Embed Size (px)
Citation preview

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA
DEPARTAMENTO DE BIOTECNOLOGIA
Dissertação de Mestrado
PRODUÇÃO DE GLICOSE-6-FOSFATO DESIDROGENASE A PARTIR DO HIDROLISADO HEMICELULÓSICO DE
PALHA DE ARROZ POR Candida guilliermondii
Tranferido da Biblioteca do DEBIQ para a Bilblioteca
Universitária em Junho/2004 Proc. nº 202/04
Lorena - SP - Brasil 2004

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA DEPARTAMENTO DE BIOTECNOLOGIA
PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INDUSTRIAL
PRODUÇÃO DE GLICOSE-6-FOSFATO DESIDROGENASE A PARTIR DO HIDROLISADO HEMICELULÓSICO DE PALHA DE
ARROZ POR Candida guil/iermondii
Dissertação de mestrado apresentada como parte das exigências para a obtenção do título de Mestre em Biotecnologia Industrial
Banca examinadora:
Ora. Inês Conceição Roberto Dr. Roberto da Silva Ora. Adriane Maria Ferreira Milagres
Estudante:
Daniela de Borba Gurpilhares
Lorena - SP - Brasil 2004
ii
1 ;
. -. - -~

iii
Agradeço a Deus, a meus pais e todas as pessoas que me apoiaram e incentivaram; tornando-se importantes para mim durante esse período.

iv
AGRADECIMENTOS
À Profa. Ora Inês Conceição Roberto, não somente pela orientação neste
trabalho, mas também pelos constantes incentivos, pela dedicação, apoio e
confiança durante a realização do trabalho;
Aos Professores Doutores João Batista, Maria das Graças Felipe e
Adriane Milagres, pelo valioso conhecimento transmitido e pela amizade;
A todos os funcionários do DEBIQ que, de um modo ou de outro, foram
responsáveis por parte deste trabalho, principalmente à Rita, ao Paulinho e
Nicamor, pelos auxílios prestados dentro e fora do laboratório como também
pelo carinho;
Aos alunos, por compartilharem o conhecimento adquirido e os bons
momentos;
À FAPESP e CAPES, pela bolsa concedida;
A todos aqueles que aqui não foram citados, mas de uma forma ou de
outra, colaboraram para a realização do meu trabalho.

V
ÍNDICE
LISTA DE TABELAS vii
LISTA DE FIGURAS viii
LISTA DE NOMENCLATURAS --------------------- X
1 - INTRODUÇÃO 1 ------------------------- 2 - REVISÃO BIBLIOGRÁFICA 4
2.1 - MATERIAIS LIGNOCELULÓSICOS 4 ------------------- 2. 1. 1 - COMPOSIÇÃO E CARACTERÍSTICAS DO MATERIAL LIGNOCELULÓSICO_ 4 2.1.2- POTENCIAIS Usos DOS MATERIAIS LIGNOCELULÓSICOS 7 2.1.3 - OBTENÇÃO DO HIDROLISADO HEM/CELULÓSICO 9
2.2 - METABOLISMO DE XILOSE EM LEVEDURAS 11 --------------- 2.3 - GLICOSE-6-FOSFATO DESIDROGENASE (G6PDH) 13
2.3.1 - IMPORTÂNCIA FISIOLÓGICA 13 2.3.2- PRINCIPAIS APLICAÇÕES 14 2.3.3 - FONTES DE OBTENÇÃO E MÉTODOS PARA LIBERAÇÃO DA ENZIMA_ 15 2.3.4 - INFLUÊNCIA DAS CONDIÇÕES DE CULTIVO 18
3 - OBJETIVOS 20
4- MATERIAIS E MÉTODOS 22
4.1 - PREPARO DA MATÉRIA PRIMA 22
4.2 - OBTENÇÃO E TRATAMENTO DO HIDROLISADO HEMICELULÓSICO 22
4.3 - MICRORGANISMO 23
4.4 - PREPARO DO INÓCULO 23
4.5 - ESTUDO DAS CONDIÇÕES DE ROMPIMENTO CELULAR 24
4.6- PROCESSO FERMENTATIVO 24 4.6.1 - MEIO DE FERMENTAÇÃO 24 4.6.2 - CONDIÇÕES DE CULTIVO 25 4.6.3-METODOLOGIA ESTATÍSTICA 25
4.7 - MÉTODOSANAlÍTICOS 27 4. 7.1 - DETERMINAÇÃO DA CONCENTRAÇÃO CELULAR 27 4.7.2 • DETERMINAÇÃO DA CONCENTRAÇÃO DE GLICOSE, XILOSE, ARABINOSE XILITOL E ÁCIDO ACÉTICO 28 4. 7.3 • DETERMINAÇÃO DA ATIVIDADE ENZIMÁTICA 28 4.7.4 • DETERMINAÇÃO DO TEOR DE PROTEÍNAS TOTAIS 29
4.8 - METODOLOGIA DE ANÁLISE DE RESULTADOS 29
5 - RESULTADOS E DISCUSSÃO 32
5.1 - OBTENÇÃO DO HIDROLISADO HEMICELULÓSICO DE PALHA DE ARROZ 32
5.2 - ESTUDO DO ROMPIMENTO DAS CÉLULAS DE CANO/DA GUILLIERMOND/1 34
5.3 - ESTUDO DA INFLUÊNCIA DE VARIÁVEIS DO PROCESSO FERMENTATIVO NA PRODUÇÃO DE GUCOSE-6-FOSFATO DESIDROGENASE 46

vi
5.4 - ANÁLISE ESTATÍSTICA DO EFEITO DA CONCENTRAÇÃO INICIAL DE XILOSE, PH E NÍVEL DE INÓCULO NA PRODUÇÃO DA ENZIMA G6PDH E DE XILITOL 53
5.5 - MODELAGEM MATEMÁTICA PARA ESTIMAR A ATIVIDADE ESPECÍFICA DE GLICOSE-6- FOSFATO DESIDROGENASE EM FUNÇÃO DAS VARIÁVEIS SIGNIFICATIVAS AO PROCESSO FERMENTATIVO 57
6 - CONCLUSÕES 62 ~~~~~~~~~~~~~~~~~~~~~~~~- 7 - PREVISÕES FUTURAS 64 8 - REFERÊNCIAS BIBLIOGRÁFICAS 65

vu
LISTA DE TABELAS
Tabela 2.1. Alguns exemplos de produtos obtidos a partir da
fermentação de xilose a Tabela 2.2. Eficiência dos métodos de rompimento celular em diferentes
tipos de células (BECKER et ai., 1996) 16
Tabela 4.1. Matriz de planejamento experimental 23, com pontos
intermediários, para investigar o efeito do pH inicial,
concentração de xilose e do nível de inóculo nos parâmetros fermentativos de produção da enzima _ 26
Tabela 4.2. Nível de probabilidade e notação a ser utilizada para cada
nível de significância, no planejamento efetuado _ 27
Tabela 5.1. Composição do hidrolisado hemicelulósico obtido por hidrólise ácida da palha de arroz _
Tabela 5.2. Valores de atividade específica de glicose-6-fosfato
desidrogenase (G6PDH) para diferentes microrganismos em
diferentes métodos de rompimento 45
Tabela 5.3. Consumo percentual dos açúcares, fator de conversão de
33
xilose em xilitol (Yp1s), e atividade específica da glicose-6-
fosfato desidrogenase (U/mgprotetna) obtidos no tempo final de
fermentação 52
Tabela 5.4. Produção máxima de xilitol cPxmto1) e atividade específica de
G6PDH (U/mgproteina) obtidas no planejamento fatorial 23 para
avaliação da influência do pH inicial, concentração de xilose e
nível de inóculo na produção da enzima glicose-6-fosfato
desidrogenase por Candida guil/iermondii cultivada em
hidrolisado hemicelulósico de palha de arroz
55 Tabela 5.5. Análise da variância para o modelo representativo da atividade
específica de G6PDH de células de Candida guil/iermondii
58

viii
LISTA DE FIGURAS
Figura 2.1. Estrutura simplificada da fibra de materiais lignocelulósicos
(SHLESER, 1999) 5
Figura 2.2. Estrutura química parcial da hemicelulose (FENGEL &
WEGENER, 1989) 6
Figura 2.3. Representação esquemática do metabolismo de xilose em
leveduras (adaptado de GÍRIO et ai., 1994) 11
Figura 2.4. Diferentes estruturas de paredes celulares de microrganismos (SCHMIDELL et ai., 2001) _
Figura 4.1. Representação esquemática da atuação da enzima G6PDH
na reação para determinação de atividade enzimática _
Figura 5.1. Porcentagem de células rompidas em função da concentração
das suspensões celulares empregando tempos de agitação em
vórtice de 1, 3 e 5 minutos. Número de células intactas(-•-) e
00600 <-•-> 37
17
28
Figura 5.2. Porcentagem de células rompidas em função do tempo de
rompimento empregando diferentes concentrações celulares _ 38
Figura 5.3. Proteínas totais (-Ã-), atividade volumétrica (-•-) e atividade
específica (-0-) de glicose-6-fosfato desidrogenase, obtidas
após o rompimento de diferentes concentrações celulares de
Candida guillierrnondii empregando o tempo de agitação em
vórtice de 1 minuto ~~~~~~~~~~~~~~~~ 41
Figura 5.4. Proteínas totais (-Ã-), atividade volumétrica (-•-) e atividade
específica (-0-) de glicose-6-fosfato desidrogenase, obtidas
após o rompimento de diferentes concentrações celulares de
Candida guillierrnondii empregando o tempo de agitação em
vórtice de 3 minutos 42
Figura 5.5. Proteínas totais (-Ã-), atividade volumétrica {-•-) e atividade
específica (-0-) de glicose-6-fosfato desidrogenase, obtidas
após o rompimento de diferentes concentrações celulares de
Candida gui/lierrnondii empregando o tempo de agitação em
vórtice de 5 minutos ~~~~~~~~~~~~~~~- 43
Figura 5.6. Consumo de D-xilose (A) e produção de células (8), xilitol {C),
glicose-6-fosfato desidrogenase (D) por Candida guil/iermondii
empregando hidrolisado hemicelulósico com concentração

ix
inicial de D-xilose de 30 g/L sob diferentes condições de
cultivo. Ensaio 1: inóculo de 1 g/L e pH inicial de 4,5 (- •-),
ensaio 2: inóculo 1 g/L e pH 7,5 (-•-). ensaio 5: inóculo 3 g/L e
pH 4,5 (- ... -), ensaio 6: inóculo 3 g/L e pH 7,5 (-T-)
47
Figura 5.7. Consumo de D-xilose (A) e produção de células (B), xilitol (C),
glicose-6-fosfato desidrogenase (D) por Candida guil/iermondii
empregando hidrolisado hemicelulósico com concentração
inicial de D-xilose de 70 gil sob diferentes condições de
cultivo. Ensaio 3: inóculo de 1 g/L e pH inicial de 4,5 (-•-).
ensaio 4: inóculo 1 g/L e pH 7,5 (-•-), ensaio 7: inóculo 3 g/L e
pH 4,5 (- ... -), ensaio 8: inóculo 3 g/L e pH 7,5 (-T-) 49
Figura 5.8. Consumo de D-xilose (A) e produção de células (B), xilitol (C),
glicose-6-fosfato desidrogenase (D) por Candida guil/iermondii
empregando hidrolisado hemicelulósico com concentração
inicial de D-xilose de 50 g/L sob diferentes condições de
cultivo. Ensaios 9, 10 e 11: inóculo de 2 g/L e pH inicial de 6,0, ensaios do ponto central(-•-).(-•-),(- ... -) _
Figura 5.9. Gráfico de Pareto para os efeitos das variáveis pH inicial,
concentração de xilose e nível de inócuto nas respostas
atividade específica da enzima G6PDH (A) e produção máxima
de xilitol (B) 56
50
Figura 5.10. Gráfico dos valores previstos pelo modelo linear em função dos
valores observados para a resposta atividade específica de
G6PDH 60 Figura 5.11. Atividade específica de glicose-6-fosfato desidrogenase em
função da concentração de xilose e do pH, usando 1 g/L de
61

Abreviaturas
Yp1s
Yxts
Ox
Qp
Pxmtol
X
G6PDH
t
rpm
ANOVA
LISTA DE NOMENCLATURAS
Fator de conversão de xilose em xilitol (g/g)
Fator de conversão de substrato em células (g/g)
Produtividade em células (g/L.h)
Produtividade em xilitol (g/L.h)
Produção máxima em xilitol (g/L)
Atividade volumétrica (U/L)
Atividade específica (U/mgproteina)
Densidade ótica a 600 nm
Concentração celular (g/L)
Glicose-6-fosfato desidrogenase
tempo
Rotações por minuto
Análise de variância

XI
RESUMO Produção de glicose-6-fosfato desidrogenase a partir de hidrolisado hemicelulósico de palha de arroz por Candida guil/iermondii. Daniela de Borba Gurpilhares. Programa de Pós-graduação em Biotecnologia Industrial, Departamento de Biotecnologia, Faculdade de Engenharia Química de Lorena. Orientador: Dra. Inês Conceição Roberto (Departamento de Biotecnologia, FAENQUIL, CP 116, 12600-000, Lorena, SP, Brasil). Co-orientador: Dr. Adalberto Pessoa Jr. (Departamento de Tecnologia Bioquímico-Farmacêutica da Faculdade de Ciências Farmacêuticas, USP). Banca examinadora: Dr. Roberto da Silva e Dra. Adriane Maria Ferreira Milagres. Janeiro de 2004.
Este projeto teve como objetivo avaliar a produção da enzima glicose-6- fosfato desidrogenase (G6PDH), por processo fermentativo, empregando a levedura éandida guil/iermondii FTl20037, em meio constituído de hidrolisado hemicelulósico de palha de arroz.
Inicialmente, foram realizados ensaios de rompimento celular empregando esferas de vidro sob agitação em vórtice, variando o tempo de rompimento e a concentração celular da suspensão de forma a se obter uma eficiente liberação da enzima. Este estudo preliminar revelou que as melhores condições de rompimento foram o tempo de 5 minutos de agitação em vórtice e concentração celular de 11,6 g/L, alcançando uma atividade específica de 0,96 U/mgproteína- Em etapa posterior, os ensaios fermentativos foram realizados com o objetivo de se avaliar o efeito do pH inicial do meio (X,) em combinação com outras variáveis do processo na produção da enzima, nomeadamente a concentração inicial de xilose (X2) e o nível de inóculo (~). Para atingir os objetivos deste projeto, foi utilizada a metodologia do planejamento experimental. Com esta metodologia foi possível estabelecer um modelo matemático para descrever a produção de G6PDH, em atividade específica, por Candida guilliermondii. O modelo previsto para esta resposta, em função das variáveis estudadas e do efeito interativo entre elas, é dado pela equação:
sendo previsto pelo modelo uma atividade específica máxima de 0,64 U/mgproteína, podendo ocorrer uma variação de 0,55 a 0, 72 U/mgprcxeína, dentro do intervalo de confiança de 95%.
Para o ensaio de validação do modelo, realizado nas condições de 70 gil de xilose, pH 7,5 e nível de inóculo de 1 g/L, obteve-se Ae=0,60 U/mgproteína, revelando que o modelo proposto é adequado para descrever essa resposta na região em estudo.
Os resultados obtidos no presente trabalho confirmam o potencial da levedura Candida guilliermondii na produção da enzima G6PDH, em substratos de baixo custo.

xii
ABSTRACT
This work was focused on the evaluation of the glucose-6-phosphate dehydrogenase (G6PDH) production, by fermentation process, employing yeast cells of Candida guilliermondii FTI 20037, in growth medium of hemicellulosic hydrolysates from rice straw.
lnitially, cell disruption assays were performed employing glass beads under vortexing agitation, varying the disruption time and the concentration of cell suspension in the way of obtaining an efficient releasing of the enzyme. This preliminary study revealed that the best disruption conditions employed were time of vortexing agitation of 5 minutes and cell concentration of 11,6 g/L, reaching a specifc activity of 0,96 U/mgprotein· At the next study, the fermentation assays were performed in shaking flasks with the aim of evaluate the effect of the initial pH of the media (X1) in combination with other process variables that could interfere in the enzyme production, namely initial xylose concentration (X2) and the levei of inoculum (X:3). To reach the aim of this project, it was used the methodology of the experimental design. This methodology enabled to find a mathematical model to describe the G6PDH production, in specific activity, by Candida guilliermondii. The model obtained for this response, as a function of the studied variables and the interactive effect between them, was as follows:
being adjusted by the model, the highest specific activity of 0,64 U/mgprotein, that could vary from 0,55 to 0,72 U/mgprotein, at the confidence levei of 95%.
To the validation of the model, an assay was performed at xylose concentration of 70 g/L, pH 7,5 and levei of inoculum of 1 g/L, reaching a specific activity of 0,60 U/mgprotein wich revealed that the proposed model is suitable to describe this response at the tested range.
The results obtained in this work confirm the potential of the yeast Candida guilliermondii in the enzyme (G6PDH) production, in low cost substrate.

1 - INTRODUÇÃO
Devido à necessidade da diminuição do impacto ambiental e ao
aproveitamento racional de fontes renováveis, como resíduos agrícolas e
florestais, estes materiais têm sido amplamente utilizados como matéria-prima
para geração de energia, ração animal e principalmente como fonte de
carboidratos para processos de bioconversão. O emprego de processos
biotecnológicos de modificação destas matérias-primas ( conversão
microbiológica ou enzimática) constituem o objetivo de estudos de vários
grupos de pesquisa no Brasil e em outros países. Entretanto, a utilização de
materiais lignocelulósicos somente terá interesse comercial se todas as suas
frações forem devidamente aproveitadas, principalmente a fração
hemicelulósica, haja vista que apenas o componente celulósico tem sido
tradicionalmente utilizado em processos industriais. A fração hemicelulósica, ao
contrário da fração celulósica, apresenta variações quanto a sua estrutura e
composição sendo composta basicamente de hexases (D-glicose, D-manose,
D-galactose), pentases (D-xilose e L-arabinose), ácidos D-glicurônicos e
grupos acetilas. A xilose, principal componente desta fração, tem sido
amplamente utilizada em processos de bioconversão visando à obtenção de
produtos como combustíveis, aditivos, ácidos orgânicos, proteína microbiana,
pigmentos e várias enzimas de interesse comercial.
Em decorrência da expansão do uso industrial de enzimas, tem surgido
grande interesse, nos últimos anos, na produção das mesmas. Recentemente,
'-.

2
tem surgido um mercado lucrativo para enzimas microbianas intracelulares,
principalmente na área da saúde como reativos em diagnóstico clínico e em
biotransformação de produtos químicos e farmacêuticos. A glicose-6-fosfato
desidrogenase é um importante exemplo de enzima microbiana utilizada em
diagnóstico clínico.
A glicose-6-fosfato desidrogenase - G6PDH (EC 1.1.1.49) desempenha
um papel fundamental na via das pentases fosfato, a qual catalisa a oxidação
da glicose 6-fosfato (G6P) em 6-fosfogliconato, gerando cofatores reduzidos
(NADPH). Esta enzima tem se mostrado importante em estudos bioquímicos e
médicos, apresentando grande interesse como reagente analítico, pois pode
ser usada em diversas análises quantitativas. Embora a G6PDH possa ser
encontrada em diversos tecidos animais e vegetais, o uso de microrganismos
como fonte de enzimas tem se tornado bastante atraente, principalmente
devido à possibilidade da utilização de matérias-primas renováveis para os
cultivos celulares.
A obtenção de produtos intracelulares, principalmente de enzimas
microbianas, tem representado um problema prático em relação aos processos
de purificação destas biomoléculas. Os processos para a separação de tais
produtos geralmente envolvem rompimento, mecânico ou químico, das células,
seguido da remoção dos fragmentos celulares e de alguns contaminantes por
centrifugação ou filtração em membrana. Além disso, a obtenção dos produtos
intracelulares pode ser influenciada de modo significativo através de variáveis
do processo fermentativo como, concentração de substrato, pH do meio, nível
de inóculo, concentração de oxigênio dissolvido, dentre outras.
Dentro deste contexto, o presente projeto visa estudar o processo de
produção da enzima glicose-6-fosfato desidrogenase (G6PDH) por processo
fermentativo, empregando a levedura Candida guilliermondii cultivadas em
hidrolisado hemicelulósico de palha de arroz.
O presente trabalho se insere no programa de pesquisa do Grupo de
Processos Fermentativos do Departamento de Biotecnologia (DEBIQ) da
Faculdade de Engenharia Química de Lorena (FAENQUIL), em cooperação
com a Universidade de São Paulo (Departamento de Tecnologia Bioquímico-
Farmacêutica da Faculdade de Ciências Farmacêuticas) que tem como objetivo
o desenvolvimento de tecnologia para o aproveitamento da fração
' - . --

3
hemicelulósica de resíduos agroindustriais visando a produção de insumos por
via biotecnológica, dando ênfase aos estudos de produção e purificação de
enzimas intracelulares, em particular, a enzima G6PDH.

4
2 - REVISÃO BIBLIOGRÁFICA
2.1 - MATERIAIS LIGNOCELULÓSICOS
2.1.1 - Composição e Características do Material Lignoce/u/ósico
Os principais componentes orgânicos dos materiais lignocelulósicos são
a celulose, a hemicelulose e a lignina, em proporções que dependem da
espécie vegetal e outras variações biológicas, como diferenças genéticas
dentro da própria espécie, e condições de crescimento (FENGEL &
WEGENER, 1989; GOLDSTEIN, 1981 ).
O termo estrutura lignocelulósica refere-se a parte do vegetal que forma
a parede celular, composta basicamente por carboidratos ( celulose e
hemicelulose) os quais estão ligados a uma estrutura contendo substâncias
aromáticas, denominada lignina. A Figura 2.1 apresenta a estrutura simplificada
da fibra de materiais lignocelulósicos.

5
Lignina
Parede celular do vegetal
Fibras de celulose
Figura 2.1. Estrutura simplificada da parede celular vegetal de materiais
lignocelulósicos (SHLESER, 1999)
A celulose é um homopolímero linear de unidades repetidas de 0-
glicose com ligações f3(1 ~4) de elevado peso molecular e alta cristalinidade.
Sua função na natureza é estrutural, sendo o principal componente da parede
do vegetal onde atua como suporte para a membrana celular (GOLDSTEIN,
1981; KUHAD & SINGH, 1993). A estrutura cristalina do material celulósico
constitui um dos principais obstáculos para a sua hidrólise, independente do
método de extração empregado (PARIS!, 1989).
A lignina constitui um polímero complexo de estrutura polifenólica e não
convertida em açúcares fermentáveis (LADISCH, 1979). Este complexo atua
como um "cimento" entre as fibras oferecendo barreira à degradação
enzimática do material celulósico (GOLDSTEIN, 1981 ).
A hemicelulose é, depois da celulose, o complexo orgânico natural de
maior ocorrência na biosfera. Este complexo está presente em todas as
camadas da parede celular do vegetal, porém concentrado nas camadas
primárias e secundárias, onde ocorre em associação com a celulose e a lignina

6
(BISARIA & GHOSE, 1981). A Figura 2.2 mostra a estrutura simplificada da
hemicelulose.
a-Arabinofuranose
Figura 2.2. Estrutura química parcial da hemicelulose (MUSSATTO, 2002)
Ao contrário da celulose, as hemiceluloses apresentam variações quanto
a sua estrutura e composição sendo constituída de polímeros heterogêneos
com cadeias lineares apresentando ramificações laterais. Estes polímeros são
compostos basicamente de hexases (D-glicose, D-manose e D-galactose),
pentases (D-xilose e L-arabinose), ácidos D-glicurônicos e grupos acetilas
(KUHAD & SINGH, 1993). As cadeias das hemiceluloses são de menores
dimensões que as da celulose apresentando grau de polimerização
normalmente menor que 200 (GONG et ai., 1981a). Além dos três
componentes majoritários, a parede celular da biomassa vegetal apresenta
outros constituintes, em menor proporção, que são classificados como
extrativos (ácidos graxos, resinas, taninos, etc.) e não-extrativos (sílica,
oxalatos, carbonatos, etc.) que variam suas composições de acordo com cada
espécie de material (KUHAD & SINGH, 1993).

7
2. 1.2 - Potenciais Usos dos Materiais Lignocelulósicos
Atualmente existe grande tendência em obter-se uma utilização mais
eficiente de resíduos agroindustriais (PANDEY et ai., 2000). O desenvolvimento
da tecnologia para o uso dessa biomassa renovável e a tentativa de diminuir o
impacto adverso sobre o meio ambiente tem-se mostrado importantes (SUN et
ai., 2000). Os materiais lignocelulósicos sob a forma de resíduos agrícolas e
florestais são acumulados anualmente, no mundo, em grandes quantidades.
Devido a sua natureza renovável e ao seu baixo custo, estes materiais têm sido
amplamente utilizados como matéria prima para geração de energia, ração
animal e principalmente como fonte de carboidratos para processos de
bioconversão (KUHAD & SINGH, 1993). De acordo com PARIS! (1989) a
utilização de materiais lignocelulósicos somente terá interesse comercial se
todas as suas frações forem devidamente aproveitadas, principalmente, a
fração hemicelulósica, haja vista que apenas o componente celulósico tem sido
tradicionalmente utilizado em processos industriais.
Segundo PARAJÓ et ai. (1994) consideráveis esforços têm sido
dispensados para o desenvolvimento de novos processos para a utilização
destes materiais. As novas tecnologias devem satisfazer a várias exigências,
tais como (i) possibilidade econômica, (ii) alta eficiência no processamento da
matéria prima, (iii) alta seletividade na separação das frações, e (iv) baixo
impacto ambiental.
Dentre os materiais lignocelulósicos considerados como potenciais
fontes alternativas para geração de diferentes produtos de interesse econômico
e social podem ser destacados o bagaço de cana de açúcar e a palha de
diversos cereais (dU TOIT et ai. 1984, WILKE et ai. 1981). Segundo GRAJEK
(1988), a conversão destes materiais em produtos de valor econômico constitui
um processo de grande importância para o futuro da humanidade.
A palha de arroz é um dos principais resíduos lignocelulósicos no
contexto brasileiro. Segundo dados da Empresa Brasileira de Pesquisa
Agropecuária (Embrapa), cerca de 14 milhões de toneladas de palha de arroz
foram produzidos na safra de 1995/96. Este resíduo contém uma fração
hemicelulósica de aproximadamente 25%, rica em xilose a qual vem sendo
utilizada principalmente para a obtenção de xilitol por via biotecnológica

(ROBERTO et ai., 2002). A palha de arroz tem sido utilizada , também, na
produção de combustíveis líquidos, fibras, ou na geração de energia (KADAM
et ai., 2000). No oriente, onde o arroz e o trigo são intensamente cultivados, a
palha é o principal recurso utilizado na alimentação animal, e na produção de
combustíveis domésticos e fibras de celulose (MISRA et ai., 1993).
Nos últimos anos, vários trabalhos vêm sendo conduzidos com o
objetivo de se obter uma separação seletiva dos componentes poliméricos dos
materiais lignocelulósicos (celulose, hemicelulose e lignina), em tratamentos
denominados de "Fracionamento". Este tipo de processo permite uma
utilização separada das frações obtidas e sua transformação em produtos de
interesse comercial por meio de processos biotecnológicos (KUHAD & SINGH,
1993)
A alta proporção de pentases, particularmente xilose, na fração
hemicelulósica, aliada a sua maior facilidade de extração do complexo
lignocelulósico, tem atraído a atenção de pesquisadores para a efetiva
utilização desta fração em processos de bioconversão. A Tabela 2.1 mostra
alguns produtos que podem ser obtidos a partir de xilose por bioconversão.
Tabela 2.1. Alguns exemplos de produtos obtidos a partir da fermentação de
xilose
Classificação Produtos Referências Combustíveis/
Solventes
Etanol, Acetona-butano!, 2,3
butanodiol
Bisaria (1998)
Magee & Kosaric (1985)
Aditivo alimentar Xilitol Roberto et ai. (1991);
Winkelhausen &.
Kusmanova ( 1998)
Acidos Orgânicos Acido lático Garde et ai. (2002)
Acido acético Brownell & Nakas (1991)
Outros Proteína microbiana
Pigmentos
Pessoa et ai. ( 1996)
Parajó et ai. ( 1998)
8

9
2.1.3 - Obtenção do Hidrolisado Hemice/ulósico
A hidrólise ácida utilizada na produção de hidrolisados hemicelulósicos
tem sido descrita como um dos métodos mais eficientes na solubilização de
açúcares provenientes da fração hemicelulósica, bem como no pré-tratamento
da biomassa visando o aumento da degradação celulósica através de enzimas
(McMILLAN, 1994). A hidrólise da fração hemicelulósica envolve solubilização
e parcial degradação dos açúcares produzidos o que demonstra ser um dos
principais problemas encontrado neste processo.
Durante a hidrólise ácida de materiais lignocelulósicos são formados
diferentes tipos de açúcares (D-glicose, D-galactose, D-manose, D-xilose, L-
arabinose) e produtos de degradação os quais são considerados tóxicos,
podendo afetar intensamente o metabolismo microbiano. Os principais
compostos tóxicos incluem hidróximetilfurfural e furfural (produtos da
degradação de açúcares), ácido acético (substância liberada da estrutura
hemicelulósica), e alguns compostos fenólicos e aromáticos (produtos da
degradação da lignina) sendo estes considerados mais tóxicos que furfural ou
hidróximetilfurfural, mesmo em baixas concentrações (PARAJÓ et ai., 1998).
Tendo em vista a necessidade de diminuição da toxicidade causada por
compostos inibidores presentes no hidrolisado, processos de destoxificação
como neutralização, evaporação, resinas de troca iônica e adsorção por carvão
ativo têm sido utilizados antes da fermentação (McMILLAN, 1994). A adsorção
com carvão ativo vem sendo utilizada como um método de destoxificação para
melhorar a capacidade de microrganismos de fermentar hidrolisados
hemicelulósicos. De acordo com MUSSATTO & ROBERTO (2001), o xilitol
produzido por Candida guilliermondii apresentou um rendimento de 0, 72 g/g e
uma produtividade volumétrica de 0,61 g/L.h após tratamento do hidrolisado de
palha de arroz com carvão ativo na razão hidrolisado: carvão de 40 g/g,
correspondendo à remoção de 27% de compostos fenólicos. Estes estudos
demonstraram ainda que esses parâmetros tiveram um aumento de 22 e 45%,
respectivamente, em relação àqueles obtidos em hidrolisado sem tratamento
com carvão.
O aumento na concentração de xilose presente nos hidrolisados
hemicelulósicos também tem demonstrado ser importante sob o ponto de vista

10
econômico, pois tende a melhorar parâmetros, como rendimento e
produtividade, na bioconversão da D-xilose em xilitol. O aumento na
concentração inicial deste açúcar aumenta a concentração do produto final,
melhorando a produtividade volumétrica e tornando o processo de separação
mais viável (MUSSATIO & ROBERTO, 2004). Em estudos realizados por
PARAJÓ et ai, (1996) a relativa baixa concentração de xilose presente no
hidrolisado hemicelulósico de madeira limitou tanto a produtividade quanto o
rendimento de xilitol por Debaryomyces hansenii. De acordo com MUSSA TIO
& ROBERTO (2004), durante a etapa de concentração do hidrolisado de palha
de arroz, o aumento dos teores de açúcares é acompanhado pelo aumento nas
quantidades de hidróximetilfurfural, ácido acético e produtos da degradação da
lignina, resultando em maiores níveis de toxicidade e inibição do metabolismo
microbiano. Segundo estes autores, a hidrólise ácida da palha de arroz
empregando O, 1 M de H2S04 na proporção líquido:sólido de 1 O: 1 (v/p) durante
27 minutos a 121ºC seguida da concentração do hidrolisado hemicelulósico
obtido produziram níveis de compostos tóxicos, tais como furfural,
hidróximetilfurfural e ácido acético, abaixo do limite de inibição preconizado na
literatura.
Outro fato importante é que a utilização de hidrolisados em processos
fermentativos implica em um efeito, nem sempre positivo, na obtenção do
produto de interesse devido à presença de diferentes tipos de açúcares como
xilose, glicose e arabinose. A capacidade da Candida guil/iermondii em
fermentar xilose, mesmo na presença de outros açúcares, foi estudada pelos
pesquisadores LEE et ai. (1996), e SUGAI & DELGENES (1995). Tais estudos
demonstraram que a glicose reprime parcialmente a indução" de aldose
redutase (xilose redutase) pela xilose e que a intensidade dessa repressão está
correlacionada a concentração de glicose no sistema de indução, ou seja, o
aumento da concentração de glicose adicionada a xilose pode anular o efeito
dessa pentase na indução da atividade da aldose redutase.

11
2.2 - METABOLISMO DE XILOSE EM LEVEDURAS
Uma vez dentro da célula da levedura, a xilose é reduzida a xilitol pela
xilose redutase (aldose redutase EC 1.1.1.21) dependente quer de NADH ou
de NADPH. O xilitol, composto relativamente estável, ou é excretado da célula
ou oxidado a xilulose pela xilitol desidrogenase (EC 1.1.1.9) dependente de
NAD ou NADP. As primeiras duas reações são consideradas limitadoras do
metabolismo de xilose. A presença quer de elevadas atividades de xilose
redutase (XR) ou baixas atividades de xilitol desidrogenase (XDH) tem sido
usada como critério para selecionar os microrganismos produtores de xilitol. A
fosforilação da D-xilulose a D-xilulose-5-fosfato é catalisada pela
xilulosequinase (WINKELHAUSEN & KUZMANOVA, 1998). A Figura 2.3
mostra um esquema simplificado do metabolismo de xilose em leveduras.
A conversão de pentoses a xilulose-5-fosfato é um pré-requisito para a
sua utilização pelas vias catabólicas centrais. A D-xilulose-5-fosfato pode
subsequentemente entrar na via das pentoses fosfato, também conhecida
como via das hexoses monofosfato (HMP) a qual possui duas etapas (oxidativa
e não oxidativa).
xy1uiOW
i
8 stnanot 4 {'\
Nf<!Í NAD>i
fn.x:tose-6-P + ·~~p
-pyruvate --+ Biomass
Figura 2.3. Representação esquemática do metabolismo de xilose em leveduras (adaptado de GÍRIO et ai., 1994)

12
A glicose-6-fosfato desidrogenase (G6PDH) é a enzima responsável
pela primeira etapa do processo oxidativo da via das pentases fosfato. Nesta
etapa, a glicose 6-fosfato é convertida em 6-fosfoglico-õ-lactona, um éster
intramolecular, que é hidrolisado para a forma ácida livre 6-fosfogliconato.
Durante esta etapa são gerados equivalentes redutores, sob a forma de
NADPH, para as reações biossintéticas redutivas dentro das células, enquanto
que na fase não oxidativa as pentases fosfato são convertidas em hexases e
triases fosfato (JEFFRIES, 1983). A via das pentases fosfato também fornece
ribose-5-fosfato usada para a síntese de ácidos nucleicos e histidina e eritrose-
4-fosfato necessária para a síntese de ácidos aromáticos.
O gliceraldeido-3-fosfato e a frutose-6-fosfato são produtos da fase não
oxidativa da via das pentases fosfato. Ambos podem ser convertidos a piruvato
na via de Embden-Meyerhof-Parnas (EMP). O piruvato pode ser
descarboxilado e reduzido a etanol ou pode entrar no ciclo dos ácidos
tricarboxílicos. A conversão de xilulose-5-fosfato em gliceraldeido-3-fosfato e
acetil fosfato pela xilulose-5-fosfato fosfocetolase apresenta uma via alternativa
para a utilização da xilulose-5-fosfato denominada desvio da fosfocetolase
(PRIOR et ai., 1989). O metabolismo da xilose em leveduras fornece uma
variedade de produtos contendo carbono que incluem, xilitol, dióxido de
carbono, etanol, ácido acético, entre outros. Os rendimentos em produto são
dependentes da regulação do fluxo de carbono através das vias metabólicas
disponíveis (SLININGER et ai., 1987). Recentemente, o metabolismo de xilose
em leveduras selvagens e recombinantes tem sido revisto (HO et ai., 1999;
JEFFRIES & SHI, 1999). Um estudo detalhado da bioquímica e fisloloqia de
leveduras capazes de metabolizar a xilose foi publicado por HAHN-HÃGERDAL
et ai., 1994.
O catabolismo de xilose está intimamente relacionado às condições
ambientais e aos microrganismos em estudo. A literatura apresenta
principalmente os fatores: suprimento de oxigênio, concentração de substrato,
pH e nível de inóculo, como as variáveis que influenciam a produção de
metabólitos no meio de cultivo (HAHN-HAGERDAL et ai., 1994;
WINKELHAUSEN & KUZMANOVA, 1998).

13
2.3 - GLICOSE-6-FOSFATO DESIDROGENASE (G6POH)
2.3.1 - Importância Fisiológica
A glicose 6-fosfato desidrogenase (G6PDH) é uma enzima
citoplasmática de grande importância para a sobrevida das células uma vez
que é responsável pela manutenção de um nível adequado da coenzima
reduzida NADPH.
Em seres humanos, a deficiência da enzima G6PDH configura uma
patologia conhecida como anemia hemolítica. Esta deficiência promove a
diminuição dos eritrócidos devido à incapacidade das células de regenerarem o
NADPH, um cofator importante nos processos biológicos oxidativos (VIEIRA-
NETO et ai. 1999). Tal patologia é resultado de mutações em diferentes pontos
no gene que codifica a enzima e é considerada a enzimopatia mais comum em
humanos pois afeta 400 milhões de pessoas no mundo. As populações de
origem Mediterrânea, Asiática e do Oriente Médio apresentam altas
freqüências de mutações desse gene. Em Hong Kong e no sul da China, 4-6%
da população masculina são deficientes em G6PDH (LUZZATIO & MEHTA,
1995).
As manifestações clínicas associadas à deficiência de G6PDH são: 1)
Anemia hemolítica induzida por drogas: antimaláricos, sulfonamidas, sulfonas e
outras drogas ou produtos químicos que estejam associados à significativa
hemólise em algumas pessoas; 2) Anemia hemolítica induzida por infecção:
numerosas infecções por bactérias ou vírus podem provocar hemólise, sendo
as mais importantes as hepatites, a pneumonia e a febre tifóide; 3) Favismo:
súbito aparecimento de anemia hemolítica aguda após 24 a 48 horas da
ingestão de fava; 4) Icterícia neonatal: surge geralmente entre 1 a 4 dias de
vida; 5) Anemia hemolítica congênita crônica: estas manifestações ocorrem
devido a um aumento de estresse oxidativo.
Em leveduras, as reações iniciais do metabolismo de xilose são
limitadas em diferentes graus em função da especificidade da enzima xilose
redutase para os cofatores reduzidos NADPH ou NADH. De acordo com
ALEXANDER et ai. (1988), em Candida shehatae o metabolismo de xilose sob
condições aeróbias é possivelmente limitado pelos níveis da enzima xilose

14
redutase ou glicose 6-fosfato desidrogenase (fonte de NADPH), enquanto que
sob anaerobiose a xilose redutase dependente de NADH é o fator limitante.
SILVA et ai. (1996) concluíram que a xilose redutase de Candida guilliermondii
requer apenas NADPH como cofator, o que demonstra a importância do fluxo
de xilose pela via das pentases para regeneração do cofator. JEPPSSON et ai.
(2002) demonstraram a importância da parte oxidativa da via das pentases
fosfato em cepas recombinates de Saccharomyces cerevisiae visando melhorar
o rendimento em etanol a partir da xilose. A cepa TMB3255, a qual teve o gene
ZWF1 rompido com conseqüente redução na produção de NADPH, apresentou
rendimentos em etanol de 0,41 g/g e em xilitol de 0,05 g/g. Porém a taxa de
consumo de xilose foi diminuída em 84% devido à limitação na redução de
xilose por NADPH. Tais resultados indicam a existência de uma estreita
conexão entre a produção de xilitol e a parte oxidativa da via das pentases
fosfato.
JEPPSSON et ai. (2003), avaliaram a influência da enzima G6PDH, em
diferentes níveis de atividade, na fermentação da D-xilose. Os diferentes niveis
na desta enzima foram obtidos através do promotor de cobre, CUP1, utilizado
para facilitar a regulação da expressão do gene ZWF1 com a mudança na
concentração de Cu2+ do meio (cepa TMB3030). A maior atividade de G6PDH,
1,567 U/mgprotetna, nessa cepa resultou em um maior rendimento de xilitol (0,33
g/g) quando comparado à cepa TMB 3001 (0,29 g/g) que continha 1,015
U/mgprotelna· Esses estudos comprovaram a existência da relação entre
atividade de G6PDH e rendimento de xilitol.
2.3.2 - Principais Aplicações
Além da importância em estudos bioquímicos e médicos (LEITE, 1989;
SEVERO, 1979), a enzima G6PDH apresenta grande interesse como reagente
analítico, pois pode ser usada em diversas análises quantitativas, incluindo
medidas de atividade da hexoquinase e creatina-quinase, concentrações de
ATP e de hexases (BERGMEYER, 1984). De acordo com CHATEL et ai.
(1996), a G6PDH pode identificar e quantificar de forma eficiente e precisa
moléculas de glicose liberadas após hidrólise do amido em sucos, mesmo na
presença de pedaços de frutas, compostos coloridos, elevadas concentrações

15
de sacarose e outros tipos de polissacarídeos. SHIROKANE et ai. (2000),
também utilizaram a G6PDH, juntamente com as enzimas maltose-1-
epimerase, maltose fosforilase e J3-fosfoglucomutase, para a determinação de
maltose.
Visando avaliar o metabolismo da fonte de carbono e sua estabilização
pelos microrganismos, presentes no solo, FREY et ai. (1999) propôs um
método simples.rápido e específico para a determinação da concentração de
glicose em solos enriquecidos, utilizando hexoquinase e glicose-6-fosfato
desidrogenase. A enzima G6PDH tem sido também empregada para a
determinação de glicose em sistema de reator em fluxo contínuo, pois o
NADPH formado pela reação enzimática pode ser facilmente detectado
espectrofotometricamente ou fluorometricamente (MORI et ai., 1999; FREY et
ai., 1999). Essa enzima pode, ainda, detectar baixíssimos níveis de
estreptavidina e biotina por ensaios de bioluminescência em reações de
hibridização de DNA, sem perda da atividade enzimática (TEROUANNE et ai.,
1989; BALAGUER et ai., 1989). Outra aplicação recente é a confecção de
biossensores, visando o rápido monitoramento da concentração de G6P no
sangue com menor custo e consumo de tempo do que os métodos tradicionais
como cromatografia e espectroscopia. Esse monitoramento é importante
porque ele pode refletir diretamente a atividade relativa da G6PDH na via
metabólica e tem sido utilizado por possibilitar o controle de G6PDH em
eritrócitos humanos e em células de fígado de ratos (BASSI et ai., 1999).
2.3.3 - Fontes de Obtenção e Métodos para Liberação da Enzima
A enzima G6PDH esta amplamente distribuída na natureza, sendo
encontrada em quase todos os tecidos de origem animal e vegetal e em células
de microrganismos. Dentre as principais fontes de obtenção são destacadas as
leveduras de panificação, Torula, Baci/lus stearothermophillus, Leuconostoc
mesenteroides, adrenais de bovinos e eritrócitos humanos (LEW, 1998).
A maioria das enzimas produzidas por microrganismos são intracelulares
e · algumas delas tem sido produzidas com sucesso em escala industrial
(ÔZBEK & ÜLGEN, 2000). O mercado para produtos microbianos intracelulares
visando aplicação industrial, principalmente na área de alimentos e médica, se

16
encontra em crescimento contínuo devido aos novos desenvolvimentos em
engenharia genética e tecnologia do DNA recombinante.
Para que ocorra a liberação dos produtos microbianos intracelulares, é necessário o uso de métodos que são capazes de romper a parede celular sem
causar inativação dos produtos biológicos. Os métodos de rompimento celular
podem ser divididos em 4 classes, como mecânicos (homogeneizador de alta
pressão, moinho de bolas, prensa francesa e ultra-som), não-mecânicos
(choque osmótico, congelamento e descongelamento, aquecimento, secagem),
químicos (álcalis, solventes, detergentes, ácidos) e enzimáticos (lise enzimática
ou inibição da síntese da parede celular). Segundo LOVITI et ai 2000, os
métodos mecânicos são geralmente os mais apropriados para rompimentos em
larga escala.
Para leveduras, o método de rompimento utilizando esferas de vidro
demonstrou ser mais conveniente e efetivo na obtenção do extrato celular. A
ação de rompimento das esferas de vidro durante agitação causa ruptura da
célula de levedura. A espessura e a rigidez da parede celular das leveduras
fazem de métodos como o ultra-som, lise osmótica e homogeneização
ineficazes no rompimento dessas células (Tabela 2.2)
Tabela 2.2. Eficiência dos métodos de rompimento celular em diferentes tipos
de células (BECKER et ai., 1996)
Métodos Bactéria Levedura Células ... Animais
Ultra-som Eficaz Ineficaz Ineficaz
Prensa Francesa Eficaz Eficaz somente a Ineficaz elevadas pressões
(20000 psi) Lise Osmótica Eficaz para Ineficaz Eficaz
liberação de enzimas
periplasmáticas Homogeneização Ineficaz Ineficaz Muito eficaz
· com pistão de vidrofTeflon Moinho com Eficaz O mais eficaz Ineficaz
esferas de vidro

17
Há diferentes estruturas de paredes celulares. As células animais
possuem membranas frágeis e fáceis de serem rompidas, enquanto que as
bactérias, leveduras e outras formas de fungos possuem paredes rígidas e que
exigem elevadas tensões de cisalhamento para o seu rompimento (Figura 2.4).
Bactérias rn:. Peptidioglicano gram (+) ! 1
l e:=:::::.-- Membrana citoplasmática
1 1 Membrana externa Bactérias '=miaiimm· gram (·) t'.:il Peptidioglicano
c:=::::::::a-- Membrana citoplasmática
Leveduras Manana I proteínas
Figura 2.4. Diferentes estruturas de paredes celulares de microrganismos
(SCHMIDELL et ai., 2001)
Segundo RICCI-SILVA et al.(2000) o procedimento escolhido deve
permitir o acoplamento de uma alta eficiência de desintegração com um curto
tempo de rompimento. A eficiência de uma técnica de rompimento particular é
usualmente avaliada em termos do grau de células rompidas e/ou do valor de
atividade específica da enzima recuperada na suspensão de células rompidas.
Os métodos físicos, destacando os homogeneizadores e os moinhos com
esferas de vidro, são os mais eficientes para o rompimento de células em
pequena e grande escala.
O rompimento envolvendo agitação rápida de uma suspensão de células
microbianas com esferas de vidro é uma excelente técnica. Este procedimento
é particularmente eficiente no rompimento de leveduras e fungos filamentosos,
os quais possuem paredes celulares rígidas constituídas principalmente por
polissacarídeos (80-90%) (KESHAVARZ et ai., 1987).

18
2.3.4 - Influência das Condições de Cultivo
Assim como para qualquer produto metabólico excretado pelos
microrganismos, fatores como pH, concentração do inóculo, concentração e
tipo da fonte de carbono, oxigênio dissolvido dentre outros podem afetar
significativamente a produção de uma enzima intracelular.
Em fermentações empregando a levedura Saccharomyces cerevisiae,
ABRAHÃO-NETO et ai. (1997) constataram que o pH do meio de cultivo
exerceu uma forte influência na atividade da G6PDH. Segundo os autores, a
atividade da enzima foi aumentada em 57% quando o pH do cultivo passou de
5,0 para pH 4,5. Neste mesmo trabalho os autores também estudaram o efeito
da concentração de oxigênio dissolvido sobre a atividade específica da enzima
sendo obtido um valor máximo de Q!~1 O U/mg quando a concentração de
oxigênio dissolvido no meio foi de 4,0 mg 02.L-1. Segundo os autores, a
diminuição da atividade enzimática observada com valores superiores (6,0 mg
02.L-1) e inferiores (0,2 mg 02.L-1) de oxigênio dissolvido foi devido à reduzida
taxa de crescimento sob estas concentrações de oxigênio. Essa observação
deve-se ao fato da G6PDH participar da via das pentoses, caminho pelo qual
as células sintetizam ribose-6-fosfato, um dos constituintes chave dos ácidos
nucleícos e nucleotídeos. Dado que a demanda por ácidos nucleícos durante a
fase de crescimento máximo é acentuada, a atividade específica da G6PDH
também aumenta.
De acordo com GÍRIO et ai (1994), a parte oxidativa da via das pentases
fosfato consiste de duas reações principais catalisadas pela glicose-6-fosfato
desidrogenase e pela 6-fosfogluconato desidrogenase dependentes de NADP,
resultando na formação de NADPH. Parte dos cofatores reduzidos são
empregados na redução da xilose, na primeira reação do metabolismo deste
substrato, e o restante é requerido pela célula em reações biossintéticas. Esses
autores estudaram o efeito da taxa de transferência de oxigênio (OTR) nos
níveis das enzimas do metabolismo de xilose em células de Debaryomyces
hansenni e observaram que a atividade de glicose-6-fosfato desidrogenase foi
estimulada pela quantidade de oxigênio disponível, aumentando em 60%
quando as condições de OTR variaram de 2,72 a 12,93 mmol 02/L.min.
Aumento na atividade de G6PDH similar foi relatado para Candida shehatae

19
cultivadas em xilose quando as condições de aeração do cultivo foram
mudadas de anaeróbio para semiaeróbio e então aeróbio (ALEXANDER et ai.,
1988). O maior valor de atividade específica de G6PDH obtido por GlRIO foi de
0,54 U/mg em meio sintético.
A atividade de várias enzimas incluindo G6PDH durante o cultivo de
Candida guilliermondii em quimiostato empregando xilose como fonte de
carbono foi determinada por GRANSTROM et ai (2001 ). Para uma taxa de
diluição de 0,097 h", a aÚvidade da enzima G6PDH decresceu cerca de 35%
(de 0,74 para 0,26 U/mg) quando a condição de oxigenação do cultivo passou
de aeróbio para oxigênio-limitado, o que demonstra que o efeito da
concentração de oxigênio é significativo na produção da enzima.
A fonte de carbono pode também influênciar a produção de glicose-6-
fosfato-desidrogenase (G6PDH) a partir de leveduras (SILVA, 2000). Em meios
contendo glicose ou glicose/sacarose a atividade da enzima foi superior
quando comparado com meios contendo apenas sacarose. Em cultivos
empregando Candida boidinii, a atividade da G6PDH também variou com a
fonte de carbono empregada (SURYADI et ai 2000). A atividade específica da
enzima decresceu de 0,51 para 0,39 U/mg quando o glicerol foi substituído pela
glicose. KATO et ai (1979), em estudos visando a purificação de glicose-6-
fosfato desidrogenase e 6-fosfogluconato desidrogenase a partir da levedura
Candida boidinii capaz de fermentar metanol, observaram que a atividade
destas enzimas foi duas vezes maior quando o metanol foi substituído pela
glicose como fonte de carbono.
De acordo com CERTIK et ai (1999), a relação carbono/nitrogênio pode
ser considerada fator extremamente importante num cultivo. Estes autores
mostraram que a atividade de G6PDH a partir do fungo Cunninghamella
echinulata aumentou de 100 para 200 nmol/min.mg quando a concentração de
nitrogênio no meio aumentou em 1,2 g/L.

20
3 - OBJETIVOS
GERAL
O presente trabalho teve como objetivo estudar a produção da enzima
glicose-6-fosfato desidrogenase (G6PDH) por processo fermentativo, sob
diferentes condições de cultivo, a partir do hidrolisado hemicelulósico de palha
de arroz utilizando a levedura Candida guilliermondii.
ESPECÍFICOS
./ Determinar as condições de rompimento de células de Candida
guilliermondii variando-se o tempo de rompimento e a concentração
celular da suspensão, visando uma eficiente liberação da enzima
G6PDH;
./ Investigar o efeito do pH inicial, concentração de xilose e do nível de
inóculo nos parâmetros fermentativos de produção da enzima, com
auxílio de um planejamento estatístico;

21
./ Determinar as melhores condições de fermentação e estabelecer um
modelo matemático que correlacione as respostas obtidas com as
variáveis significativas do processo, empregando a metodologia da
superfície de resposta.

22
4 - MATERIAIS E MÉTODOS
4.1 - PREPARO DA MATÉRIA PRIMA
A palha de arroz utilizada nos experimentos foi obtida na região de
Lorena. Após o recebimento, a palha foi seca ao sol e cominuída em moinho
tipo martelo a um tamanho de aproximadamente 1 cm de comprimento e 1 mm
de espessura.
4.2 - OBTENÇÃO E TRATAMENTO DO HIDROLISADO HEMICELULÓSICO
O hidrolisado hemicelulósico foi obtido por catálise ácida em reator
confeccionado em aço-inox AISI 316 com capacidade de 350 litros, munido de
aquecimento indireto por resistência elétrica por camisa de óleo térmico. As
reações foram realizadas a 121 ºC por 27 minutos empregando uma relação
solução ácida:palha seca de 1 Og/g. Foi utilizado o H2S04 como catalisador e a
solução ácida foi preparada para concentração final de 1 % (ROBERTO et ai,
2003). Ao final da reação, o material descarregado do reator foi centrifugado e
a fração líquida recuperada (hidrolisado hemicelulósico) foi retornada ao reator
de hidrólise para destoxificação do hidrolisado com carvão ativo nas seguintes
condições: relação carvão:hidrolisado de 3% (p/v), temperatura de 45 ºC e
tempo de agitação de 30 minutos (MUSSATIO & ROBERTO, 2001). Após o

processo de adsorção com carvão ativo, o hidrolisado foi centrifugado para
remoção de sólidos e então concentrado sob vácuo a uma temperatura de
65 ºC, visando aumentar a concentração de xilose até aproximadamente
108 g/L. O hidrolisado destoxificado e concentrado (HDC) foi estocado sob
refrigeração a 4°C, pH O, 70, para posterior utilização nos ensaios de
fermentação.
4.3 - MICRORGANISMO
Foi utilizada a levedura Candida guilliermondii FTI 20037, da coleção de
culturas do Grupo de Processos Fermentativos DEBIQ/FAENQUIL,
selecionada para produção de xilitol a partir de materiais lignocelulósicos. A
cultura foi repicada em tubos de ensaio contendo ágar malte inclinado,
incubada em estufa a 30 ºC por 24 h e então conservada em geladeira à 4ºC
para posterior preparo do inóculo.
4.4 - PREPARO DO INÓCULO
Células de Candida guilliermondii, recém repicadas em meio de
manutenção, foram transferidas em condições assépticas, com auxílio de uma
alça de platina, para tubos de ensaio contendo cerca de 5 ml de água
destilada esterilizada. Alíquotas de 1 ml desta suspensão foram transferidas
para frascos Erlenmeyer de 500 ml contendo 200 ml do meio de cultivo
constituído por hidrolisado destoxificado com pH ajustado para 6,2 e
concentração inicial de xilose de 30 g/L. Os frascos foram incubados a 30ºC
em agitador rotativo (Tecnal mod. Te 420) a 200 rpm durante 29 horas. Após
este tempo, as células foram separadas por centrifugação a 2000xg por 20
minutos e ressuspendidas em água destilada esterilizada de forma a se obter
uma suspensão celular densa. A partir desta suspensão foi calculado o volume
necessário para fornecer a concentração celular inicial desejada no meio de
fermentação, para cada ensaio.

2-4
4.5 - ESTUDO DAS CONDIÇÕES DE ROMPIMENTO CELULAR
Foi utilizado o método de rompimento com esferas de vidro ( diâmetro
médio de 0,5 mm) sob agitação em vórtice. O método consiste em adicionar
esferas de vidro a uma suspensão celular em proporções de 1: 1 (v/v) e
promover vários ciclos de agitação em vórtice alternando com períodos de
resfriamento em banho de gelo. Para este estudo, células provenientes do
cultivo do inóculo foram separadas por centrifugação e após lavagem com
água esterilizada foram ressuspendidas em tampão Tris-HCI 50 mM, pH 7,5
(adicionado de P-mercaptoetanol 10 mM, ácido aminocapróico 2 mM e EDTA
0,2 mM) de forma a se obter uma suspensão densa (da ordem de 30 g/L). A
partir desta suspensão foram preparadas várias diluições para o estudo do
rompimento celular. Volumes de 3 mi de suspensão celular e 3 ml de esferas
de vidro foram colocados em tubos de centrifuga e após agitação em vórtice
por intervalos regulares de tempo (1, 3 e 5 minutos; sendo que cada 1 min de
agitação, a suspensão foi resfriada em banho de gelo por 30 s). Os sólidos em
suspensão resultantes (fragmentos celulares e esferas) foram separados do
sobrenadante por centrifugação por 15 minutos a 4ºC e 6725g. O sobrenadante
foi então quantificado quanto ao teor de proteínas totais e atividade de G6PDH
e a eficiência do rompimento foi avaliada através da contagem de células
intactas, em câmara de contagem, e através da leitura de densidade ótica,
DO&XJ, antes e após o rompimento.
- --
4.6 - PROCESSO FERMENTATIVO
4.6.1 - Meio de Fermentação
O meio de fermentação foi preparado com o hidrolisado concentrado
conforme descrito no item 4.2. O hidrolisado foi diluído em água destilada
esterilizada de forma a se obter a concentração inicial de xilose desejada.
Antes da utilização dos hidrolisados diluídos como meio de fermentação, estes

25
tiveram os valores de pH ajustados de acordo com o planejamento fatorial
apresentado na Tabela 4.1.
4.6.2 - Condições de Cultivo
Os experimentos foram conduzidos em frascos Erlenmeyer de 500 ml
contendo 200 ml do meio de fermentação, inoculados com células cultivadas
nas condições descritas anteriormente. Os frascos foram incubados a 30ºC, em
agitador rotatório (Tecnal mod. Te 420), sob a agitação de 200 rpm. Durante as
fermentações, amostras foram retiradas, em diferentes tempos, para o
acompanhamento do consumo de xilose, glicose e arabinose, produção de
glicose-6-fosfato desidrogenase e xilitol, crescimento celular e variação do pH.
4.6.3 - Metodologia Estatística
Para avaliação da influência da concentração inicial de xilose, nível de
inóculo e pH inicial do cultivo sobre a produção da enzima G6PDH foi utilizado
um planejamento fatorial completo de dois níveis 23, conforme matriz
apresentada na Tabela 4.1.
As variáveis foram codificadas de acordo com a equação:
(1)
onde Vc -valor codificado da variável independente
VR -valor real da variável independente
V0 - valor real da variável independente no ponto central
l!J..VR - valor do passo, isto é (VRmáx-VRmín)/2

26
Tabela 4.1. Matriz de planejamento experimental com pontos
intermediários, para investigar o efeito do pH inicial,
concentração de xilose e do nível de inóculo nos parâmetros
fermentativos de produção da enzima
Ensaio Níveis originais Níveis codificados
das variáveis das variáveis
pH Xilose lnóculo pH Xilose lnóculo
inicial (g/L) (g/L) inicial (g/L) (g/L)
1 4,5 30 1 -1 -1 -1
2 7,5 30 1 +1 -1 -1
3 4,5 70 1 -1 +1 -1
4 7,5 70 1 +1 +1 -1
5 4,5 30 3 -1 -1 +1
6 7,5 30 3 +1 -1 +1
7 4,5 70 3 -1 +1 +1
8 7,5 70 3 +1 +1 +1
9 6,0 50 2 o o o 10 6,0 50 2 o o o 11 6,0 50 2 o o o
A análise estatística dos dados obtidos foi realizada utilizando-se o
programa STATGRAPHICS (versão 5.0). A significância dos fatores
experimentais foi testada pelo teste t de Student. Este teste, baseado na
hipótese de que o verdadeiro parâmetro é zero, é utilizado para elucidar a
significância dos fatores. Se o valor de t calculado (tcatc) for superior ao t tabelado tc1 _ª·À.) para um nível de significância a. e À. graus de liberdade, então o
termo em causa contribui significativamente para a resposta. Os níveis de
significância e a notação que será utilizada para denotar cada nível,
encontram-se na Tabela 4.2.

27
Tabela 4.2. Nível de probabilidade e notação a ser utilizada para cada nível de
significância, no planejamento efetuado
Nível p Notação
significância
90% p < 0,10 *
95% p < 0,05 **
99% p < 0,01 ***
Para a quantificação dos níveis das variáveis e otimização das
condições de produção da enzima foi utilizada a metodologia de superfície de
resposta.
4. 7 - MÉTODOS ANAlÍTICOS
4.7.1 - Determinação da Concentração Celular
A concentração celular foi obtida mediante uma curva de calibração que
correlaciona densidade ótica (DO) com massa seca de células. Para a
elaboração desta curva, células provenientes do cultivo do inóculo foram
centrifugadas, lavadas e ressuspendidas em água destilada de forma a se
obter uma suspensão homogênea densa (suspensão original). Esta suspensão
foi utilizada para preparar suspensões com diversas diluições (1 :50, 1 :100,
1 :125, 1 :200, 1 :250, 1 :500). Cada uma destas suspensões diluídas foram
submetida a leitura de absorbância em espectrofotômetro (Beckman DU 640) a
600 nm, utilizando-se água destilada como branco. A massa seca foi
determinada a partir da suspensão original após secagem de alíquotas com
volume conhecido, em estufa a 105 ºC por 24 horas.

28
4. 7.2 - Determinação da Concentração de Glicose, Xilose, Arabinose Xilitol e Ácido Acético
As concentrações de glicose, xilose, arabinose, ácido acético e xilitol
foram determinadas por Cromatografia Líquida de Alto Desempenho (HPLC),
em equipamento WATERS, nas seguintes condições: coluna 810-RAD Aminex
HPX-87H (300 x 7,8 mm); temperatura: 45 ºC; eluente: ácido sulfúrico 0,01 N;
fluxo 0,6 ml/min; volume de amostra: 20 µL; detector: índice de refração.
4. 7.3 - Determinação da Atividade Enzimática
A atividade de G6PDH foi determinada por análise espectrofotométrica
em aparelho Beckman 640 OU, a 340 nm e temperatura de 30 ºC de acordo
com o método descrito por BERGMEYER (1984). A atividade foi determinada
em um sistema enzimático contendo amostra, nicotinamida-adenina-
dinucleotídeo fosfato (NADP+) e glicose-6-fosfato (G6P). Nesta reação, a
glicose-6-fosfato (G6P), na presença da enzima G6PDH e do cofator NADP\ é oxidada a gliconato-6-fosfato gerando o cofator reduzido nicotinamida-adenina
dinucleotídeo fosfato (NADPH), conforme esquematizado na Figura 4.1. A
atividade da enzima G6PDH foi então determinada pela medida da velocidade
de formação de NADPH.
Glicose-6-Fosfato
;~!~::~::aro i 1 ~ Gliconato-6-Fosfato
NADPH
Figura 4.1. Representação esquemática da atuação da enzima G6PDH na
reação para determinação de atividade enzimática

29
Para a determinação da atividade da enzima G6PDH, foi utilizado 500µL
de tampão Tris-HCI (0,071 M, pH?,5), 1 OOµL de cloreto de magnésio (35mM),
5~tl NADP+ (O, 131 M), 1 OµL glicose-6-fosfato (500mM) e 1 OOµL de amostra.
Uma unidade (U) de G6PDH é definida como a quantidade da enzima capaz de
catalisar a redução de 1 µmal de NADP+/min nas condições experimentais. A
atividade volumétrica foi determinada pela seguinte equação:
~a lxl06 A\' =-x-·-x7J5xdiluição
~· 6220 .
onde:
Av = atividade volumétrica (U/L)
.1al.1t = velocidade de formação de NADPH por minuto a 340 nm
6220 = coeficiente de extinção molar ou índice de absorbância (M-1cm-1) para
NADPH
7, 15 = fator de diluição da cubeta
4.7.4 - Determinação do Teor de Proteínas Totais
O teor de proteínas totais foi determinado de acordo com o método
proposto por LOWRY. Foi utilizado como padrão a albumina de soro bovino
(BSA -"bovine serum albumin").
4.8 - METODOLOGIA DE ANÁLISE DE RESULTADOS
Para avaliação dos resultados obtidos nas diversas etapas do trabalho
foram analisados os seguintes parâmetros:
Eficiência do Rompimento Celular(%)
Nº células I mL = células,x5xl 04 xdiluição
onde:
cétútes, = número total de células contadas em Câmara de Neubauer

30
5x104 = fator de diluição da Câmara de Neubauer
/1° cé/11/asante - 11° células j . (%) = o : epors x100
/1 ce/ulas,111(es onde:
n º célulasantes = número de células antes do rompimento
n º célulasdepois = número de células depois do rompimento
Curva de Peso Seco:
DO -0,0386 [X]g / L = 600 xdiluição
1,527 onde:
[X]= concentração celular (g/L)
00600 = densidade ótica a 600 nm
(%) = [XLntes -[X]depois xlOO [XJanles
onde:
[X]antes = concentração de células antes do rompimento (00600)
[X]depois = concentração de células depois do rompimento (00600)
Fator de Conversão de Substrato em Células YXJs (g/g)
Yxis = _f1;_Y_ = (Xr - X;) ou YXJs = Coeficiente angular da -!).Sr (Sli - slf) + (S2; - S21)
equação de LiX = f(-LiST)
Fator de Conversão de Xilose em Xilitol Yp1s (g/g)
YP/s = ~ = (Pr - J:) ou Yp,s = Coeficiente angular da equação de L\P -!).S2 (S2; - S2f)
onde:
~ e Xr - Concentração inicial e final de células (g/L)
Pi e Pt - Concentração inicial e final de xilitol (g/L)
ti e tt- Tempo inicial e final da fermentação (h)

31
S1i e S1t - Concentração inicial e final de glicose (g/L)
S2i e S2t - Concentração inicial e final de xilose (g/L)
Produtividade em Xilitol Qp (g/L.h): LlP (P1 - P;)
Qp = -;;; = U1 - ti)
Produtividade em Células Qx (g/L.h)
Ox= LlX = (X1 -X;) - M (r1 -t;)
Atividade Específica de G6PDH em relação às proteínas totais (U/mg prot)
A atividade específica da enzima G6PDH foi obtida pela relação entre
atividade enzimática volumétrica (U/L) e a concentração de proteínas totais
(mg/L).

32
5 - RESULTADOS E DISCUSSÃO
5.1 - OBTENÇÃO DO HIDROLISADO HEMICELULÓSICO DE PALHA DE ARROZ
O hidrolisado hemicelulósico foi obtido através da hidrólise ácida da
palha de arroz em condições previamente determinadas por ROBERTO et ai.
(2002) conforme descrito no item 4.2. Segundo esses autores, o hidrolisado da
palha de arroz apresentou na sua composição, como componentes poliméricos
principais, glucana (43,4%), xilana (20,2%), arabinana (2,73%), lignina (17,2%)
e cinzas (11,4%).
No presente trabalho, foram obtidos 86 litros de hidrolisado tratado com
carvão ativo (3%) e 15 litros de hidrolisado concentrado a partir de 16,5 kg de
palha de arroz (peso seco).
A composição química do hidrolisado hemicelulósico da palha de arroz
após hidrólise, tratamento com carvão ativo e concentração a vácuo está
mostrada na Tabela 5.1. Verifica-se que, sob as condições de hidrólise
empregadas, o açúcar predominante no hidrolisado foi a D-xilose (18,97 g/L),
seguido de arabinose (3,29 g/L) e glicose (2,22 g/L). Concentração semelhante
de D-xilose (18,24 g/L) e teores menores de arabinose (1,71 g/L) e glicose
(1,20 g/L), foram obtidos por RODRIGUES et ai. (2001) após a hidrólise ácida
do bagaço da cana-de-açúcar empregando 100 mg de H2S04 diluído/g de
bagaço de cana (peso seco) durante 10 minutos a 121ºC. SUN et ai. (1999),
observaram que a hidrólise alcalina da palha de arroz, diferentemente da
- -~

33
hidrólise ácida, favoreceu a solubilização da O-glicose presente na fração
hemicelulósica.
Tabela 5.1. Composição do hidrolisado hemicelulósico obtido por hidrólise
ácida da palha de arroz
Componentes
(g/L)
Hidrolisado
O-glicose
D-xilose
L-arabinose
Ácido Acético
2,22
18,97
3,29
2,22
Tratado com Concentrado 6
Carvão ativo vezes -
2,17 11,28
18,37 103,1
3,20 15,29
1,69 2,84
ln Natura
Após a obtenção do hidrolisado, realizou-se o tratamento deste com
carvão ativo visando a parcial remoção de compostos tóxicos prejudiciais ao
crescimento e ao metabolismo de microrganismos. Segundo FELIPE et ai.
(1996), tais compostos devem ser removidos ou terem suas concentrações
diminuídas para uma utilização eficiente do hidrolisado hemicelulósico em
processos de bioconversão de xilose. Observa-se, na Tabela 5.1, que não
houve alteração nas concentrações de açúcares, porém ocorreu uma pequena
redução na concentração de ácido acético. De acordo com estudos realizados
por RODRIGUES et ai. (2001 ), o tratamento com carvão ativo antes da
concentração a vácuo (4 vezes) do hidrolisado de bagaço de cana em pH 0,92,
favoreceu a remoção de compostos fenólicos (98%) e apresentou uma redução
nos valores de ácido acético de 62,2%. No presente trabalho, o tratamento com
carvão ativo do hidrolisado de palha de arroz antes da concentração a vácuo
proporcionou uma redução de 24% de ácido acético. Esta diferença pode ser
atribuída aos diferentes tipos de materiais lignocelulósicos, às diferentes
condições de hidrólise empregadas além do tratamento utilizado na remoção
dos compostos tóxicos presentes no hidrolisado.
O processo de concentração a vácuo foi realizado visando o aumento
da concentração de xilose no hidrolisado, embora . tal processo fosse

acompanhado pelo aumento na quantidade de compostos tóxicos. Como pode
ser observado na Tabela 5.1, a concentração do hidrolisado produziu uma
elevação na quantidade de xilose (443%), glicose (408%) e arabinose (365%),
além do ácido acético (28%). Apesar do ácido acético ser um composto tóxico
aos microrganismos, FELIPE et a/.(1995) relataram que somente valores acima
de 3 g/L podem afetar negativamente na fermentação da xilose por Candida
g uilliermondii.
5.2 - ESTUDO DO ROMPIMENTO DAS CÉLULAS DE Candida guil/iermondii
O rompimento celular é uma operação unitária importante na obtenção
de produtos intracelulares de microrganismos, sendo os métodos físicos os
mais comumente utilizados. Em estudos da etapa de rompimento celular, a
influência de fatores como concentração celular, tempo de rompimento,
agitação, volume de esferas de vidro, pressão, potência, são freqüentemente
avaliados de acordo com o método a ser empregado. Além disso, para o
monitoramento da eficiência do rompimento celular, parâmetros como
concentração total de proteínas solúveis ou enzima intracelular liberada,
tamanho das células, número de células viáveis e concentração celular, têm
sido avaliados (CURRIE et ai., 1972; FOLLOWS et ai., 1971; AGERKVIST &
ENFORS, 1990; KULA et ai., 1990; MELENDRES et ai., 1993).
No presente trabalho, experimentos foram conduzidos visando à determinação das condições de rompimento de Candida guilliermondii,
utilizando esferas de vidro sob agitação em vórtice, variando o tempo de
rompimento e a concentração celular da suspensão para que eficiente
liberação da enzima glicose-6-fosfato desidrogenase pudesse ser alcançada. A
eficiência do método de rompimento foi estimada através da análise dos
parâmetros concentração celular medida por densidade ótica, DO&X>, e também
pelo número de células intactas, utilizando Câmara de Neubauer; além da
quantidade de proteínas solúveis e da enzima glicose-6-fosfato desidrogenase.
Tal etapa foi realizada com repetição para que a reprodutibilidade do método
também pudesse ser avaliada.

35
A Figura 5.1 apresenta a porcentagem de células rompidas em função
da concentração das suspensões celulares nos tempos de rompimento 1, 3 e 5
minutos. Nota-se que no tempo de 1 minuto, a porcentagem de células
rompidas estimada pela densidade ótica não mostrou significante variação com
a concentração celular, alcançando um valor médio de 44% de células
rompidas. De modo diferente, a porcentagem de células rompidas estimada
pelo número de células intactas neste mesmo tempo, apresentou aumento
acentuado a partir da concentração celular de 11,6 g/L, alcançando 67% de
rompimento celular com 35,4 g/L de células, o que pode ser explicado por um
possível erro experimental presente na contagem de células em Câmara de
Neubauer, uma vez que suspensão celular mais concentrada implica em maior
diluição para a leitura.
Considerando o tempo de 3 minutos, observa-se que a porcentagem de
células rompidas, estimada tanto pela densidade ótica quanto pelo número de
células intactas, apresentou comportamento parecido ao observado para o
tempo de 1 minuto. Entretanto os valores de células rompidas alcançados
foram maiores.
Para o tempo de 5 minutos de agitação em vórtice, pode-se observar
que a concentração celular não influenciou significativamente na porcentagem
de células rompidas, já que os valores de porcentagem mostraram pouca
variação quando estimados pela densidade ótica bem como pelo número de
células intactas. Vale ressaltar que os erros-padrão mostrados na Figura 5.1
para o tempo de 5 minutos foram menores se comparados aos outros tempos.
TAUBERT et ai. (2000) observaram que o aumento da concentração celular
promoveu um decréscimo na eficiência do rompimento de Ganoderma
applanatum em moinho de esferas, embora o aumento da biomassa não tenha
influenciado no rompimento de Pycnoporus cinnabarinus. No presente trabalho,
as concentrações celulares empregadas promoveram pouca variação na
porcentagem de células rompidas. Esses resultados sugerem que a influência
da concentração celular sobre a eficiência de rompimento, depende do tipo de
microrganismo estudado.
Os resultados das porcentagens de células rompidas em função do
tempo, empregando diferentes concentrações celulares, estão apresentadas na
Figura 5.2. A porcentagem de células rompidas variou com o tempo de

36
rompimento, tanto para as células rompidas estimadas por DOro:i quanto para
as estimadas pela contagem de células intactas em Câmara de Neubauer.
Nota-se, na Figura 5.2, que o aumento do tempo de agitação em vórtice
favoreceu a eficiência de rompimento, apesar da concentração celular de
5,9 g/L ter atingido um valor de porcentagem de células rompidas maior (DOEOJ)
após 3 minutos quando comparado com o tempo de 5 minutos. De um modo
geral, o tempo de 5 minutos demonstrou ser o mais adequado para a obtenção
de um eficiente rompimento celular, haja vista que os maiores valores de
porcentagens de células rompidas foram alcançados nesse tempo.
RICCI-SILVA et ai (2000) analisaram a influência da concentração
celular inicial, agitação e do tempo na porcentagem de células de
Saccharomyces cerevisiae rompidas em moinho de esferas. Segundo estes
autores, o aumento do tempo de rompimento de 3 a 15 minutos,
independentemente da concentração inicial da suspensão celular, mostrou
favorecer no rompimento dessas células até o tempo de 9 minutos, a partir do
qual a porcentagem de células rompidas permaneceu praticamente constante.
Estes autores constataram ainda que, empregando-se uma concentração
celular de 170 g/L e agitação de 2300 rpm, o tempo de 6 minutos foi suficiente
para que 83% das células fossem rompidas. No presente trabalho, a maior
porcentagem de células rompidas, 78% (em número de células intactas) e 69,5
± 8,5% (DOro:i), foi alcançada com uma concentração celular de 35,4 g/L e 5
minutos de agitação em vórtice. De acordo com MARFFY & KULA (1999),
porcentagens de rompimento variando de 60 a 80% são satisfatórias para a
obtenção de enzimas por leveduras.
RICCI-SILVA et ai. (2000), concluíram que observações feitas em
câmara de Neubauer com o auxílio do microscópio não são necessariamente
um bom critério para a contagem de células rompidas, além de não
confirmarem se a membrana citoplasmática foi afetada na mesma proporção.
KUBOI et ai (1995), durante o estudo do rompimento de células de Escherichia
coli em ultrassom visando à obtenção da enzima ~-galactosidase, avaliaram o
parâmetro concentração celular medido pela densidade ótica como uma
estimativa da eficiência do método de rompimento utilizado. Esse parâmetro

37
80 11 minuto\
70
~60 I! ~; UJ -ig 50 "ã. f7'!~i><~ E 40 o ..... ~ 30
~ 20 o .l 10
o o 5 10 15 20 25 30 35 40
80 e minutos I 70 ~- ~a) ~,V i UJ
~ 50 o.. E 40 o l ..... UJ 30 (11
::i :âj 20 o
10
o o 5 10 15 20 25 30 35 40
80 ~ minutos \
~:~~~-=-1 -ig 50 l "ã. E 40 e UJ 30 (11
's =ai 20 o
10
0-1--~~~~~~~~~~~~~~ O 5 10 15 20 25 30 35 40
Concentração celular (gil)
Figura 5.1. Porcentagem de células rompidas em função da concentração das
suspensões celulares empregando tempos de agitação em vórtice
de 1, 3 e 5 minutos. Número de células intactas (- • -) e DOEm (-•-)

38
! D Contagem D 00600rm
100 90 80 70 60 50
40 30
20 10 0+-~'-r-'~-.-~.........-~---.~~........-~
100 90 80
[ 70 :Q 60 u "ã. 50 E e 40 UI
~ 30 =o; (.) 20
10
0+----'-'-r-'-'--.-----'-........,__.__.___,~~ .......... ~
X= 1,2 g/L
3 5
100 90 80
~ 70 UI .jg 60 "ã. 50 E e 40 UI
~ 30
~ 20 10 0+-~'-r-'~-.-~.........-~---.~~........-~
X= 5,9 g/L
3 5
80 70
X - 221 gil
~ 60 ~ :Q 50 u "ã. 40 E e UI 30 1'11
~ 20 ~
-
----, e-- ~
e , ......
.·
10 o
3
Tempo (min) 5
X= 3,2 g/L
3 5
100 90 80 70 60 50
40 30
20 10 0+-~'-r-'~~~.........-~---.~~........-~
X= 11,6 gil
3
X= 35,4 g/L
5
100 90 80 70 60 50
40 30
20 10 o..-~ ........ ~-.-~....,..~~~......,~~
3
Tempo (min) 5
Figura 5.2. Porcentagem de células rompidas em função do tempo de
rompimento empregando diferentes concentrações celulares

39
provou ser eficaz na determinação da extensão do rompimento celular, sendo
uma medida simples e rápida. Entretanto, a medida da concentração celular
pela DOero (% de células rompidas) deve provar a existência de uma relação
entre densidade ótica e produto liberado (FOSTER, 1995). Com base nos
trabalhos citados anteriormente, a eficiência do método empregado foi
avaliada, em etapas posteriores, pelo parâmetro concentração celular medida
através da densidade ótica, o qual, além de ter demonstrado facilidade e
rapidez, foi semelhante à concentração celular medida pela contagem do
número de células intactas. Para que pudesse ser efetuada uma análise mais
criteriosa da porcentagem de células rompidas, medida pelo número de células
intactas, seria necessário o emprego de microscópio de contraste de fases.
Segundo FOSTER (1995), estas observações podem ser verdadeiras, porém o
rompimento das células deveria, também, ser quantificado por critérios mais
objetivos como análise do tamanho das partículas antes e após o rompimento,
atividade extracelular de uma enzima alvo, dentre outros.
Diferentes autores estudaram a eficiência do rompimento celular através
da liberação de compostos intracelulares como proteínas solúveis e enzimas
(KUBOI et ai., 1995; SONG & JACQUES, 1997; LOVITI et ai., 2000; RICCI-
SILVA et ai., 2000; TAUBERT et ai., 2000; BALASUNDARAM & PANDIT,
2001 ). No presente trabalho, o sobrenadante do extrato celular, obtido das
amostras após o rompimento e centrifugação, foi analisado com o objetivo de
se quantificar a enzima intracelular glicose-6-fosfato desidrogenase (G6PDH).
As Figuras 5.3 a 5.5 apresentam os resultados das análises relacionados às
proteínas totais e às atividades volumétricas e específicas da G6PDH nos
tempos de rompimento 1, 3 e 5 minutos, respectivamente, empregando-se
diferentes concentrações celulares.
A Figura 5.3 mostra a liberação de proteínas e da enzima G6PDH no
tempo de rompimento de 1 minuto. Observa-se que a liberação de proteínas
totais como também a da enzima G6PDH (atividade volumétrica) aumentou
com o aumento da concentração da suspensão celular proporcionando uma
maior liberação destes compostos intracelulares presentes no extrato livre de
células. Neste tempo os valores de proteínas totais e de atividade volumétrica
alcançados foram 1736 mg/L e 1022 U/L, respectivamente. Entretanto, este
comportamento não foi observado para a atividade específica desta enzima.

40
Nota-se que o aumento acentuado na atividade específica (0,47 U/mgproteína)
ocorreu até a concentração celular de 5,9 g/L, sugerindo que esse tempo de
agitação em vórtice tenha sido insuficiente para adequada liberação da mesma.
Os resultados referentes ao tempo de 3 minutos estão apresentados na
Figura 5.4. Nota-se que tanto a quantidade de proteínas totais quanto a
atividade volumétrica de G6PDH foram aumentadas até a concentração de
22, 1 g/L, atingindo um valor de 2505 mg/L e 1578 U/L, respectivamente, as
quais se mantiveram nestes níveis na concentração celular de 35,4 g/L.
Observa-se, ainda, que a concentração celular influenciou na atividade
específica da enzima somente em suspensões mais diluídas, ou seja, até 5,9
g/L, a partir da qual a atividade específica da enzima permaneceu praticamente
constante, em torno de 0,65 U/mgproteina-
Com relação ao tempo de 5 minutos (Figura 5.5), verifica-se que o
comportamento da liberação de proteínas e da atividade volumétrica foi
semelhante ao observado para o tempo de 1 minuto. Neste tempo os maiores
valores de proteínas totais, 4157 mg/L, e atividade volumétrica, 2757 U/L,
foram obtidos com a concentração celular de 35,4 g/L. Nota-se, ainda na Figura
5.5, que a atividade específica alcançou um valor máximo de 0,96 U/mQproteina
com 11,6 g/L de células, a partir da qual houve um decréscimo em cerca de
31 % quando a maior concentração celular foi empregada, 35,4 g/L. Tal fato
pode ser explicado pelo pronunciado aumento da quantidade de proteínas
totais liberadas no extrato. Estes resultados demonstram que, apesar de
maiores quantidades de proteínas terem sido solubilizadas no tempo de
agitação em vórtice de 5 minutos, esse fator promoveu um decréscimo na
atividade específica de G6PDH, sugerindo uma possível desnaturação dessa
biomolécula ou liberação de outras proteínas para esse tempo, a partir da
suspensão celular de concentração 11,6 g/L. FOLLOWS et ai (1971 ),
observaram que durante o rompimento de células de levedura por um
homogeneizador de alta pressão, o comportamento da liberação de enzimas
apresentou-se dependente de sua localização. A maior parte de glicose-6-
fosfato desidrogenase está localizada no citoplasma e sua liberação durante o
rompimento celular é instantânea. ÕZBEK & ÜLGEN (2000), em estudo da
estabilidade de enzimas após sonificação, observaram que a enzima G6PDH.
., -· ,._
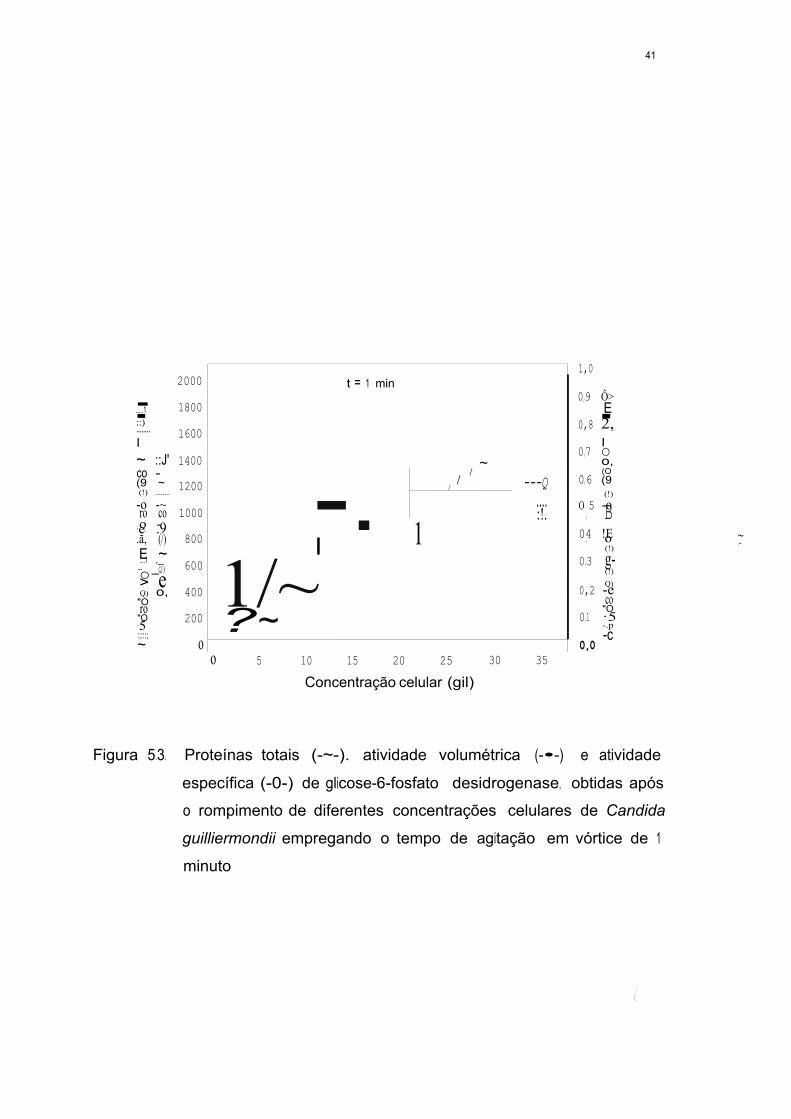
41
1,0 2000 t = 1 min -
~
0,9 Ô> .....1 1800 E - - ::) 0,8 2, ........ 1600 I I ~ ::J' 1400 0,7 O
o, co - / (O (9 ~ 1200 -. / ---Q 0,6 (9
/ (!) ........ (!) -o -~ .... O 5 -o ro co 1000 :!. . B o - 1 ·e .9 04 !E .ã, (/) 800 I . o ~ E ~ (!)
,-
::::1 ,_ 600 1/~ 0,3 g- - Q) (!) o- > e Q) Q) o, 400 0,2 -e "O co ro ?~ "O "O 200 0,1 ·5 ·5 -.p :;:::; -c ~ o º·º o 5 10 15 20 25 30 35
Concentração celular (gil)
Figura 5.3. Proteínas totais (-~-). atividade volumétrica (-•-) e atividade
específica (-0-) de glicose-6-fosfato desidrogenase, obtidas após
o rompimento de diferentes concentrações celulares de Candida
guilliermondii empregando o tempo de agitação em vórtice de 1
minuto
/ (

I o c-, a. ...J <X) õ, l9 E (l) ........,
"C .~ cu cu o +' ·e .9 :a, (/) E ~ ::J ·- - Q) o +' > e Q) a.
"C cu "C ·5 ~
42
t = 3 min 3000 ~--------------------.--1,0
900
600
300
O-+.-.::'-.---,.-~~~~-~~~-~....-~---~ ........ o.o o
2700
2400
35
,,....., 0,9 0>
.ê 0,8 2
I 0,7 O a.
(O 0,6 (.9
Q) "C
0,5 ~
04 ~ . o (l) o.
0,3 fil Q)
0,2 -o cu "C
0,1 ·5 :.::; <(
2100
1800
1500
1200
5 10 15 20 25 30
Concentração celular (gil)
Figura 5.4. Proteínas totais (-Ã-), atividade volumétrica (-•-) e atividade
específica (-0-) de glicose-6-fosfato desidrogenase, obtidas após
o rompimento de diferentes concentrações celulares de Candida
guillirmondii empregando o tempo de agitação em vórtice de 3
minutos

::J' -- ::::> .......... J: o ......... a.. _J co -- (9 E Q) .......... "O !/)
<ll g .g .9 :ã, !/)
E ~ ::J ·- - Q) o ..... > e Q) a..
"O <ll "O ·s: ~
3000 2500 2000 1500 1000 500
o -+="C...,...---,--..---,-----.---,.---~~----.--..----.------,----,--..--..,..._ o.o o
4500 t = 5 rnin
35
1.4 ......... O)
E -- 1,2 2, J: o
1,0 O.. (O (9
0,8 i s
ti= 0,6 'õ
Q) a. C/)
0.4 Q) Q) "O <ll
0,2 ~ :.:; <(
43
Figura 5.5. Proteínas totais (-À-), atividade volumétrica (-•-) e atividade
específica (-0-) de glicose-6-fosfato desidrogenase, obtidas após
o rompimento de diferentes concentrações celulares de Candida
guilliermondii empregando o tempo de agitação em vórtice de 5
minutos
4000 3500
5 10 15 20 25 30 Concentração celular (g/L)

44
apresentou menores valores de estabilidade nesse processo quando
comparada as enzimas álcool desidrogenase, maiato desidrogenase, L-lactato
desidrogenase, fosfatase alcalina e a (3-galactosidase (AP > pG > LDH > MDH
> ADH > G6PDH) sugerindo que o perfil e o grau de degradação das mesmas
variam de acordo com a enzima além da sua localização na célula.
Como discutido anteriormente, as proteínas totais liberadas mostraram-
se dependentes da concentração celular inicial da suspensão, em todos os
tempos. LOVITI et ai (2000), em estudo do rompimento de células de
Saccharomyces cerevisiae em compartimento sob pressão, também
investigaram o efeito da concentração celular na liberação de proteínas
utilizando concentrações entre 2 e 90 g/L submetidas a uma pressão de 170
MPa. Estes autores observaram que, inicialmente, até cerca de 30 g/L o
aumento da proteína liberada foi proporcional ao aumento da concentração
celular porém acima desta concentração, a quantidade de proteína não
mostrou ser proporcional, sendo que, com 90 g/L a concentração de proteínas
presentes no extrato foi 15% menor do que em suspensões celulares mais
diluídas. No presente trabalho, comportamento semelhante também foi
observado, ou seja, a relação entre proteínas liberadas foi proporcional até a
concentração celular de 35,4 g/L, em todos os tempos estudados.
Nesta etapa do trabalho, os maiores valores de atividades específicas da
G6PDH foram alcançadas no tempo de 5 minutos sendo a máxima, 0,96
U/mgprotefna, obtida na suspensão celular de concentração 11,6 g/L.
O estudo da etapa de rompimento mostrou-se importante, visto que o
mesmo possibilitou a determinação de condições adequadas a serem
aplicadas, posteriormente, no processo fermentativo visando a produção da
enzima glicose-6-fosfato desidrogenase. Porém, durante o processo
fermentativo, a baixa formação de biomassa e os pequenos volumes de
amostras retiradas com o tempo constitui um fator limitante para utilização de
uma concentração celular de 11,6 gil. Desta forma, optou-se pelo emprego da
concentração de 3,0 g/L para o estudo da influência das variáveis pH do meio,
concentração inicial de xilose e nível de inóculo sobre a produção da enzima.
Nesta concentração celular (3 g/L) obteve-se porcentagem de células rompidas
de 56,5 ± 4,5% e atividade específica de 0,72 U/mgprotelna .. Esse valor de
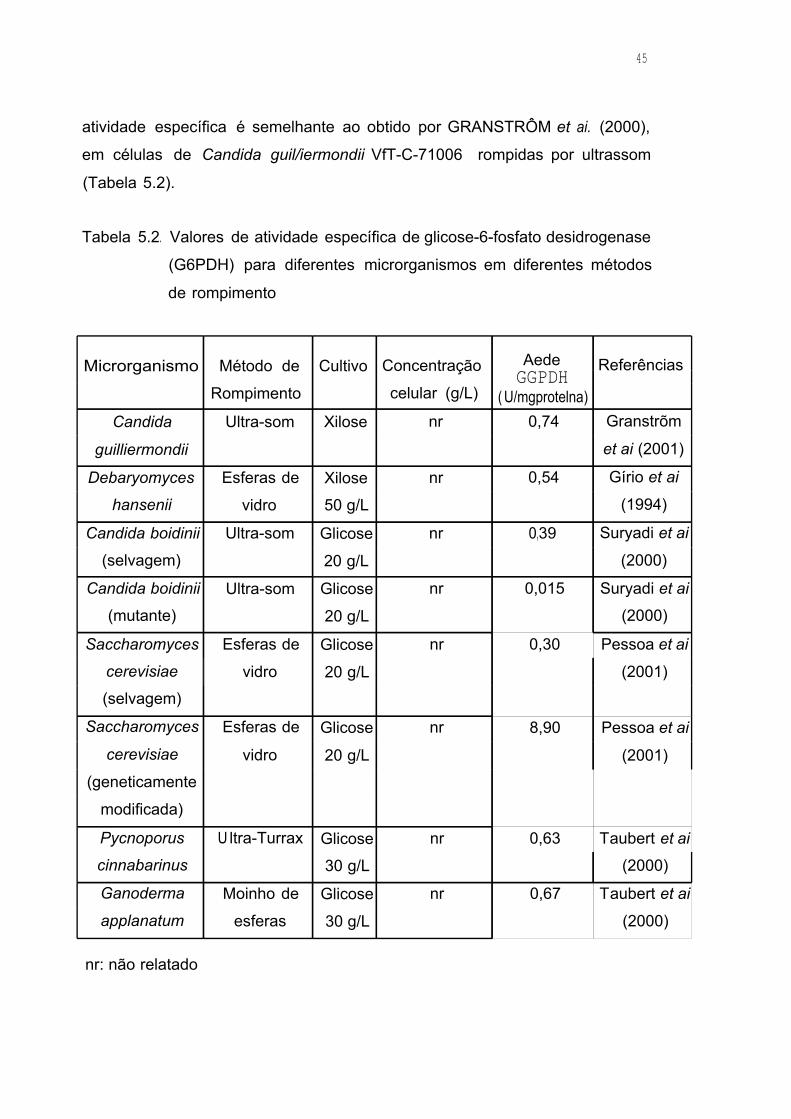
45
atividade específica é semelhante ao obtido por GRANSTRÔM et ai. (2000),
em células de Candida guil/iermondii VfT-C-71006 rompidas por ultrassom
(Tabela 5.2).
Tabela 5.2. Valores de atividade específica de glicose-6-fosfato desidrogenase
(G6PDH) para diferentes microrganismos em diferentes métodos
de rompimento
Microrganismo Método de Cultivo Concentração Aede Referências GGPDH Rompimento celular (g/L) ( U/mgprotelna)
Candida Ultra-som Xilose nr 0,74 Granstrõm
guilliermondii et ai (2001)
Debaryomyces Esferas de Xilose nr 0,54 Gírio et ai
hansenii vidro 50 g/L (1994)
Candida boidinii Ultra-som Glicose nr 0,39 Suryadi et ai
(selvagem) 20 g/L (2000)
Candida boidinii Ultra-som Glicose nr 0,015 Suryadi et ai
(mutante) 20 g/L (2000)
Saccharomyces Esferas de Glicose nr 0,30 Pessoa et ai
cerevisiae vidro 20 g/L (2001)
(selvagem)
Saccharomyces Esferas de Glicose nr 8,90 Pessoa et ai
cerevisiae vidro 20 g/L (2001) (geneticamente
modificada)
Pycnoporus U ltra-Turrax Glicose nr 0,63 T aubert et ai
cinnabarinus 30 g/L (2000)
Ganoderma Moinho de Glicose nr 0,67 T aubert et ai
applanatum esferas 30 g/L (2000)
nr: não relatado

46
5.3 - ESTUDO DA INFLUÊNCIA DE VARIÁVEIS DO PROCESSO FERMENTATIVO NA
PRODUÇÃO DE GLICOSE-6-FOSFATO DESIDROGENASE
O estudo da influência das variáveis do processo fermentativo sobre a
produção da enzima glicose-6 fosfato desidrogenase foi baseado no
planejamento experimental, conforme matriz apresentada na Tabela 4.1
presente no item 4.6.3.
O consumo de D-xilose, a concentração celular e a produção de xilitol e
glicose-6-fosfato desidrogenase, observados em diferentes condições de pH,
concentração inicial de D-xilose e nível de inóculo, nos diversos tempos de
fermentação por Candida guilliermondii estão mostrados nas Figuras 5.6 a 5.8.
A Figura 5.6 representa os ensaios com concentração inicial de D-xilose
de 30 g/L (ensaios 1, 2, 5 e 6 da matriz do planejamento fatorial 23). Como
pode ser visto na Figura 5.6 (A), a D-xilose presente no meio foi
completamente consumida ao final de 48 horas em todas as condições
estudadas. Entretanto, nas condições nível de inóculo de 3 g/L e pH 4,5 ou 7,5,
foram observadas as maiores velocidades de consumo deste açúcar, sendo
consumida cerca de 94% da xilose após 36 horas de fermentação. As
concentrações residuais de xilose nos ensaios com nível de inóculo de 1 g/L e
de 3 g/L foram da ordem de 1,5 g/L e 0,2 g/L, respectivamente, após 48 horas
de cultivo.
O crescimento celular demonstrou ser influenciado principalmente pelo
pH, sendo as maiores concentrações celulares obtidas em pH 7,5. O aumento
no nível de inóculo também favoreceu o crescimento celular quando o pH do
meio variou de 4,5 para 7,5 (Figura 5.6 B). Por outro lado, ao final da
fermentação, a concentração de xilitol presente no meio foi semelhante em
todos os ensaios independentemente das condições avaliadas. Vale ressaltar
que para as fermentações conduzidas no maior nível de inóculo a produção
máxima de xilitol foi alcançada após 36 horas de cultivo (Figura 5.6 C).
Com relação à enzima glicose-6-fosfato desidrogenase (Figura 5.6 D),
observa-se que quando o pH foi elevado de 4,5 para 7,5, nos ensaios com 1
g/L de inóculo, a atividade volumétrica de G6PDH passou de 167 para 253 U/L
(51%). Um aumento mais acentuado foi observado quando o inóculo passou de

35 A 8 7
30 6
25 5 ::::i' ::::i' § 20 ~4 (!} cn cn l\l o 15 ::i x :V 3 o
10 2
5
o o o 10 20 30 40 50 o 10 20 30 40 50
20 e D ... 300
15 250
:::]' 2' 200 § 10 2.
I o ~ 150 ~ x (O (9
5 100
50 o o o 10 20 30 40 50 o 10 20 30 40 50
Tempo (h) Tempo (h)
47
Figura 5.6. Consumo de D-xilose (A) e produção de células (8), xilitol (C),
glicose-6-fosfato desidrogenase (D) por Candida guil/iermondii
empregando hidrolisado hemicelulósico com concentração inicial
de D-xilose de 30 g/L sob diferentes condições de cultivo. Ensaio
1: inóculo de 1 g/L e pH inicial de 4,5 (-•-), ensaio 2: inóculo 1 g/L
e pH 7,5 (-0-), ensaio 5: inóculo 3 g/L e pH 4,5 (-•-), ensaio 6:
inóculo 3 g/L e pH 7,5 (-V-)

48
1 g/L para 3 g/L em pH 4,5, alcançando 90%, enquanto que um aumento de
apenas 26% foi alcançado em pH 7,5.
A Figura 5. 7 apresenta os resultados referentes aos ensaios conduzidos
com concentração inicial de 70 g/L de xilose. Com base nesta Figura (5. 7 A),
verifica-se que nas condições 3 g/L de inóculo e pH 4,5 a xilose presente no
meio de cultivo não foi totalmente consumida, apresentando um residual de
15,30 g/L, no entanto, quando o pH do cultivo passou de 4,5 para 7,5 houve um
aumento na velocidade de consumo desse açúcar além de seu esgotamento
do meio após 96 horas de fermentação. Neste mesmo tempo, os ensaios 3 e 4
apresentaram consumo percentual médio de xilose de 97%.
A produção de células (Figura 5.7 8) também foi fortemente influenciada
pela variação de pH do meio na condição de 3 g/L de inóculo, uma vez que a
elevação no pH de 4,5 para 7,5 favoreceu o aumento na concentração celular
em 54%.
Na Figura 5. 7 C nota-se que, da mesma forma que para o consumo de
xilose e a produção de células, as condições nível de inóculo de 3 g/L e pH 4,5
não foram favoráveis a produção de xilitol. É interessante observar que para o
pH 4,5, o aumento do inóculo afetou negativamente a produção de xilitol a qual
decresceu de 38,50 para 24,99 g/L (ensaios 3 e 7 da matriz do planejamento
fatorial). Tal comportamento não pode ser explicado pela inibição do
hidrolisado em baixo valor de pH conforme constatado em outros trabalhos,
uma vez que no inóculo de 1 g/L e pH 4,5 a produção de xilitol foi semelhante à de 1 g/L e pH 7,5.
Com relação a enzima glicose-6-fosfato desidrogenase (Figura 5.7 D)
verfica-se que os maiores valores de atividade volumétrica foram obtidos
quando as fermentações foram realizadas nos níveis superiores das variáveis
pH e concentração de inóculo (7,5 e 3 g/L, respectivamente). Nestes ensaios, a
variação do pH de 4,5 para 7,5 promoveu um aumento de 207% na atividade
volumétrica de G6PDH.
Na Figura 5.8 A, B, C e D, estão apresentados os ensaios conduzidos
com 50 g/L de D-xilose, 2 g/L de inóculo e pH 6,0. Nota-se que a D-xilose foi
praticamente consumida com 60 horas de fermentação. Nesses ensaios, ao
final da fermentação, obteve-se concentração média de 7 ,2 g/L em células e

80 A 10 B
60 8
,......_ ,......_ 6 ....J ~ 40 -- C) C) - - (1) (/)
(/) <ti o "S 4
>< 20 •(I) o
2 o
o o 20 40 60 80 100 o 20 40 60 80 100
50 e D 500
40 400
30 ,......_ ,......_ ....J ....J -- -- :::, 300 C) - - I - 20 =ª o o, x (O 200 o
10 100
o o
o 20 40 60 80 100 o 20 40 60 80 100
Tempo (h) Tempo (h)
49
Figura 5.7. Consumo de D-xilose (A) e produção de células (B), xilitol (C),
glicose-6-fosfato desidrogenase (D) por Candida guilliermondii
empregando hidrolisado hemicelulósico com concentração inicial
de D-xilose de 70 g/L sob diferentes condições de cultivo. Ensaio
3: inóculo de 1 g/L e pH inicial de 4, 5 (- • -), ensaio 4: inóculo 1 g/L
e pH 7,5 (-0-),ensaio 7: inóculo 3 g/L e pH 4,5 (-Ã-), ensaio 8:
inóculo 3 g/L e pH 7,5 (-V-)

60 A B
50 8
40 ,......_ ,......_ 6
_J ....J -- õ, 30 O) .._., - (!) V,
ro V, 3 o 20 x •(!) 4 o 10
2 o
o 10 20 30 40 50 60 o 10 20 30 40 50 60
30 e 400 D
25 350
20 300 ,......_
,......_ _J _J -- Ô> 15 2 250 .._.,
]: o ~ 200 ~ 10 x (O C)
150 5 100
o 50
o 10 20 30 40 50 60 o 10 20 30 40 50 60 Tempo (h) Tempo (h)
50
Figura 5.8. Consumo de 0-xilose (A) e produção de células (B}, xilitol (C),
glicose-6-fosfato desidrogenase (D) por Candida guilliermondii
empregando hidrolisado hemicelulósico com concentração inicial
de D-xilose de 50 g/L sob diferentes condições de cultivo. Ensaios
9, 1 O e 11: inóculo de 2 g/L e pH inicial de 6,0, ensaios do ponto
central(-•-}, (-0-), (-.A.-)

51
29,2 g/L em xilitol. A produção da enzima variou de 263 a 379 U/L o que pode
ser atribuído à variação na concentração inicial de inóculo nestes ensaios.
Os resultados referentes ao consumo percentual dos açúcares bem
como os parâmetros fermentativos fator de conversão de xilose em xilitol e
atividade especícica de G6PDH estão mostrados na Tabela 5.3.
Observou-se que em todos os ensaios a glicose foi totalmente
consumida antes de 12 horas de fermentação e que o consumo da arabinose
foi iniciado somente após esgotamento da xilose (Apêndice A.3)).
Vale ressaltar que o consumo de arabinose foi influenciado pelas
condições de cultivo empregadas (Tabela 5.3). Verifica-se que os maiores
percentuais de consumo deste açúcar ocorreram nos ensaios com maior nível
de inóculo, com exceção do ensaio 7 (maior concentração de xilose e menor
pH inicial), onde não se observou consumo desta pentase o que provavelmente
foi devido à presença de um grande residual de xilose no meio. O maior
consumo de arabinose (81 %) foi obtido nas condições do ensaio 5, ou seja, pH
4,5 e nível de inóculo de 3 g/L.
A Tabela 5.3 também mostra os valores dos parâmetros fator de
conversão de xilose em xilitol e de atividade específica da G6PDH. Os valores
de Y Pts nos ensaios conduzidos com 1 g/L de inóculo ( ensaios 1 a 4) variaram
de 0,59 a 0,66 g/g e de atividade específica variaram de 0,28 a 0,62
U/mgproteína- Nota-se, ainda, que nas condições do ensaio 4 (pH 7 ,5, 70 g/L de
xilose e 1 g/L de inóculo) foram obtidos os valores máximos para ambas
respostas. O mesmo comportamento foi observado para os ensaios realizados
com 3 g/L de inóculo (ensaios 5, 6, 7 e 8). Nestas fermentações os valores de
Yp1s variaram de 0,53 a 0,74 g/g e de atividade específica de 0,31 a 0,40
U/mQproteína sendo observado que o maior Yp1s e atividade específica foram obtidos no ensaio 8.
Com relação aos ensaios do ponto central (ensaios 9, 10 e 11 ), os
valores médios de Yp1s e AE foram de 0,65 g/g e 0,42 U/mQproteína, respectivamente.
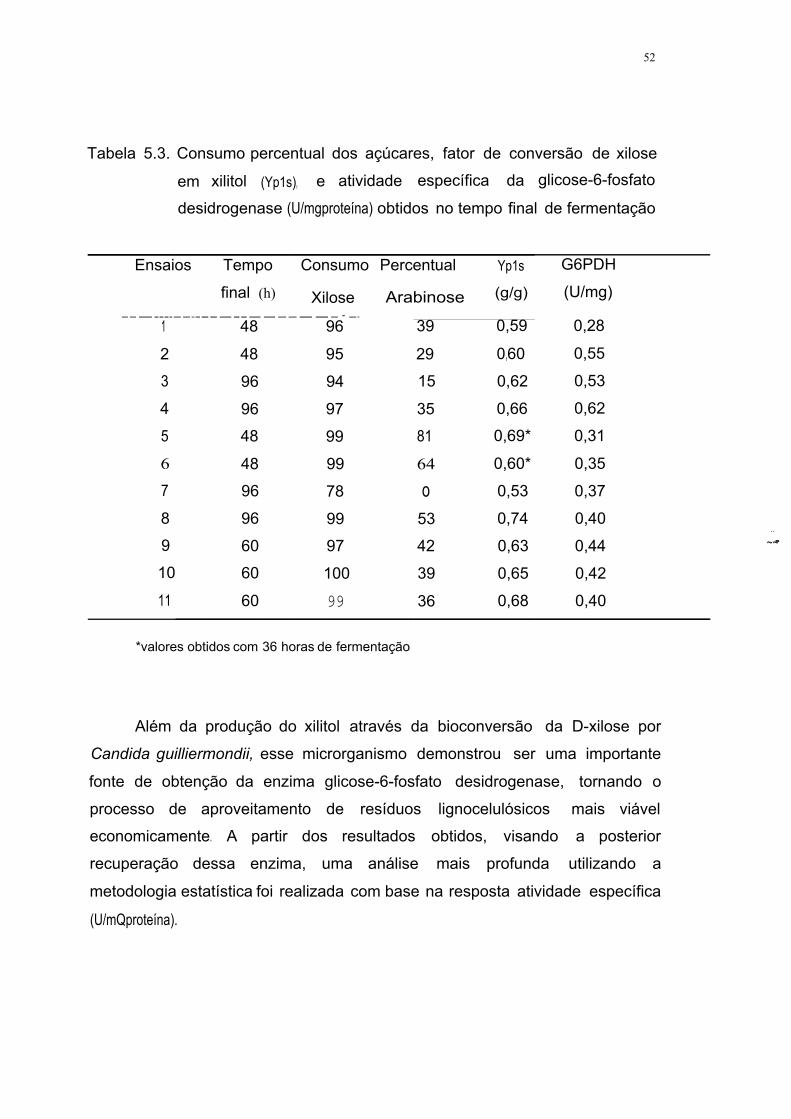
52
Tabela 5.3. Consumo percentual dos açúcares, fator de conversão de xilose
em xilitol (Yp1s), e atividade específica da glicose-6-fosfato
desidrogenase (U/mgproteína) obtidos no tempo final de fermentação
Ensaios Tempo Consumo Percentual Yp1s G6PDH
final (h) Xilose Arabinose (g/g) (U/mg) ·-----------------------·--
1 48 96 39 0,59 0,28
2 48 95 29 0,60 0,55
3 96 94 15 0,62 0,53
4 96 97 35 0,66 0,62
5 48 99 81 0,69* 0,31
6 48 99 64 0,60* 0,35
7 96 78 o 0,53 0,37
8 96 99 53 0,74 0,40 ..
9 60 97 42 0,63 0,44 - ~~
10 60 100 39 0,65 0,42
11 60 99 36 0,68 0,40
*valores obtidos com 36 horas de fermentação
Além da produção do xilitol através da bioconversão da D-xilose por
Candida guilliermondii, esse microrganismo demonstrou ser uma importante
fonte de obtenção da enzima glicose-6-fosfato desidrogenase, tornando o
processo de aproveitamento de resíduos lignocelulósicos mais viável
economicamente. A partir dos resultados obtidos, visando a posterior
recuperação dessa enzima, uma análise mais profunda utilizando a
metodologia estatística foi realizada com base na resposta atividade específica
(U/mQproteína).

53
5.4 - ANÁLISE ESTATÍSTICA DO EFEITO DA CONCENTRAÇÃO INICIAL DE XILOSE,
PH E NÍVEL DE INÓCULO NA PRODUÇÃO DA ENZIMA G6PDH E DEXILITOL
Na Tabela 5.4 estão apresentados os resultados em atividade específica
da enzima glicose-6-fosfato desidrogenase e produção máxima de xilitol
obtidos nos ensaios realizados com base no planejamento fatorial 23. De
acordo com estes resultados, pode-se observar que a menor atividade
específica, 0,28 U/mgproteína, foi obtida no ensaio 1 onde foram empregados os
menores níveis de pH inicial, concentração de xilose e nível de inóculo.
Observa-se ainda que a elevação simultânea nos níveis das variáveis pH
inicial, concentração de xilose e nível de inóculo ( ensaio 8) promoveu um
aumento de 43% nos valores de atividade específica de G6PDH e que esta se
mostrou fortemente influenciada pelo nível de inóculo, sendo que no menor
nível, 1 g/L (ensaios 2, 3 e 4), foram obtidos os maiores valores para esta
resposta, com exceção do ensaio 1. Nesta condição, a variação do pH de 4,5
para 7 ,5 promoveu um aumento de 96% quando foi empregada uma
concentração de xilose de 30 g/L e de 17% com 70 g/L de xilose. No entanto,
quando o nível de inóculo foi de 3 g/L a elevação do pH promoveu um aumento
em torno de 11 %, para ambas concentrações de xilose avaliadas. Estas
observações indicam que o pH inicial e o nível de inóculo influenciaram
fortemente na produção da glicose-6-fosfato desidrogenase.
Para a resposta produção máxima de xilitol (Tabela 5.4), observa-se que
para os ensaios conduzidos com 30 g/L de xilose (ensaios 1, 2, 5 e 6), a
concentração final do produto apresentou pouca variação (de 16,54 a 18,57
g/L). Estes resultados sugerem que, na região estudada, as variáveis pH e
nível de inóculo não influenciaram na formação do xilitol. Nos ensaios 3, 4, 7 e
8, realizados a 70 g/L de xilose, a concentração de xilitol foi de 38,50, 40, 1 O,
24,99 e 43,95 g/L, respectivamente. Nota-se que com exceção do ensaio 7, a
produção final de xilitol foi similar nas condições de pH e concentração de
inóculo avaliadas. Estes dados sugerem a existência de um efeito positivo da
variável concentração de xilose, na região investigada, sobre esta resposta.

54
A influência das variáveis investigadas, seus efeitos principais bem como
a existência de efeitos de interação entre elas, pode ser melhor avaliada
empregando-se a análise estatística.
A Figura 5.9 A e B mostram o gráfico de Pareto o qual representa os
efeitos estimados, em ordem decrescente de magnitude, das variáveis pH
inicial, concentração de xilose e nível de inóculo e suas interações nas
respostas atividade específica de G6PDH e produção máxima de xilitol. O
tamanho de cada barra é proporcional ao efeito estimado. A linha vertical é
utilizada para avaliar quais dos efeitos são estatisticamente significativos no
intervalo de confiança de 95%.

55
Tabela 5.4. Produção máxima de xilitol (Pxi1tto1) e atividade específica de G6PDH
(U/mgproteína) obtidas no planejamento fatorial 23 para avaliação da
influência do pH inicial, concentração de xilose e nível de inóculo na
produção da enzima glicose-6-fosfato desidrogenase por Candida
guilliermondii cultivada em hidrolisado hemicelulósico de palha de
arroz
Ensaio Variáveis Respostas
pH Concentração Nível de U/mgproteína Pxmtol inicial de xilose inóculo (g/L)
(g/L) (g/L)
1 4,5 30 1 0,28 16,54 2 7,5 30 1 0,55 16,73
3 4,5 70 1 0,53 38,50 4 7,5 70 1 0,62 40,10 5 4,5 30 3 0,31 18,57 6 7,5 30 3 0,35 17,07 7 4,5 70 3 0,37 24,99 8 7,5 70 3 0,40 43,95 9 6,0 50 2 0,44 29,06 10 6,0 50 2 0,42 29,04 11 6,0 50 2 0,40 29,45

p= 0,05 A
lnóculo (C) -5,79
Xilose (B)
pH (A)
A*C
B*C
A*B
1,5 2,0 2,5 3,0 3,5 4,0 4,5 5,0 5,5 6,0 6,5 Efeito estimado (Valor absoluto)
p= 0,05 B
B (Xilose) 7,60
A*B
A(pH)
A*C
B*C
e (lnóculo)
o 1 2 3 4 5 6 7 Efeito estimado (Valor absoluto)
56
8 9
Figura 5.9. Gráfico de Pareto para os efeitos das variáveis pH inicial,
concentração de xilose e nível de inóculo nas respostas atividade
específica da enzima G6PDH (A) e produção máxima de xilitol (B)

57
Como pode ser visto (Figura 5.9 A), os efeitos principais nível de inóculo,
C, concentração de xilose, 8, pH inicial, A, além da interação AC (pH e nível de
inóculo) foram significativos. O maior efeito principal observado para essa
resposta foi o da variável nível de inóculo a qual apresentou estimativa de
- 5, 79, seguido pela concentração de xilose com estimativa de + 4,53. Observa-
se ainda que o efeito interativo entre as variáveis A e C apresentou uma
estimativa de sinal negativo indicando que os resultados de atividade
específica são melhorados com a condução dos ensaios em menores valores
de concentrações de inóculo e pH mais elevado, nas respectivas regiões
estudadas. Os resultados desta análise (teste t de Student) confirmam os
efeitos sugeridos pela análise direta dos dados e também a existência de
interação entre as variáveis.
Com relação à resposta produção máxima de xilitol (Figura 5.9 8), nota-
se que apenas o efeito principal concentração de xilose, 8, mostrou-se
significativo para o intervalo de confiança de 95% na região estudada.
No entanto, como o objetivo principal deste trabalho é a produção da
enzima glicose-6-fosfato desidrogenase, seguiu-se o estudo com a resposta
atividade específica de G6PDH, visando obter um modelo matemático que
descreva a influência das variáveis do processo fermentativo sobre esta
resposta.
5.5 - MODELAGEM MATEMÁTICA PARA ESTIMAR A ATMDADE ESPECIFICA DE
GLICOSE-6-FOSFATO DESIDROGENASE EM FUNÇÃO DAS VARIÁVEIS
SIGNIFICATIVAS AO PROCESSO FERMENTATIVO
O primeiro passo para a determinação de um modelo empírico através
da metodologia de superfície de resposta é aproximar a função (f), na região
das variáveis independentes. Se a resposta for bem modelada por uma função
linear, então a função de aproximação é um modelo de primeira ordem. Se
existir curvatura no sistema ou na região de ótimo, então um polinômio de grau
superior, como um modelo de segunda ordem deverá ser utilizado para
aproximar a resposta.

58
Para a obtenção de um modelo matemático que melhor descrevesse a
relação entre as variáveis independentes (pH, nível de inóculo e concentração
inicial de xilose) e a resposta em estudo (atividade específica de glicose-6-
fosfato desidrogenase) procedeu-se uma análise de regressão múltipla, cujos
resultados estão apresentados na Tabela 5.5.
Tabela 5.5. Análise da variância para o modelo representativo da atividade
específica de G6PDH de células de Candida guilliermondii
Fonte de SQ GL MQ F p
Variação ----------·--------·-----------
Modelo 0,10 6 0,017 11,85 0,03**
Curvatura 8,SE-05 1 8,SE-05 0,06 0,83
Residual 4,4E-03 3 1,5E-03
Falta de ajuste 3,6E-03 1 3,6E-03 9,03 0,09*
Erro puro 8,0E-04 2 4,0E-04
Total 0,11 10
.. nível de confiança de 90%; .... nível de confiança de 95%
R2 = 0,959; C.V. = 9,03%
SQ = soma quadrática; GL = graus de liberdade; MQ = média quadrática
Analisando a referida Tabela, verifica-se que a regressão obtida foi
estatisticamente significativa (p< 0,05) e que o modelo não mostrou diferença
significativa para a curvatura (p>0,83) e para a falta de ajuste (p>0,09) ao nível
de 95% de confiança. Isto significa que a diferença entre os valores obtidos
experimentalmente e os valores previstos pelo modelo pode ser explicada
apenas pelo erro experimental. A validade do modelo de 1ª ordem obtido pôde
ser também comprovada pelo elevado coeficiente de determinação, R2 = 0,96,
ou seja o modelo consegue explicar 96% da variação total em torno da média,
sendo que os restantes 4% representam os resíduos. Desta forma, o modelo
matemático que descreve a produção de glicose-6-fosfato desidrogenase em

59
atividade específica, considerando a região em estudo, pode ser expresso pela
seguinte equação:
Onde: Y representa o valor previsto para a atividade específica de G6PDH
(U/mgproteína) e X1, X2, X3 representam os níveis codificados das variáveis pH
inicial, concentração de xilose e nível de inoculo, respectivamente.
A Figura 5.1 O apresenta os valores de atividade específica de G6PDH
previstos pelo modelo em função dos valores experimentais obtidos nos 11
ensaios. Observa-se que a distribuição dos pontos em torno da reta para a
resposta atividade específica de G6PDH se ajustam ao modelo linear.
A superfície de resposta obtida pela equação do modelo que representa
a atividade específica de G6PDH em função das variáveis pH inicial,
concentração de xilose, na região estudada, está mostrada na Figuras 5.11.
Nota-se uma tendência crescente da resposta avaliada com o aumento do nível
do pH e concentração inicial de xilose, empregando-se uma concentração de
inóculo de 1,0 g/L. Através da equação obtida, o modelo prevê uma atividade
específica máxima de 0,64 U/mgproteína empregando-se o hidrolisado com uma
concentração de xilose de 70 g/L, pH 7,5 e um nível de inóculo de 1 g/L,
podendo ocorrer uma variação de 0,55 a O, 72 U/mgproteína, dentro do intervalo
de confiança de 95%. Para a confirmação do modelo obtido foi realizado um
ensaio nestas condições tendo sido alcançado, neste ensaio, uma atividade
específica de 0,60 U/mgproteína- Este resultado demonstra que o modelo se
adequou aos resultados experimentais, descrevendo bem a região em estudo.

60
0,70
0,65
0,60 ,....... O> E 0,55 -- ::) ......... 0,50 (/) o
;'!:::: 0,45 "O (l> ... o. 0,40 (/) (l>
0,35 ... o (IJ > 0,30
0,25
0,20 0,20 0,25 0,30 0,35 0,40 0,45 0,50 0,55 0,60 0,65 0,70
Valores observados (U/mg)
Figura 5.1 O. Gráfico dos valores previstos pelo modelo linear em função dos
valores observados para a resposta atividade específica de
G6PDH

61
o,1
o,6
~ :ro o.s ~ A ,Z O,
:;;;-- o,S ~ ~
Figura 5.11. Atividade específica de glicose-6-fosfato desidrogenase em função
da concentração de xilose e do pH, usando 1 g/L de inóculo

62
6 - CONCLUSÕES
Os resultados obtidos no decorrer do presente trabalho, permitiram tirar
as seguintes conclusões:
../ A porcentagem de rompimento celular de Candida guilliermondii pode
ser estimada de forma simples e rápida através da leitura da densidade
ótica da suspensão, antes e após o rompimento;
../ Na etapa de rompimento, a enzima liberada mostrou ser dependente da
concentração inicial da suspensão celular e do tempo de agitação
empregados;
../ A máxima liberação da enzima G6PDH foi obtida nas condições, tempo
de agitação em vórtice de 5 minutos e concentração celular de 11,6 g/L,
alcançando uma atividade específica de 0,96 U/mgproteina· Suspensões
celulares mais concentradas, ou seja, maiores que 11,6 gil,
promoveram um declínio na atividade específica da glicose-6-fosfato
desidrogenase para este tempo;
../ A produção da glicose-6-fosfato desidrogenase, durante o processo
fermentativo, mostrou ser influenciada por todas as variáveis estudadas,
nomeadamente pH inicial, concentração de xilose e nível de inóculo. A
produção desta enzima foi favorecida com a diminuição do nível de

63
inóculo e com o aumento do pH e concentração inicial de xilose, na
região estudada;
./ Pela metodologia estatística usada foi possível estabelecer um modelo
matemático para descrever a produção de G6PDH, em atividade
específica, por Candida guilliermondii cultivadas em hidrolisado
hemicelulósico de palha de arroz. O modelo previsto para esta reposta,
em função das variáveis estudadas e do efeito interativo entre elas, é
dado pela equação:
y = 0,43 + 0,054XI + 0,054X2 -0,069X3 -0,024XIX2 -0,036XIX3 -0,026X2X3
sendo que, através desta equação, o modelo prevê uma atividade
específica máxima de 0,64 U/mQproterna, podendo ocorrer uma variação de
0,55 a 0, 72 U/mgproteína, dentro do intervalo de confiança de 95%;
./ Para o ensaio de validação do modelo, realizado nas condições de 70
g/L de xilose, pH 7,5 e nível de inóculo de 1 g/L, obteve-se AE=0,60
U/mgproteína, revelando que o modelo proposto é adequado para
descrever essa resposta na região em estudo.

64
7 - PREVISÕES FUTURAS
Para complementar estes estudos, dando continuidade ao processo
biotecnológico de produção da enzima glicose-6-fosfato desidrogenase por
Candida guilliermondii cultivadas em hidrolisado hemicelulósico de palha de
arroz; no trabalho de doutorado pretende-se purificar esta enzima através do
processo de extração líquido-líquido em sistemas de duas fases aquosas
integrado ao rompimento celular desta levedura.

65
8 - REFERÊNCIAS BIBLIOGRÁFICAS
ABRAHÃO-NETO, J.; INFANTl,P.; VITOLO, M. lnfluence of pH, Temperature and Dissolved Oxygen Concentration on the Production of Glucose-6- Phosphate Dehydrogenase and lnvertase by Saccharomyces cerevisiae. Brazilian Journal of Chemical Engineering. v.14, n.1, p.89-94, 1997.
ALEXANDER, M.A., YANG, V.W., JEFFRIES, T.W. Leveis of Pentase Phosphate Pathway Enzymes from Candida shehatae Grown in Continuous Culture. Applied Microbiology and Biotechnology, v. 29, p. 282-288, 1988.
AGERKVIST, 1., ENFORS, S. O. Characterization of E. co/i cell disintegrates from a bead mill and high pressure homogenizers. Biotechnology and Bioengineering, v. 36, p. 1083-1089, 1990.
BALAGUER, P.; TÉROUANNE, B.; ELIAOU, J. F., HUMBERT, M.; BOUSSIOUX, A M.; NICOLAS, J. C. Use of Glucose-6-Phosphate Dehydrogenase as a New Label for Nucleic Acid Hybridization Reactions. Analytical Biochemistry, v. 180, p. 50-54, 1989.
BALASUNDARAM, B.; PANDIT, A . 8. Selective release of invertase by hydrodynamic cavitation. Biochemical Engineering Journal. v. 8, p. 251-256, 2001.
BASSI, AS.; TANG, D.Q.; BERGOUGNOU, M.A. Mediated, Amperometric Biosensor for Glucose-6-Phosphate Monitoring Based on Entrapped Glucose-6- Phosphate Dehydrogenase, Mg+2 ions, Tetracyanoquinodimethane, and Nicotinamide Adenine Dinucleotide Phosphate in Carbon Paste. Analytical Biochemistry. v. 268, n. 2, p. 223-228, 1999.
BECKER, J. M., CALDWELL, G. A, ZACHGO, E .A (1996) Biotechno/ogy: A laboratory course, Academic Press, San Diego, 2. ed., 261 p.
BELFAQUIH, N., PENNINCKX, M.J. A Bifunctional f3-Xylosidase-Xylose lsomerase from Streptomyces sp. EC10. Enzyme and Microbial Technology, V. 27, p. 114-121, 2000.

66
BERGMEYER, H.U. Methods of Enzymatic Analysis. Wheinheim: Velag Chemie, 3.ed. v.2, 539p, 1984.
BISARIA, V. S. ln: Bioconversion of Waste Materiais to Industrial Products. Martin A. Eds. Ed. Chapman & Hall, Londres, 1998.
BRADFORD, M. A.; Rapid and Sensitive Method for the Quantification of Protein Utilizing the Principie of Protein-dye Binding. Analytical Biochemistry, V. 72, p. 248-254, 1976.
BROWNELL, J. E., NAKAS, J. P. Bioconversion of Acid-Hydrolyzed poplar Hemicellulose to Acetic Acid by Clostridium thermoaceticum. Journal of Industrial Microbiology, v. 7, p. 1-6, 1991.
BRUINENBERG, P. M; DIJKEN, J. P, SCHEFFERS, W. A. The role of redox balances in the anaerobic fermentation of xylose by yeasts. Applied Microbiology and Biotechnology. v. 18, p. 287-292, 1983.
CERTIK, M.; MEGOVA, J.; HORENITZKY, R.; Effect of Nitrogen Sources on the Activities of Lipogenic Enzymes in Oleaginous Fungus Cunninghamel/a echinulata. J. Gen. Appl. Microbial, v. 45, p. 289-293, 1999.
CHATEL, S., VOIRIN, A., LUCIANI, A.; ARTAUD, J. Starch ldentification and Determination in Sweetened Fruit Preparations. Journal of Agriculture and Food Chemistry, v. 44, n. 2, p. 502-506, 1996.
CURRIE; J. A, DUNNILL, P., LILL Y, M. D. Release of protein from baker's yeast by disruption in an industrial agitator mill. Biotechnology and Bioengineering, V. 14, p. 725-736, 1972.
Ou TOIT, P.J., OLIVIER, S.P., VAN , P.L. Sugar Cane Bagasse as a Possible Source of Fermentable Carbohydrates 1: Characterization of Bagasse with Regard to Monosaccharide Hemicellulose and Amino Acid Composition. Biotechnology and Bioengineering, v. 26, p. 1071-1078, 1984.
FELIPE, M. G. A.; VIEIRA, D. C.; VITOLO, M.; SILVA, S. S.; ROBERTO, 1. C.; MANCILHA, 1. M. Effect of acetic acid on xylose fermentation to xylitol by Candida guilliermondii. Joumal Basic Microbiology. v. 35, p. 171-177, 1995.
FELIPE, M. G. A.; ALVES, L. A.; SILVA, S. S.; ROBERTO, 1. C.; MANCILHA, 1. M.; ALMEIDA SILVA, J. B. Fermentation of eucalyptus hemicellulosic hydrolysate to xylitol by Candida guilliermondii. Bioresource Technology. v. 56, p.281-283, 1996.
FREY, S.D.; ELLIOTT, E.T.; PAUSTIAN, K. Application of the Hexokinase- Glucose-6-Phosphate Dehydrogenase Enzymatic Assay for Measurement of Glucose in Amended Soil. Soil Biology and Biochemistry, v. 31, n. 6, p. 933- 935, 1999.
FOLLOWS, M., HETHERINGTON, P. J., DUNNILL, P., LILL Y, M. D. Release of enzymes from baker's yeast by disruption in an industrial homogenizer. Biotechnology and Bioengineering, v.13, p. 549-560, 1971.
FOSTER, D., Optimizing recombinant product recovery through improvements in cell-disruption technologies. Current Opinion in Biotechnology, v. 6, p. 523-526, 1995.

67
GARDE, A. , JONSSON. G., SCHMIDT, A. S., AHRING, B.K. Lactic Production from Wheat Straw Hemicellulose Hydrolysate by Lactobacillus pentosus and Lactobacillus brevis. Bioresource Technology, v.81, n.3, p. 217-223, 2002.
GÍRIO, F. M.; ROSEIRO, J. C.; SÁ-MACHADO, P.; DUARTE-REIS, A R.; AMARAL-COLLAÇO, M. T. Effect of oxygen transfer rate on levels of key enzymes of xylose metabolism in Debaryomyces hansenii. Enzyme Microbiology and Technology. v. 16, p. 1074-1078, 1994.
GODFREY, T., WEST, S. Industrial Enzymology. 2nd_ Edition. New York:Stockton Press, 1996. 609p.
GRAJEK, W. Production on Protein by Thermophilic Fungi from Sugar Beet Pulp in Solid State Fermentation. Biotechnology and Bioengineering, v.32, p. 255-260, 1988.
GRÃNSTROM, T., OJAMO, H. LEISOLA, M. Chemostat Study of Xylitol Production by Candida guilliermondii. Applied Microbiology and Biotechnology. v. 55, p. 36-42, 2001.
GOLDSTEIN, I.S. Organic Chemical from Biomass. Boca Raton: CRC Press, 1981.
GONG, C. S.; CHEN, L. F.; FLICKINGER, C.; CHIANG, L. C.;TSAO, G. T. Production of ethanol from D-xylose by using D-xylose isomerase and yeasts. Applied and Environmental Microbiology. v. 41, p. 430-436, 1981.
HAHN-HÃGERDAL, B., JEPPSSON, H., SKOOG, K., PRIOR, B.A. Biochemistry and Physiology of Xylose Fermentation by Yeasts. Enzyme and Microbial Technology, v. 16, p. 933-943, 1994.
JEFFRIES, T.W. Utilization of Xylose by Bacteria, Yeasts, and Fungi. Advances in Biachem Eng, v. 27, p. 1-32, 1983.
JEPPSSON, M.; JOHANSSON, B.; HAHN-HÃGERDAL, B.; GORWA- GRAUSLUND, M. F. Reduced oxidative pentase phosphate pathway flux in recombinant xylose-utilizing Saccharomyces cerevisiae strains improves the ethanol yield from xylose. Applied and Environmental Microbiology. v. 68, p. 1604-1609, 2002.
JEPPSSON, M.; JOHANSSON, B.; JENSEN, P. R.; HAHN-HÃGERDAL, B.; GORWA-GRAUSLUND, M. F. The levei of glucose.:.S-phosphate dehydrogenase activity strongly influences xylose fermentation and inhibitor sensitivity in recombinant Saccharomyces cerevtstee strains. http:l/www.interscience.wiley.com (arquivo consultado em outubro de 2003).
KADAM, K.L., FORREST, L.H., JACOBSON, W.A. Rice Straw as a Lignocellulosic Resource: Collection, Processing, Transportation, and Environmental Aspects. Biomass and Bioenergy, v. 18, p. 369-389, 2000.
KATO, N.; SAHM, H.; SCHUTTE, H.; WAGNER, F. Purification and properties of glucose-6-phosphate dehydrogenase and 6-phophogluconate dehydrogenase from methanol-utilizing yeast, Candida boidinii. Biochemistry and Biophysis. Acta, 566, p. 1-11, 1979.

68
KUBOI, R.; UMAKOSHI, H.; TAKAGI, N.; KOMASAWA, 1. Optimal disruption methods for selective recovery of f3-galactosidase from Escherichia coli. Journal of Fermentation and Bioengineering. v. 79, p. 335-341, 1995.
KUHAD, R.C., SINGH, A. Lignocellulose Biotechnology: Current and Future Prospects. Criticai Reviews in Biotechnology, v. 13, n.2, p. 151-172, 1993.
KULA, M. R., SCHUTIE, H., VOGELS, G., FRANK, A. Cell disintegration for the purification of intracellular proteins. Fd. Biotechnology, v. 4, p. 169-183, 1990.
LANG, C.; GÔLLNITZ C.; POPOVIC, M.; STAHL, U.; Optimization of Fungai Polygalacturonase Synthesis by Saccharomyces cerevisiae in fed-batch culture. Chemical Engineering Joumal v.65, p. 219-226, 1999.
LEE, H.; SOPHER, C. R.; YAU, K. Y. F. lnduction of xylose reductase and xylitol dehydrogenase activities on mixed sugars in Candida guilliermondii. Journal Chem. Tech. Biotechnol. v. 66, p. 375-379, 1996.
LEITE, A. A. Determinação das atividades da glicose-6-fosfato- desidrogenase e 6-fosfogliconato desidrogenase em neutrófilos de eqüinos adultos puro sangue inglês. Dissertação de Mestrado. FCF/USP, 1989.
LEVY, H.R. Glucose 6-Phosphate Dehydrogenase from Leuconostoc mesenteroids. Biachem. Soe. Trans. v. 17, p. 313-315, 1988.
LILLY, M.D. Production of intracellular Microbial Enzymes. ln: WINGARD JUNIOR, L.B.; KATCHALSKI-KATZIR, E.; GOLDSTEIN, L. eds. Applied Biochemistry and Bioengineering. New York: Academic Press, v.2, 306p, 1979.
LOVITI, R.W., JONES, M., COLLINS, S.E., COSS, G.M., YAU, C.P., ATIOUCH, C. Disruption of Baker's Yeast Using a Disrupter of Simple and Novel Geometry. Process Biochemistry. v.36, p.415-421, 2000.
LUZZATIO, L., MEHTA, A. Glucose-6-phosphate dehydrogenase deficiency. ln: Scriver, C.R.; Beaudet, A.L.; Sly, W.S.; Vale, D. eds. The Metabolic and Molecular Bases of lnherited Disease, McGraw Hill, New York, 7th ed., 1995.
MAGEE, R.J., KOSARIC, N> Bioconversion of Hemicellulose. ln: Advances in Biochemical Engineering /Biotechnology, Springer-Verlag, New York, v. 32, p.60-93, 1985.
MARFY, F.; KULA, M. R. Enzyme yields from cells of brewer's yeast disrupted by treatment in horizontal disintegrator. Biotechnol. Bioeng. v. 16, p. 623-634, 1974.
McMILLAN, J. D. Pretreatment of lignocellulosic biomass. ln: Himmel, M.E.; BAKER, J.O.; OVEREND, R. P. (Eds.), Enzymatic conversion of biomass for fuels production (Symposium, Series 566). American Chemical Society, USA, p. 293-324, 1994.
MELENDRES, A. V., HONDA, H., SHIRAGAMI, N., UNNO, H. Enzyme release kinetics in a cell disruption chamber of a bead mill. J. Chem. Eng. Japan, v. 26, p. 148-152, 1993.

69
MORI, H.; ISHIDA, M.; OKAMOTO, S. Glucose determination using a flow system with enzyme reactor and application to analysis of glucose content in beverages. Journal of Health Science. v. 45, n. 3, p. 126-129, 1999.
MUSSATIO, S. I.; ROBERTO, 1. C. Hydrolysate detoxification with activated charcoal for xylitol production by Candida guil/iermondii. Biotechnology Letters. v. 23, p. 1681-1684, 2001.
MUSSATIO, S. 1. Influência do tratamento do hidrolisado hemicelulósico de palha de arroz na produção de xilitol por Candida guilliermondii. Lorena, DEBIQ/FAENQUIL, 2002. 173p.
MUSSATIO, S. I.; ROBERTO, 1. C. Optimal experimental condition for hemicellulosic hydrolysate treatment with activated charcoal for xylitol production. Biotechnology Progress. v. 20, p. 134-139, 2004.
NETO, E. C., WEBER, L., BRITES, A. Deficiência de Glicose-6- Fosfato- Desidrogenase: Qual a Incidência no Brasil? Laes Haes, ano XX, n.120, p.126- 136. 1999.
ÔZBEK, B, ÜLGEN K.Ô. The Stability of Enzymes after Sonification. Process Biochemistry. v. 35, p.1037-1043, 2000.
PANDEY, A.; SOCCOL, C. R.; NIGAM, P.; SOCCOL, V. T. Biotechnological potential of agro-industrial residues. 1: sugarcane bagasse. Bioresource Technology. v. 7 4, p. 69-80, 2000.
PARAJÓ, J.C., VAQUEZ, D., ALONSO, J.L., DOMÍNGUEZ, H. Prehydrolysis of Eucalyptus Wood With Dilute Sulphuric Acid: Operation in Autoclave. Holz ais und Werkstoff, v.52, p.102-108, 1994.
PARAJÓ, J.C., SANTOS, V., VAQUEZ, M. Production of Carotenoids by Phaffia rhodozyma Growing on Media Made from Hemicellulosic Hydrolysates of Eucaliptus globules Wood. Biotechnology and Bioengineering, v.59, n. 4, p.501-506, 1998.
PARAJÓ, J.C., DOMÍNGUEZ, H., DOMÍNGUEZ, J.M., Biotechnological Production of Xylitol. Part 1: lnterest of Xylitol and Fundamentais of its Biosynthesis. Bioresource Technology, v.65, p.191-201, 1998.
PARIS!, F. Advances in Lignocellulosics Hydrolysis and in the Utilization of the Hydrolyzates. ln: FIECHTER, A. Advances in Biochemical Engineering/Biotechnology, v.38, p.53-87, 1989.
PRIOR, B.A., KILLIAN, S.G., du PREZ, J.C. Fermentation of Xylose by the Yeasts Candida shehatae and Pichia stipitis: Prospects and Problems. Process Biochemistry, 1989.
PESSOA Jr., A., MANCILHA, 1., SATO, S. Cultivation of Candida tropicalis in Sugar Gane Hemicellulosic Hydrolysate for Microbial Protein Production. Journal of Biotechnology, v.51, p.83-88, 1996.
RICCI-SILVA, M.E., VITOLO, M., ABRAHÃO-NETO, J. Protein and Glucose 6- Phosphate Dehydrogenase Releasing from Baker's Yeast Cells Disrupted by a Vertical Bead Mill. Process Biochemistry, v.35, p.831-835, 2000.

70
RODRIGUES, R. C. L. B.; FELIPE, M. G. A.; ALMEIDA E SILVA, J. B.; VITOLO, M.; GÓMEZ, P. V. The influence of pH, temperature and hydrolysate concentration on the remova! of volatile and nonvolatile compounds from sugarcane bagasse hemicellulosic hydrolysate treated with activated charcoal before or after vacuum evaporation. Brazilian Journal of Chemical Engineering. v. 18, 2001.
ROBERTO, 1. C.; SATO, S.; MANCILHA, 1. M. Effect of inoculum level on xylitol production from rice straw hemicellulose hydrolysate by Candida guil/iermondii. Joumal lnd.Microbiol. v. 16, p. 348-350, 1996.
ROBERTO, 1. C.; MUSSATTO, S.I.; RODRIGUES, C. L. B. Dilute-acid hydrolysis for optimization of xylose recovery from rice straw in a semi-pilot reactor. Industrial Crops and Products. v. 17, p. 171-176, 2003.
ROBERTO, I.C., FELIPE, M.G.A., LACIS, L., SILVA, S.S., MANCILHA, I.M. Utilization of Sugar Gane Bagasse Hemicellulosic Hydrolisate by Candida guil/iermondii for Xylitol Production. Bioresource Technology, v.36, n.3, p.271-275, 1991.
ROBERTO, I.C. Obtenção e Fermentação do Hidrolisado Hemicelulósico de Palha de Arroz. Projeto de Pesquisa FAPESP (Processo No99/09558-5), 2001.
SEVERO, L. G. Determinação da atividade da glicose-6-fosfato desidrogenase (G-6-PD) em eritrócitos humanos. São Paulo: FCF/USP, 1979. 68p. (Dissertação de Mestrado).
SHLESER, R. Ethanol Production in Hawaii. http://www.hawaii.gov/dbedt/ert/ethanol (arquivo consultado e junho de 1999).
SHIROKANE, Y.; ICHIKAWA, K.; SUZUKI, M. A novel enzymic determination of maltose. Carbohydrate Research. v. 329, p. 699-702, 2000.
SILVA, D. P. Obtenção das Enzimas Hexoquinase e Glicose-6-fosfato Desidrogenase utilizando Saccharomyces cerevisiae. São Paulo: FCF/USP, 2000. 165p. (Dissertação de Mestrado).
SILVA, S.S., VITOLO, M., PESSOA Jr., A., FELIPE, M.G.A. Xylose Reductase and Xylitol Dehydrogenase Activities of D-Xylose-Xylitol-Fermenting Candida guilliermondii. Joumal of Basic Microbiology, v.36, n.3, p.187-191, 1996.
SLININGER, P. J.; BOLEN, P. L.; KURTZMAN, C. P. Pachysolen tannophilus: properties and process consideration for ethanol production from D-xylose. Enzyme Microb. Technol. v. 9, p. 5-15, 1987.
SONG, D. D.; JACQUES, N. A. Cell disruption of Escherichia coli by glass bead stirring for recovery of recombinant proteins. Analytical Biochemistry. v. 248, p. 300-301, 1997.
SUGAI, J. K.; DELGENES, J. P. Catabolite repression of induction of aldose reductase activity and utilization of mixed hemicellulosic sugars in Candida guil/iermondíí1 Current Microbiology. v. 31, p. 239-244, 1995.
SUN, R. C.; TOMKINSON, J.; MA, P. L.; LIANG, S. F. Comparativa study of hemicelluloses from rice straw by alkali and hydrogen peroxide treatments. Carbohydrate Polymers. v. 42, p. 111-122, 2000.

71
SURYADI, H., YOSHIDA, N., YAMADA-ONODERA, K., KATSURAGI, T., TANI, H. Characterization of a Flavinogenic Mutant of Methanol Yeast Candida boidinii and lts Extracellular Secretion of Riboflavin. Jounal of Bioscience and Bioengineering. v.90, n.1, p.52-56, 2000.
TAUBERT, J.; KRINGS, U.; BERGER, R. G. A comparative study on the disintegration of filamentous fungi. Journal of Microbiological Methods. v. 42, p. 225-232, 2000.
TEROUANNE, B.; BENCHEICK, M., BALAGUER, P.; BOUSSIOUX, A.M.; NICOLAS, I.C. Bioluminescent assays using glucose-6-phosphate dehydrogenase - Application to biotin and streptavidin detection. Anal. Biachem. v. 180, n. 1, p. 43-49, 1989.
VIEIRA-NETO, E.; PEREIRA, C. N.; FONSECA, A. A. Avaliação do Desempenho de Rastreamento Neonatal de Deficiência de Glicose-6-Fosfato Desidrogenase em Amostras de Sangue em Papel Filtro. Laes & Haes, n.119, p.98-104, 1999.
WILKE, C.R., YANG, R.D., SCIAMANNA, FREITAS, R.P. Raw Materiais Evaluation and Process Developtment Studies for Conversion of Biomass to Sugar and Ethanol. Biotechnology and Bioengineering, v. 23, p.163-183, 1981.
WINKELHAUSEN, E., KUSMANOVA, S. Microbial Conversion of D-Xylose to Xylitol. Journal of Fermentation and Bioengineering, v.86, n.1, p.1-14, 1998.
WITHAKER, J.R. Enzymes in Analytical Chemistry. ln: FOX, P.F., ed. Food Enzymology. New York: Elsevier Science Publishers, 1991. v. 2, 378pp.

APÊNDICES
APÊNDICE A.1 - CURVA DE CALIBRAÇÃO DE BIOMASSA E DE PROTEiNAS TOTAIS (LOWRY)
Encontra-se, a seguir, a curva de obtida para a calibração da concentração
celular, pelo método do peso-seco em função da absorbância a 600 nm. A
calibração foi realizada com as células crescidas em hidrolisado lavando-as duas
vezes com água destilada.
0,8
E o.r e: g 0,6 e.o ro ro 0,5 "ü e:
~ 0,4 .... o C/l ~ 0,3
ro
jº·2 // <1> 0,1 ~
• •
o.o -+--r--r--.--,r--r----.----r----r--r-.,--...---r-....--,r--r---r-r--r--.----,
0,00 0,05 0,10 0,15 0,20 0,25 0,30 0,35 0,40 0,45 0,50
Concentração Celular (g/L)
Figura A .1. Curva de calibração de biomassa em g/L
Curva de Calibração do lnóculo lavado:
D.O= 0,03857+1,52736 x Biomassa (R2=0,996)

Apresentam-se, a seguir, as curvas obtidas para a calibração das
concentrações de proteínas totais em mg/L.
0,20
0,18
~ 0,16 E e 0,14
~ 0,12
.!.'!! O 10 g ' ~ 0,08 o ~ 0,06
0,04
• 0,02 /.
Y =0,00217+6,38743E-4 X
100 150 200 250
Concentração de BSA (mg/L) 300
Figura A.2. Curva Padrão de proteínas totais utilizada nos ensaios de rompimento celular
0,9
0,8
0,7 E ~ 0,6 .... ~ 0,5
·~ 0.4
~ 0,3
j 0,2
0,1
0,8 B
0,0+-"''-.--,------------ 0 1000 200 400 6Xl 800
Concentração de 8$&. {mg/L)
0,8
0,7
E 0,6 e ~ 0,5
"' : Q.4 'g :l:1l 0,3
.§ 0,2 -c
0,1
0,7
~ 0,6
~ 0,5
.00 600 800 Concentração de BSA {mg/L)
e
0,0+-0 ~-200---400---WJ--800---1~000- Concentração de BSA (mg/L)
Figura A.3. Curva Padrão de proteínas totais utilizada nos ensaios do processo fermentativo
A)Y = -0,02861 + 6,73047E-4X (R2=0,994)
B) Y = 0,01546 + 7,49795E -4X (R2=0,999)
C)Y = 0,0177 + 7,92682E - 4X (R2=0,981)

APÊNDICEA.2 - RESULTADOS DOS ENSAIOS DE ROMPIMENTO CELULAR
Nesta seção encontram-se tabelados os valores obtidos para todos os
ensaios de rompimento celular efetuados. Foram determinados os valores de
porcentagens de células rompidas, tanto por DOaxi quanto por número de células
intactas, proteínas totais, atividades volumétrica e específica de G6PDH bem como
seus respectivos desvios-padrão.
Tabela A.1. Porcentagens de células rompidas (ensaios 1 e 2) determinadas pela contagem do
número de células em câmara de Neubauer, no tempo de 1 minuto, empregando
diferentes concentrações celulares
Concentração celular Ensaio 1
40
43
68
Média Erro-padrão
41 o 38 ±8 48 o
37,5 ±2,5
50,5 ±7,5 67 ±1
(g/L)
1,2
3,2
5,9
11,6
22,1
35,4
(%)
41
46
48
35
58
66
Ensaio 2 (%)
41
30
Tabela A.2. Porcentagens de células rompidas (ensaios 1 e 2) detemninadas pela contagem do
número de células em câmara de Neubauer, no tempo de 3 minutos, empregando
diferentes concentrações celulares
Concentração celular Ensaio 1 Ensaio 2 Média Erro-padrão
(g/L) (%) (%)
1,2 62 58,5 ±3,5
3,2 54 25 39,5 ±14,5
5,9 52 54 53 ±1
11,6 46 57 51,5 ±5,5
22,1 53 67 60 ±7
35,4 79 70 74,5 ±4,5

Tabela A.3. Porcentagens de células rompidas (ensaios 1 e 2) determinadas pela contagem do
número de células em câmara de Neubauer, no tempo de 5 minutos, empregando
diferentes concentrações celulares
--- -·-------···-··--·---·----·------ Concentração celular Ensaio 1 Ensaio 2 Média Erro-padrão
(g/L) (%) (%)
1,2 61 67 64 3,2 58 47 52,5 5,9 56 64 60 11 ,6 58 61 59,5 22,1 70 72 71 35,4 78 78 78
±3 ±5,5
±4 ±1,5 ±1 o
Nº células I mL = cétutas.x+x; 04 xdiluição
nº células j 1 - n" células, . (%) = n es aepois xl 00 nº células antes
Tabela A.4. Porcentagens de células rompidas (ensaios 1 e 2) determinadas pela 00600, no tempo de
1 minuto, empregando diferentes concentrações celulares
Concentração celular Ensaio 1 Ensaio 2 Média Erro-padrão
(g/L) (º/o) (O/o) -------H•,----------·--·--•.WU•---·------ ' ----------•OOO•o••<,O_O _
1,2 42 54 48 ±6 3,2 54 5,9 11,6
22,1 35,4
40 50 11
56
30 42 ±12 48 44 ±4 42 46 ±4 58 34,5 ±23,5 47 51,5 ±4,5

Tabela A.5. Porcentagens de células rompidas (ensaios 1 e 2) determinadas pela 00600, no tempo de
3 minutos, empregando diferentes concentrações celulares
---·--------- ·---------------------------- Concentração celular Ensaio 1 Ensaio 2 Média Erro-padrão
(g/L) (%) (%)
1,2 47 62 54,5 ±7,5 3,2 58 39 48,5 ±9,5 5,9 67 78 72,5 ±5,5 11,6 49 53 51 ±2 22,1 64 72 68 ±4 35,4 66 54 60 ±6
Tabela A.6. Porcentagens de células rompidas (ensaios 1 e 2) determinadas pela 00600, no tempo de
5 minutos, empregando diferentes concentrações celulares
Concentração celular Ensaio 1 · Ensaio 2 (g/L) (%) (%)
1,2 56 77
3,2 61 52
5,9 62 70
11,6 63 67
22,1 72 73
35,4 78 61
Média Erro-padrão
66,5 ±10,5 56,5 ±4,5 66 ±4
65 ±2 72,5 ±0,5
69,5 ±8,5
DO -0,0386 _ [ X]g I L = 600 xdiluiçao
1,527
(%) = [ XLntes - [ X]depois xlOO [Xlantes

Tabela A.7. Atividade volumétrica de glicose-6-fosfato desidrogenase (ensaios 1 e 2) no tempo de 1
minuto, em diferentes concentrações celulares ~-------------------------------------- ---
Concentração celular Av (g/L) ensaio 1
(U/L)
1,2 22
3,2 79
5,9 145
11,6 341
22,1 483
35,4 962
------·--·-·--·--·-· .. ·-·--··-·-·-·---··-····--···--·----··-···--·--··· Av Média Erro-padrão
ensaio 2 (U/L)
26
45
209
370
552
1083
24 ±2 62 ±17 177 ±32
355;5 ±14,5 517,5 ±34,5 1022,5 ±60,5 _________________________________ .. _, ., __ ,, , ,, __ ., _
Tabela A.8. Atividade volumétrica de glicose-6-fosfato desidrogenase (ensaios 1 e 2) no tempo de 3
minutos, em diferentes concentrações celulares
Concentração celular Av Av Média · · Erro-padrão
(g/L) ensaio 1 ensaio 2 .. (U/L) (U/L) --~
1,2 55 63 59 ±4 3,2 161 154 157,5 ±3,5 5,9 349 241 295 ±54
11,6 831 768 799,5 ±31,5 22,1 1614 1543 1578,5 ±35,5
35,4 2045 1404 1724,5 ±320,5. ,.,.._ .. ,.~,.L, ... , ..... o.,.LU,
Tabela A.9. Atividade volumétrica de glicose-6-fosfato desidrogenase (ensaios 1 e 2) no tempo de 5
minutos, em diferentes concentrações celulares·
Concentração celular . Av . · · Av · . Média · · . Erro-padrão . ·
(g/L) ensaio 1
(U/L)
1,2 48
3,2 239
5,9 442
11,6 1544
22,1 1740
35,4 3555
ensaio 2
(U/L)
81 64,5 ±16,5
218 228,5 ±10,5
370 406 ±36
974 1259 ±285
1807 1773,5 ±33,5 1959 2757 ±798

Tabela A.1 O. Proteínas totais (ensaios 1 e 2), no tempo de 1 minuto, em diferentes concentrações
celulares
Concentração celular Proteínas totais Média
(g/L) ensaio 1
Proteínas totais
ensaio 2 Erro-padrão
1,2 3,2 5,9 11,6 22,1 35,4
(mg/L)
188 238 344 819 771 1572
(mg/L)
98 218 403 605 1174 1901
143 ±45 228 ±10
373,5 ±29,5 712 ±107
972,5 ±201,5 1736,5 ±164,5
Tabela A.11. Proteínas totais (ensaios 1 e 2), no tempo de 3 minutos, em diferentes concentrações
celulares
Concentração celular Proteínas totais Proteínas totais Média Erro-padrão
(g/L) ensaio 1 ensaio 2
(mg/L) (mg/L)
1,2 230 103 166,5 ±63,5 3,2 298 355 326,5 ±28,5 5,9 563 373 468 ±95 11,6 1338 1113 1225,5 ±112,5 22,1 2508 2503 2505 ±2,5 35,4 2583 2635 2609 ±26
.............................. ...,~-··· .... ··~· -···~· ,..........,..,,..., ................................ --=-
Tabela A.12. Proteínas totais (ensaios 1 e 2), no tempo de 5 minutos, em diferentes concentrações
celulares
Concentração celular Proteínas totais · Proteínas totais Média · Erro-padrão .
ensaio 2 (g/L) ensaio 1 (mg/L)
1,2 196 3,2 323 5,9 498
11,6 1317 22,1 2671
(mg/L)
35,4 4586
112 315 452 1351 2530 3728
154 ±42 319 ±4 475 ±23 1334. ±17
2600,5 ±70,5 4157 ±429
-··-····--··-·-------·-··---··--·---·--·-----···--·-·--

Tabela A.13. Atividade específica de glicose-6-fosfato desidrogenase (ensaios 1 e 2) no tempo de 1
minuto, em diferentes concentrações celulares
Concentração celular
(g/L)
·--- .. -·--·-··-···--- .. - -- . Média Erro-padrão
1,2
3,2
5,9
11,6
22,1
35,4
ensaio 1 ensaio 2
(U/mgprotelna) (U/mgprotelna)
0,12 0,27 0,195 ±0,075 0,33 0,21 0,27 ±0,06 0,42 0,52 0,47 ±0,05
0,42 0,61 0,515 ±0,095 0,63 0,47 0,55 ±0,08 0,61 0,57 0,59 ±0,02
Tabela A.14. Atividade específica de glicose-6-fosfato desidrogenase (ensaios 1 e 2) no tempo de 3
minutos, em diferentes concentrações celulares ·concentração celular ·"···-A;··-····-·-·-··-··-~~~-·· ··-·····Ã~~··· ··- ····· ··- ·· ·· ·MédTa··· Erro-padrão
(g/L) ensaio 1 ensaio 2
(U/mgprotelna) (U/mgprotelna) .. - ~........,.,
1,2 0,24 0,61 0,425 ±0,185 3,2 0,54 0,43 0,485 ±0,055
5,9 0,62 0,65 0,635 ±0,015
11,6 0,62 0,69 0,655 ±0,035
22,1 0,64 0,62 0,63 ±0,01
35,4 0,79 0,53 0,66 ±0,13
Tabela A.15. Atividade específica de glicose-6-fosfato desidrogenase (ensaios 1 e 2) no tempo de 5
minutos, em diferentes concentrações celulares
Concentração celular· · AE · · AE · · . Média . · Erro-padrão .
(g/L) ensaio 1 ensaio 2
(U/mgprotefna) (U/mgprotelna)
1,2 0,24 0,72 0,48 ±0,24
3,2 0,74 0,69 0,715 ±0,025
5,9 0,89 0,82 0,855- ±0,035
11,6 1,2 0,72 0,96 ±0,24
22,1 0,65 0,71 0,68 ±0,03
35,4 0,78 0,53 0,655 ±0,125

da 106 . . _ Av(U / L) = -. x--x7,15xdrlwçao
mm 6220
Onde, AE é a atividade específica, Av atividade volumétrica e P1 as proteínas totais

APÊNDICE A. 3 - RESULTADOS DOS ENSAIOS DO PLANEJAMENTO FATORIAL 23
Nesta seção encontram-se representados os valores obtidos para todos os
ensaios do planejamento fatorial 23 efetuado. A concentração celular foi determinada
pelo método do peso-seco, enquanto a concentração de açúcares, xilitol e ácido
acético foi obtida por cromatografia líquida de alto desempenho (HPLC - WATERS).
Foram também avaliados os valores de porcentagem de rompimento por DO&XJ,
proteínas totais através da curva padrão de proteínas, atividades volumétricas e
específicas além do pH do meio, através de pHmetro.
Tabela A.16. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guil/iermondii, com 30 g/L de xilose, 1 g/L de inóculo e pH 4,5
Tempo (h) [X] antes
(g/L)
[X] depois Rompimento
(g/L) (%)
Av (U/L)
Proteínas si AE
tampão (U/mQprot)
(mg/L) ---------··---- -------------·---·····-------------------- o 3,1 1,4 55 132 353 0,37
12 2,2 1,5 32 93 224 0,42
24 2,3 1,6 30 108 194 0,57
36 2,7 1,7 37 114 291 0,39
48 3,0 2,2 27 147 530 0,28
·--~-·- ... = ........................ ... , ...... ..,......,. ... .,.......,...,...,,.,......., ... ,., ............................................ Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 33,44 3,703 5,244 0,9078 30 0,7 4,62
12 24,69 5,059 0,5391 2,918 101 2,4 4,70
24 16,13 4,930 8,418 122 2,6 5,02
36 8,467 4,631 13,47 122 2,9 4,99
48 1,489 3,180 16,54 167 3,4 4,80 _..__ ... _... .. .., .. --.. ........ -.,.. ........... "'""-"''"""' .................. - .... - ............................... ,., ....... - ...... ..,_ ..... _ .... _ . ., .......... _ .... _"'_"'"'··'·-···"'·· .. ··.oc···-···· ............ ---·--~·-·"'·· ........... ,.-" ........ ...-0-··:.-·o.-..-· .. .., ..................... _ .. ____ .. _ .. _____

Tabela A.17. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guil/iermondii, com 30 gil de xilose, 1 g/L de inóculo e pH 7,5
--------------·----- -·---·------·---·-·-··--·--·----···--·----- .. --------· Tempo (h) [X) antes [X) depois Rompimento Av Proteínas si AE
(gil) (gil) (%) (UIL) tampão (Ulmgprot)
(mgll)
o 3,1 1,4 55 132 353 0,37
12 2,5 1,5 40 137 305 0,45
24 2,8 1,8 36 134 280 0,49
36 2,5 1,8 28 145 362 0,40
48 2,7 1,8 33 167 304 0,55 __ N __________ ,. ___ .. ______________ ,,_, ______ , ____ __ .. __ ,,_ .. ___________ ,.,, ______ ., _____ ,_,,, ___
, , ,, .. ,,,,, ~.- ,0,0 ..,-,,,,, .. ., , A,•,,,0. 0, 0,,,0,<-'H,0 .. ,0,0 0,0,0,0 ,,,>,.,o,•A,0,0,0,0.0.0 0~0,< •,~ 0,0,0,0_,d,0-,0·<•0•>•••~, - •,••O,,.<•• o,,,O, ,,,,,o.- .. ,<,0 •·•·--'••~~dd .. ,M,,_.., .. ,,,o,o, .. ,,, .. ,, .. ,,,,,,.,,,,, ,, .. _.,,,,,,,0,0••••••••••"·'''•'•••• , ,.,_. ,....,_ .. ,_. ,, .. ,,,.., , ,,,0,0. •_,_..,,,,0,,,0J•
Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (gil) (gil) (gil) acético (gil) (UIL) (gil)
(gil)
o 33,78 3,685 5,315 1,091 30 0,7 7,63
12 25,67 5,197 0,8216 2,526 40 2,7 6,56
24 17,04 5,048 0,7371 7,291 144 3,0 6,59
36 8,710 4,954 0,6939 13,47 220 3,8 6,68
48 1,555 3,786 0,5561 16,73 253 4,1 6,72 .............. ~--·-- .................... , ....... L ......... , ..................... LL-< ..... <..W.1.-· ......... ,., ............................................... -- ~ .... ,J ........ ,J ..... .._. ..... ,_,,..........._.,, ..... ,., ....................... ~,.L,-.>. ................. ,....__~~- ......... ---·~----
Tabela A.18. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guil/iermondii, com 70 gil de xilose, 1 gil de inóculo e pH 4,5
Tempo (h) [X] antes [X] depois Rompimento Av . Proteínas si AE .
(gil) (gil) (%) (UIL) tampão (Ulmgprot)
(mgll)
o 2,8 1,5 46 96 303 0,32
24 2,5 1,1 56 108 238 0,45
48 2,5 1,6 36 103 195 0,53
60 3,5 2,2 37 147 246 0,60
96 2,9 1,9 34 207 393 0,53

------·--··--------------------------···------- .. --------·--···---------- . ------·-····-···-·····---····--·-···-·-····-·-·-· .. -·--··-··-··-···-····--· Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 66,08 7,309 12,69 1,670 41 1,2 4,64
24 60,37 12,65 1,446 3,951 99 2,3 4,54
48 42,87 12,05 0,5560 12,86 156 3,8 4,72
60 31,91 12,34 0,1660 20,04 160 3,8 4,88
96 3,773 10,73 38,50 350 4,9 4,75 ··--·-·····----··"·-·--·-···---···-·--·---·--------- .. ---·-··- ., __________ ................ -, ........ ·-···-··---···--·--·------- .... ,. ____ ,, _________ ,,, .. ,. __________ , ................ ................................. - ......
Tabela A.19. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guilliermondii, com 70 g/L de xilose, 1 g/L de inóculo e pH 7,5
~·-·~O,M,O,MMMM,O,o,,,0,0,0-M•-•-••••••••••••M•OM,o,,,,_.,_._. __ M ......... MOM•'•••••~- ..... ,, .. M,0,,,,,0,0,0MM,•,O,O•••••••••••••••••••-••·•"·MO,<,<M .. ,O,MOd,,M,M,,••,•M,0•• .. ••• ... , ...... ,- ..... ~>- .. ,d.,,,.,,,.,.,,,.,.,,,,,,, ... ~, ...... , ..... ~~d-·d~ .... , ..... , .. ~.,,,.0_,,, ... ,,,.,,,., ... ,-,, ... ,.,.•.•dd,•,•d,<d•'•'d,Od '•'"• .. , ... ,.,,,,,.,.,_,,,,,O,>,••••O•'••••••d•'"•~' 0 •·'
Tempo (h) [X] antes [X] depois Rompimento Av Proteínas si AE
(g/L) (g/L) (%) (U/mgprot)
o 2,8 1,5 46
24 2,8 1,7 39
48 2,9 1,3 55
60 3,3 1,8 45
96 3,0 1,5 50 -~~----~ --- ....................... ~--
(U/L) tampão
(mg/L)
96 303
136 362
136 400
174 345
208 337
0,32
0,38
0,34
0,50
0,62
- --
•••••••••••••••••••••••••••••••00••·••.,•••••••••••"••••••••••••••••••••••••••.,••••••• ...... ,,.,.,_,,.,. ... ,,.n,,,,, • .,,.,,,.u,•n•-"•"•"'-"'-"''"'·n•••"•••••~••••••••••••••••,.•••••••••••"•••••••·•••-•••--••••••••••••••••,.,.,,,.,,.,,,.,.,,,,.,,,o
Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 71,88 4,412 13,05 2,389 51 1,5 7,44
24 51,48 11,72 1,482 6,994 175 3,6 6,27
48 35,07 12,12 1,669 20,14 248 5,3 6,42
60 24,27 11,32 1,030 26,07 253 4,8 6,50
96 1,844 8,531 1,385 40,10 354 5,1 6,67

Tabela A.20. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guil/iermondii, com 30 g/L de xilose, 3 g/L de inóculo e pH 4,5
-------·-··-- ------------- ------------·----------·-----------·-·--···------ Tempo (h) [X] antes [X] depois Rompimento Av Proteínas s/ AE
(g/L) (g/L) (%) (U/L) tampão (U/mgprot)
(mg/L)
o 3,1 1,4 55 132 353 0,37
12 2,5 1,7 32 104 275 0,39
24 2,5 1,6 36 113 306 0,37
36 2,4 1,5 38 127 302 0,42
48 2,5 1,8 28 130 414 0,31 --·--------------, .. __________________ ,, ___ --·--------··-·--·- .. ----·-··-···- .. ··------·--·--- .. ---·--·--
................ ~ ............. , ... - ... , ............ -----·--·-··· ..... , ............................................. , .••.•••.•. , .• ,,.,.,,,.,,,J.•., .•. ~., ................................ , ................ _ ..... ______ ._,_,,,., .......................................... ............ ~-~--------···· .. ················--·-··"'·'·'·'·'·•··················· .. - . ._ .. .......... ,.,.,.,., .•. ,., .. .,.,.,.,,
Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 32,70 3,402 5,166 0,8264 149 3,5 4,62
12 19,88 5,056 0,4610 6,314 170 4,1 4,68
24 9,089 4,840 14,15 199 4,4 4,95
36 1,933 3,522 18,57 259 4,9 4,87
48 0,1764 0,9737 17,56 317 6,1 4,86 ~-~._._._. ....................................... ~. .J.~ .................... ,-·=_.- ........................ _. ..... w. ........ , .................................. <.&.h<-'~ ....... . ~., .......... ,w ..... ,._.._ ____ 0.0.0.•-'~•-khh~hh•·-hh~h··•••••••·h--··•-•·h-~.h•••M-h---~-
Tabela A.21. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
gui/liermondii, com 30 g/L de xilose, 3 g/L de inóculo e pH 7,5
Tempo (h) [XÍ ãntes "txJêiepÕis"-···-RÕmpimêniõ""""-'"""-· ·x:; . .. Proteínas s/
(g/L) (g/L) (%)
o 3,1 1,4 55 12 3,2 1,9 41
24 2,9 1,6 45
36 2,7 1,4 48
48 2,8 1,6 43
(U/L) tampão
(mg/L)
132 257
166 379
151 421
125 364
121 350
0,51
0,44
0,36
0,34
0,35 ··-·-----······-·--··-······-·-·-··-·---------·---------
---···-------------·-··-·--- -·---·----------··-···-··--·-··· .. ---------· .. ·-- •••••••-•••••••••••••m•••••••••-••••--•••• -·-·"··-- .. ·----·-··-····------······ Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 31,06 2,984 4,963 1,007 149 3,5 7,63
12 19,90 5, 113 0,8803 6,316 218 4,2 6,45
24 9,982 4,776 0,7370 11,79 250 4,8 6,53
36 2,082 4,157 0,6507 17,07 273 5,9 6,64
48 0,3384 1,791 0,4679 17,01 320 7,4 6,72 ··--··· .. ---····· .. ··-·-·-··-·· ··--···-···--··-·-- .. ---------·-···---· .. ·-·------------------·-··""'" ..... , ____ .. ___ ....................... _, ............................ .......................................... .................... _________
Tabela A.22. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guilliermondii, com 70 g/L de xilose, 3 g/L de inóculo e pH 4,5
Tempo (h) [X] antes . . [X] de-pois· Rõm'pimênto ······ Av··· P.rÕtê'inâs'"s/ .. . . A~---······ (g/L) (g/L) (%) (U/L)
o 3,1 1,7
24 2,7 1,5
48 2,4 1,4
60 2,4 1,3
96 2,4 1,3 -·=·~· -~-- ........... o>Jw ..
45 129
44 105
41 81
46 71
46 69
tampão (U/mgprot)
(mg/L}
369 0,35
362 0,29
218 0,37
195 0,36
188 0,37
Tempo. Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 70,40 7,712 10,83 1,508 116 2,8 4,58
24 54,68 10,98 1,663 4,397 167 4,3 4,48
48 40,19 10,91 1,820 9,947 162 4,8 4,50
60 34,32 11,13 1,600 13,45 160 5,4 4,52
96 15,30 11,26 0,8087 24,99 161 5,6 4,68 •H•u•.O••••~•-...,o. .. ,.,"'o·o;,,.,~-.-.,.__,.,_.,.,,..., .. , ....... o· .,,...,,,....,=,.--,.,.-....., •-q,.._..., ... ___ ,.,.., ___ .... __ "'"'" .. ""'"""""'-.,...,. .,_.,____ •·-·· ·=-•.u< -H--~··-- .....,.,._....,,-., _ .. .,,. ..,,., -..;,-..,._,.,< .-«,_ .. ,..,..,._...,.,.,n.•••••••4u ..... ,,.....,,.,.,. • .,.,_, ..

Tabela A.23. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guilliermondii, com 70 g/L de xilose, 3 g/L de inóculo e pH 7,5
---------··-··--·--·--·-·----- -·--··-----··------·---------------- Tempo (h) [X] antes [X] depois Rompimento Av Proteínas s/ AE
(g/L) (g/L) (%) (U/L) tampão (U/mQprot)
(mg/L)
o 3,0 1,6 47 113 328 0,34
24 3,2 1,9 41 128 384 0,33
48 2,7 1,5 44 123 342 0,36
60 2,9 1,6 45 128 398 0,32
96 2,9 1,6 45 167 423 0,40 --·----·- .. -·-·--· .. --·----·------ .... ,.,,_, _______ ···--·-··-·- .. -· ... -- .. ··-·--·----·------ .. -·- .. -·--- ----
~.,,,,,,,,,.,,,,,,, ,.,.,~,,,,,>,o,,.,,O,o,,,,,O,O,O,O,O,h•,••••~'•'•>•0,0 .. ,,.,,,,<,<,>,,,0,0,0,,,0, .. 0,0,0,>•0,00·N•,<,,.,,,,L•J,0-',~·-···-•••,0,0,,,,,o, .. ,, .. ,,ho,,,,,,,,,,.,,,,>,>,,,O,<,,,O,>,O•>,<,•-'•'•' ~,.,,O,,,O,M,,,0,>,<,>,>,0,0,0,,,0,0,0,0,0,0, .. 0,0,0•>•<•>,<,>,,,0,0,<,>,•,>,<,>,0,0,0•0,>,>, .. >,,•••-•,--l,•,,,,,>,•,><<,O,>,O,>,•,••••••'•'•''·'''·'•-•••-·••-•,•.-••,•·•·•,O,•,O·•••·•
Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 69,78 6,568 11,17 1,685 113 3,0 7,48
24 41,48 11,38 1,551 14,18 216 5,4 6,54
48 18,92 10,37 1,255 30,15 328 7,2 6,65
60 9,60 9,756 1,323 38,31 327 7,4 6,78
96 0,5946 5,195 0,9987 43,95 495 8,6 6,93 ................................................................... ......,. ...................................... _, ... _. ~.- ........... .,_............._.. ........................ __ ~ ........................ ~~·~-·- .. -·--·--·-"-'·"-""·~'- .... -~----~--- ... -- ...... , ....... _____ , .. _.,_ .. ___ .. __ .~-·---------~
Tabela A.24. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guillierrnondii, com 50 g/L de xilose, 2 g/L de inóculo e pH 6,0
Tempo (h) . [X} antes ·- -ixféfepÔis-·-·--RÕrrÍpimentéi-·--·-····-··· Av Proteínas s/ AE
(g/L) (g/L) (%) (U/L) tampão (U/mQprot)
(mg/L)
o 2,8 1,5 46 96 286 0,34
24 2,9 1,8 38 79 163 0,48
36 2,5 1,5 40 88 234 0,38
48 2,7 1,5 44 93 188 0,49
60 2,6 1,6 38 95 215 0,44

---------·-·---·-··---- ..... _,, ________________________________________________________________________ ,. _____________________________ ,,_ .. _______________________ ,,, ..... - ............... -----···--·-···--··-·····-·-·······--·- ··--···· Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 54,24 3,831 11,00 1,352 86 2,5 6,13
24 27,03 8,763 0,7655 12,51 117 4,3 5,92
36 17,48 8,582 0,6046 20,08 186 5,3 6,09
48 7,779 7,494 0,4372 24,67 234 6,8 6,13
60 1,749 6,433 0,4007 29,06 281 7,7 6,13 •••-•·---------•-•"•--•·------------•-•••-·------••••••••--••••••u•••,HM•·--H--•-·----------•••-----~·---•·•--•-- m••••••••-•••----•••,O,-HO••••••••••••••••••••••••
Tabela A.25. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guilliermondii, com 50 g/L de xilose, 2 g/L de inóculo e pH 6,0
-·····Temi>o'''fi,j'-·"-····-txj'·a-ntes·-·· '"''txfcfe.pÕis"''"''j:fo.mpime'i,iêi Av .. Proteínas si ,,.,,,.,Ã;···· '"' ''
(g/L) (g/L) (%) (U/L) tampão (U/mQprot)
(mg/L)
129 369 0,35
115 273 0,42
133 311 0,43
97 232 0,42
95 227 0,42 ____ ......_. ............................ ~~ ..........................
o 3,1 1,7 45
24 2,7 1,6 40
36 3,0 1,7 43
48 2,6 1,5 42
60 2,6 1,4 46
Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 24
36
48
60
50,85
26,31
15,50
6,246
O, 1811
5,465 7,602
7,706
7,539
7,002
4,645
1,154
0,6233
12,54
20,54
26,97
29,04
79
204
226
220
263
1,9
4,8
5,1
5,9
7,2
6,10
6,14
6,32
6,37
6,45

Tabela A.26. Ensaio fermentativo de produção de glicose-6-fosfato desidrogenase por Candida
guil/ierrnondii, com 50 g/L de xilose, 2 g/L de inóculo e pH 6,0
-----·-----·----------------·-----·----··------ ... -------·-·--·- .. ·-·-··----- Tempo (h) [X] antes [X] depois Rompimento Av
(g/L) (g/L) (%) (U/L)
o 3,1 1,7 45 129
24 2,7 1,6 40 75
36 2,6 1,6 38 92
48 2,6 1,6 38 120
60 2,4 1,4 41 136 -·---- .. ·--- .. --_ .. __ ,_,, ________ , __ ,, ________ ,, ____ ., _______________________ ,,,_, .. _ .
Proteínas s/ AE tampão (U/mQprot)
(mg/L)
369 0,35
248 0,30
221 0,42
244 0,49
341 0,40
Tempo Xilose Glicose Arabinose Acido Xilitol Av Células pH
(h) (g/L) (g/L) (g/L) acético (g/L) (U/L) (g/L)
(g/L)
o 51,49 5,134 7,733 1,150 79 1,9 6,10
24 26,69 7,510 11,92 130 4,7 6,10
36 16,52 7,563 0,7057 19,99 173 4,9 6,26 - ... ,.., 48 7,339 7,317 26,90 267 5,8 6,31
60 0,4432 4,942 29,45 379 6,7 5,42

APÊNDICE A.4 - CÁLCULO DO FATOR DE RENDIMENTO EM XILITOL, POR REGRESSÃO,
FATOR DE RENDIMENTO EM CÉLULAS, PRODUTIVIDADE EM XILITOL E PRODUTIVIDADE EM
CÉLULAS DO PLANEJAMENTO FATORIAL 23
Encontra-se nesta seção os gráficos da regressão linear dos valores de ~P
em função de (-~S), para o cálculo do fator de rendimento em xilitol em cada um dos
ensaios realizados. O valor para o fator de rendimento em xilitol, Yp1s, é fornecido
pelo coeficiente angular da reta de ~p = A+ Yp1s (-~S). O fator de rendimento em
células, YXJs, foi calculado pontualmente para cada um dos ensaios realizados. A
produtividade em xilitol (O») e a produtividade em células (Ox), também foram
calculadas. Os valores de cada parâmetro estão presentes na Tabela A.17.
18
14
.,.r,..••··•r,·,·,....,•,•o•,-, ... ..,-,,,.,.,,r,·<'•"-,-,•,-,-,•rr.,...,·~-.,.,._...., ............. ._.r•-,.r••r,-,.,,~-·,.,.,~,·rrr.-.•,-,•,•,-,•,-,•••r••••••ro~-.-·~r,•,•••
Parâmetro Valor Erro 16
o 'S 12 -c, o C: 10 o .., 8 o "" -~ 6
~ 4
YP/S 0,5962 0,5962 0,034
17,00461 0,52223
N p
a 17,00461 ---Ff. SD
0,99676 0,58804 4 0,00324
·25 -20 -15 -10 .5 -(Variação do substrato)
18
18
Parâmetro Valor Erro 14
,§ 12 -e,
l 10 o .., 8 s ~ 6
~ "
a
O,OC699 0,0354 ·
18,02981 0,56733
R SD N p
0,99661 0,6391 4 0,00339 2
o+-~~..--..--~~~~~~~~ -30 -25 -20 -15 -10 -5
-(Variação do substrato)
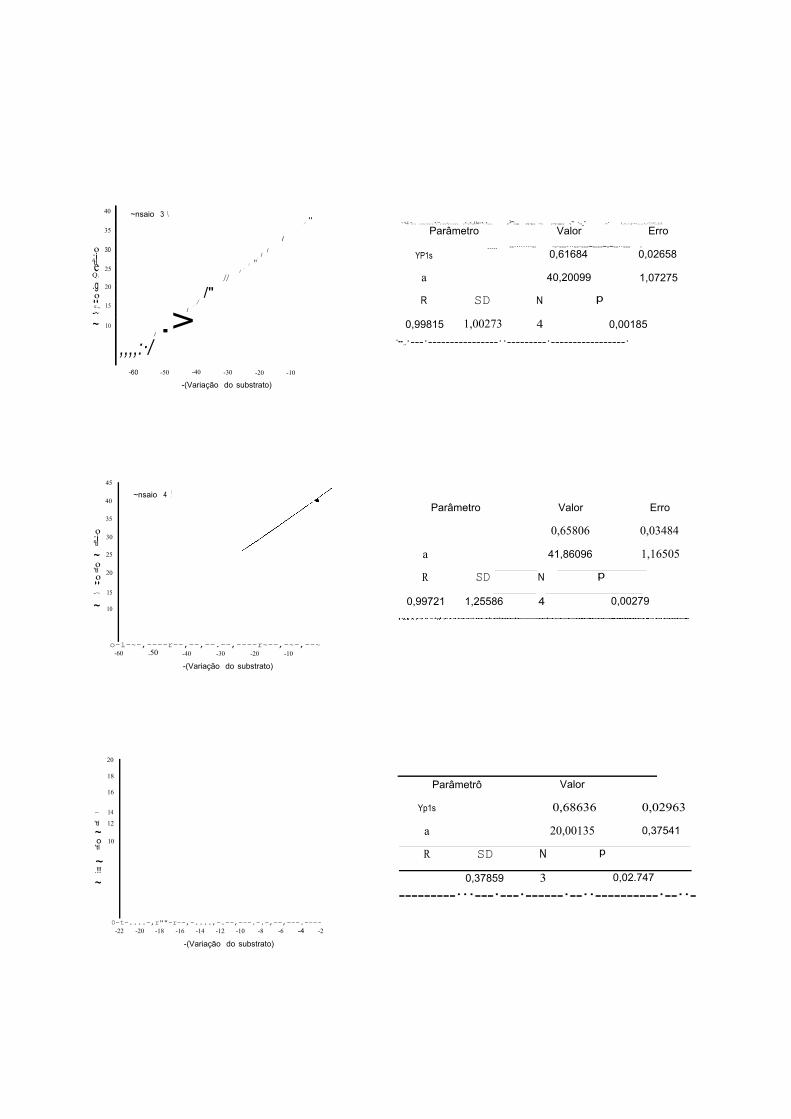
40 ~nsaio 3 \ .. /
35 /
o 30 /
/ :i /
'tl / ,,
e 25 / /
Q. // .g 20 /" o ..
/ /
e.,, 15 -~ /
~ 10 .> ,,,,:·/
/
-60 -50 -40 -30 -20 -10
-(Variação do substrato)
45
40 ~nsaio 4 !
35
o 30 :i
'tl
~ 25 o 'tl 20 o .. -~ 15
~ 10
.50 o-i-~-,----r--,--,--.--,----r~--,-~-,--~
-60 -40 -30 -20 -10
-(Variação do substrato)
20
18
16
~ 14
'tl 12
~ o 10 'tl
~ .!!!
~
0-t-....-,r""-r~-,-....,-.--,-~-.-.-,~-,-~-.-~~~ -22 -20 -18 -16 -14 -12 -10 -8 -6 -4 -2
-(Variação do substrato)
• <J ..• ,., ,.,.,.,.,., .•. ~,.,.,-,.,.,.,., ,.•.,.•,,JJo>• .. •,,., ,.>~,,,, ,.,.,,, ..., ,..,.,,,,, .,~ -...,_~ ....- L.,•.,.,~_j~•.•.,.,,.,,.,•,,.<,">.'C.,.,L.L
Parâmetro Valor Erro
YP1s ..... --········-- ·-·---···--·---~----~-~--··--- -
0,61684 0,02658
a 40,20099 1,07275
R SD N p
0,99815 1,00273 0,00185 4 ·-- .. ·---·----------------··---------·-----------------·
Parâmetro Valor Erro
a
0,65806
41,86096
0,03484
1,16505
R p SD N
0,99721 4 0,00279 1,25586
Parâmetrô Valor
Yp1s 0,68636 0,02963
a 20,00135 0,37541
R p SD N
0,37859 0,02.747 3 ---------···---·---·------·--··----------·--··-

0-t-..-,r-r-.-~.-..-,r-r-r~.-~~r-r-r~.-~~ -22 -20 -18 -18 -14 -12 -10 -S -6 -4 -2
-(Variaçao do substrato)
25
o 20 '5 "' e O.. 15 o
"' o ~ 10 .!!!
~
0-t-~~--,~r-r~-.-~~~.-~~~~~~--, -eo -50 -40 -30 -20
-(Variaçao do substrato) -10
45
~ 35
"' e o.. 30 o "' i 25
·ê ~ 20
15
-40 -30 -20 -10 -(Variaçao do substrato)
a
Parâmetro
R SD
0,99849 ···-~······-·--04·-~-·--···
0,41714
........ · ... Parãmeiro · · ·
a
R
0,99263
SD
Valor Erro
0,00139
18,13291
0,03304
0,42651
N p
3 0,03494
Valor Erro
0,52967 0,04573
32,32884 1,77345
N p
4 0,00737 1,29148
a
Parâmetro Valor Erro
0,73674 0,0224
44,64995 0,52178
R SD N p
4 9,22859E-4 0,99900 0,68123

JO 28 26 2,
o 22 ~ 20 e 1e ~ 16 ,, " .g 12 o-
·l:'i! 10 ~ 8
..JO -25 -20 -15 -10 -5
-(Variaçao do substrato)
JO 28 26 2,
o 22
~ 20 e 18
~ 16 ,, " o ca 12 o,
.S! 10
~ 8
JEnsaio 10 1 ./--;;
.> /
o+-~-.-~-...-~~-~-.-~-.--~-,-~ .JO -25 -20 -15 -10 -5
-(Variaçao do substrato)
30 :.s 26 2'
o 22
~ 20 e 18
~ 16
,, " _g 12 o-
..!!! 10
~ 8 6
• 2 O+-~-,-~-,.--,---,...-~--,-~-.-~-..~ -30
@nsaio 11 J
•
-25 -20 -15 -10 -5
-(Variaçao do substrato) o
Parâmetro
a
Valor Erro
0,63363
31,13998
0.0462
0,76ro3
·-------···-------------··-·--------------- 0,99473 0,88837 4 O,C!J527
Parâmetro
a
R SD 0,00326 1 ,05472
Valor Erro
0,64737 0,05342
31,07932 0,83254
N p
4 0,00674
Parâmetro Erro
YP/S
a
Valor ................ 0;68400 .
31,78563
0,06185
0,99698
R SD N p
0,99192 1,22031 4 O,ro!OO

Tabela A.27. Parâmetros fermentativos obtidos no planejamento fatorial 23
-- ··········-·-···-- .. ....,.,. .. , ... ,.,,,.,,., .... -= ................. -- ·---------------------------.-- ••••••••;.e.<,......m=......;.:.,.,...::,==-=.,.c,=L'-"-"-'"=.,•"•·J"I. .• ",
Ensaios Parâmetros Fermentativos
U/mQproteina Pxilitol YXJs Yp1s Qx Qp
(g/L) (g/g) (g/g) (g/L.h) (g/L.h)
1 0,28 16,54 0,076 0,59 0,056 0,34
2 0,55 16,73 0,094 0,60 0,071 0,35
3 0,53 38,50 0,050 0,62 0,038 0,40
4 0,62 40,10 0,048 0,66 0,037 0,42
5 0,31 18,57 0,041 0,69 0,039 0,52
6 0,35 17,07 0,075 0,60 0,067 0,47
7 0,37 24,99 0,044 0,53 0,029 0,26
8 0,40 43,95 0,074 0,74 0,058 0,46
9 0,44 29,06 0,092 0,63 0,087 0,48
10 0,42 29,04 0,094 0,65 0,088 0,48
11 0,40 29,45 0,085 0,68 0,080 0,49
.. _ _,/





























