
contabilizados conjuntamente com o feijão-comum (Phaseolus spp.).
A utilização do feijão-caupi se dá dos mais variados modos: na forma de pasta ou
farinha como ingredientes para alimentos e comidas regionais (Uwaegbute et al., 2000), na
forma de grãos secos ou de grãos verdes (Brasil, Senegal e sudeste dos EUA), de vagens
verdes frescas (Ásia e Caribe) e de folhas (leste da África). Há também uso como adubo
verde (Austrália e sudeste dos EUA) e na forma de forragem para animais (África
Ocidental e partes da região do Sahel) (Ehlers e Hall, 1997).
O feijão-caupi apresenta características adaptativas e nutricionais que fazem dele
uma importante fonte alimentícia em regiões pobres de clima e solo pouco favoráveis.
Entre suas características nutricionais, apresenta altos níveis de ácido fólico, baixos níveis

18
de fatores anti-nutricionais e produtores de flatulência, quando comparado com o feijão-
comum (Bressani, 1985). Além disso, é uma fonte de proteína barata e importante para a
nutrição de populações de países em desenvolvimento (Langyintuo et al., 2004),
contribuindo significativamente na agricultura de subsistência de famílias na África
Central e Oeste (Baur, 1992), bem como no Brasil.
Considerada uma planta rústica, o feijão-caupi é capaz de adaptação em diferentes
condições, desde regiões de extrema umidade (região Norte, bacia Amazônica), como em
regiões de grandes períodos de seca (região Nordeste do Brasil) ou mesmo desérticas (sul
do deserto do Sahara). Nestes locais pode produzir até uma tonelada por hectare de grãos
secos, onde outras espécies não sobreviveriam (Hall, 2004a). Além disso, devido a sua
capacidade de fixação de nitrogênio (Hall, 2004b) e eficiente simbiose com fungos
micorrízicos (Kwapata e Hall, 1985) é capaz de crescer em solos ácidos ou alcalinos (Fery,
1990), mesmo de baixa fertilidade.
Um estudo comparando a rentabilidade do cultivo da bananeira em consórcio com
feijão comum e feijão-caupi no litoral Sul da Bahia (Lima et al., 2005) mostrou uma
rentabilidade 71% superior no consórcio feijão-caupi - bananeira quando comparado com o
feijão comum - bananeira. O primeiro apresentou uma receita total de R$ 3.030,00 (receita
líquida de R$ 2.473,80) e índice de rentabilidade de 5,75 (retorno de R$ 5,75 para cada real
investido) agregando um valor significativo à cultura da bananeira, enquanto o último
apresentou índice de rentabilidade de 3,36, com retornos de R$ 3,36. De acordo com a
Companhia Nacional de Abastecimento (CONAB), entre 2005 e 2006 houve uma grande
safra do feijão-caupi no Brasil, favorecido pela regularidade climática e boa distribuição
das chuvas, principalmente nos períodos de desenvolvimento e maturação da cultura. A
grande oferta nos meses de maio e junho (período de colheita) derrubou os preços
praticados para a cultura, sendo o mesmo comercializado pelo produtor por R$ 35,00 a R$
40,00 a saca de 60 kg e, mesmo assim, foi considerado interessante para a cultura. Segundo
a CONAB (2006), para o Estado do Ceará houve um aumento de 73,7% no rendimento
(kg.ha-1), 92% na produção (toneladas) e apenas 11% na área cultivada, o que demonstra
que o aumento na produção está se dando pela melhoria do manejo e/ou utilização de
cultivares com melhores resultados agronômicos (CONAB, 2006).

19
3.1.1. Programas de melhoramento
Esforços estão sendo empregados no melhoramento do feijão-caupi no Brasil e em
países como a África e a Índia (Sanginga et al., 2000). Ehlers e Hall (1997) há 10 anos já
apontavam a necessidade de maiores estudos e investimentos na modernização do
melhoramento do feijão-caupi, e apesar das contribuições recentes, em diversas áreas,
como estresse fisiológico (Iuchi et al., 2000), resistência a doenças (Bashir e Hampton,
1996) e transgenia (Popelka et al., 2006), ainda é uma espécie com potenciais pouco
explorados quando comparado com outros grãos.
As características mais interessantes para os programas de melhoramento em V.
unguiculata são a precocidade (Ishiyaku et al., 2005), a resistência às pragas (Barreto e
Quinderé, 2000), fungos (Castro et al., 2003) e vírus (Assunção et al., 2005), a tolerância à
seca (Pimentel et al., 2002), ao calor (Hall, 2004a) e por fim, à salinidade (Wang et al.,
2003).
O Instituto Internacional de Agricultura Tropical (IITA) e o Instituto Senegalense
de Pesquisa Agrícola (ISRA) são referências mundiais no desenvolvimento de cultivares
precoces, como a cultivar “IT84S-2246” e “Mouride”, desenvolvidas pela IITA e ISRA,
respectivamente (Ehlers e Hall, 1997). A seleção de cultivares precoces é importante,
principalmente para regiões onde existe um grande período de seca. Em Sahel, região
vizinha ao deserto do Sahara, com baixa umidade, baixa taxa de precipitação e alta
evaporação, as cultivares conseguem escapar da seca devido à precocidade. Essa
precocidade ajuda a estabelecer a cultura em áreas adversas e a estabilizar a produção em
longos períodos de seca (Chittaranjan, 2007).
Insetos pragas, sejam em suas formas larvais (ex: lagartas) ou adultas (ex:
gafanhotos e afídeos), podem prejudicar o desenvolvimento de culturas. A estratégia de
controle mais difundida ainda é o uso de inseticidas, que se usados de forma inadequada
podem contaminar os solos e lençóis subterrâneos e, indiretamente, os seres humanos,
levando ao surgimento de quadros patológicos típicos de intoxicação. Logo, a resistência a
insetos também é de grande importância (Ehlers e Hall, 1997). Alguns acessos de feijão-
caupi (TVu 80016 e TVu 8381) já apresentam resistência a afídeos, enquanto que a
resistência a gorgulho tem sido incorporada a cultivares tanto pelo IITA quanto pelo ISRA,
diminuindo as perdas de grãos na estocagem e fazendo com que o tempo de
armazenamento aumentasse em dois meses, quando comparado com cultivares susceptíveis
(Roberts et al., 2004 ).

20
Perdas de produção por viroses também são preocupações no melhoramento do
feijão-caupi. Prejuízos por causas virais representam um grande problema nas plantações
nacionais, o que ressalta a importância do desenvolvimento de cultivares com resistência a
vírus fitopatogênicos. Nesse contexto, a Embrapa lançou a “BR17 Gurguéia”, uma cultivar
de V. unguiculata resistente ao Mosaico Severo, ao Mosaico Dourado e a diversos
potivírus (Ehlers e Hall, 1997). No tocante a fungos, cultivares resistentes (“CB46” e a
“CB88”) foram desenvolvidas no Sudoeste dos Estados Unidos para Fusarium
oxysporium, tendo sido considerada localmente também de alta produção (Helms et al.,
1991a; Helms et al., 1991b).
Fatores ambientais adversos são grandes fontes de estresse, como temperaturas
extremas (elevadas ou muito baixas), disponibilidade e qualidade de água, bem como solos
pouco férteis ou salinos. Nesse campo de estudos, já existem trabalhos que relatam o êxito
no desenvolvimento de cultivares tolerantes à seca (Ogbonnaya et al., 2003), calor (Ehlers
e Hall, 1998) e frio (Ismail et al., 1999).
No Brasil, o melhoramento genético do feijão-caupi é realizado pela Embrapa
(Empresa Brasileira de Pesquisa Agropecuária) Meio-Norte, localizada em Teresina, PI,
cujo programa realiza cruzamentos com aplicação de seleção fenotípica. Apesar de
lançamentos constantes de novas cultivares, material tolerante à condições salinas
nordestinas ainda não se encontram disponíveis.
3.2. Salinização
Quando os sais dissolvidos na água de reservatórios naturais ou artificiais
precipitam na superfície dos solos irrigados, acumulando-se ano após ano, dá-se início ao
processo de salinização de solos (Rai e Rai, 1999).
O processo de salinização de solos cresce a cada ano e se tornou um problema
mundial. Segundo a FAO/UNESCO, 397 milhões de hectares do globo apresentam solos
salinos, enquanto 434 milhões de hectares apresentam solos sódicos. Em 2000, o fenômeno
da salinidade já afetava 45 (19,5%) dos 230 milhões de hectares de áreas irrigadas do
globo, acarretando prejuízos na produção mundial de plantas cultivadas. As regiões áridas
e semi-áridas irrigadas do planeta são as mais afetadas, com 25% de seus solos salinizados
(FAO/AGL, 2007).
Segundo a FAO/AGL (2007) no Brasil, estima-se que 20-25% das áreas irrigadas
próximas a rios e riachos intermitentes, principalmente em solos aluviais, sofram de

21
salinidade e/ou problemas de drenagem, enquanto 2,4% do território sofre com salinização
por causas naturais. Apesar da água utilizada no Brasil apresentar, na sua maioria, bons
níveis de qualidade (como a água da chuva armazenada em açudes, água de rios e água
subterrânea de áreas sedimentares), isto nem sempre é verdadeiro para a região nordestina.
Esse mesmo estudo recolheu amostras do solo dos Estados da Bahia, Sergipe, Alagoas,
Pernambuco, Paraíba, Rio Grande do Norte e Ceará, representando um total de 110
milhões de hectares numa escala de 1:500.000. As áreas afetadas por salinidade de origem
ambiental foram estimadas em 9,1 milhões de hectares correspondendo a 9% da área de
amostragem (FAO/AGL, 2007). As principais causas da salinização no Brasil seriam
irrigações excessivas ou pouco eficientes, solos pobres com manejo de água inadequado,
sistemas de drenagens inadequados, elevação do nível do mar com invasão e acúmulo de
água salgada na superfície, solos rasos com pouca fertilidade e irrigação com água salina
sem tratamento apropriado.
A principal conseqüência da exposição de plantas a solos com excesso de sais é a
diminuição do desenvolvimento das mesmas com queda da produção agrícola (Cainnes e
Shennan, 1999). Segundo Hussain et al. (2002) a salinidades de solos é o fator ambiental
que mais limita a produtividade de plantas cultivadas. Normalmente, essa queda na
produtividade pode ser causada por um estresse osmótico resultado de um déficit hídrico
causado pela redução da absorção de água pelas raízes (Greenway e Munns, 1980). Isso
acontece no início da exposição ao sal, devido ao acúmulo de íons (Na+ e Cl-, por exemplo)
no interior de suas células, o que leva a uma diminuição do potencial hídrico no ambiente
radicular (Serrano et al., 1999; Munns, 2002). Após uma exposição mais prolongada ao
estresse salino se observa, além da diminuição da captura de água pelas raízes, os efeitos
da toxidez do acúmulo de sal, que culminam com distúrbios nutricionais e metabólicos
(Munns, 2002).
Outros fatores podem também estar relacionados à diminuição do desenvolvimento
das plantas. Stavarek e Rains (1984) indicam que o estresse salino leva a um desvio da
energia destinada ao crescimento e desenvolvimento das plantas, para processos de
regulação de íons inorgânicos e manutenção da homeostase osmótica das células. Outro
fator é a diminuição da oferta de nitrato (NO3+) nas folhas. O nitrato é um composto
indispensável para o crescimento das plantas sendo convertido em aminoácidos; seus
derivados metabólicos atuam na regulação osmótica, porém a atividade da enzima nitrato
redutase nas folhas depende do fluxo de nitrato pelas raízes, o que é drasticamente afetado
com a salinidade (Ferrario et al., 1998; Campbell, 1999).

22
Os sintomas do estresse salino são muito parecidos com os encontrados no estresse
hídrico: em baixas concentrações promove o amarelamento e queima da extremidade das
folhas, em seguida, com maior tempo de contato com o sal, apresentam coloração marrom
e apodrecimento (Rai e Rai, 1999). O estresse salino está intimamente ligado ao estresse
hídrico já que seus mecanismos para minimizar os efeitos do estresse muitas vezes se
sobrepõem, fazendo com que diversos estudos sobre sinalização de estresse hídrico sejam
focados principalmente em respostas de plantas ao sal. Essa escolha também ocorre devido
à maior precisão e facilidade de se desenhar experimentos de estresse salino em laboratório
(Hasegawa et al., 2000; Zhu, 2002).
Quando em situação de estresse, as plantas apresentam respostas que levam ao
sacrifício de parte de sua estrutura para garantir a sobrevivência do restante. Isso é
resultado de uma sinalização ativa, a qual recebe o nome de “resposta adaptativa”. A
importância de se conhecer essas vias de sinais de transdução está na possibilidade de
suprimirmos os genes envolvidos nessa resposta, aumentando a tolerância ao estresse
salino e hídrico (Zhu, 2002).
Existem, basicamente, três tipos de respostas adaptativas: a homeostase (iônica e
osmótica), a desintoxicação (controle e reparo dos danos causados pelo estresse) e o
controle do crescimento, para coordenar a divisão e a expansão celular para níveis
adequados às condições de estresse (Zhu, 2002).
Plantas capazes de desenvolvimento normal em solos com concentração maior de
sais são chamadas de halofíticas ou halotolerantes, sendo capazes de adaptação a altos
níveis de sais, restringindo a entrada de íons nas raízes, ou impedindo-os de chegarem às
folhas através do xilema (Malcom et al., 2003). Diversas espécies halofíticas burlam os
efeitos do estresse salino baixando o potencial osmótico dos seus tecidos com o acúmulo
de solutos orgânicos no citosol e inorgânicos nos vacúolos, em um processo denominado
“ajuste osmótico” (Niu et al., 1995). Esses solutos orgânicos teriam como principal função
atuar na estabilidade das proteínas e membranas, além de protegerem a célula dos efeitos
dos radicais livres de oxigênio (Bohnert e Shen, 1999).
Os genes envolvidos em processos relacionados ao estresse salino podem ser
agrupados de acordo com as vias e processos nos quais estão envolvidos: síntese de
osmólitos, proteção da integridade da célula, estresse oxidativo, homeostase de íons e
fatores de transcrição (Zhu, 2001).
Existem diversas rotas de sinalização para salinidade e seca propostas em trabalhos
científicos (Zhu, 2000), porém poucas são tão bem estabelecidas quanto a via “SOS” (Salt

23
Overly Sensitive). O excesso extracelular de Na+ é o gatilho externo para essa rota de
sinalização. O aumento de Na+ intra ou extracelular aciona um sinal Ca+ citoplasmático
que vai atuar na expressão de genes SOS e ativação de suas proteínas (Zhu, 2000). Os
resultados dessas ativações são mudanças na expressão e atividade de transportadores de
íons como Na+, K+ e H+ (Zhu, 2001). O sinal de cálcio citosólico, gerado pelo aumento de
sal no meio extra ou intracelular, é percebido por uma proteína ligadora de cálcio (SOS3)
codificada por um gene denominado sos3. Essa proteína apresenta um sítio capaz de ligá-la
a molécula de cálcio citosólico (Liu e Zhu, 1998; Ishitani et al., 2000). A proteína SOS3
ativada interage e ativa uma segunda proteína denominada SOS2 (Liu et al., 2000; Halfter
et al., 2000). O complexo SOS2-SOS3 regula a expressão de uma terceira proteína, a
SOS1, uma proteína de membrana plasmática do tipo bomba de Na+ / H+; assim como ativa
sua função transportadora que fará com que a mesma troque íons Na+ do interior da célula
por H+ oriundos do exterior. (Shi et al., 2000; Qiu et al., 2001). O transportador SOS1
apresenta uma longa cauda citoplasmática que deve funcionar como sensor para a presença
de Na+ em excesso no interior das células (Shi et al., 2000). Dessa forma SOS1 deve
controlar a ativação de SOS2 nas plantas, o que levaria a sua ativação e estimulação da
capacidade de eliminar Na+ (Zhu, 2002).
Diversos organismos foram utilizados para elucidar um pouco mais os mecanismos
envolvidos na tolerância à salinidade, como procariotos (Escherichia coli), eucariotos
unicelulares (Saccharomyces cerevisiae), plantas halófitas (Mesenbryanthemum
crystallinum) e plantas glicofíticas (arroz, tomate ou Arabdopsis thaliana) (Borsani et al.,
2003). Testes com leveduras mutantes para os genes sos2 e sos3 mostraram que a ativação
do transportador SOS1 acontece mesmo somente com a adição de SOS2 ativado, chegando
a níveis encontrados em indivíduos selvagens (Qiu et al., 2001). As proteínas SOS2 e
SOS3 também devem exercer um papel importante nos sistemas de compartimentalização
vacuolar de sal, um outro importante mecanismo para a tolerância ao estresse salino (Rus
et al., 2001). A simples presença de genes sos1 expressos confere um leve aumento na
tolerância ao sal, porém na presença de sos2 e sos3 observa-se um aumento considerável
na tolerância devido à sua ativação pelos mesmos (Shi et al. 2000).
O conhecimento desses genes não é necessário para o melhoramento convencional,
porém para as novas técnicas de melhoramento é indispensável localizar os fatores
limitantes e os processos chaves envolvidos na tolerância, assim, abordagens transgênicas
para aumentar a tolerância à salinidade, por exemplo, seriam plausíveis. Algumas fontes de
tolerância a sal foram identificadas em tomates selvagens e cultivares primitivas. Esse tipo

24
de recurso genético tem sido utilizado para o aumento da tolerância de cultivares modernas
utilizando técnicas de melhoramento genético convencional, porém os progressos são
lentos (Darlene et al., 1997).
A maior limitação do uso do melhoramento convencional se dá pelo fato da
difícil inserção de genes responsáveis pela resistência à salinidade num determinado
genoma através de cruzamentos convencionais sem afetar características multigênicas
importantes, como florescimento, qualidade do fruto ou produção de matéria seca (Flowers
et al., 2000). Porém, testes com mais cultivares se fazem necessários tendo em vista que os
mecanismos que conferem tal tolerância podem variar de espécie para espécie (Borsani et
al., 2003).
Estudos mostram que é possível induzir uma relativa tolerância em plantas
conhecidamente sensíveis. Rai e Rai (1999) conseguiram promover um aumento
significativo em Azolla pinnata R. Br. sensíveis ao sal através da pré-incubação gradual e
constante a doses não letais crescentes de NaCl. As plantas pré-incubadas apresentaram em
NaCl 60 mM o mesmo desenvolvimento que plantas não expostas previamente ao sal
cultivadas em NaCl 20 mM. O mesmo tipo de resultado foi obtido por Amzallag et al.
(1990) em Sorghum bicolor (L.) Moench. Expondo explantes de sorgo durante 20 dias a
doses não letais de NaCl (75 a 150 mol.m-3) conseguiram com que as plantas
sobrevivessem a doses letais (300 mol.m-3), condição em que plantas controles morriam
em duas semanas.
Estudo realizado em V. unguiculata por Silva et al. (2003) mostrou que plantas
crescidas em condições salinas (75 mmol NaCl) apresentaram uma forte inibição de seu
desenvolvimento ocasionada por um desbalanço hídrico intracelular. Foi observado
aumento do conteúdo de solutos inorgânicos em diversos pontos da planta pelo acúmulo de
íons Na+ e Cl-, aumento de carboidratos solúveis nas folhas e raízes e reduziu os níveis de
prolina nas raízes. Costa et al. (2003) também realizaram experimentos para entender um
pouco mais sobre as conseqüências do estresse salino na fisiologia do feijão-caupi. Nesse
estudo observaram que a presença de NaCl no ambiente radicular (75 mmol) reduziu a
produção de matéria seca e que quanto mais afetado era a cultivar, maior era o conteúdo
intracelular de Na+.

25
3.3. Cultivo de Vegetais in vitro
A idéia de regenerar plantas a partir de fragmentos de tecidos vegetais remonta ao
início do século XX. No entanto, resultados promissores só foram obtidos a partir de 1950,
com a descoberta e a subseqüente aplicação de hormônios vegetais para direcionar a
regeneração in vitro (Torres et al. 1999). No final dessa década, Tulecke e Nickell (1959)
publicaram estudo mostrando a eficácia da cultura de tecidos vegetais na produção de
grandes quantidades de plantas. Nos anos 60, George Morel (1964), ao enfrentar
problemas com vírus em cultivo de orquídeas, descobriu que ápices de brotos, uma região
de poucos milímetros e com tecido meristemático de rápido crescimento, encontravam-se
ainda livre de infecção. Após isolá-los Morel conseguiu sucesso na produção de novas
plantas livres de vírus pelo cultivo desses meristemas apicais em meio de cultura. Esse
procedimento seria adotado como técnica básica em laboratórios de cultivo e manutenção
de germoplasma.
Atualmente, plantas livres de vírus e multiplicação (clonagem) de genótipos
passaram a ser as técnicas de cultivo in vitro mais utilizadas comercialmente. Fitch et al.
(2001) provaram a eficácia da cultura de ápices caulinares na erradicação do vírus da folha
amarela em cana de açúcar (Saccharum spp.). Devido ao seu rápido crescimento e ao fato
de não apresentar sistema vascular (floema) o meristema apical pode ser retirado e
cultivado em condições estéreis livre de vírus. Acredita-se que praticamente todas as
espécies propagadas vegetativamente estejam contaminadas com um ou mais vírus, o que
leva a uma diminuição do seu desenvolvimento e produção (Manganaris, 2003).
O cultivo de suspensão celular também é uma alternativa para a obtenção de plantas
clonadas. Esta técnica pode ser resumida como a proliferação de células em meio líquido,
sob agitação contínua. É aplicada em estudos nas mais diversas áreas: bioquímica,
fisiologia vegetal, fitopatologia e genética de plantas. Exemplos podem ser vistos no
isolamento de enzimas vegetais, no estudo de fatores de crescimento, de resistências a
diferentes estresses, entre outras aplicações (Wildholm, 1972; Breindenbach e Waring,
1977).
Para o desenvolvimento de materiais tolerantes a estresses bióticos ou abióticos são
utilizadas as seguintes técnicas: fertilização in vitro, hibridização somática, identificação
de variação somaclonal, transgenia e indução de mutação.

26
A polinização e fertilização in vitro permitem estudar a fertilização e
desenvolvimento do embrião em condições controladas, vencendo barreiras reprodutivas
impostas pelo estigma, possibilitando a recuperação de híbridos específicos ou
intergenéricos (Kanta et al., 1962; Richards e Rupert, 1980).
Protoplastos, células vegetais desprovidas de parede celular obtidas pela digestão
das mesmas por enzimas específicas, são amplamente utilizados no melhoramento de
espécies de interesse pela formação de híbridos somáticos (Torres et al., 1999).
Na produção de transgênicos, o cultivo in vitro torna possível a obtenção de plantas
recombinantes pelo uso de técnicas de transformação celular associada à regeneração in
vitro. A biobalística e o cultivo na presença de Agrobacterium sp. são formas de se obter
células ou parte de tecidos transformados, porém essas células ou áreas geneticamente
modificadas precisam ser selecionadas, multiplicadas e diferenciadas em plantas (Birch,
1997). Chaleff e Parsons (1978) conseguiram selecionar sete linhagens de tabaco
(Nicotiana tabacum L.) tolerantes ao herbicida Picloram, dentre as quais quatro
regeneraram in vitro, dando origem a plantas cuja resistência segregava de forma
dominante e estável.
Para se obter rapidamente homozigose para todos os loci, o cultivo de anteras ou
ovários é empregado. Nesse caso os duplo-haplóides podem ser obtidos pelo cultivo das
células haplóides expostas a Colchicina, o que leva à duplicação do material genético,
garantindo que todos os alelos se encontrem em homozigose. Essa ferramenta pode ser
muito útil em estudos de herança de certas características em programas de melhoramento.
De Maine (1989) hibridizou fêmeas dihaplóides resistentes a podridão da folha da batata
(Phytophthora infestans) com machos tetraplóides resistentes a nematóides císticos da
batata (Globodera pallida) e obteve híbridos tetraplóides com ambas as resistências.
A maioria das técnicas apresentadas anteriormente requer que a característica alvo
exista na espécie de interesse. Porém, em alguns casos, essa característica não é observada.
Nestes casos a indução de variabilidade genética via mutagênese, é uma alternativa
recomendada (Torres et al., 1999).
Longos períodos de cultivo in vitro podem acarretar alterações fenotípicas devido a
alterações genéticas ou epigenéticas chamadas de variação somaclonal (Evans e Sharp,
1986). Essas mutações podem ser usadas na obtenção de novas variantes genéticas para
programas de melhoramento de plantas. Baer et al. (2007) provaram que a variação
somaclonal obtida pela regeneração de plantas a partir de linhagens de calos de Eleusine

27
coracana (L.) Gaertn. foi capaz de gerar características de interesse agronônico como
maior produção de grãos e biomassa, bem como a diminuição do tempo de
desenvolvimento. As novas plantas obtidas por variação somaclonal podem ser usadas
como material inicial em programas de melhoramento.
3.4 Indução de Mutação
O cientista irlandês Hugo De Vries, renomado professor de botânica em Amsterdã
já percebia a importância das mutações na evolução e especiação. Apesar de algumas de
suas idéias não serem coerentes com o conhecimento atual sobre evolução, sua visão sobre
o uso de mutantes em programas de melhoramento e indução artificial de mutantes se
encaixam perfeitamente na perspectiva atual sobre o assunto. Em seu livro “Species and
Varieties: Their Origin by Mutation” (de Vries, 1901, apud Harten, 1998) o autor comenta:
“Devemos procurar por plantas mutantes na natureza, ou devemos esperar
que algumas espécies se tornem mutáveis por métodos artificiais. A
primeira nos promete conseguir resultados mais rápidos, porém a segunda
é muito promissora e pode trazer resultados de importância muito maior.
Realmente, se em algum momento formos capazes de fazer plantas
mutarem à nossa vontade e talvez até em direções escolhidas
arbitrariamente, não existem limites para o poder que nós devemos
finalmente ganhar sobre a Natureza.”
Um ano antes do lançamento do seu livro, De Vries já se referia a novos tipos de
radiação (raios X e raios γ) descobertos por W.K. von Röntgen, H. Beccquerel, P. Curie e
M. Curie como possíveis indutores de mutação em plantas. Seu livro marcou um ponto de
início ao estudo da indução de mutação, porém somente 22 anos mais tarde foi provado
que mutações poderiam ser artificialmente induzidas (Harten, 1998).
Para fins práticos, podemos dividir as fontes ou agentes de mutação em duas
categorias, os agentes físicos (radiações) e químicos. Na categoria de agentes físicos
encontramos diversos tipos de radiação, como a radiação gama, raios x, luz ultravioleta,
elétrons, prótons e nêutrons. As radiações podem ser divididas em dois grupos de acordo
com sua forma de propagação: radiações corpusculares e radiações do espectro
eletromagnético (ondas eletromagnéticas) (Green e Roderick, 1966).

28
Para a indução de mutação são usadas radiações de alta energia, capazes de ionizar
a matéria (tecidos) pela qual atravessam. Entende-se por ionização a liberação de pequenas
quantidades de energia (eléctrons) das ondas ou partículas de radiação durante a colisão
com átomos de diferentes moléculas que compõem a matéria irradiada (Harten, 1998).
Radiações ionizantes acarretam diversos eventos físicos e químicos no DNA e
RNA e os efeitos biológicos dessa exposição vai depender da quantidade de energia
absorvida pelo tecido, mais especificamente pelos cromossomos (Harten, 1998).
Radiações corpusculares consistem nas radiações que se propagam por partículas
móveis. São exemplos de radiações corpusculares os elétrons, prótons, nêutrons, partículas
alfa e beta. Todas as radiações corpusculares são ionizantes, tanto partículas carregadas
(prótons e elétrons) como não carregadas (nêutrons e fótons). A radiação alfa é constituída
por dois prótons e dois neutrons e são emitidas de isótopos que mudam para um novo
elemento químico no momento de sua liberação. Possuem pouca velocidade e por isso não
são capazes de atravessar a pele humana ou uma folha de papel, porem podem ser
perigosas se inaladas ou ingeridas. Essa partícula é muito pouco usada em indução de
mutação. A radiação beta, por sua vez é composta de um elétron que possui efeitos
similares aos raios gama. Sua penetração é maior que a das partículas alfa, porém inferior
aos raios X e gama. É mais usada em indução de mutação e podem penetrar alguns
milímetros em tecido vegetal. Os nêutrons constituem outro exemplo de radiação
corpuscular. São partículas não carregadas que podem surgir de diversas maneiras. Uma
delas é a liberação durante uma fissão nuclear (Harten, 1998).
Radiações eletromagnéticas consistem nas radiações que se propagam através de
ondas. Radiações de ondas mais longas não são capazes de causar ionização
(infravermelho e luz visível) enquanto radiações com um comprimento de onda menor são
consideradas ionizantes (raios gama e raios X). A luz ultravioleta encontra-se numa faixa
intermediária do espectro das radiações eletromagnéticas, sendo a mais presente na
natureza. Apenas 10% da radiação ultravioleta que inside sobre um tecido vegetal é
absorvida, porém apesar de sua pouca penetrância é usada como agente mutagênico no
tratamento de pequenos alvos como grãos de pólen e suspensões de células. Funciona
aumentando a freqüência de mutações espontâneas além de ser muito menos perigosa que
outras radiações (Harten, 1998).
Amplamente usado na área médica, o raio X difere da radiação gama apenas pelo
local de origem, enquanto os raios X surgem da nuvem de elétrons que circundam o núcleo
atômico, a radiação gama surge do próprio núcleo. A radiação gama é o agente mutagênico

29
mais usado. Sua alta penetrância a torna perigosa já que pode causar muito dano ao tecido
que atravessa. Altamente ionizante, apresenta um dos menores comprimentos de onda
dentre as radiações eletromagnéticas (Segreto e Segreto, 1997).
Uma das primeiras revisões sobre cultivares induzidos por radiação foi escrita por
Singleton em 1962, na forma de uma lista, na qual nove novos cultivares propagados por
semente foram descritos (Singleton, 1964). Esse estudo incluía cv. Primex de mostarda
branca (Sinapis alba L.), Suíça (1950); cv. Schaefer`s Universal de feijão (Phaseolus
vulgaris L.), Alemanha (1950); cv. Regina II de nabo (Brassica napus L.), Suíça (1957);
cv. Salinac de feijão (P. vulgaris), Michigan, USA; cv. Pallas de feno (Hordeum vulgare
L.), Suíça (1958); cv. N.C.4X de amendoim (Arachis hypogaea L.), Carolina do Norte,
EUA (1959); cv. Florad de aveia (Avena sativa L.), Florida, EUA (1960).
Dois anos depois, durante um importante encontro sobre o uso de mutações
induzidas em programas de melhoramento de plantas, um grupo dos principais estudiosos
em mutação da época aceitou coordenar todas as atividades na área (Harten, 1998).
Naquele momento, um órgão fiscalizador dos estudos em indução de mutações em plantas
foi criado pela junção de dois outros pré-existentes (Food and Agriculture Organization of
the United Nations / International Atomic Energy Agency-FAO/IAEA).
Mas foi no ano de 1969 que a indução de mutação teve sua grande virada (Micke et
al., 1990). Durante o Simpósio “Natureza, Indução e Utilização de Plantas Mutantes” foi
apresentada uma nova lista, agora com 77 cultivares mutantes. Já em 1990, 21 anos mais
tarde, esse número subia para 1330 novos mutantes, um crescimento que refletia os
esforços investidos pelos pesquisadores da área de mutações e radiobiologia
(Sigurbjörnsson, 1991).
Segundo FAO / IAEA (2007), mais de 2.550 variedades foram oficialmente
lançadas entre 1930 a 2007, sendo que cerca de 70% dessas cultivares foram obtidas por
seleção direta de mutantes, ou seja, após seleção os mutantes foram multiplicados, testados
e introduzidos no mercado (após devido registro e liberação pela FAO/IAEA) como novas
cultivares, sem haver necessidade de novos cruzamentos. Nos últimos sete anos, 298 novas
cultivares foram geradas, formando um banco de 2550 cultivares obtidas por meio de
indução de mutação. O Brasil é responsável por 36 desse total, sendo 28 novas cultivares
de Oryza sativa L., duas cultivares de Triticum aestivum L., três de P. vulgaris e três
Chrysanthemum sp. Trinta e duas cultivares foram obtidas pelo uso de radiação gama e
apenas quatro, foram obtidas pelo cruzamento com radiomutantes (FAO / IAEA, 2007).

30
Países da Ásia e Europa são os que mais contribuem para o aumento desse banco de
cultivares, ocupando o primeiro e segundo lugar no melhoramento por mutagênese,
respectivamente. A América Latina está em quinto lugar. Em uma análise geral percebe-se
que a maioria das plantas que compõem o banco da FAO/IAEA são cultivares propagáveis
por sementes (67%) ou ornamentais (24.7%) (Tabela 1). Aproximadamente 89% das novas
cultivares/ plantas desenvolvidas nesse período (1930 - 2000) foram obtidas pelo uso de
radiação, e especificamente, 84% por radiação gama (Maluszynski et al., 2000). Bhatia et
al. (2001) relataram que até aquele ano, 265 novas cultivares de leguminosas produtoras de
grãos já haviam sido lançadas em mais de 32 países através do uso de indução por
radiação. Citaram também que a irradiação de sementes por raios gama ou X resultou na
obtenção direta de 111 cultivares/ plantas.
Tabela 1. Número de plantas de diferentes tipos agronômicos geradas a partir de indução
de mutação via radiação gama, em diferentes regiões agrícolas (Fonte: FAO/IAEA, 2007).
Tipo Agronômico Região
Legume Cereal Ornamental Propagável por sementes
Propagável vegetativamente
Total
Ásia 18 52 292 1131 75 1570
Europa 8 13 301 356 18 696
América do Norte 0 0 32 113 20 165
África 2 0 0 57 1 60
América Latina 0 0 3 43 3 49
Austrália e Pacífico 0 0 3 7 0 10
Total 28 65 631 1707 117 2550
Apesar da lista de novos mutantes continuar crescendo, não representa o número
real de novas plantas mutantes. A velocidade com que novas características surgem, bem
como a quantidade de material mutante produzido em pesquisas científicas para fins
comerciais e/ou acadêmicos dificulta o registro de todas as plantas obtidas. Apenas as
plantas selecionadas para determinada característica dentro de um programa de
melhoramento são registradas, enquanto as que apresentaram outras mudanças são

31
descartadas ou conservadas para estudos posteriores, muitas vezes não realizados (Harten,
1998).
Lyakh e Lagron (2005) irradiaram sementes de linhaça (planta cujo óleo extraído
das sementes é amplamente utilizado na indústria e alimentação humana na Europa) com
radiação gama e obteve diversas alterações de interesse agronômico-comercial. Foi
observado neste trabalho mudança na cor da corola, antera e sementes, nova forma e
tamanho das flores (forma de estrela), mudança na altura e hábito das plantas. Srivastava e
Mishra (2005) usaram radiação gama como fonte de indução de variabilidade para
conseguir flores de Hibiscus rosa-sinensis L. três vezes menores que o normal.
Além de alterações estéticas, o uso de agentes indutores de mutação pode causar
mudanças na fisiologia da planta, levando à descoberta de novas cultivares com maior
produção agrícola, e com isso aumentar o retorno do investimento no seu cultivo. Hasib e
Ganguli (2005) fizeram com que a cultivar de arroz Basmati (de alta demanda pela
população da Índia, porém de baixo desempenho) apresentasse um aumento de até 40% na
sua produção através do cruzamento de cv. Basmati com radiomutantes previamente
selecionados.
Outro caráter de interesse é a mudança no porte de plantas, tornando-as mais
compactas e possibilitando assim um melhor aproveitamento da área de cultivo. Ananda
Kumar (2005) utilizou a cultivar White Ponni, o mais popular arroz no Sul da Índia, em
estudo de redução de porte por indução com radiação gama. Ao fim do estudo conseguiu
uma redução máxima e estável de 17,2% no tamanho de algumas plantas sem alterar o seu
desempenho agronômico.
Outra alteração fisiológica de grande interesse dos agricultores é a precocidade.
Plantas precoces possibilitam ao produtor colher e oferecer seu produto antes que o
mercado sature e o preço de venda diminua. Através da técnica de cultivo de anteras
coletadas de plantas oriundas de sementes de arroz irradiadas com raios gama (450 Gy),
Myint et al. (2005) conseguiram obter mutantes com produção por hectare maior que a
cultivar original, com a vantagem de serem colhidos até 19 dias antes. Shamsuzzaman et
al. (2005) utilizando radiação gama conseguiram desenvolver um mutante de grão-de-bico
capaz de alcançar a maturidade duas semanas antes do material não tratado. Essa diferença
temporal pode ser de grande valia tendo em vista que na Índia, local onde foi desenvolvida
a pesquisa, o grão-de-bico alcança a fase adulta no período de chuvas o que leva a uma
queda drástica na sua produção.

32
Perdas de produção por fatores ambientais de estresse como seca, frio e calor
preocupam produtores e também são objetos de estudo de cientistas em todo o globo.
Gonzáles-Cepero et al. (2005) conseguiram induzir, em duas cultivares cubanas de tomate,
tolerância à seca pela irradiação de plantas na doses de 300 Gy e 500 Gy. Sementes das
plantas irradiadas foram testadas em campo e três delas mostraram uma produção até 2,6
vezes maior que o parental em condições de estresse.
Por sua vez, a indução de resistência a agentes bióticos, como bactérias e vírus,
podem interferir positivamente na produtividade das plantas. A bactéria Xanthomonas
oryzae, que pode causar perda de até 100% de cultivos de arroz cv. Basmati (Herdt, 1991)
motivou pesquisadores a desenvolver novas cultivares resistentes à podridão bacteriana.
Doze dos 16 mutantes resistentes obtidos foram gerados por mutagênese induzida por raios
gama, enquanto o restante pelo uso de metanosulfato de etila (EMS) (Agrawal et al.,
2005).
O melhoramento por indução de mutação pode aprimorar diversos parâmetros de
interesse econômico como: aumento da produção, melhoria da qualidade, redução do uso
de pesticidas, economia de água, melhor uso do solo, caracteres de interesse do
consumidor, maior valor nutritivo, maior facilidade de colheita, redução das importações e
aumento das exportações (Ahloowalia et al., 2004).
Em relação ao feijão-caupi, a lista de materiais obtidos por indução de mutação da
FAO/IAEA (2007) conta com nove cultivares, sendo sete obtidas em centros de pesquisa
na Índia e duas no Quênia. Os agentes mutagênicos mais utilizados foram a radiação gama
e DMS (Dimetil Sulfato). A maioria dos mutantes registrados apresentou melhoras no
desempenho e na produção (Tabela 2). Entretanto, nenhuma foi descrita como sendo
tolerante à salinidade presente em solos.
Tabela 2. Cultivares de V. unguiculata obtidos pelo uso de radiação gama (Fonte:
FAO/IAEA, 2007).
MUTANTE PAÍS ANO CARACTERÍSTICA Uneca-Gama Costa Rica 1998 Produção Cowpea-88 Índia 1990 Produção Co 5 Índia 1986 Valor Nutricional ICV 11 Quênia 1985 Hábito semi-ereto ICV 12 Quênia 1985 Produção

33
3.5. Seleção in vitro de Plantas
Seja selecionando mutantes induzidos por agentes físicos ou químicos, seja no
auxílio à identificação de células transgênicas em uma suspensão celular ou no isolamento
de variantes somaclonais, a seleção in vitro é uma das técnicas de cultivo de tecidos
vegetais mais simples e eficientes utilizadas no melhoramento não convencional de
plantas.
A transformação genética de plantas permite a inclusão de genes de outros
organismos em uma planta, conferindo-lhe nova(s) característica(s). Essa é uma etapa
delicada do processo de obtenção de trangênicos, porém uma seleção in vitro eficiente dará
mais credibilidade aos resultados. O sucesso de programas de transgenia em plantas
superiores depende de um protocolo de seleção e regeneração in vitro eficiente, pois as
mesmas apresentam-se notoriamente recalcitrantes, incluindo o gênero Vigna (Garcia et
al., 1987; Popelka et al., 2004). Popelka et al. (2006) não conseguiram êxito na
regeneração de embriões ou plantas a partir de calos transformados de V. unguiculata,
assim como já havia sido notificado em outros trabalhos (Anand et al.,2001; Muthukumar
et al., 1995). Apesar dos problemas com calo, Popelka et al. (2006) provaram que é
possível selecionar in vitro variantes transgênicos de caupi resistentes ao Callosobruchus
maculatus F. utilizando eixos embrionários em meio com o fitorregulador BAP
(Benzilalminopurina), para indução de regeneração direta de brotamentos múltiplos. Saini
e colaboradores (2003) transformaram eixos embrionários de Vigna mungo (L.) Hepper
utilizando Agrobacterium tumefaciens contendo genes virais do ZYMV (vírus do mosaico
amarelo), com o objetivo de produzir plantas resistentes. O processo, incluindo a obtenção
de transformantes até formação de plântulas enraizadas in vitro durou oito semanas, o que
contribuiria para a redução do tempo necessário para o melhoramento da cultivar estudada.
Considerando o caráter tolerância à salinidade (NaCl) em processos de seleção in
vitro, poucos exemplos recentes estão disponíveis na literatura internacional. Macedo et al.
(2005) demonstraram ser possível estabelecer uma dose de sal a ser usada na seleção in
vitro de gemas de bananeira Grand Naine tolerantes, pela observação dos sinais de
toxicidade atribuído ao NaCl em diferentes doses. Ulisses et al. (2000), expondo gemas de
Musa sp. cv. Nanicão a diferentes doses de NaCl (20, 40, 60, 80, 100, 120 mM) durante 60
dias, depois cultivando-as por mais 90 dias em 80 e 100 mM NaCl (após um intervalo de
30 dias em meio sem sal) conseguiram selecionar 80 plantas consideradas tolerantes a
salinidade.

34
Mutantes obtidos por variação somaclonal já foram selecionados para o caráter com
uso do cultivo in vitro. Brito et al. (2007) estudaram somaclones de abacaxizeiro
submetidos a crescentes pressões seletivas (12.5, 25 e 50 mM) com NaCl. Os autores
perceberam que a pré-inoculação em concentrações menores de NaCl não influenciava na
regeneração de plantas durante cultivo em 100 mM NaCl.
Ochatt et al. (1999) empregaram uma variante da técnica de seleção in vitro,
conhecida como seleção recorrente, para conseguir linhagens de calos de batata tolerantes
a NaCl. Intercalando subculturas de calos de Solanum tuberosum cv. Kennebec, ora em
meio contendo NaCl 150 mM, ora em meio sem pressão seletiva, conseguiram recuperar
linhagem tolerante (Clone 150) capaz de sobreviver a essas condições de estresse. No
trabalho, os autores avaliaram o efeito da concentração de NaCl na regeneração de calos e
plantas usando como material biológico material propagado de Clone 150, e tendo como
dose letal a concentração de 450 mM. Doses de 150 e 300 mM permitiram 45 e 15%,
respectivamente, de organogênese indireta (formação de ápice caulinar a partir de calo) no
material inoculado.
Gangopadhyay et al. (1997) usaram NaCl a 43 mM para pré-selecionar, em meio
semi-sólido, colônias de células de Brassica juncea (L.) Coss. oriundas de suspensão
celular. Uma linhagem pré-selecionada foi multiplicada e 20 clones de aproximadamente
20 g foram selecionados através de subcultivos (cinco por dose) em doses crescentes de
NaCl (43, 85, 128 e 171 mM). A cada repicagem para nova concentração os calos sofriam
uma redução no desenvolvimento, porém ao longo dos subcultivos naquela dose
restauravam um crescimento constante, provando sua adaptação. Células dos calos
tolerantes mostraram ter maior conteúdo em Prolina em relação ao controle, porém esse
conteúdo não aumentou significativamente com o aumento na concentração de NaCl.
Hassan et al. (2004) selecionaram in vitro explantes de V. radiata var. Kawmy-1
tolerantes ao NaCl. Cultivando cotilédones a 25, 50 100 e 150 mM de NaCl, isolaram e
aclimataram materiais tolerantes com sucesso. Os materiais aclimatados voltaram a ser
regados nas doses usadas na sua seleção in vitro, e tiveram nitrogênio total (N-total),
conteúdo protéico e nitrato redutase avaliados. Baixas doses como a de 50 mM de NaCl
aumentaram as quantidades de nitrogênio total encontradas na parte aérea e nas raízes de
V. radiata, enquanto doses superiores (100 e 150 mM) diminuíram significativamente
essas taxas. Houve diferença significativa no conteúdo total de proteínas apenas entre as
doses de 50 e 100 mM, onde a menor dose estimulou o aumento do conteúdo protéico,
enquanto doses maiores levaram a uma leve queda. Na análise do efeito da concentração

35
de NaCl na atividade da enzima nitrato redutase, foi observada uma inibição significativa
nas doses de 50 mM em diante.
Trabalhos de seleção in vitro envolvendo a espécie V. unguiculata para tolerância a
NaCl ainda não foram reportados.

36
4. REFERÊNCIAS BIBLIOGRÁFICAS
Agrawal PK, Sidu GS and Gosal SS (2005) Induction of bacterial blight resistance in elite
indiam rice (Oryza sativa L.) cultivars using gamma radiation and ethyl methane sulfonate.
Mutation and Breeding Newsletter and Reviews, 1:17–18.
Ahloowalia BS, Maluszynski M and Nichterlein K (2004) Global impact of mutation-
derived varieties. Euphytica, 135:187–204.
Amzallag GN, Lerner HR and Poljakoff-Mayber A (1990) Induction of increased salt
tolerance in Sorghum bicolor by NaCl pretreatment. J Exp Bot 41:29–34.
Anand RP, Ganapathi A, Vengadesan G, Selvaraj N, Anbazhagan VR e Kulothungan S
(2001) Plant regeneration from immature cotyledon derived callus of Vigna unguiculata
(L.) Walp. (cowpea). Current Sci, 80:671–674
Ananda Kumar CR (2005) Stable dwarf mutants of a high quality tall rice variety White
Ponni. Mutation Newsletter and Reviews, 1:15–16.
Assunção IP, Filho LRM, Resende LV, Barros MCS, Lima GSA, Coelho RSB and Lima
JAA (2005) Genes diferentes podem conferir resistência ao cowpea severe mosaic virus
em Caupi. Fitopat brasileira 30(3): 274–278.
Baer GY, Yamets AI, Stadnichuk NA, Rakhmetov DB and Blume YB (2007) Somaclonal
variability as a source for creation of new varieties of finger millet ( Eleusine coracana (L.)
Gaertn.). Cytology and Genetics 41(4):204–208.
Barreto PD and Quinderé MAW (2000) Resistência de genótipos de caupi ao caruncho.
Pesq Agropec Bras 35(4):779–785.

37
Bashir IM and Hampton RO (1996) Sources of genetic resistance in cowpea [Vigna
unguiculata (L.) Walp.] to cowpea aphid-borne mosaic potyvirus. European Journal of
Plant Pathology 102:411–419.
Baur H (1992) Farming systems in northern Ghana. Nyankpala Agricultural Experiment
Station Report. Tamale, Ghana, 140 pp.
Bhatia CR, Maluszynski M, Nichterlein K, Van Zanten L (2001) Mutation breeding
review: Grain legumes cultivars derived from induced mutation affecting nodulation. Plant
Breeding and Genetics Section, pp 44.
Birch RG (1997) Plant Transformation: Problems and Strategies for Practical Application.
Annual Review of Plant Physiology and Plant Molecular Biology, 48:297–326.
Bohnert HJ and Shen B (1999) Transformation and compatible solutes. Scientia
Horticulturae 78:237–260.
Borsani O, Valpuesta V and Botella MA (2003) Developing salt tolerant plants in a new
century: a molecular biology approach. Plant Cell Tissue and Organ Culture 73:101–115.
Bressani R (1985) Nutritive value of cowpea. In: Singh SR and Rachie KO (eds.) Cowpea
Research, Production and Utilization. John Wiley e Sons, West Sussex, UK, pp 353–359.
Briedenbach VR and Waring AJ (1977) Responses to chilling of tomato seedlings and cells
in suspension cultures. Plant Physiology 60:190–192.
Brito LKFL de, Moura GEDD, Martins CP, Aloufa MAI, Macedo CEC, Lopes DB and
Barroso PAV (2007) Cultivo in vitro de somaclones de abacaxizeiro na presença de NaCl.
Revista Brasileira de Engenharia Agrícola e Ambiental 11(3):279–283
Cainnes AM, Shennam C (1999) Interactive effects of Ca2+ and NaCl salinity on growth
of two tomato genotypes differing in Ca2+ use efficiency. Plant Physiol Biochem 37:569–
576.

38
Campbell WH (1999) Nitrato redutase structure, function and regulation: bridging the gap
between Biochemestry and Physiology. Annu Ver Plant Physiol Mol Biol 50:227–300.
Castro NR, Menezes GC and Coelho RSB (2003) Herança da resistência genética do caupi
à cercosporiose. Fitopatologia Brasileira 28:552–554.
Chaleff RS and Parsons MF (1978) Direct selection in vitro for herbicide-resistant mutants
of Nicotiana tabacum. Proc Natl Acad Sci 75(10):5104–5107.
Chittaranjan K (2007) Genome Mapping and Molecular Breeding in Plants: Sugar and
Tuber Crops. Vol 3. Springer Berlin Heidelberg, 306 pp.
CONAB (Companhia Nacional de Abastecimento)
http://www.conab.gov.br/download/safra/FeijaoTotalSerieHist.xls. (April 6, 2006)
Costa PHA, Silva JV, Bezerra MA, Filho JE, Prisco JT and Filho EG (2003) Crescimento a
níveis de solutos orgânicos e inorgânicos em cultivares de Vigna unguiculata submetidos à
salinidade. Revista Brasileira de Botânica (36) 3:289–297.
Darlene M. Lawson, China F. Lunde e Martha A. Mutschler (1997) Marker-assisted
transfer of acylsugar-mediated pest resistance from the wild tomato, Lycopersicon
pennellii, to the cultivated tomato, Lycopersicon esculentum. Molecular Breeding 3:307–
317.
De Maine MJ , Stewart HE and Philips MS (1989) The production of dihaploids and
tetraploids with combined quantitative resistance to potato cyst nematode (Globodera
pallida) and foliage blight (Phytophthora infestans). Potato Research 32:425–430.
De Vries H (1901) The mutation theory I. Leipzig, Veit & Co. pp
Ehlers JD and Hall AE (1997) Cowpea [Vigna unguiculata (L.) Walp.]. Field Crops Res
53:187–204.

39
Ehlers JD and Hall AE (1998) Heat tolerance of contrasting cowpea lines in short and long
days. Field Crops Research 55(1):11–21.
Evans DA and Sharp WR (1986) Applications of somaclonal variation. Bio/Technology
4:528–532.
FAO/AGL Global Network on Integrated Soil Management for Sustainable Use of Salt-
affected Soils http://www.fao.org/ag/AGL/agll/spush/intro.htm (Acessado em 27 de Maio
2007).
Ferrario S, Valadier M and Foyer CH (1998) Over-expression of nittrate reductase in
tabaco delays drought-induced decreases in nitrato reductase activirties and mRNA. Plant
Physiol 117:293–302.
Fery, RL (1990) The cowpea: production, utilization, and research in the United States.
Hort Rev 12:197–222.
Fitch MMM, Lehrer AT, Komor E and Moore PH (2001) Elimination of Sugarcane yellow
leaf virus from infected sugarcane plants by meristem tip culture visualized by tissue blot
immunoassay. Plant Pathology 50:676–680.
Flowers TJ, Koyama ML, Flowers SA, Chinta Sudharka KP, Shing KP and Yeo AR (2000)
QTL: their place in engineering tolerance of rice to salinity. J Exp Bot 51:99–106.
Freire-Filho FR (1988) Origem, evolução e domesticação do caupi [Vigna unguiculata (L.)
Walp.] In: ARAÚJO, J. P. P. de; WATT, E.E. Org. O Caupi no Brasil. Goiânia:
EMBRAPA-CNPAF/ Ibadan: IITA, pp 25–46.
Freire Filho FR, Ribeiro VQ and Bandeira, LMR (1997) Org. Cultivares de feijão caupi
recomendadas para plantio nas regiões Norte e Nordeste: ano agrícola 1997/98. Teresina:
EMBRAPA-CPAMN, 1997. 26p. (EMBRAPA-CPAMN. DocumentoS 22).

40
Gangopadhyay G, Basu S e Gupta S (1997) In vitro selection and physiological
characterization of NaCl- and mannitol-adapted callus lines in Brassica juncea. Plant Cell,
Tissue and Organ Culture, 50:161–169.
Garcia JA, Hillie J e Goldbach R (1987) Transformation of cowpea Vigna unguiculata
cells with a full length DNA copy of cowpea mosaic virus m-RNA. Plant Sci, 44:89–98.
Green, E. L., and Roderick, T. H. (1966). Radiation Genetics. In Biology of the Laboratory
Mouse, Green, E. L., eds. (McGraw-Hill, New York), pp. 165–185.
Greenway H and Munns R (1980) Mechanisms of salt tolerance in nonhalophytes. Annu
Ver Plant Physiol 31:149–190.
Gonzáles-Cepero MC, Mukandama JP, Fuentes JL and Alí MM (2005) Induction of
drought tolerance in tomato using 60Co gamma ray irradiation. Mutation Breeding
Newsletter and Reviews, 1:9–10.
Halfter U, Ishitani M and Zhu JK (2000). The Arabidopsis SOS2 protein kinase physically
interacts with and is activated by the calcium-binding protein SOS3. Proc Natl Acad Sci
97(37):30–34
Hall AE (2004a) Breeding for Adaptation to drought and heat. European Journal of
Agronomy 24:447–454.
Hall AE (2004b) Comparative Ecophysiology of Cowpea, Common Bean, and Peanut. In:
Nguyen HT and Blum A (eds) Physiology and Biotechnology Integration for Plant
Breeding. Technology e Industrial Arts. pp 271–325.
Harten AM van (1998) Mutation breeding: theory and practical applicantions. Cambridge
University Press, UK, 367 pp.
Hassan NM, Serag MS e El-Feky FM (2004) Changes in nitrogen content and protein
profiles following in vitro selection of NaCl resistant mung bean and tomato. Acta
Physiologiae Plantarum, 26(2):165–175.

41
Hasegawa PM, Bressan RA, Zhu JK and Bohnert HJ (2000) Plant cellular and molecular
responses to high salinity. Annu Rev Plant Physiol Plant Mol Biol 51(4):63–99.
Hasib KM and Ganguli PK (2005) High yield lins of “Basmati” rice evolved through cross
breeding with induced mutants. Mutation Breeding Newsletter and Reviews, 1:16–17.
Helms D, Panella L, Buddenhagen IW, Tucker CL and Gepts PL (1991a) Registration of
“California Blackeye 46” cowpea. Crop Sci 31:1703.
Helms D, Panella L, Buddenhagen IW, Tucker CL and Gepts PL (1991b) Registration of
“California Blackeye 88” cowpea. Crop Sci 31:1703.
Herdt RW (1991) Research priorities for rice biotechnology, In: Khush GS and
Toenniessen (eds) Rice biotechnology. UK, pp 19–54.
Hussain M, Naeem M, Ashraf MY and Iqbal Z (2002) Effects of NaCl salinity on growth
and ion partitioning in some barley varieties. Online Journ of Biol Sci 2(9)608–611.
IBGE. Instituto Brasileiro de Geografia e Estatística (1996) Anuário estatístico. Rio de
Janeiro.
Ishitani M, Liu J, Halfter U, Kim CS,Wei M, Zhu JK. 2000. SOS3 function in plant salt
tolerance requires myristoylation and calcium-binding. Plant Cell 12(16):67–77.
Ishiyaku MF, Singh BB and Craufurd PQ (2005) Inheritance of time to flowering in
cowpea [Vigna unguiculata (L.) Walp.]. Euphytica 142:291–300.
Ismail AM, Hall AE and Close TJ (1999) Purification and partial characterization of a
dehydrin involved in chilling tolerance during seedling emergence of Cowpea. Plant
Physiology 120:237–244.
Iuchi S, Kobayashi M, Yamaguchi-Shinozaki K and Shinozaki K (2000) A Stress-
Inducible Gene for 9-cis-Epoxycarotenoid Dioxygenase Involved in Abscisic Acid

42
Biosynthesis under Water Stress in Drought-Tolerant Cowpea. Plant Physiology 123:553–
562.
Kanta K, Rangaswamy NS and Maheshwari P (1962) Test-tube fertilization in a flowering
plant. Nature 194:1214–1217.
Kwapata MB and Hall AE (1985) Effects of moisture regime and phosphorus on
mycorrhizal infection, nutrient uptake, and growth of cowpea [Vigna unguiculata (L.)
Walp.]. Field Crops Res 12:241–250.
Langyintuo AS, Ntoukam G, Murdock L, Lowenberg-DeBoer J and Miller DJ (2004)
Consumer preferences for cowpea in Cameroon and Ghana
Agricultural Economics, 30(3):203–213.
Lima MB, Alves EJ, Borges AL and Nascimento FHA (2005) Efeitos das culturas de
milho (Zea mays), feijão (Phaseolus vulgaris) e caupi (Vigna unguiculata) na agregação de
valor ao cultivo da bananeira “terra”, em Teolâncdia, Litoral Sul da Bahia. Rev. Bras.
Frutic., 27(1):55–59.
Liu J, Ishitani M, Halfter U, Kim CS and Zhu JK (2000) The Arabidopsis thaliana SOS2
gene encodes a protein kinase that is required for salt tolerance. Proc Natl Acad Sci
97(37)35–40.
Liu J and Zhu JK (1998) A calcium sensor homolog required for plant salt tolerance.
Science 280(19):43–45.
Lyakh VA and Lagron VA (2005) Induced mutation variability in Linum grandiflorum
Desp. Mutation Breeding Newsletter and Reviews, 1 : 4–4.
Macedo CEC, Moura GEDD, Aloufa MAI e Barroso PAV (2005) Efeito do NaCl sobre o
crescimento e multiplicação in vitro de bananeira . Revista Brasileira de Fruticultura.,
27(2):194–197.

43
Malcolm CV, Lindley VA, O’Leary JW, Runciman HV and Barrett-Lennard EG (2003)
Halophyte and glycophyte salt tolerance at germination and the establishment of halophyte
shrubs in saline environments. Plant and Soil 253:171–185.
Maluszynski M, Nichterlein K, van Zanten L and Ahloowalia BS (2000) Officially
released mutant varieties - the FAO/IAEA Database. Mut. Breed. Rev. 12:1–84.
Manganaris GA, Economou AS, Boubourakas IN and Katis NI (2003) Elimination of PPV
and PNRSV through thermotherapy and meristem-tip culture in nectarine. Plant Cell Rep
22:195–200.
Micke A, Donini B and Maluszynski M (1990) Induced mutations for crop improvement.
Mutation Breeding Reviews, 7:1–41.
Morel G (1964) Tissue culture: A new means of clonal propagation of orchid. American
Orchid Society Bulletin 33:473–478.
Munns R (2002) Comparative physiology of salt and water stress. Plant, Cell e
Environment. 28:239–250
Muthukumar B, Mariamma M e Gnanam A (1995) Regeneration of plants fom primary
leaves of cowpea. Plant Cell, Tissue and Organ Culture, 42:153–155.
Myint KS, Aung KOA and Soe K (2005) Development of a short duration upland rice
mutant line through anther culture of gamma irradiated plants. Mutation Breeding
Newsletter and Reviews, 1:13–14.
National Plant Germplasm System; http://www.ars-grin.gov/cgi-
bin/npgs/html/taxon.pl?41647 (Acessado em 28 de Outubro de 2007)
Niu X, Bressan RA, Hasegawa PM and Pardo JM (1995) Ion homeostasis in NaCl stress
environments. Plant Physiology. 109:735–742.

44
Ochatt SJ, Marconi PL, Radice S e Caso OH (1999) In vitro recurrent selection of potato:
production and characterization of salt tolerant cell lines and plants. Plant Cell, Tissue and
Organ Culture, 55:1–8.
Ogbonnaya CI, Sarr B, Brou C, Diouf O, Diop NN and Roy-Macauley H ( 2003) Selection
of Cowpea Genotypes in Hydroponics, Pots, and Field for Drought Tolerance. Crop Sci
43:1114–1120.
Pimentel C, Abboud ACS, Roy-Macauly H, Diouf O and Sarr B (2002) Leaf protoplasmic
tolerance to drought, in two cowpea genotypes cultivated at the field. Revista Universidade
Rural, Série Ciências da Vida, 22(1):07–14.
Popelka JC, Terryn N e Higgins THV (2004) Gene technology for grain legumes: can it
contribute to the food challenge in developing countries? Plant Sci, 167:195–206.
Popelka JC, Gollasch S, Moore A, Molvig L, Higgins TJV (2006) Genetic transformation
of cowpea (Vigna unguiculata L.) and stable transmission of the transgenes to progeny.
Plant Cell Rep 25:304–312.
Qiu Q, Guo Y, Dietrich M, Schumaker KS and Zhu JK (2001) Characterization of the
plasma membrane NaC/HC exchanger in Arabidopsis thaliana. Abstr Int WorkshopPlant
Membr Biol, 12th, Madison, 235 pp.
Rai V and Rai AK (1999) Growth behaviour of Azolla pinnata at various salinity levels
and induction of high tolerance. Plant and Soul. 206:79–84.
Richards KW and Rupert EA (1980) In vitro fertilization and seed development in
Trifolium. In vitro, 21:181–184.
Roberts P, Boukar O, Cissé N and Drabo I (2004) Development of improved cowpea
cultivars with increased yield potential, tolerance to biotic and abiotic stresses, and having
grain quality traits preferred by farmers and consumers. In: Regional Partnerships to
Enhance Bean/Cowpea Consumption and Production in Africa and Latin America.
Michigan State University, USA 93pp.

45
Rus A, Yokoi S, Sharkhuu A, Reddy M and Lee BH (2001) AtHKT1 is a salt tolerance
determinant that controls Na+ entry into plant roots. Proc. Natl. Acad. Sci. 98(141)50-55.
Saini R, Jaiwal S e Jaiwal PK (2003) Stable genetic transformation of Vigna mungo L.
Hepper via Agrobacterium tumefaciens. Plant Cell Rep., 21:851–859.
Sanginga N, Lyasse O and Singh BB (2000) Phosphorus use efficiency and nitrogen
balance of cowpea breeding lines in a low P soil of the derived savanna zone in West
Africa. Plant and Soil 220:119–128.
Saini R, Jaiwal S and Jaiwal PK (2003) Stable genetic transformation of Vigna mungo L.
Herpper via Agrobacterrium tumefaciens. Plant Cell Report, 21(9):851–859.
Segreto RA and Segreto HRC (1997) Radiação ionizante e resposta celular- atualidades.
Radiologia Brasileira 30(6):301–307.
Serrano R, Mulet JM, Rios G, Marquez JA, de Larriona IF, Leube MP, Mendizabal I,
Pascual-Ahuir A, Proft MRR and Montesinos C (1999) A glimpse of the mechanism of ion
homeostasis during salt stress. J Exp Bot 50:1023-1–36.
Shamsuzzaman KM, Islam MM, Begum M and Subhan A (2005) Development of an early
maturing chickpea variety, BINASOLA-3. Mutation Breeding Newsletter and Reviews,
1:8–9.
Shi H, Ishitani M, Kim C and Zhu JK (2000) The Arabidopsis thaliana salt tolerance gene
SOS1 encodes a putative NaC/HC antiporter. Proc Natl Acad Sci 97:(6)896–901.
Silva JV, Lacerda CF, Costa PHA, Filho JE, Filho EG and Prisco JT (2003) Physiological
responses of NaCl stressed cowpea plants grown in nutrient solution supplemented with
CaCl2. Braz. J. Plant Physiol., 15(2):99–105.
Sigurbjörnsson B (1991) Opening address. In: Mutation breeding for crop improvement.
Proceedings FAO/IAEA Symposium, Viena, pp 3–5.

46
Silva PSL and Oliveira CN (1993) Rendimento de “feijão verde” e maduro de cultivares de
caupi. Horticultura Brasileira, 11(2):133–135.
Singleton WR (1964) Induction of mutations and plant breeding. In: Procedings of Third
Congress of European Association for Research on Plant Breeding. Eucarpia, Paris, pp 43–
55.
Srivastava A and Mishra R (2005) Gamma ray induced small flower mutant in Hibiscus
rosa-sinensis. Mutation Breeding Newsletter and Reviews, 1:20–21.
Stavarek SJ and Rains DW (1984) The development of tolerance to mineral stress.
HortScience 19:13–18.
Torres AC, Caldas L and Buso A (1999) Cultura de Tecidos e Transformação Genética de
Plantas. Vol. 2. Brasília: SPI/Embrapa, 354p.
Tulecke W and Nickell LG (1959) Production of Large Amounts of Plant Tissue by
Submerged Culture. Science 130(3379):863–864.
Ulisses C, Câmara TR, Willadino L, Meunier I, Rocha PSG da and Albuquerque C (2000)
Seleção in vitro de gemas de bananeira “nanicão” tolerantes à salinidade. Scientia
Agricola, 57(4):667–670.
Uwaegbute AC, Iroegbu CU and Eke O (2000). Chemical and sensory evaluation of
germinated cowpeas (Vigna unguiculata) and their products. Food Chemistry 68:141–146.
Wang W, Vinocur B and Altman A (2003) Plant responses to drought, salinity and extreme
temperatures: towards genetic engineering for stress tolerance. Planta 218:1–14.
Wildholm JM (1972) Cultured Nicotiana tabacum cells with an altered anthralinate
synthetase which is less sensitive to feedback inhibition. Biochimica et Biophysica Acta
256:52–58.
Zhu JK (2000) Genetic analysis of plant salt tolerance using Arabidopsis thaliana.
Plant Physiol. 124:941–48

47
Zhu JK (2001) Plant salt tolerance. Trends Plant Sci. 6:66–71.
Zhu JK (2002) Salt and drought stress signal transduction in plants.
Annu. Rev. Plant Biol. 53: 247–273

48
5. MANUSCRITO
5.1 Manuscrito 1
In vitro and in vivo selection of induced NaCl tolerance in
Cowpea [Vigna unguiculata (L.) Walp.]
Manuscrito a ser encaminhado à revista Genetics and Molecular Biology

49
In vitro and in vivo selection of induced NaCl tolerance
in Cowpea [Vigna unguiculata (L.) Walp.]
Luiz Rodrigo Saldanha Gazzaneo1, Laureen Michelle Houllou-Kido 2, Waldeciro
Colaço3, Gilberto Dias Alves4, Ana Maria Benko-Iseppon1 e Éderson Akio Kido1
Key words: gamma-radiation, salinity, tissue culture, abiotic stress.
* Contatos FAX: 00558134591879 E-mail: [email protected].
1 Universidade Federal de Pernambuco – UFPE, Departamento de Genética, Recife-PE. 2 Empresa Pernambucana de Pesquisa Agropecuária – IPA, Laboratório de Genoma e
Cultura de Tecido, Recife-PE.
3 Universidade Federal de Pernambuco – UFPE, Departamento de Energia Nuclear – DEN,
Recife-PE. 4 Universidade de Pernambuco – UPE, Instituto de Ciências Biológicas – ICB, Recife-PE.

50
ABSTRACT
The present study aimed to increase NaCl tolerance in cowpea [Vigna unguiculata (L.)
Walp.] cv. IPA 206 through induced mutagenesis using gamma rays followed by in vitro or
in vivo selection. The in vitro selection was performed by placing cotyledons and embryo
axis from irradiated seeds (150 Gy) in MS medium with 256 mM NaCl as selective
pressure (DL100). Salt sensibility test showed that 86 mM NaCl improved embryo axis
callus growth, while 256 mM NaCl promoted total organogenesis inhibition and explant
necrosis. Out of 3000 explants inoculated, five showed differentiated tissue development
and seven calli presented steady growth under DL100. For the in vivo selection radioseeds
from doses of 50, 100, 200, 300, 400, 600 Gy were grown in greenhouse and watered with
Hoagland nutritive solution with 60 mM NaCl. Four plants (50, 100, 300 and 600 Gy)
presented no external signs of saline stress. Nineteen semitolerant materials (from different
doses) presented great variability on seed weight, seed number per pod, pod length and
days to harvesting. Both, in vitro and in vivo approaches proved to be useful in selecting
mutation induced material.

51
INTRODUÇÃO
O feijão-caupi [Vigna unguiculata (L.) Walp.] é uma importante fonte de proteína para a
população humana de países em desenvolvimento (Prinyawiwatkul et al., 1996) e em
especial nas regiões áridas e semi-áridas do Nordeste brasileiro, onde é cultivado para
subsistência de famílias de baixa renda (Teixeira et al., 1988). Apesar de seu alto valor
nutricional e social em comparação a outras leguminosas, poucos esforços no
melhoramento genético da espécie por indução de mutação foram até agora reportados.
Apenas cinco cultivares produzidas pelo uso da radiação compõem o banco de dados da
FAO / IAEA (2007), sendo o aumento da produção, incremento no valor nutricional e
mudança do hábito da planta as características melhoradas. Mesmo com os atuais avanços
alcançados na tecnologia de produção do feijão-caupi, muitos fatores ainda contribuem
para perdas consideráveis dos investimentos no seu cultivo (Ehlers and Halls, 1997).
Estresses bióticos e abióticos são as principais causas para a restrição na sua produção.
Dentre as fontes de estresse abiótico, a pressão salina tem um considerável papel na
redução da produtividade, especialmente nos 25% da área de zonas áridas e semi-áridas do
globo afetadas pela salinização (Chhabra, 1996). África, Índia e Brasil se destacam no
melhoramento convencional do feijão-caupi, entretanto o uso de técnicas não
convencionais, como a indução por raios gama, poderia auxiliar esses programas de
melhoramento acelerando a obtenção de novas características (Ehlers e Hall 1997;
Sanginga et al. 2000; Pimentel et al. 2002). Epesar dos trabalhos realizados, não se
encontram disponíveis materiais tolerantes às condições salinas da região nordeste
brasileira. Cultivares de feijão-caupi tolerantes a concentrações crescentes de NaCl seriam
essenciais para garantir o cultivo em áreas consideradas salinizadas, gerando suprimentos e
renda para seus habitantes. O presente trabalho teve como objetivo avaliar a aplicabilidade

52
do uso da radiação gama e dos processos da seleção in vitro e in vivo no melhoramento do
feijão-caupi visando tolerância a níveis de NaCl.

53
MATERIAL E MÉTODOS
Material biológico, desinfestação de sementes e explantes para o cultivo in vitro
Sementes de feijão-caupi (Vigna unguiculata cv IPA 206), fornecidas pela Empresa
Pernambucana de Pesquisa Agropecuária (IPA), foram usadas como fontes de explantes.
Desinfestações de sementes foram feitas com uso de etanol 70% (v/v; cinco minutos),
solução de água sanitária da marca Dragão 10% (v/v) e passagens em água destilada estéril
(três vezes). Para a redução de contaminação bacteriana, sementes foram intumescidas em
solução estéril de Cefalexina (2,5 mg.mL-1) por 12 horas. Explantes para uso nos testes de
sensitividade ao NaCl, radiossensitividade e seleção in vitro foram isolados de sementes
mantidas em placas de Petri contendo papel filtro embebido em água estéril, as quais foram
seladas (filme de PVC) e mantidas no escuro (3 dias). Após germinação, as sementes
tiveram os tegumentos retirados e cotilédones e eixos embrionários isolados.
Teste de sensibilidade ao NaCl
Teste de sensibilidade ao NaCl foi feito para se determinar a dose letal de 100% (DL100).
Dez eixos embrionários por tratamento foram inoculados em meio de regeneração
constituído de micro e macronutrientes de MS (Murashigue and Skoog, 1962), 2 mg.mL-1
de BAP, vitaminas (ácido Nicotínico, Piridoxina e Tiamina), sacarose 3% (p/v) e agar
0.6% (p/v). NaCl foi adicionado ao meio visando atingir as concentrações finais de 0,5%
(86mM), 1,0% (172mM) e 1,5% (258 mM). Frascos controles (dez eixos) não tiveram
NaCl acrescidos à composição do meio. Os materiais foram cultivados em sala de
crescimento com temperatura controlada (25 ± 2oC) e fotoperíodo de 16 horas de luz Ouma
et al.2004), durante quatro semanas (Figura 1a). Ao final do período, os materiais foram
avaliados, sendo as freqüências de organogêse direta e indireta obtidas.

54
Teste de radiossensitividade in vitro
Sementes acondicionadas em envelopes (200 sementes / envelope de papel) foram
irradiadas (doses testadas: 50, 100, 200, 300, 400, 500 e 600 Grays) em irradiador
Gammacell (fonte de Co60). Sementes do grupo-controle não foram irradiadas. Sementes
irradiadas foram embebidas em água destilada estéril e germinaram em placas de Petri,
sendo cotilédones e eixos embrionários isolados e inoculados em meio de regeneração,
para identificação da dose capaz de reduzir a freqüência de regeneração dos explantes em
cerca de 35%.
Seleção in vitro
3.000 sementes irradiadas (em dose definida a partir do teste de radiossensitividade acima)
foram inoculadas em meio de regeneração. Após um mês, explantes foram transferidos
para meio de cultura acrescido de NaCl (258 mM). Plantas e calos capazes de sobreviver a
essa pressão seletiva após 60 dias de cultivo, com três transferências para novo meio (uma
a cada vinte dias) foram considerados tolerantes.
Ensaio em telado e seleção in vivo
Sementes de cv. IPA 206 irradiadas em diferentes doses (sete sementes / dose-50, 100,
200, 300, 400, 500 e 600 Gy) e sementes controles não irradiadas foram plantadas em
vasos (uma por vaso) contendo dois quilos de substrato para cultivo de hortaliças. As
plantas foram mantidas em telado até fechamento do ciclo reprodutivo e suas sementes S1
(256 irradiadas e 20 não irradiadas) foram plantadas individualmente em caselas de
madeira (10 x 10 x 20 cm) contendo saco plástico com areia fina lavada. Foram 116
sementes S1 da dose 50 Gy, 61 da dose 100 Gy, 17 da 200 Gy, 39 da 300 Gy, 23 da 400 Gy
e 10 da 600 Gy, além de 10 controles salinos (CCS) e 10 não salinos (CSS). Cada casela

55
contendo uma semente sofreu rega diária com solução nutritiva de Hoagland (50% v/v) por
três semanas, e então, NaCl até concentração final de 15 mM foi acrescido à solução de
rega realizada a cada 2 dias e durante uma semana, e assim sucessivamente com as
soluções posteriores de 30 mM, 45 mM e 60 mM de NaCl. Vagens foram coletadas após
apresentarem-se secas. O número de dias até início de colheita, comprimento de vagens,
número de sementes por vagem e peso para 100 sementes foram estimados.
Análise estatística
Tratamentos foram analisados em Teste F (ao nível de 1% de significância), sendo as
médias comparadas através de Teste de Tukey, ambos feitos com o programa Assistat –
Assistência estatística 7.4 beta (Silva 2007). Experimentos seguiram o delineamento
inteiramente ao acaso com diferentes números de repetições. Dados de organogênese direta
e indireta para eixos embrionários e cotilédones sofreram transformação segundo a fórmula
√(X+0,5).

56
RESULTADOS
A regeneração dos cotilédones e eixos embrionários é normalmente precedida pela
mudança de cor para verde, seguida de intumescimento (Figura 1b). A toxicidade pelo
NaCl pode comprometer esse processo regenerativo. Explantes em cultivo sob diferentes
concentrações de NaCl, evidenciaram quatro níveis de respostas: início de regeneração
(InR; Figura 1c), intumescimento e mudança de cor (IMc; Figura 1d), inibição do
desenvolvimento (DIn; Figura 1f) e necrose (Nec; Figure 1e). Após quatro semanas de
cultivo, 90% (18) dos eixos embrionários do tratamento controle se mostraram
intumescidos com mudança de cor e em início de regeneração (50% em IMc e 40% em
InR) e somente 10% (2) com inibição do desenvolvimento (DIn). Eixos embrionários
cultivados em NaCl 86 mM mostraram um aumento daqueles com desenvolvimento
inibido (20% em DIn); aqueles em NaCl 172 mM, além de 10% em DIn apresentaram 20%
em Nec, ao passo que 100% daqueles mantidos em NaCl 258 mM se mostraram
comprometidos (60% Nec e 40% de DIn) (Figura 2). Por inibir completamente a
regeneração de explantes cv. IPA 206, a dose de NaCl 258 mM foi considerada DL100 e a
ideal entre as como pressão seletiva na seleção in vitro para NaCl.
Organogênese direta e indireta foram observados em 10% dos eixos embrionários do
tratamento controle, em resposta ao fitorregulador presente no meio (BAP), iniciando o
processo de regeneração, enquanto que na dose de NaCl 258 mM (DL100) nenhum tipo de
organogênese foi observada. Em doses intermediárias (86 e 172 mM) a organogênese
direta se manteve constante, enquanto a indireta aumentou com a adição do sal, em relação
ao controle (Figura 3).
No tratamento controle do teste de radiossensitividade, a organogenese direta foi observada
em 44% dos eixos embrionários. Essa organogênese direta dos eixos embrionários foi

57
afetada pela radiação. A resposta desses explantes à radiação gama não foi diretamente
proporcional à dose, sendo a melhor resposta observada para a dose 100 Gy (62%). Acima
da dose 300 Gy, não se observou regeneração (Figura 4). Nos tratamentos para eixos
embrionários irradiados com alguma organogênese direta observada, as médias não
diferiram entre si ao nível estudado (1%), em relação ao controle (Figura 4).
Efeito da radiação gama sobre a organogênese direta dos cotilédones também foi
acompanhada. Em 89% dos explantes do tratamento controle, a organogênese direta foi
observada. Com o aumento da dose de radiação observou-se, em geral, uma redução da
freqüência de regeneração (Figura 4), mas ao contrário do observado para eixos
embrionários, mesmo nas doses mais elevadas (500 Gy e 600 Gy) a organogênese não foi
completamente inibida. Entretanto, as médias desses tratamentos diferiram da média do
tratamento controle (P < 0,01; Figura 4).
A freqüência de organogênese indireta observada em eixos embrionários do tratamento
controle do teste de radiossensitividade foi de 33%, enquanto que nos tratamentos
irradiados a formação de calos variou entre 20% (doses 500 e 600 Gy) a 62% (dose 200 e
400 Gy) (Figura 1f e Figura 5). As diferenças entre as médias dos tratamentos, no entanto,
não foram significativas ao nível de 1% (dados não mostrados).
Eixos embrionários expostos à radiação gama apresentaram em média freqüências de
regeneração total (organogênese direta e indireta) menor que a observada nos explantes do
tratamento controle, com exceção da dose de 100 Gy. Diferenças médias significativas
somente foram observadas entre o tratamento 100 Gy em relação a 500 ou 600 Gy (Figura
6).

58
Recuperação de plantas tolerantes após seleção in vitro
Após sobreviverem 60 dias sob pressão salina com 256 mM de NaCl (Figura 1g), cinco
explantes (três de cotilédones e dois de eixos embrionários) irradiados na dose de 150 Gy
apresentaram regeneração (organogênese direta) com formação de tecido diferenciado
(Figura 1h) e sete (um de cotilédones e seis de eixos embrionários) apresentaram
regeneração (organogênese indireta) com formação de calos, e foram consideradas
potencialmente tolerantes ao sal. Explantes apresentando início de formação de ápices
caulinares foram transferidos para meio contendo 14 μM de ácido giberélico (GA3) para
induzir o alongamento dos entrenós e permitir o isolamento, clonagem e enraizamento das
plantas (ainda em andamento).
Resultados in vivo
A maioria das plantas (182) apresentou os sinais esperados de toxicidade pelo NaCl, como
perda da pigmentação das folhas (clorose), queima das extremidade das folhas, seguido
pela queda das folhas antes da morte da planta (Figure 7a). Outras características além da
tolerância à salinidade podem ter sido afetadas pela radiação gama. No geral, em relação
ao controle não salino (CSS), as médias para algumas características agronômicas
(tamanho da vagem, número de sementes por vagem e peso estimado de 100 sementes de
plantas) das plantas sobreviventes (PS) no sistema de seleção in vivo (sob condição de
estresse) diminuíram, ao passo que o período até a colheita aumentou. (Tabela 1). Na
mesma tabela é possível verificar o número de plantas sobreviventes (pós-seleção) em
relação ao número de sementes S1 plantadas e advindas de sementes irradiadas de uma
mesma dose (SI).
Algumas plantas apresentaram sintomas moderados de toxides pelo NaCl, mas permitiram
que as mesmas características agronômicas fossem levantadas (Tabela 2). As que

59
apresentaram os maiores números de sementes em condição salina foram IIC6-100 e VD6-
400 com 13 sementes/ vagem e VC4-600, com 15 (Tabela 2). Quanto à precocidade, a
planta IIE3-100 (sob estresse salino) produziu vagem madura quatro dias antes da planta
mais precoce do controle sem sal (CSS; 65 dias) e 12 dias antes da mais precoce do
controle com sal (CCS; 73). As plantas IIIJ6-200, VG6-200 e VA1-300 também
apresentaram vagem madura em tempo igual ou inferior à das plantas CSS (65 dias).
Quanto ao peso estimado de 100 sementes, as melhores plantas em relação aos controles
CSS (18 g) foram: IIF3-50 (29 g), IJ5-50 (24 g), IIB1-100 (23 g), IID6-100 (23 g), VD5-
400 (22 g), VD2-400 (21 g), VA1-300 (20 g), VA2-600 (20 g) e IIII3-200 (20 g). Quanto
ao tamanho médio de vagens, cinco plantas [IH2-50 (20,0 cm), IA1-100 (21,0 cm), III6-50
(22,0 cm), VC5-400 (23,0 cm) e IC6-100 (23,5 cm)] apresentaram vagens maiores que a
média do controle CCS (14,2 cm) e, em alguns casos, do controle CSS (21,6 cm).
Quatro plantas (IIF4-50 - 50 Gy, VF1-300 e VE1-300 - 300 Gy; e VA6-600 - 600 Gy)
foram consideradas tolerantes por não apresentarem sintomas visíveis de toxicidade.

60
DISCUSSÃO
Os resultados indicaram que aumentos na concentração de NaCl em meio de cultura de
regeneração para feijão-caupi V. unguiculata cv. IPA 206 afetam a viabilidade e o
desenvolvimento in vitro de explantes. Baixas concentrações do sal NaCl (86 mM)
induziram a organogênese indireta de eixos embrionários, enquanto a concentração de 258
mM inibiu completamente qualquer tipo de organogênese (direta ou indireta). Resultados
similares foram descritos na literatura para banana cv. Nanicão (Alves 2000) e cv. Maçã
(Houllou-Kido 2003).
A dose de 258 mM de NaCl demonstrou ser apropriada para uso em uma rigorosa seleção
in vitro de feijão-caupi cv. IPA 206, já que ocasionou a morte em 100% de eixos
embrionários e cotilédones, sendo esta, portanto, a DL100 observada entre as testadas para
esta cultivar. Kumarv e Sharma (1989) observaram uma DL100 de 300 mM de NaCl para V.
radiata (L.) Wilczek. De acordo com Zair et al. (2003), a regeneração e formação somática
de embriões, assim como a sensibilidade ao estresse são fenômenos extremamente
dependentes do genótipo, o que justificaria as diferenças. Provavelmente o feijão-caupi cv.
IPA 206 V. unguiculata seria mais sensível ao NaCl que V. radiata (L.) Wilczek do estudo
em questão.
O uso da DL100 observada (258 mM de NaCl) foi eficiente na seleção e posterior
identificação in vitro de variantes tolerantes ao sal. Resultados similares foram descritos
por Houllou-Kido (2003), que obteve sucesso na seleção de variantes de banana (Musa
spp.) cv. Maçã tolerantes ao NaCl usando uma DL100 de 250 mM.
Após a seleção in vitro, 12 de 3.000 explantes (4 variante tolerante / 1.000 explantes)
sobreviveram à pressão seletiva (NaCl 258 mM) por 60 dias. Houllou-Kido (2003) obteve
resultados próximos utilizando também radiação gama seguida de seleção in vitro para

61
tolerância ao NaCl (DL100: 250 mM), identificando 15 variantes tolerantes entre 4.958
explantes (aprox. 3 variante tolerante / 1.000 explantes) de ápices caulinares e suspensão
de células de banana cv. Maçã.
Em geral, efeitos da radiação gama acarretaram, com o aumento da dose, uma diminuição
na organogênese direta em cotilédones. Tais resultados também seguiram os encontrados
para banana (Musa spp.) tendo gemas como explantes (Houllou-Kido, 2003). Por outro
lado, em toranja (Charbaji and Nabulsi, 1999) e batata (Li et al., 2005) houve uma melhora
na organogênese direta (aumento do número e comprimento de brotamentos e raízes) após
exposição a baixas doses de radiação gama (4 – 7 Gy). De modo similar, doses de radiação
entre 100 – 400 Gy estimularam a organogênese, agora indireta, de eixos embrionários in
vitro com feijão-caupi cv. IPA 206. Apesar desse incremento, não foram observadas
diferenças significativas ao nível estudado (1%) entre as médias dos diferentes tratamentos.
De modo similar, Brasileiro et al. (1999) não constataram diferenças significativas, no que
se refere à formação de calos (organogênese indireta), para os genótipos de tomate e doses
estudadas - 200 Gy, para sementes e 20 Gy, para botões florais. Isto é indicativo de que os
efeitos da radiação gama podem variar de acordo com o tecido, espécie vegetal e a dose
aplicada, o que reforça a realização de teste de radiossensitividade para se determinar a
melhor dose de radiação gama a ser utilizada na indução de mutação in vitro.
Considerando o fato de que a organogênese direta para eixos embrionários somente foi
observada com doses abaixo de 300 Gy e também o fato de que a organogênese direta de
cotilédones de feijão-caupi cv. IPA 206 apresentou uma redução nos níveis de regeneração
com o aumento da dose de radiação gama, em relação ao tratamento controle (89%), a dose
escolhida neste trabalho para induzir mutações em cotilédones e eixos embrionários em
feijão-caupi cv. IPA 206 deveria ficar entre 100 – 200 Gy, sendo escolhida uma dose de

62
150 Gy. Essa dose mostrou ser eficiente na geração de variabilidade, pois permitiu o
aparecimento e identificação de variantes tolerantes ao NaCl (12 plantas selecionadas).
As doses de radiação gama entre 50 - 600 Gy promoveram variabilidade em feijão-caupi
cv. IPA 206, visto que possibilitaram que indivíduos com melhor tolerância ao NaCl
fossem identificados. Aparentemente, a irradiação de sementes de feijão-caupi cv. IPA 206
proporcionou variabilidade em vários aspectos agronômicos, como precocidade, peso
estimado de 100 sementes, comprimento da vagem e número de sementes por vagem,
caracteres observados em condições de telado e sob estresse (60 mM de NaCl), além do
aparecimento de novos padrões de tamanho e cor de sementes (padrão variegado). Adu-
dapaah e Sangwan (2004) irradiaram sementes de Vigna subterranea (L.) Verdc. (150 Gy)
e obtiveram estimativas de variância genética até quatro vezes maior para peso de 100
sementes do grupo irradiado em relação ao grupo controle.
Neste estudo com a cv. IPA 206, plantas oriundas de sementes irradiadas com a dose de
500 Gy não produziram vagens e, portanto, para essa dose não havia sementes S1
disponíveis para a seleção in vivo. A dose de 500 Gy também apresentou efeito inibitório
na germinação e crescimento da planta V. radiata em estudo feito por Sangsiri et al.
(2005).
Apesar de ser amplamente reportada a inibição do desenvolvimento in vivo de feijão-caupi
pelo NaCl(Franco et al. 1999; Otoch et al. 2001; Costa et al. 2003; Silva et al. 2003; Sousa
et al. 2003), o uso da radiação gama como fonte de mutação em sementes de feijão-caupi
cv. IPA 206 resultou em quatro plantas consideradas tolerantes e 16 semi-tolerantes, essas
últimas com resultados superiores ou iguais ao controle em condições salinas. Quatro
plantas tolerantes oriundas de 162 sementes S1 provenientes de sementes irradiadas
(diferentes doses) representam uma freqüência de aproximadamente 25 variantes tolerantes
/ 1.000 sementes irradiadas, uma freqüência bem maior do que aquela in vitro (4 em

63
1.000). Mas esses variantes tolerantes, apesar de não mostrarem sinal aparente de
toxicidade pelo NaCl, apresentaram retardo no desenvolvimento quando comparados com
controles em condições não salinas principalmente, o que é de certo modo esperado, pois
estudos mostram que algumas plantas tolerantes quando expostas a condições salinas,
desaceleram seu ciclo reprodutivo devido ao redirecionamento de sua maquinaria
molecular e fisiológica para suprimir a toxicidade ao NaCl, em grande parte das vezes pela
alocação de osmólitos para recuperar a homeostase osmótica, ou pela ativação da rota SOS
para restabelecer o balanço iônico (Zhu 2001). Essas plantas, apesar do atraso, em média,
no florescimento e formação de vagens [de 22 dias quando comparado com o controle
salino (CCS; 79 dias) e de 34 dias com o controle não salino (CSS; 67 dias)], podem ser
uma opção caso se confirmem produções em solos salinos.
Apesar das abordagens in vitro e in vivo produzirem indivíduos tolerantes em feijão-caupi
cv. IPA 206, a seleção in vivo apresentou melhor eficiência, talvez devido ao
comportamento recalcitrante da espécie (Anand et al. 2001). Os procedimentos para a
seleção in vitro requerem um maior cuidado e manipulação mais minuciosa do material
biológico, além do custo com reagentes. A seleção in vivo é um procedimento
relativamente simples e não precisa de repiques constantes (transplantes) como no cultivo
in vitro. Mesmo assim, a seleção de variantes in vitro ainda tem um papel importante no
melhoramento por indução de mutação, pois é crucial na seleção de outros tipos de
explantes irradiados, como células em suspensão celular, tecidos vegetais (folhas, ápices
caulinares, calos) e órgãos (anteras) (Murakishi e Carlson 1982; Gangopadhyay et al.
1997; Houllou-Kido 2003; Hoffmann et al. 2004).
Os resultados indicaram que a indução de mutação via radiação gama seguida por seleção
in vitro ou in vivo provou ser uma metodologia eficiente na identificação de variantes para
tolerância ao NaCl em feijão-caupi V. unguiculata cv. IPA 206. A similaridade entre a

64
pressão seletiva in vitro e o estresse in vivo são os principais fatores para a identificação
exitosa de prováveis tolerantes. Entretanto, a avaliação em campo será necessária para
comprovar a viabilidade dos possíveis variantes e a estabilidade genética da característica
de interesse.

65
REFERÊNCIAS
Adu-Dapaah HK and Sangwan RS (2004) Improving bambara groundnut productivity
using gamma irradiation and in vitro techniques. African Journal of Biotechnology,
3(5):260–265.
Alves GD (2000) Indução de mutação em gemas micropropagadas de bananeira nanincão
Musa spp., grupo AAA e seleção de mutantes tolerantes a salinidade. UFRPE, Recife-PE.
Anand RP, Ganapathi A, Vengadesan G, Selvaraj N, Anbazhagan VR and Kulothungan S
(2001) Plant regeneration from immature cotyledon-derived callus of Vigna unguiculata
(L.) Walp (cowpea). Current Science, 80(5):671–674.
Brasileiro ACR, Willadino L and Guerra M (1999) Efeitos do estádio de desenvolvimento
da antera e da radiação gama na formação de calos derivados de anteras de tomate. Sci.
Agric, 56(4):835–842.
Charbaji T and Nabulsi I (1999) Effect of low doses of gamma radiation on in vitro growth
of grapevine. Plant Cell, Tissue and Organ Culture, 57(2):129–132.
Costa PHA, Silva JV, Bezerra MA, Filho JE, Prisco JT and Filho EG (2003) Crescimento e
níveis de solutos orgânicos e inorgânicos em cultivares de Vigna unguiculata submetidos à
salinidade. Revista Bresil. Bot., 26(3):289–297.

66
Ehlers JD and Hall AE (1997) Cowpea [Vigna unguiculata (L.) Walp.]. Field Crops Res
53:187–204.
Franco OL, Filho JE, Prisco JT and Filho EG (1999) Effects of CaCl2 on growth and
osmoregulator accumulation in NaCl stressed cowpea seedlings. Revista Brasil. de Fisiol.
Veget. 11(3):145–151.
Gangopadhyay G, Basu S and Gupta S (1997) In vitro selection and physiological
characterization of NaCl- and mannitol-adapted callus lines in Brassica juncea. Plant Cell,
Tissue and Organ Culture, 50(3):161–169.
Hoffmann F, Thomas E and Wenzel G (2004) Anther culture as a breeding tool in rape.
TAG Theorical and Applied Genetics, 61(3):225–232.
Houllou-Kido LM (2003) Irradiação de gemas axilares, suspensão celular e protoplastos de
bananeira cv. Maçã (Musa spp.), visando à seleção de mutantes resistentes a salinidade.
CENA/USP, São Paulo.
Kumar V and Sharma DR (1989) Isolation and characterization of sodium chloride-
resistant callus culture of Vigna radiata (L.) Wilczek var. radiate. Journal of Experimental
Botany, 40(1): 143–147.
Li HZ, Zhou WJ, Zhang ZJ, Gu HH, Takeuchi Y and Yoneyama K (2005) Effect of γ-
radiation on development, yield and quality of microtubers in vitro in Solanum tuberosum
L. Biologia Plantarum, 49(4):625–628.

67
Murakishi HH and Carlson OS (1982) In vitro selection of Nicotiana sylvestris variants
with limited resistance to TMV. Plant Cell Reports, 1(3):94–97.
Murashigue T and Skoog F (1962) A revised medium for rapid growth and biossays with
tobacco tissue cultures. Physiologia Plantarum, Kobenhawn, v. 15, p.473-497.
Oliveira AP, Sobrinho JT, Nascimento JT, Alves AU, Albuquerque IC and Bruno GB
(2002) Avaliação de linhagens e cultivares de feijão-caupi, em Areia, PB. Horticultura
Brasileira, 20(2):180–182.
Otoch MLO, Sobreira ACM, Aragão MEF, Orellano WG, Lima MGS and Melo DF (2001)
Salt modulation of vauolar H+-ATPase and H+-Pyrophosphatase activities in Vigna
unguiculata. J. Plant Physiol., 158:545–551.
Ouma JP, Young MM, Reichert NA (2004) Optimization of in vitro regeneration of
multiple shoots from hypocotyl sections of cotton (Gossypium hirsutum L.). African
Journal of Biotechnology, 3(3):169-173.
Prinyawiwatkul, W, McWatters K H, Beuchat LR and Phillips RD (1996) Cowpea flour: a
potential ingredient in food products. Crit. Rev. Food Sci. Nutr. 36(5):413-436
Sangsiri C, Sorajjapinum W and Srinives P (2005) Gamma radiation induced mutations in
mungbean. Science Asia, 31:251–255.
Segreto RA and Segreto HR (1997) Radiação ionizante e resposta celular. Radiol. Bras.
30(6):301–307.

68
Silva FAS (2007) ASSISTAT Versão 7.4 beta. Capturado em 20. mar. 2007. Online.
Disponível na internet http://www.assistatsites.uol.com.br.
Silva JV, Lacerda CF, Costa PHA, Filho JE, Filho EG and Prisco JT (2003) Physiological
responses of NaCl stressed cowpea plants grown in nutrient solution supplemented with
CaCl2. Braz. J. Plant Physiol., 15(2):99–105.
Sousa MF, Campos FAP, Prisco JT, Filho JE and Filho EG (2003) Growth and protein
pattern in cowpea seedlings subjected to salinity. Biol. Plantarum, 47(3):341–346.
Teixeira SM, May PH and Santana AC de. ( 1988) Produção e importância econômica do
caupi no Brasil. In: Araujo, JPP de and Watt EE (eds). O caupi no Brasil. Brasília :
IITA/Embrapa, pp 99–136.
Ulissues C, Camara TR, Willadino L, Meunier I, Rocha PSG and Albuquerque C (2000)
Seleção in vitro de gemas de bananeira “nanicão” tolerantes à salinidade. Scientia
Agricola, 57(4):667–670.
Zair I, Chlyah I, Sabounji K, Tittahsen M and Chlyah H (2003) Salt tolerance
improvement in some wheat cultivars after application of in vitro selection pressure. Plant
Cell, Tissue and Organ Culture, 73:237–244.
Zhu JK (2001) Plant salt tolerance. Trends in Plant Science, 26(2):66–71.

69
FIGURAS
Figura 1. Cultivo in vitro de V. unguiculata cv. IPA 206. a) explantes após inoculação; b) intumescimento em eixos embrionários; c)
aspectos da organogênese direta em cotilédones; d) cotilédones intumescidos e verdes; e) teste de salinidade (seta 1: necrose em
cotilédone; seta 2: organogênese direta em eixo embrionário); f) teste de salinidade (seta 1: cotilédones com inibição de
desenvolvimento; seta 2: organogênese indireta em eixo embrionário; g) matéria mais sensível (esquerda) e menos sensível (direita) sob
pressão seletiva; h) aspecto de material selecionado.

70
Figura 2. Efeito da concentração de NaCl na freqüência de eventos do processo de
regeneração in vitro de eixos embrionários de feijão-caupi cv. IPA206 (mudança de cor,
início de regeneração, inibição do desenvolvimento e necrose).
Figura 3 Efeito da concentração de NaCl na freqüência de organogênese direta e indireta
de eixos embrionários de feijão-caupi cv. IPA 206.

71
Figura 4. Efeito de diferentes doses de radiação gama na freqüência de organogênese
direta de cotilédones e eixos embrionários de feijão-caupi cv. IPA 206. Tratamentos com a
mesma letra não diferem entre si (Teste de Tukey à 1%).
0
10
20
30
40
50
60
Controle 50Gy 100Gy 200Gy 300Gy 400Gy 500Gy 600Gy
Dose (Gy)
Org
anog
ênes
e (%
)
Figura 5. Efeito de diferentes doses de radiação gama na freqüência da organogênese
indireta em eixos embrionários de feijão-caupi cv. IPA 206.
a a
ab abc
a
abc
c bc
ab
ab
a
ab ab
c c c

72
0
10
20
30
40
50
60
0 50 100 200 300 400 500 600
Dose (Gy)
Per
cent
ual
Figura 6. Efeito de diferentes doses de radiação gama na freqüência da organogênese total
(direta e indireta) de eixos embrionários de feijão-caupi cv. IPA 206. Tratamentos com a
mesma letra não diferem entre sim (Teste de Tukey à 1%).
ab
ab
ab ab
b b
ab
a

73
Figure 7. Seleção in vivo de feijão-caupi [Vigna unguiculata (L.) Walp.] cv. IPA 206. a)
Plantas sob pressão salina (esquerda para a direita: planta controle não irradiada, planta
irradiada semi-tolerante, planta irradiada tolerante). b) Aspecto do grid de madeira do
ensaio in vivo. c) Variação no comprimento de vagens. d) Diversidade de sementes (seta
mostra semente controle; demais: pequenas/leves e novos padrões de cores/ maiores).

74
TABELAS
Tabela 1. Valores médios das características agronômicas (tamanho da vagem, número de sementes por vagem, peso de 100 sementes, dias até colheita) de feijão-caupi cv. IPA 206 oriundas de sementes irradiadas (SI) após seleção (PS) in vivo para tolerância ao NaCl (60 mM).
TRATAMENTOS PS/SI VAGEM* (cm)
SEMENTES* (POR VAGEM)
PESO DE 100
SEMENTES* (g)
COLHEITA*(dias)
CSS 09/10 20.6 13 18.3 67 CCS 04/10 14.2 06 15.5 79 50 Gy 16/116 16.2 07 14.5 78 100 Gy 18/61 16.4 07 15.0 78 200 Gy 15/17 14.3 07 15.2 79. 300 Gy 10/39 14.6 08 14.1 78 400 Gy 16/23 16.3 08 16.3 77 600 Gy 06/10 16.1 10 16.4 77
*Resultados médios. CSS: controle sem sal; CCS: Controle com sal.
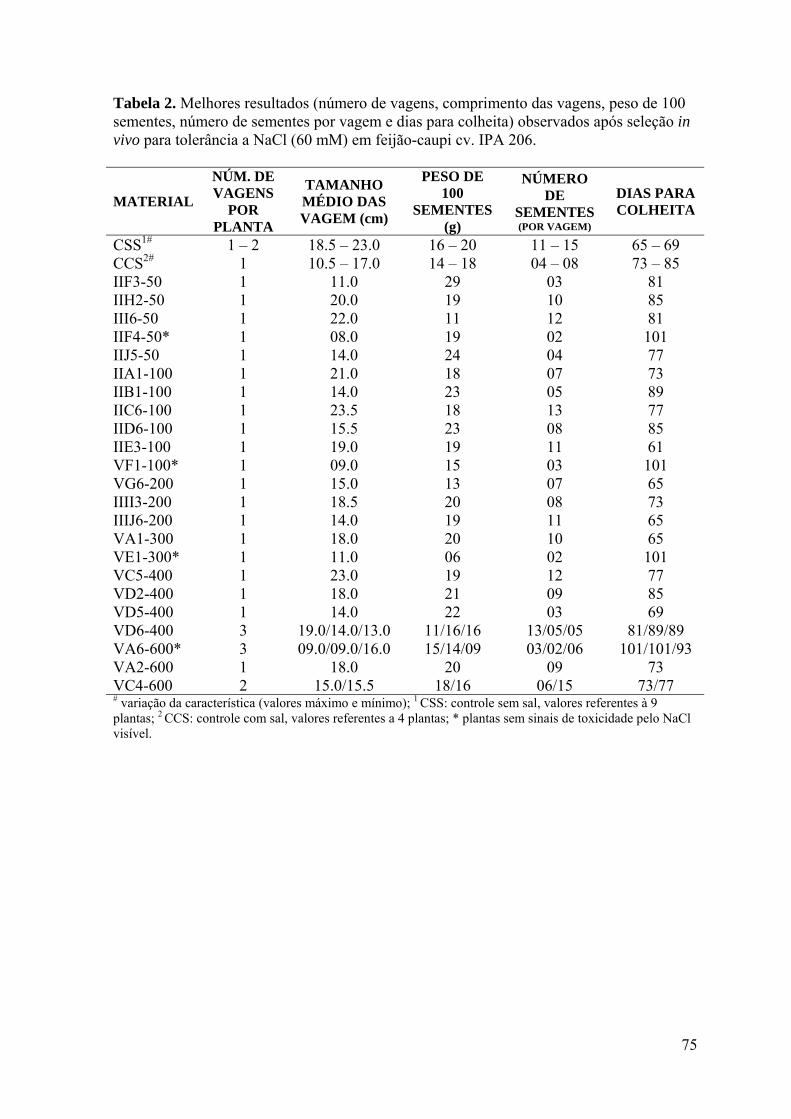
75
Tabela 2. Melhores resultados (número de vagens, comprimento das vagens, peso de 100 sementes, número de sementes por vagem e dias para colheita) observados após seleção in vivo para tolerância a NaCl (60 mM) em feijão-caupi cv. IPA 206.
MATERIAL
NÚM. DE VAGENS
POR PLANTA
TAMANHO MÉDIO DAS VAGEM (cm)
PESO DE 100
SEMENTES (g)
NÚMERO DE
SEMENTES (POR VAGEM)
DIAS PARA COLHEITA
CSS1# 1 – 2 18.5 – 23.0 16 – 20 11 – 15 65 – 69 CCS2# 1 10.5 – 17.0 14 – 18 04 – 08 73 – 85 IIF3-50 1 11.0 29 03 81 IIH2-50 1 20.0 19 10 85 III6-50 1 22.0 11 12 81 IIF4-50* 1 08.0 19 02 101 IIJ5-50 1 14.0 24 04 77 IIA1-100 1 21.0 18 07 73 IIB1-100 1 14.0 23 05 89 IIC6-100 1 23.5 18 13 77 IID6-100 1 15.5 23 08 85 IIE3-100 1 19.0 19 11 61 VF1-100* 1 09.0 15 03 101 VG6-200 1 15.0 13 07 65 IIII3-200 1 18.5 20 08 73 IIIJ6-200 1 14.0 19 11 65 VA1-300 1 18.0 20 10 65 VE1-300* 1 11.0 06 02 101 VC5-400 1 23.0 19 12 77 VD2-400 1 18.0 21 09 85 VD5-400 1 14.0 22 03 69 VD6-400 3 19.0/14.0/13.0 11/16/16 13/05/05 81/89/89 VA6-600* 3 09.0/09.0/16.0 15/14/09 03/02/06 101/101/93 VA2-600 1 18.0 20 09 73 VC4-600 2 15.0/15.5 18/16 06/15 73/77 # variação da característica (valores máximo e mínimo); 1 CSS: controle sem sal, valores referentes à 9 plantas; 2 CCS: controle com sal, valores referentes a 4 plantas; * plantas sem sinais de toxicidade pelo NaCl visível.

76
CONCLUSÕES
• A técnica de indução de mutação via radiação gama seguida da seleção in vivo ou
in vitro para a identificação de materiais com tolerância ao NaCl em feijão-caupi
[Vigna unguiculata (L.) Walp] cv. IPA 206 mostrou ser estratégia promissora para
obtenção dos variantes desejáveis.
• A escolha do explante na seleção in vitro de feijão-caupi V. unguiculata cv. IPA
206 é importante para o sucesso da técnica.
• A dose de 258 mM de NaCl demonstrou ser apropriada para uso em seleção in
vitro de feijão-caupi cv. IPA 206, correspondendo à DL100 observada para a
cultivar.
• Baixas doses de radiação gama (até 300 Gy) foram mais eficazes no aparecimento
de variantes da cv. IPA 206 oriundos de sementes irradiadas.

77
8. ANEXOS

78
Anexo 1. Dados complementares in vitro

79
Os resultados obtidos durante o teste in vitro de sensibilidade ao NaCl em eixos
embrionário de feijão-caupi cv. IPA206 são mostrados na Tabela 1. Foram avaliadas a
freqüência com que eixos embrionários cultivados em diferentes concentrações de NaCl se
desenvolveram, bem como, a presença ou ausência de organogêneses direta e/ou indireta.
Tabela 1. Resultado do teste in vitro de sensibilidade ao NaCl em eixos embrionários de feijão-caupi Vigna unguiculata (L.) Walp cv. IPA206.
Tratamento Estágio de desenvolvimento
Organogênese indireta
Organogênese Direta
Frasco 01 IMc - -Frasco 02 IMc - -Frasco 03 InR - +Frasco 04 NoD - -Frasco 05 IMc - -Frasco 06 IMc - -Frasco 07 IMc + -Frasco 08 InR - -Frasco 09 InR - -
Con
trole
Frasco 10 InR - -Frasco 01 IMc + -Frasco 02 IMc + -Frasco 03 IMc + -Frasco 04 IMc - -Frasco 05 IMc + -Frasco 06 IMc + -Frasco 07 IMc + -Frasco 08 InR - -Frasco 09 InR - +
0
,5%
NaC
l
Frasco 10 IMc + -Frasco 01 InR - -Frasco 02 InR - -Frasco 03 IMc + -Frasco 04 IMc - -Frasco 05 Nec - -Frasco 06 Nec - -Frasco 07 IMc - +Frasco 08 IMc + -Frasco 09 InR - -
1,0%
NaC
l
Frasco 10 IMc - -Frasco 01 Nec - -Frasco 02 Nec - -Frasco 03 Nec - -Frasco 04 NoD - -Frasco 05 NoD - -Frasco 06 Nec - -Frasco 07 Nec - -Frasco 08 Nec - -Frasco 09 NoD - -
1,5%
NaC
l
Frasco 10 NoD - -Nec = necrose; NoD = nenhum desenvolvimento; IMc = intumescimento e mudança de cor; InR = início de regeneração.

80
Dados para a análise do efeito da radiação gama na regeneração de feijão-caupi V.
unguiculata cv. IPA206. Ocorrências de organogênese (direta e indireta) em cotilédones
verificadas nos diferentes tratamentos são mostradas na Tabela 2.
Tabela 2. Efeito de diferentes doses de radiação gama na organogêse in vitro de cotilédones de feijão-caupi Vigna unguiculata (L.) Walp cv. IPA206.
Tratamento Organogênese indireta
Organogênese Direta
Controle 01 - + Controle 01 - + Controle 02 - + Controle 02 - + Controle 03 - + Controle 03 - + Controle 04 - + Controle 04 - + Controle 05 - + Controle 05 - - Controle 06 - + Controle 06 - + Controle 07 - + Controle 07 - - Controle 08 - + Controle 08 - + Controle 09 - + Controle 09 - + Controle 10 X X Controle 10 X X 50 Gy 01 - - 50 Gy 01 - - 50 Gy 02 - + 50 Gy 02 - + 50 Gy 03 - + 50 Gy 03 - + 50 Gy 04 - + 50 Gy 04 - + 50 Gy 05 - + 50 Gy 05 - + 50 Gy 06 - + 50 Gy 06 - + 50 Gy 07 - + 50 Gy 07 - - 50 Gy 08 - + 50 Gy 08 - - 50 Gy 09 - + 50 Gy 09 - -

81
Cont.
Tratamento Organogênese indireta
Organogênese Direta
50 Gy 10 X X 50 Gy 10 X X 100 Gy 01 - - 100 Gy 01 - - 100 Gy 02 - - 100 Gy 02 - + 100 Gy 03 - + 100 Gy 03 - + 100 Gy 04 - + 100 Gy 04 - + 100 Gy 05 - + 100 Gy 05 - - 100 Gy 06 X X 100 Gy 06 X X 100 Gy 07 - + 100 Gy 07 - + 100 Gy 08 - + 100 Gy 08 - - 100 Gy 09 - + 100 Gy 09 - - 100 Gy 10 X X 100 Gy 10 X X 200 Gy 01 - - 200 Gy 01 - - 200 Gy 02 - + 200 Gy 02 - - 200 Gy 03 - + 200 Gy 03 - + 200 Gy 04 - + 200 Gy 04 - + 200 Gy 05 X X 200 Gy 05 X X 200 Gy 06 - + 200 Gy 06 - - 200 Gy 07 - + 200 Gy 07 - - 200 Gy 08 - + 200 Gy 08 - - 200 Gy 09 - - 200 Gy 09 - - 200 Gy 10 - - 200 Gy 10 - - 300 Gy 01 - + 300 Gy 01 - - 300 Gy 02 - - 300 Gy 02 - +

82
Cont.
Tratamento Organogênese indireta
Organogênese Direta
300 Gy 03 - - 300 Gy 03 - + 300 Gy 04 - + 300 Gy 04 - + 300 Gy 05 - - 300 Gy 05 - + 300 Gy 06 - + 300 Gy 06 - - 300 Gy 07 - + 300 Gy 07 - + 300 Gy 08 - + 300 Gy 08 - + 300 Gy 09 - + 300 Gy 09 - + 300 Gy 10 - - 300 Gy 10 - + 400 Gy 01 - + 400 Gy 01 - - 400 Gy 02 - - 400 Gy 02 - - 400 Gy 03 - + 400 Gy 03 - - 400 Gy 04 - + 400 Gy 04 - + 400 Gy 05 - + 400 Gy 05 - - 400 Gy 06 - + 400 Gy 06 - - 400 Gy 07 - + 400 Gy 07 - - 400 Gy 08 - + 400 Gy 08 - - 400 Gy 09 - + 400 Gy 09 - - 400 Gy 10 - - 400 Gy 10 - - 500 Gy 01 - + 500 Gy 01 - + 500 Gy 02 - - 500 Gy 02 - - 500 Gy 03 - - 500 Gy 03 - - 500 Gy 04 - - 500 Gy 04 - - 500 Gy 05 - - 500 Gy 05 - -

83
Cont.
Tratamento Organogênese indireta
Organogênese Direta
500 Gy 06 - - 500 Gy 06 - - 500 Gy 07 - - 500 Gy 07 - - 500 Gy 08 - - 500 Gy 08 - - 500 Gy 09 - - 500 Gy 09 - - 500 Gy 10 - + 500 Gy 10 - - 600 Gy 01 - - 600 Gy 01 - - 600 Gy 02 - + 600 Gy 02 - - 600 Gy 03 - - 600 Gy 03 - - 600 Gy 04 - - 600 Gy 04 - - 600 Gy 05 - - 600 Gy 05 - - 600 Gy 06 - - 600 Gy 06 - - 600 Gy 07 - + 600 Gy 07 - - 600 Gy 08 - + 600 Gy 08 - - 600 Gy 09 - - 600 Gy 09 - - 600 Gy 10 - - 600 Gy 10 - +
O par de números iguais representa o par de cotilédones inoculados para cada semente. Os sinais “+ e -” indicam, respectivamente, presença ou ausência de organogênese. X = contaminação bacteriana e/ou fúngica.
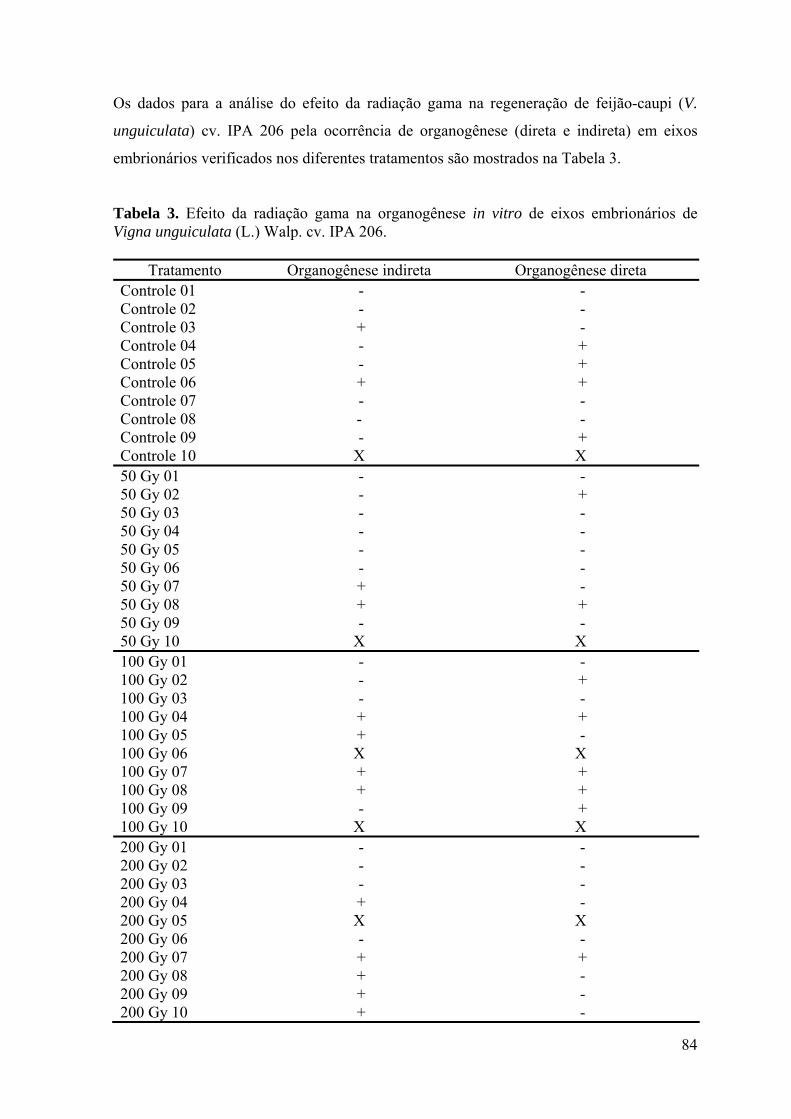
84
Os dados para a análise do efeito da radiação gama na regeneração de feijão-caupi (V.
unguiculata) cv. IPA 206 pela ocorrência de organogênese (direta e indireta) em eixos
embrionários verificados nos diferentes tratamentos são mostrados na Tabela 3.
Tabela 3. Efeito da radiação gama na organogênese in vitro de eixos embrionários de Vigna unguiculata (L.) Walp. cv. IPA 206.
Tratamento Organogênese indireta Organogênese direta Controle 01 - - Controle 02 - - Controle 03 + - Controle 04 - + Controle 05 - + Controle 06 + + Controle 07 - - Controle 08 - - Controle 09 - + Controle 10 X X 50 Gy 01 - - 50 Gy 02 - + 50 Gy 03 - - 50 Gy 04 - - 50 Gy 05 - - 50 Gy 06 - - 50 Gy 07 + - 50 Gy 08 + + 50 Gy 09 - - 50 Gy 10 X X 100 Gy 01 - - 100 Gy 02 - + 100 Gy 03 - - 100 Gy 04 + + 100 Gy 05 + - 100 Gy 06 X X 100 Gy 07 + + 100 Gy 08 + + 100 Gy 09 - + 100 Gy 10 X X 200 Gy 01 - - 200 Gy 02 - - 200 Gy 03 - - 200 Gy 04 + - 200 Gy 05 X X 200 Gy 06 - - 200 Gy 07 + + 200 Gy 08 + - 200 Gy 09 + - 200 Gy 10 + -

85
Cont.Tratamento Organogênese indireta Organogênese direta
300 Gy 02 + - 300 Gy 03 + - 300 Gy 04 - + 300 Gy 05 - + 300 Gy 06 - - 300 Gy 07 + - 300 Gy 08 - + 300 Gy 09 - - 300 Gy 10 + - 400 Gy 01 X X 400 Gy 02 - - 400 Gy 03 - - 400 Gy 04 - - 400 Gy 05 + - 400 Gy 06 + - 400 Gy 07 X X 400 Gy 08 + - 400 Gy 09 + - 400 Gy 10 + - 500 Gy 01 - - 500 Gy 02 - - 500 Gy 03 - - 500 Gy 04 + - 500 Gy 05 - - 500 Gy 06 - - 500 Gy 07 - - 500 Gy 08 - - 500 Gy 09 + - 500 Gy 10 - - 600 Gy 01 - - 600 Gy 02 - - 600 Gy 03 - - 600 Gy 04 - - 600 Gy 05 - - 600 Gy 06 + - 600 Gy 07 - - 600 Gy 08 + - 600 Gy 09 - - 600 Gy 10 - -
O par de números iguais representa o par de cotilédones inoculados para cada semente. Os sinais “+ e -” indicam, respectivamente, presença ou ausência de organogênese. X = contaminação bacteriana e/ou fúngica.

86
Anexo 2. Dados complementares in vivo

87
a) Desempenho agronômico (dados de precocidade, peso estimado de 100 sementes,
tamanho da vagem e número de sementes por vagem) das plantas durante seleção in vivo
(Tabela 1).
Tabela 1. Dados coletados das plantas durante seleção in vivo.
Variante
Número médio
de vagens
Dose
Comprimento max. e min. das vagens
(cm)
Sementes por
vagem
Peso de 100 sementes
(g)
Colheita (dias)
CSS-1 2 0 Gy 18,5/18,0 11/10 16/17 69/69CSS-2 2 0 Gy 21,5/22,0 13/15 19/19 65/65CSS-3 1 0 Gy 22,5 15 20 69CSS-4 2 0 Gy 19,0/17,0 12/9 18/19 65/69CSS-5 1 0 Gy 21,0 13 18 65CSS-6 1 0 Gy 23,0 15 20 65CSS-7 1 0 Gy 19,0 13 18 65CSS-8 2 0 Gy 19,5/19,0 12/12 17/19 65/65CSS-9 2 0 Gy 22,0 14 19 65CCS-1 1 0 Gy 13,0 04 18 73CCS-2 1 0 Gy 17,0 07 14 77CCS-3 1 0 Gy 17,0 08 16 85CCS-4 1 0 Gy 10,5 07 14 73IB1-50 1 50 Gy 19,0 09 16 85IE1-50 1 50 Gy 11,5 02 10 77IIJ1-50 2 50 Gy 18,0/17,0 09/07 12/09 77/81IB2-50 1 50 Gy 11,0 06 12 81IIE2-50 1 50 Gy 18,0 11 12 81IIF2-50 1 50 Gy 16,0 07 08 69IIH2-50 1 50 Gy 20,0 10 19 85ID2-50 2 50 Gy 16,0/15,5 07/07 12/12 69/73IB3-50 1 50 Gy 16,0 08 19 69IIF3-50 1 50 Gy 11,0 03 29 81IIJ3-50 1 50 Gy 17,0 09 13 85IIF4-50 1 50 Gy 08,0 02 19 101IIJ5-50 1 50 Gy 14,0 04 24 77III6-50 1 50 Gy 22,0 12 11 81IC2-100 2 100 Gy 10,0/16,0 04/06 18/15 69/93IC4-100 1 100 Gy 10,0 03 08 73 IC5-100 1 100 Gy 18,5 08 12 77IIA1-100 1 100 Gy 21,0 07 18 73IIA2-100 1 100 Gy 12,5 05 14 89IIA3-100 1 100 Gy 14,0 07 06 77IIB1-100 1 100 Gy 14,0 05 23 89IIC1-100 1 100 Gy 17,0 08 16 81

88
Cont.
Variante
Número médio de vagens Dose
Comprimento max. e min. da vagem
(cm)
Sementes por
vagem
Peso de 100 sementes
(g)
Colheita (dias)
IIC6-100 1 100 Gy 23,5 13 18 77IID1-100 1 100 Gy 18,0 07 14 73IID2-100 1 100 Gy 18,0 11 14 85IID6-100 1 100 Gy 15,5 08 23 85IIE1-100 1 100 Gy 13,0 05 08 69IIE3-100 1 100 Gy 19,0 11 19 61IIII4-100 1 100 Gy 15,0 07 19 89IIII5-100 1 100 Gy 15,0 08 18 69VF1-100 1 100 Gy 9,0 03 15 101IIII6-100 1 100 Gy 15,0 07 08 69IIA5-200 1 200 Gy 13,0 08 19 81IIA6-200 2 200 Gy 14,5/08,0 12/03 15/12 77/81IIII3-200 1 200 Gy 18,5 08 20 73IIIJ2-200 1 200 Gy 12,2 08 16 77IIIJ3-200 1 200 Gy 13,0 04 16 77IIIJ4-200 1 200 Gy 15,0 05 18 89IIIJ6-200 1 200 Gy 14,0 11 19 65VG3-200 1 200 Gy 15,0 06 12 77VG4-200 1 200 Gy 15,0 09 13 77VG5-200 1 200 Gy 12,0 06 07 81VG6-200 1 200 Gy 15,0 07 13 65VH2-200 1 200 Gy 14,0 06 17 69VH5-200 1 200 Gy 16,0 07 14 73VH6-200 1 200 Gy 19,0 07 17 77IF2-300 1 300 Gy 12,0 07 08 69VA1-300 1 300 Gy 18,0 10 20 65VE1-300 1 300 Gy 11,0 02 06 101VE5-300 1 300 Gy 16,0 08 17 93VF1-300 1 300 Gy 11,0 02 06 101VF2-300 1 300 Gy 15,0 07 14 77VF3-300 1 300 Gy 13,0 05 15 85VF4-300 1 300 Gy 15,0 07 13 85VF6-300 2 300 Gy 10,0/18,5 03/09 13/13 81/77VB1-400 1 400 Gy 16,0 10 16 81VB3-400 1 400 Gy 19,0 11 15 69VB6-400 2 400 Gy 16,0/13,0 09/05 16/16 69/73VC5-400 1 400 Gy 23,0 12 19 77VC6-400 1 400 Gy 18,0 10 17 73VD2-400 1 400 Gy 18,0 09 21 85VD3-400 1 400 Gy 17,0 10 18 73VD4-400 1 400 Gy 17,0 08 13 85VD5-400 1 400 Gy 14,0 03 22 69VD6-400 3 400 Gy 14,0/13,0/19,0 05/05/13 16/16/11 89/89/81VE2-400 2 400 Gy 14,0/14,0 07/05 15/13 81/77VE4-400 1 400 Gy 16,5 10 17 85

89
Cont.
Variante Número
de vagens
Dose Comprimento
da vagem (cm)
Sementes por
vagem
Peso de 100 sementes
(g)
Colheita (dias)
VA2-600 1 600 Gy 18,0 09 20 73VA6-600 3 600 Gy 09,0/16,0/09,0 03/06/02 15/09/14 101/93/101VC3-600 1 600 Gy 16,0 12 19 69VC4-600 2 600 Gy 15,0/15,5 06/15 18/16 73/77
CSS = controle sem sal; CCS = controle com sal.
b) Análise comparativa dos melhores variantes em relação às médias dos controles para
diferentes caracteres agronômicos
b.1) Precocidade
Quatro plantas apresentaram resultados interessantes para precocidade. Planta IIE3-100
produziu vagens maduras quatro dias antes do controle CSS (não salino) mais precoce e
doze dias antes do controle CCS (salino) mais precoce. IIIJ6-200, VG6-200 e VA1-300
mostraram mesmo tempo de coleta que CSS e precocidade de oito dias quando
comparados com CCS (Figura 1).
Figura 1. Plantas com melhores resultados para precocidade após seleção in vivo para tolerância a
NaCl em cv. IPA 206. CSS = controle sem sal; CCS = controle com sal.

90
b.2) Peso médio estimado para 100 sementes
b.2.1) Por dose de radiação
O peso médio de 100 sementes por tratamento (dose de radiação) não apresentou diferença
significativa quando comparado com controle salino ou não salino, porém nenhum
tratamento alcançou os valores obtidos pelo controle sem sal (Figura 2).
Figura 2. Efeito da radiação gama no peso de 100 sementes de Vigna unguiculata (L.)
Walp. cv. IPA 206 após seleção in vivo. CSS = controle sem sal; CCS = controle com sal.
b.2.2) Por variantes
Melhores variantes: IIF3-50 apresentou o melhor “peso de 100 sementes” (29g) durante a
seleção in vivo. As plantas IJ5-50 (24g), IIB1-100 (23g), IID6-100 (23g), VD5-400 (22g),
VD2-400 (21g),VA1-300 (20g), VA2-600 (20g) e IIII3-200 (20g) também mostraram
resultados acima da média CSS (Figura 3).

91
Figura 3. Plantas com melhores resultados para “peso de 100 sementes” após seleção in
vivo para tolerância a NaCl em cv. IPA 206. CSS = controle sem sal; CCS = controle com
sal.
b.3) Tamanho de vagem
b.3.1) Por dose de radiação
Controle sem sal: vagens até 4,2 cm maiores que o melhor resultado para outros
tratamentos (Figura 4). O caráter não apresentou variações significativas em Teste F.
Figura 4. Efeito da radiação gama no comprimento de vagens de Vigna unguiculata (L.)
Walp. cv. IPA 206 após seleção in vivo. Não foi encontrada diferença siginificativa (Tukey
5%). CSS = controle sem sal; CCS = controle com sal.

92
b.3.2) Por variante
Vagens com 20, 21, 22, e 23 cm foram encontradas nas plantas IH2-50, IA1-100, III6-50 e
VC5-400, respectivamente. Variante IC6-100: melhor resultado com 23.5 cm (Figura 5).
Figura 5. Plantas com melhores resultados para comprimento de vagem após seleção in
vivo para tolerância a NaCl em cv. IPA 206. CSS = controle sem sal; CCS = controle com
sal.
b.4) Número de sementes por vagem
b.4.1) Por dose de radiação
Tratamentos (exceção de 600 Gy) diferiram do controle CSS, porém não foi encontrada
diferença significativa entre os mesmos e o controle CCS (Figura 6).

93
Figura 6. Efeito da radiação gama no número de sementes por vagens de Vigna unguiculata (L.)
Walp. cv. IPA 206 após seleção in vivo. Tratamentos com a mesma letra não diferem entre si (teste
de Tukey 1%). CSS = controle sem sal; CCS = controle com sal.
b.4.2) Por variante
Três plantas se destacaram: IIC6-100 e VD6-400 com 13 sementes/vagem, e VC4-600 com
15 sementes/vagem (Figura 7).
Figura 7. Plantas com melhores resultados para número de sementes por vagem após seleção in
vivo para tolerância a NaCl em cv. IPA 206. CSS = controle sem sal; CCS = controle com sal.
a ab
b b b b b b

94
c) Teste F As tabelas de 1 a 7 representam os dados obtidos pelo Teste de F para diversas análises in
vitro e in vivo.
Tabela 1. Análise da variância para organogênese indireta de eixos embrionários de Vigna
unguiculata (L.) Walp. cultivados em diferentes doses de NaCl.
F.V. G.L. S.Q. Q.M. F
Tratamento 3 0.75429 0.25143 7.5652 **
Resíduo 36 1.19646 0.03324
Total 39 1.95075
** significativo ao nível de 1% de probabilidade (p < .01) Tabela 2. Análise da variância para organogênese direta de cotilédones de Vigna
unguiculata (L.) Walp. expostos a diferentes doses de radiação gama.
F.V. G.L. S.Q. Q.M. F
Tratamento 7 0.75429 0.25143 6.4026 **
Resíduo 142 1.19646 0.03324
Total 149 1.95075
** significativo ao nível de 1% de probabilidade (p < .01) Tabela 3. Análise da variância da freqüência média de organogênese direta de eixos
embrionários de Vigna unguiculata (L.) Walp. expostos a diferentes doses de radiação
gama.
F.V. G.L. S.Q. Q.M. F
Tratamento 7 0.85212 0.12173 3.5203 **
Resíduo 65 2.24770 0.03458
Total 72 3.09982
** significativo ao nível de 1% de probabilidade (p < .01)

95
Tabela 4. Análise da variância para organogênese total (direta e indireta) de eixos
embrionários de Vigna unguiculata (L.) Walp. expostos a diferentes doses de radiação
gama.
F.V. G.L. S.Q. Q.M. F
Tratamento 7 0.74133 0.10590 2.2004 *
Resíduo 138 6.92805 0.05020
Total 145 7.66939
* significativo ao nível de 5% de probabilidade (.01 =< p < .05) Tabela 5. Análise da variância para número de dias para a colheita (precocidade) de
vagens de plantas de Vigna unguiculata (L.) Walp. irradiadas em diferentes doses de
radiação gama.
F.V. G.L. S.Q. Q.M. F
Tratamento 7 1200.80000 171.54286 3.3546 **
Resíduo 82 4193.15556 51.13604
Total 89 5393.95556
** significativo ao nível de 1% de probabilidade (p < .01) Tabela 6. Análise da variância para tamanho de vagens de plantas de Vigna unguiculata
(L.) Walp. irradiadas em diferentes doses de radiação gama.
F.V. G.L. S.Q. Q.M. F
Tratamento 7 251.33339 35.90477 3.9346 **
Resíduo 65 593.15291 9.12543
Total 72 844.48630
* significativo ao nível de 1% de probabilidade (p < .01)

96
Tabela 7. Análise da variância para produção de sementes por vagens de plantas de
Vigna unguiculata (L.) Walp. irradiadas em diferentes doses de radiação gama.
F.V. G.L. S.Q. Q.M. F
Tratamento 7 282.38056 40.34008 4.9122 **
Resíduo 82 673.40833 8.21230
Total 89 955.78889
** significativo ao nível de 1% de probabilidade (p < .01)

97
Anexo 3. INSTRUÇÕES PARA AUTORES
Revista GENETICS AND MOLECULAR BIOLOGY
ISSN 1415-4757 Ribeirão Preto, Brasil

Gazzaneo LRS Indução de mutação e seleção em feijão-caupi [Vigna unguiculata (L.) Walp]...
98
SCOPE AND POLICY Genetics and Molecular Biology (formerly named Revista Brasileira de Genética/Brazilian Journal of Genetics - ISSN 0100-8455) is published quarterly by the Sociedade Brasileira de Genética (Brazilian Society of Genetics). The Journal considers contributions that present the results of original research in genetics, evolution and related scientific disciplines. Although Genetics and Molecular Biology is an official publication of the Brazilian Society of Genetics, contributors are not required to be members of the Society. It is a fundamental condition that submitted manuscripts have not been and will not be published elsewhere. With the acceptance of a manuscript for publication, the publishers acquire full and exclusive copyright for all languages and countries. Manuscripts considered in conformity with the scope of the journal as judged by the Editor in conjunction with the Editorial Board are reviewed by the Associate Editors and two or more external reviewers. Acceptance by the Editor is based on the quality of the work as substantial contribution to the field and on the overall presentation of the manuscript. SUBMISSION OF PAPERS 1. Manuscripts should be submitted to:
Angela M. Vianna-Morgante , Editor-in-Chief Genetics and Molecular Biology Rua Capitão Adelmio Norberto da Silva, 736 14025-670 Ribeirão Preto, SP – Brasil 2. A submission package sent to the Editorial Office must contain: a) A cover letter stating that all authors have approved the submission of the
manuscript. The letter must be signed by the Corresponding Author and must inform the e-mail addresses of all other authors so that they can be contacted by the Editorial Office for confirmation of the submission. b) An electronic copy of the text, tables and figures. Formats for text are Word or RTF, in Windows platform. Images in TIFF or JPEG formats should be sent in separate files (For Figures, see detailed instructions in 3.1.h). Mailed CD-ROMs must be labeled with the first author’s last name, platform and software (see detailed instructions below). This CD should also include files of Supplementary Material to be published online only, as well as of any unpublished or in press material cited in the text. Failure to adhere to these guidelines can delay the handling of your contribution and manuscripts may be returned before being reviewed. c) Manuscripts including photos or any other identifiable data of human subjects must be accompanied by a copy of the signed consent by the individual or his/her guardian. 3. Categories of Contribution 3.1 Research Articles Manuscripts must be written in English in double-spaced, 12-point type throughout, including the References Cited section, appendices, tables and legends; formatted to A4 paper with 2.5 cm margins; marked with consecutive page numbers, beginning with the cover page. The following elements must start on a new page and be ordered as they are listed below: a) The title page must contain: a concise and informative title; the authors’ names (first name at full length); the authors’ institutional affiliation, including department, institution, city, state or
Genetics and Molecular Biology - NOTICE TO CONTRIBUTORS

Gazzaneo LRS Indução de mutação e seleção em feijão-caupi [Vigna unguiculata (L.) Walp]...
99
province and country; different affiliations indicated with superscript numbers; a short running title of about 35 characters, including spaces; up to five key words; the corresponding author’s name, postal address, phone and fax numbers and email address. The corresponding author is the person responsible for checking the page proofs, arranging for the payment of color illustrations and author’s alteration charges. b) The Abstract must be a single paragraph that does not exceed 200 words and summarizes the main results and conclusions of the study. It should not contain references. c) The text must be as succinct as possible. Text citations: articles should be referred to by authors’ surnames and date of publication; citations with two authors must include both names; in citations with three or more authors, name the first author and use et al. List two or more references in the same citation in chronological order, separated by semi-colons. When two or more works in a citation were published in the same year, list them alphabetically by the first author surname. For two or more works by the same author(s) in a citation, list them chronologically, with the years separated by commas. (Example: Freire-Maia et al., 1966a, 1966b, 2000). Only articles that are published or in press should be cited. In the case of personal communications or unpublished results, all contributors must be listed by initials and last name (et al. should not be used). Numbers: In the text, numbers nine or less must be written out except as part of a date, a fraction or decimal, a percentage, or a unit of measurement. Use Arabic numerals for numbers larger than nine. Avoid starting a sentence with a number. Binomial Names: Latin names of genera, species and intraspecific taxa in the text must be printed in italics; names of orders
and families should appear in the Title and also when first mentioned in the text. URLs for programs, data or other sources should be listed in the Internet Resources Section, immediately after the References Section, not in the text. URLs for citations of publications in electronic journals should appear in the reference section. The text includes the following elements: Introduction – Description of the background that led to the study. Material (or Subjects) and Methods –Details relevant to the conduct of the study. Statistical methods should be explained at the end of this section. Results –Undue repetition in text and tables should be avoided. Comment on significance of results is appropriate but broader discussion should be part of the Discussion section. Discussion – The findings of the study should be placed in context of relevant published data. Ideas presented in other publications should not be discussed solely to make an exhaustive presentation. Some manuscripts may require different formats appropriate to their content. d) The Acknowledgments must be a single paragraph that immediately follows the discussion and includes references to grant support. e) The References Section: references must be ordered alphabetically by the first author surname; references with the same first author should be ordered as follows: first, as single author in chronological order; next, with only one more co-author in alphabetical order by the second author; and finally followed by references with more than two co-authors, in chronological order, independent of the second author

Gazzaneo LRS Indução de mutação e seleção em feijão-caupi [Vigna unguiculata (L.) Walp]...
100
surnames. Use standard abbreviations for journal titles. Only articles that are published or in press should be included in this section. Works submitted for publication but not yet accepted, personal communications and unpublished data must be cited within the text. “Personal communication” refers to individuals other than the authors of the manuscript being submitted; “unpublished data” refers to data produced by at least one of the authors of the manuscript being submitted. . f) Internet Resources Section this section should contain a list of URLs referring to data presented in the text, software programs and other Internet resources used during data processing. When databases are cited, date of consultation must be stated. Sample Internet Resource citation : Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/OMIM (September 4, 2005) LEM Software, http://dir.niehs.nih.gov/dirbb/weinbergfiles/hybrid_design.htm g) Tables: each table must start on a new page. A concise title should be provided above the table. Tables must be numbered consecutively in Arabic numerals. Each column must have a title in the box head. Footnotes typed directly below the table should be indicated in lowercase superscript numbers. h) Figures must be numbered consecutively in Arabic numerals. Legends should be typed on a separate sheet. Images should be in TIFF or JPEG format and provided in separate files. Figures in Word format cannot be published. Journal quality reproduction will require grayscale and color at resolution yielding 300 dpi. Authors should submit bitmapped line art at resolution yielding 600-1200 dpi. These
resolutions refer to the output size of the file; if it is anticipated that images will be enlarged or reduced, the resolutions should be adjusted accordingly. Scanned figures should not be submitted. Identify each illustration by affixing on the back a label containing: the number of the figure, the name of the first author and an arrow indicating top of illustration. Identify each illustration by the first author name and the number of the respective figure. Color illustration can be accepted, but authors may be asked to defray the cost. i) Nomenclature should adhere to current international standards. j) Sequences may appear in text or in figure. DNA, RNA and protein sequences equal to or greater than 50 units must be entered into public databases. The accession number must be provided and released to the general public together with publication of the article. Long sequences requiring more than two pages to reproduce will not be published unless the Editorial decision is that the publication is necessary. Complete mtDNA sequence will not be published. k) Data access: reference should be made to availability of detailed data and materials used for reported studies. l) Ethical issues: Reports of experiments on live vertebrates must include a brief statement that the institutional review board approved the work and the protocol number must be provided. For experiments involving human subjects, authors must also include a statement that informed consent was obtained from all subjects. If photos or any other identifiable data are included, a copy of the signed consent must accompany the manuscript. m) Supplementary Material: Data that the authors consider of importance for

Gazzaneo LRS Indução de mutação e seleção em feijão-caupi [Vigna unguiculata (L.) Walp]...
101
completeness of a study, but which are too extensive to be included in the published version, can be submitted as Supplementary Material. This material will be made available together with the electronic version. In case a manuscript contains such material, it should be appropriately identified within the text. Supplementary material in tables should be identified as Table S1, Table S2, etc., in case of figures, they should be named accordingly, Figure S1, Figure S2. For online access, Supplementary material should be in PDF, JPEG or GIFF formats In addition, a list of this material should be presented at the end of the manuscript text file, containing the following statement: Supplementary material - the following online material is available for this article: - Table S1 .... This material is available as part of the online article from http://www.scielo.br/gmb 3.2 Short Communications Present brief observations that do not warrant full-length articles. They should not be considered preliminary communications. They should be 15 or fewer typed pages in double spaced 12-point type, including literature cited. They should include an Abstract no longer than five percent of the paper’s length and no further subdivision with introduction, material and methods, results and discussion in a single section. Up to two tables and two figures may be submitted. The title page and reference section format is that of full-length article.
3.3 Letters to the Editor Relate or respond to recent published items in the journal. Discussions of political, social and ethical issues of interest to geneticists are also welcome in this form. 3.4 Review Articles Review Articles are welcome. 3.5 Book Reviews Publishers are invited to submit books on Genetics, Evolution and related disciplines, for review in the journal. Aspiring reviewers may propose writing a review. 3.6 History, Story and Memories Accounts on historical aspects of Genetics relating to Brazil. 4. Proofs and Copyright Transfer Page proofs will be sent to the corresponding author. Changes made to page proofs, apart from printer’s errors, will be charged to the authors. Notes added in proof require Editorial approval. A form of consent to publish and transfer of copyright will have to be signed by the corresponding author, also on behalf of any co-authors.




![JONNATHAN AMARAL DE FREITAS - UFC · jonnathan amaral de freitas purificaÇÃo e caracterizaÇÃo de uma peroxidase de raÍzes de feijÃo-de-corda [vigna unguiculata (l.) walp.]](https://img.document.onl/doc/110x75/601bc904fb19a36e6a00a5c6/jonnathan-amaral-de-freitas-jonnathan-amaral-de-freitas-purificafo-e-caracterizafo.jpg)





![TESE Emmanuel Jereissati - UFC · 2015. 1. 19. · 1 EMMANUEL DE SOUSA JEREISSATI Transformação genética de feijão-caupi [Vigna unguiculata (L.) Walp] e tabaco (Nicotiana tabacum)](https://img.document.onl/doc/110x75/61155d96d8bee2635d419368/tese-emmanuel-jereissati-2015-1-19-1-emmanuel-de-sousa-jereissati-transformao.jpg)


















