Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DE CAMPINAS
FACULDADE DE CIÊNCIAS MÉDICAS
TACIANE BARBOSA HENRIQUES
ESTUDO DE FREQUÊNCIA DE POLIMORFISMOS EM GENES ENVOLVIDOS NO
RISCO DE EFEITOS ADVERSOS DA RISPERIDONA EM UMA AMOSTRA DA
POPULAÇÃO BRASILEIRA
CAMPINAS
2015

TACIANE BARBOSA HENRIQUES
ESTUDO DE FREQUÊNCIA DE POLIMORFISMOS EM GENES ENVOLVIDOS NO
RISCO DE EFEITOS ADVERSOS DA RISPERIDONA EM UMA AMOSTRA DA
POPULAÇÃO BRASILEIRA
Dissertação de mestrado apresentada à Faculdade de Ciências
Médicas da Universidade Estadual de Campinas como parte dos
requisitos para obtenção do título de Mestra em Ciências Médicas,
área de concentração Ciências Biomédicas..
ORIENTADORA: MARICILDA PALANDI DE MELLO
ESTE EXEMPLAR CORRESPONDE À VERSÃO FINAL DA DISSERTAÇÃO
DEFENDIDA PELA ALUNA TACIANE BARBOSA HENRIQUES E
ORIENTADA PELA PROFa. DRa. MARICILDA PALANDI DE MELLO.
Assinatura da Orientadora
CAMPINAS
2015



AOS MEUS PAIS MAUREM E HENRIQUES,
IRMÃOS LIDIANE E AMILSON E
AO BRUNO POR TODO AMOR,
DEDICO.

AGRADECIMENTOS
Á Deus, nosso Pai, que me traz serenidade, e ilumina meu caminho.
Aos meus pais, por todo amor e apoio em todos os momentos da minha vida.
Agradeço de todo meu coração a vocês, meu porto seguro!
Aos meus irmãos Lidi por nos dar a Bia, nossa riqueza e por ser minha gêmea tão
querida. Mi, pelos conselhos, conversas e apoio.
Ao meu namorado Bruno, pelo amor, companheirismo e acima de tudo, amizade.
Foi ele que sempre me incentivou e me apoiou em todas as decisões. Como é bom tê-lo por
perto!
Á minha orientadora, Professora Doutora Maricilda Palandi de Mello, por todo
carinho, confiança, por todas as histórias de vida compartilhadas, ensinamentos e paciência.
Obrigada Professora por ter participado da minha vida em um momento tão importante como
este, sem você nada disso seria possível.
Ao Professor Doutor Gil Guerra–Júnior, pela dedicação e competência, que fez
acontecer esse projeto. Obrigada pelo presente que foi participar desse grupo de pesquisa, e
por toda ajuda que deu quando precisei.
Ao mais novo Doutor Amilton dos Santos, por toda disposição em me ajudar com
TODAS as dúvidas e inseguranças.
Á toda equipe do Ambulatório de Psiquiatria pelo trabalho realizado com as
crianças. A equipe de Pediatria por toda dedicação nas coletas dos casos e controles.
Ao Rodrigo Secolin, pela atenção e sabedoria nas estatísticas do trabalho!
Aos amigos de laboratório de Genética Molecular e Humana do CBMEG, os que
foram e os que permanecem: Débora, Dri, Helena, Regi, Flor, Zelo, Aninha, Creyto, Ana
Paula, Pamela, Pri, Marcela, Tais, Luana, Pri Jacob, Li (Amilga e Amora, que levarei para
vida, obrigada por muitas e infinitas risadas, conselhos, fofocas, sem você não seria igual foi),
Sueli, Nádia, Nathee (pela amizade, conselhos e risadas), Fábio, Carol (pelas conversas sem
fim, pela Cia nos últimos meses, obrigada “Querol”, pelo carinho imenso), Batatinha, Paulo,
Juanito e Alessandra, muito obrigada a todos por todas as risadas, conversas e cafés.
Em especial agradeço a Carol Lincoln, pelo imenso carinho que tem ao ensinar, e
pela orientação na escolha certa. A Cris, que me ensinou tudo, que me ajudou tanto. Obrigada
Cris, por toda paciência que teve. A Mara, pelo auxílio-bolsa do seu projeto, pela sabedoria,

pelos momentos “bizarros” e por todas as dicas e conversas, como sempre falo, você é
DEMAIS!
Aos funcionários do Centro de Biologia Molecular e Engenharia Genética.
Á todos que aceitaram participar desse trabalho e doaram o sangue. Sem eles esse
trabalho não seria realizado.
Ás agências financiadoras, FAPESP e CAPES pelo auxílio concedido.
Á todos que direta e indiretamente contribuíram de alguma forma para realização
desse trabalho.

“Por ai passa a escada espiral, que se abisma
e se eleva ao infinito. No vestíbulo há um
espelho, que fielmente duplica as
aparências”.
Jorge Luis Borges

RESUMO
Na infância e na adolescência, a risperidona é um medicamento de uso corrente no manejo de
diversas condições psicopatológicas. Embora eficaz, não é isenta de efeitos adversos tais
como ganho de peso, síndrome metabólica, alterações hormonais, que podem ser agravados
na interação com outras medicações. Os fatores genéticos são os agentes de risco que mais
influenciam o ganho de peso induzido por antipsicóticos e uma compreensão dos mesmos
pode permitir tratamentos adaptados a indivíduos de alto risco. Numerosos polimorfismos
genéticos vêm sendo avaliados quanto ao potencial de suscetibilidade aos efeitos adversos
relacionados ao tratamento com risperidona. No entanto, estes estudos são em geral
conduzidos em populações específicas, que não a brasileira. Neste trabalho, foram analisadas
variações nucleotídicas nos genes CYP2D6, LEP, LEPR, HTR2C, DRD2, MC4R e SCARB2,
para estimar a frequência de determinados genótipos em uma amostra da população brasileira,
comparando-as com as frequências de outras populações depositadas em bancos de dados
moleculares internacionais. A casuística foi composta por 317 indivíduos. Foram coletadas
amostras de sangue periférico total, seguida da extração de DNA genômico para determinação
das variantes nos genes descritos acima pela técnica de PCR em Tempo Real, com kits
TaqMan SNP Genotyping. Foram realizados testes como qui-quadrado para a comparação das
frequências encontradas na população em estudo com frequências em outras populações.
Testes estatísticos foram utilizados para se estimar o equilíbrio de Hardy-Weinberg através do
programa Haploview (BROAD Institute) disponível gratuitamente na WEB. Todas as single
nucleotide variations (SNVs) estudadas apresentaram distribuição em equilíbrio de Hardy-
Weinberg, exceto as rs7799039 e rs6318 nos genes LEP e HTR2C, respectivamente, não
sendo estas indicadas para estudos de associação. As frequências dos alelos menos frequentes
(MAFs) observadas, foram semelhantes às da população global constante da base de dados do
Projeto 1000 Genomas, exceto para as variantes no gene SCARB2 (rs3853188) e no DRD2
(rs6277) que, na população brasileira, se mostraram inferior e superior, respectivamente. Os
resultados descritos ressaltam a importância de estudar as frequências de variantes usadas em
estudos de associação na população brasileira, pois podem diferir de outras populações devido
à alta heterogeneidade dessa população. Futuros estudos poderão ser realizados com os dados
aqui obtidos e permitindo não só a identificação de marcadores genéticos de risco, mas
também a realização de estudos de associação desses marcadores como fatores
farmacogenéticos com os efeitos causados por risperidona, permitindo avanços na

individualização do tratamento psiquiátrico e facilitando o desenvolvimento de ferramentas
terapêuticas.
Palavras Chave: Risperidona. Polimorfismo Genético. Genótipo.

ABSTRACT
In childhood and adolescence, risperidone is a commonly used drug in the management of
several psychopathological conditions. While effective, it is not exempted of adverse effects
such as weight gain, metabolic syndrome, hormonal changes, which may be aggravated by the
interaction with other medications. Genetic factors are the most important agents of risk that
influence the weight gain induced by antipsychotic. Numerous genetic polymorphisms have
been evaluated regarding their potential susceptibility to the adverse effects to risperidone
treatment. However, these studies are generally conducted on a specific population different
from the Brazilian population. In this work, we analyzed nucleotide variations in genes such
as CYP2D6, LEP, LEPR, HTR2C, DRD2, MC4R e SCARB2, to estimate the frequency of
certain genotypes in a sample in the Brazilian population, compared with frequencies of other
populations deposited in DNA sequence databases. These data contribute to association
studies of pharmacogenetic factors, which may be used in the development of genetically
oriented personalized therapies in psychiatric practice in the future. Peripheral blood samples
of 317 individuals were collected, followed by genomic DNA extraction. To determine the
gene variants PCR Real-Time with TaqMan SNP Genotyping kits were used. X-squared test
was used to compare frequencies obtained for the Brazilian sample population here studied
with frequencies in other populations. Statistical tests using Haploview program were used to
estimate Hardy-Weinberg equilibrium for all SNVs. All SNVs were in Hardy-Weinberg
equilibrium, except the rs7799039 e rs6318 in LEP and HTR2C, respectively, indicating that
they are not suitable for association studies. The Minor Allele Frequency (MAF) observed for
each SNV was similar to the global population MAFs in the 1000 Genomes database.
However, those MAFs for SCARB2 (rs3853188) and DRD2 (rs6277) were, respectively,
lower and higher in Brazilian population when compared to the global MAFs. Results
described underscore the importance of studying the frequency of variants used in association
studies in the Brazilian population because it may differ from other populations due to its
highly heterogeneous composition. Future studies can be done with those data and will allow
improving the identification of genetic markers conferring risk, associating those markers
with pharmacogenetic factors and relating them to effects caused by medications. This
certainly will allow advances in individualized psychiatric treatment by facilitating the
development of therapeutic tools.

Keywords: Risperidone. Genetics Polymorphism. Genotype.

LISTA DE ABREVIATURAS
Arg – aminoácido arginina
CAAE- Certificado de apresentação para apreciação Ética
CYP2D6 – Gene do citocromo P450
CYP450 – citocromo P450
CBMEG – Centro de Biologia Molecular e Engenharia Genética
c. – posição do nucleotídeo no cDNA
ºC – graus Celsius
Cys – aminoácido cisteína
DNA – Ácido desoxirribonucléico
DRD2 – Gene Receptor D2 de dopamina
D2 – receptor de dopamina D2
EDTA – ácido etilenodiaminotetracético
FCM – Faculdade de Ciências Médicas
FRET – transferência de energia ressoante fluorescente
Gln – aminoácido glutamina
HDL- high density lipoprotein
HTR2C – Gene receptor 2C de serotonina
IMC – índice de massa corpórea
Kb – kilobases
LDL – Low density lipoprotein (lipoproteína de baixa densidade)
LEP – gene leptina
LEPR – gene receptor de leptina
MAF – Frequência do menor alelo
mRNA – RNA mensageiro
MC4R – gene receptor 4 de melanocortina
MgCl2 – cloreto de magnésio
mM – milimolar
MGB – minor grove-binder
mL – mililitro
µg - micrograma
µL - microlitro

pb – pares de bases
p. – posição do aminoácido na proteína
PCR – reação em cadeia da polimerase
pH – potencial hidrogeniônico
Ser – aminoácido serina
SDS – programa 7500 System Sequence Detection Software©
SM – síndrome metabólica
SNV – variação de único nucleotídeo
SCARB2 – gene receptor varredor classe B membro 2
Taq – Termus aquaticus
UV – ultravioleta
UNICAMP – Universidade Estadual de Campinas

LISTA DE FIGURAS
Figura 1: Esquema de ação de drogas atípicas e neurotransmissores. As drogas ou
neurotransmissores se ligam à superfície extracelular do receptor transmembranar (R). O
receptor acoplado a um agonista interage com a proteína G. Com a energia fornecida pela
hidrólise do GTP em GDP, a proteína G ativada então é capaz de interagir com um sistema
efetor (E) que em seguida desencadeará o efeito clínico. Adaptado de Carvalho (12).
Figura 2: Representação esquemática da ação da risperidona. Estão representados a
risperidona e seus antagonistas setoroninérgico, dopaminérgico, anti-H1 histamínicas e
antialfa-1 e alfa-2 adrenérgicas. Adaptado de Souza (17).
Figura 3: Esquema representando o metabolismo da risperidona por CYP2D6 (Isoenzima do
citocromo P450). Adaptado de http://psychopharmacologyinstitute.com/ (retirado em
10/10/2014).
Figura 4: Esquema representativo de um SNP (do inglês: single nucleotide polymorphysm)
ou SNV (do inglês: single nucleotide variation). Retirado de:
http://en.wikipedia.org/wiki/Single-nucleotide_polymorphism, em Agosto de 2014.
Figura 5: Esquema demonstrativo do processo farmacocinético e farmacodinâmico
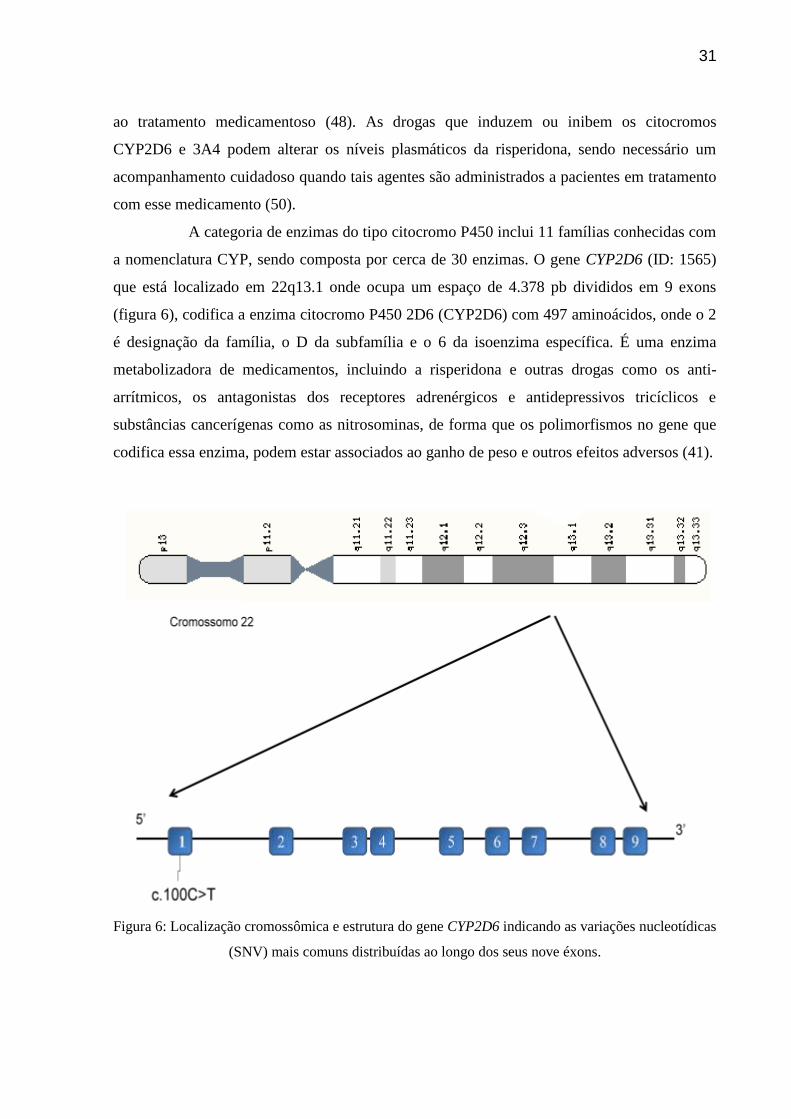
Figura 6: Localização cromossômica e estrutura do gene CYP2D6 indicando as variações
nucleotídicas (SNV) mais comuns distribuídas ao longo dos seus nove éxons.
Figura 7: Localização cromossômica e estrutura do gene do receptor D2 de dopamina
(DRD2) com a posição das variações nucleotídicas (SNVs) mais comuns.
Figura 8: Esquema dos receptores serotoninérgicos. Os segmentos retangulares verticais
representam a região hidrofóbica da proteína que ao longo da membrana forma o domínio de
ligação dos receptores. Na região citoplasmática se forma o domínio onde é acoplada a
proteína G. Adaptado de Rang et al. (78).
Figura 9: Representação esquemática do cromossomo onde o gene do receptor 2C de
serotonina (HTR2C) se localiza e sua estrutura identificando as duas variações nucleotídicas
mais estudadas.
Figura 10: Representação esquemática do cromossomo onde está o gene do receptor de
leptina (LEPR) e a estrutura do gene ressaltando as duas variações nucleotídicas mais
estudadas.
Figura 11: Possíveis funções da leptina e da via de regulação da massa de tecido adiposo.
Adaptado de He et al. (70).

Figura 12: Representação esquemática da estrutura do gene de leptina (LEP) indicando a
variação nucleotídica mais estudada e sua localização cromossômica.
Figura 13: Representação esquemática da estrutura do gene receptor 4 de melanocortina
(MC4R) indicando a região da variação nucleotídica mais estudada e o cromossomo no qual o
gene se localiza.
Figura 14: Ação do receptor varredor classe B tipo 2 (SCARB2) abundante no lisossomo e
endossomo. Adaptado de Yamayoshi et al. (116).
Figura 15: Representação esquemática da estrutura do gene receptor varredor classe B tipo 2
(SCARB2) indicando a variação nucleotídica mais estudada.
Figura 16: Esquema da reação de PCR em Tempo Real para detecção alélica.
(Disponível em:<http://www.appliedbiosystems.com/ cms_040027.pdf>. Acesso em
Fev/2014).
Figura 17: Gráfico de discriminação alélica obtido no programa 7500 Software v2.0.6, após a
reação de PCR em Tempo Real. Os homozigotos do alelo com maior frequência do SNV
rs6318 do gene HTR2C, está em azul (G/G), em verde encontram-se os heterozigotos (G/C) e
os homozigotos do alelo com menor frequência em vermelho (C/C).
Figura 18: Distribuição genotípica da população estudada e das populações depositadas no
banco do projeto 1000 Genomes para a SNV c.100C>T (rs1065852) do gene citocromo
P4502D6.
Figura 19: Distribuição genotípica da população estudada para o SNV c.-585A>G do gene do
receptor 2D de dopamina e de populações depositadas em banco de dados.
Figura 20: Distribuição genotípica da população estudada para o SNV c.957C>T do gene do
receptor 2D de dopamina e das populações depositadas em banco de dado.
Figura 21: Distribuição genotípica da população estudada para o SNV c.68G>C do gene do
receptor 2C de serotonina comparando com as populações de banco de dados (dados retirados
do Projeto 1000 Genomas).
Figura 22: Distribuição genotípica da população estudada e das populações depositadas em
banco de dados para o SNV c.-759C>T do gene do receptor 2C de serotonina.
Figura 23: Distribuição genotípica da população estudada e das populações depositadas em
banco de dados para o SNV c.668A>G do gene do receptor de leptina
Figura 24: Distribuição genotípica da população estudada e das populações depositadas em
banco de dados para o SNV c.-2548A>G do gene da leptina.

Figura 25: Distribuição genotípica da população estudada e das populações depositadas em
banco de dados para o SNV g.60183864T>C do gene receptor 4 de melanocortina.
Figura 26: Distribuição genotípica da população estudada e das populações depositadas em
banco de dados para o SNV c.275+7280A>C do gene receptor varredor classe B tipo 2.

LISTA DE TABELAS
Tabela 1. Genes e SNVs incluídos no estudo.
Tabela 2. Classificação por cor de pele dos indivíduos da casuística estudada.
Tabela 3. Distribuição genotípica dos polimorfismos dos genes HTR2C, LEP, LEPR, DRD2,
CYP2D6, SCARB2 e MC4R e os valores de MAF da população brasileira e da população
global.
Tabela 4. p value para equilíbrio de Hardy-Weinberg e frequência alélica.

SUMÁRIO
1. INTRODUÇÃO ...................................................................................................... 21
1.1 Antipsicóticos Típicos e Atípicos ............................................................... 21
1.2 Transtornos psiquiátricos ........................................................................... 22
1.3 Risperidona ................................................................................................ 23
1.4 SNV (Single Nucleotide Variations) ............................................................ 27
1.5 SNVs importantes relacionadas aos efeitos dos antipsicóticos atípicos .... 28
1.5.1 Genes da família citocromo P450 ........................................................... 30
1.5.2 Gene DRD2 ................................................................................................ 33
1.5.3 Gene HTR2C .............................................................................................. 36
1.5.4 Gene LEPR ................................................................................................ 39
1.5.5 Gene LEP .................................................................................................. 41
1.5.6 Gene MC4R ............................................................................................... 43
1.5.7 Gene SCARB2 ............................................................................................ 45
2. JUSTIFICATIVA ................................................................................................... 46
3. OBJETIVOS ........................................................................................................... 47
4. CASUÍSTICA ......................................................................................................... 48
5. MÉTODOS ............................................................................................................. 50
5.1 Genotipagem dos alelos pela Reação em Cadeia da Polimerase (PCR) em
Tempo Real ...................................................................................................... 50
5.2 Procedimentos para reação de PCR em tempo real .................................. 52
5.3 Análises estatísticas ................................................................................... 53
6. RESULTADOS E DISCUSSÃO ........................................................................... 55
6.1 CYP2D6 ..................................................................................................... 59
6.2 DRD2.......................................................................................................... 60
6.3 HTR2C ....................................................................................................... 63

6.5 LEP ............................................................................................................ 67
6.6 MC4R ......................................................................................................... 69
6.7 SCARB2 ..................................................................................................... 70
7. CONCLUSÃO ........................................................................................................ 72
8. REFERÊNCIAS ..................................................................................................... 73
9. ANEXOS ................................................................................................................. 90

21
1. INTRODUÇÃO
1.1 Antipsicóticos Típicos e Atípicos
Os antipsicóticos, também chamados de neurolépticos, são medicações que
inibem a função psicomotora; são substâncias químicas sintéticas capazes de atuar nas células
nervosas que regulam os processos psíquicos. Apesar da denominação, não têm suas ações
restritas ao tratamento das psicoses de início precoce (1). Essas medicações, que apresentam
mecanismos de ação bastante complexos no campo da psicofarmacologia clínica, são úteis,
em curto e médio prazo, no tratamento da agressividade e de outros problemas
comportamentais graves, como conduta, comportamento desafiador, autismo e outros
transtornos do desenvolvimento (1,2). São especialmente utilizados no tratamento de
transtornos de tiques, transtorno bipolar e transtorno obsessivo-compulsivo.
A história dos antipsicóticos teve início na década de 1940, com a descoberta por
acaso dos anti-histamínicos (citado em 3, 4). Em 1958 foi sintetizada a clozapina, na época o
melhor neuroléptico tricíclico caracterizado como atípico (citado em 3). No entanto, foi na
década de 1960 que se estabeleceu o importante papel do receptor de dopamina no
desenvolvimento da esquizofrenia. Assim, a maioria dos neurolépticos exerce a ação de
antagonistas de dopamina, embora este não seja único alvo possível da doença (4,5). Apenas
na década de 1970 se tornaria claro que a característica chave de todos os fármacos com
propriedades antipsicóticas era sua capacidade de bloquear receptores D2 de dopamina (2).
Mesmo sendo de indiscutível eficácia terapêutica, essa ação apresenta vários efeitos colaterais
indesejáveis (2).
A risperidona é uma droga antipsicótica classificada como atípica e utilizada para
o tratamento agudo e a longo prazo de pacientes com esquizofrenia (6). Embora seja um
antagonista potente dos receptores de dopamina e serotonina (7), pouco se conhece do
mecanismo molecular de sua ação. Além disso, o perfil farmacológico de drogas
antipsicóticas atípicas em geral sugere que tenham um mecanismo de ação mais complicado
(8). A risperidona foi introduzida em 1994 e tem sido usada para o tratamento de
esquizofrenia, de episódios maníacos de moderados a mais graves associados a distúrbio
bipolar, de comportamentos agressivos em distúrbios de conduta em crianças e adolescentes
entre outros (9).

22
Do ponto de vista clínico, um antipsicótico é definido como atípico ou de nova
geração e assim se distingue dos convencionais ou típicos, principalmente por se relacionar a
poucos sintomas extrapiramidais e/ou por possuir alguma ação terapêutica sobre sintomas
negativos (2). Os antipsicóticos atípicos, como mencionado anteriormente são antagonistas
que possuem afinidade com o receptor de dopamina e se ligam a receptores de serotonina
como agonistas inversos promovendo a estabilização do receptor na conformação inativa
(figura 1) (10,11). No Brasil dispõe-se da clozapina, risperidona, olanzapina e quetiapina.
Figura 1: Esquema de ação de drogas atípicas e neurotransmissores. As drogas ou neurotransmissores
se ligam à superfície extracelular do receptor transmembranar (R). O receptor acoplado a um agonista
interage com a proteína G. Com a energia fornecida pela hidrólise do GTP em GDP, a proteína G
ativada então é capaz de interagir com um sistema efetor (E) que em seguida desencadeará o efeito
clínico. Adaptado de Carvalho (12).
1.2 Transtornos psiquiátricos
Para se estabelecer o tratamento, na avaliação psiquiátrica de uma criança ou
adolescente, faz-se primeiramente um levantamento global de seu desenvolvimento, incluindo
dificuldades comportamentais e emocionais, passadas e atuais (13). Ao final da avaliação,
deve-se saber o quanto o desenvolvimento não satisfez as expectativas para sua idade
cronológica e é possível então se estabelecer o grau de prejuízo devido a fatores
comportamentais e emocionais (13).

23
Segundo Goodman (1), três grandes grupos de diagnósticos psicopatológicos são
especialmente relevantes para psiquiatras da infância e adolescência:
- transtornos emocionais ou internalizados (quadros ansiosos, fóbicos,
depressivos, somatizações, transtorno obsessivo-compulsivo) que causam preocupações,
medos, melancolia, dores, etc;
- transtornos do comportamento disruptivo ou externalizados (transtornos de
conduta, transtorno desafiador de oposição, hiperatividade) resultando em comportamentos
desafiadores, agressivos ou antissociais, que afetam os demais;
- transtornos do desenvolvimento (atrasos na fala e/ou leitura, retardo mental,
transtornos do espectro autista, enurese, encoprese) caracterizados por atrasos ou
anormalidades na aquisição dos marcos de funções psicomotoras, de linguagem, intelectuais e
sociais, como resultado de problemas na maturação biológica.
Goodman e Scott (14) afirmam ainda que existem transtornos como esquizofrenia
e outras psicoses de início precoce, anorexia nervosa, transtorno bipolar de humor com início
na infância ou adolescência, transtorno de vinculação com desinibição, síndrome de Gilles de
la Tourette e outros transtornos de tiques.
Nos transtornos psiquiátricos podem existir os sintomas positivos como, ilusões,
delírios, alucinações, pensamentos, fala e comportamento desorganizado e os sintomas
negativos como isolamento social, falta de interesse e déficit de atenção (2). Um antipsicótico
ideal seria aquele que, simultaneamente, tratasse os sintomas positivos referidos como
alterações de funções normais e atenuasse os sintomas negativos referidos como perda da
função normal para se evitar efeitos adversos extrapiramidais e hiperprolactinemia,
respectivamente. Os sintomas extrapiramidais ocorrem devido ao bloqueio de receptores D2
em uma de suas vias, a nigro-estriatal, ocorrendo no sistema nervoso central, onde parece
haver um balanço entre atividades dopaminérgicas e colinérgicas, os sintomas são distonia
aguda (espasmos musculares), parkinsonismo medicamentoso, acatisia (inquietação
psicomotora) e discinesia tardia (movimentos involuntários) (2,15).
1.3 Risperidona
A risperidona, como citado anteriormente, é um antagonista monoaminérgico com
alta afinidade por receptores dopaminérgico do tipo 2 (DRD2) e serotoninérgico do tipo 2 (5-

24
HT2), apesar de também ter ações sobre os receptores adrenérgicos alfa-1 e alfa-2 e
histamínico H1, como ilustrado na figura 2 (9, 16). O efeito da risperidona pode se assemelhar
aos antipsicóticos típicos quando usada em doses altas, pois pode levar a efeitos
extrapiramidais e síndrome metabólica, além de provocar elevação de prolactina, sedação e
ganho de peso (4, 16). Uma vez que alguns pacientes apresentam reações adversas, torna-se
maior o interesse em se identificar possíveis marcadores genéticos que possam direcionar
melhor o tratamento com essa medicação. Muitas variações nucleotídicas presentes em genes
que codificam as enzimas que metabolizam a droga, ou em genes que codificam os receptores
de dopamina e serotonina, ou mesmo em genes relacionados a hormônios como o da leptina,
entre outros, têm sido estudadas e avaliadas quanto à contribuição na eficácia terapêutica (4,
9).
Figura 2: Representação esquemática da ação da risperidona. Estão representados a risperidona e seus
antagonistas setoroninérgico, dopaminérgico, anti-H1 histamínicas e antialfa-1 e alfa-2 adrenérgicas.
Adaptado de Souza (17).
A risperidona é um fármaco bem absorvido pelo intestino. Em 92-94% dos
caucasianos é rapidamente metabolizada no fígado em 9-hidroxirrisperidona (9-OH-
risperidona) pelo complexo CYP2D6, como ilustrado na figura 3. Já os metabolizadores
pobres a convertem mais lentamente. A 9-OH-risperidona é o metabólito ativo na maioria dos
pacientes (18).

25
Figura 3: Esquema representando o metabolismo da risperidona por CYP2D6 (Isoenzima do
citocromo P450). Adaptado de http://psychopharmacologyinstitute.com/ (retirado em 10/10/2014).
Embora os antipsicóticos atípicos possuam vantagens em relação aos de primeira
geração, principalmente por evitarem sintomas extrapiramidais, eles também podem ocasionar
efeitos adversos potencialmente sérios (2). O ganho de peso, uma das complicações possíveis,
é um dos principais fatores de não aderência dos pacientes ao tratamento medicamentoso.
Outra preocupação relacionada à administração de risperidona diz respeito ao risco de
indução de hiperprolactinemia (19, 20).
O tratamento com risperidona está associado com ganho de peso moderado e
síndrome metabólica, em intensidade comparável à provocada por antipsicóticos típicos,
porém menor do que aquela induzida pela clozapina e pela olanzapina (21). Acredita-se que
esse ganho de peso ocorre devido ao aumento do apetite e ingestão de alimentos mediada por
receptores histamínicos e receptores de serotonina. O aumento da gordura vem acompanhado
do aumento de leptina (3). Quanto menor a idade e o peso antes do início do tratamento,
maior tende a ser o ganho de peso (21). A Síndrome metabólica (SM) é um transtorno
complexo associado a fatores de risco cardiovascular, como a resistência à insulina e a
deposição de gordura central. No gênero masculino, a eficácia, a duração do tratamento e a
dose de risperidona utilizada, além de fatores genéticos, parecem estar positivamente
associadas à intensidade do ganho de peso (21-23). Por outro lado, pacientes com
esquizofrenia e tabagistas parecem ter maior propensão ao desenvolvimento de síndrome
metabólica quando tratados com olanzapina e clozapina do que tratados com risperidona (24).
Indivíduos com síndrome metabólica possuem riscos até 3 vezes maior de morbidade

26
cardiovascular. A SM também é preditora de diabetes, e têm como sintomas a hipertensão,
níveis alterados de colesterol HDL e obesidade central (25).
Entre os antipsicóticos atípicos, a risperidona e a paliperidona são os que mais
provocam hiperprolactinemia, o que também parece se relacionar à dose (26). As alterações
podem ser verificadas em homens e mulheres, no entanto, as maiores elevações parecem
ocorrer entre as mulheres e em pacientes mais jovens (27). A prevalência de
hiperprolactinemia em doentes esquizofrênicos tratados com antipsicóticos típicos ou com
risperidona é de 42% para os homens e de 75% para as mulheres (28). Elevações nas
concentrações séricas de prolactina nem sempre se traduzem em sintomas clínicos e tendem a
aumentar naturalmente durante a puberdade (26), no entanto, os pacientes devem ser
monitorados em relação a esses efeitos, e as concentrações de prolactina devem ser aferidas,
caso os mesmos ocorram (26).
A hiperprolactinemia considerada um efeito secundário comum no tratamento
com antipsicóticos demonstra ter efeitos negativos na fertilidade, função sexual e densidade
óssea (28).
Há uma tendência maior de ocorrerem efeitos adversos extrapiramidais com a
risperidona do que com quaisquer outros antipsicóticos atípicos, mas isso pode ser
minimizado utilizando-se a menor dose com ação sobre os sintomas positivos (2). O uso
combinado de antipsicóticos típicos com risperidona aumenta o risco dos efeitos
extrapiramidais.
Devido ao bloqueio histaminérgico e adrenérgico, podem ocorrer sedação e
hipotensão. Em doses altas, podem ocorrer efeitos extrapiramidais, pois a medicação passa a
se comportar como antipsicótico típico (29).
Náuseas e vômitos, rinite, aumento de ansiedade, disfunção erétil, disfunção
orgásmica e aumento da pigmentação também podem se associar à utilização da risperidona
(2).
A risperidona pode exacerbar ou induzir sintomas de transtorno obsessivo-
compulsivo e tiques, provavelmente devido a antagonismo serotoninérgico, o que pode ser
manejado com a adição de inibidores seletivos de recaptação de serotonina (30).
Além das diferenças individuais em eficácia entre os antipsicóticos atípicos, existe
variação de indivíduo para indivíduo nos requisitos de dosagem referentes à sensibilidade aos
efeitos colaterais ou efeitos adversos. Assim, torna-se claro que as variações genéticas
influenciam na resposta individual e nos quadros de efeitos colaterais desses medicamentos

27
indicando a utilidade clínica dos testes genéticos atuais, bem como sinalizando as perspectivas
futuras de aplicação da farmacogenética e da farmacogenômica nesse tipo de tratamento (10).
1.4 SNV (Single Nucleotide Variations)
Diferentes versões de uma determinada sequência de DNA em um local
cromossômico são chamados de alelos. Quando os alelos são tão comuns que são encontrados
em mais de 1% dos cromossomos na população em geral, constituem o que é conhecido como
polimorfismo genético. Qualquer sítio no qual existam alelos múltiplos como componentes
estáveis da população é, por definição, polimórfico (31). Dentre os diferentes tipos de
polimorfismos, um tipo deles pode acontecer quando um nucleotídeo é substituído por outro
na sequência de DNA como mostra a figura 4. Este pode ser chamado de SNP (do inglês:
single nucleotide polymorphysm). Outra denominação para variações nucleotídicas com
significado mais amplo e que não se restringe à frequência ≥ 1% nas populações é a SNV (do
inglês: single nucleotide variation), (32).
Figura 4: Esquema representativo de um SNP (do inglês: single nucleotide polymorphysm) ou
SNV (do inglês: single nucleotide variation). Retirado de: http://en.wikipedia.org/wiki/Single-
nucleotide_polymorphism, em Agosto de 2014.

28
As SNVs são variações uniformemente distribuídas por todo o genoma, e
aparecem, cerca de uma vez a cada 100-300 pares de bases ao longo do genoma humano. Elas
podem ser encontradas em íntrons, éxons e sequências regulatórias no genoma (32).
A maioria das doenças é causada por interações complexas entre vários genes e
fatores ambientais. Assim, ao se estudar pequenas sequências de DNA, tem-se conseguido
associar as SNVs a determinadas características de doenças, bem como identificar genes
associados a uma determinada doença; sendo assim, é possível utilizá–las como marcadores
genéticos. Por este motivo o estudo de frequências de variações nucleotídicas, ou de
polimorfismos, em genes envolvidos no metabolismo de drogas torna-se importante, no
sentido de associá-los ao risco de efeitos adversos no uso dessas drogas, isto tornará viável o
estudo de fatores farmacogenéticos para o estabelecimento de um tratamento individualizado.
Assim, o estudo aprofundado dos SNVs se aplica não apenas ao processo de diagnóstico, mas
também à medicina preventiva (33).
Uma visão mais ampla da medicina pode ganhar nova importância à medida que
as indústrias farmacêuticas possam criar fármacos geneticamente orientados de acordo com as
necessidades de cada paciente (33,34).
1.5 SNVs importantes relacionadas aos efeitos dos antipsicóticos atípicos
A Farmacogenética é a ciência que investiga as variantes genéticas relacionadas
com a resposta dos indivíduos a determinadas medicações objetivando a prática terapêutica,
avaliando a base genética de resposta a fármacos possibilitando assim a identificação e dose
das drogas para cada indivíduo (35,36).
Com os avanços na área de pesquisa ao longo de 50 anos na tentativa de
individualização terapêutica, a farmacogenética evoluiu para farmacogenômica, que por sua
vez estuda como os marcadores genéticos podem ser usados nas respostas ao tratamento,
visando a descoberta de novos compostos com potencial de aplicação em tratamentos
individuais (37). Outro aspecto importante que diz respeito a farmacogenômica, é a
possibilidade de usar técnicas genômicas a fim de identificar novos genes regulados por
drogas que serão alvos terapêuticos para desenvolvimento de novos fármacos com menores
efeitos adversos e maior tolerabilidade (38).

29
Os fatores genéticos são os agentes de risco que mais influenciam no ganho de
peso induzido por antipsicóticos e uma melhor compreensão dos mesmos pode permitir
tratamentos adaptados para indivíduos que apresentem genótipos de maior risco (39).
Entretanto, não está totalmente claro se concentrações plasmáticas dos antipsicóticos de
segunda geração correlacionam-se com o ganho de peso (40,41).
A seleção dos genes para estudos de farmacogenética depende das proteínas que
eles codificam, sendo estas associadas a efeitos funcionais e processos fisiológicos da
medicação (42).
Os genes envolvidos com processos farmacocinéticos são aqueles onde os
polimorfismos podem estar associados com a resposta a drogas no metabolismo, bem como
no seu transporte, podendo levar a alterações na eficácia do tratamento, diminuindo ou
aumentando o nível plasmático. Um caso típico é o do gene CYP2D6, que tem sido um dos
candidatos mais prováveis por determinar traços mais simples que aqueles influenciados por
genes envolvidos em processos farmacodinâmicos, que estuda os mecanismos de ação da
droga, interagindo com os receptores, estabelecendo uma resposta clínica através do seu efeito
farmacológico. Assim, há sempre o interesse em se estudar os polimorfismos em
determinados genes envolvidos nos processos farmacogenéticos e associá-los aos resultados
terapêuticos ou aos efeitos colaterais de uma determinada droga. Além do CYP2D6, podem-se
citar os genes DRD2 e HTR2C como genes bastante estudados e envolvidos em processos
farmacodinâmicos (43, 44). Fatores genéticos podem modificar os efeitos farmacocinéticos e
dinâmicos dos fármacos desde a sua administração até sua interação fármaco-alvo como
ilustrado na figura 5.

30
Figura 5: Esquema demonstrativo do processo farmacocinético e farmacodinâmico.
1.5.1 Genes da família citocromo P450
O caminho de uma determinada droga no organismo depende de alguns fatores
como absorção, ligação a proteínas, distribuição, interação dos receptores e excreção, sendo
que cada um desses processos ocorre em regiões específicas (45). Esses processos, por sua
vez, são catalizados por enzimas, portanto os polimorfismos nos genes que produzem essas
enzimas podem influenciar diretamente a biotransformação das drogas (46). Quando ocorre
uma anormalidade em algum desses genes, podem ocorrer alterações na resposta ao processo
terapêutico chegando até à toxicidade, sendo a genética a base para analisar as variações
observadas na prática clínica do tratamento (47-49).
A maioria dos antipsicóticos atípicos são metabolizados pelas isoenzimas de fase I
do citocromo P450 (39). A CYP2D6 presente no fígado humano metaboliza 25% de todos os
medicamentos. Os estudos de farmacocinética têm como hipótese a associação da baixa
atividade do CYP2D6 com o aumento nos níveis séricos de antipsicóticos, o que pode levar
ao aumento de peso (41). O sistema citocromo P450 é responsável pelas reações de oxidação
de inúmeras drogas, sendo mais de 65% dos fármacos comuns metabolizados por CYP2D6.
Tendo esse papel fundamental na biotransformação, é provável que as variações no gene
CYP2D6 que levam a alterações na atividade da proteína possam causar mudanças na resposta
31
ao tratamento medicamentoso (48). As drogas que induzem ou inibem os citocromos
CYP2D6 e 3A4 podem alterar os níveis plasmáticos da risperidona, sendo necessário um
acompanhamento cuidadoso quando tais agentes são administrados a pacientes em tratamento
com esse medicamento (50).
A categoria de enzimas do tipo citocromo P450 inclui 11 famílias conhecidas com
a nomenclatura CYP, sendo composta por cerca de 30 enzimas. O gene CYP2D6 (ID: 1565)
que está localizado em 22q13.1 onde ocupa um espaço de 4.378 pb divididos em 9 exons
(figura 6), codifica a enzima citocromo P450 2D6 (CYP2D6) com 497 aminoácidos, onde o 2
é designação da família, o D da subfamília e o 6 da isoenzima específica. É uma enzima
metabolizadora de medicamentos, incluindo a risperidona e outras drogas como os anti-
arrítmicos, os antagonistas dos receptores adrenérgicos e antidepressivos tricíclicos e
substâncias cancerígenas como as nitrosominas, de forma que os polimorfismos no gene que
codifica essa enzima, podem estar associados ao ganho de peso e outros efeitos adversos (41).
Figura 6: Localização cromossômica e estrutura do gene CYP2D6 indicando as variações nucleotídicas
(SNV) mais comuns distribuídas ao longo dos seus nove éxons.

32
Segundo o Human Cytochrome P450 (CYP) Allele Nomenclature Committee
(2010) (http://www.cypalleles.ki.se/cyp2d6.htm), o gene CYP2D6 apresenta mais de 90
variantes alélicas, sendo que algumas das quais podem afetar significativamente a atividade
enzimática. Assim, essas variações se agrupam de acordo com a atividade enzimática de cada
isoforma de CYP2D6. De uma forma geral, quatro grupos são formados: 1) o de isoformas
não funcionais; 2) o de isoformas com função reduzida; 3) o de isoformas funcionais e, 4) o
de isoformas com função aumentada. Portanto, os riscos de efeitos colaterais decorrentes da
administração de drogas podem se correlacionar a diferentes genótipos, tais como os
portadores de dois alelos não funcionais, que são metabolizadores pobres; os portadores de
um alelo com função normal e o outro não funcional ou com os dois alelos com funções
reduzidas, sendo metabolizadores intermediários; os com dois alelos funcionais que
apresentam metabolização extensiva; e, os que apresentam alelos com aumento de atividade
enzimática podendo, nesse caso, ocorrer a falta de resposta à terapia, devido ao seu
metabolismo muito rápido (51, 52).
Tem-se especulado que metabolizadores pobres, na presença de nitrosaminas, que
são xenobióticos constituintes da fumaça de cigarro, são mais propensos ao câncer de pulmão
em comparação com metabolizadores extensivos (53). Em outro estudo mostrou-se que
pacientes com náuseas e vômitos em tratamento quimioterápico possuíam metabolizador
ultra-rápido (54). Esses estudos levam a concluir que a genotipagem pode ajudar no sentido
de se estabelecer a medicação adequada para os diferentes genótipos encontrados.
Alelos variantes de CYP2D6 apresentam-se amplamente distribuídos entre as
populações. Em particular, o alelo CYP2D6*10 é bastante frequente entre Asiáticos e
Caucasianos e representa a maior causa de baixa atividade enzimática em Orientais (55). Este
alelo se caracteriza pela troca c.100C>T (anteriormente designada de C188T), cujo registro no
banco de SNPs é rs1065852 (http://www.ncbi.nlm.nih.gov/projects/SNP). Essa variação
nucleotídica ocorre no exon 1 do gene CYP2D6 e leva à troca p.Pro34Ser formando uma
enzima instável com menor atividade (56,57).
Quando tratados com risperidona, os portadores do genótipo homozigoto da
variante menos frequente (T/T), que são metabolizadores pobres, apresentam maior taxa de
eventos adversos do que os metabolizadores extensivos cujo genótipo é C/C (58). Em
pacientes tratados com risperidona e portadores do alelo T, a atividade reduzida do CYP2D6
foi associada à maior probabilidade de ganho de peso induzida pelo antipsicótico.

33
Variantes de CYP2D6 podem também influenciar os níveis plasmáticos de
metabólitos ativos de drogas específicas, como por exemplo, os níveis plasmáticos do
endoxifeno que é produto da conversão de tamoxifeno mediada por CYP2D6 (51,59). Sobre a
variação interindividual observada no aumento de prolactina induzido pela risperidona,
podem-se também considerar explicações a partir de diferenças genéticas em seu metabolismo
(60). Os níveis de prolactina parecem estar correlacionados aos níveis do metabólito ativo 9-
OH-risperidona, mas não aos níveis séricos da risperidona em si (60, 61). Este achado pode
implicar que pacientes com deficiência de CYP2D6 apresentem menores níveis de
prolactinemia que metabolizadores normais ou ultrarrápidos, mas são necessários mais
estudos nesse sentido (60).
1.5.2 Gene DRD2
A dopamina é responsável pelo controle da secreção de prolactina, envolvida na
regulação do apetite, atividade motora e cognição. O sistema dopaminérgico parece ser o
maior componente nos mecanismos da variação do uso abusivo de substâncias. A função da
dopamina é mediada por dois grupos de receptores, D1 e D2.
Embora o antagonismo dopaminérgico seja parte integral da eficácia antipsicótica,
foi mais recentemente que se registraram evidências de que também pudesse estar envolvido
nos efeitos adversos relacionados ao ganho de peso (41). A disponibilidade de receptores D2
dopaminérgicos estriatais é significativamente menor em indivíduos obesos (62). Ademais, o
não funcionamento do gene DRD2 que codifica o receptor de dopamina é associado à
obesidade, conferindo nestes casos mais vulnerabilidade a dependências (39).
O gene do receptor D2 de dopamina (ID: 1813), localizado em 11q23.2, possui 8
éxons distribuídos em 66 kb dos quais cerca de 50 kb correspondem ao íntron que separa o
primeiro éxon dos demais exons codificantes. Sete éxons codificam uma proteína com 443
aminoácidos do receptor de dopamina D2 (figura 7). Este gene possui 339 variações
nucleotídicas descritas (http://www.ensembl.org/Homo_sapiens/Transcript/ProtVariations?
db=core;g=ENSG00000149295;r=11:113409615-113475691;t=ENST00000362072).
O DRD2 apresenta duas isoformas uma longa e outra curta as quais são geradas
por um splicing alternativo do exon 6 no mRNA. Ambas mostram funções distintas bem
como uma capacidade diferencial para aumentar a concentração de DRD2. A isoforma curta é
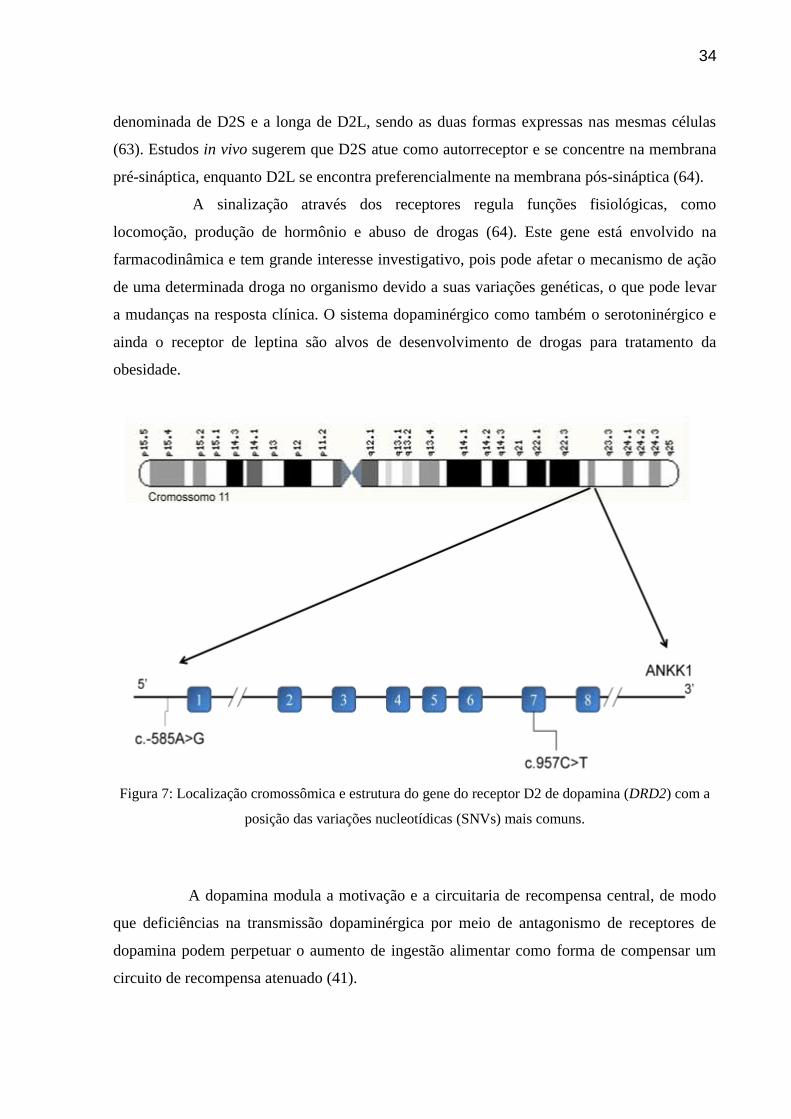
34
denominada de D2S e a longa de D2L, sendo as duas formas expressas nas mesmas células
(63). Estudos in vivo sugerem que D2S atue como autorreceptor e se concentre na membrana
pré-sináptica, enquanto D2L se encontra preferencialmente na membrana pós-sináptica (64).
A sinalização através dos receptores regula funções fisiológicas, como
locomoção, produção de hormônio e abuso de drogas (64). Este gene está envolvido na
farmacodinâmica e tem grande interesse investigativo, pois pode afetar o mecanismo de ação
de uma determinada droga no organismo devido a suas variações genéticas, o que pode levar
a mudanças na resposta clínica. O sistema dopaminérgico como também o serotoninérgico e
ainda o receptor de leptina são alvos de desenvolvimento de drogas para tratamento da
obesidade.
Figura 7: Localização cromossômica e estrutura do gene do receptor D2 de dopamina (DRD2) com a
posição das variações nucleotídicas (SNVs) mais comuns.
A dopamina modula a motivação e a circuitaria de recompensa central, de modo
que deficiências na transmissão dopaminérgica por meio de antagonismo de receptores de
dopamina podem perpetuar o aumento de ingestão alimentar como forma de compensar um
circuito de recompensa atenuado (41).

35
Estudos recentes usando variações nucleotídicas como alvos apontaram que
determinadas variantes de DRD2 estão envolvidas no ganho de peso induzido por
antipsicóticos, ou seja, variações genéticas podem levar a alterações na expressão dos
receptores dopaminérgicos, modificando a resposta clínica dos pacientes em uso de
antipsicóticos, causando, portanto, efeitos colaterais indesejáveis.
A variação c.-585A>G (rs1799978) tem sido associada a uma resposta mais
rápida ao tratamento com antipsicóticos em adultos com esquizofrenia (65). Este achado pode
ser visto como consistente com os resultados de que uma menor densidade de DRD2 poderia
estar associada a uma ocupação relativamente maior de receptores, resultando em uma melhor
resposta ao tratamento, porém poderia também se correlacionar a uma maior concentração de
prolactina, uma vez que o bloqueio dopaminérgico aumenta a sua liberação (65, 66). De
acordo com um estudo sobre variantes do receptor D2 e indução de hiperprolactinemia em
crianças e adolescentes, o genótipo A/A desta variante foi associado a um efeito protetor forte
contra a elevação da prolactina (26). O alelo A tem resposta mais rápida ao tratamento com
risperidona (65). A dopamina inibe a secreção de prolactina ao atuar nos receptores D2,
portanto drogas que bloqueiam esses receptores aumentam níveis de prolactina.
O polimorfismo c.957C>T (rs6277), localizado no sétimo éxon do gene,
representa uma mudança silenciosa no códon 319. O alelo T está associado com a redução e
estabilidade do mRNA (67) e com uma baixa disponibilidade de DRD2 no estriado em
indivíduos saudáveis (68). Esta mesma variante encontra-se em menor frequência em
indivíduos esquizofrênicos possuindo um efeito protetor contra esquizofrenia, comparado
com a variante C, associada à maior expressão de receptores DRD2 (69). O genótipo T/T foi
encontrado ainda em maior quantidade em indivíduos com sintomas de depressão (70). Este
SNV encontra-se também, associado com consumo de álcool (71,72). Em outro estudo com
pacientes com doença de Parkinson, em tratamento com Levodopa, encontrou-se uma
associação deste marcador genético com discinesia (73).
Os neurotransmissores de dopamina têm tido um papel chave nos estudos dos
sintomas da esquizofrenia, pois a ativação de receptores D2 por uma variedade de agonistas
provoca aumento da atividade motora e comportamental, agravando a esquizofrenia em
humanos (37, 65).

36
1.5.3 Gene HTR2C
Um dos neurotransmissores mais importantes que regula o balanço energético é a
serotonina. Os antipsicóticos atípicos associados ao maior risco de ganho de peso e de
aumento de prolactina possuem um antagonismo importante com receptores serotoninérgicos
(41) e variantes genéticas no gene codificante de receptores serotoninérgicos têm sido ligadas
à resposta terapêutica à clozapina e à risperidona (21, 74, 75). O aumento de peso ocorre em
pacientes submetidos ao uso de medicamento antipsicótico por terem seus receptores
bloqueados. Os antipsicóticos mostram alta propensão para induzir o ganho de peso, devido à
alta afinidade com o receptor de serotonina envolvidos na regulação de energia e massa
corpórea e por atuarem nas vias de sinalização de leptina e melanocortina, influenciando as
vias centrais que afetam os sistemas controladores de apetite e saciedade (41, 76).
Os receptores serotoninérgicos fazem parte de sete famílias acopladas à proteína
G (figura 8), dentre esses receptores, o HTR2C tem sido o mais estudado em relação ao ganho
de peso induzido por antipsicóticos, por se tratar de um importante receptor na regulação do
apetite (41, 77). Esse também está envolvido na regulação da ansiedade, locomoção e balanço
energético (77).
Figura 8: Esquema dos receptores serotoninérgicos. Os segmentos retangulares verticais representam a
região hidrofóbica da proteína que ao longo da membrana forma o domínio de ligação dos receptores.
Na região citoplasmática se forma o domínio onde é acoplada a proteína G. Adaptado de Rang (78).

37
O gene do receptor 2C de serotonina (ID: 3358), localizado na banda
cromossômica Xq24-q28, codifica o receptor 2C de 5-hidroxitriptamina com 458
aminoácidos. O gene se estende por 326 kb de DNA e possui 6 éxons sendo que a transcrição
se inicia no final do éxon 3. Cerca de 231 variações nucleotídicas já foram identificadas nesse
gene(http://www.ensembl.org/Homo_sapiens/Transcript/ProtVariations?db=core;g=ENSG00
000147246;r=X:114584078-114910061;t=ENST00000276198). Os receptores 5-HT2C são
densamente expressos em múltiplas regiões do cérebro e se mostraram envolvidos na
modulação dos efeitos de estímulos discriminativos provocados pela cocaína (79,80). Por
estar localizado no cromossomo X, os genótipos para variações no gene variam entre
mulheres, que podem apresentar genótipos homozigotos da variante mais frequente,
homozigotos da variante menos frequente ou heterozigotos, e homens que serão sempre
hemizigotos.
Figura 9: Representação esquemática do cromossomo onde o gene do receptor 2C de serotonina
(HTR2C) se localiza e sua estrutura identificando as duas variações nucleotídicas mais estudadas.
Lappalainen et al. (81) descreveram a variação c.68G>C (rs6318) que leva à
mutação p.Cys23Ser e é frequente na população caucasiana. Essa transversão ocorre numa
região do gene que codifica a parte hidrofóbica da proteína. Esta variação afeta a estrutura da
proteína receptora sendo que a substituição ocorre na região N-terminal extracelular mostrada
na figura 9 (42). O interesse nesta variação cresceu quando se observou que a substituição por

38
serina no resíduo 23 estava associada com o metabólito de degradação da norpinefrina, 3-
metoxi-4-hidroxifenilglicol (MHPG), tanto em indivíduos violentos com dependência de
álcool quanto na população controle.
Em um estudo de associação de genes candidatos à eficácia e a reações adversas à
risperidona, realizado em pacientes autistas, mulheres homozigotas ou homens hemizigotos
para a variante G tiveram um menor aumento de prolactina, quando comparados a indivíduos
homozigotos ou hemizigotos para o alelo C (82). Segundo Yuan et al. (83) essa variação não
está associada à obesidade, pois no seu estudo o SNV foi mais comum em não obesos e não
diabéticos.
A variação c.68G>C tem sido estudada também em relação a respostas à
clozapina, e se sugere que o alelo C pode ser indicativo de resposta positiva para tratamentos
com antipsicóticos. Estudos envolvendo pacientes com esquizofrenia indicam ainda que este
alelo possa estar de alguma forma envolvido na resposta clínica ou na expressão fenotípica da
doença, pois os pacientes com essa variação passam maior tempo hospitalizados (42).
A hiperprolactinemia induzida pela risperidona é principalmente derivada do
bloqueio de receptores D2, o que abole o efeito inibitório da dopamina sobre a liberação de
prolactina (2, 82, 84). Ao contrário da dopamina, a serotonina estimula a secreção de
prolactina e seus antagonistas reduzem a concentração de prolactina (84). Receptores 5HT2A
e 2C estão envolvidos na liberação de prolactina, e agonistas desses receptores aumentam a
secreção desse hormônio (85). O efeito antagonista exercido pela risperidona sobre esses
receptores inibe a secreção de prolactina, e dessa forma, compensando o aumento de
prolactina induzido pela risperidona ao se ligar a receptores D2 (82, 84).
A variação nucleotídica c.-759C>T (rs3813929) na região 5’ do gene HTR2C, é
um dos marcadores mais consistentes em relação à associação ao ganho de peso induzido por
antipsicóticos, apesar de haver resultados conflitantes entre os trabalhos na literatura (41, 83,
86-88).
No local dessa variação há a ligação a fatores de transcrição que regulam os níveis
de produção do receptor e mudanças nessa expressão podem mudar a regulação neuronal de
muitos processos fisiológicos, incluindo o apetite (89).
Yuan et al. (83) e Hill e Reynolds (86), mostraram que o alelo T encontra-se
associado com maior atividade promotora que o alelo C, sendo que este último apresentou
maior risco de ganho de peso em pacientes esquizofrênicos, podendo ser resultado da
diminuição da expressão neuronal do receptor. A variação C associou-se ainda a diabetes

39
melittus tipo 2 em um estudo com a população Asiática. Pooley et al. (90) encontraram uma
maior frequência do alelo C em grupos de mulheres obesas que foi associada com a
resistência à perda de peso quando em heterozigose.
Em um estudo com Chineses encontrou-se associação deste polimorfismo com o
ganho de peso em pacientes sob uso de risperidona. Como alelo T dessa variação encontra-se
associado com uma maior expressão do gene, aparentemente exerce um efeito protetor ao
ganho de peso em homens hemizigotos e mulheres heterozigotas, e exercendo também
resistência a Diabetes melittus tipo 2 (43, 75, 91) e em pacientes com esquizofrenia recebendo
risperidona (86) ou clozapina (92). Este efeito protetor pode estar relacionado ao maior nível
de transcrição deste gene, resultando em maior resistência à obesidade (83). Este mesmo alelo
demonstra-se associado com o aumento da tendência para discinesias (42). Já o alelo C dessa
variante está relacionado com uma menor taxa de transcrição, tornando os indivíduos mais
suscetíveis ao aumento de peso, pois a resposta do hipotálamo à leptina encontra-se
diminuída, logo, à saciedade também, o que pode levar ao desenvolvimento de diabetes
melitus tipo 2 (33). Há estudos, ainda, que mostram que pode estar ou não associado com a
síndrome metabólica, embora em um estudo com pacientes em uso de Clozapina que
apresentavam esta variante alélica a associação à síndrome metabólica não ficou clara (93),
mas houve influência no ganho de peso em pacientes com esquizofrenia (92).
1.5.4 Gene LEPR
O gene do receptor de leptina, LEPR (ID: 3953), está localizado em 1p31.3 ao
longo de cerca de 221 kb, com 20 éxons dos quais 18 são codificantes. Registram-se 660
variações nucleotídicas para esse gene
(http://www.ensembl.org/Homo_sapiens/Transcript/ProtVariations?db=core;
g=ENSG00000116678;r=1:65420652-65641559;t=ENST00000349533). Esse gene codifica o
receptor de leptina com 1165 aminoácidos cuja estrutura apresenta um único domínio
transmembranar. O receptor de leptina, da família de receptores de citocinas, está envolvido
na regulação do metabolismo da gordura e, participa de uma via hematopoiética. A leptina
atua através dos receptores, que funciona como parte de uma via de sinalização que pode
inibir a ingestão de alimento e/ou regular o gasto de energia, possuindo ainda várias funções
endócrinas. Mutações neste gene e no gene de leptina têm sido descritas e podem levar à

40
obesidade, hiperfagia, resistência à insulina e distúrbios de função imune em animais e
obesidade monogenética humana (94).
Figura 10: Representação esquemática do cromossomo onde está o gene do receptor de leptina (LEPR)
e a estrutura do gene ressaltando as duas variações nucleotídicas mais estudadas.
Algumas variações alélicas deste gene vêm sendo estudadas para avaliação de
ligação com obesidade, sendo que a variante c.668A>G (rs1137101), localizada no éxon 6 do
gene, que faz a troca p.Gln223Arg numa região extracelular, demonstrou-se associada a este
fenótipo em um estudo no Brasil (95). O alelo G deve predispor indivíduos saudáveis a
desenvolver distúrbios metabólicos. Segundo Yiannakouris et al. (96), houve prevalência do
alelo G na forma homozigótica entre os indivíduos na população da Grécia com sobrepeso e
obesos comparados com indivíduos com peso normal, suportando assim a hipótese de que
este alelo está associado à obesidade. Em um estudo com a população Mediterrânea, o alelo G
foi também mais frequente em obesos do que não obesos, apresentando maior nível de leptina
e maior Índice de massa corporal (IMC) do que os encontrados nos indivíduos com o alelo A.
Essa variante pode levar a alterações na via de sinalização e predispõem a um estado de
resistência a leptina. Segundo o autor, o genótipo A/A é um preditor significativo do índice de
massa corpórea.

41
1.5.5 Gene LEP
A leptina é um hormônio peptídico secretado exclusivamente pelo tecido adiposo,
que age no hipotálamo após se ligar a seus receptores e ativar uma cascata de sinais
secundários (41), como ilustrado na figura 11. Estes receptores inibem a ingestão alimentar e
aumentam o gasto energético (97). Altos níveis séricos de leptina inibem a expressão do RNA
mensageiro do neuropeptídeo Y (NPY) (98) e estimulam a liberação do hormônio estimulante
alfa-melanocítico (-MCH) (94). Este efeito leva à supressão do apetite, armazenamento de
energia e secreção de insulina (41). A falta de leptina leva ao aumento do apetite, obesidade e
disfunção neuroendócrina. A LEP é responsável pelo controle da ingestão alimentar atuando
no sistema nervoso central.
Figura 11: Possíveis funções da leptina e da via de regulação da massa de tecido adiposo. Adaptado de
He et al. (70).
O gene da leptina, LEP (ID: 3952) está localizado em 7q32.1 onde ocupa pouco
mais que 16 kb. Produz um mRNA de aproximadamente 3,5 kb em três éxons, com uma
sequência aberta de leitura correspondendo a 167 aminoácidos altamente conservados (figura
12). Registram-se para esse gene 83 variações nucleotídicas
(http://www.ensembl.org/Homo_sapiens/Transcript/ProtVariations?db=core;g=ENSG000001
74697;r=7:128241284-128257628;t=ENST00000308868). A proteína resultante possui um

42
papel importante na regulação do peso corporal. Esta proteína, que atua por meio do seu
receptor, funciona como parte de uma via de sinalização que pode inibir a ingestão de
alimentos e/ou regular o gasto de energia para manter a constância da massa adiposa. Ela está
envolvida na regulação de respostas imunes e inflamatórias. As mutações neste gene e/ou nas
suas regiões reguladoras podem causar obesidade. Este gene também tem sido associado ao
desenvolvimento de diabetes mellitus tipo 2 (http://ghr.nlm.nih.gov/gene/LEP). Variações no
gene LEP podem explicar a diferença interindividual nos níveis de leptina em indivíduos com
maior gordura corporal. O ganho de peso provocado por antipsicóticos ocorre mesmo na
presença de níveis altos de leptina.
Figura 12: Representação esquemática da estrutura do gene de leptina (LEP) indicando a variação
nucleotídica mais estudada e sua localização cromossômica.
Os níveis baixos de leptina estão envolvidos com a depressão. Pacientes
depressivos e ansiosos apresentam níveis mais baixos de leptina quando comparados com
indivíduos saudáveis. Alguns estudos afirmam que os níveis baixos de leptina têm sido
correlacionados com a tendência ao suicídio (99, 100).
A variação c.-2548A>G (rs7799039) na região promotora do gene LEP pode
afetar a expressão gênica no processo de transcrição o que levaria a uma alteração na secreção

43
desse hormônio no tecido adiposo (101). Há indicação de que o genótipo G/G da variação c.-
2548A>G possa ser importante para a regulação e manutenção dos níveis de leptina. Já o
genótipo A/A pode conduzir a uma maior expressão do mRNA e aumento dos níveis
plasmáticos de leptina. Em um estudo chinês o genótipo A/A foi associado com maior ganho
de peso e maior taxa de IMC, em pacientes sob uso da risperidona (102). Em mulheres esta
mesma variante foi associada com a obesidade extrema (103). Porém indivíduos com alelo G
são mais sensíveis ao ganho de peso devido à redução da sinalização da leptina no sistema
nervoso central (101). Para uma interação gene-gene, pode haver uma associação na maior
taxa de IMC em portadores do alelo C para o SNV rs3813929 do gene HTR2C, com os
genótipos G/G e A/G desta variante (93 ).
1.5.6 Gene MC4R
O receptor 4 de melanocortina (MC4R) é uma proteína G transmembranar
expressa no sistema nervoso central, primariamente no hipotálamo (41, 104). O MCR4 é um
regulador chave do controle leptinérgico da homeostase energética. Mutações heterozigóticas
no gene MC4R são encontradas em 0,5 - 6,3% em populações de obesos ou de início precoce,
sendo a causa mais frequente das formas monogênicas de obesidade humana (94, 105, 106).
A genética não está relacionada somente com a variação fenotípica, mas também
influencia no comportamento alimentar. Muitos genes codificam proteínas com papéis
fundamentais na ingestão alimentar e na homeostase energética, como é o caso do MC4R.
O gene MC4R (ID: 4160) possui somente um éxon com 1.438 pb e está localizado
na região18q22 onde ocupa cerca 1670 pb (figura 13). Este gene codifica uma proteína de 332
aminoácidos, o receptor 4 de melanocortina. Há aproximadamente 300 variações registradas
para esse gene (http://www.ensembl.org/Homo_sapiens/Transcript/
ProtVariations?db=core;g=ENSG00000166603;r=18:6037111060372775;t=ENST000002997
66).

44
Figura 13: Representação esquemática da estrutura do gene receptor 4 de melanocortina (MC4R)
indicando a região da variação nucleotídica mais estudada e o cromossomo no qual o gene se localiza.
A variação genética g.60183864T>C (rs17782313), cuja posição se baseia na
sequência NC_000018.10, está localizada 188 kb além do gene MC4R na direção 3’ e tem
sido associada com aumento de massa gordurosa, peso e risco de obesidade (107,108), sendo
também relacionada com maior ingestão de gorduras alimentares e de energia total, resultando
em maior aumento de peso a longo prazo em populações saudáveis (41). Uma região que se
sobrepõe ao MC4R foi identificada em um estudo genético amplo sobre obesidade em uma
população pediátrica livre de tratamentos medicamentosos. Esse resultado altamente
significativo repetiu-se em duas amostras independentes de adultos recebendo medicamentos
antipsicóticos em um estudo conduzido por Lett et al. (41). Estes estudos indicam que a
região 3’ do gene MC4R é uma forte candidata a estar envolvida no ganho de peso induzido
por antipsicóticos, sem desprezar a ação potencial de outros genes ainda não identificados
presentes nessa região (109).
Um estudo em chineses e europeus mostrou associação da variação com o índice
de massa corporal e risco de obesidade (110, 111). O genótipo C/C foi associado com o
aumento de consumo em grandes quantidades de alimentos em crianças da população

45
europeia (112), relatado também em mulheres com maior consumo de energia total e
gordurosa (113). Em um outro estudo foi encontrada uma maior frequência do alelo C em
crianças obesas comparados com não obesas (114).
1.5.7 Gene SCARB2
Apolipoproteínas são proteínas que se ligam a lipídios para facilitar seu transporte
e regular seu metabolismo (23). O receptor varredor classe B (SCARB), que possui os
membros 1 e 2, medeia a retirada seletiva de ésteres de colesteril das partículas de HDL do
colesterol (115). É uma glicoproteina tipo III, localizada nas limitações das membranas de
lisossomos e endossomos (figura 14) e possui um domínio transmembranar com N- e C-
terminal. Um estudo de análise fisiogenômica, numa abordagem previamente utilizada para
estudar a genética de drogas e respostas dietéticas, foi usado para comparar associações
genéticas com perfis de massa corporal observados em pacientes tratados com olanzapina e
risperidona, como uma tentativa de desvendar mecanismos contrastantes nas características
relacionadas ao ganho de peso de ambas as drogas (23). Perfis de ganho de peso em pacientes
tratados com risperidona foram nominalmente associados ao gene do SCARB2 (23, 41).
Figura 14: Ação do receptor varredor classe B tipo 2 (SCARB2) abundante no lisossomo e
endossomo. Adaptado de Yamayoshi et al. (116).

46
O gene do receptor varredor classe B tipo 2, SCARB2 (ID: 950) localiza-se em
4q21.1 ocupando pouco mais que 55 kb de DNA. Produz um mRNA de 4.763 pb em 12 éxons
e codifica a proteína de membrana lisossomo 2 que possui 478 aminoácidos (figura 15). Já se
encontram registradas cerca de 135 variações nucleotídicas para esse gene
(http://www.ensembl.org/Homo_sapiens/Transcript/ProtVariations?db=core;g=ENSG000001
38760;r=4:76158737-76213893;t=ENST00000264896). O SCARB2 é expresso em vários de
tecidos tais como cérebro e rins. Acredita-se que este gene desempenhe um papel na
biogênese e manutenção do compartimento endossomal e lisossomal
(http://www.ncbi.nlm.nih.gov/gene/950). Atua como um receptor lisossômico. Variantes
genéticas podem atuar como modificadores da expressão fenotípica e da gravidade da doença
de Gaucher. Outra doença que se acredita estar associada a esse gene é a Epilepsia Mioclônica
Progressiva 4 com ou sem insuficiência renal (117)
Uma maior expressão deste gene provoca alargamento de endossomo e prejudica
o tráfico de membrana para fora do compartimento.
Figura 15: Representação esquemática da estrutura do gene receptor varredor classe B tipo 2
(SCARB2) indicando a variação nucleotídica mais estudada.
A variação c.275+7280A>C (rs3853188) neste gene dentro do intron 2 tem sido
incluída em estudos para se avaliar os efeitos do tratamento com risperidona (23).

47
2. JUSTIFICATIVA
A risperidona é o antipsicótico atípico mais utilizado em psiquiatria infantil e
juvenil, sendo prescrita para uma grande variedade de condições psicopatológicas (2). Embora
os antipsicóticos atípicos possuam vantagens em relação aos de primeira geração,
principalmente por evitarem sintomas extrapiramidais, eles também podem ocasionar efeitos
adversos potencialmente sérios (2). O ganho de peso, uma das complicações possíveis, é um
dos principais fatores de não aderência dos pacientes ao tratamento medicamentoso. Outra
preocupação relacionada à administração de risperidona concerne ao risco de indução de
hiperprolactinemia (19, 20, 60).
Este trabalho faz parte de um estudo de pesquisa com possíveis fatores
farmacogenéticos que exerçam influência na ocorrência dos efeitos adversos no uso de
risperidona. Para que esse estudo de associação seja viável é necessário o conhecimento das
frequências alélicas e genotípicas, em uma amostra da população brasileira, dos marcadores
presentes nos genes envolvidos nos mecanismos de ação da risperidona. Uma vez que as
frequências disponíveis como referência são as descritas para outras populações podem não se
aplicar à população no Brasil dada a grande diversidade e alto grau de miscigenação da
referida população.
Assim, esse trabalho se fundamenta no conhecimento da distribuição em uma
amostra na população brasileira de variantes conhecidas e que podem estar associadas às
respostas clínicas de indivíduos em uso de medicamentos. É por meio dessa distribuição e
pelas associações que porventura vierem a ser demonstradas que os médicos poderão
estabelecer um tratamento, buscando dados estatísticos de frequências alélicas e genotípicas
em principais genes envolvidos com os efeitos de drogas, contribuindo também para o
desenvolvimento de práticas terapêuticas e psiquiátricas personalizadas, geneticamente
orientadas, que possam ter aplicações futuras diretas no tratamento medicamentoso de
indivíduos com transtornos mentais graves.

48
3. OBJETIVOS
Em indivíduos da população brasileira, obter as frequências de variantes
alélicas e as frequências genotípicas de SNVs (single nucleotide variations) que possam estar
associadas aos efeitos adversos da risperidona.
Comparar as frequências obtidas com as populações asiática, africana, europeia
e ameríndios depositadas nos bancos de dados do Projeto 1000 genomas e HapMap.

49
4. CASUÍSTICA
A casuística foi composta por 317 indivíduos, entre eles crianças, adolescentes e
jovens adultos com idades de 10 á 25 anos, de uma amostra da população brasileira recrutados
no Hospital de Clínicas da Unicamp. . Esses indivíduos são familiares de pacientes, pacientes,
estudantes e trabalhadores da Unicamp. Todos assinaram o termo de consentimento livre e
esclarecido (em anexo).
A pesquisa tem como objetivo avaliar as frequências de polimorfismos em genes
que não estão envolvidos no desenvolvimento de doença psiquiátrica visando a distribuição
genotípica em amostra população brasileira aleatoriamente recrutados, sendo assim indivíduos
portadores de transtornos psiquiátricos ou em uso de medicação não foram excluídos, visto
que a doença não interfere nos resultados da pesquisa.
Este trabalho foi conduzido sob aprovação do Comitê de Ética em Pesquisa da
Faculdade de Ciências Médicas-Unicamp, sob o número, CAAE: 14693513.5.0000.5404.

50
5. MÉTODOS
As amostras de DNA genômico foram obtidas a partir de 8 mL de sangue total
periférico colhidos em tubo com o anticoagulante ácido etilenodiamino tetra-acético (EDTA)
0,5 M pH 8,0. A extração do DNA genômico foi realizada pela técnica padronizada no
laboratório de Genética Molecular Humana-CBMEG (118) que resumidamente se dá através
de lise com proteinase K (Boehringer Mannhein, Alemanha) seguida de extrações com fenol
saturado com tampão Tris-HCl pH 8,0, com fenol/clorofórmio/álcool isoamílico na proporção
25:24:1 v/v e com clorofórmio/álcool isoamílico na proporção 24:1 v/v e, finalmente da
precipitação do DNA com etanol absoluto gelado.
As concentrações e a pureza (razão 260/280 >= 1,8 e razão 260/230 = 2,0) das
amostras extraídas foram medidas no equipamento Nanodrop 8000 - Multi-Sample Micro-
Volume UV-Vis Spectrophotometer (Thermo Scientific, USA), posteriormente testadas em gel
de agarose para verificar a fragmentação. O marcador de peso molecular utilizado foi o DNA
ladder de 1 Kb Plus (Invitrogen Corporation, Estados Unidos) em concentração de 0,15 ug/ul.
O gel foi imerso em solução diluída de brometo de etídio (0,5 μL/mL em água destilada)
durante 15 minutos, sendo visualizado em transluminador de luz ultravioleta e digitalizado,
utilizando o MiniBis Pro Bio-Imaging Systems (Uniscience).
5.1 Genotipagem dos alelos pela Reação em Cadeia da Polimerase (PCR) em Tempo
Real
Para genotipagem dos SNVs descritos na tabela 1, foi utilizado o ensaio de
discriminação alélica TaqMan® (Applied Biosystems, Foster City, CA - USA). Este ensaio,
além das frequências alélicas, determina se as amostras são homozigotas do alelo comum,
homozigotas do alelo raro ou heterozigotas.
51
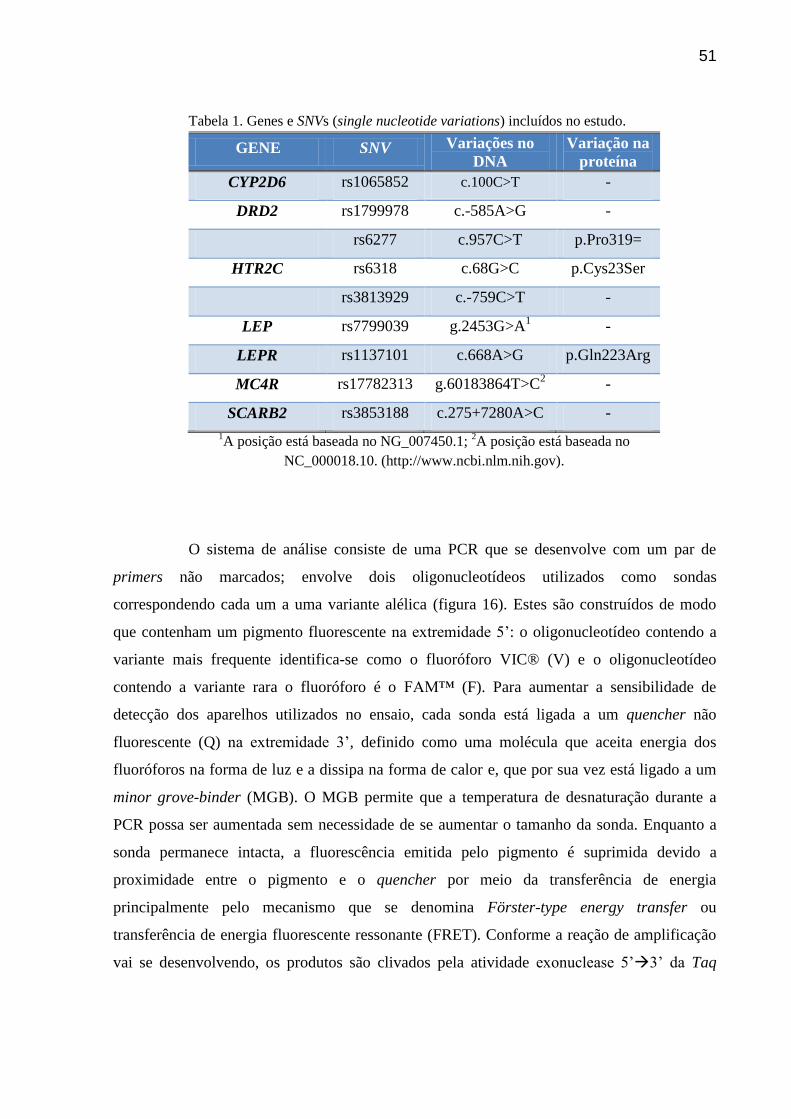
Tabela 1. Genes e SNVs (single nucleotide variations) incluídos no estudo.
GENE SNV Variações no
DNA
Variação na
proteína
CYP2D6 rs1065852 c.100C>T -
DRD2 rs1799978 c.-585A>G -
rs6277 c.957C>T p.Pro319=
HTR2C rs6318 c.68G>C p.Cys23Ser
rs3813929 c.-759C>T -
LEP rs7799039 g.2453G>A1 -
LEPR rs1137101 c.668A>G p.Gln223Arg
MC4R rs17782313 g.60183864T>C2 -
SCARB2 rs3853188 c.275+7280A>C -
1A posição está baseada no NG_007450.1;
2A posição está baseada no
NC_000018.10. (http://www.ncbi.nlm.nih.gov).
O sistema de análise consiste de uma PCR que se desenvolve com um par de
primers não marcados; envolve dois oligonucleotídeos utilizados como sondas
correspondendo cada um a uma variante alélica (figura 16). Estes são construídos de modo
que contenham um pigmento fluorescente na extremidade 5’: o oligonucleotídeo contendo a
variante mais frequente identifica-se como o fluoróforo VIC® (V) e o oligonucleotídeo
contendo a variante rara o fluoróforo é o FAM™ (F). Para aumentar a sensibilidade de
detecção dos aparelhos utilizados no ensaio, cada sonda está ligada a um quencher não
fluorescente (Q) na extremidade 3’, definido como uma molécula que aceita energia dos
fluoróforos na forma de luz e a dissipa na forma de calor e, que por sua vez está ligado a um
minor grove-binder (MGB). O MGB permite que a temperatura de desnaturação durante a
PCR possa ser aumentada sem necessidade de se aumentar o tamanho da sonda. Enquanto a
sonda permanece intacta, a fluorescência emitida pelo pigmento é suprimida devido a
proximidade entre o pigmento e o quencher por meio da transferência de energia
principalmente pelo mecanismo que se denomina Förster-type energy transfer ou
transferência de energia fluorescente ressonante (FRET). Conforme a reação de amplificação
vai se desenvolvendo, os produtos são clivados pela atividade exonuclease 5’3’ da Taq
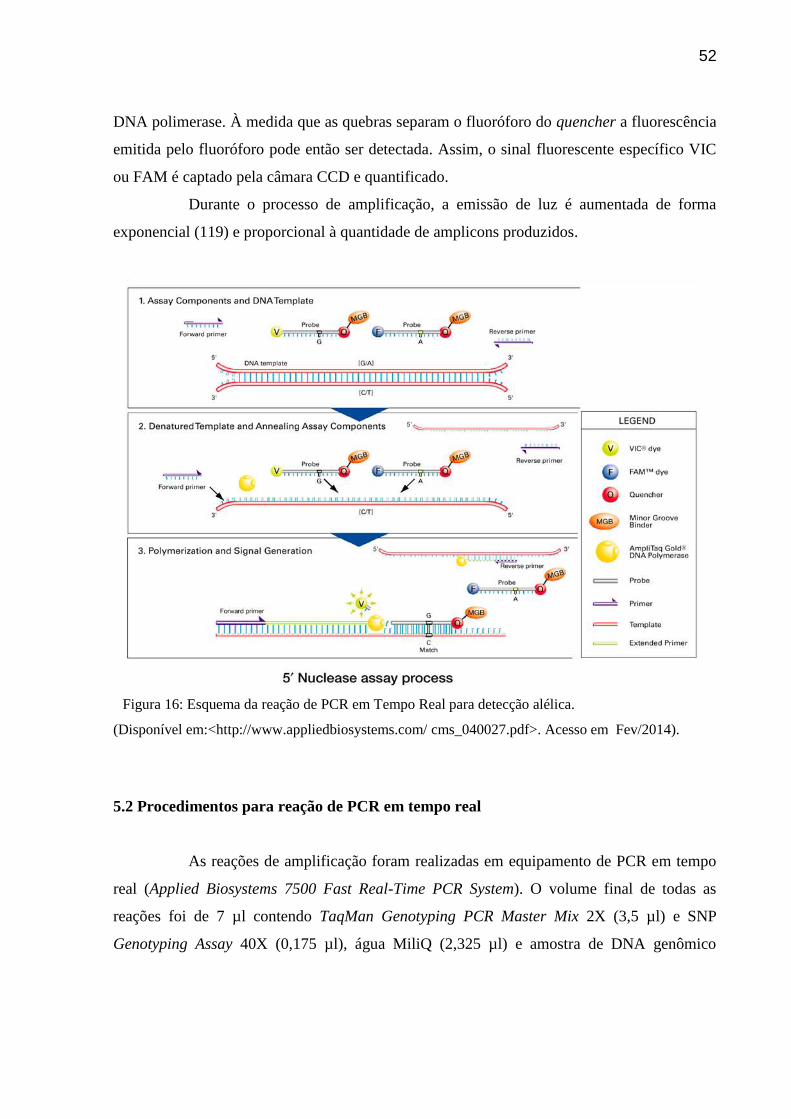
52
DNA polimerase. À medida que as quebras separam o fluoróforo do quencher a fluorescência
emitida pelo fluoróforo pode então ser detectada. Assim, o sinal fluorescente específico VIC
ou FAM é captado pela câmara CCD e quantificado.
Durante o processo de amplificação, a emissão de luz é aumentada de forma
exponencial (119) e proporcional à quantidade de amplicons produzidos.
Figura 16: Esquema da reação de PCR em Tempo Real para detecção alélica.
(Disponível em:<http://www.appliedbiosystems.com/ cms_040027.pdf>. Acesso em Fev/2014).
5.2 Procedimentos para reação de PCR em tempo real
As reações de amplificação foram realizadas em equipamento de PCR em tempo
real (Applied Biosystems 7500 Fast Real-Time PCR System). O volume final de todas as
reações foi de 7 µl contendo TaqMan Genotyping PCR Master Mix 2X (3,5 µl) e SNP
Genotyping Assay 40X (0,175 µl), água MiliQ (2,325 µl) e amostra de DNA genômico

53
(1µl/10ng). Este ensaio consiste em se amplificar um fragmento onde o SNV esteja contido e
ao mesmo tempo há a hibridização alelo-específica para cada variante alélica.
O Genotyping Master Mix possui todos os componentes para realizar o ensaio
com as sondas TaqMan® (Applied Biosystems), e contém 250 U de Amplitaq Gold® DNA
Polymerase, 100 U de AmpErase® UNG, dNTP, 10X TaqMan® Buffer A, 25 mM de MgCl2
em solução. O SNP Genotyping Assay contém um par de primers (sense e antisense), uma
sonda marcada com pigmento VIC® e uma sonda marcada com pigmento FAM™.
A concentração de DNA genômico foi de 10 ng/µl. Para genotipagem, as reações
foram realizadas em placa ótica de 96 poços (MicroAmp 0,1 mL, Applied Biosystems) de
acordo com as seguintes temperaturas e ciclos: desnaturação de 10 minutos a 95°C, seguida
de 40 ciclos a 95°C por 15 segundos e finalizando com uma extensão a 60°C por um minuto.
Os dados foram gravados e analisados utilizando o programa 7500 System
Sequence Detection Software© (SDS), no qual mostram os resultados num histograma dos
genótipos homozigotos X (eixo horizontal) versos os homozigotos Y (eixo vertical), ou
heterozigotos (na diagonal).
Essa técnica é muito indicada para análise de polimorfismo, pois permite a
quantificação do produto final amplificado, ao contrário da PCR convencional que é
qualitativo e estima os resultados por meio de corrida eletroforética. A técnica de PCR em
tempo real permite aumento no rendimento e diminui risco, possuindo elevada especificidade
e uma grande precisão. O tempo de reação é uma vantagem pois é menor quando comparado
com a PCR convencional, gerando o resultado através do gráfico de discriminação alélica, o
qual é facilmente interpretado. Portanto, podemos concluir em três dias os resultados da
genotipagem de até 95 pacientes para uma SNV em uma única placa. Dois dias são utilizados
para a extração de DNA separada em duas etapas, sendo necessários dois dias para sua
realização.
5.3 Análises estatísticas
Todos os dados dos polimorfismos foram arquivados em formato de tabela no
programa Excel 2007. O equilíbrio de Hardy–Weinberg (limite do valor de p = 0.01) dos
SNVs foram avaliados pelo software Haploview (BROAD Institute) para determinar se

54
determinados SNVs são viáveis para estudos de associação na população brasileira. Teste de
associação como qui-quadrado foi utilizado para comparar a frequência dos genótipos das
populações disponibilizadas em bancos de sequência de DNA com a população do presente
estudo, pelo ambiente de estatística R i386 3.1.1, versão (2014). Para os SNVs em
desequilíbrio de Hardy-Weinberg, verificou-se a distribuição em outras populações pelo
programa Haploview, para que se pudesse estabelecer as comparações.

55
6. RESULTADOS E DISCUSSÃO
As análises foram realizadas em 317 indivíduos participantes, dos quais 176 eram
homens e 141 mulheres. Esse número representa uma amostra da população brasileira, de
regiões que incluem o interior do estado de São Paulo e Sul de Minas Gerais. Estudos com um
tamanho amostral maior e de diferentes regiões do país, são necessários, a fim de se
determinar um valor significativo que represente a população brasileira em geral. Além disso,
a ampliação do número amostral também será mais significativo para demonstrar diferenças
nas frequências desses polimorfismos devido à miscigenação presente nessa população.
Na população brasileira, a heterogeneidade pode representar um problema para os
estudos de polimorfismos que possuem alguma relação com a raça (120, 121). Por esse
motivo estudá-la é importante para verificar as diferenças que ocorrem nas frequências
genotípicas quando comparadas com as outras populações.
O Brasil é um país onde há uma variada composição genética entre seus
habitantes, e essa diversidade resulta em uma população miscigenada composta por
contribuições de três grupos principais, os caucasianos, africanos e ameríndios. Nesse estudo
utilizamos o meio de classificação por cor de pele segundo o Instituto Brasileiro de Geografia
e Estatística (IBGE) (122), com a finalidade de melhor representar a população da região em
estudo. Essa classificação foi realizada por meio de auto-denominação, na qual os próprios
indivíduos declaram sua cor de pele. São elas: negra, parda, amarela e branca, como mostra na
tabela 2.
Tabela 2. Classificação por cor de pele dos indivíduos da casuística estudada.
COR INDIVÍDUOS (%)
Branco 254 (81%)
Negro 13 (4%)
Pardo 33 (10%)
Amarelo 17 (5%)

56
Utilizou-se o modo de discriminação alélica para verificação dos resultados dos
polimorfismos, conforme a Figura 17. Foram genotipados 317 indivíduos para 9 SNVs em
locos de 7 genes diferentes.
Figura 17: Gráfico de discriminação alélica obtido no programa 7500 Software v2.0.6, após a reação
de PCR em Tempo Real. Os homozigotos do alelo com maior frequência da variação nucleotídica do
rs6318 do gene receptor 2C de serotonina (HTR2C), está em azul (G/G), em verde encontram-se os
heterozigotos (G/C) e os homozigotos do alelo com menor frequência em vermelho (C/C).
Os resultados das frequências genotípicas observadas para as SNVs na população
brasileira estão representadas na tabela 3, juntamente com a comparação entre as frequências
do menor alelo (MAF) obtido para população estudada e a frequência correspondente a
população global obtida no site do projeto 1000 Genomas.
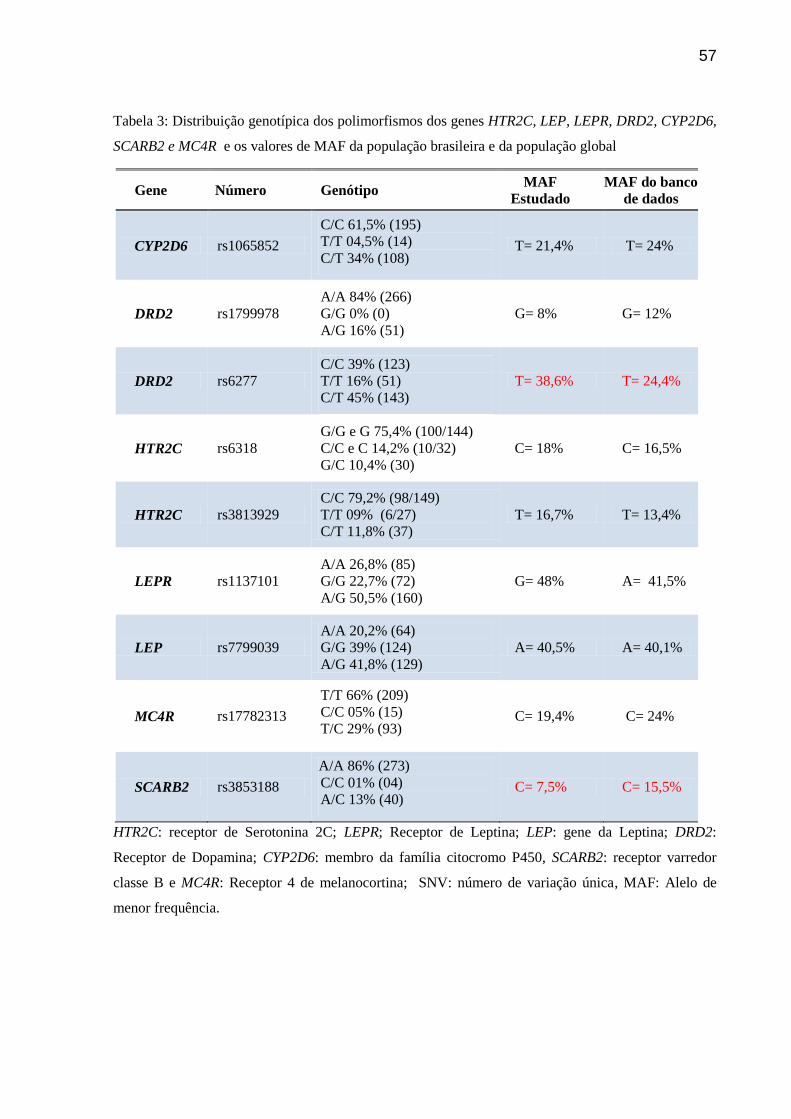
57
Tabela 3: Distribuição genotípica dos polimorfismos dos genes HTR2C, LEP, LEPR, DRD2, CYP2D6,
SCARB2 e MC4R e os valores de MAF da população brasileira e da população global
HTR2C: receptor de Serotonina 2C; LEPR; Receptor de Leptina; LEP: gene da Leptina; DRD2:
Receptor de Dopamina; CYP2D6: membro da família citocromo P450, SCARB2: receptor varredor
classe B e MC4R: Receptor 4 de melanocortina; SNV: número de variação única, MAF: Alelo de
menor frequência.
Gene Número Genótipo MAF
Estudado
MAF do banco
de dados
CYP2D6 rs1065852
C/C 61,5% (195)
T/T 04,5% (14)
C/T 34% (108) T= 21,4% T= 24%
DRD2 rs1799978
A/A 84% (266)
G/G 0% (0)
A/G 16% (51)
G= 8% G= 12%
DRD2 rs6277
C/C 39% (123)
T/T 16% (51)
C/T 45% (143)
T= 38,6% T= 24,4%
HTR2C rs6318
G/G e G 75,4% (100/144)
C/C e C 14,2% (10/32)
G/C 10,4% (30)
C= 18% C= 16,5%
HTR2C rs3813929
C/C 79,2% (98/149)
T/T 09% (6/27)
C/T 11,8% (37)
T= 16,7% T= 13,4%
LEPR rs1137101
A/A 26,8% (85)
G/G 22,7% (72)
A/G 50,5% (160)
G= 48% A= 41,5%
LEP rs7799039
A/A 20,2% (64)
G/G 39% (124)
A/G 41,8% (129)
A= 40,5% A= 40,1%
MC4R rs17782313
T/T 66% (209)
C/C 05% (15)
T/C 29% (93) C= 19,4% C= 24%
SCARB2 rs3853188
A/A 86% (273)
C/C 01% (04)
A/C 13% (40) C= 7,5% C= 15,5%
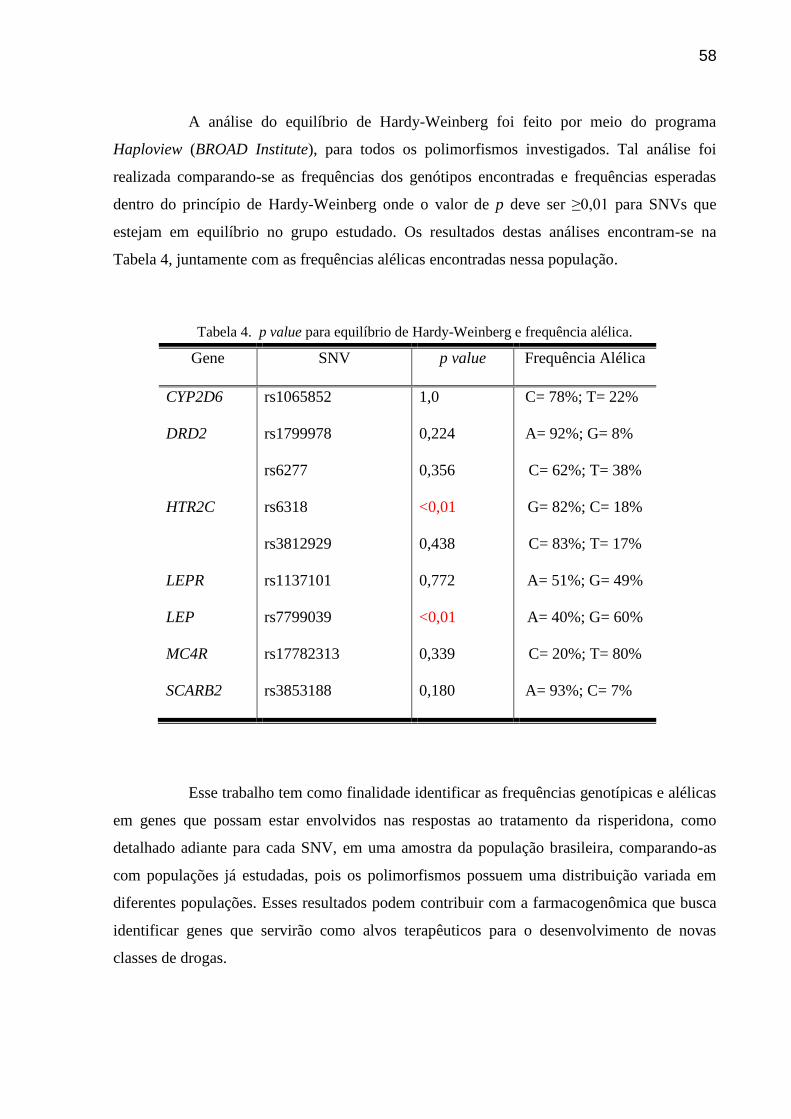
58
A análise do equilíbrio de Hardy-Weinberg foi feito por meio do programa
Haploview (BROAD Institute), para todos os polimorfismos investigados. Tal análise foi
realizada comparando-se as frequências dos genótipos encontradas e frequências esperadas
dentro do princípio de Hardy-Weinberg onde o valor de p deve ser ≥0,01 para SNVs que
estejam em equilíbrio no grupo estudado. Os resultados destas análises encontram-se na
Tabela 4, juntamente com as frequências alélicas encontradas nessa população.
Tabela 4. p value para equilíbrio de Hardy-Weinberg e frequência alélica.
Esse trabalho tem como finalidade identificar as frequências genotípicas e alélicas
em genes que possam estar envolvidos nas respostas ao tratamento da risperidona, como
detalhado adiante para cada SNV, em uma amostra da população brasileira, comparando-as
com populações já estudadas, pois os polimorfismos possuem uma distribuição variada em
diferentes populações. Esses resultados podem contribuir com a farmacogenômica que busca
identificar genes que servirão como alvos terapêuticos para o desenvolvimento de novas
classes de drogas.
Gene SNV p value Frequência Alélica
CYP2D6
DRD2
HTR2C
LEPR
LEP
MC4R
SCARB2
rs1065852
rs1799978
rs6277
rs6318
rs3812929
rs1137101
rs7799039
rs17782313
rs3853188
1,0
0,224
0,356
<0,01
0,438
0,772
<0,01
0,339
0,180
C= 78%; T= 22%
A= 92%; G= 8%
C= 62%; T= 38%
G= 82%; C= 18%
C= 83%; T= 17%
A= 51%; G= 49%
A= 40%; G= 60%
C= 20%; T= 80%
A= 93%; C= 7%

59
6.1 CYP2D6
A SNV c.100C>T rs1065852 que leva à troca p.Pro34Ser forma uma enzima
instável com menor atividade (56,57). O genótipo de maior frequência para essa SNV foi o
C/C com 61,5% (tabela 3), para o qual a atividade enzimática é considerada normal, sendo
denominados metabolizadores extensivos os indivíduos portadores desse genótipo. A
frequência do genótipo homozigoto T/T foi de 4,5%, sendo que estes são considerados
metabolizadores pobres. Uma vez que há a perda da atividade enzimática nesses casos,
haverá, portanto, maior risco de desenvolvimento de efeitos secundários, como o ganho de
peso quando expostos ao tratamento com risperidona. A frequência do alelo menos frequente
(MAF) foi 0, 2145 indicando uma frequência alélica semelhante à global descrita no banco de
referência do projeto 1000 Genomes
(http://www.ncbi.nlm.nih.gov/projects/SNP/snp_ref.cgi?rs=1065852) que é de 0,2380.
Dentre os dados disponíveis para várias populações, o da população asiática se
destaca por ser o alelo T o de maior frequência sendo este maior que 50%. Esta população foi
diferente quando comparada com esse estudo (p> 0,01). Se compararmos a frequência para
esse alelo obtida no presente trabalho com as de populações individuais, verificamos que a da
população brasileira aqui apresentada está mais próxima à da europeia (p = 0,747), do que as
das ameríndias (p = 0,055), e africanas (p > 0,01). Assim, pode-se dizer que a população
asiática está em maior risco de desenvolver efeitos adversos quando em uso de antipsicóticos
do que a população brasileira e latino-americana de origem hispânica (58).
Vários trabalhos avaliam a correlação da presença do alelo c.100T no genótipo
com a eficácia de tratamentos ou riscos de efeitos colaterais. Um estudo com a população do
Sri Lanka mostrou que 39% apresentaram o alelo com perda parcial da atividade enzimática
de CYP2D6, comparados a 10% dos Indianos e 40% nos Japoneses, confirmando, portanto,
uma maior frequência T na população do Sri Lanka e do Japão quando comparados com as
outras populações (123). No presente estudo a frequência de heterozigotos para esta variante
foi de 34% o que indica uma semelhança de frequência com as populações européias e norte-
americanas (figura 18).
Metabolizadores ultra-rápidos, que apresentam a amplificação da expressão do
gene, foram identificados com frequência maior que 7% na Espanha, provavelmente devido à
imigração Asiática, já no norte da Europa a prevalência é baixa (124-127).
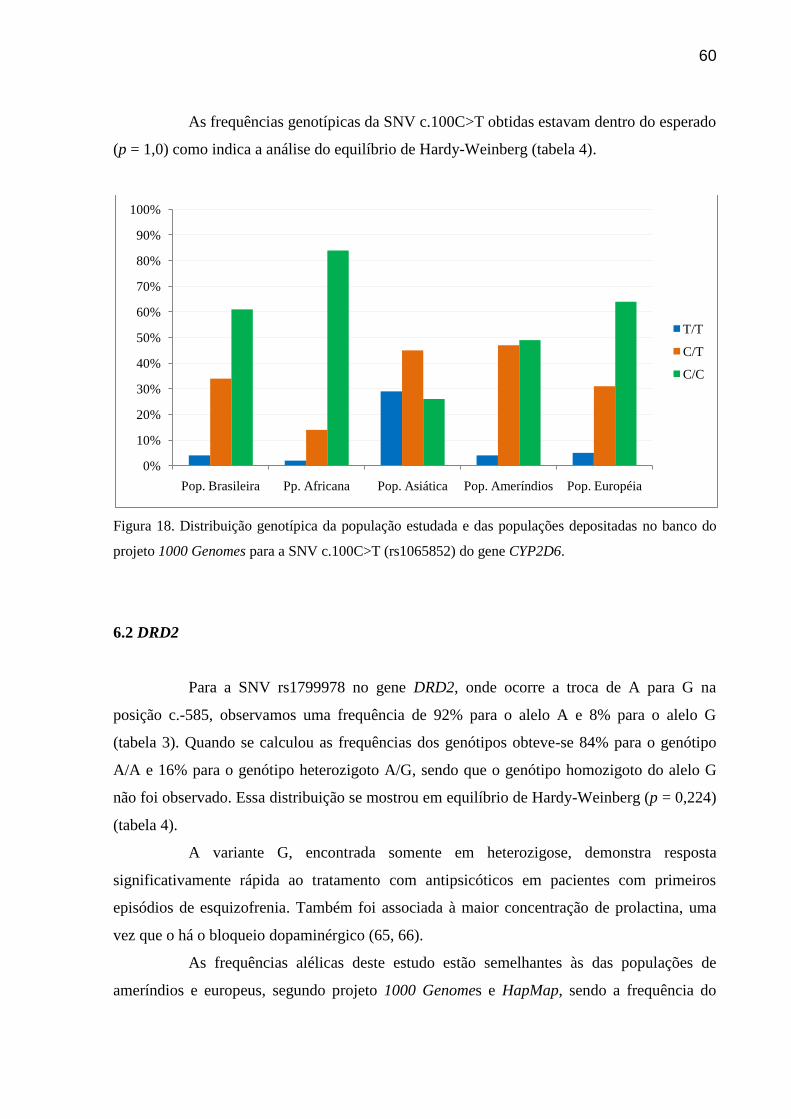
60
As frequências genotípicas da SNV c.100C>T obtidas estavam dentro do esperado
(p = 1,0) como indica a análise do equilíbrio de Hardy-Weinberg (tabela 4).
Figura 18. Distribuição genotípica da população estudada e das populações depositadas no banco do
projeto 1000 Genomes para a SNV c.100C>T (rs1065852) do gene CYP2D6.
6.2 DRD2
Para a SNV rs1799978 no gene DRD2, onde ocorre a troca de A para G na
posição c.-585, observamos uma frequência de 92% para o alelo A e 8% para o alelo G
(tabela 3). Quando se calculou as frequências dos genótipos obteve-se 84% para o genótipo
A/A e 16% para o genótipo heterozigoto A/G, sendo que o genótipo homozigoto do alelo G
não foi observado. Essa distribuição se mostrou em equilíbrio de Hardy-Weinberg (p = 0,224)
(tabela 4).
A variante G, encontrada somente em heterozigose, demonstra resposta
significativamente rápida ao tratamento com antipsicóticos em pacientes com primeiros
episódios de esquizofrenia. Também foi associada à maior concentração de prolactina, uma
vez que o há o bloqueio dopaminérgico (65, 66).
As frequências alélicas deste estudo estão semelhantes às das populações de
ameríndios e europeus, segundo projeto 1000 Genomes e HapMap, sendo a frequência do
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pp. Africana Pop. Asiática Pop. Ameríndios Pop. Européia
T/T
C/T
C/C
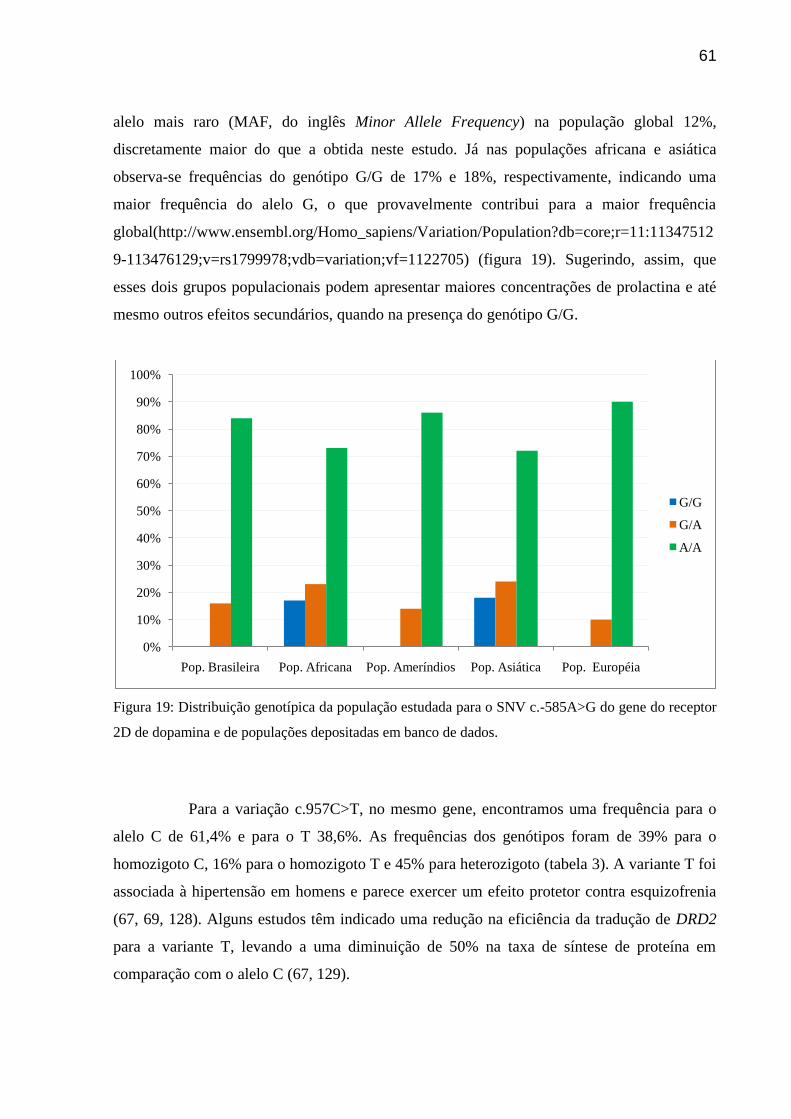
61
alelo mais raro (MAF, do inglês Minor Allele Frequency) na população global 12%,
discretamente maior do que a obtida neste estudo. Já nas populações africana e asiática
observa-se frequências do genótipo G/G de 17% e 18%, respectivamente, indicando uma
maior frequência do alelo G, o que provavelmente contribui para a maior frequência
global(http://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=11:11347512
9-113476129;v=rs1799978;vdb=variation;vf=1122705) (figura 19). Sugerindo, assim, que
esses dois grupos populacionais podem apresentar maiores concentrações de prolactina e até
mesmo outros efeitos secundários, quando na presença do genótipo G/G.
Figura 19: Distribuição genotípica da população estudada para o SNV c.-585A>G do gene do receptor
2D de dopamina e de populações depositadas em banco de dados.
Para a variação c.957C>T, no mesmo gene, encontramos uma frequência para o
alelo C de 61,4% e para o T 38,6%. As frequências dos genótipos foram de 39% para o
homozigoto C, 16% para o homozigoto T e 45% para heterozigoto (tabela 3). A variante T foi
associada à hipertensão em homens e parece exercer um efeito protetor contra esquizofrenia
(67, 69, 128). Alguns estudos têm indicado uma redução na eficiência da tradução de DRD2
para a variante T, levando a uma diminuição de 50% na taxa de síntese de proteína em
comparação com o alelo C (67, 129).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pop. Africana Pop. Ameríndios Pop. Asiática Pop. Européia
G/G
G/A
A/A
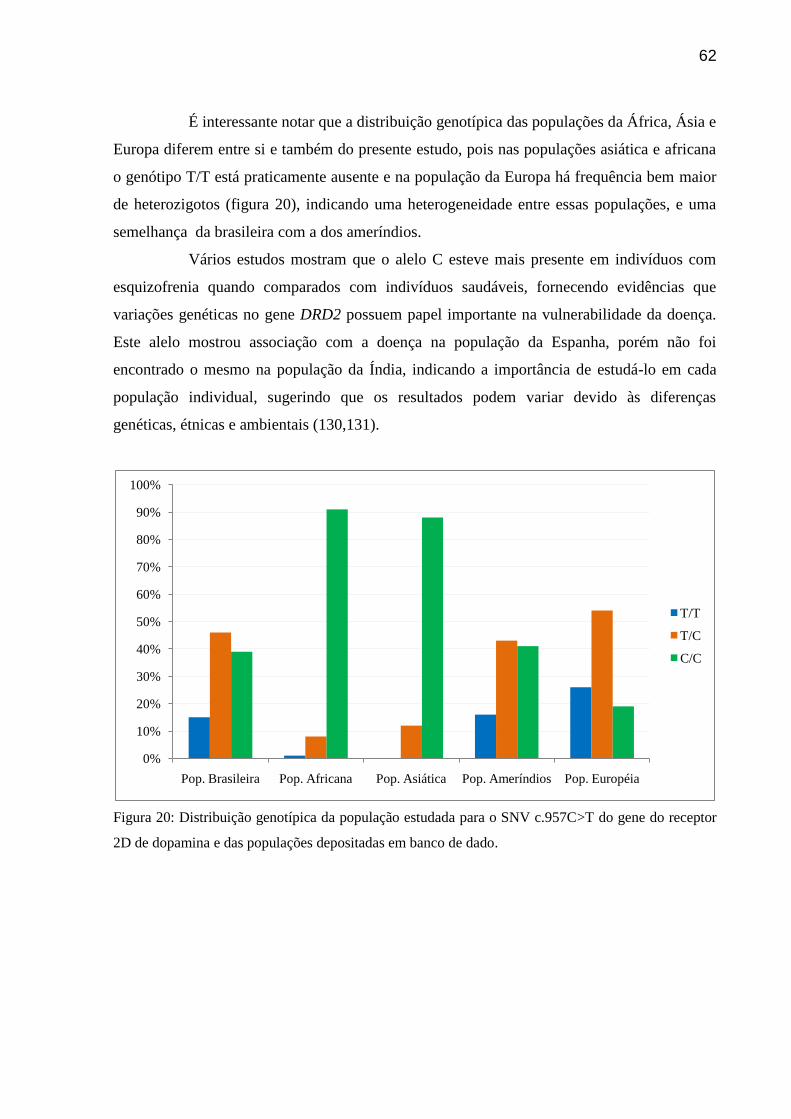
62
É interessante notar que a distribuição genotípica das populações da África, Ásia e
Europa diferem entre si e também do presente estudo, pois nas populações asiática e africana
o genótipo T/T está praticamente ausente e na população da Europa há frequência bem maior
de heterozigotos (figura 20), indicando uma heterogeneidade entre essas populações, e uma
semelhança da brasileira com a dos ameríndios.
Vários estudos mostram que o alelo C esteve mais presente em indivíduos com
esquizofrenia quando comparados com indivíduos saudáveis, fornecendo evidências que
variações genéticas no gene DRD2 possuem papel importante na vulnerabilidade da doença.
Este alelo mostrou associação com a doença na população da Espanha, porém não foi
encontrado o mesmo na população da Índia, indicando a importância de estudá-lo em cada
população individual, sugerindo que os resultados podem variar devido às diferenças
genéticas, étnicas e ambientais (130,131).
Figura 20: Distribuição genotípica da população estudada para o SNV c.957C>T do gene do receptor
2D de dopamina e das populações depositadas em banco de dado.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pop. Africana Pop. Asiática Pop. Ameríndios Pop. Européia
T/T
T/C
C/C

63
6.3 HTR2C
Duas SNVs presentes no gene HTR2C foram avaliadas neste estudo. Como o gene
está mapeado no cromossomo X, para os cálculos das frequências alélicas e genotípicas foram
levados em consideração os números de homens e mulheres que compuseram a amostra.
A SNV c.68G>C (rs6318) está localizada no gene HTR2C. Corresponde ao
resíduo 23 no primeiro domínio hidrofóbico da proteína e faz a troca do aminoácido serina
por cisteina.
As frequências estabelecidas para os indivíduos foram de 31, 5% para G/G e 45,
4% para 0/G, 3, 5% para C/C e 10,1% para 0/C e, 9, 5% para G/C, conforme indicado na
tabela 3. A MAF encontrada nesse estudo foi semelhante à da população global, mostrando
uma distribuição alélica dentro da esperada. Porém essa SNV não se encontra em equilíbrio
de Hardy-Weinberg (tabela 3), sugerindo que esta não seria adequada para estudos de
associação na população brasileira.
Segundo Reynolds et al. (42), o alelo C, em homens pode estar mais associado a
níveis elevados de cortisol e insulina do que o G. Este mesmo alelo associou-se também a
hiperprolactinemia em pacientes com uso de risperidona (132) e pode estar envolvido na
resposta clínica ou expressão fenotípica da esquizofrenia (40). Este SNV está envolvido com
distúrbios alimentares como a anorexia nervosa e influencia sintomas psicóticos e respostas a
drogas.
Em outras populações como africana, americana, europeia e asiática, a frequência
de G é mais elevada do que o alelo C, porém na população africana a diferença entre as duas
variantes é pequena, sendo 41% e 59% para C e G, respectivamente. Na população em geral a
frequência alélica de G é de 83% e a de C é de 17%. A frequência na população em estudo foi
de 80% para o alelo G e 20% para o alelo C, aproximando-se da frequência encontrada nas
populações europeia e americana. Todas as populações mantiveram o alelo G mais frequente,
no entanto todas com a presença de genótipos C/C, exceto a população asiática que apresenta
uma frequência de 1% para o alelo C, o qual os efeitos adversos estão associados. A figura 21
mostra a comparação das frequências genotípicas para as populações, ressaltando a
semelhança da amostra estudada com a população europeia para o SNV rs6318.
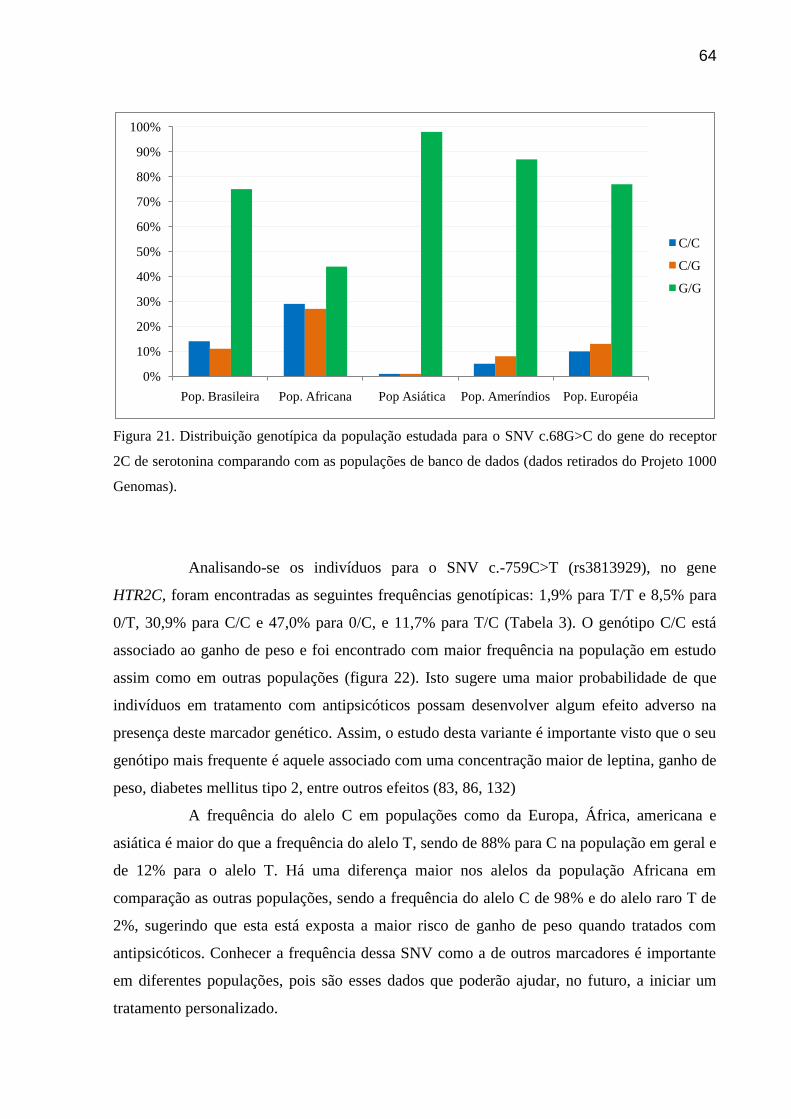
64
Figura 21. Distribuição genotípica da população estudada para o SNV c.68G>C do gene do receptor
2C de serotonina comparando com as populações de banco de dados (dados retirados do Projeto 1000
Genomas).
Analisando-se os indivíduos para o SNV c.-759C>T (rs3813929), no gene
HTR2C, foram encontradas as seguintes frequências genotípicas: 1,9% para T/T e 8,5% para
0/T, 30,9% para C/C e 47,0% para 0/C, e 11,7% para T/C (Tabela 3). O genótipo C/C está
associado ao ganho de peso e foi encontrado com maior frequência na população em estudo
assim como em outras populações (figura 22). Isto sugere uma maior probabilidade de que
indivíduos em tratamento com antipsicóticos possam desenvolver algum efeito adverso na
presença deste marcador genético. Assim, o estudo desta variante é importante visto que o seu
genótipo mais frequente é aquele associado com uma concentração maior de leptina, ganho de
peso, diabetes mellitus tipo 2, entre outros efeitos (83, 86, 132)
A frequência do alelo C em populações como da Europa, África, americana e
asiática é maior do que a frequência do alelo T, sendo de 88% para C na população em geral e
de 12% para o alelo T. Há uma diferença maior nos alelos da população Africana em
comparação as outras populações, sendo a frequência do alelo C de 98% e do alelo raro T de
2%, sugerindo que esta está exposta a maior risco de ganho de peso quando tratados com
antipsicóticos. Conhecer a frequência dessa SNV como a de outros marcadores é importante
em diferentes populações, pois são esses dados que poderão ajudar, no futuro, a iniciar um
tratamento personalizado.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pop. Africana Pop Asiática Pop. Ameríndios Pop. Européia
C/C
C/G
G/G
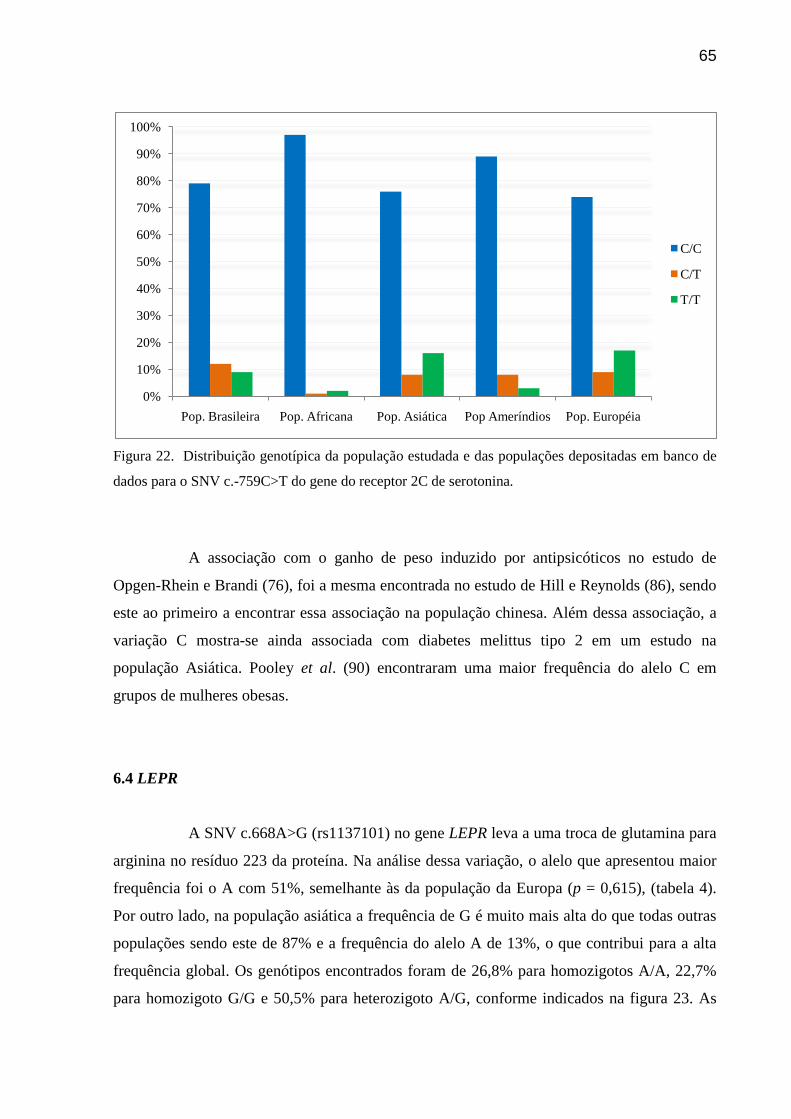
65
Figura 22. Distribuição genotípica da população estudada e das populações depositadas em banco de
dados para o SNV c.-759C>T do gene do receptor 2C de serotonina.
A associação com o ganho de peso induzido por antipsicóticos no estudo de
Opgen-Rhein e Brandi (76), foi a mesma encontrada no estudo de Hill e Reynolds (86), sendo
este ao primeiro a encontrar essa associação na população chinesa. Além dessa associação, a
variação C mostra-se ainda associada com diabetes melittus tipo 2 em um estudo na
população Asiática. Pooley et al. (90) encontraram uma maior frequência do alelo C em
grupos de mulheres obesas.
6.4 LEPR
A SNV c.668A>G (rs1137101) no gene LEPR leva a uma troca de glutamina para
arginina no resíduo 223 da proteína. Na análise dessa variação, o alelo que apresentou maior
frequência foi o A com 51%, semelhante às da população da Europa (p = 0,615), (tabela 4).
Por outro lado, na população asiática a frequência de G é muito mais alta do que todas outras
populações sendo este de 87% e a frequência do alelo A de 13%, o que contribui para a alta
frequência global. Os genótipos encontrados foram de 26,8% para homozigotos A/A, 22,7%
para homozigoto G/G e 50,5% para heterozigoto A/G, conforme indicados na figura 23. As
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pop. Africana Pop. Asiática Pop Ameríndios Pop. Européia
C/C
C/T
T/T
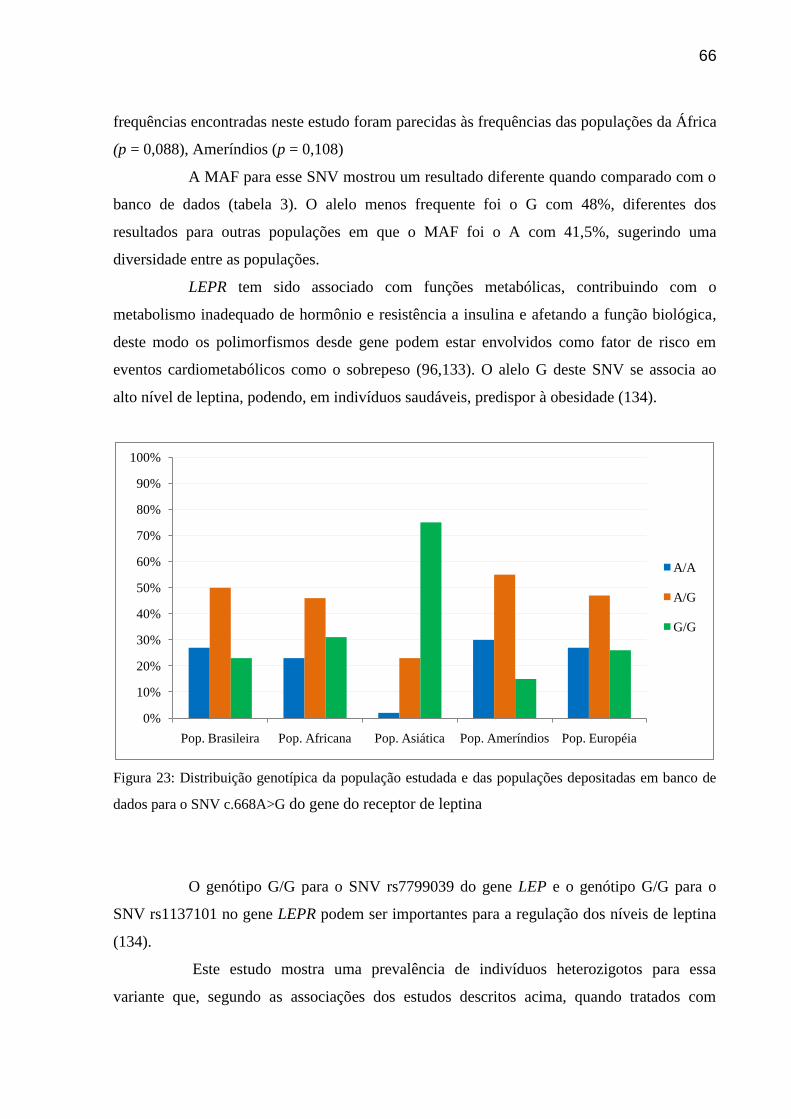
66
frequências encontradas neste estudo foram parecidas às frequências das populações da África
(p = 0,088), Ameríndios (p = 0,108)
A MAF para esse SNV mostrou um resultado diferente quando comparado com o
banco de dados (tabela 3). O alelo menos frequente foi o G com 48%, diferentes dos
resultados para outras populações em que o MAF foi o A com 41,5%, sugerindo uma
diversidade entre as populações.
LEPR tem sido associado com funções metabólicas, contribuindo com o
metabolismo inadequado de hormônio e resistência a insulina e afetando a função biológica,
deste modo os polimorfismos desde gene podem estar envolvidos como fator de risco em
eventos cardiometabólicos como o sobrepeso (96,133). O alelo G deste SNV se associa ao
alto nível de leptina, podendo, em indivíduos saudáveis, predispor à obesidade (134).
Figura 23: Distribuição genotípica da população estudada e das populações depositadas em banco de
dados para o SNV c.668A>G do gene do receptor de leptina
O genótipo G/G para o SNV rs7799039 do gene LEP e o genótipo G/G para o
SNV rs1137101 no gene LEPR podem ser importantes para a regulação dos níveis de leptina
(134).
Este estudo mostra uma prevalência de indivíduos heterozigotos para essa
variante que, segundo as associações dos estudos descritos acima, quando tratados com
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pop. Africana Pop. Asiática Pop. Ameríndios Pop. Européia
A/A
A/G
G/G

67
antipsicóticos, podem apresentar efeitos colaterais como o ganho de peso. Na população
asiática devido a prevalência de G, sugere-se que os indivíduos tenham risco de obesidade
maior do que nas outras populações.
6.5 LEP
Para o SNV c.-2548A>G (rs7799039) do gene LEP foi encontrada uma frequência
genotípica de 20,2% A/A, 39% G/G e de 41, 8% A/G (tabela 3). O alelo com maior
frequência no presente estudo foi o G com 60% enquanto o alelo com menor frequência foi de
40%. Na população geral, segundo os projetos HapMap e 1000 Genomes, o alelo A tem uma
frequência de 43% e o alelo G de 57%, sendo diferente somente na população asiática, com
75% para A e 25% para G. Este SNV mostrou-se em desequilíbrio de Hardy-Weinberg
(p<0,01), portanto não é recomendável de ser utilizado como marcador genético para estudos
de associação. Uma possível SNV que esteja em equilíbrio de ligação com este no mesmo
gene, seria o marcador ideal para o estudo de associação. Diferente dos resultados obtidos no
equilíbrio de Hardy-Weinberg para essa SNV, o MAF foi o mesmo encontrado em outras
populações, sendo o alelo A com 40% para ambos.
Em um estudo com a população da Espanha sugeriu que este polimorfismo pode
estar associado a outro polimorfismo funcional que realmente afete o fenótipo, com graus
diferentes de desequilíbrio de ligação entre as populações. A variante estudada não está em
uma região conservada, sendo sua importância funcional especulada. Nesse mesmo estudo
esta SNV mostrou-se em equilíbrio de Hardy-Weinberg (135).
A frequência genotípica encontrada neste trabalho manteve-se semelhante
somente quando comparada com a população de ameríndios (p= 0,864) do banco de dados do
projeto 1000 Genomes e HapMap (figura 24). No entanto, quando realizamos as estimativas
através do programa Haploview para a população de ameríndios, esta se manteve em
equilíbrio de Hardy-Weinberg (p=0,167). Todas as populações apresentadas na figura 24
mantiveram-se em equilíbrio de Hardy-Weinberg, diferentemente da população estudada.
Portanto podemos dizer que esse desequilíbrio se deve aos modificadores de efeito, entre eles
os fatores ambientais e a etnia da população, que é bastante miscigenada.
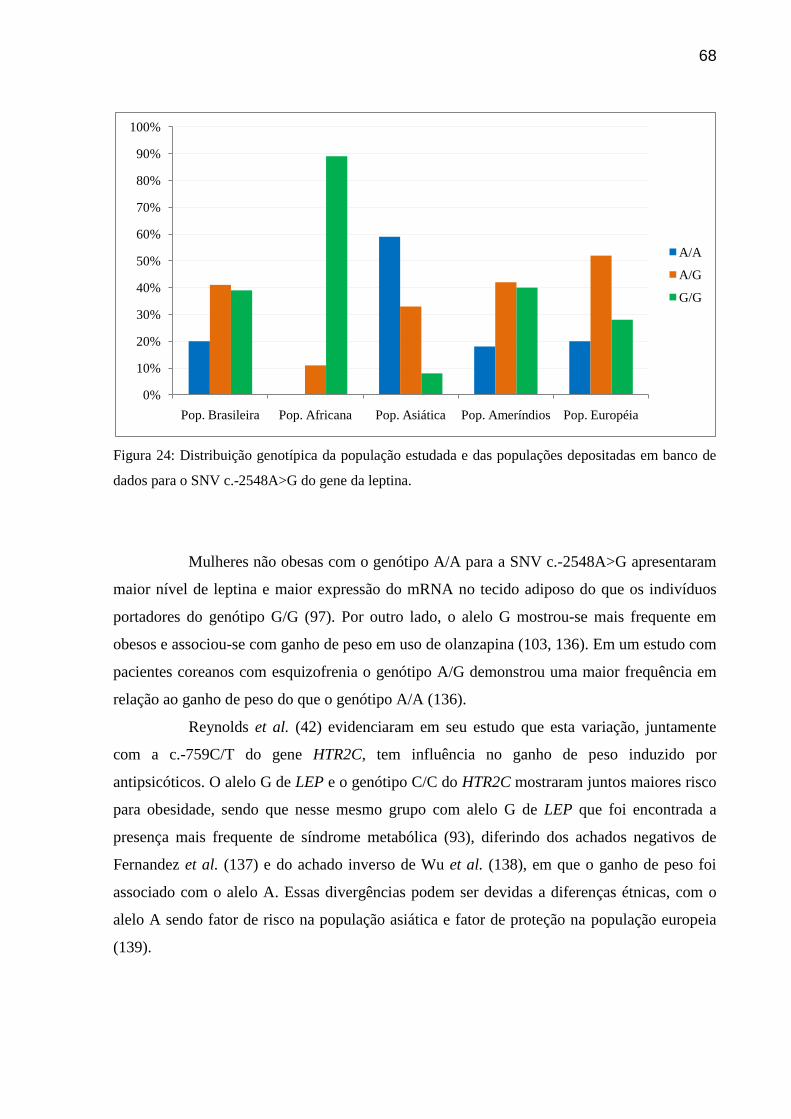
68
Figura 24: Distribuição genotípica da população estudada e das populações depositadas em banco de
dados para o SNV c.-2548A>G do gene da leptina.
Mulheres não obesas com o genótipo A/A para a SNV c.-2548A>G apresentaram
maior nível de leptina e maior expressão do mRNA no tecido adiposo do que os indivíduos
portadores do genótipo G/G (97). Por outro lado, o alelo G mostrou-se mais frequente em
obesos e associou-se com ganho de peso em uso de olanzapina (103, 136). Em um estudo com
pacientes coreanos com esquizofrenia o genótipo A/G demonstrou uma maior frequência em
relação ao ganho de peso do que o genótipo A/A (136).
Reynolds et al. (42) evidenciaram em seu estudo que esta variação, juntamente
com a c.-759C/T do gene HTR2C, tem influência no ganho de peso induzido por
antipsicóticos. O alelo G de LEP e o genótipo C/C do HTR2C mostraram juntos maiores risco
para obesidade, sendo que nesse mesmo grupo com alelo G de LEP que foi encontrada a
presença mais frequente de síndrome metabólica (93), diferindo dos achados negativos de
Fernandez et al. (137) e do achado inverso de Wu et al. (138), em que o ganho de peso foi
associado com o alelo A. Essas divergências podem ser devidas a diferenças étnicas, com o
alelo A sendo fator de risco na população asiática e fator de proteção na população europeia
(139).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pop. Africana Pop. Asiática Pop. Ameríndios Pop. Européia
A/A
A/G
G/G
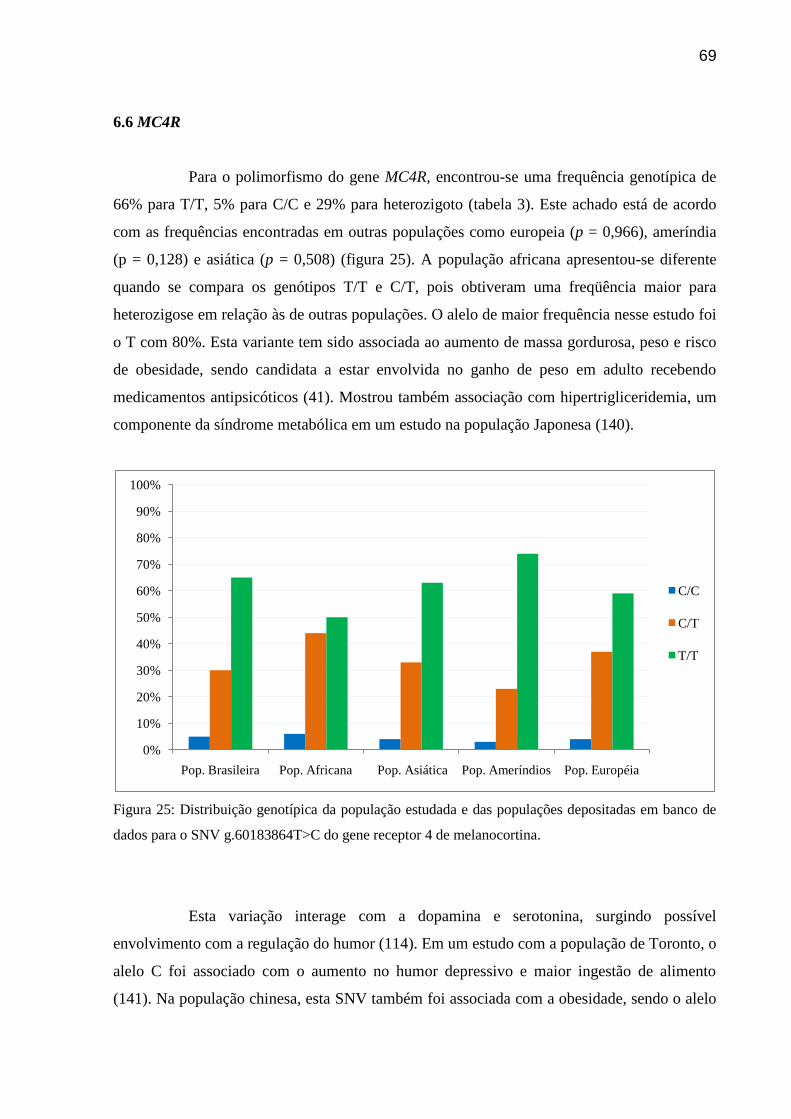
69
6.6 MC4R
Para o polimorfismo do gene MC4R, encontrou-se uma frequência genotípica de
66% para T/T, 5% para C/C e 29% para heterozigoto (tabela 3). Este achado está de acordo
com as frequências encontradas em outras populações como europeia (p = 0,966), ameríndia
(p = 0,128) e asiática (p = 0,508) (figura 25). A população africana apresentou-se diferente
quando se compara os genótipos T/T e C/T, pois obtiveram uma freqüência maior para
heterozigose em relação às de outras populações. O alelo de maior frequência nesse estudo foi
o T com 80%. Esta variante tem sido associada ao aumento de massa gordurosa, peso e risco
de obesidade, sendo candidata a estar envolvida no ganho de peso em adulto recebendo
medicamentos antipsicóticos (41). Mostrou também associação com hipertrigliceridemia, um
componente da síndrome metabólica em um estudo na população Japonesa (140).
Figura 25: Distribuição genotípica da população estudada e das populações depositadas em banco de
dados para o SNV g.60183864T>C do gene receptor 4 de melanocortina.
Esta variação interage com a dopamina e serotonina, surgindo possível
envolvimento com a regulação do humor (114). Em um estudo com a população de Toronto, o
alelo C foi associado com o aumento no humor depressivo e maior ingestão de alimento
(141). Na população chinesa, esta SNV também foi associada com a obesidade, sendo o alelo
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pop. Africana Pop. Asiática Pop. Ameríndios Pop. Européia
C/C
C/T
T/T

70
C o fator de risco (111). Os genótipos C/C e C/T foram associados com aumento de risco de
diabetes tipo 2 em mulheres caucasianas de descendência europeia, sendo também associados
à compulsão alimentar, fome excessiva e hiperfagia (113).
6.7 SCARB2
A variação c.275+7280A>C no gene SCARB2, dentro do íntron 2, tem sido
incluída em estudos para se avaliar os efeitos decorrentes do tratamento com risperidona, pois
perfis de ganho de peso em pacientes tratados com risperidona foram associados ao gene (23).
As frequências observadas para esta SNV foram de 86% para o genótipo A/A, 1% para C/C e
13% para o heterozigoto (tabela 3). Frequências semelhantes às deste estudo para os alelos A
e C foram encontradas na população europeia, sendo A 93% e C 7%. As frequências
observadas em nosso estudo também não diferiram muito da observada para a população
ameríndia, mas as populações da Ásia e da África apresentaram maior frequência do genótipo
heterozigoto (figura 26).
Este receptor participa de processos de mediação da retirada seletiva de ésteres de
colesterol das partículas de high density lipoprotein (HDL) colesterol, por isso estudá-lo é
importante, pois polimorfismos desse gene podem estar envolvidos na ocorrência de
problemas no metabolismo de lipídeos (41, 115).

71
Figura 26: Distribuição genotípica da população estudada e das populações depositadas em banco de
dados para o SNV c.275+7280A>C do gene receptor varredor classe B tipo 2.
A necessidade de fazer testes genéticos para determinação da conduta terapêutica
abrirá uma série imensa de problemas éticos, legais, sociais e econômicos. A todas as
especialidades da medicina, dentre elas a psiquiatria, deverão se manter a par dos avanços da
farmacogenômica, pois essa não será só um avanço científico mas deverá estar junto da
prática médica no que diz respeito a condutas de tratamento. Todo corpo clínico terá que ter
conhecimento dessa área para poder prescrever tratamentos adaptados (38).
A técnica utilizada é rápida e de custo relativamente médio podendo ser acessível
a todos. Para que isso se torne possível os médicos deverão estar diretamente ligados a
laboratórios de análises moleculares para realização desses testes, bem como ter à disposição
bancos de dados de frequências populacionais de polimorfismos relevantes para diferentes
áreas de pesquisa com diferentes drogas e tratamentos. Por isso, a importância deste trabalho
na contribuição para futuros estudos com indivíduos de diferentes regiões da população
brasileira a fim de estabelecer dados representativos de variações genotípicas em genes que
possam estar associados às respostas ao tratamento da risperidona, e por fim colaborando com
a farmacogenômica na individualização do tratamento.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Pop. Brasileira Pop. Africana Pop. Asiática Pop. Ameríndios Pop. Européia
C/C
A/C
A/A

72
7. CONCLUSÃO
As frequências alélicas obtidas para os SNVs de uma forma geral não diferiram
das frequências registradas para população global de referência exceto as das variações
rs6277 no gene DRD2 e rs3853188 no gene SCARB2.
A distribuição genotípica se mostrou em equilíbrio de Hardy-Weinberg para a
maioria das SNVs, sendo portanto bons marcadores para estudo de associação, exceto para a
rs7799039 no gene da leptina e rs6318 no gene HTR2C.
Por fim, esse trabalho contribui para estudos de farmacogenética futuros que
poderão ser realizados com base nos resultados aqui descritos e permitirá uma melhor
identificação de marcadores genéticos de risco principalmente para indivíduos em uso de
risperidona.

73
8. REFERÊNCIAS
1. Goodman R, Simonoff E, Stevenson J. The impact of child IQ, parent IQ and sibling IQ
on child behavioural deviance scores. J Child Psychol Psychiatr 1995; 36: 409-425.
2. Stahl SM. Essential Psychopharmacology. In: Neuroscientific Basis and Pratical
Applications. Cambridge University Press, New York, 1996.
3. Hood DS, Orr KGD, Nutt D. Antipsychotics. Elsevier. 2007; 6(7): 295-300.
4. Tsermpini EE, Assimakopoulos K, Bartsakoulia M, Iconomou G, Merkouri E,
Konstantinos P, et al. Individualizin clozapine and risperidone treatment for
schizophrenia patients. Pharmacogenomics. 2014; 15(1): 95-110.
5. Hjerde E, Dahl SG, Sylte I. Atypical and typical antipsychotic drug interactions with the
dopamine D2 receptor. Eur. J. Med. Chemistry. 2005; 40: 185-194.
6. Pajonk FG. Risperidone in acute and long-term therapy of schizophrenia a clinical
profile. Prog. Neuropsy. Biol Psychiatry. 2004; 28(1): 15-23.
7. Janssen PA, Niemegeers CJ, Awouters F, Schellekens KH, Megens AA, Meert TF.
Pharmacology of risperidone (R64 766), a new antipsychotic with serotonin-s2 and
dopamine-D2 antagonistic properties. J. Pharmacol. Exp. Ther. 1988; 244(2): 685-693.
8. Mas s, Gassó P, Bernardo M, Lafuente A. Functional analysis of gene expression in
risperidone trated cells provide new insights in molecular mechanism and new candidate
genes for pharmacogenetic. Eur. Neuropsyc. 2013: 23: 329-337.
9. Llerena A, BereczR, Peñas-Lledó E,Suveges A, Fariñas H. Farmacogenetics of clinical
response to risperidone. Pharmacogenomics. 2013; 14(2): 177-194.
10. Brennan MD Pharmacogenetics of second-generation antipsychotics. Future Medicine,
Pharmacogenomics. 2014; 15: 869–884.

74
11. Meltzer HY. Serotonergic mechanisms as targets for existing and novel antipsychotics.
Handb. Exp. Pharmacol. 2012; 212: 87–124.
12. Carvalho WA, Carvalho RDS, Medrado VC, Vianna PTG. Biologia Molecular dos
receptores farmacológicos e seus sistemas efetores de interesse em anestesiologia. Rev.
Bras. Anestesiol. 1997; 47(2): 152-167.
13. Sadock BJ, Sadock VA. Ciências do comportamento e psiquiatria clínica. In: Compêndio
de Psiquiatria. 9a ed. Porto Alegre: Artmed; 2007.
14. Goodman R, Scott S. Psiquiatria infantil. 1a ed. São Paulo: Roca, 2004.
15. Lesch KP. Schizophrenia. Weird world inside the brains. The Lancet. 2001; 358.
16. Moore TR, Hill AM, Panguluri SK. PharmacogenomicsinPsychiatry: implications for
practice. Recent Patents on Biotechnology. 2014; 8(2): 152-159.
17. Souza A. Comparando o uso da olanzapina com antipsicóticos típicos e atípicos no
tratamento da psicose esquizofrênica e em quadros agudos de agitação psicomotora em
diversas etiologias. RBM Especial Neuropsiquiatria. 2012; 69: 3-8.
18. Huang ML, Van Peer A, Woestenborghs R, De Costger R, Hevkantas J, Jansen AAI, et
al. Pharmacokinetics of the novel antipsychotic agent risperidone and the prolactin
response in healthy subjects. Clin Pharmacol Ther. 1993; 54: 247-268.
19. Haddad PM, Wieck A. Antipsychotic-induced hyperprolactinaemia: Mechanisms, clinical
features and management. Drugs. 2004; 6: 2291-22314.
20. Pappagallo M, Silva R. The effect of atypical antipsychotic agents on prolactin levels in
children and adolescents. J Child Adolesc Psychopharmacol. 2004; 14: 359-371.

75
21. Lane HY, Liu YC, Huang.L, Chang YC, Wu PL, Lu CT, et al. Risperidone-related
weight gain: Genetic and nongenetic predictors. J Clin Psychopharmacol, 2006; 26: 128-
134.
22. Hong CJ, Liou YJ, Bai YM, Chen TT, Wang YC, Tsai SJ. Dopamine receptor D2 gene
is associated with weight gain in schizophrenic patients under long-term atypical
antipsychotic treatment. Pharmacogenetics Genomics.2010; 20: 359-366.
23. Ruaño G, Goethe JW, Caley C, Woolley S, Holford TR, Kocherla M, et al.
Physiogenomic comparison of weight profiles of olanzapine and risperidone-treated
patients. Mol Psychiatry, 2007; 12: 474–482.
24. Yevtushenko OO, Cooper SJ, O’neill R, Doherty JK, Woodside JV, Reynolds GP.
Influence of 5-HT2C receptor and leptin gene polymorphisms, smoking and drug
treatment on metabolic disturbances in patients with schizophrenia. Br J Psychiatry,
2008; 192: 424–428.
25. Ribeiro Filho FF, Mariosa LS, Ferreira SRG, Zanella MT. Gordura Visceral e Síndrome
Metabólica, Arq Bras Endocrinol Metab 2006; 50(2).
26. Calarge CA, Ellingrod VL, Acion L, Miller DD, Moline J, Tansey MJ, et al. Variants of
the dopamine D2 receptor gene and risperidone-induced hyperprolactinemia in children
and adolescents. Pharmacogenet Genomics, 2009; 19: 73-82.
27. Yasui-Furukori N, Saito M, Tsuchimine S, Nakagami T, Sato Y, Sugawara N, et al.
Association between dopamine-related polymorphisms and plasma concentrations of
prolactin during risperidone treatment in schizophrenic patients. 2008; 32(6): 1491-495.
28. Carvalho MM, Góis C. Hiperprolactinémia em psiquiatria, Acta Med Port.2011; 24(6):
1005-1012.

76
29. Haddad PM, Das A, Keyhani S, Chaudhry IB. Antipsychotic drugs and extrapyramidal
side effects in first episode psychosis: A systematic review of head-head comparisons. J
Psychopharmacol, 2012; 26: 15-26.
30. Marsanic VB, Aukst-Margetic B, Grgic N, Kusmic E. Misdiagnosis and exacerbation of
unusual obsessive-compulsive disorder presentation with risperidone and clozapine in an
adolescent girl - A case report. Coll Antropol, 2011; 35 (1): 293-296.
31. Thompson & Thompson. Genética Médica, 6° edição: Editora Guanabara Koogan. Rio
de Janeiro, 2002, p. 76-82.
32. Lewin B. Genes VII. Editora ARTMED. São Paulo, 2001: 39-42.
33. Silva De Sá SM. Importância dos polimorfismos do metabolismo da Serotonina e da
Catecol O- metiltransferase na suscetibilidade para a Biabetes Mellitus tipo 2.
[Dissertação de mestrado], Universidade de Lisboa, 2011.
34. Shastry BS. Pharmacogenetics and the concept of individualized medicine. The
Pharmacogenomic Journal.2006; 6: 16-21.
35. Lindpaintner K. Pharmacogenetics and the future of medical practice. BR J Clin
Pharmacol.2002; 54(2): 221 – 230.
36. Hiemke C, Hartter S. Pharmacokinetics of selective serotonin reuptake inhibitors.
Pharmacol. Ther.2000; 85(1): 11-28.
37. Cordeiro Q, Shavin RG, Cappi C, Sampaio AS, Morais IA, Hounie AG, et al.
Pharmacogenomics and psychiatry – Review; Rev. Fac. Ciênc. Méd. Sorocaba. 2009;
11(1): 4-10.
38. Licinio J. Farmacogenômica: oportunidades e desafios. Rev. Bras. Psiquiatria. 2001;
23(3).

77
39. Müller DJ, Kennedy JL. Genetics of antipsychotic treatment emergent weight gain in
schizophrenia. Pharmacogenomics.J.2006; 7: 863–87.
40. Gentile S. Contributing factors to weight gain during long-term treatment with second-
generation antipsychotics. A systematic appraisal and clinical implications. Obes
Rev.2009; 1: 527-542.
41. Lett TAP, Wallace TJM, Chowdhury NI, Tiwaru AK, Kennedy JL, Muller DJ,
Pharmacogenetics of antipsychotic-induced weight gain: review and clinical
implications. Mol Psychiatry.2012: 1-25..
42. Reynolds GP, Templeman LA, Zhang .J. The role of 5-HT2C receptor polymorphisms in
the pharmacogenetics of antipsychotic drug treatment. Progress in Neuro-
Psychopharmacology & Biological Psychiatry.2005; 29: 1021–1028.
43. De Leon, J. Pharmacogenomics: the promise of personalized medicine for CNS
disorders. Neuropsychopharmacology. 2009; 34: 159-172.
44. Nebert DW, Zhang G, Vesell ES. From human genetics and genomics to
pharmacogenetics and pharmacogenomics: past lessons, future directions. Drug Metab
Ver. 2008; 40: 187–224..
45. Thompson, JS, Thompson M W. Genética bioquímica humana: farmacogenética. In:
Genética médica. 4.ed. Rio de Janeiro: Guanabara Koogan, 1990: 124-133.
46. Nora JJ, Fraser C. Farmacogenética. In: Genética médica. 2.ed. Rio de Janeiro:
Guanabara Koogan, 1985, p. 354-360.
47. Beiguelman B. Farmacogenética. In: (Org.) Farmacogenética e sistemas sangüíneos
eritrocitários. Rio de Janeiro: Ed. Guanabara Kogan-FUNCAMP, 1983: 2-78.

78
48. de Lucia R, Oliveira-Filho RM, Valle LBS, OGA S. Farmacocinética: Eliminação
Metabólica de Fármacos. In: Farmacologia integrada: princípios básicos. Rio de Janeiro:
Ateheneu, 1988; 1: 102-112.
49. Frota-Pessoa O, Otto PA. Os genes recessivos: farmacogenética. In: Genética clínica.
Rio de Janeiro: Francisco Alves, 1984: 74-79.
50. Bazire S. Psychotropic Drug Directory. London: Health Comm; 2008.
51. Beverage JN, Sissung TM, Sion AM, Danesi R, Figg WD. CYP2D6 polymorphisms and
the impact on tamoxifen therapy. J Pharm Sci.2007; 96: 2224-2231.
52. Blazquez A, Mas S, Plana MT, Lafuente A, Lázaro L. Fluoxetine pharmacogenetics in
child and adult populations – Review. Eur Child Adolesc. Psychiatry, 2012.
53. Wolf CR, Smith G. Cytochrome P450 CYP2D6. Ch. 18. IARC Sci Publ. 1999; 148:
209–229.
54. Kaiser R, Sezer O, Papies A, Bauer S, Schelenz C, Tremblay P, et al. Patient-tailored
antiemetic treatment with 5-hydroxytryptamine type 3 receptor antagonists according to
cytochrome P-450 2D6 genotypes. J Clin Oncol, 2002; 20: 2805-2811.
55. Bertilsson L. Geographical/interracial differences in polymorphic drug oxidation.
Current state of knowledge of cytochromes P450 (CYP) 2D6 and 2C19. Clin
Pharmacokinet,v.29, p. 192–209, 1995.
56. Cai WM, Chen B, Zhang WX. Frequency of CYP2D6*10 and *14 alleles and their
influence on the metabolic activity of CYP2D6 in a healthy Chinese population. Clin
Pharmacol Ther. 2007; 81: 95-98.
57. Wang G, Zhang H, He F, Fang X. Effect of the CYP2D6*10 C188T polymorphism on
postoperative tramadol analgesia in a Chinese population. Eur J Clin Pharmacol, 2006;
62: 927-931.

79
58. Orengo-Mercado C, Nieves BB, López L, Vallés-Ortiz N, Renta JR, Santiago-Borrero
PJ, et al. Frequencies of Functional Polymorphisms in Three Pharmacokinetic Genes of
Clinical Interest within the Admixed Puerto Rican Population. J Pharmacogenomics
Pharmacoproteomics. 2013; 4(113).
59. Souza RDM, Martins DMF, Chein MBC, Brito LMO. Importance of CYP2D6 in users
of tamoxifen in breast cancer. Femina, 2011; 39(5).
60. Troost PW, Lahuis BR, Hemans MH, Buitelaar JK, Engeland H, Scahill L, et al.
Prolactin release in children treated with risperidone: Impact and role of CYP2D6
metabolism. J Clin Psychopharmacol, 2007; 27: 52-57.
61. Knegtering R, Baselmans P, Castelein S, et al. Predominant role of the 9-hydroxy
metabolite of risperidone in elevating blood prolactin levels. Am J Psychiatry.2005; 162:
1010-1012.
62. Wang GJ, Volkow ND, Logan J, PappaS NR, Wong CT, Zhu W, et al. Brain dopamine
and obesity. Lancet, 2001; 357: 354–357.
63. Picetti R, Saiardi A, Samad TA, Bozzi Y, Baik JH, Borrelli E. Dopamine D2 receptors in
signal transduction and behavior. Crit. Rev. Neurobiol, 1997; 11: 121-142..
64. Usiello A, Baik JH, Rougé-Pont F, Picetti R, Dierich A, Lemeur M, et al. Distinct
functions of the two isoforms od dopamine D2 recetors. Nature, 2000; 408: 199-203.
65. Lencz T, Robinson DG, Xu K, Ekholm J, Sevy S, Gunduz-Bruce H, et al. DRD2
promoter region variation as a predictor of sustained response to antipsychotic
medication in first-episode schizophrenia patients. Am J Psychiatry, 2006; 163: 163-529.
66. Mihara K, Kondo T, Suziki A, Yasui N, Nagashima U, Ono S, et al. Prolactin response
to nemonapride, a selective antagonist for D2 like dopamine receptors, in schizophrenic

80
patients in relation to Taq1A polymorphism of DRD2 gene. Psychopharmacology (Berl).
2000; 149: 246–50.
67. Duan J, Wainwright MS, Comeron JM, SAITOU N, SANDERS A.R, GELERNTER J,
et al. Synonymous mutations in the human dopamine receptor D2 (DRD2) affect mRNA
stability and synthesis of the receptor. Human Molecular Genetics, v. 12, n. 3, p. 205-
216, 2003.
68. Hirnoven M, Laakso A, Nagren K, Rinne JO, Pohjalainen T, Hietala J. C957T
polymorphism of the dopamine D2 receptor (DRD2) gene affects striatal DRD2
availability in vivo. Molecular Psychiatric. 2004; 9: 1060-1066.
69. Lawford BR, Young RM, Swagell CD, Barnes M, Burton S.C, Ward WK, et al. The C/C
genotype of the C957T polymorphism of the dopamine D2 receptor is associated with
schizophrenia. Elsevier, 2005; 73: 31-37.
70. He M, Hong Y, Duan ZX, Qu W, Gong HY, Fan ZL, et al. Genetic distribution and
association analysis of DRD2 gene polymorphisms with major depressive disorder in the
Chinese Han population. Int J Clin Exp Pathol. 2013;.6: 1142-1149.
71. Hill SY. Dopaminergic mutations: within-family association and linkage in multiplex
alcohol dependence families. AM J Med Genet B Neuropsychiatric Genet, 2008; 10:
107-113.
72. Yang BZ, Kranzler HR, Zhao H, Gruen JR, Luo X, Gelernter J. Haplotypic variants in
DRD2, ANKK1, TTC12 and NCAM1 are associated with comorbid alcohol and drug
dependence. Alcohol Clin Exp Res, 2008; 32: 2117-2127.
73. Rieck M, Schumacher-Schuh AF, Altmann V, Francisconi CLM, Fagundes PTB, Monte
TL, et al. DRD2 haplotype is associated with dyskinesia induced by levodopatherapy in
Parkinson’s disease patients. Pharmacogenomics, 2012; 13: 1701-1710.

81
74. de Luca V, Muuller DJ, Hwang R, Lieberman JA, Volavka J, Meltzer HY, et al. HTR2C
haplotypes and antipsychotics induced weight gain: X-linked multimarker analysis. Hum
Psychopharmacol. 2007; 22: 463-467.
75. Mulder H, Franke B, Van Der-Beek, Van Der AA, Arends J, Wilmink FW, et al. The
association between HTR2C polymorphisms and obesity in psychiatric patients using
antipsychotics: a cross-sectional study. Pharmacogenomics J. 2007; 7. 318-124.
76. Opgen-Rhein C, Brandi EJ, Muller DJ, Neuhaus AH, Tiwari AK, Sander T et al.
Association of HTR2C, but not LEP or INSIG2, genes with antipsychotic-induced
weight gain in a German Sample. Future Medicine, Pharmacogenomics, 2010; 11(6):
773-780.
77. Berger M, Gray JA, Roth BL. The expanded Biology of serotonin. Annual Reviews
Med. 2009; 60: 355 – 366.
78. Rang H., Dale MM. Farmacologia, Rio de Janeiro, 2ª Ed. Guanabara Koogan, p.431-435, 1993.
79. Callahan PM, Cunningham KA. Modulation of the discriminative stimulus properties of
cocaine by 5-HT1B and 5-HT2C receptors. J Pharmacol Exp Ther, 1995; 274(3): 1414–
1424.
80. Filip M, Faron-Gorecka A, Kusmider M, Golda A, Frankowska M, Dziedzicka-
Wasylewska M. Alterations in BDNF and trkB mRNAs following acute or sensitizing
cocaine treatments and withdrawal. Brain Res. 2006; 1071(1): 218–225.
81. Lappalainen J, Zhang L, Dean M, et al. Identification, expression, and pharmacology of
a Cys23- Ser23 substitution in the human 5-HT2c receptor gene (HTR2C). Genomics,
1995; 27(2): 274-279.
82. Correia CT, Almeida JP, Santos PE, Sequeira AF, Marques CE, Miguel TS, et al.
Pharmacogenetics of risperidone therapy in autism: association analysis of eight

82
candidate genes with drug efficacy and adverse drug reactions. Pharmacogenomics J.
2010; 10(5): 418-430.
83. Yuan X, Yamada K, Ishiyama-Shigemoto S, Koyama W, Nonaka K. Identification of
polymorphic loci in the promoter region of the serotonin 5-HT2C receptor gene and their
association with obesity and type II diabetes. Diabetologia, 2000; 43: 373–376.
84. Torre DL, Falorni A. Pharmacological causes of hyperprolactinemia. Ther Clin Risk
Manag, 2007; 3: 929–951.
85. Van De Kar LD, Javeda, Zhang Y, Serres F, Raap DK, Gray TS. 5-HT2A receptors
stimulate ACTH, corticosterone, oxytocin, renin, and prolactin release and activate
hypothalamic CRF and oxytocin-expressing cells. J Neurosci, 2001; 21: 3572–3579.
86. Hill MJ, Reynolds GP. 5-HT2C receptor gene polymorphisms associated with
antipsychotic drug action alter promoter activity. Brain Res, 2007; 1149: 14–17.
87. Mccarthy S, Mottagui-Tabar S, Mizuno Y, Sennblad B, Hoffstedt J, Arner P, et al.
Complex HTR2C linkage disequilibrium and promoter associations with body mass
index and serum leptin. Human Genetics, 2005; 117: 545–57.
88. Buckland PR, Hoogendoom B, Guy CA, Smith SK, Coleman SL, O’donovan MC. Low
gene expression conferred by association of an allele of the 5-HT2C receptor gene with
antipsychotic induced weight gain. Am J Psychiatry. 2005; 162: 613-615.
89. Iordanidou M, Tavridou A, Vasiliadis M.V, Arvanitidis K.I, Petridis J, Christakidis D, et al. The
-759C/T polymorphism of the 5-HT2C receptor is associated with type 2 diabetes in male and
female Caucasians. Pharmacogenetics and Genomics. 2008; 18; 153-159.
90. Pooley EC, Fairburn CG, Cooper Z, Sodhi MS, Cowen PJ, Harrison PJ. 5-HT2C
receptor promoter polymorphism (HTR2C-759C/T) is associated with obesity in women,
and with resistance to weight loss in heterozygotes. Am J Med Genet. 2004; 126: 124–
127.

83
91. Ferrari M, Bolla E, Bortolaso P, Callegari C, Poloni N, Lecchini S, et al. Association
between CYP1A2 polymorphisms and clozapine-induced adverse reactions in patients
with schizophrenia. Psychiatry Res, 2012; 200(2–3): 1014–1017.
92. Reynolds GP, Zhang Z, Zhang X. Polymorphism of the Promoter Region of the
Serotonin 5-HT2C Receptor Gene and Clozapine- Induced Weight Gain. Am J
Psychiatry. 2003; 160.
93. Yevtushenko OO, Cooper SJ, O’neill R, Doherty JK, Woodside JV, Reynolds GP.
Influence of 5-HT2C receptor and leptin gene polymorphisms, smoking and drug
treatment on metabolic disturbances in patients with schizophrenia. Br J Psychiatry,
2008; 192: 424–428.
94. Farooqi IS, O’rahilly S. Monogenic obesity in humans. Annu Rev Med, 2005; 56: 443-
458.
95. Duarte SF, Francischetti EA, Genelhu-Abreu V, Barroso SG, Braga JU, Cabello PH, et
al. p.Q223R leptin receptor polymorphism associated with obesity in Brazilian
multiethnic subjects. Am J Hum Biol. 2006; 18(4): 448-453.
96. Yiannakouris N, Yannakoulia M, Melistas L, Chan JI, Klimis-Zacas D, MANTZOROS
CS. The Q223R polymorphism of the leptin receptor gene is significantly associated
with obesity and predicts a small percentage of bodyweight and body composition
variability. J Clin Endocrinol Metab, 2001; 86: 4434-4439.
97. Pi-Sunyer FX, Laferrere B. Metabolic abnormalities and the role of leptin in human
obesity. J Clin Endocrinol Metab, 1999; 84: 3-7.
98. Sentissi O, Epelbaum J, Olie JP, Poirier MF. Leptin and ghrelin levels in patients with
schizophrenia during different antipsychotics treatment: A review. Schizophr Bull. 2008;
34: 1189–99.

84
99. Westling S, Ahrén B, Traskman-Bendz L, Westrin A. Low csf leptin in female suicide
attempters with major depression. J Affect Disord, 2004; 81(1): 41–48.
100. Atmaca M, Kugoglu M, Tezcan E, Parmaksiz S, Saribas M, Ozler S. et al. Serum
leptin and cholesterol values in suicide attempters. Neuropsychobiology, 2002; 45 (3):
124–7.
101. Hoffstedt J, Eriksson P, Mottagui-Tabar S, Arner P. A Polymorphism in the Leptin
Promoter Region (-2548 G/A) Influence Gene Expression and Adipose Tissue Scretion
of leptin. Horm Metab Res, 2002; 34(7): 355-359.
102. Zhang ZJ, Yao ZJ, Mou XD, Chrn JF, Zhu RX, Liu W, et al. Association of -
2548G/A functional polymorphism in the promoter region of leptin gene with
antipsychotic agent-induced weight gain. Zhonghua Yi Xue Za Zhi, 2003; 83: 2119–
2123.
103. Li WD, Reed DR, Lee JH, Xu W, Kilker RL, Sodam BR, et al. Sequence variants in
the 5′ flanking region of the leptin gene are associated with obesity in women. Ann Hum
Genet.1999; 63: 227-234.
104. List JF, Habener JF. Defective melanocortin 4 receptors in hyperphagia and morbid
obesity. N Engl J Med. 2003; 348: 1160-1163.
105. Dubern B, Clement K, Pelloux V, Froguel P, Girardet JP, Guy-Grand B, et al.
Mutational analysis of melanocortin-4 receptor, agoutirelated protein, and alpha-
melanocyte-stimulating hormone genes in severely obese children. J Pediatr, 2001; 139:.
204-209.
106. Lubrano-Berthelier C, Cavazos M, Dubern B, Shapiro A, Stunff CL, Zhang S, et al.
Molecular genetics of human obesity associated MC4R mutations. Ann NY Acad Sci.
2003; 994: 49–57.

85
107. Loos RJ, Lindgren CM, Li S, Wheeler E, Zhao JH, Prokopenko I, et al. Common
variants near MC4R are associated with fat mass, weight and risk of obesity. Nat Genet.
2008; 40: 768–75.
108. Meyre D, Delplanque J, Chevre JC, Lecoeur C, Lobbens S, Gallina S, et al. Genome-
wide association study for early-onset and morbid adult obesity identifies three new risk
loci in European populations. Nat Genet., 2009; 41: 157–9.
109. Kapur S, Seeman P. Does fast dissociation from the dopamine d(2) receptor explain
the action of atypical antipsychotics?: a new hypothesis. A. J Psychiatry. 2001; 158:
158-360.
110. Shi J, Long J, Gao YT, Lu W, Cai Q, Wen W, et al. Evaluation of genetic
susceptibility loci for obesity in Chinese women. Am J Epidemiol. 2010; 172: 244–254.
111. Cheung C, Tso AWK, Cheung BMY, Xu A, Ong KL, Fong CHY, et al. Obesity
susceptibility genetic variants identified from recent genome-wide association studies:
implications in a Chinese population. J Clin Endocrinol Metab, 2010; 95: 1395-1403.
112. Stutzmann F, Cauchi S, Durand E, Calvacanti-Proença C, Pigeure M, Hartikainen AL,
et al. Common genetic variation near MC4R is associated with eating behavior patterns
in European populations. Int J Obes, 2009; 33: 373-378.
113. Qi L, Kraft P, Hunter DJ, Hu FB. The common obesity variant near MC4R gene is
associated with higher intakes of total energy and dietary fat, weight change and diabetes
risk in women. Hum Mol Genet.2008; 17: 3502-3508.
114. Ho-Urriola J, Guzmán-Guzmán IP, Smalley SV, González A, Weisstaub G,
Dominguez-Vásquez P, et al. Melanocortina- 4 receptor polymorphism rs 17782313:
Association with obesity and eating in the absence of hunger in Chilean children.
Nutrition, 2014; 30: 145-149.

86
115. Eckhardt ER, Cai L, Sun B, Webb NR, Van Der Westhuyzen DR. High density
lipoprotein uptake by scavenger receptor SR-BII. J Biol Chem, 2004; 279: 14372-14381.
116. Yamayoshi S, Fujii K, Koikes. Receptor for enterovirus 71 Open. Emergin Microbes &
Infections. 2014; 3.
117. Berkovic SF, Dibbens LM, Oshlack A, Silver JD, Katerelos M, Vears DF. et al.
Array-bases gene discovery with three unrelated subjects shows SCARB2/LIMP-2
deficiency causes myoclonus epilepsy and glomerulosclerosis. Am. J. Hum. Genet.
2008; 82: 673-684.
118. De Araújo M. Estudo da organização molecular do “cluster” gênico CYP21 e C4 em
famílias com a forma clássica de deficiência da 21-hidroxilase no Brasil [tese de
mestrado]. Campinas: Instituto de Biologia; Universidade Estadual de Campinas, 1996.
119. Heid CA, Stevens J, Livak KJ, William PM. Real time quantitative PCR. Genome
Res.1996; 6(10): 986 – 994.
120. Osorio RG. O sistema classificatório de “cor ou raça” do IBGE. ISSN 1415-4765,
Instituto de Pesquisa Econômica Aplicada. Brasília, 2003.
121. Pena SJ, Di Pietro G, Fuchshuber-Moares M, Genro JP, Hutz MH, Gomes FS, et al.
The Genomic ancestry of individuals from different geographical regions of Brazil is
more uniform than expected. Plos One; 2011 6: E17063.
122. IBGE – Instituto brasileiro de geografia e estatística. Característicos étnicos- raciais da
população. Coordenação de População e Indicadores Sociais, 2008.
123. Tharanga TDP, Jinadasa CM, Risama MF, Galappatthy P, Jayakody RL,
Dissanayakevh Genetic variants in the cytochrome P450 2D6 gene in the Sri Lankan
population. Indian J Hum Genet. 2013; 19: 392-396.

87
124. Dahl ML, Johansson I, Bertilsson L, Ingelman-Sundberg M, Sjoqvist F. Ultrarapid
hydroxylation of debrisoquine in a Swedish population. Analysis of the molecular
genetic basis. J Pharmacol Exp Ther, 1995; 274: 516-520.
125. Agundez JA, Ledesma MC, Ladero JM, Benitez J. Prevalence of CYP2D6 gene
duplication and its repercussion on the oxidative phenotype in a white population. Clin
Pharmacol Ther. 1995; 57: 265-269.
126. Aklillu E, Persson I, Bertilsson L, Johansson I, Rodrigues F, Ingelman-Sundberg M.
Frequent distribution of ultrarapid metabolizers of debrisoquine in an ethiopian
population carrying duplicated and multiduplicated functional CYP2D6 alleles. J
Pharmacol Exp Ther.1996; 278: 441–6.
127. Gaedigk A, Bradford LD, Marcucci KA, Leeder JS. Unique CYP2D6 activity
distribution and genotype–phenotype discordance in black Americans. Clin. Pharmacol.
Ther, 2002; 72 (1): 76–89.
128. Rosmond R, Rankinen T, Chagnon M, Pérusse L, Chagnon YC, Bouchard C, et al.
Polymorphism in exon 6 of the dopamine D(2) receptor gene (DRD2) is associated with
elevated blood pressure and personality disorders in men. J Hum Hypertens, 2001; 15:
553-558.
129. Hanninen K, Katila H, Kampman O, Anttila S, Lli A, Rontu R, et al. Association
between the C957T polymorphism of the dopamine D2 receptor gene and schizophrenia.
Neurosci Lett. 2006; 407: 195-198..
130. Hoenick J, Araqués M, Rodríquez-Jiménez R, Ponce G, Martínez I, Rubio G, et al.
C957T DRD2 polymorphism is associated with schizophrenia in Spanish patients. Acta
Psychiatr Scand, 2006; 114(6): 435-438.
131. Kukreti R, Tripathi S, Bhatnagar P et al. Association of DRD2 gene variant with
schizophrenia. Neurosci Lett, 2006; 392: 68-71.

88
132. Santos-Junior A. Farmacogenética dos efeitos adversos da risperidona em crianças e
adolescentes Tese - Doutorado. Campinas/SP. Universidade Estadual de Campinas,
2015.
133. Tritos NA, Mantzoros CS. Leptin: its role in obesity and beyond, Diabetologia, 1997;
40(12): 1371–1379.
134. Constantin A, Costache G, Sima AV, Glavce CS, Vladica M, Popov DL. Leptin G-
2548A and leptin receptor Q223R gene polymorphisms are not associated with obesity
in Romanian subjects. Biochem Biophys Res Commun, 2010.; 391(1): 283-286.
135. Portolés O, Sorlí JV, Francés F, Coltell O, González JI, Sáiz C, Carella D. Effect of
genetic variation in the leptin gene promoter and the leptin receptor gene on obesity risk
in a population-based case-control study in Spain. European Journal of Epidemiology,
2006; 21: 605-612.
136. Kang SG, Heon-Jeong L, Young-Min P, Jung-Eun C, Changsu H, Young-Ku K, et al.
Possible association between the -2548A/G polymorphism of the leptin gene and
olanzapine-induced weight gain. Progress in: Neuro-Psychopharmacology & Biological
Psychiatry, 2008; 32: 160–163.
137. Fernández E, Carrizo E, Fernández V, Connell L, Sandia I, Prieto D, et al.
Polymorphisms of the LEP- and LEPR genes, metabolic profile after prolonged
clozapine administration and response to the antidiabetic metformin. Schizophr Res,
2010; 121: 213-217.
138. Wu R, Zhao J, Shao P, Ou J, Chang M. Genetic predictors of antipsychotic-induced
weight gain: a case-matched multi-gene study. Zhong Nan Da Xue Xue Bao Yi Xue
Ban, 2011; 36: 720-723.
139. Shen J, Ge W, Zhang J, Zhu HJ, Fang Y. Leptin -2548g/a gene polymorphism in
association with antipsychotic-induced weight gain: a meta-analysis study. Psychiatr
Danub. 2014; 26: 145-151.

89
140. Katsuura-Kamanos, Uemura H, Arisawa K, Yamaguchi M, Hamajima N, Wakai K, et
al. A polymorphism near MC4R gene (rs17782313) is associated with serum triglyceride
levels in the general Japanese population the J-MICC Study. Endocr. Genetic/
epigenetics, 2014.
141. Yilmaz Z, Davis C, Loxton NJ, Kaplan AS, Levitan RD, Carter RD Kennedy JL.
Association between MC4R rs17782313 polymorphism and overeating behaviors.
International Journal of Obesity. 2014: 1-7.
142. Gonzalez A, Valeiras M, Sidransky E, Tayebi N. Lysosomal integral membrane
protein-2: A new player in Lysosome-related pathology. Mol Genet Metab, 2014; 111:
84-91.

90
9. ANEXOS
Termo de Consentimento Livre e Esclarecido
PROJETO: Farmacogenética da resposta da risperidona em uso isolado ou não no
tratamento de transtornos mentais entre 10 e 20 anos de idade
PESQUISADORES ENVOLVIDOS:
Adriana Perez - Aluna de Iniciação Científica do curso de Graduação em Medicina - FCM -
UNICAMP; Luiz Fernando Longuim Pegoraro - Psicólogo, Mestre em Saúde da Criança e do
Adolescente - FCM - UNICAMP; Maricilda Palandi de Mello - Química, Livre-Docente,
Centro de Biologia Molecular e Engenharia Genética (CBMEG) - UNICAMP; Taciane
Barbosa Henriques – Aluna de Mestrado em Ciências Médicas – FCM – UNICAMP; Prof.
Dr. Stephen Hyslop - Bioquímico - Professor Doutor – Departamento de Farmacologia - FCM
- UNICAMP; Prof. Dr. Paulo Dalgalarrondo – Médico Psiquiatra, Professor Titular -
Departamento de Psicologia Médica e Psiquiatria - FCM - UNICAMP; Profa. Dra. Eloisa
Helena Rubello Valler Celeri - Médica Psiquiatra, Professora Doutora - Departamento de
Psicologia Médica e Psiquiatria – FCM UNICAMP; Prof. Dr. Gil Guerra-Júnior - Médico
Endocrinologista Pediátrico Professor Titular - Departamento de Pediatria - FCM –
UNICAMP
PESQUISADOR RESPONSÁVEL:
Amilton dos Santos Júnior (Médico Psiquiatra, Mestre em Saúde da Criança e do Adolescente
- FCM - UNICAMP)
ENDEREÇO:
Departamentos de Pediatria e Psicologia Médica e Psiquiatria - FCM – UNICAMP Cidade
Universitária - Campinas - SP - CEP 13081-970
Fone: 19-3521-7206 e-mail: [email protected]
IDENTIFICAÇÃO DO(A) SUJEITO DE PESQUISA:
Nome:
Endereço:
Bairro: Cidade:
UF: CEP: Fone:
OBJETIVO DA PESQUISA:
Eu __________________________________________________________________,
R.G.: ________________________, entendo que fui convidado(a) a participar de um projeto
de pesquisa, aprovado pelo Comitê de Ética da FCM-UNICAMP, envolvendo pacientes que

91
fazem uso de risperidona. O sigilo será mantido por meio da identificação das pacientes por
um código.
PROCEDIMENTO:
Eu entendo que, se concordar em participar desse estudo serei submetido(a) à coleta de
sangue para extração de DNA para análise dos seguintes polimorfismos: gene CYP2D6: SNV rs72552269, c.100C>T, p.P34S (alelo CYP2D6*10)
gene HTR2C: SNVs rs6318, c.68G>C; p.Cys23Ser e rs3813929, c.-759C>T
gene DRD2: SNVs rs1799732, c.-141ins_delC e rs1799978, c.-241A>G
gene LEPR: SNVs rs7799039, -2548A>G e rs1137101, c.668A>G, p.Q223R
gene MC4R: SNV rs17782313 na região 3’ do gene
gene SCARB2: SNV rs3853188, c.275+7280A>C
O estudo dos polimorfismos será realizado para análise das frequências dos genótipos desses
genes na população. Tal procedimento não apresenta risco importante.
RISCO E DESCONFORTO:
Para a realização dos exames de sangue serão necessários cerca de 08 ml de sangue venoso,
obtidos em uma única coleta. Os riscos associados a esse procedimentos são mínimos,
podendo ocorrer dor e/ou manchas rochas (equimoses) no local da coleta de sangue. O
desconforto será mínimo, pois, em geral, essa coleta será realizada de veia do braço, por
profissional treinado e devidamente habilitado para realizá-la
VANTAGENS:
Entendo que não terei benefício direto com a participação no estudo, a não ser o de conhecer
os polimorfismos nos genes estudados. Trata-se de estudo experimental testando a hipótese de
que pacientes com transtorno mental em uso de risperidona (isolada ou não) podem apresentar
a curto e longo prazo efeitos colaterais das medicações e que estes efeitos podem estar
relacionados com a presença de determinados polimorfismos genéticos. Entendo que somente
no final do estudo poderei saber sobre a presença de algum outro benefício ligado a esta
avaliação.
SIGILO:
Eu entendo que toda informação será submetida aos regulamentos do Hospital, referentes ao
sigilo da informação médica. Se os resultados ou informações fornecidas forem utilizados
para fins de publicação científica, nenhum nome será mencionado.
FORNECIMENTO DE INFORMAÇÃO ADICIONAL:
Eu entendo que posso requisitar informações adicionais referentes ao estudo a qualquer
momento. Em caso de alguma dúvida sobre a pesquisa o médico responsável, Amilton dos
Santos Júnior, fone (19)35217514, estará disponível para responder às minhas questões e
preocupações.
Deseja ser informado sobre os resultados do estudo das análises dos polimorfismos descritos?
____________________________.

92
RECUSA OU DESCONTINUAÇÃO DA PARTICIPAÇÃO:
Eu entendo que minha participação nesse projeto de pesquisa é voluntária e que posso recusar
ou retirar meu consentimento a qualquer momento (incluindo a retirada da amostra de
sangue). Eu reconheço também que o médico Amilton dos Santos Júnior pode interromper
minha participação nesse estudo a qualquer momento que julgar apropriado.
Se você tiver alguma denúncia ou reclamação a respeito dos aspectos éticos do projeto entre
em contato com o Comitê de Ética em Pesquisa da FCM - UNICAMP, e-mail:
[email protected] Telefone: 19-3521-8936, Rua: Tessália Vieira de Camargo, 126 Distrito
de Barão Geraldo Campinas – SP CEP: 13083-887.
Nome do (a) participante:__________________________________________________
Assinatura do(a) participante:_______________________________________________
Nome do(a) responsável:__________________________________________________
Assinatura do(a) responsável:______________________________________________
Local e Data:___________________________________________________________
RESPONSABILIDADE DO PESQUISADOR:
Eu expliquei à _________________________________________________ o objetivo do
estudo, os procedimentos requeridos e os possíveis riscos e vantagens que poderão advir do
estudo. Eu me comprometo e fornecer uma cópia desse formulário de consentimento à
participante.
Nome do pesquisador: Amilton dos Santos Júnior
Assinatura do pesquisador:________________________________________________
Local e Data: __________________________________________________________

93
Aprovação do Comitê de ética





























