Embed Size (px)
Citation preview

1
UNIVERSIDADE ESTADUAL DO CEARÁ
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS
VETERINÁRIAS
LUCELINA DA SILVA ARAÚJO
ESTUDO DO POTENCIAL CITOTÓXICO DO VENENO DA SERPENTE Crotalus durissus cascavella EM ADENOCARCINOMA
OVARIANO-OVCAR-8.
FORTALEZA
2012

2
LUCELINA DA SILVA ARAÚJO
ESTUDO DO POTENCIAL CITOTÓXICO DO VENENO DA SERPENTE Crotalus durissus cascavella EM ADENOCARCINOMA
OVARIANO-OVCAR-8.
Dissertação apresentada ao Programa de Pós-Graduação em Ciências Veterinárias da Faculdade de Veterinária da Universidade Estadual do Ceará, como requisito parcial para a obtenção do grau de Mestre em Ciências Veterinárias. Área de Concentração: Reprodução e Sanidade Animal. Linha de Pesquisa: Reprodução e sanidade de carnívoros,onívoros, herbívoros e aves. Orientadora: Profª. Dra. Janaina Serra Azul Monteiro Evangelista
FORTALEZA
2012

3
Dados Internacionais de Catalogação na Publicação
Universidade Estadual do Ceará
Biblioteca Central Prof. Antônio Martins Filho
Bibliotecário(a) Responsável – Giordana Nascimento de Freitas CRB-3/1070
A663e Araújo, Lucelina da Silva
Estudo do potencial citotóxico do veneno da serpente Crotalus Durissus Cascavella em adenocarcinoma ovariano – OVCAR - 8 / Lucelina da Silva Araújo. — 2012.
CD-ROM. 71f. il. (algumas color) ; 4 ¾ pol.
“CD-ROM contendo o arquivo no formato PDF do trabalho acadêmico, acondicionado em caixa de DVD Slin (19 x 14 cm x 7 mm)”.
Dissertação (mestrado) – Universidade Estadual do Ceará, Faculdade de Veterinária, Programa de Pós-graduação em Ciências Veterinárias, Fortaleza, 2012.
Área de concentração: Reprodução e Sanidade animal.
Orientação: Profa. Dra. Janaina Serra Azul Monteiro.
1. Crotalus Durissus Cascavella. 2. Veneno. 3. Citotóxidade. 4. Apoptose. I. Título.
CDD: 636.089

4

5
Dedico,
Ao meu esposo, Hudson Rocha por todo apoio, amor e paciência.

6
AGRADECIMENTOS
Agradeço a Deus e a Nossa Senhora por estarem presentes em cada segundo da
minha vida me dando força, proteção e coragem para continuar.
À Universidade Estadual do Ceará (UECE) e ao Programa de Pós-graduação
em Ciências Veterinárias (PPGCV) e a Faculdade de Veterinária (FAVET) pela
capacitação profissional que me proporcionaram.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
pelo incentivo concedido na forma de bolsa de estudo, ao Conselho Nacional de
Desenvolvimento Científico e Tecnológico (CNPq) e à Fundação Cearense de Apoio ao
Desenvolvimento Científico e Tecnológico (FUNCAP) pelo auxílio financeiro dessa
pesquisa.
À Universidade Federal do Ceará (UFC), em especial ao Laboratório de
Oncologia Experimental (LOE) por ter disponibilizado a sua estrutura física para a
realização da parte experimental deste projeto. Em especial, a Professora Dra. Letícia
Veras Costa Lotufo por autorizar a realização de nossos experimentos, ao Dr. Danilo
Rocha, que me orientou de forma brilhante durante a realização dos testes laboratoriais
e por todos os ensinamentos transmitidos nesse período. Ao professor Dr. Diego Wilke
pela iniciativa e auxilio no início da execução dos experimentos e a todos os que fazem
parte desta equipe que de alguma forma contribuíram para o desenvolvimento deste
projeto.
Ao Dr. Daniel Viana, pelo auxílio na leitura das lâminas e belíssimo laudo
emitido após sua atenção e disponibilidade.
À Dra. Diva Nojosa, do Núcleo Regional de Ofiologia da Universidade Federal
do Ceará – NUROF-UFC por ter gentilmente cedido o veneno para realização dos
experimentos.
À minha querida orientadora, Dra. Janaina Serra Azul Monteiro Evangelista,
por ter me acolhido e me orientado, sempre me fornecendo apoio acadêmico e moral,
por todos os conhecimentos transmitidos e por toda a ajuda concedida em todos os
momentos que precisei. Muito obrigada pela amizade e pela confiança que depositou
em mim, que seja o início de uma longa caminhada!

7
Aos mestres que passaram por mim ao longo do meu crescimento acadêmico,
pelos ensinamentos não só para minha vida profissional, mas para o meu dia a dia, em
especial a minha eterna orientadora Dra. Nilza Dutra Alves.
À professora Dra. Lúcia de Fátima (querida professora Lucinha) pelos
ensinamentos, pelo carinho, atenção e principalmente pelas palavras de apoio e
reconhecimento.
Aos colegas e amigos de laboratório que me receberam no HISTOVESP,
Magna Matos, Duanny Murinelly, André Figueiredo, Júnior Félix, Renato Pinheiro,
Deborah Queiroz, Karen Denise, Jorge Luis, Raquel Colares, Keyttyanne Sampaio,
Amanda Brito, Jéssica Nóbrega, Celito Filho, Davi Rocha, Glayciane Bezerra, Douglas
Varela, D. Zirlene, em especial a minha querida Anna Sérgia, meu braço direito durante
esses dois anos de trabalho, a Nathalia Santiago e a Wanessa Rodrigues pela amizade e
palavras de conforto e amor.
Aos professores do PPGCV, pelo conhecimento e experiência compartilhados.
Aos colegas do curso de mestrado por todos os momentos divertidos e por todo
o apoio que tive de vocês.
Aos funcionários do programa, Adriana Albuquerque e aos demais
funcionários por toda a ajuda e suporte em todos os procedimentos acadêmicos.
Aos meus amigos que não pertencem ao “mundo acadêmico”, pelos momentos
de descontração e por todo o apoio que obtive de vocês, principalmente, nos períodos
mais estressantes.
A minha família que me apoiou ao longo destes anos, compreendendo as
minhas ausências e me dando força para continuar. Principalmente ao meu esposo
Hudson Rocha pela paciência, pelo amor, dedicação e confiança. Aos meus pais, Angela
Maria e Paulo Araújo pelo amor incondicional e educação transmitidos a mim. Ao meu
irmão amado Paulo Filho, pelo companheirismo e brincadeiras.
Finalmente, agradeço a todos que aqui não foram citados, mas ajudaram direta
ou indiretamente para que essa dissertação fosse feita.

8
RESUMO
Nas últimas décadas o câncer alcançou grande dimensão, sendo considerado um
problema de saúde publica a nível mundial, representando uma das principais causas de
morte no mundo. As serpentes do gênero Crotalus vêm ganhando espaço significativo
no campo científico, pois pesquisas realizadas com seu veneno estão demostrando que
este apresenta mecanismos envolvidos em atividades citotóxicas em linhagens tumorais.
Logo, o objetivo deste trabalho foi analisar o efeito citotóxico do veneno bruto da
serpente Crotalus durissus cascavella (Cdcasca), in vitro, em linhagens tumorais, bem
como, os mecanismos de morte celular induzidos por este veneno. Os efeitos citotóxicos
da Cdcasca foram avaliados nas linhagens de carcinoma ovariano, carcinoma de
próstata, carcinoma de mama e glioblastoma. Foram realizados também avaliação de
alterações no DNA, integridade da membrana celular, ciclo celular e atividade das
Caspases 3 e 7. Os resultados demostraram relevante citotoxicidade apresentando
valores de CI50 de 2.4 até 6.7 µg/mL, para as cinco linhagens testadas. As células
tratadas com o Cdcasca demostraram alterações morfológicas de forma dose-
dependente, apresentando redução no volume celular, fragmentação nuclear e formação
de corpos apoptóticos. A atividade anti-proliferativa e integridade de membrana celular
foi confirmada, com uma significante redução do número de células para a concentração
do veneno de 4 μg/mL. A ocorrência de apoptose foi confirmada através da ativação de
Caspases 3 e 7. Em conclusão, o veneno da Crotalus durissus cascavella demostrou
efeito citotóxico dose-dependente em diversas linhagens tumorais, demonstrando
diversas alterações morfológicas em especial as células de linhagem de carcinoma
ovariano, causando morte celular.
Palavras chave: Crotalus durissus cascavella. Veneno. Citotoxicidade. Apoptose.

9
ABSTRACT
In the last decades cancer has reached a great dimension being considered a worldwide
public health problem, representing a major cause of death in world. Snakes from the
genus Crotalus has gained significant space in the scientific field, since researches with
their venom show that its mechanisms are involved in the cytotoxic activity.Therefore,
the aim of the present study was to analyze the crude venom of Crotalus durissus
cascavella (Cdcasca) cytotoxic activity in vitro against tumor cell lines, as well as, the
mechanism of cell death induced by this venom. . The cytotoxic effects of Cdcasca were
evaluated on ovarian cell carcinoma, prostate carcinoma, breast carcinoma and
glioblastoma. Evaluations to DNA fragmentation, cell membrane integrity, cell cycle
and the activity of Caspases 3 and 7 were also present in this study. Results showed
relevant cytotoxicity presenting values of CI50 of 2.4 to 6.7 µg/mL on the five lineages
tested. The cells treated with Cdcasca showed morphologic alterations in a dose-
dependent way, presenting reduction on the cell volume, nuclear fragmentation and
presence of apoptotic bodies. The antiproliferactive activity and the membrane integrity
were confirmed, with a significant reduction of the number of cells to the concentration
of the venom of μg/mL. The occurrence of apoptosis was confirmed trough the
activation of Caspases 3 and 7. In conclusion, the venom of Crotalus durissus cascavella
showed a dose-dependent cytotoxic effect in several tumor cell lines, presenting several
morphological alterations, especially to the ovarian cell carcinoma, causing cell death.
Keywords: Crotalus durissus cascavella. Venom. Cytotoxicity. Apoptosis.

10
LISTA DE FIGURAS
Revisão de literatura
Figura 1. Serpente Crotalus durissus cascavella............................................................ 19
Figura 2. Morte celular por apoptose, imagem adaptada............................................... 27
Figura 3. Fotomicrografia de adenocarcinoma ovariano. Evidenciando formação dos
corpos apoptóticos (setas), H.E, aumento de 400x, Motic BA310, Moticam
2000................................................................................................................................ 28
Figura 4. Fotomicrografia de adenocarcinoma ovariano. Evidenciando (A) Cariorrexis;
(B) Mitoses atípicas; (C) Sincícios, picnocse. Coloração H.E, aumento de 400x, Motic
BA310, Moticam 2000................................................................................................... 31
Figura 5. Imunologia da morte celular por necrose........................................................ 32
Figura 6. Diagrama representativo dos produtos da autofagia e metabolismo de célula
cancerígena..................................................................................................................... 33
Capitulo 1
Figure 1. Photomicrographs of OVCAR-8 cells stained with Hematoxilin/Eosin, 400×.
Cells were incubated with Cdcasca for 24 h. A: Control; B: Doxorubicin 0.5 μM; C:
Cdcasca 1.0 μg/mL; D: Cdcasca 2.0 μg/mL; E: Cdcasca 4.0 μg/mL. Black arrow: cell
membrane blebbing; White arrow: atypical mitosis; Black dashed arrow: Nucleus
fragmentation; White dashed arrow: membrane damage and pyknosis………………. 55
Figure 2. Confocal (Zeiss LSM 710) images of OVCAR-8 cells stained with DAPI,
400×. Cells were incubated with Cdcasca for 24 h. A: Control; B: Doxorubicin 0.5 μM;
C: Cdcasca 1.0 μg/mL; D: Cdcasca 2.0 μg/mL; E: Cdcasca 4.0 μg/mL. White arrow:
atypical mitosis; White dashed arrow: Nucleus fragmentation……….......................... 56
Figure 3. Effect of Cdcasca on OVCAR-8 cell membrane integrity (A) and cell number
(B) determined by flow cytometry, using propidium iodide after 24 h incubation. The
negative control was treated with the vehicle saline (Control). Paclitaxel (Ptx) at 0.05

11
µM was used as the positive control. Data are presented as mean values ± S.E.M. of
three independent experiments performed in triplicate. *p<0.05 compared to the
negative control by ANOVA followed by Dunnett’s test.............................................. 57
Figure 4. Effect of Cdcasca on caspases 3,7 determined by flow cytometry, after 24h
incubation. The negative control was treated with the vehicle saline (Control).
Doxorubicin (Dox) at 0.5 µM was used as the positive control. Data are presented as
mean values ± S.E.M. of three independent experiments performed in triplicate.
*p<0.05 compared to the negative control by ANOVA followed by Dunnett’s test…. 58

12
LISTA DE TABELAS
Revisão de literatura
Tabela 1: Estimativas para o ano de 2012 das taxas brutas de incidência por 100 mil
habitantes e de número de casos novos por câncer, segundo sexo e localização primária*
no Brasil.......................................................................................................................... 36
Tabela 2: Distribuição proporcional dos dez tipos de câncer mais incidentes estimados
para 2012 por sexo, exceto pele não melanoma* no Brasil............................................ 36
Capitulo 1
Table 1. Cytotoxic activity of Cdcasca on tumor cell lines…........................................ 59

13
LISTA DE ABREVIATURAS E SIGLAS
AA Ácido araquidônico
ATP Adenosina trifosfato
Bcl-2 Família de proteínas envolvidas no processo de morte celular
Cdcasca Crotalus durissus cascavella
cFLA2 Enzimas citosólicas como as fosfolipases A2
CHO-K1 Linhagem celular de carcinoma ováriano
Crotoxina A Fração ácida da Crotoxina (Crotapotina)
Crotoxina B Fração básica (Fosfolipase A2)
Cvxα e Cvxβ Convulxina tipo α e β.
DAPI Dihidrocloreto de 4',6-diamidino-2-fenilindole
DMSO Dimethyl sulfoxide (Dimetil sulfóxido)
GH3 Linhagem celular de adenoma benigno de pituitária
H.E. Corante hematoxilina e eosina.
HL-60 Linhagem celular de leucemia promielocítica humano
IAP Inibidor da proteína de apoptose
CI50 Concentração Inibitória Média capaz de provocar 50% do efeito
máximo
IL-6 Interleucina-6
INCA Instituto Nacional do Câncer
LAO L-amino-oxidase
LOE Laboratório de Oncologia Experimental

14
MCF-7 Linhagem celular de carcinoma mamário humano
MTT Sal 3-(4,5-dimetil-2-tiazol)-2,5-difenil-2-H-brometo de tetrazolium
MX-1 Linhagem celular de carcinoma mamário humano
NaCl Cloreto de sódio
NaHCO3 Bicarbonato de sódio
NCI National Cancer Institute
OMS Organização Mundial da Saúde
OVCAR-8 Linhagem celular de adenocarcinoma ovariano humano
PC-3M Linhagem celular de carcinoma de próstata humano
PI Iodeto de Propídio
PLA2 Fosfolipase A2
ROS Espécies reativas de oxigênio
RT2 Linhagem celular de glioblastoma humano
SF-268 Linhagem celular de glioblastoma humano
SKOV3 Linhagen celular de carcinoma ovariano humano
TBS Tampão Tris
TNF-α Fator de necrose tumoral- α
Tumor Ehrlich Adenocacinoma mamário
TXA2 Tromboxano A2
WHO World Health Organization

15
SUMÁRIO
1. INTRODUÇÃO........................................................................................................ 16
2. REVISÃO DE LITERATURA................................................................................ 18
2.1. Aspectos Gerais e Epidemiológicos do Gênero Crotalus............................ 18
2.2. Constituintes do veneno da espécie Crotalus durissus................................ 19
2.2.1. Crotoxina e suas subunidades....................................................... 19
2.2.2. Crotamina..................................................................................... 21
2.2.3. Convulxina.................................................................................... 22
2.2.4. Giroxina........................................................................................ 22
2.2.5. Peptídio natriurético..................................................................... 23
2.3. Estudo da atividade citotóxica do veneno do Gênero Crotalus e suas
frações................................................................................................................. 24
2.4. Morte celular............................................................................................... 26
2.4.1. Apoptose ...................................................................................... 27
2.4.2. Necrose ........................................................................................ 30
2.4.3. Autofagia...................................................................................... 32
2.5. A importância do estudo sobre o câncer...................................................... 33
3. JUSTIFICATIVA.................................................................................................... 38
4. HIPÓTESE CIENTÍFICA....................................................................................... 39
5. OBJETIVOS............................................................................................................. 40
5.1. Objetivo geral.............................................................................................. 40
5.2. Objetivos específicos……………………………………………………... 40
6. CAPÍTULO I - Cytotoxic effect of Crotalus durissus cascavella venom in tumor
cell lines.…………………………….…………………..…………………………….. 41
7. CONSIDERAÇÕES FINAIS................................................................................... 60
8. PERSPECTIVAS...................................................................................................... 61
REFERÊNCIAS BIBLIOGRÁFICAS....................................................................... 62

16
1. INTRODUÇÃO
As serpentes do Gênero Crotalus tem ganhado importante espaço no campo
científico, visto que em algumas décadas seu veneno vem sendo alvo de pesquisadores
devido sua constituição e o efeito que essas substâncias podem causar. No Brasil elas
são representadas por apenas uma espécie, a Crotalus durissus, e classificadas nas
subespécies Crotalus durissus terrificus, Crotalus durissus collilineatus, Crotalus
durissus cascavella, Crotalus durissus ruruima, Crotalus durissus marajoensis,
observadas de forma específica em diferentes localidades do país (PINHO & PEREIRA,
2001). Alguns autores reconhecem uma sexta subespécie no território brasileiro, a
Crotalus durissus trigonicus, encontrada em algumas regiões de Roraima e na savana de
Rupununi na Guiana (CAMILLO, 1998; AUTO, 1999; RATELSLANGEN, 2010).
O veneno de diversas espécies exercem efeitos em quase todas as células e
tecidos do organismo humano e suas propriedades farmacológicas podem ser
determinadas de acordo com a quantidade de constituintes específicos e biologicamente
ativos, presentes nessas substâncias. A natureza proteinácea desses venenos foi
estabelecida em 1843 por Lucien Bonaparte (BON, 1997). Atualmente considera-se que
cerca de 90 a 95 % do peso seco dos venenos ofídicos têm propriedade protéica, e são
estas proteínas as responsáveis por quase a totalidade dos efeitos biológicos
encontrados.
A peçonha da Crotalus apresenta-se como um complexo tóxico-enzimático,
onde são encontradas as enzimas fosfodiesterase, L-amino-oxidase, 5-nucleotidase e
toxinas como a crotoxina, convulxina, crotamina e giroxina (RANGEL-SANTOS et al.,
2004).
Estudos realizados por Soares et al (2010) e Tamieti et al. (2007) testaram o
veneno da espécie Crotalus durissus em linhagens tumorais, verificando sua ação
citotóxica e atividade antitumoral. As neoplasias malignas expressam características
anormais devido a padrões alterados de expressão gênica nas células cancerosas em
decorrência de alterações genéticas, Da Silva e colaboradores (2002) observaram a ação
do veneno da Crotalus durissus terrificus e da Bothrops jararaca diretamente sobre as
células do tumor de Ehrlich (Adenocacinoma mamário).

17
Grandes avanços nas pesquisas com toxinas e enzimas ofídicas desenvolveram-
se nos últimos trinta anos com o advento das análises da estrutura bioquímica,
principalmente dos venenos elapídicos, em consequência de sua maior letalidade.
O veneno da espécie Crotalus durissus tem demonstrado atividade tanto em
linhagens tumorais, como em atividade antitumoral (in vitro e in vivo), sendo testado em
sua forma original, veneno bruto, bem como em suas frações isoladas (SOARES et al.,
2010). A crotoxina está presente como principal componente do veneno do gênero
Crotalus, sendo necessário ressaltar que de acordo com as subespécies, contidas neste
gênero, existem diferenças no potencial de atividade do veneno. De acordo com Rangel
e colaboradores (2004) mesmo com um perfil eletroforético semelhante nas subespécies
Crotalus durissus cascavella, Crotalus durissus collilineatus e Crotalus durissus
terrificus, existem diferenças em suas atividades, talvez relacionadas à existência de
isoformas dessa proteína.
Em vista disso, torna-se relevante estudar os efeitos citotóxicos do veneno da
subespécie Crotalus durissus cascavella, frente às linhagens tumorais direcionando-se a
entender seus efeitos biológicos e mecanismo de ação.

18
2. REVISÃO DE LITERATURA
2.1. Aspectos Gerais e Epidemiológicos do Gênero Crotalus
As serpentes possuem grande importância tanto na cadeia ecológica, como na
área biomédica. Pesquisas realizadas com os venenos por elas produzidos têm se
intensificado, uma vez que suas frações enzimáticas-protéicas vêm sendo usadas na
fabricação de medicamentos. Destacando a anatomofisiologia da família Viperidae,
estas serpentes apresentam grande especialização no crânio, o que justifica a extrema
mobilidade da maxila e outros ossos da cabeça, conformação que favorece a efetividade
nos ataques desses animais e possibilita a inoculação do veneno durante seus botes . O
veneno dessas serpentes é produzido por duas glândulas especiais localizadas na cabeça,
atrás e abaixo dos olhos, que são glândulas salivares modificadas, onde a saliva é a
própria toxina. Elas possuem reservas para vários botes seguidos, sendo que, uma vez
totalmente extraído o veneno ou secreção, as glândulas só voltarão a estar totalmente
cheias após duas semanas (SANTOS, 1995).
As serpentes do gênero Crotalus tem a seguinte distribuição e são classificadas
nas seguintes subespécies: Crotalus durissus terrificus, encontrada nas regiões sul
oriental e meridional; Crotalus durissus collilineatus, distribuídas nas regiões secas do
Centro-Oeste, em Minas Gerais e no norte de São Paulo; Crotalus durissus cascavella
(Figura 01), encontrada nas áreas de caatinga da região Nordeste; Crotalus durissus
ruruima, observada na região Norte; Crotalus durissus marajoensis, observada na Ilha
de Marajó; e a Crotalus durissus trigonicus, encontrada em algumas regiões de Roraima
(PINHO; PEREIRA, 2001; RATELSLANGEN, 2010).
O veneno crotálico apresenta-se como um complexo tóxico-enzimático de várias
substâncias, dentre as enzimas podemos destacar fosfodiesterase, L-amino oxidase, 5-
nucleotidase e o fracionamento destas revela as seguintes toxinas: crotoxina, crotamina,
giroxina, convulxina e proteinases séricas, como crotalocitina, trombocitina e enzimas
trombina-like (identificadas como proteases séricas são capazes de degradar cadeias de
fibrinogênio, induzindo a agregação plaquetária). (RANGEL-SANTOS et al., 2004;
FONSECA, et al., 2006; SPINOSA, 2008)

19
Figura 01: Imagem ilustrativa da serpente Crotalus durissus cascavella. Fonte: http://www.biologados.com.br/especiais/instituto_butantan_turismo 2.2. Constituintes do veneno da espécie Crotalus durissus
2.2.1. Crotoxina e suas subunidades
A Crotoxina principal polipeptídeo presente no veneno crotálico foi isolada
inicialmente por Silva (1982) na subespécie Crotalus durissus terrificus, este autor
destacou que este componente correspondia a 68% do peso total do veneno. De Acordo
com Hendon (1971) a crotoxina é um β-neurotoxina que tem duas subunidades, uma
ácida, a crotapotina (crotoxina A) e outra básica, a fosfolipase A2 (crotoxina B).
A crotapotina tem sua acidez justificada pela composição de aminoácidos: 12
resíduos de aspartato/asparagina e 14 de glutamato/glutamina versus 2 resíduos de lisina
e 2 de arginina. A fosfolipase A2 crotálica contém 11 resíduos de aspartato/asparagina e
10 de glutamato/glutamina versus 11 resíduos de lisina e 12 de arginina, caracterizando
seu caráter básico. A crotapotina e a fosfolipase A2 são ricas em cistina, contendo 7 e 8
pontes dissulfídicas respectivamente. As duas subunidades formam a crotoxina, um
complexo molar de proporção 1:1 (BREITHAUPT et al., 1974; SPINOSA, 2008).
Variações interespecíficas e intraespecíficas foram descritas para o complexo
crotoxina. As variações interespecíficas estão relacionadas com diferenças desse
componente nas diversas subespécies das serpentes e as variações intraespecíficas
diferem na combinação das isoformas das subunidades A e B, as quais possuem
potência e atividade biológica diferentes em uma subespécie específica (PEREAÑEZ et
al., 2009).

20
A crotapotina (crotoxina A) não possui a toxicidade e a atividade enzimática da
crotoxina, mas potencializa a sua toxicidade. Essa potencialização é explicada pelo fato
de que, quando associado ao componente B (PLA2), o componente A (crotapotina)
previne a ligação da PLA2 a sítios de baixas especificidades e afinidade, direcionando a
PLA2 a se ligar a sítios de receptores de membrana (pós-sináptica) de alta
especificidade e alta afinidade, o que potencializa a toxicidade da crotoxina. Esses sítios
de alta especificidade podem ser receptores colinérgicos ou componentes da membrana
fortemente associados a estes (BON et al., 1979). Esses receptores foram denominados
como tipo N (neuronal) e M (muscular). Os receptores do tipo N foram os primeiros a
serem identificados e estão presentes em grande quantidade nas membranas de cérebro
de ratos. Tais receptores ligam-se com alta afinidade às PLA2s neurotóxicas. Os
receptores do tipo M foram identificados em músculo esquelético de coelhos
(MATSUBARA, 2009).
A crotapotina demonstrou outros efeitos como, potencializar edema inflamatório
(induzido por carragenina), possuir efeito antimicrobiano contra, principalmente,
bactérias Gram-negativas e causar alterações renais, tais como, aumentar a pressão de
perfusão renal e a resistência vascular renal em rins perfundidos sem, entretanto, alterar
o fluxo urinário e a filtração glomerular, ao contrário da crotoxina (OLIVEIRA, et al.,
2003).
O veneno da Crotalus durissus cascavella contém pelo menos 4 isoformas de
crotoxinas formadas por diferentes combinações de crotapotinas com uma única
isoforma de PLA2. Ao contrário do veneno de Crotalus durissus terrificus, que contém
3 ou 4 isoformas de PLA2 (BEGHINI et al., 2000).
Os dados supracitados podem servir como indicio para sugerir as diferenças na
intensidade de atuação do veneno das diferentes subespécies.
A crotoxina é uma β-neurotoxina que possui um efeito tóxico bloqueador pós-
sináptico, diminuindo a ação da acetilcolina e impedindo a despolarização da
membrana, e pré-sináptico da liberação de acetilcolina, ou seja, ela bloqueia a
transmissão neuromuscular, impedindo a contração muscular estriada esquelética. Ela
pode causar uma paralisia dos músculos respiratórios, culminando na morte do
envenenado. (BON et al., 1979; GOPALAKRISHNAKONE et al., 1984; RANGEL-
SANTOS et al., 2004; SPINOSA, 2008; MATSUBARA, 2009). O veneno da Crotalus

21
durrissus terrificus e a crotoxina B induzem a expressão protéica de cicloxigenase-2
pelas células endoteliais (MATSUBARA, 2009).
A superfamília de PLA2 consiste de um amplo espectro de enzimas definidas
pela sua habilidade de catalisar a hidrólise da ligação do centro estérico (posição sn-2)
de fosfolipídios (SIX; DENNIS, 2000). Elas conferem à crotoxina uma ação miotóxica
por atuarem clivando fosfolipídios em sítios específicos de membranas de fibras
musculares estriadas esqueléticas sensíveis à PLA2, causando mionecrose, com perda
de estriação das fibras e degeneração hialina (4-6 horas após a injeção da crotoxina)
(GOPALAKRISHNAKONE et al., 1984).
As PLA2s induzem a um extravasamento de plasma, sendo capazes de aumentar
a permeabilidade microvascular da pele na região dorsal de ratos. Os mecanismos de
formação de edema induzido pelas PLA2s envolvem a ativação de fibras sensoriais C,
as quais liberam neuropeptídeos, como a substância P, que medeiam efeitos
neurogênicos inflamatórios locais (respostas vasoativas) em tecidos inervados e de
mastócitos locais (pela substância P), os quais liberam histamina e serotonina
(CÂMARA et al., 2003).
Fonseca et al. (2006) afirmaram que a crotoxina apresenta duas diferentes
atividades enzimáticas (PLA2 e da protease de serina) e dois tipos diferentes de ações
biológicas (neurotóxica e coagulante).
2.2.2. Crotamina
A crotamina é um polipeptídeo básico, pertencente à família das miotoxinas
polipetitídicas básicas, as quais possuem a capacidade de penetração intracelular,
através de mecanismos independentes de gasto energético, por interação com
proteoglicanos (MATSUBARA, 2009).
O efeito miotóxico também é causado pela crotamina, esta fração apresenta
atividade sinérgica à da crotoxina, ela atua causando principalmente contrações
musculares, dependentes da despolarização da membrana das células musculares
estriadas esqueléticas. Esta ação provavelmente é exercida sobre os canais de sódio,
pela indução do influxo de cálcio (LOMONTE et al., 2003). Também foi observado que
a crotamina interage ativamente com as membranas lipídicas das células, formando
vacúolos e demonstrando uma atividade mionecrótica.

22
2.2.3. Convulxina
A convulxina é uma toxina de alto peso molecular, pertencente à família das
lectinas do tipo C, formada por duas cadeias polipeptídicas (Cvxα e Cvxβ)
covalentemente associadas em uma estrutura trimérica (αβ)3. Esta fração já foi isolada
das subespécies Crotalus durissus terrificus, Crotalus durissus cascavella e Crotalus
durissus collilineatus e consiste em um potente ativador plaquetário. A indução
plaquetária provocada por esta toxina pode está relacionada com uma reação
dependente de Ca+, iniciada pela ligação da convulxina à glicoproteína Ib (GPIb)
(JANDROT-PERRUS, 1997; POLGAR et al., 1997; KANAJI et al., 2003). Matsubara
(2009) também descreveu que esta toxina é capaz de agregar e lisar plaquetas, ligando-
se com alta afinidade a um pequeno número de sítios plaquetários, por mecanismo
dependente de cálcio, fibrinogênio e adenosina difosfato. De acordo com Lima et al.
(2005) além de induzir a agregação plaquetária ao ligar-se ao receptor GPV1 das
plaquetas, ela causa convulsões, alterações respiratórias e circulatórias.
Leduc e Bon (1998) clonaram e sequenciaram as duas cadeias da convulxina de
Crotalus durissus terrificus. Neste estudo, observou-se uma alta similaridade entre as
cadeias α e β, com uma média de porcentagem de identidade de 74%. O
sequenciamento das subunidades resultou em cadeias compreendendo 158 e 148
resíduos de aminoácidos para Cvxα e Cvxβ, respectivamente e um peptídeo sinal de 23
resíduos de aminoácidos idênticos para as duas subunidades. Toyama e colaboradores
(2001) isolaram e caracterizaram uma proteína semelhante à convulxina da peçonha de
Crotalus durissus collilineatus com alta similaridade em relação à convulxina da
Crotalus durissus terrificus.
2.2.4. Giroxina
A giroxina pertence ao grupo do gênero trombina e foi isolado a partir do veneno
da serpente Crotalus durissus terrificus por Raw em 1986, por meio de três passos
cromatográficos de purificação (precipitação com sulfato de amônio, cromatografia em
Sephadex G-75 e Sepharose-1, 4-butanodiol diglycyl-p-aminobenzamidina). Após
análise eletroforética decobriu-se que a giroxina possuia um peso molecular de 34 kDa e

23
um pH 8,0, ótimo para a coagulação do sangue humano (RAW et al., 1986). A giroxina
converte o fibrinogênio em fibrina e assim aumenta o tempo de coagulação do sangue
(SANO-MARTINS et al., 2001). Além de atuar na coagulação, a giroxina apresenta
uma excelente atividade neurotóxica (BARROS et al., 2011). Fonseca e colabradores
(2006) descreveram a giroxina apresentando duas frações principais: atividade de
trombina-like e L-amino-oxidase (LAO). A LAO é a única oxidase dependente de FAD
(Flavina adenina dinucleotídeo) encontrada em veneno de serpentes e sua toxicidade,
possivelmente, envolve a geração de peróxido de hidrogênio, produto final da oxidação
de L-aminoácidos. A LAO também induz a agregação plaquetária, envolvendo a
ativação da PLA2 (TOYAMA et al., 2006). Em estudos mais recentes Barros et al.
(2011) verificaram que a giroxina demonstrou um elevado nível de atividade
coagulante, com uma dose mínima entre 0,015-0,037µg/µL. Outras atividades da
giroxina são a amidásica e a esterásica. (MARTINS et al., 2002)
Prado-Franceschi (1990) sugere que esta toxina produz uma síndrome
convulsiva peculiar em camundongos, caracterizada por movimentos rápidos de rotação
do corpo em torno de seu eixo longitudinal. Estudos posteriores confirmaram sua ação
no sistema nervoso central, causando a síndrome labiríntica em camundongos,
corroborando os achados anteriores (MATSUBARA, 2009). O estudo desta enzima
poderá servir como um modelo molecular interessante para o desenvolvimento de novos
fármacos ou agentes terapêuticos (MORAES et al., 2004; FERRARO et al., 2005),
principalmente devido a sua resistência aos inibidores de proteinases fisiológicas
(MATSUI et al., 2000; SERRANO et al., 2005).
2.2.5. Peptídio natriurético
Outro componente importante do veneno é o peptídio natriurético que ao ser
isolado da Crotalus durissus cascavella foi capaz de aumentar a pressão de perfusão e a
resistência vascular renal em rins perfundidos de ratos. Este componente demonstrou ser
capaz de aumentar o fluxo urinário e a filtração tubular renal e diminuir o transporte
tubular de sódio e a pressão arterial sistêmica. EVANGELISTA et al. (2008)
observaram que este componente do veneno crotálico apresentou efeitos renais e
vasculares, com consequente efeito diurético e hipotensor.

24
2.3. Estudo da atividade citotóxica do veneno do Gênero Crotalus e suas
frações
A diversidade de toxinas e enzimas que constituem os venenos de serpentes
pode conferir a estas substâncias a habilidade de interagir com múltiplas integrinas,
podendo resultar na inibição da agregação celular, inibição da angiogênese, bem como
indução da morte das células tumorais, seja por vias apoptóticas ou necróticas. Russel
(1983) em um trabalho de revisão mostra os efeitos de toxinas de veneno de serpentes
que se ligam a integrinas e provocam efeitos antiangiogênicos e antitumorais.
Células de mamíferos em cultura são importantes ferramentas para avaliar a
atividade citotóxica de substâncias com potencial terapêutico (PARKIN et al., 2001)
Diversos estudos vêm sendo realizados testando o veneno da espécie Crotalus
durissus em linhagens tumorais, testando seu efeito citotóxico, bem como sua atividade
antitumoral. Em 2010, Soares et al. verificaram em seus experimentos que células das
linhagens de glioblastoma (RT2) e adenoma benigno de pituitária (GH3) quando
expostas ao veneno da serpente Crotalus durissus terrificus apresentaram alterações
morfológicas como irregularidade de membrana celular, encolhimento celular,
condensação nuclear, fragmentação de DNA e formação de corpos apoptóticos,
características típicas de apoptose,.
Em um outro estudo realizado por Tamieti et al. (2007) o veneno da cascavel
induziu apoptose em células tumorais de carcinoma de ováriano de hamster (CHO-K1).
Yan e colaboradores (2006) testaram a crotoxina, principal fração do veneno
crotálico e demonstram a ocorrência de autofagia e apoptose celular em linhagem de
leucemia mieloide crônica (K562), a toxina diminuiu a viabilidade das células,
estimulando a autofagia, e também foi constatado alterações da membrana mitocondrial
com a liberação de citocromo-c e ativação da caspase-3. No ano de 2007 os mesmos
autores repetiram os estudos em outra linhagem celular (MCF-7) de câncer mamário
humano, deficientes em caspase-3. Desta vez, o estimulo da autofagia ainda foi
observado, assim como a apoptose pela condensação e fragmentação nuclear. A
viabilidade celular foi inibida de maneira dose-dependente de acordo com o tempo de
exposição.
No ensaio realizado por Soares et al. (2010) os testes citotóxicos com a principal
fração do veneno, a crotoxina, demonstrou que as linhagens RT2 e GH3, apresentaram

25
alterações morfológicas e degradação de DNA, só que em um menor intensidade. O
efeito citotóxico da crotoxina, neste caso, não foi tão intenso como quando comparado
ao veneno total, o que levou os autores a acreditarem que a crotoxina pode não ser o
único componente envolvido na citotoxicidade do veneno para estas linhagens.
Outro componente que vem ganhando destaque nas pesquisas é a PLA2
crotálica. Estudos foram realizados para verificar seus efeitos em varios tipos celulares,
como em macrófagos, linfócitos, plaquetas e células neoplásicas (NEWMAN et al.,
1993, DONATO et al., 1996, SAMPAIO et al. 2005, YAN et al., 2006 e 2007).
Fonseca et al. (2006) testaram tanto a crotoxina como suas subunidades, PLA2 e
crotapotina, em plasma humano rico em plaquetas e puderam verificar que na
concentração de 20µL a crotoxina demonstrou potente ação para agregação plaquetária,
enquanto que suas principais subunidades não foram capazes de apresentar o mesmo
potencial ao serem testadas isoladamente. A interação dos dois componentes pode ser
um possível indício para este evento ter ocorrido, visto que estudos anteriores relataram
que a crotapotina pode agir potencializando a atividade da PLA2.
Newman et al. (1993) verificaram uma eficácia na ação antitumoral in vivo,
usando uma administração diária de Crotoxina, demonstrando inibição no crescimento
em 83% no carcinoma de pulmão de Lewis e uma inibição de 69% em carcinoma
mamário humano (MX-1). Uma menor atividade foi observada em células de leucemia
(HL-60), inibindo o crescimento em apenas 44%, o que sugere que a Crotoxina pode
ter, certa, especificidade para tumores sólidos.
Estudos de citotoxicidade de toxinas do veneno da cascavel têm fornecido dados
promissores para uma futura utilização destas no tratamento de câncer. Como exemplo
pode-se citar o produto natural, VRCTC-310-ONCO, resultante da combinação de duas
toxinas da Crotalus durissus terrificus, que se encontra em fase de investigação clínica
como um potente agente antitumorigênico (STANCHI et al., 2002).
Estudos toxicológicos demonstraram que a administração da concentração
antitumoral do veneno da Crotalus durissus terrificus, calculada in vitro (0,016 gμ/mm2
de massa tumoral), em um tumor induzido, não provocou efeitos tóxicos agudos e
nenhuma alteração funcional ou histopatológica nos animais tratados (SOARES et al.,
2010).
Apesar da enorme variabilidade de cânceres, evidências demonstraram que a
resistência à apoptose é uma das características mais marcantes da maioria dos tumores

26
malignos (OKADA et al. 2004), o que torna ainda mais importante estudos com estas
substâncias, visto que dados revelaram essa possível interferência sobre as vias
apoptóticas ou mesmo quando agem ativando outras vias de morte celular, seja por
meio da autofagia ou mesmo necrose, como já citado anteriormente.
Diversos estudos vêm sendo realizados com o veneno da espécie Crotalus
durissus em linhagens tumorais, in vitro e na atividade antitumoral, in vivo,
principalmente com a fração crotoxina. Em vista disso, é relevante estudar os efeitos
citotóxicos do veneno da subespécie Crotalus durissus cascavella, serpente encontrada
nas áreas de caatinga da região Nordeste do Brasil, frente aos efeitos citotóxicos em
linhagens tumorais.
2.4. Morte celular
O desenvolvimento e a manutenção dos organismos dependem de uma interação
entre as células que o compõe. Por exemplo, no desenvolvimento embrionário, muitas
células produzidas em excesso são levadas à morte, contribuindo para a formação dos
órgãos e tecidos (MEIER et al., 2000).
Durante muito tempo, a morte celular foi considerada um processo passivo de
caráter degenerativo, ocorrendo em casos de infecções, lesões celulares e ausência de
fatores de crescimento e como consequência, a célula altera a integridade da membrana
plasmática, aumentando o seu volume e perde suas funções metabólicas (YU et al.,
2000).
Entretanto, nem todos os eventos de morte celular são processos passivos.
Organismos multicelulares são capazes de induzir a morte celular programada como
resposta a estímulos intracelulares ou extracelulares (HENGARTNER, 2002).
Esses processos de morte celular podem ser classificados de acordo com suas
características morfológicas e bioquímicas em: necrose, apoptose e autofagia
(CASTEDO et al., 2004; OKADA, 2004). Alterações na coordenação desses tipos de
morte celular estão implicadas na tumorigênese (RICCI, 2006).

27
2.4.1. Apoptose
A apoptose é um tipo de morte celular regulada de forma ordenada por uma série
de eventos em cascatas de sinalização; É um mecanismo que desempenha papel
essencial na regulação do crescimento, desenvolvimento, manutenção da quantidade de
células ou atividade sobre células anormais nos organismos; Promove uma eliminção
seletiva de células danificadas, com o intuito de manter a integridade do organismo e a
manutenção da vida; É um processo que requer gasto de energia e tem por característica
morfológica principal a diminuição do volume citoplasmático, condensação da
cromatina, degradação do DNA e formação de corpos apoptóticos (REED, 2000;
DAMIANI, 2004; DANIAL e KORSMEYER, 2004; RELLO et al., 2005) (Figura 2).
Figura 2: Esquema da fisiologia de morte celular por apoptose, imagem adaptada. Fonte: http://www.utm.utoronto.ca/~w3cellan/apoptosis.html

28
A figura 3 demonstra uma alteração marcante presente em células com
característica de apoptose, que é a formação dos corpos apoptóticos.
Figura 3: Fotomicrografia de adenocarcinoma ovariano. Evidenciando formação dos
corpos apoptóticos (setas), H.E, aumento de 400x, Motic BA310, Moticam 2000. Fonte: Arquivo pessoal, 2012.
Diferente da morte por necrose, que tem por característica provocar resposta
inflamatória potencialmente prejudicial aos tecidos, a apoptose causa alterações na
superfície da célula que promovem a fagocitose por macrófagos, antes de ocorrer a
liberação do seu conteúdo citoplasmático sem causar inflamação (ZIEGLER et al.,
2004).
Genes que controlam o ciclo celular, como P53, c-MYC e RB, estão envolvidos
no processo de apoptose, bem como, proteínas da família BCL-2 (Bcl-2, Bax, Bad, Bid)
também desempenham papel importante no processo de morte celular programada
(RICCI et al., 2006).
A morte por apoptose pode ser regulada por duas vias: via de receptores de
morte ou extrínseca e via mitocondrial ou intrínseca (FAN et al., 2005). Estas vias
ativam caspases iniciadoras ou pró-caspases e ambas convergem para ativação das
caspases efetoras. A indução de apoptose e execução requer a cooperação de uma série
de moléculas, incluindo sinal molecular, receptores, enzimas e os genes reguladores.
Entre eles, podemos destacar outros constituintes reguladores epecíficos da cascata, tais
como o inibidor da proteína de apoptose (IAP), Bcl-2 família de proteínas e calpaína, é

29
essencial no processo de apoptose (LAUNAY et al., 2005) As caspases iniciadoras
correspondem as caspase-8 e caspase-9, estas ativam caspases efetoras pela clivagem
proteolítica de pró-caspase, como exemplo a caspase-3, após ativação pelas caspases
iniciadoras, agem sobre substratos específicos, como inibidores de apoptose (Bcl-2),
proteínas regulatórias do citoesqueleto, moléculas que mantém estoques de ATP (PARP
– Polimerase poli ADP- ribose), fator de fragmentação de DNA (DFF) entre outros,
como já descrito anteriormente. (STRASSER et al., 2000).
A via intrínseca é caracterizada por falta de fatores de crescimento para
sobrevivência das células no ambiente em que se encontram. A ausência desses fatores
levaria a ativação da via mitocondrial de várias moléculas, tais como: Bcl-2, Bid
caspase 9 e por último a caspase 3. Já a via extrínseca a apoptose é induzida via Fas e
ativa caspase 8, que por sua vez ativa caspase 3, sem que ocorra dano na mitocôndria
(LOGUE e MARTIN, 2008).
A apoptose também pode ser induzida por enzimas citosólicas como as
fosfolipases A2 (cFLA2) (ATSUMI et al., 2000). A FLA2 é uma classe diversa de
enzimas que catalisam a hidrólise de fosfolipídios e ácidos graxos da membrana
plasmática, desempenhando papel importante em vários eventos biológicos
(HIRABAYASHI, 2004). A hidrólise de fosfolipídios pela FLA2 gera ácido
araquidônico (AA) e lisofosfolipídios. São os ácidos araquidônicos que, mediados por
enzimas cicloxigenases e lipoxigenases, geram as ROS (espécies reativas de oxigênio),
as quais causam o rompimento da membrana mitocondrial, permitindo a liberação de
citocromo c e, consequentemente, induzindo a apoptose. O ácido araquidônico também
tem sido indicado como mediador da formação de ceramidas através da
esfingomielinase (ZHAO et al., 2002). Defeitos na via apoptótica são bastante comuns
no processo de formação de tumores, pois permite que células danificadas continuem
proliferando. Além disso, a apoptose desregulada afeta a quimio e radio resitências,
aumentando o limiar para que a morte celular seja inicializada.
Mesmo o câncer exibindo características celulares heterogêneas, os tumores
malignos adquiriram a propriedade de crescer além dos limites considerados para um
tipo celular normal. A expansão clonal de uma célula transformada depende de um
descontrole da sua capacidade proliferativa e de uma crescente incapacidade de morrer
por apoptose. Apesar da enorme variabilidade do câncer, evidências demonstram que a

30
resistência à apoptose é uma das características mais marcantes da maioria dos tumores
malignos (OKADA E MAK, 2004).
2.4.2. Necrose
A necrose refere-se a um espectro de alterações morfológicas que se seguem à
morte celular, ela é resultante em grande parte, da ação progressiva de enzimas sobre a
célula, letalmente lesada, que se caracteriza pela morte celular causada por injúrias no
tecido, constituindo-se num processo não mediado por ATP, levando a ruptura da célula
e gerando inflamação tecidual (FADEEL; ORRENIUS, 2005).
As características mais comuns da necrose são: ruptura da membrana
citoplasmática e extravasamento de material antigênico, o que induz à reação
inflamatória, intensificando assim o dano tecidual. O padrão morfológico é decorrente
do resultado de dois processos: digestão enzimática que se traduz microscopicamente
como uma necrose de liquefação e a desnaturação de proteínas que origina a necrose de
coagulação. Na necrose de coagulação os contornos celulares são preservados por
algum tempo enquanto que na necrose de liquefação a célula é completamente digerida
(ZIEGLER; GROSCURTH, 2004).
As alterações nucleares podem apresentar-se sob três padrões: a basofilia da
cromatina pode diminuir (cariólise), uma alteração que supostamente reflete na
atividade da DNAse; a picnose, caracterizada pela retração nuclear e aumento da
basofilia. Neste caso o DNA tem uma aparência condensada, como uma massa sólida,
basofílica. Por fim um padrão, conhecido como cariorrexe, o núcleo picnótico ou
parcialmente picnótico sofre fragmentação (COTRANS et al., 2000; BOHM, SCHILD,
2003) (Figura 4).
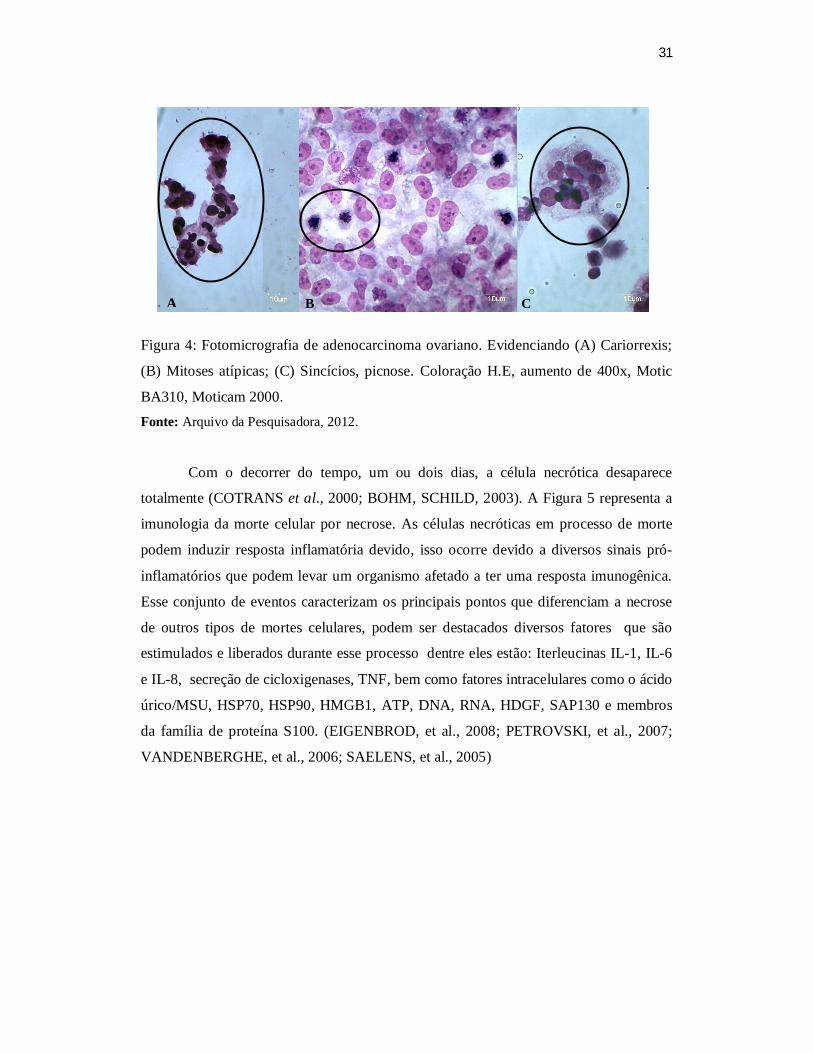
31
Figura 4: Fotomicrografia de adenocarcinoma ovariano. Evidenciando (A) Cariorrexis;
(B) Mitoses atípicas; (C) Sincícios, picnose. Coloração H.E, aumento de 400x, Motic
BA310, Moticam 2000. Fonte: Arquivo da Pesquisadora, 2012.
Com o decorrer do tempo, um ou dois dias, a célula necrótica desaparece
totalmente (COTRANS et al., 2000; BOHM, SCHILD, 2003). A Figura 5 representa a
imunologia da morte celular por necrose. As células necróticas em processo de morte
podem induzir resposta inflamatória devido, isso ocorre devido a diversos sinais pró-
inflamatórios que podem levar um organismo afetado a ter uma resposta imunogênica.
Esse conjunto de eventos caracterizam os principais pontos que diferenciam a necrose
de outros tipos de mortes celulares, podem ser destacados diversos fatores que são
estimulados e liberados durante esse processo dentre eles estão: Iterleucinas IL-1, IL-6
e IL-8, secreção de cicloxigenases, TNF, bem como fatores intracelulares como o ácido
úrico/MSU, HSP70, HSP90, HMGB1, ATP, DNA, RNA, HDGF, SAP130 e membros
da família de proteína S100. (EIGENBROD, et al., 2008; PETROVSKI, et al., 2007;
VANDENBERGHE, et al., 2006; SAELENS, et al., 2005)
A B C

32
Figura 5: Esquema ilustrativo da imunologia da morte celular por necrose. Células
necróticas liberaram organelas intracelulares, bem como fatores intracelulares como o
ácido úrico/MSU, HSP70, HSP90, HMGB1, ATP, DNA, RNA, HDGF, SAP130 e
membros da família de proteína S100. Além disso, as células necróticas secretam IL-1α
e IL-6 de modo passivo e ativo, respectivamente. Imagem adaptada de Garg et al., 2010.
2.4.3. Autofagia
A autofagia é um processo adaptativo conservado evolutivamente e controlado
geneticamente. Ela ocorre em resposta a um estresse metabólico que resulta na
degradação de componentes celulares (DANIAL, 2004; LUM, 2005). Durante a
autofagia, porções do citoplasma são encapsuladas por membrana, originando estruturas
denominadas autofagossomos (Figura 06). Estes irão se fusionar com os lisossomos e
posteriormente o conteúdo dos autofagossomos será degradado pelas hidrolases
lisossomais (KELEKAR, 2005).

33
Figura 6: Diagrama representativo dos produtos da autofagia e metabolismo de célula
cancerígena. Imagem adaptada de Lozy & Karantza, 2012.
2.5. A importância do estudo sobre o câncer
A importância do câncer na área de Saúde Pública ganha destaque à medida que
ocorre o controle de outras doenças, fazendo com que haja um interesse especial em
formas de controle para uma enfermidade que aflige a humanidade. O avanço da ciência
e da tecnologia tem possibilitado a melhoria dos meios de diagnóstico e de tratamento,
contribuindo para um declínio das taxas de mortalidade por enfermidades controláveis,
tais como a tuberculose, a desnutrição, o diabetes mellitus e outras afecções. A
urbanização, a industrialização e a maior expectativa de vida da população são os
principais fatores que contribuem para o aumento da incidência das doenças crônico
degenerativas, dentre elas, o câncer.
O câncer é um termo genérico para um amplo grupo de doenças que podem
afetar qualquer parte do corpo. Outros termos utilizados são tumores e neoplasias
malignas. Uma característica marcante do câncer é a formação rápida de células

34
anormais que podem invadir tecidos próximos, bem como se disseminar para outros
tecidos e órgãos (OMS, 2012). As neoplasias podem diferir de acordo com o tipo de
célula, tecido ou órgão em que se desenvolvem. Geralmente as células neoplásicas
levam a formação de uma massa tumoral, podendo esta ser benigna ou maligna (INCA,
2008).
A presença de células em crescimento contínuo, com propriedades de invasão e
destruição do tecido adjacente, bem como, de crescimento em outros sítios diferentes do
tumor primário (metastização), pode determinar a severidade da doença.
Essas propriedades normalmente são ocasionadas por um acúmulo de mutações
nos oncogenes, nos genes supressores e nos genes reparadores de DNA, o que
caracteriza o câncer como doença genética. (FOSTER, 2008)
Os oncogenes promovem a proliferação celular de forma ordenada, enquanto os
genes supressores mantêm essa proliferação sob controle, restringindo assim o
crescimento celular. Os genes reparadores de danos do DNA têm por função refazer as
moléculas que sofrem mutação. Quando ocorre o mau funcionamento dos mecanismos
de regulação do ciclo celular, permitindo a passagem das células mutadas pelo ciclo, o
acumulo dessas mutações, acaba por contribuir no surgimento das características do
tumor maligno (VERMEULEN; VAN BOCKSTAELE; BERNEMAN, 2003).
Para o estabelecimento de medidas efetivas de controle do câncer fazem-se
necessárias informações de qualidade sobre sua distribuição de incidência e
mortalidade, o que possibilita melhor compreensão sobre a doença e seus determinantes;
avaliação dos avanços tecnológicos aplicados à prevenção e tratamento, bem como a
efetividade na atenção à saúde, perspectiva descrita pelo Instituto Nacional do Câncer -
INCA. Ao ser considerado um problema de saúde pública mundial faz-se necessário
estudos aprofundados acerca desta patologia. A Organização Mundial da Saúde (OMS)
estimou que, para o ano 2030, podem-se esperar 27 milhões de novos casos de câncer,
17 milhões de mortes por câncer e 75 milhões de pessoas vivas, anualmente, com
câncer. (OMS, 2012)
No Brasil o problema do câncer destaca-se devido à forma com que seu perfil
epidemiológico vem se apresentando, e, com isso, o tema vem conquistando espaço nas
prioridades do governo federal, que investe em centros específicos direcionados a
tratamentos e apoio a pacientes acometidos por esta patologia. O conhecimento sobre a
situação dessa doença permite estabelecer prioridades e alocar recursos de forma

35
direcionada para a modificação positiva desse cenário. As estimativas para o ano de
2012 serão válidas também para o ano de 2013 e apontam a ocorrência de
aproximadamente 518.510 novos casos de câncer, incluindo os casos de pele não
melanoma, o que reforça a magnitude do problema do câncer no país. (INCA, 2012)
O câncer no Brasil é destacado como a segunda causa de morte em números
absolutos, subsequentes às doenças cardiovasculares, quando não são considerados os
óbitos por causas externas. Os tumores malignos são responsáveis por um número
expressivo e crescente de pacientes em todo o mundo, também representando a segunda
causa de morte da população mundial. (SRIVASTAVA et al., 2005) A análise da
tendência temporal mostra um aumento significativo de sua participação percentual
dentre todos os óbitos durante as últimas décadas, fazendo com que as neoplasias
malignas ocupem, progressivamente, maior dimensão em termos de mortalidade.
O câncer afeta, portanto, a população de um modo geral, incluindo pessoas de
ambos os sexos e de todas as idades.
Nas tabelas 1 e 2, estão descritas as principais localizações primárias de
neoplasias no organismo, bem como a distribuição proporcional dos dez tipos de câncer
mais incidentes estimados para 2012 por sexo, no Brasil.

36
Tabela 1: Estimativas para o ano de 2012 das taxas brutas de incidência por 100 mil
habitantes e de número de casos novos por câncer, segundo sexo e localização primária*
no Brasil.
Tabela 2: Distribuição proporcional dos dez tipos de câncer mais incidentes estimados
para 2012 por sexo, exceto pele não melanoma* no Brasil.
*Números arredondados para 10 ou múltiplos de 10
Fonte: INCA, 2011.
*Números arredondados para 10 ou múltiplos de 10. Fonte: INCA, 2011.

37
A terapêutica do câncer baseia-se, de forma geral, na associação de
procedimentos cirúrgicos, tratamento radioterápico e quimioterapia desses tumores. Ao
longo dos anos as formas de abordagem para o tratamento sofreram alterações
significativas após o inicio da utilização do tratamento adjuvante. Os protocolos pós-
cirúrgicos, juntamente com o uso da quimioterapia com diferentes mecanismos de ação
e a radioterapia, associados a outros utensílios vem melhorando os resultados dos
tratamentos associados a alguns tipos de câncer. Infelizmente, muitos tumores ainda
apresentam respostas modestas aos protocolos clínicos, limitando a indicação e a
eficácia do tratamento adjuvante tanto para os tumores primários quanto para as
metástases (KUMMAR et al., 2004).
Portanto torna-se necessário encontrar e desenvolver modalidades terapêuticas
mais eficientes, oferecendo então aos pacientes com câncer oportunidades reais de
controle da doença.
Pesquisas utilizando produtos naturais vêm aumentando ao longo das décadas,
especialmente estudos relacionados à ação de componentes de plantas, microrganismos,
toxinas ou mesmo substâncias oriundas de animais, como os venenos e mesmo a própria
saliva. O avanço no estudo desses componentes vem interferindo diretamente na área da
oncologia, propiciando a descoberta de diversas substâncias utilizadas na terapêutica
antineoplásica. Existem diversas fontes naturais disponíveis e estas oferecem
possibilidades de encontrar substâncias de grande interesse para medicina. De acordo
com Harvey (2008), mais de uma centena de compostos derivados de produtos naturais
estão em fase de testes clínicos, principalmente para o tratamento do câncer e de
doenças infecciosas. Visto que a morbidade associada aos quimioterápicos ainda é um
obstáculo, a descoberta de novos fármacos antineoplásicos de fácil administração e com
mínimos ou nulos efeitos adversos é de extrema importância.
Várias pesquisas identificaram fármacos com atividade específica contra alguns
mecanismos metabólicos da célula tumoral, bem como fármacos que inibem a
neovascularização tumoral, que induzem a rediferenciação celular ou que levem a célula
tumoral à apoptose. Desta forma, as moléculas e substâncias a serem pesquisadas
precisam ser testadas de forma a favorecer a descoberta de novas ferramentas
terapêuticas.

38
1. JUSTIFICATIVA
O câncer apresenta-se como um problema de saúde publica, seu impacto na
sociedade e o aumento em número de ocorrências preocupa a sociedade brasileira. No
Brasil a doença se torna relevante devido seu perfil epidemiológico, o que requisita cada
vez mais investimentos voltados à descoberta de novas ferramentas terapêuticas que
possam agir de forma mais efetiva e menos agressiva aos pacientes. O conhecimento
sobre a situação atual destas patologias permite aos órgãos responsáveis e grupos de
pesquisas estabelecer prioridades e alocar recursos de forma direcionada para a
modificação positiva desse cenário. O estudo da citotoxicidade sobre linhagens tumorais
utilizando veneno de serpentes vem aumentando de forma significante. O que garante
este aumento são os resultados promissores que vem sendo obtidos por diversos
pesquisadores (ANIM et al., 1997; TAMIETI et al., 2007; GALÁN et al., 2008;
SOARES et al., 2010). Diversas vias de morte celular vêm sendo estudadas tentando
determinar qual a forma de atuação do veneno da espécie Crotalus durissus, em
linhagens tumorais, seja pela apoptose, necrose ou mesmo autofagia (YAN et al., 2007;
SOARES et al., 2010). Estudos sobre o veneno da Crotalus durissus cascavella são
indispensáveis, desta forma, é interessante isolar substâncias constituintes do veneno e
testar sua atividade específica sobre a linhagem OVCAR-8, bem como em outras
linhagens, e assim aplicar testes citotóxicos que favoreçam identificar qual ou em quais
vias de morte celular este veneno pode atuar.

39
2. HIPÓTESE CIENTÍFICA
Diante do exposto formulou-se a seguinte hipótese:
1) O veneno da serpente Crotalus durissus cascavella apresenta potencial
citotóxico sobre diversas linhagens tumorais, em específico, ação em linhagem
de carcinoma ovariano (OVCAR-8).
2) O veneno pode está relacionado com a ativação do mecanismo de apoptose, pela
ativação das caspases, bem como ação via necrose.

40
3. OBJETIVOS
5.1. Objetivo geral
Estudar a atividade citotóxica do veneno da serpente Crotalus durissus
cascavella em linhagens tumorais.
5.2. Objetivos específicos
Estudar a atividade citotóxica do veneno da Crotalus durissus cascavella em
linhagens de células tumorais de OVCAR-8 e SKOV3 (carcinoma ovariano), PC-3M
(carcinoma prostático), MCF-7 (carcinoma mamário), e SF-268 (glioblastoma).
Analisar a integridade de membrana, viabilidade celular e alterações na
morfologia celular.
Determinar a ativação de caspases efetoras (3 e 7), na linhagem tumoral de
adenocarcinoma ovariano (OVCAR-8), após tratamento com veneno da serpente
Crotalus durissus cascavella.

41
6. CAPÍTULO I
Estudo do potencial citotóxico do veneno da Crotalus durissus cascavella em
linhagens tumorais
(Study of the cytotoxic potential of the venom of Crotalus durissus cascavella in tumor
cell lines)
Periódico: Toxicon
(Submetido em, 20 de outubro de 2012)

42
Elsevier Editorial System(tm) for Toxicon Manuscript Draft
Manuscript Number: TOXCON-D-12-00452 Article Type: Research Paper
Cytotoxic effect of Crotalus durissus cascavella venom in tumor cell lines.
L.S. Araújoa, J.S.A.M. Evangelistaa, D.D. Rochab, L.V.C. Lotufob, M.O. Moraesb, D.V.
Wilkeb, D.M.Borges-Nojosa, C. O’Pessoab, D.A. Vianac, N.S.C. Rosasa.
aVeterinary Faculty, State University of Ceará, Fortaleza, Ceará Brazil. bDepartment of Physiology and Pharmacology, Federal University of Ceará. cDepartment of Biology, Federal University of Ceará. dRenorbio, State University of Ceará, Fortaleza, Ceará Brazil.
* Corresponding author. Graduate Program in Veterinary Sciences, Veterinary Faculty,
State University of Ceará, Fortaleza, Ceará Brazil, Av. Dedé Brasil, 1700 – Campus
Itaperi, 60740-903 – Fortaleza, Ceará, Brazil. Tel.: 055-(85)31019889; fax: 055-
(85)31019850. E-mail address: [email protected] (L.S. Araújo),
[email protected] (J.S.A.M. Evangelista).

43
Abstract
In the last decades cancer has reached a bigger dimension being considered a worldwide
public health problem, representing a major cause of death in world. Snakes from the
genus Crotalus has gained significant space in the scientific field in a few decades,
since their venom is being researched target because of its mechanisms involved in the
cytotoxic activity.Therefore, the aim of this present study was to analyze the crude
venom of Crotalus durissus cascavella (Cdcasca), a typical snake form the northeast of
Brazil, for its in vitro cytotoxic activity against tumor cell lines, as well as, the
mechanism of cell death induced by this venom. The cytotoxic effects of Cdcasca was
evaluated by using the MTT (3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium
bromide) assay in the following cell lines: OVCAR-8 and SKOV3 (ovarian carcinoma),
PC-3M (prostate carcinoma), MCF-7 (breast carcinoma) and SF-268 (glioblastoma ). In
order to investigate DNA alterations cells were incubated with 4’,6-diamidine-2’-
phenylindole dihydrochloride (DAPI). We analysed in the flow cytometry the cell
membrane integrity, internucleosomal DNA fragmentation, cell cycle and Caspases 3
and 7 activities. Data were presented as mean values ± S.E.M. of three independent
experiments performed in triplicate. *p<0.05 compared to the negative control by
ANOVA followed by Dunnett’s test, then further analyzed in the Graphpad Prism 5.
The results had shown that the IC50 values ranged from 2.4 to 6.7 µg/mL between the
five cell lines tested. Cells treated with Cdcasca displayed morphological alterations in
dose-dependent effect, presenting reduction in cell volume, nuclear fragmentation and
the formation of apoptotic bodies. After that, cells were also stained with DAPI to better
evaluate the morphology of the nucleus after treatment with Cdcasca. The
antiproliferative activity of the venom was confirmed by flow cytometry, with a
significant reduction of cell number at 4 μg/mL. We analyzed the integrity of cell
membrane with the highest concentration of Cdcasca, which reduced cell membrane
integrity. The occurrence of apoptosis was later confirmed through the activation of
caspases 3 and 7. We concluded that Cdcasca showed dose-dependent cytotoxic effects
against several tumor cell lines, which demonstrated that, the cell death activation
occured by necrosis and apoptosis.
Keywords: Crotalus durissus; venom; cytotoxicity; apoptosis

44
1. Introduction
In the last decades cancer has reached a bigger dimension being considered a
worldwide public health problem, representing a leading cause of death (Srivastava et
al., 2005). According to the World Health Organization (WHO), in the year of 2030, an
incidence of 27 million cases of cancer are expected, 17 million cancer deaths and
around 75 million people living with cancer (WHO, 2012).
There are several sources of natural products available and these offer a great
opportunity to find interest medical substances and according to Harvey (2008), over
one hundred compounds derived from natural sources are undergoing clinical trials,
particularly for treating cancer and infectious diseases (Harvey, 2008). Since the
morbidity associated with chemotherapy is still an obstacle, the discovery of new
anticancer drugs is of utmost importance and therefore it is necessary to search for new
alternatives to improve the drug efficacy to neoplastic diseases.
Snakes from the genus Crotalus has gained significant space in the scientific
field in a few decades, since their venom is being researched target because of its
constituents and the effect that these substances can cause. In Brazil they can be found
in different parts of the country, but they are represented by only one species, the
Crotalus durissus (Cd) which can be classified into subspecies Cd terrificus, Cd
collilineatus, Cd cascavella, Cd ruruima and Cd marajoensis (Pinho & Pereira, 2001).
Most poison exerts its effects in almost all cells and tissues and their
pharmacological properties are determined by the amount of specific and biologically
active substances. Between 90 and 95% of the dry weight of snake venoms have
ownership protein, and these proteins are responsible for almost all of the biological
effects attributed to snake venoms (Bon, 1997).
The venom of Crotalus presents itself as a complex-toxic enzyme, which are
found phosphodiesterase enzymes, amino oxidase l, 5-nucleotidase and toxins such as,
crotoxin, convulxin, and crotamine gyroxin (Rangel et al., 2004). The crotoxin is
considered as the major component of the venom from this genus, and depending on the
snake subspecies, differences in this protein isoform may be found as well as difference
in their activities (Rangel et al., 2004).

45
A few studies have been conducted by testing the crude venom and some
isolated fractions of Crotalus durissus species in tumor cell lines, to access their
cytotoxic and antitumor activity (Soares et al., 2010; Tamieti et al., 2007). Therefore,
the aim of this present study was to analyze the crude venom of Crotalus durissus
cascavella (Cdcasca), a typical snake form the northeast of Brazil, for its in vitro
cytotoxic activity against tumor cell lines, as well as, the mechanism of cell death
induced by this venom.
2. Material and Methods
2.1. Venom obtaining
The venom of the snake Crotalus durissus cascavella was gently provided by
the Ofiology Regional Center, Federal University of Ceará.
2.2. Cell lines
In the present study, we used OVCAR-8 and SKOV3 (ovarian carcinoma), PC-
3M (prostate carcinoma), MCF-7 (breast carcinoma), and SF-268 (glioblastoma ) cell
lines, kindly provided by the National Cancer Institute U.S. (Bethesda, MD). Cells were
maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum, 2 mM
glutamine, 100 μg/mL streptomycin and 100 U/mL penicillin and incubated at 37 °C
under a 5% CO2 atmosphere.
2.3. Cytotoxicity assay
Cytotoxicity was determined using the MTT (3-(4,5-dimethyl-2-thiazolyl)-2,5-
diphenyl-2H-tetrazolium bromide; Sigma-Aldrich Co., St. Louis, MO, USA) assay
developed by Mosmann (1983). For that, cells were seeded at 40x103 cells/mL in 96-
well plates 24 h prior to addition of test substances , then incubated with Cdcasca (0.19
- 25 ug/mL) or vehicle control (saline) for 72 h. Three hours before the end of the
incubation, 150 μL of a MTT stock solution (0.5 mg/mL) was added to each well. Later
the formazan product was dissolved in 150 μL of DMSO, absorbance was measured at

46
570 nm using a multiplate reader (DTX 880 Multimode Detector, Beckman Coulter,
Inc. Fullerton, CA, USA) and IC50 values determined by nonlinear regression using the
software GraphPad® Prism 5.0 (Intuitive Software for Science, San Diego, CA).
2.4. Analysis of mechanisms involved in the cytotoxic activity of Cdcasca
For the next set of experiments OVCAR-8 cells were seeded at a density of
40x103 cells/mL and treated with Cdcasca at concentrations of 1, 2 and 4 μg/mL for 24
h. The vehicle (saline) and paclitaxel (0.05 μM) or doxorubicin (0.5 μM) was used as
negative and positive controls, respectively.
2.4.1. Hematoxilin-Eosin stain
After the incubation period OVCAR-8 cells were examined for morphological
changes by light microscopy (Motic BA310, Moticam 2000). To evaluate cell
morphology, after the incubation period cells were harvested, transferred to cytospin
slides, fixed with methanol for 1 min and stained with hematoxilin and eosin.
2.4.2. Nucleus stain
In order to investigate DNA alterations characteristic of apoptotic cells were
incubated with 4’,6-diamidine-2’-phenylindole dihydrochloride (DAPI). Briefly, after
the incubation period cells were fixed with 3.7 % formaldehyde and incubated with
DAPI. After extensive washing with PBS, images were captured by confocal
microscopy (LSM 710 Zeiss).
2.4.3. Flow cytometry analysis
Five thousand events were analyzed for each replicate in three independent
experiments and cellular debris was omitted from the analysis. OVCAR-8 cells
fluorescence was then determined by flow cytometry in a Guava EasyCyte Mine using
Guava Express Plus software.

47
2.4.3.1. Cell membrane integrity
The cell membrane integrity was evaluated by the exclusion of propidium iodide
(Sigma Aldrich Co., St. Louis, MO/USA). Briefly, 100 μL suspension of treated and
untreated OVCAR-8 cells was incubated with 50 μg/mL propidium iodide for 10 min.
Fluorescence was measured and analyzed for cell number and membrane integrity.
2.4.3.2 Internucleosomal DNA fragmentation and cell cycle analysis.
DNA fragmentation and cell cycle distribution was analyzed by flow cytometry
after DNA staining with propidium iodide. Briefly, 100 μL of treated and untreated
OVCAR-8 cells were incubated for 30 min, in the dark, with hypotonic solution
containing 50 μg/mL propidium iodide, 0.1% sodium citrate, and 0.1% Triton X-100.
Fluorescence was measured and DNA fragmentation and cell cycle distribution were
analyzed.
2.4.3.3. Caspases activity
Caspase 3,7 activities were analyzed by flow cytometry, using Guava®
EasyCyte Caspase Kit, after 24 h of incubation. OVCAR-8 cells (40x103 cells/mL) were
incubated with Fluorescent Labeled Inhibitor of Caspases (FLICA) and maintained for 1
h at 37º C and 5% CO2. After a sequence of washing and spinning, the cells were
resuspended in the working solution (7-AAD 1:200 in 1× washing buffer) and analyzed
immediately using flow cytometry.
2.4.3.4. Statistical analysis
Data are presented as mean values ± S.E.M. of three independent experiments
performed in triplicate. *p<0.05 compared to the negative control by ANOVA followed
by Dunnett’s test. Data was further analyzed in the Graphpad Prism 5.

48
2.5. Ethical aspects
This project was submitted to the Ethics Committee on Animal Research at the
State University of Ceará (number of the submission process: 11516651-3). All care
were taken to preserve the health of the researchers and other professionals involved in
this work and the environment in the areas where it conducts the research project.

49
Results and Discussion
Snake venoms are recognized for its biochemical complexity, being composed
of a mixture of toxins, enzymes, growth factors and other proteins with different
biological activities (Tanjoni et al., 2003; Braga et al., 2007). In the last decades snake
venoms has attracted the interest of many researchers looking to characterize the
components of these venoms as well as identify their biological properties (Bon, 2000).
Thinking about that, we then decided to study the in vitro effects of the crude venom
from the snake Crotalus durissus cascavella (Cdcasca) in tumor cell lines.
First, the cytotoxicity of the Cdcasca venom was evaluated against five tumor
cell lines of different histotypes using the MTT assay (Mosmann, 1983). As shown in
Table 1, the IC50 values ranged from 2.4 to 6.9 µg/mL between the five cell lines tested.
The prostate cancer cell line PC-3M was the most sensitive cell line with IC50 of 2.4
µg/mL, followed by the ovarian carcinoma cell line OVCAR-8, with IC50 value of 2.7
µg/mL. Interestingly, SKOV-3 cells, which is also an ovarian carcinoma cell line,
showed to be the less sensitive cell line (IC50 = 6.9 µg/mL) among all five tested,
showing the non-specificity of Cdcasca towards any cell line type.
Until today there are not many studies reporting the cytotoxicity of Cdcasca,
however there are several studies which have demonstrated the cytotoxic activity of the
crude venom of different species of snakes in other tumor cell lines (Das et al., 2011;
Gomes et al., 2010 e Yan et al., 2007), which is in agreement with the results found here
for Cdcasca.
Soares (2010) verified on it’s experiments that glioblastoma tumor lines (RT2)
and benign pituitary adenoma (GH3) when treated with the Crotalus durissus terrificus
venom showed morphologic changes such as, cell wall irregularity, cellular shrink,
nucleus condensation, DNA fragmentation and development of apoptotic bodies, typical
characters of apoptosis. Such characteristics of apoptosis were also showed by Tamieti
(2007), that exposed hamster ovarian carcinoma cells (CHO-K1) to crotalic venom.
Treating cells with cytotoxic substances usually leads them to cell death, which
may be by apoptosis, necrosis, autophagy and mitotic catastrophe, among others (Okada
and Mak, 2004). Despite the recognition by some authors that the distinction between
all those types of cell death is difficult and controversial, there are at least some well-
defined characteristics to distinguish principally the cell death by apoptosis and necrosis

50
Cells undergoing necrosis typically exhibit rapid swelling and loss of membrane
integrity which ends up releasing their cytoplasmic content into the environment, as for
cells undergoing apoptosis the typical morphological changes are cytoplasmic
shrinkage, plasma membrane blebbing, chromatin condensation and DNA
fragmentation (Kanduc et al., 2002).
Cells treated with Cdcasca at the concentration of 1.0 and 2.0 μg/mL displayed
some of those morphological features including reduction in cell volume, nuclear
fragmentation and the formation of apoptotic bodies, suggesting the induction of
apoptosis in OVCAR-8 cells (Figures 1C and D). On the other hand, at 4 μg/mL, the
effects of Cdcasca were more intense and some characteristics of necrosis could also be
seen (Figure 1E). After that, cells were also stained with DAPI to better evaluate the
morphology of the nucleus after treatment with Cdcasca. As shown in Figure 2,
negative control cells show a round nucleus typical of interphase, while in doxorubicin
treated cells the nucleus appears to be already fragmenting. At the lowest concentration
tested of Cdcasca the nucleus of the cells looks similar to the negative control cells,
being considered viable, and as the concentration increases the number of cells with
fragmented DNA starts to show (Figures 2C, 2D and 2E).
Considering the observed morphological features, we then decided to analyze
some cellular and biochemical events to assess the mechanisms involved in cell death
induced by Cdcasca. The antiproliferative activity of the venom was confirmed by flow
cytometry (Figure 3B), with a significant reduction of cell number at 4 μg/mL. The
inhibition of cell proliferation can be associated with loss of cell membrane integrity,
with only 64 % of cells presenting intact membrane in the presence of Cdcasca at 4
μg/mL (Figure 3A), corroborating data from morphological analysis. Cell membrane
integrity serves as a parameter of the viability of cells and at the initial stages of
apoptosis the cell shrinks while its membrane remains intact, however, necrotic cell
swelling occurs as a result of the early failure of cell membrane integrity (Ziegler and
Groscurth, 2004), thus we might say that at lower concentrations Cdcasca (shows an
antiproliferative activity and becomes cytotoxic at higher concentrations.
As the venom at lower concentrations showed no cytotoxicity, we decided to
analyze whether the cell cycle was being affected. However, we found that Cdcasca was
not interfering in the cell cycle distribution at all (Data not shown) and the only

51
alteration observed was a slight increase in the sub G0/G1 phase of the cell cycle, which
again is an indicative of DNA fragmentation and consequently apoptosis.
The occurrence of apoptosis was later confirmed through the activation of
caspases 3 and 7 by Cdcasca, especially at the concentration of 4 µg/mL, where 32 % of
the cells were showing caspases activation (Figure 4). The caspases are shown to be one
of the important executors of apoptosis, being generally activated by several anti-tumor
drugs. This activation involves mainly two signaling pathways, one is the mitochondrial
and the other is the cell death receptor pathway, which consecutively activates caspases
9 and 8 and these both ends up by activating caspases 3 and 7 (Hengartner, 2000;
Mehmet, 2002).
Yan et al. (2006) had already shown that crotoxin, the major component of the
venom from Crotalus durissus terrificus, acts through caspase 3 activation and although
this is in accordance with our results, further studies are needed to elucidate the exact
mechanism involved in the cytotoxicity of Cdcasca.
The present paper showed that the Crotalus durissus cascavella venom had a
cytotoxic action on a variety of tumor cell lines, specially in ovarian carcinoma line
(OVCAR-8). The Cdcasca demonstrated an efficient activity, causing considerable
cytotoxic effects, such as, morphological changes and activating cell death, by necrosis
and apoptosis. More studies are necessaries through specific assays, to determine on
wich pathway this venom is more efficient on the cell death process.
Acknowledgments
We wish to thank CNPq, CAPES, FUNCAP, FINEP, BNB/FUNDECI and
PRONEX for the financial support in the form of grants and fellowship awards.
Conflict of interest
The authors of this article declare that there are no potential conflicts of
interest including employment, consultancies, stock ownership, honoraria, paid expert
testimony and patent applications/registrations related to the current manuscript

52
References
Bon, C., 1997. Snake venom and pharmacopoeia. In: Bauchot, R. (edit) Snakes a
Natural History. New York: Sterling Publishing Co., 194-209.
Bon, C., 2000. The natural toxins. Biochimie. 82, 791-792.
Braga, M. D. M., Martins, A. M. C., Amora, D. N., Menezes D. B., Toyama, M. H.,
Toyama D. O., Marangoni, S., Alves, C. D., Barbosa, P. S. F., Alves, R. S., Fonteles,
M. C., Monteiro, H. S. A., 2007. Purification and biological effects of L- amino acid
oxidase isolated from Bothrops insularis venom. Toxicon 51, 199-207.
Das, T., Bhattacharya, S., Halder, B., Biswas, A., Das Gupta, S., Gomes, A., & Gomes,
A. 2011. Cytotoxic and antioxidant property of a purified fraction (NN-32) of Indian
Naja naja venom on Ehrlich ascites carcinoma in BALB/c mice. Toxicon : official
journal of the International Society on Toxinology 57(7-8), 1065–72.
Gomes, A., Bhattacharjee, P., Mishra, R., Biswas, A. K., Dasgupta, S. C., & Giri, B.
2010. Anticancer potential of animal venoms and toxins. Indian journal of experimental
biology 48(2), 93–103.
Harvey, A. L., 2008. Drug Discov. Today, 13, 894.
Hengartner, M.O., 2000. The biochemistry of apoptosis. Nature 407, 770–776.
Kanduc, D., Mittelman, A., Serpico, R., Sinigaglia, E., Sinha, A.A., Natale, C.,
Santacroce, R., Di Corcia, M.G., Lucchese, A., Dini, L., Pani, P., Santacroce, S.,
Simone, S., Bucci, R., Farber, E., 2002. Cell death: apoptosis versus necrosis. Int J
Oncol. 21(1), 165-70.
Mehmet, H., 2002. Apoptosis: caspase find a new place to hide. Nature 403, 29–30.

53
Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival:
application to proliferation and cytotoxicity assays. J. Immunol. Methods. 65(2), 55-63.
Okada, H. and Mak, T.W. 2004. Pathways of apoptotic and non-apoptotic death in
tumour cells. Nat. Rev. Cancer 4, 592–603.
World health Organization, 2012.
Pinho, F. M. O., Pereira, I. D., 2001. Ofidismo. Ver. Ass. Méd. Brás. 47, 1, 24-29.
Rangel, A. S., Dos-Santos, E. C., Lopes-Ferreira, M., Lima, C., Cardoso, D. F., Mota, I.
2004. A comparative study of biological activities of crotoxina and CB fraction of
venoms from Crotalus durissus terrificus, Crotalus durissus cascavella and
Crotalus durissus collilineatus. Toxicon 43 (7), 801-810.
Soares M.A., Pujatti P.B., Fortes-Dias CL, Antonelli L, S. R., 2010. Crotalus durissus
terrificus venom as a source of antitumoral agents. The Journal of Venomous Animals
and Toxins including Tropical Diseases 16 (3), 480–492.
Srivastava, V., Negi, J. K., Gupta, M., Khanuja, S. P. S., 2005. Bioorg. Med. Chem. 13,
58-92.
Tamieti B. P., Damatta R. A., Cogo J. C., Da Silva N. S., Mittmann J., P.-S. C., 2007.
Cytoskeleton, endoplasmic reticulum and nucleus alterations in cho-k1 cell line after
Crotalus durissus terrificus (South American Rattlesnake) venom treatment. Journal of
Venomous Animals and Toxins Including Tropical Diseases 13, 56–58.
Tanjoni, I., Butera, D., Bento, L., Della-Casa, M. S., Marques-Porto, R., Takehara,
H. A., Gutierrez, J. M., Fernandes, I., Moura-da-Silva, A. M., 2003. Snake venom
metalloproteinases: structure/function relationships studies using monoclonal
antibodies. Toxicon 42(7), 801-808.

54
Yan, C., Liang, Z., Gu, Z., Yang, Y., Reid, P., & Qin, Z. 2006. Contributions of
autophagic and apoptotic mechanisms to CrTX-induced death of K562 cells. Toxicon
47, 521–530.
Yan, C., Yang, Y., Qin, Z., Gu, Z., Reid, P., & Liang, Z., 2007. Autophagy is involved
in cytotoxic effects of crotoxin in human breast cancer cell line MCF-7 cells. Acta
Pharmacologica Sinica 28(4), 540–8.
Ziegler, U., Groscurth, P., 2004. Morphological features of cell death. News Physiol.
Sci. 19, 124–128

55
ANNEX
Figures
Figure 1. Photomicrographs of OVCAR-8 cells stained with Hematoxilin/Eosin, 400×.
Cells were incubated with Cdcasca for 24 h. A: Control; B: Doxorubicin 0.5 μM; C:
Cdcasca 1.0 μg/mL; D: Cdcasca 2.0 μg/mL; E: Cdcasca 4.0 μg/mL. Black arrow: cell
membrane blebbing; White arrow: atypical mitosis; Black dashed arrow: Nucleus
fragmentation; White dashed arrow: membrane damage and pyknosis.

56
Figure 2. Confocal (Zeiss LSM 710) images of OVCAR-8 cells stained with DAPI,
400×. Cells were incubated with Cdcasca for 24 h. A: Control; B: Doxorubicin 0.5 μM;
C: Cdcasca 1.0 μg/mL; D: Cdcasca 2.0 μg/mL; E: Cdcasca 4.0 μg/mL. White arrow:
atypical mitosis; White dashed arrow: Nucleus fragmentation.

57
Figure 3. Effect of Cdcasca on OVCAR-8 cell membrane integrity (A) and cell number
(B) determined by flow cytometry, using propidium iodide after 24 h incubation. The
negative control was treated with the vehicle saline (Control). Paclitaxel (Ptx) at 0.05
µM was used as the positive control. Data are presented as mean values ± S.E.M. of
three independent experiments performed in triplicate. *p<0.05 compared to the
negative control by ANOVA followed by Dunnett’s test.

58
Figure 4. Effect of Cdcasca on caspases 3,7 determined by flow cytometry, after 24 h
incubation. The negative control was treated with the vehicle saline (Control).
Doxorubicin (Dox) at 0.5 µM was used as the positive control. Data are presented as
mean values ± S.E.M. of three independent experiments performed in triplicate.
*p<0.05 compared to the negative control by ANOVA followed by Dunnett’s test

59
Tables
Table 1. Cytotoxic activity of Cdcasca on tumor cell lines.
Cell lines
OVCAR-8 PC-3M MCF-7 SF-268 SKOV3
Cdcasca 2.7
(2.6 – 2.9)
2.4
(2.2 – 2.6)
4.9
(4.2 – 5.8)
4.1
(3.5 – 4.7)
6.9
(5.9 – 8.0)
Doxorubicin 0.3
(0.1 – 0.4)
0.8
(0,7 – 0.9)
0.4
(0.3 – 0.6)
0.6
(0.4 – 0.9) -
Data are presented as IC50 values in μg/mL and as the 95% confidence interval obtained by
nonlinear regression for all of the cell lines from two independent experiments, performed in
duplicate, after 72 h of incubation.

60
7. CONSIDERAÇÕES FINAIS
O presente trabalho demonstrou que o veneno da serpente Crotalus durissus
cascavella (Cdcasca) apresentou atividade citotóxica para as linhagens tumorais
testadas.
Os resultados de CI50 apresentados confirmaram sua citotoxicidade em
linhagens de células tumorais de carcinoma ovariano (OVCAR-8 e SKOV3), carcinoma
prostático (PC-3M), carcinoma mamário (MCF-7) e glioblastoma (SF-268), sendo
importante ressaltar que devido sua excelente resposta ao ensaio citotóxico e sua
disponibilidade a linhagem OVCAR-8 foi estudada de forma mais aprofundada.
O Cdcasca demonstrou potencial de atividade eficiente em linhagem OVCAR-8,
provocando consideráveis alterações morfológicas nas células, dentre elas destacam-se:
problemas na integridade de membrana, alterações nucleares, formação de vacúolos
citoplasmáticos/nucleares, diminuição da viabilidade celular e por consequência morte
celular, seja pela via necrótica ou apoptótica.
A ativação das caspases efetoras (3 e 7) ocorreu em células tratadas com a
concentração mais elevada, o que pode sugerir uma ação do veneno na ativação desta
via de morte celular, tornando então fundamental que mais estudos sejam realizados
visando determinar por qual via este veneno é mais eficiente no processo de morte
celular.

61
8. PERSPECTIVAS
O estudo da utilização de substâncias naturais bioativas é uma excelente opção
para o surgimento de novas ferramentas terapêuticas, independente de qual aplicação na
área da saúde. O veneno de serpentes e seus componentes, um exemplo desse tipo de
substância, são largamente testados, atualmente já é confirmado seu potencial de ação
sobre diversas linhagens tumorais, seja alterando e inibindo sua capacidade de
proliferação celular, induzindo morte celular, ou mesmo sua atividade antiangiogênica.
Outras atividades descobertas são: sua ação bactericida, leishmanicida, ativação de
processos de fagocitose em infecções virais, ação sobre processos hematológicos,
influência no sistema nervoso, bem como, ativação das vias apoptóticas, necróticas ou
fagocitose celular. Os bons resultados obtidos neste trabalho, utilizando o veneno da
serpente Crotalus durissus cascavella, subespécie característica da Região Nordeste, é
de grande valia para a elaboração de linhas de pesquisas direcionadas e aprofundadas
sobre sua ação citotóxica. Isto ocorrendo poderá então permitir a descoberta e produção
de uma nova ferramenta terapêutica, que venha a servir de apoio para melhorar o
cenário, atual, do câncer na saúde pública. Entretanto, sua ação ainda não está
compreendida, a interação entre seus componentes e seu verdadeiro potencial de
atividade são uma incógnita, o que torna necessária a purificação desta substância para
então serem realizados estudos mais específicos.

62
9. REFERÊNCIAS BIBLIOGRÁFICAS
AUTO, H. J. F., Animais peçonhentos. Maceió: EDUFAL, p.114, 1999.
BARROS, L C., SOARES, A.M., COSTA, F.L., RODRIGUES, V.M., FULY, A.L.,
GIGLIO, J.R., GALLACCI, M., THOMAZINI-SANTOS, I.A., BARRAVIEIRA,
S.R.C.S., FERREIRA JUNIOR, R. S., Biochemical and biological evaluation of gyroxin
isolated from Crotalus durissus terrificus venom. The Journal of Venomous Animals
and Toxins including Tropical Diseases, v.17, n.1, p.23–33, 2011.
BEGHINI, D. G.; TOYAMA, M H; HYSLOP, S.; SODEK, L.; NOVELLO, J. C.;
MARANGONI1, S., 5 Enzymatic characterization of a novel phospholipase A2 from
Crotalus durissus cascavella rattlesnake (Maracambóia) venom. Journal of protein
chemistry, v. 19, n. 8, p. 679-84, 2000.
BOHN, I.; SCHILD, H., Apoptosis: the complex scenario for a silent cell death.
Molecular Imaging and Biology, v.5, n.1, 2-14, 2003.
BON, C., Snake venom and pharmacopoeia. In: Bauchot, R. (edit) Snakes a Natural
History. New York: Sterling Publishing Company, p.194-209, 1997.
BREITHAUPT, H.; RÜBSAMEN, K.; HABERMANN, E., Biochemistry and
pharmacology of the crotoxin complex. Biochemical analysis of crotapotin and the
basic Crotalus phospholipase A. European journal of biochemistry / FEBS, v. 49, n. 2,
p. 333-45, 1974.
CÂMARA, P.; ESQUISATTO, L.; CAMARGO, E.; RIBELA, M.; TOYAMA, M.;
MARANGONI, S.; DE NUCCI, G.; ANTUNES, E., Inflammatory edema induced by
phospholipases A2 isolated from Crotalus durissus sp. in the rat dorsal skin: A role for
mast cells and sensory c-fibers. Toxicon, v.41, p.823-829, 2003.
CAMILLO, M.A.P., Contribuição ao estudo das giroxinas (enzimas semelhantes à
trombina) dos venenos das serpentes brasileiras Lachesis muta muta e Crotalus durissus

63
terrificus. 1998. 75 f.Tese (Doutorado) – Instituto de Pesquisas Energéticas e Nucleares,
São Paulo, São Paulo, 1998.
CASTEDO, M.; PERFETTINI, J.L.; ROUMIER, T.; ANDREAU, K.; MEDEMA, R.,
KROEMER, G., Cell death by mitotic catastrophe: a molecular definition. Oncogene,
v.23, p.2825-837, 2004.
COTRANS, R.S.; KUMAR, V.; COLLINS, T., Robbins patologia estrutural e
funcional. 6.ed. Rio de Janeiro: Guanabara Koogan; 2000; p.14-22; p.233-93.
DA SILVA, R. J.; DA SILVA, M. G.; VILELA, L. C.; FECCHIO, D.Antitumor effect
of Bothrops jararaca venom. Mediators Inflammation, v.11, n.2, p.99-104, 2002.
DANIAL, N.N.; KORSMEYER, S.J., Cell death: critical control points. Cell.,v.116,
p.205-19, 2004.
DAMIANI, D., Mecanismos da Apoptose. Manual de Patologia e Citopatologia
Oncológica, 2004.
DONATO, N.J.; MARTIN, C.A.; PEREZ, M.; NEWMAN, R.A.; VIDAL, J.C.;
ETCHEVERRY, M.A., Regulation of epidermal growth factor receptor activity by
crotoxin, a snake venom phospholipase A2 toxin. Biochemical Pharmacology. v. 51,
p.1535–1543, 1996.
EIGENBROD, T., PARK, J.H., HARDER, J., IWAKURA, Y., NUNEZ, G., Cutting
edge: critical role for mesothelial cells in necrosis-induced inflammation through the
recognition of IL-1 alpha released from dying cells, The Journal of Immunology. v.181,
p.8194–8198, 2008.
EVANGELISTA, J. S. A M.; MARTINS, A. M. C.; NASCIMENTO, N. R. F.; SOUSA,
C.M.; ALVES, R.S.; TOYAMA, D.O.; TOYAMA, M.H.; EVANGELISTA, J.J.F.;
MENEZES, D.M.; MANASSÉS, C.F.; MORAES, M.E.A.; MONTEIRO, H.S.A., Renal
and vascular effects of the natriuretic peptide isolated from Crotalus durissus cascavella

64
venom. Toxicon : official journal of the International Society on Toxinology, v.52, n.7,
p.737-44, 2008.
FAN, T.J.; HAN, L.H.; CONG, R.S.; LIANG, J., Caspase Family Proteases and
Apoptosis. Acta Biochimica et Biophysica Sinica, v.37, n.11, p.719-727, 2005.
FADEEL, B., ORRENIUS, S. Apoptosis: a basic biological phenomenon with wide-
ranging implications in human disease. Journal of Internal Medicine, v.258, p.479-517,
2005.
FERRARO, G.C.; MORAES, J.R.E.; SHIMANO, A.C.; PEREIRA, G.T.; MORAES,
F.R.; BUENO DE CAMARGO, M.H., Efect of snake venom derived fbrin glue on the
tendon healing in dogs. Clinical and biomechanical study. Journal Venom Animals
Toxins including Tropical Diseases, v.11, n.3, p.261-74, 2005.
FONSECA, F. V.; ANTUNES, E.; MORGANTI, R. P.; MONTEIRO, H. S. A.;
MARTINS, A M. C.; TOYAMA, D. O.; MARANGONI, S., Characterization of a new
platelet aggregating factor from crotoxin Crotalus durissus cascavella venom. The
protein journal, v. 25, n. 3, p.183–92, 2006.
FOSTER, I., Cancer: A cell cycle defect. Radiography, v.14 (2), p.144–149, 2008.
GARG, A.D.; NOWIS, D.; GOLAB, J.; VANDENABEELE, P.; KRYSKO, D.V. &
AGOSTINIS, P., Immunogenic cell death, DAMPs and anticancer therapeutics: an
emerging amalgamation. Biochimica et biophysica acta, v.1805, n.1, p.5-371, 2010.
GOPALAKRISHNAKONE, P.; DEMPSTER, D. W.; HAWGOOD, B. J.; ELDER, H.
Y. Cellular and mitochondrial changes induced in the structure of murine skeletal
muscle by crotoxin, a neurotoxic phospholipase A2 complex. Toxicon : official journal
of the International Society on Toxinology, v. 22, n. 1, p. 85-98, 1984.
HARVEY, A. L., Natural products in drug discovery. Drug Discovery Today, v.13,
n.19/20, p.894-901, 2008.

65
HENDON, R.A., FRAENKEL-CONRAT, H., Biological role of the two components of
CrTX. Proceedings of the National Academy of Sciences. USA, v. 68, n. 7, 1560–1563.
1971.
HENGARTNER, M.O., The biochemistry of apoptosis. Nature., v.407, p.770-76, 2000.
HIRABAYASHI, T.; MURAYAMA, T.; SHIMIZU, T., Regulatory mechanism and
physiological role of cytosolic phospholipase A2. Biological and Pharmaceutical
Bulletin. v.27, p. 1168- 1173, 2004.
JANDROT-PERRUS, M.; LAGRUE, A. H.; OKUMA, M.; BON C. Adhesion and
activation of human platelets induced by convulxin involves glycoprotein VI and
integrin alpha2beta1. Journal of Biological Chemistry. V. 272, n. 43, p. 27035-27041,
1997.
KANAJI, S.; KANAJI, T.; FURIHATA, K.; KATO, K.; WARE, J. L.; KUNICLI, T. J.
Convulxin binds to native, human glycoprotein IBα. The Journal of Biological
Quemistry. V. 278, n. 41, p. 39452-39460, 2003.
KELEKAR, A., Autophagy. Annals of the New York Academy of Sciences, 1066,
p.259-71, 2005.
KUMMAR, V.; ABBAS, A.; FAUSTO, N.; ROBBIN; COTRAN – Pathology Basis of
Disease, 7a. ed., WB Saunders: China, 2004.
LAUNAY, S.; HERMINE, O.; FONTENAY, M.; KROEMER, G.; SOLARY, E.;
GARRIDO, C., Vital functions for lethal caspases. Oncogene, v.24, p.5137-5148, 2005
LEDUC, M. & BON, C. Cloning of subunits of convulxin, a collagen-like platelet-
aggregating protein from Crotalus durissus terrificus venom. Biochem, J. v. 333, p.
389-393, 1998.

66
LIMA, D.C.; ABREU, P. A.; FREITAS, C.C.; SANTOS, D.O.; BORGES, R.O.;
SANTOS, T.C.; CABRAL, L.M.; RODRIGUES, C.R.; CASTRO, H.C., Snake Venom:
Any Clue for Antibiotics and CAM? e CAM., v.2,n.1, p. 39-47, 2005.
LOGUE, S.E.; MARTIN, S.J., Caspase activation cascades in apoptosis. Biochemical
Society Transactions, v. 36, p. 1-9, 2008.
LOMONTE, B.; ANGULO, Y.; CALDERÓN, L., An overview of lysine-49
phospholipase A2 myotoxin from crotalid snake venoms and their structural
determinants of myotoxic action. Toxicon., v.42, p. 885-901, 2003.
LOZY, F.,; KARANTZA, V., Autophagy and cancer cell metabolism. Seminars in cell
& developmental biology, v.23, n.4, p.395–401, 2012.
LUM, J.J.; DEBERARDINIS, R.J.; THOMPSON, C.B., Autophagy in metazoans: cell
survival in the land of plenty. Nature Reviews Molecular Cell Biology, v.6, p.439-48,
2005.
MARTINS, A. M. C.; TOYAMA, M.H.; HAVT. A.; NOVELLO, J.C.; MARANGONI,
S.F; MANASSÉS, C.F.; MONTEIRO, H.S.A., Determination of Crotalus durissus
cascavella venom components that induce renal toxicity in isolated rat kidneys.
Toxicon : official journal of the International Society on Toxinology, v. 40, n. 8, p.
1165-171, 2002.
MATSUBARA, M. H. Efeitos do veneno de Crotalus durissus terrificus, da crotoxina e
de suas subunidades fosfolipase A2 e crotapotina em monocamadas de células
endoteliais em cultura. 2009. 106f. Dissertação (Mestrado em Biotecnologia) -
Interunidades em Biotecnologia, USP/Instituto Butantan/IPT, São Paulo, 2009.
MATSUI, T.; FUJIMURA, Y.; TITANI K., Snake venom proteases afecting hemostasis
and thrombosis. Biochim Biophys Acta.;1477(1-2):146-56, 2000.

67
MEIER, P.; FINCH, A.; EVAN, G., Apoptosis in development. Nature, v.407, p.796-
801, 2000.
MINISTÉRIO DA SAÚDE. Instituto Nacional de Câncer - INCA. Disponível em:
<http://www.inca.gov.br>, Acesso em Setembro de 2012.
MORAES, J.R.E.; CORREA, P.H.A.; CAMPLESI, A.C.; MORAES, F.R.,
Experimental use of fbrin glue derived from snake venom in non-pregnant canine
uterus. Journal Venom Animals Toxins including Tropical Diseases., v.10 (2):133-143,
2004.
NEWMAN, R.A.; VIDAL, J.C.; VISKATIS, L.J.; JOHNSON, J.I.; ETCHEVERRY,
M.A., VRCTC–310 - a novel compound of purified animal toxins separates antitumor
efficacy from neurotoxicity. Invest New Drugs, v.11, p.151–9, 1993.
NICASTRO, G.; FRANZONI, L.; DE CHIARA, C.; MANCIN, A. C.; GIGLIO, J. R. &
SPISNI, A., "Solution structure of crotamine, a Na+ channel affecting toxin from
Crotalus durissus terrificus venom." Eur J Biochem, v.270 (9): 1969-1979, 2003.
OKADA, H.; MAK, T.W., Pathways of apoptotic and non-apoptotic death in tumour
cells. Nature Reviews Cancer, v.4, p.592–603, 2004.
OLIVEIRA, D. G. DE; TOYAMA, MARCOS H; MARTINS, A. M.; HAVT, A.;
NOBRE, A.C.L; MARANGONI, S.; CÂMARA, P.R.; ANTUNES, E.; DE NUCCI, G.;
BELIAM, L.O.S; FONTELES, M.C.; MONTEIRO, HELENA S.A., Structural and
biological characterization of a crotapotin isoform isolated from Crotalus durissus
cascavella venom. Toxicon, v. 42, n. 1, p. 53-62, 2003.
Organização Mundial da Saúde - OMS (WHO). Disponível em:
<http://www.who.int/countries/bra/es/>, Acesso em Outubro de 2012.
PARKIN, D.M.; BRAY, F.; FERLAY, J.; PISANI, P., Estimating the world cancer
burden: Globocan 2000. International Journal of Cancer, v.94, n.2, p.153-156, 2001.

68
PEREAÑEZ, J.A.; NÚÑEZ, V.; HUANCAHUIRE-VEGA, S.; MARANGONI,
SERGIO; PONCE-SOTO, L. A., Biochemical and biological characterization of a
PLAR (2R) from crotoxin complex of Crotalus durissus cumanensis. Toxicon : official
journal of the International Society on Toxinology, v. 53, p. 534-542, 2009.
PETROVSKI, G., ZAHUCZKY, G., MAJAI, G., FESUS, L., Phagocytosis of cells
dying through autophagy evokes a pro-inflammatory response in macrophages,
Autophagy, v.3, p.509–511, 2007.
PINHO, F. M. O.; PEREIRA, I. D., Ofidismo. Revista da Associação Médica Brasileira,
v.47, n.1, p. 24-29, 2001.
POLGÁR, J.; CLEMETSON, J. M.; KEHREL, B. E.; WIEDEMANN, M.;
MAGNENAT, E. M.; WELLS, T. N.; CLEMETSON, H. J., Platelet activation and
signal trans by convulxin, a C-type lectin from Crotalus durissus terrificus (tropical
rattlesnake) venom via the p62/GPVI collagen receptor. J. Biol. Chem. v. 272, n. 21, p.
13576-13583, 1997.
PRADO-FRANCESCHI, J., On the pharmacology of convulxin and giroxin. Memórias
do Instituto Butantan, v.52, p.25-6, 1990. Suplemento.
RANGEL-SANTOS, A.; DOS-SANTOS, E. C.; LOPES-FERREIRA, M.; LIMA, C.;
CARDOSO, D. F.; MOTA, I., A comparative study of biological activities of crotoxina
and CB fraction of venoms from Crotalus durissus terrificus, Crotalus durissus
cascavella and Crotalus durissus collilineatus. Toxicon., v.43, n.7, p. 801-810, 2004.
RATELSLANGEN. Crotalus durissus - South American rattlesnake. [2012].
Disponível em: http://www.ratelslangen.nl/gallery/Crotalus_durissus_South_
American_rattlesnake/. Acesso em novembro de 2012.
RAW I.; ROCHA M.C.; ESTEVES M.I.; KAMIGUTI A.S., Isolation and
characterization of a thrombin-like enzyme from the venom of Crotalus durissus

69
terrifcus. Brazilian Journal of Medical and Biological Research., v.19, n.3, p.333-338,
1986.
REED, CJ. Apoptosis and cancer: strategies for integrating programmed cell death.
Semin. Hemtol. v. 37, p.9-16, 2000.
RELLO, S.; STOCKERT, J.C.; MORENO, V.; GÁMEZ, A.; PACHECO, M.;
JUARRANZ, A.; CAÑETE, M. AND VILLANUEVA. Morphological criteria to
distinguís cell death induced by apoptotic and necrotic treatments. Apoptosis. v.10,
p.201-208, 2005.
RICCI, M.S.; ZONG, W.X., Chemotherapeutic approaches for targeting cell death
pathways. Oncologist, v.11, p.342-57, 2006.
RUSSEL, F.E., Snake Venom Poisoning. New York: Scholium International Grat Neck,
1983.
SAELENS, X., FESTJENS, N., PARTHOENS, E., VANOVERBERGHE, I., KALAI,
M., VAN KUPPEVELD, F., VANDENABEELE, P., Protein synthesis persists during
necrotic cell death, The Journal of Cell Biology, v.168, p.545–551, 2005.
SAMPAIO, S. C.; RANGEL-SANTOS, A.C.; PERES, C.M.; CURI, Y. Inhibitory effect
of phospholipase A2 isloated from Crotalus durissus terrificus venom on maccrophage
function. Toxicon, v.45, p.671-676, 2005.
SANO-MARTINS, I.S.; TOMY, S.C.; CAMPOLINA, D.; DIAS, M.B.; DE CASTRO,
S.C.B.; SOUSA-E-SILVA, M.C.C.;AMARAL, C.F.S.; REZENDE, M.A.; KAMIGUTI,
A.S.; WARRELL, D.A.; THEAKSTON, R.D.G. Coagulopathy following lethal and
non-lethal envenoming of humans by the South American rattlesnake in Brazil. Q J M,
v.94, n.10, p.551-559, 2001.
SANTOS, E., Anfíbios e répteis. 3. ed., Belo Horizonte, Itatiaia,1995, p.263.

70
SERRANO, S.M.; MAROUN, R.C., Snake venom serine proteinases: sequence
homology vs. substrate specifcity, a paradox to be solved. Toxicon., v.45(8), p.1115-32,
2005.
SILVA, M. G. Câncer em Fortaleza 1978-1980. Instituto e Registro de câncer do Ceará.
Fortaleza, p.135, 1982.
SIX, D. A.; DENNIS, E. A., The expanding superfamily of phospholipase A 2
enzymes : classi ¢ cation and characterization. Group, v. 1488, 2000.
SOARES M.A.; PUJATTI P.B.; FORTES-DIAS C.L.; ANTONELLI L, S. R., Crotalus
durissus terrificus venom as a source of antitumoral agents. The Journal of Venomous
Animals and Toxins including Tropical Diseases, v.16, n.3, p.480–492, 2010.
SPINOSA, H. S.; GÓRNIAK, S. L.; PALERMO-NETO, J., Toxicologia aplicada à
Medicina Veterinária. 1.ed. Barueri: Manole, 2008. 942p.
SRIVASTAVA V.; ARVIND S.N.; KUMAR J.K.; GUPTA M.M.; SUMAN P.S., Plant-
based anticancer molecules: A chemical and biological profile of some important leads.
Bioorganic & Medicinal Chemistry, v.3, n.908, p.58-92, 2005.
STANCHI, N.O.; ARIAS, D.; MARTINO, P.E.; DIEZ, R.A.; COSTA, L.A., 30-day
intravenous administration of VRCTC-310-ONCO in rabbits. Farmaco, v. 57, n.2,
p.167-70, 2002.
STRASSER, A.; O´CONNOR L.; DIXIT V.M., Apoptosis signaling. Annual Review of
Biochemistry, v.69, p. 217-245, 2000.
TAMIETI B. P.; DAMATTA R. A.; COGO J. C.; DA SILVA N. S.; MITTMANN J. P.-
S. C., Cytoskeleton, endoplasmic reticulum and nucleus alterations in cho-k1 cell line
after Crotalus durissus terrificus (south american rattlesnake) venom treatment. Journal
of Venomous Animals and Toxins Including Tropical Diseases, v.13, p.56–58, 2007.

71
TOYAMA, M. H. ; CARNEIRO, Everardo M. ; MARANGONI, Sergio ; AMARAL,
M. E. C. ; VELLOSO, L. A. ; BOSCHERO, A. C. . Isolation and characterization of a
Convulxin like protein from Crotalus durisssus collilineatus venom. Journal of Protein
Chemistry, NY, USA, v. 20, n.7, p. 585-591, 2001.
TOYAMA, M.H.; TOYAMA, D.D.O.; PASSERO, L.F.D.; LAURENTI, M.D.;
CORBETT, C.E.; TOMOKANE, T.Y.; FONSECA, F.V.; ANTUNES, E.; JOAZEIRO,
P.P.; BERIAM, L.O.S.; MARTINS, M.A.C.; MONTEIRO, H.S.A; FONTELES, M.C.,
Isolation of a new L-amino acid oxidase from Crotalus durissus cascavella venom.
Toxicon : official journal of the International Society on Toxinology, v. 47, n. 1, p. 47-
57, 2006.
VANDENBERGHE, T., KALAI, M.,. DENECKER, G., MEEUS, A., SAELENS, X.,
VANDENABEELE, P., Necrosis is associated with IL-6 production but apoptosis is
not, Cellular Signalling, v.18, p.328–335, 2006.
VERMEULEN, K.; VAN BOCKSTAELE, D. R.; BERNEMAN, Z. N., The cell cycle:
a review of regulation, deregulation and therapeutic targets in cancer, Cell
Proliferation., v.36, p.131-149, 2003.
YAN, C.; LIANG, Z.; GU, Z.; YANG, Y.; REID, P.; QIN, Z., Contributions of
autophagic and apoptotic mechanisms to CrTX-induced death of K562 cells. Toxicon
47, 521–530, 2006.
YAN, C.; YANG, Y.; QIN, Z.; GU, Z.; REID, P.; LIANG, Z., Autophagy is involved in
cytotoxic effects of crotoxin in human breast cancer cell line MCF-7 cells. Acta
pharmacologica Sinica, v.28, n.4, p.540–8, 2007.
YU, S.P.; CHOI, D.W., Ions, cell volume, and apoptosis. Proceedings of the National
Academy of Sciences USA. V. 97, 9 360-362, 2000.

72
ZHAO, S.; DU, X.; CHAI M.; CHEN, J.; ZHOU, Y.; SONG, J., Secretory
phospholipase A2 induces apoptosis via a mechanism involving ceramide generation.
Biochimica et. Biophysica Acta. v.1581, p.75-88, 2002.
ZIEGLER, U.; GROSCURTH, P., Morphological features of cell death. News Physiol.
Sci, v.19, p.124-128, 2004.





























