Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE ENGENHARIA FLORESTAL
Programa de Pós-Graduação em Ciências Florestais e Ambientais
REGENERAÇÃO in vitro DE Eucalyptus cloeziana F. Muell.
ALEX ZICHNER ZORZ
Cuiabá – MT 2016

ALEX ZICHNER ZORZ
REGENERAÇÃO in vitro DE Eucalyptus cloeziana F. Muell.
Orientador: Dr. GILVANO EBLING BRONDANI
Co-orientadores: Dr. LEANDRO SILVA DE OLIVEIRA
Dr. ANDRÉ LUIS LOPES DA SILVA
Dissertação apresentada à Faculdade de
Engenharia Florestal da Universidade
Federal de Mato Grosso, como parte das
exigências do Curso de Pós-Graduação
em Ciências Florestais e Ambientais, para
obtenção do título de mestre.
Cuiabá – MT 2016


Agradecimentos
À minha família, pelo apoio constante e a fortaleza que representam para
minha vida.
À OEA e o Grupo Coimbra, à Universidade Federal de Mato Grosso e ao
“Programa de Pós-graduação em Ciências Florestais e Ambientais”, pela
oportunidade da realização do mestrado.
À CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior), pela concessão da bolsa de estudo.
Ao Professor Dr. Gilvano E. Brondani, pela orientação académica e
profissional e também pela amizade construída nesses anos.
Ao Professor Dr. Leandro Silva de Oliveira, pela amizade e ajuda na
condução dos experimentos.
Aos colegas do mestrado, essencialmente a Fernanda Furlan e Natália
Gavilan, pelo apoio, ajuda incondicional, amizade construída e pela
atmosfera de confiança que ofereceram para mim desde o primeiro dia do
curso.
À família do “Laboratório de Cultura de Tecidos Vegetais” e à grande família
dos Bolsistas Estrangeiros que também contribuíram direta ou
indiretamente na realização do trabalho.

SUMÁRIO
Lista de tabelas ........................................................................................ i
Lista de figuras ...................................................................................... iii
Resume .................................................................................................... v
Abstract ................................................................................................. vii
Resumen ............................................................................................... viii
1 INTRODUÇÃO ........................................................................................ 1
2 REVISÃO BIBLIOGRAFICA ................................................................. 4
2.1 Eucalyptus sp. ................................................................................. 4
2.2 Eucalyptus cloeziana ...................................................................... 5
2.3 Propagação de Eucalyptus cloeziana ........................................... 8
2.4 Micropropagação de Eucalyptus sp. ............................................. 9
2.5 Organogênese ................................................................................ 10
2.6 Fatores que influenciam a organogênese in vitro ..................... 12
2.6.1 Tipo de explante ......................................................................... 12
2.6.2 Reguladores de crescimento ..................................................... 14
3 MATERIAL E MÉTODOS .................................................................... 17
3.1 Local dos experimentos ................................................................ 17
3.2 Material vegetal ............................................................................. 17
3.3 Desinfestação das sementes e germinação in vitro .................. 17
3.4 Preparação do meio de cultura e condições experimentais ..... 17
3.5 Cultura organogênica .................................................................... 19
3.5.1 Fonte de explantes e tratamentos ........................................... 19
3.5.2 Regeneração in vitro ............................................................... 22
3.5.3 Multiplicação e alongamento in vitro ...................................... 22
3.5.4 Enraizamento ex vitro e aclimatização .................................. 23
3.6 Análise histológica ....................................................................... 23
3.7 Análise estatística dos dados ....................................................... 24
4 RESULTADOS E DISCUSSÃO ............................................................ 25
5 CONCLUSÕES ..................................................................................... 49
6 CONSIDERAÇÕES FINAIS E RECOMENDAÇÕES ............................ 50
7 REFERENCIAS BIBLIOGRAFICAS ................................................... 51

i
Lista de Tabelas
Tabela 1. Composição básica do meio de cultura WPM utilizado para a indução organogênica de Eucalyptus cloeziana ...................................... 18 Tabela 2. Descrição das combinações de fito-reguladores empregados na cultura calogênica de tecidos de Eucalyptus cloeziana inoculados in vitro ................................................................................................................. 20 Tabela 3. Intensidade de calejamento dos diferentes tecidos de Eucalyptus cloeziana inoculados in vitro aos 30 dias ................................................. 21 Tabela 4. Combinação dos fito-reguladores BAP e AIB na etapa de multiplicação e alongamento in vitro de explantes de Eucalyptus cloeziana ................................................................................................................. 22 Tabela 5. Resumo da análise de variância para a porcentagem de manifestação bacteriana (BAC), oxidação (OXI), calo (CAL) e regeneração dos brotos (REG) in vitro de Eucalyptus cloeziana em relação aos tratamentos testados aos 30 dias ............................................................ 25 Tabela 6. Porcentagem de explantes de Eucalyptus cloeziana que apresentaram manifestação bacteriana in vitro em relação aos diferentes tecidos e combinações de fito-reguladores na indução à calogênese, aos 30 dias após a instalação do experiment ................................................. 28 Tabela 7. Porcentagem de explantes de Eucalyptus cloeziana que apresentaram oxidação in vitro em relação aos diferentes tecidos e combinações de fito-reguladores na indução à calogênese, aos 30 dias após a instalação do experiment ............................................................. 30 Tabela 8. Porcentagem de explantes de Eucalyptus cloeziana que apresentaram calo in vitro em relação aos diferentes tecidos e combinações de fito-reguladores na indução à calogênese, aos 30 dias após a instalação do experiment .......................................................................................... 32 Tabela 9. Resumo da intensidade calogênica in vitro nos diferentes tratamentos .............................................................................................. 33 Tabela 10. Resumo das características anatômicas identificadas em calos de Eucalyptus cloeziana relacionados aos tratamentos na fase de indução calogênica ................................................................................................ 34 Tabela 11. Porcentagem de explantes de Eucalyptus cloeziana que apresentaram regeneração adventícia in vitro no meio de indução à organogênese, aos 30 dias ...................................................................... 40

ii
Tabela 12. Descrição dos tratamentos que apresentaram formação de brotos adventícios na etapa de regeneração ........................................... 41 Tabela 13. Resumo da análise de variância do número de brotos (NB) por calo regenerado in vitro de Eucalyptus cloeziana na etapa de regeneração de brotos em relação aos tratamentos, aos 90 dias ................................ 41 Tabela 14. Número de brotos (NB) por calo regenerado in vitro de Eucalyptus cloeziana em relação aos diferentes tecidos de origem e as combinações de fito-reguladores na etapa de regeneração .................... 43 Tabela 15. Resumo da análise de variância para a porcentagem de sobrevivência (SOB%) e enraizamento (ENR%) ex vitro de brotações de Eucalyptus cloeziana aos 20 dias ............................................................ 45 Tabela 16. Porcentagem de sobrevivência (SOB%) ex vitro de microestacas de Eucalyptus cloeziana segundo a origem e os meios de multiplicação e alongamento in vitro (MMA), aos 20 dias ........................ 46 Tabela 17. Porcentagem de enraizamento (ENR%) ex vitro de microestacas de Eucalyptus cloeziana segundo a origem e os meios de multiplicação e alongamento in vitro (MMA), aos 20 dias ........................ 47

iii
Lista de figuras Figura 1. Ocorrência natural de Eucalyptus cloeziana .............................. 6 Figura 2. Tecidos de Eucalyptus cloeziana inoculados in vitro. (A) Hipocótilo; (B) cotilédone; (C) raiz. Barra = 0,25 cm ................................ 19 Figura 3. Manifestação de colônias bacterianas e oxidação in vitro em explantes de Eucalyptus cloeziana, aos 30 dias após a inoculação. (A) Tecido do hipocótilo apresentando manifestação de bactérias (setas) e (D) oxidação. (B) Bactérias presentes (setas) no tecido cotiledonar e (E) oxidação do tecido. (C) Segmentos de raiz apresentando manifestação de bactérias (setas) e (F) oxidação do tecido. Barra = 0,5 cm ...................... 27 Figura 4. Histologia de calos de Eucalyptus cloeziana. (A) Formação de calo (setas) na epiderme do explante. (B) Detalhe de estruturas meristemáticas (EM) induzidas na epiderme do explante. (C) Raízes adventícias (Ra) geradas a partir do calo (C). (E) Calos apresentando área com características meristemáticas e (D) formação de meristemoides (m). Barra = 100 µm ........................................................................................ 36 Figura 5. Calos embriogênicos de Eucalyptus cloeziana. (A e C) Detalhe de embriões somáticos de origem multicelular (ES) na fase globular desenvolvidos diretamente do explante inicial e (E) indiretamente através do calo. Barra = 100 µm. (B e D) Calos apresentando células pró- embrionárias (CPE). Barra = 25 µm. S: suspensor. Pt: Protoderme ........ 37 Figura 6. Histologia de calos organogênicos de Eucalyptus cloeziana obtidos do tratamento constituído pela combinação 7 (2,0 mg L-1 de TDZ) e o tecido do hipocótilo, evidenciando a regeneração de brotos. (A) Detalhe de regiões de atividade meristemática (setas vermelhas) e (A e B) brotações adventícias (setas amarelas) apresentando conexão vascular com a massa celular de origem (seta preta). C= Calo; B= Broto; F= Folha; MA= Meristema apical ............................................................................. 39 Figura 7. Detalhe da síntese de antocianina (setas) na superfície de calos organogênicos no 1o subcultivo (30 dias) dos explantes no meio de regeneração de brotos. Calos advindos tratamentos de indução calogênica da combinação 7 (2,0 mg L-1 de TDZ) nos tecidos do (A) hipocótilo e (B) cotilédone. Barra = 1 cm .......................................................................... 39 Figura 8. Organogênese in vitro, enraizamento ex vitro e aclimatização de microestacas de Eucalyptus cloeziana. (A e B) Calo apresentando regeneração de brotos adventícios in vitro no meio de regeneração, aos 90 dias (barra = 1 cm); (C) Calo regenerado apresentando brotos alongados ao terceiro subcultivo (90 dias) (barra = 0,5 cm); (D) Broto coletado para enraizamento ex vitro (barra = 1 cm); (E e F) Enraizamento ex vitro debrotos aos 20 dias (barra = 1 cm); (G) Enraizamento in vitro e formação de micro-

iv
cepa (barra = 1cm); (H) Mudas aclimatizadas e alocadas em casa de sombra aos 150 dias ................................................................................ 42 Figura 9. Detalhe do tufo padrão utilizado para a multiplicação e alongamento dos brotos. Corte longitudinal do calo organogênico procedente do hipocótilo induzido pela combinação 7 (2,0 mg L -1 de TDZ), conservando na base o tecido de origem. C= calo .................................. 44

v
RESUMO
ZICHNER, A. Z. Regeneração in vitro de Eucalyptus cloeziana F. Muell. 2015. (Mestrado em Ciências Florestais e Ambientais) – Universidade Federal de Mato Grosso, Cuiabá-MT. Orientador: Dr. Gilvano Ebling Brondani. Co-orientadores: Dr. Leandro Silva de Oliveira; Dr. André Luís Lopes da Silva.
Dentre as espécies de Eucalyptus plantadas no Brasil, o Eucalyptus cloeziana F. Muell tem papel destacado no setor florestal em razão das características da madeira, especialmente nos setores industriais energéticos e moveleiros. Atualmente, as instalações de plantios comerciais da espécie se reduzem a materiais genéticos por via seminal, resultando povoamentos de alta heterogeneidade com baixos rendimentos econômicos. A dificuldade de promoção rizogênica de propágulos representa o principal limitante na reprodução clonal de genótipos selecionados, na qual a cultura de tecidos, através da técnica de micropropagação, apresenta-se como uma opção atrativa e amplamente reconhecida pelas vantagens que oferece no intuito de propagar espécies e genótipos de alto valor genético. Considerando os escassos estudos direcionados ao desenvolvimento de metodologias de reprodução vegetativa de E. cloeziana, o presente trabalho teve por objetivo principal desenvolver um protocolo efetivo de micropropagação através da organogênese indireta, avaliando a interação de fito-reguladores em tecidos juvenis nas distintas etapas estabelecidas. Hipocótilo, cotilédone e raiz, foram inoculados in vitro e submetidos a 27 combinações de fito-
reguladores, compostas por concentrações entre 0, 2,0 e 4,0 mg L-1 do ácido naftalenoacético (ANA) e 0, 1,0 e 2,0 mg L-1 do thidiazuron (TDZ) e do ácido diclorofenoxiacético (2,4-D) respectivamente, sendo estes os principais tratamentos nas fases de calogênese, regeneração adventícia de brotos, multiplicação e alongamento das brotações in vitro e sua competência no enraizamento e aclimatização ex vitro. Os resultados foram avaliados mediante a análise de variância (p<0,01 e p<0,05) e a comparação de médias pelo teste de Duncan (p<0,05). A resposta à desdiferenciação celular e formação de calo foi eficiente na maioria dos tratamentos, destacando as elevadas intensidades calogênicas fornecidas
pelas concentrações de 1,0 mg L-1 de 2,4-D e 2,0 mg L-1 de TDZ nos tecidos utilizados. Analises histológicas favoreceram a identificação de características morfogênicas relacionadas à formação de estruturas pro embriogênicas e embriôes somáticos na fase globular destacados nas combinações dos reguladores ANA e 2,4-D. Todos os tecidos apresentando calo foram transferidos no meio de regeneração constituído
pela concentração de 1,0 mg L-1 de benzilaminopurina (BAP) para estimular a competência dos calos à formação de brotações adventícias destacando- se, principalmente, o tratamento composto pelo tecido do hipocótilo na
concentração de 2,0 mg L-1 do TDZ do meio de indução, o qual forneceu uma média de 41,5 brotos por calo organogênico. Na fase de enraizamento e aclimatização ex vitro, o mesmo tratamento foi destacado nas variáveis de porcentagem de sobrevivência (80 e 100%), e enraizamento (55,6 e 80%) das brotações advindas dos meios de cultura de multiplicação e

vi
alongamento. O sistema de miniestufa foi adequado para a aclimatização inicial e posteriormente as mudas foram rustificadas em casa de sombra (50%). Dessa forma, comprovou-se a eficiência da cultura de tecidos através da organogênese in vitro e sua possível implementação para o desenvolvimento de novas metodologias visando a produção de mudas de Eucalyptus cloeziana.
Palavras-chave: Eucalipto, organogênese, calo, enraizamento adventício.

vii
ABSTRACT
ZICHNER, A. Z. In vitro regeneration of Eucalyptus cloeziana F. Muell. 2015. 2015. Dissertation (Master in Forestry and Environmental Sciences) - Federal University of Mato Grosso, Cuiabá. Adviser: Dr. Gilvano Ebling Brondani. Co-advisers: Dr. Leandro Silva de Oliveira; Dr. André Luís Lopes da Silva. Among the species of Eucalyptus planted in Brazil, Eucalyptus cloeziana F. Muell has a prominent role in the forestry sector due to the wood characteristics, especially in the energy and furniture industrial sectors. Currently, the commercial growing of plants of the species are reduced to genetic material by seminal seedlings, resulting stands high heterogeneity with low economic returns. The difficulty of rizogenic seedlings promotion is the main limiting the clonal reproduction of selected genotypes, in which tissue culture through micropropagation technique presents itself as an attractive option and widely recognized by the advantages it offers in order to propagate species and high genetic value genotypes. Considering the few studies directed to the development of vegetative reproduction methods of E. cloeziana, the present work had as main objective to develop an effective micropropagation protocol by indirect organogenesis, assessing the interaction of phyto-regulators in juvenile tissues in different stages established. Hypocotyl, cotyledon and root were inoculated in vitro and submitted to 27 combinations of phyto-regulators, concentrations
comprised between 0, 2.0 and 4.0 mg L-1 naphthalene acetic acid (NAA) and 0, 1.0 and 2.0 mg L -1 thidiazuron (TDZ) and dichlorophenoxyacetic acid (2,4-D) respectively, which are the main treatments in phases of callus formation, adventitious bud regeneration, multiplication and elongation of shoots in vitro and competence rooting and acclimatization ex vitro. The results were evaluated by analysis of variance (p<0.01 and p<0.05) and comparison of means by Duncan test (p<0.05). The response to cell dedifferentiation and callus formation was efficient most of the treatments
highlighting the high calogenic intensities provided by 2.0 mg L-1 of TDZ concentration and 1.0 mg L-1 of 2,4-D on all tissues tested. Histological analysis facilitated the identification of morphogenesis features related to the formation of pro embryogenic structures and somatic embryos in globular stage highlighting combinations of NAA and 2,4-D regulators. All callus tissue was exposed to the effect of the benzylaminopurine (BAP) concentration of 1.0 mg L-1 to enhance the competence of the callus regeneration of adventitious shoots highlighting mainly the treatment
composed by the hypocotyl tissue in the TDZ concentration of 1.0 mg L-1
induction medium, which provided an average of 41.5 shoots per organogenic callus. In the ex vitro acclimatization phase, the same treatment was highlighted in survival percentage variables (80 and 100%), and rooting (55.6 and 80%) of shoots arising from multiplying and stretching culture medium. The mini-incubator system was adequate for the initial acclimatization and later the seedlings were rustic in shade house (50%). Thus, it proved the efficiency of tissue culture through organogenesis in vitro and possible implementation for the development of new methods aiming at

viii
the production of Eucalyptus cloeziana plants.
Keywords: Eucalypt, organogenesis, calli, adventitious rooting

ix
RESUMEN
ZICHNER, A. Z. Regeneración in vitro de Eucalyptus cloeziana F. Muell. 2015. (Maestría en Ciencias Forestales y Ambientales) – Universidad Federal de Mato Grosso, Cuiabá-MT. Orientador: Dr. Gilvano Ebling Brondani. Coorietadores: Dr. Leandro Silva de Oliveira; Dr. André Luís Lopes da Silva.
Entre las especies de eucalipto plantados en Brasil, Eucalyptus cloeziana F. Muell tiene un papel destacado en el sector forestal, debido a las características de la madera, especialmente en los sectores energéticos e fabricantes de muebles. Actualmente, el cultivo comercial de plantas de la especie se reduce a material genético por vía seminal, lo que demuestra alta heterogeneidad con retornos económicos bajos. La dificultad de la promoción rizogénica de las plántulas es la principal limitación de la reproducción clonal de genotipos seleccionados, en los que el cultivo de tejidos a través de la técnica de micropropagación, representa como una opción atractiva y ampliamente reconocida por las ventajas que ofrece para propagar especies y genotipos de altos valor genético. Teniendo en cuenta los pocos estudios dirigidos al desarrollo de métodos de reproducción vegetativa de E. cloeziana, el presente trabajo tuvo como objetivo principal el desarrollo de un protocolo efectivo de micropropagación E. cloeziana por organogénesis indirecta, mediante la evaluación de la interacción de los fito-reguladores en los tejidos juveniles en las diferentes etapas establecidas. Hipocótilo, cotiledón y segmentos de raíz fueron inoculados in vitro y sometidos a 27 combinaciones de fito-reguladores, con concentraciones comprendidas entre 0, 2,0 y 4,0 mg L-1 ácido acético naftaleno (ANA) y 0, 1,0 y 2,0 mg L-1 thidiazurón (TDZ) y ácido diclorofenoxiacético (2,4-D), respectivamente, fueron los principales tratamientos en las fases de formación de callo, la regeneración yemas adventicias y la competencia enraizamiento ex vitro, evaluados mediante análisis de la varianza (p<0,01 y p<0,05) y la comparación de medias mediante el test de Duncan (p <0,05). La respuesta a la desdiferenciación celular y la formación de callo fue eficiente para la mayoría de los tratamientos destacando altas intensidades de calogênicas proporcionadas por las concentraciones de 2,0 mg L-1 de TDZ y 1,0 mg L-1 2,4-D en los tejidos testados. Análisis histológicos detectaron características morfogénicas relacionadas a la formación de estructuras pre embriogénicas y embriones somáticos en fase globular, destacados en las combinaciones de los reguladores ANA y 2,4-D. Todos los callos han sido expuestos al efecto de la concentración 1,0 mg L-1 de la citocinina bencilaminopurina (BAP) para inducir a la regeneración de brotes adventicios, en los cuales se destacó el tratamiento que comprende el tejido de hipocótilo y la concentración de 1,0 mg L-1 de TDZ del medio de inducción, que proporcionó un promedio de 41,5 brotes por callo organogénico. En la fase de aclimatación ex vitro, el mismo tratamiento se destacó en las variables de porcentaje de supervivencia (80 y 100%), y enraizamiento (55,6 y 80%) de los brotes resultantes de los medios de multiplicación y alongamiento in vitro. El sistema miniestufa fue adecuado para la aclimatación inicial y más tarde las plántulas fueron rustificadas en casa de sombra (50%). Por lo

x
tanto, se demostró la eficacia de cultivo de tejidos a través de organogénesis in vitro y su posible aplicación para el desarrollo de nuevos métodos con el objetivo de la producción satisfactoria de plantas de Eucalyptus cloeziana. Palabras claves: Eucalipto, organogénesis, callos, enraizamiento adventicio.

1
1 INTRODUÇÃO
Dentro das espécies exóticas mais cultivadas no setor florestal
brasileiro destacam-se as espécies do gênero Eucalyptus; reconhecidas
por suas características silviculturais, rápido crescimento, grande
plasticidade ecológica e propriedades na madeira, as quais tornam à árvore
atrativa para atividades comerciais e de pesquisa.
A maioria dessas espécies se desenvolve em regiões tropicais e
subtropicais, assim como no Brasil onde é considerado o gênero florestal
mais importante. Diversas espécies vêm sendo avaliadas para o
estabelecimento de plantações comerciais devido à grande demanda de
produtos florestais. Dentre as espécies promissoras destaca-se o
Eucalyptus cloeziana pela aptidão e competência na silvicultura. Ademais,
características tecnológicas da sua madeira como alta densidade, grande
durabilidade e excelente qualidade para serrarias, produção de carvão e
uso na construção civil apresentam-se atrativas para o setor florestal.
O melhoramento genético é uma realidade de grande
importância no setor florestal atual, a qual se fundamenta na seleção de
espécies e indivíduos com características determinadas por fatores
genéticos e na evolução das ferramentas para obter produtos madeireiros
de maior qualidade, num espaço e tempo menor, para diversos ramos
industriais que expressam constante crescimento.
Associada às técnicas de propagação clonal destes indivíduos
selecionados, a biotecnologia também tem avançado com os programas de
melhoramento das espécies florestais, fornecendo metodologias para a
produção massiva de mudas através da clonagem de árvores. Como forma
de propagação vegetativa, baseada em técnicas de cultura de tecidos in
vitro, a micropropagação torna-se uma alternativa para a regeneração de
plantas que apresentam dificuldades de reprodução natural e também onde
os métodos convencionais de propagação clonal não se tornam viáveis
(THORPE et al., 1991).
Uma das metodologias atuais adotadas para a obtenção de
mudas de espécies de Eucalyptus constitui a organogênese, atualmente
escolhida como alternativa para micropropagar espécies lenhosas em

2
grande escala. Ademais atende trabalhos de transformação genética que
estão sendo gerados com a tentativa de inserir caracteres genéticos que
poderiam promover propriedades de interesse econômico, não apenas à
produtividade, mas também à qualidade de madeira, resistência a
herbicidas, pragas e doenças (GIRIJASHANKAR, 2011).
A realização de análises histológicas em trabalhos de
organogênese in vitro é fundamental para a confirmação da origem dos
novos órgãos formados em cultivo, excluindo-se a possibilidade de
interpretação equivocada dos resultados advindos da presença de gemas
pré-formadas no explante original (ALVES et al., 2004b).
Apesar dos grandes avanços na silvicultura clonal das espécies
de eucalipto, a propagação vegetativa de Eucalyptus cloeziana constitui
ainda um fator limitante para a produção de mudas em escala comercial,
sendo a espécie considerada, por muitos autores, como de difícil
enraizamento em comparação às demais espécies do gênero (ALFENAS
et al., 2004).
Por tanto, a importância de desenvolver novas metodologias e
protocolos efetivos de regeneração in vitro, especificamente para espécie
Eucalyptus cloeziana são atrativas, visando conhecer as diversas
respostas envolvidas no processo da micropropagação, as quais estão
associadas não apenas à resposta da espécie também as respostas
envolvidas na idade fisiológica dos explantes e a resposta aos reguladores
de crescimento.
1.1 Objetivos
Objetivo geral
Estabelecer um protocolo efetivo para a micropropagação de
Eucalyptus cloeziana através da organogênese indireta.
Objetivos específicos
1. Avaliar o efeito das combinações de fito-reguladores (ácido
naftalenoacético (ANA), thidiazuron (TDZ) e ácido diclorofenoxiacético (2,4-
D)) na indução de calos em tecidos juvenis (hipocótilo, cotilédone e raiz).
2. Avaliar a competência morfogênica dos calos na regeneração
de órgãos adventícios.

3
3. Avaliar a composição da origem dos brotos na resposta à
promoção de raízes adventícias.
4. Identificar padrões de diferenciação histológica e a origem
do desenvolvimento organogênico e/ou embriogênico nos explantes.

4
2 REVISÃO BIBLIOGRAFICA
2.1 Eucalyptus sp.
O gênero Eucalyptus engloba aproximadamente 700 espécies
arbóreas que pertencem à família Myrtaceae (BOLAND et al., 2006) onde
naturalmente ocorrem nos territórios da Austrália, Filipinas, Ilhas Timor e
Nova Guiné e representam, provavelmente, as árvores mais conhecidas
(ALVES, 2007), com maior quantidade de investigações e mais utilizadas
para plantações florestais (ALVES et al., 2004).
As principais características destas árvores se referem ao rápido
crescimento, a plasticidade e adaptabilidade às diferentes condições edafo-
climáticas que permite inúmeras utilizações como matéria-prima, onde sua
madeira é aproveitada para a construção civil, carvão para energia,
produção de papel, e as folhas para a extração de óleos essenciais (TIBOK
et al., 1995) e, atualmente, escolhidas também por sua capacidade de
oferecer serviços ambientais, tais como a captura de carbono atmosférico
e recuperação de áreas degradadas.
Muitas espécies de Eucalyptus, e seus híbridos, são utilizados
amplamente como espécies exóticas para plantações em muitas áreas
tropicais e subtropicais do mundo (POTTS e DUNGEY, 2004; PEREIRA et
al., 2000). Os principais híbridos (incluindo os recíprocos) utilizados
comercialmente são o E. grandis × E. urophylla, E. grandis × E.
camaldulensis incluindo as espécies E. saligna, E. pellita, E. exserta e E.
tereticornis (DUNGEY e NIKLES 2000). Embora no Brasil e no Congo são
plantados em grande escala, consideráveis plantações também ocorrem na
China, Indonésia e África do Sul. Há também pequenas áreas destes
híbridos e espécies de eucalipto em outros países da Ásia (Filipinas, Vietnã,
Tailândia e Malásia) e América do Sul (Chile, Uruguai, Argentina, Paraguai)
(DUNGEY e NIKLES, 2000).
Nas áreas de florestas plantadas no Brasil com Eucalyptus,
predomina a espécie E. grandis, que ocupa 55% da área total, seguido de
17% com E. saligna, 11% com híbridos de E. grandis × E. urophylla, 9%
com E. urophylla, 2% com E. viminalis, e 6% com outras espécies
(CAMPOS et al., 2011), onde as espécies de E. cloeziana e E. dunnii são

5
consideradas promissoras para as regiões central e sul, respectivamente,
e o E. benthamii, vem se destacando nas regiões frias do sul do país
(HIGASHI et al., 2000).
Segundo o Anuário Estatístico da Associação Brasileira de
Produtores de Florestas Plantadas (ABRAF, 2013) em 2012, a área de
plantios de Eucalyptus no Brasil totalizou 5.102.030 ha, representando
crescimento de 4,5% (228.078 ha) frente ao indicador de 2011. Tratando-
se de produtos de florestas plantadas, o Brasil figura como o maior
exportador mundial de celulose e fibra de eucalipto (SARTORETTO et al.,
2008). O principal fator que alavancou esse crescimento foi o
estabelecimento de novos plantios com base na biotecnologia, frente à
demanda futura dos projetos industriais, principalmente do segmento de
papel e celulose.
2.2 Eucalyptus cloeziana
Natural da Austrália, o Eucalyptus cloeziana F. Muell também é
denominado como Gympie messmate, e representa a única espécie do
subgenero Idiogenes, onde ocorre ao leste do Estado de Queensland, no
distrito de Gympie no sudeste, com extensas ocorrências em Mundubbera
ao oeste de Springsure, e alguns pomares isolados no norte e oeste de
Townsville (Figura 1). Apresenta ocorrências descontínuas ao norte da
região de Townsville e ao oeste de Cooktown (BOLAND et al., 2006).
Na região de origem, o clima é quente e sub-úmido a úmido, com
temperatura média do mês mais quente entre 29 e 34ºC, e a média das
mínimas do mês mais frio entre 5 e 18ºC, podendo ocorrer fracas geadas
em até 5 dias por ano. A precipitação pluviométrica média anual varia entre
550 e 2300 mm (IPEF, 2015). O melhor desenvolvimento da espécie ocorre
nos solos bem drenados, ácidos e de baixa a média fertilidade natural, ou
seja, situações associadas a áreas de depósito de sedimentos ou de
misturas calcário e argila de origem vulcânica, de profundidade moderada
(BOLAND et al., 2006).
Eucalyptus cloeziana é uma espécie rústica e de rápido
crescimento, adequada para reflorestamentos destinados à produção de
madeira para diversos usos.

6
No Brasil, é uma espécie considerada de recente introdução e
tem-se observado bom desempenho de diferentes procedências em
regiões tropicais como no Cerrado (savana) onde, segundo Moura e
Guimarães (1988) foram registrados crescimentos em volume iguais a 35
m3/ha/ano, destacando-se materiais procedentes de Kennedy, assim como
também na região nordeste do pais onde plantios das procedências de
Gympie e Paluma tiveram desenvolvimento esperados (MOURA e
GUIMARÃES, 1988). Em regiões subtropicais também apresentaram
desenvolvimento adequado, principalmente no sudeste e o Sul (MOURA,
2003; LORENZI et al., 2003).
Figura 1. Ocorrência natural de Eucalyptus cloeziana (BOLAND et
al., 2006)
Segundo Moura (2003), existem registros de plantações
estabelecidas com essa espécie em outras regiões do mundo, onde é
considerado importante para reflorestamentos destinados para suportes de
linhas telefônicas e transmissão de energia, especialmente em países no
sul da África como Congo, Quênia, Malawi, Nigéria, Moçambique, África do
Sul, Zimbábue e Uganda e Zâmbia. Os plantios são realizados,
principalmente nas regiões de chuvas de verão com precipitação anual de
1000 a 1500 mm e uma estação seca de 4 a 5 meses (HOPEWELL et al.,
2008).

7
Hopewell et al. (2008) e Chen et al. (2010) consideram a
Eucalyptus cloeziana como uma das espécies candidatas para o
estabelecimento de plantios comerciais com objetivo de produção de
madeira sólida, devido a sua excepcional forma e retidão do fuste
(BLEAKLEY e CANT, 1985), o que garante a presença de toras com alto
aproveitamento no processamento mecânico.
Além disso, a alta densidade (0,855 a 1,140 g/cm3) e a durabilidade
natural da madeira, mesmo sem tratamento químico, são, sem dúvida, as
principais qualidades desejadas para a construção civil, produção de
escoras, caibros e mourões (PEREIRA et al., 2000; PALUDZYSZYN FILHO
et al., 2006), além de oferecer matéria-prima para atender às exigências da
indústria moveleira e de laminados (GONZALEZ et al., 2006;
INTERAMNENSE, 1998).
Eucalyptus cloeziana, em conjunto com outras espécies de
eucalipto, tais como E. camaldulensis, E. citriodora e E. deglupta,
representa uma das espécies escolhidas para a instalação de plantações
para fins energéticos (COUTO et al., 2011). Em programas de
melhoramento genético são selecionados espécies e clones que
apresentem resistência a doenças e alto conteúdo de carbono fixo e lignina
na madeira, desejado para a produção de carvão (AMS, 2004),
características que diferem em projetos para plantios direcionados às
indústrias de celulose e papel.
Entre as limitações silviculturais importantes da espécie, E.
cloeziana apresenta susceptibilidade às geadas fortes (CLARKE et al.,
2009) e não se adapta bem em regiões com deficiência hídrica severa
(NGUGI et al., 2004).
Em relação à instalação de plantações, esporadicamente esta
espécie é afetada pelo lepidóptero desfolhador Thyrintheina arnobia em
solos arenosos e de baixa fertilidade (MOURA, 2001). Também se destaca
a susceptibilidade à ferrugem causada por Puccinia psidii (FERREIRA.
1989; ALFENAS et al., 2004).
A incidência severa de ferrugem causada por este fungo foi
verificada nos plantios de E. cloeziana no sul da Bahia, com até seis a oito
meses de idade (MORAES et al., 1982). Esta doença incide sobre partes

8
jovens da planta, tais como primórdios foliares, pecíolos, ápices dos ramos
e na haste principal de mudas no viveiro; e ataca da mesma forma,
brotações de cepas (RUIZ et al., 1987), limitando o seu plantio em regiões
sujeitas a esta enfermidade.
2.3 Propagação de Eucalyptus cloeziana
Escassos são os estudos científicos que envolvem a produção
de mudas de Eucalyptus cloeziana o qual necessita, todavia, de maior
desenvolvimento e pesquisa que possam consolidar metodologias de
propagação, tanto seminal como clonal, que possam apoiar programas de
melhoramento genético e o estabelecimento de plantios comerciais.
A metodologia de propagação dessa espécie ainda é restrita
pela obtenção de mudas por sementes, mesmo que a espécie apresente
altos índices de produção de sementes, todavia há baixa porcentagem de
germinação e dificuldade na repicagem em condições de viveiro (MOURA
e GUIMARÃES, 2003). Em condições in vitro Cid et al. (1997) relataram a
dificuldade de germinação e estabelecimento de sementes de E. cloeziana
devido, principalmente, a manifestação de fungos e a presença de
bactérias no meio de cultura.
Atualmente, os avanços na silvicultura clonal de diferentes
espécies do gênero Eucalyptus são relevantes na comunidade científica e
empresarial. Contudo, são poucos os estudos que envolvem a propagação
vegetativa de Eucalyptus cloeziana em escala comercial, considerando que
o enraizamento adventício em estacas e miniestacas apresenta-se como
fator limitante para a produção de mudas clonais (ALFENAS et al., 2004).
Técnicas de propagação amplamente utilizadas como estaquia
e miniestaquia foram desenvolvidas por Almeida et al. (2007a) e Almeida
et al. (2007b) respectivamente, os quais verificaram altas porcentagens de
enraizamento dos propágulos, mesmo assim destacaram a variabilidade
apresentada dos resultados dependendo dos materiais genéticos e da fonte
de fornecimento dos propágulos.
Trueman et al. (2013) também avaliaram a porcentagem de
enraizamento de miniestacas de E. cloeziana frente a condições de
temperatura nas quais as mini-cepas foram submetidas, concluindo que

9
elevando a temperatura, em torno de 33 °C, foi eficiente para promover a
produção de propágulos e aumentar a porcentagem de enraizamento.
Atualmente, visando o estabelecimento de protocolos efetivos
de propagação de espécies de Eucalyptus que apresentam dificuldades ao
enraizamento de propágulos, são utilizadas estratégias mediante a cultura
in vitro, que é amplamente descrito na literatura (SHARMA e
RAMAMURTHY, 2000; BORGES et al., 2011, OLIVEIRA et al., 2012;
BRONDANI et al., 2012a), em situações em que as metodologias
convencionais não foram satisfatórias para a promoção da juvenilidade dos
tecidos ou mesmo para aumentar a multiplicação e produção de mudas
clonais (CHAPERON, 1987).
Baseado nesse contexto, Oliveira (2014) estabeleceu
protocolos pioneiros de micropropagação por meio das técnicas de
organogênese indireta e a proliferação de gemas axilares de Eucalyptus
cloeziana, evidenciando os procedimentos como efetivos para a
propagação extensiva da espécie. Assim também, Oliveira et al. (2015)
avaliaram o processo da micropropagação de material adulto de árvores
selecionadas de Eucalyptus cloeziana promovendo a reversão do estado
juvenil e em consequência, o enraizamento efetivo das micro-estacas,
fornecendo resultados promissores para a propagação clonal da espécie.
2.4 Micropropagação de Eucalyptus sp.
A micropropagação é a aplicação mais prática da cultura de
tecidos de plantas, sendo uma das técnicas de propagação de maior
impacto e mais difundida nos últimos anos (GRATTAPAGLIA e MACHADO,
1998; OLIVEIRA et al., 2013). Atualmente se encontra embutida no setor
florestal, principalmente no que diz respeito aos programas de
melhoramento genético.
A clonagem de Eucalyptus no Brasil passou por inúmeras
transformações, tendo iniciado com a técnica de estaquia que foi
implementada em escala comercial no final da década de 70 (XAVIER et
al., 2013).
Devido às dificuldades de enraizamento encontradas em
algumas espécies e híbridos de Eucalyptus na propagação clonal por

10
estaquia, principalmente no que envolve material adulto, o
desenvolvimento de técnicas visando a reversão à juvenilidade e o vigor
dos propágulos, como a mini-estaquia seriada (WENDLING e XAVIER,
2005) e da micro-estaquia (XAVIER e COMÉRIO, 1996; ASSIS, 1997) têm
permitido avanços consideráveis na propagação clonal de Eucalyptus. Em
clones com maior dificuldade de enraizar, existe maior eficiência de
enraizamento em resposta ao rejuvenescimento in vitro pelo uso da micro-
estaquia (SANTOS et al., 2005; TITON et al., 2006; OLIVEIRA et al., 2012),
além de favorecer a maximização do processo de produção de mudas
micropropagadas, através do enraizamento ex vitro.
Dentre outras vantagens são descritos a maximização ou a
manutenção do valor genético do clone a ser propagado (DUTRA et al.,
2009), preservação de germoplasma, produção de plantas livres de
doenças (XAVIER et al., 2013) e constituir uma base para o emprego de
outras técnicas biotecnológicas, como a transformação genética
(GONZALEZ, 2002).
Conforme o explante utilizado e sua subsequente manipulação,
a micropropagação pode ser conduzida por uma das três maneiras:
mediante a multiplicação por meio da proliferação de gemas axilares;
mediante a indução de gemas adventícias por organogênese direta ou
indireta ou mediante embriogênese somática (THORPE et al., 1991;
BONGA e VON ADERKAS, 1992; LAKSHMI SITA, 1993; GRATTAPAGLIA
e MACHADO, 1998), onde a primeira corresponde ao principal método de
cultivo in vitro do Eucalyptus, com diversos trabalhos relatando o uso dessa
metodologia para a clonagem de espécies deste gênero (XAVIER e SILVA,
2010), sendo este sistema mais facilmente controlado e apresenta
fidelidade genética muito alta, em que as gemas pré-formadas possuem
determinação para o crescimento vegetativo, as quais irão se desenvolver
naturalmente em plantas completas (GRATTAPAGLIA e MACHADO,
1998).
2.5 Organogênese
O processo de organogênese in vitro é considerado um evento
complexo (XAVIER e OTONI, 2009). Este dá-se mediante a

11
desdiferenciação e diferenciação celular, retomada da atividade
meristemática em células maduras diferenciadas ou em um tecido
calogênico desorganizado (ALVES et al., 2004a). Em outras palavras, pela
indução de gemas adventícias diretamente sobre os explantes ou sobre
uma massa de células não-organizadas, denominada calo (THORPE et al.,
1991; BONGA e VON ADERKAS, 1992).
Quando as gemas se formam diretamente sobre os explantes,
a organogênese é denominada direta, ocorrendo em tecidos que
apresentam potencial morfogênico. Em certas espécies, gemas
adventícias podem surgir diretamente do tecido do explante, não havendo
a formação prévia de calos, o que proporciona fidelidade na propagação
(XAVIER et al., 2013).
A organogênese é denominada indireta quando formação da
gema é precedida pela formação de calo, onde o processo se produz
previamente a desdiferenciação e diferenciação celular e surgem gemas
que crescem e se desenvolvem em novas partes aéreas (VIJAYA e GIRI,
2003).
Dentre a principais utilidades dessa técnica, além de oferecer
uma alta taxa de multiplicação e regeneração, reproduzindo grandes
quantidades de plantas (THORPE et al., 1991). Também é adotado quando
se deseja produzir células para manipulações genéticas, como
poliploidizações e transformações genéticas (VENTURIERI e
VENTURIERI, 2004). Por tanto, a fidelidade das características genéticas
se vê comprometida em protocolos de propagação clonal que induzem a
formação de calo, possivelmente na promoção de variações somaclonais
(GIRIJASHANKAR, 2012), as quais não são facilmente controladas.
As multiplicações sucessivas podem dar-se pela subdivisão do
calo e manutenção de um sistema adventício, ou pela alteração do
processo para proliferação axilar (GRATTAPAGLIA e MACHADO, 1998).
A regeneração in vitro de espécies de Eucalyptus através da
organogênese tem sido relatada para algumas espécies, tais como o E.
tereticornis (SUBBAIAH e MINOCHA, 1990; AGGARWAL et al., 2010), E.
grandis (BRONDANI et al., 2012b; ARENHART e ZAFFARI, 2008;
PEREIRA et al. 2013), E. grandis × E. urophylla (ALCANTARA et al., 2011;

12
BATISTA, 2012; HAJARI et al., 2006), E. nitens (BANDYOPADHYAY et al.,
1999), E. saligna (DIBAX et al., 2010a), E. globulus (NUGENT et al., 2001),
camaldulensis (MURALIDHARAN e MASCARENHAS, 1987; DIBAX et al.,
2010b; GIRIJASHANKAR, 2012), E. benthamii × E. dunnii (CAUDURO,
2013) e E. gunnii (HERVÉ et al., 2001).
Apesar dos estudos demonstrarem a viabilidade e relativa
facilidade da multiplicação in vitro de diferentes espécies de eucalipto via
organogênese é preciso levar em consideração que o protocolo de
regeneração deve ser adaptado para cada espécie, pois embora um
protocolo seja estabelecido para uma espécie, ou até mesmo para um
clone, normalmente pode não ser eficiente para outra espécie
(ALCANTARA, 2008). Alguns estudos demonstraram que diferentes
espécies submetidas às mesmas condições de cultivo, resultaram com
desempenho produtivo divergente (NASCIMENTO, 2013).
2.6 Fatores que influenciam a regeneração in vitro
Na obtenção da regeneração in vitro são testados múltiplos
fatores externos e internos, envolvendo interação entre o meio de cultura,
fatores do ambiente de incubação (PERES, 2002; BORGES et al., 2012),
ação de reguladores de crescimento, em particular a relação entre auxinas
e citocininas (LEMOS, 2010) como também da habilidade ou competência
dos tecidos para responder às mudanças hormonais durante o período de
cultivo (GEORGE et al., 2008; PIERIK, 1997).
2.6.1 Tipo de explante
A partir da seleção da espécie que se deseja micropropagar,
identifica-se a fonte de explante que possa apresentar melhores condições
de resposta ao cultivo in vitro (CID e TEIXEIRA. 2010). Qualquer parte do
tecido extraído da planta destinada ao uso in vitro denomina-se explante.
A lista de possíveis explantes é longa e variada, mas não todos
os explantes reagem da mesma forma a uma determinada condição in vitro.
Assim, é importante a identificação de partes da planta que possam
responder satisfatoriamente à propagação vegetativa, mas haverá maior
sucesso se forem utilizados tecidos jovens, os quais possuem maior
competência organogênica (XAVIER et al., 2013; WENDLING et al., 2014).

13
O sucesso no cultivo do material vegetal in vitro é influenciado
pela idade do tecido ou órgão que será utilizado como explante inicial.
Assim, os requerimentos na formação do calo frequentemente diferem de
acordo com a idade do explante (GEORGE et al., 2008). Ainda,
considerando o tipo de material inicial, juvenil ou adulto, assim como a
posição original do explante na planta refletem os níveis hormonais
endógenos das células do tecido utilizado, podendo ter importante
influência sobre cada processo como o da divisão celular nos processos de
regeneração in vitro, tanto na organogênese e a embriogênese (PIERIK,
1997).
Na regeneração in vitro, tanto para processos organogênicos,
como embriogênicos em espécies de Eucalyptus, são utilizados uma ampla
gama de explantes dependendo da rota morfogênica desejável, como
embriões zigóticos (PINTO et al., 2002; PRAKASH e GURUMURTHI,
2010), cotilédones (BANDYOPADHYAY et al., 1999; DIBAX et al., 2010b;
DIBAX et al., 2005), hipocótilos (SUBBAIAH e MINOCHA, 1990; HUANG et
al., 2010), ápices meristemáticos (GLOCKE et al., 2006) segmentos nodais
(GIRIJASHANKAR, 2012; BRONDANI et al., 2012b).
Embora segmentos de raiz apresentam capacidade
morfogênica (KERBAUY, 1998), estudos em processos orgenogênicos de
Eucalyptus este tipo de explante não são escolhidos por apresentar
grande sensibilidade ao estresse oxidativo.
A escolha do explante dependerá da viabilidade para a sua
utilização, sobretudo pela capacidade de resposta morfogênica em relação
aos estímulos induzidos no cultivo in vitro (LEMOS, 2010). Dessa forma,
a opção por explantes que apresentem um menor grau de diferenciação
reflete em uma maior probabilidade de sucesso quanto à regeneração de
gemas adventícias ou embriões nos explantes. Embora explantes adultos
coletados de materiais selecionados são poucos relatados na literatura
devido a sua dificuldade na expressão de respostas in vitro, mas através
de culturas sucessivas pode-se reverter o estado juvenil, favorecendo
assim altos índices de regeneração.

14
2.6.2 Reguladores de crescimento
Reguladores de crescimento ou fito-hormônios vegetais são
aqueles produzidos pela própria planta; que em baixas concentrações, são
biologicamente ativos, podendo promover, inibir ou modificar o
crescimento, geralmente em um local diferente daquele onde foi produzido,
mas também esses podem ser sintetizadas produzindo efeitos
semelhantes aos hormônios naturais (LEMOS, 2010).
A escolha do fito-hormônio a ser utilizado na cultura in vitro
dependerá do tipo de morfogênese desejada; de seu nível endógeno no
explante no momento da excisão; da capacidade do tecido sintetizar o
regulador durante o período da cultura e da possível interação entre os fito-
hormônios endógenos e aqueles adicionados ao meio (SANTOS, 2003).
Cid e Teixeira (2010) salientam que na cultura de tecidos as
auxinas são utilizadas no enraizamento de brotos, e combinações com
citocininas são frequentemente usadas na organogênese e/ou
embriogênese favorecendo à indução do calo a partir de um explante.
Combinações de auxinas, como o ácido naftalenoacético (ANA)
e o ácido diclorofenildioxiacético (2,4-D), com as citocininas,
benzilaminopurina (BAP) e o thidiazuron (TDZ), são amplamente utilizadas
nas fases de indução à formação de calo e formação de brotos, destacando
que alguns reguladores de crescimento têm uma posição ambígua, pois
atuam como auxina ou como citocinina. Esse é o caso do Thidiazuron ou
TDZ (LU, 1993), que grande parte dos trabalhos com espécies de
Eucalyptus relataram o uso do TDZ em concentrações que variam de 0,01
mg L-1 a 10 mg L-1 para a indução de calos in vitro (CID et al., 1999; ALVES
et al., 2004a, b).
Embora sejam destacadas as vantagens do uso do TDZ,
Huetteman e Preece, (1993) e Lu (1993) reportam dificuldades na
conversão das brotações induzidas por esse regulador em plantas inteiras,
reduzidos índices nos processos de alongamento e enraizamento, os quais
recomendam a adequação de concentrações efetivas nos processos de
regeneração in vitro e evitar prolongadas exposições nos meios de cultura
onde concentrações de TDZ foram adicionados.

15
Calogênese
Arenhart e Zaffari (2008) avaliaram diferentes combinações dos
reguladores 2,4-D, TDZ, ANA e AIA (ácido indolilacético) na formação de
calos, registrando porcentagens reduzidas e nulas nas combinações entre
2,4-D e TDZ, mas nos tratamentos de concentrações isoladas responderam
efetivamente. Segundo George et al. (2008), o 2,4-D representa a auxina
mais usada na indução de calo em estudos de organogênese indireta.
Apesar disso, o 2,4-D é pouco usado na cultura de Eucalyptus, mas
atualmente vem sendo avaliado nos processos de embriogênese, em
combinação com outras auxinas (PINTO et al. 2002).
Oliveira (2014) destacou a efetividade do uso de concentrações
isoladas de TDZ na formação calos organogênicos de Eucalyptus
cloeziana, em comparação às concentrações de ANA, sendo esta última
não favorável. Contudo, as experiências de Alves et al. (2004b) e Dibax et
al. (2010b) indicam melhores resultados de calejamento de tecidos nos
tratamentos com a combinação dos reguladores de crescimento desses
reguladores (TDZ e ANA) no hibrido Eucalyptus grandis × Eucalyptus
urophylla e em Eucalyptus saligna, respectivamente.
Em explantes de hipocótilo de plântulas com idade de 14 dias de
Eucalyptus urophylla em meio de cultura com TDZ mais ANA, além de que
induziram tanto à organogênese quanto a formação de estruturas
semelhantes a embriões no estádio cordiforme (TIBOK et al., 1995).
Dibax et al. (2005) também testaram a relação auxina/citocininas
na organogênese de E. camaldulensis mediante diferentes combinações
de ANA e BAP durante a fase de calogênese, e observaram que em
concentrações estabelecidas de BAP, a formação de calos foi favorecida
com a presença de maiores concentrações de ANA.
Diferenciação celular e formação de brotos
Na etapa de indução à diferenciação celular e adoção à
competência organogênica para a formação de multibrotações adventícias
em calos, as citocininas são utilizadas, dentre as quais comumente em
cultura de tecidos como ZEA (zeatina), a 2iP (isopententenil adenina), KIN
(cinetina) e a BAP (benzilaminopurina). Esta última é reportada como a

16
principal citocinina utilizada na micropropagação de Eucalyptus (BUNN,
2005).
O BAP favoreceu a geração de calos organogênicos em cultivos
que contiveram a concentração isolada da citocinina, sendo pouco efetiva
a combinação com concentrações de ANA, sendo essa combinação de
reguladores amplamente utilizada nas fases de multiplicação in vitro de
explantes de diversas espécies de eucalipto, sendo demonstrado sua
efetividade na regeneração de calos de Eucalyptus cloeziana (OLIVEIRA,
2014).
Os estudos citados anteriormente descrevem os protocolos de
calogênese e regeneração sujeitos ao material genético e à resposta
dependente das concentrações, tipos e combinações de reguladores para
cada espécie a ser testada. Contudo, muitos dos estudos são restritos nas
fases iniciais e também pouco aprofundados no sentido de
acompanhamento do processo organogênico. Este processo é de grande
importância uma vez que podem auxiliar em futuros estudos no sentido de
aprimoramento de técnicas de multiplicação.

17
3 MATERIAL E MÉTODOS
3.1 Local do experimento
O presente experimento foi conduzido no Laboratório de Cultura
de Tecidos Vegetais da Faculdade de Engenharia Florestal, Universidade
Federal de Mato Grosso, campus Cuiabá.
3.2 Material vegetal
Foram utilizados explantes oriundos de plântulas germinadas in
vitro de Eucalyptus cloeziana F. Muell cuja procedência das sementes foi
de uma Área Produtora de Sementes (APS) localizada em Anhembi,
Estado do São Paulo, determinado pelo cultivar LCFA026 do Instituto de
pesquisas e estudos florestais (IPEF).
3.3 Desinfestação das sementes e germinação in vitro
As sementes de Eucalyptus cloeziana foram pré-tratadas
mediante embebição em água destilada por 24h. Posteriormente, as
sementes foram lavadas com água corrente por cinco minutos, sendo
desinfestadas em uma solução de álcool 70% por 90 segundos e em
seguida em hipoclorito de sódio 2,5% (v/v) contendo uma gota (0,05 mL)
de Tween-20 durante 25 minutos.
Após o processo de desinfestação, as sementes foram lavadas
três vezes em água destilada autoclavada. As sementes selecionadas para
o cultivo foram aquelas que apresentaram estado de turgência e coloração
branca, as quais foram inoculadas in vitro em tubos de ensaio contendo
meio de cultura constituído por água destilada e uma concentração de ágar
de 6,0 gL-1.As sementes foram mantidas em sala de crescimento com
temperatura de 25 ± 2ºC, fotoperíodo de 16h de luz e irradiância de 32 µmol
m-2s-1.
3.4 Preparação do meio de cultura e condições experimentais
O meio de cultura utilizado, para o cultivo das sementes assim como
para a indução calogênica e organogênica dos explantes, foi composto pelos sais
básicos do meio de cultura Woody Plant Medium (WPM) (LLOYD e McCOWN,
1980), utilizando água deionizada, o valor do pH foi para 5,8 mediante uso
de NaOH (0,1 mol L-1) ou HCl (0,1 mol L-1), antes da adição do ágar e da

18
Tabela 1. Composição básica do meio de cultura WPM utilizado para a
indução organogênica de Eucalyptus cloeziana
Nutriente (mg.L-1)
NO3- 135,89
NH4+ 69,97
P 38,69
Ca 120,54
K 493,11
S 237,67
Mg 36,49
B 1,08
Cu 0,064
Fe 5,58
Mo 0,10
Mn 5,49
Zn 1,96
Fonte de Macro e Micronutrientes FQ / PM (mg.L-1)
Nitrato de amônio NH4NO3 / 80,04 400,00
Cloreto de cálcio CaCl2.2H2O / 147,02 96,00
Nitrato de cálcio Ca(NO3)2.4H2O / 236,15 556,00
Fosfato de potássio monobásico KH2PO4 / 136,09 170,00
Sulfato de potássio K2SO4 / 174,26 990,00
Sulfato de magnésio MgSO4.7H2O / 246,48 370,00
Sódio - EDTA Na2-EDTA.2H2O / 372,24 37,30
Sulfato de ferro FeSO4.7H2O / 278,02 27,80
Sulfato de manganês MnSO4.H2O / 169,01 16,90
Sulfato de zinco ZnSO4.7H2O / 287,54 8,60
Ácido bórico H3BO3 / 61,83 6,20
Sulfato de cobre CuSO4.5H2O / 249,68 0,25
Molibdato de sódio Na2MoO4.2H2O / 241,95 0,25
Fonte de Vitamina FQ / PM (mg.L-1)
Tiamina-HCl C12H18Cl2N4O5.xH2O/337,27 1,00
Piridoxina-HCl C8H12ClNO3 / 205,64 1,00
Ácido nicotínico C6H5NO2 / 123,11 1,00
Pantotenato Ca2+ C18H32CaN2O10 / 476,54 1,00
Biotina C10H16N2O3S / 244,30 1,00
Fonte de Aminoácido FQ / PM (mg.L-1)
Glicina H2NCH2COOH / 75,07 1,00
Fonte de Carbono FQ / PM (mg.L-1)
Mio-inositol C6H12O6 / 180,20 100,00
Sacarose C12H22O11 / 342,30 30.000,00
Agente polimerizador (mg.L-1)
Ágar 6.000,00

19
autoclavagem. O meio de cultura foi autoclavado na temperatura de
121ºC, que corresponde a pressão aproximada de 1 kgf cm-2, durante 15
minutos (Tabela 1).
3.5 Cultura organogênica
3.5.1 Fonte de explantes e tratamentos
Plântulas obtidas de sementes germinadas in vitro com 20 dias
de idade foram utilizadas como fonte dos explantes. Segmentos de raízes
e hipocótilo, medindo 0,5 cm de comprimento, e cotilédones seccionados
à metade contendo o pecíolo (Figura 2 A-C) foram excisados e inoculados
individualmente em tubos de ensaio de vidro contendo 6,0 ml de meio de
cultura WPM, suplementados com diferentes combinações e
concentrações de fito-reguladores.
Figura 2. Tecidos de Eucalyptus cloeziana inoculados in vitro. (A) Hipocótilo; (B) cotilédone; (C) raiz. Barra = 0,25 cm.
Após a inoculação, os explantes foram mantidos em sala de
crescimento na ausência de luminosidade (escuro) e temperatura de 25 ±
2ºC por 30 dias.
Os fatores que constituíram os tratamentos na cultura calogênica
foram os tipos de explantes e as combinações de diferentes concentrações
de fito-reguladores. Concentrações de 0,0; 1,0 e 2,0 mg L-1 foram
estabelecidos para o thidiazuron (TDZ) e para o ácido diclorofenoxiacético
(2,4-D), e 0,0; 2,0 e 4,0 mg L-1 para o ácido naftaleno acético (ANA). Cada
um dos tratamentos foi constituído por 10 repetições (10 tubos de ensaio),
contendo 1 explante em cada repetição, as quais foram dispostas em um
delineamento inteiramente causalizado em arranjo fatorial (3x27),
avaliando a interação de 3 tipos de tecidos (explantes) e 27 combinações
A B C

20
de fito-reguladores (Tabela 2).
Após 30 dias da aplicação dos tratamentos foram avaliados o
aspecto visual dos tecidos (visando verificar alguma anormalidade), além
da porcentagem de manifestação bacteriana, oxidação e tecidos que
apresentaram calogênese. Para os tecidos que apresentam calogênese
foram adotadas referências de classificação: 1 para presença e 0 para
ausência de calo. Considerou-se contaminação as colônias bacterianas
que surgiram no início do estabelecimento in vitro dos explantes,
principalmente na superfície do meio de cultura.
Tabela 2. Descrição das combinações de fito-reguladores empregados na cultura calogênica de tecidos de Eucalyptus cloeziana inoculados in vitro.
Combinações de
Fito-reguladores (mg L-1)
Nº ANA TDZ 2,4-D
1 0,0 0,0 0,0 2 0,0 0,0 1,0 3 0,0 0,0 2,0 4 0,0 1,0 0,0 5 0,0 1,0 1,0 6 0,0 1,0 2,0 7 0,0 2,0 0,0 8 0,0 2,0 1,0 9 0,0 2,0 2,0 10 2,0 0,0 0,0 11 2,0 0,0 1,0 12 2,0 0,0 2,0 13 2,0 1,0 0,0 14 2,0 1,0 1,0 15 2,0 1,0 2,0 16 2,0 2,0 0,0 17 2,0 2,0 1,0 18 2,0 2,0 2,0 19 4,0 0,0 0,0 20 4,0 0,0 1,0 21 4,0 0,0 2,0 22 4,0 1,0 0,0 23 4,0 1,0 1,0 24 4,0 1,0 2,0 25 4,0 2,0 0,0 26 4,0 2,0 1,0
27 4,0 2,0 2,0

21
Para os tratamentos que apresentaram calo foram
determinadas categorias de intensidade de calejamento dos tecidos. As
categorias foram utilizadas para estabelecer a porcentagem da superfície
do explante original que formou calo (Tabela 3).
Tabela 3. Intensidade de calejamento dos diferentes tecidos de Eucalyptus cloeziana inoculados in vitro aos 30 dias
Intensidade do calo Hipocótilo Cotilédone Raiz
1
(x ≤ 25%)
2
(25% < x < 50%)
3 (x = 50%)
4
(50% < x < 75%)
5
(x ≥ 100%)

22
3.5.2 Regeneração in vitro
Explantes iniciais da cultura calogênica, que apresentaram ou
não calos, foram transferidos para o meio de regeneração organogênica,
constituído pelo meio básico WPM suplementado com 1,0 mg L-1 de BAP
(6-benzilamino purina).
Subcultivos foram realizados a cada 30 dias, com a intenção de
renovar a concentração nutritiva do meio de cultura, tanto dos sais WPM
quanto do fito-regulador. Foram avaliados os tratamentos; constituídos
pelas culturas calogênicas (tecidos originais mais as combinações de fito-
reguladores); que apresentaram regeneração organogênica (broto e/ou
raiz). Também foi contabilizada a quantidade de brotações adventícias
induzidas de cada tecido regenerado após o 3o subcultivo (90 dias).
3.5.3 Multiplicação e alongamento in vitro
Os calos regenerados foram divididos em explantes padrões
(tufos ou aglomerado de brotações) contendo, aproximadamente, 3
brotações de comprimento superior a 1 cm, mantendo a identificação do
tratamento que deu origem ao calo inicial. Os explantes foram sub-
cultivados a cada 30 dias e sub-divididos conforme a necessidade, com o
intuito de oferecer melhores condições para seu desenvolvimento in vitro
e manter o vigor vegetativo.
O meio de cultivo para a etapa de multiplicação in vitro foi
constituído pelo meio básico (WPM) suplementado com BAP e ácido indol-
butírico (AIB) (Tabela 4).
Tabela 4. Combinação dos fito-reguladores BAP e AIB na etapa de multiplicação e alongamento in vitro de explantes de Eucalyptus cloeziana.
Meios de multiplicação
e alongamento BAP AIB
1 1,0 mg L-1 0,5 mg L-1
2 0,5 mg L-1 1,0 mg L-1
Nesta etapa objetivou-se manter os tufos sob condições
hormonais diferentes visando à produção de material vegetativo e a futura

23
coleta de brotações alongadas para a instalação de
ensaios de enraizamento ex vitro.
3.5.4 Enraizamento ex vitro e aclimatização
Brotações alongadas, possuindo comprimento superior ou igual
a 1 cm foram coletadas e inoculadas ex vitro no sistema de mini-estufa, a
qual foi constituída de uma estrutura plástica de cor transparente, a fim de
favorecer a luminosidade e evitar variação da umidade relativa do ar. O
substrato utilizado para promover o enraizamento das micro-estacas foi
constituído por substrato orgânico comercial adicionado com argila sintética
vermiculita, na proporção igual a 1:1 (v/v).
As condições ambientais dos primeiros 20 dias de incubação ex
vitro foram iguais as condições à sala de crescimento das culturas in vitro
(temperatura média de 25±2 ºC e o fotoperíodo de 16h).
Variáveis como a porcentagem de sobrevivência, enraizamento
adventício das microestacas foram avaliadas aos 20 dias de incubação ex
vitro.
Posteriormente, as mudas foram retiradas da miniestufa e
subcultivadas em recipientes individuais de volume maior (250 ml)
contendo a mesma proporção do substrato utilizado e fornecidos de uma
solução nutritiva composta por o 50% das concentrações dos sais macro e
micronutrientes correspondentes ao meio MS (Murashige e Skoog) a fim de
fornecer uma fonte nutritiva e promover o continuo desenvolvimento das
mudas.
Para completar a fase de aclimatização, as mudas
micropropagadas foram transplantadas para uma casa de sombrite (50%)
e transferidas para copos plásticos de maior dimensão (500 ml).
3.6 Análise histológica
Amostras do corte transversal e longitudinal da região da gema
axilar foram fixadas em solução de formaldeído e glutaraldeído (Karnovsky,
1965) modificado (glutaraldeído 1%; paraformaldeído 4% em tampão
fostato de sódio (NaH2PO4.H2O à 0,1 M; pH 7,2) e submetidas a seis séries
de vácuo (-600 mmHg) por 30 minutos cada. Posteriormente, as amostras
foram armazenadas durante 30 dias a 4°C. Em seguida, as amostras foram

24
desidratadas por meio de série alcoólica-etílica em concentrações
crescentes (10, 20, 30, 40, 50, 60, 70, 80, 90 e 100%, v/v), permanecendo
em cada solução por 15 minutos. Em seguida as amostras foram imersas
em meio de infiltração (Historesina, Leica) durante 24 horas e após, foram
alocadas na porção inferior do recipiente de moldura. Em seguida as
amostras foram emblocadas em Historesina (hidroxietil metacrilato, Leica)
com endurecedor conforme recomendação do fabricante onde
permaneceram durante 28 dias à temperatura de 24°C. Os blocos contendo
as amostras foram seccionados longitudinalmente ou transversalmente à
5 μm de espessura com o uso de micrótomo rotativo automático Microm
HM 355S (Thermo Scientific). Os cortes obtidos foram corados com azul
de toluidina (0,05%, v/v) em tampão fostato de sódio e ácido cítrico (Sakai,
1973) durante 30 minutos e montados em lâminas histológicas com resina
sintética (Entellan). As lâminas histológicas foram analisadas e
fotomicrografadas em microscópio de luz (Opton) sendo as imagens
capturadas em escala micrométrica.
3.7 Análise estatística dos dados
Os dados mensurados de todos os experimentos foram
submetidos ao teste de Hartley (p<0,05) e Shapiro-Wilk (p<0,05), a fim de
verificar a homogeneidade das variâncias e a distribuição normal dos
dados, respectivamente. Os dados foram transformados conforme a
necessidade pelo teste de Box-Cox. Em seguida, foi realizada a análise
de variância (ANOVA, p<0,01 e p<0,05). De acordo com a significância da
ANOVA, os dados dos fatores qualitativos foram comparados pelo teste
de Duncan (p<0,01 e p<0,05). Para a análise estatística dos dados foi
utilizado os programas computacionais R (R Development Core Team,
2012) e SOC (Embrapa, 1990).

25
4 RESULTADOS E DISCUSSÃO
4.1 Cultura calogênica e regeneração in vitro
De acordo com a análise de variância foram detectadas
diferenças significativas (p<0,01) em relação a interação dos fatores
testados, tanto nas variáveis da fase de indução à calogênese
(Manifestação de bactérias (BAC), oxidação (OXI) e presença de calos
(CAL)), como também na regeneração das brotações (REG) (Tabela 5).
Tabela 5. Resumo da análise de variância para a porcentagem de manifestação bacteriana (BAC), oxidação (OXI), calo (CAL) e regeneração dos brotos (REG) in vitro de Eucalyptus cloeziana em relação aos tratamentos testados aos 30 dias.
Fonte de variação GL Quadrados médios
BAC(%)(1) OXI(%)(1)
CAL(%)(1) REG(%)(1)
COMB 26 1,13** 0,71** 5,62** 0,035** TEC 2 0,62** 1,56** 17,18** 0,033**
COMB × TEC 52 0,012** 0,19** 1,42** 0,018** Resíduo 697 0,026 0,025 0,198 0,004 Média - 23,5 19,9 78,4 1,5
CVexp(%) - 19,2 18,2 18,8 6,4
** Valor significativo ao nível de 1% de probabilidade de erro, respectivamente pelo teste F. (1) Dados transformados por 1/exp[(n+0,5)/100] , onde n = dado amostrado. GL = graus de liberdade, CVexp. = coeficiente de variação experimental. COMB = combinação de fito- regulador, TEC = tipo de tecido.
Sementes de Eucalyptus cloeziana foram inoculadas in vitro
para a obtenção de plântulas como fonte de explantes, dentre as quais
foram selecionados aqueles cultivos que não apresentaram manifestação
de patógenos (bactérias ou fungos), geralmente encontrados em explantes
introduzidos in vitro.
4.1.1 Manifestação de bactérias
Apesar da metodologia de desinfestação das sementes tenha
controlado o desenvolvimento de micro-organismos na maioria dos cultivos
seminais, tecidos utilizados na indução à calogênese apresentaram
manifestação de bactérias (Figura 3 A, B e C), atingindo uma média geral
de 23,5%, incluindo o tratamento da testemunha. Com base nesse
resultado, pode-se supor que nos tecidos desta espécie encontram-se
bactérias endógenas que podem comprometer o sucesso do cultivo in vitro.

26
A presença de manifestação bacteriana foi avaliada dentre os
tipos de tecidos testados, apresentando porcentagens entre 0 a 100% para
os explantes de hipocótilos e raiz, e para o cotilédone porcentagens de 0
a 70%.
Ressalta-se que os tratamentos referentes as combinações de
fito-reguladores que apresentaram a maior concentração de ANA (4,0 mg
L-1), destacando os tratamentos das combinações 25 (4,0 mgL-1 ANA + 2,0
mgL-1 TDZ) e 26 (4,0 mgL-1 ANA + 2,0 mgL-1 TDZ + 1,0 mgL-1 2,4-D) nas
quais se registraram maiores porcentagens de manifestação bacteriana
nos três tecidos; 100%, 60-70% e 100% para hipocótilo, cotilédone e raiz,
respectivamente. Além disso, os tratamentos das combinações 2, 6, 13, 20,
22, 23 e 27 apresentaram elevada manifestação de bactérias nos diferentes
tecidos (Tabela 6).
A contaminação dos tecidos por bactérias endógenas é um
problema relativamente comum nos procedimentos de micropropagação,
devido à dificuldade de eliminação durante o processo de desinfestação,
representando problemas para o estabelecimento das culturas (PIERIK,
1997).
Todavia, se o procedimento de desinfestação dos explantes for
efetivo, a manifestação endofítica in vitro pode ocorrer em condições de
qualquer tipo e/ou intensidade de estresse abiótico, que eventualmente
possa ocorrer durante as diferentes fases de micropropagação (ALMEIDA
et al., 2005). Dessa forma, as concentrações de fito-reguladores podem ter
causado estresse fisiológico nos explantes, especificamente nos
tratamentos que apresentaram a concentração mais elevada (4,0 mg L-1)
do ANA, acarretando a manifestação de bactérias, efeito também
registrado em cultura in vitro de Eucalyptus citriodora (ALMEIDA, 2012).
Em brotações epicórmicas de Eucalyptus cloeziana coletadas
para introdução in vitro, Oliveira (2014) observou porcentagens de
manifestação bacteriana similares ao presente trabalho, entre 26,4 a 35,8%
sujeitas à época de coleta, com reduzida manifestação durante o período
de baixa precipitação pluviométrica.

27
Figura 3. Manifestação de colônias bacterianas e oxidação in vitro em explantes de Eucalyptus cloeziana, aos 30 dias após a inoculação. (A) Tecido do hipocótilo apresentando manifestação de bactérias (setas) e (D) oxidação. (B) Bactérias presentes (setas) no tecido cotiledonar e (E) oxidação do tecido. (C) Segmentos de raiz apresentando manifestação de bactérias (setas) e (F) oxidação do tecido. Barra = 0,5 cm
Além disso, o mesmo autor registrou diferenças nas taxas de
multiplicação dentre vários materiais genéticos, considerando que a
presença de bactérias endofíticas podem ter influenciado o
desenvolvimento de explantes, principalmente ao considerar a
competição pelos nutrientes do meio de cultura.
Porém, alguns autores (ESPOSITO-POLESI, 2011; ALMEIDA
et al., 2005) não consideram a presença de microrganismos endofíticos
como uma barreira nos procedimentos da cultura in vitro, sendo descrita a
relação planta/microrganismos como benéfica na micropropagação de
clones de Eucalyptus benthamii, relatado por Piotto (2013), o qual
evidenciou essa interação no crescimento dos índices de massa seca das
brotações e na promoção do alongamento das mesmas, favorecendo a
posterior aclimatização dos materiais genéticos.
E F
B A

28
Tabela 6. Porcentagem de explantes de Eucalyptus cloeziana que apresentaram manifestação bacteriana in vitro em relação aos diferentes tecidos e combinações de fito-reguladores na indução à calogênese, aos 30 dias após a instalação do experimento.
Combinações de fito- reguladores (mg L-1)
Tecido de origem
Nº ANA TDZ 2,4-D Hipocótilo Cotilédone Raiz
1 0,0 0,0 0,0 33,3±16,7Db 55,6±17,6ABa
37,5±18,3DEb
2 0,0 0,0 1,0 50,0±16,7Ca 0,0±0,0Eb
50,0±18,9CDa
3 0,0 0,0 2,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
4 0,0 1,0 0,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
5 0,0 1,0 1,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
6 0,0 1,0 2,0 40,0±16,3CDa 44,4±17,6BCa
33,3±16,7Eb
7 0,0 2,0 0,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
8 0,0 2,0 1,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
9 0,0 2,0 2,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
10 2,0 0,0 0,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
11 2,0 0,0 1,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
12 2,0 0,0 2,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
13 2,0 1,0 0,0 100,0±0,0Aa 20,0±13,3Db
100,0±0,0Aa
14 2,0 1,0 1,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
15 2,0 1,0 2,0 0,0±0,0Eb 0,0±0,0Eb
10,0±10Fa
16 2,0 2,0 0,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
17 2,0 2,0 1,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
18 2,0 2,0 2,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
19 4,0 0,0 0,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
20 4,0 0,0 1,0 50,0±16,7Cb 0,0±0,0Ec
100,0±0,0Aa
21 4,0 0,0 2,0 0,0±0,0Eb 0,0±0,0Eb
80,0±13,3Ba
22 4,0 1,0 0,0 90,0±10Aa 40,0±16,3Cc
60,0±16,3Cb
23 4,0 1,0 1,0 40,0±16,3CDc 60,0±16,3Aa
50,0±16,7CDb
24 4,0 1,0 2,0 0,0±0,0Ea 0,0±0,0Ea
0,0±0,0Fa
25 4,0 2,0 0,0 100,0±0,0Aa 70,0±15,3Ab
100,0±0,0Aa
26 4,0 2,0 1,0 100,0±0,0Aa 60,0±16,3Ab
100,0±0,0Aa
27 4,0 2,0 2,0 70,0±15,3Bb 60,0±16,3Ac
90,0±10ABa
Nas colunas, médias seguidas por mesma letra maiúscula e, nas linhas, médias seguidas por mesma letra minúscula não diferem significativamente pelo teste de Duncan ao nível de 5% de probabilidade de erro. Dados apresentados como: média ± erro padrão.
4.1.2 Oxidação dos tecidos
Os tecidos cultivados in vitro também apresentaram
porcentagem de oxidação, com valor de 19,9% entre todos os tratamentos,
porém o valor pode ser considerado reduzido, pelo fato que este representa

29
uma limitação na cultura de tecidos vegetais podendo dificultar o
estabelecimento do experimento (GRATTAPAGLIA e MACHADO, 1998)
considerando que os explantes foram cultivados em condições de escuro
como estratégia para a prevenção da oxidação dos tecidos.
A oxidação foi registrada mediante avaliação visual dos tecidos
que apresentaram necrose (Figura 3 D, E e F), dentre os quais o tecido
cotiledonar se destacou apresentou menor quantidade de tratamentos com
explantes oxidados, favorecido pelo maior grau de diferenciação celular em
comparação aos outros utilizados (hipocótilo e raiz).
Os valores da porcentagem de oxidação (Tabela 7) foram
semelhantes à manifestação bacteriana entre os diferentes tipos de
tecidos; 0-100%, 0-60% e 0-100%, respectivamente para hipocótilo,
cotilédone e raiz. A raiz foi o tecido mais susceptível, registrando os maiores
valores de manifestação bacteriana e oxidação (90 - 100%) nas
combinações 13 (2,0 mg L-1 ANA + 1,0 mg L-1 TDZ), 20 (4,0 mg L-1 ANA +
1,0 mg L-1 2,4-D), 25 (4,0 mg L-1 ANA + 2,0 mg L-1 TDZ), 26 (4,0 mg L-1
ANA + 2,0 mg L-1 TDZ + 1,0 mg L-1 2,4-D) e 27 (4,0 mg L-1 ANA + 2,0 mg
L-1 TDZ + 2,0 mg L-1 2,4-D).
Cabe ressaltar que, semelhante ao tecido radicular, o hipocótilo
também demonstrou elevados valores de manifestação bacteriana nas
mesmas combinações (13, 20, 25, 26 e 27), mas diferiu em relação a
porcentagem de oxidação, apresentando valores porcentuais menores (20
– 100%). Esse resultado pode estar relacionado ao fato que estes
explantes responderam mais rapidamente ao estímulo produzido pelos
reguladores de crescimento, situação similar que também foi relatada na
etapa de calogênese em tecidos de Eucalyptus benthamii x E. dunnii
(CAUDURO, 2011).
Reguladores de crescimento também podem intensificar o
estres oxidativo desencadeando o desequilíbrio hormonal devido à
natureza, combinação e/ou concentração dos reguladores adicionados ao
meio de cultura, ou estar relacionados à síntese de outros compostos que
promovem a oxidação celular como o caso do TDZ, o qual está relacionado
com a liberação de etileno que induz a senescência e necrose dos
explantes cultivados in vitro (CARVALHO et al., 2011).

30
Tabela 7. Porcentagem de explantes de Eucalyptus cloeziana que apresentaram oxidação in vitro em relação aos diferentes tecidos e combinações de fito-reguladores na indução à calogênese, aos 30 dias após a instalação do experimento.
Combinações de fito- reguladores (mg L-1)
Tecidos de origem
Nº ANA TDZ 2,4-D Hipocótilo Cotilédone Raiz
1 0,0 0,0 0,0 100,0±0,0Aa 33,3±16,7BCb
25,0±16,4Ec
2 0,0 0,0 1,0 50,0±16,7CDa 11,1±11,1Dc
37,5±18,3DEb
3 0,0 0,0 2,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
4 0,0 1,0 0,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
5 0,0 1,0 1,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
6 0,0 1,0 2,0 40,0±16,3Db 44,4±17,6Ba
33,3±16,7Ec
7 0,0 2,0 0,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
8 0,0 2,0 1,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
9 0,0 2,0 2,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
10 2,0 0,0 0,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
11 2,0 0,0 1,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
12 2,0 0,0 2,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
13 2,0 1,0 0,0 20,0±13,3Eb 0,0±0,0Dc
100±0,0Aa
14 2,0 1,0 1,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
15 2,0 1,0 2,0 10,0±10,0EFb 0,0±0,0Dc
80,0±13,3Ba
16 2,0 2,0 0,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
17 2,0 2,0 1,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
18 2,0 2,0 2,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
19 4,0 0,0 0,0 0,0±0,0Fa 0,0±0,0Da
0,0±0,0Fa
20 4,0 0,0 1,0 50,0±16,7CDb 0,0±0,0Dc
100,0±0,0Aa
21 4,0 0,0 2,0 10,0±10,0EFc 40,0±16,3BCb
80,0±13,3Ba
22 4,0 1,0 0,0 10,0±10,0EFb 0,0±0,0Dc
60,0±16,3Ca
23 4,0 1,0 1,0 10,0±10,0EFb 0,0±0,0Dc
50,0±16,7CDa
24 4,0 1,0 2,0 10,0±10,0EFa 0,0±0,0Db
0,0±0,0Fb
25 4,0 2,0 0,0 60,0±16,3BCb 0,0±0,0Dc
100,0±0,0Aa
26 4,0 2,0 1,0 100,0±0,0Aa 30,0±15,3Cb
100,0±0,0Aa
27 4,0 2,0 2,0 70,0±15,3Bb 60,0±16,3Ac
90,0±10,0ABa
Nas colunas, médias seguidas por mesma letra maiúscula e, nas linhas, médias seguidas por mesma letra minúscula não diferem significativamente pelo teste de Duncan ao nível de 5% de probabilidade de erro. Dados apresentados como: média ± erro padrão.
4.1.3 Indução da calogênese e intensidade calogênica
A indução a calogênese foi considerada elevada em todos os
tratamentos, atingindo média geral de 78,4%, demonstrando adequação
dos fito-reguladores testados e a competência dos tecidos em relação a
indução de estruturas celulares desdiferenciadas (Figura 4 A e B), os quais

31
registraram porcentagens iguais dentre 0-100% para o hipocótilo e tecido
radicular, e 40-100% para o cotilédone (Tabela 8).
A calogênese também foi categorizada por meio da intensidade
de calo induzido na área do tecido em 30 dias de cultivo in vitro (Tabela 9).
As categorias de calejamento 4 (50%≥75% de calejamento) e 5 (100% de
calejamento) foram adotadas para determinar os maiores índices de
calogênese registrados nos tratamentos de indução (tecidos e
combinações de fito-reguladores).
Tratamentos que apresentaram quantidades percentuais iguais
ou superiores a 50% de tecidos apresentando calo nas categorias
superiores de calejamento (4 e 5), foram aquelas combinações que
apresentaram apenas um regulador. As diferentes combinações
estabelecidas entre os fito-reguladores ANA, TDZ e 2,4-D não favoreceram
a indução de elevadas intensidades calogênicas, com uma única exceção
dos tratamentos constituído pelo tecido cotiledonar nas combinações 12
(2,0 mg L-1 de ANA + 2,0 mgL-1 de 2,4-D) e 27 (4,0 mg L-1 de ANA + 2,0
mgL-1 de TDZ + 2,0 mgL-1 de 2,4-D)
Os tratamentos que registraram maior quantidade explantes de
elevada intensidade de calejamento foram destacados os tratamentos das
combinações 7 (2,0 mg L-1 do TDZ), o qual apresentou 88,9% e 77,8% para
hipocótilo e raiz respectivamente, e da combinação 2 (2,0 mgL-1 de 2,4-D)
que apresentou 62.5% no tecido cotiledonar. Assim como também
ressaltaram as combinações 3 (2,0 mg L-1 de 2,4-D) e 19 (4,0 mg L-1 de
ANA) nos tecidos cotiledonar e hipocótilo respectivamente. (Tabela 9).
Em termos gerais, os tecidos do hipocótilo (25,5%) e cotiledonar
(25,0%) apresentaram destaque em relação às intensidades superiores de
calejamento. O tecido radicular apresentou a menor quantidade de calos
nas categorias 4 e 5 (11,6%), evidenciando a reduzida aptidão à
desdiferenciação celular.
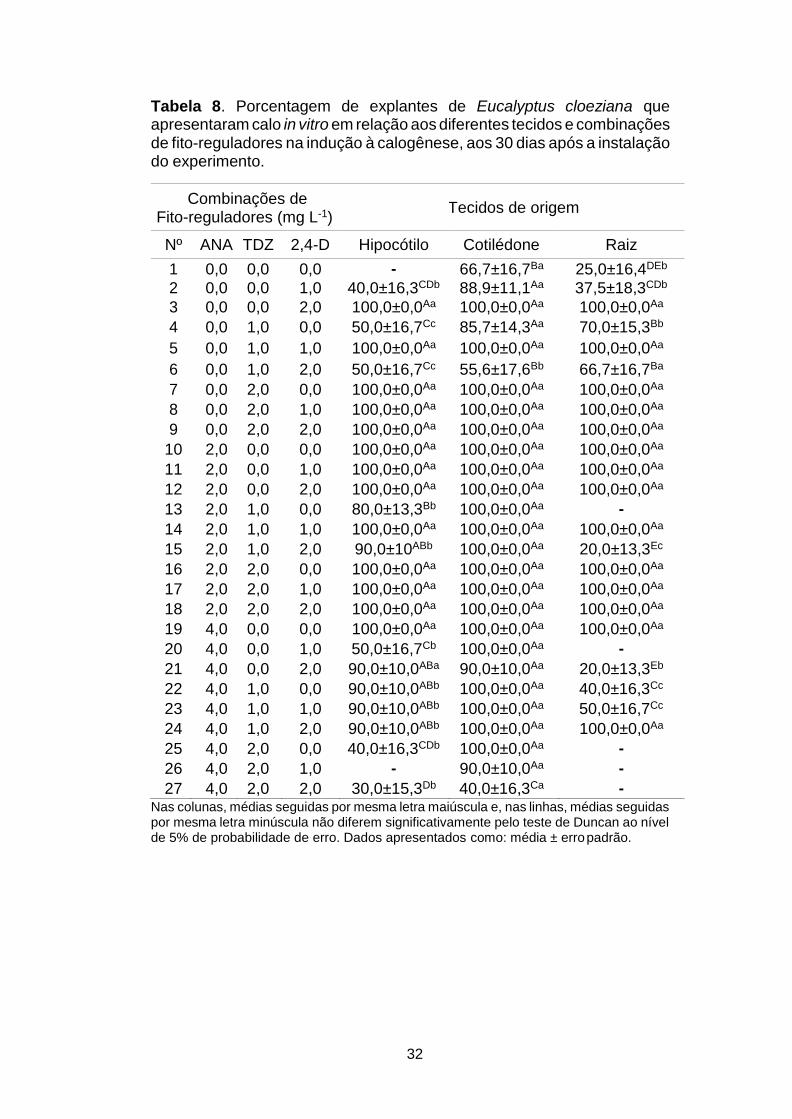
32
Tabela 8. Porcentagem de explantes de Eucalyptus cloeziana que apresentaram calo in vitro em relação aos diferentes tecidos e combinações de fito-reguladores na indução à calogênese, aos 30 dias após a instalação do experimento.
Combinações de Fito-reguladores (mg L-1)
Tecidos de origem
Nº ANA TDZ 2,4-D Hipocótilo Cotilédone Raiz
1 0,0 0,0 0,0 - 66,7±16,7Ba 25,0±16,4DEb
2 0,0 0,0 1,0 40,0±16,3CDb 88,9±11,1Aa
37,5±18,3CDb
3 0,0 0,0 2,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
4 0,0 1,0 0,0 50,0±16,7Cc 85,7±14,3Aa
70,0±15,3Bb
5 0,0 1,0 1,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
6 0,0 1,0 2,0 50,0±16,7Cc 55,6±17,6Bb
66,7±16,7Ba
7 0,0 2,0 0,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
8 0,0 2,0 1,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
9 0,0 2,0 2,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
10 2,0 0,0 0,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
11 2,0 0,0 1,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
12 2,0 0,0 2,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
13 2,0 1,0 0,0 80,0±13,3Bb 100,0±0,0Aa
-
14 2,0 1,0 1,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
15 2,0 1,0 2,0 90,0±10ABb 100,0±0,0Aa
20,0±13,3Ec
16 2,0 2,0 0,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
17 2,0 2,0 1,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
18 2,0 2,0 2,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
19 4,0 0,0 0,0 100,0±0,0Aa 100,0±0,0Aa
100,0±0,0Aa
20 4,0 0,0 1,0 50,0±16,7Cb 100,0±0,0Aa
-
21 4,0 0,0 2,0 90,0±10,0ABa 90,0±10,0Aa
20,0±13,3Eb
22 4,0 1,0 0,0 90,0±10,0ABb 100,0±0,0Aa
40,0±16,3Cc
23 4,0 1,0 1,0 90,0±10,0ABb 100,0±0,0Aa
50,0±16,7Cc
24 4,0 1,0 2,0 90,0±10,0ABb 100,0±0,0Aa
100,0±0,0Aa
25 4,0 2,0 0,0 40,0±16,3CDb 100,0±0,0Aa
-
26 4,0 2,0 1,0 - 90,0±10,0Aa -
27 4,0 2,0 2,0 30,0±15,3Db 40,0±16,3Ca
- Nas colunas, médias seguidas por mesma letra maiúscula e, nas linhas, médias seguidas por mesma letra minúscula não diferem significativamente pelo teste de Duncan ao nível de 5% de probabilidade de erro. Dados apresentados como: média ± erro padrão.

33
Tabela 9. Resumo da intensidade calogênica in vitro nos diferentes tratamentos.
Nº ANA TDZ 2,4-D 1(1) 2 3 4 5 1 2 3 4 5 1 2 3 4 5
1 0,0 0,0 0,0 - - - - - 100,0 - - - - 100,0 - - - -
2 0,0 0,0 1,0 - - 25,0 25,0 50,0 - 12,5 12,5 12,5 62,5 - 33,3 - - 66,7
3 0,0 0,0 2,0 - 11,1 11,1 22,2 55,6 - 11,1 11,1 22,2 55,6 100,0 - - - -
4 0,0 1,0 0,0 40,0 - 20,0 40,0 - - 16,7 33,3 33,3 16,7 42,8 28,6 14,3 - 14,3
5 0,0 1,0 1,0 - 66,7 11,1 22,2 - 11,1 44,4 33,4 11,1 - 55,6 33,3 11,1 - -
6 0,0 1,0 2,0 60,0 40,0 - - - 40,0 40,0 - 20,0 - 100,0 - - - -
7 0,0 2,0 0,0 - - - 11,1 88,9 - 22,2 11,1 66,7 - 11,1 - - 11,1 77,8
8 0,0 2,0 1,0 - 50,0 20,0 - 30,0 60,0 30,0 - 10 - 40,0 20,0 - 10,0 30,0
9 0,0 2,0 2,0 77,8 22,2 - - - 100,0 - - - - 100,0 - - - -
10 2,0 0,0 0,0 - 33,3 33,3 11,1 22,3 - 33,3 44,4 22,3 - 12,5 50,0 - - 37,5
11 2,0 0,0 1,0 70,0 - - 10,0 20,0 10,0 40,0 40,0 10,0 - 80,0 20,0 - - -
12 2,0 0,0 2,0 100,0 - - - - 30,0 20,0 - - 50 100,0 - - - -
13 2,0 1,0 0,0 75,0 12,5 - - 12,5 70,0 20,0 10,0 - - - - - - -
14 2,0 1,0 1,0 40,0 50,0 - 10,0 - 50,0 50,0 - - - 100,0 - - - -
15 2,0 1,0 2,0 11,1 55,6 33,3 - - 70,0 20,0 10,0 - - 50,0 50,0 - - -
16 2,0 2,0 0,0 40,0 20,0 10,0 10,0 20,0 20,0 40,0 10,0 30,0 - 62,5 25,0 12,5 - -
17 2,0 2,0 1,0 10,0 80,0 10,0 - - 70,0 30,0 - - - 70,0 30,0 - - -
18 2,0 2,0 2,0 90,0 10,0 - - - 70,0 - 10,0 - 20,0 100 - - - -
19 4,0 0,0 0,0 - - 20,0 50,0 30,0 - 10,0 30,0 20,0 40,0 33,4 44,4 11,1 - 11,1
20 4,0 0,0 1,0 - 40,0 20,0 20,0 20,0 - 10,0 20,0 30,0 40,0 - - - - -
21 4,0 0,0 2,0 - - 44,4 33,4 22,2 11,1 - 22,2 22,2 44,5 - 100,0 - - -
22 4,0 1,0 0,0 77,8 11,1 11,1 - - 50,0 10,0 10,0 20,0 10,0 100,0 - - - -
23 4,0 1,0 1,0 100,0 - - - - 90,0 10,0 - - - 100,0 - - - -
24 4,0 1,0 2,0 66,7 33,3 - - - 90,0 10,0 - - - 90,0 10,0 - - -
25 4,0 2,0 0,0 100,0 - - - - 80,0 10,0 10,0 - - - - - - -
26 4,0 2,0 1,0 - - - - - 77,8 22,2 - - - - - - - -
27 4,0 2,0 2,0 100,0 - - - - 50,0 - - - 50,0 - - - - -
Média 40,69 22,55 10,78 9,80 15,69 43,33 20,00 11,67 11,25 13,75 70,73 15,24 3,05 2,44 9,15 (1) Parâmetros estabelecidos para a avaliação da intensidade calogênica descritas na Tabela 3.

34
Com o intuito de avaliar o desempenho morfogênico a nível
celular, foram coletados explantes calogênicos de diferentes tratamentos
e foram identificadas características anatômicas relacionadas com a
competência organogênica e/ou embriogênica dos tecidos induzidos pelas
combinações de fito-reguladores utilizados (Tabela 10).
Tabela 10. Resumo das características anatômicas identificadas em calos de Eucalyptus cloeziana relacionados aos tratamentos na fase de indução calogênica.
Combinações de Fito-reguladores (mg L-1)
Tecidos de origem
Nº ANA TDZ 2,4-D Hipocótilo Cotilédone Raiz
1 0,0 0,0 0,0 - - - 2 0,0 0,0 1,0 Ra CPE CPE, RM, ESg 3 0,0 0,0 2,0 C CPE, ESg - 4 0,0 1,0 0,0 CPE RM CPE, ESg 5 0,0 1,0 1,0 C - CPE 6 0,0 1,0 2,0 CPE, m - ESg 7 0,0 2,0 0,0 - CPE, RM m 8 0,0 2,0 1,0 m - C 9 0,0 2,0 2,0 - - -
10 2,0 0,0 0,0 RM, CPE CPE, m, RM CPE 11 2,0 0,0 1,0 RM ESg, CPE C 12 2,0 0,0 2,0 ESg, RM - - 13 2,0 1,0 0,0 - - - 14 2,0 1,0 1,0 - C - 15 2,0 1,0 2,0 - RM - 16 2,0 2,0 0,0 - - - 17 2,0 2,0 1,0 - C C 18 2,0 2,0 2,0 - RM, CPE - 19 4,0 0,0 0,0 - ESg, RM - 20 4,0 0,0 1,0 - CPE, ESg - 21 4,0 0,0 2,0 ESg, CPE, RM ESg, Ra ESg 22 4,0 1,0 0,0 - - RM 23 4,0 1,0 1,0 - - - 24 4,0 1,0 2,0 - - - 25 4,0 2,0 0,0 - m - 26 4,0 2,0 1,0 - - - 27 4,0 2,0 2,0 - m - Ra: raiz adventícia; CPE: Células pro-embrionarias; RM: Região meristemática; ESg: Embrião somático no estado globular; C: calo; m: Meristemoide.
Dentre as características mais relevantes foram identificadas
áreas celulares de competência meristemática, destacados pelo tamanho
reduzido das células, isodiamétricas e de relação núcleo/plasmática densa

35
(Figura 4 E) as quais promovem a formação de meristemoides (Figura 4 D)
e, em consequência, a geração de novos órgãos de origem adventício
(THORPE e KUMAR, 1993), dos quais os tratamentos das combinações
2 (1,0 mg L-1 2,4-D) e 21 (4,0 mg L-1 ANA + 2,0 mg L-1 2,4-D) forneceram
o balanço hormonal efetivo para a proliferação inicial de raízes adventícias
a partir do calo nos tecidos do hipocótilo e do cotilédone respetivamente.
(Figura 4 C).
Outras características morfogênicas encontradas nas amostras
se diferenciaram em estruturas ou células pró embriogênicas (Figura 5 B
e D) e embriões somáticos, as quais se destacaram os tratamentos que
apresentaram concentrações, tanto isoladas como combinadas, de ANA e
2,4-D na formação de embriões em estágios iniciais (globular) (Figura 5 A,
B, C e E) essencialmente os tratamentos da combinação 21 constituído de
concentrações de 4,0 mg L-1 ANA e 2,0 mg L-1 de 2,4-D em todos os tecidos
testados.
A efetividade desses reguladores também foi relatada por
Muralidharan e Mascarenhas (1987; 1995) na embriogênese somática de
E. citriodora a partir de embriões zigoticos utilizando 3,5 mg L-1 de ANA.
Em E. dunnii, Termignoni et al. (1996) constataram a formação de setores
embriogênicos em explantes cultivados em meio de cultura contendo ANA
ou a combinação de ANA e 2,4-D, nas proporções de 1:1 e 3:1. Pinto et al.
(2002) também observaram a embriogênese somática em E. globulus em
calos derivados de embriões zigóticos maduros inteiros na presença de 3-
15 mg L-1 de ANA isoladamente ou na combinação de 1 mg L-1 de ANA com
1 mg L-1 de 2,4-D.
No entanto, a maioria dos trabalhos relata a inexistência de um
protocolo eficiente de regeneração de Eucalyptus por embriogênese
somática, uma vez que ainda existem problemas básicos a serem
solucionados como metodologias para maturação dos embriões, o número
reduzido de embriões obtidos e a baixa taxa de conversão dos embriões
em plantas.

36
Figura 4. Histologia de calos de Eucalyptus cloeziana. (A) Formação de calo (setas) na epiderme do explante. (B) Detalhe da estrutura celular desdiferenciada (calo) e acúmulo de amido. (C) Raízes adventícias (Ra) geradas a partir do calo (c). (E) Calos apresentando área com características meristemáticas e (D) formação de meristemoides (m). Barra= 100 µm
A
C
EI
B
Ra
Ra
D
E

37
Figura 5. Calos embriogênicos de Eucalyptus cloeziana. (A, C e E) Embriões somáticos (ES) na fase globular. Barra = 100 µm (B) Imagem aproximada do embrião somático presente em A. Barra = 25 µm. (D) Calo apresentando células pró-embrionárias (CPE). Barra = 25 µm. S: suspensor. Pt: Protoderme
A
ES ES
ES ES
ES ES
ES
ES ES
ES
ES ES
B
Pt
CPE
S
ES
ES E

38
4.1.4 Regeneração adventícia
Todos os tecidos de Eucalyptus cloeziana que formaram calo
foram transferidos para meio de cultura, o qual foi suplementado com 1,0
mg L-1 de BAP para todos os tratamentos, visando a regeneração de gemas
e brotos. Considerando a avaliação histológica dos explantes calosos de
alguns tratamentos na formação de estruturas embriogênicas, o meio de
regeneração não apresentou eficiência na promoção da diferenciação dos
embriões nas fases de maturação (coração, torpedo e cotiledonar).
A diferenciação dos tecidos e a formação de órgãos (brotações
adventícias) foram consideradas como reduzidas. Em termos gerais,
observou-se que cerca de 1,5% dos calos demonstraram competência
organogênica (Figura 8 A e B), dos quais sobressaíram os tratamentos
constituídos pelas combinações 4 (1,0 mg L-1 de TDZ), 7 (2,0 mg L-1 de
TDZ) e 16 (2,0 mg L-1 de ANA + 2,0 mg L-1 de TDZ). Os tecidos com
potencial morfogênico ressaltaram o hipocótilo e o cotilédone.
Calos regenerados a partir de tecidos do hipocótilo não
apresentaram diferença significativa nas combinações 7 (2,0 mg L-1 de
TDZ) e 16 (2,0 mg L-1 de ANA + 2,0 mg L-1 de TDZ), os quais resultaram
valores médios de regeneração de brotos de 22,2% e 20,0%,
respectivamente. Por outro lado, dentre os calos obtidos de origem
cotiledonar, houve diferença significativa entre os tratamentos que foram
constituídos pelas combinações 4 e 7, constituídas pelas concentrações de
1,0 mg L-1 e 2,0 mg L-1 de TDZ, respectivamente, sendo este último mais
responsivo ao meio de regeneração, atingindo uma porcentagem igual a
55,6% de calos que regeneraram brotos (Tabela 11).
Diante dos cortes histológicos foi confirmada a regeneração de
brotações adventícias, as quais foram originadas de calos que
apresentaram competência organogênica, evidenciando regiões
meristemáticas (Figura 6 A) e a conexão vascular das brotações com a
massa celular de origem (Figura 6 B).

39
A B
Figura 6. Histologia de calos organogênicos de Eucalyptus cloeziana
obtidos do tratamento constituído pela combinação 7 (2,0 mg L-1 de TDZ) e o tecido do hipocótilo, evidenciando a regeneração de brotos. (A) Detalhe de regiões de atividade meristemática (setas vermelhas) e (A e B) brotações adventícias (setas amarelas) apresentando conexão vascular com a massa celular de origem (seta preta). C= Calo; B= Broto; F= Folha; MA= Meristema apical.
Apesar da elevada intensidade calogênica de todos tecidos por
meio da aplicação de concentrações de ANA e 2,4-D, estes não
apresentaram competência organogênica ao serem transferidos para o
meio de regeneração. O tecido radicular não apresentou competência para
a indução de órgãos adventícios no meio de regeneração em nenhuma das
combinações de reguladores devido à alta determinação epigenética das
células deste tipo de tecido.
Figura 7. Detalhe da síntese de antocianina (setas) na superfície de calos
organogênicos no 1o subcultivo (30 dias) dos explantes no meio de regeneração de brotos. Calos advindos tratamentos de indução calogênica
da combinação 7 (2,0 mg L-1 de TDZ) nos tecidos do (A) hipocótilo e (B) cotilédone. Barra = 1 cm.
A
C
B
B
F
MA
B

40
Além disso, os calos oriundos dos tecidos radiculares foram os
únicos que não apresentaram pigmentação avermelhada, característica
própria da síntese de antocianinas que responde à continua divisão celular
e a possível formação de pontos meristemáticos (Figura 7 A e B), relação
detalhada em diversos trabalhos na regeneração in vitro de espécies de
eucalipto (MURALIDHARAN e MASCARENHAS, 1987; SUBBAIAH e
MINOCHA, 1990; HERVÉ et al., 2001; ALVES et al., 2004a; DIBAX et al.,
2005).
Tabela 11. Porcentagem de explantes de Eucalyptus cloeziana que apresentaram regeneração adventícia in vitro no meio de indução à organogênese, aos 30 dias.
Nas colunas, médias seguidas por mesma letra maiúscula e, nas linhas, médias seguidas por mesma letra minúscula não diferem significativamente pelo teste de Duncan ao nível de 5% de probabilidade de erro. Dados apresentados como: média ± erro padrão.
Combinações de Fito-reguladores (mg L-1) Tecidos de origem
Nº ANA TDZ 2,4-D Hipocótilo Cotilédone Raiz
1 0,0 0,0 0,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
2 0,0 0,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
3 0,0 0,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
4 0,0 1,0 0,0 0,0±0,0Bb 42,8±20,2Ba
0,0±0,0Ab
5 0,0 1,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
6 0,0 1,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
7 0,0 2,0 0,0 22,2±14,7Ab 55,5±17,5Aa
0,0±0,0Ac
8 0,0 2,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
9 0,0 2,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
10 2,0 0,0 0,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
11 2,0 0,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
12 2,0 0,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
13 2,0 1,0 0,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
14 2,0 1,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
15 2,0 1,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
16 2,0 2,0 0,0 20,0±13,3Aa 0,0±0,0Cb
0,0±0,0Ab
17 2,0 2,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
18 2,0 2,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
19 4,0 0,0 0,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
20 4,0 0,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
21 4,0 0,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
22 4,0 1,0 0,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
23 4,0 1,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
24 4,0 1,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
25 4,0 2,0 0,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
26 4,0 2,0 1,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa
27 4,0 2,0 2,0 0,0±0,0Ba 0,0±0,0Ca
0,0±0,0Aa

41
4.1.5 Número de brotos
Dentre os calos regenerados foi avaliado o número de brotações
adventícias (NB) no meio de regeneração ao final da 3o subcultivo (90 dias
de cultivo in vitro). Os tratamentos foram constituídos pelos tecidos e
combinações de fito-reguladores que deram origem aos calos
organogênicos (Tabela 12).
Tabela 12. Descrição dos tratamentos que apresentaram formação de
brotos adventícios na etapa de regeneração.
C = cotilédone; H = hipocótilo.
Por meio da análise de variância (Tabela 13) foi constatado o
efeito significativo dos tratamentos testados (p<0,05) inoculados no meio
de regeneração.
Tabela 13. Resumo da análise de variância do número de brotos (NB) por calo regenerado in vitro de Eucalyptus cloeziana na etapa de regeneração de brotos em relação aos tratamentos, aos 90 dias.
Fonte de variância
GL
Quadrado médio
NB (1)
Tratamento 3 0,17*
Resíduo 8 0,038
Média - 18,5
CVexp(%) - 16,52
* Valor significativo ao nível de 5% de probabilidade de erro, respectivamente pelo teste F. (1) Dados transformados por log10 (n+0,5), onde n = dado amostrado. GL = graus de liberdade, CVexp. = coeficiente de variação experimental.
Tratamentos Fito-reguladores (mg L-1) e tecido de origem
ANA TDZ 2,4-D Tecido
1 0,0 1,0 0,0 C
2 0,0 2,0 0,0 H
3 0,0 2,0 0,0 C
4 2,0 2,0 0,0 H

42
Figura 8. Organogênese in vitro, enraizamento ex vitro e aclimatização de
microestacas de Eucalyptus cloeziana. (A e B) Calo apresentando
regeneração de brotos adventícios in vitro no meio de regeneração, aos 90
dias (barra = 1 cm); (C) Broto coletado para enraizamento ex vitro (barra
= 1 cm); (D) Calo regenerado apresentando brotos alongados ao terceiro
subcultivo (90 dias) (barra = 0,5 cm); (E) Enraizamento in vitro e formação
de micro-cepa (barra = 1cm); (G) Mudas aclimatizadas e alocadas em casa
de sombra aos 150 dias; (F e H) Enraizamento ex vitro de brotos aos 20
dias (barra = 1 cm).
O tratamento 2 o qual foi constituído pelo calo regenerado da
combinação 2,0 mg L-1 de TDZ e o tecido do hipocótilo, apresentou maior
quantidade de brotações adventícias no meio de regeneração (Figura 8 B),
F
G
A B
E

43
estabelecendo uma média geral de 41,5 brotações por calo, diferindo
estatisticamente dos demais tratamentos (Tabela 14).
Considerando que os calos procedentes deste tratamento
apresentaram índices elevados na intensidade calogênica (100%) o que
pode ter favorecido a competência organogênica, resultando calos com
regiões de alta atividade meristemática e promovendo quantidades
superiores de brotos adventícios. Esta relação pode ter estimulado as
diferenças quanto à regeneração entre os tecidos de hipocótilo em
comparação ao tecido cotiledonar (Tratamento 3), nos quais os dos
tratamentos foram induzidos na mesma concentração de TDZ (2,0 mg L-1).
Tabela 14. Número de brotos (NB) por calo regenerado in vitro de Eucalyptus cloeziana em relação aos diferentes tecidos de origem e as combinações de fito-reguladores na etapa de regeneração.
Tratamento
Fito-regulador (mg L-1)
e tecido de origem NB
ANA TDZ 2,4-D Tecido
1 0,0 1,0 0,0 C 20,0±3,5b
2 0,0 2,0 0,0 H 41,5±16,5a
3 0,0 2,0 0,0 C 12,0±2,9b
4 2,0 2,0 0,0 H 9,5±3,5b
Médias seguidas por mesma letra minúscula não diferem significativamente pelo teste de Duncan ao nível de 5% de probabilidade de erro. Dados apresentados como: média ± erro padrão. C = cotilédone; H = hipocótilo.
Comparando os tratamentos 2 e 4, os quais possuíram
características similares, tais como o tecido de origem (hipocótilo), e a
combinação do meio de indução calogênica (2,0 mg L-1 de TDZ e 2,0 mg L-
1 de ANA + 2,0 mg L-1 de TDZ, respectivamente), foi observada diferença
significativa em relação ao número de brotações regeneradas (Tabela 14).
A concentração da auxina ANA contida nos calos do tratamento 4 pode ter
apresentado um efeito desfavorável na adoção à competência celular na
formação dos brotos, considerando que este grupo de fito-reguladores
inibem a formação de brotações aéreas (TAIZ e ZIEGER, 2004).
Todos os calos regenerados foram subdivididos em explantes
padrão (tufos) contendo como mínimo 3 brotações aéreas (Figura 9),

44
conservando a base calogênica que deu origem, e cultivadas em 2 meios
de multiplicação e alongamento (Tabela 4) constituídas por concentrações
diferentes dos fito-reguladores BAP e AIB (Figura 8 C e D), como também
se objetivou induzir a competência rizogênica das bases calosas para o
estabelecimento de micro-cepas in vitro, o qual foi apenas eficiente para
um explante, procedente do tecido cotiledonar e do meio de indução 4 (1,0
mgL-1 de TDZ), inoculado no meio de multiplicação e alongamento 2 (0,5
mgL-1 de BAP + 1,0 mgL-1 de AIB) (Figura 8 E).
Teoricamente, em uma estrutura celular desdiferenciada (calo)
depois de ter-se diferenciado e regenerado em um tipo de órgão (brotos),
a retomada de uma rota morfogênica e formação de um órgão diferente é
pouco provável devido a que, comumente, o processo organogênico
apresenta-se como unipolar, quer dizer, a formação de uma brotação aérea
ou raiz em uma mesma massa celular, ou no caso de se formar os dois
órgãos, estes não apresentarão conexão vascular (Peres, 2002).
Figura 9. Detalhe do tufo padrão utilizado para a multiplicação e alongamento dos brotos. Corte longitudinal do calo organogênico
procedente do hipocótilo induzido pela combinação 7 (2,0 mg L -1 de TDZ), conservando na base o tecido de origem. C= calo 4.2 Sobrevivência e enraizamento ex vitro de micro-estacas
A sobrevivência e enraizamento das micro-estacas foram
avaliados aos 20 dias de incubação no sistema de miniestufa, alocados na
sala de crescimento, as quais apresentaram respostas significativas
descritas através da análise de variância (Tabela 15).
O sistema de mini-estufa foi adotado por meio das vantagens
destacadas em trabalhos de aclimatização ex vitro, principalmente, de
Eucalyptus benthamii relatados por Brondani et al. (2012a) e Baccarin et al.
C

45
(2015) e Eucalyptus cloeziana por Oliveira et al. (2015).
Tabela 15. Resumo da análise de variância para a porcentagem de sobrevivência (SOB%) e enraizamento (ENR%) ex vitro de brotações de Eucalyptus cloeziana aos 20 dias.
Fonte de variância gl Quadrado meio
SOB(%) ENR(%)
Fito-reguladores e Tecidos de origem
(FTO)
2
3,51**
4,62**
Meio de cultura (ME)
1
0,0006ns
1,28ns
FTOxME 2 1,027ns 2,61**
Resíduo 48 0,534 0,47
Média - 68,51 38,9
CVexp(%) - 33,4 40,87
ns Valor não significativo ao nível de 5% de probabilidade de erro pelo teste F. * e ** Valor significativo ao nível de 5% e 1% de probabilidade de erro, respectivamente pelo teste F. Dados transformados por exp(n/100) , onde n = dado amostrado. GL = graus de liberdade, CVexp. = coeficiente de variação experimental.
Esse sistema, além de favorecer condições adequadas de
umidade relativa do ar e temperatura também incrementou o sucesso da
aclimatização dos explantes, evitando fatores relacionados ao estresse no
momento de transferências de plantas com raízes induzidas in vitro no
substrato nas condições ex vitro, como também os danos possivelmente
provocados nas operações dificultando a aclimatização e crescimento das
mudas.
Considerado que isso foi satisfatório para a aclimatização e
acondicionamento ex vitro das microestacas de Eucalyptus cloeziana, no
qual a porcentagem de sobrevivência esteve diretamente influenciada
(p<0,01) pelo tecido e o meio de indução que deram origem aos propágulos
utilizados, atingindo uma porcentagem média do 68,51% de explantes
vivos.
O sistema de mini-estufa permitiu controle fitossanitário e
desenvolvimento individual das micro-estacas, permitindo a mínima
influência das condições externas e atuando como barreira para

46
disseminação diversos agentes patogênicos, podendo inferir que nas
principais etapas para a otimização do processo rizogênico é fundamental
a mínima interferência ambiental para o sucesso da aclimatização e
enraizamento ex vitro das micro-estacas.
No entanto, porcentagens de sobrevivência ex vitro
apresentaram diferencias significativas entre os tratamentos que deram
origem às micro-estacas, destacando novamente o tratamento constituído
pelo tecido do hipocótilo + 2,0 mg L-1 de TDZ que apresentou melhores
porcentagens de sobrevivência dos propágulos, dentre 80 e 100% de
acordo com os MMA 1 e MMA2 respectivamente (Tabela 16).
Tabela 16. Porcentagem de sobrevivência (SOB%) ex vitro de microestacas de Eucalyptus cloeziana segundo a origem e os meios de multiplicação e alongamento in vitro (MMA), aos 20 dias.
Fito-regulador (mg L-1) e
tecido de origem MMA
Combinação
ANA TDZ 2,4-D Tecido 1 2
1 0,0 1,0 0,0 C 66,7±14,2a 66,7±12,6b
2 0,0 2,0 0,0 H 80,0±13,3a 100,0±0,0a
3 2,0 2,0 0,0 H 50,0±28,9a 0,0±0,0c
Médias seguidas por mesma letra minúscula não diferem significativamente na coluna, pelo teste de Duncan ao nível de 5% de probabilidade de erro. Dados apresentados como: média ± erro padrão. C = cotilédone; H = hipocótilo.
Micro-estacas provenientes da combinação 3 (2,0 mg L-1 de
ANA + 2,0 mg L-1 de TDZ), coletados do MMA2 (0,5 mg L-1 de BAP + 1,0
mg L-1 de AIB) apresentaram dificuldade na aclimatização ex vitro. Esse
efeito pode estar relacionado ao comprimento reduzido das brotações
alongadas (comprimento médio= 1,2 cm), as quais apresentaram estresse
durante a transferência para a mini-estufa.
A etapa de sobrevivência de propágulos ex vitro, reflete a
adequação do sistema de aclimatização e enraizamento, visando a
maximização dos processos de produção de mudas.
A porcentagem de enraizamento ex vitro das micro-estacas de
Eucalyptus cloeziana (Figura 8 F e H) a presentou média de 38,9%, a qual

47
pode ser considerada como resultado prometedor para a espécie, a qual
se caracteriza pela dificuldade de enraizamento de propágulos (ALFENAS
et al., 2004). Discriminando entre os diferentes tratamentos, foram
encontradas diferencias significativas dependendo da origem das micro-
estacas (tecidos e combinações de fito-reguladores) e os meios de
multiplicação e alongamento nos quais estiveram cultivados no ambiente in
vitro.
Tabela 17. Porcentagem de enraizamento (ENR%) ex vitro de microestacas de Eucalyptus cloeziana segundo a origem e os meios de multiplicação e alongamento in vitro (MMA), aos 20 dias.
Fito-regulador (mg L-1) e
tecido de origem MMA
Combinação
ANA TDZ 2,4-D Tecido 1 2
1 0,0 1,0 0,0 C 0,0Bb 53,3±13,33Aa
2 0,0 2,0 0,0 H 80,0±13,33Aa 55,6±17,57Ab
3 2,0 2,0 0,0 H 0,0Ba 0,0Ba
Nas colunas, médias seguidas por mesma letra maiúscula e, nas linhas, médias seguidas por mesma letra minúscula não diferem significativamente pelo teste de Duncan ao nível de 5% de probabilidade de erro. Dados apresentados como: média ± erro padrão. C = cotilédone; H = hipocótilo
Apesar da média geral de enraizamento das micro-estacas
tinha sido considerada reduzida, os diferentes tratamentos apresentaram
respostas divergentes, porém promissoras para a produção clonal da
espécie, sendo registrado o porcentual maior de brotos enraizados (80%)
em brotações procedentes da combinação 2 (hipocótilo + 2, mg L-1 de TDZ)
cultivados no meio MMA1 (1,0 mg L-1 de BAP + 0,5 mg L-1 de AIB) (Figura
8 F) que foram suplementadas ao meio de cultura (Tabela 17).
Cabe salientar a relação entre os dois reguladores como pouco
comuns na estimulação a competência à rizogênese de propágulos pelo
fato de conter a concentração de citocinina superior, as quais são definidas
como inibidores na promoção de raízes adventícias, em comparação às
auxinas utilizadas no meio de cultura (TAIZ e ZEIGER 2004).
Concentrações de AIB em torno de 0,5 mg L-1 poderiam
promover a predisposição ao enraizamento em brotos alongados,

48
diferindo estatisticamente em comparação com os propágulos da
mesma combinação de origem cultivamos no meio MMA2 (0,5 mg L-1 de
BAP + 1,0 mg L-1 de AIB), que teoricamente, pelo fato de conter a
concentração superior do regulador AIB (1,0 mg L-1), deveriam ter
fornecido maiores porcentagens dos propágulos, os quais forneceram
valor igual a 55,6%.
Porcentagens similares aos registrados neste trabalho foram
mencionados por Oliveira et al. (2015), os quais observaram porcentagens
de enraizamento de micro-estacas da mesma espécie, apresentando
elevada resposta quanto ao tipo do material genético avaliado. Os valores
de enraizamento ex vitro das micro-estacas variaram de 25% e 55%,
respectivamente para matrizes adultas e rejuvenescidas por meio da micro-
estaquia.
Cabe destacar o tempo de cultivo in vitro, desde a etapa de
calogênese (30 dias), regeneração de brotações (90 dias), multiplicação e
alongamento de brotos (90 dias), o enraizamento ex vitro (20 dias) e a
aclimatização e rustificação (Figura 8 G) das mudas (100 dias) o protocolo
apresentado (330 dias) pode ser considerado como uma alternativa rápida
e efetiva de propagação da espécie.
A metodologia desenvolvida pode ser testada em outras
espécies que apresentam dificuldade de enraizamento, levando em
consideração que nos processos atuais de rejuvenescimento in vitro
comumente são considerados dentre 12 a 19 sub-cultivos sucessivos (360
a 523 dias) visando a juvenilidade/revigoramento dos tecidos e
consequentemente o aumento na competência rizogênica de propágulos.
Cabe salientar que o protocolo foi desenvolvido para explantes coletados
de plântulas obtidas pela germinação in vitro, sendo necessário testar, para
plantas adultas selecionadas, a viabilidade da aplicação da técnica.

49
5 CONCLUSÕES
1. A competência dos tecidos e as concentrações dos fito-
reguladores foram efetivas à formação de calo.
2. As combinações 2 (1,0 mg L-1 de 2,4-D) e 7 (2,0 mg L-1 de TDZ)
induziram elevadas intensidades calogênicas nos tecidos testados.
3. A presença de estruturas pró embrionárias e embriões somáticos no
estágio globular foram promovidas principalmente nas combinações de
ANA e 2,4-D, destacando os tratamentos da combinação 21 (4,0 mg L-1 de
ANA + 2,0 mg L-1 de 2,4-D) nos tecidos utilizados.
4. O BAP no meio de regeneração adventícia dos explantes promoveu
a diferenciação celular e a formação de brotos aéreos, mas não foi efetivo
na diferenciação de embriões somáticos em estágios maduros.
5. O tratamento constituído pelo tecido cotiledonar induzido na
combinação 7 (2,0 mg L-1 de TDZ) destacou na promoção dos calos
organogênicos (55,6%), ademais das variáveis quantidade de brotos (41,5
brotos/explante), porcentagens de sobrevivência (80 - 100%) e
enraizamento ex vitro (55,6 – 80%).
6. Mediante o sistema de mini-estufa e a seguinte transferência a casa
se sombra forneceram condições ótimas para aclimatização e rustificação
das microestacas, respectivamente.

50
6 CONSIDERAÇÕES FINAIS E RECOMENDAÇÕES
Porcentagens de manifestação bacteriana e a ocorrência da
oxidação nos tecidos apresentaram-se interligadas nos primeiros estágios
da cultura calogênica, mas estas não constituíram uma problemática para
o desenvolvimento do experimento.
Tecidos juvenis apresentaram resposta eficiente ao estimulo do
fito-regulador TDZ principalmente ás concentrações isoladas de 1,0 e 2,0
mg L-1 destacando-se nas diferentes etapas in vitro e ex vitro, podendo ser
utilizado para o estabelecimento de protocolos de organogênese indireta
mediante a utilização de explantes maduros provenientes de matrizes
selecionadas a campo.
A utilização de combinações de BAP e AIB foram adequadas
para a multiplicação e alongamento das brotações, inclusive foi possível
combinar as duas etapas no mesmo período de cultivo, mas a identificação
de concentrações que promovam maiores taxas de multiplicação e índices
de alongamento das brotações favoreceriam na eficiência da metodologia.
Protocolos efetivos para a formação de micro-cepas in vitro e/ou
mini-cepas ex vitro de Eucalyptus cloeziana poderiam beneficiar à
constituição de bancos de matrizes para o fornecimento de brotações com
elevada competência ao enraizamento.
Metodologias para o desenvolvimento efetivo e obtenção de
embriões somáticos maduros in vitro poderiam favorecer ao
estabelecimento do protocolo de embriogênese somática de Eucalyptus
cloeziana para estratégias para a obtenção de sementes artificiais e
propagação clonal da espécie.

51
7 REFERENCIAS BIBLIOGRAFICAS
AGGARWAL, D.; KUMAR, A.; REDDY, M. S. Shoot organogenesis from elite plants of Eucalyptus tereticornis. Plant Cell Tissue and Organ Culture, Dordrecht, v. 102, n. 1, p. 45–52, 2010. ALCANTARA, G. B. Transformação genética de Eucalyptus grandis x E. urophylla com o gene gfp sob controle de promotores cbfs induzidos. Curitiba. Tese (Doutorado), Universidade Federal de Paraná. 2008. 133 p. ALCANTARA, G. B.; BESPALHOK FILHO, J. C.; QUOIRIN, M. Organogenesis and transient genetic transformation of the hybrid Eucalyptus grandis × Eucalyptus urophylla. Sci. Agric., v.68, n.2, 2011, p.246-251.
ALFENAS, A. C.; ZAUZA, E. A. V.; MAFIA, R. G.; ASSIS, T. F. Clonagem e doenças do eucalipto. Viçosa: UFV, 2004. 442 p. ALVES, E. C. S. C.; XAVIER, A.; OTONI, W. C. Organogênese in vitro a partir de explante caulinar na regeneração de clones de Eucalyptus grandis W. Hill ex Maiden x E. urophylla S. T. Blake. Revista Árvore, Viçosa, v. 28, n. 5, pp. 643-653, 2004a
. Organogênese de explante foliar de clones de Eucalyptus grandis x E. urophylla. Pesquisa Agropecuária Brasileira, Rio de Janeiro, v. 39, n. 5, p. 421-430, 2004b
ALVES, E. C. S. C. Antioxidantes, reguladores de crescimento e estresse térmico na embriogênese somática de Eucalyptus spp. Viçosa. Tese (Doutorado). Universidade Federal de Viçosa. 2007. 102 p.
ALMEIDA, C.V.; YARA, R.; ALMEIDA, M. Fungos endofíticos isolados de ápices caulinares de pupunheira cultivada in vivo e in vitro. Pesquisa Agropecuária Brasileira, Brasília, v. 40, p. 467-470, 2005.
ALMEIDA, F.D., XAVIER, A. DIAS, J.M.M. Vegetative propagation of selected Eucalyptus cloeziana F. Muell. trees through cutting technique. Revista Árvore 31, 445–453, 2007a
ALMEIDA, F.D.; XAVIER, A.; DIAS, J.M.M.; PAIVA, H.N. Eficiência das auxinas (AIB e ANA) no enraizamento de miniestacas de clones de Eucalyptus cloeziana F. Muell. Revista Árvore, Viçosa, v. 31, n. 3, p. 455–463, 2007b
ALMEIDA, L.V. Técnicas para a otimização da multiplicação in vitro de
brotações de Eucalyptus citriodora (Hook) K. D. Hill & L. A. Johnson.
2012. 107 p. (Mestrado em Recursos Florestais) - Escola Superior de
Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 2012.

52
AMS: Associação Mineira de Silvicultura. Silvicultura: desenvolvimento social, econômico e ambiental com plantios florestais. Belo Horizonte: Associação Mineira de Silvicultura, 18p, 2004
ASSIS, T. F. Propagação vegetativa de Eucalyptus por microestaquia. In: IUFRO CONFERENCE ON SILVICULTURE AND IMPROVEMENT OF EUCALYPTS, 1997, Salvador. Proceedings... Colombo: EMBRAPA, 1997. v. 1, p. 300-304
ARENHART, R. A.; ZAFFARI, G. R. Otimização do protocolo de micropropagação por organogênese indireta de Eucalyptus grandis. Revista de Ciências Agroveterinárias. Lages, v.7, n.1, p. 16-22, 2008
ASSOCIAÇÃO BRASILEIRA DE PRODUTORES DE FLORESTAS PLANTADAS (ABRAF). Anuário estatístico da ABRAF: ano base 2012. Brasília, 2013. 140 p BACCARIN, F. J. B.; BRONDANI, G. E.; ALMEIDA, L. V.; VIEIRA, I. G.; OLIVEIRA, L. S.; ALMEIDA, M. Vegetative rescue and cloning of Eucalyptus benthamii selected adult trees, New Forests. V. 46, n. 4, pp 465-483, 2015 BANDYOPADHYAY, S.; CANE, K.; RASMUSSEN, G.; HAMILL, J. Efficient plant regeneration from seedling explants of two commercially important temperate eucalypt species – Eucalyptus nitens and E. globulus. Plant Science, New York, v.140, n. 2, p. 189- 198, 1999 BATISTA, T. R. Organogênese e embriogênese somática de híbrido de Eucalyptus grandis. Lavras. Tese (Mestrado). Universidade Federal de Lavras. 2012. 73 p. BLEAKLEY, S.; CANT, R.T. Australian species in Zimbabwe: existing role and potential. In: WORKSHOP ON SEED HANDLING AND EUCALYPT TAXONOMY, 1984, Harare. Proceedings... Harare: [s.n.], 1985. p. 246-252. BOLAND, D.J.; BROOKER, M.I.H.; CHIPPENDALE, G.M.; HALL, N.; HYLAND, B.P.M.; JOHNSTON, R.D.; KLEINIG, D.A.; MCDONALD, M.W.; TURNER, J.D. Forest trees of Australia. 5ª ed. CSIRO. p. 736. 2006. BONGA, J. M.; VON ADERKAS, P. In vitro culture of trees. Netherlands: Kluwer Academic Publishers, 1992. 236 p. BORGES, S.R.; XAVIER, A.; OLIVEIRA, L.S.; LOPES. A.P.; OTONI, W.C; TAKAHASHI, E.K.; MELO, L.A. Estabelecimento in vitro de clones híbridos de Eucalyptus globulus. Ciência Florestal, Santa Maria, v. 22, n. 3, p. 605-616, 2012.

53
BORGES, S.R.; XAVIER, A.; OLIVEIRA, L.S.; LOPES, A.P.; OTONI, W.C. Multiplicação in vitro de clones híbridos de Eucalyptus globulus. Revista Árvore, Viçosa, v. 35, n. 2, p. 173-182, 2011 BRONDANI, G.E.; ONDAS, H.W.W.; BACCARIN, F.J.B.; GONÇALVES, A.N.; ALMEIDA, M. Micropropagation of Eucalyptus benthamii to form a clonal micro-garden. In Vitro Cellular Development and Biology-Plant, Columbia, v. 48, n. 5, p. 478-487, 2012a BRONDANI, G. E.; ARAUJO, M. A. de; ALCÂNTARA, B. K. de; CARVALHO, J. G. de; GONÇALVES, A. N.; ALMEIDA, M. de. Efeitos do boro e cálcio na organogênese in vitro de Eucalyptus grandis. Acta Scientiarum. Agronomy, 34(4), 403-411, 2012b.
BUNN, E. Development of in vitro methods for ex situ conservation of Eucalyptus impensa, an endangered mallee from southwest Western Australia.Plant Cell, Tissue and Organ Culture, Dordrecht, v. 83, n. 1, p. 97–102, 2005. CHEN, S.X.; WU, Z.H.; LI, Z.H.; XIE, Y.J.; LI, T.H.; ZHOU, Q.Y.; ARNOLD, R. Selection of species for solid wood production in southern China. Journal of Tropical Forest Science, Kepong, v. 22, n. 3, p. 308-316, 2010 CID, L.P. B.; TEIXEIRA, J. B. Explante, meio de cultivo, luz e temperatura. In: CID, L. P. B. Cultivo in vitro de plantas. Brasilia DF. Embrapa. 103-128 p. 2010 CID, L. P. B.; MACHADO, A. C. M.; CARVALHEIRA, S. R. C.; BRASILEIRO, A. C. M. In vitro contamination and germination of Eucalyptus cloeziana seeds. Proceedings of the IUFRO conference on silviculture and improvement of eucalypts, Salvador, Brazil. EMBRAPA, Colombo, pp. 14–34. 1997
. Plant regeneration from seedling explants of Eucalyptus grandis x E. urophylla. Plant Cell Tissue and Organ Culture, Dordrecht, v. 56, n. 1, p. 17–23, 1999 CAMPOS, D. T. S.; SILVA, M. C. S.; DA LUZ, J. M. R.; TELESFORA, R. J.; KASUYA, M. C. M. Colonização micorrízica em plantios de eucalipto. Revista Árvore, v. 35, n. 5, p. 965- 974, 2011. CARVALHO, D.C.; SILVA, A.L.L.; TANNO, G.N.; PURCINO, M.; BIASI, L.A. Organogênese a partir de segmentos foliares e internodais de videira cv. Merlot. Ciênc. agrotec., v. 35. n. 1. 2011. 108-114. CAUDURO, Y. O. Organogênese indireta em Eucalyptus benthamii x Eucalyptus dunnii e transformação genética de Eucalyptus saligna E Nicotiana tabacum. Curitiba. Universidade Federal de Paraná. 2013. 151p.

54
CHAPERON, H. Vegetative propagation of Eucalyptus. In: Symposio sobre Silvicultura y Mejoramiento Genético de Especies Forestales, 1987, Buenos Aires, Argentina. Anales... (s.l): AFOCEL, 1987. p. 215-232. CLARKE, B.; MCLEOD, I.; VERCOE, T. Trees for farm forestry: 22 promising species. Canberra: Rural Industries Research and Development Corporation, 2009. 239 p. COUTO, L.; NICHOLAS, I.; WRIGHT, L. Short rotation eucalypt plantations for energy in Brazil. IEA Bioenergy, Washington, v. 43, n. 1, p.17, 2011 DIBAX, R.; EISFELD, C. L; CUQUEL, F. L.; KOEHLER. H.; QUOIRIN, M. Plant regeneration from cotyledonary explants of Eucalyptus camaldulensis. Scientia Agricola, v. 62, n. 4, p. 406-412, 2005
DIBAX, R.; DESCHAMPS, C.; BESPALHOK FILHO, J, C.; VIEIRA, L. G. E.; MOLINARI, H. B. C.; CAMPOS, M. K. F. DE; QUOIRIN, M. Organogenesis and Agrobacterium tumefaciens mediated transformation of Eucalyptus saligna with P5CS gene. Biologia Plantarum, v. 54, n. 1, p. 6-12. 2010a.
DIBAX, R.; QUISEN, R. C.; BONA, C.; QUOIRIN, M. Plant regeneration from cotyledonary explants of Eucalyptus camaldulensis Dehn and histological study of organogenesis in vitro. Brazilian archives of Biology and technology, Vol. 53, n. 2: pp.311-318, 2010b. DUNGEY, H.S.; NIKLES D.G. An international survey of interspecific hybrids in forestry. In: DUNGEY H.S.; DIETERS M.J.; NIKLES D.G. (eds), Hybrid Breeding and Genetics of Forest Trees. Proceedings of QFRI/CRC-SPF Symposium, 9–14th April 2000 Noosa, Queensland, Australia. Department of Primary Industries, Brisbane, pp. 419–42
DUTRA, L. F.; WENDLING, I.; BRONDANI, G. E. A micropropagação de eucalipto. Pesquisa Florestal Brasileira, Colombo, n. 58, p.49-59, jan./jun. 2009
EMBRAPA-Empresa Brasileira de Pesquisa Agropecuária. Programa SOC - software científico: versão 2.1. Campinas: Embrapa Informática Agropecuária, 1990.
FERREIRA, F. A. Patologia florestal: principais doenças florestais do Brasil. Viçosa: Sociedade de Investigações Florestais, 1989.
GEORGE, E. F.; HALL, M. A.; KLERK, G. J. Plant propagation by tissue culture. 3rd ed. Dordrecht: The Backgrouns, v. 1. 2008.
GIRIJASHANKAR, V. Genetic transformation of eucalyptus. Physiology and Molecular Biology of Plants. vol. 17, n. 1, p. 9–23. 2011.
. In vitro regeneration of Eucalyptus camaldulensis. Physiology and Molecular Biology of Plants, v. 18, n. 1, p. 79–87, 2012.

55
GLOCKE, P.; COLLINS, G.; SEDGLEY, M. 6-Benzylamino purine stimulates in vitro shoot organogenesis in Eucalyptus erythronema, E. stricklandii and their interspecific hybrids. Scientia Horticulturae, Amsterdan, v.109, n. 4, p. 339–344, 2006
GONZALEZ, E. R. Transformação genética de Eucalyptus grandis e do híbrido E. grandis x E. urophylla via Agrobacterium. Piracicaba, Tese (Doutorado em Genética e Melhoramento de Plantas) - Escola Superior de Agricultura “Luís de Queiroz”, Piracicaba, 2002, 93 p. GONÇALEZ, J. C.; BREDA, L. de C. S.; BARROS, J. F. M.; MACEDO, D. G.; JANIN, G.; COSTA, A. F. da; VALE, A. T. do. Características tecnológicas das madeiras de Eucalyptus grandis W.Hill ex Maiden e Eucalyptus cloeziana F. Muell visando ao seu aproveitamento na indústria moveleira. Ciência Florestal, v. 16, n. 3, p. 329-341, 2006 GRATTAPAGLIA, D.; MACHADO, M. A. Micropropagação. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/CNPH, p.183-260,1998 HAJARI, E.; WATT, M.P.; MYCOCK, D.J.; MCALISTER, B. Plant regeneration from induced callus of improved Eucalyptus clones. South African Journal of Botany, Pretoria, v. 72, p. 195- 201, 2006.
HERVÉ, P.; JAUNEAU, A.; PÂQUES, M.; MARIEN, J.; BOUDET, A. M.; TEULIÈRES, C. A procedure for shoot organogenesis in vitro from leaves and nodes of an elite Eucalyptus gunnii clone: comparative histology. Plant Science, Limerick, v. 161, p. 645-653, 2001 HIGASHI, E.N.; SILVEIRA, R.L.V.A.; GONÇALVES, A.N., Propagação vegetativa de Eucalyptus: princípios básicos e a sua evolução no Brasil. Circular Técnica IPEF, Piracicaba, n. 192, p. 11, 2000.
HOPEWELL, G. P.; ATYEO, W.J.; MCGAVIN, R.L. Evaluation of wood characteristics of tropical post-mid rotation plantation Eucalyptus cloeziana and E. pellita: Part (d) Veneer and plywood potential. Victoria: Forest and Wood Products Australia, 2008, 47p HUANG, Z. C.; ZENG, F. H.; LU, X. Y. Efficient regeneration of Eucalyptus urophylla from seedling-derived hypocotyls. Biologia Plantarum, v. 54, n.1, p. 131-134, 2010.
HUETTEMAN, C.A.; PREECE, J.E. Thidiazuron: a potent cytokinin for wood plant tissue culture. Plant Cell, Tissue and Organ Culture, Notherlands, v.33, n.2, p.105-119, 1993.
INTERAMNENSE, M.T. Utilização das madeiras de Eucalyptus cloeziana (F. Muell), Eucalyptus maculata (Hook) e Eucalyptus punctata DC var. punctata para produção de painéis compensados. 1998. 81 p. Dissertação (Mestrado em Engenharia Florestal), Universidade Federal do Paraná, Curitiba,1998.

56
INSTITUTO DE PESQUISAS E ESTUDOS FLORESTAIS. Identificação de espécies florestais, Eucalyptus cloeziana F. Muell. Disponível em: <http://www.ipef.br/identificacao/cief/especies/cloeziana>. Acesso em: 22 setembro de 2015. KARNOVSKY, M.J. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. Journal Cellular Biology, New York, v. 27, n. 15, p. 137–138, 1965
KERBAUY, G, B. Cultura de raízes e regeneração de plantas. In: TORRES, A.C.; CALDAS, L.S.; BUSO, J.A. Cultura de tecidos e transformação genética de plantas. Brasília: Embrapa/CNPH, p.183-260,1998
LAKSHMI SITA, G. Micropropagation of Eucalyptus. In: AHUJA, M. R. (Ed.). Micropropagation of woody plants. Boston: Kluwer Academic Publishers, p.263-280. 1993.
LEMOS, E. E. P. Organogênese. In: CID, L. P. B. Cultivo in vitro de plantas. Brasilia DF. Embrapa. 103-128 p. 2010 LLOYD, G.; MCCOWN, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. International Plant Propagation Society Proceedings, v. 30, p. 421-427, 1980. LORENZI, H.; SOUZA, H.M.; TORRES, M. A. V.; BACHER, L. B. Árvores exóticas no Brasil. Nova Odessa, São Paulo: Ed. Instituto Plantarum.384p. 2003
LU, C. The use of thidiazuron in tissue culture. In Vitro Cell and Development Biology, v. 29, p: 92-96, 1993
MORAES, T. S. A.; GONÇALVES, E. L.; RESENDE, G. C.; MENDES, J.; SUITER, F. W. Evolução da ferrugem causada pela Puccinia psidii WINTER, em Eucalyptus spp. Dados preliminares. Piracicaba: IPEF, 1982. 12p. (Circular Técnica, 144)
MOURA, V. P. G. Seleção de espécies de Eucalyptus e Corymbia de acordo com locais e usos. Brasília: Embrapa Recursos Genéticos e Biotecnologia, 2001. 32p. (Embrapa Recursos Genéticos e Biotecnologia. Documentos, 68).
. O germoplasma de Eucalyptus cloeziana F. Muell no Brasil. Brasília: Embrapa Recursos Genéticos e Biotecnologia; CNPF, 2003. 9p. (EMBRAPA. Comunicado Técnico 102).
MOURA, V.P.G.; GUIMARÃES, D.P. Uma análise da atividade florestal nos Cerrados. In: SIMPÓSIO SOBRE CERRADO, Brasília. Anais... Planaltina: EMBRAPA, CPAC, 1988. p. 853-870.
. Produção de mudas de Eucalyptus para o estabelecimento de plantios florestais. Brasília: Embrapa Recursos Genéticos e Biotecnologia; CNPF. 2003. 9 p. (EMBRAPA. Comunicado Técnico 85).

57
MURALIDHARAN, E. M.; MASCARENHAS, A. F. Somatic embryogenesis in Eucalyptus. In: JAIN, S. M.; GUPTA, P. K.; NEWTON, R. J. (Ed.). Somatic embryogenesis in woody plants. Dordrecht: Kluwer Academic Publishers, v.2. p.23-40, 1995. . In vitro plantlet formation by organogenesis in Eucalyptus camaldulensis and by somatic embryogenesis in Eucalyptus citriodora. Plant Cell Reports, Berlin, v. 6, p. 256-259, 1987.
NASCIMENTO, M. S. do. Organogênese a partir de segmentos cotiledonares e calogênese de anteras de pinhão-manso (Jatropha curcas L.), Lavras. Tese (Mestrado), Universidade Federal de Lavras. 2012. 89 p.
NGUGI, M.R.; DOLEY, D.; HUNT, M.A.; RYAN, P.; DART, P. Physiological responses to water stress in Eucalyptus cloeziana and E. argophloia seedlings. Trees, Berlin, v. 18, n. 4, p.381-389, 2004
NUGENT, G.; CHANDLER, S.F.; WHITEMAN, P.; STEVENSON, T.W. Adventitious bud induction in Eucalyptus globulus Labill. In Vitro Cellular Development and Biology-Plant, Columbia, v. 37, n. 3, p. 388–391, 2001. OLIVEIRA, L.S.; XAVIER, A.; DIAS, P.C.; CORREIA, A.C.G.; BORGES, S.R.; TAKAHASHI, E.K.; PAIVA, H.N. Enraizamento de miniestacas e microestacas de clones de Eucalyptus urophylla x E. globulus e de Eucalyptus grandis x E. globulus. Scientia Forestalis, Piracicaba, v. 40, n.96, p. 507-516, 2012. OLIVEIRA. L. S.; DIAS P. C.; BRONDANI, G. E. Micropropagação de espécies florestais brasileiras. Pesquisa Florestal Brasileira, Colombo, v. 33, n. 76, 2013: p. 439-53. OLIVEIRA, L. S. Propagação de Eucalyptus cloeziana F. Muell. Piracicaba, Tese (Doutorado). Escola Superior de Agricultura “Luiz de Queiroz”, 2014. 144 p.
OLIVEIRA, L. S.; BRONDANI, G. E.; PIOTTO, K. D. B.; CALSAVARA, R.; GONÇALVES, A. N.; ALMEIDA, M. Micropropagation of Eucalyptus cloeziana mature trees. Australian Forestry. 2015. DOI:
10.1080/00049158.2015.107321
PALUDZYSZYN FILHO, E.; SANTOS, P. E. T. dos; FERREIRA, C. A. Eucaliptos indicados para plantio no Estado do Paraná. Colombo: Embrapa Florestas - CNPF, 2006. 45 p. (Documentos, 129). PRAKASH, M. G.; GURUMURTHI, K. S. Effects of type of explant and age, plant growth regulators and medium strength on somatic embryogenesis and plant regeneration in Eucalyptus camaldulensis. Plant Cell, Tissue and Organ Culture, v. 100, p. 13–20, 2010

58
PEREIRA, R. P. W.; MONTEIRO, M. B. O.; ABREU, H. S. Callogenesis in stem explants of Eucalyptus grandis (Hill ex Miden) showing resilience in oxide calluses. Floresta e Ambiente. 20(3): 407- 416 p. 2013. PEREIRA, J. C. D.; STURION, J. A.; HIGA, A. R.; HIGA, C. V.; SHIMIZU, J.Y. Características da madeira de algumas espécies de eucalipto plantadas no Brasil. Colombo: Embrapa Florestas, 2000. 113p. (Embrapa Florestas. Documentos, 38). PERES, L. E. P. Bases fisiológicas e genéticas da regeneração de plantas in vitro. Biotecnologia Ciência e Desenvolvimento, v. 4, n. 25, p. 44-48,2002. PIERIK, R.L.M. In vitro culture of higher plants. Dordrecht: Kluwer Academic, 1997. 348p
PINTO, G.; SANTOS, C.; NEVES, L.; ARAÚJO, C. Somatic embryogenesis and plant regeneration in Eucalyptus globulus Labill. Plant Cell Reports, v.21, n.3, p.208-213, 2002.
PIOTTO, K. D. B. Atuação da manifestação bacteriana no desenvolvimento in vitro de clones de Eucalyptus benthamii Maiden & Cambage. 168 p. Tese (Doutorado em Fisiologia e Bioquímica de Plantas) - Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 2013
POTTS, B. M.; DUNGEY, H. S. Interspecific hybridization of Eucalyptus: key issues for breeders and geneticists. New Forests, v. 27, n. 2, pp 115-138, 2004 R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R- project.org/. 2015 RUIZ, R. A.; ALFENAS, A. C.; FERREIRA, F. A.; ZAMBOLIM, L. Fungicidas protetores e sistêmicos para o controle da ferrugem do eucalipto, causada por Puccinia psidii. Revista Árvore, v. 11, n.1, p. 56-65, 1987
SAKAI, W. S. Simple method for differential staining of paraffin embedded plant material using toluidine blue O. Biotechnic and Histochemistry, Baltimore, v. 48, n. 5, p. 247–249, 1973
SANTOS, E. K. dos. Totipotência celular e cultura de tecidos vegetais. In: FREITAS, L. B.; BERED, F. Genética e evolução vegetal. Porto Alegre: Editora de UFRGS, 2003. p.415-444. SANTOS, A. P. D.; XAVIER A.; OLIVEIRA M. L.; REIS. G. G. Efeito da estaquia, miniestaquia, microestaquia e micropropagação no desempenho silvicultural de clones de Eucalyptus grandis. Scientia Forestalis n. 68, p.29-38, ago. 2005

59
SARTORETTO, L. M. Transformação genética: estratégias e aplicações para o melhoramento genético de espécies florestais. Ciência Rural, Santa Maria, v. 38, n. 3, p. 861-871, 2008. SHARMA, S.K.; RAMAMURTHY, V. Micropropagation of 4-year-old elite Eucalyptus tereticornis trees. Plant Cell Reports, Heidelberg, v. 19, n. 5, p.511–518, 2000
SUBBAIAH, M. M.; MINOCHA, S. C. Shoot regeneration from stem and leaf callus of Eucalyptus tereticornis. Plant Cell Reports, Berlin, v. 9, p. 370-373, 1990.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 3ª Ed. Artmed: 677 p. 2006
TERMIGNONI, R. R.; WANG, P.; HU, C. Somatic embryo induction in Eucalyptus dunnii. Plant Cell, Tissue and Organ Culture, v.45, p.129-132,1996. TIBOK, A.; N.W. BLACKHALL, J.B. POWER, M.R. DAVEY. Optimized plant regeneration from callus derived from seedling hypocotyls of Eucalyptus urophylla. Plant Science, 110. 139-145. 1995.
TITON, M.; XAVIER, A.; OTONI, W. C. Clonal propagation of Eucalyptus grandis using the mini-cutting and micro-cutting techniques. Scientia Forestalis, n. 71, p. 109-117, 2006.
TRUEMAN, S.J.; MCMAHON, T.V.; BRISTOW, M. Production of Eucalyptus cloeziana cuttings in response to stock plant temperature. Journal of Tropical Forest Science, Kuala Lumpur, v. 25, n. 1, p. 60–69,2013
THORPE, T. A.; KUMAR, P.P. Cellular control of morphogenesis. In: AHUJA, M.R. Micropropagation of wood plants. Dordrecht: Kluwer Academic, 1993. p. 11-29.
THORPE, T. A.; HARRY, I. S.; KUMAR, P. P. Application of micropropagation to forestry. In: DEBERGH, P. C.; ZIMMERMAN, R. H. Micropropagation: technology and aplication. Dordrecht: Kluwer Academic Press, 1991. p. 311 - 336.
VENTURIERI, G. A.; VENTURIERI, G. C. Calogênese do híbrido Theobroma randiflorum x T. obovatum (Sterculiaceae). Acta amazônica. v. 34(4) 2004: p. 507 – 511 VIJAYA, G.L.; GIRI, C.C. Plant regeneration via organogenesis from shoot base-derived callus of Arachis stenosperma and A. villosa. Current Science, 85(11). 2003:1624-1629. WATT, M. P; BLAKEWAY, F.; CRESSWELL, C. F.; HERMAN, B. Somatic embryogenesis in Eucalyptus grandis. South African Forestry Journal, v.157, p.59-65, 1991.

60
WENDLING. I.; XAVIER, A. influência da miniestaquia seriada no vigor radicular de clones de Eucalyptus grandis. R. Árvore, Viçosa-MG, v.29, n.5, p.681-689, 2005 WENDLING, I., TRUEMAN, S.J.; XAVIER, A. Maturation and related aspects in clonal forestry—part II: reinvigoration, rejuvenation and juvenility maintenance. New Forests. v. 45, n. 4. p.473–486, 2014 XAVIER, A.; COMÉRIO, J. Microestaquia: uma maximização da micropropagação de Eucalyptus. Revista Árvore, v. 20, n. 1, p. 9-16, 1996 XAVIER, A.; OTONI, W.C. Aplicações da micropropagação na clonagem de Eucalyptus no Brasil. Agronomía Costarricense, San Jose, v. 33, n. 2, p. 303-307, 2009 XAVIER, A.; SILVA, R.L. Evolução da silvicultura clonal de Eucalyptus no Brasil. Agronomía Costarricense, San Jose, v. 34, n. 1, p. 93-98, 2010. XAVIER, A.; WENDLING, I.; SILVA, R.L. Silvicultura clonal: princípios e técnicas. 2da ed. Viçosa: Editora UFV, 2013. 279 p.





























