Embed Size (px)
Citation preview

UNIVERSIDADE CIDADE DE SÃO PAULO
PROGRAMA DE MESTRADO EM FISIOTERAPIA
SAMIRA SALAME
NEUROPLASTICIDADE INDUZIDA POR
DIFERENTES PERÍODOS DE TREINAMENTO DE
EXERCÍCIO ACROBÁTICO
SÃO PAULO
2013

SAMIRA SALAME
NEUROPLASTICIDADE INDUZIDA POR
DIFERENTES PERÍODOS DE TREINAMENTO DE
EXERCÍCIO ACROBÁTICO
Dissertação apresentada ao Programa de Mestrado em Fisioterapia da Universidade Cidade de São Paulo, como requisito para obtenção do título de Mestre, sob orientação da Profa. Drª. Raquel Simoni Pires .
SÃO PAULO
2013

Ficha Elaborada pela Biblioteca Prof. Lúcio de Souza. UNICID
S159n
Salame, Samira
Neuroplasticidade induzida por diferentes períodos de treinamento de exercício acrobático. / Samira Salame. --- São Paulo, 2013.
89 p.
Bibliografia
Dissertação (Mestrado) – Universidade Cidade de São Paulo - Orientadora: Profa. Dra. Raquel Simoni Pires
1. Plasticidade neural. 2. Proteína 1 de resposta de crescimento precoce. 3. Proteínas do tecido nervoso. 4. Exercícios. I. Pires, Raquel Simoni, oriente. II. Título.
CDD 615.82

SAMIRA SALAME
Neuroplasticidade induzida por diferentes períodos de
treinamento de exercício acrobático
Dissertação apresentada ao
Programa de Mestrado em
Fisioterapia da Universidade
Cidade de São Paulo, como
requisito para obtenção do
título de Mestre
A Comissão Julgadora dos trabalhos de Defesa da Tese de Mestrado, em
sessão pública realizada a ......../........../............., considerou
( ) Aprovada ( ) Reprovada
Examinadora:.........................................................................................................
Profª.Drª Rosana Macher Teodori
Universidade Metodista de Piracicaba
Examinadora:.........................................................................................................
Profª.Drª Sandra Regina Alouche
Universidade Cidade de São Paulo
Presidente:.............................................................................................................
Profª.Drª Raquel Simoni Pires
Universidade Cidade de São Paulo

Dedico primeiramente à Deus,
e a minha querida Família.

Agradecimentos
Agradeço primeiramente a Deus por conceder-me a vida, por ser a razão do meu viver, por andar sempre comigo mostrando-me o melhor caminho a seguir, por me abençoar e ter dado sabedoria para que eu pudesse alcançar aquilo que almejei. Pois sei, que este sonho já estava no coração Dele. Agradeço a todos da minha família por serem meus maiores incentivadores e por estarem comigo sempre. Aos meus Pais, meus fãs número um, que vibram com minhas vitórias, que estão sempre comigo durante as dificuldades, que me colocam para cima quando me faltam forças e que sempre me fazem sentir a melhor pessoa do mundo! Agradeço também aos meus irmãos Jihad e Ramza, os mais lindos que eu pude ter. Eu os amo e sou eternamente grata, pelo simples fato deles existirem em minha vida e torná-la mais completa e feliz. Agradeço também a minha Tia Walide por ser minha segunda mãe, acompanhando-me sempre em todas as etapas da minha vida e dando todo o apoio quando foi preciso, além de todo o incentivo necessário na minha educação com seu jeito forte e marcante! E a minha querida Vó Luiza, por seu imensurável amor, carinho e suas orações incessantes por mim, elas funcionaram, deu tudo certo Vózinha! (risos) Agradeço ao Pr. Leonilton, que já faz parte da família, pelas orações e apoio sempre!! Agradeço a minha orientadora, “Prof” (como costumo chamar) Raquel pelos ensinamentos valiosos, incentivo, pelas palavras de apoio e ânimo, por acreditar sempre em mim, pela amizade e por toda ajuda concedida. Agradeço minha companheira de trabalho e maninha Juliana (Ju), por tudo! Pelos experimentos que realizamos juntas, pela força que sempre me deu, pelas palavras amigas na hora do sufoco e desânimo. Mas também, por todas as risadas e brincadeiras de todos os dias! Sentirei falta!

Agradeço ao Prof. Britto, chefe do Laboratório USP por conceder o espaço para a realização da pesquisa, e por todo o ensinamento e disposição! Agradeço também as colegas da USP pelas colaborações, sugestões, ensinamentos, por estarem sempre dispostas a me ajudar: Carol, Priscila e Ana! Muito Obrigada! Agradeço ao Adilson, técnico do Laboratório por toda a ajuda com os experimentos, pela paciência e alegria sempre!! Obrigada!! Agradeço a Prof.ª Andréia por permitir que eu usufruir-se dos equipamentos do seu laboratório quando foi preciso! Agradeço aos amigos da USP pelas brincadeiras, pelo acolhimento e pela amizade: Gabi, Marina, Lucíla, Francis, Mauro, Jáfia, Cecília, Vivian, Taísa, Erika, Kallene, Cleyton e em especial ao André e Danilo, a dupla dinâmica, pelas risadas, pela ajuda, pelas distrações, amizade e incentivo sempre, muito obrigada! Aos amigos da Unicid: Andressa, Indiara, Naiane, Flávia Manfredi, Diego, Anderson, Quilza e Wellington pela amizade e apoio sempre! Aos colegas e Professores (Profª Sandra Freitas, Profª Mônica, Profª Sandra Alouche e Profª Raquel) do grupo de Neuro da Unicid pelos ensinamentos e contribuições essenciais para o meu crescimento! À minha Fonoaudióloga Cláudia por toda ajuda, paciência e carinho em todo esse tempo! Às minhas amigas da Faculdade: Renata Kalil e Renata Hernandes que sempre farão parte da minha vida! Obrigada por toda força mesmo de longe! E aos meus amigos: Marluci, Karen, Dri, Mayara, Alê Ataíde, Ellys, Arly , Alex, Polly, Samantha e Samuel por de alguma forma me ajudarem a vencer! Alguns pela distância eu nunca vejo, mas são sempre muito presentes na minha vida! Agradeço a FAPESP pelo auxílio financeiro! Obrigada...

SUMÁRIO
LISTA DE ILUSTRAÇÕES ................................................................................................................ 16
LISTA DE ABREVIATURAS E SIGLAS .............................................................................................. 18
RESUMO ...................................................................................................................................... 20
ABSTRACT .................................................................................................................................... 22
1. INTRODUÇÃO ........................................................................................................................ 7
1.1. Neuroplasticidade x Exercício ................................................................................................ 7
1.1.1. Modalidade do Exercício Físico ........................................................................................... 8
1.1.2. Períodos de treinamento: Curto e Longo prazo ............................................................... 11
1.2. Neurogênese Hipocampal adulta ......................................................................................... 13
1.3. Neuroanatomia Funcional .................................................................................................... 14
1.3.1. Córtex Motor ..................................................................................................................... 15
1.3.2. Estriado ............................................................................................................................. 16
1.3.3. Cerebelo ............................................................................................................................ 17
1.3.4. Hipocampo ........................................................................................................................ 20
1.4. Marcadores de Plasticidade ................................................................................................. 20
1.4.1. Proteínas Sinápticas e Proteínas Estruturais ..................................................................... 21
1.4.2. Gene Egr1 .......................................................................................................................... 23
2. JUSTIFICATIVA ..................................................................................................................... 25
3. OBJETIVOS ........................................................................................................................... 25
3.1. Objetivo Geral ...................................................................................................................... 25
3.2. Objetivos Específicos ............................................................................................................ 25
4. MATERIAL E MÉTODO ......................................................................................................... 26
4.1. Animais ................................................................................................................................. 26
4.2. Desenho experimental ......................................................................................................... 26
4.2.1. Protocolo de Exercício Acrobático .................................................................................... 26
4.3. Protocolo de Imuno‐histoquímica ........................................................................................ 27

4.4. Protocolo de Avaliação da Neurogênese pelo “Doublecortin” (DCX) .................................. 30
4.5. Protocolo de “Immunoblotting” .......................................................................................... 31
4.6. Análise Estatística ................................................................................................................. 32
5. RESULTADOS ........................................................................................................................... 33
5.1. Desempenho Motor ............................................................................................................. 33
5.2. Expressão de Proteínas Estruturais e Sinápticas .................................................................. 34
5.2.1. Córtex Motor ..................................................................................................................... 36
5.2.2. Estriado ............................................................................................................................. 40
5.2.3. Cerebelo ............................................................................................................................ 44
5.3. Expressão de Egr1 ................................................................................................................ 52
5.3.1. Córtex Motor ..................................................................................................................... 52
5.3.2. Estriado ............................................................................................................................. 52
5.3.3. Cerebelo ............................................................................................................................ 52
5.4.1 Hipocampo ......................................................................................................................... 56
6. DISCUSSÃO .............................................................................................................................. 57
6.1. Comportamento do Desempenho Motor ............................................................................ 57
6.2. Efeitos dos Diferentes Tempos de Treinamento em Regiões Motoras ............................... 58
6.2.1. Córtex Motor ..................................................................................................................... 58
6.2.2. Estriado ............................................................................................................................. 61
6.2.3. Cerebelo ............................................................................................................................ 63
6.2.4. Hipocampo ........................................................................................................................ 66
7. CONCLUSÕES ........................................................................................................................... 67
8. REFERÊNCIAS ........................................................................................................................... 68
ANEXO I ....................................................................................................................................... 78

LISTA DE ILUSTRAÇÕES
Figura 1 – Imagens digitais dos obstáculos que compõem o circuito para as
atividades acrobáticas.......................................................................................27
Figura 2 - Desempenho dos ratos em diferentes períodos de treinamento
acrobático avaliado pela média do tempo gasto para realização das seis tarefas
presentes no circuito por semana......................................................................33
Tabela 1 – Níveis de proteínas estruturais e sinápticas nos diferentes períodos
de treinamento de exercício acrobático.............................................................35
Figura 3 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína associada ao microtúbulo (MAP2) no córtex
motor..................................................................................................................37
Figura 4 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a expressão de neurofilamento (NF) no córtex motor...........38
Figura 5 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína sinapsina I (SYS) no córtex motor........................39
Figura 6 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína sinaptofisina (SYP) no córtex motor......................41
Figura 7 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína associada ao microtúbulo (MAP2) no
estriado..............................................................................................................42
Figura 8 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a expressão de neurofilamento (NF) no estriado...................43
Figura 9 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína sinapsina I (SYS) no estriado................................45

Figura 10 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína sinaptofisina (SYP) no estriado.............................46
Figura 11 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína associada ao microtúbulo (MAP2) no
cerebelo.............................................................................................................47
Figura 12 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a expressão de neurofilamento (NF) no cerebelo..................49
Figura 13 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína sinapsina I (SYS) no cerebelo...............................50
Figura 14 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre a proteína sinaptofisina (SYP) no cerebelo............................51
Figura 15 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre o gene de resposta precoce ao crescimento (Egr1) no córtex
motor..................................................................................................................53
Figura 16 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre o gene de resposta precoce ao crescimento (Egr1) no
estriado..............................................................................................................54
Figura 17 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre o gene de resposta precoce ao crescimento (Egr1) no
cerebelo.............................................................................................................55
Figura 18 – Efeitos dos diferentes períodos de treinamento de exercício
acrobático sobre o “doublecortin” (DCX) no hipocampo....................................56

LISTA DE ABREVIATURAS E SIGLAS
- AMS: Área Motora Complementar
- AC: Exercício Acrobático
- AC1: Treinamento de Exercício Acrobático por 1 semana
- AC2: Treinamento de Exercício Acrobático por 2 semanas
- AC4: Treinamento de Exercício Acrobático por 4 semanas
- AE: Ambiente Enriquecido
- BDNF: Fator Neurotrófico Derivado do Encéfalo
- Cb: Cerebelo
- CG: Camada Granular
- CM: Camada Molecular
- CaM-K: Cálcio calmodulina/quinase
- DCX: “Doublecortin”
- DG: Giro Denteado
- DLS: Estriado Dorsolateral
- DMS: Estriado Dorsomedial
- EE: Exercício em Esteira
- EG: Externamente Guiado
- Egr1: Gene de Resposta Precoce ao Crescimento
- EV: Exercício Voluntário
- GluR: Receptor de Glutamato
- GPe: Globo Pálido Externo
- GPi: Globo Pálido Interno
- IF: Filamentos Intermediários
- IG: Internamente Guiado
- LTP: Potenciação a Longo Prazo
- MAP: Proteína Associado ao Microtúbulo
- MF: Microfilamento de Actina
- MT: Microtúbulo
- NB: Núcleos da Base
- NF: Neurofilamento

- NT: Neurotransmissor
- SED: Sedentário
- SGV: Zona Subgranular
- SNC: Sistema Nervoso Central
- SYP: Sinaptofisina
- SYS: Sinapsina
- SVZ: Zona Subventricular

RESUMO
Exercícios realizados a curto e a longo prazo podem induzir mudanças
funcionais e estruturais distintas nas áreas encefálicas envolvidas com controle
motor. Portanto, o estudo visou avaliar a expressão das proteínas sinapsina I
(SYS), sinaptofisina (SYP), MAP2 e neurofilamento de pequeno peso molecular
(NF68) além do gene Egr1 nas regiões do córtex motor, estriado e cerebelo e
níveis de neurogênese no hipocampo de ratos adultos jovens submetidos a
diferentes períodos de treinamento de exercícios acrobáticos. O desempenho
motor também foi avaliado. Para tal, utilizamos 76 ratos (2 meses), Wistar,
aproximadamente 250g, divididos aleatoriamente em 2 grupos: exercício
acrobático (AC=57) e controle-sedentário (SED=19). O grupo submetido ao
exercício acrobático foi subdividido em 3 grupos com diferentes períodos de
treinamento: 1semana (AC1), 2 semanas (AC2) e 4 semanas (AC4), com 5
passagens pelo circuito acrobático, 3 vezes na semana. Para analisar a
expressão de proteínas nas áreas motoras envolvidas com a coordenação,
equilíbrio e planejamento foi utilizada a técnica de “immunoblotting” e imuno-
histoquímica. Para o desempenho motor mediu-se o tempo na execução de
cada tentativa. As comparações entre os grupos foram conduzidas com o teste
ANOVA para medidas repetidas, seguida do pós-teste de Tukey quando a
análise de variância apresentou diferença estatisticamente significante
(p≤0,05). Observou-se na análise do desempenho motor que os animais
pertencentes aos grupos AC1, AC2 e AC4 mostraram melhora do desempenho
motor com a diminuição do tempo na execução do exercício acrobático a partir
da segunda semana. Em nossos resultados, na região do córtex motor, a curto
prazo de treinamento houve diminuição de MAP2 e aumento de SYP e a longo
prazo houve aumento de MAP2 e SYP com aumento progressivo de células
Egr1-positivas ao longo do período de treinamento. No estriado, o treinamento
a curto prazo de atividades motoras complexas induziu aumento de NF, SYP e
SYS na região dorsomedial e aumento de MAP2, SYP e SYS na região
dorsolateral e a longo período de treinamento aumento de MAP2, SYP e SYS
na região dorsomedial e somente de MAP2 na região dorsolateral dos animais
treinados. Juntamente com maior marcação de celulas Egr1-positivas na região

dorsomedial com pico na segunda semana de treinamento (AC2) e retornando
aos níveis basais. E no cerebelo, a curto prazo houve aumento de NFs nas
duas camadas estudadas e de SYP na camada CM e a longo prazo aumento
somente de NFs juntamente com o aumento de marcação de células
granulares Egr1-positivas com pico na primeira semana de treinamento (AC1).
Não houve alteração significativa na contagem de células DCX-positivas no
hipocampo. Desta forma, nosso estudo sugere que os diferentes tempos de
treinamento modulam proteínas sinápticas e estruturais e o gene Egr1 de forma
distinta nas áreas encefálicas, desempenhando um importante papel na
plasticidade tempo-dependente em regiões motoras.
Palavras chave: Neuroplasticidade, gene Egr1, proteínas estruturais, proteínas
sinápticas, exercício acrobático

ABSTRACT
Exercises performed in the short and long term can induce functional and
structural changes in distinct brain areas involved in motor control. Therefore,
this study aimed to evaluate the expression of proteins synapsin I (SYS),
synaptophysin (SYP), MAP2 and low molecular weight neurofilament (NF68)
beyond the Egr1 gene regions of the motor cortex, striatum and cerebellum and
levels of neurogenesis in the hippocampus of young adult rats submitted to
different periods of training of acrobatic exercises. Motor performance was also
evaluated. Therefore, we used 76 rats (2 months), Wistar approximately 250g
were randomly divided into 2 groups: exercise Acrobatic (AC = 57) and control-
sedentary (SED = 19). The group submitted the acrobatic exercise was divided
into 3 groups with different training times: 1week (AC1), 2 weeks (AC2) and 4
weeks (AC4), with 5 passes through the circuit acrobatic, 3 times a week. To
analyze the expression of proteins involved in motor areas with balance,
coordination and planning technique was used for "immunoblotting" and
immunohistochemistry. To motor performance was measured execution time in
each trial. Comparisons between groups were performed with ANOVA for
repeated measures not followed by the Tukey post-test when the analysis of
variance showed a statistically significant difference (p ≤ 0.05). It was observed
in engine performance analysis that the animals belonging to the groups AC1,
AC2 and AC4 showed increased engine performance with reduced time in
acrobatic performance of the exercise after the second week. In our results, the
region of the motor cortex, short-term training of decreased MAP2 and SYP
increase and long-term increased by MAP2 and SYP with progressive increase
in Egr1-positive cells throughout the training period. In the striatum, the short-
term training of complex motor activities induced increase in NF, SYP and SYS
in the region dorsomedial and increased MAP2, SYP and SYS in dorsolateral
and long training period increased MAP2, SYP and SYS in the region and
dorsomedial MAP2 only in the dorsolateral region of the trained animals. Along
with higher markup Egr1-positive cells in the dorsomedial region with a peak in
the second week of training (AC2) and returning to baseline levels. And in the
cerebellum, increased short term NFS studied in two layers and SYP layer CM

and long-term increase only NFs along with the increase in labeling of cells with
granular Egr1 positive peak during the first week of training (AC1) . There was
no significant change in cell count DCX-positive in the hippocampus. Thus, our
study suggests that the different training times and modulate synaptic proteins
and structural gene Egr1 differently in brain areas play an important role in time-
dependent plasticity in motor regions.
Keywords: Neuroplasticity, Egr1 gene, Structural Proteins, Synaptic Proteins,
Acrobatic exercise

7
1. INTRODUÇÃO
1.1. Neuroplasticidade x Exercício
Durante muito tempo, acreditou-se que o sistema nervoso central (SNC),
após seu desenvolvimento, tornava-se uma estrutura rígida, que não poderia
ser modificada, e que lesões neste seriam permanentes, pois suas células não
poderiam ser reconstituídas ou reorganizadas. Hoje sabemos da capacidade
de modificação do SNC e da grande adaptabilidade em função de suas
experiências ou como tentativa de regeneração1-3.
A plasticidade neural refere-se à capacidade que o SNC possui em
modificar algumas das suas propriedades morfológicas e funcionais em
resposta às alterações do ambiente. Na presença de lesões, o SNC utiliza-se
desta capacidade na tentativa de recuperar funções perdidas3-5 e/ou,
principalmente, fortalecer funções similares relacionadas às originais.
Nas últimas décadas alguns estudos também têm se dedicado ao
entendimento das bases neurobiológicas do exercício físico para manter e
melhorar as funções cerebrais. Evidências mostram que exercícios físicos
trazem inúmeros benefícios ao sistema nervoso de humanos6,7 e animais8-11,
com consequentes melhoras no aprendizado, na memória e na plasticidade do
sistema nervoso como resposta ao exercício 9,12-14.
O exercício físico acelera cascatas celulares e moleculares que
aumentam e mantém a plasticidade cerebral, induz expressão de genes
associados à plasticidade, pode promover neurogênese e aumento da
vascularização e metabolismo cerebral9,15,16.
As adaptações que o exercício físico causa no sistema nervoso têm
implicações na prevenção e tratamento de obesidade, câncer, depressão,
declínio cognitivo associado ao envelhecimento, mostrando que em animais
envelhecidos há melhora da memória espacial, aumentando a neurogênese e
suprimindo a apoptose no giro denteado17,18, e ameniza os distúrbios
neurológicos gerados pela doença de Parkinson, doença de Alzheimer,
acidente vascular isquêmico e lesões medulares ou encefálicas19,20.

8
1.1.1. Modalidade do Exercício Físico
Diversos estudos têm mostrado que diferentes modalidades de exercício
físico geram um mecanismo de neuroproteção, promovendo redução da
apoptose no hipocampo21,22 e aumento da sinaptogênese no córtex motor 23 de
ratos pós lesão encefálica, além de facilitar a recuperação funcional pós lesão
medular11,24.
Os roedores são os principais modelos animais de estudo para os
paradigmas do exercício físico nas funções cerebrais e seus mecanismos.
Vários modelos de exercícios vêm sendo desenvolvidos a fim de simular os
exercícios físicos realizados por seres humanos9,12,25-27. Esses modelos de
exercícios físicos em animais têm protocolos que incluem diferentes
intensidades, frequência de treinamento contínua e intermitente, e realizados
em um período de curta e longa duração25.
Os principais modelos de atividade física estudados são: exercício
voluntário28-31, ambientes enriquecidos4,32,33, exercícios forçados como natação 34-36e esteira26,37-40, e exercício acrobático40-43. O tipo de atividade ou tarefa, e a
forma como esta é realizada tem diferentes efeitos sobre o encéfalo44.
No exercício voluntário (EV) os animais executam a atividade com
tempo, frequência e intensidade definidos por eles mesmos. Por exemplo, ao
correr em uma roda dentro do seu ambiente, podemos observar que alguns
ratos correm vários quilômetros por dia, enquanto outros pouco se envolvem na
atividade. É um exercício realizado geralmente de forma intermitente, com
curtos períodos de alta a baixa intensidade durante um período de 12h
noturnas25,28.
Vários estudos que envolvem o EV avaliam a plasticidade neural em
áreas relacionadas a aprendizagem motora e memória. O aumento do fator
neurotrófico derivado do cérebro (BDNF), de genes associados à plasticidade
sináptica, excitabilidade celular e de MAP Kinase foram observados no
hipocampo e suas subáreas9,45-47. Vaynman e colaboradores38 demonstraram
que esse tipo de atividade promove um aumento dos níveis de proteínas
sinápticas (sinapsina I e sinaptofisina) sendo que a sinapsina I mostrou uma
correlação positiva com a quantidade de exercícios executados. E mais, que

9
essas respostas das proteínas sinápticas frente ao EV são controladas pelo
BDNF.
O EV mostrou ser capaz de aumentar a proliferação celular no giro
denteado do hipocampo e a capacidade de induzir a potenciação à longo prazo
(LTP)48, além de alterar a atividade de neurotransmissores nos encéfalos de
ratos. Soares e colaboradores49 observaram aumento de noradrenalina no
córtex frontal de ratos adultos após 4 a 5 semanas de treinamento, ressaltando
a importância deste tipo de exercício em áreas relacionadas à memória.
Vários estudos em mamíferos tiveram foco nas respostas encefálicas
frente à experiência ambiental. Um dos paradigmas mais utilizados para
demonstrar esse fenômeno é o ambiente enriquecido (AE)4,12. O AE
tipicamente envolve a exposição de roedores a uma variedade de estímulos,
tais como brinquedos, túneis, rodas de correr e interações sociais com outros
roedores. Estes estudos têm fornecido informações importantes sobre os
mecanismos envolvidos com aprendizagem e memória, visto que o ambiente
enriquecido pode produzir uma grande variedade de alterações morfológicas e
funcionais nas regiões específicas do cérebro, tal como o hipocampo e o
neocórtex12.
O enriquecimento ambiental aumenta as ramificações dendríticas,
contatos sinápticos, neurotransmissão e o tamanho dos neurônios na área do
neocórtex, além de favorecer a potenciação à longo prazo (LTP)12 no
hipocampo, a neurogênese4,50, os níveis de neurotrofinas e sinaptofisina, e o
gene de expressão CREB12,51.
Outro modelo de exercício físico realizado em ratos é a natação. Apesar
das críticas quanto à temperatura, ao estresse do esforço forçado e ao tempo
possível de submersão dos animais durante o exercício, a natação tem sido
muito empregada, pois é uma habilidade inata aos ratos e os estudos
realizados utilizando esse modelo revelam a ocorrência de adaptações ao
treinamento físico semelhante àquelas observadas em humanos52. No entanto,
uma limitação do modelo é a dificuldade de precisão da sobrecarga de
esforço53.
No exercício físico forçado em esteira (EE) os animais o executam de
acordo com as determinações do pesquisador, que estabelece a velocidade,

10
inclinação e tempo de treinamento25. Este exercício é capaz de promover
mudanças no SNC como neurogênese e supressão da apoptose no hipocampo
de ratos idosos, prevenindo assim o déficit de memória ocorrida com o
envelhecimento17,18. O exercício em esteira a curto e a longo prazo induz
mudanças distintas na expressão dos receptores de glutamato do tipo AMPA
(GluR1-3) nas diferentes estruturas encefálicas estudadas quando comparado
o grupo controle sedentário39. Carro e colaboradores54, observaram que o
exercício em esteira protege contra agentes excitotóxicos, prevenindo dano e
perda neuronal, promovendo um efeito neuroprotetor.
Outros estudos concentraram-se na aprendizagem de habilidade motora
associada ao treinamento acrobático, em oposição ao exercício físico repetido
como uma única variável enriquecedora. O exercício acrobático (AC) consiste
no treinamento de novas habilidades motoras utilizando uma sequência de
tarefas destinadas a incentivar a resolução de problemas e coordenação. O AC
promove alterações em proteínas neuronais como MAP2 e sinaptofisina55,
aumento de BDNF e seu receptor 56, sinaptogênse em regiões do cerebelo e
córtex motor32,41,43,57 além de alterações comportamentais, como redução do
tempo de realização das tarefas motoras, indicando um aumento substancial
na aquisição de habilidades motoras32.
Este tipo de aprendizagem motora (em oposição à atividade motora
envolvida no exercício voluntário ou forçado) aumenta a formação de sinapses
no córtex cerebelar42,43, enquanto que o exercício físico repetitivo aumenta a
densidade dos vasos sanguíneos cerebelares58, sugerindo que os elementos
individuais do ambiente enriquecido podem ter efeitos diferentes sobre os
componentes específicos da plasticidade neural. O treinamento acrobático em
ratos também aumenta sinaptogênese no córtex motor e coordenação motora
após lesão no córtex sensório-motor23.
Garcia e colaboradores40 ao comparar diferentes modalidades de
exercício físico (exercício acrobático e exercício em esteira) observou que no
grupo treinado em esteira a área do córtex motor apresentou aumento
significativo das proteínas sinapsina I (SYS) e sinaptofisina (SYP) e diminuição
de neurofilamento (NF), no estriado houve um aumento de neurofilamento
(NF68), SYS e SYP, e no cerebelo aumento de MAP2 e neurofilamento (NF).

11
Por outro lado, o grupo do exercício acrobático mostrou um aumento
significativo de MAP2 e SYP no córtex motor, aumento de MAP2, SYS, SYP e
NF68 no estriado e aumento de SYS no cerebelo sugerindo que o exercício
promove aumento das proteínas estruturais e sinápticas no córtex motor e
estriado, suportando o envolvimento dessas regiões no processo de
aprendizagem de novas habilidades motoras enquanto que o exercício em
esteira promoveu alterações mais evidentes no córtex motor e cerebelo.
Com intuito de compreender a contribuição dos mais variados estímulos
ambientais sobre funções cognitivas, Lambert e colaboradores12 observaram
que cada elemento individual do ambiente enriquecido pode gerar alterações
neurobiológicas específicas e contribuir de forma diferente na memória espacial
e alterações sinápticas. O exercício voluntário foi capaz de melhorar a memória
espacial comparada ao controle, enquanto que a estimulação cognitiva e o
treinamento acrobático não. Apesar disso, o exercício e a estimulação cognitiva
induziram um aumento da expressão da sinaptofisina no neocórtex e
hipocampo.
1.1.2. Períodos de treinamento: Curto e Longo prazo
Além do tipo de atividade, o que se tem estudado são respostas a
diferentes períodos de treinamento. Porém, a determinação da duração do
exercício nos estudos que avaliam seu potencial terapêutico é uma dúvida que
persiste.
Exercícios tanto a curto quanto a longo prazo foram descritos como tendo
um grande impacto no encéfalo e no comportamento de animais. A maioria
desses estudos analisa a neuroplasticidade ocorrida em áreas relacionadas à
aprendizagem e memória12,40,47,49,56,59.
Em 2002, Molteni e colaboradores9, demonstraram no hipocampo que
existem respostas diferentes na expressão de genes após o exercício em
esteira agudo e crônico, porém o fator neurotrófico derivado do encéfalo
(BNDF) apresentou acréscimo em todos os tempos estudados (3, 7 e 28 dias).
A proteína dependente de cálcio/calmodulina quinase (CaM-K) é ativada
durante os períodos de exercício a curto prazo e a longo prazo, após ocorrer

12
aumento da transmissão glutamatérgica, enquanto MAP-K/ERK é mais ativada
durante o exercício à longo prazo. Já os subtipos NR2A e NR2B do receptor de
glutamato do tipo NMDA apresentou pico de expressão gênica após 3 dias de
corrida, sofrendo decréscimo gradativo após 7 e 28 dias, evidenciando o
envolvimento do exercício físico na neuroplasticidade. O perfil temporal da
expressão do gene parece delinear um mecanismo no qual vias moleculares
específicas são ativadas após o desempenho do exercício9.
O exercício voluntário aumenta a expressão de muitos genes associados
à função sináptica9. Por exemplo, a sinapsina I é aumentada
predominantemente em curtos períodos de exercício voluntário (3 e 7 dias) no
hipocampo, consistente com seu papel na liberação de vesículas sinápticas.
Em adição à sinapsina I, o exercício voluntário aumenta os níveis de mRNA
para sintaxina e sinaptogamina60.
A expressão de proteínas envolvidas na plasticidade neuronal como
MAP2 e sinaptofisina, foi analisada em ratos expostos ao exercício AC a curto
prazo, revelando um aumento na expressão de sinaptofisina no córtex motor,
que parece estar relacionada com os primeiros cinco dias de aprendizagem da
habilidade motora55. Entretanto, a MAP2 não parece aumentar de acordo com
a dificuldade da tarefa e nem com a duração do treinamento55.
Estudos revelaram mudanças na expressão do BDNF e seu receptor
frente a diferentes tipos de treinamento físico56. Nos grupos acrobáticos e
exercício forçado houve um aumento na expressão de BDNF e de seu receptor
TrkB na camada molecular do cerebelo nos primeiros sete dias de treinamento.
Após o décimo quarto dia de treinamento, o BDNF permaneceu elevado
apenas no grupo acrobático, enquanto o TrkB retornou aos níveis basais em
ambos os grupos, no exercício acrobático e exercício forçado. No córtex motor,
houve um aumento significativo tanto da proteína BDNF quanto do seu receptor
TrkB após 14 dias de treinamento acrobático. No grupo exercício voluntário
houve apenas o aumento da expressão da proteína BDNF e não de seu
receptor56. Real e colaboradores39 mostraram que o exercício forçado a curto e
a longo prazo induz mudanças diferentes na expressão dos receptores de
glutamato do tipo AMPA (GluR1-3) nas diferentes estruturas encefálicas
estudadas quando comparado ao grupo controle sedentário. O córtex sensório-

13
motor apresentou resposta bifásica, revelando uma diminuição da expressão
de GluRs na fase inicial e aumento na tardia. Já no cerebelo foi observado uma
diminuição de GluRs na fase inicial, enquanto no estriado houve um aumento
somente na fase tardia.
Diferentes tempos de treinamento de ratos em esteira mostraram um
aumento de sinapsina I no estriado após 3 e 7 dias de treino retornando a
níveis basais no décimo quinto dia, o córtex motor não revelou alteração na
expressão de proteínas sinápticas mas redução do nível da expressão de NFs
após um curto prazo de treinamento (3 dias), demonstrando que o exercício em
esteira de intensidade moderada é capaz de promover mudanças distintas nas
proteínas estruturais e sinápticas em diversas regiões do encéfalo de ratos a
curto e longo período de treinamento26.
Estudos avaliando a influência dos diferentes períodos de treinamento
do exercício acrobático sobre os níveis de proteínas estruturais e sinápticas, a
resposta do gene Egr1 e os níveis de neurogênese são escassos.
1.2. Neurogênese Hipocampal adulta
A descoberta de neurogênese no encéfalo de mamíferos adultos
derrubou o dogma de que o encéfalo adulto não tem a capacidade de gerar
novos neurônios. Porém, hoje em dia já é bem aceito que neurônios são
continuamente adicionados em poucas regiões do encéfalo durante toda a vida
adulta61.
A neurogênese adulta originam-se de células progenitoras neurais que
tem sido constantemente observadas em duas regiões no cérebro adulto: a
zona subventricular (SVZ) do ventrículo lateral e na zona subgranular (SGZ) do
giro denteado (DG) do hipocampo. Neurônios nascidos na SVZ migram através
da corrente migratória rostral e se tornam neurônios granulares e
periglomerulares no bulbo olfatório. Neurônios nascidos na SGZ se diferenciam
e integram-se na rede local de neurônios como células granulares do DG61.

14
No cérebro adulto, o hipocampo é uma estrutura crucial para a formação
de certos tipos de memória, como a memória temporal e espacial. Por meio de
suas interações com estruturas associadas do cérebro62.
Muitos estudos demonstraram que a atividade comportamental como o
enriquecimento ambiental4,63, o exercício físico48, e as tarefas específicas
dependente da aprendizagem64,65 podem regular a atividade neurogênica no
DG do hipocampo61.
O exercício voluntário tem mostrado aumentar a neurogênese e
melhorar o desempenho cognitivo66,67, aumentando a proliferação e
sobrevivência de células14,48,68,69. Este aumento da neurogênese, contudo, é
dependente da fase circadiana e do volume diário de exercício. Foi observado
que camundongos tiveram níveis de neurogênese aumentados apenas quando
faziam exercício em roda de correr por 3 horas e no meio do período ativo
(ciclo escuro)70.
Vários protocolos de exercício também têm demonstrado um aumento
da neurogênese hipocampal de animais adultos revelados pelo número de
células “doublecortin”–positivas (DCX) na SGZ do giro denteado do
hipocampo17,71,72.
Estudo como o de Ferreira e colaboradores71 mostraram que o exercício
em esteira induziu um aumento da proliferação celular e neurogênese na SGZ
do giro denteado do hipocampo após todos os períodos de treinamento (3, 7 e
15 dias) e revelou que 3 dias de exercício em esteira de moderada intensidade
foram suficientes para induzir estas alterações.
É importante ressaltar que o efeito benéfico do exercício sobre a
neurogênese hipocampal adulta é observado tanto em animais jovens, quanto
em animais idosos14. Há, no entanto, a hipótese de que não é simplesmente o
exercício que promove benefícios para o encéfalo, mas sim o exercício com
desafios cognitivos66.
1.3. Neuroanatomia Funcional
Nos próximos tópicos serão abordadas as estruturas do Sistema
Nervoso que foram estudadas neste trabalho, permitindo um melhor

15
entendimento de sua composição e função na atividade motora e aprendizado
motor. Estas regiões motoras foram escolhidas por serem frequentemente
afetadas por doenças neurodegerativas ou por lesões, por estarem envolvidas
direta ou indiretamente com o planejamento e/ou execução do movimento, ou
por sofrerem influência direta dos efeitos do exercício físico.
1.3.1. Córtex Motor
Muitas áreas do córtex cerebral estão envolvidas principalmente com o
processamento da informação sensorial ou com o planejamento e execução
dos comandos motores73. Essas áreas são subdividas em motora primária e
em várias áreas pré-motoras (áreas motoras secundárias ou de associação). A
região motora primária é responsável por executar o movimento, participando
de padrões de movimentos mais complexos74. O córtex cerebral esta
organizado em camadas celulares. O neocórtex contem seis camadas
numeradas de I-VI, como descrito por Lorente de Nó em 194275, numeradas da
superfície externa (pia máter) do córtex para a substância branca73.
Embora cada camada do córtex cerebral seja definida primariamente
pela presença ou ausência de corpos celulares de neurônios, cada camada
contém também elementos adicionais. Nas camadas I, II e III encontramos
dendritos apicais de neurônios que tem seus corpos celulares nas camadas V e
VI, enquanto as camadas V e VI contem os dendritos basais de neurônios com
corpos celulares nas camadas III e IV. Quanto a camada IV estão as fibras
aferentes e na V temos células que dão origem às fibras eferentes, chamadas
também de células de Betz na área motora, isto é, que deixam o córtex
cerebral indo para o tronco encefálico, medula espinal e núcleos da base , e na
VI temos as fibras que partem desta zona em direção ao tálamo73.
Vários estudos mostram mudanças no número de sinapses32,41 e da
eficiência sináptica76 no córtex motor em associação com as mudanças nas
áreas corticais relacionadas ao movimento32,77. Outros autores têm
demonstrado que o exercício acrobático após uma lesão cerebral promove o
aumento da sinaptogênese, isto é, mais sinapses por neurônios na camada V
do córtex cerebral quando comparado aos outros grupos do estudo23.

16
1.3.2. Estriado
Quatro estruturas subcorticais interconectadas compõem os núcleos da
base (NB), com projeções principais para o córtex cerebral, tálamo e certos
núcleos do tronco encefálico73. Estas estruturas controlam a atividade motora
por meio da regulação de impulsos neuromotores que facilitam sua atividade
tônica, auxiliando o planejamento e a execução de movimentos
sequenciados78,79. A principal função destas é manter a eficácia dos neurônios
corticais, sobretudo na área motora suplementar (AMS), afim de organizar e
ativar seqüências de movimentos ou programas motores, em tempos
adequados dentro de uma seqüência de movimentos auto-gerados73. Outra
função importante é a de ativar e finalizar programas motores que sejam
adequados para a aquisição de uma meta, como por exemplo, alcançar e
segurar um objeto. Seu papel funcional também está relacionado à
manutenção desse movimento durante sua execução e que pode ser
observado pelas variações na velocidade e na amplitude do movimento80,81.
Além disso, os NB possuem importante participação na formação da memória e
aprendizado82 83.
Os núcleos da base são constituídos pelo: estriado, globo pálido (interno
- GPi e externo - GPe), substância negra (parte compacta e reticulada) e o
núcleo subtalâmico. O estriado por sua vez, consiste de três importantes
subdivisões: o núcleo caudado, o putâmen e o estriado ventral (o qual inclui o
núcleo acumbens)73.
Embora o estriado possua vários tipos diferentes de células, 90-95%
delas são neurônios com espinhos dendriticos e projeções GABA-érgicas.
Essas células são alvos importantes da aferência cortical84.
Os núcleos da base auxiliam o córtex motor no controle de movimentos
através de duas vias eferentes do estriado, sendo uma direta e outra indireta. A
vida direta é composta pelo globo pálido interno e pela substância negra(parte
reticulata), ambos compostos, em parte, por neurônios GABA-érgicos e a via
indireta inclui conexões inicialmente com o globo pálido externo e o núcleo
subtalâmico, sendo essa uma via puramente GABA-érgica85.

17
Em linhas gerais, a organização funcional dos núcleos da base se inicia
com uma aferência cortical ao corpo estriado. A alça direta retira a inibição que
o globo pálido teria sobre o núcleo ventrolateral do tálamo, permitindo então
que este estimule o córtex cerebral (liberando o movimento). Já a alça indireta,
da qual participa o núcleo subtalâmico, produz uma inibição do tálamo e
consequentemente uma estimulação reduzida do córtex (inibindo o
movimento)86. A substância negra aferenta então o estriado fazendo sinapses
dopaminérgicas com neurônios que contêm receptores D1 para formar a alça
direta, facilitando a via tálamo-cortical e com neurônios que contêm receptores
D2 para formar a alça indireta, inibindo a inibição do tálamo. Em resumo, a
ativação da via nigroestriatal resultará na facilitação do movimento.
Essas duas vias têm funções distintas para o controle de movimentos. A
indireta está envolvida com a iniciação e/ou com a finalização de movimentos e
a direta, além da iniciação, é responsável pela manutenção do programa motor
durante a ação87 88.
Alguns estudos sugerem que durante as fases precoces e tardias de
aprendizagem de habilidade, os circuitos estriatais e os processos envolvidos
podem ser distintos. O estriado dorsomedial ou associativo (DMS – homólogo
ao caudado em primatas) que recebe aferências principalmente da área motora
suplementar e córtex pré-frontal, parece estar envolvido nas fases iniciais de
aprendizagem visuomotora. De outra forma, o estriado dorsolateral ou
sensório-motor (DLS – homólogo ao putâmen em primatas), recebe aferências
do córtex sensório-motor e esta relacionado as aquisições de comportamento
habituais e automáticas89.
As tarefas internamente guiadas (IG), ou seja, movimento iniciados
voluntariamente, são principalmente processadas pelo circuito NB-tálamo-
cortical com recrutamento do circuito cerebelo-tálamo-cortical90.
1.3.3. Cerebelo
O cerebelo foi escolhido para ser analisado neste estudo, uma vez que
desempenha funções importantes no controle motor. Ele ajusta respostas
motoras por meio de uma comparação do resultado pretendido com o

18
movimento executado. Além disso, ele modula a força e a amplitude dos
movimentos e está envolvido na aprendizagem motora91,92.
O cerebelo de mamíferos é formado pelo córtex cerebelar (substância
cinzenta externa), substância branca interna e por núcleos cerebelares
profundos, que representam suas principais estruturas de saída. Ele pode ser
dividido de acordo com o desenvolvimento evolutivo ou sua funcionalidade93,94.
O sistema nervoso utiliza o cerebelo para coordenar as funções de
controle motor em três níveis funcionais: vestíbulocerebelo, constituído
principalmente pelos lobos floculonodulares e porções adjacentes do vermis,
responsável pela manutenção da postura e do equilíbrio. O espinocerebelo
formado pela maior parte do vermis do cerebelo posterior e anterior e os lobos
intermediários, que contém circuitos para coordenar, sobretudo, os movimentos
das porções distais dos membros, especialmente mãos e dedos, além de
regular o tônus muscular e a postura. E o cerebrocerebelo, composto pelas
zonas laterais dos hemisférios cerebelares, responsáveis pelo planejamento
dos movimentos voluntários sequenciais73,95.
O córtex cerebelar, é formado por três camadas distintas: molecular, das
células de Purkinje e granular. A disposição celular em cada uma dessas
camadas é a mesma em todos os lobos, diferindo somente na procedência das
vias aferentes e no destino das vias eferentes86.
A camada mais externa ou camada molecular do córtex cerebelar é
basicamente uma área de sinapses e nela encontram-se os corpos celulares
de dois tipos de interneurônios inibitórios, estrelados e em cesto, dispersos
entre as fibras paralelas, originárias das células granulares (excitatórias) e os
dendritos das células de Purkinje (inibitórias), os quais encontram-se
orientados perpendicularmente a esses axônios73,86.
Abaixo da camada molecular está a camada das células de Purkinje,
que consiste em uma única camada de corpos celulares grandes e
arborizações dendríticas em forma de leque que se estendem até a camada
molecular. Os axônios das células de Purkinje projetam-se para os núcleos
cerebelares profundos ou para os núcleos vestibulares, constituindo assim a
via de saída do córtex cerebelar. Estas células constituem a eferência do córtex
cerebelar73,86.

19
E a camada mais interna ou camada granular contém um grande
número de células granulares e um pequeno número de interneurônios de
Golgi. As fibras musgosas, a maior fonte de aferências ao cerebelo, terminam
nessa camada73,86,96.
O cerebelo recebe dois tipos principais de aferências excitatórias: as
fibras musgosas e fibras trepadeiras. Ambos os grupos de fibras fazem
sinapses excitatórias com neurônios cerebelares, mas os dois grupos terminam
diferentemente no córtex cerebelar e produzem diferentes padrões de disparos
nas células de Purkinje73.
As fibras musgosas originam-se de núcleos do tronco encefálico e
neurônios medulares que dão origem ao trato espinocerebelar e conduzem
informações sensórias da periferia, bem como informações do córtex cerebral.
Essas fibras fazem sinapses excitatórias sobre os dendritos das células
granulares na camada granular e indiretamente sobre as células de Purkinje via
fibras paralelas73,93 .
As fibras trepadeiras originam-se do núcleo olivar inferior e formação
reticular medial, que recebem aferências do córtex cerebral, núcleo rubro e
medula espinal e conduzem informações somatossensórias e/ou visuais. Essas
fibras fazem sinapses diretamente com o soma e os dendritos das células de
Purkinje93.
De uma forma geral, essas informações processadas no córtex
cerebelar saem pelos axônios das células de Purkinje direcionados para os
núcleos cerebelares profundos e daí para o tálamo formando assim o circuito
cerebelo-tálamo-cortical (Cb-tálamo-cortical)97. Esse circuito fornece ao córtex
motor informações sobre os movimentos em andamento, além da velocidade e
da força desse movimento44.
Enquanto os circuitos NB-tálamo-corticais são principalmente ativados em
tarefas que são IG, as tarefas externamente guiadas (EG), ou seja, tarefas
orientadas por estímulos visuais ou auditivas são principalmente processadas
nos circuitos Cb-tálamo-corticais, com recrutamento do circuito NB-tálamo-
cortical90.

20
1.3.4. Hipocampo
O hipocampo desempenha importante papel em algumas formas de
aprendizado e memória98. É uma região do lobo temporal do cérebro,
responsável por aspectos da neurobiologia da memória emocional e declarativa
(memória de fatos e eventos). Processa a informação recentemente adquirida
por um período de dias, semanas ou até meses, e depois a transfere para
áreas importantes do córtex cerebral para um armazenamento mais
prolongado73,99,100.
É composto por duas áreas principais: corno de Amon (subdividido em
quatro campos – CA1, CA2, CA3, CA4), e o giro denteado (GD)101,102. CA2 é
pequeno e indistinto em algumas espécies, por este motivo ele é
frequentemente incluído em CA1101,103.O termo “formação hipocampal” inclui o
GD, o subículo, o hipocampo propriamente dito, além dos córtices entorrinal,
perirrinal e paraipocampal102.
O giro denteado é composto por três camadas de células: a camada
molecular (relativamente desprovida de células e ocupada basicamente pelos
dendritos das células granulares, em cesto e polimórficas), a camada de
células granulares (principal camada de células que apresenta empacotamento
denso de células granulares e uma população de células em cesto na sua
camada mais profunda) e a camada de células polimórficas (também chamada
de hilo e onde se encontram as células musgosas)85.
Um aspecto que confere extrema importância a região do hipocampo
seria a população de células tronco neurais nela encontrada. Além da zona
subventricular dos ventrículos laterais, a zona subgranular do giro denteado do
hipocampo é a única outra área onde foram identificadas estas células
progenitoras no encéfalo de mamíferos adultos98.
1.4. Marcadores de Plasticidade
Estudos demonstram evidências sobre neurogênese e plasticidade
cerebral104,105 induzidas especificamente por famílias de moléculas
neutrotróficas106,107. Durante o processo de aprendizagem, há modificações nas

21
estruturas e funcionamento das células neurais e de suas conexões, ou seja, o
aprendizado promove modificações plásticas, como crescimento de novas
terminações e botões sinápticos, crescimento de espículas dendríticas,
aumento das áreas sinápticas funcionais42, estreitamento da fenda sináptica,
mudanças de conformação de proteínas receptoras, incremento de
neurotransmissores108.
No contexto da plasticidade algumas ferramentas são frequentemente
usadas para a verificação e avaliação de possíveis alterações morfológicas
plásticas, como elementos do citoesqueleto e de densidades sinápticas109.
A seguir faremos uma breve descrição dos marcadores de plasticidade aqui
estudados.
1.4.1. Proteínas Sinápticas e Proteínas Estruturais
Atualmente, pesquisas propiciaram uma sequência de estudos
analisando a implicação de proteínas de vesículas sinápticas, como
sinapsinas8,11,60,110-112 e sinaptofisina10,12,113 na neuroplasticidade frente ao
exercício físico. A avaliação de elementos estruturais também tem mostrado
efeitos plásticos de intervenções como o exercício e/ou ambientes
enriquecidos, além de mostrar os efeitos da falta destes, como a redução do
número de sinapses e de espinhos dendríticos em regiões envolvidas com a
cognição114. Pode haver também em decorrência do exercício mudanças na
atividade e concentração de determinados subtipos de receptores39,115.
A Sinapsina I (SYS) é membro da família de fosfoproteínas expressas
principalmente em neurônios, envolvidas no ancoramento e liberação
de vesículas sinápticas, que medeia a transmissão sináptica116. Acredita-se
que a proteína sinapsina I, a mais abundante de todas as fosfoproteinas
neuronais, é mediadora de interações de vesículas sinápticas com o
citoesqueleto. Esta proteína está envolvida com a regulação da formação de
sinapses em vários estágios do desenvolvimento neuronal, regulando a
maturação estrutural e funcional das sinapses, exercendo papel importante na
organização da estrutura do terminal nervoso117,118.

22
Outra proteína sináptica é a sinaptofisina (SYP), uma glicoproteína
associada a vesículas que é fosforilada em tirosina. Responsável pela
biogênese e ancoragem das vesículas sinápticas no terminal sináptico e
resgate de proteínas dessas vesículas por endocitose, o que é importante para
uma neurotransmissão rápida e eficiente38. Aumento na imunorreatividade de
sinaptofisina parece refletir um aumento dos terminais pré-sinápticos119-121
importante para a biogênese das vesículas sinápticas e endocitose122,123.
Os microtúbulos e suas proteínas associadas (MAP2) têm um papel
fundamental no desenvolvimento de axônios e dendritos. É uma proteína que
faz parte de um grupo de componentes do citoesqueleto, predominantemente
expressos nos neurônios e servem como substrato para a maioria das
proteínas quinases e fosfatases presentes em neurônios. A MAP2 pode
desempenhar múltiplas funções, incluindo a nucleação e estabilização dos
microtúbulos e possíveis microfilamentos, o transporte de organelas dentro de
axônios e dendritos, bem como a fixação de proteínas reguladoras, tais como
proteínas quinases que podem ser importantes para a transdução de sinal.
Além disso, participam do processo de crescimento e arborização dendrítica,
remodelamento dendríticos pós-lesão, estabelecimento e manutenção da
sinaptogênese124.
E por fim, o citoesqueleto neuronal é composto por três filamentos
interconectados: os microfilamentos de actina, os microtúbulos e os filamentos
intermediários125. Os neurofilamentos(NF), filamentos intermediários
específicos de neurônios, constituem o principal tipo de filamento em neurônios
adultos105. São essenciais para a manutenção da forma neuronal, da
arborização dendrítica e para o crescimento axonal, e transporte de moléculas
e organelas no sistema nervoso126.
Existem três tipos diferentes de neurofilamentos que diferem entre si
pelo peso molecular: as de baixo peso molecular, NF-L (68kDa), as de peso
molecular intermediário, NF-M (160kDa) e as de alto peso molecular, NF-H
(200kDa), que formam heteropolímeros entre si, necessitando de co-
polimerização do NF68 com pelo menos uma das outras subunidades126.
Durante o estágio de morfogênese neuronal os componentes do
citoesqueleto auxiliam nos processos de crescimento neurítico e estabilização

23
de axônios e dendritos formados127. Mutações em genes que codificam
neurofilamentos ou suas proteínas ligantes podem causar inibição no
transporte axonal. Adicionalmente, o acúmulo anormal de proteínas dos
neurofilamentos é uma característica comum a muitas doenças
neurodegenerativas128.
1.4.2. Gene Egr1
O Gene Egr1 (gene de resposta precoce ao crescimento 1) é membro de
uma família Egr, caracterizada pela presença de três proteínas zinc-fingers e
outras proteínas DNA-ligantes, que juntas participam da regulação da
expressão de genes alvo, importantes para os efeitos a longo prazo no
crescimento e diferenção celular129,130. O Egr1 recebeu denominações (Zif,
NGFI-A, Krox-24, TIS8) na medida em que foi definido por sua rápida
expressão em diferentes tecidos, como cérebro, pulmão e gânglio cervical
superior, após exposição desses tecidos ao fator de crescimento neural
(NGF)131.
O Egr1 é induzido em neurônios após estimulação extracelular por
substâncias tróficas e neurotransmissores, dentre eles o glutamato, bem como
seus agonistas específicos, que ativam diversas classes de receptores
glutamatérgicos, como kainato, NMDA e AMPA. Em 1997, Beckmann e
colaboradores132 destacaram o aumento da atividade do Egr1 no córtex
cerebral e hipocampo de ratos após a administração de NMDA e kainato, em
compensação, um pré-tratamento com um antagonista do receptor NMDA (MK-
801) não afetou a expressão dos genes imediatos (incluindo Egr1) no sistema
límbico, sugerindo que a interação de Egr1 com os receptores de glutamato
depende da região do sistema nervoso central observada133.
Devido a essa interação entre Egr1 e os fatores de transcrição
desencadeando a expressão de genes de resposta tardia, deve-se considerar
um importante papel funcional deste gene no contexto da plasticidade e do
aprendizado134.
A expressão de Egr1 apresentou-se aumentada após indução da
potenciação de longo prazo (LTP) no giro denteado de ratos, superando a

24
expressão dos demais genes imediatos pesquisados, c-Fos, c-jun e jun-B135. É
importante a pesquisa do gene Egr1 em regiões do encéfalo como cerebelo,
estriado e córtex motor devido a sua relação com a LTP e aprendizado. O
treinamento acrobático em animais promove sinaptogênese em regiões do
cerebelo e córtex motor32,41,43, além de alterações comportamentais, como
redução do tempo de realização das tarefas, indicando um aumento
substancial na aquisição das habilidades motoras32.
Entretanto os mecanismos intrínsecos a essa plasticidade que o
treinamento acrobático gera durante um treino a curto e a longo prazo ainda
não são claros. No presente estudo, buscamos associar aos mecanismos
plásticos observados pela expressão de proteínas estruturais e sinápticas a
expressão de Egr1 nessas regiões ao realizar o treinamento de ratos adultos
jovens em um circuito acrobático, traçando um perfil do comportamento do
gene relacionado ao protocolo proposto.

25
2. JUSTIFICATIVA
A neuroplasticidade que ocorre no sistema nervoso central de animais e
humanos através do exercício físico, independentemente da modalidade, a
curto ou a longo prazo foi descrita por diversos autores. Entretanto, evidências
demonstrando resposta de proteínas estruturais, sinápticas e genes de
expressão precoce após exercícios acrobáticos com diferentes tempos de
treinamento são escassas.
3. OBJETIVOS
3.1. Objetivo Geral
Este estudo visa observar o desempenho motor e mensurar a
neuroplasticidade induzida por diferentes tempos de treinamento de exercício
acrobático nas áreas motoras do encéfalo de ratos.
3.2. Objetivos Específicos
3.2.1. Analisar a performance motora dos ratos submetidos a curto (
uma e duas semanas), e longo (quatro semanas) períodos de treinamento
acrobático.
3.2.2. Avaliar a expressão das proteínas sinápticas (sinapsina I e
sinaptofisina) e do citoesqueleto (microtúbulos e neurofilamentos) no córtex
motor (primário e secundário), estriado e cerebelo de ratos adultos submetidos
a diferentes períodos de treinamento de exercício acrobático.
3.2.3. Quantificar a expressão de Egr1 nas áreas motoras do encéfalo
de ratos adultos envolvidas com o planejamento e aprendizado motor,
submetidos a períodos distintos de treinamento de exercício acrobático.
3.2.4. Quantificar a expressão de DCX na região hipocampal (giro
denteado) do encéfalo de ratos adultos treinados durante diferentes períodos
de treinamento de exercício acrobático.

26
4. MATERIAL E MÉTODO
4.1. Animais
Neste estudo foram utilizados 76 ratos machos da linhagem Wistar
fornecidos pelo Biotério Central do Instituto de Ciências Biomédicas da USP.
Os animais foram mantidos em uma sala com temperatura constante de 23 ºC
e ciclo claro/escuro artificialmente controlado de 12/12 h, tendo livre acesso à
alimentação e água ad libitum. Antes do início do treinamento físico os animais
ficaram 15 dias na sala para adaptação de ciclo invertido 36 136. Este estudo foi
conduzido de acordo com os Princípios Éticos de Experimentação Animal
adotado pelo Colégio Brasileiro de Experimentação Animal (COBEA) e foi
aprovado pela Comissão Ética em Experimentação Animal (CEEA) do ICB/USP
em 21/05/2010, protocolo registrado sob nº 152 nas folhas 95 do livro 2 para
uso de animais de experimentação (anexo I).
4.2. Desenho experimental
Os animais foram divididos aleatoriamente em 2 grupos: acrobático
(n=57) e controle-sedentário (SED, n=19). O grupo acrobático passou por um
período de adaptação à tarefa motora passando 2 vezes por todo o circuito em
2 dias numa semana. Posteriormente, este grupo foi subdividido em 3 com
diferentes períodos de treinamento: 1semana (AC1, n=19), 2 semanas (AC2,
n=19) e 4 semanas (AC4, n=19). Os animais sedentários permaneceram no
mesmo ambiente que os treinados. Os encéfalos foram analisados pelas
técnicas de imuno-histoquímica e “immunoblotting”.
4.2.1. Protocolo de Exercício Acrobático
O treinamento acrobático foi feito no período ativo do animal70 e
consistiu em atravessar um circuito constituído pelos seguintes obstáculos:
gangorra, barreiras verticais, ponte de cordas paralelas, traves paralelas de
madeira roliça, pontes de madeira roliça e corda, como mostra a Figura 1. O

27
circuito foi baseado no utilizado por Kleim e colaboradores32 e localizado a uma
altura de 1,50m do chão. O treinamento foi realizado em dias alternados, com
frequência de três vezes por semana e com 5 passagens nesse circuito por dia
até completar o tempo de treinamento de cada grupo. Os animais receberam
estímulos manuais leves no trem posterior quando necessário como forma de
estímulo para percorrer todo o caminho. Foi registrado o tempo necessário
para cada rato percorrer cada passagem pelo circuito41,58. Após o término das 5
passagens de cada animal pelo circuito, todos os obstáculos e plataformas
foram limpos com etanol 70%.
Figura 1: Imagens digitais dos obstáculos que compõem o circuito de treinamento de exercícios acrobáticos40.
4.3. Protocolo de Imuno-histoquímica
Os animais treinados (n=5 de cada grupo) e os controles (n=5) foram
anestesiados com ketamina (100mg/kg) e xilazina (25mg/kg) por via
intraperitonial e submetidos à perfusão transcardíaca, com solução salina
0,9%, seguida de solução fixadora constituída de paraformaldeído 2%
dissolvido em tampão fosfato 0,1 M (PB, pH 7,4). Os animais foram submetidos
a esse procedimento após uma hora do término do treinamento. Após a
perfusão, os encéfalos foram coletados e armazenados em paraformaldeído
2%, durante 4 horas. Após este período, o material foi transferido para uma
solução crioprotetora de sacarose a 30% em PB 0,1M, armazenados a 4oC.
Após 36 horas, foram feitos cortes frontais dos encéfalos em uma espessura de
30 m em um micrótomo deslizante de congelamento (Leica SM 2000 R).

28
Os cortes histológicos foram colocados em placa de cultivo, em solução
crioprotetora, e mantidos em freezer a -20ºC até o momento do procedimento
de imuno-histoquímica. Os cortes contendo o córtex motor, estriado e o
cerebelo foram selecionados137 e submetidos à metodologia de imuno-
histoquímica com anticorpos primários específicos para detecção de Egr1
(anticorpo policlonal obtido em coelho dirigido contra o gene Egr1, Santa Cruz
Biotechnology, Inc. CA, USA), na concentração 1:100, sinapsina I (anticorpo
policlonal obtido em coelho dirigido contra proteína SYS – Chemicon ,
Temecula, CA, EUA) na concentração 1:1000, sinaptofisina (anticorpo
policlonal obtido em coelho dirigido contra proteína SYP – DakoCytomation,
Glostrup, Dinamarca) na concentração 1:1000, MAP2 (anticorpo monoclonal
obtido em camundongo dirigido contra a proteína MAP2 – Chemicon,
Temecula, CA, EUA) na concentração 1:1000 e neurofilamentos (anticorpo
monoclonal obtido em camundongo dirigido contra os neurofilamentos – Zymed
Laboratories, San Francisco, CA, EUA) na concentração 1:2000. O anticorpo
PAN reconhece uma região homóloga nos três neurofilamentos (NF68, NF160
e NF200), o que torna impossível distinguí-los com a técnica de imuno-
histoquímica, mas apenas quando analisados pela técnica de “immunoblotting”.
A imuno-histoquímica foi processada da seguinte forma: os cortes
selecionados foram lavados em tampão fosfato (PB 0,1M) por três vezes de 10
minutos, em seguida foram incubados com anticorpos primários específicos
para os anticorpos citados acima, diluídos em PB 0,1M com 0,3% de Triton X-
100 e 5% de soro normal do animal em que foi feito o anticorpo secundário e
mantido por um período de 14 a 18 horas à temperatura ambiente (24ºC).
Os cortes foram então novamente lavados em PB 0,1M em temperatura
ambiente e incubados por duas horas com o anticorpo secundário biotinilado
(1:200) contra as imunoglobulinas do animal no qual foi feito o anticorpo
primário. Após nova série de lavagens à temperatura ambiente, os cortes foram
colocados por duas horas numa solução de Triton-X-100 0,3 % em tampão
fosfato 0,1M com 0,4 M de NaCl, onde se incluiu o complexo avidina-biotina-
peroxidase (ABC ELITE kit, Vector Labs.). Após nova série de lavagens, os
cortes foram imersos num meio contendo 3-3’diaminobenzidina (DAB- Sigma-
Aldrich) 0,05% em PB 0,1M por cerca de 5 minutos e posteriormente,

29
adicionado em torno de 3 ml de solução de H2O2 a 0,3% em água destilada,
mantendo-se os cortes neste banho até que a reação fosse evidenciada.
Atingida a imunorreatividade desejada com o desenvolvimento de coloração
marrom clara, os cortes foram removidos da solução com DAB e imersos em
tampão PB 0,1M. Depois de nova série de lavagens em PB 0,1M com o
objetivo de remoção do excesso de reagente, os cortes foram colocados sobre
lâminas de vidro gelatinizadas e colocadas em placa quente. Após secarem
foram hidratadas em água destilada por 1 minuto, banhadas em solução de
tetróxido de ósmio 0,1% por 15–30 segundos para intensificar a coloração,
desidratadas por uma série de álcoois em concentrações crescentes, clareadas
com Hemo-De (Fisher) e cobertas com lamínulas, tendo como meio de
montagem o Permount (Sigma). A análise qualitativa do material e as
quantificações por densitometria foram realizadas com a utilização do
microscópio óptico (E1000, Nikon) acoplado à câmera digital e programa Nikon
Imaging Software ACT-U.
A análise semi-quantitativa da densidade óptica relativa foi realizada em
4 animais de cada grupo. Para cada animal analisamos 5 cortes dos quais
foram capturados 8 campos tanto do córtex motor como do cerebelo, e 12
campos do estriado. No córtex foram selecionadas as áreas motoras primária
(M1) e secundária (M2), por serem áreas envolvidas diretamente com a
execução e o planejamento motor, respectivamente, sendo analisada a
camada V e suas imediações, pois esta camada contém células eferentes do
córtex. No estriado foram analisados campos da região dorsomedial e
dorsolateral em toda extensão rostro-caudal. E por fim, foi selecionada a
região paramediana do cerebelo, e analisado as camadas molecular e granular
do córtex cerebelar exceto com relação ao anticorpo Egr1 no qual foi analisado
somente a camada granular do córtex cerebelar.
As regiões de interesse foram identificadas baseado no atlas Stereotaxic
Coordinates137. Para o córtex motor foi analisado entre 2.52-1.56 da área de
bregma. Para o estriado foi analisado entre 1.80-0.36 da área de bregma, e
para o cerebelo analisamos o córtex cerebelar (lóbulo paramediano) entre
14.08-12.30 da área de bregma. As imagens foram analisadas utilizando o
programa Image J (NIH).

30
4.4. Protocolo de Avaliação da Neurogênese pelo “Doublecortin”
(DCX)
Animais do grupo sedentário (SED, n=4) e grupo treinado (AC1, AC2 e
AC4, n=12) foram analisados pela técnica de imuno-histoquímica com algumas
especificidades: a solução fixadora constituiu-se de paraformaldeído 4%
dissolvido em tampão fosfato 0,1 M (PB, pH 7,4). Os animais foram submetidos
a esse procedimento após uma hora do término do treinamento. Após a
perfusão, os encéfalos foram coletados e armazenados em paraformaldeído
4%, durante 4 horas. Após este período, o material foi transferido para uma
solução crioprotetora de sacarose a 30% em PB 0,1M, armazenados a 4oC.
Após 16 horas aproximadamente, foram feitos cortes frontais dos encéfalos em
uma espessura de 30 m em um micrótomo deslizante de congelamento (Leica
SM 2000 R).
Os cortes histológicos foram colocados em placa de cultivo, em solução
PB 0,2M e submetidos no mesmo dia ao procedimento de imuno-histoquímica.
Os cortes contendo áreas da região hipocampal foram selecionados e
submetidos à metodologia de imuno-histoquímica com anticorpos primários
específicos para detecção de DCX (anticorpo policlonal obtido em coelho
dirigido contra o gene DCX, Santa Cruz Biotechnology, Inc. CA, USA), como
citado acima.
A análise qualitativa do material e as quantificações das células DCX-
positivas foram realizadas com a utilização do microscópio óptico (E1000,
Nikon) acoplado à câmera digital e programa Nikon Imaging Software ACT-U.
A contagem das células DCX-positivas foi realizada em 4 animais de
cada grupo. Para cada animal analisamos 5 cortes dos quais foram capturadas
4 áreas do hipocampo, na região do giro denteado (GD) especificamente na
zona subgranular (ZSG), entre 3 e 4mm da área de bregma137,por estar
envolvido com a neurogênese adulta61. As imagens foram analisadas utilizando
o programa Image J (NIH).

31
4.5. Protocolo de “Immunoblotting”
Após o treino físico, 10 animais de cada um dos 4 grupos foram
decapitados, e o córtex motor (primário e secundário), cerebelo e estriado
foram rapidamente coletados, e congelados em nitrogênio líquido e
armazenadas a -80ºC até o uso. Para iniciar a extração, as amostras foram
homogenizadas a 4ºC em tampão de extração (Tris pH 7,4 1000 mM; EDTA
10mM; PMSF 2mM aprotinina 0,01 mg/ml) com um homogenizador do tipo
Turratec, modelo MA-102/mini (Marconi; São Paulo, Brasil). Os homogenatos
foram centrifugados a 12.000 rpm, por 20 minutos a 4ºC em uma centrífuga
modelo CT14000 DR (Cientec, São Paulo, Brasil). O sobrenadante foi
separado do “pellet” e uma pequena amostra deste foi utilizada para
determinação do conteúdo proteico, e o restante tratado com tampão Laemmli
(Tris/HCL 125mM, pH 6,8, contendo 2,5% (p/v) de SDS, 2,5% de 2-
mercaptoetanol (2-ME), 4mM de EDTA e 0,05% de azul de bromofenol)138,
contendo DTT 100nM 138, fervido em banho-maria por 5 minutos e
armazenado. O conteúdo protéico do material isolado dos diferentes grupos de
animais foi dosado pelo método de Bradford (Amresco, U.S.A) 139. Cerca de
100 µg de proteína destas amostras armazenadas em Laemmli foram
submetidas a separação por eletroforese com corrente constante de 25mA em
géis de acrilamida de 6,5% e 8% contendo dodecil sulfato de sódio (SDS, Bio-
Rad, EUA) utilizando uma cuba para mini-gel (Mini-Protean 3; Bio-Rad). Após a
separação eletroforética, as proteínas foram eletro-transferidas para a
membrana de nitrocelulose (Millipore, 0,2um de diâmetro) utilizando-se um
sistema de transferência (Trans-Blot cell system; BioRad), em tampão
contendo SDS, de acordo com a técnica descrita por Towbin et al., 1979. Após
a transferência, as membranas foram incubadas em solução de bloqueio com
leite desnatado (Molico, Nestlé) a 5% em tampão Tris-Salina (Tris 10mM e
NaCl 0,15M, pH 7,5) por 2 horas em temperatura ambiente sob agitação leve.
Após este período, as membranas foram lavadas por três vezes de 10 minutos
com Tris-Salina e incubadas com os mesmos anticorpos utilizados na imuno-
histoquímica, além do anticorpo contra β-actina (Sigma, St.Louis, MO, EUA ,
concentração 1:10.000) por 18 horas a 4ºC, sob agitação leve. Em seguida as

32
membranas foram lavadas novamente e incubadas por 2 horas com um
anticorpo secundário marcado com peroxidase (Amersham Biosciences),
diluído a 1:10000 em solução bloqueadora. O excesso de conjugado foi
removido com mais um ciclo de lavagens e as membranas foram reveladas
utilizando o Kit ECL (Amersham Biosciences) de quimioluminescência. Logo
após a revelação as membranas foram então incubadas com solução
“stripping” por 20 minutos à temperatura ambiente para retirar o anticorpo da
membrana e reincubar a membrana com novo anticorpo. Finalmente as bandas
obtidas nos filmes foram escaneadas e analisadas quanto a densidade óptica
da imunorreatividade usando o programa Image J (National Institutes of
Health/USA). Dos dados obtidos realizou-se uma razão entre a densidade
óptica de cada uma das proteínas estruturais e sinápticas com a β-actina que
funcionou como controle interno de pipetagem, e estes foram então submetidos
às análises estatísticas com o programa GraphPad Prism 5. Os dados
representados nos gráficos foram normalizados pelo grupo controle/sedentário
que assumimos ser 1, ou seja 100%.
4.6. Análise Estatística
Os dados de densidade óptica relativa, níveis relativos de proteína e
densidade média de células Egr1 e DCX-positivas foram submetidos à análise
estatística com o uso do software GraphPad Prism 5. As comparações entre os
grupos foram conduzidas a partir do teste ANOVA para medidas repetidas,
seguida do pós-teste de Tukey quando a análise de variância apresentou
diferença estatisticamente significante. Na análise dos dados utilizou-se Média
+ Erro Padrão. Para todos os testes, o nível de significância assumido foi de
p≤0,05.
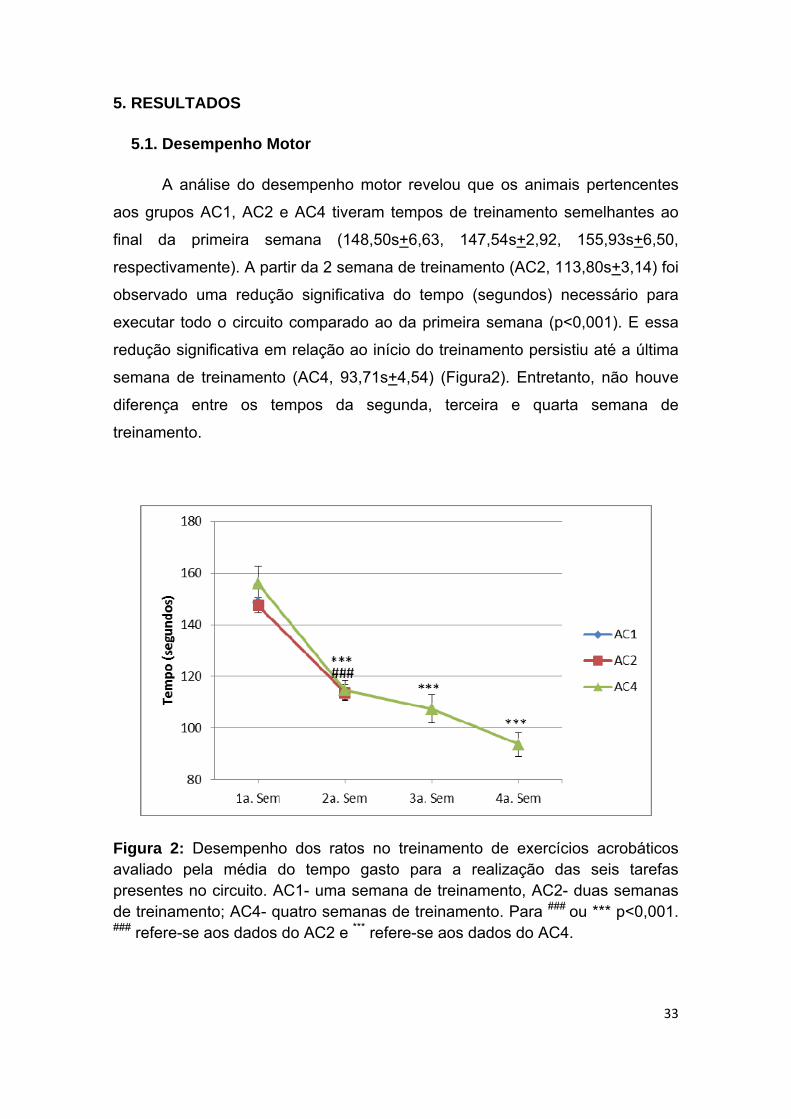
33
5. RESULTADOS
5.1. Desempenho Motor
A análise do desempenho motor revelou que os animais pertencentes
aos grupos AC1, AC2 e AC4 tiveram tempos de treinamento semelhantes ao
final da primeira semana (148,50s+6,63, 147,54s+2,92, 155,93s+6,50,
respectivamente). A partir da 2 semana de treinamento (AC2, 113,80s+3,14) foi
observado uma redução significativa do tempo (segundos) necessário para
executar todo o circuito comparado ao da primeira semana (p<0,001). E essa
redução significativa em relação ao início do treinamento persistiu até a última
semana de treinamento (AC4, 93,71s+4,54) (Figura2). Entretanto, não houve
diferença entre os tempos da segunda, terceira e quarta semana de
treinamento.
Figura 2: Desempenho dos ratos no treinamento de exercícios acrobáticos avaliado pela média do tempo gasto para a realização das seis tarefas presentes no circuito. AC1- uma semana de treinamento, AC2- duas semanas de treinamento; AC4- quatro semanas de treinamento. Para ### ou *** p<0,001. ### refere-se aos dados do AC2 e *** refere-se aos dados do AC4.

34
5.2. Expressão de Proteínas Estruturais e Sinápticas
Diferentes tempos de treinamento acrobático induziram mudanças
significativas na expressão de proteínas nas áreas motoras encefálicas
reveladas pelos métodos de “immunoblotting” e imuno-histoquímica (Tabela 1).
Entretanto, alguns dados foram discordantes entre as técnicas empregadas,
visto que a técnica de imuno-histoquímica nos permite avaliar os tipos e
localização das estruturas imunorreativas ao marcador plástico em camadas
específicas do córtex motor, região lateral e medial do estriado e área
paramediana do cerebelo, enquanto a técnica de “immunoblotting” avalia o
conteúdo proteico da estrutura toda retirada do encéfalo, podendo assim
mascarar as mudanças alcançadas em uma área específica de uma
determinada estrutura.
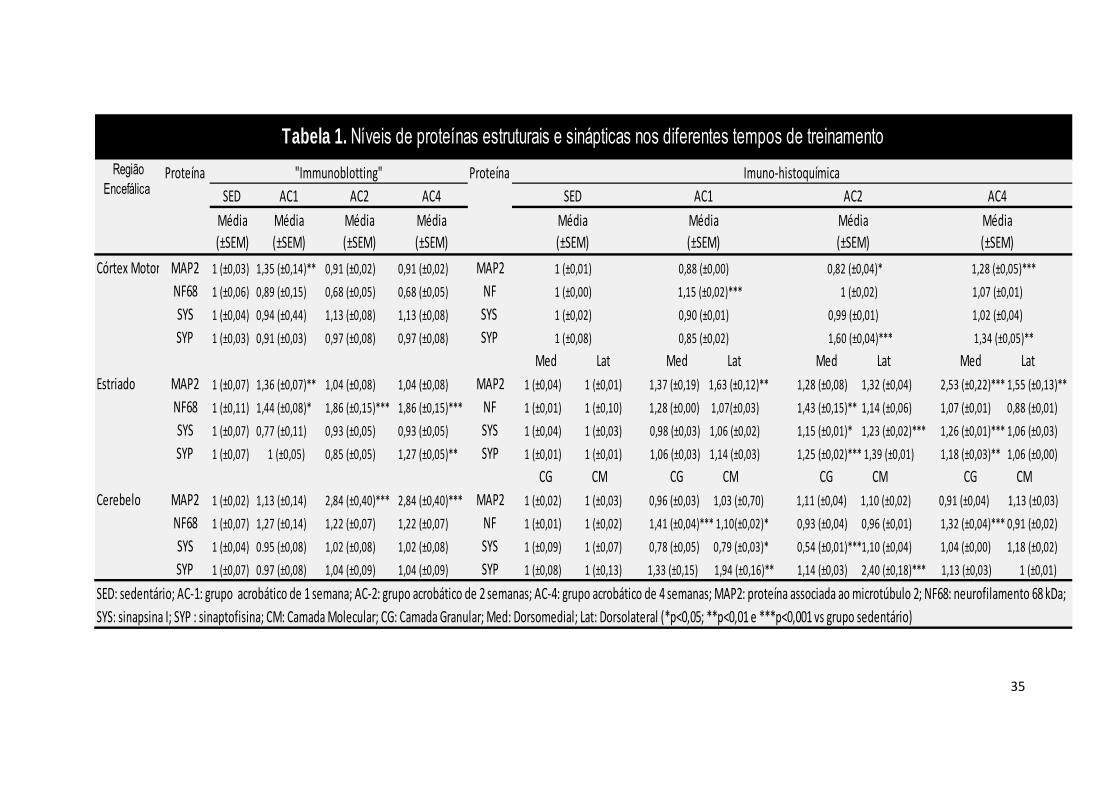
35
SED AC1 AC2 AC4 SED AC1 AC2 AC4
Média Média Média Média Média Média Média Média
(±SEM) (±SEM) (±SEM) (±SEM) (±SEM) (±SEM) (±SEM) (±SEM)
Córtex Motor MAP2 1 (±0,03) 1,35 (±0,14)** 0,91 (±0,02) 0,91 (±0,02) MAP2 1 (±0,01) 0,88 (±0,00) 0,82 (±0,04)* 1,28 (±0,05)***
NF68 1 (±0,06) 0,89 (±0,15) 0,68 (±0,05) 0,68 (±0,05) NF 1 (±0,00) 1,15 (±0,02)*** 1 (±0,02) 1,07 (±0,01)
SYS 1 (±0,04) 0,94 (±0,44) 1,13 (±0,08) 1,13 (±0,08) SYS 1 (±0,02) 0,90 (±0,01) 0,99 (±0,01) 1,02 (±0,04)
SYP 1 (±0,03) 0,91 (±0,03) 0,97 (±0,08) 0,97 (±0,08) SYP 1 (±0,08) 0,85 (±0,02) 1,60 (±0,04)*** 1,34 (±0,05)**
Med Lat Med Lat Med Lat Med Lat
Estriado MAP2 1 (±0,07) 1,36 (±0,07)** 1,04 (±0,08) 1,04 (±0,08) MAP2 1 (±0,04) 1 (±0,01) 1,37 (±0,19) 1,63 (±0,12)** 1,28 (±0,08) 1,32 (±0,04) 2,53 (±0,22)*** 1,55 (±0,13)**
NF68 1 (±0,11) 1,44 (±0,08)* 1,86 (±0,15)*** 1,86 (±0,15)*** NF 1 (±0,01) 1 (±0,10) 1,28 (±0,00) 1,07(±0,03) 1,43 (±0,15)** 1,14 (±0,06) 1,07 (±0,01) 0,88 (±0,01)
SYS 1 (±0,07) 0,77 (±0,11) 0,93 (±0,05) 0,93 (±0,05) SYS 1 (±0,04) 1 (±0,03) 0,98 (±0,03) 1,06 (±0,02) 1,15 (±0,01)* 1,23 (±0,02)*** 1,26 (±0,01)*** 1,06 (±0,03)
SYP 1 (±0,07) 1 (±0,05) 0,85 (±0,05) 1,27 (±0,05)** SYP 1 (±0,01) 1 (±0,01) 1,06 (±0,03) 1,14 (±0,03) 1,25 (±0,02)*** 1,39 (±0,01) 1,18 (±0,03)** 1,06 (±0,00)
CG CM CG CM CG CM CG CM
Cerebelo MAP2 1 (±0,02) 1,13 (±0,14) 2,84 (±0,40)*** 2,84 (±0,40)*** MAP2 1 (±0,02) 1 (±0,03) 0,96 (±0,03) 1,03 (±0,70) 1,11 (±0,04) 1,10 (±0,02) 0,91 (±0,04) 1,13 (±0,03)
NF68 1 (±0,07) 1,27 (±0,14) 1,22 (±0,07) 1,22 (±0,07) NF 1 (±0,01) 1 (±0,02) 1,41 (±0,04)*** 1,10(±0,02)* 0,93 (±0,04) 0,96 (±0,01) 1,32 (±0,04)*** 0,91 (±0,02)
SYS 1 (±0,04) 0.95 (±0,08) 1,02 (±0,08) 1,02 (±0,08) SYS 1 (±0,09) 1 (±0,07) 0,78 (±0,05) 0,79 (±0,03)* 0,54 (±0,01)***1,10 (±0,04) 1,04 (±0,00) 1,18 (±0,02)
SYP 1 (±0,07) 0.97 (±0,08) 1,04 (±0,09) 1,04 (±0,09) SYP 1 (±0,08) 1 (±0,13) 1,33 (±0,15) 1,94 (±0,16)** 1,14 (±0,03) 2,40 (±0,18)*** 1,13 (±0,03) 1 (±0,01)
SED: sedentário; AC‐1: grupo acrobático de 1 semana; AC‐2: grupo acrobático de 2 semanas; AC‐4: grupo acrobático de 4 semanas; MAP2: proteína associada ao microtúbulo 2; NF68: neurofilamento 68 kDa;
SYS: sinapsina I; SYP : sinaptofisina; CM: Camada Molecular; CG: Camada Granular; Med: Dorsomedial; Lat: Dorsolateral (*p<0,05; **p<0,01 e ***p<0,001 vs grupo sedentário)
Tabela 1. Níveis de proteínas estruturais e sinápticas nos diferentes tempos de treinamento
Região Encefálica
Proteína "Immunoblotting" Proteína Imuno‐histoquímica

36
5.2.1. Córtex Motor
Células das várias camadas do córtex motor primário e secundário (M1 e
M2) apresentaram-se imunorreativas aos anticorpos contra MAP2, NFs,
sinapsina I e sinaptofisina em todos os grupos estudados.
O exercício acrobático induziu mudanças distintas na expressão de
proteínas estruturais e sinápticas no córtex motor de ratos no decorrer de um
mês de treinamento.
Quanto às proteínas estruturais, a imunorreatividade ao anticorpo MAP2
mostrou-se ao longo das neurópilas, por toda a região da camada V do córtex
motor primário e secundário, nos 4 grupos estudados (Figura 3). Análise da
densidade óptica relativa nessa região cortical revelou que duas semanas de
treinamento acrobático induziu uma diminuição estatisticamente significante de
da proteína MAP2 (AC2, aprox. 18%, p≤0,05) e aumento significativo na quarta
semana de treinamento (AC4, aprox. 28%, p≤0,05) quando comparado ao
grupo controle (SED) (Figura 3). Os dados de “immunoblotting” corroboram
com esses resultados mostrando um aumento dos níveis proteicos somente no
grupo AC4 (aprox. 40%, p≤0,05) comparado ao SED, AC1 e AC2 (Figura 3).
Nesta região, a expressão do NF foi observada em processos e corpos
celulares, sendo identificado um aumento estatisticamente significativo da
densidade óptica relativa no grupo AC1 (aprox. 15%, p≤0,05) em relação ao
grupo controle (SED) (Figura 4). Os dados de “immunoblotting” não revelaram
alteração significativa no nível de expressão de NF68 entre os grupos
estudados (Figura 4).
As proteínas sinápticas SYS e SYP foram expressas de forma
puntiforme e difusa (Figuras 5 e 6). A densidade óptica relativa de SYS não
revelou mudanças significativas frente aos diferentes tempos de treinamento
acrobático (Figura 5). Entretanto, nos dados de “immunoblotting” foi observado
uma diminuição de SYS (aprox. 32%, p≤0,001) no grupo AC2 com retorno aos
níveis do controle no grupo AC4 (Figura 5).
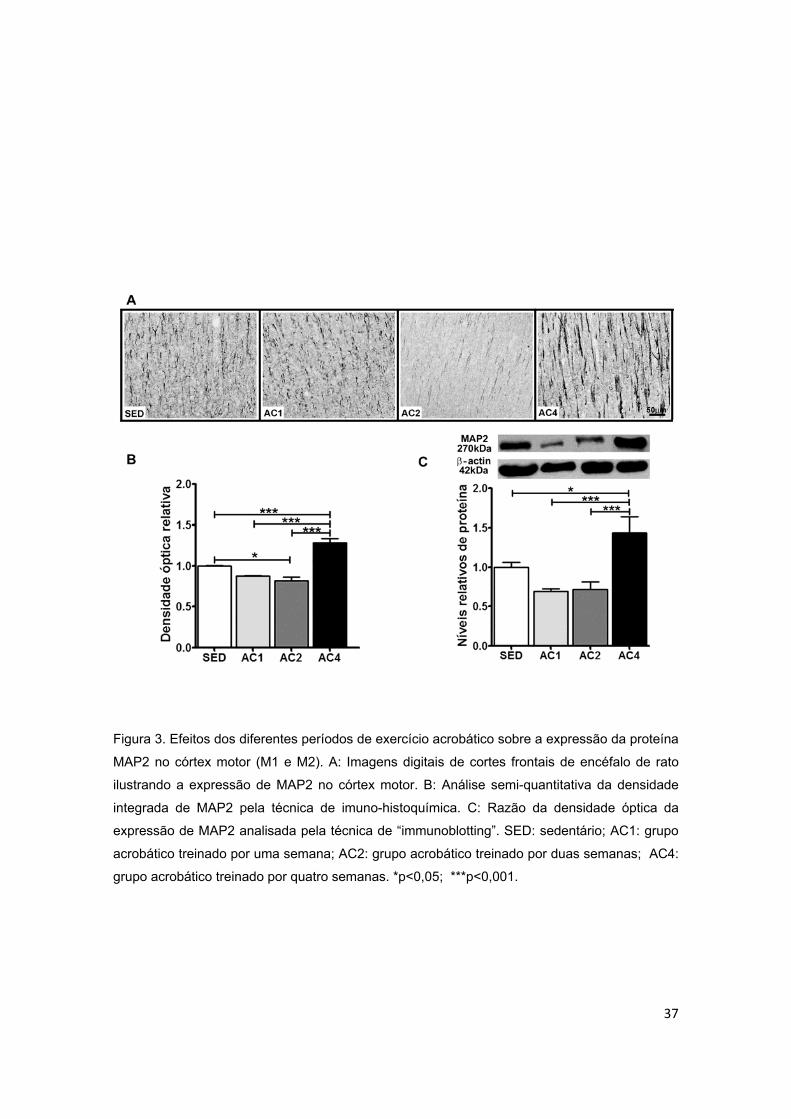
37
Figura 3. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da proteína
MAP2 no córtex motor (M1 e M2). A: Imagens digitais de cortes frontais de encéfalo de rato
ilustrando a expressão de MAP2 no córtex motor. B: Análise semi-quantitativa da densidade
integrada de MAP2 pela técnica de imuno-histoquímica. C: Razão da densidade óptica da
expressão de MAP2 analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo
acrobático treinado por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4:
grupo acrobático treinado por quatro semanas. *p<0,05; ***p<0,001.
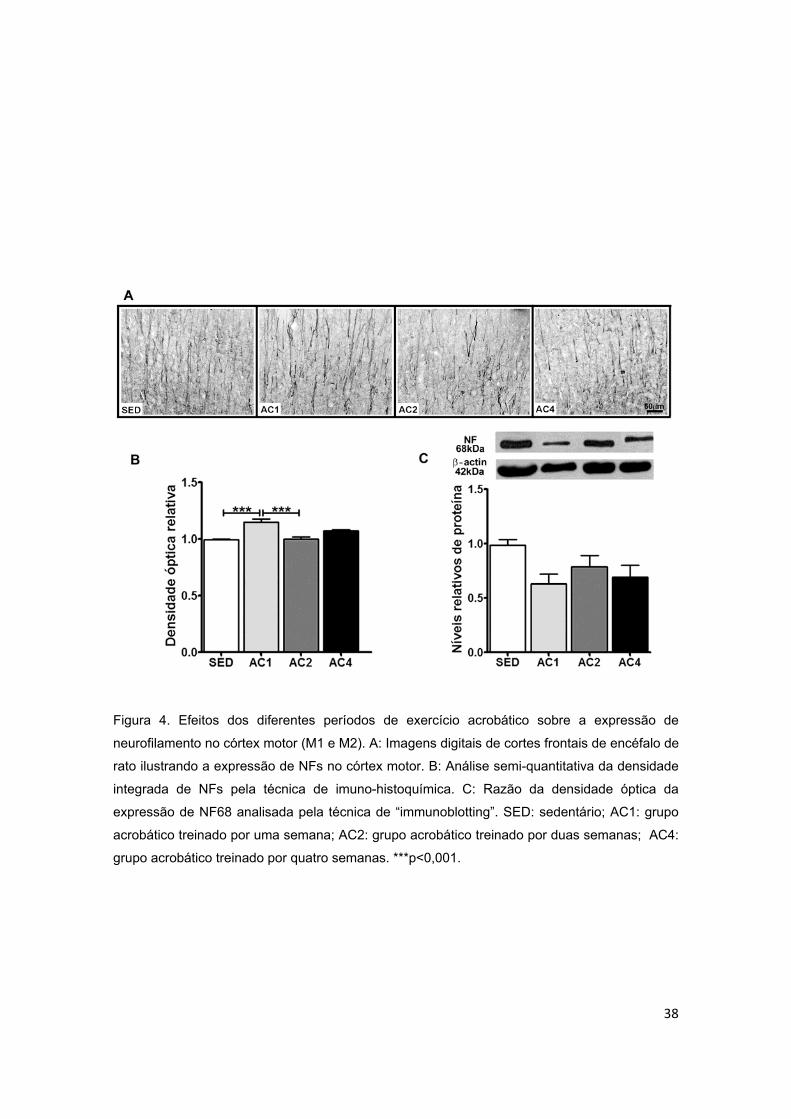
38
Figura 4. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão de
neurofilamento no córtex motor (M1 e M2). A: Imagens digitais de cortes frontais de encéfalo de
rato ilustrando a expressão de NFs no córtex motor. B: Análise semi-quantitativa da densidade
integrada de NFs pela técnica de imuno-histoquímica. C: Razão da densidade óptica da
expressão de NF68 analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo
acrobático treinado por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4:
grupo acrobático treinado por quatro semanas. ***p<0,001.
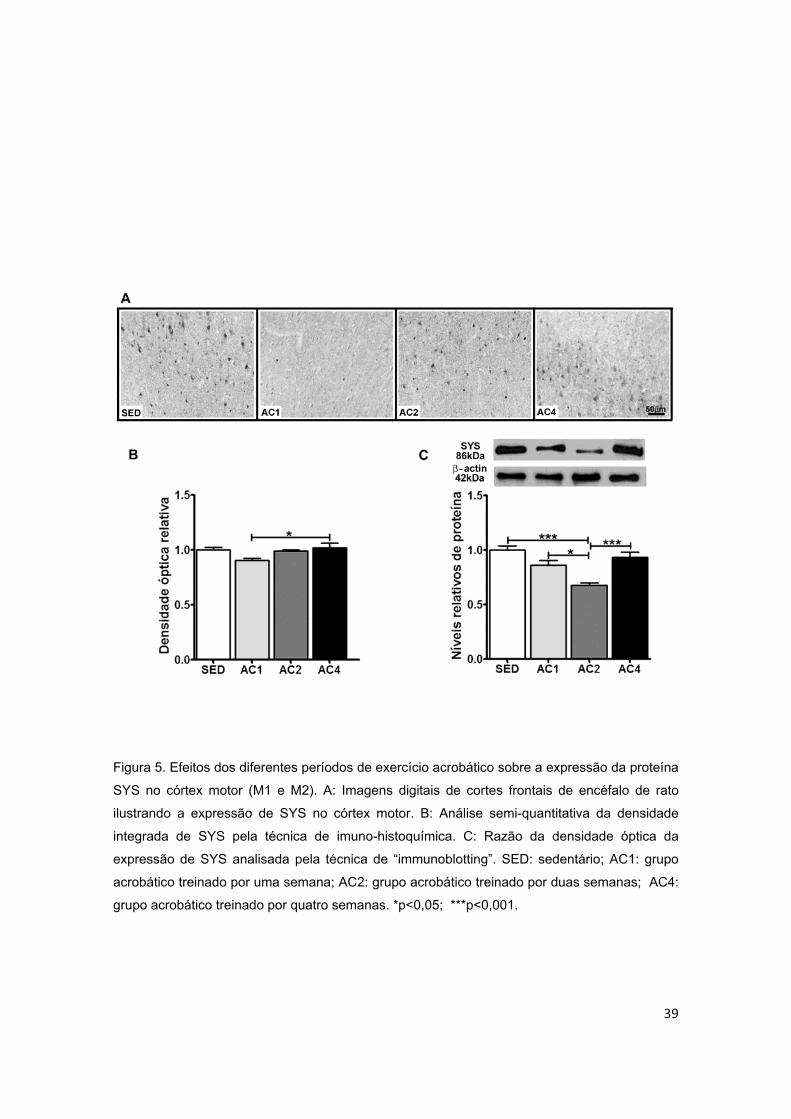
39
Figura 5. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da proteína
SYS no córtex motor (M1 e M2). A: Imagens digitais de cortes frontais de encéfalo de rato
ilustrando a expressão de SYS no córtex motor. B: Análise semi-quantitativa da densidade
integrada de SYS pela técnica de imuno-histoquímica. C: Razão da densidade óptica da
expressão de SYS analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo
acrobático treinado por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4:
grupo acrobático treinado por quatro semanas. *p<0,05; ***p<0,001.

40
O exercício acrobático por duas semanas (AC2) promoveu um aumento
estatisticamente significante da densidade óptica relativa de SYP (aprox. 60%,
p≤0,05) em relação ao grupo controle, que persistiu no AC4 (aprox. 34%,
p≤0,05) (Figura 6). Já os dados de “immunoblotting” revelaram que o exercício
acrobático promoveu uma diminuição da expressão de SYP (aprox. 19%,
p≤0,05) nas duas primeiras semanas, retornando aos níveis do controle nos
ratos do grupo AC4 (Figura 6).
5.2.2. Estriado
No estriado, o exercício acrobático induziu mudanças distintas na
expressão de proteínas estruturais e sinápticas em relação aos diferentes
tempos de treinamento de exercício acrobático.
Nos quatro grupos estudados, a proteína MAP2 foi expressa ao longo da
neurópila de forma difusa por toda extensão rostro-caudal e médio-lateral do
estriado (Figura 7). A densidade óptica relativa de MAP2 no estriado
apresentou um aumento significativo na região dorsomedial do grupo AC4 em
relação aos outros grupos estudados, e na região dorsolateral foram os grupos
AC1 e AC4 que demonstraram um aumento estatisticamente significativo
quando comparado ao grupo sedentário (Figura 7). Entretanto, os dados de
“immunoblotting” revelaram um aumento significante da expressão de MAP2 no
grupo AC2 em relação ao grupo controle (aprox. 55%, p≤0,05) com retorno ao
basal no grupo AC4 (Figura 7).
A imunorreatividade contra o anticorpo para NF revelou uma marcação
puntiforme e agrupada em regiões onde possivelmente há a passagem de
feixes nervosos (Figura 8). A densidade óptica relativa de NF na região
dorsolateral mostrou-se similar nos diferentes períodos de treinamento
estudados. Entretanto, na região dorsomedial houve um aumento significativo
(aprox. 43%, p≤0,05) do grupo AC2 quando comparado ao controle, retornando
aos níveis do controle na quarta semana de treinamento (Figura 8).
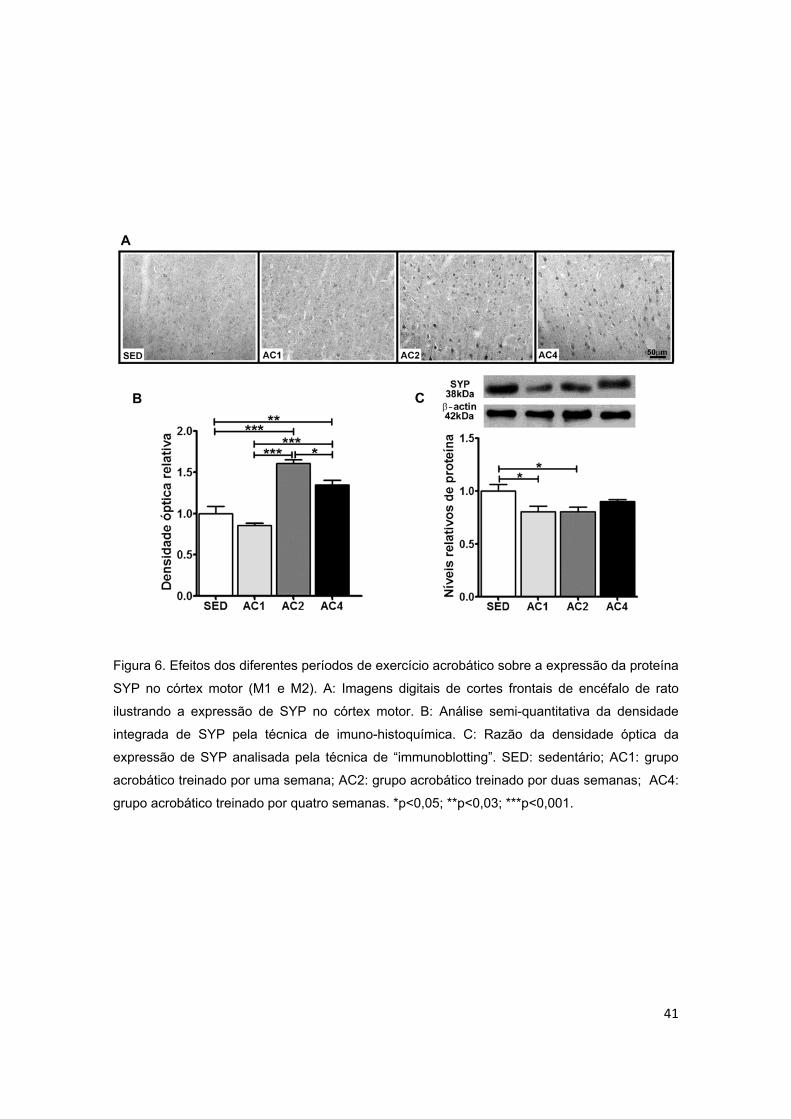
41
Figura 6. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da proteína
SYP no córtex motor (M1 e M2). A: Imagens digitais de cortes frontais de encéfalo de rato
ilustrando a expressão de SYP no córtex motor. B: Análise semi-quantitativa da densidade
integrada de SYP pela técnica de imuno-histoquímica. C: Razão da densidade óptica da
expressão de SYP analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo
acrobático treinado por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4:
grupo acrobático treinado por quatro semanas. *p<0,05; **p<0,03; ***p<0,001.
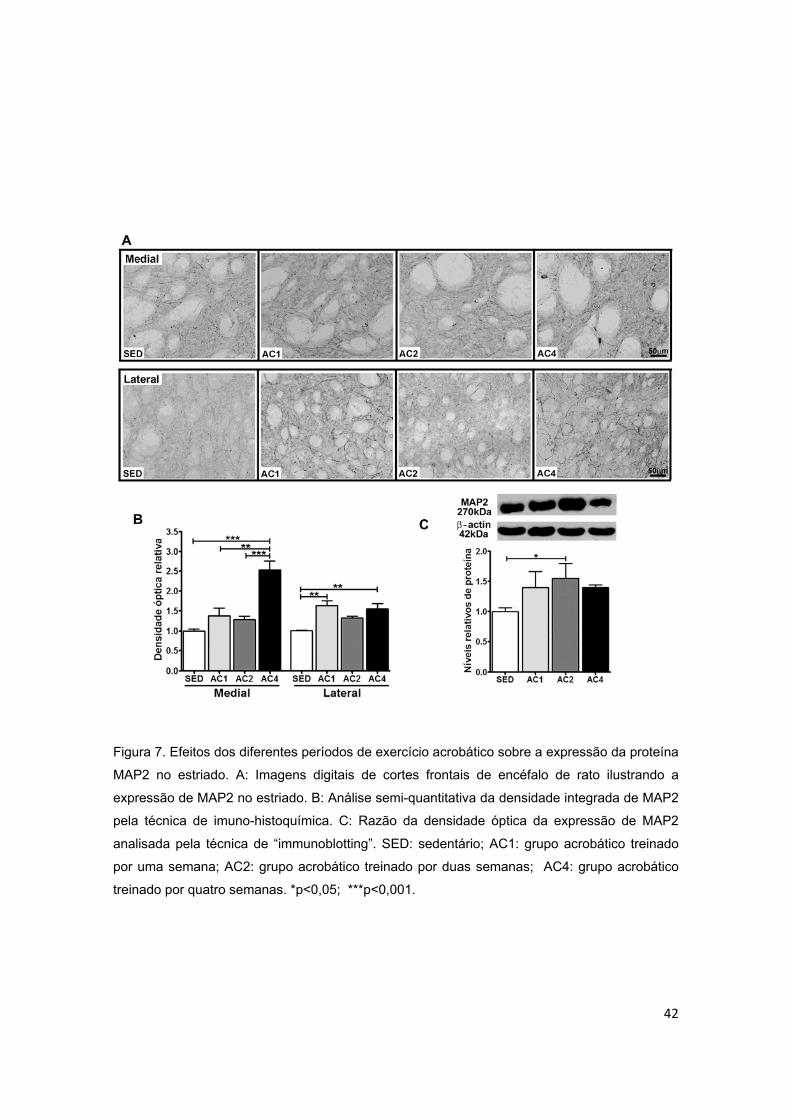
42
Figura 7. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da proteína
MAP2 no estriado. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando a
expressão de MAP2 no estriado. B: Análise semi-quantitativa da densidade integrada de MAP2
pela técnica de imuno-histoquímica. C: Razão da densidade óptica da expressão de MAP2
analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas. *p<0,05; ***p<0,001.
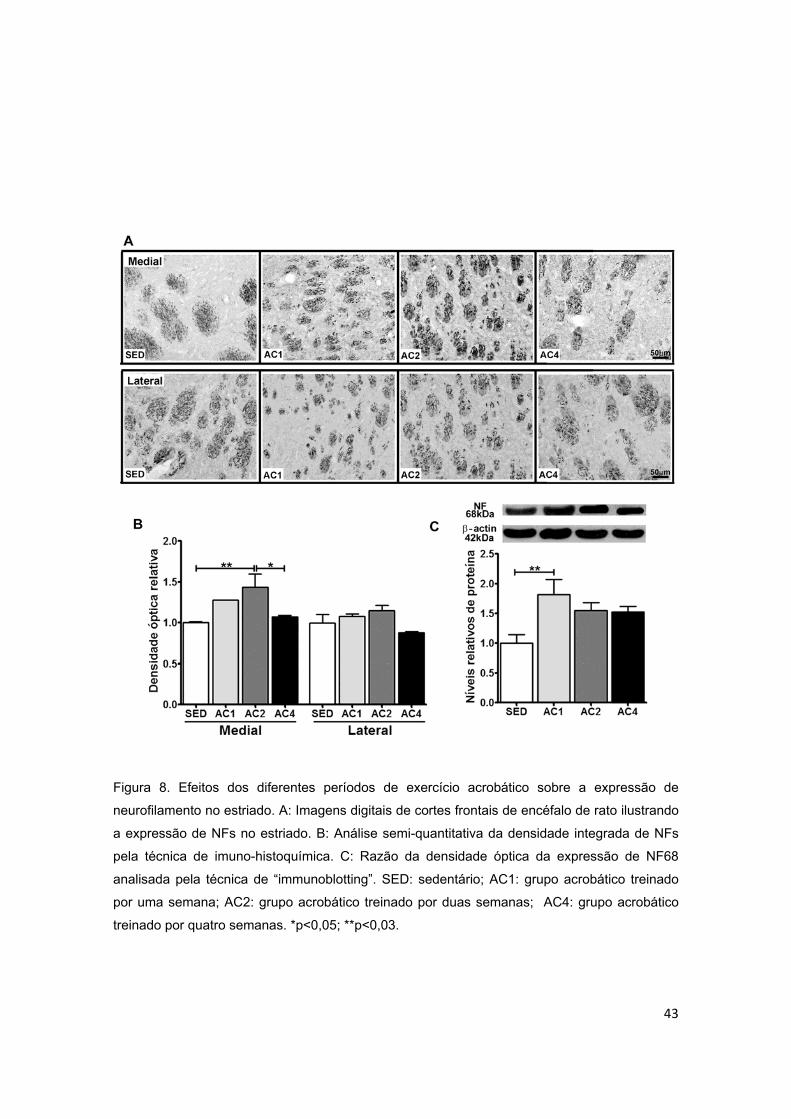
43
Figura 8. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão de
neurofilamento no estriado. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando
a expressão de NFs no estriado. B: Análise semi-quantitativa da densidade integrada de NFs
pela técnica de imuno-histoquímica. C: Razão da densidade óptica da expressão de NF68
analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas. *p<0,05; **p<0,03.

44
Todavia, os dados do “immunoblotting” demonstraram um aumento de
NF68 no grupo AC1 (aprox. 82%, p≤0,01), retornando aos níveis do controle na
segunda semana de treinamento (Figura 8).
As proteínas sinápticas SYS e SYP nesta região revelaram uma
imunorreatividade de padrão puntiforme e difusa por todo o estriado (Figuras 9
e 10). A densidade óptica relativa de SYS mostrou-se significativamente
aumentada na região dorsomedial após duas e quatro semanas de treinamento
acrobático (AC2 e AC4) comparada ao grupo controle. Enquanto na região
dorsolateral, o aumento estatisticamente significativo (aprox. 23%, p≤0,05)
ocorreu somente no grupo AC2 retornando ao nível do controle na quarta
semana de treinamento (Figura 9). Já os dados de “immunoblotting” não
revelaram mudanças significativas com relação ao grupo controle (Figura 9).
A quantificação da imunorreativade para SYP na região dorsomedial do
estriado revelou que os grupos AC2 e AC4 induziram um aumento
estatisticamente significante em relação ao grupo controle e na região
dorsolateral esse aumento foi observado nos grupos AC1 e AC2 comparado ao
controle (Figura 10). Os dados de “immunoblotting” do estriado revelaram que o
treinamento acrobático induziu um aumento da proteína SYP (aprox. 27%,
p≤0,01) no grupo AC4 quando comparado aos outros grupos estudados (Figura
10).
5.2.3. Cerebelo
No cerebelo dos quatro grupos de animais estudados foi observado
processos axonais e dendríticos ao longo das camadas granular e molecular
imunorreativos a MAP2 e NF (Figuras 11 e 12). A imunorreatividade para essas
proteínas estruturais apresentaram um padrão citoplasmático de marcação nas
células de Purkinje. Análises da proteína MAP2 realizadas pela imuno-
histoquímica na camada granular e molecular do cerebelo e pelo
“immunoblotting” de todo o cerebelo revelaram que os diferentes tempos de
treinamento acrobático não foi capaz de produzir mudanças estatisticamente
significativas quando comparado ao controle (Figura 11).
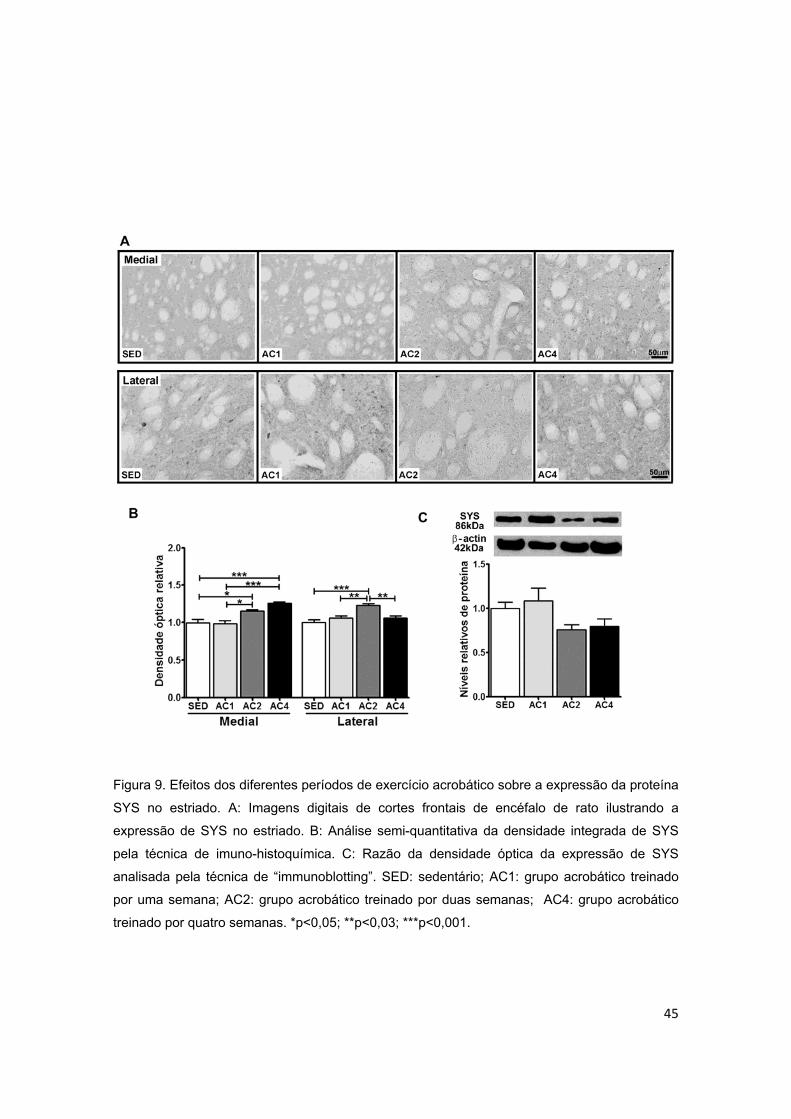
45
Figura 9. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da proteína
SYS no estriado. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando a
expressão de SYS no estriado. B: Análise semi-quantitativa da densidade integrada de SYS
pela técnica de imuno-histoquímica. C: Razão da densidade óptica da expressão de SYS
analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas. *p<0,05; **p<0,03; ***p<0,001.
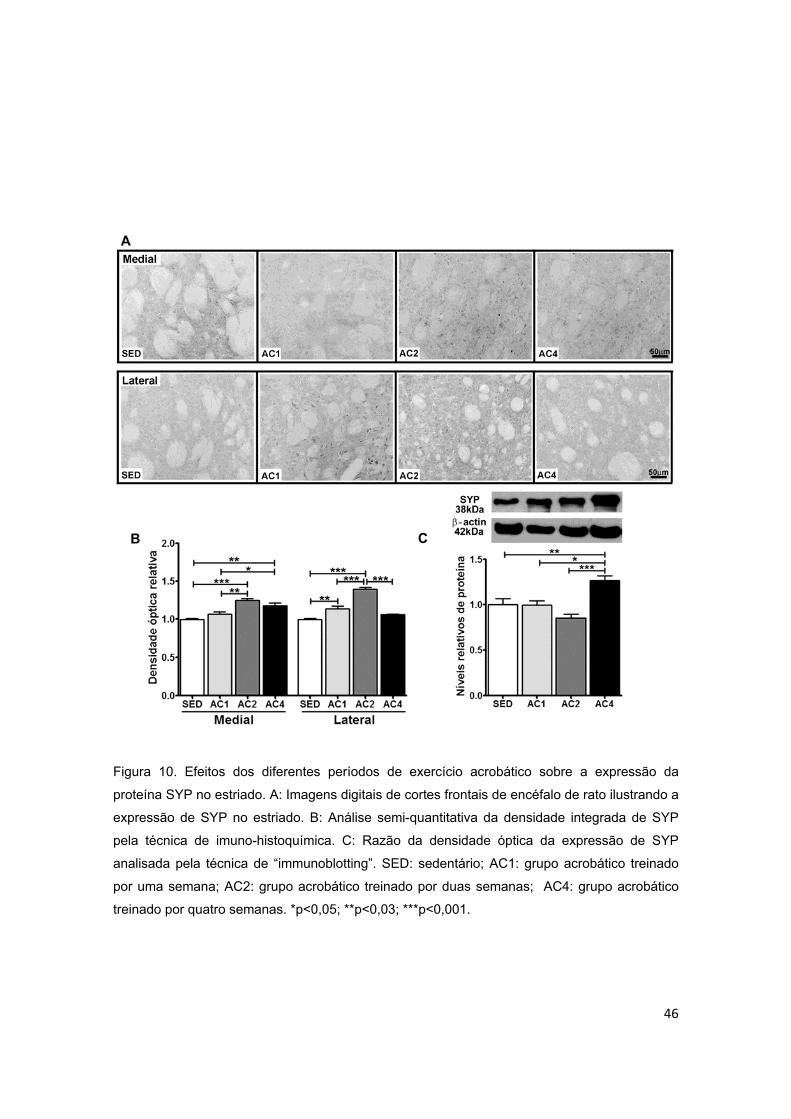
46
Figura 10. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da
proteína SYP no estriado. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando a
expressão de SYP no estriado. B: Análise semi-quantitativa da densidade integrada de SYP
pela técnica de imuno-histoquímica. C: Razão da densidade óptica da expressão de SYP
analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas. *p<0,05; **p<0,03; ***p<0,001.
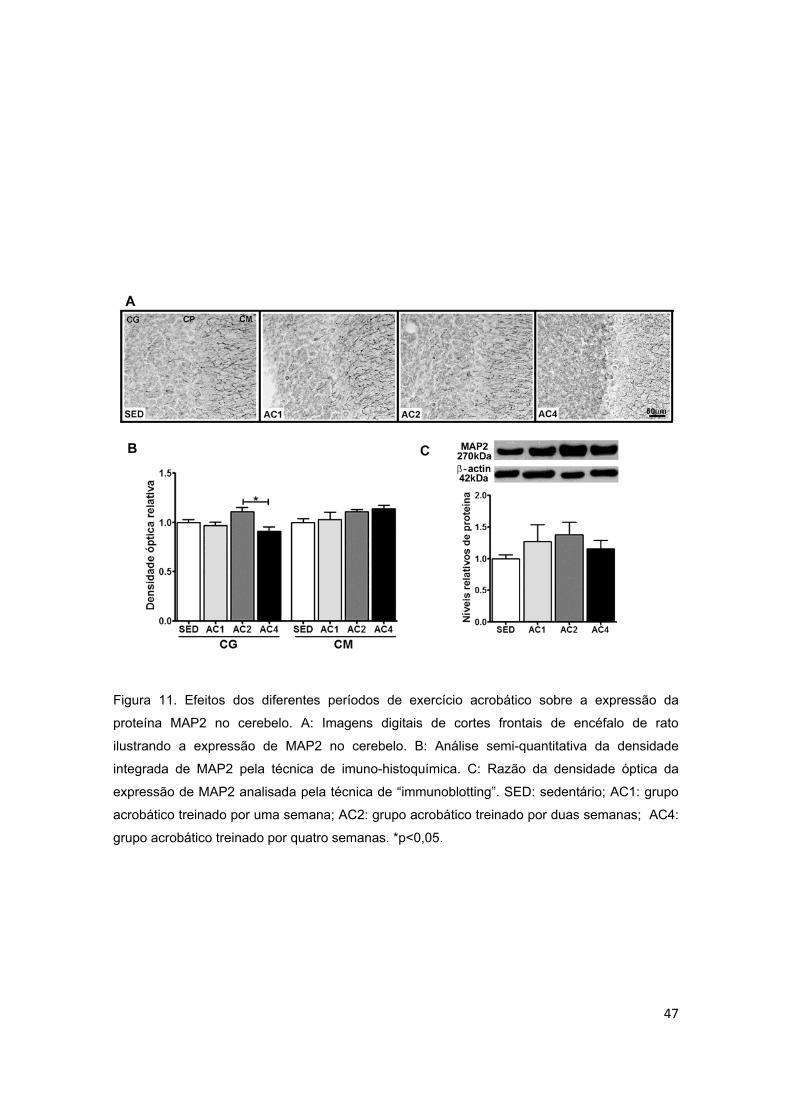
47
Figura 11. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da
proteína MAP2 no cerebelo. A: Imagens digitais de cortes frontais de encéfalo de rato
ilustrando a expressão de MAP2 no cerebelo. B: Análise semi-quantitativa da densidade
integrada de MAP2 pela técnica de imuno-histoquímica. C: Razão da densidade óptica da
expressão de MAP2 analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo
acrobático treinado por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4:
grupo acrobático treinado por quatro semanas. *p<0,05.

48
A proteína NF nos grupos treinados AC1 e AC4 mostrou um aumento
significativo da densidade óptica relativa na camada granular em relação ao
grupo controle, enquanto na camada molecular esse aumento foi produzido
somente pelo grupo AC1 (Figura 12). Dados de “immunoblotting” mostraram
que o exercício acrobático induziu um aumento significativo de NF68 (aprox.
48%, p≤0,05) no grupo AC2 comparado ao SED, com retorno a níveis do
controle na 4ª semana (Figura 12).
Quanto a expressão das proteínas sinápticas SYS e SYP no cerebelo
dos grupos estudados foi observado imunorreatividade de características
puntiforme entre as células de Purkinje e em aglomerados difusos na camada
molecular e esparsos na camada granular (Figuras 13 e 14). O treinamento
acrobático promoveu uma diminuição estatisticamente significante da
intensidade de marcação de SYS na camada granular após duas semanas
(AC2) e na camada molecular após uma semana de treinamento (AC1) em
relação ao grupo controle, retornando ao nível do controle após esses períodos
(Figura 13). Dados de “immunoblotting” não revelaram alterações significativas
de SYS entre os quatro grupos estudados (Figura 13).
O exercício acrobático induziu mudanças na imunorreatividade para SYP
somente na camada molecular, onde observamos um aumento da densidade
óptica relativa nos grupos AC1 e AC2 comparado ao controle, retornando ao
nível basal na quarta semana de treinamento (Figura 14). Dados de
“immunoblotting” não revelaram alterações significativas desta proteína entre
os 4 grupos estudados (Figura 14).
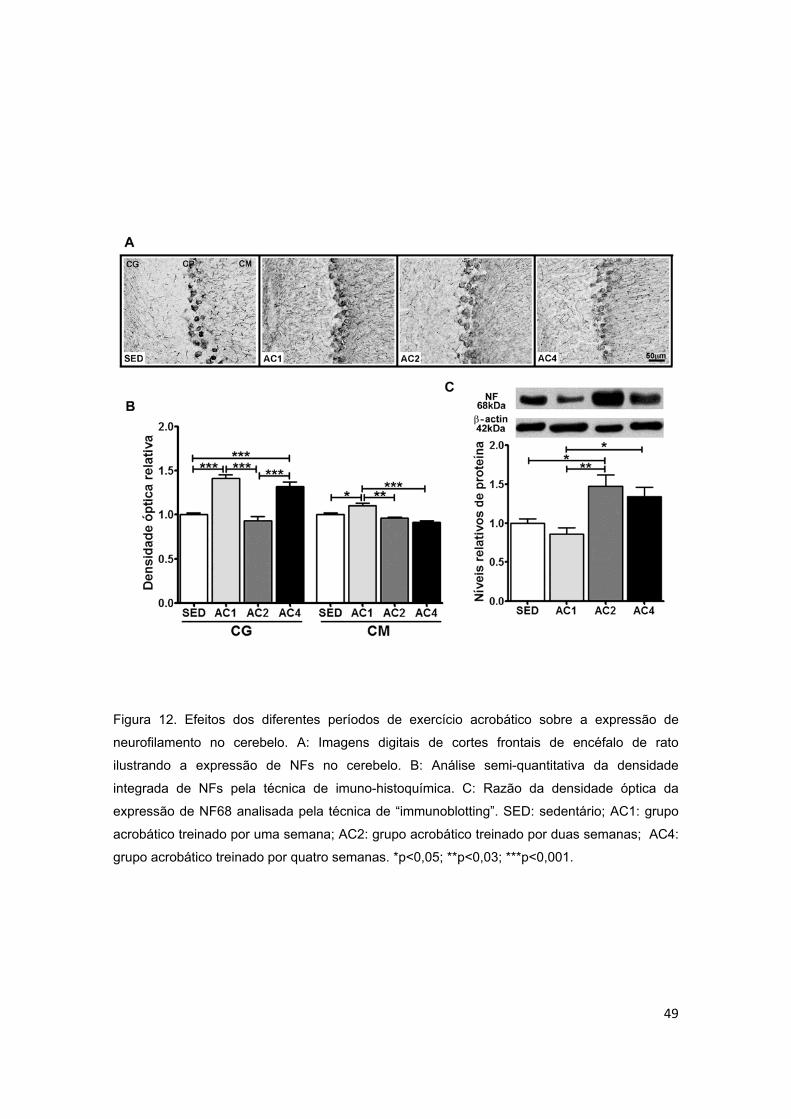
49
Figura 12. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão de
neurofilamento no cerebelo. A: Imagens digitais de cortes frontais de encéfalo de rato
ilustrando a expressão de NFs no cerebelo. B: Análise semi-quantitativa da densidade
integrada de NFs pela técnica de imuno-histoquímica. C: Razão da densidade óptica da
expressão de NF68 analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo
acrobático treinado por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4:
grupo acrobático treinado por quatro semanas. *p<0,05; **p<0,03; ***p<0,001.
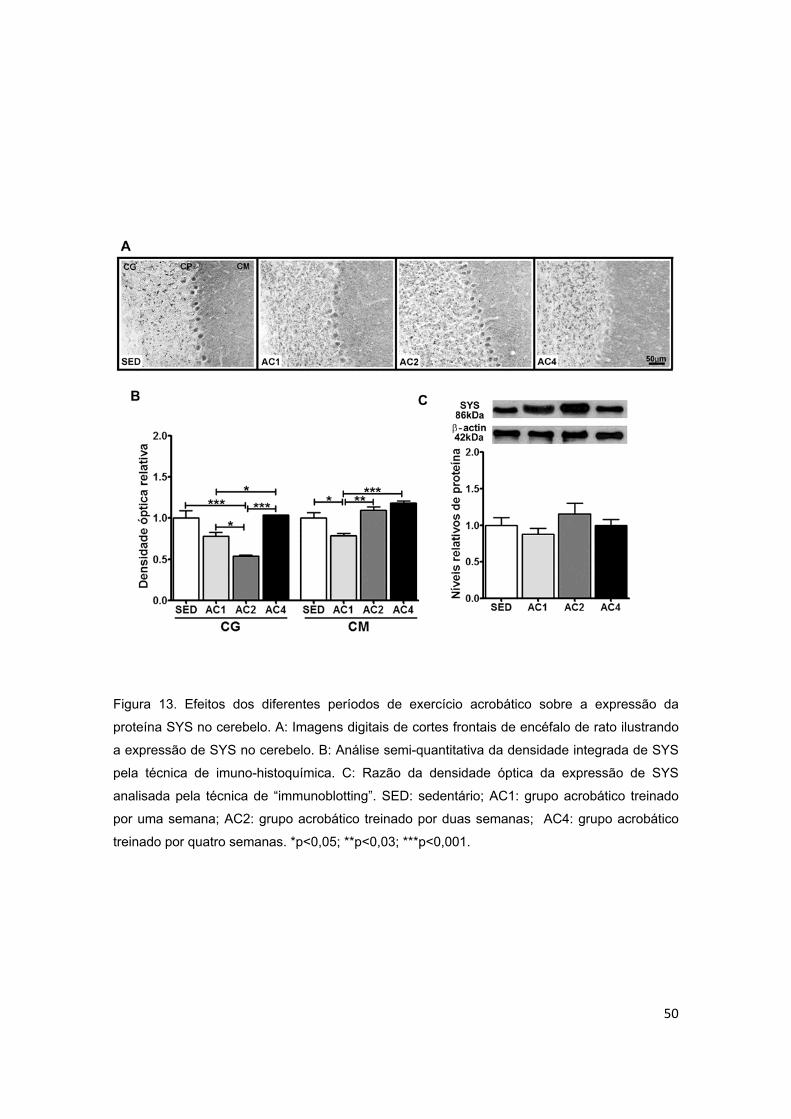
50
Figura 13. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da
proteína SYS no cerebelo. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando
a expressão de SYS no cerebelo. B: Análise semi-quantitativa da densidade integrada de SYS
pela técnica de imuno-histoquímica. C: Razão da densidade óptica da expressão de SYS
analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas. *p<0,05; **p<0,03; ***p<0,001.
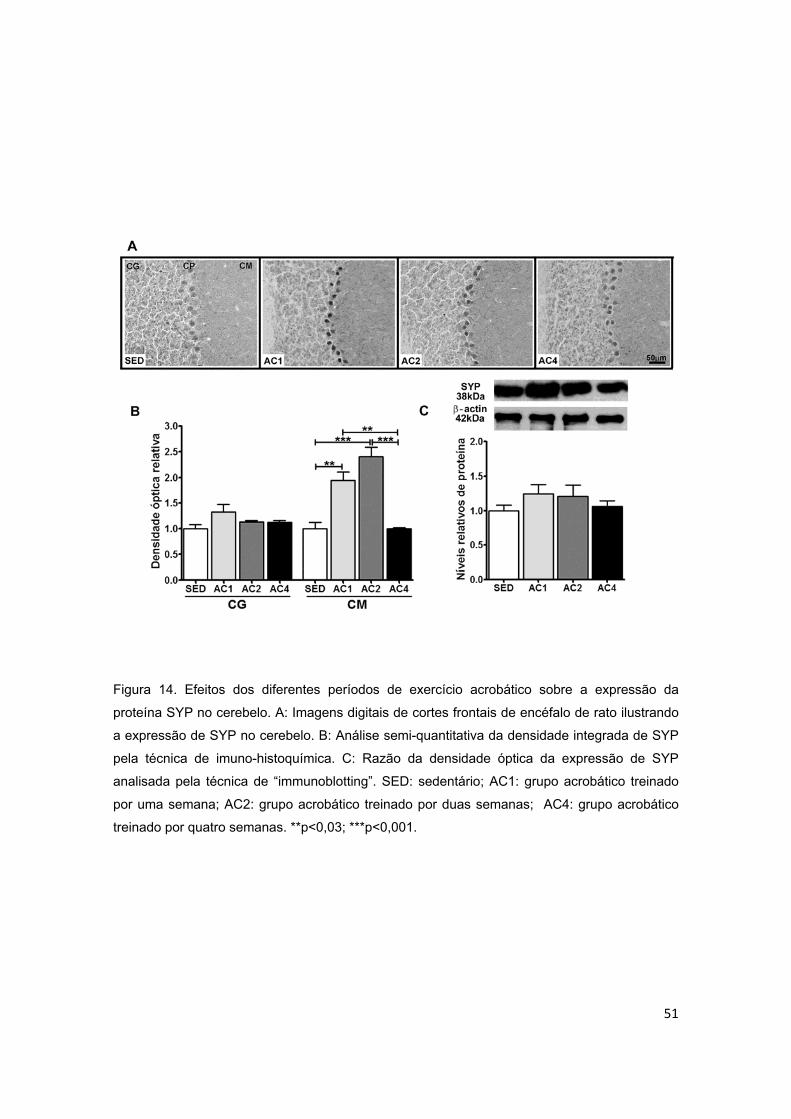
51
Figura 14. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão da
proteína SYP no cerebelo. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando
a expressão de SYP no cerebelo. B: Análise semi-quantitativa da densidade integrada de SYP
pela técnica de imuno-histoquímica. C: Razão da densidade óptica da expressão de SYP
analisada pela técnica de “immunoblotting”. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas. **p<0,03; ***p<0,001.

52
5.3. Expressão de Egr1
5.3.1. Córtex Motor
No córtex motor (M1 e M2) dos quatro grupos de animais estudados a
imunorreatividade contra o gene Egr1 se apresentou de forma difusa com
marcações no núcleo da célula (Figura 15).
A análise da densidade média de células Egr1-positivas nessa região
cortical revelou um crescente aumento significativo de células Egr1-positiva nos
três grupos treinados em relação ao grupo controle (Figura 15).
5.3.2. Estriado
No estriado dos quatro grupos estudados, o gene Egr1 foi expresso no
núcleo da célula de forma difusa por toda a extensão rostro-caudal e médio-
lateral (Figura 16).
A análise quantitativa mostrou que o AC2 induziu um aumento
estatisticamente significante da densidade média de células Egr1-positivas na
região dorsomedial em relação ao grupo controle e grupo AC1, retornando ao
nível do controle no AC4 (Figura 16). Já na região dorsolateral não houve
diferença entre os grupos experimentais e controle (Figura 16). Entretanto, foi
observado um aumento de células Egr1-positivas no grupo AC2 comparado ao
grupo AC1 (Figura 16).
5.3.3. Cerebelo
No cerebelo dos quatro grupos estudados, pode-se observar
imunorreatividade contra o gene Egr1 de forma difusa por toda a camada
granular com marcação no núcleo das células (Figura 17).
A análise da camada granular mostrou que o exercício acrobático
induziu um aumento estatisticamente significante de células Egr1-positivas
comparado ao controle nos três diferentes tempos de treinamento. Embora
esse aumento foi mais acentuado no grupo AC1(Figura 17).
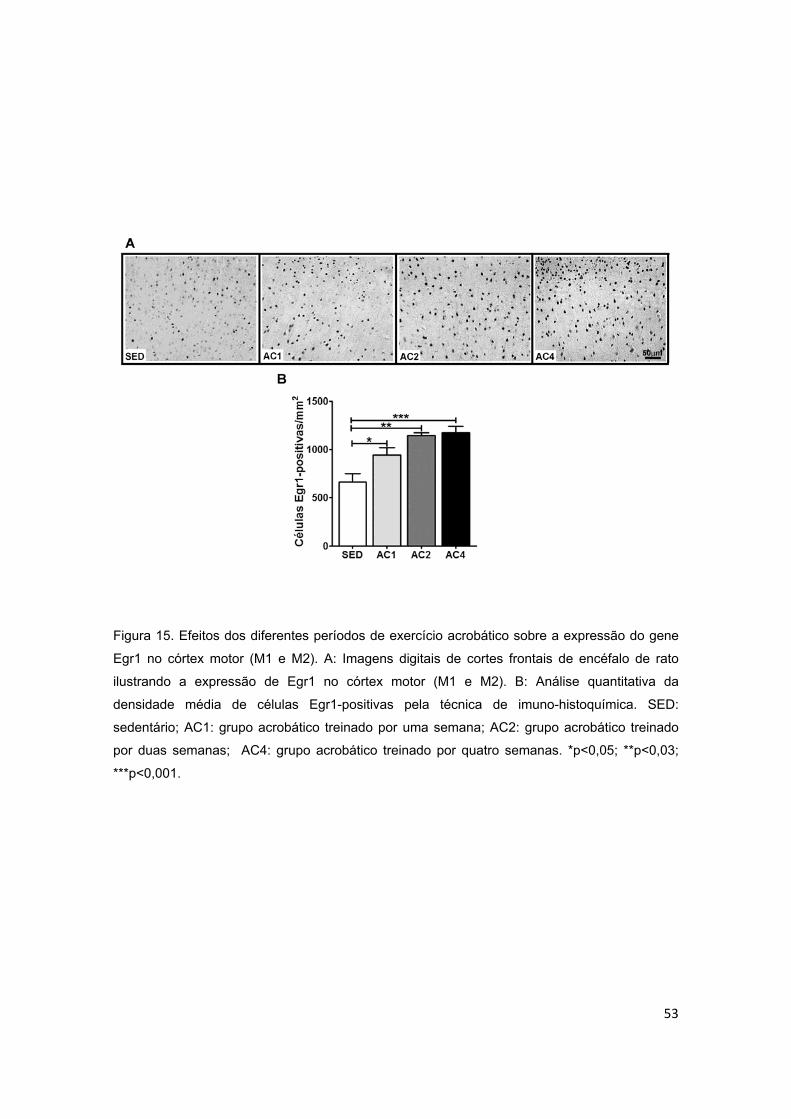
53
Figura 15. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão do gene
Egr1 no córtex motor (M1 e M2). A: Imagens digitais de cortes frontais de encéfalo de rato
ilustrando a expressão de Egr1 no córtex motor (M1 e M2). B: Análise quantitativa da
densidade média de células Egr1-positivas pela técnica de imuno-histoquímica. SED:
sedentário; AC1: grupo acrobático treinado por uma semana; AC2: grupo acrobático treinado
por duas semanas; AC4: grupo acrobático treinado por quatro semanas. *p<0,05; **p<0,03;
***p<0,001.
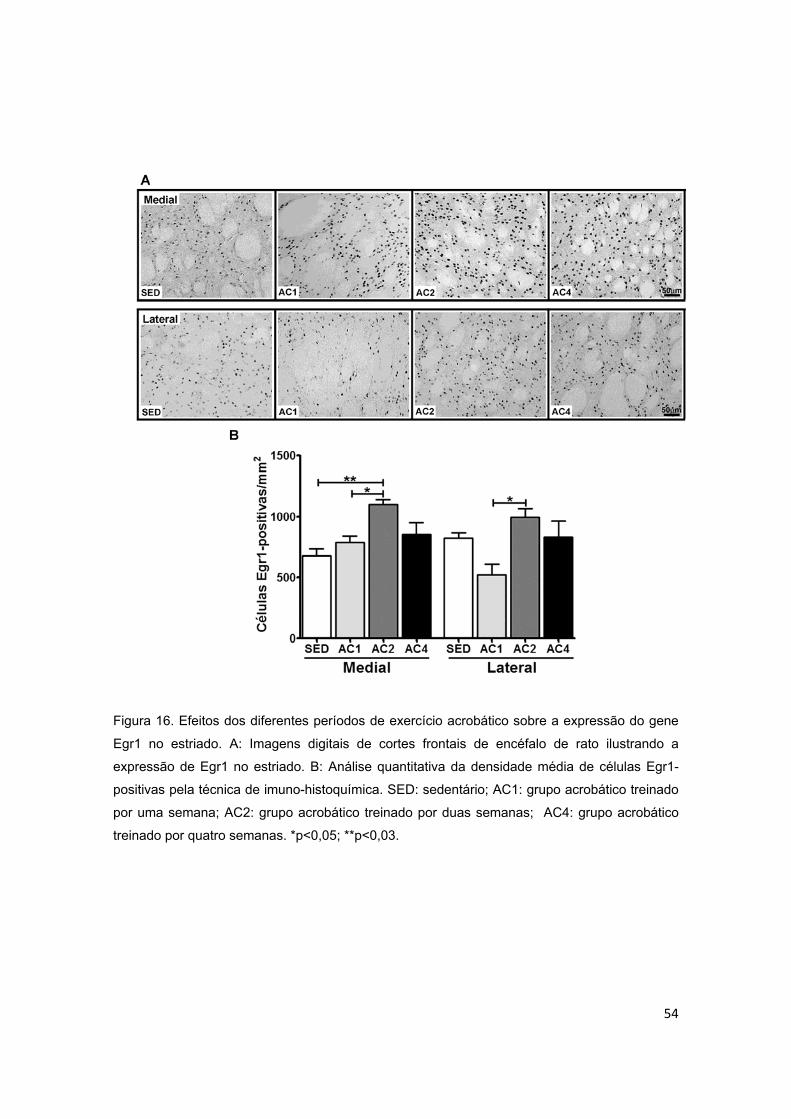
54
Figura 16. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão do gene
Egr1 no estriado. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando a
expressão de Egr1 no estriado. B: Análise quantitativa da densidade média de células Egr1-
positivas pela técnica de imuno-histoquímica. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas. *p<0,05; **p<0,03.
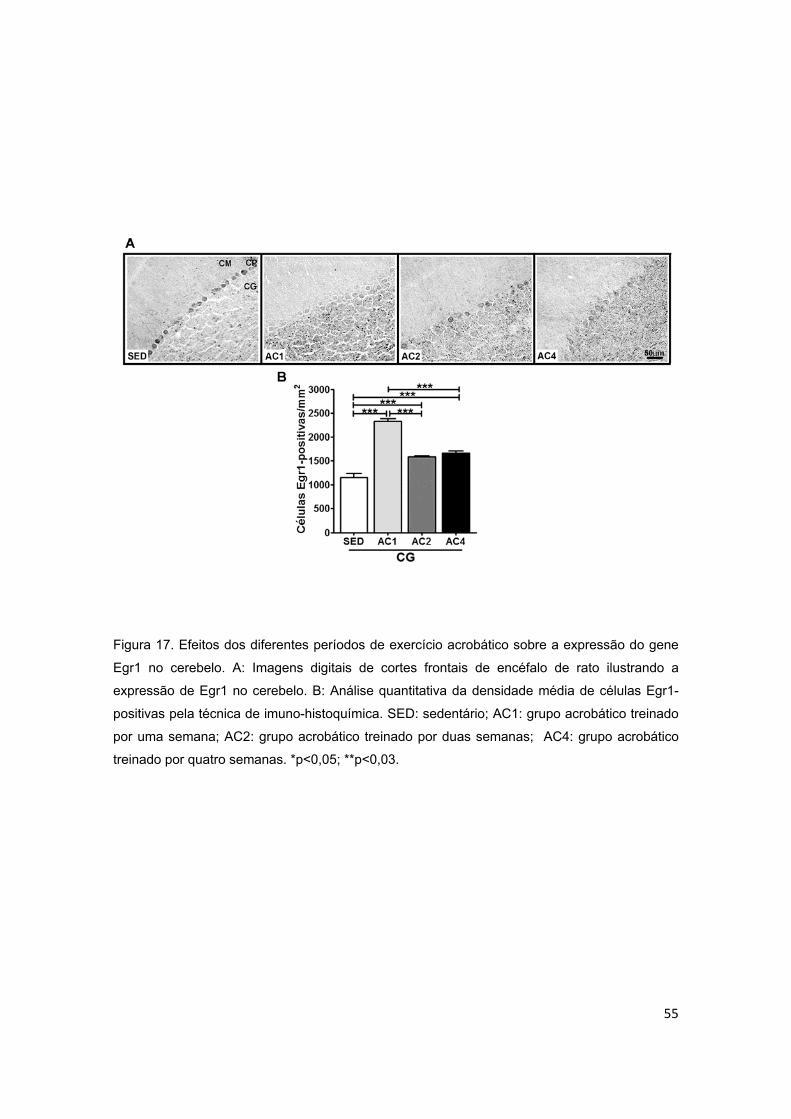
55
Figura 17. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão do gene
Egr1 no cerebelo. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando a
expressão de Egr1 no cerebelo. B: Análise quantitativa da densidade média de células Egr1-
positivas pela técnica de imuno-histoquímica. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas. *p<0,05; **p<0,03.
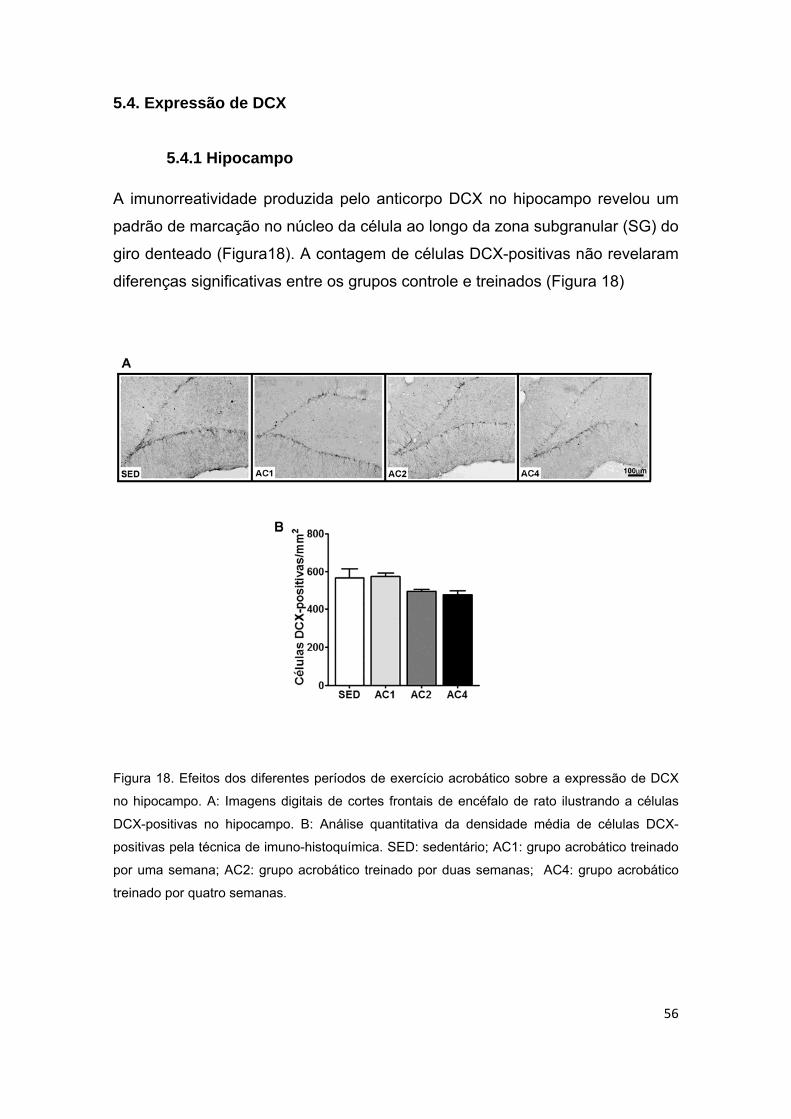
56
5.4. Expressão de DCX
5.4.1 Hipocampo
A imunorreatividade produzida pelo anticorpo DCX no hipocampo revelou um
padrão de marcação no núcleo da célula ao longo da zona subgranular (SG) do
giro denteado (Figura18). A contagem de células DCX-positivas não revelaram
diferenças significativas entre os grupos controle e treinados (Figura 18)
Figura 18. Efeitos dos diferentes períodos de exercício acrobático sobre a expressão de DCX
no hipocampo. A: Imagens digitais de cortes frontais de encéfalo de rato ilustrando a células
DCX-positivas no hipocampo. B: Análise quantitativa da densidade média de células DCX-
positivas pela técnica de imuno-histoquímica. SED: sedentário; AC1: grupo acrobático treinado
por uma semana; AC2: grupo acrobático treinado por duas semanas; AC4: grupo acrobático
treinado por quatro semanas.

57
6. DISCUSSÃO
6.1. Comportamento do Desempenho Motor
Aprendizagem de uma habilidade motora é caracterizada pela melhora
do desempenho do comportamento aprendido.55 Essa melhora do desempenho
motor consiste de duas fases distintas do aprendizado. A primeira fase consiste
de um período de rápida melhora do desempenho que é observado após
poucas sessões de treinamento, e a segunda fase é caracterizada por um
ganho lentificado do desempenho motor que ocorre ao longo de várias sessões
de treinamento.41 Essas duas fases são chamadas de aquisição e
consolidação, respectivamente. Os nossos dados revelaram que a partir da 2ª
semana de treinamento acrobático (AC2) foi observado uma redução
significativa do tempo necessário para executar todo o circuito comparado ao
da primeira semana. E essa redução significativa em relação ao início do
treinamento persistiu na terceira e quarta semana de treinamento (AC4)
apresentando uma crescente queda no tempo necessário para a realização das
tarefas motoras.
Vários estudos com diferentes protocolos de treinamento e exigências
motoras distintas revelaram uma redução do tempo de passagem pelo circuito
que variou entre o segundo, quinto, sexto e décimo dia de treinamento
dependendo do tipo de atividade exigida e o número de repetições do
circuito.40-43,55,56 E mais, tanto o nosso estudo como os citados anteriormente40-
43,55,56 revelaram reduções do tempo com padrões distintos ao longo do
treinamento destacando-se as duas fases do aprendizado. Além disso, esses
estudos demonstraram que quanto maior foi os desafios motores presentes nos
circuitos de treinamento maior será a redução de tempo comparado ao início
do treinamento sugerindo um importante melhora do desempenho motor nesta
fase inicial de aquisição, enquanto aquelas tarefas com pouca exigência
motora não revelaram reduções significativas no tempo de realização da tarefa
durante as fases de aquisição e consolidação do aprendizado.

58
Os dados do nosso estudo sugerem que a fase de aquisição ocorreu nas
duas primeiras semanas de treinamento, grupos AC1 e AC2, e a de
consolidação do aprendizado foi observada no grupo AC4.
A diminuição do tempo gasto para percorrer o circuito observado no
nosso estudo e nos citados acima foi decorrente de mudanças
comportamentais, tais como diminuição de erros, hesitações e dos estímulos
manuais.
6.2. Efeitos dos Diferentes Tempos de Treinamento em Regiões
Motoras
Com o objetivo de estudar proteínas estruturais e sinápticas em regiões
motoras do encéfalo, observamos que os diferentes períodos de treinamento
de exercício acrobático utilizado induziu respostas distintas tanto nos níveis das
proteínas estudadas (NFs, MAP2, SYS e SYP) quanto na densidade óptica
relativa do gene Egr1.
O presente estudo demonstrou que algumas proteínas envolvidas na
estrutura e na função neuronal são dinamicamente alteradas durante o
processo de aquisição e consolidação da aprendizagem motora enquanto
outros seguem um padrão de alteração de expressão mais previsível.
Portanto, nos tópicos a seguir, cada região motora será discutida
separadamente com o intuito de identificar efeitos específicos dos períodos de
treinamento sobre essas regiões envolvidas no controle e desempenho motor.
6.2.1. Córtex Motor
Os nossos dados revelaram que diferentes tempos de treinamento de
exercício acrobático induziram mudanças distintas na expressão de proteínas
estruturais e sinápticas no córtex motor de ratos adultos.
A expressão dessas proteínas foi diferente diante das técnicas de
“immunoblotiing” e imuno-histoquímica, provavelmente devido a diferença na
quantia e no tipo de tecido examinado com essas duas técnicas. Os dados da
imuno-histoquímica resultam da análise de uma discreta área do córtex motor

59
correspondente a camada V enquanto os obtidos pela técnica de
“immunoblotting” são oriundos da análise do córtex motor como um todo.
Quanto as proteínas estruturais, o exercício acrobático a curto prazo
induziu diminuição da expressão de MAP2 e aumento de NF68 e a longo prazo
somente aumento do nível proteico de MAP2 quando comparado ao grupo
sedentário, revelando um remodelamento dendrítico e sinaptogênese de
maneira distinta ao longo dos períodos de treinamento.
Aumento da arborização dendrítica e sinaptogênese no córtex motor
após treinamento de uma tarefa complexa tem sido bastante observada em
alguns estudos relacionados com a aprendizagem motora23,41,140,141 A
sinaptogênese resultante da aprendizagem motora tem sido demonstrada nos
neurônios piramidais do córtex motor durante a fase de manutenção ou
consolidação do aprendizado de habilidade motora, não sendo evidente na
fase de aquisição.41 Além disso, Kleim e colaboradores141 investigando a
relação temporal entre sinaptogênese e reorganização do mapa motor cortical
associado com aprendizagem de habilidade motora em ratos treinados por 3, 7
ou 10 revelaram que o aumento significativo no número de sinapses por
neurônio não foram observado até o sétimo dia de treinamento e que a
reorganização do mapa motor só foi evidenciado após 10 dias de treinamento.
Garcia e colaboradores40 observaram que animais treinados por 4 semanas de
exercício acrobático, 3 vezes/semana com 5 passagens/dia induziu um
aumento na expressão da proteína MAP2 e SYP, corroborando nossos dados
que apresentaram resultados semelhantes.
Estudo com mapeamento cortical demonstrou que após treinamentos de
novas habilidades motoras em ratos ocorreu uma expansão da área cortical
envolvida com os movimentos utilizados para a realização dessas tarefas.77
Portanto, os nossos dados corroboram com as citados acima, revelando que
aumentos na expressão de MAP2 após quatro semanas de treinamento AC
demonstram uma crescimento da árvore dendrítica e estabelecimento de novas
sinapses, resultando num aumento da área cortical envolvida no controle dessa
atividade motora e consequente reorganização do mapa cortical juntamente
com a fase de consolidação observada pela melhora comportamental do
desempenho motor.

60
Entretanto, nossos dados revelaram que curtos períodos de treinamento
acrobático induziu uma diminuição da proteína MAP2 e um aumento do NF,
sugerindo um remodelamento axonal favorecendo aumento do calibre axonal e
consequente aumento da velocidade de condução do impulso nervoso.
A expressão das proteínas sinápticas SYS e SYP no córtex motor dos
animais AC tanto a curto como a longo prazo foi diferente diante das duas
técnicas de análise “immunoblotting” e imuno-histoquímica. Discrepâncias
como essas são encontradas na literatura, e a explicação mais plausível estão
nas diferentes características das metodologias empregadas. Curto período de
treinamento (AC1 e AC2) promoveu uma diminuição dos níveis proteicos de
SYS e SYP no córtex motor como um todo, revelado pela técnica de
“immunoblotting”; entretanto, quando analisadas pelo método de imuno-
histoquímica, observamos aumento significativo de SYP tanto a curto como a
longo prazo do treinamento acrobático.
Os nossos dados de imuno-histoquímica, revelando aumento da
expressão de SYP no córtex motor dos animais treinados, foram obtidos com
análises de densidade óptica relativa na camada V. Esses dados corroboram
com vários estudos que mostraram um aumento de sinapses por neurônio na
camada V em ratos que realizavam exercício acrobático comparados aos
animais controle,23,32,77,141 sugerindo que o treinamento acrobático a curto
prazo pode favorecer o fortalecimento das sinapses já existentes140 ou
aumentar a excitabilidade neuronal142. Já o aumento de SYP após longo prazo
de treinamento acrobático aparece juntamente com os achados de
sinaptogênese e potenciação sináptica em áreas corticais envolvidas com o
planejamento dessa atividade motora ao longo da fase de consolidação do
aprendizado de habilidades motoras. Entretanto, existe um outro estudo que
revelou que o treinamento acrobático diário por 6 semanas não afetou a
expressão dos níveis de SYP no córtex motor de ratos adultos analisado pelo
método de ELISA.55 Uma explicação para a diferença entre os nossos dados é
que eles analisaram uma área cortical maior e diferente da que nós estudamos,
. Considerando nossos dados citados acima, constatamos que o córtex
motor está ativo tanto durante a fase de aquisição como na fase de
consolidação da aprendizagem de habilidade motora. Nossos dados de

61
densidade de células Egr1-positivas na camada V do córtex motor revelaram
um aumento progressivo de células ativas ao longo do período de treinamento.
Avaliar a expressão do gene Egr1 nessa região cortical ao longo dos
diferentes períodos de treinamento acrobático é importante devido a sua
relação com o mecanismo de potenciação de longo prazo (LTP) e aprendizado.
A expressão de Egr1 pode ser induzida, de forma rápida e transitória, por
moléculas sinalizadoras externas como neurotransmissores e fatores de
crescimento, que se ligam a receptores de glutamato e ativam uma cascata de
eventos, levando a expressão deste gene.
A crescente ativação de Egr-1 encontrada no nosso trabalho corrobora
com os nossos achados de mudanças na expressão de proteínas estruturais e
sinápticas juntamente com os dados da literatura citados acima que revelaram
uma remodelamento dendrítico, sinaptogênese e reorganização do mapa
cortical juntamente com a melhora do desempenho motor ao longo da
aprendizagem de habilidade motora, refletindo possivelmente os mecanismos
intrínsecos envolvidos no contexto da plasticidade sináptica.
6.2.2. Estriado
Os diferentes períodos de treinamento (AC1, AC2 e AC4) promoveram
alterações na expressão das proteínas estruturais e sinápticas no estriado.
Quanto as proteínas estruturais, observamos aumento da expressão das
proteínas MAP2 e NF68 a curto prazo de treinamento do exercício acrobático
enquanto que a longo prazo não houve alterações. A ausência de mudanças
na expressão de proteínas estruturais no estriado a longo prazo de treinamento
é discordante do obtido por Garcia e colaboradores40, que demonstraram um
aumento de MAP2 e NF68 após 4 semanas de treinamento. Possivelmente
essa discrepância dos resultados se deu em virtude da diferença existente
entre as amostras utilizadas, pois em nosso estudo comparamos 3 grupos com
períodos de treinamento distintos, enquanto que nos estudo citado foi utilizado
2 grupos com diferentes tipos de exercícios

62
O estriado é uma estrutura encefálica envolvida tanto com a fase inicial
do aprendizado de novas habilidades e sequências motoras, como com a
consolidação dessas habilidades motoras numa fase mais tardia. 44,143. Sendo
assim, esse padrão distinto do aumento das proteínas estruturais entre os
diferentes períodos de treinamento parece apoiar as diferentes funções do
estriado na aquisição de uma habilidade motora complexa e consolidação da
aprendizagem motora.89
Com relação a expressão das proteínas sinápticas no decorrer do
período de treinamento obtidos pelo método de “immunoblotting”, observamos
mudanças na expressão da SYP a longo prazo de treinamento, sem alterações
na expressão da proteína SYS. Este aumento da expressão da proteína SYP
no estriado pode ter ocorrido devido a uma liberação aumentada de glutamato
que está diretamente ligada à fosforilação da SYP, sugerindo uma possível
potenciação de longo prazo (LTP).144
A imuno-histoquímica revelou um aumento de SYP e SYS a curto prazo
de treinamento nas regiões dorsolateral, e ao longo do período de treinamento
na região dorsomedial dos animais treinados, diferentemente dos dados de
“immunoblotting”. Essa diferença pode ser devido às variadas formas de
reconhecimento e ligação dos anticorpos aos seus epítopos. Contudo, esse
aumento da eficiência sináptica nessas regiões do estriado induzida pelos
diferentes períodos de treinamento estão de acordo com o aumento do
potencial excitatório pós-sináptico observados nos registros eletrofisológicos do
estriado de camundongos submetidos igualmente a realização de habilidades
motoras.89
Os dados de imuno-histoquímica obtidos em nosso estudo sugerem uma
reorganização estrutural e funcional do estriado dorsal dependente do tempo
de treinamento e da atividade motora complexa. Estudos anteriores com
roedores demonstraram o envolvimento de diferentes circuitos do estriado
durante a fase inicial e tardia do aprendizado de uma habilidade motora e
também de respostas motoras habituais e automáticas.1,40 No nosso estudo
observamos que o treinamento a curto prazo de atividades motoras complexas
induziu aumento de PAN, SYP e SYS na região dorsomedial e aumento de
MAP, SYP e SYS na região dorsolateral enquanto que a longo período de

63
treinamento podemos observar aumento de MAP2, SYP e SYS na região
dorsomedial e somente de MAP2 na região dorsolateral. Sugerindo um
remodelamento dendrítico e axonal e maior eficiência sináptica na região
dorsomedial ao longo do período de treinamento. Juntamente com nossos
dados de Egr1-positivo que mostraram uma ativação maior da região
dorsomedial tendo seu pico na segunda semana de treinamento (AC2) e
retornando aos níveis basais.
Nossos dados corroboram com estudos que observaram que a região
dorsomedial parece estar mais envolvida com a aquisição de habilidades
motoras , enquanto que a região dorsolateral parece estar envolvido além das
respostas habituais e automáticas com a consolidação da aprendizagem
motora. 40,44,143 Observamos em nosso estudo que na fase de aquisição, há
uma melhora rápida do desempenho motor revelado pela diminuição do tempo
necessário para realização daquela tarefa, e tanto a fase de aquisição como a
de consolidação parecem ser acompanhadas de respostas plásticas distintas,
que podem ser tanto em regiões diferentes como em relação a expressão de
proteínas. Podemos sugerir também que essa melhora rápida do desempenho
motor pode ter sido proporcionado pelo aumento da eficiência sináptica tanto
da região dorsomedial quanto da região dorsolateral.
6.2.3. Cerebelo
Os nossos dados revelaram que as mudanças na expressão das
proteínas estruturais e sinápticas no cerebelo ocorreram preferencialmente
após curto período de treinamento acrobático, retornando ao nível do controle
na quarta semana de treinamento. Ao analisarmos o cerebelo como um todo,
através da técnica de “immunoblotting”, observarmos mudanças somente a
curto prazo de treinamento na expressão da proteína NF68. Porém, quando
analisamos especificamente a região do lóbulo paramediano do cerebelo, tanto
na camada molecular quanto na camada granular observamos mudanças
tempo-dependente distintas na expressão das proteínas sinápticas e
estruturais.

64
Quanto as proteínas estruturais, observamos que curto período de
exercício acrobático promoveu aumento da proteína NF68 revelado pela
técnica de “immunoblotting” e quando analisadas separadamente pela técnica
de imuno-histoquímica observamos um aumento da expressão da proteína
PAN nas camadas granular e molecular após curto período de treinamento e
após longo período somente na camada granular. Além disso, nossos dados
revelaram ausência de alterações na expressão da proteína MAP2 em ambas
as técnicas utilizadas. Esses achados de expressão de NF e MAP2 após longo
período de treinamento (AC4) corroboram com os de Garcia e colaboradores40
que não revelaram mudanças após quatro semanas de treinamento. Mas o
aumento da expressão de NF nas camadas granular e molecular frente a uma
semana de treinamento acrobático sugere uma importante reestruturação
axonal com possível aumento no diâmetro e/ou extensão dos axônios
favorecendo uma maior velocidade de condução do impulso nervoso nessas
camadas do córtex cerebelar. Vale ressaltar que as fibras nervosas
encontradas nessas camadas são axônios das aferências, fibras musgosas e
trepadeiras, parte dos axônios das células granulares e as fibras paralelas.
Considerando que a atividade acrobática requer coordenação e equilíbrio, esse
aumento de NF, principalmente na fase inicial do treinamento, decorre da
intensa reaferentação proprioceptiva e cortical.
Essa mudança da expressão de NF está acompanhada de aumento de
expressão da proteína sináptica SYP na camada molecular e diminuição da
expressão da proteína SYS na camada molecular e granular após curtos
períodos de treinamento acrobático. O aumento da SYP concomitante ao do
NF sugere incrementos da eficiência sináptica juntamente com aumento da
velocidade de condução dos impulsos nervosos na camada molecular que é
confirmado pelo aumento da densidade de células granulares Egr1-positivas ao
longo do período de treinamento com pico na primeira semana de treinamento
(AC1).
O cerebelo participa da integração somatosensorial145, do fornecimento
de informação do movimento ao córtex146 e na aquisição de coordenação
motora e aprendizado espacial147. O exercício acrobático é uma tarefa motora
complexa que exige coordenação e equilíbrio do animal para executar todo o

65
circuito. Com base nisso, podemos sugerir que na primeira semana de
treinamento quando encontramos o pico de ativação das células granulares se
deveu ao aumento da demanda proprioceptiva através da via espinocerebelar
que adentra ao cerebelo como fibras musgosas. Essa demanda aumentada
pode gerar uma intensa estimulação induzindo aumento de NF e consequente
remodelamento axonal, além do aumento da formação de vesículas e
recaptação através do aumento de SYP que parece dar suporte ao aumento da
eficiência sináptica na camada molecular onde se encontram as arborizações
dendríticas das células de Purkinje em resposta ao exercício acrobático. Além
de que esse aumento de SYP no cerebelo pode ser devido ao aumento de
liberação de glutamato que está diretamente relacionado à fosforilação da SYP,
sugerindo uma possível potenciação de longo prazo (LTP)144.
A longo prazo de treinamento, nossos dados não mostraram um
aumento da densidade óptica relativa de SYP e SYS. No entanto, Kleim e
colaboradores43 mostraram que o exercício acrobático é capaz de promover
sinaptogênese nos animais submetidos a 30 dias consecutivos de acrobacias
visto que apresentaram um aumento do número de sinapses por célula de
Purkinje. Uma possível explicação para isto é que a frequência de treinamento
utilizada por Kleim e colaboradores24 foi de 30 dias consecutivos, enquanto que
do nosso estudo foi 3 vezes por semana durante 30 dias. Um tempo tão
prolongado de treinamento pode favorecer os mecanismos plásticos da fase de
consolidação do aprendizado de uma nova habilidade motora gerando um
automatismo motor. Já foi muito bem descrito a importância do circuito
cerebelo-tálamo-cortical nesse tipo de atividade motora44.
Em contrapartida, nossos dados revelaram também que na fase inicial
de treinamento houve uma diminuição da SYS na camada molecular e
granular. Porém estudos anteriores já mostraram que o exercício acrobático foi
capaz de aumentar significativamente a expressão de SYS na camada
molecular a longo prazo, ou seja após 4 semanas treinamento de exercício
acrobático.40 A associação desses dois dados sugerem que o exercício
acrobático parece induzir respostas distintas na expressão da proteína SYS
frente a diferentes períodos de treinamento, revelando assim uma mudança
plástica tempo-dependente.

66
6.2.4. Hipocampo
O diferentes períodos de treinamento de exercício acrobático não
revelaram mudanças na contagem de células DCX-positivas na zona
subgranular (SGV) do giro denteado do hipocampo entre os grupos treinados.
Lambert e colaboradores12 ao analisar ratos submetidos ao treinamento de
exercício voluntário (EV), exercício acrobático (AC) e exposição ao ambiente
enriquecido (AE) por 6 semanas, 7 vezes na semana, observaram aumento da
proteína SYP na região do hipocampo em ratos submetidos ao EV e AE, sem
alterações no grupo AC. Sugerindo menor ativação hipocampal através do
exercício acrobático. Isso levanta a hipótese de que a falta de mudança na
eficiência sináptica no hipocampo observada pelo estudo do Lambert e
colaboradores pode dar suporte a falta de neurôgenese induzida pelo exercício
acrobático. Porém precisa de mais análises afim de maior esclarecimento.

67
7. CONCLUSÕES
- Com base nos resultados obtidos nesse estudo, podemos concluir que
os diferentes períodos de treinamento de exercício acrobático promoveram
melhora acentuada do desempenho motor dos ratos ao final da segunda
semana, e ao mesmo tempo que geraram alterações nas proteínas
sinápticas e estruturais e no gene Egr1 de forma distinta nas áreas
encefálicas. Além disso, o exercício acrobático não promoveu alterações
nos níveis de neurogênese observado pelo gene DCX no hipocampo.
- No córtex motor, o AC promoveu mudanças na expressão de proteínas
MAP2 e SYP após duas e quatro semanas de treinamento, coincidindo com
aumento de células Egr1-positivas,
- O estriado respondeu com aumento de proteínas estruturais MAP2 e
NFs principalmente após uma a duas semanas de treinamento precedendo
o aumento das proteínas sinápticas SYP e SYS que aconteceram na
segunda e quarta semana.
- No cerebelo, o aumento da proteína estrutural NF e sináptica SYP
aconteceram após curto período de treinamento acrobático (AC1 e AC2)
retornando ao nível controle na quarta semana.
- O conhecimento sobre as respostas plásticas induzidas por diferentes
tempos de treinamento do exercício acrobático e respectivas mudanças no
desempenho motor podem facilitar a escolha da duração do emprego de
determinadas intervenções terapêuticas e contribuir para um melhor
planejamento do protocolo de exercício escolhido.

68
8. REFERÊNCIAS
1. Mora F, Segovia G, del Arco A. Aging, plasticity and environmental enrichment: structural changes and neurotransmitter dynamics in several areas of the brain. Brain research reviews. 2007;55(1):78-88. 2. Rosenzweig MR, Bennett EL. Psychobiology of plasticity: effects of training and experience on brain and behavior. Behavioural brain research. 1996;78(1):57-65. 3. Stein DG, Brailowsky S, Will B. Brain repair. New York ; Oxford: Oxford University Press; 1995. x, 156 p. p. 4. Kempermann G, Kuhn HG, Gage FH. More hippocampal neurons in adult mice living in an enriched environment. Nature. 1997;386(6624):493-5. 5. Azizi SA, Vendrame M. Exercise: a workout for neuroregeneration. Neuroscience letters. 2007;418(3):211-2. 6. Dustman RE, Emmerson RY, Ruhling RO, Shearer DE, Steinhaus LA, Johnson SC, et al. Age and fitness effects on EEG, ERPs, visual sensitivity, and cognition. Neurobiology of aging. 1990;11(3):193-200. 7. Lupinacci NS, Rikli RE, Jones CJ, Ross D. Age and physical activity effects on reaction time and digit symbol substitution performance in cognitively active adults. Research quarterly for exercise and sport. 1993;64(2):144-50. 8. Gomez-Pinilla F, Ying Z, Roy RR, Molteni R, Edgerton VR. Voluntary exercise induces a BDNF-mediated mechanism that promotes neuroplasticity. Journal of neurophysiology. 2002;88(5):2187-95. 9. Molteni R, Ying Z, Gomez-Pinilla F. Differential effects of acute and chronic exercise on plasticity-related genes in the rat hippocampus revealed by microarray. The European journal of neuroscience. 2002;16(6):1107-16. 10. Marin R, Williams A, Hale S, Burge B, Mense M, Bauman R, et al. The effect of voluntary exercise exposure on histological and neurobehavioral outcomes after ischemic brain injury in the rat. Physiology & behavior. 2003;80(2-3):167-75. 11. Ying Z, Roy RR, Edgerton VR, Gomez-Pinilla F. Exercise restores levels of neurotrophins and synaptic plasticity following spinal cord injury. Experimental neurology. 2005;193(2):411-9. 12. Lambert TJ, Fernandez SM, Frick KM. Different types of environmental enrichment have discrepant effects on spatial memory and synaptophysin levels in female mice. Neurobiology of learning and memory. 2005;83(3):206-16. 13. Neeper SA, Gomez-Pinilla F, Choi J, Cotman C. Exercise and brain neurotrophins. Nature. 1995;373(6510):109. 14. van Praag H, Shubert T, Zhao C, Gage FH. Exercise enhances learning and hippocampal neurogenesis in aged mice. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2005;25(38):8680-5. 15. Liepert J, Miltner WH, Bauder H, Sommer M, Dettmers C, Taub E, et al. Motor cortex plasticity during constraint-induced movement therapy in stroke patients. Neuroscience letters. 1998;250(1):5-8. 16. Kramer AF, Hahn S, Cohen NJ, Banich MT, McAuley E, Harrison CR, et al. Ageing, fitness and neurocognitive function. Nature. 1999;400(6743):418-9. 17. Kim DH, Ko IG, Kim BK, Kim TW, Kim SE, Shin MS, et al. Treadmill exercise inhibits traumatic brain injury-induced hippocampal apoptosis. Physiology & behavior. 2010;101(5):660-5.

69
18. Kim YP, Kim H, Shin MS, Chang HK, Jang MH, Shin MC, et al. Age-dependence of the effect of treadmill exercise on cell proliferation in the dentate gyrus of rats. Neuroscience letters. 2004;355(1-2):152-4. 19. Dishman RK, Berthoud HR, Booth FW, Cotman CW, Edgerton VR, Fleshner MR, et al. Neurobiology of exercise. Obesity. 2006;14(3):345-56. 20. Sutoo D, Akiyama K. Regulation of brain function by exercise. Neurobiology of disease. 2003;13(1):1-14. 21. Itoh T, Imano M, Nishida S, Tsubaki M, Hashimoto S, Ito A, et al. Exercise inhibits neuronal apoptosis and improves cerebral function following rat traumatic brain injury. Journal of neural transmission. 2011;118(9):1263-72. 22. Sim YJ, Kim SS, Kim JY, Shin MS, Kim CJ. Treadmill exercise improves short-term memory by suppressing ischemia-induced apoptosis of neuronal cells in gerbils. Neuroscience letters. 2004;372(3):256-61. 23. Jones TA, Chu CJ, Grande LA, Gregory AD. Motor skills training enhances lesion-induced structural plasticity in the motor cortex of adult rats. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1999;19(22):10153-63. 24. Ahmed Z, Wagdy M, Benjamin M, Mohamed S, Mohamed H, Ahmed S, et al. Therapeutic effects of acrobatic exercise and magnetic field exposure on functional recovery after spinal cord injury in mice. Bioelectromagnetics. 2011;32(1):49-57. 25. Arida RM, Scorza FA, Gomes da Silva S, Cysneiros RM, Cavalheiro EA. Exercise paradigms to study brain injury recovery in rodents. American journal of physical medicine & rehabilitation / Association of Academic Physiatrists. 2011;90(6):452-65. 26. Ferreira AF, Real CC, Rodrigues AC, Alves AS, Britto LR. Moderate exercise changes synaptic and cytoskeletal proteins in motor regions of the rat brain. Brain research. 2010;1361:31-42. 27. Leasure JL, Jones M. Forced and voluntary exercise differentially affect brain and behavior. Neuroscience. 2008;156(3):456-65. 28. Sherwin CM. Voluntary wheel running: a review and novel interpretation. Animal behaviour. 1998;56(1):11-27. 29. Tong L, Shen H, Perreau VM, Balazs R, Cotman CW. Effects of exercise on gene-expression profile in the rat hippocampus. Neurobiology of disease. 2001;8(6):1046-56. 30. Pang TY, Stam NC, Nithianantharajah J, Howard ML, Hannan AJ. Differential effects of voluntary physical exercise on behavioral and brain-derived neurotrophic factor expression deficits in Huntington's disease transgenic mice. Neuroscience. 2006;141(2):569-84. 31. Zheng H, Liu Y, Li W, Yang B, Chen D, Wang X, et al. Beneficial effects of exercise and its molecular mechanisms on depression in rats. Behavioural brain research. 2006;168(1):47-55. 32. Kleim JA, Barbay S, Cooper NR, Hogg TM, Reidel CN, Remple MS, et al. Motor learning-dependent synaptogenesis is localized to functionally reorganized motor cortex. Neurobiology of learning and memory. 2002;77(1):63-77. 33. Lewis MH. Environmental complexity and central nervous system development and function. Mental retardation and developmental disabilities research reviews. 2004;10(2):91-5.

70
34. Inal M, Akyuz F, Turgut A, Getsfrid WM. Effect of aerobic and anaerobic metabolism on free radical generation swimmers. Medicine and science in sports and exercise. 2001;33(4):564-7. 35. Ding Q, Vaynman S, Akhavan M, Ying Z, Gomez-Pinilla F. Insulin-like growth factor I interfaces with brain-derived neurotrophic factor-mediated synaptic plasticity to modulate aspects of exercise-induced cognitive function. Neuroscience. 2006;140(3):823-33. 36. Salgado-Delgado R, Angeles-Castellanos M, Buijs MR, Escobar C. Internal desynchronization in a model of night-work by forced activity in rats. Neuroscience. 2008;154(3):922-31. 37. Ang ET, Dawe GS, Wong PT, Moochhala S, Ng YK. Alterations in spatial learning and memory after forced exercise. Brain research. 2006;1113(1):186-93. 38. Vaynman SS, Ying Z, Yin D, Gomez-Pinilla F. Exercise differentially regulates synaptic proteins associated to the function of BDNF. Brain research. 2006;1070(1):124-30. 39. Real CC, Ferreira AF, Hernandes MS, Britto LR, Pires RS. Exercise-induced plasticity of AMPA-type glutamate receptor subunits in the rat brain. Brain research. 2010;1363:63-71. 40. Garcia PC, Real CC, Ferreira AF, Alouche SR, Britto LR, Pires RS. Different protocols of physical exercise produce different effects on synaptic and structural proteins in motor areas of the rat brain. Brain research. 2012;1456:36-48. 41. Kleim JA, Lussnig E, Schwarz ER, Comery TA, Greenough WT. Synaptogenesis and Fos expression in the motor cortex of the adult rat after motor skill learning. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1996;16(14):4529-35. 42. Kleim JA, Vij K, Ballard DH, Greenough WT. Learning-dependent synaptic modifications in the cerebellar cortex of the adult rat persist for at least four weeks. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1997;17(2):717-21. 43. Kleim JA, Swain RA, Armstrong KA, Napper RM, Jones TA, Greenough WT. Selective synaptic plasticity within the cerebellar cortex following complex motor skill learning. Neurobiology of learning and memory. 1998;69(3):274-89. 44. Holschneider DP, Yang J, Guo Y, Maarek JM. Reorganization of functional brain maps after exercise training: Importance of cerebellar-thalamic-cortical pathway. Brain research. 2007;1184:96-107. 45. Neeper SA, Gomez-Pinilla F, Choi J, Cotman CW. Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain research. 1996;726(1-2):49-56. 46. Vaynman S, Ying Z, Gomez-Pinilla F. The select action of hippocampal calcium calmodulin protein kinase II in mediating exercise-enhanced cognitive function. Neuroscience. 2007;144(3):825-33. 47. Stranahan AM, Lee K, Becker KG, Zhang Y, Maudsley S, Martin B, et al. Hippocampal gene expression patterns underlying the enhancement of memory by running in aged mice. Neurobiology of aging. 2010;31(11):1937-49. 48. van Praag H, Christie BR, Sejnowski TJ, Gage FH. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proceedings of the

71
National Academy of Sciences of the United States of America. 1999;96(23):13427-31. 49. Soares J, Holmes PV, Renner KJ, Edwards GL, Bunnell BN, Dishman RK. Brain noradrenergic responses to footshock after chronic activity-wheel running. Behavioral neuroscience. 1999;113(3):558-66. 50. Nilsson M, Perfilieva E, Johansson U, Orwar O, Eriksson PS. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. Journal of neurobiology. 1999;39(4):569-78. 51. Williams BM, Luo Y, Ward C, Redd K, Gibson R, Kuczaj SA, et al. Environmental enrichment: effects on spatial memory and hippocampal CREB immunoreactivity. Physiology & behavior. 2001;73(4):649-58. 52. Gobatto CA, Sibuya CY, Azevedo JRM, Luciano E, Kokubun E, Mello MAR. Caracterização da intensidade de exercício e do efeito de treinamento físico no modelo de natação de ratos Wistar. Revista Motriz. 2001; 7:57-62. 53. Galdino RS, Souza CCA, Luciano E, Mello MAR. Protein calorie malnutritio does not impair glucose metabolism adaptations to physical exercise. Nutrition. 2000;20:527-35. 54. Carro E, Trejo JL, Busiguina S, Torres-Aleman I. Circulating insulin-like growth factor I mediates the protective effects of physical exercise against brain insults of different etiology and anatomy. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2001;21(15):5678-84. 55. Derksen MJ, Ward NL, Hartle KD, Ivanco TL. MAP2 and synaptophysin protein expression following motor learning suggests dynamic regulation and distinct alterations coinciding with synaptogenesis. Neurobiology of learning and memory. 2007;87(3):404-15. 56. Klintsova AY, Dickson E, Yoshida R, Greenough WT. Altered expression of BDNF and its high-affinity receptor TrkB in response to complex motor learning and moderate exercise. Brain research. 2004;1028(1):92-104. 57. Kleim JA, Freeman JH, Jr., Bruneau R, Nolan BC, Cooper NR, Zook A, et al. Synapse formation is associated with memory storage in the cerebellum. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(20):13228-31. 58. Black JE, Isaacs KR, Anderson BJ, Alcantara AA, Greenough WT. Learning causes synaptogenesis, whereas motor activity causes angiogenesis, in cerebellar cortex of adult rats. Proceedings of the National Academy of Sciences of the United States of America. 1990;87(14):5568-72. 59. Buitrago MM, Schulz JB, Dichgans J, Luft AR. Short and long-term motor skill learning in an accelerated rotarod training paradigm. Neurobiology of learning and memory. 2004;81(3):211-6. 60. Vaynman S, Ying Z, Gomez-Pinilla F. Exercise induces BDNF and synapsin I to specific hippocampal subfields. Journal of neuroscience research. 2004;76(3):356-62. 61. Deng W, Aimone JB, Gage FH. New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nature reviews Neuroscience. 2010;11(5):339-50. 62. Squire LR. Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychological review. 1992;99(2):195-231. 63. Bruel-Jungerman E, Laroche S, Rampon C. New neurons in the dentate gyrus are involved in the expression of enhanced long-term memory following

72
environmental enrichment. The European journal of neuroscience. 2005;21(2):513-21. 64. Snyder JS, Hong NS, McDonald RJ, Wojtowicz JM. A role for adult neurogenesis in spatial long-term memory. Neuroscience. 2005;130(4):843-52. 65. Gould E, Beylin A, Tanapat P, Reeves A, Shors TJ. Learning enhances adult neurogenesis in the hippocampal formation. Nature neuroscience. 1999;2(3):260-5. 66. Fabel K, Kempermann G. Physical activity and the regulation of neurogenesis in the adult and aging brain. Neuromolecular medicine. 2008;10(2):59-66. 67. van Praag H. Neurogenesis and exercise: past and future directions. Neuromolecular medicine. 2008;10(2):128-40. 68. Ehninger D, Kempermann G. Regional effects of wheel running and environmental enrichment on cell genesis and microglia proliferation in the adult murine neocortex. Cerebral cortex. 2003;13(8):845-51. 69. van Praag H, Kempermann G, Gage FH. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nature neuroscience. 1999;2(3):266-70. 70. Holmes MM, Galea LA, Mistlberger RE, Kempermann G. Adult hippocampal neurogenesis and voluntary running activity: circadian and dose-dependent effects. Journal of neuroscience research. 2004;76(2):216-22. 71. Ferreira AF, Real CC, Rodrigues AC, Alves AS, Britto LR. Short-term, moderate exercise is capable of inducing structural, BDNF-independent hippocampal plasticity. Brain research. 2011;1425:111-22. 72. Uda M, Ishido M, Kami K, Masuhara M. Effects of chronic treadmill running on neurogenesis in the dentate gyrus of the hippocampus of adult rat. Brain research. 2006;1104(1):64-72. 73. Kandel ER, Schwartz JH, Jessell TM. Mecanismos Celulares do Aprendizado e da Memória. Fundamentos da Neurociência e do Comportamento. 4 ed. California: McGraw-Hill; 2000. 74. Schwartz JH, Jessell TM, Kandel ER. Princípios Da Neurociência. Barueri: Manole; 2003. 75. R. LdN. Cerebral cortex: architecture, intercortical conections, motor projection. . Physiology of the nervous system. ed. New York: Oxford University Press; 1942. 76. Teskey GC, Monfils MH, VandenBerg PM, Kleim JA. Motor map expansion following repeated cortical and limbic seizures is related to synaptic potentiation. Cerebral cortex. 2002;12(1):98-105. 77. Kleim JA, Barbay S, Nudo RJ. Functional reorganization of the rat motor cortex following motor skill learning. Journal of neurophysiology. 1998;80(6):3321-5. 78. Côté L, Crutcher MD. The basal ganglia. In: KANDEL, E.R.; SCHWARTZ S.H.; JESSEL, T.M. Principles of neural Science. ganglia Tb, editor. Norwalk: Apleton & Lange; 1991. 79. Freund HJ. Mechanisms of voluntary movements. Parkinsonism & related disorders. 2002;9(1):55-9. 80. Gentilucci M, Negrotti A. The control of an action in Parkinson's disease. Experimental brain research Experimentelle Hirnforschung Experimentation cerebrale. 1999;129(2):269-77.

73
81. Gentilucci M, Bertolani L, Benuzzi F, Negrotti A, Pavesi G, Gangitano M. Impaired control of an action after supplementary motor area lesion: a case study. Neuropsychologia. 2000;38(10):1398-404. 82. Mink JW. Fundamental Neuroscience. San Diego: Academic Press; 1999. 83. Fearnley JM, Lees AJ. Ageing and Parkinson's disease: substantia nigra regional selectivity. Brain : a journal of neurology. 1991;114 ( Pt 5):2283-301. 84. Gerfen CR. Synaptic organization of the striatum. Journal of electron microscopy technique. 1988;10(3):265-81. 85. Paxinos G. The Rat Nervous System. 3 ed. Amsterdam: Elsevier Academic Press; 2004. 86. Kiernan J. Neuroanatomia Humana de Barr. 7 ed. Barueri: Manole; 2003. 87. Onla-or S, Winstein CJ. Function of the 'direct' and 'indirect' pathways of the basal ganglia motor loop: evidence from reciprocal aiming movements in Parkinson's disease. Brain research Cognitive brain research. 2001;10(3):329-32. 88. Grillner S, Hellgren J, Menard A, Saitoh K, Wikstrom MA. Mechanisms for selection of basic motor programs--roles for the striatum and pallidum. Trends in neurosciences. 2005;28(7):364-70. 89. Yin HH. The sensorimotor striatum is necessary for serial order learning. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2010;30(44):14719-23. 90. Lewis MM, Slagle CG, Smith AB, Truong Y, Bai P, McKeown MJ, et al. Task specific influences of Parkinson's disease on the striato-thalamo-cortical and cerebello-thalamo-cortical motor circuitries. Neuroscience. 2007;147(1):224-35. 91. Machado A. Neuroanatomia Funcional. Rio de Janeiro: Atheneu; 1993. 92. Lundy-Ekman L. Neurociência fundamentos para a reabilitação. São Paulo: Guanabara Koogan; 2000. 93. Delgado-Garcia JM. [Structure and function of the cerebellum]. Revista de neurologia. 2001;33(7):635-42. Estructura y funcion del cerebelo. 94. Sarna JR, Hawkes R. Patterned Purkinje cell death in the cerebellum. Progress in neurobiology. 2003;70(6):473-507. 95. Guyton AC, Hall JE. Tratado de fisiologia médica. 9 ed. Rio de Janeiro: Guanabara Koogan; 1997. 96. Eccles JC. Neurogenesis and morphogenesis in the cerebellar cortex. Proceedings of the National Academy of Sciences of the United States of America. 1970;66(2):294-301. 97. Ghez C, Thach WT. O Cerebelo. In: Kandel ER, Schwartz JH, Jessell TM, editors. Princípios da Neurociência: Manole; 2003. 98. Zhao C, Deng W, Gage FH. Mechanisms and functional implications of adult neurogenesis. Cell. 2008;132(4):645-60. 99. Kempermann G, Gage FH. Genetic determinants of adult hippocampal neurogenesis correlate with acquisition, but not probe trial performance, in the water maze task. The European journal of neuroscience. 2002;16(1):129-36. 100. Larry RS, Kandel ER. Um Mecanismo de Armazenamento Sináptico para Memória Declarativa. Porto Alegre: Artmed; 2003.

74
101. van Groen T, Miettinen P, Kadish I. The entorhinal cortex of the mouse: organization of the projection to the hippocampal formation. Hippocampus. 2003;13(1):133-49. 102. Shepherd GM. The Synaptic organization of the brain. 3rd ed. New York: Oxford University Press; 1990. vii, 561 p. p. 103. Blackstad TW. Commissural connections of the hippocampal region in the rat, with special reference to their mode of termination. The Journal of comparative neurology. 1956;105(3):417-537. 104. Yu LC, Zheng EM, Lundeberg T. Calcitonin gene-related peptide 8-37 inhibits the evoked discharge frequency of wide dynamic range neurons in dorsal horn of the spinal cord in rats. Regulatory peptides. 1999;83(1):21-4. 105. Lee MK, Cleveland DW. Neuronal intermediate filaments. Annual review of neuroscience. 1996;19:187-217. 106. Briones TL. Environment, physical activity, and neurogenesis: implications for prevention and treatment of Alzhemier's disease. Current Alzheimer research. 2006;3(1):49-54. 107. Redila VA, Christie BR. Exercise-induced changes in dendritic structure and complexity in the adult hippocampal dentate gyrus. Neuroscience. 2006;137(4):1299-307. 108. Arnstein PM. The neuroplastic phenomenon: a physiologic link between chronic pain and learning. The Journal of neuroscience nursing : journal of the American Association of Neuroscience Nurses. 1997;29(3):179-86. 109. Tagliaferro P, Javier Ramos A, Onaivi ES, Evrard SG, Lujilde J, Brusco A. Neuronal cytoskeleton and synaptic densities are altered after a chronic treatment with the cannabinoid receptor agonist WIN 55,212-2. Brain research. 2006;1085(1):163-76. 110. Griesbach GS, Gomez-Pinilla F, Hovda DA. The upregulation of plasticity-related proteins following TBI is disrupted with acute voluntary exercise. Brain research. 2004;1016(2):154-62. 111. Griesbach GS, Hovda DA, Molteni R, Wu A, Gomez-Pinilla F. Voluntary exercise following traumatic brain injury: brain-derived neurotrophic factor upregulation and recovery of function. Neuroscience. 2004;125(1):129-39. 112. Molteni R, Wu A, Vaynman S, Ying Z, Barnard RJ, Gomez-Pinilla F. Exercise reverses the harmful effects of consumption of a high-fat diet on synaptic and behavioral plasticity associated to the action of brain-derived neurotrophic factor. Neuroscience. 2004;123(2):429-40. 113. Ding Y, Li J, Lai Q, Azam S, Rafols JA, Diaz FG. Functional improvement after motor training is correlated with synaptic plasticity in rat thalamus. Neurological research. 2002;24(8):829-36. 114. Kozorovitskiy Y, Gross CG, Kopil C, Battaglia L, McBreen M, Stranahan AM, et al. Experience induces structural and biochemical changes in the adult primate brain. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(48):17478-82. 115. Sarbadhikari SN, Saha AK. Moderate exercise and chronic stress produce counteractive effects on different areas of the brain by acting through various neurotransmitter receptor subtypes: a hypothesis. Theoretical biology & medical modelling. 2006;3:33. 116. Jovanovic JN, Benfenati F, Siow YL, Sihra TS, Sanghera JS, Pelech SL, et al. Neurotrophins stimulate phosphorylation of synapsin I by MAP kinase and

75
regulate synapsin I-actin interactions. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(8):3679-83. 117. Chin LS, Li L, Ferreira A, Kosik KS, Greengard P. Impairment of axonal development and of synaptogenesis in hippocampal neurons of synapsin I-deficient mice. Proceedings of the National Academy of Sciences of the United States of America. 1995;92(20):9230-4. 118. Fornasiero EF, Bonanomi D, Benfenati F, Valtorta F. The role of synapsins in neuronal development. Cellular and molecular life sciences : CMLS. 2010;67(9):1383-96. 119. Masliah E, Terry RD, Alford M, DeTeresa R. Quantitative immunohistochemistry of synaptophysin in human neocortex: an alternative method to estimate density of presynaptic terminals in paraffin sections. The journal of histochemistry and cytochemistry : official journal of the Histochemistry Society. 1990;38(6):837-44. 120. Eastwood SL, Burnet PW, McDonald B, Clinton J, Harrison PJ. Synaptophysin gene expression in human brain: a quantitative in situ hybridization and immunocytochemical study. Neuroscience. 1994;59(4):881-92. 121. Chen KS, Masliah E, Mallory M, Gage FH. Synaptic loss in cognitively impaired aged rats is ameliorated by chronic human nerve growth factor infusion. Neuroscience. 1995;68(1):19-27. 122. Daly C, Sugimori M, Moreira JE, Ziff EB, Llinas R. Synaptophysin regulates clathrin-independent endocytosis of synaptic vesicles. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(11):6120-5. 123. Tartaglia N, Du J, Tyler WJ, Neale E, Pozzo-Miller L, Lu B. Protein synthesis-dependent and -independent regulation of hippocampal synapses by brain-derived neurotrophic factor. The Journal of biological chemistry. 2001;276(40):37585-93. 124. Sanchez C, Diaz-Nido J, Avila J. Phosphorylation of microtubule-associated protein 2 (MAP2) and its relevance for the regulation of the neuronal cytoskeleton function. Progress in neurobiology. 2000;61(2):133-68. 125. Julien JP. Neurofilament functions in health and disease. Current opinion in neurobiology. 1999;9(5):554-60. 126. Perrot R, Berges R, Bocquet A, Eyer J. Review of the multiple aspects of neurofilament functions, and their possible contribution to neurodegeneration. Molecular neurobiology. 2008;38(1):27-65. 127. Hoffman PN, Cleveland DW, Griffin JW, Landes PW, Cowan NJ, Price DL. Neurofilament gene expression: a major determinant of axonal caliber. Proceedings of the National Academy of Sciences of the United States of America. 1987;84(10):3472-6. 128. Gama Sosa MA, Friedrich VL, Jr., DeGasperi R, Kelley K, Wen PH, Senturk E, et al. Human midsized neurofilament subunit induces motor neuron disease in transgenic mice. Experimental neurology. 2003;184(1):408-19. 129. Sheng M, Greenberg ME. The regulation and function of c-fos and other immediate early genes in the nervous system. Neuron. 1990;4(4):477-85. 130. O'Donovan KJ, Tourtellotte WG, Millbrandt J, Baraban JM. The EGR family of transcription-regulatory factors: progress at the interface of molecular and systems neuroscience. Trends in neurosciences. 1999;22(4):167-73.

76
131. Milbrandt J. A nerve growth factor-induced gene encodes a possible transcriptional regulatory factor. Science. 1987;238(4828):797-9. 132. Beckmann AM, Davidson MS, Goodenough S, Wilce PA. Differential expression of Egr-1-like DNA-binding activities in the naive rat brain and after excitatory stimulation. Journal of neurochemistry. 1997;69(6):2227-37. 133. Gass P, Herdegen T, Bravo R, Kiessling M. Spatiotemporal induction of immediate early genes in the rat brain after limbic seizures: effects of NMDA receptor antagonist MK-801. The European journal of neuroscience. 1993;5(7):933-43. 134. Kaczmarek L. Gene expression in learning processes. Acta neurobiologiae experimentalis. 2000;60(3):419-24. 135. Cole AJ, Saffen DW, Baraban JM, Worley PF. Rapid increase of an immediate early gene messenger RNA in hippocampal neurons by synaptic NMDA receptor activation. Nature. 1989;340(6233):474-6. 136. Campos LA, Plehm R, Cipolla-Neto J, Bader M, Baltatu OC. Altered circadian rhythm reentrainment to light phase shifts in rats with low levels of brain angiotensinogen. American journal of physiology Regulatory, integrative and comparative physiology. 2006;290(4):R1122-7. 137. Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. Burlington: Elsevier Academic Press; 2005. 138. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680-5. 139. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry. 1976;72:248-54. 140. Rioult-Pedotti MS, Friedman D, Hess G, Donoghue JP. Strengthening of horizontal cortical connections following skill learning. Nature neuroscience. 1998;1(3):230-4. 141. Kleim JA, Hogg TM, VandenBerg PM, Cooper NR, Bruneau R, Remple M. Cortical synaptogenesis and motor map reorganization occur during late, but not early, phase of motor skill learning. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2004;24(3):628-33. 142. Aou S, Woody CD, Birt D. Increases in excitability of neurons of the motor cortex of cats after rapid acquisition of eye blink conditioning. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1992;12(2):560-9. 143. Doyon J, Bellec P, Amsel R, Penhune V, Monchi O, Carrier J, et al. Contributions of the basal ganglia and functionally related brain structures to motor learning. Behavioural brain research. 2009;199(1):61-75. 144. Evans GJ, Cousin MA. Tyrosine phosphorylation of synaptophysin in synaptic vesicle recycling. Biochemical Society transactions. 2005;33(Pt 6):1350-3. 145. Manzoni D. The cerebellum and sensorimotor coupling: looking at the problem from the perspective of vestibular reflexes. Cerebellum. 2007;6(1):24-37. 146. Bonnefoi-Kyriacou B, Legallet E, Lee RG, Trouche E. Spatio-temporal and kinematic analysis of pointing movements performed by cerebellar patients with limb ataxia. Experimental brain research Experimentelle Hirnforschung Experimentation cerebrale. 1998;119(4):460-6.

77
147. Joyal CC, Meyer C, Jacquart G, Mahler P, Caston J, Lalonde R. Effects of midline and lateral cerebellar lesions on motor coordination and spatial orientation. Brain research. 1996;739(1-2):1-11.

78
ANEXO I

79


![neuropatia periférica induzida pela quimioterapia (npiq) · 26 On 34 > [jan-jun 2017] neuropatia periférica induzida pela quimioterapia (npiq) resumo: a neuropatia Periférica Induzida](https://img.document.onl/doc/110x75/5fad5917dd23b224ef7f5622/neuropatia-perifrica-induzida-pela-quimioterapia-npiq-26-on-34-jan-jun.jpg)
![ARTI SÃO Neuroplasticidade e a Recuperação da Função após ... · SOBRINHO, ].BR e col. -Neuroplasticidade e a Recuperação da Função após Lesões Cerebrais Acta Fisiátrica](https://img.document.onl/doc/110x75/5fea19200cc2930796337610/arti-sfo-neuroplasticidade-e-a-recuperao-da-funo-aps-sobrinho-br.jpg)


















![Neuroplasticidade [em portugues] (por: carlitosrangel)](https://img.document.onl/doc/110x75/557df8b2d8b42ab3268b4c77/neuroplasticidade-em-portugues-por-carlitosrangel.jpg)






