Embed Size (px)
Citation preview

UNIVERSIDADE DO ESTADO DO AMAZONAS FUNDAÇÃO DE MEDICINA TROPICAL DR. HEITOR VIEIRA DOURADO
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL DOUTORADO EM DOENÇAS TROPICAIS E INFECCIOSAS
CARACTERIZAÇÃO GENÉTICA E SENSIBILIDADE MICROBIANA DE BACTÉRIAS AERÓBICAS ISOLADAS DE PROCESSOS INFECCIOSOS DE
PACIENTES COM DOENÇAS HEMATOLÓGICAS DA FUNDAÇÃO HEMOAM.
CRISTINA MOTTA FERREIRA
MANAUS 2011

i
CRISTINA MOTTA FERREIRA
CARACTERIZAÇÃO GENÉTICA E SENSIBILIDADE MICROBIANA DE BACTÉRIAS AERÓBICAS ISOLADAS DE PROCESSOS INFECCIOSOS DE
PACIENTES COM DOENÇAS HEMATOLÓGICAS DA FUNDAÇÃO HEMOAM
Tese apresentada ao Programa de Pós-Graduação em Medicina Tropical da Universidade do Estado do Amazonas em Convênio com a Fundação de Medicina Tropical Dr. Heitor Vieira Dourado, para obtenção do grau de Doutor em Doenças Tropicais e Infecciosas.
Orientador (a): Profª Dra. Maria das Graças do Vale Barbosa Co-orientador: Dr. Felipe Gomes Naveca
MANAUS 2011

Ficha Catalográfica
F383c Ferreira, Cristina Motta. Caracterização genética e sensibilidade microbiana das
bactérias aeróbicas isoladas de processos infecciosos de pacientes com doenças hematológicas da Fundação HEMOAM / Cristina Motta Ferreira. -- Manaus : Universidade do Estado do Amazonas, 2011.
208 f. : il.
Tese de Doutorado do Programa de Pós-Graduação em Medicina Tropical da Universidade do Estado do Amazonas em parceria com a Fundação de Medicina Tropical do Amazonas – UEA/FMT, 2011.
Orientadora: Profª. Dra. Maria das Graças do Vale Barbosa. 1. Bactérias aeróbicas. 2. ESBL. 3. Beta-lactamase. 4.
MRSA. 5. SCCmec. I. Título. CDU: 579.84

ii
FOLHA DE JULGAMENTO
CARACTERIZAÇÃO GENÉTICA E SENSIBILIDADE MICROBIANA DE BACTÉRIAS AERÓBICAS ISOLADAS DE PROCESSOS
INFECCIOSOS DE PACIENTES COM DOENÇAS HEMATOLÓGICAS DA FUNDAÇÃO HEMOAM.
CRISTINA MOTTA FERREIRA
“Esta Tese foi julgada adequada para obtenção do Título de Doutor em Doenças Tropicais e Infecciosas, aprovada em sua forma final pelo programa de Pós-Graduação em Medicina Tropical da Universidade do Estado do Amazonas, em convênio com a Fundação de Medicina Tropical Dr. Heitor Vieira Dourado”.
Banca Julgadora:
___________________________________________ Profª. Maria das Graças Vale Barbosa, Dra.
Presidente
_________________________________________ Prof. Marcelo Cordeiro dos Santos, Dr.
Membro
________________________________________________ Prof. Henrique Manuel Condinho da Silveira, Dr.
Membro
____________________________________ Profª. Ivete de Araújo Rolland, Dra.
Membro
____________________________________
Prof. João Vicente Braga de Souza, Dr. Membro

iii
DEDICATÓRIA
Dedico esta grande conquista primeiramente a DEUS Pai, por seu amor incondicional por mim. A Minha família: William, Guilherme e Gabriel. Aos meus pais: Cauby Braga Motta e Ana Leite Motta que sempre me incentivaram.

iv
AGRADECIMENTOS
Universidade do Estado do Amazonas e Fundação de Medicina Tropical Dr. Heitor
Vieira Dourado.
A SUFRAMA pelo suporte financeiro para custear as despesas do Programa de Pós-
Graduação.
A Fundação de Amparo a Pesquisa do Estado do Amazonas- FAPEAM.
A todos os pacientes que participaram desta pesquisa.
Aos servidores da Fundação HEMOAM.
Aos meus orientadores Maria das Graças Vale Barbosa e Felipe Gomes Naveca.
A Dra. Cíntia Mara Costa de Oliveira e equipe.
As minhas alunas do PAIC Mirela Andrade Neves e Andrea Laureano de Carvalho.
A minha equipe de plantão da Maternidade Moura Tapajós, por sua compreensão.
Aos servidores da Fundação de Dermatologia e Venereologia Alfredo da Matta.
A Dra. Leny Nascimento da Motta Passos, Dra. Nazaré Saunier, Dr. Rodrigo Leitão,
Fundação Sangue Nativo e todos os seus funcionários.
Aos meus coordenadores da Universidade Nilton Lins Maria das Graças da Costa
Alecrim e Raphael Borges.
A Conceição Tufic, Secretária do PPGMT, pelo seu carinho e paciência nesta longa
jornada.
A todos os professores e funcionários do curso de Doutorado em Doenças Tropicais
e Infecciosas. Obrigada pela paciência e esforço dedicado.

v
EPÍGRAFE
“O grande barato da vida é olhar para trás e sentir orgulho da sua própria história”.
Carlos Drummond de Andrade

vi
RESUMO
Introdução: Espécies de estafilococos resistentes a meticilina, mundialmente disseminados tanto pelos hospitais como nas comunidades, e de bactérias gram-negativas resistentes aos beta-lactâmicos, ainda permanecem como um severo problema de saúde pública. Objetivo: Descrever a freqüência, sensibilidade a antibióticos e caracterização genética de bactérias patogênicas isoladas de processos infecciosos de pacientes com doenças hematológicas da Fundação HEMOAM. Metodologia: Estudo transversal, descritivo, incluindo-se pacientes de ambos os sexos, qualquer faixa etária, admitidos na Fundação, dos quais foram coletadas amostras biológicas (urina, sangue, escarro, fezes, sangue de cateter, aspirado de medula, abscesso ocular, perianal, secreção de orofaringe, de nasofaringe, feridas e lesão de pele) no período de julho/2007 a agosto/2008. As espécies bacterianas foram identificadas através de testes bioquímicos padronizados de rotina, confirmadas pelo 16S rRNA e seqüenciamento. Nas bactérias gram-positivas a resistência a meticilina foi determinada pelos testes PBP2a e epsilométrico com cefoxitina e oxacilina. A detecção da presença do gene de resistência (MecA, SCCmec), origem dos isolados (comunitários ou hospitalares) e clones estafilocócicos foi realizada através de PCR e genotipagem. Nas bactérias gram-negativas, a determinação do fenótipo ESBL foi realizada através do teste epsilométrico. Os genes envolvidos na produção das ESBLs (TEM-, OXA-, SHV-, CTX-M-) foram identificados através de PCR. Resultados: De 146 amostras biológicas coletadas, foram isoladas 44 espécies bacterianas, sendo 17 gram-negativas e 27 gram-positivas. A doença hematológica mais freqüente foi leucemia linfocítica aguda e leucemia mielóide aguda. O protocolo molecular do 16S rRNA identificou como espécies bacterianas mais freqüentes Staphylococcus epidermidis, S. intermedius, S. aureus, Staphylococcus sp., E. coli, Serratia sp., Serratia liquefaciens e B. cepacia. Das 17 bactérias gram-negativas 12 foram produtoras da enzima ESBL e carrearam os genes blaTEM, blaSHV, blaCTX-M and blaOXA. Das 27 bactérias gram-positivas, dez apresentaram perfil de resistência a meticilina, sendo nove estafilococos coagulase negativos e 01 S. aureus. Os cassetes cromossômicos mec (SCCmec) mais freqüentes foram os dos tipos I, III, e V . O MLST revelou a presença dos seguintes clones: S. aureus ST243 carreando o SCCmec tipo I, III, V; S. epidermidis ST2 carreando o SCCmec tipo II, III, V e de um novo clone de S. epidermidis ST365 carreando o SCCmec tipo I, III, V. Conclusão: A identificação dos clones de S.aureus ST243, S. epidermidis ST2 e ST365, nas espécies de estafilococos meticilina resistentes, e dos genes blaTEM, blaSHV, blaCTX-M e blaOXA nos plasmídeos de cepas produtoras de ESBL, comprovam a presença de espécies bacterianas implicadas em processos infecciosos hospitalares. Estes achados servem como alerta para que sejam introduzidas ações para a redução da disseminação dessas linhagens clonais patogênicas, a fim de se evitar surtos de infecções hospitalares e óbitos, principalmente em pacientes imunodebilitados. Palavras-chave: ESBL, mecA, beta-lactamase, MRSA, SCCmec, ST365, ST243,S. epidermidis, S. aureus, infecção hospitalar.

vii
ABSTRACT
Introduction: Methicillin-resistant Staphylococci and Gram-negative bacteria, being spread worldwide, by both hospitals and in communities still remain as a severe public health problem. Objective: To describe the frequency, sensitivity to antibiotics and genetic characterization of pathogenic bacteria isolated from hematologic patients with infectious processes at the HEMOAM Foundation. Methods: We conducted a descriptive cross-sectional study including patients of both sexes at any age who had been admitted to the HEMOAM Foundation. A total of 146 biological samples such as of urine, blood, sputum, feces, catheter tip, bone marrow aspirate, and abscess fluid along with oropharyngeal, ocular, perianal, nasopharynx, open wound, and skin lesion secretions were collected in the period from July 2007 to August 2008. Bacterial strains identified by standard biochemical tests of laboratorial routine had their species confirmed by 16S rRNA protocols and sequencing. Resistance to methicillin for gram-positive strains was determined by PBP2a and epsolometric tests with cefoxitin/oxacillin. The presence of the resistance gene (mecA and SCCmec), origin of isolates (community/ hospital) and staphylococci clones was performed by molecular tests such as PCR and genotyping. The identification of the ESBL phenotype in gram-negative bacteria was performed by epsolometric test. The identification of the genes involved in the production of ESBLs (TEM-, OXA-, SHV-, and CTX-M-) was performed by molecular tests such as PCR. Bacterial resistance to antimicrobials used in routine therapy was performed using the E-test. Results: We isolated 44 bacterial strains from 146 biological samples of which, 17 were gram-negative and 27 gram-positive. The most frequent hematologic disease was acute lymphocytic leukemia and acute myeloid leukemia. Biochemical tests and 16S rRNA sequencing detected S. epidermidis, S. intermedius, S. aureus, Staphylococcus spp., E. coli, Serratia sp., Serratia liquefaciens and B. cepacia as the most frequent bacteria. 12 of the 17gram-negative bacteria were ESBL-producing and carried blaTEM, blaSHV, blaCTX-M and blaOXA genes. 10 of the 27 gram-positive bacteria were methicillin-resistant of which, nine were coagulase-negative staphylococci and one S. aureus. The most frequent cromossomal cassettes were SCCmec types I, III, V. MLST revealed the presence of the clones: S. aureus ST243 (SCCmec type I, III, V) , S. epidermidis ST2 (SCCmec tipo II, III, V) and new clone of the S. epidermidis ST365(SCCmec type I, III, V). Conclusion: The identification of S. aureus ST243 (SCCmec type I, III, V) clone, S. epidermidis ST2 (SCCmec type II, III, V), ST365 (SCCmec type I, III, V) clones and blaTEM, blaSHV, blaCTX-M , blaOXA genes in Gram-negative bacteria plasmids, proving the presence of pathogenic species implied in hospital infectious processes. These findings appear as an alert to introduce actions to reduce the spread of those pathogenic lineages of S. aureus and S. epidermidis to prevent hospital infection outbreaks as well as mortality risks, mainly for immune compromised patients. Keywords: ESBL, mecA, beta-lactamase, MRSA, SCCmec, ST365, ST243,S. epidermidis, S. aureus, nosocomial infection.

viii
LISTA DE FIGURAS- TESE
Figura 1 Árvore filogenética da vida de organismos com destaque em azul para o grupo das bactérias.......................................................................................... 1
Figura 2 Desenho esquemático da parede celular de Bactéria Gram-positiva.............. 3 Figura 3 Desenho esquemático da parede celular de Bactéria Gram-negativa............. 3 Figura 4 Desenho esquemático de gene mecA............................................................. 16 Figura 5 Classificação dos principais tipos de cassetes................................................ 17 Figura 6 Fluxograma dos procedimentos executados na realização do estudo............ 33
Figura 7 Imagem das fitas de cefinase plus utilizadas na realização do teste da beta-lactamase......................................................................................................... 37
Figura 8 Desenho esquemático das fitas utilizadas na realização do Etest ESBL com os gradientes de concentrações dos antibióticos CT e TZ e em associação com o inibidor, o ácido clavulânico.................................................................. 38
Figura 9 Imagem de detecção do fenótipo ESBL pelo método E-teste Fita com os gradientes de concentrações dos antibióticos TZ e em associação com o inibidor, o ácido clavulânico............................................................................. 39
Figura 10 Imagem do resultado do teste de suscetibilidade método E-teste para a detecção da MIC (concentração inibitória mínima).......................................... 40
Figura 11 Protocolos laboratoriais- Figura1 e 2- Imagem da fita de E-teste para ESBL com a escala em MIC aplicadas no meio de cultura....................................... 167
Figura 12 Protocolos laboratoriais- Padrões de inibição do crescimento- Fig 4 –corte ESBL positiva................................................................................................... 172
Figura 13 Protocolos laboratoriais- Padrões de inibição do crescimento- Fig 5 –zona fantasma.......................................................................................................... 172
Figura 14 Protocolos laboratoriais- Padrões de inibição do crescimento- Fig 6 –de formação da inibição da elipse......................................................................... 172
Figura 15 Protocolos laboratoriais- Padrões de inibição do crescimento- Fig 7 –resultado não determinado.............................................................................. 172

ix
LISTA DE TABELAS-TESE
Tabela 1 Meios de culturas de triagem para semeadura primária........................... 105 Tabela 2 Divisão dos cocos Gram positivos pela prova da catalase....................... 119
Tabela 3 Provas bioquímicas para identificação dos gêneros de Cocos Gram-positivos.................................................................................................... 113
Tabela 4 Identificação das espécies de cocos Gram-positivos catalase positivos.. 113 Tabela 5 Esquema para diferenciação de espécies de Cocos Gram-positivos....... 114 Tabela 6 Identificação de gênero e espécie bacteriana.......................................... 120 Tabela 7 Diferenciação das enterobactérias pelo Enterokit C................................ 124 Tabela 8 Características dos enteropatógenos da família Enterobacteriaceae nos
meios EPM-MILi........................................................................................ 128 Tabela 9 Glicose O/F............................................................................................... 130 Tabela 10 Valores das provas de oxidase e motilidade............................................ 133 Tabela 11 Valores das provas de OF/MC e morfologia............................................ 133 Tabela 12 Valores dos testes.................................................................................... 134 Tabela 13 Identificação de isolados Não fermentadores........................................... 135 Tabela 14 Valores das provas bioquímicas............................................................... 137 Tabela 15 Leitura e interpretação dos resultados da PBP 2a................................... 144 Tabela 16 Tipos de reação de aglutinação................................................................ 144 Tabela 17 Resultados do teste com 201 isolados clínicos frescos de S aureus de
quatro laboratórios.................................................................................... 146 Tabela 18 Guia para interpretação do Eteste ESBL.................................................. 156

x
LISTA DE TABELAS- ARTIGO 1
Tabela 1 Frequency of hematologic diseases diagnosed in this study…………………. 58 Tabela 2 Primer sequence and PCR conditions…………………………………………… 57 Tabela 3 Gram-negative strains, ESBL phenotypes and genes detected from patients
with infections………………………………………………………………………. 60 Tabela 4 Antimicrobial Susceptibility test (E-test) for 17Gram-negative bacteria
isolated from patients with bacterial infection…………………………………… 61 Tabela 5 Strains isolated from different specimen type detected from patients with
infections……………………………………………………………………………. 58

xi
LISTA DE TABELAS- ARTIGO 2
Tabela 1 Strains isolated from patients with hematologic disease……………………… 74
Tabela 2 Phenotypes and genotypes profiles of the Staphylococcal strains isolated from patients with hematologic diseases……………………………………… 75

xii
LISTA DE QUADROS
Quadro 1 Mecanismo de ação das drogas antimicrobianas......................................... 10 Quadro 2 Características diferenciais entre HA-MRSA e CA-MRSA........................... 22
Quadro 3 Protocolo de interpretação do Etest® ESBL............................................ 38 Quadro 4 Iniciadores mecA e SCCmec utilizados na reação.............................. 44 Quadro 5 Iniciadores das toxinas bacterianas utilizados na reação................... 46 Quadro 6 Iniciadores das ESBLs ........................................................................ 46

xiii
ABREVIATURAS ATCC American Type Culture Collection CA-MRSA Community acquired methicillin resistant Staphylococcus aureus CLSI Clinical and Laboratorial Standard Institute CVI Complexo violeta-iodo ET-A Esfoliatin Toxin A ET-B Esfoliatin Toxin B ESBL Extended spectrum beta –lactamase HA- MRSA Hospital acquired methicillin resistant Staphylococcus aureus HA- MR-SCoN Hospital acquired methicillin resistant Staphylococcus sp. LOS Lipooligossacarídeo LUKPV ou PVL Panton Valentine Leucocidin MHC II Complexo de histocompatibilidade tipo II MIC Minimal inhibitory concentration Meio de cultura MIO Motilidade, Indol, Ornitina MRSA Methicillin- or oxacillin-resistant S. aureus MR-SCoN Methicillin- or oxacillin-resistant Staphylococcus sp. MSSA Methicillin- or oxacillin-sensitive S. aureus PBP 2a ou PBP 2' Penicillin-binding protein 2a or 2’ PCR Polymerase Chain reaction PFGE Pulsed-field gel electrophoresis Phantom zone Zona fantasma SCCmec Staphylococcal chromosomal cassette mec SCoN Coagulase negative Staphylococci TSI Tríplice Açúcar Ferro

xiv
SUMÁRIO
1 INTRODUÇÃO .................................................................................................... 1 1.1 Histórico .............................................................................................................. 1 1.2 Morfologia bacteriana........................................................................................... 2 1.3 Parede celular de bactéria gram-positiva............................................................. 2 1.3.1 Parede celular de bactéria Gram-negativa.......................................................... 3 1.4 Relações microbiota normal/parasita/hospedeiro................................................ 4 1.5 Patogenicidade e virulência bacteriana............................................................... 5 1.5.1 Patogenicidade de cocos Gram-positivos............................................................ 6 1.5.2 Patogenicidade de bastonetes Gram-negativos.................................................. 7 1.6 Drogas antimicrobianas....................................................................................... 8 1.6.1 Mecanismo de ação das drogas antimicrobianas................................................ 9 1.7 Resistência bacteriana aos antimicrobianos........................................................ 9 1.7.1 Resistência dos S aureus e Staphylococcus spp aos ß-lactâmicos.................... 12 1.8 Gene mecA......................................................................................................... 14 1.8.1 Complexo genético mec....................................................................................... 15 1.8.2 Complexo genético ccr e principais tipos de SCCmec........................................ 15 1.9 Staphylococcus spp hospitalares e comunitários................................................ 17 1.9.1 Prevalência dos estafilococos resistentes a meticilina........................................ 18 1.9.2 Processos infecciosos ocasionados por estafilococos resistentes a meticilina... 18 1.9.3 Cassetes cromossômicos e clones estafilocócicos............................................. 19 1.9.4 Características diferenciais entre HA-MRSA e CA-MRSA................................... 20 1.10 Prevalência de bactérias Gram-negativas produtoras de ESBLs........................ 21 1.10.1 Enzima Beta-Lactamase...................................................................................... 23 1.10.2 Classificação das Enzimas Beta-Lactamases..................................................... 24 1.10.3 Beta-Lactamase de espectro estendido (ESBL).................................................. 25 1.10.4 Subtipos de enzimas (ESBL)............................................................................... 26 1.10.5 Importância Clínica das ESBL............................................................................. 27 1.10.6 Preocupações relativas às infecções bacterianas............................................... 28 2 OBJETIVOS......................................................................................................... 31 2.1 Geral..................................................................................................................... 31 2.2 Específicos........................................................................................................... 31 3 MATERIAL E MÉTODOS.................................................................................... 32 3.1 Delineamentos...................................................................................................... 32 3.2 População de estudo............................................................................................ 32 3.3 Amostragem......................................................................................................... 32 3.4 Critérios de inclusão............................................................................................. 34 3.5 Critérios de exclusão............................................................................................ 34 3.6 Solicitação de exame bacteriológico.................................................................... 34 3.6.1 Pacientes hematológicos imunodeprimidos......................................................... 34 3.6.2 Fatores de baixo risco para complicação............................................................. 35 3.7 Coleta de amostra biológica................................................................................. 35 3.8 Cultura, isolamento e identificação de bactérias isoladas.................................... 35 3.9 Teste da beta-lactamase...................................................................................... 36 3.9.1 Teste da beta-lactamase de espectro estendido (ESBL)..................................... 36

xv
3.9.2 Teste de suscetibilidade antimicrobiana............................................................... 39 3.9.3 Teste da PBP 2a.................................................................................................. 40 3.9.4 Caracterizações moleculares............................................................................... 40 3.9.4.1 Extração do DNA Total Bacteriano e Quantificação........................................... 40 3.9.4.2 Extração do DNA plasmidial................................................................................ 41 3.9.4.3 Reação em cadeia da polimerase (PCR)............................................................ 42 3.9.4.3.1 Para o gene 16S rRNA........................................................................................ 42 3.9.4.3.2 PCR em tempo real (q-PCR) para o gene mecA................................................ 44 3.9.4.3.3 Gene mecA........................................................................................................... 43 3.9.4.3.4 Tipos de SCCmec................................................................................................ 45 3.9.4.3.5 Genes seh, etd, arcA e LUKPV das toxinas bacterianas.................................... 45 3.9.4.3.6 Genes das Beta-lactamases de espectro estendido (ESBLs)............................. 46 3.9.4.3.7 Genotipagem para identificação dos clones bacterianos..................................... 47 3.10 Purificação pré-sequenciamento.......................................................................... 48 3.11 Precipitação do DNA com Isopropanol/etanol...................................................... 48 3.12 Sequenciamento................................................................................................... 49 3.13 Controle de Qualidade.......................................................................................... 49 3.14 Armazenamentos de amostras............................................................................. 50 3.15 Análise de Dados................................................................................................. 50 3.16 Fontes de financiamento (Nacionais e/ou Internacionais)................................... 50 4. RESULTADOS..................................................................................................... 51 4.1 Artigo 1................................................................................................................. 52 4.2 Artigo 2................................................................................................................. 71 5. DISCUSSÃO........................................................................................................ 80 6 CONCLUSÃO...................................................................................................... 83 7 REFERÊNCIAS.................................................................................................... 85 8 ANEXOS............................................................................................................... 103 8.1 Anexo 1-TCLE...................................................................................................... 103 8.2 Anexo 2- Protocolo médico/laboratorial................................................................ 105 8.3 Anexo 3- Protocolos laboratoriais......................................................................... 107

1
1 INTRODUÇÃO 1.1 Histórico
As bactérias são os organismos mais antigos, procariontes, constituídos por
uma célula, sem organelas e núcleo celular, encontradas na forma isolada ou em
colônias, pertencentes ao Domínio homônimo Bactéria. Foram descobertas por
Antoni van Leeuwenhoek em 1683 após observar resíduos de seus próprios dentes
e de outros materiais ao microscópio (3 ,5 ,59, 122).
No final do século XIX, as bactérias passaram a despertar o interesse de
muitos cientistas após Louis Pasteur e Robert Koch descreverem o seu papel como
vetores de várias doenças e a demonstração da origem bacteriana de diversas
doenças como o tifo, lepra e gonorréia (3 ,5 ,98, 122).
Atualmente as bactérias compõem um dos três domínios do sistema de
classificação cladístico. Inicialmente foram classificadas entre as plantas por Lineu e
agrupadas como fungos. Em 1866, Ernst Haeckel as inseriu no reino Protista (Figura
1). Em 1969 foram incluídas entre os procariotas no reino Monera por Whittaker. Em
1977, Carl Woese dividiu os procariotas em dois grupos, com base nas seqüências
“16S” do rRNA, que denominou de Eubacteria e Archaebacteria, e mais tarde,
renomeados pelo mesmo para Bacteria e Archaea devido as mesmas formarem
domínios separados com origem e evolução separadas a partir de um organismo
primordial (3 ,5 ,59, 122, 132,143).
Fonte: http://nai.arc.nasa.gov/news_stories/news_detail.cfm?ID=274.
Figura 1 – Árvore filogenética da vida de organismos, com destaque em azul para o
grupo das bactérias

2
1.2 Morfologia Bacteriana Morfologicamente as bactérias estão classificadas de acordo com a forma da
célula em cocos, bacilos, vibriões, espirilos, espiroquetas e, quanto ao grau de
agregação, os cocos podem se apresentar de forma isolada, aos pares (diplococos),
ligados em cadeias (colar), forma de cachos, cúbica (sarcina), bacilos aos pares e
alinhados em cadeia. Já os bacilos podem se dividr ao longo de seu eixo curto;
assim, existem menos grupamento de bacilos que de cocos. A maioria deles se
apresenta isolados. Os diplobacilos aparecem após a divisão e os estreptobacilos
ocorrem em cadeias (127,142).
1.3 Parede celular de Bactéria Gram-positiva Na maioria das bactérias Gram-positivas a parede celular é constituída por uma
espessa camada de peptideoglicano, ácido teicóico que se divide em ácido
lipoteicóico, atravessa a camada de peptideoglicano e se liga a membrana
plasmática, e o ácido teicóico da parede celular, ligado à camada de peptideoglicano
(Figura 2) (18, 129,142).
O principal componente da parede é o peptideoglicano, um polímero de longas
cadeias de glicídio com ligações cruzadas flexíveis de peptídeos formando uma
estrutura forte, em forma de rede ao redor da bactéria. Esta rede é formada por
dissacarídeo denominado de N-acetilglucosamina e ácido N-acetil murâmico, ligado
ao peptídeo L-lisina,D-alanina, L-alanina, D-glutamina, que permite a ligação
cruzada com um grupamento de pentaciclinas. Reação de transpeptidação une os
peptídeos enquanto que os dissacarídeos são polimerizados ao longo da cadeia de
glicano por meio de reação de transglicolização. O processo da ligação cruzada é
catalisado pelas enzimas transpeptidases, as proteínas ligadoras de penicilina
PBPs(18, 129, 142).

3
Fonte: http://vsites.unb.br/ib/cel/microbiologia/morfologia1/pcgpos.gif
Figura 2 – Desenho esquemático da parede celular de Bactéria Gram-positiva
1.3.1 Parede celular de Bactéria Gram-negativa
Consiste de uma fina camada de peptideoglicano e membrana externa. O
peptideoglicano está ligado a lipoproteínas e fosfolipídeos na membrana externa e
localiza-se no periplasma, um espaço fluido entre a membrana externa e membrana
plasmática (Figura 3) (129, 142).
A membrana externa possui o lipopolissacarídeo (LPS) formado por uma porção
lipídica denominada de lipídeo A (endotoxina) e pelo açúcar polissacarídeo “0” que
atua como antígeno (121,142). Algumas bactérias Gram-negativas possuem lipo-
oligossacarídeos (LOS) e não LPS em suas paredes celulares (46, 147). O LOS contêm
lipídeo A e um oligossacarídeo central sem antígeno O, polissacarídeo de cadeia
longa como o encontrado no LPS de bactérias entéricas (22).
Figura 3 – Desenho esquemático da parede celular de bactéria Gram-negativa

4
A técnica de coloração do Gram desenvolvida por Hans Christian Joacquin Gram
em 1884, beneficiou a microbiologia, pois classificou as bactérias em dois grandes
grupos: Gram-positivas e Gram-negativas (78,142). Esta técnica baseia-se na
composição química e integridade da parede celular das duas bactérias e,
dependendo da cor que adquirirem, são classificadas em Gram-positivas (coradas
em roxo) e Gram-negativas (vermelho) (78,142).
As características estruturais da parede bacteriana compõem a base da técnica
que funciona da seguinte forma: Ao se adicionar o corante primário cristal violeta e o
mordente lugol, ambos se combinam no citoplasma de cada bactéria e as coram de
violeta escura ou púrpura, formando um complexo grande denominado de complexo
violeta-iodo (CV-I) ou iodopararrosanilina (78, 127, 142).
Ao se proceder a descoloração com álcool-acetona, devido ao fato de a parede
celular das Gram-positivas possuírem uma espessa camada de peptideoglicano, o
complexo CV-I não é arrastado e permanece intacto na mesma. Já nas Gram-
negativas, devido o peptideoglicano ser constituído por apenas uma fina camada, o
complexo CV-I é arrastado tornando a bactéria incolor (78,127,142).
Procede-se então a coloração de fundo com o contracorante safranina para que
possa tingir as estruturas incolores, permanecendo as já coradas da mesma cor. As
bactérias descoradas absorvem a safranina tornando-se assim vermelhas. Por isso,
as bactérias Gram-negativas coram-se em vermelho e as Gram-positivas em azuis (78,127,142).
1.4 Relações microbiota normal/parasita/hospedeiro
Os agentes etiológicos de doenças infecciosas humanas pertencem a cinco
grupos principais de organismos: bactérias, fungos, protozoários, helmintos e vírus,
os quais possuem características fenotípicas e genotípicas essenciais que facilitam
sua identificação (21, 36, 110, 127, 132). Esses agentes podem co-existir em conjunto com
o hospedeiro (127) beneficiando-o, impedindo assim o crescimento de microrganismos
nocivos. Tal relação é denominada de antagonismo microbiano, pois envolve uma

5
competição entre as bactérias da microbiota normal do corpo humano e os
microrganismos nocivos (138, 142).
A microbiota normal, cuja função é a manutenção da saúde corporal do
hospedeiro, compete com tais microrganismos por nutrientes, produzindo
substâncias nocivas aos mesmos, afetando o pH da pele, produzindo bacteriocinas
e alterando, até mesmo o oxigênio disponível, objetivando assim evitar a
colonização por agentes patogênicos. A microbiota normal do hospedeiro coloniza
os olhos, pele, trato gastrointestinal, trato genito-urinário e nariz de ambos os sexos.
Isto pode ocorrer de forma saprofítica, comensal, mutualística ou parasitária,
podendo haver uma relação de cooperação entre os microrganismos ( 78, 127, 142).
Em algumas situações esta cooperação pode ocasionar doenças ao hospedeiro.
Certas espécies de estreptococos patogênicos utilizam os seus receptores de
adesão não para se fixar aos dentes, mas sim em outros estreptococos orais,
ocasionado assim processos infecciosos como gengivites e doenças periodônticas
ao hospedeiro (78, 142).
1.5 Patogenicidade e virulência bacteriana
A virulência dos microrganismos não é atribuída somente a um fator único,
mas sim a vários fatores que estão relacionados ao organismo, hospedeiro e
interação entre os mesmos. A virulência envolve a infectividade do microrganismo e
a gravidade da condição produzida. A capacidade de virulência entre as cepas pode
ocorrer numa mesma espécie ou em um determinado grupo de microrganismos
considerados patogênicos. A capacidade de um microrganismo de causar doença
envolve tanto fatores microbianos como do hospedeiro. Os microrganismos
desenvolvem respostas adaptativas geneticamente modificadas a fim de lhes
proporcionar versatilidade, sobrevivência e desenvolvimento em ambientes hostis (127).
As bactérias desenvolveram métodos que as auxiliaram a captar alterações
no ambiente externo e a expressar seus genes envolvidos nas respostas
adaptativas. Quando a população atinge um determinado limiar, há um auto-indutor

6
suficiente que ativa os fatores de transcrição na célula e expressam os genes de
fatores de virulência, mecanismo este denominado de “sensores em quórum” (47,
127, 100).
Os fatores de virulência possibilitam as bactérias causar danos ao
hospedeiro. Alguns deles estão associados a células ou podem ser extracelulares,
ou então, fazerem parte da composição anatômica ou fisiológica da célula. Dentre os
diversos fatores de virulência das bactérias podem-se citar as adesinas (auxiliam na
adesão às células do hospedeiro como as proteínas ligadoras de colágeno, de
aderência extracelular, de ligação a fibronectina e fator de aglutinação), agressinas
(incluem cápsulas, substâncias viscosas extracelulares, proteínas, carboidratos de
superfície e outras pequenas moléculas) (78,127,142), toxinas (síndrome do choque
tóxico estafilocócico-TSST-1(19,20), enterotoxina, toxina da síndrome da pele
escaldada (125), endotoxinas, exotoxinas (78,127,142) e enzimas (hialuronidase,
estafiloquinase, estreptoquinase(109), coagulase (54,133), lipase, hemolisinas (40),
proteases, leucocidinas como a Panton Valentine(71), determinantes de superfícies e
de resistência, responsáveis pela patologia ocasionada pelas mesmas.
1.5.1 Patogenicidade de Cocos Gram-positivos
O gênero Staphylococcus é composto por diversas espécies, muitas das
quais podem ser encontradas em amostras biológicas humanas. Dentre elas a mais
importante é o Staphylococcus aureus. Apesar de fazerem parte da microbiota
normal humana, as espécies de estafilococos podem produzir infecções oportunistas
significativas em condições apropriadas (78,127,142). Estas espécies possuem vários
fatores de virulência como polissacarídeos capsulares, peptideoglicano, ácidos
teicóicos, proteína A, hemolisinas como a Panton Valentine (71), toxinas (esfoliatina
(ET-A e ET-B) (136), enterotoxinas (A, até E, H e I) (69) e superantígenos bacterianos
(enterotoxinas estafilocócicas, TSST-1,exotoxinas pirogênicas estafilocócicas (SPE
A, B, C, F, G, H e J) e principalmente biofilmes (19, 78,142).
A superantigenicidade se refere à capacidade das toxinas de estimularem a
proliferação maciça de células T, sem relação com a especificidade antigênica das
mesmas. Ocorre uma indução da proliferação de células T policlonais após a ligação

7
do MHC II de uma célula apresentadora de antígeno com os receptores de células T
e a porção variável da cadeia ß (beta) do receptor de antígenos das células T.
Ocorre então uma liberação maciça de citocinas produzindo os sintomas da TSST-1
que incluem febre, exantema, descamação, hipotensão, comprometimento
gastrintestinal, muscular, das mucosas, renal, hepático, hematológico e sistema
nervoso central. Algumas toxinas como a Panton-Valentine e a TSST-1 também
podem ser produzidas pelos estafilococos coagulase negativos (127,142).
A ação da Panton-Valentine, leucotoxina formadora de poro de membrana,
inicialmente conhecida como substância leucocidina por Van deVelde em 1894, por
sua capacidade de lisar leucócitos, ocorre através da liberação de dois componentes
pertencentes as classes proteína S (LukS-PV) e proteína F (LukF-PV); antes de se
tornarem um heptâmero formador de poros de membrana nos leucócitos
polimorfonucleares (PMNs). Quando o LukS-PV se liga aos PMNs sofre fosforilação
pela proteína quinase do hospedeiro (A ou C) através da indução dos canais de íons
cálcio. Ocorre a indução do sinal de eventos de transdução que levam a liberação de
interleucinas e mediadores inflamatórios. Dependendo da concentração da PVL,
pode ocorrer lise dos PMNs ou apoptose, sendo que esta última pode envolver a
formação de poros na membrana mitocondrial (21).
Tanto os Staphylococcus aureus quanto os Staphylococcus spp. podem
ocasionar uma variedade de processos patogênicos como furúnculos, carbúnculos,
impetigo, mastite, hidradenite supurativa, infecções de feridas, meningite,
pericardite, osteomielite, piomiosite, intoxicação alimentar, síndrome da pele
escaldada, síndrome do choque tóxico, bacteremia, endocardite, infecções
pulmonares e artrite séptica (127,142).
1.5.2 Patogenicidade de Bastonetes Gram-negativos
As bactérias Gram-negativas pertencentes às Enterobacteriaceae estão
amplamente distribuídas na natureza, sendo encontradas no solo, na água, em
vegetais e no trato intestinal humano. Os membros da família Enterobacteriaceae
podem ser agentes patológicos de qualquer tipo de doença infecciosa e isolados de

8
qualquer tipo de amostra recebida no laboratório. Os pacientes debilitados ou
imunocomprometidos são altamente suscetíveis às infecções hospitalares após
serem colonizados por cepas ambientais, ou por processos invasivos como
cateterismo ou biópsias cirúrgicas após as mucosas serem traumatizadas (128,142).
Quando os mecanismos de defesa do hospedeiro são anulados devido a
causas não-infecciosas subjacentes ou então devido a intervenções diversas como
neoplasisas, terapia imunossupressora ou diabetes, os microrganismos da própria
flora (endógenos) podem ocasionar processos patológicos como uma nova resposta
adaptativa ao seu hospedeiro (127,142).
Estas espécies podem produzir vários fatores de virulência como as
endotoxinas (lipopolissacarídeo-LPS e lipooligossacarídeos-LOS), exotoxinas
(exotoxina A), toxinas (enterotoxinas termolábel ou termoestável), viscosidade
extracelular, leucocidina, proteases (IgA), fosfolipases, enzimas, fímbrias (Pili) (1),
PorI-PorA/PorB, proteínas Opa (proteína II) (53), proteína Rmp, cápsulas, plasmídeos (6), hemaglutininas (127, 94).
As bactérias Gram-negativas podem ocasionar uma série de doenças
dependendo do tipo de tecido que podem infectar como feridas, infecções urinárias,
infecções de garganta, meninigite, infecções de ouvido, septicemias, disenterias,
gastrenterites, síndrome similar ao choque tóxico estafilocócico e bacteremias (127,142).
1.6 Drogas antimicrobianas Os antimicrobianos conhecidos como antibióticos, são produtos químicos
produzidos por microrganismos ou sinteticamente, podendo ter diferentes espectros
de ação. Os antibióticos de estreito espectro, são aqueles que agem somente
contra determinados tipos de bactérias (penicilina G, glicopeptídeos, macrolídeos,
nitrofurantoína, metronidazol, aztreonam, ácido nalidíxico e fosfomicina), enquanto
que os de largo espectro, afetam amplo número de bactérias como as Gram-
positivas e Gram-negativas (carbapenêmicos,cefalosporinas,ß-lactâmicos e

9
combinações de inibidores de ß-lactâmicos e fluoroquinolonas). Estes antibióticos
geralmente são utilizados num processo infeccioso no qual o patógeno não é
imediatamente conhecido. Os antibióticos também podem se classificar como
bactericidas (destroem os patógenos) ou bacteriostáticos (inibem o crescimento
do patógeno) (95, 109, 127,142).
1.6.1 Mecanismo de ação das drogas antimicrobianas
As drogas antimicrobianas agem através de diferentes mecanismos de ação:
inibição da síntese da parede celular, inibição das β-lactamases bacterianas, dano
a membrana citoplasmática, inibição da síntese protéica, inibição da síntese de
ácido nucléico e inibição da síntese de metabólitos essenciais (quadro 1) (95, 96, 126,
142).
1.7 Resistência Bacteriana aos Antimicrobianos
As bactérias existem há milhões de anos e os mecanismos de defesa
desenvolvidos pelas mesmas, contra o ataque de outras bactérias, também são
utilizados para os antibióticos. Os mecanismos de resistência microbiana são
complexos, variados, não completamente conhecidos, que existem antes mesmo da
introdução dos antibióticos. Surgiram devido a mutações dos genes microbianos e
pela aquisição de genes oriundos de fontes externas. O surgimento de múltiplas
espécies bacterianas resistentes aos agentes antimicrobianos ainda constitui um
sério problema para os clínicos. Muitos fatores contribuíram para o aparecimento de
isolados multiresistentes, dentre eles o uso indiscriminado de antibióticos, elevação
da população de risco, pacientes imunodebilitados, tempo de internação prolongado
de pacientes portadores de doenças crônicas debilitantes nos hospitais, enfermarias,
Unidade de Terapia Intensiva (UTI), falhas nas práticas de controle de infecção e
creches (34, 44).

10
Quadro1. Mecanismo de ação das drogas antimicrobianas
Agente antimicrobiano Mecanismo de ação
β-lactâmicos (penicilinas) e β-lactâmicos
semi-sintéticos (meticilina,nafcilina,
oxacilina), cefalosporinas, imipenem,
aztreonam, vancomicina,bacitracina.
Ácido clavulânico (adicionado ao β-
lactâmico amoxacilina)
Inibição da síntese da parede celular Inibe na parede celular a síntese
do peptideoglicano e mureína.
Inibição das β-lactamases (penicilinases) Inibe as enzimas β-lactamases bacterianas
cloranfenicol, clindamicina, eritromicina
aminoglicosídeos e tetraciclinas
Inibição da síntese protéica Age na subunidade ribosomal 50S
Age na subunidade ribosomal 30S
Quinolona, ciprofloxacina, rifampicina Inibição da síntese de ácidos nucléicos Inibe a síntese de DNA
Inibe a síntese de mRNA
Análogo ao fator de crescimento
(sulfanilamida, acetil sulfisoxazol,
trimetoprima
Inibição da síntese de metabólitos essenciais Inibe o metabolismo do ácido fólico (anti-
folato)
Polimixina Dano a membrana citoplasmática Fonte: (Tenover 2006 (126), Winn et al. 2008 (142))
A resistência bacteriana pode se expressar de forma constitutiva, quando é
produzida pela bactéria independentemente da presença da droga, ou ser induzida,
quando é produzida após a exposição da bactéria a droga, como por exemplo, a
enzima beta-lactamase que hidrolisa o antibiótico. A resistência também pode ser
adquirida. Neste caso podem ocorrer alterações mutacionais no genoma bacteriano
ou serem adquiridos materiais genéticos codificados transferidos por conjugação
como plasmídeo, transposon ou seqüências de inserção (31, 44, 127).
Os plasmídeos contêm genes de resistência a uma ampla variedade de
antibióticos, portanto, a sua transferência horizontal (ou lateral) entre bactérias não
importando a espécie, pode acarretar na aquisição de vários determinantes de

11
resistência tornando a espécie resistente a vários tipos de antimicrobianos. A
transferência horizontal é responsável pela disseminação de numerosos
determinantes de resistência antimicrobiana entre as bactérias, o que justifica a
rápida disseminação dos genes de resistência (31, 44). Ela pode ocorrer por:
a) conjugação- requer contato entre as células, sendo um processo no qual
os plasmídeos ou transposons são trasnferidos do doador para o receptor.
b) Transformação- envolve a captura e incorporação do DNA nú.
c) Transdução- quando o DNA do hospedeiro é transferido via bacteriófago,
que age como vetor para sua injeção na célula receptora. A transdução é
um mecanismo extremamente importante de transferência genética entre
as bactérias resistentes aos antibióticos (142).
Os mecanismos de resistência residem em três categorias: a) inativação
enzimática (beta-lactamases); b) receptores alterados (alterações nos ribossomas,
na DNA girase ou enzimas bacterianas alteradas); c) alteração do transporte de
antibióticos (diminuição da força motriz da proteína, alterações nas proteínas da
membrana externa (porinas) e o transporte ativo por células bacterianas) (31,143).
Dentre os citados, o mecanismo de modificação do alvo é utilizado por um grande
número de espécies bacterianas contra uma ampla variedade de antibióticos, sendo
o de indução ou inativação enzimática o mais predominante (31).
Como as drogas β-lactâmicas são amplamente utilizadas devido a sua
elevada efetividade, baixo custo e mínimo efeito colateral, o alvo dos antibióticos β-
lactâmicos nas espécies de estafilococos reside nas quatro proteínas nativas PBP,
envolvidas na síntese da parede celular bacteriana. As bactérias resistentes inativam
os β-lactâmicos por três mecanismos primários (16, 126, 141):
1˚ Pelas β-lactamases, que são enzimas produzidas por um largo grupo de
bactérias Gram-positivas e negativas, que inativam os β-lactâmicos pela
hidrólise do anel β-lactâmico.
2˚ Pela aquisição da proteína ligadora de penicilina 2a (PBP2a) de baixa
afinidade ou alteração do alvo do antibiótico.
3˚ Evitando o acesso do antibiótico ao seu alvo pela alteração da
permeabilidade da parede ou por efluxo.

12
Os mecanismos 1 e 2 estão relacionados entre si e podem co-existir na
mesma célula (16, 80).
1.7.1 Resistência dos S. aureus e Staphylococcus spp. aos ß-lactâmicos A resistência bacteriana dos estafilococos aos antibióticos ainda continua
sendo um desafio para a terapia, tanto em infecções nosocomiais quanto as
adquiridas na comunidade. Nos Estados Unidos, dois milhões de pessoas são
acometidas por infecções e 60% delas, são ocasionadas por bactérias resistentes a
antibióticos. A resistência dos estafilococos a oxacilina, antibiótico atualmente
utilizado como representante da classe PPR (oxacilina, meticilina, nafcilina,
cloxacilina, dicloxacilina), deve-se a expressão da proteína PBP 2a ou PBP 2’,
ausente nos Staphylococcus spp. sensíveis a oxacilina. O outro mecanismo de
resistência é pela produção da enzima beta-lactamase (β-lactamase) que hidrolisa o
antibiótico (108, 115).
Os Staphylococcus spp. sensíveis a meticilina, possuem cinco PBPs, que são
enzimas que se localizam na parede celular bacteriana e catalisam a etapa final de
formação da mesma. As proteínas ligadoras de penicilina 1, 2, 3, 3’,4 tem afinidade
por antibióticos β-lactâmicos. A resistência a meticilina é derivada da produção de
uma PBP adicional denominada de PBP 2a ou PBP 2’, que possui baixa afinidade
por drogas β-lactâmicas, sendo codificada pelo gene mecA (115).
Além da alteração na PBP 2a, os Staphylococcus spp podem apresentar mais
dois mecanismos interagindo entre si, como a hiperprodução de β-lactamases
(BORSA) e modificações na capacidade de ligação das PBPs (MODSA). Entretanto,
o fato de portar o gene não significa que a bactéria seja resistente, pois ele pode ter
um baixo nível de expressão e assim, o desenvolvimento da resistência bacteriana,
fica dependente do modo e do nível de expressão desse gene (42, 81, 105, 108, 115).
As β-lactamases dos estafilococos são enzimas codificadas por plasmídeos
extracelulares, que podem ser dividas em quatro tipos, de A até D. Diferem das β-
lactamases das bactérias Gram-negativas por serem expressas em altos níveis na

13
presença dos indutores, exceto o tipo D, como penicilina ou seus análogos. A
expressão do gene blaZ das β-lactamases está sob o controle de dois genes
regulatórios adjacentes, o indutor blaRI e o gene repressor blaI. A inibição ocorre
quando o produto do gene blaI liga-se ao operador do gene estrutural blaZ,
ocorrendo então o bloqueio da transcrição do gene. A transcrição ocorre quando o
produto de clivagem do BLAR1 cliva a proteína blaI permitindo que o gene seja
transcrito (112, 113).
Alguns genes independentes do lócus mec, denominados de fem (fatores
essenciais para a expressão de resistência) contribuem para a resistência aos β-
lactâmicos. Estão presentes no cromossomo dos Staphylococcus spp. resistentes a
meticilina. Dois genes, femA e femB, estão envolvidos na formação da ligação
cruzada dos precursores da cadeia de petideoglicano. A disfunção de um dos dois
diminui o conteúdo de glicina nos precursores do peptideoglicano e confere
suscetibilidade a é necessária para elevar a atividade autolítica nos isolados que
possuem o gene mecA (57, 67, 130). meticilina. A presença do femA
Os Staphylococcus spp muitas vezes expressam resistência heterogênea,
significando que seu crescimento em presença dos antibióticos β-lactâmicos,
seleciona subclones altamente resistentes em uma população com MICs baixos
para meticilina. A causa da heteroresistência são fatores cromossômicos cuja
atividade afeta o nível da resistência. Isto pode ocasionar erros laboratoriais nos
testes convencionais de cultura, podendo as mesmas serem diagnosticadas como
isolados MSSA e assim, tornarem-se um reservatório nos ambientes hospitalares (39,
42, 107, 115, 117).
Nos S. aureus e Staphylococus coagulase negativa (SCoN), o gene de
resistência a meticilina denominado de mecA, localiza-se em um elemento genético
móvel denominado de cassete cromossômico estafilocócico mec (SCCmec) (42, 91, 93,
99, 106, 111), e nos S. aureus, está integrado em seu cromossomo em um único sítio
(attBscc) próximo a origem de replicação (55), possuindo baixa afinidade por β-
lactâmicos e sem relação com as β-lactamases (81).

14
1.8 Gene mecA O gene mecA é altamente preservado entre as espécies de estafilococos. O
mecanismo responsável pela sua transferência horizontal do S. epidermidis para o
S. aureus ou entre as espécies Gram-positivas ainda é desconhecido (55). O mesmo
pode ter se originado do Staphylococcus sciuri, possivelmente uma espécie
ancestral, e posteriormente ter sido transferido para as outras espécies (S.
epidermidis e S. aureus). O gene mecA pode ser um gene nativo no S.sciuri que
possivelmente originou o mecA no S. aureus. Evidências genéticas demonstram que
a pbpD do S.sciuri foi o precursor evolucionário do gene mecA do S.aureus. A
proteína codificada pela pbpD tornou-se conhecida como PBP4, uma das seis PBP’s
detectadas no S.sciuri. Esta proteína possui algumas propriedades bioquímicas
semelhantes a PBP2a do S. aureus relacionada com a resistência aos antibióticos ß-
lactâmicos (7, 33,146).
A expressão gênica do mecA pode ter dois fenótipos: induzível ou
constitutivo. O fenótipo induzível possui um lócus regulatório com os genes mecI e
mecRI, homólogos aos genes reguladores blaI e blaRI que regula a produção das β-
lactamases. Os genes reguladores bla ou mec regulam a produção da PBP 2a e β-
lactamase devido ao alto grau de homologia dos dois sistemas (28,85).
Os isolados com disfunção na região regulatória dos genes mecI e mecRI ,
expressam o mecA constitutivamente, pois o BlaR1 é um forte indutor do mecA e
BlaI fraco repressor. Entretanto, nos isolados clínicos a regulação é feita pelo gene
bla devido a deleções e mutações que ocorrem no gene mecI responsável pela
repressão. Cepas com estas características são muito importantes porque são
desreprimidas lentamente dificultando a detecção da resistência, necessitando até
de 48hs para se expressar totalmente. O BlaR1 e o BlaI são capazes de controlar a
expressão da PBP2a quando os genes mecR1 e o mecI estão desativados. As
classes mec são definidas pelo rearranjo dos genes adjacentes ao mecA (16,59, 78).
Igualmente ao S. aureus, a resistência a oxacilina no S.epidermidis é derivada
da produção de uma PBP 2a alternativa, codificada pelo gene mecA (120), que possui

15
homologia entre os MRSA e MR-SCoN, indicando que o mesmo pode ser
proveniente dos Staphylococcus sp. (28, 65). O gene mecI reprime o mecA e auto-
reprime a transcrição do mecR1 e mecI, através da ligação do promotor-operador
(OP). A repressão da transcrição do mecA pelo mecI, faz com que a cepa torne-se
suscetível a drogas ß-lactâmicas, devido a ausência da produção de PBP 2a. A
inativação por deleção, faz com que a mesma expresse altos níveis de resistência
(65, 140).
1.8.1 Complexo genético mec Três classes principais do complexo genético mec foram identificadas. O
protótipo mec (classe A) é composto pelo mecA, seus genes reguladores
mecR1(gene do sinal transdutor), mecI (gene repressor) (120) intactos e a seqüência
de inserção IS431: classe A- IS431- mecI- mecR1- mecA- IS431. No complexo
classe B o domínio PB do mecR1 e o gene completo mecI estão truncados por uma
cópia parcial de IS272: classe B- IS272- ΔmecR1- mecA- IS431. Já o complexo C
possui duas variantes, C1 e C2. Na classe C1 o domínio PB do mecR1 e todo o
mecI estão truncados pelo IS431, e na classe C2, os domínios MS e PB do mecR1
e do mecI estão truncados por IS431: classe C- IS431- ΔmecR1- mecA-IS431. Na
classe D o mecI está deletado e o domínio PB do mecR1 truncado:Classe D- ΔmecR1- mecA- IS431 (65, 78). O complexo mec mais prevalente e disseminado
possui a estrutura mecI-mecR1-mecA-IS431R (IS431mec) (68, 140).
1.8.2 Complexo genético ccr e principais tipos de SCCmec O complexo genético ccr é composto por um ou dois genes recombinases
sítios-específicos, responsáveis pela mobilidade do SCCmec. O SCCmec é
integrado no sítio genômico attBscc localizado na sequência aberta de leitura orfX
(open reading frame) com função desconhecida (17). O gene ccr catalisa a excisão e
integração do elemento SCCmec. Atualmente três foram identificados: ccrA, ccrB,
ccrC (Figura 4) (17, 64).

16
Fonte: Hussain et al. 2001.
Figura 4 – Desenho esquemático de gene mecA
As enzimas ccrA, ccrB estão classificadas em quatro alótipos: ccrA1 e ccrB1
(carreados pelo SCCmec tipo-I), ccrA2 e ccrB2 (carreado pelo SCCmec tipo-II e
SCCmec tipo-IV), ccrA3 e ccrB3 (carreados pelo SCCmec tipo-III), ccrA4 e ccrB4
(carreado pelo SCCmec tipo-VI). O gene ccrC foi identificado pelo SCCmec tipo-V
ou elemento SCC (17, 64).
O cassete estafilocócico cromossômico (SCCmec) classifica-se conforme a
combinação do complexo genético mecA e as recombinases do cassete
cromossômico ccr (17, 29, 83). O complexo genético ccr codifica as recombinases
responsáveis pela mobilidade do SCCmec (17, 98).
As partes remanescentes do SCCmec são denominadas de região J (região
J1, J2, J3), componentes não essenciais do cassete que podem carrear
determinantes de resistência aos antibióticos. Variações nessas cadeias são
utilizadas para definir os subtipos de SCCmec (17, 87). A estrutura do SCCmec pode
ser resumida em: J1-ccr-J2-mec-J3 (87). Até o presente momento seis tipos de
SCCmec foram descritos (Figura 5).
Isolados sensíveis a meticilina (MSSA) tornam-se resistentes (MRSA ou MR-
SCoN) através da aquisição do SCCmec, elemento genético móvel que carreia o
gene mecA encontrado nas espécies de estafilococos (23, 29, 64). O mesmo é um
veículo de troca de genes de resistência a meticilina e está amplamente distribuído
entre os S.aureus e Staphylococcus sp. (83).

17
Figura 5. Classificação dos principais tipos de Cassetes
1.9 Staphylococcus spp. hospitalares e comunitários
Com o advento da penicilina na década de quarenta, houve redução na taxa
de mortalidade dos pacientes portadores de infecções ocasionadas por tais
espécies. Porém, surgiram isolados produtores de enzima beta-lactamase que se
disseminaram pelos hospitais. Nos anos 50, surgiu uma nova droga β-lactâmica anti-
estafilocócica (meticilina), resistente a ação de β-lactamases produzidas pelas
bactérias, para fins de tratamento de infecções ocasionadas por espécies de
estafilococos resistentes à penicilina (21,132). Novamente os estafilococos
desenvolveram resistência cromossômica a meticilina, a partir de um mecanismo
que altera as proteínas ligadoras de penicilina (PBP) e confere resistência cruzada a
todos os β-lactâmicos (21,132).

18
1.9.1 Prevalência de estafilococos resistentes a meticilina
A prevalência de isolados MRSA varia em diferentes regiões do planeta. Na
Suíça, o percentual é de 85,7% de HA-MRSA e 14,3% de CA-MRSA (124). Na
Finlândia, 80% de HA-MRSA(63) e nos EUA 55% de HA-MRSA (73). No Japão 67% e
na Austrália 23%(26). Na Argentina a prevalência de HA-MRSA é de 51%, enquanto
que na Bolívia é de 55%, no Paraguai 30%, Uruguai 24% e Venezuela 27%(13). Para
o Brasil, em 2006, a Associação Pan-Americana de Doenças Infecciosas registrou
que 54% das infecções comunitárias e hospitalares eram ocasionadas por isolados
MRSA(13). Especificamente no Rio de Janeiro, a taxa foi de 56% (27), em Porto Alegre
100% de estafilococos coagulase negativos (83) e 40,5% de S.aureus resistentes a
meticilina (104) , enquanto que na cidade de Manaus, no ano de 2003, Egido et al.
detectaram 90% de S. aureus sensíveis a meticilina (MSSA). No de 2009, na mesma
cidade, Ferreira et al. detectaram 15% de MRSA em um estudo realizado na
Fundação Afredo da Matta sem identificar a origem dos isolados (44).
1.9.2 Processos infecciosos ocasionados por estafilococos resistentes a meticilina
Devido a sua ampla disseminação nos hospitais em todo o mundo, o S.aureus
(HA-MRSA) continua sendo o agente responsável por inúmeros processos
infecciosos nosocomiais (138), como infecções de pele e tecido mole (123), pneumonia,
artrite séptica, endocardite, osteomielite, septicemias e infecções intravasculares (138). Com o surgimento de casos na comunidade (CA-MRSA), passou a afetar
também indivíduos saudáveis (atletas, presidiários, soldados em treinamento nas
bases) sem fatores de risco associados (contato direto, compartilhamento de itens
pessoais sem higiene freqüente, colonização nasal) (124,138). Assim como os
S.aureus, os coagulase negativos podem também causar diferentes processos
patológicos, principalmente em pacientes imunodebilitados, como bacteremia,
endocardite, infecções urinárias e relacionadas com cateteres e próteses. Portanto,
a importância deles, baseia-se no fato de estarem se tornando prevalentes e
associados a processos infecciosos de elevada morbidade e taxas de mortalidade
que podem atingir até 14% das infecções hospitalares (41, 135).

19
1.9.3 Cassetes cromossômicos e Clones estafilocócicos
Apesar dos estafilococos resistentes a meticilina possuirem o gene mecA
localizado no SCCmec (63, 124) apresentarem resistência aos antibióticos beta-
lactâmicos, possuem outros determinantes comuns de resistência que contribuem
para caracterizar os diferentes tipos de SCCmec, além dos complexos essenciais
mec e ccr (54, 63, 97, 104), resultando assim na identificação de diferentes tipos de
cassetes cromossômicos (tipo I até V) (63, 85, 97) relatados em vários países (80), cada
qual possuindo tamanho e características diferentes, dispersos em vários
continentes, constituindo clones de estafilococos responsáveis por infecção
comunitárias (CA-MRSA) e hospitalares (HA-MRSA), como o ST8 (spa tipo t008,
USA 300),ST1 (spa tipo t127, USA400) (110) e tipo 80 (ST80; spa tipo t044, clone
europeu) (110), sendo altamente virulentos para os pacientes internados em hospitais,
e se tornando um veículo de troca de genes de resistência entre os estafilococos (63).
Esses clones estafilocócicos diferem entre si devido a organização genômica
e estrutural, sendo esta refletida pela distribuição no SCCmec, nos determinantes de
toxinas estafilocócicas como a enterotoxina, exfoliatina D, e leucocidina Panton
Valentine (LuKPV), que ocorrem também em isolados de estafilococos coagulase
negativa, demonstrando que podem ser passíveis de transferência horizontal,
apesar do mecanismo ainda não ser bem conhecido (54, 82, 110). Sabe-se atualmente,
que cepas identificadas como sensíveis (MSSA) LuKPV negativas, podem ser
reservatórios comunitários aguardando apenas a aquisição do SCCmec, LuKPV e
outros determinantes de virulência e resistência para se tornarem clones CA-MRSA
virulentos (63, 82, 110). Portanto, a LuKPV é utilizada mundialmente nos testes
laboratoriais como marcador para determinação de isolados comunitários (CA-
MRSA), por estar envolvida em infecções invasivas como pneumonia necrotizante (24, 38, 82), pois geralmente os estafilococos pvl-positivos, estão associados com
infecções cutâneas piogênicas necrotizantes (abcessos e furúnculos) e menos
frequentemente com celulite e necrose tecidual (103), apesar de já existirem relatos
de S. aureus pvl-positivos isolados de pacientes portadores de artrite, bacteremia (71)
e pneumonia necrotizante (49, 74).

20
Atualmente a nomenclatura dos estafilococos meticilina resistentes baseia-se
na sequência de tipagem multilocus (ST) dos fragmentos de sete genes
denominados “housekeeping”, que referem a linhagem da bactéria, no caso S.
aureus e S. epidermidis, resultando em um perfil alélico designado de ST e o tipo do
elemento SCCmec. A detecção de linhagens divergentes através de técnicas
moleculares como o MLST, sugere que o estafilococo resistente a meticilina surgiu
pela introdução do SCCmec em uma linhagem de cepa sensível a essa droga(26).
Atualmente existe uma diversidade de clones de S. aureus e de S.
epidermidis dispersos pelo mundo. Na China os clones mais prevalentes são o
ST239 e ST1018(138). Na Oceania, o ST30 (11). No continente europeu, a Romênia
apresenta os clones ST239,ST5,ST80,ST1,ST254 (75) como os mais prevalentes,
enquanto que nos demais países prevalece o ST80(11). No Kuwait prevalece o ST80 (131a,131b). Nos Estados Unidos os clones prevalentes são USA300 (ST8) e USA 400
(ST1)(11,131a,131b), enquanto que no Brasil, especificamente nas cidades de Porto
Alegre e Rio de Janeiro, foram identificados três complexos clonais de S. aureus
como ST8,ST30 e ST1, correspondendo aos clones USA300, clone do Oceano
Pacífico Sul e USA400 (26, 116). Em relação ao S. epidermidis, dados publicados
demonstram a prevalência dos clones ST2 e ST22 na Grécia (76), ST2 na Suiça,
Finlândia e China (61, 77, 139), e nos Estados Unidos, os clones ST2, ST5,ST6, ST22,
ST23, ST87, ST185,ST186 (88,145). No Brasil os clones conhecidos são os
ST2,ST231 e o ST263 (www.mlst.net).
1.9.4 Características diferenciais entre HA-MRSA e CA-MRSA
Vários fatores de risco dentre eles hospitalização, antibióticoterapia,
internação em enfermarias, contato direto com o corpo clínico, contribuem para
infecção ou colonização dos pacientes por isolados de estafilococos coagulase
negativos e S.aureus resistentes a meticilina. Entretanto, deve-se ressaltar que os
CA-MRSA e HA-MRSA são tanto genética como fenotipicamente distintos (110).
Isolados HA-MRSA apresentam resistência a múltiplos antibióticos beta-
lactâmicos (penicilinas lábeis, estáveis, penicilina G, amoxacilina, carbapenêmicos,

21
cefalosporinas de 1º, 2º, 3º e 4º geração) e, em especial, a fluoroquinolona. Não
estão associados ao gene codificador da Leucocidina Panton-Valentine (PVL) e
carreiam um cassete SCCmec largo do tipo I, II, III, raramente o IV (21, 24, 45, 47, 63). Os
CA-MRSA carreiam um cassete SCCmec menor tipo IV, V ou VI, que os torna
suscetíveis a antibióticos não beta-lactâmicos como clindamicina, tetraciclina, e
também a quinolona, sulfametoxazol/trimetoprima e doxiciclina. Freqüentemente
estão associados com a presença da toxina Leucocidina Panton Valentine (PVL),
localizada no interior do bacteriófago phiSLT, e apresentam múltiplas características
no pulsed-field gel electrophoresis (PFGE) (21, 24, 45, 47, 63). São também mais
virulentos e ocasionam doenças altamente invasivas e progressivas (pneumonia
necrotizante), geralmente em indivíduos saudáveis, jovens, e sem exposição a
fatores de risco hospitalar (124), devendo-se verificar se o tipo do SCCmec possui ou
não importância nessas infecções (Quadro 2) (21, 49, 124).
1.10 Prevalência de bactérias Gram-negativas produtoras de ESBLs
Os processos patológicos que acometem pacientes infectados por bactérias
Gram-positivas, também podem ocorrer em bactérias Gram-negativas, e a grande
preocupação clínica está relacionada com sua capacidade de adquirir resistência
aos antibióticos, que as torna capazes de produzir a enzima beta-lactamase de
espectro estendido (ESBL), que hidrolisa as oxiamino-cefalosporinas e aztreonam
dificultando assim a escolha terapêutica, o que afeta os pacientes, principalmente
idosos, imunocomprometidos e debilitados, já que esses antibióticos são utilizados
diariamente na rotina hospitalar e comunitária (48, 109, 130, 134) .
Dentre as bactérias não-fermentadoras, a Pseudomonas aeruginosa é a que
apresenta índice de mortalidade mais elevado (5%) (32), tendo sido isolada de 10,7% (32) das amostras biológicas coletadas de pacientes sob antibióticoterapia profilática,
seguida posteriormente por outras espécies bacterianas como E. coli e Salmonella
sp.(46, 48). Já nos pacientes internados portadores de doenças hematológicas, como
lúpus, anemia hemolítica, aplasia de medula, linfoma, doença maligna dentre outras,
a imunodeficiência ocasionada pela doença de base os torna muito mais vulneráveis
a infecções bacterianas por espécies Gram-negativas (32), principalmente se

22
estiverem utilizando acesso venoso central como cateteres (136). Relatos de
bacteremia nesses pacientes assim como infecções hospitalares ocasionadas por
E.coli, principalmente em pacientes imunocomprometidos, já foram registrados na
literatura (79, 136).
Quadro 2. Características diferenciais entre HA-MRSA e CA-MRSA
Aquisição HA-MRSA CA-MRSA
Hospitalar Comunitário
Infecções clínicas Pacientes imunodebilitados, contato
direto e.
Indivíduos jovens, saudáveis, sem
fatores de risco associados e.
Fatores de risco
Internação prolongada, uso prolongado
de antibióticos, enfermarias, diabetes,
dispositivos intravasculares, doença
pulmonar crônica b.
Contato físico, compartilhamento de
itens pessoais, lesões de pele, uso
de drogas, prisão, atletas b
Fenótipo Resistência a múltiplos antibióticos beta-
lactâmicos e em especial a
fluoroquinolona a, f.
Suscetíveis a antibióticos não beta-
lactâmicos como clindamicina,
tetraciclina,quinolona,sulfametoxazol
/trimetoprima e doxiciclina
(multidroga) a, f.
Marcadores genéticos
Negativo para o gene codificador da
Leucocidina Panton-Valentine (PVL).
Carreiam um cassete SCCmec largo,
do tipo I,II, III, e raramente o IV d, f.
Gene codificador da (PVL) c
Carreiam um cassete SCCmec
menor, tipo IV ou V. Múltiplas
características no PFGE d, f.
Fonte: Fey et al. 2003 a (45); Huang et al. 2006b (60); Ma et al. 2008c (82), Bouchami et al. 2009d (23) , Strandén et al. 2009e (124),
Boyle-Vavra et al. 2007f (21).
A prevalência de bactérias produtoras de ESBLs oscila entre as instituições
de saúde e países. Na Suíça é de 8%, Portugal 34%, Ásia 20% (92), Turquia 58% (105)
e África do Sul 20% (93). Nos EUA a média anual é de 3% (14), e nas regiões da
América Latina 60%. No Brasil, a prevalência registrada para espécies como a
Klebsiella pneumoniae (K. pneumoniae) é de 45,4% enquanto que para a
Escherichia coli (E. coli) 6% (10). Na região Norte, e especificamente no Amazonas,
ainda não há registros publicados sobre infecções ocasionadas por bactérias
produtoras de ESBL.

23
1.10.1 Enzima Beta-Lactamase As ß-lactamases são enzimas que se localizam no espaço periplasmático nas
bactérias Gram-negativas ou extracelularmente nas Gram-positivas. A informação
genética para sua síntese pode estar contida no plasmídeo ou cromossomo
bacteriano (25). No caso do plasmídeo, o mesmo também pode ser incorporado no
cromossomo bacteriano (25) desenvolvendo-se assim a resistência cromossômica.
Esta resistência muitas vezes não é detectada em condições normais de
crescimento do isolado, surgindo apenas quando na presença de antibióticos
indutores como os ß-lactâmicos (25).
Os plasmídeos ou transposons contém os genes da ß-lactamase que são
transferíveis entre as bactérias não importando a espécie (109), e que atualmente
estão disseminados no mundo inteiro, sendo encontrados em diferentes espécies
das famílias Enterobacteriaceae, Pseudomonas aeruginosa, Haemophilus influenzae
e Neisseria gonorrhoeae (14, 103).
A beta-lactamase SHV-1 (variável sulfidril), codificada cromossomicamente na
maioria dos isolados de K. pneumoniae, é freqüentemente mediada por plasmídeo
em E. coli. Assim, devido a essa forma de disseminação dos genes, nos últimos 20
anos, novos antibióticos surgiram e com eles também novas beta-lactamases, que
induziram o surgimento de cepas resistentes (14).
As ß-lactamases destroem os antibióticos ß-lactâmicos como penicilinas e
cefalosporinas através da hidrólise do anel beta-lactâmico (25), mas podem ser
inibidas por inibidores de beta-lactamases como o clavulanato, sulbactam e
tazobactam, ineficazes contra a classe “C” destas enzimas (36).
Mais de 340 ß-lactamases já foram identificadas estando classificadas em
quatro classes de A à D conforme sua estrutura primária e características funcionais
e bioquímicas (grupo I à IV) (37, 43). As ESBLs clássicas são comumente
transmitidas/codificadas por plasmídeo como o da família TEM-, SHV- e OXA-. No
decorrer dos anos, dentro de cada família, ocorreram mutações e novas variantes

24
surgiram como por exemplo, TEM-6, CTX-M-8, CTX-M-2, PER, VEB, GES, e TLA (36,
89, 102, 108, 134).
A complexidade das ß-lactamases é acentuada por vários fatores dentre eles
o fato de uma mesma cepa de bactéria expressar múltiplas ß-lactamases de tipos
variados, podendo ocasionar diferentes implicações clínicas e dificultar a análise
laboratorial de seu perfil enzimático. Outro fator relacionado é o “efeito do inóculo”
que dificulta a detecção de resistência clinicamente relevante. As bactérias
produtoras de ESBLs, demonstram resistência cada vez mais elevada as
cefalosporinas de terceira geração com o aumento do inóculo bacteriano (efeito do
inóculo), fenômeno ocasionado pela incapacidade dos antibióticos de destruírem
ou inbirem as bactérias(142).
Outro exemplo importante relacionado a essa complexidade, é o fato dessas
enzimas apresentarem afinidade por diferentes antibióticos ß-lactâmicos. Porém,
sua hiperprodução pode inativar a eficácia desses antibióticos, apesar de terem sido
classificadas como ativas contra todo o antibiótico ß-lactâmico. Não é raro também,
a presença de mecanismos redundantes de resistência na mesma cepa bacteriana,
a qual pode albergar uma ß-lactamase combinada com uma mutação na porina,
tornando-a multiressitente, ou então portar transposons contendo genes de
resistência a múltiplos antibióticos além dos ß-lactâmicos, como fluoroquinolonas,
aminoglicosídeos, trimetoprim-sulfametoxazol(142).
1.10.2 Classificação das Enzimas Beta-Lactamases
As enzimas da classe A ou grupo II incluem as famílias blaTEM e blaSHV (110)
que hidrolisam as penicilinas e cefalosporinas; classe B ou grupo III,
carbapenêmicos; classe C ou grupo I, cefalosporinas; classe D, penicilinas e
cloxacilina; e grupo IV (peniclinases) penicilina(36). As enzimas pertencentes à classe
A, geralmente são encontradas em plasmídeo ou transposon sendo consideradas as
mais importantes. A SHV, por exemplo, é freqüentemente encontrada em isolados
clínicos (36, 43).

25
Posteriormente Ambler classificou as ESBLs em 4 classes evolucionárias (A,
B, C, D). As enzimas SHV- e TEM- pertencem à classe A e possuem a serina como
seu principal resíduo catalítico, enquanto que as OXA- pertencem à classe D
(oxacilinases). O esquema de Bush classifica as ESBLs de acordo com o perfil do
substrato e características físicas como ponto isoelétrico e peso molecular. Neste
esquema as ESBLs estão divididas em dois subgrupos: 2be (TEM- e SHV-) e 2d
(OXA-) inibidos pelo clavulanato. Em 1995 Bush-Jacoby-Medeiros apresentaram um
novo esquema no qual as enzimas foram divididas em quatro grupos (1-4) e
subgrupos (a-f) (36).
O grupo 1 é formado por cefalosporinases que não são inibidas pelo ácido
clavulânico e pertencem a classe molecular C. O grupo 2 são penicilinases,
cefalosporinases ou ambas, que são inibidas pelo ácido clavulânico e pertencem a
classe molecular A e D. Devido ao surgimento de vários mutantes ESBLs foi criada
duas subclasses 2a (penicilinases) e 2b (ESBLs de amplo espectro).
Posteriormente foram criados subgrupos 2be (inativam cefalosporinas de terceira
geração), 2br (ESBLs resistentes aos inibidores de beta-lactamases), 2c (inativam a
carbenicilina), 2d (inativam a cloxacilina e são fracamente inibidas pelos inibidores
de ß-lactamases) e 2f (serina-beta-lactamases). O grupo 3 possuem enzimas que
necessitam de zinco para para exercerem seu efeito sendo denominadas de zinco-
beta-lactamases ou metallo-beta-lactamases e correspondem a classe molecular B.
No grupo 4 estão as penicilinases não inibidas pelo ácido clavulânico (36).
1.10.3 Beta-Lactamase de espectro estendido (ESBL)
A primeira descrição de ESBL mediada por plasmídeo denominado TEM-1, foi
feita em 1960. Esta enzima foi encontrada em cepa de E. coli isolada da
hemocultura de uma paciente denominada Temoniera, na Grécia, daí a origem do
nome (14, 35, 43). Em 1983 foi identificado um novo grupo de enzima beta-lactamase
em isolados clínicos de Serratia marcescens e Klebsiella pneumoniae. Esta enzima
passou a denominar-se de Beta-lactamase de espectro estendido ou ampliado
(ESBLs) (2, 35).

26
Posteriormente, as ESBLs foram classificadas em dois grandes grupos sem
relação entre si, apesar de algumas enzimas agirem sobre o mesmo substrato (ß-
lactâmicos) ligando-se a sítios diferentes (35). Assim sua ação ampliou-se também
sobre as oxiamino-cefalosporinas e monobactans, reduzindo as opções terapêuticas
da classe dos ß-lactâmicos para os carbapenêmicos, elevando os custos do
tratamento e risco para os pacientes devido a longos períodos de hospitalização e
antibióticoterapia (2, 35, 102). No caso dos pacientes portadores de doenças
hematológicas, os mesmos são frequentemente submetidos à quimioterapia, o que
ocasiona imunodeficiência deixando-os susceptíveis a uma ampla variedade de
infecções, principalmente as hospitalares, e especialmente aquelas ocasionadas por
bactérias produtoras de ESBL, contribuindo assim para a elevação do risco de
morbi-mortalidade (113, 117).
As ESBLs são produtos derivados de mutações pontuais no sítio ativo da
enzima TEM e SHV (104), codificadas por plasmídeos conjugativos transferíveis entre
as bactérias Gram-negativas (9, 128), que freqüentemente produzem determinantes de
resistência a outras classes de antibióticos (65). As bactérias que comumente
produzem esse tipo de enzima são E. coli e K. pneumoniae (2, 10, 65, 69, 102, 108),
podendo também ser elaboradas por outras espécies como Enterobacter,
Salmonella, Proteus, Citrobacter, Morganella morganii, Serratia marcescens,
Shigella dysenteriae, Pseudomonas aeruginosa, Burkholderia cepacia (4, 10, 69).
1.10.4 Subtipos das enzimas Beta-Lactamases de espectro estendido (ESBLs)
Vários são os subtipos de ESBL. Em E. coli a maioria das ß-lactamases
pertencem a classe A de Ambler e podem ser divididas em ß-lactamases de
pequeno espectro como a TEM-1,TEM-2 e SHV, e as de amplo espectro, como
TEM-3, SHV-5 e CTX-M-like. As do tipo CTX-M são de largo espectro e derivadas
das ß-lactamases codificadas cromossomicamente pelo gênero Kluyvera. Foram
isolados mais de 70 tipos, divididos em cinco clones, que possuem similaridades em
sua sequência de aminoácidos conhecidos como CTX-M-1, CTX-M-2, CTX-M-8,
CTX-M-9, e CTX-M-25 (89,109). Estudos realizados nos genes codificadores de ESBL

27
revelaram que a maioria dos isolados produtores carreiam os genes TEM-1 (88%),
CTX-M (20%), SHV (8%), e OXA (5%), sendo o CTX-M o mais prevalente e disperso
pelo mundo (79, 110). Nas espécies de E. coli, os subtipos SHV, TEM e CTX-M-15 são
os mais prevalentes (108), já em Pseudomonas aeruginosa, é o GES- 1 e GES-5 (86).
Bactérias produtoras de enzimas mediadoras de resistência aos
carbapenêmicos estão descritas mundialmente. As carbapenamases são
representadas por 3 classes moleculares de ß-lactamases: A,B e D. A mais
conhecida é a K. pneumoniae (KPC); enterobactéria produtora de carbapenamase,
responsável por vários surtos hospitalares. A classe B são as metalo-
betalactamases (MBL) que apresentam um amplo espectro de substrato. As
carbapenamases da classe D tipo OXA, são mais frequentemente detectadas nas
bactérias não fermentadoras, exceto a OXA-48, detectada somente em
enterobactérias e que possui uma notável atividade de hidrolisar os carbapenêmicos (35, 90).
1.10.5 Importância Clínica das ESBL
As mutações genéticas pontuais que ocorrem nos isolados produtores de
ESBL, torna-os capazes de sintetizar as ESBLs que atuam hidrolisando antibióticos
de amplo espectro como as cefalosporinas (ceftazidima, ceftriaxona, cefotaxima),
monobactans (aztreonam) (4, 36, 43, 66, 105), e, como os plasmídeos muitas vezes
albergam genes codificam as ESBLs e outros que codificam resistência aos
antibióticos não beta-lactâmicos, quando a bactéria é estimulada para um gene de
determinado antibiótico, pode-se tornar concomitantemente, resistente aos outros
antibióticos também, podendo ampliar assim sua resistência contra outros
antimicrobianos não ß-lactâmicos como aminoglicosídeos, fluoroquinolonas,
tetraciclina, cloranfenicol e sulfonamidas (36, 93).
Outro aspecto importante é que esses genes, localizados no mesmo
plasmídeo conjugativo, podem ser transmitidos conjuntamente de um isolado para
outro não importando o gênero bacteriano, ou então serem estimulados
separadamente por pressão seletiva, tornando assim mais difícil a escolha

28
terapêutica nesses casos, principalmente, quando da ausência do diagnóstico
laboratorial desse fenótipo (84, 102)
Portanto, torna-se importante a realização dos testes de suscetibilidade de
acordo com os critérios determinados pelo CLSI 2010, pois até 2009 a
recomendação era de se realizar a detecção de cepas produtoras de ESBL em duas
etapas, screening e testes confirmatórios. Entretanto, em 2010 (manual M100-S20)
esta orientação deixou de existir como condição única, sendo incluídas as
orientações posológicas mínimas além dos pontos de corte modificados para
interpretar os resultados obtidos (halo de inibição e concentração inibitória
mínima)(30).
1.10.6 Preocupações relativas às infecções bacterianas
Isolados resistentes a antimicrobianos como os HA-MRSA, CA-MRSA e
ESBLs, ocasionam quadros graves de infecções muitas vezes difíceis de tratar
devido à limitada opção terapêutica, elevando assim o percentual de morbi-
mortalidade e os custos do tratamento decorrentes da internação prolongada nos
hospitais. (85, 93). Outro fator importante é a dificuldade para detecção desses
isolados nos testes laboratoriais de suscetibilidade a antimicrobianos que, apesar
das normas do CLSI 2010 (manual M100-S20), para algumas espécies ainda é
necessário teste confirmatório para este tipo de fenótipo, devido ao surgimento de
novos mutantes bacterianos produtores de ESBL e novas espécies de estafilococos
coagulase negativos resistentes a meticilina, para as quais o CLSI ainda não possui
padronização da concentração inibitória mínima (MIC) aos antimicrobianos utilizados
na rotina hospitalar (85).
A importância desses patógenos é tão relevante para a saúde pública, que
relatos de doenças causadas por bactérias Gram-positivas resistentes a meticilina
(MRSA), Gram-negativas produtoras de beta lactamase de espectro estendido
(ESBL) incluindo aquelas que ampliaram seu espectro de resistência aos
carbapenêmicos (ampC), estão amplamente descritos e discutidos na literatura
científica nacional e internacional (115,117,138,139). Portanto, a resistência microbiana

29
aos antimicrobianios utilizados no tratamento, ainda é um grande desafio tanto nas
infecções comprovadamente hospitalares, quanto nas adquiridas na comunidade,
em todo o mundo. Em razão dos pacientes internados nos hospitais estarem mais
expostos aos antimicrobianos de largo espectro (que muitas vezes debilitam a
barreira química natural do organismo ou ocasionam alteração da resposta imune),
ao contato freqüente entre profissional da saúde-paciente (possibilitando a
transmissão cruzada de patógenos), a ruptura da barreira tecidual por processos
invasivos, como cirurgia, inserção de sondas, cateteres, e ao déficit nutricional
devido à dificuldade de ingestão, a probabilidade de ocorrerem processos
infecciosos graves e até mesmo fatais nos mesmos, ocasionados por bactérias
resistentes presentes no ambiente hospitalar, é elevadíssima(114,115,117).
Atualmente observa-se que os fatores de risco para que um paciente
desenvolva algum processo infeccioso bacteriano, de origem hospitalar ou
comunitária, tanto em hospitais como em ambulatórios já é uma realidade, e a
Fundação de Hematologia e Hemoterapia do Amazonas (FHEMOAM), como um
centro que promove assistência ambulatorial e internações a pacientes com doenças
hematológicas (hemofilia, talassemia, anemia hemolítica, leucemia,etc) muitas vezes
graves, não é uma exceção no processo. Muitos dos usuários assistidos requerem
tratamentos que incluem seções de quimioterapia que, às vezes, causam efeitos
colaterais como imunodeficiência, deixando-os mais suscetíveis a infecções
bacterianas graves que, quando tratadas de modo ineficaz, podem levar a estado de
morbidez prolongado podendo ocasionar a morte. Nestes casos, a resistência
microbiana aos antibióticos utilizados no tratamento, ainda é um desafio tanto nas
infecções comprovadamente hospitalares, quanto nas adquiridas na comunidade.
Pesquisas científicas nesse campo são fundamentais para a saúde pública e
bem estar dos indivíduos e das comunidades, pois as questões que envolvem a
saúde pública são complexas e seus problemas devem ser confrontados em todos
os aspectos, dos mais abrangentes aos mais localizados, e o estudo científico pode
contribuir sobremaneira para o equacionamento e solução dos mesmos.

30
Pesquisas direcionadas para os mecanismos de resistência bacteriana aos
antibióticos, utilizando-se protocolos moleculares(110,130) que possibilitem visualizar
se os genes específicos e suas possíveis mutações são comuns ou exclusivas,
como as descritas por pesquisadores em outras regiões(132,135), auxiliarão
diretamente no desenvolvimento de novos projetos envolvendo outras instituições de
saúde ligadas ao SUS, de diferentes níveis de complexidade, o que ampliará a visão
epidemiológica sobre a fisiopatogenia das doenças ocasionadas por esses agentes,
o monitoramento dos mecanismos de resistência dos mesmos, a importância da
pressão seletiva relacionada ao uso de antimicrobianos, os mecanismos de infecção
cruzada bem como os de disseminação, auxiliando sobremaneira no controle das
infecções, das decisões sobre a terapia antimicrobiana e das políticas de saúde a
serem adotadas para a prevenção de surtos hospitalares ou comunitários.

31
2 OBJETIVOS 2.1 Geral
Descrever a freqüência, sensibilidade aos antimicrobianos e caracterizar as
alterações genético-moleculares das bactérias patogênicas isoladas de processos
infecciosos de pacientes com doenças hematológicas da Fundação HEMOAM 2.2 Específicos
Identificar a freqüência dos gêneros e espécies bacterianas.
Identificar os fenótipos de sensibilidade, sensibilidade reduzida e a resistência
dessas bactérias aos antibióticos indicados no tratamento.
Identificar as alterações genético-moleculares de resistência envolvidas.

32
3 MATERIAL E MÉTODOS 3.1 Delineamento Estudo transversal, descritivo, com componentes analíticos, realizado a a
partir da demanda espontânea de pacientes atendidos na Fundação de Hematologia
e Hemoterapia do Amazonas/FHEMOAM, autarquia do governo do Estado do
Amazonas, localizada na zona centro oeste de Manaus, onde são atendidos
anualmente 6.820 pacientes com 49.333 doações de sangue entre Capital e Interior
do Estado. 3.2 População de estudo Pacientes de ambos os sexos, de qualquer faixa etária, portadores de
doenças hematológicas, com suspeita clínica de processo infeccioso
bacteriano, internados na FHEMOAM.
3.3 Amostragem Foram incluídos todos os casos de pacientes com doença hematológica
apresentando quadro de infecção bacteriana durante o período de julho/2007 a
agosto/2008. O estudo foi aprovado pelo comitê de ética da FHEMOAM sob número
170 CAAE- 0005.0.112.000-06- versão 003/06.
A abordagem do paciente foi realizada após consulta médica na qual foi
detectada provável infecção. Foram realizados esclarecimentos quanto aos objetivos
e métodos a serem utilizados, apresentando ao sujeito da pesquisa o Termo de
Consentimento Livre Esclarecido adequado para adultos e crianças (anexo 1).
Aqueles que não concordaram em participar do estudo não tiveram nenhum tipo de
prejuízo no atendimento e receberam o mesmo tratamento oferecido pela
FHEMOAM aos demais pacientes. Após esclarecimento e aceite dos pacientes, as
amostras foram coletadas, as informações referentes ao cadastro e resultado dos
testes foram incluídas no anexo 2, e os testes foram realizados conforme protocolos
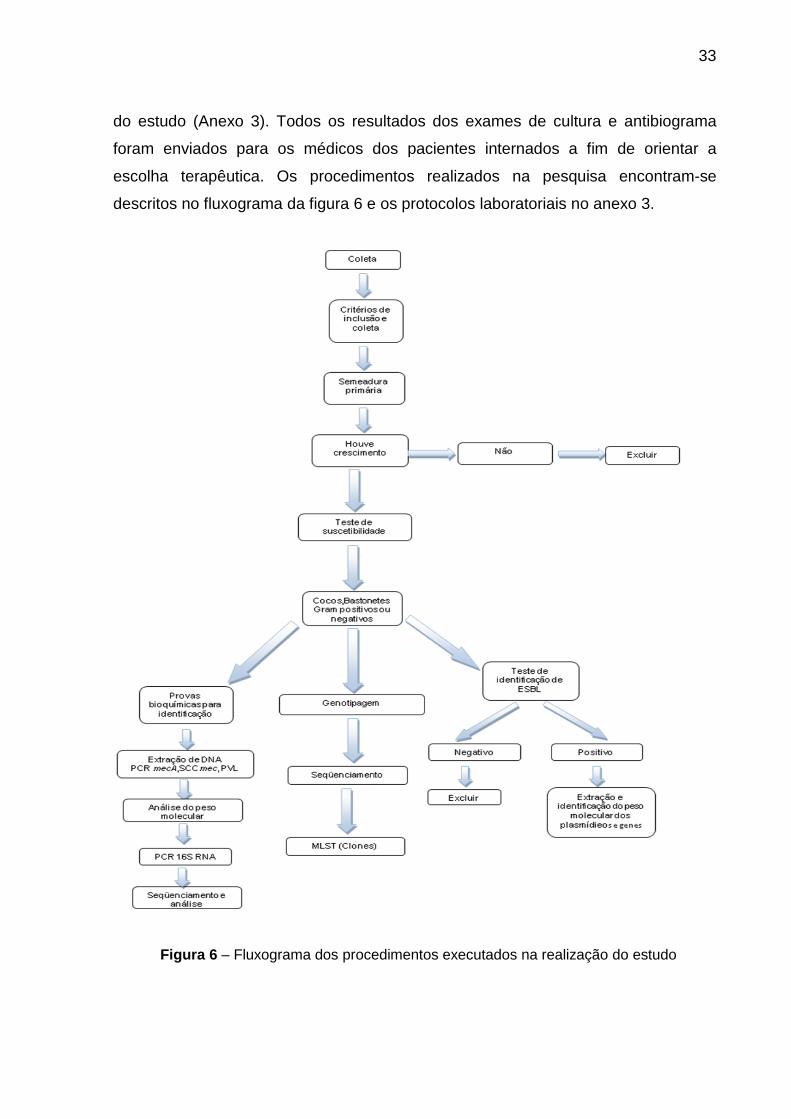
33
do estudo (Anexo 3). Todos os resultados dos exames de cultura e antibiograma
foram enviados para os médicos dos pacientes internados a fim de orientar a
escolha terapêutica. Os procedimentos realizados na pesquisa encontram-se
descritos no fluxograma da figura 6 e os protocolos laboratoriais no anexo 3.
Figura 6 – Fluxograma dos procedimentos executados na realização do estudo

34
3.4 Critérios de inclusão
a) Pacientes com diagnóstico prévio de doença hematológica
b) Pacientes atendidos no ambulatório ou internado na FHEMOAM
c) Pacientes apresentando sinais e sintomas clínicos de processo infeccioso
bacteriano
d) Pacientes utilizando ou não antibióticos de amplo espectro a partir de
qualquer via de administração
e) Pacientes neutropênicos (número de neutrófilos inferior a 1000 mm3 de
sangue) ou que apresentarem quadro febril durante o período de
internação.
3.5 Critérios de exclusão a) Pacientes cujas culturas apresentaram contaminação por outras
bactérias durante o processo de manipulação
b) Pacientes portadores de outras doenças que não as hematológicas.
c) Culturas que apresentaram crescimento microbiano de Micrococcus sp.
3.6 Solicitações do Exame Bacteriológico O exame laboratorial foi realizado após solicitação médica seguindo o
protocolo interno padronizado da FHEMOAM, conforme normas técnicas
padronizadas (Pop/HEMOAM/Goldin 2001), adequadas para diferentes tipos de
infecções bacterianas. O protocolo clínico (Pop/FHEMOAM) interno específico
para pacientes hematológicos imunodeprimidos define como critério de infecção:
3.6.1 Pacientes Hematológicos Imunodeprimidos: Foram considerados pacientes portadores de doenças hematológicas
imunodeprimidos aqueles que apresentaram:
1- Neutropenia- quando a contagem de neutrófilo está abaixo de 500/células
por mm3 ou quando inferior a 1000/células por mm3 após quimioterapia.
2- Quebra de barreiras naturais: lesão de pele, mucosa, quimioterapia

35
3- Febre durante neutropenia (temperatura acima de 38,3°C ou acima de
38°C mais de uma hora). 3.6.2 Fatores de baixo risco para complicação:
Os pacientes hematológicos que apresentaram um dos sintomas abaixo
relacionados foram considerados como de baixo risco para complicação, mas
com probabilidade de desenvolver infecção bacteriana.
1- Contagem absoluta de neutrófilos acima de 100 células, monócitos
acima de 100 células raios X de tórax e função renal normal .
2- Leucopenia com menos de sete dias.
3- Ausência de infecção na corrente sanguínea e no cateter.
4- Recuperação precoce de produção celular pela medula óssea.
5- Remissão de malignidade.
6- Pico de temperatura abaixo de 39°C, ausência de rebaixamento de
alteração neurológica (ausência de dor abdominal e co-morbidade,
choque, pneumonia, vômito, diarréia).
3.7 Coleta de amostra biológica
A coleta de material biológico foi realizada seguindo as recomendações
de Winn et al., 2008 (142) e Manual Anvisa Módulo III (8,9), sendo transportados
imediatamente para o laboratório. Material de ponta de cateter foi coletado por
médico especializado e enviado imediatamente para o laboratório (frasco
estéril sem conservante) (8,9,142).
3.8 Cultura, Isolamento e Identificação das bactérias isoladas Para o isolamento primário de bactérias aeróbicas, foram utilizados meios
de cultura como ágar sangue de carneiro a 5% (Himedia Hexasystens-Mumbai,
India), ágar manitol salgado (Himedia), ágar eosina azul de metileno (EMB)
(Himedia), ágar Mac Conkey (Himedia), Mueller Hinton (Himedia).

36
Após coleta, as amostras foram semeadas e incubadas em estufa
bacteriológica à temperatura de 35,6ºC, por um período de 24/48 horas. Culturas
que apresentaram crescimento foram submetidas à reação de Gram (Newprov). As
colônias Gram-positivas foram submetidas aos testes de identificação de gênero e
espécie como, catalase, coagulase (Newprov), fermentação do manitol (Himedia),
teste do L-pyrrolidonyl-beta-naphtylamide-PYR-Test (Probac do Brasil), testes com
monodiscos de antibióticos como novobiocina 5µg (Laborclin), bacitracina 0,04UI
(Newprov), polimixina B-Pol 300 (Newprov), furazolidona-100mcg (Cefar).
As bactérias identificadas como Gram-negativas, foram submetidas a testes
de identificação de gênero e espécie como oxidase, Kit EPM-MILi-Citrato (Probac do
Brasil), Enterokit B (Probac do Brasil), Enterokit C (Probac do Brasil), Kit NFII
(Probac do Brasil), testes bioquímicos adicionais com os meios de Mac Conkey
(Himedia), ágar uréia (Himedia), meio de MIO (Himedia), caldo Lisina (Himedia),
meio de TSI (Himedia), açúcares como glicose (Himedia), maltose (Himedia), lactose
(Himedia), sacarose (Himedia), realizados conforme protocolos padronizados (142).
Os protcolos laboratoriais encontram-se descritos no anexo 3.
3.9 Teste da Beta-Lactamase (cefinase) Foi realizado o teste da beta-lactamase tanto nas bactérias Gram-positivas
quanto Gram negativas, a partir de fita impregnada com reagente contendo
cefalosporina cromogênica (Cefinase plus), segundo recomendações do fabricante
Beckton-Dickson (BD Diagnostic System, Sparks, MD). A detecção da enzima
sugere que a espécie bacteriana produz a enzima beta-lactamase sendo resistentes
aos antibióticos beta-lactâmicos (12).
3.9.1 Teste da Beta-Lactamase de espectro estendido (ESBL) O teste aplicado para detecção do fenótipo ESBL foi o epsilométrico (E-
teste®) utilizando-se os antibióticos cefotaxima (0.25–16 μg/mL)/cefotaxime (0.016–1
μg/mL) mais 4 μg/mL de ácido clavulânico; ceftazidima (0.5–32 μg/mL) /ceftazidima
(0.064–4 μg/mL) mais 4 μg/mL de ácido clavulânico, segundo recomendações do

37
fabricante (AB-Biodisk-Sweden®).
Fonte: William Antunes Ferreira
Figura 7: Imagem das fitas de cefinase plus utilizadas na realização do teste da beta-
lactamase.
Os organismos testes foram suspensos em solução salina a 0.8%, padrão de
turbidez número 0.5 da escala de McFarland e inoculados no meio de ágar Mueller-
Hinton (Himedia), seguidas por uma incubação a 37°C por 18–24h. A interpretação
do testes foi realizada segundo o manual técnico do Clinical and Laboratory
Standards (CLSI; Wayne, Pennsylvania, USA)(29), o qual fornece o ponto de corte
(break point) correspondente ao diâmetro da zona de inibição. Cepas de E. coli
ATCC 25922 e K. pneumoniae ATCC 700603 foram utilizadas como controle de
qualidade para os meios de cultura e testes de suscetibilidade.
O teste epsilométrico E-teste® consiste na aplicação de fitas plásticas, não-
porosas com cinco milímetros (mm) de altura e 60 mm de comprimento contendo em
um dos lados o MIC (Concentração Inibitória Mínima) num gradiente de µg/mL para
os antibióticos cefotaxime (CT) e ceftazidime (TZ) com escalas de 0,25-16 µg/mL e
0,50-32µg/mL, respectivamente. No outro apresenta dois gradientes predefinidos de
CTL (cefotaxime/ácido clavulânico) e TZL (ceftazidime/ácido clavulânico) com
escalas de 0,016-1 µg/mL mais 4 µg/mL de ácido clavulânico, e 0,064-4 g/mL mais 4
µg/mL de ácido clavulânico (Figura 7). Após o cultivo e isolamento, a cultura pura foi
armazenada em freezer a -70ºC segundo CLSI, 2005(29). Posteriormente, as cepas
resistentes foram submetidas a estudos moleculares para determinação genética de
sua resistência e os resultados armazenados em banco de dados apropriado.

38
Fonte: AB BIODISK.
Figura 8: Desenho esquemático das fitas utilizadas na realização do Etest® ESBL com os
gradientes de concentrações dos antibióticos CT e TZ e em associação com o inibidor, o
ácido clavulânico.
A produção de ESBL (figura 8) foi determinada a partir do aparecimento de
uma zona fantasma (phantom zone), deformação da elipse do lado contendo o CT
ou TZ, ou quando o MIC de CT ou TZ se apresentassem reduzidos a ≥ 3 log2 de
diluições na presença do ácido clavulânico (Quadro 3).
Quadro 3. Protocolo de interpretação do Etest® ESBL, conforme recomendações do fabricante AB BIODISK
ESBL MIC Descrição
Positivo
CT > 0,5 e CT/CTL > 8
ou
TZ > 1 e TZ/TZL > 8
ou
Zona fantasma ou deformação
da elipse no CT ou TZ
Produção de ESBL e resistência
a todos as penicilinas,
cefalosporinas e aztreonam
(CLSI)
Negativo CT < 0,5 ou CT/CTL < 8
e
TZ < 1 ou TZ/TZL < 8
Não produtora de ESBL
Não determinada
CT > 16 e CTL > 1
e
Quando uma fita é negativa
para ESBL e a outra não
determinada
ESBL não determinada. Se a
ESBL é suscetível, confirmar o
resultado com a genotipagem.
Fonte: (Etest® ESBL, AB BIODISK)

39
Fonte: Cristina Motta Ferreira
Figura 9: Imagem de detecção do fenótipo ESBL pelo método epsilométrico E-teste®.
Fita com os gradientes de concentrações dos antibióticos TZ e em associação com o
inibidor, o ácido clavulânico.
3.9.2 Teste de Suscetibilidade Antimicrobiana
O teste antimicrobiano epsilométrico E-teste® (figura 9), para a detecção da
MIC (concentração inibitótria mínima), foi realizado utilizando-se o meio de cultura
ágar Mueller Hinton (Himedia), conforme recomendações do fabricante AB Biodisk
(Solna, Sweden). Os critérios de interpretação utilizados seguiram as
recomendações do Institute for Clinical and Laboratory Standards CLSI (CDC-
Atlanta, USA, 2005)(29) . Os antibióticos amicacina, oxacilina, cefoxitina, penicilina G,
ciprofloxacina, cefepime, vancomicina, ceftazidime, cloranfenicol, imipenem e
tetraciclina foram escolhidos em comum acordo com o corpo clínico do HEMOAM.
As fitas de E-test® contendo os antibióticos utilizados para tratamento de
infecções bacterianas Gram-positivas foram: oxacilina, cefoxitina, vancomicina,
tetraciclina, ceftazidima, cloranfenicol, amicacina, cefepime (0.016-256.0 µg/mL),
benzilpenicillina, ciprofloxacina, imipenem (0.002-32.00 µg/mL). Para as bactérias
Gram-negativas foram: tetraciclina, ceftazidima, cefoxitina, cloranfenicol, cefepime,
amicacina (0.016–256.0 µg/mL), ciprofloxacina e imipenem (0.002–32.00 µg/mL).
Os organismos testes foram suspensos em solução salina a 0.8%, padrão
número 0.5 da escala de McFarland e inoculadas no meio de ágar Mueller-Hinton,
seguidas por uma incubação a 37°C por 18–24h. A interpretação do teste foi
realizada segundo o manual técnico do Clinical and Laboratory Standards (CLSI;

40
Wayne, Pennsylvania, USA)(29), o qual fornece o ponto de corte (break point),
correspondente ao diâmetro da zona de inibição. Cepa de S. aureus ATCC 25923 e
E. coli ATCC 25922 foram utilizadas com controle de qualidade para os meios de
cultura e testes de suscetibilidade.
Fonte: William Antunes Ferreira
Figura 10: Imagem do resultado do teste de suscetibilidade método
epsilométrico E-teste® para a detecção da MIC (concentração inibitótria
mínima).
3.9.3 Teste da PBP 2a
Os S. aureus com resistência no MIC/E-test maior ou igual a quatro
(>/=4µg/mL) para oxacilina e cefoxitina, e os SCoN com resistência no MIC/E-teste
maior ou igual a cinco (>/=0,5µg/mL) (CLSI) para oxacilina e cefoxitina, foram
submetidos ao teste da proteína ligadora de penicilina (PBP 2a ou PBP2’) conforme
protocolo do kit “PBP 2’Test Kit” (Oxoid,Japan). Este kit detecta os (MRSA) e MR-
SCoN, pois a PBP2’ é uma proteína presente na parede celular bacteriana
responsável pela resistência cromossômica a meticilina (oxacilina) nesta espécie.
3.9.4 Caracterizações Moleculares 3.9.4.1 Extração do DNA Total Bacteriano e Quantificação
Todas as espécies bacterianas identificadas através de metodologia
padronizada foram semeadas em meio sólido de Luria Bertani (LB) (Promega,
Madison, WI, USA) e incubadas a 37°C por 24hs. Após 24hs, retirou-se 01 colônia

41
pura para inoculação em 5ml de meio líquido de cultura LB com nova incubação a
37°C por uma noite, sob agitação, sendo a extração realizada conforme protocolo do
kit Easy DNA for genomic DNA isolation (Invitrogen, Carlsbad, CA, USA) com
adaptações em relação ao tempo, temperatura de incubação das amostras e
concentração enzimática, utilizando-se previamente tratamento com lisozima 50µl
(10mg/mL) para permitir a lise bacteriana.
O princípio do kit baseou-se na adição da solução A (solução de lise) ao pellet
bacteriano resuspenso em solução tampão PBS seguida de incubação a 65°C. As
proteínas e os lipídios das células foram precipitados e extraídos pela adição da
solução B (solução de precipitação) e clorofórmio. A fase de centrifugação teve por
objetivo separar a solução em duas fases com uma interface sólida entre as
mesmas. O DNA ficou na fase aquosa clara de cima e as proteínas e os lipídios na
interface sólida. O clorofórmio forma a fase abaixo da nterface sólida. O DNA foi
então removido, precipitado com etanol e ressuspenso com tampão TE (Invitrogen).
Após a extração o DNA foi submetido à eletroforese em gel de agarose a
0,8% com SYBR safe DNA gel stain 10.000X concentrate in DMSO (Invitrogen) por
uma hora e comparado com padrão de peso molecular 100bp DNA Ladder (2-
2,072bp) (Invitrogen). O gel foi examinado em transiluminador ultravioleta a 300nm e
posteriormente fotografado em sistema apropriado de fotodocumentação. O DNA foi
quantificado em aparelho UV Spectrophotometer 50 Conc Varian e diluído para uma
concentração final de 5ng/µl sendo as amostras armazenadas a -70ºC.
3.9.4.2 Extração do DNA plasmidial
As bactérias Gram-negativas produtoras de ESBL, foram semeadas em meio
sólido de Luria Bertani (LB) com ampicilina (50mg/ml) (Promega) e incubadas a
37°C por 16hs. Após 16hs, retirou-se 01 colônia pura para inoculação em 5ml de
meio líquido de cultura LB com nova incubação a 37°C 16-18hs sob agitação sendo
a extração realizada conforme protocolo do kit Wizard® Plus SV Minipreps-DNA
purification System (Promega® , Madison, WI, USA).

42
A técnica baseou-se no rompimento da parede celular para se obter um
lisado, separando o DNA solúvel dos restos celulares e outros materiais insolúveis,
purificando-o de proteínas solúveis e outros ácidos nucléicos. O rompimento da
célula foi feito utilizando-se detergentes ou desnaturação alcalina. O lisado foi
clareado por centrifugação, filtração ou clareamento magnético. O DNA foi então
purificado da parte solúvel do lisado. Quando as matrizes de sílica foram utilizadas,
o DNA foi eluído em água livre de nuclease. Obteve-se então um material de alta
qualidade e pronto para ser utilizado em várias aplicações moleculares (Promega®).
Após a extração, o DNA plasmidial foi submetido à eletroforese em gel de
agarose a 0,8%, com SYBR safe DNA gel stain 10.000X concentrate in DMSO
(Invitrogen) por uma hora e vinte minutos, e comparado com padrão de peso
molecular 2-16Kb DNA Ladder (Promega). O gel foi examinado em transiluminador
ultravioleta a 300nm e posteriormente fotografado em sistema apropriado de
fotodocumentação. A avaliação e classificação do plasmídio foi realizada a partir da
análise comparativa dos padrões de peso molecular do DNA plasmidial das cepas
isoladas resistentes aos antibióticos.
3.9.4.3 Reação em cadeia da polimerase (PCR) 3.9.4.3.1 Para o gene 16S rRNA
Realizou-se o PCR para amplificação do DNA genômico das bactérias Gram-
positivas e Gram-negativas. O protocolo para o 16S rRNA utilizando o primer
bacteriano universal (Invitrogen) com um produto de PCR de aproximadamente 900
bp, foram realizados para confirmar o gênero e espécie, conforme protocolo de
Miyoshi et al.(2005)(91). A seqüência do ciclo de amplificação da reação foi de 35
ciclos de 95ºC por 2 minutos (min.), 94ºC por 1 min., 58ºC por 1 min., 72ºC por 1
min., 72ºC por 5 min., e 4ºC forever totalizando um tempo de 2,5hs.
A análise eletroforética foi realizada em gel de agarose a 1%, corado com
Sybr safe DNA gel stain 10.000X concentrate in DMSO (Invitrogen) em 80v, por 50
minutos. Um marcador de peso molecular 100bp DNA Ladder (Invitrogen) foi
aplicado para se estimar o tamanho das amostras. O gel foi examinado em

43
transiluminador ultravioleta a 300nm e posteriormente fotografado em sistema
apropriado de fotodocumentação.
A sequência de similaridade da região 16S rRNA foi comparada com os
dados do Genbank utilizando-se a ferramenta BLAST com o algoritmo MegaBLAST.
Os Primers universais utilizados na reação foram 27F-5′-
AGAGTTTGATCCTGGCTCA G-3′; 149 R -5′-GGTTA CCTTGTTACGACTT-3′ (91) .
3.9.4.3.2 PCR em tempo real (q-PCR) para o gene mecA
Para a amplificação do gene mecA e Nuc utilizou-se 17µl de mix para reação
(330µl de Power sybr, 66µl de primers mix, 165µl de água mili-Q) e 3µl da amostra.
A seqüência do ciclo de amplificação da reação foi de 95º C por 10 minutos, 45
ciclos de 95ºC por 10 segundos, 50ºC e 60ºC por 15 segundos, e a curva de
desnaturação (melt) padrão.
3.9.4.3.3 Gene mecA
Os primers mecA (mecA1 e mecA2), tipos de SCCmec (CIF2, ccrC, RIF5,
SCCmecV, SCCmecIII, dcs, ccrB2, kdp, mecIP2, mecIP3, mecAP4 e mecAP7),
toxinas bacterianas (seh, arcA, etd, LUKPV) foram obtidos de acordo com o
protocolo descrito por Jonas et al. (2002) (66), Thomas et al. (2007) (130), Milheiriço et
al. (2007) (87), e Strommenger et al., (2008)(110). Cepas S. aureus ATCC (33591) e
S. epidermidis (ATCC12228) foram utilizadas como controle positivo e negativo
(quadro 4).
Para uma reação com volume final de 25µl, utilizou-se 20µl de mix para a
reação (14,45µl de água ultrapura, 2,5µl de tampão 10X (Invitrogen), 1,25µl de
MgCl2 (50µm) (Invitrogen), 0,5µl dNTP (10µm) (Invitrogen), 1,0µl do total de primers
(forward e reverse), 0,3µl de DNA Taq polimerase (5U/ml) (Invitrogen) e 5µl de DNA
cromossomal (5ng). A seqüência do ciclo de amplificação da reação foi de 30 ciclos
de 94º C por 4 minutos (min.), 94ºC por 45seg., 50ºC por 45seg., 72ºC por 60 seg.,
72ºC por 2min., e temperatura final de 4ºC, totalizando um tempo de 2,25hs.
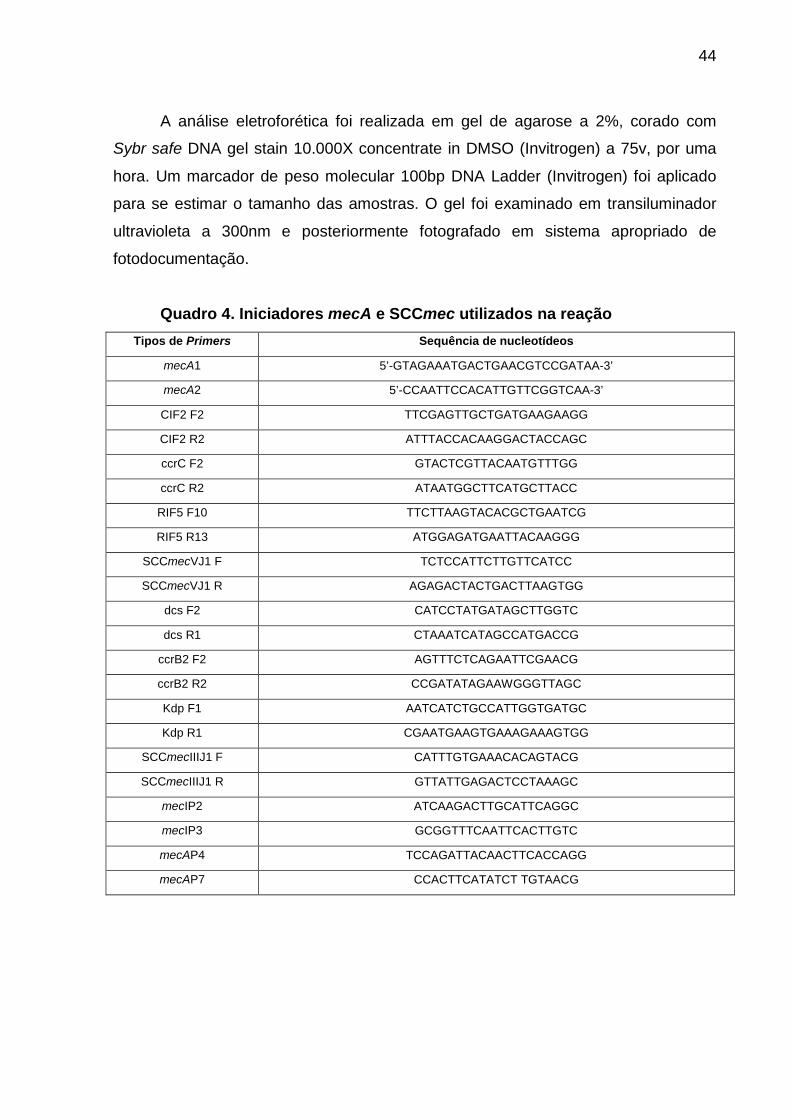
44
A análise eletroforética foi realizada em gel de agarose a 2%, corado com
Sybr safe DNA gel stain 10.000X concentrate in DMSO (Invitrogen) a 75v, por uma
hora. Um marcador de peso molecular 100bp DNA Ladder (Invitrogen) foi aplicado
para se estimar o tamanho das amostras. O gel foi examinado em transiluminador
ultravioleta a 300nm e posteriormente fotografado em sistema apropriado de
fotodocumentação.
Quadro 4. Iniciadores mecA e SCCmec utilizados na reação Tipos de Primers Sequência de nucleotídeos
mecA1 5’-GTAGAAATGACTGAACGTCCGATAA-3’
mecA2 5’-CCAATTCCACATTGTTCGGTCAA-3’
CIF2 F2 TTCGAGTTGCTGATGAAGAAGG
CIF2 R2 ATTTACCACAAGGACTACCAGC
ccrC F2 GTACTCGTTACAATGTTTGG
ccrC R2 ATAATGGCTTCATGCTTACC
RIF5 F10 TTCTTAAGTACACGCTGAATCG
RIF5 R13 ATGGAGATGAATTACAAGGG
SCCmecVJ1 F TCTCCATTCTTGTTCATCC
SCCmecVJ1 R AGAGACTACTGACTTAAGTGG
dcs F2 CATCCTATGATAGCTTGGTC
dcs R1 CTAAATCATAGCCATGACCG
ccrB2 F2 AGTTTCTCAGAATTCGAACG
ccrB2 R2 CCGATATAGAAWGGGTTAGC
Kdp F1 AATCATCTGCCATTGGTGATGC
Kdp R1 CGAATGAAGTGAAAGAAAGTGG
SCCmecIIIJ1 F CATTTGTGAAACACAGTACG
SCCmecIIIJ1 R GTTATTGAGACTCCTAAAGC
mecIP2 ATCAAGACTTGCATTCAGGC
mecIP3 GCGGTTTCAATTCACTTGTC
mecAP4 TCCAGATTACAACTTCACCAGG
mecAP7 CCACTTCATATCT TGTAACG

45
3.9.4.3.4 Tipos de SCCmec
Para uma reação com volume final de 25µl, utilizou-se 20µl de mix para a
reação (13,5µl de água ultrapura, 2,5µl de tampão 10X (Invitrogen), 0,75µl de MgCl2
(50µm) (Invitrogen), 0,5µl dNTP (10µm) (Invitrogen), 1,25µl do total de primers
(forward e reverse), 0,25µl de DNA Taq polimerase (5U/ml) (Invitrogen) e 5µl de
DNA cromossomal (5ng). A seqüência do ciclo de amplificação da reação foi de 30
ciclos de 94º C por 4 minutos (min.), 94ºC por 30seg., 53ºC por 30seg., 72ºC por 1
min., 72ºC por 4 min., e temperatura final de 4ºC, totalizando um tempo de 2,25hs.
Cepas S. aureus ATCC (33591) e S. epidermidis (ATCC12228) foram utilizadas
como controle positivo e negativo
A análise eletroforética foi realizada em gel de agarose a 3%, corado com
Sybr safe DNA gel stain 10.000X concentrate in DMSO (Invitrogen) a 75v, por uma
hora. Um marcador de peso molecular 100bp DNA Ladder (Invitrogen) foi aplicado
para se estimar o tamanho das amostras. O gel foi examinado em transiluminador
ultravioleta a 300nm e posteriormente fotografado em sistema apropriado de
fotodocumentação.
3.9.4.3.5 Genes seh, etd, arcA e lukPV das toxinas bacterianas
Para a reação com volume final de 25µl, utilizou 20µl de mix para a reação
(12,95µl de água ultrapura, 2,5µl de tampão 10X (Invitrogen), 1,25µl de MgCl2
(50µm), 0,5µl de dNTPs (10µm) (Invitrogen), 2,5 µl do mix de todos os primers
(forward e reverse), 0,3µl de DNA Taq polimerase (Invitrogen) e 5µl de DNA
cromossomal (5ng). A seqüência do ciclo de amplificação da reação foi de 30 ciclos
de 94ºC a 4 min., 94ºC por 45seg., 50ºC 45 seg., 72 ºC 60seg., 72 ºC 2min., e
temperatura final de 4ºC, perfazendo um total de 2,25horas. Os primers seh, etd,
arcA, lukPV foram obtidos de acordo com o protocolo descrito por Strommenger et
al. (2008)(110). Cepas S. aureus ATCC (29213) S. epidermidis (ATCC 12228) foram
utilizadas como controle positivo (quadro 5).

46
A análise eletroforética foi realizada em gel de agarose a 2%, corado com
SYBR safe DNA gel stain 10.000X concentrate in DMSO (Invitrogen) a 75v, por uma
hora. Um marcador de peso molecular 100bp DNA Ladder (Invitrogen) foi aplicado
para se estimar o tamanho das amostras. O gel foi examinado em transiluminador
ultravioleta a 300nm e posteriormente fotografado em sistema apropriado de
fotodocumentação.
Quadro 5. Iniciadores das toxinas bacterianas utilizados na reação Tipos de primers Sequência de nucleotídeos
Seh f CAACTGCT GATTTAGCTCAG
Seh r GTCGAATGAGTAATCTCTAGG
arcA f TTGCTCAAACTTTGAGAGATGAA
arcA r TTACGTACGCCAGCCATGAT
Setd f CCCGTTGATTAGTCATGCAG
Setd r CCAGAATTTCCCGACTCAG
lukPV f ATCATTAGGTAAAATGTCTGGACATGATCCA
lukPV r GCATCAAGTGTATTGGATAGCAA AAGC
3.9.4.3.6 Genes das Beta-lactamases de espectro estendido (ESBLs)
A detecção dos genes bla (beta-lactamase) foi realizada através da
amplificação por PCR com os iniciadores para os genes blaTEM, blaSHV, blaCTX-M e
blaOXA, respectivamente (quadro 6).
Quadro 6. Iniciadores das ESBLs
Genes Tipos de primers Sequência de nucleotídeos
blaTEM TEM-F 5’ – ATGAGTATTCAACATTTCCG – 3’
TEM-R 5’ – CTGACAGTTACCAATGCTTA – 3’
blaSHV SHV-F 5’ – GGTTATGCGTTATATTCGCC – 3’
SHV-R 5’ – TTAGCGTTGCCAGTGCTC – 3’
blaCTX-M CTX-F 5’ – ATGTGCAGYACCAGTAARGT – 3’
CTX-R 5’ – TGGGTRAARTARGTSACCAGA – 3’
blaOXA OXA-F 5’ – ACACAATACATATCAACTTCGC – 3’
OXA-R 5’ – AGTGTGTTTAGAATGGTGATC – 3’
Legenda: Y = C ou T; R = A ou G; S = G ou C.

47
Para uma reação com volume final de 25µl, utilizou-se 5µL de tampão 5X;
2,5µL de MgCl2 (25mM); 0,5µL de dNTP (10µM); 1µL de cada iniciador (10µM);
0,2µL de GoTaq™ DNA Polymerase, 5µL de DNA cromossomal (5ng) e 1µL para
DNA plasmidial (1:10). A seqüência do ciclo de amplificação foi de 96ºC por 5 min;
35 ciclos de 96ºC por 1 min, 58ºC(TEM) / 60ºC (SHV) por 1 min e 72ºC por 1 min.
Após a finalização dos ciclos, houve uma extensão final de 72ºC por 10 min. Para o
gene blaCTX-M: desnaturação inicial a 94ºC por 7 min; seguida de 35 ciclos de 50 seg
a 94ºC, 40 seg a 50ºC, 1 min a 72ºC; e extensão final de 5 min a 72ºC. Para blaOXA:
desnaturação inicial de 5 min a 96ºC; seguido de 35 ciclos de 1 min a 96ºC, 1 min a
60ºC, 2 min a 72ºC; e extensão final de 10 min a 72ºC.
A análise eletroforética foi realizada em gel de agarose a 1%, corado com
Sybr safe DNA gel stain 10.000X concentrate in DMSO (Invitrogen) a 75v, por uma
hora. Um marcador de peso molecular 100bp DNA Ladder (Invitrogen) foi aplicado
para se estimar o tamanho das amostras. O gel foi examinado em transiluminador
ultravioleta a 300nm e posteriormente fotografado em sistema apropriado de
fotodocumentação. Foi utilizado como controle positivo para o gene blaSHV cepa de
K. pneumoniae ATCC 700603 tanto para o plasmídeo como para o cromossomo.
3.9.4.3.7 Genotipagem para identificação dos clones bacterianos Os primers referentes aos sete loci dos genes housekeeping e os protocolos
para realização das PCRs para o clone do S. aureus: arc (Carbamato quinase), aro
(Siquimato deidrogenase), glp (Glicerol quinase), gmk (Guanilato quinase), pta (Fosfato acetiltransferase), tpi (Triosefosfato isomerase), yqi (Acetil coenzima A
acetiltransferase) e dos clones dos S. epidermidis: arcC (Carbamato quinase),
aroE (Siquimato deidrogenase), gtr (Transportador ABC), mutS (Proteína de reparo
do DNA), pyrR (Proteína operon reguladora da pirimidina), tpiA (Triosefosfato
isomerase), yqiL (Acetil coenzima A acetiltransferase), foram obtidos de acordo
com os protocolos descritos no site (http://www.mlst.net/) com adequações nas
concentrações dos reagentes e na temperatura do ciclo da reação (anexo 3).

48
A análise eletroforética foi realizada em gel de agarose a 2%, corado com
SYBR safe DNA gel stain 10.000X concentrate in DMSO (Invitrogen) a 75v, por uma
hora. Um marcador de peso molecular 100bp DNA Ladder (Invitrogen) foi aplicado
para se estimar o tamanho das amostras. O gel foi examinado em transiluminador
ultravioleta a 300nm e posteriormente fotografado em sistema apropriado de
fotodocumentação.
3.10 Purificação pré-sequenciamento
A purificação foi realizada conforme recomendação do fabricante do kit illustra TM GFX TM PCR DNA and Gel Band Purification (GE Healthcare, Buckinghamshire,
UK). A análise eletroforética foi realizada em gel de agarose a 1%, corado com
SYBR safe DNA gel stain 10.000X concentrate in DMSO (Invitrogen) a 80 v, por uma
hora. Um marcador de peso molecular 100bp DNA Ladder (Invitrogen) foi aplicado
para se estimar o tamanho das amostras. O gel foi examinado em transiluminador
ultravioleta a 300nm e posteriormente fotografado em sistema apropriado de
fotodocumentação. O seqüenciamento de todas as amostras foi realizado no
aparelho ABI Sequence (Applied Biosystems) para se detectar a seqüência
nucleotídica e posterior comparação com o banco de dados público BLAST e
GenbanK databases. Para os genes pesquisados no MLST (Multi Locus Sequence
Typing) a purificação foi realizada conforme protocolo do Big Dye® X terminatorTM
purification kit (Applied Biosystems) sendo posteriormente sequenciados.
3.11 Precipitação do DNA com Isopropanol/etanol
Para a reação com volume final de 20µl, utilizou 19µl de mix para a reação de
pré-sequencimamento (12,0µl de água ultrapura, 3,0µl de tampão 5X (Invitrogen),
1,5µl de cada primer separadamente, 2,5µl de Big Dye (invitrogen) e 1,0µl de DNA
amplificado (forward e reverse separadamente) com Big Dye. A seqüência do ciclo
de amplificação da reação foi de 30 ciclos de 96 ºC a 1 min., 96ºC por 15seg., 50ºC
15 seg., 60ºC 4 min., 4ºC forever, perfazendo um total de 2,30horas.

49
Posteriormente as amostras foram precipitadas adicionando-se 80µl de
isopropanol a 75% e submetidas ao vortex. A placa foi selada com papel colante e
deixada a T.A. (temperatura ambiente) por 15min. protegida da luz. Em seguida a
mesma foi centrifugada por 20min. a 14.000 rpm descartando-se o sobrenadante por
inversão sobre um papel absorvente. Aplicou-se um pulso de rotação da centrífuga
com a placa invertida sobre o papel absorvente. Adicionou-se 20µl de etanol a 70%
em cada amostra e selou-se a placa novamente, submetendo a mesma a uma nova
centrifugação a 14000 rpm a T.A. Posteriormente, descartou-se o sobrenadante e
repetiu-se a centrifugação com a placa invertida sobre o papel absorvente. O pellet
preso no fundo do poço ficou a temperatura ambiente para secar, protegido da luz, e
a placa foi preparada para o seqüenciamento.
3.12 Seqüenciamento
Para a ressuspensão da amostra foi utilizada 10 µl formamida Hi-Di (applied
Biosystems P/N 4311320) em cada uma, sendo posteriormente tampadas com papel
laminado. A palca foi selada, submetida ao vortex, uma rotação na centrifuga e
desanturada no termociclador a 95ºC por 2 min. Posteriormente a placa foi colocada
no gelo seco por 2 min. e submetida a uma nova rotação na centrífuga para
remoção de bolhas de ar. Removeu-se o adesivo selante da placa. Posteriormente,
ambas placa, bandeja e tampa adaptativa foram inseridas no autosampler do
aparelho de seqüenciamento 3130/genetic analyser (applied Biosystems Hitachi) e
procedida à reação de seqüenciamento.
3.13 Controle de Qualidade Dos meios de cultura e do antibiótico utilizado no teste de suscetibilidade,
foram utilizados as cepas de: S. aureus ATCC 25923, S. aureus ATCC 33591, S.
aureus ATCC 29213, E. coli ATCC 25922, S. saprophyticus ATCC 15305, S.
epidermidis ATCC 12228, Pseudomonas aeruginosa ATCC 27853, S. pyogenes
ATCC 19615, K. pneumoniae ATCC 13883; , K. pneumoniae ATCC 700603; E.
aerogenes ATCC 13048; E. cloacae ATCC13047 (American Type Culture and
Collection), recomendado pelo Institute for Clinical and Laboratory Standards (CLSI,

50
2005) (29). Para os testes moleculares utilizou-se os padrões de peso molecular
Supercoiled DNA Ladder de 2- 10Kb (Promega), DNA Ladder de 100bp e 123bp
(Invitrogen).
3.14 Armazenamentos de amostras As amostras de bactérias isoladas foram armazenadas em meio de cistina
tripticase soja com 10% de glicerol em freezer a -70ºc (CLSI, 2005) (29).
3.15 Análise de Dados Os dados coletados foram armazenados em planilha do Excel e os
resutados descritos e demonstrados através de tabelas de freqüência e
gráficos. 3.16 Fontes de financiamento (Nacionais e/ou Internacionais)
Projeto submetido e aprovado pelo Edital MS/CNPq/FAPEAM N. 014/2006,
Programa de pesquisa para o SUS: gestão compartilhada em saúde.

51
4 RESULTADOS Os resultados estão apresentados na forma de artigos originais.

52
4.1 Artigo 1 Extended-spectrum beta-lactamase-producing bacteria isolated from hematologic patients in Manaus, State of Amazonas, Brazil
Cristina Motta Ferreira1*, William Antunes Ferreira2, Nayanne Cristina Oliveira da
Silva Almeida3, Felipe Gomes Naveca4, Maria das Graças Vale Barbosa5
1Universidade Estadual do Amazonas (UEA); Fundação de Medicina Tropical do
Amazonas (FMT-AM); Fundação de Hematologia e Hemoterapia do Amazonas
(HEMOAM), Manaus, State of Amazonas, Brazil. 2Fundação de Dermatologia Tropical e Venereologia Alfredo da Matta (FUAM). 3Universidade Federal do Amazonas (UFAM), Manaus, State of Amazonas, Brazil. 4Leônidas and Maria Deane Institute/FIOCRUZ, Manaus, State of Amazonas, Brazil. 5The Amazon Tropical Medicine Foundation FMT-AM; Universidade Estadual do
Amazonas (UEA), Manaus, State of Amazonas, Brazil.
*Corresponding Author Cristina Motta Ferreira
Fundação de Hematologia e Hemoterapia do Amazonas (HEMOAM)
Address: Avenida Constantino Nery, 2533 Apto. 704, Bloco B, Residencial.
Maria da Fé. CEP: 69050-001, Manaus, Amazonas, Brazil.
Telephone: (55)-92-3233-30-89; Email: [email protected]
Abstract Antibiotic therapy in hematologic patients, often weak and susceptible to a
wide range of infections, particularly nosocomial infections derived from long
hospitalization periods, is a challenging issue. This paper presents ESBL-producing
strains isolated from such hematologic patients treated at the Amazon Hematology
and Hemotherapy Foundation (HEMOAM) in the Brazilian Amazon Region to identify
the ESBL genes carried by them as well as the susceptibility to 11 antimicrobial
agents using the E-test method. A total of 146 clinical samples were obtained from
July 2007 to August 2008, when 17 gram-negative strains were isolated in our

53
institution. The most frequent isolates confirmed by biochemical tests and 16S rRNA
sequencing were E. coli (8/17), Serratia spp. (3/17) and B.cepacia (2/17).
All gram-negative strains were tested for extended-spectrum-beta-lactamases
(ESBLs), where: (12/17) strains carried ESBL; among these, (8/12) isolates carried
blaTEM, blaCTX-M, blaOXA , blaSHV genes, (1/12) blaTEM gene and (3/12) blaTEM, blaCTX-M,
blaOXA genes. Antibiotic resistance was found in (15/17) of the isolates for
tetracycline, (12/17) for ciprofloxacin, (1/17) resistance for cefoxitin and
chloramphenicol, (1/17) for amikacin and (3/17) cefepime. This research showed the
presence of gram-negative ESBL-producing bacteria infecting hematologic patients
in HEMOAM. These strains carried the blaTEM, blaSHV, blaCTX-M and blaOXA genes
and were resistant to different antibiotics used in the treatment. This finding was
based on a period of 13 months, during which clinical samples from specific
populations were obtained. Therefore, caution is required when generalizing the
results that must be based on posological orientations and new breakpoints for disk
diffusion and microdilution published by CLSI 2010.
Key words ESBL, beta-lactams, nosocomial infection, TEM, SHV, OXA, CTX-M
Running Title ESBL-producing bacteria from hematologic patients
Introduction
Extended-spectrum beta-lactamases (ESBL) are enzymes produced by gram-
negative bacteria such as Klebsiella pneumoniae and Escherichia coli (24) as well as
by species from other genera, such as Enterobacter sp., Salmonella sp., Proteus sp.,
Serratia marcescens, Shigella dysenteriae, Pseudomonas aeruginosa, and
Burkholderia cepacia (2, 3, 31, 26).
ESBLs originate from punctual genetic mutations in the active sites of the
TEM-1, TEM-2, and SHV-1 enzymes (3, 10, 12, 31, 19, 23, 26, 34), which enable the

54
hydrolysis of extended-spectrum cephalosporins, monobactams, penicillins. (2, 3, 5,
9, 11, 12, 27, 30). However, these plasmid mediated enzymes have their activity
inhibited by clavulanic acid, tazobactam and sulbactam. Another important
epidemiological characteristic is the possibility of these enzymes being transferred to
other species via plasmids or transposons (2, 3, 5, 6, 9, 11, 12, 21, 27, 28, 30, 34).
The emergence of ESBL-producing bacteria, responsible for infectious
processes with high levels of morbidity and mortality has complicated the therapeutic
options available for elderly, immunocompromised, and debilitated individuals (4, 31).
The prevalence of ESBLs among clinical isolates varies according to countries and
institutions (31). In the United States, the presence of these enzymes among
Enterobacteriaceae has increased from 0% to 25%, with a national average of 3%,
whereas the percentage of Klebsiella sp. has demonstrated an increased resistance
from 5% to 10% to ceftazidime (6).
In other countries, such as Switzerland, the frequency of ESBL-producing
bacteria varies from 3% to 8%, whereas it is 34% in Portugal, 4.8% in Korea, 8.5% in
Taiwan, 12% in Hong Kong, and 58% in Turkey. In Latin America, studies have
reported frequencies ranging from 30% to 60% (6, 21). The prevalence of ESBL
resistance to antibiotics in Brazil has reached 45.4%, 65% of which stand for K.
pneumoniae and E. coli, which is considered one of the highest levels in the world.
For the city of Manaus, in the State of Amazonas, Brazil, no published data is found
regarding the prevalence of such bacteria (3, 5).
ESBL-producing bacteria are responsible for serious infectious processes,
commonly associated with outbreaks of nosocomial infections. This situation is
relevant as most patients with hematologic diseases undergo chemotherapy that can
lead to a deficient immune response. These medical conditions can lead to
prolonged hospitalization, elevation of the risks of mortality and morbidity, and
increased treatment costs.
This study therefore aimed to detect ESBL-producing strains isolated from
such hematologic patients treated at the HEMOAM in the Brazilian Amazon region to

55
identify the ESBL genes carried by them and susceptibility to 11 antimicrobial agents
using the E-test method. Materials and Methods The study was approved by the HEMOAM Ethics Committee (Approval
number 170 CAAE- 0005.0.112.000-06 -Version 003/06). An informed consent was
obtained from each patient before specimens were collected and the results were
used for the management of each respective patient.
We conducted a descriptive cross-sectional study including patients of both
sexes at any age who had been admitted to the HEMOAM Foundation in the city of
Manaus, State of Amazonas, with prior diagnosis of hematologic disease, presenting
clinical signs and symptoms suggestive of acute gram-negative bacterial infection,
whether feverish or not, or having developed fever during hospital stay in the period
from July 2007 to August 2008.
Gram-negative bacteria strains were identified using colonial morphology on
blood, MacConkey, Eosin Methylene Blue (EMB), and Mueller-Hinton agar plates
(Himedia- Hexasystems, Mumbai, India), followed by biochemical panels like NF II
Kit, enterokit C, and enterokit B (Probac, São Paulo, Brazil). Other tests, such as
urease, oxidase, glucose, mannose, lactose, and maltose fermentation, were also
performed. The 16S rRNA protocol with the bacterial primers 27F (5′-
GAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTAC GACTT-3′)
(Invitrogen, Carlsbad, CA, USA) with a product size around 900 bp, were performed
to confirm genus and species (17).
The sequence analysis of the 16S rRNA region was compared with GenBank
data using BLAST search with the MegaBLAST algorithm for identifying high-
similarity sequences. The sequences with the highest levels of similarity were
considered as being consistent for species identified by the biochemical tests.
The antibiotics E-test® strips representing common drugs used in the

56
treatment of suspected gram-negative bacterial infections in our hospital were
chosen for antimicrobial testing. Strips included tetracycline, ceftazidime, cefoxitin,
chloramphenicol, cefepime, and amikacin (0.016–256.0 µg/mL), and ciprofloxacin
and imipenem (0.002–32.00 µg/mL).
Test organisms were suspended in 0.8% saline to 0.5 McFarland standards
and then inoculated on Mueller-Hinton agar plates, followed by overnight incubation
at 37°C for 18–24 hours. Interpretation was performed using guidelines laid down in
the Clinical and Laboratory Standards (CLSI; Wayne, Pennsylvania, USA) manual,
which provides break points corresponding to the diameter of the zone of inhibition.
The isolates were screened for the production of ESBL by the E-test® method
with the antibiotics cefotaxime (0.25–16 μg/mL)/cefotaxime (0.016–1 μg/mL) plus 4
μg/mL clavulanic acid, ceftazidime (0.5–32 μg/mL) /ceftazidime (0.064–4 μg/mL) plus
4 μg/mL clavulanic acid, following manufacturer’s recommendations (7). Strains of E.
coli ATCC 25922 and K. pneumoniae ATCC 700603 were used as quality controls for
culture media, susceptibility tests, PCR and the ESBL-production E-test®.
The bacteria deemed as positive for ESBL-production were inoculated in 5 ml
of Luria-Bertani (LB) broth with ampicillin (50 µg/mL) and incubated at 37°C for 16
hours for plasmid extraction following the protocol described in the Kit Wizard® Plus
SV Minipreps-DNA purification System (Promega®; São Paulo, Brazil).
Electrophoresis was performed in 0.8% agarose gel stained with SYBR Safe
(Invitrogen, Carlsbad, CA, USA) for the identification of plasmid sizes. The
Supercoiled Loading Dye Ladder 2-16 Kb (Invitrogen) was used as a standard
molecular weight. Data were filed by using Excel programs. ESBL isolates were
preserved for future molecular analysis. PCR amplification for the detection of
chromosomal beta-lactamase resistance AmpC type in Pseudomonas spp. isolates
was not included in our study.
The Polimerase Chain Reaction (PCR) assay to amplification and detection of
the blaTEM, blaSHV, blaCTX-M and blaOXA was carried out in a final volume reaction of

57
25µl using 10X buffer, MgCl2(50mµ), dNTP (10mµ), DNA Taq polimerase (5U/mL)
(Invitrogen), 1,25µl of each primer TEM-F, TEM-R, SHV-F, SHV-R, blaCTX-M-F,
blaCTX-M-R, blaOXA-F, blaOXA-R and 5µl of plasmidial DNA. Cycling parameters were
previously described (Table 2). The amplified products obtained have not been
sequenced up to the present moment.
Results
A total of 146 clinical samples were obtained from 69 patients over a period of
thirteen months. These samples included the following: urine, pus, blood, marrow
aspirate, sputum, feces, abscesses, and oropharyngeal and nasopharyngeal swabs.
The strains isolated from different specimen type are presented in Table 5.
Table 2. Primer sequence and PCR conditions
Primers Oligonucleotide sequence
(5’to 3’) PCR conditions Reference
Expected size (bp)
TEM-F ATGAGTATTCAACATTTCCG 1 cycle of 5 min at 96oC; 35 cycles
of 1min at 96ºC; 1min at 58 ºC;
1min at 72 ºC ; 1 cycle of 10 min at
72oC
14 867 TEM-R CTGACAGTTACCAATGCTTA
SHV-F GGTTATGCGTTATATTCGCC 1 cycle of 5min at 96 ºC; 35 cycles
of 1min at 96 ºC, 1min at 60oC,
1min at 72 ºC; 1 cycle of 10 min at
72 ºC
14 867 SHV-R TTAGCGTTGCCAGTGCTC
CTX-M-F ATGTGCAGYACCAGTAARGT 1 cycle of 7min at 94oC; 35 cycles
of 50 sec at 94 ºC, 40 sec at 50 ºC,
1 min at 72 ºC; 1 cycle of 5min at
72 ºC.
14 593 CTX-M-
R TGGGTRAARTARGTSACCAGA
OXA- F ACACAATACATATCAACTTCGC 1 cycle of 5min at 96oC; 35 cycles
of 1min at 96 ºC, 1min at 60 ºC, 2
min at 72 ºC; 1 cycle of 10 min at
72 ºC.
14 885 OXA- R AGTGTGTTTAGAATGGTGATC

58
Table 5. Strains isolated from different specimen type detected from patients with
infections
Specimen types Isolates
Blood
K. pneumoniae, E. coli, Pseudomonas sp., P.
stutzery, Serratia sp., B. cepacia
Urine E. coli
feces E. coli
Eye secretion Pseudomonas aeruginosa
In relation to gender, (35/69) were male and (34/69) were female. Among the
collected samples, we isolated 44 bacterial strains, 17 of which were gram-negative.
The most frequent hematological diseases in the patients were acute lymphocytic
leukemia (36/69) and acute myeloid leukemia (6/69) (Table 1).
Table 1. Frequency of hematologic diseases diagnosed in this study
Hematologic disease N
Acute lymphocytic leukemia 36
Acute myeloid leukemia 6
Sickle cell anemia 4
Pancytopenia 4
Chronic myeloid leukemia 3
Chronic lymphocytic leukemia 3
Hemophilia 2
Multiple myeloma 2
Others● 9
Total 69 ● Hypofibrinogenemia + hemophilia (2/9), Thalassemia (1/9), aplastic anemia (1/9),
thrombocytopenic idiopathic purpura (1/9), acute hemolytic anemia (1/9), lymphoma (1/9),
unidentified leucopenic disease (1/9), myelodysplastic syndrome (1/9)

59
Biochemical tests and 16S rRNA sequencing detected E. coli (8/17), followed
by Serratia sp. (2/17), Serratia liquefaciens (1/17) and B. cepacia (2/17) as the most
frequent gram-negative strains. All the strains were tested for the production of ESBL
and the E-test showed that (12/17) were ESBL producers (Table 3).
All of them had their plasmid extracted and the molecular weights were 3.5, 7
and >15 Kb. The PCR assay showed that that (8/12) isolates carried blaTEM, blaCTX-M,
blaOXA , blaSHV genes, (1/12) blaTEM gene and (3/12) blaTEM, blaCTX-M, blaOXA genes.
As presented in Table 4, the results from antimicrobial susceptibility tests
demonstrated antibiotic resistance in (15/17) of the isolates for tetracycline, (12/17)
for ciprofloxacin, (1/17) resistance for cefoxitin/chloramphenicol, and (1/17) for
amikacin and (3/17) for cefepime.
Data regarding the molecular epidemiology of ESBL clonality and PCR
amplification for the detection of chromosomal beta-lactamase resistance AmpC type
in Pseudomonas spp. isolates have not been presented so far, but we believe that
this will not expressively impact on this study as it was aimed to detect ESBL-
producing strains in hematologic patients only. Our finding was based on a period of
13 months, during which clinical samples were obtained from a specific population.
Therefore, caution is required when results are generalized as they must be based
on posological orientations and new breakpoints for disk diffusion and microdilution according to the CLSI 2010 manual M100-S20.
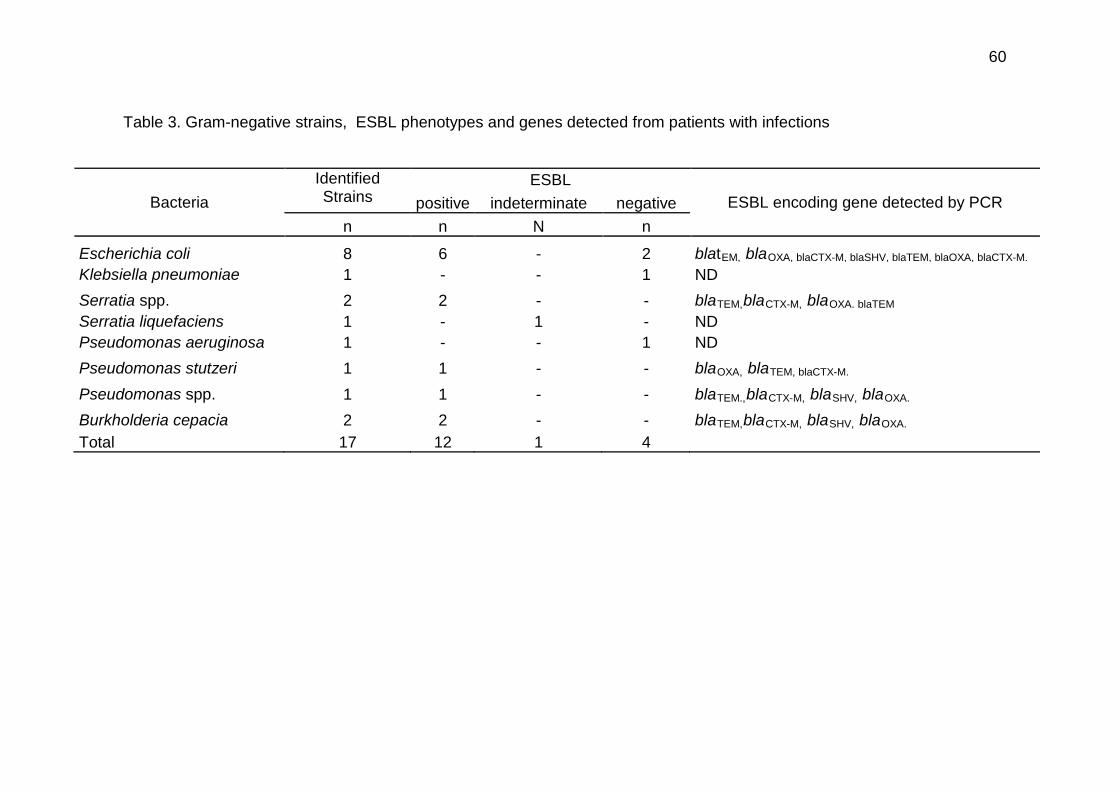
60
Table 3. Gram-negative strains, ESBL phenotypes and genes detected from patients with infections
Bacteria Identified Strains
ESBL ESBL encoding gene detected by PCR positive indeterminate negative
n n N n
Escherichia coli 8 6 - 2 blatEM, blaOXA, blaCTX-M, blaSHV, blaTEM, blaOXA, blaCTX-M. Klebsiella pneumoniae 1 - - 1 ND Serratia spp. 2 2 - - blaTEM,blaCTX-M, blaOXA. blaTEM Serratia liquefaciens 1 - 1 - ND Pseudomonas aeruginosa 1 - - 1 ND Pseudomonas stutzeri 1 1 - - blaOXA, blaTEM, blaCTX-M. Pseudomonas spp. 1 1 - - blaTEM.,blaCTX-M, blaSHV, blaOXA. Burkholderia cepacia 2 2 - - blaTEM,blaCTX-M, blaSHV, blaOXA. Total 17 12 1 4

61
Table 4. Antimicrobial Susceptibility test (E-test) for 17 Gram-negative bacteria isolated from patients with
bacterial infection
Antibiotics
Bacteria
E. coli K.
pneumoniae
Serratia
spp.
S.
liquefaciens
P.
aeruginosa P. stutzeri
Pseudomonas
spp.
B.
cepacia
S I R S I R S I R S I R S I R S I R S I R S I R
Tetracycline - - 8 - - 1 1 - 1 - - 1 1 - - - - 1 - - 1 - - 2
Ciprofloxacin 1 - 7 1 - - 1 - 1 1 - - 1 - - - - 1 - - 1 - - 2
Ceftazidime 2 6 - 1 - - 1 1 - 1 - - 1 - - 1 - - - 1 - 1 1 -
Cefoxitin 5 2 1 1 - - 1 1 - 1 - - - 1 - 1 - - - 1 - 2 - -
Chloramphenicol 6 1 1 1 - - 2 - - 1 - - 1 - - 1 - - 1 - - 2 - -
Amikacin 7 - 1 1 - - 2 - - 1 - - 1 - - 1 - - 1 - - 2 - -
Imipenem 8 - - 1 - - 2 - - 1 - - 1 - - 1 - - 1 - - 2 - -
Cefepime 3 3 2 1 - - 1 - 1 1 - - 1 - - 1 - - - 1 - 1 1 -
S: susceptible; I: intermediate; R: resistant

62
Discussion
ESBLs are enzymes that act by inhibiting the action of beta-lactam antibiotics
(1, 25). Concerns regarding the clinical and epidemiological aspects of such ESBL-
producing pathogens are related to the use of extended-spectrum antimicrobials
such as carbapenems for their treatment (12, 22), thus increasing treatment costs as
well as patient risks due to longer hospital stay and prolonged therapy (1, 29).
Furthermore, hematologic patients often undergo chemotherapy, which can
lead to an immunodeficient status and leave these patients susceptible to a wide
range of infections (32, 33). This is particularly important with respect to nosocomial
infection in the hospital environment, which can elevate the risks of mortality and
morbidity, mainly in the case of infections caused by ESBL-producing bacteria.
In our study, the most frequent ESBLs-producing bacteria were E. coli, B.
cepacia, Serratia spp., and Pseudomonas spp. In addition, ESBL-producing strains
of E. coli and K. pneumoniae have also been isolated in other studies from patients
with outbreaks of bacterial peritonitis (32), bloodstream infections (33) in ICU (1) from
different clinical samples from patients in Mwanza (Tanzania) (20), in Enugu (Nigeria)
(31), and in France, Portugal (9), Brazil (33), as well as several other countries (2,
13, 16, 19, 37).
These E. coli isolated from samples of the patients point to the possibility of a
hospital-acquired infection probably due to inadequate antibiotic therapy,
inappropriate prescriptions without culture results and susceptibility testing and
cross-infections that contributed to increase the number of these resistant isolates (1,
10, 25, 27, 40).
The detection of ESBL enzymes is very important, because according to CLSI
M100-S17 and M100-S18 (25, 27) criteria, these organisms should be reported as
resistant to all extended-spectrum beta-lactam antibiotics, regardless of their
susceptibility test results. Currently, according to CLSI M100-S20, new interpretive
criteria are available for routine ESBL testing (1, 25, 27).

63
The reduced sensitivity to cephalosporins detected in E. coli strains in this
study might be due to the variations in substratum preference, the differing accuracy
between tests for enzyme detection, weak inhibition by clavulanic acid, or different
combined resistance mechanisms that led either to reduced or no sensitivity (26, 35).
This type of phenotype could be demonstrated in other multicenter study developed
in Norway in 2003, in which 60% of ESBL-producing isolates showed reduced
sensitivity to extended-spectrum cephalosporin, possibly indicating an increased
level of resistance (26, 36).
Our findings demonstrated that all the ESBL-producing Enterobacteriaceae
were sensitive to imipenem, while 94.1% were sensitive to amikacin, indicating that
these agents can still be used in the treatment of infections caused by these
pathogens in HEMOAM. Similar results have been observed by Tumbarello et al.
(37) in a teaching hospital in Rome (Italy).
The existence of ESBL-producing Pseudomonas spp. is of serious concern as
it stands for a pathogen with the capacity to acquire resistance to a wide range of
clinically important antimicrobials and it can lead to numerous infectious processes.
In this study, we detected 33.4% of the isolates to be resistant to tetracycline and
ciprofloxacin (12, 25, 29). Similar results have been observed in India, Iran, Spain,
Portugal, Canada, and Brazil. However, in Norway, positive CTX-M isolates showed
reduced sensitivity only to ciprofloxacin (12, 16, 24, 36).
In Europe, 20% of hyperproductive ESBL isolates have demonstrated
resistance to ceftriaxone and aztreonam, while in Latin America, reports show that
the resistance rate is higher in relation to other regions of the world (12, 25). In
Manaus (Brazil), our study provides the first data regarding the existence of ESBLs in
clinical samples from patients. Thus, it is not possible to compare our results
regarding the spectrum of antimicrobial resistance.
The therapeutic alternative for treating infections caused by ESBL-positive
pathogens such as Pseudomonas spp. lies in the use of imipenem or meropenem;
however, this may stimulate the appearance of multiresistant strains in hospitals,

64
which could lead to additional serious problems for patients as well as difficulties for
physicians regarding viable therapeutic options (12, 16, 25, 39).
The control of bacterial resistance is directly related to the detection of this
mechanism through laboratory and molecular tests as well as the implementation of
rational use of antibiotics in hospitals (26, 39). We documented the presence of
blaTEM, blaSHV, blaCTX-M and blaOXA genes in E.coli, Serratia spp., Pseudomonas spp.,
and B.cepacia strains (Table 3).
Similar results were detected in other studies such as the blaCTX-M gene
detected in E.coli isolates in Cambodia, Canada, United Kingdom (28) and in clinical
outbreaks of CTX-M-15-type in India, Spain, and Portugal (9), considered as the
most prevalent ESBL-encoding gene worldwide, now replacing TEM and SHV as the
predominant ESBL isolate in European countries (14).
The prevalence of ESBL production in Enterobacteriaceae in France (18)
belonged to CTX-M variants such as CTX-M-I, and CTX-M-9. In Spain and Portugal,
they frequently detected TEM-24, CTX-M-15, CTX-M-32 and SHV-12. In Bulgaria,
CTX-M-3, CTX-M-15 and SHV-12 in S.marcescens isolates (18). In Amsterdam,
CTX-M-1, SHV-1, TEM-116 in P. aeruginosa and CTX-M-1, SHV-1 in S. maltophilia
(22). In Brazil, CTX-M-14, TEM-1, CTX-M-15, OXA-1, OXA-4 and OXA-7 (7, 22, 23).
The detection of the blaTEM, blaSHV, blaCTX-M and blaOXA genes in gram-
negative isolates in this study allows us to consider the possibility of plasmidial
resistance. Both the genes that encode the ESBLs and those that encode resistance
to non-beta-lactam antibiotics are frequently located in the same conjugative plasmid,
transmitted conjunctively from one isolate to another, showing the ability of being
separated by selective pressure due to the use of multiple antibiotics under effective
control (16, 26).
The early identification of infectious processes linked to the use of appropriate
antimicrobials based on microbiological diagnosis, rational prescription of antibiotics,
and effective control can help avoid serious hospital infections. Most significantly, it is
important to consider the inclusion of new antibiotics as therapeutic choices. These

65
measures will help the implementation of effective public policies for epidemiological
surveillance and control by Brazilian health authorities in the State of Amazonas.
This research showed the presence of gram-negative ESBL-producing
bacteria infecting hematologic patients in HEMOAM. These strains carried the blaTEM,
blaSHV, blaCTX-M and blaOXA genes and were resistant to different antibiotics used in
the treatment. It is necessary to perform screening and confirmatory test for
phenotypic detection of these strains in a routine laboratory to avoid health care
failure.
Acknowledgements
The authors are grateful to acknowledge the support provided by the
Amazonas State Research Foundation (Fundação de Amparo a Pesquisa do Estado
do Amazonas - FAPEAM). Inputs to carry out this research derived from a project
approved by the Research Program of SUS (the Brazilian National Health Care
System): Shared Health Management - PPSUS- Posting 014/2006-FAPEAM
Manaus, State of Amazonas, Brazil. This paper stems from the preliminary findings
reported by the author’s doctoral thesis at the Amazon State University (UEA) and
the Amazon Tropical Medicine Foundation (FMT-AM) in the field of clinical
bacteriology.
In addition, the Amazon Hematology and Hemotherapy Foundation
(HEMOAM), the Tropical Dermatology and Veneorology Foundation Alfredo da Matta
(FUAM), and the Graduate Program of Biotechnology (PPGBiotec).
Disclosure Statement No competing financial interests exist.
References 1. Ahmed, S.H., Daef, E.A. , Badary, M.S., Mahmoud, M.A., Elsayed, A.A.A. 2009.
Nosocomial blood stream infection in intensive care units at Assiut University
Hospitals (Upper Egypt) with special reference to extended spectrum b-lactamase
producing organisms. BMC Research Notes. 2:76.

66
2. Aladag, M.O., Durak, Y. 2009. Investigating Some Antibiotics, Plasmids Profile
and ESBL Characteristic of Klebsiella pneumoniae Isolated from Urinary System
Infection. World Applied Sciences Journal. 6:630-636 (In Turkish).
3. Al-Zarouni, M., Senok, A., Rashid, F., Al-Jesmi, S.M., Panigrahi, D.M. 2008.
Prevalence and antimicrobial susceptibility pattern of extended-spectrum beta-
lactamase-producing Enterobacteriaceae in the United Arab Emirates. Medical
Principles and Practices. 17:32-36.
4. Atifah, M.N., Loo, H.K.C., Subramaniam, G., Wong, E.H., Selvi, P., Ho, S.,
Kamarulzaman, A., Parasakthi, N.2005. Faecal prevalence of extended-spectrum
ß-lactamase (ESBL)-producing coliforms in a geriatric population and among
haematology patients. Malaysian J Pathol. 27:75-81.
5. Augusti, G.R., Superti, S., Zavascki, A.P. 2007. Prevalence of beta-lactamase
production of extended-spectrum bacteremia in Klebsiella pneumoniae and
Escherichia coli. Scientia Medica. 17:192-196 (In Portuguese).
6. Bradford, P.A. 2001. Extended-Spectrum-Lactamases in the 21st Century:
Characterization, Epidemiology, and Detection of This Important Resistance
Threat. Clin Microbiol Rev. 14:933-951.
7. Chmelnitsky, I., Carmeli,Y., Leavitt,A.,Schwaber,M.,Venezia, S.N. 2005. CTX-M-2
and new CTX-M-39 enzyme are the major extended-spectrum beta-lacamase in
multiple Escherichia coli clones isolated in Tel Aviv, Israel. Antimicrob Agents
Chemother.49 (11): 4745-4750.
8. Clinical and Laboratory Standards Institute. Performance Standards for
Antimicrobial Susceptibility Testing; Twentieth Informational Supplement.
Document M100-S20. CLSI, Wayne PA, 2010.
9. Cormican, M.G., Marshall, S.A., Jones, R.N. 1996. Detection of extended-
spectrum β-lactamase (ESBL)-producing strains by the Etest ESBL screen. J Clin
Microbiol. 34:1880-1884.
10. Coque, T.M., Novais, A., Carattoli, A., Poirel, L., Pitout, J., Peixe, L., Baquero, F.,
Cantón, R., Nordmann, D.P. 2008. Dissemination of clonally related Escherichia
coli strains expressing extended-spectrum β-lactamase CTX-M-15. Emerg Infect
Dis. 14:195-200.
11. Dalmarco, E.M., Blatt, S.L., Córdova, C.M.M. 2006. Laboratory identification of
beta-lactamase extended-spectrum (ESBLs) - Review. RBAC. 38:171-177(In
Portuguese).

67
12. Habeeb, K.C., Surekha, S., Lakshmi, S., Narasimha, G. 2007. Multidrug
resistance and Beta-lactamase production by Klebsiella pneumoniae. African
Journal of Biotechnology. 6:1791-1793.
13. Hujer, A.M., Kslar, K.S., Dietenberger, N.J., Bethel, C.R., Endimiani, A., Bonomo,
R.A.. 2009. Detection of SHV β-lactamases in Gram-negative bacilli using
fluorescein-labeled antibodies. BMC Microbiology. 9:1-4.
14. Lim, K.T., Yasin, R., Yeo, C.C., Puthucheary,S., Thong, K.L. 2009.
Characterization of Multidrug Resistant ESBL-Producing Escherichia coli Isolates
from Hospitals in Malaysia. Journal of Biomedicine and Biotechnology.1-10.
15. Macedo, M.L.A.P., Cartaxo, R.S., Almeida, T.C.C., Souza, L.B.S., Santana, W.J.,
Coutinho, H.D.M. 2005. Mechanisms of resistance and detection of beta-
lactamases. Ciênc Biol Saúde. 7:59-63 (In Portuguese).
16. Mansouri, M., Ramazanzadeh, R. 2009. Spread of extended-spectrum beta-
lactamase producing Escherichia coli clinical isolates in Sanandaj hospitals. J Biol
Sci. 9:362-366.
17. Miyoshi, T., Iwatsuki, T., Naganuma, T. 2005. Phylogenetic characterization of
16S rRNA gene clones from deep-groundwater microorganisms that pass through
0.2-micrometer-pore-size filters. Appl Environ Microbiol. 71:1084-1088.
18. Mohammad, M. F., Somayeh, D., Nafiseh, R., Araz, M., Marzieh, Aligholi.,
Fereshteh, S., Mahmood, P., Davud, Y. 2009. Distribution of blaTEM, blaSHV,
blaCTX-M Genes Among Clinical Isolates of Klebsiella pneumoniae at
Labbafinejad Hospital, Tehran, Iran. MDR 00 (00): 1-6.
19. Monstein, H.J., Tärnberg, M., Nilsson, L.E. 2009. Molecular identification of CTX-
M and blaOXY/K1 β-lactamase genes in Enterobacteriaceae by sequencing of
universal M13-sequence tagged PCR-amplicons. BMC Infect Dis 9:7. http://
www.biomedcentral.com/1471-2334/9/7.
20. Mshana, S.E., Kamugisha, E., Mirambo, M., Chakraborty, T., Lyamuya, E.F.
2009. Prevalence of multiresistant gram-negative organisms in a tertiary hospital
in Mwanza, Tanzânia. BMC Res Notes. 2:49. Available from: http://www.
biomedcentral.com/1756-0500/2/49.
21. Mulvey, R.M., Bryce, E., Boyd, D., Agostini, M.O., Christianson, S., Simor, A.E.,
Paton, S. 2004. Ambler Class A extended-spectrum beta-lactamase-producing
Escherichia coli and Klebsiella spp. in Canadian hospitals. Antimicrob Agents
Chemother. 48:1204-1214.

68
22. Naiemi, N., Duim, B., Bart, A. 2006.A CTX-M extended-spectrum beta-lactamase
in Pseudomonas aeruginosa and Stenotrophomonas maltophilia. JMM 55: 1607-
1608.
23. Novella, M.C.C., Guth, B.E.C., Castanheira, M., Carmo, M.S., Pignatari, A.C.C.
2010. First description of blaCTX-M-14 and blaCTX-M-15–producing Escherichia
coli isolates in Brazil. MDR 16(3): 177-184 (In Portuguese).
24. Pasta, A.A.C., Fração, F.H.A., Magalhães, G.L.G., Quesada, R.M.B. 2008.
Prevalence and antimicrobial susceptibility test in strains of extended-spectrum
beta-lactamase-producing (ESBL) Klebsiella pneumoniae isolated from patients in
University Hospital /UEL. RBAC. 40:137-141 (In Portuguese).
25. Perez, F., Endimiani, A., Hujer, K.M., Bonomo, R.A. 2007. The continuing
challenge of ESBLs. Curr Opin Pharmacol. 7:459-469.
26. Picão, R.C., Gales, A.C. 2007. Extended-spectrum beta-lactamase-producing
(ESBL) Pseudomonas aeruginosa: Nightmare or imagination? Prática Hospitalar.
49:79-84 (In Portuguese).
27. Romanus, I.I., Egwu, O.A., Ngozi, A.T., Chidiebube, N.A., Chika, E.P. 2009.
Extended-spectrum β-lactamase (ESBL)-mediated resistance to antibiotics
among Klebsiella Pneumoniae in Enugu Metropolis. Maced J Med Sci. 2:1-4 (In
Nigerian).
28. Ruppé, E., Hem, S., Lath, S., Gautier, V., Ariey, F., Sarthou, J.L., Monchy, D.,
Arlet, G. 2009. CTX-M β-lactamases in Escherichia coli from community-acquired
urinary tract infections, Cambodia. Emerg Infect Dis. 15:741-748.
29. Schmitt, J., Jacobs, E., Schmidt, H. 2007. Molecular characterization of extended-
spectrum beta-lactamases in Enterobacteriaceae from patients of two hospitals in
Saxony, Germany. J Clin Microbiol. 56:241-249.
30. Seok, H.J., Kwon, B., Jung, H.L., Seung, G.S., Geun, H.K., Ghil, J.J., Young,
H.K., Byeong, C.J., Sang, H.L. 2004. Molecular characterization of extended-
spectrum beta-lactamases produced by clinical isolates of Klebsiella pneumoniae
and Escherichia coli from a Korean Nationwide Survey. J Clin Microbiol. 42:2902-
2906.
31. Slama, T.G. 2008. Gram-negative antibiotic resistance: there is a price to pay.
Review. Critical Care. 12:4. http://ccforum.com/content/12/S4/S4.
32. Song, K.H., Jeonn, J.H., Park, W.B., Park, S.W., Kim, H.B., Oh, M.D., Lee, H.S.,
Kim, N.J., Choe, K.W. 2009. Clinical outcomes of spontaneous bacterial

69
peritonitis due to extended-spectrum beta-lactamase-producing Escherichia coli
and Klebsiella species: A retrospective matched case-control study. BMC Infect
Dis 9:41. http://www.biomecentral.com/1471-2334/9/41.
33. Superti, S.V., Augusti, G., Zavascki, A.P. 2009. Risk factors for and mortality of
extended-spectrum-b-lactamase-producing Klebsiella pneumoniae and
Escherichia coli nosocomial bloodstream infections. Rev Inst Med Trop. S. Paulo.
51:211-216 (In Portuguese).
34. Thierry, F., Guillaume, A., Valerie, G., Talarmin A., Raymond, B. 2006.
Extended-spectrum-lactamase-producing Enterobacteriaceae, Central African
Republic. Emerg Infect Dis. 12:863-865.
35. Thomson, KS. 2001. Controversies about extended-spectrum and ampc beta-
lactamases. Emerg Infect Dis. 7:333.
36. Tofteland, S., Haldorsen, B., Dahl, K.H., Simonsen, G.S., Steinbakk, M.,
Walsh, T.R., Sundsfjord, A. 2007. Effects of phenotype and genotype on
methods for detection of extended-spectrum-lactamase-producing clinical
isolates of Escherichia coli and Klebsiella pneumoniae in Norway. J Clin
Microbiol. 45:199-205.
37. Tumbarello, M., Sali, M., Trecarichi, E.M., Leone, F., Rossi, M., Fiori, B., De
Pascale, G., D’Inzeo, T., Sanguinetti, M., Fadda, G., Cauda, R., Spanu, T. 2008.
Bloodstream infections caused by extended-spectrum-lactamase-producing
Escherichia coli: Risk factors for inadequate initial antimicrobial therapy.
Antimicrob Agents Chemother. 52:3244-3252.
38. Venkatachalam, K.V., Wanzhi, H., Mark, L.R., Timothy, P. 1994.
Characterization of TEM-1 B-lactamase mutants from position 238 to 241 with
increased catalytic efficiency for ceftazidime. J Biol Chem. 269:23444-23450.
39. Winn, J.W.C., Allen, S.D., Janda, W.M., Koneman, E.W., Procop, G.W.,
Schreckenberger, P.C., Woods, G.L. 2008. Microbiological diagnosis. Color Atlas
and Text. 6th ed. Guanabara Koogan, Rio de Janeiro.
40. Smith, B.A., Gabasan, A., Zivin, T., Sordillo, E.M., Polsky, B. 2009. Management
of an outbreak due to ESBL-positive Klebsiella pneumoniae (ESBL+KP) in an
ICU: Focusing on the basics. AJIC 33(5): e121-e122.

70
4.2 Artigo 2 Novel Methicillin resistant coagulase-negative Staphylococci clone isolated from patients with hematological diseases at the Blood Bank Center of Amazon, Brazil Cristina Motta Ferreira1*, Felipe Gomes Naveca2*, William Antunes Ferreira3*, Cíntia
Mara Costa de Oliveira4, Maria das Graças Vale Barbosa5.
Authors affiliations: 1 Universidade do Estado do Amazonas ; Fundação de Medicina Tropical Dr. Heitor
Vieira Dourado; Fundação de Hematologia e Hemoterapia do Amazonas-HEMOAM,
Manaus-Amazon-Brazil. Manaus-Amazon-Brazil. PhD student in Infectious and
Tropical Diseases. 2Instituto Leônidas e Maria Deane/FIOCRUZ. PhD in Microbiology. Manaus-Amazon-
Brazil. 3 Fundação de Dermatologia Tropical e Venereologia Alfredo da Matta, Manaus-
Amazon-Brazil; Universidade do Estado do Amazonas, Fundação de Medicina
Tropical Dr. Heitor Vieira Dourado. PhD student in Infectious and Tropical Diseases.
Manaus-Amazon-Brazil. 4,5 Fundação de Medicina Tropical Dr. Heitor Vieira Dourado. Manaus-Amazon-
Brazil. 4PhD in Biotecnhology; 5PhD in Biological Sciences.
*These authors contributed equally to this work Financial support: This Research Project (Nr. 3098766263) was funded by
Fundação de Amparo a Pesquisa do Estado do Amazonas-FAPEAM. Manaus,
Amazonas, Brazil, through a Grant Proposal approved for the SUS Research
Program (called “Gestão Compartilhada em Saúde-PPSUS-014/2006”).

71
Corresponding author: Dr. Cristina Motta Ferreira. Fundação de Hematologia e Hemoterapia do Amazonas-
HEMOAM, 4397 Constantino Nery Avenue, Chapada. Laboratory of Bacteriology,
CEP: 69050-002, Manaus, Amazonas, Brazil. Telephone: (55)-92-3655-01-00;
E-mail: [email protected]
Running title: Methicillin resistant Staphylococcus in Manaus Keywords: SCCmec, ST clones, coagulase-negative Staphylococcus, S. aureus,
methicillin resistant, nosocomial infection, blood infection, hospital, hematologic
disease.
Abstract Methicillin-resistant Staphylococcus remains as a severe public health problem
worldwide. We conducted MLST analysis that revealed three different sequence
types isolated from patients with hematological diseases: S. aureus ST243, S.
epidermidis ST2, and a new clone of S. epidermidis ST365. Those findings reinforce
the potential of dissemination presented by multiresistant Staphylococcus.
Methicillin-resistant Staphylococci stand as main pathogens responsible for
high levels of infectious process worldwide. The methicillin resistance has to do with
penicillin binding protein (PBP 2a) encoded by the mecA gene, located at the
staphylococcal cassette chromosome mec (SCCmec). In S. aureus types I and VI
are noticed, whereas types IV and V are found in S. epidermidis and S. haemolyticus
(1).
The genotype distribution of Staphylococci differs in different geographical
regions. USA300 CAMRSA clone ST8 is dominant in the United States and the ST80
and ST30 in Europe, Kuwait, Singapore, the Southwest Pacific islands and, New
Zealand (2). In relation to S. epidermidis, the ST69 clone, is predominant in Greece;
ST57, ST88 in Portugal, ST2 in China, ST2, ST22, ST61, ST71 in Mexico and ST63
in Argentina (4).
The risk for immunocompromised patients can be significant if they catch
infections caused by those species in hospitals. The presence of

72
immunocompromised patients at the hospital and blood bank where this research
took place is common due to chemotherapy applied against leukemia, myeloma,
hemolytic anemia, hemophilia, etc. This can increase morbimortality risks due to
prolonged hospitalization, spread of multi-resistant strains, and rise in treatment
costs.
Therefore, the resistance of these bacteria against antibiotics, their respective
SCCmec and ST was identified. The knowledge of geographic distribution,
transmission mechanisms, factors that contribute to the appearance and spread of
these genotypes will help control hospital infection outbreaks and dissemination, and
implement programs to supervise the resistance presented by these pathogens. The Study
In the period from July 2007 to August 2008, we conducted a descriptive
cross-sectional study including patients of both sexes, admitted to the HEMOAM
Foundation, who had a previous diagnosis of hematologic disease, presenting clinical
signs and symptoms suggestive of acute bacterial infection, whether feverish or not,
or having developed fever during hospital stay. The HEMOAM Foundation is a high
complexity hospital for hematological diseases that supplies blood and
hemoderivatives for the city.
Each patient provided more than one sample: urine, blood, sputum, feces,
catheter tip, bone marrow aspirate, and abscess fluid along with oropharyngeal,
ocular, perianal, nasopharynx, open wound, and skin lesion secretions were
collected. Biochemical tests and nucleotide sequencing of the 16S rRNA region
presented Staphylococcus epidermidis, S. intermedius, S. aureus and
Staphylococcus sp. as the most frequent species. Other isolated species were S.
hyicus, S. haemolyticus, S. simulans, S. intermedius, S. lugdunensis (Table 1).

73
Table 1. Strains isolated from patients with hematologic disease
Strains n°
Micrococcus sp 1
Staphylococcus aureus 4
Staphylococcus hyicus 1
Staphylococcus sp 5
Staphylococcus haemolyticus 1
Staphylococcus epidermidis 8
Staphylococcus simulans 1
Staphylococcus intermedius 4
Staphylococcus lugdunensis 2
TOTAL 27
The extraction of the genomic DNA of the 26 Gram-positive bacteria isolated
from the 146 biological samples collected from 69 patients, and the PCR protocol for
the identification of the mecA gene, SCCmec, toxins (seh, arcA, Setd) and the
Panton-Valentine leukocidin gene was performed as described by Jonas et al. (2002)
(5), Milheiriço et al. (2007) (6), and Strommenger et al. (2008) (7), confirming the
presence of the mecA gene and the absence of the lukPV gene.
The PCR for MLST was made as described by Thomas et al. (2007) (8) and
Enright et al. (2000) (9), disclosing three types of different sequences typing (ST): S.
aureus ST243, one of S. epidermidis ST2, and two newly isolates of S. epidermidis
ST365 clone. The ST243 strain harboured the SCCmec type I, III, V, ST2 the
SCCmec type II, III, V, and ST365 the SCCmec type I, III, V (Table 2). The
sequence of genes of this study is found at GenBank with the following access
numbers: JN398483-07.

74
Table 2. Phenotypes and genotypes profiles of the Staphylococcal strains isolated from patients with hematologic diseases
*Gold standard; ǂ S. epidermdis: arcC, aroE, gtr,
mutS, pyrR, tpiA, yqiL; S. aureus: arcC, aroE, glpF,
gmk,pta, tpi, yqiL ;
† not determined (there is no MLST protocols for those
species yet).
Strains
Phenotypes tests Genotypes tests
Oxac Cefox PBP2a mecA* SCCmec types
Allelic profileǂ MLST
S. epidermidis R S + + I,III,V 3 25 5 5 3 4 4 365
S. epidermidis R S + + I,III,V 3 25 5 5 3 4 4 365
S. epidermidis R R + + II,III,V 7 1 2 2 4 1 1 2
S. aureus R R + + I,III,V 2 2 5 2 6 3 2 243
S. intermedius R S + + I,III,V NT† NT NT NT NT NT NT NT
S. lugdunensis S S + + III NT NT NT NT NT NT NT NT
Staphylococcus sp R S + + I,III,V NT NT NT NT NT NT NT NT
Staphylococcus sp R S + + I,III,V NT NT NT NT NT NT NT NT
Staphylococcus sp. R R + + I NT NT NT NT NT NT NT NT
S. hyicus R R + + I,II,IV NT NT NT NT NT NT NT NT

75
The S. epidermidis ST 365 clone was isolated from the peripheral blood of two
male and female patients (9 and 2 years old) presenting acute lymphocytic leukemia.
The susceptibility tests showed that both presented resistance against methicillin,
tetracycline, penicillin, oxacillin, and chloramphenicol; reduced sensitivity to
ceftazidime and sensitivity to vancomycin, cefoxitin, ciprofloxacin, amikacin,
imipenem and cefepime.
The S. epidermidis ST2 clone isolated from blood infection from a 24-year-
old patient with chronic myeloid leukemia presented resistance against methicillin,
cefoxitin, oxacillin, amikacin, ceftazidime, ciprofloxacin, imipenem, cefepime; reduced
sensitivity to chloramphenicol, and sensitivity to vancomycin. Its presence in our
hospital may be due to colonized patients (reservoirs) or colonized patient,
contaminated healthcare workers-to-patient transmission, resistance profile of the
isolate in relation to antimicrobial agents, high formation of biofilm, recognized as a
critical factor for the for the colonization of medical devices, heterogeneous
expression of the gene and the SCCmec found in disease-associated isolates, ica
gene cluster, insertion sequence element IS256, and the presence of SCCmec
present in disease-associated strains. This reflects highly ability to adjust through
genetic exchange to environmental changing and antibiotics therapy (10, 11).
ST2, founder of the CC2 clonal complex, is considered as the prevalent
international cause of most S. epidermidis hospital infections, also detected as an
epidemic hospital pathogen worldwide and in different European hospitals (10, 11).
The detection of resistant methicillin strains belonging to this virulent clonal
lineage in hospitalized - mainly immunocompromised - patients, points to a possible
hospital infection, as they had been in hospital for over 72 hours, did not come from
other hospitals, received no prolonged antibiotic therapy at the moment of collection,
and presented frequent use of catheter because of the treatment.
The S. aureus ST243 clone isolated from a 30-year-old hemophilic patient with
blood infection presented resistance to methicillin, penicillin, tetracycline, cefoxitin,
oxacillin, and ceftazidime; sensitivity to ciprofloxacin, chloramphenicol, vancomycin,

76
and amikacin. Despite the in vitro sensitivity against imipenem and cefepime, CLSI
M-100 S20 recommendations suggest that they should be reported as resistant due
to low treatment effectiveness.
The presence of ST243 in our hospital may have come from patient/patient or
colonized patient/ contaminated healthcare workers- transmission and prolonged
antibiotic therapy. Other risk factors that can be considered for hematogenous
infection are nasal carriers, and skin/soft tissue infections (11, 12). This clone is
spread over the USA, Thailand and Japan (data available at www.mlst.net), though
only the Thailand clone is methicillin-resistant.
The presence of the SCCmec III cassette in S. epidermidis, with no clinical
importance in infections, must be revised, because the horizontal transfer of a large
number of resistance genes between species opens way to methicillin resistance
(13,14), which induces to the use of different antibiotics as a therapeutic option
(mainly vancomycin).
Another fact observed was the presence of SCCmec type V in S. epidermidis.
Types IV and V are small, structurally similar elements that carry the mecA gene as a
unique antibiotic resistance encoder. As they have a high competitive transfer
capacity, this can explain the presence of SCCmec type V in relation to type IV (15).
Another important factor detected was the existence of various SCCmec types. This
may suggest that multiple introductions are occurring. And their presence in the
same ST may suggest a possible horizontal transfer between the species (9).
Conclusions A horizontal transfer of the mec gene may be occurring, which favors the
appearance of methicillin resistance between the species. Microbial resistance
monitoring programs are important to avoid hospital infection outbreaks and the
spread of pathogenic clonal lines.
Acknowledgements We are grateful to acknowledge the support provided by Fundação de Amparo a
Pesquisa do Estado do Amazonas - FAPEAM. In addition, the Superintendent of the
State of the Amazonas (SUFRAMA).

77
Conflict of Interest The authors have no conflict of interest related to the development of the present
study.
Biographical Sketch Cristina Motta Ferreira, Biochemistry, MSc in Tropical Pathology, PhD student in
Infectious and Tropical Diseases, professor of clinical bacteriology in the Nilton Lins
University. The author develops activity, with emphasis on methicillin resistant
staphylococci and beta-lactamase-producing bacteria (ESBL), at the Fundação de
Hematologia e Hemoterapia do Amazonas-HEMOAM, in clinical bacteriology
laboratory.
References: 1. Strandén A M, Frei R, Adler H, Flückiger U, Widmer AF. Emergence of SCCmec
type IV as the most common type of methicillin-resistant Staphylococcus aureus in a
university hospital. Infection . 2009; 37: 44-48.
2. Udo EE, O’Brien FG, Al-Sweih N, Noronha B, Matthew B, Grubb WB. Genetic
Lineages of Community-Associated Methicillin-Resistant Staphylococcus aureus in
Kuwait Hospitals. J.Clin Microbiol. 2008; 46: 3514–16.
3. Liakopoulos A, Spiliopoulou I, Damani A, Kanellopoulou M, Schoina S, Papafragas
E, et al. Dissemination of two international linezolid-resistant Staphylococcus
epidermidis clones in Greek hospitals. J Antimicrob Chemother. 2010; 65: 1070–77.
4. Miragaia M, Lencastre H, Perdreau-Remington F, Chambers HF, Higashi J,
Sullam PM, et al. Genetic Diversity of Arginine Catabolic Mobile Element in
Staphylococcus epidermidis. Plos one. 2009; 4: e7722.
Doi:10.1371/journal.pone.0007722
5. Jonas D, Speck M, Daschner FD, Grundmann H. Rapide PCR-based
identification of methicillin- resistant Staphylococcus aureus from screening swabs. J
Clin Microbiol. 2002; 40: 1821-23.
6. Milheiriço C, Oliveira DC, de Lencastre H. Update to the multiplex strategy for
assignment of mec element types in Staphylococcus aureus. Antimicrob Agents
Chemother. 2007; 51: 3374-77.
7. Strommenger B, Braulke C, Pasemann B, Schmidt C, Witte W. Multiplex PCR for

78
rapid detection of Staphylococcus aureus isolates suspected to represent
community-acquired strains. J Clin Microbiol . 2008; 46: 582-87.
8. Thomas LC, Gidding HF, Ginn AN, Olma T, Iredell J. Development of a real-time
Staphylococcus aureus and MRSA (SAM-) PCR for routine blood culture. J.
micmeth.2007; 68: 296-302.
9. Enright, MC, Nicholas PJD, Davies CE, Peacock SJ, Spratt B. Multilocus
Sequence Typing for Characterization of Methicillin- Resistant and Methicillin-
Susceptible Clones of Staphylococcus aureus. J Clin Microbiol. 2000; 38: 1008-15.
10.Weisser M, Schoenfelder SMK, Orasch C, Arber C, Gratwohl A, Frei R, et al.
Hypervariability of Biofilm Formation and Oxacillin Resistance in a Staphylococcus
epidermidis Strain Causing Persistent Severe Infection in an Immunocompromised
Patient. J Clin Microbiol. 2010; 48: 2407–12.
11. Widerström M, Wiström J, Sjöstedt A, Monsen T. Coagulase-negative
staphylococci: update on the molecular epidemiology and clinical presentation, with a
focus on Staphylococcus epidermidis and Staphylococcus saprophyticus. Eur J Clin
Microbiol Infect Dis. 2011. DOI 10.1007/s10096-011-1270-6.
12.Tatsuo Yamamoto T, Nishiyama A, Takano T, Yabe S, Higuchi W, Razvina O, et
al. Community-acquired methicillin-resistant Staphylococcus aureus: community
transmission, pathogenesis, and drug resistance. J Infect Chemother. 2010; 16: 225–
54.
13. Sousa FCJ, Nunes EWF, Nascimento ED, Oliveira SM, Melo MCN,Fernandes
MJBC. Prevalência de Staphylococcus spp resistentes à meticilina isolados em uma
maternidade escola da Cidade de Natal, Estado do Rio Grande do Norte. Rev Soc
Bras Med Trop. 2009; 42: 179-82.
14. Barbier F, Ruppe´E, Hernandez D, Lebeaux D, Francois P, Felix B, et al. Methicillin-Resistant Coagulase-Negative Staphylococci in the Community: High
Homology of SCCmec IVa between Staphylococcus epidermidis and Major Clones of
Methicillin-Resistant Staphylococcus aureus. JID. 2010; 202: 270-81.
15. Ito T, Ma XX, Takeuchi F, Okuma K, Yuzawa H, Hiramatsu K. Novel Type V
Staphylococcal Cassette Chromosome mec Driven by a Novel Cassette
Chromosome Recombinase, ccrC. Antimicrob Agents Chemother . 2004;48:2637–51

79
5 DISCUSSÃO
5.1 Freqüência dos gêneros e espécies bacterianas.
Neste estudo as bactérias mais freqüentes produtoras de ESBL foram a E.
coli, B. cepacia, Serratia sp. e Pseudomonas aeruginosa. As E. coli produtoras de
ESBL identificadas indicam uma possível infecção hospitalar, devido a uma
inadequada antibióticoterapia, sem exame de cultura e teste de suscetibilidade e
infecções cruzadas que contribuíram para a elevação do número desses isolados
resistentes (1, 106, 142). Bactérias gram-negativas como K. pneumoniae, E. coli assim
como outras espécies de enterobactérias tais como Salmonella sp., Serratia
marcescens, Proteus sp., Shigella dysenteriae, Pseudomonas aeruginosa e B.
cepacia, podem produzir uma enzima denominada de beta-lactamase de espectro
estendido (ESBL) (2, 102) .
As ESBLS são enzimas que agem inibindo a ação de antibióticos beta-
lactâmicos. A grande preocupação clínica em relação a esses isolados, reside no
uso de outras opções terapêuticas como os carbapenêmicos e ao elevado tempo de
internação dos pacientes. Na FHEMOAM , os pacientes hematológicos são
frequentemente submetidos a quimioterapia apresentando baixa em sua imunidade,
tornando-se assim suscetíveis a uma ampla gama de infecção, principalmente as
hospitalares, elevando o risco de mortalidade (114).
5.2 Identificação dos fenótipos de sensibilidade, sensibilidade reduzida e a
resistência dessas bactérias aos antibióticos indicados no tratamento.
Com relação a sensibilidade e resistência bacterianas, foram identificadas
bactérias Gram-negativas produtoras da enzima ESBL resistentes as drogas beta-
lactâmicas, e espécies de estafilococos (MRSA e MR-SCoN) resistentes aos
antibióticos beta-lactâmicos, com a presença do gene mecA, localizado no SCCmec.
A sensibilidade reduzida detectada nas E. coli pode ser devida a variação no
substrato preferencial de ação, na fraca acurácia dos testes de detecção desta
enzima, fraca inibição pelo ácido clavulânico ou diferentes mecanismos combinados
de resistência bacteriana. Este tipo de fenótipo também foi detectado em outros

80
estudos. As enterobactérias produtoras de ESBL identificadas, demonstraram
sensibilidade ao imipenem e amicacina, indicando que estes antimicrobianos ainda
podem ser utilizados na rotina de tratamento (102, 118).
A presença de Pseudomonas sp produtoras de ESBLs é preocupante, pois
apresentam elevada capacidade de adquirir resistência a vários tipos de antibióticos.
Neste estudo, 33,4% das mesmas apresentaram resistência a tetraciclina e
ciprofloxacina. A opção terapêutica nesses casos reside no uso de meropenem e
imipenem. Entretanto, o uso freqüente desses antimicrobianos pode estimular o
surgimento de isolados mutiresistentes, o que pode prejudicar os pacientes (84, 118).
A detecção de enzimas ESBL é muito importante pois, conforme os critérios
do CLSI M100-S17 e M100-S18, devem ser reportadas como resistentes a todos os
antibióticos beta-lactâmicos, independentemente do resultado do teste de
suscetibilidade. O surgimento de bactérias produtoras de ESBL, responsáveis por
processos infecciosos com elevada morbi-mortalidade, tem dificultado a opção
terapêutica (29).
Os genes detectados nos isolados produtores de ESBL como a E. coli,
Serratia sp. Pseudomonas sp., e B. cepacia foram blaTEM, blaSHV, blaCTX-M e blaOXA ,
indicando uma possível resistência plasmidial. Os genes que codificam as ESBLs e
aqueles que codificam a resistência aos antibióticos não beta-lactâmicos, estão
localizados no mesmo plasmído conjugativo, sendo transmitidos conjuntamente
entre as bactérias, podendo ser separados por pressão seletiva devido ao uso de
múltiplos antibióticos (84, 102).
Nas bactérias Gram-positivas, a resistência a meticilina nos estafilococos
ainda continua sendo um desafio para a saúde pública em diversos hospitais do
mundo. Ela é expressa devido a produção da proteína ligadora de penicilina,
codificada pelo gene mecA, localizado no cassete cromossômico estafilocócico mec,
sendo o mais freqüente o tipo I e VI nos S. aureus e, nos estafilococos coagulase
negativos o tipo IV e V. Esses patógenos representam um risco potencial para os
pacientes, principalmente os imunodebiltados. Na FHEMOAM, a presença de
pacientes imunodebilitados é comum devido a quimioterapia a que são submetidos

81
para tratamento da doença de base como leucemia, mieloma,anemia hemolítica
dentre outras, contribuindo assim para a elevação do risco de morbi-mortalidade,
prolongamento no tempo de internação e disseminação de isolados resistentes entre
os pacientes (124).
5.3 Identificar as alterações genético-moleculares de resistência envolvidas.
O clone do estafilococo coagulase negativo ST2, isolado do sangue periférico
dos pacientes, apresentou resistência a meticilina. Sua presença pode estar
relacionada com a formação de biofilme, reconhecido como fator crítico para
colonização de equipamentos hospitalares, perfil de resistência do isolado aos
antibióticos utilizados na rotina hospitalar, expressão heterogênea do gene mec,
clone do gene ica, sequência do elemento de inserção IS256 e ao SCCmec presente
em isolados associados a doença. O clone ST2 é reconhecido internacionalmente
como o agente responsável por inúmeros casos de infecções em vários hospitais da
Europa. A detecção de isolados resistentes a meticilina pertencentes a esse clone
virulento, sugere uma possível infecção hospitalar, pois os pacientes também
utilizavam cateteres, estavam internados há mais de 72 horas, não eram
procedentes de outros hospitais e nem submetidos a antibióticoterapia prolongada (139).
A presença do clone de S. aureus ST243, isolado de infecção hematogênica
dos pacientes, pode ser devida a transmissão por contato direto entre pacientes,
antibióticoterapia prolongada ou pacientes colonizados/ corpo clínico contaminado.
Outros fatores importantes que podem contribuir para o desenvolvimento de
infecção sanguínea são o portador nasal e as infecções de tecido mole. O clone
ST243 apresentou resistência a meticilina sendo detectado apenas nos Estados
Unidos, Japão e Tailânda. Somente o clone da Tailândia apresentou resistência a
meticilina, os demais foram sensíveis (139).
Em relação ao cassete cromossômico, o tipo III detectado no S. epidermidis,
considerado anteriormente como agente sem importância clínica, deve ser revisto,
pois a transferência de vários genes de resistência, pode induzir o desenvolvimento
da resistência a meticilina, tendo como conseqüência o uso de diferentes antibióticos

82
como opção terapêutica (principalmente a vancomicina). A presença do SCCmec V
em relação ao tipo IV no S. epidermidis, pode ser explicada pela elevada
competitividade entre ambos no momento da transferência entre os isolados. Outro
fator importante observado foi a presença de varios tipos de SCCmec. Isto sugere
que múltiplas introduções de cassetes podem estar ocorrendo, e a presença de um
mesmo tipo de SCCmec em um único clone, pode indicar também transferência
horizontal do gene mec entre as espécies (11, 64) .

83
6 CONCLUSÃO
Freqüência dos gêneros e espécies bacterianas
1. Identificaram-se pacientes hematológicos com infecção ocasionada por bactérias
Gram-positivas e negativas resistentes a diferentes grupos de antibióticos. As
bactérias Gram-positivas mais frequentes foram os Staphylococcus epidermidis,
Staphylococcus intermedius, Staphylococcus aureus e Staphylococcus sp.,
enquanto que as Gram-negativas, foram a Escherichia coli, Serratia sp., Serratia
liquefaciens e Burkholderia cepacia.
Identificação dos fenótipos de sensibilidade, sensibilidade reduzida e a resistência dessas bactérias aos antibióticos indicados no tratamento.
2. Com relação a sensibilidade e resistência bacterianas, foram identificadas
bactérias Gram-negativas produtoras da enzima ESBL resistentes as drogas
beta-lactâmicas, e espécies de estafilococos (MRSA e MR-SCoN) resistentes
aos antibióticos beta-lactâmicos, com a presença do gene mecA, localizado no
SCCmec.
Identificar as alterações genético-moleculares de resistência envolvidas.
Os genes detectados nos plasmídeos isolados das bactérias Gram-negativas foram
blaTEM, blaSHV, blaCTX-M e blaOXA comprovando a presença de isolados implicados
em processos infecciosos hospitalares.
Os clones das espécies de estafilococos detectados foram:
- S. aureus ST 243 (SCCmec I, III, V) (4133 MLST database).
- S. epidermidis ST2 (SCCmec II, III, IV,V) (529 MLST database).
- Dois isolados pertencentes ao novo clone de S. epidermidis - ST 365
(SCCmec I, III, V) (527,528 MLST database).
A identificação do S. aureus ST 243 (SCCmec I, III, V, S. epidermidis ST2 (SCCmec
II, III, IV,V), e do novo clone S. epidermidis ST 365 (SCCmec I, III, V), nos

84
estafilococos meticilina resistentes, comprovam a presença de linhagens
patogênicas implicadas em processos infecciosos hospitalares.
A presença dos diferentes tipos de cassetes nos isolados meticilina resistentes
sugere múltipla introdução e, sua presença no mesmo ST, transferência horizontal
do gene mec.

85
7 REFERÊNCIAS 1-Ahmed K. Fimbriae of Branhamella catarrhalis as possible mediators of adherence
to pharyngeal epithelial cells. APMIS 100, 1066-1072. 1992.
2-Aladag MO, Y D. Investigating Some Antibiotics, Plasmids Profile and ESBL
Characteristic of Klebsiella pneumoniae Isolated from Urinary System Infection.
World Applied Sciences Journal 6, 630-363. 2009.
3- Alcamo IE. Fundamentals of Microbiology. 5th ed ed. California: 1997.
4-Al-Zarouni M, Senok A, Rashid F, Al-Jesmi SM, Panigrahi DM. Prevalence and
Antimicrobial susceptibility pattern of extended-spectrum beta-lactamase-producing
Enterobacteriaceae in the United Arab Emirates. Medical Principles and Practices
17, 32-36. 2008.
5-Amabis JM, Martho GR. Biologia 2. 2 ed. Rio de Janeiro: 2004.
6-Anderson B, Albrritton W.I, Biddle J, Johnson SR. Common ß-lactamase specifying
plasmid in Haemophilus ducreyi and Neisseria gonorrhoeae. Antimicrob Agents
Chemother 25, 296-297. 1984.
7-Antignac A, Tomasz A. Reconstruction of the phenotypes of methicillin resistant
Staphylococcus aureus by replacement of the staphylococcal cassette THL –
Research 35/2010 104 Methicillin Resistance in Staphylococci: Horizontal Transfer of
Mobile Genetic Element (SCCmec) between Staphylococcal Species chromosome
mec with a plasmid-borne copy of Staphylococcus sciuri pbpD gene. Antimicrob
Agents Chemother. 53[2], 435-441. 2009.
8-ANVISA. Detecção e Identificação de Bactérias de Importância Médica. Brasília:
2005
9-ANVISA. Procedimentos Laboratoriais: da Requisição do Exame à Análise
Microbiológica. Brasília: 2010.
10- Augusti GR, Superti S, Zavascki AP. Prevalência de produção de beta-

86
lactamases de espectro estendido em bacteremias por Klebsiella pneumoniae e
Escherichia coli. Scientia Medica 17[4], 192-196. 2007.
11-Barbier F, Ruppé E, Hernandez D, Lebeaux D, Francois P, Felix B, Desprez A,
Maiga A, Woerther PL, Gaillard K, Jeanrot C, Wolff M, Schrenzel J, Andremont A,
Ruimy R. Methicillin-Resistant Coagulase-Negative Staphylococci in the Community:
High Homology of SCCmec IVa between Staphylococcus epidermidis and Major
Clones of Methicillin-Resistant Staphylococcus aureus. The Journal of Infectious
Diseases 202[2], 270-281. 2010.
12-BD Diagnostic System. BBL Paper Discs for the Detection of â-Lactamase
Enzymes- Cefinase Disc. USA: 2010.
13- Blanco MG, Mejía C, Isturiz R, Alvarez C, Gotuzzo LBE, Labarca J, Luna CM,
Rodríguez-Noriega E, Salles MJC, Zurita J, Seas C. Epidemiology of methicillin-
resistant Syaphylococcus aureus (MRSA) in Latin America. Int J Antimicrob Agents
34, 304-308. 2009.
14-Bradford PA. Extended- Spectrum- Lactamases in the 21st Century:
Characterization , Epidemiology, and Detection of This Important Resistance Threat.
Clin Microbiol Rev 14[4], 933-951. 2001.
15-Berger-Bachi B. Resistance mechanisms of gram-positive bacteria. Int J Med
Microbiol. 292[1], 27-35.2002.
16-Berglund C, Ito T, Ikeda M, Ma XX, Soderquist B, Hiramatsu K. Novel Type of
Staphylococcal Cassette Chromosome mec in a Methicillin-Resistant Staphylococcus
aureus Strain Isolated in Sweden. Antimicrob Agents Chemother 52[10], 3512-3516.
2008.
17-Berglund C, Ito T, Ma XX, Ikeda M, Watanabe S, Soderquist B, et al. Genetic
diversity of methicillin-resistant Staphylococcus aureus carrying type IV SCCmec in
Orebro Country and the western region of Sweden. Journal of Antimicrobial
Chemotherapy 63, 32-41. 2008.

87
18-Beveridge TJ. Ultraestructure, chemistry, and function of the bacterial wall. Int
Rev Cytol 12, 229-317. 1981.
19-Bohach GA, Fast DJ, Nelson RD, Schlievert PM. Staphylococcal and
Streptococcal pyrogenic toxins involved in toxic shock syndrome and related
illnesses. Crit Rev Microbiol 17, 251-272. 1990.
20-Boyle-Vavra S, Daum RS. Community-acquired methicillin-resistant
Staphylococcus aureus: the role of Panton–Valentine leukocidin. Laboratory
Investigation 87, 03-09. 2007.
21-Brodeur BR, Martin D. Antigenic analysis of the saccharide moiety of the
lipooligosaccharide of Bordetella pertussis. Springer Semin Immunopathol 15, 205-
215. 1993.
22-Bouchami O, Achour W, Hassen AB. Typing of Staphylococcal Cassette
Chromosome mec Encoding Methicillin Resistance in Staphylococcus aureus Strains
Isolated at the Bone Marrow Transplant Centre of Tunisia. Curr Microbiol 59, 380-
385. 2009.
23-Bouguessa NR, Bes M, Meugnier H, Forey F, Reverdy ME, Lina G. Detection of
Methicillin-Resistant Staphylococcus aureus Strains Resistant to Multiple Antibiotics
and Carrying the Panton-Valentine Leukocidin Genes in an Algiers Hospital.
Antimicrob Agents Chemother 50[3], 1083-1085l. 2006.
24-Bush K. ß-Lactamase Inhibitors from Laboratory to Clinic. Clin Microbiol Rev 1[1],
109-123. 1988.
25-Carvalho MCS, Bonelli RR, Souza RR, Moreira S, Santos LCG, Conceição MS, et
al. Emergence of multiresistant variants of the community-acquired methicillin-
resistant Staphylococcus aureus lineage ST1-SCCmecIV in 2 hospitals in Rio de
Janeiro, Brazil. Diagn Microbiol Infect Dis. 65[3], 300-305. 2009.

88
26-Carvalho KS, Mamizuka EM, Gontijo FP. Methicillin/Oxacillin-resistant
Staphylococcus aureus as a hospital and public health threat in Brazil. Braz J Infect
Dis 14[1], 71-76. 2010.
27-Chambers HF. Methicillin resistance in staphylococci: molecular and biochemical
basis and clinical implications. Clin Microbiol Rev 10[4], 781-791. 1997.
28-Chongtrakool P, Ito T, Ma XX, Kondo Y, Trakulsomboon S, Tiensasitorn C, et al.
Staphylococcal Cassette Chromosome mec (SCCmec) Typing of Methicillin-
Resistant Staphylococcus aureus Strains Isolated in 11 Asian Countries: a Proposal
for a New Nomenclature for SCCmec Elements. Antimicrob Agents Chemother 50[3],
1001-1012. 2006.
29-Clinical and Laboratory Standards Institute (CLSI). Performance Standards for
Antimicrobial Susceptibility Testing; Twentieth Informational Supplement. Document
M100-S20. Wayne PA: 2010.
30- Columbus C. Antimicrobial resistance in bacterial pathogens. BUMC Proceedings
11, 183-185. 1998.
31-Cordonnier C, Herbrecht R, Buzyn A, Leverger G, Leclercq R, Nitenberg G, et al.
Risk factors for Gram-negative bacterial infections in febrile neutropenia.
Haematologica 90[8], 102-109. 2005.
32-Couto I, Sanches IS, Sa-Leao R, de Lencastre H. Molecular characterization of
Staphylococcus sciuri strains isolated from humans. J Clin Microbiol. 38[3], 1136–
1143. 2000.
33-Cristie C. Antimicrobial resistance in bacterial pathogens. BUMC Proceedings 11,
183-185. 1998.
34-Cunha MLRS, Lopes CAM, Rugolo LMSS, Chalita LV. Significância clínica de
estafilococos coagulase-negativa isolados de recém-nascidos. Arch Pediatr Urug
75[2], 183-194. 2004.
35-Chu YW, Tung VWN, Cheung TKM, Chu MY, Cheng N, Lai C, Tsang DNC,Lo
JYC. Carbapenemases in Enterobacteria, Hong Kong,China, 2009. Emerg Infect Dis
17[1],130-132. 2011.

89
36-Dalmarco EM, Blatt SL, Córdova CM. Identificação Laboratorial de ß-Lactamases
de Espectro Estendido (ESBLs) – Revisão. RBAC 38[3], 171-177. 2006.
37-D'Azevedo PA, Inoue FM, Andrade SS, Tranchesi R, Pignatari ACC. Pneumonia
necrotizante por Staphylococcus aureus resistente à meticilina.Rev. Soc. Bras.Med.
Trop 42[4],461-462. 2009.
38-Dickinson TM, Archer GL. Expression of Oxacillin Resistance in Staphylococcus
epidermidis: Roles of mecA Transcriptional Regulation and Resistant-Subpopulation
Selection. Antimicrob Agents Chemother 44[6], 1616-1623. 2000.
39-Dinges MM, Orwin PM, Schlievert PM, Schmidt C. Exotoxins of Staphylococus
aureus. Clin Microbiol Rev 13, 16-34. 2000.
40-Edwards KJ, Kaufmann ME, Saunders NA. Rapid and Accurate Identification of
Coagulase-Negative Staphylococci by Real-Time PCR. J Clin Microbiol 39[9], 3047-
3051. 2001.
41-Ellis M, Zwaan F, Hedström U, Poynton C, Kristensen J, Jumaa P, et al.
Recombinant human interleukin 11 and bacterial infection in patients with [correction
of] haematological malignant disease undergoing chemotherapy: a double-blind
placebo-controlled randomised trial. Lancet 361[9361], 972. 2003.
42-Feizabadi MM, Delfani S, Raji N, Majnooni A, Aligholi M, Shahcheraghi F, et al.
Distribution of blaTEM, blaSHV, blaCTX-M Genes Among Clinical Isolates of
Klebsiella pneumoniae at Labbafinejad Hospital, Tehran, Ira. MDR 00[00], 1-5. 2009.
43-File Jr. TM. Overview of Resistance in the 1990s. Chest 115[3], 3S-8S. 1999.
44-Ferreira WA, Vasconcelos WS, Ferreira CM, Silva MFP, Gomes JS, Alecrim
MGC. Prevalência de Staphylococcus aureus meticilina resistente (MRSA) em
pacientes atendidos em ambulatório de dermatologia geral em Manaus-Amazonas.
Rev Pat trop 38[20], 83-92. 2009.

90
45-Fey PD, Salim BS, Rupp ME, Hinrichs SH, Boxrud DJ, Davis CC, et al.
Comparative Molecular Analysis of Community- or Hospital-Acquired Methicillin-
Resistant Staphylococcus aureus. Antimicrob Agents Chemother 47[1], 196-203.
2003.
46-Gambello MJ, Kaye S, Iglewski BH. LasR of Pseudomonas aeruginosa is a
transcriptional activator of the alkaline protease gene (apr) and an enhancer of
exotoxin A expression. Infect Immun 61, 1180-1184. 1993.
47-Ganga R, Riederer K, Sharma M, Fakih MG, Johnson LB, Shemes S, et al. Role
of SCCmec Type in Outcome of Staphylococcus aureus Bacteremia in a Single
Medical Center. J Clin Microbiol 47[3], 590-595. 2009.
48-Gomez J, Alcantara M, Simarro E, Martinez B, Ruiz J, Guerra B. Pseudomonas
aeruginosa bacteremia: clinical and epidemiological patterns and treatment. Rev Esp
Quimioter 15[4], 360-365. 2002.
49-Gillet Y, Issartel B, Vanhems P, Fournet JC, Lina G, Bes M, et al. Association
between Staphylococcus aureus strains carrying gene for Panton-Valentine
leukocidin and highly lethal necrotising pneumonia in young immunocompetent
patients. Lancet 359[9308], 753–759.
50-Griffiss JM, Schneider H, Mandrell RE, Yamasaki R, Jarvis GA, Kim JJ, et al.
Lipooligosaccharides: the principal glycolipids of the neisserial outer membrane. Rev
Infect Dis 10, S87-S95. 1988.
51-Hackbarth CJ, Chambers HF. blaI and blaR1 regulate beta-lactamase and PBP2a
production in methicillin-resistant Staphylococcus aureus. Antimicrob Agents
Chemother. 37[5], 1144-1149.1993.
52-Hackbarth CJ, Miick C, Chambers HF. Altered production of penicillinbinding
protein 2a can affect phenotypic expression of methicillin resistance in
Staphylococcus aureus. Antimicrob Agents Chemother. 38[11], 2568-2571.1994.

91
53-Hajek V. Staphylococcus intermedius, a new species isolated from animals. Int J
Syst Bacteriol 26, 401-408. 1976.
54-Hanssen AM, Kjeldsen G, Sollid JUE. Local Variants of Staphylococcal Cassette
Chromosome mec in Sporadic Methicillin-Resistant Staphylococcus aureus and
Methicillin-Resistant Coagulase-Negative Staphylococci: Evidence of Horizontal
Gene Transfer?. Antimicrob Agents Chemother 48[1], 285-296. 2004.
55-Hauck CR, Meyer TF. Small Talk Opa proteins as mediators of Neisseria host cell
communication. Curr Opin Microbiol [6], 43-49. 2003.
56-Henze U, Sidow T, Wecke J, Labischinski H, Berger-Bachi B. Influence of femB
on methicillin resistanceand peptidoglycan metabolism in Staphylococcus aureus. J
Bacteriol 175[6], 1612-1620. 1993.
57-Higuchi W, Takano T, Teng LJ, Yamamoto T. Structure and specific detection of
staphylococcal cassette chromosome mec type VII. Biochem Biophys Res Commun.
377[3], 752-756. 2008.
58-Hiramatsu K. Molecular evolution of MRSA. Microbiol Immunol. 39[8],531-
543.1995.
59-Holt JG. Bergey's Manual of Determinative Bacteriology. 9th ed ed. Baltimore,
Maryland: 1994.
60-Huang H, Flynn NM, King JH, Monchaud C, Morita M, Cohen SH. Comparisons of
Community-Associated Methicillin-Resistant Staphylococcus aureus (MRSA) and
Hospital-Associated MSRA Infections in Sacramento, California. J Clin Microbiol
44[7], 2423-2427. 2006.
61-Ibrahem S, Salmenlinna S, Virolainen A, Kerttula AM, Lyytikainen O, Jagerroos H,
et al. Carriage of Methicillin-Resistant Staphylococci and Their SCCmec Types in a
Long-Term-Care Facility. J Clin Microbiol 41[1], 32-37. 2009.

92
62-Ibrahem S, Salmenlinna S, Lyytikainen O, Vaara M, Varkila JV. Molecular
characterization of methicillin-resistant Staphylococcus epidermidis strains from
bacteraemic patients. Clin Microbiol Infect 14, 1020-1027. 2008.
63-International Working Group on the Classification of Staphylococcal Cassette
Chromosome Elements (IWG-SCC). Classification of Staphylococcal Cassette
Chromosome mec (SCCmec): Guidelines for Reporting Novel SCCmec Elements.
Antimicrob Agents Chemother 53[12], 4961-4967. 2009.
64-Ito T, Okuma K, Ma XX, Yuzawa H, Hiramatsu K. Insights on antibiotic resistance
of Staphylococcus aureus from its whole genome: genomic island SCC. Drug Resist
Updat 6, 41-52. 2003.
65-Jeong SH, Bae IK, Lee JH, Sohn SG, Kang GH, Jeon GJ, et al. Molecular
Characterization of Extended-Spectrum Beta-Lactamases Produced by Clinical
Isolates of Klebsiella pneumoniae and Escherichia coli from a Korean Nationwide
Survey. J Clin Microbiol 42[7], 2902-2906. 2004.
66-Jonas D, Speck M, Daschner FD, Grundmann H. Rapid PCR-Based Identification
of Methicillin-Resistant Staphylococcus aureus from Screening Swabs. J Clin
Microbiol 40[5], 1821-1823. 2002.
67-Katayama Y, Ito T, Hiramatsu K. Genetic organization of the chromosome region
surrounding mecA in clinical staphylococcal strains: role of IS431-mediated mecI
deletion in expression of resistance in mecA-carrying, low-level methicillin-resistant
Staphylococcus haemolyticus. Antimicrob Agents Chemother 45[7], 1955-1963.
2001.
68-Kent TH. Staphylococcal enterotoxin gastroenteritis in rhesus monkeys. Am J
Pathol 48, 387-407. 1966.
69-Kenneth ST. Controversies about extended-spectrum and ampc beta-lactamases.
Emerg Infect Dis 7, 333. 2001.

93
70-Konig B, Prevost G, Konig W. Composition of Staphylococcal bi-component toxins
determines pathophysiological reactions. J Med Microbiol 46, 479-485. 1993.
71-Kravitz GR, Dries DJ, Peterson ML, Schlievert PM. Purpura fulminans due to
Staphylococcus aureus. Clin Infect Dis. 40[7], 941–947. 2005.
72-Kumar SM, Haccuria K, Michiels M, Ieven M, Poyart C, Hryniewicz W, et al.
Current Trends in Rapid Diagnostics for Methicillin-Resistant Staphylococcus aureus
and Glycopeptide-Resistant Enterococcus Species. J Clin Microbiol 46[5], 1577-
1587. 2008.
73-Labandeira-Rey M, Couzon F, Boisset S, Brown EL, Bes M, Benito Y, et al.
Staphylococcus aureus Panton-Valentine leukocidin causes necrotizing pneumonia.
Science. 315[5815], 1130-1133. 2007.
74-Levinson W,Jawetz E. Microbiologia. 5th ed. Rio Porto Alegre: 2005.
75-Lonescu R, Mediavilla JR, Chen L, Grigorescu DO,Idomir M, Kreiswirth N,Roberts
RB. Molecular Characterization and Antibiotic Susceptibility of Staphylococcus
aureus from a Multidisciplinary Hospital in Romania. Microbial Drug Resistance 16
[4], 263-272. 2010.
76-Liakopoulos A, Spiliopoulou I, Damani A, Kanellopoulou M, Schoina S,
Papafragas E, Marangos M, Fligou F, Zakynthinos E, Makris D, Protonotariou E,
Tsiapara F, Filos K, Diza E, Anastassiou ED, Petinaki E. Dissemination of two
international linezolid- resistant Staphylococcus epidermidis clones in Greek
hospitals. J Antimicrob Chemother 65, 1070-1077. 2010.
77-Li M, Xing Wang, Qian Gao,Yuan. Molecular characterization of Staphylococcus
epidermidis strains isolated from a teaching hospital in Shanghai, China. Journal of
Medical Microbiology 58, 456-461. 2009
78-Lim TT, Coombs GW, Grubb WB. Genetic organization of mecA and mecA-
regulatory genes in epidemic methicillin-resistant Staphylococcus aureus from
Australia and England. J Antimicrob Chemoter 50[6], 819-824. 2002.

94
79-Lim KT, Yasin R, Yeo CC, Puthucheary S, Thong KL. Characterization of
Multidrug Resistant ESBL-Producing Escherichia coli Isolates from Hospitals in
Malaysia. Journal of Biomedicine and Biotechnology 2009, 1-10. 2009.
80-Lyon BR, Skurray R. Antimicrobial resistance of Staphylococcus aureus: genetic
basis. Microbiol Rev 51[1], 88-134. 1987.
81-Ma XX, Ito T, Tiensasitorn C, Jamklang M, Chongtrakool P, Boyle-Vavra S, et al.
Novel Type of Staphylococcal Cassette Chromosome mec Identified in Community-
Acquired Methicillin-Resistant Staphylococcus aureus Strains. Antimicrob agents and
chemother 46[4], 1147-1152. 2002.
82-Ma XX, Ito T, Kondo Y, Cho M, Yoshizawa Y, Kaneko J, et al. Two Different
Panton-Valentine Leukocidin Phage Lineages Predominate in Japan. J Clin Microbiol
46[10], 3246-3258. 2008.
83-Machado ABMP, Reiter KC, Paiva RM, Barth AL. Distribution of staphylococcal
cassette chromosome mec (SCCmec) types I, II, III and IV in coagulase-negative
staphylococci from patients attending a tertiary hospital in southern Brazil. J Med
Microbiol 56, 1328-1333. 2007.
84-Mansouri M, Ramazanzadeh R. Spread of extended-spectrum beta-lactamase
producing Escherichia coli clinical isolates in Sanandaj hospitals. J Biol Sci 9, 362-
366. 2009.
85-Martins A, Cunha MLRS. Methicillin Resistance in Staphylococcus aureus and
coagulase-negative Staphylococci: Epidemiological and Molecular Aspects.
Microbiol.Immunol 51[9], 787-795. 2007.
86-Menezes EA, Alencar AM, Cunha FA, Ângelo MRF, Salviano MNC, Oliveira IRN.
Freqüência de cepas produtoras de enzima beta lactamase de espectro expandido
(ESBL) e perfil de susceptibilidade de Klebsiella pneumoniae em hemoculturas no
berçário de um hospital de Fortaleza. RBAC [40], 1-7. 2008.

95
87- Milheiriço C, Oliveira DC, De Lencastre H. Update to the Multiplex PCR Strategy
for Assignment of mec Element Types in Staphylococcus aureus. Antimicrob Agents
Chemother 51[9], 3374-3377. 2007.
88-Monk AB, Boundy S, Chu VH, Bettinger JC, Robles JR, Fowler Jr.VG, Archer GL.
Analysis of the Genotype and Virulence of Staphylococcus epidermidis Isolates from
Patients with Infective Endocarditis. Infection and immunity 76[11], 5127-5132. 2008.
89-Monstein HJTM, Nilsson LE. Molecular identification of CTX-M and blaOXY/K1 ß-
lactamase genes in Enterobacteriaceae by sequencing of universal M13-sequence
tagged PCR-amplicons. BMC Infectious Diseases 9, 7. 2009. 15-8-2009.
90-Moquet O, Bouchiat C, Kinana A, Seck A, Arouna O, Bercion R, Breurec
S,Moquet BG. Class D OXA-48 Carbapenemase in Multidrug-Resistant
Enterobacteria, Senegal. Emerg Infect Dis 17[1],143-144.2011.
91-Miyoshi T, Iwatsuki T, Naganuma T. Phylogenetic Characterization of 16S rRNA
Gene Clones from Deep-Groundwater Microorganisms That Pass through 0.2-
Micrometer-Pore-Size Filters. Applied and Environmental Microbiology 71[2], 1084-
1088. 2005.
92-Mshana SE, Kamugisha E, Mirambo M, Chakraborty T, Lyamuya EF. Prevalence
of multiresistant gram-negative organisms in a tertiary hospital in Mwanza, Tanzania.
BMC Research Notes 2, 49. 2009.
93-Mulvey MR, Bryce E, BD, Agostini MO, Christianson S, Simor AE, et al. Ambler
Class A Extended-Spectrum Beta-Lactamase-Producing Escherichia coli and
Klebsiella spp. in Canadian Hospitals. Antimicrob Agents Chemother 48[4], 1204-
1214. 2004.
94-Murphy S, Fitzgerald M, Mulcahy R, Keane C, Coakley D, Scott T. Studies on
haemagglutination and serum resistance status of strains of Moraxella catarrhalis
isolated from elderly. Gerontology 43, 277-282. 1997.

96
95-Nathwani D, Wood MJ. Penicillins. A current review of their clinical pharmacology
and therapeutic use. Drugs 45[6], 866-894.1993.
96-Neu HC. The crisis in antibiotic resistance. Science 257[5073], 1064-1073.1992.
97-O'Brien FG, Coombs GW, Pearson JP, Christiansen KJ, Grubb WB. Type V
Staphylococcal Cassette Chromosome mec in Community Staphylococci from
Australia. Antimicrob agents and chemother 49[12], 5129-5132. 2005.
98-Oplustil CA, Zoccoli CM, Tobouti NR, Sinto SI. Procedimentos básicos em
Microbiologia Clínica. São Paulo: 2000. 108- ANVISA. Detecção e Identificação de
Bactérias de Importância Médica. Brasília: 2005.
99-Park C, ee DG, Choi SM, Park SH, Choi JH, oo JH, et al. A Case of Perianal
Abscess due to Panton-Valentine Leukocidin Positive Community-Associated
Methicillin-Resistant Staphylococcus aureus: Report in Korea and Literature Review
from the Far East. Infection and Chemotherapy 40[2], 121-126. 2008.
100-Pearson JP, Passador L, Iglewski BH, Greenberg EP. A second acyl-
homoserine lactone signal produced by Pseudomonas aeruginosa. Proc Natl Acad
Sci USA 92, 1490-1494. 1995.
101-Prevost G, Mourey L, Colin DA, Menestrina G. Staphylococcal pore-forming
toxins. Curr Top Microbiol Immunol 257, 53-83. .2001.
102-Picão RC, Gales AC. Extended-spectrum beta-lactamase-producing (ESBL)
Pseudomonas aeruginosa: Nightmare or imagination? Prática Hospitalar 49, 79-84.
2007.
103-Reichert B, Birrell G, Bignardi G. Severe non-pneumonic necrotising infections in
children caused by Panton-Valentine leukocidin producing Staphylococcus aureus
strains. J Infect 50[5], 438-442. 2005.
104-Reiter KC. Distribuição dos SCCmec tipos I, II, III e IV em Staphylococcus

97
aureus meticilina-resistente isolados de pacientes do Hospital de Clínicas de Porto
Alegre. LUME. Repositório Digital. Universidade Federal do Rio Grande do Sul.
LUME- Repositório Digital-UFRGS . 2009. 10-11-2009.
105-Remonatto G, Bolzan V, Zanchi AC, D'azevedo PA. Detecção Molecular da
Resistência Bacteriana- Ênfase para Enterococcus e Streptococcus. Newslab 70,
100-112. 2005.
106-Romanus II, Egwu OA, Ngozi AT, Chidiebube NA, Chika EP. Extended
Spectrum Beta –Lactamase (ESBL) Mediated Resistance to Antibiotics Among
Klebsiella Pneumoniae in Enugu Metropolis. Macedonian Journal of Medical
Sciences 2[3], 1-4. 2009. 15-8-2009.
107-Rosenstein R, Gotz F. Staphylococcal lipases:biochemical and molecular
characterization. Biochemie 82, 1005-1014. 2000.
108-Rossi F, Andreazzi DB. Resistência Bacteriana. Interpretando o antibiograma.
São Paulo: 2005.
109-Ruppé E, Sopheak H, Sovannarith L, Gautier V, Ariey F, Sarthou JL, et al. CTX-
M Beta-Lactamases in Escherichia coli from Community-acquired Urinary Tract
Infections, Cambodia. Emerg Infect Dis 15[5], 741-748. 2009.
110-Strommenger B, Braulke C, Pasemann B, Schmidt C, Witte W. 2008. Multiplex
PCR for Rapid Detection of Staphylococcus aureus Isolates Suspected to Represent
Community-Acquired. J Clinl Microbiol 46[2], 582-587. 2008.
111-Sabra AH, Araj GF, Kattar MM, Abi-Rached RY, Khairallah MT, Lena JD, et al.
Molecular characterization of ESBL-producing Shigella sonnei isolates from patients
with bacilliary dysentery in Lebanon. J Infect Dev Ctries 3[4], 300-305. 2009.
112-Safo MK, Zhao Q, Ko T, Musayev FN, Robinson H, Scarsdale N, et al. Crystal
Structures of the BlaI Repressor from Staphylococcus aureus and Its Complex with
DNA: Insights into Transcriptional Regulation of the bla and mec Operon. Journal of

98
Bacteriology 187[5], 1833-1844. 2005.
113-Safo MK, Ko T, MFN, Zhao Q, Wang AHJ, Arche GL. Structure of the MecI
repressor from Staphylococcus aureus in complex with the cognate DNA operator of
mec. Acta Crystallogr Sect F Struct Biol Cryst Commun 62[Pt(4)], 320-324. 2006.
114-Song KH, Jeonn JH, Park WB, Park SW, Kim HB, Oh MD, et al. Clinical
outcomes of spontaneous bacterial peritonitis due to extended-spectrum beta-
lactamase-producing Escherichia coli and Klebsiella species: A retrospective
matched case-control study. BMC Infectious Diseases 9, 41. 2009.
115-Souza MV, Reis C, Pimenta FC. Revisão sobre a aquisição gradual de
resistência de Staphylococcus aureus aos antimicrobianos. Rev Pat trop 34[1], 27-
36. 2005.
116-Silva-Carvalho MC, Bonelli RR,Souza RR,Moreira S,dos Santos LC,de Souza
Conceição M,de Mello Junior SJ,Carballido JM,Rito PN,Vieira VV,Teixeira LA, Sá
Figueiredo AM. Emergence of multiresistant variants of the community-acquired
methicillin-resistant Staphylococcus aureus lineage ST1-SCCmecIV in 2 hospitals in
Rio de Janeiro, Brazil. Diagn Microbiol Infect Dis. 65[3], 300-305. 2009.
117-Suarez GJ, Krsnik I, Reyes E, De MD, Hernanz N, Barr-Ali M, et al. Elderly
haematological patients with chemotherapy-induced febrile neutropenia have similar
rates of infection and outcome to younger adults: a prospective study of risk-adapted
therapy. Br J Haematol 120[2], 209-216. 2003.
118-Superti SV, Augusti G, Zavascki AP. Risk factors for and mortality of extended-
spectrum-beta-lactamase-producing Klebsiella pneumoniae and Escherichia coli
nosocomial bloodstream infections. Rev Inst Med trop.S.Paulo 51, 211-216. 2009.
119-Shore A, Rossney AS, Keane CT, Enright MC, Coleman DC. Seven novel
variants of the staphylococcal chromosomal cassette mec in methicillin-resistant
Staphylococcus aureus isolates from Ireland. Antimicrob Agents Chemother 49[5],
2070-2083. 2005.

99
120-Shukla SK, Ramaswamy SV, Conradt J, Stemper ME, Reich R, Reed KD, et al.
Novel Polymorphisms in mec Genes and a New mec Complex Type in Methicillin-
Resistant Staphylococcus aureus Isolates Obtained in Rural Wisconsin. Antimicrob
Agents Chemother 48[8], 3080-3085. 2004.
121-Slom SJ, Biech-Andersen A, Henrichsen J. Ultrastructural localization of
capsules, cell wall polysaccharide, cell wall proteins, and F antigen in penumococci.
Infect Immun 56, 1980-1986. 1988.
122-Stanier RY, Ingraham JL, Wheelis ML, ainter PR. General Microbiology. Upper
Saddle River, New Jersey: 1986.
123-Stevens AM, Hennessy T, Baggett HC, Bruden DPD, Klejka J. Methicillin-
Resistant Staphylococcus aureus Carriage and Risk Factors for Skin Infections,
Southwestern Alaska, USA. Emerg Infect Dis 16[5], 797-803. 2010.
124-Strandén AM, Frei R, Adler H, Flückiger U, Widmer AF. Emergence of SCCmec
Type IV as the Most Common Type of Methicillin-Resistant Staphylococcus aureus in
a University Hospital. Infection 37, 44-48. 2009.
125-Takagi Y, Futurama S, Asada Y. Action site of exfoliative toxin on keratinocytes.
J Invest Dermatol [94], 52-54. 1990.
126-Tenover FC. Mechanisms of antimicrobial resistance in bacteria. Am J Infect
Control 34(5 Suppl 1):S3-10; discussion S64–73. 2006.
127-Tortora G, Funke BR, Case CL. Microbiologia. 8th ed. Rio Porto Alegre:2005.
128-Thierry F, Arlet G, Gautier V, Talarmin. A, Bercion R. Extended-spectrum-
Lactamase producing Enterobacteriaceae, Central African Republic. Emerg Infect
Dis 12[5], 863-865. 2006.
129-Thomasz A, Fischetti VANRPFJJ. The staphylococcal cell wall. In: ASM Press,
editor. Gram Positive Pahogens.Washington, DC: 2000. p. 351-60.

100
130-Thomas LC, Gidding HF, Ginn AN, Olma T, Iredell T. Development of a real-time
Staphylococcus aureus and MRSA(SAM-)PCR for routine blood culture. Journal of
Microbiological Methods 68, 296-302. 2007.
1301a-Udo EE, Sarkhoo E. The dissemination of ST80-SCCmec-IV community-
associated methicillin resistant Staphylococcus aureus clone in Kuwait hospitals.
Annals of Clinical Microbiology and Antimicrobials 9[31], 1-7. 2010.
131b-Udo EE, Sarkhoo E. Genetic analysis of high-level mupirocin resistance in the
ST80 clone of community-associated meticillin-resistant Staphylococcus aureus.
Journal of Medical Microbiology 59, 193-199. 2010.
132-Ueno M, Jorge AOC. Caracterização de Staphylococcus aureus resistentes à
meticilina, envolvidos em infecções nosocomiais, por meio de técnicas fenotípicas e
análise de perfil plasmidial. Rev.biociênc.Taubaté 7[2], 15-22. 2001.
133-Varaldo PE, Kilpper-Balz R, Biavasco F, Satta G, Schleifer HK. Staphylococcus
delphini SP. Nov., a coagulase-positive species isolated from dolphins. Int J Syst
Bacteriol 38, 436-439. 1998.
134-Venkatachalam KV, Huang W, LaRoccos M, Palzkill T. Characterization of
TEM-1 B-Lactamase Mutants from Position 238 to 241 with Increased Catalytic
Efficiency for Ceftazidime. The Journal of Biological Chemistry 269[38], 23444-
23450. 1994.
135-Vermont CL, Hartwig NG, Fleer A, Man P, Verbrugh H, Groot R, et al.
Persistence of Clones of Coagulase-Negative Staphylococci among Premature
Neonates in Neonatal Intensive Care Units: Two-Center Study of Bacterial
Genotyping and Patient Risk Factors. J Clinl Microbiol 36[9], 2485-2490. 1998.
136-Vidal F, Mensa J, Almela M, Olona M, Martinez JA, Marco F, et al. Bacteraemia
in adults due to glucose non-fermentative Gram-negative bacilli other than P.
aeruginosa. QJ Med 96[3], 227-234. 2003.

101
137-Vignaroli C, Varaldo PE, Camporese A. Methicillin-Resistant Staphylococcus
aureus USA400 Clone, Italy. v. Emerg Infect Dis 15[6], 995. 2009.
138-Yao D, Yu FY, Qin ZG, Chen C, He SS, Chen ZG, et al. Molecular
characterization of Staphylococcus aureus isolates causing skin and soft tissue
infections (SSTIs). BMC Infectious Diseases 10[133], 3-5. 2010.
139-Weisser M,Schoenfelder SMK,Orasch C,Arber C,Gratwohl A,Frei R,Eckart
M,Fluckiger U,Ziebuhr W. Hypervariability of Biofilm Formation and Oxacillin
Resistance in a Staphylococcus epidermidis Strain Causing Persistent Severe
Infection in an Immunocompromised Patient. Journal of Clinical Microbiology 48[7],
2407-2412. 2010.
140-Weller TMA. The distribution of mecA, mecRI and mecI and sequence analysis
of mecI and the mec promoter region in staphylococci expressing resistance to
methicillin. J Antimicrob Chemoter 43, 15-22. 1999.
141-Wilke MS, Lovering AL, Strynadka NC. Beta-lactam antibiotic resistance: a
current structural perspective. Curr Opin Microbiol 8[5], 525-533. 2005.
142-Winn jr WC, Allen SD, Janda WM, Konemam EW, Procop GW, Schreckenberger
PC, et al. Diagnóstico Microbiológico. Texto e Atlas colorido. 6th ed ed. Rio de
Janeiro: 2008.
143-Witzany G. Bio-Communication of Bacteria and their Evolutionary Roots in
Natural Genome Editing Competences of Viruses. Open Evolution Journal 2,44-54.
2008.
144-Wright GD. Bacterial resistance to antibiotics: Enzymatic degradation and
modification. Advanced Drug Delivery. Advanced Drug Delivery Reviews 57, 1451-
1470. 2005.
145-Wong A, Reddy SP, Smyth DS, Rosenfeld MEA, Sakoulas G, Robinson DA.

102
Polyphyletic Emergence of Linezolid-Resistant Staphylococci in the United States.
Antimicrobial Agents and Chemotherapy 54[2], 742-748. 2010
146-Wu SW, de Lencastre H, Tomasz A. Recruitment of the mecA gene homologue
of Staphylococcus sciuri into a resistance determinant and expression of the resistant
phenotype in Staphylococcus aureus. J Bacteriol 183[8], 2417-2424.2001.
147-Zamze SE, Moxon ER. Composition of the lipopolysaccharide from different
capsular serotype strains of Haemophilus influenza. J Gen Microbiol 133, 1443-1451.
1987.
148-Zhang K, McClure JA, Elsayed S, Conly JM. Novel staphylococcal cassette
chromosome mec type, tentatively designated type VIII, harboring class A mec and
type 4 ccr gene complexes in a Canadian epidemic strain of methicillin-resistant
Staphylococcus aureus. Antimicrob Agents Chemother 53[2], 531-540.2009.

103
8 Anexos 8.1 Termo de Consentimento Livre esclarecido
Prevalência e sensibilidade a antibióticos das bactérias aeróbicas isoladas de processos
infecciosos de pacientes com doenças hematológicas da Fundação HEMOAM
Este é um trabalho de pesquisa que tem como finalidade realizar um exame do micróbio que esteja
provavelmente causando uma infecção em seu corpo, e o resultado do exame poderá trazer boas
coisas para a sua saúde, pois será encaminhado para o médico que fará o tratamento adequado.
Para realizar o teste será necessária a coleta do seu sangue com agulha descartável, onde você
sentirá um leve aperto em seu braço e um pouco de dor no momento da picada da agulha, mas nada
disto comprometerá sua saúde ou seu bem estar físico. Poderá sentir também, dependendo do tipo
de exame e a solicitação do médico, uma sensação de um swab (cotonete grande) que será passado
em alguma parte de seu corpo (garganta, nariz, ferida), ou a urina que tiver que coletar. Não se
preocupe com seu sangue, pois logo após termos feito os exames ele será desprezado e somente
será guardado num freezer o germe que foi identificado como o responsável pela sua doença
infecciosa. Caso o(a) Sr. (a) ou seu filho concorde em participar, estarão nos ajudando a ter
informações sobre sua saúde e também como prevenir algumas doenças graves. O projeto poderá
ser encerrado se ocorrerem problemas além dos mencionados acima que possam prejudicá-lo. O Sr
(a) ou seu filho poderão ser ressarcidos (as) pelos integrantes da pesquisa, caso haja algum tipo de
prejuízo físico extremo, durante a coleta do exame para a pesquisa, mas não se preocupe pois para
este projeto vocês não correm esse risco. Todos os resultados dos exames feitos serão mantidos em
segredo e somente o (a) Sr.(a), e/ou seu filho e/ou o responsável pelo projeto poderão ver os
resultados. Também queremos lhe dizer que eles poderão ser publicados em uma revista médica
para que outros colegas da nossa profissão possam tomar conhecimento e tentar ajudar outras
pessoas. O (a) Sr. (a) ou seu filho terão liberdade a qualquer momento de sair do estudo sem que
isso lhe traga algum prejuízo ao seu modo de vida e qualquer dúvida poderá ser tirada com a
responsável que é a Drª Cristina Motta Ferreira pelo telefone 3655-01-00 ramal 0109 ou na Fundação
HEMOAM, Laboratório de Microbiologia pelo período da tarde.
Eu,.............................................................................................................................................................
................................recebi a explicação de que serei um dos participantes dessa pesquisa. Se eu não
souber ler ou escrever, uma pessoa de minha confiança lerá este documento para mim e depois
escreverá nesta página o meu nome. Por estar devidamente informado (a) e esclarecido (a) sobre o
este termo, livremente, sem qualquer pressão por parte dos pesquisadores, expresso meu
consentimento para a minha inclusão.
Eu,...............................................................................................responsável pelo paciente menor de
idade ................................................................................................recebi a explicação de que o
mesmo será um dos participantes dessa pesquisa. Se eu não souber ler ou escrever, uma pessoa de
minha confiança lerá este documento para mim e depois escreverá nesta página o meu nome. Por

104
estar devidamente informado (a) e esclarecido (a) sobre o este termo, livremente, sem qualquer
pressão por parte dos pesquisadores, expresso meu consentimento para a minha inclusão.
......................................................................
......../......./.........
Assinatura do paciente Data / /
Impressão do polegar direito do paciente,
caso este não saiba escrever seu nome.
............................................................................................
Assinatura do pesquisador que conversou com o paciente

105
8.2 Anexo 2 PROTOCOLO MÉDICO
Prevalência e sensibilidade a antibióticos das bactérias aeróbicas isoladas de processos infecciosos de pacientes com doenças hematológicas da Fundação HEMOAM
Nome do paciente:_______________________ Registro:___________ Data do exame:_____
D. Nasc:_______________ Idade:__________ Procedência________________________
Clínica ( ) Leito ( ) nº ___ Ambulatório ( )
Dados Clínicos: Provável origem do processo infeccioso: comunitário ( ) hospitalar ( )
Relacionado a procedimento invasivo? Sim ( ) Não ( )
Cirurgia ? Qual? ___________________________________________
Doença Hematológica: Anemia aplástica ( ) Leucemia (LLA) ( ) A.falciforme ( )
Leucemia (LMC) ( ) Leucemia (LLC) ( ) LMA( )
Mieloma Múltiplo( ) Linfoma ( ) Hemofilia ( )
Foco infeccioso- Descrição da Amostra:
Trato respiratório:
Ouvido: Otite ( ) Seios da face: Sinusite ( )
Boca: Abcesso dentário ( ) Úlcera ( ) Faringe( ) amigdala ( )
Pulmão: tuberculose ( ) pneumonia ( ) escarrro( )
Coração: endocardite ( )
Trato urinário: infecção urinária ( )
Sintomático ( ) assintomático ( ) jato médio ( )
sonda de alívio ( ) punção suprapúbica ( ) saco coletor( )
Trato intestinal: diarréia ( ) tiflite ( )
Pele: ferida cutânea ( ) abcessos ( ) úlceras ( ) foliculite ( ) sepse( ) ponta
de cateter ( ) foco de origem desconhecida ( ) próteses ( ) outros ( )
Hemocultura: sangue periférico ( ) sangue colhido de cateter ( )
Topografia(local da coleta) _________________________________
nível de coleta : superficial ( ) profunda ( )
Via de obtenção de material: punção ( ) swab ( ) raspado ( ) drenagem ou fístula ( )
dreno ( ) outros : _______________
Fez uso nos últimos dias de antibiótico ? Sim ( ) não ( )
Quais ?______________________________________________
Existe infecção em outra topografia ? Sim ( ) não ( )
Existe comprometimento imunológico ? Sim ( ) não ( )
É paciente transferido de outro hospital ? Sim ( ) não ( )

106
É portador de bactérias multirresistentes? Sim ( ) não ( )
Condição clínica do paciente: febre antes da internação ( ) febre no decorrer da internação ( )
neutropenia febril ( ) neutropenia afebril ( ) febre após alta da internação ( )
Data do pedido _/___/___
Médico solicitante (Nome legível) e CRM : ____________________________

107
8.3 Anexo 3 PROTOCOLO LABORATORIAL
Nome:______________________________Registro:________________ Procedência_______________
Data:_______ Clínica ( ) Leito ( ) Ambulatório ( )
Local do Nascimento____________________ Médico:_____________ Amostra biológica
coletada:________________________________ Data da coleta ___/___/___
Responsável pela coleta _______________________________________
Natureza do teste solicitado: teste direto ( ) Gram ( ) Ziehl ( )
Rotina bacteriológica (Gram+cultura) ( )
Resultados: Método de Gram:
BGN ( ) BGP ( ) CGP ( ) CGN ( )
Testes Identificação de cocos (Micrococos e estafilococos):
Catalase pos ( ) negativa( ) Ferment. Manitol_______
Coagulase pos ( ) negativa( )
Novobiocina halo ___mmm –sensível ( ) halo ___mmm –resistente ( )
Furazolidona halo ___mmm –sensível ( ) halo ___mmm –resistente ( )
Bacitracina halo ___mmm –sensível ( ) halo ___mmm –resistente ( )
polimixina |B halo ___mmm –sensível ( ) DNAse _________ PYR ______
Testes Identificação de Estreptococos:
Catalase pos ( ) negativa( ) Hemólise alfa (parcial) ( ) beta (total) ( ) gama (ausência) ( )
Bacitracina halo ___mmm –sensível ( ) halo ___mmm –resistente ( )
Optoquina halo ___mmm –sensível ( ) halo ___mmm –resistente ( )
STX halo ___mmm –sensível ( ) halo ___mmm –resistente ( )
Cresc e em NaCl 6,5%_______ PYR ______Bile-esculina ____CAMP _____
Testes Bioquímicos:
Oxidase______ Glicose (gás)_____Sacarose _____ Maltose ________ Lactose ______H2S ______
Oxidase_______ Citrato ______ Uréia _____ Manitol ______ TSI_______Motilidade _____
Lisina _____ Arginina________Ornitina_______ Indol_____ Triptofano desaminase_____
gelatina_______ Beta lactamase________
Microorganismo identificado:_________________________
Resultado do Teste de Suscetibilidade (difusão com disco): Microrganismo Gram-positivo ( )
Eritromicina ( ) S ( )R Sulfazotrim ( )S ( )R
Oxacilina ( )S ( )R Rifampicina ( )S ( )R
Gentamicina ( )S ( )R Tetraciclina ( )S ( )R
Cefepime ( )S ( )R Vancomicina ( )S ( )R
Penicilina G ( )S ( )R Cloranfenicol ( )S ( )R
Clindamicina ( )S ( )R Ciprofloxacin ( )S ( )R
Microrganismo Gram-negativo ( )
Ampicilina ( )S ( )R Amicacina ( )S ( )R
Sulfazotrim ( )S ( )R Gentamicina ( )S ( )R
Imipenem ( )S ( )R Amoxicilina + Clavulanato ( )S ( ) R
108
Ceftazidima ( )S ( )R Ciprofloxacin ( )S ( )R
Cefalotina ( )S ( )R Cefuroxima ( )S ( )R
Cefepime ( )S ( )R Cefoxitina ( )S ( )R
Urinocultura ( ) sim ( ) Não Contagem de colônias.................................
Ácido Nalidixico ( )S ( )R Gatifloxacin ( )S ( )R
Ampicilina ( )S ( )R Norfloxacina ( )S ( )R
Gentamicina ( )S ( )R Nitrofurantoína ( )S ( )R
Tetraciclina ( )S ( )R Cefalotina ( )S ( )R
Cefepime ( )S ( )R Sulfazotrim ( )S ( )R
Cloranfenicol ( )S ( )R Ciprofloxacin ( )S ( )R
Técnico responsável pelos resultados de exame:_________________

109
8.4 Anexo 4
Protocolos Laboratoriais
Coloração pelo método de Gram Fonte: Winn et al. 2008.
1 - Solução de violeta de metila Solução 1: Violeta de metila P.A. (Neon)............................................. 2,0g
Álcool etílico 99,5GL P.A. (Labsynth) .............................. 100,0mL
Álcool metílico 99,5GL P.A. (Granquimica) ...................... 100,0mL
Solução 2: Oxalato de amônia P.A. (Reagen) ................................... 4.0g
Água deionizada .............................................................. 400,0mL Obs: Misturar as soluções 1 e 2, deixando em repouso por 24 horas ao abrigo da luz
filtrando em papel de filtro comum, estocando em frasco âmbar ao abrigo da luz.
2 - Preparo da solução de lugol (estoque) Iodeto de potássio P.A. ............................................................ 4,5g
Iodo metálico P.A. .................................................................. 3,0g
Água deionizada .................................................................. 450,0mL
2.1 - Preparo da solução de lugol diluído 1/20 (uso) Em frasco apropriado, juntar 2ml de lugol (estoque) com 38ml de água
deionizada. Obs: Misturar o Iodeto de Potássio (Merck) na água, até a completa dissolução.
Acrescentar o Iodo metálico (Cinética Química) até completa dissolução com água
destilada. Estocar em frasco âmbar ao abrigo da luz.

110
3 - Solução de safranina Safranina O P.A. ............................................................ 2,5g
Água deionizada ............................................................ 500mL
Obs: Misturar a safranina O (Vetec) na água até completa dissolução. Estocar em
frasco âmbar ao abrigo da luz. 4 - Solução descorante
Álcool etílico a 95,5° GL.
5 - Técnica de coloração
a) Cobrir o esfregaço com violeta de metila (Vetec, lt. 25) por 15seg;
b) Adicionar água deionizada sobre a lâmina com violeta de metila por 45seg;
c) Escorrer o corante e lavar em água corrente;
d) Cobrir a lâmina com lugol diluído 1/20 (uso), por aproximadamente 1
minuto;
e) Escorrer o lugol e lavar em água corrente;
f) Descorar o esfregaço adicionando álcool etílico 95,5° GL até que não haja
mais o desprendimento de corante;
g) Lavar em água corrente;
h) Cobrir a lâmina com safranina O por aproximadamente 30seg;
i) Lavar em água corrente;
j) Deixar secar a temperatura ambiente antes de examinar ao microscópio.

111
Protocolo de Coleta de material Biológico Fonte: Manual Anvisa módulo III (116).
Instrução para coleta
Hemocultura: A hemocultura é o exame definitivo para o diagnostico de invasão de
microorganismos na corrente sanguínea.
Definição de hemocultura: Coleta de um par de frascos, num mesmo momento e
num único local (frasco aeróbio e anaeróbio ou dois frascos aeróbios, por exemplo)
Obs.: A coleta de mais de 2 frascos (3,4,5 ...) num mesmo momento e de um único
local deve ser interpretada como uma hemocultura única.
Técnica: 1-Colher antes da administração de antibióticos (quando possível) ou na troca dos
mesmos;
2-Lavar as mãos e seca-las;
3-Usar luvas estéreis;
4-Remover o selo da tampa do frasco de hemocultura, e fazer assepsia previa nas
tampas com álcool a 70 %.
5-Garrotear o braço do paciente e selecionar uma veia adequada. Esta área não
deverá mais ser tocada com os dedos. Fazer a anti-sepsia com álcool a 70 % de
forma circular e de dentro para fora, concetricamente.
6-Aplicar solução de iodo (tintura de iodo 1% a 2% ou PVPI 10%) também com
movimento em espiral de dentro para fora. Para ação adequada do iodo, deixar
secar por 1 a 2 minutos antes de efetuar a coleta. Para pacientes alérgicos ao iodo,
passar novamente o álcool a 70 %.
7-Venopunção: coletar a quantidade de sangue e o numero de amostras
recomendadas de acordo com as orientações descritas ou discriminadas no pedido
medico.
8-Não é necessária a troca de agulhas para inoculação nos frascos.

112
9-Misturar o conteúdo nos frascos por inversão (método manual).
10-Limpar o local da venopunção com álcool a 70% removendo todo o iodo;
11-Identificar cada frasco conforme a padronização e enviar ao laboratório
juntamente com a solicitação médica devidamente preenchida, em caso de frasco
automatizado não rasurar o código de barra.
Observações: Não é recomendada a coleta de hemocultura através de cateteres ou cânula, por
demonstrarem nível significativo de contaminação quando comparada à veia
periférica.
Preferir punções venosas, pois as artérias não trazem beneficio em relação a
recuperação de microorganismos.
Método de coleta do sangue e o volume coletado influenciam diretamente no
sucesso de recuperação de microorganismos e uma interpretação adequada dos
resultados.
O melhor momento da coleta:
1-Antes de instituir a terapêutica antimicrobiana.
2-Antes do pico febril ou da ascenção da temperatura (30 a 90 minutos: bacteremia
máxima).
3-Se não for possível detectar a ascensão da temperatura, coletar durante ou
imediatamente ao pico febril.
Volume e número de amostras:
A sensibilidade aumenta 3% a cada ml de sangue coletado. Trabalhos recentes
mostram que, quanto maior o volume de sangue, por amostra, melhor a recuperação
do microorganismo(respeitando a porção sangue/ meio), sendo que o volume ideal
corresponde a 10 % do volume total do frasco de coleta. Os frascos de sistemas
automatizados permitem a coleta de até 10 ml de sangue. O número de frascos

113
deverá ser coletado de acordo com as condições clinicas do paciente. Preconiza-se
para o paciente adulto a coleta de 20 ml de sangue a cada hemocultura, ou seja, 02
frascos de hemocultura a cada coleta. Os frascos do sistema manual com 45 ml de
meio, coletar 5 ml de sangue. Frascos pediátricos de sistemas automatizados
permitem a coleta de 1 a 4 ml de sangue.No caso de recém - nascidos e crianças
pequenas, coletas apenas um frasco de hemocultura. Os frascos de sistema manual
com 9 ml de meio , coletar 1 ml de sangue. Um total de 3 culturas em 24 horas
costuma ser suficiente para descartar bacteremia ou endocardite. Coletas acima de
quatro amostras não trouxeram maior índice de recuperação microbiana.
Obs.: O uso de frascos específicos para cultura de anaeróbios e micobactérias,
deve ser considerado apenas em situação especificas onde ser pode esperar
participação de anaeróbios e micobacterias, e, deverá ser solicitado para tal. Ou
ainda, nos quadros clínicos cirúrgicos envolvendo o trato digestivo, infecções
pélvicas e outros, coletar sistematicamente um par de frascos: aeróbio e anaeróbio.
Nunca resfriar o frasco e enviar o mais rápido possível ao laboratório
Condições clínicas especificas Infecções Sistêmicas e localizadas (sepse, osteomielite, meningite): Duas amostras sanguíneas de punções venosas diferentes (braço direito e
esquerdo) com intervalo de, no máximo 5 minutos , entre as punções. Se possível
10 a 20 ml por amostra.
Bacteremia ou febre de origem desconhecida (abscessos, febre tifóide, brucelose): Duas as três amostras coletadas no primeiro dia de punções venosas diferentes com
intervalos superiores a uma hora . Se negativo, 24 ou 36 horas após, colher mais
duas amostras.
Endocardite: Aguda: três amostras sanguíneas de punções venosas diferentes (braço esquerdo e
direito) nas 1º a 2 º horas com intervalos mínimos de 20 a 30 minutos.

114
Subaguda: três amostras no primeiro dia (15 min. Ou mais de intervalo). Se o
resultado for negativo, após 24 horas obter mais duas amostras.
Pacientes com picos febris regulares, coletar duas amostras antes do inicio da febre
(1 hora), evitar o pico febril.
Crianças: duas culturas são recomendadas para diagnostico de
bacteremias em recém-nascidos. Pacientes em uso de antimicrobianos, coletar duas
hemoculturas separadamente, em cada dia, durante 3 dias consecutivos , se as
primeiras negativas.
Trato respiratório:
Escarro: 1-Lembrar que este material não é considerado ideal para avaliação microbiológica,
do trato respiratório.
2-A melhor coleta é feita com a supervisão direta do pessoal de enfermagem
treinado ou fisioterapeuta.
3-Orientar o paciente sobre a importância da coleta do escarro e não da saliva.
4-Colher o escarro através da tosse, de modo que o material venha do “peito” e não
da garganta.
5-Colher somente uma amostra por dia, se possível, o primeiro escarro da manhã,
antes da ingestão de alimentos, em recipiente estéril da boca larga e enviar o mais
rapidamente possível ao laboratório.
6-Orientar o paciente para escovar os dentes somente com água, e gargarejar varias
vezes a boca com água. Não usar pasta de dente ou colutórios.
Obs.: Caso o paciente tenha dificuldade de escarrar, esta amostra poderá se
induzida por inalação de 20 ml de 3% a 10% de NaCl 0,85% ou ser realizada coleta
por aspiração transtraqueal.

115
Secreção de orofaringe: 1-O sucesso de detecção do germe depende da amostra coletada exatamente na
área inflamada, evitando outros sítios da cavidade oral.
2-Solicitar ao paciente que abra bem a boca.
3-Usando o abaixador de língua e swab estéril, fazer esfregaço sobre as amídalas e
faringe posterior, evitando tocar na língua e bochechas.
4-Passar o swab nas áreas hiperemiadas (faringe posterior, pilare D e E, ou abaixo
das amigdalas), próximo aos pontos de supuração ou abaixo das placas ou
membranas.
5-Coletar preferencialmente dois swabs e colocá-los em meio transporte quando
houver.
6-Enviar imediatamente ao laboratório para evitar a secagem do marterial, quando
não houver meio de transporte.
Secreção de nasofaringe: 1-A coleta deve ser feita antes do uso de produtos tópicos.
2-Introduzir em swab flexível e estéril, pelo meato nasal, paralelo ao palato superior,
buscando atingir o orifício posterior das fossas nasais, tentando evitar tocar o swab
na mucosa da narina, fazer um discreto movimento circular e retirá- lo.
3-Recolocar o swab no tubo com meio de transporte e enviar ao laboratório
imediatamente.
URINA:
1- A coleta deve ser feita pela manhã, preferencialmente da primeira micção do dia,
ou então após retenção vesical de duas a três horas.
Crianças: 1-Realizar assepsia rigorosa prévia dos genitais com água e sabão neutro e
posterior secagem com gaze estéril.

116
2-Ideal: jato médio espontâneo.
3-Em lactentes em que não se consegue colher através do jato médio, pode se usar
o saco coletor de urina, porém, a troca deve ser realizada de 30 em 30 minutos e ao
trocar o coletor, refazer a assepsia.
4-Casos especiais (RN, lactentes de baixo peso, resultados repetidamente
duvidosos) indicar punção vesical suprapúbica, que deverá ser realizada por médico.
Adultos do sexo feminino: 1-A coleta deve ser supervisionada pessoalmente por uma enfermeira ou auxiliar
treinada.
2-Orientar a paciente que ao sentar no vaso sanitário, deverá afastar os grandes
lábios com uma das mãos e com a outra lavar as genitálias de frente para trás,
utilizando gaze estéril.
3-Continuar afastando os grandes lábios para urinar, o primeiro jato de urina deve
ser desprezado no vaso sanitário. Colher o jato médio urinário no frasco fornecido
pela enfermagem, evitando encher o frasco.
4-Fechar bem o frasco e entregar ao pessoal de enfermagem.
5-Enviar ao laboratório até 30 minutos, caso não seja possível , as amostras
deverão ser resfriadas até 2 horas.
Adultos do sexo masculino: 1-A coleta deve ser feita pela manhã, preferencialmente da primeira micção do dia,
ou então após retenção vesical de duas a três horas.
2- Realizar a higiene local da genitália utilizando uma gaze estéril embebida com
água e sabão neutro e gaze estéril.
3-Coletar jato médio em um frasco estéril descartável e levar ao laboratório
Pacientes cateterizados com sistema de drenagem fechada:
1-Colher a urina puncionando- se o cateter na proximidade da junção com o tudo de
drenagem. Não Colher urina da bolsa coletora.
2-No pedido laboratorial deverá constar que o paciente está cateterizado.

117
Ponta de sonda vesical:
1-Não realizar cultura da ponta da sonda vesical, porque o crescimento bacteriano
representa a flora de uretra distal.
2-Recomenda-se cultura de urina após 48 horas da retirada da sonda na
monitoração de processos infecciosos.
3-Urocultura realizadas antes deste período podem fornecer resultados positivos
sem que eles estejam, necessariamente, associadas à infecção. Fezes:
1-Devem ser coletadas no inicio ou fase aguda da doença, quando os patógenos
estão usualmente presentes em maior número e, preferencialmente, antes da
antibióticoterapia, e quatro dias após o uso de contrastes radiológicos.
2-Coletar as fezes e colocar em um frasco contendo o meio para transporte (Cary
Blair ou salina glicerinada tamponada), em quantidade equivalente a uma colher de
sobremesa. Preferir sempre as porções mucosas e sanguinolentas.
3-Fechar bem o frasco e agitar o material.
4-Se a amostra não for entregue no laboratório em uma hora, conservar na geladeira
a 4ºC, no máximo por um período de 12 horas. Marcar o horário da coleta.
Swab retal: 1-Usar swab de algodão.
2-Umedecer o swab em solução salina estéril (não usar gel lubrificante) e inserir no
esfíncter retal, fazendo movimentos rotatórios.
3-Ao retirar certifique-se que existe coloração fecal no algodão.
4-Enviar ao laboratório no intervalo de 30 minutos ou utilizar o meio de transporte
Cary Blair.
Lesões abertas úmidas e úlceras: 1-Esta coleta deve ser realizada pelo médico assistente.

118
2-A área ao redor da lesão deverá ser limpa com gaze embebida em álcool a 70%.
3-Remover crostas e excesso de secreção com solução salina e gaze estéril.
4-Passar o swab na base e nas bordas da lesão, colocar o swab no meio de
transporte e enviar o mais rápido possível para o laboratório, sem refrigerar.
Observações: Não coletar o pus emergente. O material das margens da lesão e a parte mais
profunda do sítio escolhido são mais representativos e possuem maior viabilidade de
microrganismos. A cultura de lesões secas e crostas não são recomendadas, a
menos que a obtenção de exsudato não seja possível. A coleta de ferida de
queimadura deve ser realizada após extensa limpeza e debridamento da lesão.
Biopsia de pele é a técnica mais recomendada.
Espécimes cirúrgicos – biópsias e secreções: 1-Esta coleta é realizada pelo médico assistente.
2-Verificar, no momento do recebimento do material, se as instruções de coleta
foram seguidas.
3-Colher o material por aspiração com seringa.
4-Técnica de coleta asséptica.
Secreção conjuntival (Ocular): 1-A coleta deverá ser realizada antes do uso de qualquer produto tópico.
2-Retirar o excesso de secreção purulenta com gaze estéril.
3-Com uma das mãos afastar as pálpebras superior e inferior, e com a outra passar
o swab na mucosa conjuntival.
3-Colocá-lo no meio de transporte e enviar diretamente ao laboratório.
Semeadura em meios de cultura
Fonte: Manual Anvisa módulo V (110) ; Oplustil et al. 2000(109)

119
Tabela 1- Meios de culturas de triagem para semeadura primária Meios de cultura Meios utilizados
Urina Laminocultivos ( CLED/MacConkey), Ágar EMB + CLED
Fezes Selenito (Meio Líquido), ágar EMB e Agar SS
HEMOCULTURAS Frascos de Hemocultura
Secreções Ágar sangue ou chocolate BEM
Ponta de cateter Ágar sangue
Líquidos Ágar sangue (EMB), frascos de hemocultura
Fragmentos Ágar sangue meio líquido TSB (frascos de hemocultura)
Lavabo Brônquico Ágar sangue, Ágar chocolate EMB.
Preparo dos materiais clinicos e semeadura inicial
Amostras recebidas em “swab”:
Amostras colhidas com “swab” são utilizadas para a coleta de diversos materiais
clínicos, mas seu uso deve ser evitado quando outras metodologias podem ser
empregadas. Quando as amostras forem coletadas em “swab” duplo utilizar um dos
“swabs” para fazer o esfregaço em uma lâmina, e o outro, para colocar no meio de
transporte (caso não seja semeado na mesma hora).
Quando somente um “swab” for coletado, suspender o swab em um pequeno
volume de solução fisiológica estéril (esta na geladeira), agitar (vortex ou centrifuga)
2000 por dois comprimir o swab na parede do tubo e utilizar esta suspensão para o
preparo do esfregaço e semeadura nos meios aquáticos. Semear por esgotamento
nos meios apropriados, colocar na estufa a 35º C.
OBS.: Não deixar o “swab dentro do tubo sem meio de transporte, ou colocar na
geladeira ou estufa para ser semeado no outro dia”. Sempre executar semeadura
primaria.
Cultura de urina:

120
1-Abrir o lâminocultivo e submergir a lâmina contendo e meio de cultura na urina
recém-coletada. Ambas as superfícies dos meios de cultura devem ser
completamente umedecidas pela urina.
2-Retirar o excesso de urina deixando-a escorrer junto à borda do recipiente.
3-Fechar o tubo e encubar ente 35º C+- 1ºC em posição vertical durante 18 a 24
horas.
Obs.: Sempre fazer uma lâmina a fresco e observar a presença de leucócitos,
eritrócitos e bactérias.
Cultura de fezes:
Amostra aceitável: 1-Fezes sem conservante devem ser encaminhadas e processadas no máximo
dentro de 1 hora após a coleta.
2-Fezes com conservante podem ser mantidas á temperatura ambiente por ate 24
horas.
Amostra não aceitável: Amostras sem conservantes que foram obtidas há mais de três horas
Várias amostras colhidas no mesmo dia de urina e fezes
Fezes liquidam colidas ou enviadas em fraldas.
Procedimento 1-Semear três a quatro alçadas de fezes no tubo contendo meio selenito (meio
enriquecida).
2-Colocar o tubo na estufa por 12 a 18 horas.
3-Semear depois para os meios selenito, ágar SS e ágae BEM.
4-Incubar por mais de 18 a 24 horas.

121
Conservantes mais comuns: Glicerina tamponada
Cary-blair (meio de transporte para fezes)
Swab com meio de transporte Stuart
Obs.: Se houver suspeita de vibrio spp, a amostra de fezes ou “swab” retal deve ser
encaminhada em cary-blair à temperatura ambiente.
Hemocultura: Procedimento
1- Identificar o frasco com: nome do paciente, data e hora da coleta e numero da
amostra.
2-Levantar a parte central da tampa de alumínio com o auxilio de uma pinça.
3-Fazer assepsia da tampa de borracha com álcool 70% e deixar secar e introduzir o
sangue: 5ml no frasco para adulto e 1ml no frasco pediátrico.
4-Incubar a 35ºC com agulha estéril para realizar a incubação em aerobiose.
5-Se possível coletar 02 amostras.
6- Sepse, pneumonia, meningite, artritre e pielonefrite: 2 a 3 hemoculturas antes
do início da terapia. Coletar sem necessidade de intervalo, intercalando os sítios da
punção.
7-FOI, endocardite ou outra bacteriemia ou fungemia contínua: 2 a 3
hemoculturas com 20 min de intervalo entre as coletas. Se forem negativas nas
primeiras 24hs, repetir o procedimento.
8-Paciente em uso de terapia antimicrobiana- utilizar frasco com inibidores de
antibióticos e coletar antes da próxima dose do antibiótico.
Técnica para processamento de hemocultura manual Frasco de hemocultura- incubar a 35ºC em C02. Após 24hs (1º dia)-- semear em
ágar: sangue, chocolate, Mac Conkey e realizar o teste de Gram (sugerir

122
agrupamento).
Resultados: Positivo- sem crescimento em ágar sangue e chocolate com bactérias no Gram,
semear em anaerobiose. Relatar cultura positiva para anaeróbio. Não realizamos
este tipo de cultura.
Negativo- emitir resultado parcial – reincubar.
Reincubar a 35ºC. Após 48hs(2º dia) – semear em Agar sangue, chocolate, Mac
Conkey e BEM. Realizar o Gram.
Resultados:
Positivo- emitir resultado parcial- identificação e TSA (cocos ou bastonetes)
Negativo- emitir resultado parcial – reincubar.
Reincubar 35ºC. Após 120hs (5º dia) – semear em Agar: Chocolate, Mac Conkey e
EMB. Realizar o Gram.
Resultados:
Positivo- emitir resultado parcial- identificação e TSA (cocos ou bastonetes).
Negativo- emitir resultado parcial – reincubar.
Reincubar 35ºC. Após o 7º dia: Semear uma gota em ágar chocolate.
Resultados:
Positivo- emitir resultado parcial- identificação e TSA (cocos ou bastonetes).
Negativo- resultado neg, desprezar o frasco. —FUNGOS????

123
OBS: No resultado emitir o sítio da coleta, horário e temperatura do paciente no momento da coleta.
Microrganismos mais freqüentes em hemocultura: S. aureus, Staphylococcus
coagulase negativa, Enterococcus spp., Streptococcus spp., mais raramente Listeria
spp..
Entre os fungos temos Cândida spp., Crytococcus neoformans.
Entre os gram-negativos: E. coli, Klebsiella pneumoniae, Salmonella spp.,
Enterobacter spp., Pseudomonas aeruginosa, Haemophilus influenzae, N.
meningitidis e outros. Em crianças ocorrem também os mesmos que nos adultos,
mas há maior prevalência de bacteremia por estafilococos, estreptococos, (S.
pneumoniae), meningococos, hemófilos.
Cultura de secreções: Material clínico: Amostras de exsuadato de lesão:
Superficial
Profunda
Operatória
Obs.: Quando for solicitada bacterioscopia conjuntamente com a cultura, é
recomendado coletar dois “swabs”. Um dos “swabs” deve ser utilizado
imediatamente após a coleta para confeccionar uma lâmina e outro ser colocado em
meio de transporte.
Procedimento Incubação nos meios de cultura: Amostra não coletada com “swab”: Ágar sangue, chocolate (opcional para feridas profundas) e EMB.

124
“Swab” sem meio de transporte:
Ressuspender a amostra colhida com “swab” em volume pequeno de caldo TSB (o
mesmo de hemocultura) ou solução fisiológica estéril. Homogeneizar vigorosamente
a solução para inocular em ágar sangue e ágar EMB e fazer uma lâmina para a
coloração de gram.
“Swab” como meio de transporte:
Semear em ágar sangue e ágar EMB.
Seringa:
Inocular diretamente em ágar sangue e ágar EMB.
Protocolo para Identificação de Cocos Gram-positivos Fonte: Manual Anvisa módulo V (110).
Tabela 2- Divisão dos cocos Gram positivos pela prova da catalase
Staphylococcus spp Enterococcus spp Micrococcus spp. Streptococcus spp. Planococcus spp Aerococcus spp Stomatococcus spp. Gemella spp., Leuconostoc spp. Lactococcus spp., Stomatococcus spp.

125
1-Teste da catalase
1- Reagentes a) Peróxido de hidrogênio a 3 ou 30% armazenado em frasco escuro, sob
refrigeração.
b) Cultura pura (24h) do isolado bacteriano.
2 - Controle de qualidade a) Devem-se realizar os testes de controle com microorganismos positivos (S.
aureus) e negativos (Streptococcus spp.), diariamente ou antes de se testar a
bactéria em estudo.
3 – Procedimento técnico: a) Com agulha de inoculação ou aplicador de madeira estéril, transferir coletar uma
colônia para a superfície de uma lâmina de vidro.
b) Acrescentar uma gota de peróxido de hidrogênio a 3% ou 30%.
4 – Interpretação
O aparecimento rápido e prolongado de bolhas ou de efervescência significa que o
teste é positivo.
2-Teste da citocromo oxidase (oxidase) 1- Reagentes a) Diidrocloreto de tretrametil-p-fenilenodiamina, 1% (reagente de Kovac)
b) Diidrocloreto de dimetilp-fenilenodiamina, 1% (reagente de Gordon e Mcleod)
Reativo: (Merck) Diidrocloreto de tetrametil ou dimetil-p-fenilenodiamina P. A. ......... 1g
Água deionizada ............................................................................... 10mL

126
2 - Controle de qualidade a) Devem-se realizar testes de controle com bactérias que apresentam reações
positivas (E. coli) e negativas (P. aeruginosa), rotineiramente.
3 – Procedimento a) Adicionar duas a três gotas diretamente na colônia bacteriana a ser estada
b) Procedimento direto em tira de papel.
OBS: Algumas gotas do reagente são adicionadas em papel de filtro ou em discos
de tiras de papel comerciais impregnados com reagente seco. Em qualquer dos
métodos, utiliza-se alça de platina ou aplicador de madeira impregnada com a
colônia suspeita na área do papel de filtro.
4 – Interpretação
O aparecimento de cor azul-escura no local da inoculação no intervalo de 10
segundos significa que a colônia é positiva para produção de citocromo oxidase.
3-Diferenciação de gêneros e espécies Fonte: Manual Anvisa módulo V (110).
Tabela 3 - Provas bioquímicas para identificação dos gêneros de Cocos Gram-positivos catalase positivos
Gênero Motilidade NaCl 5% Oxidase Bacitracina Tétrade Manitol
Staphylococcus Neg + Neg Resist até o
disco
variável Cresce
fermenta
Planococcus + + Neg variável
Micrococcus Neg + + Sens
>=10mm
+ Cresce não
fermenta
Stomatococcus Neg Neg Neg variável
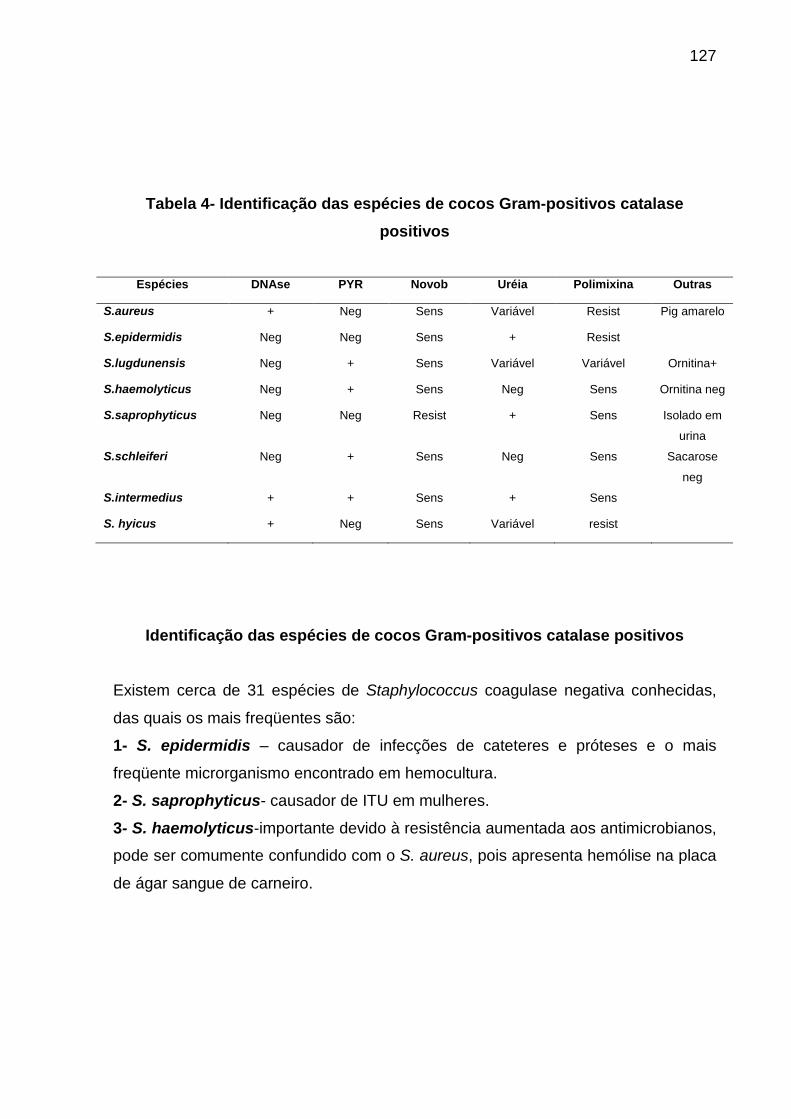
127
Tabela 4- Identificação das espécies de cocos Gram-positivos catalase positivos
Espécies DNAse PYR Novob Uréia Polimixina Outras
S.aureus + Neg Sens Variável Resist Pig amarelo
S.epidermidis Neg Neg Sens + Resist
S.lugdunensis Neg + Sens Variável Variável Ornitina+
S.haemolyticus Neg + Sens Neg Sens Ornitina neg
S.saprophyticus Neg Neg Resist + Sens Isolado em
urina
S.schleiferi Neg + Sens Neg Sens Sacarose
neg
S.intermedius + + Sens + Sens
S. hyicus + Neg Sens Variável resist
Identificação das espécies de cocos Gram-positivos catalase positivos
Existem cerca de 31 espécies de Staphylococcus coagulase negativa conhecidas,
das quais os mais freqüentes são:
1- S. epidermidis – causador de infecções de cateteres e próteses e o mais
freqüente microrganismo encontrado em hemocultura.
2- S. saprophyticus- causador de ITU em mulheres.
3- S. haemolyticus-importante devido à resistência aumentada aos antimicrobianos,
pode ser comumente confundido com o S. aureus, pois apresenta hemólise na placa
de ágar sangue de carneiro.

128
Tabela 5- Esquema para diferenciação de espécies de Cocos Gram-positivos catalase negativa sensíveis a vancomicina
Espécie Hemólise Bacitracina Optoquina SXT NaCl 6,5%
CAMP Bile
esculina
S. pneumoniae α- hemólise Variável Sens Sens - - -
S. pyogenes β- hemólise Sens Resist Resist - - -
S. agalactiae Novobiocina -
sensível
Gama
hemólise Resist Resist Resist - + -
Enterococcus
gama ou α-
hemólise Resist Resist Resist + - +
Grupo Viridans α- hemólise - - -
S. bovis Gama ou α-
hemólise - - +
PYR Test Fonte: PROBAC DO BRASIL
Indicações: Baseado na hidrólise enzimática da L-pyrrolidonyl-beta-naphtylamide, o PYR TEST
destina-se à identificação dos Streptococcus spp, Staphylococcus spp e bacilos
Gram negativos não fermentadores de glicose.
A hidrólise do L-pyrrolidonyl-beta-naphtylamide através da enzima L-pyroglutamyl-
aminopeptidase é verificada através da reação do PYR Reagente com a beta-
naphtylamide livre, que forma uma Base de Schiff de coloração vermelha.
Procedimento: 1-Com o auxílio de uma pinça previamente flambada, retirar um disco de PYR do
frasco.

129
2-Utilizando uma pipeta ou um frasco conta-gotas, impregnar o disco de PYR com
água destilada estéril ou água de torneira* Depositá-lo sobre uma lâmina ou placa
de Petri.
3-Com o auxílio de uma alça previamente flambada, faça um esfregaço da bactéria
recém isolada a ser identificada no disco de PYR umedecido.
4-Aguardar 5 minutos, a temperatura ambiente, e colocar uma gota do PYR
reagente. As reações positivas ocorrem em até 1 minuto.
* O uso de salina torna a reação mais lenta e menos intensa.
Leitura: PYR positivo: O desenvolvimento de uma cor vermelho-cereja indica resultado
positivo.
PYR negativo: Uma coloração amarela ou alaranjada indica resultado negativo.
Observação: Dos Streptococcus spp (cocos Gram positivos, catalase negativa) o Streptococcus
pyogenes e o Enterococcus spp são PYR positivo. Nos casos de amigdalite
bacteriana, com isolamento de estreptococos beta hemolíticos
a prova rápida de PYR é extremamente útil para o diagnóstico do S.pyogenes e o
tratamento precoce do paciente.
Precauções: Após leitura os discos devem ser descartados conforme as recomendações vigentes
para resíduos de serviços de saúde.
Apresentação: Caixa com 24 discos de PYR e 1 frasco com 2 mL de PYR
Reagente.
Conservação: Manter em geladeira (2º a 8º C), ao abrigo da luz.
Validade: 3 meses.

130
Protocolo para Identificação de Bastonetes Gram-negativos
Principais provas para a identificação das enterobactérias de importância clínica
ENTEROKIT B Fonte: PROBAC DO BRASIL
1. Fermentação da glicose
2. Fermentação da lactose
3. Motilidade
4. Utilização de citrato
5. Descarboxilação da lisina
6. Produção de sulfeto de hidrogênio (H2S)
7. Produção de gás (CO2)
8. Oxidase
9. Produção de indol
10. Produção de urease
11. Produção de fenilalanina desaminase ou opção triptofanase
12. Produção de gelatinase ou opção DNAse
Provas complementares de Identificação 1- Fermentação de outros carboidratos: sacarose, maltose, arabinose, salicina,
dulcitol, manitol, etc.
2- Utilização de aminoácidos: arginina e ornitina
3-Hidrólise da esculina, etc.
4- ONPG
5-Utilização de acetato
6-Provas úteis, mas pouco utilizadas: vermelho de metila, voges-proskauer,
crescimento em KCN, tartarato de jordan e lipase.
Os esquemas de identificação baseiam-se na determinação dos gêneros e
espécies mais isolados na clínica, e nas provas mais características de cada gênero

131
e espécie, baseado em alguns critérios como: facilidade de execução, facilidade de
interpretação, custo, rapidez para leitura, etc.
ENTEROKIT B Fonte: PROBAC DO BRASIL
Indicações: O Enterokit B consiste dos seguintes meios: EPM, MILi e Citrato de Simmons. O
meio EPM contém os testes de fermentação e produção de gás em glicose,
produção de H2S, hidrólise da uréia e desaminação do triptofano.
O meio MlLi contém os testes de motilidade, indol e descarboxilação de lisina. O
Citrato de Simmons oferece o teste de utilização do citrato como única fonte de
carbono. Os três meios totalizam 8 testes que somados ao da fermentação da
lactose na placa de isolamento, permitem identificar com fidelidade a grande maioria
das enterobactérias isoladas de espécimes clínicos.
Procedimento: 1-Tocar a colônia com agulha de platina e semear os 3 meios na seguinte ordem:
Citrato, EPM e MlLi.
2-Para inocular o meio de Citrato deslizar a agulha pelo centro estriando toda a
superfície inclinada. No meio
3-EPM introduzir a agulha até o fundo do tubo e ao retirá-la semear a superfície do
meio enquanto o MILi deve ser semeado por picada central que deve atingir o fundo
do tubo.
4-Incubar o Enterokit a 35°C ± 2°C com as tampas semi-rosqueadas e fazer a leitura
após 18 horas a 24 horas.
Leitura e interpretação:
1. Meio EPM Produção de gás: Aparecimento de bolhas ou deslocamento do meio de cultura do
fundo do tubo.
Produção de H2S: Enegrecimento do meio em qualquer intensidade.
Hidrólise da uréia: Aparecimento de cor azul ou verde azulada (reação fraca) que
se estende para a base do meio, envolvendo-a totalmente ou não.

132
Desaminação do triptofano (LTD): Aparecimento de cor verde-garrafa na
superfície do meio.
2. Meio MlLi Motilidade (MOT): A bactéria móvel cresce além da linha de picada. A imóvel
somente nesta linha.
Descarboxilação da lisina: Quando a lisina é descarboxilada o meio adquire cor
púrpura acentuada ou discreta. Quando o aminoácido não é utilizado, o meio
adquire cor amarelada nos seus 2/3 inferiores. Considerar o teste positivo sempre
que o meio não estiver amarelo.
Produção de indol: Adicionar 3 a 4 gotas do reativo de Kovacs à superfície do meio
e agitar levemente. Quando a bactéria produz indol, o reativo adquire cor rosa ou
vermelha. Quando não produz, o reativo mantém sua cor inalterada.
3. Meio Citrato de Simmons
A utilização do citrato revela-se pelo aparecimento de cor azul na superfície
do meio. O teste deve ser considerado negativo quando a cor do meio não sofre
alteração.
Precauções: Após o uso, o produto deverá ser descartado conforme as recomendações vigentes
para resíduos de serviços de saúde.
Apresentação: Caixa com 16 conjuntos e um frasco de Reativo de Kovacs.
Conservação: Manter em temperatura ambiente entre 10º e 25ºC (local fresco).
Validade: 6 meses.

133
Tabela 6- Identificação de gênero e espécie bacteriana
NÚMERO ESPÉCIE BACTERIANA
000 Shigella
004 Shigella spp - Yersinia enterocolitica - Escherichia coli
006 Escherichia coli
011 Citrobacter freundii
012 Hafnia alvei
013 Serratia marcescens - Hafnia alvei
014 Escherichia coli
015 Citrobacter diversus
016 Escherichia coli
035 Providencia stuartii - Providencia alcalifaciens
040 Yersinia pseudotuberculosis - Yersinia enterocolitica
044 Yersinia enterocolitica
051 Citrobacter freundi
053 Serratia marcescens
055 Citrobacter diversus
070 Proteus penneri
074 Morganella morganii
075 Providencia stuartii - Providencia rettgerii
111 Citrobacter freundii
112 Salmonella Typhi
113 Salmonella spp
151 Citrobacter freundii
170 Proteus mirabilis - Proteus penneri
171 Proteus mirabilis
174 Proteus vulgaris
175 Proteus vulgaris
202 Hafnia alvei
204 Escherichia coli
206 Escherichia coli
210 Salmonella Paratyphi A - Serratia liquefaciens
211 Citrobacter freundii - Serratia liquefaciens
212 Hafnia alvei - Salmonella Choleraesuis - Serratia liquefaciens
213 Serratia marcescens - Salmonella Choleraesuis - Serratia liquefaciens
214 Escherichia coli

134
Tabela 6- Identificação de gênero e espécie bacteriana NÚMERO ESPÉCIE BACTERIANA
215 Citrobacter diversus
216 Escherichia coli
235 Providencia alcalifaciens
251 Citrobacter freundii
253 Serratia marcescens
255 Citrobacter diversus
270 Proteus penneri
274 Morganella morganii
310 Salmonella Paratyph A
311 Citrobacter freundii
312 Salmonella Choleraesuis
313 Salmonella Choleraesuis - Salmonella spp
316 Edwardsiella tarda
351 Citrobacter freundii
370 Proteus mirabillis - Proteus penneri
371 Proteus mirabillis
374 Proteus vulgaris
375 Proteus vulgaris
404 Escherichia coli
406 Escherichia coli
410 Serratia liquefaciens
411 Citrobacter freundii - Enterobacter cloacae - Serratia sp
412 Serratia liquefaciens
413 Serratia sp.
414 Escherichia coli
415 Citrobacter diversus
416 Escherichia coli
417 Serratia odorífera
443 Klebsiella pneumoniae
447 Klebsiella oxytoca
451 Citrobacter freundii - Enterobacter cloacae
455 Citrobacter diversus
511 Citrobacter freundii
551 Citrobacter freundii

135
Tabela 6- Identificação de gênero e espécie bacteriana
NÚMERO ESPÉCIE BACTERIANA
604 Escherichia coli
606 Escherichia coli
607 Klebsiella oxytoca
610 Serratia liquefaciens
611 Citrobacter freundii - Enterobacter cloacae - Serratia sp
612 Serratia liquefaciens
613 Enterobacter aerogenes - Serratia SP
614 Escherichia coli
615 Citrobacter diversusi
616 Escherichia coli
643 Klebsiella pneumoniae
647 Klebsiella oxytoca
651 Citrobacter freundii - Enterobacter cloacae
655 Citrobacter diversus
711 Citrobacter freundii
751 Citrobacter freundii
Referências bibliográficas: 1. Ewing, W. H., 1986 - Edwards and Ewing’s Identification of Enterobacteriaceae,
4th edition. Elsevier Science Publishing Co., Inc., N. York.
2. Toledo, M. R. F.; Fontes, C. F. & Trabulsi, L. R., 1982 - EPM - Uma modificação
do meio de Rugai e Araújo, para a realização simultânea dos testes de produção de
gás a partir de glicose, H2S, urease e triptofano desaminase. Rev. Microbiol., 13:
309 - 315.
3. Toledo, M. R. F.; Fontes, C. F. & Trabulsi, L. R., 1982 - MILi - Um meio para a
realização dos testes de motilidade, indol e lisina descarboxilase. Rev. Microbiol., 13:
230 - 235.
4. Farmer III, J. J. e cols. 1985 - Biochemical Identification of new species and
biogroups of Enterobacteriaceae isolated from clinical specimens. J. Clin. Microbiol.,
21: 46 - 76.

136
ENTEROKIT C Fonte: PROBAC DO BRASIL
Indicações: O ENTEROKIT C consiste nos testes de descarboxilação da lisina, arginina e
ornitina (base Moeller), fermentação de adonitol e arabinose e liquefação da
gelatina. Sua finalidade principal é complementar o Enterokit B na identificação das
espécies mais importantes de Klebsiella, Enterobacter, Hafnia e Serratia. Assim
todas as vezes que o Enterokit B não identificar o microrganismo com precisão,
recorre-se ao Enterokit C.
A soma dos testes do Enterokit B e C permite diagnosticar com segurança as
espécies mais importantes de Klebsiella, Enterobacter, Hafnia e Serratia. O Enterokit
C também é extremamente útil na identificação de amostras atípicas pertencentes
aos outros gêneros de enterobactéria.
Composição/Conjunto: Meios Descarboxilase de Moeller: Controle, Lisina, Arginina e
Ornitina...................................................................................................4 mL/tubo
Meio de Adonitol....................................................................................4 mL/tubo
Meio de Arabinose................................................................................. 4 mL/tubo
Meio de Gelatina.................................................................................... 1 fita/tubo
Procedimento: 1-Inocular os meios de Moeller (lisina, arginina, ornitina e controle) da maneira usual,
e os meios de adonitol e arabinose por picada central, introduzindo a agulha até o
fundo do tubo
2-Pingar 10 gotas de salina estéril, no tubo identificado como Gelatina, com o auxílio
do conta gotas, que acompanha o produto.
3-Com o auxílio de uma alça, turvar a salina (inóculo pesado).
4-Incubar a 35º ± 2°C durante 24 a 48 hs.

137
Leitura e interpretação: Descarboxilação de lisina, arginina e ornitina: quando positiva, o meio adquire
cor púrpura discreta ou intensa; se negativa, a cor do meio permanece inalterada.
Fermentação de adonitol e arabinose: quando positiva, o meio se torna rosa
discreto ou intenso; se negativa, a cor do meio permanece inalterada.
Liquefação da gelatina: Gelatinase +: utilização da gelatina impregnada na fita, com alteração da cor cinza-
esverdeada opaca para azul transparente.
Gelatinase -: a cor da fita de gelatina permanece inalterada.
Na tabela, são apresentadas as características das principais espécies de
enterobactérias.
Apresentação: Caixa com 6 conjuntos.
Conservação: Manter em temperatura ambiente (entre 10° e 25°C)
Validade: 6 meses.
Tabela 7- Diferenciação das enterobactérias pelo Enterokit C
Bactérias % de Amostras positivas
Enterob. Lisina Argin. Ornitina Gelat. Adon. Arabinose
E. coli 90 17 65 0 5 99
Shigella sp. 0 5 1 0 0 60
S. sonnei 0 2 98 0 0 95
E. tarda 100 0 100 0 0 9
Salmonella sp 98 70 97 0 0 99
S. typhi 98 3 0 0 0 2
S. paratyphi A 0 15 95 0 0 100
C. freundii 0 65 20 0 0 100
C. diversus 0 65 99 0 98 100
K. pneumoniae 98 0 0 0 90 99
K. oxytoca 99 0 0 0 99 98
E. aerogenes 98 0 8 0 98 100
E. cloacae 0 97 96 0 25 100
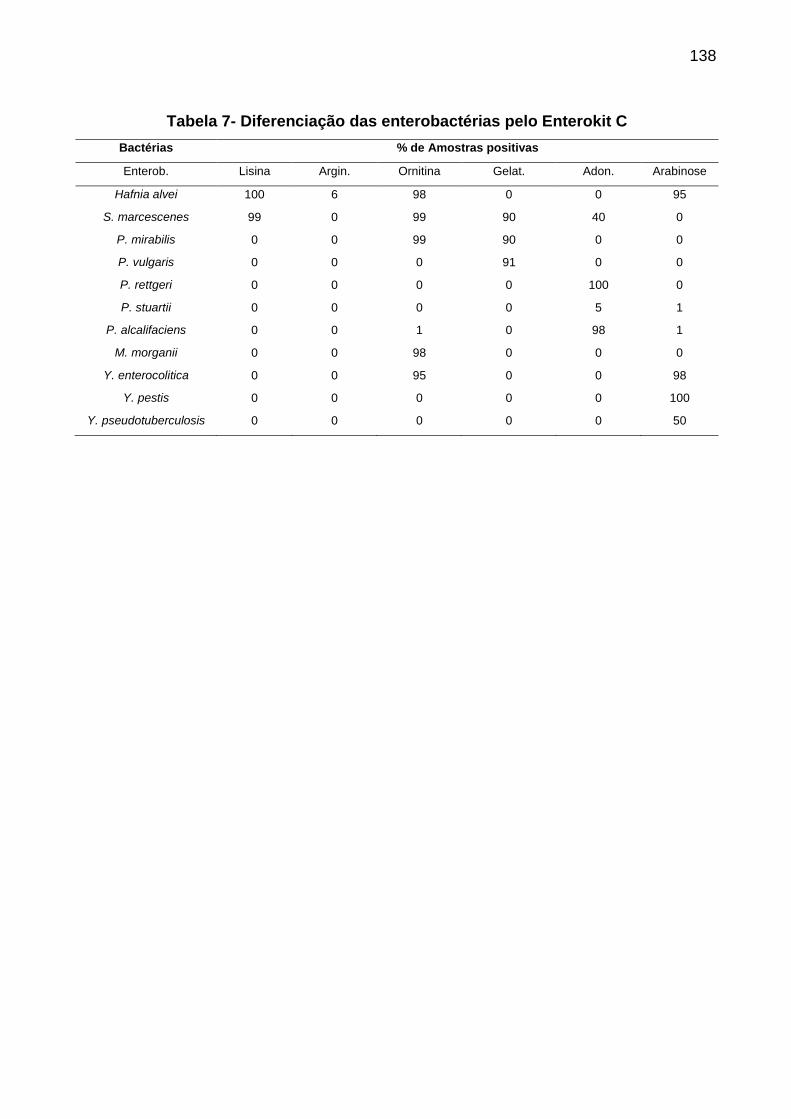
138
Tabela 7- Diferenciação das enterobactérias pelo Enterokit C Bactérias % de Amostras positivas
Enterob. Lisina Argin. Ornitina Gelat. Adon. Arabinose
Hafnia alvei 100 6 98 0 0 95
S. marcescenes 99 0 99 90 40 0
P. mirabilis 0 0 99 90 0 0
P. vulgaris 0 0 0 91 0 0
P. rettgeri 0 0 0 0 100 0
P. stuartii 0 0 0 0 5 1
P. alcalifaciens 0 0 1 0 98 1
M. morganii 0 0 98 0 0 0
Y. enterocolitica 0 0 95 0 0 98
Y. pestis 0 0 0 0 0 100
Y. pseudotuberculosis 0 0 0 0 0 50

139
EPM-MILi
Fonte: PROBAC DO BRASIL
Indicações: Identificação bioquímica das bactérias da Família Enterobacteriaceae.
Composição: Meio EPM: Este meio é uma modificação do meio de Rugai e Araújo e contém os
seguintes testes: produção de gás por fermentação de glicose, produção de H2S,
hidrólise de uréia e desaminação do triptofano.
Meio MILi: Este meio é utilizado para avaliar a motilidade, produção de indol e
descarboxilação da lisina.
Os sete testes, quando considerados com os resultados da reação de fermentação
da lactose observada nas placas de isolamento (lactose positiva ou negativa),
permitem identificar presuntivamente as seguintes enterobactérias: Shigella,
Salmonella, E. coli e Y.enterocolitica.
Procedimento:
1. Semeadura
Tocar com agulha de platina uma colônia bem isolada e inocular os 2 meios com o
mesmo inóculo. Para inocular o meio EPM, introduzir a agulha até o fundo do tubo e
ao retirá-la semear a superfície do meio. A inoculação do MILi deve ser feita por
picada central, introduzindo a agulha até o fundo do tubo.
Incubar entre 35°C a 37°C com as tampas semi-rosqueadas e fazer a leitura após 18
a 24 horas.
2. Leitura e interpretação
Meio EPM Produção de gás: Aparecimento de bolhas ou deslocamento do meio do fundo do
tubo.
Produção de H2S: Enegrecimento do meio em qualquer intensidade.

140
Hidrólise da uréia: Aparecimento de cor azul ou verde-azulada (reação fraca) que
se estende para a base do meio, envolvendo-a totalmente ou não.
Desaminação do triptofano: Aparecimento de cor verde-garrafa na superfície do
meio. Quando a reação é negativa a superfície do meio adquire cor azul ou
raramente amarela.
Meio MILi Motilidade: A bactéria móvel cresce além da picada turvando parcial ou totalmente
o meio. A bactéria imóvel cresce somente na picada.
Descarboxilação da lisina: Quando a lisina é descarboxilada, o meio adquire cor
púrpura. Quando o aminoácido não é utilizado, o meio adquire cor amarela nítida
nos seus 2/3 inferiores.
Produção do indol: Após leitura dos testes de motilidade e lisina, adicionar 3 a 4
gotas do Reativo de Kovacs a superfície do meio e agitar levemente. Quando a
bactéria produz indol, o reativo adquire cor rosa ou vermelha. Quando não produz, o
reativo mantém sua cor inalterada.
A tabela mostra as características dos enteropatógenos nos meios EPM e MILi, bem
como os resultados da reação de fermentação da lactose observados nos meios de
isolamento (lactose positiva ou negativa). A identificação complementar destas
bactérias pode ser feita por meio de testes de aglutinação em lâmina com anti-soros
correspondentes a cada um deles (ver linha de soros Probac do Brasil).
Precauções:
Após o uso o produto deverá ser descartado seguindo-se as normas vigentes de
resíduos de serviços de saúde. Não utilizar o produto em caso de turvação do meio
líquido ou na presença de outros sinais de contaminação.
Conservação:
Manter em temperatura ambiente entre 10° a 25°C (local fresco).

141
Validade: 6 meses.
Tabela 8- Características dos enteropatógenos da família Enterobacteriaceae nos meios EPM-MILi.
Lactose
(placa de
isolamento)
Meio EPM Meio MILi
Aspecto Geral Gás H2S U LTD Mot I Lis
Shigella
dysenteriae
flexneri
boydii
- B/Am e S/Az
Au/b(a) e Au/e - a - - - - +/- -
Shigella
sonnei -
B/Am e S/Az
Au/b e Au/e - - - - - - -
E.coli
invasora +/-
B/Am e S/Az
AP/b e Au/e +/- - - - - + -
E.coli
outras +
B/Am e S/Az
G/b e Au/e +/- - - - - + -
Salmonella -b B/Am e S/Az
P/b e P/e + + - - + - +
Yersinia
enterocoliti
ca
- B/Am e S/Az
Au/b e Au/e - - + - - -/+ -
U: urease, LTD: L-triptofano desaminase, Mot: motilidade, I: indol, Lis: lisina, B/Am: Base Amarela, Au/b: Ausência de bolhas de gás, Au/e: Ausência de enegrecimento, B/Az: Base Azul, P/b: Presença de bolhas de
gás, P/e: Presença de enegrecimento, S/Az: Superfície Azul, G/b: Geralmente com bolhas de gás e AP/b:
Ausência ou presença de bolhas de gás
a) Alguns sorotipos de Shigella flexneri e Shigella boydii produzem gás.
b) A Salmonella Typhimurium lactose + é raramente isolada atualmente.
Referências bibliográficas: 1. Almeida, P. C. A. & Trabulsi, L. R. - Características culturais, bioquímicas,
sorológicas e virulência de amostras de Salmonella typhimurium fermentadoras de
lactose. Rev. Microbiol., 5:27 - 35, 1974.
2. Edwards, P. R. & Ewing, W. H. - Identification of Enterobacteriaceae Burgess
Publishing Company, Minneapolis, Minnesota, 1972.

142
3. Toledo, M. R. F.; Fontes, C. F. & Trabulsi, L. R., 1982 - MILi - Um meio para a
realização dos testes de motilidade, indol e lisina descarboxilase. Rev. Microbiol., 13:
230 - 235.
4. Toledo, M. R. F.; Fontes, C. F. & Trabulsi, L. R., 1982 - EPM - Uma modificação
do meio de Rugai e Araújo, para a realização simultânea dos testes de produção de
gás a partir de glicose, H2S, urease e triptofano desaminase. Rev. Microbiol., 13: 09
- 315.

143
KIT NF Fonte: PROBAC DO BRASIL
Indicações: O KIT NF da PROBAC DO BRASIL é constituído pelos testes de oxidase, utilização
de glicose em meio base OF, descarboxilação de lisina e arginina (base Moeller),
liquefação da gelatina, hidrólise da uréia, DNAse, e sensibilidade a polimixina. Os
resultados destes testes permitem identificar a maioria dos bacilos Gram negativos
não fermentadores da glicose isolados na rotina laboratorial.
Características dos componentes: Meios OF glicose com óleo e sem óleo............................................ 4 ml/tubo.
Meios Descarboxilase de Moeller: controle, lisina e arginina .............2 ml/tubo.
Meio de Gelatina ................................................................................ 1fita/tubo.
Meio DNAse..........................................................................................6 ml/placa.
Uréia......................................................................................................3 ml/tubo
Polimixina..............................................................................................6 discos/frasco
Fitas para determinação de oxidase.....................................................6 fitas/frasco
Procedimento: Os testes devem ser realizados a partir de colônias isoladas e puras com 24 horas de crescimento. 1-Oxidase: com auxílio de alça de platina, bastão de vidro ou madeira, faça um
esfregaço da colônia na fita de oxidase.
-Não utilize qualquer tipo de alça ou bastão que contenha vestígios de ferro, pois
este poderá provocar uma reação falso-positiva.
-Não utilize colônias de meios com indicadores, pois poderão apresentar resultados
falso-positivos.
-Meios de cultura: inocular densamente todos os meios do KIT NF, a partir do
crescimento bacteriano no meio EPM ou agar inclinado:

144
2-OF glicose com óleo e sem óleo: inocular por picada central, introduzindo a
agulha até o fundo.
-Controle, Lisina e Arginina: inocular com alça ou agulha, depositando o inóculo na
parede do tubo, abaixo do óleo mineral.
-Gelatina: - pingar 10 gotas de salina estéril, no tubo identificado como GEL, com o
auxílio do conta gotas, que acompanha o produto. Com o auxílio de uma alça turvar
a salina (inóculo pesado).
-Uréia: semear com inóculo denso na superfície do meio.
-DNAse: semear 2-3 colônias em spot ( círculo de 0,5 cm de diaâmetro) deixando
bastante espaço entre os teste para avaliar a presença de halo.
-Polimixina: fazer antibiograma em M.Hinton e colocar o disco.
-Incubar os meios de cultura a 37ºC durante 24-48 h; quando houver dúvidas quanto
a interpretação dos testes, reincubar por mais 24 a 48 h e fazer nova leitura.
Interpretação do resultado: A) Teste de Oxidase: a leitura é feita em poucos segundos:
Oxidase + : o esfregaço bacteriano na fita apresenta coloração rosa intenso, que
após alguns segundos pode mudar para preta.
Oxidase -: o esfregaço bacteriano não apresenta alteração de cor.
A cor rosa intensa aparece rapidamente (15-20 segundos). Após este intervalo,
cores: rosa, violeta pálido são falsos positivos. A B. cepacia pode dar reação fraca.
B) Tabela 9- Glicose O/F: OF glicose
Tubos c/ óleo s/ óleo
Oxidação do açúcar (Oxidativo) Inalterado amarelo só na superfície
Não utilização do açúcar (Inerte ou alcalino) 1 Inalterado inalterado ou azulado
Fermentação da glicose (Fermentador ou Ácido)2 amarelo amarelo
1. Aguardar no mínimo 72 h para definir como inerte, pois pode ocorrer a oxidação
tardia ou lenta. 2. Quando a bactéria fermenta a glicose, não pode ser identificada pelo KIT NF II.
Deve ser repicada em ENTEROKIT B para diagnóstico.

145
C) Descarboxilação de aminoácidos: Lisina e Arginina:
-Cor púrpura do meio mais acentuado que a do tubo controle: utilização do
aminoácido.
-Cor do meio igual ou mais amarelada que a do tubo controle: não utilização do
aminoácido.
Se negativo aguardar até 5 dias. Apenas uma prova deve ser positiva, exceto a
B.cepacia que pode ser ornitina + e lisina +.
D) Gelatina: Gelatinase (+): Utilização da gelatina impregnada na fita, com alteração da cor de
cinza metálica para azul transparente.
Gelatinase (-): A cor da fita permanece inalterada.
Obs: antes de fazer a leitura, agitar suavemente o tubo.
E) Uréia: a cor rosa forte aparece em parte do ápice ou em todo o meio após 24-72
h de incubação: ligeira mudança de cor rósea no ápice que não progride com maior
incubação é considerada negativa. A Bordetella bronchiseptica pode dar reação
positiva em 4 h.
F) DNAse: adicionar ácido clorídrico 1N e aguardar 5 minutos. Observar a presença
de halo transparente em volta do inoculo e o restante do meio fica leitoso. Se
negativo repetir o teste com leitura em 48 h. Sempre usar uma cepa controle positivo
e negativo.
G) Polimixina: qualquer halo em torno do disco significa sensibilidade.
Provas adicionais não incluídas: -Crescimento em Mac Conkey: Semear com alça uma colônia isolada o crescimento
deverá ocorrer entre 1 e 3 dias.
-Motilidade: Se a motilidade em tubo for negativa, recomenda-se a confirmação do
resultado com o teste em lâmina.
-Teste de motilidade em lâmina: Inocular 2 colônias em um caldo TSB e incubá-lo pó
18-24 h á 37ºC.
Agitar o tubo e com uma pipeta ou alça estéril retirar uma gota e depositar sobre
uma lâmina. Sobre a gota depositar uma lamínula e observar com aumento de 400

146
vezes. A presença de muitas ou poucas bactérias cruzando o campo é significativa
de motilidade positiva. Movimentos vibratórios fracos são considerados negativos. A
motilidade deve
ser observada em 37º e 20ºC.
-Coloração de Gram: Realizar a coloração de Gram para a observação da
morfologia bacteriana
-Crescimento a 42º C: semear 2 colônias no meio. Incubar a 42º C. A prova será
positiva quando ocorrer com a turbidez do meio ou for nítido o aumento da
densidade bacteriana. O ideal é comparar com um controle mantido a temperatura
ambiente.
A interpretação dos resultados obtidos no KIT NF deve ser realizada de acordo com
o Sistema Numérico que acompanha o produto. Quando necessário, outros testes
podem ser realizados para confirmação do diagnóstico.
Apresentação: Caixa com 6 conjuntos.
Validade: 6 meses.
Conservação: Manter os meios de cultura em temperatura ambiente 10º- 25ºC
(local fresco). Manter as Fitas
de Oxidase, os discos de polimixina e as placas de DNAse em geladeira (2 a 8º C).
Precauções: Após o uso, o produto deverá ser descartado conforme as
recomendações vigentes para resíduos de serviços de saúde.

147
ESQUEMA NÚMERICO PARA IDENTIFICAÇÃO EM BACTÉRIAS NÃO FERMENTADORAS
Etapa 1 Fazer as seguintes provas:
• Oxidase
• Motilidade
• O/F Glicose / Controle
• Crescimento em Mac Conkey
• Gram
• Gelatina
• DNAse
• Uréia
• Lisina
• Arginina
• Crescimento a 42ºC
• Teste de sensibilidade a polimixina
Tabela 10- Valores das provas de Oxidase e Motilidade VALORES
PROVAS Provas positivas Provas negativas
Oxidase 4 0
Motilidade 2 0
Somar valores obtidos para as provas de oxidase e motilidade= A
Tabela 11- Valores das provas de OF, MC e Morfologia VALORES
PROVAS Provas positivas Provas negativas
OF Oxidativo=4 Inerte ou alcalino=0
Mac Conkey Crescimento=2 Não crescimento=0
Morfologia (Gram) Bacilo=1 Coco/cocobacilo=0
Somar valores obtidos para as provas de OF glicose + Crescimento em Mac Conkey + Gram= B

148
Tabela 12- Valores dos testes bioquímicos
Leitura AB C D E Leitura ABCDE
42º C
4 Gel 2
DNA 1
URE 4
LIS 2
ARG 1
PMX 1
00 * *
01 * *
02 * *
03 * *
06 * *
07 * *
22 * * *
26 * * * * * *
40 * * *
41 * * * *
42 * * *
43 * * *
45 * * *
47 * * *
63 * * * * *
65 * *
67 * *
*Campos preenchidos não considerar a leitura da prova
Etapa 3 Provas complementares Estas provas poderão ser indicadas para caracterizar alguns não fermentadores ou
diferenciar outros cujas provas empregadas apresentaram padrão semelhante.
1. Citrato
2. Crescimento em NaCl 6,5%
3. Esculina
4. Indol
5. PYR
Etapa 4 Localizar o número com 5 algarismos obtido e a correspondente identificação na
coluna da esquerda.

149
A tabela de freqüência mostra para alguns microrganismos a freqüência que o
conjunto de provas pode ocorrer.
Frequências baixas (<10% ) sugerem necessidade de revisão das provas (p. ex.
aguardar mais tempo para interpretar provas negativas ou repetição de todas elas).
Números não encontrados sugerem bactéria com padrão não comum, falha de
interpretação de uma ou mais provas ou bactéria rara.

150
Tabela 13- Identificação de isolados Não fermentadores A B C D E Freq.
% Provas complementares
Acinetobacter iwofii 0 0 0 0 0 80 Citrato negativo
Acinetobacter iwofii 0 0 0 0 0 10 Citrato negativo
A.haemolyticus 0 2 2 1 0 45 Citrato negativo
A.haemolyticus 0 2 2 1 0 ˂10 Citrato positivo
A.calcoaceticus 0 3 0 1 0 90 Citrato pos. hemólise em
Agar sangue pos. A.baumannii 0 6 4 1 0 90 Citrato positivo
A.baumannii 0 6 4 5 0 ˂10 Citrato positivo
A.calcoaceticus 0 6 0 1 0 ˂10 Citrato positivo
A.baumannii 0 7 4 1 0 ˂10 Citrato positivo
A.baumannii 0 7 4 5 4 ˂10 Citrato positivo
A.haemolyticus 0 6 2 1 0 45 Citrato positivo
A.haemolyticus 0 7 2 1 0 ˂10 Citrato pos. hemólise em
Agar sangue pos. Acinetobacter spp 0 0/1/2/3/
4/5/6/7
2/0 2/0 0 ˂10 Citrato pos. hemólise em
Agar sangue pos
S.maltophilia 2 2 1 2 0 15
S.maltophilia 2 6 1 2 0 56
P.oryzihabitans 2 6 0 0 0 86
B. cepacia 2 6 0 2 1 80
P. luteola 2 6 0 1 0 99 Esculina pos. e PYR pos
P.oryzihabitans 2 6 0 0 0 14 Esculina neg. e PYR pos
B.cepacia 2 2 0 2 1 80 PYR negativo
S.maltophilia 2 6 0 1 0 29 PYR negativo
Moraxella spp 4 0 0 0 0 50
Moraxella catarrhalis 4 0 1 0 0 99
Moraxella lacunata 4 0 2 0 0 99
C.indologenes 4 1 2 0 1 ˂10

151
Tabela 13- Identificação de isolados Não fermentadores A B C D E Freq.
% Provas complementares
C.meningosepticum 4 1 3 0 1 ˂10
Moraxella spp 4 2 0 0 0 50
M/Ureolytica 4 2 0 4 0 99 M.canis 4 2 1 0 0 99
C.Indologenes 4 3 2 0 1 ˂10 indol pos C.Meningosepticum 4 3 3 0 1 ˂10 indol pos
S.paucimobilis 4 5 0 0 0 indol neg
Sphingobacterium 4 5 0 4 1 indol neg
Sphingobacterium 4 5 1 4 1 indol neg
C.Indologenes 4 5 2 0 1 90 indol pos
C.Meningosepticum 4 5 3 0 1 45 indol pos
Sphingobacterium 4 7 0 4 1 indol neg Sphingobacterium 4 7 1 4 1 indol neg
C.Indologenes 4 7 2 4 1 ˂10 indol pos
C.Meningosepticum 4 7 3 4 1 45 indol pos
A.Denitrificans 6 3 0 0 0 NaCl 6,5% neg. e PYR pos
A.faecalis 6 3 0 0 2 NaCl 6,5% pos.e PYR neg
B. bronchiseptica 6 3 0 4 0 Uréia pos. ++ e PYR neg.
A. Denitrificans 6 3 0 4 0 Uréia Fraca e PYR pos
S. Paucimobilis 6 5 0 0 0
S. Paucimobilis 6 5 4 0 0
S. Paucimobilis 6 5 1 0 0
S. Paucimobilis 6 5 5 0 0
A. xilosoxidans 6 7 0 0 0 07 Esculinaneg., NaCl6,5%neg.
S.Paucimobilis 6 7 0 0 0 Esculina positiva
P.Stutzeri 6 7 0 0 0
B.cepacia 6 7 0 0 1 03
A. xilosoxidans 6 7 4 0 0 43 Esculina neg.,NaCl 6,5%
S.paucimobilis 6 7 4 0 0 Esculina positiva
P.stutzeri 6 7 4 0 0
B.cepacia 6 7 4 0 1 13 Esculinaneg., NaCl6,5%pos.
P. putida 6 7 0 1 0
P.aeruginosa 6 7 4 1 0 18 NaCl 6,5%=Variável P.mendocina 6 7 4 1 0 90 NaCl 6,5%=positiva
B.pseudomallei 6 7 4 1 1 21
A.Xilosoxidans 6 7 0 2 0 07

152
Tabela 13- Identificação de isolados Não fermentadores A B C D E Freq.
% Provas complementares
B. cepacia 6 7 0 2 1 13
A.Xilosoxidans 6 7 4 2 0 43
B.cepacia 6 7 4 2 1 51
S.paucimobilis 6 7 1 0 0 Esculina positiva
S.putrefaciens 6 7 1 0 0 H2S pos.,no TSI e Esculina
S.Paucimobilis 6 7 5 0 0
B.cepacia 6 7 2 0 1 01
B.cepacia 6 7 6 0 10 3
P.fluorescens 6 7 2 1 0
P.aeruginosa 6 7 6 1 0 82
B. pseudomallei 6 7 6 1 1 79
B. cepacia 6 7 2 2 1 03
B. cepacia 6 7 6 2 1 13
S. putrefaciens 6 7 3 0 0 H2S positivo no TSI
S. putrefaciens 6 7 5 0 0 H2S positivo no TSI
S. putrefaciens 6 7 7 0 0 H2S positivo no TSI
Tabela 14- Valores das provas bioquímicas Colônias com Pigmento rosa
Microrganismo A B C D E Microrganismo
Methylobacterium spp 6/4/2/0 5/1 0 4 0
Coral e seca
Roseomonas spp 6/4/2/0 7/6/3/2 0 4 4
Rosada e mucóide

153
Penicillin-binding protein (PBP2’) Teste de Aglutinação em Látex
Fonte: Kit PBP 2’- Oxoid Brasil
Introdução Este teste é um ensaio de aglutinação rápida em látex, para a detecção de PBP2’
(também chamado de PBP2a), em Staphylococcus spp. Isolados, como uma ajuda a
identificação de Staphylococcus aureus meticilina resistente (MRSA) e
Staphylococcus coagulase negativa meticilina resistente.
Princípio do teste
Os Staphylococcus spp. são a principal causa de infecções nosocomiais e de
infecções comunitárias adquiridas no mundo. Em muitas instituições
aproximadamente 25% a 50% das cepas de S. aureus e 75% das cepas de
Staphylococcus coagulase (CoNS) são resistentes a meticilina. Os MRSA podem ser
vistos como um problema particular pela facilidade com que certas cepas
epidêmicas se difundem e colonizam pacientes debilitados. O tratamento de cepas
sensíveis com penicilinas resistentes a penicilinase (PRP) é recomendável, uma vez
que as drogas ß-lactâmicas são facilmente absorvidas pelos líquidos corpóreos e
tecidos, causam menores complicações durante o tratamento e não selecionam os
organismos vancomicina-resistentes. A identificação segura de meticilina resistente
é, portanto importante.
Cepas de S. aureus com reduzida sensibilidade para PRP estão divididas nas
seguintes categorias:
(I) S. aureus meticilina resistente (MRSA), que possui uma baixa afinidade a
proteína de ligação à penicilina PBP2’, codificada pelo gen mecA;
(II) S. aureus “bordeline” meticilina resistente (BORSA), geralmente assim
considerado por possuir ums hiperprodução de beta lactamase – tipo A; e
(III) Cepas com PBPs modificado devido à capacidade de alteração da
penicilina “binding” ou a hiperprodução de PBPs (MODSA). MODSA são
raramente isolados e sua resposta clínica à terapia aos beta-lactâmicos
não tem sido bem estudada. Desse modo, para propostas clínicas, com

154
raras exceções, a presença de PBP2’ é responsável pela meticilina
resistência nos tratamentos de infecções com S. aureus e CoNS.
O fenótipo de meticilina resistente pode ser altamente heterogêneo,
dificultando a detecção pelos métodos convencionais dos testes de sensibilidade a
antimicrobianos, como a concentração mínima inibitória (MIC) ou a triagem em disco
e Agar. A precisão deste método é afetada pelo tamanho do inoculo, tempo e
temperatura de incubação, meio, pH, concentração de sal e outros fatores. Em
adição, estes métodos de cultura requerem 24 horas de incubação para resultados
precisos. CoNS com freqüência produzem os mais baixos níveis de PBP2’ e
requerem indução por exposição de um PRPS para produzir suficiente produto a ser
detectado. A detecção do gen mecA, tem sido considerado o padrão (“gold
standard”) para a determinação de meticilina resistente devido a sua precisão, mas o
método é extremamente trabalhoso e de alto custo.
O teste de Látex Oxoid PBP2’ possui a vantagem de detectar diretamente a
proteína do PBP2’ num tempo rápido e com o mínimo de trabalho. O teste tem
potencial para ser até mais preciso que a detecção pelo gen mecA, uma vez que
resultados falso positivos não ocorrem com cepas que apresentam mecA, mas são
incapazes de produzir o produto de proteína do gene. Em adição, o teste não
detecta cepas que são hiperprodutoras de beta lactamase ou PBPs.
O teste Oxoid PBP2’ foi previamente avaliado mundialmente, demonstrando
possuir uma alta sensibilidade e especificidade. Partículas de látex sensibilizadas
com anticorpo monoclonal contra PBP2’ reagem especificamente com os
Staphylococcus meticilina resistentes causando aglutinação visível a olho nu.
Componentes do kit - DR901 Látex Teste – sensibilizado com anticorpo monoclonal contra PBP2’.
- DR902 Látex Controle – sensibilizado com anticorpo monoclonal da mesma
subclasse de IgG mostrando não reatividade com as proteínas de S. aureus.
- DR903 Reagente de Extração 1
- DR904 Regente de Extração 2
Cartões de Teste
Bastões homogenizadores
Folheto de instrução

155
Materiais necessários, mas não fornecidos - Micropipetas e ponteiras (50µl)
- Alças microbiológicas (5µl/1µl)
- Banho-maria ou banho aquecido
- Centrífuga (1500 x g)
- Tubos de microcentrífuga (tampa de segurança)
- Desinfetante apropriado para laboratório
Precauções
Este produto se destina exclusivamente ao uso diagnóstico in vitro. O tempo
de aquecimento deve ser de três minutos. Aquecimento por mais de cinco minutos
pode causar diminuição da sensibilidade. Aquecimento somente por um minuto ou
menos, pode causar uma aglutinação não específica.
Ao remover o sobrenadante a ser utilizado pelo teste após a centrifugação,
retirara a pipeta cuidadosamente para evitar que o material sólido do fundo do tubo
não seja aspirado. A utilização de material sólido pode causar aglutinação não
específica. Misturar bem os reagentes (Látex) antes se sua utilização, para a
obtenção de uma suspensão homogênea.
Os reagentes contém como preservante azida sódica a 0,095%. A azida sódica é
tóxica e pode reagir com encanamentos de chumbo ou cobre produzindo metais de
azida que são explosivos por contato. Para prevenir o acúmulo de azida nos
encanamentos, lavar abundantemente com água imediatamente após despejar os
reagentes no esgoto.
Como as amostras podem conter organismos patogênicos, manipular utilizando
precauções apropriadas. Os procedimentos de extração podem não matar as
bactérias; portanto a extração deve ser realizada segundo as mesmas precauções.
Os reagentes de extração 1 e 2 contém uma leve solução irritante e um fraco
ácido. Evitar contato direto através do uso de equipamentos de proteção
convenientes. Caso o material entrar em contato com a pele, membranas mucosas
ou olhos, lavar imediatamente o local com água em abundância.
Armazenagem
Armazenar o kit entre 2° e 8°C. Nestas condições os reagentes manterão sua

156
atividade até a data de validade mostrada na caixa.
Procedimento para controles Para cada novo lote do kit e semanalmente, os seguintes procedimentos para
controle deverão ser realizados.
1. Controle Positivo – Utilizar uma cepa MRSA conhecida como ATCC®
43300 (Culti-Loops Oxoid® C9022L). Seguir o método informado no
procedimento do teste. Assegurar que a aglutinação ocorra dentro de 3
minutos.
2. Controle Negativo – Utilizar uma cepa conhecida de Staphylococcus
aureus Meticilina Sensível (MRSA) como a ATCC® 25933 ou ATCC® 29213
(Culti-Loops Oxoid® C7010L ou C7011L). Seguir o método informado no
procedimento do teste. Assegurar que a aglutinação não ocorra dentro de 3
minutos. Não utilizar o teste se as reações observadas com os organismos
controles estiverem incorretas. 3. Não utilizar o kit após a data de validade.
Importante nota de procedimento Não encostar o conta gotas no cartão de teste onde se encontra a amostra para
evitar a contaminação dos reagentes. Assegurar que as tampas dos reagentes
estejam fechadas após a sua utilização evitando assim uma contaminação ou
ressecamento dos mesmos. Após o uso, retornar o kit ao refrigerador assegurando
que os frascos estejam na posição correta.
Preparação da cultura As colônias poderão ser testadas a partir dos seguintes meios de cultura:
Tryptone Soya Agar (TSA) com 55 de sangue de carneiro, Muller-Hinton Agar. O uso
de culturas recentes (18 A 24 hs) é recomendado. Entretanto, para garantir
crescimento suficiente, culturas com crescimento entre 24 e 48 horas poderão ser
testadas se necessário. O desempenho verificado nos dados constantes nesta bula
foi gerado utilizando estes meios como parte do processo submetido ao FDA (North
American Food and Drugs Administration). Os dados demonstram que o teste
também é efetivo com colônias de S. aureus provenientes de Tryptone Soya Agar e

157
Columbia Agar com 5% de sangue de cavalo, Iso-Sensitest Agar e DSTA. Estes
meios são comumente utilizados na Europa, mas não nos EUA, e, portanto, não
foram utilizados nos testes que geram os dados mostrados na seção
“Características de Desempenho”.
Requisito especial O teste PBP2’deve ser realizado somente com as espécies de Staphylococcus spp.
(cocos Gram positivos). O teste de coagulase ou equivalente deve ser realizado para
determinar se as cepas isoladas são Staphylococcus aureus ou outras espécies de
estafilococos.
Preparação para inoculo Para os Staphylococcus aureus, o teste pode ser realizado a partir de colônias em
placa de isolamento primário, se o crescimento for suficiente, ou a partir de
subculturas do isolado. Outras espécies de bactérias não devem estar presentes na
placa para não interferir no teste. Para Staphylococcus coagulase negativa, uma
indução é necessária para a produção de PBP2’ suficiente.
1. Preparar uma suspensão em caldo do microrganismo no padrão da escala
de McFarland 1 e semear em uma placa de TSA com sangue, Columbia
Sangue Agar ou Mueller-Hinton Agar. Alternativamente inocular a placa de
ágar com várias colônias e semear em quatro quadrantes.
2. Colocar um disco de oxacilina (1µg/ml) sobre o microrganismo semeado ou
no principal quadrante inoculado.
3. Incubar a 37°C por não menos de 24hs e não mais de 48hs.
Cuidado: Para evitar resultados falsos negativos, não realizar o teste se não
houver inóculo suficiente disponível.

158
Método do teste Procedimento de Extração do PBP2’
1. Adicionar 4 gotas do Reagente 1 em um tudo de microcentrífuga.
2. Aproximadamente 1.5 x 109 (3-5 µl) células devem ser testadas. Isto pode ser
conseguido usando uma alça estéril de 5µl para remover suficiente
quantidade de células e completar o diâmetro interno da alça.
Alternativamente, uma alça estéril de 1 µl pode ser utilizada para remover
várias colônias num tamanho equivalente de 1 µl ou 1mm de altura cobrindo
todo o diâmetro caso grumos estejam presentes. Uma suspensão muito turva
deve ser observada.
3. Colocar o tubo em um Banho-Maria ou em um bloco aquecido (SUPERIOR A
95°C) e aquecer Por 3 minutos.
4. Retirar o tubo de microcentrífuga e deixar atingir a temperatura ambiente.
5. Adicionar 1 gota do Reagente 2 no tubo e misturar.
6. Centrifugar a 1500 x g por 5 minutos, ou seja, 3.000 rpm para um raio de
rotação de 15 cm ou 4500rpm para um raio de rotação de 4,5cm. Utilizar o
sobrenadante para o teste.
Procedimento de aglutinação em Látex
1. Para cada sobrenadante a ser testado, utilizar um círculo do cartão de teste
para testar com o Látex Reagente e outro para testar com Látex Controle.
2. Homogenizar os reagentes de Látex por inversão por várias vezes, e
adicionar uma gota de Látex Teste ou Látex Controle em cada um dos
círculos.
3. Colocar 50µl do sobrenadante no círculo teste e no círculo controle, com
cuidado para evitar a formação de bolhas. Misturar o látex e o
sobrenadante em cada círculo utilizando os bastões homogenizadores.
4. Imprimir ao cartão teste movimentos rotatórios por três minutos e observar
a aglutinação sob condições normais de luminosidade. Anotar os
resultados observados no teste e controle.

159
5. Descartar o cartão teste com segurança em recipiente contendo solução
desinfetante ou em descarte de materiais infecciosos.
Tabela 15 - Leitura e interpretação dos resultados da PBP 2a
Aglutinação verificada com Látex teste, mas não com o Látex controle em 3 minutos PBP2’Positivo (MRSA)
Nenhuma aglutinação verificada com o Látex Teste em 3 minutos PBP2’Negativo (MRSA)
Aglutinação verificada com o Látex Controle em 3 minutos Indeterminado
Aglutinação verificada com Látex teste, mas não com o Látex controle em 3 minutos PBP2’Positivo (MRSA)
Tabela 16- Tipos de reação de aglutinação
Negativa (-) Uma suspensão de partículas homogênea sem grupos visíveis.
Fracamente Positiva (+) Grupos pequenos, mas definidos contra o fundo turvo
Fortemente Positiva (+) Grupos grandes e pequenos contra um fundo ligeiramente turvo ou grandes grumos
contra um fundo muito claro
Obs: Ocasionalmente, reações negativas podem apresentar uma aparência granular
fina ou reações podem ser observadas. Nestes casos, a visualização do fundo pode
ser utilizada para interpretar o resultado. Um fundo opaco indica um resultado
negativo e um fundo claro pode ser interpretado como resultado positivo.
Limitações
Resultados indeterminados devem ser retestados com um material extraído
recentemente. Se, sob reteste o resultado se mantiver indeterminado, a resistência a
meticilina deverá ser determinada por outro método.
Resultados falsos negativos podem ocorrer se for utilizado no teste material

160
insuficiente. Neste caso, o teste deverá ser repetido com a quantidade suficiente de
material. Verdadeiros resultados positivos geralmente apresentam fortes reações.
Reações falso positivas ocorrem raramente, e são limitadas geralmente a
fracas reações. Estes resultados podem ser verificados por retestes em culturas
recentes. Staphylococcus aureus hiperprodutores de PBP ou chamados de S.
aureus modificados (MODSA) e cepas S. aureus bordeline resistente (BORSA) não
possuel PBP2’e não se espera reação neste teste.
Cepas de alguns microrganismos podem ter baixo nível de meticilina
resistência ou em raros caos, produzirem PBP2’em baixas quantidades e
apresentarem resultados falsos negativos.
Devido às limitações de sensibilidade e especificidade dos testes de
sensibilidade para Staphylococcus coagulase negativa, determinados pelo NCCLS,
particularmente para outras cepas que não o Staphylococcus epidermidis, os
resultados do ensaio de PBP2’ não devem concordar com os resultados dos testes
de sensibilidade. Cepas com MICs > 4 g/ml de
resistentes, apesar do resultado no teste de PBP2’.
Características de Desempenho
O teste de aglutinação em látex Oxid Penicillin-Binding protein foi avaliado em
4 laboratórios geograficamente diversos com isolados clínicos frescos de S. aureus.
201 isolados foram testados com os métodos do NCCLS e com o teste Oxid PBP2’
para cada três meios. Uma fraca reação falso positiva (negativa na repetição) em
TSA com sangue foi encontrada no látex Oxoid. Todas as reações positivas foram
fortes, exceto três reações fracas (mas positivas) com ágar Mueller-Hinton.
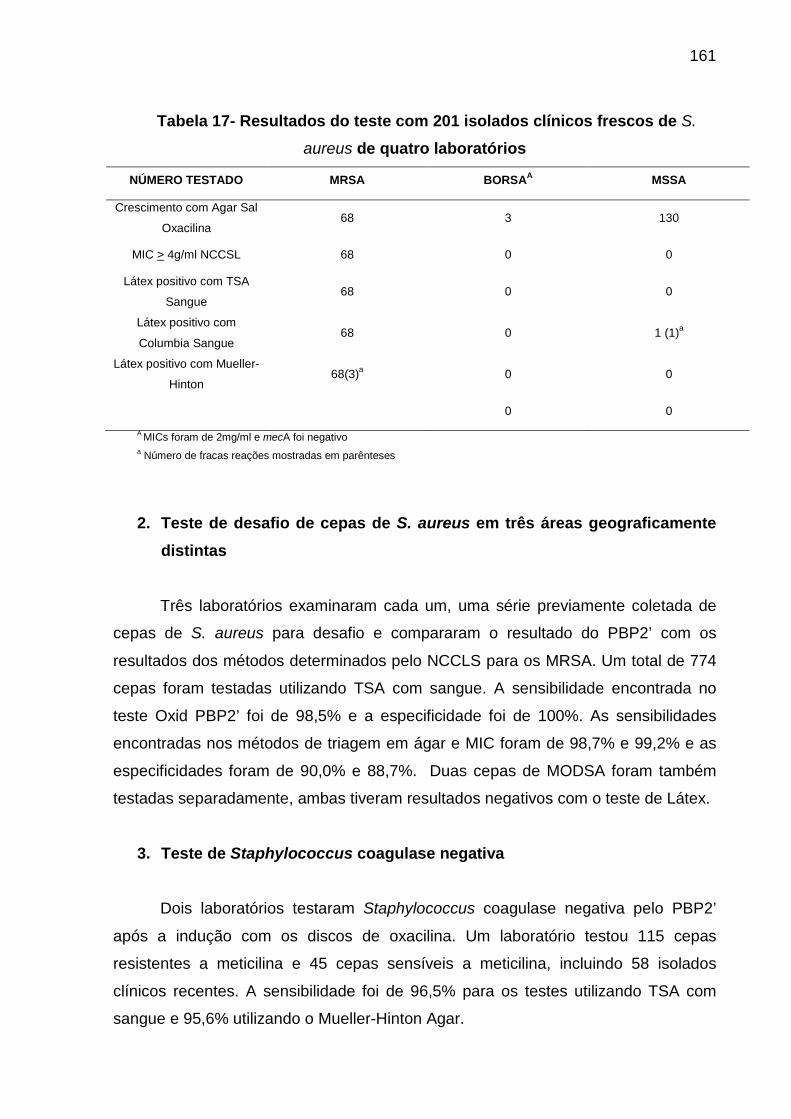
161
Tabela 17- Resultados do teste com 201 isolados clínicos frescos de S.
aureus de quatro laboratórios
NÚMERO TESTADO MRSA BORSAA MSSA
Crescimento com Agar Sal
Oxacilina 68 3 130
MIC > 4g/ml NCCSL 68 0 0
Látex positivo com TSA
Sangue 68 0 0
Látex positivo com
Columbia Sangue 68 0 1 (1)a
Látex positivo com Mueller-
Hinton 68(3)a 0 0
0 0
A MICs foram de 2mg/ml e mecA foi negativo a Número de fracas reações mostradas em parênteses
2. Teste de desafio de cepas de S. aureus em três áreas geograficamente distintas Três laboratórios examinaram cada um, uma série previamente coletada de
cepas de S. aureus para desafio e compararam o resultado do PBP2’ com os
resultados dos métodos determinados pelo NCCLS para os MRSA. Um total de 774
cepas foram testadas utilizando TSA com sangue. A sensibilidade encontrada no
teste Oxid PBP2’ foi de 98,5% e a especificidade foi de 100%. As sensibilidades
encontradas nos métodos de triagem em ágar e MIC foram de 98,7% e 99,2% e as
especificidades foram de 90,0% e 88,7%. Duas cepas de MODSA foram também
testadas separadamente, ambas tiveram resultados negativos com o teste de Látex.
3. Teste de Staphylococcus coagulase negativa Dois laboratórios testaram Staphylococcus coagulase negativa pelo PBP2’
após a indução com os discos de oxacilina. Um laboratório testou 115 cepas
resistentes a meticilina e 45 cepas sensíveis a meticilina, incluindo 58 isolados
clínicos recentes. A sensibilidade foi de 96,5% para os testes utilizando TSA com
sangue e 95,6% utilizando o Mueller-Hinton Agar.

162
A especificidade foi de 100% utilizando TSA com sangue e 98% utilizando o
Mueller-Hinton Agar. A obtenção de um bom inóculo foi mais difícil com Mueller-
Hinton Agar. O segundo laboratório testou 212 cepas de meticilina resistente e 203
cepas de meticilina sensível obtendo uma sensibilidade de 99,5% e uma
especificidade de 99,5% utilizando o inóculo crescido em Columbia Agar.
4. Reprodutibilidade Dez diferentes cepas bem caracterizadas de Staphylococcus aureus (3MRSA,
3MSSA, 3 BORSA e MODSA) foram enviadas a três laboratórios geograficamente
diversos com cada cepa submetida por 5 vezes em códigos cegos. Todos os 150
resultados obtidos concordaram com os resultados esperados com 100% de
reprodutividade. As três cepas mecA positivas foram positivas todas as vezes que
foram testadas (45/45 testes). As três cepas MSSA e as três cepas BORSA
apresentaram resultados negativos toda vez que o teste foi realizado (90/90 testes).
A MODSA obteve um MIC de 16µg/ml para a oxacilina e um resultado negativo no
Oxoid Latex Teste, como esperado, toda vez que os teste foi realizado (15/15
testes).

163
Teste de suscetibilidade pelo método ETEST
Fonte: Etest- PROBAC DO BRASIL
1 - Utilização
O Etest é uma técnica quantitativa, que permite a determinação da
concentração mínima inibitória (MIC) dos agentes antimicrobianos e a detecção dos
mecanismos de resistência para uma variedade de microrganismos.
2 - Fundamento
O Etest consiste numa tira de plástico inerte, não poroso (5 x 57 mm). A
superfície superior da tira contém uma escala que expressa a CMI em μg/mL e o
código identificador do agente antimicrobiano ou reagente (figura 1).
Na superfície inferior da tira está um gradiente de concentrações,
correspondentes a 15 diluições duplas, do antibiótico pré-definido baseado no
método convencional para a determinação da MIC. Quando a tira de Etest é
aplicada à superfície, previamente inoculada, do meio, o gradiente do antimicrobiano
e imediatamente transferido para a matriz da gelose. O gradiente contínuo de
concentrações, estabelecido ao longo da tira, mantém-se estável durante um longo
período de tempo, cobrindo os tempos críticos para uma grande variedade de
microrganismos.
Após 24 horas ou mais de incubação, forma-se uma elipse de inibição,
simétrica e centrada ao longo da tira. O ponto de intercepção da elipse com a tira
corresponde ao valor da MIC em μg/mL, que é lido diretamente na escala e
posteriormente relacionado à tabela padrão para indises de resistência,
sensibilidade reduzida ou sensibilidade. 3 - Conservação
A embalagem selada deve ser conservada, de acordo com as condições
especificadas na carteira de folha de alumínio até à expiração do prazo de validade.
As tiras Etest que permanecem numa embalagem aberta, devem ser mantidas secas, conservadas num contentor estanque ao ar ou num tubo de conservação
com dessecante ou re-selando a carteira com uma mola e utilizá-la até à expiração

164
do prazo de validade. Quando não estão em utilização, as tiras Etest devem ser
protegidas da humidade e da luz.
4 – Manipulação A menos que seja conservada numa sala de temperatura controlada (18 a
22°C), a embalagem deve atingir a temperatura ambiente antes de ser aberta. Abra
a carteira de folha de alumínio e retire as tiras manualmente com os dedos, pinça ou
com aplicador específico. Não toque no lado da tira, que tem os reagentes, isto é, no
lado oposto da escala MIC.
5 – Método
Meio de cultura Seleccione os suplementos adequados ao microrganismo a ser ensaiado.
Assegure-se que o meio cumpre as especificações do controlo de qualidade,
profundidade da gelose de 4.0 ± 0.5 mm. Preparação do Inóculo
Emulsione várias colónias isoladas, a partir de uma cultura pura “overnight”,
para um meio de suspensão adequado. Os organismos fastidiosos devem ser
suspensos num caldo e utilizados no período de 15 minutos. Compare a turvação
com o padrão McFarland adequado.
Inoculação
Mergulhe uma zaragatoa de algodão estéril na suspensão de inóculo e
pressione contra a parede interna do tubo, para remover o excesso de líquido.
Espalhe cuidadosamente em toda a superfície do meio gelosado, em três direcções,
também pode ser utilizado o Retro C80™ (inoculador rotativo). Para testes com
antifúngicos, abra 2 sulcos na placa, mergulhando a ansa no inoculo, entre eles.
Deixe que o excesso de humidade seja completamente absorvido e que a
superfície esteja completamente seca, antes de aplicar as tiras Etest.

165
Aplicação das tiras
Aplique a tira de Etest à superfície do meio de cultura com a escala de CMI
de frente e virada para cima (para a abertura da placa). Tal pode ser feito com pinça,
com um aplicador manual, Nema C88™ (caneta de vácuo) ou Simplex C76™
(aplicador automático). Assegure-se que a tira fica totalmente em contacto com a
superfície do meio de cultura e sem bolhas de ar. Uma vez aplicada, não remova a
tira (figura 2).
Incubação
As placas são colocadas na estufa de incubação, na posição invertida (com a
tampa para baixo); as temperaturas e os tempos de incubação são aplicados de
acordo com os microrganismos em estudo.
Leitura
O valor de MIC é lido no ponto onde a elipse de inibição intersecta a tira. No
endereço www.abbiodisk.com estão disponíveis vários padrões de
crescimento/inibição para diferentes microrganismos, assim como oprocedimento
correto para a leitura dos resultados.
Figura 1 Figura 2
Figura 1 e 2 – Imagem da fita de E-teste para ESBL com a escala em MIC
aplicadas no meio de cultura. Fonte: AB Bidisk

166
Teste de Suscetibilidade pelo método Eteste para confirmação in vitro de ESBL
cefotaxime/cefotaxime + clavulanic acid ceftazidime/ceftazidime + clavulanic acid
Fonte: Etest para ESBL- PROBAC DO BRASIL
Reagents
100 ou 30 unidades de fitas E-teste reagents de cada CT/CTL ou TZ/TZL
Armazenamento
Todos os pacotes com fitas não abertas ou não utilizadas devem ser
armazenados a -20°C ou na temperatura sigerida na bula do kit até a data de
expiração do mesmo. Fitas não utilizadas devem ser armazenadas em um frasco
contendo sílica colorida para retirar a umidade. Proteger as fitas Etestes da
exposição a misturas, calor e luz direta forte em qualquer hora.
Evitar a penetração de umidade dentro do pacote ou no frasco. As fitas devem ser mantidas secas. Manuseio
Antes de utilizar as fitas Eteste de um pacote fechado, inspecionar
visualmente para verificar se está intacto. Não utilizar as fitas se o pacote estiver
danificado.
Ao retirar do freezer a -20°C, deixar o pacote ou container atingir a
temperatura ambiente por 30 minutos. A umidade condensada na superfície de fora
deve evaporar antes de abrir o pacote. Abra o pacote de acordo com as instruções.
Ao segurar as fitas de Eteste, pegue apenas a fita na área de Eteste ESBL. Não
toque na superfície da fita com o antibiotic, por exemplo, o lado oposto da escala. As
fitas podem ser colocadas utilizando-se um aplicador (figura 2).

167
Precauções
• O Eteste ESBL é somente para uso diagnóstco in vitro.
• O uso apropriado do sistema requer o conhecimento de pessoal terinado em
microbiologia e teste antimicrobiano.
• O Eteste ESBL deve ser utilizado de acordo com os procedimentos descritos.
Procedimentos assépticos devem ser realizados todas às vezes que se utilizar
espécimes bacterianas e medidas de precaução devem ser tomadas contra danos
microbiológicos. As placas de Agar devem ser esterilizadas após uso, antes do
descarte.
• Ocasionalmente a estática pode ocasionar a aderência de uma ou mais fitas.
Tenha certeza que você separou as fitas e as appliqué somente uma vez na
superfície do ágar.
• Devido à rápida difusão do antibiotico, as fitas Eteste ESBL não podem se
removidas uma vez em contato com a superfície do ágar.
• Consultar o guia técnico do Eteste (www.abbiodisk.com), e ler a bula do mesmo
antes da utilização. Procedimento
Materiais necessários que não acompanham as fitas:
• Placas de ágar Mueller Hinton (depth of 4 ± 0.5 mm)
• Salina estéril (0.85% NaCl)
• lupa estéril, swabs, tubos, pipetas e tesouras
• Aplicador manual de Eteste
• Turbidez número 0.5 da escala padrão de McFarland
• Incubatora (35 ± 2 °C)
• Cepas de controle de qualidade
• Frasco de estocagem com sílica
• Informação técnica adicional www.abbiodisk.com/Etest Technical Manual

168
Meio de ágar
Tenha certeza de que a espessura do ágar seja 4.0 ± 0.5 mm com pH 7.3 ± 0.1 .
Preparação do inóculo
Emulsionar várias colônias recém isoladas de uma cultura de 24hs em meio
sólido, em salina até uma turbidez de 0.5 da escala padrão de McFarland. Se o
inoculo estiver correto, uma camda confluente será obtida após a incubação. Faça
uma contagem de colônias para verificar se o seu procedimento atingiu uma densidade correta do inóculo no que se refere à UFC/mL
Nota:
Como as ESBL são dependentes do inóculo, uma quantidade muito pesada
ou leve da mesma afeta diretamente o resultado. O excesso da enzima pode
interferir no componente do ácido clavulânico no teste e reduzir potencialmente o
raio do MIC da CT/CTL ou TZ/TZL e fornecer um resultado falso negativo. Ao
contrário, pouca enzima pode fornecer um MIC baixo para a CT ou TZ, e reduzir o
raio da CT/CTL e TZ/TZL.
Inoculação
Inserir um swab estéril, não tóxico na suspensão do inóculo. Remover o
excesso de fluido pressionando o swab contra a parede interna do tubo. Passar o
swab na superfície do ágar por 3 vezes, rodando a placa aproximadamente 60º cada
vez, para assegurar uma boa distribuição. Permitir que a umidade seja absorvida por
aproximadamente 15 minutos de modo que a superfície esteja completamente seca antes de aplicar as fitas de ESBL.

169
Aplicação
Verificar se a superfície do ágar inoculado esteja completamente seca antes
de aplicar as fitas. Abra o pacote e aplique às fitas conforme descrito abaixo.
Manuseio
As fitas de Eteste ESBL podem ser aplicadas na superfície do ágar inoculado
com uma pinça, um aplicador de Eteste manual, ou Nema C88. Sempre coloque a
fita no ágar com a escala do MIC para cima (na direção da placa aberta), e o
gradiente do antibiótico na superfície do ágar. Se for colocada de modo errado, não
será formada a zona de elipse devido ao antibiótico não ter se difundido pela fita
pástica não porosa. Tenha certeza de que toda a fita esteja em completo contato
com a superfície do ágar. Se necessário, remover o ar das mesmas pressionando-as
gentilmente com a pinça, sempre movendo da mais baixa concentração até a mais
alta. Pequenas bolhas abaixo das fitas não afetam o resultado. Uma vez aplicada, a
fita não pode ser movida pois o antibiótico é rapidamente liberado na superfície do
ágar.
Incubação
Incubar as places de agar numa posição invertida a 35 ± 2 °C por 16-20 horas
em temperatua ambiente.
Leitura Quando o crecimento bacteriano estiver visível, ler os valores do MIC para
CT, CTL, TZ e TZL, onde as respectivas elipses inibitórias se encontram com a fita
(figura 4). Crecimento em todo o gradiente sem zona de inibição indica que o MIC é
maior ou igual (≥) ao maior valor na escala de leitura. Uma elipse inibitória abaixo do
gradiente indica que o MIC é menor (<) do que o menor valor na escala. Quando
colônias mutantes estiverempresentes na elipse de inibição, ler o MIC onde estas
colônias estiverem completamente inibidas. Para valor de MIC em alta escala, elipse
de inibição pode ser muito pequena ou não claramente discernível. Ocasionalmente,
uma zona fantasma (phantom zone) pode ser vidualizada abaixo do gradiente de
CTL ou TZL e a elipse pode ou não ser vista em prózimo ao fim da fita de CT ou TZ
(Figura 5). Elipse de inibição de CT ou TZ pode também ser deformada no final da
fita (Figure 6). A presença de uma zona fantasma ou deformação da elipse é uma
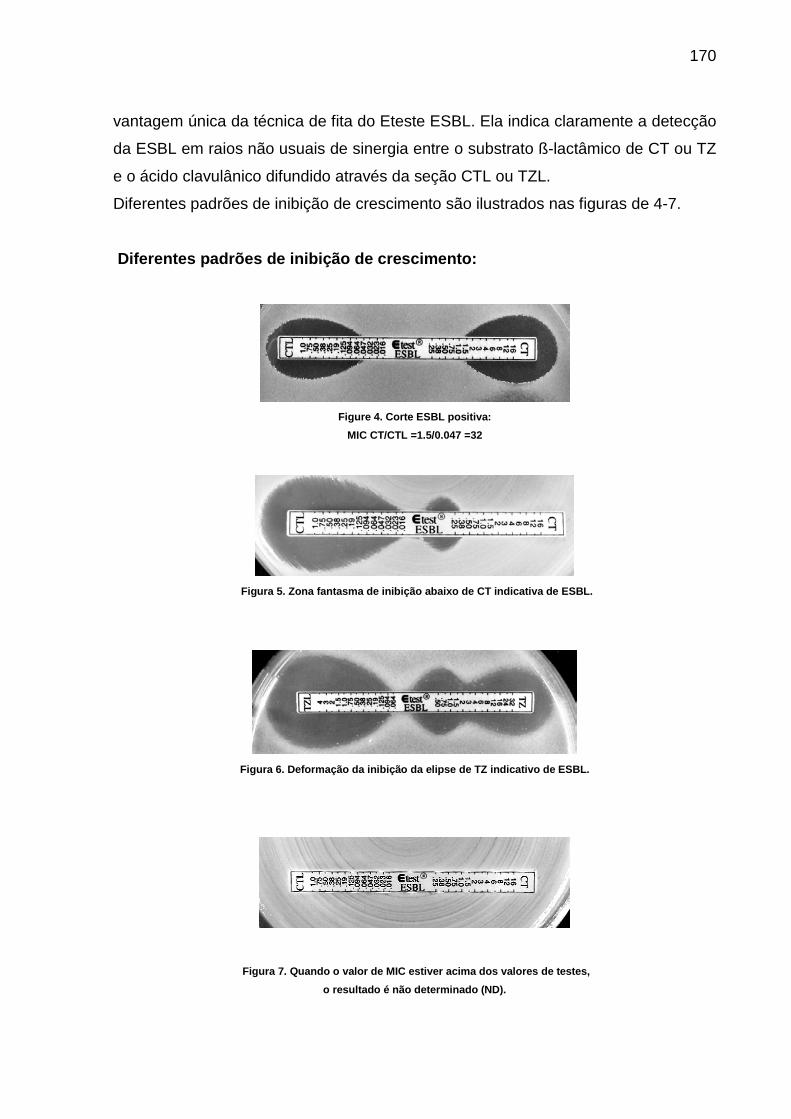
170
vantagem única da técnica de fita do Eteste ESBL. Ela indica claramente a detecção
da ESBL em raios não usuais de sinergia entre o substrato ß-lactâmico de CT ou TZ
e o ácido clavulânico difundido através da seção CTL ou TZL.
Diferentes padrões de inibição de crescimento são ilustrados nas figuras de 4-7.
Diferentes padrões de inibição de crescimento:
Figure 4. Corte ESBL positiva:
MIC CT/CTL =1.5/0.047 =32
Figura 5. Zona fantasma de inibição abaixo de CT indicativa de ESBL.
Figura 6. Deformação da inibição da elipse de TZ indicativo de ESBL.
Figura 7. Quando o valor de MIC estiver acima dos valores de testes, o resultado é não determinado (ND).

171
Tabela 18 - Guia para interpretação do Eteste ESBL
ESBL MIC Raio do MIC Reporting
Positivo
CT > 0,5 e CT/CTL > 8
ou
TZ > 1 e TZ/TZL > 8
ou
Zona fantasma ou deformação da
eclipse no CT ou TZ
Produção de ESBL e resistência a
todos as penicilinas, cefalosporinas e
aztreonam (CLSI)
Negativo CT < 0,5 ou CT/CTL < 8
e
TZ < 1 ou TZ/TZL < 8
Não produtora de ESBL
Non-determinado (ND)
CT > 16 e CTL > 1
e
Quando uma fita é negativa para
ESBL e a outra não determinada
ESBL não determinada. Se a ESBL é
suscetível, confirmar o resultado com
a genotipagem.
Exemplos de como interpretar os raios de MIC: CT/CTL 8/0.125 = 64 = ESBL +
TZ/TZL >32/<0.064 = >500 = ESBL +
CT/CTL 1/<0.016 = >62 = ESBL +
CT/CTL 4/>1 = <4 = ESBL -
TZ/TZL 0.5/0.25 = 2 = ESBL -
CT/CTL 0.25/0.19 = 1.3 = ESBL -
TZ/TZL 1/41) = 0.25 = ESBL -
TZ/TZL >32/>4 = out of range = ND 2)
CT/CTL ESBL negative and TZ/TZL ND = ND 3)
Notas:
1) Quando os MICs de CTL ou TZL forem mais elevados do que CT ou TZ
respectivamente, pode refletir a indução da produção de ß-lactamase pelo ácido
clavulânico.
2) Quando ambos os valores do MIC estiverem acima da escala do teste, o
resultado é ND= não-determinado. Isto pode sugerir a presença de um inibidor

172
resistente TEM ou enzima AmpC ou que o valor do MIC está fora do alcance do
valor do teste.
3) Quando um resultado for ESBL negativo e o outro ND, a interpretação deve ser
ND.
Controle de qualidade
Cepas de controle de qualidade devem ser testadas pelo método descrito em
procedimentos para verificar a qualidade dos reagents e a abilidade do teste em
confirmer a produçõ de ESBL. E. coli ATCC® 35218 produz ß-lactamase TEM-1
(não ESBL) e serve como controle negative para o teste. K. pneumoniae ATCC
700603,um genótipo positive para ESBL, fornece um valor de MIC dentro da escala
e serve como controle positive, ambos pelo critério do raio de MIC ou pelo fenômeno
de deformação da elipse. Atenção cuidadosa deve ser dada para a mautenção e
armazenamento de K. pneumoniae ATCC 700603 pois a perda espontânea do
plasmídeo codificador de ESBL tem sido documentada e pode dar resultados de
controle de qualidade fora dos limites aceitáveis. Valor de MIC para CTL ou TZL
acima das especificações pode indicar degradação do ácido clavulâncio ou um
inóculo excessivamente pesado. Verificar o armazenamento e manuseio das fitas e
repetir o teste utilizando o inoculo de forma correta.

173
Protocolo de extração plasmidial KIT PROMEGA Wizard® Plus SV Minipreps DNA Purification System
1 - Centrifugar de 1 a 10 ml de cultura em meio líquido crescido overnight em tubo
de eppendorf de contendo 1,5ml por 5minutos.
2 - Dispensar o sobrenadante e retirar o excesso com pipeta de 200μl. Em seguida,
adicionar 250μl da solução de ressuspensão (Tris-HCl, EDTA e RNase A). 3- Desprender o pellet com o auxílio de ponteira esterilizada.
4 - Adicionar 250μl da solução de lise (NaOH e SDS - 2), invertendo 4 vezes para
misturar.
5 - Adicionar 10μl da solução de protease alcalina, invertendo 4 vezes para
misturar. Incubar por 5 minutos em temperatura ambiente.
6- Adicionar 350μl de solução de neutralização (hidrocloridrato de guanidina, Acetato de potássio e Ác. acético glacial - 4), invertendo 4 vezes para misturar.
7-Centrifugar a 12.000 rpm (velocidade máxima), por 10 minutos a temperatura
ambiente.
Obtenção do DNA plasmidial 8 – Centrifugar a 12.000 rpm (velocidade máxima), por 1 minuto a temperatura
ambiente dispensando o decantado e voltando a inserir a coluna Spin no mesmo
tubo de coleta.
Lavagem 1-Inserir a coluna Spin no tubo de coleta.
2-Decantar o lisado na coluna Spin retirando com cuidado (pipeta de 1000μl - vol. de
750μl). 3-Deixar 1 minuto a T.A.
4- Centrifugar a 12.000 rpm (velocidade máxima), por 1 minuto a temperatura
ambiente dispensando o decantado e voltando a inserir a coluna Spin no mesmo
tubo de coleta.
Lavagem: 1-Adicionar 750μl de solução de lavagem (com etanol adicionado). Centrifugar a
12.000 rpm (velocidade máxima), por 1 minuto a temperatura ambiente
dispensando o decantado e voltando a inserir a coluna Spin no mesmo tubo de

174
coleta.
2-Repetir os passo 1 com 250μl de solução de lavagem.
3-Centrifugar a 12.000 rpm (velocidade máxima), por 2 minutos a temperatura
ambiente por 2 vezes.
Eluição: 1-Transferir a coluna Spin num microtubo de 1,5ml, tendo o cuidado de não
transferir a solução de lavagem junto. Se por acaso estiver solução de lavagem,
centrifugar novamente por 1 minuto a velocidade máxima, e então, transferir em um
novo tubo estéril de 1,5ml.
2-Adicionar 100μl de água livre de nuclease na coluna spin. Centrifugar na
velocidade máxima por 1 minuto a T.A.
3-Descartar a coluna e guardar o DNA a -20ºC ou temperatura menos.
4-Correr em gel de agarose a 0.8% 1μl de cada amostra com 1μl de Ladder por 50
minutos registrando o resultado em fotodocumentação.
Observação: Frasco 1- (Sol. de resuspensão): Tris-HCl → Atua como tampão.
EDTA :Age como quelante sequestrando os íons Ca2+ e Mg2-, desestabilizando a
membrana da célula, causando o inchamento devido a entrada da glicose.
RNase: Degrada o RNA presente, evitando que este possa prejudicar a extração.
Frasco 2 (Sol. de lise): NaOH: Auxilia na formação dos poros devido a elevação do pH o que desestrutura a
membrana.
SDS: Detergente aniônico que atua rompendo as ligações covalentes da membrana,
retirando lipídios e levando a formação de pequenos poros que possibilitam a
passagem dos plasmídios para o meio extracelular sem permitir a passagem do
DNA cromossomal.
Frasco 3 (Protease alcalina): Inativa endonucleases e outras proteínas liberadas durante o processo de lise que
podem afetar a qualidade do DNA purificado.
Frasco 4 (Sol. de neutralização): Atua retirando as proteínas que envolvem o plasmídio e elevando o pH da solução
para próximo de 8.

175
Protocolo de extração de DNA total
Kit Invitrogentm - Easy-DNAtm Kit for Genomic DNA Isolation
Adaptações realizadas por Cristina Ferreira, Nayanne Oliveira e William Ferreira
Repicar uma colônia da bactéria em LB sólido por 24hs a 35 ºC. Pescar com a
haste do swab estéril 01 colônia da bactéria recém-crescida e inseri-la no tubo
baquelite de rosca contendo o LB líquido, deixando a tampa frouxa, em shaker por
25hs na estufa a 35 ºC.
Após 24hs, se o mesmo turvar, retirar da estufa e do shaker. Levar para
capela de fluxo laminar classe II de extração de DNA, colocar um pouco no tubo de
eppendorf de 1,5mL, centrifugar por 5min. Na centrífuga de microtubo a 20 ºC
(deixar o cool ligado). Após centrifugação, decantar por inversão o sobrenadante no
descartex, e acrescentar mais meio ao tubo centrifugando novamente até acabar
todo o LB líquido e ficar somente o pellet bacteriano no eppendorf.
Retirar todo o sobrenadante que restar após inversão do tubo com pipeta de
20 µl. Ressuspender o pellet com 200 µl de tampão PBS 1X. Soltar o pellet com
muito cuidado.
Isolamento do DNA 1-Adicionar 350µl da Solução A na suspensão de célula e homogeneizar por
inversão ou com ponteira com muito cuidado.
2- incubar em banho-maria a 65ºC por 20min.
3-Adicionar 150µl da Solução B e homogeneizar por inversão o tubo até que a
amostra esteja uniformemente viscosa (10seg. a 1 minuto).
4-Adicionar 500µl de clorofórmio (PA) e agitar até diminuir a viscosidade e a mistura
estar homogênea.
5-Centrifugar com velocidade máxima (12.000rpm) por 15min. a 4ºC para separar
as fases. Retirar com pipeta o sobrenadante e colocá-lo em um novo tubo de
eppendorff de 1,5ml desprezando o usado.
6-Iniciar a precipitação do DNA.

176
Precipitação do DNA 10- A solução de DNA, adicionar 1mL de etanol a 100% (PA), refrigerado
(congelador) a -20 ºC e homogeneizar por inversão o tubo.
11- Incubar por 1 hora e 30 minutos ou mais em gelo (congelador e mergulhar o tubo
no gelo)
12- Centrifugar com velocidade máxima (12.000 rpm) por 12min. a 4ºC , removendo
o etanol por inversão do tubo e retirando excesso com pipeta de 200µl.
13-Adicionar 500µl de etanol a 80% (contendo 80 mL de etanol + 20 mL de água)
refrigerado a -20 ºC e homogeneizar por inversão por 20 vezes.
14- Centrifugar com velocidade máxima (12.000 rpm) por 5min. a 4ºC, retirar o
excesso por inversão do tubo.
15- Centrifugar com velocidade máxima (12.000 rpm) por 3min. Retirar o etanol
residual com a pipeta de 20 µl. Deixar secar na capela por 10-15min. Cuidado ao
retirar para não desprender o pellet que fica no fundo do tubo ou parede .
16-Ressuspender o pellet com 50µl de tampão TE, com a ponteira com muito
cuidado. Adicionar 2µl de RNase a 2mg/mL para ficar a uma concentração de
40µl/mL. Retirar do congelador a RNase somente na hora em que for colocá-la,
descongelar na mão, usar e congelar novamente.
17-Incubar em banho-maria a 37ºC por 30 min. O DNA está pronto para futuros
experimentos. Guardar a 4ºC.
18- Correr em gel de agarose a 0.8% 1μl de cada amostra com 1μl de Ladder por 50
minutos registrando o resultado em fotodocumentação.

177
Protocolo adaptado para PCR 16S rRNA
Fonte: Myoshi et al. 2005
Utilizar como controle o S. aureus (ATCC 25923) e o S. epidermidis (ATCC12228)
Preparação do Master Mix 16S no DNA Workstation Reagentes Para 1 reação (µl)
Água ultrapura 13,45
Tampão 10X * 2,5
MgCl2 (50 mM)* 1,25
dNTP (10 mM) 0,5
Primer F (10 mM) 0,5
Primer R (10 mM) 0,5
Taq polimerase (5U/mL) 0,3
20
* Obs: Descongelar bem o tampão e MgCl2 em vórtex, para não atrapalhar a reação.
Acrescentar em cada tubo de eppendorff de 0,2ml, 20 µl do máster mix + 5 µl do
DNA lisado (5ng/ µl) (da extração, quantificado e diluído com água Q). Fazer na
capela de fluxo laminar classe IIA. O volume final da reação será de 25 µl.
Utilizar o termociclador com o programa 16S
Temp tempo 95ºC 2min
94ºC 1min
35 Ciclos 58ºC 1min 72ºC 1min
72ºC 5 min
4ºC forever
Total 2,5horas
Correr em gel de agarose a 1% 5µl do produto do PCR (DNA). Preparar o gel antes
e deixar esfriar. Aplicar 18 bolinhas de 3µl de corante xilenocianol. Pipetar 5 µl do
DNA lisado, misturar com o corante e depois 1µl do ladder com o xilenocianol e
aplicar nos poços do gel . Congelar o restante a -20C. Utilizar o Ladder de 100bp.

178
Purificar antes do seqüenciamento com o Kit GE. Esperar bandas de amplicon de aproximadamente 900bp. Caso necessite preparar o dNTP 100 µl. na (10 mµ) misturar: 10 µl dATP, 10 µl
dttp , 10 µl dctp, 10 µl dgtp + 60 µl de água Q . Colocar em um único tubo.

179
Protocolo adaptado para PCR mecA1 e mecA2 femB1 e femB2
Fonte: Jonas et al, 2002.
Diluir os primers liofilizados obtidos por compra.
Preparação do Master Mix mecA1 e A2 e femB1 e B2 no DNA Workstation Reagentes Para 1 reação (µl)
Água ultrapura 13,45
Tampão 10X * 2,5
MgCl2 (50 mM)* 1,25
dNTP (10 mM) 0,5
Primer mecA1 e A2(5 mM) 1,0
Primer FemB1 e B2(5 mM) 1,0
Taq polimerase (5U/mL) 0,3
20
Utilizar o termociclador com o programa Jonas.
Temp tempo 94ºC 45seg
50ºC 45seg
30 Ciclos 72ºC 60seg
72ºC 2 min
4ºC forever
Total 2,25horas
Correr em gel de agarose a 1% 5 µl do produto do PCR (DNA). Deixar correr 1hora a
75 volts. Utilizar o Ladder 100bp. Esperar para o mecA1 e mecA2 bandas de amplicon em torno de 310bp. Esperar para o femB1 e femB2 bandas de amplicon de aproximadamente de 651bp.

180
Protocolo adaptado de PCR para PCR mecP4 e P7
Fonte: Milheiriço et al. 2007.
Diluir os primers liofilizados obtidos por compra. O mecP4 e mecP7 são controles
internos positivos da Milheiriço.
Preparação do Master Mix mecP4 e mecP7 no DNA Workstation Reagentes Para 1 reação (µl)
Água ultrapura 13,45
Tampão 10X * 2,5
MgCl2 (50 mM)* 1,25
dNTP (10 mM) 0,5
Primer mecP4 (5 mM) 1,0
Primer MEC P7 (5 mM) 1,0
Taq polimerase (5U/mL) 0,3
20
* Obs: Acrescentar em cada tubo de eppendorff de 0,2ml, 20 µl do máster mix + 5 µl
do DNA lisado (5ng/ µl) (extraído, quantificado e diluído com água Q). Fazer na
capela de fluxo laminar classe IIA. O volume final da reação será de 25 µl.
Utilizar o termociclador com o programa denominado de Jonas.
Temp tempo 94ºC 4 min
94ºC 45seg
30 Ciclos 50ºC 45seg
72ºC 60 seg
72ºC 2min
4ºC Forever
Total 2,25horas
Correr em gel de agarose a 1% 5 µl do produto do PCR (DNA). Pipetar 5 µl do DNA
lisado, misturar com 3µl do corante. Depois misturar 1µl do ladder 100bp com o

181
xilenocianol e aplicar nos poços do gel. Deixar correr 1hora a 75 volts.
Congelar o restante a -20C. Descongelar e purificar antes do seqüenciamento com o
Kit GE. Esperar para o mecP41 e mecP7 duas bandas de amplicon de aproximadamente 162bp.

182
Protocolo adaptado de PCR para toxinas estafilocócicas
Fonte: Strommmenger et al., 2008.
Diluir os primers liofilizados obtidos por compra. O primer mãe está a uma
concentração de 100 mµ. Diluir para uma concentração de (10 µm) e, depois, (5µm).
Cepa controle do PCR é a ATCC 29213 (MSSA PVL neg). Zhang et al.,2008.
A partir do mix de cada par de (5µm), retirar de cada um deles:
Setd F+ Setd R 1- retirar 25µl
ArcA F+ ArcA R 2 - retirar 25µl
Seh F+ seh R 3- retirar 25µl
luKPV F + lukPV R 4- retirar 25µl
Acrescentar mais 25µl de água Q ou de injeção. Teremos um único mix denominado
de mix toxina que terá 125 µl a uma concentração de 1 mµ. Retirar 1/10 do volume
total da reação (2,5 µl).
Preparação do Master Mix Toxinas no DNA Workstation Reagentes Para 1 reação (µl)
Água ultrapura 12,95
Tampão 10X * 2,5
MgCl2 (50 mM)* 1,25
dNTP (10 mM) 0,5
Primers setdF/R, ArcAF/R, 2,5
sehF/R, lukPV F/R (5 mM).
Taq polimerase (5U/mL) 0,3
20
* Obs: Acrescentar em cada tubo de eppendorff de 0,2ml, 20 µl do master mix + 5 µl
do DNA lisado(5ng/ µl) (extraído, quantificado e diluído com água Q). Fazer na
capela de fluxo laminar classe IIA. O volume final da reação será de 25 µl.
Utilizar o termociclador com o programa denominado de Toxinas.

183
Temp tempo 94ºC 4 min
94ºC 45seg
30 Ciclos 50ºC 45seg
72ºC 60 seg
72ºC 2min
4ºC Forever
Total 2,25horas
Correr em gel de agarose a 2% 5 µl do produto do PCR (DNA). Pipetar 5 µl do DNA
lisado, misturar com 3 µl do corante. Depois misturar 1µl do ladder 100bp com o
xilenocianol e aplicar nos poços do gel. Deixar correr 1hora a 75 volts.
Congelar o restante a -20C. Descongelar e purificar antes do seqüenciamento com o
Kit GE.
Esperar para a toxina bandas de amplicon entre 200 a 600bp.
Setd ........................607 bp arcA...........................215bp
Seh............................358 bp
lukPV ........................432 bp

184
Protocolo apatado de PCR para cassetes cromossômicos Fonte: Milheiriço et al. , 2007.
Diluir os primers liofilizados obtidos por compra. O primer mãe está a uma
concentração de 100 mµ. Diluir depois para uma concentração de (10µm)= 10µl do
primer mãe +90 µl de água Q ou injeção) e após retirar 50µl de cada par de primer,
misturar os mesmos e ficará a uma concentração de (5µm).
Primes controles internos positivos ctles mecA P4 mecA P7 - Milheiriço et al. ,
2008.
Cepas controles ATCC 12228 S. epidermidis (ctle negativo) e ATCC 33591 S. aureus (ctle pos) Machado et al,2007. Utilizar 4 primers (2 F+R) mais os 2 ctles internos positivos e proceder o PCR.
CIF2 F+ CIF2 R e os ccrC F2+ ccrC R2 com os primers ctles mecA P4 e mecA P7
RIF5 F10+ RIF5 R13 e os SCCmec V J1 F+ SCCmec V J1 R com os primers ctles
mecA P4 mecA P7 dcs F2 + dcs R1 e os ccrB2 F2+ccrB2 R2 com os primers ctles mecA P4 mecA P7 Kdp F1 + Kdp R1 e os SCCmec III J1 F+ SCCmec III J1 R os primers ctles mecA P4 mecA P7 mecI P2+ mecI P3 e os primers ctles mecA P4 e mecA P7 .
Preparação do Master Mix Cassete cromossômico no DNA Workstation Reagentes Para 1 reação (µl)
Água ultrapura 13,5
Tampão 10X * 2,5
MgCl2 (50 mM)* 0,75
dNTP (10 mM) 0,5
Primers 2 pares- 2 + 2ctles 1,25
Taq polimerase (5U/mL) 0,25
50

185
* Obs: Acrescentar em cada tubo de eppendorff de 0,2ml, 20 µl do master mix
+ 5 µl do DNA lisado(5ng/ µl) (extraído, quantificado e diluído com água Q). Fazer na
capela de fluxo laminar classe IIA. O volume final da reação será de 25 µl.
Utilizar o termociclador com o programa denominado de Cassete.
Temp tempo
94ºC 4 min
94ºC 30seg
30 Ciclos 53ºC 30seg
72ºC 1min
72ºC 4min
4ºC Forever
Correr em gel de agarose a 3% 5 µl do produto do PCR (DNA). Pipetar 5 µl do DNA
lisado, misturar com 3µl do corante. Depois misturar 1µl do ladder 100bp com o
xilenocianol e aplicar nos poços do gel. Deixar correr 1hora a 75 volts.
Congelar o restante a -20C. Descongelar e purificar antes do seqüenciamento com o
Kit GE. Esperar para o cassete produtos com amplicon de: CIF2 ..........................495 bp
ccrC ...........................449 bp
FIF5 ...........................4141 bp
SCCmec………….…377 bp
dcs ……..........……....342 bp
ccrB2 ……..................311 bp
Kdp ………………..…284 bp
SCCmec ………….....243 bp
mecI ………................209 bp
mecA P4 eP7 ............162 bp

186
Protocolo adpatado de PCR para Genotipagem dos clones estafilocócicos
Fonte: http://www.mlst.net/
Para o S. aureus: Diluir os primers liofilizados obtidos por compra. O primer mãe está a uma
concentração de 100 mM. Diluir depois para uma concentração de (10mM)= 10µl do
primer mãe +90 µl de água Q ou injeção) e após retirar 50µl de cada par de primer,
misturar os mesmos e ficará a uma concentração de (5mM).Cepas controles ATCC 12228 S. epidermidis (ctle negativo) e ATCC 33591 S. aureus (ctle pos). Realizar adaptações na temperatura de hibridização, se necessário. Preparação do Master Mix no DNA Workstation Reagentes Para 1 reação (µl)
Água ultrapura 15,0
Tampão 10X * 2,5
MgCl2 [50mM]* 0,75
dNTP [10 mM] 0,5
Mix genes [5mM] 1,0
Taq polimerase [5U/mL] 0,25
20
* Obs: Acrescentar em cada tubo de eppendorff de 0,2ml, 20 µl do master mix + 5 µl
do DNA a 5ng/ µl. Fazer na capela de fluxo laminar classe IIA. O volume final da
reação será de 25 µl. Utilizar o termociclador com o programa denominado de
MLST-SA.
Temp tempo 95ºC 5 min
95ºC 1 min
57ºC 1 min
30 Ciclos 72ºC 1 min
72ºC 5 min
4ºC forever
4ºC Forever

187
Correr em gel de agarose a 0,7% 5 µl do produto do PCR (DNA) misturado com 3µl
do corante Load Blue. Depois homogeneizar 1µl do ladder 100bp com o Load Blue e
aplicar nos poços do gel. Deixar correr 1hora a 75 volts.
Congelar o restante a -20C. Descongelar e purificar antes do seqüenciamento com o
Kit GE. Esperar produtos com amplicon de aproximadamente 500bp para os genes: arc, aro, glp,gmk,pta,tpi,yqi.
Para o S. epidermidis Realizar adaptações na temperatura de hibridização, se necessário.
Preparação do Master Mix no DNA Workstation Reagentes Para 1 reação (µl)
Água ultrapura 15,0
Tampão 10X * 2,5
MgCl2 [50mM]* 0,75
dNTP [10 mM] 0,5
Mix genes [5mM] 1,0
Taq polimerase [5U/mL] 0,25
20
* Obs: Acrescentar em cada tubo de eppendorff de 0,2ml, 20 µl do master mix
+ 5 µl do DNA a 5ng/µl. Fazer na capela de fluxo laminar classe IIA. O volume final
da reação será de 25 µl.
Utilizar o termociclador com o programa denominado de MLST-SE.
Temp tempo 95ºC 3 min
95ºC 30 seg
50ºC 1 min
34 Ciclos 72ºC 1 min
72ºC 10 min
4ºC forever
4ºC Forever

188
Correr em gel de agarose a 0,7% 5 µl do produto do PCR (DNA) misturado com 3µl
do corante Load Blue. Depois homogeneizar 1µl do ladder 100bp com o Load Blue e
aplicar nos poços do gel. Deixar correr 1hora a 75 volts.
Congelar o restante a -20C. Descongelar e purificar antes do seqüenciamento com o
Kit GE. Esperar para os genes descritos abaixo produtos com amplicon de: arcC............................................465bp
aroE............................................420bp
gtr................................................438bp
mutS...........................................412bp
pyr..............................................428bp
tpi................................................424bp
yqiL.............................................416bp

189
Protocolo para preparação de gel agarose
Fonte: Pop de Preparação de gel de agarose-Laboratório de Biologia Molecular- Fundação Alfredo da Matta
Descrição
Preparar o gel agarose a uma concentração de 0,8%, 1% e 2%
Produto Agarose NA frs 500 ou 100U invitrogen.
Protocolo
Observações: Esta preparação é para ser utilizada em cuba de eletroforese para 50ml de
gel. A proporção é de Massa/volume. O tampão diluidor é o TAE 1X. Caso não
tenha, pegar o frasco grande com o tampão TAE50X e seguir a bula para fazer a
diluição para 1X utilizando a água Milli Q ou deionizada.
Pesar em papel laminado na balança analítica:
Gel a 0,8%. Pesar 0,2g de agarose para 25mL de tampão TAE
Gel a 1%. Pesar 1,5g de agarose para 150ml de tampão TAE
Gel a 2% Pesar 0,6g de agarose para 30 de tampão TAE
Gel a 3% Pesar 1,5g de agarose para 50mL de tampão TAE

190
Técnica 1. Colocar o gel em pó no Erlenmeyer, diluir com tampão TAE, homogeneizar e levar
para o microondas durante 40 segundos. Quando faltarem 7 segundos para terminar
o ciclo no microondas (33seg), desligá-lo, observar se houve dissolução (isto é para
evitar que entre em ebulição).
2.Levar o erlenmeyer para a torneira e ficar molhando o vidro para acelerar o
resfriamento. Assim que a temperatura estiver morna (tipo mamadeira na mão),
adicionar o Syber safe e homogeneizar bem.
3. Despejar e caso fiquem bolhas, perfurá-las com agulha estéril. Colocar os pentes
e deixar gelificar cobrindo a cuba com papel alumínio (evita ação da luz). Após
gelificação, retirar os pentes com cuidado. Retirar o gel e encaixá-lo na cuba de
corrida. Acrescentar o tampão TAE 1X até mergulhar todo o gel. Caso ele esteja a
60 mA, deve ser trocado por um novo.
4-Sempre que correr tampar com papel alumínio. Um pente de 10 corre 45minutos.
Caso a coloração do gel não fique boa, pode-se acrescentar o TAE 1X mais 5ul de
Syber e deixar no aparelho Hibrid capture para homogeneizar.
8. Para observar a corrida do gel, só uma pessoa com óculos vê. Deve-se limpar a
placa com gaze e água deionizada.

191
Preparo das amostras para aplicação no gel
Fonte: Pop Preparação de gel de agarose-Laboratório de Biologia Molecular-Fundação Alfredo da Matta
1- Cortar um segmento de PARAFILM, adicionando 1,5μl de SYBR safe (Invitrogen),
sobre a superfície do mesmo, ou o igual volume de xilenocianol.
2- Pipetar 8μl do DNA extraído, misturando com o SYBR depositado no PARAFILM.
3- Misturar 2μl do BLUE ORANGE 6X LOADING DYE (Promega®) em PARAFILM e
1μl do Super Coiled DNA ladder 2-16kb (Promega®), misturando e adicionando no
poço do GEL destinado ao MARCADOR DE DNA. 3-Ligar a cuba de eletroforese, calibrando a corrida para 75 volts por 1 hora de 20min. Cobrir o GEL com papel alumínio. (preto -) → (vermelho +).
Obs: Blue/Orange Loading Dye, 6X – Marcador contendo 0.4% de orange G,
0.03% azul de bomofenol, 0.03% xileno cianolc FF, 15% Ficoll® 400, 10mM Tris-HCl
(pH 7.5) e 50mM EDTA (pH 8.0). Utilizado para carrear e monitorar a migração das
em eletroforese em gel de poços.

192
Procedimento para acionamento da fonte de eletroforese
Fonte: Pop Preparação de gel de agarose-Laboratório de Biologia Molecular-Fundação Alfredo da Matta
1-Ligar a cuba de eletroforese na tomada de 220v.
2- Colocar os cabos nos seus respectivos eletrodos. Ligar o FIO PRETO no pólo
POSITIVO e o VERMELHO no NEGATIVO (o DNA migra do pólo negativo (cátodo
– fio preto) para o positivo (ânodo – fio vermelho).
3- Depois de ligada, acionar o botão SET – AMPERAGEM: NÃO ALTERAR.
4- Apertar SET novamente: PROGRAMA: NÃO ALTERAR.
5- Apertar SET novamente: TEMPO: ACIONAR AS SETAS PARA ALTERAR.
6-- Apertar SET novamente. A palavar OFF aparecerá na tela.
7-Apertar o run.

193
Protocolo para purificação de DNA de solução ou de reação enzimática.
KIT ILLUSTRATM GFXTM PCR DNA AND GEL BAND PURIFICATION KIT FOR
THE PURIFICATION AND CONCENTRATION OF DNA
Captura da amostra
1-Adicionar 100µL de tampão de captura 2 (tampa azul) em 20µL de amostra.
2- Misture completamente.
Ligação da amostra 1- Centrifugar o mix tampão de captura 2 + amostra brevemente (30 seg) para
coletar o líquido do fundo do tubo.
2- Transferir o centrifugado x na coluna e no tubo coletor.
3- Centrifugar a 12.000 rpm por 30 seg.
4- Descartar o sobrenadante e recolocar a coluna no tubo coletor.
Lavagem e secagem 1- Adicionar 100µL de tampão de lavagem tipo 1 (tampa amarela) na microcoluna
Spin.
2- Centrifuguar a 12.000 rpm por 30 seg.
3- Descartar o tubo coletor e transferir a microcoluna spin para um tubo eppendorff
de 1,5mL.
Eluição 1-Adicionar 20µL do tampão de eluição tipo 6 (tampa rosa) ao CENTRO da
membrana da microcoluna ou ÁGUA ULTRA-PURA.
2- Incubar por 1 min a temperatura ambiente.
3-Centrifugar a 12.00 rpm por 1 min para recuperar o DNA.
4- Armazenar o DNA purificado a -20°C.
5-Correr em gel de agarose a 1% por 30min e registrar o resultado

194
Reação de PCR para seqüenciamento
Fonte: Manual da Applied Biosystems aparelho 3130/genetic analyser- Hitachi.
Mix para Reação de Seqüenciamento
A reação deve ser feita com os primers Forward e reverse separadamente.
Fazer um mix para cada um deles se for o caso.
A ordem a ser acrescentada no tubo de eppendorf deve ser: Água mili-Q, Tampão
5X do Big Dye, primer específico.
CUIDADO! O DYE É SENSÍVEL À LUZ. DEVE SER ADICIONADO POR ÚLTIMO A REAÇÃO E COM A LUZ DA CAPELA DO DNA WORKSTATION APAGADA. APÓS ADIÇÃO DO DYE AO MIX, COBRIR O MIX COM PAPEL ALUMÍNIO. DISPENSAR O MIX NOS POCINHOS SEM DEMORA. Cálculo das amostras para microplaca: PCR (n=1) Mix Pgem H2O mili-Q__________5,0µl Big Dye_____2,0µl
Tampão 5X_________2,0 µl pgem ______2,0 µl
Big Dye____________1,0 µl M13________3 µl
Primer (1:3)_________ 1,0 µl H20 mili Q ___3 µl
Amostra ___ ________ 1,0 µl (pode variar- 1,5 µl).
Levar ao termociclador: Programa de 25 ciclos por 2,10hs:
96ºC ___1 min
96ºC ____15seg
50ºC ____15 seg
60ºC ____4 min
60ºC ____1min

195
4ºC _____forever.
Protocolo para precipitação de DNA com Isopropanol/Etanol
Fonte: Manual da Applied Biosystems aparelho 3130/genetic analyser- Hitachi.
1-Utilizar placa com 96 poços, o papel selante do próprio fabricante
2-Para a reação de 20µl, adicionar 80µl de isopropanol a 75% e submetê-las ao
vortex.
3- Selar a placa com papel selante e deixar a T.A. (temperatura ambiente) por
15min. protegida da luz.
4- Centrifugar a placa por 20min. a 14.000 rpm e descartado o sobrenadante
invertendo a placa sobre um papel absorvente.
5- Aplicar um pulso de rotação na centrífuga com a placa invertida sobre o papel
absorvente.
6- Adicionar 20µl de etanol a 70% em cada amostra e selar a placa novamente.
7- Centrifugar a placa por 10min. a 14.000 rpm e descartado o sobrenadante
invertendo a placa sobre um papel absorvente.
8-Repetir os passos 5 a 7 por mais uma vez, só que ao descartar o sobrendante
inverter a placa com foca na pia e depois dar umas batidas sobre o papel
absorvente até estar mauis seca.
9-Deixar o pellet secando a T.A. protegido da luz com papel laminado.
10-A placa com o pellet seco pode ser armazenada no freezer até estar pronta para
ser seqüenciada.

196
Protocolo de preparo de amostras para seqüenciamento com Big-Dye X-Terminator
Fonte: Manual da Applied Biosystems aparelho 3130/genetic analyser- Hitachi.
1- Para reação de 10 µl utilizar 45 µl do SAM solution.
2-Ajustar a pipeta multicanal para 45 µl e pipetar a SAM solution acrescentando
nos poços.
3-Acrescentar para reação de 10 µl, 10 µl de Big Dye X-terminator solution.
4-Homogeneizar sempre que possível no vortex o Big Dye X-terminator, pois a
solução é difícil de pipetar e acrescentar em cada poço.
5-Selar bem a placa com o selo adesivo (optical adhesive covers).
6-Levar para o homogeneizador (vortex) por 1hora e 30 minutos.
7-Retirar do vortex e levar para o aparelho seqüenciador
8-Iniciar o procedimento de seqüenciamento.

197
Protocolo para preparo de amostras para seqüenciamento
Fonte: Manual da Applied Biosystems aparelho 3130/genetic analyser- Hitachi.
1-Para ressuspender a amostra, utilizar formamida Hi-Di (applied Biosystems P/N
4311320). Ela vem congelada e deve ser aliquotada em tubos de 1,5mL para não
degradar.
2-Adicionar 10µl de formamida Hi-Di em cada amostra. Tampar com papel
laminado.
3-Selar a placa, submeter ao vortex e aplicar um pulso na centrífuga.
4-Marcar o lado adesivo plástico que está usando.
5-Desnaturar no termociclador a 95ºC por 2 min.
6-Colocar a placa no gelo por 2min.
7-Aplicar um pulso na centrífuga para remover bolhas de ar.
8-Remover o adesivo e colocar a septa (aparador do aparelho) na placa.
9-Colocar a placa com a bandeja e tampa adaptativa no “autosampler”.
10-As amostras com formamida devem ser corridas em até 48hs para evitar a
degradação da fluoresceína e da formamida.
11- Iniciar o seqüenciamento no aparelho seqüenciador da Applied Biosystems
3130/genetic analyser- Hitachi.





























