Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE ESTUDOS COSTEIROS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL
MESTRADO EM ECOLOGIA DE ECOSSISTEMAS COSTEIROS E
ESTUARINOS
ECOLOGIA TRÓFICA DE SEIS ESPÉCIES DE ANUROS EM UMA
ÁREA DOMINADA POR FLORESTAS DE MANGUE NA AMAZÔNIA
BRASILEIRA
ARTHUR LUIZ RAIOL DE BRITO
Orientador: Prof. Dr. Marcus Emanuel Barroncas Fernandes
Bragança
2012

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE ESTUDOS COSTEIROS
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL
MESTRADO EM ECOLOGIA DE ECOSSISTEMAS COSTEIROS E
ESTUARINOS
ECOLOGIA TRÓFICA DE SEIS ESPÉCIES DE ANUROS EM UMA
ÁREA DOMINADA POR FLORESTAS DE MANGUE NA AMAZÔNIA
BRASILEIRA
ARTHUR LUIZ RAIOL DE BRITO
Dissertação de mestrado apresentada ao
Programa de Pós-Graduação em Biologia
Ambiental, do Instituto de Estudos
Costeiros, da Universidade Federal do
Pará, como parte dos requisitos
necessários para a obtenção do Grau de
Mestre em Biologia Ambiental.
Prof. Dr. Marcus E. B. Fernandes Orientador
Bragança
2012

ARTHUR LUIZ RAIOL DE BRITO
ECOLOGIA TRÓFICA DE SEIS ESPÉCIES DE ANUROS EM UMA
ÁREA DOMINADA POR FLORESTAS DE MANGUE NA AMAZÔNIA
BRASILEIRA
Dissertação de mestrado apresentada ao
Programa de Pós-Graduação em Biologia
Ambiental, do Instituto de Estudos
Costeiros, da Universidade Federal do
Pará, como parte dos requisitos
necessários para a obtenção do Grau de
Mestre em Biologia Ambiental.
Prof. Dr. Marcus E. B. Fernandes
Orientador:
BANCA EXAMINADORA
---------------------------------------------------
Prof. Dr. Marcus Emanuel Barroncas Fernandes (Orientador)
UFPA - Campus de Bragança
______________________________________________
Profa. Dra. Maria Cristina dos Santos Costa (UFPA - ICB) (Titular)
_______________________________________
Prof. Dr Gleomar Fabiano Maschio (UFPA - ICB) (Titular)
_______________________________________
Profa. Dra. Rita de Cássia Oliveira Santos (Titular)
__________________________________________________
Prof. Dr. José Antonio Renan Bernardi (Instituto Federal do Pará – IFPA) (Suplente)

AGRADECIMENTOS
Agradeço a FAPESPA pela bolsa fornecida durante o curso.
Ao Programa de Pós-Graduação em Biologia Ambiental, IECOS, Campus de Bragança.
Ao Laboratório de Ecologia de Manguezal.
Ao Dr. Marcus Fernandes, que se dispôs a me orientar e acreditar na viabilidade do
trabalho, bem como por estar aberto a trabalhar com um grupo diferenciado de sua linha
de pesquisa.
A minha família, Ruth Helena, Luiz Alberto e Aimée que sempre esteve perto pra me
apoiar a ajudar de todas as formas possíveis e impossíveis.
A Tayana Maria pelo seu companheirismo durante quase 7 anos me aturando e me
dando forças, em cada uma das batalhas, que juntos tivemos que superar.
A Dra. Ilmarina por fazer parte da minha vida, me dando e recebendo muito amor e
carinho.
Aos amigos Rosivan, Kelle, Folha, André Palmeira, Dani, Cinthya, Adriano, Dante,
Denis, Kelli, Nara, Cristovam e Célio agradeço pela ajuda durante esta jornada, tendo
enfrentado juntos as adversidades e alegrias. Certamente não conseguirei me lembrar de
todos. Para aqueles aqui não citados, minhas sinceras desculpas.
Agradeço também ao João e o Eliphas, que me ajudaram muito nos trabalhos de
laboratório, se tornando grandes amigos.
Enfim, a todos que ajudaram nessa jornada.

i
Sumário
CAPÍTULO 1 ................................................................................................................................ 1
INTRODUÇÃO GERAL .............................................................................................................. 1
ESTUDOS DE ECOLOGIA ALIMENTAR ............................................................................. 2
FATORES QUE INFLUENCIAM AS RELAÇÕES TRÓFICAS ........................................... 2
A DIETA DOS ANUROS......................................................................................................... 3
ESTRATÉGIAS DE FORRAGEIO DOS ANUROS ............................................................... 4
OS ANUROS ANALISADOS .................................................................................................. 5
REFERÊNCIAS BIBLIOGRÁFICAS ...................................................................................... 7
CAPÍTULO 2 .............................................................................................................................. 11
ECOLOGIA TRÓFICA DE SEIS ESPÉCIES DE ANURAS EM UMA ÁREA DOMINADA
POR FLORESTAS DE MANGUE NA AMAZÔNIA BRASILEIRA ....................................... 11
ABSTRAT ................................................................................................................................. 12
RESUMO ................................................................................................................................... 13
INTRODUÇÃO ........................................................................................................................... 14
MATERIAL E MÉTODOS ........................................................................................................... 15
Área de Estudo .................................................................................................................... 15
Espécies de Anuros.............................................................................................................. 16
Composição da Dieta .......................................................................................................... 16
Análise Estatística ............................................................................................................... 18
RESULTADO ............................................................................................................................. 19
Composição da Dieta .......................................................................................................... 19
Sobreposição e Índices de Diversidade............................................................................... 23
Morfologia e Dieta .............................................................................................................. 25
DISCUSSÃO .............................................................................................................................. 26
Partição de Recursos .......................................................................................................... 26
Dieta e Forrageio ................................................................................................................ 28
LITERATURA CITADA ............................................................................................................... 31

1
CAPÍTULO 1
INTRODUÇÃO GERAL
Capitulo escrito segundo as normas da ABNT (autor-data).

2
ESTUDOS DE ECOLOGIA ALIMENTAR
Em geral, um dos principais objetivos em estudos de ecologia animal é avaliar os tipos de
alimento e a variedade de hábitat que os animais utilizam (Johnson, 1980), sendo a ideia de teia
alimentar central nesses estudos. Tais relações tróficas atuam na história de vida, fisiologia,
comportamento e regulação populacional em resposta às adaptações a competidores intra e
interespecíficos, à defesa de suas presas e aos próprios inimigos naturais (Wilbur, 1997).
Deste modo, as relações tróficas atuam como um importante componente que possibilita um
melhor entendimento da estrutura de comunidade animais, desempenhando grande papel na
dinâmica de suas populações. Seu estudo ainda possibilita a identificação de possíveis
competidores, a definição do período em que estão ativos e as interações predatórias em que
estão envolvidos (Caldwell, 1996; Peltzer e Lajmanovich, 1999).
Na ecologia trófica de uma dada assembleia, a amplitude (diversidade das presas consumidas)
e a sobreposição (similaridade da dieta) do nicho trófico são medidas importantes para o
entendimento da dinâmica das populações estudadas (Pianka, 1974). Estes índices têm sido
amplamente utilizados em estudos da dieta de anfíbios que vivem em simpatria (Lima e
Magnusson, 1998; Santos, Almeida e Vasconcelos, 2004; Menin, Rossa-Feres e Giaretta, 2005;
Piatti e Souza, 2011).
FATORES QUE INFLUENCIAM AS RELAÇÕES TRÓFICAS
Os ecólogos também têm apontado para o fato de que a dieta dos anuros é fortemente
influenciada por interações interespecíficas, relacionados a predação e a competição (Gordon,
2000). Nesse sentido varios estudos tem correlacionado a dieta dos anfíbios com fatores, como:
tamanho da presa e do predador, número e volume das presas consumidas e mudanças
ontogenéticas (Lima e Moreira, 1993; Lima e Magnusson, 1998; Menin, Rossa-Feres e Giaretta,

3
2005; Batista et al., 2011; Piatti e Souza, 2011). Deste modo, a dieta pode afetar a organização de
uma dada comunidade, podendo determinar assim, a existência ou co-existência de uma
determinada espécie, onde a utilização de vários itens alimentares é de extrema importância
nessas relações, diminuindo a competição entre as espécies (Caldwell, 1996). Além do mais,
estudos recentes sugerem ainda que a mirmecofagia e a toxicidade são traços coevolutivos em
algumas famílias de anuros (Caldwell, 1996; Darst et al., 2005).
Tais mecanismo que permite que as diferentes espécies utilizem os recursos (ex. espaço,
alimento e tempo) disponíveis de forma diferenciada, foi o que Schoener (1989) chamou de
partição de recursos. Essa partição é resultado de dois ou mais fatores e ocorre independente de
cada espécie, sendo essencial para as interações de uma comunidade, determinando a diversidade,
distribuição e abundância dessas espécies (Pianka, 1974; Toft, 1985). De maneira geral, os
estudos realizados sobre a partição de recursos assumem que a similaridade morfológica entre as
espécies conduz ao uso de alguns recursos de modo similar (MacArthur e Levins, 1967) e que
para acessar a dieta de diferentes espécies dentro de uma determinada assembleia é importante o
melhor entendimento das relações entre essas espécies (Duré, Kehr e Schaefer, 2009).
A DIETA DOS ANUROS
Os anfíbios são, excepcionalmente, carnívoros quando adultos, alimentando-se principalmente
de invertebrados, tais como: insetos, crustáceos e moluscos (Beebee, 1996). Alguns espécies de
maior porte podem chegar a consumir uma diversidade de vertebrados, como doninhas (Beringer
e Johnson, 1995), aves (Wu et al., 2005), roedores (Quiroga, Sanabria e Acosta, 2009) e,
eventualmente, morcegos (Silva, Santos e Amorim, 2010). Outras podem consumir anuros, como
por exemplo, as rãs neotropicais do gênero Leptodactylus (Teixeira e Vrcibradic, 2003; Brito,

4
2011; Heitor et al., 2012). Como exceção ao grupo dos anuros carnívoros podemos mencionar a
espécie Rana hexadactyla, cujos indivíduos podem consumir insetos na fase de girino e
macrófitas aquáticas na fase adulta (Das, 1996), além de Xenohyla truncata, cujos espécimes
incluem frutas em sua dieta (Da Silva e De Britto-Pereira, 2006).
Apesar do grande número de estudos que trata sobre a dieta de anfíbios, ainda são poucos
aqueles que enfocam a partição de recursos em nível interespecífico (Toft, 1980; 1981; Rosa et
al., 2002; Santos, Almeida e Vasconcelos, 2004; Vignoli e Luiselli, 2012). A maioria investiga a
dieta de uma espécie ou grupos taxonômicos (Hirai e Matsui, 2002; Duré, Kehr e Schaefer, 2009;
Quiroga, Sanabria e Acosta, 2009; Solé et al., 2009; Valderrama-Vernaza, Ramírez-Pinilla e
Serrano-Cardozo, 2009; Paula-Lima, Roedder e Solé, 2010).
ESTRATÉGIAS DE FORRAGEIO DOS ANUROS
Muitos herpetólogos consideram os anuros como sendo consumidores generalistas, entretanto
diversos estudos têm revelado a existência de várias guildas alimentares (Parmelee, 1999).
Existem duas estratégias de forrageio relacionadas aos anfíbios anuros: i) o forrageio senta-e-
espera e ii) o forrageio por busca ativa. Na primeira estratégia o predador espera imóvel a presa
passar. Suas presas são relativamente grandes, de alta mobilidade, o número de captura por dia é
baixo, gasta-se pouca energia para a captura, utilizando-se principalmente a visão. As espécies de
sapos são geralmente robustas, de coloração críptica, apresentam baixas taxas metabólicas e boca
larga. Esse é um tipo de forrageamento utilizado por diversas espécies de leptodactilídeos e
hilídeos (Toft, 1981; Perry e Pianka, 1997). Na segunda estratégia, o predador procura ativamente
suas presas, as quais são geralmente menores, de baixa mobilidade e maior abundância,
utilizando o olfato, além da visão, para efeito de captura. São animais geralmente menores

5
quando comparados àqueles que usam a técnica senta-e-espera. Muitas espécies possuem toxinas
na pele, como os dendrobatídeos e bufonídeos, além de apresentarem altas taxas metabólicas,
bem como altas taxas de capturas de presas por dia (Toft, 1981; Perry e Pianka, 1997).
Ambas as estratégias alimentares podem estar relacionadas a fatores externos (ex.
disponibilidade de presas, presença ou ausência de competidores e risco de predação) e fatores
intrínsecos (ex. idade, sexo e estado reprodutivo), além da preferência da dieta e dos nutrientes
que cada espécie necessita (Perry e Pianka, 1997). Por último, fatores como as limitações
sensoriais, características morfológicas e parâmetros fisiológicos (Perry e Pianka, 1997).
OS ANUROS ANALISADOS
O presente trabalho analisou a dieta de seis espécies de anuros pertencentes a três famílias:
Hylidae, Leptodactylidae e Leiuperidae, ocorrentes na península de Ajuruteua, em Bragança,
Pará. Das espécies estudadas, quatro pertencem à família Hylidae:
Hypsiboas raniceps Cope, 1862 – um hilídeo, amplamente distribuídos na
Amazônia brasileira, sendo registrado em vários tipos de hábitats. Em áreas de floresta essa
espécie pode ser encontrada em lagos ou na vegetação de entorno, podendo ser encontrada
também nas savanas amazônicas, áreas urbanas e hábitats degradados (La Marca, 2004; Zina, De
Sá e Prado, 2010).
Phyllomedusa hypochondrialis (Daudin, 1800) - uma espécie noturna que pode ser
encontrada em grande parte da bacia amazônica brasileira. É facilmente encontrada em arbustos e
herbáceas de savanas inundadas e de hábitats florestais, bem como em hábitats degradados e
áreas urbanas (Azevedo-Ramos et al., 2010).

6
Scinax x-signatus (Spix, 1824) - espécie facilmente adaptável à perturbação
humana, ocorrendo em savanas tropicais e bordas de floresta da Venezuela até o Suriname e em
boa parte do Brasil (Miquel, Ulisses e Abraham, 2010). Este hilídeo é muito comum próximo às
habitações humanas, apresentando hábitos noturnos e arborícolas (Galatti et al., 2007).
Dendropsophus nanus (Boulenger, 1889) – Espécie de hilídeo que ocorre em
quase todo o território brasileiro, em vegetações herbáceas na beira da água parada, podendo ser
encontrada em florestas e savanas, além de áreas abertas e perturbadas (Reichle et al., 2004).
A família Leptodactylidae está representada, no presente trabalho, por apenas uma espécie:
Leptodactylus macrosternum Miranda-Ribeiro, 1926 - uma rã que pode ser
encontrada em uma grande variedade de hábitats, tanto em florestas tropicais úmidas quanto em
áreas abertas, como os ambientes savânicos, campos e áreas secas (Heyer et al., 2008). A espécie
L. macrosternum é amplamente distribuída na América do Sul, a leste dos Andes.
A família Leiuperidae também está representada por apenas uma espécie:
Pseudopaludicola cf. canga (Giaretta & Kokubum, 2003) - uma rã classificada
como endêmica da região de Carajás (Giaretta e Kokubum, 2003), porém estudos recentes têm
demonstrado que a distribuição geográfica dessa espécie pode ser maior do que o conhecido
(Duarte et al., 2010; Pansonato, Morais e Ávila, 2012).
O manguezal é um ecossistema que apresenta condições ambientais (ex. solo lamoso, inundação
diária e salinidade da água) que pode limitar a ocorrência e o estabelecimento de muitas espécies de
anuros. Considerando que as diferenças na ecologia alimentar são decorrentes do mecanismo de
partição dos recursos disponíveis entre espécies que vivem em simpátria, espera-se que o grau de
sobreposição dessas espécies seja maior entre aquelas taxonomicamente semelhantes. Assim, o

7
presente estudo tem como objetivo maior descrever as relações tróficas de espécies de anuros
presentes em áreas de manguezal.
REFERÊNCIAS BIBLIOGRÁFICAS
AZEVEDO-RAMOS, C. et al. Phyllomedusa hypochondrialis. IUCN Red List of Threatened
Species. Version 2013.1.: IUCN 2013. < www.iucnredlist.org>. 2010.
BATISTA, R. C. et al. Diet of Rhinella schneideri (Werner, 1894) (Anura: Bufonidae) in the
Cerrado, Central Brazil. Herpetology Notes, v. 4, p. 17-21, 2011.
BEEBEE, T. J. C. Ecology and Conservation of amphibians. London, UK: Chapmam Hall,
1996.
BERINGER, J.; JOHNSON, T. R. Rana catesbeiana (Bullfrog). Diet. Herpetological Review, p.
26:98, 1995.
BRITO, A. L. R. Dieta de Leptodactylus macrosternum (Anura-Leptodactylidae) em áreas de
restinga e manguezal na península de Ajuruteua, Bragança, Pará, Brasil. 2011. Trabalho de
Conclusão de Curso (Graduação em Ciências Biológicas). Faculdade de Biologia, Universidade
Federal do Pará, Bragança, Pará.
CALDWELL, J. P. The evolution of myrmecophagy and its correlates in poison frogs (Family
Dendrobatidae). Journal of Zoology, v. 240, p. 75-101, 1996.
DARST, C. R. et al. Evolution of dietary specialization and chemical defense in poison frogs
(Dendrobatidae): A comparative analysis. The American Naturalist, v. 165, n. 1, p. 56-69,
2005.
DAS, I. Folivory and seasonal changes in diet in Rana hexadactyla (Anura: Ranidae). Journal of
Zoology, v. 238, n. 4, p. 785-794, 1996.
DA SILVA, H. R.; DE BRITTO-PEREIRA, M. C. How much fruit do fruit-eating frogs eat? An
investigation on the diet of Xenohyla truncata (Lissamphibia: Anura: Hylidae). Journal of
Zoology, v. 270, n. 4, p. 692-698, 2006.
DUARTE, T. C. et al. Chromosome analysis in Pseudopaludicola (Anura, Leiuperidae), with
description of sex chromosomes XX/XY in P. saltica. Hereditas, v. 147, n. 2, p. 43-52, 2010.
DURÉ, M. I.; KEHR, A. I.; SCHAEFER, E. F. Niche overlap and resorce partitioning among
five sympatric bofonids (Anura - Bufonidae) from northeastern Argentina Phyllomedusa, v. 8, n.
1, p. 27-39, 2009.

8
GALATTI, U. et al. Anfíbios da Área de Pesquisa Ecológica do Guamá (APEG) e Região de
Belém. In: GOMES, J. I.;MARTINS-DA-SILVA, M., et al (Ed.). Mocambo: Diversidade e
Dinâmica Biológica da Área de Pesquisa Ecológica do Guamá (APEG). Belém: MCT/Museu
Paraense Emílio Goeldi, 2007. p.75-95.
GIARETTA, A. A.; KOKUBUM, M. N. C. A new species of Pseudopaludicola (Anura,
Leptodactylidae) from northern Brazil. Zootaxa, v. 383, p. 1–8, 2003.
GORDON, C. E. The coexistence of species La coexistencia de especies. Revista chilena de
Historia natural, v. 73, n. 1, p. 175-198, 2000.
HEITOR, R. C. et al. Predation of Hypsiboas pardalis (Anura, Hylidae) by the butter frog
Leptodactylus cf. latrans (Anura, Leptodactylidae), in municipality of Espera Feliz, State of
Minas Gerais, southeastern Brazil. Herpetology Notes, v. 5, p. 23-25, 2012.
HEYER, R. et al. Leptodactylus ocellatus. IUCN Red List of Threatened Species. Version
2010.2.: IUCN 2010. < www.iucnredlist.org>. 2008.
HIRAI, T.; MATSUI, M. Feeding relationships between Hyla japonica and Rana nigromaculata
in rice fields of Japan. Journal of Herpetology, p. 662-667, 2002.
JOHNSON, H. E. The comparison of use and availability measurements for evaluating resource
preference. Ecology, v. 61, n. 1, p. 65-71, 1980.
LAMARCA, E. et al. Hypsiboas raniceps. IUCN Red List of Threatened Species. Version
2013.2.: IUCN 2013 < www.iucnredlist.org> 2004.
LIMA, A. P.; MAGNUSSON, W. E. Partitioning seasonal time: interactions among size,
foraging activity and diet in leaf-litter frogs. Oecologia, v. 116, p. 259-266, 1998.
LIMA, A. P.; MOREIRA, G. Effects of prey size and foraging mode on the ontogenetic change
in feeding niche of Colostethus stepheni (Anura: Dendrobatidae). Oecologia, v. 95, n. 1, p. 93-
102, 1993.
MACARTHUR, R. H.; LEVINS, R. The limiting similarity, convergence and divergence of
coexisting species. The American Naturalist v. 101, p. 377-385, 1967.
MENIN, M.; ROSSA-FERES, D. C.; GIARETTA, A. A. Resource use and coexistence of two
syntopic hylid frogs (Anura, Hylidae). Revista Brasileira de Zoologia, v. 21, n. 1, p. 61-72,
2005.
MIGUEL, T. R.; ULISSES, C.; ABRAHAM, M. Scinax x-signatus. IUCN Red List of
Threatened Species. Version 2013.1.: IUCN 2013 2010.
PANSONATO, A.; MORAIS, D. H.; ÁVILA, R. W. A new species of Pseudopaludicola
Miranda-Ribeiro, 1926 (Anura: Leiuperidae) from the state of Mato Grosso, Brazil, with

9
comments on the geographic distribution of Pseudopaludicola canga Giaretta & Kokubum, 2003.
Zootaxa, v. 3523, p. 49-58, 2012.
PARMELEE J. R. Trophic ecology of a tropical anuran assemblage. Natural History Museum,
University of Kansas, 11, 1-59.
PAULA-LIMA, J. E.; ROEDDER, D.; SOLÉ, M. Diet of two sympatric Phyllomedusa (Anura:
Hylidae) species from a cacao plantation in southern Bahia, Brazil. North-Western Journal of
Zoology, v. 6, n. 1, p. 13-24, 2010.
PELTZER, P. M.; LAJMANOVICH, R. C. Análisis trófico en dos poblaciones de Scinax nasicus
(Anura, Hylidae) de Argentina. Alytes, v. 16, p. 84-96, 1999.
PERRY, G.; PIANKA, E. R. Animal foraging: past, present and future Tree, v. 12, n. 9, p. 360-
364, 1997.
PIANKA, E. R. Niche overlap and diffuse competition. Proceedings of the National Academy
of Science, v. 71, n. 5, p. 2141-2145, 1974.
PIATTI, L.; SOUZA, F. L. Diet and resource partitioning among anurans in irrigated rice fields
in Pantanal, Brazil. Brazilian Journal of Biology, v. 71, n. 3, p. 653-661, 2011.
QUIROGA, L. B.; SANABRIA, E. A.; ACOSTA, J. C. Size- and Sex-Dependent Variation in
Diet of Rhinella arenarum (Anura: Bufonidae) in a Wetland of San Juan, Argentina Journal of
Herpetology, v. 43, n. 2, p. 311-317, 2009.
REICHLE, S. et al. Dendropsophus nanus. IUCN Red List of Threatened Species. Version
2013.1.: IUCN 2013 2004.
ROSA, I. et al. Diet of four sympatric anuran species in a temperate environment. Boletín de la
Sociedad Zoológica del Uruguay, v. 13, p. 12-20, 2002.
SANTOS, E. M.; ALMEIDA, A. V.; VASCONCELOS, S. D. Feeding habits of six anuran
(Amphibia: Anura) species in a rainforest fragment in Northeastern Brazil. Iheringia. Série
Zoologia, v. 94, n. 4, p. 433-438, 2004.
SCHOENER, T. W. Resource partitioning in ecological communities. Science, v. 185, n. 4145,
p. 27-39, 1974.
SILVA, L. A. M. D.; SANTOS, E. M. D.; AMORIM, F. O. D. Predação oportunística de
Molossus molossus (Pallas, 1766) (Chiroptera: Molossidae) por Rhinella jimi (Stevaux, 2002)
(Anura: Bufonidae) na Caatinga, Pernambuco, Brasil. Biotemas, v. 23, n. 2, p. 215-218, 2010.
SOLÉ, M. et al. Diet of Leptodactylus ocellatus (Anura: Leptodactylidae) from a cacao
plantation in southern Bahia, Brazil. Herpetology Notes, v. 2, p. 9-15, 2009

10
TEIXEIRA, R. L.; VRCIBRADIC, D. Diet of Leptodactylus ocellatus (Anura; Leptodactylidae)
from coastal lagoons of southeastern Brazil. Cuadernos de Herpetologia, v. 17, p. 113-120,
2003.
TOFT, C. A. Feeding ecology of thirteen syntopic species of anurans in a seasonal tropical
environment. Oecologia, v. 45, n. 1, p. 131-141, 1980.
______. Feeding ecology of panamanian litter anurans: patterns in diet and foraging mode.
Journal of Herpetology, v. 15, p. 139-144, 1981.
______. Resource partitioning in amphibians and reptiles. Copeia, v. 1985, n. 1-21, 1985.
VALDERRAMA-VERNAZA, M.; RAMÍREZ-PINILLA, M. P.; SERRANO-CARDOZO, V. H.
Diet of the Andean frog Ranitomeya virolinensis (Athesphatanura: Dendrobatidae). Journal of
Herpetology, v. 43, n. 1, p. 114-123, 2009.
VIGNOLI, L.; LUISELLI, L. Dietary relationships among coexisting anuran amphibians: a
WILBUR, H. M. Experimental ecology of food webs: complex systems in temporary ponds.
Ecology, v. 78, n. 8, p. 2279-2302, 1997.
WRIGHT, P. et al. The crab‐eating frog, Rana cancrivora, up‐regulates hepatic carbamoyl
phosphate synthetase I activity and tissue osmolyte levels in response to increased salinity.
Journal of Experimental Zoology Part A: Comparative Experimental Biology, v. 301, n. 7,
p. 559-568, 2004.
WU, Z. et al. Diet of Introduced Bullfrogs (Rana catesbeiana): Predation on and Diet Overlap
with Native Frogs on Daishan Island, China. Journal of Herpetology, v. 39, n. 4, p. 668-674,
2005/12/01 2005.
ZINA, J.; DESÁ, F. P.; PRADO, C. A. P. Amphibia, Anura, Hylidae, Hypsiboas raniceps Cope,
1862: Distribution extension. Check List, v. 6, n. 2, 2010.

11
CAPÍTULO 2
ECOLOGIA TRÓFICA DE SEIS ESPÉCIES DE ANURAS EM UMA ÁREA
DOMINADA POR FLORESTAS DE MANGUE NA AMAZÔNIA
BRASILEIRA
Artigo a ser submetido à revista Herpetologica

12
ABSTRAT.- Most anurans are regarded as generalist predator because of its diet reflect the availability
of prey. However, the feeding of the animals may also be influenced by environmental and
morphological factors. We evaluate trophic relationships in order to describe the dietary composition of
six sympatric species of frogs and verify the effect of breadth and niche overlap on this resource
partitioning in areas dominated by mangrove forests. We observed that the highest values of overlap
were recorded between taxonomically similar species, however this trend was not true for all species.
Our results showed that among the six species studied three are ant specialists (H. Raniceps, L.
macrosternum, and P. cf. canga) two are not ant specialists (P. hypocondrialis and D nanus) and one is
generalist (S. x-signatus). However, other tests involving the collection of more detailed information
about prey electivity can improve the understanding of how frogs use the available resources in their
environment, besides better determine trends that nocturnal anurans present about their foraging way.
Keywords: diet, mangrove, anurans assemblage, Pará, Brazil

13
RESUMO.- A maioria dos anuros é considerada como predador generalista, devido a sua dieta refletir a
disponibilidade das presas. Entretanto, sua alimentação pode ser influenciada também por fatores
ecológicos e morfológicos. Aqui nós avaliamos as relações tróficas no intuito de descrever a
composição da dieta de seis espécies de anuros que vivem em simpatria e verificar o efeito da
amplitude e sobreposição de nicho sobre a partição desse recurso em áreas dominadas por florestas de
mangue. Observamos que os maiores valores de sobreposição foram entre espécies taxonomicamente
semelhantes, entretanto essa tendência não foi verdadeira para todas as espécies. Nossos resultados
mostraram que entre as seis espécies estudadas, três são especialistas em formiga (H. raniceps, L.
macrosternum e P. cf. canga) duas são não especialistas em formiga (P. hipocondrialis e D. nanus) e
uma generalista (S. x-signatus). Entretanto, outros testes que envolvem a coleta de informações mais
detalhadas sobre a eletividade da dieta podem melhorar o entendimento de como os anuros usam os
recursos disponíveis em seu ambiente, além de melhor determinar as tendências que os anuros noturnos
apresentam quanto ao modo de forrageio.
Palavras-chave: dieta, manguezal, assembleia de anuros, Pará, Brasil

14
INTRODUÇÃO
Estudos têm demonstrado que os animais partilham os recursos disponíveis no ambiente de três
formas básicas: eles diferem quanto ao tempo de atividade, quanto ao espaço que exploram e quanto
aos alimentos que ingerem (Pianka, 1973; Schoener, 1974). Além desses tipos de partilha de recursos,
tradicionalmente os anuros também diferem quanto suas estratégias de forrageio. Eles podem ser do
tipo de predador que procura ativamente suas presas (forrageador ativo), ou podem ser predadores por
emboscada, que esperam suas presas (forrageador senta-e-espera) (Perry e Pianka, 1997). Tais
diferenças tanto no uso dos recursos como nas estratégias de forrageio reduzem a competição, pois lhes
permitem capturar uma gama variada de presas, além de permitir uma coexistência de uma variedade
de espécies (Duré e Kehr, 2004).
A maioria dos anuros tem uma alimentação considerada oportunista, já que sua dieta é altamente
influenciada pela disponibilidade de alimentos (Parmelee, 1999). Entretanto, trabalhos em laboratório
têm revelado que algumas espécies são seletivas em sua alimentação, principalmente das famílias
Bufonidae e Dendrobatidae (Drewes e Roth, 1981; Simon e Toft, 1991). Outros estudos ainda apontam
para uma especialização a nível individual (Bolnick et al., 2003; Araújo et al., 2009), onde cada
indivíduo utiliza apenas um pequeno subconjunto do seu nicho populacional.
Fatores como a plasticidade da dieta e a habilidade de explorar vários tipos de presas são essenciais
para o sucesso das espécies de anuros em ambientes inóspitos, como o manguezal. Globalmente, pouco
se sabe sobre a fauna de anuros no ecossistema de manguezal (Kathiresan e Bingham, 2001).
Adicionalmente, a maioria dos trabalhos se limita apenas a reportar a ocorrência de certas espécies
nesse ambiente (Kathiresan e Rajendran, 2004; Rao, 2004; Satheeshkumar, 2011; Dev Roy e
Sivaperuman, 2012; Jena et. al., 2013), sendo escassos os trabalhos que tratam de ecologia trófica
(Glorioso, et al., 2012).

15
De fato, as relações tróficas apresentam-se como uma ferramenta importante que melhora o
entendimento da história de vida dos anuros (Vignoli e Luiselli, 2012). Tais relações ajudam a
compreender o modo de forrageio, a forma de utilização do habitat, além do comportamento de
regulação populacional, em resposta às adaptações a competidores e à defesa de suas presas (Wilbur,
1997; Parmelee, 1999). Partindo desse pressuposto, nós avaliamos as relações tróficas no intuito de
descrever a composição da dieta de seis espécies de anuros que vivem em simpatria e verificar o efeito
tanto da amplitude quanto da sobreposição de nicho sobre a partição de recursos em áreas dominadas
por florestas de mangue.
MATERIAL E MÉTODOS
Área de Estudo
A área de estudo está localizada na península de Ajuruteua (46º50’– 46º38’W e 00º05’–01º00’S),
nos limites da Reserva Extrativista Marinha Caeté-Taperaçú, município de Bragança, nordeste do
Estado do Pará. Essa região é entrecortada por canais-de-maré que ligam as florestas de mangue ao
estuário (Fernandes, 2003) possuindo um regime de macromarés semidiurnas (DHN, 2011). Para o
ano amostrado, os níveis de umidade relativa do ar ultrapassam 90%, com taxa de precipitação
pluviométrica anual de 3.613 mm e temperatura do ar média anual de 26°C. Duas estações foram
definidas de acordo com a pluviosidade mensal: uma estação seca, que abrangeu o período de agosto
a dezembro/2008 (precipitação <100 mm), e uma estação chuvosa, compreendendo o período
representado pelos meses de julho/2008 e de janeiro a junho/2009 (precipitação >100 mm). Dados
foram obtidos da Estação meteorológica automática (A226), localizada no município de Bragança-
PA.
A vegetação de mangue ocupa a maior parte da Planície Costeira Bragantina, distribuindo-se ao
longo dos canais-de-maré em direção ao continente (Souza-Filho e El-Robrini, 2000). Ao longo da

16
península também ocorrem manchas de vegetação de campo natural, as quais são alagadas
periodicamente, sob influência de águas pluviais e estuarinas (Souza-Filho e El-Robrini, 2000).
O local onde os anuros foram coletados é uma área dominada por florestas de mangue, contígua às
zonas de marismas de água doce (freshwater marshes), e é conhecida localmente como “Furo do
Taici”. Os manguezais são dominados por Avicennia germinans (L.) Stearn, com ocorrência de
Rhizophora mangle L. e Laguncularia racemosa (L.) Gaertn f., sendo esta última encontrada
principalmente nas bordas da floresta. Apenas as marés equinociais inundam essa área de manguezal, e
por esse motivo o chão da floresta permanece a maior parte do tempo consolidado, facilitando a entrada
e o forrageio de espécies de anuros terrícolas nesse ambiente. As áreas de marisma contíguas ao
manguezal acumulam água doce no período chuvoso, secando totalmente no período seco. Espécies
herbáceas como Heliconia sp. e Eleucharias sp. são abundantes nesse ambiente .
Espécies de Anuros
Todos os espécimes analisados pertencem à Coleção de Zoologia de Bragança (CZB), da
Universidade Federal do Pará (UFPA), e foram cedidos para a produção deste trabalho. Seis espécies
de anuros foram selecionadas, por serem as espécies mais representativas na área do Taici. As espécies
analisadas pertencem a três famílias, sendo a família Hylidae composta por anuros arborícolas
[Hypsiboas raniceps Cope, 1862; Phyllomedusa hypochondrialis (Daudin, 1800); Scinax x-signatus
(Spix, 1824) e Dendropsophus nanus (Boulenger, 1889)], enquanto Leptodactylidae (Leptodactylus
macrosternum Miranda-Ribeiro, 1926) e Leiuperidae (Pseudopaludicola cf. canga Giaretta &
Kokubum, 2003) são famílias representadas por anuros terrestres.
Composição da Dieta
Medimos o comprimento rostro-cloacal (CRC) e a largura da boca (LB) de cada espécime com
ajuda de um paquímetro digital (precisão 0,01mm). O sexo de cada indivíduo foi determinado através
da presença do saco vocal, ou pela observação direta das gônadas. Indivíduos nos quais as gônadas

17
observadas não foram diferenciadas, tais como testículos ou ovários, foram classificados como
imaturos. Investigamos o conteúdo gastrointestinal, haja vista que presas pequenas passariam mais
rapidamente para a porção intestinal do tubo digestivo, não sendo contabilizadas no cômputo total dos
itens alimentares (Schoener, 1989).
Considerando apenas as presas inteiras, ou completas, e não levando em conta antenas e apêndices
locomotores, medimos o maior comprimento e largura, para calcularmos o volume (mm³) usando a
fórmula do ovóide-esferóide:
𝑉 = 43𝜋⁄ . 𝐶
2⁄ . (𝐿2⁄ )² (1)
onde V é o volume, C é o comprimento e L a largura da presa (Magnusson et al., 2003). Acessamos o
Índice de Importância Relativa (IR) de cada categoria de presa usando a fórmula:
IR = (N% + F% + V%) 3⁄ (2)
onde N% é o percentual numérico, F% é o percentual da frequência e V% é o percentual volumétrico
(Biavati et al., 2004).
Para investigar a diversidade de itens alimentares, nós usamos o índice de Shannon (H’):
(3)
onde pi é a proporção dos recursos, ln é p logaritmo natural e s o número de presas (Shannon e Weaver,
1949). A fim de fazer uma comparação mais precisa usamos o índice de Shannon padronizado proposto
por (Krebs, 1999):
Hstd = H’/ln(s) (4)
onde H’ é o índice de Shannon. Para estima a amplitude do nicho trófico de cada espécie nós usamos o
Índice de Levins (1968):
(5)

18
onde j é a categoria do recurso utilizado e p é a proporção da categoria j utilizada por cada espécie.
O índice de Levins foi padronizado em uma escala que varia de 0 a 1 seguindo Hulbert (1978):
(6)
onde B é o índice de Levins e N o número de possíveis presas. Para esse cálculo usamos tanto dados
numéricos como volumétricos.
Análise Estatística
Nós estimamos o valor de similaridade entre as dietas das espécies de anuros, com o auxílio da
rotina SIMPER (Similarity Percentage). Desta forma, identificamos as diferentes presas que
apresentavam maior contribuição na dieta. Realizamos análises comparativas (ANOSIM-um fator) da
dieta entre as espécies e entre os grupos de anuros terrestre e arborícola, utilizando os dados de
composição numérica da dieta, a partir de uma matriz de similaridade utilizando-se o software
PRIMER v6, PML (Clarke e Gorley, 2006). Adicionalmente, verificamos possíveis diferenças na dieta
entre os sexos, para cada espécie, através do teste não paramétrico de Kruskal-Wallis (U), com o
auxílio do pacote estatístico Bioestat 5.0, IDSM (Ayres et al., 2007).
A fim de avaliar a sobreposição da dieta entre as espécies, usamos o índice de sobreposição de
Pianka (Ojk) (Pianka, 1974), com a ajuda do software EcoSim v7, Intelligence Inc. (Gotelli e
Entsminger, 2001). Este programa realiza permutações para criar “pseudo-comunidades” (Pianka
1974), e compara estatisticamente os padrões dessas “pseudo-comunidades” com a matriz original.
Para essas análises foram feitas 1.000 permutações com base na matriz de dados, usando a opção do
algoritmo RA3 do EcoSim (Gotelli e Entsminger, 2001).
Conduzimos uma Análise de Correspondência (AC), no intuito de avaliar as associações entre as
espécies de predadores e os itens de presas, utilizando o software Statistica v6.0, StatSoft (StatSoft,
2001).

19
Após a transformação normalizando os dados originais, as diferenças no CRC entre as espécies de
anuros foram testadas através da análise de variância (ANOVA-um fator). Da mesma forma, os dados
transformados foram utilizados para a análise de regressão linear entre as variáveis morfométricas CRC
e LB, para cada espécie. Para verificar possíveis relações entre essas duas variáveis com os volumes
médios e máximos das presas ingeridas, nós usamos a correlação de Pearson. Todos os testes acima
foram rodados com o auxílio do pacote estatístico Bioestat 5.0 (Ayres et al., 2007).
RESULTADO
Composição da Dieta
Analisamos um total de 246 indivíduos (Tabela 1), onde 12% desses indivíduos não apresentavam
nenhuma presa no seu trato digestório. A mesma tabela mostra que L. macrosternum foi a espécie que
apresentou o maior número de presas (19,4 ± 36,7) e volume (67,3 ± 121,1) por indivíduo. Scinax x-
signatus foi a espécie que apresentou o menor valor de presas por indivíduo (2,4 ± 2,1), enquanto P. cf.
canga apresentou os menores volumes (0,4 ± 0,6).
TABELA 1.—Valores (Média ± Desvio Padrão) dos dados biométricos e tróficos das seis espécies de anuros em áreas
dominadas por manguezais da costa amazônica brasileira. CRC= Comprimento Rostro-Cloacal, LB= Largura da Boca
Hypsiboas
raniceps
Phyllomedusa
hypochondrialis
Scinax
x-signatus
Dendropsophus
nanus
Leptodactylus
macrosternum
Pseudopaludicola
cf. canga
Número de
indivíduos (n=246) 46 45 50 32 23 50
CRC (mm) 47,0 ± 12,7 34,9 ± 4,0 33,4 ± 26,3 16,1 ± 1,1 47,8 ± 9,8 14,1 ± 1,2
LB (mm) 15,0 ± 3,9 11,3 ± 1,1 10,1 ± 1,7 5,6 ± 0,3 15,8 ± 2,8 4,7 ± 0,4
Número de presas por
indivíduo 12,0 ± 60,9 5,3 ± 6,1 2,4 ± 2,1 3,4 ± 5,0 19,4 ± 36,7 12,3 ± 13,2
Volume das presas por
indivíduo (mm²) 21,3 ± 76,5 4,9 ± 12,8 32,3 ± 59,5 2,2 ± 3,2 67,3 ± 121,1 0,4 ± 0,6
Número de táxon por
indivíduo 1,8 ± 1,2 2,5 ± 1,4 1,4 ± 1,1 1,5 ± 1,5 2,5 ± 2,0 3,8 ± 2,0
Hábito/Hábitat Noturno/Arbóreo Noturno/Arbóreo Noturno/Arbóreo Noturno/Arbóreo Noturno/Terrestre Noturno/Terrestre
20
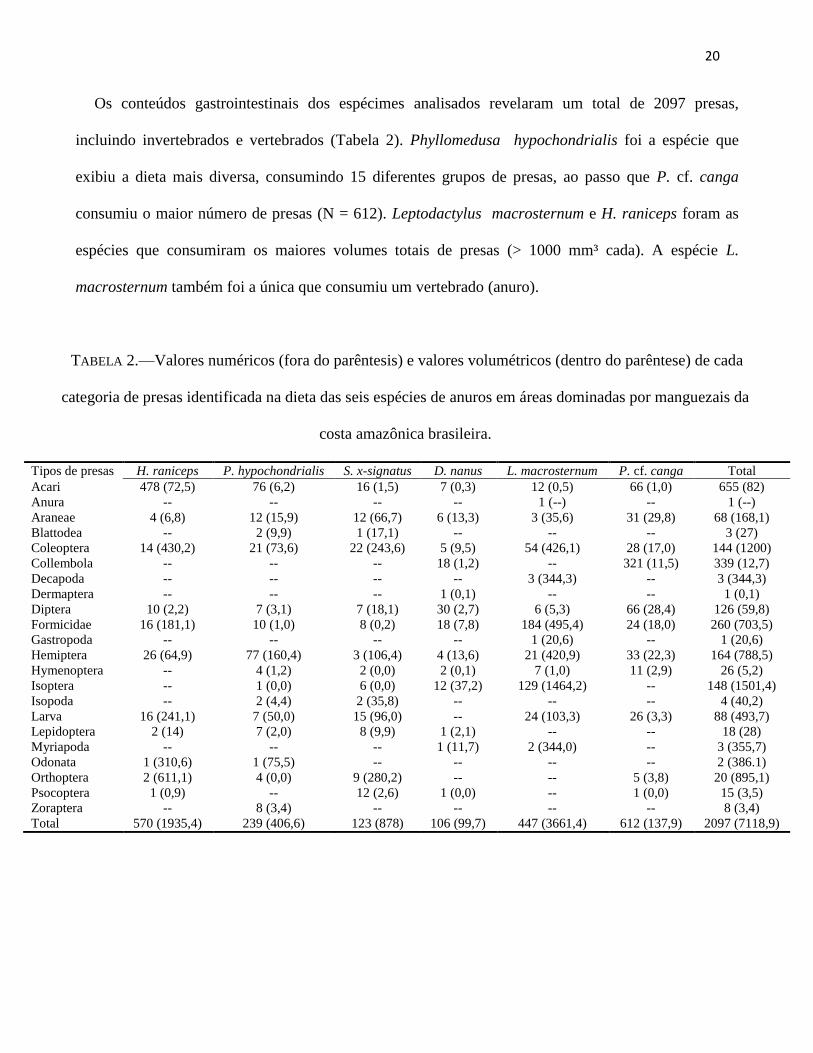
Os conteúdos gastrointestinais dos espécimes analisados revelaram um total de 2097 presas,
incluindo invertebrados e vertebrados (Tabela 2). Phyllomedusa hypochondrialis foi a espécie que
exibiu a dieta mais diversa, consumindo 15 diferentes grupos de presas, ao passo que P. cf. canga
consumiu o maior número de presas (N = 612). Leptodactylus macrosternum e H. raniceps foram as
espécies que consumiram os maiores volumes totais de presas (> 1000 mm³ cada). A espécie L.
macrosternum também foi a única que consumiu um vertebrado (anuro).
TABELA 2.—Valores numéricos (fora do parêntesis) e valores volumétricos (dentro do parêntese) de cada
categoria de presas identificada na dieta das seis espécies de anuros em áreas dominadas por manguezais da
costa amazônica brasileira.
Tipos de presas H. raniceps P. hypochondrialis S. x-signatus D. nanus L. macrosternum P. cf. canga Total
Acari 478 (72,5) 76 (6,2) 16 (1,5) 7 (0,3) 12 (0,5) 66 (1,0) 655 (82)
Anura -- -- -- -- 1 (--) -- 1 (--)
Araneae 4 (6,8) 12 (15,9) 12 (66,7) 6 (13,3) 3 (35,6) 31 (29,8) 68 (168,1)
Blattodea -- 2 (9,9) 1 (17,1) -- -- -- 3 (27)
Coleoptera 14 (430,2) 21 (73,6) 22 (243,6) 5 (9,5) 54 (426,1) 28 (17,0) 144 (1200)
Collembola -- -- -- 18 (1,2) -- 321 (11,5) 339 (12,7)
Decapoda -- -- -- -- 3 (344,3) -- 3 (344,3)
Dermaptera -- -- -- 1 (0,1) -- -- 1 (0,1)
Diptera 10 (2,2) 7 (3,1) 7 (18,1) 30 (2,7) 6 (5,3) 66 (28,4) 126 (59,8)
Formicidae 16 (181,1) 10 (1,0) 8 (0,2) 18 (7,8) 184 (495,4) 24 (18,0) 260 (703,5)
Gastropoda -- -- -- -- 1 (20,6) -- 1 (20,6)
Hemiptera 26 (64,9) 77 (160,4) 3 (106,4) 4 (13,6) 21 (420,9) 33 (22,3) 164 (788,5)
Hymenoptera -- 4 (1,2) 2 (0,0) 2 (0,1) 7 (1,0) 11 (2,9) 26 (5,2)
Isoptera -- 1 (0,0) 6 (0,0) 12 (37,2) 129 (1464,2) -- 148 (1501,4)
Isopoda -- 2 (4,4) 2 (35,8) -- -- -- 4 (40,2)
Larva 16 (241,1) 7 (50,0) 15 (96,0) -- 24 (103,3) 26 (3,3) 88 (493,7)
Lepidoptera 2 (14) 7 (2,0) 8 (9,9) 1 (2,1) -- -- 18 (28)
Myriapoda -- -- -- 1 (11,7) 2 (344,0) -- 3 (355,7)
Odonata 1 (310,6) 1 (75,5) -- -- -- -- 2 (386.1)
Orthoptera 2 (611,1) 4 (0,0) 9 (280,2) -- -- 5 (3,8) 20 (895,1)
Psocoptera 1 (0,9) -- 12 (2,6) 1 (0,0) -- 1 (0,0) 15 (3,5)
Zoraptera -- 8 (3,4) -- -- -- -- 8 (3,4)
Total 570 (1935,4) 239 (406,6) 123 (878) 106 (99,7) 447 (3661,4) 612 (137,9) 2097 (7118,9)

21
Os maiores valores do Índice de Importância Relativa (IR) das presas consumidas diferem entre
todas as espécies de anuros estudadas (Tabela 3). A mesma tabela mostra que a ordem Acari
apresentou o maior valor de IR dentre todas as presas, na dieta de H. raniceps (IR=37,4), enquanto que
as ordens Acari, Coleoptera, Hemiptera e a família Formicidae foram os tipos de presa que
apresentaram as maiores médias desse índice (IR=13,5; 12,6; 12,5 e 10,4, respectivamente).
TABELA 3.—Valores do Índice de Importância Relativa (IR) de cada presa consumida pelas seis espécies
de anuros em áreas dominadas por manguezais da costa amazônica brasileira.
Tipos de
presa H. raniceps P. hypochondrialis S. x-gnatus D. nanus L. macrosternum P. cf. canga Média (%)
Acari 37,4 16,6 8,4 5,2 3,8 9,3 13,5
Anuro - - - - 0,7 - 0,1
Araneae 2,1 6,1 9,4 10,0 2,3 11,0 6,8
Blattodea - 1,7 1,4 - - - 0,5
Coleoptera 13,4 12,4 19,7 6,2 15,5 8,6 12,6
Collembola - - - 8,3 - 25,5 5,6
Decapoda - - - - 4,5 - 0,8
Dermaptera - - - 1,1 - - 0,2
Diptera 4,1 2,7 4,4 21,2 1,7 16,2 8,4
Formicidae 7,5 4,4 4,9 11,2 27,0 7,6 10,4
Gastropoda - - - - 0,8 - 0,1
Hemiptera 8,3 32,2 6,2 8,7 8,9 10,5 12,5
Hymenoptera - 1,9 1,4 1,4 1,1 2,9 1,5
Isoptera - 0,4 3,0 19,1 24,1 - 7,8
Isopoda - 1,2 2,8 - - - 0,7
Larva 8,1 6,8 11,3 - 5,1 6,1 6,2
Lepidoptera 1,2 2,9 4,8 1,7 - - 1,8
Myriapoda - - - - 4,5 - 0,8
Odonata 5,8 6,6 - - - - 2,1
Orthoptera 11,5 1,4 16,7 5,0 - 2,1 6,1
Psocoptera 0,5 - 5,6 1,0 - 0,2 1,2
Zoraptera - 3,1 - - - - 0,5
Total (%) 100 100 100 100 100 100 -
22
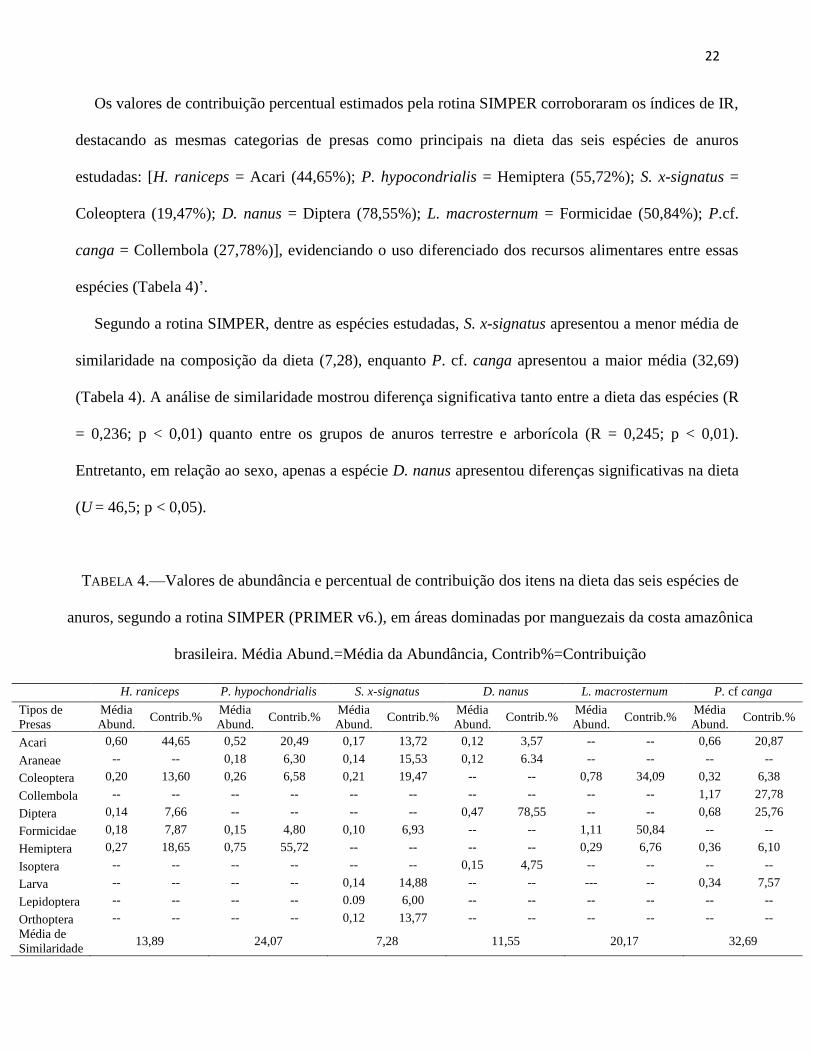
Os valores de contribuição percentual estimados pela rotina SIMPER corroboraram os índices de IR,
destacando as mesmas categorias de presas como principais na dieta das seis espécies de anuros
estudadas: [H. raniceps = Acari (44,65%); P. hypocondrialis = Hemiptera (55,72%); S. x-signatus =
Coleoptera (19,47%); D. nanus = Diptera (78,55%); L. macrosternum = Formicidae (50,84%); P.cf.
canga = Collembola (27,78%)], evidenciando o uso diferenciado dos recursos alimentares entre essas
espécies (Tabela 4)’.
Segundo a rotina SIMPER, dentre as espécies estudadas, S. x-signatus apresentou a menor média de
similaridade na composição da dieta (7,28), enquanto P. cf. canga apresentou a maior média (32,69)
(Tabela 4). A análise de similaridade mostrou diferença significativa tanto entre a dieta das espécies (R
= 0,236; p < 0,01) quanto entre os grupos de anuros terrestre e arborícola (R = 0,245; p < 0,01).
Entretanto, em relação ao sexo, apenas a espécie D. nanus apresentou diferenças significativas na dieta
(U = 46,5; p < 0,05).
TABELA 4.—Valores de abundância e percentual de contribuição dos itens na dieta das seis espécies de
anuros, segundo a rotina SIMPER (PRIMER v6.), em áreas dominadas por manguezais da costa amazônica
brasileira. Média Abund.=Média da Abundância, Contrib%=Contribuição
H. raniceps P. hypochondrialis S. x-signatus D. nanus L. macrosternum P. cf canga
Tipos de
Presas
Média
Abund. Contrib.%
Média
Abund. Contrib.%
Média
Abund. Contrib.%
Média
Abund. Contrib.%
Média
Abund. Contrib.%
Média
Abund. Contrib.%
Acari 0,60 44,65 0,52 20,49 0,17 13,72 0,12 3,57 -- -- 0,66 20,87
Araneae -- -- 0,18 6,30 0,14 15,53 0,12 6.34 -- -- -- --
Coleoptera 0,20 13,60 0,26 6,58 0,21 19,47 -- -- 0,78 34,09 0,32 6,38
Collembola -- -- -- -- -- -- -- -- -- -- 1,17 27,78
Diptera 0,14 7,66 -- -- -- -- 0,47 78,55 -- -- 0,68 25,76
Formicidae 0,18 7,87 0,15 4,80 0,10 6,93 -- -- 1,11 50,84 -- --
Hemiptera 0,27 18,65 0,75 55,72 -- -- -- -- 0,29 6,76 0,36 6,10
Isoptera -- -- -- -- -- -- 0,15 4,75 -- -- -- --
Larva -- -- -- -- 0,14 14,88 -- -- --- -- 0,34 7,57
Lepidoptera -- -- -- -- 0.09 6,00 -- -- -- -- -- --
Orthoptera -- -- -- -- 0,12 13,77 -- -- -- -- -- --
Média de
Similaridade 13,89 24,07 7,28 11,55 20,17 32,69

23
Sobreposição e Índices de Diversidade
A média observada de sobreposição de nicho trófico, com dados nos valores numéricos, entre as
espécies foi baixa (Ojk = 0,36), mas foi maior que a média esperada ao acaso, sendo a diferença entre
elas significativa (Ojk = 0,19; p < 0,01). O maior valor de sobreposição (Ojk = 0,72) foi entre H.
raniceps e P. hipocondrialis, ao passo que o menor (Ojk = 0,09) foi entre L. macrosternum e H.
raniceps (Tabela 5). Já com relação ao volume, a média observada (Ojk = 0,45) também foi maior que
a esperada (Ojk = 0,22), entretanto, nenhuma diferença significativa foi observada entre elas. Ao passo
que, o maior valor de sobreposição volumétrica foi entre as espécies H. raniceps e S. x-signatus (Ojk =
0,91), e o menor entre S. x-signatus e L. macrosternum (Ojk = 0,25).
TABELA 5.—Valores de sobreposição de nicho trófico, calculado pelo índice de Pianka, com a ajuda do
programa EcoSim 7.0, das seis espécies de anuros em áreas dominadas por manguezais da costa amazônica
brasileira.
H. raniceps P. hypochondrialis S. x-signatus D. nanus L. macrosternum P. cf. canga
H. raniceps --
P. hypochondrialis 0,723 (0,646) --
S. x-sgnatus 0,451 (0,911) 0,539 (0,663) --
D. nanus 0,210 (0,261) 0,356 (0,272) 0,621 (0,339) --
L. macrosternum 0,093 (0,270) 0,224 (0,340) 0,447 (0,255) 0,621 (0,865) --
P. cf. canga 0,210 (0,343) 0,247 (0,506) 0,242 (0,446) 0,366 (0,433) 0,108 (0,304) --

24
A ordenação da Análise de Correspondência confirma alguns dos resultados de sobreposição de
nicho trófico (Fig. 1). A dimensão 1 claramente separa L. macrosternum, que é mais relacionado com
os grupos Isoptera e Formicidae do que as outras espécies de anuros. Por outro lado, a dimensão 2
separa as espécies de menor tamanho como P. cf. canga e D. nanus dos outros anuros, relacionado-os
com os grupos de presas que apresentam pequenos volumes, como aquelas pertencentes às ordens
Collembola e Diptera.
FIG. 1.—Grupos das espécies de predador e itens de presas em duas dimensões. Hyp =
Hypsiboas raniceps, Phy = Phyllomedusa hipochondrialis, Sci = Scinax x-signatus,
Den = Dendropsophus nanus, Lep = Leptodactylus macrosternum e Pse =
Pseudopaludicola cf. canga

25
A Tabela 6 apresenta os índices de amplitude de nicho e de diversidade de Shannon para as seis
espécies de anuros em manguezal. Scinax x-signatus foi a espécie que apresentou o maior valor de
amplitude de nicho numérico 9,66 (padronizado = 0,41) e de diversidade de Shannon 1,04
(padronizado = 0,49). Levando em consideração dados volumétricos, Pseudopaludicula sp. apresentou
o maior valor de amplitude 7,64 (BA = 0,31), enquanto todas as outras espécies apresentaram valores
similares.
TABELA 6.—Valores dos índices de diversidade de presas e amplitude de nicho da dieta
dos anuros em áreas dominadas por manguezais da costa amazônica brasileira. Para a
coluna de amplitude de nicho os valores fora dos parêntesis representam os dados
numéricos e dentro dos parêntesis os dados volumétricos.
Morfologia e Dieta
Nós encontramos diferenças significativas no CRC das seis espécies de anuros investigadas
(ANOVA, F(1,34) = 305,7; p < 0,001). O teste post hoc de Tukey mostrou que o único par de espécies
que não apresentou diferença significativa foi L. macrosternum e H. raniceps. Todas as espécies
apresentaram correlação positiva e significativa entre os seus valores de CRC e LB. Contudo, quando
comparamos as médias dos volumes das presas com o valor de CRC, apenas o táxon Pseudopaludicula
cf. canga mostrou correlação significativa (r = 0,32; n = 42; p < 0,05), enquanto o volume máximo das
Diversidade de presas
(Índice de Shannon - Hstd)
Amplitude do nicho
(Índice de Levins - BA)
Hypsiboas raniceps 0,11 0,01 (0,16) Phyllomedusa hypochondrialis 0,34 0,16 (0,15) Scinax x-signatus 0,49 0,41 (0,17) Dendropsophus nanus 0,44 0,24 (0,18) Leptodactylus macrosternum 0,26 0,12 (0,18) Pseudopaludicola cf. canga 0,25 0,10 (0,31)

26
presas não mostrou relação significativa com nenhuma das espécies. Assim como Da mesma forma,
nenhuma relação significativa foi encontrada entre LB e os volumes das presas.
DISCUSSÃO
Partição de Recursos
Nossos resultados mostram que a sobreposição de nicho trófico entre as espécies de anuros
estudadas em área de manguezal foi maior do que o esperado por acaso. Entretanto, em comunidades
reais, seria esperada uma menor sobreposição, do que se espera no caso nulo (Schoener, 1974). Para
esse resultado são conhecidas duas interpretações: i) que a alta sobreposição de nicho é resultado de
uma disponibilidade de recursos no hábitat suficiente pra satisfazer as espécies (Jiménez e Bolaños,
2012) ou ii) que os altos valores de sobreposição estão atuando como uma força seletiva, porém sem
expressar divergência na dieta das espécies (Connel, 1980; Piatti e Souza, 2011).
Encontramos também grandes diferenças na composição e quantidade das presas, entre a dieta dos
anuros estudados. Isto certamente reflete a ampla variedade de presas registradas, cuja diferença é
expressa de várias formas, tais como: tamanho (ex. de microhimenópteros a libélulas), grau de
esclerotização do exoesqueleto (ex. colêmbolas a crustáceos), e até com relação ao estágio de
desenvolvimento (ex. de larvas a adultos). Além disso, cada uma das seis espécies de anuros apresentou
um grupo de presa típico e com maior representatividade em sua dieta, o que pode funcionar como uma
estratégia de coexistência, que reflete o grau de diferenciação no uso do nicho entre essas espécies
(Begon et al., 2007).
De acordo com Zimmerman e Simberloff (1996), a sobreposição da dieta é maior entre espécies
taxonomicamente semelhantes, devido estas compartilharem traços comportamentais, fisiológicos e
morfológicos. Nossos resultados corroboram essa teoria mostrando que os maiores valores de
sobreposição foram registrados entre espécies da mesma família (Hylidae) e menores valores entre

27
espécies de famílias diferentes (Tabela 5). Contudo, L. macrosternum e D. nanus apresentaram um
valor de sobreposição elevado em relação a média, mesmo sendo de famílias distintas e possuindo
tamanhos diferentes. Isto pode estar associado ao fato de essas duas espécies terem apresentado
elevado consumo de indivíduos da ordem Isoptera e da família Formicidae. As formigas, por exemplo,
figuram entre os insetos mais importantes para o manguezal, tanto pela sua grande abundância quanto
por suas múltiplas interações com o sistema (Cannicci et al., 2008). É importante ressaltar que amplas
sobreposições de nichos nem sempre indicam alta competição, já que a abundância do recurso no
ambiente resulta em partição do mesmo sem prejuízos ou perdas para as espécies consumidoras
(Jiménez e Bolaños, 2012).
O alto valor de sobreposição de nicho também foi registrado entre H. raniceps e P. hypochondrialis.
Neste caso, a sobreposição pode ser resultado não apenas da proximidade taxonômica, mas também da
distribuição vertical dos seus indivíduos (Menin et al., 2005), cuja variação é muito pequena nos
estratos verticais que ocupam, como foi relatado para essas espécies em um trabalho feito nessa mesma
área (Silva, 2010). Outro resultado que pode estar demonstrando essa relação com a distribuição
vertical é a sobreposição entre as espécies H. raniceps e D. nanus, que apresentaram o menor valor de
sobreposição entre os hilídeos, e possuem a maior variação no uso dos estratos verticais (Silva, 2010).
No caso das duas espécies terrestres, L. macrosternum e P. cf. canga, essas apresentaram um baixo
valor de sobreposição, que pode ser uma mera consequência da grande diferença no tamanho do corpo
entre essas espécies. Contudo, em assembleias de anuros, os padrões de uso dos recursos alimentares
estão relacionados, principalmente, às habilidades comuns de utilização de recursos, que são herdadas
de suas respectivas linhagens e que surgiram concomitantemente com a divergência das famílias (Toft,
1980), podendo ter coevoluído com outras características, como a defesa contra predadores (Caldwell,
1996).

28
Dieta e Forrageio
Hypsiboas raniceps apresentou grande número de presas por indivíduo, além de uma baixa
diversidade de presas e de amplitude de nicho, indicando uma dieta especializada, onde há o consumo
de uma grande quantidade de itens taxonomicamente limitados. Observamos que nas áreas estudadas,
H. raniceps alimentou-se principalmente de ácaros, sendo este resultado diferente daqueles já
reportados por outros autores, os quais indicam maiores valores de insetos na dieta dessa espécie
(Peltzer e Lajmanovich 2001; Vaz-Silva, 2004). Simon e Toft (1991) também não encontraram
nenhuma especialização das espécies da família Hylidae em direção ao grupo Acari. Adicionalmente,
alguns trabalhos têm demonstrado que especialistas tendem a usar a estratégia de forrageio ativo para
capturar suas presas, de preferência sedentárias, irregularmente distribuídas e de corpo duro, como
formigas, besouros e ácaros (Toft, 1980; Huey e Pianka, 1981).
Já a espécie P. hypochondrialis consumiu principalmente Hemiptera, seguido de Acari e Coleoptera.
Esta espécie de anuro mostrou uma dieta generalista, apresentando números intermediários de captura
de presa e de amplitude de nicho (Toft, 1980, 1981). Esses resultados se mostraram semelhantes aos
descritos por Silva-Filho (2011), em áreas de terra firme no nordeste do Pará, onde Phyllomedusa
hypochondrialis se alimentou principalmente de Hemiptera. Esse resultado pode estar relacionado com
a influência do hábitat sobre a disponibilidade de presas. Esse tipo de dieta está relacionado com
forrageio tipo senta-e-espera, onde suas presas são caracterizadas por artrópodes mais ativos e de corpo
mole, como ortopteras, hemipteras e aranhas (Perry e Pianka, 1997). Segundo Toft (1981), existem
mais especialistas, de qualquer tipo, do que espécies generalistas.
Scinax x-signatus alimentou-se principalmente de coleoptera, aranhas, larvas e ortopteros, que
juntos contribuíram com cerca de 60% da dieta. Nesse sentido, S. x-signatus apresentou os maiores
valores de diversidade de presas e amplitude de nicho, e os menores valores para a média de presas por
individuo, indicando assim uma dieta não especialista. Adicionalmente, S. x-signatus apresentou o

29
menor valor de similaridade com base na rotina SIMPER, o que indica que esta espécie utiliza de forma
mais igualitária seu espectro de presas. Nenhuma referência a dieta dessa espécie foi encontrada na
literatura.
A dieta de D. nanus em nosso trabalho foi composta principalmente de Diptera, seguido de Araneae
e Isoptera. A espécie teve um número baixo de presas por individuo, porém um alto valor de
diversidade de presas, mostrando uma dieta não especialista em formiga (Toft, 1980; 1981). Nosso
resultado de dieta foi similar ao reportado por Menin et al. (2005), onde Diptera foi o grupo de presa
mais abundante na dieta da espécie.
Leptodactylus macrosternum teve uma dieta bastante diversificada, entretanto, com altos valores de
contribuição para apenas dois grupos de presas (formigas e besouros). Por outro lado, obteve valores de
diversidade e amplitude de nicho relativamente baixos. Também apresentou altas taxas de capturas de
presas por individuo, o que ligado ao fato da desproporcionalidade de algumas presas, nos faz pensar
que esta espécie tem um comportamento alimentar especialista, que consome outros tipos de presas por
oportunismo (Sabagh e Carvelho-e-Silva, 2008). Esses altos valores de Formicidae e Coleoptera já
foram reportados para a espécie em outro trabalho (Pereira, 2012), entretanto, o autor trata a espécie
como generalista.
A família Leptodactylidae sempre foi reconhecida por apresentar forrageador do tipo generalista,
com estratégia senta-e-espera (Duré e Kehr, 2004). Entretanto, exceções já foram registradas, como a
espécie Eleutherodactylus vocator, que utiliza formigas em proporções similares à especialistas em
formiga (Toft, 1980).
A rã Pseudopaludicola cf. canga utilizou principalmente itens menores de presas dos grupos
Collembola, Diptera e Acari. A taxa de presas por individuo nesta espécie foi relativamente alta, com
baixos valores de diversidade e amplitude de nicho, indicando uma dieta aparentemente especialista. A
ordem Collembola é amplamente distribuída, estando entre os artrópodes terrestres mais comuns e

30
abundantes, apresentando um comportamento agregado e irregular, com altas densidades por m²
(Hopkin, 1997; Culik et al., 2006), sendo portanto muitas vezes relacionados a dietas de especialistas e
forrageio tipo ativo (Perry e Pianka, 1997; Sabagh e Carvalho-e-Silva, 2008).
Muitos trabalhos têm demonstrado correlações significativas entre o tamanho do predador (ou
largura da boca) e o volume da presa (Quiroga et al., 2009; Piatti e Souza, 2011; Batista et al., 2011).
No entanto, em nosso estudo apenas a espécie P. cf canga mostrou essa relação entre CRC e o volume
máximo das presas, além de ser a espécie com maior amplitude volumétrica. Este resultado pode estar
relacionado com grande o número de presas inteiras em vários espécimes de P. cf. canga, o que
aumentou e melhorou a amostragem.
Os padrões dentro das famílias de anuros apresentam provas de que as especializações em formigas
derivam de um modo de forrageio não especializado ou generalista (Toft, 1981), como por exemplo, os
dendrobatídeos, que surgiram na América do Sul, e podem ter divergido de leptodactilídeos (Linch,
1971; Savage, 1973; Toft, 1981). Por exemplo, espécies do gênero Colostethus, que são considerados
primitivos, estão entre os dendrobatídeos em que a especialização em formiga é menos evidente (Darst
et al., 2005). No entanto, alguns modelos preveem que o modo de forrageio, pode mudar em resposta
da disponibilidade de presas (Dullman e Trueb, 1994).
Contudo, os estudos de modo de forrageio têm sido restrito a anuros diurnos de serapilheira (Toft,
1981), existindo ainda uma insuficiência de dados sobre o comportamento de forrageio de anuros
noturnos, tanto arborícola, terrestres ou aquáticos. Entretanto, estudos de anuros de hábito noturno
indicam que eles apresentam padrões semelhantes de especialização de dieta (Duellman, 1978). As
evidências apresentadas em nosso estudo sugerem que semelhanças taxonômicas aumentam a
sobreposição da dieta, mas não necessariamente a competição interespecífica. Contudo, nem todas as
evidências aqui apresentadas são consistentes com esta hipótese. Outros testes que envolvem a coleta
de informações mais detalhadas sobre a eletividade da dieta podem melhorar o entendimento de como

31
os anuros usam os recursos disponíveis em seu ambiente, além de melhor determinar as tendências que
os anuros noturnos apresentam quanto ao modo de forrageio.
LITERATURA CITADA
Araújo, M.S., D.I. Bolnick, L.A. Martinelli, A.A. Giaretta e S.F. Dos Reis. 2009. Individual-level
diet variation in four species of Brazilian frogs. Journal of Animal Ecology 78:(4), 848-856.
Ayres, M., A.J. M, D.L. Ayres e A.S. Santos. 2007. BioEstat: Aplicações Estatísticas nas Áreas
das Ciências Bio-Medicas. Versão 5.0. Instituto de Desenvolvimento Sustentável Mamirauá,
IDSM-MCT-CNPq, Brasil.
Batista, R.C., C.B. De-Carvalho, E.B. Freitas, S.C. Franco, C.C. Batista, W.A. Coelho e R.G.
Faria. 2011. Diet of Rhinella schneideri (Werner, 1894) (Anura: Bufonidae) in the Cerrado,
Central Brazil. Herpetology Notes 4: 17-21.
Begon, M., C.R. Townsend e J.L. Harper. 2007. Ecologia: de indivíduos a ecossistemas, Artmed.
Biavati, G.M., H.C. Wiederhecker e G.R. Colli. 2004. Diet of Epipedobates flavopictus (Anura:
Dendrobatidae) in a Neotropical Savanna. Journal of Herpetology 38: 510-518.
Bolnick, D.I., R. Svanbäck, J.A. Fordyce, L.H. Yang, J.M. Davis, C.D. Hulsey e M. L. Forister.
2003. The Ecology of Individuals: Incidence and Implications of Individual Specialization.
The American Naturalist 161:(1) 1-28.
Caldwell, J.P. 1996. The evolution of myrmecophagy and its correlates in poison frogs (Family
Dendrobatidae). Journal of Zoology 240: 75-101.
Cannicci, S., D. Burrows, S. Fratini, T.J.I. Smith, J. Offenberg e F. Dahdouh-Guebas. 2008.
Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review.
Aquatic Botany 89: 186-200.

32
Clarke, K.R. e R.N. Gorley. 2006. PRIMER: User Manual/ Tutorial. Versão 6.0. PRIMER-E
Plymouth, United Kingdom.
Connell, J.H. 1975. Some mechanisms producing structure in natural communities. Pp. 460–491
in Cody, M.L. e J.M. Diamond (Eds.), Ecology and evolution of communities. Harvard
University Press,
Connell, J.H. 1980. Diversity and the coevolution of competitors, or the ghost of competition
past. Oikos 131-138.
Culik, M.P., D.d.S. Martins e J.A. Ventura. 2006. Collembola (Arthropoda: Hexapoda)
communities in the soil of papaya orchards managed with conventional and integrated
production in Espírito Santo, Brazil. Biota Neotropica 6: 2-10.
Darst, C.R., P.A. Menéndez-Guerrero, L.A. Coloma e D.C. Cannatella. 2005. Evolution of
dietary specialization and chemical defense in poison frogs (Dendrobatidae): A comparative
analysis. The American Naturalist 165: 56-69.
Dev Roy, M.K. e C. Sivaperuman. 2012. Fauna of Ecosystems of India-Mangroves. Zoological
Society of India 1-11.
Drewes, R.C., and B. Roth. 1981. Snail-eating frogs from the Ethiopian highlands: a new anuran
specialization. Zoological Journal Linnean Society 73: 267-287.
DHN. 2012. Diretoria de Hidrografia e Navegação. Disponivel em
http://www.mar.mil.br/dhn/chm/tabuas/index.htm. Diretoria de Hidrografia e Navegação,
Brasil. Arquivado por WebCite em http://www.webcitation.org/6LUbSS5a0 em 23 de
Novembro 2013.
Duellman, W.E. 1978. The biology of an equatorial herpetofauna in Amazonian Ecuador.
University of Kansas Museum of Natural History Miscellaneous Publications, Kansas.

33
Duellman, W.E. e L. Trueb. 1994. Biology of Amphibians. The Johns Hopkins University Press,
USA.
Duré, M.I. 1999. Phyllomedusa hypocondrialis. Diet. Herpetological Review 30: 92.
Duré, M.I. e A.I. Kehr. 2004. Influenci of microhabitat on the trophic ecology of two
Leptodactylids from northeastern Argentina. Herpetologica 60: 295-303.
Fernandes, M.E.B. 2003. Os Manguezais da Costa Norte Brasileira. Fundação Rio Bacanga, São
Luiz.
Glorioso, B.M., J.H. Waddle, M.E. Crockett, K.G. Rice e H.F. Percival. 2012. Diet of the
invasive Cuban Treefrog (Osteopilus septentrionalis) in pine rockland and mangrove habitats
in South Florida. Caribbean Journal of Science 46:(2-3) 346-355.
Gotelli, N.J. e G. Entsminger. 2001. EcoSim: Null models software for ecology. Versão 7.0.
Adiquirido por Intelligence Inc. and KeseyBear, USA.
Hopkin, S.P. 1997. Biology of the Springtails:(Insecta: Collembola):(Insecta: Collembola).
Oxford University Press,
Huey, R.B. e E.R. Pianka. 1981. Ecological Consequences of Foraging Mode. Ecology 62: 991-
999.
Hulbert, S.H. 1978. The measurement of niche overlap and some relatives. Ecology 59: 67-77.
Jena, S.C., S.K. Palita, M.K. Mahapatra. 2013. Anurans of Bhitarkanika mangroves, Odisha, east
coast of India. Check List 9:(2), 400-404.
Jiménez, R. e F. Bolaños. 2012. Use of food and spatial resources by two frogs of the genus
Dendropsophus (Anura: Hylidae) from La Selva, Costa Rica. Phyllomedusa: Journal of
Herpetology 11: 51-62.
Kathiresan, K., e B.L. Bingham. 2001. Biology of Mangroves and Mangrove Ecosystems.
Advances in marine biology 40: 81-251.

34
Kathiresan, K. e N. Rajendran. 2004. Mangrove ecosystem of India Ocean region. Indian Journal
of Marine Sciences 34:(1) 104-113.
Krebs, C.J. 1999. Ecological methodology. Benjamin/Cummings Menlo Park, USA.
Levins, R. 1968. Evolution in Changing environments: Some theoretical explorations. Princeton
University Press, USA.
Lynch, J.D. 1971. Evolutionary relationships, osteology, and zoogeography of leptodactyloid
frogs.
Magnusson, W.E., A.P. Lima, W.A. da Silva e M.C. Araújo. 2003. Use of Geometric Forms to
Estimate Volume of Invertebrates in Ecological Studies of Dietary Overlap. Copeia 1: 13-19.
Menin, M., D.C. Rossa-Feres e A.A. Giaretta. 2005. Resource use and coexistence of two
syntopic hylid frogs (Anura, Hylidae). Revista Brasileira de Zoologia 21: 61-72.
Parmelee, J.R. 1999. Trophic ecology of a tropical anuran assemblage. Scientific Papers Natural
History Museum, University of Kansas 11: 1-59.
Peltzer, P.M. e R.C. Lajmanovich. 2001. Hyla raniceps (NCN). Diet. Herp. Rev 32: 247-248.
Pereira, T.C.S.O. 2012. Ecologia trófica em uma taxocenose de anfíbios: estrutura, filogenia e
especialização individual. Dissertação de Mestrado, Universidade Federal do Rio Grande do
Norte, Natal, Brasil.
Perry, G. e E.R. Pianka. 1997. Animal foraging: past, present and future. Tree 12: 360-364.
Pianka, E.R. 1973. The structure of lizard communities. Annual Review of Ecology and
Systematics 4: 53-74.
Pianka, E.R. 1974. Niche overlap and diffuse competition. Proceedings of the National Academy
of Science 71: 2141-2145.
Piatti, L. e F.L. Souza. 2011. Diet and resource partitioning among anurans in irrigated rice fields
in Pantanal, Brazil. Brazilian Journal of Biology 71: 653-661.

35
Quiroga, L.B., E.A. Sanabria e J.C. Acosta. 2009. Size- and Sex-Dependent Variation in Diet of
Rhinella arenarum (Anura: Bufonidae) in a Wetland of San Juan, Argentina Journal of
Herpetology 43: 311-317.
Rao, C.A.N. 2004. Faunal Diversity: Estuarine Ecosystem. Environment Protection Training
Research Institute-Environmental Information System (EPTRI-ENVIS) Newsletter (3) 8-11.
Sabagh, L.T. e A.M.P.T. Carvalho-e-Silva. 2008. Feeding overlap in two sympatric species of
Rhinella (Anura: Bufonidae) of the Atlantic Rain Forest. Revista Brasileira de Zoologia 25:
247-253.
Satheeshkumar, P. 2011. First record of a mangrove frog Fejervarya cancrivora (Amphibia:
Ranidae) in the Pondicherry Mangroves, Bay of Bengal-India. World Journal of Zoology 6:(3)
328-330.
Savage, J.M. 1973. The geographic distribution of frogs: patterns and predictions. Pp. 351–445 in
Via, J.L. (Eds.), Evolutionary biology of the Anurans: contemporary research on major
problems. University of Missouri Press, USA.
Schoener, T.W. 1974. Resource partitioning in ecological communities. Science 185: 27-39.
Schoener, T.W. 1989. Should hingut contents be included in lizard dietary compilations? Journal
of Herpetology 23: 455-458.
Shannon, C.E. e W. Weaver. 1949. The mathematical theory of communication (Urbana, IL.
University of Illinois Press 19: 1.
Silva-Filho, H.F. 2011. Ecologia trófica e reprodutiva de Phyllomedusa hypochondrialis (Daudin,
1802)(Hylidae, Phyllomedusinae) na Amazônia Oriental, Pará, Brasil. Universidade Federal
do Pará, Pará, Brasil.

36
Silva, R.P. 2010. A herpetofauna associada à zona costeira da Amazônia brasileira e o padrão de
distribuição espaço temporal de anuros na península de Ajuruteua, Bragança-Pa. Dissertação
de Mestrado, Universidade Federal do Pará, Brasil.
Simon, M.P. e C.A. Toft. 1991. Diet specialization in small vertebrates: mite-eating in frogs.
Oikos 263-278.
Souza-Filho, P.W.M. e M. El-Robrini. 2000. Geomorphology of the Bragança Coastal zone,
northeastern Pará state. Brazilian Journal of Geology 30: 522-526.
StatSoft. 2001. Statistica: Data Analysis Software System. Versão 6.0. StatSoft, USA.
Toft, C.A. 1980. Feeding ecology of thirteen syntopic species of anurans in a seasonal tropical
environment. Oecologia 45: 131-141.
Toft, C.A. 1981. Feeding ecology of panamanian litter anurans: patterns in diet and foraging
mode. Journal of Herpetology 15: 139-144.
Vaz-Silva, W. 2004. Hyla raniceps (Chaco Treefrog). Diet. Herp. Rev. 35: 159.
Vignoli, L. e L. Luiselli. 2012. Dietary relationships among coexisting anuran amphibians: a
worldwide quantitative review. Oecologia 169: 499-509.
Wilbur, H.M. 1997. Experimental ecology of food webs: complex systems in temporary ponds.
Ecology 78: 2279-2302.
Zimmerman, B.L. e D. Simberloff. 1996. An historical interpretation of habitat use by frogs in a
Central Amazonian Forest. Journal of Biogeography 23: 27-46.





























