Embed Size (px)
Citation preview

RAFAELA GAGETI BULGARELLI
RESPOSTA AO BAIXO FÓSFORO EM SOJA NODULADA:
CONTRIBUIÇÃO DA MICORRIZA ARBUSCULAR
RESPONSE TO PHOSPHORUS STARVATION IN NODULATED
SOYBEANS: CONTRIBUTION OF ARBUSCULAR MYCORRHIZA
CAMPINAS
2017
UNIVERSIDADE ESTADUAL DE CAMPINAS
Instituto de Biologia

RAFAELA GAGETI BULGARELLI
RESPOSTA AO BAIXO FÓSFORO EM SOJA NODULADA:
CONTRIBUIÇÃO DA MICORRIZA ARBUSCULAR
RESPONSE TO PHOSPHORUS STARVATION IN NODULATED
SOYBEANS: CONTRIBUTION OF ARBUSCULAR MYCORRHIZA
Dissertação apresentada ao Instituto
de Biologia da Universidade Estadual
de Campinas como parte dos
requisitos para a obtenção do Título
de Mestra em BIOLOGIA VEGETAL.
Dissertation presented to the Institute
of Biology in partial fulfillment of the
requirements for the degree of Master
in PLANT BIOLOGY
Orientadora: SARA ADRIÁN LÓPEZ DE ANDRADE
ESTE ARQUIVO DIGITAL CORRESPONDE À VERSÃO
FINAL DA TESE DEFENDIDA PELA ALUNA
RAFAELA GAGETI BULGARELLI, E ORIENTADA PELA
PROFA. DRA. SARA ADRIÁN LÓPEZ DE ANDRADE
CAMPINAS
2017


Campinas, 17 de fevereiro de 2017
COMISSÃO EXAMINADORA
Prof. Dra. Sara Adrián López Andrade (Orientador)
Prof. Dr. Ladaslav Sodek
Dr. Tiago Tezotto
Os membros da Comissão Examinadora acima assinaram a Ata de Defesa, que se encontra no
processo de vida acadêmica do aluno.

Agradecimentos
À Universidade Estadual de Campinas, ao Departamento de Biologia Vegetal, e ao
Programa de Pós-Graduação em biologia vegetal pela oportunidade de realização do curso.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela
concessão da bolsa de estudos, fundamental para a realização deste curso.
À minha orientadora Professora Doutora Sara Adrián López Andrade, por acreditar em
mim e pela valiosa oportunidade de orientação, amizade e confiança. Agradeço também por
sua gentileza, dedicação, determinação e muitos outros adjetivos que fazem com que seja,
para mim, um exemplo de pessoa e pesquisadora.
Ao professor Paulo Mazzafera, pela ajuda, confiança e pela disponibilidade do
laboratório para realização do projeto.
A todos os professores que compartilharam seus conhecimentos, especialmente aos
professores do departamento Rafael e Sodek que sempre estiveram dispostos a ajudar e a
sanar qualquer dúvida.
Aos colegas do departamento que sempre estiveram presentes e me ajudaram diversas
vezes a tomar decisões mais sensatas, principalmente a Flávia, Eduardo, Sarah, Simone,
Luciano, Nathalia, Alexandre e Pedro. Agradeço a todos pela troca de conhecimento, e
também pelas discussões sobre diversos assuntos. Obrigada pelo apoio, incentivo, pela alegria
de todos os cafés e no dia a dia, sendo assim fundamentais nesse processo.
Aos meus amigos e familiares principalmente aos meus pais Joaquim e Fátima pelo
amor incondicional, pelo apoio, pelo exemplo, e por sempre estarem presentes.
Ao meu noivo Raphael pelo amor e companheirismo, por estar sempre presente
partilhando os bons e maus momentos. Pelo imenso apoio e incentivo que me deu desde o
começo dessa trajetória que foi fundamental.
A todos que de qualquer forma fizeram parte desse processo meu sincero
agradecimento.

RESUMO
O nitrogênio (N) e o fósforo (P) são nutrientes que podem limitar o desempenho das
plantas. Atualmente, há crescente conscientização da importância das interações planta-
microrganismo na aquisição de nutrientes e na promoção do crescimento de plantas,
especialmente sob estresse. A micorriza arbuscular (MA) é uma das simbioses mais úbiquas
entre raízes e fungos do solo, com reconhecido papel na nutrição mineral e na atenuação de
diversos estresses em plantas. O objetivo foi avaliar os efeitos da simbiose micorrízica
arbuscular (MA) sob aspectos fisiológicos e moleculares de soja nodulada exposta à
deficiência de P, com especial interesse na capacidade de nodulação e no transporte e
absorção de P. Realizaram-se dois experimentos em sistema hidropônico, usando vermiculita
como substrato, nos quais plantas de soja (Glycine max [L.] Merrill) mantidas sob baixo nível
de P (50 μM) e noduladas com Bradyrhizobium elkanii foram inoculadas (+MA) ou não (-
MA) com o fungo MA Glomus macrocarpum. Plantas não inoculadas mantidas em condições
suficientes de P (500 μM) constituiram o tratamento denominado -MA+P. A coleta das
plantas foi realizada no início da floração e durante enchimento da semente. No primeiro
experimento os resultados mostraram que a MA influenciou positivamente o crescimento e a
nodulação em ambos os estádios estudados em relação às plantas -MA. Embora a
micorrização não tenha compensado completamente a demanda por P quando comparada a
planta de soja bem nutrida, a simbiose aumentou a atividade da nitrogenase na floração e
levou a menor limitação metabólica da fotossíntese em condições de escassez de P, quando
comparada às plantas não micorrizadas. Essas respostas poderiam ser resultantes da
contribuição da micorriza na absorção de P, estímulo da fixação simbiótica de nitrogênio
(FSN) e possivelmente do estímulo da fotossíntese pelo aumento da força dreno. No segundo
experimento, os resultados foram diferentes ao primeiro quanto ao beneficio na nodulação e
no crescimento, o que pode ter sido efeito de fatores climáticos pouco favoráveis ou da
diminuição da capacidade colonizadora do fungo MA, uma vez que os níveis de colonização
micorrízica neste experimento foram menores aos do primeiro experimento. Considerando o
crescimento das plantas, a soja nodulada não se beneficiou de forma significativa da simbiose
MA em condições de limitação de P. No entanto, foi possível verificar a influência da MA em
vários aspectos fisiológicos e moleculares como atividade fotoquímica, metabolismo do N,
padrão de expressão de transportadores de P e de genes relacionados com o metabolismo de
N. O perfil de expressão de genes da família de transportadores PHT1 foi significativamente
alterado pela simbiose, o que sugere diferentes vias de aquisição de P em plantas micorrizadas
e não micorrizadas durante a deficiência em P.
Palavra Chave: Glycine max, Glomus macrocarpum, Bradyrhizobium elkanii, deficiência em
fósforo, fixação simbiótica de nitrogênio, fotossíntese, transportadores de fosfato

ABSTRACT
Nitrogen (N) and phosphorus (P) are nutrients that can limit plant performance.
Currently, there is growing awareness of the importance of plant-microorganism
interactions in nutrient acquisition and in promoting plant growth, especially under
stress. Arbuscular mycorrhiza (AM) is one of the most ubiquitous symbioses between
soil roots and fungi, with a recognized role in mineral nutrition and the attenuation of
several plant stresses. The objective was to evaluate the effects of arbuscular
mycorrhizal (AM) symbiosis on physiological and molecular aspects of nodulated
soybean exposed to P deficiency, with special interest in nodulation capacity and
transport and absorption of P. Two experiments were carried out in hydroponic system,
using vermiculite as substrate, which soybean plants (Glycine max [L.] Merrill) was
kept under low P levels (50 μM) and nodulated with Bradyrhizobium elkanii were
inoculated (+AM) or not (-AM) with the fungus AM Glomus macrocarpum. Non-
inoculated plants maintained under conditions of sufficient P (500 μM) constituted the
treatment denominated - AM + P. Harvest were performed at the beginning of flowering
and during filling of the seed. In the first experiment the results showed that AM
positively influenced growth and nodulation in both studied stages when compared to
the - AM plants. Although the symbiosis did not fully compensate for the demand of P
when compared to the well-nourished soybean plant, symbiosis increased nitrogenase
activity at flowering and presented less metabolic limitation of photosynthesis in
conditions of P shortage than non-mycorrhizal ones. These responses could result from
the contribution of mycorrhizae to P uptake, Nitrogen fixation (NF) stimulus and
increased drain strength. In the second experiment, the results were different to the first
one regarding the nodulation and growth benefit, which may have been due to
unfavourable climatic factors or the decrease in the colonizing capacity of the fungus
AM, since the levels of mycorrhizal colonization in this experiment were significantly
lower than in the first experiment. Considering plant growth, nodulated soybean did not
benefit significantly from the AM symbiosis under conditions of P limitation. However,
it was possible to verify the influence of AM in several physiological and molecular
aspects such as photochemical activity, N metabolism, expression of P transporters and
genes related to N metabolism. The gene expression profile of the PHT1 transporters
family was significantly altered by symbiosis, suggesting different pathways of Pi
acquisition in mycorrhized and non-mycorrhized plants during P starvation.
Keywords: Glycine max, Glomus macrocarpum, Bradyrhizobium elkanii, phosphorus-
starvation, nitrogen fixation, photosynthesis, phosphate transporter

SUMÁRIO
Resumo....................................................................................................06
Abstract...................................................................................................07
Introdução.........................................................................................10
Soja e fixação simbiótica de nitrogênio……….........…….…....10
Fósforo: importância, disponibilidade e absorção…......…..….13
Transportadores de fosfato em soja……………........…......…..14
Resposta das plantas à baixa disponibilidade de fósforo...........17
Simbiose micorrízica arbuscular………………………............19
Hipóteses e objetivos………………………………………......….…...23
Capítulo 1:
Mycorrhiza enhances nitrogen fixation and photosynthesis in
phosphorus-starved soybean plants
Abstract………………………………………………………..……….25
Introduction………………………………………………..…...………26
Material and methods
Experimental design and soil preparation……………………..29
Biological material and plant growth conditions……………....29
Measurements and analytical determinations……….………....30
Statistical analysis……………..…………………..…………...31
Results
Plant growth, nodulation and mycorrhizal colonization……….33
Nutrient concentration, N and P contents and use efficiencies and
nitrogenase activity….…..……………………………………..34
Photosynthesis………….……………………….….……….….37
Discussion……………………………..…....…………………….........39
Conclusion…………………………………..……….………………....44
Acknowledgements…………………………………..………...………44
References……………………………………………...…...………….45
Capítulo 2:
Influência da simbiose micorrízica na expressão de transportadores
de fosfato em raízes e nódulos de soja cultivada em deficiência de
fósforo

Introdução............................................................................................50
Material e Métodos
Delineamento experimental………....…….……………............54
Materiais biológicos, plantio e condições de crescimento..........54
Preparo da solução nutritiva…………………....…….........….55
Preparo da cultura de rizóbio……………………..….........…..55
Inóculo de fungos micorrízicos arbusculares e avaliação da
colonização micorrízica ………………...…………............…..56
Matéria seca de parte aérea, raízes e nódulos…………............56
Determinação da concentração de nutrientes……….........…....56
Determinação da concentração de fosforo inorgânico…...........56
Atividade fotoquímica..................................................................57
Determinação da concentração de aminoácidos e ureídeos.......57
Extração de RNA e síntese de cDNA……………………….......58
Obtenção das sequências e primers……………………….…...58
Análise de expressão por PCR quantitativa em
tempo real (RT-qPCR) ……………............................................61
Análises estatísticas………………….........………..………......61
Resultados
Crescimento, nodulação e colonização micorrízica……….......62
Concentração de nutrientes..…………………....….………......63
Atividade fotoquímica..........…………………………….….......68
Conteúdo de aminoácidos e ureídeos totais…………….……...68
Expressão gênica em raízes e nódulos da soja:
metabolismo de N e transportadores de P…………....…..…....69
Discussão.................................................................................................75
Conclusão................................................................................................81
Considerações finais................................................................................82
Referências..............................................................................................84
Anexo I..........................................................................................94
Anexo II.........................................................................................95

10
Introdução
Soja e fixação simbiótica de nitrogênio
A soja, Glycine max (L.) Merrill, é uma leguminosa da subfamília Faboideae,
oriunda do Leste da Ásia, é cultivada em grande quantidade nas Américas,
principalmente nos Estados Unidos e no Brasil, maiores produtores desta cultura que
hoje movimenta milhões no mercado de grãos (Ministério da Agricultura
http://www.agricultura.gov.br/vegetal/culturas/soja). Na safra 2015/2016, o Brasil foi o
segundo maior produtor de grãos desta leguminosa atrás apenas dos Estados Unidos. Da
semente da soja se obtém diversos produtos principalmente farinha e óleo. Por possuir
alto teor de proteínas, o grão da soja é muito utilizado na alimentação humana e animal.
A planta pode ser utilizada como forragem animal e como adubo verde quando plantada
em consórcio com outras espécies. A soja é capaz de se associar com pelo menos cinco
espécies de bactérias fixadoras de dinitrogênio (N2), do filo das Proteobactérias, dos
gêneros Bradyrhizobium e Sinorhizobium: Bradyrhizobium japonicum, B. elkanii, B.
liaoningense, B. yuanmingense e Sinorhizobium fredii. A simbiose da soja com
bactérias do gênero Bradyrhizobium é uma das mais eficientes no que se refere ao
aporte de nitrogênio (N) para a planta. No Brasil, a fixação simbiótica de N2 (FSN) na
cultura da soja é altamente eficiente e traz importantes benefícios econômicos
(Döbereiner, 1997; Hungria et al., 2001), já que a soja nodulada não precisa ou tem a
necessidade de adubação nitrogenada reduzida, podendo fixar de 109 a 250 kg/ha ano
de N2, o que representa entre 70% e 85% do N total acumulado pela planta (Hungria et
al., 2001).
Na FSN, o N2 atmosférico é convertido em amônia a qual é posteriormente
incorporada em aminoácidos e/ou ureídeos, que são transportados à parte aérea via
xilema (Schubert, 1986). As raízes da soja são capazes de formar nódulos por
estabelecerem associação simbiótica com bactérias fixadoras de N2 do grupo “rizóbia”
(Balestrasse et al., 2006; Udvardi et al., 1988). De forma prévia ao estabelecimento da
simbiose ocorre o processo de reconhecimento entre as raízes e as bactérias por meio de
uma complexa troca de sinais químicos e moleculares, na qual as raízes liberam
flavonóides, os quais induzem a expressão de genes bacterianos conhecidos como
fatores nod (D’Haeze & Holsters, 2002). Esses fatores de nodulação são essenciais
durante o processo de colonização dos rizóbios através dos pelos radiculares e na

11
indução da divisão das células do córtex da raiz, onde se inicia a organogênese do
nódulo (Broughton et al., 2003; Dakora, 2000; Geurts & Bisseling, 2002). Dentro do
nódulo as bactérias se dividem e passam por um processo de transformação dentro das
membranas peribacteróides, formando os chamados bacteróides; o conjunto de
bacteróides envolto por membrana vegetal forma o denominado simbiosoma (Gage,
2004).
As células infectadas estão rodeadas por tecido radicular não infectado que
contém tecido vascular (xilema e floema), o qual mantém o fluxo de metabólitos entre
nódulo, raiz e parte aérea (Atkins, 1987). A enzima nitrogenase, responsável pela
atividade de fixação de N2, se encontra no interior dos bacteróides, em um ambiente
com baixa concentração de O2, uma vez que a atividade desta enzima é sensível à
presença deste elemento inibindo-a (Luciñski et al., 2002). Esta condição de baixa
disponibilidade de O2 é obtida por meio da presença de uma camada de células que
forma uma barreira à difusão do O2 (Minchin, 1997) e também devido à presença da
proteína leghemoglobina que possui alta afinidade pelo O2 e realiza o transporte do
mesmo dentro das células reduzindo a concentração de O2 livre (Appleby, 1984). Na
FSN o N2 atmosférico é fixado em amônia pela ação da nitrogenase, a qual é um
complexo enzimático formado por duas subunidades: o componente I, denominado
proteína ferro-molibdênio (FeMo) e que catalisa a redução do N2, e o componente II, a
dinitrogênio redutase, denominada Fe-proteína e que transfere os elétrons para a
proteína FeMo. A nitrogenase somente é ativa quando as duas subunidades estão
associadas (Andrews et al., 2009). As subunidades α e β do componente I são produtos
dos genes nifD e nifK, respectivamente. A Fe-proteína ou componente II é um
homodímero que se liga a um centro de ferro-enxofre e é codificada pelo gene nifH. A
nitrogenase clássica é, portanto, codificada pelo complexo gênico nifHDK (Andrews et
al., 2009).
A reação catalisada pela nitrogenase quebra a tripla ligação do N2 reduzindo-o à
amônia e consumindo energia metabólica na forma de ATP, na seguinte reação:
N2 + 8e- + 8 H
+ + 16 ATP → 2NH3 + H2 + 16ADP + 16 Pi
A amônia produzida é então assimilada em glutamina (Gln) pela enzima
glutamina sintetase (GS), a qual pode ser seguida da formação de glutamato (Glu) pela
enzima glutamato sintase (GOGAT) (Lam et al., 1996). A GS catalisa a aminação do

12
Glu produzindo Gln e esta pode ser transportada para a parte aérea via xilema, ou pode
ser canalizada para a síntese de outros aminoácidos ou de ureídeos dentro do nódulo
(Forde & Lea, 2007) (Figura 1). A GOGAT transfere o grupo amido da Gln para o 2-
oxoglutarato produzindo duas moléculas de Glu (Andrews et al., 2009), que podem ser
utilizadas também na síntese de outros aminoácidos (Forde & Lea, 2007). Em plantas
fixadoras das regiões tropicais, como a soja, os ureídeos alantoína e alantoato são as
principais moléculas nitrogenadas transportadas para a parte aérea (Atkins, 1987). Estas
moléculas apresentam proporção N:C de 1:1, e uma vez que são sintetizadas nos
nódulos podem ser consideradas boas indicadoras do metabolismo do nódulo e da FSN
(McClure et al., 1980).
A biossíntese de ureídeos é complexa, começa a partir da inosina monofosfato
(IMP) e de guanina proveniente da degradação das purinas (Figura 1). A IMP é
convertida em xantina, que é então convertida em ácido úrico, pela xantina oxidase. O
ácido úrico formado é convertido pela enzima uricase, ou oxidase do ácido úrico, em
alantoína, e esta pode ser transformada a alantoato pela alantoinase (Figura 1) (Smith &
Atkins, 2002; Zrenner et al., 2006). Sabe-se que a soja nodulada transporta mais de 80%
dos compostos nitrogenados na forma de ureídeos (Schubert et al., 1995), enquanto que
a não nodulada transporta principalmente aminoácidos como a asparagina (Asn) e
nitrato (Amarante et al., 2006). Apesar da predominância dos ureídeos na soja
simbiótica quantidade significativa de aminoácidos no xilema tem sido observada
(Amarante & Sodek, 2006).
Figura 1. Esquema mostrando o metabolismo de fixação de N2, assimilação de N, síntese de
ureideos e transporte em nódulos de soja. (Sym, simbiosoma; Bac, bacteróide; Glu, glutamato;
GS, glutamina sintetase; Gln, glutamina, UA, ácido úrico; Aln, alantoína; Alc, ácido alantóico;
CS; estria de Caspary) (Carter & Tegeder, 2016).

13
Por outro lado, a sacarose proveniente da fotossíntese é a fonte primaria de
carbono reduzido para o metabolismo do nódulo, a qual é clivada pela sacarose sintase e
metabolizada na via glicolítica, principalmente no citoplasma de células vegetais na área
infectada. Esta via glicolítica está regulada positivamente no nível transcricional durante
o desenvolvimento do nódulo (Udvardi et al., 1988). Sabe-se que mutantes com
atividade fortemente reduzida da enzima sacarose sintase são defectivos na FSN
(Gordon et al., 1999). Nos nódulos as atividades fosfoenolpiruvato carboxilase
(PEPCase) e malato desidrogenase (MDH) divergem o fluxo de carbono da glicólise
para a formação de malato, o qual é considerado a principal fonte de carbono
transportada ao bacteroide. Numerosos estudos mostram a regulação transcricional
positiva dos genes da PEPCase e da MDH, assim como daqueles para a anidrase
carbônica, que converte o CO2 em ácido carbônico, para o seu uso pela PEPCase
durante o desenvolvimento do nódulo. Evidências indicam que os ácidos dicarboxílicos,
principalmente o malato, são as principais fontes de carbono fornecidas pela planta ao
bacteróide e usadas durante a FSN. Alguns estudos mostram que os genes codificadores
de proteínas responsáveis pelo o transporte e o metabolismo de ácidos dicarboxílicos
são indispensáveis para a FSN (Udvardi et al., 1988), evidenciando importância dos
ácidos dicarboxílicos no metabolismo primário dos bacteróides. A membrana do
simbiosoma de todas as leguminosas estudadas até o momento possuem transportadores
de dicarboxilato com suficiente atividade para comportar as taxas de FSN registradas
em plantas (Jeong et al., 2004). Dada a alta demanda por energia metabólica durante o
processo de fixação, um dos nutrientes mais importantes para a FSN é o fósforo.
Fósforo: importância, disponibilidade e absorção
O fósforo (P) é componente de muitas moléculas com função biológica, como
por exemplo, DNA, RNA, proteínas, lipídios, açúcares, ATP, ADP e NAD(P)H,
envolvidas em processos como a transferência de informação e de energia, a
fotossíntese, a sinalização intracelular, replicação e expressão de genes. O que definea
essencialidade do P para as plantas (Marschner, 2011). Depois do N, o P é o
macronutriente que mais limita o crescimento e desenvolvimento das plantas. Os solos
contem várias formas inorgânicas deste nutriente que variam conforme o pH do solo,
incluindo as formas inorgânicas PO43-
, HPO42-
, H2PO4-
. O P é absorvido pelas plantas
na forma de ortofosfato (H2PO4-), forma de P inorgânico (Pi) predominante em solos
com pH entre 5 e 6 (Schachtman et al., 1998). O P do solo também pode estar na forma

14
orgânica como ésteres fosfatos, entre os quais os inositois fosfatos, ácidos nucleicos e
fosfolipídios são os mais frequentes no solo (Nussaume et al., 2011). Ainda, o P é
considerado um elemento de baixa mobilidade no solo, devido a sua lenta difusão no
solo (10-12
a 10-15
m2 s
-1) e à sua alta adsorção às partículas do solo (óxidos e hidróxidos
de Fe e Al, cátions e matéria orgânica) (Schachtman et al., 1998). A absorção de Pi
pelas raízes das plantas cria uma área de depleção ao redor da superfície da raiz, motivo
pelo qual o P pode ser considerado um dos macronutrientes menos disponíveis para a
planta na solução do solo, em concentrações entre 0,5 e 10 μM. Assim, a aquisição e
utilização de P pelas plantas são fatores determinantes na produção agrícola (Zhang et
al., 2014).
O sistema radicular da planta tem a função de explorar o ambiente do solo e
absorver quantidades adequadas de nutrientes para o crescimento e o desenvolvimento
(Zhang et al., 2014). Na agricultura tradicional os agricultores dependem da inerente
fertilidade do solo ou da adição de fertilizantes fosfatados para fornecer o P demandado
pela planta. Contudo, a intensificação da agricultura resultou na dependência de
aplicação de fertilizantes para aumentar a produção agrícola. O fosfato usado nos
fertilizantes é originado principalmente de rochas fosfatadas, única reserva global
significante de Pi, e um recurso considerado não-renovável e finito. Alguns estudos
mostram que está havendo uma diminuição contínua na qualidade e quantidade das
rochas fosfatadas, pois as reservas de alto grau de qualidade vêm sendo esgotadas (Van
Kauwenbergh, 2010). Portanto, é de extrema importância entender a resposta das
plantas à escassez de P com o fim de desenvolver estratégias que possam reduzir a
dependência de adição de fertilizantes à produção agrícola (Jez et al., 2016). Um fator
agravante é que o aumento na aplicação de fertilizantes fosfatados não é sempre eficaz
na melhoria da produção vegetal, pois apenas de 10% a 45% do P aplicado é absorvido
pelas culturas (Adesemoye & Kloepper, 2009), diminuindo a eficiência econômica da
produção agrícola e aumentando o risco de poluição de águas e outros recursos
ambientais (Jez et al., 2016).
Transportadores de fosfato em soja
As plantas absorvem nutrientes do solo principalmente através de proteínas
transportadoras localizadas na membrana plasmática das células epidérmicas e pelos
radiculares. O Pi é absorvido do solo, transportado através das células do córtex e da

15
endoderme da raiz até chegar ao xilema e transportado para as células e tecidos dos
diversos órgãos da planta através de diferentes classes de proteínas transportadoras de
fosfato (Nussaume et al., 2011). Em plantas, os transportadores de Pi pertencem a
quatro famílias: PHT1, PHT2, PHT3 e PHT4. Os transportadores envolvidos na
aquisição de Pi são codificados pela família de genes PHT1, são predominantemente
expressos nas células epidérmicas e na região externa do córtex dos pelos radiculares e
tem sido indicados como responsáveis pela absorção de Pi na interface solo-planta. Em
contraste com os genes Pht1, membros das famílias PHT2, PHT3 e PHT4 têm sido
associados principalmente com a distribuição de Pi entre compartimentos subcelulares, e
seus produtos gênicos estão localizados no retículo endoplasmático (PHT2), membranas
de plastídios (PHT3), membrana mitocondrial interna e aparato de Golgi (PHT4)
(Lopez-Arredondo et al., 2014).
As proteínas PHT1 transportam Pi através do mecanismo de co-transporte, tipo
simporte com íons de hidrogênio (Ullrich-Eberius et al., 1981). Como ocorre para
outros nutrientes, o Pi é, na maioria das vezes, absorvido seletivamente pela raiz por
meio de transportadores PHT1 contra um gradiente eletroquímico, dada a acentuada
variação no gradiente de concentração entre o meio externo e o intracelular. Esse
processo de absorção é energeticamente mediado pela atividade de proteínas de
membrana específicas: ATPases do tipo P, que unidirecionalmente transportam os
prótons através da membrana celular às custas da hidrólise de ATP. O gradiente de
concentração de prótons e o potencial de membrana gerado formam uma força próton-
motriz que facilita a passagem de Pi através das PHT1 contra seu gradiente de
concentração (Karandashov & Bucher, 2005). A via direta de absorção de Pi consiste na
absorção de P através da epiderme e pelos radiculares; sendo que para melhorar a
eficiência de absorção de Pi em condições de baixa disponibilidade as plantas podem
regular a arquitetura da raiz com o fim de aumentar a relação superfície da raiz/volume
do solo, por meio da emissão de pelos radiculares ou de raízes secundárias,
principalmente na região mais superficial do solo (Smith et al., 2003).
No genoma da soja foram identificados 14 genes da família de transportadores
PHT1, distribuídos em oito cromossomos, e mais um pseudogene (Fan et al., 2013), os
quais se originaram de quatro antecessores comuns antes do evento de triplicação do
genoma (Schlueter et al., 2007; Schmutz et al., 2010) que ocorreu durante a história
evolutiva da soja no período entre 130 e 240 milhões de anos atrás. Esses

16
transportadores antecessores foram denominados: GmPHT1 A, B, C e D e deram
origem a sete pares de genes parálogos e um gene singleton. Esses sete pares de genes
parálogos PHT1 sofreram sub-funcionalização e tiveram conservada a sua função na
absorção de Pi, mesmo que com diferentes padrões de expressão espaço-temporal e/ou
com diferentes afinidades pelo Pi. Já o gene parálogo GmPHT1;4 passou por processo
de pseudogenização, processo pelo qual uma das cópias de genes que passaram por
processo de duplicação é perdida (Moore & Purugganan, 2005). Os genes PHT1
mostram maior expressão em raízes sob deficiência de Pi, mas em diferentes graus
dependendo do transportador, sendo que a maioria das proteínas PHT1 tem localização
subcelular similar na membrana plasmática. Exceção para GmPHT1;8 que se localiza
no reticulo endoplasmático, e o GmPHT1;10, que além de se expressar na membrana
plasmática também foi localizado nas bandas de Hechtian, entre a membrana plasmática
e a parede celular (Fan et al., 2013). Dos 14 genes da família de transportadores de Pi no
genoma da soja, três GmPT7, GmPT10 e GmPT11 foram descritos como induzidos
especificamente pela presença de micorriza arbuscular MA (Tamura et al., 2012). Pouco
se sabe sobre como ocorre o transporte de Pi até os nódulos de soja simbiótica e é
praticamente desconhecido o papel da MA na modulação ou a extensão do aporte de Pi
a este órgão. Em leguminosas noduladas sabe-se que a epiderme dos nódulos possui
capacidade de absorver Pi e que este pode chegar até o nódulo pelo tecido vascular
(Carter & Tegeder, 2016). Trabalhos recentes têm identificado o transportador GmPT5
como chave no controle do transporte de Pi entre a raiz e o nódulo, particularmente sob
condições de deficiência de P (Qin et al., 2012).
A maioria dos genes de PHT1 são induzidos pela privação de Pi ou pela
simbiose com fungos micorrízicos arbusculares (FMAs) nas raízes, indicando que a
absorção de Pi é fortemente dependente de PHT1 (Nussaume et al., 2011). A família de
genes codificando proteínas transportadoras PHT1 é monofilética (Karandashov &
Bucher, 2005) e nas angiospermas pode ser agrupada em 4 subfamílias, a subfamília I é
composta por genes PHT1 induzidos por FMAs, a subfamília II por genes de mono e
eudicotiledôneas, subfamília III são exclusivos de eudicotiledôneas, e a subfamília IV
são exclusivas de monocotiledôneas (Loth-Pereda et al., 2011; Tamura et al., 2012).
Antes da ocorrência das primeiras plantas terrestres, há aproximadamente 475 milhões
de anos, a subfamília II das PHT1 divergiu, indicando que o mecanismo de absorção
direta de Pi foi a principal via de aquisição do nutriente pelas plantas mais basais

17
(Wellman et al., 2003). Cerca de 460 milhões de anos atrás surgiram também os FMAs
que podem ter desempenhado um papel crucial na facilitação da colonização do meio
terrestre pelas primeiras plantas que provavelmente eram briófitas (Redecker et al.,
2000).
Resposta das plantas à baixa disponibilidade de fósforo
Em plantas a deficiência em P pode gerar um conjunto diverso de respostas
denominadas PSR (Phosphorus Starving Response), que na parte aérea da planta
incluem o aumento da reutilização e remobilização de Pi, a redução da taxa
fotossintética, o acúmulo de antocianinas, aumento na concentração de açúcares, a
remodelação de lipídeos e a diminuição no crescimento da mesma, o que resulta na
perda de produtividade das culturas (Zhang et al., 2014). Já as raízes em resposta à
baixa disponibilidade de P apresentam aumento na capacidade de absorção, transporte e
translocação de Pi, alterações na arquitetura do sistema radicular (ASR), aumento na
exsudação de ácidos orgânicos e secreção de fosfatases ácidas, mudanças em rotas
metabólicas, associação com fungos micorrízicos arbusculares (FMAs) e também a
substituição de lipídeos, fosfolipídeos por sulfolipídeos ou galactolipídeos (Zhang et al.,
2014). O status de Pi no tecido vegetal influencia diretamente os processos metabólicos
fotossintéticos de transferência de energia (geração de ATP e NADPH), a regeneração
de substratos e utilização de fotoassimilados e a difusão de CO2 nas folhas (Singh et al.,
2013). Devido à regulação negativa da fotossíntese durante a deficiência em Pi as
plantas podem se ver privadas de carboidratos levando a diminuição do crescimento e
reduzindo a produtividade primária e o rendimento agrícola (Høgh Jensen et al., 2002).
Para se adaptar à variação de nutrientes no solo as plantas desenvolveram
mecanismos complexos para integrar a percepção e resposta sistêmica de sinais e
manter a homeostase celular de nutrientes em todos os órgãos. As raízes ao perceberem
alterações na disponibilidade de nutrientes enviam sinais para a parte aérea através do
xilema. A parte aérea percebendo os sinais enviados se comunica com o ápice dos
ramos e raízes através do floema para ajustar os processos de desenvolvimento e
absorção de nutrientes (Liu et al., 2009). Estudos em Arabidopsis mostraram que a
deficiência em P é percebida pela região da ponta da raiz, incluindo a região
meristemática e a coifa (Svistoonoff et al., 2007). As mudanças externas de Pi são
percebidas e sinais moleculares como açúcares, hormônios e microRNA (miRNA) são

18
enviados. Estes sinais funcionam como sinais locais ou sistêmicos para induzir respostas
à distância. Sabe-se que o próprio Pi pode atuar como sinalizador pois o aumento da
concentração interna de Pi reprime diversos genes da PSR (Varadarajan et al., 2002). O
microRNA miR399 funciona como um sinal sistêmico, móvel, quando as plantas
experimentam deficiência de Pi. A expressão do miR399 é induzida, este miRNA liga-
se à ligase E2 PHO2 a qual é suprimida, levando à indução de genes transportadores de
Pi e à ativação da absorção de Pi. Isto ocorre porque PHO2 regula negativamente a
expressão de PHT1 por ubiquitinização (Aung et al., 2006). Análises de promotor-
repórter sugerem que miR399 e PHO2 são expressos predominantemente em tecidos
vasculares (Aung et al., 2006). Alguns fitohormônios também atuam como sinais e
alterações na disponibilidade de Pi podem afetar a produção, a sensibilidade e o
transporte de hormônios como a auxina e o etileno, que estão associados a sinalização
para modificação da ASR em resposta à deficiência em P (Al‐Ghazi et al., 2003; Ma et
al., 2003). Devido às diversas funções exercidas pelo Pi nas vias de sinalização
interligadas, a resposta à baixa disponibilidade de P geralmente resulta da combinação
de várias rotas de sinalização (Chiou & Lin, 2011).
Em situações de escassez de P as plantas podem presentar respostas específicas
que procurem aumentar tanto a capacidade de absorção quanto a eficiência do uso
interno de P. As características estruturais do sistema radicular que estão relacionadas à
PSR são o aumento na relação raiz: parte aérea e mudanças na arquitetura do sistema
radicular, como aumento na produção de raízes laterais, aumento no comprimento e no
número de pelos ou produção de raízes nas camadas mais superficiais do solo (van de
Wiel et al., 2016) e estabelecimento de simbioses micorrízicas, as quais contribuem
para a maior absorção e/ou mobilização de P no solo. Outra característica fisiológica
que tem sido observada é a exsudação radicular, a qual pode contribuir na mobilização
de P no solo. Os exsudatos radiculares incluem desde prótons, ácidos orgânicos, tais
como o ácido cítrico, malato e oxalato, e compostos fenólicos, até enzimas hidrolíticas
de formas orgânicas de P no solo, como as fosfatases ácidas e nucleases, como as
ribonucleases. Para a eficiente absorção do Pi liberado ou mobilizado, a expressão de
transportadores de fosfato de alta afinidade deve ser induzida em baixas concentrações
de P, aumentando a eficiência de absorção de P (Plaxton & Tran, 2011). Um aspecto
importante é a efetiva reciclagem de P de tecidos maduros ou senescentes à tecidos em
crescimento ativo ou o reuso de P armazenado nos vacúolos. As fosfatases não estão

19
somente envolvidas na mobilização do P do solo, mas também na realocação do mesmo
na planta, envolvendo grupos específicos de fosfatases (Duff et al., 1994), como as
fosfatases ácidas e as fosfatases ácidas purpúreas. Outros mecanismos relacionados à
PSR podem ser considerados importantes no aumento da eficiência do uso de P (EUP)
durante a deficiência de P, incluindo mudanças metabólicas que reduzam a necessidade
de P, como a remodelação de membranas celulares pela substituição de fosfolipídios por
sulfolipídeos, galactolipídeos ou outros glicolipídeos (Siebers et al., 2015), a redução na
quantidade de RNA ribossomal, o uso de vias metabólicas que evitam o uso de Pi e
adenilatos fosfatados (ATP), acúmulo de açúcares e amido ou de aminoácidos e
metabólitos secundários como fenilpropanóides, flavonóides ou glucosinolatos
(Scheible & Rojas-Triana, 2015).
Simbiose micorrízica arbuscular
Estima-se que mais de 80% das espécies vegetais terrestres sejam capazes de se
associar com fungos do solo do filo Glomeromycota (Schüßler et al., 2001), formando a
simbiose conhecida como micorriza arbuscular (MA), provavelmente a ubíqua das
simbioses terrestres. O desenvolvimento da MA resulta na colonização do córtex da raiz
pelo FMA e na formação de estruturas intracelulares específicas chamadas de
arbúsculos, onde acontece a troca de nutrientes entre ambos os simbiontes. A rede de
hifas extra-radiculares se estende desde a raiz no solo e conecta a planta com regiões
além da área de depleção de nutrientes como o P, aumentando, normalmente, a sua
absorção. Assim, plantas associadas a FMA apresentam frequentemente maior vigor e
maior conteúdo de P, o que pode resultar em maior produção de biomassa. Em troca, os
FMAs associados utilizam carboidratos oriundos da atividade fotossintética da planta
hospedeira.
O reconhecimento entre FMAs e as raízes hospedeiras ocorre, como no caso do
rizóbio com leguminosas, através de sinais moleculares presentes nos exsudatos
radiculares antes de qualquer contato físico entre raiz e fungo. O sinal que induz a
ramificação e crescimento das hifas do FMA são flavonóides ou sesquiterpenos
produzidos por plantas, especificamente hormônios do grupo das estrigolactonas. Por
outro lado, moléculas produzidas pelo fungo, lipo-quito-oligossacarídeos, induzem a
ativação de diferentes genes na planta que controlam a colonização do fungo pela raiz
(Kosuta et al., 2003). Assim, o FMA entra em contato com a raiz e coloniza

20
extensivamente células epidérmicas e corticais da raiz hospedeira. Esta interação entre
plantas e FMAs demanda importantes mudanças no metabolismo da raiz, entre elas a
reorganização e metabolismo de plastídeos e mitocôndrias das células colonizadas
(Lohse et al., 2005). Mas, ao contrário do que ocorre nas simbioses leguminosa-rizóbio,
a associação MA é essencialmente não específica, podendo mais de uma espécie de
FMA se associar de forma mutualística com variadas espécies vegetais e mais de uma
espécie de FMA colonizar a raiz de uma mesma planta.
Na via direta de absorção de P, atuam membros da família PHT1
desempenhando um importante papel na retirada de Pi da solução do solo e exibindo
uma forte expressão na raiz (Smith et al., 2003). A colonização por FMAs abre uma via
eficaz para adquirir Pi e que não envolve a via direta de absorção de Pi, a captação
simbiótica de Pi através da interação de plantas com fungos micorrízicos aumenta a
capacidade de adquirir P do solo aumentando a área de absorção radicular por meio das
hifas extra-radiculares, dissolvendo Pi insolúvel, e promovendo a mineralização
biológica de P (Richardson & Simpson, 2011); esta rota de absorção de P tem sido
denominada via micorrízica. Nesta rota, o Pi é transportado via hifas rapidamente para
células corticais da raiz, evitando o lento movimento impulsionado por difusão de Pi da
solução do solo à superfície da raiz. Além disso, a simbiose com FMAs ativa a
expressão de uma série de genes induzidos pela deficiência de Pi que tem como objetivo
aumentar a eficiência de aquisição de P pelo sistema radicular (Smith et al., 2011).
Trabalhos anteriores mostram que existem genes induzidos pela associação de
FMAs, que codificam transportadores de P em plantas. Esses incluem MtPHT1;4 em
Medicago truncatula (Harrison, 2002), OsPHT1;11 e OsPHT1;13 em arroz (Glassop et
al., 2007), StPHT1;3, StPHT1;4 e StPHT1;5 em batata (Rausch et al., 2001),
GmPHT1;7, GmPHT1;10 e GmPHT1;11 em soja (Tamura et al., 2012), AsPHT1;1 em
Astragalus sinicus (Xie et al., 2013), ZmPHT1;6 em milho (Willmann et al., 2013),
SlPHT1;3, SlPHT1;4 e SlPHT1;5 em tomate (Chen et al., 2007), e BdPHT1;3,
BdPHT1;7, BdPHT1;12 e BdPHT1;13 em Brachypodium distachyon (Hong et al.,
2012). Leguminosas como soja, alfafa, ervilha e feijão são capazes de estabelecer dupla
simbiose com FMAs e rizóbios. A capacidade da MA no incremento da absorção de P
pela planta, contribui para a absorção deste elemento pela planta em solos com baixa
fertilidade, superando desta forma a deficiência de P e conduzindo ao incremento da
nodulação, FSN e produção de biomassa vegetal. A MA também pode aumentar a

21
absorção de micronutrientes como o Cu e o Zn, modula a distribuição de
fotoassimilados e o balanço hormonal em leguminosas (van der Heijden et al., 2008). A
simbiose dupla entre leguminosas e rizóbios é crucial na formação e composição das
comunidades vegetais (van der Heijden et al., 2008). A MA pode, dependendo das
espécies envolvidas e das condições de crescimento, promover a FSN pelo aporte de P e
outros nutrientes que são essenciais para o processo de fixação (Xavier & Germida,
2002), com casos de redução ou inibição da FSN em leguminosas crescendo em solo
com baixa fertilidade e na ausência da associação com FMAs (Asimi et al., 1980;
Azcón et al., 1991).
A associação tripartite, rizóbio-FMA-leguminosa, tem mostrado na maioria das
vezes interação sinérgica ou aditiva dos benefícios dos simbiontes microbianos,
resultando na promoção da produtividade da leguminosa hospedeira (Andrade et al.,
2004; Azcón et al., 1992; Barea & Azcon-Aguilar, 1983). Os FMAs aumentam a
produção de biomassa e a eficiência fotossintética pela diminuição na razão N:P, sendo
a maior absorção de P frequentemente associada ao aumento no acúmulo de N e da
produtividade de leguminosas (Gray, 2010). A eficiência do uso de N na fotossíntese é
normalmente aumentada pelo suprimento de P pela MA. Tanto as taxas de fotossíntese
saturadas por luz e o rendimento quântico aumentam em resposta ao suprimento de N
por parte da simbiose com rizóbios. Portanto, a relação N:P controla o aumento da
resposta fotossintética e indica que a relação N:P:C possui papel fundamental nas
associações tripartites, determinando a produção de biomassa das plantas (Gray, 2010).
Em leguminosas, a aquisição de recursos e sua alocação são dependentes do
conjunto de complexas mudanças dos três membros da simbiose tripartite. É bem
conhecido que a FSN diminui em plantas deficientes em P, o que tem sido explicado
pelo efeito do baixo suprimento de P no crescimento da planta hospedeira, no
crescimento e funcionamento do nódulo ou em ambos, planta e nódulo (Israel, 1987;
Robson et al., 1981). Alguns estudos sugerem que a baixa disponibilidade de P reduz o
crescimento da parte aérea e, em consequência, a menor taxa de fotossíntese limita o
suprimento de C aos nódulos, reduzindo a FSN (Israel, 1987). O processo de redução de
N2 requer ao redor de 2,9 mg de C por miligrama de N fixado, e esta demanda pode
equivaler a até 32% da taxa fotossintética das plantas. Estudo sobre os efeitos da
deficiência de P e o suprimento de fotoassimilados da parte aérea para o nódulo tem
função importante na FBN. As leguminosas que adquirem N por meio da FSN possuem

22
geralmente um maior requerimento de P e outros nutrientes para manutenção do
adequado metabolismo do nódulo (Sulieman et al., 2013). A escassez ou deficiência de
P podem afetar diretamente a FSN, modulando a formação, crescimento e
funcionamento do nódulo. Tem sido sugerido que à alta demanda de ATP para o
funcionamento da nitrogenase e a disponibilidade de P são fatores críticos para a ótima
atividade do nódulo (Al-Niemi et al., 1997).

23
Hipóteses e objetivos
Diante do relatado na literatura levantam-se as seguintes hipóteses:
- O fósforo absorvido pela MA pode atender a demanda de nódulos sob escassez
de P, promovendo a nodulação e a FSN estimulando o desenvolvimento e atenuando os
efeitos de deficiência de P em plantas de soja.
- A MA modula a expressão de proteínas transportadoras de P, contribuindo com
o aporte de Pi ao tecido do nódulo influenciando a fixação de N2.
- O Pi absorvido pela MA e o estimulo da fotossintese devida a dupla simbiose,
pode influenciar de forma positiva a fotossintese.
Portanto o objetivo deste trabalho foi avaliar os efeitos da simbiose micorrizica
arbuscular (MA) em aspectos fisiólogicos e moleculares de soja nodulada exposta à
deficiencia de P, com especial interesse na capacidade de nodulação, transporte e
absorção de P.
Para isso foi realizado um estudo composto de dois experimentos com igual
delineamento experimental os quais são apresentados em dois capítulos. No primeiro
foram analisados diferentes parâmetros de crescimento, a nodulação, concentração de
nutrientes nos tecidos vegetais, a atividade da nitrogenase e as trocas gasosas. No
segundo, além das analises de crescimento, nodulação e concentração de nutrientes, foi
avaliada a atividade fotoquímica, conteudo de aminoácidos e uréideos totais nas folhas
e, a expressão de genes relacionados com o metabolismo de N e com transportadores de
P nas raízes e nódulos. Finalmente, é apresentada uma seção de considerações finais que
discute e comenta o conjunto de resultados de ambos os experimentos realizados.

24
CAPÍTULO 1
MYCORRHIZA ENHANCES NITROGEN FIXATION AND
PHOTOSYNTHESIS IN PHOSPHORUS-STARVED SOYBEAN PLANTS

25
Mycorrhizae enhance nitrogen fixation and photosynthesis in phosphorus-starved
soybean plants
Rafaela Gageti Bulgarelli, Fernanda Castro Correia Marcos, Rafael Vasconcelos
Ribeiro, Sara Adrián López Andrade*
Department of Plant Biology, Institute of Biology, Institute of Biology, P.O. Box: 6109,
University of Campinas – UNICAMP 13083-970, Campinas, SP, Brazil
*Corresponding author: [email protected]
Abstract
Nitrogen (N) and phosphorus (P) are nutrients that frequently limit plant
performance and crop yield. Currently, awareness of the importance of plant-microbe
interactions in plant nutrient acquisition and growth promotion persist, particularly
under stress challenges, such as those resulting from P scarcity. The aim of the present
study was to evaluate the effects of P starvation on the influence of arbuscular
mycorrhiza (AM) symbiosis on N2-fixing capacities, plant growth, nutrient uptake and
photosynthetic performance of soybean plants at flowering and maturity. To this end,
soybean (Glycine max) plants were nodulated with Bradyrhizobium elkanii and
inoculated or not with the AM fungus Glomus macrocarpum. The +AM and –AM
plants were grown under low P conditions (50 µM), and a control treatment with –AM
plants under sufficient P concentration (500 µM) was also performed (-AM+P). Plants
were grown until the beginning of the flowering and another set till the grain filling
stage. The results showed that mycorrhization improved soybean plant performance
under P scarcity compared with non-AM associated plants under the same conditions.
AM positively influenced growth and plant nodulation at both the flowering and grain-
filling stages, without significant changes in plant P contents. Under P starvation AM
symbiosis increased nitrogenase activity and N contents, the maximum carboxylation
rate of the Rubisco and reduced CO2 compensation point and the metabolic limitation of
photosynthesis, which indicated the AM stimulus to symbiotic N2 fixation (SNF) and
photosynthesis. Nonetheless, under these conditions, mycorrhizal symbiosis could not
completely meet soybean P demand compared with well-nourished soybean plants,
which produced higher plant and nodules biomass. In conclusion, AM association in
nodulated soybean plants is likely an important strategy for maintaining SNF

26
functioning, conferring increased plant growth and better N status under P scarcity
conditions and significantly attenuating negative effects of low Pi in nodulated-soybean
plants.
Keywords: dual inoculation, Bradyrhizobium elkanii, Glomus macrocarpum, Glycine
max, nitrogenase activity, nitrogen fixation, phosphorus-starvation
Introduction
Soybean (Glycine max (L.) Merrill) is a major oilseed crop that obtains nitrogen (N)
through symbiotic N2-fixation (SNF) by establishing an association with specific
diazotrophic proteobacteria collectively known as rhizobia (Whitehead & Day, 1997).
In Brazil, soybean inoculation with N2-fixing rhizobia provides important economic
benefits, significantly reducing N fertilization (Kaschuk et al., 2009). Accordingly,
estimates suggest that nodulated soybean can fix approximately 109-250 kg/ha of N per
year through SNF, representing up to 85% of the total N accumulated by the plant
(Hungria et al., 2001). Atmospheric N2 is converted through SNF to ammonia in a
reaction catalyzed by the prokaryotic enzyme nitrogenase, which in N2-fixing root
nodules are located inside the symbiosome and consumes high amounts of metabolic
energy (Schubert et al., 1995). Subsequently, NH3 is incorporated through the GS-
GOGAT cycle into amino acids or metabolized to ureides in tropical legumes, such as
soybeans (Lam et al., 1996). Subsequently, organic N is transported via xylem to the
shoots, where several important physiological processes occur, such as photosynthesis
(Schubert et al., 1995). Furthermore, sucrose formed during photosynthetic CO2
fixation is the primary source of reduced carbon for nodule metabolism. Inside the
nodules, phosphoenolpyruvate carboxylase and malate dehydrogenase form malate,
considered the main carbon source respired by the bacteroid to support nitrogenase
activity (Udvardi et al., 1988).
As a large proportion of the terrestrial plant species form mutualistic associations
with soil fungi of the sub-phylum Glomeromycotina (Spatafora et al., 2016), arbuscular
mycorrhiza (AM) symbiosis is the most widespread symbiosis in terrestrial ecosystems
(Schüßler et al., 2002). AM establishment results in the fungal colonization of the root
cortex and the formation of specific intracellular structures called arbuscules, where
nutrient exchange between symonbionts occurs. Fungal hyphae grow out of the roots,
acting as bridges between the roots and soil or among roots of different plants, with

27
strong implications in plant physiology (Lynch, 2011). Thus, AM association is
considered a plant nutritional strategy within a set of plant responses to low P
availability. AM fungi use plant photoassimilates as the sole source of carbon,
consuming up to 16% of recent photoassimilates (Kaschuk et al., 2009).
Legumes such as soybean establish dual symbiosis with rhizobia and AM fungi and
both symbionts function as carbon sinks, which depending on the conditions may
stimulate photosynthesis in the host plant (Kaschuk et al., 2009). AM may also enhance
photosynthesis through their influence on phytohormone balance and nutrient
acquisition (Huang et al. 1985). Nodule metabolism requires high amounts of P, such as
ATP, for use during N2 reduction (Xavier & Germida, 2002). Increases in net
photosynthetic rates resulting from the augmented carbon sink of dual symbiosis have
previously been observed in soybean (Harris et al., 1985; Kaschuk et al., 2009; Kucey
& Paul, 1982). However, under limiting conditions, such as those imposed by P or light
scarcity, the beneficial effects of the symbiosis can be neutralized or even eliminated,
causing negative effects on leguminous plants (Ballhorn et al., 2016; Pang & Paul,
1980). AM fungi can also increase the absorption of other nutrients, such as Cu and Zn,
and alter the distribution of photoassimilates and the hormonal balance in legumes (van
der Heijden et al., 2008). The overall modulation of plant physiology acts
synergistically or additively in rhizobia-AMF-leguminous symbiosis, increasing host
productivity, as shown for soybean, peanut and Hedysarum coronarium (Andrade et al.,
2004; Azcón et al., 1992; Barea & Azcon-Aguilar, 1983; Ibrahim et al., 1995).
Phosphorus plays an important role in the composition of chloroplasts and in
photosynthesis, with a regulatory role in ribulose-1,5-bisphosphate carboxylase-
oxygenase (Rubisco) activation and in the ratio of phosphorylated intermediates of the
Calvin cycle. Under severe P limitation, net CO2 assimilation may be reduced,
reflecting a limitation in Rubisco efficiency and activity and the inactivation of the
enzymes involved in ribulose-1,5-bisphosphate (RuBP) regeneration (Brooks et al.,
1988; Fredeen et al., 1990). ADP phosphorylation is also inhibited in chloroplast stroma
under Pi scarcity, limiting RuBP regeneration. In addition, the low demand of NADPH,
ATP and reduced ferredoxin by impaired activity of the Calvin cycle may lead to low
PSII efficiency and photo-oxidative stress (Hernández & Munné-Bosch, 2015). As a
result of photosynthesis impairment, plants may show symptoms of P deficiency, such

28
as chlorophyll loss, anthocyanin accumulation and growth reduction (Hernández &
Munné-Bosch, 2015).
Nonetheless, low P availability is a major constraint for SNF and crop production
worldwide (Tesfaye et al., 2007). P is considered an element of low mobility in soil
(Nussaume et al., 2011; Schachtman et al., 1998), reflecting its low diffusion coefficient
in soil solution (10-12
to 10-15
m2 s
-1) and its high adsorption to soil particles, such as to
Fe and Al hydroxides and sesqui-oxides, and organic matter. As inorganic P (Pi) is
taken up by plant roots, a depletion area is formed surrounding the roots (Lynch, 2011).
Thus, P is one of the less available nutrients to plants, which also vary depending on
soil pH. Pi concentration in soil solution are commonly lower than 10 μM, making P
uptake and use efficiency determinant factors for plant growth (Zhang et al., 2014).
Soils with high P retention capacities are common worldwide, constraining crop yield
and enhancing P fertilizer dependence to maintain or increase productivity. The current
intensification of agriculture has led to the increasing dependence of P-fertilizers
obtained primarily from phosphate rocks, the most significant global P reserve and non-
renewable resource that can be depleted in the next century (Van Kauwenbergh, 2010).
However, the increasing application of P fertilizers is not always effective in improving
crop yields, as only 10% to 45% of applied P is readily used by crops (Adesemoye &
Kloepper, 2009). Thus, plants with higher Pi uptake efficiency under low Pi conditions
may play an important role in increasing productivity and contributing to a more
sustainable agriculture.
Herein, we hypothesized that under Pi scarcity AM enhance P uptake and stimulates
photosynthesis, promoting nodulation and symbiotic nitrogen fixation (SNF) by
stimulating photosynthesis and alleviating P deficiency effects in soybean plants. To
this end, the impact of AM symbiosis on N2-fixing capacity, plant nutrient uptake and
photosynthesis in soybean plants under low Pi conditions at two different developmental
stages (flowering and grain filling) was evaluated.

29
Materials and Methods
Experimental design and substrate preparation
To evaluate the impact of the double symbiosis on SNF, nutrient uptake and
photosynthesis, soybean (Glycine max [L.] Merrill) plants nodulated with
Bradyrhizobium elkanii (SEMIA 5019) were inoculated (+AM) or not (-AM) with the
AM fungus Glomus macrocarpum and grown under low Pi conditions (50 µM). In
addition, a control treatment with -AM plants under sufficient Pi concentration (500
µM) was also included (-AM+P). These Pi concentrations were established based in
previous studies with soybean (Li et al., 2012). Half of the plants from each treatment
were grown until the beginning of flowering (R1-R2 stages, approximately 60 days after
germination), and the remaining plants were grown until the grain filling stage (R5-R6
stages), approximately 100 days after germination, close to the final period of the
reproductive stage (Fehr et al., 1971). The experiment was in a completely randomized
design and for each of the three treatments (+AM, -AM and -AM+P) twenty-four
replications were performed, twelve for flowering and twelve for grain filling stages.
Biological material and plant growth conditions
Soybean seeds of the cultivar “Foscarin IAC-31” were used, which were kindly
provided by the Center for Research and Development of Grains and Fibers, Agronomic
Institute (IAC), Campinas, Brazil. Seeds, previously surface-sterilized (5 min, 3%
NaClO), were immersed for 20 min in liquid inoculum containing approximately 108
cells mL-1
of B. elkanii (SEMIA 5019), grown at 26°C in yeast-mannitol broth (YMB)
for 7 days until the exponential phase of growth (when the optical density at 600 nm
was between 0.6 and 1.0). The AM fungus G. macrocarpum was used as inocula and
provided (treatment +AM) as a mixed sand-soil-inoculum (sand-soil with colonized
roots, spores, and hyphae), obtained from the Centre for Research and Development in
Soil and Environmental Resources, IAC. Each pot received 20 mL of sand-soil-
inoculum with approximately 1,300 spores at the time of sowing. Four seeds were sown
per pot (2 L), previously filled with sterilized fine-grained vermiculite. Non-mycorrhizal
treatments received washings of the soil–inoculum mixture filtered through Whatman
No. 42 filter paper. After cotyledon emergence, the seedlings were thinned to one plant
per pot.

30
The plants were irrigated two times per week with 200 mL of modified Hoagland &
Arnon (1950) nutrient solution without mineral source of N and with P concentration
modified according to the treatment: 50 μM for +AM and -AM and 500 μM for –
AM+P, P was added as KH2PO4. The plants were irrigated with distilled water when
necessary and grown for seven weeks until harvest at the beginning of the flowering and
for 12 weeks for the second harvest at the beginning of the grain filling stage. The
experiment was performed under greenhouse conditions, between September and
November 2015, when maximum and minimum temperatures ranged between 41 and
19°C and day/night duration ranged between 12/12 h and 13/11 h. During the
experiment, maximum photosynthetic photon flux density was around 1,800 μmol m- 2
s- 1
.
Measurements and analytical determinations
At harvest, the shoots and roots were separated, the roots were washed, and the
nodules were detached and counted. Approximately 1 g of fresh root was stored in 50%
ethanol for determining mycorrhizal colonization. The remaining plant material was air-
dried at 60°C, and shoots and roots were weighed and ground. The contents of N, P, K,
Ca, Mg, S, Cu, Mn, Zn, and B were determined in shoots and roots using inductively
coupled plasma-optical emission spectroscopy (ICP-OES; Varian Vista MPX, Palo
Alto, CA, USA) after HNO3–HClO4 digestion. Total N was determined using the
Kjeldahl analysis (Bremner, 1965) after sulfuric acid digestion of plant samples.
Mycorrhizal colonization was evaluated using the slide method (Giovannetti &
Mosse, 1980) in four replicates per treatment after clearing the roots with 2.5% KOH
and staining with 0.05% trypan blue, and it was estimated according to Trouvelot
(1986). For each replicate, 30 root segments of approximately 1 cm were analyzed, and
the presence of intraradical, extraradical hyphae and arbuscules was recorded, and
mycorrhizal colonization intensity (M%), frequency of mycorrhiza (F%) and arbuscule
abundance (A%) were calculated using the MYCOCALC program
(http://www.dijon.inra.fr/mychintec/Mycocalc-prg/download.html).
In vivo nitrogenase activity (NA) was evaluated in nodulated plants (four per
treatment) at the flowering stage by measuring H2 evolution in a synthetic air mixture
using the Qubit™ Open Flow Gas Exchange System (Qubit Systems Inc., Kingston,
Ontario, Canada) according to Justino & Sodek (2013). Qubit™ pots were used, and

31
each pot contained a mixture of gravel and vermiculite (2:1, v:v) as substrate to
facilitate gas flow around the root system. A whole plant was carefully transferred to the
Qubit™ pots and maintained for 1 day under these conditions for acclimation prior to
measuring NA. NA was expressed as the rate of H2 evolution based on the plant nodule
fresh mass: μmol H2 h-1
plant-1
. We considered that H2 evolution is closely equivalent to
the rate of N2 reduction to ammonia, assuming that one mole of N2 is reduced for each
mole of H2 formed.
One week prior to harvest at the flowering stage, leaf gas exchange was evaluated
on the central leaflet of the second fully expanded trifoliate leaf using an infrared gas
analyzer (Li-6400, Licor, Lincoln NE, USA), with a leaf chamber of 6 cm2. The short-
term response of leaf CO2 assimilation (An) to increasing air CO2 concentration was
examined. The response of An to the intercellular CO2 partial pressure (Ci) was
determined after leaf tissue was briefly acclimated under a photosynthetic photon flux
density (Q) of 2,000 μmol m−2
s−1
for 5 min. After temporal stability at an air
temperature of 27.4 ± 1.0°C, the air CO2 partial pressure (Ca) inside the leaf chamber
varied between 5 and 200 Pa in 12 steps: 38, 25, 12.5, 10, 7.5, 5, 4, 38, 60, 80, 130 and
200 Pa. During the measurements, the leaf temperature was 29.3 ± 1.3°C and the leaf-
to-air vapor pressure difference was 1.1 ± 0.4 kPa. Measurements were recorded when
the total coefficient of variation was lower than 5% and under temporal stability (on
average, 3 min after the beginning of each step). The following biochemical parameters
were calculated from the analytical solutions according to Sharkey et al. (2007):
maximum carboxylation rate of Rubisco (Vc,max), maximum electron transport rate
(Jmax), dark respiration (Rd), CO2 compensation point (Г) and CO2 assimilation rate (A)
at 380 µmol mol–1
CO2 (A380). The metabolic limitation (Lm) of photosynthesis of
mycorrhizal and non-AM plants was calculated as Lm=[(AUL−ALL)/AUL]*100 (Lawlor,
2002), where AUL is the leaf CO2 assimilation of –MA+P plants and ALL is the leaf CO2
assimilation of +AM or -AM plants, both under air CO2 saturation.
Statistical analysis
For growth, nutrient contents, gas exchange, and NA data, two-way ANOVA was
used, and the means were compared using Tukey’s test (P<0.05). Significant differences
in mycorrhizal colonization parameters among plants of different developmental stages
were analyzed using Student’s t-test (P<0.05). Data expressed as percentages and data

32
expressed as relative to counts were arcsine-square-root and log(x+1) transformed,
respectively, prior to statistical analyses.
33
Results
Plant growth, nodulation, and mycorrhizal colonization
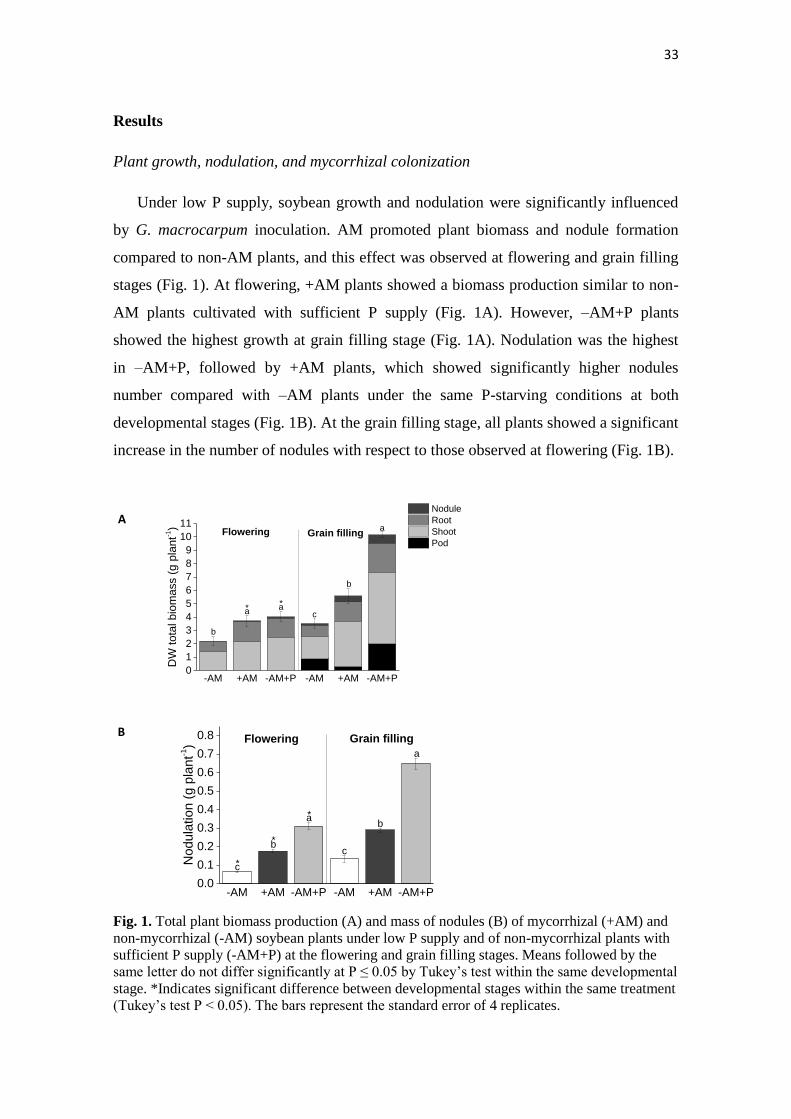
Under low P supply, soybean growth and nodulation were significantly influenced
by G. macrocarpum inoculation. AM promoted plant biomass and nodule formation
compared to non-AM plants, and this effect was observed at flowering and grain filling
stages (Fig. 1). At flowering, +AM plants showed a biomass production similar to non-
AM plants cultivated with sufficient P supply (Fig. 1A). However, –AM+P plants
showed the highest growth at grain filling stage (Fig. 1A). Nodulation was the highest
in –AM+P, followed by +AM plants, which showed significantly higher nodules
number compared with –AM plants under the same P-starving conditions at both
developmental stages (Fig. 1B). At the grain filling stage, all plants showed a significant
increase in the number of nodules with respect to those observed at flowering (Fig. 1B).
Fig. 1. Total plant biomass production (A) and mass of nodules (B) of mycorrhizal (+AM) and
non-mycorrhizal (-AM) soybean plants under low P supply and of non-mycorrhizal plants with
sufficient P supply (-AM+P) at the flowering and grain filling stages. Means followed by the
same letter do not differ significantly at P ≤ 0.05 by Tukey’s test within the same developmental
stage. *Indicates significant difference between developmental stages within the same treatment
(Tukey’s test P < 0.05). The bars represent the standard error of 4 replicates.
-AM +AM -AM+P -AM +AM -AM+P0
1
2
3
4
5
6
7
8
9
10
11
*
b
DW
tota
l bio
mass (
g p
lant-1
)
Nodule
Root
Shoot
Pod
Flowering Grain filling
aa
c
b
a
*
-AM +AM -AM+P -AM +AM -AM+P0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
*
*
Nodula
tion (
g p
lant-1
)
c
b
ab
c
a
Flowering Grain filling
*
A
B

34
Roots of inoculated soybean plants were highly colonized, with high abundance
of arbuscules, and no significant differences were observed between developmental
stages (Table 1). Roots of plants without AM fungus inoculation did not show
mycorrhizal structures.
Table 1. Mycorrhizal colonization intensity
(M%), frequency of mycorrhiza (F%) and
arbuscule abundance (A%) of soybean plants
inoculated with Glomus macrocarpum at the
flowering and grain filling stages. Different
letters indicate significant differences among
developmental stages using Student’s t-test
at P < 0.05.
Parameter Flowering Grain filling
M% 58.4 a 70.8 a
F% 94.2 a 95.8 a
A% 50.1 a 45.3 a
Nutrient concentration, N and P contents and nitrogenase activity
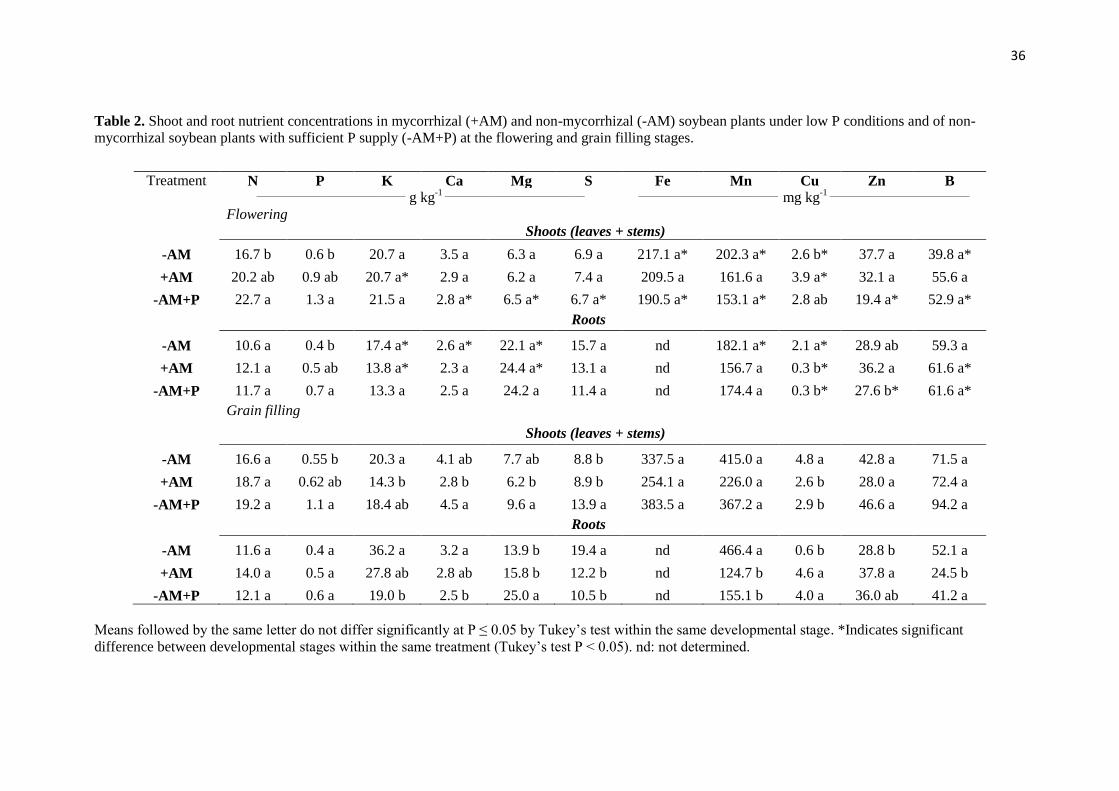
The nutrient concentrations in the shoots and roots differed depending on the AM
and P treatment and on the developmental stage of the plant (Table 2). At flowering, the
concentrations of K, Ca, Mg, S, Fe, Mn, Zn and B in the shoots were similar among
treatments (Table 2). However, the Cu concentration was significantly higher in the
+AM plants than in the –AM plants, and none of these plants showed differences from
the Cu concentration in –AM+P plants. Shoot N and P concentrations were higher in the
–AM+P plants than in the –AM plants under low P conditions; +AM plants showed
intermediate concentrations of these elements that did not significantly differ from the
plants with the non-AM treatments, regardless of P supply (Table 2). In the roots, the
concentrations of N, K, Ca, Mg, S, Mn, and B were similar between treatments.
However, root P concentration was higher in –AM+P plants and lower in –AM plants;
root P concentration in +AM plants showed non-significant differences compared to
other treatments. The Cu concentration in roots was significantly lower in +AM and –
AM+ P than in –AM plants under low Pi conditions. AM plants showed the highest Zn
concentrations in the roots (Table 2).
At the grain filling stage, the concentrations of N, Fe, Mn, Zn and B in the shoots
were similar among treatments (Table 2). However, shoot Ca and Mg concentrations
were lower in +AM than in –AM plants. Under P-deficient conditions, shoot K and Cu
concentrations were higher in –AM than in +AM plants. At this developmental stage,
the shoot S concentration was lower in plants under low P conditions, irrespective of

35
mycorrhizal status. Root N and P concentrations were similar among treatments, and the
root concentrations of K and Ca were significantly higher in -AM plants than in those
from –AM+P treatment. –AM+P plants showed higher root Mg concentrations than the
roots of plants under low P availability, whereas S and Mn concentrations were higher
in roots of –AM. Cu and Zn concentrations were significantly higher in the roots of
+AM and –AM+P plants than in -AM plants, and the root B concentration was lower in
+AM than in –AM plants.
The concentration of some nutrients significantly changed between the flowering
and grain filling stages. At grain filling, -AM plants showed increases in shoot Fe, Mn,
Cu and B concentrations and in root K, Ca and Mn concentrations and decreases in the
root concentrations of Mg and Cu compared to the flowering period. From flowering to
grain filling, the K and Cu concentrations in the shoot diminished and increased in the
roots, while the root concentrations of Mg and B decreased. At the grain filling stage, –
AM+P plants showed an increase in shoot Ca, Mg, S, Fe, Mn, Zn and B concentrations
with respect to flowering (Table 2).
AM symbiosis increased plant N content in relation to non-AM under low P
conditions in both stages of development (Fig. 2A). N contents in +AM plants were
similar to those in non-AM plants grown with sufficient P supply at flowering. The P
content of AM plants at flowering was not significantly different from the other two
treatments and was lower than that in plants with sufficient P supply (Fig. 2B). At
flowering, NUE was higher in –AM plants than in –AM+P or +AM plants (Fig. 3). The
PUE values in +AM plants were similar to those in –AM plants, which showed higher
PUE than –AM+P plants (Fig. 3B). At grain filling, there were no significant
differences in either PUE or NUE among treatments (Fig. 3). Under low P conditions,
NA was higher in +AM than in –AM plants, and the highest NA was observed in –
AM+P plants (Fig. 4).
36
Table 2. Shoot and root nutrient concentrations in mycorrhizal (+AM) and non-mycorrhizal (-AM) soybean plants under low P conditions and of non-
mycorrhizal soybean plants with sufficient P supply (-AM+P) at the flowering and grain filling stages.
Treatment N P K Ca Mg S Fe Mn Cu Zn B
________________________________
g kg-1 ______________________________
______________________________
mg kg-1 ______________________________
Flowering
Shoots (leaves + stems)
-AM 16.7 b 0.6 b 20.7 a 3.5 a 6.3 a 6.9 a 217.1 a* 202.3 a* 2.6 b* 37.7 a 39.8 a*
+AM 20.2 ab 0.9 ab 20.7 a* 2.9 a 6.2 a 7.4 a 209.5 a 161.6 a 3.9 a* 32.1 a 55.6 a
-AM+P 22.7 a 1.3 a 21.5 a 2.8 a* 6.5 a* 6.7 a* 190.5 a* 153.1 a* 2.8 ab 19.4 a* 52.9 a*
Roots
-AM 10.6 a 0.4 b 17.4 a* 2.6 a* 22.1 a* 15.7 a nd 182.1 a* 2.1 a* 28.9 ab 59.3 a
+AM 12.1 a 0.5 ab 13.8 a* 2.3 a 24.4 a* 13.1 a nd 156.7 a 0.3 b* 36.2 a 61.6 a*
-AM+P 11.7 a 0.7 a 13.3 a 2.5 a 24.2 a 11.4 a nd 174.4 a 0.3 b* 27.6 b* 61.6 a*
Grain filling
Shoots (leaves + stems)
-AM 16.6 a 0.55 b 20.3 a 4.1 ab 7.7 ab 8.8 b 337.5 a 415.0 a 4.8 a 42.8 a 71.5 a
+AM 18.7 a 0.62 ab 14.3 b 2.8 b 6.2 b 8.9 b 254.1 a 226.0 a 2.6 b 28.0 a 72.4 a
-AM+P 19.2 a 1.1 a 18.4 ab 4.5 a 9.6 a 13.9 a 383.5 a 367.2 a 2.9 b 46.6 a 94.2 a
Roots
-AM 11.6 a 0.4 a 36.2 a 3.2 a 13.9 b 19.4 a nd 466.4 a 0.6 b 28.8 b 52.1 a
+AM 14.0 a 0.5 a 27.8 ab 2.8 ab 15.8 b 12.2 b nd 124.7 b 4.6 a 37.8 a 24.5 b
-AM+P 12.1 a 0.6 a 19.0 b 2.5 b 25.0 a 10.5 b nd 155.1 b 4.0 a 36.0 ab 41.2 a
Means followed by the same letter do not differ significantly at P ≤ 0.05 by Tukey’s test within the same developmental stage. *Indicates significant
difference between developmental stages within the same treatment (Tukey’s test P < 0.05). nd: not determined.
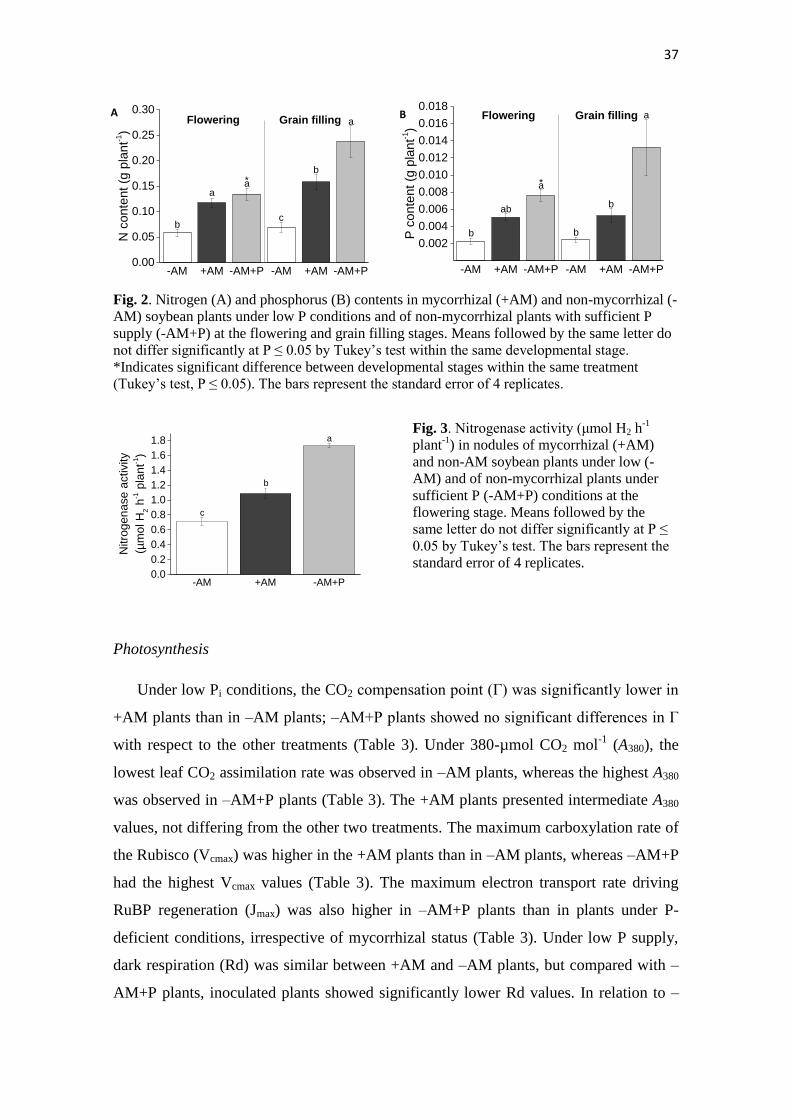
37
Fig. 2. Nitrogen (A) and phosphorus (B) contents in mycorrhizal (+AM) and non-mycorrhizal (-
AM) soybean plants under low P conditions and of non-mycorrhizal plants with sufficient P
supply (-AM+P) at the flowering and grain filling stages. Means followed by the same letter do
not differ significantly at P ≤ 0.05 by Tukey’s test within the same developmental stage.
*Indicates significant difference between developmental stages within the same treatment
(Tukey’s test, P ≤ 0.05). The bars represent the standard error of 4 replicates.
Fig. 3. Nitrogenase activity (μmol H2 h-1
plant-1
) in nodules of mycorrhizal (+AM)
and non-AM soybean plants under low (-
AM) and of non-mycorrhizal plants under
sufficient P (-AM+P) conditions at the
flowering stage. Means followed by the
same letter do not differ significantly at P ≤
0.05 by Tukey’s test. The bars represent the
standard error of 4 replicates.
Photosynthesis
Under low Pi conditions, the CO2 compensation point (Г) was significantly lower in
+AM plants than in –AM plants; –AM+P plants showed no significant differences in Г
with respect to the other treatments (Table 3). Under 380-µmol CO2 mol-1
(A380), the
lowest leaf CO2 assimilation rate was observed in –AM plants, whereas the highest A380
was observed in –AM+P plants (Table 3). The +AM plants presented intermediate A380
values, not differing from the other two treatments. The maximum carboxylation rate of
the Rubisco (Vcmax) was higher in the +AM plants than in –AM plants, whereas –AM+P
had the highest Vcmax values (Table 3). The maximum electron transport rate driving
RuBP regeneration (Jmax) was also higher in –AM+P plants than in plants under P-
deficient conditions, irrespective of mycorrhizal status (Table 3). Under low P supply,
dark respiration (Rd) was similar between +AM and –AM plants, but compared with –
AM+P plants, inoculated plants showed significantly lower Rd values. In relation to –
-AM +AM -AM+P -AM +AM -AM+P0.00
0.05
0.10
0.15
0.20
0.25
0.30N
conte
nt (g
pla
nt-1
)Flowering Grain filling
b
aa
c
b
a
*
-AM +AM -AM+P -AM +AM -AM+P
0.002
0.004
0.006
0.008
0.010
0.012
0.014
0.016
0.018
b
P c
onte
nt (g
pla
nt-1
)
Flowering Grain filling
b
ab
a
b
a
*
-AM +AM -AM+P0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
Nitro
genase a
ctivity
(µm
ol H
2 h
-1 p
lant-1
)
c
b
a
A B
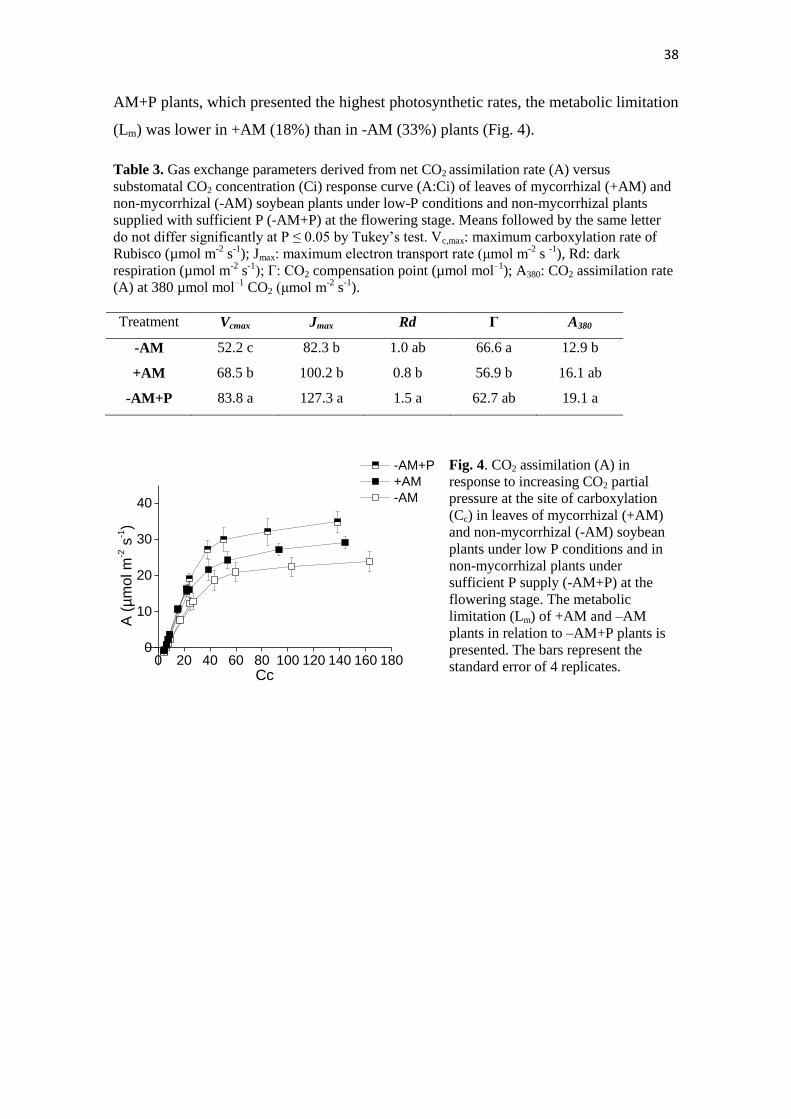
38
AM+P plants, which presented the highest photosynthetic rates, the metabolic limitation
(Lm) was lower in +AM (18%) than in -AM (33%) plants (Fig. 4).
Table 3. Gas exchange parameters derived from net CO2 assimilation rate (A) versus
substomatal CO2 concentration (Ci) response curve (A:Ci) of leaves of mycorrhizal (+AM) and
non-mycorrhizal (-AM) soybean plants under low-P conditions and non-mycorrhizal plants
supplied with sufficient P (-AM+P) at the flowering stage. Means followed by the same letter
do not differ significantly at P ≤ 0.05 by Tukey’s test. Vc,max: maximum carboxylation rate of
Rubisco (µmol m-2
s-1
); Jmax: maximum electron transport rate (μmol m-2
s -1
), Rd: dark
respiration (µmol m-2
s-1
); Г: CO2 compensation point (µmol mol–1
); A380: CO2 assimilation rate
(A) at 380 µmol mol–1
CO2 (μmol m-2
s-1
).
Treatment Vcmax Jmax Rd Г A380
-AM 52.2 c 82.3 b 1.0 ab 66.6 a 12.9 b
+AM 68.5 b 100.2 b 0.8 b 56.9 b 16.1 ab
-AM+P 83.8 a 127.3 a 1.5 a 62.7 ab 19.1 a
Fig. 4. CO2 assimilation (A) in
response to increasing CO2 partial
pressure at the site of carboxylation
(Cc) in leaves of mycorrhizal (+AM)
and non-mycorrhizal (-AM) soybean
plants under low P conditions and in
non-mycorrhizal plants under
sufficient P supply (-AM+P) at the
flowering stage. The metabolic
limitation (Lm) of +AM and –AM
plants in relation to –AM+P plants is
presented. The bars represent the
standard error of 4 replicates.
0 20 40 60 80 100 120 140 160 1800
10
20
30
40
A (
µm
ol m
-2 s
-1)
Cc
-AM+P
+AM
-AM

39
Discussion
Nitrogen and phosphorus are nutrients that may often limit plant growth and
performance under agricultural and natural systems. Currently, there is a growing
awareness of the importance of plant-microbe interactions in plant performance,
particularly under stress challenges and in view of the necessity to reduce nutrient
inputs and minimize the environmental impacts of mineral fertilizers (Jez et al., 2016).
From this perspective, the aim of the present study was to evaluate the effects of P
starvation on AM symbiosis and determine how this stressful condition influences N2-
fixing capacities, plant growth, nutrient uptake and photosynthetic performance of
soybean plants.
The results clearly showed that the soybean association with the AM fungus G.
macrocarpum improved plant performance under P scarcity compared with non-AM
plants under the same conditions. AM positively influenced growth and plant
nodulation at both the flowering and grain filling stages (Fig. 1). Nonetheless,
mycorrhizal symbiosis could not completely compensate for soybean full P demand
when compared with plants grown with sufficient Pi; as AM soybean plants showed
lower biomass production and nodulation (Fig. 1), particularly at plant maturity when
seed formation is a strong nutrient sink (Bender et al., 2015). In fact, the maximum rate
of nutrient uptake in legume crops such as soybean is expected at flowering, whereas
seeds become the main nutrient sink at the early reproductive stage, when nutrient
accumulation in vegetative tissues is reduced (Bender et al., 2015).
Depending on the AM fungus-rhizobium species combination and the
environmental conditions, AM symbioses may promote SNF and plant growth by
increasing Pi uptake, as demonstrated in alfalfa, lentil, Amorpha canescens and lima
bean (Azcón et al., 1991; Ballhorn et al., 2016; Larimer et al., 2014; Xavier & Germida,
2002). Nonetheless, plants may experience growth reduction, reflecting the high
demand for photoassimilates from both symbionts under imbalanced symbiotic reward
host (Koide & Elliott, 1989). In addition, stress conditions, such as those resulting from
P limitation, can neutralize the beneficial effect of the AM symbiosis or even eliminate
this relationship and cause negative effects on the host plant (Ballhorn et al., 2016; Pang
& Paul, 1980). This fact may in part explain some results in which the increased benefit
of mycorrhiza might have been limited by the severe Pi scarcity imposed to plants

40
during the experiment. AM plants under low P did not perform as well as plants grown
with sufficient P (Fig. 1 and 2), suggesting that AM association had and extra-cost
limiting biomass production or that under the P concentration used in this experiment
the acquisition of P via AM symbiosis did not fully satisfy plant demand. The high
values of AM root colonization indicated a high fungal biomass and carbon flux toward
the fungus (Table 1). In addition, soybean plants grown under sufficient P supply more
than doubled their biomass production and nodules number between flowering and
grain filling stages, whereas the same did not occur in mycorrhizal plants under Pi
scarcity. Accordingly, some studies have addressed reduced plant benefits of dual root
symbioses, AM and rhizobia, under limiting resources, such as light and Pi availability
(Ballhorn et al., 2016; Ronsheim, 2012).
In addition to the high colonization and efficient association of the AM fungus G.
macrocarpum, the strain of B. elkanii used in the present study also efficiently
nodulated the soybean cultivar Foscarin IAC-31 (Table 1, Fig. 1); nodule formation and
growth were maintained until the maturation stage with active N2 fixation, once the
nutrient solution had no mineral N source and plant increased the number and mass of
nodules and N contents until the grain filling stage (Fig. 1 and 2). Under P scarcity, AM
plants exhibited significantly higher biomass production and greater nodulation than
non-AM plants, likely reflecting the stimulation of photosynthesis (Table 3), higher
SNF and increased nitrogenase activity (Fig. 1 and 3), which contributed to greater P
uptake and accumulation (Fig. 2 and Table 2). Increased nodulation by the same AM
fungus G. macrocarpum in soybean was already reported (Andrade et al., 2004). Thus,
AM soybean plants formed more and bigger nodules than non-AM plants under P
scarcity, evidencing the positive effect of AM symbiosis on SNF, which resulted in
higher N contents in AM plants (Fig. 2). Conversely, the formation of fewer and smaller
nodules in non-AM plants under P scarcity may reflect lower SNF efficiency, in which
the smaller nodules with higher oxygen diffusion through a greater specific area may
have impaired N2-fixation capacities (Esfahani et al., 2016), as suggested by the low
nitrogenase activity of non-AM and +AM plants under low P conditions (Fig. 3). Higher
SNF efficiency has been associated with the rate of P allocation to the nodules and
higher nitrogenase activities, as reported here in soybeans grown with sufficient P
supply (Fig. 1 and 3). Soybeans can be more efficient in P partitioning than other

41
legumes, such as Vigna radiata, indicating a good level of adaptability to low P
conditions (Sulieman & Tran, 2015).
Once inorganic P is taken from the soil, either through a direct plant pathway or the
mycorrhizal pathway, this nutrient is transported to sink organs, where the metabolic
demand may be high. SNF is one of these P-demanding processes, constituting a strong
sink not only for Pi but also for carbon in the nodules (Kaschuk et al., 2009). During P
scarcity, nodule functioning can be impaired due to the negative effects of Pi-deficiency
on photosynthesis, which ultimately limits carbohydrate loading into the phloem and
import into the nodules for metabolism (Sulieman & Tran, 2015), and reflects the
reduced P itself for N2 fixation reactions. In the present study, the P and N
concentrations in AM plants did not significantly differ from non-AM plants regardless
of the P supply, showing in general intermediate P and N concentrations in both shoots
and roots at both plant stages (Table 2). In terms of content per plant, AM plants
showed 50% higher shoot P and N contents than non-AM plants under the same low P
conditions and at both developmental stages, indicating more effective P uptake and
SNF under AM symbiosis. This finding is supported by the high nitrogenase activity of
the nodules of AM plants, which are entirely responsible for the plant N contents.
Nodular tissue is a strong sink for P, even in P-starved legumes (Sa & Israel, 1991;
Sulieman et al., 2013), and some authors have estimated that up to 20% of the plant P
can be allocated to the nodules, with the nodular tissue becoming increasing P sinks
under P deficiency (Sulieman & Tran, 2015). The AM association with nodulated
legumes can enhance Pi uptake and transfer via root vascular tissues (Tajini et al.,
2009), although some P can also be directly absorbed independently through the
epidermis of the nodules (Sulieman & Tran, 2015). Although the root P concentrations
between non-AM and AM plants were not very different, the P concentration in the
nodules was not determined here, and this topic should be addressed in future studies to
explain nutrient partitioning in legume plants.
AM symbiosis may have contributed to the metal demand of nodules, as for
example, the Zn concentration was higher in mycorrhizal than in non-AM roots at both
developmental stages under low P supply (Table 2). This finding may reflect Zn
allocation to nodular tissues, where this nutrient acts as a cofactor for several enzymes
and as a regulator of plant gene expression (Moreau et al., 2002). Zn and P homeostasis
in plants is tightly and complexly linked (Briat et al., 2015; Saenchai et al., 2016), and

42
Zn may also control P status in nodules, playing a critical role in nodulation (Moreau et
al., 2002). Some high-affinity phosphate transporters are regulated by Zn (Huang et al.,
2000), and the mycorrhizal pathway may contribute to plant micronutrient acquisition.
Some estimations have established that between 20 to 50% of the absorbed metal
nutrients Cu, Zn, or Fe, for example (Lehmann et al., 2014), can be delivered through
the symbiotic pathway, with contributions depending on host and fungal species and on
the soil conditions (González-Guerrero et al., 2016). Other studies have shown that the
Zn contents significantly increased in mycorrhizal tomato plants and fruits under
sufficient Zn concentrations (Cavagnaro et al., 2006), whereas under excessive Zn
levels, AM symbiosis may either increase or reduce Zn entrance into the plant (Andrade
et al., 2010; Wong et al., 2007).
SNF in the soybean-rhizobia association contributes to N nutrition, and AM fungi
association can contribute to the uptake of several nutrients, mainly P, often resulting in
positive effects and increased photosynthetic capacity (Kaschuk et al., 2009). However,
dual symbiosis may outbalance carbon costs and constrain mutualistic benefits under
limiting conditions, such as those resulting from severe nutrient scarcity or low light
conditions (Harris et al., 1985; Koide & Elliott, 1989). In the present study, AM
symbiosis clearly alleviated the effects of P deficiency, stimulating N2-fixation activity
and enhancing plant N status, as the biochemical reactions of N2-fixation demand large
amounts of photoassimilates, and P, that are used during nodules metabolism (Xavier &
Germida, 2002). Similarly, AM fungi use plant photoassimilates as the sole source of
fungal carbon, and if the energy invested in the symbioses is not sufficiently
compensated by enhanced photosynthetic rates and nutrient acquisition, then the host
plant may show lower growth than non-symbiotic plants (Kaschuk et al., 2009). When
comparing the photosynthetic performance of soybean plants grown with sufficient P
with plants under P-starvation, a negative effect of Pi starvation on the biochemical
reactions of photosynthesis related to Rubisco activity and RuBP regeneration was
observed (Table 3), as maximum rates of carboxylation (Vcmax) and CO2 assimilation
rates (A380) decreased. These findings may reflect the central role of P in energy transfer
as ATP and NADPH in the regeneration of photosynthetic intermediaries in the
utilization of photoassimilates and in CO2 diffusion within the leaves (Singh & Reddy,
2016). In addition, increases in CO2 compensation point (Г) have been related to the
response to different stress (Lauer et al., 1989; Zlatev sand Yordanov, 2004). The higher

43
Г of soybean leaves under low Pi conditions were related to high respiration rates in
soybean under low Pi conditions (Lauer et al., 1989), particularly to increases in the
alternative oxidase pathway in relation to the cytochrome oxidase pathway of
mitochondrial respiration, as the former bypass adenylates and Pi control (Vanlerberghe,
2013). In this study, and under Pi starving conditions, non-AM plants showed
significantly higher Г than AM plants (Table 3), suggesting that low Pi stress may has
led to higher respiration rates probably through the involvement of alternative oxidase
pathway, related to lower ATP yields, which may negatively impact growth. AM plants
showed better Pi-status and were able to maintain lower Г under Pi deficiency (Table 3)
and higher maximum rates of carboxylation (Vcmax) than that in non-AM associated
plants, probably contributing to the observed growth and SNF stimulation.
The results of the present study showed that relative to plants with sufficient P
supply, there was a lower metabolic limitation of photosynthesis in +AM plants than in
-AM plants, and photosynthesis was less sensitive to P starvation in +AM plants (Fig.
4). This effect is a likely consequence of increased P uptake and improved N status
through enhanced SNF (Fig. 2). These results are consistent with a previous study in
which a lower metabolic limitation of photosynthesis was observed in watermelon
plants associated with AM and grown under drought (Mo et al., 2016).

44
Conclusions
In view of the perspective for the necessity of more efficient use of scarce P
resources by legumes, this study showed that AM association can be considered an
important plant strategy for maintaining SNF functioning and improving plant growth,
even when P availability is low. Under Pi starvation, AM symbiosis increased plant
biomass, nodulation, and nitrogenase activity in relation to non-AM plants at both
flowering and grain filling stages, despite the costs to maintain the dual symbiosis.
Although, mycorrhizal symbiosis could not completely compensate for soybean P
demand when compared with well-nourished plants under Pi starvation, AM
significantly attenuated the negative effects of low Pi in nodulated-soybean plants.
Acknowledgements
This work was financially supported by a grant from the São Paulo Research
Foundation FAPESP (Grant 2016/06396-1). The authors would like to thank the
National Council for Scientific and Technological Development (CNPq, Brazil) for
studentship (RGB) and fellowship (RVR) grants.

45
References
Adesemoye AO, Kloepper JW (2009) Plant-microbes interactions in enhanced fertilizer-
use efficiency. Appl Microbiol Biotechnol 85,1-12.
Andrade SA, Silveira AP, Mazzafera P (2010) Arbuscular mycorrhiza alters metal
uptake and the physiological response of Coffea arabica seedlings to increasing
Zn and Cu concentrations in soil. Sci Total Environ 408,5381-5391.
Andrade SAL, Abreu CA, de Abreu MF, Silveira APD (2004) Influence of lead
additions on arbuscular mycorrhiza and Rhizobium symbioses under soybean
plants. Appl Soil Ecol 26,123-131.
Azcón R, Gomez M, Tobar R (1992) Effects of nitrogen source on growth, nutrition,
photosynthetic rate and nitrogen metabolism of mycorrhizal and phosphorus-
fertilized plants of Lactuca sativa L. . New Phytol 121,227–234.
Azcón R, Rubio R, Barea J (1991) Selective interactions between different species of
mycorrhizal fungi and Rhizobium meliloti strains, and their effects on growth,
N2‐fixation (15N) and nutrition of Medicago sativa L. New Phytol 117,399-404.
Ballhorn DJ, Schadler M, Elias JD, Millar JA, Kautz S (2016) Friend or Foe-Light
Availability Determines the Relationship between Mycorrhizal Fungi, Rhizobia
and Lima Bean (Phaseolus lunatus L.). PLoS One 11,e0154116.
Barea J, Azcon-Aguilar C (1983) Mycorrhizas and their significance in nodulating
nitrogen-fixing plants vol 36. Advances in agronomy, vol 4.
Bender RR, Haegele JW, Below FE (2015) Nutrient uptake, partitioning, and
remobilization in modern soybean varieties. Agron J 107,563-573.
Bremner J (1965) Inorganic forms of nitrogen. Methods of soil analysis Part 2
Chemical and microbiological properties,1179-1237.
Briat JF, Rouached H, Tissot N, Gaymard F, Dubos C (2015) Integration of P, S, Fe,
and Zn nutrition signals in Arabidopsis thaliana: potential involvement of
PHOSPHATE STARVATION RESPONSE 1 (PHR1). Front Plant Sci 6,290.
Brooks A, Woo KC, Wong SC (1988) Effects of phosphorus nutrition on the response
of photosynthesis to CO2 and O2, activation of ribulose bisphosphate
carboxylase and amounts of ribulose bisphosphate and 3-phosphoglycerate in
spinach leaves. Photosynth Res 15,133-141.
Cavagnaro TR, Jackson LE, Six J, Ferris H, Goyal S, Asami D, Scow KM (2006)
Arbuscular mycorrhizas, microbial communities, nutrient availability, and soil
aggregates in organic tomato production. Plant Soil 282,209-225.
Esfahani MN et al. (2016) Adaptation of the symbiotic Mesorhizobium–chickpea
relationship to phosphate deficiency relies on reprogramming of whole-plant
metabolism. Proceedings of the National Academy of Sciences 113,E4610-
E4619.

46
Fehr W, Caviness C, Burmood D, Pennington J (1971) Stage of development
descriptions for soybeans, Glycine max (L.) Merrill. Crop science 11,929-931.
Fredeen AL, Raab TK, Rao IM, Terry N (1990) Effects of phosphorus nutrition on
photosynthesis in Glycine max (L.) Merr. Planta 181,399-405.
Giovannetti M, Mosse B (1980) An evaluation of techniques for measuring vesicular
arbuscular mycorrhizal infection in roots. New Phytol 84,489-500.
González-Guerrero M, Escudero V, Saéz Á, Tejada-Jiménez M (2016) Transition Metal
Transport in Plants and Associated Endosymbionts: Arbuscular Mycorrhizal
Fungi and Rhizobia. Front Plant Sci 7.
Harris D, Pacovsky R, Paul E (1985) Carbon economy of soybean–Rhizobium–Glomus
associations. New Phytol 101,427-440.
Hernández I, Munné-Bosch S (2015) Linking phosphorus availability with photo-
oxidative stress in plants. J Exp Bot 66,2889-2900.
Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without
soil. California Agricultural Experiment Station Circular 347.
Huang C, Barker SJ, Langridge P, Smith FW, Graham RD (2000) Zinc deficiency up-
regulates expression of high-affinity phosphate transporter genes in both
phosphate-sufficient and-deficient barley roots. Plant Physiol 124,415-422.
Hungria M, Campo RJ, MENDES IdC Fixação biológica do nitrogênio na cultura da
soja. In, 2001. Embrapa Soja; Brasília, DF: Embrapa Cerrados,
Ibrahim KK, Arunachalam V, Rao PSK, Tilak K (1995) Seasonal response of groundnut
genotypes to arbuscular mycorrhiza—Bradyrhizobium inoculation. Microbiol
Res 150,218-224.
Jez JM, Lee SG, Sherp AM (2016) The next green movement: Plant biology for the
environment and sustainability. Science 353,1241-1244.
Justino GC, Sodek L (2013) Recovery of nitrogen fixation after short-term flooding of
the nodulated root system of soybean. J Plant Physiol 170,235-241.
Kaschuk G, Kuyper TW, Leffelaar PA, Hungria M, Giller KE (2009) Are the rates of
photosynthesis stimulated by the carbon sink strength of rhizobial and
arbuscular mycorrhizal symbioses? Soil Biol Biochem 41,1233-1244.
Koide R, Elliott G (1989) Cost, benefit and efficiency of the vesicular-arbuscular
mycorrhizal symbiosis vol 3. Functional Ecology. British Ecological Society.
Kucey R, Paul E (1982) Carbon flow, photosynthesis, and N2 fixation in mycorrhizal
and nodulated faba beans (Vicia faba L.). Soil Biol Biochem 14,407-412.
Lam H-M, Coschigano K, Oliveira I, Melo-Oliveira R, Coruzzi G (1996) The
molecular-genetics of nitrogen assimilation into amino acids in higher plants.
Annu Rev Plant Physiol Plant Mol Biol 47,569-593.

47
Larimer AL, Clay K, Bever JD (2014) Synergism and context dependency of
interactions between arbuscular mycorrhizal fungi and rhizobia with a prairie
legume. Ecology 95,1045-1054.
Lawlor DW (2002) Limitation to Photosynthesis in Water‐stressed Leaves: Stomata vs.
Metabolism and the Role of ATP. Ann Bot 89,871-885.
Lehmann A, Veresoglou SD, Leifheit EF, Rillig MC (2014) Arbuscular mycorrhizal
influence on zinc nutrition in crop plants–A meta-analysis. Soil Biol Biochem
69,123-131.
Li C, Gui S, Yang T, Walk T, Wang X, Liao H (2012) Identification of soybean purple
acid phosphatase genes and their expression responses to phosphorus availability
and symbiosis. Ann Bot 109,275-285.
Lynch JP (2011) Root phenes for enhanced soil exploration and phosphorus acquisition:
tools for future crops. Plant Physiol 156,1041-1049.
Mo Y et al. (2016) Regulation of Plant Growth, Photosynthesis, Antioxidation and
Osmosis by an Arbuscular Mycorrhizal Fungus in Watermelon Seedlings under
Well-Watered and Drought Conditions. Front Plant Sci 7.
Moreau RA, Whitaker BD, Hicks KB (2002) Phytosterols, phytostanols, and their
conjugates in foods: structural diversity, quantitative analysis, and health-
promoting uses. Prog Lipid Res 41,457-500.
Nussaume L et al. (2011) Phosphate import in plants: Focus on the PHT1 transporters.
Front Plant Sci 2,83.
Pang P, Paul E (1980) Effects of vesicular-arbuscular mycorrhiza on 14C and 15N
distribution in nodulated fababeans. Can J Soil Sci 60,241-250.
Ronsheim ML (2012) The effect of mycorrhizae on plant growth and reproduction
varies with soil phosphorus and developmental stage. Am Midl Nat 167,28-39.
Sa T-M, Israel DW (1991) Energy status and functioning of phosphorus-deficient
soybean nodules. Plant Physiol 97,928-935.
Saenchai C, Bouain N, Kisko M (2016) The Involvement of OsPHO1; 1 in the
Regulation of Iron Transport Through Integration of Phosphate and Zinc
Deficiency Signaling. Front Plant Sci 7.
Schachtman DP, Reid RJ, Ayling SM (1998) Phosphorus Uptake by Plants: From Soil
to Cell. Plant Physiol 116,447–453.
Schubert S, Serraj R, Plies-Balzer E, Mengel K (1995) Effect of drought stress on
growth, sugar concentrations and amino acid accumulation in N2-fixing Alfalfa
(Medicago sativa). J Plant Physiol 146,541-546.
Schüßler A, Schwarzott D, Walker C (2002) A new fungal phylum, the
Glomeromycota: phylogeny and evolution. Mycological Research 105,1413-
1421.

48
Sharkey TD, Bernacchi CJ, Farquhar GD, Singsaas EL (2007) Fitting photosynthetic
carbon dioxide response curves for C(3) leaves. Plant Cell Environ 30,1035-
1040.
Singh SK, Reddy VR (2016) Methods of mesophyll conductance estimation: its impact
on key biochemical parameters and photosynthetic limitations in phosphorus-
stressed soybean across CO2. Physiol Plant 157,234-254.
Spatafora JW et al. (2016) A phylum-level phylogenetic classification of zygomycete
fungi based on genome-scale data. Mycologia 108,1028-1046.
Sulieman S, Tran L-SP (2015) Phosphorus homeostasis in legume nodules as an
adaptive strategy to phosphorus deficiency. Plant Science 239,36-43.
Sulieman S, Van Ha C, Schulze J, Tran L-SP (2013) Growth and nodulation of
symbiotic Medicago truncatula at different levels of phosphorus availability. J
Exp Bot,ert122.
Tajini F, Suriyakup P, Vailhe H, Jansa J, Drevon J-J (2009) Assess suitability of
hydroaeroponic culture to establish tripartite symbiosis between different AMF
species, beans, and rhizobia. BMC Plant Biology 9,1.
Tesfaye M, Liu J, Allan DL, Vance CP (2007) Genomic and genetic control of
phosphate stress in legumes. Plant Physiol 144,594-603.
Trouvelot A (1986) Mesure du taux de mycorhization VA d'un systeme radiculaire.
Recherche de methodes d'estimation ayant une significantion fonctionnelle.
Mycorrhizae: physiology and genetics,217-221.
Udvardi MK, Price GD, Gresshoff PM, Day DA (1988) A dicarboxylate transporter on
the peribacteroid membrane of soybean nodules. FEBS letters 231,36-40.
van der Heijden MG, Bardgett RD, van Straalen NM (2008) The unseen majority: soil
microbes as drivers of plant diversity and productivity in terrestrial ecosystems.
Ecol Lett 11,296-310.
Van Kauwenbergh SJ (2010) World phosphate rock reserves and resources.
Whitehead LF, Day DA (1997) The peribacteroid membrane. Physiol Plantarum
100,30-44.
Wong CC, Wu SC, Kuek C, Khan AG, Wong MH (2007) The Role of Mycorrhizae
Associated with Vetiver Grown in Pb‐/Zn‐Contaminated Soils: Greenhouse
Study. Restoe Ecol 15,60-67.
Xavier L, Germida J (2002) Response of lentil under controlled conditions to co-
inoculation with arbuscular mycorrhizal fungi and rhizobia varying in efficacy.
Soil Biol and Biochem 34,181-188.
Zhang Z, Liao H, Lucas WJ (2014) Molecular mechanisms underlying phosphate
sensing, signaling, and adaptation in plants. J Integr Plant Biol 56,192-220.

49
CAPÍTULO 2
A MICORRIZA ARBUSCULAR ALTERA EXPRESSÃO DE
TRANSPORTADORES DE FOSFATO EM RAÍZES E NÓDULOS DE SOJA
SEM ALTERAR O CONTÉUDO DE P DA PLANTA

50
A micorriza arbuscular altera expressão de transportadores de fosfato em raízes e
nódulos de soja sem alterar o conteúdo de P da planta
Introdução
O nitrogênio (N) e o fósforo (P) são os nutrientes que mais limitam o
crescimento de plantas e a produção das culturas. As leguminosas são capazes de se
associar a bactérias fixadoras de dinitrogênio (N2) e, através da fixação simbiótica de N2
(FSN) suprir suas necessidades mesmo em solos deficientes neste nutriente (Vance et
al., 2003). O P, no entanto, é absorvido pelas plantas na forma de ortofosfato (H2PO4-),
forma de P inorgânico (Pi) predominante em solos com pH entre 5 e 6 (Schachtman et
al., 1998). Porém, o P é considerado um elemento de baixa mobilidade no solo, devido
a sua baixa mobilidade no solo e à sua alta capacidade de adsorção a partículas do solo
(Schachtman et al., 1998). A absorção de Pi pelas plantas cria uma área de depleção ao
redor da superfície da raiz, motivo que limita ainda mais taxa de difusão e torna o P um
dos nutrientes menos disponíveis. Na solução do solo o Pi encontra-se em concentrações
de 0,5 a 10 μM, e assim, a aquisição e utilização de P pelas plantas são fatores
determinantes para o desenvolvimento (Zhang et al., 2014).
Em situações de escassez de P as plantas podem apresentar respostas específicas
para aumentar tanto a capacidade de absorção quanto a eficiência do uso interno de P,
denominadas em conjunto como Phosphorus Starving Response (PSR) (Zhang et al.,
2014). Algumas características relacionadas à PSR são aumento na relação raiz:parte
aérea e mudanças na arquitetura do sistema radicular, com aumento na produção de
raízes laterais, aumento no comprimento e número de pelos ou produção de raízes nas
camadas mais superficiais do solo, por exemplo (van de Wiel et al., 2016), ou o
estabelecimento de simbioses micorrízicas e exsudação radicular as quais contribuem
para a maior absorção e/ou mobilização de P no solo (Li et al., 2012). Para a eficiente
absorção do Pi do solo, a expressão de transportadores de fosfato de alta afinidade deve
ser induzida em baixas concentrações de P, aumentando assim a eficiência de absorção
e uso de P (Plaxton & Tran, 2011).
Como uma das estratégias para superar a limitação na disponibilidade de P no
ambiente as plantas podem se associar simbioticamente com fungos micorrízicos
arbusculares (FMA), pertencentes ao sub-phylum Glomeromycotina (Spatafora et al.,
2016). O desenvolvimento da micorriza arbuscular (MA) resulta na colonização do

51
córtex da raiz pelo FMA e na formação de arbúsculos, onde acontece a troca de
nutrientes entre ambos os simbiontes. A rede de hifas extrarradiculares se estende no
solo desde a raiz e conecta a planta com regiões que podem ir além da área de depleção,
contribuindo para a absorção do nutriente em solos com baixa disponibilidade de P
(Oldroyd, 2013). Esta maior absorção de P faz com que plantas com MA apresentem
frequentemente maior crescimento. Em troca, o FMA associado utiliza quantidades
significativas (até 20%) de carboidratos oriundos da atividade fotossintética do
hospedeiro (Adesemoye & Kloepper, 2009).
Leguminosas como a soja são capazes de estabelecer a dupla simbiose com
FMAs e rizóbios e se beneficiar da FSN e do aumento na absorção de P pela MA
(Azcón et al., 1991; Ballhorn et al., 2016; Larimer et al., 2014; Xavier & Germida,
2002). A MA pode, dependendo das espécies envolvidas na tripla simbiose, planta-
rizóbio-FMA, e das condições de crescimento, promover a FSN, o que tem sido
relacionado ao aporte de P e outros nutrientes essenciais para o processo de fixação de
nitrogênio (Xavier & Germida, 2002). A dupla simbiose também funciona como dreno
de carbono e pode estimular a fotossíntese (Kaschuk et al., 2009), fenômeno reportado
em soja e fava-comum (Harris et al., 1985; Kaschuk et al., 2009; Kucey & Paul, 1982).
Por outro lado, há casos de redução ou inibição da FSN em leguminosas crescendo em
solos com baixa fertilidade e ausência da associação com FMAs (Asimi et al., 1980;
Azcón et al., 1991; Ballhorn et al., 2016). O nódulo que é responsável pela fixação
requer alta quantidade de P para manter a atividade de fixação de N2 e a escassez ou
deficiência de P pode afetar diretamente a FSN e a nodulação (Sulieman et al., 2013).
O status de Pi no tecido foliar influencia diretamente os processos fotossintéticos
de transferência de energia, a regeneração de substratos, utilização de fotoassimilados e
a difusão de CO2 nas folhas (Singh et al., 2013). Devido à inibição da fotossíntese
durante a deficiência em Pi as plantas podem ser privadas de carboidratos, levando a
diminuição do crescimento e menor produção das culturas (Høgh Jensen et al., 2002).
Condições severas de deficiência de P podem levar a alterações do aparato
fotossintético incluindo reduções na assimilação de CO2, redução na expressão de genes
relacionados à fotossíntese e fotoinibição do fotossistema II, causando estresse
oxidativo e danos a lipídios, proteínas e ácidos nucleicos (Hernández & Munné-Bosch,
2015; Singh et al., 2013; Zhang et al., 2014).

52
As plantas absorvem Pi do solo principalmente através de proteínas
transportadoras localizadas na membrana plasmática de células epidérmicas e de pêlos
radiculares (Nussaume et al., 2011). Esses transportadores de Pi, codificados pela
família de genes PHT1, são predominantemente expressos na epiderme e na região
externa do córtex da raiz. Nos pelos radiculares, eles têm sido indicados como
responsáveis pela absorção de Pi na interface solo-planta (Lopez-Arredondo et al.,
2014) e também pelo transporte de Pi para o xilema, afetando o movimento de P da raiz
à parte aérea (Lapis-Gaza et al., 2014). No genoma da soja foram identificados 14 genes
da família de transportadores PHT1, distribuídos em oito cromossomos, e mais um
pseudogene (Fan et al., 2013), sendo a maioria deles induzida pela privação de Pi ou
pela simbiose com FMAs (Nussaume et al., 2011). O transportador GmPHT1;8
localiza-se no retículo endoplasmático diferentemente dos demais que estão localizados
na região subcelular da membrana plasmática (Fan et al., 2013). Transportadores de Pi
induzidos por MA tem sido identificados em várias espécies (Chen et al., 2007; Glassop
et al., 2007; Harrison, 2002; Hong et al., 2012; Rausch et al., 2001; Tamura et al., 2012;
Willmann et al., 2013; Xie et al., 2013) e em alguns casos a expressão destes
transportadores de Pi foi associada com a absorção de P por raízes colonizadas
(Fellbaum et al., 2014). Qin et al. (2012) identificaram o transportador GmPT5 como
chave no controle do transporte de Pi entre a raiz e o nódulo, particularmente sob
condições de deficiência de P. No entanto, na literatura encontram-se resultados
contraditórios e nem sempre os níveis de expressão de genes PHT1 relacionaram-se
com benefício micorrízico na absorção de P (Grace et al., 2009; Sisaphaithong et al.,
2012).
O entendimento dos mecanismos de absorção, uso e remobilização de P é
essencial no intuito de aumentar a eficiência do uso do P (EUP). No entanto, ainda são
escassos os estudos que avaliem mecanismos moleculares relacionados à maior EUP em
plantas micorrizadas. Também pouco se sabe sobre o transporte de Pi aos nódulos de soja
simbiótica e é praticamente desconhecido o papel da micorriza arbuscular na modulação
ou a extensão do aporte de Pi a este tecido nodular. Assim, o objetivo deste trabalho foi
avaliar como a soja nodulada e associada a MA responde a escassez de P. Neste sentido,
hipotetizamos que o fósforo absorvido pela MA pode atender a demanda de nódulos sob
escassez de P, promovendo a nodulação e a FSN estimulando o desenvolvimento
aliviando os efeitos de deficiência de P em plantas de soja. Para isso foi realizado um

53
experimento para avaliar o impacto da simbiose MA sobre o padrão de expressão de
genes de transportadores de Pi na raiz e no nodulo e relaciona-los com o crescimento,
nodulação atividade fotoquimica, nutrição, expressão de genes e metabolitos do
metabolismo nitrogenado das plantas de soja em condições de P baixas em dois estádios
de desenvolvimento diferentes (floração e enchimento de grãos).

54
Materiais e métodos
Delineamento experimental
Foi realizado um experimento em vasos, em casa de vegetação com condições
naturais de luz e temperatura, na qual plantas de soja [Glycine max (L.) cv. IAC
Foscarin-31] foram cultivadas. O experimento consistiu de seis tratamentos: plantas
com baixo suprimento de P (50 μM) e inoculadas (+MA) ou não (-MA) com FMA;
plantas com suprimento suficiente de P (500 μM) e sem inoculação de FMA (-MA+P),
com duas épocas de coleta: florescimento e formação da semente. O delineamento
experimental foi totalmente ao acaso e com 12 repetições para cada tratamento,
totalizando 72 parcelas experimentais.
Materiais biológicos, plantio e condições de crescimento
Sementes de soja [G. max (L.) cv. IAC Foscarin-31], cedidas pela Seção de
Grãos e Fibras do Instituto Agronômico (IAC), Campinas, SP, foram previamente
esterilizadas com solução de hipoclorito 3% por 5 min, e semeadas diretamente em
vasos plásticos de 3 L de capacidade contendo vermiculita de granulação fina como
substrato, também previamente esterilizado por autoclavagem por 1 h a 121°C e 1 atm.
Imediatamente antes da semeadura, as sementes foram mergulhadas por 20 min em
solução aquosa com meio de cultura YMB (Yeast Mannitol Broth) contendo
Bradyrhizobium elkanii estirpe SEMIA 5019. Na semeadura, as sementes do tratamento
+MA foram inoculados com 20 mL de inóculo misto de Glomus macrocarpum
contendo aproximadamente 1.360 esporos por vaso. Foram colocadas 3 sementes por
vaso. Após aproximadamente uma semana ocorreu a germinação e 15 dias após a
semeadura foi feito desbaste mantendo uma planta por vaso. Após o surgimento do
primeiro par de folhas unifoliada foi realizada uma reinoculação do rizóbio, com 10 mL
de meio de cultura YMB contendo B. elkanii.
Desde o início da germinação, as plantas foram irrigadas com 100 mL de água
destilada quando necessário, a partir do surgimento da folha unifoliada foram irrigadas
com 100 mL de solução de nutritiva ½ força e quando completaram o desenvolvimento
da primeira folha trifoliada passaram a ser irrigadas duas vezes na semana com 300 mL
de solução de nutritiva e com 100 mL de uma a três vezes por semana de acordo com a
necessidade da planta. A solução nutritiva utilizada foi a solução de Hoagland & Arnon

55
(1950) modificada, sem fonte de N e com a concentração de P alterada de acordo com o
tratamento: 50 μM para os tratamentos +MA e -MA e 500 μM para o tratamento -
MA+P.
As plantas foram coletadas no florescimento, entre os estádios R1 e R2,
aproximadamente 60 após a germinação ou no estádio de formação da semente, entre
R4 e R5, aproximadamente 100 dias após a germinação. O experimento foi conduzido
no período de fevereiro a abril de 2016, quando a temperatura média mínima foi de
20°C e a média da máxima foi de 40°C.Cabe ressaltar que os meses de fevereiro e
março foram de muita precipitação e alta cobertura de nuvens durante o período diurno
em quanto o mês de abril foi quente e sem apenas cobertura de nuvens, dados que
podem ser acessados no arquivo meteorológico disponível no site:
www.meteoblue.com/pt/tempo/previsao/archive/campinas.
Preparo da solução nutritiva
A solução de Hoagland foi modificada para atender as concentrações de P
estabelecidas para os tratamentos, contendo: MgSO4.7H2O (2 mM), KH2PO4 (0,05 mM
para os tratamentos +MA e –MA e 0,5 mM para o tratamento –MA+P), CaSO4.2H2O (2
mM), K2SO4 (2 mM), H3BO3 (0,046 mM), MnCl2.4H2O (9,1 μM), ZnSO4.7H2O (0,765
μM), CuSO4.5H2O (0,32 μM) e H2MoO4 (0,56 μM). O ferro foi acrescentado apenas no
momento da irrigação, na quantidade de 1 mL L-1
de solução, na forma de solução de
Fe-EDTA contendo Na2-EDTA (33,2 g L-1
), FeSO4.7H2O (25 g L-1
) e NaOH (3,65 g L-
1).
Preparo da cultura de rizóbio
As culturas de B. elkanii, estirpe SEMIA 5019 utilizadas no experimento foram
replicadas e mantidas em meio de cultura líquido YMB segundo proposto por Norris &
Date (1976), com pH ajustado entre 6,8 e 7,0. A formulação do meio foi a seguinte:
K2HPO4 (0,5 g L-1
), MgSO4.7H2O (0,8 g L-1
), NaCl (0,1 g L-1
), FeCl3.6H2O (0,01 g L-1
),
extrato de levedura (0,8 g L-1
), manitol (10 g L-1
) e 5 mL L-1
de solução de azul de
bromotimol 0,5% (m/v) em etanol. Os meios foram autoclavados a 120ºC por 20 min.
inoculados com a estirpe utilizada em câmara de fluxo laminar e incubados à 25ºC, sob
agitação, por um período de 7 dias até a fase exponencial de crescimento, quando a

56
densidade óptica a 600 nm estava entre 0,6 e 1. Antes de proceder com a inoculação, a
cultura no meio líquido foi diluída 10 vezes em água destilada.
Inóculo de fungos micorrízicos arbusculares e avaliação da colonização micorrízica
A espécie de FMA utilizada foi Glomus macrocarpum, cedido pela Seção de
Microbiologia do Solo do IAC e proveniente de vasos de multiplicação em Brachiaria
brizantha. A colonização micorrízica foi avaliada usando o método de lâmina proposto
por Giovannetti & Mosse (1980), em 4 replicatas biológicas por tratamento. Após as
raízes sem clareadas com 2,5% de KOH e coradas com azul de tripan 0,05%, a
colonização foi estimada como descrito por Trouvelot (1986). Para cada replicata, 20
segmentos de raiz de aproximadamente 1 cm foram avaliados quanto à presença de
colonização, atribuindo uma nota para classificar a porcentagem de colonização de
acordo com Trouvelot (1986).
Matéria seca de parte aérea, raízes e nódulos
Na coleta quatro plantas de cada tratamento foram separadas em parte aérea e
raiz. As raízes foram lavadas em água de torneira para retirada da vermiculita, os
nódulos e vagens foram destacados e contados e todo o material foi acondicionado em
sacos de papel e liofilizado. Posteriormente cada parte foi pesada para determinação da
matéria seca de caule e folhas (MSPA), raízes (MSR), vagens (MSV) e nódulos (MSN).
Determinação da concentração de nutrientes
Após a liofilização e pesagem, a parte aérea e a raiz foram moídas e
encaminhadas para análise da concentração de nutrientes no Centro de Solos e Recursos
Agroambientais do IAC. As concentrações de P, K, Ca, Mg, S, Mn, Fe, Cu, Zn e B
foram determinadas por espectrometria de emissão óptica com plasma (ICP-OES;
JobinYvon, JY50P Longjumeau, France), após digestão nítrico-perclórica (HNO3–
HClO4). O nitrogênio foi determinado pelo método Kjeldahl após digestão sulfúrica
(Bremner, 1965). O conteúdo de P ou N foi definido como sendo a quantidade total de P
ou N presente na parte aérea e raiz da planta (g planta-1
) (Li et al., 2016).
Determinação da concentração de fósforo inorgânico
Para a determinação da concentração de P inorgânico (Pi) foram utilizados
diferentes materiais vegetais: nódulos, folha nova, folha velha e raiz. Consideramos

57
folhas novas as duas folhas trifoliadas mais jovens completamente expandidas e folhas
velhas as duas folhas trifoliadas em fase de senescência há mais tempo. O material
vegetal foi imediatamente congelado a -80°C após a coleta das plantas. Cem miligramas
de matéria fresca foram macerados e colocados em 1 mL de ácido acético 1% (v:v), o
homogenato foi agitado por 4 min, seguido de centrifugação a 8000 g por 5 min a 4°C.
O material foi analisado segundo o método proposto por Ames (1966) e modificado por
Chiou et al. (2005). Em 100 µL do extrato foram adicionados 700 µL de solução de
reação e 200 µL de água. A reação foi incubada por 20 min a 42°C e realizamos a
leitura em espectrofotômetro a 820 nm. A solução de reação consistiu em uma mistura
(1:6, v:v) das soluções A (ácido ascórbico 10%) e B (0,42% de molibdato de amônia em
H2SO4 1 N).
Atividade fotoquímica
A avaliação da fluorescência da clorofila a foi realizada no segundo par de
folhas mais novas completamente expandidas, utilizando um fluorômetro modelo
FluorPen FP100 (Photon Systems Instruments, Czech Republic) utilizando sinais
registados antes e depois dos impulsos de saturação [λ < 710 nm, PPFD ~ 10,000 µ mol
(photon) m–2
s–1
, 0.7 s] após excitação do PSI com um fonte longa de luz [λ = 735 nm,
PPFD < 15 W m–2
, 3.0 s]. Algumas variáveis fotoquímicas foram estimadas: em tecidos
escuros adaptados (30 min), como os sinais de fluorescência máxima (Fm), mínima (F0)
e variável (Fv = Fm-F0) e eficiência quântica potencial (Fv / Fm); e nos tecidos adaptados
à luz, tais como o rendimento quântico efetivo do fotossistema II (ФPSII = Fm' - Fs), o
quenching fotoquímico e não-fotoquímico (NPQ) da fluorescência.
Determinação da concentração de aminoácidos e ureídeos
O tecido foliar foi coletado, congelado imediatamente em nitrogênio líquido e
liofilizado. Uma amostra de 0,01 mg de folha macerada foi colocada em 1 mL de
metanol 80% em água (v:v), agitada por 2 min, seguido de sonicação por 15 min e o
extrato centrifugado a 8000 g por 10 min (Mokochinski et al., 2013). Os extratos foram
acondicionados em tubos vedados e armazenados a -40°C até as análises. Deste extrato
foram analisados os aminoácidos livres e os ureídeos totais.
A concentração de aminoácidos totais foi determinada com ninhidrina pelo
método descrito por Yemm et al. (1955), usando leucina como padrão. Para a análise

58
foram utilizados 200 µL do extrato metanólico, a leitura foi realizada em
espectrofotômetro a 570 nm. A quantificação de ureídeos totais foi feita utilizando 250
µL do extrato de acordo com o método proposto por Vogels & Van der Drift (1970),
utilizando alantoína como padrão, e a leitura realizada em espectrofotômetro a 535 nm.
Extração de RNA e síntese de cDNA
O RNA total foi extraído de amostras de raiz e nódulos utilizando o reagente
Tri-Phasis (BioAgency) e quantificado em espectrofotômetro de microvolumes
(Nanodrop, Thermo Scientific, Waltham, US). A qualidade e integridade do RNA foram
verificadas em eletroforese em gel de agarose a 1% com brometo de etídeo. Todas as
amostras de RNA foram tratadas com DNase I, RNase-free (Thermo Scientific), a fim
de remover quaisquer resquício de DNA genômico. A primeira fita de cDNA foi
sintetizada utilizando a polimerase de transcrição reversa a partir de 2 μg de RNA de
acordo com o protocolo do kit da EasyScript Strand cDNA Synthesis SuperMix (Trans).
Obtenção das sequências e primers
Foram utilizados primers específicos para análise da expressão na raiz e nos
nódulos dos genes transportadores de fosfato PHT1, GmPHT1; 1/14 (Qin et al., 2012) e
genes de sacarose sintase GmSS1 (Chiasson et al., 2014). Ainda, de forma
complementar, foi analisada a expressão de genes ligados ao metabolismo de N, como
os genes que codificam a enzima asparagina sintetase GmAS, três isoformas: GmAS1,
GmAS2 e GmAS3, a glutamina sintetase citosólica GmGS1e plastídica GmGS2 (Souza et
al., 2016) e a glutamato sintase GmGOGAT e ureato oxidase GmUO1 (Chiasson et al.,
2014), envolvido na síntese de ureídeos. Referente a genes bacterianos, foi analisada a
expressão de BeNifH e BeNifD (Souza et al., 2016), que codificam a subunidade α dos
componentes I e II da nitrogenase, respectivamente.
A busca das sequências dos genes de estudo foi realizada ou verificada no banco
de dados do NCBI (National Center for Biotechnology Information) e/ou no
Phythozome (http://www.phytozome.net/). Para o desenho dos primers para PCR
quantitativa em tempo real (RT-qPCR) foi utilizado o programa Primer3 Plus®
(http://frodo.wi.mit.edu/primer3/), considerando as características especiais da PCR
quantitativa, como o tamanho do amplicon, menor que 150 pares de base (pb), primers
com 19-20 pb, temperatura de anelamento entre 60 e 90°C, entre outros.

59
Como genes constitutivos foram utilizados os primers do gene GmTefS1 (Qin et
al., 2012), GmAct11 e GmEF1b (Jian et al., 2008); e como constitutivo para genes
bacterianos foram testados os genes BeGapA (glyceraldeído-3-fosfato desidrogenase) e
o gene Be16S que codifica a subunidade 16S ribossomal (Chueire et al., 2003). As
sequências específicas utilizadas neste estudo estão listadas na Tabela 4.

60
Tabela 4. Sequências utilizadas como primers para analise da expressão gênica por RT-qPCR.
Primer (5’- 3’)
Nome do gene GenBank (NCBI) Locus name (Phytozome) CDS F R
Amplicon size (bp)
Eficiência (%)
GmPHT1;1 FJ797401 Glyma10g33030 1611 TGACCACAAGTACGATCTTCC CGCCAATAGTAGGTAAGAGCA 129 108
GmPHT1;2 FJ797402 Glyma03g31950 1620 GACATAGCGCGAAATCTGTC CAAACACGGCCGCAATGAAG 150 93
GmPHT1;3 FJ797403 Glyma10g00950 1602 AGGTGCACCAAAGCCGGGAACT TGGCCATGACACCCTCTGCA 215 95
GmPHT1;4 FJ797404 Glyma10g04230 1566 GAACACTTTCAGGGCAACTC GTCATCACAGTCTTTGCATCG 145 91
GmPHT1;5 FJ797405 Glyma20g34610 1611 GAGGGGCATTCATTGCTGCA AGCGAATCCACCTTCGAACCT 151 102
GmPHT1;6 FJ797406 Glyma10g33020 1509 CTGCTCACATACTATTGGCGT GTCCAACAGGAACCAAGTAAC 249 114
GmPHT1;7 FJ797407 Glyma19g34710 1620 GAGCACTCCCAGCTGCATTG GGCGACTGAGGAAGTCCTTG 204 84
GmPHT1;8 FJ797408 Glyma20g02660 1545 GGACCACCAAGGAACATCATAA CCCAGAAATGCCATGACAAC 155 94
GmPHT1;9 FJ797409 Glyma07g34870 1536 ACAAGAAGACAAGAGGGTCG AACCGAGCATGAGAATCAAC 162 92
GmPHT1;10 FJ797410 Glyma20g34620 1584 GAGCAATTGGACACAAGAAG TCCAACAGGAACCAAGTAGT 110 110
GmPHT1;11 FJ797411 Glyma14g36650 1674 GGACTCCCGAATGAATGCTA AGCTGCAGTCAACTCCCCTA 218 -
GmPHT1;12 FJ797412 Glyma14g28780 1578 ATGTTTAACTGTGGGCGGCG CCCTATTATTGGGCGTCGGT 182 78
GmPHT1;13 FJ797413 Glyma13g08720 1632 TCATTTTCGCGGGTTTAGTC GCTTGCTTCACGTTTCCTTC 232 85
GmPHT1;14 FJ797414 Glyma02g00840 1602 CAGGTTCTGGCTAGGGTTTG ACATAGTCAAATGCGGGGTC 243 107
GmSS1 XM_006606164 Glyma20g31730 1668 TTGGTCTTGCGAGAGCGGGC CGGCAACCATGGGGTGAGGC 120 89
GmAS1 NM_001248809.1 1740 GAGGGCCTTTGATGATGAAG CCATCAATCCAACCATAGCC 102 71
GmAS2 NM_001248792.1 1746 AAAGCTGTTGAGTGGGATGC GCTGAAATGTGCACTCCAAG 80 83
GmAS3 XM_003552013.2 1680 TAAGGCCTGATCTTGGAAGG CCAACCCCATCACTGAATTG 124 100
GmGS1 NM_001255403.2 Glyma18g04660.1 1071 CCATTGAGAAGTTGGGGAAG TGCAACTCCCCATAAGAAGG 122 105
GmGS2 XM_006597503.1 Glyma.15G102000 1299 CGCCACAAAGATCACATCAG TGCTGTCTCATGCTTTCCAG 72 98
GmGOGAT-Fd1 XM_006576724.1 Glyma.03G128300 4881 GCAACTGAAGAGCCTGATTG GGAACCAGCTGCCAAAATAG 111 97
GmGOGAT-Fd2 XR_419957.1 Glyma.19G130800 4746 AACCGCCTCCATAATTCCTC CAATGAATCCAACCCCACAG 124 97
GmUO1 D86929 Glyma10g23790 930 AGTTCGAGCAGAGGCACGGGA CCCACGCGCCACTCCACAAT 94 101
BeNifH Contig GCATCCTGAAATACGCCAAC AGATCAGCTGAGTGCCCAAC 126 114
BeNifD Contig ATTGCGGAGTCAAGTCCAAC TATGGACCATGTCCTTGATCG 114 112
GmTefS1 X56856 1979 TGCAAAGGAGGCTGCTAACT CAGCATCACCGTTCTTCAAA 200 98
GmAct11 BW652479 Glyma18g52780 1134 CGGTGGTTCTATCTTGGCATC GTCTTTCGCTTCAATAACCCTA 141 83
GmEF1b EV279336 Glyma02g44460 672 GTTGAAAAGCCAGGGGACA TCTTACCCCTTGAGCGTGG 102 89
BeGapA AM295406 798 AGATCAACGAGGCGATGAAG TGGTTGAAGTCGATCGAGAC 94 100
Be16S AF237422 1411 CTACAATGGCGGTGACAATG CGAACTGAGACGGCTTTTTG 70 65

61
Análise de expressão por PCR quantitativa em tempo real (RT-qPCR)
A expressão dos níveis de transcritos dos genes de interesse foi analisada por
RT-qPCR em raízes e nódulos das plantas de soja no florescimento. As análises por RT-
qPCR foram realizadas no equipamento StepOne Plus (Applied Biosystems, Carlsbad,
USA) usando QuantiFast SYBR Green PCR Kit (Qiagen) para acompanhar a síntese de
dsDNA. As condições dos PCR foram: 95°C por 3 min, seguido por 40 ciclos de 95°C
por 10 s e 60°C por 30 s. A confirmação de especificidade do amplicon foi baseada na
curva de dissociação no final de cada corrida. Ainda foi realizada uma curva de
eficiência de amplificação para cada par de primers, na qual foi verificada eficiência e
especificidade analisada com base na curva de dissociação para todos os primers. Para
normalização interna adequada dos genes de soja, foi utilizada a expressão dos genes de
referencia anteriormente citados e avaliados como estáveis pelo geNorm
exclusivamente para este experimento (valor de estabilidade M<0,15 e coeficiente de
variação<0,05) (Vandesompele et al., 2002). A expressão relativa foi normalizada pela
expressão da amostra de menor valor e os dados foram exportados do qBasePLUS
(Derveaux et al., 2010; Hellemans et al., 2007) para o Excel, para análise estatística.
A quantificação relativa da expressão dos genes de B. elkanii foi determinada
comparando a expressão transcricional entre genes alvo e referencia pelo método 2-ΔCt
,
onde: 2 representa a eficiência do primer do gene alvo (eficiência é calculada como a %
de eficiência X 0,01+1); ΔCt= (Ctalvo – Ctref). Onde, Ct: ciclo limiar, alvo: gene alvo, e
ref: gene referência (Livak & Schmittgen, 2001).
Análises estatísticas
Os dados foram submetidos à análise de variância (ANOVA) e as médias foram
comparadas pelo teste de Tukey (P<0,05), usando o software estatístico Sisvar, versão
5.6 (Ferreira, 2011). Os dados de contagem e os expressos em porcentagem foram
transformados em log(x+1) e arcoseno(√𝑥/100), respectivamente, antes da analise
estatística. Os gráficos foram feitos no programa OriginPro, versão 9.1 para Windows.
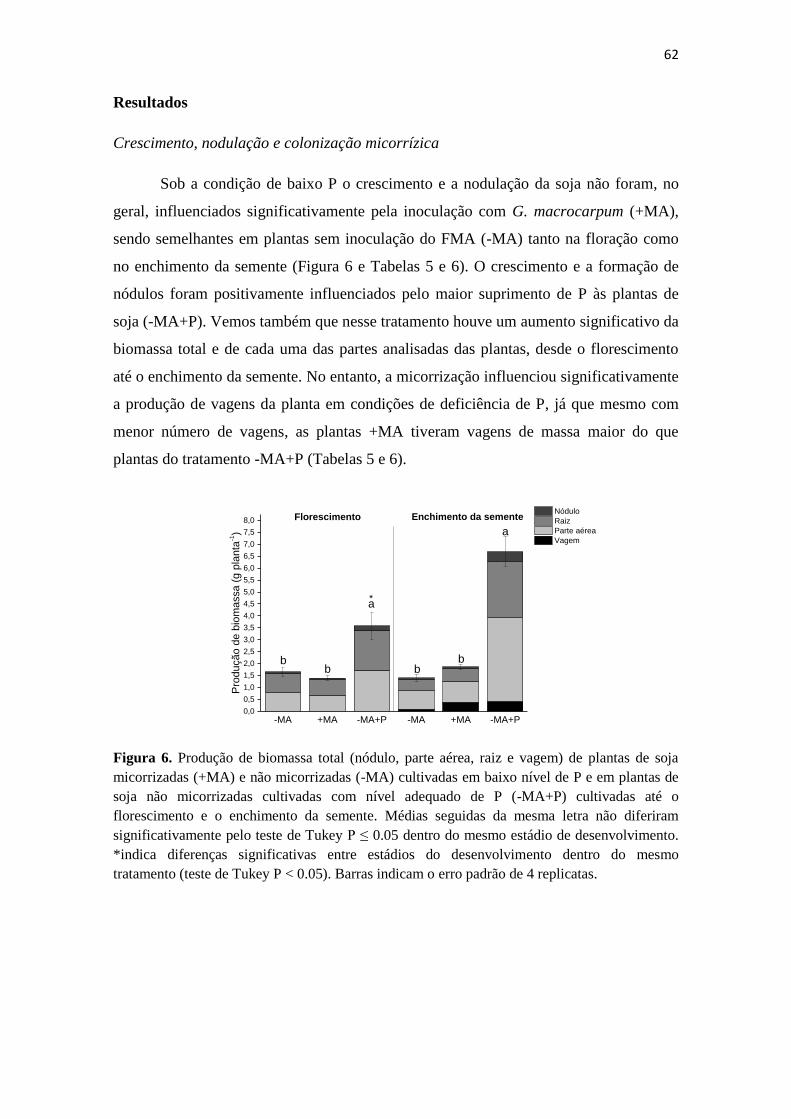
62
Resultados
Crescimento, nodulação e colonização micorrízica
Sob a condição de baixo P o crescimento e a nodulação da soja não foram, no
geral, influenciados significativamente pela inoculação com G. macrocarpum (+MA),
sendo semelhantes em plantas sem inoculação do FMA (-MA) tanto na floração como
no enchimento da semente (Figura 6 e Tabelas 5 e 6). O crescimento e a formação de
nódulos foram positivamente influenciados pelo maior suprimento de P às plantas de
soja (-MA+P). Vemos também que nesse tratamento houve um aumento significativo da
biomassa total e de cada uma das partes analisadas das plantas, desde o florescimento
até o enchimento da semente. No entanto, a micorrização influenciou significativamente
a produção de vagens da planta em condições de deficiência de P, já que mesmo com
menor número de vagens, as plantas +MA tiveram vagens de massa maior do que
plantas do tratamento -MA+P (Tabelas 5 e 6).
Figura 6. Produção de biomassa total (nódulo, parte aérea, raiz e vagem) de plantas de soja
micorrizadas (+MA) e não micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de
soja não micorrizadas cultivadas com nível adequado de P (-MA+P) cultivadas até o
florescimento e o enchimento da semente. Médias seguidas da mesma letra não diferiram
significativamente pelo teste de Tukey P ≤ 0.05 dentro do mesmo estádio de desenvolvimento.
*indica diferenças significativas entre estádios do desenvolvimento dentro do mesmo
tratamento (teste de Tukey P < 0.05). Barras indicam o erro padrão de 4 replicatas.
-MA +MA -MA+P -MA +MA -MA+P0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
5,0
5,5
6,0
6,5
7,0
7,5
8,0
Pro
duçã
o d
e b
iom
assa
(g
pla
nta
-1)
Nódulo
Raiz
Parte aérea
Vagem
bb
a
bb
a
Florescimento Enchimento da semente
*

63
Tabela 5. Produção de biomassa de nódulos, parte aérea, raiz e vagens de plantas de soja
micorrizadas (+MA) e não micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de
soja não micorrizadas cultivadas com nível adequado de P (-MA+P) até o florescimento e o
enchimento da semente, respectivamente. Médias seguidas da mesma letra não diferiram
significativamente pelo teste de Tukey P ≤ 0.05 dentro do mesmo estádio de desenvolvimento.
*indica diferenças significativas entre estádios do desenvolvimento dentro do mesmo
tratamento (teste de Tukey P < 0.05).
Florescimento Enchimento da semente
Parte aérea Raiz Nódulo Vagem Parte aérea Raiz Nódulo
g planta-1
-MA 0,78 b 0,82 b 0,05 b 0,09 b 0,77 b 0,49 b 0,05 b
+MA 0,68 b 0,67 b 0,05 b 0,38 a 0,88 b 0,53 b 0,07 b
-MA+P 1,73 a* 1,64 a* 0,21 a* 0,42 a 3,50 a 2,36 a 0,41 a
Tabela 6. Número de nódulos e de vagens de plantas de soja micorrizadas (+MA) e não
micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de soja não micorrizadas
cultivadas com nível adequado de P (-MA+P) cultivadas até florescimento e o enchimento da
semente. Médias seguidas da mesma letra não diferiram significativamente pelo teste de Tukey
P ≤ 0.05 dentro do mesmo estádio de desenvolvimento. *indica diferenças significativas entre
estádios do desenvolvimento dentro do mesmo tratamento (teste de Tukey P < 0.05).
Número de nódulos Número de vagens
Florescimento Enchimento da semente Enchimento da semente
-MA 30 b 33 b 4 b
+MA 33 b 46 b 4 b
-MA+P 57 a 69 a 10 a
As raízes de soja inoculadas com o FMA estavam no geral pouco colonizadas e
apresentaram colonização micorrízica entre 5% e 10% em ambas as fases de
desenvolvimento, não havendo diferença significativa entre os florescimento e
enchimento da semente. Baixo número de arbúsculos e vesículas também foram
observados (dados não mostrados).
Concentração de nutrientes
No florescimento a concentração de Ca, Mg, S, Mn, Cu, Zn e B na parte aérea
não foi alterada pelos tratamentos estudados (Tabela 7). Já a deficiência em P diminuiu
a concentração de N, P e K na parte aérea da soja, sendo a concentração destes três
elementos similar na parte aérea de plantas +MA e –MA crescidas com baixo
suprimento de P (Tabela 7). Na raiz somente a concentração de P e Mn em plantas não
inoculadas foi influenciada pela deficiência em P, com plantas -MA+P apresentando

64
maiores concentrações. A inoculação do FMA aumentou a concentração de N, Zn e B
nas raízes da soja em condição de deficiência de P (Tabela 7).
No estádio de enchimento da semente as plantas com suprimento adequado de P
apresentaram concentrações de N, P e B na parte aérea maiores do que plantas –MA
(Tabela 7). A concentração de P e Mn em plantas +MA não diferiu das observadas em
plantas sem inoculação (–MA ou –MA+P). A concentração de Mn na raiz foi maior em
plantas –MA do que nas inoculadas +MA ou com aporte suficiente de P (-MA+P). A
concentração de Zn na parte aérea foi maior em plantas +MA do que em plantas não
inoculadas. Nas raízes de plantas –MA+P a concentração de N foi inferior do que me
plantas -MA e sem diferenças significativas com plantas +MA (Tabela 7).
A concentração de alguns nutrientes em raízes e parte aérea mudou
significativamente entre os dois estádios reprodutivos analisados. No enchimento da
semente as plantas apresentaram diminuição na concentração de K, Mn e Cu na parte
aérea com relação ao observado no florescimento (Tabela 7). Já em plantas –MA+P a
concentração de B de parte aérea e de N na raiz aumentou no estádio de enchimento da
semente. A concentração de K na raiz de plantas crescidas em baixo nível de P
aumentou na maturidade. Em relação ao florescimento, a concentração de Ca, Mg, Mn,
Cu, Zn e B aumentou de forma significativa somente em raízes de plantas –MA no
enchimento da semente, havendo redução na concentração de S (Tabela 7).
A concentração de Fe na parte aérea e raízes foi determinada, sendo
extremamente alta nas raízes. Talvez esse fato esteja associado à contaminação da raiz
com resíduos de vermiculita usada como substrato para o crescimento da planta, uma
vez que vermiculita é constituída de silicatos de alumino-ferro-magnésio. Por este
motivo decidimos não apresentar esses dados.
O conteúdo de N e P acumulado por planta, obtido da multiplicação da
concentração de N e P pela biomassa, não foi influenciado pela simbiose em nenhum
dos estádios analisados (Figura 7A e B). Plantas crescidas com maior concentração de P
tiveram maiores conteúdos de N e P e ambos aumentaram do florescimento ao
enchimento da semente (Figura 7A e B).
A concentração de Pi foi maior na condição –MA+P em todas as partes da planta
e em ambos os estádios analisados, com exceção do Pi na raiz de plantas coletadas no

65
florescimento que foi igual para plantas dos três tratamentos (Tabela 8). Entre o
florescimento e o enchimento de grão, a concentração de Pi em plantas –MA+P
aumentou na raiz e diminuiu em folhas novas e velhas (Tabela 8). Em plantas crescendo
em baixo nível de P, folhas e nódulos tiveram concentrações de Pi similares entre si e
maiores do que aquelas em raízes. Já em plantas crescidas com níveis suficientes de P, a
concentração de Pi foi maior em folhas do que em nódulos e raízes, as quais
apresentaram as menores concentrações de Pi (Tabela 8).
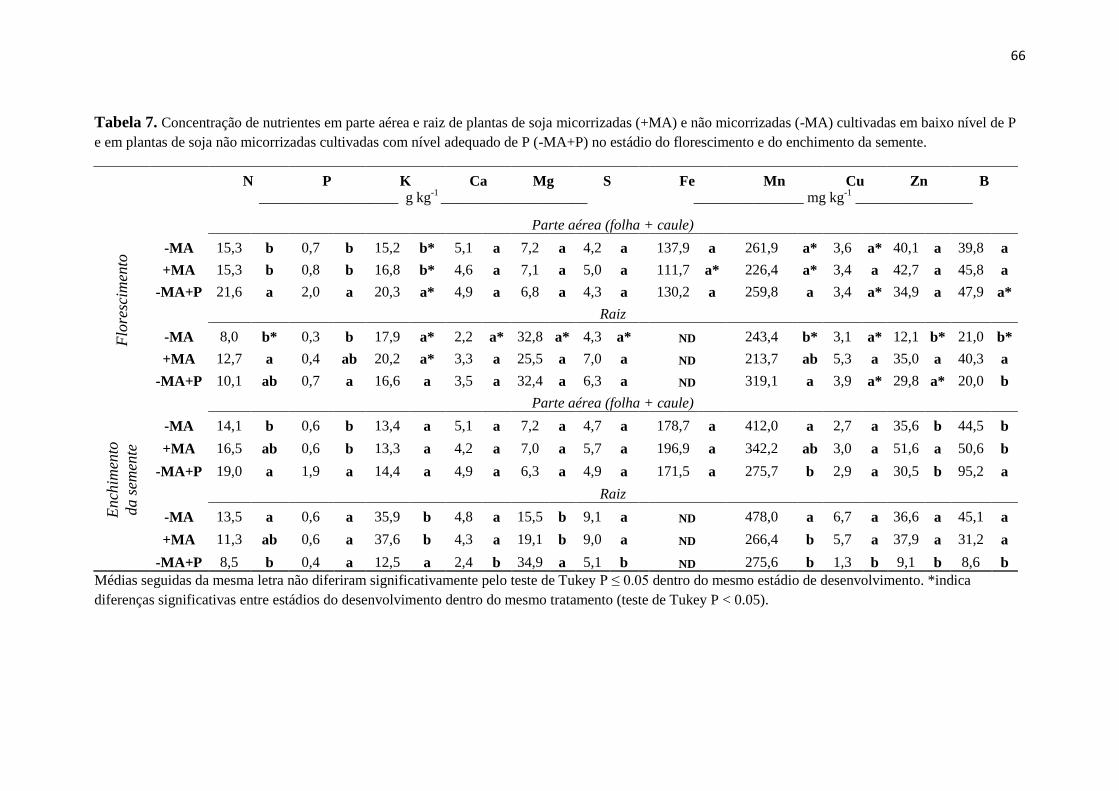
66
Tabela 7. Concentração de nutrientes em parte aérea e raiz de plantas de soja micorrizadas (+MA) e não micorrizadas (-MA) cultivadas em baixo nível de P
e em plantas de soja não micorrizadas cultivadas com nível adequado de P (-MA+P) no estádio do florescimento e do enchimento da semente.
N P K Ca Mg S Fe Mn Cu Zn B
___________________ g kg-1
____________________ _______________ mg kg-1
________________
Flo
resc
imen
to
Parte aérea (folha + caule)
-MA 15,3 b 0,7 b 15,2 b* 5,1 a 7,2 a 4,2 a 137,9 a 261,9 a* 3,6 a* 40,1 a 39,8 a
+MA 15,3 b 0,8 b 16,8 b* 4,6 a 7,1 a 5,0 a 111,7 a* 226,4 a* 3,4 a 42,7 a 45,8 a
-MA+P 21,6 a 2,0 a 20,3 a* 4,9 a 6,8 a 4,3 a 130,2 a 259,8 a 3,4 a* 34,9 a 47,9 a*
Raiz
-MA 8,0 b* 0,3 b 17,9 a* 2,2 a* 32,8 a* 4,3 a* ND 243,4 b* 3,1 a* 12,1 b* 21,0 b*
+MA 12,7 a 0,4 ab 20,2 a* 3,3 a 25,5 a 7,0 a ND 213,7 ab 5,3 a 35,0 a 40,3 a
-MA+P 10,1 ab 0,7 a 16,6 a 3,5 a 32,4 a 6,3 a ND 319,1 a 3,9 a* 29,8 a* 20,0 b
Ench
imen
to
da s
emen
te
Parte aérea (folha + caule)
-MA 14,1 b 0,6 b 13,4 a 5,1 a 7,2 a 4,7 a 178,7 a 412,0 a 2,7 a 35,6 b 44,5 b
+MA 16,5 ab 0,6 b 13,3 a 4,2 a 7,0 a 5,7 a 196,9 a 342,2 ab 3,0 a 51,6 a 50,6 b
-MA+P 19,0 a 1,9 a 14,4 a 4,9 a 6,3 a 4,9 a 171,5 a 275,7 b 2,9 a 30,5 b 95,2 a
Raiz
-MA 13,5 a 0,6 a 35,9 b 4,8 a 15,5 b 9,1 a ND 478,0 a 6,7 a 36,6 a 45,1 a
+MA 11,3 ab 0,6 a 37,6 b 4,3 a 19,1 b 9,0 a ND 266,4 b 5,7 a 37,9 a 31,2 a
-MA+P 8,5 b 0,4 a 12,5 a 2,4 b 34,9 a 5,1 b ND 275,6 b 1,3 b 9,1 b 8,6 b
Médias seguidas da mesma letra não diferiram significativamente pelo teste de Tukey P ≤ 0.05 dentro do mesmo estádio de desenvolvimento. *indica
diferenças significativas entre estádios do desenvolvimento dentro do mesmo tratamento (teste de Tukey P < 0.05).
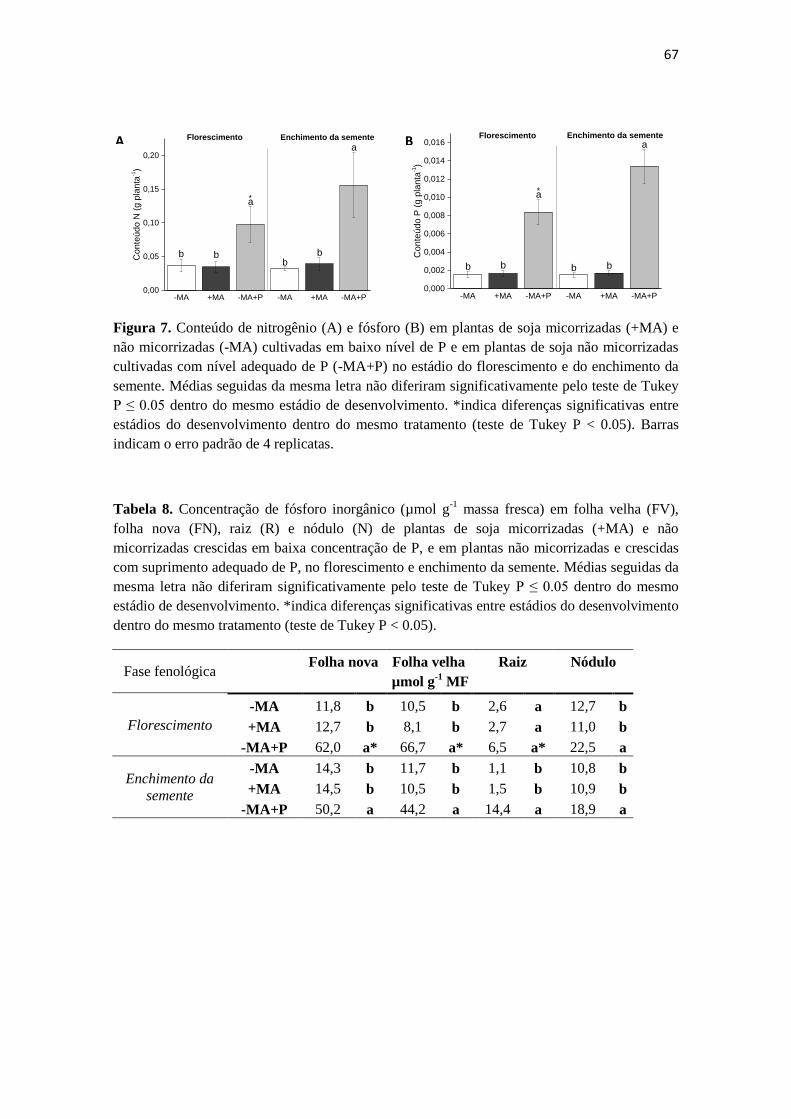
67
Figura 7. Conteúdo de nitrogênio (A) e fósforo (B) em plantas de soja micorrizadas (+MA) e
não micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de soja não micorrizadas
cultivadas com nível adequado de P (-MA+P) no estádio do florescimento e do enchimento da
semente. Médias seguidas da mesma letra não diferiram significativamente pelo teste de Tukey
P ≤ 0.05 dentro do mesmo estádio de desenvolvimento. *indica diferenças significativas entre
estádios do desenvolvimento dentro do mesmo tratamento (teste de Tukey P < 0.05). Barras
indicam o erro padrão de 4 replicatas.
Tabela 8. Concentração de fósforo inorgânico (µmol g-1
massa fresca) em folha velha (FV),
folha nova (FN), raiz (R) e nódulo (N) de plantas de soja micorrizadas (+MA) e não
micorrizadas crescidas em baixa concentração de P, e em plantas não micorrizadas e crescidas
com suprimento adequado de P, no florescimento e enchimento da semente. Médias seguidas da
mesma letra não diferiram significativamente pelo teste de Tukey P ≤ 0.05 dentro do mesmo
estádio de desenvolvimento. *indica diferenças significativas entre estádios do desenvolvimento
dentro do mesmo tratamento (teste de Tukey P < 0.05).
Fase fenológica
Folha nova Folha velha Raiz Nódulo
µmol g-1
MF
Florescimento
-MA 11,8 b 10,5 b 2,6 a 12,7 b
+MA 12,7 b 8,1 b 2,7 a 11,0 b
-MA+P 62,0 a* 66,7 a* 6,5 a* 22,5 a
Enchimento da
semente
-MA 14,3 b 11,7 b 1,1 b 10,8 b
+MA 14,5 b 10,5 b 1,5 b 10,9 b
-MA+P 50,2 a 44,2 a 14,4 a 18,9 a
-MA +MA -MA+P -MA +MA -MA+P0,00
0,05
0,10
0,15
0,20a
bb
bConte
údo N
(g p
lanta
-1)
b
a
Enchimento da sementeFlorescimento
*
-MA +MA -MA+P -MA +MA -MA+P0,000
0,002
0,004
0,006
0,008
0,010
0,012
0,014
0,016 a
a
bbb
Con
teú
do P
(g
pla
nta
-1)
b
Florescimento Enchimento da semente
*
A B

68
Atividade fotoquímica
As eficiências quânticas potencial (Fv/Fm) e efetiva do fotossistema II (ΦPSII)
foram similares nos tratamentos –MA e +MA, porém foram significativamente maiores
em plantas –MA+P (Tabela 9). O coeficiente de extinção não fotoquímica (NPQ) foi
significativamente maior em folhas de plantas com baixo suprimento de P quando
comparadas àquelas crescidas com suficiência de P (Tabela 9). O coeficiente de
extinção fotoquímica (Qp) não foi afetado pelos tratamentos (Tabela 9).
Tabela 9. Atividade fotoquímica em folhas de plantas de soja micorrizadas (+MA) e não
micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de soja não micorrizadas
cultivadas com nível adequado de P (-MA+P) no estádio do florescimento. Fv/Fm: eficiência
quântica potencial, ФPSII: rendimento quântico efetivo do fotossistema II, Qp: quenching
fotoquímico, NPQ: quenching não-fotoquímico. Médias seguidas de letras diferentes indicam
diferenças significativas entre os tratamentos no florescimento (teste de Tukey P < 0.05).
Fv/Fm ΦPSII NPQ Qp
-MA 0,69 b 0,14 b 5,50 a 0,36 a
+MA 0,73 b 0,18 b 5,33 a 0,41 a
-MA+P 0,78 a 0,25 a 3,02 b 0,42 a
Conteúdo de aminoácidos e ureídeos totais
No florescimento o conteúdo de aminoácidos livres totais nas folhas não foi
afetado pelos tratamentos. Já no enchimento da semente houve maior concentração em
folhas de plantas –MA+P, sendo que plantas micorrizadas apresentaram a menor
concentração (Tabela 10). A concentração de ureídeos de plantas no florescimento foi
significativamente menor em +MA e maior em –MA+P (Tabela 10). Na época do
enchimento de semente, plantas –MA+P continuaram a apresentar maior teor de
ureídeos totais do que plantas +MA, com plantas não inoculadas e sob deficiência de P
apresentando teores foliares intermediários (Tabela 10).

69
Tabela 10. Conteúdo de aminoácidos livres e ureídeos totais (µmol g-1
massa seca) em folhas de
soja micorrizadas (+MA) e não micorrizadas (-MA) cultivadas em baixo nível de P e em plantas
de soja não micorrizadas cultivadas com nível adequado de P (-MA+P), no estádio do
florescimento e do enchimento da semente. Médias seguidas da mesma letra não diferiram
significativamente pelo teste de Tukey P ≤ 0.05 dentro do mesmo estádio de desenvolvimento.
*indica diferenças significativas entre estádios do desenvolvimento dentro do mesmo
tratamento (teste de Tukey P < 0.05).
Fase
fenológica Aminoácidos Ureídeos
µmol g-1
MS
Florescimento
-MA 7,54 a 3,97 b
+MA 6,14 a 4,50 ab
-MA+P 8,09 a 5,23 a
Enchimento
da semente
-MA 8,60 ab 4,83 ab
+MA 6,85 b 4,27 b
-MA+P 9,03 a 5,76 a
Expressão gênica em raízes e nódulos da soja: metabolismo de N e transportadores de
P
A expressão de alguns genes relacionados com o metabolismo do N foi
analisada nos nódulos de plantas coletadas no florescimento (Figura 9). Observou-se
que o nível de expressão dos genes da glutamato sintase GmGOGAT1 e GmGOGAT2,
ureato oxidase GmUO1 e asparagina sintetase GmAS1 e GmAS2 não tiveram alteração
significativa nas condições estudadas (Figura 9). Já o gene da isoforma 3 da asparagina
sintetase GmAS3 apresentou maior nível de expressão em nódulos de plantas -MA,
seguido por nódulos de plantas +MA e com menor expressão nos nódulos de plantas -
MA+P (Figura 9). O gene da glutamina sintetase citoplasmática GmGS1 e cloroplástica
GmGS2 apresentou maior expressão em nódulos de plantas com maior disponibilidade
de P (-MA+P), sendo menor nos nódulos de planta micorrizadas (+MA). Em nódulos de
plantas -MA o nível de expressão do gene GmGS2 foi intermediária, não diferindo do
observado em nódulos de plantas de outros tratamentos (Figura 9).
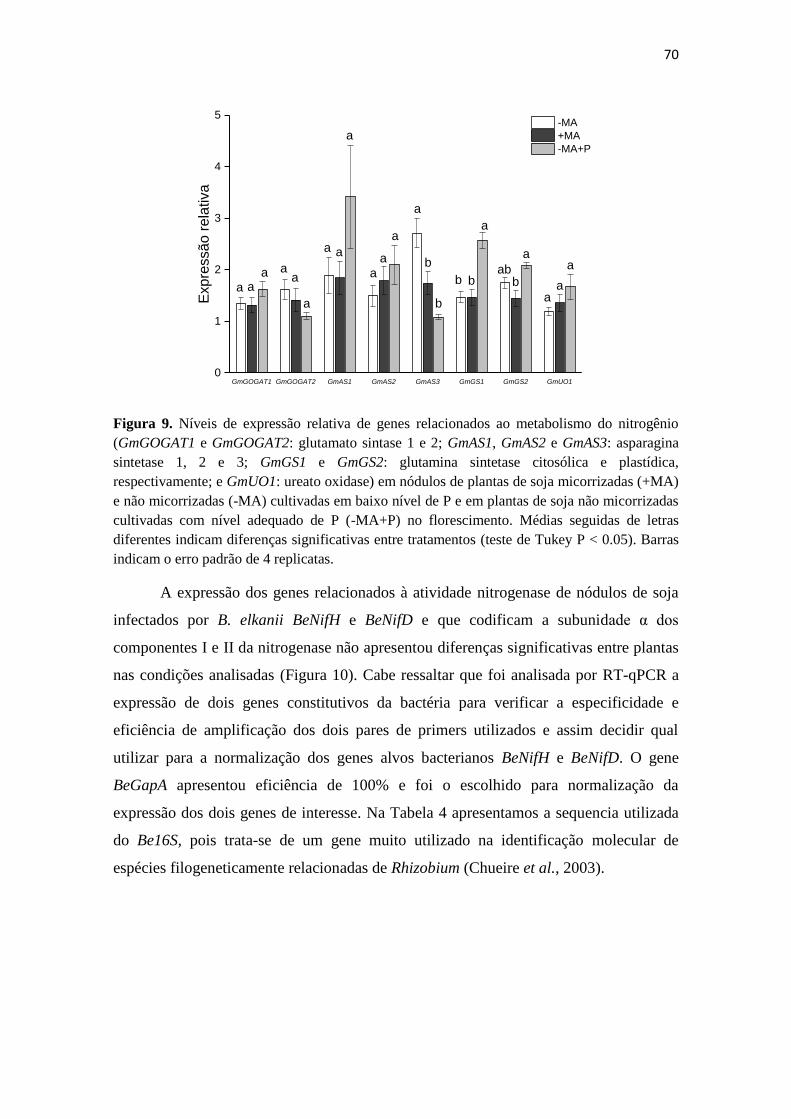
70
Figura 9. Níveis de expressão relativa de genes relacionados ao metabolismo do nitrogênio
(GmGOGAT1 e GmGOGAT2: glutamato sintase 1 e 2; GmAS1, GmAS2 e GmAS3: asparagina
sintetase 1, 2 e 3; GmGS1 e GmGS2: glutamina sintetase citosólica e plastídica,
respectivamente; e GmUO1: ureato oxidase) em nódulos de plantas de soja micorrizadas (+MA)
e não micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de soja não micorrizadas
cultivadas com nível adequado de P (-MA+P) no florescimento. Médias seguidas de letras
diferentes indicam diferenças significativas entre tratamentos (teste de Tukey P < 0.05). Barras
indicam o erro padrão de 4 replicatas.
A expressão dos genes relacionados à atividade nitrogenase de nódulos de soja
infectados por B. elkanii BeNifH e BeNifD e que codificam a subunidade α dos
componentes I e II da nitrogenase não apresentou diferenças significativas entre plantas
nas condições analisadas (Figura 10). Cabe ressaltar que foi analisada por RT-qPCR a
expressão de dois genes constitutivos da bactéria para verificar a especificidade e
eficiência de amplificação dos dois pares de primers utilizados e assim decidir qual
utilizar para a normalização dos genes alvos bacterianos BeNifH e BeNifD. O gene
BeGapA apresentou eficiência de 100% e foi o escolhido para normalização da
expressão dos dois genes de interesse. Na Tabela 4 apresentamos a sequencia utilizada
do Be16S, pois trata-se de um gene muito utilizado na identificação molecular de
espécies filogeneticamente relacionadas de Rhizobium (Chueire et al., 2003).
GmGOGAT1 GmGOGAT2 GmAS1 GmAS2 GmAS3 GmGS1 GmGS2 GmUO10
1
2
3
4
5
bbb
a
ab
a
b
b
a
a
a
a
aa
aa
a
a
aaaa
Exp
ressã
o r
ela
tiva
-MA
+MA
-MA+P
a
a
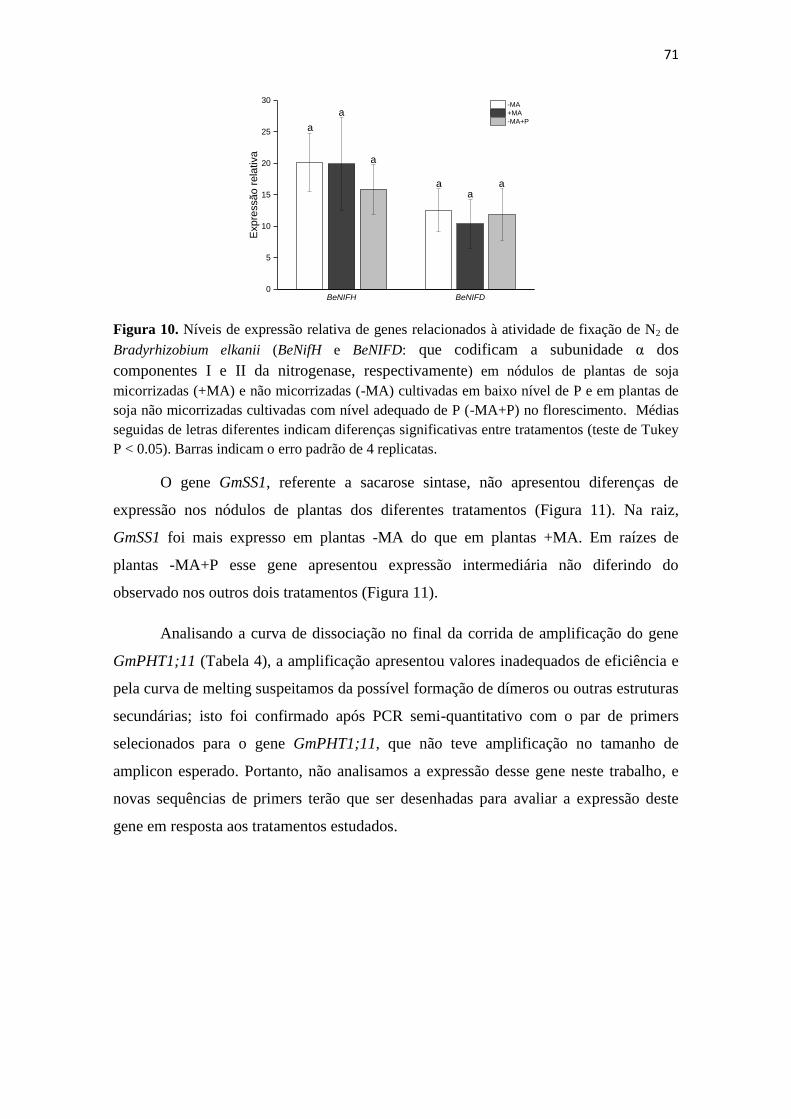
71
Figura 10. Níveis de expressão relativa de genes relacionados à atividade de fixação de N2 de
Bradyrhizobium elkanii (BeNifH e BeNIFD: que codificam a subunidade α dos
componentes I e II da nitrogenase, respectivamente) em nódulos de plantas de soja
micorrizadas (+MA) e não micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de
soja não micorrizadas cultivadas com nível adequado de P (-MA+P) no florescimento. Médias
seguidas de letras diferentes indicam diferenças significativas entre tratamentos (teste de Tukey
P < 0.05). Barras indicam o erro padrão de 4 replicatas.
O gene GmSS1, referente a sacarose sintase, não apresentou diferenças de
expressão nos nódulos de plantas dos diferentes tratamentos (Figura 11). Na raiz,
GmSS1 foi mais expresso em plantas -MA do que em plantas +MA. Em raízes de
plantas -MA+P esse gene apresentou expressão intermediária não diferindo do
observado nos outros dois tratamentos (Figura 11).
Analisando a curva de dissociação no final da corrida de amplificação do gene
GmPHT1;11 (Tabela 4), a amplificação apresentou valores inadequados de eficiência e
pela curva de melting suspeitamos da possível formação de dímeros ou outras estruturas
secundárias; isto foi confirmado após PCR semi-quantitativo com o par de primers
selecionados para o gene GmPHT1;11, que não teve amplificação no tamanho de
amplicon esperado. Portanto, não analisamos a expressão desse gene neste trabalho, e
novas sequências de primers terão que ser desenhadas para avaliar a expressão deste
gene em resposta aos tratamentos estudados.
BeNIFH BeNIFD0
5
10
15
20
25
30
aa
a
a
a
Exp
ressã
o r
ela
tiva
-MA
+MA
-MA+Pa
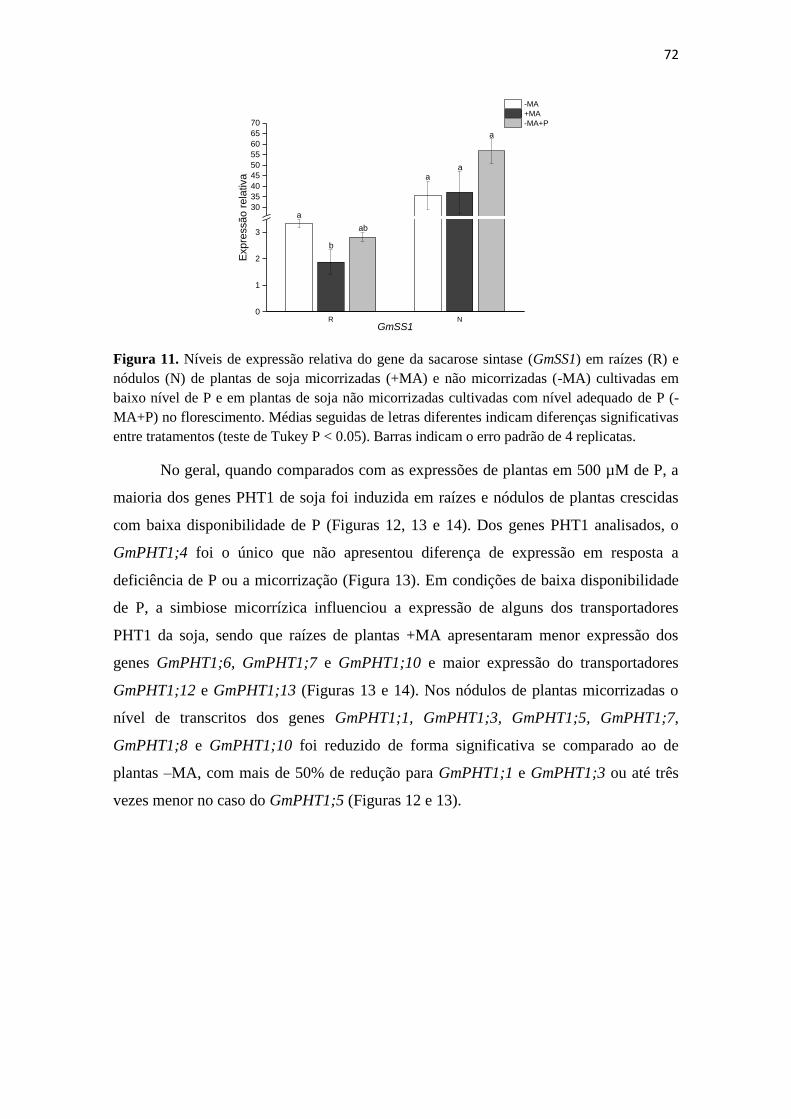
72
Figura 11. Níveis de expressão relativa do gene da sacarose sintase (GmSS1) em raízes (R) e
nódulos (N) de plantas de soja micorrizadas (+MA) e não micorrizadas (-MA) cultivadas em
baixo nível de P e em plantas de soja não micorrizadas cultivadas com nível adequado de P (-
MA+P) no florescimento. Médias seguidas de letras diferentes indicam diferenças significativas
entre tratamentos (teste de Tukey P < 0.05). Barras indicam o erro padrão de 4 replicatas.
No geral, quando comparados com as expressões de plantas em 500 µM de P, a
maioria dos genes PHT1 de soja foi induzida em raízes e nódulos de plantas crescidas
com baixa disponibilidade de P (Figuras 12, 13 e 14). Dos genes PHT1 analisados, o
GmPHT1;4 foi o único que não apresentou diferença de expressão em resposta a
deficiência de P ou a micorrização (Figura 13). Em condições de baixa disponibilidade
de P, a simbiose micorrízica influenciou a expressão de alguns dos transportadores
PHT1 da soja, sendo que raízes de plantas +MA apresentaram menor expressão dos
genes GmPHT1;6, GmPHT1;7 e GmPHT1;10 e maior expressão do transportadores
GmPHT1;12 e GmPHT1;13 (Figuras 13 e 14). Nos nódulos de plantas micorrizadas o
nível de transcritos dos genes GmPHT1;1, GmPHT1;3, GmPHT1;5, GmPHT1;7,
GmPHT1;8 e GmPHT1;10 foi reduzido de forma significativa se comparado ao de
plantas –MA, com mais de 50% de redução para GmPHT1;1 e GmPHT1;3 ou até três
vezes menor no caso do GmPHT1;5 (Figuras 12 e 13).
R N0
1
2
3
30
35
40
45
50
55
60
65
70
b
a
aa
ab
Expre
ssã
o r
ela
tiva
GmSS1
-MA
+MA
-MA+P
a
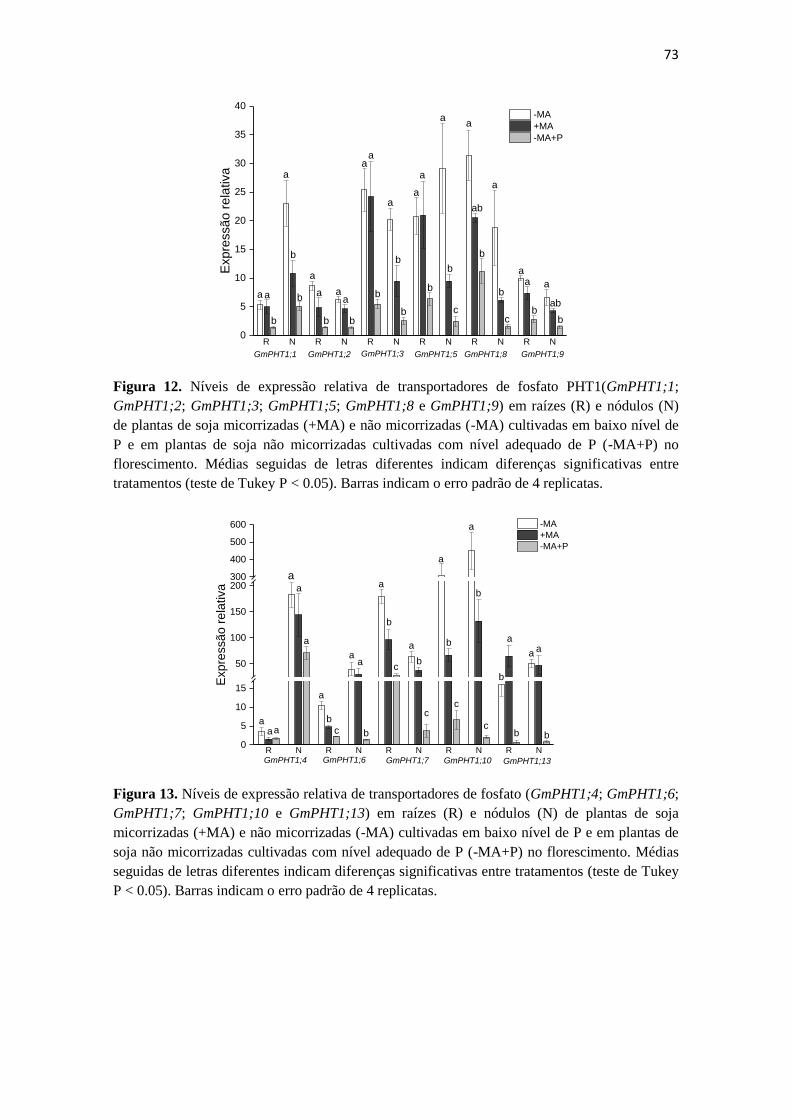
73
Figura 12. Níveis de expressão relativa de transportadores de fosfato PHT1(GmPHT1;1;
GmPHT1;2; GmPHT1;3; GmPHT1;5; GmPHT1;8 e GmPHT1;9) em raízes (R) e nódulos (N)
de plantas de soja micorrizadas (+MA) e não micorrizadas (-MA) cultivadas em baixo nível de
P e em plantas de soja não micorrizadas cultivadas com nível adequado de P (-MA+P) no
florescimento. Médias seguidas de letras diferentes indicam diferenças significativas entre
tratamentos (teste de Tukey P < 0.05). Barras indicam o erro padrão de 4 replicatas.
Figura 13. Níveis de expressão relativa de transportadores de fosfato (GmPHT1;4; GmPHT1;6;
GmPHT1;7; GmPHT1;10 e GmPHT1;13) em raízes (R) e nódulos (N) de plantas de soja
micorrizadas (+MA) e não micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de
soja não micorrizadas cultivadas com nível adequado de P (-MA+P) no florescimento. Médias
seguidas de letras diferentes indicam diferenças significativas entre tratamentos (teste de Tukey
P < 0.05). Barras indicam o erro padrão de 4 replicatas.
R N R N R N R N R N R N0
5
10
15
20
25
30
35
40
aa
a
ab
a
b
abb
c
b
b
b
b
b
b
bb
b
b
bc
a
a
aa
a
aa
a
a
a
a
a
a
GmPHT1;2 GmPHT1;3 GmPHT1;5 GmPHT1;8 GmPHT1;9
Expre
ssã
o r
ela
tiva
-MA
+MA
-MA+P
GmPHT1;1
a
b
R N R N R N R N R N0
5
10
15
50
100
150
200300
400
500
600
bb
c
c
b
b
a
a
c
b
a aa
a
bc
ba
b
a
c
b
a
a
a
a
aa
a
GmPHT1;6 GmPHT1;7 GmPHT1;10 GmPHT1;13
Exp
ressã
o r
ela
tiva
-MA
+MA
-MA+P
GmPHT1;4
a
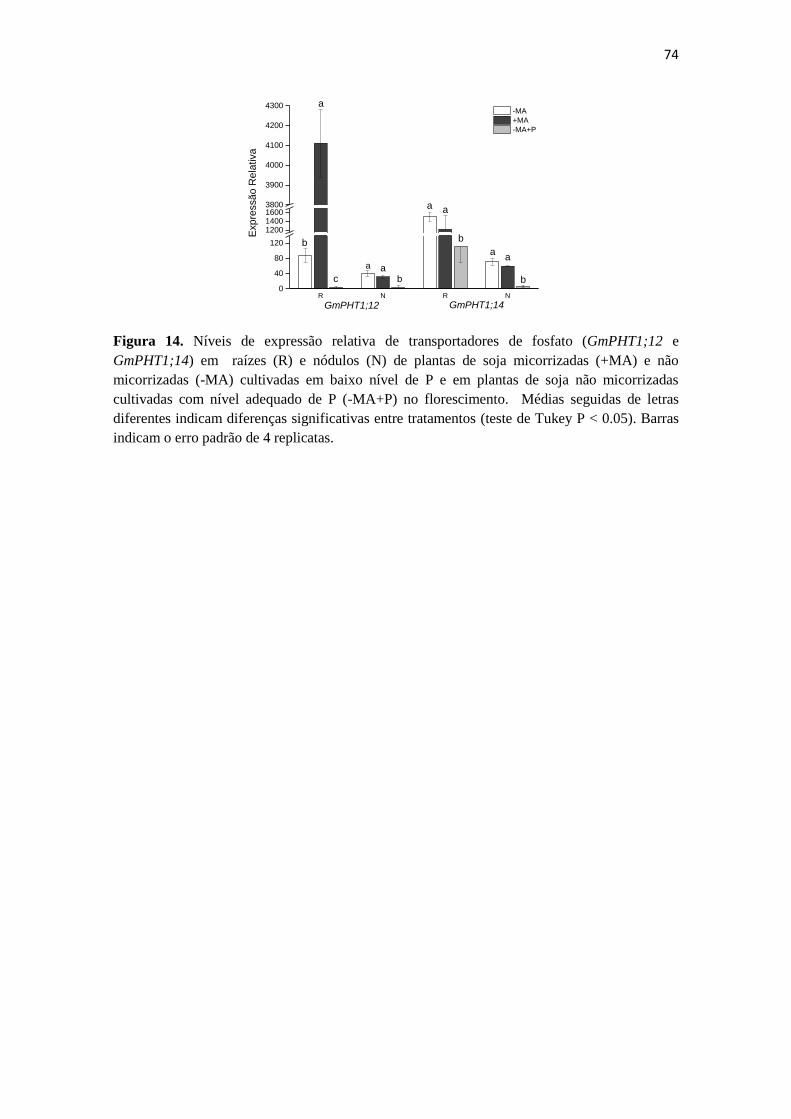
74
Figura 14. Níveis de expressão relativa de transportadores de fosfato (GmPHT1;12 e
GmPHT1;14) em raízes (R) e nódulos (N) de plantas de soja micorrizadas (+MA) e não
micorrizadas (-MA) cultivadas em baixo nível de P e em plantas de soja não micorrizadas
cultivadas com nível adequado de P (-MA+P) no florescimento. Médias seguidas de letras
diferentes indicam diferenças significativas entre tratamentos (teste de Tukey P < 0.05). Barras
indicam o erro padrão de 4 replicatas.
R N R N0
40
80
120
1200140016003800
3900
4000
4100
4200
4300
b b
aa
aa
b
a
a
GmPHT1;14
Expre
ssã
o R
ela
tiva
-MA
+MA
-MA+P
GmPHT1;12
b
a
c

75
Discussão
A baixa disponibilidade de P é especialmente limitante para o crescimento de
plantas, especialmente para leguminosas que realizam fixação simbiótica de N2 (FSN),
afetando o processo de fixação e a nodulação. Portanto, entender os mecanismos de
transporte de P e a influência da micorriza arbuscular (MA) na nutrição fosfatada em
condições de escassez de P é de grande importância para contribuir com o
desenvolvimento de abordagens que possam incrementar a eficiência de absorção e uso
do P e, consequentemente, a eficiência FSN. Assim, nesse estudo avaliamos a absorção
e a partição de P em plantas de soja com dupla associação micorriza arbuscular-rizóbio.
Em condições de baixa disponibilidade de P, a micorrização da soja ao FMA G.
macrocarpum não resultou, no geral, em alterações de crescimento de raízes e parte
aérea ou na nodulação (Figura 6 e Tabelas 5 e 6). A produção de biomassa total da
planta foi similar em plantas micorrizadas e não micorrizadas (Figura 6). Porém, a
biomassa produzida pelas vagens de plantas micorrizadas foi significativamente maior
no estádio de enchimento da semente (Tabela 5), apesar da quantidade de vagens entre
os tratamentos (Tabela 6), o que indica que a micorriza contribuiu para a maior alocação
de recursos para os frutos. A presença da micorriza pode ter contribuído para a maior
absorção de P (Oldroyd, 2013) na fase de enchimento da semente, nutriente que pode ter
sido preferencialmente alocado para as vagens quando comparadas à parte aérea e raiz
das plantas.
A deficiência de P tanto de plantas micorrizadas quanto daquelas não
micorrizadas causou importante redução na produção de biomassa da planta, o que
poderia ser explicado pela redução na assimilação de CO2 e da expansão foliar (Fredeen
et al., 1989). Neste estudo, a simbiose não conseguiu compensar a demanda por P na
condição de deficiência do nutriente. Sabe-se que a simbiose micorrízica pode causar
respostas desde altamente benéficas a prejudiciais para a planta, o denominado
continuum mutualismo parasitismo (Smith & Smith, 2013). Em determinadas situações
como níveis excessivos de P ou o estresse causado pela limitação de P os efeitos
positivos da simbiose micorrízica podem ser neutralizados ou mesmo eliminados e
provocar efeitos negativos para a planta hospedeira (Ballhorn et al., 2016). Portanto, a
severa limitação de P imposta à planta durante o nosso experimento pode ter mitigado
os efeitos benéficos da micorriza, sugerindo que a associação aumentou o custo em

76
termos de carbono para a planta e limitou o crescimento ou que a aquisição de P pela
via micorrízica não foi suficiente para promover maior produção de biomassa pela
planta. Plantas que cresceram com níveis suficientes de P dobraram a produção de
biomassa entre o florescimento e o enchimento da semente, o que não ocorreu em
plantas sob a deficiência de P, independente da micorrização (Tabela 5).
A deficiência em P é um dos principais fatores nutricionais que limitam a
fotossíntese, isto pelo seu envolvimento direto na transferência de energia e no
metabolismo de carboidratos nos cloroplastos (Fredeen et al., 1989; Hernández &
Munné-Bosch, 2015). A fotossíntese em plantas de soja é severamente reduzida pela
deficiência de P afetando o crescimento e a sua produção (Qiu & Israel, 1992). Durante
as reações dependentes de luz da fotossíntese, grande parte da energia de excitação é
convertida em energia química através do transporte linear de elétrons nas membranas
dos tilacóides, levando à produção de ATP e NADPH. Como esperado, a deficiência em
P causou uma redução significativa da eficiência fotoquímica das plantas de soja
(Tabela 9), o que está associado à escassez de P e possivelmente à menor concentração
de clorofila (Hernández & Munné-Bosch, 2015). Tanto plantas micorrizadas como não
micorrizadas tiveram um aumento do quenching não-fotoquímico (NPQ) em condições
de baixa disponibilidade de P (Tabela 9), o que mostra que plantas deficientes em P
devem ativar mecanismos de dissipação do excesso de energia para evitar o dano foto-
oxidativo (Hernández & Munné-Bosch, 2015). Durante a deficiência em P a redução na
assimilação de CO2 reduz o consumo de energia química e promove-se um
desequilíbrio entre a taxa de transporte de elétrons e consequente produção de ATP e
NADPH e a assimilação de carbono e consumo dos produtos da atividade fotoquímica.
Ainda, a produção de ATP é afetada uma vez que fotofosforilação do ADP é inibida
durante a deficiência de Pi no estroma, o que em consequência limita a regeneração de
RuBP (Heber et al., 1989). Em nosso experimento os níveis de Pi de folhas velhas e
novas de plantas de soja crescida com baixo P foi muito reduzido se comparado com
plantas em condições suficientes de P (Tabela 8), o que mostra a limitação de Pi para
processos que ocorrem durante a fotossíntese e o efeito negativo observado na eficiência
fotoquímica (Tabela 9).
Neste trabalho, curiosamente os níveis de Pi na raiz foram semelhantes em
plantas sob baixa e adequada disponibilidade de P. Nestas condições, o conteúdo de Pi
de nódulos e folhas foi similar e significativamente maior do que o observado na raiz

77
(Tabela 8). A este respeito observou-se que plantas coletadas durante o estádio de
enchimento da semente tiveram menores concentrações de Pi nas folhas, velhas e novas,
se comparado ao florescimento (Tabela 8), o que seria uma provável consequência da
remobilização de Pi para o fruto.
O conteúdo de aminoácidos livres nas folhas e no xilema podem ser indicadores
da assimilação de N na raiz (Justino & Sodek, 2013; Pate, 1973), e o seu conteúdo
também pode ser alterado pelo desenvolvimento do rizóbio. A presença da dupla
simbiose e a condição de escassez de P no geral diminuiu o conteúdo de aminoácidos e
ureídeos totais nas plantas micorrizadas em comparação com as plantas não
micorrizadas com condições adequadas de P (Tabela 10) provavelmente devido à menor
quantidade de N fixado em plantas deficientes de P e ao próprio dreno energético e
nutricional exercido pelos simbiontes. Teria sido interessante analisar o conteúdo de
aminoácidos e ureídeos totais no nódulo para poder correlacionar o conteúdo destes
compostos nitrogenados com a expressão de genes do metabolismo de N neste tecido,
infelizmente a massa coletada dos nódulos não foi suficiente, especialmente no caso do
tratamento –MA. Teria sido igualmente importante a avaliação da atividade da
nitrogenase com o fim de quantificar de forma mais direta a FSN.
No entanto, vários genes relacionados com o metabolismo de N tiveram sua
expressão analisada para avaliar o efeito da micorrização e da deficiência em P nos
nódulos da soja. No geral, houve pouca alteração na expressão dos genes estudados em
resposta à deficiência de P ou à micorrização (Figura 8). Nódulos de plantas deficientes
em P apresentaram menor expressão da isoforma citoplasmática da glutamina sintetase
ou GS1 do que nódulos de plantas crescidas com níveis suficientes de P (Figura 8)
assim como expressão da isoforma cloroplastídica, GS2, dos nódulos das plantas
micorrizadas. Esses resultados indicam que a amônia produzida durante a FSN deve
estar sendo ativamente assimilada no aminoácido Gln em nódulos de plantas com
suficiente Pi (Schulze et al., 2015). A expressão de genes da GOGAT, no entanto, não
teve alterações significativas, assim como o gene da uricase (GmUO1) envolvido na
síntese de ureídeos (Figura 8). No entanto, a expressão de um dos três genes de
asparagina sintetase analisados, o GmAS3, foi induzida em nódulos de plantas não
micorrizadas sob deficiência de P (Figura 8). Esta resposta não seria esperada uma vez
que alguns estudos mostram repressão da asparagina sintetase em condições de
deficiência de P (Keller et al., 2002). Sabe-se que a redução do crescimento em plantas

78
deficientes em P determina uma menor demanda de N (Høgh Jensen et al., 2002)
causando um mecanismo de feedback de N sobre a FSN, devido à essa menor demanda
de N, substâncias nitrogenadas deixam de ser transportadas das raízes ao nódulo via
floema alterando assim a fixação de N2 (Schulze et al., 2015). Tal suposição é reforçada
pelo fato da relação N/P em soja deficiente ser significativamente maior do que em
suficiência de P, corroborando com o obtido em outros trabalhos com alfafa e trevo
(Høgh Jensen et al., 2002; Schulze et al., 2015). Tem sido sugerido que em condições
de baixo P compostos nitrogenados importados pelo nódulo, tal como Asn, podem
controlar a atividade do nódulo e reprimir a expressão da asparagina sintetase (Keller et
al., 2002). O perfil de aminoácidos em nódulos ou a concentração de aminoácidos totais
neste tecido ajudaria a testar esta hipótese e deve ser tema de pesquisa futura.
Mesmo com as alterações observadas no metabolismo de N nos nódulos, a
expressão dos genes BeNifH e BeNifD não foi alterada em resposta à micorriza e ao
baixo P (Figura 9). Mudanças na expressão desses genes que codificam componentes da
nitrogenase podem ser um indicativo da atividade de fixação da bactéria. Assim, a
atividade de FSN poderia não ter sido alterada pela deficiência de P nos nódulos. Mas
quando analisamos o conteúdo de N na parte aérea e raiz, observamos que o conteúdo
de N foi maior nas plantas em suficiência de P do que em deficiência (Figura 7),
sugerindo que a regulação na expressão de genes relacionados à atividade nitrogenase
foi similar entre os tratamentos. Dado o maior número e tamanho dos nódulos de
plantas sob deficiência de P (Tabelas 5 e 6) a fixação de N foi maior para plantas com P
suficiente P, e provavelmente pelo efeito positivo do estímulo da FSN pela demanda de
N para a parte aérea e a menor relação N/P destas plantas. O nódulo possui alta
demanda de P sendo considerado um forte dreno deste nutriente (Sulieman et al., 2013),
já que a fixação e o metabolismo de N requer gastos significativos de energia os quais
são afetados pela deficiência de P (Sa & Israel, 1991). Isto corrobora-se dadas as altas
concentrações de Pi nos nódulos de plantas deficientes em P se comparadas às da raiz
sob condições de baixo P (Tabela 8) (Schulze et al., 2006), no entanto a concentração de
P total não foi analisada nos nódulos, apenas na raiz e parte aérea.
A sacarose não é utilizada diretamente como substrato para a maioria dos
processos envolvidos no crescimento, desenvolvimento e armazenamento, tanto na
fonte como no dreno, sendo o destino metabólico da sacarose mediado principalmente
pelas enzimas sacarose sintetase e as diferentes invertases intra e extracelulares (Sung et

79
al., 1989). Em nódulos a sacarose importada via floema é metabolizada
preferencialmente pela sacarose sintetase (SS) e não pela ação de invertases (Koch,
2004), mas vemos que no nódulo não houve diferenças significativas na expressão da
enzima entre os tratamentos (Figura 9). Em raiz de plantas micorrizadas, a expressão da
SS foi menor que em plantas não micorrizadas (Figura 9). Teria sido interessante avaliar
teor de amido e sacarose nos nódulos e raízes para relaciona-los aos dados de expressão
da SS, a analise da sacarose invertase poderia também contribuir para um melhor
entendimento do metabolismo da sacarose na presença da micorriza e durante a escassez
de P.
A raiz é obviamente o principal órgão responsável pela manutenção da
homeostase de Pi nas folhas, possuindo mecanismos de sinalização altamente regulados
e que inclui sinais hormonais, como citocininas e strigolactonas, translocados à parte
aérea via xilema (Zhang et al., 2014). Uma das respostas à limitação de P é a indução de
transportadores de Pi de alta afinidade na raiz com o objetivo de aumentar a capacidade
de absorção. Em soja foram identificados 14 genes da família PHT1 (Fan et al., 2013) e
nossos resultados mostram que a maioria deles respondeu a deficiência de P, como foi o
caso do GmPHT1;3, 5 e 9 na raiz, GmPHT1;6 e 12 no nódulo e GmPHT1;2, 12 e 14 na
raiz e nódulo (Figuras 11 a 13), sugerindo uma tentativa de aumentar a eficiência de
absorção de Pi do solo (Zhang et al., 2014). Já a expressão dos transportadores
GmPHT1;6, 7 e 10 na raiz e GmPHT1;3, 5, 7, 8, e 10 no nódulo (Figuras 11 e 12) foi
reprimida na presença da simbiose MA, o que estaria associado à via de aquisição de P
em plantas associadas ao FMA. Outros trabalhos demostraram que alguns
transportadores de P envolvidos na absorção de Pi pela via direta são reprimidos na
presença de micorriza (Chiou et al., 2001; Grunwald et al., 2009; Tamura et al., 2012;
Wang et al., 2016). O transportador GmPHT1;4 não apresentou diferença de expressão
em raízes e nódulos de plantas sob as diferentes condições deste trabalho e foi altamente
expresso em nódulos (Figura 12). Qin et al. (2012) demonstraram que esse
transportador (GmPT5) coordena o transporte de Pi da raiz para o nódulo, especialmente
sob condições limitadas de P, sendo fundamental na manutenção da homeostase de P
nos nódulos, tecido com alta demanda neste nutriente. Outros transportadores de Pi
como GmPHT1;11, 12 e 13 em soja são induzidos em raízes micorrizadas (Tamura et
al., 2012) e isto foi de fato observado no nosso experimento. Os genes GmPHT1;12 e
13 apresentaram alta expressão em raízes associadas ao FMA G. macrocarpum (Figuras

80
12 e 13), infelizmente não foi possível, até o momento, a analise do gene GmPHT1;11
neste trabalho. Este transportador PHT1;11 é o que tem sido mais estreitamente
relacionado com os benefícios da simbiose na nutrição fosfatada (Wang et al., 2016),
sugerindo estar envolvido na transferência de P na interface micorrízica (Wang et al.,
2016), isto pode explicar a relação entre falta de expressão significativa de este gene
neste experimento dada a baixa colonização radicular e os poucos arbúsculos
observados nas raízes de plantas inoculadas.

81
Conclusão
A soja nodulada não se beneficiou de forma significativa da simbiose MA em
condições de limitação de P, ao menos em termos de crescimento, nodulação e nutrição.
A severa limitação de P imposta à planta durante o experimento possivelmente mitigou
os efeitos benéficos da micorriza sugerindo que a simbiose não conseguiu compensar a
demanda por P na condição de escassez deste nutriente, porém não foi prejudicial ou
causou efeitos deletérios já que as respostas fisiológicas como crescimento, absorção de
nutrientes, atividade fotoquímica, metabolismo de N de plantas micorrizadas foram
similares às das plantas não micorrizadas nessa mesma condição. No entanto, foi
possível verificar a influencia da micorriza no aspecto molecular já que o perfil de
expressão de genes da família PHT1 foi significativamente alterado pela simbiose e
corroborando a existência de diferentes vias de aquisição de Pi em plantas micorrizadas
e não micorrizadas, mesmo na ausência de benefício na quantidade de P absorvida.

82
CONSIDERAÇÕES FINAIS
Os dois experimentos realizados neste estudo e abordados individualmente em
cada um dos capítulos desta dissertação apresentaram respostas distintas, mesmo que o
delineamento experimental foi o mesmo em ambos. No primeiro, os resultados
mostraram que a MA influenciou positivamente o crescimento e a nodulação em ambos
os estádios estudados em relação às plantas não micorrizadas e sob deficiência de P.
Embora a micorriza não tenha promovido o crescimento da planta de forma mais
próxima ao observado em plantas bem nutridas, a simbiose aumentou a atividade da
nitrogenase e levou a menor limitação metabólica da fotossíntese em condições de
escassez de P se comparado às plantas não micorrizadas na mesma condição. Isto
mostra que as respostas poderiam ser resultantes da contribuição da micorriza na
absorção de P, estimulando a FSN e aumentando o conteúdo de N na planta.
Outras análises moleculares e bioquímicas (expressão de transportadores de P,
enzimas do metabolismo de N, teor de amido e sacarose) dessas mesmas plantas do
primeiro experimento possivelmente teriam contribuído no entendimento da resposta da
soja nodulada e micorrizada à deficiência em P. Porém devido a um incidente com o
ultrafreezer onde o material vegetal coletado foi armazenado, o mesmo foi perdido antes
das análises serem feitas. Por esse motivo o experimento foi repetido, porém em época
diferente do ano ao realizado para o primeiro experimento desta dissertação.
No segundo experimento as plantas expostas as mesmas condições de limitação
de P responderam de forma diferente quanto ao beneficio na nodulação e no
crescimento, o que pode ter sido efeito de fatores climáticos pouco favoráveis durante a
condução deste experimento: período muito chuvoso com maior cobertura de nuvens.
Outro fator a ser considerado, é que no segundo experimentos houve uma diminuição da
capacidade colonizadora do fungo MA utilizado como inóculo, uma vez que os níveis
de colonização micorrízica foram significativamente menores aos observados em
plantas do primeiro experimento. No entanto, a soja nodulada mesmo não se
beneficiando de forma significativa da simbiose MA em condições de limitação de P
não foi prejudicial ou deletéria, pois plantas associadas à micorriza se desenvolveram de
forma semelhante a plantas não micorrizadas. As respostas bioquímicas e respostas
moleculares dos genes do metabolismo de N também não foram diferentes para plantas

83
micorrizadas e não micorrizadas, apenas a expressão de alguns transportadores de Pi foi
significativamente alterado pela simbiose.
A simbiose micorrízica, dentro do continuum mutualismo-prasatismo pode
causar respostas desde altamente benéficas a prejudiciais para a planta (Smith & Smith,
2013). Em determinadas situações de estresse os efeitos positivos da simbiose
micorrízica podem provocar efeitos negativos para a planta hospedeira (Ballhorn et al.,
2016), porem, no presente estudo mesmo na condição de estresse a simbiose não causou
redução no crescimento. A intensidade de colonização do FMA foi baixa e
possivelmente se tivesse sido maior poderia ter causado um maior custo em termos de
carbono causando uma redução no crescimento da planta ou mesmo estimulando a
fotossíntese, resultando em efeitos parecidos ao do primeiro experimento. Futuros
estudos serão necessários para compreender melhor a resposta de soja nodulada à
deficiência de P e à micorrização, o que sem dúvida será de importância desde uma
perspectiva atual e mais holística que procura entender a planta em interação com seus
simbiontes e seus microrganismos associados.

84
Referências
Adesemoye AO, Kloepper JW (2009) Plant-microbes interactions in enhanced fertilizer-
use efficiency. Appl Microbiol Biotechnol 85,1-12.
Al-Niemi TS, Kahn ML, McDermott TR (1997) P metabolism in the bean-Rhizobium
tropici symbiosis. Plant Physiol 113,1233-1242.
Al‐Ghazi Y et al. (2003) Temporal responses of Arabidopsis root architecture to
phosphate starvation: evidence for the involvement of auxin signalling. Plant
Cell Environ 26,1053-1066.
Amarante Ld, Lima J, Sodek L (2006) Growth and stress conditions cause similar
changes in xylem amino acids for different legume species. Environ Exp Bot
58,123-129.
Amarante Ld, Sodek L (2006) Waterlogging effect on xylem sap glutamine of
nodulated soybean. Biol plantarum 50,405-410.
Ames BN (1966) Assay of inorganic phosphate, total phosphate and phosphatases. In:
Methods in enzymology, vol 8. pp 115-118.
Andrade SAL, Abreu CA, de Abreu MF, Silveira APD (2004) Influence of lead
additions on arbuscular mycorrhiza and Rhizobium symbioses under soybean
plants. Appl Soil Ecol 26,123-131.
Andrews M, Lea PJ, Raven JA, Azevedo RA (2009) Nitrogen use efficiency. 3.
Nitrogen fixation: genes and costs. Annals of Applied Biology 155,1-13.
Appleby CA (1984) Leghemoglobin and Rhizobium respiration. Ann Rev Plant Physiol
35,443-478.
Asimi S, Gianinazzi-Pearson V, Gianinazzi S (1980) Influence of increasing soil
phosphorus levels on interactions between vesicular-arbuscular mycorrhizae and
Rhizobium in soybeans. Can J Bot 58,2200-2205.
Atkins C (1987) Metabolism and translocation of fixed nitrogen in the nodulated
legume. Plant Soil 100,157-169.
Aung K, Lin S-I, Wu C-C, Huang Y-T, Su C-l, Chiou T-J (2006) pho2, a phosphate
overaccumulator, is caused by a nonsense mutation in a microRNA399 target
gene. Plant Physiol 141,1000-1011.
Azcón R, Gomez M, Tobar R (1992) Effects of nitrogen source on growth, nutrition,
photosynthetic rate and nitrogen metabolism of mycorrhizal and phosphorus-
fertilized plants of Lactuca sativa L. . New Phytol 121,227–234.
Azcón R, Rubio R, Barea J (1991) Selective interactions between different species of
mycorrhizal fungi and Rhizobium meliloti strains, and their effects on growth,
N2‐fixation (15N) and nutrition of Medicago sativa L. New Phytol 117,399-404.

85
Balestrasse KB, Gallego SM, Tomaro ML (2006) Aluminium stress affects nitrogen
fixation and assimilation in soybean (Glycine max L.). Plant Growth Regul
48,271-281.
Ballhorn DJ, Schadler M, Elias JD, Millar JA, Kautz S (2016) Friend or Foe-Light
Availability Determines the Relationship between Mycorrhizal Fungi, Rhizobia
and Lima Bean (Phaseolus lunatus L.). PLoS One 11,e0154116.
Barea J, Azcon-Aguilar C (1983) Mycorrhizas and their significance in nodulating
nitrogen-fixing plants vol 36. Advances in agronomy, vol 4.
Bremner J (1965) Inorganic forms of nitrogen. Methods of soil analysis Part 2
Chemical and microbiological properties,1179-1237.
Broughton WJ, Zhang F, Perret X, Staehelin C (2003) Signals exchanged between
legumes and Rhizobium: agricultural uses and perspectives. Plant Soil 252,129-
137.
Carter AM, Tegeder M (2016) Increasing Nitrogen Fixation and Seed Development in
Soybean Requires Complex Adjustments of Nodule Nitrogen Metabolism and
Partitioning Processes. Curr Biol 26,2044-2051.
Chen A, Hu J, Sun S, Xu G (2007) Conservation and divergence of both phosphate- and
mycorrhiza-regulated physiological responses and expression patterns of
phosphate transporters in solanaceous species. New Phytol 173,817-831.
Chiasson DM et al. (2014) Soybean SAT1 (Symbiotic Ammonium Transporter 1)
encodes a bHLH transcription factor involved in nodule growth and NH4+
transport. P Natl Acad Sci U S A 111,4814-4819.
Chiou TJ, Aung K, Lin SI, Wu CC, Chiang SF, Su CL (2005) Regulation of phosphate
homeostasis by MicroRNA in Arabidopsis. Plant Cell 18,412-421.
Chiou TJ, Lin SI (2011) Signaling network in sensing phosphate availability in plants.
Annu Rev Plant Biol 62,185-206.
Chiou TJ, Liu H, Harrison MJ (2001) The spatial expression patterns of a phosphate
transporter (MtPT1) from Medicago truncatula indicate a role in phosphate
transport at the root/soil interface. Plant J 25,281-293.
Chueire L, Bangel E, Mostasso F, Campo R, Pedrosa F, Hungria M (2003) Taxonomic
classification of rhizobial strains recommended for soybean and common bean
crops in Brazil based on the sequencing of the 16s rRNA gene. Rev Bras Ciênc
Solo 27,833-840.
D’Haeze W, Holsters M (2002) Nod factor structures, responses, and perception during
initiation of nodule development. Glycobiology 12,79R-105R.
Dakora FD (2000) Commonality of root nodulation signals and nitrogen assimilation in
tropical grain legumes belonging to the tribe Phaseoleae. Functional Plant
Biology 27,885-892.

86
Derveaux S, Vandesompele J, Hellemans J (2010) How to do successful gene
expression analysis using real-time PCR. Methods 50,227-230.
Döbereiner J (1997) Biological nitrogen fixation in the tropics: social and economic
contributions. Soil Biol Biochem 29,771-774.
Duff SM, Sarath G, Plaxton WC (1994) The role of acid phosphatases in plant
phosphorus metabolism. Physiol Plantarum 90,791-800.
Fan C et al. (2013) The pattern of Phosphate transporter 1 genes evolutionary
divergence in Glycine max L. BMC Plant Biology 13,48.
Fellbaum CR, Mensah JA, Cloos AJ, Strahan GE, Pfeffer PE, Kiers ET, Bücking H
(2014) Fungal nutrient allocation in common mycorrhizal networks is regulated
by the carbon source strength of individual host plants. New Phytol 203,646-
656.
Ferreira DF (2011) Sisvar: a computer statistical analysis system. Ciência e
agrotecnologia 35,1039-1042.
Forde BG, Lea PJ (2007) Glutamate in plants: metabolism, regulation, and signalling. J
Exp Bot 58,2339-2358.
Fredeen AL, Rao IM, Terry N (1989) Influence of phosphorus nutrition on growth and
carbon partitioning in Glycine max. Plant Physiol 89,225-230.
Gage DJ (2004) Infection and invasion of roots by symbiotic, nitrogen-fixing rhizobia
during nodulation of temperate legumes. Microbiol Mol Biol Rev 68,280-300.
Geurts R, Bisseling T (2002) Rhizobium Nod factor perception and signalling. Plant
Cell 14,S239-S249.
Giovannetti M, Mosse B (1980) An evaluation of techniques for measuring vesicular
arbuscular mycorrhizal infection in roots. New Phytol 84,489-500.
Glassop D, Godwin RM, Smith SE, Smith FW (2007) Rice phosphate transporters
associated with phosphate uptake in rice roots colonised with arbuscular
mycorrhizal fungi. Can J Bot 85,644-651.
Gordon AJ, Minchin FR, James CL, Komina O (1999) Sucrose synthase in legume
nodules is essential for nitrogen fixation. Plant Physiol 120,867-878.
Grace E, Cotsaftis O, Tester M, Smith F, Smith S (2009) Arbuscular mycorrhizal
inhibition of growth in barley cannot be attributed to extent of colonization,
fungal phosphorus uptake or effects on expression of plant phosphate transporter
genes. New Phytol 181,938-949.
Gray VM (2010) The role of the C:N:P stoichiometry in the carbon balance dynamics of
the legume–AMF–Rhizobium tripartite symbiotic association. In, vol 18. pp
387-416.

87
Grunwald U et al. (2009) Overlapping expression patterns and differential transcript
levels of phosphate transporter genes in arbuscular mycorrhizal, Pi-fertilised and
phytohormone-treated Medicago truncatula roots. Planta 229,1023-1034.
Harris D, Pacovsky R, Paul E (1985) Carbon economy of soybean–Rhizobium–Glomus
associations. New Phytol 101,427-440.
Harrison MJ (2002) A phosphate transporter from Medicago truncatula involved in the
acquisition of phosphate released by Arbuscular Mycorrhizal Fungi. The Plant
Cell Online 14,2413-2429.
Heber U, Viil J, Neimanis S, Mimura T, Dietz KJ (1989) Photoinhibitory damage to
chloroplasts under phosphate deficiency and alleviation of deficiency and
damage by photorespiratory reactions. Z Naturforsch C 44,524-536.
Hellemans J, Mortier G, De Paepe A, Speleman F, Vandesompele J (2007) qBase
relative quantification framework and software for management and automated
analysis of real-time quantitative PCR data. Genome biology 8,1.
Hernández I, Munné-Bosch S (2015) Linking phosphorus availability with photo-
oxidative stress in plants. J Exp Bot 66,2889-2900.
Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without
soil. California Agricultural Experiment Station Circular 347.
Høgh Jensen H, Schjoerring JK, SOUSSANA JF (2002) The influence of phosphorus
deficiency on growth and nitrogen fixation of white clover plants. Ann Bot
90,745-753.
Hong JJ, Park YS, Bravo A, Bhattarai KK, Daniels DA, Harrison MJ (2012) Diversity
of morphology and function in arbuscular mycorrhizal symbioses in
Brachypodium distachyon. Planta 236,851-865.
Hungria M, Campo RJ, MENDES IdC Fixação biológica do nitrogênio na cultura da
soja. In, 2001. Embrapa Soja; Brasília, DF: Embrapa Cerrados,
Israel DW (1987) Investigation of the role of phosphorus in symbiotic dinitrogen
fixation. Plant Physiol 84,835-840.
Jeong J et al. (2004) A nodule-specific dicarboxylate transporter from alder is a member
of the peptide transporter family. Plant Physiol 134,969-978.
Jez JM, Lee SG, Sherp AM (2016) The next green movement: Plant biology for the
environment and sustainability. Science 353,1241-1244.
Jian B, Liu B, Bi Y, Hou W, Wu C, Han T (2008) Validation of internal control for
gene expression study in soybean by quantitative real-time PCR. BMC Mol Biol
9,59.
Justino GC, Sodek L (2013) Recovery of nitrogen fixation after short-term flooding of
the nodulated root system of soybean. J Plant Physiol 170,235-241.

88
Karandashov V, Bucher M (2005) Symbiotic phosphate transport in arbuscular
mycorrhizas. Trends Plant Sci 10,22-29.
Kaschuk G, Kuyper TW, Leffelaar PA, Hungria M, Giller KE (2009) Are the rates of
photosynthesis stimulated by the carbon sink strength of rhizobial and
arbuscular mycorrhizal symbioses? Soil Biol Biochem 41,1233-1244.
Keller S et al. (2002) Nitrogen feedback regulation in phosphorus deficient Lotus
japonicus plants. In: Nitrogen Fixation: Global Perspectives. CABI Publishing,
p 433.
Koch K (2004) Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar
sensing and plant development. Curr Opin Plant Biol 7,235-246.
Kosuta S, Chabaud M, Lougnon G, Gough C, Dénarié J, Barker DG, Bécard G (2003)
A Diffusible Factor from Arbuscular Mycorrhizal Fungi Induces Symbiosis-
Specific MtENOD11 Expression in Roots of Medicago truncatula. Plant Physiol
131,952-962.
Kucey R, Paul E (1982) Carbon flow, photosynthesis, and N2 fixation in mycorrhizal
and nodulated faba beans (Vicia faba L.). Soil Biol Biochem 14,407-412.
Lam H-M, Coschigano K, Oliveira I, Melo-Oliveira R, Coruzzi G (1996) The
molecular-genetics of nitrogen assimilation into amino acids in higher plants.
Annu Rev Plant Physiol Plant Mol Biol 47,569-593.
Lapis-Gaza HR, Jost R, Finnegan PM (2014) Arabidopsis PHOSPHATE
TRANSPORTER1 genes PHT1; 8 and PHT1; 9 are involved in root-to-shoot
translocation of orthophosphate. BMC Plant Biology 14,1.
Larimer AL, Clay K, Bever JD (2014) Synergism and context dependency of
interactions between arbuscular mycorrhizal fungi and rhizobia with a prairie
legume. Ecology 95,1045-1054.
Li C, Gui S, Yang T, Walk T, Wang X, Liao H (2012) Identification of soybean purple
acid phosphatase genes and their expression responses to phosphorus availability
and symbiosis. Ann Bot 109,275-285.
Li H et al. (2016) A Genetic Relationship between Phosphorus Efficiency and
Photosynthetic Traits in Soybean As Revealed by QTL Analysis Using a High-
Density Genetic Map. Front Plant Sci 7,924.
Liu T-Y, Chang C-Y, Chiou T-J (2009) The long-distance signaling of mineral
macronutrients. Curr Opin Plant Biol 12,312-319.
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-
time quantitative PCR and the 2− ΔΔCT
method. Methods 25,402-408.
Lohse S, Schliemann W, Ammer C, Kopka J, Strack D, Fester T (2005) Organization
and metabolism of plastids and mitochondria in arbuscular mycorrhizal roots of
Medicago truncatula. Plant Physiol 139,329-340.

89
Lopez-Arredondo DL, Leyva-Gonzalez MA, Gonzalez-Morales SI, Lopez-Bucio J,
Herrera-Estrella L (2014) Phosphate nutrition: improving low-phosphate
tolerance in crops. Annu Rev Plant Biol 65,95-123.
Loth-Pereda V et al. (2011) Structure and expression profile of the phosphate Pht1
transporter gene family in mycorrhizal Populus trichocarpa. Plant Physiol
156,2141-2154.
Luciñski R, Polcyn W, Ratajczak L (2002) Nitrate reduction and nitrogen fixation in
symbiotic association Rhizobium-legumes. Acta Biochim Pol 49,537-546.
Ma Z, Baskin TI, Brown KM, Lynch JP (2003) Regulation of root elongation under
phosphorus stress involves changes in ethylene responsiveness. Plant Physiol
131,1381-1390.
Marschner H (2011) Marschner's mineral nutrition of higher plants. Academic press.
McClure PR, Israel DW, Volk RJ (1980) Evaluation of the relative ureide content of
xylem sap as an indicator of N2 fixation in Soybeans. Plant Physiol 66,720-725.
Minchin F (1997) Regulation of oxygen diffusion in legume nodules. Soil Biol Biochem
29,881-888.
Mokochinski JB, Soares DX, Bruns RE, Mazzafera P, Sawaya A Optimization of
extraction conditions of free amino acids in plants by factorial design. In:
Congresso Brasileiro de Espectrometria de Massas, 2013. pp 7-11
Moore RC, Purugganan MD (2005) The evolutionary dynamics of plant duplicate
genes. Curr Opin Plant Biol 8,122-128.
Norris D, Date R (1976) Legume bacteriology. Tropical Pastures Research; Principles
and Methods. Comonwealth Bureal of pastures and Field Crops, , Bull. 51
Hurley, England.
Nussaume L et al. (2011) Phosphate import in plants: Focus on the PHT1 transporters.
Front Plant Sci 2,83.
Oldroyd GE (2013) Speak, friend, and enter: signalling systems that promote beneficial
symbiotic associations in plants. Nat Rev Microbiol 11,252-263.
Pate J (1973) Uptake, assimilation and transport of nitrogen compounds by plants. Soil
Biology and Biochemistry 5,109-119.
Plaxton WC, Tran HT (2011) Metabolic adaptations of phosphate-starved plants. Plant
Physiol 156,1006-1015.
Qin L et al. (2012) The high-affinity phosphate transporter GmPT5 regulates phosphate
transport to nodules and nodulation in soybean. Plant Physiol 159,1634-1643.
Qiu J, Israel DW (1992) Diurnal starch accumulation and utilization in phosphorus-
deficient soybean plants. Plant Physiol 98,316-323.

90
Rausch C et al. (2001) A phosphate transporter expressed in arbuscule-containing cells
in potato. Nature 414,462–470.
Redecker D, Kodner R, Graham LE (2000) Glomalean fungi from the Ordovician.
Science 289,1920–1921.
Richardson AE, Simpson RJ (2011) Soil microorganisms mediating phosphorus
availability update on microbial phosphorus. Plant Physiol 156,989-996.
Robson A, O'hara G, Abbott L (1981) Involvement of phosphorus in nitrogen fixation
by subterranean clover (Trifolium subterraneum L.). Funct Plant Biol 8,427-
436.
Sa T-M, Israel DW (1991) Energy status and functioning of phosphorus-deficient
soybean nodules. Plant Physiol 97,928-935.
Schachtman DP, Reid RJ, Ayling SM (1998) Phosphorus Uptake by Plants: From Soil
to Cell. Plant Physiol 116,447–453.
Scheible W-R, Rojas-Triana M (2015) Sensing, signaLling, and CONTROL of
phosphate starvation in plants: molecular players and applications. In: Annual
Plant Reviews, Phosphorus Metabolism in Plants, vol 48. p 25.
Schlueter JA et al. (2007) Gene duplication and paleopolyploidy in soybean and the
implications for whole genome sequencing. BMC Genomics 8,330.
Schmutz J et al. (2010) Genome sequence of the palaeopolyploid soybean. Nature
463,178-183.
Schubert KR (1986) Products of biological nitrogen fixation in higher plants: synthesis,
transport, and metabolism. Ann Rev Plant Physiol 37,539-574.
Schubert S, Serraj R, Plies-Balzer E, Mengel K (1995) Effect of drought stress on
growth, sugar concentrations and amino acid accumulation in N2-fixing Alfalfa
(Medicago sativa). J Plant Physiol 146,541-546.
Schulze J, Sulieman S, Fischinger S (2015) N-Feedback Regulation of N2 Fixation In
Medicago Truncatula Under P-Deficiency. Gen.Appl. Plant Physiology 34, 33-54.
Schulze J, Temple G, Temple SJ, Beschow H, Vance CP (2006) Nitrogen fixation by
white lupin under phosphorus deficiency. Ann Bot 98,731-740.
Schüßler A, Schwarzott D, Walker C (2001) A new fungal phylum, the
Glomeromycota: phylogeny and evolution. Mycol Res 105,1413-1421.
Siebers M, Dormann P, Holzl G (2015) Membrane Remodelling in Phosphorus-
deficient Plants. In: Annual Plant Reviews, Phosphorus Metabolism in Plants,
vol 48. Annu Plant Rev, pp 237-263.
Singh SK, Badgujar G, Reddy VR, Fleisher DH, Bunce JA (2013) Carbon dioxide
diffusion across stomata and mesophyll and photo-biochemical processes as

91
affected by growth CO2 and phosphorus nutrition in cotton. J Plant Physiol
170,801-813.
Sisaphaithong T, Kondo D, Matsunaga H, Kobae Y, Hata S (2012) Expression of plant
genes for arbuscular mycorrhiza-inducible phosphate transporters and fungal
vesicle formation in sorghum, barley, and wheat roots. Biosci Biotechnol
Biochem 76,2364-2367
Smith FA, Smith SE (2013) How useful is the mutualism-parasitism continuum of
arbuscular mycorrhizal functioning? Plant and Soil 363,7-18
Smith FW, Mudge SR, Rae AL, Glassop D (2003) Phosphate transport in plants. Plant
Soil 248,71–83.
Smith PM, Atkins CA (2002) Purine biosynthesis. Big in cell division, even bigger in
nitrogen assimilation. Plant Physiol 128,793-802.
Smith SE, Jakobsen I, Gronlund M, Smith FA (2011) Roles of arbuscular mycorrhizas
in plant phosphorus nutrition: interactions between pathways of phosphorus
uptake in arbuscular mycorrhizal roots have important implications for
understanding and manipulating plant phosphorus acquisition. Plant Physiol
156,1050-1057.
Souza SC, Mazzafera P, Sodek L (2016) Flooding of the root system in soybean:
biochemical and molecular aspects of N metabolism in the nodule during stress
and recovery. Amino Acids.
Spatafora JW et al. (2016) A phylum-level phylogenetic classification of zygomycete
fungi based on genome-scale data. Mycologia 108,1028-1046.
Sulieman S, Van Ha C, Schulze J, Tran L-SP (2013) Growth and nodulation of
symbiotic Medicago truncatula at different levels of phosphorus availability. J
Exp Bot,ert122.
Sung S-JS, Xu D-P, Black CC (1989) Identification of actively filling sucrose sinks.
Plant Physiol 89,1117-1121.
Svistoonoff S et al. (2007) Root tip contact with low-phosphate media reprograms plant
root architecture. Nat Genet 39,792-796.
Tamura Y, Kobae Y, Mizuno T, Hata S (2012) Identification and expression analysis of
arbuscular mycorrhiza-inducible phosphate transporter genes of soybean. Biosci
Biotechnol Biochem 76,309-313.
Trouvelot A (1986) Mesure du taux de mycorhization VA d'un systeme radiculaire.
Recherche de methodes d'estimation ayant une significantion fonctionnelle.
Mycorrhizae: physiology and genetics,217-221.
Udvardi MK, Price GD, Gresshoff PM, Day DA (1988) A dicarboxylate transporter on
the peribacteroid membrane of soybean nodules. FEBS letters 231,36-40.

92
Ullrich-Eberius CI, Novacky A, Fischer E, Lüttge U (1981) Relationship between
energy-dependent phosphate uptake and the electrical membrane potential in
Lemna gibba G1. Plant Physiol 67,797-801.
van de Wiel CCM, van der Linden CG, Scholten OE (2016) Improving phosphorus use
efficiency in agriculture: opportunities for breeding. Euphytica 207,1-22.
van der Heijden MG, Bardgett RD, van Straalen NM (2008) The unseen majority: soil
microbes as drivers of plant diversity and productivity in terrestrial ecosystems.
Ecol Lett 11,296-310.
Van Kauwenbergh SJ (2010) World phosphate rock reserves and resources.
Vance CP, Uhde-Stone C, Allan DL (2003) Phosphorus acquisition and use: critical
adaptations by plants for securing a nonrenewable resource. New Phytol
157,423-447.
Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F
(2002) Accurate normalization of real-time quantitative RT-PCR data by
geometric averaging of multiple internal control genes. Genome biology 3,1.
Varadarajan DK, Karthikeyan AS, Matilda PD, Raghothama KG (2002) Phosphite, an
analog of phosphate, suppresses the coordinated expression of genes under
phosphate starvation. Plant Physiol 129,1232-1240
Vogels G, Van der Drift C (1970) Differential analyses of glyoxylate derivatives. Anal
Biochem 33,143-157.
Wang X, Zhao S, Bucking H (2016) Arbuscular mycorrhizal growth responses are
fungal specific but do not differ between soybean genotypes with different
phosphate efficiency. Ann Bot 118,11-21.
Wellman CH, Osterloff PL, Mohiuddin U (2003) Fragments of the earliest land plants.
Nature 425,282–285.
Willmann M et al. (2013) Mycorrhizal phosphate uptake pathway in maize: vital for
growth and cob development on nutrient poor agricultural and greenhouse soils.
Front Plant Sci 4,533.
Xavier L, Germida J (2002) Response of lentil under controlled conditions to co-
inoculation with arbuscular mycorrhizal fungi and rhizobia varying in efficacy.
Soil Biol and Biochem 34,181-188.
Xie X, Huang W, Liu F, Tang N, Liu Y, Lin H, Zhao B (2013) Functional analysis of
the novel mycorrhiza-specific phosphate transporter AsPT1 and PHT1 family
from Astragalus sinicus during the arbuscular mycorrhizal symbiosis. New
Phytol 198,836-852.
Yemm E, Cocking E, Ricketts R (1955) The determination of amino-acids with
ninhydrin. Analyst 80,209-214.

93
Zhang Z, Liao H, Lucas WJ (2014) Molecular mechanisms underlying phosphate
sensing, signaling, and adaptation in plants. J Integr Plant Biol 56,192-220.
Zrenner R, Stitt M, Sonnewald U, Boldt R (2006) Pyrimidine and purine biosynthesis
and degradation in plants. Annu Rev Plant Biol 57,805-836.

94
Anexo I

95
Anexo II