Embed Size (px)
Citation preview

UNIVERSIDADE DE SÃO PAULO
INSTITUTO DE PSIQUIATRIA
JESSYKA MARIA DE FRANÇA BRAM
Biomarcadores em líquor e em leucócitos no transtorno depressivo
maior, comprometimento cognitivo leve e doença de Alzheimer
(Versão corrigida. Resolução CoPGr 6018/11, de 1 de novembro de 2011. A versão original está disponível na Biblioteca da FMUSP)
São Paulo
2017

JESSYKA MARIA DE FRANÇA BRAM
Biomarcadores em líquor e em leucócitos no transtorno depressivo
maior, comprometimento cognitivo leve e doença de Alzheimer
Dissertação apresentada à Faculdade de
Medicina da Universidade de São Paulo para a
obtenção do título de Mestre em Ciências
Programa de Psiquiatria
Orientador: Prof. Dr. Orestes Vicente Forlenza
(Versão corrigida. Resolução CoPGr 6018/11, de 1 de novembro de 2011. A versão original está disponível na Biblioteca da FMUSP)
São Paulo
2017

Bram J.M.F. Biomarcadores em líquor e em leucócitos no Transtorno
Depressivo Maior, Comprometimento Cognitivo Leve e doença de Alzheimer
[Dissertação]. São Paulo: Faculdade de Medicina, Universidade de São Paulo;
2017.
Aprovado em:
Banca Examinadora
Prof. Dr. ______________________________________________________ Instituição: ______________________________________________________ Julgamento: _______________________________________________________ Prof. Dr. ______________________________________________________ Instituição: ______________________________________________________ Julgamento: _______________________________________________________ Prof. Dr. ______________________________________________________ Instituição: ______________________________________________________ Julgamento: _______________________________________________________ Prof. Dr. ______________________________________________________ Instituição: ______________________________________________________ Julgamento: _______________________________________________________

Aos meus pais, Roberto e Maria, pelo incentivo
e apoio em todas as minhas escolhas e
decisões.
Aos meus irmãos e sobrinhos, pelo
companheirismo e pela compreensão nos
momentos ausentes.
Ao meu noivo, Thiago, pela complacência e por
estar ao meu lado durante todo o
desenvolvimento dessa Dissertação.
Aos idosos que consentiram em participar
desse estudo.

Agradecimentos
Agradeço primeiramente a Deus, por estar sempre ao meu lado. À
minha família, por me acompanhar sempre e confiar em mim e ao meu futuro
marido Thiago, por ser meu companheiro de todas as horas e por me aguentar
nos momentos mais importantes dessa trajetória.
Ao Dr. Orestes Vicente Forlenza, pela orientação e por acreditar em
mim.
À Dra. Leda Leme Talib, por estar sempre ao meu lado, pela dedicação,
carinho, esperança, honestidade, sinceridade e por ter me recebido e ajudado
assiduamente na elaboração desse trabalho de maneira profissional e materna.
À Mestre e doutoranda Helena Passarelli Giroud Joaquim Nadal, que me
incentiva a continuar sempre, mesmo com as dificuldades encontradas pelo
caminho, mostrando-se presente e disponível todo o tempo. Agradeço pela
amizade e companheirismo.
Agradeço também a essas mentoras por terem paciência em me ensinar
toda a parte de bancada, por ajudarem a me organizar conforme a prioridade e
a escrever de modo mais coerente e objetivo, pois assim, permitiram que eu
adquirisse maturidade científica.
Às minhas amigas Leda (Chó), Helena (Lê), Bruna (Bu), Eliane (Beide),
Alana (Alhana), Tamires (Tami), Mislene (Mis) e Ana (Namir) pelo
companheirismo, desabafos, risadas, ajuda sempre que necessário, conselhos,
brincadeiras e muito mais. Sou muito grata por tornarem todos meus dias mais
alegres e divertidos! Levo vocês comigo sempre!
Às funcionárias do LIM-27, Luciana e Sandra, por realizarem a coleta
dos pacientes do estudo e também pela paciência em ensinar-me a preparar a
amostra coletada. Agradeço à Dona Edivani pela dedicação, por todo material

do laboratório sempre limpo e organizado, pelo companheirismo e pelos
deliciosos cafés. Agradeço também pelo carinho, pelas conversas, pelas
risadas e por todo o aprendizado.
Ao Laboratório de Neurociências – LIM-27, como um todo, por confiar
em mim e abrir as portas para que eu pudesse desenvolver o Projeto de
Mestrado ora proposto.
A todos que de alguma forma contribuíram para o desenvolvimento
desse projeto e aos profissionais do Ambulatório de Psiquiatria Geriátrica do
LIM-27 do Instituto de Psiquiatria do Hospital das Clínicas da Faculdade de
Medicina da Universidade de São Paulo (IPq-HCFMUSP), pois sem eles o
recrutamento e a realização de avaliações neuropsicológicas, bem como
diagnóstico dos mesmos, simplesmente não teriam ocorrido.
Agradeço também à Eliza e Isabel do Programa de Pós-Graduação do
Instituto de Psiquiatria, por sempre torcerem por mim, por se preocuparem com
os prazos e me alertarem sobre os mesmos, pelo carinho com que me tratam e
por estarem sempre presentes durante o desenvolvimento desse Mestrado.
À Zelinda, por ser solicita e estar sempre presente.
À Denise, por ter sido essencial no processo de preparação e envio dos
documentos necessários para a emissão de um parecer pelo Comitê de Ética,
em todas as instâncias. Obrigada pelo carisma, paciência, disponibilidade e
preocupação.
Ao Instituto de Psiquiatria (IPq) pela oportunidade de realizar esse
mestrado pelo Programa de Pós-Graduação.
Ao Laboratório de pesquisa Básica em Doenças Renais, LIM-12, em
especial à Mestre Ivone Braga de Oliveira, pela parceria, pela atenção e

cuidado, e pela colaboração colocando à disposição o fotodocumentador
utilizado para conclusão do método de Western Blotting.
À Associação Beneficiente Alzira Denize Hertzog da Silva (ABADHS)
pelo financiamento e apoio à pesquisa.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
(CAPES), agência de fomento que financiou parte da pesquisa realizada,
possibilitando condições propícias para a realização do projeto.
À Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP),
processo nº 2014/21946-6, agência de fomento que disponibilizou recursos
para o desenvolvimento desse mestrado e para a divulgação do mesmo por
meio de Congressos.

“Mesmo quando tudo parece desabar, cabe a
mim decidir entre rir ou chorar, ir ou ficar,
desistir ou lutar; porque descobri, no caminho
incerto da vida, que o mais importante é
decidir!”
Cora Coralina

Normalização Adotada
Esta dissertação está de acordo com as seguintes normas, em vigor no
momento desta publicação:
Referências: adaptado de International Committee of Medical Journal
Editors (Vancouver).
Universidade de São Paulo. Sistema Integrado de Bibliotecas. Diretrizes
para apresentação de dissertações e teses da USP – Parte IV (Vancouver).
Elaborado por Vânia Martins Bueno de Oliveira Funaro (coordenadora), Maria
Cláudia Pestana, Maria Cristina Cavarette Dziabas, Eliana Maria Garcia, Maria
Fátima dos Santos, Maria Marta Nascimento e Suely Campos Cardoso. 3ª ed
revisada, ampliada e modificada. São Paulo: Sistema Integrado de Bibliotecas;
2016.
Abreviaturas dos títulos dos periódicos de acordo com o Listo f Journals
Indexed in Index Medicus.

SUMÁRIO
1 INTRODUÇÃO ......................................................................................................................... 1
1.1 Transtorno Depressivo ................................................................................................ 2
1.2 Comprometimento Cognitivo Leve ............................................................................ 4
1.3 Doença de Alzheimer .................................................................................................. 5
1.5 Possíveis biomarcadores para diagnóstico precoce da Doença de Alzheimer . 9
1.5.1 A Disintegrin And Metallopeptidase 10 ............................................................... 10
1.5.2 β-site APP-cleaving enzyme 1.............................................................................. 11
1.5.3 Presenilina 1 ............................................................................................................ 12
1.5.4 Peptídeo Aβ ............................................................................................................. 12
1.6 Justificativa .................................................................................................................. 13
2 OBJETIVOS ........................................................................................................................... 14
2.1 Objetivo geral .............................................................................................................. 14
2.2 Objetivos específicos ................................................................................................. 14
3 MATERIAL E MÉTODOS .................................................................................................... 15
3.1 Casuística .................................................................................................................... 15
3.2 Matriz leucocitária ...................................................................................................... 18
3.2.1 Obtenção e preparo de leucócitos ....................................................................... 18
3.2.2 Determinação de BACE1, ADAM10 e PSEN1 ................................................... 19
3.2.3 Determinação do peptídeo Aβ .............................................................................. 21
3.3 Matriz liquórica............................................................................................................ 22
3.3.1 Obtenção e preparo de líquor ............................................................................... 22
3.3.2 Determinação de BACE1, ADAM10 e PSEN1 ................................................... 23
3.3.3 Determinação do peptídeo Aβ42 ........................................................................... 23
3.4 Análise estatística ...................................................................................................... 24
4 RESULTADOS ...................................................................................................................... 26
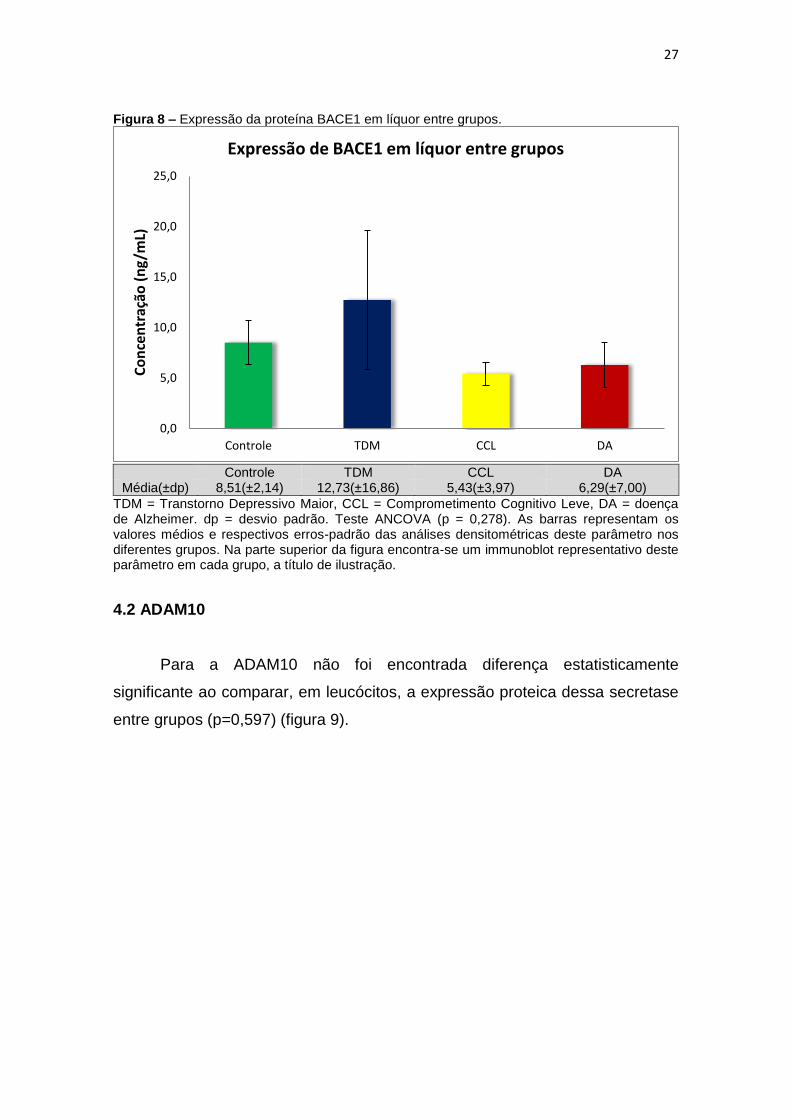
4.1 BACE1 ......................................................................................................................... 26
4.2 ADAM10 ...................................................................................................................... 27
4.3 PSEN1 ......................................................................................................................... 29
4.4 Peptídeo Aβ ................................................................................................................ 30
4.5 Análise de correlação entre variáveis ..................................................................... 32

5 DISCUSSÃO .......................................................................................................................... 38
6 CONSIDERAÇÕES FINAIS ................................................................................................ 44
REFERÊNCIAS BIBLIOGRÁFICAS ...................................................................................... 45
APÊNDICES .............................................................................................................................. 60
ANEXOS .................................................................................................................................... 81

LISTA DE FIGURAS
Figura 1 – Marcos patológicos da doença de Alzheimer. ................................................... 6
Figura 2 – Placas Senis ............................................................................................................ 7
Figura 3 – Processamento proteolítico da APP. ................................................................... 8
Figura 4 – Casuística conforme grupo e matriz biológica. ................................................ 17
Figura 5 – Método de ELISA sanduíche. ............................................................................. 22
Figura 6 – Metodologia Luminex. .......................................................................................... 24
Figura 7 – Expressão da proteína BACE1 em leucócitos e representação do
Immunoblotting entre grupos. ................................................................................................. 26
Figura 8 – Expressão da proteína BACE1 em líquor entre grupos. ................................ 27
Figura 9 – Expressão da proteína ADAM10 em leucócitos e representação do Immunoblotting entre grupos. ................................................................................................. 28
Figura 10 – Expressão da proteína ADAM10 em líquor entre grupos. ........................... 29
Figura 11 – Expressão da proteína PSEN1 em leucócitos e representação do
Immunoblotting entre grupos. ................................................................................................. 30
Figura 12 – Expressão do peptídeo Aβ em leucócitos entre grupos. ............................. 31
Figura 13 – Expressão do peptídeo Aβ em líquor entre grupos. ..................................... 32
Figura 14 – Correlação entre BACE1 expressa em leucócitos e BACE1 expressa em
líquor. .......................................................................................................................................... 33
Figura 15 – Correlação entre as proteínas e peptídeo expressos em leucócitos. ........ 34
Figura 16 – Correlação entre BACE1 e ADAM10 no líquor. ............................................. 35
Figura 17 – Correlação entre expressão de BACE1 no líquor e idade. .......................... 35
Figura 18 – Correlação entre expressão proteica e resultados obtidos nas avaliações
neuropsicológicas. .................................................................................................................... 37

LISTA DE TABELAS
Tabela 1 – Prevalência de transtornos mentais em idosos de comunidades urbanas brasileiras. ............................................................................................ 2
Tabela 2 – Perfil demográfico dos indivíduos recrutados. ............................... 18
Tabela 3 – Diluição e tempo de incubação dos anticorpos das secretases e do controle endógeno. ........................................................................................... 21

LISTA DE SIGLAS
ADAM10 A Desintegrin And Metallopeptidase 10
ADAMs A Desintegrin And Metallopeptidase
ADRDA Associação da Doença de Alzheimer e Doenças
Relacionadas
ANCOVA Análise de Variância
APH-1 Anterior pharynx-defective
ApoE4 Apolipoproteína E alelo ԑ4
APP Proteína Precursora Amilóide
Aβ Peptídeo β-Amilóide
BACE1 β-site APP-cleaving enzyme 1
BDNF Fator Neurotrófico Derivado do Cérebro
BSA Albumina de Soro Bovino
CAMCOG Teste Cognitivo de Cambridge
CCL Comprometimento Cognitivo Leve
DA Doença de Alzheimer
DSM-IV Manual Diagnóstico e Estatístico de Transtornos Mentais –
Quarta Edição
DSM-V Manual Diagnóstico e Estatístico de Transtornos Mentais –
Quinta Edição
EGTA Ácido etileno glicol-bis N,N,N9,N9-tetra-acético
ELISA Enzyme Linked Immuno Sorbent Assay
ENFs Emaranhados Neurofibrilares Intracelulares
HAM-D 21 Escala de Depressão de Hamilton – 21 itens
IL Interleucinas
IPq – FMUSP Instituto de Psiquiatria da Faculdade de Medicina da
Universidade de São Paulo
kDa Kilodalton
KS Kolmogorov-Smirnov
LIM-16 Laboratório de Fisiopatologia Renal
LIM-27 Laboratório de Neurociências
mA Miliampéres

MEEM Mini-Exame do Estado Mental
NCT Niscastrin
NIA National Institute on Aging
NINCDS Instituto Nacional de Doenças Neurológicas e Comunicativas
e Acidente Vascular Cerebral
OMS Organização Mundial da Saúde
PEN-2 Presenilin enhancer 2
PMSF Fluoreto de fenilmetilsulfonil
PSEN Presenilin
PSEN1 Presenilina 1
RIPA Radio-Immunopreciptation Assay
SCID – I/P Entrevista Clínica Estruturada para o Diagnóstico de
Transtornos Mentais do Eixo I
SPSS Statistical Package for Social Science
TBS -Tween Tris-buffered saline – Tween
TCLE Termo de Consentimento Livre e Esclarecido
TDM Transtorno Depressivo Maior
TNCm Transtorno Neurocognitivo Menor
αAPPs Fragmento solúvel de APP de 130kDa
βAPPs Fragmento solúvel de APP de 110kDa
LISTA DE SÍMBOLOS
α Alfa
β Beta
γ Gama
r Coeficiente de Correlação de Pearson

RESUMO
Bram JMF. Biomarcadores em líquor e em leucócitos no transtorno depressivo
maior, comprometimento cognitivo leve e doença de Alzheimer [Dissertação].
São Paulo: Faculdade de Medicina, Universidade de São Paulo; 2017.
O acelerado processo de envelhecimento tem trazido não só mudanças
no perfil demográfico, mas também no perfil epidemiológico da população.
Entre os problemas de saúde encontrados em idosos, merecem destaque os
transtornos mentais, visto que acometem cerca de um terço dessa população,
sendo os transtornos depressivos e a demência os problemas de saúde mental
mais prevalentes. O transtorno depressivo maior (TDM), bem como o
comprometimento cognitivo leve (CCL), tem sido associado prejuízo das
funções cognitivas, além de aumentar o risco de desenvolvimento de
demência, principalmente doença de Alzheimer (DA). Sendo assim, é
importante que haja identificação preventiva de pessoas com maior risco para o
desenvolvimento de DA a fim de proporcionar um tratamento precoce. Dessa
maneira este estudo objetivou comparar os marcadores biológicos envolvidos
na cascata amiloide, expressos em leucócitos e líquor, nas condições clínicas
TDM, CCL, DA e controles saudáveis. Para tanto, foi determinada a expressão
proteica das secretases ADAM10, BACE1 e PSEN1 e do peptídeo Aβ em
leucócitos e líquor pelos métodos de Western Blotting, ELISA e Luminex. Ao
analisarmos a expressão das secretases e do peptídeo Aβ em leucócitos não
observamos diferenças estatisticamente significantes entre os grupos, exceto
pela expressão da proteína PSEN1, que apresentou menores níveis em DA em
relação à TDM. Em relação ao líquor, observamos uma significativa diminuição
do peptídeo Aβ no grupo DA quando comparado aos grupos CCL, TDM e
também a controles saudáveis; porém não foram observadas diferenças de
expressão desse peptídeo entre CCL, TDM e controles saudáveis. Esses
resultados sugerem que a matriz leucocitária, apesar de expressar os
componentes desta via, sugerindo dispor da maquinaria enzimática necessária
para o processamento da proteína precursora do amiloide (APP), não é a ideal
para estudos sobre a cascata amiloide. Além disso, os resultados obtidos em

amostras de líquor, de acordo com a casuística analisada, reforçam os dados
da literatura que estabelecem a redução da concentração do peptídio Aβ no
líquor como um biomarcador da DA, sem termos detectado alterações nos
demais grupos estudados. Nossos resultados (negativos) em relação à
expressão das APP-secretases no líquor acrescentam dados a este domínio
ainda controverso da literatura. Assim sendo, mais estudos devem ser
realizados para que possamos compreender melhor as vias relacionadas ao
desenvolvimento da DA.
Descritores: transtorno depressivo maior; comprometimento cognitivo leve;
doença de Alzheimer; ADAM10; Presenilina-1; Peptídeos beta-amiloides;
leucócitos; líquor.

ABSTRACT
Bram J.M.F. Biomarkers in cerebrospinal fluid and leukocytes in Major
Depressive Disorder, Mild Cognitive Impairment and Alzheimer's disease
[dissertation]. São Paulo: “Faculdade de Medicina, Universidade de São Paulo”;
2017.
The accelerated aging process has brought not only changes in the
demographic profile, but also in the epidemiological profile of the population.
Among the health problems found in the elderly, mental disorders are worth
mentioning, since they affect about a third of this population, being the
depressive disorders and dementia the most prevalent mental health problems.
Major depressive disorder (MDD), as well as mild cognitive impairment (MCI),
have been associated with higher levels of cognitive symptoms and increased
risk of developing dementia, especially Alzheimer's disease (AD). Therefore, it
is important to identify individuals at greater risk for the development of AD in
order to provide treatment at early stages of the disease process. In this way,
this study aimed to compare the biological markers involved in the amyloid
cascade under the clinical conditions TDM, CCL, DA and healthy controls
expressed in leukocytes and CSF. We determined the protein expression of
ADAM10, BACE1 and PSEN1 and Aβ peptides in leucocytes and CSF using
the Western Blotting, ELISA and Luminex methods. When analyzing the
expression of the secretases and the Aβ peptide in leukocytes, we did not
observe statistically significant differences between the groups, except for the
expression of the PSEN1 protein, which presented lower levels in AD in relation
to MDD. In relation to CSF, we observed a significant reduction of the Aβ
peptide in AD when compared to MCI, TDM and also to healthy controls, but no
differences in expression of this peptide were observed between MCI, MDD and
healthy controls. These results suggest that the leukocytes, although
expressing the key components and the enzymatic machinery necessary for the
proteolytic processing of the amyloid precursor protein (APP), may not
represent an adequate biological matrix for studies addessing the amyloid
cascade. In addition, results obtained in CSF samples, according to the

analyzed series, reinforce the literature data that establish the reduction of Aβ
peptide concentration in CSF as a biomarker of AD, without detecting
alterations in the other groups studied. Our (negative) results in relation to the
expression of APP-secretases in CSF add data to this still controversial domain
of the literature. Therefore, more studies should be done so that we can better
understand the pathways related to the development of AD.
Descriptors: Major Depressive Disorder, Mild Cognitive Impairment, Alzheimer's
Disease, ADAM10, BACE1, PSEN1, Aβ Peptide, Leukocytes and CSF.

1
1 INTRODUÇÃO
Chegar à velhice não é mais um privilégio de poucos, visto que a
população mundial está envelhecendo rapidamente. A Organização Mundial da
Saúde (OMS) revela que, até 2050, a proporção de pessoas com mais de 60
anos de idade irá corresponder a 22% da população mundial, representando
aproximadamente dois bilhões de pessoas idosas no mundo (World Health
Organization, 2014).
Esses dados são de extrema importância dado que o processo de
envelhecimento é responsável por mudanças morfofuncionais que afetam
todos os sistemas fisiológicos principais de forma variável e causam maior
vulnerabilidade do organismo às agressões externas e internas (Moraes et al,
2010). Além disso, o eminente envelhecimento populacional vem
acompanhado pelo aumento da prevalência de doenças crônicas não
transmissíveis que podem gerar um processo incapacitante, dificultando ou
impedindo o desempenho na realização de atividades cotidianas. Todas essas
condições acima citadas podem culminar no comprometimento significativo da
qualidade de vida dos idosos, trazendo consigo desafios tanto para a saúde
física quanto mental dos mesmos (Ministério da Saúde, 2007; Clemente et al,
2011).
Dos problemas de saúde que mais afetam os idosos merecem ênfase os
transtornos neuropsiquiátricos, visto que acometem atualmente mais de 20%
dos idosos com 60 anos ou mais, sendo responsáveis por 6,6% dos casos de
incapacidade nessa população. Dentre eles, os transtornos depressivos e
demência apresentam-se como os mais comuns para essa faixa etária (World
Health Organization, 2016).
Esse cenário não se difere no Brasil. Os idosos representam o segmento
populacional que mais têm aumentado na população brasileira, com
estimativas de incremento médio para os próximos dez anos de mais de 1
milhão de idosos ao ano (Ervatti et al., 2015), o que é preocupante pois dados
epidemiológicos demonstram que cerca de um terço dos idosos brasileiros são

2
acometidos por transtornos neuropsiquiátricos, em especial por transtornos
depressivos e demência (tabela 1), não se distanciando da realidade do
contexto mundial (Garrido, Menezes, 2002; Clemente et al., 2011).
Tabela 1 – Prevalência de transtornos mentais em idosos de comunidades urbanas brasileiras.
PROBLEMA DE SAÚDE MENTAL NO IDOSO PREVALÊNCIA (%)
Transtornos Neuropsiquiátricos (Ramos et al., 1993; Coelho Filho, Ramos, 1999; Maragno et al., 2006)
26,4 a 33,6
Transtorno Depressivo (Castro-Costa et al., 2008; Lima et al., 2009)
19,8 a 38,5
Demência (Scazufca, 2002)
4,2 a 7,2
A atenção a esses transtornos se faz necessária à medida que trazem
importantes impactos sociais e econômicos para a sociedade. Além de haver
considerável piora da qualidade de vida do idoso e de seus familiares e/ou
cuidadores, há uma crescente demanda por recursos públicos, aumentando os
custos na área da saúde, principalmente nas assistências de média e alta
complexidade. Nesse sentido, o desenvolvimento de pesquisas na área é de
grande valia e pode proporcionar não só uma redução de gastos públicos, mas
também mais anos de vida com autonomia e independência.
1.1 Transtorno Depressivo
O transtorno depressivo é uma das principais causas de incapacidade
no mundo, afetando atualmente cerca de 350 milhões de pessoas, sendo o
gênero feminino o mais atingido (World Health Organization, 2016). Dentre a
população idosa, a prevalência global desse transtorno pode variar de 10 a
20%, dependendo da situação cultural (Rangaswamy, 2001; Archana, Nisha,
2009). É importante ressaltar que essa condição não é inerente ao processo de
envelhecimento, porém há muitos fatores que aumentam o risco desse
problema de saúde mental em idosos (Jeste et al., 2010).
O Manual Diagnóstico e Estatístico de Transtornos Mentais – Quinta
Edição (DSM-V) inclui em seu capítulo sobre transtornos depressivos oito
diferentes subtipos que apresentam em comum “presença de humor triste,
vazio ou irritável, acompanhado de alterações somáticas e cognitivas que

3
afetam significativamente a capacidade de funcionamento do indivíduo”,
diferindo-os pelos aspectos de duração, momento ou etiologia presumida
(American Psychiatric Association, 2013).
Dentre esses transtornos depressivos, o Transtorno Depressivo Maior
(TDM) é o que representa a condição mais clássica, sendo caracterizado por
alterações no humor e cognição com episódios distintos de pelo menos duas
semanas de duração (Sierksma et al., 2010; American Psychiatric Association,
2013). No idoso sua apresentação sintomatológica manifesta-se
diferentemente quando comparada a outras faixas etárias (Martins, 2008),
expressando-se de maneira mais heterogênea e menos estereotipada que nos
adultos, havendo uma tendência na manifestação de maior quantidade de
sintomas somáticos, como dor crônica, fadiga, alterações de sono e apetite, em
relação a sintomas psíquicos, como a tristeza (Diniz; Forlenza, 2008). Além
disso, o TDM tem sido associado a altos níveis de morbimortalidade, piora da
qualidade de vida, maiores níveis de comprometimento cognitivo e funcional
(Alexopoulos et al., 2002; Andrei et al., 2007; Chachamovic et al., 2008; Ávila et
al., 2009), além de aumentar o risco de desenvolvimento de demência (Ownby
et al., 2006; Diniz et al., 2013).
A principal teoria relacionada às bases biológicas do transtorno
depressivo envolve neurotransmissores cerebrais e é denominada hipótese das
monoaminas (Bahls, 1999; Wuwongse, Chang, Law, 2010). Essa hipótese
fundamenta-se no conceito de deficiência das aminas biogênicas serotonina,
noradrenalina e dopamina como etiologia desse transtorno (Bahls, 1999).
Tais neurotransmissores, juntamente com a acetilcolina são
responsáveis pela modulação e integração sobre outras atividades corticais e
subcorticais, além de estarem envolvidos na regulação de apetite, sono,
atividade psicomotora e, possivelmente, humor (Bandeira, 2012).
Alterações em cascatas inflamatórias e neurotróficas também têm sido
estudadas e relacionadas à fisiopatologia do transtorno depressivo,
principalmente do subtipo TDM. Estudos com pacientes deprimidos
demonstraram elevação de interleucinas (IL) pró inflamatórias e diminuição das
IL anti-inflamatórias (Maes et al., 2009). Já com relação ao suporte
neurotrófico, uma pesquisa desenvolvida com modelos animais transgênicos

4
para o transtorno depressivo evidenciou redução significativa da síntese de
fator neurotrófico derivado do cérebro (BDNF), além disso, observou que o
restabelecimento de níveis de BDNF ocasionou melhora do comportamento
depressivo nestes modelos (Duman e Monteggia, 2006).
1.2 Comprometimento Cognitivo Leve
Outra condição clínica que têm sido alvo de muitos estudos diz respeito
ao conceito de comprometimento cognitivo leve (CCL), atualmente conhecido
como transtorno neurocognitivo menor, segundo DSM-V (American Psychiatric
Association, 2013). Ele tem sido utilizado na pesquisa clínica para identificar
indivíduos que apresentam um grande risco de conversão para a doença de
Alzheimer (DA), pois a taxa de conversão do CCL para DA é de
aproximadamente 10 a 15% ao ano (Petersen et al., 2001a; 2001b; Hamdan,
2008).
O CCL caracteriza-se pela queixa de memória corroborada por um
informante, pelo comprometimento da memória quando comparado à idade e
nível de escolaridade, pela função cognitiva geral preservada, pelas atividades
de vida diárias intactas e pela ausência de demência (Petersen et al., 2001a;
2001b).
O CCL pode ser classificado em subtipo amnéstico ou não-amnéstico
(Petersen, 2004). O primeiro é o mais conhecido e faz referência a indivíduos
que apresentam comprometimento da memória, podendo existir de maneira
isolada (CCL amnéstico único domínio) ou ser acompanhado por outros déficits
cognitivos (CCL amnéstico múltiplos domínios). Já no CCL não-amnéstico a
memória encontra-se preservada, podendo haver um ou mais domínios
comprometidos (CCL não-amnéstico único domínio e CCL não-amnéstico
múltiplos domínios) (Winblad et al., 2004).
Como o CCL é um fator de risco para o desenvolvimento de demência,
principalmente para a DA, muitas pesquisas têm sido desenvolvidas a fim de
compreender os processos metabólicos que ocorrem no cérebro, e que estão
associados ao comprometimento cognitivo.

5
Estudos longitudinais revelaram que idosos com CCL com altas taxas de
conversão para DA já apresentavam um aumento nos níveis da proteína Tau
hiperfosforilada no líquor (Sunderland et al., 1999). Maruyama et al. (2001)
quantificaram em líquor níveis de peptídeo β-amilóide (Aβ) e proteína Tau em
pacientes com CCL e com DA em diversos estágios da doença. Os níveis de
Aβ não apresentaram diferenças significantes entre CCL e controles, porém
esses valores diminuíram significantemente na medida em que a DA tornou-se
clinicamente evidente. Já estudos recentes sobre placas amilóides têm
identificado que a deposição do Aβ, assim como o hipometabolismo, está
associada ao declínio cognitivo (Storandt et al., 2009; Villemagne et al., 2011) e
à conversão (Okello et al., 2009; Jack et al., 2010) de CCL para DA, além de
preceder disfunções sinápticas e neuronais (Landau et al., 2012).
1.3 Doença de Alzheimer
Dentre as demências, merece destaque a DA, por ser considerada não
só a etiologia mais comum de demência entre idosos com 65 anos ou mais,
mas também o principal problema de saúde pública entre idosos (Rodgers,
2008), tanto pela prevalência quanto pelas projeções para o futuro e pelas
consequências desta doença (Zigman et al., 1997; Helmer et al., 2006).
A DA é compreendida como uma doença neurodegenerativa
progressiva, heterogênea em seus aspectos etiológico, clínico e patológico
(Machado, 2002), e há evidências de que seu processo degenerativo se inicie
de vinte a trinta anos antes do aparecimento clínico da doença (Catricala; Torti;
Ricevuti, 2012). Atualmente seu diagnóstico é baseado em uma combinação de
critérios clínicos que incluem histórico familiar, exame neurológico, avaliações
neuropsicológicas e exames de imagem (McKhann et al, 2011).
Os principais marcos biológicos da doença são os emaranhados
neurofibrilares intracelulares (ENFs) e as placas amilóides (neuríticas ou senis),
juntamente com a perda progressiva de neurônios no cérebro (figura 1). Em
algumas áreas cerebrais, a perda neuronal pode chegar a 90% (Bussière et al.,
2003; Qin et al., 2009).
6
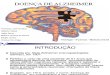
Figura 1 - Marcos patológicos da doença de Alzheimer.
A figura representa os marcos patológicos da DA: à esquerda a via amiloidogênica, em que o processamento proteolítico da APP ocorre por meio da β- e γ-secretase, respectivamente, levando à formação de placas senis; a desintegração dos microtúbulos do citoesqueleto neural, levando à formação de emaranhados neurofibrilares (ENFs); e a morte neuronal ocasionada pelos processos anteriores. À direita está representada a via fisiológica. Fonte: Adaptado de National Institute on Aging (NIA), s/a.
Os ENFs formam-se a partir do colapso do citoesqueleto neuronal, que
ocorre através da hiperfosforilação da proteína Tau (Brion et al.,
1985; Grundke-Iqbal et al., 1986; Kosik et al., 1986; Wood et al., 1986). Nos
tecidos cerebrais a fosforilação da proteína Tau é resultado da ação conjunta
de fosfatases e quinases (De Paula et al., 2009). Assim sendo, a inibição da
hiperfosforilação da Tau pela inibição de quinases torna-se uma via importante
para desenvolvimento de planos terapêuticos na DA (Forlenza et al., 2000).
A formação de placas neuríticas senis ou placas amiloides, outro marco
patológico da doença, resultam da deposição e agregação, em meio
extracelular, de Aβ (figura 2), sendo a Proteína Precursora Amilóide (APP) um
elemento chave neste processo.

7
Figura 2 - Placas Senis
A) Neurônios saudáveis; B) Formação de placas senis, por meio da agregação e deposição de Aβ nos espaços interneurais. Fonte: Adaptado de Alzheimer’s Association, 2017.
Na DA, a clivagem da APP por proteases encontra-se alterada,
favorecendo a via amiloidogênica, ao aumentar a clivagem da APP por β-
secretase, que gera a liberação de um derivado conhecido como βAPP solúvel
(βAPPs). Em seguida a APP é clivada, em sua outra extremidade, por -
secretase. Isso gera a liberação do Aβ que, por consequência, acumula-se em
agregados extracelulares nos neurônios (Laferla, 2002; Zinseret al., 2007;
Rodgers, 2008).
Já em condições fisiologicamente normais a clivagem de APP ocorre
pela via não-amiloidogênica, em que a APP é clivada por α-secretase entre os
resíduos de aminoácidos 612 e 613 (lisina 16 e leucina 17, respectivamente),
ou seja, dentro da sequência do Aβ, evitando assim a formação das placas
amilóides (Hooper, Turner, 2002; Gonçalves, 2012). Nesta via, a clivagem por
α-secretase produz αAPP solúvel (αAPPs) e um fragmento residual p3, o qual
possui propriedades neuroprotetoras (Rodgers, 2008; Kreutz, 2010). Após esta
clivagem inicial a APP é clivada também por -secretase (figura 3). Em
pacientes com DA, o processamento da APP via α-secretase encontra-se
diminuído (Colciaghi et al., 2004).
A B

8
Figura 3 - Processamento proteolítico da APP.
A clivagem da APP pode ocorrer mediante a via não-amiloidogênica ou secretória (à direita), por α- e γ-secretase, ou via amiloidogênica (à esquerda), por β- e γsecretase, favorecendo a liberação de Aβ. Fonte: Adaptado de Wang et al., 2012.
Pesquisas têm evidenciado relações entre transtorno depressivo e
demência, sendo aquele considerado um fator de risco para o desenvolvimento
de demência, em particular para a DA. Além disso, processos fisiopatológicos
comuns entre ambas têm sido identificados (Caraci, 2010).
Idosos com TDM, tanto de início precoce quanto tardio, apresentam
comprometimentos cognitivos significativos que podem não ser completamente
recuperados após tratamento com antidepressivos. Diniz et al. (2013) ao
realizarem uma meta-análise de estudos de coorte e caso-controle
encontraram achados que sustentam a hipótese de que idosos com TDM de
início tardio apresentam um maior risco para desenvolvimento de demência,
em particular para DA e Demência Vascular, quando comparados a idosos
saudáveis. Estes resultados foram semelhantes ao estudo cujo achado
evidenciou uma associação entre TDM de início tardio e DA (Ownby et al.,
2006).
O TDM de início tardio apresenta algumas características relevantes
para o desenvolvimento de DA, sendo elas, déficits de memória, função
executiva e diminuição da velocidade de processamento (Gallassi et al., 2006;
Sheline et al., 2006). Além disso, este TDM é frequentemente associado a

9
sintomas que refletem hipofrontalidade e apresenta uma associação com
fatores de risco cardiovasculares, marcadores inflamatórios e da
Apolipoproteína E alelo ԑ4 (ApoE4) (Gallarda, 1999). Tais características
também podem ser observadas nos primeiros sinais de demência.
Wuwongse et al. (2010) apresentaram em seus estudos semelhanças e
diferenças entre fatores fisiopatológicos tanto do TD quanto da DA, destacando
fatores inflamatórios, fatores neurotróficos e marcadores apoptóticos e
concluíram que estas duas condições patológicas parecem compartilhar fatores
comuns como processos neuroinflamatórios com aumento de citocinas pró-
inflamatórias e diminuição das anti-inflamatórias, diminuição de fatores
neurotróficos e degeneração sináptica. Já com relação ao CCL, estudos
constataram que a persistência de sintomas depressivos nessa condição
clínica ao longo de dois ou três anos pode estar intimamente relacionada à
conversão para DA, representando um risco duas vezes maior do
desenvolvimento de DA quando comparado a casos de CCL sem depressão
(Modrego e Ferrández, 2004; Houde et al., 2008).
Visto que o TDM evidencia-se como fator de risco para o
desenvolvimento de DA, assim como o CCL, e que apresenta alguns processos
fisiopatológicos comuns à DA, faz-se importante a busca por possíveis
marcadores biológicos, envolvidos na neurobiologia da DA, em particular na
formação de placas amiloides, que possam corroborar os achados na literatura.
1.5 Possíveis biomarcadores para diagnóstico precoce da Doença de
Alzheimer
A detecção de indicadores do processo patogênico da DA antes do
desenvolvimento de danos significativos aos tecidos neurais e,
consequentemente, antes do aparecimento das manifestações clínicas da
doença, por meio de biomarcadores com alta sensibilidade e especificidade,
podem contribuir para o diagnóstico precoce, com implicações sobre a
abordagem terapêutica e até mesmo a eventual prevenção da demência
(Ravid, 2009; Hampel et al., 2008). O potencial de aplicação dos
biomarcadores tem gerado muita expectativa na área biomédica, sendo que

10
muitos têm sido propostos para avaliar os riscos de desenvolvimento da DA,
bem como identificá-la precocemente (Di Luca et.al., 2005).
Por definição, biomarcadores são características objetivas e
quantificáveis dos processos biológicos (Strimbu e Tavel, 2010), e para serem
considerados ideais devem ser capazes de detectar uma característica
fundamental da doença, ser validado em casos de DA comprovada por
autopsia, ser reprodutível, não requerer procedimentos invasivos e de elevado
custo financeiro e possuir especificidade superior a 75% e sensibilidade
superior a 85% (Borroni et al., 2006).
Muitos alvos que possuem marcadores bioquímicos ante-mortem, têm
sido investigados para possível diagnóstico precoce da doença. Além disso,
vêm sendo investigadas proteínas que reflitam in vivo os processos que levam
a formação de achados histopatológicos e que possam ter seus níveis
mensurados em fluidos biológicos (Ravid, 2009; Hampel et al., 2008).
Enzimas e produtos de clivagem proteolítica da APP, relacionadas à
cascata do amiloide, têm sido amplamente estudadas por serem associadas
diretamente às atividades das secretases α-, β- e γ, respectivamente
identificadas como A Desintegrin And Metallopeptidase 10 (ADAM10), β-site
APP-cleaving enzyme 1 (BACE1), e presinilina 1 (PSEN1). As mesmas serão
foco desse estudo e serão descritas adiante, juntamente com o peptídeo Aβ.
1.5.1 A Disintegrin And Metallopeptidase 10
Alguns membros das A Disintegrin And Metallopeptidase (ADAMs)
podem atuar como α-secretases (Cole e Vassar, 2007a; Cole e Vassar, 2007b;
Cole e Vassar, 2008), sendo eles ADAM9, 10 e 17 (Asai et al., 2003; Deuss et
al., 2008). ADAM corresponde a uma família de proteínas multi-modulares
(White et al., 2003; Moss e Lambert, 2002), sendo muitas delas identificadas
como proteínas transmembrana do tipo I que se ancoram devido à presença do
domínio transmembrana próximo à região C-terminal.
A região N-terminal das ADAMs possui um peptídeo sinal que a
direciona à via secretória e um pró-domínio que possui função na maturação,
pois sua presença mantém o domínio metalopeptidase inativo bloqueando a

11
atividade proteolítica da proteína. Após a remoção do pró-domínio o domínio
metalopeptidase torna-se ativo e é capaz de realizar suas funções catalíticas
(Seals e Courtneidge, 2003).
A ADAM10 pertence à superfamília das ADAMs e é expressa em
oligodendrócitos, em subconjunto de neurônios em desenvolvimento, em
núcleos cerebrais e na diferenciação da massa cinzenta (Lin et al., 2008).
Também é provável que seja responsável pela clivagem de várias proteínas da
superfície das células do cérebro, tais como efrinas (Hattori et al., 2000; Janes
et al., 2005), APP (Hooper e Turner, 2002), moléculas de direcionamento
axonal como, por exemplo, a molécula de adesão neuronal (Hinkle et al.,
2006), entre outras.
Essa enzima tem sido associada diretamente com a atividade de α-
secretase (Vingtdeux e Marambaud, 2012). Em estudos com camundongos
transgênicos para a DA, observaram que a superexpressão desta proteína,
considerada a principal α-secretase do cérebro (Fahrenholz et al., 2000;
Postina, 2008; Postina, 2012), resultou na diminuição da deposição de
peptídeos Aβ e dos déficits cognitivos (Postina et al., 2004).
1.5.2 β-site APP-cleaving enzyme 1
A BACE1 apresenta todas as características previstas para uma β-
secretase e está diretamente relacionada à DA (Vassar et al., 1999; Vassar et
al., 2011). Essa proteína caracteriza-se por ser uma proteína transmembrana
tipo 1, que possui 501 aminoácidos em sua cadeia e tem atividade de protease
do tipo aspartil. É largamente expressa no cérebro e está menos presente em
outros órgãos, localizando-se predominantemente em compartimentos
intracelulares ácidos, com seu centro ativo no lúmen da vesícula (Vassar et al.,
1999; Marcinkiewicz, Seidah, 2000; Yang; Vassar, 2014).
Ela apresenta outros substratos além da APP, sugerindo uma variedade
de funções fisiológicas, sendo a identificação da BACE1 importante para a
concepção de inibidores potentes e seletivos (Vassar e Kandalepas, 2011),
visto que essa proteína tem se mostrado como a principal responsável pela
formação de Aβ no cérebro por meio da via amiloidogênica (Gonçalves, 2012).

12
1.5.3 Presenilina 1
A PSEN1 é um dos quatro principais componentes do complexo de
multiproteínas -secretase: presenilin (PSEN), niscastrin (NCT), anterior
pharynx-defective (APH-1) e presenilin enhancer 2 (PEN-2), sendo responsável
pela geração de fragmentos resultantes da clivagem da APP por α e β-
secretase (Capell et al., 2005).
A PSEN1 é sintetizada como um polipeptídio de 50 kDa, mas pode
sofrer clivagem endoproteolítica gerando derivados estáveis C e N-terminais de
20 e 29 kDa, respectivamente (Doan et al., 1996), e localiza-se na membrana
plasmática, onde se liga diretamente a complexos de caderina e catenina (Baki
et al., 2001). Determinadas mutações nos genes que codificam a PSEN1,
assim como nos que codificam a PSEN2, levam a um aumento da
produção/libertação e secreção de Aβ (Martins, 2010).
Estudos recentes realizados por García-Ayllón et al. (2013), ao
analisarem o líquor de pacientes com DA leve a moderada e compará-los com
controles saudáveis, observaram a presença de complexos de PSEN1 em
ambos os grupos, sendo que o grupo com DA apresentou maiores níveis totais
de PSEN1 quando comparados aos controles, sugerindo assim que a mesma
possa ser usada como possível biomarcador para a DA.
1.5.4 Peptídeo Aβ
O Aβ é um peptídeo produzido por seres humanos e outros mamíferos,
estando presente em fluidos biológicos e parênquima cerebral (Seubert et al.,
1992; Tabaton et al., 1994; Ghiso et al., 1997). Na senescência, ele permanece
solúvel no cérebro, sendo progressivamente degradado e/ou removido. Já em
pacientes com DA, o mesmo se agrega em meio extracelular constituindo as
placas senis (Stromer e Serpell, 2005).
A produção do peptídeo Aβ é proveniente do processamento proteolítico
da APP e é liberado por clivagens sequenciais de β e -secretases. Como o
ponto de clivagem do domínio transmembrana da APP é variável, os peptídeos

13
Aβ liberados no cérebro podem apresentar diferentes tamanhos, variando de
38 a 43 aminoácidos (Schoonenboom et al., 2005). Dessas isoformas, somente
os peptídeos Aβ1-40, Aβ1-42 e Aβ1-43 são encontrados nas placas neuríticas
senis, sendo o Aβ1-42 a mais prevalente nas lesões (Jarrett et al., 1993).
1.6 Justificativa
Segundo Richard et al. (2013), o TD pode representar um fator de risco
para demência ou sintoma de fator de risco relacionado, bem como ser um
sintoma de demência precoce ou até mesmo uma reação à incapacidade
cognitiva e funcional. Seus estudos sobre possíveis relações entre TDM de
início tardio, demência e CCL sugerem que a depressão esteja relacionada a
um maior risco de desenvolvimento de DA e de progressão de CCL para DA.
Pelo fato da DA e dos transtornos depressivos serem as principais
causas de problemas de saúde mental em idosos e visto que a depressão
geriátrica está associada a maiores níveis de comprometimento cognitivo e
aumento do risco de desenvolvimento de DA, assim como o CCL, eles têm sido
alvo de muitos estudos, principalmente àqueles relacionados a possíveis
biomarcadores.
Estudar e compreender, principalmente in vivo, as alterações fisiológicas
no cérebro, tem incentivado a busca por marcadores biológicos periféricos que
espelhem como tais processos metabólicos ocorrem neste órgão (Busnello et
al., 2008). Neste sentido, a determinação destes possíveis biomarcadores no
sangue, por meio de leucócitos, ou no líquor pode representar um avanço no
diagnóstico e em intervenções terapêuticas, além de permitir uma detecção
precoce de possíveis evoluções clínicas para DA (Busnello et al., 2006).

14
2 OBJETIVOS
2.1 Objetivo geral
Comparar um conjunto de marcadores biológicos envolvidos na cascata
do amiloide expressos em leucócitos e líquor nas condições clínicas TDM,
CCL, DA e controles saudáveis.
2.2 Objetivos específicos
Determinar os níveis de expressão proteica das secretases ADAM10,
BACE1, PSEN1 e do peptídeo Aβ em leucócitos de pacientes idosos
com TDM, CCL e DA e compará-la com controles saudáveis;
Determinar os níveis de expressão proteica das secretases ADAM10,
BACE1, PSEN1 e do peptídeo Aβ em líquor de pacientes idosos com
TDM, CCL e DA e compará-la com controles saudáveis;
Correlacionar a expressão das proteínas acima e do peptídeo Aβ
quantificadas nas duas matrizes biológicas.

15
3 MATERIAL E MÉTODOS
3.1 Casuística
Trata-se de um estudo transversal, aprovado pelo Comitê de Ética em
Pesquisa – Nº 982.945/2015 (Anexo 1), em que foram recrutados 80
indivíduos, com 60 anos ou mais, dentre aqueles admitidos no Ambulatório de
Psiquiatria Geriátrica do Laboratório de Neurociênicas (LIM-27), localizado no
Instituto de Psiquiatria do Hospital das Clínicas da Faculdade de Medicina da
Universidade de São Paulo (IPq – HCFMUSP), coordenado pelo Professor
Doutor Orestes Vicente Forlenza. Cada grupo foi constituído por 20 indivíduos,
por meio dos critérios de inclusão:
Grupo 1. Idosos diagnosticados com TDM de início tardio (episódio único ou
recorrente), com definição do episódio realizada por meio da versão brasileira
da Entrevista Clínica Estruturada para o Diagnóstico de Transtornos Mentais do
Eixo I (SCID – I/P) do Manual Diagnóstico e Estatístico de Transtornos Mentais
– Quarta Edição (DSM-IV) (First et al., 2002), sendo a gravidade dos sintomas
depressivos avaliada por meio da Escala de Depressão de Hamilton – 21 itens
(HAM-D 21) (Hamilton, 1960).
Grupo 2. Idosos diagnosticados com CCL conforme a presença de queixas
consistentes de memória corroboradas por um informante; comprometimento
objetivo da memória ao executar testes cognitivos, porém não sendo grave o
suficiente para atender a critérios de demência; função intelectual global
preservada; execução de atividades de vida diária preservada ou com
dificuldades mínimas; sem diagnóstico de demência (Petersen et al., 2014;
Winblad et al., 2004).
Grupo 3. Idosos diagnosticados com DA em estágio leve a moderado,
conforme critérios estabelecidos pelo Instituto Nacional de Doenças
Neurológicas e Comunicativas e Associação da Doença de Alzheimer e
Doenças Relacionadas (NINCDS-ADRDA) (McKhann et al., 1984).

16
Grupo Controle. Idosos saudáveis participantes como voluntários das rotinas
de avaliação da clínica de memória oferecida pelo Ambulatório de Psiquiatria
Geriátrica do LIM-27 (IPq – HCFMUSP), com função cognitiva
comprovadamente normal segundo avaliações neuropsicológicas, e que se
mantiveram estáveis ao longo do seguimento; e ausência de história atual ou
pregressa de depressão ou em outros transtornos psiquiátricos, confirmados
pelo SCID I/P e escore no HAM-D 21 entre 0 e 7.
Vale salientar que todos os pacientes e controles realizaram uma
avaliação neuropsiquiátrica completa e avaliação neuropsicológica antes do
estabelecimento do diagnóstico cognitivo e que o teste cognitivo de Cambridge
(CAMCOG) (Roth et al., 1986; Nunes et al., 2008) e o Mini-Exame do Estado
Mental (MEEM) (Folstein et al., 1975) foram utilizados como medida do
desempenho cognitivo global.
Foram excluídos do estudo todos aqueles que (1) apresentaram
deficiências sensoriais e/ou intelectuais que os impedisse de desempenhar
adequadamente os testes necessários para avaliação das funções cognitivas;
(2) comorbidades neuropsiquiátricas; (3) doenças clínicas ou neurológicas com
impacto sobre a capacidade cognitiva; (4) diagnóstico de outras doenças
neurodegenerativas; (5) doenças clínicas sem controle clínico adequado ou
indivíduos com saúde frágil no momento da avaliação inicial; (6) outros
transtornos neuropsiquiátricos maiores, tais como esquizofrenia e transtorno
bipolar; (7) história ou evidência de alcoolismo ou abuso de substâncias
depressoras do sistema nervoso central; (8) estar em uso atual de medicações
antidepressivas, anticonvulsivantes, antipsicóticas, carbonato de lítio ou
qualquer medicação que sabidamente esteja associada aos mecanismos a
serem investigados neste estudo; (9) se recusaram a assinar o Termo de
Consentimento Livre e Esclarecido (TCLE) (Apêndice A).
Após a alocação dos participantes nos respectivos grupos, mediante
reuniões de consenso multidisciplinar para estabelecimento dos diagnósticos,
estes foram submetidos à coleta de sangue para preparo dos leucócitos e uma
parte deles (n= 37) realizou ainda, a coleta de líquor (figura 4).

17
Figura 4 – Casuística conforme grupo e matriz biológica.
Não foram observadas diferenças estatisticamente significantes na
distribuição por gênero entre os grupos ao realizar a análise estatística do perfil
demográfico dos indivíduos recrutados (tabela 2), diferentemente das variáveis
idade, escolaridade e escores nos testes cognitivos (MEEM e CAMCOG), onde
observamos diferenças estatisticamente significantes, indicando que os
indivíduos do grupo DA são discretamente mais velhos, têm escolaridade mais
baixa e, como esperado, têm pior desempenho cognitivo global.
18
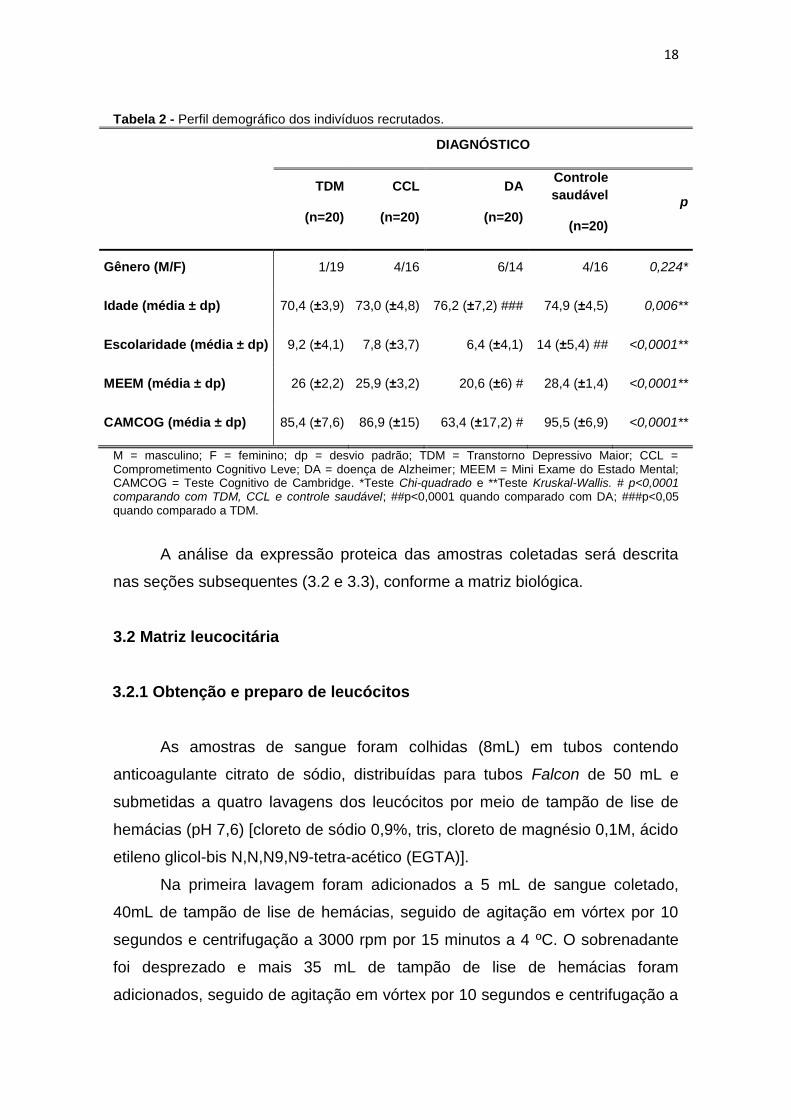
Tabela 2 - Perfil demográfico dos indivíduos recrutados.
DIAGNÓSTICO
TDM
(n=20)
CCL
(n=20)
DA
(n=20)
Controle
saudável
(n=20)
p
Gênero (M/F) 1/19 4/16 6/14 4/16 0,224*
Idade (média ± dp) 70,4 (±3,9) 73,0 (±4,8) 76,2 (±7,2) ### 74,9 (±4,5) 0,006**
Escolaridade (média ± dp) 9,2 (±4,1) 7,8 (±3,7) 6,4 (±4,1) 14 (±5,4) ## <0,0001**
MEEM (média ± dp) 26 (±2,2) 25,9 (±3,2) 20,6 (±6) # 28,4 (±1,4) <0,0001**
CAMCOG (média ± dp) 85,4 (±7,6) 86,9 (±15) 63,4 (±17,2) # 95,5 (±6,9) <0,0001**
M = masculino; F = feminino; dp = desvio padrão; TDM = Transtorno Depressivo Maior; CCL = Comprometimento Cognitivo Leve; DA = doença de Alzheimer; MEEM = Mini Exame do Estado Mental; CAMCOG = Teste Cognitivo de Cambridge. *Teste Chi-quadrado e **Teste Kruskal-Wallis. # p<0,0001 comparando com TDM, CCL e controle saudável; ##p<0,0001 quando comparado com DA; ###p<0,05 quando comparado a TDM.
A análise da expressão proteica das amostras coletadas será descrita
nas seções subsequentes (3.2 e 3.3), conforme a matriz biológica.
3.2 Matriz leucocitária
3.2.1 Obtenção e preparo de leucócitos
As amostras de sangue foram colhidas (8mL) em tubos contendo
anticoagulante citrato de sódio, distribuídas para tubos Falcon de 50 mL e
submetidas a quatro lavagens dos leucócitos por meio de tampão de lise de
hemácias (pH 7,6) [cloreto de sódio 0,9%, tris, cloreto de magnésio 0,1M, ácido
etileno glicol-bis N,N,N9,N9-tetra-acético (EGTA)].
Na primeira lavagem foram adicionados a 5 mL de sangue coletado,
40mL de tampão de lise de hemácias, seguido de agitação em vórtex por 10
segundos e centrifugação a 3000 rpm por 15 minutos a 4 ºC. O sobrenadante
foi desprezado e mais 35 mL de tampão de lise de hemácias foram
adicionados, seguido de agitação em vórtex por 10 segundos e centrifugação a

19
1500 rpm por 10 minutos a 4ºC, efetivando dessa maneira a segunda lavagem.
Novamente o sobrenadante foi desprezado e o segundo procedimento de
lavagem foi repetido. Na quarta lavagem, o sobrenadante foi desprezado e 1
mL de tampão de lise de hemácias foi adicionado ao Falcon, juntamente com
10 µL de fluoreto de fenilmetilsulfonil (PMSF), seguido de agitação em vórtex
para homogeneizar. Cada tubo de Falcon originou 2 alíquotas de, no mínimo,
0,5 mL cada, sendo estas armazenadas em freezer (-80ºC) para posterior
análise.
3.2.1.1 Extração de proteínas totais de leucócitos
Para realizar a extração de proteínas de leucócitos, as amostras
congeladas em freezer (-80ºC) foram descongeladas em gelo (uma alíquota) e
imediatamente submetidas à centrifugação a 1500 rpm por 15 minutos a 4 ºC.
O sobrenadante foi desprezado e o pellet foi ressuspendido com Radio-
Immunopreciptation Assay (RIPA) buffer, sendo este um tampão de lise [tris
25mM, ácido clorídrico (pH 7,6), cloreto de sódio 150mM, Nonidet P-40 1%,
desoxicolato de sódio 1%, dodecil sulfato de sódio 0,1%]. Após ressuspensão,
a amostra foi centrifugada novamente a 1500 rpm por 15 minutos a 4 ºC e o
sobrenadante (aproximadamente 0,5 mL) foi armazenado.
Em seguida, uma alíquota de 10μL de amostra/paciente foi destinada à
quantificação de proteínas pelo método de Lowry, ensaio colorimétrico para
determinação de proteínas totais (Lowry et al., 1951), por meio do kit Bio-Rad
DC Protein assay (Bio-Rad Hercules). A determinação da concentração de
proteínas totais foi obtida por meio de uma curva padrão de proteínas,
empregando-se albumina de soro bovino (BSA; Sigma-Adrich) nas seguintes
concentrações: 0,1 mg/mL, 0,2 mg/mL, 0,5 mg/mL, 1,0 mg/mL e 1,5 mg/mL.
3.2.2 Determinação de BACE1, ADAM10 e PSEN1
Para a separação das proteínas por peso molecular foi utilizado o
método semi-quantitativo Western Blotting (Anexo 1). As amostras foram
diluídas em tampão de amostra [tris 0,8M, dodecil sulfato de sódio 8%, glicerol

20
10%, β-mercaptoetanol 12%, azul de bromofenol 0,02%] (Laemmli, 1970), e
aquecidas a 100ºC por 5 minutos em banho seco (DRI-BLOCK DB-3, Techne),
desnaturando as proteínas, desenovelando-as completamente, a fim de facilitar
a separação durante a corrida eletroforética. No gel de poliacrilamida foram
aplicados 25µg de proteínas totais/amostra, além do marcador padrão de
massa molecular Precision Plus Protein™ Dual Xtra Standards (Bio-Rad).
A corrida eletroforética se desenvolveu em meio de tampão de corrida
[tris 25mM, glicina 160mM, dodecil sulfato de sódio 0,1%, água destilada], por
120 minutos a 120V. Logo em seguida, o gel foi submetido à transferência das
proteínas para membrana de nitrocelulose Amersham Hybond ECL
Nitrocellulose Membrane (GE Healthcare). A transferência se deu no sistema
Mini-Protean® II Cell (Bio-Rad) em meio a tampão de transferência [tris 25mM,
glicina 192mM, metanol 20%, água destilada], à amperagem de 200
miliampéres (mA), por 90 minutos.
A transferência das proteínas para membrana de nitrocelulose
possibilitou posterior ensaio imunoenzimático (immunoblotting), por meio da
reação com anticorpos específicos. Para a realização do immunoblotting, as
membranas foram bloqueadas com leite desnatado 5% em Tris-buffered saline
– Tween (TBS-Tween) 0,2% [tris 250mM, cloreto de sódio 1,4M, cloreto de
potássio 50mM, Tween 20, água destilada], a fim de evitar interações
inespecíficas do anticorpo primário. Após o bloqueio, a membrana foi incubada
com anticorpo primário e em seguida com o anticorpo secundário. Entre o
bloqueio e as incubações com os anticorpos, as membranas foram lavadas
quatro vezes, por 5 minutos cada, com TBS-Tween 0,2%.
Vale salientar que, para corrigir possíveis variações analíticas inerentes
à técnica, foi utilizada a β-actina (Abcam) como controle endógeno. O protocolo
para análise da mesma seguiu a mesma lógica daquele utilizado para as
proteínas (Apêndice B). A média das densitometrias das amostras foi dividida
pela média da densitometria da β-actina correspondente à membrana em
análise.
Tanto o bloqueio com leite quanto as incubações com anticorpos foram
realizados sobre o agitador Platform Vari Mix (Barnstead/Thermolyne) (tabela
3). Já as lavagens foram realizadas à temperatura ambiente em agitadores

21
Roto Mix-50800 (Barnstead/Thermolyne).
Em seguida à reação com anticorpo secundário e lavagem, foi
adicionado sobre a membrana, em local escuro, o reagente Amersham™
ECL™ Select Western Blotting Detection Reagent (GE Healthcare - Life
Sciences), que ao reagir com a peroxidase conjugada ao anticorpo secundário
por, no mínimo 5 minutos, emite luz, sendo esta captada por equipamento
específico. Para adquirir a imagem da membrana, foi utilizado o
Fotodocumentador Alliance 4.7 (UVITEC Cambridge), em parceria com o LIM-
16, denominado Laboratório de Fisiopatologia Renal, da Faculdade de
Medicina da Universidade de São Paulo.
Após a aquisição da imagem, foi realizada a densitometria das bandas
(Apêndice C), bem como a análise estatística por meio de softwares
adequados.
Tabela 3 – Diluição e tempo de incubação dos anticorpos das secretases e do controle endógeno.
PROTEÍNA BLOQUEIO INCUBAÇÃO
Anticorpo Primário Anticorpo Secundário
BACE1 1 hora 1:5000 / Overnight
Anti-BACE1 antibody (Abcam)
1:20000 / 1 hora Anti-Rabbit IgG –
conjugado à peroxidase (GE Healthcare Life
Sciences)
PSEN1 Overnight 1:10000 / 1 hora
Anti-PSEN1 antibody (Abcam)
1:10000 / 1 hora Anti-Rabbit IgG –
conjugado à peroxidase (GE Healthcare Life
Sciences)
ADAM10 Overnight 1:15000 / 1 hora
Anti-ADAM10 antibody (Abcam)
1:30000 / 1 hora Anti-Rabbit IgG –
conjugado à peroxidase (GE Healthcare Life
Sciences)
β-ACTINA Overnight 1:6000 / 1 hora
Anti-Beta Actin Antibody (Abcam)
1:2000 / 1 hora Anti-Mouse IGG (whole molecule) – Peroxidase
Antibody (SIGMA)
3.2.3 Determinação do peptídeo Aβ
As amostras foram descongeladas em gelo, seguido da dosagem de

22
proteínas totais por Lowry (Lowry et al., 1951). Posteriormente, deu-se
prosseguimento ao desenvolvimento do método. Para a análise do peptideo Aβ
por meio do kit comercial Human Aβ42 Enzyme Linked Immuno Sorbent Assay
(ELISA) kit (Invitrogem, Thermo Fisher Scientific). Foram analisadas 39
amostras em duplicata.
ELISA é um ensaio imunoenzimático que se baseia na interação
antígeno-anticorpo, permitindo a mensuração dos mesmos (Arosa; Cardoso,
2012). Existem algumas variantes do método, sendo o ELISA sanduíche o
utilizado em nosso estudo (figura 5). A leitura foi realizada em
espectrofotômetro (HIDEX®) a um comprimento de onda de 450nm.
Figura 5 – Método de ELISA sanduíche.
Nesse método, (1) aplica-se na placa de poliestireno sensibilizada com anticorpo primário a amostra com o antígeno a ser analisado. (2) Após a inserção da amostra, adiciona-se um segundo anticorpo específico para o antígeno capaz de reconhecê-lo em um epítopo diferente daquele reconhecido pelo primeiro anticorpo. (3) Em seguida adiciona-se o anticorpo secundário conjugado e, por fim, (4) insere-se o cromógeno, que, ao ser clivado emite luz, sendo sua intensidade diretamente proporcional à quantidade de antígeno identificado. Vale salientar que são realizadas lavagens entre as etapas supracitadas.
3.3 Matriz liquórica
3.3.1 Obtenção e preparo de líquor
As amostras de líquor foram obtidas por meio de punção lombar nos
espaços intervertebrais L3/L4 ou L4/L5, com uma agulha de calibre 23. As

23
coletas foram realizadas por médico neurologista, no período matutino, em
tubos de polipropileno com capacidade para 12-15mL. Em seguida, as mesmas
foram centrifugadas a 3200 rpm por 10 minutos a 4ºC, separadas em alíquotas
(0,5mL cada) e armazenadas em freezer (-80ºC) para posterior análise.
3.3.2 Determinação de BACE1, ADAM10 e PSEN1
Para análise das proteínas BACE1, ADAM10 e PSEN1 no líquor foram
utilizados os kits comerciais de ELISA sanduíche Human Beta-secretase 1
ELISA kit (Thermo Fisher Scientific), Human A Desintegrin And Metalloprotease
10 (ADAM10) ELISA kit (Abbexa), Human Presenilin 1 ELISA kit (RayBiotech),
respectivamente. Vale ressaltar que os ensaios foram realizados em duplicata.
Todas as leituras foram realizadas no equipamento HIDEX, a um
comprimento de onda de 450nm. Para o kit Human Beta-secretase 1 ELISA kit
(Thermo Fisher Scientific) foram feitas duas leituras, uma a 450nm e outra a
550nm, sendo calculado o resultado por meio da subtração dos valores obtidos
nos comprimentos de onda (450nm – 550nm) conforme preconiza o kit. As
análises gráficas puderam ser geradas por meio de regressão linear para o kit
de ADAM10 e da curva de quatro parâmetros logísticos para os kits de BACE1
e PSEN1.
3.3.3 Determinação do peptídeo Aβ42
As concentrações do peptídeo Aβ42 foram determinadas pelo
imunoensaio fluorimétrico INNO-BIA AlzBio3 (Innogenetics) e mensuradas pela
plataforma Luminex® xMAP®.
A tecnologia Luminex xMAP® (MAP: Multiple Analyte Profiling) envolve
microesferas de látex com dois fluoróforos. Utilizando proporções precisas de
dois fluoróforos, podem ser criados 100 conjuntos diferentes de microesferas –
cada uma delas com uma assinatura baseada em “códigos de cores” e que
podem ser identificadas pelo instrumento Luminex (Talib et al., 2011).
Os kits Multiplex utilizam estas microesferas como base do imunoensaio
ligando anticorpos diferentes a esferas de cores diferentes. Os ensaios se

24
fundamentam na metodologia “sanduíche” convencional de dois sítios.
Anticorpos de captura específicos para cada analito estão imobilizados às
microesferas por ligações covalentes não reversíveis (figura 6) (Talib et al.,
2011).
Figura 6 – Metodologia Luminex.
A) Microesfera acoplada com anticorpo de captura específico. B) O anticorpo de captura se liga ao analito específico. C) o anticorpo de detecção biotinilado se liga ao analito específico. D) Estreptavidina-ficoeritrina (SA-PE) se liga ao receptor biotinilado e emite sinal fluorescente. Fonte: Extraído de Talib et al., 2011.
A próxima etapa é a adição do anticorpo de detecção biotinilado. Depois
que o analito (proteína de interesse) se liga aos anticorpos de captura
localizados na superfície das microesferas (figura 3B), a detecção final é feita
através de um terceiro marcador fluorescente, ficoeritrina (PE) ligada ao
anticorpo de detecção (figura 3C e D) (Talib et al., 2011).
O equipamento Luminex movimenta estas esferas em uma fila única
através de feixes de dois lasers diferentes em um citômetro de fluxo. O primeiro
feixe de laser classifica o código de cor para o ensaio e o segundo laser
quantifica o sinal de reporte em cada microesfera (Talib et al., 2011).
3.4 Análise estatística
A análise estatística foi realizada por meio do programa estatístico
Statistical Package for Social Science (SPSS) versão 14, para Windows e a
significância estatística para todas as análises foi estabelecida para valores de
p<0,05 (α = 95%).
Para verificar a normalidade dos dados obtidos foi utilizado o teste
Kolmogorov-Smirnov (KS). Esse teste fornece o nível descritivo que pode ser
interpretado de maneira a aceitar ou não a hipótese nula.

25
A partir do KS, foi possível identificar que o perfil dos dados do estudo
não apresenta distribuição normal. Tendo em vista esse resultado, foi
necessária a transformação dos dados em paramétricos para a realização da
Análise de Covariância (ANCOVA), a fim de avaliar o potencial de interação
entre diferentes fatores nos grupos de estudo. Dados referentes à idade e
escolaridade foram determinados como co-variáveis, permitindo que os
mesmos não interferissem na análise da relação entre as variáveis
independentes. A ANCOVA foi realizada com post-hoc de Bonferroni para
identificar quais pares de grupo diferem.
O teste Chi-quadrado, por sua vez, foi utilizado a fim de encontrar um
valor de dispersão para duas variáveis nominais, avaliando a associação
existente entre elas.
Por fim, para análise de correlação, utilizamos o Coeficiente de
Correlação de Pearson (r), para refletir a intensidade de uma relação linear
entre dois conjuntos de dados.
26
4 RESULTADOS
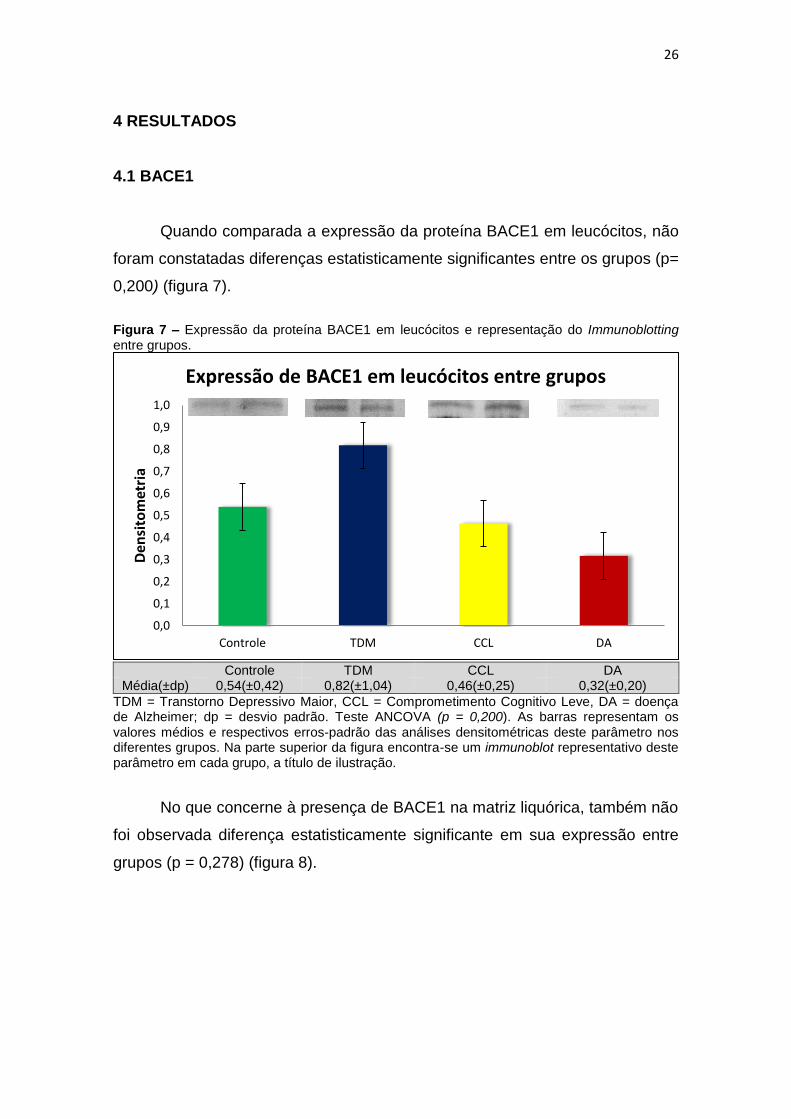
4.1 BACE1
Quando comparada a expressão da proteína BACE1 em leucócitos, não
foram constatadas diferenças estatisticamente significantes entre os grupos (p=
0,200) (figura 7).
Figura 7 – Expressão da proteína BACE1 em leucócitos e representação do Immunoblotting entre grupos.
Controle TDM CCL DA
Média(±dp) 0,54(±0,42) 0,82(±1,04) 0,46(±0,25) 0,32(±0,20)
TDM = Transtorno Depressivo Maior, CCL = Comprometimento Cognitivo Leve, DA = doença de Alzheimer; dp = desvio padrão. Teste ANCOVA (p = 0,200). As barras representam os valores médios e respectivos erros-padrão das análises densitométricas deste parâmetro nos diferentes grupos. Na parte superior da figura encontra-se um immunoblot representativo deste parâmetro em cada grupo, a título de ilustração.
No que concerne à presença de BACE1 na matriz liquórica, também não
foi observada diferença estatisticamente significante em sua expressão entre
grupos (p = 0,278) (figura 8).
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
Controle TDM CCL DA
Den
sito
met
ria
Expressão de BACE1 em leucócitos entre grupos
27
Figura 8 – Expressão da proteína BACE1 em líquor entre grupos.
Controle TDM CCL DA
Média(±dp) 8,51(±2,14) 12,73(±16,86) 5,43(±3,97) 6,29(±7,00)
TDM = Transtorno Depressivo Maior, CCL = Comprometimento Cognitivo Leve, DA = doença de Alzheimer. dp = desvio padrão. Teste ANCOVA (p = 0,278). As barras representam os valores médios e respectivos erros-padrão das análises densitométricas deste parâmetro nos diferentes grupos. Na parte superior da figura encontra-se um immunoblot representativo deste parâmetro em cada grupo, a título de ilustração.
4.2 ADAM10
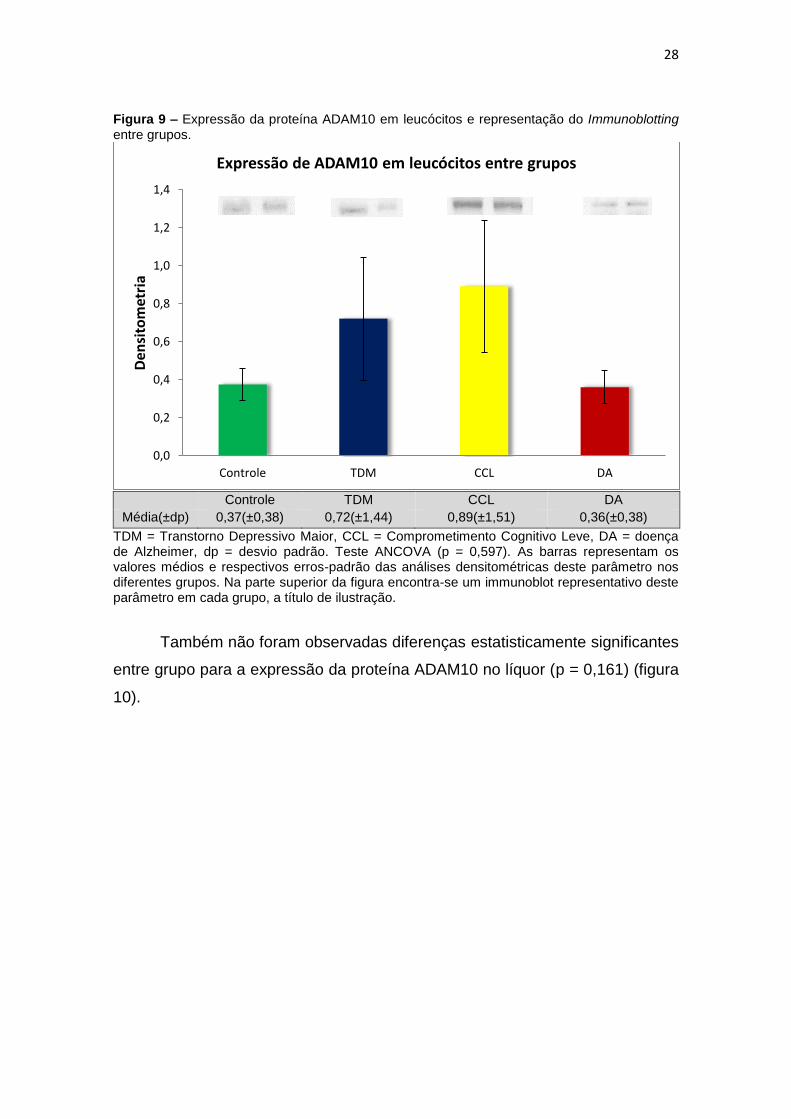
Para a ADAM10 não foi encontrada diferença estatisticamente
significante ao comparar, em leucócitos, a expressão proteica dessa secretase
entre grupos (p=0,597) (figura 9).
0,0
5,0
10,0
15,0
20,0
25,0
Controle TDM CCL DA
Co
nce
ntr
ação
(n
g/m
L)
Expressão de BACE1 em líquor entre grupos
28
Figura 9 – Expressão da proteína ADAM10 em leucócitos e representação do Immunoblotting entre grupos.
Controle TDM CCL DA
Média(±dp) 0,37(±0,38) 0,72(±1,44) 0,89(±1,51) 0,36(±0,38)
TDM = Transtorno Depressivo Maior, CCL = Comprometimento Cognitivo Leve, DA = doença de Alzheimer, dp = desvio padrão. Teste ANCOVA (p = 0,597). As barras representam os valores médios e respectivos erros-padrão das análises densitométricas deste parâmetro nos diferentes grupos. Na parte superior da figura encontra-se um immunoblot representativo deste parâmetro em cada grupo, a título de ilustração.
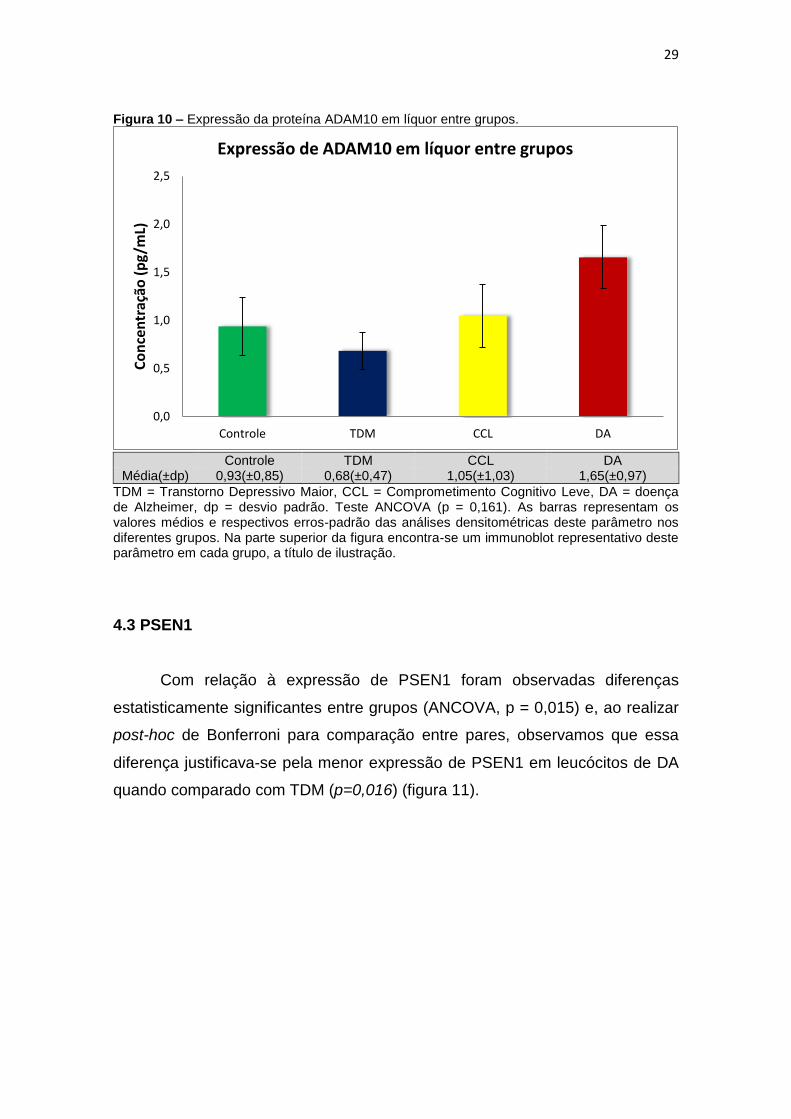
Também não foram observadas diferenças estatisticamente significantes
entre grupo para a expressão da proteína ADAM10 no líquor (p = 0,161) (figura
10).
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Controle TDM CCL DA
Den
sito
met
ria
Expressão de ADAM10 em leucócitos entre grupos
29
Figura 10 – Expressão da proteína ADAM10 em líquor entre grupos.
Controle TDM CCL DA
Média(±dp) 0,93(±0,85) 0,68(±0,47) 1,05(±1,03) 1,65(±0,97)
TDM = Transtorno Depressivo Maior, CCL = Comprometimento Cognitivo Leve, DA = doença de Alzheimer, dp = desvio padrão. Teste ANCOVA (p = 0,161). As barras representam os valores médios e respectivos erros-padrão das análises densitométricas deste parâmetro nos diferentes grupos. Na parte superior da figura encontra-se um immunoblot representativo deste parâmetro em cada grupo, a título de ilustração.
4.3 PSEN1
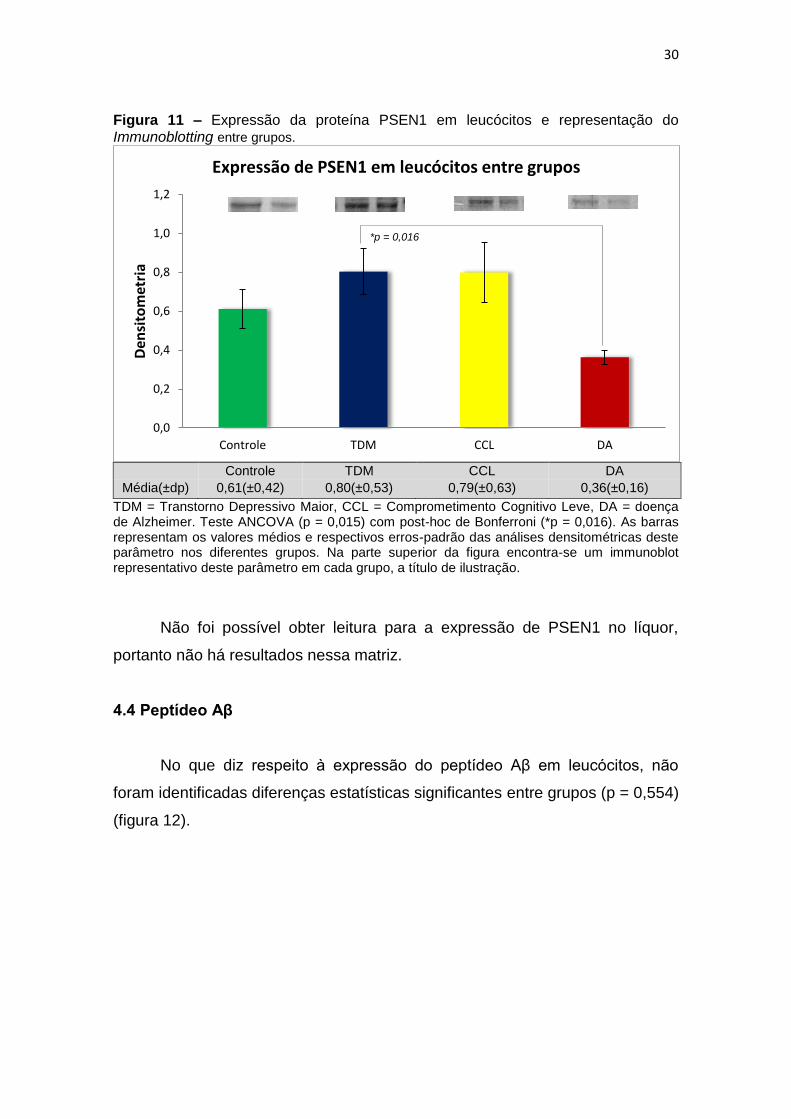
Com relação à expressão de PSEN1 foram observadas diferenças
estatisticamente significantes entre grupos (ANCOVA, p = 0,015) e, ao realizar
post-hoc de Bonferroni para comparação entre pares, observamos que essa
diferença justificava-se pela menor expressão de PSEN1 em leucócitos de DA
quando comparado com TDM (p=0,016) (figura 11).
0,0
0,5
1,0
1,5
2,0
2,5
Controle TDM CCL DA
Co
nce
ntr
ação
(p
g/m
L)
Expressão de ADAM10 em líquor entre grupos
30
Figura 11 – Expressão da proteína PSEN1 em leucócitos e representação do Immunoblotting entre grupos.
Controle TDM CCL DA
Média(±dp) 0,61(±0,42) 0,80(±0,53) 0,79(±0,63) 0,36(±0,16)
TDM = Transtorno Depressivo Maior, CCL = Comprometimento Cognitivo Leve, DA = doença de Alzheimer. Teste ANCOVA (p = 0,015) com post-hoc de Bonferroni (*p = 0,016). As barras representam os valores médios e respectivos erros-padrão das análises densitométricas deste parâmetro nos diferentes grupos. Na parte superior da figura encontra-se um immunoblot representativo deste parâmetro em cada grupo, a título de ilustração.
Não foi possível obter leitura para a expressão de PSEN1 no líquor,
portanto não há resultados nessa matriz.
4.4 Peptídeo Aβ
No que diz respeito à expressão do peptídeo Aβ em leucócitos, não
foram identificadas diferenças estatísticas significantes entre grupos (p = 0,554)
(figura 12).
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Controle TDM CCL DA
Den
sito
met
ria
Expressão de PSEN1 em leucócitos entre grupos
*p = 0,016
31
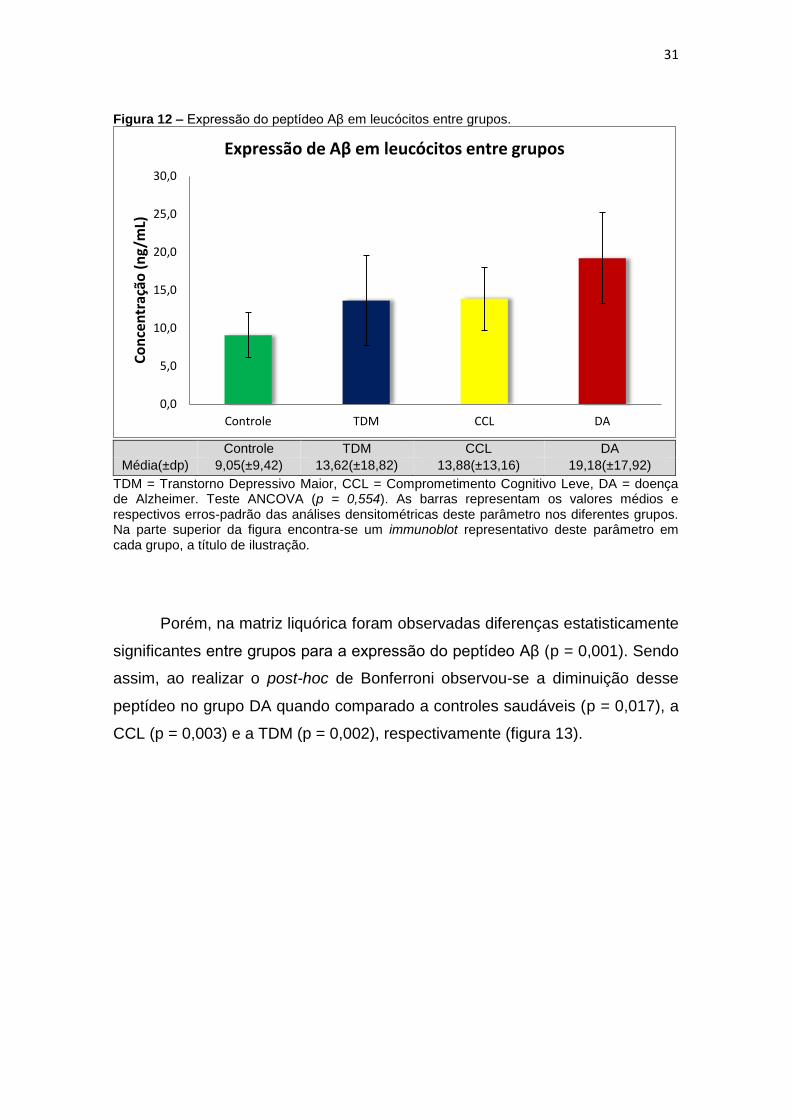
Figura 12 – Expressão do peptídeo Aβ em leucócitos entre grupos.
Controle TDM CCL DA
Média(±dp) 9,05(±9,42) 13,62(±18,82) 13,88(±13,16) 19,18(±17,92)
TDM = Transtorno Depressivo Maior, CCL = Comprometimento Cognitivo Leve, DA = doença de Alzheimer. Teste ANCOVA (p = 0,554). As barras representam os valores médios e respectivos erros-padrão das análises densitométricas deste parâmetro nos diferentes grupos. Na parte superior da figura encontra-se um immunoblot representativo deste parâmetro em cada grupo, a título de ilustração.
Porém, na matriz liquórica foram observadas diferenças estatisticamente
significantes entre grupos para a expressão do peptídeo Aβ (p = 0,001). Sendo
assim, ao realizar o post-hoc de Bonferroni observou-se a diminuição desse
peptídeo no grupo DA quando comparado a controles saudáveis (p = 0,017), a
CCL (p = 0,003) e a TDM (p = 0,002), respectivamente (figura 13).
0,0
5,0
10,0
15,0
20,0
25,0
30,0
Controle TDM CCL DA
Co
nce
ntr
ação
(n
g/m
L)
Expressão de Aβ em leucócitos entre grupos
32
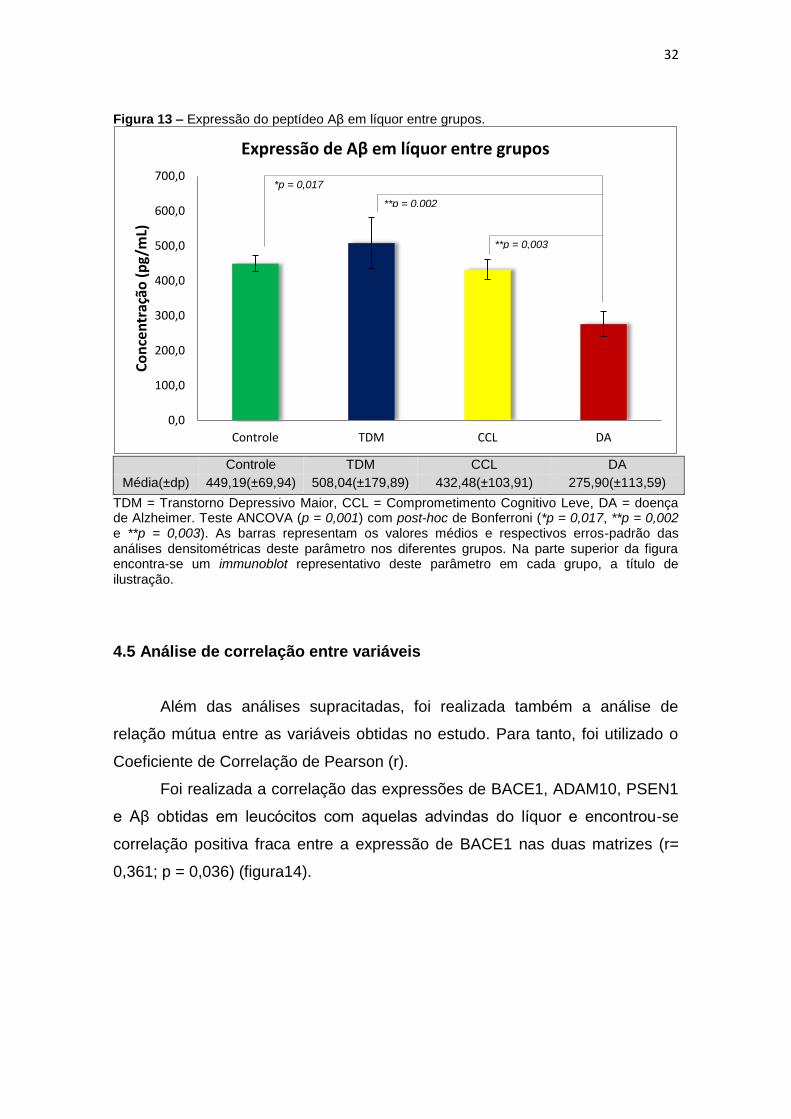
Figura 13 – Expressão do peptídeo Aβ em líquor entre grupos.
Controle TDM CCL DA
Média(±dp) 449,19(±69,94) 508,04(±179,89) 432,48(±103,91) 275,90(±113,59)
TDM = Transtorno Depressivo Maior, CCL = Comprometimento Cognitivo Leve, DA = doença de Alzheimer. Teste ANCOVA (p = 0,001) com post-hoc de Bonferroni (*p = 0,017, **p = 0,002 e **p = 0,003). As barras representam os valores médios e respectivos erros-padrão das análises densitométricas deste parâmetro nos diferentes grupos. Na parte superior da figura encontra-se um immunoblot representativo deste parâmetro em cada grupo, a título de ilustração.
4.5 Análise de correlação entre variáveis
Além das análises supracitadas, foi realizada também a análise de
relação mútua entre as variáveis obtidas no estudo. Para tanto, foi utilizado o
Coeficiente de Correlação de Pearson (r).
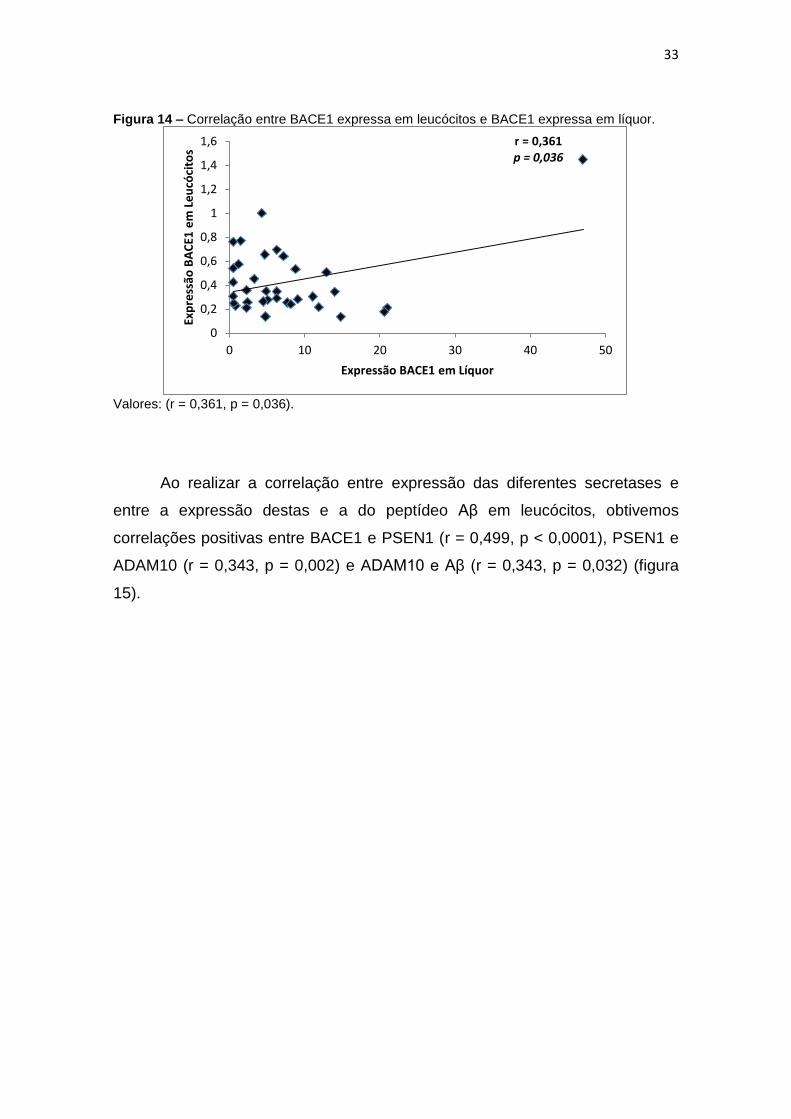
Foi realizada a correlação das expressões de BACE1, ADAM10, PSEN1
e Aβ obtidas em leucócitos com aquelas advindas do líquor e encontrou-se
correlação positiva fraca entre a expressão de BACE1 nas duas matrizes (r=
0,361; p = 0,036) (figura14).
0,0
100,0
200,0
300,0
400,0
500,0
600,0
700,0
Controle TDM CCL DA
Co
nce
ntr
ação
(p
g/m
L)
Expressão de Aβ em líquor entre grupos
*p = 0,017
**p = 0,002
**p = 0,003
33
Figura 14 – Correlação entre BACE1 expressa em leucócitos e BACE1 expressa em líquor.
Valores: (r = 0,361, p = 0,036).
Ao realizar a correlação entre expressão das diferentes secretases e
entre a expressão destas e a do peptídeo Aβ em leucócitos, obtivemos
correlações positivas entre BACE1 e PSEN1 (r = 0,499, p < 0,0001), PSEN1 e
ADAM10 (r = 0,343, p = 0,002) e ADAM10 e Aβ (r = 0,343, p = 0,032) (figura
15).
r = 0,361 p = 0,036
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
0 10 20 30 40 50
Exp
ress
ão B
AC
E1 e
m L
eu
cóci
tos
Expressão BACE1 em Líquor
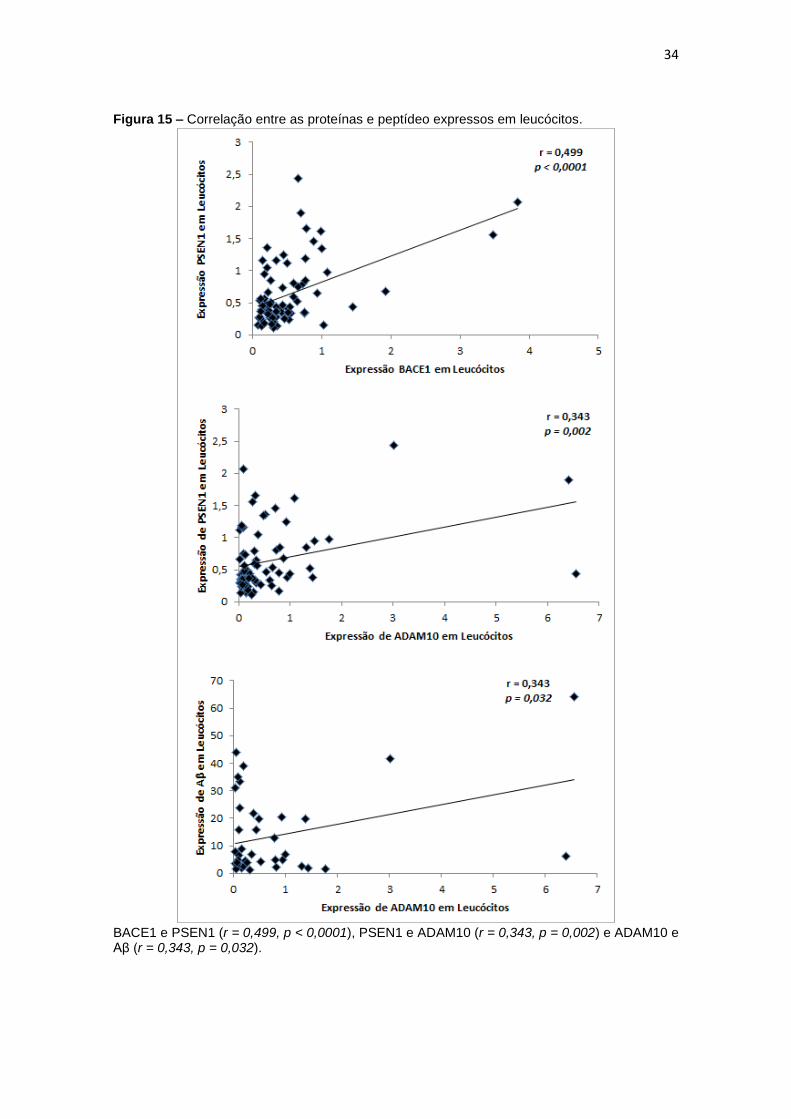
34
Figura 15 – Correlação entre as proteínas e peptídeo expressos em leucócitos.
BACE1 e PSEN1 (r = 0,499, p < 0,0001), PSEN1 e ADAM10 (r = 0,343, p = 0,002) e ADAM10 e Aβ (r = 0,343, p = 0,032).
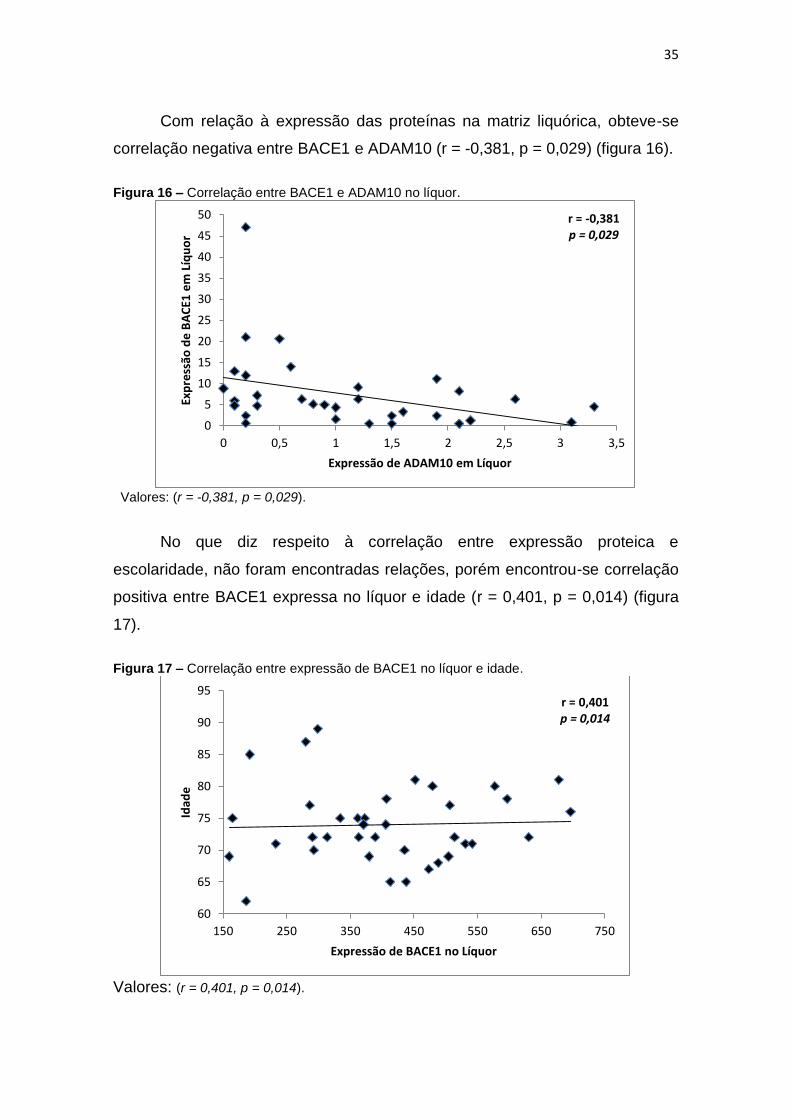
35
Com relação à expressão das proteínas na matriz liquórica, obteve-se
correlação negativa entre BACE1 e ADAM10 (r = -0,381, p = 0,029) (figura 16).
Figura 16 – Correlação entre BACE1 e ADAM10 no líquor.
Valores: (r = -0,381, p = 0,029).
No que diz respeito à correlação entre expressão proteica e
escolaridade, não foram encontradas relações, porém encontrou-se correlação
positiva entre BACE1 expressa no líquor e idade (r = 0,401, p = 0,014) (figura
17).
Figura 17 – Correlação entre expressão de BACE1 no líquor e idade.
Valores: (r = 0,401, p = 0,014).
r = -0,381 p = 0,029
0
5
10
15
20
25
30
35
40
45
50
0 0,5 1 1,5 2 2,5 3 3,5
Exp
ress
ão d
e B
AC
E1 e
m L
íqu
or
Expressão de ADAM10 em Líquor
r = 0,401 p = 0,014
60
65
70
75
80
85
90
95
150 250 350 450 550 650 750
Idad
e
Expressão de BACE1 no Líquor

36
Quanto à correlação entre expressão proteica e resultados obtidos nas
avaliações neuropsicológicas, tem-se correlação positiva entre Aβ no líquor e
as avaliações MEEM (r = 0,504, p = 0,002) e CAMCOG (r = 0,572, p < 0,0001),
e correlação negativa entre ADAM10 no líquor e CAMCOG (r = -0,408, p =
0,023) (figura 18).
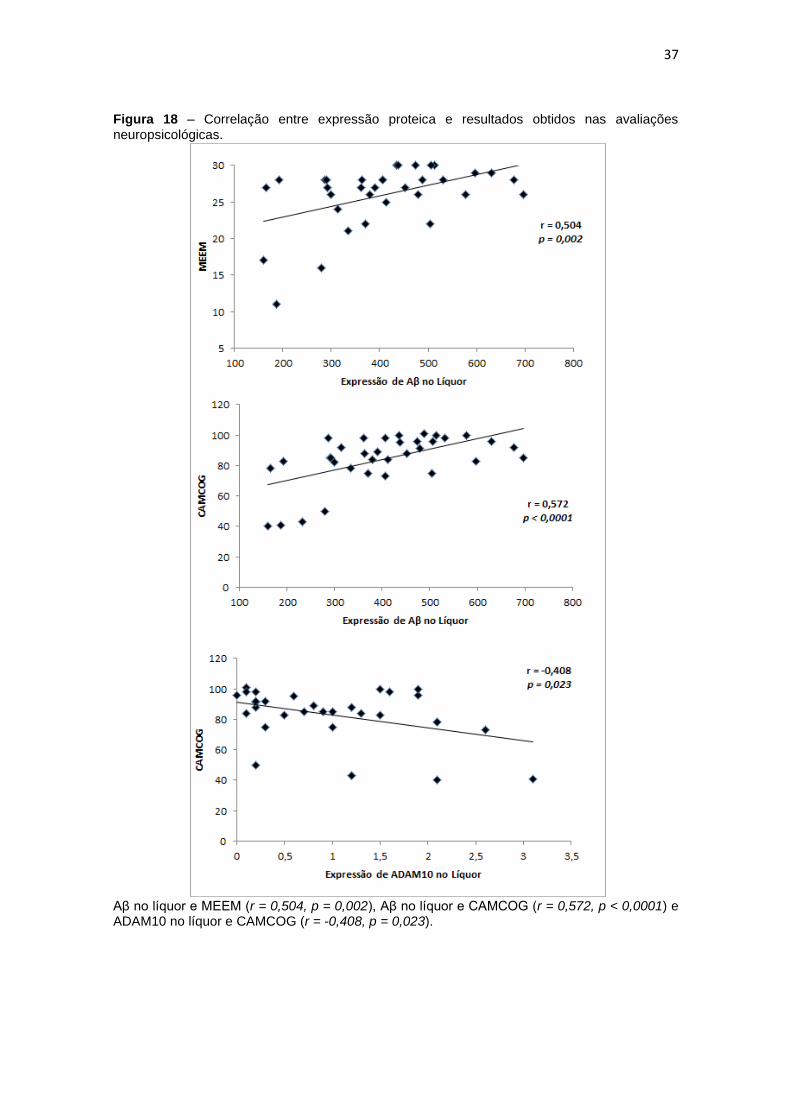
37
Figura 18 – Correlação entre expressão proteica e resultados obtidos nas avaliações neuropsicológicas.
Aβ no líquor e MEEM (r = 0,504, p = 0,002), Aβ no líquor e CAMCOG (r = 0,572, p < 0,0001) e ADAM10 no líquor e CAMCOG (r = -0,408, p = 0,023).

38
5 DISCUSSÃO
Nesse estudo comparamos a expressão de BACE1, ADAM10, PSEN1 e
Aβ tanto em leucócitos quanto em líquor de indivíduos com TDM, CCL, DA e
controles saudáveis e correlacionamos a expressão das secretases e do
peptídeo Aβ nas duas matrizes biológicas. Nossos achados demonstraram a
presença das três secretases e do Aβ em leucócitos assim como de BACE1,
ADAM10 e Aβ no líquor, corroborando os dados da literatura (Colciaghi et al.,
2002; Blennow e Hampel, 2003; Carmona et al., 2012; García-Ayllón et al.,
2013; Delvaux et al., 2013; Schuck et al., 2016).
Há poucos relatos sobre a presença de BACE1, PSEN1, ADAM10 e Aβ
em leucócitos bem como sua função biológica nesta matriz. Expressões dessas
secretases já foram descritas em leucócitos de sangue periférico, apesar de
terem sido observados padrões diferentes daqueles investigados no cérebro,
dependendo da metodologia aplicada (Delvaux et al., 2013). Quanto ao Aβ, sua
expressão também já foi descrita na matriz leucocitária (Carmona et al., 2012).
No líquor podemos observar maior variedade de estudos que têm por
objetivo identificar tanto a expressão dessas proteínas quanto sua atividade,
visto que o líquor possui uma relação direta com o sistema nervoso central e
que as secretases em questão estão envolvidas na cascata amilóide. O
peptídeo Aβ também tem sido amplamente investigado no líquor (Motter et al.,
1995; Blennow e Hampel, 2003; Buchhave et al., 2012; van Harten et al., 2013;
Blennow et al., 2015), inclusive pelo nosso grupo (Diniz et al., 2008; Forlenza et
al., 2015a; Forlenza et al., 2015b).
Não encontramos diferença de expressão proteica de BACE1 em
leucócitos ao realizarmos a comparação entre todos os grupos desse estudo. A
BACE1, por ser caracterizada como a principal responsável pela formação de
Aβ no cérebro por meio da via amiloidogênica (Gonçalves, 2012), tem sido
muito estudada tanto no líquor quanto em matrizes periféricas. Nossos
achados de expressão proteica corroboram dados já descritos na literatura
referentes à expressão gênica dessa proteína em leucócitos de humanos
(Herrera-Rivero et al., 2013; Herrera-Rivero et al., 2015). No líquor também não
descrito um aumento da expressão da BACE1 no líquor de idosos com CCL

39
quando comparados a pacientes com DA e também a controles saudáveis,
porém quando compararam DA com controles não observaram diferença
(Zhong et al., 2007; Ewers et al., 2008). Há também na literatura relatos de
aumento dos níveis de BACE1 em pacientes com DA nas regiões corticais
(Holsinger et al., 2002; Yang et al., 2003; Fukumoto et al., 2002; Ahmed et
al., 2010), em plaquetas (Marksteiner e Humpel, 2013), bem como aumento da
atividade e expressão dessa proteína em líquor de pacientes com DA e até
mesmo CCL (Holsinger et al., 2006; Verheijen et al., 2006; Zhong et al., 2007;
Zetterberg et al., 2008, Mulder et al., 2010). Porém, também há pesquisas que
mostram níveis diminuídos (Decourt et al., 2013); ou até mesmo um estudo em
que não foram encontradas diferenças tanto na atividade dessa proteína,
quanto na expressão do fragmento βAPPs no líquor (Perneczky et al., 2014)
quando comparados os grupos DA, CCL e controles saudáveis.
O fato de não encontrarmos em nossa casuística diferença de expressão
proteica de BACE1 entre os grupos na matriz leucocitária, pode significar que
essa proteína seja capaz de desempenhar em leucócitos outras funções que
não a de β-secretase, questionando o papel desta matriz para pesquisas sobre
a via que representa a cascata amiloide. No que concerne ao líquor, é razoável
pensar no aumento da BACE1 uma vez que essa matriz está em contato direto
com o SNC, porém nossos dados não demonstraram diferença entre grupos, o
que pode ser devido ao tamanho relativamente pequeno da amostra para os
métodos aplicados. Contudo, como dito acima, há controvérsias na literatura
sobre a magnitude (e o sentido) da alteração da expressão proteica de BACE1
no LCR.
Ao analisarmos a expressão de ADAM10 na matriz leucocitária, proteína
associada diretamente à atividade de α-secretase (Vingtdeux e Marambaud,
2012), não observamos diferença ao compararmos os grupos diagnósticos.
Resultados semelhantes foram obtidos no líquor, em que não encontramos
diferença na expressão entre grupos. Não há estudos que descrevam a
expressão da proteína ADAM10 em leucócitos ou líquor. Já foi descrita
diminuição dos níveis de αAPPs, também no líquor (Sennvik et al., 2000;
Colciaghi et al., 2002, Fellgiebel et al., 2009), o que provavelmente ocorre
como resultado de uma redução de expressão ou atividade da ADAM10.

40
(Endres e Fahrenholz, 2012). Além disso, em plaquetas já foi demonstrada
diminuição significativa dos níveis de ADAM10 em pacientes com DA quando
comparado a controles (Colciaghi et al., 2002; Tang et al., 2006; Manzine et al.,
2013), bem como uma diminuição dos níveis de αAPPs (Colciaghi et al., 2002).
No entanto, há na literatura resultados que não demonstraram significância na
comparação da expressão desse fragmento (Olsson et al., 2003), corroborando
nossos achados.
No que diz respeito à expressão proteica de PSEN1 (De Strooper et al.,
1998; Capell et al., 2005), enzima responsável pela liberação do produto da
clivagem da APP por α- ou β-secretase, encontramos diferença ao
compararmos os grupos diagnósticos. Na análise entre pares, observamos
diminuição dos níveis de PSEN1 em DA quando comparado à TDM. Estudos
sobre a expressão proteica de PSEN1 e suas respectivas funções têm sido
abordados na literatura tanto no tecido cerebral (Mathews et al., 2000;
Davidsson et al., 2001; Verdile et al., 2004; Borghi et al., 2010; Kakuda et al.,
2012) quanto em outras matrizes, tais como plaquetas e megacariócitos (Vidal
et al., 1996), plasma (García-Ayllón et al., 2013) e leucócitos (Rogaev et al.,
1997). Em plaquetas já foi descrito um aumento da expressão proteica de
PSEN1 em pacientes com DA quando comparado a controles, contudo não
foram observadas alterações quando realizada a comparação entre CCL e
controles (Bermejo-Bescós et al., 2013). Com relação à expressão proteica de
PSEN1 no tecido cerebral de pacientes com DA, os estudos têm mostrado
níveis de PSEN1 inalterados (Mathews et al., 2000), aumentados (Borghi et al.,
2010; Kakuda et al., 2012) e até mesmo diminuídos (Davidsson et al., 2001;
Verdile et al., 2004). Apesar de dados comparativos sobre a expressão proteica
de PSEN1 em leucócitos entre TDM e DA não terem sido encontrados na
literatura, estudos já mencionaram a relação dessa proteína em processos
inflamatórios (Laky e Fowlkes, 2007; Saura, 2010), inclusive foi demonstrado
por Jiang et al. (2009) que camundongos knock-out para PSEN1 e PSEN2
apresentaram resposta inflamatória robusta tanto no cérebro quanto na
periferia. Neste contexto, faz sentido pensarmos em níveis diminuídos de
PSEN1 em DA quando comparados a TDM, uma vez que na DA os processos
inflamatórios encontram-se mais exacerbados em comparação com TDM,

41
devido ao estímulo de vias de sinalização pró-inflamatórias pelo peptídeo Aβ,
que por sua vez contribui para o aumento da inflamação sistêmica (Goldeck et
al., 2016; Herbert e Lucassen, 2016).
A PSEN1 também já foi identificada no líquor de indivíduos idosos,
porém foi relatado que a mesma, por possuir muitas regiões hidrofóbicas,
encontra-se instável nessa matriz (García-Ayllón, 2013), o que pode justificar
não termos observado a presença dessa proteína em nossa casuística.
Ao compararmos os grupos não observamos diferenças nos níveis de
peptídeo Aβ em leucócitos. Ao analisarmos a matriz liquórica, encontramos
menor expressão de Aβ em pacientes com DA quando comparados a CCL,
TDM e controles. Porém, não foi encontrada diferença nos níveis de Aβ entre
os grupos TDM, CCL e controles.
Apesar de não encontrarmos diferença na expressão do Aβ em
leucócitos, já foi descrito aumento da porcentagem das estruturas beta-folha do
peptídeo Aβ (conformação assumida pelo peptídeo à medida que se agrega em
forma de oligômeros) em leucócitos mononucleares de pacientes com DA
quando comparados a controles (Carmona et al., 2012). Em contrapartida,
outro estudo realizado em plasma revelou diminuição desse peptídeo em DA
quando comparados a CCL e controles, apresentando correlações positivas
com os achados em líquor (Janelidze et al., 2016).
Os níveis diminuídos de Aβ no líquor de indivíduos em DA, quando
comparado a controles, CCL e TDM encontrados em nosso estudo corroboram
outros trabalhos já desenvolvidos pelo nosso grupo e também àqueles
descritos na literatura (Motter et al., 1995; Buchhave et al., 2012; Reis et al.,
2012; van Harten et al., 2013; Blennow et al., 2015, Forlenza et al., 2015b).
Quanto às correlações, ao analisarmos as mesmas proteínas em
diferentes matrizes, observamos uma correlação positiva entre BACE1
expressa em leucócitos com aquela expressa no líquor, corroborando o estudo
realizado em plasma (Janelidze et al., 2016). Para as outras proteínas não
houve correlação da expressão nas diferentes matrizes analisadas.
Em leucócitos, as correlações positivas obtidas entre BACE1 e PSEN1
e ADAM10 e PSEN1 podem ser explicadas pelo fato das duas vias
necessitarem da ação da γ-secretase para a liberação do fragmento gerado a

42
partir da clivagem da APP por meio das vias não-amiloidogênica (ADAM10) e
amiloidogênica (BACE1). Se mantivermos esse raciocínio, a correlação positiva
apresentada entre ADAM10 e o Aβ mostra-se controversa, visto que sua
formação é inversamente proporcional à expressão de ADAM10.
Ao verificar as correlações no líquor, pudemos identificar uma correlação
negativa entre as proteínas BACE1 e ADAM10, o que é relevante se
pensarmos na fisiopatologia da DA, pois há um aumento da clivagem da APP
por BACE1 em detrimento da ADAM10, proporcionando maior formação de Aβ.
Houve também correlação positiva entre BACE1 e idade, o que pode explicar o
fato de haver na senescência a formação do peptídeo em sua forma solúvel
(Stromer e Serpell, 2005).
Considerando as varáveis clínicas obtidas por meio das avaliações
MEEM e CAMCOG verificamos uma correlação positiva com a expressão do
peptídeo Aβ no líquor. Os escores do MEEM podem variar 0 a 30 pontos
(Folstein et al., 1975; Brucki et al. 2003), enquanto o CAMCOG pode
apresentar o valor 105 como pontuação máxima (Roth et al., 1986; Nunes et
al., 2008). Para ambas as avaliações, quanto menor a pontuação maior o grau
de comprometimento cognitivo dos indivíduos. Visto que na DA há a diminuição
do peptídeo Aβ no líquor e que as pontuações nos instrumentos tendem a
diminuir com o declínio cognitivo, essas pontuações demonstram-se coerentes
com o que é observado tanto na clínica quanto na fisiopatologia da doença.
Tendo em vista que nosso estudo não demonstrou diferença na
expressão de Aβ em leucócitos e que também não houve diferenças na
expressão de BACE1 e ADAM10, podemos sugerir que essa matriz não reflita
os processos fisiopatológicos da DA apesar de conter a maquinaria necessária
para tal. Com exceção dos resultados sobre PSEN1, não pudemos observar
diferenças entre a casuística estudada, sugerindo que a expressão dessas
proteínas em leucócitos possa estar ligada a outras funções biológicas, tais
como ativação de células T envolvidas na resposta imune, adesão celular,
desenvolvimento do timo, adesão e migração de leucócitos, dentre outras (Tian
et al., 2008; Dreymueller et al., 2015; Universal Protein Resource, 2017).
Já com relação ao líquor, pudemos identificar que o mesmo apresentou
níveis diminuídos de Aβ, corroborando dados já descritos na literatura. Estudos

43
têm demonstrado que a combinação dos valores do peptídeo Aβ1-42 e das
proteínas Tau total (T-total) e Tau fosforilada (fosfo-Tau) têm apresentado boa
sensibilidade e especificidade (Forlenza et al., 2015b), porém ainda não são
capazes de determinar a conversão para a DA. Ao buscar a relação das
proteínas relacionadas à cascata amiloide nessa matriz, não percebemos
diferença entre os grupos estudados, o que pode ser atribuído ao pequeno
tamanho da amostra. No entanto, ao olharmos para a literatura, notamos
divergência nos resultados sobre essas proteínas no líquor, sugerindo a
necessidade de mais estudos na área para podermos compreender melhor as
vias relacionadas ao desenvolvimento da doença.

44
6 CONSIDERAÇÕES FINAIS
Os níveis de BACE1, ADAM10, PSEN1 e Aβ encontrados em leucócitos
sugerem que essa matriz, apesar de expressar a maquinaria necessária para o
processamento proteolítico da APP, não é a ideal para estudos sobre a cascata
amiloide. Além disso, os resultados obtidos em amostras de líquor, de acordo
com a casuística analisada, reforçam os dados da literatura que estabelecem a
redução da concentração do peptídio Aβ no líquor como um biomarcador da
DA, sem termos detectado alterações nos demais grupos estudados. Nossos
resultados (negativos) em relação à expressão das APP-secretases no líquor
acrescentam dados a este domínio ainda controverso da literatura. Assim
sendo, mais estudos devem ser realizados para que possamos compreender
melhor as vias relacionadas ao desenvolvimento da DA.

45
REFERÊNCIAS BIBLIOGRÁFICAS
Ahmed RR, Holler CJ, Webb RL, Li F, Beckett TL, Murphy MP. BACE1 and BACE2 enzymatic activities in Alzheimer’s disease. J Neurochem 2010; 112:1045–53.
Alexopoulos GS, Borson S, Cuthbert BN, Devanand DP, Mulsant BH, Olin JT, et al. Assessment of late life depression. Biol Psychiatry 2002;52:164-74.
Alzheimer’s Association. A doença de Alzheimer e o cérebro. Disponível em: < http://www.alz.org/brain_portuguese/10.asp>. Acesso em: 24 de janeiro de 2017.
American Psychiatric Association.Diagnostic and Statistical Manual of Mental Disorders – Fifht Edition. Arlington, V.A., American Psychiatric Association, 2013.
Andrei AM, Fraguas R, Telles RM, Alves TC, Strunz CM, Nussbacher A, et al. Major depressive disorder and inflammatory markers in elderly patients with heart failure. Psychosomatics 2007;48:319-24.
Archana Singh, Nisha Misra. A study on Loneliness, Depression and Sociability. Industrial Psychiatry journal. 2009; 18(1): 51-52.
Asai M, Hattori C, Szabó B, Sasagawa N, Maruyama K, Tanuma S. Putative function of ADAM9, ADAM10, and ADAM17 as APP alpha-secretase. Biochem Biophys Res Commun 2003 Jan;301(1):231-5.
Ávila R, Moscoso MA, Ribeiz S, Arrais J, Jaluul O, Bottino CM. Influence of education and depressive symptoms on cognitive function in elderly. Int Psychogeriatr 2009 Jun;21(3):560-7.
Bahls CS. Depressão: uma breve revisão dos fundamentos biológicos e cognitivos. Interação 1999 Jan-Dec;3:49-60.
Baki L, Marambaud P, Efthimiopoulos S, Georgakopoulos A, Wen P, Cui W et al. Presenilin-1 binds cytoplasmic epithelial cadherin, inhibits cadherin/p120 association, and regulates stability and function of the cadherin/catenin adhesion complex. Proc Natl Acad Sci USA 2001 Feb;98(5):2381–6.
Bandeira CB. Prevalência de depressão em idosos institucionalizados. Fortaleza/CE. Dissertação [Mestrado em Saúde Coletiva] – Universidade de Fortaleza; 2012.
Bermejo-Bescós P, Martín-Aragón S, Jiménez-Aliaga K, Benedí J, Felici E, Gil P et al. Processing of the Platelet Amyloid Precursor Protein in the Mild Cognitive Impairment (MCI) Neurochem Res 2013;38:1415–1423.

46
Blennow K, Dubois B, Fagan AM, Lewczuk P, de Leon MJ, Hampel H. Clinical utility of cerebrospinal fluid biomarkers in the diagnosis of early Alzheimer’s disease. Alzheimers Dement 2015;11(1):58-69.
Blennow K, Hampel H. CSF markers for incipient Alzheimer’s disease. Lancet Neurol 2003;2:605–613.
Borghi R, Piccini A, Barini E, Cirmena G, Guglielmotto M, Tamagno E, et al. Upregulation of presenilin 1 in brains of sporadic, late-onset Alzheimer’s disease. J Alzheimers Dis 2010;22:771–775.
Borroni B, Di Luca M, Padovani A. Predicting Alzheimer dementia in mild cognitive impairment patients. Are biomarkers useful? Eur J Pharmacol 2006;545(1):73-80.
Brasil. Ministério da Saúde. Secretaria de Atenção à Saúde. Departamento de Atenção Básica. Envelhecimento e saúde da pessoa idosa. Brasília (DF): Ministério da Saúde; 2007.
Brion JP, Couck AM, Passareiro E, Flament-Durand J. Neurofibrillary tangles of Alzheimer’s disease: An immunohistochemical study. J Submicrosc Cytol 1985;17: 89–96.
Brucki SMD, Nitrini R, Caramelli P, Bertolucci PHF, Okamoto IH. Sugestóes para o uso do mini-exame do estado mental no Brasil. Arq Neuro-Psiquiatr 2003;61(3):777-81.
Buchhave P, Minthon L, Zetterberg H, Wallin AK, Blennow K, Hansson O. Cerebrospinal fluid levels of β-amyloid 1-42, but not of tau, are fully changed already 5 to 10 years before the onset of Alzheimer dementia. Arch Gen Psychiatry 2012;69:98–106.
Busnello JV, Leke R, Oses JP, Feier G, Bruch RS, Quevedo J et al. Acute and chronic electroconvulsive shock in rats: Effects on peripheral markers of neuronal injury and glial activity. Life Sciences 2006;78(26): 3013-3017.
Busnello JV, Oses JP, da Silva RS, Feier G, Barichello T, Quevedo J, et al. Peripheral nucleotide hydrolysis in rats submitted to a model ofelectroconvulsive therapy. Prog Neuropsychopharmacology Biol Psychiatry, 2008 Dec;32(8):1829-33.
Bussière T, Giannakopoulos P, Bouras C, Perl DP, Morrison JH, Hof PR. Progressive degeneration of nonphosphorylated neurofilament protein-enriched pyramidal neurons predicts cognitive impairment in Alzheimer's disease: stereologic analysis of prefrontal cortex area 9. The Journal of Compatative Neurology 2003 Aug;463(3):281-302.

47
Capell A, Beher D, Prokop S, Steiner H, Kaether C, Shearman MS, et al.-Secretase complex assembly within the early secretory pathway. The J Biol Chem 2005 Feb;280(8):6471-8.
Caraci F, Copani A, Nicoletti F, Drago F. Depression and Alzheimer’s disease: Neurobiological links and common pharmacological targets. Eur J Pharmacol 2010 Jan;626(1):64-71.
Carmona P, Molina M, Calero M, Bermejo-Pareja F, Martinéz-Martín P, Alvarez I, et al. Infrared spectroscopic analysis of mononuclear leukocytes in peripheral blood from Alzheimer’s disease patients. Annal Bioanal Chem 2012;402(6):2015-21.
Castro-Costa E, Lima-Costa MF, Carvalhais S, Firmo JO, Uchoa E. Factors associated with depressive sypmtoms measured by the 12-item General Health Questionnaire in community-dwelling older adults (The Bambuí Health Aging Study). Rev Bras Psiquiatr. 2008 Jun;30(2):104-9.
Catricala S, Torti M, Ricevuti G. Alzheimer disease and platelets: how’s that relevant. Immun Ageing 2012 Sep; 9(1):20.
Chachamovich E, Fleck M, Laidlaw K, Power M. Impact of major depression and subsyndromal symptoms on quality of life and attitudes toward aging in an international sample of older adults. The Gerontologist 2008;48:593-602.
Clemente AS, Filho AIL, Firmo JOA. Concepções sobre os transtornos mentais e seu tratamento entre idosos atendidos em um serviço público de saúde mental. Cad Saúde Pública. 2011 Mar;27(3):555-64.
Coelho Filho JM, Ramos LR. Epidemiologia do envelhecimento no Nordeste do Brasil: resultados de inquérito domiciliar. Rev Saúde Pública. 1999;33:445-53.
Colciaghi F, Borroni B, Pastorino L, Marcello E, Zimmermann M, Cattabeni F et al. [alpha]-Secretase ADAM10 as Well as [alpha]APPs Is Reduced in Platelets and CSF of Alzheimer Disease Patients. Molecular Medicine 2002;8(2):67–74.
Colciaghi F, Borroni B, Zimmermann M, Bellone C, Longhi A, Padovani A, et al. Amyloid precursor protein metabolism is regulated toward alpha-secretase pathway by Ginkgo biloba extracts. Neurobiology of Disease 2004a Jul;16(2):454-60.
Cole SL, Vassar R. BACE1 structure and function in health and Alzheimer’s disease. Current Alzheimer Research 2008 Apr; 5(2):100-20.
Cole SL, Vassar R. The Alzheimer's disease beta-secretase enzyme, BACE1. Mol Neurodegener 2007a Nov;2:22.
Cole SL, Vassar R. The Basic Biology of BACE1: A Key Therapeutic Target for Alzheimer's Disease. Curr Genomics 2007b Dec;8(8):509-30.

48
Courtneidge SA, Seals DF. The ADAMs family of metalloproteases: multidomain proteins functions. Genes Dev 2003 Jan;17(1):7-30.
Davidsson P, Bogdanovic N, Lannfelt L, Blennow K. Reduced expression of amyloid precursor protein, presenilin-1 and rab3a in cortical brain regions in Alzheimer’s disease. Dement Geriatr Cogn Disord 2001;12:243–250.
De Paula VJR, Guimarães FM, Forlenza OV. Papel da proteína Tau na fisiopatologia da demência frontotemporal. Rev Psiquiatr Clín 2009 Mar;36(5):197-202.
De Strooper B, Saftig P, Craessaerts K, Vanderstichele H, Guhde G, Annaert W, et al. Deficiency of presenilin-1 inhibits the normal cleavage of amyloid precursor protein. Nature 1998;391(6665):387–90.
Decourt B, Walker A, Gonzales A, Malek-Ahmadi M, Liesback C, Davis KJ et al. Can platelet BACE1 levels be used as a biomarker for Alzheimer’s disease? Proof-of-concept study. Platelets 2013;24(3):235–238.
Delvaux E, Bentley K, Stubbs V, Sabbagh M, Coleman PD. Differential processing of amyloid precursor protein in brain and in peripheral blood leukocytes. Neurobiol Aging 2013 Jun;34(6):1680-6.
Deuss M, Reiss K, Hartmann D. Part-time alpha-secretases: the functional biology of ADAM 9, 10 and 17. Current Alzheimer Research 2008 Apr;5(2):187-201.
Di Luca M, Grossi E, Borroni B, Zimmermann M, Marcello E, Colciaghi F, et al. Artificial neural networks allow the use of simultaneous measurements of Alzheimer Disease markers for early detection of the disease. J Transl Med 2005 Jul;23(3).
Diniz BS, Butters MA, Albert SM, Dew MA, Reynolds CF 3rd. Late-life depression and risk of vascular dementia and Alzheimer’s disease: systematic review and meta-analysis of community-based cohort studies. Br J Psychiatry 2013;202:329-35.
Diniz BS, Pinto Junior JA, Forlenza OV. Do CSF total tau, phosphorylated tau, and beta amyloid 42 help to predict progression of mild cognitive impairment to Alzheimer1s disease? A systematic review and meta-analysis of the literature. World J Biol Psychiatry 2008;9(3):172-82.
Diniz BSO,Forlenza OV. Depressão Geriátrica. In: De Lacerda ALT, Quarantini LC, Miranda-Scippa AMA, Del Porto JA, editores. Depressão: do neurônio ao funcionamento social. 1. ed. São Paulo: Artmed, 2008. p. 85-98.
Doan A, Thinakaran G, Borchelt DR, Slunt HH, Ratovitsky T, Podlisny M, et al. Protein Topology of Presenilin 1. Neuron 1996 Nov;17:1023-30.

49
Dreymueller D, Uhlig S, Ludwig A. ADAM-family metalloproteinases in lung inflammation: potential therapeutic targets. Am J Physiol Lung Cell Mol Phisiol 2015;308(4):L325-43.
Duman RS, Monteggia LM.A neurotrophic model for stress-related mood disorders. Biol Psychiatry 2006 Jun;59(12):1116-27.
Endres K, Fahrenholz F. Regulation of alpha-secretase ADAM10 expression and activity. Exp Brain Res 2012;217:343-52.
Ervatti LR, Borges GM, Jardim AP (orgs). Mudança demográfica no Brasil no início do século XXI: subsídios para as projeções da população. Rio de Janeiro: Instituto Brasileiro de Geografia e Estatística, 2015.
Ewers M, Zhong Z, Bürger K, Wallin A, Blennow K, Teipel SJ, et al. Increased CSF-BACE1 activity is associated with ApoE-ε4 genotype in subjects with mild cognitive impairment and Alzheimer’s disease. Brain 2008 May;131(Pt5):1252-8.
Fahrenholz F, Gilbert S, Kojro E, Lammich S, Postina R. Alpha-secretase activity of the disintegrinmetalloprotease ADAM 10. Influences of domain structure. Ann N Y. AcadSci 2000;920:215–22.
Fellgiebel A, Kojro E, Müller MJ, Scheurich LG, Fahrenholz F. CSF APPs alpha and phosphorylated tau protein levels in mild cognitive impairment and dementia of Alzheimer’s type. J Geriatr Psychiatry Neurol 2009;22(1):3-9.
First MB, Splitzer RL, Gibbon M, Williams JBW. Structured Clinical Interview for DSM-IV Axis I Disorders, Research Version, Patient Edition (SCID-I/P) New York, NY: Biometrics Research, New York State Psychiatric Institute; 2002.
Folstein MF, Folstein SE, McHugh PR. "Mini-mental state". A practical method for grading the cognitive state of patients for the clinician. J Psychiatr Res 1975 Nov;12(3):189-98.
Forlenza OV, Diniz BS, Teixeira AL, Radanovic M, Talib LL, Rocha NP, et al. Lower cerebrospinal fluid concentration of Brain-Derived Neurotruphic Factor Predicts Progression from Mild Cognitive Impairment to Alzheimer’s disease. Neuromolecular Med 2015a;17(3):326-32.
Forlenza OV, Radanovic M, Talib LL, Aprahamian I, Diniz BS, Zetterberg H et al. Cerebrospinal fluid biomarkers in Alzheimer’s disease: diagnostic accuracy and prediction of dementia. Alzheimers Dement (Amst) 2015b;1(4):455-63.
Forlenza OV, Spink JM, Dayanandan R, Anderton BH, Olesen OF, Lovestone S. Muscarinic Agonists Reduce tau Phosphorylation in Non-neuronal Cells Via GSK-3B Inhibition and in Neurons. J Neural Transm 2000;107(10):1201-12.

50
Fukumoto H, Cheung BS, Hyman BT, Irizarry MC. Beta-secretase protein and activity are increased in the neocortex in Alzheimer disease. Arch Neurol 2002; 59:1381–9.
Gallarda T. Alzheimer’s disease and depression. Encephale 1999 Nov;25(5):14-7.
Gallassi R, Di Sarro R, Morreale A, Amore M. Memory impairment in patients with late-onset major depression: The effect of antidepressant therapy. J Affect Disord 2006 Apr;91(2-3):243-50.
García-Ayllón MS, Campanari ML, Brinkmalm G, Rábano A, Alom J, Saura CA, et al. CSF Presenilin-1 complexes are increased in Alzheimer’s disease. Acta Neuropathol Commun 2013 Aug;1(46).
Garrido R, Menezes PR. O Brasil está envelhecendo: boas e más notícias por uma perspectiva epidemiológica. Rev Bras Psiquiatr. 2002 Apr;24(Suppl1):3-6.
Ghiso J, Calero M, Matsubara E, Governale S, Chuba J, Beavis R, et al. Alzheimer's soluble amyloid beta is a normal component of human urine. FEBS Lett 1997 May;408(1):105-8.
Goldeck D, Witkowski JM, Fülop T, Pawelec G. Peripheral immune signatures in Alzheimer disease. Curr Alzheimer Res 2016;13(7):739-49.
Gonçalves RGJ. Nicotina modula o processamento da proteína precursora amilóide e a plasticidade no sistema retinocolicular. Dissertação [Mestrado apresentado à Universidade Federal Fluminense] – Universidade Federal Fluminense; 2012.
Grundke-Iqbal I, Iqbal K, Tung YC, Quinlan M, Wisniewski HM, Binder LI. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc Natl Acad Sci 1986;83: 4913–4917.
Hamdan, AC. Avaliação neuropsicológica na doença de Alzheimer e no Comprometimento Cognitivo Leve. Psicol Argum 2008 Jul-Set;26(54):183-92.
Hamiltom M. A rating scale for depression. Journal Neurol Neurosurg Psychiatry 1960;23:56-62.
Hampel H, Bürger K, Teipel SJ, Bokde AL, Zetterberg H, Blennow K. Core candidate neurochemical and imaging biomarkers of Alzheimer’s disease. Alzheimers Dement 2008 Jan;4(1):38-48.
Hattori M, Osterfield M, Flanagan JG. Regulated cleavage of a contact-mediated axon repellent. Science 2000 Aug;289(5483):1360-5.
Helmer C, Pasquier F, Dartigues JF. Epidemiology of Alzheimer disease and related disorders. Med Sci (Paris) 2006 Mar;22(3):288-96.

51
Herbert J, Lucassen PJ. Depression as a risk factor for Alzheimer’s disease: genes, steroids, cytokines and neurogenesis – what do you need to know? Front Neuroendocrinol 2016;41:153-71.
Herrera-Rivero M, Hernández-Aguilar MH, Aranda-Abreu GE. A strategy focused on MAPT, APP, NCSTN and BACE1 to build blood classifiers fo Alzheimer’s disease. J Theor Biol 2015;7(376):32-8.
Herrera-Rivero M, Soto-Cid A, Hernández ME, Aranda-Abreu GE. Tau, APP, NCT and BACE1 in lymphocytes through cognitively normal ageing and neuropathology. An Acad Bras Cienc 2013;85(4):1489-96.
Hinkle CL, Diestel S, Lieberman J, Maness PF.Metalloprotease induced ectodomain shedding of neural cell adhesion molecule (NCAM). J Neurobiol 2006 Oct;66(12):1378 –95.
Holsinger RM, Lee JS, Boyd A, Masters CL, Collins SJ. CSF BACE1 activity is increased in CJD and Alzheimer disease versus [corrected] other dementias. Neurology 2006; 67:710–2.
Holsinger RM, McLean CA, Beyreuther K, Masters CL, Evin G. Increased expression of the amyloid precursor beta-secretase in Alzheimer’s disease. Ann Neurol 2002; 51:783–6.
Hooper NM, Turner AJ. The search for alpha-secretase and its potential as a therapeutic approach to Alzheimer s disease. Curr Med Chem 2002 Jun;9(11):1107-19.
Houde M, Bergman H, Whitehead V, Chertkow H. A predictive depression pattern in mild cognitive impairment. Int J Geriatr Psychopharmacol 2008; 23:1028–33.
Jack Junior CRJ, Wiste HJ, Vemuri P, Weigand SD, Senjem ML, Zeng G, et al. Brain β-amyloid measures and magnetic resonance imaging atrophy both predict time-to-progression from mild cognitive impairment to Alzheimer’s disease. Brain 2010Nov;133(11):3336-48.
Janelidze D, Stomrud E, Palmqvist S, Zettemberg H, van Westen D, Jeromin A, et al. Plasma β-amyloid in Alzheimer’s disease and vascular disease. Sci Rep 2016;6:26801.
Janes PW, Saha N, Barton WA, Kolev MV, Wimmer-Kleikamp SH, Nievergall E, et al. Adam meets Eph: an ADAM substrate recognition module acts as a molecular switch for ephrin cleavage in trans. Cell 2005 Oct;123(2):291–304.
Jarret JT, Berger EP, Lansbury PT Jr. The carboxy terminus of the beta amyloid protein is critical for the seeding of amyloid formation: implication for the pathogenesis of Alzheimer’s disease. Biochemistry 1993;32(18):4693-7.

52
Jeste DV, Depp CA, Vahia IV. Successful cognitive and emotional aging. World psychiatry. 2010;9(2):78-84.
Jiang X, Zhang D, Shi J, Chen Y, Zhang P, Mei B. Increased inflammatory response both in brain and in periphery in presenilin 1 and presenilin 2 conditional double knock-out mice. J Alzheimers Dis 2009;18(15):515-23.
Kakuda N, Shoji M, Arai H, Furukawa K, Ikeuchi T, Akazawa K, et al. Altered γ-secretase activity in mild cognitive impairment and Alzheimer’s disease. EMBO Mol Med 2012;4:344–352.
Kosik KS, Joachim CL, Selkoe DJ. Microtubule-associated protein tau (tau) is a major antigenic component of paired helical filaments in Alzheimer disease. Proc Natl Acad Sci 1986;83: 4044–4048.
Kreutz, F. Efeito do peptídeo beta amilóide sobre a biossíntese de gangliosídios e avaliação da atividade neuroprotetora do GM1. Dissertação [Mestrado apresentado ao Programa de Pós-Graduação em Ciências Biológicas: Bioquímica como requisito parcial à obtenção do título de Mestre] – Universidade Federal do Rio Grande do Sul; 2010.
Laemmli U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970 Aug;227(5259):680-5.
Laferla EM. Calcium dyshomeostasis and intracellular signaling in Alzheimer’s disease. Nat Rev Neurosci 2002 Nov;3(11):862-72.
Laky K, Fowlkes BJ. Presenilins regulate αβ T cell development by modulating TCR signaling. J Exp Med 2007;204(9):2115-29.
Landau SM, Mintun MA, Joshi AD, Koeppe RA, Petersen RC, Aisen PS, et al. Amyloid Deposition, Hypometabolism, and Longitudinal Cognitive Decline. Ann Neurol 2012 Oct;72(4):578-86.
Lima MTR, Silva RS, Ramos LR. Fatores associados à sintomatologia depressiva numa coorte urbana de idosos. J Bras Psiquiatr. 2009 Jan;58:1-7.
Lin J, Luo J, Redies C. Differential expression of five members of the ADAM family in the developing chicken brain. Neuroscience 2008 Nov;157(2):360–75.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem 1951 Nov;193(1):265-75.
Machado JCB. Doença de Alzheimer.In: Freitas EV, PY L, Neri AL,CançadoFAX,DollJ,Gorzoni M, organizadores. Tratado de Geriatria e Gerontologia. 1. ed. Rio de Janeiro: Guanabara Koogan; 2002. p. 133-14.
Maes M, Yirmyia R, Noraberg J, Brene S, Hibbeln J, Perini G, et al. The inflammatory & neurodegenerative hypothesis of depression: leads for future

53
research and new drug developments in depression. Metab Brain Dis 2009 Mar;24(1):27-53.
Manzine PR, de França Bram JM, Barham EJ, Vale FAC, Selistre-de-Araújo HS, Cominetti MR et al. ADAM10 as a biomarker for Alzheimer’s disease: a study with brazilian elderly. Dement Geriatr Cogn Disord 2013;35:58-66.
Maragno L, Goldbaum M, Gianini RJ, Novaes HMD, César CLG. Prevalência de transtornos mentais comuns em populações atendidas pelo Programa Saúde da Família (QUALIS) no Município de São Paulo, Brasil. Cad Saúde Pública. 2006;22(8):1639-48.
Marcinkiewicz M, Seidah NG. Coordinated expression of beta-amyloid precursor protein and the putative beta-secretase BACE and alpha-secretase ADAM10 in mouse and human brain.J Neurochem 2000 Nov;75(5):2133-43.
Marksteiner J, Humpel C. Platelet-Derived Secreted Amyloid-Precursor Protein-β as a Marker for Diagnosing Alzheimer’s Disease. Curr Neurovasc Res 2013 November;10(4): 297–303.
Martins PAG. Abordagem farmacogenómica na doença de Alzheimer: variação genética associada a CYP2D6. Dissertação [Mestrado apresentado à medicina legal e Ciências Forenses] – Faculdade de Medicina da Universidade de Coimbra; 2010.
Martins RM. A depressão no idoso. Millenium 2008 Apr;(34)119-23.
Maruyama M, Arai H, Sugita M, Tanji H, Higuchi M, Okamura N, et al. Cerebrospinal fluid amyloid beta(1-42) levels in the mild cognitive impairment stage of Alzheimer's disease. Exp Neurol 2001 Dec;172(2):433-6.
Mathews PM, Cataldo AM, Kao BH, Rudnicki AG, Qin X, Yang JL, et al. Brain expression of presenilins in sporadic and early-onset, familial Alzheimer’s disease. Mol Med 2000;6:878–891.
McKhann G, Drachman D, Folstein M, Katzman R, Price D, Stadlan EM. Clinical diagnosis of Alzheimer’s disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984 July;34(7939):939-44.
McKhann GM, Knopman DS, Chertkow H, Hyman BT, Jack CR Jr, Kawas CH, et al.The diagnosis of dementia due to Alzheimer’s disease: recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 2011 May;7(3):263–9.
Modrego PJ, Ferrández J. Depression in patients with mild cognitive impairment increases the risk of developing dementia of Alzheimer type: a prospective cohort study. Arch Neurol 2004;61:1290–93.

54
Moraes EM, Moraes FL, Lima SPP. Características biológicas e psicológicas do envelhecimento. Rev Med Minas Gerais 2010 Feb;20(1):67-73.
Moss ML, Lambert MH. Shedding of membrane proteins by ADAM family proteases. Essays Biochem 2002;38:141-53.
Motter R, Vigo-Pelfrey C, Kholodenko D, Barbour R, Johson-Wood K, Galasko D. Reduction of beta-amyloid peptide42 in the cerebrospinal fluid of patients with Alzheimer's disease. Ann Neurol 1995;38:643–8.
Mulder SD, Van der Flier WM, Verheijen JH, Mulder C, Scheltens P, Blankenstein MA, et al. BACE1 activity in cerebrospinal fluid and its relation to markers os AD pathology. J Alzheimers Dis 2010;20:253-60.
National Institutes of Health (NIH): National Institute on Aging (NIA). Scientific images. Disponível em: <https://www.nia.nih.gov/alzheimers/scientific-images>. Acesso em: 27 de janeiro de2017.
Nunes PV, Diniz BS, Radanovic M, Abreu ID, Borelli DT, Yassuda MS, et al. CAMcog as a screening tool for diagnosis of mild cognitive impairment and dementia in a Brazilian clinical sample of moderate to high education. Int J Geriatr Psychiatry 2008 Nov;23(11):1127-33.
Okello A, Koivunen J, Edison P, Archer HA, Turkheimer FE, Någren K, et al.Conversion of amyloid positive and negative MCI to AD over 3 years: an 11C-PIB PET study. Neurology 2009 Sep; 73(10):754-60.
Olsson A, Höglund K, Sjögren M, Andreasen N, Minthon L, Lannfelt L, et al. Measurement of alpha- and beta-secretase cleaved amyloid precursor protein in cerebrospinal fluid from Alzheimer patients. Expo Neurol 2003;183(1):74-80.
Ownby RL, Crocco E, Acevedo A, John V, Loeweinstein D. Depression and Risk for Alzheimer Disease: Systematic Review, Meta-analysis, and Metaregression Analysis. Arch Gen Psychiatry 2006 May;63(5):530–8.
Perneczky R, Alexopoulos P, Alzheimer’s disease euroimaging Initiative. Cerebrospinal fluid BACE1 activity and markers of amyloid precursor protein metabolism and axonal degeneration in Alzheimer’s disease. Alzheimers Dement 2014;10(5 Suppl):S425-29.
Petersen RC, Caracciolo B, Brayne C, Gauthier S, Jelic V, Fratiglioni L. Mild cognitive impairment: a concept in evolution. J Intern Med 2014 mar;275(3):214-28.
Petersen RC, Doody R, Kurz A, Mohs RC, Morris JC, Rabins PV, et al.Currentconcepts in mildcognitiveimpairment. Arch Neurol 2001a Dec;58(12):1985-92.

55
Petersen RC, Stevens JC, Ganguli M, Tangalos EG, Cummings JL, DeKosky ST. Practice parameter: early detection of dementia: mild cognitive impairment (an evidence-based review). Report of the Quality Standards Subcommittee of the American Academy of Neurology. Neurology 2001b May;56(9):1133-42.
Petersen RC. Mild cognitive impairment as a diagnostic entity. J Intern Med 2004 Sep;256(3):183–94.
Postina R, Schroeder A, Dewachter I, Bohl J, Schmitt U, Kojro E, et al. A disintegrin-metalloproteinase prevents amyloid plaque formation and hippocampal defects in an Alzheimer disease mouse model. J Clin Invest 2004 May;113(10):1456-64.
Postina R. A closer look at alpha-secretase. Curr Alzheimer Res 2008 Apr;5(2):179-86.
Postina R. Activation of alpha-secretase cleavage. J Neurochem 2012 Jan;120(Suppl 1):46-54.
Qin W,Ho L, Wang J, Peskind E, Pasinetti GM.S100A7, a novel Alzheimer's disease biomarker with non-amyloidogenic alpha-secretase activity acts via selective promotion of ADAM-10. PLoS One 2009;4(1):e4183.
Ramos, L.R. e col. Perfil do idoso em área metropolitana na região Sudeste do Brasil: resultados de inquérito domiciliar. Rev Saúde Pública. 1993;27(2):87-94.
Rangaswamy SM. Geneva, Switzerland: The World Health Organization; 2001. World Health Report on Mental Health: New understanding New Hope. Indian J Psychiatry. 2001; 43(1): 1–4.
Ravid R. Biobanks for biomarkers in neurological disorders: the Da vinci bridge for optimal clinico-pathological connection. J Neurol Sci 2009 Aug;286(1-2):119-27.
Reis T, Brandão CO, Coutinho ESF, Engelhardt E, Laks J. Cerebrospinal Fluid Biomarkers in Alzheimer’s Disease and Geriatric Depression: Preliminary Findings from Brazil. CNS Neurosci Ther 2012 Jul;18(7):524-9.
Richard E, Reitz C, Honig LS, Schupf N, Tang MX, Manly JJ, et al. Late-Life Depression, Mild Cognitive Impairment and Dementia. JAMA Neurol 2013 Mar;70(3):374-82.
Rodgers AB. Alzheimer's disease: unraveling the mystery. Maryland: NIH; 2008.
Rogaev EI, Sherrington R, Wu C, Levesque G, Liang Y Rogaeva EA, et al. Analysis of the 59 sequences, genomic structure, and alternative splicing of the presenilin-1 gene (PSEN1) associated with early onset Alzheimer’s disease. Genomics 1997;40:415-24.

56
Roth M, Tym E, Mountjoy CO, Huppert FA, Hendrie H, Verma S, et al. CAMDEX: a standardized instrument for the diagnosis of mental disorders in the elderly with special reference to the early detection of dementia. Br J Psychiatry 1986;149:698-709.
Saura CA. Presenilin/γ-secretase and inflammation. Front Aging Neurosci 2010;2:16.
Scazufca M, Cerqueira ATAR, Menezes PR, Prince M, Vallada HP, Miyazaki MCOS, et al. Investigações epidemiológicas sobre demência nos países em desenvolvimento. Rev Saúde Pública. 2002 Dec;36(6):773-8.
Schoonenboom NS, Mulder C, Van Kamp GJ, Mehta SP, Scheltens P, Blankenstein MA et al. Amyloid beta 38, 40, and 42 species in cerebrospinal fluid: more of the same? Ann Neurol 2005 Jul;58(1): 139-42.
Schuck F, Wolf D, Fellgiebel A, Endres K. Increase of α-secretase ADAM10 in platelets along cognitively healthy aging. J Alzheimers Dis 2016;50(3):817-26.
Seals DF, Courtneidge SA. The ADAMs family of metalloproteases: multidomain proteins with multiple functions. Genes Dev 2003 Jan;17(1):7-30.
Sennvik K, Fastbom J, Blomberg M, Wahlund L-O, Winblad B, Benedikz E. Levels of α- and β-secretase cleaved amyloid precursor protein in the cerebrospinal fluid of Alzheimer’s disease patients. Neurosci Lett 2000;278(3):168-72.
Seubert P, Vigo-Pelfrey C, Esch F, Lee M, Dovey H, Davis D, et al. Isolation and quantification of soluble Alzheimer's beta-peptide from biological fluids. Nature 1992 Sep;359(6393): 325-7.
Sheline YI, Barch MD, Garcia K, Gersing K, Pieper C, Welsh-Bohmer K, et al. Cognitive Function in Late Life Depression: Relationships to Depression Severity, Cerebrovascular Risk Factors and Processing Speed. Biol Psychiatry 2006 Jul;60(1):58-65.
Sierksma A, Van Den Hove DL, Steinbusch HW, Prickaerts J. Major depression, cognitive dysfunction and Alzheimer’s disease: is there a link? Eur J Pharmacol 2010 Jan;626(1):72-82.
Storandt M, Mintun MA, Head D, Morris JC. Cognitive decline and brain volume loss as signatures of cerebral amyloid-β peptide deposition identified with Pittsburgh compound B: cognitive decline associated with Abeta deposition. Arch Neurol 2009 Dec;66(12):1476-81.
Strimbu K, Tavel JA. What are biomarkers? Curr Opin HIV AIDS 2010;5(6): 463-6.

57
Stromer T, Serpell LC. Structure and morphology of the Alzheimer's amyloid fibril. Microsc Res Tech 2005 Jul;67(3-4):210-7.
Sunderland T, Wolozin B, Galasko D, Levy J, Dukoff R, Bahro M, et al. Longitudinal Stability of CSF TAU Levels. Biol Psychiatry 1999 Sep;46(6):750-5.
Tabaton M, Nunzi MG, Xue R, Usiak M, Autilio-Gambetti L, Gambetti P. Soluble amyloid beta-protein is a marker of Alzheimer amyloid in brain but not in cerebrospinal fluid. Biochem Biophys Res Commun 1994 May;200(3):1598-1603.
Talib LL, Joaquim HPG, de Paula VJR. Metodologia em neuroquímica. In: Miguel EC, Gentil V, Gattaz WF. Clínica Psiquiátrica: a visão do departamento e do Instituto de Psiquiatria do HCFMUSP. São Paulo: Manole; 2011. Cap. 151.
Tang K, Hynan LS, Baskin F, Rosenberg RN. Platelet amyloid protein processing: a bio-marker for Alzheimer’s disease. J Neurol Sci 2006;240(1-2):53-8.
Tian L, Wu X, Chi C, Han M, Xu T and Zhuang Y. ADAM10 is essential for proteolytic activation of Notch during thymocyte development. Int Immunol 2008;20(8):1181-7.
Universal Protein Resource – UniProt. UniProtKB – P49768 (PSEN1_HUMAN). Disponível em: <http://www.uniprot.org/uniprot/P49768>. Acesso em: 13 de Fevereiro de 2017.
van Harten AC, Visser PJ, Pijnenburg YA, Teunissen CE, Blankenstein MA, Scheltens P. Cerebrospinal fluid Aβ42 is the best predictor of clinical progression in patients with subjective complaints. Alzheimers Dement 2013;9:481–7.
Vassar R Bennett BD, Babu-Khan S, Kahn S, Mendiaz EA, Denis P, et al. Beta-secretase cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE. Science 1999 Oct;286(5440):735–41.
Vassar R, Kandalepas PC. The β-secretase enzyme BACE1 as a therapeutic target for Alzheimer’s disease. Alzheimers Res Ther 2011 May;3(3):20.
Verdile G, Gnjec A, Miklossy J, Fonte J, Veurink G, Bates K, et al. Protein markers for Alzheimer disease in the frontal cortex and cerebellum. Neurology 2004;63:1385–1392.
Verheijen JH, Huisman LG, van Lent N, Neumann U, Paganetti P, Hack CE, et al. Detection of a soluble form of BACE-1 in human cerebrospinal fluid by a sensitive activity assay. Clin Chem 2006; 52:1168–74.

58
Vidal R, Ghiso J, Wisniewski T, Frangione B. Alzheimer’s presenilin 1 gene expression in platelets and megakaryocytes. Identification of a novel splice variant. FEBS Lett 1996;393:19–23.
Villemagne VL, Pike KE, Chételat G, Ellis KA, Mulligan RS, Bourgeat P, et al. Longitudinal assessment of Aβ and cognition in aging and Alzheimer disease. Ann Neurol 2011 Jan;69(1):181-92.
Vingtdeux V, Marambaud P. Identification and biology of α-secretase. J Neurochem 2012 Jan;120(Suppl 1):34-45.
Wang H, Megill A, Kaiwen H, Kirkwood A, Hey-Kyoung L. Consequences of inihibiting amyloid precursor protein processing enzymes on synaptic function and plasticity. Neural Plast 2012; 272374:24p.
White JM. ADAMs: modulators of cell-cell and cell-matrix interactions. Curr Opin Cell Biol 2003 Oct;15(5):598-606.
Winblad B, Palmer K, Kivipelto M, Jelic V, Fratiglioni L, Wahlund LO, et al. Mild cognitive impairment-beyond controversies, towards a consensus: report of the International Working Group on Mild Cognitive Impairment. J Intern Med 2004 Sep;256(3):240-6.
Wood JG, Mirra SS, Pollock NJ, Binder LI. Neurofibrillary tangles of Alzheimer disease share antigenic determinants with the axonal microtubule-associated protein tau (tau). Proc Natl Acad Sci 1986;83: 4040–4043.
World Health Organization. Ageing and Life Course: Facts about ageing, 2014. Disponível em: <http://www.who.int/ageing/about/facts/en/>. Acesso em: 17 de Agosto de 2015.
World Health Organization. Mental disorders, 2016. Disponível em: <http://www.who.int/mediacentre/factsheets/fs396/en/>. Acesso em: 31 de Maio de 2016.
World Health Organization. Mental health and older adults, 2016. Disponível em: <http://www.who.int/mediacentre/factsheets/fs381/en/>. Acesso em: 18 de Maio de 2016.
Wuwongse S, Chuen-Chung Chang R, Law ACK. The putative neurodegenerative links between depression and Alzheimer’s disease. Prog Neurobiol 2010 Aug;91(4):362-75.
Yang LB, Lindholm K, Yan R, Citron M, Xia W, Yang XL, et al. Elevated beta-secretase expression and enzymatic activity detected in sporadic Alzheimer disease. Nat Med 2003; 9:3–4.
Yang R, Vassar R. Targeting the β secretase BACE1 for Alzheimer’s disease therapy. Lancet Neurol 2014 Mar;13(3):319-29.

59
Zetterberg H, Andreasson U, Hansson O, Wu G, Sankaranarayanan S, Andersson ME, et al. Elevated cerebrospinal fluid BACE1 activity in incipient Alzheimer disease. Arch Neurol 2008; 65:1102–7.
Zhong Z, Ewers M, Teipel S, Burger K, Wallin A, Blennow K, et al. Levels of beta-secretase (BACE1) in cerebrospinal fluid as a predictor of risk in mild cognitive impairment. Arch Gen Psychiatry 2007; 64:718–26.
Zigman W, Schupf N, Haveman M, Silverman W. The epidemiology of Alzheimer disease in intellectual disability: results and recommendations from an international conference. J Intellect Disabil Res 1997 Feb; 41(Pt 1):76-80.
Zinser EV, Hartmann T, Grimm MO. Amyloid beta-protein and lipid metabolism. Biochim Biophys Acta 2007 Aug;1768(8):1991–2001.

60
APÊNDICES
APÊNDICE A – Termo de Consentimento Livre e Esclarecido
HOSPITAL DAS CLÍNICAS DA FACULDADE DE MEDICINA DA
UNIVERSIDADE DE SÃO PAULO-HCFMUSP
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO
_________________________________________________________________ DADOS DE IDENTIFICAÇÃO DO SUJEITO DA PESQUISA OU RESPONSÁVEL
LEGAL 1. NOME:_______________________________________________________________
DOCUMENTO DE IDENTIDADE Nº:____________________ SEXO: M □ F □
DATA NASCIMENTO:___/____/____ ENDEREÇO:___________________________________________________________ Nº:_________APTO:__________________BAIRRO:____________________________ CIDADE:_______________________________CEP:____________________________ TELEFONE: DDD (___)____________________________
2. RESPONSÁVEL LEGAL:_________________________________________________
NATUREZA (grau de parentesco, tutor, curador etc.)____________________________
DOCUMENTO DE IDENTIDADE:_______________________ SEXO: M □ F □
DATA NASCIMENTO:___/____/____ ENDEREÇO:___________________________________________________________ Nº:_________APTO:__________________BAIRRO:____________________________ CIDADE:_______________________________CEP:____________________________ TELEFONE: DDD (___)____________________________
_______________________________________________________________________________________
DADOS SOBRE A PESQUISA
1. TÍTULO DO PROTOCOLO DE PESQUISA: Biomarcadores em líquor e em leucócitos no Transtorno
Depressivo Maior, Comprometimento Cognitivo Leve e doença de Alzheimer.
PESQUISADOR: Prof. Dr. Orestes Vicente Forlenza
CARGO/FUNÇÃO: Professor Associado do Departamento de Psiquiatria/Psiquiatra
INSCRIÇÃO CONSELHO REGIONAL Nº 65590
UNIDADE DO HCFMUSP: Laboratório de Neurociências (LIM-27), Departamento de Psiquiatria.
2. AVALIAÇÃO DO RISCO DA PESQUISA:
RISCO MÍNIMO □ RISCO MÉDIO □

61
RISCO BAIXO ■ RISCO MAIOR □
3. DURAÇÃO DA PESQUISA: 24 meses
HOSPITAL DAS CLÍNICAS DA FACULDADE DE MEDICINA DA UNIVERSIDADE DE SÃO PAULO-HCFMUSP
1- Desenho e objetivos do estudo: Essas informações estão sendo fornecidas para
sua participação voluntária neste estudo, que visa o estudo de biomarcadores e suas
alterações em indivíduos portadores de Transtorno Depressivo Maior (TDM),
Comprometimento Cognitivo Leve (CCL) e doença de Alzheimer (DA), estabelecendo-
se possíveis correlações entre as doenças em estudo. Pelo fato da DA e dos
transtornos depressivos serem as principais causas de problemas de saúde mental em
idosos e visto que a depressão geriátrica está associada a maiores níveis de
comprometimento cognitivo e aumento do risco de desenvolvimento de DA, assim
como o CCL, eles têm sido alvo de muitos estudos, principalmente àqueles
relacionados a possíveis biomarcadores.
2- Descrição dos procedimentos que serão realizados, com seus propósitos e
identificação dos que forem experimentais e não rotineiros: Os participantes da
pesquisa realizarão entrevista médica com profissional treinado, visando
preenchimento de ficha de histórico e avaliação clínica (psiquiátrica e cognitiva) e,
posteriormente passarão por dois procedimentos realizados por profissionais treinados
e qualificados, sendo eles: coleta de 10-40 mL (equivalente a 3 colheres de sopa) de
sangue por punção periférica da veia do antebraço e coleta de 12-15mL (equivalente a
1 colher de sopa) de líquor, no período matutino, por punção lombar nos espaços
intervertebrais L3/L4 ou L4/L5, com uma agulha de calibre 23. As amostras serão
utilizadas para as determinações bioquímicas. Todo material será codificado de modo
que a identidade dos participantes não seja revelada.
3- Desconfortos e riscos esperados: Os desconfortos e riscos esperados são
mínimos e são decorrentes do processo da coleta de sangue e de coleta de líquor.
Você poderá apresentar dor, desconforto ou hematoma quando o sangue for retirado.
Com relação à coleta de líquor, você poderá apresentar dor de cabeça ao levantar,
sensações cutâneas subjetivas (ex., frio, calor, formigamento, pressão etc.), dor local e

62
infecção. Entretanto, a incidência destas complicações é muito pequena (em torno de
4% frente a todas as coletas realizadas até hoje por nosso grupo) e essa porcentagem
torna-se ainda menor no que se refere à cefaleia pós-raquianestesia em idosos.
4- Potenciais benefícios para o participante: Os participantes poderão receber
orientações sobre acesso ao tratamento na rede de saúde, e os resultados dos
exames realizados poderão auxiliar seu médico na condução do caso. Não há
benefício direto para o participante; trata-se de estudo experimental testando a
existência de variantes bioquímicas nos transtornos depressivos maiores e
neurocognitivos menores associadas ao desenvolvimento da DA. É possível, mas não
garantido, que os pacientes com estas doenças possam ser beneficiados pelos
resultados deste estudo no futuro, bem como seus familiares. O estudo visa maior
esclarecimento dos mecanismos biológicos responsáveis pelo aparecimento da DA.
De posse destes conhecimentos, espera-se conseguir, no futuro, melhores estratégias
para o tratamento do referido transtorno.
5- Garantia de acesso: Em qualquer etapa do estudo, você terá acesso aos
profissionais responsáveis pela pesquisa para esclarecimento de eventuais dúvidas. O
principal investigador é o Dr. Wagner Farid Gattaz, que pode ser encontrado no
endereço Rua Dr. Ovídio Pires de Campos, 785, Instituto de Psiquiatria, 1º andar, CEP
05403-010, São Paulo (SP), Telefone (11) 2661-7283. Se você tiver alguma
consideração ou dúvida sobre a ética da pesquisa, entre em contato com o Comitê de
Ética em Pesquisa (CEP) – Rua Ovídio Pires de Campos, 225 – Prédio da
Administração, 5º andar – Tel: 2661-7585 ou 2661-6442 (ramais 16 e 17), e-mail:
6- É garantida a liberdade da retirada de consentimento a qualquer momento e
deixar de participar do estudo, sem qualquer prejuízo à continuidade de seu
tratamento na Instituição;
7- Direito de confidencialidade: As informações obtidas serão analisadas em
conjunto com outros pacientes, não sendo divulgada a identificação de nenhum
paciente.

63
9- Direito de ser mantido atualizado sobre os resultados parciais das pesquisas,
quando em estudos abertos, ou de resultados que sejam do conhecimento dos
pesquisadores;
10- Despesas e compensações: Não há despesas pessoais para o participante em
qualquer fase do estudo, incluindo exames e consultas. Também não há
compensação financeira relacionada à sua participação. Se existir qualquer despesa
adicional, ela será absorvida pelo orçamento da pesquisa.
11- Compromisso do pesquisador de utilizar os dados e o material coletado
somente para esta pesquisa.
Acredito ter sido suficientemente informado a respeito das informações que li
ou que foram lidas para mim, descrevendo o estudo “Biomarcadores em líquor e em
leucócitos no Transtorno Depressivo Maior, Comprometimento Cognitivo Leve e
doença de Alzheimer”.
Eu discuti com o Dr. Orestes Vicente Forlenza sobre a minha decisão em
participar nesse estudo. Ficaram claros para mim quais são os propósitos do estudo,
os procedimentos a serem realizados, seus desconfortos e riscos, as garantias de
confidencialidade e de esclarecimentos permanentes. Ficou claro também que minha
participação é isenta de despesas e que tenho garantia do acesso a tratamento
hospitalar quando necessário. Concordo voluntariamente em participar deste estudo e
poderei retirar o meu consentimento a qualquer momento, antes ou durante o mesmo,
sem penalidades ou prejuízo ou perda de qualquer benefício que eu possa ter
adquirido, ou no meu atendimento neste Serviço.
__________________________________________ Data: ___/___/___ Assinatura do paciente/representante legal __________________________________________ Data: ___/___/___ Assinatura da testemunha para casos de pacientes menores de 18 anos, analfabetos, semi-analfabetos ou portadores de deficiência auditiva ou visual. (Somente para o responsável do estudo) Declaro que obtive de forma apropriada e voluntária o Consentimento Livre e Esclarecido deste paciente ou representante legal para a participação neste estudo. ____________________________________________ Data: ___/___/___ Assinatura do responsável pelo estudo

64
APÊNDICE B – Padronização método Western Blotting
Para efetuar a padronização do método Western Blotting para
determinação das proteínas BACE1, PSEN1 e ADAM10 foi usado um pool de
leucócitos de indivíduos saudáveis, seguindo o mesmo protocolo do preparo de
amostra (figuras 1,2 e 3).
Figura 1 - Western blotting da BACE1 em diferentes condições para immunoblotting.
A) Bloqueio com 5% de leite desnatado por 1 hora, anticorpo primário 1:1000 overnight e anticorpo secundário 1:20000 por 1 hora. B) Bloqueio com 5% de leite desnatado por 1 hora, anticorpo primário 1:5000 overnight e anticorpo secundário 1:20000 por 1 hora.
Figura 2 – Western blotting da PSEN1 em diferentes condições para immunoblotting.
A) Bloqueio com 5% de leite desnatado overnight, anticorpo primário 1:10000 por 1 hora e anticorpo secundário 1:10000 por 1 hora; B) Bloqueio com 3% de soro de albumina bovina (BSA) overnight, anticorpo primário 1:7000 (com 3% de BSA) por 1 hora e anticorpo secundário 1:20000 por 1 hora.
Figura 3 – Western blotting da ADAM10 em diferentes condições para immunoblotting.

65
A) Bloqueio com 5% de leite desnatado overnight, anticorpo primário 1:10000 por 1 hora e anticorpo secundário 1:30000 por 1 hora; B) Bloqueio com 5% de leite desnatado overnight, anticorpo primário 1:15000 por 1 hora e anticorpo secundário 1:30000 por 1 hora.
Com base nos resultados obtidos e acima relatados, padronizou-se o
immunoblotting para cada uma das proteínas de interesse, bem como para o
controle endógeno (β-actina), que utilizou protocolo semelhante.
66
APÊNDICE C – Densitometrias
Abaixo estão representadas as densitometrias das proteínas BACE1 (tabela 1), ADAM10 (tabela 2) e PSEN1 (tabela 3).
Tabela 1 – Densitometria da Proteína BACE1 (continua)
AMOSTRAS INTEGRATED DENSITY VALUES
BACE1 β-ACTINA BACE1/β-ACTINA (BACE1/β-ACTINA)/2
1612 25056 35931 0,697336562
0,7700 1612
29232 34692 0,842615012
964 11484 17346 0,662054652
0,6400 964
11484 18585 0,617917676
1248 28188 91686 0,307440613
0,2764 1248
24012 97881 0,245318295
1708 7308 64428 0,113428944
0,1343 1708
7308 47082 0,155218555
984 30276 45843 0,660427982
0,7613 984
45936 53277 0,86221071
2606 34452 76818 0,448488635
0,4232 2606
35496 89208 0,397901533
2340 16704 95403 0,175088834
0,2412 2340 32364 105315 0,307306651
1063 28188 117705 0,239480056
0,2231 1063 27144 131334 0,206679154
1604 68904 101598 0,678202327
0,7290 1604 64728 83013 0,779733295
1415 31320 156114 0,200622622
0,2561 1415 45936 147441 0,311555131
1099 50112 128856 0,388899236
0,4414 1099 53244 107793 0,493946731
1864 61596 91686 0,671814672
0,6651 1864 52200 79296 0,658292978
2390 22968 142485 0,161195915
0,2257 2390 32364 111510 0,29023406
1899 51156 120183 0,425650882
0,4235 1899 42804 101598 0,421307506
1942 59508 100359 0,592951305
0,5126 1942 61596 142485 0,432298137
2397 33408 102837 0,324863619
0,5394 2397 35496 47082 0,753918695
67
Tabela 1 – Densitometria da Proteína BACE1 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
BACE1 β-ACTINA BACE1/β-ACTINA (BACE1/β-ACTINA)/2
1944 34452 49560 0,695157385 0,5904
1944 35496 73101 0,485574753
1104 26100 78057 0,334371037 0,3459
1104 29232 81774 0,357473035
2148 56376 69384 0,812521619 0,7574
2148 52200 74340 0,702179177
1103 43848 76818 0,570803718 0,5344
1103 40716 81774 0,497908871
1995 143028 74340 1,923970944 0,9620
1995 -- 70623 --
2345 32364 70623 0,458264305 0,4341
2345 18792 45843 0,409920817
2086 12528 50799 0,246619028 0,2607
2086 15660 56994 0,274765765
2089 22968 81774 0,280871671 0,2549
2089 22968 100359 0,228858398
2027 24012 94164 0,255001912 0,5956
2027 52200 55755 0,936238902
1112 59508 43365 1,372258734 1,0787
1112 42804 54516 0,785163989
2118 56376 54516 1,034118424 0,9910
2118 65772 69384 0,947941889
2490 10440 151158 0,069066804 0,0929
2490 9396 80535 0,116669771
1125 4176 29736 0,140435835 0,1435
1125 4176 28497 0,146541741
2120 9396 85491 0,109906306 0,1316
2120 8352 54516 0,153202729
2501 37584 76818 0,48926033 0,5117
2501 46980 87969 0,534051768
2088 74124 97881 0,75728691 0,7482
2088 52200 70623 0,739135976
1131 49068 12390 3,960290557 3,8409
1131 55332 14868 3,721549637
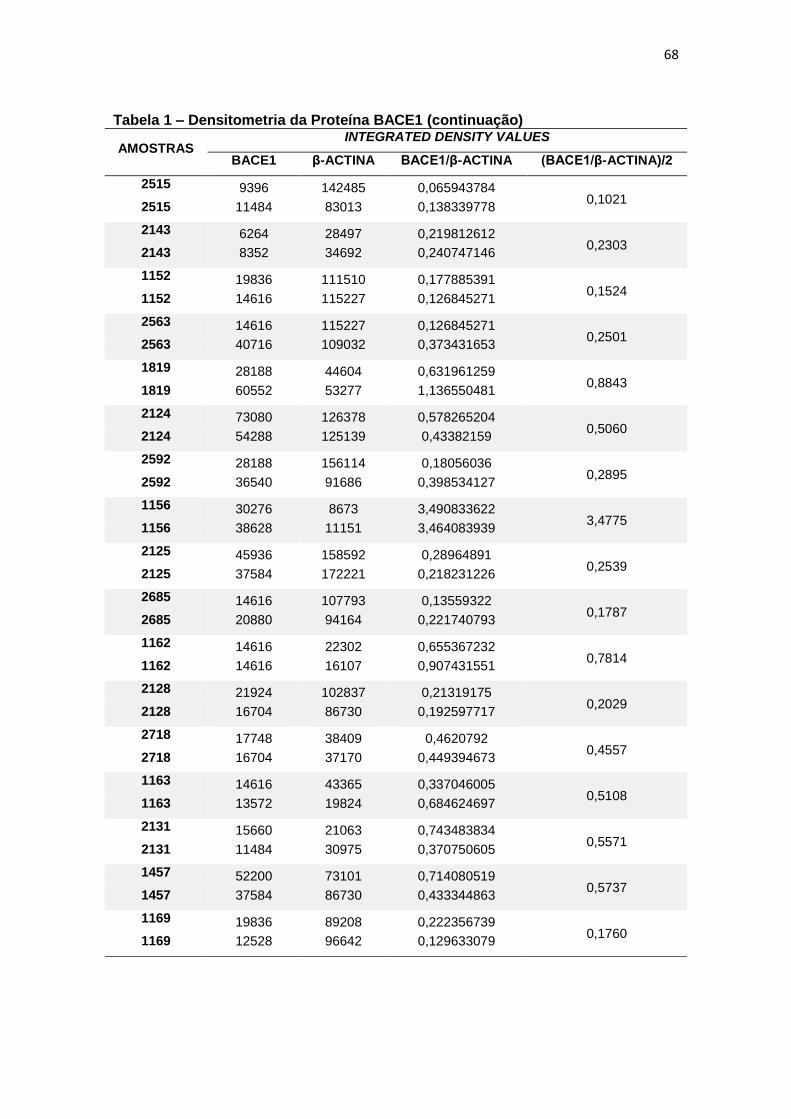
68
Tabela 1 – Densitometria da Proteína BACE1 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
BACE1 β-ACTINA BACE1/β-ACTINA (BACE1/β-ACTINA)/2
2515 9396 142485 0,065943784 0,1021
2515 11484 83013 0,138339778
2143 6264 28497 0,219812612 0,2303
2143 8352 34692 0,240747146
1152 19836 111510 0,177885391 0,1524
1152 14616 115227 0,126845271
2563 14616 115227 0,126845271 0,2501
2563 40716 109032 0,373431653
1819 28188 44604 0,631961259 0,8843
1819 60552 53277 1,136550481
2124 73080 126378 0,578265204 0,5060
2124 54288 125139 0,43382159
2592 28188 156114 0,18056036 0,2895
2592 36540 91686 0,398534127
1156 30276 8673 3,490833622 3,4775
1156 38628 11151 3,464083939
2125 45936 158592 0,28964891 0,2539
2125 37584 172221 0,218231226
2685 14616 107793 0,13559322 0,1787
2685 20880 94164 0,221740793
1162 14616 22302 0,655367232 0,7814
1162 14616 16107 0,907431551
2128 21924 102837 0,21319175 0,2029
2128 16704 86730 0,192597717
2718 17748 38409 0,4620792 0,4557
2718 16704 37170 0,449394673
1163 14616 43365 0,337046005 0,5108
1163 13572 19824 0,684624697
2131 15660 21063 0,743483834 0,5571
2131 11484 30975 0,370750605
1457 52200 73101 0,714080519 0,5737
1457 37584 86730 0,433344863
1169 19836 89208 0,222356739 0,1760
1169 12528 96642 0,129633079
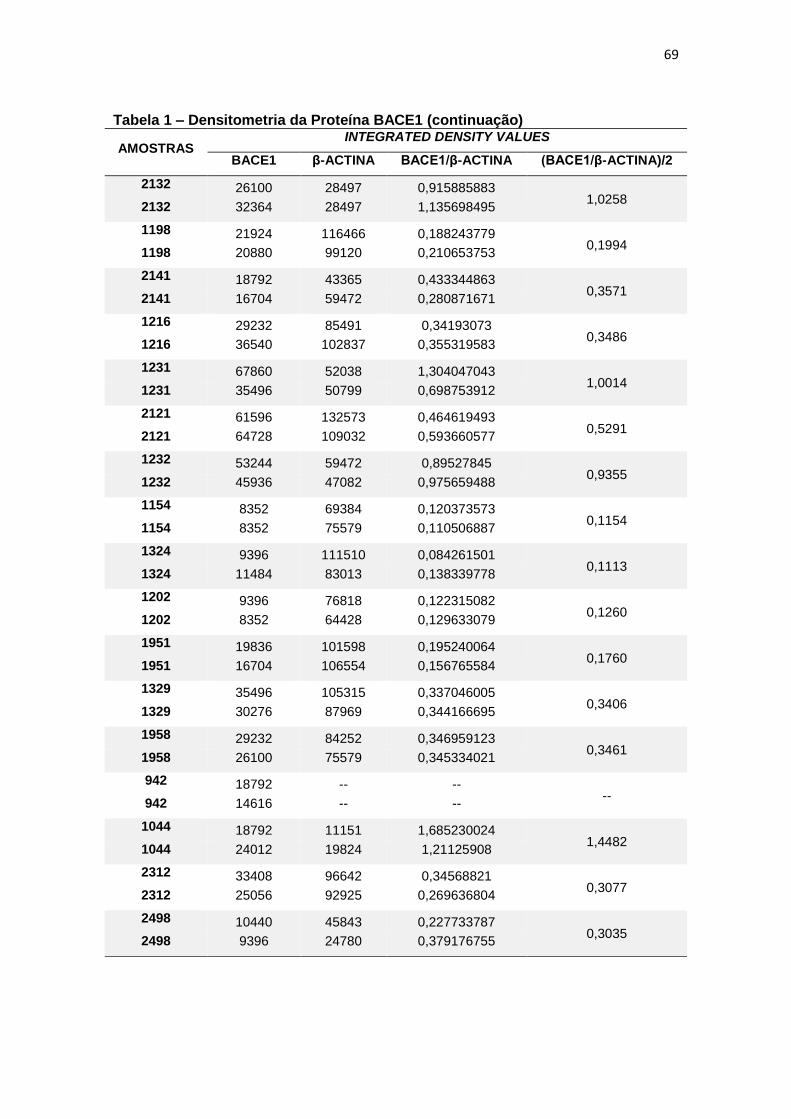
69
Tabela 1 – Densitometria da Proteína BACE1 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
BACE1 β-ACTINA BACE1/β-ACTINA (BACE1/β-ACTINA)/2
2132 26100 28497 0,915885883 1,0258
2132 32364 28497 1,135698495
1198 21924 116466 0,188243779 0,1994
1198 20880 99120 0,210653753
2141 18792 43365 0,433344863 0,3571
2141 16704 59472 0,280871671
1216 29232 85491 0,34193073 0,3486
1216 36540 102837 0,355319583
1231 67860 52038 1,304047043 1,0014
1231 35496 50799 0,698753912
2121 61596 132573 0,464619493 0,5291
2121 64728 109032 0,593660577
1232 53244 59472 0,89527845 0,9355
1232 45936 47082 0,975659488
1154 8352 69384 0,120373573 0,1154
1154 8352 75579 0,110506887
1324 9396 111510 0,084261501 0,1113
1324 11484 83013 0,138339778
1202 9396 76818 0,122315082 0,1260
1202 8352 64428 0,129633079
1951 19836 101598 0,195240064 0,1760
1951 16704 106554 0,156765584
1329 35496 105315 0,337046005 0,3406
1329 30276 87969 0,344166695
1958 29232 84252 0,346959123 0,3461
1958 26100 75579 0,345334021
942 18792 -- -- --
942 14616 -- --
1044 18792 11151 1,685230024 1,4482
1044 24012 19824 1,21125908
2312 33408 96642 0,34568821 0,3077
2312 25056 92925 0,269636804
2498 10440 45843 0,227733787 0,3035
2498 9396 24780 0,379176755
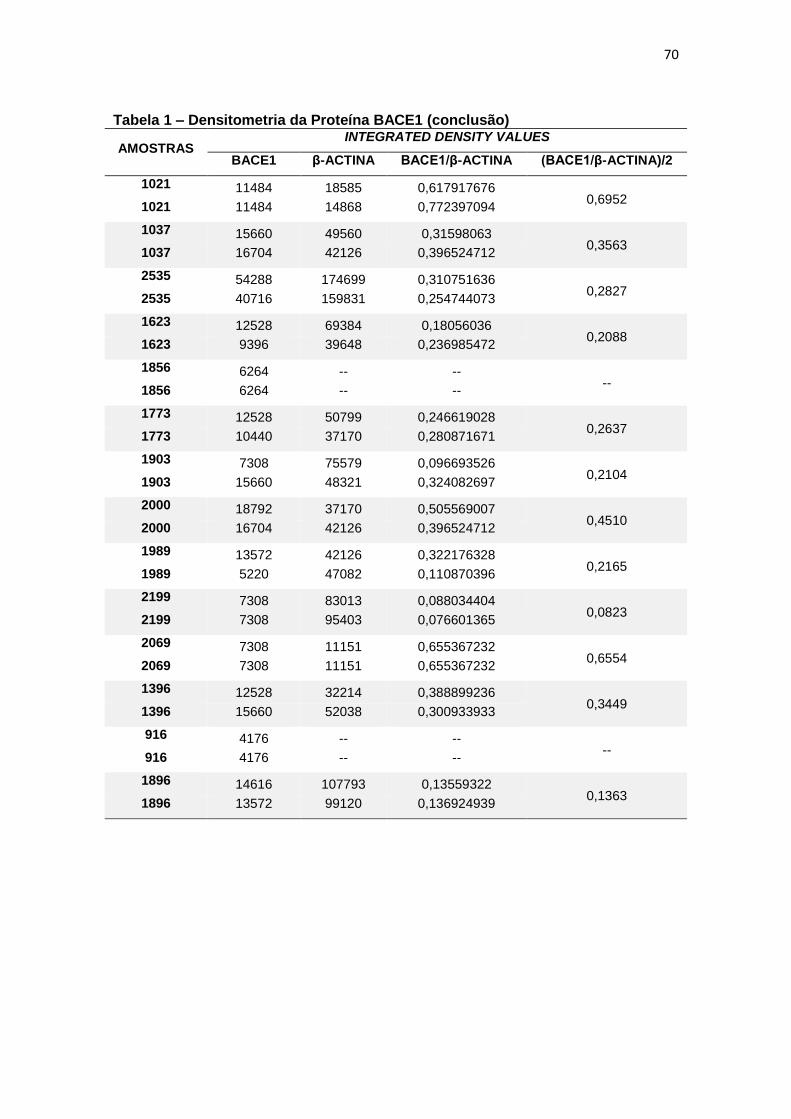
70
Tabela 1 – Densitometria da Proteína BACE1 (conclusão)
AMOSTRAS INTEGRATED DENSITY VALUES
BACE1 β-ACTINA BACE1/β-ACTINA (BACE1/β-ACTINA)/2
1021 11484 18585 0,617917676 0,6952
1021 11484 14868 0,772397094
1037 15660 49560 0,31598063 0,3563
1037 16704 42126 0,396524712
2535 54288 174699 0,310751636 0,2827
2535 40716 159831 0,254744073
1623 12528 69384 0,18056036 0,2088
1623 9396 39648 0,236985472
1856 6264 -- -- --
1856 6264 -- --
1773 12528 50799 0,246619028 0,2637
1773 10440 37170 0,280871671
1903 7308 75579 0,096693526 0,2104
1903 15660 48321 0,324082697
2000 18792 37170 0,505569007 0,4510
2000 16704 42126 0,396524712
1989 13572 42126 0,322176328 0,2165
1989 5220 47082 0,110870396
2199 7308 83013 0,088034404 0,0823
2199 7308 95403 0,076601365
2069 7308 11151 0,655367232 0,6554
2069 7308 11151 0,655367232
1396 12528 32214 0,388899236 0,3449
1396 15660 52038 0,300933933
916 4176 -- -- --
916 4176 -- --
1896 14616 107793 0,13559322 0,1363
1896 13572 99120 0,136924939
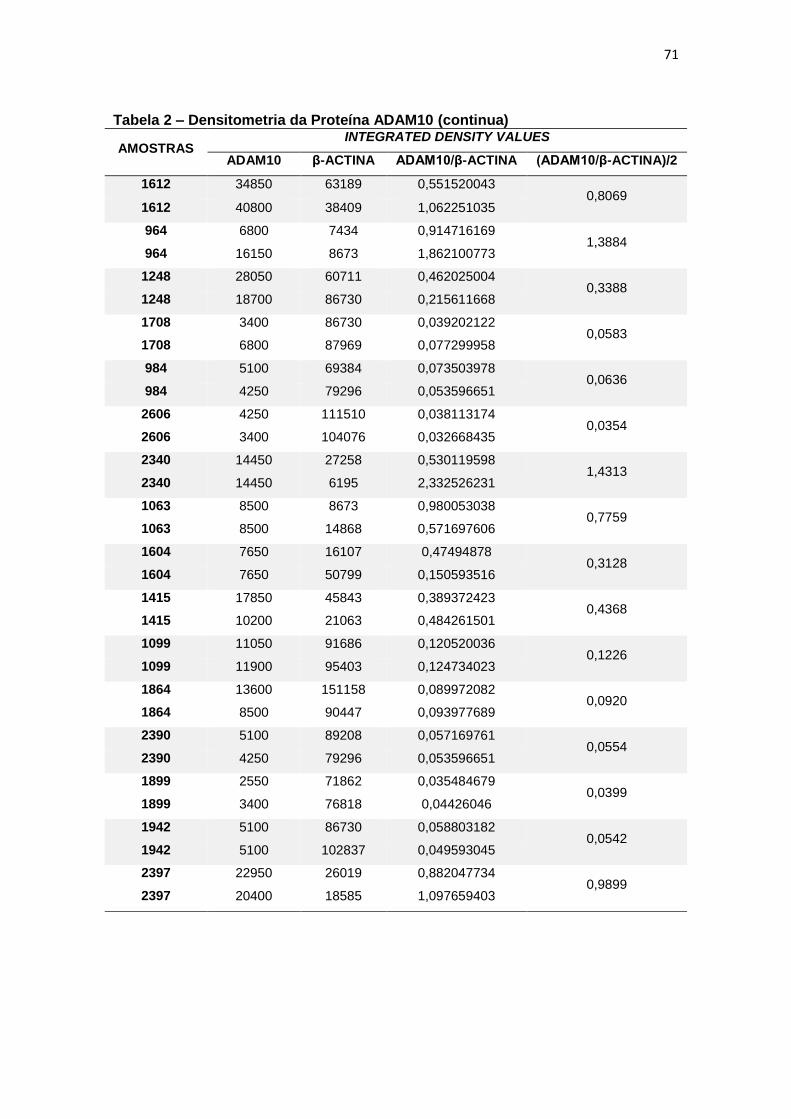
71
Tabela 2 – Densitometria da Proteína ADAM10 (continua)
AMOSTRAS INTEGRATED DENSITY VALUES
ADAM10 β-ACTINA ADAM10/β-ACTINA (ADAM10/β-ACTINA)/2
1612 34850 63189 0,551520043 0,8069
1612 40800 38409 1,062251035
964 6800 7434 0,914716169 1,3884
964 16150 8673 1,862100773
1248 28050 60711 0,462025004 0,3388
1248 18700 86730 0,215611668
1708 3400 86730 0,039202122 0,0583
1708 6800 87969 0,077299958
984 5100 69384 0,073503978 0,0636
984 4250 79296 0,053596651
2606 4250 111510 0,038113174 0,0354
2606 3400 104076 0,032668435
2340 14450 27258 0,530119598 1,4313
2340 14450 6195 2,332526231
1063 8500 8673 0,980053038 0,7759
1063 8500 14868 0,571697606
1604 7650 16107 0,47494878 0,3128
1604 7650 50799 0,150593516
1415 17850 45843 0,389372423 0,4368
1415 10200 21063 0,484261501
1099 11050 91686 0,120520036 0,1226
1099 11900 95403 0,124734023
1864 13600 151158 0,089972082 0,0920
1864 8500 90447 0,093977689
2390 5100 89208 0,057169761 0,0554
2390 4250 79296 0,053596651
1899 2550 71862 0,035484679 0,0399
1899 3400 76818 0,04426046
1942 5100 86730 0,058803182 0,0542
1942 5100 102837 0,049593045
2397 22950 26019 0,882047734 0,9899
2397 20400 18585 1,097659403
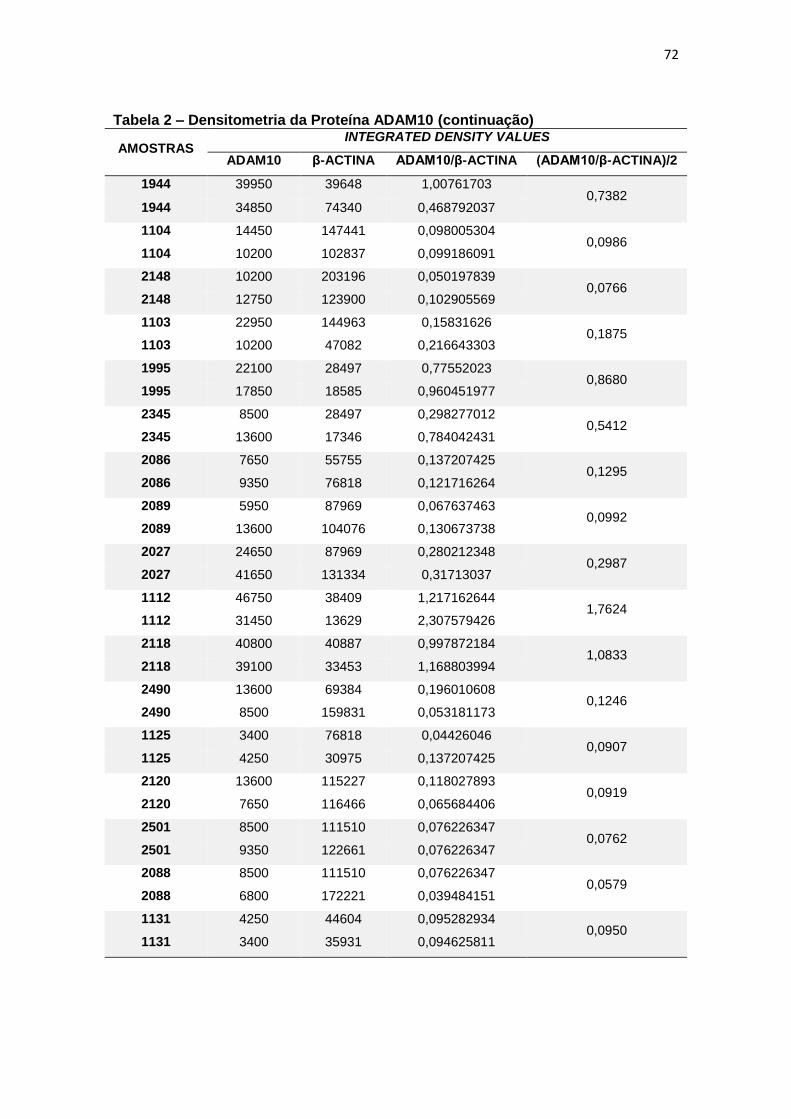
72
Tabela 2 – Densitometria da Proteína ADAM10 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
ADAM10 β-ACTINA ADAM10/β-ACTINA (ADAM10/β-ACTINA)/2
1944 39950 39648 1,00761703 0,7382
1944 34850 74340 0,468792037
1104 14450 147441 0,098005304 0,0986
1104 10200 102837 0,099186091
2148 10200 203196 0,050197839 0,0766
2148 12750 123900 0,102905569
1103 22950 144963 0,15831626 0,1875
1103 10200 47082 0,216643303
1995 22100 28497 0,77552023 0,8680
1995 17850 18585 0,960451977
2345 8500 28497 0,298277012 0,5412
2345 13600 17346 0,784042431
2086 7650 55755 0,137207425 0,1295
2086 9350 76818 0,121716264
2089 5950 87969 0,067637463 0,0992
2089 13600 104076 0,130673738
2027 24650 87969 0,280212348 0,2987
2027 41650 131334 0,31713037
1112 46750 38409 1,217162644 1,7624
1112 31450 13629 2,307579426
2118 40800 40887 0,997872184 1,0833
2118 39100 33453 1,168803994
2490 13600 69384 0,196010608 0,1246
2490 8500 159831 0,053181173
1125 3400 76818 0,04426046 0,0907
1125 4250 30975 0,137207425
2120 13600 115227 0,118027893 0,0919
2120 7650 116466 0,065684406
2501 8500 111510 0,076226347 0,0762
2501 9350 122661 0,076226347
2088 8500 111510 0,076226347 0,0579
2088 6800 172221 0,039484151
1131 4250 44604 0,095282934 0,0950
1131 3400 35931 0,094625811
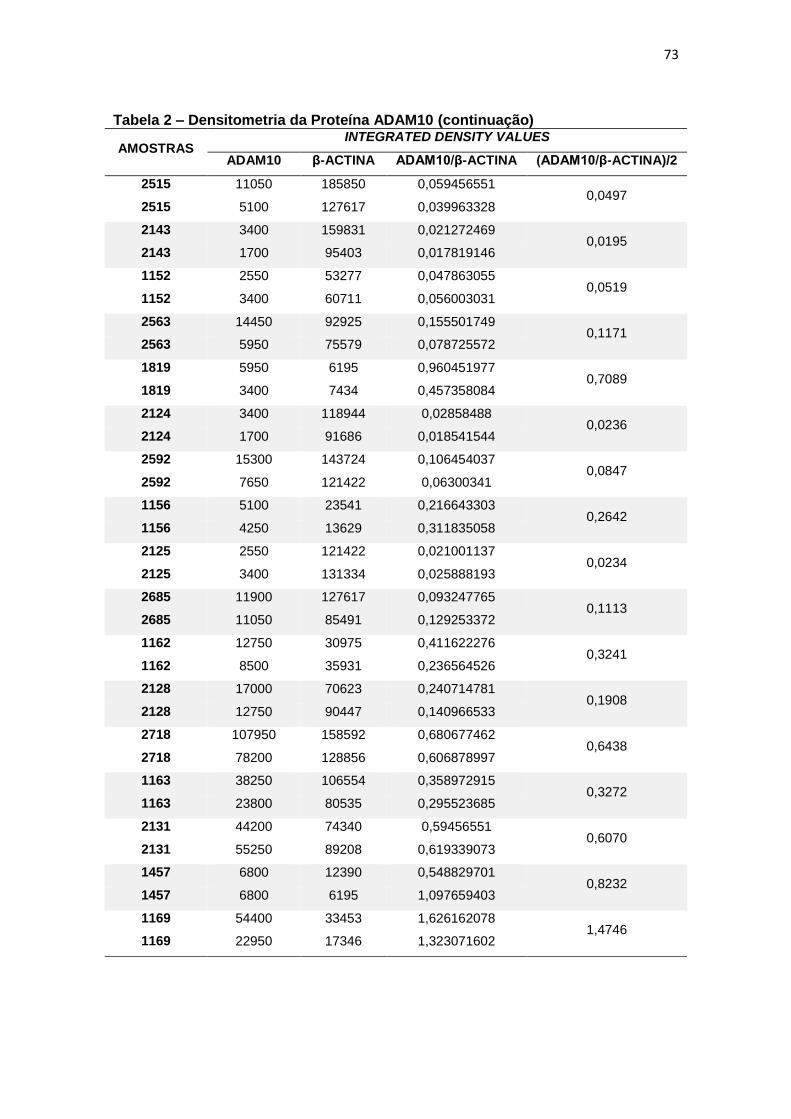
73
Tabela 2 – Densitometria da Proteína ADAM10 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
ADAM10 β-ACTINA ADAM10/β-ACTINA (ADAM10/β-ACTINA)/2
2515 11050 185850 0,059456551 0,0497
2515 5100 127617 0,039963328
2143 3400 159831 0,021272469 0,0195
2143 1700 95403 0,017819146
1152 2550 53277 0,047863055 0,0519
1152 3400 60711 0,056003031
2563 14450 92925 0,155501749 0,1171
2563 5950 75579 0,078725572
1819 5950 6195 0,960451977 0,7089
1819 3400 7434 0,457358084
2124 3400 118944 0,02858488 0,0236
2124 1700 91686 0,018541544
2592 15300 143724 0,106454037 0,0847
2592 7650 121422 0,06300341
1156 5100 23541 0,216643303 0,2642
1156 4250 13629 0,311835058
2125 2550 121422 0,021001137 0,0234
2125 3400 131334 0,025888193
2685 11900 127617 0,093247765 0,1113
2685 11050 85491 0,129253372
1162 12750 30975 0,411622276 0,3241
1162 8500 35931 0,236564526
2128 17000 70623 0,240714781 0,1908
2128 12750 90447 0,140966533
2718 107950 158592 0,680677462 0,6438
2718 78200 128856 0,606878997
1163 38250 106554 0,358972915 0,3272
1163 23800 80535 0,295523685
2131 44200 74340 0,59456551 0,6070
2131 55250 89208 0,619339073
1457 6800 12390 0,548829701 0,8232
1457 6800 6195 1,097659403
1169 54400 33453 1,626162078 1,4746
1169 22950 17346 1,323071602
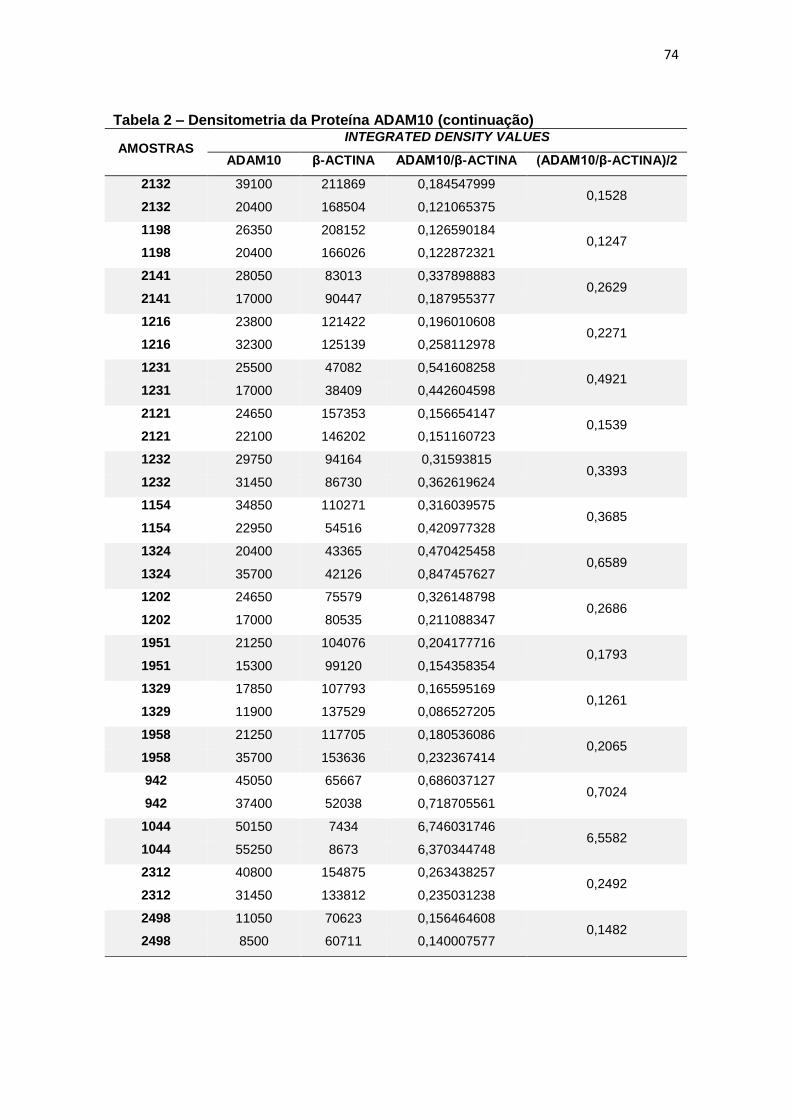
74
Tabela 2 – Densitometria da Proteína ADAM10 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
ADAM10 β-ACTINA ADAM10/β-ACTINA (ADAM10/β-ACTINA)/2
2132 39100 211869 0,184547999 0,1528
2132 20400 168504 0,121065375
1198 26350 208152 0,126590184 0,1247
1198 20400 166026 0,122872321
2141 28050 83013 0,337898883 0,2629
2141 17000 90447 0,187955377
1216 23800 121422 0,196010608 0,2271
1216 32300 125139 0,258112978
1231 25500 47082 0,541608258 0,4921
1231 17000 38409 0,442604598
2121 24650 157353 0,156654147 0,1539
2121 22100 146202 0,151160723
1232 29750 94164 0,31593815 0,3393
1232 31450 86730 0,362619624
1154 34850 110271 0,316039575 0,3685
1154 22950 54516 0,420977328
1324 20400 43365 0,470425458 0,6589
1324 35700 42126 0,847457627
1202 24650 75579 0,326148798 0,2686
1202 17000 80535 0,211088347
1951 21250 104076 0,204177716 0,1793
1951 15300 99120 0,154358354
1329 17850 107793 0,165595169 0,1261
1329 11900 137529 0,086527205
1958 21250 117705 0,180536086 0,2065
1958 35700 153636 0,232367414
942 45050 65667 0,686037127 0,7024
942 37400 52038 0,718705561
1044 50150 7434 6,746031746 6,5582
1044 55250 8673 6,370344748
2312 40800 154875 0,263438257 0,2492
2312 31450 133812 0,235031238
2498 11050 70623 0,156464608 0,1482
2498 8500 60711 0,140007577
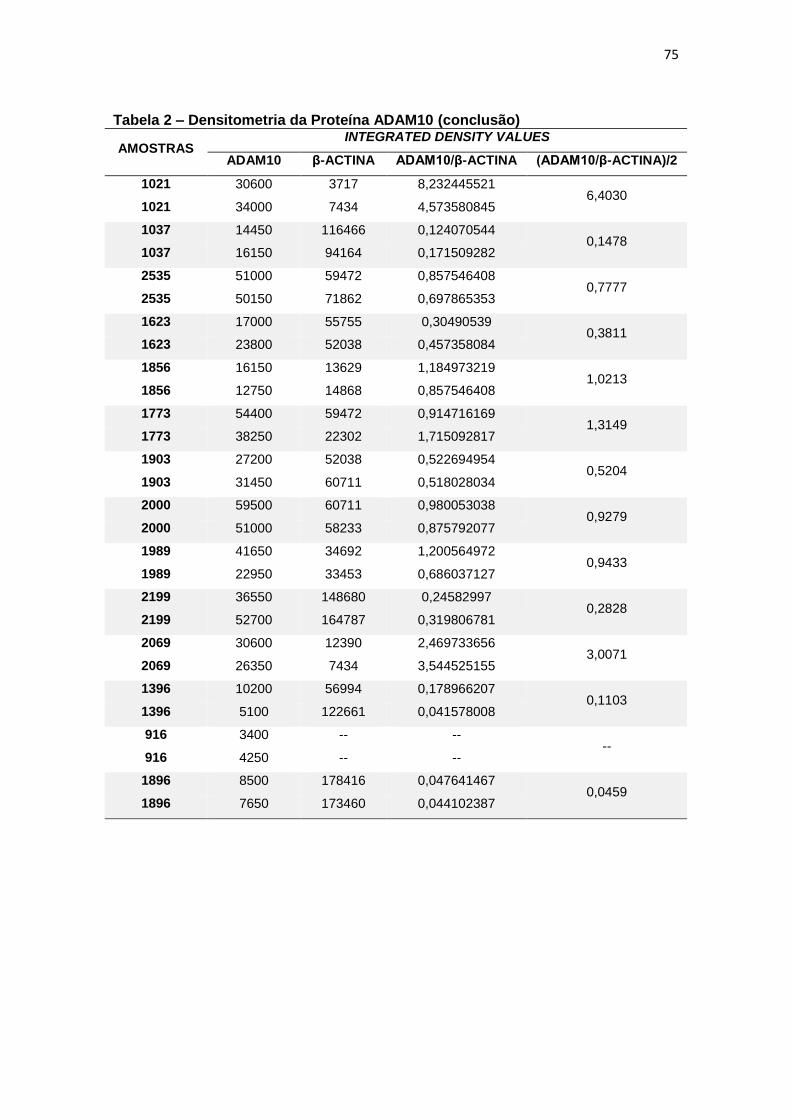
75
Tabela 2 – Densitometria da Proteína ADAM10 (conclusão)
AMOSTRAS INTEGRATED DENSITY VALUES
ADAM10 β-ACTINA ADAM10/β-ACTINA (ADAM10/β-ACTINA)/2
1021 30600 3717 8,232445521 6,4030
1021 34000 7434 4,573580845
1037 14450 116466 0,124070544 0,1478
1037 16150 94164 0,171509282
2535 51000 59472 0,857546408 0,7777
2535 50150 71862 0,697865353
1623 17000 55755 0,30490539 0,3811
1623 23800 52038 0,457358084
1856 16150 13629 1,184973219 1,0213
1856 12750 14868 0,857546408
1773 54400 59472 0,914716169 1,3149
1773 38250 22302 1,715092817
1903 27200 52038 0,522694954 0,5204
1903 31450 60711 0,518028034
2000 59500 60711 0,980053038 0,9279
2000 51000 58233 0,875792077
1989 41650 34692 1,200564972 0,9433
1989 22950 33453 0,686037127
2199 36550 148680 0,24582997 0,2828
2199 52700 164787 0,319806781
2069 30600 12390 2,469733656 3,0071
2069 26350 7434 3,544525155
1396 10200 56994 0,178966207 0,1103
1396 5100 122661 0,041578008
916 3400 -- -- --
916 4250 -- --
1896 8500 178416 0,047641467 0,0459
1896 7650 173460 0,044102387
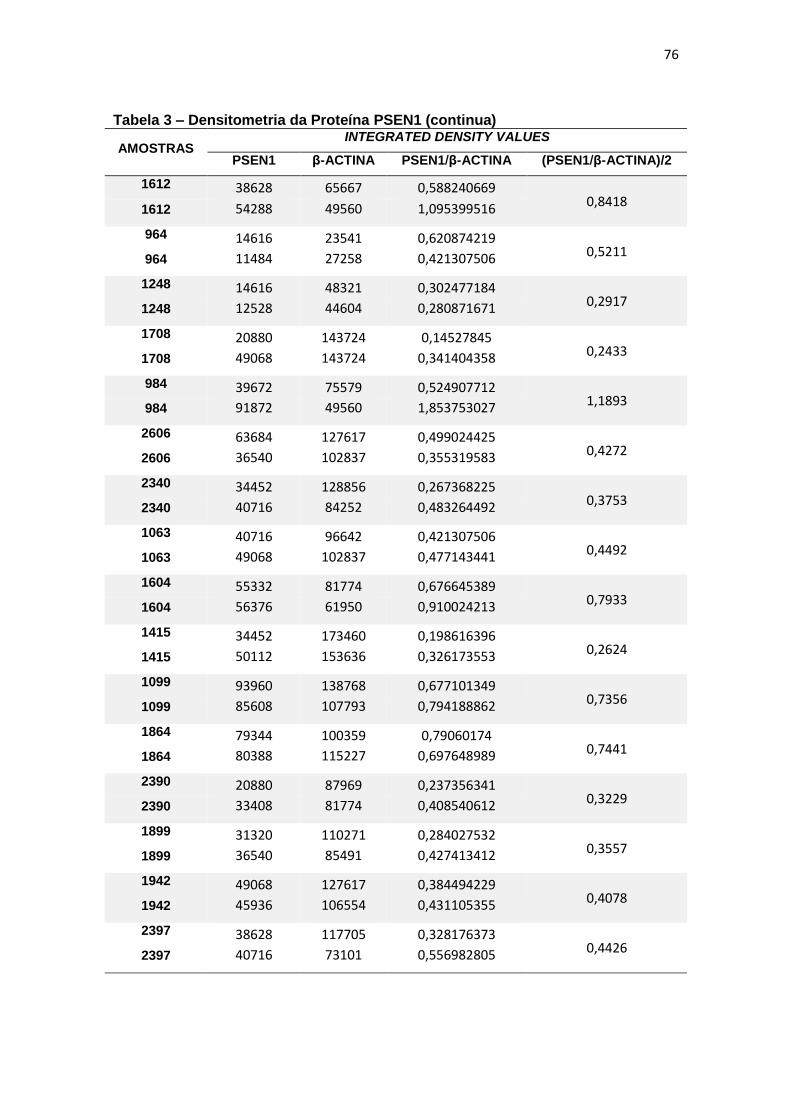
76
Tabela 3 – Densitometria da Proteína PSEN1 (continua)
AMOSTRAS INTEGRATED DENSITY VALUES
PSEN1 β-ACTINA PSEN1/β-ACTINA (PSEN1/β-ACTINA)/2
1612 38628 65667 0,588240669 0,8418
1612 54288 49560 1,095399516
964 14616 23541 0,620874219 0,5211
964 11484 27258 0,421307506
1248 14616 48321 0,302477184 0,2917
1248 12528 44604 0,280871671
1708 20880 143724 0,14527845 0,2433
1708 49068 143724 0,341404358
984 39672 75579 0,524907712 1,1893
984 91872 49560 1,853753027
2606 63684 127617 0,499024425 0,4272
2606 36540 102837 0,355319583
2340 34452 128856 0,267368225 0,3753
2340 40716 84252 0,483264492
1063 40716 96642 0,421307506 0,4492
1063 49068 102837 0,477143441
1604 55332 81774 0,676645389 0,7933
1604 56376 61950 0,910024213
1415 34452 173460 0,198616396 0,2624
1415 50112 153636 0,326173553
1099 93960 138768 0,677101349 0,7356
1099 85608 107793 0,794188862
1864 79344 100359 0,79060174 0,7441
1864 80388 115227 0,697648989
2390 20880 87969 0,237356341 0,3229
2390 33408 81774 0,408540612
1899 31320 110271 0,284027532 0,3557
1899 36540 85491 0,427413412
1942 49068 127617 0,384494229 0,4078
1942 45936 106554 0,431105355
2397 38628 117705 0,328176373 0,4426
2397 40716 73101 0,556982805
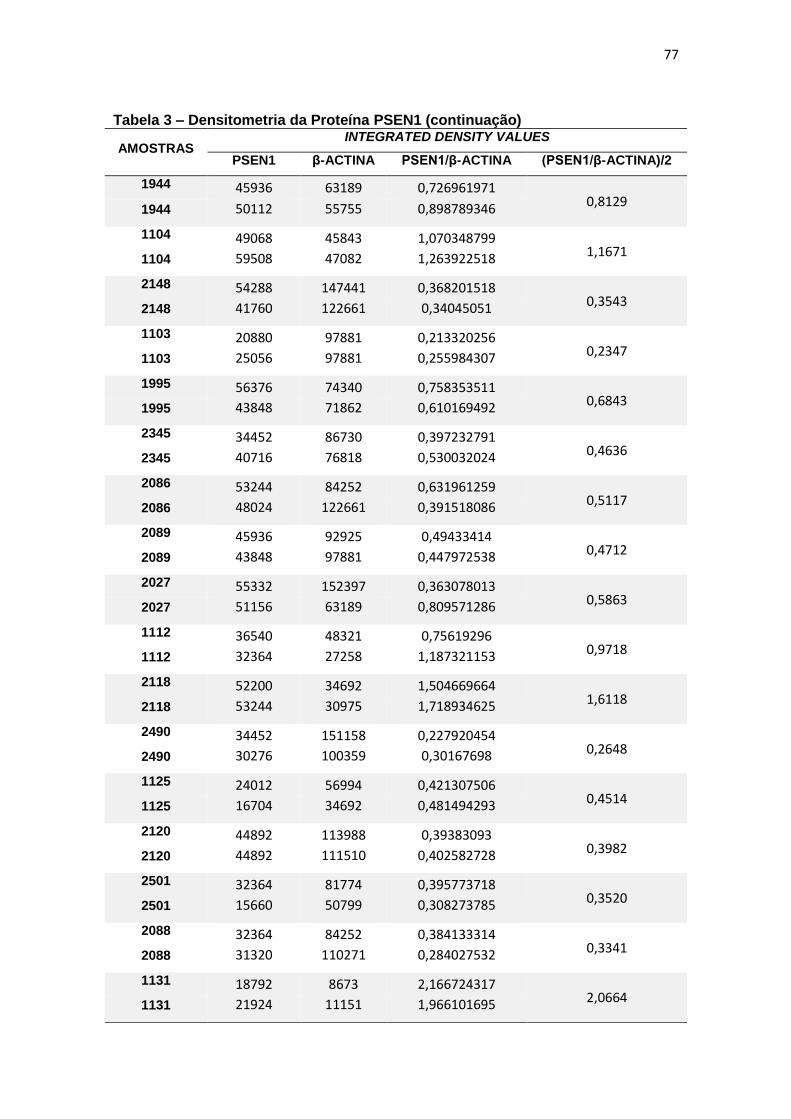
77
Tabela 3 – Densitometria da Proteína PSEN1 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
PSEN1 β-ACTINA PSEN1/β-ACTINA (PSEN1/β-ACTINA)/2
1944 45936 63189 0,726961971 0,8129
1944 50112 55755 0,898789346
1104 49068 45843 1,070348799 1,1671
1104 59508 47082 1,263922518
2148 54288 147441 0,368201518 0,3543
2148 41760 122661 0,34045051
1103 20880 97881 0,213320256 0,2347
1103 25056 97881 0,255984307
1995 56376 74340 0,758353511 0,6843
1995 43848 71862 0,610169492
2345 34452 86730 0,397232791 0,4636
2345 40716 76818 0,530032024
2086 53244 84252 0,631961259 0,5117
2086 48024 122661 0,391518086
2089 45936 92925 0,49433414 0,4712
2089 43848 97881 0,447972538
2027 55332 152397 0,363078013 0,5863
2027 51156 63189 0,809571286
1112 36540 48321 0,75619296 0,9718
1112 32364 27258 1,187321153
2118 52200 34692 1,504669664 1,6118
2118 53244 30975 1,718934625
2490 34452 151158 0,227920454 0,2648
2490 30276 100359 0,30167698
1125 24012 56994 0,421307506 0,4514
1125 16704 34692 0,481494293
2120 44892 113988 0,39383093 0,3982
2120 44892 111510 0,402582728
2501 32364 81774 0,395773718 0,3520
2501 15660 50799 0,308273785
2088 32364 84252 0,384133314 0,3341
2088 31320 110271 0,284027532
1131 18792 8673 2,166724317 2,0664
1131 21924 11151 1,966101695
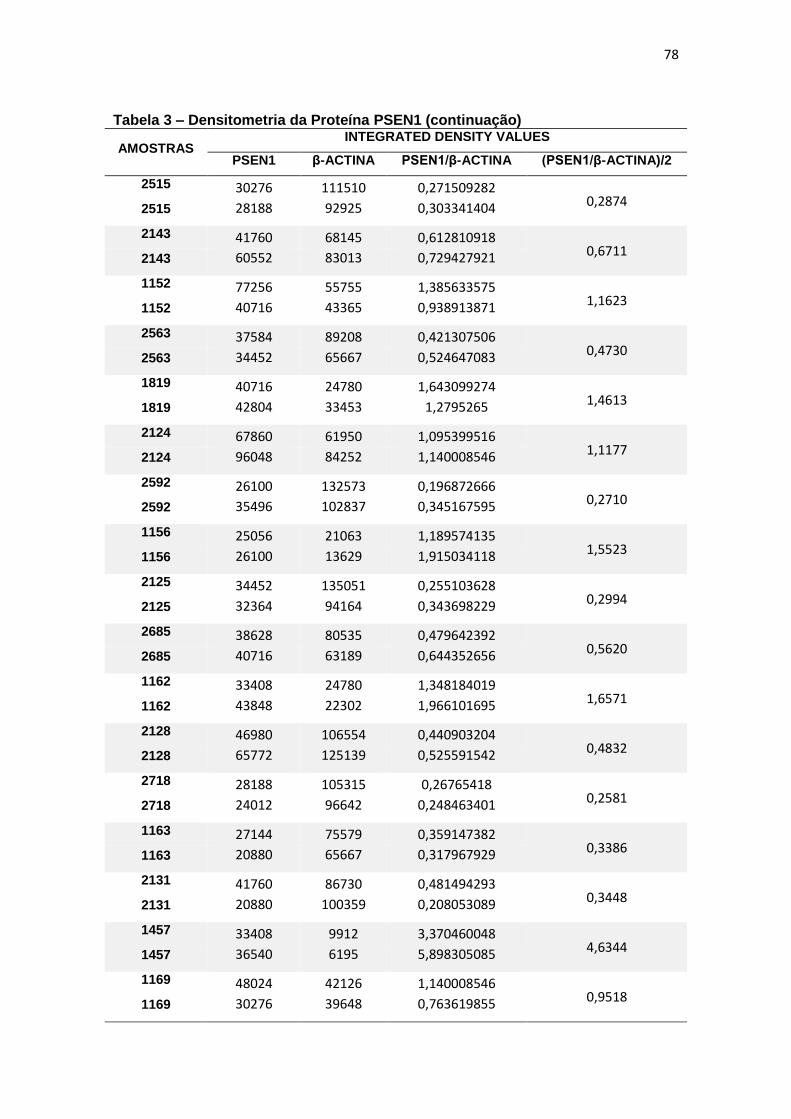
78
Tabela 3 – Densitometria da Proteína PSEN1 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
PSEN1 β-ACTINA PSEN1/β-ACTINA (PSEN1/β-ACTINA)/2
2515 30276 111510 0,271509282 0,2874
2515 28188 92925 0,303341404
2143 41760 68145 0,612810918 0,6711
2143 60552 83013 0,729427921
1152 77256 55755 1,385633575 1,1623
1152 40716 43365 0,938913871
2563 37584 89208 0,421307506 0,4730
2563 34452 65667 0,524647083
1819 40716 24780 1,643099274 1,4613
1819 42804 33453 1,2795265
2124 67860 61950 1,095399516 1,1177
2124 96048 84252 1,140008546
2592 26100 132573 0,196872666 0,2710
2592 35496 102837 0,345167595
1156 25056 21063 1,189574135 1,5523
1156 26100 13629 1,915034118
2125 34452 135051 0,255103628 0,2994
2125 32364 94164 0,343698229
2685 38628 80535 0,479642392 0,5620
2685 40716 63189 0,644352656
1162 33408 24780 1,348184019 1,6571
1162 43848 22302 1,966101695
2128 46980 106554 0,440903204 0,4832
2128 65772 125139 0,525591542
2718 28188 105315 0,26765418 0,2581
2718 24012 96642 0,248463401
1163 27144 75579 0,359147382 0,3386
1163 20880 65667 0,317967929
2131 41760 86730 0,481494293 0,3448
2131 20880 100359 0,208053089
1457 33408 9912 3,370460048 4,6344
1457 36540 6195 5,898305085
1169 48024 42126 1,140008546 0,9518
1169 30276 39648 0,763619855
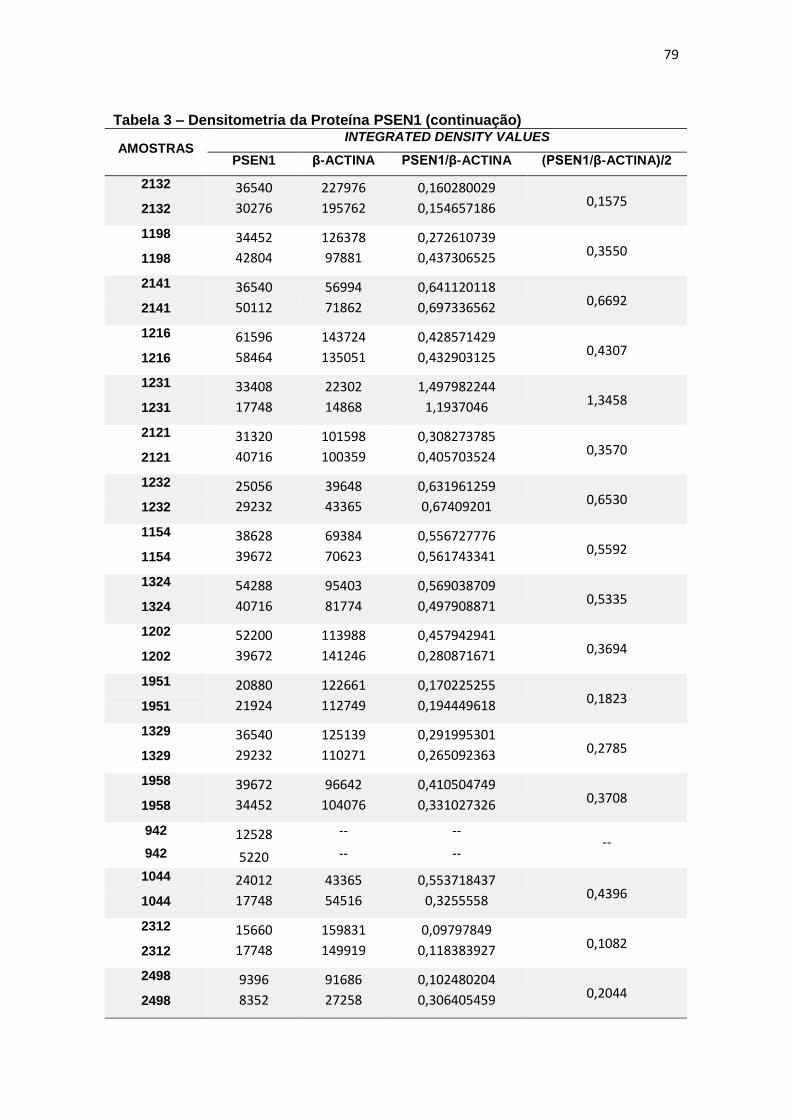
79
Tabela 3 – Densitometria da Proteína PSEN1 (continuação)
AMOSTRAS INTEGRATED DENSITY VALUES
PSEN1 β-ACTINA PSEN1/β-ACTINA (PSEN1/β-ACTINA)/2
2132 36540 227976 0,160280029 0,1575
2132 30276 195762 0,154657186
1198 34452 126378 0,272610739 0,3550
1198 42804 97881 0,437306525
2141 36540 56994 0,641120118 0,6692
2141 50112 71862 0,697336562
1216 61596 143724 0,428571429 0,4307
1216 58464 135051 0,432903125
1231 33408 22302 1,497982244 1,3458
1231 17748 14868 1,1937046
2121 31320 101598 0,308273785 0,3570
2121 40716 100359 0,405703524
1232 25056 39648 0,631961259 0,6530
1232 29232 43365 0,67409201
1154 38628 69384 0,556727776 0,5592
1154 39672 70623 0,561743341
1324 54288 95403 0,569038709 0,5335
1324 40716 81774 0,497908871
1202 52200 113988 0,457942941 0,3694
1202 39672 141246 0,280871671
1951 20880 122661 0,170225255 0,1823
1951 21924 112749 0,194449618
1329 36540 125139 0,291995301 0,2785
1329 29232 110271 0,265092363
1958 39672 96642 0,410504749 0,3708
1958 34452 104076 0,331027326
942 12528 -- -- --
942 5220 -- --
1044 24012 43365 0,553718437 0,4396
1044 17748 54516 0,3255558
2312 15660 159831 0,09797849 0,1082
2312 17748 149919 0,118383927
2498 9396 91686 0,102480204 0,2044
2498 8352 27258 0,306405459
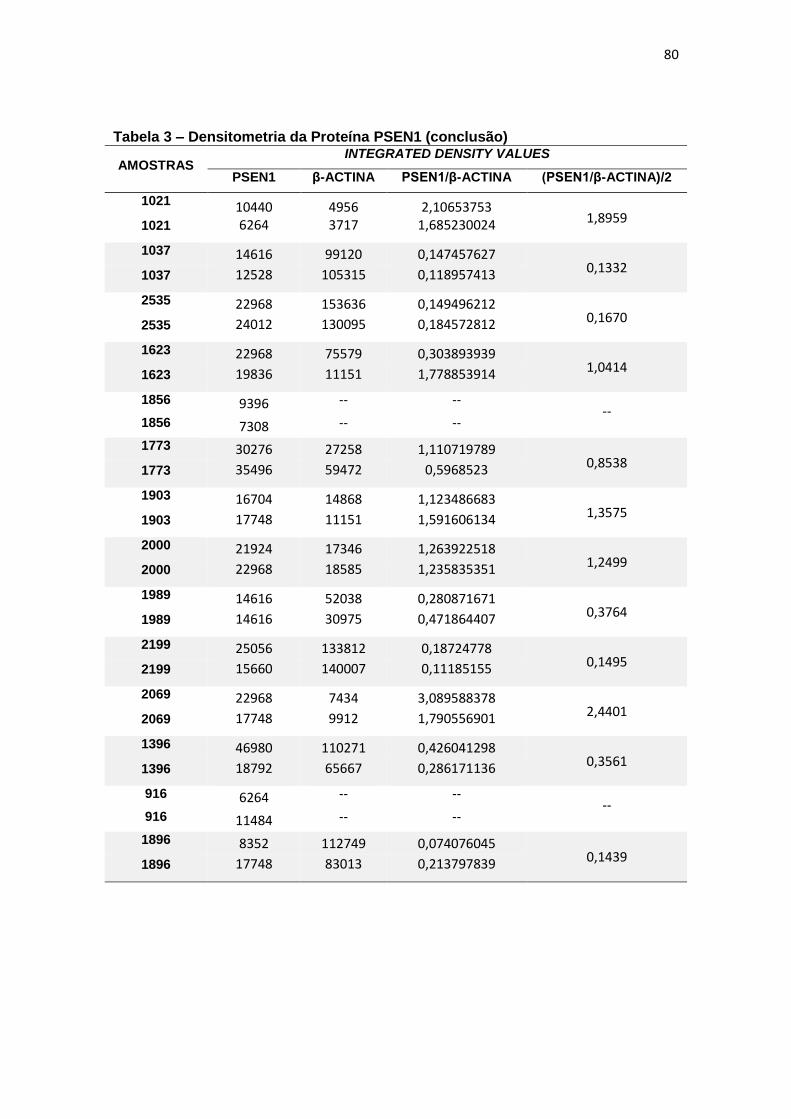
80
Tabela 3 – Densitometria da Proteína PSEN1 (conclusão)
AMOSTRAS INTEGRATED DENSITY VALUES
PSEN1 β-ACTINA PSEN1/β-ACTINA (PSEN1/β-ACTINA)/2
1021 10440 4956 2,10653753 1,8959
1021 6264 3717 1,685230024
1037 14616 99120 0,147457627 0,1332
1037 12528 105315 0,118957413
2535 22968 153636 0,149496212 0,1670
2535 24012 130095 0,184572812
1623 22968 75579 0,303893939 1,0414
1623 19836 11151 1,778853914
1856 9396 -- -- --
1856 7308 -- --
1773 30276 27258 1,110719789 0,8538
1773 35496 59472 0,5968523
1903 16704 14868 1,123486683 1,3575
1903 17748 11151 1,591606134
2000 21924 17346 1,263922518 1,2499
2000 22968 18585 1,235835351
1989 14616 52038 0,280871671 0,3764
1989 14616 30975 0,471864407
2199 25056 133812 0,18724778 0,1495
2199 15660 140007 0,11185155
2069 22968 7434 3,089588378 2,4401
2069 17748 9912 1,790556901
1396 46980 110271 0,426041298 0,3561
1396 18792 65667 0,286171136
916 6264 -- -- --
916 11484 -- --
1896 8352 112749 0,074076045 0,1439
1896 17748 83013 0,213797839

81
ANEXOS
ANEXO A – Parecer Consubstanciado do Comitê de Ética em Pesquisa