Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO CEARÁ
INSTITUTO DE CIÊNCIA DO MAR
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS MARINHAS TROPICAIS
WALLACE ALVES DE SOUSA
PADRÕES DA DIVERSIDADE BETA EM ASSEMBLEIAS DE PEIXES DE
ESTUÁRIOS DO SEMIÁRIDO BRASILEIRO
FORTALEZA
2019

WALLACE ALVES DE SOUSA
PADRÕES DA DIVERSIDADE BETA EM ASSEMBLEIAS DE PEIXES DE
ESTUÁRIOS DO SEMIÁRIDO BRASILEIRO
Dissertação apresentada ao programa de Pós-
Graduação em Ciências Marinhas Tropicais do
Instituto de Ciências do Mar da Universidade
Federal do Ceará, como requisito parcial à
obtenção do título de Mestre em Ciências
Marinhas Tropicais. Área de concentração:
Utilização e manejo de ecossistemas marinhos
e estuarinos.
Orientador: Prof. Dr. Jorge Iván Sánchez
Botero.
FORTALEZA
2019


PADRÕES DA DIVERSIDADE BETA EM ASSEMBLEIAS DE PEIXES DE
ESTUÁRIOS DO SEMIÁRIDO BRASILEIRO
Dissertação apresentada ao programa de Pós-
Graduação em Ciências Marinhas Tropicais do
Instituto de Ciências do Mar da Universidade
Federal do Ceará, como requisito parcial à
obtenção do título de Mestre em Ciências
Marinhas Tropicais. Área de concentração:
Utilização e manejo de ecossistemas marinhos
e estuarinos.
Aprovada em: ___/___/______.
BANCA EXAMINADORA
________________________________________
Prof. Dr. Jorge Iván Sánchez Botero
(Orientador)
Universidade Federal do Ceará (UFC)
_________________________________________
Prof. Dr. Marcelo de Oliveira Soares
Universidade Federal do Ceará (UFC)
_________________________________________
Dra. Cristiane Xerez Barroso
Universidade Federal do Ceará (UFC)

A todas as manifestações da Vida.
Aos meus pais, Aila e Francisco.

AGRADECIMENTOS
À minha família, destacando minha mãe Maria Aila, por todo o sentimento
contínuo de amor em todas as etapas de minha vida.
Ao meu orientador, Jorge Iván Sánchez Botero, pelo o acompanhamento, respeito
e pelas oportunidades criadas durante minha trajetória acadêmica como aluno da Universidade
Federal do Ceará.
À Viviana Pittalis, por todo companheirismo, paciência, amizade e amor
compartilhado durante nossa trajetória (insieme).
Aos amigos e colegas (“as rachas”) dos laboratórios de Ecologia Aquática e
Conservação (LEAC) no campus do Pici e Ecologia Pesqueira (LEP) no Labomar que de
muitos modos contribuíram com aquisição de informações e sugestões para o
desenvolvimento desse e de outros estudos. São eles: Ronaldo Gurgel, Leonardo Mesquita,
Gabriela Valentim, Grazielly Matias, Victória Maria, Yasmim Vieira, Carlos Alberto, Eliêta
Alves, Felipe Braga, Thaís Chaves, Guilherme Scheidt, Henrique Fonseca e Lucas Peixoto.
Ao colega e amigo Ronaldo Gurgel (Ronaaldoo fenômeno da Biologia) por todo o
auxílio em todas as etapas da realização desse estudo.
Aos pescadores, sem os quais não seria possível o acesso e nem as capturas de
peixes em áreas estuarinas mais distantes da foz.
À Secretária do Meio Ambiente do Estado do Ceará (SEMA) por disponibilizar
infraestrutura de lancha, piloto (Tenente Araújo) e segurança policial durante todas as coletas
realizadas no estuário do rio Cocó em Fortaleza.
Ao CNPq pelo apoio financeiro através da concessão de bolsa de estudos,
auxiliando durante todo o período do curso Mestrado, e a aos membros do quadro docente e
administrativo do Programa de Pós-Graduação em Ciências Marinhas Tropicais pelas
disciplinas ofertadas e pela excelente gerência dos assuntos de interesse do quadro discente.
E finalmente aos membros da banca de defesa de dissertação, Marcelo Oliveira
Soares (Universidade Federal do Ceará – UFC/LABOMAR) e Cristiane Xerez Barroso
(Universidade Federal do Ceará) pelas críticas, sugestões e tempo dedicado na correção do
texto. E aos professores Vicente Vieira Faria (Universidade Federal do Ceará – UFC) e
Danielle Garcez (Universidade Federal do Ceará - UFC/LABOMAR) pela colaboração e no
apoio com as correções e sugestões para a melhoria do texto da dissertação.

RESUMO
A diversidade beta descreve mudanças na composição de espécies espacial e temporalmente,
apresentando, caso existam, padrões ecológicos nas assembleias. As componentes turnover
(ganho e perda simultâneos de espécies entre locais) e nestedness (diferença de riqueza de
espécies entre locais) fornecem um modo de avaliar a composição de espécies em diferentes
habitats. Dessa forma, este estudo avalia a beta diversidade da assembleia de peixes ao longo
de cinco estuários (Choró, Cocó, Curu, Pacoti,Pirangi no Estado do Ceará, Brasil) e compara
a dissimilaridade entre as zonas inferiores, intermediárias e superiores destes ecossistemas.
Para tal, considerou-se a densidade do número de indivíduos (N/100m²) de espécies de peixes
capturadas em coletas padronizadas utilizando arrasto de praia em diversos pontos dos canais
estuarinos em zonas inferiores (próximas ao mar adjacente), intermediárias e superiores (a
montante do mar adjacente). A diversidade beta, calculada pelo índice de Bray-Curtis, foi
dividida nas componentes turnover e nestedness avaliando as contribuições de cada, em
relação à heterogeneidade total. A análise espacial dos estuários revelou que a componente
turnover explicou em maior percentual a β – diversidade; enquanto que a análise temporal
mostrou um aumento da explicação da β – diversidade pela componente nestedness nos meses
com maior precipitação. As altas taxas de turnover observadas refletem mudanças
relacionadas à abundância e composição de espécies de peixes ao longo dos gradientes
estuarinos avaliados. A dominância dessa componente sugere que vários locais desses
estuários devem ser adequadamente conservados, por abrigar alta diversidade e/ou passam por
um processo de dano ambiental, acarretando em perda de espécies de peixes. Espacialmente,
estuários que experimentam maior permanência sob condições de influência dulcícola ou de
hipersalinidade têm maiores β-diversidades em suas zonas superiores. Temporalmente, o
período chuvoso diminuiu as áreas de dispersão para a maioria dos peixes nos estuários
avaliados, possibilitando aumento da explicação da β-diversidades por nestedness
(aninhamento).
Palavras-chave: Distribuição. Ictiofauna. Ecossistemas litorâneos. Semiárido. Brasil.

ABSTRACT
Beta diversity describes spatial temporary changes in species composition, showing eventual
assemblage’s ecological standards. Turnover (the simultaneous / concurrent replacement of
species from site to site) and nestedness (difference on species abundance from site to site)
components provide us with an instrument to estimate species composition in different
habitats. Thus, this study describes fish assemblage’s beta diversity in five estuaries (Choró,
Cocó, Curu, Pacoti e Pirangi) and compares dissimilarities in between inferior, intermediate
and superior zones of these ecosystems. For this purpose it has been considered individuals’
number density (N/100m²) of fish species captured in standardized gatherings by using beach
seines in different points of estuarine channels in inferior (near to the adjacent sea),
intermediate and superior zones. Beta diversity, measured through the Bray-Curtis index, has
been divided in turnover and nestedness components, assessing each contribution compared
to total heterogeneity. The estuaries spatial analysis showed that turnover component
explains β – diversity with a larger percentage. On the other side, the temporal shows that
nestedness component has a better explanation of β – diversity during the rainiest months. The
high index of turnover observed reflects changes in fish species’ abundance and composition
along the estuarine gradients analyzed. The dominance of these components suggests that
many sites of the estuaries have to be correctly preserved, since they host high diversity
and/or get through a process of environmental damage that causes a loss of fish species.
Spacially, estuaries with higher permanence under freshwater or hypersalinity have highest β
– diversity in their superior zones. Temporally, the rainy period has reduced dispersion areas
for the largest amount of fishes that composed fish assemblages of the estuaries analyzed,
allowing for a higher explanation of β – diversity through nestedness.
Keywords: Distribution. Ichthyofaunal. Coastal Ecosystems. Semiarid. Brazil.

LISTA DE FIGURA
Figura 1 Localização geográfica das áreas de ocorrência e coleta dos estuários dos
rios Choró, Cocó, Curu, Pacoti e Pirangi ....................................................... 5
Figura 2 Rank de espécies e suas respectivas densidades (indivíduos /100 m²) para os
estuários dos rios Choró, Cocó, Curu, Pacoti e Pirangi .................................. 17
Figura 3 Riqueza de espécies de peixes nos estuários dos rios (I) - Choró (Ch), (II) -
Cocó (Co), (III) - Curu (Cr), (IV) - Pacoti (Pa) e (V) - Pirangi (Pi), nos
períodos de seca (Se) e chuva (C), com base na composição das guildas de
peixes das zonas inferior (ZI), intermediária (ZM) e superior (ZS) de cada
ecossistema ................................................................................................... 20
Figura 4 Valores da β-diversidade média e suas componentes turnover e nestedness
para as zonas inferiores (ZI), intermediárias (ZM) e superiores (ZS) os
estuários dos rios Choró, Cocó, Curu, Pacoti e Pirangi .................................. 23
Figura 5 Valores da β-diversidade total, turnover e nestedness para os estuários dos
rios Cocó (Co), Pirangi (Pi), Curu (Cr), Choró (Ch) e Pacoti (Pa) por
coleta................................................................................................................ 24
Figura 6 Relação entre densidade média (D_med) de espécies de peixes, riqueza de
espécies (R), beta diversidade média (β-div) e salinidades médias referentes
a todas as coletas realizada no estuário dos rios Choró (I), Cocó (II), Curu
(III), Pacoti (IV) e Pirangi (V) nas zonas inferiores (ZI), intermediárias (ZM)
e superiores (ZS) ................................................................................... 25
Figura 7 Valores da β-diversidade, turnover e nestedness para a ictiofauna dos
estuários dos rios Cocó (Co), Pirangi (Pi), Curu (Cr), Choró (Ch) e Pacoti
(Pa) nos meses de seca e chuva..................................................................... 29
Figura 8 Valores da β-diversidade média e suas componentes turnover e nestedness
para as zonas inferiores (ZI), intermediárias (ZM) e superiores (ZS) nos
estuários dos rios Choró, Cocó, Curu, Pacoti e Pirangi para os períodos
chuvoso (I) e seco (II)................................................................................ 29
Figura 9 Escalonamento multidimensional não métrico (“NMDS”) das
dissimilaridades entre as zonas para os períodos de seca (S) e chuva (C) nos
estuários dos rios Choró (I), Cocó (II), Curu (III), Pacoti (IV) e Pirangi

(V)................................................................................................................ 31
Figura 10 Análise SIMPER das contribuições percentuais das espécies de peixes nos
períodos de chuva e seca nos estuários dos rios Choró (I), Cocó (II), Curu
(III), Pacoti (IV) e Pirangi (V)...................................................................... 34

LISTA DE TABELAS
Tabela 1 Áreas de coleta pertencente a cada zona nos estuários dos rios Choró (Ch),
Cocó (Co), Curu (Cr), Pacoti (Pa) e Pirangi (Pi) .......................................... 6
Tabela 2 Lista de espécies de peixes, guildas estuarinas (G) (MS – Marinha –
visitante; MO – Marinha estuarina oportunista; MD – Marinha estuarino
dependente; E – Estuarina; FO – Dulcícola estuarino oportunista; D –
Dulcícola e estuário de ocorrência (X) - Choró (Ch), Cocó (Co), Curu
(Cu), Pacoti (Pa) e Pirangi (Pi). Tombo indicado pela iniciais UFRN e seu
respetivo número na LISA............................................................................. 10
Tabela 3 Número de indivíduos, densidade total para todas as coletas e riqueza de
espécies de peixes registradas nos estuários dos rios Choró, Cocó, Curu,
Pacoti e Pirangi nos períodos de chuva e seca.............................................. 16
Tabela 4 Valores absolutos e percentuais da β-diversidade e de suas componentes
turnover e nestedness para os estuários dos rios Choró, Cocó, Curu, Pacoti
e Pirangi......................................................................................................... 23

SUMÁRIO
1 INTRODUÇÃO 1
2 OBJETIVOS 3
2.1 Objetivos específicos 3
3 MATERIAL E MÉTODOS 4
3.1 Área de estudo 4
3.2 Amostragem 5
3.3 Análise de dados 8
4 RESULTADOS 9
4.1 Estrutura da assembleia de peixes
9
4.2 Dissimilaridades da composição de peixes nos estuários 22
4.2.1 Beta diversidade espacial 22
4.2.2 Beta diversidade temporal
28
4.2.3 Dissimilaridade da ictiofauna entre zonas
30
4.2.4 Espécies de peixes com maiores influências na β - diversidade
33
5 DISCUSSÃO 36
6 CONCLUSÕES 41
REFERÊNCIAS 42

1
1. INTRODUÇÃO
A diversidade biológica pode ser expressa como uma composição das
diversidades alfa (α), gama (γ) e beta (β). A α – diversidade ou a riqueza de espécies é a
diversidade em uma escala, a γ - diversidade compreende todo ecossistema ou região de
interesse de um estudo e a β - diversidade é a variação na composição de espécies entre locais,
mostrando padrões ecológicos (BARROS et al., 2014; LEGENDRE, 2005). Existem vários
modos de avaliar a beta diversidade (ANDERSON et al., 2011). Independente da variante que
esteja sendo utilizada, dois processos distintos moldam as comunidades e as suas diferenças:
substituição e perda ou ganho de espécies (BASELGA, 2010, 2017).
A substituição ou turnover indica uma alteração de espécies de um local para
outro, enquanto a perda (ou ganho) resulta em diferenças de riqueza entre os locais
(CARVALHO et al., 2012). Quando ao longo de uma gradiente há locais mais pobres que são
subconjuntos de locais mais ricos em espécies revela-se um padrão aninhado (BASELGA,
2013). Dessa forma, o aninhamento (nestedness) se caracteriza por ser um caso especial, o
qual revela um padrão ordenado de diferença na riqueza de espécies (CARVALHO et al.,
2012). A substituição e o aninhamento de espécies são formas marcadamente diferentes de
gerar dissimilaridade. Assim as contribuições dessas duas componentes da β – diversidade
indicam diferentes processos ecológicos e são fundamentais para entender influências nas
assembleias pelas mudanças espaciais, ambientais e temporais (LEGENDRE, 2014;
BASELGA, 2013; CARVALHO et al., 2012).
A abundância de indivíduos também fornece informações sobre o funcionamento
do ecossistema, e seu monitoramento pode indicar respostas a diferentes distúrbios (BALATA
et al., 2007). Dados de abundância são claramente mais ricos em informações do que os
dados de presença-ausência, podendo mudar a forma como interpretamos a variação espacial
na estrutura da assembléia (CASSEY et al.,2008). Medidas baseadas na abundância detectam
variações mais sutis. Por exemplo, podemos observar todas as mesmas espécies em dois
locais, no entanto, estas podem apresentar diferentes valores de abundância (as espécies mais
comuns em um local podem ser raras em outro local e vice-versa) (BARWELL et al., 2015).
Ainda conforme os mesmos autores, quando os ranks de espécies são iguais, a uniformidade
das abundâncias pode variar, ou seja, espécies consideradas comuns podem ser mais ou
menos dominantes.
Embora a informação sobre abundância torne as nossas inferências sobre a β -
diversidade mais significativas (BARWELL et al., 2015), esta pode aumentar com o

2
incremento da heterogeneidade ambiental e da produtividade (ASTORGA et al., 2014;
CHASE, 2010). Assim, diferentes espécies de uma assembleia podem ter sucesso ecológico
sob condições de alta produtividade, resultando numa alta β - diversidade (CESCHIN et al.,
2017). No entanto, uma diminuição na β - diversidade pode ocorrer em resposta à
eutrofização, tal cenário poderá implicar a dominância de algumas espécies com maior
tolerância associadas à extinção local de espécies mais sensíveis (ZORZAL-ALMEIDA et al.,
2017).
Dada a importância sócio-ambiental dos estuários em todo o mundo, poucos
estudos avaliaram padrões de β - diversidade (BARROS et al., 2014), necessários para
entender os processos que moldam e mantêm a biodiversidade , assim como orientar esforços
de conservação (TEICHERT et al., 2018). Assim, o presente estudo utiliza peixes como
modelo para avaliar os padrões de beta-diversidade em ambientes estuarinos do semiárido
brasileiro.
Os peixes são um importante recurso biológico que sustentam pescarias
comerciais e comunidadades litorâneas de pescadores. Em escalas locais, os padrões que
influenciam a riqueza de espécies de peixes em estuários são: tamanho do estuário, fluxo do
rio, largura da boca e proporção da área entremarés (PASQUAUD et al., 2015). Diante disso,
avaliar se as assembleias de peixes em estuários dominam padrões de substituição ou
aninhamento, auxilia estratégias para conservação (SOCOLAR et al., 2016). Por exemplo,
uma alta substituição (turnover) pode indicar que os locais diferem em características
ambientais, selecionando determinado grupo de espécies. Portanto, uma alta diversidade de
espécies pode ser preservada quando a heterogeneidade ambiental é mantida (PELÁEZ et al.,
2017). Por outro lado, um acentuado aninhamento pode indicar que locais com alta riqueza de
espécies (α – diversidade) devem ser prioridade para a conservação (GIANUCA et al., 2017).
No nordeste do Brasil, entre os estados do Ceará e Pernambuco, estuários com
quadros de hipersalinidade (SOUSA BARROSO et al. 2018; LACERDA et al., 2006)
apresentam um reduzido aporte hídrico dos rios e estão sujeitos a altas taxas de evaporação,
maior penetração da água marinha e gradiente salino a montante (POTTER et al., 2010).
Assim, potencialmente, estes estuários com alta variabilidade nas condições ambientais,
principalmente relacionados à salinidade, devem possuir alta substituição de espécies espacial
e temporal (VILLEGER et al., 2012). No entanto, sabendo que os estuários avaliados neste
estudo apresentam quadro de hipersalinidade (Choró, Curu, Pacoti e Pirangi) ou são destino
do escoamento de águas urbanas (Pacoti e Cocó), indaga-se sobre a existência de um padrão
comum nas distribuições de espécies de peixes nestes ecossistemas. Dessa forma, este estudo

3
tem por objetivo descrever padrões da β – diversidade da assembleia de peixes em cinco
estuários do semiárido brasileiro.
2. OBJETIVOS
2.1. Objetivo Geral
Caracterizar a β – diversidade da assembleia de peixes em estuários do semiárido
brasileiro.
2.2. Objetivos específicos
Caracterizar a β – diversidade espacial e temporalmente em cada um dos
estuários.
Avaliar a relação da salinidade com a β – diversidade avaliada em estuários do
semiárido.
Avaliar a dissimilaridade da comunidade de peixes entre as zonas inferiores,
intermediárias e superiores em estuários da região semiárida.

4
3 MATERIAL E MÉTODOS
3.1 Área de estudo
Os estuários Choró, Cocó, Curu, Pacoti e Pirangi, localizados no estado do Ceará,
estão inseridos dentro da região nordeste do Brasil (entre as coordenadas 4° 24’2, 86” S 37°
49’13. 61” O e 3° 24’40,74” S 39° 3’ 52,44”) que possui clima semiárido, com um período
seco longo e outro úmido curto e irregular (Figura 1). O regime de chuvas dessa região é
controlado pela Zona de Convergência Intertropical (ZCIT). A precipitação acontece de modo
irregular, principalmente durante o primeiro semestre do ano e correspondem a 91% do total
anual. Mais de 60% dessa precipitação ocorre em três meses do ano (fevereiro/março/abril ou
março/abril/maio). Pela proximidade do território cearense com a linha do equador, dominam
temperaturas altas com amplitudes reduzidas (máximas – 29,4 °C março e 30,7 °C novembro;
mínimas – 21,2 °C – julho e 23,7 °C entre janeiro a março). A insolação e a evaporação são
altas com maiores valores no segundo semestre do ano. As bacias hidrográficas, que contém
os rios que drenam os estuários avaliados neste estudo, desembocam no mar (exorréicos) com
regime de escoamento intermitente sazonal. Também possuem amplo número de barramentos,
construídos para abastecimento humano e atividades agropecuárias (MORAIS et al., 2006).
Dessa forma, a zona litorânea na qual os estuários estão localizados sofrem diversos
distúrbios como: diminuição da função de espigão hidráulico, aceleração dos processos
morfodinâmicos na foz, ampliação longitudinal da influência da maré salina, hipersalinização
sazonal e diminuição do aporte sedimentar argilo – siltoso na planície flúvio – marinha
(MORAIS & PINHEIRO, 2011; MORAIS & PINHEIRO, 2010; MORAIS et al., 2006;
MOLISANI et al., 2006).
O espaço territorial onde os estuários estão inseridos é caracterizado pela presença
de florestas de mangues fragmentadas, com acentuada ocupação urbana e atividades de
carcinicultura que causam a perda dessa vegetação e afetam serviços ambientais fornecidos
por esse ecossistema (MEIRELES et al., 2007). Os estuários dos rios Choró, Curu e Pirangi
apresentam em suas margens fazendas de criação de camarão, enquanto que Cocó e Pacoti
possuem ampla ocupação urbana em seu entorno. Todos os estuários em questão são afetados
por emissões antrópicas de nitrogênio (N) e fósforo (P), com excedentes que comprometem a
qualidade das águas (SOUSA BARROSO et al., 2016; LACERDA et al., 2008; MEIRELES
et al., 2007).
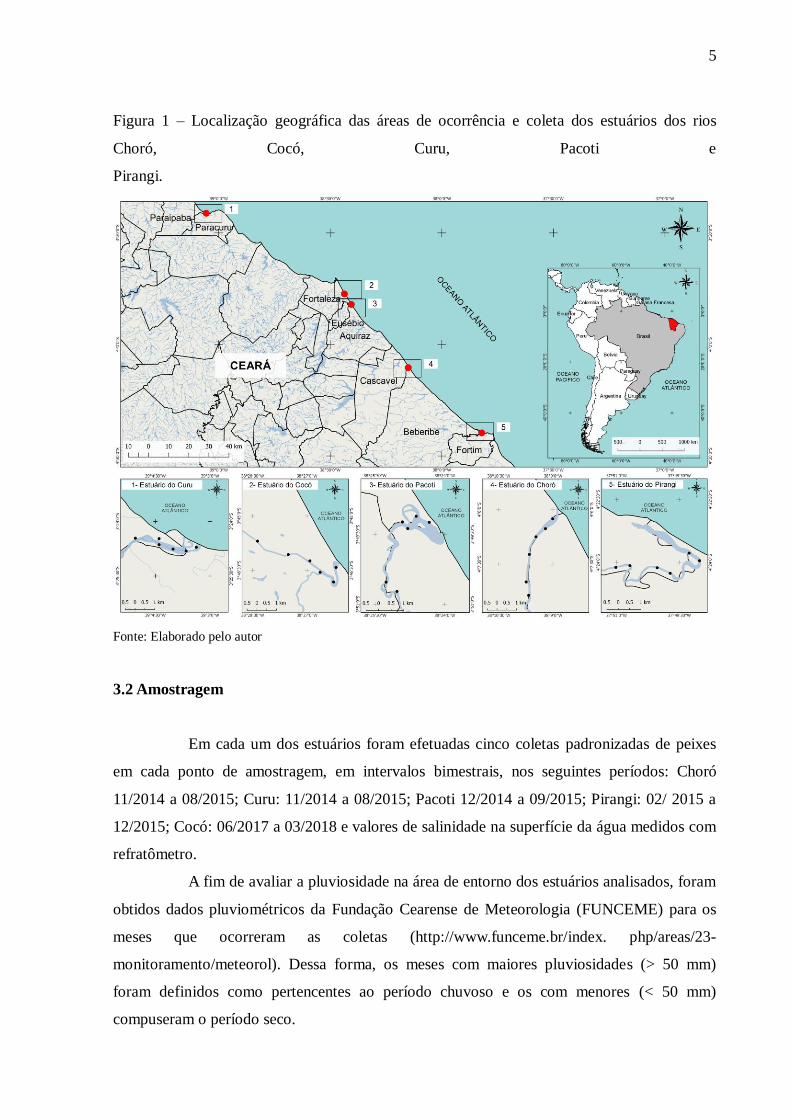
5
Figura 1 – Localização geográfica das áreas de ocorrência e coleta dos estuários dos rios
Choró, Cocó, Curu, Pacoti e
Pirangi.
Fonte: Elaborado pelo autor
3.2 Amostragem
Em cada um dos estuários foram efetuadas cinco coletas padronizadas de peixes
em cada ponto de amostragem, em intervalos bimestrais, nos seguintes períodos: Choró
11/2014 a 08/2015; Curu: 11/2014 a 08/2015; Pacoti 12/2014 a 09/2015; Pirangi: 02/ 2015 a
12/2015; Cocó: 06/2017 a 03/2018 e valores de salinidade na superfície da água medidos com
refratômetro.
A fim de avaliar a pluviosidade na área de entorno dos estuários analisados, foram
obtidos dados pluviométricos da Fundação Cearense de Meteorologia (FUNCEME) para os
meses que ocorreram as coletas (http://www.funceme.br/index. php/areas/23-
monitoramento/meteorol). Dessa forma, os meses com maiores pluviosidades (> 50 mm)
foram definidos como pertencentes ao período chuvoso e os com menores (< 50 mm)
compuseram o período seco.

6
Os pontos de amostragem nos estuários avaliados foram divididos em zonas,
usando como critério os valores de salinidade obtidos durante o período chuvoso (fevereiro –
maio), as porcentagens de cada tipo de substrato e largura do rio. A zona inferior (ZI) perfez
áreas mais próximas ao ambiente marinho com substratos areno-siltosos, e as maiores larguras
dos rios (entre 300 m a 500 m). A zona superior (ZS) (a montante da foz) teve a porção do rio
com menor salinidade durante o período chuvoso (fevereiro – maio) (valores menores que: 15
– Curu; 9,4 – Pirangi; 1 - Choró, Cocó e Pacoti) (Tabela 1), substratos siltosos com maior
percentual de argila e menor largura (entre 50 m a 65 m). A zona intermediária (ZM) foi
definida entre a zona inferior e superior, teve a contribuição mais equilibrada dos três tipos de
substratos (arenoso, siltoso e argila) e larguras entre 75 m e 160 m. A zonas indicadas são
representadas por pontos na figura 1 sendo os dois primeiros, mais próximos ao mar
adjacente, indicando a zona inferior (ZI), os dois pontos seguintes, à zona intermediária (ZM)
e os dois últimos, a montante, representando a zona superior (ZS).
Tabela 1 – Áreas de coleta pertencente a cada zona nos estuários dos rio Choró (Ch),
Cocó (Co), Curu (Cr), Pacoti (Pa) e Pirangi (Pi).
Estuário Zona Coordenadas
Choró
Zona Inferior (ZI) 4°06'15.08"S 38°09'00.53"O
-
4°06'31.31"S 38°09'12.54"O
Zona Intermediária (ZM) 4°06'44.93"S 38°09'28.25"O
-
4°07'14.10"S 38°09'39.25"O
Zona Superior (ZS) 4°07'59.73"S 38°09'44.63"O
-
4°08'22.00"S 38°09'40.47"O
Cocó
Zona Inferior (ZI) 3°46'25.19"S 38°26'12.13”O
-
3°46'49.12"S 38°26'16.10”O
Zona Intermediária (ZM) 3°46'33.08"S 38°26'38.30”O
-
3°46'14.26"S 38°26'55.19”O
Zona Superior (ZS) 3°46'03.82"S 38°27'31.00”O
-
3°45'41.60"S 38°28'21.87”O
Curu
Zona Inferior (ZI) 3°24'42.65"S 39°03'22.52"O
-
3°24'48.81"S 39°03'39.35"O
Zona Intermediária (ZM) 3°24'45.63"S 39°04'00.88"O

7
-
3°24'38.34"S 39°04'23.96"O
Zona Superior (ZS) 3°24'38.34"S 39°04'23.96"O
-
3°24'27.62"S 39°05'03.08"O
Pacoti
Zona Inferior (ZI) 3°49'14.36"S 38°24'20.07"O
-
3°49'06.49"S 38°24'36.93"O
Zona Intermediária (ZM) 3°49'22.34"S 38°24'47.86"O
-
3°49'13.84"S 38°24'54.83"O
Zona Superior (ZS) 3°50'31.61"S 38°25'15.87"O
-
3°50'43.63"S 38°25'01.74"O
Pirangi
Zona Inferior (ZI) 4°23'46.77"S 37°49'16.13”O
-
4°23'57.65"S 37°49'1.20”O
Zona Intermediária (ZM) 4°24'15.65"S 37°49'43.15”O
-
4°24'7.75"S 37°50'1.72"O
Zona Superior (ZS) 4°24'4.74"S 37°50'40.78”O
-
4°24'7.16"S 37°51'0.91”O
Fonte: Elaborado pelo autor.
Para a captura das espécies de peixes, foi utilizado arrasto de praia de 25 m de
comprimento, 2 m de altura e 12 mm de malha entre nós. Em cada ponto de coleta, foram
realizados três arrastos, em áreas de 200 m2, com profundidades entre 0,3 a 1,5m, durante os
períodos de maré baixa nos intervalos de vazante (2,0 a 0,0) ou de enchente (0,0 a 2,0) e,
durante horários diurnos (07:00 as 17:00h). Cada arrasto perfez uma extensão de
aproximadamente 200 m² de área estuarina obtida a partir da formula A = D x L, onde A é a
área, D é a distância partindo da margem (10 m) e L é o comprimento real da rede que foi
empregado (20 m).
Os peixes capturados foram crioanestesiados com eugenol, condicionados em
sacos plásticos e, em seguida, fixados em formol 10 %. Após período de 48 horas, eram
preservados em álcool etílico 70%. Todas as coletas foram autorizadas pelo ICMBio/SISBio
sob as Licenças nº 43014-3 para os estuários do Choró, Curu, Pacoti e Pirangi e nº 57780
para o estuário do Cocó.

8
No Laboratório de Ecologia Aquática e Conservação (LEAC) do Departamento de
Biologia da Universidade Federal do Ceará, os peixes foram identificados ao nível de espécie
usando os estudos realizados por Araújo et al. (2004); Carpenter (2002a); Carpenter (2002b),
Fischer et al., (2011), Garcia Jr. et al., (2010), Marceniuk (2005), Marceniuk et al., (2012),
Menezes et al., (2015) e Moura e Lindeman (2007). Posteriormente, exemplares de cada
espécie foram tombados na coleção ictiológica do Laboratório de Ictiologia Sistemática e
Evolutiva (LISE) da Universidade Federal do Rio Grande Norte (UFRN).
3.3 Análise dos dados
Para a caracterização da assembleia de peixes, os indivíduos coletados foram
identificados até o mínimo táxon possível e contabilizados. A partir das informações obtidas
com os arrastos, a unidade de densidade escolhida para a apresentação e uso dos dados de
espécies foi a de número de indivíduos /100 m².
Com base em Potter et al. (2015), as espécies foram classificadas pela guilda de
uso do ambiente estuarino em: marinho – visitante (MS); marinhas estuarino – oportunistas
(MO); marinhas estuarinas dependentes (MD), estuarinas (E), dulcícolas (D) e em dulcícolas
estuarino oportunistas (FO).
Para obter as dissimilaridades na composição de espécies de peixes entre as zonas
inferior (ZI), intermediária (ZM) e superior (ZS) para cada estuário, assim como, as
dissimilaridades de peixes entre os períodos de seca e chuva, foram calculadas as
dissimilaridades entre múltiplos locais usando o coeficiente de Bray – Curtis.
O referido coeficiente foi escolhido, pois as amostras de peixes foram obtidas a
partir de espaços de dimensões físicas iguais, ou seja, abrangeram áreas iguais. Além disso, o
índice é sensível a diferenças na abundância entre espécies, onde espécies abundantes
assumem maiores importâncias do que espécies raras (RICOTTA & PODANI 2017).
Os cálculos das dissimilaridades foram realizados com o uso do software R a
partir da extensão betapart (BASELGA, 2016). Dentre as funções presentes nessa extensão,
duas foram usadas para obter tanto os valores totais quanto entre zonas das componentes
turnover (substituição) e nestedness (aninhamento). São elas: beta.multi.abund e
beta.pair.abund.
A primeira função (beta.multi.abund) gera três valores que são (i) variação
balanceada de abundância, que é similar ao turnover, (ii) gradientes de abundância similiar ao
nestedness e (iii) a dissimilaridade total (Bray-Curtis), que é a soma de ambas as componentes

9
para todos os locais da tabela de abundância de espécies. A segunda (beta. pair.abund) calcula
as mesmas três dissimilaridades da primeira função; no entanto, gera três matrizes contendo
os valores entre locais de cada componente de dissimilaridade (BASELGA, 2016). Estas
análises separam as componentes resultantes da β – diversidade em turnover (substituição) e
nestedness (aninhamento) (BASELGA, 2013, 2016), gerando o coeficiente valores que
variam entre 0 a 1, quanto mais se aproxima do 1 maior é a diferença entre os locais ou zonas.
A fim de obter os gráficos com a β – diversidade e suas componentes turnover e
nestedness para cada um dos estuários, foram considerados a média e o desvio padrão do
valor da dissimilaridade que cada zona apresentou.
Para analisar espacialmente a salinidade e relacioná-la com a densidade de
espécies e a β – diversidade, foram consideradas as médias dos valores da salinidade obtidos
em cada local onde foram realizadas as coletas.
Para avaliar as diferenças na β – diversidade entre as zonas de cada estuário
temporalmente, foram confeccionados gráficos de escalonamento multidimensional não
métrico (nMDS) usando Bray_Curtis como índice de dissimilaridade.
Por último, para verificar quais as espécies de peixes contribuem com um maior
percentual para gerar as dissimilaridades entre zonas de cada estuário analisado nos períodos
de seca e chuva, foi realizada uma análise de similaridade de porcentagens (SIMPER).
4 RESULTADOS
4.1 Estrutura da assembleia de peixes
Nos cinco estuários avaliados, foram capturados um total de 23108 peixes de 16
ordens, 35 famílias e 85 espécies. Todos os estuários apresentaram ocorrências únicas,
totalizando 24 espécies de peixes no total. O estuário do rio Pacoti teve o maior número de
ocorrências únicas, oito espécies. Quanto para os outros estuários foram: seis espécies para
Choró, quatro espécies para Curu e Cocó e duas espécies no Pirangi (Tabela 2).
Foram registradas 12 espécies de peixes comuns aos cinco estuários: Lycengraulis
grossidens (Spix & Agassiz, 1829), Lile piquitinga Schreiner & Miranda Ribeiro, 1903),
Mugil curema Valenciennes, 1836, Atherinella brasiliensis (Quoy & Gaimard, 1825),
Strongylura timucu (Walbaum, 1792), Caranx latus Agassiz, 1831, Lutjanus griseus
(Linnaeus, 1758), Diapterus auratus Ranzani, 1842 , Eucinostomus argenteus Baird & Girard,
1855, Eucinostomus melanopterus (Bleeker, 1863), Sphoeroides testudineus (Linnaeus, 1758)

10
e Hyporhamphus unifasciatus (Razani, 1841). A ordem com maior número de famílias foi
Perciforme (14). Dentro desta ordem, as famílias mais representativas foram Gerreidae (8
espécies), Engraulidae e Gobiidae (6 espécies), Lutjanidae e Carangidae (5 espécies) com
cinco cada e Mugilidae (4 espécies).
Seis guildas de uso do ambiente estuarino contemplaram todas as espécies de
peixes que ocorreram nos estuários analisados. As guildas com maior número de espécies
foram marinho estuarino dependente (39 espécies), marinho estuarino oportunista (19
espécies) e estuarino (16 espécies). Enquanto as demais guildas apresentaram número
reduzido de espécies: dulcícola (6), marinha visitante (4) e dulcícola estuarino oportunista (1)
(Tabela 2).
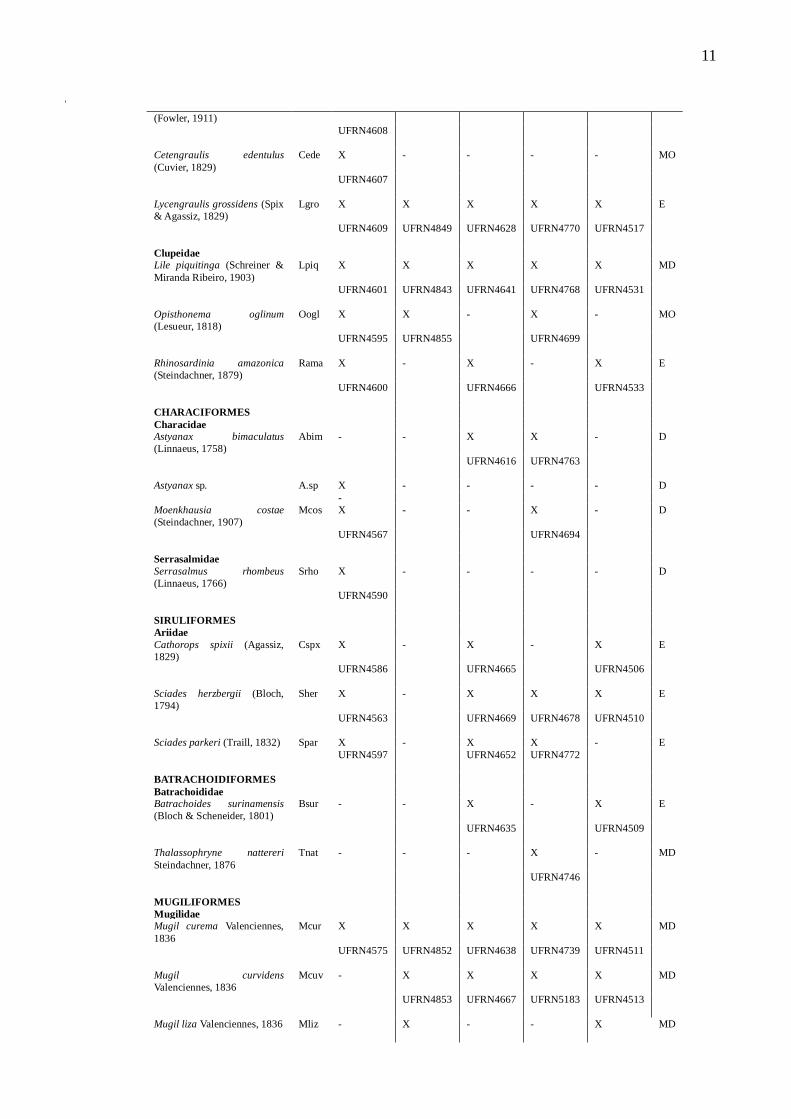
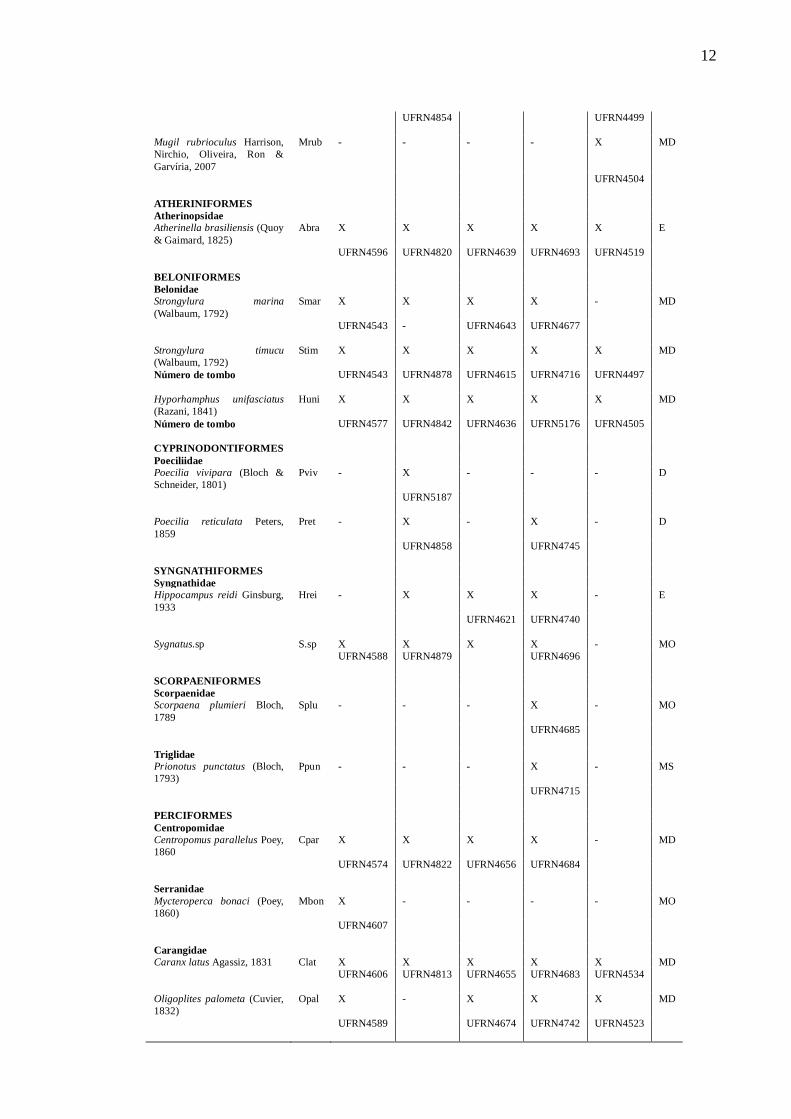
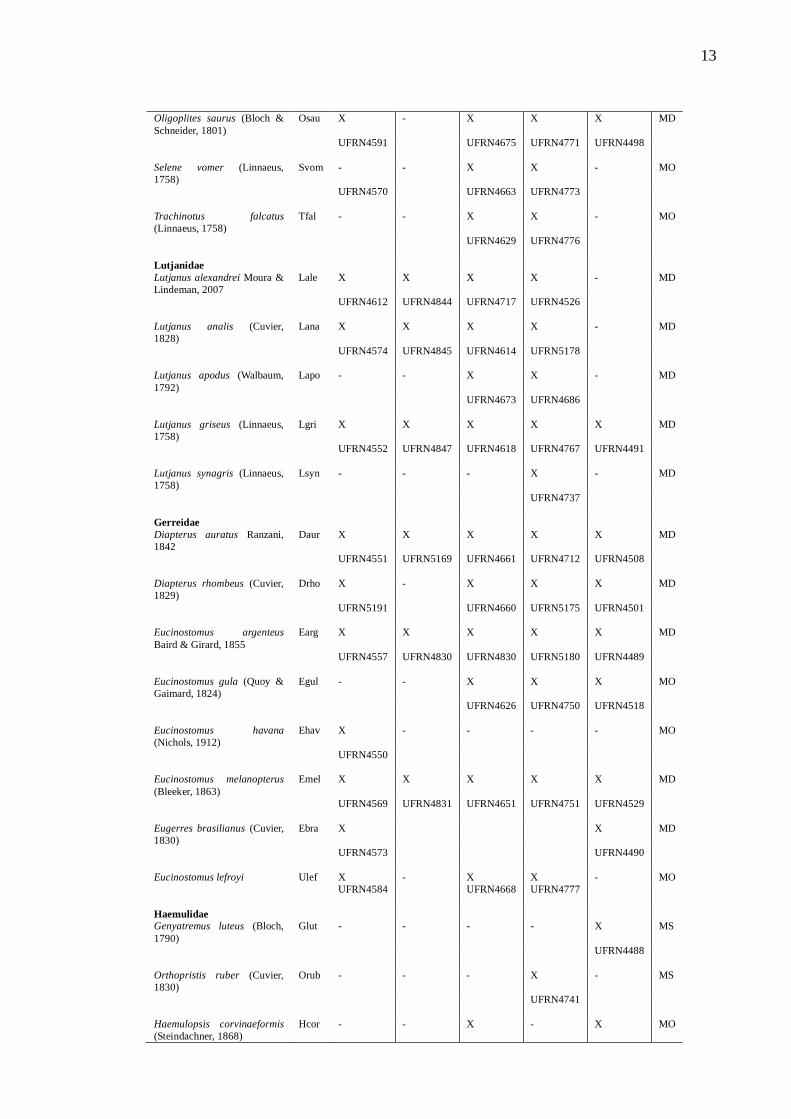
Tabela 2 – Lista de espécies de peixes, guildas estuarinas (G) (MS – Marinha –
visitante; MO – Marinha estuarina oportunista; MD – Marinha estuarino dependente;
E – Estuarina; FO – Dulcícola estuarino oportunista; D – Dulcícola e estuário de
ocorrência (X) - Choró (Ch), Cocó (Co), Curu (Cu), Pacoti (Pa) e Pirangi (Pi). Tombo
indicado pela iniciais UFRN e seu respetivo número na LISA.
Táxon Cód. Ch Co Cu Pa Pi G
ELOPIFORMES
Elopidae
Elops saurus Linnaeus, 1766 Esau X - - X - MD
UFRN4545
UFRN4697
Megalopidae
Megalops atlanticus
Valenciennes, 1847
Matl - X - - - MO
UFRN4850
ALBULIFORMES
Albulidae
Albula vulpes (Linnaeus,
1758)
Avul - - X - - MO
UFRN4671
ANGUILLIFORMES
Ophichthidae
Myrichthys ocellatus
(Lesueur, 1825)
Moce X - X X - MO
UFRN4547
UFRN4637
UFRN4695
CLUPEIFORMES
Engraulidae
Anchoa hepsetus (Linnaeus,
1758)
Ahep X - - - - MO
UFRN4603
Anchoa spinifer
(Valenciennes, 1848)
Aspi X - X - - MO
UFRN4608
UFRN4649
Anchovia clupeoides
(Swainson, 1839)
Aclu X - X X X MD
UFRN4598
UFRN4625
UFRN4762
UFRN4525
Anchoviella lepidentostole Alep X - - - - MO
11
(Fowler, 1911)
UFRN4608
Cetengraulis edentulus
(Cuvier, 1829)
Cede X - - - - MO
UFRN4607
Lycengraulis grossidens (Spix
& Agassiz, 1829)
Lgro X X X X X E
UFRN4609
UFRN4849
UFRN4628
UFRN4770
UFRN4517
Clupeidae
Lile piquitinga (Schreiner &
Miranda Ribeiro, 1903)
Lpiq X X X X X MD
UFRN4601
UFRN4843
UFRN4641
UFRN4768
UFRN4531
Opisthonema oglinum
(Lesueur, 1818)
Oogl X X - X - MO
UFRN4595
UFRN4855
UFRN4699
Rhinosardinia amazonica
(Steindachner, 1879)
Rama X - X - X E
UFRN4600
UFRN4666
UFRN4533
CHARACIFORMES
Characidae
Astyanax bimaculatus
(Linnaeus, 1758)
Abim - - X X - D
UFRN4616
UFRN4763
Astyanax sp. A.sp X - - - - D
-
Moenkhausia costae
(Steindachner, 1907)
Mcos X - - X - D
UFRN4567
UFRN4694
Serrasalmidae
Serrasalmus rhombeus
(Linnaeus, 1766)
Srho X - - - - D
UFRN4590
SIRULIFORMES
Ariidae
Cathorops spixii (Agassiz,
1829)
Cspx X - X - X E
UFRN4586
UFRN4665
UFRN4506
Sciades herzbergii (Bloch,
1794)
Sher X - X X X E
UFRN4563
UFRN4669
UFRN4678
UFRN4510
Sciades parkeri (Traill, 1832) Spar X - X X - E
UFRN4597
UFRN4652
UFRN4772
BATRACHOIDIFORMES
Batrachoididae
Batrachoides surinamensis
(Bloch & Scheneider, 1801)
Bsur - - X - X E
UFRN4635
UFRN4509
Thalassophryne nattereri
Steindachner, 1876
Tnat - - - X - MD
UFRN4746
MUGILIFORMES
Mugilidae
Mugil curema Valenciennes,
1836
Mcur X X X X X MD
UFRN4575
UFRN4852
UFRN4638
UFRN4739
UFRN4511
Mugil curvidens
Valenciennes, 1836
Mcuv - X X X X MD
UFRN4853
UFRN4667
UFRN5183
UFRN4513
Mugil liza Valenciennes, 1836 Mliz - X - - X MD
12
UFRN4854
UFRN4499
Mugil rubrioculus Harrison,
Nirchio, Oliveira, Ron &
Garvíria, 2007
Mrub - - - - X MD
UFRN4504
ATHERINIFORMES
Atherinopsidae
Atherinella brasiliensis (Quoy
& Gaimard, 1825)
Abra X X X X X E
UFRN4596
UFRN4820
UFRN4639
UFRN4693
UFRN4519
BELONIFORMES
Belonidae
Strongylura marina
(Walbaum, 1792)
Smar X X X X - MD
UFRN4543
- UFRN4643
UFRN4677
Strongylura timucu
(Walbaum, 1792)
Stim X X X X X MD
Número de tombo UFRN4543
UFRN4878
UFRN4615
UFRN4716
UFRN4497
Hyporhamphus unifasciatus
(Razani, 1841)
Huni X X X X X MD
Número de tombo UFRN4577
UFRN4842
UFRN4636
UFRN5176
UFRN4505
CYPRINODONTIFORMES
Poeciliidae
Poecilia vivipara (Bloch &
Schneider, 1801)
Pviv - X - - - D
UFRN5187
Poecilia reticulata Peters,
1859
Pret - X - X - D
UFRN4858
UFRN4745
SYNGNATHIFORMES
Syngnathidae
Hippocampus reidi Ginsburg,
1933
Hrei - X X X - E
UFRN4621
UFRN4740
Sygnatus.sp S.sp X X X X - MO
UFRN4588
UFRN4879
UFRN4696
SCORPAENIFORMES
Scorpaenidae
Scorpaena plumieri Bloch,
1789
Splu - - - X - MO
UFRN4685
Triglidae
Prionotus punctatus (Bloch,
1793)
Ppun - - - X - MS
UFRN4715
PERCIFORMES
Centropomidae
Centropomus parallelus Poey,
1860
Cpar X X X X - MD
UFRN4574
UFRN4822
UFRN4656
UFRN4684
Serranidae
Mycteroperca bonaci (Poey,
1860)
Mbon X - - - - MO
UFRN4607
Carangidae
Caranx latus Agassiz, 1831 Clat X X X X X MD
UFRN4606
UFRN4813
UFRN4655
UFRN4683
UFRN4534
Oligoplites palometa (Cuvier,
1832)
Opal X - X X X MD
UFRN4589
UFRN4674
UFRN4742
UFRN4523
13
Oligoplites saurus (Bloch &
Schneider, 1801)
Osau X - X X X MD
UFRN4591
UFRN4675
UFRN4771
UFRN4498
Selene vomer (Linnaeus,
1758)
Svom - - X X - MO
UFRN4570
UFRN4663
UFRN4773
Trachinotus falcatus
(Linnaeus, 1758)
Tfal - - X X - MO
UFRN4629
UFRN4776
Lutjanidae
Lutjanus alexandrei Moura &
Lindeman, 2007
Lale X X X X - MD
UFRN4612
UFRN4844
UFRN4717
UFRN4526
Lutjanus analis (Cuvier,
1828)
Lana X X X X - MD
UFRN4574
UFRN4845
UFRN4614
UFRN5178
Lutjanus apodus (Walbaum,
1792)
Lapo - - X X - MD
UFRN4673
UFRN4686
Lutjanus griseus (Linnaeus,
1758)
Lgri X X X X X MD
UFRN4552
UFRN4847
UFRN4618
UFRN4767
UFRN4491
Lutjanus synagris (Linnaeus,
1758)
Lsyn - - - X - MD
UFRN4737
Gerreidae
Diapterus auratus Ranzani,
1842
Daur X X X X X MD
UFRN4551
UFRN5169
UFRN4661
UFRN4712
UFRN4508
Diapterus rhombeus (Cuvier,
1829)
Drho X - X X X MD
UFRN5191
UFRN4660
UFRN5175
UFRN4501
Eucinostomus argenteus
Baird & Girard, 1855
Earg X X X X X MD
UFRN4557
UFRN4830
UFRN4830
UFRN5180
UFRN4489
Eucinostomus gula (Quoy &
Gaimard, 1824)
Egul - - X X X MO
UFRN4626
UFRN4750
UFRN4518
Eucinostomus havana
(Nichols, 1912)
Ehav X - - - - MO
UFRN4550
Eucinostomus melanopterus
(Bleeker, 1863)
Emel X X X X X MD
UFRN4569
UFRN4831
UFRN4651
UFRN4751
UFRN4529
Eugerres brasilianus (Cuvier,
1830)
Ebra X X MD
UFRN4573
UFRN4490
Eucinostomus lefroyi Ulef X - X X - MO
UFRN4584
UFRN4668
UFRN4777
Haemulidae
Genyatremus luteus (Bloch,
1790)
Glut - - - - X MS
UFRN4488
Orthopristis ruber (Cuvier,
1830)
Orub - - - X - MS
UFRN4741
Haemulopsis corvinaeformis
(Steindachner, 1868)
Hcor - - X - X MO

14
UFRN4645
UFRN4520
Sparidae
Archosargus probatocephalus
(Walbaum, 1792)
Apro - - X - - MS
UFRN4672
Archosargus rhomboidalis
(Linnaeus, 1758)
Arho X - - X - MO
UFRN4599
UFRN4747
Sciaenidae
Cynoscion acoupa (Lacepède,
1801)
Caco - - - X - MD
UFRN4751
Micropogonias furnieri
(Desmarest, 1823)
Mfur - - X X X MD
UFRN4664
UFRN4743
UFRN4495
Stellifer naso (Jordan, 1889) Snas - - - X - MD
UFRN4691
Cichlidae
Oreochromis niloticus
(Linnaeus, 1758)
Onil - X - - X FO
UFRN4856
UFRN4492
Scaridae
Sparisoma radians
(Valenciennes, 1840)
Srad X - X X - MO
UFRN4579
UFRN4630
UFRN4774
Eleotridae
Eleotris pisonis (Gmelin,
1789)
Epis - X - - - E
UFRN4824
Gobiidae
Bathygobius soporator
(Valenciennes, 1837)
Bsop X - X X X E
UFRN4565
UFRN4627
UFRN4764
UFRN4540
Ctenogobius boleosoma
(Jordan & Gilbert, 1882)
Cbol X X X X - MD
UFRN4613
UFRN4826
UFRN4659
UFRN4766
Ctenogobius smaragdus
(Valenciennes, 1837)
Csma - - X X X MD
UFRN4658
UFRN4767
UFRN4539
Evorthodus lyricus (Girard,
1858)
Elyr - - - X - MD
UFRN4744
Gobionellus oceanicus
(Pallas, 1770)
Goce X - X X X E
UFRN4572
UFRN4633
UFRN4682
UFRN4528
Gobionellus stomatus Starks,
1913
Gsto X - X X - E
UFRN4558
UFRN4642
UFRN4695
Ephippidae
Chaetodipterus faber
(Broussonet, 1782)
Cfab X - X X X MD
UFRN4583
UFRN4640
UFRN4765
UFRN4494
Sphyraenidae
Sphyraena barracuda
(Edwards, 1771)
Sbar - X X X - MD
UFRN4863
UFRN4634
UFRN4736
Sphyraena guachancho
Cuvier, 1829
Sgua - X - - - MD
UFRN4864
15
PLEURONECTIFORMES
Paralichthyidae
Citharichthys arenaceus
Evermann & Marsh, 1900
Care - X - - X MD
UFRN4818
UFRN4522
Citharichthys spilopterus
Günther, 1862
Cspi X - X - X E
UFRN4562
UFRN4617
UFRN4486
Paralichthys brasiliensis
(Ranzani, 1842)
Pbra - - - - X MD
-
Achiridae
Achirus achirus (Linnaeus,
1758)
Aach X - X X X MD
UFRN4582
UFRN4654
UFRN4761
UFRN4421
Achirus lineatus (Linnaeus,
1758)
Alin X - X X X MD
UFRN4582
UFRN4646
UFRN4713
UFRN4515
Trinectes paulistanus
(Miranda Ribeiro, 1915)
Tpau - X X - X MD
UFRN5171
UFRN4648
UFRN4542
Cynoglossidae
Symphurus tessellatus (Quoy
& Gaimard, 1824)
Stss - - - X - E
UFRN4690
TETRAODONTIFORMES
Tetraodontidae
Sphoeroides greeleyi Gilbert,
1900
Sgre X X X X - E
UFRN4568
UFRN4861
UFRN4631
UFRN4678
Sphoeroides testudineus
(Linnaeus, 1758)
Stes X X X X X E
UFRN4564
UFRN4862
UFRN4632
UFRN4676
UFRN4493
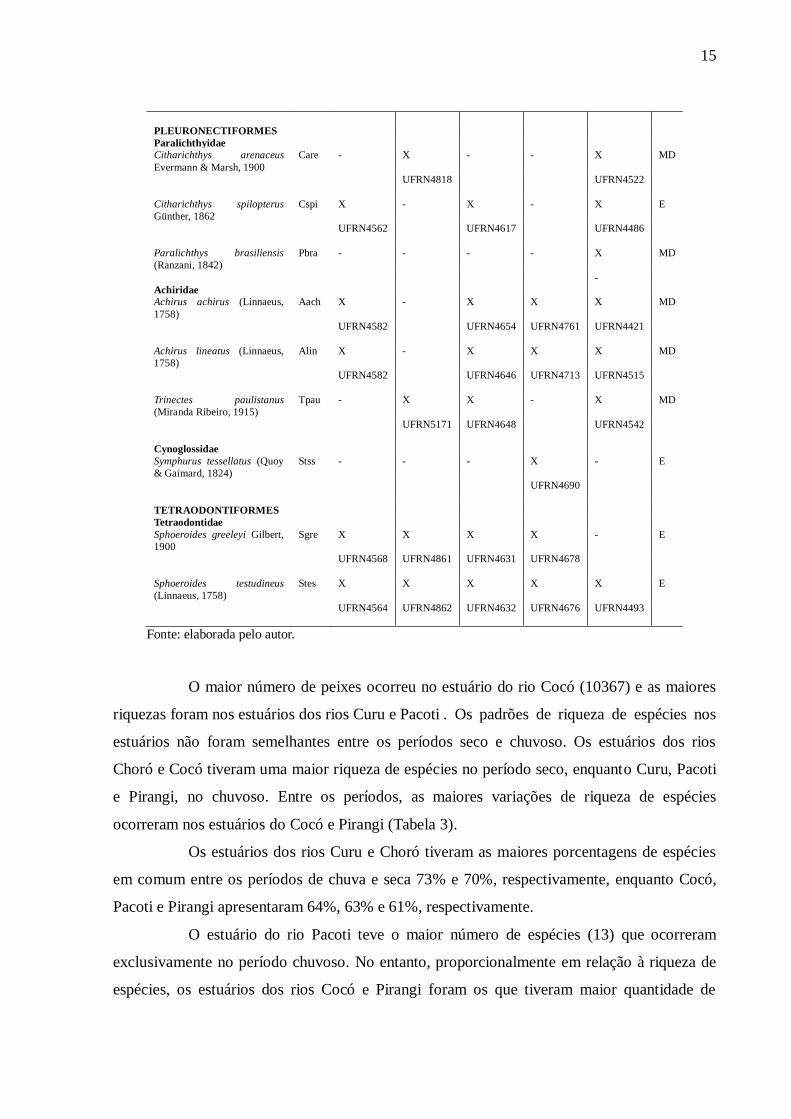
Fonte: elaborada pelo autor.
O maior número de peixes ocorreu no estuário do rio Cocó (10367) e as maiores
riquezas foram nos estuários dos rios Curu e Pacoti . Os padrões de riqueza de espécies nos
estuários não foram semelhantes entre os períodos seco e chuvoso. Os estuários dos rios
Choró e Cocó tiveram uma maior riqueza de espécies no período seco, enquanto Curu, Pacoti
e Pirangi, no chuvoso. Entre os períodos, as maiores variações de riqueza de espécies
ocorreram nos estuários do Cocó e Pirangi (Tabela 3).
Os estuários dos rios Curu e Choró tiveram as maiores porcentagens de espécies
em comum entre os períodos de chuva e seca 73% e 70%, respectivamente, enquanto Cocó,
Pacoti e Pirangi apresentaram 64%, 63% e 61%, respectivamente.
O estuário do rio Pacoti teve o maior número de espécies (13) que ocorreram
exclusivamente no período chuvoso. No entanto, proporcionalmente em relação à riqueza de
espécies, os estuários dos rios Cocó e Pirangi foram os que tiveram maior quantidade de

16
espécies exclusivas temporalmente, sendo 10 espécies nos meses secos no primeiro estuário e
12 no período chuvoso no segundo.
Tabela 3 – Número de indivíduos, densidade total para todas as coletas e riqueza de
espécies de peixes registradas nos estuários dos rios Choró, Cocó, Curu, Pacoti e
Pirangi nos períodos de chuva e seca.
Estuário
Número de indivíduos
Riqueza de espécies Riqueza de espécies
Chuva Seca
Choró
2881
50
42 45
Cocó
10367
36
26 33
Curu
4176
53
48 47
Pacoti
4196
59
50 45
Pirangi
1488
39
36 27
Total 23108
Fonte: elaborado pelo autor.
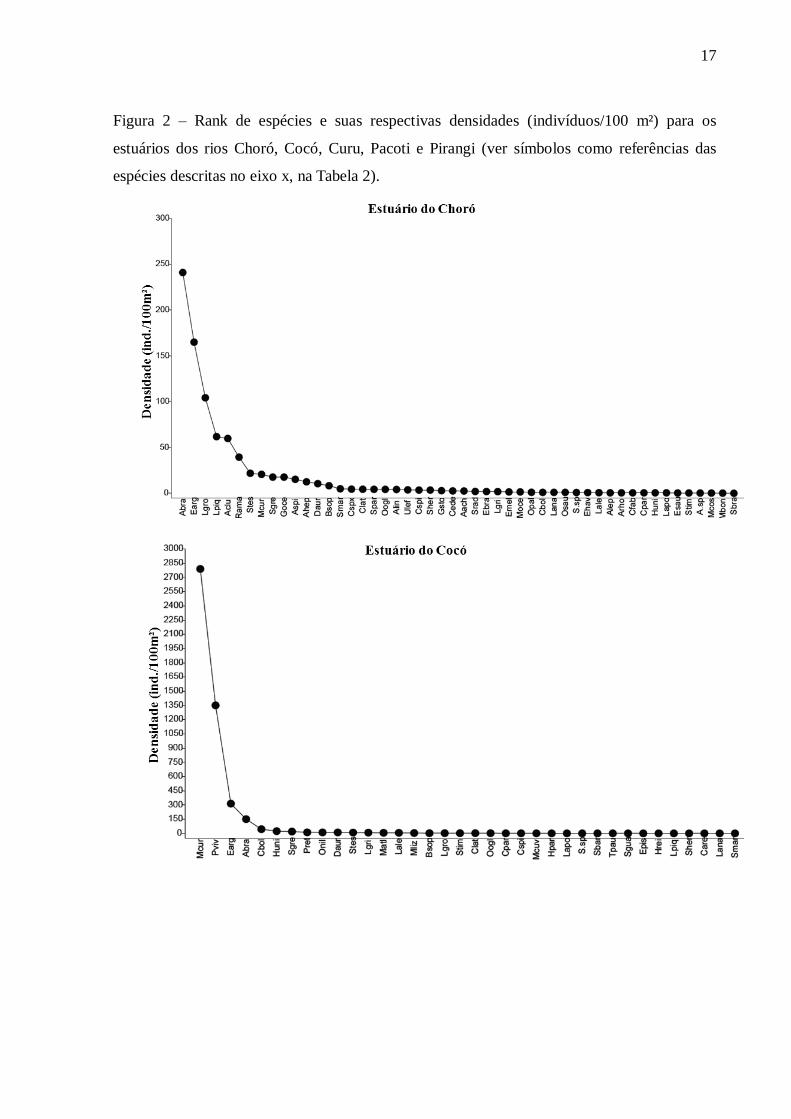
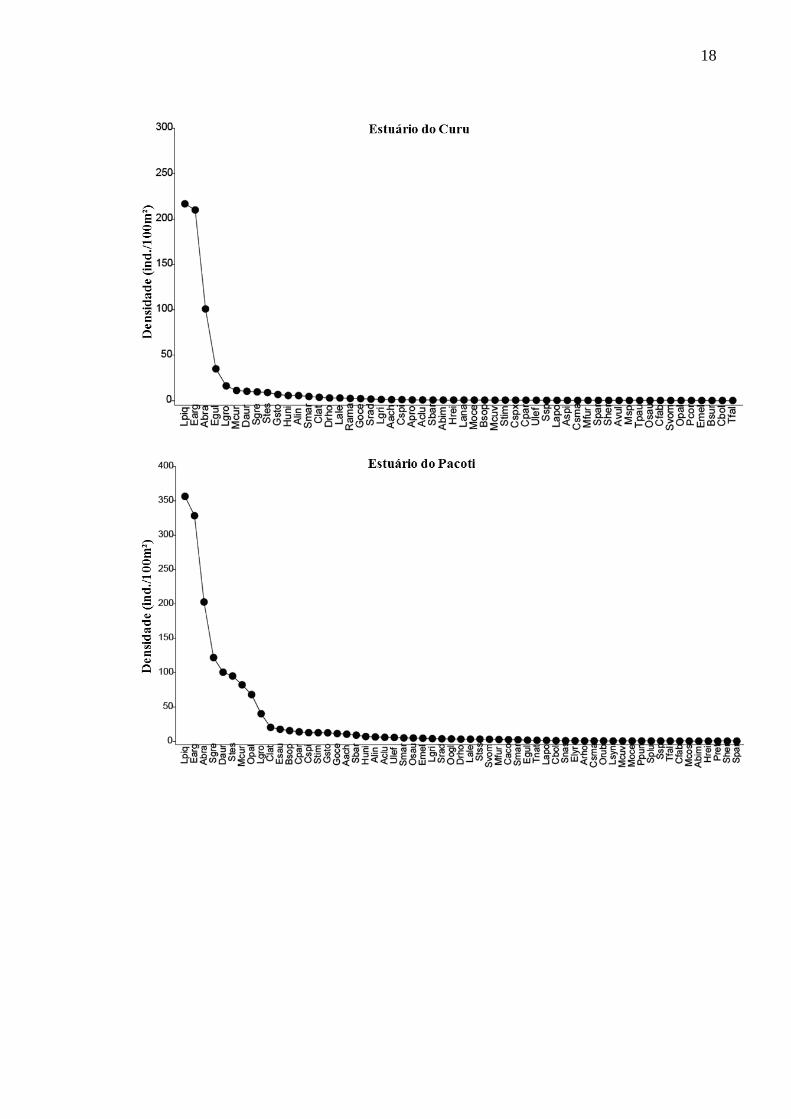
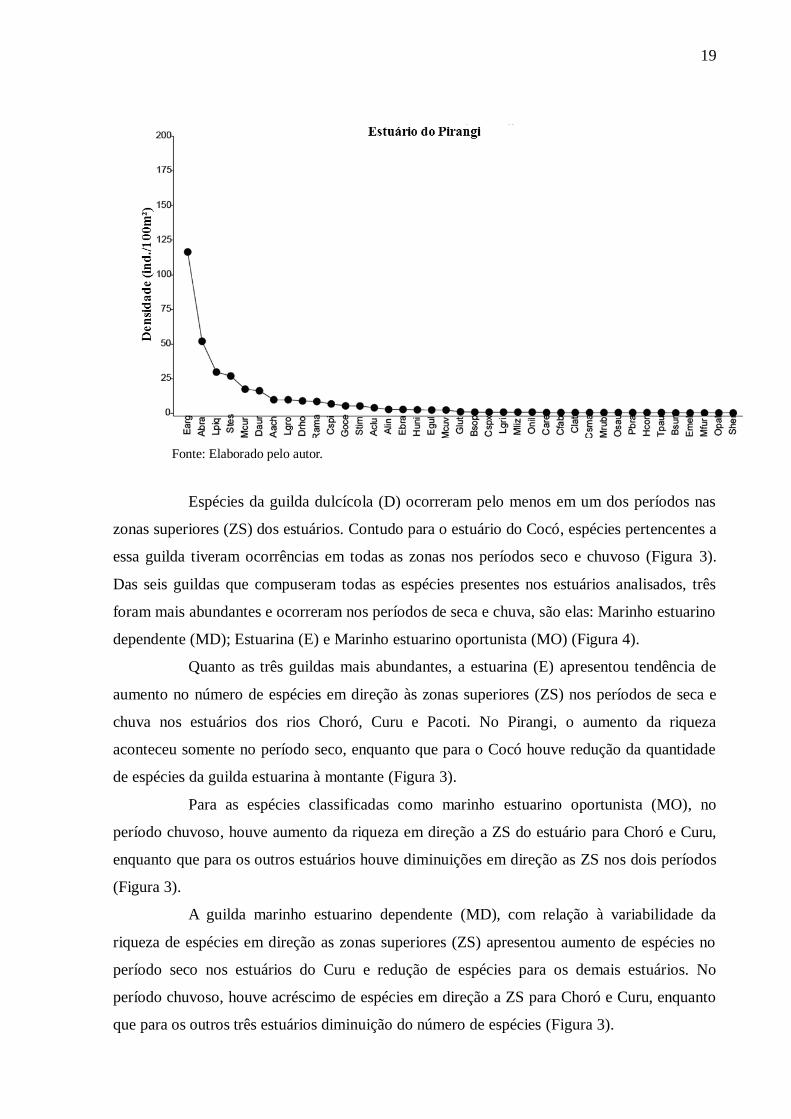
Cinco espécies de peixes correspondem entre 69% e 97% da densidade total em
cada estuário. Para os estuários dos rios Pacoti, Pirangi e Choró, as cinco espécies de peixes
com maior densidade compuseram 69%, 71,2% e 73,4% da densidade total, respectivamente.
No Curu e Cocó, os valores foram 86,4% e 97%. Dentre as cinco espécies com as maiores
densidades em cada estuário, três são comuns a todos os estuários, são elas: Eucinostomus
argenteus, Atherinella brasiliensis e Lile piquitinga. No rio Cocó, a espécie dulcícola Poecilia
vivipara representou 28,2% da densidade total, contrastando com os demais estuários onde as
maiores densidades foram de espécies marinhas ou estuarinas (Figura 2).
17
Figura 2 – Rank de espécies e suas respectivas densidades (indivíduos/100 m²) para os
estuários dos rios Choró, Cocó, Curu, Pacoti e Pirangi (ver símbolos como referências das
espécies descritas no eixo x, na Tabela 2).
18
19
Fonte: Elaborado pelo autor.
Espécies da guilda dulcícola (D) ocorreram pelo menos em um dos períodos nas
zonas superiores (ZS) dos estuários. Contudo para o estuário do Cocó, espécies pertencentes a
essa guilda tiveram ocorrências em todas as zonas nos períodos seco e chuvoso (Figura 3).
Das seis guildas que compuseram todas as espécies presentes nos estuários analisados, três
foram mais abundantes e ocorreram nos períodos de seca e chuva, são elas: Marinho estuarino
dependente (MD); Estuarina (E) e Marinho estuarino oportunista (MO) (Figura 4).
Quanto as três guildas mais abundantes, a estuarina (E) apresentou tendência de
aumento no número de espécies em direção às zonas superiores (ZS) nos períodos de seca e
chuva nos estuários dos rios Choró, Curu e Pacoti. No Pirangi, o aumento da riqueza
aconteceu somente no período seco, enquanto que para o Cocó houve redução da quantidade
de espécies da guilda estuarina à montante (Figura 3).
Para as espécies classificadas como marinho estuarino oportunista (MO), no
período chuvoso, houve aumento da riqueza em direção a ZS do estuário para Choró e Curu,
enquanto que para os outros estuários houve diminuições em direção as ZS nos dois períodos
(Figura 3).
A guilda marinho estuarino dependente (MD), com relação à variabilidade da
riqueza de espécies em direção as zonas superiores (ZS) apresentou aumento de espécies no
período seco nos estuários do Curu e redução de espécies para os demais estuários. No
período chuvoso, houve acréscimo de espécies em direção a ZS para Choró e Curu, enquanto
que para os outros três estuários diminuição do número de espécies (Figura 3).
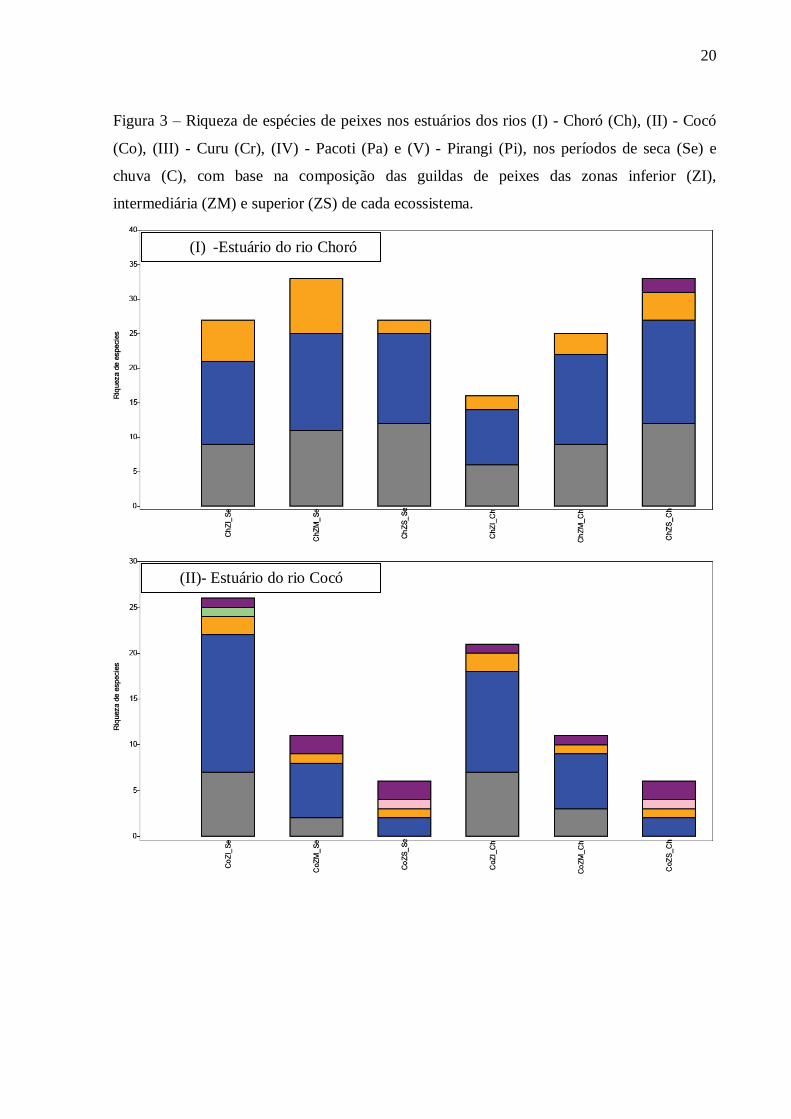
20
Figura 3 – Riqueza de espécies de peixes nos estuários dos rios (I) - Choró (Ch), (II) - Cocó
(Co), (III) - Curu (Cr), (IV) - Pacoti (Pa) e (V) - Pirangi (Pi), nos períodos de seca (Se) e
chuva (C), com base na composição das guildas de peixes das zonas inferior (ZI),
intermediária (ZM) e superior (ZS) de cada ecossistema.
(I) -Estuário do rio Choró
(II)- Estuário do rio Cocó
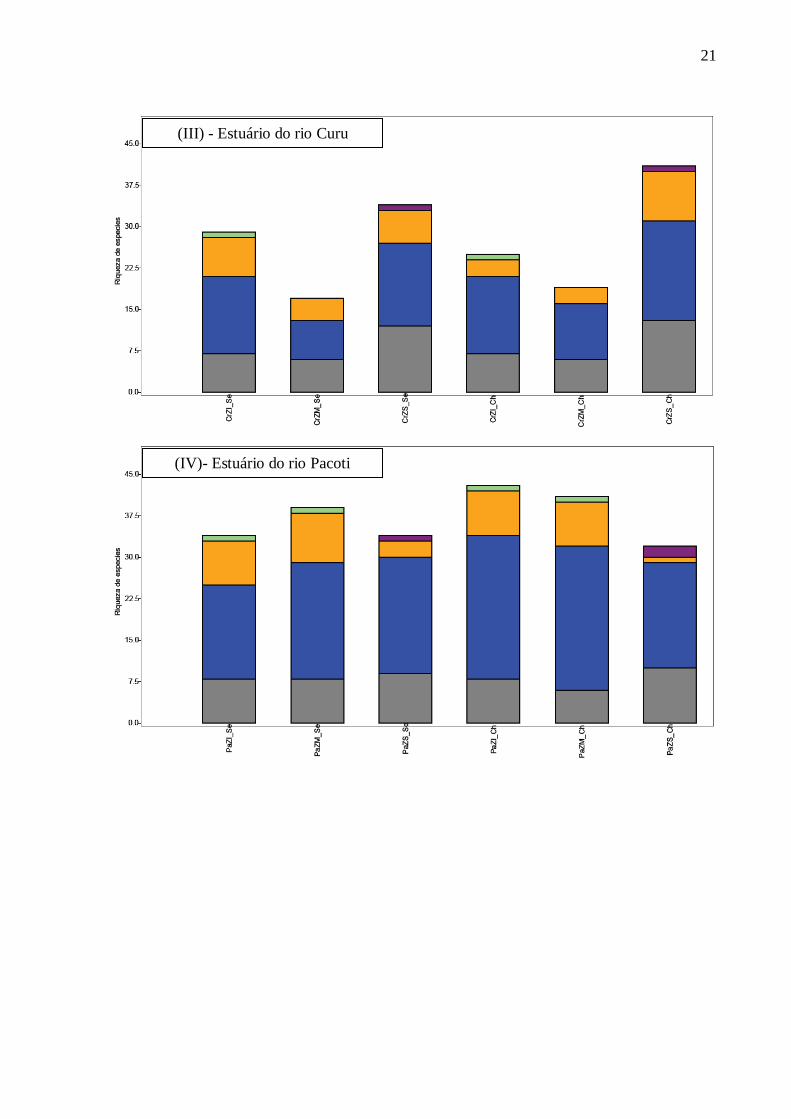
21
(III) - Estuário do rio Curu
(IV)- Estuário do rio Pacoti
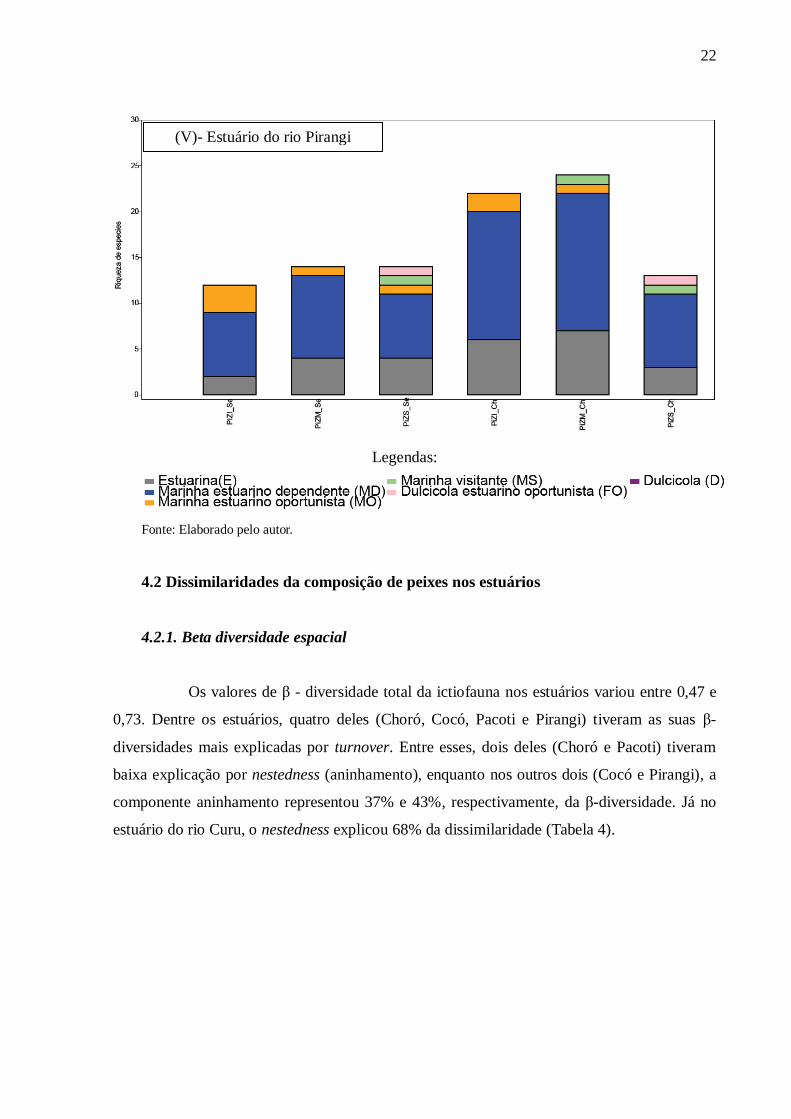
22
Fonte: Elaborado pelo autor.
4.2 Dissimilaridades da composição de peixes nos estuários
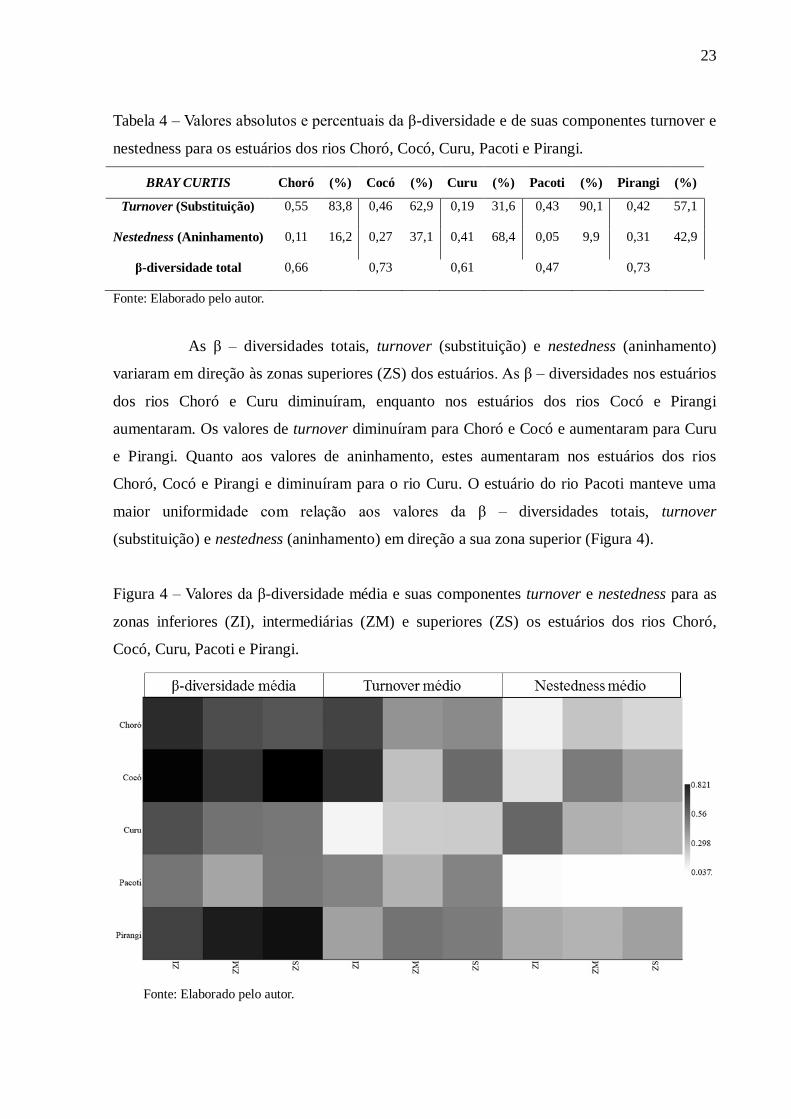
4.2.1. Beta diversidade espacial
Os valores de β - diversidade total da ictiofauna nos estuários variou entre 0,47 e
0,73. Dentre os estuários, quatro deles (Choró, Cocó, Pacoti e Pirangi) tiveram as suas β-
diversidades mais explicadas por turnover. Entre esses, dois deles (Choró e Pacoti) tiveram
baixa explicação por nestedness (aninhamento), enquanto nos outros dois (Cocó e Pirangi), a
componente aninhamento representou 37% e 43%, respectivamente, da β-diversidade. Já no
estuário do rio Curu, o nestedness explicou 68% da dissimilaridade (Tabela 4).
(V)- Estuário do rio Pirangi
Legendas:
23
Tabela 4 – Valores absolutos e percentuais da β-diversidade e de suas componentes turnover e
nestedness para os estuários dos rios Choró, Cocó, Curu, Pacoti e Pirangi.
BRAY CURTIS Choró (%) Cocó (%) Curu (%) Pacoti (%) Pirangi (%)
Turnover (Substituição) 0,55
83,8
0,46
62,9
0,19
31,6
0,43
90,1
0,42
57,1
Nestedness (Aninhamento) 0,11
16,2
0,27
37,1
0,41
68,4
0,05
9,9
0,31
42,9
β-diversidade total 0,66
0,73
0,61
0,47
0,73
Fonte: Elaborado pelo autor.
As β – diversidades totais, turnover (substituição) e nestedness (aninhamento)
variaram em direção às zonas superiores (ZS) dos estuários. As β – diversidades nos estuários
dos rios Choró e Curu diminuíram, enquanto nos estuários dos rios Cocó e Pirangi
aumentaram. Os valores de turnover diminuíram para Choró e Cocó e aumentaram para Curu
e Pirangi. Quanto aos valores de aninhamento, estes aumentaram nos estuários dos rios
Choró, Cocó e Pirangi e diminuíram para o rio Curu. O estuário do rio Pacoti manteve uma
maior uniformidade com relação aos valores da β – diversidades totais, turnover
(substituição) e nestedness (aninhamento) em direção a sua zona superior (Figura 4).
Figura 4 – Valores da β-diversidade média e suas componentes turnover e nestedness para as
zonas inferiores (ZI), intermediárias (ZM) e superiores (ZS) os estuários dos rios Choró,
Cocó, Curu, Pacoti e Pirangi.
Fonte: Elaborado pelo autor.
24
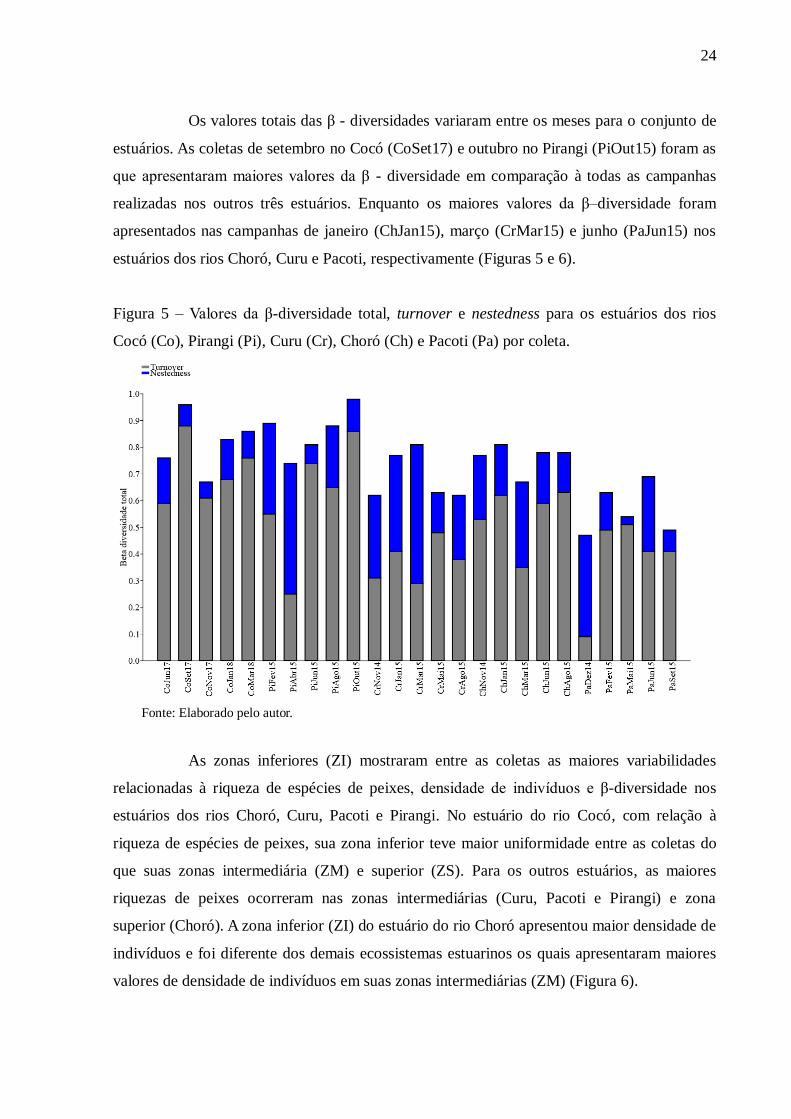
Os valores totais das β - diversidades variaram entre os meses para o conjunto de
estuários. As coletas de setembro no Cocó (CoSet17) e outubro no Pirangi (PiOut15) foram as
que apresentaram maiores valores da β - diversidade em comparação à todas as campanhas
realizadas nos outros três estuários. Enquanto os maiores valores da β–diversidade foram
apresentados nas campanhas de janeiro (ChJan15), março (CrMar15) e junho (PaJun15) nos
estuários dos rios Choró, Curu e Pacoti, respectivamente (Figuras 5 e 6).
Figura 5 – Valores da β-diversidade total, turnover e nestedness para os estuários dos rios
Cocó (Co), Pirangi (Pi), Curu (Cr), Choró (Ch) e Pacoti (Pa) por coleta.
Fonte: Elaborado pelo autor.
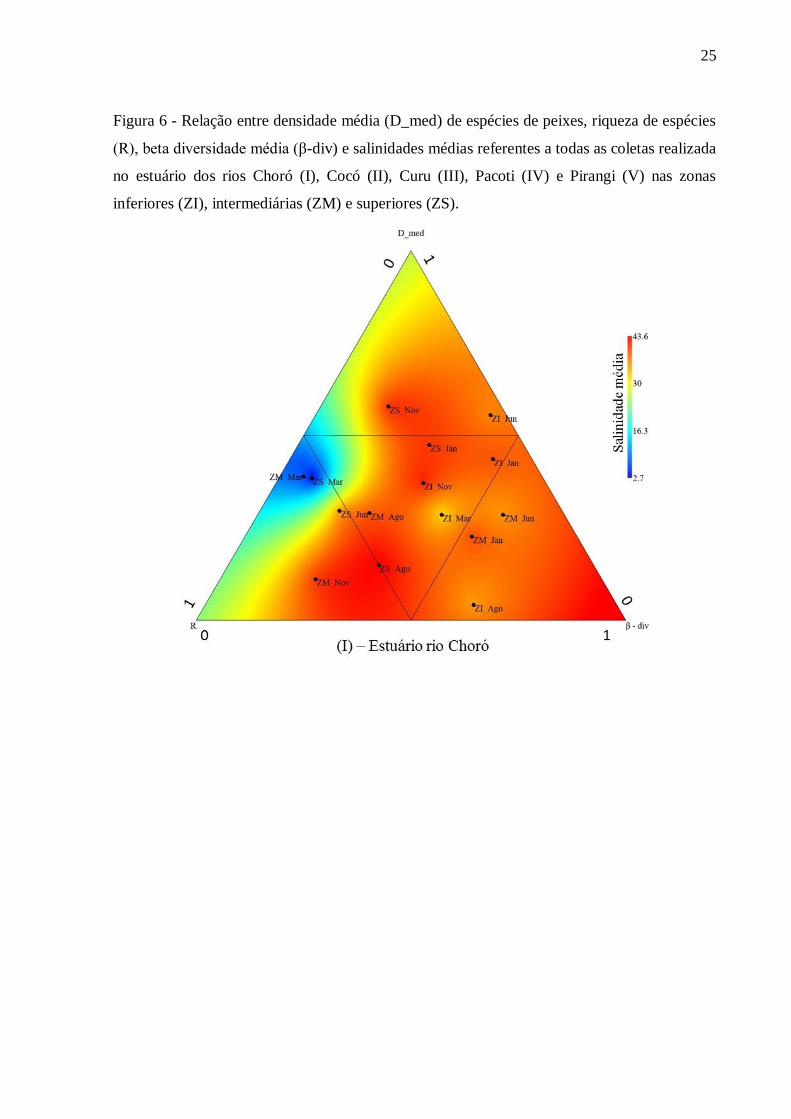
As zonas inferiores (ZI) mostraram entre as coletas as maiores variabilidades
relacionadas à riqueza de espécies de peixes, densidade de indivíduos e β-diversidade nos
estuários dos rios Choró, Curu, Pacoti e Pirangi. No estuário do rio Cocó, com relação à
riqueza de espécies de peixes, sua zona inferior teve maior uniformidade entre as coletas do
que suas zonas intermediária (ZM) e superior (ZS). Para os outros estuários, as maiores
riquezas de peixes ocorreram nas zonas intermediárias (Curu, Pacoti e Pirangi) e zona
superior (Choró). A zona inferior (ZI) do estuário do rio Choró apresentou maior densidade de
indivíduos e foi diferente dos demais ecossistemas estuarinos os quais apresentaram maiores
valores de densidade de indivíduos em suas zonas intermediárias (ZM) (Figura 6).
25
Figura 6 - Relação entre densidade média (D_med) de espécies de peixes, riqueza de espécies
(R), beta diversidade média (β-div) e salinidades médias referentes a todas as coletas realizada
no estuário dos rios Choró (I), Cocó (II), Curu (III), Pacoti (IV) e Pirangi (V) nas zonas
inferiores (ZI), intermediárias (ZM) e superiores (ZS).
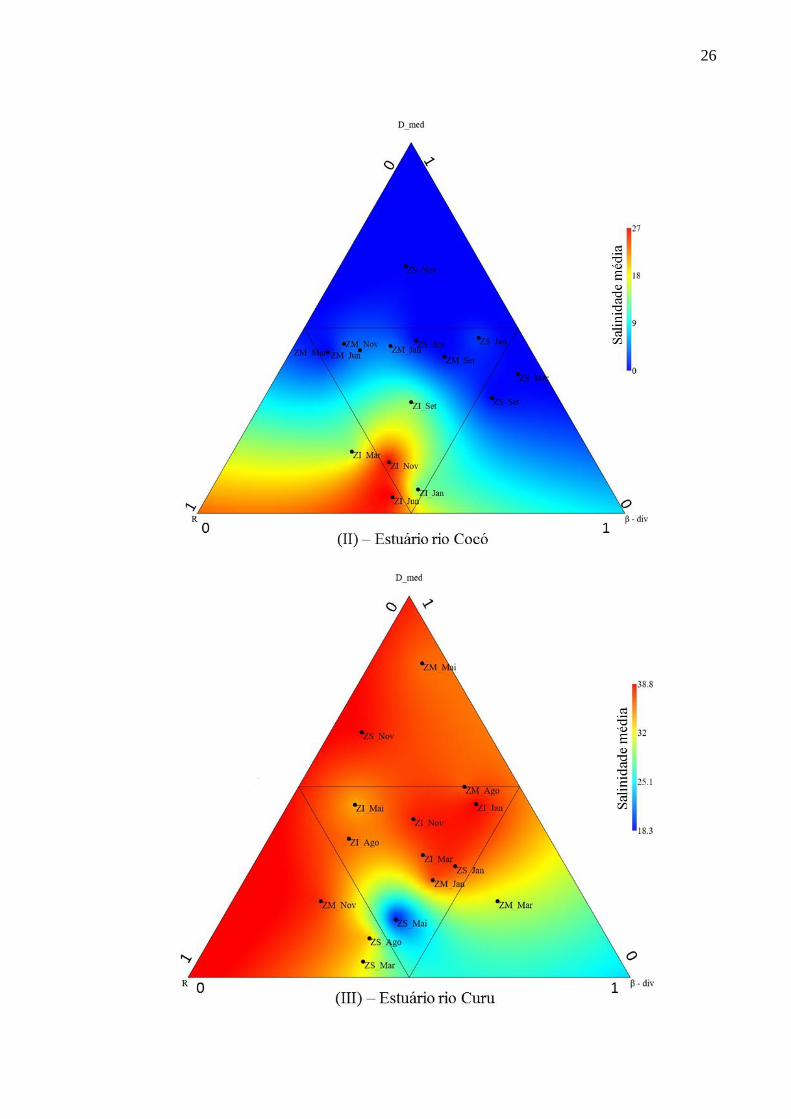
26
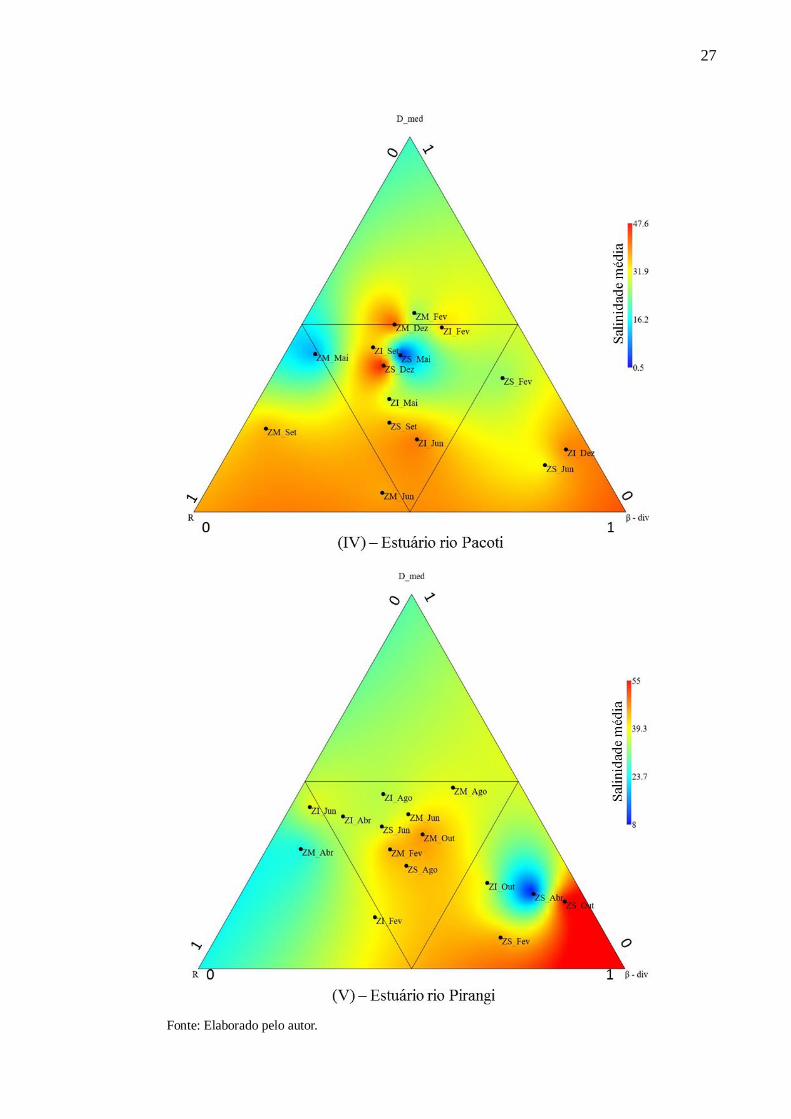
27
Fonte: Elaborado pelo autor.

28
4.2.2. Beta diversidade temporal
Nos períodos de chuva e seca a β-diversidade da ictiofauna é explicada pela
componente turnover (substituição) para os estuários dos rios Choró, Cocó, Pacoti e Pirangi.
No estuário do rio Curu, a β-diversidade continuou sendo em maior parte explicada por
nestedness (aninhamento) para ambos os períodos. No entanto, na estação chuvosa apesar do
turnover explicar em maior parte a β-diversidade, a componente aninhamento (nestedness)
teve maiores valores que no período seco em todos os estuários avaliados (Figura 7).
Os valores da β-diversidade da ictiofauna nos estuários apresentaram
variabilidade em direção às zonas superiores (ZS) em ambos os períodos. No período
chuvoso, diminuíram nos estuários dos rios Cocó, Curu e Pacoti, mantiveram-se constantes e
aumentaram para Choró e Pirangi, respectivamente (Figura 8-I). No período seco, diminuíram
para Choró, Cocó e Pacoti, foram uniformes para o estuário do rio Curu e aumentaram no
estuário do Pirangi, respectivamente (Figura 8-II).
Os valores da componente turnover, no período chuvoso, diminuíram em direção
as zonas superiores para os estuários Choró e Cocó e aumentaram para estuários Curu, Pacoti
e Pirangi (Figura 8-I). Na estação seca, os valores de turnover (substituição) reduziram para
os estuários dos rios Choró, Cocó e Pacoti e aumentaram para os estuários dos rios Curu e
Pirangi (Figura 8-II).
Os valores de nestedness aumentaram em direção as zonas superiores no período
chuvoso para quatro estuários e mantiveram uniformidade no Pacoti (Figura 8-I). Enquanto
que no período seco diminuíram no estuário do rio Curu, aumentaram os valores para Pirangi
e Cocó e mantiveram baixa variabilidade dos valores de nestedness (aninhamento) para Choró
e Pacoti (Figura 8-II).
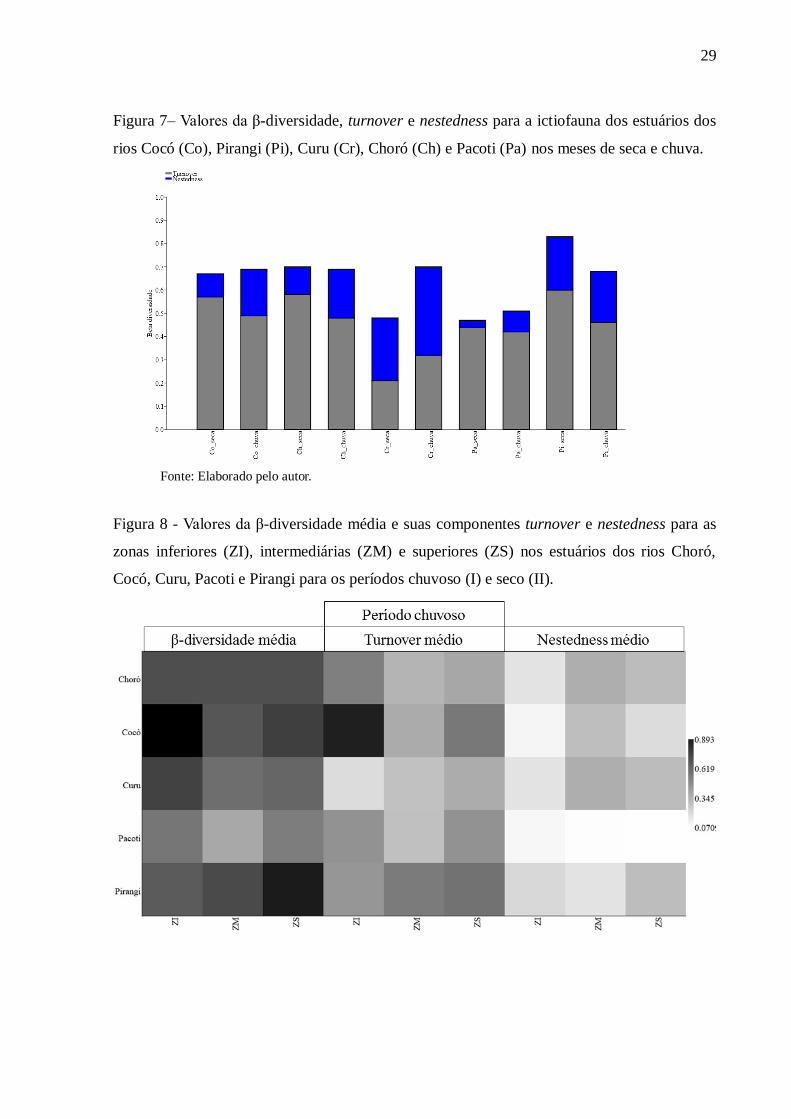
29
Figura 7– Valores da β-diversidade, turnover e nestedness para a ictiofauna dos estuários dos
rios Cocó (Co), Pirangi (Pi), Curu (Cr), Choró (Ch) e Pacoti (Pa) nos meses de seca e chuva.
Fonte: Elaborado pelo autor.
Figura 8 - Valores da β-diversidade média e suas componentes turnover e nestedness para as
zonas inferiores (ZI), intermediárias (ZM) e superiores (ZS) nos estuários dos rios Choró,
Cocó, Curu, Pacoti e Pirangi para os períodos chuvoso (I) e seco (II).
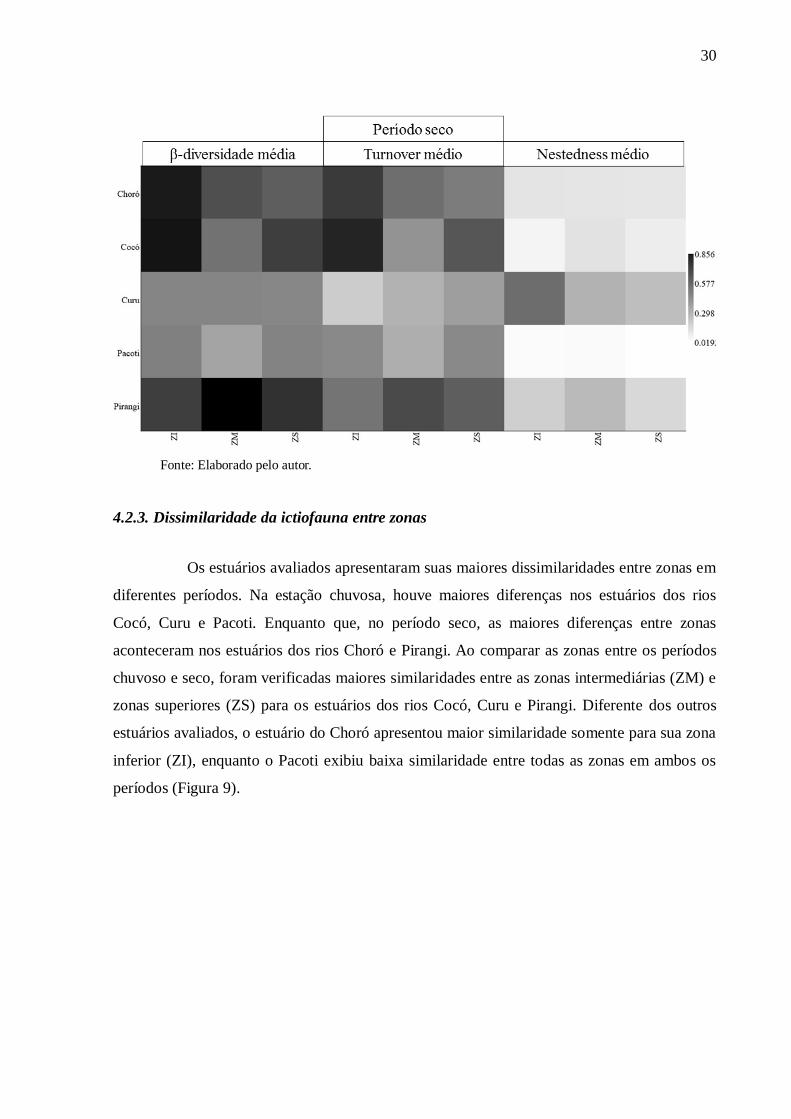
30
Fonte: Elaborado pelo autor.
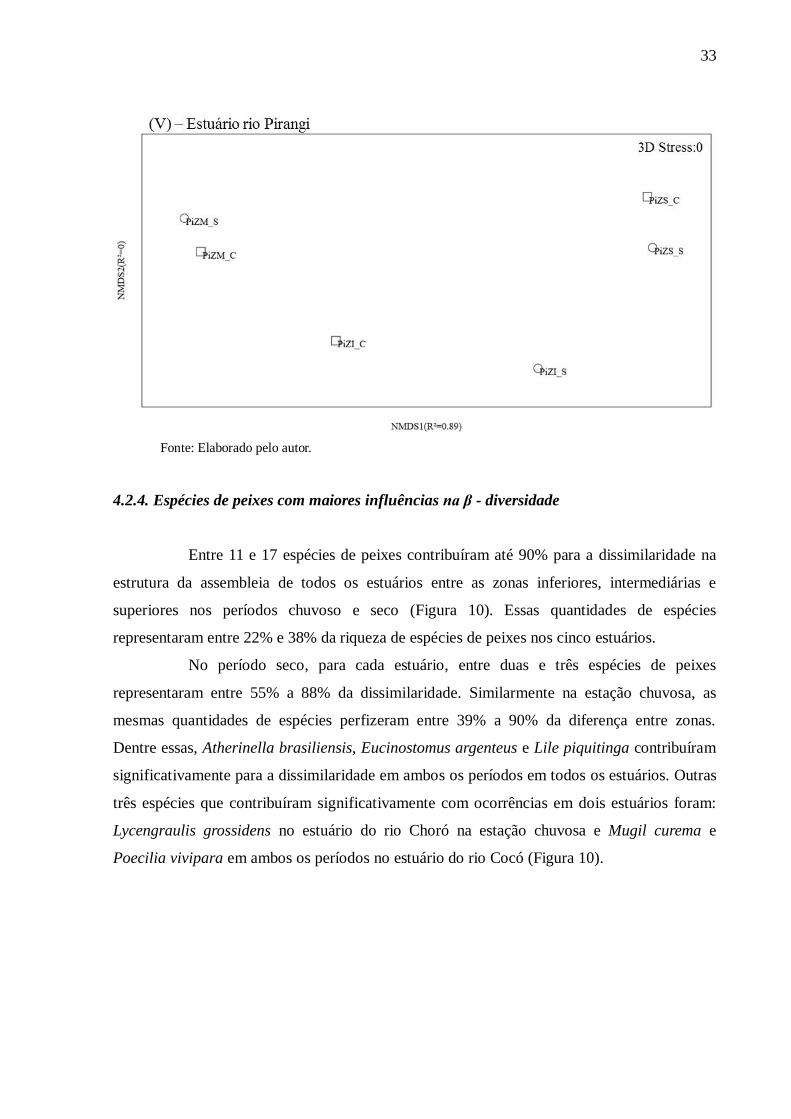
4.2.3. Dissimilaridade da ictiofauna entre zonas
Os estuários avaliados apresentaram suas maiores dissimilaridades entre zonas em
diferentes períodos. Na estação chuvosa, houve maiores diferenças nos estuários dos rios
Cocó, Curu e Pacoti. Enquanto que, no período seco, as maiores diferenças entre zonas
aconteceram nos estuários dos rios Choró e Pirangi. Ao comparar as zonas entre os períodos
chuvoso e seco, foram verificadas maiores similaridades entre as zonas intermediárias (ZM) e
zonas superiores (ZS) para os estuários dos rios Cocó, Curu e Pirangi. Diferente dos outros
estuários avaliados, o estuário do Choró apresentou maior similaridade somente para sua zona
inferior (ZI), enquanto o Pacoti exibiu baixa similaridade entre todas as zonas em ambos os
períodos (Figura 9).

31
Figura 9 – Escalonamento multidimensional não métrico (“NMDS”) mostrando as
dissimilaridades entre as zonas para os períodos de seca (S) e chuva (C) nos estuários dos rios
Choró (I), Cocó (II), Curu (III), Pacoti (IV) e Pirangi (V).

32
33
Fonte: Elaborado pelo autor.
4.2.4. Espécies de peixes com maiores influências na β - diversidade
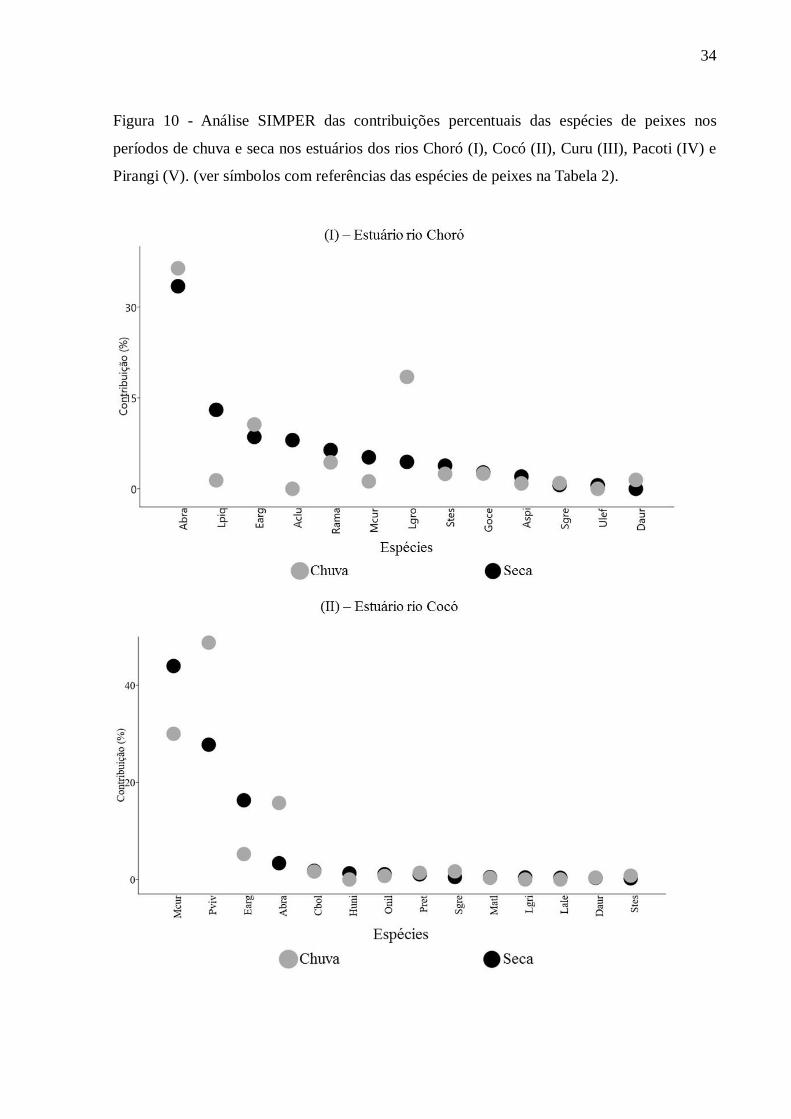
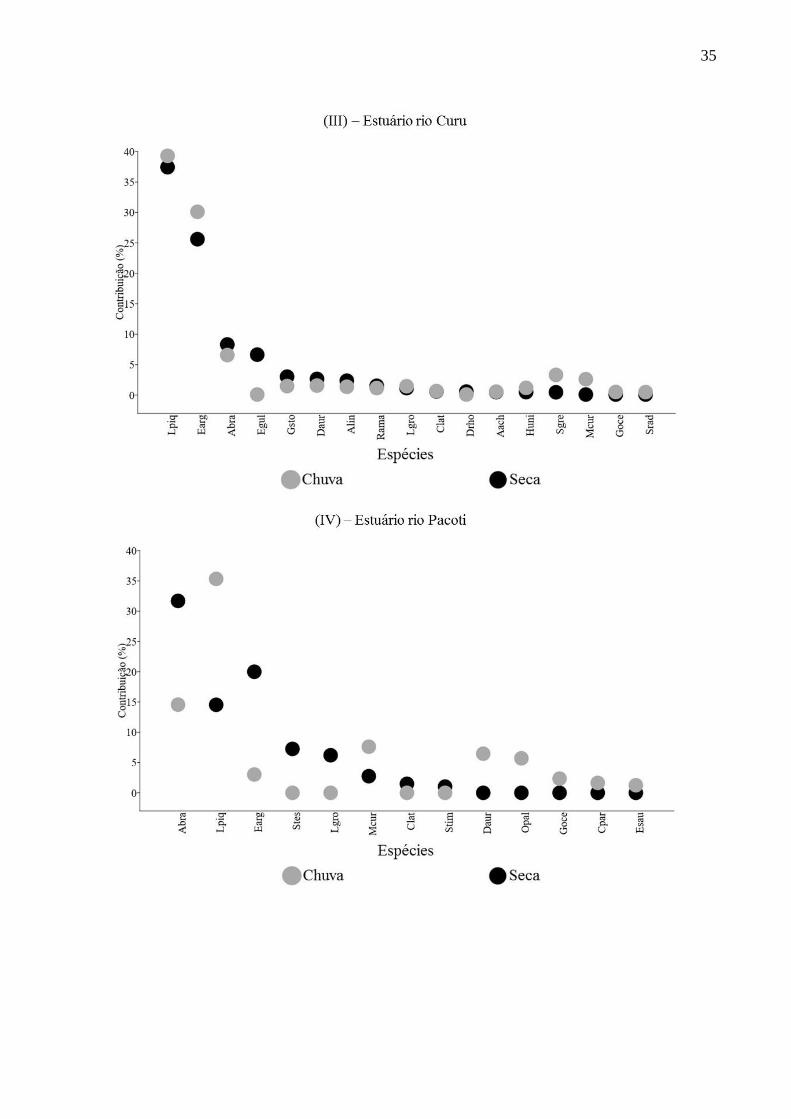
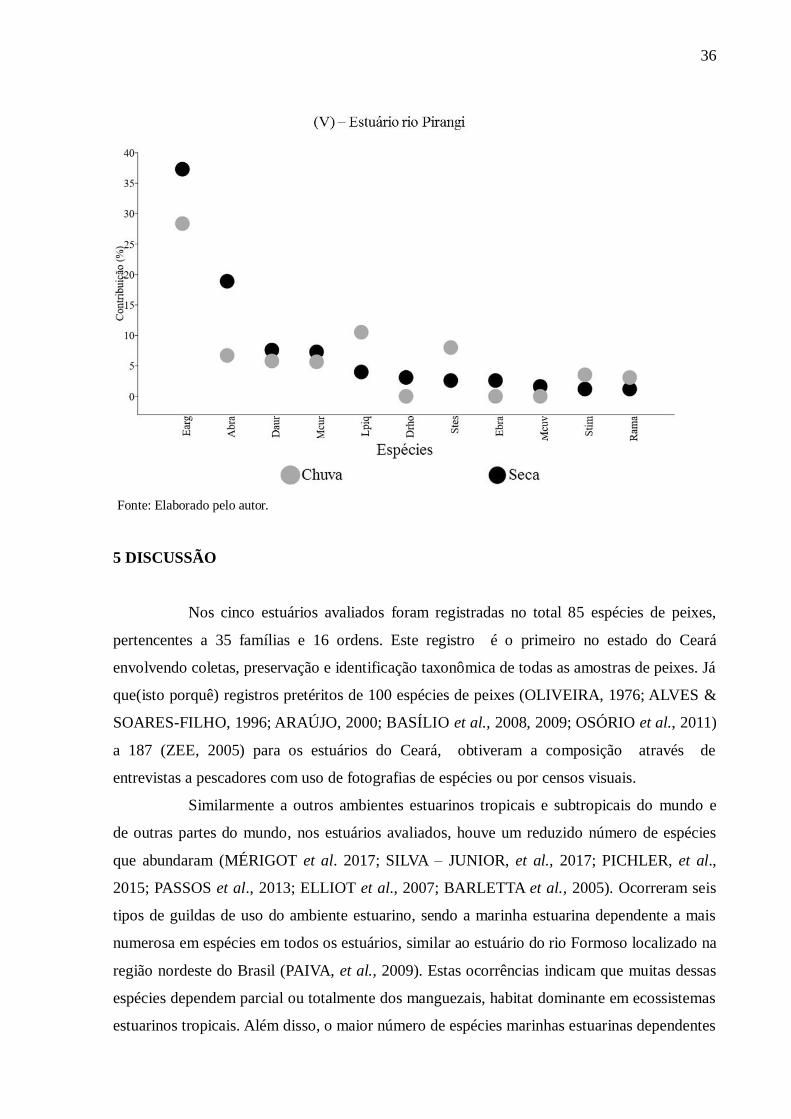
Entre 11 e 17 espécies de peixes contribuíram até 90% para a dissimilaridade na
estrutura da assembleia de todos os estuários entre as zonas inferiores, intermediárias e
superiores nos períodos chuvoso e seco (Figura 10). Essas quantidades de espécies
representaram entre 22% e 38% da riqueza de espécies de peixes nos cinco estuários.
No período seco, para cada estuário, entre duas e três espécies de peixes
representaram entre 55% a 88% da dissimilaridade. Similarmente na estação chuvosa, as
mesmas quantidades de espécies perfizeram entre 39% a 90% da diferença entre zonas.
Dentre essas, Atherinella brasiliensis, Eucinostomus argenteus e Lile piquitinga contribuíram
significativamente para a dissimilaridade em ambos os períodos em todos os estuários. Outras
três espécies que contribuíram significativamente com ocorrências em dois estuários foram:
Lycengraulis grossidens no estuário do rio Choró na estação chuvosa e Mugil curema e
Poecilia vivipara em ambos os períodos no estuário do rio Cocó (Figura 10).
34
Figura 10 - Análise SIMPER das contribuições percentuais das espécies de peixes nos
períodos de chuva e seca nos estuários dos rios Choró (I), Cocó (II), Curu (III), Pacoti (IV) e
Pirangi (V). (ver símbolos com referências das espécies de peixes na Tabela 2).
35
36
Fonte: Elaborado pelo autor.
5 DISCUSSÃO
Nos cinco estuários avaliados foram registradas no total 85 espécies de peixes,
pertencentes a 35 famílias e 16 ordens. Este registro é o primeiro no estado do Ceará
envolvendo coletas, preservação e identificação taxonômica de todas as amostras de peixes. Já
que(isto porquê) registros pretéritos de 100 espécies de peixes (OLIVEIRA, 1976; ALVES &
SOARES-FILHO, 1996; ARAÚJO, 2000; BASÍLIO et al., 2008, 2009; OSÓRIO et al., 2011)
a 187 (ZEE, 2005) para os estuários do Ceará, obtiveram a composição através de
entrevistas a pescadores com uso de fotografias de espécies ou por censos visuais.
Similarmente a outros ambientes estuarinos tropicais e subtropicais do mundo e
de outras partes do mundo, nos estuários avaliados, houve um reduzido número de espécies
que abundaram (MÉRIGOT et al. 2017; SILVA – JUNIOR, et al., 2017; PICHLER, et al.,
2015; PASSOS et al., 2013; ELLIOT et al., 2007; BARLETTA et al., 2005). Ocorreram seis
tipos de guildas de uso do ambiente estuarino, sendo a marinha estuarina dependente a mais
numerosa em espécies em todos os estuários, similar ao estuário do rio Formoso localizado na
região nordeste do Brasil (PAIVA, et al., 2009). Estas ocorrências indicam que muitas dessas
espécies dependem parcial ou totalmente dos manguezais, habitat dominante em ecossistemas
estuarinos tropicais. Além disso, o maior número de espécies marinhas estuarinas dependentes

37
destaca a alta conectividade do estuário com o ambiente marinho, confirmando-o como
ecossistema de berçário, alimentação e refúgio, sendo necessário evitar a perda de manguezais
e a pressão pesqueira como medida para uma melhor gestão da pesca marinha (MÉRIGOT et
al. 2017; LESLIE, et al., 2017; SELLESLAGH & AMARA, 2008).
Nos cinco estuários avaliados neste estudo, seis famílias (Gerreidae, Engraulidae,
Gobiidae, Lutjanidae, Carangidade e Mugilidae) compõem 32 espécies de peixes. Espécies
destas famílias ocorrem com alta frequência e em alguns casos em altas densidades em
ecossistemas estuarinos tropicais e subtropicais (BLABER, 2000).
A espécie Eucinostomus argenteus da família Gerreidae apresentou maiores
densidades em todos os estuários avaliados. Essa família caracteriza-se pela alta plasticidade
alimentar, sendo os bivalves os itens principais de suas dietas, e por uma ampla tolerância as
variações de salinidade, permitindo-as ocupar ambientes eurihalinos, estuarinos e marinhos
(ARAÚJO, et al., 2016; BLABER, 2000).
A espécie Lycengraulis grossidens da família Engraulidae também esteve entre as
maiores densidades nos estuários analisados. Indivíduos dessa família quando adultos formam
agregações, assumindo importância comercial na atividade pesqueira e são um importante elo
trófico nos ambientes costeiros, devido a sua função na conversão da biomassa planctônica
em forrageira para peixes piscívoros (ARAÚJO et al., 2008).
As espécies da família Gobiidae tiveram altas frequências com baixa densidade.
Indivíduos dessa família têm preferencia por fundos moles (lamosos), são onívoros, desovam
no ambiente estuarino e representam grande parte da assembleia de larvas de peixes em
muitos estuários tropicais (MÉRIGOT et al. 2017; BLABER, 2000). Essas características
destacam a importância dessa família para a manutenção da estrutura trófica dos estuários.
Na família Mugilidae, Mugil curema apresenta alta densidade nos estuários, com
destaque para o rio Cocó. Espécies dessa família se alimentam de organismos através da
ingestão do substrato (iliófagos), suas presas são abundantes em águas rasas com baixa
hidrodinâmica ou sobre substratos com alto conteúdo orgânico (BLABER, 2000). Tais
condições são encontradas principalmente em estuários onde têm alta entrada de detritos
orgânicos tornando essas espécies dependentes desses ecossistemas. Portanto ações de
manutenção da área de manguezal, principal fonte de detritos orgânicos para o estuário, são
importantes para sustentar populações de espécies da família Mugilidae, as quais representam
o principal recurso pesqueiro estuarino do nordeste do Brasil.

38
As famílias Carangidae e Lutjanidade apresentaram alta frequência nas coletas,
indivíduos jovens das espécies ocorrem nos estuários atrelados às raízes dos mangues
(OSÓRIO et al., 2011; BLABER, 2000) e compõem, quando adultas, o estoque pesqueiro
costeiro.
Em média, 30% das espécies de peixes que ocorreram nos estuários analisados
explicaram 90% da β-diversidade. Essa baixa contribuição é explicada pela alta densidade de
indivíduos de poucas espécies de peixes que ocorrem em todas as assembleias estuarinas.
Espécies como Eucinostomus argenteus, Atherinella brasiliensis, Lile piquitinga,
Lycengraulis grossidens, Mugil curema e Poecilia vivipara apresentaram as maiores
densidades de indivíduos em todos os ecossistemas avaliados nos períodos de chuva e seca.
Atherinella brasiliensis, pertencente à guilda estuarina, teve alta densidade junto com
Eucinostomus argenteus nas zonas inferiores dos estuários. Essa espécie geralmente habita em
estuários formando cardumes, possui dieta generalista que varia de copépodos a detritos,
apresenta alta plasticidade trófica, rápido crescimento e curto ciclo de vida (PASSOS et al.,
2013; CONTENTE et al., 2011). A espécie Lile piquitinga possui baixo valor comercial é
encontrada frequentemente em estuários tropicais, com ciclo de vida de curta duração e
produz poucos ovos, porém com desova múltipla durante um período prolongado (BLABER,
2000). Entre as espécies que dominaram as assembleias estuarinas, a única da guilda dulcícola
ocorreu estuário do rio Cocó, Poecilia vivipara que além de ocorrer em água doce, podem
ocorrer em águas salobras e águas costeiras marinhas, sendo utilizada no controle de
mosquitos e com potencial para colonizar habitats com baixa qualidade de água (NELSON,
2016; HRBEK et al., 2007). Assim, os estuários avaliados apresentaram baixa diversidade da
ictiofauna, altas abundâncias de poucas espécies que dominam nestes ambientes, com amplos
limites de tolerância às condições físicas - químicas nos ecossistemas estuarinos, variadas
estratégias reprodutivas e plasticidade trófica.
Os estuários dos rios Choró, Cocó, Curu, Pacoti e Pirangi tiveram suas β-
diversidades explicadas entre as componentes turnover (substituição) e nestedness
(aninhamento), as quais dependeram tanto da área de dispersão de espécies de peixes em
direção as zonas superiores (ZS) quanto da heterogeneidade ambiental controlada pelos
valores de salinidade ao longo do ano. Assim, valores de salinidade atenuaram a
heterogeneidade desses ecossistemas possibilitando maiores ou menores áreas para a
dispersão das espécies de peixes das guildas marinho estuarino dependente (MD), marinho
estuarino oportunista (MO) e estuarino (E) em função do período chuvoso ou seco.

39
Nos estuários dos rios Choró e Pacoti, a salinidade similar ao do ambiente
marinho em todas as zonas aumentando a área de dispersão aos peixes das guildas marinho
estuarino dependente, marinho estuarino visitante e estuarina. Quanto aos estuários dos rios
Cocó e Pirangi, a respectiva influência dulcícola para o primeiro e condição hipersalina em
direção as zonas superiores para o segundo, reduzem as áreas de dispersão para as guildas de
peixes com maior número de espécies. O estuário do Curu apresenta características marinhas
relacionadas à salinidade ampliando a área de dispersão aos peixes das guildas marinho
estuarino dependente, marinho estuarino oportunista e estuarina; no entanto, tem a menor
distância entre sua zona inferior e superior quando comparado aos outros estuários avaliados.
No esquema conceitual das relações esperadas entre heterogeneidade ambiental,
áreas de dispersão e dissimilaridade (turnover e nestedness) espera-se que a componente
nestedness (aninhamento) diminua com a ampliação da área de dispersão de espécies,
enquanto a componente turnover (substituição) aumente (GIANUCA et al., 2017). Assim, os
resultados do presente estudo apresentaram similaridade ao esquema conceitual anterior,
sendo as β-diversidades mais explicadas por turnover nos estuários dos rios Choró e Pacoti.
Nos estuários do Cocó e Pirangi, os valores de turnover foram dominantes a explicar a β-
diversidade, porém houve explicações por aninhamento elevadas, enquanto que para o
estuário do rio Curu uma menor distância entre a zona inferior e superior contribuiu para uma
menor área de dispersão de espécies de peixes e consequentemente para um maior valor de
aninhamento explicando a β-diversidade. Assim, houve um padrão relacionado às áreas de
dispersão as quais foram dependentes da salinidade, e mostram que áreas menores apresentam
maiores valores de aninhamento e áreas maiores menores valores desta componente.
A estruturação de assembleias estuarinas de peixes por turnover relacionando à
riqueza de espécies e a densidades de indivíduos entre locais também foi relatada em estuários
para invertebrados bentônicos e peixes (TEICHERT et al., 2018; MEDEIROS et al., 2016;
GUTIÉRREZ – CÁNOVAS et al., 2013; VILLEGER et al., 2012). Estes resultados indicaram
que, independente do táxon animal, ambientes estuarinos mostram substituição de espécies ao
longo do gradiente salino, com espécies apresentando alto grau de adaptação às variações da
salinidade (BARROS et al., 2014; 2012). No entanto, os resultados deste estudo destacam que
em estuários com forte influência dulcícola (Cocó) ou com características hipersalinas
constantes (Pirangi) apresentam uma diminuição do número de espécies e uma baixa
densidade de indivíduos em direção as suas zonas superiores.
Na estação chuvosa, houve maiores valores da componente aninhamento da β-
diversidade para todos os estuários avaliados. Isto indica que os incrementos de água doce

40
nestes ecossistemas contribuem para as flutuações de variáveis ambientais estuarinas, como
salinidade, matéria orgânica, turbidez, concentração de nutrientes e carga de sedimentos,
fundamentais para a sobrevivência da flora e fauna (STEICHEN & QUIGG, 2018). De fato,
como no modelo conceitual de Gianuca (GIANUCA et al., 2017), a maior influência dulcícola
reduziu a área de dispersão da assembleia de peixes estuarinas, diminuindo os valores de
turnover (substituições).
Padrões de aninhamento nos estuários avaliados indicam que há estressores de
origem natural ou antrópica ao longo do gradiente estuarino (BARROS et al., 2014;
GUTIÉRREZ – CÁNOVAS et al., 2013). Estudos anteriores indicaram que estes ecossistemas
sofrem, em proporções diversas, modificações ambientais (GODOY et al., 2018;
QUINTELA-FALCÃO et al., 2011; CAVALCANTE et al.,2009; MOLISANI et al., 2006),
que tendem a torná-los homogêneos, promovendo a perda de espécies de peixes com menores
substituições de espécies, domínio de generalistas, espécies com maior amplitude de nicho
(WHITFIELD et al., 2012; ELLIOT & WHITFIELD, 2011 ), e consequentemente menores β-
diversidades (SOCOLAR et al., 2016).
A riqueza, a densidade e a beta diversidade da ictiofauna dos estuários avaliados
na região semiárida do nordeste do Brasil são influenciadas pelas variações da salinidade. A
beta diversidade é modelada principalmente pelas densidades das espécies Atherinella
brasiliensis, Eucinostomus argenteus e Lile piquitinga da guildas estuarino e marinho
estuarino dependente. Já as assembleias de peixes são estruturadas por turnover (substituição
de espécies), havendo domínio da ordenação nestedness nos meses com maiores
precipitações. Isto indica que vários locais desses estuários devem ser adequadamente
conservados, por abrigar uma ampla diversidade e/ou passam por sofrer amplo processo de
degradação ambiental, acarretando em perda de espécies de peixes.

41
6 CONCLUSÕES
As β-diversidades da ictiofauna nos estuários avaliados apresentam padrões
distintos, tanto a nível espacial como temporal. Espacialmente, estuários do semiárido que
experimentam maior permanência sob condições de influência dulcícola ou de hipersalinidade
têm maiores β-diversidades em suas zonas superiores. Temporalmente, o período chuvoso nos
estuários do semiárido diminui as áreas de dispersão aos peixes nos estuários avaliados,
possibilitando aumento da explicação da β-diversidades por nestedness (aninhamento).

42
REFERÊNCIAS
ALVES, M. I. M.; SOARES-FILHO, A. A. Peixes do estuário do rio Jaguaribe (Ceará Brasil):
aspectos fisioecológicos. Ciência Agronômica, v. 27, n. 1/2, p. 5 -16, 1996.
ANDERSON, M.; CRIST, T; CHASE, J.; VELLEND, M.; INOUYE, B.; FREESTONE, A.;
SANDERS, N.; CORNELL, H.; COMITA, L.; DAVIES, K.; HARRISON, S.; KRAFT, N.;
STEGEN, J.; SWENSON, N. Navigating the multiple meanings of b diversity: a roadmap for
the practicing ecologist. Ecology Letters, v. 14, p. 19–28, 2011.
ARAÚJO, A. L. F.; DANTAS, R. P.; PESSANHA, A. L. M. Feeding ecology of three juvenile
mojarras (Gerreidae) in a tropical estuary of northeastern Brazil. Neotropical Ichthyology, v.
14, n. 1, ´p. 1 – 10, 2016.
ARAÚJO, M. E.; TEIXEIRA, J. M. C.; OLIVEIRA, A. M. E. Peixes estuarinos marinhos
do Nordeste brasileiro. Fortaleza: Edições UFC, 2004.
ARAÚJO, M. E.; TEIXEIRA, J. M.; OLIVEIRA, A. M. E. Ictiofauna marinha do Estado do
Ceará, Brasil: III. Actinopterygii de estuários. Arquivos de Ciências do Mar, v. 33, p. 139-
142, 2000.
ASTORGA, A.; DEATH, R.; DEATH, F.; PAAVOLA, R.; CHAKRABORTY, M.; MUOTKA,
T. Habitat heterogeneity drives the geographical distribution of beta diversity: the case of
New Zealand stream invertebrates. Ecology and Evolution, v. 4, n. 13, p. 2693– 2702, 2014.
BALATA, D.; PIAZZI, L.; BENEDETTI - CECCHI, L. Sediment disturbance and loss of beta
diversity on sub tidal rocky reefs. Ecology, v. 88, n. 10, p. 2455–2461, 2007.
BARLETTA, M.; BARLETTA-BERGAN, A.; SAINT-PAUL, U.; HUBOLD, G. The role of
salinity in structuring the fish assemblages in a tropical estuary. Journal of Fish Biology, v.
66, p. 45-72, 2005.
BARROS, F.; BLANCHET, H.; HAMMERSTROM, K.; SAURIAU, P-G.; OLIVER, J. A
framework for investigating general patterns of benthic β-diversity along estuaries.
Estuarine, Coastal and Shelf Science, v. 149, p. 223 – 231, 2014
BARROS, F.; CARVALHO, G. C.; COSTA, Y.; HATJE, V. Subtidal benthic macroinfaunal
assemblages in tropical estuaries: Generality amongst highly variable gradients. Marine
Environmental Research, v. 81, p. 43 – 52, 2012.
BARROS, F; BLANCHET, H; HAMMERSTON, K; SAURIAU, P; OLIVER, J. A framework
for investigating general patterns of benthic b-diversity along estuaries. Estuarine, Coastal
and Shelf Science, v. 149, p. 223 – 231, 2014.
BARWELL, L.; ISAAC, N.; KUNIN, W. Measuring b-diversity with species abundance data.
Journal of Animal Ecology, v. 84, p. 1112–1122, 2015.

43
BASELGA, A. Partitioning abundance-based multiple-site dissimilarity into components:
balanced variation in abundance and abundance gradients. Methods in Ecology and
Evolution, v. 8, p. 799 – 808, 2017.
BASELGA, A. Partitioning the turnover and nestedness components of beta diversity. Global
Ecology and Biogeography, v. 19, p. 134 – 143, 2010.
BASELGA, A. Separating the two components of abundance-based dissimilarity: balanced
changes in abundance vs. abundance gradients. Methods in Ecology and Evolution, v. 4, p.
552 – 557,2013.
BASÍLIO, T. H.; FARIA V. V.; FURTADO-NETO, M. A. A. Fauna de Elasmobrânquios do
estuário do rio Curu, Ceará,Brasil. Arquivos de Ciências do Mar, v. 41, n. 2, p. 65 – 72,
2008.
BASÍLIO, T. H.; GODINHO, W. O.; ARAÚJO, M. E.; FURTADO-NETO, M. A.; FARIA, V.
V. Ictiofauna do estuário do rio Curu, Ceará, Brasil. Arquivos de Ciências do Mar, v. 42, n.
2, p. 81-88, 2009.
BLABER, S. J. M. Tropical Estuarine Fishes: Ecology, Exploitation and Conservation.
London, Oxford: Blackwell Science, 2000.
CARPENTER, K. E. (Ed.) The living marine resources of the Western Central Atlantic.
Volume 2: Bony fishes part 1 (Acipenseridae to Grammatidae). FAO Species
Identification Guide for Fishery Purposes and American Society of Ichthyologists and
Herpetologists Special Publication No. 5.Rome: FAO. 2002a.
CARPENTER, K. E. (Ed.) The living marine resources of the Western Central Atlantic.
Volume 2: Bony fishes part 2 (Acipenseridae to Grammatidae). FAO Species
Identification Guide for Fishery Purposes and American Society of Ichthyologists and
Herpetologists Special Publication No. 5.Rome: FAO. 2002b.
CARVALHO, J.; CARDOSO, P.; GOMES, P. Determining the relative roles of species
replacement and species richness differences in generating beta diversity patterns. Global
Ecology and Biogeography, v. 21, p. 760–771, 2012.
CASSEY, P.; LOCKWOOD, J.; OLDEN, J.; BLACKBURN, T. The varying role of
populationabundance in structuring indices of biotic homogenization. Journal of
Biogeography, v. 35, p. 884–892, 2008.
CAVALCANTE, R.; SOUSA, F.; NASCIMENTO, R.; SILVEIRA, E.; FREIRE, S. S. G. The
impact of urbanization on tropical mangroves (Fortaleza, Brazil): Evidence from PAH
distribution in sediments. Journal of Environmental Management, v. 91, p. 328–335, 2009.
CESCHIN, F.; BINI, L.; PADIAL, A. Correlates of fish and aquatic macrophyte beta diversity
in the Upper Parana River floodplain. Hydrobiologia, v. 805, p. 377 – 389, 2017.
CHASE, J. M. Stochastic Community AssemblyCauses Higher Biodiversity in More
Productive Environments. Science, v. 328, p. 1388 – 1391, Jun. 2010.

44
CONTENTE, R. F.; STEFANONI, M. F.; SPACH, H. L. Fish assemblage structure in an
estuary of the Atlantic Forest biodiversity hotspot (southern Brazil). Ichthyological
Research, v. 58, p. 38 – 50, 2011.
ELLIOT, M.; WHITFIELD, A. K. Challenging paradigms in estuarine ecology and
management. Estuarine, Coastal and Shelf Science, v. 94, p. 306 – 314, 2011.
ELLIOTT, M.; WHITFIELD, A. K.; POTTER, I. C.; BLABER, S. J. M.; CYRUS, D. P.;
NORDLIE, F. G.; HARRISON, T. D. The guild approach to categorizing estuarine fish
assemblages: a global review. Fish and Fisheries, v. 8, p. 241-268, 2007.
FISCHER, L. G.; PEREIRA, L. E. D.; VIEIRA, J. P. Peixes estuarinos e costeiros. 2. ed. Rio
Grande: Luciano Gomes Fischer, 2011.
GIANUCA, A.; DECLERCK, S.; LEMMENS, P.; MEESTER, L. Effects of dispersal and
environmental heterogeneity on the replacement and nestedness components of β-diversity.
Ecology, v.98, n. 2, p. 525–533, 2017.
GODOY, D. P. M.; MEIRELES, A. J.; LACERDA, L. D. Mangrove Response to Land Use
Change in Estuaries along the Semiarid Coast of Ceará, Brazil. Journal of Coastal Research,
v. 34, n. 3, p.524-533, 2018.
GUTIÉRREZ – CÁNOVAS, C.; MILLÁN, A.; VELASCO, J.; VAUGHAN, P. I.;
ORMEROD, J. S. Contrasting effects of natural and anthropogenic stressors on beta diversity
in river organisms. Global Ecology and Biogeography, v. 22, p. 796–805, 2013.
HRBECK, T.; SECKINGER, J.; MEYER, A. A phylogenetic and biogeographic perspective
on the evolution of poeciliid fishes. Molecular Phylogenetics and Evolution, v. 43, p. 986 –
998, 2007.
LACERDA, D. L.; MOLISANI, M.; SENA, D.; MAIA, P. L. Estimating the importance of
natural and anthropogenic sources on N and P emission to estuaries along the Ceará State
Coast NE Brazil. Environmental Monitoring and Assessment, v. 141, p. 149 – 164, 2008.
LACERDA, L.; MAIA, L.; SOUZA, G.; BEZERRA, L.; MENEZES, M. Manguezais do
nordeste e mudanças ambientais. Ciência Hoje, v. 39, n. 229, p. 24 – 29, 2006.
LEGENDRE, P. Interpreting the replacement and richness difference components of beta
diversity. Global Ecology and Biogeography, v. 23, p. 1324–1334. 2005.
LEGENDRE, P. Interpreting the replacement and richness difference components of beta
diversity. Global Ecology and Biogeography, v. 23, p. 1324 – 1334, 2014.
LESLIE, T.; JAMES, N. C.; POTTS, W. M.; RAJKARAN, A. The relationship between
habitat complexity and nursery provision for an estuarine dependent fish species in a
permanently open South African estuary. Estuarine, Coastal and Shelf Science, v. 17, p. 1 –
32, 2017.
MARCENIUK, A. P. Chave para identificação das espécies de bagres marinhos (Siluriformes,
Ariidae) da costa brasileira. Boletim do Instituto de Pesca, v. 31, n. 2, p. 89-101, 2005.

45
MARCENIUK, A. P.; BETANCUR-R.; ACERO, A. P.; MURIEL-CUNHA, J. Review of the
Genus Cathorops (Siluriformes: Ariidae) from the Caribbean and Atlantic South America,
with Description of a New Species. Copeia, n. 1, 77-97, 2012.
MEDEIROS, R. C.; HEPP, U. L.; PATRÍCIO, J.; MOLOZZI, J. Tropical estuarine
macrobenthic communities are structured by turnover rather than nestedness. PLoS ONE, v.
11, n. 9, 2016.
MEIRELES, J. A. A.; CASSOLA, R.; TUPINAMBÁ, S.; QUEIROZ, L. Impactos ambientais
decorrentes das atividades da carcinicultura ao longo do litoral cearense, nordeste do Brasil.
Mercator - Revista de Geografia da UFC, v. 6, n. 12, 2007.
MENEZES, N. A.; NIRCHIO, M.; OLIVEIRA, C.; SICCHARAMIREZ, R. Taxonomic
review of the species of Mugil (Teleostei: Perciformes: Mugilidae) from the Atlantic South
Caribbean and South America, with integration of morphological, cytogenetic and molecular
data. Zootaxa, v. 3918, n. 1, p. 1-38, 2015.
MÉRIGOT, B.; FREDOU, F. L.; PONTES VIANA, P. A.; PADOVANI FERREIRA, B.; DO
NASCIMENTO COSTA JUNIOR, E.; BESERRA DA SILVA JÚNIOR, C.A.; FREDOU, T.
Ocean & Coastal Management, v. 143, p. 175- 183, 2017.
MOLISANI, M. M.; CRUZ, A. L. V.; MAIA, L. P. Estimativa da descarga fluvial para os
estuários do Estado do Ceará, Brasil. Arquivo de Ciências do Mar, v.39, p. 53-60, 2006.
MORAIS, J. O.; PINHEIRO, L. S. Interferências de barramentos no regime hidrológico do
Estuário do Rio Catú – Ceará – Nordeste do Brasil. Sociedade & Natureza, v. 22, n. 2, p.
237-250, Ago. 2010.
MORAIS, J. O.; PINHEIRO, L. S. The effect of semi-aridity and damming on sedimentary
dynamics in estuaries -Northeastern region of Brazil. Journal of Coastal Research, v. 64, p.
1540 – 1544, 2011.
MORAIS, J. O.; SÁ FREIRE, G. S. PINHEIRO, L. S.; SOUZA, M. J. N.; CARVALHO, A.
M.; PESSOA, P. R. S. OLIVEIRA, S. H. M. EROSÃO e progradação do litoral
brasileiro/CEARÁ In: Erosão e progradação do litoral brasileiro. Brasília: Ministério do
Meio Ambiente, 2006. 475 p.
MOURA, R. L.; LINDEMAN, K. C. A new species of snapper (Perciformes: Lutjanidae)
from Brazil, with comments on the distribution of Lutjanus griseus and Lutjanus apodus.
Zootaxa, v. 1422, p. 31-43, 2007.
NELSON, Joseph S.; GRANDE, Terry C.; WILSON, Mark V. H. Fish of the world. 5 ed.
New Jersey, USA: John Wiley & Sons, 2016.
OLIVEIRA, A. M. E. Composição e distribuição da ictiofauna, nas águas estuarinas do rio
Jaguaribe, Jaguaribe (Ceará - Brasil). Arquivos de Ciências do Mar, v. 16, n. 1, p. 9- 18,
1976.

46
OSÓRIO, F. M.; GODINHO, W. O.; LOTUFO, T. M. C. Ictiofauna associada às raízes de
mangue do estuário do Rio Pacoti -CE, Brasil. Biota Neotropica, v. 11, p. 415-420, 2011.
PAIVA, A. C. G.; LIMA, M. F. V.; SOUZA, J. R. B.; ARAÚJO, M. E. Spatial distribution of
the estuarine ichthyofauna of the Rio Formoso (Pernambuco, Brazil), with emphasis on reef
fish. Zoologia, v. 26, n. 2, p. 266-278, 2009.
PASQUAUD, S.; VASCONCELOS, R.; FRANÇA, S.; HENRIQUES, S.; COSTA, M.;
CABRAL, H. Worldwide patterns of fish biodiversity in estuaries: Effect of global vs. local
factors. Estuarine, Coastal and Shelf Science, v.154, p. 122 – 128, 2015.
PASSOS, A. C.; CONTENTE, R. F.; ABBATE PAULO, F. V.; SPACH, H. L.; VILAR, C. C.;
JOYEUX, J. C.; CARTAGENA. B. F. C.; FÁVARO, L. F. Analysis of fish assemblages in
sectors along a salinity gradient based on species, families and functional groups.
BrazilianJournal of Oceanography, v. 61, n. 4, p. 251-264, 2013.
PELÁEZ, O.; AZEVEDO, F.; PAVANELLI, C. Environmental heterogeneity explains
species turnover but not nestedness in fish assemblages of a Neotropical basin. Acta
Limnologica Brasiliensia, v. 29, n. e117, 2017.
PICHLER, H. A.; SPACH, H. L.; GRAY, C. A.; BROADHURST, M. K.; SCHWARZ JR. R.;
OLIVEIRA-NETO, J. Environmental influences on resident and transient fishes across
shallow estuarine beaches and tidal flats in a Brazilian World Heritage area. Estuarine,
Coastal and Shelf Science, v. 164, p. 482-492, 2015.
POTTER, I. C.; CHUWEN, B. M.; HOEKSEMA, S. D.; ELLIOT, M. The concept of an
estuary: a definition that incorporates systems which can become closed to the ocean and
hypersaline. Estuarine, Coastal and Shelf Science, v. 87, p. 497–500, 2010.
POTTER, I. C.; TWEEDLEY, J. R.; ELLIOTT, M.; WHITFIELD, A. K. The ways in which
fish use estuaries: a refinement and expansion of the guild approach. Fish and Fisheries, v.
16, n. 2, p. 230 – 239, 2015.
QUINTELA – FALCÃO, T. O.; de MORAIS, J. O.; PINHEIRO, L. S. Morphodynamics of
the Curu estuary inlet – Brazil. Journal of Coastal Research, v. 64, p. 805 – 808, 2011.
SELLESLAGH J.; AMARA R. Environmental factors structuring fish composition and
assemblages in a small macrotidal estuary (eastern English Channel). Estuarine, Coastal and
Shelf Science, v. 79, p. 507-517, 2008.
SILVA – JÚNIOR, C. A. B.; MÉRIGOT, B.; LUCENA – FRÉDOU, F.; FERREIRA, B. P.;
COXEY, M. S.; REZENDE, S.M.; FRÉDOU, T. Functional diversity of fish in tropical
estuaries: A traits-based approach of communities in Pernambuco, Brazil. Estuarine, Coastal
and Shelf Science, v. 198, p. 413 – 420, 2017.
SOCOLAR, J.; GILROY, J.; KUNIN, W.; EDWARDS, D. How should beta-diversity inform
biodiversity conservation? Trends in Ecology & Evolution, v. 31, n. 1, p. 67 – 80, Jan. 2016.

47
SOUSA BARROSO, H.; BECKER, H.; MELO, M. V. Influence of river discharge on
phytoplankton structure and nutrient concentrations in four tropical semiarid estuaries.
Brazilian journal of oceanography, v.64, n. 1, p. 37 – 48, 2016.
SOUSA BARROSO, H.; LOPES TAVARES, T. C.; de OLIVEIRA SOARES, M.; GARCIA,
T. M.; MELO, B. R.; CAVALCANTE VIEIRA, A. S.; VIANA, P. B.; TAVARES FERREIRA,
T. J.; FILHO, J. P.; FRANÇA SCHETTINI, C. A.; SANTAELLA, S. T. Intra-annual
variability of phytoplankton biomass and nutrients in a tropical estuary during a severe
drought. Estuarine, coastal and shelf science, 2018.
TEICHERT, N.; LEPAGE, M.; CHEVILLOT, X.; LOBRY, J. Environmental drivers of
taxonomic, functional and phylogenetic diversity (alpha, beta and gamma components) in
estuarine fish communities. Journal of Biogeography, v. 45, p. 406 – 417, 2018.
VILLÉGER, S.; MIRANDA, J.; HERNANDEZ, D.; MOUILLOT, D. Low functional β-
Diversity despite high taxonomic β - Diversity among Tropical Estuarine Fish Communities.
PLoS ONE, v. 7, p. 1 - 10, 2012.
WHITFIELD, A. K.; ELLIOT, M.; BASSET, A.; BLABER, S. J. M.; WEST, R. J. Paradigms
in estuarine ecology - A review of the Remane diagram with a suggested revised model for
estuaries. Estuarine, Coastal and Shelf Science, v. 97, p. 78 – 90, 2012.
ZEE –Zoneamento Ecológico e Econômico da Zona Costeira do Estado do Ceará:
ictiofauna dos estuários do estado do Ceará. Fortaleza, Governo do Estado do Ceará:
UFC/SEMACE/LABOMAR, 2005.
ZORZAL-ALMEIDA, S.; BINI, L.; BICUDO, D. Beta diversity of diatoms is driven by
environmental heterogeneity, spatial extent and productivity. Hydrobiologia, v. 800, p. 7 –
16, 2017.





























