Embed Size (px)
DESCRIPTION
Dissertação de Mestrado apresentado ao Programa de pós graduação em Geologia da UFRJ
Citation preview

UFRJ Rio de Janeiro
2008
Rommulo Mendes Carvalho Barreiro
MACROESTRUTURAS BIOEROSIVAS EM CONCHAS DE
MOLUSCOS DO PÂNTANO DA MALHADA (HOLOCENO), CABO
FRIO, RIO DE JANEIRO
Dissertação de Mestrado (Geologia)

UFRJ
Rio de Janeiro Fevereiro 2008
Rommulo Mendes Carvalho Barreiro
MACROESTRUTURAS BIOEROSIVAS EM CONCHAS DE
MOLUSCOS DO PÂNTANO DA MALHADA (HOLOCENO), CABO
FRIO, RIO DE JANEIRO
Dissertação de Mestrado submetida ao Programa de Pós-graduação em Geologia, Instituto de Geociências, da Universidade Federal do Rio de Janeiro – UFRJ, como requisito necessário à obtenção do grau de Mestre em Ciências (Geologia).
Área de concentração:
Paleontologia e Estratigrafia
Orientadoras:
Drª. Claudia Gutteres Vilela Drª. Maria Célia Elias Senra

BARREIRO, Rommulo Mendes Carvalho
Macroestruturas bioerosivas em conchas de moluscos do Pântano da Malhada (Holoceno), Cabo Frio, Rio de Janeiro / Rommulo Mendes Carvalho Barreiro - - Rio de Janeiro: UFRJ / IGeo, 2008.
xv, 85f. : 25il., 1 apênd.;
Dissertação (Mestrado em Geologia) – Universidade Federal do Rio de Janeiro, Instituto de Geociências, Programa de Pós-graduação em Geologia, 2008.
Orientadoras: Claudia Guterres Vilela, Maria Célia Elias Senra
1. Geologia. 2. Paleontologia e Estratigrafia – Dissertação de Mestrado. I. Vilela, Claudia Gutterres. II. Universidade Federal do Rio de Janeiro, Instituto de Geociências, Programa de Pós-graduação em Geologia. III.. Macroestruturas bioerosivas em conchas de moluscos do Pântano da Malhada (Holoceno), Cabo Frio, Rio de Janeiro

Rommulo Mendes Carvalho Barreiro
MACROESTRUTURAS BIOEROSIVAS EM CONCHAS DE
MOLUSCOS DO PÂNTANO DA MALHADA (HOLOCENO), CABO
FRIO, RIO DE JANEIRO
Dissertação de Mestrado submetida ao Programa de Pós-graduação em Geologia, Instituto de Geociências, da Universidade Federal do Rio de Janeiro – UFRJ, como requisito necessário à obtenção do grau de Mestre em Ciências (Geologia).
Área de concentração:
Paleontologia e Estratigrafia
Orientadoras:
Drª. Claudia Gutterres Vilela Drª. Maria Célia Elias Senra
Aprovada em: 26/02/2008
Por:
_____________________________________ Presidente: Dr. Ismar de Souza Carvalho, IGEO/UFRJ
_____________________________________ Dr. Antônio Carlos Sequeira Fernandes, MN/UFRJ
_____________________________________ Drª. Rita de Cassia Tardin Cassab, DNPM

vi
Dedico este trabalho a Maria da
Conceição, minha avó, que esteve
ao meu lado por todo este período.

vii
Agradecimentos
À professora Doutora Maria Célia Elias Senra (UNIRIO) pela orientação do trabalho realizado e dedicação demonstrada durante minha vida acadêmica. À professora Doutora Claudia Gutterres Vilela pela orientação acadêmica do mestrado. Aos professores Doutor Antonio Carlos Sequeira Fernandes (MN/UFRJ) e Doutora Assunta D´Alessandro (Universitat di Bari/ Itália) por ceder parte da bibliografia utilizada. Aos amigos Márcio Aguiar da Costa, Natalia Pereira Benaim, Thatyanna Ramos Barreira, Diana Mariani e Marcela da Silva Rosa, pelas revisões gramaticais e contextuais e à equipe do Núcleo de Estudos Tafonômicos (NEST/UNIRIO) pelo auxílio nas coletas realizadas. Ao Programa de Pós-graduação em Geologia do Instituto de Geociências da Universidade Federal do Rio de Janeiro e à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pelo suporte financeiro.

viii
Resumo
BARREIRO, Rommulo Mendes Carvalho. Macroestruturas bioerosivas do Pântano da Malhada (Holoceno), Cabo Frio, Rio de Janeiro. Rio de Janeiro, 2008. xv, 85f. Dissertação (Mestrado em Geologia) – Programa de Pós-graduação em Geologia, Instituto de Geociências, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2008. Bioerosão é a denominação dada aos processos de degradação de substratos litificados por ação biológica. Seu estudo em substratos bioclásticos vem adquirindo notabilidade nas últimas décadas, contribuindo de forma significativa para ciências como a Icnologia, a Ecologia das interações e a Paleontologia. A atividade bioerosiva, um dos mais importantes processos ocorrentes no limiar Geologia-Biologia, por ser de natureza física, química ou ambas, resulta na produção de sedimentos em ambientes marinhos. Pesquisados em detalhe, estes elementos ampliam o horizonte de conhecimento acerca dos organismos, com difícil preservação, que habitaram ambientes pretéritos, bem como a variedade das estruturas por eles produzidas. Caracteriza-se por ser uma ferramenta fundamental na compreensão de processos evolutivos e paleoecológicos ocorridos no período que vai desde o Cambriano até o Holoceno. Este trabalho se propõe ao estudo icnotaxonômico e a análise quantitativa das macroestruturas bioerosivas encontradas em conchas de Anomalocardia brasiliana, Crassostrea rhizophorae e Ostrea cf. puelchana provenientes das coquinas da planície costeira de Cabo Frio (Região do Pântano da Malhada). São identificadas seis icnoespécies: Entobia laquea, E. paradoxa, Caulostrepsis taeniola, Maeandropolydora aff. sulcans, Oichnus simplex e O. paraboloides. Estruturas estas produzidas por esponjas clionaídeas, poliquetas espionídeos e gastrópodos naticídeos e muricídeos. São interpretados padrões de simbiose entre organismo invasor e hospedeiro, estudo geométrico das estruturas e deduções paleoambientais. Palavras-chave: Bioerosão, Holoceno, Icnologia.

ix
Abstract
BARREIRO, Rommulo Mendes Carvalho. Macroestruturas bioerosivas do Pântano da Malhada (Holoceno), Cabo Frio, Rio de Janeiro [Macrobioerosion structures from Pântano da Malhada (Holocene), Cabo Frio, Rio de Janeiro] Rio de Janeiro, 2008. xv, 85f. Dissertação (Mestrado em Geologia) – Programa de Pós-graduação em Geologia, Instituto de Geociências, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2008. Bioerosion is the name given to the processes of hard substrate degradation by biological means. Its study in bioclastic substrate becomes in the last few decades more notable and has been contributing in a significant way for sciences as Ichnology, Ecology of the interactions and Paleontology. The bioerosive activity is one of the most important common processes in the Geology-Biology threshold. It results in the production of sediments in marine environments through physical and/or chemical means. These traces widen the horizon of knowledge concerning the organisms with difficult preservation that had inhabited paleoenvironments, as well as the variety structures produced by them. It is characterized for being a basic tool in the understanding of evolutionary and paleoecologic processes, in particular of the occurrences in the period between Cambrian and Holocene. This work focuses icnotaxonomy study and the quantitative analysis of the bioerosive macrostructures present in shells of Anomalocardia brasiliana, Crassostrea rhizophorae and Ostrea cf. puelchana from the Cabo Frio coastal plain shellbed (Pântano da Malhada region). They are identified in six icnoespecies: Entobia laquea, E. paradoxa, Caulostrepsis taeniola, Maeandropolydora aff. sulcans, Oichnus simplex and O. paraboloides. These structures were produced by clionaid sponges, spionid polychaetes and naticid and muricid gastropods. This work interpreted standards of symbiosis between invading organism and host, geometric study of the bioerosive structures and paleoenviromental deductions.
Key-words: Bioerosion, Holocene, Ichnology

x
Lista de figuras
Figura 1: Esquema representativo das classes etológicas atuais. 6
Figura 2: Esquema da perfuração produzida por esponjas clinaídeas. 12
Figura 3: Esquema apresentando as fases de crescimento de A a E. Escalas
com valor de 0,5 mm. 14
Figura 4: Esquema do padrão geral de perfurações produzidas por poliquetas. 18
Figura 5: Esquemas da geometria transversal das perfurações completas e
incompletas de muricídeos e naticídeos (B). 21
Figura 6: Mapa de localização da área de estudo (Pântano da Malhada). 24
Figura 7: Perfil sedimentar da área de estudo. 25
Figura 8: Imagens da área de estudo. 27
Figura 9: Esquemas de mediação dos eixos de A. brasiliana e ostreóideos. 29
Figura 10: Diagrama de setores para as valvas de (a) A. brasiliana e (b)
ostreóideos. 30
Figura 11: Esquema de um indivíduo de A. brasiliana em posição de vida no
sedimento. As setas simbolizam as correntes hídricas criadas pelos sifões
inalante e exalante. 31
Figura 12: Esquema de um indivíduo de C. rhizophorae em posição de vida. As
setas escuras simbolizam os movimentos da valva superior, as setas claras
simbolizam as correntes hídricas criadas pelo manto. 31
Figura 13: Histogramas de distribuição das medidas do eixo antero-posterior de
A. brasiliana nas três localidades. 41
Figura 14: Freqüências das estruturas bioerosivas de cada produtor, em A.
brasiliana bioerodidas por localidade de coleta. 42
Figura 15: Freqüências das estruturas bioerosivas de cada produtor em valvas
bioerodidas de ostreóideos por localidade de coleta. Legenda: Cr: C.
rhizophorae, Op: O. cf. puelchana. 42
Figura 16: Boxplot’s de distribuição das valvas de A. brasiliana nas localidades
agrupadas segundo presença e ausência bioerosão de esponja e poliquetas. 43
Figura 17: Boxplot’s de distribuição das valvas direitas e esquerdas de C.
rhizophorae de #SP agrupadas por bioerosões de esponja e poliquetas e em 44

xi
ausente e presentes.
Figura 18: Freqüências dos setores com estruturas bioerosivas de poliquetas
em A. brasiliana. 45
Figura 19: Freqüências das conchas com estruturas bioerosivas de poliquetas
nos setores central e marginal, tanto no periostraco quanto no hipostraco em C.
rhizophorae e O. cf. puelchana. 46
Figura 20: Freqüências das bioerosões por poliquetas na face externa
(periostraco) nas valvas de ostreóideos de acordo com a quantidade de
estruturas encontradas (raro, moderado e abundante). 47
Figura 21: Freqüências das bioerosões por poliquetas na face interna
(hipostraco) nas valvas de ostreóideos de acordo com a quantidade de
estruturas encontradas (raro, moderado e abundante). 47
Estampa 1 63
Estampa 2 64
Estampa 3 65
Estampa 4 66
Estampa 5 67

xii
Lista de tabelas
Tabela 1: Dados quantitativos de exemplares encontrados nas amostras
estudadas 68
Tabela 2: Valores absolutos e freqüências de valvas de ostreóideos
bioerodidos por esponjas. 68
Tabela 3: Freqüências das estruturas bioerosivas em valvas de A. brasiliana. 69
Tabela 4: Freqüências das estruturas bioerosivas de poliquetas em valvas de
ostreóideos. 70

xiii
Lista de quadros
Quadro 1: Alguns exemplos de icnotáxons produzidos por bioerosão e suas
respectivas classificações etológicas. 7

xiv
Sumário Agradecimentos .................................................................................................. vii
Resumo .............................................................................................................. viii
Abstract ........................................................................................ ...................... ix
Lista de figuras ........................................................................................ ............. x
Lista de tabelas ........................................................................................ ........... xii
Lista de quadros ................................................................................................ xiii
1. INTRODUÇÃO ................................................................................................. 1
1.1. Objetivos ....................................................................................................... 4
2. ICNOLOGIA E BIOEROSÃO .......................................................................... 5
2.1. Bioclausuras ................................................................................................ 8
3. AGENTES E PROCESSOS ............................................................................ 9
3.1. Esponjas ...................................................................................................... 10
3.2. Poliquetas .................................................................................................... 15
3.3. Gastrópodos ................................................................................ ................. 18
4. ÁREA DE ESTUDO ......................................................................................... 22
4.1. LOCALIDADES DE COLETA ...................................................................... 25
4.1.1. Reserva Tauá............................................................................................ 25
4.1.2. Sítio da Paz .............................................................................................. 26
4.1.3. Fazenda Junqueira ................................................................................... 26
5. MATERIAL E MÉTODOS ............................................................................... 28
5.1. Coleta do material ....................................................................................... 28
5.2. Tratamento laboratorial ................................................................................ 28
5.3. Tratamento estatístico .................................................................................. 32
6. RESULTADOS ................................................................................................ 33
6.1. Icnotaxonomia .............................................................................................. 33
6.2. Análise quantitativa das populações de moluscos representadas nas
coquinas.............................................................................................................. 41
6.3. Análise quantitativa das estruturas bioerosivas .......................................... 43
6.4. Distribuição espacial da bioerosão produzida por poliquetas .................... 45
7. DISCUSSÃO ................................................................................................... 48

xv
8. CONCLUSÕES ............................................................................................... 53
Referências bibliográficas ................................................................................... 55
Glossário ............................................................................................................. 62
Apêndice 1 – Tabelas ......................................................................................... 68

1
1. INTRODUÇÃO
O estudo de estruturas bioerosivas em substratos rochosos e bioclásticos vem
adquirindo notabilidade nas últimas décadas devido a contribuições à Icnologia,
Ecologia das interações e Paleontologia. Esta realidade, entretanto, não se aplica ao
Brasil, onde o estudo ainda é incipiente.
As estruturas bioerosivas são resultantes de processos biológicos em
substratos litificados ou lignificados, nos quais ocorre dissolução ou raspagem, com
posterior remoção do material por organismos vivos, representando atividades de
predação e habitação. São denominadas bioerosões, termo proposto por Neumann
em 1966, porém seu conhecimento vem desde meados do século XIX, quando Lyell
(1842) denomina de lithophagi e lithodomi os moluscos que perfuram o substrato
para própria habitação.
A atividade bioerosiva, por ser de natureza física, química ou ambas, resulta
na produção de sedimentos em ambientes marinhos, a partir da degradação de
esqueletos carbonáticos. Acker & Risk (1985) mostram que qualquer tratamento de
sedimentologia carbonática ou modelo de fácies que ignora o efeito da bioerosão
corre o grave risco de estar incompleto. Dentro de uma concepção tafonômica, a
bioerosão é um dos mais importantes processos ocorrentes no limiar Geologia-
Biologia.
Estudadas dentro da Icnologia como icnofósseis, as bioerosões são
importantes registros que auxiliam o estudo geológico e paleontológico, ampliando
nossa imagem dos organismos com difícil preservação que habitaram ambientes
pretéritos (Ekdale, 1978).
O estudo em bioerosão pode ser utilizado como ferramenta em análises
paleoecológicas (Boekschoten, 1966, 1967), reconstruções paleoambientais
(Boeckshoten, 1967; Arua, 1982; Bromley & D’Alessandro, 1987; Lorenzo & Verde,
2004), processos tafonômicos (Walker & Goldstein, 1999; Pickerril et al., 2002;
Farinati et al., 2005), interpretações paleobatimétricas (Bromley, 1978; Nielsen et
al.,2003; Försterra et al., 2005), bem como na compreensão de processos evolutivos
(Baumiller et al., 1999; Harper, 2005; Ekdale, 2006; Tapanila, 2006a) e de interações

2
predador-presa (Kowaleski et al.,1998; Roopnarine & Beussink, 1999; Dietl &
Alexander, 2000).
Seus processos de formação são estudados em programas de monitoramento
ambiental (Rützler, 1975, 2002; Silina, 2003; Ward-Paige et al., 2005), pesquisa de
“patologias” causadas por infestações em conchas de moluscos cultivados para o
comércio (Handley & Bergquist, 1997; Mortensen et al., 2000; Boscolo & Giovanardi,
2002; Silina, 2006) e padrões de sedimentologia (Rützler, 1975; Acker & Risk,1985).
A bioerosão é o principal processo ecológico no ciclo dos carbonatos, tanto
nas áreas temperadas, quanto nas tropicais (Hutchings, 1986 apud Calcinai et al.,
2003).
As estruturas bioerosivas podem ser produzidas por moluscos, anelídeos,
esponjas, algas, entre outros, sendo registradas desde o Cambriano até o Holoceno
(Taylor & Wilson, 2003). Além dos organismos que produzem furos distintos, a
bioerosão é complementada por seres que formam ninhos, arranham, raspam,
mordem ou mesmo causam atrito no fundo marinho ou em materiais a eles
associados. Engloba relações simbióticas e de predação (Martin & Britayev, 1998)
com ampla aplicação ambiental.
Os fatores ambientais com maior relevância em cenários marinhos, que
influenciam na composição das assembléias de traços fósseis são: batimetria,
substrato, oxigênio e hidrodinâmica (Ekdale,1985); em comunidades atuais, Ward-
Paige et al. (2005) citam ainda a influência do aporte de detritos e nutrientes
terrestres (poluição) no aumento da população de esponjas clionaídeas perfurantes,
aumentando, assim, a bioerosão em substratos carbonáticos localmente.
As macroperfurações são estruturas com dimensões transversais maiores que
um milímetro (Ekdale, 2006). São produzidas quase exclusivamente por bioerosões
de domichnia, servindo de habitação ao ocupante por toda a vida.
Uma das principais características da malacofauna do Pântano da Malhada é
a predominância numérica de conchas de moluscos das seguintes espécies:
Anomalocardia brasiliana (Gmelin, 1791), Anadara cheminitzi (Philippi, 1851),
Anadara brasiliana (Lamarck, 1819), Anadara ovalis Brugière, 1789, Crassostrea
rhizophorae (Guilding, 1828), Dendostrea frons (Linnaeus, 1758), Lucina pectinata
(Gmelin, 1791), Ostrea cf. puelchana d´Orbigny, 1842, Protothaca antiqua (King &

3
Broderip, 1835), Tellina sp. Linnaeus 1758, Acteocina candei (d’Orbigny, 1841),
Bulla striata Bruguière, 1792, Cerithium atratum (Born, 1778), Mitrella albovittata
Lopes,Coelho & Cardoso, 1965, Nassarius vibex (Say, 1822), Neritina virginea
(Linnaeus, 1758), Odostomia canaliculata Adams, 1850, Antalis disparile (d’Orbigny,
1842). Destas destacam-se Anomalocardia brasiliana, Crassostrea rhizophorae e
Ostrea puelchama com estruturas bioerosivas preservadas (Barreiro & Senra, 2005,
2006; Barreiro et al., 2006).
O Pântano da Malhada, localizado na Região dos Lagos do estado do Rio de
Janeiro, apresenta níveis de coquinas com extensão em área de aproximadamente
60.000 m2 e alto grau de preservação. Estes depósitos, inicialmente estudados por
Senra et al. (2002, 2003, 2004), têm excelente exposição na Reserva Tauá,
atualmente um Sítio Paleobiológico (Castro et al., 2006) cadastrado na SIGEP
(Comissão Brasileira de Sítios Geológicos e Paleobiológicos).
As coquinas estão relacionadas à última transgressão marinha ocorrida
durante o Holoceno na região (5.100 anos AP), que propiciou a formação de um
corpo lagunar onde desaguavam os rios da região, sendo depois ressecado após a
regressão marinha subseqüente (4.900 anos AP) (Castro et al., 2006).
A pesquisa em bioerosão dispõe de grande potencial para o
conhecimento macroevolutivo e macroecológico de organismos perfuradores e suas
estratégias comportamentais (Tapanila, 2006a). a morfologia das perfurações é,
contudo, muito variável e, em muitos casos, extremamente simples, raramente
permitindo a determinação precisa do organismo produtor (Bromley, 1978). Apesar
do amplo conhecimento destas estruturas, ainda se faz necessário um detalhamento
dos padrões de perfuração relacionados ao tipo de ambiente e às respostas do
hospedeiro ao agente, bem como os mecanismos fisiológicos envolvidos, além de
estabelecer modelos quantitativos das icnotramas em relação aos diversos táxons
providos de exoesqueleto carbonático. Essa pesquisa, portanto, pode contribuir com
o melhor entendimento das relações ecológicas entre organismos diferentes
(sinecologia) ou na relação organismo/substrato (autoecologia do organismo
perfurador). Poderá fornecer informações relativas à fidelidade do registro fossilífero,
uma vez que parte dos esqueletos é enfraquecida e se torna suscetível aos agentes
hidrodinâmicos e biológicos.

4
As perfurações provocadas por organismos endolíticos possuem dois
atributos que os transformam em potenciais ferramentas para a interpretação
paleoambiental. (1) As perfurações são, de certa forma, diageneticamente
indestrutíveis. (2) Elas são preservadas genuinamente in situ em relação ao seu
substrato (concha). (3) Em coquinas de natureza autóctone, podem adquirir valor
estratigráfico.
1.2. Objetivos
O presente trabalho tem por objetivo o estudo icnotaxonômico das
macroestruturas bioerosivas encontradas em conchas do Pântano da Malhada, com
indicação de seus produtores, relações ecológicas e paleoambientais.

5
2. Icnologia e Bioerosão
Icnologia é a ciência que se ocupa do estudo dos vestígios resultantes das
atividades de organismos em substratos consolidados (litificados e lignificados) ou
inconsolidados (sedimentos). Pode apresentar duas divisões: a Paleoicnologia, que
estuda os icnitos do passado (icnofósseis ou traços fósseis), e a Neoicnologia,
estudo dos icnitos recentes.
Icnitos ou traços são estruturas biogênicas recentes ou fósseis que refletem o
comportamento do organismo. Estão, de certa forma, relacionadas à morfologia de
seu produtor, tais como pegadas, pistas, escavações e perfurações, incluindo ainda
coprólitos, gastrólitos, regurgitólitos, ninhos, teias de aranha, casulos (tecidos ou
feitos por cimentação de sedimentos), marcas de mordidas, entre outras (Fernandes
et al., 2002; Bertling et al., 2006).
A definição mais atual de traço fóssil, proposta por Bertling et al. (2006), o
aponta como estrutura morfologicamente recorrente resultante da atividade vital de
um organismo individual (ou grupo monoespecífico de organismos) que modifica o
substrato.
Os traços fósseis devem ser vistos como resultantes da combinação de três
parâmetros intimamente ligados: substrato, organismo produtor e comportamento
(Gibert et al., 2004). Segundo Ekdale (1985), os icnofósseis podem fornecer
informações das populações como abundância, diversidade e densidade, dados
relevantes na interpretação das estratégias populacionais (espécies oportunistas ou
de equilíbrio).
Apresentam uma parataxonomia (icnotaxonomia) regida pelo Código
Internacional de Nomenclatura Zoológica que os define como “trabalho fossilizado
de organismos” (“fossilized work of organisms”) (ICZN, 1999, Art. 1.2.1). A
icnotaxonomia apresenta os níveis de icnoespécie e icnogênero. Subespécie e
subgênero são também aceitos na sistemática icnológica, apesar de raramente
usados (Rindsberg, 1990 apud Bertling et al., 2006). O ICZN (1999, Art. 10.3)
permite o uso de icnofamílias para que sejam aplicadas do mesmo modo que os

6
níveis inferiores. Estas deveriam, contudo, ser baseadas somente na morfologia das
estruturas e não na biologia do produtor (Bromley, 1996 apud Bertling et al., 2006).
Além da classificação icnotaxonômica, há a classificação etológica. Proposta
primeiramente por Seilacher (1953 apud Gibert et al., 2004), desde então se tornou
o meio mais robusto e completo de agrupar os traços fósseis em um nível acima do
icnogenérico.
O primeiro sistema de classificação etológico agrupava cinco categorias de
estruturas. Após contribuições mais recentes, se encontra atualmente com doze
categorias (Figura 1), assim denominadas: Repichnia, estruturas de locomoção
direta; Pascichnia, estruturas de locomoção com alimentação; Fodinichnia,
estruturas de habitação com alimentação; Domichnia, estruturas de habitação;
Cubichnia, estruturas de repouso; Fugichnia, estruturas de fuga; Agrichnia,
estruturas de armadilha e cultivo; Praedichnia, marcas de predação; Aedifichnia,
estruturas de construção sobre o substrato; Equilibrichnia, estruturas de reajuste
gradual; Calichnia, estruturas de nidificação; e Fixichnia, marcas de ancoragem e
fixação (Fernandes et al., 2002; Gibert et al., 2004).
Figura 1: Esquema representativo das classes etológicas atuais (fonte: Gibert et al., 2004).
As estruturas bioerosivas, portanto, se enquadram em cinco categorias: duas
exclusivamente bioerosivas (Praedichnia e Fixichnia) e três que englobam também
estruturas de bioturbação (Domichnia, Equilibrichnia e Pascichnia). Alguns exemplos
podem ser vistos no quadro a seguir.
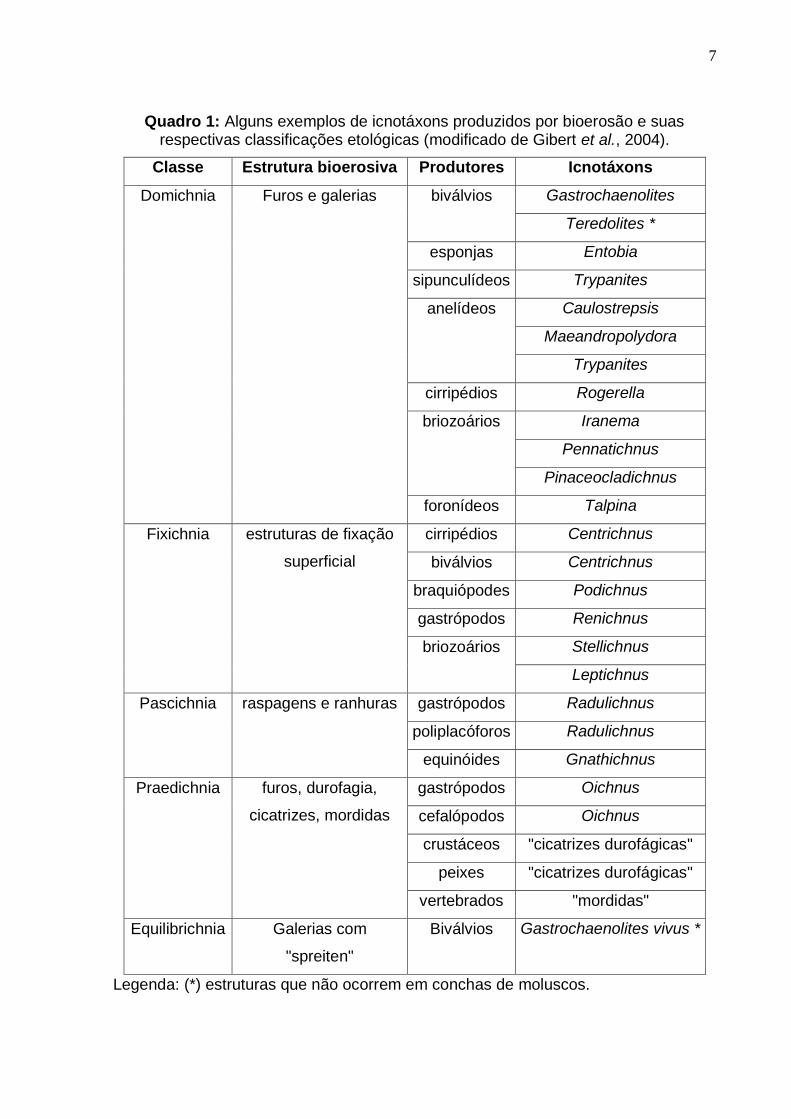
7
Quadro 1: Alguns exemplos de icnotáxons produzidos por bioerosão e suas respectivas classificações etológicas (modificado de Gibert et al., 2004).
Classe Estrutura bioerosiva Produtores Icnotáxons
Domichnia Furos e galerias biválvios Gastrochaenolites
Teredolites *
esponjas Entobia
sipunculídeos Trypanites
anelídeos Caulostrepsis
Maeandropolydora
Trypanites
cirripédios Rogerella
briozoários Iranema
Pennatichnus
Pinaceocladichnus
foronídeos Talpina
Fixichnia estruturas de fixação
superficial
cirripédios Centrichnus
biválvios Centrichnus
braquiópodes Podichnus
gastrópodos Renichnus
briozoários Stellichnus
Leptichnus
Pascichnia raspagens e ranhuras gastrópodos Radulichnus
poliplacóforos Radulichnus
equinóides Gnathichnus
Praedichnia furos, durofagia,
cicatrizes, mordidas
gastrópodos Oichnus
cefalópodos Oichnus
crustáceos "cicatrizes durofágicas"
peixes "cicatrizes durofágicas"
vertebrados "mordidas"
Equilibrichnia Galerias com
"spreiten"
Biválvios Gastrochaenolites vivus *
Legenda: (*) estruturas que não ocorrem em conchas de moluscos.

8
2.1. Bioclausuras
Para alguns autores (eg.: Palmer & Wilson, 1988; Taylor, 1990; Tapanila,
2005), há ainda um grupo distinto de traços fósseis, intimamente ligado aos
processos de bioerosão produzidos por organismos endossimbiontes.
Quando um endossimbionte penetra o esqueleto de seu hospedeiro, este, em
alguns casos, reage à invasão e cria uma estrutura calcificada que isola o organismo
invasor. Este processo pode ser observado em corais e em moluscos produtores de
concha. Nestes, há a formação de uma estrutura denominada ampola nacarada
(Boscolo & Giovanardi, 2002).
Palmer & Wilson (1988) dão a estas estruturas a denominação de
bioclausuras, que seriam caracterizadas pelo aprisionamento de um organismo
infestante (de corpo mole) por crescimento esquelético do hospedeiro. Taylor (1990)
as classifica como bioimuração, por se tratarem da preservação como molde da
morfologia de um organismo em uma estrutura esquelética externa não própria.
Para Tapanila (2005), elas são classificadas como traços fósseis e
compreendem uma nova classificação etológica, denominada Impedichnia
(estruturas isolamento ao organismo invasor). Representam, no registro fossilífero,
antigas interações simbióticas (hospedeiro invasor), configurando-se como
deformações no esqueleto do hospedeiro.
Porém, Bertling et al. (2006) não consideram a bioclausura uma estrutura
abrangida pelo conceito de traço fóssil, pois, assim como a pérola, não se trata de
modificação no crescimento padrão do organismo, o que dispensa a existência da
classe Impedichnia. Para estes autores, a bioclausura deve ser estudada dentro da
Paleopatologia.

9
3. AGENTES E PROCESSOS
Grupos de invertebrados marinhos desprovidos de defesas corporais contra
predadores desenvolveram, ao longo da evolução, o hábito endolítico. Vivendo
dentro de perfurações, produzidas para sua proteção, adquirem uma estrutura
externa protetora e obtêm a vantagem de ocupar um espaço inapropriado para a
maioria dos seres vivos (Zundelevich et al., 2007).
A escavação de um “lar” em um substrato duro não é uma tarefa fácil para um
invertebrado simples. Penetração e remoção de materiais mineralizados requer uma
anatomia especializada, habilidade comportamental variada e um considerável gasto
de energia (Ekdale, 2006).
Além das perfurações de habitação, existem aquelas que representam o
hábito alimentar durófago, ou seja, pequenos orifícios em carapaças carbonáticas
que auxiliam na retirada da carne da presa.
Segundo Warme (1975), pelo menos doze filos de animais estão envolvidos
nos processos bioerosivos, além de alguns grupos de plantas e protozoários. Dentre
estes, os mais óbvios e provavelmente os mais importantes organismos perfurantes
são: algas e fungos microscópicos, esponjas clionídeas, sipunculídeos, anelídeos
poliquetas, biválvios foladídeos, gastrópodos, crustáceos, equinóides, peixes e
briozoários (Taylor et al., 1999).
Estudos de laboratório confirmaram que fatores físicos e químicos de
diferentes origens interferem no recrutamento larvar de organismos dependentes do
substrato duro (Sebesvari et al., 2006).
Perry (2000) mostra que em corais tanto recentes quanto fósseis, as
bioerosões de maior expressão são aquelas ocasionadas por esponjas, sendo as
estruturas produzidas por poliquetas um componente secundário. E que a
abundância relativa de estruturas produzidas por vermes em relação àquelas
produzidas por esponjas sugere que o material seja de um ambiente de águas
rasas, favoráveis para a proliferação de poliquetas.

10
3.1. Esponjas
Dentre os numerosos organismos com habilidade de perfurar rochas
carbonáticas, as esponjas, principalmente da família Clionaidae, são de especial
interesse, devido a seu modo eficiente de penetração no substrato e por produzirem
grãos carbonáticos de formato típico (“chips”) que podem ser facilmente identificados
na fração lamosa dos sedimentos marinhos (Rützler, 1975). Grandes quantidades de
sedimento são produzidas por estas esponjas e alguns substratos são dominados
por seus fragmentos (Acker & Risk, 1985).
Considerando que muitas esponjas apresentam defesas químicas e/ou
mecânicas contra predadores, aquelas que não as têm podem apresentar a
vantagem competitiva de usar elemento esquelético como proteção. Tal estratégia
se baseia em perfurar estruturas sólidas de composição carbonática, que estão
inacessíveis para a maioria dos predadores e obter o benefício adicional de usar um
espaço não disponível a seus concorrentes (Zundelevich et al., 2007).
A erosão por esponja produz uma série de câmaras e galerias conectadas
dentro do substrato calcário onde a esponja vive, o organismo mantém contato com
o exterior através de papilas inalantes e exalantes ou, em algumas vezes, por meio
de um tecido incrustante (Calcinai et al., 2003).
As esponjas perfurantes desenvolveram um meio peculiar de penetração no
substrato. Ao se fixarem, usam células cauterizantes (corrosivas) que separam
fragmentos de CaCO3 (com tamanho variando entre 15-85m) da rocha,
transformando-os em grãos de calcário. Uma pequena depressão é produzida
quimicamente, com a utilização de anidrase carbônica e ácido fosfatase, em torno do
grão, isolando-o do substrato, que depois é transferido para fora do corpo da
esponja pelos canais de circulação de água e pelo ósculo. Esses dois processos de
corrosão e transporte dos grãos constituem a perfuração química e mecânica
respectivamente (Zundelevich et al., 2007; Calcinai et al., 2003).
As taxas de bioerosão dependem de alguns fatores bióticos e abióticos, que
incluem: disponibilidade de nutrientes e alimentos (Holmes et al., 2000), estado
fisiológico do organismo (Rützler, 1975), densidade e tipo de substrato onde o
organismo perfura (Hutchings, 1986).

11
Estímulo mecânico, alta intensidade luminosa, fortes correntes e,
possivelmente, baixa temperatura parecem acelerar o processo escavador em
ambientes coralíneos (Rützler, 1975).
Zundelevich et al. (2007), em laboratório, estimaram uma taxa total de
bioerosão do carbonato de cálcio em 340170g/m² por ano, sendo 260g/m²
dissolvidos quimicamente e o restante mecanicamente. Este estudo mostrou que a
taxa de bioerosão química é cerca de três vezes maior que a mecânica,
confrontando os dados de Rützler (1975), que estimou, em observações in situ, a
bioerosão total em 256g/m² por ano, estipulando que somente de 2 a 3% do
substrato seriam dissolvidos.
Esponjas perfurantes provavelmente datam do Devoniano (Tapanila, 2006b) e
as morfologias das perfurações, similares às formas modernas, são abundantes em
rochas cretáceas e mais recentes (Acker & Risk, 1985).
Suas estruturas bioerosivas são classificadas sob o icnogênero Entobia
Bronn, 1838, uma perfuração complexa com múltiplas aberturas e câmaras (Figura
2). Apresenta dezoito icnoespécies, que foram descritas com base nas
configurações tridimensionais (Gibert et al., 1998). Este icnotáxon é conhecido
desde pelo menos o Devoniano Médio (Tapanila, 2006b).
Calcinai et al. (2003) estudaram (em microscopia eletrônica de varredura) os
diferentes padrões estruturais das paredes internas das perfurações produzidas por
dois gêneros diferentes de esponjas clionaídeas. Eles consideram as variedades
estruturais um efeito diferenciado da morfologia dos pseudópodos de cada gênero,
mostrando, com isso, que as microesculturas internas das perfurações são
idiomórficas, ou seja, são específicas para cada grupo perfurador.

12
Figura 2: Esquema da perfuração produzida por esponjas clinaídeas (adaptado de Bromley & D’Alessandro, 1984)
Para Bromley & D’Alessandro (1984) as morfologias das estruturas (Entobia)
são influenciadas por fatores bióticos e abióticos, entre eles:
a) Natureza e estrutura do substrato: esponjas bioerosivas são restritas a substratos
calcários totalmente litificados (calcita e aragonita). Contudo, se o substrato possui
pequena área para fixação (e.g.: conchas de moluscos), a perfuração se torna
estenomórfica. Além disso, as heterogeneidades do substrato podem influenciar
fortemente a morfologia da perfuração.
b) Natureza do ambiente: gradiente fótico, hidrodinâmica, presença de incrustantes e
predadores. Em áreas propícias, as esponjas podem completar seu crescimento
ontogenético rapidamente, ao passo que esponjas mal localizadas podem usar sua
energia na construção de longos canais exploratórios na procura de melhores
condições de habitação. Em conchas parcialmente enterradas, as esponjas
cessam abruptamente seu crescimento abaixo da linha do sedimento.

13
c) Proximidade de outros organismos endolíticos: nos locais onde outras perfurações
(como: Trypanites e Caulostrepsis) estão ainda ocupadas por seus produtores, a
esponja evita o contato com estes e, onde estas estruturas são abundantes, a
esponja competidora é forçada a escavar um sistema estenomórfico entre e ao redor
de tais estruturas. Quando duas esponjas ─ da mesma espécie ou de espécies
diferentes ─ ocupam um mesmo substrato simultaneamente, ocorre um crescimento
lateral de cada uma, por canais exploratórios finos e curtos, que não chegam a
intercomunicar os dois indivíduos na região de proximidade, modificando, assim, a
morfologia drasticamente (Bromley & Tendal, 1973).
d) Tipos de esponjas perfurantes: cada espécie de esponja perfurante reflete
diferentes padrões anatômicos durante a escavação das galerias no substrato,
resultando em perfurações idiomórficas.
De acordo com Bromley & D’Alessandro (1984), o crescimento (ontogenia)
das esponjas dentro do substrato apresenta fases distintas de desenvolvimento e
maturação (Figura 3):
Fase de crescimento A: Imediatamente após o recrutamento, a esponja jovem
penetra no substrato produzindo finos canais exploratórios que se ramificam e
espalham radialmente a partir do ponto de abertura. Assim sendo, na zona mais
jovem de uma perfuração madura, onde o crescimento linear está ainda ativo, a
morfologia se apresenta como finos canais exploratórios.
Fase de crescimento B: Atrás da zona de crescimento linear, o diâmetro do furo é
aumentado por crescimento lateral. Normalmente neste estágio, formas nodulares
(“cameradas”) começam a se dilatar localmente formando as câmaras propriamente
ditas, ao passo que as formas não cameradas apresentam um formato de galhos de
árvore.

14
Fase de crescimento C: Nesta fase é alcançada uma morfologia relativamente
estável, representada por grandes áreas da perfuração tendo características
peculiares, como o tamanho da câmara, dimensões dos canais intercamerais, etc. O
formato radial original do sistema pode ainda ser visível. Muitas esponjas perfurantes
não ultrapassam essa fase.
Fase de crescimento D: O crescimento lateral sucessivo continua a ampliar as
câmaras e ramificações, permanecendo apenas um pequeno resíduo de substrato
entre as câmaras. Novas ramificações são introduzidas na área entre os ramos
existentes, até que o formato radial original seja mascarado. Em algumas formas
nodulares, ocorre a fusão de câmaras vizinhas, produzindo cavidades largas e
compostas.
Fase de crescimento E: Algumas esponjas continuam a crescer lateralmente. A
fusão associada destrói todas as características iniciais da perfuração e produz uma
câmara única ou poucas cavidades ou, ainda, um sistema de túneis não nodulares
de grandes extensões.
Figura 3: Esquema apresentando as fases de crescimento de A a E. Escalas com valor de 0,5mm (adaptado de Bromley & D’Alessandro, 1984).

15
3.2. Poliquetas
Os poliquetas são invertebrados bentônicos que desenvolveram uma série de
estratégias de vida para interagir com outros organismos. A escavação de substratos
carbonáticos, ligada à sobrevivência de formas imaturas, é um importante
mecanismo responsável pela produção de galerias em conchas.
A família Spionidae reúne espécies, dentro dos gêneros Polydora, Dipolydora
e Crazziella, com a habilidade de escavar substratos compostos por carbonato de
cálcio, tais como rochas carbonáticas, corais, algas calcárias coralinas incrustantes,
e também conchas de biválvios e gastrópodos. São animais sedentários que
apresentam dois palpos longos, finos e protráteis, com corpo apresentando mais de
180 segmentos (Boscolo & Giovanardi, 2002).
Em gastrópodos e biválvios é maior o grau de invasão por Polydora (29% e
23%, respectivamente) e, por isso, estão envolvidos na maioria das relações
parasitárias por poliquetas (Martin & Britayev,1998).
Os estudos iniciais apontavam que as perfurações eram produzidas
mecanicamente. Entretanto, evidências mais recentes indicam desgaste químico,
com secreção de ácido acético, associado à escavação mecânica, com raspagem
dos cristais de calcita da concha pelo quinto segmento modificado para abrasão
(McDiarmid et al., 2004; Silina, 2006; Wisshak & Neumann, 2006).
Tinoco-Orta & Cáceres-Martínez (2003) desenvolveram estudo detalhado
sobre a atividade bioerosiva de Polydora, gênero que tem como hábito viver no
interior de galerias em forma de U perfurados na concha do hospedeiro,
apresentando duas aberturas fundidas em forma de 8 (oito). A parte anterior do
anelídeo emerge da galeria e se alimenta de partículas tiradas do sedimento
superficial ou do material em suspensão na coluna d’água com o auxílio dos palpos
(Blake & Evans, 1973).
Os tubos não são produzidos por polidorídeos adultos, mas por suas larvas
quando estas se fixam, bem como por indivíduos pós-larvais durante o estágio de
recrutamento (Liu & Hsieh, 2000).

16
Nos polidorídeos, o recrutamento e a fixação ao substrato estão relacionados
ao acúmulo de argila na superfície da concha. A turbulência da água pode ter um
papel negativo no processo de infestação, pois além de comprometer a fixação,
remove larvas recém-fixadas. De acordo com Mortensen et al. (2000), a presença de
sedimentação lamosa pode causar um ambiente favorável à proliferação de larvas
de Polydora.
Diferentes parâmetros foram identificados como positivos e negativos no
processo de fixação de larvas, pós-larvas e formas jovens. São eles: conteúdo
orgânico do sedimento, distribuição do tamanho do grão, presença de formas jovens
e adultos coespecíficos, metabólitos haloaromáticos de organismos simpátricos e a
presença de bactérias (Sebesvari et al., 2006).
Silina (2003) mostra que o aumento no gradiente de bioerosão das conchas
está ligado a um aumento gradual no conteúdo de matéria orgânica nos sedimentos
de fundo. Este é um fator favorável ao desenvolvimento de bactérias e fitoplâncton,
que é uma fonte de alimentos para poliquetas. Bactérias e diatomáceas são os itens
de maior importância na dieta de polidorídeos que escavam conchas (Silina, 2006).
Galerias de Polydora em ostras estão relacionadas ao aumento da salinidade da
água (Almeida et al., 1997).
As larvas de poliquetas geralmente se fixam e melhor sobrevivem na margem
de crescimento da valva superior de ostreóideos ─ onde o suprimento de nutrientes
e oxigênio para a larva é mais favorável devido às correntes hídricas ─ e,
preferencialmente, nas fendas formadas pelas lamelas de crescimento, que são
locais propícios ao acúmulo de argila, apresentando, assim, um microclima ótimo
para seu assentamento. Perfurações nas fendas indicam taxas de sedimentação
pelítica indispensáveis ao recrutamento larvar (Silina, 2006).
Nos substratos ocupados por poliquetas, é comum a abundância de
indivíduos. A infestação é mais freqüente em moluscos previamente infestados do
que naqueles livres de vermes, mostrando, assim, uma preferência na invasão em
moluscos pré-infestados. Adultos preexistentes produzem um tipo de estímulo
químico à fixação de larvas, como ocorre em outros invertebrados (Tinoco-Orta &
Cáceres-Martínez, 2003; Silina, 2006).

17
Em biválvios semiendobentônicos a fixação de larvas de Polydora se dá em
torno da abertura sifonal, posição de alojamento vantajosa em relação a outras
regiões da superfície da concha, pois permite que o verme se alimente de partículas
que estão sendo inaladas ou expelidas pelo molusco (Tinoco-Orta & Cáceres-
Martínez, 2003). Tal preferência pode ser vista em conchas de A. brasiliana, onde as
perfurações encontram-se com maior freqüência na região posterior, local onde se
formam as correntes inalantes (Barreiro & Senra, 2006).
Quando a infestação é grande, a concha é enfraquecida e torna o molusco
mais suscetível à predação por crustáceos, além de potencializar a fragmentação
sob transporte hidráulico acentuado (Bower & McGladdery, 2003).
Segundo Blake & Evans (1973), as perfurações não entram em contato com a
parte mole do animal, pois o mesmo secreta camadas de conchiolina que são, em
seguida, cobertas por nácar, uma proteção contra a invasão do poliqueta, formando
uma estrutura denominada ampola nacarada. As galerias (Figura 4) podem se
apresentar no formato em U (mais comum), formato em Y, em forma de pêra,
complexos ramificados e depressões rasas, com ou sem ampola nacarada.
Em conchas de mexilhões, as ampolas nacaradas se desenvolvem mais
freqüentemente na valva direita (superior) que na esquerda (inferior), devido a
exposição relacionada à luminosidade e acúmulo de sedimento fino próprios para a
fixação larvar. O percentual de mexilhões com ampolas aumenta na mesma
proporção que a quantidade de pelitos nos sedimentos de fundo e a idade dos
moluscos. Isso se deve ao maior conteúdo de carbono orgânico em sedimento
lamoso que em sedimento arenoso (Silina, 2006).
As perfurações provenientes da ação de polidorídeos são classificadas nos
icnogêneros Caulostrepsis Clarke, 1908, contendo seis icnoespécies, e
Maeandropolydora Voigt, 1965, com cinco icnoespécies.

18
Figura 4: Esquema do padrão geral de perfurações produzidas por poliquetas. Icnogêneros (A) Caulostrepsis e (B) Maeandropolydora.
3.3. Gastrópodos
Exemplos clássicos de interações que deixam traços fósseis são as
perfurações de natureza predatória em esqueletos mineralizados de invertebrados.
Geralmente, podem ser distinguidas facilmente de furos de origem não biogênica.
Perfurações biogênicas aparecem em vários táxons paleozóicos, mais comuns em
braquiópodes do que em moluscos. Estruturas bioerosivas ligadas ao hábito
alimentar de gastrópodos carnívoros se apresentam como um único furo circular (ou
subcircular), na superfície externa da concha, atravessando-a perpendicularmente
em toda a espessura e se afunilando na extremidade interna (Baumiller et al., 1999).
A curiosidade sobre as perfurações em conchas e carapaças antecede muitos
séculos, pois desde a época de Aristóteles, é sabido que alguns gastrópodos
marinhos apresentam a capacidade de perfurar buracos em conchas de biválvios,
cracas, etc., para o consumo dos tecidos moles internos (Clelland, 1999).
A presença de furos nos esqueletos mineralizados de invertebrados marinhos
tem uma história geológica que remonta ao Pré-Cambriano Superior (Baumiller,
1996).
Abertura

19
Kowalewski et al. (1998) interpretaram a história evolutiva das perfurações
predatórias em três fases. A fase paleozóica, que se estende do Pré-Cambriano
Superior ao Carbonífero, é ilustrada pela aparição de predadores com uma
estratégia perfurante relativamente bem-sucedida (0,3-0,5 ocorrências por milhão de
anos) em braquiópodes, pequenos fósseis com conchas, moluscos bentônicos, entre
outros. No final desta fase (Devoniano ao Carbonífero), os predadores perfuravam
principalmente braquiópodes articulados, enquanto gastrópodos platiceratídeos se
tornavam importantes parasitas de equinodermas sésseis. Durante a fase mesozóica
(Permiano ao Cretáceo Inferior), predadores perfurantes eram tão raros que seu
registro não passa de poucas perfurações isoladas (< 0,1 ocorrência por milhão de
anos). Esta fase pode ser interpretada como o intervalo em que os perfuradores
possuíam uma adaptação perfurante latente. Por algum motivo, o grupo não se
tornou bem-sucedido e difundido, ou esta característica era facultativa para o hábito
alimentar. A fase cenozóica (Cretáceo Superior ao Holoceno) representa uma
segunda expansão dos perfurantes, associados a uma maior diversificação dos
predadores durófagos. Os perfuradores, especialmente os gastrópodos
prosobrânquios que se alimentam de moluscos, tornaram-se cada vez mais comuns
(> quatro ocorrências por milhão de anos).
Gastrópodos perfurantes atuais ocorrem nos Prosobranchia, ordem
Mesogastropoda (família Capulidae e superfamílias Naticoidea e Tonnoidea), e
ordem Neogastropoda (superfamília Muricoidea), nos Opistobranchia, ordem
Nudibranchia (família Vayssiereidae) (Bromley, 1981; Clelland, 1999). Estão
presentes desde a zona entre-marés até 270 m de profundidade e a maioria é
encontrada em águas rasas costeiras (Carriker, 1981 apud Clelland, 1999).
Segundo Cintra-Buenrostro et al. (2005), a maioria das perfurações presentes
em moluscos biválvios é resultado da predação por gastrópodos naticídeos e
muricídeos. O tamanho, forma e localização das perfurações são evidências para a
identidade, tamanho, comportamento e preferência por presa de predadores
perfurantes.
Para Roopnarine & Beussink (1999), a morfologia da presa é também um
importante componente no exame da dinâmica predatória no registro fossilífero.
Os gastrópodos predadores possuem uma pequena glândula especializada
(~1,00-2,5 mm de diâmetro), denominada órgão acessório perfurador (accessory

20
boring organ – sistema ABO), essencial no processo de perfuração. O sistema ABO
está localizado na extremidade da probóscide dos naticídeos e na sola do pé de
muricídeos e sua habilidade na dissolução do substrato depende da composição da
concha, temperatura e composição iônica do meio aquoso circundante (Clelland,
1999).
Duarte & Holler (1986), em estudo sobre a preferência de presas pelo
gastrópodo muricóideo Thais haemastoma (Linnaeus, 1767), observaram que sua
dieta está relacionada à disponibilidade de presas e à relação de tamanho predador-
presa.
O tamanho da presa é determinado por exame tátil da concha pela
probóscide e região anterior do pé (Carriker & Van Zandt, 1972 apud Clelland, 1999)
e a localização das presas é dada por quimiopercepção, porém Lorenzo & Verde
(2004) apontam que este método não é totalmente eficaz. Os autores observaram,
em seu material de estudo, uma perfuração na face interna da concha, indicando
que o gastrópodo atacou a presa morta.
O processo de perfuração consiste na dissolução química associada à
raspagem mecânica pela rádula da seguinte maneira: o periostraco é removido com
movimentos circulares pela rádula, expondo a concha calcária; em seguida, o
sistema ABO é posicionado e secreta substâncias ácidas (enzima ácido fosfatase,
ácido clorídrico e agente quelante de cálcio) que amolecem a concha; a rádula,
então, raspa o material que foi dissociado, aprofundando o furo; este processo de
alternância de dissolução e raspagem se repete novamente até que o furo se
complete e a parte carnosa do animal seja atingida. Quando o furo atravessa a
concha, a probóscide do gastrópodo, então, é evertida para dentro do orifício
produzido e começa a consumir a presa. Os fragmentos da concha resultantes do
processo de escavação são engolidos pelo gastrópodo (Clelland, 1999).
O mecanismo de raspagem em muricídeos e naticídeos é diferente. Nos
primeiros o odontóforo (e a rádula) são capazes de rotaciornar 180° enquanto, nos
últimos, a rotação é de somente 90° (tornando-se necessário um maior número de
repetições da raspagem durante o processo). As taxas de perfuração giram em torno
de 0,33 mm por dia para muricídeos e 0,54 mm por dia para naticídeos, variando de
acordo com a espécie de gastrópodo. Estas baixas taxas fazem com que um furo

21
leve em torno de três dias para ser completado, dependendo da espessura da
concha a ser perfurada (Clelland, 1999).
Essas peculiaridades se refletem nas morfologias das estruturas resultantes
(Figura 5): perfurações feitas por muricídeos apresentam um formato de cone
invertido, com a base chata, pois o material é removido uniformemente durante o
processo. Já as perfurações de naticídeos, quando incompletas, possuem uma
elevação central, pois o material é removido por partes (Bromley, 1981; Clelland,
1999).
Figura 5: Esquemas da geometria transversal das perfurações completas (esquerda) e incompletas (direita) de muricídeos (A) e naticídeos (B).
Perfurações por predação podem enfraquecer as conchas dos biválvios,
submetendo-as a tendências tafonômicas. A dimensão do furo, o número de
perfurações e ainda a sua ocorrência na valva direita ou esquerda afetam, também,
a hidrodinâmica do bioclasto. Conchas perfuradas, com densidade menor, podem
ser preferencialmente transportadas e soterradas (Walker & Goldstein, 1999).
Segundo Lorenzo & Verde (2004), esta bioerosão não é indicativa de
ambientes específicos, apenas evidencia aspectos comportamentais dos
gastrópodos predadores e registram relações interespecíficas entre estes e suas
presas. Contudo, não havendo registro do fóssil corporal do predador, a perfuração
delineia um retrato mais próximo da biodiversidade de uma assembléia.
Estas estruturas correspondem ao icnogênero Oichnus Bromley, 1981.
Podem estar presentes em biválvios, gastrópodos e outros grupos, como cirripédios
e poliquetas coloniais (sabelídeos) (Duarte & Holler, 1986).

22
4. ÁREA DE ESTUDO
As planícies costeiras são superfícies geomorfológicas deposicionais de baixo
gradiente, formadas por sedimentação predominantemente subaquosa, que
margeiam corpos de água de grandes dimensões, como o mar ou oceano,
representadas comumente por faixas de terrenos recentemente (em termos
geológicos) emersos e compostos por sedimentos marinhos, continentais, flúvio-
marinhos, lagunares, paludiais, em geral de idade quaternária (Suguio, 2003).
Na região costeira do estado do Rio de Janeiro ocorre uma série de
ambientes de sedimentação quaternária gerados em sistemas deposicionais de
origem continental e transicional/marinha. Tais depósitos são mais desenvolvidos na
porção que vai de Niterói até o limite com o estado do Espírito Santo do que no setor
de Paraty até Mangaratiba. Contém sedimentos sílticos e/ou areno-argilosos ricos
em matéria orgânica, separando terraços arenosos pleistocênicos, podendo
freqüentemente conter grande quantidade de conchas de moluscos de ambientes
lagunares (Silva & Cunha, 2001).
O desenvolvimento desta planície costeira foi, em grande parte, guiado pelas
direções estruturais do embasamento, que exerceram controle sobre a formação de
baías e sobre a disposição dos remanescentes rochosos interiores às antigas baías
que passaram a receber sedimentos provenientes das terras altas (Silva & Cunha,
2001).
Neste contexto, localiza-se a região compreendida pelo Pântano da Malhada
(22°44’48”/22°47’07” S – 42°01’14”/41°59’52” W – entre os municípios de Cabo Frio
e Armação dos Búzios), uma área de quase 60.000 m² (Figuras 6, 7 e 8), que
apresenta um expressivo nível conchífero subordinado ao intervalo pelítico (Senra et
al., 2003; Senra & Silva e Silva, 2005). Este intervalo que se destaca pela sua
singularidade geológica e pelo estado de preservação ambiental resultante de um
evento transgressivo-regressivo marinho ocorrido no Holoceno (Castro et al., 2006).
Estes depósitos foram originados na formação de um grande sistema lagunar,
por ocasião da fase de submersão, que antecedeu o nível máximo de 5.100 anos
A.P.. O litoral foi invadido pelo mar, formando sistemas lagunares, que se

23
estabeleceram por detrás de ilhas-barreiras como na desembocadura do rio Paraíba
do Sul ou nos cursos inferiores dos rios Itabapoana, Macaé e São João.
Alguns pontos representam depósitos provenientes de manguezais, pois são
caracterizados por sedimentos, em geral pelíticos, localmente arenosos e, quase
sempre, ricos em matéria orgânica, podendo conter fragmentos de madeira e
conchas de moluscos (Martin et al., 1997). Durante esse período, o nível relativo do
mar na região do Pântano da Malhada situava-se em torno de 5,0 m acima do nível
atual no centro da paleolaguna e 0,5 m na borda (Castro et al., 2006).
Análises anteriores efetuadas no Pântano da Malhada registraram sedimentos
estuarinos, que correspondem parcialmente à atual planície litorânea de Cabo Frio,
acumulados diretamente sobre rochas do embasamento cristalino ou do Grupo
Barreiras (Senra et al., 2002). A biodiversidade das assembléias e sua análise
tafonômica mostram que pertencem a sub-ambientes relacionados aos diferentes
eventos de inundação marinha (Senra et al., 2005).
A seqüência sedimentar na área da planície litorânea de Cabo Frio contém
intervalos pelíticos, arenosos e bioclastos intercalados (Figura 7). O ambiente
deposicional foi interpretado como um ambiente estuarino com manguezal a
hipersalino, e uma laguna hipohalina e fluvial com base na microfauna de
foraminíferos obtidos em duas localidades (Rodrigues & Carvalho, 1980). Senra et
al. (2002) sugerem ambiente lagunar e, posteriormente, Bernardes & Senra (2006)
reconheceram condições estuarinas conforme a distribuição da malacofauna.
Embora não existam dados concretos sobre a causa da mortandade
generalizada das espécies de conchas, atribui-se a mesma a uma rápida regressão
ocorrida por volta de 4.900 anos A.P., gerando possivelmente condições de
hipersalinidade. O depósito de coquina situado a aproximadamente 4,0 m acima do
nível do mar atual representa evidência inquestionável de nível marinho pretérito
mais elevado (Castro et al., 2006).

24
Figura 6: Mapa de localização da área de estudo (Pântano da Malhada). Fonte: CPRM, 2001 (adaptado).
Falha Contracional

25
Figura 7: Perfil sedimentar da área de estudo.
4.1. LOCALIDADES DE COLETA
4.1.1. Reserva Tauá (22°45’45” S, 41°59’52” W)
O intervalo sedimentar é composto por argila cinza na base, seguido de um
depósito bioclasto-suportado (Figura 8B e C), densamente empacotado, a uma
profundidade de 40 cm, sobreposto por argila castanho-avermelhada, seguida uma
camada de turfosa no topo.
Neste local foram identificados os biválvios Anomalocardia brasiliana,
Dendostrea frons, Tellina sp., Ostrea cf. puelchama; os gastrópodos Bulla striata,
Nassarius vibex, Neritina virginea, Odostomia canaliculata, Acteocina candeii, e o
escafópodo Antalis disparile.

26
4.1.2. Sítio da Paz (22°45’31” S, 41°59’50” W)
Na localidade o perfil sedimentar amostrado apresenta 1,80 m de argila cinza
com o nível de bioclastos intercalado a uma profundidade de 0,40 m. A
concentração de conchas é matriz-suportada argilosa, fortemente empacotada
(Figura 8D e F). Das três localidades é a que apresentou menor diversidade nas
amostras obtidas das coletas mecânicas, enquanto a coleta manual evidenciou
espécies ainda não registradas na região.
Foram identificadas nesta localidade os biválvios Anomalocardia brasiliana,
Tellina sp., Protothaca antiqua, Lucina pectinata, Ostrea cf. puelchana e Crassostrea
rhizophorae; os gastrópodos Bulla striata e Mitrella albovittata e o escafópodo Antalis
disparile.
4.1.3. Fazenda Junqueira (22°44’55” S, 42°01’20” W)
A concentração de conchas é moderadamente empacotada, bioclasto
suportada, em matriz areno-argilosa, onde a fração areia está presente em cerca de
50% da matriz (Figura 8E). Nela foram identificados os biválvios Anomalocardia
brasiliana, Crassostrea rhizophorae, Ostrea cf. puelchana, Lucina pectinata, Tellina
sp., Protothaca antiqua, Anadara cheminitz, A. brasiliana, A. ovalis, os gastrópodos
Bulla striata, Cerithium atratum, Neritina virginea e o escafópodo Antalis disparile.

27
Figura 8: Imagens da área de estudo. (A) Vista geral do Pântano da Malhada; (B) afloramento na Reserva Tauá; (C) seção da coquina na Reserva Tauá; (D) seção da coquina no sítio da Paz; (E) Fazenda Junqueira, ponto de coleta próximo ao canal de drenagem do rio Uma; (F) ponto de coleta do sítio da Paz próximo ao canal de drenagem.

28
5. MATERIAIS E MÉTODOS
5.1. Coleta do material
As coletas foram realizadas durante as campanhas bianuais do Núcleo de
Estudos Tafonômicos-NEST/UNIRIO, entre outubro de 2002 e setembro de 2006,
fornecendo um total de oito amostras das seguintes localidades: Reserva Tauá, sítio
da Paz e fazenda Junqueira. Os procedimentos de campo incluíram medições dos
intervalos sedimentares, verificação de coordenadas geográficas (GPR) e análise
das propriedades tafonômicas.
Para designação das localidades, adotaram-se os sinais: #RT: Reserva Tauá,
#SP: sítio da Paz, #FJ: fazenda Junqueira.
Nas coletas da Reserva Tauá e Fazenda Junqueira, foram empregadas
cavadeiras manuais e o material foi acondicionado em embalagens plásticas
rotuladas. No Sítio da Paz, a coleta foi realizada com uma retroescavadeira para
retirar amostras de 68 dm3 de sedimento, das quais foram analisados 2 kg de cada.
5.2. Tratamento laboratorial
O tratamento laboratorial para todas as amostras consistiu de lavagem em
água corrente, peneiramento, triagem manual e identificação dos bioclastos até o
nível de espécie (quando possível) e análise de feições tafonômicas bioerosivas.
Foram considerados apenas os exemplares com umbo e eixo antero-posterior
completos medindo no mínimo 6,00 mm. Como o grau de desarticulação dos
biválvios não foi considerado, os termos valva e concha são empregados como
sinônimos, embora na prática tenham significados diferentes.
A análise das bioerosões nas conchas de A. brasiliana englobou: (a) medida
em milímetros do eixo antero-posterior, utilizando um paquímetro com erro de leitura
de 0,05 mm; (b) valva direita ou esquerda; (c) localização da bioerosão de acordo no
diagrama por setores (Figura 9A); e (d) identificação do tipo de perfuração (furos

29
isolados, canais em forma de U ou galerias cameradas). Já nas conchas dos
ostreóideos, consideraram-se as seguintes características: (a) medida do eixo umbo-
palial; (b) valva direita (superior) ou esquerda (inferior); (c) localização da bioerosão
de acordo com o diagrama setorizado (Figura 9B); (d) presença ou ausência de
bioerosão nas faces externa (periostraco) e interna (hipostraco) da valva; (e)
identificação do tipo de perfuração.
Figura 9: Esquemas de mediação dos eixos de (a) A. brasiliana e (b) ostreóideos. Legenda:EAP- eixo antero-posterior, EUP- eixo umbo-palial.
Para a localização das perfurações ocasionadas por poliquetas e
gastrópodos, foi elaborado um diagrama de setorização da superfície das valvas.
Para A. brasiliana, estipularam-se nove setores (Figura 10A), assim denominados:
antero-dorsal (AD), dorsal (D), postero-dorsal (PD), anterior (A), central (C), posterior
(P), antero-ventral (AV), ventral (V), postero-ventral (PV). Nos ostreóideos dois
setores (Figura 10B): central, englobando o centro da valva e seu umbo (C), e
marginal, periferia da valva (M).
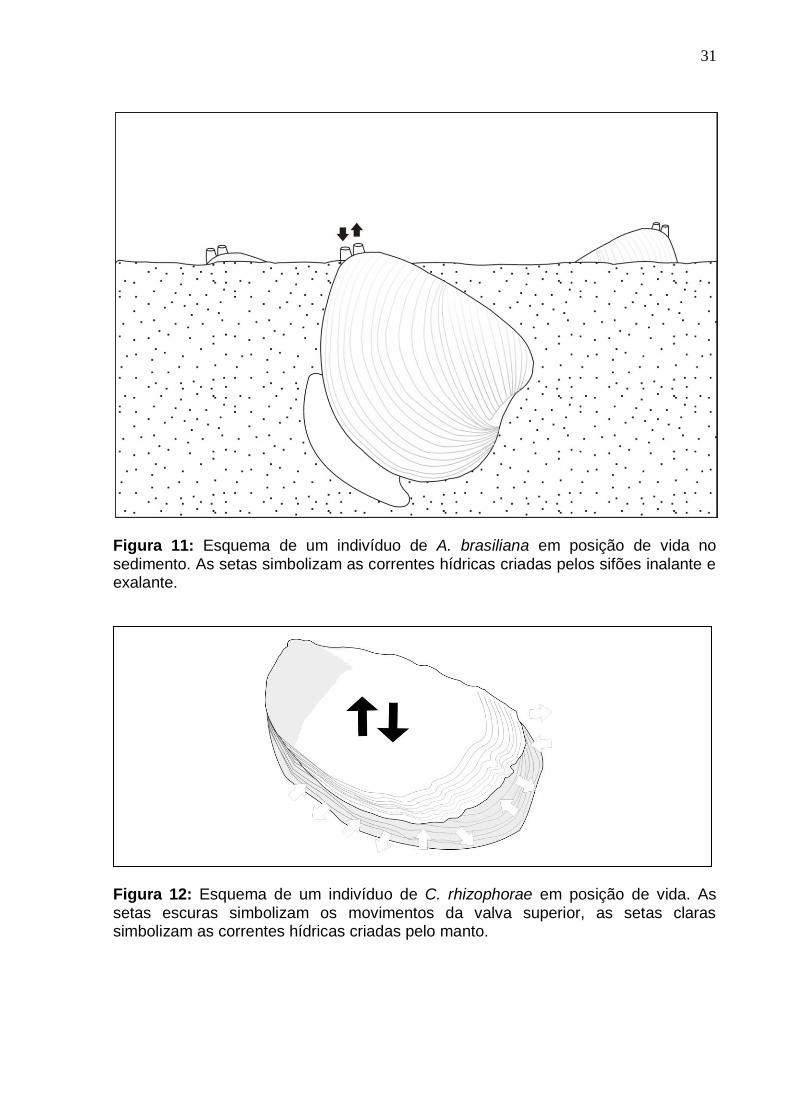
A diferença nos diagramas de setorização se deve à morfologia funcional de
cada grupo estudado (Figuras 11 e 12). Os biválvios venerídeos, ao qual pertence
à espécie A. brasiliana, apresentam hábito endobentônico semi-endobentônico,
projetando sua região posterior, onde estão posicionados seus sifões inalante e
exalante, para fora do sedimento. Já os biválvios ostreóideos possuem hábito
epibentônico, com a valva esquerda (inferior) se fixando ao substrato e a valva
A B

30
direita (superior) livre. Provocam, a partir de movimentos de abrir e fechar da valva
superior, a criação de uma bolsa dentro da cavidade do manto, onde ocorre a
movimentação da água, que se faz ao redor de toda a margem da valva.
Figura 10: Diagrama de setores para as valvas de (a) A. brasiliana e (b) ostreóideos (abreviações no texto).
Devido também à grande plasticidade morfológica das conchas de
ostreóideos, a medida do eixo antero-posterior não foi considerada, por não
representar com fidelidade a idade do indivíduo.
As ampolas nacaradas e perfurações de poliquetas foram quantificadas de
acordo com os seguintes parâmetros: raro, para conchas com infestação/perfuração
<10%; moderado, entre >10% <40%; e abundante, >40%.
A geometria das galerias produzidas por poliquetas foi classificada em
formato em U, ramificado ou linear, bem como entre transfixantes ou não.
A identificação das estruturas bioerosivas dentro da icnologia se deu através
da comparação dos dados morfológicos analisados com bibliografia específica de
descrição dos icnogêneros e icnoespécies (Häntzschel, 1975; Bromley, 1981;
Bromley & D’Alessandro, 1983, 1984).
A B
31
Figura 11: Esquema de um indivíduo de A. brasiliana em posição de vida no sedimento. As setas simbolizam as correntes hídricas criadas pelos sifões inalante e exalante.
Figura 12: Esquema de um indivíduo de C. rhizophorae em posição de vida. As setas escuras simbolizam os movimentos da valva superior, as setas claras simbolizam as correntes hídricas criadas pelo manto.

32
As estruturas bioerosivas representativas foram fotografadas em microscópio
estereoscópio da marca Olympus (modelo XTZ-5) com o auxílio do programa Image-
Pro Plus® versão 5.1.1.38, que também auxiliou nas medições digitais das
estruturas.
5.3. Tratamento estatístico
Os dados quantitativos e qualitativos das bioerosões inseridos no Microsoft
Excel serviram de base para a confecção de gráficos no programa RGui de
estatística, que possibilitou a construção de histogramas e boxplot’s, bem como
gráficos no próprio Microsoft Excel.
Os histogramas mostram como se comportam variáveis numéricas contínuas
(medidas dos eixos) e os boxplot´s mostram como essas dados se distribuem em
grupos definidos por variáveis qualitativas (e.g.: presença e ausência de bioerosão).
Além do cálculo das freqüências da bioerosão, utilizou-se teste de hipótese
onde se verificou a diferença entre duas amostras de uma mesma população. Para
isto, nos baseamos em duas possibilidades: a hipótese nula (H0), que considera não
haver diferença entre as amostras, e a hipótese alternativa (H1), que julga as duas
amostras diferentes. Quando p > 0,10, é possível afirmar que as amostras são iguais
e aceitar H0. Do contrário se p < 0,10, as amostras têm uma diferença significativa. O
valor p representa a possibilidade de se estar errando ou não na escolha das
hipóteses. Para o teste das hipóteses, foram utilizados os testes Kruskal-Wallis.
O teste Kruskal-Wallis verifica a diferença entre amostras a partir de um valor
estatístico calculado com a ordenação dos valores das amostras. Este teste é
utilizado quando a população é considerada não paramétrica, ou seja, não segue um
parâmetro definido.

33
6. Resultados
O exame das estruturas bioerosivas com base na geometria estrutural e
morfologia evidenciou seis icnoespécies dentro de quatro icnogêneros a seguir
descritos.
6.1. Icnotaxonomia
Icnogênero Entobia Bronn, 1837
Icnoespécie-tipo: Entobia cretacea Portlock, 1843
Classificação etológica: Domichnia.
Distribuição estratigráfica: Devoniano ao Holoceno.
Diagnose: Perfuração em substratos carbonáticos, consistindo de câmaras únicas
ou cadeias/redes de galerias conectadas a superfície do substrato por poucas ou
numerosas aberturas. A morfologia é fortemente modificada pela sua ontogenia. As
galerias apresentam um aumento progressivo no diâmetro durante o crescimento,
em algumas formas, as dilatações, a distâncias mais ou menos regulares, produzem
um sistema de câmaras próximas e interconectadas (“cameração”), porém em
algumas formas estruturais esta característica não se faz presente. A superfície
interna da perfuração possui uma microestrutura cuspidada que pode ser perdida em
espécimes mais maduros. Finas apófises podem aparecer em todas ou na maioria
das superfícies do sistema (Bromley & D’Alessandro, 1984).

34
Entobia paradoxa (Fischer, 1868)
Estampa 01: Fig. C, D, E e F, Estampa 04: Fig. E e F.
Diagnose: Perfuração camerada, composta nos estágios mais maduros por uma
rede de câmaras muito irregulares, com uma morfologia amebóide, com arranjo em
duas séries paralelas a superfície do substrato. Todas as câmaras são
interconectadas e o formato se torna extremamente irregular devido ao afunilamento,
como pescoços de diversos comprimentos nos espaços de substrato que separam
cada câmara da sua vizinha. Em formas mais maduras, uma fusão parcial das
câmaras permite a formação de galerias não cameradas, usualmente variável em
diâmetro e ausente de características diagnósticas. Possui aberturas com formas
circulares, geralmente com baixa densidade na superfície (Bromley & D’Alessandro,
1984).
Descrição: Câmaras de geometria muito irregular, dispostas de forma a ocupar o
máximo de espaço no substrato. Canais intercamerais reduzidos a orifícios que
comunicam a câmaras vizinhas, ou, algumas vezes, modificados em pescoços
alargados das câmaras. Nem todas as câmaras se conectam à superfície, as que
possuem conexão para o exterior a fazem por orifícios ou, com menor freqüência
através de um curto canal de abertura. Algumas câmaras estão fusionadas gerando
galerias de geometria amebóide.
Comentários: Os espécimes de A. brasiliana apresentaram estruturas do tipo E.
paradoxa, com câmaras irregulares distribuindo-se pelo interior da concha, em
estágios ontogenéticos, que segundo a classificação de Bromley & D’Alessandro
(1984) variaram da Fase A até a Fase D, não ocorrendo a Fase E. Ao microscópio
estereoscópio as faces internas das perfurações, mostraram uma microescultura em
forma de pequenas fossas semicirculares justapostas, de superfície polida com raio
(r) entre 37,97 m e 56,96 m e área média de 7.862,63 m² (Estampa 01: Figura
E).

35
Entobia laquea Bromley & D’Alessandro, 1984
Estampa 01: Fig. A e B; Estampa 02: Fig. A; Estampa 04: Fig. C e D.
Diagnose: Perfuração camerada composta, nos estágios maduros, por redes e
tramas de pequenas câmaras dispostas em séries subparalelas à superfície do
substrato. As câmaras, variáveis em formato, se afunilam abruptamente perto das
constrições que as separam das outras câmaras vizinhas. Elas estão organizadas
em cadeias curtas, mais ou menos arqueadas, que circundam pequenos espaços de
uma forma que lembram pequenos laços. As aberturas, de formatos circulares,
raramente fusionadas e ovaladas, são pequenas, numerosas e com distribuição
irregular, abrindo-se diretamente na câmara ou em um diminuto canal de abertura
(Bromley & D’Alessandro, 1984).
Descrição: Perfuração com câmaras de geometria esférica a subesférica,
distribuídas em séries que lembram cordões de contas dispostas paralelamente e a
superfície do substrato com sinuosidade dentro do substrato. Aberturas pequenas e
circulares, dispostas aleatoriamente na superfície. Canais de abertura muito curtos
ou inexistentes. Câmaras muito próximas umas das outras, fazendo com que os
canais intercamerais sejam curtos, ou mesmo, se apresentem como uma pequena
constrição entre duas câmaras vizinhas. Em alguns pontos ocorre a fusão de duas
câmaras devido ao alargamento das mesmas.
Comentários: Estrutura encontrada em conchas de C. rhizopharae e O. cf.
puelchana. A visualização da parte interna da perfuração foi facilitada pelo grau de
preservação dos bioclastos que apresenta lamelas que se desprendem umas das
outras expondo as camadas mais internas da concha. As faces internas das
câmaras se apresentaram de forma polida, sem a presença de uma microescultura
característica. As câmaras possuíam diâmetros variando entre 0,21 mm e 0,78 mm
(média 0,46 mm) com volume médio de 0,23 mm³ e suas aberturas (para o exterior)
possuíam raio variando entre 34,3 m e 240,5 m, com média de 182,0 m. Entobia

36
laquea é facilmente confundida com E. geometrica, se diferenciando da segunda
apenas pelo seu menor tamanho das câmaras. As conchas de C. rhizophorae
provenientes da fazenda Junqueira não possuem estruturas bioerosivas produzidas
por esponjas.
Icnogênero Caulostrepsis Clarke, 1908
Icnoespécie-tipo: Caulostrepsis taeniola Clarke, 1908
Classificação etológica: Domichnia.
Distribuição estratigráfica: Devoniano Inferior ao Holoceno.
Diagnose: Estruturas perfurantes tubulares, totalmente inseridas no substrato com
formato de bolsa ou orelha, produzidas por uma galeria moldada em U, possuindo
abertura única para o exterior. A superfície interna da perfuração se mostra polida,
sem qualquer ornamentação. Estruturas mais complexas podem ser produzidas pelo
desenvolvimento de múltiplos lóbulos no plano básico do U. Os ramos da galeria
podem ser distinguíveis através de toda sua extensão e separados por um tabique
central ou podem se fusionar formando uma bolsa oval ou achatada sem separação
central. Podem ocorrer estados intermediários envolvendo uma depressão axial. Na
porção distal a largura é no mínimo o dobro da espessura, o corte transversal pode
mostrar uma geometria oval-comprimida, elíptica, formato em oito ou semelhante a
um halter. Na abertura a largura é notavelmente menor que na porção distal, mas o
corte transversal deve ter o mesmo formato do corte transversal da porção proximal.
Ela pode ser modificada por ramificações superficiais ou depressões em forma da
calha, normalmente de dois a quatro divergindo da abertura (Bromley &
D’Alessandro, 1983).
Comentários: Caulostrepsis engloba seis icnoespécies e se diferencia de
Trypanites por apresentar dupla abertura ou seção transversal achatada, e de

37
Maeandropolydora pela ausência de galerias cilíndricas bem desenvolvidas e por
possuir os ramos paralelos ao longo de toda sua extensão.
Caulostrepsis taeniola, Clarke 1908
Estampa 02: Fig. C; Estampa 04: Fig. A, B e C; Estampa 05: Fig. A, B, C, D e E.
Diagnose: Galeria cilíndrica, disposta em formato de U estreito que algumas vezes
se alarga tomando um aspecto de língua. As faces internas do centro da perfuração
estão sempre interconectadas por um tabique. As galerias se fusionam próximas à
abertura. Corte transversal mediano em formato de halter e da abertura em formato
de 8 (Bromley & D’Alessandro, 1983).
Descrição: Estrutura perfurante composta por uma galeria cilíndrica dobrada ao
meio formando um U, podendo ás vezes se mostrar sinuosa ao longo do interior do
substrato. Corre paralela ao substrato em extensões variáveis, podendo às vezes
mudar de direção e atravessar a concha perpendicularmente (transfixante), entrando
em contato com sua superfície interna. As faces internas interiores da estrutura se
conectam por um pequeno espaço, formando, assim, um tabique (e não um septo
completo) que separa os dois ramos da perfuração. O corte transversal ao longo do
corpo da estrutura bem como da abertura se mostra com forma de 8 ou halter, na
região distal (base do U) sempre em forma de halter, onde há a formação de uma
pequena bolsa. A abertura pode ser perpendicular ao substrato ou formar um ângulo
inclinado em relação a ele, dando origem a pequenas calhas antes que se aprofunde
no substrato.
Comentários: Estrutura encontrada nas três espécies de moluscos estudadas. Sua
extensão e sinuosidade estão relacionadas ao tamanho da concha e disponibilidade
de substrato para a perfuração. Em alguns casos estava associado a estruturas
denominadas ampola nacarada, estrutura esta que é a resposta do molusco ao
ataque do organismo perfurador invasor. As galerias apresentaram largura média de
0,38mm (variando entre 0,23 mm e 0,53 mm) com o tabique central tendo largura
média de 0,26 mm (variando entre 0,12 mm e 0,34 mm).

38
Icnogênero Maeandropolydora Voigt, 1965
Icnoespécie-tipo: Maeandropolydora decipiens Voigt, 1965
Classificação etológica: Domichnia.
Distribuição estratigráfica: Cretáceo ao Holoceno.
Diagnose: Galerias longas e cilíndricas contendo duas ou mais aberturas, entrando
através do substrato sinuosamente, ou em contorções irregulares. As galerias
podem estar em contanto, paralelas entre si em pares, com ou sem fusão. Podem
ocorrer pequenos ou grandes giros, os ramos podem estar separados pelo tabique
ou formar uma bolsa (Bromley & D’Alessandro, 1983).
Comentários: Maeandropolydora se diferencia de Caulostrepsis por apresentar
duas aberturas distintas em posições não fusionadas.
Maeandropolydora aff. sulcans Voigt, 1965
Estampa 02: Fig. F; Estampa 05: Fig. C e D.
Diagnose: Galeria cilíndrica apresentando pelo menos duas aberturas,
irregularmente contorcida, geralmente disposta em curvas, nunca apresentando
fusão onde as paredes exibem contato mútuo; tabique ausente (Bromley &
D’Alessandro, 1983).
Descrição: Estruturas perfurantes em forma de calha, como a metade de um cilindro
esculpida no substrato formando curvas e voltas assimétricas. Diâmetro constante,

39
direção constante paralela à superfície, comprimento variável. Profundidades
variáveis, desde pequenas ranhuras no substrato até fossas moderadamente
profundas bem delimitadas, em alguns casos a perfuração entra no substrato
chegando a formar galerias internas.
Comentários: Diferentemente das estruturas descritas em Bromley & D’Alessandro
(1983), as presentes no Pântano da Malhada se apresentam pequenas e algumas
vezes associadas a perfurações do tipo Caulostrepsis, indicando que talvez sejam
perfurações incompletas deste icnogênero. Possuem pouca sinuosidade e larguras
que variavam entre 0,37 mm e 0,30 mm com média de 0,34 mm.
Icnogênero Oichnus Bromley, 1981
Icnoespécie-tipo: Oichnus simplex Bromley, 1981
Classificação etológica: Praedichnia.
Distribuição estratigráfica: Cambriano ao Holoceno.
Diagnose: Furos com geometria de circular a subcircular, de origem biogênica em
substratos duros. A perfuração pode atravessar o substrato penetrando-o, ou
simplesmente terminar dentro dele como uma depressão rasa ou profunda (Bromley,
1981).
Comentários: Por se tratarem de estruturas de origem predatória ou parasitária
(Bromley, 1981; Baumiller, 1996; Nielsen et al., 2003), são restritas a substratos
carbonáticos esqueletais, como conchas de moluscos e braquiópodes, tecas de
foraminíferos e carapaças de equinodermos.

40
Oichnus simplex Bromley, 1981
Estampa 03: Fig. B, D e E.
Diagnose: Oichnus possuindo forma circular ou subcircular simples, eixo mais ou
menos perpendicular a superfície do substrato. Quando esta estrutura não penetra
por completo o substrato o final distal é achatada e hemisférica (Bromley, 1981).
Descrição: Furos circulares, cônicos, perpendiculares à superfície da concha,
atravessando-a de um lado ao outro. Diâmetro do furo na face externa da concha
(periostraco) sempre maior que o diâmetro na face interna (hipostraco). Paredes
lisas sem ornamentações, ou evidenciando a microestrutura da concha.
Comentários: Estruturas presentes em A. brasiliana, C. rhizophorae e O. cf.
puelchana. No material estudado ocorre sempre de maneira individual e completa
(com sucesso), nunca em duplicidade, preferencialmente no centro das valvas de
venerídeos e em ostreóideos nas proximidades das margens. O diâmetro da
circunferência externa apresentou medidas entre 2,24 mm e 3,08 mm e o diâmetro
da circunferência interna apresentou medidas entre 1,68 mm e 2,40 mm.
Oichnus paraboloides Bromley, 1981
Estampa 03: Fig. A, C e F.
Diagnose: Oichnus tendo um formato esférico parabolóide, truncado, nos casos em
que a perfuração atravessa diretamente o substrato (Bromley, 1981).
Descrição: Furos circulares e semicirculares, parabólicos ou esféricos,
perpendiculares à superfície da concha, atravessando-a de um lado ao outro, ou

41
terminado em fundo cego com pequena elevação central. Diâmetro do furo na face
externa da concha (periostraco) sempre maior que o diâmetro do final da perfuração.
Paredes parabólicas, lisas sem ornamentações, ou evidenciando a ultraestrutura da
concha.
Comentários: Estruturas presentes nas três espécies estudadas. Ocorrem sempre
de maneira única, nunca em duplicidade e distribuição nas conchas similar a O.
simplex. Dois exemplares representavam tentativa de predação sem sucesso, pois o
furo não chegava a atravessar a concha por completo (Estampa 03: Figuras C e D).
Diâmetro externo com tamanho máximo de 3,48 mm e mínimo de 2,12 mm ,
diâmetro interno (nos furos completos) com medida máxima de 1,74 mm e 2,26 mm.
6.2. Análise quantitativa das populações de moluscos representadas nas
coquinas
As amostras provenientes das localidades forneceram exemplares (#RT:
n=1216, #SP: n=308, #FJ: n=172) de A. brasiliana, cujas medidas do EAP
demonstraram a estrutura etária da população estudada (Figura 13).
Figura 13: Histogramas de distribuição das medidas do eixo antero-posterior de A. brasiliana nas três localidades.
Para o total de estrutura bioerosivas encontradas em A. brasiliana a
perfuração por esponja apresentou 18,75%, perfuração por poliquetas 79,75% e
predação por gastrópodo 1,50%.

42
Figura 14: Freqüências das estruturas bioerosivas de cada produtor, em A. brasiliana bioerodidas por localidade de coleta (abreviações no texto).
Figura 15: Freqüências das estruturas bioerosivas de cada produtor em valvas bioerodidas de ostreóideos por localidade de coleta. Legenda: Cr: C. rhizophorae, Op: O. cf. puelchana (demais abreviações no texto).
Para os ostreóideos as estruturas bioerosivas se comportaram da seguinte
forma: 3,45%; 71,43% e 25,125 de esponjas, poliquetas e gastrópodos
respectivamente.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
#RT #SP #FJ Total
Esponja
Poliqueta
Gastrópodo
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Cr #SP Cr #FJ Op #FJ Total
Esponja
Poliqueta
Gastrópodo

43
6.3. Análise quantitativa das estruturas bioerosivas
A partir das medições obtidas do eixos EAP (de A. brasiliana nas três
localidades) e EUP (C. rhizophorae de #SP), e do registro dos traços de perfuração
obtiveram-se os diagramas boxplot´s indicando as diferenças nos subgrupos
ausente e presente (Figuras 16 e 17).
Figura 16: Boxplot’s de distribuição das valvas de A. brasiliana nas localidades agrupadas segundo presença e ausência bioerosão de esponja e poliquetas.

44
As diferenças entre os grupos ausente e presente para as bioerosões de
esponjas em A. brasiliana foram corroborados pelo teste Krukal-Wallis que apontou
os seguintes valores de p: 1,72∙10-4 e 0,028 para #RT e #FJ respectivamente. Este
mesmo teste desconsiderou a diferença entre os grupos ausente e presente de
perfuração de esponjas em #SP com valor de p igual a 0,84.
Para as bioerosões de poliquetas em A. brasiliana o teste apontou os
seguintes valores de p: 2,47∙10-6; 9,7∙10-3 e 0,009 em #RT, #SP e #FJ
respectivamente para os grupos ausente e presente.
Figura 17: Boxplot’s de distribuição das valvas direitas e esquerdas de C. rhizophorae de #SP agrupadas por bioerosões de esponja e poliquetas e em ausente e presentes.
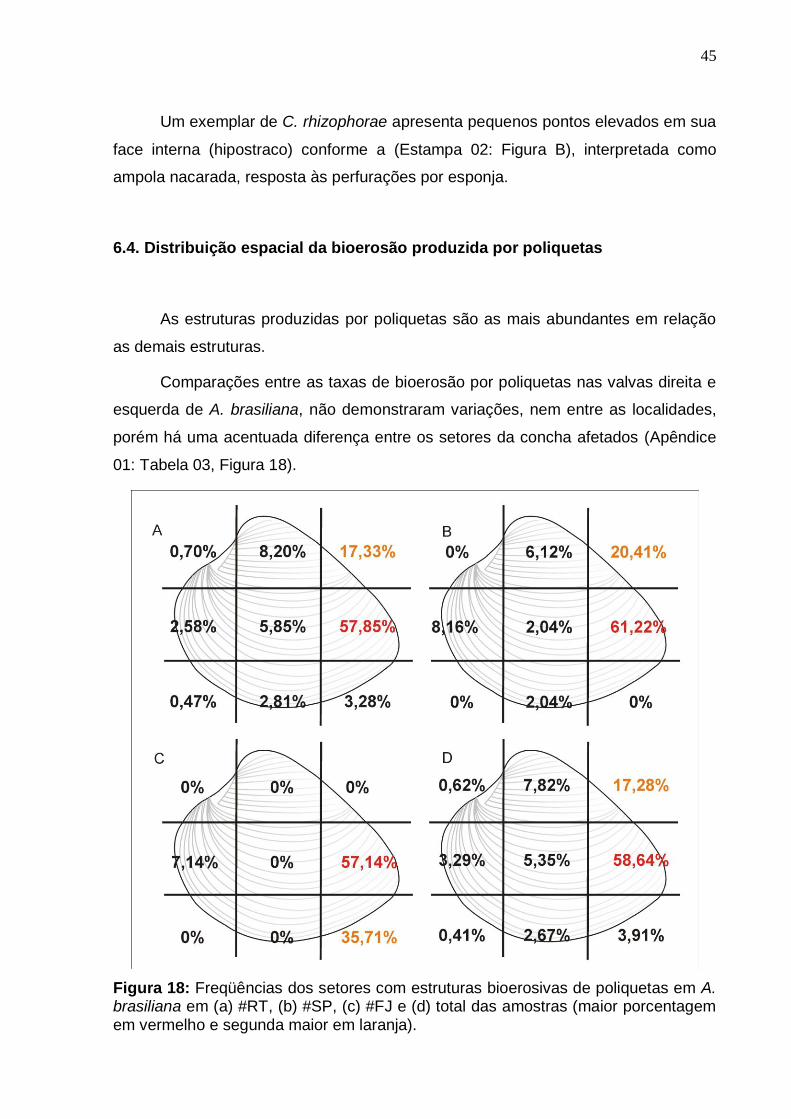
45
Um exemplar de C. rhizophorae apresenta pequenos pontos elevados em sua
face interna (hipostraco) conforme a (Estampa 02: Figura B), interpretada como
ampola nacarada, resposta às perfurações por esponja.
6.4. Distribuição espacial da bioerosão produzida por poliquetas
As estruturas produzidas por poliquetas são as mais abundantes em relação
as demais estruturas.
Comparações entre as taxas de bioerosão por poliquetas nas valvas direita e
esquerda de A. brasiliana, não demonstraram variações, nem entre as localidades,
porém há uma acentuada diferença entre os setores da concha afetados (Apêndice
01: Tabela 03, Figura 18).
Figura 18: Freqüências dos setores com estruturas bioerosivas de poliquetas em A. brasiliana em (a) #RT, (b) #SP, (c) #FJ e (d) total das amostras (maior porcentagem em vermelho e segunda maior em laranja).
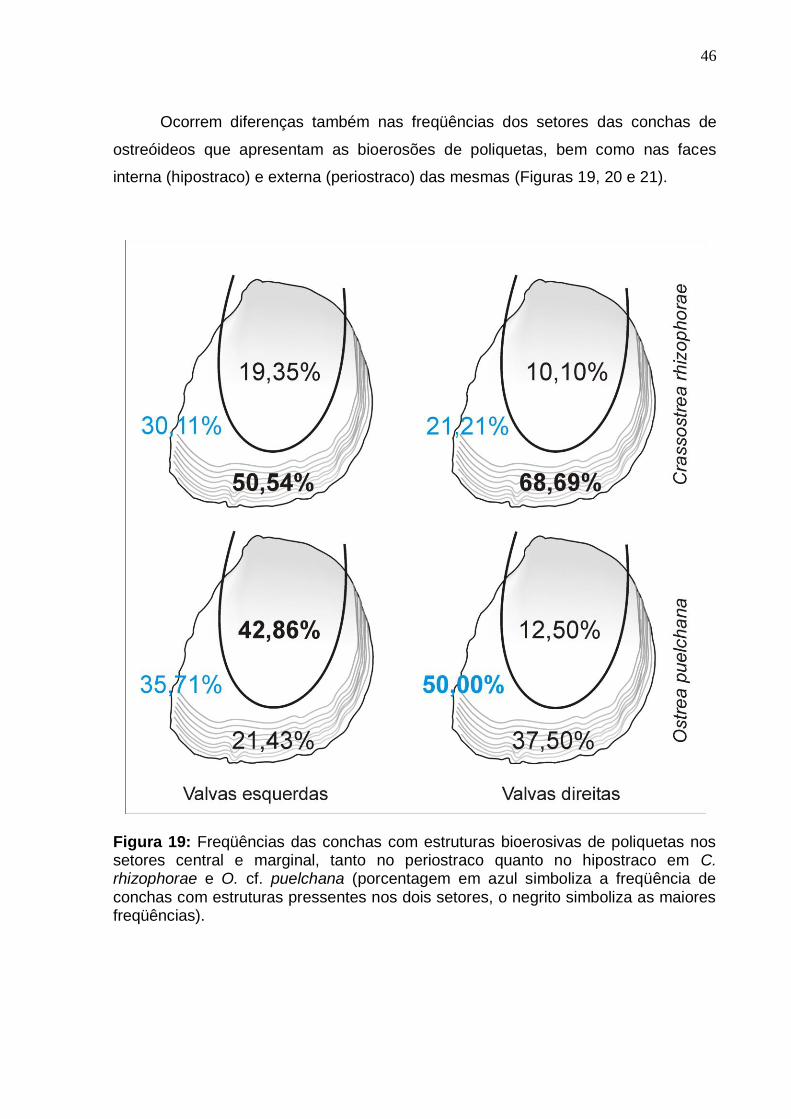
46
Ocorrem diferenças também nas freqüências dos setores das conchas de
ostreóideos que apresentam as bioerosões de poliquetas, bem como nas faces
interna (hipostraco) e externa (periostraco) das mesmas (Figuras 19, 20 e 21).
Figura 19: Freqüências das conchas com estruturas bioerosivas de poliquetas nos setores central e marginal, tanto no periostraco quanto no hipostraco em C. rhizophorae e O. cf. puelchana (porcentagem em azul simboliza a freqüência de conchas com estruturas pressentes nos dois setores, o negrito simboliza as maiores freqüências).

47
Figura 20: Freqüências das bioerosões por poliquetas na face externa (periostraco) nas valvas de ostreóideos de acordo com a quantidade de estruturas encontradas (raro, moderado e abundante).
Figura 21: Freqüências das bioerosões por poliquetas na face interna (hipostraco) nas valvas de ostreóideos de acordo com a quantidade de estruturas encontradas (raro, moderado e abundante).
0%
20%
40%
60%
80%
100%
direita esquerda direita esquerda
C. rhizophorae O. cf. puelchana
raro
moderado
abundante
0%
20%
40%
60%
80%
100%
direita esquerda direita esquerda
C. rhizophorae O. cf. puelchana
raro
moderado
abundante

48
7. DISCUSSÃO
A distribuição das medidas do eixo antero-posterior dos exemplares de A.
brasiliana convertida aos histogramas demonstra padrão populacional normal da
espécie onde predomina o número de formas jovens, reportando ainda o episódio de
morte não seletiva ou catastrófica. A representação para #FJ é invertida, com
predomínio de formas adultas, fato que pode ser explicado pelo tipo de material
coletado no entorno de tocas de crustáceos decápodes. Estes artrópodes
remobilizam bioclastos do nível de conchas até 20 cm abaixo da superfície, como
registrado na planície costeira do Paraná (Ângulo et al., 2002). Tal fato, não invalida
o estudo das perfurações tendo em vista que há representatividade de todas as
classes de tamanho e o material provém das coquinas, ou seja, têm a mesma
origem que os demais, considerando sua ampla distribuição horizontal.
Conchas de A. brasiliana de #RT com medidas do eixo antero-posterior entre
6,00 mm e 34,15 mm, média de 14,51 mm e mediana de 12,00 mm (Apêndice 01:
Tabela 01). Apresentam 4,84% bioerosão por esponjas, 22,91% bioerosão por
poliquetas e 0,16% predação por gastrópodo.
Em #SP as valvas de A. brasiliana têm as medidas do eixo antero-posterior
entre 6,00 mm e 31,00 mm, média de 14,55 mm e mediana de 13,00 mm; e 0,65%
possuíam bioerosão por esponjas, 9,42% bioerosão por poliquetas e 0,32%
predação por gastrópodo. Conchas de C. rhizophorae têm as medidas do eixo
umbo-palial entre 21,00 mm e 133,00 mm, com média de 57,08 mm e mediana de
57,00 mm (Apêndice 01: Tabela 01), 11,68% registraram bioerosão por esponjas,
33,23% bioerosão por poliquetas e 1,20% predação por gastrópodo.
Na localidade de #FJ as valvas de A. brasiliana têm as medidas do eixo
antero-posterior variando entre 7,00 mm e 32,05 mm, média de 20,45 mm e mediana
de 22,00 mm (Apêndice 01: Tabela 01), das quais 8,14% apresentavam bioerosão
por esponjas, 6,40% bioerosão por poliquetas e 1,74% predação por gastrópodo. As
conchas de C. rhizophorae apresentam as medidas do eixo umbo-palial entre 11,00
mm e 68,00 mm, média de 38,95 mm e mediana de 40,00 mm (Apêndice 01: Tabela
01), 63,16% possuíam bioerosão por poliquetas. As valvas de O. cf. puelchana

49
apresentam as medidas do eixo umbo-palial entre 9,00 mm e 33,00 mm, com média
de 17,92 mm e mediana de 17,00 mm (Apêndice 01: Tabela 01), 24,49% tem
bioerosão por esponjas, 44,90% por poliquetas e 6,12% predação por gastrópodo.
A bioerosão por esponja teve maior expressão nas valvas esquerdas do que
nas direitas (Apêndice 01: Tabela 02), tanto para a espécie C. rhizophorae (16,46%
contra 7,06%, respectivamente), quanto para O. cf. puelchana (33,33% contra
10,53%, respectivamente).
Os estágios são comuns a todas as esponjas com hábito perfurador, como
apontado por Bromley & D’Alessandro (1984) que designaram cinco fases de
crescimento, as iniciais são definidas por finos canais exploratórios radiais e a última
representada por fusão das câmaras ou sistema de túneis extensos. Em razão disso
a icnotaxonomia fica prejudicada nas duas fases sendo a classificação
icnoespecífica de Entobia possível nas fases B, C e D.
A microescultura observada nas faces internas das câmaras de Entobia
paradoxa apresenta um padrão estrutural semelhante à microescultura produzida
pela esponja perfurante do gênero Cliona durante o ato escavador, conforme
demonstrado por Calcinai et al. (2003).
A ausência da microescultura das câmaras de Entobia nas valvas de
ostreídeos, deve-se provavelmente ao padrão lamelar de crescimento da concha,
que dificulta sua preservação.
Em A. brasiliana a região posterior das valvas apresenta maior freqüência de
aberturas de tubos de poliquetas: 58,64%, no setor posterior; 17,28%, no setor
postero-dorsal e 7,82%, no setor dorsal. Enquanto que a região ventral apresenta
baixa freqüência de perfurações: 2,67%, setor anterior; 0,41%, setor antero-ventral e
3,29%, setor ventral. Estruturas de ampola nacarada foram observadas em 7,92%
das valvas com bioerosão por poliqueta. E perfurações transfixantes foram
observadas em 33,96% dos bioclastos (Apêndice 01: Tabela 03).
Já em C. rhizophorae foi observada uma diferença significativa nas
freqüências das valvas esquerdas e direitas afetadas pela bioerosão de poliqueta
(Apêndice 01: Tabela 04). Na localidade #SP há uma preferência por valvas direitas
(54,12%) em relação às esquerdas (11,59%), ao contrário de #FJ, que apresenta
preferência por valvas esquerdas (70,0%) em relação às valvas direitas (55,56%).

50
Porém quando observamos o total de valvas analisadas desta espécie, vimos que a
preferência é por valvas direitas (54,19%) em relação às valvas esquerdas (14,92%).
Em O. cf. puelchana a diferença entre as valvas bioerodidas mostrou-se irrelevante
(42,11% das direitas e 46,67% das esquerdas).
Estas bioerosões em C. rhizophorae apresentam localização com maior
freqüência na região periférica de ambas as valvas (68,69% para valvas direitas e
50,54% para as esquerdas); e na face externa, periostraco (95,96% para direitas e
89,25% para esquerdas), em relação à face interna, hipostraco (30,30% para as
direitas e 39,78% para as esquerdas) (Apêndice 01: Tabela 04). Estruturas de
ampola nacarada estão presentes em 41,41% das valvas direitas e 21,51% das
valvas esquerdas.
Foi observada diferença nos tamanhos das valvas de A. brasiliana utilizadas
como substrato pelas esponjas e poliquetas perfurantes, como evidenciam os
boxplot´s (Figura 15) montados a partir das medidas do eixo antero-posterior das
conchas com e sem a presença das perfurações. Fato este corroborado pelo teste
Kruskal-Wallis com valores abaixo de p abaixo de 0,10, confirmando a preferência
de valvas maiores pelos poliquetas perfurantes.
Em virtude da grande variedade morfológica apresentada por C. rhizophorae
e O. cf. puelchana causado pelo crescimento não isométrico, associado à pequena
quantidade amostral, os ostreóideos provenientes da #FJ não puderam ser
estudados estatisticamente na correlação entre estruturas bioerosivas e tamanho
das valvas.
Não foi observada diferença entre os tamanhos de conchas de C. rhizophorae
com e sem bioerosão de esponjas (Figura 16).
Em relação às demais espécies estudadas os ostreóideos mostram maiores
taxas de bioerosão provavelmente por se tratarem de espécies com hábito
epibentônico, sendo assim mais suscetíveis ao recrutamento das larvas planctônicas
de esponjas e poliquetas perfurantes.
Nas conchas de O. cf. puelchana a localização da bioerosão teve maior
freqüência na região periférica 37,50% para valvas direitas e região central 42,86%
para as esquerdas (Figura 18). A face externa (periostraco) apresentou maior
freqüência de estruturas bioerosivas tanto nas valvas direitas (100%) quanto nas

51
esquerdas (78,55%) em relação à face interna (hipostraco) (Figuras 19 e 20).
Estruturas de ampola nacarada estão presentes em 12,50% das valvas direitas e
7,16% das valvas esquerdas.
As conchas de O. cf. puelchana são planas ou até mesmo convexas o que
não permite acumulo de matéria orgânica, indispensável ao recrutamento larvar.
A preferência pela região posterior das valvas de A. brasiliana e marginal de
C. rhizophorae e O. cf. puelchana corroboram Tinoco-Orta & Cáceres-Martinez
(2003) que destacam a freqüência do poliqueta invasor por regiões da concha onde
se encontram as correntes hídricas produzidas pelos sifões ou manto.
No Sul e Sudeste brasileiro são conhecidas dez famílias de poliquetas.
Destas, a família Spionidae é representada por dez gêneros e quinze espécies, das
quais Polydora neocaeca Williams & Radashevsky, 1999 e P. cornuta Bosc, 1802
são as únicas encontradas no inteiro de galerias por ela perfuradas (Amaral et al.,
2006). As medidas do diâmetro e comprimento corporais destas espécies são
compatíveis com as medições encontras em Caulostrepsis e Maeandropolydora.
Perfurações por poliquetas geradoras de fraturas nas conchas (Estampa 05:
Figura E) não foram quantificadas, porém pôde-se observar que estas potencializam
a fragmentação dos bioclastos devido ao enfraquecimento da estrutura cristalina
original.
A presença de ampolas nacaradas representam o ataque do organismo
invasor quando o hospedeiro ainda se encontrava vivo, enquanto que perfurações
transfixantes representam o ataque quando o hospedeiro já se encontrava morto,
descaracterizando a exigência primordial de correntes hídricas ricas em nutrientes,
que explica a localização preferencial das perfurações.
A presença de pequenas elevações arredondadas (ampolas) na superfície
interna da valva de C. rhizophorae (Estampa 02: Figura B) com presença de Entobia
é interpretada como a resposta do organismo vivo ao ataque da esponja perfurante.
Ao atravessar a concha, a esponja entra em contato com o epitélio do manto do
molusco e o induz a secretar uma fina camada de conchiolina e carbonato de cálcio
para se isolar e proteger do organismo invasor, num processo parecido com aquele
formador de pérolas.

52
Não se encontrou literatura de ampolas nacaradas referentes ao ataque de
esponjas em conchas de moluscos, possivelmente devido a maioria do estudos
sobre bioerosão por esponjas abordarem substratos coralíneos.
As perfurações atribuídas à atividade predatória de gastrópodos muricídeos
(Oichnus simplex) foram provavelmente produzidas pela espécie Stramonita
haemastoma (Linnaeus, 1767), gastrópodo carnívoro pertencente à família
Muricidae, que apresenta registro nos sedimentos provenientes do Pântano da
Malhada.
Em A. brasiliana as perfurações se localizavam preferencialmente na região
central da valva, com um exemplar ocorrendo na região dorsal (umbonal). Possuíam
raio da circunferência externa máximo de 1,72 mm e mínimo de 1,13 mm, e da
circunferência interna máximo de 1,20 mm e mínimo de 0,84 mm. Todas as
estruturas estavam completas, e a razão entre os raios externo e interno girou em
torno de 1,38.
Já em C. rhizophorae e O. cf. puelchana estas estruturas ocorrerem sempre
na valva direita (superior), preferencialmente, na região marginal. Possuem raio
máximo externo de 1,16 mm e mínimo de 1,09 mm, o raio da circunferência interno,
quando o furo era completo, apresentava medida máxima de 0,92 mm e mínima de
0,87 mm, com razão média de 1,28 entre os raios da circunferência externa e
interna.
O. cf. puelchana é uma ostra com pequenas dimensões e relativamente
delgada, e os exemplares de Stramonita haemastoma, seu possível predador são
também pequenos, o que explica as maiores taxas de predação, nesse biválvio.
O possível produtor das estruturas O. parabolóides não pôde ser deduzido,
porém acredita-se que seja pertencente à família Naticidae, sem representante nos
sedimentos das localidades por se tratar de uma espécie de corpo mole.

53
8 -CONCLUSÕES
A bioerosão com maior expressão observada nas amostras foi produzida por
poliquetas, seguida das produzidas por esponjas e por último, a predação por
gastrópodos.
Há uma preferência pelo local de assentamento de larvas de poliquetas na região
posterior de Anomalocardia brasiliana e marginais de Crassostrea rhizophorae e
Ostrea cf. puelchana, regiões onde se formam as correntes exalantes e inalantes
das espécies.
Valvas de Anomalocardia brasiliana adultas são selecionadas para a fixação das
larvas de polidorídeos, fato este corroborado pelos testes estatísticos,
possibilitando afirmar que os poliquetas são substrato-seletivos, ao menos em
relação ao tamanho do bioclasto.
Existem variações na morfologia de Entobia nas espécies, devido à microestrutura
das conchas.
A despeito das restrições quanto à natureza do ambiente para perfurações por
esponja, estas ocorrem associadas a poliquetas que necessitam de condições
opostas.
Conchas com traços de poliquetas e esponjas concomitantes podem sugerir que
as últimas são posteriores à presença das primeiras.
As estruturas bioerosivas do icnogênero Entobia foram produzidas pelo gênero
Cliona (Demospongiae: Hadromerida: Clionaidae), gênero que não deixou registro
microscópico nos depósitos estudados.
O padrão de perfuração de Caulostrepsis e Maeandropolydora sugere que o
organismo produtor é um polidorídeo.
Há o registro indireto de gastrópodos naticídeos representadas pelas estruturas
de Oichnus paraboloides.
A identificação das icnoespécies englobadas pelo icnogênero Entobia é
complexa, pois algumas delas apresentam características muito sutis, que podem

54
facilmente ser mascaradas pela ontogenia da perfuração e pelo estenomorfismo
imposto pelo substrato.
Ampolas nacaradas não devem ser consideradas como icnofósseis, mas sim
como estruturas representativas de paleopatologias.
Este estudo é o primeiro no registro de ampolas nacaradas como reação ao
ataque de esponjas perfurantes.
Com este estudo pode-se observar que as estruturas bioerosivas por si só não
são suficientes para uma interpretação paleoambiental precisa, porém deve-se
ressaltar a sua importância como ferramenta acessória em conjunto com outros
dados (sedimentológicos, geoquímicos, etc.) para uma reconstrução mais próxima
da realidade.
A relação entre as taxas de bioerosão de poliquetas e esponjas indica um
ambiente de águas rasas, com aporte de sedimentos finos constante e altas
concentrações de matéria orgânica, durante o Holoceno da Planície Costeira de
Cabo Frio.

55
Referências Bibliográficas
ACKER, K.L.; RISK, M.J. Substrate destruction and sediment production by the boring sponge Cliona caribbea on Grand Cayman Island. Journal of sedimentary petrology, v.55, n.5, p.705-711, 1985. ALMEIDA, M.J.; MACHADO, J.; COIMBRA, J. Growth and biochemical composition of Crassostrea gigas (Thunberg) at three fishfarm earthen ponds. Journal of Shellfish Research, v.16, p.455-462, 1997. AMARAL, A.C.Z.; RIZZO, A.E.; ARRUDA, E. Manual de identificação dos invertebrados marinhos da Região Sudeste e Sul do Brasil. São Paulo: Edusp, 2006, 288p. ANGULO, R.J.; PESSENDA, L.C.R.; SOUZA, M.C. O significado das datações ao C14 na reconstrução de paleoníveis marinhos e na evolução das barreiras quaternárias do litoral paranaense. Revista Brasileira de Geociências, v.32, p.95-106, 2002. ARUA, I. Borings and shell damage in eocene gastropoda: Southern Nigeria. Palaeogeography, Palaeoclimatology, Palaeoecology, v.38, p.269-282. 1982. BARREIRO, R.M.C.; SENRA, M.C.E. Contribuição ao conhecimento da fidelidade do registro fossilífero em conchas de moluscos. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, XIX, 2005, Aracajú. CD de Resumos, Aracajú: Sociedade Brasileira de Paleontologia. BARREIRO, R.M.C.; SENRA, M.C.E. Estudo de estruturas bioerosivas produzidas por poliquetas em valvas de Anomalocadia brasiliana (Gmelin, 1789). In: JORNADA DE INICIAÇÃO CIENTÍFICA DA UNIRIO, 5, 2006, Rio de Janeiro. Resumos, Rio de Janeiro: UNIRIO, 2006. p.40-41. BARREIRO, R.M.C.; LOPES, G.N.; SENRA, M.C.E. Estudo da distribuição espacial de feições bioerosivas em conchas de Crassostrea rhizophorae (Guilding, 1828). Paleontologia em Destaque, v.53, p.32-33, 2006. BAUMILLER, T.K. Boreholes in the Middle Devonian blastoid Heteroschisma and their implications for gastropod drilling. Palaeogeography, Palaeoclimatology, Palaeoecology, v.123, p.343-351, 1996. BAUMILLER, T.K.; LEIGHTON, L.R.; THOMPSON, D.L. Boreholes in Mississippian spiriferide brachiopods and their implications for Paleozoic gastropod drilling. Palaeogeography, Palaeoclimatology, Palaeoecology, v.147, p.283-289, 1999. BERNARDES, A.P.; SENRA, M.C.E. Caracterização paleoambiental dos bioclastos holocênicos de Cabo Frio. In: Jornada de Iniciação Cientifica-UNIRIO, 5, 2006, Rio de Janeiro. Resumos, Rio de Janeiro: UNIRIO, 2006. p.1. BERTLING, M.; BRADDY, S.J.; BROMLEY, R.G.; DEMATHIEU, G.R.; GENISE, J.; MIKULAS, R.; NIELSEN, J. K.; NIELSEN, K.S.S.; RINDSBERG, A.K.; SCHLIRF, M.;

56
UCHMAN, A. Names for trace fossils: a uniform approach. Lethaia, v.39, p.265-286, 2006. BLAKE, J. A.; EVANS, J. W. Polydora and related genera as borers in mollusk shells and other calcareous substrates. The Veliger, v.15, n.3, p.235-249, 1973. BOSCOLO, R. & GIOVANARDI, O. Polydora ciliata shell infestation in Tapes philippinarum Manila clam held out of the substrate in the Adriatic sea, Italy. Journal of Invertebrate Pathology, v.79, p.97-198, 2002. BOEKSCHOTEN, G. J. Shell borings of sessile epibiontic organisms as palaeoecological guides (with) examples from the Dutch coast). Palaeogeography, Palaeoclimatology, Palaeoecology, v.2, p.333-379, 1966. BOEKSCHOTEN, G. J. Palaeoecology of some Mollusca from the Tielrolde Sands (Pliocene, Belgium). Palaeogeography, Palaeoclimatology, Palaeoecology, v.3, p.311-362, 1967. BOWER, S.M. & MCGLADDERY, S.E. Synopsis of infectious diseases and parasites of commercially exploited shellfish. Disponível em: < http://www-sci.pac.dfo-mpo.gc.ca/shelldis/title_e.htm >, acessado em: 01 de fevereiro de 2008, 2003. BROMLEY, R.C. Bioerosion of the Bermuda Reefs. Palaeogeography, Palaeoclimatology, Palaeoecology, v.23, p.169-197, 1978. BROMLEY, R.C. Concepts in ichnotaxonomy illustrated by small round holes in shells. Acta Geològica Hispànica, v.16, p.55-64, 1981. BROMLEY, R.C.; D’ALESSANDRO, A. Bioerosion in the Pleistocene of Southern Italy: ichnogenera Caulostrepsis and Maeandropolydora. Rivista Italiana di Paleontologia e Stratigrafia, v.89, n.2, p.283-309, 1983. BROMLEY, R.C.; D’ALESSANDRO, A. The Ichnogenus Entobia from the Miocene, Pliocene and Pleistocene of Southern Italy. Rivista Italiana di Paleontologia e Stratigrafia, v.90, n.2, p.227-296, 1984. BROMLEY, R.C.; D’ALESSANDRO, A. Bioerosion of the Plio-Pleistocene transgression of Southern Italy. Rivista Italiana di Paleontologia e Stratigrafia, v.93, n.3, p379-442, 1987. BROMLEY, R.C.; TENDAL, O.S. Example of substrate competition and phobotropism between two clionid sponges. Journal Zoological, v.169, p.151-155, 1973. CALCINAI, B.; ARILLO, A.; CERRANO, C.; BAVESTRELLO, G. Taxonomy-related differences in the excavating micro-patterns of boring sponges. Journal of the Marine Biological Association of the United Kingdom, v.83. p.37-39, 2003. CASTRO, J.W.A.; SENRA, M.C.E.; RAMOS,R.R.C. Coquinas da Paleolaguna da Reserva Tauá-Pântano da Malhada, RJ - Um registro de optimum climático

57
holocênico. In: WINGE,M.; SCHOBBENHAUS,C.; BERBERT-BORN,M.; QUEIROZ,E.T.; CAMPOS,D.A.; SOUZA,C.R.G.; FERNANDES,A.C.S. (Edit.), Sítios Geológicos e Paleontológicos do Brasil, 2006. Publicado na Internet em 13/09/2006 no endereço: http://www.unb.br/ig/sigep/sitio004/sitio004.pdf CINTRA-BUENROSTRO, C.E.; FLESSA, K.W.; ÁVILA-SERRANO, G. Who cares about a vanish Clam? Trophic importance of Mulinia coloradoensis inferres from predatory damage. Palaios, v.20, p.296-302, 2005. CLELLAND, E.S. Vacuolar-type ATPase in the accessory boring organ of Nucella lamellose (Mollusca: Gastropoda): role in shell penetration. 1999. 73f. Dissertação (Mestrado em Biologia) Programa de Graduação em Biologia, York University, Toronto, Ontário. CPRM. SERVIÇO GEOLÓGICO DO BRASIL. Geologia do Estado do Rio de Janeiro: Mapas. Rio de Janeiro, 2001, CD-ROM. DIETL, G.P.; ALEXANDER, R.R. Post-Miocene Shift in Stereotypic Naticid Predation on Confamilial Prey from the Mid-Atlantic Shelf: Coevolution with Dangerous Prey. Palaios, v.15, p.414-429, 2000. DUARTE, L.F.L., HOLLER, M.T. Estudos preliminares sobre a preferência de Thais haemastoma (L.) (Gastropodoa-Prosobranquia) por diferentes espécies de presas. In: SIMPÓSIO DE ECOSSISTEMAS DA COSTA SUL E SUDESTE BRASILEIRAS, 1986, Porto Alegre, p. 192-200. EKDALE, A.A. Preface. Palaeogeography, Palaeoclimatology, Palaeoecology, v.23, p.167-168, 1978. EKDALE, A.A. Paleoecology of the marine endobenthos. Palaeogeography, Palaeoclimatology, Palaecology, v.50, p.63-81. EKDALE, A.A. History of hard habitat homes: The role of bivalves in the evolution of the macroboring niche. In: INTERNATIONAL BIOEROSION WORKSHOP, 5, 2006, Erlangen, Alemanha. Programme & Abstracts, Erlangen, Alemanha, p. 10-11. FARINATI, E. A.; SPAGNUOLO, J.; ALIOTTA, S. Tafonomía comparativa de moluscos del estuario de Bahía Blanca, Argentina. In: INTERNATIONAL MEETING TAPHOS´05, 2, Barcelona. Resumos, Barcelona, p.105-106. FERNANDES, A.C.S.; BORGHI, L.; CARVALHO, I.S.; ABREU, C.J. Guia dos icnofósseis de invertebrados do Brasil. Rio de Janeiro: Editora Interciência, 2002, 260p. FÖRSTERRA, G.; BEUCK, L.; HÄUSSERMANN, V. & FREIWALD, A. Shallow-water Desmophyllum dianthus (Scleractinia) from Chile: characteristics of the biocoenoses, the bioeroding community, heterotrophic interactions and (paleo)-bathymetric implications. In: COLD-WATER CORALS AND ECOSYSTEMS. Freiwald A, Roberts JM (eds): Springer-Verlag, Berlin Heidelberg, pp 937-977, 2005.

58
GIBERT, J.M; MARTINELL, J.; DOMÈNECH, J.M. Entobia ichnofacies in fossil rocky shores, Lower Pliocene, northwestern Mediterranean. Palaios, v.13, p.476-487, 1998. GIBERT, J.M; DOMÈNECH, R.; MARTINELL, J. An ethological framework for animal bioerosion trace fossils upon mineral substrates. Palaios, v.13, p.476-487, 2004. HANDLEY, S. J.; BERGQUIST, P. R. Spionid polychaete infestations of intertidal pacific oysters Crassostrea gigas (Thunberg), Mahurangi Harbour, northern New Zealand. Aquaculture, v.153, p.191-205, 1997. HÄNTZSCHEL, W. Trace Fossils and Problematica. In: TEICHERT, R.C. (ed.), Treatise on Invertebrate Paleontology. Boulder: Geological Society of America & University of Kansas, Part W, Miscellanea, suppl. 1, 269p, 1975. HARPER, E.M. Foosils explained 50- Recognizing predator-prey interactions in the fossil record. Geology Today, v.21, n.5, p.191-196, 2005. Holmes, K.E.; Edinger, E.N.; Harijadi Limmon, G.V. & Risk, M.J. 2000. Bioerosion of live massive corals and branching coral rubble on Indonesian cral reefs. Marine Pollution Bulletin, 40:606-617. HUTCHINGS, P.A. Biological destruction of coral reefs: a review. Coral Reefs, v.4, p.329-252, 1986. ICZN. International Comission for Zoological Nomenclature. International Code of Zoological Nomenclature, adopted by the International Union of Biological Sciences, 4a ed., Londres, 1999. Disponível em: <http://www.iczn.org/iczn/index.jsp>. Acessado em: 02 de fevereiro de 2008. KOWALEWSKI, M.; DULAI, A.; FÜRSICH, F.T. The Phanerozoic history of drilling predation. Geology, v.26, n.12, p.1091-1094, 1998. LIU, P. J.; HSIEH, H. L. Burrow architecture of the spionid polychaete Polydora villosa in the corals Montipora and Porites. Zoological Studies, v.39, n.1, p.47-54, 2000. LORENZO, N.; VERDE, M. Estructuras de bioerosión em moluscos marinos de la formación Villa Soriano (Pleistoceno Tardío-Holoceno) de Uruguay. Revista Brasileira de Paleontologia, v.7, n.3, p.319-328, 2004. LYELL. C. Principles of geology: or, the modern changes of the Earth and its inhabitants, considered as illustrative of geology. Boston: Hilliard, Gray & Co. v.3. 475p., 1842. MCDIARMID, H.; DAY, R.; WILSON, R. The ecology of polychaetes that infest abalone shells in Victoria, Australia. Journal of Shellfisheries Research, v.23, n.4, p.1179-1188, 2004.

59
MARTIN, D.; BRITAYEV, T. A. Simbiotic polychaetes: review of known species. Oceanography and Marine Biology Annual Review, v.36, p.217-340, 1998. MARTIN, L.; SUGUIO, K.; DOMINGUEZ, J.M.L.; FLEXOR, J-M. Geologia do Quaternário Costeiro do Litoral Norte do Rio de Janeiro e do Espírito Santo. Belo Horizonte: CPRM. 112pp.:II, mapas, 1997. MORTENSEN, S.; VAN DER MEEREN, T.; FOSSHAGEN, A.; HERNAR, I.; HARKESTAD, L.; TORKILDSEN, L. & BERGH, O. Mortality of scallop spat in cultivation, infested with tube dwelling bristle worms, Polydora sp. Aquaculture international, v.8, p.267-271, 2000. NEUMANN, A.C. Observations on coastal erosion in Bermuda and measurements of the boring rate of the esponge, Cliona lampa. Limnolgy and Oceanography, v.11, n.1, p.92-108, 1966. NIELSEN, K.S.S.; NIELSEN, J.K.; BROMLEY, R.G. Palaeoecological and ichnological significance of microborings in Quaternary Foraminifera. Palaentologia Electronica, v.6, n.2, 13p. 2003. PALMER, T.J.; WILSON, M.A. Parasitism of ordovician Bryozoans and the origin of pseudoborings. Palaeontology, v.31, n.4, p.939-949, 1988. PERRY, C.T. Macroborings of Pleistocene coral communities, Falmouth Formation, Jamaica. Palaios, v.15, p.483-491, 2000. PICKERILL, R.K., DONOVAN, S.K; PORTELL, R.W. Bioerosional Trace Fossils from the Miocene of Cariacou, Lesser Antilles. Caribbean Journal of Science, v.38, p.106-117, 2002. RODRIGUES, M.A.C.; CARVALHO, M.G.P. Interpretação paleoecológica da planície costeira do Rio São João, RJ, com base em foraminíferos. Anais da Academia Brasileira de Ciências, v.52, v.4, p.763-771, 1980. ROOPNARINE, P.D.; BEUSSINK, A. Extiction and naticid predation of the bivalve Chione von Mühlfeld in the Late Neogene of Florida. Palaeontogia Electronica, v.2, n.1, 1999. url: http://palaeo-electronica.org/1999_1/bivalve/issue1_99.htm. Acessado em julho de 2007. RÜTZLER, K. The role of burrowing sponges in bioerosion. Oecologia,v.19, p.203-216, 1975. RÜTZLER, K. Impact of Crustose Clionid Sponges on Caribbean Reef Corals. Acta Geològica Hispànica, v.37, n.1, p.61-72, 2002. SEBESVARI, Z.; ESSER, F.; HARDER, T. Sediment-associated cues for larval settlement of the infaunal spionid polychaetes Polydora cornuta and Streblospio benedicti. Journal of experimental marine biology and ecology, v.337, p.109-120, 2006.

60
SENRA, M.C.E.; MELLO, K.; JUSTO, R.; BATISTA, D.S.; ASSIS, C.M. & MARCELLA, C. Composição biótica dos depósitos holocênicos da Praia Rasa-Reserva Tauá, Municípios de Armação do Búzios – Cabo Frio. Estado do Rio de Janeiro. Paleontologia em Destaque, v.40, p.33, 2002. SENRA, M.C.E.; BARREIRO, R.M.C.; ASSIS, C.M.; BATISTA, D.S.; SILVA E SILVA, L.H. Tafonomia e paleoambientes do Pântano da Malhada – Reserva Tauá, planície costeira de Armação dos Búzios/Cabo Frio (Quaternário Superior) estado do rio de Janeiro, Brasil. In: CONGRESSO DA ASSOCIAÇÃO BRASILEIRA DE ESTUDOS DO QUATERNÁRIO, IX, Recife, 2003, CD de Trabalhos Completos, Recife, 2003. SENRA, M.C.E.; SILVA E SILVA, L.H.; BATISTA, D.S.; BARREIRO, R.M.C.; ASSIS, C.M. Assossiações fauníticas, ficoflora e sedimentos em depósitos holocênicos da baixada litorânea do Estado do Rio de Janeiro. In: JORNADA FLUMINENSE DE PALEONTOLOGIA, I, Rio de Janeiro, 2004. Paleonotícias, Rio de Janeiro, v.36, p.15-18, 2004. SENRA, M.C.E.; SILVA E SILVA, L.H. Taphonomy of Quaternary molluscan assemblages, Reserva Tauá, Rio de Janeiro, Brazil. In: 2nd International Meeting TAPHO’S 05/ 4a Reunión de Tafonomia y Fossilización, Barcelona, 2005. Abstract Volume, Barcelona, p.139-140, 2005. SILINA, A. V. Long-term variation of Polychaete bioerosion of the scallop Mizuhopecten yessoesis from the northwest part of the Sea of Japan. Russian Journal of Marine Biology, v.29, n.6, p.390-394, 2003. SILINA, A. V. Tumor-like formations on the shells of Japanese scallops Patinopecten yessoesis (Jay). Marine Biology, v.148, p.833-840, 2006. SILVA, L.C.; CUNHA, H.C.S. Geologia do Estado do Rio de Janeiro: texto explicativo do mapa geológico do Estado do Rio de Janeiro, Brasília: CPRM, 2001, CD-ROM. SUGUIO, K. Tópicos de geociências para o desenvolvimento sustentável: as regiões litorâneas. Revista do Instituto de Geociências da USP, v. 1, p. 1-40, 2003. TAPANILA, L. Palaeocology and diversity of endosymbionts in Paleozoic marine invertabrates: Trace fossil evidence. Lethaia, v.38, p.89-99, 2005. TAPANILA, L. Discussion session: using bioerosion trace fossils in evolutionary paleoecology. In: INTERNATIONAL BIOEROSION WORKSHOP, 5, Erlangen, 2006. Programme & Abstracts, Erlangen, Alemanha, p. 12-13, 2006a. TAPANILA, L. Devonian Entobia borings from Nevada, with a revision of Topsentopsis. Journal of Paleontology, v.80, p.760-767, 2006b. TAYLOR, P.D. Preservation of soft-bodied and other organisms by bioimmuration – a review. Palaeontology, v.33, n.1, p.1-17, 1990.

61
TAYLOR, P.D.; WILSON, M.A. Palaeoecology and evolution of marine hard substrate communities. Earth Science Reviews, v.62, p.1-103, 2003. TAYLOR, P.D.; WILSON, M.A.; BROMLEY, R.G. A new Ichnogenus for etchings made by cheilostome bryozoans into calcareous substrates. Palaeontology, v.42, n.4, p.595-604, 1999. TINOCO-ORTA, G. D.; CÁCERES-MARTÍNEZ, J. Infestation of the clam Chione fluctifraga by the burrowing worm Polydora sp. nov. in laboratory conditions. Journal of Invertebrate Pathology, v.83, p.196-205, 2003. WALKER, S.E.; GOLDSTEIN, S.T. Taphonomic tiering: experimental field taphonomy of mollusks and foraminifera above and below the sediment – water interface. Palaeogeography, Palaeoclimatology, Palaeoecology, v.149, p.227-244, 1999. WARD-PAIGE, C.A.; RISK, M.J.; SHERWOOD, O.A.; JAAP, W.C. Clionid sponge surveys on the Florida Reef Tract suggest land-based nutrient inputs. Marine Pollution Bulletin, v.51, p.570-579, 2005. WARME, J.E. Borings as trace fossils, and the processes of marine bioerosion. In: FREY, R.W. (ed.), The Study of Trace Fossils. New York: Springer-Verlag. p.181-227, 1975. WISSHAK, M.; NEUMANN, C. A symbiotic association of a boring polychaete and an echinoid from the Late Cretaceous of Germany. Acta Palaeontologica Polonica, v.51, n.3, p.589–597, 2006. ZUNDELEVICH, A. LAZAR, B.; ILAN, M. Chemical versus mechanical bioerosion of coral reefs by boring sponges – lessons from Pione cf. vastifica. The Journal of Experimental Biology, v.210, p. 91-96, 2007.

62
GLOSSÁRIO
Abertura: abertura na superfície do substrato que conecta o lúmen da perfuração com o exterior. No icnogênerio Entobia representa o ósculo da esponja vivente.
Apófises: pequenas projeções em formato de agulha que se extendem de todas as superfícies da perfuração de esponjas. Câmaras: porções dilatadas, globulares ou fusiformes, nos sistemas de perfurações por esponjas. Camerada: morfologia das perfurações de esponjas que apresentam partes alargadas ao longo dos canais, lembrando um cordão de contas, que formarão mais tarde as câmaras propriamente ditas. Canal de entrada: canais subcilíndricos ou suavemente cônicos que conectam as câmaras ou galerias com as aberturas na superfície do substrato. Canal intercameral: canais subcilíndricos que interconectam as câmaras. Canal exploratório: a porção mais juvenil da perfuração de esponja, onde predomina o crescimento linear, consistindo de canais finos e ramificados. Endolítico: qualidade do organismo que vive dentro de substratos litificados (rochas, conchas, carapaças, etc.) e que apresenta habilidade perfuradora. Estenomórfico: morfologia da perfuração que é restringida pelo espaço disponível no substrato, por exemplo conchas de moluscos. Galeria: passagem principal da perfuração, onde vive o organismo endolítico. Idiomórfico: característica peculiar a perfuração representada pelas diferenças nos grupos produtores. Microescultura: características morfológicas microscópicas da superfície da perfuração. Tabique: região central elevada em forma de mureta que divide os canais e câmaras das pefurações. Endosimbionte: organismo que vive dentro de um hospedeiro ou de sua carapaça, sem, contudo, ser prejudicial a ele. Bioimuração: do inglês bioimmuration, se trata de um modo de preservação que resulta do crescimento orgânico sobre um organismo séssil. Organismos de corpo mole (ou com pouquíssima mineralização) podem se preservar quando um outro organismo séssil (de carapaça mineralizada) cresce sobre seu corpo, formando assim um molde externo do organismo substrato.
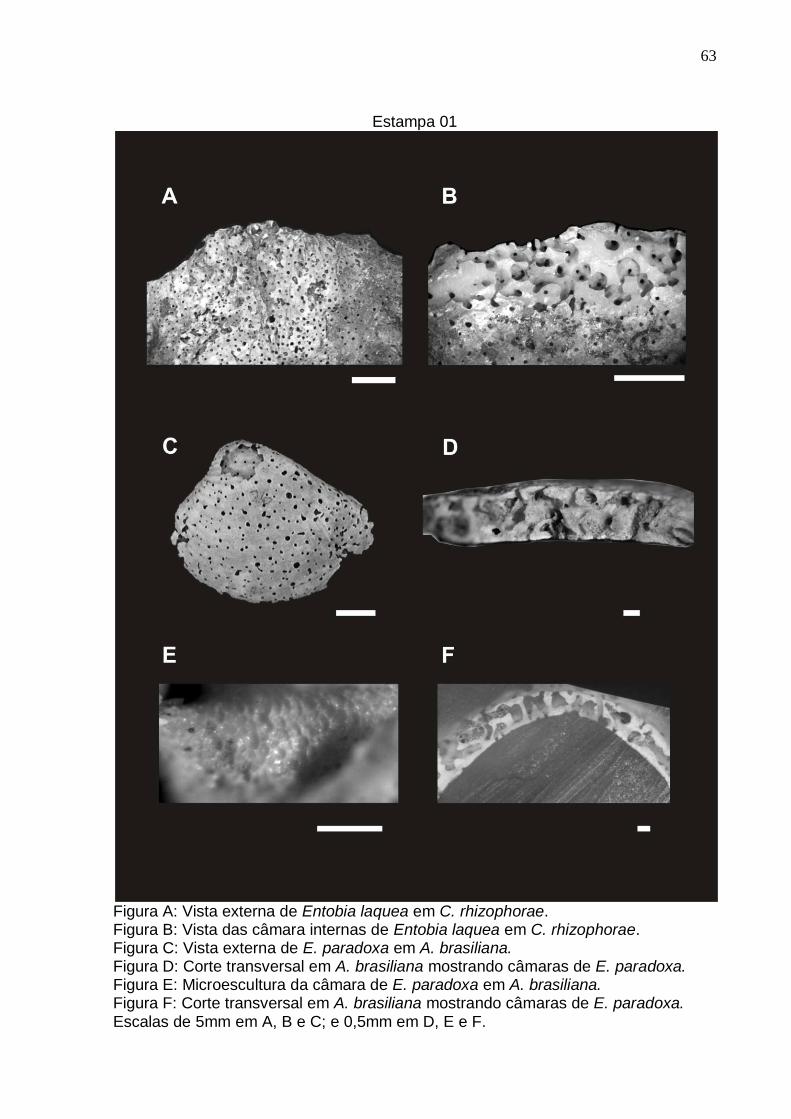
63
Estampa 01
Figura A: Vista externa de Entobia laquea em C. rhizophorae. Figura B: Vista das câmara internas de Entobia laquea em C. rhizophorae. Figura C: Vista externa de E. paradoxa em A. brasiliana. Figura D: Corte transversal em A. brasiliana mostrando câmaras de E. paradoxa. Figura E: Microescultura da câmara de E. paradoxa em A. brasiliana. Figura F: Corte transversal em A. brasiliana mostrando câmaras de E. paradoxa. Escalas de 5mm em A, B e C; e 0,5mm em D, E e F.
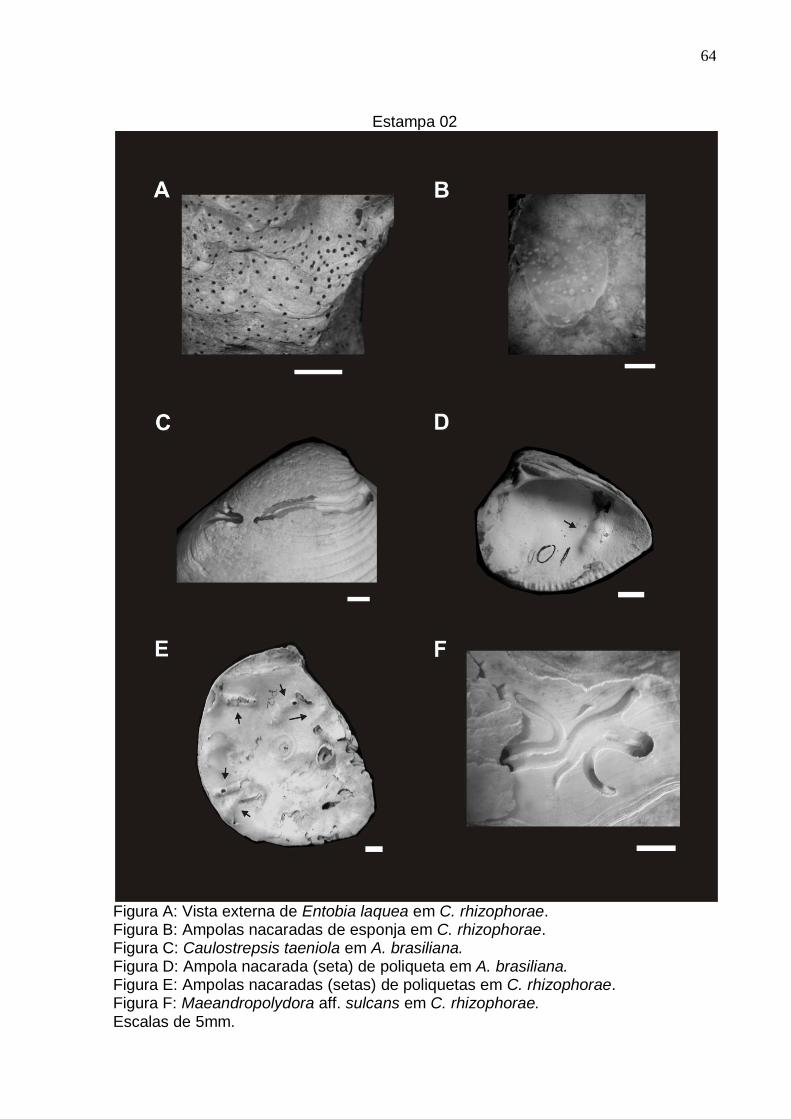
64
Estampa 02
Figura A: Vista externa de Entobia laquea em C. rhizophorae. Figura B: Ampolas nacaradas de esponja em C. rhizophorae. Figura C: Caulostrepsis taeniola em A. brasiliana. Figura D: Ampola nacarada (seta) de poliqueta em A. brasiliana. Figura E: Ampolas nacaradas (setas) de poliquetas em C. rhizophorae. Figura F: Maeandropolydora aff. sulcans em C. rhizophorae. Escalas de 5mm.
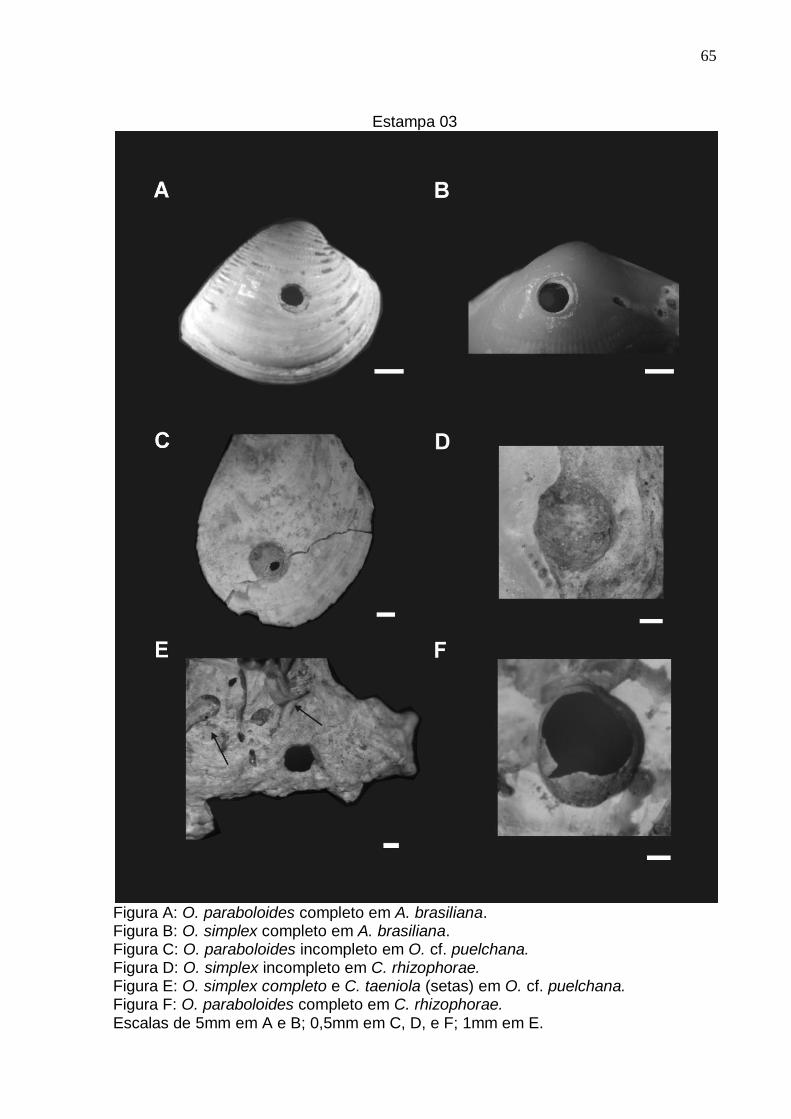
65
Estampa 03
Figura A: O. paraboloides completo em A. brasiliana. Figura B: O. simplex completo em A. brasiliana. Figura C: O. paraboloides incompleto em O. cf. puelchana. Figura D: O. simplex incompleto em C. rhizophorae. Figura E: O. simplex completo e C. taeniola (setas) em O. cf. puelchana. Figura F: O. paraboloides completo em C. rhizophorae. Escalas de 5mm em A e B; 0,5mm em C, D, e F; 1mm em E.

66
Estampa 04
Figura A: Aberturas de C. taeniola em C. rhizophorae. Figura B: C. taeniola em O. cf. puelchana. Figura C: C. taeniola associado a E. laquea em C. rhizophorae. Figura D: Fase B de E. laquea em C. rhizophorae. Figura E: Corte transversal de câmara de E. paradoxa em A. brasiliana. Figura F: E. paradoxa em A. brasiliana. Escalas de 5mm em A, B e F; 1mm em C e D; 0,5mm em E.

67
Estampa 05
Figura A: Aberturas de C. taeniola na região posteriro de A. brasiliana. Figura B: Detalhe Aberturas de C. taeniola em A. brasiliana. Figura C: A. brasiliana com C. taeniola e M. aff. sulcans. Figura D: M. aff. sulcans na região posteiror A. brasiliana. Figura E: Perfuração transfixante de poliqueta em A. brasiliana. Figura F: C. taeniola no hipostraco de A. brasiliana. Escalas de 5mm em A, B, C e D; 1cm em E e F; 0,5mm em E.
A B
C D
E F
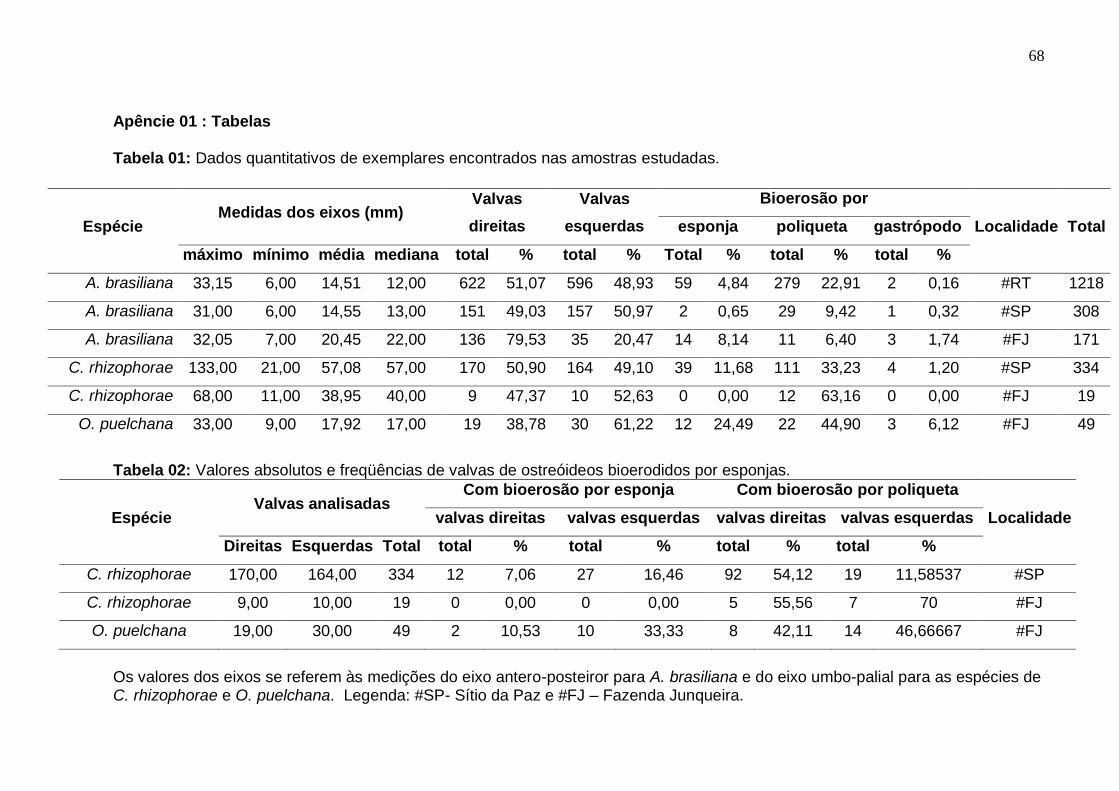
68
Apêncie 01 : Tabelas Tabela 01: Dados quantitativos de exemplares encontrados nas amostras estudadas.
Tabela 02: Valores absolutos e freqüências de valvas de ostreóideos bioerodidos por esponjas.
Espécie Valvas analisadas
Com bioerosão por esponja Com bioerosão por poliqueta
Localidade valvas direitas valvas esquerdas valvas direitas valvas esquerdas
Direitas Esquerdas Total total % total % total % total %
C. rhizophorae 170,00 164,00 334 12 7,06 27 16,46 92 54,12 19 11,58537 #SP
C. rhizophorae 9,00 10,00 19 0 0,00 0 0,00 5 55,56 7 70 #FJ
O. puelchana 19,00 30,00 49 2 10,53 10 33,33 8 42,11 14 46,66667 #FJ
Os valores dos eixos se referem às medições do eixo antero-posteiror para A. brasiliana e do eixo umbo-palial para as espécies de C. rhizophorae e O. puelchana. Legenda: #SP- Sítio da Paz e #FJ – Fazenda Junqueira.
Espécie Medidas dos eixos (mm)
Valvas
direitas
Valvas
esquerdas
Bioerosão por
Localidade Total esponja poliqueta gastrópodo
máximo mínimo média mediana total % total % Total % total % total %
A. brasiliana 33,15 6,00 14,51 12,00 622 51,07 596 48,93 59 4,84 279 22,91 2 0,16 #RT 1218
A. brasiliana 31,00 6,00 14,55 13,00 151 49,03 157 50,97 2 0,65 29 9,42 1 0,32 #SP 308
A. brasiliana 32,05 7,00 20,45 22,00 136 79,53 35 20,47 14 8,14 11 6,40 3 1,74 #FJ 171
C. rhizophorae 133,00 21,00 57,08 57,00 170 50,90 164 49,10 39 11,68 111 33,23 4 1,20 #SP 334
C. rhizophorae 68,00 11,00 38,95 40,00 9 47,37 10 52,63 0 0,00 12 63,16 0 0,00 #FJ 19
O. puelchana 33,00 9,00 17,92 17,00 19 38,78 30 61,22 12 24,49 22 44,90 3 6,12 #FJ 49
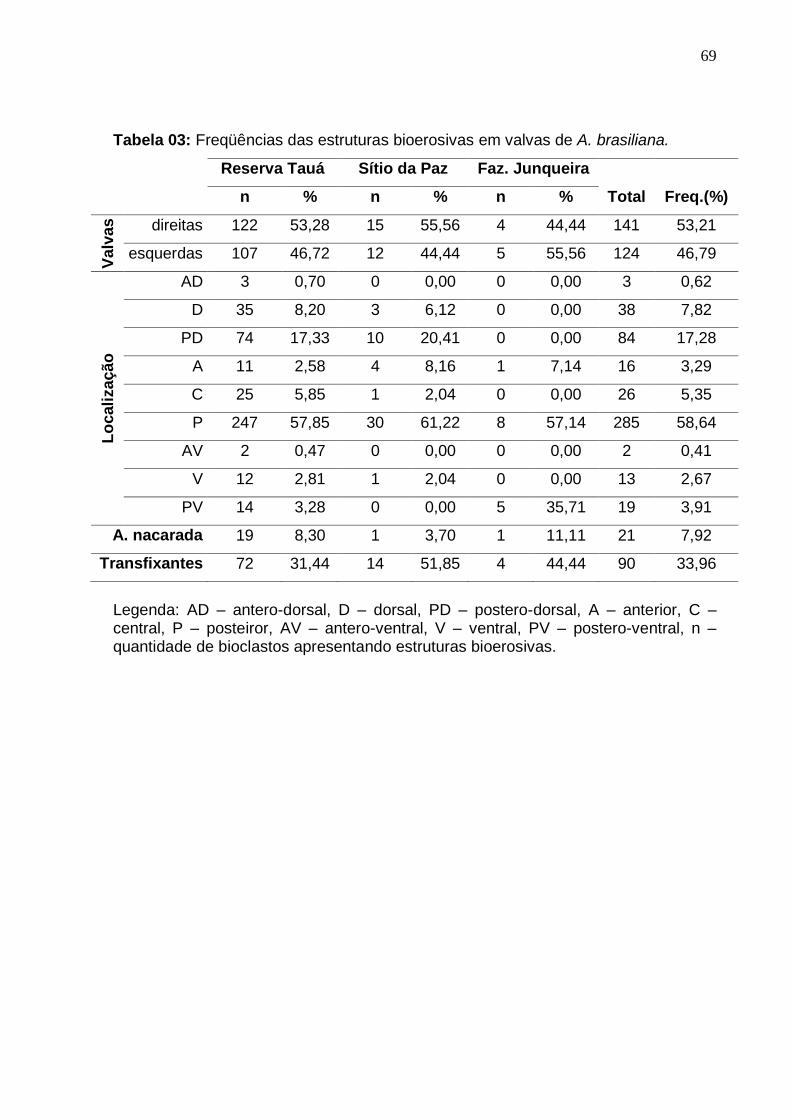
69
Tabela 03: Freqüências das estruturas bioerosivas em valvas de A. brasiliana.
Legenda: AD – antero-dorsal, D – dorsal, PD – postero-dorsal, A – anterior, C – central, P – posteiror, AV – antero-ventral, V – ventral, PV – postero-ventral, n – quantidade de bioclastos apresentando estruturas bioerosivas.
Reserva Tauá Sítio da Paz Faz. Junqueira
Total Freq.(%) n % n % n %
Valv
as
direitas 122 53,28 15 55,56 4 44,44 141 53,21
esquerdas 107 46,72 12 44,44 5 55,56 124 46,79
Lo
cali
zação
AD 3 0,70 0 0,00 0 0,00 3 0,62
D 35 8,20 3 6,12 0 0,00 38 7,82
PD 74 17,33 10 20,41 0 0,00 84 17,28
A 11 2,58 4 8,16 1 7,14 16 3,29
C 25 5,85 1 2,04 0 0,00 26 5,35
P 247 57,85 30 61,22 8 57,14 285 58,64
AV 2 0,47 0 0,00 0 0,00 2 0,41
V 12 2,81 1 2,04 0 0,00 13 2,67
PV 14 3,28 0 0,00 5 35,71 19 3,91
A. nacarada 19 8,30 1 3,70 1 11,11 21 7,92
Transfixantes 72 31,44 14 51,85 4 44,44 90 33,96

70
Tabela 04 : Freqüências das estruturas bioerosivas de poliquetas em valvas de ostreóideos.
Crassostrea rhizophorae Ostrea puelchana
direita % esquerda % direita % esquerda %
Localização
central 10 10,10 18 19,35 1 12,50 6 42,86
periferico 68 68,69 47 50,54 3 37,50 3 21,43
ambas 21 21,21 28 30,11 4 50,00 5 35,71
Periostraco
abundante 13 13,13 11 11,83 4 50,00 7 50,04
moderado 25 25,25 20 21,51 4 50,00 2 14,22
raro 57 57,58 52 55,91 0 0,00 2 14,29
total 95 95,96 83 89,25 8 100,00 11 78,55
Hipostraco
abundante 0 0,00 4 4,30 0 0,00 5 35,67
moderado 4 4,04 5 5,38 0 0,00 0 0,00
raro 26 26,26 28 30,11 0 0,00 0 0,00
total 30 30,30 37 39,78 0 0,00 5 35,67
Ampola
nacarada
abundante 4 4,04 1 1,08 0 0,00 0 0,00
moderado 9 9,09 3 3,23 0 0,00 0 0,00
raro 28 28,28 16 17,20 1 12,50 1 7,16
total 41 41,41 20 21,51 1 12,50 1 7,16





























