Embed Size (px)
Citation preview

Espermatófitas (Fanerógamas)
Em 1960, Beck, baseado em um fóssil de Archaeopteris (gênero com fronde de samambaia) em conexão com Callixylon (gênero que apresentava crescimento secundário característico de gymnospermas), reconheceu um novo grupo, denominando-o Progymnosperma. Posteriormente, descobriu-se que, em vez de folhas compostas, a suposta fronde representava ramos e as folhas eram simples e laminares. Essas plantas tinham cerca de 8,5 m alt. e eram caducifólias, ocupando ambientes mais sazonais, possivelmente próximos a rios. Com raízes mais desenvolvidas, as progimnospermas intensificaram o desgaste das rochas, aumentando a profundidade e a extensão dos solos. O cálcio e o magnésio do solo reagiam quimicamente com o dióxido de carbono formando carbonatos solúveis que eram carregados pelos rios até o oceano, onde precipitavam. A transferência do carbono da atmosfera para as rochas diminuiu o efeito estufa e foi responsável pelo declínio da temperatura global no fim do Permiano
Progimnosperma. (note o hábito arbóreo e os ramos simulando folhas compostas).
A partir desse grupo de plantas do
Devoniano, com crescimento secundário e produção de heterósporos liberados pela planta mãe, teriam derivado as plantas com semente. Nas espermatófitas (ou fanerógama), os
megasporângio (nucelo), retido no esporófito e protegidos por um tegumento; nas chamadas criptógamas, os megásporos são liberados. O conjunto formado pelo nucelo e pelo tegumento é denominado óvulo. O tegumento surgiu a partir de redução e fusão de ramos arranjados ao redor do nucelo. Esse arranjo aumentaria a captação de pólen e auxiliariam a dispersão da semente. Nas plantas atuais, com exceção de uma abertura chamada micrópila, o tegumento engloba todo o nucelo, provendo ao gametófito e ao embrião maior proteção contra a dessecação e a herbivoria.
O
megásporos foram reduzidos a um único por
gametófito feminino tornou-se complet
ação do zigo
ro e a formação da seme
amente dependente do esporófito, nutrindo-se a partir do nucelo. Os gametófitos masculinos, ou grãos de pólen, são liberados no ambiente e devem alcançar a micropila, penetrando no óvulo até alcançarem o gametófito feminino para reprodução. O transporte do pólen de uma estrutura masculina para uma feminina, chamado de polinização, é uma inovação do ciclo reprodutivo das espermatófitas. Apenas após a polinização o gametófito masculino completa sua maturação, com o crescimento do tubo polínico e liberação de gametas masculinos próximo à oosfera. O desenvolvimento de ambos os gametófitos vai variar conforme o grupo, sendo bastante reduzido nas angiospermas.
Mesmo após a fecundação e a formto, a embriogênese da nova geração
esporofítica ocorre com o gametófito ainda ligado ao esporófito-mãe. A semente é, portanto, resultante do óvulo após a fecundação do gameta feminino, incluindo o tecido de três gerações: uma testa derivada do tegumento diplóide de origem esporofítica, um endosperma haplóide de origem gametofítica (triplóide, derivada da dupla fecundação nas angiospermas), e um embrião diplóide, resultado do desenvolvimento do zigoto formado pela fusão de gametas.
A retenção do megásponte trouxeram uma enorme vantagem
adaptativa para as espermatófitas. A fecundação do gametófito feminino tornou-se menos dependente de água. Nas primeiras espermatófitas, ocorria a zooidogamia; os grãos de pólen entravam pela micrópila, uma abertura no tegumento do megasporângio, chegando até a câmara polínica. O gametófito masculino crescia nutrindo-se do nucelo até alcançar a câmara arquegonial, onde lançava dois gametas
33
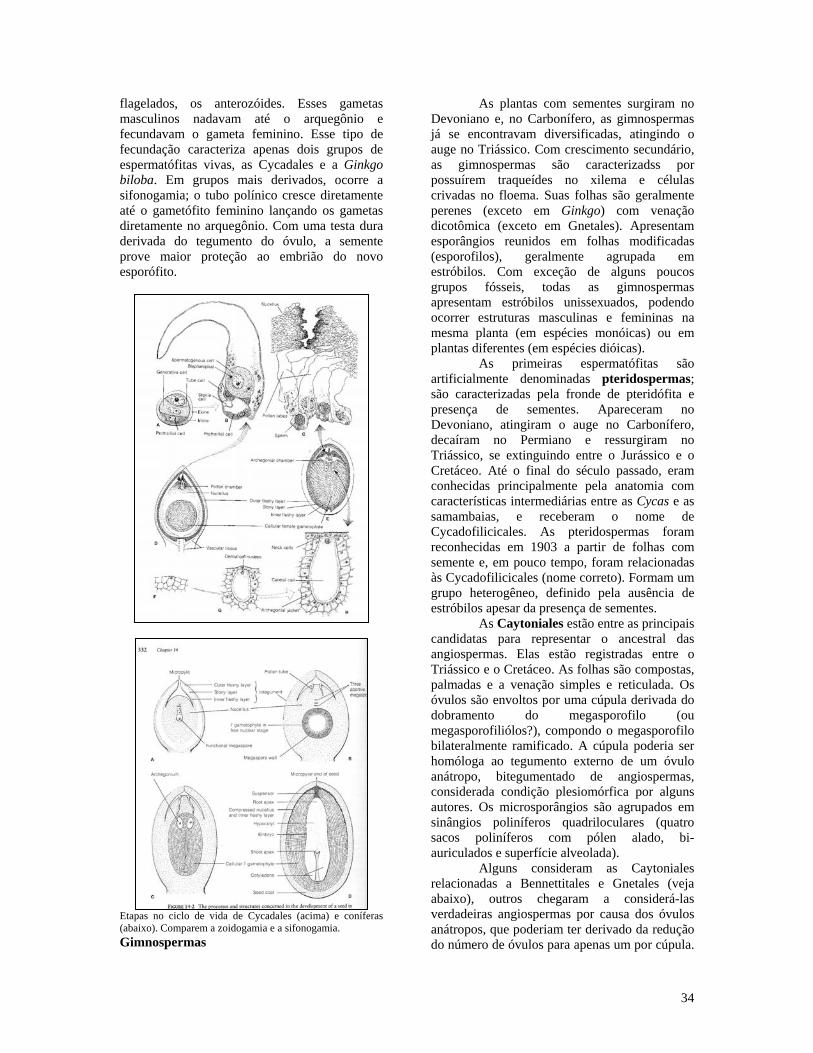
flagelados, os anterozóides. Esses gametas masculinos nadavam até o arquegônio e fecundavam o gameta feminino. Esse tipo de fecundação caracteriza apenas dois grupos de espermatófitas vivas, as Cycadales e a Ginkgo biloba. Em grupos mais derivados, ocorre a sifonogamia; o tubo polínico cresce diretamente até o gametófito feminino lançando os gametas diretamente no arquegônio. Com uma testa dura derivada do tegumento do óvulo, a semente prove maior proteção ao embrião do novo esporófito.
mentes surgiram no evonia
s são artificial
cipais candidat
Caytoniales relacion
Etapas no ciclo de vida de Cycadales (acima) e coníferas (abaixo). Comparem a zoidogamia e a sifonogamia. Gimnospermas
As plantas com seD no e, no Carbonífero, as gimnospermas já se encontravam diversificadas, atingindo o auge no Triássico. Com crescimento secundário, as gimnospermas são caracterizadss por possuírem traqueídes no xilema e células crivadas no floema. Suas folhas são geralmente perenes (exceto em Ginkgo) com venação dicotômica (exceto em Gnetales). Apresentam esporângios reunidos em folhas modificadas (esporofilos), geralmente agrupada em estróbilos. Com exceção de alguns poucos grupos fósseis, todas as gimnospermas apresentam estróbilos unissexuados, podendo ocorrer estruturas masculinas e femininas na mesma planta (em espécies monóicas) ou em plantas diferentes (em espécies dióicas).
As primeiras espermatófitamente denominadas pteridospermas;
são caracterizadas pela fronde de pteridófita e presença de sementes. Apareceram no Devoniano, atingiram o auge no Carbonífero, decaíram no Permiano e ressurgiram no Triássico, se extinguindo entre o Jurássico e o Cretáceo. Até o final do século passado, eram conhecidas principalmente pela anatomia com características intermediárias entre as Cycas e as samambaias, e receberam o nome de Cycadofilicicales. As pteridospermas foram reconhecidas em 1903 a partir de folhas com semente e, em pouco tempo, foram relacionadas às Cycadofilicicales (nome correto). Formam um grupo heterogêneo, definido pela ausência de estróbilos apesar da presença de sementes.
As Caytoniales estão entre as prinas para representar o ancestral das
angiospermas. Elas estão registradas entre o Triássico e o Cretáceo. As folhas são compostas, palmadas e a venação simples e reticulada. Os óvulos são envoltos por uma cúpula derivada do dobramento do megasporofilo (ou megasporofiliólos?), compondo o megasporofilo bilateralmente ramificado. A cúpula poderia ser homóloga ao tegumento externo de um óvulo anátropo, bitegumentado de angiospermas, considerada condição plesiomórfica por alguns autores. Os microsporângios são agrupados em sinângios poliníferos quadriloculares (quatro sacos poliníferos com pólen alado, bi-auriculados e superfície alveolada).
Alguns consideram as adas a Bennettitales e Gnetales (veja
abaixo), outros chegaram a considerá-las verdadeiras angiospermas por causa dos óvulos anátropos, que poderiam ter derivado da redução do número de óvulos para apenas um por cúpula.
34

Outros autores desvinculam essa relação e acreditam num paralelismo.
As Coritospermales existiram no Gondwa
ospermas recentes abrangem 80 gêneros
YCADALES dióicas; caule não ramíficado,
(-18) m
rdem é sustentada pela capacidade form
(1990) dividiu a ordem em
As Cycadales lembram palmeiras
na, durante o Triássico. Eram plantas lenhosas caracterizadas por folhas em forma de samambaias e megasporofilos bipinados organizados dorsiventralmente. A redução do número de óvulos por cúpula para um ajudaria a corroborar a hipótese de que o segundo tegumento dos óvulos bitegumentados de angiospermas poderia ter derivado da cúpula das pteridospermas.
As gimne 870 espécies, encontradas
principalmente nas regiões temperadas. Alguns grupos dominam grandes áreas florestais como é o caso das coníferas na taiga, enquanto outros são representados por populações isoladas nas regiões tropical e subtropical, como é o caso da maioria das Cycas. Algums grupos são extremamente localizados como é o caso da Gingko biloba, considerada um fóssil vivo, a única remanescente de uma linhagem, atualmente encontrada apenas em florestas remotas da China, ou a Welwitschia mirabilis, espécie pitoresca, encontrada somente no deserto da Namíbia, no sudeste da África. C Plantas 10 alt., ou subterrâneo e tuberoso. Folhas pinadas, raramente bipinadas, persistentes, até 1,5 m compr., formando uma coroa no ápice do caule, geralmente a base persiste no caule após a queda. Estróbilo 1(-3), terminal; o masculino (microstróbilo) com numerosos esporofilos espiraladamente dispostos ao longo de um eixo central, cada qual com 1-milhares de microsporângios radialmente agrupados na face abaxial; o feminino (megastróbilo) com megasporofilos compostos por uma estipe, um escudo distal e 2 óvulos protegidos internamente (exceto Cycas, 1-6 óvulos marginais cada, e megasporofilos foliáceos dispersos no ápice da coroa de folhas, não formando megastróbilo). Sementes subglobosas, 0,5-6 cm compr., de cores vivas. A ode ar nódulos de cianobactérias do gênero Anabaena que atuam como fixadoras de nitrogênio nas raízes, o que possibilita a ocupação de regiões com solos pobres, e pela produção de cicasina (Stevenson 1990). Está distribuída em regiões tropicais e subtropicais da África, incluindo Madagascar, Austrália e ilhas do Pacífico, e nas Américas incluindo ilhas do
Caribe, ocorrendo desde matas úmidas até mangues e dunas. Stevensontrês famílias: Cycadaceae, incluindo apenas Cycas, Stangeriaceae, com Stangeria e Bowenia, e Zamiaceae. Nos Neotrópicos, estão representados Zamia (cerca de 45 espécies), Z. ulei e Z. lecointei chegando até o Brasil, Chigua (1) descrita recentemente para a Colômbia, Ceratozamia (11) e Dioon (9) endêmicas do México e Mycrocycas (1) em Cuba.
Exemplo de Cycadales: A. hábito; B. microstróbilo
B
megasporofilo; D. conjunto de megasporofilos em C(note que nesse gênero não há formação de megastróbilo pteridófitas, com caule não ramificeventualmente subterrâneo e uma coroa de fogeralmente pinadas no ápice. O caulemanoxílico; o tecido parenquimatoso é bastdesenvolvido, armazenando água na formamucilagem e toxinas, em detrimento do tecvascular, que forma um lenho fino. polinizadas principalmente por besouros. megastróbilos, que podem ir de 2 a 80 compr. e alcançar até 40 kg, podem chegaproduzir até 500 sementes. As sementes
A
ou
; C.
D
C
ycas ).
ado, lhas é
ante de ido São Os cm r a são
35

esponjosas em Cycas, o que possibilita sua flutuação e conseqüentemente dispersão pela água, mas na maioria dos casos as sementes são vistosas atraindo animais, principalmente aves que as dispersam. Apesar de terem dominado a Terra
rante
as espécies serem
INKGOALES óicas, até 30 m alt. e 0,5 m
iâm.; tr
nas por inkgo
INALES vores até 100 m alt., dióicas ou
onóica
ic
Gynkgo biloba: inverno e verão e verão (acima). Folhas e
rrespondem às coníferas ortado
du a maior parte do Mesozóico, a ordem conta atualmente com apenas cerca de 150 espécies, freqüentemente representadas por populações raras, com poucos indivíduos. Devido a sua toxidade, muitas foram sacrificadas na Austrália por causarem prejuízos ao gado. Houve um grande declínio demográfico das populações de várias espécies de Cycadales no século passado e muitas espécies encontram-se atualmente ameaçadas de extinção, podendo ser consideradas fósseis vivos. Apesar de algumutilizadas na alimentação de populações indígenas e outras possuírem usos na medicina popular, a maior importância econômica das Cycadales é a utilização ornamental de espécies como Cycas revoluta e C. circinalis. G Árvores did onco ramificado, simpodial; mucilagem presente na raiz e nas folhas. Folhas flabeliformes, freqüentemente lobadas, venação paralelo-dicotômica, espiraladamente dispostas ou agrupadas em braquiblastos, decíduas no inverno. Estróbilos axilares, em braquiblastos; o masculino composto por vários microsporangióforos, cada qual com 2 sacos polínicos; o feminino composto por um pedúnculo com 2 óvulos divergentes no ápice (1 geralmente aborta). Semente verde, globosa, desenvolvendo externamente um tegumento carnoso quase livre do resto da semente. É representada atualmente apeG biloba. A espécie foi preservada graças ao seu cultivo em templos budistas da China e do Japão, mas a existência de populações naturais em florestas chinesas não foi completamente descartada. É utilizada na urbanização, mas as sementes produzem odor desagradável de manteiga rançosa. A amêndoa da semente é utilizada há mais de 300 anos no oriente para diversos tratamentos. P Árm s; tronco com crescimento monopodial, geralmente com anéis de crescimento visíveis em corte transversal e canais resiníferos presentes. Folhas simples, geralmente espiraladas ou
agrupadas em braquiblastos aciculares, lineares ou escamiformes. Estróbilos masculinos geralmente constituídos por m rosporofilos simples, microsporângios abaxiais. Cones femininos geralmente compostos; escamas ovulíferas e bracteais com diferentes graus de modificação e fusão, raramente óvulo no ápice de um ramo lateral. Sementes raramente envoltas em um arilo.
ramo com semente (abaixo).
As Pinales co(p ras de cones). Os cones que produzem pólen são estróbilos simples formados por um eixo central rodeado de microsporofilos. Os cones femininos, no entanto, são compostos por escamas ovulíferas derivadas da redução de um ramo (braquiblasto) na axila de uma bráctea (escama bracteal) e não são homólogos aos estróbilos masculinos. Taxaceae parece ser uma exceção. Nessa família, não existem evidências de uma bráctea e a semente surge no ápice de um ramo lateral. São reconhecidas sete famílias e cerca de 600 espécies, um terço delas em Pinaceae, família característica do hemisfério norte. No Brasil, ocorrem apenas duas famílias. Araucariaceae, que inclui Araucaria e Agathis e 32 espécies, está distribuída exclusivamente no hemisfério sul. Apenas Araucaria angustifolia (o pinheiro-do-paraná) está representada no Brasil. Os cones femininos (chamados de pinhões) produzem um único óvulo por escama
36

ovulífera, que geralmente se transformam em sementes recheadas de tecido nutritivo bastante calórico, geralmente consumidas em festas juninas regadas a quentão. Podocarpaceae abrange 105 espécies em sete gêneros e recentemente tem sido subdividida em quatro famílias. Podem apresentar uma a muitas escamas, cada qual com um óvulo, subtendidas por uma bráctea, mas geralmente apenas um óvulo se desenvolve. As escamas podem estar
Exemplo de conífera. A. hábi B. cone; C. conjunt
carnoso
; microstróbilos (note a quantidade de pólen).
fundidas e formar um receptáculobase da semente. Produzem uma grande quantidade
len e pó são polinizadas pelo vento. A dispedas sementes geralmente também é feita pvento, embora existam grupos nos quaitegumento da semente torna-se carnosocolorido, atraindo pássaros como dispers(p.ex. Juniperus). Enquanto algumas espé
formam florestas de grandes extensões, outras espécies estão reduzidas a populações localizadas, como as sequóias gigantes (Sequoiadendron gigantea) protegidas em parques na Califórnia. As coníferas estão entre as maiores plantas do mundo, e as mais longevas, alguns espécimes de Lagarastrobus franklinii, espécie de Podocarpaceae endêmica da Tasmânia, chegam a viver 11.000 anos. São utilizadas principalmente como fonte de celulose e pela industria madeireira. A resina de espécies de Pinus é utilizada na indústria química.
A
de
NETALES bustos, trepadeiras, raramente
rvores,
Exemplo de conífera: semente de Podocarpaceae (note as brácteas suculentas formando um receptáculo vistoso). G Subará geralmente dióicos. Folhas opostas ou verticiladas, de forma variada. Estróbilos
toB
o Cna
de rsão elo
s e e
ores cies
funcionalmente unissexuados, subtendidos por brácteas decussadas, arranjados em um estróbilo composto; os masculinos com microsporângios pedunculados (esporangióforos) subtendidos por pares de brácteas decussadas; os femininos com dois óvulos incluídos em um par de brácteas; óvulos com tegumento rostrado na região da micrópila. Sementes globosas, bacáceas e coloridas ou secas e aladas.
37

Exemplo de conífera: Taxodiaceae.
neros muito intos
amo com microstróbilos; E. ramo vegetativo (note as folhas reticuladas semelhantes às encontradas em angiospermas).
Gnetales reúne três gêdis entre si, cada um constituindo uma família própria. Ephedra (Ephedraceae) conta com aproximadamente 50 espécies. São geralmente arbustos cespitosos com folhas reduzidas encontrados em solos alcalinos de desertos e montanhas rochosas da Ásia, norte da África, Europa, América do Norte e América do Sul. Gnetum (Gnetaceae) abrange 40 espécies tropicais. São geralmente lianas com folhas opostas, peninérveas, semelhantes às de angiospermas. Os estróbilos são agrupados em anéis derivados da fusão das brácteas, formando uma espiga. Welwitschia (Welwitschiaceae) é representada apenas por W. mirabilis, uma espécie bastante bizarra, restrita aos desertos da Namíbia, no sudoeste da África. Possuem um caule tuberoso de onde partem duas folhas de crescimento indeterminado e que podem ultrapassar 6 m compr. e 1,8 cm larg., chegando a viver mais de 2.000 anos.
t
Exemplos de Gnetales: A-C. Ephedra. A hábito; B. megastróbilo (note o tegumento rostrado); C. microstróbilo (note os andróforos). D-E. Gnetum. D. r
DB
A EC
38

Welwitchia mirabilis (à esquerda). Distribuição geográfica dos gêneros de Gentales (à direita). Os óvulos são polinizados por insetos atraídos por um líquido açucarado secretado pela micrópila. Plantas produtoras de pólen podem também possuir estróbilos portando óvulos não funcionais. O par de brácteas que precede o óvulo se desenvolve com a semente. Em algumas espécies, elas formam alas, auxiliando na dispersão pela água, enquanto em outras, elas são vistosas, atraindo animais que podem atuar na dispersão das sementes.
Espécies de Ephedra são utilizadas para a produção do estimulante efedrina. Welwitchia é utilizada como combustível e algumas espécies de Gnetum possuem sementes comestíveis, e produzem casca utilizada para a produção de papel, corda e rede de pesca. Filogenia
Ao avaliar as relações entre as plantas terrestres, o ictiologista Parenti (1980) realizou o primeiro estudo filogenético com dados morfológicos incluindo as espermatófitas. Foi Crane (1985), entretanto, que introduziu o método cladístico de maneira detalhada na botânica. Ele utilizou Gnetales como estudo de caso, incorporando fósseis na análise e discutindo problemas e dificuldades do método, bem como a fragilidade de seus resultados.
Estudos subseqüentes, incluindo (e.g. Doyle & Donoghue 1987, 1992) ou não (e.g. Loconte & Stevenson 1990) fósseis reavaliaram a definição dos caracteres e seus estados e aplicaram novas ferramentas na análise dos dados. As árvores mais parcimoniosas na análise de Doyle & Donoghue (1992) indicavam as angiospermas como grupo irmão de um clado formado por Bennettitales e Gnetales. No entanto, cladogramas com um passo a mais, resultavam em uma hipótese distinta: as Gnetales como grupo irmão de um clado formado por angiosperma e Bennettitales. Apesar de ambas as topologias colocarem as Gnetales como grupo
irmão vivo das angiospermas, a interpretação para a evolução dos caracteres a partir dessas duas topologias é completamente distinta (Doyle 1998).
Até meados de 1990, os estudos morfológicos mostravam quase invariavelmente as Gnetales como grupo irmão das angiospermas, formando o clado das antófitas, que incluía também os grupos fósseis Bennettitales e Pentoxylales. Essa relação entre Gnetales e angiospermas era geralmente bem sustentada, mesmo em estudos combinando dados morfológicos e moleculares (e.g. Doyle et al. 1994, Doyle 1996; mas veja Nixon et al. 1994 para uma análise indicando Gnetales parafilética em relação às angiospermas).
A consagração da antófitas dominou a sistemática até meados da década de 1990, rendendo às gimnospermas a condição de grupo parafiético. Carquist (1996) foi uma exceção nesse contexto. Baseado na natureza da perfuração dos vasos de Gnetales e de angiospermas, ele revitalizou a posição de Bailey (1944, 1953) associando Gnetales às coníferas, contestando assim a proximidade filogenética de Gnetales às angiospermas. Diferente das análises morfológicas, as análises moleculares indicavam freqüentemente um clado formado pelas gimnospermas atuais, não confirmando as antófitas. A falta de congruência entre os estudos moleculares, no entanto, gerava desconfiança. Ao avaliarem a relação entre angiospermas e Gnetales utilizando rbcL, Goremykin et al. (1996) detectaram um grande número de substituições na terceira base. Essas taxas elevadas de substituição sugeriam que essa posição estaria saturada e sua inclusão poderia levar a desvios nos resultados. De fato, partições das posições dos codóns em análises com regiões de plastídeo (rbcL, Chaw et al. 2000; psaA e psbB, Magallón & Sanderson 2002)
G
39

Comparação entre cladogramas obtidos com dados morfológicos e moleculares (note a variação na posição de Gnetales) demonstraram que a terceira posição da seqüência sustenta um resultado inconsistente àquele obtido a partir da primeira e da segunda posições, distorcendo o resultado geral da análise. A partir do final dos anos de 1990, a reconstrução filogenética dos cinco grandes grupos de espermatófitas (Cycadales, Ginkgo, coníferas, Gnetales e angiospermas) passou a ser monopolizado por análises filogenéticas exclusivamente moleculares. A grande maioria dos estudos baseados em regiões de plastídeo (ITScp, Goremykin et al. 1996), nuclear (18S DNAr, Chaw et al. 1997) e mitocondrial (cox1, Bowe & de Pamphilis 1997) indicavam uma única origem para as gimnospermas atuais. A baixa sustentação para esse clado (ITScp e DNAr 18S) e a possibilidade da influência de ramos longos (cox1), entretanto, estimularam a ampliação dos dados (Hansen et al. 1999, cerca de 9.000 pares de base), com a implementação de análises combinadas incluindo regiões dos três gênomas, associadas à aplicação de vários algoritmos de busca, diferentes modelos
evolutivos e programas para avaliação do sinal filogenético (rbcL e as subunidades pequenas nuclear e mitocondrial do DNAr, Chaw et al. 2000; rbcL, DNAr 18S, cox1 e atpA, Bowe et al. 2000). Esses estudos multigênicos não estabeleceram ainda uma topologia confiável para as relações internas em gimnospermas. Mesmo com oito regiões (ca. 15,8 Kpb; Soltis et al. 2002), a posição de Ginkgo e a inclusão ou não de Gnetales em coníferas são discutíveis. Entretanto, a natureza monofilética das gimnospermas tem sido altamente sustentada nas análises mais recentes, confirmando a dicotomia entre as plantas atuais com sementes nuas, as gimnospermas, e com sementes inclusas no fruto, as angiospermas.
40
























![Darlan Lima - Alexandre Arcanjo e Orlando Nascimento - Teologia Sistemtica [Apostila]](https://img.document.onl/doc/110x75/55cf9cde550346d033ab56f8/darlan-lima-alexandre-arcanjo-e-orlando-nascimento-teologia-sistemtica.jpg)



