Embed Size (px)
Citation preview

837
Braz J Med Biol Res 38(6) 2005
HLA-A typing by PCR-RFLP
Typing class I HLA-A gene usinga nested PCR-RFLP procedure
1TOXICAN, Departamento de Patologia, Faculdade de Medicina,Universidade Estadual Paulista, Botucatu, SP, Brasil2Departamento de Genética, Faculdade de Medicina de Ribeirão Preto,Universidade de São Paulo, Ribeirão Preto, SP, Brasil
E.C. Castelli1,D.S. Gil1,
L.C.S. Veiga2 andJ.L.V. de Camargo1
Abstract
In order to detect several new HLA-A class I alleles that have beendescribed since 1998, the original PCR-RFLP method developed toidentify the 78 alleles recognized at that time at high resolution levelwas adapted by us for low and medium resolution levels using a nestedPCR-RFLP approach. The results obtained from blood samples of 23subjects using both the PCR-RFLP method and a commercial kit(MicroSSP1A®, One Lambda Inc.) showed an agreement higher than95%. The PCR-RFLP adapted method was effective in low andmedium resolution histocompatibility evaluations.
CorrespondenceE.C. Castelli
TOXICAN
Departamento de Patologia
FMB, UNESP
18618-000 Botucatu, SP
Brasil
E-mail: [email protected]
Research supported by FAPESP
(Nos. 01/08139-0 and 01/9448-7),
and CNPq (No. 302361/2003-0).
Received April 30, 2004
Accepted February 10, 2005
Key words• Human leukocyte antigen-A• HLA-A• Typing• RFLP• Polymorphism• Major histocompatibility
complex
The human major histocompatibility com-plex, also known as the human leukocyteantigen (HLA) complex, is a 4-Mb high-density and polymorphic region located at6p21.3 with more than 200 genes and whichrepresents about 2.5% of the human chromo-some 6. These genes belong to three maingroups, class I, II and III, which are structur-ally and functionally different (1-3). TheHLA class I molecules are glycoproteinsexpressed in almost all nucleated cells thatcollect intracellular peptide fragments andtransport them to the cell surface, where theHLA-peptide combination is presented toCD8+ T cells. Class II molecules are ex-pressed in antigen-presenting cells such as Blymphocytes and dendritic cells, and presentpeptides to CD4+ T cells. Class III genesencode soluble proteins of the complementsystem and cytokines like tumor necrosisfactor (TNA-α and LTA-α) (1-3).
High polymorphism is an important char-acteristic of the HLA complex. By Septem-
ber 2004, 1114 alleles had been describedfor class I genes: 325 for HLA-A, 592 forHLA-B, 175 for HLA-C, 6 for HLA-E, 2 forHLA-F, and 15 for HLA-G. For class II, 707alleles had been described: 3 for DRA, 458for DRB, 28 for DQA1, 56 for DQB1, 20 forDPA1, 110 for DPB1, 4 for DMA, 6 forDMB, 8 for DOA, and 8 for DOB (4; http://www.ebi.ac.uk/imgt/hla/stats.html). Thecompatibility of these polymorphisms be-tween transplant recipients and organ donorsis necessary for graft acceptance and, there-fore, detailed histocompatibility evaluationis required to reduce or avoid tissue rejection(5).
The homology among class I genes is alimiting factor for several HLA typing meth-ods and justifies molecular instead of sero-logical approaches (6). Sequence-based meth-ods involving sequence-specific primers (6-8), sequence-specific oligonucleotide probes(9) and sequencing-based techniques (10)are powerful tools used to assign HLA poly-
Brazilian Journal of Medical and Biological Research (2005) 38: 837-842ISSN 0100-879X Short Communication

838
Braz J Med Biol Res 38(6) 2005
E.C. Castelli et al.
morphisms. Their specific uses depend onindividual case needs, resolution level, thenumber of analyses to be performed, andavailable funds. Laboratories that do nothave sequencing facilities or sufficient fi-nancial support for expensive kits may ben-efit from alternative tools for HLA typing.
We present here an alternative procedurefor HLA-A polymorphism assignment basedon a PCR-RFLP methodology originally usedby Mitsunaga et al. (11) to analyze 78 HLA-A class I alleles. In order to validate thisalternative procedure at low (allele family)and medium (allele family/allele) resolutionlevels, the present study was carried out todetect all of the 274 alleles described in theliterature up to January 2003 (12). The Hos-pital Ethics Committee at this Medical Schoolapproved this study protocol.
A nested PCR approach with genericprimers was the first step for specific ampli-fication of exons 2 and 3, where the poly-morphisms within the HLA class I genes aremainly located. However, due to the greatsimilarity of class I genes and the presence ofpseudogenes in the HLA complex, the PCRprimer used in the original RFLP methodol-ogy (11), ASP5 - GCCCCGAACCCTCSTCCTGCTA/ASP3 - CCGTGGCCCCTGGTACCCGT, frequently also amplify other classI genes. The HLA-B and -C loci have onlyone or two mismatches in the region of theoriginal ASP5, depending on the polymor-phism of each individual. As a consequence,these loci usually provide PCR products ofthe same size (not distinguished by commonelectrophoresis) and provide the same RFLPpatterns for most endonucleases applied,confusing the analysis.
Therefore, a new forward primer(ASPTBE) with 3 terminal mismatches wasdeveloped in our laboratory to be used withthe original ASP3 primer. Initially, amplifi-cation of genomic DNA with generic prim-ers (ASPTBE, CAGACSCCGAGGATGGCC/ASP3 - CCGTGGCCCCTGGTACCCGT) was performed to obtain a 1017-bp
sequence including exon 1 to intron 3 of theHLA-A gene. This PCR product was diluted1:1000 in TE buffer (10 mM Tris-HCl, pH7.5, and 1 mM EDTA) and used as templatefor two further PCR procedures to amplifyseparately exon 2 (NA23 - GKCCTCGCTCTGGTTGTAGTAGC/NA25 - CAGGCTCYCACTCCATGAGGTATTTC primers) (11)and exon 3 (NA33 - CGTCTCCTTCCCGTTCTCCAGGT/NA35 - GTCSGGGCCAGGTTCTCACAC primers) (11). DMSO was in-cluded in all reactions to increase PCR speci-ficity. Restriction analyses were performedfor exons 2 and 3 with endonucleases thatwere chosen on the basis of the resolutionlevel needed (see below).
In order to obtain a low resolution level,the PCR-RFLP methodology was adapted topermit the separate evaluation of each HLA-A allele family. After amplification of exons2 and 3, RFLP analyses were performed.Endonucleases Bsp1286I, BsrI, BstNI, MnlI,HinfI, MspI, SacII, and PstI were used forexon 2, and endonucleases BslI, Fnu4HI,BsrI, Bsp1286I, HaeIII, NlaIII, HhaI, andMspI were used for exon 3, for a total of 16RFLP analyses (with 13 different endonu-cleases). Tables with the RFLP patterns foreach allele family are available from thecorresponding author.
To obtain a level of medium resolution,the endonucleases originally proposed (11),BsaJI, Bsp1286I, BsrI, BstNI, HinfI, MspI,MnlI, and SacII for exon 2, and BsaAI, BslI,BsoFI, Bsp1286I, BsrI, HaeIII, HgaI, HhaI,MspI, MnlI, and NlaIII for exon 3, are notsuitable because they do not permit the de-tection of the recently described and specifichigh frequency HLA-A alleles. Therefore,other restriction endonucleases were addedin order to increase the resolution and/or tofacilitate the detection of heterozygous com-binations. For exon 2 analysis, 12 enzymeswere added: PstI and NlaIV for standarddigestion and TspRI, AvaII, BslI, HhaI, NlaIII,HgaI, BsaHI, HaeIII, FokI, and HphI foradditional digestion. For exon 3, seven en-
839
Braz J Med Biol Res 38(6) 2005
HLA-A typing by PCR-RFLP
donucleases were added: TspRI and NlaIVfor standard digestion and MspA1, BsaJI,BstNI, SacII, and HphI for additional diges-tion. The RFLP pattern for each HLA-Aallele and the endonucleases proposed forthe medium resolution level are available inPDF format (contact the corresponding au-thor). Currently, 325 alleles for the HLA-Alocus are known (4) and new ones are ex-pected to be continuously described; all ofthese new alleles can be adequately insertedinto the technical flowchart presented.
The procedure is as follows: genomicDNA was obtained using the GFX® Genom-ic Blood DNA Purification kit (AmershamPharmacia Biotech Inc., Piscataway, NJ,USA) following manufacturer recommen-dations. The genomic DNA was amplifiedby PCR using the ASPTBE/ASP3 primers.PCR was performed in a final volume of 20µl containing 4% DMSO, 0.3 mM of eachdNTP, 2.0 mM magnesium chloride, 0.30µM of each primer, and 1 unit of DNApolymerase (Invitrogen Corporation, Carls-bad, CA, USA). The mixture was heated to94ºC for 6 min for DNA denaturation fol-lowed by 34 cycles of 94ºC (1 min), 64ºC (1min), and 72ºC (1 min) each. An 8-µl aliquotof each reaction was checked on 2% agarosegel stained with ethidium bromide. The PCRproduct was diluted in TE buffer or ultrapurewater at 1:1000 and used as template (usu-ally 2 µl) to amplify the HLA-A gene exons2 (NA23/NA25) and 3 (NA33/NA35). Re-actions for exons 2 and 3 were performed in100 µl and 120 µl final volumes, respective-ly, containing 3.5% DMSO, 0.25 mM ofeach dNTP, 2.0 mM magnesium chloride,0.30 µM of each primer, and 1.5 unit ofDNA polymerase (Invitrogen). The reactionmixtures were heated to 64ºC for 3 min forDNA denaturation followed by 34 cycles of94ºC (1 min), 65ºC (1 min), and 72ºC (1min) each. After the reaction, 5 µl of eachreaction mixture was checked on 2% aga-rose gel stained with ethidium bromide.
Aliquots of 7 µl of each PCR product
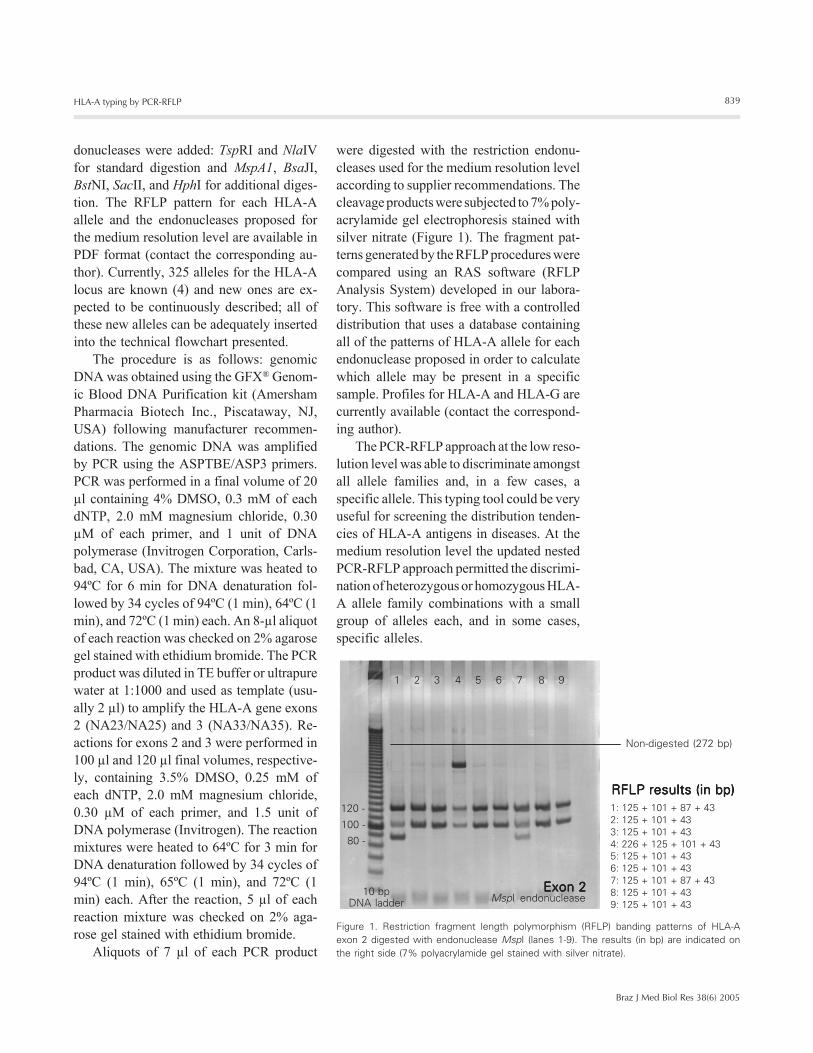
were digested with the restriction endonu-cleases used for the medium resolution levelaccording to supplier recommendations. Thecleavage products were subjected to 7% poly-acrylamide gel electrophoresis stained withsilver nitrate (Figure 1). The fragment pat-terns generated by the RFLP procedures werecompared using an RAS software (RFLPAnalysis System) developed in our labora-tory. This software is free with a controlleddistribution that uses a database containingall of the patterns of HLA-A allele for eachendonuclease proposed in order to calculatewhich allele may be present in a specificsample. Profiles for HLA-A and HLA-G arecurrently available (contact the correspond-ing author).
The PCR-RFLP approach at the low reso-lution level was able to discriminate amongstall allele families and, in a few cases, aspecific allele. This typing tool could be veryuseful for screening the distribution tenden-cies of HLA-A antigens in diseases. At themedium resolution level the updated nestedPCR-RFLP approach permitted the discrimi-nation of heterozygous or homozygous HLA-A allele family combinations with a smallgroup of alleles each, and in some cases,specific alleles.
1 2 3 4 5 6 7 8 9
Non-digested (272 bp)
RFLP results (in bp)RFLP results (in bp)RFLP results (in bp)RFLP results (in bp)RFLP results (in bp)1: 125 + 101 + 87 + 432: 125 + 101 + 433: 125 + 101 + 434: 226 + 125 + 101 + 435: 125 + 101 + 436: 125 + 101 + 437: 125 + 101 + 87 + 438: 125 + 101 + 439: 125 + 101 + 43
Exon 2Exon 2Exon 2Exon 2Exon 2MspI endonuclease
10 bpDNA ladder
120 -
100 -
80 -
Figure 1. Restriction fragment length polymorphism (RFLP) banding patterns of HLA-Aexon 2 digested with endonuclease MspI (lanes 1-9). The results (in bp) are indicated onthe right side (7% polyacrylamide gel stained with silver nitrate).

840
Braz J Med Biol Res 38(6) 2005
E.C. Castelli et al.
To test the efficiency of the nested PCR-RFLP methodology at a medium resolutionlevel, DNA from blood samples of 23 volun-teers was evaluated using the proposedmethod and a commercial kit (MicroSSP1A®,One Lambda Inc., Canoga Park, CA, USA),widely employed in histocompatibility testsfor organ transplantation. All analyses per-
formed using the nested PCR-RFLP meth-odology and the commercial kit were com-patible, assigning either homozygous or het-erozygous combinations (Table 1). Twoevaluations using the commercial kit did notdetect one of the alleles, probably due to theuse of samples under conditions which dif-fered from the kit recommendations, leading
Table 1. Comparison between nested PCR-RFLP and a commercial kit (MicroSSP1A®): results of 23 blood samples from normal subjects. Theresults of the analyses agreed, except for 2 cases for which only one allele family was detected by the commercial kit (weak amplificationprobably due to low quality DNA).

841
Braz J Med Biol Res 38(6) 2005
HLA-A typing by PCR-RFLP
to weak amplifications. However, the RFLPmethodology detected both alleles (allelefamilies) in these two samples, in which thesingle allele assigned by the kit was alsodetected by RFLP. Although only 23 indi-viduals were tested, the more than 95% agree-ment obtained between the methodologiessuggests that the PCR-RFLP technique forHLA-A typing is as sensitive as the commer-cial kit and shows a high level of reliability.Rare alleles are theoretically distinguishableusing this approach since RFLP is based onspecific endonucleases that recognize spe-cific DNA sequences, but this could not beconfirmed due to the absence of samplescarrying these alleles.
In addition, blood samples from 70 ran-domly chosen individuals were evaluated bythe PCR-RFLP methodology to compare theirHLA-A distribution with the previously re-ported distribution within the Brazilian popu-lation (13-15). The most frequent HLA-Aallele family in Brazil is the A2 group, butrepresentative frequencies also occur for theA1, A3, A24, and A68 groups (13-15). Inprevious reports of the HLA-A frequency in3 geographically separate Brazilian subpopu-lations, Paraná (Southern Brazil), Pernam-buco (Northeastern Brazil) and Minas Gerais(Southeastern Brazil), the frequencies werearound 8-10% for HLA-A1, 26-29% forHLA-A2, 8-14% for HLA-A3, 10-14% forHLA-A24, and 5-9% for HLA-A68. In thepresent study, the frequencies of these alle-les were 10% for A1, 27% for A2, 8.6% for
A3, 7% for A24, and 8% for A68, showingorders of magnitude similar to those of thefrequencies reported (13-15). Therefore, themore representative alleles reported for ahighly heterogeneous population like theBrazilian one demonstrated a good resolu-tion with the nested PCR-RFLP methodol-ogy and a high reliability using the typingprocedures reported in the present study.
The alternative methodology for HLA-Apolymorphism assignment proposed here wasshown to be adequate for low or mediumresolution levels. Furthermore, it also per-mitted the positive discrimination of het-erozygous combinations, as do other typingmethods (7,16). This may be very usefulsince the use of commercial kits is relativelyexpensive for each individual sample. Al-though the nested PCR-RFLP methodologyis more laborious, with a large initial invest-ment in endonucleases, HLA-A typing usingthis procedure is cheaper since the endonu-cleases can be used for several analyses, andare at least as sensitive, if not more so, thangeneric commercial kits.
Acknowledgments
The authors would like to thank the blooddonors, Drs. Elida Benquique Ojopi andMaria Inês de Campos Pardini for helpfulsuggestions and discussions, Maria LuizaAndanaz Falaguera for blood collection, andBruna Liboni for invaluable technical help.
References
1. Doherty PC & Zinkernagel RM (1975). A biological role for the majorhistocompatibility antigens. Lancet, 1: 1406-1409.
2. Klein J & Sato A (2000). The HLA system. First of two parts. NewEngland Journal of Medicine, 343: 702-709.
3. Undlien DE, Lie BA & Thorsby E (2001). HLA complex genes in type1 diabetes and other autoimmune diseases. Which genes are in-volved? Trends in Genetics, 17: 93-100.
4. Robinson J, Waller MJ, Parham P, de Groot N, Bontrop R, KennedyLJ, Stoehr P & Marsh SGE (2003). IMGT/HLA and IMGT/MHC:sequence databases for the study of the major histocompatibility
complex. Nucleic Acids Research, 31: 311-314.5. Fleischhauer K, Kernan NA, O’Reilly RJ, Dupont B & Yang SY (1990).
Bone marrow-allograft rejection by T lymphocytes recognizing asingle amino acid difference in HLA-B44. New England Journal ofMedicine, 323: 1818-1822.
6. Schaffer M & Olerup O (2001). HLA-AB typing by polymerase-chainreaction with sequence-specific primers: more accurate, less er-rors, and increased resolution compared to serological typing. Tis-sue Antigens, 58: 299-307.
7. Mytilineos J, Lempert M, Scherer S, Schwarz V & Opelz G (1998).

842
Braz J Med Biol Res 38(6) 2005
E.C. Castelli et al.
Comparison of serological and DNA PCR-SSP typing results forHLA-A and HLA-B in 421 Black individuals: a Collaborative Trans-plant Study report. Human Immunology, 59: 512-517.
8. Chen DF, Seibert I, Chen HY, Herbst-Kiene I, Pastucha LT & StangelW (1998). Improvement of HLA class I and class II PCR-SSP typingby using timed-release activity of DNA polymerase. Tissue Anti-gens, 51: 645-648.
9. Williams F, Meenagh A, Maxwell AP & Middleton D (1999). Alleleresolution of HLA-A using oligonucleotide probes in a two-stagetyping strategy. Tissue Antigens, 54: 59-68.
10. Scheltinga SA, Johnston-Dow LA, White CB, van der ZwA, BakemaJE, Rozemuller EH, van den TwJ, Kronick MN & Tilanus MG (1997).A generic sequencing based typing approach for the identificationof HLA-A diversity. Human Immunology, 57: 120-128.
11. Mitsunaga S, Tokunaga K, Kashiwase K, Akaza T, Tadokoro K & JujiT (1998). A nested PCR-RFLP method for high-resolution typing ofHLA-A alleles. European Journal of Immunogenetics, 25: 15-27.
12. Marsh SG, Albert ED, Bodmer WF et al. (2002). Nomenclature forfactors of the HLA system. Tissue Antigens, 60: 407-464.
13. Middleton D, Williams F, Meenagh A, Daar AS, Gorodezky C,Hammond M, Nascimento E, Briceno I & Perez MP (2000). Analysisof the distribution of HLA-A alleles in populations from five conti-nents. Human Immunology, 61: 1048-1052.
14. Nigam P, Dellalibera E, Maurício-da-Silva L, Donadi EA & Silva RS(2004). Polymorphism of HLA class I genes in the Brazilian popula-tion from the Northeastern State of Pernambuco corroborates an-thropological evidence of its origin. Tissue Antigens, 64: 204-209.
15. Braun-Prado K, Mion ALV, Pereira NF, Culpi L & Petzl-Erler M (2000).HLA class I polymorphism, as characterized by PCR-SSOP, in aBrazilian exogamic population. Tissue Antigens, 56: 417-427.
16. Moribe T, Kaneshige T & Inoko H (1997). Complete HLA-A DNAtyping using the PCR-RFLP method combined with allele group- andsequence-specific amplification. Tissue Antigens, 50: 535-545.





























