Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CÂMPUS DE BOTUCATU
Avaliações ambiental e nutricional da produção de microcepas e
microestacas de Eucalyptus grandis x E. urophylla em sistema hidropônico
em casa-de-vegetação.
AGNALDO SCARASSATI
Tese apresentada à Faculdade de Ciências Agronômicas da UNESP – Câmpus de Botucatu, para obtenção do título de Doutor em Agronomia - Área de Concentração em Energia na Agricultura.
BOTUCATU – SP Dezembro – 2003

UNIVERSIDADE ESTADUAL PAULISTA “JÚLIO DE MESQUITA FILHO”
FACULDADE DE CIÊNCIAS AGRONÔMICAS
CÂMPUS DE BOTUCATU
Avaliações ambiental e nutricional da produção de microcepas e
microestacas de Eucalyptus grandis x E. urophylla em sistema hidropônico
em casa-de-vegetação
AGNALDO SCARASSATI
Orientador: Prof. Dr. Iraê Amaral Guerrini
Tese apresentada à Faculdade de Ciências Agronômicas da UNESP – Câmpus de Botucatu, para obtenção do título de Doutor em Agronomia - Área de Concentração em Energia na Agricultura.
BOTUCATU – SP Dezembro – 2003

iv
BIOGRAFIA DO AUTOR
AGNALDO SCARASSATI nascido em Piracicaba, estado de São Paulo, Brasil, no
dia 17 de julho de 1962 e formado pela Escola Superior de Agricultura Luiz de Queiroz –
ESALQ –USP, em Engenharia Florestal; Pesquisador Rhae-CNPq (Recursos Humanos em
Áreas Estratégicas do Governo Federal) na iniciação científica; Mestrado em Ciências
Florestais (ESALQ-USP) com financiamento do projeto pela FAPESP (Fundação de
Amparo à Pesquisa do Estado de São Paulo); Professor na Faculdade de Agronomia Dr.
Francisco Maeda, em Ituverava , São Paulo, durante dez anos; Consultor/Professor na
Cooperrual-Oisca-Brasil, Organization for Industrial, Spiritual, Cultural and Advancement
(ONG com status consultivo no Conselho Econômico e Social da ONU); Consultor do BID
– FOMIN (Banco Interamericano de Desenvolvimento – Fundo Multilateral de
Investimento); Pesquisador Rhae-DTI-CNPq (Desenvolvimento Técnico Industrial) na
International Paper do Brasil; Contratado pela International Paper do Brasil de dezembro de
2001 a janeiro de 2003, para as funções de pesquisador em Biotecnologia e Produção
Clonal (área de concentração: nutrição e fisiologia vegetal); Atualmente é Docente na
Faculdade Cantareira de São Paulo – Capital – acumulando as funções de Coordenador
Geral dos Cursos de Pós-Graduação Lato Sensu na IES (2004).

v
AGRADECIMENTOS Ao Prof. Dr. Iraê Amaral Guerrini pela orientação, paciência, disponibilidade e acima
de tudo, pela confiança durante o desenvolvimento deste trabalho.
Ao Departamento de Engenharia Rural da Faculdade de Ciências Agronômicas da
UNESP – Câmpus de Botucatu.
Ao Programa de Pós-Graduação em Agronomia, Área de Concentração em Energia na
Agricultura, por ter aceitado-me como discente.
Aos membros da banca de qualificação e defesa por ter emprestado um pouco do seu
conhecimento para melhoria deste trabalho.
A Faculdade Cantareira na pessoa do Professor Paulo Meinberg, que dedicou a sua
vida inteira a educação, por ter apoiado-me durante o tempo que permaneci afastado da
instituição.
A International Paper do Brasil pela concessão das instalações, material genético e
cobertura dos custos de análises laboratoriais.
Ao Prof. Dr. José Iran de Oliveira e a sua equipe da ESALQ/USP, pelo apoio nas
análises dos parâmetros climáticos e ao colega Carlos Henrique Garcia pelas análises
estatísticas.

vi
OFEREÇO A todos os funcionários da International Paper do Brasil, Chamflora e, em especial ao
Departamento de Tecnologia Florestal, na pessoa de seus funcionários, dos mais simples, aos
engenheiros e diretores, sem os quais seria impossível a realização desta tese, preferindo não
relatar nomes para não deixar, por puro esquecimento, pessoas importantes e que me
auxiliaram muito, fora da relação, mas tenham a certeza que jamais esquecerei a amizade e o
carinho de todos os companheiros de trabalho.
DEDICO
A minha filha, razão dos meus dias.
As minhas irmãs, sobrinhos e sobrinhas.
A minha mãe e meu pai (em memória) , pela educação e dedicação,
pois ninguém é alguém na vida sem a presença da família.
DEDICAÇÃO ESPECIAL
Existem pessoas que passam pela vida e deixam caminhos e marcas para que outras
possam seguir. Conheci uma destas pessoas na International Paper do Brasil, Chamflora, e tive
o prazer de conviver com ela e ver que a força de vontade pode, apesar das adversidades da
vida, suplantar todos os obstáculos. A você, Aparecia R. Faquinete, ou simplesmente, Dona
Cida, dedico este trabalho e sei que com Deus certamente está.

vii
SUMÁRIO
Página
LISTA DE TABELAS..................................................................................................... ix
LISTA DE FIGURAS...................................................................................................... xv
RESUMO......................................................................................................................... xvii
SUMMARY..................................................................................................................... xix
1 INTRODUÇÃO............................................................................................................ 1
2 OBJETIVOS................................................................................................................. 5
3 REVISÃO DE LITERAURA....................................................................................... 6
3.1 Sistemas hidropônicos...................................................................................... 6
3.2 Os nutrientes minerais e funções na planta...................................................... 8
3.3 Ação do pH....................................................................................................... 19
3.4 Parâmetros ambientais...................................................................................... 19
4 MATERIAIS E MÉTODOS......................................................................................... 22
4.1 Determinações realizadas................................................................................. 31
5 RESULTADOS E DISCUSSÃO.................................................................................. 33
5.1 Análise geral dos nutrientes presentes na solução nutritiva, substrato,
microestacas, broto e caule...........................................................................
34
5.2 Análise por coleta dos nutrientes presentes na solução nutritiva, substrato,
microestacas (broto) e caule..........................................................................
37
5.3 Análises dos nutrientes presentes nas brotações das microcepas, de maneira
geral, para identificação da produtividade das microestacas........................
45
5.4 Análises dos nutrientes presentes nas brotações das microcepas, por coleta
de microestacas, para identificação da produtividade...................................
47
5.5 Análises da produção de microestacas em função dos nutrientes presentes
nas soluções consideradas.............................................................................
54
5.6 Análises da produção de microestacas em função dos nutrientes presentes
nas soluções, analisando-se todas as coletas simultaneamente.....................
62

viii
5.7 Análises da produção de microestacas em função dos nutrientes presentes
nas soluções, analisando-se separadamente o efeito da produção por
coleta.............................................................................................................
63
5.8 Análises da produção de microestacas em função dos parâmetros
ambientais.....................................................................................................
70
5.9 Análise conjunta das informações tabeladas..................................................... 77
6 CONCLUSÕES............................................................................................................ 80
REFERÊNCIAS BIBLIOGRÁFICAS........................................................................... 81
APÊNDICE 1................................................................................................................... 93
APÊNDICE 2................................................................................................................... 131

ix
LISTA DE TABELAS
Página
Tabela 1. Parâmetros analisados e unidades fornecidas pelos laboratórios nas análises
do substrato, material vegetal e solução nutritiva...........................................
26
Tabela 2. Concentração de macronutrientes (g L-1) e micronutrientes (mg L-1)
presentes nas soluções nutritivas utilizadas durante o teste de omissão de
nutrientes em microcepas de Eucalyptus grandis x E.
urophylla.........................................................................................................
28
Tabela 3. Cronograma de coleta de amostras em material vegetal (folha, caule raiz e
brotação), solução hidropônica e substrato, para análise de nutrientes, no
teste de omissão, em mudas de Eucalyptus grandis x E. urophylla,
mantidas em sistema hidropônico..................................................................
30
Tabela 4. Análise de regressão para produção de microestaca de Eucalyptus grandis x
E. urophylla em função dos elementos analisados na solução, raiz, broto,
caule e substrato, geral, em função da Tabela 3, mantida em sistema
hidropônico, com omissão de nutriente e
completa..........................................................................................................
35
Tabela 5. Resumo do Stepwise Procedure for Dependent Variable para produção de
microestaca do Eucalyptus grandis x E. urophylla realizado no SAS,
referente à equação de regressão polinomial quadrática (01).........................
36
Tabela 6. Análise de regressão quadrática para produção de microestaca de
Eucalyptus grandis x E. urophylla em função dos elementos analisados na
folha, caule, raiz, broto, solução e substrato, por coleta, em função da
Tabela 3, mantida em sistema hidropônico com omissão de nutriente e
completa..........................................................................................................
38
Tabela 7. Resumo do Stepwise Procedure for Dependent Variable para microestaca
de Eucalyptus grandis x E. urophylla, realizado no SAS, referente às
equações de regressão quadrática de 02 a 09.................................................
38

x
Tabela 8. Análise de regressão quadrática para produção de microestaca de
Eucalyptus grandis x E. urophylla em função dos elementos analisados nas
brotações, geral, conforme a Tabela 3, mantida em sistema hidropônico,
com omissão de nutriente e completa.............................................................
46
Tabela 9. Resumo do Stepwise Procedure for Dependent Variable para microestaca
de Eucalyptus grandis x E. urophylla em função dos elementos analisados
nas brotações, geral, de acordo com as Tabelas 3, mantida em sistema
hidropônico com omissão de nutriente e completa.......................................
46
Tabela 10. Análise de regressão quadrática para produção de microestaca de
Eucalyptus grandis x E. urophylla em função dos elementos analisados nas
brotações, por coleta, em função da Tabela 3, mantida em sistema
hidropônico, com omissão de nutriente e completa.......................................
47
Tabela 11. Resumo do Stepwise Procedure for Dependent Variable para produção
microestacas de Eucalyptus grandis x E. urophylla em função dos
elementos analisados nas brotações, por coleta, de acordo com a Tabela 3,
mantida em sistema hidropônico, com omissão de nutriente e
completa..........................................................................................................
48
Tabela 12. Análise de regressão quadrática para produção de microestaca de
Eucalyptus grandis x E. urophylla em função da solução considerada, de
acordo com a Tabela 3, mantida em sistema hidropônico,com omissão de
nutriente e completa........................................................................................
54
Tabela 13. Resumo do Stepwise Procedure for Dependent Variable para microestaca
de Eucalyptus grandis x E. urophylla em função da solução considerada,
de acordo com a Tabela 3, mantida em sistema hidropônico, com omissão
de nutriente e completa...................................................................................
55
Tabela 14. Análise de regressão quadrática para produção de microestacas de
Eucalyptus grandis x E. urophylla, analisando-se apenas o efeito das
soluções, para todas as coletas, de acordo com a Tabela 3, mantida em
sistema hidropônico, com omissão de nutriente e
completa.........................................................................................................
62

xi
Tabela 15. Resumo do Stepwise Procedure for Dependent Variable para microestacas
de Eucalyptus grandys x E. urophylla, realizado no SAS, referente à
equação de regressão quadrática (27).............................................................
Tabela 16. Análise de regressão quadrática para produção de microestacas de
Eucalyptus grandis x E. urophylla em função dos elementos analisados na
solução, por coleta, de acordo com a Tabela 3, mantida em sistema
hidropônico, com omissão de nutriente e completa......................................
63
64
Tabela 17. Resumo do Stepwise Procedure for Dependent Variable para produção de
microestacas de Eucalyptus grandis x E. urophylla, realizado no SAS,
referente às equações de regressões quadráticas de 28 a 35...........................
64
Tabela 18. Teor de nitrogênio (g kg-1) presente na análise foliar da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
93
Tabela 19. Teor de fósforo (g kg-1) presente na análise foliar da cepa de Eucalyptus
grandis x E. urophylla, em função da coleta e da omissão de nutrientes.......
94
Tabela 20. Teor de potássio (g kg-1) presente na análise foliar da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
95
Tabela 21. Teor de cálcio (g kg-1) presente na análise foliar da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
96
Tabela 22. Teor de magnésio (g kg-1) presente na análise foliar da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
97
Tabela 23. Teor de ferro (mg kg-1) presente na análise foliar da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
98

xii
Tabela 24. Teor de boro (mg kg-1) presente na análise foliar da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
99
Tabela 25. Quantidade de microestacas de Eucalyptus grandis x E. urophylla, em
função da coleta e da omissão de nutrientes...................................................
Tabela 26. Teor de nitrogênio (g kg-1) presente em microestacas de Eucalyptus
grandis x urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
100
101
Tabela 27. Teor de fósforo (g kg-1) presente em microestacas de Eucalyptus grandis
x E. urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
Tabela 28. Teor de potássio (g kg-1) presente em microestacas de Eucalyptus grandis
x urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
102
103
Tabela 29. Teor de Cálcio (g kg-1) presente em microestacas de Eucalyptus grandis x
E. urophylla, em função da coleta e da omissão de nutrientes.......................
104
Tabela 30. Teor de magnésio (g kg-1) presente em microestacas de Eucalyptus
grandis x E. urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
105
Tabela 31. Teor de ferro (mg kg-1) presente em microestacas de Eucalyptus grandis x
E. urophylla, em função da coleta e da omissão de nutrientes.......................
106
Tabela 32. Teor de boro (mg kg-1) presente em microestacas de Eucalyptus grandis x
E. urophylla, em função da coleta e da omissão de nutrientes......................
107
Tabela 33. Teor de nitrogênio (g kg-1) presente na análise da raiz da cepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
108
Tabela 34. Teor de fósforo (g kg-1) presente na análise da raiz da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
109

xiii
Tabela 35. Teor de potássio (g kg-1) presente na análise da raiz da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
110
Tabela 36. Teor de cálcio (g kg-1) presente na análise da raiz da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
111
Tabela 37. Teor de magnésio (g kg-1) presente na análise da raiz da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
112
Tabela 38. Teor de ferro (mg kg-1) presente na análise da raiz da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
113
Tabela 39. Teor de boro (mg kg-1) presente na análise da raiz da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
114
Tabela 40. Teor de fósforo (mg dm-3) presente na análise do substrato da microcepa
de Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
115
Tabela 41. Teor potássio (mmolc dm-3) presente na análise do substrato da microcepa
de Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
116
Tabela 42. Teor de cálcio (mmolc dm-3) presente na análise do substrato da microcepa
de Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes......................................................................................
117
Tabela 43. Teor de magnésio (mmolc dm-3) presente na análise do substrato da
microcepa de Eucalyptus grandis x E. urophylla, em função da coleta e da
omissão de nutrientes......................................................................................
118
Tabela 44. Teor de ferro presente (mg dm-3) na análise do substrato da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes....................................................................................................
119

xiv
Tabela 45. Teor de boro (mg dm-3) presente na análise do substrato da microcepa de
Eucalyptus grandis x urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
120
Tabela 46. Valores de pH da solução da microcepa de Eucalyptus grandis x E.
urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
121
Tabela 47. Teor de NH+4 (mg L-1) presente na análise da solução da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
122
Tabela 48. Teor de NO-3 (mg L-1) presente na análise da solução da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
123
Tabela 49. Teor de fósforo (mg L-1) presente na análise da solução da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
124
Tabela 50. Teor de potássio (mg L-1) presente na análise da solução da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
125
Tabela 51. Teor de cálcio (mg L-1) presente na análise da solução da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes.......................................................................................................
126
Tabela 52. Teor de magnésio (mg L-1) presente na análise da solução da microcepa
de Eucalyptus grandis x E. urophylla, em função da coleta e da omissão
de nutrientes...................................................................................................
127
Tabela 53. Teor de ferro (mg L-1) presente na análise da solução da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
128
Tabela 54. Teor de boro (mg L-1) presente na análise da solução da microcepa de
Eucalyptus grandis x E. urophylla, em função da coleta e da omissão de
nutrientes........................................................................................................
129

xv
LISTA DE FIGURAS
Figura 1. Metodologia utilizada para produção de mudas a partir de matrizes
selecionadas, cultivadas in vitro e sistema hidropônico...............................
25
Figura 2. Etapas e equipamentos utilizados para realização do experimento. “A” e
“B”: limpeza das cubas de recebimento da solução; “C”: material vegetal
em água; “D”: material vegetal coletado para análise laboratorial; “E”:
tanques de solução; “F”: retirada de amostras para análises “G”;
automatização do sistema hidropônico em “H” e “I” e sensores de
radiação global “J”, radiação PAR “L” e temperatura e umidade
“M”.Metodologia utilizada para produção de mudas a partir de matrizes
selecionadas, cultivo in vitro e sistema hidrôponico.....................................
27
Figura 3. Efeito da umidade relativa externa e interna máximas sobre a produção de
microestacas de Eucalyptus grandis x E. urophylla, em solução completa
e com omissão de nutrientes, mantidas em sistema hidropônico, em casa-
de-vegetação, durante o período de 23 de maio a 19 de setembro de 2002.
72
Figura 4. Efeito da umidade relativa externa e interna médias sobre a produção de
microestacas de Eucalyptus grandis x E. urophylla, em solução completa e
com omissão de nutrientes, mantidas em sistema hidropônico em casa-de-
vegetação, durante o período de 23 de maio a 19 de setembro de 2002.......
72
Figura 5. Efeito da umidade relativa externa e interna mínimas sobre a produção de
microestacas de Eucalyptus grandis x E. urophylla, em solução completa e
com omissão de nutrientes, mantidas em sistema hidropônico em casa-de-
vegetação, durante o período de 23 de maio a 19 de setembro de 2002........
73
Figura 6. Efeito da temperatura externa e interna máximas sobre a produção de
microestacas de Eucalyptus grandis x E. urophylla, em solução completa e
com omissão de nutrientes, mantidas em sistema hidropônico em casa-de-
vegetação, durante o período de 23 de maio a 19 de setembro de 2002.......
74
Figura 7. Efeito da radiação global externa e interna máximas sobre a produção de
microestacas de Eucalyptus grandis x E. urophylla, em solução completa e
com omissão de nutrientes, mantidas em sistema hidropônico, em casa-de-

xvi
vegetação,durante o período de 23 de maio a 19 de setembro de 2002........ 75
Figura 8. Efeito da radiação fotossintéticamente ativa, externa e interna máximas
sobre a produção de microestacas de Eucalyptus grandis x E. urophylla,
em solução completa e com omissão de nutrientes, mantidas sistema
hidropônico, em casa-de-vegetação, durante o período de 23 de maio a 19
de setembro de 2002.....................................................................................
75
Figura 9. Efeito da radiação fotossintéticamente ativa, externa e interna máximas
sobre a produção de microestacas de Eucalyptus grandis x E. urophylla,
em solução completa e com omissão de nutrientes, mantidas em sistema
hidropônico, em casa-de-vegetação, durante o período de 23 de maio a 19
de setembro de 2002....................................................................................
76
Figura 10. Sintomas de deficiência de nitrogênio e de fósforo em híbrido de
Eucalyptus grandis x E. urophylla, comparados à testemunha (solução
completa), 79 dias após o momento em que as mudas receberam solução
nutritiva (M.0.)...............................................................................................
132
Figura 11. Sintomas de deficiência de potássio e de cálcio apresentados pelo híbrido
de Eucalyptus grandis x E. urophylla, comparados à testemunha (solução
completa), 79 dias após o momento em que as mudas receberam solução
nutritiva (M.0.)..............................................................................................
133
Figura 12. Sintomas de deficiência de magnésio e boro apresentados pelo híbrido de
Eucalyptus grandis x E. urophylla, comparados à testemunha (solução
completa), 79 dias após o momento em que as mudas receberam solução
nutritiva (M.0.)..............................................................................................
134
Figura 13. Sintomas de deficiência de ferro apresentados pelo híbrido de Eucalyptus
grandis x E. urophylla, comparados à testemunha (solução completa), 79
dias após o momento em que as mudas receberam solução nutritiva
(M.0.)............................................................................................................
135

xvii
AVALIAÇÕES AMBIENTAL E NUTRICIONAL DA PRODUÇÃO DE MICROCEPAS E
MICROESTACAS DE Eucalyptus grandis x E. urophylla EM SISTEMA
HIDROPÔNICO EM CASA-DE-VEGETAÇÃO. Botucatu, 2003. 135 p. Doutorado em
Agronomia – Área de Concentração em Energia na Agricultura. Faculdade de Ciências
Agronômicas, Universidade Estadual Paulista.
Autor: AGNALDO SCARASSATI
Orientador: IRAÊ AMARAL GUERRINI
RESUMO
Este trabalho foi realizado na International Paper do Brasil,
localizada no município de Mogi-Guaçu, SP. O objetivo foi verificar através da omissão de
nutrientes minerais nas soluções nutritivas, a produção de microestacas das microcepas,
mantidas em casa-de-vegetação, através de sistemas hidropônicos em Eucalyptus grandis x E.
urophylla, acompanhando as interrelações entre os nutrientes, além de caracterizar os
parâmetros ambientais que poderiam estar influenciando no processo. O delineamento
experimental utilizado foi inteiramente casualizado, com três repetições para a solução
hidropônica, substrato e material vegetal. Os resultados obtidos permitiram verificar que para
a otimização do sistema de produção de microestacas são necessárias análises laboratoriais
para os nutrientes presentes na solução nutritiva, substrato e material vegetal (brotação, caule e
folha) e que há modificações estatísticas em relação ao valor que cada nutriente apresentado
tem, em relação à época e fase de desenvolvimento do material. Em relação aos parâmetros
ambientais: temperatura, umidade relativa, radiação global e PAR, pode-se afirmar que a
produção de microestacas oscila em relação à umidade relativa na casa-de-vegetação em torno
de 91,7% para valores máximos, de 79% a 81% para valores médios e de 32% a 51,4% para
valores mínimos. As temperaturas máximas internas que permitiram maiores produtividades
estão entre 26oC a 32oC. A radiação global adequada para maximizar a produção variou de
336,7 W/m2 a 416,9 W/m2 dentro da casa-de-vegetação, enquanto que a PAR (V/Wm2) variou

xviii
de 357,7 V/Wm2 a 1446,2 V/Wm2, com maiores valores de produção de microestacas a 722
V/Wm2.
Palavras-Chaves: Eucalipto, hidroponia, nutrição mineral, microestaquia.

xix
NUTRICIONAL EVALUATION OF MICROHADGE AND MICROCUTTING OF
Eucalyptus grandis x E. urophylla IN HYDROPONIC SYSTEM IN GREENHOUSES.
Botucatu, 2003. 135 p. Doutorado em Agronomia/Energia na Agricultura. Faculdade de
Ciências Agronômicas, Universidade Estadual Paulista.
Author: AGNALDO SCARASSATI
Adviser: IRAÊ AMARAL GUERRINI
SUMMARY
This work was realized at International Paper of Brazil located
in the city of Mogi-Guaçu, state of São Paulo. The aim of the study was to verify, through the
omission of mineral nutrients in nutritive solutions, the production of microcuttings of the
minicuttings, kept in greenhouses, through hydroponics systems in Eucalyptus grandis x E.
urophylla, clones. Following the inter relations among the nutrients, besides characterizing the
environmental parameters that could be influencing the process. The experimental line used
ANOVA model and regression analysis was entirely, with three repetitions for the
hydroponics solution, substrate and biomass. The results obtained allowed to verify that, for
the optimization of the production system of micro stakes, laboratory analysis are necessary
for the nutrients that are present in the nutritive solution, substrate and biomass (sprout, stalk
and leaf) and that are statistical changes related to the participation that each nutrient shows,
related to the cycle and development stage of the material. Regarding environmental
parameters: temperature, relative moisture, global radiation and PAR, we can say that the
highest productions of micro cuttings oscillate. The relative moisture in the greenhouse was
around 91,7% maximum: from 79% to 81% medium values and between 32% and 51,4%
minimum values. Regarding maximum internal temperatures that allow the highest
productivities, they range from 26ºC to 32ºC. The adequate global radiation to maximize the
production is 336,7 W/m2 to 416,9 W/m2 inside the greenhouse and the radiation PAR
(V/Wm2) varied a lot (357,7 V/Wm2 at 1446,2 V/Wm2) but the highest productions were
obtained at 722 V/Wm2.
_________________________________________
Key words: Eucalyptus, hydroponics culture, mineral nutrition, soil less culture, micro cutting

1
1 INTRODUÇÃO
O eucalipto é a mais importante espécie nas plantações comerciais no Brasil com
finalidade de produção de pasta celulósica, papéis, lenha, carvão-vegetal e mais recentemente,
está sendo utilizado para substituição de outras madeiras na confecção de móveis.
São milhões de hectares plantados em todo o país. A sua produção sustentada requer
pesquisas com a espécie, principalmente em relação ao teor de celulose, potencial calorímetro
(no caso da produção de carvão-vegetal) e estudos das movimentações dimensionais para a
sua utilização na indústria moveleira.

2
Porém, nenhumas destas características poderiam ser aprimoradas e reproduzidas se não
houvesse um processo competente de melhoramento genético e de propagação. Em função
desta importância, o gênero Eucalyptus, no atual cenário da silvicultura, teve aperfeiçoado a
técnica de estaquia com o advento da miniestaquia e microestaquia (Assis, 2002).
Atualmente esse sistema de clonagem torna possível a continuidade genética do material
e a sua estabilidade nos viveiros florestais, e os sistemas de produções hidropônicos
maximizam a produção de mudas.
Esta técnica de propagação é utilizada há mais de dez anos, mas em alguns casos,
porém, para que o processo possa ser mantido dentro de padrões médios de produtividade e
homogeneidade do material, otimizando a produção em escala, é necessário que ocorra a
reversão à juvenilidade das matrizes, conseguida através de processos da micropropagação in
vitro e constante substituição, pelo menos uma vez a cada seis meses, das microcepas que
estão em sistema hidropônico.
O progresso desta técnica de cultivo de plantas está ligado diretamente aos
conhecimentos obtidos com a fisiologia e a nutrição da espécie, com especial estudo para os
processos fotossintéticos, transpiração foliar, parâmetros de conforto térmico vegetal, diagnose
nutricional, juntamente com o uso de estruturas protegidas, equipadas com sensores diversos,
desde os mais simples, como os eletrocondutivimetros (para medição da concentração de sais
da solução - EC), ao sensor PAR, instalados em estações metereológicas para medição da
fotossíntese ativa.
A nutrição e as condições ambientais de produção são parâmetros importantes para se
entender e otimizar o sistema. Marschner (1995) salienta que a nutrição das plantas deve ser
estudada em relação à função exercida pelo elemento mineral na planta e, uma das maneiras

3
de avaliar sua ação, segundo Kramer & Kozlowski (1960 e 1979), é através do processo de
indução de deficiências, quer dos assim chamados macronutrientes ou através da indução de
deficiências dos micronutrientes, com a verificação da ação deste processo no metabolismo.
Se há um papel específico e essencial dependente da presença de um nutriente, ao
retirá-lo da nutrição da planta, tem-se essa função explicitada, com algum grau de
especificidade conhecida, embora, em termos químicos, a maneira pela qual o elemento está
implicado no processo possa ser complexa e merecedora de estudos mais detalhados (Epstein,
1975). Hoje já se sabe que alterações nos níveis de carboidratos e proteínas têm relação com
os parâmetros nutricionais.
Experimentos realizados com eucalipto, segundo Neves (1983), tem-se mostrado
algumas correlações entre a nutrição deste em casa de vegetação e no campo. Alta tolerância
ao alumínio, preferencial absorção por amônio em relação a nitrato e baixa concentração
crítica de cálcio e magnésio, têm-se observado em espécies de eucalipto tanto em casa-de-
vegetação quanto no campo (Barros & Silva, 1990).
Barros & Silva (1990) também verificaram que a limitação para o crescimento do
eucalipto em solos de cerrado era devido ao pouco enxofre existente. Este fato foi inicialmente
observado em casa de vegetação e a adição de enxofre em plantações comerciais é atualmente
prática rotineira.
De um modo geral, os sintomas de deficiência ou excesso de um determinado elemento
mineral é semelhante em todas as espécies de plantas. E o motivo pelo qual o sintoma é típico
do elemento, deve-se ao fato de que ele exerce sempre a mesma função, qualquer que seja o
vegetal (Meyer et al., 1983).

4
Entretanto, existem respostas peculiares entre e dentro das espécies, como resultado da
expressão genética, influenciando a distribuição dos elementos ou a sensibilidade de sistemas
metabólicos (Vose, 1963).

5
2 OBJETIVOS
a) Verificar a interrelação entre nutrientes, utilizando a omissão de alguns, como fonte
principal de informação do efeito sobre as microcepas (parte aérea e sistema radicular)
e microestacas de Eucalyptus grandis x E. urophylla, substrato e soluções nutritivas
utilizadas;
b) Acompanhar a produtividade das microcepas, em solução completa e com a omissão
de nutrientes, em relação às microestacas de Eucalyptus grandis x E. urophylla,
cultivadas em sistema hidropônico;
c) Caracterizar os parâmetros ambientais que poderiam estar influenciando na produção
de biomassa vegetal.

6
3 REVISÃO DE LITERATURA
3.1. Sistemas hidropônicos
A hidroponia, termo derivado de duas palavras de origem grega “hydro”, que significa
água e “ponos”, que significa trabalho, foi proposto inicialmente por Willian F. Gericke, nos
EUA, por volta de 1930, quando utilizou esta técnica de cultivo em escala comercial (Bliska &
Honório, 1996). Atualmente está se desenvolvendo rapidamente como meio de produção
vegetal (Furlani, 1995 e 1998), sendo utilizada na área florestal para o estabelecimento do
jardim miniclonal (Xavier, 2002; Assis, 2002).
De todos os métodos de cultivo sem solo, o cultivo em água, por definição, é o
autêntico cultivo hidropônico.

7
Existem várias maneiras de se cultivar plantas em sistemas hidropônicos de acordo
com Ikeda & Osawa (1980) e Resh (1992), porém os mais utilizados são nutrient film
technique (NFT) ou técnica do fluxo laminar de nutrientes, deep film technique (DFT) ou
cultivo na água ou floating, e técnicas de cultivo com substrato.
No caso do deep film technique (DFT), a solução nutritiva forma uma lâmina profunda
(5 a 20 cm), onde as raízes ficam submersas. Não existem canais e sim uma mesa plana onde
circula a solução, através de um sistema de entrada e drenagem característica (Furlani, 1998).
Pode-se misturar neste sistema, vasos, como os tubetes, que são preenchidos com materiais
diversos como areia, vermiculitas, perlita, lã de rocha, espuma fenólica, espuma de
poliuretano, fibra de coco ou misturas destes, além de outros compostos como palha de arroz
carbonizada e turfas.
Alburuqerque & Dechen (2000) utilizaram o sistema hidropônico para avaliar a
capacidade de absorção de macronutrientes nos porta-enxertos de videiras, concluíndo que há
absorção seletiva de nutrientes em função dos cultivares analisados.
O método de cultivo em solução hidropônica ou “soilless culture” permite a
observação mais apurada dos efeitos fisiológicos, nutricionais e anatômicos de plantas
mantidas neste sistema. Souza (2001) trabalhando com soja nos sistemas com solo e em
solução hidropônica, comprovou que os experimentos em hidroponia e em solo são
igualmente eficientes no caso de um projeto de seleção de soja tolerante ao alumínio e
evidenciou que o sistema hidropônico permitiu melhor controle das variáveis não
experimentais, o que aumenta a precisão dos testes.
Garland et al. (1990) encontraram correlação entre o comprimento das raízes de
plântulas de soja cultivadas em sistema de hidroponia e o peso seco das plantas, aferindo a

8
este sistema a capacidade de imitar uma possível situação de campo, como mencionado
anteriormente.
Outra vantagem ao se trabalhar com soluções nutritivas é poder correlacionar melhor o
efeito de um nutriente em outro como, por exemplo, o efeito do zinco e do fósforo na absorção
e na produtividade da alface (Lactuca sativa) (Moreira et al. 2001).
Além das interações apresentadas, quer para seleção do material, quer para o estudo
individualizado de nutrientes ou suas interações, os sistemas hidropônicos são mais produtivos
para algumas espécies. Maia (1998) trabalhando com um sistema automático de fornecimento
de solução nutritiva para cultivo hidropônico de plantas em vasos, comprovou que esse
sistema, em relação ao convencional, foi três vezes mais produtivo para o cultivo de hortelã
(Mentha crispa).
3.2. Os nutrientes minerais e as suas funções na planta
O atual estágio de produção de mudas de eucalipto, criou um sistema conhecido como
jardim miniclonal, em que as minicepas ou microcepas (explantes vegetais produzidos “in
vitro” e formadores das cepas), podem ser conduzidas diretamente em tubetes, por sistema
hidropônico (Wendling et al. 2000).
A nutrição mineral utilizada no jardim miniclonal é, normalmente, composta por macro
e micronutrientes, sendo vartiável. O fator material genético (clone), ambiente e sistema de
condução das mini ou microcepas, são determinantes para a melhor formulação e balanço dos
elementos que compõem tal solução nutritiva (Assis et al., 1990).

9
O método apresenta como vantagem à rapidez e não ser destrutivo, na maioria das
vezes, ou permitir uma quantidade maior de repetições em espaços reduzidos, aumentando a
confiabilidade estatística dos resultados (Souza, 2001).
O atual estágio de desenvolvimento das técnicas de cultivo, permite melhor
detalhamento das condições de cultivo, por utilizarem métodos de propagação assexuada e
casa-de-vegetação com sensores diversos instalados.
Portanto, as informações básicas sobre a nutrição mineral assumem grande
importância, principalmente quando se visa a utilização mais adequada dos fertilizantes em
função das exigências nutricionais dos clones ou classes de clones (Sgarbi et al., 1999).
Não obstante os estudos sobre o crescimento do eucalipto, em condições de carências
nutricionais, foram realizados para diferentes espécies, a partir de mudas propagadas por
sementes (Rocha Filho et al., 1979).
Segundo Maffeis et al. (2000) em trabalho realizado com a omissão de N, P, K, Ca, S,
Mg, B, em Eucalyptus citriodora, somente nas ausências de N, K e B foram visualizados
sintomas de carência nutricional. Na falta de nitrogênio, inicialmente as folhas mais velhas
apresentaram uma coloração verde-clara, que com o passar do tempo progredia para um
amarelecimento generalizado, proporcionando rápida senescência das folhas. Segunda os
autores, este sintoma deve-se ao decréscimo no teor de clorofila. O cultivo das plantas em
solução completa durante os três meses iniciais forneceu quantidade suficiente de P, Ca, Mg e
S para atender à demanda até a idade de 11 meses. Observaram que os tratamentos, em que se
omitiu N, Ca, Mg, S e B apresentaram menor crescimento em altura quando comparado a
solução completa.

10
As concentrações foliares de N, P, Mg, S e B, obtidas nos tratamentos, segundo os
mesmos autores (Maffeis et al. 2000), estavam abaixo da faixa proposta como adequada para o
gênero Eucalyptus ssp. por Malavolta (1987) e, para E. maculata por Dell & Robinson (1993).
Nos tratamentos com omissão de K e Ca as concentrações situaram-se dentro da faixa
proposta como adequada por Malavolta (1987). Segundo também Malavolta & Crocomo
(1982) e Malavolta et al. (1997) maiores concentrações foliares de potássio ocorreram quando
o magnésio estava ausente na solução, pois esses macronutrientes competem pelo mesmo sítio
de absorção.
A competição entre magnésio e potássio ocorre durante o processo de absorção
radicular, uma vez que utilizam os mesmos sítios de absorção. Entretanto, o efeito negativo do
desbalanço de nutrientes somente é detectado a partir do incremento na produção de matéria
seca, florescimento ou frutificação da planta, através da determinação do acúmulo de
nutrientes. Porém a correção do problema, dependendo do estágio de desenvolvimento do
material, torna-se difícil ou a custos elevados. Mortvedt & Khasawneh (1986) atribuíram ao
balanço de nutrientes no solo, a responsabilidade por limitações no crescimento das plantas.
Os efeitos interiônicos entre K, Ca e Mg ocorrem na forma de inibição competitiva,
normalmente ao nível de membrana celular (Epstein, 1975). Segundo Malavolta (1980) esse
processo ocorre quando dois elementos se combinam pelo mesmo sítio ativo do carregador.
Um exemplo clássico é dado pelas altas doses de potássio no meio, inibindo a absorção de Ca
e Mg, chegando muitas vezes a causar deficiência desses dois nutrientes com queda de
produção. Marschner (1995) também relata que cátions como o potássio pode atravessar a
membrana plasmática com maior velocidade, deprimindo a absorção de cátions mais lentos

11
como Ca e Mg. A absorção preferencial do íon K+ ocorre por ser monovalente e de menor
grau de hidratação quando comparado aos divalentes (Kabata-Pendias & Pendias, 1984).
3.2.1 Nitrogênio
O nitrogênio é um dos elementos essenciais para o desenvolvimento e produção das
plantas. Não há, entretanto, um consenso a respeito do nível ótimo de adubação nitrogenada
para se obter o máximo rendimento em diversas culturas e mesmo em sistemas hidropônicos.
Aparentemente, a resposta das culturas a diferentes níveis de N depende do cultivar e das
condições ambientais.
A redução do crescimento proporcionada pela deficiência de nitrogênio é resultado das
funções que o nutriente desempenha na planta. Normalmente, o N é o nutriente mais exigido
pelas culturas, uma vez que atua como estrutural nas moléculas dos aminoácidos, proteínas,
enzimas, pigmentos e produtos secundários (Marschner, 1995; Malavolta et al. 1997).
Carelli et al. (1996) investigaram a atividade da redutase de nitrato in vivo, a
fotossíntese líquida, as trocas gasosas e o crescimento em plantas de girassol (Helianthus
annuus L., cv. Contisol), cultivadas em vasos com areia lavada e irrigadas com soluções
nutritivas com níveis de nitrato de 3,75; 7,5; 15 e 30 mM. A fotossíntese, a transpiração e o
teor de N total das folhas aumentaram com o nível de nitrato das soluções nutritivas até um
valor máximo, que variou de 21 mM a 26 mM de nitrato, decrescendo a seguir. A resistência à
difusão diminuiu com o aumento da concentração de nitrato até o nível de 22 mM. A atividade
da redutase de nitrato aumentou linearmente com os níveis de nitrato das soluções nutritivas.
O crescimento da parte aérea das plantas avaliadas pela massa seca das folhas e hastes, área

12
foliar, número de folhas e da altura das plantas, apresentou resposta quadrática ao nível de
nitrato, atingindo o valor máximo entre 20 e 22 mM. O crescimento das raízes não foi alterado
pelo fornecimento de nitrato. A deficiência em nitrogênio provocou decréscimos na
fotossíntese, na atividade da redutase de nitrato, na transpiração e no crescimento, e aumento
na resistência a difusão.
A produtividade das culturas é grandemente determinada pela interação entre o
metabolismo do carbono e do nitrogênio. Estes estão estreitamente interligados uma vez que a
energia necessária para a assimilação do nitrogênio deriva direta ou indiretamente da
fotossíntese. Por sua vez, a capacidade fotossintética depende do suprimento de nitrogênio,
pois grande parte deste nas folhas está alocada nas proteínas, envolvidas no processo
fotossintético, principalmente na enzima Rubisco no caso das plantas C3. De fato, numerosos
trabalhos em diversas espécies de plantas mostraram acentuada e positiva relação entre o teor
de nitrogênio na folha e a capacidade fotossintética (Malavolta, 1980).
Bugarím et al. (1998), em estudo da relação entre o nitrato (N-N0-3) e a amônia (N-
NH+4) em crisântemo var. Puma, observaram as diferentes pressões osmóticas exercidas por
combinações variadas destes elementos na extração de nutrientes, cultivada esta espécie em
um sistema hidropônico por subirrigação. Com o incremento da pressão osmótica de 0,54 a
0,90 atm, diminuiu a concentração de Ca e Mg nas folhas; os demais nutrientes não foram
afetados pela concentração osmótica da solução.
Segundo Steiner (1984 e 1985) quando a pressão osmótica da solução nutritiva é maior
em relação á requerida pela espécie, os processos antes utilizados de absorção se invertem e a
absorção de nutrientes por fluxo de massa diminui, aumentando a absorção por difusão. No
caso do P, Ca e Mg, à explicação das alterações na absorção, estão ligadas à acidificação

13
decorrente de maiores quantidades de NH+4 na solução nutritiva. Conforme o pH da solução se
acidifica, aumenta a absorção destes elementos.
Silveira et al. (1996) constataram que a omissão de nitrogênio e boro foram limitantes
ao crescimento de híbrido de E. grandis x E. urophylla.
Pode haver também interações no valor do pH da solução em combinações com baixas
concentrações de nitrogênio, induzindo a deficiências de Mn (Savvas et al., 2003)
3.2.2 Fósforo
O papel central que o fosfato desempenha na energética do metabolismo e nas reações
biossintéticas sugere que sua deficiência dificilmente seria menos desastrosa do que a do
nitrogênio, o que de fato acontece. O fosfato é necessário para a síntese de trifosfato de
adenosina (ATP) e de numerosos outros compostos fosforilados e sua carência, por
conseguinte causa transtornos imediatos e severos no metabolismo e no desenvolvimento
(Epstein, 1975).
Plantas deficientes em fósforo caracterizam por possuir, crescimento lento,
freqüentemente coloração avermelhada devido ao aumento da formação de antocianinas;
folhas com uma coloração mais escura quando comparadas com folhas de plantas com
nutrição completa e redução de muitos processos metabólicos, incluindo divisão e expansão
celular, respiração e fotossíntese (Marschner, 1995).
As diagnoses de fertilidade de solo e problemas nutricionais para crescimento de
eucalipto no Brasil são baseadas em análises de alta drenagem e geralmente são utilizados

14
solos pobres para o cultivo de eucalipto. Tem-se focado mais na planta como uma ferramenta
para a diagnose de limitações nutricionais.
Fósforo é certamente o principal nutriente nos solos pobres e de baixa fertilidade da
região do cerrado e muito se tem pesquisado sobre métodos de acesso, avaliação e
interpretação de resultados. A concentração critica de fósforo disponível para o crescimento de
mudas de E. grandis pode variar dependo do solo e dependendo da solução de extração
(Neves, 1983).
A grande demanda inicial de fósforo por eucaliptos justifica a grande quantidade de
fertilização fosfatada recomendada no momento do transplante. (Barros & Silva, 1990).
Porém, a omissão de P, segundo Sgarbi et al. (1999) não proporcionou redução de
biomassa do eucalipto, quando comparado ao tratamento completo. Rocha Filho et al. (1979)
encontraram quedas de 69% na produção de biomassa de mudas de E. urophylla, quando
cultivadas sob carência de P em relação à solução completa.
3.2.3 Potássio
Já o potássio é altamente móvel no floema. Sua utilização é por isso eficiente no
sentido de ser prontamente redistribuído das folhas mais velhas para os órgãos mais novos em
crescimento. Como conseqüência disso os sintomas de deficiência aparecem em primeiro
lugar nas folhas mais velhas, que ficam com coloração nas bordas que progride em direção ao
centro das folhas (Epstein, 1975; Muniz et al., 2001).
A diminuição na proporção de ramos e caule na omissão de potássio é dada pela menor
migração de fotoassimilados através de vasos condutores, relacionada com o processo de

15
fotofosforilação, pois quando o teor de potássio é grande, ocorre estímulo na produção de ATP
(Malavolta & Crocomo, 1982).
Em plantas carentes de potássio, ocorrem danos nas atividades estomáticas, pois esse
íon quando em teor adequado reduz a taxa de transpiração, regulando o potencial osmótico do
mesófilo celular (Mengel & Kirkby, 1979 e 1982).
3.2.4 Magnésio
No caso do magnésio, ele é componente da clorofila e ativador de numerosas enzimas
que afetam a transferência do fosfato e é um elemento cuja deficiência afeta fortemente o
metabolismo. A clorose é um sintoma inicial, seguido da diminuição da fotossíntese. Os
caminhos biossintéticos são desarranjados como conseqüência da inibição em transforilações
enzimáticas essenciais (Epstein, 1975).
O teor de magnésio nos órgãos vegetais é menor quando há deficiência do mesmo
(Camargos, 1999). Epstein (1975) evidenciou inibição competitiva do cálcio e do potássio na
absorção do magnésio.
Segundo trabalho executado por Sgarbi et al. (1999), plantas cultivadas na ausência de
K e de Mg não apresentam redução significativa na produção de matéria seca quando
comparadas com as do tratamento completo. Admite-se, segundo os mesmos autores, que o
clone de E. grandis x E. urophylla, utilizado no estudo, é pouco exigente em K, haja vista os
baixos teores foliares encontrados no tratamento com omissão deste macronutriente e a
elevada produção de matéria seca. O E. grandis x E. urophylla apresentou teores foliares
adequados de Mg, de acordo com os valores de Dell et al. (1995), mesmo quando cultivado na

16
ausência deste macronutriente. Este fato pode ter ocorrido em função do Mg fornecido até os 9
meses de idade ter sido suficiente para garantir o crescimento normal das plantas até a idade
de 26 meses, segundo citam os autores.
3.2.5 Cálcio
Outro elemento químico de relevância é o cálcio. Sendo um elemento imóvel no
floema, os desarranjos devido à deficiência, podem ser muito localizados, e a raiz é o órgão
afetado mais severamente, ele cessa de crescer, torna - se desorganizado e escuro e às vezes
até morre. Antes que isso aconteça, o processo de divisão mitótica da célula é desarranjado
podendo haver núcleos poliplóides, células binucleadas, núcleos constritos e divisões
amitóticas (Epstein, 1975).
Kaul et al. (1968), Rocha Filho et al. (1978) observaram, a omissão de cálcio na
solução não reduziu a produção de biomassa do E. grandis x E. urophylla. Esses resultados
indicam que a exigência nutricional do Eucalyptus em relação ao Ca é baixa quando
comparada com outras culturas, conforme relatado por Barros & Silva (1990).
3.2.6 Boro
Quanto ao boro, Valência (1964) salientou a necessidade de aplicações de B no solo
para prevenir o aparecimento de sintomas, como a morte de gemas, usando doses de 20 g de
borax®/planta/ano. Apesar de ser o nutriente cuja participação no metabolismo vegetal é
menos compreendida, a visão global de trabalhos com este elemento permite que hipóteses

17
sejam aventadas para explicar as mudanças fisiológicas, anatômicas ou morfológicas que
ocorrem quando há deficiência no substrato para suprir as necessidades do metabolismo da
planta.
A redução de altura na carência de B e Ca deve-se ao fato desses nutrientes atuarem no
crescimento meristemático das plantas (Marschner, 1995; Malavolta et al. 1997). As plantas
de E. citriodora deficientes, principalmente em boro, apresentaram morte das gemas apicais,
com posterior brotação das gemas axilares, resultando na paralisação do crescimento em
altura. Em relação ao diâmetro, verificou-se redução e o significativo crescimento apenas na
omissão de N. Para a produção de folhas, o comportamento dos tratamentos foi semelhante ao
obtido para o crescimento em altura, onde a carência de N e B diminuiu drasticamente a
biomassa foliar.
A omissão de boro na solução resultou em maiores concentrações de N, P, Ca e S nas
folhas, provavelmente devido ao menor crescimento das plantas, provocando menor efeito de
diluição dos nutrientes. Verificou-se que as concentrações de Cu, Fe e Zn nas folhas foram
mais elevadas nas plantas cultivadas na ausência de boro. O mesmo não ocorreu em relação às
concentrações foliares de Mn, uma vez que não se verificaram diferenças significativas entre
os tratamentos. Em todos os tratamentos, exceto na omissão de boro, ocorreram baixas
concentrações de Cu, podendo estas ser consideradas deficientes de acordo com Malavolta
(1987), Dell et al. (1995) e Silveira (2000).
Segundo Silveira (2000) no mesmo trabalho, as concentrações de Fe nas folhas para
todos os tratamentos foram consideradas acima da faixa adequada (Malavolta, 1987; Dell et
al., 1995). Em relação ao Mn, as concentrações obtidas para todos os tratamentos estavam
dentro da faixa adequada. As concentrações foliares de Zn foram consideradas adequadas com

18
base em Malavolta (1987), exceto na omissão de boro, cujos valores foram em torno de duas
vezes maiores que dos demais tratamentos (Silveira, 2000).
Em trabalho sob diferentes doses de boro em Eucalyptus citriodora, Maffeis et al.
(2000) observaram redução drástica no crescimento das plantas, quando comparados aos
demais tratamentos que apresentação boro na solução. Nas doses entre 0,125 a 4,0 mg L-1 não
houve registro de diferenças significativas. Segundo os mesmos autores, a deficiência de boro
proporcionou menor concentração de cálcio nas folhas e quando os valores estavam abaixo de
31 mg kg-1 nas folhas, a planta apresentava valores reduzidos de crescimento.
3.2.7 Ferro
Em termos de broto, o Fe é particularmente importante na síntese de clorofila e
estabilização de tilacóides. A conversão de energia da fotossíntese é largamente afetada
através de cloroses de Fe e pode ser avaliada através de parâmetros de fluorescência (Dechen
et al., 1991).
Mengel & Kirkby (1982), Ingestad (1982) e Malavolta (1980), descrevem de maneira
similar às deficiências de ferro, sempre com início nas folhas novas, cloroses internervais e
pontos avermelhados na folha. A explicação está na falta da ferrodoxina, que atua em
processos metabólicos ligados ao NADP+, ou seja, diretamente na taxa fotossintética.
Altas concentrações de zinco na solução podem alterar a absorção de ferro, como
observado por Soares et al. (2000) em E. urophylla, induzindo deficiência na planta.

19
3.3 Ação do pH
Mudanças no pH da solução nutritiva durante experimentações podem explicar, a partir
dos princípios de eletrocondutividade das células das plantas; alterações na carga total de
cátions e de ânions na solução. Quando um cátion é absorvido pela raiz, um ânion também é
absorvido e um cátion é liberado; assim, a absorção de NH+4
pode ser acompanhada da
absorção de H+ e afetar com isso o pH da vizinhança da raiz. A absorção de NO-3 é do tipo
ativo, e seu transporte no interior da célula reduz consideravelmente quando é inibida a síntese
de ATP e, seu aceptor, que é o NO-3, é co-transportado com H+ (Maldonado, 1987) uma
estequiometria de 2H+:1NO-3, isto explica a alcalinização do meio externo quando as plantas
absorvem NO-3. O incremento do pH da solução nutritiva se deve a absorção de NO-
3 que se
acompanha da absorção de H+ e excreção de OH- para manter o balanço de cargas. A
magnitude da redução do pH da solução pode explicar em certa medida a redução da menor
produção de matéria seca da parte aérea e das raízes das plantas (Gallegos-Vásquez et al.
2000).
Valores de pH na rizosfera podem restringir a absorção de Cu, Mn e Zn, indicado em
análise e visualmente por deficiência de Cu e Mn (Savvas et al., 2003).
3.4 Parâmetros Ambientais
Segundo Engel & Poggiani (1991) a eficiência fotossintética está ligada ao teor de
clorofila das plantas, afetando o crescimento e influenciando a adaptabilidade das mesmas aos
diversos ambientes. Em presença da luz a substância é constantemente sintetizada e destruída

20
pelo processo da foto-oxidação, sendo que a velocidade de decomposição é diretamente
proporcional à intensidade luminosa muito alta, causando prejuízos para a fotossíntese. De
acordo com Lee (1988) pesquisas evidenciaram que o teor de clorofila varia muito entre as
espécies, assim como entre genótipos de mesma espécie solar.
Dentre os fatores que controlam a formação de raízes em segmentos do caule e a
produção de biomassa, estão àqueles inerentes às plantas utilizadas como fontes de
propágulos, como o ambiente onde estão crescendo. Embora alguns aspectos do seu
mecanismo de ação não estejam ainda completamente elucidados, a influência do genótipo,
estado nutricional, estágio fisiológico, substâncias de reserva, temperatura, sazonalidade,
turgidez, substâncias de crescimento endógenas e exógenas, umidade relativa do ar,
quantidade de luz e evapotranspiração, tem sido amplamente reconhecida como interferentes
no processo (Hansen, 1987; Xavier et al., 2001).
Em virtude da grande influência que exerce no enraizamento, merece destaque as
condições de luminosidade nos locais onde são produzidas as estacas, sendo que as respostas
mais positivas estão freqüentemente associadas à redução (estiolamento) (Assis et al., 1990).
Apesar dos fatores ambientais não atuarem isoladamente sobre as plantas, a luz é
fundamental, como fonte direta de energia para o desenvolvimento de todos os vegetais (Engel
& Poggiani, 1991).
Cada espécie tem exigências próprias para seu desenvolvimento e, a intensidade de luz
que chega ao indivíduo é especialmente importante para seu crescimento e desenvolvimento
(Poggiani et al., 1990).
A luminosidade também exerce influência sobre todos os estágios de crescimento dos
vegetais, existindo um ponto ótimo para cada fase (Drumond & Lima, 1993).

21
Embora sejam variáveis as exigências das diferentes espécies, as temperaturas do leito
de enraizamento variando de 21 a 27°C durante o dia, e ao redor de 15°C durante a noite, são
satisfatórias para espécies de eucaliptos. Temperaturas do ar elevadas devem ser evitadas, pois
o aumento do metabolismo, além de estimular o desenvolvimento das raízes, pode favorecer a
perda de água pelas folhas, levando as estacas ao dessecamento, tendo em vista que a perda de
água é sempre mais rápida que a sua absorção. Portanto, deve-se induzir primeiro a iniciação
radicular através de um meio artificial, onde se mantenha a temperatura do substrato mais alta
do que a do ar garantindo a absorção (Hartman et al., 1997).
A perda de água de uma cultura não só esta ligada ao estágio de crescimento e
desenvolvimento, mas também a sua condição de sanidade, representada pela infestação de
pragas e doenças. O conhecimento da evapotranspiração real é de grande valor, pois com ela é
possível estabelecer a relação com o potencial de água na planta (índice de rendimento
vegetativo). Quanto mais próximo da unidade for este índice, tanto mais próximo das
condições ideais de crescimento e desenvolvimento se encontra a planta (Ometto, 1981).
Alguns processos fisiológicos são fortemente influenciados pela umidade relativa do
ar, como a fotossíntese que tende a diminuir com a diminuição desse parâmetro, em
decorrência ao fechamento dos estômatos e do aumento da resistência estomática à difusão de
CO2 (Horie, 1979). Na transpiração, o aumento no déficit de saturação causa aumento na
resistência foliar à difusão do vapor d’água, fazendo com que o aumento da transpiração seja
inferior ao aumento no déficit de pressão de vapor (Whitehead et al., 1981).

22
4 MATERIAL E MÉTODOS
As mudas de Eucalyptus grandis x E. urophylla utilizadas neste trabalho foram
coletadas no mês de maio de 2002 na área de pleno sol do viveiro de produção de mudas
florestais da International Paper, em Mogi-Guaçu, estado de São Paulo – Brasil, estando com
90 dias de idade, contados a partir de sua entrada em casa-de-vegetação para enraizamento,
possuindo 60 cm de altura, diâmetro de colo de 2 mm e com 3 pares de folha. A metodologia
de produção destas mudas está especificada na Figura 2.
Por serem plantas originárias de uma área de produção, poderia conter resíduos da
adubação de base utilizada normalmente (4,0 kg de Fosmag® + 10,0 Kg de NPK 20-00-20

23
para cada m3 de substrato com 50% de vermiculita fina; e 50% de casca de arroz carbonizada),
o que interferiria na obtenção de alguns resultados almejados. Para solucionar este problema,
as plantas foram mantidas no viveiro experimental da empresa, na mesma região, sob sistema
de irrigação forçado, com 100 mm/dia de água, quatorze dias, totalizando 1400 mm de água de
irrigação, sem a qualquer fertilizante adicionado a esta.
Posteriormente as mudas foram levadas para os canaletes com água corrente por sete
dias, com altura de inundação de 5 cm e tempo de 15 minutos com água e uma hora sem.
Este procedimento adotado permitiu que na análise do material vegetal e substrato, os
nutrientes estivessem em concentração constante e somente em 23 de maio de 2002 as plantas
permaneceram em soluções nutritivas diversas em função da omissão de nutrientes e solução
completa, sendo que o período de inundação foi de 15 minutos com solução por uma hora sem
esta, até 19 de setembro do mesmo ano,
A altura de poda das mudas variou de 8 a 10 cm, permanecendo com um par de folhas..
Isto permitiu a formação das microcepas, sendo que as microestacas retiradas destas e
consideradas para contagem de progresso do sistema, apresentavam de 4cm de altura na
solução completa, sendo este procedimento padrão da empresa para formação de microcepas.
Para a execução deste experimento, foi(ram) utilizada(s) (Figura 1):
a) Oito canaletes para o crescimento das microcepas, produzidos em fibra de vidro,
pintados no fundo com tinta azul, coberto com isopor a 3 cm de altura e plástico
preto pelo lado de dentro e branco pelo lado de fora, de modo que as mudas se
estabeleceram em células recortadas em função do diâmetro do tubete e, em cada
canalete continha um tratamento ;

24
b) Oito reservatórios para as soluções nutritivas, cobertos e colocados pelo lado de
dentro da casa-de-vegetação; oito moto-bombas para o bombeamento das soluções
nutritivas;
c) Casa-de-vegetação com cobertura plástica e lateral abertas, sem presença de
exaustores de ar e
d) Fertilizantes para a composição das soluções nutritivas, sendo nitrato de amônia,
nitrato de potássio, nitrato de cálcio, cloreto de potássio, sulfato de magnésio,
sulfato de manganês, ácido fosfórico, ácido bórico, sulfato de cobre, sulfato de
zinco, molibdato de sódio, cloreto de cálcio, cloreto de manganês, ferro quelatado
e fosfato de potássio.
e) Duas estações metereológicas estacionadas, uma dentro da casa-de-vegetação e
outra fora desta, da marca SKUITTER®, para mensurações de quinze em quinze
minutos da radiação global, PAR, umidade relativa e temperatura.
Durante o período que permaneceram na solução nutritiva, as plantas foi submetido às
omissões (Tabela 2) de ferro (tratamento1), boro (tratamento 2), magnésio (tratamento 3),
cálcio (tratamento 4), potássio (tratamento 5), fósforo (tratamento 6), nitrogênio (tratamento
7). O tratamento 8 corresponde à solução completa.

25
A B C D
E F G H
I J L
M
Figura 1. Etapas e equipamentos utilizados para realização do experimento. “A e B”:limpeza das cubas de recebimento da solução nutritiva; “C”: material vegetal em água; “D”: material vegetal sendo coletado para análise laboratorial; “E”: tanquesde solução; “F”: retirada de amostras para análises e colocação em frascos “G”;automatização do sistema hidropônico em “H e I” e sensores de radiação global “J”, radiação PAR “L” e temperatura e umidade “M”.

26
Tabela 1. Parâmetros analisados e unidades fornecidas pelos laboratórios nas análises do
substrato, material vegetal e solução nutritiva.
Solução Nutritiva
mg L-1
Material Vegetal Substrato
NH+4 N (g kg-1) M.O. (g kg-1)
NO-3
P P (g kg-1) P em Mehlich (mg dm-3)
P em Resina (mg dm-3)
K K (g kg-1) K (mmolc dm-3)
Ca Ca (g kg-1) Ca (mmolc dm-3)
Mg Mg (g kg-1) Mg (mmolc dm-3)
S S (g kg-1) S (mg dm-3)
Fe Fe (mg kg-1) Fe (mg m-3)
Mn Mn (mg kg-1) Mn (mg dm-3)
Cu Cu (mg kg-1) Cu (mg dm-3)
Zn Zn (mg kg-1) Zn (mg dm-3)
Na Na (mg kg-1) Na (mg dm-3)
B B (mg kg-1) B (mg dm-3)
Co (mg kg-1)
Mo Mo (mg kg-1)
pH pH em água

Mat
riz
Sele
cion
ada
Res
gate
de
Bro
tos
Enr
aiza
men
toda
m
acro
esta
cas
Intr
oduç
ãoM
ultip
licaç
ão
Alo
ngam
ento
Form
ação
da
Mic
roce
pa
Mic
roce
pa n
o H
idro
jard
im
Hid
roja
rdim
Clo
nal
Col
eta
de
Mic
roes
taca
s
Con
fecç
ão d
e M
inic
roes
taca
sE
nrai
zam
ento
das
Min
iest
acas
Mud
a Pr
onta
Flor
esta
Clo
nal
Prod
ução
de
mac
roes
taca
spar
a o
labo
rató
rio
Áre
a de
Rus
tific
ação
Man
ipul
ação
da
mic
roce
pas
Figu
ra 2
. M
etod
olog
ia u
tiliz
ada
para
pro
duçã
o de
mud
as a
par
tirde
mat
rizes
sele
cion
adas
, cul
tivo
in v
itro
e si
stem
a hi
drôp
onic
o.
27

28
Tabela 2. Concentração de macronutrientes (g L-1) e micronutrientes (mg L-1) presentes nas
soluções nutritivas utilizadas durante o teste de omissão de nutrientes em
microcepas Eucalyptus grandis x E. urophylla. Nutrientes
N P K Ca Mg S B Cu Zn Mn Mo Fe Na Cl EDTA
-N 0,00 56,70 231,75 77,22 52,00 57,58 0,52 0,10 0,34 1,00 0,20 8,10 7,01 334,69 43,96
-P 104,19 0,00 231,26 77,60 52,00 57,58 0,52 0,10 0,34 1,00 0,20 8,10 7,01 69,42 43,96
-K 104,82 56,90 0,00 77,60 52,00 57,58 0,52 0,10 0,34 1,00 0,20 8,10 7,01 0,00 43,96
-Ca 104,31 56,75 230,01 0,00 52,00 57,58 0,52 0,10 0,34 1,00 0,20 8,10 7,01 0,00 43,96
-Mg 105,43 56,70 229,47 77,60 0,00 57,81 0,52 0,10 0,34 1,00 0,20 8,10 7,01 142,65 43,96
-B 104,19 56,75 230,01 77,60 52,00 57,58 0,00 0,10 0,34 1,00 0,20 8,10 7,01 0,00 43,96
-Fe 104,19 56,75 230,01 77,60 52,00 52,92 0,52 0,10 0,34 1,00 0,20 0,00 0,10 0,00 0,00
Completa 104,19 56,75 230,01 77,60 52,00 57,58 0,52 0,10 0,34 1,00 0,20 8,10 7,01 0,00 43,96
As análises laboratoriais para os materiais vegetais e substrato, foram realizados na
empresa Unithal® ,Campinas (SP), e as análises das soluções nutritivas foram realizadas no
laboratório da empresa Dessa Consult em Holambra (SP), através de procedimentos indicados
pelo IAC. Os parâmetros analisados assim como as suas unidades estão citadas na Tabela 1.
As amostras do material vegetal, num total de quinze, divididas em três repetições,
foram secas e moídas e submetidas às digestões nítrico-perclórica e sulfúrica para a obtenção
dos extratos, visando à determinação dos macros e micronutrientes, conforme metodologia
descrita por Malavolta et al. (1997). A determinação do nitrogênio foi realizada através do
método micro Kjeldahl. A determinação do P foi realizada pelo método da colorimetria de
molibdato-vanadato; do K por fotometria de chama; do Ca, Mg, Cu, Fe, Mn e Zn por
espectrometria de absorção atômica; do S por turbidimetria de suspensão de sulfato de bário e

29
do B por colorimetria de azometina-H. O cronograma total das atividades é apresentado na
Tabela 3.

30
Tabe
la 3
. Cro
nogr
ama
de c
olet
a de
am
ostra
s em
mat
eria
l veg
etal
(fol
ha, c
aule
, rai
z e
brot
ação
), so
luçã
o hi
drop
ônic
a e
subs
trato
par
a an
ális
e de
nut
rient
es, n
o te
ste
de o
mis
são,
em
mud
as d
e Eu
caly
ptus
gra
ndis
x E
. uro
phyl
la, m
antid
os e
m si
stem
a hi
drop
ônic
o.
Folh
a R
aiz
Cau
le
Bro
taçõ
esSo
luçã
o m
iner
alSu
bstra
to
T
reze
dia
s ant
es d
as m
udas
rec
eber
em so
luçã
o nu
triti
va
T -
13d
T -
13d
T -
13d
T -
13d
Seis
dia
s ant
es d
as m
udas
rece
bere
m so
luçã
o nu
tritiv
a
T - 0
6d T
- 06
dT
- 06d
T - 0
7d
Mom
ento
em
que
as m
udas
rec
ebem
solu
ções
nut
ritiv
as
M0
M0
M0
M
0 M
0 D
uas h
oras
apó
s o re
cebi
men
to d
e so
luçã
o nu
tritiv
a T0
2h
T02h
T0
2h
T0
2h
T02h
Q
uatro
hor
as a
pós o
rece
bim
ento
de
solu
ção
nutri
tiva
T04h
T0
4h
T04h
T0
4h
Sete
hor
as e
mei
a ap
ós o
rece
bim
ento
de
solu
ção
nutri
tiva
T7,5
h T7
,5h
T7,5
h
T7
,5h
Vin
te e
dua
s hor
as a
pós o
rece
bim
ento
de
solu
ção
nutri
tiva
T22h
T2
2h
T22h
T22h
T2
2h
Qua
rent
a e
seis
hor
as a
pós o
rece
bim
ento
de
solu
ção
nutri
tiva
T46h
T4
6h
T46h
T4
6h
Trez
e di
as a
pós M
0 T1
3d
T13d
T1
3d
T1
3d
T13d
T
rint
a e
cinc
o di
as a
pós M
0 (s
egun
da c
olet
a de
mic
roes
taca
s)
T35d
T3
5d
T35d
T
35d
T35d
T3
5d
Qua
rent
a e
cinc
o di
as a
pós M
0 T4
5d
T45d
T4
5d
T45d
T4
5d
T45d
Se
ssen
ta e
um
dia
s apó
s M0
T61d
T6
1d
T61d
T6
1d
T61d
T6
1d
Sete
nta
e no
ve d
ias a
pós M
0 (f
inal
da
pres
ença
de
folh
as)
T79
d T7
9d
T79d
T7
9d
T79d
T7
9d
Nov
enta
e se
te d
ias a
pós M
0
T97D
T9
7D
T97D
T9
7D
T97D
C
ento
e se
te d
ias a
pós M
0
T107
DT1
07D
T107
D
T107
D
T107
D
Cen
to e
qui
nze
dias
apó
s M0
T1
15D
T115
DT1
15D
T1
15D
T1
15D
C
ento
e v
inte
e q
uatro
dia
s apó
s M0
T1
24D
T124
DT1
24D
T1
24D
T1
24D
*O
s neg
ritos
no
text
o re
fere
m-s
e, re
spec
tivam
ente
aos
tem
pos d
e in
ício
do
expe
rimen
to (1
3d);
mom
ento
em
que
as m
udas
com
eçam
a re
cebe
r a so
luçã
o nu
tritiv
a (M
0); i
níci
o da
col
eta
das m
icro
cepa
s (T3
5d) e
fina
l da
pres
ença
de
folh
as n
as m
icro
touç
as (T
79d)
.

31
A proposta do trabalho, como definida nos objetivos foi, através da estatística,
verificar o efeito da omissão de alguns elementos químicos nas soluções administradas às
microcepas sobre a produção destas e efeitos na microtouça, além de avaliações ambientais da
área de produção. O delineamento experimental foi inteiramente casualizado sendo que nas
Tabelas de 4 a 17 são apresentadas a ANOVA e Equações de Regressão, optando-se pelas
quadráticas em função de sua maior acuidade, indicada pelos valores de R2, utilizando o
software estatístico SAS© com valores de significância de 15%, padrão para este tipo de
experimentação (Pimentel-Gomes & Garcia, 2003).
São apresentadas no Apêndice 1, através das Tabelas de 18 a 54, as análises dos teores
de nutrientes presentes nas partes vegetais, substrato e solução, além da produção de
microestacas, com e sem transformação das médias das partes consideradas, através do teste
de Tukey a 5% de probabilidade (Pimentel-Gomes & Garcia, 2003). Apenas alguns resultados
foram utilizados nas discussões dos experimentos em função de não explicarem de maneira
clara e objetiva os fenômenos observados na área experimental, porém foram deixadas em
apêndice por conterem informações relativas as produções de microestacas e aos teores de
nutrientes nas partes analisadas importantes para os pesquisadores da área.
4.1. Determinações realizadas
• Análise laboratorial para avaliação dos teores de nutrientes presentes no substrato: Ca,
Mg, K, P em extrator Menlich-1, P em Resina trocadora de íons, M.O., S, Na, B, Fe,
Mn, Cu, Zn; na folha: N, P, Ca, Mg, S, Fe, Mn, Cu, Zn, Na, B, Co, Mo; no broto: N, P,
K, Ca, Mg, S, Fe, Mn, Cu, Zn, Na, B, Co, Mo; no caule: N, P, K, Ca, Mg, S, Fe, Mn,
Cu, Zn, Na, B, Co, Mo e na raiz: N, P, K, Ca, Mg, S, Fe, Mn, Cu, Zn, Na, B, Co, Mo.

32
Para cada tratamento, incluindo-se a testemunha, quinze plantas inteiras foram
coletadas aleatoriamente e posteriormente divididas em três blocos com cinco plantas,
foram encaminhadas para análises dos teores de nutrientes ;
• A solução nutritiva utilizada no teste foi analisada quimicamente para determinação
das concentrações dos seguintes teores de nutrientes: NH+4, NO-
3, Na, K, Ca, Mg, S,
P, Cl, Fe, Mn, Cu, Zn, Be Mo; sendo retiradas três amostras de 250 mL por
tratamento, no momento da troca da solução (solução velha), em função da presença
de microestacas maiores ou iguais a 4 cm na solução completa;
• Foram obtidas informações também em relação ao pH e eletrocondutividade da
solução e
• As estações metereológicas colocada dentro da casa-de-vegetação e fora desta,
realizaram a verificação de umidade relativa do ar (%), temperatura (oC), radiação
global (W/m2) e radiação PAR (V/ Wm2).
Foi acrescentada, diariamente, na caixa de solução nutritiva, água destilada para
completar o consumo em função da evapotranspiração, o que permitiu maior confiabilidade
dos resultados obtidos.

33
5 RESULTADOS E DISCUSSÃO
As recomendações de nutrientes em uma solução nutritiva, assim como em talhões
florestais, seguem mesmo princípio designado por Ingestad & Ägren (1995), ou seja, por
produtividade nutricional Pn.
Como se sabe, o processo fotossintético permite a produção de biomassa mediante a
presença de macro e micronutrientes que podem ser administrados a planta de diversas
maneiras e formulações, acumulando-se em diversas partes da mesma, formando os “pools”
metabólicos (Cromer et al., 1993).

34
A máxima desta produtividade vegetal é obtida quando a planta está em condições
fisiológicas estabilizadas, “steady-state”, ou seja, a quantidade de nutrientes internas se
estabiliza, situação em que a quantidade de biomassa e a absorção de nutrientes estão
dinamicamente equilibradas (Ingestad & Ägren, 1995). Outras condições interferem no
sistema com as condições bioclimáticas, que alteram esta estabilização/equilíbrio e podem
modificar os níveis de absorção, velocidade, relação e formação dos “pools” de nutrientes
(Fyles et al., 1994).
Desta forma, podemos notar que para cada uma das análises estatísticas realizadas há
comportamentos explicitados nas análises de variância e equações de regressão que indicam
estas alterações, nas fases de desenvolvimento do material (tempo), condições ambientais e
“pools” de nutrientes acumulados, além das alterações do pH da solução, avaliados neste
experimento e, temperaturas, não mencionadas no trabalho.
5.1. Análise geral dos nutrientes presentes na solução nutritiva, substrato, microestacas
(broto) e caule.
Pode-se observar que na equação de regressão quadrática 1, as variáveis que mais
contribuíram para a produção de microestacas foram o magnésio, o molibdênio, o cobre, o
potássio, o enxofre e o sódio. A retirando do potássio, magnésio e enxofre provocaram
incremento na produção de microestacas, e o aumento do molibdênio, cobre e sódio, agiram
da mesma forma.
As partes da planta onde podemos notar, através da equação de regressão, incremento
na produção de microestacas são o broto e o caule, incluindo-se substrato e solução nutritiva.

35
Para cada uma destas partes há pelo menos um elemento nutricional que deve ser considerado,
indicando que apenas análises de partes da planta, ou mesmo solução e outras, não são
suficientes para o entendimento da produção das microestacas.
Há um conjunto de fatores que deve ser observado e que está relacionado na equação
de regressão quadrática 1 e que explicam este processo fisiológico com 38,44 % de exatidão
(Tabela 5) e com “R2” (coeficiente de determinação) de 0,65364897 (Tabela 4).
Os valores em negrito, nas equações de regressão quadrática, referem-se às variáveis
de maior importância para a função “produção de microestacas”, resultado da interpretação
dos valores parciais encontrados no Stepwise Procedure for Dependent Variable.
Tabela 4. Análise de regressão quadrática para produção de microestaca de Eucalyptus
grandis x E. urophylla em função dos elementos analisados na solução, raiz, broto,
caule e substrato, geral, em função da Tabela 3, mantida em sistema hidropônico,
com omissão de nutriente e completa.
R2 = 0,65364897
DF SQ QM F Prob>F Regressão 22 36827,72335871 1673,98742540 14,50 0,0001 Erro 169 19514,02143295 115,46758244 Total 191 56341,74479167
Equação de regressão quadrática (01).
Produção de Microestacas = + (20,29) – (22,56 K do broto) + (9,75 Mo do broto) – (53,97
Mg do broto) + (0,198 Cu do caule) + (0,0299 Na do caule) + (0,0069 Fe da raiz) + (0,082
Mn da raiz) – (0,085 B da raiz) – (5,42 K do substrato) – (0,0613 Cl da solução) + (5,13 K2
do broto) – (233,51 S2 do caule) – (0,00000043 Fe2 da raiz) + (0,00000721 Cu2 da raiz) –
(0,3649 Co2 da raiz) + (0,4772 Mo2 da raiz) + (0,021 NH4(2) da solução) + (0,00344 Na2
solução) + ( 0,08009 Mn2 da solução).

36
Tabela 5. Resumo do Stepwise Procedure for Dependent Variable para produção de
microestaca do Eucalyptus grandis x E. urophylla realizado no SAS, referente à
equação de regressão quadrática (01).
Variação Número Parcial Acumulada R2 R2 C(p) F Prob>F
Mg do broto 1 0,0654 0,0654 288,3743 13,2905 0,0003 Mo do broto 2 0,0569 0,1222 261,3965 12,2415 0,0006 Cu do caule 3 0,0511 0,1733 237,3646 11,6147 0,0008 K do substrato 4 0,0693 0,2426 204,0623 17,0996 0,0001 S2 do caule 5 0,0564 0,2989 177,3230 14,9599 0,0002 Na2 da solução 6 0,0362 0,3352 160,8574 10,0813 0,0018 B da raiz 7 0,0247 0,3598 150,2891 7,0875 0,0084 Mo2 do broto 8 0,0218 0,3817 141,1561 6,4645 0,0118 Mo2 da raiz 9 0,0206 0,4023 132,6593 6,2707 0,0132 K2 do broto 10 0,0151 0,4174 126,9424 4,7038 0,0314 K do broto 11 0,0247 0,4421 116,3572 7,9665 0,0053 Mn2 da solução 12 0,0171 0,4592 109,6212 5,6735 0,0183 Na2 do broto 13 0,0134 0,4727 104,7677 4,5390 0,0345 Na do caule 14 0,0492 0,5219 81,6796 18,2231 0,0001 Fe da raiz 15 0,0309 0,5528 67,9293 12,1619 0,0006 Cu2 da raiz 16 0,0173 0,5701 61,0998 7,0523 0,0086 Zn do caule 17 0,0157 0,5858 55,1102 6,5852 0,0111 Co2 da raiz 18 0,0136 0,5994 50,1692 5,8813 0,0163 Mg do broto 17 0,0029 0,5965 49,6576 1,2612 0,2630 NH4
(2) da solução 18 0,0085 0,6050 47,3497 3,7014 0,0560 Cl da solução 19 0,0107 0,6157 43,8802 4,8027 0,0298 Mn da raiz 20 0,0155 0,6312 37,9914 7,1758 0,0081 Fe2 da raiz 21 0,0115 0,6426 34,1470 5,4547 0,0207 Mg do caule 22 0,0110 0,6536 30,5340 5,3734 0,0216

37
5.2. Análise por coleta dos nutrientes presentes na solução nutritiva, substrato,
microestacas (broto) e caule.
5.2.1. Segunda coleta.
A primeira coleta de material das ainda mudas crescidas no viveiro foi descartada nas
análises em função das possíveis interferências que o explante poderia ter nos resultados.
As Tabelas 6 e 7 indicam a atuação dos elementos nutricionais em cada momento da
coleta nas microcepas em relação à produção de microestacas e a parte da planta coletada,
assim como a influência dos nutrientes presentes na solução e no substrato.
Considerou-se que quando a microcepa que estava em solução completa apresentasse
microestacas acima de 4cm de comprimento, seria realizada coleta geral de material dos outros
tratamentos. Desta forma, observa-se que todas as Tabelas fazem referência a segunda coleta,
como sendo inicial.
Nesta segunda coleta (Tabela 6), nota-se menor necessidade de cobalto pelo sistema
radicular, assim como a presença do fósforo, no mesmo órgão e que pode aumentar a
produção de microestacas. De fato, a importância da presença ou ausência destes elementos na
raiz pode ser explicada tendo em vista que, por estarem em soluções com deficiências
induzidas, a raiz parece ser o primeiro órgão a sentir este processo de ausência de nutrientes.
Há que se fazer referência que não foi retirado cobalto da solução.

38
Tabela 6. Análise de regressão quadrática para produção de microestaca de Eucalyptus
grandis x E. urophylla em função dos elementos analisados na folha, caule, raiz,
broto, solução e substrato, por coleta, em função da Tabela 3, mantida em sistema
hidropônico com omissão de nutriente e completa.
Segunda Coleta R2 = 0,85574900 DF SQ QM F Prob>F Regressão 5 7475,78758148 1495,15751630 21,36 0,0001 Error 18 1260,17075186 70,00948621 Total 23 8735,95833333 Equação de regressão quadrática (02).
Produção de microestacas = (-16,7180) + (55,85 P da raiz) + (0,0010 Zn2 do caule) + (96,71
S2 da raiz) – (1,5143 Co2 da raiz) + (7,2513 Ca2 da folha).
Tabela 7. Resumo do Stepwise Procedure for Dependent Variable para microestaca de
Eucalyptus grandys x E. urophylla, realizado no SAS, referente às equações de
regressão polinomial quadrática de 02 a 09.
Equação de Regressão 02:
Variação Número Parcial Acumulada F Prob>F R2 R2 Co2 da raiz 1 0,3016 0,3016 9,5008 0,0054 P da raiz 2 0,2227 0,5243 9,8325 0,0050 Ca2 da folha 3 0,1790 0,7033 12,0675 0,0024 S2 da raiz 4 0,0948 0,7982 8,9271 0,0076 Zn2 do caule 5 0,0576 0,8557 7,1860 0,0153
5.2.2. Terceira coleta.
Já na terceira coleta de material ocorreu maior necessidade de manganês e cobre na
solução (equação de regressão quadrática 03) que, em conjunto, contribuíram com 35,25%
para a explicação neste momento do processo de produção de microestacas (Tabela 7a).

39
Tabela 6a. Continuação da Tabela 6. Terceira Coleta R2 = 0,92658863 DF SQ QM F Prob>F Regressão 10 5242,59987440 524,25998744 16,41 0,0001 Erro 13 415,35845893 31,95065069 Total 23 5657,95833333 Equação de regressão quadrática (03).
Produção de microestacas = (173,48) – (1,85 Fe do broto) – (0,065 Na do broto) – (64,45 Ca
da raiz) + (6,3851 Mn da solução) + (74,60 Ca2 do broto) + (0,0055 Fe2 do broto) + 68,08
(Mg2 da raiz) + (0,00019 Zn2 da raiz) – (0,00806 P2 da solução) + (2,1731 Cu2 da solução).
Tabela 7a. Continuação da Tabela 7. Equação de regressão 03: Variação Número Parcial Acumulada F Prob>F R2 R2 Mn da solução 1 0,2433 0,2433 7,0748 0,0143 Cu2 da solução 2 0,1092 0,3525 3,5413 0,0738 Mg2 da raiz 3 0,0847 0,4373 3,0119 0,0980 Fe2 do broto 4 0,0650 0,5023 2,4812 0,1317 Na do broto 5 0,0669 0,5692 2,7966 0,1118 Ca da raiz 6 0,0960 0,6652 4,8726 0,0413 Fe do broto 7 0,0613 0,7265 3,5854 0,0765 Zn2 da raiz 8 0,0773 0,8038 5,9090 0,0281 P2 da solução 9 0,0649 0,8687 6,9239 0,0197 Ca2 do broto 10 0,0579 0,9266 10,2509 0,0069
5.2.3. Quarta coleta.
Nesta quarta coleta, as manifestações do efeito das deficiências já aparecem na
microcepa, através da importância do enxofre no caule e nas brotações, ambos favorecendo o
processo de produção (Tabela 6b). Neste momento respondem por 73,48% da resposta de

40
produção de microestacas (Tabela 7b). Há também que se considerar que o enxofre e o
manganês não foram retirados das soluções.
Tabela 6b. Continuação da Tabela 6. Quarta Coleta R2 = 0,89603856 DF SQ QM F Prob>F Regressão 4 5374,10325809 1343,52581452 40,94 0,0001 Erro 19 623,52174191 32,81693378 Total 23 5997,62500000 Equação de regressão quadrática (04).
Produção de Microestacas = - (8,1212) + (0,05667 Mn do caule) + (478,36 S2 do broto) –
(0,0066 B2 do broto) + (0,012 B2 do caule).
Tabela 7b. Continuação da Tabela 7. Equação de regressão 04: Variação Número Parcial Acumulada F Prob>F R2 R2 S2 do broto 1 0,4822 0,4822 20,4846 0,0002 Mn do caule 2 0,2526 0,7348 19,9980 0,0002 B2 do broto 3 0,1047 0,8394 13,0359 0,0017 B2 do caule 4 0,0566 0,8960 10,3476 0,0045
5.2.4. Quinta coleta.
A quinta coleta de microestacas apresentou no caule a menor necessidade do
molibdênio para o aumento de produção destas (Tabela 6c). Também o elemento citado não
foi retirado da solução nutritiva e sua contribuição para o processo nesta fase foi de 57,17%
(Tabela 7c).

41
Tabela 6c. Continuação da Tabela 6 Quinta Coleta R2 = 0,86952465 DF SQ QM F Prob>F Regressão 5 5856,97311065 1171,39462213 23,99 0,0001 Erro 18 878,86022269 48,82556793 Total 23 6735,83333333 Equação de regressão quadrática (05).
Produção de microestacas = + (285,02) – (23,64 K do broto) – (1,44 P do substrato em resina)
+ (65,93 P2 do broto) – (2,27 Mo2 do caule) + (0,0001 Cu2 da raiz).
Tabela 7c. Continuação da Tabela 7. Equação de regressão 05
Variação Número Parcial Acumulada F Prob>F R2 R2 Mo2 do caule 1 0,5717 0,5717 29,3699 0,0001 P em resina 2 0,1545 0,7262 11,8487 0,0024 K no broto 3 0,0362 0,7624 3,0430 0,0964 Cu2 da raiz 4 0,0525 0,8148 5,3821 0,0316 P2 do broto 5 0,0547 0,8695 7,5467 0,0133
5.2.5. Sexta coleta.
Nesta sexta coleta, o molibdênio volta a ter a sua importância na análise do caule,
contribuindo com 32,94% do resultado, assim como o cobre na brotação. O potássio que foi
retirado de uma das soluções, aparece como elemento que contribui com 30,64%, quando
analisamos as brotações. Nota-se que a ausência deste último elemento é favorável ao
processo de produção de microestacas e que outras partes da planta começam a contribuição
para a explicação do processo fisiológico da produção (Tabelas 6d e 7d).

42
Tabela 6d. Continuação da Tabela 6. Sexta Coleta R2 = 0,90778182 DF SQ QM F Prob>F Regressão 6 3793,92283476 632,32047246 27,89 0,0001 Erro 17 385,41049857 22,67120580 Total 23 4179,33333333 Equação de regressão quadrática (06).
Produção de microestacas = - (9,8427) + (1,2515 Cu do broto) + (8,4739 Mo do caule)
-(4,9878 K2 do broto) + (0,0027 P2 do substrato em MEL.) + (0,001071 S2 do substrato) +
(0,0022 Mg2 da solução).
Tabela 7d. Continuação da Tabela 7. Equação de regressão 06 Variação Número Parcial Acumulada F Prob>F R2 R2 Mo do caule 1 0,3294 0,3294 10,8051 0,0034 K2 do broto 2 0,3064 0,6358 17,6646 0,0004 Cu do broto 3 0,1678 0,8035 17,0773 0,0005 P2 em MELS 4 0,0433 0,8468 5,3678 0,0318 Mg2 em solução 5 0,0378 0,8846 5,9008 0,0258 S2 no substrato 6 0,0232 0,9078 4,2686 0,0544
5.2.6. Sétima coleta.
A análise da equação de regressão quadrática 07, permite verificar que a planta já
sente, de maneira pronunciada a ausência de ferro, sendo neste momento, o elemento mais
exigido para que haja aumento de produção. Na Tabela 23, (Apêndice 1) pode-se verificar que
as menores produções de microestacas ocorrem na ausência deste elemento, principalmente ao
correr do tempo de cultivo.

43
Tabela 6e. Continuação da Tabela 6. Sétima Coleta R2 = 0,97351039 DF SQ QM F Prob>F Regressão 11 3420,75325082 310,97756826 40,09 0,0001 Erro 12 93,08008251 7,75667354 Total 23 3513,83333333 Equação de regressão quadrática (07).
Produção de microestacas = + (32,4454) + (7,74 N do caule) + (0,05 Zn do caule) – (8,22 Mg
do substrato) – (0,2384 Ca da solução) + (0,211 Mg da solução) + (0,1243 Fe da solução) +
(4,93 Mo2 do broto) + (67,39 P2 do caule) – (0,001 B2 do caule) + (0,0008 Ca2 da solução) –
(135,17 Mo2 da solução).
Tabela 7e. Continuação da Tabela 7. Equação de regressão 07 Variação Número Parcial Acumulada F Prob>F R2 R2 Fe da solução 1 0,3493 0,3493 11,8094 0,0024 Zn do caule 2 0,1748 0,5241 7,7139 0,0113 Ca2 da solução 3 0,1563 0,6804 9,7805 0,0053 N do caule 4 0,0933 0,7737 7,8382 0,0114 Mg do substrato 5 0,0538 0,8275 5,6084 0,0293 Mo2 do broto 6 0,0420 0,8695 5,4656 0,0319 B2 do caule 7 0,0425 0,9120 7,7282 0,0134 Mg da solução 8 0,0181 0,9301 3,8804 0,0676 P2 do caule 9 0,0160 0,9460 4,1435 0,0612 Mo2 da solução 10 0,0112 0,9572 3,3944 0,0883 Ca da solução 11 0,0163 0,9735 7,3839 0,0187
5.2.7. Oitava coleta.
Analisando-se a equação de regressão quadrática 08 pode-se notar a importância da
presença do fósforo na raiz, aparecendo como indicativo positivo de produtividade. Observa-
se também que grande quantidade de matéria orgânica, originária do substrato utilizado,
interfere negativamente no processo, talvez pelo fato de reter nutrientes, influenciar na relação

44
C/N, que pode estar escondendo a retirada de nitrogênio da solução, em detrimento para a
planta ou decompondo-se , liberando nutrientes que não estão, neste momento do processo,
contribuindo para a produção de micorestacas.
Tabela 6f. Continuação da Tabela 6. Oitava Coleta R2 = 0,80957190 DF SQ QM F Prob>F Regressão 5 3014,84574901 602,96914980 15,30 0,0001 Erro 18 709,15425099 39,39745839 Total 23 3724,00000000 Equação de regressão quadrática (08).
Produção de microestacas = + (33,77) + (73,47 P da raiz) – (0,135 B da raiz) – (0,2088 Na do
substrato) – (20,32 Ca2 do broto) – (5,5172 M.O.2 do substrato).
Tabela 7f. Continuação da Tabela 7. Equação de regressão 08 Variação Número Parcial Acumulada F Prob>F R2 R2 P da raiz 1 0,2573 0,2573 7,6225 0,0114 M.O.2 do substrato 2 0,2912 0,5485 13,5468 0,0014 Ca2do broto 3 0,1510 0,6995 10,0505 0,0048 Na do substrato 4 0,0594 0,7590 4,6846 0,0434 B da raiz 5 0,0506 0,8096 4,7835 0,0422
5.2.8. Nona coleta.
Na última coleta realizada a retirada do fósforo da solução influencia de maneira
positiva a produção de microestacas, sendo este responsável por 53,62% (Tabela 7g) da
contribuição para explicar o processo fisiológico.

45
Tabela 6g. Continuação da Tabela 6 Nona Coleta R2 = 0,82031524 DF SQ QM F Prob>F Regressão 5 6985,80462385 1397,16092477 16,44 0,0001 Erro 18 1530,19537615 85,01085423 Total 23 8516,00000000 Equação de regressão quadrática (09)
Produção de microestacas = +(17,092) + (6,27 Ca do substrato) – (4,47 K do substrato) +
(0,000047 Mn2 do caule) + (13,075 N2 da raiz) – (0,0088147 P2 da solução).
Tabela 7g. Continuação da Tabela 7.
Equação de regressão 09 Variação Número Parcial Acumulada F Prob>F R2 R2 P2 da solução 1 0,5362 0,5362 25,4334 0,0001 Ca do substrato 2 0,1224 0,6586 7,5281 0,0122 N2 da raiz 3 0,0503 0,7089 3,4558 0,0778 Mn2 do caule 4 0,0714 0,7803 6,1766 0,0224 K do substrato 5 0,0400 0,8203 4,0081 0,0606
5.3. Análises dos nutrientes presentes nas brotações das microcepas, de maneira geral,
para identificação da produtividade das microestacas.
Através da observação realizada na equação de regressão quadrática 10, os elementos
presentes nas brotações da microcepas que influenciaram o processo de produção de
microestacas foram o magnésio, o cobre e o molibdênio. Porém juntos, responderam por
apenas 18,13% para o entendimento do processo fisiológico, indicando que este tipo de
material não é o mais adequado para obtermos respostas em relação à produção.
Na análise dos elementos de maior influência, dentro deste processo, nota-se que a
ausência do magnésio da brotação e o aumento da concentração do molibdênio no mesmo

46
explante, favorecem o processo de produção de microestacas, porém, ambos respondem por
apenas 12,22% da resposta da resposta obtida.
Tabela 8 Análise de regressão quadrática para produção de microestaca de Eucalyptus grandis
x E. urophylla em função dos elementos analisados nas brotações, geral, conforme a
Tabela 3, mantida em sistema hidropônico, com omissão de nutriente e completa.
Todas as Coletas R2 = 0,18126134 DF SQ QM F Prob>F Regressão 4 10212,58029526 2553,145073 10,35 0,0001 Erro 187 46129,16449640 246,680024 Total 191 Equação de regressão quadrática (10).
Produção de microestacas = + (27,4823) – (77,020 Mg do broto) + (0,2790 Cu do broto) +
(16,73 Mo do broto) – (1,9539 Mo2 do broto).
Tabela 9 Resumo do Stepwise Procedure for Dependent Variable para microestaca de
Eucalyptus grandis x E. urophylla em função dos elementos analisados nas
brotações, geral, de acordo com as Tabelas 3, mantidas em sistema hidropônico com
omissão de nutriente e completa.
Todas as coletas Variação Número Parcial Acumulada R2 R2 F Prob>F Mg do broto 1 0,0654 0,0654 13,2905 0,0003 Mo do broto 2 0,0569 0,1222 12,2415 0,0006 Mo2 do broto 3 0,0291 0,1513 6,4478 0,0119 Cu do broto 4 0,0299 0,1813 6,8348 0,0097

47
5.4. Análises dos nutrientes presentes nas brotações das microcepas, por coleta de
microestacas, para identificação da produtividade.
Na seqüência das análises, foram verificados os elementos que interferiram no
processo de produção de microestacas de maneira separada, por coleta.
Na segunda coleta do material ocorre menor necessidade de potássio na brotação da
microcepas. Isto pode ser observado na Tabela 20 (Apêndice 1), principalmente ao final do
processo de 124 dias. Ele foi responsável por 39,03% da resposta de produção (Tabela 11).
Tabela 10. Análise de regressão quadrática para produção de microestaca de Eucalyptus
grandis x E. urophylla em função dos elementos analisados nas brotações, por coleta,
em função da Tabela 3, mantida em sistema hidropônico, com omissão de nutriente e
completa.
Segunda Coleta R2= 0,45377660 DF SQ QM F Prob>F Regressão 3 3964,17348975 1321,39116 5,54 0,0062 Erro 20 Total 23 Equação de regressão quadrática (11).
Produção de microestacas = + (24,91) – (25,99 K do broto) + (5,27 K2 do broto) +
(0,0001010 Na2 do broto).

48
Tabela 11. Resumo do Stepwise Procedure for Dependent Variable para produção
microestacas de Eucalyptus grandis x E. urophylla em função dos elementos
analisados nas brotações, por coleta, de acordo com a Tabela 3, mantida em sistema
hidropônico, com omissão de nutriente e completa.
Segunda Coleta
Variação Número Parcial Acumulada R2 R2 F Prob>F
K2 do broto 1 0,2905 0,2905 9,0071 0,0066 K do broto 2 0,0999 0,3903 3,4396 0,0777 Na2 do broto 3 0,0634 0,4538 2,3227 0,1432
Na terceira coleta há efeito negativo do acúmulo de sódio nas brotações em relação à
produção, talvez resultante de um processo de acúmulo de sais que carrearam consigo o Na,
tendo em vista a sua grande permeabilidade celular. No caso do ferro, repete o comportamento
anteriormente observado. A sua presença na solução e conseqüentemente na planta é de
fundamental importância para o seu desenvolvimento e produção, de acordo com a equação de
regressão quadrática 12.
Tabela 10a .Continuação da Tabela 10. Terceira Coleta R2= 0,053519837 DF SQ QM F Prob>F Regressão 5 3028,13000 605,6260 4,15 0,0111 Error 18 2629,8282 146,1015 Total 23 5657,9584 Equação de regressão quadrática (12).
Produção de Microestacas = + (29,67) + (29,78 Co do broto) + (0,0011 Fe2 do broto) –
(0,0448 Cu2 do broto) + (0,00557 Zn2 do broto) – (0,000098 Na2 do broto).

49
Tabela 11a. Continuação da Tabela 11. Terceira Coleta Variação Número Parcial Acumulada R2 R2 F Prob>F
Fe2 do broto 1 0,1397 0,1397 3,5729 0,0720 Co do broto 2 0,0934 0,2331 2,5578 0,1247 Na2 do broto 3 0,1591 0,3922 5,2347 0,0332 Zn2 do broto 4 0,0822 0,4744 2,9734 0,1009 Cu2 do broto 5 0,0608 0,5352 2,3526 0,1425
Apesar do enxofre não ter sido retirado de nenhuma das soluções, a sua presença no
broto aparece com importante e está respondendo por 48,22% do resultado alcançado na
produção de microestaca nesta fase do processo, de acordo com a Tabela 11b.
Tabela 10b. Continuação da Tabela 10. Quarta Coleta R2 = 0,45377660 DF SQ QM F Prob>F Regressão 3 3964,17348975 1321,39116325 5,54 0,0062 Erro 20 4771,78484359 238,58924218 Total 23 8735,95833333 Equação de regressão quadrática (13).
Produção de microestacas = + (3,532) + (0,023 Mn do broto) + (476,44 S2 do broto) –
(0,00568 B2 do broto).
Tabela 11b. Continuação da Tabela 11. Quarta Coleta Variação Número Parcial Acumulada R2 R2 F Prob>F
S2 do broto 1 0,4822 0,4822 20,4846 0,0002 Mn do broto 2 0,1867 0,6688 11,8386 0,0025 B2 do broto 3 0,1241 0,7929 11,9872 0,0025

50
Observações realizadas “in loco” davam conta do resultado obtido pela equação de
regressão quadrática 14. A ausência de potássio nas brotações permitiu uma maior quantidade
de microestacas produzidas, assim como veremos em outras equações, a ausência do magnésio
também produziu o mesmo efeito. A sua contribuição para o processo foi de 39,78%.
Tabela 10c. Continuação da Tabela 10. Quinta Coleta R2 = 0,79361677 DF SQ QM F Prob>F Regressão 5 5345,67030669 1069,13406134 13,84 0,0001 Erro 18 1390,16302665 77,23127926 Total 23 6735,83333333 Equação de regressão quadrática (14).
Produção de microestacas = - (71,74) + (48,43 N do broto) + (1,48 Cu do broto) – (6,83 N2 do
broto) – (4,186 K2 do broto) + (6,78 Co2 do broto).
Tabela 11c. Continuação da Tabela 11 Quinta Coleta Variação Número Parcial Acumulada R2 R2 F Prob>F K2 do broto 1 0,3978 0,3978 14,5323 0,0010 Co2 do broto 2 0,1435 0,5413 6,5705 0,0181 Cu do broto 3 0,0894 0,6308 4,8449 0,0396 N do broto 4 0,1112 0,7420 8,1922 0,0100 N2 do broto 5 0,0516 0,7936 4,5020 0,0480
A retirada do magnésio da solução e a sua ausência nas brotações aumentaram a
produção das microestacas, como pode ser observado na equação de regressão quadrática 15.
Este elemento respondeu por 21,04% (Tabela 11d) pelo resultado obtido. No caso do enxofre,
não houve solução em que tenha sido retirado e a sua presença no broto de maneira positiva,
permite dizer que este aumenta, até níveis a serem testados, a produção de microestacas. Este

51
elemento respondeu por 30,53% dos resultados obtidos. Pode-se também notar que na equação
polinomial citada, o enxofre interfere positivamente na produção e, ambos, magnésio e
enxofre, respondem por 51,57% para explicar a produção nesta sexta coleta de microestacas.
Tabela 10d. Continuação da Tabela 10. Sexta Coleta R2 = 0,64374198 DF SQ QM F Prob>F Regressão 3 2690,41229639 896,80409880 12,05 0,0001 Erro 20 1488,92103694 74,44605185 Total 23 4179,33333333 Equação de regressão quadrática (15).
Produção de Microestacas = +(34,57) + (0,9171 Cu do broto) – (176,21 Mg2 do broto) +
(298,64 S2 do broto).
Tabela 11d. Continuação da Tabela 11. Sexta Coleta Variação Número Parcial Acumulada R2 R2 F Prob>F
S2 do broto 1 0,3053 0,3053 9,6689 0,0051 Mg2 do broto 2 0,2104 0,5157 9,1223 0,0065 Cu do broto 3 0,1280 0,6437 7,1885 0,0144
Há uma inversão de ordem no caso do magnésio, em relação a sua necessidade nas
brotações, talvez causada pela diminuição muito grande nas quantidades do magnésio nas
brotações, em função do período de tempo em que estas permaneceram em solução com
omissão deste nutriente. A produção de microestacas foi mais adequada quando da presença
deste elemento na brotação, assim como o cálcio, muito embora percentualmente, eles tenham
respondido, em conjunto por apenas 33,35% dos resultados conseguidos (Tabela 11e).

52
Tabela 10e. Continuação da Tabela 10. Sétima Coleta R2 = 0,69373349 DF SQ QM F Prob>F Regressão 6 2437,66386555 406,27731092 6,42 0,0011 Erro 17 1076,16946778 63,30408634 Total 23 3513,83333333 Equação de regressão quadrática (16).
Produção de microestacas = +(245,43) – (1308,77 Mg do broto) – (0,0368 Mn do broto) +
(63,142 Ca2 do broto) + (1845,92 Mg2 do broto) + (0,00012 Fe2 do broto) + (0,00001247
Mn2 do broto).
Tabela 11e. Continuação da Tabela 11. Sétima Coleta Variação Número Parcial Acumulada R2 R2 F Prob>F
Ca2 do broto 1 0,2074 0,2074 5,7563 0,0253 Fe2 do broto 2 0,1083 0,3157 3,3248 0,0825 Mn2 do broto 3 0,0869 0,4026 2,9090 0,1036 Mg do broto 4 0,0933 0,4960 3,5184 0,0761 Mg2 do broto 5 0,1261 0,6221 6,0081 0,0247 Mn do broto 6 0,0716 0,6937 3,9766 0,0624
A oitava e nona coleta respondeu de maneira diferenciada a ausência de nutrientes. No
caso da oitava coleta, apenas ao boro parece estar legando algum efeito sobre a produção de
microestacas, fato este não esperado, tendo em vista que a ausência de boro na brotação, leva a
um desenvolvimento inadequado e baixa produção de microestacas, como observado “in
loco”. Porém este efeito responde por apenas 12,88% da resposta fisiológica que, em termos
estatísticos é muito baixo.

53
Tabela 10f. Continuação da Tabela 10. Oitava Coleta R2 = 0,12882901 DF SQ QM F Prob>F Regressão 1 479,75923143 479,75923143 3,25 0,0850 Erro 22 3244,24076857 147,46548948 Total 23 3724,00000000 Equação de regressão quadrática (17). Produção de microestacas = + (44,26) – (0,0016 B2 do broto)
Tabela 11f. Continuação da Tabela 11. Oitava Coleta Variação Número Parcial Acumulada R2 R2 F Prob>F B2 do broto 1 0,1288 0,1288 3,2534 0,0850
No caso da nona coleta, novamente os resultados matemáticos confirmam a realidade.
A ausência de magnésio nas brotações aumenta a produção de microestacas (equação de
regressão quadrática 18). O efeito deste nos resultados de produção é de 33,57% (Tabela 11g).
Tabela 10g. Continuação da Tabela 10. Nona Coleta R2 = 0,3567903 DF SQ QM F Prob>F Regressão 1 2858,64261191 2858,64261191 11,12 0,0030 Erro 22 5657,35738809 257,15260855 Total 23 8516,00000000 Equação de regressão polinomial quadrática (18).
Produção de microestacas = + (4,5664) + (0,0398 Na do broto) – (351,088 Mg2 do broto).

54
Tabela 11g. Continuação da Tabela 11. Nona Coleta Variação Número Parcial Acumulada R2 R2 F Prob>F
Mg2 do broto 1 0,3357 0,3357 11,1165 0,0030 Na do broto 2 0,2104 0,5461 9,7361 0,0052
5.5. Análises da produção de microestacas em função dos nutrientes presentes nas
soluções consideradas.
Na solução completa a diminuição dos níveis de potássio e o acréscimo nos de boro na
brotação, assim como a maior quantidade de magnésio na raiz, auxiliam a produção de
microestacas (equação de regressão 19)
Estes teores dos elementos na solução completa (Tabela 13), em resultados parciais,
responderam a 22,96% (boro na brotação), 18,57% (potássio na brotação) e 15,97%
(manganês na raiz) e no total a 57,50% (soma das porcentagens descriminadas).
Tabela 12. Análise de regressão quadrática para produção de microestaca de Eucalyptus
grandis x E. urophylla em função da solução considerada, de acordo com a Tabela 3,
mantida em sistema hidropônico, com omissão nutriente e completa.
Solução Completa R2 = 0,81861203
DF SQ QM F Prob>F Regressão 6 2607,61628840 434,60271473 10,71 0,0001 Erro 17 689,71704493 40,57159088 Total 23 3297,33333333 Equação de regressão quadrática (19).
Produção de microestacas = + (19,532) – (73 P do broto) + (54,37 S do broto) + (0,1365 Mn
da raiz) – (1,331 K2 do broto) + (0,000045 Mn2 do broto) + (0,0032 B2 do broto).
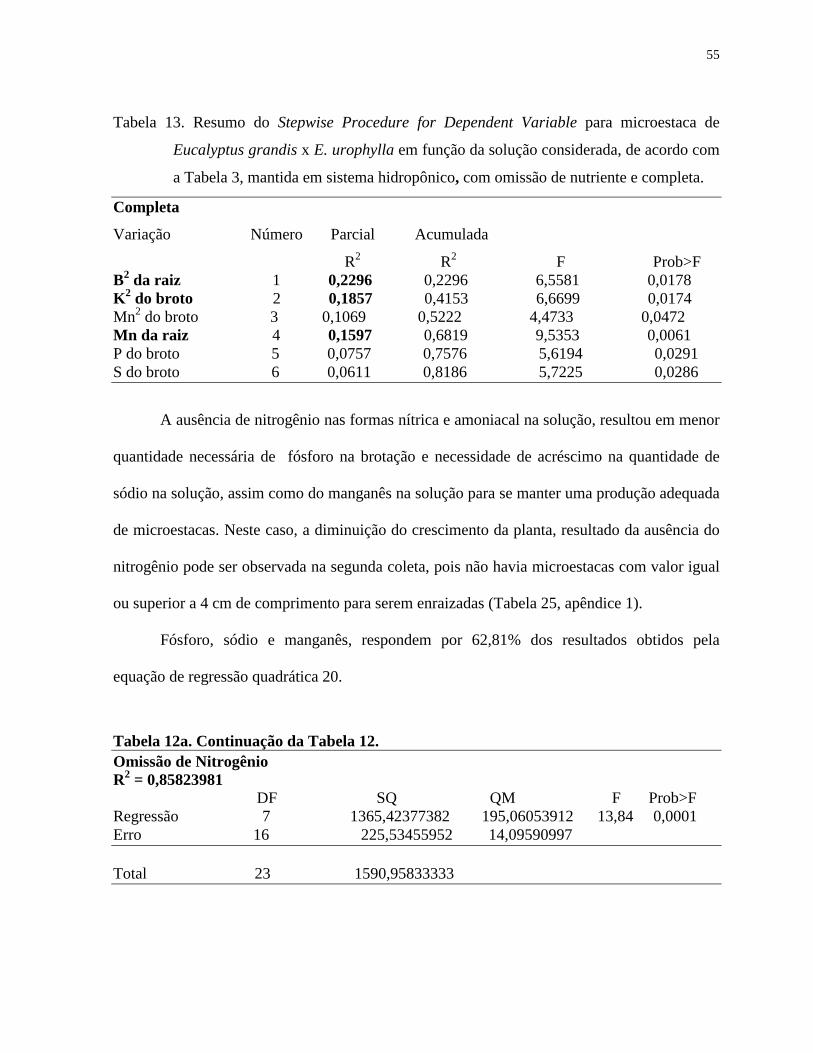
55
Tabela 13. Resumo do Stepwise Procedure for Dependent Variable para microestaca de
Eucalyptus grandis x E. urophylla em função da solução considerada, de acordo com
a Tabela 3, mantida em sistema hidropônico, com omissão de nutriente e completa.
Completa
Variação Número Parcial Acumulada
R2 R2 F Prob>F B2 da raiz 1 0,2296 0,2296 6,5581 0,0178 K2 do broto 2 0,1857 0,4153 6,6699 0,0174 Mn2 do broto 3 0,1069 0,5222 4,4733 0,0472 Mn da raiz 4 0,1597 0,6819 9,5353 0,0061 P do broto 5 0,0757 0,7576 5,6194 0,0291 S do broto 6 0,0611 0,8186 5,7225 0,0286
A ausência de nitrogênio nas formas nítrica e amoniacal na solução, resultou em menor
quantidade necessária de fósforo na brotação e necessidade de acréscimo na quantidade de
sódio na solução, assim como do manganês na solução para se manter uma produção adequada
de microestacas. Neste caso, a diminuição do crescimento da planta, resultado da ausência do
nitrogênio pode ser observada na segunda coleta, pois não havia microestacas com valor igual
ou superior a 4 cm de comprimento para serem enraizadas (Tabela 25, apêndice 1).
Fósforo, sódio e manganês, respondem por 62,81% dos resultados obtidos pela
equação de regressão quadrática 20.
Tabela 12a. Continuação da Tabela 12. Omissão de Nitrogênio R2 = 0,85823981 DF SQ QM F Prob>F Regressão 7 1365,42377382 195,06053912 13,84 0,0001 Erro 16 225,53455952 14,09590997 Total 23 1590,95833333
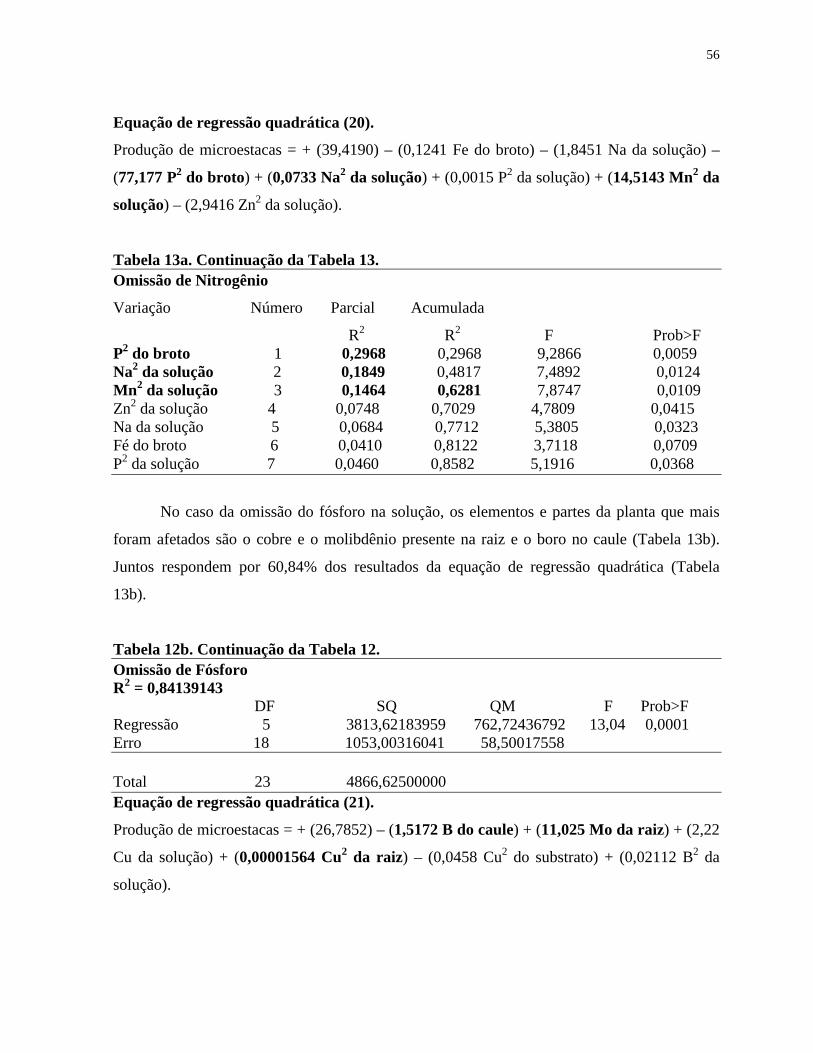
56
Equação de regressão quadrática (20).
Produção de microestacas = + (39,4190) – (0,1241 Fe do broto) – (1,8451 Na da solução) –
(77,177 P2 do broto) + (0,0733 Na2 da solução) + (0,0015 P2 da solução) + (14,5143 Mn2 da
solução) – (2,9416 Zn2 da solução).
Tabela 13a. Continuação da Tabela 13. Omissão de Nitrogênio
Variação Número Parcial Acumulada
R2 R2 F Prob>F P2 do broto 1 0,2968 0,2968 9,2866 0,0059 Na2 da solução 2 0,1849 0,4817 7,4892 0,0124 Mn2 da solução 3 0,1464 0,6281 7,8747 0,0109 Zn2 da solução 4 0,0748 0,7029 4,7809 0,0415 Na da solução 5 0,0684 0,7712 5,3805 0,0323 Fé do broto 6 0,0410 0,8122 3,7118 0,0709 P2 da solução 7 0,0460 0,8582 5,1916 0,0368
No caso da omissão do fósforo na solução, os elementos e partes da planta que mais
foram afetados são o cobre e o molibdênio presente na raiz e o boro no caule (Tabela 13b).
Juntos respondem por 60,84% dos resultados da equação de regressão quadrática (Tabela
13b).
Tabela 12b. Continuação da Tabela 12. Omissão de Fósforo R2 = 0,84139143
DF SQ QM F Prob>F Regressão 5 3813,62183959 762,72436792 13,04 0,0001 Erro 18 1053,00316041 58,50017558 Total 23 4866,62500000 Equação de regressão quadrática (21).
Produção de microestacas = + (26,7852) – (1,5172 B do caule) + (11,025 Mo da raiz) + (2,22
Cu da solução) + (0,00001564 Cu2 da raiz) – (0,0458 Cu2 do substrato) + (0,02112 B2 da
solução).

57
Tabela 13b. Continuação da Tabela13. Omissão de Fósforo
Variação Número Parcial Acumulada R2 R2 F Prob>F Mo da raiz 1 0,2405 0,2405 6,9674 0,0150 Cu2 da raiz 2 0,1593 0,3998 5,5748 0,0280 B do caule 3 0,2086 0,6084 10,6543 0,0039 Cu2 do substrato 4 0,0999 0,7083 6,5040 0,0195 (continuação da Tabela13b). B2 da solução 5 0,0753 0,7836 6,2671 0,0222 Cu da solução 6 0,0578 0,8414 6,1912 0,0235
A omissão de potássio na solução levou a efeito direto na sua concentração no
substrato e também a necessidade de diminuição do cloro na solução para manutenção de
produção de microestacas, tendo em vista que o sal utilizado na solução completa é o cloreto
de potássio. Apesar de ter sido omitido, o potássio ainda aparece no substrato, talvez fruto do
processo de decomposição deste e há aumento do cloro na solução, que provavelmente na
ausência do elemento não é carreado para o substrato ou planta (Tabela 12c).
Os elementos em suas posições na solução e substrato, respondem por 60,87% (Tabela
13c), dos resultados esperados pela equação de regressão quadrática 22, da Tabela 12c.
Tabela 12c. Continuação da Tabela12. Omissão de Potássio R2 = 0,86954883
DF SQ QM F Prob>F Regressão 5 4662,19473111 932,43894622 24,00 0,0001 Erro 18 699,43026889 38,85723716 Total 23 5361,62500000 Equação de regressão quadrática (22).
Produção de microestacas = + (169,8864) – (11,52 K do substrato) – (0,7978 P em MELLS)
– (0,5336 Cl em solução) + (6,9542 N2 da raiz).

58
Tabela 13c. Continuação da Tabela 13. Omissão de Potássio
Variação Número Parcial Acumulada R2 R2 F Prob>F K do substrato 1 0,4133 0,4133 15,4998 0,0007 Cl da solução 2 0,1954 0,6087 10,4850 0,0039 N2 da raiz 3 0,0732 0,6819 4,6051 0,0443 P em MELS 4 0,1255 0,8074 12,3829 0,0023 Ca da solução 5 0,0621 0,8695 8,5704 0,0090
A retirada do cálcio na solução levou a uma menor necessidade de zinco na brotação e
aumento na necessidade de manganês na raiz (equação de regressão quadrática 23), indicando
que estes elementos podem estar associados no transporte dentro da planta, uma vez que o
cálcio desloca-se por fluxo de massa e não havendo este na solução, o zinco pode ter o seu
deslocamento comprometido.
Tanto o manganês da raiz como o zinco da brotação, respondem em conjunto por
69,88% da resposta de produção (Tabela 13d), representada pela equação de regressão citada.
Tabela 12d. Continuação da Tabela 12. Omissão de Cálcio R2 = 0,76720635
DF SQ QM F Prob>F Regressão 3 3080,30152883 1026,76717628 21,97 0,0001 Erro 20 934,65680450 46,73284023 Total 23 4014,95833333 Equação de regressão quadrática (23).
Produção de microestacas = + (21,3541) + (0,2061 Mn da raiz) – (0,08454 Na do substrato) –
(0,0007 Zn2 do broto).

59
Tabela 13d. Continuação da Tabela 13. Omissão de cálcio
Variação Número Parcial Acumulada R2 R2 F Prob>F Zn2 do broto 1 0,4651 0,4651 19,1287 0,0002 Mn da raiz 2 0,2337 0,6988 16,2964 0,0006 Na do substrato 3 0,0684 0,7672 5,8755 0,0250
A omissão de magnésio na solução levou a necessidade de aumento nas concentrações
de matéria orgânica no substrato, representada pela casca de arroz (equação de regressão
quadrática 24). Nestas condições de omissão de magnésio, a colocação de maior quantidade de
matéria orgânica, pode aumentar a produção de microestacas, talvez pela liberação de
magnésio pelo substrato (vermiculita).
A equação de regressão citada responde por 46,29% da resposta de produção de
microestacas na ausência de magnésio.
Tabela 12e. Continuação da Tabela 12. Omissão de Magnésio R2 = 0,81448992
DF SQ QM F Prob>F Regressão 4 5017,53 1254,3839 14,04 0,0001 Erro 19 1697,42 89,33803 Total 23 6714,95 Equação de regressão quadrática (24).
Produção de microestacas = + (70,063) – (12,98 N do broto) + (6,6487 M.O.substrato) +
(0,00010865 Mn2 do caule) + (2,87648 Co2 do caule).

60
Tabela 13e. Continuação da Tabela 13. Omissão de Magnésio
Variação Número Parcial Acumulada R2 R2 F Prob>F M.O. substrato 1 0,4629 0,4629 18,9628 0,0003 N do broto 2 0,1376 0,6006 7,2359 0,0137 Mn2 do caule 3 0,0951 0,6957 6,2512 0,0212 Co2 do caule 4 0,0515 0,7472 3,8738 0,0638
A omissão de ferro na solução tem uma correlação direta com o sódio, tendo em vista
que o ferro utilizado é quelatado com Na.EDTA. Desta forma, a retirada do ferro faz com que
a solução exija maiores quantidades de sódio como pode ser observada na equação de
regressão quadrática 25.
Tabela 12f. Continuação da Tabela 12. Omissão de Ferro R = 0,83441085
DF SQ QM F Prob>F Regressão 6 3689,45187609 614,90864601 14,28 0,0001 Erro 17 732,17312391 43,06900729 Total 23 4421,62500000 Equação de regressão quadrática (25).
Produção de microestacas = + (45,9579) + (0,4810 Mg da solução) – (463,95 S2 do broto) –
(0,0008 Na2 do substrato) – (0,0022 B2 do substrato) – (0,0621 Cu2 do substrato) + (0,03856
Na2 da solução).
Tabela 13f. Continuação da Tabela 13. Omissão de Ferro
Variação Número Parcial Acumulada
R2 R2 F Prob>F Na2 da solução 1 0,3870 0,3870 13,8886 0,0012 B2 do substrato 2 0,1663 0,5533 7,8200 0,0108 S2 do broto 3 0,1084 0,6617 6,4104 0,0198

61
(continuação da Tabela 13f) Cu2 do substrato 4 0,0586 0,7204 3,9848 0,0604 Na2 do substrato 5 0,0527 0,7731 4,1807 0,0558 Mg da solução 6 0,0613 0,8344 6,2956 0,0225
A retirada do boro da solução, fez com que o cloro tivesse importância sobre a
produção de microestacas, evidenciada na equação de regressão quadrática 26, sendo que a
resposta do cloro na solução em função da produção de microestacas é de 23,81%, de acordo
com a Tabela 13g.
Tabela 12g. Continuação da Tabela 12. Omissão de Boro R2 = 0,45355134
DF SQ QM F Prob>F Regressão 3 1468,12679849 489,37559950 5,53 0,0062 Erro 20 1768,83153484 88,44157674 Total 23 3236,95833333 Equação de regressão quadrática (26). Produção de microestacas = + (26,0271) + (0,09953 Mn da raiz) – (0,00094 Zn2 do broto) – (0,000227 Cl2 da solução).
Tabela 13g. Continuação da Tabela 13. Omissão de Boro
Variação Número Parcial Acumulada R2 R2 F Prob>F Cl2 da solução 1 0,2381 0,2381 6,8733 0,0156 Zn2 do broto 2 0,1362 0,3742 4,5694 0,0445 Mn da raiz 3 0,0793 0,4536 2,9036 0,1039

62
5.6. Análises da produção de microestacas em função dos nutrientes presentes nas
soluções, analisando-se todas as coletas simultaneamente.
Como se esperava, em função das observações “in loco”, a retirada do potássio da
solução aumentou a produtividade de microestacas, porém o fato a se estranhar é que também
a retirada do cálcio produziu o mesmo efeito (equação de regressão quadrática 27). O
manganês, por sua vez, é um desconhecido neste processo de produção. Sua ação pode estar
ligada à ocupação de espaços iônicos liberados pelo cálcio e o magnésio. Porém, os três
elementos juntos responderam por apenas 15,47% dos resultados de produção de microestacas
(Tabela 15) e o grau de confiabilidade da equação foi de apenas 0,266.
Tabela 14. Análise de regressão quadrática para produção de microestacas de Eucalyptus
grandis x E. urophylla, analisando-se apenas o efeito das soluções, para todas as
coletas, de acordo com a Tabela 3, mantida em sistema hidropônico, com omissão
de nutriente e completa.
Todas as Coletas R2 = 0,26693371 DF SQ QM F Prob>F Regressão 8 15039,5107 1879,9388 8,33 0,0001 Erro 183 41302,2340 225,6952 Total 191 56341,7447 Equação de regressão quadrática (27).
Produção de microestacas = +(41,90) + (2,65 pH da solução) – (0,7963 NH4 da solução) –
(0,2072 K da solução) – (0,0672 Ca da solução) + (1,8568 Mn da solução) + (0,988 Zn da
solução) + (0,04114 NH4(2) da solução) – (0,00006039 S2 da solução).
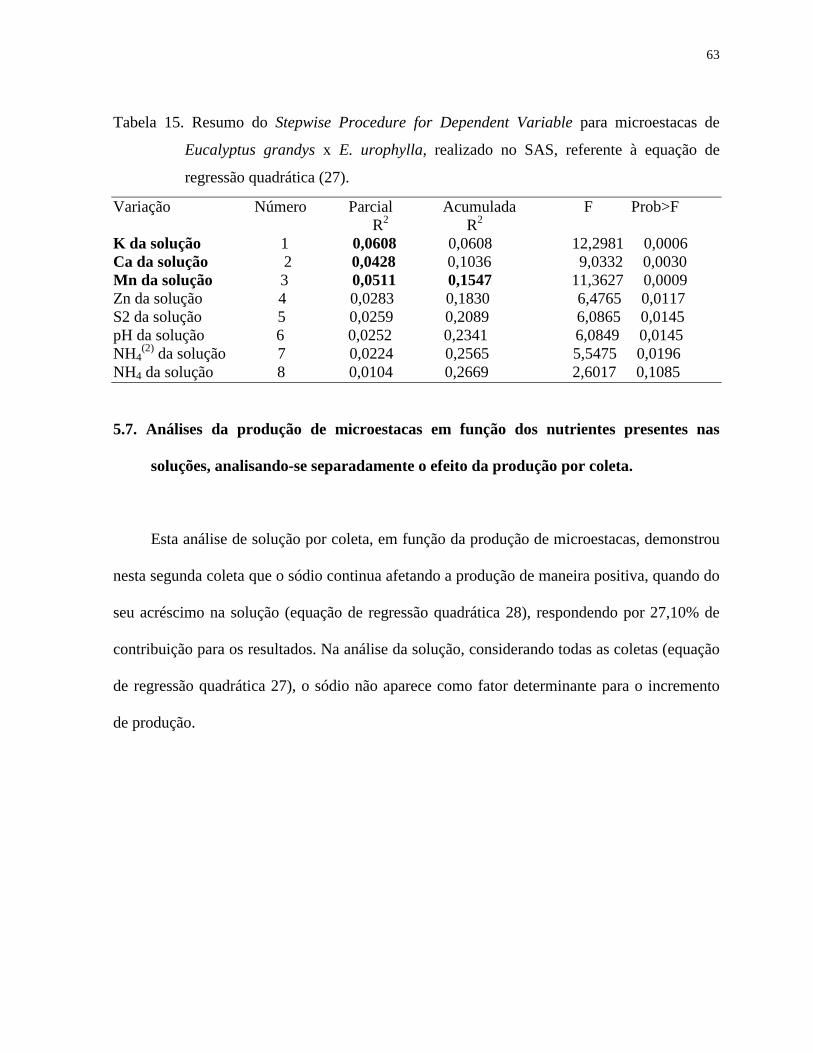
63
Tabela 15. Resumo do Stepwise Procedure for Dependent Variable para microestacas de
Eucalyptus grandys x E. urophylla, realizado no SAS, referente à equação de
regressão quadrática (27).
Variação Número Parcial Acumulada F Prob>F R2 R2 K da solução 1 0,0608 0,0608 12,2981 0,0006 Ca da solução 2 0,0428 0,1036 9,0332 0,0030 Mn da solução 3 0,0511 0,1547 11,3627 0,0009 Zn da solução 4 0,0283 0,1830 6,4765 0,0117 S2 da solução 5 0,0259 0,2089 6,0865 0,0145 pH da solução 6 0,0252 0,2341 6,0849 0,0145 NH4
(2) da solução 7 0,0224 0,2565 5,5475 0,0196 NH4 da solução 8 0,0104 0,2669 2,6017 0,1085
5.7. Análises da produção de microestacas em função dos nutrientes presentes nas
soluções, analisando-se separadamente o efeito da produção por coleta.
Esta análise de solução por coleta, em função da produção de microestacas, demonstrou
nesta segunda coleta que o sódio continua afetando a produção de maneira positiva, quando do
seu acréscimo na solução (equação de regressão quadrática 28), respondendo por 27,10% de
contribuição para os resultados. Na análise da solução, considerando todas as coletas (equação
de regressão quadrática 27), o sódio não aparece como fator determinante para o incremento
de produção.

64
Tabela 16. Análise de regressão quadrática para produção de microestacas de Eucalyptus
grandis x E. urophylla em função dos elementos analisados na solução, por coleta,
de acordo com a Tabela 3, mantida em sistema hidropônico, com omissão de
nutriente e completa.
Segunda Coleta R2 = 0,67005917 DF SQ QM F Prob>F Regressão 1 2367,7107 2367,7107 8,18 0,0091 Erro 22 6368,2476 289,4658 Total 23 8735,958 Equação de regressão quadrática (28).
Produção de microestacas = - (26,5747) + (4,0240 P da solução) + (0,0257 Na2 da solução) –
(0,0585 P2 da solução) + (1,8726 Mn2 da solução).
Tabela 17. Resumo do Stepwise Procedure for Dependent Variable para produção
microestacas de Eucalyptus grandis x E. urophylla, realizado no SAS, referente às
equações de regressão quadráticas de 28 a 35.
Segunda Coleta Variação Número Parcial Acumulada F Prob>F R2 R2 Na2 da solução 1 0,2710 0,2710 8,1796 0,0091 Mn2 da solução 2 0,0812 0,3522 2,6312 0,1197 P2 da solução 3 0,1812 0,5334 7,7686 0,0114 P da solução 4 0,1366 0,6701 7,8681 0,0113
Nesta terceira coleta volta a evidenciar a necessidade de manganês na solução, agora
atuando de maneira positiva no processo, ou seja, em função das omissões de outros
elementos, o Mn na solução adiciona produção às microestacas (equação de regressão
quadrática 29). A sua contribuição para o resultado da equação é de 24,33%, sendo que o
coeficiente de determinação (R2) é de 0,3525 (Tabela 16a).

65
Tabela 16 a. Continuação da Tabela 16. Terceira Coleta R2 = 0,35251781 DF SQ QM F Prob>F Regressão 2 1994,5310 997,2655 5,72 0,0104 Erro 21 3663,4272 174,4489 Total 23 5657,9583 Equação de regressão quadrática (29). Produção de microestacas = +(27,2810) + (2,4134 Mn da solução) + (1,7855 Cu2 da solução).
Tabela 17a. Continuação da Tabela 17. Terceira Coleta Variação Número Parcial Acumulada F Prob>F R2 R2 Mn da solução 1 0,2433 0,2433 7,0748 0,0143 Cu2 da solução 2 0,1092 0,3525 3,5413 0,0738
Comprovando que há interações que devem ser estudadas de maneira mais profunda,
nesta coleta inverte-se o papel do sódio, sendo considerada a sua presença desnecessária neste
momento (equação de regressão quadrática 30) e a presença de maiores quantidades de
enxofre como benéfica no processo fisiológico. O coeficiente da equação citada é de 0,7138
(Tabela 16b) e o grau de interferência destes elementos é de 44,92% (Tabela 17b).
Tabela 16b. Continuação da Tabela 16. Quarta Coleta R2 = 0,71388214 DF SQ QM F Prob>F Regressão 4 4281,5973 1070,3993 11,85 0,0001 Erro 19 1716,0276 90,3172 Total 23 5997,625 Equação de regressão quadrática (30). Produção de microestacas = +(81,9183) – (2,8843 Na da solução) – (0,2891 Ca da solução) + (0,0833 S da solução) – (1,6977 Zn da solução).
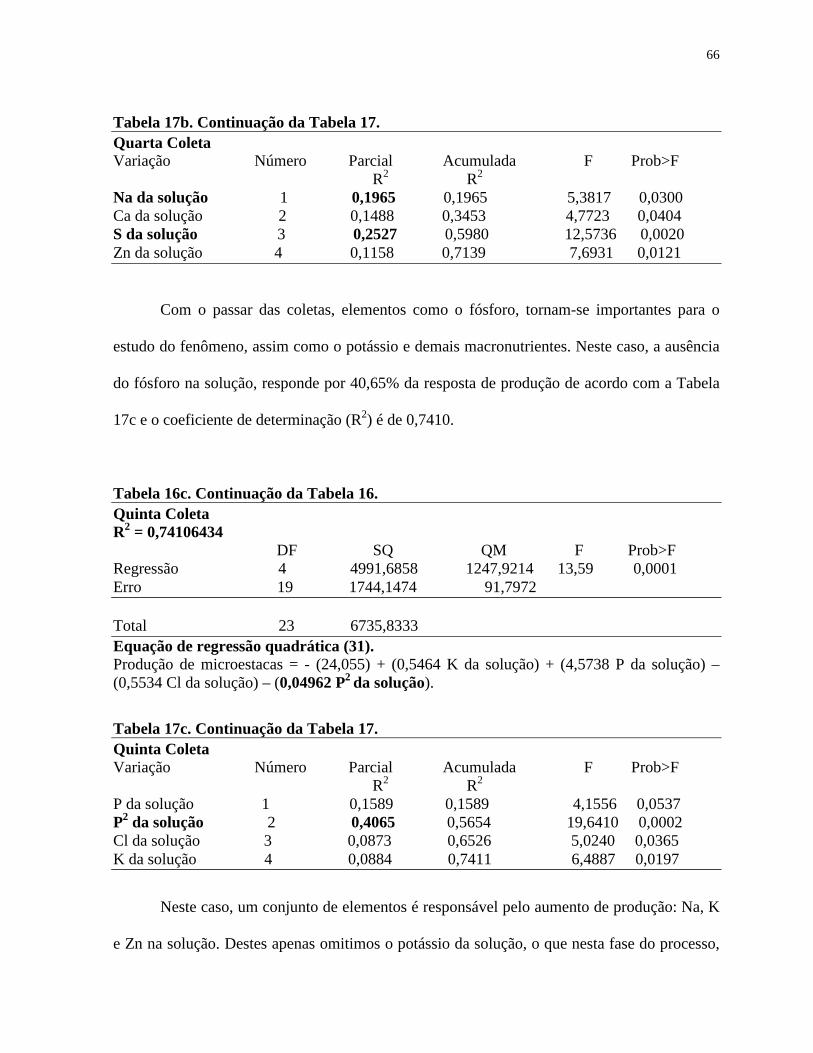
66
Tabela 17b. Continuação da Tabela 17. Quarta Coleta Variação Número Parcial Acumulada F Prob>F R2 R2 Na da solução 1 0,1965 0,1965 5,3817 0,0300 Ca da solução 2 0,1488 0,3453 4,7723 0,0404 S da solução 3 0,2527 0,5980 12,5736 0,0020 Zn da solução 4 0,1158 0,7139 7,6931 0,0121
Com o passar das coletas, elementos como o fósforo, tornam-se importantes para o
estudo do fenômeno, assim como o potássio e demais macronutrientes. Neste caso, a ausência
do fósforo na solução, responde por 40,65% da resposta de produção de acordo com a Tabela
17c e o coeficiente de determinação (R2) é de 0,7410.
Tabela 16c. Continuação da Tabela 16. Quinta Coleta R2 = 0,74106434 DF SQ QM F Prob>F Regressão 4 4991,6858 1247,9214 13,59 0,0001 Erro 19 1744,1474 91,7972 Total 23 6735,8333 Equação de regressão quadrática (31). Produção de microestacas = - (24,055) + (0,5464 K da solução) + (4,5738 P da solução) – (0,5534 Cl da solução) – (0,04962 P2 da solução).
Tabela 17c. Continuação da Tabela 17. Quinta Coleta Variação Número Parcial Acumulada F Prob>F R2 R2 P da solução 1 0,1589 0,1589 4,1556 0,0537 P2 da solução 2 0,4065 0,5654 19,6410 0,0002 Cl da solução 3 0,0873 0,6526 5,0240 0,0365 K da solução 4 0,0884 0,7411 6,4887 0,0197
Neste caso, um conjunto de elementos é responsável pelo aumento de produção: Na, K
e Zn na solução. Destes apenas omitimos o potássio da solução, o que nesta fase do processo,

67
demandou uma necessidade de aumento nas concentrações destes elementos (Equação de
regressão quadrática 32), sendo que o coeficiente de determinação (R2) é de 0,7119 (Tabela
16d).
A soma das influências destes elementos na equação é de 54,85% (Tabela.17d).
Tabela 16d. Continuação da Tabela 16. Sexta Coleta R2 = 0,71194579 DF SQ QM F Prob>F Regressão 5 2975,4587 595,0917 8,90 0,0002 Erro 18 1203,8745 66,8819 Total 23 4179,3333 Equação de regressão quadrática (32). Produção de microestacas = + (68,4322) – (0,1364 NO3 da solução) – (0,4412 Cl da solução) + (0,0079 Na2 da solução) + (0,0016 K2 da solução) + (0,2635 Zn2 da solução). Tabela 17d. Continuação da Tabela 17. Sexta Coleta Variação Número Parcial Acumulada F Prob>F R2 R2 Cl da solução 1 0,0933 0,0933 2,2634 0,1467 Zn2 da solução 2 0,1504 0,2437 4,1762 0,0537 Na2 da solução 3 0,1622 0,4059 5,4610 0,0300 NO3 da solução 4 0,0701 0,4761 2,5437 0,1272 K2 da solução 5 0,2359 0,7119 14,7405 0,0012
A necessidade da presença de ferro na solução para aumento na produção de
microestacas, pode estar ligada a menores quantidades do elemento na planta, aparecendo
como reflexo, a sua necessidade de incremento na solução (equação de regressão quadrática
33), Ele é responsável por 34,93% da resposta de produção da equação citada (Tabela 17e),
sendo que o coeficiente de determinação da equação (R2) é de 0,5405 (Tabela 16e).

68
Tabela 16e. Continuação da Tabela 16. Sétima Coleta R2 = 0,54053459 DF SQ QM F Prob>F Regressão 3 1899,3484 633,1161 7,84 0,0012 Erro 20 1614,4848 80,7242 Total 23 3513,8333 Equação de regressão polinomial quadrática (33). Produção de microestacas = + (20,6571) + (0,1420 Fe da solução) + (0,0007246 Ec2 da solução) – (0,0007627 Ca2 da solução).
Tabela 17e. Continuação da Tabela 17. Sétima Coleta Variação Número Parcial Acumulada F Prob>F R2 R2 Fe da solução 1 0,3493 0,3493 11,8094 0,0024 Ca2 da solução 2 0,0877 0,4370 3,2697 0,0849 Ec2 da solução 3 0,1036 0,5405 4,5086 0,0464
A partir da oitava coleta, elementos como o amônio podem sobrar na solução (equação
de regressão quadrática 34), fruto de processos de decomposição do substrato, muito embora,
em trabalhos realizados em cultura in vitro, haja evidências de que este elemento é utilizado de
maneira preferencial pelas plantas, ao contrário do nitrato.
No caso do cloro, ele é bem absorvido pela plantas, podendo estar ocorrendo excesso
deste no material vegetal, o que pode evidenciar excesso na solução, de acordo com a mesma
equação de regressão.
Ambos respondem por 24,41% da resposta de produtividade de microestacas
identificadas pela equação 34, sendo que o coeficiente de determinação (R2) desta é de 0,45
(Tabela 16f).

69
Tabela 16f. Continuação da Tabela 16. Oitava Coleta R2 = 0,45056241 DF SQ QM F Prob>F Regressão 4 1677,8944 419,4736 3,90 0,0179 Erro 19 2046,1055 107,6897 Total 23 3724,000 Equação de regressão quadrática (34). Produção de microestacas = + (51,015) – (3,2328 NH4 da solução) – (0,1041 Cl da solução) + (1,0919 Zn da solução) – (0,0001637 NO3
(2) da solução).
Tabela 17f. Continuação da Tabela 17. Oitava Coleta Variação Número Parcial Acumulada F Prob>F R2 R2 NO3
2 da solução 1 0,1037 0,1037 2,5460 0,1248 Zn da solução 2 0,1028 0,2065 2,7208 0,1139 Cl da solução 3 0,1177 0,3242 3,4824 0,0768 NH4 da solução 4 0,1264 0,4506 4,3698 0,0503
Na última coleta aparecem resultados semelhantes à quinta coleta, em relação ao
elemento de maior peso no resultado, o fósforo, com 53,62%, Tabela 17g e equação de
regressão quadrática 35. Sabe-se da sua importância no desenvolvimento do sistema radicular,
além de fazer parte do ATP (adenosina-trifosfato), fonte de energia para a planta, na sua
transformação em ADP (adenosina-difosfato).
A confiabilidade da equação é evidenciada na Tabela 16g, com “R2” igual a 0,6197.
Tabela 16g. Continuação da Tabela 16. Nona Coleta R2 = 0,61979141 DF SQ QM F Prob>F Regressão 2 5278,1436 2639,0718 17,12 0,0001 Erro 21 3237,8563 154,1836 Total 23 8516,0000

70
Equação de regressão quadrática (35). Produção de microestacas = + (73,6948) – (0,2216 Cl da solução) – (0,01093 P2 da solução).
Tabela 17g. Continuação da Tabela 17. Nona Coleta Variação Número Parcial Acumulada F Prob>F R2 R2 P2 da solução 1 0,5362 0,5362 25,4334 0,0001 Cl da solução 2 0,0836 0,6198 4,6175 0,0435
5.8. Análises da produção de microestacas em função dos parâmetros ambientais.
5.8.1 Umidade Relativa
As Figuras 3 e 4 são representativas de médias de coletas durante o tempo de 124 dias
em que o material se manteve em solução nutritiva completa e com omissão de nutrientes.
As avaliações foram realizadas internamente na área coberta e externamente, Os
valores apresentados, assim como as curvas de tendências, são as mais representativas deste
processo.
Pode-se observar, pela Figura 3, que mede a umidade relativa interna máxima, que há
uma maior tendência de produção de microestacas quando a umidade interna atinge 91,7%,
declinando a valores de 92,5%. Há que se observar que em condições como as descritas, pode
haver deficiência de cálcio no material vegetal, tendo em vista que o seu transporte é feito por
fluxo de massa.
A umidade relativa máxima externa flutua muito, como já era de se esperar, tendo em
vista que não há um controle como na área coberta (estufa aberta em suas laterais). Qualquer

71
fator como aumento da evapotranspiração da grama e ventos podem alterar a umidade medida
pelo equipamento.
Analisando a umidade relativa média, medida pelo equipamento na área externa e
interna (Figura 4), pode-se notar uma correlação de aferições, começando com valores em
torno de 64% de umidade e tende uma maior convergência de produção de microestacas ao
redor de 79 a 81% de umidade relativa.
A mesma situação de correlação de resultados podem ser observadas na Figura 5 que
mede a umidade relativa interna e externa mínima, variando entre um mínimo de 31 a 32% de
mínimas a 51,4% de máximas.

72
Produção de microestacas em função da umidade relativa
y = -13,23x2 + 2430,2x - 111556R2 = 0,9182 y = -14,149x3 + 4083,1x2 - 392752x + 1E+07
R2 = 0,5517
05
101520253035404550
90 91 92 93 94 95 96 97 98
Umidade relativa em %
Núm
ero
méd
io d
e m
icro
esta
cas*
Umidade relativa externa máxima Umidade relativa interna máxima
Figura 3. Efeito da umidade relativa externa e interna máximas sobre a produção de microestacas de Eucalyptus grandis x E. urophylla, em solução completa e com omissão de nutrientes, mantidas sistema hidropônico, em casa-de-vegetação, durante o período de 23 de maio a 19 de setembro de 2002. *Refere-se ao número médio de microestacas, por coleta, de 15 plantas.
Produção de microestacas em função da umidade relativa média
y = -0,0221x2 + 4,5139x - 178,45R2 = 0,8727
y = -0,0288x2 + 5,6793x - 223,77R2 = 0,9002
0
5
10
15
20
25
30
35
40
45
50
60 65 70 75 80 85
Umidade relativa em %
Núm
ero
méd
io d
e m
icro
esta
cas*
Umidade relativa externa média Umidade relativa interna média
Figura 4. Efeito da umidade relativa externa e interna médias sobre a produção de microestacas de Eucalyptus grandis x E. urophylla, em solução completa e com omissão de nutrientes, mantidas em sistema hidropônico em casa-de-vegetação, durante o período de 23 de maio a 19 de setembro de 2002. *Refere-se ao número médio de microestacas, por coleta, de 15 plantas.

73
Produção de microestacas em função da umidade relativa
y = -0,0358x2 + 4,1959x - 76,605R2 = 0,9134
y = -0,0347x2 + 3,937x - 65,071R2 = 0,9052
05
101520253035404550
25 30 35 40 45 50 55
Umidade relativa em %
Núm
ero
méd
io d
e m
icro
esta
cas*
Umidade relativa mínima externa Umidade relativa mínima interna
Figura 5. Efeito da umidade relativa externa e interna mínimas sobre a produção de microestacas de Eucalyptus grandis x E.urophylla, em solução completa e com omissão de nutrientes, mantidas em sistema hidropônica em casa-de-vegetação, durante o período de 23 de maio a 19 de setembro de 2002. *Refere-se ao número médio de microestacas, por coleta, de 15 plantas.
5.8.2 Temperatura, radiação global e radiação PAR
Em relação à temperatura externa máxima, Figura 6, varia entre 24,4oC a 29,8oC,
enquanto a interna de 26oC a 32,7oC, percebendo-se uma tendência de queda na produtividade
quando as temperaturas máximas internas sobem.
A Figura 7 mede a radiação global externa e interna. Trata-se da quantidade de energia
que chega ao solo, medida em W/m2. Esta identificação pode determinar a média adequada em
que as produções de microestacas podem ser maximizadas, ou seja, há uma flutuação de
valores em torno de 336,7 W/m2 a 416,9 W/m2. Esta referência pode ser utilizada para cobrir
ou permitir a penetração de luz em área de produção de microestacas, indicando que as
microcepas necessitam destes valores de radiação global para melhor funcionamento
fisiológico.

74
As diferenças entre as curvas de tendência e valores das radiações globais máximas
externas e internas, referem-se a cobertura na estufa de produção.
Em relação à radiação PAR (V/Wm2) média, que mede a radiação fotossintéticamente
ativa, podemos notar que ela varia entre 357,7V/Wm2 com maiores produções de microestacas
a 722V/Wm2, quando a produção decai, flutuando os valores em torno de 589V/Wm2.
Produção de microestacas em função da temperatura interna máxima
y = -0,8965x3 + 71,687x2 - 1907,6x + 16931R2 = 0,8214
y = -0,3659x3 + 31,376x2 - 895,25x + 8540R2 = 0,8476
05
101520253035404550
20 22 24 26 28 30 32 34
Temperatura em oC
Núm
ero
méd
io d
e m
icro
esta
cas*
Temperetura externa máxima Temperatura interna máxima
Figura 6. Efeito da temperatura externa e interna máximas sobre a produção de microestacas de Eucalyptus grandis x E. urophylla, em solução completa e com omissão de nutrientes, mantidas em sistema hidropônico em casa-de-vegetação, durante o período de 23 de maio a 19 de setembro de 2002. *Refere-se ao número médio de microestacas, por coleta, de 15 plantas.

75
Produção de microestacas (x10) em função da radiação global máxima
420450
360
430400
220
340
440420
450
360
430400
220
340
440
y = -0,0018x2 + 0,5115x + 468,49R2 = 0,8358
y = -0,0023x2 + 2,4581x - 240,04R2 = 0,6727
0
50
100
150
200
250
300
350
400
450
500
0 100 200 300 400 500 600 700 800 900
W/m2
Núm
ero
méd
io d
e m
icro
esta
cas
(x10
)*
Radiação Global externa máxima Radiação global interna máxima
Figura 7. Efeito da radiação global externa e interna máximas sobre a produção de microestacas de Eucalyptus grandis x .E. urophylla, em solução completa e com omissão de nutrientes, mantidas sistema hidropônico, em casa-de-vegetação, durante o período de 23 de maio a 19 de setembro de 2002. *Refere-se ao número médio de microestacas, por coleta, de 15 plantas.
Produção de microestacas (x10) em função da radiação PAR máxima
y = -0,0009x2 + 1,7019x - 373,67R2 = 0,7936
y = -0,0005x2 + 1,0492x - 127,44R2 = 0,6715
0
50
100
150
200
250
300
350
400
450
500
0 200 400 600 800 1000 1200 1400 1600 1800
Valores de PAR em V/Wm2
Núm
ero
méd
io d
e m
icro
esta
cas
(x10
)*
Radiação PAR externa máxima Radiação PAR interna máxima
Figura 8. Efeito da radiação fotossintéticamente ativa, externa e interna máximas sobre a produção de microestacas de Eucalyptus grandis x E. urophylla, em solução completa e com omissão de nutrientes, mantidas no sistema hidropônico, em casa-de-vegetação, durante o período de 23 de maio a 19 de setembro de 2002. *Refere-se ao número médio de microestacas, por coleta, de 15 plantas.

76
Produção de microestacas (x10) em função dda radiação PAR média
y = -0,0029x2 + 2,6686x - 154,15R2 = 0,7591
y = -0,0017x2 + 2,0507x - 169,32R2 = 0,6542
050
100150200250300350400450500
0 100 200 300 400 500 600 700 800 900 1000
V/Wm2
Núm
ero
méd
io d
e m
icro
esta
cas
(x10
)*
Radiação PAR externa média Radiação PAR média internaPolinômio (Radiação PAR média interna) Polinômio (Radiação PAR externa média)
Figura 9. Efeito da radiação fotossinteticamente ativa, externa e interna máximas sobre a produção de
microestacas de Eucalyptus grandis x E. urophylla, em solução completa e com omissão de nutrientes, mantidas no sistema hidropônico, em casa-de-vegetação, durante o período de 23 de maio a 19 de setembro de 2002. *Refere-se ao número médio de microestacas, por coleta, de 15 plantas.

77
5.9 Análise conjunta das informações
Seria de certa forma inadmissível que um estudo mais aprofundado do sistema não
fosse realizado, apesar do processo ter se iniciado em 1997 e ter mostrado às empresas que o
enraizamento e a produtividade das mini ou microestacas duplicavam, poucos estudos foram
publicados sobre a questão, permitindo compreensão mais adequada do sistema de produção.
Campinhos et al. (2000) no trabalho intitulado “Hidrojardim Clonal Champion: uma
otimização na produção de mudas de eucalipto”, salientaram que a produtividade de
microestacas/microcepas/mês sobe de 3 para 6 no viveiro de mudas de São Paulo e, que o
enraizamento das microestacas no viveiro do Mato-Grosso-do-Sul aumentou de 50% para
80%.
Porém, quanto mais delicado o sistema, maior a possibilidade de erros que podem ser
cometidos e maior a necessidade de pesquisas. É o que ficou evidenciado entre 2001 e 2002 na
International Paper do Brasil (Mogi-Guaçu, São Paulo), quando houve perda de estabilidade
fisiológica das miniestacas utilizadas no processo de produção de mudas, em função do fato
do mini jardim clonal hidropônico ter deixado de ser renovado por microestacas originárias do
laboratório de cultura de tecidos. Isto se deveu ao fato das novas mudas introduzidas no
hidrojardim clonal não terem a necessária reversão a juvenilidade requisitada para melhoria na
produção e estabilidade do clone. Na realidade as touças do hidrojardim eram substituídas
quando apresentavam baixa produtividade, por mudas do viveiro comercial, ou seja, mudas
originárias de miniestacas, parte das próprias touças que estavam sendo substituídas e que
forneceram as miniestacas.

78
Clones consagrados pela empresa tiveram queda de enraizamento e aproveitamento
finais (informações pessoais) o que reabilitou o laboratório em 2002 e possibilitou um melhor
aproveitamento do sistema de produção.
Através dos resultados obtidos neste trabalho, nota-se também que existem algumas
incongruências em relação ao monitoramento das condições nutricionais das microcepas e
minicepas, assim como da solução nutritiva, tão importante para a produção das mudas, a se
destacar:
A) Na Tabela 4, em que todos os teores nutricionais foram analisados conjuntamente
(macro e micronutrientes), assim como as origens destes (material vegetal, solução e
substrato) na equação de regressão quadrática (01), observa-se que várias partes da
planta, além das já tradicionalmente analisadas, tiveram importância no resultado final
da regressão, como o substrato e o caule da microcepa, além do fato de mostrar que
existem relações que estão sendo desconsideradas quando do ajuste do processo de
produção, pois o molibdênio do broto da microcepa contribuiu com 5,69% do resultado
expresso na equação de regressão; o cobre e o sódio do caule da microcepa
contribuíram, em conjunto, com 10,03% e segundo a equação de regressão, na solução
existia a necessidade da colocação de maiores quantidades de sódio (Tabela 5). Estes
elementos (Cu, Mo e Na) não foram alterados em seus teores no preparo das soluções
nutritivas, e mesmo assim apareceram nas equações de regressão, de forma
determinante, demonstrando terem ações fisiológicas que precisam ser melhores
estudadas;
B) Analisando-se os teores dos nutrientes de maior importância (superior a 3% nos
resultados parciais), a Tabela 5 apresentou resultado interessante pois a resposta da

79
equação de regressão (01) ao processo de produção de microestacas em relação aos
elementos em negrito correspondeu a 38,44% e, pesar do grande número de interações,
o coeficiente de determinação foi de 65,36% (Tabela 4);
C) Na Tabela 9, o resultado com apenas brotações, retirado das microcepas, e que são
comuns de análises de seus teores de nutrientes para se verificar o conteúdo de macro e
micro elementos para ajuste da solução nutritiva e comparações com os índices de
enraizamento da microestacas, contribuiu com apenas 12,22% dos teores nutricionais
envolvidos (resultados parciais) contra 38,44% da Tabela 5, sendo que o coeficiente de
determinação foi de 18,12% (Tabela 8) contra 65,36% indicados na Tabela 4,
demonstrando que a retirada deste material (brotações das microcepas), por si só, não é
suficiente para tomadas de decisão em relação às melhorias no processo de produção
de microestacas e;
D) Outro fator a se considerar na análise das tabelas é que, conforme o resultado da coleta
ou da análise individual das soluções em função das suas omissões, podem ocorrer
diversas formas de interpretação das equações de regressão e acuidade de R2. Isto já
era previsto por Ingestad (1982) e Ingestad & Ägren (1992 e 1995), Fyles et al., 1994 e
Cromer et al. (1993), quando tratam do “steady-state” e dos “polls” de armazenamento,
uma vez que as necessidades dos nutrientes mudam em função do tempo de
crescimento (∆T), da parte do vegetal analisada e do estágio de desenvolvimento do
vegetal (ontogênese).

80
6 CONCLUSÕES
1. Para o conjunto de fatores analisados em relação à parte nutricional e suas
implicações no processo de produção de microestacas, é de fundamental
importância a análise da planta como um todo, solução nutritiva e substrato;
2. O processo de produção é dinâmico, alterando-se a cada coleta, sendo um ou
outro elemento nutricional de fundamental importância naquele momento do
processo fisiológico, importando ao pesquisador a interpretação pontual para
decisão de quando recomendar alguma intervenção nutricional;
3. O entendimento dos parâmetros climáticos, em especial a radiação global e
PAR é fundamental para o aumento da produtividade das microestacas e;
4. Os valores apresentados neste trabalho permitirão interpretar de maneira mais
adequada as temperaturas internas das áreas de produção, assim como as
umidades relativas, evitando-se perdas de produção de microestacas por falta
de conhecimento destes fatores ambientais.

81
REFERÊNCIAS BIBLIOGRÁFICAS
ASSIS, T.F. Evolution of Technology for Cloning Eucalyptus in Large Scale. Guaíba
Klabin, 2002. 11 p. (Comunicação Interna).
ASSIS, T.F. et al. Efeito da redução da luz em jardins clonais sobre o enraizamento de estacas
de E. urophylla x E. grandis. In: CONGRSSO FLORESTAL BRASILEIRO, 6, 1990, Campos
de Jordão. Anais... São Paulo:SBS-SBEF, 1990. p. 454-455.

82
ALBUQUERQUE, T. C. S.; DECHEN, A.R. Absorção de macronutrientes por porta-enxertos
e cultivares de videira em hidroponia. Scientia Agrícola, Piracicaba, v.57, n.1, p. 45-50, 2000.
BARROS, N. F.; SILVA, A. P. Condução de touças de Eucalyptus: resultados preliminares.
Piracicaba: IPEF, 1990. 20 p.
BLISKA, A. Jr.; HONÓRIO, S.L. Cartilha tecnológica de hidroponia. Campinas: Faculdade
de Engenharia Agrícola, 1996. 50 p.
BUGARÍN, R. M. et al. Amonio / nitrato y concentración iónica total de la solución nutritiva
en Crisantemo. II. Extracción nutrimental de hojas. Terra, Bucuresti, RO, v.16, p.113-124,
1998.
CAMARGOS, S.L. Diagnose de deficiência, teor e acúmulo de nutrientes em
Castanheira-do-Brasil. 90f. 1999. Tese (Doutoramento em Ciências: área de concentração
em Energia Nuclear na Agricultura) – Centro de Energia Nuclear na Agricultura - Escola
Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba. 1998.
CAMPINHOS, E. N. et al. Hidrojardim clonal Champion: Uma otimização na produção de
mudas de eucaliptos. Silvicultura, São Paulo, v.80, p.42-46, 2000.
CARELLI, M. L. C. et al. Níveis de nitrogênio, metabolismo, crescimento e produção de
girassol. Revista Brasileira de Fisiologia Vegetal, Londrina, v.8, n.2, p.123-130, 1996.

83
CROMER, R.N.; CAMERON, D.M.; RANCE, S.J. Response to nutrients in Eucalyptus
grandis: 2. Nitrogen accumulation. Forest Ecology and Management, Amsterdam, v.62,
p.231-243, 1993.
DECHEN, A. R.; HAAG, H. P.; CARMELO, Q. A. de C. Funções dos micronutrientes nas
plantas. In: FERREIRA, M.E.; CRUZ, M.C.P. (Ed.) Micronutrientes na agricultura.
Piracicaba: POTAPÓS/CNPq, 1991. p. 65-78.
DELL, B.; ROBINSON, J.M. Symptoms of mineral nutrient deficiency and nutrients
concentration ranges in seedlings of Eucalyptus maculata Hook. Plant and Soil, Dordrecht,
v.155/156, p.255-261, 1993.
DELL, B.; MALAJCZUK, N.; GROVE, T.S. Nutrient disorders in plantation eucalypts.
Canberra: Australian Centre for International Agricultural Research, 1995. 104 p.
DRUMOND, M. A.; LIMA, P. C. F. Sombreamento e produção de mudas de leucena e
cumaru. In: CONGRESSO FLORESTAL BRASILEIRO, 1993, Curitiba. Anais... São
Paulo:SBS-SBEF, 1993. p.309-311.
ENGEL, V. L.; POGGIANI, F. Estudo da concentração de clorofila nas folhas e seu espectro
de absorção de luz em função do sombreamento em mudas de quatro espécies florestais
nativas. Revista Brasileira de Fisiologia Vegetal, Londrina, v.3, p.39-45, 1991.

84
EPSTEIN, E. Nutrição mineral de plantas: princípios e perspectivas. Rio de Janeiro: Livros
Técnicos e Científicos, 1975. 341 p.
FYLES, J. W.; CÔTE, B.; COURCHESNE, F. Effects of base cation fertilization on soil and
foliage nutrient concentrations and litter-fall and throghfall nutrien fluxes in a sugar maple
forest. Canadian Journal of Forest Researches, Ottawa, v.24, p.542-549, 1994.
FURLANI, P.R. Cultivo de alface pela técnica de hidroponia - NFT. Campinas: Instituto
Agronômico de Campinas, 1995. 18 p. (Documentos, 55).
FURLANI, P.R. Instruções para o cultivo de hortaliças de folha pela técnica de
hidroponia – NFT. Campinas: Instituto Agronômico de Campinas, 30 p. 1998.
GALLEGOS-VÁSQUEZ, C. et al. Absorción de nitrato y amonio por plantas de nopal en
hidroponía: Nitrate and ammonium Uptake by Cactus Pear in hidroponics. Terra, Bucuresti,
RO, v.18, n.2, 2000.
GARLAND, M. L.; CAMPBELL, K. A.; CARTER JÚNIOR, T. E. Aluminum tolerance in
soybean: genotypic correlation and repeatability of solution culture and greenhouse screening
methods. Crop Science, Madison, v.30, p.1049-1054, 1990.

85
HANSEN, J. Stock plant lighting and adventitious root formation. HortScience, v.22, n.5,
p.746-749, 1987.
HARTMANN, H.T.; KESTER,D.E.; DAVIS JR., F.T. Plant propagation: principles and
practices. 6. ed. New York: Englewood Clipps/ Prentice Hall, 1997. 770 p.
HORIE, T. Studies on photosynthesis and primary production of rice plants in relation to
meteorological environmental. Agricultural and Forest Meteorology, Davis, v. 35, p.1-12,
1979.
INGESTAD, T.; ÄGREN, G.I. Plant nutrition and growth: Basic principles. Plant and Soil,
Dordrecht, v.168-169, p.15-20, 1995.
INGESTAD, T.; ÄGREN, G.I. Theory for growth of plants and methods. Physiologia
Plantarum, Kobenhavn, v. 84, n.1, p. 177-84, 1992.
INGESTAD, T. Relative addition rate external concentration: driving variables used nutrition
research. Plant Cell Environmental, Oxford, v. 5, p. 443-53, 1982.
IKEDA, H.; OSAWA, T. Lettuce growth as influenced by N source and temperature of the
nutrient solution. In: INTERNATIONAL SOCIETY on SOLILESS CULTURE, n.5, 1980,
Wageningen. Proceedings…Wageningen: ISOSC, 1980. p.273-284.

86
KABATA-PENDIAS, A.; PENDIAS, H. Trace elements in soils and plants, Boca Raton
Press, 1984. 315 p.
KAUL, O. N.; SRIVASTAVA, P. B. L.; TANDON, V. N. Nutrition studies on Eucalyptus: 3-
diagnosis of mineral deficiencies in Eucalyptus grandis seedlings. Indian Forester, Calcutta,
v.94, n.11, p.831-834, 1968.
KRAMER P. J.; KOZLOWSKI, T.T. Physiology of trees. New York: McGraw –Hill Book,
1960. 642 p.
KRAMER, P. J.; KOZLOWSKI, T. T. Physiology of woody plants. New York: Academic
Press, 1979. 811 p.
LEE, S.G.; ANOROFF, S Boron in plants: A biochemical role. Science, Washinghton, v.158,
p.798-99, 1967.
LEE, D. W. Simulating forest shade to study the development ecology of tropical plants:
juvenile growth in three vines in India. Journal of Tropical Ecology, Cambridge, v.4, p.281-
292, 1988.
MAFFEIS, A.R.; SILVEIRA, R.L.V.A.; BRITO J.O. Reflexos das deficiências de
macronutrientes e boro no crescimento de plantas, produção e qualidade de óleo essencial em
Eucalyptus citriodora. Scientia Forestalis, Piracicaba, n. 57, p.87-98, jun. 2000.

87
MAIA, N.B. Produção e qualidade do óleo essencial de duas espécies de menta cultivadas
em soluções nutritivas - 1998. 105f. Tese (Doutorado em Agronomia – área de concentração
em Solos e Nutrição de Plantas) - Escola Superior de Agricultura “Luiz de Queiroz”,
Universidade de São Paulo, Piracicaba, 1998.
MALAVOLTA, E. Elementos da nutrição mineral das plantas. São Paulo: Agronômica
Ceres, 1980. 251 p.
MALAVOLTA, E. Essências florestais: eucalipto e pinus. In: MALAVOLTA, E. Manual de
calagem e adubação das principais culturas. São Paulo: Ceres, 1987. cap.10, p. 376-396.
MALAVOLTA, E.; CROCOMO, O. O potássio e planta. In: TSUIOSHI YAMADA. Potássio
na agricultura brasileira. Londrina: Fundação IAPAR, 1982. 555p.
MALAVOLTA, E.; VITTI, G. C.; OLIVEIRA, S. A. Avaliação do estado nutricional das
plantas: princípios e aplicações. 2. ed. Piracicaba: Associação Brasileira para a Pesquisa da
Potassa e do Fosfato, 1997. 319 p.
MALDONADO, J.M.. Asimilación del nitrógeno y del azufre. In: AZCON, J; TALON. B. M.
Fisiología y bioquímica vegetal: manual de calagem e adubação das principais culturas: São
Paulo, Ceres, 1987. p.215-236.

88
MARSCHNER, H. Mineral nutrition of higher plants. 2. ed. London: Academic Press,
1995. 889 p.
MEYER, B.; et al. Introdução à fisiologia vegetal. 2. ed. Lisboa: Fundação Calouste
Gulbenkian, 1983. 710 p.
MENGEL, K.; KIRKBY, E. A. Principles of plant nutrition. 2.ed. Switzerland: Worblanfen-
Bern: International Potash Institute, 1979. 593 p.
MENGEL, K.; KIRKBY, E.A. Principles of plant nutrition. 3. ed. Bern: International Potash
Institute, 1982. 655 p.
MOREIRA, M. A., FONTES, P. C. R.; CAMARGOS, M. I. de. Interação zinco e fósforo em
solução nutritiva influenciando o crescimento e a produtividade da alface. Pesquisa.
Agropecuária. Brasileira, Brasília, v. 36, n. 6, p. 903-909, jun. 2001.
MORTVEDT, J.J.; KHASAWNEH, F.E. Effects of growth responses on cationic relationships
in plants. Soil Science, New Brunswick, v.141, p.200-207, 1986.
MUNIZ, M.R.A ; SGARBI, F.; HIGASHI, E.N.; SILVEIRA, R.L.V.A. Seja doutor do seu
eucalipto. Arquivo agronômico, Piracicaba, n.12, 32p. 2001.

89
NEVES, J. C. L. Aspectos nutricionais em mudas de Eucalyptus ssp. – tolerância ao
alumínio e níveis críticos de fósforo no solo. Viçosa – MG: Universidade Federal de Viçosa,
1983. 34p.
OMETTO, J.C. Bioclimatologia vegetal. São Paulo. 1981. 440 p.
PIMENTEL-GOMES; GARCIA. Estatística aplicada a experimentos agronômicos e
florestais: exposição com exemplos e orientações para uso de aplicativos. Piracicaba:
FEALQ, 2002. 309 p.
POGGIANI, F.; BRUNI, S.; BARBOSA, E. S. Q. Efeito do sombreamento das mudas de três
espécies florestais. Revista do Instituto Florestal de São Paulo, São Paulo, v.4, n.2, p.564 –
569. 1990.
RESH, H.M. Cultivos hidropônicos. nuevas técnicas de producción. 2.ed. Madri: Mundi –
Prensa, 1992. 318 p.
ROCHA FILHO, J.V.; HAAG, H.P; OLIVEIRA, G.D.; SARRUGE, J.R. Influência do B no
crescimento e na composição química do Eucalyptus grandis. Brasil Florestal, Brasília, v.39,
p.29-33, 1979.

90
SAVVAS, D. et al. Interactions between ammonium and pH of the nutrient solution supplied
to gerbera (Gerbera jamesonii) grown in pumice. Plant and Soil, Dordrecht, v. 254, n. 2,
p.393-402, 2003.
SGARBI, F. et al. Crescimento e produção de biomassa de clone de Eucalyptus grandis x
Eucalyptus urophylla em condições de deficiência de macronutrientes, B e Zn. Scientia
Forestalis, Piracicaba, n. 56, p. 69-82, dez. 1999.
SILVEIRA, R. L. V. A:. Efeito do potássio no crescimento, nas concentrações dos
nutrientes e nas características da madeira juvenil de progênies de Eucalyptus grandis
W. Hill ex Maiden cultivadas em solução nutritiva. 2000. 151f. Tese (Doutorado em
Agronomia – área de concentração em Solos e Nutrição de Plantas) - Escola Superior de
Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 2000.
SILVEIRA, R. L A. et al. Sintomas de deficiência de macronutrientes e boro em híbridos de
Eucalyptus grandis x Eucalyptus urophylla em solução nutritiva (compact disc). In:
CONGRESSO LATINO AMERICANO de CIÊNCIAS do SOLO, 13 ,1996, Águas de
Lindóia. Anais.... Águas de Lindóia: SBCS/SLCS, 1996. 96p.
SOARES, C. R. F. S. et al. Crescimento e nutrição mineral de Eucalyptus maculata e
Eucalyptus urophylla em solução nutritiva com concentração crescente de cobre. Revista.
Brasileira de Fisiologia Vegetal, Londrina, v.12, n.3, p.213-225, 2000.

91
SOUZA, L. A. C. Reação de genótipos de soja ao alumínio em hidroponia e no solo Pesquisa
Agropecuária Brasileira, Brasília, v. 36, n. 10, p. 1255-1260, out. 2001.
STEINER, A. The universal nutrient solution. In: I INTERNATIONAL SOCIETY on
SOILLESS CULTURE, 6, 1984. Proceedings. Lunteren: ISOC, 1984. p. 633-649.
STEINER, A. The history of mineral plant nutrition about 1860 as source of tree origin of
soilless culture methods. Soilless Culture, v.1, n.1, p. 7-23, 1985.
VALÊNCIA, G. La deficiencia de boro en el cafeto y su control. Cenicafé, Chinchina, v.15,
p.115-25, 1964.
VOSE, P.B. Varietal differences in plant nutrition. Herbage Abstracts, Farnham Royal, v.33,
p. 1 – 13, 1963.
KAUL, O.N.; SRIVASTAVA, P.B.L.; TANDON, V.N. Nutrition studies on Eucalyptus: 3-
diagnosis of mineral deficiencies in Eucalyptus grandis seedlings. Indian Forester, Calcutta,
v.94, n.11, p.831-834, 1968.
XAVIER, A. Silvicultura clonal I – Princípios e Técnicas de Propagação Vegetativa. Viçosa:
Universidade Federal de Viçosa, 2002. 60 p. (Caderno Didático).

92
XAVIER, A. et al. Desempenho do enraizamento de microestacas e miniestacas de clones
híbridos de Eucalyptus grandis. Revista Árvore, Viçosa, v.25, n.4, p.403-411, 2001.
WENDLING, I. et al. Propagação clonal de híbridos de Eucalyptus spp. por miniestaquia.
Revista Árvore, Viçosa, v.24, n.2, p.181-186, 2000.
WHITEHEAD, D.;OKALI, D.U.U.; FASEHUN, F.E. Stomatal response to environmental
variables in two tropical forest species during the dry season in Nigerian. Journal of
Application in Ecology, v.18, p.571-587, 1981.

93
APÊNDICE 1

94
Tab
ela
18. T
eor d
e ni
trogê
nio
(g k
g-1) p
rese
nte
na a
nális
e fo
liar d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
C
ompl
eta
18,0
a (2
2,6)
17
,6a
(21,
1)
16,6
a (1
7,9)
17
,2a
(19,
7)
17,2
a (1
9,7)
20
,2ab
(3
1,1)
17
,8a
(21,
9)
18,2
a (2
3,5)
14
,4c
(22,
1)
18,0
a (2
2,9)
O
mis
são
de N
17
,6a
(21,
1)
18,2
a (2
3,2)
19
,0a
(26,
3)
16,5
a (1
7,5)
17
,7a
(21,
6)
17,1
c (1
9,6)
15
,9a
(15,
5)
16,1
a (1
6,2)
17
,8b
(10,
8)
16,4
a (1
6,9)
O
mis
são
de P
18
,0a
(22,
6)
17,6
a (2
1,2)
17
,9a
(22,
3)
17,9
a (2
2,4)
18
,5a
(24,
3)
17,4
bc
(20,
5)
18,6
a (2
5,0)
18
,6a
(25,
1)
18,0
b (2
2,6)
17
,1a
(19,
5)
Om
issã
o de
K
17,5
a (2
0,9)
18
,6a
(24,
8)
18,2
a (2
3,3)
17
,6a
(21,
1)
16,4
a (1
7,1)
18
,7ab
c (2
5,0)
17
,7a
(21,
5)
18,1
a (2
3,2)
21
,3a
(35,
5)
19,3
a (2
7,4)
O
mis
são
de
Ca
17,2
a (1
9,9)
17
,9a
(22,
1)
17,7
a (2
1,7)
18
,5a
(24,
5)
18,2
a (2
3,5)
18
,6ab
c (2
4,8)
17
,6a
(21,
2)
17,6
a (2
1,2)
18
,3b
(23,
8)
17,5
a (2
0,9)
O
mis
são
de
Mg
17,1
a (1
9,4)
16
,1a
(16,
1)
18,0
a (2
2,6)
17
,6a
(21,
5)
17,7
a (2
1,4)
19
,9ab
c (3
0,0)
18
,7a
(25,
2)
17,4
a (2
0,6)
20
,3ab
(3
0,7)
17
,9a
(22,
2)
Om
issã
o de
B
17,5
a (2
0,9)
16
,8a
(18,
6)
19,1
a (2
6,6)
17
,1a
(19,
5)
18,3
a (2
3,7)
21
,2a
(35,
0)
18,6
a (2
5,0)
17
,1a
(19,
4)
18,0
b (2
2,6)
17
,6a
(21,
1)
Om
issã
o de
Fe
17
,8a
(22,
0)
18,5
a (2
4,6)
18
,5a
(24,
6)
16,5
a (1
7,5)
16
,8a
(18,
6)
17,1
c (1
9,4)
17
,3a
(20,
2)
17,8
a (2
2,0)
18
,0b
(25,
0)
17,6
a (2
1,3)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,24
61
0,27
75
0,31
93
0,31
27
0,19
81
0,31
01
0,39
04
0,27
88
0,24
81
0,33
63
CV
(%)
4,93
5,
54
6,21
6,
35
3,96
5,
83
7,75
5,
57
4,77
6,
71
Tra
nsfo
rmaç
ão
RQ
(y+1
) R
Q (y
+1)
RQ
(y+1
) R
Q (y
+1)
RQ
(y+1
) R
Q (y
+1)
RQ
(y+1
) R
Q (y
+1)
RQ
(y+1
) R
Q (y
+1)
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
sess
refe
rent
es à
s méd
ias s
em tr
ansf
orm
ação
.

95
Tab
ela
19. T
eor d
e fó
sfor
o (g
kg-1
) pre
sent
e na
aná
lise
folia
r da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
C
ompl
eta
11,2
a (2
,6)
11,1
b (2
,4)
10,9
a (2
,0)
11,1
ab
(2,4
) 11
,1 a
(2,3
) 11
,4 a
(3,1
) 11
,3c
(3,5
) 11
,6cd
(3
,7)
11,7
c (3
,8)
11,6
b (3
,6)
Om
issã
o de
N
11,2
a (2
,7)
11,1
b (2
,5)
11,1
a
(2,3
) 11
,1ab
(2
,4)
11,1
a (2
,5)
11,2
a (2
,6)
12,3
a (5
,1)
12,6
a (6
,0)
12,7
a (6
,1)
12,6
a (6
,1)
Om
issã
o de
P
11,2
a (2
,7)
11,3
ab
(2,9
) 11
,3 a
(2,8
) 11
,2ab
(2
,6)
11,1
a (2
,5)
11,4
a (3
,0)
11,4
bc
(3,1
) 11
,2d
(2,7
) 11
,1d
(2,5
) 11
,4b
(3,1
) O
mis
são
de K
11
,2a
(2,6
) 11
,7a
(3,8
) 11
,1 a
(2,4
) 11
,2ab
(2
,6)
11,1
a (2
,3)
11,4
a (3
,1)
12,0
a (4
,6)
12,1
b (4
,8)
12,2
b (5
,0)
12,0
ab
(4,5
) O
mis
são
de
Ca
11,4
a (3
,2)
11,3
ab
(3,0
) 11
,0 a
(2,3
) 11
,2ab
(2
,6)
11,2
a (2
,6)
11,4
a (3
,1)
11,8
ab
(4,0
) 11
,8bc
(4
,1)
11,8
bc
(4,0
) 11
,8ab
(4
,0)
Om
issã
o de
M
g 11
,3a
(2,9
) 11
,2b
(2,6
) 11
,0 a
(2,3
) 11
,2a
(2,7
) 11
,1 a
(2,4
) 11
,3 a
(2,9
) 12
,0 a
(4,5
) 11
,9bc
(4
,2)
11,8
bc
(4,0
) 12
,1ab
(4
,7)
Om
issã
o de
B
11,3
a (2
,9)
11,3
ab
(2,8
) 11
,1 a
(2,5
) 11
,1ab
(2
,3)
11,2
a (2
,6)
11,4
a (3
,0)
11,4
bc
(3,0
) 11
,5cd
(3
,4)
11,5
cd
(3,2
) 11
,6b
(3,6
) O
mis
são
de F
e 11
,2a
(2,6
) 11
,3ab
(2
,8)
11,2
a (2
,8)
11,0
b (2
,2)
11,1
a (2
,4)
11,4
a (3
,0)
11,4
bc
(3,1
) 11
,3d
(2,8
) 11
,6c
(3,6
) 11
,4b
(3,2
)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,03
5 0,
0426
0,
0493
0,
0224
0,
0132
0,
0403
0,
0474
0,
0443
0,
0443
0,
0935
C
V(%
) 1,
09
1,33
1,
56
0,71
0,
41
1,25
1,
42
1,32
1,
32
2,78
T
rans
form
ação
R
Q (y
+1)
RQ
(y+1
) R
Q (y
+1)
RQ
(y+1
) R
Q (y
+1)
RQ
(y+1
) R
Q (y
+1)
RQ
(y+1
) R
Q (y
+1)
RQ
(y+1
) *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
sess
refe
rent
es à
s méd
ias s
em tr
ansf
orm
ação
.

96
Tab
ela
20. T
eor d
e po
táss
io (g
kg-1
) pre
sent
e na
aná
lise
folia
r da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
C
ompl
eta
34,0
ab
(15,
8)
33,7
c (1
3,7)
33
,2a
(10,
5)
33,8
ab
(14,
5)
33,9
a (1
5,3)
34
,5a
(19,
2)
33,8
ab
(14,
9)
33,6
ab
(14,
7)
33,9
abc
(15,
6)
33,5
bcd
(12,
7)
Om
issã
o de
N
34
,1ab
(1
6,3)
33
,8bc
(1
4,3)
33
,2a
(10,
4)
33,6
b (1
3,6)
33
,9a
(15,
5)
34,1
ab
(16,
8)
34,2
a (1
7,4)
33
,9a
(15,
0)
34,2
a (1
7,4)
33
,4cd
(1
1,9)
O
mis
são
de P
34
,0ab
(1
5,8)
33
,9bc
(1
5,4)
33
,4a
(11,
9)
33,9
ab
(15,
2)
33,7
a (1
3,7)
34
,3ab
(1
8,1)
34
,1ab
(1
6,5)
33
,9a
(14,
2)
34,1
ab
(16,
6)
34,1
ab
(16,
4)
Om
issã
o de
K
33
,8ab
(1
4,8)
34
,1ab
c (1
6,8)
33
,2a
(10,
3)
34,0
a (1
6,0)
33
,7a
(14,
1)
33,9
b (1
5,6)
33
,2b
(10,
5)
33,1
b (1
5,2)
32
,8e
(7,8
) 32
,8e
(7,6
) O
mis
são
de
Ca
34,3
ab
(17,
7)
34,9
a (2
2,1)
33
,0a
(9,5
) 33
,7ab
(1
4,2)
33
,7a
(14,
0)
34,3
ab
(17,
8)
33,9
ab
(15,
2)
33,9
a (9
,6)
33,6
cd
(13,
1)
33,2
de
(10,
3)
Om
issã
o de
M
g 35
,1a
(23,
8)
34,6
ab
(19,
7)
33,4
a (1
2,1)
33
,8ab
(1
4,4)
33
,7a
(13,
8)
34,2
ab
(17,
0)
34,5
a (1
9,1)
33
,7a
(15,
4)
33,4
d (1
2,0)
33
,2de
(1
0,4)
O
mis
são
de B
33
,9ab
(1
5,5)
34
,5ab
c (1
9,5)
33
,3a
(11,
2)
33,6
b (1
3,4)
33
,8a
(14,
5)
34,3
ab
(18,
1)
34,1
ab
(16,
6)
33,9
a (1
5,4)
34
,2a
(17,
3)
33,9
abc
(15,
4)
Om
issã
o de
Fe
33
,6b
(13,
6)
34,1
abc
(16,
6)
33,6
a (1
3,5)
33
,6b
(13,
6)
33,7
a (1
4,2)
34
,3ab
(1
8,0)
33
,9ab
(1
5,3)
33
,8a
(13,
1)
33,8
bc
(14,
6)
34,2
a
(17,
0)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
1365
0,
0864
0,
1076
0,
0359
0,
0541
0,
0506
0,
092
0,06
28
0,03
83
0,06
32
CV
(%)
1,41
0,
89
1,14
0,
36
0,56
0,
52
0,95
0,
65
0,40
0,
66
Tra
nsfo
rmaç
ão
RQ
(y+1
0)
RQ
(y+1
0)R
Q (y
+10)
RQ
(y+1
0)R
Q (y
+10)
RQ
(y+1
0)
RQ
(y+1
0)R
Q (y
+10)
RQ
(y+1
0)R
Q (y
+10)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
ss re
fere
ntes
às m
édia
s sem
tran
sfor
maç
ão.

97
Tab
ela
21. T
eor d
e cá
lcio
(g k
g-1) p
rese
nte
na a
nális
e fo
liar d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
C
ompl
eta
13,2
ab
(7,6
) 13
,8ab
(9
,8)
13,4
a (8
,5)
13,6
a (9
,4)
13,5
a (8
,7)
14,1
a (1
1,3)
12
,1d
(3,6
) 12
,5b
(5,2
) 12
,8b
(6,2
) 13
,9ab
c (1
0,4)
O
mis
são
de
N
13,0
b (7
,0)
13,2
c (7
,6)
13,1
a (7
,1)
13,3
ab
(8,0
) 13
,7a
(9,4
) 13
,7c
(9,5
) 12
,4cd
(4
,6)
12,7
ab
(6,0
) 13
,0ab
(7
,0)
14,1
ab
(11,
1)
Om
issã
o de
P
13,2
ab
(7,7
) 13
,3bc
(8
,0)
13,1
a (7
,1)
13,6
ab
(9,3
) 13
,6a
(9,0
) 13
,7bc
(9
,7)
12,3
d (4
,3)
12,8
ab
(6,1
) 12
,8ab
(6
,3)
13,4
ab
(8,5
) O
mis
são
de
K
13,8
a (9
,8)
14,0
a (1
0,9)
13
,4a
(8,4
) 13
,5ab
(8
,7)
13,6
a (9
,3)
14,0
ab
(10,
9)
13,2
a (7
,5)
13,5
a (8
,7)
13,6
a (9
,1)
14,3
a (1
2,0)
O
mis
são
de
Ca
13,1
ab
(7,1
) 13
,6ab
c (9
,2)
13,4
a (8
,0)
13,2
ab
(7,8
) 13
,4a
(8,4
) 13
,5c
(8,9
) 12
,6bc
(5
,5)
12,9
ab
(6,5
) 12
,9ab
(6
,4)
13,5
bc
(8,9
) O
mis
são
de
Mg
13,0
b (7
,0)
13,2
c (7
,7)
13,4
a (8
,3)
13,0
b (6
,9)
13,3
a (8
,0)
13,7
c (9
,5)
12,7
b (5
,8)
13,2
ab
(7,8
) 12
,7b
(5,8
) 14
,0ab
c (1
0,9)
O
mis
são
de B
13
,1ab
(7
,3)
13,3
bc
(8,0
) 13
,3a
(8,1
) 13
,1ab
(7
,2)
13,2
a (7
,8)
13,6
c (9
,3)
12,7
a (5
,8)
12,7
b (5
,7)
12,7
b (5
,8)
13,4
c (8
,3)
Om
issã
o de
Fe
13
,0b
(6,7
) 13
,2c
(7,8
) 13
,4a
(8,4
) 13
,1ab
(7
,1)
13,1
a (7
,2)
13,5
c (8
,8)
12,7
b (5
,8)
12,6
b (5
,4)
12,8
b (6
,1)
13,4
c (8
,2)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
0691
0,
0511
0,
0887
0,
063
0,05
94
0,03
55
0,03
12
0,07
61
0,07
53
0,07
09
CV
(%)
1,84
1,
33
2,35
1,
66
1,56
0,
91
0,95
2,
08
2,05
1,
81
Tra
nsfo
rmaç
ão
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
ss re
fere
ntes
às m
édia
s sem
tran
sfor
maç
ão.

98
Tab
ela
22. T
eor d
e m
agné
sio
(g k
g-1) p
rese
nte
na a
nális
e fo
liar d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
Col
etas
T
rata
men
tos
T02h
T0
4h
T7,5
h T2
2h
T46h
T1
3d
T35d
T4
5d
T61d
T7
9d
Com
plet
a 12
,2a
(4,0
) 12
,2ab
(4
,0)
12,2
a (4
,0)
12,2
a (4
,0)
12,0
bc
(3,4
) 12
,3a
(4,4
) 11
,8b
(2,7
) 11
,8a
(2,7
) 11
,8a
(2,8
) 11
,8ab
(2
,6)
Om
issã
o de
N
12
,1a
(3,8
) 12
,1b
(3,6
) 12
,2a
(4,0
) 12
,0ab
(3
,5)
12,1
abc
(3,8
) 12
,2a
(4,0
) 11
,8ab
(2
,9)
11,9
a (3
,1)
11,9
a (3
,0)
11,9
a (3
,1)
Om
issã
o de
P
12,1
a (3
,9)
12,1
ab
(3,9
) 12
,2a
(4,1
) 12
,1ab
(3
,7)
12,2
abc
(3,9
) 12
,3a
(4,3
) 11
,8b
(2,7
) 11
,9a
(3,0
) 11
,9a
(2,9
) 11
,9a
(3,0
) O
mis
são
de
K
12,1
a (3
,7)
12,2
ab
(4,1
) 12
,2a
(4,2
) 12
,1ab
(3
,6)
12,2
ab
(4,2
) 12
,2a
(4,1
) 12
,1a
(3,9
) 12
,0a
(3,3
) 11
,9a
(2,9
) 11
,8ab
(2
,7)
Om
issã
o de
C
a 12
,0a
(3,4
) 12
,4a
(4,7
) 12
,0a
(3,3
) 11
,9b
(3,1
) 12
,2a
(4,2
) 12
,2a
(4,1
) 11
,9ab
(2
,9)
12,0
a (3
,2)
11,9
a (3
,0)
11,8
ab
(2,8
) O
mis
são
de
Mg
12,1
a (3
,9)
12,2
ab
(3,9
) 12
,1a
(3,6
) 12
,0ab
(3
,5)
12,1
abc
(3,9
) 12
,2a
(3,9
) 12
,0ab
(3
,2)
11,9
a (2
,9)
11,7
a (2
,3)
11,7
b (2
,3)
Om
issã
o de
B
12,0
a (3
,4)
12,2
ab
(4,2
) 12
,1a
(3,8
) 12
,0ab
(3
,3)
12,0
abc
(3,5
) 12
,2a
(3,9
) 12
,0ab
(3
,5)
11,9
a (3
,2)
11,9
a (3
,0)
11,8
ab
(2,7
) O
mis
são
de
Fe
12,0
a (3
,3)
12,2
ab
(4,2
) 12
,1a
(3,7
) 11
,9b
(3,2
) 12
,0c
(3,3
) 12
,2a
(4,2
) 11
,9ab
(3
,0)
11,8
a (2
,9)
11,8
a (2
,8)
11,9
a (3
,1)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
0326
0,
7595
0,
0403
0,
0218
0,
0223
0,
0157
0,
0313
0,
0318
0,
0232
0,
0207
C
V (%
) 0,
95
4,82
1,
17
0,63
0,
64
0,45
0,
92
0,94
0,
68
0,61
T
rans
form
ação
Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
sess
refe
rent
es à
s méd
ias s
em tr
ansf
orm
ação
.

99
Tab
ela
23. T
eor d
e fe
rro
(mg
kg-1
) pre
sent
e na
aná
lise
folia
r da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
C
ompl
eta
5,73
a (3
06,3
) 5,
36b
(213
,3)
5,94
a (3
79,0
) 5,
34a
(209
,6)
5,28
a (2
05,6
) 5,
38a
(219
,1)
4,96
a (1
43,3
) 5,
26ab
(1
96,1
) 5,
37ab
(2
15,0
) 5,
52ab
(2
51,3
) O
mis
são
de
N
5,59
a (2
65,0
) 5,
34b
(209
,0)
5,83
a (3
43,3
) 5,
13a
(167
,7)
5,22
a (1
88,3
) 5,
22a
(185
,1)
5,05
a (1
56,6
) 5,
17ab
c (1
79,3
) 4,
97b
(143
,3)
5,44
ab
(230
,6)
Om
issã
o de
P
6,16
a (8
36,0
) 5,
36b
(215
,6)
5,97
a (3
91,6
) 5,
13a
(169
,4)
5,23
a (1
87,1
) 5,
20a
(181
,8)
5,06
a (1
61,8
) 5,
00ab
c (1
50,3
) 5,
07ab
(1
61,0
) 5,
31ab
(2
02,0
) O
mis
são
de
K
5,49
a (2
40,2
) 5,
06b
(157
,5)
5,59
a (2
70,0
) 5,
14a
(171
,6)
5,21
a (1
82,8
) 5,
21a
(183
,0)
5,17
a (1
76,6
) 5,
23ab
(1
86,6
) 5,
31ab
(1
81,3
) 5,
48ab
(2
40,1
) O
mis
são
de
Ca
5,32
a (2
04,0
) 5,
72ab
(3
06,5
) 5,
62a
(278
,3)
5,07
a (1
59,6
) 5,
27a
(194
,5)
5,34
a (2
10,1
) 4,
98a
(145
,7)
5,18
abc
(177
,0)
5,19
ab
(239
,6)
5,41
ab
(225
,0)
Om
issã
o de
M
g 5,
33a
(206
,6)
6,21
a (5
94,9
) 5,
59a
(270
,0)
5,21
a (1
88,6
) 5,
38a
(226
,5)
5,31
a (2
02,6
) 5,
28a
(197
,2)
5,41
a (2
28,0
) 5,
46a
(239
,6)
5,67
a (2
91,6
) O
mis
são
de B
5,
54a
(253
,3)
5,73
ab
(309
,8)
5,65
a (2
91,6
) 4,
99a
(146
,6)
5,08
a (1
60,2
) 5,
19a
(180
,0)
5,16
a (2
18,1
) 4,
81bc
(1
23,6
) 5,
22ab
(1
88,3
) 5,
37ab
(2
20,0
) O
mis
são
de
Fe
5,36
a (2
13,3
) 5,
73ab
(3
08,1
) 5,
55a
(265
,0)
5,01
a (1
51,6
) 5,
04a
(155
,2)
5,19
a (1
80,0
) 5,
00a
(148
,5)
4,67
c (1
06,6
) 5,
12ab
(1
69,3
) 5,
23b
(187
,3)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
1,
3616
0,
7595
0,
4533
0,
415
0,19
91
0,88
23
0,88
23
0,53
57
0,47
84
0,37
32
CV
(%)
8,65
4,
82
2,80
2,
85
1,33
6,
13
6,13
3,
71
3,24
2,
43
Tra
nsfo
rmaç
ão
Log
(y+4
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
Log
(y+3
) Lo
g (y
+3)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
ss re
fere
ntes
às m
édia
s sem
tran
sfor
maç
ão.

100
Tab
ela
24. T
eor d
e bo
ro (m
g kg
-1) p
rese
nte
na a
nális
e fo
liar d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
Col
etas
T
rata
men
tos
T02h
T0
4h
T7,5
h T2
2h
T46h
T1
3d
T35d
T4
5d
T61d
T7
9d
Com
plet
a 3,
79a
(45,
4)
3,67
a (3
8,3)
3,
72a
(40,
8)
3,45
ab
(31,
4)
3,32
b (2
6,8)
3,
95a
(51,
5)
3,89
c (4
8,2)
4,
55ab
c (9
4,4)
4,
50ab
(8
9,3)
4,
77ab
(1
18,5
) O
mis
são
de
N
3,62
a (3
6,5)
3,
53a
(34,
1)
3,82
a (4
5,4)
3,
73ab
(4
1,4)
3,
54ab
(3
3,6)
3,
60bc
(3
6,0)
4,
34b
(76,
6)
4,70
abc
(109
,4)
4,64
ab
(102
,7)
4,67
ab
(106
,2)
Om
issã
o de
P
3,68
a (3
9,7)
3,
73a
(41,
5)
3,59
a (3
5,5)
3,
66ab
(3
8,7)
3,
64a
(37,
9)
3,69
b (3
9,3)
4,
37b
(78,
6)
4,71
ab
(111
,3)
4,57
ab
(96,
1)
4,65
ab
(104
,43)
O
mis
são
de
K
3,88
a (4
7,8)
3,
61a
(36,
3)
3,60
a (3
5,8)
3,
28b
(25,
7)
3,53
ab
(33,
3)
3,79
ab
(43,
9)
4,60
a (9
9,1)
4,
86a
(129
,3)
4,45
ab
(84,
8)
4,60
ab
(99,
30)
Om
issã
o de
C
a 3,
60a
(35,
7)
3,71
a (4
0,2)
3,
67a
(38,
6)
3,81
a (4
4,4)
3,
59a
(35,
3)
3,73
ab
(41,
0)
4,34
b (7
6,4)
4,
71ab
(1
11,0
) 4,
72a
(114
,7)
4,70
ab
(109
,3)
Om
issã
o de
M
g 3,
48a
(32,
3)
3,71
a (4
0,1)
3,
70a
(40,
1)
3,66
ab
(38,
4)
3,53
ab
(33,
3)
3,57
bc
(34,
8)
4,31
b (7
3,7)
4,
45bc
(8
4,7)
4,
35b
(76,
8)
4,53
b (9
5,3)
O
mis
são
de B
3,
60a
(35,
9)
3,49
a (3
2,1)
3,
69a
(40,
2)
3,33
b (2
7,7)
3,
29b
(25,
9)
3,38
c (2
8,7)
3,
33d
(27,
1)
4,39
c (2
9,8)
3,
32c
(26,
7)
3,57
c (3
4,7)
O
mis
são
de
Fe
3,56
a (3
4,3)
3,
65a
(37,
6)
3,73
a (4
1,3)
3,
40ab
(2
9,3)
3,
39ab
(2
8,8)
3,
81ab
(4
4,2)
4,
39b
(80,
0)
3,42
d (8
0,7)
4,
63ab
(1
01,8
) 4,
81a
(122
,7)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
5527
0,
4052
0,
4971
0,
4525
0,
2574
0,
2604
0,
2018
0,
3136
0,
2994
0,
2502
C
V(%
) 5,
34
3,93
4,
75
4,51
2,
61
2,49
1,
69
2,47
2,
40
1,94
T
rans
form
ação
Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
sess
refe
rent
es à
s méd
ias s
em tr
ansf
orm
ação
.

101
Tab
ela
25. M
édia
de
prod
ução
de
mic
roes
taca
s de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m se
tent
a e
cinc
o m
icro
cepa
s, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d T
rata
men
to
Prod
ução
Pr
oduç
ão
Prod
ução
Pr
oduç
ão
Prod
ução
Pr
oduç
ão
Prod
ução
Pr
oduç
ão
Com
plet
a 3,
43b
(31)
3,
88a
(49)
3,
32bc
(2
9)
3,71
a (4
3)
3,82
a (4
6)
3,63
a (2
9)
3,63
ab
(38)
4,
15a
(54)
O
mis
são
de N
- -
3,16
a (2
4)
2,98
c (2
0)
1,99
b (9
) 2,
48b
(12)
2,
85a
(9)
2,73
b (1
6)
3,21
b (1
6)
Om
issã
o de
P
3,69
ab
(41)
3,
65a
(44)
3,
42ab
c (3
1)
3,73
a (4
2)
3,83
a (4
7)
3,35
a (1
9)
3,60
ab
(41)
4,
02ab
(4
7)
Om
issã
o de
K
4,22
a (6
9)
3,87
a (5
1)
4,14
a (6
3)
3,81
a (4
6)
3,88
a (4
9)
3,85
a (3
8)
3,37
ab
(31)
4,
20a
(57)
O
mis
são
de
Ca
3,77
ab
(44)
3,
77a
(45)
3,
55ab
c (3
6)
3,84
a (4
8)
3,82
a (4
6)
3,16
a (1
6)
3,82
a (4
6)
3,87
ab
(39)
O
mis
são
de
Mg
3,77
ab
(44)
3,
92a
(53)
4,
01ab
(5
5)
4,15
a (6
5)
3,85
a (4
8)
3,49
a (2
4)
3,56
ab
(37)
4,
35a
(69)
O
mis
são
de B
3,
45b
(33)
3,
73a
(42)
3,
54ab
c (3
5)
3,76
a (4
4)
3,57
a (3
6)
3,38
a (2
5)
3,39
ab
(30)
3,
57ab
(2
9)
Om
issã
o de
Fe
3,
43b
(33)
3,
99a
(55)
2,
93c
(21)
3,
78a
(44)
3,
49a
(33)
3,
28a
(17)
3,
37ab
(3
0)
3,81
ab
(36)
Tes
te F
4,
829
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,65
25
1,02
78
0,81
92
0,98
17
0,45
58
1,10
3 0,
9603
0,
8324
C
V (%
) 6,
41
9,69
8,
30
9,64
4,
48
11,5
5 9,
87
7,54
T
rans
form
ação
Lo
g (y
) Lo
g (y
) Lo
g (y
) Lo
g (y
) Lo
g (y
) Lo
g (y
+10)
Lo
g (y
) Lo
g (y
+10)
*M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
ss re
fere
ntes
às m
édia
s sem
tran
sfor
maç
ão.
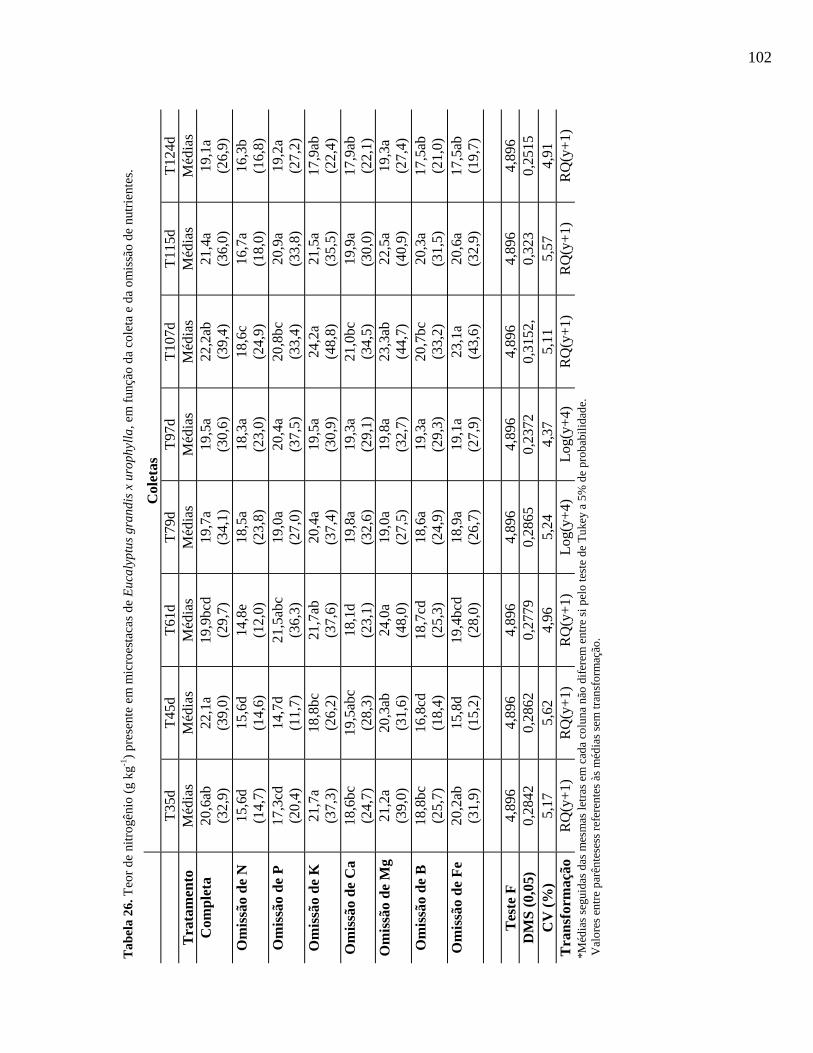
102
Tab
ela
26. T
eor d
e ni
trogê
nio
(g k
g-1) p
rese
nte
em m
icro
esta
cas d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d T
rata
men
to
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Com
plet
a 20
,6ab
(3
2,9)
22
,1a
(39,
0)
19,9
bcd
(29,
7)
19,7
a (3
4,1)
19
,5a
(30,
6)
22,2
ab
(39,
4)
21,4
a (3
6,0)
19
,1a
(26,
9)
Om
issã
o de
N
15,6
d (1
4,7)
15
,6d
(14,
6)
14,8
e (1
2,0)
18
,5a
(23,
8)
18,3
a (2
3,0)
18
,6c
(24,
9)
16,7
a (1
8,0)
16
,3b
(16,
8)
Om
issã
o de
P
17,3
cd
(20,
4)
14,7
d (1
1,7)
21
,5ab
c (3
6,3)
19
,0a
(27,
0)
20,4
a (3
7,5)
20
,8bc
(3
3,4)
20
,9a
(33,
8)
19,2
a (2
7,2)
O
mis
são
de K
21
,7a
(37,
3)
18,8
bc
(26,
2)
21,7
ab
(37,
6)
20,4
a (3
7,4)
19
,5a
(30,
9)
24,2
a (4
8,8)
21
,5a
(35,
5)
17,9
ab
(22,
4)
Om
issã
o de
Ca
18,6
bc
(24,
7)
19,5
abc
(28,
3)
18,1
d (2
3,1)
19
,8a
(32,
6)
19,3
a (2
9,1)
21
,0bc
(3
4,5)
19
,9a
(30,
0)
17,9
ab
(22,
1)
Om
issã
o de
Mg
21,2
a (3
9,0)
20
,3ab
(3
1,6)
24
,0a
(48,
0)
19,0
a (2
7,5)
19
,8a
(32,
7)
23,3
ab
(44,
7)
22,5
a (4
0,9)
19
,3a
(27,
4)
Om
issã
o de
B
18,8
bc
(25,
7)
16,8
cd
(18,
4)
18,7
cd
(25,
3)
18,6
a (2
4,9)
19
,3a
(29,
3)
20,7
bc
(33,
2)
20,3
a (3
1,5)
17
,5ab
(2
1,0)
O
mis
são
de F
e 20
,2ab
(3
1,9)
15
,8d
(15,
2)
19,4
bcd
(28,
0)
18,9
a (2
6,7)
19
,1a
(27,
9)
23,1
a (4
3,6)
20
,6a
(32,
9)
17,5
ab
(19,
7)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
2842
0,
2862
0,
2779
0,
2865
0,
2372
0,
3152
, 0,
323
0,25
15
CV
(%)
5,17
5,
62
4,96
5,
24
4,37
5,
11
5,57
4,
91
Tra
nsfo
rmaç
ão
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
) Lo
g(y+
4)
Log(
y+4)
R
Q(y
+1)
RQ
(y+1
) R
Q(y
+1)
*Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
sess
refe
rent
es à
s méd
ias s
em tr
ansf
orm
ação
.

103
Tab
ela
27. T
eor d
e fó
sfor
o (g
kg-1
) pre
sent
e em
mic
roes
taca
s de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d T
rata
men
to
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Com
plet
a 12
,1a
(4,9
) 12
,2ab
(4
,9)
12,1
a (4
,8)
12,2
a (5
,1)
11,9
ab
(4,2
) 12
,4a
(5,4
) 11
,8ab
(4
,1)
11,5
ab
(3,4
) O
mis
são
de N
12
,6a
(6,1
) 12
,1ab
(4
,8)
11,3
b (2
,9)
12,1
a (4
,9)
12,0
ab
(4,5
) 12
,3ab
(5
,3)
12,0
ab
(4,5
) 11
,8a
(4,0
) O
mis
são
de P
12
,3a
(5,4
) 11
,9bc
(4
,2)
12,2
a (5
,0)
12,0
a (4
,4)
11,7
b (3
,8)
11,9
b (4
,4)
11,5
c (3
,4)
11,4
b (3
,1)
Om
issã
o de
K
12,5
a (5
,6)
12,5
a (5
,8)
12,5
a (5
,0)
12,0
a (4
,5)
12,1
a (4
,9)
12,5
a (5
,7)
11,9
ab
(4,3
) 11
,4b
(3,2
) O
mis
são
de C
a 12
,5a
(5,7
) 12
,2ab
(5
,0)
12,2
a (5
,0)
12,1
a (4
,8)
11,9
ab
(4,3
) 12
,4a
(5,4
) 12
,0ab
(4
,4)
11,7
ab
(3,8
) O
mis
são
de M
g 12
,7a
(6,3
) 12
,2ab
(5
,1)
12,6
a (6
,0)
12,3
a (5
,3)
12,1
a (4
,8)
12,6
a (6
,0)
12,1
a (4
,7)
11,8
a (4
,1)
Om
issã
o de
B
12,4
a (5
,4)
11,4
c (3
,0)
11,9
ab
(4,4
) 11
,9a
(4,4
) 11
,9ab
(4
,3)
12,3
ab
(5,3
) 11
,8bc
(4
,0)
11,4
b (3
,1)
Om
issã
o de
Fe
12,5
a (5
,7)
11,8
bc
(4,0
) 12
,3a
(5,1
) 12
,1a
(4,8
) 11
,9ab
(4
,2)
12,3
ab
(5,2
) 12
,0ab
(4
,5)
11,7
a (3
,7)
T
este
F
4,89
6 4,
896
4,89
6a 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
0583
0,
0539
0,
0669
0,
1021
0,
0355
0,
0415
0,
0279
0,
0365
C
V (%
) 1,
64
1,57
1,
94
2,97
1,
04
1,18
0,
82
1,10
T
rans
form
ação
R
Q(y
+1)
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
) *M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
ss re
fere
ntes
às m
édia
s sem
tran
sfor
maç
ão.

104
Tab
ela
28. T
eor d
e po
táss
io (g
kg-1
) pre
sent
e em
mic
roes
taca
s de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d T
rata
men
to
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Com
plet
a 35
,3bc
(2
5,3)
35
,0ab
(2
2,7)
35
,1a
(23,
0)
35,0
bc
(23,
1)
35,4
ab
(25,
7)
36,1
abc
(30,
4)
36,6
a (3
4,4)
35
,1a
(23,
3)
Om
issã
o de
N
33,6
d (1
3,2)
34
,5ab
c (1
9,3)
34
,2a
(17,
3)
34,8
bc
(21,
3)
34,9
bc
(22,
4)
35,8
abc
(28,
6)
35,4
bc
(25,
9)
34,5
a (1
9,4)
O
mis
são
de P
35
,1bc
(2
3,9)
35
,2a
(24,
0)
35,2
a (2
4,1)
35
,5ab
(2
6,5)
35
,2ab
(2
4,3)
36
,2ab
(3
1,6)
36
,2a
(31,
1)
34,8
a (2
1,1)
O
mis
são
de K
34
,2cd
(1
7,4)
34
,2bc
(1
7,5)
34
,6a
(20,
0)
34,4
c (1
8,5)
33
,7d
(13,
9)
33,8
d (1
4,7)
34
,2d
(17,
3)
33,6
b (1
3,4)
O
mis
são
de C
a 35
,3bc
(2
5,1)
35
,3a
(25,
1)
34,7
a (2
0,9)
34
,4c
(18,
9)
34,7
c (2
0,8)
35
,3bc
(2
5,2)
36
,2a
(31,
5)
35,0
a (2
2,5)
O
mis
são
de M
g 38
,5a
(48,
6)
35,2
a (2
4,1)
35
,6a
(27,
4)
35,0
bc
(23,
2)
34,7
c (2
0,5)
35
,2c
(24,
4)
35,3
a (2
5,0)
34
,7a
(20,
8)
Om
issã
o de
B
36,0
b (3
0,1)
33
,8c
(14,
7)
35,2
a (2
4,4)
35
,3ab
(2
5,3)
35
,5ab
(2
6,7)
35
,7ab
c (2
7,7)
36
,3a
(32,
4)
35,0
a (2
2,8)
O
mis
são
de F
e 35
,5bc
(2
6,1)
34
,7ab
c (2
0,5)
35
,6a
(27,
2)
35,9
a (2
9,0)
35
,6a
(27,
4)
36,6
a (3
4,0)
36
,7a
(35,
1)
35,0
a (2
2,9)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,13
61
0,08
36
0,15
71
0,08
02
0,06
99
0,09
3 0,
0726
0,
0676
C
V (%
) 1,
35
0,85
1,
58
0,80
0,
67
0,92
0,
71
0,68
T
rans
form
ação
R
Q(y
+10)
R
Q(y
+10)
R
Q(y
+10)
R
Q(y
+10)
R
Q(y
+10)
R
Q(y
+10)
R
Q(y
+10)
R
Q(y
+10)
*
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.
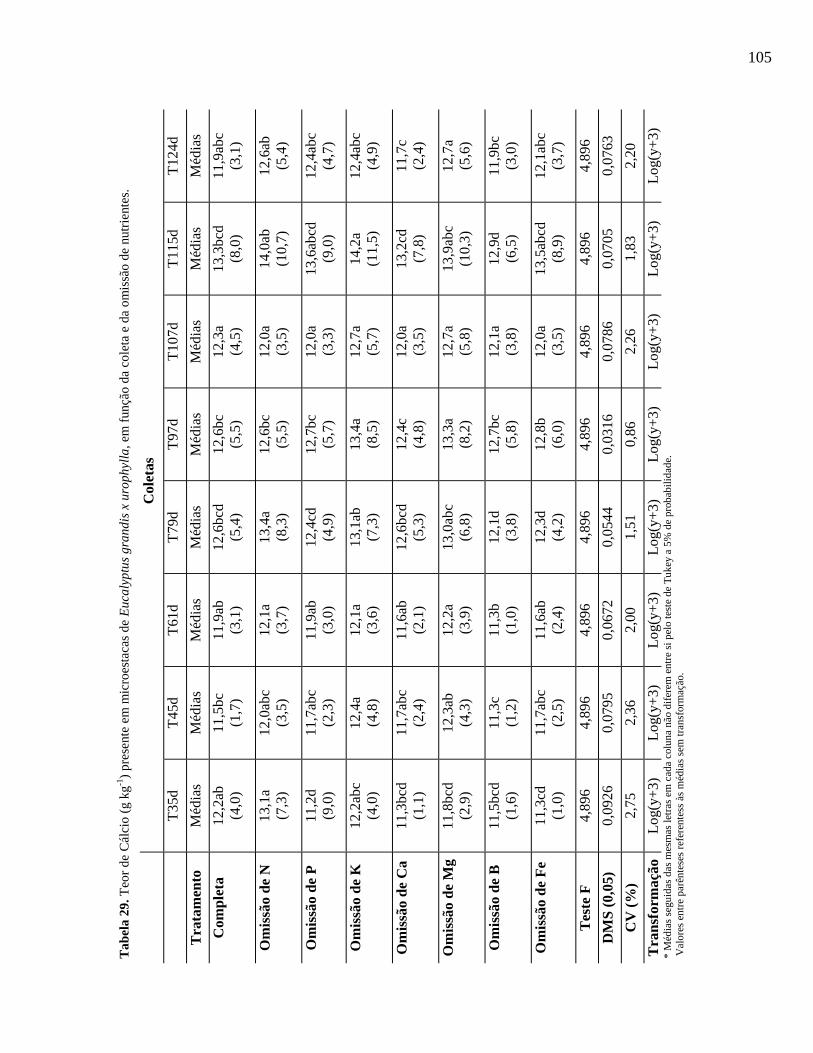
105
Tab
ela
29. T
eor d
e C
álci
o (g
kg-1
) pre
sent
e em
mic
roes
taca
s de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d T
rata
men
to
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Com
plet
a 12
,2ab
(4
,0)
11,5
bc
(1,7
) 11
,9ab
(3
,1)
12,6
bcd
(5,4
) 12
,6bc
(5
,5)
12,3
a (4
,5)
13,3
bcd
(8,0
) 11
,9ab
c (3
,1)
Om
issã
o de
N
13,1
a (7
,3)
12,0
abc
(3,5
) 12
,1a
(3,7
) 13
,4a
(8,3
) 12
,6bc
(5
,5)
12,0
a (3
,5)
14,0
ab
(10,
7)
12,6
ab
(5,4
)
Om
issã
o de
P
11,2
d (9
,0)
11,7
abc
(2,3
) 11
,9ab
(3
,0)
12,4
cd
(4,9
) 12
,7bc
(5
,7)
12,0
a (3
,3)
13,6
abcd
(9
,0)
12,4
abc
(4,7
)
Om
issã
o de
K
12,2
abc
(4,0
) 12
,4a
(4,8
) 12
,1a
(3,6
) 13
,1ab
(7
,3)
13,4
a (8
,5)
12,7
a (5
,7)
14,2
a (1
1,5)
12
,4ab
c (4
,9)
Om
issã
o de
Ca
11,3
bcd
(1,1
) 11
,7ab
c (2
,4)
11,6
ab
(2,1
) 12
,6bc
d (5
,3)
12,4
c (4
,8)
12,0
a (3
,5)
13,2
cd
(7,8
) 11
,7c
(2,4
)
Om
issã
o de
Mg
11,8
bcd
(2,9
) 12
,3ab
(4
,3)
12,2
a (3
,9)
13,0
abc
(6,8
) 13
,3a
(8,2
) 12
,7a
(5,8
) 13
,9ab
c (1
0,3)
12
,7a
(5,6
)
Om
issã
o de
B
11,5
bcd
(1,6
) 11
,3c
(1,2
) 11
,3b
(1,0
) 12
,1d
(3,8
) 12
,7bc
(5
,8)
12,1
a (3
,8)
12,9
d (6
,5)
11,9
bc
(3,0
)
Om
issã
o de
Fe
11,3
cd
(1,0
) 11
,7ab
c (2
,5)
11,6
ab
(2,4
) 12
,3d
(4,2
) 12
,8b
(6,0
) 12
,0a
(3,5
) 13
,5ab
cd
(8,9
) 12
,1ab
c (3
,7)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,09
26
0,07
95
0,06
72
0,05
44
0,03
16
0,07
86
0,07
05
0,07
63
CV
(%)
2,75
2,
36
2,00
1,
51
0,86
2,
26
1,83
2,
20
Tra
nsfo
rmaç
ão
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tess
às m
édia
s sem
tran
sfor
maç
ão.

106
Tab
ela
30. T
eor d
e m
agné
sio
(g k
g-1) p
rese
nte
em m
icro
esta
cas d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d T
rata
men
to
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Com
plet
a 12
,1ab
(3
,7)
11,9
abc
(3,0
) 11
,9a
(2,9
) 11
,8ab
(2
,8)
12,1
ab
(3,7
) 12
,1a
(3,7
) 12
,0a
(3,5
) 11
,9a
(3,1
) O
mis
são
de N
12
,5a
(5,1
) 12
,0ab
c (3
,3)
11,8
a (2
,6)
11,9
ab
(3,1
) 12
,1ab
(3
,7)
12,1
a (3
,6)
12,2
a (4
,1)
12,0
a (3
,5)
Om
issã
o de
P
11,9
b (3
,0)
12,1
ab
(3,8
) 11
,9a
(3,0
) 12
,0ab
(3
,3)
11,9
bcd
(3,1
) 11
,9a
(3,2
) 12
,0a
(3,3
) 11
,9a
(3,1
) O
mis
são
de K
12
,1ab
(3
,8)
12,1
ab
(3,6
) 11
,9a
(3,0
) 11
,9ab
(3
,2)
12,1
ab
(3,7
) 12
,1a
(3,7
) 12
,1a
(3,6
) 11
,7a
(2,5
) O
mis
são
de C
a 12
,0b
(3,3
) 12
,2a
(4,0
) 11
,8a
(2,8
) 12
,0a
(3,4
) 11
,9cd
(3
,0)
12,0
a (3
,5)
12,1
a
(3,6
) 11
,9a
(2,9
) O
mis
são
de M
g 12
,0b
(3,3
) 11
,7c
(2,5
) 11
,8a
(2,6
) 11
,7b
(2,4
) 11
,7d
(2,5
) 11
,9a
(3,0
) 11
,7a
(2,4
) 11
,7a
(2,5
) O
mis
são
de B
12
,1ab
(3
,8)
11,9
bc
(2,9
) 11
,9a
(2,9
) 11
,9ab
(2
,9)
12,0
bc
(3,5
) 11
,9a
(3,2
) 12
,1a
(3,6
) 11
,9a
(2,9
) O
mis
são
de F
e 12
,0b
(3,4
) 12
,1ab
(3
,6)
12,0
a (3
,5)
12,0
a (3
,5)
12,3
a (4
,3)
12,1
a (3
,6)
12,1
a (3
,7)
11,9
a (3
,0)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
0425
0,
0315
0,
044
0,02
79
0,02
15
0,03
4 0,
0282
0,
0325
C
V (%
) 1,
23
0,94
1,
30
0,82
0,
63
1,00
0,
82
0,96
T
rans
form
ação
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
*
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

107
Tab
ela
31. T
eor d
e fe
rro
(mg
kg-1
) pre
sent
e em
mic
roes
taca
s de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d T
rata
men
to
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Com
plet
a 4,
34b
(76,
6)
4,82
ab
(124
,7)
4,72
ab
(112
,6)
5,09
a (1
65,6
) 4,
92a
(138
,3)
5,50
ab
(272
,0)
4,68
a (1
09,0
) 4,
69ab
(1
11,0
) O
mis
são
de N
5,
73a
(311
,4)
4,78
ab
(118
,3)
4,73
ab
(114
,6)
5,21
a (1
84,0
) 5,
35a
(218
,3)
4,99
ab
(148
,0)
4,81
a (1
26,6
) 4,
93a
(145
,0)
Om
issã
o de
P
4,81
b (1
25,1
) 4,
72ab
(1
12,0
) 4,
87ab
(1
31,0
) 4,
80a
(125
,0)
5,25
a (1
96,6
) 5,
24ab
(1
92,0
) 4,
82a
(123
,6)
4,80
ab
(121
,6)
Om
issã
o de
K
4,91
ab
(143
,0)
5,11
a (1
75,3
) 4,
90ab
(1
34,0
) 5,
28a
(197
,3)
5,18
a (1
80,0
) 5,
38ab
(2
17,3
) 4,
91a
(135
,6)
4,52
ab
(91,
1)
Om
issã
o de
Ca
4,79
b (1
10,0
) 4,
90ab
(1
34,0
) 4,
64b
(103
,3)
4,99
a (1
47,6
) 4,
96a
(144
,0)
5,73
a (3
20,0
) 4,
97a
(145
,0)
4,80
ab
(121
,6)
Om
issã
o de
Mg
4,90
ab
(135
,0)
4,89
ab
(134
,0)
5,05
a
(156
,6)
5,19
a (1
87,6
) 5,
12a
(168
,3)
5,34
ab
(208
,3)
5,08
a (1
61,3
) 4,
93a
(138
,5)
Om
issã
o de
B
5,02
ab
(178
,9)
4,42
b (8
3,3)
4,
63b
(101
,6)
4,95
a (1
44,0
) 4,
93a
(145
,0)
5,22
ab
(184
,3)
4,67
a (1
06,6
) 4,
35b
(76,
7)
Om
issã
o de
Fe
4,36
b (7
7,9)
4,
56b
(96,
0)
4,61
b (1
00,0
) 4,
70a
(110
,0)
4,71
a (1
12,0
) 4,
96b
(144
,3)
4,84
a (1
28,0
) 4,
36b
(77,
9)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
874
0,53
48
0,34
3 0,
6308
0,
677
0,74
0,
4524
0,
4992
C
V (%
) 6,
57
3,95
2,
54
4,43
4,
73
4,94
3,
29
3,77
T
rans
form
ação
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
*
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

108
Tab
ela
32. T
eor d
e bo
ro (m
g kg
-1) p
rese
nte
em m
icro
esta
cas d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d T
rata
men
to
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Méd
ias
Com
plet
a 3,
74b
(41,
7)
4,07
b (5
7,6)
3,
87ab
(4
7,4)
4,
12ab
(6
1,5)
4,
17a
(64,
1)
4,30
ab
(73,
3)
4,55
a (9
4,2)
4,
13a
(61,
2)
Om
issã
o de
N
4,36
a (7
7,6)
4,
53a
(92,
5)
3,75
abc
(42,
0)
4,20
ab
(66,
3)
4,19
a (6
5,7)
4,
36ab
(7
7,3)
4,
46ab
(8
6,2)
4,
00ab
(5
4,9)
O
mis
são
de P
4,
12ab
(6
1,7)
4,
20ab
(6
5,8)
4,
13a
(61,
5)
4,40
(8
3,3)
4,
18a
(65,
0)
4,50
a (8
9,3)
4,
50ab
(8
9,7)
3,
86ab
(4
6,8)
O
mis
são
de K
4,
10ab
(6
0,4)
4,
31ab
(7
4,5)
3,
92a
(50,
4)
3,92
b (5
0,0)
4,
11a
(60,
4)
3,93
bc
(53,
5)
4,18
c (6
5,0)
3,
68b
(38,
8)
Om
issã
o de
Ca
4,12
ab
(61,
0)
4,02
b (5
6,3)
3,
51bc
(3
2,9)
4,
26ab
(7
0,5)
4,
13a
(62,
0)
3,46
cd
(30,
4)
4,39
b (7
9,9)
4,
10a
(59,
4)
Om
issã
o de
Mg
3,98
ab
(52,
9)
4,02
b (5
5,7)
4,
09a
(59,
3)
3,93
b (5
0,3)
4,
12a
(61,
2)
4,25
ab
(69,
4)
4,21
c (6
6,7)
3,
99ab
(5
3,4)
O
mis
são
de B
3,
97ab
(5
4,8)
3,
43c
(30,
0)
3,43
c (3
0,0)
3,
31c
(26,
6)
3,08
b (2
1,0)
3,
41d
(29,
5)
3,48
d (3
1,8)
3,
19c
(23,
5)
Om
issã
o de
Fe
4,19
ab
(65,
7)
3,34
c (2
7,3)
3,
96a
(51,
9)
4,22
ab
(67,
3)
4,28
a (3
71,7
) 4,
46a
(85,
9)
4,58
a
(77,
1)
4,08
a (5
9,1)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,57
1 0,
3321
0,
401
0,41
79
0,27
18
0,50
06
0,17
59
0,38
26
CV
(%)
4,95
2,
94
3,69
3,
64
2,38
4,
33
1,44
3,
48
Tra
nsfo
rmaç
ão
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

109
Tab
ela
33. T
eor d
e ni
trogê
nio
(g k
g-1) p
rese
nte
na a
nális
e da
raiz
da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
15,1
a (1
2,9)
15
,5a
(14,
1)
15,8
a (1
5,0)
16
,7a
(18,
3)
16,6
a (1
8,0)
16
,2a
(16,
5)
15,2
a (1
3,4)
14
,8ab
(1
2,0)
15
,8b
(15,
0)
16,7
a (1
7,9)
15
,3a
(16,
5)
15,7
a (1
8,4)
15
,4ab
(1
3,8)
15
,6a
(14,
5)
Om
issã
o de
N
15,7
a (1
4,8)
15
,6a
(14,
5)
15,5
a (1
4,1)
16
,3a
(16,
7)
15,2
a (1
3,3)
14
,1b
(10,
1)
14,9
a (1
2,4)
13
,7b
(9,5
) 13
,7c
(8,8
) 14
,2b
(10,
3)
13,5
a (8
,6)
13,9
a (1
0,3)
14
,2b
(10,
3)
13,5
b (8
,4)
Om
issã
o de
P
14,9
a (1
2,2)
16
,3a
(16,
9)
15,3
a (1
3,6)
16
,4a
(17,
2)
15,9
a (1
5,4)
17
,1a
(19,
3)
15,6
a (1
4,7)
17
,4a
(20,
5)
16,2
ab
(16,
3)
15,8
ab
(15,
2)
15,5
a (1
7,2)
17
,0a
(26,
4)
16,1
ab
(16,
2)
15,2
ab
(13,
2)
Om
issã
o de
K
16,1
a (1
6,1)
15
,9a
(15,
6)
16,4
a (1
7,2)
16
,2a
(16,
5)
16,7
a (1
8,0)
17
,1a
(19,
3)
15,9
a (1
5,6)
16
,1ab
(1
6,2)
17
,9a
(22,
4)
16,5
a
(17,
6)
14,6
a (1
3,2)
15
,7a
(18,
1)
16,4
a (1
7,2)
14
,6ab
(1
1,5)
O
mis
são
de C
a 15
,7a
(14,
9)
15,1
ab
(13,
0)
15,3
a (1
3,4)
16
,1a
(15,
9)
16,7
a (1
7,9)
16
,3a
(16,
6)
16,6
a (1
7,8)
15
,6ab
(1
4,7)
15
,6bc
(1
4,5)
16
,0ab
(1
5,6)
14
,4a
(12,
7)
15,8
a (1
9,1)
16
,1ab
(1
6,1)
15
,4ab
(1
3,8)
O
mis
são
de M
g 15
,0a
(12,
8)
13,2
b (7
,7)
16,0
a (1
5,9)
15
,2a
(13,
2)
17,3
a (2
0,0)
16
,7a
(18,
2)
16,5
a (1
7,6)
16
,6ab
(1
8,0)
15
,9ab
(1
5,4)
16
,1ab
(1
6,2)
14
,5a
(12,
7)
15,3
a (1
6,4)
17
,0a
(19,
4)
15,1
ab
(12,
9)
Om
issã
o de
B
15,0
a (1
2,7)
15
,8a
(15,
2)
16,1
a (1
6,0)
16
,2a
(16,
4)
16,5
a (1
7,3)
15
,5ab
(1
4,2)
15
,4a
(14,
1)
16,5
ab
(17,
4)
16,0
ab
(15,
7)
16,4
a (1
7,2)
14
,8a
(14,
3)
15,2
a (1
6,1)
16
,5a
(17,
4)
15,7
a (1
4,9)
O
mis
são
de F
e 15
,3a
(13,
6)
15,9
a (1
5,4)
15
,9a
(15,
4)
17,1
a (1
9,5)
16
,9a
(18,
9)
15,8
ab
(15,
3)
16,4
a (1
7,3)
16
,3ab
(1
6,6)
15
,3bc
(1
3,6)
16
,6a
(17,
8)
15,2
a (1
5,9)
16
,1a
(20,
3)
16,5
a (1
7,5)
14
,6ab
(1
1,4)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,17
64
0,21
87
0,23
8 0,
2561
0,
2693
0,
1947
0,
3421
0,
3204
0,
205
0,20
77
0,23
17
0,33
13
0,21
81
0,18
81
CV
(%)
4,04
4,
99
5,32
5,
54
5,76
4,
26
7,61
7,
10
4,58
4,
56
5,55
7,
49
4,7
4,43
T
rans
form
ação
R
Q(y
+1)
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
)Lo
g(y+
3)
Log(
y+3
) R
Q(y
+1)
RQ
(y+1
)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

110
Tab
ela
34. T
eor d
e fó
sfor
o (g
kg
-1) p
rese
nte
na a
nális
e da
raiz
da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
10,8
a (1
,8)
11,1
a (2
,4)
11,1
a (2
,3)
11,0
b (2
,1)
11,2
a (2
,7)
11,6
a (3
,5)
11,6
d (3
,5)
13,4
ab
(8,2
) 13
,3a
(7,9
) 12
,5a
(5,7
) 12
,6ab
(5
,9)
13,0
a (7
,2)
12,9
ab
(6,7
) 13
,5ab
(8
,2)
Om
issã
o de
N
10,8
a (1
,8)
10,9
a (2
,0)
11,0
a (2
,3)
11,1
ab
(2,4
) 11
,2a
(2,6
) 11
,4a
(3,1
) 12
,0bc
d(4
,5)
12,4
bc
(5,5
) 12
,3a
(5,3
) 12
,8a
(6,5
) 12
,4ab
(5
,6)
12,7
a (6
,3)
12,7
b (6
,2)
12,5
b (5
,7)
Om
issã
o de
P
10,9
a (2
,0)
11,3
a (2
,8)
11,0
a (2
,1)
11,1
ab
(2,5
) 11
,3a
(2,9
) 11
,4a
(3,0
) 11
,8cd
(4
,0)
12,2
c (4
,9)
12,3
a (5
,2)
12,8
a (6
,6)
12,1
ab
(4,8
) 12
,1a
(4,9
) 12
,9ab
(6
,8)
12,1
b (4
,8)
Om
issã
o de
K
10,9
a (2
,0)
11,0
a (2
,2)
11,1
a (2
,5)
11,2
ab
(2,6
) 11
,3a
(2,9
) 11
,7a
(3,7
) 13
,1a
(7,3
) 13
,8a
(9,2
) 13
,3a
(7,8
) 12
,8a
(6,6
) 11
,9a
(4,3
) 13
,2a
(7,5
) 13
,3ab
(7
,8)
12,9
ab
(6,7
) O
mis
são
de
Ca
11,0
a (2
,3)
11,3
a (2
,9)
11,2
a (2
,7)
11,3
a (2
,8)
11,5
a (3
,3)
11,6
a (3
,6)
12,9
ab
(6,9
) 12
,5bc
(5
,9)
12,2
a (5
,0)
12,6
a (6
,1)
12,6
ab
(5,9
) 12
,7a
(6,4
) 13
,7a
(9,0
) 14
,1a
(9,9
) O
mis
são
de
Mg
10,9
a (1
,9)
11,3
a (2
,8)
11,2
a (2
,6)
11,0
b (2
,2)
11,4
a (3
,0)
11,7
a (3
,8)
12,4
abcd
(5,4
) 12
,6ab
c(6
,1)
12,8
a (6
,6)
12,6
a (6
,0)
13,2
a (7
,5)
13,4
a (8
,7)
13,5
ab
(8,5
) 14
,2a
(10,
2)
Om
issã
o de
B
10,8
a (1
,7)
11,1
a (2
,3)
11,0
a (2
,2)
11,0
ab
(2,2
) 11
,3a
(2,9
) 11
,7a
(3,7
) 12
,8ab
(6
,6)
12,7
abc
(6,2
) 12
,8a
(6,5
) 12
,7a
(6,2
) 12
,5ab
(5
,8)
13,6
a (8
,7)
13,5
ab
(8,4
) 12
,9ab
(6
,9)
Om
issã
o de
Fe
11
,0a
(2,1
) 11
,1a
(2,5
) 11
,2a
(2,7
) 11
,2ab
(2
,7)
11,3
a (2
,9)
11,7
a (3
,7)
12,6
ab
(6,1
) 12
,3bc
(5
,3)
13,0
a (7
,1)
12,5
a (5
,7)
12,0
ab
(4,5
) 12
,7a
(6,1
) 12
,7ab
(6
,2)
12,9
ab
(6,8
)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,03
2 0,
043
0,05
24
0,02
96
0,03
64
0,04
0,
01
0,12
06
0,18
27
0,12
04
0,12
33
0,15
95
0,10
35
0,13
59
CV
(%)
1,03
1,
36
1,66
0,
93
1,13
1,
21
2,84
3,
33
5,05
3,
34
3,50
4,
34
2,77
3,
65
Tra
nsfo
rmaç
ão
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

111
Tab
ela
35. T
eor d
e po
táss
io (g
kg-1
) pre
sent
e na
aná
lise
da ra
iz d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
Col
etas
T
rata
men
tos
T02h
T0
4h
T7,5
h T2
2h
T46h
T1
3d
T35d
T4
5d
T61d
T7
9d
T97d
T1
07d
T115
d T1
24d
Com
plet
a 32
,6a
(6,6
) 33
,0a
(9,0
) 32
,5a
(6,2
) 32
,9a
(8,6
) 33
,1a
(10,
1)
33,7
a (1
3,7)
33
,4a
(12,
0)
33,5
a (1
2,8)
33
,1b
(9,7
) 33
,2ab
c (1
2,2)
33
,1ab
(1
0,1)
32
,9a
(8,8
) 33
,6a
(13,
3)
33,4
a (1
1,8)
O
mis
são
de
N
32,7
a (7
,4)
32,7
a (7
,6)
32,6
a (6
,5)
33,0
a (9
,0)
33,2
a (1
0,6)
33
,8a
(14,
7)
33,5
a (1
2,3)
33
,6a
(13,
2)
33,2
ab
(10,
5)
33,0
bc
(11,
8)
33,3
a (1
1,1)
32
,8a
(8,1
) 33
,3ab
(1
1,2)
32
,8bc
d (7
8)
Om
issã
o de
P
32,6
a (6
,6)
33,2
a (1
0,8)
32
,6a
(6,7
) 33
,0a
(9,1
) 33
,2a
(10,
4)
33,9
a (1
5,5)
33
,7a
(13,
6)
33,7
a (1
3,8)
33
,7a
(14,
1)
33,3
ab
(8,3
) 33
,2ab
(1
0,5)
32
,8a
(8,1
) 33
,7a
(13,
8)
33,1
ab
(10,
1)
Om
issã
o de
K
32
,5a
(6,0
) 33
,1a
(9,6
) 32
,7a
(7,5
) 32
,9a
(8,3
) 32
,8a
(8,0
) 33
,0a
(9,5
) 32
,7a
(7,2
) 32
,7a
(7,3
) 32
,3c
(4,8
) 32
,4d
(8,2
) 32
,3c
(4,4
) 31
,9b
(2,0
) 32
,5c
(5,9
) 32
,4d
(5,1
) O
mis
são
de
Ca
33,0
a (9
,3)
33,6
a (1
2,9)
32
,6a
(6,7
) 33
,0a
(8,9
) 33
,0a
(8,9
) 33
,7a
(13,
8)
33,5
a (1
2,5)
33
,4a
(11,
6)
32,9
b (8
,8)
32,8
c (5
,1)
32,7
bc
(7,3
) 32
,6a
(6,3
) 33
,5ab
(1
2,4)
33
,2ab
(1
0,2)
O
mis
são
de
Mg
33,3
a (1
0,9)
33
,2a
(10,
8)
32,7
a (7
,5)
32,6
a (6
,6)
32,6
a (9
,4)
33,7
a (1
3,7)
33
,5a
(12,
5)
33,4
a (1
2,2)
33
,2ab
(1
0,8)
32
,9c
(11,
5)
32,7
bc
(7,5
) 32
,6a
(6,5
) 33
,0b
(9,5
) 32
,7cd
(7
,1)
Om
issã
o de
B
32,5
a (6
,0)
32,7
a (7
,1)
32,6
a (6
,6)
32,8
a (7
,7)
32,8
a (1
0,5)
33
,5a
(12,
6)
33,4
a (1
2,0)
33
,5a
(12,
5)
33,3
ab
(11,
2)
33,4
ab
(90)
33
,3a
(11,
0)
32,9
a (8
,9)
33,5
ab
(12,
3)
32,9
bc
(8,8
) O
mis
são
de
Fe
33,0
a (9
,1)
32,6
a (6
,7)
33,3
a (1
1,4)
32
,8a
(8,2
) 32
,8a
(10,
0)
33,8
a (1
4,3)
33
,6a
(13,
3)
33,5
a (1
2,6)
33
,5a
(12,
5)
33,4
a
(10,
4)
33,2
ab
(10,
5)
33,6
a (8
,5)
33,6
ab
(13,
1)
33,1
ab
(10,
1)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
0864
0,
1022
0,
0848
0,
0506
0,
0581
0,
0441
0,
0279
0,
0398
0,
0606
0,
0429
0,
501
0,04
57
0,04
69
0,04
45
CV
(%)
0,93
1,
09
0,91
0,
54
0,62
0,
46
0,29
0,
42
0,64
0,
45
0,53
0,
49
0,49
0,
47
Tra
nsfo
rmaç
ão
RQ
(y+
10)
RQ
(y+
10)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

112
Tab
ela
36. T
eor d
e cá
lcio
(g k
g-1) p
rese
nte
na a
nális
e da
raiz
da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
12,5
ab
(4,9
) 12
,9a
(6,5
) 12
,6ab
c(5
,5)
12,8
ab
(6,1
) 12
,9a
(6,4
) 13
5a
(8,6
) 11
,7b
(2,3
) 13
,6a
(9,2
) 14
,0a
(11,
0)
13,9
ab
(10,
5)
14,1
a (1
1,4)
13
,8b
(10,
0)
15,0
ab
(15,
2)
14,7
a (1
3,9)
O
mis
são
de N
12
,4ab
(4
,7)
12,5
a (5
,0)
12,4
bc
(4,8
) 12
,9ab
(6
,5)
12,8
a (6
,2)
12,9
b (6
,4)
12,0
b (3
,5)
1,30
ab
(6,9
) 13
,2a
(7,6
) 13
,6ab
(9
,1)
14,0
a (1
1,0)
13
,2b
(7,6
) 14
,5ab
(1
2,9)
12
,9b
(6,6
) O
mis
são
de P
12
,4ab
(4
,8)
12,6
a (5
,5)
12,2
c (4
,1)
13,0
a (6
,8)
13,3
a (7
,8)
13,1
ab
(7,4
) 12
,3ab
(4
,5)
13,3
ab
(8,0
) 13
,9a
(10,
5)
14,4
a (1
2,5)
14
,2a
(11,
7)
13,7
b (9
,7)
16,0
a (1
9,6)
13
,4ab
(8
,6)
Om
issã
o de
K
12,8
a (6
,3)
12,8
a (6
,1)
13,1
a (7
,3)
12,5
ab
(5,1
) 13
,0a
(6,7
) 13
,2ab
(7
,6)
1,27
a (5
,9)
1,25
ab
(5,0
) 12
,8a
(6,2
) 13
,4ab
(8
,5)
13,0
a (6
,8)
13,3
b (8
,1)
14,6
ab
(13,
2)
13,0
b (6
,7)
Om
issã
o de
C
a 12
,3b
(4,2
) 12
,7a
(5,7
) 12
,9ab
(6
,5)
12,6
ab
(5,5
) 12
,8a
(6,1
) 12
,9b
(6,4
) 12
,9ab
(6
,6)
12,4
ab
(4,3
) 12
,7a
(6,0
) 13
,2ab
(7
,6)
13,1
a (7
,2)
12,7
b (5
,9)
15,5
ab
(17,
7)
13,6
ab
(9,2
) O
mis
são
de
Mg
12,2
b (3
,9)
12,7
a (5
,8)
12,6
abc
(5,6
) 12
,4b
(4,8
) 12
,8a
(6,2
) 13
,0b
(6,9
) 12
,1ab
(3
,7)
12,3
b (4
,9)
12,6
a (5
,5)
13,0
b (6
,9)
13,3
a (8
,0)
13,4
b (8
,3)
14,2
b (1
1,6)
13
,2b
(7,5
) O
mis
são
de B
12
,3b
(4,3
) 12
,5a
(5,0
) 12
,7ab
c(5
,7)
12,6
ab
(5,2
) 12
,8a
(6,3
) 13
,1ab
(7
,1)
1,36
a (9
,3)
13,3
ab
(7,9
) 13
,5a
(8,8
) 13
,9ab
(1
0,3)
14
,5a
(13,
1)
15,4
a (1
7,1)
15
,8a
(18,
8)
13,5
ab
(8,9
) O
mis
são
de
Fe
12,5
ab
(5,2
) 12
,7a
(5,8
) 12
,9ab
(6
,4)
12,6
ab
(5,3
) 12
,9a
(6,5
) 12
,8b
(6,0
) 13
,4a
(8,5
) 12
,6ab
(5
,4)
13,6
a (9
,4)
13,5
ab
(8,9
) 13
,6a
(9,3
) 13
,3b
(8,2
) 14
,7ab
(1
3,8)
13
,8ab
(1
0,1)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,05
3 0,
0667
0,
063
0,05
37
0,05
3 0,
0428
0,
1303
0,
1242
0,
2093
0,
1323
0,
1801
0,
1612
0,
1485
0,
1379
C
V(%
) 1,
50
1,85
1,
75
1,49
1,
44
1,15
3,
64
3,40
5,
54
3,42
4,
62
4,17
3,
48
3,59
T
rans
form
ação
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

113
Tab
ela
37 T
eor d
e m
agné
sio
(g k
g-1) p
rese
nte
na a
nális
e da
raiz
da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
12,2
a (3
,9)
13,0
a (6
,9)
13,2
a (7
,8)
12,6
a (5
,5)
12,4
b (4
,6)
12,3
a (4
,2)
11,6
d (2
,2)
12,7
a (6
,0)
13,2
ab
(7,7
) 12
,9ab
(6
,7)
13,4
a (8
,4)
13,0
a (7
,1)
13,1
c (7
,1)
16,7
a (2
3,7)
O
mis
são
de N
12
,1a
(3,7
) 12
,4a
(4,8
) 13
,1a
(7,4
) 12
,9a
(6,5
) 12
,8ab
(6
,0)
12,1
a (3
,8)
12,3
abc
(4,3
) 12
,6a
(5,4
) 13
,3ab
(8
,2)
12,5
ab
(5,0
) 13
,6a
(9,2
) 12
,6a
(5,5
) 12
,8c
(6,0
) 12
,5b
(5,2
) O
mis
são
de P
12
,1a
(3,7
) 12
,7a
(5,8
) 12
,9a
(6,7
) 12
,9a
(6,6
) 13
,2a
(7,8
) 12
,3a
(4,3
) 12
,1bc
d(3
,7)
12,8
a (6
,2)
13,5
ab
(8,9
) 13
,4a
(8,6
) 12
,9a
(6,4
) 12
,7a
(5,7
) 16
,7a
(23,
5)
16,0
a (2
0,0)
O
mis
são
de K
12
,2a
(4,1
) 12
,5a
(5,0
) 12
,7a
(5,8
) 12
,6a
(5,4
) 12
,8ab
(6
,2)
12,2
a (3
,9)
12,8
a (6
,1)
12,6
a (5
,4)
13,3
ab
(8,0
) 12
,1ab
(3
,6)
12,5
a (5
,1)
12,7
a (5
,8)
13,4
bc
(8,6
) 12
,8b
(6,1
) O
mis
são
de
Ca
12,2
a (4
,0)
13,0
a (6
,8)
13,4
a (8
,4)
13,2
a (7
,5)
13,2
a (7
,7)
12,1
a (3
,6)
12,8
a (6
,2)
12,5
a (5
,0)
12,6
ab
(5,5
) 12
,8ab
(6
,3)
12,8
a (6
,3)
12,6
a (5
,4)
16,1
a (2
0,6)
16
,0a
(20,
1)
Om
issã
o de
M
g 12
,0a
(3,3
) 13
,2a
(7,6
) 12
,9a
(6,6
) 12
,6a
(5,5
) 12
,7ab
(5
,9)
12,2
a (3
,9)
11,9
cd
(3,2
) 12
,4a
(4,8
) 13
,0ab
(6
,8)
11,9
b (3
,1)
12,9
a (6
,3)
12,4
a (4
,8)
13,5
bc
(9,1
) 12
,3b
(4,4
) O
mis
são
de B
12
,1a
(3,7
) 12
,9a
(6,7
) 13
,2a
(7,5
) 12
,9a
(6,6
) 12
,7ab
(5
,9)
12,4
a (4
,9)
12,7
ab
(5,8
) 12
,6a
(5,5
) 13
,1ab
(7
,2)
12,7
ab
(5,8
) 1,
32a
(7,6
) 13
,8a
(10,
8)
15,4
ab
(17,
3)
12,8
b (6
,2)
Om
issã
o de
Fe
12
,4a
(4,6
) 13
,4a
(8,3
) 13
,2a
(7,7
) 13
,3a
(7,8
) 12
,7ab
(5
,8)
12,3
a (4
,3)
12,7
ab
(5,7
) 1,
25a
(5,2
) 13
,8a
(10,
0)
12,8
ab
(6,1
) 13
,2a
(7,6
) 12
,9a
(6,4
) 1,
28c
(6,3
) 15
,2a
(16,
0)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
0468
0,
1156
0,
1504
0,
098
0,07
87
0,05
74
0,06
27
0,07
69
0,11
4 0,
1433
0,
1424
0,
1399
0,
2225
0,
2131
C
V(%
) 1,
35
3,15
4,
04
2,68
2,
16
1,65
1,
78
2,14
3,
03
3,99
3,
84
3,84
5,
50
5,25
T
rans
form
ação
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
Lo
g(y+
3)
Log(
y+3)
*
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

114
Tab
ela
38. T
eor d
e fe
rro
(mg
kg-1
) pre
sent
e na
aná
lise
da ra
iz d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
6,72
a (8
36,9
) 7,
82a
(280
3,3)
7,
99a
(343
1,6)
7,51
a (1
985,
0)7,
15
(129
8,8)
6,68
a (8
08,3
) 5,
14b
(175
,9)
8,33
a (4
243,
3)
8,20
ab
(365
3,3)
8,54
a (5
390,
0)8,
73a
(626
6,6)
8,61
ab
(573
3,3)
8,59
a (5
466,
6)8,
75ab
(6
416,
6)
Om
issã
o de
N
6,67
a (8
41,0
) 7,
30a
(149
5,0)
7,
75a
(278
8,3)
7,89
a (2
790,
0)7,
42a
(169
1,6)
6,48
a (6
58,3
) 7,
32a
(156
7,3)
8,02
abc
(306
6,6)
8,
25ab
(3
858,
3)8,
60a
(560
0,0)
8,63
a (5
816,
6)8,
40ab
(4
566,
6)8,
51a
(523
3,3)
8,13
c (3
446,
6)
Om
issã
o de
P
6,76
a (8
67,5
) 7,
39a
(165
5,0)
7,
61a
(223
1,6)
7,80
a (2
580,
0)7,
33a
(186
5,0)
6,81
a (9
38,4
) 6,
73ab
(1
414,
1)7,
92bc
(2
783,
3)
8,21
ab
(372
5,0)
8,90
a (7
483,
3)8,
50a
(520
0,0)
8,62
ab
(576
6,6)
9,15
a (9
466,
6)8,
18c
(363
6,6)
Om
issã
o de
K
6,88
a (9
95,0
) 7,
17a
(131
5,0)
7,
03a
(186
6,0)
7,79
a (2
606,
6)7,
51a
(199
8,2)
6,70
a (8
18,1
) 7,
77a
(238
0,0)
8,15
ab
(348
3,3)
8,
81a
(678
3,3)
8,82
a (6
950,
0)8,
48a
(490
0,0)
8,88
a (7
400,
0)9,
10a
(900
0,0)
8,63
abc
(576
6,6)
Om
issã
o de
C
a 6,
78a
(899
,9)
8,28
a (3
966,
6)
8,66
a (5
833,
3)8,
08a
(323
5,0)
7,79
a (2
576,
6)6,
52a
(686
,6)
7,70
a (2
216,
6)7,
68c
(216
6,6)
7,
59b
(199
0,0)
8,62
a (5
620,
0)8,
49a
(508
3,0)
8,46
ab
(475
0,0)
8,84
a (7
000,
0)8,
83a
(690
0,0)
Om
issã
o de
M
g 6,
47a
(651
,7)
8,10
a (3
330,
0)
7,84
a (2
774,
0)7,
55a
(212
3,3)
7,56
a (1
995,
0)6,
80a
(902
,0)
7,54
a (1
893,
3)8,
06ab
(3
166,
6)
8,58
a (5
733,
3)8,
91a
(748
3,3)
9,15
a (9
450,
0)9,
05a
(863
3,3)
9,11
a (9
133,
3)9,
00a
(816
6,6)
Om
issã
o de
B
7,05
a (1
284,
7)
7,89
a (3
281,
6)
8,24
a (4
233,
3)7,
75a
(257
3,3)
7,50
a (1
900,
0)6,
90a
(106
1,6)
7,66
a (2
123,
3)7,
65c
(216
6,6)
8,
11ab
(3
583,
3)8,
51a
(512
6,6)
8,65
a (6
233,
3)8,
65ab
(5
900,
0)8,
88a
(723
3,3)
8,61
abc
(559
6,6)
Om
issã
o de
Fe
6,
98a
(109
8,9)
8,
00a
(328
3,3)
8,
11a
(335
0,0)
8,08
a (3
250,
0)7,
48a
(190
8,3)
6,48
a (6
59,1
) 7,
19a
(142
3,7)
7,18
d (1
316,
6)
8,36
a (4
300,
0)7,
75b
(240
0,0)
8,39
a (5
320,
0)7,
94b
(288
6,6)
7,37
b (1
738,
3)8,
26bc
(3
931,
6)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
8304
1,
1758
1,
9554
1,
1929
1,
2692
0,
6402
1,
6652
0,
3735
0,
7061
0,
7496
1,
1114
0,
7598
0,
7598
0,
5086
C
V(%
) 4,
32
5,36
8,
75
5,40
6,
00
3,39
8,
25
1,67
3,
02
3,08
4,
55
3,13
3,
13
2,10
T
rans
form
açã
o lo
g(y+
1)
log(
y+1
) Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
*
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

115
Tab
ela
39. T
eor d
e bo
ro (m
g kg
-1) p
rese
nte
na a
nális
e da
raiz
da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T04h
T7
,5h
T22h
T4
6h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
3,82
a (4
4,86
) 4,
12ab
(6
0,90
) 4,
37ab
(7
9,70
) 3,
90a
(51,
03)
3,90
a (4
9,06
) 3,
93a
(50,
53)
3,48
c (3
2,10
) 4,
40ab
(8
3,33
) 4,
13ab
(6
2,33
) 4,
20bc
(6
6,57
) 4,
31a
(73,
87)
4,24
ab
(69,
07)
4,60
a (9
9,57
) 4,
54a
(93,
40)
Om
issã
o de
N
3,84
a (4
7,16
) 4,
02b
(55,
20)
4,45
a (8
5,80
) 4,
09a
(59,
33)
3,87
a (4
7,33
) 3,
76a
(42,
63)
3,93
b (5
0,83
) 4,
27ab
(7
0,83
) 4,
02ab
(5
5,33
) 4,
04cd
(5
6,20
) 3,
83bc
(4
6,27
) 4,
00b
(53,
77)
4,00
b (5
4,07
) 3,
85bc
(4
6,50
)
Om
issã
o de
P
3,88
a
(48,
13)
4,16
ab
(64,
56)
4,12
ab
(61,
23)
4,14
a (6
2,43
) 3,
91a
(49,
43)
3,64
ab
(37,
53)
4,30
a (7
2,77
) 4,
60a
(99,
00)
4,18
a (6
5,57
) 4,
33ab
(7
5,37
) 4,
26a
(70,
93)
4,44
a (8
4,10
) 4,
53a
(92,
37)
4,24
ab
(69,
70)
Om
issã
o de
K
3,47
ab
(31,
46)
4,01
b (5
4,80
) 4,
13ab
(6
3,46
) 3,
70a
(46,
73)
3,82
a (4
5,63
) 3,
93a
(51,
00)
4,33
a (7
5,87
) 4,
43ab
(8
3,50
) 3,
76b
(42,
70)
3,95
cd
(51,
73)
3,58
c (3
5,33
) 4,
04ab
(5
7,77
) 4,
03b
(55,
50)
3,74
c (4
1,40
)
Om
issã
o de
Ca
3,65
a (3
8,20
) 4,
31ab
(7
3,86
) 3,
90b
(49,
67)
4,17
a (6
3,80
) 3,
83a
(45,
60)
3,84
a (4
6,10
) 4,
19ab
(6
5,37
) 4,
28ab
(7
2,00
) 4,
03ab
56
,40)
4,
20bc
(6
6,43
) 3,
96ab
c (5
2,13
) 4,
04b
(56,
40)
4,40
a (8
0,83
) 4,
20ab
(6
6,47
)
Om
issã
o de
M
g 3,
13b
(21,
95)
4,19
ab
(66,
00)
4,06
ab
(57,
30)
4,32
a (7
6,43
) 3,
93a
(50,
43)
3,78
a (4
3,30
) 4,
15ab
(6
2,50
) 4,
14b
(62,
07)
3,94
ab
(50,
73)
3,91
d (4
9,17
) 3,
89ab
c (4
8,40
) 4,
11ab
(6
0,50
) 4,
00b
(53,
97)
3,75
c (4
1,67
)
Om
issã
o de
B
3,52
ab
(33,
06)
4,45
a (8
5,40
) 4,
24ab
(6
9,93
) 3,
37a
(28,
93)
3,69
a (3
9,13
) 3,
30b
(26,
13)
3,30
c (2
6,27
) 3,
32c
(27,
10)
3,01
c (1
9,40
) 3,
42e
(29,
70)
3,08
d (2
0,80
) 3,
27c
(26,
10)
3,45
c (3
1,30
) 3,
26d
(25,
43)
Om
issã
o de
Fe
3,48
ab
(31,
60)
4,09
b (5
9,16
) 4,
24ab
(6
8,60
) 3,
45a
(30,
83)
3,76
a (4
2,43
) 3,
82a
(45,
00)
4,44
a (8
3,90
) 4,
55ab
(9
4,30
) 4,
00ab
(5
4,17
) 4,
58a
(96,
90)
4,27
a (7
0,80
) 4,
20ab
(6
6,30
) 4,
50a
(89,
80)
4,29
a
(73,
27)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
4628
0,
3576
0,
5385
0,
9599
0,
3601
0,
3528
0,
3632
0,
4169
0,
4088
0,
27
0,43
74
0,39
6 0,
3288
0,
3972
C
V(%
) 4,
54
3,03
4,
54
8,75
3,
31
3,32
3,
19
3,46
3,
71
2,33
3,
90
3,45
2,
77
3,52
T
rans
form
ação
Lo
g(y+
1)
Log(
y+1)
Lo
g(y+
1)
Log(
y+1)
Log(
y+1)
Lo
g(y+
1)Lo
g(y+
1)Lo
g(y+
1)
Log(
y+1)
Log(
y+1)
Log(
y+1)
Lo
g(y+
1)Lo
g(y+
1)Lo
g(y+
1)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

116
Tab
ela
40. T
eor d
e fó
sfor
o (m
g dm
-3) p
rese
nte
na a
nális
e do
subs
trato
da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T7
,5h
T22h
T7
0h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d
Com
plet
a 4,
10a
(59,
66)
4,17
a (6
4,33
) 4,
16a
(63,
33)
4,56
a (9
5,00
) 45
,7a
(96,
00)
4,66
a (1
05,0
0)
4,62
ab
(101
,00)
4,
75ab
c (1
15,0
0)
4,73
a (1
13,3
3)
4,63
ab
(102
,66)
4,
75a
(115
,00)
4,
58ab
c (9
6,66
)
Om
issã
o de
N
3,90
a (4
9,66
) 3,
96a
(52,
33)
4,05
a (5
7,33
) 4,
47ab
(5
6,67
) 45
,4a
(93,
33)
4,61
a (1
00,0
0)
4,49
b (8
8,66
) 4,
72bc
d (1
11,6
6)
4,64
ab
(103
,33)
4,
61ab
(1
00,0
0)
4,84
a (1
26,6
6)
4,53
bcd
(92,
33)
Om
issã
o de
P
4,09
a (6
0,50
) 3,
96a
(52,
00)
3,96
a (5
2,66
) 4,
41b
(82,
00)
4,45
a (8
4,66
) 4,
58a
(97,
33)
4,54
ab
(93,
33)
4,66
e (1
05,0
0)
4,47
c (8
6,66
) 4,
55b
(94,
00)
4,72
a (1
11,6
6)
4,51
cd
(90,
33)
Om
issã
o de
K
4,15
a (6
3,33
) 3,
89a
(50,
00)
4,20
a (6
6,00
) 4,
56a
(95,
00)
4,57
a (9
6,00
) 4,
66a
(105
,00)
4,
58ab
(9
7,00
) 4,
69de
(1
08,3
3)
4,55
bc
(93,
66)
4,62
ab
(100
,66)
4,
73a
(113
,33)
4,
45d
(85,
33)
Om
issã
o de
C
a 4,
16a
(63,
66)
4,14
a (6
2,33
) 4,
14a
(62,
33)
4,54
ab
(93,
00)
4,56
a (9
5,00
) 4,
61a
(100
,00)
4,
67a
(106
,66)
4,
78a
(118
,33)
4,
75a
(115
,00)
4,
70a
(110
,00)
4,
80a
(121
,66)
4,
66a
(105
,00)
O
mis
são
de
Mg
3,99
a (5
3,66
) 4,
11a
(60,
33)
4,15
a (6
3,33
) 4,
55a
(94,
33)
4,57
a (9
6,00
) 4,
64a
(102
,66)
4,
69a
(108
,33)
4,
76ab
(1
16,6
6)
4,58
bc
(96,
67)
4,63
ab
(101
,66)
4,
79a
(120
,00)
4,
59ab
c (9
8,00
)
Om
issã
o de
B
4,15
a (6
3,00
) 4,
12a
(61,
00)
4,15
a (6
3,00
) 4,
53ab
(9
2,67
) 4,
58a
(96,
67)
4,64
a (1
03,3
3)
4,67
a (1
06,6
6)
4,75
abc
(115
,00)
4,
58bc
(9
7,33
) 4,
62ab
(1
00,6
6)
4,76
a (1
16,6
6)
4,60
ab
(99,
00)
Om
issã
o de
Fe
4,05
a (5
6,67
) 3,
98a
(53,
67)
4,32
a (7
5,33
) 4,
50ab
(8
9,33
) 4,
34a
(80,
33)
4,59
a (9
8,00
) 4,
62a
(101
,66)
4,
70cd
e (1
10,0
0)
4,57
bc
(95,
66)
4,69
a (1
08,3
3)
4,76
a (1
16,6
6)
4,62
a (1
00,6
6)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
3808
0,
4990
0,
423
0,13
55
0,41
91
0,07
71
0,16
76
0,05
04
0,14
56
0,10
62
0,17
25
0,08
44
CV
(%)
3,30
4,
36
3,60
1,
06
3,27
0,
58
1,28
0,
37
1,11
0,
81
1,27
0,
65
Tra
nsfo
rmaç
ão
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

117
Tab
ela
41. T
eor
de p
otás
sio
(mm
olc
dm-3
) pr
esen
te n
a an
ális
e do
sub
stra
to d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m f
unçã
o da
col
eta
e da
om
issã
o de
nu
trien
tes.
C
olet
as
Tra
tam
ento
s T7
,5h
T22h
T7
0h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
9,6a
(6
,3)
9,3a
(5
,5)
10,0
a (7
,4)
11,9
a (1
3,0)
10
,1a
(7,7
) 14
,3ab
(2
1,8)
10
,3ab
(8
,0)
16,0
ab
(29,
8)
14,1
b (2
1,3)
15
,7b
(28,
3)
15,4
c (2
6,8)
15
,1b
(25,
6)
Om
issã
o de
N
9,4a
(5
,8)
9,5a
(5
,9)
9,7a
b (6
,4)
11,4
ab
(11,
2)
10,1
a (7
,6)
15,2
a (2
6,0)
10
,1ab
(7
,7)
15,6
abc
(27,
6)
14,2
b (2
1,1)
15
,2bc
(2
6,0)
15
,9bc
(2
9,5)
14
,6b
(23,
1)
Om
issã
o de
P
9,8a
(6
,5)
9,3a
(5
,6)
9,8a
b (6
,7)
10,6
bc
(8,7
) 10
,4a
(8,4
) 14
,5ab
(2
2,8)
10
,3ab
(8
,2)
16,2
a (3
1,0)
14
,0b
(20,
8)
15,4
bc
(27,
0)
15,8
bc
(28,
6)
14,9
b (2
4,8)
O
mis
são
de K
9,
6a
(6,3
) 9,
1a
(4,9
) 8,
9c
(4,4
) 8,
4d
(3,4
) 8,
7b
(3,9
) 8,
3c
(3,1
) 8,
8c
(4,3
) 8,
9e
(4,3
) 8,
3c
(02,
9)
8,7d
(3
,9)
8,1d
(2
,7)
9,3c
(5
,4)
Om
issã
o de
C
a 9,
5a
(5,9
) 9,
2a
(5,3
) 9,
4bc
(5,7
) 10
,2c
(8,1
) 10
,4a
(8,4
) 13
,5b
(18,
8)
10,0
b (7
,3)
14,9
c (2
4,5)
14
,2c
(21,
3)
14,5
bc
(22,
8)
15,2
c (2
6,1)
15
,2b
(26,
1)
Om
issã
o de
M
g 9,
4a
(5,6
) 9,
4a
(5,7
) 9,
6ab
(6,1
) 10
,4c
(8,3
) 10
,2a
(7,9
) 14
,3ab
(2
2,1)
10
,0b
(7,2
) 15
,3bc
d (2
6,3)
13
,5b
(18,
8)
14,2
c (2
1,8)
15
,8bc
(2
8,8)
15
,2b
(26,
1)
Om
issã
o de
B
9,6a
(6
,5)
9,4a
(5
,6)
9,7a
b (6
,4)
11,9
a (1
2,7)
10
,4a
(8,5
) 14
,7ab
(2
3,5)
10
,4ab
(8
,5)
15,5
abcd
(2
7,1)
16
,3a
(31,
5)
17,1
a (3
5,5)
17
,6a
(38,
6)
18,3
a (4
3,1)
O
mis
são
de F
e 10
,0a
(7,0
) 9,
4a
(5,8
) 10
,0a
(7,3
) 11
,8a
(12,
5)
10,4
a (8
,3)
14,6
ab
(23,
3)
10,5
a (8
,7)
14,6
d (2
3,3)
15
,9a
(29,
3)
15,0
bc
(25,
0)
16,5
b (3
2,3)
18
,4a
(43,
6)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,06
96
0,05
3 0,
0538
0,
0878
0,
0508
0,
1145
0,
0447
0,
0862
0,
1607
0,
135
0,11
07
0,22
83
CV
(%)
2,54
1,
99
1,97
2,
85
1,77
2,
95
1,56
2,
07
4,09
3,
28
2,59
5,
31
Tra
nsfo
rmaç
ão
Log
(y+2
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

118
Tab
ela
42. T
eor d
e cá
lcio
(mm
olc d
m-3
) pre
sent
e na
aná
lise
do su
bstra
to d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
Tra
tam
ento
s T7
,5h
T22h
T7
0h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
21,3
a (7
5,0)
20
,4a
(67,
7)
20,5
a (6
8,3)
19
,0a
(57,
3)
19,6
a (6
1,3)
19
,1ab
(5
8,3)
20
,7a
(69,
3)
20,4
a (6
7,0)
15
,8ab
c (5
6,0)
20
,3a
(66,
3)
20,7
ab
(69,
6)
19,4
ab
(60,
0)
Om
issã
o de
N
19,4
a (6
0,3)
20
,1a
(65,
3)
20,0
a (6
4,6)
20
,0a
(64,
7)
18,5
a (5
36)
19,8
ab
(62,
6)
21,2
a (7
3,6)
19
,3a
(59,
0)
17,0
ac
(46,
0)
18,5
a (5
4,0)
19
,0b
(57,
0)
16,5
c (4
2,3)
O
mis
são
de P
20
,7a
(71,
0)
21,9
a (8
0,0)
21
,4a
(75,
3)
20,6
a (6
9,0)
19
,2a
(58,
6)
20,5
ab
(68,
3)
19,2
ab
(59,
3)
20,9
a (7
1,6)
20
,1ab
(6
5,3)
20
,9a
(71,
0)
20,9
ab
(71,
0)
18,1
bc
(51,
3)
Om
issã
o de
K
21,1
a (7
3,3)
20
,4a
(67,
7)
21,2
a (7
4,0)
20
,4a
(67,
7)
20,5
a (6
8,3)
21
,2a
(73,
6)
19,7
ab
(62,
3)
20,5
a (6
8,3)
18
,3bc
(5
2,6)
20
,5a
(68,
3)
20,5
ab
(68,
3)
19,5
ab
(60,
6)
Om
issã
o de
C
a 21
,1a
(73,
6)
21,4
a (7
6,0)
20
,1a
(65,
0)
19,0
a (5
7,3)
17
,7a
(49,
0)
17,5
b (4
8,0)
17
,5b
(48,
0)
15,3
b (3
6,3)
14
,8d
(34,
3)
14,4
a (3
2,0)
14
,2c
(32,
0)
13,5
d (2
8,6)
O
mis
são
de
Mg
19,1
a (5
8,6)
20
,8a
(70,
7)
21,0
a (7
2,0)
20
,7a
(70,
3)
20,7
a (7
0,0)
20
,7a
(70,
0)
20,0
a (6
4,6)
20
,6a
(69,
0)
20,6
a (6
9,3)
20
,0a
(64,
3)
22,1
a (8
1,3)
20
,8a
(70,
6)
Om
issã
o de
B
20,1
a (6
5,0)
20
,2a
(65,
7)
20,6
a (6
9,0)
20
,8a
(70,
3)
19,4
a (6
0,3)
19
,1ab
(5
8,3)
20
,2a
(66,
0)
19,1
a (5
7,6)
18
,2bc
(5
2,3)
19
,5a
(61,
3)
19,9
ab
(64,
3)
18,7
ab
(55,
3)
Om
issã
o de
Fe
21,3
a (7
5,0)
19
,6a
(58,
3)
21,0
a (7
2,0)
18
,8a
(56,
3)
19,2
a (5
8,6)
20
,8a
(72,
3)
20,8
a (7
1,0)
18
,8a
(55,
6)
18,7
abc
(55,
3)
19,9
a (6
3,6)
20
,3ab
(6
7,0)
19
,2ab
(5
8,6)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,29
8 0,
2779
0,
1955
0,
3167
0,
3125
0,
3066
0,
2507
0,
2355
0,
2244
0,
2705
0,
2672
0,
2427
C
V (%
) 5,
12
4,77
3,
32
5,61
5,
70
5,44
4,
44
4,29
4,
31
4,96
4,
78
4,70
T
rans
form
ação
Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) Lo
g (y
+1)
Log
(y+1
) *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

119
Tab
ela
43. T
eor d
e m
agné
sio
(mm
olc d
m-3
) pre
sent
e na
aná
lise
do su
bstra
to d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de
nutri
ente
s.
Col
etas
T
rata
men
tos
T7,5
h T2
2h
T70h
T1
3d
T35d
T4
5d
T61d
T7
9d
T97d
T1
07d
T115
d T1
24d
Com
plet
a 48
,0 a
(48,
0)
45,0
a (4
5,0)
52
,6ab
(5
2,6)
24
,2 a
(48,
7)
35,0
a (3
5,0)
23
,8ab
(4
6,6)
22
,5 a
(40,
6)
31,6
ab
(90,
0)
24,8
cd
(52,
0)
25,5
a (5
5,3)
26
,1 a
(58,
3)
23,0
ab
(43,
0)
Om
issã
o de
N
50
,6 a
(50,
6)
54,0
a (5
4,0)
51
,3ab
(5
1,3)
24
,7 a
(51,
7)
36,6
a (3
6,6)
25
,7 a
(56,
3)
22,3
a (4
0,0)
32
,5a
(96,
0)
26,4
ab
(60,
0)
26,7
a (6
1,3)
26
,8 a
(62,
3)
22,7
ab
(41,
6)
Om
issã
o de
P
51,6
a (5
1,6)
55
,3 a
(55,
3)
47,0
ab
(47,
0)
24,0
a (4
8,0)
33
,6 a
(33,
6)
23,6
ab
(46,
0)
22,3
a (4
0,0)
29
,6ab
(7
8,3)
25
,6ab
c (5
6,0)
25
,4 a
(55,
0)
25,5
a (5
5,3)
22
,7ab
(4
1,6)
O
mis
são
de
K
52,0
a (5
2,0)
51
,6 a
(51,
7)
44,6
ab
(44,
6)
23,9
a (4
7,7)
36
,6 a
(36,
6)
23,6
ab
(46,
0)
22,2
a (3
9,6)
27
,3ab
(6
5,0)
22
,8e
(42,
3)
24,8
ab
(52,
0)
25,3
a (5
4,3)
22
,2ab
(3
9,6)
O
mis
são
de
Ca
47,6
a (4
7,6)
50
,6 a
(50,
7)
54,6
a (5
4,6)
24
,0 a
(48,
0)
35,0
a (3
5,0)
25
,2ab
(5
4,0)
22
,6 a
(41,
3)
28,7
ab
(73,
3)
27,0
a (6
3,3)
26
,1 a
(58,
6)
26,0
a (5
8,0)
24
,0a
(48,
0)
Om
issã
o de
M
g 48
,0 a
(48,
0)
48,6
a (4
8,7)
47
,6ab
(4
7,6)
26
,2 a
(59,
0)
35,0
a (3
5,0)
23
,3b
(44,
6)
22,0
a (3
8,6)
26
,8b
(62,
6)
23,7
de
(46,
6)
23,1
b (4
3,6)
24
,7 a
(51,
3)
19,7
b (2
9,6)
O
mis
são
de
B
49,0
a (4
9,0)
50
,6 a
(50,
7)
49,0
ab
(49,
0)
26,5
a (6
0,3)
36
,3 a
(36,
3)
24,1
ab
(48,
3)
22,2
a (3
9,6)
27
,9ab
(6
8,3)
24
,9cd
(5
2,3)
25
,2ab
(5
3,6)
25
,4 a
(55,
0)
23,4
ab
(45,
0)
Om
issã
o de
Fe
56
,0 a
(56,
0)
50,0
a (5
0,0)
44
,3b
(44,
3)
24,0
a (4
7,7)
36
,6 a
(36,
6)
24,6
ab
(50,
6)
22,1
a (3
9,0)
28
,5ab
(7
1,6)
25
,1bc
d (5
3,3)
26
,1 a
(58,
3)
25,1
a (5
8,3)
22
,9ab
(4
2,6)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,86
55
1,11
1,
00
0,34
03
0,58
84
0,21
83
0,10
1 0,
5257
0,
1404
0,
2354
0,
2176
0,
3697
C
V (%
) 6,
07
7,77
7,
29
4,86
5,
84
3,17
1,
60
6,37
1,
97
3,27
2,
99
5,78
T
rans
form
ação
-
- -
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
)R
Q(y
+1)
RQ
(y+1
)*
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

120
Tab
ela
44. T
eor d
e fe
rro
(mg
dm-3
) pre
sent
e na
aná
lise
do su
bstra
to d
a m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
Tra
tam
ento
s T7
,5h
T22h
T7
0h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
11,5
4a
(132
,3)
11,9
9a
(143
,0)
12,2
6a
(149
,3)
4,64
a (1
01,6
) 4,
48ab
(8
6,8)
4,
80bc
(1
20,0
) 4,
94a
(144
,1)
5,39
b (2
17,6
) 5,
32bc
(2
03,0
) 5,
56c
(258
,0)
5,37
a (2
15,6
) 5,
53a
(253
,0)
Om
issã
o de
N
11,7
8a
(138
,0)
11,9
4a
(142
,0)
10,5
7c
(111
,0)
4,65
a (1
03,7
) 4,
53ab
(9
2,0)
5,
19a
(180
,8)
4,97
a (1
45,5
) 5,
50ab
(2
43,0
) 5,
40ab
c (2
24,1
) 5,
69ab
c (2
96,6
) 5,
52a
(250
,0)
5,39
ab
(219
,0)
Om
issã
o de
P
11,2
8a
(126
,3)
11,4
3a
(129
,8)
11,5
9ab
(133
,5)
4,69
a (1
07,3
) 4,
52ab
(9
0,0)
5,
06ab
(1
56,1
) 4,
84a
(124
,6)
5,49
ab
(241
,0)
5,41
abc
(223
,6)
5,78
ab
(323
,0)
5,41
a (2
22,8
) 5,
40ab
(2
22,3
) O
mis
são
de K
9,
89b
(97,
0)
11,5
0a
(131
,6)
11,5
5ab
(132
,5)
4,10
a (8
1,6)
4,
60a
(98,
3)
5,06
ab
(156
,1)
4,83
a (1
23,6
) 5,
45ab
(2
31,3
) 5,
44ab
c (2
30,6
) 5,
65ab
c (2
83,6
) 5,
34a
(206
,6)
5,44
a (2
29,3
) O
mis
são
de
Ca
10,1
3b
(101
,8)
12,3
6a
(151
,8)
10,7
4bc
(114
,6)
4,18
a (1
06,5
) 4,
62a
(100
,7)
4,98
ab
(144
,6)
5,13
a (1
72,0
) 5,
56ab
(2
59,3
) 5,
67a
(291
,0)
5,83
a (3
42,6
) 5,
54a
(254
,1)
5,53
a (2
50,6
) O
mis
são
de
Mg
10,1
2b
(101
,6)
11,5
9a
(133
,5)
11,6
6a
(135
,1)
4,72
a (1
10,6
) 4,
51ab
(9
0,0)
5,
10a
(163
,8)
5,11
a (1
68,3
) 5,
67a
(292
,6)
5,55
ab
(257
,0)
5,71
abc
(301
,3)
5,48
a (2
38,6
) 5,
59a
(268
,6)
Om
issã
o de
B
10,4
1b
(107
,5)
11,4
9a
(131
,1)
11,9
6a
(142
,1)
4,74
a (1
13,6
) 4,
42ab
(8
2,2)
4,
97ab
c (1
42,6
) 4,
93a
(137
,5)
5,67
a (2
90,0
) 5,
47ab
c (2
37,3
) 5,
61bc
(2
73,3
) 5,
43a
(228
,3)
5,46
a (2
35,3
) O
mis
são
de F
e 10
,07b
(1
00,5
) 11
,77a
(1
37,6
) 12
,37a
(1
52,1
) 4,
67a
(105
,5)
4,27
b (6
9,6)
4,
73c
(111
,5)
4,78
a (1
18,3
) 4,
91c
(135
,0)
5,23
c (1
86,0
) 4,
79d
(118
,5)
4,76
b (1
15,3
) 5,
20b
(179
,3)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
7813
1,
1585
0,
8499
1,
1643
0,
2892
0,
3012
0,
5575
0,
246
0,27
63
0,22
28
0,22
46
0,22
98
CV
(%)
2,59
3,
48
2,59
8,
92
5,23
2,
13
3,98
1,
59
1,79
1,
41
1,48
1,
49
Tra
nsfo
rmaç
ão
RQ
(y+1
) R
Q(y
+1)
RQ
(y+1
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
Log
(y+2
) Lo
g (y
+2)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

121
Tab
ela
45. T
eor d
e bo
ro (m
g dm
-3) p
rese
nte
na a
nális
e do
subs
trato
da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T7
,5h
T22h
T7
0h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
1,09
a (0
,20)
1,
11b
(0,2
3)
1,09
ab
(0,2
0)
1,06
a (0
,13)
1,
09ab
(0
,20)
1,
15a
(0,3
3)
1,11
b (0
,23)
1,
14ab
(0
,30)
1,
07a
(0,1
6)
1,16
a (0
,36)
1,
21a
(0,4
6)
1,14
a (0
,30)
O
mis
são
de
N
1,12
a (0
,26)
1,
09b
(0,2
0)
1,11
ab
(0,2
3)
1,07
a (0
,17)
1,
04c
(0,1
0)
1,18
a (0
,40)
1,
09b
(0,2
0)
1,14
ab
(0,3
0)
1,07
a (0
,16)
1,
15ab
(0
,33)
1,
16a
(0,3
6)
1,14
a (0
,30)
O
mis
são
de
P 1,
09a
(0,2
0)
1,09
b (0
,17)
1,
09ab
(0
,20)
1,
06a
(0,1
3)
1,11
ab
(0,2
3)
1,14
a (0
,30)
1,
09b
(0,2
0)
1,16
a (0
,36)
1,
04a
(0,1
0)
1,16
a (0
,36)
1,
09b
(0,2
0)
1,11
a (0
,23)
O
mis
são
de
K
1,07
a (0
,16)
1,
09b
(0,1
7)
1,09
ab
(0,2
0)
1,07
a (0
,17)
1,
09ab
(0
,20)
1,
12a
(0,2
6)
1,11
b (0
,23)
1,
11bc
(0
,23)
1,
06a
(0,1
3)
1,16
a (0
,36)
1,
04b
(0,1
0)
1,09
a (0
,20)
O
mis
são
de
Ca
1,11
a (0
,23)
1,
06b
(0,1
3)
1,07
b (0
,16)
1,
07a
(0,1
7)
1,09
ab
(0,2
0)
1,12
a (0
,26)
1,
33a
(0,8
0)
1,12
ab
(0,2
6)
1,09
a (0
,20)
1,
16a
(0,3
6)
1,07
b (0
,16)
1,
14a
(0,3
0)
Om
issã
o de
M
g 1,
07a
(0,1
6)
1,09
b (0
,20)
1,
14a
(0,3
0)
1,06
a (0
,13)
1,
07bc
(0
,16)
1,
16a
(0,3
6)
1,09
b (0
,20)
1,
14ab
(0
,30)
1,
04a
(0,1
0)
1,12
ab
(0,2
6)
1,04
b (0
,16)
1,
15a
(0,3
3)
Om
issã
o de
B
1,
07a
(0,1
6)
1,18
a (0
,40)
1,
09ab
(0
,20)
1,
09a
(020
) 1,
09ab
(0
,20)
1,
18a
(0,4
0)
1,11
b (0
,23)
1,
06c
(0,1
3)
1,06
a (0
,13)
1,
09b
(0,2
0)
1,04
b (0
,10)
1,
09a
(0,2
0)
Om
issã
o de
Fe
1,
12a
(0,2
6)
1,09
b (0
,20)
1,
07b
(0,1
6)
1,04
a (0
,10)
1,
12a
(0,2
6)
1,09
a (0
,20)
1,
14b
(0,3
0)
1,14
ab
(0,3
0)
1,06
a (0
,13)
1,
12ab
(0
,26)
1,
06b
(0,1
3)
1,12
a (0
,26)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
89
DM
S (0
,05)
0,
0646
0,
0625
0,
046
0,08
0,
0453
0,
1091
0,
147
0,05
17
0,08
79
0,08
79
0,07
38
0,06
35
CV
(%)
2,07
2,
00
1,48
2,
63
1,46
3,
31
4,57
1,
62
2,91
2,
04
2,37
1,
99
Tra
nsfo
rmaç
ão
Rq(
y+1)
R
q(y+
1)
Rq(
y+1)
Rq(
y+1)
Rq(
y+1)
Rq(
y+1)
Rq(
y+1)
R
q(y+
1)R
q(y+
1)R
q(y+
1)R
q(y+
1)R
q(y+
1)*
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

122
Tab
ela
46.V
alor
es d
e pH
pre
sent
e na
aná
lise
da so
luçã
o da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T22h
T1
3d
T35d
T4
5d
T61d
T7
9d
T97d
T1
07d
T115
d T1
24d
Com
plet
a 5,
73b
(5,7
3)
6,00
b (6
,00)
4,
30e
(4,3
0)
4,26
bc
(4,2
6)
3,93
bc
(3,9
3)
4,00
c (4
,00)
2,
65ab
(4
,07)
3,
90c
(3,9
0)
4,33
b (4
,33)
4,
86ab
(4
,86)
5,
30b
(5,3
0)
Om
issã
o de
N
4,
10d
(4,1
0)
4,63
d (4
,63)
5,
26c
(5,2
6)
5,60
a (5
,60)
5,
50a
(5,5
0)
5,30
a (5
,30)
2,
80a
(4,8
7)
5,83
a (5
,83)
5,
66a
(5,6
6)
5,00
a (5
,00)
5,
30b
(5,3
0)
Om
issã
o de
P
5,33
c (5
,33)
5,
90bc
(5
,90)
5,
06d
(5,0
6)
5,46
a (5
,46)
3,
93bc
(3
,93)
4,
13b
(4,1
3)
2,71
ab
(4,4
0)
3,90
c (3
,90)
4,
00c
(4,0
0)
4,16
b (4
,13)
4,
96cd
(4
,96)
O
mis
são
de
K
3,56
e (3
,56)
3,
43e
(3,4
3)
3,46
g (3
,46)
3,
26d
(3,2
6)
3,00
d (3
,00)
5,
26a
(5,2
6)
2,65
ab
(4,0
3)
4,16
b (4
,16)
4,
03c
(4,0
3)
4,60
ab
(4,6
0)
5,93
a (5
,93)
O
mis
são
de
Ca
6,30
a (6
,30)
6,
46a
(6,4
6)
5,40
bc
(5,4
0)
5,66
a (5
,66)
4,
10b
(4,1
0)
3,50
f (3
,50)
2,
72ab
(4
,43)
4,
00bc
(4
,00)
3,
43e
(3,4
3)
3,00
c (3
,00)
3,
40e
(3,4
0)
Om
issã
o de
M
g 4,
13d
(4,1
3)
4,56
d (4
,56)
4,
06f
(4,0
6)
2,90
c (2
,90)
2,
93d
(2,9
3)
3,40
g (3
,40)
2,
63ab
(3
,97)
3,
36e
(3,3
6)
3,40
e (3
,40)
2,
73c
(2,7
3)
3,03
f (3
,03)
O
mis
são
de B
5,
90b
(5,9
0)
5,96
b (5
,96)
5,
50b
(5,5
0)
4,03
c (4
,03)
3,
86c
(3,8
6)
3,63
e (3
,63)
2,
56b
(3,5
7)
3,56
d (3
,56)
4,
10c
(4,1
0)
4,93
ab
(4,9
3)
5,20
bc
(5,2
0)
Om
issã
o de
Fe
5,
73b
(5,7
3)
5,76
(5
,76)
6,
13a
(6,1
3)
4,46
b (4
,46)
3,
90d
(3,9
0)
3,90
d (3
,90)
2,
65ab
(4
,07)
3,
66e
(3,6
6)
3,76
d (3
,76)
4,
26ab
(4
,26)
4,
86d
(4,8
6)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
0,
2379
0,
1999
0,
1914
0,
2998
0,
1825
0,
0999
0,
2219
0,
1914
0,
2159
0,
0820
0,
2706
C
V (%
) 1,
65
1,32
1,
38
2,37
1,
65
0,85
2,
93
1,67
1,
86
6,83
2,
01
Tra
nsfo
rmaç
ão
- -
- -
- -
RQ
(y +
3)
- -
- -
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

123
Tab
ela
47. T
eor d
e N
H+ 4 (
mg
L-1) p
rese
nte
na a
nális
e da
solu
ção
da m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
Col
etas
T
rata
men
tos
T02h
T2
2h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
4,04
abc
(56,
0)
3,33
c (2
7,50
) 3,
18ab
(1
8,0)
2,
48a
(10,
8)
3,28
d (2
5,6)
2,
52bc
(1
1,5)
2,
45c
(8,6
) 1,
49de
(1
,97)
3,
16c
(0,0
) 3,
17a
(0,0
9)
3,17
b (0
,09)
O
mis
são
de N
2,
49d
(11,
5)
1,11
d (2
,33)
1,
79c
(3,6
7)
0,59
b (1
,07)
0,
09e
(0,1
0)
0,18
e (0
,17)
1,
19e
(0,3
3)
1,32
de
(0,7
5)
3,17
c (0
,1)
3,17
a (0
,09)
3,
20ab
(0
,30)
O
mis
são
de P
3,
89bc
(4
7,6)
3,
83ab
c (4
5,6)
3,
47ab
(2
9,3)
2,
68a
(14,
17)
3,45
c (3
0,7)
2,
85ab
(1
6,4)
3,
42b
(28,
3)
4,23
b (2
2,3)
3,
16c
(0,0
) 3,
17a
(0,0
9)
3,18
ab
(0,1
3)
Om
issã
o de
K
3,68
c (3
9,1)
3,
67
(38,
3)
3,08
b (2
0,0)
2,
77a
(14,
8)
3,38
cd
(28,
5)
2,30
c (9
,0)
2,61
c (1
0,6)
2,
26c
(6,6
) 4,
53b
(10,
6)
3,23
a (0
,47)
3,
21a
(0,3
6)
Om
issã
o de
Ca
4,53
a (9
2,3)
4,
19ab
(6
5,3)
3,
86a
(44,
8)
3,04
a (1
9,5)
3,
93a
(50,
0)
3,16
a (2
2,6)
4,
10a
(57,
6)
4,02
a (5
3,0)
4,
56b
(10,
8)
3,17
a (0
,09)
3,
17b
(0,0
9)
Om
issã
o de
M
g 4,
35ab
(7
6,8)
4,
27a
(70,
8)
3,91
a (4
7,3)
3,
10a
(21,
0)
3,65
b (3
7,6)
1,
49d
(3,5
) 1,
09e
(0,0
) 2,
86b
(14,
5)
7,10
a (4
0,5)
3,
22a
(0,4
0)
3,17
b (0
,09)
O
mis
são
de B
4,
04ab
c (5
6,5)
3,
61ab
c (3
6,1)
3,
39ab
(2
7,0)
2,
61a
(12,
5)
3,63
b (3
6,8)
2,
54bc
(1
2,2)
1,
84d
(3,3
) 1,
09e
(0,0
) 3,
20c
(0,2
6)
3,17
a (0
,09)
3,
17b
(0,0
9)
Om
issã
o de
Fe
3,68
c (3
9,0)
3,
44bc
(3
0,5)
3,
57ab
(3
2,8)
2,
81a
(15,
5)
3,36
cd
(28,
0)
2,81
a (1
5,7)
1,
09e
(0,0
) 1,
85cd
(3
,42)
3,
17c
(0,1
) 3,
17a
(0,0
9)
3,17
b (0
,09)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
D
MS
(0,0
5)
0,53
31
0,80
24
0,73
88
0,66
2 0,
1305
0,
4381
0,
3012
0,
5917
0,
2156
0,
134
0,04
1 C
V (%
) 4,
90
8,25
7,
95
9,31
1,
48
6,94
4,
77
9,22
1,
90
1,48
0,
45
Tra
nsfo
rmaç
ão
Log
(y +
1)
Log
(y +
1)
Log
(y +
3)
Log
(y +
1)
Log
(y +
1)
Log
(y +
1)
Log
(y +
3)
Log
(y +
3)
RQ
(y+1
0)
RQ
(y+1
0)
RQ
(y+1
0)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

124
Tab
ela
48. T
eor d
e N
O- 3 (
mg
L-1) p
rese
nte
na a
nális
e da
solu
ção
da m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
Col
etas
T
rata
men
tos
T02h
T2
2h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
236,
33e
(236
,30)
19
9,00
d (1
99,0
0)
5,53
b (2
50,5
0)
250,
33c
(250
,13)
30
2,60
c (3
02,6
0)
274,
33d
(274
,23)
38
0,00
b (3
80,1
0)
521,
33 a
(521
,30)
6,
16ab
(4
74,1
0)
5,78
d (3
23,1
3)
5,66
b (2
87,3
3)
Om
issã
o de
N
14,0
0g
(13,
70)
6,00
f (6
,07)
8,
68a
(6,0
0)
9,67
f (9
,63)
8,
26g
(8,2
7)
5,00
f (4
,97)
8,
33f
(8,5
0)
8,33
e (8
,30)
2,
36e
(9,6
0)
2,07
f (6
,66)
2,
32e
(9,3
0)
Om
issã
o de
P
245,
33e
(245
,33)
19
0,00
d (1
89,9
0)
5,28
c (1
93,7
0)
231,
33d
(231
,13)
29
2,60
c (2
92,2
0)
291,
33b
(291
,17)
37
7,33
b (3
77,1
3)
485,
67 a
(485
,60)
6,
17ab
(4
77,1
7)
6,11
b (4
53,1
6)
5,88
a (3
57,0
0)
Om
issã
o de
K
369,
33a
(369
,37)
35
1,67
a (3
51,5
3)
5,57
b (2
59,5
3)
268,
67b
(268
,50)
37
8,26
a (3
78,2
7)
53,0
0e
(53,
07)
204,
67c
(204
,33)
15
7,33
d (1
57,3
0)
5,12
d (1
66,4
) 4,
48e
(87,
90)
4,13
d (6
0,90
) O
mis
são
de C
a 11
5,67
f (1
15,7
7)
85,3
3e
(85,
,47)
4,
64d
(100
,69)
12
4,33
e (1
24,3
0)
175,
63f
(175
,63)
28
0,33
c (2
80,3
0)
164,
33d
(164
,33)
19
6,00
d (1
96,0
3)
5,87
c (3
38,7
3)
5,87
c (3
53,6
0)
5,25
c (1
90,0
3)
Om
issã
o de
M
g 29
9,67
b (2
99,5
7)
280,
33b
(280
,33)
5,
38bc
(2
15,1
0)
243,
67c
(243
,60)
27
9,27
e (2
79,2
7)
276,
00cd
(2
76,0
7)
165,
33d
(165
,27)
32
4,00
bc
(324
,10)
5,
82c
(338
,73)
5,
78d
(323
,30)
5,
62b
(277
,07)
O
mis
são
de B
26
8,33
d (2
68,1
7)
256,
67c
(256
,67)
5,
44bc
(2
27,9
3)
278,
33 a
(278
,47)
33
0,00
b (3
30,6
0)
325,
00 a
(324
,90)
44
3,00
a (4
43,1
0)
352,
67b
(352
,87)
6,
05b
(426
,97)
5,
81d
(334
,66)
5,
59b
(268
,33)
O
mis
são
de F
e 28
2,00
c (2
81,7
7)
267,
67bc
(2
67,6
7)
5,35
bc
(208
,23)
27
6,67
a (2
76,6
0)
305,
00c
(305
,60)
29
5,00
b (2
95,4
7)
141,
00e
(141
,00)
28
1,00
c (2
80,7
0)
6,28
a (5
34,7
7)
6,16
a (4
75,2
3)
5,88
a (3
59,8
3)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
9,
7412
14
,333
0,
2229
7,
4567
5,
2831
5,
683
4,43
22
69,2
62
0,15
05
0,03
56
0,14
93
CV
(%)
1,50
2,
47
1,37
1,
25
0,72
0,
89
0,66
8,
42
1,37
1,
62
1,46
T
rans
form
ação
-
- Lo
g (y
+ 3
)-
- -
- -
Log
(y +
1)
Log
(y +
1)
Log
(y +
1)
* M
édia
s seg
uida
s das
mes
mas
letra
s em
cad
a co
luna
não
dife
rem
ent
re si
pel
o te
ste
de T
ukey
a 5
% d
e pr
obab
ilida
de.
Val
ores
ent
re p
arên
tese
s ref
eren
tes à
s méd
ias s
em tr
ansf
orm
ação
.

125
Tab
ela
49. T
eor d
e fó
sfor
o (m
g L-1
) pre
sent
e na
aná
lise
da so
luçã
o da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T22h
T1
3d
T35d
T4
5d
T61d
T7
9d
T97d
T1
07d
T115
d T1
24d
Com
plet
a 40
,33d
(4
0,33
) 42
,67b
(4
2,67
) 55
,67a
(5
5,67
) 49
,67b
(4
9,67
) 57
,00a
(5
7,00
) 59
,66b
(5
9,67
) 75
,33a
(7
5,33
) 62
,00c
(6
2,00
) 50
,33e
(5
0,33
) 59
,00c
(5
9,00
) 44
,33c
(4
4,33
) O
mis
são
de
N
50,0
0a
(50,
00)
56,0
0a
(56,
00)
53,0
0b
(53,
00)
52,3
3b
(52,
33)
52,3
3b
(52,
33)
63,6
7a
(63,
67)
72,0
0b
(72,
00)
78,6
7b
(78,
67)
69,0
0a
(69,
00)
77,3
3b
(77,
33)
64,3
3b
(64,
33)
Om
issã
o de
P
7,67
e (7
,67)
8,
00d
(8,0
0)
25,3
3c
(25,
33)
18,6
7f
(18,
67)
21,3
3e
(21,
33)
31,6
7e
(31,
47)
41,3
3d
(41,
33)
57,0
0d
(57,
00)
30,0
0f
(30,
00)
34,3
3e
(34,
33)
27,0
0e
(27,
00)
Om
issã
o de
K
45
,33b
c (4
5,33
) 43
,00b
(4
3,00
) 27
,67c
(2
7,67
) 26
,00e
(2
6,00
) 47
,33c
(4
7,33
) 25
,33d
(2
5,33
) 39
,67e
(3
9,67
) 30
,00f
(3
0,00
) 30
,67f
(3
0,66
) 16
,33f
(1
6,33
) 15
,66f
(1
5,67
) O
mis
são
de
Ca
49,0
0ab
(49,
00)
42,3
3b
(42,
33)
54,3
3ab
(54,
33)
36,3
3d
(36,
33)
56,6
7a
(56,
67)
38,6
6d
(38,
67)
64,3
3c
(64,
33)
92,3
3a
(92,
33)
59,6
6c
(59,
66)
73,3
3b
(73,
33)
37,3
3d
(37,
33)
Om
issã
o de
M
g 44
,67c
(4
4,67
) 43
,33b
(4
3,33
) 56
,00a
(5
6,00
) 44
,33c
(4
4,33
) 34
,33d
(3
4,33
) 18
,33g
(1
8,33
) 14
,33g
(1
4,33
) 47
,33e
(4
7,33
) 53
,67d
(5
3,67
) 18
,33f
(1
8,33
) 2,
00g
(2,0
0)
Om
issã
o de
B
44
,00c
d (4
4,00
) 40
,00d
(4
0,00
) 56
,67a
(5
6,67
) 51
,33b
(5
1,33
) 58
,33a
(5
8,33
) 57
,00c
(5
7,00
) 72
,67b
(7
2,67
) 81
,33b
(8
1,33
) 63
,00b
(6
3,00
) 54
,33d
(5
4,33
) 45
,33c
(4
5,33
) O
mis
são
de
Fe
42,6
7cd
(42,
67)
43,3
3b
(43,
33)
56,6
7a
(56,
67)
57,3
3a
(57,
33)
58,0
0a
(57,
33)
61,3
3b
(61,
33)
18,6
7f
(18,
67)
80,0
0b
(80,
00)
62,3
3b
(62,
33)
88,0
0a
(88,
00)
68,3
3a
(68,
33)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
3,
6947
2,
3081
2,
44
2,88
51
2,82
68
2,08
05
1,52
67
2,88
51
2,15
9 4,
3564
2,
2348
C
V (%
) 3,
23
2,04
1,
79
2,43
2,
07
1,65
1,
08
1,54
1,
45
2,92
2,
07
Tra
nsfo
rmaç
ão
- -
- -
- -
- -
- -
- *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

126
Tab
ela
50. T
eor d
e po
táss
io (m
g L-1
) pre
sent
e na
aná
lise
da so
luçã
o da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T22h
T1
3d
T35d
T4
5d
T61d
T7
9d
T97d
T1
07d
T115
d T1
24d
Com
plet
a 15
0,67
c (1
50,6
7)
149,
33d
(149
,33)
12
0,67
ab
(120
,67)
12
0,33
a (1
20,3
3)
122,
00bc
(1
22,0
0)
143,
00ab
(1
43,0
0)
158,
67b
(158
,67)
17
7,33
b (1
77,3
3)
147,
33b
(147
,33)
11
8,67
cd
(118
,66)
8,
67b
(98,
33)
Om
issã
o de
N
15
8,00
bc
(158
,00)
15
5,00
cd
(155
,00)
11
2,33
c (1
12,3
) 11
0,67
a (1
10,6
7)
110,
67c
(110
,67)
15
2,33
a (1
52,3
3)
156,
67b
(156
,67)
11
8,33
de
(118
,33)
13
4,00
c (1
34,0
0)
127,
33bc
(1
27,3
3)
12,3
3a
(116
,67)
O
mis
são
de
P 16
1,33
abc
(161
,33)
17
0,00
ab
(170
,00)
12
3,67
a (1
23,6
7)
119,
33a
(119
,33)
12
2,33
bc
(122
,33)
15
2,33
a (1
52,3
3)
181,
00a
(181
,00)
20
2,33
(2
02,3
3)
148,
33b
(148
,33)
15
3,33
a (1
53,3
3)
12,0
0a
(132
,00)
O
mis
são
de
K
3,00
d (3
,00)
3,
00e
(3,0
0)
2,00
d (2
,00)
14
,67c
(1
4,67
) 2,
00d
(2,0
0)
45,0
0d
(45,
00)
4,00
e (4
,00)
2,
33g
(2,3
3)
4,00
d (4
,00)
2,
00f
(2,0
0)
5,00
d (1
,00)
O
mis
são
de
Ca
162,
00ab
c (1
62,0
0)
165,
67b
(165
,67)
12
0,00
ab
(120
,00)
73
,67b
(7
3,67
) 11
9,00
c (1
19,0
0)
8,33
e (8
,33)
11
3,33
c (1
13,3
3)
137,
00c
(137
,00)
13
3,33
c (1
33,3
3)
131,
66b
(131
,66)
7,
67bc
(7
8,33
) O
mis
são
de
Mg
156,
00c
(156
,00)
16
1,67
bc
(161
,67)
11
4,00
bc
(114
,00)
11
1,67
a (1
11,6
7)
118,
33c
(118
,33)
12
5,00
c (1
25,0
0)
58,6
7d
(58,
67)
52,3
3f
(52,
33)
128,
33c
(128
,33)
10
3,33
e (1
03,3
3)
11,3
3a
(100
,33)
O
mis
são
de
B
171,
67a
(171
,67)
16
8,00
ab
(168
,00)
12
4,33
a (1
24,3
3)
115,
00a
(115
,00)
14
2,00
a (1
42,0
0)
137,
00bc
(1
37,0
0)
159,
33b
(159
,33)
12
7,67
cd
(127
,67)
13
2,67
c (1
32,6
7)
111,
67de
(1
11,6
7)
8,00
b (8
8,00
) O
mis
são
de
Fe
169,
67ab
(1
69,6
7)
178,
33a
(178
,33)
12
5,33
a (1
25,3
3)
118,
67a
(118
,67)
13
2,67
ab
(132
,67)
14
8,33
ab
(148
,33)
49
,67d
(4
9,67
) 11
5,00
e (1
15,0
0)
163,
37a
(163
,33)
15
8,33
a
(158
,33)
6,
33cd
(1
19,6
7)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
12
,576
10
,64
7,45
67
15,1
02
12,3
76
15,1
68
15,7
71
10,7
18
11,7
41
11,5
4 1,
6321
C
V (%
) 3,
14
2,61
2,
50
5,45
4,
03
4,71
5,
06
3,25
3,
35
3,60
3,
23
Tra
nsfo
rmaç
ão
- -
- -
- -
- -
- -
- *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

127
Tab
ela
51. T
eor d
e cá
lcio
(mg
L-1) p
rese
nte
na a
nális
e da
solu
ção
da m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
Tra
tam
ento
s T0
2h
T22h
T1
3d
T35d
T4
5d
T61d
T7
9d
T97d
T1
07d
T115
d T1
24d
Com
plet
a 48
,33b
c (4
8,33
) 52
,00c
(5
2,00
) 41
,67a
(4
1,67
) 6,
32a
(37,
33)
7,13
a (4
8,00
) 48
,33a
b (4
8,33
) 54
,33a
(5
4,33
) 61
,00b
(6
1,00
) 8,
46a
(67,
66)
43,0
0d
(43,
00)
12,3
3c
(12,
33)
Om
issã
o de
N
44,6
7c
(44,
67)
50,0
0c
(50,
00)
32,6
7c
(32,
67)
5,38
b (2
6,00
) 7,
07a
(47,
33)
41,0
0b
(41,
00)
43,0
0b
(43,
00)
16,6
6e
(16,
67)
7,57
a (5
3,33
) 41
,00d
(4
1,00
) 13
,00c
(1
3,00
) O
mis
são
de P
55
,33a
b (5
5,33
) 58
,33a
b (5
8,33
) 36
,33b
(3
6,33
) 6,
10a
(34,
33)
7,39
a (5
1,67
) 49
,67a
(4
9,67
) 58
,67a
(5
8,67
) 78
,67a
(7
8,67
) 8,
30
(65,
00)
55,3
3b
(55,
33)
25,0
0ab
(25,
00)
Om
issã
o de
K
50,0
0bc
(50,
00)
56,3
3b
(56,
33)
43,0
0a
(43,
00)
6,75
a (4
2,67
) 8,
01a
(61,
33)
12,0
0c
(12,
00)
21,6
8d
(21,
67)
38,3
3d
(38,
33)
8,07
a (6
3,66
) 20
,00e
(2
0,00
) 1,
00e
(1,0
0)
Om
issã
o de
Ca
7,00
d (7
,00)
6,
33d
(6,3
3)
4,33
d (4
,33)
2,
82c
(5,0
0)
3,58
b (1
0,00
) 42
,33a
b (4
2,33
) 4,
67e
(4,6
7)
6,67
f (6
,67)
5,
06b
(21,
66)
12,3
3f
(12,
33)
2,00
e (2
,00)
O
mis
são
de
Mg
51,3
3abc
(5
1,33
) 60
,67a
(6
0,67
) 42
,33a
(4
2,33
) 6,
73a
(42,
33)
7,57
a (5
4,67
) 44
,00a
b (4
4,00
) 16
,67d
(1
6,67
) 51
,33c
(5
1,33
) 8,
19a
(63,
33)
50,3
3c
(50,
33)
22,3
3b
(22,
33)
Om
issã
o de
B
57,6
7a
(57,
67)
56,3
3b
(56,
33)
39,6
7ab
(39,
67)
6,73
a (4
2,33
) 7,
37a
(51,
33)
46,6
7ab
(46,
67)
54,0
0a
(54,
00)
52,3
3c
(52,
33)
7,76
a (5
6,33
) 44
,33d
(4
4,33
) 7,
67d
(7,6
7)
Om
issã
o de
Fe
58,3
3a
(58,
33)
60,3
3a
(60,
33)
42,6
7a
(42,
67)
6,77
a (4
3,00
) 7,
10a
(48,
67)
48,0
0ab
(48,
00)
30,0
0c
(30,
00)
48,0
0c
(48,
00)
8,52
a 68
,66)
62
,67a
(6
2,66
) 27
,66a
(2
7,67
)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
7,20
7 3,
1605
3,
6494
0,
8863
1,
0972
7,
4344
7,
72
7,61
2,
00
3,69
47
3,0-
533
CV
(%)
5,47
2,
23
3,65
5,
26
5,61
6,
33
7,72
6,
10
9,60
3,
17
7,78
T
rans
form
ação
-
- -
RQ
(y +
3)
RQ
(y +
3)
- -
- R
Q (y
+ 4
)-
- *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

128
Tab
ela
52. T
eor d
e m
agné
sio
(mg
L-1) p
rese
nte
na a
nális
e da
solu
ção
da m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
Col
etas
T
rata
men
tos
T02h
T2
2h
T13d
T3
5d
T45d
T6
1d
T79d
T9
7d
T107
d T1
15d
T124
d C
ompl
eta
40,6
7b
(40,
67)
40,3
3d
(40,
33)
37,0
a (3
7,00
) 5,
67a
(29,
33)
36,3
3b
(36,
33)
39,6
7bc
(39,
67)
42,3
3b
(42,
33)
40,6
7b
(40,
67)
38,6
7a
(38,
66)
24,3
3c
(24,
33)
20,6
7d
(20,
67)
Om
issã
o de
N
41
,00b
(4
1,00
) 42
,67c
(4
2,67
) 32
,33c
(3
2,33
) 5,
32ab
(2
5,33
) 38
,67a
b (3
8,67
) 37
,00c
d (3
7,00
) 39
,67b
(3
9,67
) 29
,00e
(2
9,00
) 38
,00a
(3
8,00
) 27
,67b
(2
7,67
) 26
,00b
(2
6,00
) O
mis
são
de P
43
,33a
b (4
3,33
) 43
,67b
c (4
3,67
) 37
,0a
(37,
00)
5,53
a (2
7,67
) 38
,33a
b (3
8,33
) 42
,66b
(4
2,67
) 47
,67a
(4
7,67
) 52
,33a
(5
2,33
) 40
,00a
(4
0,00
) 31
,33a
(3
1,33
) 28
,67a
(2
8,67
) O
mis
são
de
K
40,6
7b
(40,
67)
42,3
3cd
(42,
33)
35,0
0ab
(35,
00)
5,80
a (3
0,67
) 44
,33a
(4
4,33
) 21
5,67
a (2
15,6
7)
24,0
0d
(24,
00)
24,3
3f
(24,
33)
31,0
0b
(31,
00)
9,33
d (9
,33)
6,
33g
(6,3
3)
Om
issã
o de
C
a 42
,00a
b (4
2,00
) 43
,00c
(4
3,00
) 35
,00a
b (3
5,00
) 4,
76b
(19,
67)
35,0
0b
(35,
00)
33,6
7cd
(33,
67)
33,3
3c
(33,
33)
33,3
3cd
(33,
33)
35,6
7ab
(35,
67)
27,3
3b
(27,
33)
17,0
0f
(17,
00)
Om
issã
o de
M
g 3,
67c
(3,6
7)
4,67
e (4
,67)
5,
33d
(5,3
3)
2,81
c (5
,00)
5,
33c
(5,3
3)
4,00
e (4
,00)
3,
33f
(3,3
3)
5,00
g (5
,00)
18
,00c
(1
8,00
) 4,
00e
(4,0
0)
4,00
h (4
,00)
O
mis
são
de B
45
,33a
(4
5,33
) 45
,33a
b (4
5,33
) 35
,00a
b (3
5,00
) 5,
59a
(28,
33)
38,0
0ab
(38,
00)
32,6
7d
(32,
67)
40,3
3b
(40,
33)
34,3
3c
(34,
33)
35,3
3ab
(35,
33)
23,0
0c
(23,
00)
18,0
0e
(18,
00)
Om
issã
o de
Fe
45
,33a
(4
5,33
) 47
,00a
(4
7,00
) 34
,67b
(3
4,67
) 5,
44a
(26,
67)
33,6
7b
(33,
67)
33,3
3cd
(33,
33)
17,3
3e
(17,
33)
30,6
7de
(30,
67)
39,6
7a
(39,
66)
32,0
0a
(32,
00)
25,0
0c
(25,
00)
Tes
te F
4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
DM
S (0
,05)
3,
8708
2,
2348
2,
2348
0,
6517
7,
8695
6,
6295
5,
0303
3,
3147
6,
1879
1,
6321
0,
9994
C
V (%
) 3,
62
2,04
2,
51
4,50
8,
25
4,27
5,
74
3,75
6,
33
2,58
1,
94
Tra
nsfo
rmaç
ão
- -
- R
Q (y
+3)
- -
- -
- -
- *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

129
Tab
ela
53. T
eor d
e fe
rro
(mg
L-1) p
rese
nte
na a
nális
e da
solu
ção
da m
icro
cepa
de
Euca
lypt
us g
rand
is x
uro
phyl
la, e
m fu
nção
da
cole
ta e
da
omis
são
de n
utrie
ntes
.
C
olet
as
Tra
tam
ento
s T0
2h
T22h
T1
3d
T35d
T4
5d
T61d
T7
9d
T97d
T1
07d
T115
d T1
24d
Com
plet
a 6,
13d
(6,1
3)
6,03
b (6
,03)
3,
90d
(3,9
0)
2,44
bc
(2,9
7)
7,10
d (7
,10)
6,
00b
(6,0
0)
7,10
b (7
,10)
3,
22b
(7,4
3)
1,00
d (0
,730
0,
50e
(0,5
0)
0,09
f (0
,09)
O
mis
são
de N
6,
73c
(6,7
3)
7,00
a (7
,00)
4,
30c
(4,3
0)
2,50
b (3
,27)
6,
80d
(6,8
0)
5,76
b (5
,77)
0,
90f
(0,9
0)
2,35
c (2
,53)
1,
07d
(0,9
6)
8,33
b (8
,3)
5,00
b (5
,00)
O
mis
são
de P
7,
63ab
(7
,63)
7,
00a
(7,0
0)
3,67
de
(3,6
7)
2,23
d (2
,03)
13
,93a
(1
3,93
) 6,
26b
(6,2
7)
7,16
b (7
,17)
3,
76a
(11,
20)
1,97
b (5
,20)
11
,66
(11,
66)
6,33
a
(6,3
3)
Om
issã
o de
K
8,03
a (8
,03)
7,
26a
(7,2
7)
5,50
a (5
,50)
2,
76a
(4,6
7)
9,67
b (9
,77)
3,
16c
(3,1
7)
4,70
c (7
,17)
2,
24c
(2,0
7)
1,09
d (1
,00)
2,
26d
(2,2
6)
0,76
e (0
,77)
O
mis
são
de C
a 6,
97c
(6,9
7)
6,13
b (6
,13)
4,
46c
(4,4
6)
2,28
cd
(2,2
3)
7,66
c (7
,67)
7,
16a
(7,1
7)
4,06
d (4
,07)
3,
07b
(6,4
7)
2,15
b (6
,73)
11
,67a
(1
1,67
) 3,
53c
(3,5
3)
Om
issã
o de
M
g 7,
13bc
(7
,13)
6,
76a
(6,7
7)
4,90
b (4
,90)
2,
69a
(4,2
7)
7,93
c (7
,93)
6,
23b
(6,2
3)
2,76
e (2
,77)
3,
28b
(7,7
7)
2,64
a (1
2,13
) 5,
83c
(5,8
3)
2,33
d (2
,33)
O
mis
são
de B
6,
63cd
(6
,63)
6,
20b
(6,2
0)
3,50
e (3
,50)
2,
35bc
d (2
,53)
4,
56e
(4,5
7)
6,20
b (6
,20)
10
,13a
(1
0,13
) 3,
43ab
(8
,80)
1,
45c
(2,3
0)
0,30
e (0
,30)
0,
09e
(0,0
9)
Om
issã
o de
Fe
0,87
c (0
,87)
1,
16c
(1,1
7)
0,73
f (0
,73)
1,
80e
(0,2
7)
0,16
f (0
,17)
1,
20d
(1,2
0)
1,11
d (1
,10)
1,
84d
(0,4
0)
0,84
d (0
,33)
0,
12e
(0,1
2)
0,09
e (0
,09)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,51
61
0,56
24
0,31
6 0,
1773
0,
4356
0,
5161
0,
2194
0,
3678
0,
2615
0,
9164
0,
532
CV
(%)
2,91
3,
34
2,88
2,
62
2,12
3,
97
1,49
4,
48
9,71
6,
37
8,25
T
rans
form
ação
-
- -
RQ
(y +
3)
- -
- R
Q (y
+ 3
) Lo
g (y
+ 2
) -
- *
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

130
Tab
ela
54. T
eor d
e bo
ro (m
g L-1
) pre
sent
e na
aná
lise
da so
luçã
o da
mic
roce
pa d
e Eu
caly
ptus
gra
ndis
x u
roph
ylla
, em
funç
ão d
a co
leta
e d
a om
issã
o de
nut
rient
es.
C
olet
as
Tra
tam
ento
s T0
2h
T22h
T1
3d
T35d
T4
5d
T61d
T7
9d
T97d
T1
07d
T115
d T1
24d
Com
plet
a 0,
83a
(0,7
0)
0,81
ab
(0,6
7)
1,78
ab
(0,2
0)
0,54
a (0
,30)
0,
54a
(0,3
0)
1,80
a (0
,27)
1,
81ab
c (0
,30)
0,
63b
(0,4
0)
1,14
ab
(0,3
0)
1,12
a (0
,26)
1,
14a
(0,3
0)
Om
issã
o de
N
0,70
abc
(0,5
0)
0,74
ab
(0,5
7)
1,78
ab
(0,2
0)
0,51
a (0
,27)
0,
54a
(0,3
0)
1,79
ab
(0,2
3)
1,81
abc
(0,3
0)
0,54
c (0
,30)
1,
15ab
(0
,33)
1,
11ab
(0
,23)
1,
14a
(0,3
0)
Om
issã
o de
P
0,77
ab
(0,6
0)
0,89
a (0
,80)
1,
77ab
(0
,17)
0,
54a
(0,3
0)
0,51
ab
(0,2
7)
1,81
a (0
,30)
1,
84ab
(0
,40)
0,
70a
(0,5
0)
1,14
ab
(0,3
0)
1,14
a (0
,30)
1,
14a
(0,3
0)
Om
issã
o de
K
0,63
bc
(0,4
0)
0,77
ab
(0,6
0)
1,80
a (0
,27)
0,
54a
(0,3
0)
0,54
a (0
,30)
1,
77bc
(0
,13)
1,
80ab
c (0
,27)
0,
57c
(0,3
3)
1,11
b (0
,23)
1,
04b
(0,1
0)
1,09
a (0
,20)
O
mis
são
de C
a 0,
71ab
c (0
,53)
0,
74ab
(0
,54)
1,
80a
(0,2
7)
0,54
a (0
,30)
0,
54a
(0,3
0)
1,81
a (0
,30)
1,
86a
(0,4
7)
0,54
c (0
,30)
1,
15ab
(0
,33)
1,
12a
(0,2
6)
1,14
a (0
,30)
O
mis
são
de
Mg
0,72
abc
(0,5
3)
0,76
ab
(0,6
0)
1,78
ab
(0,2
0)
0,54
a (0
,30)
0,
48ab
(0
,23)
1,
78ab
c (0
,20)
1,
78bc
(0
,20)
0,
54c
(0,3
0)
1,14
ab
(0,3
0)
1,12
a (0
,26)
1,
14a
(0,3
0)
Om
issã
o de
B
0,54
c (0
,30)
0,
48c
(0,2
3)
1,76
b (0
,10)
0,
44b
(0,2
0)
0,30
c (0
,09)
1,
76c
(0,1
0)
1,77
c (0
,13)
0,
31d
(0,1
0)
1,04
c (0
,10)
1,
04b
(0,0
9)
1,09
b (0
,20)
O
mis
são
de F
e 0,
63bc
(0
,40)
0,
68bc
(0
,47)
1,
78ab
(0
,20)
0,
54a
(0,3
0)
0,44
b (0
,20)
1,
80a
(0,2
7)
1,77
c (0
,17)
0,
54c
(0,3
0)
1,16
a (0
,36)
1,
14a
(0,3
0)
1,15
a (0
,33)
T
este
F
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 4,
896
4,89
6 D
MS
(0,0
5)
0,18
24
0,20
69
0,03
91
0,05
8 0,
082
0,03
21
0,06
22
0,04
89
0,05
02
0,06
76
0,02
48
CV
(%)
9,25
9,
98
0,77
3,
86
5,90
0,
63
1,21
3,
12
6,92
2,
16
3,28
T
rans
form
ação
R
Q (y
) R
Q (y
) R
Q (y
+ 3
)R
Q (y
) R
Q (y
) R
Q (y
+ 3
)R
Q (y
+ 3
)R
Q (y
) R
Q (y
+ 1
)R
Q (y
+ 1
)R
Q (y
+ 1
)*
Méd
ias s
egui
das d
as m
esm
as le
tras e
m c
ada
colu
na n
ão d
ifere
m e
ntre
si p
elo
test
e de
Tuk
ey a
5%
de
prob
abili
dade
. V
alor
es e
ntre
par
ênte
ses r
efer
ente
s às m
édia
s sem
tran
sfor
maç
ão.

131
APÊNDICE 2

Com
plet
a-N
itrog
ênio
-Fós
foro
Folh
as v
erde
s e se
m m
anch
as.
Folh
as a
mar
elad
as e
com
m
anch
as v
erm
elha
s.Pr
esen
ça d
e lim
bo fo
liar m
aior
qu
e o
norm
al.
Figu
ra 1
0. S
into
mas
de
defic
iênc
ia d
e ni
trogê
nio
e de
fósf
oro
apre
sent
ados
pel
o hí
brid
o de
Euc
alyp
tus g
rand
isx
E. u
roph
ylla
, co
mpa
rado
s à te
stem
unha
(sol
ução
com
plet
a), 7
9 di
as a
pós o
mom
ento
em
que
as m
udas
rece
bera
m so
luçã
o nu
tritiv
a (M
.0.)
132

Com
plet
a-P
otás
sio
-Cál
cio
Folh
as v
erde
s e se
m m
anch
as.
Figu
ra 1
1. S
into
mas
de
defic
iênc
ia d
e po
táss
io e
de
cálc
io a
pres
enta
dos p
elo
híbr
ido
de E
ucal
yptu
s gra
ndis
x E.
uro
phyl
la,
com
para
dos à
test
emun
ha (s
oluç
ão c
ompl
eta)
, 79
dias
apó
s o m
omen
to e
m q
ue a
s mud
as re
cebe
ram
solu
ção
nutri
tiva
(M.0
.).
Lim
bo fo
liar m
ais a
long
ado
Folh
as n
ovas
enc
arqu
ilhad
as
e m
al fo
rmad
as
133

Com
plet
a-M
agné
sio
-Bor
o
Folh
as v
erde
s e se
m m
anch
as.
Figu
ra 1
2. S
into
mas
de
defic
iênc
ia d
e m
agné
sio
e de
bor
o ap
rese
ntad
os p
elo
híbr
ido
de E
ucal
yptu
s gra
ndis
x E.
uro
phyl
la,
com
para
dos à
test
emun
ha (s
oluç
ão c
ompl
eta)
, 79
dia
s apó
s o m
omen
to e
m q
ue a
s mud
as re
cebe
ram
solu
ção
nutri
tiva
(M.0
.).
Mai
or q
uant
idad
e de
mas
sa
folia
r pro
duzi
da.
Folh
as m
al fo
rmad
as e
av
erm
elha
das.
134

Com
plet
a-F
erro
-Fer
ro
Folh
as v
erde
s e se
m m
anch
as.
Figu
ra 1
3. S
into
mas
de
defic
iênc
ia d
e fe
rro
apre
sent
ado
pelo
híb
rido
de E
ucal
yptu
s gra
ndis
x E.
uro
phyl
la,
com
para
dos à
test
emun
ha (s
oluç
ão c
ompl
eta)
, 79
dias
apó
s o m
omen
to e
m q
ue a
s mud
as re
cebe
ram
a
solu
ção
nutri
tiva
(M.0
.).
Pres
ença
de
man
chas
nec
rótic
as.
Pont
os a
verm
elha
dos n
a fo
lha.
135





























