Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DE OURO PRETO
INSTITUTO DE CIENCIAS EXATAS E BIOLÓGICAS DEPARTAMENTO DE BIODIVERSIDADE, EVOLUÇÃO E MEIO AMBIENTE
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA DE BIOMAS TROPICAIS
Dissertação de Mestrado
Ecologia, Diversidade e Conservação dos Lagartos da Serra de Ouro Branco, Minas
Gerais
António Jorge do Rosário Cruz
Ouro Preto, Março de 2012

Ecologia, Diversidade e Conservação dos Lagartos da Serra de Ouro
Branco, Minas Gerais
Autor : António Jorge do Rosário Cruz
Dissertação apresentada ao Programa de Pós-
Graduação em Ecologia de Biomas Tropicais, do
Instituto de Ciências Exatas e Biológicas da
Universidade Federal de Ouro Preto, como parte
dos requisitos para obtenção do título de Mestre em
Ecologia.
Ouro Preto, Março de 2012

Catalogação: [email protected]
C957e Cruz, António Jorge do Rosário. Ecologia, diversidade e conservação dos lagartos da Serra de Ouro
Branco, Minas Gerais [manuscrito] / António Jorge do Rosário Cruz. - 2012.
106f.: il., color; grafs.; tabs.; mapas. Orientadora: Profª Drª Maria Rita Silvério Pires. Dissertação (Mestrado) - Universidade Federal de Ouro Preto. Instituto de Ciências Exatas e Biológicas. Departamento de Biodiversidade, Evolução e Meio Ambiente. Programa de Pós-Graduação em Ecologia de
Biomas Tropicais. Área de concentração: Ecologia.
1. Espinhaço, Serra do (MG e BA) - Teses. 2. Répteis - Teses. 3. Lagarto - Teses. 4. Habitat - Teses. 5. Florestas - Efeito de borda - Teses. I. Pires, Maria Rita Silvério. II. Universidade Federal de Ouro Preto. III. Título.
CDU: 598.112:630*15(815.1)


Ao Brasil, pela oportunidade de aprender e
construir uma África melhor!

AGRADECIMENTOS À minha família, principalmente a minha Mãe, Ana Manuela, por acreditar sempre em mim, nos meus sonhos e que mesmo de longe, andou sempre ao meu lado. Obrigado por tudo! DOD NA BÔ. Ao Brasil, mais uma vez, pela recepção, pelo Samba, pelo futebol, pelos abraços, pelo aprendizado e por me permitir com intimidade chamar seus filhos de meus irmãos! Senti-me em casa! Àprofessora Maria Rita, pela simpatia, disponibilidade, orientação e paciência em todos os momentos. Serei sempre grato! À Adriele Prisca, vulgo Adrianely, hehehe... pela amizade, pela companhia, pelos momentos mais engraçados de trabalhos árduos do campo em Ouro Branco! Dri, amo Você muito! À Núbia, quer dizer, Nubão, pela irmandade, pelas rizadas, pelo café da tarde e horas de papo no quintal, pela paciência, pela alegria, simpatia e amizade. Obrigado, você tem influência direta nesta conquista. Ao Leandro Drummond, vulgo Difs, por ser um professor nato e me ajudar em todos os momentos de dúvidas, pelos dias de campo, pelas rizadas e principalmente pela herpetologia! Aprendi e continuo aprendendo muito com você! Um obrigado! À Caryne Braga, pelo convite e sugestão ambiciosa de trabalharmos no mesmo projeto. Você tinha razão, foi muito bom! Também não poderia esquecer-me de agradecer pelos rangos do campo em Itatiaia. Dá saudade! Ao Hudson, pela amizade, pelo companheirismo, pelas trocas de ideia, pelas alegrias e como não poderia faltar, pela ajuda na análise de dados. Valeu aí, mano! À Michelle e ao Alexandre, pelos momentos de compadres. Gosto muuuuuuito de vcs! À Valéria Braga, pelo ajuda no campo e no laboratório, pelo papo e pelos momentos legais no LZV. Ao Raphael, pela ajuda no campo, pelas comédias e rizadas, pelo papo cabeça e pela cumplicidade na hora de tramar alguém! Ao Rubens, pela simpatia, e pela disponibilidade 1000. Muito obrigado por tudo! À Jaqueline Rolin, pela ajuda no campo e no laboratório. Pela vontade e disponibilidade nas horas em que eu precisava de mão de obra solidária. Hehehehehe... Valeu! À Lorena, pelos momentos no campo em Itatiaia e no laboratório. Ta vendo, deu tudo certo! Obrigada pelo apoio.

Ao pessoal do LZV: Lívia, Ana Clara, Ceres, Krisna, Vivi, Thales e Dany, pela ajuda e pelos momentos no campo e no laboratório. Valeu aí galera! Muito obrigado! Aos professores Yasmine Antonini, Sérvio Pontes, Eneida, Alessandra, Júlio e ao Professor Hilde pelos ensinamentos em salas de aulas e pelos momentos no Colegiado. Foram momentos muito importantes na minha vida! Ao Henrique C. Costa, pela ajuda, vontade e disponibilidade á qualquer momento. À poluta casa Republica Calamidade Pública, Minha Escola e minha casa! Lugar a onde eu aprendi a ver, e a questionar os lados de uma mesma moeda. Lugar onde eu aprendi a escutar e a respeitar as ideias ou visões dos outros. Mas não poderia me esquecer da Irmandade, dos rocks, do companheirismo e pela Biritagem! Deus é pai, e a cachaça vai! À Dona Maria, Mãe do mundo e de quem quiser! Bondade amor tem de sobra! À república Tropicália, pela amizade e bons momentos republicanos! As amigas da Republica Bico Doce pelos momentos inesquecíveis. Tchanas, muito obrigado pelo café, pelo violão, pela cachaça, pela cerveja e pelos momentos de amizade! Sentirei saudades de vocês, bicudas! Ao Flavinho e a Naiara, pelos momentos Happy Family. Foram momentos ótimos e muito especiais para mim. Ao Vinícius Terror, pelas aulas de Samba e de fotografia, pelos papos de final de tarde e pelos conselhos! Valeu aí Terror! À Luana e ao Rodolfinho, pelos momentos muito legais nos sambas e em Ouro Preto. Ao Pedro, pela irmandade! Ao Filipe, pela amizade, simplicidade e pela bagunça! Obrigado por tudo mano! Ao Elias e a Yara, pela vizinhança amiga e simpática. Valeu aí pelas horas de papo, pelo café e pela Música, sem falar nos almoços! Muito obrigado por tudo! As republicas amigas: Taranóia, Partenon, 171, Masmorra, Maria Maria, Favinho, Colmeia, Damas de Ouro, Santuário, Mandala, Lua Azul, Tira Burca, Quase Normal, Arriba entre outras... Valeu gente! À FAPEMIG pelo financiamento do projeto. Ao Programa de Pós graduação em Ecologia de Biomas Tropicais pela oportunidade.

À Universidade Federal de Ouro Preto, pela oportunidade e pela Bolsa de estudos. À Ouro Preto, pela hospitalidade, pelo aprendizado e pela vida! Amo demais, cada pedaço de calçada desta cidade! Sentirei saudades! Obrigado! À Minas Gerais. É verdade, quem te conhece, não esquece jamais.

1
RESUMO A Serra de Ouro Branco, porção sul da cadeia do Espinhaço, situada entre os limites da Mata
Atlântica com o Cerrado, abriga uma expressiva diversidade faunística. No entanto, faltam
informações sobre os lagartos. Com a utilização de diferentes metodologias de campo, o
presente trabalho apresenta o registro de 15 espécies de lagartos distribuídos em oito famílias
para essa região. Esse resultado amplia a distribuição conhecida de várias espécies, tanto para
os limites de cada bioma, como também para os limites de altitude. No segundo capítulo deste
trabalho foi analisada a utilização de hábitats florestais pelos lagartos da serapilheira em áreas
de mata impactadas. Nesse sentido, foram montadas armadilhas no interior de fragmentos
florestais e em bordas delimitadas por áreas utilizadas para agricultura e por estradas. Esses
dois tipos de áreas são consideradas impactantes para a diversidade faunística, uma vez que
constituem matrizes de permeabilidade variável para diferentes espécies ou grupos
faunísticos. Entre elas, foi inicialmente proposto que nas regiões onde a floresta está próxima
da borda com a estrada seria encontrada menor riqueza e diversidade de lagartos, em relação à
às regiões florestais com borda para áreas de agricultura. Variáveis ambientais foram
mensuradas como forma de caracterização do hábitat, as análises dessas variáveis foram
associadas à distribuição dos lagartos nas respectivas áreas. Os resultados indicaram que os
lagartos responderam mais às variações ambientais relacionadas à vegetação do que às
variações quanto aos tipos de bordas entre os ambientes amostrados. Alguns lagartos se
mostraram muito correlacionados a alguns fatores ambientais, como áreas sombreadas e de
maior complexidade estrutural. No entanto, todas as espécies da área ocorreram em bordas de
mata, onde, segundo a literatura, corre maior risco de extinção. Estes fatores se juntam ao fato
de que as populações de lagartos típicos de áreas florestadas são bem reduzidas na área de
estudo, o que pode ser ao estado de regeneração da floresta, que por sua vez pode ter
favorecido espécies mais generalistas quanto ao hábitat, levando à dominância de Enyalius
perditus. Felizmente, a região conta com duas Unidades de Conservação e pequenas
comunidades humanas no seu entorno, que constituem peça chave na preservação desse grupo
animal.
Espinhaço; Ouro Branco; Repteis, Lagartos; Matriz; Efeito de bordas; Estrutura do habitat.

2
ABSTRACT
Ouro Branco’s ridge, at the south of Espinhaço’s range, is located between the Atlantic Forest
and the Cerrado, and housesan expressive faunal diversity. Although, some information about
lizards is missing. Through different field methodologies, this work presents 15 lizard species,
among eight families in this region. This result amplifies the known distribution of several
species, both for the biome’s limits as for the altitude limits. The second chapter of this work
analyses the use of forests’ habitats by litter lizards in highly damaged wood areas. To do so,
traps were set inside forest fragments and in borders, whose limits were defined by
agricultural areas and roads. These two areas are considered to cause an impact in the faunal
diversity, since they make variable permeability matrixes for different species or faunal
groups. Between, it was initially proposed that, in regions where the forest is next to the road,
smaller richness and diversity of lizards would be found, related to the forest regions next to
agricultural fields. We measured some environmental variables as a way to characterize the
habitat, and these analyses were associated to the lizards’ distribution in each area. The results
show that the lizards were more responsive to the environmental changes related to vegetation
than to the type of border in each environment. Some lizards showed themselves highly
correlated to some environmental factors, such as shadow areas and higher structural
complexity. Although, some species of the area occurred in wood borders, where, according
to the literature, they have a higher risk of extinction. These factors add themselves to the fact
that the woodlands typical populations of lizards are very reduced in the analyzed area, which
might be attributed to the forest regeneration state, which could have favored more generalist
species, related to habitat, which leads to a dominance of Enyalius perditus. Happily, the
region comprehends two Conservational Units and small human communities in its
surrounding, both constituting a key element in this animal group’s preservation.
Espinhaço; Ouro Branco; Reptiles; Lizards; Matrix; Border Effect; Habitat’s structure

3
SUMÁRIO
INTRODUÇÃO GERAL ........................................................................................................................................ 5 BIBLIOGRAFIA ................................................................................................................................................... 9 CAPÍTULO 1: FAUNA DE LAGARTOS (SQUAMATA, “SAURIA”) DA SERRA DE OURO BRANCO, PORÇÃO SUL DA SERRA DO ESPINHAÇO, MINAS GERAIS .................................................................. 18
RESUMO: ........................................................................................................................................................ 19 ABSTRACT: .................................................................................................................................................... 20 INTRODUÇÃO ............................................................................................................................................... 21 MATERIAIS E MÉTODOS ........................................................................................................................... 23
Área de estudos ............................................................................................................................................ 23 Coleta dos dados .......................................................................................................................................... 25 Armadilhas de interceptação e queda ........................................................................................................ 25
Encontros casuais ou ocasionais ..................................................................................................................... 27 RESULTADOS ................................................................................................................................................ 27 DISCUSSÃO .................................................................................................................................................... 32 LISTA COMENTADA DOS LAGARTOS REGISTRADOS PARA A SERRA DO OURO BRANCO, MINAS GERAIS ............................................................................................................................................. 36
Família Anguidae ........................................................................................................................................ 36 Família Gekkonidae .................................................................................................................................... 36 Família Gymnophthalmidae ....................................................................................................................... 37 Família Leiosauridae ................................................................................................................................... 39 Família Polychrotidae ................................................................................................................................. 40 Família Scincidae ......................................................................................................................................... 41 Família Teiidae ............................................................................................................................................ 42 Família Tropiduridae .................................................................................................................................. 43
CHAVE ARTIFICIAL PARA IDENTIFICAÇÃO DAS ESPÉCIES DE LAGARTOS DA SERRA DE OURO BRANCO, OURO BRANCO, PORÇÃO SUL DO ESPINHAÇO MINAS GERAIS. .................. 44 BIBLIOGRAFIA ............................................................................................................................................. 46
CAPITULO 2: INFLUÊNCIA DE BORDAS E DAS VARIÁVEIS AMBIENTAIS NA ESTRUTURAÇÃO DA COMUNIDADE DE LAGARTOS EM FRAGMENTOS DE MATA EM UMA ÁREA DE TRANSIÇÃO MATA ATLÂNTICA- CERRADO ........................................................................................... 60
RESUMO ......................................................................................................................................................... 61 ABSTRACT ..................................................................................................................................................... 62 INTRODUÇÃO ............................................................................................................................................... 63 METODOLOGIAS ......................................................................................................................................... 67
Área de estudo ............................................................................................................................................. 67 Desenho amostral ........................................................................................................................................ 68 Características ambientais .......................................................................................................................... 70 Análise dos dados ......................................................................................................................................... 71
RESULTADOS ................................................................................................................................................ 72 DISCUSSÃO .................................................................................................................................................... 79 CONCLUSÃO ................................................................................................................................................. 85 BIBLIOGRAFIA ............................................................................................................................................. 86
4
CONSIDERAÇÕES FINAIS .............................................................................................................................. 93 BIBLIOGRAFIA ................................................................................................................................................. 96 ANEXO I ............................................................................................................................................................ 105

5
INTRODUÇÃO GERAL
Os lagartos são considerados bons modelos para a pesquisa ecológica, e excelentes
organismos experimentais. Informações extraídas através de análises de suas comunidades
como riqueza e abundância de espécies, uso de habitat, dieta e comportamentos ligados a
reprodução e predação servem como base para o entendimento de padrões ecológicos em
diferentes ecossistemas (Pianka, 1967, 1969, 1971; Pianka & Vitt, 2003). No entanto,
populações de lagartos no mundo todo tem sofrido um declínio preocupante (Gibbons et al.,
2000). Problemas como a perda de habitat, introdução de espécies exóticas, poluição
ambiental, doenças além das mudanças climáticas são apontados como os principais
causadores dessa perda, em diferentes ecossistemas (Gibbons et al., 2000).
Considerada um dos maiores hotspots da biodiversidade (Myers et al., 2000), a Mata
Atlântica está atualmente reduzida a menos de 10% do seu tamanho original, devido
principalmente à pressão sofrida pelo crescimento e expansão dos grandes centros urbanos
(Morellato & Haddad, 2000). A herpetofauna de lagartos neste bioma se mostra
extremamente rica apresentando diversos endemismos (Rodrigues, 2005). Estudos sobre a
história natural de algumas espécies (Sturaro & Silva, 2010; Barros & Teixeira, 2007; Sousa
& Cruz, 2008; Zamprogno et al., 2001; Van Sluys et al., 2004), inventários e análise de
materiais em coleções (Dixo & Verdade, 2006; Feio & Caramashi, 2002; Cicchi et al., 2009;
Condez et al., 2009; Costa et al., 2009; Forlani et al., 2010; Marques et al., 2010; Sousa et al.,
2010; Zaher et al., 2011), fazem parte de esforços recentes direcionados a preservação do
grupo neste bioma. Entretanto, esses trabalhos são ainda insuficientes frente à velocidade com
que os ambientes naturais são modificados.

6
O Cerrado é também uma das regiões prioritárias para a conservação da biodiversidade,
apresentando vários endemismos (Maury, 2002). Esse bioma é uma das regiões Neotropicais
menos conhecidas (Vanzolini, 1988), sendo que sua fauna de lagartos se mostra
extremamente rica, apresentando comunidades com até 25 espécies (Nogueira, 2006).
Estudos com os lagartos neste bioma têm produzido trabalhos relevantes, principalmente
sobre a ecologia (Vitt, 1991; Mesquita & Colli, 2006; Colli et al., 2002; Werneck & Colli,
2006). No entanto esses trabalhos mostram ser bem recentes, sendo que poucos abordam a
problemática da destruição de habitat e a resposta da comunidade de lagartos.
A Serra do Espinhaço é considerada um acidente geográfico de grande importância biológica.
Este figura-se como um ecossistema formada por um conjunto de diversos ambientes como o
Cerrado, fragmentos de Mata Atlântica e Caatinga, além de apresentar os Campos Rupestres
em suas áreas de maior altitude. Por deter uma elevada riqueza biológica, com vários
endemismos e sofrer uma elevada pressão antrópica, várias espécies se encontram ameaçadas
neste ecossistema, sendo considerada uma região prioritária para a conservação da
biodiversidade (UNESCO, 2005).
A fauna de lagartos assim como de vários outros grupos zoológicos na Serra do Espinhaço
ainda são pouco exploradas. A falta de conhecimento aliada à urgência e gravidade de
ameaças na serra do Espinhaço, levou a criação de estratégias como a criação do Plano de
Ação Nacional para a conservação dos repteis ameaçados de extinção no Espinhaço (PAN-
Herpetofauna do Espinhaço) pela portaria N° 24 de 17 de fevereiro de 2012, com objetivos de
conhecer e proteger inúmeras espécies de repteis e anfíbios. Apesar dessas atitudes e
esforções recentes, permanecem ainda diversas lacunas, principalmente as ligadas à
distribuição geográfica, ecologia e história natural, o que torna difícil a tomada de decisões

7
mais coerentes, visto que inúmeras espécies são consideradas deficientes de dados (PAN-
Herpetofauna do Espinhaço, 2012).
A serra de Ouro Branco, porção sul do Espinhaço, apresenta uma alta riqueza de espécies de
anfíbios (Leite et al., 2008; São Pedro & Feio, 2010; 2011) e de serpentes (São Pedro & Pires,
2009; Silveira et al., 2010). Trabalhos referentes ao conhecimento dessa diversidade pelos
moradores da região foram recentemente desenvolvidos (Pires et al., 2010; Mateus et al.,
2011 e Pinto, 2011), demonstrando a necessidade de envolvimento das comunidades humanas
no entorno das Unidades de Conservação na preservação da fauna local. No entanto faltam
estudos envolvendo os lagartos da região.
Sabendo-se da carência de dados e trabalhos sobre a fauna de lagartos na serra do espinhaço, e
indo de encontro com os mesmo objetivos do PAN- herpetofauna,o primeiro capítulo desta
dissertação tem como objetivo ampliar o conhecimento sobre a diversidade de lagartos na
serra de Ouro Branco, Porção sul da Serra do Espinhaço e zona de transição entre Mata
Atlântica e Cerrado. Com isso o presente trabalho apresenta uma lista de todas as espécies
com ocorrência registrada, reunindo observações referentes à ecologia, história natural e
distribuição geográfica, além de fotos e uma chave de identificação simplificada para os
lagartos da serra de Ouro Branco.
Entre os fatores que mais causam impactos negativos as comunidade naturais, a construção de
estradas tem sido uma das mais preocupantes, principalmente porque altera tanto a paisagem
como também direciona e potencializa ações que causam perdas de habitats e,
consequentemente, perda de diversidade biológica (Trombulak & Frissell, 2000). Este tipo de
infraestrutura facilita o acesso ao desmatamento e a práticas de caça (Peres & Lake, 2003),
problemas de atropelamentos da fauna silvestre (Melo & Santos-Filho, 2007; Hartmann et al.,

8
2011), contaminação por poluição e incêndios (Santos & Tabarelli, 2002; Skole & Tucker,
1993).
A agricultura é uma das atividades humanas mais agressivas ao meio ambiente. A sua prática,
tem impactos severos como a destruição ou fragmentação de habitat, poluição e contaminação
da fauna silvestre por uso de pesticidas, além de facilitar a introdução de espécies exóticas,
alterando assim comunidades de todo o ecossistema (Fistzon, 2003; Broadbent et al., 2008).
Apesar disso, dependendo do tipo de cultivo, ela pode servir como matriz, favorecendo
algumas espécies na comunidade, permitindo a circulação e realização de ações básicas como
termoregulação, forrageio e predação.
A estrutura do habitat costuma ser uma das primeiras alterações decorrentes do
desmatamento. Com as mudanças decorrentes na estrutura vegetal, diversas características no
meio também sofrem modificações (Ries et al., 2004; Harper et al., 2005; Murcia, 1995;
Jellinek et al., 2004), criando condições antes desconhecidas e pouco favoráveis para algumas
espécies, principalmente as estritamente florestais. No entanto, essas mudanças nem sempre
causam perdas da biodiversidade. Algumas alterações inclusive, podem até tornar o habitat
mais heterogêneo, oferecendo espaços antes não ocupados e aumentando a riqueza local. Mas
mudanças de grande envergadura e prolongadas por muito tempo, podem causar
desiquilíbrios sérios na comunidade, favorecendo o aumento de alguma população, que por
vezes pode ser interpretado como uma diminuição da riqueza local (Vitt & Carvalho, 1995).
O segundo capítulo desta dissertação intitulado tenta entender como a comunidade de lagartos
de serapilheira da serra de Ouro Branco se comporta perante diferentes mosaicos, constituídos
por áreas agrícolas, estradas e florestas secundárias, através da interpretação de dados
referentes à abundância relativa, riqueza e uso de habitat.

9
BIBLIOGRAFIA
AB'SABER, A. N. 1977. Os domínios morfoclimáticos da América do Sul. Primeira
aproximação. Geomorfologia 52: 1-21.
AL-JOHANY, A. M. 1995. The ecology of Agama yemenesis Klausewitz (Lacertilia:
Agamidae) in South-western Arabia. Jornal of Arid Environments. 29: 495-503.
ÁVILA-PIRES, T.C.S. 1995. Lizards of Brazilian Amazonia (Reptilia: Squamata). Zool.
Verh. Leiden. 299: 1- 706.
BARROS, E. H. & TEIXEIRA, R. L. 2007.Diet and fecundity of the Glass-lizard, Ophiodes
striatus (Sauria, Anguidae) from the Atlantic Forest in southeastern Brazil.Bol. Mus.
Biol. Mello Leitão (n. sér.) 22:11-23.
BÉRNILS, R. S.; NOGUEIRA, C. & SILVA, V. X. 2009. Biota minas -
diagnóstico do conhecimento de vertebrados: Répteis. In Biota minas - diagnóstico do
conhecimento sobre a biodiversidade no estado de Minas Gerais (G.M. DRUMMOND,
C.S. MARTINS, M.S. GRECO & F. VIEIRA, eds.), pp. 252-278, Belo Horizonte, MG.
BIERREGAARD JR, R. O.; C. GASCON, T.E. LOVEJOY & R. MESQUITA. 2001.Lessons
from Amazonia: the ecology and conservation of a fragmented forest. Yale University
Press, New Haven, EUA.
BROADBENT, E. N.; ASNER, G. P.; KELLER, M.; KNAPP, D. E.; OLIVEIRA, P. J. C. &
SILVA, J. N. 2008.Forest fragmentation and edge effects from deforestation and
selective logging in the Brazilian amazon.Biological Conservation 141. 1745–1757.
CARVALHO, A. L. G. & ARAÚJO, A. F. B. 2004. Ecologia dos lagartos da ilha de
Marambaia, RJ. Rev. Univ. Rural, Sér. Ci. Vida. Seropédica, RJ, EDUR, v. 24, n.2, p.
159-165.
CASTRO, E. R. & GALETTI, M. 2004. Frugivoria e dispersão de sementes pelo lagarto teiú
Tupinambis merianae (Reptilia: Teiidae). Papéis Avulsos de Zoologia, 44(6): 91-97.

10
CECCHI, P. J. P.; SERAFIM, H.; SENA, M. A; CENTENO, F. C. & JIM, J. 2009. Atlantic
Rainforest herpetofauna of Ilha Anchieta, an island on municipality of Ubatuba,
southeastern Brazil.Biota Neotrop., 9(2):
http://www.biotaneotropica.org.br/v9n2/en/abstract?inventory+bn0100902.
COLLI, G. R.; BASTOS, R. P, & ARAÚJO, A. F. B. 2002. The character and dynamics of
the Cerrado herpetofauna.In: Oliveira, p.s. & Marquis, R. L (eds.) The Cerrados of
Brazil: ecology and natural history of a Neotropical savana, p.223-241. New York,
Columbia University Press.
CONDEZ, T. H.; SAWAYA, R. J. & DIXO, M. 2009.Herpetofauna of the Atlantic Forest
remnants of Tapiraí and Piedade region, São Paulo state, southeastern Brazil.Biota
Neotrop.9(1):http://www.biotaneotropica.org.br/v9n1/en/abstract?inventory+bn0180901
2009.
COSTA, H. C.; FERNANDES, V. D.; RODRIGUES, A. C.; FEIO, R. N. 2009. Lizards and
amphisbaenians, municipality of Viçosa, State of Minas Gerais, southeastern
Brazil.Check List. 5(3): 732-745.
COSTA, H. C.; FERNANDES, V. D.; VRCIBRADIC, D.; FEIO, R. N. 2008. Reptilia,
Scincidae, Mabuya frenata: Distribution Extension. Check List 4(1): 86–88.
DIXO, M & MARTINS, M. 2008. Are leaf-litter frogs and lizards affected by edge effects
due to forest fragmentation in Brazilian Atlantic forest? Journal of Tropical Ecology.
24:551–554.
DIXO, M. & METZGER, J. P. 2009.Are corridors fragment size and forest structure
important for leaf-litter lizards in fragmented landscape? Fauna & Flora International,
Oryx, 43(3), 435-442.
DIXO, M. & VERDADE, V. K. 2006. Herpetofauna de serrapilheira da Reserva Florestal de
Morro Grande, Cotia (SP). Biota Neotrop. vol. 6 no. 2,
http://www.biotaneotropica.org.br/v6n2/pt/abstract?article+bn00806022006. ISSN1676-
0603.
FEIO, R. N. & CARAMASCHI, U. 2002. Contribuição ao conhecimento da herpetofauna do
nordeste do estado de Minas Gerais, Brasil. Phyllomedusa 1(2):105-111.

11
FEIO, R. N. & U. CARAMASCHI. 2002. Contribuição ao conhecimento da herpetofauna do
nordeste do estado de Minas Gerais, Brasil. Phyllomedusa 1(2):105-111.
FISZON, J.T.; MARCHIORO, N. DE P.X.; BRITEZ, R.M. DE; CABRAL, D. DE C.;
CAMELY, N.C.; CANAVESI, V.; CASTELLA, P.R.; CASTRO, E.B.V. DE; CULLEN
JR., L.; CUNHA, M.B.S.; FIGUEIREDO, E.O.; FRANKE, I.L.; GOMES, H.; GOMES,
L.J.; HREISEMNOU, V.H.V.; LANDAU, E.C.; LIMA, S.M.F.; LOPES, A.T.L.;
NETO, E.M.; MELLO, A.L. DE; OLIVEIRA, A.L.DE; ONO, K.Y.; PEREIRA,
N.W.V.; RODRIGUES, A. DOS S.; RODRIGUES, A.A.F.; RUIZ, C.R.; SANTOS,
L.F.G.L. DOS; SMITH, W.S. &C.R. DE SOUZA. 2003. Causas Antrópicas. In:
Rambaldi, D.M. and D.A.S. de Oliveira (orgs.). Fragmentação de ecossistemas: causas,
efeitos sobre a biodiversidade e recomendações de políticas públicas, pp. 66 – 99.
Brasília: MMA/SBF.
FORLANI, M. C.; BERNARDO, P.H.; HADDAD, C.B.F. & ZAHER, H. 2010. Herpetofauna
of the Carlos Botelho State Park, São Paulo State, Brazil. Biota Neotrop. 10(3):
http://www.biotaneotropica.org.br/v10n3/en/abstract?inventory+bn0021003.
GASCON, C.; LOVEJOY,T. E.; BIERREGAARD JR, R. O.; MALCOLM, J. R.;
STOUFFER, P. C.; VASCONCELOS, H.; LAURANCE W.F.; ZIMMERMAN, B.;
TOCHER, M. & BORGES, S. 1999. Matrix habitat and species persistence in tropical
forest remnants. Biological Conservation 91: 223-230.
GIBBONS, J. W.; SCOTT, D. E.; RYAN, T. J.; BUHLMANN, K. A.; TUBERVILLE, T. D.;
METTS, B. S.; GREENE, J. L.; MILLS, T.; LEIDEN, Y.; POPPY, S.; WINNE, C. T.
2000. The global decline of reptiles, deja-vu amphibians. Bioscience 50, 653–667.
HARPER, K. A., MACDONALD, S. E., BURTON, P., CHEN, J., BROSOFSKY, K. D.,
SAUNDERS, S., EUSKIRCHEN, E. S., ROBERTS, D., JAITEH, M. & ESSEEN, P. A.
2005. Edge influence on forest structure and composition in fragmented
landscapes.ConservationBiology19:768–782.
HARTMANN, P. A.; HARTMANN, M. & MARTINS, M. 2011.Snake road mortality in a
protected area in the Atlantic Forest of Southeastern Brazil. South American Journal of
Herpetoloy, 6(1), 35-42.

12
JELLINEK, S.; DRISCOLL, D. A. & KIRKPATRIK, J. B. 2004. Environmental and
vegetation variables have a greater influence than habitat fragmentation in structuring
lizard communities in remnant urban bushland. Austral Ecology. 29, 294-304.
LEITE, F. S. F.; JUNCÁ, F. A. & ETEROVICK, P. C. 2008. Status do conhecimento,
endemismo e conservação de anfíbios anuros da Serra do Espinhaço, Brasil.
Megadiversidade.
MARQUES, O. A. V; NOGUEIRA, C; MARTINS, M & SAWAYA, R. J. 2010. Impactos
potenciais das mudanças propostas no Código Florestal Brasileiro sobre os répteis
brasileiros. Biota Neotrop.,vol. 10, no. 4.
MATEUS, M. B.; PINTO, L. C. L.; MOURA, M. R. & PIRES, M. R. S. 2011. A Cobra-de –
duas –cabeças na percepção dos moradores do povoado de Itatiaia, Minas Gerais.
Biotemas. 24(3): 111-117.
MELO, E. S. & SANTOS-FILHO, M. 2007. Efeitos da BR-070 na Província Serrana de
Cáceres, Mato Grosso, sobre a comunidade de vertebrados silvestres. Revista Brasileira
de zoociências 9(2): 185-192.
MESQUITA, D. O. & COLLI, G. R. 2003. The Ecology of Cnemidophorus ocellifer
(Squamata, Teiidae) in a Neotropical Savanna. Journal of Herpetology, Vol. 37, No. 3,
pp. 498–509.
MESQUITA, D.O.; COLLI, G.R.; FRANÇA, F.G.R. & VITT, L.J. 2006.Ecology of a
Cerrado lizard assemblage in the Jalapão region of Brazil. Copeia 2006(3): 460-471.
MORELLATO, L. P. C.; HADDAD, C. F. B. 2000.Introduction: The Brazilian Atlantic
Forest. Biotropica, v. 32(4b), p. 786-792.
MURCIA, C. 1995. Edges effects in fragmented forest: Implications for conservation. Tree,
Vol: 10. no. 2.
MYERS, N.; MITTERMEIER, R. A.; MITTERMEIER, C. G.; DA FONSECA, G. A. B. &
KENT, J. 2000. Biodiversity hotspots for conservation priorities. Nature 430: 853-858.

13
NOGUEIRA, C. C. 2006. Diversidade e padrões de distribuição da fauna de lagartos do
Cerrado. Tese de Doutorado. USP-Instituto de Biociências São Paulo.
NOGUEIRA, K. O. P.C.; RODRIGUES, S. S.; ARAÚJO, V.A. & NEVES, C. A. 2011.
Oviductal Structure and Ultrastructure of the Oviparous Gecko, Hemidactylus Mabouia
(Moreau De Jonne` s, 1818).The anatomical record294:883–892.
PAN-HERPETOFAUNA DO ESPINHAÇO. 2012. Portaria N° 24 de 17 de fevereiro de
2012.
PERES, C. A. & LAKE, I. R. 2003. Extent of nontimber resource extraction in tropical
forests: Accessibility to game vertebrates by hunters in the Amazon basin. Conserv.
Biol. 17: 521–535.
PIANKA, E. R. & VITT, L. J. 2003. Lizards: Windows to the Evolution of Diversity.
University of California Press, Berkeley.
PIANKA, E. R. & R. B. HUEY.1978. Comparative ecology, niche segregation, and resource
utilization among gekkonid lizards in the southern Kalahari. Copeia 1978: 691- 701.
PIANKA, E. R. 1967. On lizard species diversity: North American flatland deserts. Ecology
48: 333-351.
PIANKA, E. R. 1969. Habitat specificity, speciation, and species density in Australian desert
lizards. Ecology 50: 498-502.
PIANKA, E. R. 1971. Species diversity. Chapter in Topics in the Study of Life: The Bio
Source Book, pp. 401-406. Harper and Row, New York.
PINTO, L. C. L. 2011.Etnozoologia e conservação de biodiversidade em comunidades rurais
da Serra do Ouro Branco. Dissertação de Mestrado. Universidade Federal de Ouro
Preto, Minas Gerais.
POUGH, F. H.; JANIS, C. M. & HEISER, J. B. 2008.A vida dos vertebrados. 4 ed. São
Paulo: Atheneu Editora,. 684 p. I.
POUGH, F.H; JANIS, C. M. & HEISER, J. B. 2003.A Vida dos Vertebrados.3ª edição.
Atheneu, São Paulo, 699p.

14
RAMOS, L. O. & SILVEIRA, A. L. 2011. New Record and distribuition mape of Ecpleopus
gaudichaudii (Reptilia, Squamata, Gymnophthalmidae) in southeastern
Brazil.Herpetology Notes. 4. 173- 176.
RIES, L.; FLETCHER, R. J. J.; BATTIN, J. & SISK, T. D. 2004. Ecological responses to
habitat edges: mechanisms, models and variability explained. Annual Review of
Ecology, Evolution and Systematics 35:491–522.
RODRIGUES, M.T. 2005. Conservação dos répteis brasileiros: os desafios para um país
megadiverso. Megadiversidade. 1(1): 87-94.
RODRIGUES, M. T. 1987, Sistemática, ecologia e zoogeografia dos Tropidurus do grupo
torquatus ao sul do Rio Amazonas (Sauria, Iguanidae). Arq. Zool., São Paulo, 31:105-
230.
RODRIGUES, M. T.; FREIRE, E. M. X.; PELLEGRINO, K. C. M. & SITES Jr, J. W. 2005.
Phylogenetic relationships of a new genus and species of microteiid lizard from the
Atlantic forest of north-eastern Brazil (Squamata, Gymnophthalmidae).Zoological
Journal of the Linnean Society. 144, 543–557.
RODRIGUES, M.T.; DIXO, M.; PAVAN, D. & VERDADE, V.K. 2002.A new species of
Leposoma (Squamata, Gymnophthalmidae) from the remnant Atlantic forests of the
state of Bahia, Brazil.Pap. Avulsos Zool. (São Paulo) 42:335-350.
RODRIGUES, M.T.;FREITAS, M. A.; SILVA, T. F. S. & BERTOLOTTO, C. E. V. 2006. A
new species of lizard genus Enyalius (Squamata, Leiosauridae) from the highlands of
Chapada Diamantina, state of Bahia, Brazil, with a key to species. Phyllomedusa
5(1):11-24.
SANTOS, A. M. & TABARELLI, M. 2002. Distance from roads and cities as a predictor of
habits loss and fragmentation in the Caatinga dry forest of Brasil. Revista Brasileira de
Botânica. 62: 897-905.
SANTOS, R.C.; LUCAS, P. S.; SOUSA, B. M. & NOVELLI, I. A. 2009. Reptilia, Squamata,
Leiosauridae, Urostrophus vautieri: Distribution extensionand geographic distribution
map. Check List 5: 533-536.

15
SÃO PEDRO, V. A & FEIO, R. N. 2010. Distribuição espacial e sazonal de Anuros em três
ambientes na Serra de Ouro Branco, extremo sul da Cadeia do Espinhaço, Minas
Gerais, Brasil. Biotemas, 23 (1): 143-154.
SÃO PEDRO, V. A & FEIO, R. N. 2011. Anuran species composition from Serra do Ouro
Branco, southernmost Espinhaço Mountain Range, state of Minas Gerais, Brazil. Check
List,Vol.7. (5).
SÃO PEDRO, V. A & PIRES, M. R. S. 2009. As Serpentes da região de Ouro Branco,
Extremo Sul da Cadeia do Espinhaço, Minas Gerais. Ceres 56 (2): 166-171.
SAZIMA, I.; SAZIMA, C.& SAZIMA, M. 2005. Little dragons prefer flowers to maidens: a
lizard that laps nectar and pollinates trees. Biota Neotropica v5(n1)
http://www.biotaneotropica.org.br/v5n1/pt/abstract?shortcommunication+BN0080501.
SILVA, T. F.; ANDRADE, B. F. E.; TEIXEIRA, R. L. & GIOVANELLI, M. 2003. Ecologia
de Ameiva ameiva ( sauria, Teiidae), na Restinga do Guriri, São Mateus, Espirito Santo,
sudeste do Brasil. Boletim Museu de Biologia Mello Leitão (N. Sér.)15:5-15.
SILVEIRA, A. L.; PIRES, M. R. & COTTA, G.A. 2010. Serpentes de uma área de transição
entre o Cerrado e a Mata Atlântica, no sudeste do Brasil. Arquivos do Museu Nacional,
Rio de Janeiro.
SKOLE, D. & TUCKER, C. 1993. Tropical deforestation and habitat fragmentation in the
Amazon: satellite data from 1978 to 1988. Science 260: 1905-1910.
SOCIEDADE Brasileira de Herpetologia. 2012. List of Brazilian reptile species.
http://www.sbherpetologia.org.br (último acesso em 05/01/2012).
SOUSA, B. M. & CRUZ, C. A. G. 2008. Hábitos alimentares em Enyalius perditus
(Squamata, Leiosauridae) no Parque Estadual do Ibitipoca, Minas Gerais. Iheringia
98(2): 260-265.
SOUSA, B. M.; NASCIMENTO, A. E. R.; GOMIDES, S. C.; VARELA RIOS, C. H.;
HUDSON, A. H. & NOVELLI, I. A. 2010. Reptiles in fragments of Cerrado and
Atlantic Forest at the Campo das Vertentes, MinasGerais State, Southeastern

16
Brazil.BiotaNeotrop.10(2):http://www.biotaneotropica.org.br/v10n2/en/abstract?article+
bn0351002.
STURARO, M. J. & ÁVILA-PIRES, T. C. S. 2011. Taxonomic revision of the geckos of the
Gonatodes concinnatus complex (Squamata: Sphaerodactylidae), with description of
two new species. Zootaxa 2869: 1-36.
STURARO, M. J. & SILVA, V. X. 2010.Natural history of the lizard Enyalius perditus
(Squamata: Leiosauridae) from an Atlantic forest remnant in southeastern Brazil.
Journal of Natural History 44 (19-20): 1225-1238.
TEIXEIRA, R. L. & GIOVANELLI, M. 1999. Ecologia de Tropidurus torquatus
(Sauria:Tropiduridae) da Restinga de Guriri, São Mateus, ES. Rev. Brasil. Biol., 59(1):
11-18.
TORRES-CARVAJAL, O. 2005.A new species of Stenocercus (Squamata, Iguanidae) from
central-western Brazil with a key to Brazilian Stenocercus.Phyllomedusa 4:123-132.
TROMBULAK , S. C. & FRISSELL, C. A. 2000. Review of ecological effects of roads on
terrestrial and aquatic communities. Conservation Biology.p 18-30. Vol. 14, No. 1.
VAN SLUYS, M.; FERREIRA, V. M. & ROCHA, C.F. D. 2004.Natural history of the lizard
Enyalius brasiliensis (LESSON,1828) (Leiosauridae) from Atlântic Forest of
southeastern Brazil. Brazilian j. Biol., 64(2): 353-356.
VANZOLINI, P. E. 1988. Distributional patterns of South American lizards. In:
VANZOLINI, P.E. & HEYER, W.R. (Eds.) Proceedings of a Workshop on Neotropical
Distribution Patterns.
VITT, L. J. 1991. An introduction to the ecology of Cerrado lizards. Journal Herpetol., 25:79-
90.
VITT, L. J. & CARVALHO C. M. 1995.Niche partitioning in a tropical wet season: lizards in
the lavrado area of northern Brazil.Copeia 2, 305-29.
WERNECK, F. P & COLLI, G. R. 2006.The lizard assemblage from Seasonally Dry Tropical
Forest enclaves in the Cerrado biome, Brazil, and its association with the Pleistocenic
Arc. J. Biogeogr. 33:1983-1992.

17
ZAHER, H.; BARBO, F. E.; MARTÍNEZ, P. S; NOGUEIRA, C.; RODRIGUES, M. T. &
SAWAYA R. J. 2011. Reptiles from São Paulo State: current knowledge and
perspectives. Biota Neotrop.11(1a): http://www.biotaneotropica.
org.br/v11n1a/en/abstract?inventory+bn0051101a.
ZAMPROGNO, C.; ZAMPROGNO, M. das G. F. & TEIXEIRA, R. L. 2001. Evidence of
terrestrial feeding in the arboreal lizard Enyalius bilineatus (Sauria, Polychrotidae) of
South-Eastern Brazil.Rev. Brasil. Biol., 61(1): 91-94.

18
CAPÍTULO 1: FAUNA DE LAGARTOS (SQUAMATA, “SAURIA”) DA SERRA DE OURO BRANCO, PORÇÃO SUL DA SERRA DO ESPINHAÇO, MINAS GERAIS

19
RESUMO:
O presente estudo avaliou a fauna de lagartos da erra de Ouro Branco, Minas Gerais, Brasil,
uma área de transição entre a Mata Atlântica e o Cerrado. Os dados foram coletados com o
uso de armadilhas de interceptação e queda (pitfall traps), procura ativa, encontros ocasionais,
além de informações de outras coleções e análise de material disponível na literatura. As
amostragens em campo foram realizadas em dois estágios em um total de 36 meses: de
dezembro de 2006 a dezembro de 2008, e de janeiro a dezembro de 2010. A área de estudos
apresentou 15 espécies de lagartos distribuídas por oito famílias: Anguidae, Gekkonidae,
Gymnophthalmidae, Leiosauridae, Polychrotidae, Mabuyidae, Teiidae e Tropiduridae. Os
lagartos encontrados neste estudo representam um número relevante de espécies para este
grupo zoológico que habitam áreas de transição entre a Mata Atlântica e o Cerrado. A área
também apresenta espécies exclusivamente florestais e endêmicas da Mata Atlântica, como
Ecpleopus gaudichaudii e Enyalius perditus, para além de espécies típicas do Cerrado e
comum aos dois biomas.
Palavras chave: Lagartos; Serra de Ouro Branco; Cadeia do Espinhaço, Ecótone.

20
ABSTRACT:
The present study evaluated the lizard fauna in Serra do Ouro Branco, Minas Gerais, Brazil, a
transition area between the Atlantic Forest and Cerrado. Data was collected using pitfall traps,
active and occasional encounters, and through information from zoological collections and the
literature. Field sampling was performed in two stages over a period of 36 months: from
December 2006 to December 2008, and from January to December 2010. The study area is
home to 15 species belonging to eight families: Anguidae, Gekkonidae, Gymnophthalmidae,
Leiosauridae, Polychrotidae, Mabuyidae, Teiidae, and Tropiduridae. Lizard species evaluated
in this study represent a relevant asset for this zoological group inhabiting the transitional
areas between the Atlantic Forest and Cerrado. They include strictly forest species and
endemic Atlantic Forest species such as Ecpleopus gaudichaudii, and Enyalius perditus, in
addition to their typical Cerrado forms, however, common to both biomes.
Keywords:Lizards; Serra de Ouro Branco; Cadeia do Espinhaço, Ecótone.

21
INTRODUÇÃO
Os lagartos são répteis Lepidosauria e constituem um dos grupos mais diversificados entre os
Amniota com cerca de 4.800 espécies (Pough et al., 2008). A diversidade de formas,
tamanhos, cores, comportamentos incluindo estratégias reprodutivas e de forrageio, uso de
habitats, e uma grande variedade de itens alimentares utilizados na dieta, faz com que este
grupo ocorra em quase todos os ecossistemas globais, com exceção das regiões polares
(Pough et al., 2008), sendo registradas 248 espécies para o Brasil, um dos países mais
biodiversos quanto ao grupo (Bérnils & Costa, 2011). Entre os vertebrados, os lagartos são
considerados organismos modelos. A análise da composição de suas comunidades, quer no
estudo de impactos antrópicos na fauna (Jellinek et al., 2004; Kitchener et al., 1980, Gibbons
et al., 2000; Gardner et al., 2007; Dixo & Martins, 2008; Lehtinen et al., 2003; Bell &
Donnelly, 2006; Dixo & Metzger, 2009), ou na interpretação de dados sobre padrões de
distribuição e endemismo de espécies entre ecossistemas (Pianka, 1973; Vanzolini et al.,
1980; Huey et al., 1983; Rodrigues, 1987; 2005; Recorder & Nogueira, 2007), são exemplos
de contribuições científicas que servem de base para políticas e ações voltadas à conservação
da biodiversidade.
Estudos recentes sobre a composição de espécies de Squamata no estado de Minas Gerais se
concentram principalmente em trabalhos sobre serpentes (Brites & Bauab, 1988; Bertoluci,
1998; Assis, 1999; Palmuti et al., 2009; São Pedro & Pires, 2009; Costa et al., 2010; Silveira
et al., 2010), havendo menor número de levantamentos citando a ocorrência de lagartos (Feio
& Caramashi, 2002; Recorder & Nogueira, 2007; Bertoluci et al., 2009; Costa et al., 2009;
Sousa et al., 2010). A diversidade de ecossistemas que o estado de Minas Gerais apresenta,
como o Cerrado, a Mata Atlântica e a Caatinga, além das áreas de altitude, tal como a Serra
do Espinhaço, permitem a sobrevivência de aproximadamente 60 espécies de lagartos (Bérnils

22
et al., 2009), o que representa cerca de 23% da fauna de lagartos do Brasil (Bérnils & Costa,
2011). Apesar disso, ainda existem grandes lacunas de conhecimento, devido a ausência de
informações sobre distribuição, ocorrência, ecologia e história natural para a maioria das
espécies.
A Cadeia do Espinhaço é uma formação montanhosa que se estende por cerca de 1000 km do
estado de Minas Gerais até a Bahia. Reconhecida como uma região prioritária para a
conservação de flora e fauna, esse conjunto de serras foi decretado pela UNESCO, em 2005,
Reserva da Biosfera, devido ao alto grau de endemismo de fauna e flora, associadas a
aspectos humanos ligados à cultura. Entretanto, grande parte da sua diversidade permanecem
ainda pouco conhecidas (Drummond et al., 2005; Costa et al., 1998), sendo que a maior parte
dos estudos são concentrados em poucas áreas (Silva et al. 2008).
A serra do Ouro Branco, localizada no extremo sul do Espinhaço, passou a contar a partir de
2009, com duas Unidades de Conservação: o Parque Estadual da Serra de Ouro Branco e o
Monumento Natural Estadual de Itatiaia todos no município de Ouro Branco, Minas Gerais.
Estas unidades devem proteger toda a biodiversidade das áreas de Campo Rupestre, Cerrado,
Mata Atlântica e da zona de transição entre esses biomas. Em esforços recentes, estudos
relacionados à herpetofauna da região, nomeadamente sobre serpentes (São Pedro & Pires,
2009; Silveira et al., 2010), anfíbios (Leite et al., 2008; São Pedro & Feio, 2010; 2011) e a
relação e percepção dos povoados em torno das Unidades de Conservação sobre a fauna local
foram desenvolvidos (Pires et al., 2010; Mateus et al. 2011; Pinto, 2011), contribuindo para o
conhecimento da fauna e das relações humanas com a biodiversidade.

23
Visando o maior conhecimento da biodiversidade da serra do Ouro Branco, porção sul da
Cadeia do Espinhaço, o presente estudo apresenta uma lista de espécies de lagartos da área,
com comentários quanto à história natural, ecologia, além de uma chave taxonômica artificial
para essas espécies.
MATERIAIS E MÉTODOS
Área de estudos
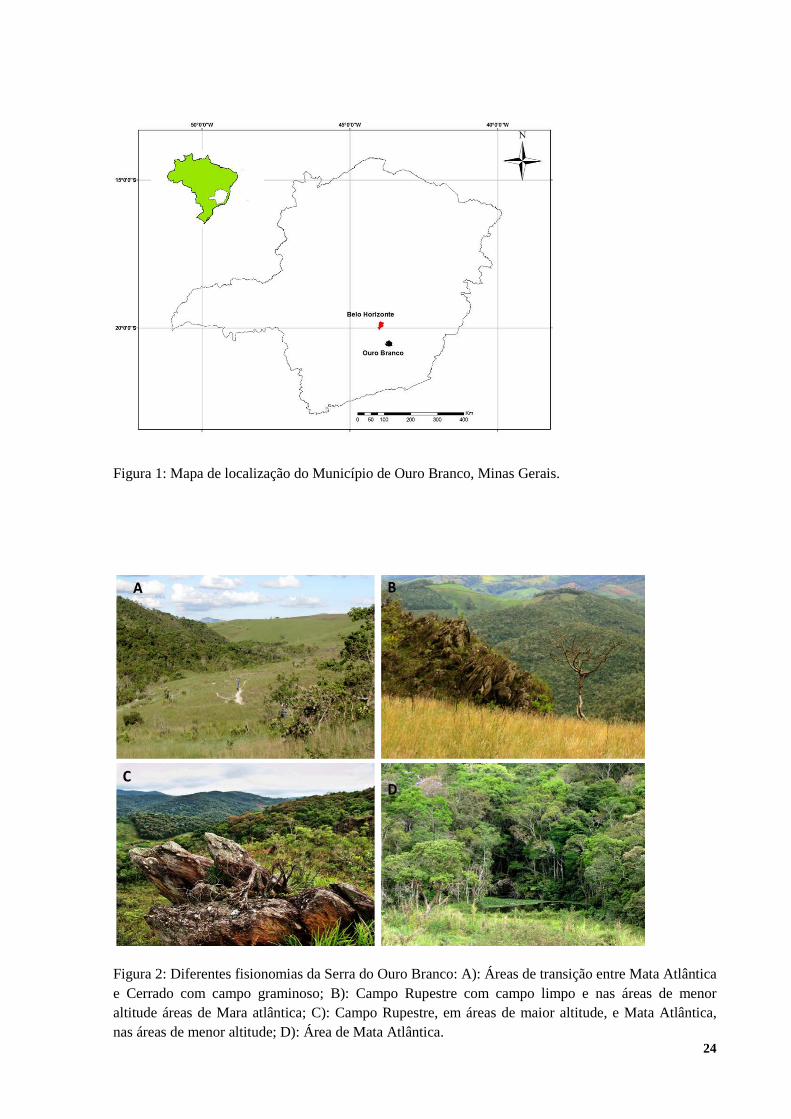
O estudo foi realizado na serra do Ouro Branco, situada no município de Ouro Branco, a 96
km de Belo Horizonte e a 20 km de Ouro Preto, MG. A área de estudo está inserida ao sul do
Quadrilátero Ferrífero e da Serra do Espinhaço, sendo o setor mais meridional destas duas
delimitações geográficas (figura 1). As formações florestais podem ocupar áreas transitórias
entre os campos rupestres, onde o solo permite a ocupação por elementos arbóreos, sendo
classificadas como Florestas Estacionais Semideciduais (Veloso et al., 1991). A altitude da
região varia entre 1250 e 1568 metros (Paula, 2003), o clima predominante é mesotérmico
(Cwb, segundo a classificação de Köppen, 1936). A temperatura média anual entre os anos de
1997 a 2010, foi de 20,7ºC e a precipitação média anual no mesmo período foi de 1321,07
mm, sendo que as chuvas se distribuíram principalmente entre os meses de novembro a
fevereiro, com uma estação seca no inverno (fonte Estação Meteorológica da Gerdau
Açominas, localizada nas proximidades dos locais de coletas).
24
Figura 1: Mapa de localização do Município de Ouro Branco, Minas Gerais.
Figura 2: Diferentes fisionomias da Serra do Ouro Branco: A): Áreas de transição entre Mata Atlântica e Cerrado com campo graminoso; B): Campo Rupestre com campo limpo e nas áreas de menor altitude áreas de Mara atlântica; C): Campo Rupestre, em áreas de maior altitude, e Mata Atlântica, nas áreas de menor altitude; D): Área de Mata Atlântica.

25
Coleta dos dados
O presente trabalho é resultado de três anos de coletas na serra do Ouro Branco, sendo que os
dados foram obtidos em duas etapas: a primeira de dezembro de 2006 a dezembro de 2008 e a
segunda, de janeiro a dezembro de 2010. Devido aos diferentes ambientes que compõem a
serra do Ouro Branco e a variedade de hábitats de ocorrência de lagartos, foram utilizados três
métodos de coleta de forma combinada, sendo eles, armadilha de interceptação e queda ou
pitfall traps (Cechin & Martins, 2000), procura ativa (Crump & Scott Jr., 1994) e encontros
ocasionais. Para complementar o inventário de espécies foram examinados espécimes
depositados na coleção de Zoologia dos Vertebrados da Universidade Federal de Ouro Preto,
(LZV-UFOP) e na coleção de lagartos do Museu de Zoologia João Moojen da Universidade
Federal de Viçosa (MZUFV), Minas Gerais, além de consultas à literatura disponível.
Armadilhas de interceptação e queda
Na primeira etapa foram instaladas em cinco áreas florestadas distintas (20°23’45’’S
43°30’34’’O; 20°29’45’’S 43°37’31’’O;20°29’58’’S 43°37’43’’O; 20°30’00’’S 43°42’48’’O;
20°29’59’’S 43°42’56’’O) armadilhas do tipo pitfall contendo cada uma 10 baldes de 60
litros. Os baldes foram enterrados até o nível do solo, a intervalos de quatro metros de
distância e interligados por cercas direcionadoras de aproximadamente 100 cm de altura, que
foram confeccionadas com lona plástica e sustentadas por estacas de madeira. As armadilhas
permaneceram abertas por cinco dias consecutivos a cada mês, totalizando um esforço
amostral de 50 baldes por dia, 250 por mês, e 3000 baldes dia por ano.

26
Na segunda etapa também foram instaladas armadilhas do tipo pitfall, com cercas
direcionadoras incluindo novas áreas de amostragem e um maior número de baldes dispostos
diferentemente dos transectos anteriores. Cada armadilha foi composta por quatro baldes
dispostos em formato de ‘Y’, ou seja, três baldes periféricos interligados ao balde central
através de uma cerca direcionadora, com dimensões de 1 metro de altura e 5 metros de
comprimento. Foram selecionadas nove áreas de amostragem, e em cada uma foram
instaladas três armadilhas, distanciadas a 50 metros entre si. Nesta metodologia só foram
georreferenciadas as armadilhas centrais de cada área sendo elas: 20°29'43.64"S
43°35'49.71"O; 20°29'45.00"S 43°35'48.53"O; 20°29'44.56"S 43°35'46.53"O; 20°29'35.13"S
43°36'24.17"O; 20°29'37.08"S 43°36'25.55"O; 20°29'39.07"S 43°36'20.88"O; 20°29'50.11"S
43°37'24.93"O; 20°29'58.98"S 43°37'29.60"O; 20°30'7.35"S 43°37'34.39"O.
Em todas as etapas deste trabalho as armadilhas do tipo pitfall permaneceram abertas por
cinco dias/mês, totalizando um esforço amostral de 108 baldes por dia, 540 por mês e 6400 no
ano. Este tipo de armadilha foi utilizado exclusivamente nos ambientes de mata, onde foi
possível encontrar solos suficientemente profundos para a sua instalação. Cabe ainda ressaltar
que, tanto na primeira como na segunda etapa, os baldes foram inspecionados no período de
manhã, para evitar predação ou morte de indivíduos capturados.
Pelo método de procura ativa (Crump & Scott Jr., 1994), os répteis foram amostrados através
de vasculha nos ambientes mais propícios ao seu encontro (Vanzolini & Papavero, 1967,
Forlani et al., 2010). Para esse método de coleta, foram vasculhadas tanto as áreas de mata,
como áreas de campo, contemplando diferentes fisionomias vegetais, tais como, campo
graminoso, campo rupestre, vegetação de velózias, matas de galeria e áreas de florestas. Os
lagartos encontrados por meio de procura ativa foram capturados à mão, sempre que possível.
Eventualmente, somente anotações, descrições e registros fotográficos puderam ser efetuados
para confirmação posterior da identificação. Para os indivíduos capturados ou avistados,

27
foram anotados em caderno de campo informações como: espécie, data e horário de captura,
tipo de ambiente (floresta, campo rupestre, clareiras e bordas e áreas urbanas/antropizadas),
micro hábitats (vegetação, tronco, folhiço, interior de cupinzeiros, reentrâncias nas rochas,
tocas de mamíferos, entre outros). Essa atividade foi praticada em campanhas diurnas (09h00-
15h00) e noturnas (18h00- 22h00) não consecutivas, por dois ou mais pesquisadores que se
deslocavam a pé.
Encontros casuais ou ocasionais
Este método de coleta consiste no registro ocasional de lagartos em áreas naturais ou urbanas
como estradas, muros, cercas e interior de habitações humanas, segundo São Pedro & Pires
(2009). O registro ocasional também ocorreu ao longo do percurso nas trilhas de acesso às
áreas de armadilhas de interceptação e queda. Para os indivíduos coletados ou registrados por
este método foram seguidos os mesmos procedimentos de descrição mencionados
anteriormente para a procura ativa.
Após o término de cada coleta, alguns espécimes foram sacrificados com o uso de xilocaína
0,5%. Amostras de tecido foram extraídas e conservadas no banco de tecido de répteis do
LZV-UFOP. Em seguida os espécimes foram fixados com formol a 10%, preservadas em
álcool a 70% e depositados na coleção herpetológica do mesmo Laboratório.
RESULTADOS
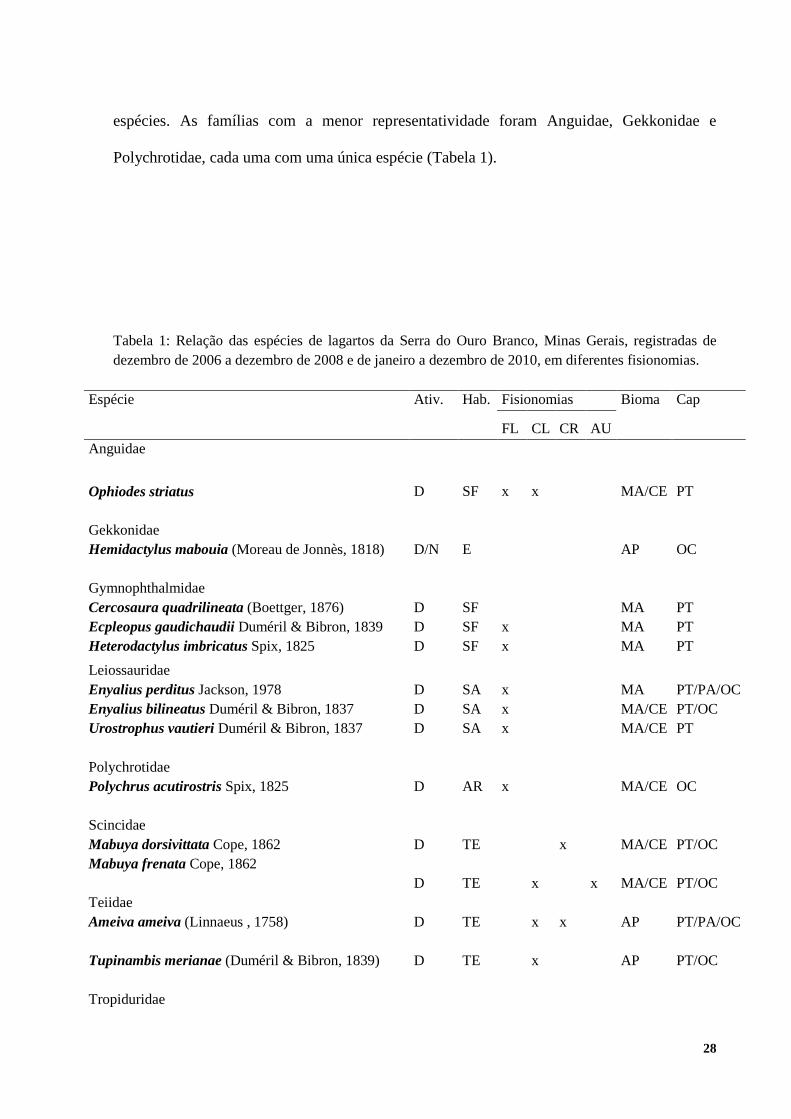
Foram registradas para Serra do Ouro Branco 15 espécies de lagartos, distribuídas em oito
famílias. As famílias com maior representatividade foram Gymnophthalmidae e Leiosauridae,
com três espécies cada uma, seguidas de Scincidae, Teiidae, e Tropiduridae com duas
28
espécies. As famílias com a menor representatividade foram Anguidae, Gekkonidae e
Polychrotidae, cada uma com uma única espécie (Tabela 1).
Tabela 1: Relação das espécies de lagartos da Serra do Ouro Branco, Minas Gerais, registradas de dezembro de 2006 a dezembro de 2008 e de janeiro a dezembro de 2010, em diferentes fisionomias.
Espécie Ativ. Hab. Fisionomias Bioma Cap
FL CL CR AU Anguidae
Ophiodes striatus D SF x x
MA/CE
PT
Gekkonidae Hemidactylus mabouia (Moreau de Jonnès, 1818) D/N E
AP OC
Gymnophthalmidae Cercosaura quadrilineata (Boettger, 1876) D SF
MA PT
Ecpleopus gaudichaudii Duméril & Bibron, 1839 D SF x
MA PT Heterodactylus imbricatus Spix, 1825 D SF x
MA PT
Leiossauridae Enyalius perditus Jackson, 1978 D SA x
MA PT/PA/OC
Enyalius bilineatus Duméril & Bibron, 1837 D SA x
MA/CE PT/OC Urostrophus vautieri Duméril & Bibron, 1837 D SA x
MA/CE PT
Polychrotidae Polychrus acutirostris Spix, 1825 D AR x
MA/CE OC
Scincidae Mabuya dorsivittata Cope, 1862 D TE
x
MA/CE PT/OC
Mabuya frenata Cope, 1862 D TE
x
x MA/CE PT/OC
Teiidae Ameiva ameiva (Linnaeus , 1758) D TE
x x
AP PT/PA/OC
Tupinambis merianae (Duméril & Bibron, 1839) D TE
x
AP PT/OC
Tropiduridae

29
Tropidurus itambere Rodrigues, 1987 D TE
x
MA/CE PA Tropidurus torquatus Wiegmann, 1934 D TE
x x x MA/CE PA/OC
Legendas: FL- florestais, CL- clareiras, CR- Campo Rupestre, AU- áreas urbanas, Distribuição dessas espécies entre os Biomas: MA - Mata Atlântica, CE- Cerrado, AP- ampla distribuição; Ativ-Atividade, Hab-hábito, D – diurna, N – noturna, Hábitos: SF - semi fossorial, TR- terrícola, SA- semi- arborícola, AR- arborícola, E- escalador; Modo de Captura: Cap- modo de captura, PT- pitfall, PA - procura ativa, OC - encontro ocasional.
Como pode se observar na figura 1, com exceção do geconídeoHemidactylus mabouia, todas
as espécies registradas apresentam atividade diurna. Os indivíduos desta espécie foram
registrados a noite em paredes próximas a fontes de luz, possivelmente predando pequenos
invertebrados. Quanto ao hábitat, 11 espécies pertencentes a seis famílias foram encontradas
próximas a áreas de borda de mata e clareiras, sendo elas: Ophiodes striatus, Enyalius
perditus, Enyalius bilineatus, U. vautieri, Mabuya dorsivittata, Mabuya frenata, E.
gaudichaudii, H. imbricatus, Ameiva ameiva, Tupinambis merianae, e ainda Tropidurus
torquatus. T. merianae e A. ameiva também foram observados em pequenas clareiras de mata,
resultantes do corte de árvores próximos aos locais de coleta.
Todos os lagartos das famílias Gymnophthalmidae, Leiosauridae e Polychrotidae registradas
para a região, ou seja, 47% do total das espécies são consideradas estritamente florestais. Para
as áreas urbanas, foram registradas três espécies, sendo elas, H. mabouia, M. frenata, e T.
torquatus. Esta última foi a espécie mais generalista, ocupando todos os tipos de hábitat,
exceto o interior de floresta, enquanto que H. Mabouia foiparticularmente associada aos
domicílios. Para as áreas de Campo Rupestre, foram registradas quatro espécies: M.
dorsivittata, T. itambere, A. ameiva e T. torquatus, sendo que as duas primeiras só ocorreram
neste hábitat.
As espécies M. frenata, M. dosivittata, A. ameiva, T. merianae, T. torquatus e T. itambere,
correspondendo a cerca de 40% da riqueza amostrada, apresentaram hábito terrícola,

30
emboraos tropidurídeos também foram observados sobre troncos de árvores em bordas
florestais. Todas as espécies pertencentes à família Gymnophthalmidae foram encontradas
somente em ambientes florestais, assim como o anguídeo Ophiodes striatus. As espécies
pertencentes à família Leiosauridae apresentaram hábitos arborícola e semi-arborícolas.

31
Figura 2. Lagartos registrados na Serra do Ouro Branco, porção sul da Cadeia do Espinhaço, Minas Gerais, Brasil. a) Ophiodes striatus; b) Hemidatylus mabouia; c) Cercosaura quadrilineata; d) Ecpleopus gaudichaudii; e) Heterodactylus imbricatus(foto: Leandro Drummmond); f) Enyalius bilineatus; g) Enyalius perditus(foto: Leandro Drummmond); h) Urostrophus vautieri(foto: Leandro Drummmond); i) Polychrus acutirostris(foto: Pedro Bernardo); j) Mabuya frenata(foto: Leandro Drummmond); k) Mabuya dorsivittata(foto: Henrique C. Costa); l)Ameiva ameiva(foto: Pedro Bernardo); m) Tupinambis merianae; n) Tropidurus torquatus; o) Tropidurus itambere(foto: Leandro Drummmond).

32
A espécie M. dorsivittata foi registrada para a região a partir um indivíduo coletado na serra
do Ouro Branco e tombado no MZUFV, não tendo sido observada no presente estudo. Onze
espécies foram capturadas com a utilização de armadilhas de interceptação e queda, sendo que
os representantes da família Gymnophthalmidae, bem como O. striatus e U. vautieri, só
foram capturadas nas armadilhas de interceptação e queda. Quatro espécies foram registradas
por procura ativa, sendo elas: E. perditus, A. ameiva, além das duas espécies da família
Tropiduridae, sendo que T. itambere foi coletado exclusivamente por meio deste método de
captura. Nove espécies foram encontradas ocasionalmente durante a execução dos trabalhos
de campo, sendo que duas espécies, H. mabouya e P. acutirostris, foram capturadas
exclusivamente por este método.
DISCUSSÃO
As 15 espécies de lagartos registradas para a Serra do Ouro Branco correspondem a
aproximadamente 26, 31 % da fauna de lagartos registradas para Minas Gerais (Bérnils et al.,
2009) e 6% das espécies de lagartos brasileiros (Bérnils & Costa, 2011). Colli et al. (2002) e
Nogueira, (2006) afirmam que em localidades bem amostradas do Cerrado, a estimativa de
riqueza de lagartos é de 13 a 28 espécies, sendo essa relação desconhecida para a Mata
Atlântica e áreas de transição entre esses dois biomas. De fato, as comparações não são
conclusivas, visto que diferenças na extensão das áreas de amostragem e as metodologias
utilizadas em cada bioma e locais de estudo podem influenciar diretamente na relação de
espécies (Rodrigues, 2005). No entanto segundo Rodrigues (2005), a fauna de lagartos em
conjunto com os anfisbenídeos é estimada em 70 espécies para todo o Cerrado, 67 para a
Mata Atlântica e 12 para áreas de transição, sendo os Campos Rupestres inclusos em áreas de

33
transição pelo autor. No presente trabalho foram encontradas 13 espécies já registradas na
Mata Atlântica, sendo quatro delas endêmicas desse bioma, oito espécies para o Cerrado, e
nove já registrados em regiões de transição entre esses dois biomas.
De acordo com a literatura disponível, a maior parte dos lagartos coletados na Serra do Ouro
Branco, cerca de 60%, partilham tanto áreas do domínio da Mata Atlântica, como também do
Cerrado. Porém, a Mata Atlântica foi o bioma com maior número de espécies registradas,
apresentando inclusive quatro espécies consideradas endêmicas desse domínio como C.
quadrilineatus, E. gaudichadi, H. Imbicatus, e E. perditus. Nenhuma espécie registrada é
endêmica do Cerrado. As espéciesA. ameiva, H. mabouia, e T. merianae apresentam ampla
ocupação e distribuição geográfica.
Em Minas Gerais, próximo a divisa com os estados de Goiás e da Bahia, Recorder &
Nogueira (2007) registraram 25 espécies de lagartos em uma região de Cerrado, com clima e
topografia bem distintos de uma região de altitude, como é caso a Serra do Ouro Branco.
Entretanto todas as espécies das famílias Tropiduridae, Scincidae e Anguidae e a espécie A.
ameiva, encontradas em Ouro Branco também foram reportadas por Recorder & Nogueira
(2007). Uma explicação para essa semelhança faunística pode ser a preferência dessas
espécies por áreas abertas. Todas essas espécies em comum preferem áreas abertas, sendo
essas mais comuns no Cerrado. Os Teiideos, por exemplo, ocorrem em áreas abertas pela ação
humana (Sartorius et al., 1999), o que na região do presente estudo foi frequentemente
observado. Além de um exemplar muito jovem coletado no pitfall, essa espécie só foi avistada
em áreas permanentemente abertas para a passagem de linhas de transmissão de energia
elétrica e locais com cortes recentes de árvores.
Para o município de Viçosa, Minas Gerais, nos domínios da Mata Atlântica, foram reportadas
12 espécies de lagartos, a partir da coleção do Museu de Zoologia João Moojen (Costa et al.,

34
2009). Dentre as espécies encontradas no município de Viçosa, O. cf striatus, H. mabouia, E.
gaudichaudii, E. bilineatus, M. dorsivittata, A. ameiva, T. merianae e T. torquatus, também
foram encontradas na área do presente estudo. Todavia, exceto E. gaudichaudii, endêmica da
Mata Atlântica, e H. mabouia, que prefere ambientes antropizados (Benesi, 2007; Costa et al.,
2009; Forlani et al., 2010), todas as espécies encontradas em Viçosa também foram
reportadas nos domínios do Cerrado (Recorder & Nogueira, 2007) em uma área de
preservação. Isso mostra a flexibilidade que algumas espécies de lagartos podem exibir, sendo
as suas distribuições registradas em dois biomas tão distintos como a Mata Atlântica e o
Cerrado. No entanto, verificou-se também que as espécies em comum preferem ambientes
abertos aos ambientes de mata.
Quanto a trabalhos realizados em áreas de transição (Bertoluci et al., 2009; Sousa et al.,
2010), o presente estudo apresentou maior riqueza de espécies de lagartos. Todas as espécies
de lagartos registrados por Bertoluci et al. (2009), na Estação Ambiental de Peti, Minas
Gerais, nomeadamente: Enyalius cf bilineatus, H. mabouia, A. ameiva, T. merianae e T.
torquatus exceto Ophiodes sp.foram amostrados em Ouro Branco. No entanto, Bertoluci et al.
(2009) não utilizou armadilhas do tipo pitfall, podendo a comunidade de lagartos da
serapilheira ter sido sub amostrada, nomeadamente os pertencentes a família
Gymnophthalmidae. Esses lagartos apresentam em geral um tamanho muito reduzido, sendo
de difícil captura e visualização em meio à serapilheira.
Segundo Sousa et al. (2010), no Campo das Vertentes em Minas Gerais, que também
constitui uma área de transição, vivem em simpatria nove espécies de lagartos distribuídas em
oito famílias. Dessas, oito espécies foram comuns ao presente trabalho. Essa similaridade
pode ser explicada tanto pela presença de espécies com ampla distribuição nos dois biomas,
como também pela similaridade topográfica entre as duas áreas. Tanto o campo das Vertentes,

35
quanto a Serra do Ouro Branco são áreas de altitude elevada compostas por fragmentos
florestais de Mata Atlântica e áreas de Cerrado.
Toda a semelhança da fauna de lagartos encontrada nas diversas regiões demonstra realmente
que a área do presente estudo reúne uma fauna de lagartos com espécies representativas dos
dois biomas, e destaca a importância das áreas de transição para o entendimento de como e
quais espécies avançam para as fronteiras entre o Cerrado e a Mata Atlântica, o que pode
fornecer respostas importantes para a preservação desse grupo animal.
Algumas espécies como T. torquatus, T. merianae e A. ameiva, aparentemente parecem se
beneficiar de intervenções antrópicas, principalmente aquelas relacionadas à abertura de
clareiras nas florestas. No entanto, as espécies das famílias Gymnophthalmidae, Leiosauridae
e Polychrotidae, ou seja, 47% do total das espécies do presente estudo são consideradas
estritamente florestais (Jackson, 1978; Eisemberg et al., 2004; Dixo & Verdade, 2006). Esses
lagartos são sensíveis a mudanças no microclima e problemas associados à mudança na
vegetação podem causar alterações ou mesmo extinção de suas populações (Barlow & Peres
2004; Román-Cuesta & Martínez-Vivalta, 2006).
Corroborando com Cechin & Martins (2000), as armadilhas de interceptação foram eficientes
na representação da comunidade dos lagartos, capturando cerca de 70 % da riqueza de
espécies registradas para a Serra do Ouro Branco. Essas armadilhas foram mais eficientes na
captura de espécies de serapilheira, mas por meio delas foi possível a captura de espécies de
hábitos arborícolas e semi-arborícolas.
Nenhum lagarto da Serra do Ouro Branco está incluído na lista de espécies ameaçadas para o
Estado de Minas Gerais. No entanto, Urostrophus vautieri consta como vulnerável, na lista
das espécies da fauna ameaçada de extinção do Estado do Rio Grande do Sul (Marques et al.,
2002) e E. perditus possivelmente ameaçada no estado do Rio de Janeiro (Rocha et al., 2000).
Vale lembrar que estudos envolvendo a fauna de lagartos estão ainda em fase exploratória, e

36
nenhum grupo deve ser considerado como sendo de pouca importância biológica, sem
exaustivos inventários e pesquisas de campo (Rodrigues, 2005).
LISTA COMENTADA DOS LAGARTOS REGISTRADOS PARA A SERRA DO OURO BRANCO, MINAS GERAIS
Família Anguidae
Ophiodes striatus (Spix, 1825) apresenta corpo cilíndrico e alongado, membros anteriores
ausentes e membros posteriores vestigiais. É um animal denominado popularmente por
‘cobra de vidro’ devido à aparência serpentiforme e à facilidade com que realiza a autotomia
caudal quando manipulado. Apresenta hábito diurno e reprodução vivípara (Bernardo & Pires,
2006). Sua alimentação consiste basicamente de artrópodes, sendo Blatodea, Aranae e
Orthoptera os itens mais frequentes em sua dieta, podendo também consumir pequenos
lagartos e serpentes (Barros & Teixeira, 2007). É uma espécie semi-fossorial e pode ser
encontrada na Argentina, Uruguai e Brasil, onde ocorre principalmente em áreas de altitude
(Vaz-Ferreira & Soriano, 1960; Gallardo, 1966; Cunha, 1961; Peters et al., 1970). Até o
presente momento, o gênero Ophiodes inclui O. striatus e mais duas espécies. Entretanto
Borges-Martins (1998), mencionou que ainda restam algumas espécies a serem descritas.
Família Gekkonidae
Hemidactylus mabouia Moreau de Jonnès, 1818 são animais de pequeno porte, ovíparos,
sendo que as fêmeas podem apresentar estocagem e armazenamento de esperma no tubo
uterino (Nogueira et al., 2011), aspecto que certamente contribuiu para o sucesso da
introdução dessa espécie nas zonas tropicais. É uma espécie exótica originada do continente
africano (Carranza & Arnold, 2006; Vanzolini et al., 1980; Ávila-Pires, 1995), sendo

37
presente também em ilhas oceânicas (Carranza & Arnold, 2006; Carvalho et al., 2007). A
espécie apresenta atividade noturna. No entanto é possível encontrá-los em plena atividade
durante do dia (Carvalho et al. 2007). Indivíduos dessa espécie são encontrados
principalmente nos ambientes urbanos e até mesmo edifícios (Benesi, 2007; Costa et al.,
2009; Forlani et al., 2010), onde se alimentam de pequenos invertebrados atraídos pela luz
artificial.
Família Gymnophthalmidae
Cercosaura quadrilineata Doan, 2003 apresenta pequeno tamanho corpóreo, com membros
bem desenvolvidos e cauda longa. O gênero Cercosaura se distribui em praticamente todos os
países da América Latina, exceto no Chile. Entre as espécies representadas no Brasil, C.
quadrilineatas se diferencia das outras cinco por suas escamas dorsais dispostas em séries
transversais e oblíquas e ventrais em quatro linhas longitudinais, além de apresentar 9 a 10
poros femorais (Doan, 2003; Vanzolini, 1948). Essa espécie é deficiente em informação sobre
distribuição geográfica e dados ecológicos, sendo uma das espécies menos representadas no
presente estudo.
Ecpleopus gaudichaudii Duméril & Bibron, 1839 é a menor espécie de lagartos da região da
serra do Ouro Branco, endêmica da Mata Atlântica e única do gênero presente no Brasil.
Caracteriza-se pelo porte pequeno, cauda comprida, membros curtos e modo de vida
associado à serapilheira de áreas florestadas, alimentando-se basicamente de pequenos
invertebrados (Eisemberg et al., 2004). A reprodução é ovípara com ninhadas de um a dois
ovos (Perini & Butti, 2008). Segundo Uzzell (1969), E. gaudichaudii ocorre em altitudes não
muito elevadas, podendo chegar até os 1000m acima do nível do mar. Contudo, esta espécie
foi encontrada no presente estudo em altitudes que variam entre 1250 e 1568 metros de

38
altitude ampliando assim a sua distribuição e ocorrência, quanto a área e altitude. Apesar de
ser considera restrita de áreas preservadas, esta espécie já foi capturada em locais
antropizados por Dixo & Metzger (2009) e Costa et al. (2010), assim como no presente
trabalho, onde foi encontrada em matas secundárias próximas a estrada. Esse animal não é de
fácil captura por sua coloração críptica e tamanho reduzido. Graças a estudos recentes com
utilização de armadilhas do tipo pitfall e análises de materiais em coleções zoológicas, sua
distribuição geográfica conhecida tem aumentado consideravelmente (Costa et al., 2010; Dixo
& Metzger, 2009; Dixo & Verdade, 2006; Ramos & Silveira, 2011).
Heterodactylus imbricatus Spix, 1825 é um lagarto de pequeno porte e cauda muito longa,
podendo ser maior que o próprio corpo. É frequente o encontro de indivíduos com sinais de
autotomia caudal. Essa espécie é conhecida na região como “Cobra-de-patas”. Além de H.
imbricatus, ocorrem outras duas espécies do mesmo gênero no Brasil, H. Lundii e H.
Septentrionalis. A espécie da área do presente estudo se diferencia destas duas principalmente
por ter maior porte; pela presença de uma sexta escama supra labial e por apresentar o quinto
dedo vestigial (Rodrigues et al., 2009). A espécie não apresenta dimorfismo sexual aparente.
É encontrada na serapilheira de áreas florestadas montanhosas da Mata Atlântica do sudeste e
do sul do Brasil (Dixo & Verdade, 2006; Rodrigues et al., 2007, 2009). Recentemente, foi
reportada a ocorrência dessa espécie em ambientes ripários associados ao bioma Cerrado em
Minas Gerais (Novelli et al., 2011). Existem poucas informações sobre a reprodução e
alimentação de H. imbricatus.

39
Família Leiosauridae
Enyalius bilineatus Duméril & Bibron, 1837 é uma espécie diurna, passando a maior parte do
tempo imóvel (Jackson, 1978), apresenta corpo cilíndrico e delgado com membros e cauda
longos. O dorso é marrom ou cinza e seu ventre é branco ou acinzentado. Esse lagarto ocorre
apenas no Brasil, habitando áreas florestadas dos estados do Rio de Janeiro, Espírito Santo e
Minas Gerais. É encontrado em matas de galeria e em áreas de Cerrado com mata, onde se
alimenta de artrópodes. É considerado um forrageador do tipo senta-e-espera, predando
principalmente no solo (Zamprogno et al., 2001).
Enyalius perditus Jackson, 1978 é um lagarto de hábito semi-arborícola, que durante o dia é
encontrado em atividade no chão ou sobre vegetação, repousando nos ramos e folhas largas à
noite. Apresenta dimorfismo sexual, sendo que as fêmeas adultas geralmente apresentam
maior largura corporal e coloração do corpo marrom. Os machos adultos são mais esguios e
de coloração verde. Juvenis desta espécie se assemelham às fêmeas quanto à coloração,
demonstrando variação ontogênica na coloração (Sturaro & Silva, 2010). A espécie se
alimenta principalmente de artrópodes (Sousa & Cruz, 2008; Sturaro & Silva, 2010).
Segundo Sousa & Cruz (2008), os machos desta espécie são forrageadores ativos, enquanto
que as fêmeas utilizam a estratégia de senta-e-espera. A espécie é exclusiva de áreas com
mata, sendo sensível a fragmentação de hábitat (Dixo & Metzger, 2009). É uma espécie
endêmica da Mata Atlântica (Jackson,1978), sendo dessa forma considerada como
possivelmente ameaçada no estado do Rio de Janeiro (Rocha et al., 2000). Segundo Sturaro et
al. (2010), a espécie apresenta um ciclo reprodutivo por ano, com início em novembro e final
em março. Esta espécie ocorre com relativa abundância nas áreas florestadas da serra do Ouro
Branco.

40
Urostrophus vautieri Duméril & Bibron, 1837 é um lagarto de corpo pequeno, cilíndrico e
delgado, com membros e cauda compridos, apresenta aspecto críptico, confundindo-se com o
substrato. Essa espécie é encontrada na Argentina, Bolívia e no Brasil, onde é conhecida a sua
ocorrência em áreas de Mata Atlântica nos estados: Espírito Santo, Minas Gerais, Rio de
Janeiro, São Paulo, Paraná, Santa Catarina e Rio Grande do Sul (Etheridge & Williams, 1991;
Sazima & Haddad, 1992; Santos et al., 2009; Gasparini et al., 2010). No entanto,
recentemente, essa espécie foi encontrada nos domínios do Cerrado, em florestas ripárias na
Reserva de Boquirão, município de Ingaí, Minas Gerais (Santos et al., 2009). Trata-se da
única espécie do gênero presente no Brasil (Bérnils & Costa, 2011).
Família Polychrotidae
Polychrus acutirostris Spix, 1825: é uma espécie diurna de hábito arborícola, mas também
com registros de ocorrência no solo (Vanzolini, 1983). É um animal de pequeno porte,
apresenta corpo e cauda longos e os membros curtos. Alimenta-se de artrópodes arbóreos e de
vegetais (Vitt & Lacher-Jr, 1981). O período reprodutivo ocorre durante a estação chuvosa,
sendo reportada a postura de uma única ninhada, contendo de sete a 31 ovos (Vit & Lacher-Jr,
1981). O comportamento de camuflagem dessa espécie é bem desenvolvido, o que dificulta o
seu encontro em trabalhos de levantamento de espécies e estudos de comunidade. No Brasil,
ocorrem outras duas espécies do mesmo gênero: P. liogaster e P. marmuratus (Bérnils &
Costa, 2011). A distribuição geográfica de P. acutirostris é bastante extensa na América do
Sul ocorrendo também na Argentina, Bolívia, Paraguai e Uruguai (Burt & Burt 1931;
Vanzolini et al., 1980; Vanzolini, 1983). No Brasil, essa espécie ocorre no sul do Pará, nos
estados das regiões Nordeste e Centro Oeste, além de Minas Gerais, São Paulo, na região
Sudeste. A espécie costuma ocupar vegetações abertas e está presente no Cerrado, Caatinga,

41
Chaco, Pantanal (Vanzolini et al., 1980; Vanzolini, 1983; Kawashita-Ribeiro & Ávila, 2008)
e no presente estudo, nos limites da Mata Atlântica com o Cerrado.
Família Scincidae
Mabuya dorsivittata Cope 1862 é uma espécie de pequeno porte, corpo cilíndrico, com os
membros bem desenvolvidos e cauda longa. Vive em simpatria com M. frenata na região do
presente estudo, se diferenciando morfologicamente desta quanto ao número de escamas
frontoparientais. Alimenta-se de pequenos invertebrados e vertebrados e apresenta
dimorfismo sexual, sendo as fêmeas adultas maiores que os machos. Ocorre no Cerrado e em
matas de galeria nos estados de Minas Gerais (Recorder & Nogueira, 2007), nas dunas
costeiras, no estado do Rio Grande do Sul (Quintela et al., 2006) e em campos de altitude no
Rio de Janeiro (Rocha et al., 2003). Um único exemplar coletado para a serra do Ouro Branco
está tombada na coleção de répteis do MZUFV(MZUFV-530).
Mabuya frenata (Cope, 1862) é uma espécie diurna, heliófila que ocorre em áreas abertas,
onde seu micro hábitat preferencial são as rochas (Vrcibradic & Rocha, 1998). A dieta dessa
espécie inclui um leque variado de artrópodes, sendo os cupins os itens alimentares
preferenciais. A espécie apresenta pequeno porte, corpo cilíndrico e cauda longa e apresenta
dimorfismo sexual, com fêmeas de tamanho corpóreo maior (Vrcibradic & Rocha, 1998).
Essa espécie ocorre no sudeste e centro-oeste do Brasil, além de estar presente na Argentina,
Bolívia e Paraguai (Peters et al., 1970). Em Minas Gerais, M. frenata é registrada nas regiões
do Cerrado, Mata Atlântica, e em ambientes de transição (Recorder & Nogueira, 2007; Costa
et al., 2008; Sousa et al., 2010). No presente trabalho, foram coletados indivíduos tanto em
ambientes de mata, como também em áreas urbanas.

42
Família Teiidae
Ameiva ameiva (Linnaeus, 1758) é uma espécie de lagarto terrícola de porte médio. É um
forrageador ativo e sua dieta é composta por artrópodes, pequenos vertebrados e materiais
vegetais (Vanzolini et al., 1980; Colli et al., 1997; Teixeira, 2001). A espécie apresenta
dimorfismo sexual, sendo os machos maiores e com mais massa corporal do que as fêmeas
(Rocha, 2008). As fêmeas produzem várias ninhadas, com uma média de cinco a seis ovos por
ano (Vanzolini et al., 1980). É a única espécie do gênero descrita no Brasil e ocorre em áreas
abertas em praticamente todo o seu território (Vanzolini, 1986; Vitt & Colli, 1994). Esse
lagarto apresenta ampla distribuição geográfica, e se encontra também representado no
Panamá, norte da Argentina e leste dos Andes, ocorrendo principalmente em áreas abertas,
ecótones e áreas perturbadas (Vanzolini, 1986; Vitt & Colli, 1994; Sartorius et al., 1999).
Costuma ser uma espécie abundante no Cerrado, onde ocupa áreas abertas e bordas de matas
(Recorder & Nogueira, 2007). Neste presente trabalho a espécie foi registrada em áreas de
borda de estrada e no campo rupestre.
Tupinambis merianae (Duméril & Bibron, 1839) é uma das espécies com maior distribuição
geográfica no Novo Mundo sendo a espécie de maior porte entre os Teiídeos (Fitzgerald et
al., 1993). No Brasil, T. merianae ocorre na maioria das florestas da região sudeste
forrageando em áreas abertas de Cerrado, na Caatinga e em bordas de matas de galeria
(Sazima & Haddad, 1992; Vanzolini et al., 1980). Sua dieta é onívora, composta por
invertebrados, pequenos vertebrados, ovos, carniça e frutos (Kiefer & Sazima, 2002; Castro &
Galleti, 2004; Ávila- Pires, 1995; Colli et al., 1998), podendo atuar como dispersor de
sementes (Castro & Galetti, 2004). Este lagarto merece uma atenção na área do presente
estudo, visto que a sua carne é apreciada como recurso alimentar pelos habitantes da região
(Pinto, 2011).

43
Família Tropiduridae
Tropidurus torquatus (Wied, 1820) é um lagarto de pequeno porte, comumente encontrada
em áreas abertas, sendo vista sobre rochas, cupinzeiros, troncos caídos e no chão. É comum
em áreas antropizadas, podendo ser observada com facilidade sobre muros e cercas de casas e
ruas da área do presente estudo. Sua dieta consiste basicamente de artrópodes. No entanto,
podem ingerir partes de material vegetal e outros pequenos vertebrados (Teixeira &
Giovanelli, 1999). Sua reprodução é cíclica, ocorrendo quase que inteiramente na estação
chuvosa. Cada fêmea pode depositar mais de uma ninhada durante o ano, contendo entre
quatro a 14 ovos, dependendo do tamanho da fêmea (Vanzolini et al., 1980). Esse lagarto é
encontrado na Argentina, Uruguai, Paraguai e nas regiões centro-oeste, sudeste e sul do Brasil
(Rodrigues, 1987). Trata-se de uma das espécies mais abundantes na região, ocupando
cupinzeiros, tronco de árvores grossas, bordas de mata, muros de domicílios, em solos
arenosos próximos a área urbana e em rochas, sempre em áreas abertas.
Tropidurus itambere Rodrigues, 1987 é um animal de pequeno porte, corpo robusto e
deprimido, membros longos e cauda curta. Apresenta atividade diurna sendo observado em
atividades de forrageio e termorregulação nas horas mais quentes do dia. Sua dieta consiste
basicamente de tecidos vegetais e artrópodes (Van Sluys, 1992). Este lagarto é endêmico do
Brasil, ocorrendo nos estados de Minas Gerais, São Paulo, Goiás, Mato Grosso, Mato Grosso
do Sul e Tocantins, sendo encontrada em áreas de Cerrado pouco perturbadas, especialmente
com abundância de rochas (Rodrigues, 1987).

44
CHAVE ARTIFICIAL PARA IDENTIFICAÇÃO DAS ESPÉCIES DE LAGARTOS DA SERRA DE OURO BRANCO, OURO BRANCO, PORÇÃO SUL DO ESPINHAÇO MINAS GERAIS.
1. A: Olho com formato cônico e saliente; corpo e cauda compridos; presença de poros
femorais.............................................................................Polychrus acutirostris (fig.: 3. i).
1. B: Olho sem o formato descrito acima........................................................................2.
2. A: Membros anteriores ausentes.............................................Ophiodes striatus (fig.: 3. a).
2. B: Membros anteriores presentes..........................................................................................3.
3. A: Escamas dorsais hexagonais, quilhadas e alongadas.......................................................4.
3. B: Escamas dorsais não descritas como acima.....................................................................6.
4. A: Membros anteriores com quatro dígitos bem reduzidos, com cauda e corpo bem
alongados........................................................................Heterodactylus imbricatus (fig.: 3. e).
4. B: Membros anteriores com cinco dígitos............................................................................5.
5. A: Com 09 a 10 poros femorais..................................Cercosaura quadrilineatus (fig.: 3. c).
5. B: Poros femorais ausentes ............................................Ecpleopus gaudichaudii (fig.: 3. d).
6. A: Escamas dorsais e ventrais ciclóides................................................................................7.
6. B: Escamas dorsais não como descritos acima....................................................................8.
7. A: Uma frontoparietal..................................................................Mabuya frenata (fig.: 3. j).
7. B: Duas fronto-parietais.......................................................Mabuya dorsivittata (fig.: 3. k).

45
8. A: Bolsas ou dobras de ácaros presentes....................................................................9.
8. B: Bolsas e dobras de ácaros ausentes............................................................................10.
9. A: Bolsa de ácaros comprida e profunda nos lados do pescoço; axila sem bolsa de
ácaros...............................................................................Tropidurus itambere (fig.: 3. o).
9.B: Duas bolsas de ácaros rasas nos lados do pescoço e preenchidas por
grânulos............................................................................Tropidurus torquatus (fig.: 3. n).
10. A: Escamas ventrais quadrangulares, dispostas em fileiras regulares longitudinais e
transversais. Escamas dorsais menores que ventrais frequentemente granulares e poros
femorais presentes..............................................................................................11.
10. B: Escamas ventrais não descritas como acima e pequenas; escamas granulares ao longo
do corpo.............................................................................................................................12.
11. A: Doze ou menos ventrais em fileira transversal no meio do corpo; poros femorais
presentes..................................................................................Ameiva ameiva ( fig.: 3. i).
11. B: Quinze ou mais ventrais em fileira transversal no meio do corpo; dorso com faixas
transversais negras ou quase totalmente negras; poros femorais
presentes......................................................................Tupinambis merianae (fig.: 3. m).
12. A: Dígitos curtos com lamelas subdigitais bem desenvolvidas; corpo com pequenas
projeções ou tubérculos de escamas que se tornam mais salientes na região posterior do corpo
e na cauda, lembrando espinhos; pálpebra ausente.......Hemidactylus mabouia (fig.: 3. b).
12. B: Dígitos não descritos como acima; corpo e caudas esguios e
alongados..............................13.

46
13. A. Fileira ou crista de escamas vertebrais aumentadas e destacadas das
demais.........................................................................................................................14.
13. B. Sem distinção de escamas vertebrais....................................Urostrophus vautieri (fig.:
3. h ).
14. A: A Escama da sub-ocular bem distinta e alongada em relação as
demais.................................................................................Enyalius bilineatus (fig.: 3. f).
14. B: Escama da Sub-ocular pequena e uniforme, sendo que os machos adultos da espécie
apresentam coloração esverdeada, e as fêmeas coloração
marrom..................................................................................Enyalius perditus (fig.: 3. g).
BIBLIOGRAFIA
ASSIS, V.B. 1999. Introdução às serpentes da “Serra do Cipó” (municípios de Santana do
Riacho e Jaboticatubas). Bios. 7(7):69-71.
ÁVILA-PIRES, T.C.S. 1995. Lizards of Brazilian Amazonia (Reptilia: Squamata). Zool.
Verh. Leiden. 299: 1- 706.
BARLOW, J. & PERES, C.A. 2004.Ecological responses to El Nino induced surface fires in
central Amazonia: management implications for flammable tropical forests.
Philosophical Transactions of the Royal Society of London 359: 367-380.
BARROS, E.H. & TEIXEIRA, R.L. 2007.Diet and fecundity of the Glass-lizard, Ophiodes
striatus (Sauria, Anguidae) from the Atlantic Forest in southeastern Brazil.In Anais do
VIII Congresso de Ecologia do Brasil, 23 a 28 de setembro de 2007, Caxambu – MG.
BELL, K. E. & DONNELLY, M. A. 2006.Influence of Forest Fragmentation on Community
Structure of Frogs and Lizards in Northeastern Costa Rica.Conservation Biology.
Vol.20, No. 6, 1750–1760.

47
BENESI, R.G.Q. 2007. Répteis do Município de São Paulo. In Fauna Silvestre: Quem são e
onde vivem os animais na metrópole Paulistana. Secretaria Municipal do Verde e Meio
Ambiente, São Paulo, p. 106-147.
BERNARDO, P. H. & PIRES, M. R. S. 2006. Ophiodes striatus (glass Snake).
Preproduction.Herpetological review.37 (4). p. 469-470.
BÉRNILS, R. S. & COSTA, H. C.C. 2011.Brazilian reptiles: list of species. http://www.sbherpetologia.org.br/ (último acesso em 12/12/2011).
BÉRNILS, R.S., NOGUEIRA, C. & SILVA, V.X. 2009. Biota minas - diagnóstico do
conhecimento de vertebrados: Répteis. In Biota minas - diagnóstico do conhecimento
sobre a biodiversidade no estado de Minas Gerais (G.M. DRUMMOND, C.S.
MARTINS, M.S. GRECO & F. VIEIRA, eds.), pp. 252-278, Belo Horizonte, MG.
BERTOLUCI, J. 1998. Serpentes. In Livro Vermelho das Espécies Ameaçadas de Extinção
da Fauna de Minas Gerais (A.B.M. Machado, G.A. Fonseca, L.M.S. Aguiar & L.V.
Lins, eds.). Fundação Biodiversitas, Belo Horizonte, p.417.
BERTOLUCI, J. CANELAS, M.A.S., EISEMBERG, C.C., PALMUTI C.F.S. &
MONTINGELLI G.G. 2009. Herpetofauna of Estação Ambiental de Peti, an Atlantic
Rainforest fragment of Minas Gerais State, southeastern Brazil. Biota Neotrop., 9(1):
http://www.biotaneotropica.org.br/v9n1/en/abstract?inventory+bn0140901.
BORGES-MARTINS, M. 1998. Revisão taxonômica e sistemática filogenética do
gênero Ophiodes Wagler, 1828 (Sauria, Anguidae, Diploglossinae). Tese de Doutorado,
Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre.
BORGES-NOJOSA & SANTOS, E.M. 2005. Herpetofauna da área de Betânia e Floresta,
Pernambuco. In: Análise das variações da biodiversidade do bioma Caatinga: Suporte a
estratégias regionais de conservação/ Brasília: Ministério do Meio Ambiente. 278-291.

48
BRITES, V.L.C. & BAUAB, F.A. 1988. Fauna ofidiana do município de Uberlândia, Minas
Gerais - Brasil. I. Ocorrência na área urbana. Rev.Cent. Cienc. Biomed. Univ. Fed.
Uberlandia 4(1): 3-8.
BURT, C. E & BURT, M. D. 1931.South American Lizards in the collection of American
Museum of Natural History.Bull. Amer. Mus. Nat. Hist., 61, 7: 227-395.
CAPOBIANCO, J.P.R. 2002. Metodologia. In: Ministério do Meio Ambiente. Biodiversidade
Brasileira: avaliação e identificação de áreas e ações prioritárias para a conservação,
utilização sustentável e repartição de benefícios da biodiversidade brasileira. Brasília:
MMA/SBF. p. 5-18.
CARRANZA, S. & ARNOLD, E. N.2006. Systematics, biogeography, and evolution of
Hemidactylus geckos (Reptilia: Gekkonidae) elucidated using mitochondrial DNA
sequences. Molecular Phylogenetics and Evolution 38(2006): 531–545.
CARVALHO, A.L.G., ARAÚJO, A.F.B. & SILVA, H.R. 2007.Lizards of Marambaia, an
insular remnant of Restinga and Atlantic Forest in the State of Rio de Janeiro,
Brasil.Biota Neotrop. Vol.7, no.2.
CASSIMIRO, J & BERTOLUCI, J. 2002. Répteis da Estação Ecológica de Caratinga, um
fragmento de Mata Atlântica do estado de Minas Gerais. XXIV Congresso Brasileiro de
Zoologia: A Zoologia e os Ecossistemas Costeiros, SBZ, Itajaí, SC. p. 455.
CASTRO, E.R. & GALETTI, M. 2004. Frugivoria e dispersão de sementes pelo lagarto teiú
Tupinambis merianae (Reptilia: Teiidae). Papéis Avulsos de Zoologia, 44(6): 91-97.
CECHIN, S.Z. & MARTINS, M. 2000. Eficiência de armadilhas de queda (pitfall traps) em
amostragens de anfíbios e répteis no Brasil. Revista Brasileira de Zoologia 17 (3): 729-
740.
COLLI, G. R. 2011. Chave artificial para os lagartos do Distrito Federal. Disponível em
http://vsites.unb.br/ib/zoo/grcolli/guia/chave.htm. (Último acesso em Setembro de
2011).

49
COLLI, G. R.,PÉRES JR,A. K. &CUNHA,H. J. 1998. A new species of Tupinambis
(Squamata: Teiidae) from Central Brazil, with an analysis of morphological and genetic
variation in the genus. Herpetologica 54(4): 477-492.
COLLI, G. R., BASTOS, R.P, & ARAÚJO, A.F.B. 2002. The character and dynamics of the
Cerrado herpetofauna. In: Oliveira, p.s. & Marquis, R.L (eds.) The Cerrados of Brazil:
ecology and natural history of a neotropical savana, p.223-241. New York, Columbia
University Press.
COLLI, G. R., PÉRES-JR, A.K., ZATZ, M.G. & PINTO, A.C.S.1997.Estratégias de
forrageamento e dieta em lagartos do Cerrado e Savanas Amazônicas. In, LEITE,L. L.
& SAITO, C. H. (org.).Contribuição ao conhecimento ecológico do Cerrado- Trabalhos
selecionados no 3° Congresso de ecologia do Brasil ( Brasília 6-11/10/96). Brasília,
Dept. Ecologia- Universidade de Brasília.
COSTA, M. R.C.,HERRMANN,G., MARTINS,C.S.,LINS,L. V. &LAMAS,I.R. (Orgs.).
1998. Biodiversidade em Minas Gerais – Um Atlas para sua conservação. Belo
Horizonte: Fundação Biodiversitas.
COSTA, H. C., FERNANDES, V. D., VRCIBRADIC, D., FEIO, R. N. 2008. Reptilia,
Scincidae, Mabuya frenata: Distribution Extension. Check List 4(1): 86–88.
COSTA, H.C., PANTOJA, D.L., PONTES, J.L. & FEIO, R.N.2010. Snakes of the
Municipality of Viçosa, Atlantic Forest of Southeastern Brazil.Biota Neotrop. 10(3):
http://www.biotaneotropica.org.br/v10n3/en/abstract?inventory+bn0361003.
COSTA, H. C., FERNANDES, V.D., RODRIGUES, A. C., FEIO, R. N. 2009. Lizards and
amphisbaenians, municipality of Viçosa, State of Minas Gerais, southeastern Brazil.
Check List. 5(3): 732-745.
CRUMP, M.L. & SCOTT JR, N.J. 1994. Visual encounter surveys. In Standard methods for
amphibians (W.R. Heyer, M.A. Donnely, R.W. Mc Diarmid, L.A.C. Hayek & M.S.
Foster, eds). Smithsonian Institution Press, Washington, p. 84-92.

50
CUNHA, O.R. 1961. II. Lacertílios da Amazônia. Bol. Mus. Para. Emílio Goeldi, Nova Sér.,
Zool., Belém 39: 1-189.
DIXO, M & MARTINS, M. 2008.Are leaf-litter frogs and lizards affected by edge effects due
to forest fragmentation in Brazilian Atlantic forest? Journal of Tropical Ecology.
24:551–554.
DIXO, M. & METZGER, J. P. 2009.Are corridors fragment size and forest structure
important for leaf-litter lizards in fragmented landscape? Fauna & Flora International,
Oryx, 43(3), 435-442.
DIXO, M. & METZGER, J. P. 2009.Are corridors fragment size and forest structure
important for leaf-litter lizards in fragmented landscape? Fauna & Flora International,
Oryx, 43(3), 435-442.
DIXO, M. & VERDADE, V.K. Herpetofauna de serrapilheira da Reserva Florestal de Morro
Grande, Cotia (SP). Biota Neotrop.May/Aug 2006 vol. 6 no. 2,
http://www.biotaneotropica.org.br/v6n2/pt/abstract?article+bn00806022006.ISSN1676-
0603.
DOAN, T. M. 2003. A new phylogenetic classitication for the Gymnophthalmid genera
Cercosaura, Pantadactylus and Prinodactylus ( Reptilia: Squamata). Zoological Jornal of
Linnean Society, 137, 101-115.
EISEMBERG, C. C.; CASSIMIRO, J. & BERTOLUCI, J. 2004.Notes on the diet of the rare
Gymnophthalmid lizard Ecpleopus gaudichaudii from Southeastern Brazil.
Herpetological Review, 35(4): 336-337.
ETHERIDGE, R. &WILLIAMS, E.E. 1991. A review of the South American lizard genera
Urostrophus and Anisolepis (Squamata: Iguania: Polychridae). Bulletin of the Museum
of Camparative Zoology 152: 317-361.
FEIO, R.N. & CARAMASCHI, U. 2002. Contribuição ao conhecimento da herpetofauna do
nordeste do estado de Minas Gerais, Brasil. Phyllomedusa 1(2):105-111.

51
FITZGERALD L. A; CRUZ, F. B & PEROTTI, G. 1993 The reproductive cycle and the size
at maturity of Tupinambis rufescens (Sauria:Teiidae) in the dry Chaco of Argentina.
Journal of Herpetology 27:70-78.
FORLANI, M.C., BERNARDO, P.H., HADDAD, C.B.F. & ZAHER, H. 2010. Herpetofauna
of the Carlos Botelho State Park, São Paulo State, Brazil.Biota Neotrop. 10(3):
http://www.biotaneotropica.org.br/v10n3/en/abstract?inventory+bn00210032010.
FROST, D.R., ETHERIDGE, R., JANIES, D. & TITUS, T.A. 2001. Total evidence, sequence
alignment, evolution of polychrotid lizards, and a reclassification of the Iguania
(Squamata, Iguania) Am. Mus. Novitates, 3343:1-38.
GALLARDO, J.M. 1966. Las espécies argentinas del género Ophiodes Wagler (Anguidae,
Sauria). Rev. Mus. Arg. Ci. Nat. "Bernardino Rivadavia", Zool.,9(6):122-46.
GARDNER, T. A., BARLOW, J. & PERES, C. A. 2007.Paradox, presumption and pitfalls in
conservation biology:The importance of habitat change for amphibians and reptiles.
Biological Conservation: 138. 166–179.
GASPARINI, J. L., KOSKI, D. A. &PELOSO, P.L.V. 2010.Reptilia, Squamata,
Leiosauridae, Urostrophus vautieri Duméril and Bribon, 1837: Distribution extension,
new state record, and geographic distribution map. Check List. Vol.6. Issue 3
GIBBONS, J.W., SCOTT, D.E., RYAN, T.J., BUHLMANN, K.A., TUBERVILLE, T.D.,
METTS, B.S., GREENE, J.L., MILLS, T., LEIDEN, Y., POPPY, S. & WINNE, C.T.
2000. The global decline of reptiles, deja-vu amphibians. Bioscience 50, 653–667.
GREENBERG, C.H., NEARY, D.G. & HARRIS.L.D. 1994. A comparison of herpetofaunal
sampling effectiveness of pitfall, single-ended, and double-ended funnel traps used with
drift fences. Journal of Herpetology, 28(3): 319-324.
HUEY, R. B.,PIANKA, E. R. & SCHOENER,T. W. 1983. Introduction (pp. 1-6) in R. B.
HUEY, E. R. PIANKA, & T. W. SCHOENER.(eds.) Lizard Ecology: Studies of a
Model Organism. Harvard University Press.

52
IBAMA, 2007. Lista nacional das espécies da fauna brasileira ameaçada de extinção.
Disponível em <http://www.mma.gov.br/port/sbf/fauna/index.cfm> (último acesso em
outubro de 2007).
JACKSON, J.F. 1978. Differentiation in the genera Enyalius and Strobilurus (Iguanidae):
implications for Pleistocene climatic changes in eastern Brazil. Arquivos de Zoologia,
30: 1-79.
JELLINEK, S., DRISCOLL, D. A. & KIRKPATRIK, J. B. 2004. Environmental and
vegetation variables have a greater influence than habitat fragmentation in structuring
lizard communities in remnant urban bushland. Austral Ecology. 29, 294-304.
KAWASHITA-RIBEIRO, R.A., ÁVILA, R.W. 2008. Reptilia, Squamata, Polychrus spp.:
New record, range extensions, and distribution map in the state of Mato Grosso, Brazil.
Check List 4(3): 362–365.
KIEFER, M. C. & SAZIMA, I. 2002. Diet of juvenile tegu lizard, Tupinambis merianae
(Teiidae) in Southeastern Brazil.Anphibia-Reptilia. 23: 105-108.
KITCHENER, D. J., CHAPMAN, A., DELL, J.; MUIR, B. G. & PALMER, M. 1980. Lizard
assemblages and reserve size and structure in the Western Australian wheatbelt – some
implications for conservation. Biol. Conserv. 17, 25–62.
LEHTINEN, R. M.; RAMANAMANJATO, J-B.& RAVELOARISON, J. B. 2003.Edge
effects and extinction proneness in a herpetofauna from Madagascar. Biodiversity and
Conservation 12: 1357–1370
LEITE, F. S. F.; JUNCÁ, F. A. & ETEROVICK, P. C. 2008.Status do conhecimento,
endemismo e conservação de anfíbios anuros da Serra do Espinhaço, Brasil.
Megadiversidade.
MARQUES, A.A. B., FONTANA, C. S., VÉLEZ, E., BENCKE, G. A., SCHNEIDER, M. &
REIS, R.S. 2002. Lista de Referência da Fauna Ameaçada de Extinção no Rio Grande
do Sul. Decreto no. 41.672, 11 de junho de 2002. Porto Alegre: FZB/MCT–
PUCRS/PANGEA, 52p.

53
MESQUITA, D. O., COLLI, G. R., FRANÇA, F.G.R. & VITT, L.J. 2006. Ecology of a
Cerrado lizard assemblage in the Jalapão region of Brazil. Copeia 2006(3): 460-471.
MYERS, N., MITTERMEIER, R. A., MITTERMEIER, C. G., FONSECA, G.A.B. & KENT,
J. 2000. Biodiversity hotspots for conservation priorities. Nature 430: 853-858.
NOGUEIRA, C. C. 2006. Diversidade e padrões de distribuição da fauna de lagartos do
Cerrado. Tese de Doutorado. USP-Instituto de Biociências São Paulo.
NOGUEIRA, K. O. P.C., RODRIGUES, S. S.; ARAÚJO, V.A. & NEVES, C. A. 2011.
Oviductal Structure and Ultrastructure of the Oviparous Gecko, Hemidactylus mabouia
(Moreau De Jonne` s, 1818).The anatomical record 294:883–892.
NOVAES-SILVA, V. & ARAÚJO, A.F.B. 2008. Ecologia dos lagartos brasileiros. Technical
Books editora. Rio de janeiro.
NOVELLI, I. A., LUCAS, P. S. & SANTOS, R. C. 2011. Reptila, Squamata,
Gymnophthalmidae, Heterodactylus imbricatus Spix, 1825: Filling gaps in the state of
Minas Gerais. Check List. Vol. 7. 1.
PALMUTI, C.F.S., CASSIMIRO, J. & BERTOLUCCI, J. 2009.Food habits of snakes from
the RPPN Feliciano Miguel Abdala, an Atlantic Forest fragment of southeastern Brazil.
Biot Neotrop. 9(1):http://www.biotaneotropica. org.br/v9n1/en/abstract?short-
communication+BN0 2209012009 (ultimo acesso em 22/04/2010).
PAULA, C.C. 2003. Flora fanerogâmica da serra do Ouro Branco. In: 54º Congresso
Nacional de Botânica, 2003, Belém, PA. Livro de resumos. UFRA (FCAP)/MPEG/
Embrapa Amazônia Oriental.
PERINI, F. A. & BUTTI, M. 2008. Ecpleopus gaudichaudii (NCN).Reproduction.
Herpetological Review 39(2): 222.

54
PETERS, J. A. & DONOSO-BARROS, R. 1970. Catalogue of the Neotropical Squamata.Part
II.Lizards and Amphisbaenians. U. S. Natl. Mus. Bull. (297):viii+293.
PIANKA, E. R. & VITT, L. J. 2006. Lizards: Windows to the Evolution of Diversity.
University of California Press, Berkeley.
PIANKA, E. R. 1973. The structure of lizards communities. Anual review of ecology and
systematics.4. 53-74.
PINTO, L. C. L. 2011.Etnozoologia e conservação de biodiversidade em comunidades rurais
da Serra do Ouro Branco. Dissertação de Mestrado. Universidade Federal de Ouro
Preto, Minas Gerais.
PIRES, M.R.S.; PINTO, L.C.L.; MATEUS, M.B. 2010. A etnozoologia como instrumento para a
conservação da fauna da Serra do Ouro Branco. InALVES R.R.N.; SOUTO, W.M.S.; MOURÃO,
J.S.A Etnozoologia no Brasil: Importância, Status atual e Perspectivas, vol 7. Estudos & Avanços, 1
ed. NUPEEA, Recife, PE, Brazil., Cap. 21, pag. 471- 493.
POUGH, F. H., JANIS, C. M. & HEISER, J. B. 2008.A vida dos vertebrados. 4 ed. São
Paulo: Atheneu Editora. 684 p. I.
QUINTELA, F. M., LOEBMANN, D.& GIANUCA, N.M. 2006.Répteis continentais do
Município de Rio Grande, Rio Grande do Sul, Brasil. Biociências, Porto Alegre, v. 14,
n. 2, p. 180-188.
RAMOS, L. O. & SILVEIRA, A. L. 2011. New Record and distribuition mape of Ecpleopus
gaudichaudii (Reptilia, Squamata, Gymnophthalmidae) in southeastern
Brazil.Herpetology Notes. 4. 173- 176.
RECODER, R. & NOGUEIRA, C. 2007. Composição e diversidade de répteis na região sul
do Parque Nacional Grande Sertão Veredas, Brasil Central. Biota Neotrop. 7(3):
http://www.biotaneotropica.org.br/v7n3/pt/abstract?inventory+BN01107032007.

55
ROCHA, C. F. D., VRCIBRADIC, D., VICENTE, J. J.& CUNHA-BARROS, M. 2003.
Helminths infecting Mabuya dorsivittata (Lacertilia, Scincidae) From a high-altitude
habitat in Itatiaia National Park, Rio de Janeiro State, Southeastern Brazil. Braz. J.
Biol., 63(1): 129-132.
ROCHA, C.F.D. 2008. Body size, female reproduction and sexual dimorphism in the lizard
Ameiva ameiva (Teiidae) in a restinga of southeastern Brazil.Revista Brasileira de
Zoologia 25 (2): 370–372.
ROCHA, C.F.D., VAN-SLUYS, M.,PUORTO,G. R., FERNANDES, J. D., BARROS-
FILHO, R. R. E., SILVA, F. A., NEO, E. A. & MELGAREJO. 2000. Répteis; p. 79-87
In BERGALLO,H. G.;ROCHA,C. F. D.;ALVES,M. A. S. &VAN SLUYS,M. (ed.). A
Fauna Ameaçada de Extinção do Estado do Rio de Janeiro. Rio de Janeiro: Editora
UERJ.
RODRIGUES, M.T. 2005. Conservação dos répteis brasileiros: os desafios para um país
megadiverso. Megadiversidade. 1(1): 87-94.
RODRIGUES, M. T. 1987. Sistemática, ecologia e zoogeografia dos Tropidurus do grupo
torquatus ao sul do Rio Amazonas (Sauria, Iguanidae). Arq. Zool., São Paulo, 31:105-
230.
RODRIGUES, M.T., DIXO, M., PAVAN, D.& VERDADE, V.K. 2002. A new species of
Leposoma (Squamata, Gymnophthalmidae) from the remnant Atlantic forests of the
state of Bahia, Brazil.Pap. Avulsos Zool. (São Paulo) 42:335-350.
RODRIGUES, M.T., FREITAS, M.A., SILVA, T.F.S. & BERTOLOTTO, C.E.V. 2006. A
new species of lizard genus Enyalius (Squamata, Leiosauridae) from the highlands of
Chapada Diamantina, state of Bahia, Brazil, with a key to species. Phyllomedusa
5(1):11-24.
RODRIGUES, M. T.,FREITAS, M.A. &SILVA, T.F. S. 2009. New Species of earless lizard
genus Heterodactylus (Squamata: Gymnophthalmidae) from the highlands of Chapada
Diamantina, State of Bahia, Brazil. Journal of Herpetology 43(4): 605-611.

56
RODRIGUES, M.T., PELLEGRINO, K.C.M., DIXO, M., VERDADE, V.K., PAVAN, D.,
ARGOLO, A.J.S. & SITES JR, J. W.2007.A new genus of Microteiid Lizard from the
Atlantic Forests of State Bahia, Brazil, with a New Generic Name for Colobosaura
mentalis mentalis, and a Discussion of Relationships Among the Heterodactylini
(Squamata, Gymnophthalmidae). American Museum Novitates 3565: 1-27.
ROMÁN-CUESTA, R. M. & MARTINEZ-VILALTA, J. 2006. Effectiveness of protected
areas in mitigating firewith in their boundaries: Case study of Chiapas, Mexico.
Conservation Biology Volume 20, No. 4.
SANTOS, R.C.; LUCAS, P. S.; SOUSA, B. M. & NOVELLI, I. A. 2009. Reptilia, Squamata,
Leiosauridae, Urostrophus vautieri: Distribution extensionand geographic distribution
map. Check List 5: 533-536.
SÃO PEDRO, V. A & FEIO, R. N. 2010. Distribuição espacial e sazonal de Anuros em três
ambientes na Serra de Ouro Branco, extremo sul da Cadeia do Espinhaço, Minas
Gerais, Brasil.Biotemas, 23 (1): 143-154,
SÃO PEDRO, V. A & FEIO, R. N. 2011. Anuran species composition from Serra do Ouro
Branco, southernmost Espinhaço Mountain Range, state of Minas Gerais, Brazil. Check
List,Vol.7. (5).
SÃO PEDRO, V. A &PIRES, M. R. S. 2009. As Serpentes da região de Ouro Branco,
Extremo Sul da Cadeia do Espinhaço, Minas Gerais. Ceres 56 (2): 166-171.
SARTORIUS, S.S., VITT,L.J. &COLLI,G.R. 1999.Use of naturally and anthropogenically
disturbed habitats in Amazonian rainforests by the teiid lizard Ameiva
ameiva.Biological Conservation 90(2): 91-101.
SAZIMA, I. &HADDAD,C. F. B. 1992. Répteis da Serra do Japi: notas sobre História
Natural; p. 212-236 In: L. P. C. Morellato (ed.). História Natural da Serra do Japi:
ecologia e preservação de uma área florestal do sudeste do Brasil. Campinas: Editora da
UNICAMP/FAPESP.

57
SILVA, J. A., MACHADO, R. B., AZEVEDO, A. A., DRUMOND, G. M., FONSECA, R.
L., GOULART, M.F., JÚNIOR, E.A.M.; MARTINS,C.S. &NETO, M. B.R. 2008.
Identificação de áreas insubstituíveis para conservação da Cadeia do Espinhaço, Estado
de Minas Gerais e Bahia, Brasil. Megadiversidade, vol. 4 N:1-2.
SOUSA, B.M. & C.A.G. CRUZ. 2008. Hábitos alimentares em Enyalius perditus (Squamata,
Leiosauridae) no Parque Estadual do Ibitipoca, Minas Gerais. Iheringia 98(2): 260-265.
SOUSA, B.M., NASCIMENTO, A.E.R., GOMIDES, S.C., VARELA RIOS, C.H.,
HUDSON, A.H. & NOVELLI, I.A. 2010. Reptiles in fragments of Cerrado and Atlantic
Forest at the Campo das Vertentes, MinasGerais State, Southeastern Brazil.Biota
Neotrop.10(2):http://www.biotaneotropica.org.br/v10n2/en/abstract?article+bn0351002.
STURARO, M.J. & SILVA, V.X. 2010. Natural history of the lizard Enyalius perditus
(Squamata: Leiosauridae) from an Atlantic forest remnant in southeastern Brazil.
Journal of Natural History 44 (19-20): 1225-1238.
TEIXEIRA, R. L. & GIOVANELLI, M. 1999. Ecologia de Tropidurus torquatus
(Sauria:Tropiduridae) da Restinga de Guriri, São Mateus, ES. Rev. Brasil. Biol., 59(1):
11-18.
TEIXEIRA, R.L. 2001. Comunidade de lagartos da restinga de Guriri, São Mateus- ES,
Sudeste do Brasil. Atlântica, Rio Grande, 23: 77-84.
TORRES-CARVAJAL, O. 2005.A new species of Stenocercus (Squamata, Iguania) from
central-western Brazil with a key to Brazilian Stenocercus.Phyllomedusa.4(2):123-132.
UZZELL, T. 1969. The status of the genera Ecpleopus, Arthroseps, and Aspidolaemus
(Sauria, Teiidae).Postilla, 135: 1-23.
VAN SLUYS, M. 1992. Aspectos da ecologia do lagarto Tropidurus itambere (Tropiduridae),
em uma área do sudeste do Brasil. Rev. Bras. Biol. 52: 181-185.

58
VANZOLINI, P. E. 1948. Sobre um novo Pantadactylus do Estado de Minas Gerais (Sauria,
Teeidae). Papeis a vulsos do Departamento de Zoologia, Secretaria da Agricultura, São
Paulo, Brasil, Vol. VIII,No. 27. Pp.337-340.
VANZOLINI, P. E. 1983. Guiano-Brasilian Polychrus: distribution and speciation (Sauria:
Iguanidae). In: A. G. J. Rhodin e K. Miyata (eds.), Advances in Herpetology and
Evolutionary Biology. Museum of Comparative Zoology, Cambridge. pp. 118-131.
VANZOLINI, P. E.,RAMOS-COSTA,A. M. &VITT,L. J. 1980. Répteis das Caatingas. Rio
de Janeiro: Academia Brasileira de Ciências. 161 p.
VANZOLINI, P. E. & PAPAVERO, N. (Org.) 1967. Manual de coleta e preparação de
animais terrestres e de água doce. Departamento de Zoologia da Universidade de São
Paulo e Secretaria da Agricultura do Estado de São Paulo, São Paulo, Brasil, 222pp.
VANZOLINI, P.E. 1986. Addenda and corrigenda to the Catalogue of Neotropical
Squamata.Smithsonian Herpetological Information Service 70: 1-25.
VANZOLINI, P. E., RAMOS COSTA, A.M.M. & VITT, L.J. 1980. Répteis das Caatingas.
Academia Brasileira de Ciências, Rio de Janeiro.
VAZ-FERREIRA, R. & SIERRA DE SORIANO, B. 1960. Notas sobre los reptiles del
Uruguay. Rev. Fac. Human. y Ci. (18): 133-206.
VELOSO, H.P.; RANGEL-FILHO, A.L.R. & LIMA, J.C.A.1991. Classificação da vegetação
brasileira adaptada a um sistema universal. Rio de Janeiro: Instituto Brasileiro de
Geografia e Estatística (IBGE).
VITT, E. R. & CARVALHO C. M. 1995. Niche partitioning in a tropical wet season: lizards
in the lavrado area of northern Brazil.Copeia 2, 305-29.
VITT, L.J. & COLLI, G.R. 1994. Geographical ecology of a Neotropical lizard: Ameiva
ameiva (Teiidae) in Brazil. Canadian Journal of Zoology 72: 1986-2008.

59
VITT, L.J. & LARCHER JR, T. E. 1981. Behavior, habitat, diet, and reprodutction of the
iganid lizard Polychrus acutirostris in the Caatinga of northesastern Brazil.
Herpetologica, 37 (1), 53-63.
VITT, L.J. 1991. An introduction to the ecology of Cerrado lizards. Journal Herpetol., 25:79-
90.
VRCIBRADIC, D. & ROCHA, C.F.D. 1998.Ecology of the skink Mabuya frenata in an area
of rock outcrops in southeastern Brazil. Journal of Herpetology, 32(2): 229-237.
ZAMPROGNO, C., ZAMPROGNO, M., DAS G. F. & TEIXEIRA, R. L. 2001.Evidence of
terrestrial feeding in the arboreal lizard Enyalius bilineatus (Sauria, Polychrotidae) of
South-Eastern Brazil.Rev. Brasil. Biol., 61(1): 91-94

60
CAPITULO 2: INFLUÊNCIA DE BORDAS E DAS VARIÁVEIS AMBIENTAIS NA ESTRUTURAÇÃO DA COMUNIDADE DE LAGARTOS EM FRAGMENTOS DE MATA EM UMA ÁREA DE TRANSIÇÃO MATA ATLÂNTICA- CERRADO

61
RESUMO
A estrutura do hábitat é considerada um dos fatores primordiais no estudo da ecologia de
qualquer espécie. Sua alteração provoca mudanças na composição da comunidade, podendo
favorecer, ou levar a extinção de populações ou espécies. Este trabalho teve como objetivo de
analisar a influência da estrutura do hábitat e efeito de bordas na organização da comunidade
dos lagartos de serapilheira em fragmentos de matas situadas em uma zona de transição entre
a Mata Atlântica e o Cerrado. Para tanto foram escolhidas três áreas de mata secundárias e em
processo de regeneração denominadas como A, B e C na serra de Ouro Branco. Cada área foi
amostrada em três ambientes diferentes: borda com a estrada, borda com ambientes de prática
agrícola e interior do ambiente de mata. Em cada um desses ambientes foram instaladas
armadilhas de interceptação e queda as quais foram vistoriadas ao longo de um ano, entre
janeiro e dezembro de 2010, com um esforço amostral de 5184 dias-balde e 1728 dias-baldes
por área. Foram coletadas oito espécies, distribuídas em cinco famílias: Anguidae,
Gymnophthalmidae, Leiosauridae, Scincidae e Teiidae, totalizando 101 indivíduos. E.
perditus foi a espécie mais abundante na região, seguida de E. gaudichaudii e H. imbricatus.
Quanto a riqueza, as áreas A e B se mostraram iguais e maiores que a área C. Não foi
encontrada uma diferença significativa na riqueza de lagartos entre os diferentes ambientes
amostrados. A abundância de lagartos não variou entre as áreas e nem entre os ambientes. A
similaridade na composição da fauna de lagartos de serapilheira pode ser devido à estrutura
vegetacional similar entre os ambientes. Tal fato pode estar também relacionado à baixa
abundância de lagartos coletados. No entanto, em matas secundárias, as variáveis ambientais e
vegetacionais parecem ter exercido maior influência na estruturação da comunidade de
lagartos do que os efeitos das bordas. A partir dos dados do presente trabalho, foi possível
verificar que os ambientes de borda de matas secundárias comportam tanto as espécies
consideradas estritamente florestais e mais sensíveis a alterações no habitat, como também
espécies de áreas abertas. Considerando que espécies que ocorrem em bordas florestais estão
sujeitas aos mais variados riscos, torna-se necessário dar maior importância a conservação e
restauração de áreas perturbadas.
Palavras chave: Uso de hábitat; Fragmentação; Répteis; Ambientes perturbados.

62
ABSTRACT
Habitat structure is considered to be one of the most important factor in the study of any
species’ ecology. Its alteration incurs in changes in the composition of the community, either
favoring or extinguishing populations or species. With the purpose of analyzing the influence
of the structure of habitat and the border effects in the organization of litter lizards
communities in wood fragments located in a transition zone between the Atlantic Forest and
the Cerrado. For that, three secondary woodland areas in regeneration process were chosen,
from now on called A, B and C. In each area, three different environments were analyzed:
borders with roads, borders with agricultural fields and woodland interior. In each of these
spots, were installed interception and fall traps, which were monitored for over a year, from
January 2010 to December 2010, with a sample-effort of 5184 bucket-days and 1728 bucket-
days per area. Eight species were collected, distributed amongst five families: Anguidae,
Gymnophthalmidae, Leiosauridae, Scincidae and Teiidae, with a total of 101 individuals. E.
perditus was the most abundant species in the region, followed by E. gaudichaudii and H.
imbricatus. Areas A and B showed the same richness, higher than area C. There was no
significant difference in the richness of lizards in the analyzed regions. Their abundance
showed no signs of variation whatsoever, either between the areas or between the
environments. The similarity in the composition of litter lizards’ fauna could be attributed to
the similar vegetation structure between the environments. But in secondary forests, the
environmental and vegetation variables seem to have exerted higher influence in the structure
of the communities of lizards than the border effects. Given the present data collected by this
study, it was possible to infer that the secondary forests environments hold species considered
strictly forest related and more sensitive to habitat alterations, as species of open areas.
Considering the fact that species that occur in forest borders are subjected to more risks, it is
necessary to give a greater importance to the conservation and restoration of disturbed areas.
Keywords: Habitat using; Fragmentation; Reptiles; Disturbed Environments

63
INTRODUÇÃO
A estrutura do hábitat é considerada um dos fatores primordiais a serem levados em conta no
estudo da ecologia de qualquer grupo biológico. Alterações no hábitat provocam mudanças na
composição das espécies de uma comunidade, influenciando no sucesso dos indivíduos
quanto à capacidade de aproveitamento dos recursos disponíveis, podendo favorecer ou levar
a extinção de populações ou espécies (Murcia, 1995; Tabarelli & Gascon, 2005; Mendonça et
al., 2009; Alford & Richards, 1999; Jellinek et al., 2004; Gascon et al., 2000; Sala et al.,
2000; Fahrig, 2003).
A fragmentação de habitat, tendo como objetivo a urbanização, prática de agricultura ou
formação de estradas, levam a criação de bordas que constitui grande ameaça a biodiversidade
(Bierregaard et al., 2001; Tabarelli et al., 1999; Broadbent et al., 2008). A prática da
agricultura, além de provocar a perda de habitat, isola fragmentos de matas, facilita a entrada
de espécies exóticas e aumenta os riscos de incêndios e contaminação por agrotóxicos para as
espécies nativas (Fiszon et al., 2003; Broadbent et al., 2008). A criação de estradas, por sua
vez, tem um importante papel no direcionamento da perda de hábitats naturais e no aumento
de vias que causam impactos, como atropelamentos de animais silvestres, riscos de
contaminação por poluição, incêndios e práticas de caça, resultando assim na diminuição da
diversidade de organismos nas comunidades (Trombulak & Frissell, 2000;Cerqueira et al.,
2003; Skole & tocker, 1993; Santos & Tabarelli,2002; Fiszon et al., 2003; Peres & Lake,
2003; Tabarelli & Gascon, 2005; Hartmann et al., 2011). Esses riscos dependem muito do
ambiente que circunda o fragmento, ou seja, a “matriz” (Gascon et al., 1999; Bierregaard et
al., 2001).

64
Estudos para entender como espécies, populações ou comunidades reagem aos efeitos de
bordas são cruciais na construção de idéias e reflexões conservacionistas. A rápida destruição
de ecossistemas em todo mundo exige decisões planejadas e um conhecimento profundo das
consequências de alterações. Inúmeros estudos reúnem evidências dos impactos dessas
alterações em diversos grupos biológicos, tais como plantas (Bierregaard et al., 2001;
Laurence et al., 2000;Tabarelli et al., 1999), invertebrados (Kruess & Tscharntke, 1994),
anfíbios (Collins & Storfer, 2003; Hazell, 2003; Lehtinen et al., 2003; Gardner et al., 2007;
Bell & Donnelly, 2006; Dixo & Martins, 2008), répteis (Kitchener et al., 1980; Gibbons et al.,
2000; Gardner et al., 2007; Dixo & Martins, 2008; Lehtinen et al., 2003; Bell & Donnelly,
2006; Dixo & Metzger, 2009), aves (Mendonça et al., 2009) e mamíferos (Adler, 1996;
Pardini, 2004). Apesar dos cientistas terem apresentado um número relativamente grande de
trabalhos relacionados às consequências deste tipo de distúrbio e da identificação das ações
prioritárias, esses riscos ainda permanecem (Tabarelli & Gascon, 2005; Gardner et al., 2007).
O novo código florestal em tramitação no Brasil, um dos países mais biodiversos do mundo, é
a mais pura demonstração desta realidade, dado que estudos que apontam a perda de hábitat
como o principal causador da perda da riqueza biológica foram ignorados (Ab’Sáber, 2010).
Os lagartos ocupam diversos ambientes e sãoconsiderados particularmente sensíveis a
alterações no meio, o que os torna muito útil em estudos de qualidade ambiental (Vitt et al.,
1998; Jellinek et al.,2004; Dixo & Martins, 2008; Dixo & Metzger, 2009), sendo
considerados excelentes modelos para o entendimento de processos ecológicos que envolvem
alterações no hábitat (Kitchener et al., 1980; Lehtinen et al., 2003; Gardner et al., 2007).
Populações deste grupo vem sofrendo um declínio global, devido principalmente à perda e
alteração de habitats, introdução de espécies exóticas, poluição ambiental, doenças, além das
mudanças climáticas (Gibbons et al., 2000).

65
Devido à elevada pressão antrópica, riqueza e endemismos da flora e fauna, além de aspectos
da riqueza cultural, a Serra do Espinhaço foi elevada em 2005 a categoria de Reserva
daBiosfera pela UNESCO, pelo programa o Homem e a Biosfera (MAB). Este ambiente
singular representa um dos poucos ecossistemas montanhosos no Brasil, sendo considerado o
divisor de dois hotspots extremamente ameaçados: Mata Atlântica e a Cerrado (Myers et al.,
2000). Suas características geológicas e geográficas são argumentos interessantes para estudos
de caráter ecológico e biogeográfico que participam na geração da diversidade.
O presente trabalho foi realizado na serra do Ouro Branco, localizada na porção sul da Cadeia
do Espinhaço. A comunidade de lagartos em ambientes de interior de matas secundárias e de
bordas criadas por ações antrópicas tais como: estradas e áreas de prática agrícola foram
analisadas, incluindo elementos da estrutura do hábitat ou variáveis ambientais. Este trabalho
teve como objetivo contribuir para o entendimento dos processos ecológicos que atuam na
estruturação de comunidades em paisagens formadas por um mosaico de ambientes
perturbados. Para isso, foram propostas as seguintes questões: Como é composta a
comunidade de lagartos em diferentes áreas dos fragmentos florestais de mata secundária? Há
diferença na riqueza, abundância e composição de espécies em ambientes de interior ou
ambiente de bordas florestais? As variáveis ambientais mensuradas no trabalho influenciam
na distribuição dos lagartos no fragmento? Se sim, que variáveis ambientais mais afetam na
distribuição dos lagartos no fragmento? Para responder essas questões foram apresentadas as
seguintes hipóteses:
1) Ambientes de borda terão uma composição, riqueza e abundância diferentes dos
ambientes de interior de mata. Apesar de todos os ambientes apresentarem sinais
claros de interferência antrópica, é esperado que os ambientes de borda

66
apresentemalterações no microclima (vento, temperatura, umidade) mais bruscas do
que os ambientes de interior (Murcia, 1995), os quais oferecem condições mais
estáveis, além de apresentarem menos distúrbios, tais como, poluição e presença
antrópica.
2) Ambientes de borda com áreas de agricultura ou com estrada terão composição,
riqueza e abundância de lagartos diferentes entre si. As matrizes que circundam os
fragmentos de mata nos dois casos, são de diferentes composições, qualidade e
permeabilidade, podendo atuar como barreira ou filtro para algumas espécies
(Gascon et al.,1999). Os ambientes utilizados para a prática agrícola podem ser
utilizados pelos lagartos, constituindo uma matriz permeável para algumas espécies,
enquanto que ambientes de estrada ou rodovia apresentam-se totalmente desprovido
de vegetação e serapilheira, além do fluxo de automóveis, aumentando riscos como
atropelamentos.
3) Variáveis ambientais vegetacionais como número e circunferência do tronco das
árvores, cobertura do dossel, cobertura do sub-bosque e biomassa da serapilheira
estão correlacionados com a distribuição dos lagartos no fragmento. Uma das
razões para algumas espécies conviverem dentro da mesma comunidade está
relacionada a diferentes estratégias de uso de hábitats. Assim, a qualidade de hábitat
pode influenciar até mais do que o próprio efeito de bordas em fragmentos
perturbados (Jellinek et al., 2004). A complexidade estrutural pode estimular o
aumento do número de espécies na comunidade, sendo esperado que hábitats
estruturalmente mais simples contenham menos espécies devido à menor
disponibilidade de microambientes (Pianka, 1973).

67
METODOLOGIAS
Área de estudo
O estudo foi realizado na Serra do Ouro Branco, situada no município de Ouro Branco, Minas
Gerais, Brasil (figura 1). A área de estudo está inserida ao sul do Quadrilátero Ferrífero e da
Cadeia do Espinhaço, sendo considerada uma área de transição entre os biomas Mata
Atlântica e Cerrado. Esta região é caracterizada por matas secundárias, resultantes do
processo exploratório intenso iniciado no período de colonização do Brasil.
As áreas de mata da região de estudo são classificadas como Florestas Estacionais
Semideciduais (Veloso, 1991), ocorrendo em cotas de até 1.400 metros e em áreas transitórias
entre os Campos Rupestres, onde o solo permite a ocupação por elementos arbóreos (Paula,
2003). As chuvas se distribuem principalmente entre os meses de novembro a fevereiro e a
temperatura média anual e precipitação média anual de 20,7º e 1188,2 mm sucessivamente de
acordo com os dados relativos aos anos 1998-2010, fornecidos pela Estação Meteorológica da
Gerdau Açominas.

68
Figura 3: Localização da serra do Ouro Branco, Minas Gerais, Brasil e delimitação das duas Unidades
de Conservação da região em um mapa de altitudes.
Desenho amostral
Para o estudo foram selecionadas três áreas de florestas (A, B e C) na Serra do Ouro Branco.
Em cada área foram instaladas nove armadilhas de interceptação e queda (Cechin & Martins,
2000), sendo três em ambientes de interior da mata; três em ambientes de borda da mata com
área de agricultura, e três nas bordas da mata com a estrada (figura 2). Considerando a
distribuição das armadilhas entre os ambientes, foram nove armadilhas localizadas no interior
da mata, nove em bordas de mata com a agricultura e nove nas bordas de mata com a estrada.

69
Assim, foram considerados ambientes de interior, aqueles localizados a mais de 50 metros das
bordas de mata, enquanto que as armadilhas das bordas foram instaladas bem próximas aos
seus respectivos fatores impactantes, ou seja, estrada ou plantação de milho, não excedendo
10 metros.
As armadilhas de interceptação e queda foram dispostas em cada ambiente em forma de ‘Y’,
ou seja, cada uma foi constituída por quatro baldas de 60 Litros, distanciados a cinco metros
do balde central, e interligados por cercas guias de aproximadamente um metro de altura
(figura 3). Cada armadilha se distanciava 20 metros da próxima. Resumindo, foram utilizados
quatro baldes por cada armadilha de interceptação e queda, 12 em cada ambiente e 36 baldes
por área e 108 baldes em todo experimento.
Figura 4: Desenho amostral demonstrando a disposição das armadilhas em relação a cada área
(A, B e C) e os ambientes interior e bordas com estradas e agricultura.

70
Figura 3: Desenho esquemático demonstrando a disposição das armadilhas de interceptação e
queda em cada ambiente, com as distâncias entre os baldes e as armadilhas.
As coletas ocorreram de janeiro a dezembro de 2010, sendo que, em cada mês as armadilhas
foram abertas por um período de quatro noites consecutivas e vistoriadas sempre pela manhã,
entre às 09h00min e 11h00min. Com isso, o esforço total de amostragem correspondeu a
5.184 dias-balde e 1.728 dias-balde por área. Os lagartos coletados foram depositados na
Coleção Herpetológica do Laboratório de Zoologia da Universidade Federal de Ouro Preto
(LZV-UFOP).
Características ambientais
Foram tomadas medidas das variáveis ambientais para a descrição da estrutura da vegetação e
verificação de diferença estrutural entre os ambientes amostrados e, ainda, para testar a
correlação entre essas variáveis e a ocorrências de lagartos. Neste sentido, ao redor da
extremidade de cada armadilha, foram mensuradas 75 m² divididos em três quadrados iguais
(5 X 5 metros). Todas as árvores dentro desses quadrados, com circunferência do tronco na
altura do peito acima ou igual a cinco cm, foram contabilizadas e sua suas circunferências
anotadas. Desta forma, um total de 675 metros quadrados foram mesurados em cada área, e
2.025 metros quadrados em todo o experimento. Outras medidas como conectividade de
dossel e conectividade de sub-bosque foram anotadas categoricamente, sendo o valor zero (0)

71
atribuído para a ausência da variável; um (1), para baixa; dois (2), para média e três (3) para
alta ocorrência da variável.
Outro fator ambiental considerado foi à produção de serapilheira entre as áreas. Por ser
composto por folhas, troncos, caules, raízes, sementes, flores, frutos e resíduos animais, a
serapilheira constitui um micro-hábitat para muitas espécies em comunidades florestais, sendo
particularmente ocupada por lagartos (Dixo & Martins, 2008; Dixo & Metzger, 2009).
Durante 12 meses de amostragem, foi calculada a taxa de produção da serapilheira próximo a
cada armadilha. Para isso, foram confeccionados coletores de serapilheira, formando
quadrados de 50 cm X 50 cm e de 20 cm de altura com tela do tipo mosquiteiro de nylon 1.0
mm. Dois coletores foram colocados próximos a cada armadilha, distribuídos alternadamente
entre o lado direito e esquerdo, totalizando 18 quadrados por área e 54 coletores em todo o
experimento. Esses coletores permaneciam abertos durante 30 dias. O material recolhido foi
levado ao laboratório e colocado na estufa a 70º C durante 48 horas. Após esse procedimento,
a massa seca de serapilheira foi pesada em balança eletrônica (Shimadzu BL 3200 de peso
máximo 3200 g e peso mínimo 0,5 g).
Análise dos dados
Para a análise da riqueza de lagartos entre as diferentes áreas e ambientes, foi efetuada uma
ANOVA de duas vias, com post hoc/ Tukey. Para as análises da abundância de cada espécie
em cada área e ambiente só foram consideradas as espécies com mais de cinco indivíduos
coletados. Para as análises da abundância, foi utilizado Generalized linear mixed model
(GLMM), com distribuição quase Poisson. Para verificar a similaridade entre as diferentes
áreas e ambientes, usou-se o índice de Bray-Curtis, e para examinar as características

72
ambientais nas diferentes áreas e suas relações com as diferentes espécies florestais, foi
efetuada uma Análise de Correspondência Canônica (CCA) (Hill, 1973).
RESULTADOS
Ao longo do período de amostragem foram registrados 101 exemplares de lagartos
distribuídos em oito espécies e cinco famílias: Anguidae, Gymnophthalmidae, Leiosauridae,
Scincidae e Teiidae (tabela 2). A maioria das espécies coletadas são descritas na literatura
como semi-fossoriais, semi-arborícolas e terrícolas, ocorrendo na serapilheira de áreas de
mata.
Tabela 1: Famílias e espécies de lagartos coletados na serra do Ouro Branco nos diferentes ambientes.
Ambiente
Famílias e espécies Hábito E I A Total
Anguidae Ophiodes Cf. Striatus Spix, 1825 SF 2 0 0 2
Gymnophthalmidae Ecpleopus gaudichaudii Duméril & Bibron,
1839 SF 12 7 10 29 Heterodactylus imbricatus Spix, 1825 SF 4 4 2 10
Leiosauridae Enyalius bilineatus Duméril & Bibron, 1837 SA 2 0 2 4
Enyalius perditus Jackson, 1978 SA 16 17 19 52 Urostrophus vautieri Duméril & Bibron, 1837 SA 0 0 1 1
Scincidae Mabuya frenataCope,1862 T 1 0 1 2
Teiidae Tupinambis merianae (Duméril & Bibron,
1839) T 1 0 0 1 Total 38 28 35 101
Abreviaturas: E: estrada; I: interior; A: agricultura; AS: semi –arborícola; SF: semi-fossorial; T:
terrícola.
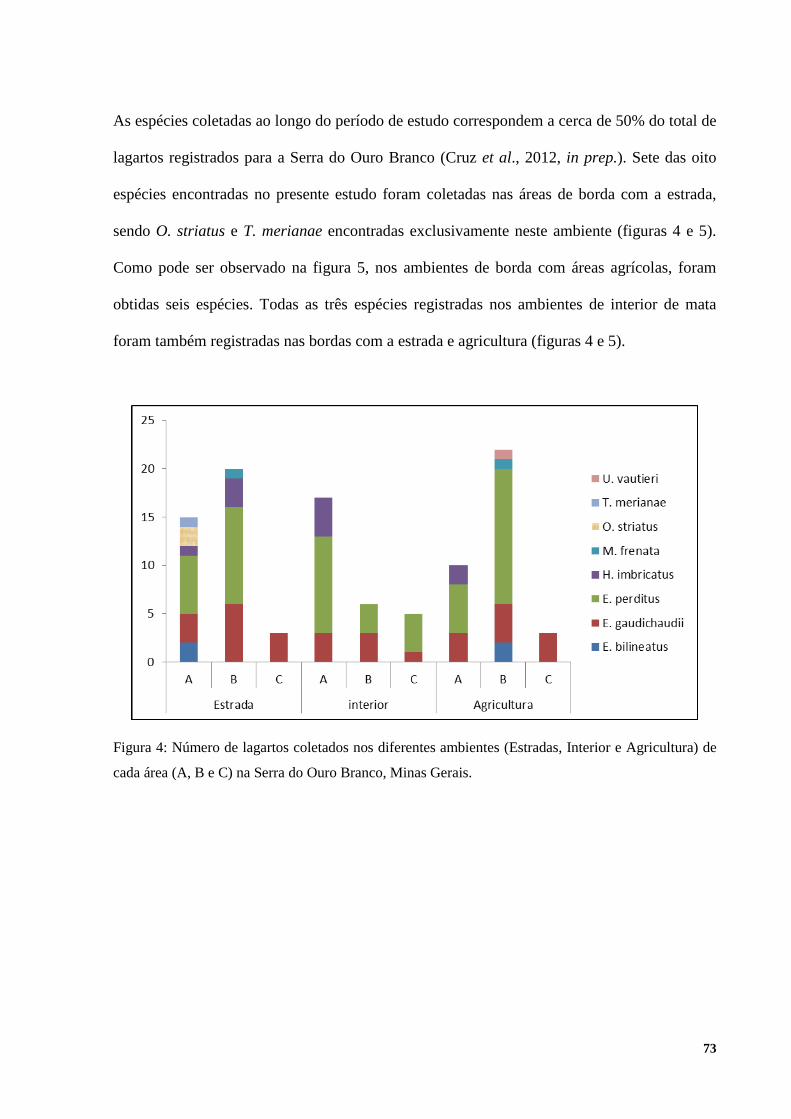
73
As espécies coletadas ao longo do período de estudo correspondem a cerca de 50% do total de
lagartos registrados para a Serra do Ouro Branco (Cruz et al., 2012, in prep.). Sete das oito
espécies encontradas no presente estudo foram coletadas nas áreas de borda com a estrada,
sendo O. striatus e T. merianae encontradas exclusivamente neste ambiente (figuras 4 e 5).
Como pode ser observado na figura 5, nos ambientes de borda com áreas agrícolas, foram
obtidas seis espécies. Todas as três espécies registradas nos ambientes de interior de mata
foram também registradas nas bordas com a estrada e agricultura (figuras 4 e 5).
Figura 4: Número de lagartos coletados nos diferentes ambientes (Estradas, Interior e Agricultura) de
cada área (A, B e C) na Serra do Ouro Branco, Minas Gerais.
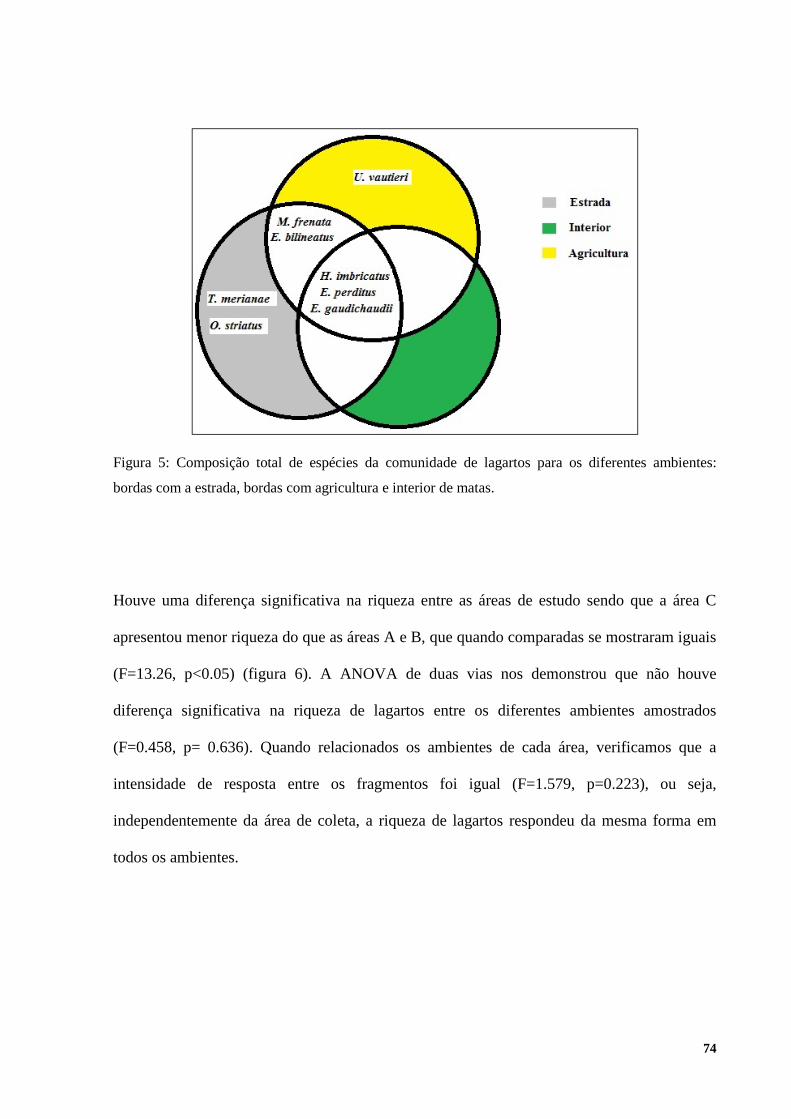
74
Figura 5: Composição total de espécies da comunidade de lagartos para os diferentes ambientes:
bordas com a estrada, bordas com agricultura e interior de matas.
Houve uma diferença significativa na riqueza entre as áreas de estudo sendo que a área C
apresentou menor riqueza do que as áreas A e B, que quando comparadas se mostraram iguais
(F=13.26, p<0.05) (figura 6). A ANOVA de duas vias nos demonstrou que não houve
diferença significativa na riqueza de lagartos entre os diferentes ambientes amostrados
(F=0.458, p= 0.636). Quando relacionados os ambientes de cada área, verificamos que a
intensidade de resposta entre os fragmentos foi igual (F=1.579, p=0.223), ou seja,
independentemente da área de coleta, a riqueza de lagartos respondeu da mesma forma em
todos os ambientes.
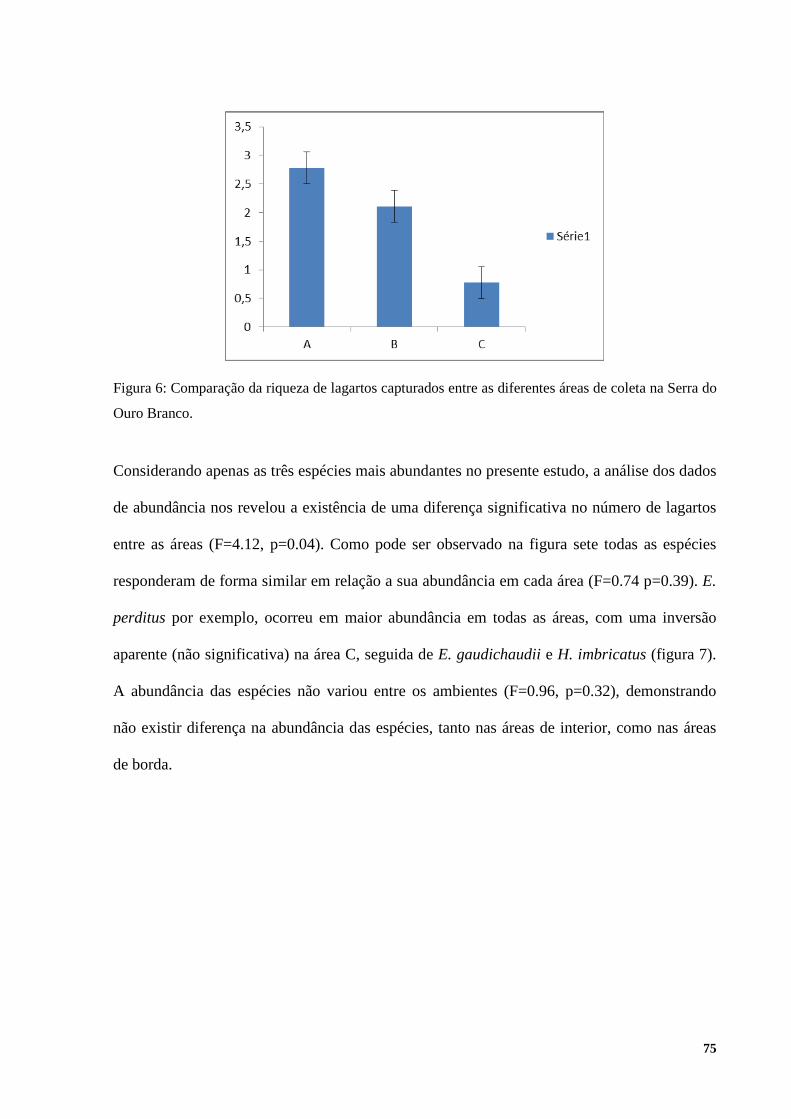
75
Figura 6: Comparação da riqueza de lagartos capturados entre as diferentes áreas de coleta na Serra do
Ouro Branco.
Considerando apenas as três espécies mais abundantes no presente estudo, a análise dos dados
de abundância nos revelou a existência de uma diferença significativa no número de lagartos
entre as áreas (F=4.12, p=0.04). Como pode ser observado na figura sete todas as espécies
responderam de forma similar em relação a sua abundância em cada área (F=0.74 p=0.39). E.
perditus por exemplo, ocorreu em maior abundância em todas as áreas, com uma inversão
aparente (não significativa) na área C, seguida de E. gaudichaudii e H. imbricatus (figura 7).
A abundância das espécies não variou entre os ambientes (F=0.96, p=0.32), demonstrando
não existir diferença na abundância das espécies, tanto nas áreas de interior, como nas áreas
de borda.
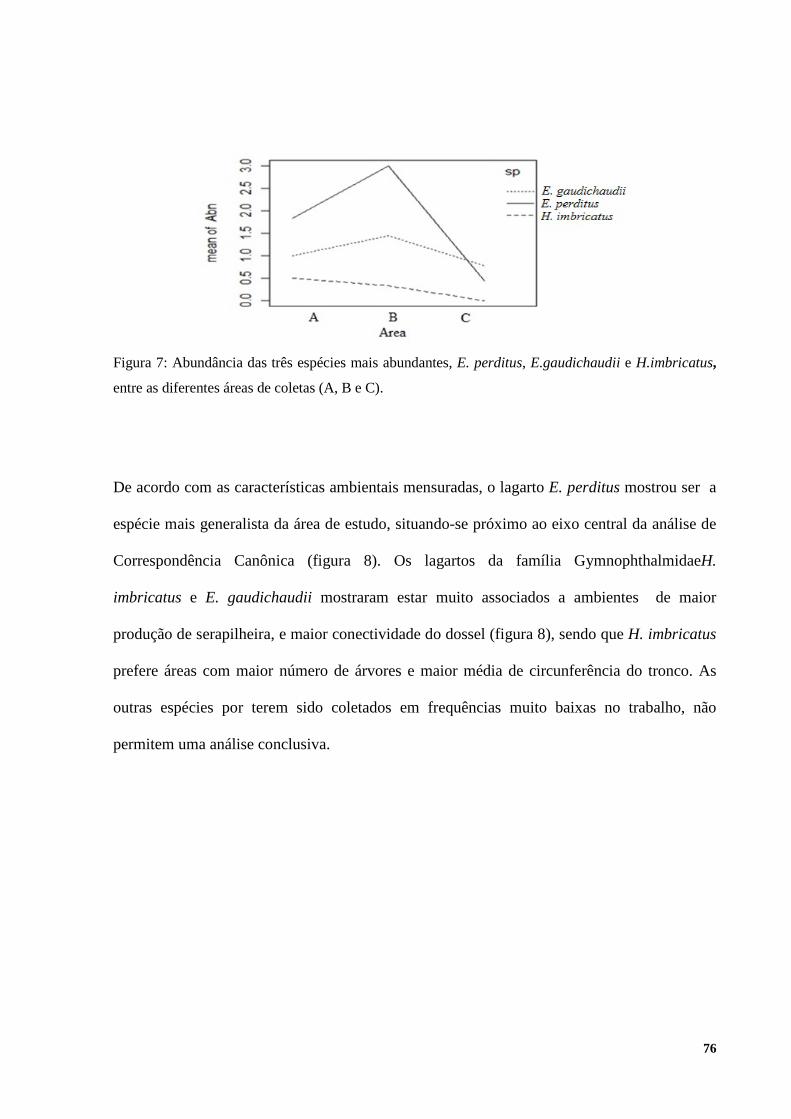
76
Figura 7: Abundância das três espécies mais abundantes, E. perditus, E.gaudichaudii e H.imbricatus,
entre as diferentes áreas de coletas (A, B e C).
De acordo com as características ambientais mensuradas, o lagarto E. perditus mostrou ser a
espécie mais generalista da área de estudo, situando-se próximo ao eixo central da análise de
Correspondência Canônica (figura 8). Os lagartos da família GymnophthalmidaeH.
imbricatus e E. gaudichaudii mostraram estar muito associados a ambientes de maior
produção de serapilheira, e maior conectividade do dossel (figura 8), sendo que H. imbricatus
prefere áreas com maior número de árvores e maior média de circunferência do tronco. As
outras espécies por terem sido coletados em frequências muito baixas no trabalho, não
permitem uma análise conclusiva.

77
Figura 8: Análise da distribuição das espécies de acordo com as variáveis ambientais encontradas na
mata secundária, na serra do Ouro Branco, Minas Gerais.
Para a caracterização das áreas de estudos e correlação das espécies com as diferentes
variáveis ambientais os autovalores da CCA dos dois primeiros eixos explicam 88,16 % da
variância dos dados, sendo que o primeiro eixo apresentou o autovalor de 0,15 (53,13%) e o
segundo eixo 0,10 (35,03%).
A análise de similaridade com o índice de Bray-Curtis (figura 8) demonstra que apesar de
formarem alguns grupos com características similares, os ambientes de borda se mostraram
similares a ambientes de interior e vice-versa. A área A é a que mais se diferenciou das outras,
principalmente devido à média da circunferência do tronco e o número de árvores (tabela 2 e
figura 5). No entanto, assim como as outras áreas, os ambientes de borda se mostraram
bastante similares, com ambientes de interior vice-versa (figura 8 e 9).
Tabela 2: Localização e caracterização das áreas e ambientes de coleta, com as médias das
diferentesvariáveis ambientais na serra do Ouro Branco, Minas Gerais. Média do número de árvores:
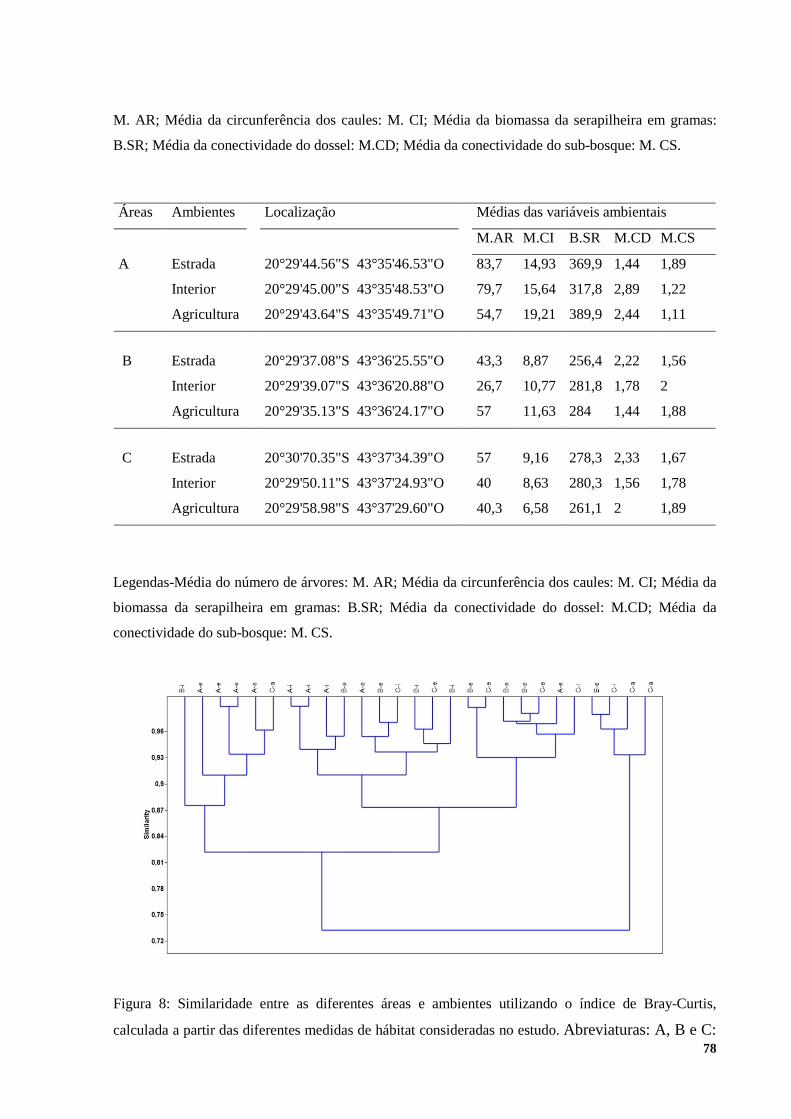
78
M. AR; Média da circunferência dos caules: M. CI; Média da biomassa da serapilheira em gramas:
B.SR; Média da conectividade do dossel: M.CD; Média da conectividade do sub-bosque: M. CS.
Áreas Ambientes
Localização
Médias das variáveis ambientais
M.AR M.CI B.SR M.CD M.CS
A Estrada
20°29'44.56"S 43°35'46.53"O
83,7 14,93 369,9 1,44 1,89
Interior
20°29'45.00"S 43°35'48.53"O
79,7 15,64 317,8 2,89 1,22
Agricultura 20°29'43.64"S 43°35'49.71"O 54,7 19,21 389,9 2,44 1,11
B Estrada
20°29'37.08"S 43°36'25.55"O
43,3 8,87 256,4 2,22 1,56
Interior
20°29'39.07"S 43°36'20.88"O
26,7 10,77 281,8 1,78 2
Agricultura 20°29'35.13"S 43°36'24.17"O 57 11,63 284 1,44 1,88
C Estrada
20°30'70.35"S 43°37'34.39"O
57 9,16 278,3 2,33 1,67
Interior
20°29'50.11"S 43°37'24.93"O
40 8,63 280,3 1,56 1,78
Agricultura 20°29'58.98"S 43°37'29.60"O 40,3 6,58 261,1 2 1,89
Legendas-Média do número de árvores: M. AR; Média da circunferência dos caules: M. CI; Média da
biomassa da serapilheira em gramas: B.SR; Média da conectividade do dossel: M.CD; Média da
conectividade do sub-bosque: M. CS.
Figura 8: Similaridade entre as diferentes áreas e ambientes utilizando o índice de Bray-Curtis,
calculada a partir das diferentes medidas de hábitat consideradas no estudo. Abreviaturas: A, B e C:

79
Áreas de coletas. As letras minúsculas: a: Bordas com agricultura; e: Bordas com estradas; i:
Interior de áreas de Mata.
Figura 9: Análise de Correspondência Canônica demonstrando o agrupamento dos diferentes
ambientes entre as diferentes áreas. Áreas de coletas: A, B e C. As letras minúsculas: a: Bordas com
agricultura; e: Bordas com estradas; i: Interior de áreas de Mata.
DISCUSSÃO
A composição de lagartos de serapilheira da serra do Ouro Branco se mostrou similar a outros
estudos realizados em fragmentos de Mata Atlântica, utilizando o mesmo método de
amostragem, ou seja, com armadilhas do tipo pitfall (Dixo & verdade, 2006; Dixo & Metzger,
2009). Espécies consideradas endêmicas da Mata Atlântica: E. perditus, E. gaudichaudii e H.
imbricatus foram registradas nesses trabalhos, sendo essas duas primeiras espécies
particularmente as mais abundantes em todos os estudos, inclusive no presente trabalho.
Exceto T. merianae registrada por Dixo & Metzger (2009), as quatro espécies restantes que

80
compõem a comunidade de lagartos serapilheira de áreas de mata representados na Serra do
Ouro Branco, não foram encontradas nos estudos citados acima.
Quando comparamos a abundância de espécies da comunidade de lagartos de serapilheira da
serra do Ouro Branco, com outras comunidades de lagartos em remanescentes da Mata
Atlântica, ela se mostrou semelhante à Dixo & Verdade (2006) e menor do que Dixo &
Martins (2008) eDixo & Metzger (2009). Entretanto, outros fatores biológicos, bem como
detalhes na metodologia de coleta podem tanto afetar a riqueza, como a abundância de
indivíduos coletados. Dixo & Martins (2008), capturaram 302 lagartos, pertencentes a 13
espécies. Entretanto, no referido trabalho, foi efetuado um esforço amostral de 6.612 baldes
por dia, distribuídos em uma única estação chuvosa, de outubro a fevereiro. Além de esse
esforço ser ligeiramente superior ao apresentado no presente estudo (5184 dias balde), ele
também foi mais concentrado, aproveitando dos padrões de sazonalidade da precipitação
pluvial, época de maior abundância relativa para os lagartos de regiões tropicais, devido a
maior disponibilidade de alimento e taxa reprodutiva (Fitch, 1982). Dixo & Verdade (2006)
também concentraram as suas coletas nos períodos chuvosos. Contudo, este apresentou um
esforço amostral bem menor em todo experimento com 2.112 baldes dia.
Na reserva do Morro Grande, São Paulo, Dixo & Metzger (2009) capturaram 228 lagartos,
divididos em cinco espécies. Para este trabalho, achamos que a qualidade do habitat pode
estar interferindo na abundância de lagartos. A área de estudos desses autores também é uma
região com uma altitude considerável (870-1030 m), e características climáticas (precipitação)
próximas às condições encontradas em Ouro Branco, se diferenciando apenas na estrutura da
mata, apresentando inclusive áreas de matas mais maduras, fato que aconteceu também no
estudo de Dixo & Martins (2008).

81
Assim como no presente estudo, E. gaudicahudii e E. perditus foram também as espécies
mais abundantes no trabalho de Dixo & Metzger (2009), apesar das populações em Ouro
Branco serem menores para ambas espécies. Essas diferenças registradas na abundância
dessas populações na comunidade podem também depender, entre outros, de inúmeros fatores
envolvidos, no histórico de desmatamento em cada uma dessas áreas. Dixo & Martins (2008)
realizaram o estudo em florestas com diferentes estágios sucessionais e em áreas de florestas
maduras, que oferecem maior complexidade de hábitat. Por outro lado, a área do presente
estudo, apresenta características marcantes de florestas em estágio inicial de regeneração.
Segundo Pianka (1973), hábitats mais complexos tendem a apresentar maior diversidade de
lagartos, devido à variedade de abrigos disponíveis, permitindo a convivência de uma maior
diversidade de espécies na comunidade, com comportamentos, formas e tamanhos diferentes.
Assim, a pouca heterogeneidade no hábitat, caracterizada pela estrutura vegetacional pode
explicar a baixa abundância e riqueza dos lagartos de serapilheira na serra do Ouro Branco.
A pouca representação das espécies O. striatus, T. merianae, E. bilineatus, U. vautieri e M.
frenata no estudo provavelmente se devem ao uso de hábitat dessas espécies. Algumas dessas
espécies, como por exemploO. striatus, T. merianae e M. frenata por preferirem áreas abertas,
costumam ser encontradas em atividade em ambientes de bordas de mata (Vanzolini et al.,
1980; Nogueira, 2006; Vrcibradic & Rocha, 1998; Costa et al., 2009).
A pouca abundância das espécies E. bilineatuse U. vautieri pode estar, tanto ligado ao
método de amostragem quanto a possibilidade de suas respectivas populações serem de
tamanho reduzido na área de estudo. Apesar de arborícolas, estas espécies também são
encontradas no chão de áreas florestadas e, portanto susceptíveis às armadilhas do tipo pitfall.
Segundo Zamprogno et al., (2001), E. bilineatus baseia sua dieta em itens que ocorrem no

82
chão das florestas. No entanto, essa espécie apresentou uma ocorrência muito menor na
serapilheira do que E. perditus, outra espécie do mesmo gênero e também considerada semi-
arboricola. Por serem espécies próximas filogeneticamente, a possibilidade da segregação de
hábitats pode ser outro fator interferindo nesta diferença. Segundo Pinka (1973), é comum que
o espaço seja um dos eixos fundamentais da segregação quando espécies aparentadas se
encontram em simpatria. Como só capturamos os lagartos na serapilheira, não sabemos
realmente qual é a representação real de lagartos de outros micro-habitats da área de estudo e,
assim, afirmar a ocorrência da segregação.
Assim como encontrado em Jellinek et al. (2004) e Dixo & Martins (2008), os ambientes de
borda não diferiram dos ambientes do interior em termos de abundância e riqueza de lagartos,
ou seja, a comunidade de lagartos não respondeu como esperado aos efeitos das bordas. No
nosso trabalho, verificamos ainda que não houve diferença na distribuição dos lagartos nos
dois tipos de ambientes de borda analisados, refutando a predição de que bordas das estradas
seriam menos ricas. Atribuímos este resultado ao estado de regeneração da mata, e à pouca
heterogeneidade entre os ambientes. Gascon (1993) e Dixo & Martins (2008) sugerem que
esta falta de resposta às bordas pode ser a resposta dos lagartos a qualidade da matriz e das
áreas do entorno. Como foi verificado no nosso estudo, apesar das áreas demonstrarem
alguma diferença estrutural entre si, os ambientes amostrados, borda e interior não diferiram
estruturalmente entre si.
Segundo Kitchner et al. (1980), os lagartos podem resistir a certos impactos da fragmentação,
sendo por vezes menos afetados que outros grupos biológicos. Hutchinson et al. (2001)
sugerem que espécies generalistas conseguem aproveitar de diferentes recursos no
ecossistema, em diferentes hábitats como remanescentes, interiores e bordas de mata, além da
matriz urbana. No entanto, alguns lagartos estritamente florestais, apesar de estarem
condenados a pequenos fragmentos de mata, por vezes bem deterioradas e sem a

83
conectividade, conseguem escolher pequenas ilhas de condições ótimas ou micro-hábitats em
boas condições, o que nem sempre corresponde a uma condição próxima à qualidade do
micro-hábitat original. Assim a capacidade de resiliência não deve ser considerada como uma
tendência a ser generalista.
Segundo Dixo & Metzeger (2009), a espécie E.gaudichauddi apresenta alguma resiliência às
perturbações e à fragmentação de hábitat, sendo encontrada inclusive em ambientes
antropizados (Costa et al., 2010). No entanto, no nosso estudo, apesar de E. gaudichaudii e H.
imbricatus serem amostradas em bordas de matas e interiores, elas mostraram ser exigentes
quanto ao hábitat. Essas espécies foram relacionadas principalmente a ambientes com maior
conectividade do dossel, portanto, menor entrada de Luz, e maior biomassa da serapilheira,
que pode significar também a existência de maior número de hábitas e maior complexidade de
microhábitat para as espécies que vivem no folhiço. Este resultado corrobora com Jellinek et
al. (2004), que demonstrou em um estudo de comunidades de lagartos na Austrália que
diversas variáveis vegetacionais podem influenciar muito mais na distribuição e ocorrência de
espécies na comunidade do que as mudanças proporcionadas em função das bordas. Dixo &
Verdade (2006) acreditam que outras características, como pequeno tamanho corporal,
associado às baixas exigências energéticas podem ser outros fatores envolvidos na resiliência
dos lagartos a fragmentação.
Pela maior abundância relativa, ocorrência em todas as áreas e ambientes amostrados, E.
perditus se comportou como uma espécie generalista na área de estudos. Entretanto, este
lagarto é considerado sensível a fragmentação de habitat (Dixo & Metzger, 2009), constando
inclusive na lista de espécies possivelmente ameaçadas no estado do Rio de Janeiro (Rocha et
al., 2000). E. perditus tem sido reportada com relativa abundância em alguns estudos em
áreas de mata secundária, sobre tudo fragmentados (Condez et al., 2009; Sturaro et al., 2010;

84
Dixo & Verdade, 2006), como é o caso do presente estudo. O micro-habitat pode ser um dos
principais responsáveis pela abundância relativa de E. perditus em áreas de matas
perturbadas. Trata-se de uma espécie restrita a áreas florestadas na Mata Atlântica que utiliza
pequenos galhos, ramos de arbustos e troncos finos, não muito acima do solo (Liou, 2008;
Sturaro et al., 2010). Esses microhábitats são de certa forma, bem abundantes em florestas
que passaram por cortes recentes e se encontram em processo de regeneração, como é o caso
dos fragmentos florestais da Serra do Ouro Branco. Além desse motivo, a espécie não
mostrou estar correlacionada as variáveis ambientais, demonstrando pouca seletividade
referentes aos fatores mensurados no estudo.
A abundância de E. perditus, em ambientes secundários ou fragmentados pode ser favorecida
pela sua estratégia de forrageio, que é do tipo senta-e-espera. Huey & Pianka, (1981)
apontaram que espécies do tipo senta-e-espera apresentam grande tamanho de ninhadas, uso
da camuflagem como defesa e pouca movimentação, o que acarreta muita economia
energética. Essas características de certa forma podem facilitar a sua sobrevivência em
fragmentos de diversos tamanhos. De fato, E. perditus, apresenta a maior ninhada de todas as
espécies coletadas no estudo, se camufla muito bem, tanto na serapilheira como em pequenos
troncos e se mantem imóvel por muito tempo. Esse predomínio e abundância de uma espécie
na comunidade pode indicar um empobrecimento na fauna local de lagartos (Vitt & Carvalho,
1995).
H. imbricatus foi à terceira espécie mais abundante no estudo, com 10 indivíduos. Apesar de
haver poucos estudos sobre esta espécie, sabemos que ela depende de áreas de mata, tendo
sido recentemente encontrada em ambientes ripários associados ao bioma do Cerrado em
Minas Gerais, Brasil (Novelli et al., 2011). No presente estudo, sua ocorrência esteve ligada

85
aos maiores valores de biomassa da serapilheira, número e circunferência das árvores, além da
sua associação a ambientes com alta conectividade do dossel florestal, demonstrando ser
exigente quanto a características no hábitat. Dessa forma, alterações nestes ambientes podem
causar efeitos negativos sobre sua população, o que demonstra a necessidade de se ter mais
estudos sobre esta espécie.
Segundo Lehtinen et al. (2003), espécies que circulam pelas bordas de mata correm maior
risco de extinção do que as espécies que habitam o interior das matas. Os ambientes de borda
de mata comportam tanto as espécies consideradas estritamente florestais e mais sensíveis às
alterações no hábitat, como também espécies de áreas abertas. Dessa forma, esses são
ambientes importantes para a conservação da fauna de lagartos, visto que acabam sendo áreas
de convergência de recursos biológicos, tanto para espécies com hábito florestal, como
também para espécies de áreas abertas.
CONCLUSÃO
A comunidade de lagartos de serapilheira do presente trabalho apresentou espécies mais
associadas ao bioma Mata Atlântica. Algumas espécies da comunidade se mostraram
resilientes aos impactos das bordas, apesar de ocorrem na serra do Ouro Branco em
populações extremamente reduzidas.
Em matas secundárias, as variáveis ambientais e vegetacionais exercem maior influência na
estruturação da comunidade de lagartos do que os efeitos das bordas. A serapilheira, por
exemplo, demonstrou ser um importante componente do ecossistema florestal para os

86
lagartos, principalmente para os representantes da família Gymnophthalmidae, que se
mostraram correlacionados positivamente com este material.
Espécies que circulam pelas bordas de mata correm maior risco de extinção do que as
espécies que habitam o interior das matas (Lehtinen et al., 2003). No caso da serra do Ouro
Branco, todas as espécies ocorrem nas bordas e estão sujeitas aos mais variados riscos, o que
aponta para a necessidade de atribuir maior importância a conservação e restauração de áreas
perturbadas.
Preservar as paisagens perturbadas como é o caso das florestas da Serra do Ouro Branco são
de extrema importância para a conservação. Isto porque grande parte da área utilizada no
estudo está inserida em uma das mais novas áreas de proteção ambiental do Espinhaço,
podendo em estudos futuros e novos esforços, fornecer subsídios sobre a dinâmica temporal
da estrutura da comunidade dentro do fragmento de matas em diferentes estágios, envolvendo
as mesmas áreas e ambientes.
BIBLIOGRAFIA
AB’SÁBER, A. N. 2010.Do Código Florestal para o Código da Biodiversidade. Biota
Neotrop.,vol. 10, no. 4.
ADLER, G. H. 1996. The island syndrome in isolated populations of a tropical Forest
rodent.Oecologia, 108, pp.694-700.
ALFORD, R. A. & RICHARDS, S. J. 1999. Global amphibian declines: a problem in applied
ecology. Annual Review of Ecology and Systematics 30, 133–165.
BELL, K. E. & DONNELLY, M. A. 2006.Influence of Forest Fragmentation on Community
Structure of Frogs and Lizards in Northeastern Costa Rica.Conservation Biology.
Vol.20, No. 6, 1750–1760.

87
BIERREGAARD JR, R. O.; GASCON, C.; LOVEJOY, T. E. & MESQUITA, R. 2001.
Lessons from Amazonia: the ecology and conservation of a fragmented forest. Yale
University Press, New Haven, EUA.
BROADBENT, E. N.; ASNER, G. P.; KELLER, M.; KNAPP, D. E.; OLIVEIRA, P. J. C. &
SILVA, J. N. 2008.forest fragmentation and edge effects from deforestation and
selective logging in the Brazilian amazon. Biological Conservation 141. 1745–1757.
CECHIN, S. Z. & MARTINS, M. 2000. Eficiência de armadilhas de queda (pitfall traps) em
amostragens de anfíbios e répteis no Brasil. Revista Brasileira de Zoologia 17 (3): 729-
740.
CECHIN, S. Z. & MARTINS, M. 2000. Eficiência de armadilhas de queda (pitfall traps) em
amostragens de anfíbios e répteis no Brasil. Revista Brasileira de Zoologia 17 (3): 729-
740.
COLLINS, J.P. & STORFER, A. 2003. Global amphibian declines: sorting the hypotheses.
Diversity and Distributions.9, 89–98.
CONDEZ, T. H; SAWAYA, R. J & DIXO, M. 2009.Herpetofauna dos remanescentes de
Mata Atlântica da região de Tapiraí e Piedade, SP, sudeste do Brasil.Biota Neotrop.,vol.
9, no. 1.
CONDEZ, T. H; SAWAYA, R. J. & DIXO, M. 2009. Herpetofauna of the Atlantic Forest
remnants of Tapiraí and Piedade region, São Paulo state, southeastern Brazil.Biota
Neotrop.9(1):http://www.biotaneotropica.org.br/v9n1/en/abstract?inventory+bn0180901
2009.
COSTA, H. C.; FERNANDES, V. D.; RODRIGUES, A.C.; FEIO, R. N. 2009. Lizards and
amphisbaenians, municipality of Viçosa, State of Minas Gerais, southeastern
Brazil.Check List. 5(3): 732-745.
COSTA, H.C.; PANTOJA, D. L.; PONTES, J. L. & FEIO, R. N. 2010. Snakes of the
Municipality of Viçosa, Atlantic Forest of Southeastern Brazil.Biota Neotrop. 10(3):
http://www.biotaneotropica.org.br/v10n3/en/abstract?inventory+bn0361003.

88
CRUZ, A. J. R. 2011. Fauna de Lagartos da Serra de Ouro Branco, porção sul da Serra do
Espinhaço, Minas Gerais, In prep.
DIXO, M & MARTINS, M. 2008.Are leaf-litter frogs and lizards affected by edge effects due
to forest fragmentation in Brazilian Atlantic forest? Journal of Tropical Ecology.
24:551–554.
DIXO, M. & METZGER, J. P. 2009.Are corridors fragment size and forest structure
important for leaf-litter lizards in fragmented landscape? Fauna & Flora International,
Oryx, 43(3), 435-442.
FAHRIG, L. 2003. Effects of habitat fragmentation on biodiversity. Annual Reviews of
Ecology and Systematics 34: 487-515.
FISZON, J.T.; MARCHIORO, N. DE P.X.; BRITEZ, R.M. DE; CABRAL, D. DE C.;
CAMELY, N.C.; CANAVESI, V.; CASTELLA, P.R.; CASTRO, E.B.V. DE; CULLEN
JR., L.; CUNHA, M.B.S.; FIGUEIREDO, E.O.; FRANKE, I.L.; GOMES, H.; GOMES,
L.J.; HREISEMNOU, V.H.V.; LANDAU, E.C.; LIMA, S.M.F.; LOPES, A.T.L.;
NETO, E.M.; MELLO, A.L. DE; OLIVEIRA, A.L. DE; ONO, K.Y.; PEREIRA,
N.W.V.; RODRIGUES, A. DOS S.; RODRIGUES, A.A.F.; RUIZ, C.R.; SANTOS,
L.F.G.L. DOS; SMITH, W.S. & C.R. DE SOUZA. 2003. Causas Antrópicas. In:
Rambaldi, D.M. and D.A.S. de Oliveira (orgs.). Fragmentação de ecossistemas: causas,
efeitos sobre a biodiversidade e recomendações de políticas públicas, pp. 66 – 99.
Brasília: MMA/SBF.
FITCH, H.S.1982. Reproductive cycles in tropical reptiles.The University of Kansas Museum
of Natural History Occasional Papers. 96, 1-53.
GARDNER, T. A., BARLOW, J. AND PERES, C. A. 2007.Paradox, presumption and pitfalls
in conservation biology:The importance of habitat change for amphibians and reptiles.
Biological Conservation: 138. 166–179.
GASCON, C. 1993. Breeding-habitat use by five Amazonian frogs at forest edge.Biodiversity
and Conservation 2:438–444.

89
GASCON, C.; WILLIAMSON, B. G. & DA FONSECA, G.A.B., 2000.Receding forest edges
and vanishing reserves. Science 288, 1356–1358.
GIBBONS, J. W., SCOTT, D. E., RYAN, T. J., BUHLMANN, K. A., TUBERVILLE, T. D.,
METTS, B.S., GREENE, J.L., MILLS, T., LEIDEN, Y., POPPY, S., WINNE, C.T.
2000. The global decline of reptiles, deja-vu amphibians. Bioscience 50, 653–667.
HARTMANN, P. A., HARTMANN, M. & MARTINS, M. 2011.Snake road mortality in a
protected area in the Atlantic Forest of Southeastern Brazil. South American Journal of
Herpetoloy, 6(1), 35-42.
HAZELL, D., 2003. Frog ecology in modified Australian landscapes: a review. Wildlife
Research 30, 193–205.
HUEY, R. B. & PIANKA, E. R. 1981.Ecological consequences of foraging mode. Ecology
62: 991-999.
HUTCHINSON M., SWAIN R., DRIESSEN M. (2001) Snakes and Lizards of Tasmania.
Nature Conservation Branch, Hobart.
JELLINEK, S.; DRISCOLL, D. A. & KIRKPATRIK, J. B. 2004. Environmental and
vegetation variables have a greater influence than habitat fragmentation in structuring
lizard communities in remnant urban bushland. Austral Ecology. 29, 294-304.
KITCHENER, D. J.; CHAPMAN A., DELL, J.; MUIR, B. G. AND PALMER, M. 1980.
Lizard assemblages and reserve size and structure in the Western Australian wheatbelt –
some implications for conservation. Biol. Conserv. 17, 25–62.
KRUESS, A., TSCHARNTKE, T. 1994. Habitat fragmentation, species loss,and biological
control.Science 264, 1581±1584.
LAURANCE, W. F.; DELAMÔNICA, P.; LAURANCE,S. G.; VASCONCELOS, H .L. &
LOVEJOY , T. E. 2000. Rainforest fragmentation kills big trees. Nature 404: 836.
LEHTINEN, R. M.; RAMANAMANJATO, J-B.& RAVELOARISON, J. B. 2003.Edge
effects and extinction proneness in a herpetofauna from Madagascar.Biodiversity and
Conservation 12: 1357–1370.

90
LIOU, N. S. 2008. História Natural de duas espécies simpátricas de Enyalius (Squamata,
Leiosauridae) na Mata Atlãntica do sudeste brasileiro. MSc thesis. University of São
Paulo, São Paulo, Brasil.
MARQUES, O. A. V; NOGUEIRA, C; MARTINS, M & SAWAYA, R. J. 2010.Impactos
potenciais das mudanças propostas no Código Florestal Brasileiro sobre os répteis
brasileiros. Biota Neotrop.,vol. 10, no. 4.
MENDONÇA, L. B.; LOPES, E. V. & ANJOS, L. 2009. On the possible extinction of bird
species in the Upper Paraná River floodplain, Brazil. Braz. J. Biol., 69(2, Suppl.): 747-
755.
MURCIA, C. 1995. Edges effects in fragmented forest: Implications for conservation. Tree,
Vol: 10. no. 2.
MYERS, N.; MITTERMEIER, R. A.; MITTERMEIER, C. G.; DA FONSECA, G. A. B. &
KENT, J. 2000. Biodiversity hotspots for conservation priorities. Nature 430: 853-858.
MYERS, N.; MITTERMEIER, R. A.; MITTERMEIER, C. G.; FONSECA, G. A. B. &
KENT, J. 2000. Biodiversity hotspots for conservation priorities. Nature430: 853-858.
NOGUEIRA, C. 2006. Diversidade e padrões de distribuição da fauna de lagartos do Cerrado.
Tese de Doutorado, Universidade de São Paulo, São Paulo.
NOVELLI, I. A.; LUCAS, P. S. & SANTOS, R. C. 2011. Reptila, Squamata,
Gymnophthalmidae, Heterodactylus imbricatus Spix, 1825: Filling gaps in the state of
Minas Gerais. Check List. Vol. 7. 1.
NOVELLI, I. A.; LUCAS, P. S. & SANTOS, R. C. 2011.Reptila, Squamata,
Gymnophthalmidae, Heterodactylus imbricatus Spix, 1825: Filling gaps in the state of
Minas Gerais. Check List. Vol. 7. 1.
PARDINI, R. 2004. Effects of forest fragmentation on small mammals in an Atlantic Forest
landscape.Biodiversity and Conservation, 13, pp.2567-2586. Available at:
http://www.springerlink.com/openurl.asp?id=doi:10.1023/B:BIOC.0000048452.18878.
2d.

91
PAULA, C. C. 2003. Flora fanerogâmica da Serra de Ouro Branco. In: 54º Congresso
Nacional de Botânica, 2003, Belém, PA. Livro de resumos. UFRA (FCAP)/MPEG/
Embrapa Amazônia Oriental.
PIANKA, E. R. & VITT, L. J. 2003. Lizards: Windows to the Evolution of Diversity.
University of California Press, Berkeley.
PIANKA, E. R. 1973. The structure of lizards communities. Anual review of ecology and
systematics.4. 53-74.
POUGH, F. H., JANIS, C. M. & HEISER, J. B. A vida dos vertebrados. 4 ed. São Paulo:
Atheneu Editora, 2008. 684 p. I.
ROCHA, C.F.D.; VAN-SLUYS, M.; PUORTO G.; FERNANDES, R.; BARROS-FILHO J.
D.; SILVA, R. R. E.; NEO, F. A. & MELGAREJO E. A. 2000. Répteis; p. 79-87 In H.
G. BERGALLO, C. F. D ROCHA, M. A. S. ALVES E M. VAN-SLUYS (ed.). A Fauna
Ameaçada de Extinção do Estado do Rio de Janeiro. Rio de Janeiro: Editora UERJ.
RODRIGUES, M. T. 1987, Sistemática, ecologia e zoogeografia dos Tropidurus do grupo
torquatus ao sul do Rio Amazonas (Sauria, Iguanidae). Arq. Zool., São Paulo, 31:105-
230.
SALA, O. E.; STUART CHAPIN III, F.; ARMESTO, J. J.; BERLOW, E.; BLOOMÞELD,
J. DIRZO, R.; HUBER-SANWALD, E.; HUENNEKE, L. F.; JACKSON, R. B.;
KINZIG, A.; LEEMANS, R.; LODGE, D. M.; MOONEY, H. A.; OESTERHELD, M.;
POFF, N. L.; SYKES, M. T.; WALKER , B. H.; WALKER, M. & WALL, D. H. 2000.
Global Biodiversity Scenarios for the Year 2100.Science’s Compass. Vol: 287. 1770-
1774.
SKOLE, D. & TUCKER, C. 1993. Tropical deforestation and habitat fragmentation in the
Amazon: satellite data from 1978 to 1988. Science 260: 1905-1910.
STURARO, M.J. & SILVA, V. X. 2010.Natural history of the lizard Enyalius perditus
(Squamata: Leiosauridae) from an Atlantic forest remnant in southeastern Brazil.
Journal of Natural History 44 (19-20): 1225-1238.

92
TABARELLI, M. and GASCON, C., 2005. Lessons from fragmentation research: improving
management and policy guidelines for biodiversity conservation. Conservation Biology,
vol. 19, no. 3, p. 734-739.
TABARELLI, M.; MANTOVANI, W. & PERES, C. A. 1999.Effects of habitat fragmentation
on plant guild structure in the montane Atlantic forest of southeastern Brazil. Biological
Conservation 91: 119-127.
TABARELLI, M.; MANTOVANI, W. & PERES, C. A. 1999.Effects of habitat fragmentation
on plant guild structure in the montane Atlantic forest of southeastern Brazil. Biological
Conservation 91: 119-127.
TROMBULAK , S. C. & FRISSELL, C. A. 2000. Review of ecological effects of roads on
terrestrial and aquatic communities. Conservation Biology. p 18-30. Vol. 14, No. 1.
VANZOLINI, P. E.; RAMOS COSTA, A. M. M. & VITT, L. J. 1980. Répteis das Caatingas.
Academia Brasileira de Ciências, Rio de Janeiro.
VELOSO, H. P.; RANGEL-FILHO, A. L. R.; LIMA, J. C. A. 1991. Classificação da
vegetação brasileira adaptada a um sistema universal. Rio de Janeiro: Instituto
Brasileiro de Geografia e Estatística (IBGE).
VITT, L. J. & CARVALHO C. M. 1995. Niche partitioning in a tropical wet season: lizards
in the lavrado area of northern Brazil.Copeia 2, 305-29.
VITT, L. J.; AVILA-PIRES, T. C.; CALDWELL, J. P. & OLIVEIRA, V. R. L. 1998. The
impact of individual tree harvesting on thermal environments of lizards in Amazonian
rain forest.Conservation Biology 12, 1±12.
VRCIBRADIC, D. & ROCHA, C. F. D. 1998. The ecology of the skink Mabuya frenata in an
area of rock outcrops in southeastern Brazil. J. Herpetol. 32: 229-237.
ZAMPROGNO, C.; ZAMPROGNO, M. das G. F. & TEIXEIRA, R. L. 2001. Evidence of
terrestrial feeding in the arboreal lizard Enyalius bilineatus (Sauria, Polychrotidae) of
South-Eastern Brazil.Rev. Brasil. Biol., 61(1): 91-94

93
CONSIDERAÇÕES FINAIS
Obedecendo as prioridades estabelecidas em Bérnils et al., 2009, o presente trabalho
apresenta uma lista de 15 espécies pertencentes a oito famílias de lagartos com ocorrência na
serra de Ouro Branco. Apesar de todas as espécies já serem encontradas em outras regiões do
Estado de Minas Gerais, o presente estudo amplia a distribuição para a região de várias
espécies, tanto para os limites de cada bioma, como também para os limites de altitude. No
entanto cabe ressaltar que excetoE. perditus, E. gaudichaudii e Tropidurus torquatus, todas as
populações de lagartos ocorrem em pequenas populações na região.
Além de fazer o uso de materiais acondicionados em coleções, o presente trabalho acrescenta
para a coleção de repteis do Laboratório de Zoologia dos Vertebrados da Universidade
Federal de Ouro Preto (LZV-UFOP) cerca de 100 espécimes de lagartos. Esta ação coloca a
Universidade de Ouro Preto como detentora de um importante acervo de repteis para o Estado
de Minas Gerais, preservando inclusive um banco de tecidos, contento material genético
extraído dos lagartos podendo fomentar pesquisas futuras com filogeografia, genética e
taxonomia.
Os dados ecológicos sobre o hábitatindicam que as áreas de mata da serra de Ouro Branco,
estão em fase de regeneração, o que de pode ter favorecido E. perditus, resultando na maior
abundância relativa dessa espécies e na diminuição da abundância de outras populações na
comunidade. Verificamos que os lagartos de serapilheira de áreas de mata são muito afetados
pelas variáveis ambientais derivados da alteração na vegetação, corroborando com os estudos

94
de Jellinek et al. (2004) e Dixo & Martins (2008), principalmente os da família
Gymnophthalmidae que se correlacionaram positivamente com a quantidade de serapilheira e
áreas sombreadas. Este resultado ressalta a importância de variáveis associados à serapilheira,
como determinantes na estrutura da comunidade de lagartos em ecossistemas florestais.
Na área de estudos ocorrem inúmeras clareiras abertas ilegalmente no interior dos fragmentos
para a exploração das maiores árvores, podendo modificar drasticamente o micro-clima da
floresta. Como consequência as espécies estritamente florestais e exigentes quanto a hábitat,
principalmente os Gymnophthalmidae, que preferem áreas sombreadas e com boa cobertura
de serapilheira, correm sérios riscos de extinção.
Trabalhos utilizando as armadilhas do tipo pitfallem áreas de mata secundária indicam que H.
imbricatus pode ocorrer em pequenas populações, o que se torna mais preocupante, quando se
tem muitas ameaças no entorno (Dixo &Verdade, 2006; Dixo & Metzeger, 2009). Vale
ressaltar queestaé uma das espécies mais carentes de informação encontrados na Serra de
Ouro Branco e na serra do Espinhaço (PAN-Herpetofauna Espinhaço).
A espécie C. quadrilineataé também pouco conhecida quanto à história Natural e ecologia.
Durante as coletas do presente trabalho foi encontrado um único indivíduo em áreas de mata.
Contudo, informações por enquanto são inconclusivas, já que pouco se sabe sobre este
pequeno Gymnophthalmidae, e estranhamos inclusive, a sua ausência em listas de espécies
com carência de informações em Minas Gerais.
Quando analisamos os dados referentes à utilização de ambientes de borda e de interior
verificamos que, além da riqueza,a abundância dos lagartos se mostrou similar entre os
diferentes ambientes. No entanto, quase 75% do total da abundância de lagartos coletados,
ocorreram em áreas de bordas. Segundo Lehtinen et al. (2003), espécies que circulam pelas
bordas de mata correm maior risco de extinção do que as espécies que habitam o interior das
matas. Os dados do presente estudo apontam sérios riscos à biodiversidade de lagartos

95
naregião, pois, esses seres vivem tanto em ambientes de interior como também nas bordas,
estando sujeitas a riscos como: perda de hábitat, entrada de espécies exóticas, aumento na
frequência de incêndios, contaminação por agrotóxicos e poluentes, atropelamentos e práticas
de caça, resultando assim na diminuição da diversidade de organismos na comunidade
(Bierregaard et al., 2001; Tabarelli et al., 1999; Broadbent et al., 2008; Trombulak &
Christopher, 2000; Cerqueira et al., 2003; Skole & Tocker, 1993; Santos & Tabarelli,2002;
Fiston et al., 2003; Peres & Lake, 2003; Tabarelli & Gascon, 2005; Hartmann et al., 2011).
Apesar de terem sido serem criadas Unidades de Conservação próximas à área de estudo,
relações conflituosas entre os moradores e a fauna ainda são frequentes (Pinto, 2011).
Segundo Pinto, 2011, o lagarto T. merianae é o que mais chama atenção dos moradores, por
ser predador de ovos invadindo galinheiros, ter uma carne apreciada na região e partes do seu
corpo ser utilizado em crenças como a cura de dores. Por meio de esforços contínuos em
educação ambiental, junto aos agricultores e proprietários rurais, achamos ser possível
diminuir esses riscos. Incentivos encorajando a popularização dessas espécies e diminuição
do uso de pesticidas e incêndios florestais,podem ser atitudes importantes para permanência
desses lagartos na região.
Para um melhor convívio entre os lagartos e os seres humanos na matriz que circula as
Unidades de conservação da Serra de Ouro Branco, recentemente com os frutos deste
trabalho, foi apresentado a eles em forma de palestras educativas todos os lagartos com
registros para a região, enfocando a importância ecológica, biológica, assim como
curiosidades e superstições ligadas aos lagartos. Esperamos, por meio da educação ambiental,
conseguir um maior envolvimento por parte dos moradores nativos da região, aumentando a
estima, engajando-os em ações que visam à conservação deste grupo.

96
Trabalhos futuros, enfocando aspectos da história natural como dieta, razão sexual, período de
reprodução, número de ninhadas e sazonalidade das espécies mais abundantes, como E.
perditus, E. gaudichaudii e H. imbricatus estão em andamento, sendo desenvolvidas graças a
inúmeros dados ecológicos fornecidos pelos exemplares coletados. Esses estudos juntamente
com o presente trabalho, com certeza nos fornecerão mais algumas dicas de como se
comportam essas comunidades em ambientes perturbados.
BIBLIOGRAFIA
AB'SABER, A. N. 1977. Os domínios morfoclimáticos da América do Sul. Primeira
aproximação. Geomorfologia 52: 1-21.
AL-JOHANY, A. M. 1995. The ecology of Agama yemenesis Klausewitz (Lacertilia:
Agamidae) in South-western Arabia. Jornal of Arid Environments. 29: 495-503.
ÁVILA-PIRES, T.C.S. 1995. Lizards of Brazilian Amazonia (Reptilia: Squamata). Zool.
Verh. Leiden. 299: 1- 706.
BARROS, E. H. & TEIXEIRA, R. L. 2007.Diet and fecundity of the Glass-lizard, Ophiodes
striatus (Sauria, Anguidae) from the Atlantic Forest in southeastern Brazil.Bol. Mus.
Biol. Mello Leitão (n. sér.) 22:11-23.
BÉRNILS, R. S.; NOGUEIRA, C. & SILVA, V. X. 2009. Biota minas -
diagnóstico do conhecimento de vertebrados: Répteis. In Biota minas - diagnóstico do
conhecimento sobre a biodiversidade no estado de Minas Gerais (G.M. DRUMMOND,
C.S. MARTINS, M.S. GRECO & F. VIEIRA, eds.), pp. 252-278, Belo Horizonte, MG.
BIERREGAARD JR, R. O.; C. GASCON, T.E. LOVEJOY & R. MESQUITA. 2001. Lessons
from Amazonia: the ecology and conservation of a fragmented forest. Yale University
Press, New Haven, EUA.

97
BROADBENT, E. N.; ASNER, G. P.; KELLER, M.; KNAPP, D. E.; OLIVEIRA, P. J. C. &
SILVA, J. N. 2008.forest fragmentation and edge effects from deforestation and
selective logging in the Brazilian amazon. Biological Conservation 141. 1745–1757.
CARVALHO, A. L. G. & ARAÚJO, A. F. B. 2004. Ecologia dos lagartos da ilha de
Marambaia, RJ. Rev. Univ. Rural, Sér. Ci. Vida. Seropédica, RJ, EDUR, v. 24, n.2, p.
159-165.
CASTRO, E. R. & GALETTI, M. 2004. Frugivoria e dispersão de sementes pelo lagarto teiú
Tupinambis merianae (Reptilia: Teiidae). Papéis Avulsos de Zoologia, 44(6): 91-97.
CERQUEIRA, R.; BRANT, A.; NASCIMENTO, M. T. & PARDINI, R. 2003. Fragmentação:
alguns conceitos. In RAMBALDI, D. M. & OLIVEIRA, D. A. S. (orgs.). Fragmentação de
ecossistemas: causas, efeitos sobre a biodiversidade e recomendações de políticas públicas.
Brasília: MMA-SBF. p 23-40.
CICCHI, P. J. P.; SERAFIM, H.; SENA, M. A; CENTENO, F. C. & JIM, J. 2009. Atlantic
Rainforest herpetofauna of Ilha Anchieta, an island on municipality of Ubatuba,
southeastern Brazil.Biota Neotrop., 9(2):
http://www.biotaneotropica.org.br/v9n2/en/abstract?inventory+bn0100902.
COLLI, G. R.; BASTOS, R. P, & ARAÚJO, A. F. B. 2002. The character and dynamics of
the Cerrado herpetofauna.In: Oliveira, p.s. & Marquis, R. L (eds.) The Cerrados of
Brazil: ecology and natural history of a Neotropical savana, p.223-241. New York,
Columbia University Press.
CONDEZ, T. H.; SAWAYA, R. J. & DIXO, M. 2009.Herpetofauna of the Atlantic Forest
remnants of Tapiraí and Piedade region, São Paulo state, southeastern Brazil.Biota
Neotrop.9(1):http://www.biotaneotropica.org.br/v9n1/en/abstract?inventory+bn0180901
2009.
COSTA, H. C.; FERNANDES, V. D.; RODRIGUES, A. C.; FEIO, R. N. 2009. Lizards and
amphisbaenians, municipality of Viçosa, State of Minas Gerais, southeastern
Brazil.Check List. 5(3): 732-745.
COSTA, H. C.; FERNANDES, V. D.; VRCIBRADIC, D.; FEIO, R. N. 2008. Reptilia,
Scincidae, Mabuya frenata: Distribution Extension. Check List 4(1): 86–88.

98
DIXO, M & MARTINS, M. 2008. Are leaf-litter frogs and lizards affected by edge effects
due to forest fragmentation in Brazilian Atlantic forest? Journal of Tropical Ecology.
24:551–554.
DIXO, M. & METZGER, J. P. 2009.Are corridors fragment size and forest structure
important for leaf-litter lizards in fragmented landscape? Fauna & Flora International,
Oryx, 43(3), 435-442.
DIXO, M. & VERDADE, V. K. 2006. Herpetofauna de serrapilheira da Reserva Florestal de
Morro Grande, Cotia (SP). Biota Neotrop. vol. 6 no. 2,
http://www.biotaneotropica.org.br/v6n2/pt/abstract?article+bn00806022006. ISSN1676-
0603.
FEIO, R. N. & CARAMASCHI, U. 2002. Contribuição ao conhecimento da herpetofauna do
nordeste do estado de Minas Gerais, Brasil. Phyllomedusa 1(2):105-111.
FEIO, R. N. & U. CARAMASCHI. 2002. Contribuição ao conhecimento da herpetofauna do
nordeste do estado de Minas Gerais, Brasil. Phyllomedusa 1(2):105-111.
FISZTON, J.T.; MARCHIORO, N. DE P.X.; BRITEZ, R.M. DE; CABRAL, D. DE C.;
CAMELY, N.C.; CANAVESI, V.; CASTELLA, P.R.; CASTRO, E.B.V. DE; CULLEN
JR., L.; CUNHA, M.B.S.; FIGUEIREDO, E.O.; FRANKE, I.L.; GOMES, H.; GOMES,
L.J.; HREISEMNOU, V.H.V.; LANDAU, E.C.; LIMA, S.M.F.; LOPES, A.T.L.;
NETO, E.M.; MELLO, A.L. DE; OLIVEIRA, A.L.DE; ONO, K.Y.; PEREIRA,
N.W.V.; RODRIGUES, A. DOS S.; RODRIGUES, A.A.F.; RUIZ, C.R.; SANTOS,
L.F.G.L. DOS; SMITH, W.S. & C.R. DE SOUZA. 2003. Causas Antrópicas. In:
Rambaldi, D.M. and D.A.S. de Oliveira (orgs.). Fragmentação de ecossistemas: causas,
efeitos sobre a biodiversidade e recomendações de políticas públicas, pp. 66 – 99.
Brasília: MMA/SBF.
FORLANI, M. C.; BERNARDO, P.H.; HADDAD, C.B.F. & ZAHER, H. 2010. Herpetofauna
of the Carlos Botelho State Park, São Paulo State, Brazil. Biota Neotrop. 10(3):
http://www.biotaneotropica.org.br/v10n3/en/abstract?inventory+bn0021003.
GASCON, C.; LOVEJOY,T. E.; BIERREGAARD JR, R. O.; MALCOLM, J. R.;
STOUFFER, P. C.; VASCONCELOS, H.; LAURANCE W.F.; ZIMMERMAN, B.;

99
TOCHER, M. & BORGES, S. 1999. Matrix habitat and species persistence in tropical
forest remnants. Biological Conservation 91: 223-230.
GIBBONS, J. W.; SCOTT, D. E.; RYAN, T. J.; BUHLMANN, K. A.; TUBERVILLE, T. D.;
METTS, B. S.; GREENE, J. L.; MILLS, T.; LEIDEN, Y.; POPPY, S.; WINNE, C. T.
2000. The global decline of reptiles, deja-vu amphibians. Bioscience 50, 653–667.
HARPER, K. A., MACDONALD, S. E., BURTON, P., CHEN, J., BROSOFSKY, K. D.,
SAUNDERS, S., EUSKIRCHEN, E. S., ROBERTS, D., JAITEH, M. & ESSEEN, P. A.
2005. Edge influence on forest structure and composition in fragmented
landscapes.ConservationBiology19:768–782.
HARTMANN, P. A.; HARTMANN, M. & MARTINS, M. 2011.Snake road mortality in a
protected area in the Atlantic Forest of Southeastern Brazil. South American Journal of
Herpetoloy, 6(1), 35-42.
JELLINEK, S.; DRISCOLL, D. A. & KIRKPATRIK, J. B. 2004. Environmental and
vegetation variables have a greater influence than habitat fragmentation in structuring
lizard communities in remnant urban bushland. Austral Ecology. 29, 294-304.
LEITE, D. L. P. 2007. Efeitos do fogo sobre a taxocenose de lagartos em áreas de Cerrado
Sensu stricto no Brasil central. Dissertação de Mestrado. Universidade de Brasília.
LEITE, F. S. F.; JUNCÁ, F. A. & ETEROVICK, P. C. 2008. Status do conhecimento,
endemismo e conservação de anfíbios anuros da Serra do Espinhaço, Brasil.
Megadiversidade.
MARQUES, O. A. V; NOGUEIRA, C; MARTINS, M & SAWAYA, R. J. 2010. Impactos
potenciais das mudanças propostas no Código Florestal Brasileiro sobre os répteis
brasileiros. Biota Neotrop.,vol. 10, no. 4.
MELO, E. S. & SANTOS-FILHO, M. 2007. Efeitos da BR-070 na Província Serrana de
Cáceres, Mato Grosso, sobre a comunidade de vertebrados silvestres. Revista Brasileira
de zoociências 9(2): 185-192.

100
MESQUITA, D. O. & COLLI, G. R. 2003. The Ecology of Cnemidophorus ocellifer
(Squamata, Teiidae) in a Neotropical Savanna. Journal of Herpetology, Vol. 37, No. 3,
pp. 498–509.
MESQUITA, D.O.; COLLI, G.R.; FRANÇA, F.G.R. & VITT, L.J. 2006.Ecology of a
Cerrado lizard assemblage in the Jalapão region of Brazil. Copeia 2006(3): 460-471.
MURCIA, C. 1995. Edges effects in fragmented forest: Implications for conservation. Tree,
Vol: 10. no. 2.
MYERS, N.; MITTERMEIER, R. A.; MITTERMEIER, C. G.; DA FONSECA, G. A. B. &
KENT, J. 2000. Biodiversity hotspots for conservation priorities. Nature 430: 853-858.
NOGUEIRA, K. O. P.C.; RODRIGUES, S. S.; ARAÚJO, V.A.; NEVES, C. A. 2011.
Oviductal Structure and Ultrastructure of the Oviparous Gecko, Hemidactylus Mabouia
(Moreau De Jonne` s, 1818).The anatomical record294:883–892.
PERES, C. A. & LAKE, I. R. 2003.Extent of Nomtimber Resource Extration in Tropical Forest:
Accessibility to Game Vertebrates by Hunters in the Amazon Basin. Conservation
Biology.17(2), 521-533(15).
PERES, C. A. & LAKE, I. R. 2003. Extent of nontimber resource extraction in tropical
forests: Accessibility to game vertebrates by hunters in the Amazon basin. Conserv.
Biol. 17: 521–535.
PETERS, J. A.; OREJAS-MIRANDA, B. 1970.Catalogue of the Neotropical Squamata: Part I
– Snakes, Part II- Lizards and Amphisbaenians. Washington, D. C.; London:
Smithsonian Institution Press.
PIANKA, E. R. & VITT, L. J. 2003. Lizards: Windows to the Evolution of Diversity.
University of California Press, Berkeley.
PIANKA, E. R. & R. B. HUEY.1978. Comparative ecology, niche segregation, and resource
utilization among gekkonid lizards in the southern Kalahari. Copeia 1978: 691- 701.
PIANKA, E. R. 1967. On lizard species diversity: North American flatland deserts. Ecology
48: 333-351.

101
PIANKA, E. R. 1969. Habitat specificity, speciation, and species density in Australian desert
lizards. Ecology 50: 498-502.
PIANKA, E. R. 1971. Species diversity. Chapter in Topics in the Study of Life: The Bio
Source Book, pp. 401-406. Harper and Row, New York.
PINTO, L. C. L. 2011.Etnozoologia e conservação de biodiversidade em comunidades rurais
da Serra do Ouro Branco. Dissertação de Mestrado. Universidade Federal de Ouro
Preto, Minas Gerais.
POUGH, F. H.; JANIS, C. M. & HEISER, J. B. A vida dos vertebrados. 4 ed. São Paulo:
Atheneu Editora, 2008. 684 p. I.
POUGH, F.H; JANIS, C. M. & HEISER, J. B. 2003.A Vida dos Vertebrados.3ª edição.
Atheneu, São Paulo, 699p.
RAMOS, L. O. & SILVEIRA, A. L. 2011. New Record and distribuition mape of Ecpleopus
gaudichaudii (Reptilia, Squamata, Gymnophthalmidae) in southeastern
Brazil.Herpetology Notes. 4. 173- 176.
RIES, L.; FLETCHER, R. J. J.; BATTIN, J. & SISK, T. D. 2004. Ecological responses to
habitat edges: mechanisms, models and variability explained. Annual Review of
Ecology, Evolution and Systematics 35:491–522.
RODRIGUES, M. T. 1987, Sistemática, ecologia e zoogeografia dos Tropidurus do grupo
torquatus ao sul do Rio Amazonas (Sauria, Iguanidae). Arq. Zool., São Paulo, 31:105-
230.
RODRIGUES, M. T.; FREIRE, E. M. X.; PELLEGRINO, K. C. M. & SITES Jr, J. W. 2005.
Phylogenetic relationships of a new genus and species of microteiid lizard from the
Atlantic forest of north-eastern Brazil (Squamata, Gymnophthalmidae).Zoological
Journal of the Linnean Society. 144, 543–557.
RODRIGUES, M.T.; DIXO, M.; PAVAN, D. & VERDADE, V.K. 2002.A new species of
Leposoma (Squamata, Gymnophthalmidae) from the remnant Atlantic forests of the
state of Bahia, Brazil.Pap. Avulsos Zool. (São Paulo) 42:335-350.

102
RODRIGUES, M.T.;FREITAS, M. A.; SILVA, T. F. S. & BERTOLOTTO, C. E. V. 2006. A
new species of lizard genus Enyalius (Squamata, Leiosauridae) from the highlands of
Chapada Diamantina, state of Bahia, Brazil, with a key to species. Phyllomedusa
5(1):11-24.
SANTOS, A. M. & TABARELLI, M. 2002. Distance from roads and cities as a predictor of
habits loss and fragmentation in the Caatinga dry forest of Brasil. Revista Brasileira de
Botânica. 62: 897-905.
SANTOS, A. M. & TABARELLI, M. 2002. Distance from roads and Cities as a predictor of habitat
loss and fragmentation in the caatinga vegetation of Brasil. Brasilian Jornal of Biology.
62(4B): 897-905.
SANTOS, R.C.; LUCAS, P. S.; SOUSA, B. M. & NOVELLI, I. A. 2009. Reptilia, Squamata,
Leiosauridae, Urostrophus vautieri: Distribution extensionand geographic distribution
map. Check List 5: 533-536.
SÃO PEDRO, V. A & FEIO, R. N. 2010. Distribuição espacial e sazonal de Anuros em três
ambientes na Serra de Ouro Branco, extremo sul da Cadeia do Espinhaço, Minas
Gerais, Brasil. Biotemas, 23 (1): 143-154.
SÃO PEDRO, V. A & FEIO, R. N. 2011. Anuran species composition from Serra do Ouro
Branco, southernmost Espinhaço Mountain Range, state of Minas Gerais, Brazil. Check
List,Vol.7. (5).
SÃO PEDRO, V. A & Pires, M. R. S. 2009. As Serpentes da região de Ouro Branco, Extremo
Sul da Cadeia do Espinhaço, Minas Gerais. Ceres 56 (2): 166-171.
SAZIMA, I.; SAZIMA, C.& SAZIMA, M. 2005. Little dragons prefer flowers to maidens: a
lizard that laps nectar and pollinates trees. Biota Neotropica v5(n1)
http://www.biotaneotropica.org.br/v5n1/pt/abstract?shortcommunication+BN0080501.
SILVA, T. F.; ANDRADE, B. F. E.; TEIXEIRA, R. L. & GIOVANELLI, M. 2003. Ecologia
de Ameiva ameiva ( sauria, Teiidae), na Restinga do Guriri, São Mateus, Espirito Santo,
sudeste do Brasil. Boletim Museu de Biologia Mello Leitão (N. Sér.)15:5-15.

103
SKOLE, D. & TUCKER, C. 1993. Tropical deforestation and habitat fragmentation in the
Amazon: satellite data from 1978 to 1988. Science 260: 1905-1910.
SOCIEDADE Brasileira de Herpetologia. 2012. List of Brazilian reptile species.
http://www.sbherpetologia.org.br (último acesso em 05/01/2012).
SOUSA, B. M. & CRUZ, C. A. G. 2008. Hábitos alimentares em Enyalius perditus
(Squamata, Leiosauridae) no Parque Estadual do Ibitipoca, Minas Gerais. Iheringia
98(2): 260-265.
SOUSA, B. M.; NASCIMENTO, A. E. R.; GOMIDES, S. C.; VARELA RIOS, C. H.;
HUDSON, A. H. & NOVELLI, I. A. 2010. Reptiles in fragments of Cerrado and
Atlantic Forest at the Campo das Vertentes, MinasGerais State, Southeastern
Brazil.BiotaNeotrop.10(2):http://www.biotaneotropica.org.br/v10n2/en/abstract?article+
bn0351002.
STURARO, M. J. & ÁVILA-PIRES, T. C. S. 2011. Taxonomic revision of the geckos of the
Gonatodes concinnatus complex (Squamata: Sphaerodactylidae), with description of
two new species. Zootaxa 2869: 1-36.
STURARO, M. J. & SILVA, V. X. 2010.Natural history of the lizard Enyalius perditus
(Squamata: Leiosauridae) from an Atlantic forest remnant in southeastern Brazil.
Journal of Natural History 44 (19-20): 1225-1238.
TEIXEIRA, R. L. & GIOVANELLI, M. 1999. Ecologia de Tropidurus torquatus
(Sauria:Tropiduridae) da Restinga de Guriri, São Mateus, ES. Rev. Brasil. Biol., 59(1):
11-18.
TORRES-CARVAJAL, O. 2005.A new species of Stenocercus (Squamata, Iguanidae) from
central-western Brazil with a key to Brazilian Stenocercus. Phyllomedusa 4:123-132.
TROMBULAK , S. C. & FRISSELL, C. A. 2000. Review of ecological effects of roads on
terrestrial and aquatic communities. Conservation Biology.p 18-30. Vol. 14, No. 1.
VANZOLINI, P. E. 1988. Distributional patterns of South American lizards. In:
VANZOLINI, P.E. & HEYER, W.R. (Eds.) Proceedings of a Workshop on Neotropical

104
Distribution Patterns. MORELLATO, L.P.C. & HADDAD, C.F.B. 2000. Introduction:
The Brazilian Atlantic Forest. Biotropica 32: 786-792.
VITT, L. J. 1991. An introduction to the ecology of Cerrado lizards. Journal Herpetol., 25:79-
90.
WERNECK, F. P & COLLI, G. R. 2006.The lizard assemblage from Seasonally Dry Tropical
Forest enclaves in the Cerrado biome, Brazil, and its association with the Pleistocenic
Arc. J. Biogeogr. 33:1983-1992.
ZAHER, H.; BARBO, F. E.; MARTÍNEZ, P. S; NOGUEIRA, C.; RODRIGUES, M. T. &
SAWAYA R. J. 2011. Reptiles from São Paulo State: current knowledge and
perspectives. Biota Neotrop.11(1a): http://www.biotaneotropica.
org.br/v11n1a/en/abstract?inventory+bn0051101a.
ZAMPROGNO, C.; ZAMPROGNO, M. das G. F. & TEIXEIRA, R. L. 2001. Evidence of
terrestrial feeding in the arboreal lizard Enyalius bilineatus (Sauria, Polychrotidae) of
South-Eastern Brazil.Rev. Brasil. Biol., 61(1): 91-94.

105
ANEXO I
Exemplares coletados e/ou verificados:
Ameiva ameiva:LZV 494S
Cercosaura quadrilineatus: LZV 003S-L.
Ecpleopus gaudichaudii: LZV 973S, 991S, 992S, 1000S, 1006S, 1007S, 1008S, 1009S,
1010S, 1024S, 1041S, 1044S, 1049S, 1050S, 1051S, 1063S, 1064S, 1068S, 1069S, 1090S,
1091S, 1101S, 1111S, 1112S, 1113S, 1114S, 1115S, 1116S, 1148S.
Enyalius bilineatus: LZV 943S, 971S, 1060S, 1065S.
Enyalius perditus: LZV 939S, 940S, 942S, 944S, 966S, 967S, 968S, 976S, 977S, 978S,
993S, 995S, 996S, 997S, 1005S, 1038S, 1039S, 1040S, 1042S, 1043S, 1045S, 1047S, 1048S,
1057S, 1059S, 1062S, 1067S, 1070S, 1071S, 1075S, 1076S, 1077S, 1079S, 1080S, 1081S,
1082S, 1083S, 1085S, 1086S, 1087S, 1098S, 1099S, 1117S, 1118S, 1109S, 1110S.
Hemidactylus mabouia: LZV 207S
Heterodactylus imbricatus: LZV 941S, 974S, 990S, 999S, 1017S, 1019S, 1052S, 1058S,
1119S 1100S.

106
Mabuya dorsivittata: MZUFV-530.
Mabuya frenata: LZV 1061S, 1066S, 002S-L.
Ophiodes striatus: LZV 937S, 938S.
Polychrus acutirostris: LZV 028S-L.
Tropidurus itambere: LZV 873S
Tropidurus torquatus: LZV.232S
Tupinambis merianae: LZV 994S.
Urostrophus vautieri: LZV 1046S.





























