Embed Size (px)
Citation preview

Metabolismo dos Lipídeos
Coordenação do Curso de QuímicaCurso de Licenciatura Plena em Química
Disciplina: BioquímicaProfessor: Francisca das Chagas
Por: Carlos Átila Ézio Raul Francisco Jônatas Josivaldo Mendes Letícia Cabral Robert Ribeiro

DIGESTÃO, ABSORÇÃO E TRANSPORTE DE LIPÍDIOS
• Os triacilgliceróis constituem aproximadamente 90% dos lipídios da dieta e são a principal forma de armazenamento de energia metabólica nos seres humanos.
• Os triacilgliceróis são constituídos por triésteres de glicerol e ácidos graxos.

DIGESTÃO E ABSORÇÃO
• A taxa de digestão dos triacilgliceróis depende da área de superfície da interface, a qual é bastante aumentada pelos movimentos peristáuticos do intestino combinados à ação emulsificante dos ácidos biliares.
• Os ácidos biliares são moléculas anfipáticas do tipo detergente que atuam na solubilização dos glóbulos de gordura.
• São derivados do colesterol, sintetizados pelo fígado e secretados na vesícula biliar como conjugados de glicina ou de taurina para armazenamento. Dali são secretados para o intestino delgado, onde ocorre digestão e absorção dos lipídios.
• Os ácidos biliares não apenas auxiliam a digestão de lipídios, são fundamentais para a absorção dos produtos digeridos.

TRANSPORTE DE LIPÍDEOS
• Os ácidos produzidos pela digestão de lipídeos que são absorvidos pela mucosa intestinal se distribuem para outros tecidos para catabolismo ou armazenamento.
• Como são pouco solúveis em soluções aquosas, são transportados pela circulação na forma de complexos de proteínas.

TRANSPORTE DE LIPÍDEOS
• As lipoproteínas são partículas globulares semelhantes a micelas que consistem em um núcleo apolar de triacilgliceróis e ésteres de colesterol envoltas por uma cobertura anfifílica de proteínas, fosfolipídeos e colesterol.
• São lipoproteínas conhecidas: VLDL, IDL, LDL e HDL.

TRANSPORTE DE LIPÍDEOS
• LDL – as células animais podem obter colesterol, um componente essencial das membranas celulares, tanto por meio de síntese interna ou por captação de LDL, que são ricas em colesterol e éteres de colesteril.
• Os receptores LDL agrupam-se em cavidades recobertas por clatrina, as quais reúnem os receptores da superfície celular destinadas a endocifose, à proporção que exclui outras proteínas da superfície da célula.

TRANSPORTE DE LIPÍDEOS
• HDL – desempenha essencialmente a função oposta da LDL: remove colesterol dos tecidos. A HDL é montada no plasma a partir de componentes na sua maioria obtidos pela degradação de outras lipoproteínas. A HDL circulante provavelmente adquire seu colesterol extraindo-o das membranas da superfície celular. O colesterol é então convertido em éster de colesteril pela enzima associada à HDL, funciona como um removedor de colesterol.

Ácidos Graxos
Definição: são ácidos monocarboxílicos de longas cadeias de hidrocarbonetos acíclicos.
Propriedades Gerais
Quanto aos tipos de ligação podem ser:
saturados, monoinsaturados e poliinsaturados.
Os mais abundantes contém C16 e C18 átomos.

Ácidos Graxos
o ponto de fusão depende de fatores:• Tamanho da cadeia carbônica.• Numero de ligações duplas Aqueles com maior cadeia carbônica
são sólidos a temperatura ambiente.

Nomenclatura dos Ácidos Graxos.
• os átomos são numerados a partir do carbono da carboxila.
• Os carbonos C2 e C3 são designados α e β.
• Para representar uma ligação dupla • Exemplo:

Oxidação de Ácidos Graxos
• Ativação de Ácidos graxos.• Reação de acilação de dependente
de ATP para formar acil-CoA graxa.• Ácido graxo + CoA + ATP → acil−CoA + AMP +
PPi
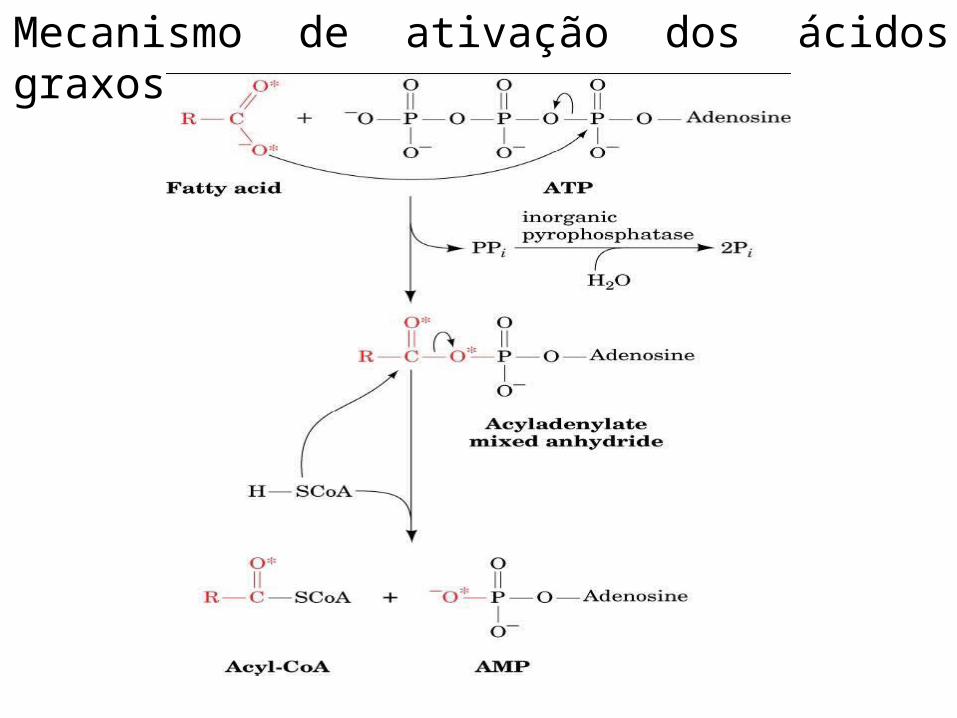
Mecanismo de ativação dos ácidos graxos
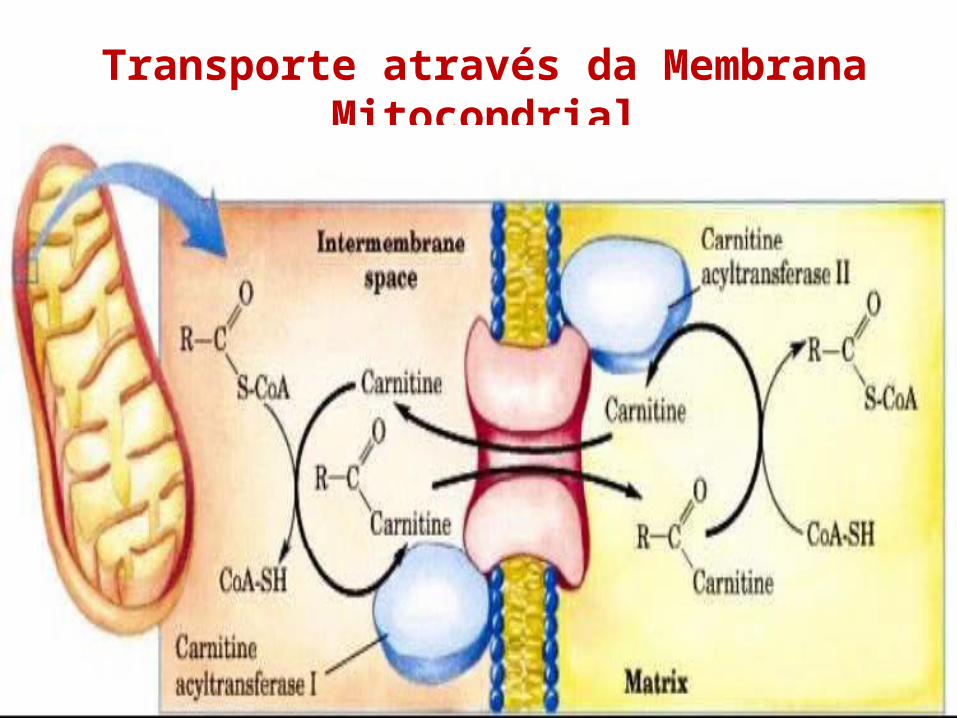
Transporte através da Membrana Mitocondrial

Oxidação.
Observe o esquema abaixo.
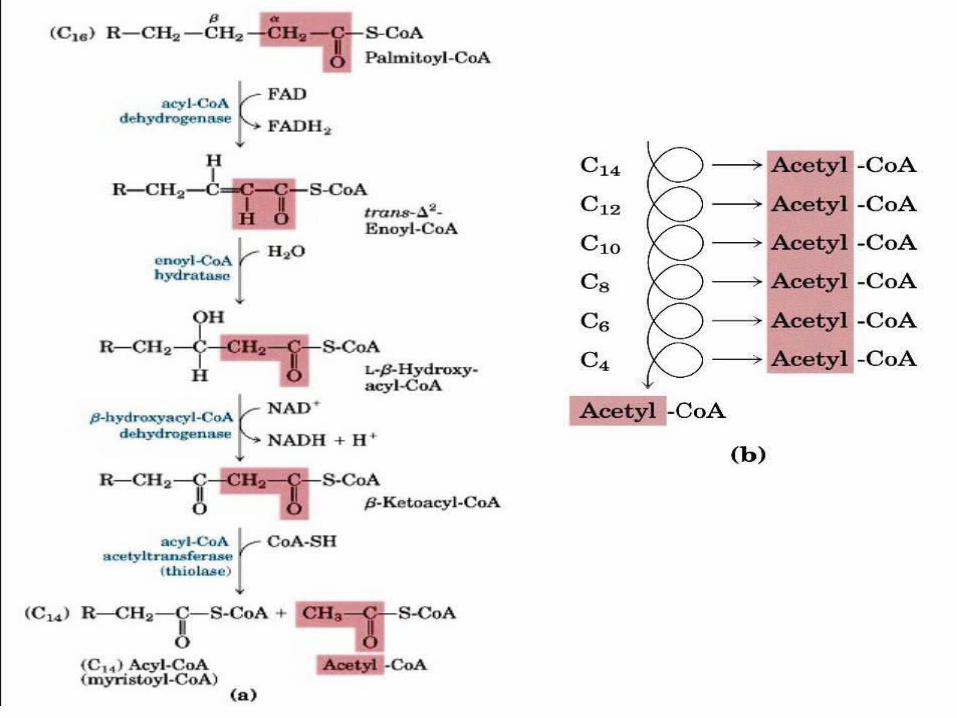

1. A formação da dupla trans α e β por meio da desidrogenarão a acil-Coa-desidrogenase.
2. Hidratação da dupla pela enoil-CoA-hidratase formando 3-L-hidroaxil-CoA
3. Desidrogenação dependente de NAD +desta β –hidroxiacil-CoA pela 3-L-hidroxiacil-CoA
4. Clivagem da ligação Cα –Cβ em uma reaçõa de tiólise com CoA catalisada pela β-cetoacil-CoA-tiolase, formand0 acetil-CoA e acil-CoA.

-Oxidação de ácidos graxos de cadeia ímpar
• A maioria do ácidos graxos possui um numero par de carbonos, sendo, portanto convertidos por completo em acetil-CoA. Algumas plantas e organismos marinhos, sintetizam ácidos graxos com um numero impar de carbonos.

• Ácidos graxos de cadeias ímpares são raros nos tecidos da maioria dos mamíferos. Em ruminantes, entretanto, podem aparecer em porcentagem considerável.
• Após os ciclos de beta-oxidação forma-se, além das várias moléculas de acetil-CoA, uma de Propionil-CoA, um acil-CoA de três carbonos que também é um produto da degradação dos aminoácidos valina, isoleucina e metoonina, sendo por esta via convertido a succinil-CoA, para entrar no ciclo do ácido citrico intermediário do Ciclo de Krebs.

• Apesar de produzirem um acetil-coA a menos em relação a um ácido graxo com um número a menos de carbono, o propionil-CoA aumenta o número de Ciclos de Krebs funcionantes, portanto, embora produzam menos energia diretamente, estes ácidos graxos contribuem na utilização da energia proveniente dos demais acetil-coA.

-Oxidação no Peroxissomo.
• Nas células de mamíferos, a maior parte da -oxidação de ácidos graxos ocorre na mitocôndria, mas os peroxissomos também oxidam os ácidos graxos com cadeias muito longas ou ramificadas.
• A -oxidação peroxissomal que difere pouco da mitocondrial encurta ácidos graxos com cadeias muito longas, as quais são então totalmente degradados na mitocôndria

Essas alterações químicas requerem apenas três enzimas:
1- A Acil-CoA-oxidase catalisa a reação Acil-CoA graxa + O2 (trans-D2-enoil-CoA + H2O2)
Essa enzima usa o co-fator FAD, mas os elétrons removidos são transferidos diretamente ao O2.
2- As atividades da enoil-CoA-hidratase peroxissomal e da 3-L-hidroxiacil-CoA-desidrogenase ocorrem em um único polipeptideo.
3- A tiolase peroxissomal catalisa a etapa final da oxidação.


Oxidação dos ácidos graxos
BALANÇO ENERGÉTICO=> em cada volta da espira => 4 ATP
ÞCada acetil-coA oxidado no Ciclo de Krebs => 10 ATP
Þ rendimento energético por cada átomo de carbono => 7ATP e como triglicéridos possuem muito mais átomos de carbono por unidade de peso em relação aos glúcidos => constituição de reservas energéticas com massa muito menor
CONCLUSÃO: Nos animais superiores e nos microorganismos a oxidação dos ácidos gordos fornece grandes quantidades de energia (NADH, ATP e GTP) para a neoglucogénese
O facto de as enzimas que intervêm na betaoxidação estar , nos animais, localizadas nas mitocôndrias, permite a conjugação, com elevado rendimento, dos processos de oxidação dos compostos energéticos coma a produção de ATP

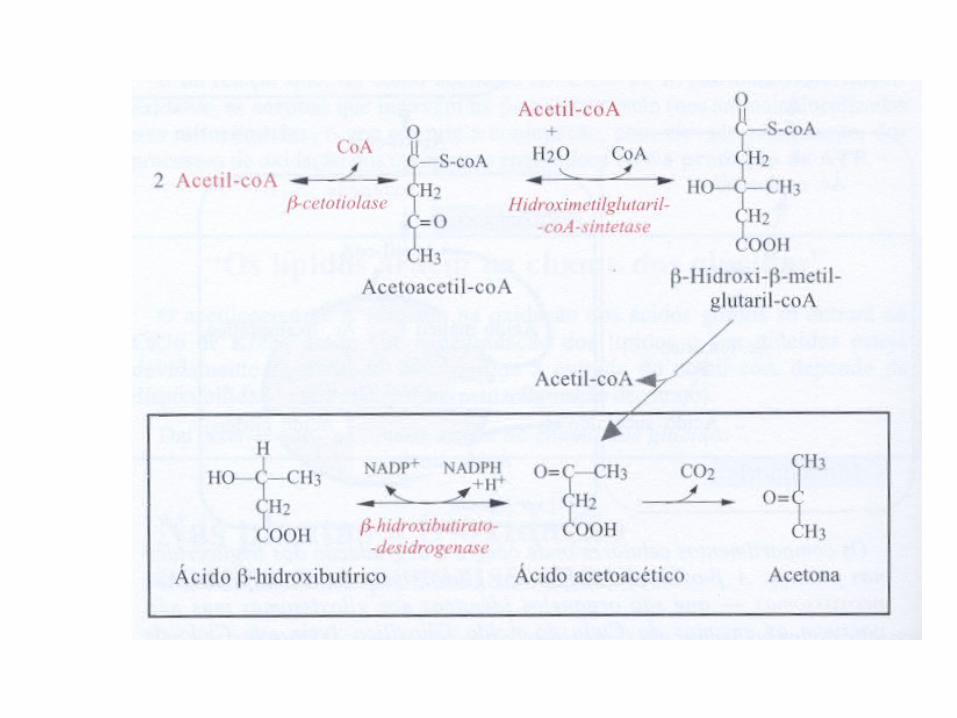
Corpos Cetônicos• São combustíveis metabólicos importantes para vários
tecido periféricos em particular para o coração e para os músculos esqueléticos;
• Quando predominar a oxidação dos lípidos sobre o catabolismo glucídico => baixa a concentração de ácido oxaloacético e o acetilco-A, não pode entrar no ciclo de Krebs => corpos cetónicos (ácido acetoacético, ácido betahidroxibutírico e acetona)
• Nos mamíferos, a formação de corpos cetónicos (ou cetogénese) ocorre no fígado e a sua acumulação dá-se no sangue (cetonemia) e na urina (cetourina)
• Os ácidos ácido acetoacético e betahidroxibutírico são indispensáveis para o organismo humano porque depois de transportados no sangue são oxidados no ciclo de Krebs e fornecem energia ao cérebro, músculo cardíaco e ao córtex renal

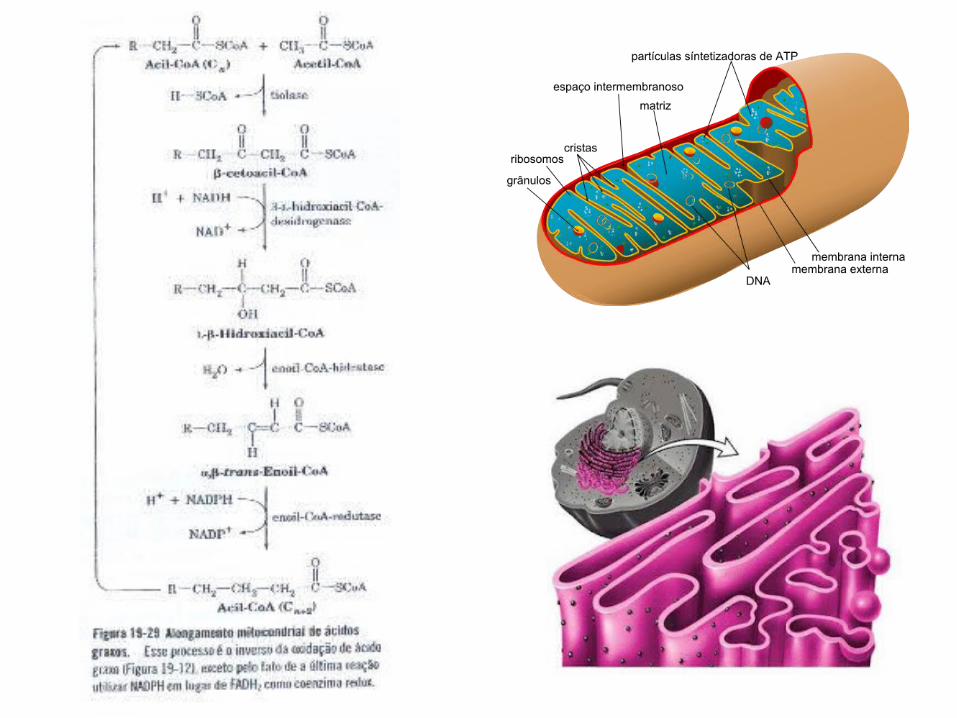
D. Alongases e Dessaturases
• As alongases estão presentes na mitocôndria e no retículo endoplasmático, mas os mecanismos de alongamento nos dois locais são diferentes;
• O alongamento mitocondrial; • O alongamento no retículo
endoplasmático;
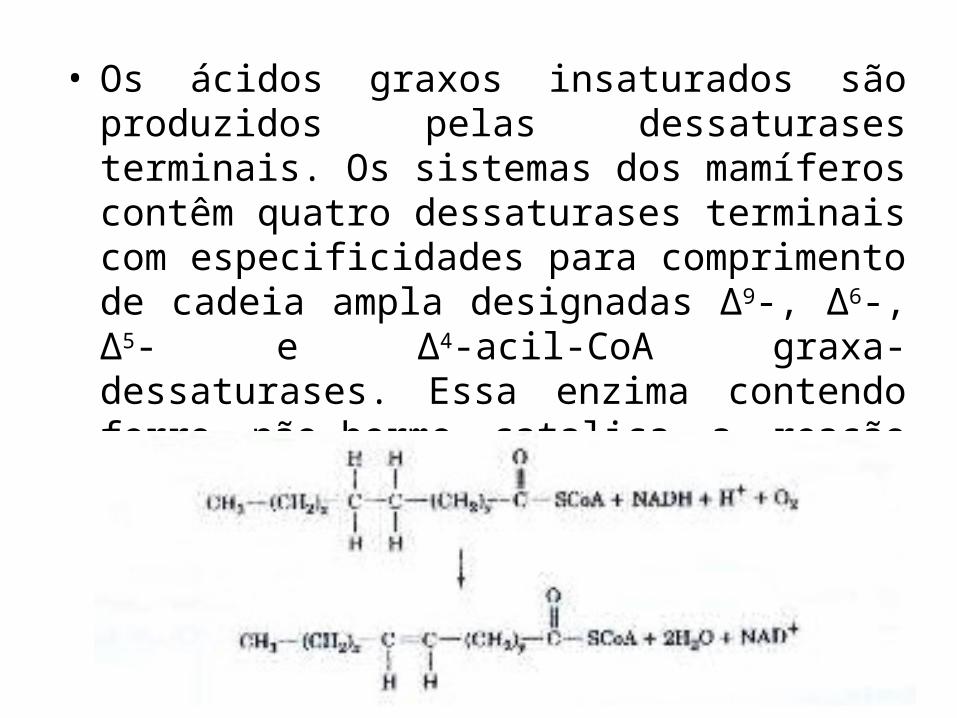
• Os ácidos graxos insaturados são produzidos pelas dessaturases terminais. Os sistemas dos mamíferos contêm quatro dessaturases terminais com especificidades para comprimento de cadeia ampla designadas ∆9-, ∆6-, ∆5- e ∆4-acil-CoA graxa-dessaturases. Essa enzima contendo ferro não-herme catalisa a reação geral:

Determinados Ácidos Graxos Poli-insaturados Devem
Ser Obtidos da Dieta
• Uma grande variedade de ácidos graxos insaturados pode ser sintetizada por combinações de reações de alongamento e dessaturação;
• O ácido linoléico é um importante constituinte dos esfingolipídeos da epiderme, que funcionam como uma barreira de impermeabilidade da pele, sendo esse um ácido graxo essencial.

E. Síntese de Triacilgliceróis
• Os triacilgliceróis são sintetizados a partir de ésteres de acil-CoA graxa e de glicerol-3-fosfato ou diidroxiacetona-fosfato;
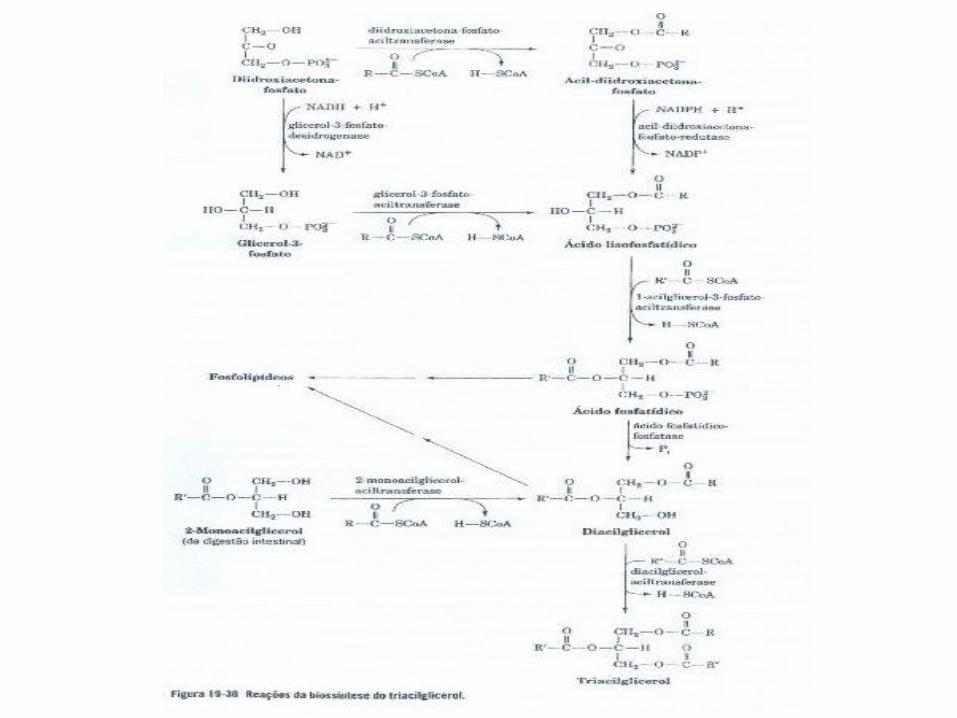

A Gliceroneogênese é Importante para a Biossíntese do Triacilglicerol
•A Gliceroneogênese é necessária em períodos de jejum, uma vez que aproximadamente 30% dos ácidos graxos que chegam ao fígado durante uma refeição são reesterificados a triacilglicerol e exportados em VLDL;
•Os adipócitos
Enzima gliconeogênica fosfoenolpiruvato-carboxicinase (PEPCK).

Regulação do Metabolismo de Ácidos Graxos
• As discussões sobre controle metabólico normalmente tratam da regulação do fluxo do metabólico através de uma rota em resposta às necessidades energéticas e aos estados alimentares distintos de um organismo;
• No metabolismo de lipídeos, hormônios como a insulina e o glucagon regulam as taxas das rotas opostas;

• A oxidação de ácidos graxos
A ação da triacilglicerol-lipase sensível a hormônio.
• A fosforilação ativa a lipase sensível a hormônios, elevando a concentração plasmática de ácidos graxos, ativando a rota de β-oxidação em músculo e fígado;
• A proporção glucagon:insulina determina a taxa e a direção do metabolismo de ácidos graxos;
• A regulação de longo prazo do metabolismo de lipídeos inclui a estimulação por insulina da síntese da acetil-CoA-carboxilase e do ácido graxo-sintase e a inibição delas por jejum prolongado.

BIOSSÍTESE DE ÁCIDOS GRAXOS
• A biossíntese de ácidos graxos ocorre por meio da condensação de unidades C2, o inverso do processo de beta oxidação.
• Uma investigação mostrou que tanto acetil-CoA como bicarbonato são necessários, e que um aunidade C3, a manolil-CoA, é um intermediário na biossíntese de ácidos graxos.

BIOSSÍTESE DE ÁCIDOS GRAXOS
• A acetil - CoA, material de partida para a síntese de ácidos graxos, é produzida na mitocôndria pela descarboxilação oxidativa de piruvato catalisada pela piruvato -desidrogenase e também pela oxidação de ácidos graxos.

BIOSSÍTESE DE ÁCIDOS GRAXOS
• Quando a demanda de ATP é baixa, de modo que a oxidação fosforilativa são mínimas, essa acetil – CoA mitocondrial pode ser armazenada para uso futuro como gordura. Entretanto a biossíntese de ácidos graxos ocorre no citosol, e a membrana mitocondrial é essencialmente impermeável à acetil – CoA.
• A acetil – CoA entra no citosol sob a forma de citrato pelo sistema de transporte de tricarboxilato. Então, a ATP – citrato – liase catalisa a reação.
• Citrato + CoA + ATP acetil-CoA + oxaloacetato + ADP + Pi

BIOSSÍTESE DE ÁCIDOS GRAXOS
• A acetil-CoA-carboxilase (ACC) catalisa a primeira etapa comprometida com a biossíntese de ácidos graxos, sendo uma das etapas controladoras da velocidade. O mecanismo dessa enzima dependente de biotina é similar aos da propionil-CoA-carboxilase e da piruvato-carboxilase.

BIOSSÍTESE DE ÁCIDOS GRAXOS
• O resultado é um grupo de três carbonos (manoil) ligado na forma de tioéster à CoA.
• A acetil-CoA-carboxilase de mamíferos está sujeita a controle alostérico e hormonal. Por exemplo, o citrato estimula a acetil-CoA-carboxilase, e as acil-CoA graxas de cadeia longa são inibidoras de enzima. O ajuste fino da atividade enzimática é realizado por modificação covalente.

BIOSSÍTESE DE ÁCIDOS GRAXOS
• Existem duas isoformas princcipais de ACC. A α ACC ocorre no tecido adiposo, e a β ACC ocore em tecidos que oxidam ácidos graxos, mas não os sintetizam, como o músculo cardíaco. Os tecidos que sintetizam e oxidam ácidos graxos, como o fígado, possuem ambas as isoformas.

BIOSSÍTESE DE ÁCIDOS GRAXOS
• A síntese de ácidos graxos, em especial do ácido palmíticco, a partir de acetil-CoA e de malonil-CoA envolve sete reações enzimáticas. Tais reações foram primeiramente estudadas em extratos e E. coli livres de células, nos quais as reações eram catalisadas por enzimas independenetes.
• Enzimas individuais com essas atividades também existem nos cloroplastos.

SÍNTESE DE
OUTROS LIPÍDEOS
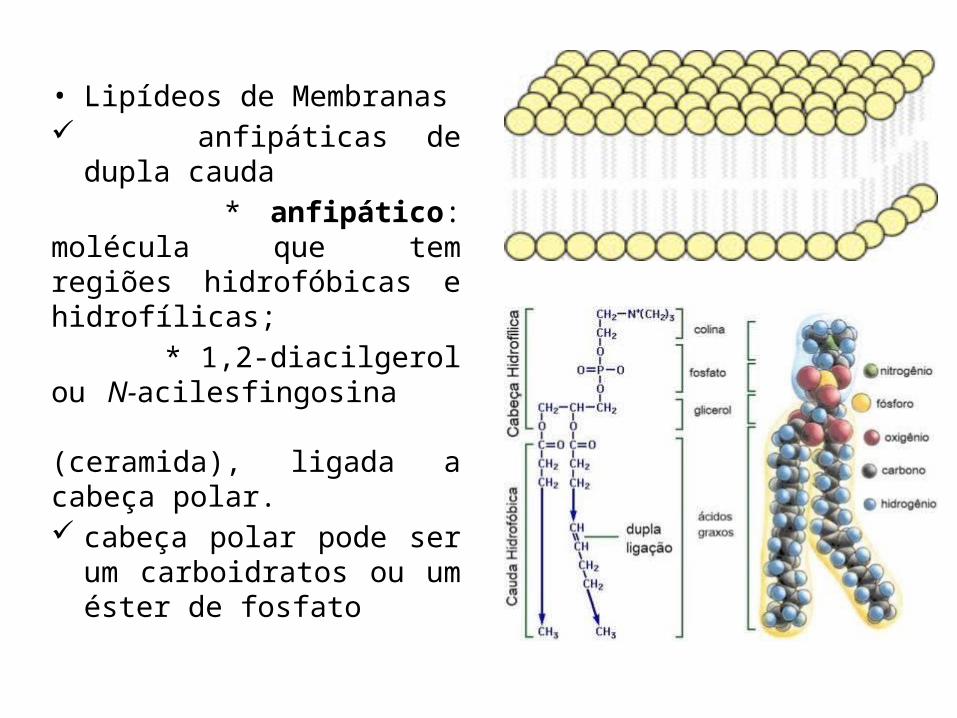
• Lipídeos de Membranas anfipáticas de dupla cauda * anfipático: molécula que tem regiões hidrofóbicas e hidrofílicas; * 1,2-diacilgerol ou N-acilesfingosina (ceramida), ligada a cabeça polar. cabeça polar pode ser um
carboidratos ou um éster de fosfato

GLICEROFOSFOLIPÍDEOS
• A síntese dos TRIACIGLICERÓIS e os GLICEROFOSFOLIPÍDEOS (lipídeos estruturais de membranas) são sintetizados a partir de precursores comuns
glicerol-3-fosfato e acil CoA-graxo e vários passos enzimáticos.
são sintetizados nas mambranas, principalmente na face citosólica do retículo endoplasmatico, sendo a partir daí transportados em vesiculas aos seus destinos celulares.

GLICEROFOSFOLIPÍDEOS
• DiacilglicerolfosfolipídeosPrecusores: 1,2-diacil e o ácido
fosfatídicoOs grupos cabeça polar estão ligados
ao C3 do glicerol por meio de uma ligação fosfodiéster;
Nos mamíferos os grupos da cabeça são etanolamina e colina são ativados antes de serem ligados ao lipídeos.

Ativação da colina e etanolaminaCTP: fosfoetanolamina-citidiltransferaseOu fosfocolina-citidiltransferase
CDP- colina: 1,2 diacilglicerol-fosfocolina-transferase
Fígado: sítio primário da síntese de triacilglicerol
Glicose
Glicólise
Dihidroxiacetona fosfato
NADH
NAD+
Glicerol 3- fosfato desidrogenase
ADP ATP
GlicerolGlicerol quinase
Glicerol fosfato
Tecido adiposo
Glicose
Glicólise
Dihidroxiacetona fosfato
NADH
NAD+
Glicerol 3-fosfato desidrogenase
Glicerol fosfato
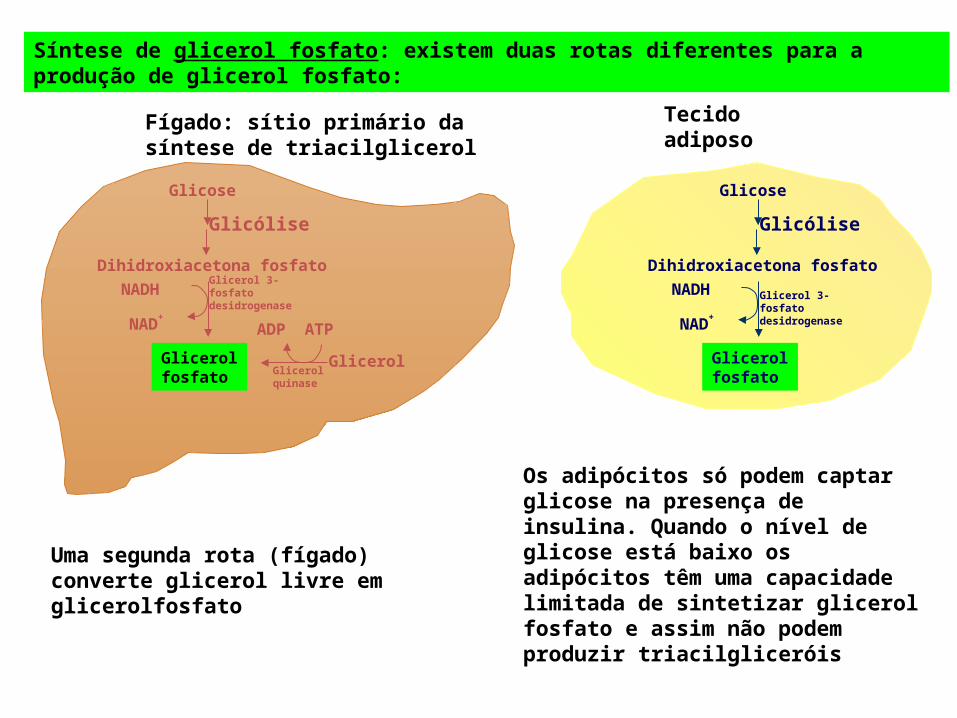
Síntese de glicerol fosfato: existem duas rotas diferentes para a produção de glicerol fosfato:
Uma segunda rota (fígado) converte glicerol livre em glicerolfosfato
Os adipócitos só podem captar glicose na presença de insulina. Quando o nível de glicose está baixo os adipócitos têm uma capacidade limitada de sintetizar glicerol fosfato e assim não podem produzir triacilgliceróis
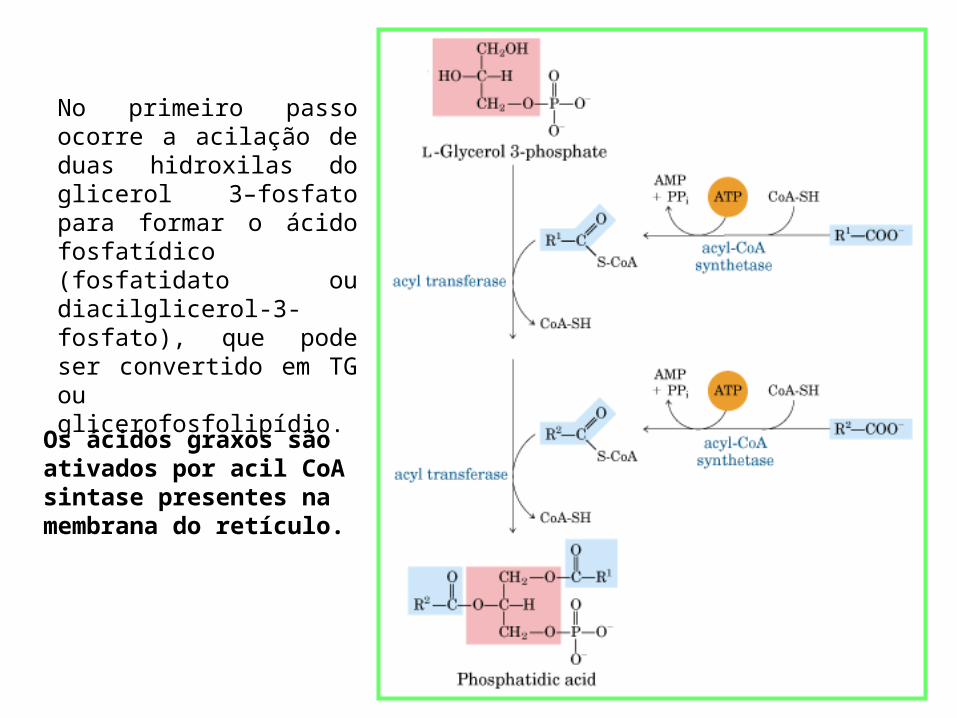
No primeiro passo ocorre a acilação de duas hidroxilas do glicerol 3–fosfato para formar o ácido fosfatídico (fosfatidato ou diacilglicerol-3-fosfato), que pode ser convertido em TG ou glicerofosfolipídio.
Os ácidos graxos são ativados por acil CoA sintase presentes na membrana do retículo.
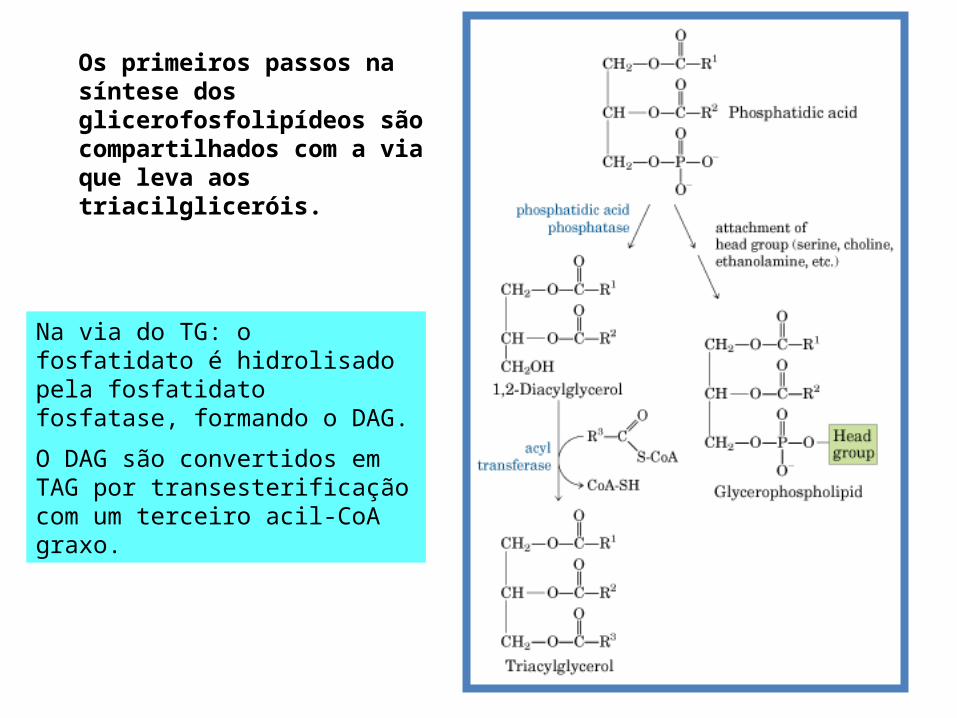
Os primeiros passos na síntese dos glicerofosfolipídeos são compartilhados com a via que leva aos triacilgliceróis.
Na via do TG: o fosfatidato é hidrolisado pela fosfatidato fosfatase, formando o DAG.
O DAG são convertidos em TAG por transesterificação com um terceiro acil-CoA graxo.
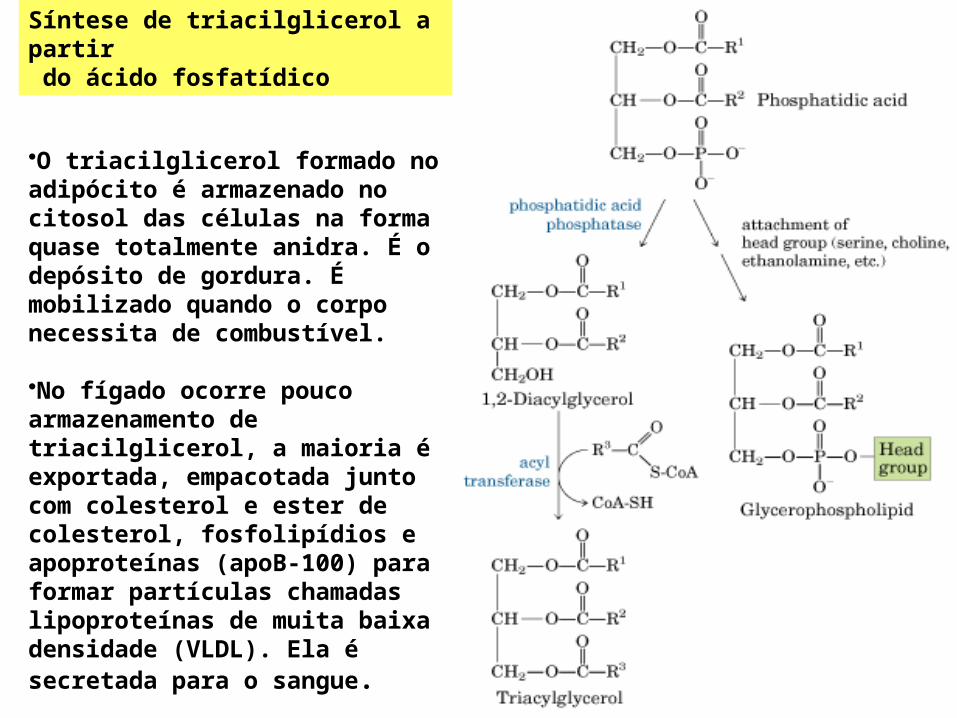
Síntese de triacilglicerol a partir do ácido fosfatídico
•O triacilglicerol formado no adipócito é armazenado no citosol das células na forma quase totalmente anidra. É o depósito de gordura. É mobilizado quando o corpo necessita de combustível.
•No fígado ocorre pouco armazenamento de triacilglicerol, a maioria é exportada, empacotada junto com colesterol e ester de colesterol, fosfolipídios e apoproteínas (apoB-100) para formar partículas chamadas lipoproteínas de muita baixa densidade (VLDL). Ela é secretada para o sangue.

Nas células eucarióticas a síntese de fosfolipídeos ocorre principalmente na face citosólica do retículo endoplasmático liso. A partir daí são transportados em vesículas ao seu destino final.
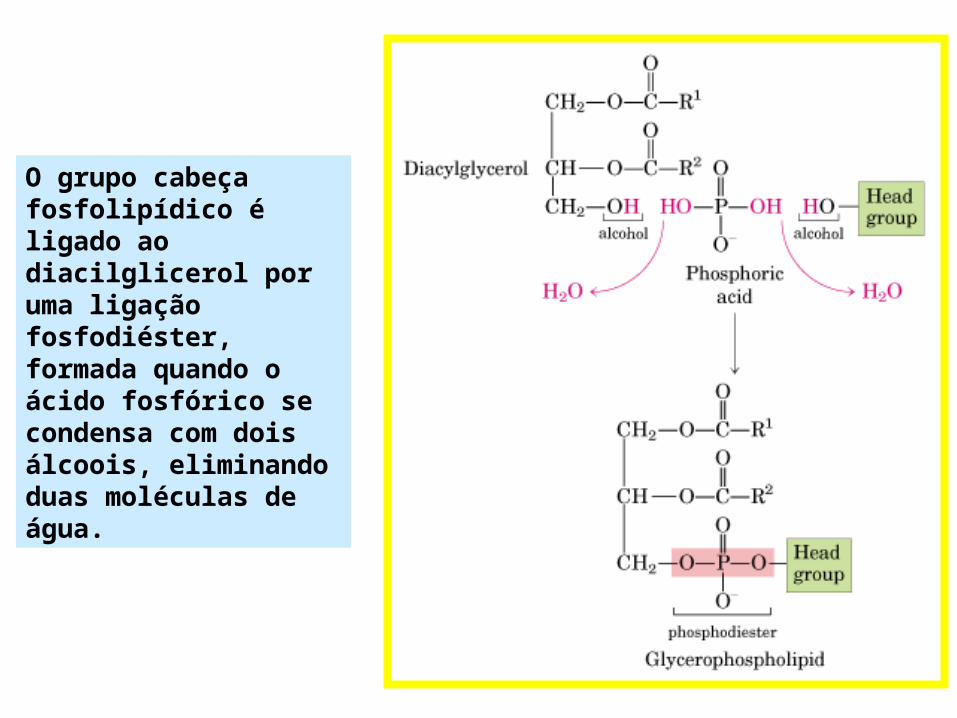
O grupo cabeça fosfolipídico é ligado ao diacilglicerol por uma ligação fosfodiéster, formada quando o ácido fosfórico se condensa com dois álcoois, eliminando duas moléculas de água.

ESFINGOLIPÍDEOS• Esfingolipídeos ou
esfingomielinas são Lipídeos conjugados presentes entre as células nervosas, os neurônios - e funcionam como uma espécie de "lubrificante", acelerando o processo de transmissão de impulso nervoso
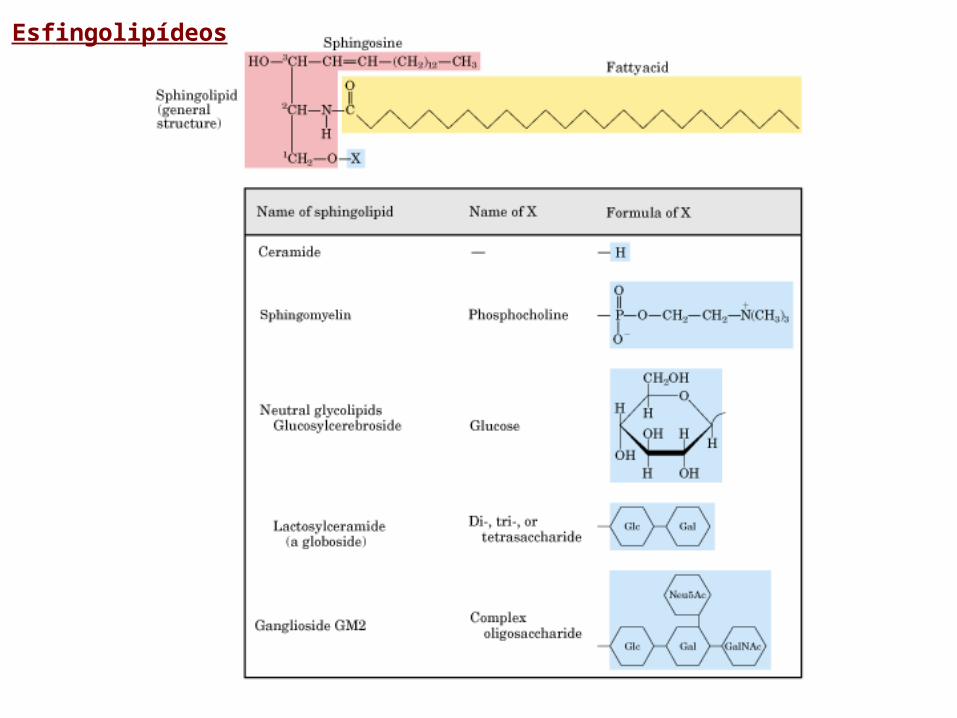
Esfingolipídeos

• Na sua maioria esfingoglicolipídeos; isto é seu grupo da cabaça consiste em unidades de carboidratos
• A principal diferença entre os esfingolipídios e os fosfolipídios é o álcool no qual estes se baseiam

ESFINGOLIPÍDEOS
Existem 3 subclasses : Esfingomielinas = Possuem a fosfocolina ou a
fosfoetanolamina como cabeça polar alcoólica;
Os Cerebrosídeos = Não possuem fosfato, e sim, um açúcar simples como álcool polar - são glicoesfingolipídios, ou glicolipídios;
Gângliosídeos = Possuem estrutura complexa, com cabeças polares muito grandes formadas por várias unidades de açúcar como por exemplo, o ácido siálico - ceramidas

ESFINGOLIPÍDEOS
• Ceramidas ( N-acilesfingosina)
Metabolismo do Colesterol

Biossíntese do Colesterol
Síntese do HMG-CoA (β-hidróxi-β-
metilglutaril-CoA) a partir de acetil-
CoA;
Conversão do HMG-CoA a esqualeno;
Conversão do esqualeno em
colesterol.

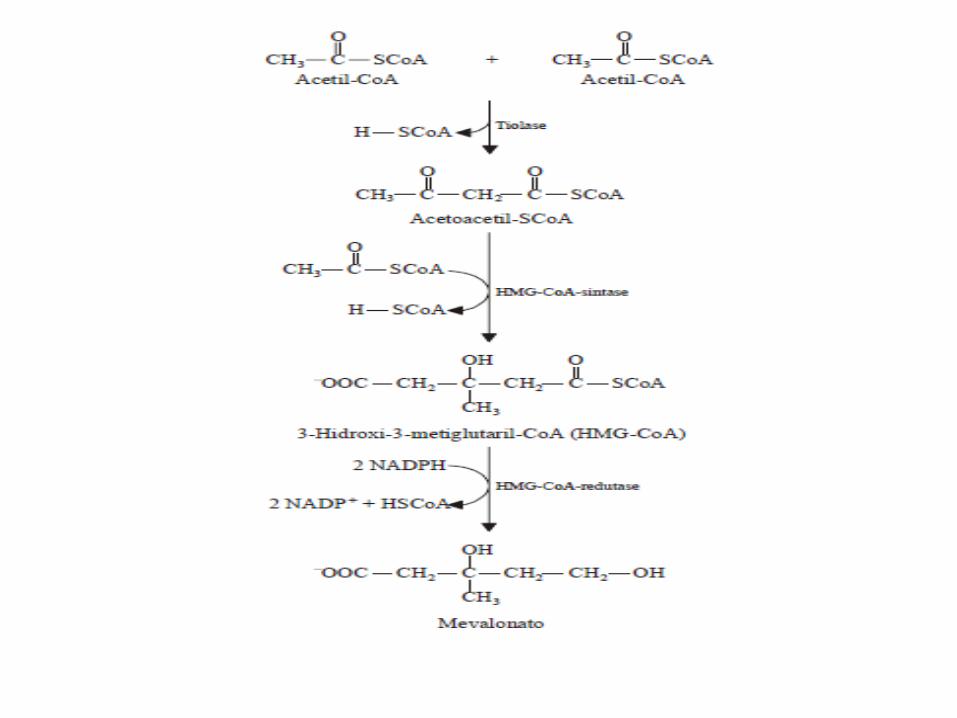
1. Síntese do HMG-CoA a partir de acetil-CoA
• Condensação de duas moléculas de acetil-CoA para formar acetoacetil-CoA, em reação catalisada pela tiolase;
• Na reação seguinte, o acetoacetil-CoA condensa com uma terceira molécula de acetil-CoA pra formar β-hidróxi-β-metilglutaril-CoA, reação sintetizada pela HMG-CoA-sintase

2. Redução do HMG-CoA a mevalonato
• A HMG-CoA é reduzida por 2 NADPH originando mevalonato;
• Esta reação é catalisada pela HMG-CoA redutase, localizada na membrana do retículo endoplasmático, e é a reação limitante da síntese de colesterol.

3ª. Etapa
O mevalonato sofre duas fosforilações, à custa de 3 ATP, e uma descarboxilação, originando isopentenil-pirofosfato (5C).
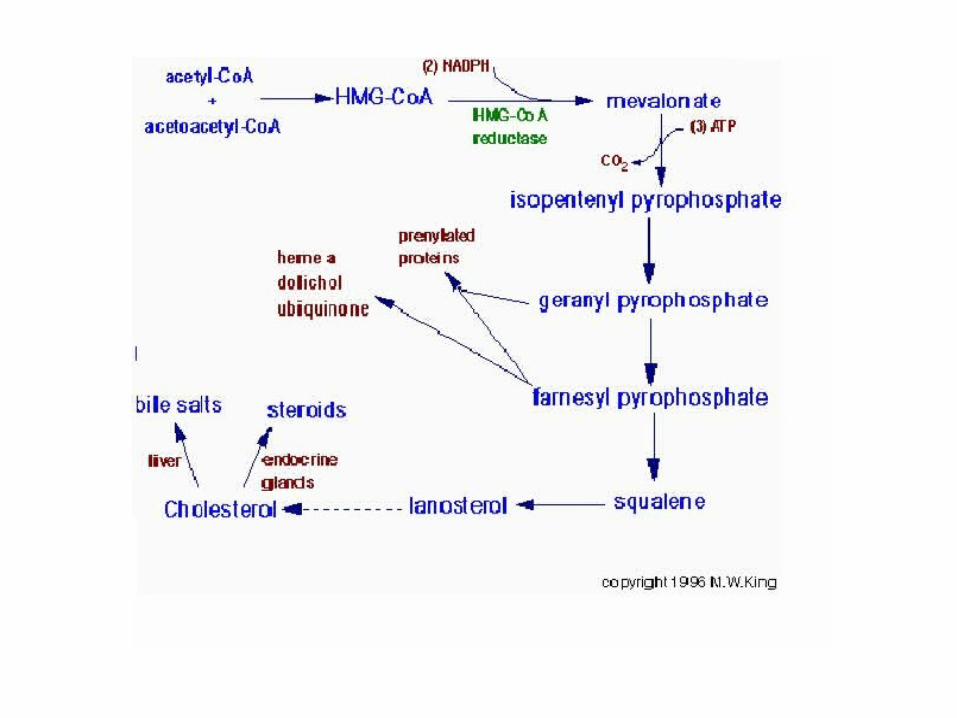

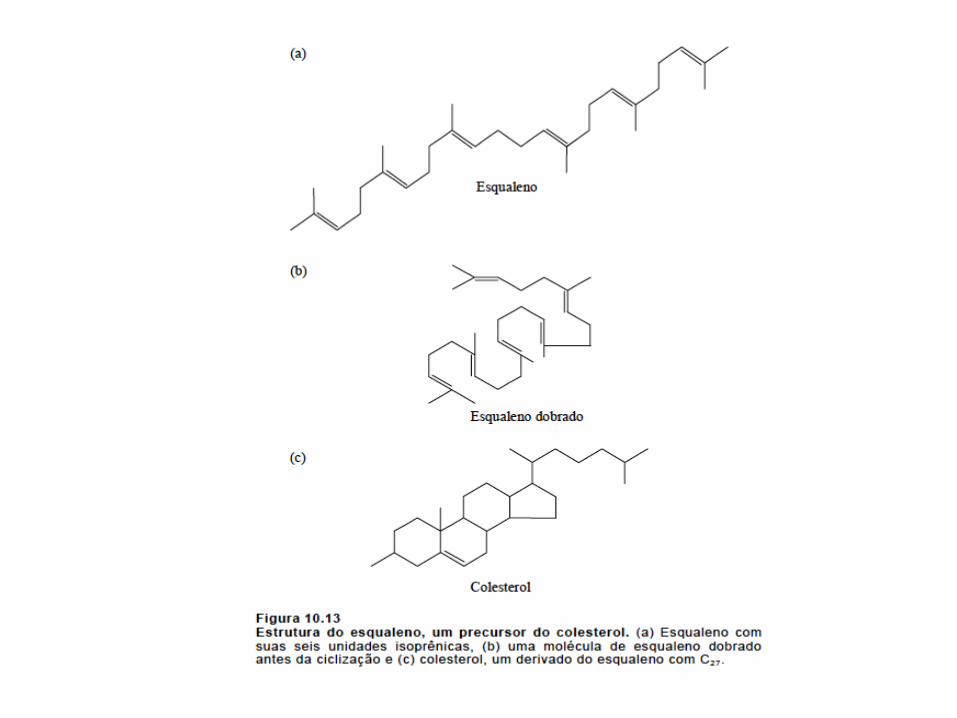
4.Conversão do Esqualeno a Colesterol
• A esqualeno-monoxigenase acrescenta um átomo de oxigênio na extremidade da cadeia, produzindo um epóxido e a subsequente formação do lanosterol;(requer NADPH e FAD)
• O lanosterol liga-se a uma segunda proteína e no fim é convertido a 7-diidrocolesterol, que é reduzido pelo NADPH para formar o colesterol;

Regulação da Síntese de Colesterol
• A enzima (HMG-CoA-redutase) esta sujeita à regulação de curto prazo por inibição competitiva, efeitos alostéricos e modificação covalente envolvendo fosforilação competitiva;
• Controle de longo prazo, por retroalimentação, da quantidade de enzima presente na célula;

Obrigada...
Boa Noite!